/
































































































































































































































Author: Туманов И.Л.
Tags: carnivora хищные млекопитающие зоология биология млекопитающие
ISBN: 5-02-026201-3
Year: 2003




Text
ЛОГИЧЕСКИЕ
ХИЩНЫХ
млекопитающих
Друзьям и близким - спутникам
по охотничьим тропам и дорогам жизни
I. L. TUMANOV
BIOLOGICAL
CHARACTERISTICS
OF CARNIVORES
mammals
OF RUSSIA
Saint-Petersburg
«NAUKA»
2003
И. Л. ТУМАНОВ
БИОЛОГИЧЕСКИЕ
ОСОБЕННОСТИ
ХИЩНЫХ
млекопитающих
РОССИИ
Санкт-Петербург
«НАУКА»
2003
УДК 599.742.4 : 591.1/.4/.5
ББК 28.673; 28.693.36
Т 78
Туманов И. Л. Биологические особенности хищных млекопитающих России. — СПб.: Наука,
2003. — 448 с.
ISBN 5-02-026201-3
На материалах многолетних полевых и экспериментальных исследований в монографии
рассматриваются адаптивные особенности хищных млекопитающих России. Подробно освещаются
вопросы пищевой специализации и сезонной смены кормов, суточной активности и перемещения
животных, сроков спаривания и плодовитости, развития и скорости роста молодняка исследуемых
видов. Особое внимание уделено анализу эколого-физиологических и морфологических адаптаций
систем органов, а также зависимости функционального состояния хищников и их популяций от
уровня заражения гельминтами. На основе изучения специфики энергетического обмена, хими-
ческой, физической, экологической терморегуляции и связанных с ними закономерностей струк-
турных преобразований показаны пути приспособления хищных млекопитающих к разным
условиям существования. Книга представляет интерес для экологов, зоологов, эколого-физиологов,
а также широкого круга специалистов в области рационального природопользования и охотничьего
хозяйства. Библиогр. 737 назв. Табл. 158. Илл. 101, в том числе 32 на вкл.
Tumanov I. L. Biological characteristics of carnivores mammals of Russia. — Saint-Petersburg:
Nauka, 2003. — 448 p.
Adaptive characteristics of predatory mammals of Russia are considered on the basis of long-term
field and experimental research. Food specialization and seasonal change of foods, daily activity and
moving, terms of coupling and fertility, development and growth rate of juveniles of predatory species
are elucidated in detail. Particular emphasis is placed on the analysis of the ecological-physiological
and morphological adaptations of systems of organs and also relationship of functional state of predators
and their populations on helminth infection. The ways of adaptation of predatory mammals to different
environmental conditions are shown on the basis of study of specificity of energy exchange, thermo-
regulations (chemical, physical, and ecological) and structural transformation patterns related to them.
The book is intended for ecologists, zoologists, ecologists-physiologists and also specialists in rational
nature management and game husbandry. References 737 titles. Tables 158. Ill. 101.
Рецензенты:
член-корреспондент РАН, проф. Э. В. ИВАНТЕР,
доктор биологических наук, проф. П. И. ДАНИЛОВ
Без объявления
ISBN 5-02-026201-3
© И. Л. Туманов, 2003
© Издательство «Наука», оформление, 2003
ВВЕДЕНИЕ
Любые изменения внешней среды вызывают в живом организме
соответствующие адаптивные реакции, которые нередко имеют достаточно
сложный характер. Они могут выражаться в изменении форм поведения и различных
функциональных систем, определяя их качественно новую структуру. В результате
изменившихся условий существования или под действием внешних раздражителей
в организме вырабатываются адекватные компенсаторные реакции, обеспе-
чивающие нормальную его жизнедеятельность и общий гомеостаз.
Однако адаптивные реакции не следует рассматривать только как быстрое
реагирование организма на кратковременные воздействия отдельных
факторов. В основном они формировались в процессе эволюции, многократно
закрепляясь в последующих поколениях, способствуя сохранению вида.
Всестороннее эколого-морфологическое и физиологическое изучение
особенностей организма может служить ключом к пониманию общего
направления приспособления видов к изменившимся условиям существования
и объяснить некоторые поведенческие реакции животных с позиции
эволюционной экологии. Полученные материалы способствуют разработке
ряда природоохранных мероприятий и служат необходимой предпосылкой для
научно-обоснованного управления популяциями промысловых видов.
Быстрое развитие биологической науки делает необходимым применение
различных методов исследования, часто заимствованных из смежных областей
знаний и способствующих более полной характеристике видовых, региональных и
популяционных особенностей экологии животных, в том числе и хищных
млекопитающих. На основании последовательного и углубленного изучения
их кормовой специализации, двигательных реакций, ряда эколого-морфо-
логических и физиологических показателей можно выявить адаптивные
механизмы, определяющие динамику численности видов, и разработать
способы контроля и управления популяциями. Эти положения определили
выбор как методических подходов, так и общих направлений данной работы.
Приспособительные реакции хищных млекопитающих, характеризу-
ющихся высокой степенью экологической радиации, носят комплексный
характер и ведут к сокращению энергетических затрат, сохранению гомеостаза
организма и поддержанию высокого репродуктивного потенциала. Стратегия
реализации этих тенденций определяется, с одной стороны, размерами
животных и, следовательно, уровнем метаболизма, а с другой - принадлеж-
ностью к определенному адаптивному типу.
Конкретный характер морфофункциональных адаптаций связан с образом
жизни видов и особенностями поведения животных. Если типичные норники
5
приспосабливаются к условиям существования посредствам адаптивных
структур, обеспечивающих работу мощного двигательного аппарата, а
специализированные полуводные виды - за счет соответствующей перестройки
сердечно-сосудистой, дыхательной и некоторых других систем, то остальные
хищники формируют универсальные приспособления, направленные на
под держание оптимального энергетического баланса в условиях неустойчивой
среды обитания.
Особое место в адаптивной стратегии млекопитающих занимает
реализация функций, связанных с сезонной периодикой факторов среды и
обеспечивающих им успешное переживание наиболее трудных переходных
периодов года. Изучение биологических циклов и закономерностей
приспособления животных к сезонным факторам до настоящего времени
остается актуальным направлением современной экологии (Яблоков, 1966,
1987; Шварц, 1968, 1980; Пианка, 1981; Шилов, 1982, 1985; Бигон и др.,
1989; Macdonald, 1995; Gittleman, 1996).
Следует также отметить, что виды, характеризующиеся низкой индиви-
дуальной резистентностью, повышенной смертностью и коротким жизненным
циклом, компенсируют данные особенности реализацией комплекса специальных
экологических адаптаций. Это - прежде всего потенциальная полиэстричность,
сокращение эмбрионального периода, увеличение плодовитости, высокая скорость
роста молодняка, раннее половое созревание, специфические возрастные
изменения соотношения полов, устойчивость к гельминтам и др. Изучение
приспособительных реакций, с разной степенью интенсивности проявляющихся в
той или иной системе органов, даёт возможность выявить их компенсаторный
характер и общую направленность для обеспечения высокой численности животных
в изменяющихся условиях среды. Определение доверительных границ эколого-
морфологических и физиологических параметров у хищных млекопитающих из
естественных местообитаний позволяет использовать эти данные в качестве
индикаторов нормального состояния организма, что необходимо для успешного
клеточного разведения пушных видов и расселения некоторых из них в природных
резерватах.
Изменяющаяся экологическая обстановка в биоценозах, связанная с
постоянно усиливающимся антропогенным прессом, оказывает негативное
воздействие на биосферу. В данных условиях особое значение имеет выявление
путей адаптаций животных к антропогенным воздействиям, что позволит
усовершенствовать стратегию и тактику природоохранной работы, направленной
на увеличение численности популяций и сохранение исчезающих видов. Крайне
важно также разработать рациональные методы и способы промышленного
разведения в неволе новых видов промысловых млекопитающих, среди которых
многие хищники определяют уровень общего объёма пушных заготовок.
Сравнительный анализ морфофункциональных преобразований различных
органов близких в систематическом отношении видов позволяет проследить
основные направления их адаптивной радиации. В этой связи цель выполненного
исследования заключалась в выяснении видоспецифических эколого-морфо-
6
логических и физиологических аспектов адаптаций хищных млекопитающих
к условиям существования для разработки биологических основ их хозяй-
ственного разведения, охраны и рационального освоения ресурсов. Важной
особенностью данной работы следует признать мониторинговый характер
оригинального материала и его однородность. В течение многих лет наши
данные по большинству видов хищных млекопитающих (прежде всего по
представителям семейств куньих и собачьих) собирались по единой
унифицированной методике, в одно время и на постоянных участках.
Исследования адаптивных биологических особенностей животных позволяют
выявить пути приспособления их организма к изменяющимся условиям среды
и играют важную роль в понимании многих общебиологических законо-
мерностей. В этом плане особый интерес представляют хищники из семейства
куньих, характеризующиеся видовым разнообразием, а некоторые из них - и
многочисленностью. В наземных экосистемах они представляют собой одно
из основных звеньев трофических связей, определяя ее развитие, целостность и
биологическую продуктивность, поэтому в нашей работе этим видам уделено
основное внимание.
В предлагаемой монографии представлены результаты многолетних
исследований, выполненных автором в Западном зональном отделении ВНИИ
охотничьего хозяйства и звероводства им. проф. Б.М. Житкова (ВНИИОЗ), а также
в Арктическом и Антарктическом НИИ (ААНИИ). Она написана главным образом
на оригинальных материалах, собранных во время экспед иций, полевых выездов и
при проведении экспериментальных работ.
При составлении программ исследований, обсуждении рабочих гипотез и
анализе собранных материалов были учтены ценные предложения и замечания
сотрудников ВНИИОЗ им. проф. Б.М. Житкова (г. Киров), его зональных отделений,
атакже коллег и друзей: Э.В. Ивантера, П.И. Данилова, О.С. Русакова, В.В. Рожнова.
Значительную техническую помощь на разных этапах работы оказали мои
постоянные помощники: Н.Л. Коровник, В. А. Смелов, Е.А. Сорина и В.В. Кожечкин.
Интересные сведения и первичные материалы наблюдений были получены от
сотрудников Ленинградского (Санкт-Петербургского) зоопарка В.С.
Андреевской, Л.М. Бородулиной и И.В. Корнеева. В сборе данных по
возрастной изменчивости физиологических параметров у куньих оказали
помощь Д.В. и Ю.Г. Терновские.
Всем лицам, оказавшим посильное содействие в выполнении данных
исследований и подготовке рукописи, автор выражает свою искреннюю
признательность.
МАТЕРИАЛ И МЕТОДИКИ
В работе обобщены материалы, собранные за 35-летний период (1966-2000
гг.) в процессе стационарных и полевых исследований. Экологические наблю-
дения проводили на северо-западе европейской части России, Северном
Таймыре, в Краснодарском крае, Болыпеземельской тундре, а также в горных
районах Иркутской области (Восточные Саяны), Бурятии, Дагестана и
Азербайджана.
Лабораторные и экспериментальные исследования выполнялись в
Ленинградском (Санкт-Петербургском), Архангельском и Красноярском
зональных отделениях ВНИИОЗ им. проф. Б.М. Житкова, в Зоологическом
институте РАН, Ленинградском (Санкт-Петербургском) зоопарке и на
экспериментальной базе Института биологии СО РАН (г. Новосибирск).
Основной материал по питанию и суточной активности хищных млеко-
питающих был собран в северо-западном регионе европейской части России.
Особенности их пищевого рациона изучали путём разбора содержимого
желудочно-кишечного тракта, экскрементов и разовых остатков добычи, каждое
из которых условно принимали за одну пробу. Всего просмотрено около 3.3 тыс.
проб по мелким и более 1 тыс. - по крупным хищникам. При их анализе
устанавливали частоту встречаемости различных компонентов корма и объемное
соотношение видов пищи во всей её массе. Встречаемость рассчитывали как
отношение частоты обнаружения одного вида добычи ко всему количеству
проб (%), а интенсивность потребления отдельных пищевых компонентов - как
процентное соотношение их объёма к общему условному объёму биопроб.
Калорийность основных объектов питания мелких хищников определяли
совместно с В.А. Смеловым путем сжигания в калориметрической бомбе
дозированных навесок (290-660 мг) с образцами, помещенными в крахмальные
капсулы (Туманов, Смелов, 1980).
Сезонную динамику массы тела и суточной потребности в пище изучали
на группах животных, содержащихся в виварии Западного зонального
отделения ВНИИОЗ и Ленинградском (Санкт-Петербургском) зоопарке.
Продолжительность контрольных кормлений при наличии избытка кормов в
условиях эксперимента составляла 3-5 дней. Дополнительные сведения по
питанию исследуемых видов, предпромысловой их численности, .величине
выводков и смертности молодняка за гнездовой период получены при анализе
более 7 тыс. сообщений, поступивших от охотников-корреспондентов
ВНИИОЗ. Продолжительность суточного хода мелких хищников и характер
их охотничьей деятельности устанавливали в природных условиях методом
троплений, а распределение периодов активности и отдыха в течение суток - в
основном путём круглосуточных наблюдений за животными, отловленными в
природе и содержащимися в вольерах. Суточную активность их основных
жертв (мышевидных грызунов) изучали в летнее время при естественной смене
освещения с помощью актографа с подвижным полом в кормовой камере
8
(Калабухова, 1951). При расшифровке актограмм условно за «ночной» период
принималось время с 19 до 5 ч утра (Туманов, 1968,1969). Всего за период
исследований было проведено 187 осенне-зимних троплений куньих, 28
наблюдений за их суточной активностью в условиях неволи и записано 56
актограмм мышевидных грызунов. По следам других хищников с целью
изучения различных форм их охотничьей деятельности пройдено в общей
сложности около 160 км.
Для изучения особенностей питания, размножения и возрастной
изменчивости соотношения полов в биопробах было собрано, вскрыто и
проанализировано более 1800 тушек различных видов хищников, добытых
во время полевых выездов и поступавших от охотников. Дополнительный
материал по демографической структуре популяций получен в 1975-1984 гг.
на пушно-меховом холодильнике (Санкт-Петербург), где было просмотрено
около 24 тыс. шкурок от животных северных регионов России.
При анализе биологических проб молодые особи (до 1 года) отделялись от
взрослых по комплексу морфологических признаков (Тимофеев, Надеев, 1955;
Новиков, 1956). Возраст взрослых хищных млекопитающих определяли гисто-
логическим методом (более 4 тыс. препаратов) путем регистрации числа ростовых
слоёв в цементе корневой части клыка (Клевезаль, Клейненберг, 1967). Упитанность
устанавливали либо визуально - по наличию подкожных и внутренних жировых
отложений, либо по весу жира из паховой области (генитальное депо), индекс
которого (%<>) служит хорошим её показателем.
Систематическое положение видов выясняли на основании морфоло-
гического и краниометрического изучения добытых животных, а также осмотра
их шкурок. При определении вида норок по шкуркам за основной
диагностический признак были приняты их цвет и наличие или отсутствие
белого пятна на губах. Видовую принадлежность норок по тушкам
устанавливали на основании краниологических различий. При этом главными
критериями служили форма слуховых капсул и количество корней на первых
по счёту верхних предкоренных зубах: двух у американской и одного у
европейской норок (Петров, 1958; Данилов, Туманов, 1976а). В настоящей
работе русские названия видов даны согласно номенклатуре, предложенной
Г.А. Новиковым (1971), а латинские - по В.Г. Гептнеру и др. (1967).
В процессе изучения размножения куньих у самок мелких и средних по
размеру видов, содержащихся в вольерах, исследовали вагинальные мазки
(1.6 тыс. проб), отбор которых проводили во все месяцы года. Размножение
хищников из природных популяций изучали как обычными макрометодами
(измерение и взвешивание репродуктивных органов, констатация
беременности, количества и величины эмбрионов и т.д.), так и путём
изготовления и анализа гистологических препаратов (Заварзин, Румянцев,
1946; Кацнельсон, Рихтер, 1959). У самцов брали семенники с придатками,
у самок - все элементы гениталий. Материал фиксировали в 10%-ном
нейтральном формалине или жидкости Буэна. Последующая обработка
состояла из обезвоживания в спиртах возрастающих концентраций и
9
хлороформе, пропитки и заливки препаратов в парафин. Толщина срезов -
7-10 мкм, окраска гематоксилин-эозином. Срезы семенников и придатков
делали совмещенными. У самцов измеряли диаметры семенных канальцев,
у самок - фолликул и желтых тел. Всего было изготовлено и просмотрено
более 2 тыс. гистологических препаратов (Данилов, Туманов, 1972, 1975а,
б; Данилов, Русаков, Туманов, 1979).
Плодовитость самок определяли по наличию желтых тел в яичниках,
количеству эмбрионов в рогах матки или детенышей в помётах. У самок
куниц и соболей из репродуктивных органов вымывали бластоцисты по
методике, разработанной нами (Туманов, 1988а, б). Примитивный способ
определения плодовитости самок, когда «отпрепарированную матку
смачивают водой, расправляют на предметном стекле и просматривают на
просвет при электрическом свете», рекомендуемый Д.В. и Ю.Г. Терновскими
(1994), не выдерживает никакой критики. Он совершенно неприемлем для
работы с соболями, куницами и более крупными хищниками, имеющими
толстостенные малопрозрачные рога матки.
При выяснении вопроса о соотношении полов в выводках самок разных
видов и возрастов кроме наблюдений за животными, содержащимися в
вольерах, использовали ведомственные данные, накопленные в Ленин-
градском (Санкт-Петербургском) зоопарке и зверосовхозе «Заря» (Ленин-
градская обл.). В последнем в 1976-1980 гг. был просмотрен и проанали-
зирован материал по рождению молодняка у самок американской норки
стандартного окраса (1382 выводка, 8730 щенков) и соболя (154 выводка,
435 детенышей). Закономерности роста и развития молодняка изучали на
животных, родившихся в виварии Западного зонального отделения
ВНИИОЗ или в указанном выше зоопарке. В процессе роста и развития
детенышей их периодически осматривали, взвешивали и измеряли.
Органометрические исследования проводили на свежих тушках зверей.
Все индексы рассчитывали по отношению к массе тела животных в шкурке.
Общее количество использованного для этих целей материала представлено в
соответствующих таблицах.
Особенности строения кровеносной системы хищных млекопитающих
выясняли с применением метода инъекции сосудов рентгеноконтрастными
массами с последующей тотальной рентгенографией и препарированием. Наливку
венозных сосудов осуществляли свинцовыми белилами через общую под-
вздошную вену, а артериальных - через дугу аорты свинцовым суриком,
разведенным на глицерине и воде. При этом тонкую канюлю выводили через
левый желудочек в аорту, где и фиксировали наложенной лигатурой (Туманов,
1974; Э. Ивантер, Т. Ивантер, И. Туманов, 1985). Всего было инъецировано
контрастными массами 75 животных, рентгенографию которых проводили
аппаратом РУМ-4 на базе Зоологического института РАН. Кроме того, на
свежих тушках было отпрепарировано 170 различных отделов основных
кровеносных магистралей, отражающих характер ветвления сосудистого русла
в связи с особенностями образа жизни исследуемых хищников.
10
При анализе микростроения дуги аорты и надпочечной железы было
изготовлено и просмотрено более 1.2 тыс. гистологических препаратов.
Морфологические исследования желудочно-кишечного тракта осуществляли
совместно с В. А. Смеловым на 210 тушках животных в возрасте старше года.
Границу между толстым и тонким отделами кишечника устанавливали по
линии окончания ворсинчатой зоны, которая отсутствует в толстом отделе, и
по размещению подвздошно-ободочной артерии (Михеичев, 1955, 1960;
Халилов, 1955; Drescher, 1976).
Основные физиологические исследования проводили на особях, отлов-
ленных в природе и находившихся в условиях вольерного содержания.
Динамику сердечного ритма и термометрических показателей в постнатальном
онтогенезе мелких хищников определяли на экспериментальной базе Института
биологии СО РАН (г. Новосибирск, 1975-1978 гг.).
Ректальную температуру измеряли медицинским электротермометром
«Темп-60». Интенсивность потребления кислорода изучали в специальных
респираторных камерах при температуре среды 20-22°С (Калабухов, 1951). Частоту
дыхания устанавливали визуально, на спящих зверьках, путем подсчёта количества
экскурсий грудной клетки в единицу времени. В ряде случаев частоту дыхательного
акта записывали на ленте двухканального электрокардиографа с помощью
микротермосопротивлений и измерительного моста от медицинского электро-
термометра, улавливающего перепады температур при дыхании.
Количество эритроцитов в крови животных учитывали в счетной камере
Горяева, а содержание гемоглобина - по гемометру Сали. Кровь для анализов
брали из хвоста. Всего за годы работы снято около 500 показателей по
респираторной деятельности и 380 - по газообмену изучаемых хищников, а
также проведено более 1.2 тыс. термометрических и 86 гематологических
измерений.
В электрокардиографических (ЭКГ) исследованиях использовали
одноканальный (реже - двухканальный) чернильнопишущий кардиограф типа
ЭКПСЧ и ЭЛКАР-6 со скоростью лентопротяжного механизма 50 и 100 мм/с.
Режим работы аппаратов -1 мв = 10 мм. ЭКГ снимали без применения наркоза
на иммобилизированных животных в 3 стандартных отведениях от конечностей.
Для регистрации ЭКГ применяли тонкие пластинчатые электроды контактного
действия. Всего преимущественно в летнее время было записано 680 ЭКГ от
хищников разного пола и возраста.
При расшифровке ЭКГ мы руководствовались работами Н.Г. Никулина
(1956) и М.В. Рощевского (1972), а при исследованиях зараженности животных
нематодами-монографиями А.М. Петрова (1941) и В.Л. Контримавичуса(1969).
Характер динамики численности разных видов хищников оценивали по
многолетним учетным данным, полученным на стационарах, и при анализе
анкетного материала, поступавшего преимущественно из северо-западных
областей России.
Глава 1. СРАВНИТЕЛЬНО-ЭКОЛОГИЧЕСКАЯ
ХАРАКТЕРИСТИКА ПИТАНИЯ И
ОСОБЕННОСТИ СТРОЕНИЯ
ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
При сравнительном анализе рациона хищных млекопитающих Северо-Запада
России, выделяются виды как с ярко выраженной пищевой специализацией, так и
с более широким кормовым спектром (Г. Новиков, 1939; Насимович, 1948,1949;
Ивантер, 1963,1967;Русаков, 1963; Данилов, 1968,1969; Новиков и др., 1970; Туманов,
Смелов, 1978,1980; Данилов и др., 1979,1985,1993). Причём эти особенности
исследуемых видов прослеживаются в разных частях их ареалов (Терновский, 1958,
1977; Абеленцев, 1968; Гептнеридр., 1967; Вайсфельд, 1977,1993; Граков, 1973,
1981;Руковский, 1985;Пажетнов, 1990; Сидорович, 1991; Б. Новиков, 1993; Полежаев,
1998;Lockie, 1961;Haglund, 1965; Erling, 1967,1981; Mason, Macdonald, 1986;Kruuk,
1989, 1995; Dunston, 1993, и др.).
Семейство куньих (Mustelidae)
В пищевом рационе куньих представлены различные группы кормов: мелкие
млекопитающие, птицы, пресмыкающиеся, земноводные, рыбы, насекомые,
моллюски, ракообразные и части растений (табл. 1,2). Тем не менее набор основных
пищевых компонентов у них заметно ограничен. Главные объекты питания - это
мелкие грызуны и земноводные. Реже, но также регулярно все они добывают птиц
и их яйца, а выдра {Lutra lutra L.), европейская и американская норки {Mustela
lutreola L. и М. vison Schreb.) - ещё и рыбу. В летнее время молодые животные
охотно потребляют водных и почвенных насекомых. Европейский барсук {Meles
meles L.) и лесная куница {Martes martes L.), кроме того, - еще и некоторые
корма растительного происхождения.
В северных областях европейской части России (Граков, 1973) осенью в рационе
лесной куницы значительное место занимают ягоды (особенно рябины) и некоторые
плоды. Ягоды брусники, черники и голубики с выпадением снега становятся
труднодоступными, и питание ими не может компенсировать энергетических затрат
зверька. Судя по зимним сборам, самки значительно чаще самцов поедают лягушек
и насекомых, тогда как остатки зайца-беляка и белки в питании обоих полов
встречаются примерно одинаково часто. Более существенны различия в зимнем
питании сеголеток и взрослых куниц. Первые, возможно, несколько чаще, чем
вторые, голодают, активнее поедают плоды и ягоды, ящериц и насекомых, но
реже - птиц и их яйца.
В южных регионах страны (особенно на Кавказе) одна из наиболее характерных
особенностей питания и лесной, и каменной куниц - это максимальное
использование доступного им урожая растительных кормов. Здесь эти хищники,
таким образом, выступают в роли не только полифагов, но и (в определенные
12
периоды года) преимущественно фитофагов, что специфично для большин-ства
представителей poj\a.Martes (Бакеев, 1973). У просмотренных нами 38 лесных
куниц, добытых в конце 60 - начале 70 годов в Дагестане и горном
Азербайджане, даже в январе-феврале в желудках обнаруживались преиму-
щественно остатки ягод и плодов, встречаемость которых составляла 77%.
На Кавказе в питании лесной куницы (Бакеев, 1973) можно выделить три
резко отличающихся друг от друга периода: весенне-летний (май-июль),
Таблица 1/Table 1
Сравнительная характеристика питания мелких видов куньих из Ленинградской
и Псковской областей (% встречаемости)
Comparative characteristics of food of small species of mustelids (Mustelidae) from
Leningrad and Pskov Provinces (% of occurrence).
Вид корма/ Food component Ласка обыквовев./ Mustela nivalis (88) Горностай/ Mustela enninea (471) Хорёк лесной/ Mustela putorius . (309) Норка европейск. /Mustela lutreola (767) Норка америк. /Mustela vison (539)
Млекопигаюшие/Mammals: 96.6±2.9 95.8±0.9 30.1±2.6 155±1.3 26.8±1.9
овдатра/muskrat - 0.2±0.2 0.3±0.3 1.6+0.4 1.9±0.6
мелкие грызуны/small rodents 89.8±3.7 93.4±1.1 29.4±2.6 13.3±1.2 23.0+1.8
насекомоядные/insectivora 6.8+5 2.2+1.2 04+0.4 0.6±0.3 1.9+0.6
Птицы/Birds: 5.7±4.2 2.3+O.7 2.5±0.9 8.6±1 8.5±1.2
утиные/Anatidae - - - 2.0+0.5 3.0+0.7
тетеревиные/Tetraonidae - 0.4±0.3 0.3±0.3 0.3±0.2 2.4±0.7
воробьинообразные/Passeriformes 5.7+4.2 1.9+0.6 0.6±0.4 4.6±0.8 2.4±0.7
прочие/other - - 1.6±0.7 1.7±0.5 0.7±0.4
Яйца птиц/Eggs of birds - 0.4±0.3 0.3±0.3 0.3±0.2 -
Пресмыкающиеся/Reptiles - - 0.6±0.4 - -
Земноводные/Amphibians - 0.6+0.4 71.8±2.6 58.0+1.8 36.4±2.1
Рыбы/Fishs - - 1.3±0.6 18.8±1.4 28.2±1.9
Насекомые/Insects: - 0.2±0.2 2.3±0.9 19.2±1.4 14.5+1.5
жуки и их личинки/ beetles and their larves - 0.2±0.2 1.6±0.7 19.2±1.4 8.2+1.2
Ракообразные/Crustaceans - - - 0.8±0.3 2.4±0.7
Моллюски/Molluscs Растительные отстатки/ - - - 0.9±0.3 -
Carcass of plants - 0.8±0.4 - 0.8±0.3 2.1±0.6
Падаль/Carrion - 1.1 ±0.5 2.9±0.9 0.5±0.2 0.7+0.4
Примечание. Здесь и далее в скобках указано количество просмотренных проб.
13
позднеосенний-раннезимний (август-декабрь) и зимне-весенний (январь-
апрель). В первый из них она питается преимущественно грызунами, птицами
и насекомыми. Осенью и в начале зимы она поедает преимущественно плоды и
ягоды, а зимой-ранней весной основу рациона хищника составляют
кустарниковая полевка, лесная и желтогорлая мыши. Растительные корма в
это время имеют второстепенное значение.
Таблица 2/ТаЫе 2
Сравнительная характеристика питания куницы лесной, барсука и выдры
из Ленинградской и Псковской областей (% встречаемости)
Comparative characteristics of food of the pine marten {Martes martes), badger {Meles
meles) and otter {Lutra lutra) in Leningrad and Pskov Provinces (% of occurrence)
Вид корма/ Куница лесная/ Барсук/ Выдра/
Martes martes Meles meles Lutra lutra
Food component (229) (102) (759)
Млекопитающие/Mammals: 60.3±3.2 56.9±4.9 3.7+0.7
ондатра/muskrat 0.4±0.4 - 0.8±0.3
мелкие грызуны/small rodents 42.8±3.3 52.0+4.9 2.9±0.6
насекомоядны e/insectivora 7.4±1.7 4.9±2.1 -
прочие/other 9.7±2 - -
Птицы/Birds: 24.8±2.8 9.8±2.8 2.5±0.6
утиные/Anatidae 0.4±0.4 - 0.9±0.3
тетеревиные/Tetraonidae 14.4±2.3 2.0+1.4 -
воробьинообразные/Passeriformes 9.6±2 7.8±2.7 1.2±0.4
прочие/other 0.4±0.4 - 0.4±0.2
Яйца птиц/Eggs of birds 0.9±0.6 l±0.5 -
Пресмыкающиеся/Reptiles 0.9+0.6 7.8+2.7 - '
Земноводные/Amphibians 13.5±2.3 16.7+3.7 79.8±1.5
Рыбы/Fishs - - 20.0+1.5
Насекомые/Insects: 6.1±1.6 54.9+4.9 3.3±0.6
жуки и их личинки/ 1.7±0.8 47.1+4.9 3.3±0.6
beetles and their larves
Ракообразные/ Crustaceans - 2.0±1.4 4.1±0.7
Моллюски/Molluscs - 3.9±1.9 0.3±0.2
Растительные остатки/ 8.3±1.2 13.7±3.6 0.1±0.1
Carcass of plants
Падаль/Carrion 5.7±1.5 - -
14
Следует отметить, что в отдельные годы в северных регионах значительную
роль в рационе лесной куницы имеют птицы (воробьинообразные, тетеревиные,
дятловые) и белка (Язан, 1962; Граков, 1962 а, 1965; Морозов, 1963,1976;Руковский,
1974), причём.для данного хищника характерно уменьшение плотоядности от
северных районов обитания к южным (Юргенсон, 1951). В соответствии с этой
закономерностью снижается и значение птиц в питании вида. В северной части его
ареала в разных регионах прослеживается более частое потребление тетеревиных
птиц зимой (20-33% встреч), чем летом (1.5-9%), когда они поедают в основном
мышевидных грызунов, насекомых и ягоды (Насимович, 1948; Данилов, Ивантер,
1967; Руковский, 1974). Остальные корма (пресмыкающиеся, ракообразные,
моллюски, черви, падаль) используются большинством куньих нерегулярно и имеют
второстепенное значение.
Среди всех изучаемых хищников особый интерес представляют трофические
связи близких видов с широким кормовым спектром - таких как лесной хорёк (Л/.
putorius L.) и норки, которые часто обитают в одних биотопах.
Анализ материалов (табл. 1), собранных в северо-западных областях России,
показал, что в целом за год по большей встречаемости в пищевом рационе остатков
мелких млекопитающих лесной хорёк достоверно отличается от европейской норки
(р=99.9%), но находится в конкурентных отношениях с американской (р=0). В то же
время полуводные хищники потребляют грызунов и землероек с различной
степенью интенсивности (р=99.9%), что свидетельствует о б отсутствии здесь острой
конкуренции между ними. Птицы, прежде всего водоплавающие и мелкие
воробьиные, также составляют основу пищевого рациона изучаемых хищников.
Оказалось, что леснойхорёкдобывает их достоверно меиьше(р=99.9%), чем норки,
в питании которых птицы представлены равными долями.
Во все сезоны земноводные служат наиболее важным кормом зверьков. При
этом лесной хорёк поедает их заметно чаще, чем аборигенная (р=99.9%) или
интродуцированная (/2=99.9%) норки. Среди них особи американского вида заметно
интенсивнее потребляют рыбу (р=99.9%). Хитиновый покров водных и
почвенных насекомых в остатках пищи лесного хорька встречается достоверно
реже, чем у норок обоих видов (р=99.9%).
Таким образом, кормовой спектр рассматриваемых видов оказывается
различным. Если основу питания хорька составляют земноводные и мышевидные
грызуны, то рацион околоводных видов намного разнообразнее и включает, кроме
того, рыб, насекомых и птиц. Американская норка по сравнению с европейской
представляет собой трофически более пластичный вид. Она успешнее осваивает
основные пищевые ресурсы: мелких млекопитающих, водоплавающих птиц, рыбу
и ракообразных, что позволяет ей широко расселяться по различным водоёмам и
быть более конкурентоспособной.
Из всех видов куньих, обитающих в исследуемом регионе, обыкновенная ласка
(М.nivalis L.) является наиболее узкоспециализированным хищником, который
во все времена года в основном питается мелкими грызунами (табл. 1). Близко
к ней по интенсивности потребления мышей и полёвок стоит горностай
(М. erminea L.). Сравнительно широкий ассортимент кормов использует лесная
15
куница, чем, вероятно, обусловливается стабильность её численности в
малоопромышляемых угодьях. В рационе выдры чаще преобладали земно-
водные и рыбы, а в периоды активной жизни барсука - мелкие млекопитающие,
лягушки и насекомые (табл.2).
В большинстве регионов России рыба играет основную роль в питании
выдры, из которой она поедает несколько десятков видов. В тех частях ареала
хищника, где ихтиофауна богаче, разнообразие ее видового состава
возрастает. В основном выдра добывает мелкую, непромысловую рыбу, но на
Дальнем Востоке и особенно на Сахалине роль лососевых в ее питании
возрастает от осени (встречаемость 26.6%) к зиме (46.6) и особенно к весеннему
периоду (53%). Летом лососевые полностью выпадают из рациона данного
амфибионта. Обычно выдра поедает только отнерестившихся и снулых лососей,
доставая их со дна реки. Трупы этих рыб, вынесенных течением на берег, она
потребляет редко (Вшивцев, 1972; Вайсфельд, 1977).
Если лягушек в разных частях ареала вида можно отнести к основным
компонентам питания выдры, то мелких млекопитающих и птиц она потребляет
относительно нечасто. На водотоках, богатых ракообразными, выдра
потребляет их с большой охотой, однако это обстоятельство не оказывает
существенного влияния на состояние запасов речного рака (Erlinge, 1967).
Как сообщает Вайсфельд (1977), из насекомых данный околоводный хищник
чаще всего поедает виды, связанные с водой - жуков-плавунцов, водолюбов,
ручейников, стрекоз и др. Поскольку размеры насекомых невелики, а общее
количество их остатков в пробах по питанию зверя незначительно, то их роль
в кормовом рационе выдры в целом невелика. Часть беспозвоночных она
заглатывает пассивно, вместе с желудками добываемых ею жертв, а часть
разыскивает активно и ловит самостоятельно при случайных встречах.
Суточный рацион хищника обычно не превышает 1 кг рыбы, а частота
кормежки в основном зависит от размеров единовременно пойманной добычи.
Таким образом, несмотря на заметную специализацию в питании рыбой и
земноводными, выдра все же достаточно пластична в поиске и использовании
других кормов.
Близкий родственник выдры - калан (Enhydra lutris L.) относится к
стенобионтным видам. Обитая на морском побережье Курильских и
Командорских островов, а также на юге Камчатки, звери питаются в основном
морскими ежами, рыбой, двустворчатыми моллюсками и крабами. Главная
особенность питания каланов в районе Командорских островов состоит в том,
что в осенне-зимний период резко падает удельный вес крабов (с 24 до 4%) и
рыбы (с 22 до 6%), уходящих в это время от берегов на большие глубины. Доля
моллюсков в их рационе снижается немного (с 50 до 45%). Они и морские ежи
(98-100% встреч) остаются основными пищевыми компонентами рассма-
триваемого околоводного хищника (Гентнер и др., 1967).
У островов Курильской гряды в феврале-марте каланы на берегу Охот-
ского моря кормятся прежде всего морскими ежами, разными моллюсками,
затем ракообразными, морскими звездами и рыбой. На восточном побережье
16
островов первое место в питании занимают в это время ракообразные, второе -
различные моллюски и третье - морские ежи и даже некоторые водоросли
(Николаев, 1958; Гептнеридр., 1967).
Суточная норма пищи для калана (Ильина, 1950) в среднем составляет
6.5-7.5 ккал, а для крупных особей увеличивается до 10 ккал. Поскольку
калорийность морских ежей, которых он может употребить в пищу за день,
составляет не более 3 ккал, то, по мнению автора, большое значение в рационе
зверя должна играть и рыба. При содержании каланов в неволе их рацион
состоял на 67% из рыбы и только на 33% - из морских ежей.
Как и на Северо-Западе России (Ивантер, 1973), в других регионах
основными группами кормов у европейского барсука {Meles meles) в разные
сезоны служат мелкие млекопитающие, насекомые и земноводные, реже птицы.
Интересные данные по сезонной изменчивости элементов питания этого
хищника из Татарстана приводит П.К. Горшков (1997). Здесь барсук,
вышедший в апреле из норы, поедает прежде всего грызунов и лягушек, а в
конце месяца - еще и некоторых насекомых. В мае значение земноводных,
мышей и полевок заметно снижается, а рацион становится разнообразнее
прежде всего за счет ящериц, ужей, личинок и взрослых майских хрущей, а
также корешков некоторых растений. В июне доля взрослых хрущей среди
кормовых объектов падает, а их личинок и взрослых жуков других видов
(например, лесной навозник) - возрастает. В этот период происходит массовый
выход из водоемов лягушек, которые становятся его постоянной добычей.
Таблица З/ТаЫе 3
Основные корма барсука в Райфском лесу (Татарстан) в разные месяцы
(по: Горшков, 1997)
The main foods of the badger {Meles meles) in the Raifskii forest
(Tatarstan) in various months
Вид корма/ Food component Апрель/ April (44) Май/ May (593) Июнь/ June (204) Июль/ July (158) Август/ August (38) Сентябрь/ September (197) Октябрь/ October (23)
Млекопитающие/ Mammals: 34.1 9.5 27.0 23.4 60.5 22.3 8.7
мыши/ mouses 22.7 2.7 2.9 3.1 28.9 5.1 -
полевки/ Micro tinae 29.5 6.6 20.1 18.9 21.0 14.7 -
Птицы/ Birds - 0.5 - - 2.6 - -
Пресмыкающиеся/ Reptiles - 8.4 4.4 - 7.9 0.5 -
Земноводные/ Amphibians 18.2 11.8 14.2 24.6 28.9 14.7 4.3
Насекомые/ Insects 59.1 91.6 96.1 96.2 92.1 96.4 95.6
Ягоды/ Berries - - 2.5 36.3 34.2 5.1 -
Грибы/ Mushrooms - - - 14.6 - 6.6 -
17
В июне-июле добычей зверя чаще становятся птицы, плоды и ягоды
растений, прежде всего земляники и малины. В августе-сентябре барсук
интенсивно питается, накапливая жировые отложения. В эти месяцы наряду с
насекомыми он много поедает мелких грызунов, численность которых
увеличивается вследствие летнего размножения. В октябре зверь становится
малоактивным, между тем еще выходит из нор и продолжает питаться
разнообразным, наиболее доступным в это время года кормом (табл. 3).
Сезонная смена основных пищевых компонентов у норок и лесного хорька
прослеживается достаточно отчётливо (табл. 4). Весной они с одинаковой энергией
добывают лягушек, грызунов и птиц, поэтому при неблагоприятных кормовых
условиях трофическая конкуренция между хищниками может усиливаться.
Летом и осенью эти виды имеют наиболее богатый кормовой рацион. В июне-
августе американская норка чаще других потребляет мышей и полёвок, однако
статистически значимых различий по этому признаку между нею и хорьком (р=68%)
или европейской норкой (р=82%) не установлено. Заметно активнее их она собирает
на отмелях и водных насекомых (соответственнор=56% ир=70%). Кроме того, в
летний период рацион обеих норок заметно дополняют водоплавающие птицы и
рыбы, которых они отлавливают одинаково интенсивно. Различия между
видами в количестве потребляемых земноводных хотя и реальны, но надежность
вывода оказалась невысокой (/>=70%). В то же время лесной хорёк поедает
летом лягушек очень интенсивно. В его рационе они присутствуют достоверно
чаще, чем у европейской (/>=90%) и американской (/>=99.9%) норок, поэтому в
июне-августе лесной хорек не составляет серьёзной конкуренции норкам даже
при совместном обитании у водоёмов.
Осенью в питании изучаемых хищников большое значение имеют земноводные
и мелкие млекопитающие. Последних американская норка добывает активнее, чем
европейская (/>=93%) и лесной хорёк (/>=95%), которые в эти месяцы охотнее
потребляют амфибий. Птиц и насекомых все эти виды отлавливают нечасто и
в небольшом количестве, обычно в теплое время года. Следует отметить, что
насекомые попадают в желудочно-кишечный тракт хищников не только «попутно
в виде примесей из желудков поедаемых ими рыб», как ошибочно считают Д.В. и
Ю Г. Терновские (1994). В природе молодые зверьки активно собирают их в весенне-
летний период, причем не только представители околоводных видов, но также
лесного хорька, а иногда и горностая, которые на рыбу не охотятся. О регулярной
встречаемости насекомых и моллюсков в желудках выдры, норок и лесного хорька
неоднократно сообщает и В.Е. Сидорович (Сидорович, 1991; Sidorovich, 2000).
По данным автора, встречаемость насекомых в желудках этих видов в летнее
время достигает 5.4-9.6%. Доля рыбы в питании норок постепенно уменьшается
в направлении от летних к зимним месяцам. В то же время следует отметить,
что данный корм в жизни американской норки играет важную роль во все сезоны.
По-видимому, высокий уровень термоадаптивных реакций меньше ограничивает
её доступ к воде в холодное время года по сравнению с аборигенным видом.
Зимой лесной хорёк интенсивнее норок (р=92%) добывает мышей и полёвок,
но в остальном их рацион примерно одинаков, поэтому в холодное время года
18
Таблица 4/Table 4
Сезонные изменения интенсивности потребления мелкими куньими основных
кормов в Ленинградской и Псковской областях (% от общего объёма проб в сезоне)
Seasonal changes of intensity of main foods consumption in small mustelids in the Leningrad
and Pskov Provinces (% from total volume of samples in the season)
Сезон, вид корма/ Season, food component Ласка/ Mustela nivalis Горностай/ Mustela erminea Лесной хорёк/ Mustela putorius Европейск. норка/ Mustela lutreola Американск. норка/ Mustela vison
BECHA/SPRING (16) (391) (15) (16) (18)
Млекопитающие/ Mammals 93.8±7.3 98.5±0.6 33.3±12.2 25.0±10.8 33.3±11.1
Птицы/ 6.2+4.3 1.4±0.6 6.7±6.5 12.5±8.3 11.1±7.8
Земноводные/ Amphibians - - 60.0±12.б 62.5+12.1 50.0+11.8
Рыбы/ Fishs Насекомые/ - 0.1±0.2 - - 5.6+5.4
Insects - - - -
ЛЕТО/SUMMER (10) (9) (16) (9) (14)
Млекопитающие/ Mammals 77.8±13.1 77.8±13.8 18.8±9.8 11.1+±10.5 35.7±12.8
Птицы/ Birds 22.2±13.1 11.1±10.5 - 6.7±3.3 7.2±6.9
Земноводные/ Amphibians - 11.1±10.5 78.7±10.2 44.4±16.6 21.4±10.4
Рыбы/ - - - 33.4±15.7 18.6±10.4
Насекомые/ - - 2.5±1.7 4.4±3.8 17.1±10.1
ОСЕНЬ/AUTUMN (19) (15) (36) (64) (59)
Млекопитающие/ Mammals 100 100 18.3+6.4 10.9±3.9 39.2±6.4
Птицы/ Birds - - 2.8±2.7 3.1±2.2 8.1 ±3.6
Земноводные/ 78.3+6.9 79.7+5 34.8+6.2
Amphibians Рыбы/ - - - 5.2+2.8 14.9±4.б
Насекомые/ 0.6+1.3 1.1+1.3 3+2.2
Insects ЗИМА/WINTER (28) (49) (20) (16) (33)
Млекопитающие/ 100 88.4±4.6 40±11.1 18.8±9.8 18.2+6.8
Птицы/ Birds - 8.6±4 5±4.9 6.2±6 3±3
Земноводные/ Amphibians - 3.1±2.5 55±11.1 75±10.8 66.7±8.2
Рыбы/ Fishs Насекомые/ - - - - 12.1+5.7
Insects
19
Таблица 5/Table 5
Сезонные изменения интенсивности потребления основных кормов лесной
куницей, барсуком и выдрой в Ленинградской и Псковской областях
(% от общего объёма проб в сезоне)
Seasonal changes of intensity in main foods consumption in the common marten (Martes
martes), badger (Meles meles) and otter (Lutra lutra) in Leningrad and Pskov Provinces
(% of total sample size per season)
Сезон, вид корма/ Season, food component Лесная куница/ Martes martes Барсук/ Meles meles Выдра/ Lutra lutra
BECHA/SPRING (23) (31) (14)
Млекопитающие/ 58.7+10.3 60.6+8.8 5.0+5.8
Mammals Птицы/ Birds 28.3±9.4 2.6±2.6 2.1±1.8
Земноводные/ Amphibians Рыбы/ 4.3±4.2 26.8+8.0 78.6+11 14.3±9.4
- -
Fishs
Насекомые/ Insects 8.7±5.9 10+5.4 -
ЛЕТО/SUMMER (12) (12) (38)
Млекопитающие/ Mammals 40.8±14.2 41.6±14.2 8.4±4.5
Птицы/ Birds 20+11.5 8.3±8 2.7±2.6
Земноводные/ Amphibians 28.3+13 23.4+12.2 41.6±8
Рыбы/ Fishs - - 43.1±8
Насекомые/ Insects 10.9±9 26.7±12.8 4.2±3.3
ОСЕНЬ/AUTUMN (38) (16) (66)
Млекопитающие/ 40+7.9 50+12.5 4.6+2.6
Mammals Птицы/ Birds 24.2±6.9 - 3.0±2.1
Земноводные/ 31.6+7.5 25.0+10.8 77.1+5.2
Amphibians Рыбы/ Fishs - 12.3±4
Насекомые/ Insects 4.2±3.2 25.0+10.8 3.0±2.1
ЗИМА/WINTER (62) - (44)
Млекопитающие/ Mammals 65.2±6 - 3.2±2.6
Птицы/ Birds 23.4±5.4 - 0.7±0.3
Земноводные/ Amphibians 6.6±3.1 - 77.5±6.3
Рыбы/ Fishs Насекомые/ 4.8+2.7 - 15.4+5.4 3.2±2.6
Insects -
20
(зима-начало весны), при недостатке пищи или ограниченных возможностях
её использования, сходство рациона может определять конкурентные
отношения между видами (Pulliainen, 1984; Palazon, Ruiz-Olmo, 1992;
Сидорович и др., 1997). Вместе с тем широкий спектр кормов и их обилие,
обычно наблюдаемое у водоёмов, заметно смягчают трофическую конкурен-
цию лесного хорька и норок, оказывая слабое воздействие на их численность.
Необходимо отметить, что в местах исследований, так же как и на соседних
территориях, основу пищевого рациона околоводных и некоторых других
видов куньих составляют амфибии (табл. 5). Они равномерно и устойчиво
освоили разнотипные водоёмы и прибрежные биотопы, во многом обеспечивая
возможность постоянного заселения их хищниками (Пикулик, Сидорович, 1991;
Ruiz-Olmo, Delibes, 1998). Следовательно, регулярное питание околоводных
животных земноводными представляет собой важную трофическую адаптацию.
Она позволяет им широко расселяться по угодьям, продуктивно их осваивать
и создавать необходимые жировые резервы в организме, что способствует
успешной зимовке и готовности зверьков к гону.
Одним из достаточно редких и интересных представителей семейства куньих
является росомаха (Gulo gulo L.), которая потребляет всё, что хоть как-то можно
использовать в пищу. Она часто питается остатками добычи других хищников, но
может и сама успешно нападать на копытных животных, добывать мелких грызунов,
зайцев, куропаток и иных птиц, выкапывать из-под снега падаль, обгрызать
найденные, уже позеленевшие кости животных. В северных территориях росомаха
«пасёт» табунки диких или стада домашних оленей, нападая на ослабленных и
больных особей, а также отставший молодняк.
В Ловозерских тундрах Кольского полуострова в зимне-весенний период
стада домашних оленей постоянно сопровождают 1-3 росомахи. Длительные
кочёвки копытных на большие расстояния (400-800 км) нередко вызывают
естественный падёж оленей, остатками которых или последом отелившихся
важенок питаются эти хищники. На европейском Севере встречаемость
основных кормов в остатках пищи росомахи была следующей: олень северный
23.7-81.6%, лось 7.9-36.9, заяц-беляк 3.9-13.2, тетеревиные птицы 5.7-19.5,
грызуны 7.9-14.5 и растительные корма 0.9-5.2 % (Б. Новиков, 1993).
Значительную долю в кормовом рационе росомахи зимой занимают мелкие
животные, попадающие в охотничьи самоловы. Так, в Красноярском крае в
отдельные годы появившиеся на участках некоторых охотников росомахи
вытаскивали из капканов и уносили от 4 до 20 соболей (табл.6).
Следует отметить, что обитающую на промысловом участке росомаху
интересует не только мелкая дичь, попавшая в самоловы, но и любая более крупная
добыча, которую можно использовать в пищу. Так, по имеющимся сведениям, в
горах Хамар-Дабана она утащила ночью крупную голову кабана, которая была
оставлена удачливыми охотниками до утра после разделки и выноса основной
части туши. В другом случае росомаха вышла на попавшую в капкан и замерзшую
в нем рысь. Объев голову, она уволокла её остатки метров за 100 и прикопала
в снег. Не брезгует росомаха и падалью: в 1989 г. в Курагинской тайге
21
Таблица 6/Table 6
Количество соболей, утащенных росомахой из капканов охотников-
промысловиков в Красноярском крае (по: Туманов и др., 1999)
The number of sables taken by wolverine (Gulo gulo) from the traps of hunters in the
Krasnoyarskii Territory
Охотучасток в долине реки/ Hunting site in the river valley Сезон охоты, год/ Hunting season, year Утащено соболей/ The number of taken sables
всего/total % от общего кол-ва/ % of total
Kac/Kas 1982/83 4 4.7
1983/84 16 18.6
1986/87 18 19.5
Кия-Северная/ 1986/87 20 16.3
Kiya-Sevemaya 1989/90 17 15.4
Тым/Тут 1988/89 12 12.7
1991/92 8 9.3
Северная Комса/ 1988/89 6 8.3
Severnaya Komsa 1989/90 7 8.8
Bax/Vakh 1989/90 12 15.8
Kypena/Kurepa 1991/92 8 12.5
Тис/Tis 1987/88 19 14.8
(Западные Саяны) росомаха подошла к избушке промысловика, где лежал
труп собаки, утащила, а затем съела его. По сообщению охотника С. Писарева,
промышлявшего в Западном Саяне по р. Она, росомаха, опередив его на
несколько минут, выхватила из капкана тушку выдры и только после
преследования и выстрелов в воздух бросила её. Интересно, что хищник
практически не реагирует на мелкую приваду в виде крылышек или лапок
птиц, подвешенных у самоловов, зато попавшую в них сойку, кедровку или
сову съедает полностью (Туманов и др.,1999).
Среди представителей семейства куньих, обитающих в России, кроме
барсука и лесной куницы к типичным эврифагам следует отнести соболя
(Martes zibellina L.) и каменную куницу (M.foina Erxl.). Потребность соболей
в животной или растительной пище в разные сезоны года определяется их
физиологическим состоянием. В определенные периоды, независимо от обилия
животной пищи, в природе эти хищники почти полностью переключаются на
питание растительными кормами: ягодами, плодами лесных растений,
орешками кедра и кедрового стланника. По мере созревания голубики,
брусники, рябины соболь регулярно и в большом количестве поедает эти ягоды.
В годы обильного урожая рябина и орешки кедра часто служат его основной
пищей (Монахов, Бакеев, 1981).
22
Таблица7/ТаЫе 7
Возрастные различия в питании соболя, встречаемость в % к количеству желудков
с пищей (по: Борзых, 2001)
Age differences in the feeding of sable (Martes zibellina), occurrence in % to the number of
stomachs with food
1995 1996
Вид корма/ ru. 2+ и старше/ 2+ и старше/
Food component 2+ and older 2+ and older
(12) (19) (23) (И) (18) (35)
Млекопитающие/ Mammals: 42 68 78 54 50 54
полевки/ Microtinae 33 42 52 36 38 37
Птицы/ Birds 0 21 26 0 0 25
Насекомые/ 16
Insects /
Кедровые орехи/ Siberian pine nuts 50 89 100 64 83 94
По нашим наблюдениям, в Восточных Саянах в осенне-зимнем сезоне
соболь регулярно питается растительными кормами и в частности кедровыми
орешками. Их доля в общем количестве просмотренных желудков (я=85)
составляла осенью и в начале зимы 70-80%, а встречаемость животной пищи в
биопробах не превышала 26-30%. В основном она была представлена красной
и красно-серой полёвками, пищухой, белкой, тетеревиными и птицами из отряда
воробьинообразных.
В 118 желудках соболей, добытых осенью 1995 и 1996 гг. в районе Обь-
Енисейского междуречья (Борзых, 2001), обнаружены остатки мелких млеко-
питающих и зайца-беляка (64.3%), птиц(14.4%), насекомых (13.4%) и растительных
кормов (93.9%). Основными кормами в этот период были кедровые орехи и полевки.
Большая доля содержания орехов в желудках зверьков объясняется хорошим
урожаем кедра в годы работы и длительным отсутствием глубокого снегового
покрова, позволяющим долгое время использовать семена кедра. Отмечено, что
до начала зимы самки несколько чаще используют в пищу животные корма, в то
время как самцы-растительные. По наблюдениям автора (табл. 7), молодые
особи в это время не добывали в пищу птиц, заметно реже питались семенами
кедра, но чаще поедали насекомых.
Нередко добычей хищника становятся и более крупные млекопитаю-
щие - заяц-беляк или кабарга (Зырянов, Туманов, Кожечкин, 1997, 2001).
Хищничество соболя в отношении кабарги тесно связано с зимним (чаще
многоснежным) периодом. По Красноярскому краю нам известно 25 случаев
гибели кабарги в результате нападения на нее соболя. Это происходит прежде
всего в многоснежные годы при повышенном дефиците кормов. В такие сезоны
быстрое нарастание высоты снежного покрова и уплотнение его верхнего слоя
23
создают неодинаковые условия для передвижения хищника и жертвы: они
становятся более благоприятными для первого. По нашему мнению,
элиминирующее воздействие соболя на кабаргу может приводить к ее
исчезновению на окраинах ареала вида не только в вертикальных (горах), но
и в широтных (на севере) пределах распространения вида (Зырянов и др., 2001).
Влияние других, более крупных хищников (росомаха, рысь) из-за их
малочисленности не играет здесь существенной роли (Хлебников, 1977).
Состав пищи каменной куницы во многом сходен с таковым лесной (Г.
Новиков, 1956; Гептнер и др., 1967). Основу её рациона составляют мелкие
млекопитающие, птицы, насекомые, ягоды и плоды фруктовых деревьев. По
нашим наблюдениям, на Кавказе (Дагестан, Азербайджан) в осеннее время
встречаемость растительных остатков в пробах (и=83) равна 56-63%, хотя даже в
этот сезон доля мышевидных грызунов в рационе достигала 60-80%. Эти данные
близки к материалам, полученным А.Л. Мамедовым (1972) в горных районах
Азербайджана. По сведениям автора, каменная куница (п= 160) питается главным
образом млекопитающими (81.9% встреч), птицами (46.2%), насекомыми (80%) и
частями растений (72.5%). В рационе лесной куницы (и=150) доля этих кормов
была соответственно следующей: 74, 34, 78 и 68%. О значительной роли
растительных кормов для каменной куницы сообщает также А.И. Соков (1972а).
По материалам этого исследователя в Таджикистане в сентябре-декабре она
особенно охотно поедает яблоки, груши, алычу, жёлтоплодную боярку, шиповник,
ежевику. Встречаемость грызунов в её питании (и=52) составляет 44.2%, птиц
7.7, насекомых 21.1, а плодов фруктовых деревьев 38.4%.
Наблюдения, проведенные в Ставропольском крае (Бакеев, 1973), показали,
что в зимнее время каменные куницы добывали преимущественно мышевидных
грызунов и мелких воробьиных птиц. В августе-сентябре они переходили на
питание растительными кормами даже в те годы, когда численность лесных и
полевых мышей, обыкновенных и кустарниковых полевок в угодьях была очень
высокой. В весенне-летний период рацион каменных куниц (164 пробы) состоял в
основном из кротов (74% встреч), мышевидных грызунов (43%), птиц (19%) и
насекомых (15%). В Краснодарском крае основу их питания в разные сезоны
составляли мышевидные грызуны и птицы, хотя в значительном количестве
хищники потребляли и корма растительного происхождения. В отдельные годы
встречаемость в рационе зверьков плодов и ягод достигала 31-48%. В Дагестане
(Гинеев, 1991) зимой плоды терна составляли 40%, боярышника и груши - по 27%
всех встреч. Мелкие млекопитающие были отмечены в 40%, птицы - в 27,
насекомые - в 13 и падаль - в 20% случаев. В пойменных (часто островных) лесах,
а в горных районах - по опушкам леса и каменистым осыпям каменная куница во
все сезоны года находит себе достаточное количество корма, что и определяет ее
тяготение к этим стациям.
Харза (Martesflavigula Boddaert) по характеру питания - типичный хищник.
Из всех куниц это наиболее крупное и сильное животное, способное быстро и
подолгу преследовать избранную жертву или же подстерегать ее из засады.
Харзы часто охотятся группами от 3 до 5 особей, чего не делает больше никто
24
из других куньих. Явную специализацию они проявляют при преследовании
кабарги, которую стараются выгнать на речную наледь, где быстро и
настигают скользящую жертву (Бромлей, 1973).
Использование различных компонентов пищи подчинено определенной
сезонности. С телятами крупных копытных и поросятами кабанов харза может
справиться только весной, когда жертва весит меньше 10 кг. Более крупных копытных
ей задушить, видимо, не удается. Бурундуков, мышей и полевок она ловит
больше в теплое время года. Зимой разнообразие спектра животных кормов
{п-185) несколько уменьшается, и их основу составляют кабарга (57.3%), белка
обыкновенная (7.6%) и летяга (2.7%), заяц маньчжурский (4.3%) и беляк (2.7%).
Растительная пища, по-видимому, используется редко и в основном молодыми
особями в раннеосеннюю пору или же совсем старыми животными. То же
можно сказать и о поедании беспозвоночных и отнерестившейся рыбы (по 1.6%).
В целом эти корма вместе с растительной пищей составляли лишь 5.4% от
общей их встречаемости в пробах. Специфика питания харзы заключается в
потреблении лишь свежей пищи, т.е. недавно пойманных животных. Падаль
или сильно промерзшие остатки даже своей прежней добычи она обычно не
трогает (Бромлей, 1973).
Рацион колонка {Mustela sibirica Pall.) состоит преимущественно из
теплокровных животных. Основной его корм - мышевидные грызуны. В своих
местообитаниях он, как и большинство других мелких хищников, предпочитает
охотиться за полевками и мышами, часто встречающимися и более доступными.
В Западной Сибири среди его кормов преобладают водяная полевка и мышевид ные
грызуны, в поймах крупных рек в рационе заметно увеличивается доля мелкой
рыбы и падали. Растительные корма он потребляет в незначительном количестве
(Войлочников, 1977). Молодые зверьки, видимо, опасаются нападать на животных
крупнее их по размеру. Так, в Бурятии нами было отловлено 15 колонков (10
сеголеток и 5 взрослых особей). В пищеварительном тракте только у 4 зверьков
старше года находились остатки рябчика или ондатры. В желудках всех остальных
зверьков были обнаружены шерсть и кости мышевидных грызунов.
В осенне-зимнем питании колонка лесостепного Забайкалья наибольшее
значение имеют даурская пищуха, остатки которой отмечены в 32.5% данных по
питанию вида, и мышевидные грызуны, преимущественно стадная полевка (10.6%).
На долю птиц приходится 13.6%, в том числе на тетеревиных - 7.8%. В горно-
таежных лесах Южного и Юго-Западного Предбайкалья основной пищей колонка
служат грызуны (67.2%), ондатра и птицы (по 14.9%), а также землеройки (8.9%).
В местах с высокой плотностью населения ондатры колонок может специализиро-
ваться на ее добыче. В таких угодьях ее остатки встречаются более чем в 50%
осмотренных желудков добытых хищников (Фетисов, 1942; Войлочников, 1977).
Солонгой {Mustela altaica Pall.) по характеристике питания близок к
горностаю. Основу его рациона составляют крупные по размеру грызуны и
птицы, реже насекомые и ящерицы. В Западном Забайкалье в его зимней
добыче (Гептнер и др., 1967) доля грызунов достигает 73.2%, птиц 22.7 и
землероек 3.1% (по 94 пробам). По частоте встречаемости среди них первое
25
Таблица 8/ТаЫе 8
Сезонная изменчивость массы тела (г) и калорийности (ккал)
основных кормов куньих
Seasonal variation of the body mass (g) and calorific value (kcal) of main foods in mustelids
Вид корма/ Food component 11 Показатели/ Parameter Весна/ Spring Лето/ Summer Осень/ Autumn Зима/ Winter
Рыжая полевка/ 20 Вес тела/ 16.5 16.8 17.1 16.7
Clethrionomys Weight
glareolus Калорийность/ Caloricity:
общая/total 17.2 27.1 22.8 19.9
на 1 г веса/ 1 1.6 1.3 1.3
per Igbody mass
Обыкновенная 19 Вес тела/ 21.5 20.5 25.3 22.2
полёвка/ Weight
Microtus arvalis Калорийность/ Caloricity:
общая/ total 26.2 33.6 27.6 25.4
на 1 г веса/ per 1g body mass 1.2 1.6 1.1 1.1
Большая синица/ 15 Вес тела/ 14.8 14.0 15.2 14.2
Parus major Weight Калорийность/ Caloricity:
общая/ total на 1 г веса/ 20.7 1.4 15.9 1.1 22.1 1.5 19.7 1.4
per lg body mass
Травяная 25 Вес тела/ 26.5 24.5 21.3 19
лягушка/ Weight
Rana temporaria Калорийность/ Caloricity:
общая/ total 16 22.7 26.0 14.3
на 1 г веса/ per lg body mass 0.6 0.9 1.2 6.8
Обыкновенный окунь/ 17 Вес тела/ Weight 33.9 43.6 45.0 42.6
Perea fluviatilis Калорийность/ Caloricity:
общая/ total 31.9 44.5 63.7 48.6
на 1 г веса/ 0.9 1 1.4 1.1
per lg body mass
Жук плавунец/ 19 Вес тела/ 1.2 1.8
Dytiscus Weight
marginalis Калорийность/ Caloricity:
общая/ total - 2.1 3.8 -
на 1 г веса/ per lg body mass - 1.8 2.1 -
26
место занимает сеноставка даурская (34.2%). В Прибалхашье в рационе
солонгоя во все сезоны также преобладают мышевидные грызуны, а летом
спектр его кормов заметно увеличивается за счет ящериц (22.5% встреч) и
насекомых (8.3%). В дельте р. Или и на побережье Балхаша (Злобин, 1972)
важным кормовым объектом солонгоя оказывается ондатра (встречаемость в
пробах 10.3%). Самцы хищника, будучи в 2.5 раза крупнее самок, преследуют
жертву заметно интенсивнее. В их рационе ондатра встречается в 3 раза чаще,
чем у самок. Вообще, солонгои, обитающие по побережью Балхаша, видимо,
больше адаптированы к добыванию этого околоводного грызуна, чем особи
из других мест. В их питании ондатра имеет заметно большее значение, чем у
хищников верхней и нижней части дельты р. Или, где в отдельные годы она
также бывает многочисленна.
В горах Средней Азии солонгой типичен для высот от 1400 до 3000 м над
ур. м., но в горных районах Памира его встречали и на высотах 2710-3700 м
над ур. м. Здесь солонгой питается в основном серым хомячком, большеухой
песчанкой, серебристой полевкой, молодняком зайца-толая, реже воробьиными
птицами. В долинах рек в зимний период он добывает мышей и мелких полевок
(24.2%), ондатру (14.7%), водяную полевку (5.8%). С апреля по октябрь кроме
грызунов (66.6%) значительное место в питании (22.8%) занимают ящерицы
(Соков, 1972 б).
В Западной Сибири, на юго-западной оконечности Алтайских гор,
солонгой часто обитает в одних стациях с соболем и горностаем, где их
основной пищей служат мышевидные грызуны, однако обнаружить следы
антагонистических отношений между ними даже в местах совместного
обитания зверьков не удавалось. Каждый вид придерживается свойственной
только ему экологической ниши, что уменьшает напряженность конкуренции
(Терновский, Терновская, 1994).
Наличие и доступность различных групп основных кормов во многом
определяют физиологическое состояние хищников, их перемещения, воспроизвод-
ственный потенциал, а в конечном итоге и численность всей популяции. В этой
связи заметный интерес представляют данные о качественном составе добычи,
который влияет на величину суточной потребности в пище и продолжительность
охотничьего поиска куньих. Исследования показали, что наибольший процент
сухого вещества (жиров, протеинов, углеводов, минеральных веществ и других
соединений) в корме и его калорийность характерны для насекомых, грызунов
и птиц. У земноводных и рыб она заметно ниже (Туманов, Смелов, 1980).
Качественный состав пищи хищников существенно изменяется по сезонам года.
Так, полёвки достигают максимальной калорийности летом, что совпадает с
данными А. Гурецки (Gorechki, 1965), а остальные компоненты корма - осенью.
Причём у них прослеживается сезонная изменчивость не только массы тела, но
и её калорийности (табл. 8).
Таким образом, к концу лета-осени, когда в организме куньих создаются
жировые резервы, необходимые для зимовки, калорийность основных компонентов
их рациона существенно возрастает.
27
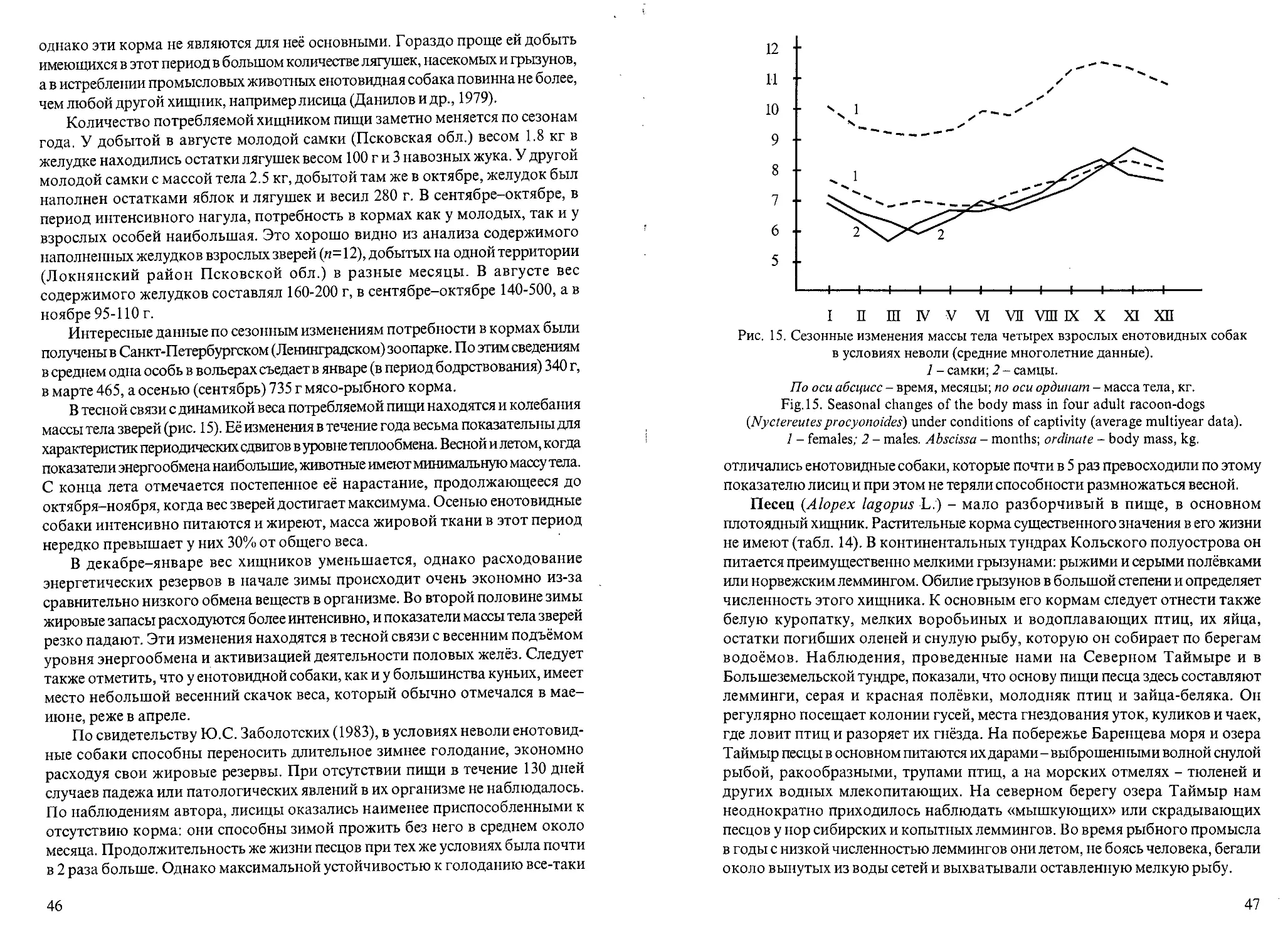
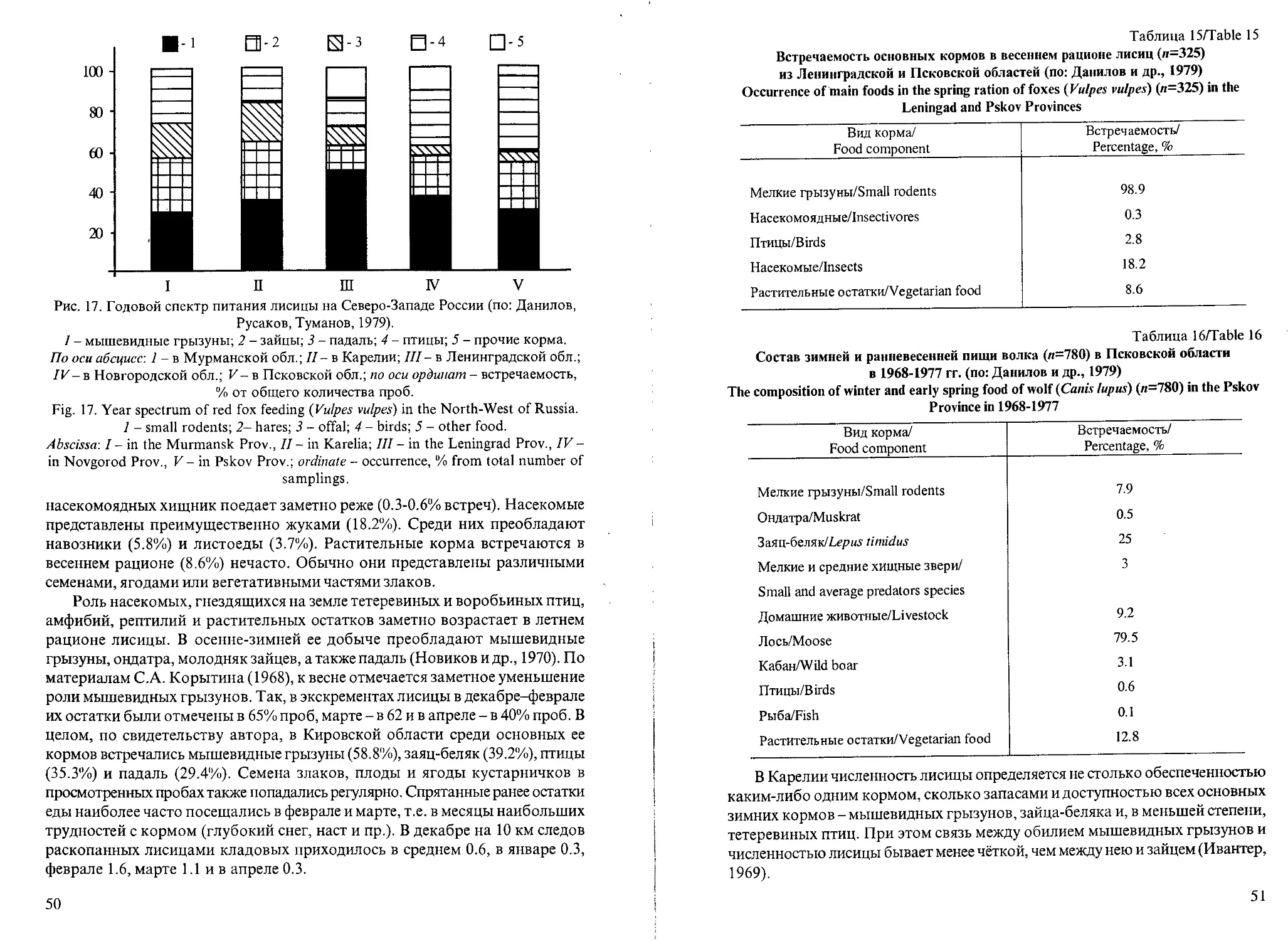
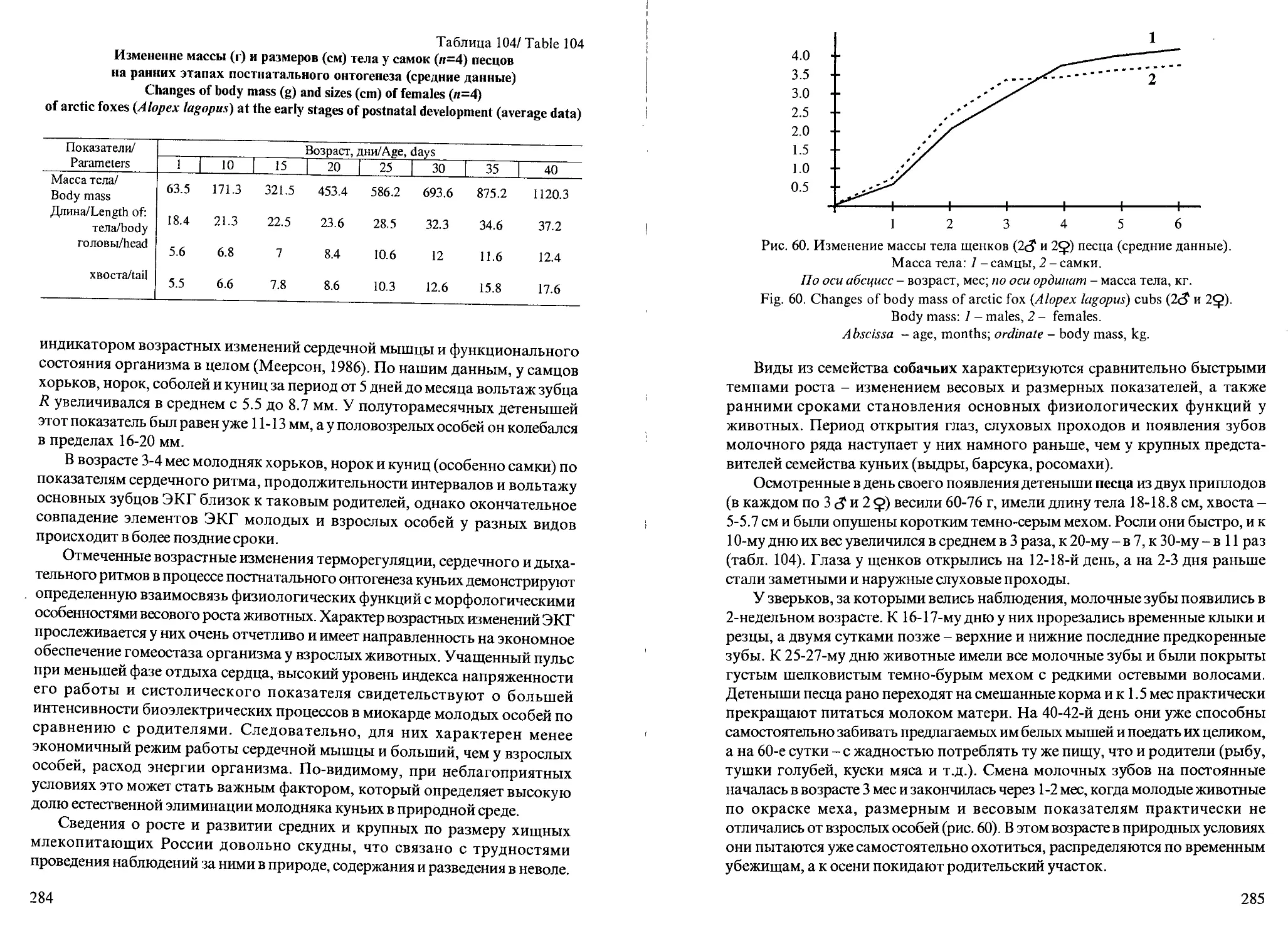
Рис. 1. Динамика массы тела самцов мелких видов куньих в течение года.
1 - американская норка; 2 - европейская норка; 3 - лесной хорек; 4 - ласка.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, г.
Fig. 1. Dynamics of the body mass in males of small mustelids species during the year.
1 - American mink (Mustela visori), 2 - European mink (Mustela lutreola), 3 - polecat
(Mustelaputorius), 4 - weasel (Mustela nivalis).
Abscissa - months; ordinate - body mass, g.
Наблюдения за разными представителями семейства куньих, содержа-
щимися в вольерах, свидетельствуют о том, что суточная потребность в пище
и масса тела заметно изменяются по сезонам года (Туманов, 1976а; Туманов,
Сорина, 1997а, б, 1999). При этом сезонные флуктуации массы тела зверьков
весьма показательны как признак периодических сдвигов в уровне их
энергетического обмена.
У всех самцов мелких хищников в конце лета-осенью масса тела
увеличивалась очень заметно (рис.1). Это согласуется с интенсивным
потреблением в данный период кормов и накоплением энергетических
жировых резервов, необходимых для существования в холодное время года.
Например, осенью по сравнению с летними месяцами масса тела
увеличивалась (в среднем) у ласки на 17%, у норок обоих видов - на 21% и
у лесного хорька - на 34%. Таким образом, в осеннее время наиболее
интенсивное жиронакопление прослеживалось у лесного хорька. Это
следует рассматривать как видовое приспособление терморегуляции,
которое обеспечивает зимой снижение теплоотдачи и расхода энергии. В
холодное время года у всех куньих жировые резервы постепенно
уменьшались, вес падал, а весной, перед началом линьки и сезона
28
Рис. 2. Динамика массы тела самца европейской норки в течение четырех лет.
1 - 1986; 2 - 1987; 3 - 1988; 4 - 1989.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, г.
Fig. 2. Multiyear dynamics of the body mass in the male of European mink (Mustela
lutreola).
1 - 1986; 2- 1987; 3- 1988; 4- 1989.
Abscissa - months; ordinate - body mass, g.
Рис. 3. Динамика массы тела самца лесного хорька в течение четырех лет.
Условные обозначения как на рис. 2.
Fig. 3. Multiyear dynamics of the body mass in the male of polecat
(Mustela putorius).
1 - 1986; 2 - 1987; 3 - 1988; 4- 1989.
Abscissa - months; ordinate - body mass, g.
29
размножения, величина этих показателей вновь возрастала. В конце весны-
летом они имели наименьшие массу тела и жировой резерв, причём данные
изменения отчётливо прослеживались во все годы наблюдений (рис. 2-3).
У самок сезонные изменения массы тела также прослеживались достаточно
отчётливо. Здесь следует отметить, что в природе динамика их веса имеет меньшую
амплитуду колебаний по сравнению с самцами. Данная особенность, по-видимому,
объясняется тем, что за время гона, родов и лактации самки заметно теряют в весе.
За короткий период осени они не успевают накопить достаточное количество
жировых резервов, что и определяет малый диапазон флуктуаций их массы и
большую по сравнению с самцами долю элиминации в природе (Tumanov, 1993а).
В то же время необходимо отметить, что при содержании в неволе не
участвовавшие в размножении самки также имели меньшую по сравнению с
самцами сезонную амплитуду колебаний массы тела. Отмеченные половые
различия, по-видимому, следует рассматривать как наследственно закреплён-
ную и притом достаточно консервативную реакцию организма самцов и самок,
обусловленную соответствующей разницей в уровне их обменных процессов.
Исследования показали, что масса тела самок интенсивно нарастала в осенний
сезон за счёт образования в организме жировых (или энергетических) ресурсов,
необходимых для нормального хода линьки и существования в холодное время
года. Например, за период с сентября по декабрь увеличение массы тела
зверьков (по отношению к июню-июлю) составляло в среднем для ласки 4.8%,
норки европейской 3.9, американской 5.6, куницы 10.4, горностая 14.8 и лесного
хорька 20.2%.
В холодное время года жировые резервы самок постепенно расходовались, и
масса тела уменьшалась. В конце зимы-ранней весной уровень этих показателей у
них вновь заметно возрастал (рис. 4-5). После окончания весенней линьки, а у
многих видов - и сезона размножения, самки были малоупитанными. Летом
они всегда имели наименьшие массу тела и жировой запас (рис. 6-7).
Материалы проведенных исследований свидетельствуют о том, что у куньих
сезонная динамика массы тела тесно связана с характером изменений потребности
в кормах. Самцы, активно потребляющие пищу зимой, летом в жару ели
сравнительно мало (рис. 8). Существенное уменьшение их суточной нормы
становилось заметным у хорей и ласок в июне, а у норок - в июле. Примерно с
середины августа количество съедаемой за сутки пищи начинало резко возрастать,
а в сентябре потребность в кормах у животных была максимальной. Так, в сентябре
этот показатель возрастал в среднем у самцов ласки на 49%, американской нор-
ки - на 50% и у европейской - на 52% по сравнению с летним периодом года.
Наибольшая потребность в корме наблюдалась у лесного хорька, у которого
суточное потребление пищи в сентябре (по сравнению с летом) возрастало в 2.2
раза. Отмеченная особенность у представителей данного вида чётко согласуется
и с увеличением их массы тела осенью за счёт депонирования в организме
значительных жировых резервов. В холодное время года у самцов всех видов
куньих потребность в высококалорийной пище заметно снижалась, что следует
рассматривать как стойкую адаптивную реакцию организма, направленную
30
950
i nmivvvivnvfflixxxixn
Рис. 4. Динамика массы тела самок норок и ласки в течение года.
/ - американская норка; 2 - европейская норка; 3 - ласка.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, г.
Fig. 4. Dynamics of body mass in females of weasel (Mustela nivalis) and minks
(Mustela lutreola, Mustela vison) during the year.
1 - American mink (Mustela vison); 2 - European mink (Mustela lutreola); 3 - weasel
(Mustela nivalis). Abscissa - months; ordinate - body mass, g.
I П Ш IV V VI VIII К X XI ХП
Рис. 5. Динамика массы тела самок лесной куницы, хорька и горностая,
в течение года.
1 - лесная куница; 2 - лесной хорек; 3 - горностай.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, г.
Fig. 5. Dynamics of body mass in females of ermine (Mustela erminea), pine marten
(Martes martes) and polecat (Mustela putorius) during the year.
I - pine marten (Martes martes); 2 - polecat (Mustela putorius); 3 - ermine
(Mustela erminea). Abscissa - months; ordinate - body mass, g.
31
-1 B-2 S-3 В-4
Рис. 6. Многолетняя динамика массы тела самки лесного хорька.
1 - 1986; 2- 1987; 5- 1988; 4- 1989.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, г.
Fig. 6. Multiyear dynamics of the body mass in the female of polecat {Mustela
putorius).
1 - 1986; 2-1987; 3- 1988; 4-1989.
Abscissa - months; ordinate - body mass, g.
Рис. 7. Многолетняя динамика массы тела самки европейской норки.
Условные обозначения как на рис. 6.
Fig. 7. Multiyear dynamics of the body mass in the female of the European mink
{Mustela lutreoki).
1 - 1986; 2-1987; 3-1988; 4-1989.
Abscissa - months; ordinate - body mass, g.
32
300
i nmivvvivnvinixxxixn
Рис. 8. Суточное потребление корма самцами из семейства
куньих в разные месяцы.
I - американская норка; 2 - европейская норка; 3 - лесной хорек; 4 - ласка.
По оси абсцисс - время, месяцы; по оси ординат - масса потребляемого корма, г.
Fig. 8. Circadian consumption of food by mustelids males in different months.
1 - American mink {Mustela vison); 2 - European mink {Mustela lutreoldy, 3 - polecat
(Mustela putorius); 4 - weasel {Mustela nivalis).
Abscissa - months; ordinate - consumption of food mass, g.
Рис. 9. Изменение суточного потребления корма самцом европейской норки
по годам.
1 - 1986; 2- \М>1-,3- 1988; 4- 1989.
По оси абсцисс - время, месяцы; по оси ординат - масса потребляемого корма, г.
Fig. 9. Changes in the circadian food consumption in the male of European mink
{Mustela lutreola) by year.
1- 1986; 2- 1987; 3- 1988; 4- 1989.
Abscissa - months; ordinate - consumption of food mass, g.
2 Зак. № 4309
33
Рис. 10. Изменение суточного потребления корма самцом лесного хорька
по годам.
Условные обозначения как на рис. 9.
Fig. 10. Changes in the circadian food consumption in the polecat (Mustela putorius)
male by year.
1 - 1986; 2-1987; 3-1988; 4-1989.
Abscissa - months; ordinate - consumption of food mass, g.
на более экономное расходование созданных осенью энергозапасов. При
падении зимой уровня обменных процессов потребность в большом количестве
добываемой пищи и энергозатратах у хищников снижается, что и определяет
благополучный исход их зимовки (Слоним, 1952; Tumanov, 1993а). Весной,
перед линькой и гоном, суточное потребление корма у самцов во все годы
исследований вновь возрастало в определенные для видов сроки (рис. 9-10).
Динамика суточной потребности в пище самок изучаемых мелких видов
куньих по периодам года имеет сходный характер с таковой у самцов, хотя и
разнится по массе поедаемого корма (рис. 11-12). В летний сезон, при высокой
температуре наружного воздуха, они ели сравнительно мало. Как и у самцов,
заметное снижение уровня суточного рациона отмечалось у них в июне или
июле. Затем в начале августа потребность норок и хорьков в пище начинала
возрастать, достигая своего максимума в сентябре-октябре, а у ласки и
горностая - в ноябре. По сравнению с летним периодом (июнь-июль) в
сентябре-декабре суточное потребление пищи в среднем повышалось у ласки
на 2.3%, у норки американской - на 1'2.5 и у европейской - на 20.1%. У самок
других видов оно было ещё выше, достигая у горностая 23.4%, лесной куницы
26.1 и у хорька 30.4%, причём характер флуктуаций массы пищи, съедаемой
хищниками за сутки, в разные годы был примерно одинаков (рис. 13-14). Таким
образом, осенью, когда зверьки начинали интенсивно питаться, их жировой
запас (или энергетический потенциал) резко возрастал. По-видимому,
выживаемость мелких хищников зимой в естественных местообитаниях
определяется наличием и доступностью высококалорийных кормов в осеннее
время (сентябрь-ноябрь), когда потребность в них наиболее высока. Особенно
нуждаются в жировых резервах самки, которым необходимо возместить
энергетические затраты за период репродуктивного сезона.
Зимой зверькам требовалось заметно меньше пищи, что обеспечивало им
экономное расходование созданных осенью жировых резервов и лучшую
34
200
180 1
160 /. . >-< X Z
140‘
120
Рис. 11. Изменение суточного потребления корма самками норок и ласки
в течение года.
1 - американская норка; 2 - европейская норка; 3 - ласка.
По оси абсцисс - время, месяцы; по оси ординат - масса потребляемого корма, г.
Fig. 11. Changes in the circadian food consumption in females of weasel and minks
during the year.
1 - American mink {Mustela visoii); 2 - European mink {Mustela lutreola)', 3 - weasel
{Mustela nivalis).
Abscissa - months; ordinate - consumption of food mass, g.
выживаемость в холодное время года. Весной, перед линькой и началом сезона
размножения, необходимость в высококалорийных кормах вновь возрастала
и сопровождалась увеличением массы тела животных, а далее весь цикл
повторялся. У барсуков суточный рацион увеличивался на протяжении всего
активного периода, от весны к осени.
Наши материалы в общих чертах совпадают с данными Н.Н. Гракова (1966)
по динамике массы тела и кормов, потребляемых лесной куницей и соболем при их
вольерном содержании. По свидетельству автора, все животные имели два периода,
когда их масса тела увеличивалась - весенний и особенно осенний. Летом у
большинства особей вес был минимальный. Максимальное количество корма они
поедали в августе-сентябре, а минимальное - в декабре. Осеннее увеличение
потребности в пище совпадало с периодом интенсивного роста зимнего волоса и
жиронакопления: в его начале масса съедаемой за сутки пищи была наиболее
35
300
Рис. 12. Изменение суточного потребления корма самками лесной куницы, хорька
и горностая в течение года.
1 - лесная куница; 2 - лесной хорек; 3 - горностай.
По оси абсцисс - время, месяцы; по оси ординат - масса потребляемого корма, г.
Fig. 12. Changes in the circadian food consumption in females of pine marten (Martes
martes), polecat (Mustela putorius) and ermine (Mustela erminea) during the year.
1 - pine marten (Martes martes); 2 - polecat (Mustela putorius); 3 - ermine (Mustela
erminea).
Abscissa - months; ordinate - consumption of food mass, g.
высокой, но потом резко снижалась. Конечная стадия осенней линьки сопро-
вождалась падением массы тела и необходимого для жизнеобеспечения корма. В
среднем в январе при вольерном содержании самец лесной куницы потреблял 258.5 г,
а самка 133.2 г полноценного корма в сутки. Таким образом, добытую белку
голодный хищник может съесть за сутки, а рябчика - максимум за двое.
В природных условиях изменение массы тела взрослых куниц, соболей и
других мелких куньих в осенне-зимний период происходит аналогично тому,
что наблюдается в вольерах. Некоторый спад упитанности зверьков в декабре-
январе характерен и для молодых особей - сеголеток. По-видимому, в данном
случае имеют место адаптации, связанные с сезонной спецификой физио-
логических функций организма. Сезонная изменчивость пищевого рациона и
его общего веса у представителей семейства куньих отмечается многими
исследователями (Fairley, 1972; Moors, 1975; Erlinge, 1975, 1981; Blandford,
36
250 л
- 1 В-2 В-3 В-4
Рис. 13. Изменение суточного потребления корма самкой лесного хорька
по годам.
I- 1986; 2- 1987; 3- 1988; 4- 1989.
По оси абсцисс - время, месяцы; по оси ординат - масса потребляемого корма, г.
Fig. 13. Changes in the circadian food consumption in the female polecat
{Mustela putorius) by years.
1 - 1986; 2- 1987; 5- 1988; 4- 1989.
Abscissa - months; ordinate - consumption of food mass, g.
Рис. 14. Изменение суточного потребления корма самкой европейской норки
по годам.
Условные обозначения как на рис. 13.
Fig. 14. Changes in the circadian food consumption in the female of European mink
(Mustela lutreola) by year.
1 -1986; 2-1987; 3-1988; 4- 1989.
Abscissa - months; ordinate - consumption of food mass, g.
1987; Kruuk, 1989, 1995; Mason, Macdonald, 1986; Brzezinski et al., 1993;
Сидорович и др., 1997).
На основании данных о потребности исследуемых хищников в пище и её
калорийности нами была рассчитана энергия их кормового рациона (Туманов,
Смелов, 1980, 1982). Материалы показали, что она тесно связана с массой
тела животных. У самых мелких представителей семейства (ласки и горностая)
удельная величина энергии потребляемого корма оказалась в 4-6 раз больше,
37
чем у выдры и барсука. Так, у обыкновенной ласки (независимо от пола) на
100 г веса она составляла 50.8-69.9 ккал, у горностая 62.3-64.1, у выдры
13.8-14.5 и у барсука 8.5-8.6 ккал в сутки.
У разных видов куньих, близких по массе тела, размеры этого показателя
во многом определяются уровнем локомоторной деятельности и теплообмена
животных. Например, у лесной куницы энергия суточного рациона,
рассчитанная на 100 г веса, была выше (26.6-29.7 ккал), чем у норок и лесного
хорька со сходной массой тела (20.1-23.6 ккал).
Собранные материалы позволили также рассчитать показатель, отражающий
сезонную динамику энергетической ёмкости рациона куньих в природных условиях
(табл. 9), и выразить его в процентном отношении к максимальному уровню,
принятому за 100%. Для этого были использованы материалы по объёмному
соотношению основных групп потребляемых в природе кормов, их калорийности
и результатам контрольных кормлений зверьков в неволе. Оказалось, что у всех
видов потребность в высококалорийной пище существенно возрастает осенью, а
Таблица 9/Table 9
Динамика суточной потребности организма куньих в калорийности кормов в
природных условиях (по: Туманов, Смелов, 1980)
Dynamics of daily demands of mustelids (Mustelidae) in the calorific value of foods in
natural conditions
Показатели/ Parameters Ласка/ Mustela nivalis Г орностай/ Mustela erminea Лесной хорёк/ Mustela putorius Европ. норка/ Mustela lutreola Америк, норка/ Mustela vison Лесная куница/ Martes tnartes Выдра/ Lutra lutra Барсук/ Meles tneles
Ср. масса тела, г/ Average body 66 161 790 776 806 930 6400 8400
mass, g Весна, ккал/ Spring, kcal. 40.5 89 155.6 141.7 173.6 268.1 661.1 900.3
% 81 84 46 62 58 80 43 36
Лето, ккал/ Summer, kcal. 50.1 101.2 161.8 179 249.8 264.4 885.3 1416.5
% 100 96 48 78 83 79 57 57
Осень, ккал/ Autumn, kcal. 49.7 105.7 339.5 230.3 300.2 335.3 1551.4 2502.2
% 99 100 100 100 100 100 100 100
Зима, ккал/ Winter, kcal. 35.2 88.6 134 130.9 154.3 271.7 676.0 -
% 70 84 39 57 51 81 44 -
В среднем за сутки, ккал/ On the average per day, kcal В пересчёте на 43.9 96.1 197.7 170.5 219.5 284.9 943.5 1606.3
100 г массы тела/ Recalculated per 100 g of the body 66.5 59.7 25 22 27.2 30.6 14.7 19.1
mass
Примечание. Процент рассчитывали относительно максимального уровня калорийности
корма для данного вида, принятого за 100%.
38
зимой, наоборот, снижается. Эти изменения наиболее отчётливо проявляются у
лесного хорька, норок и выдры. Их следует трактовать как стойкую адаптивную
реакцию, направленную на экономное расходование созданных в организме
значительных энергозапасов. При падении уровня обмена веществ в холодный
период года резко снижается потребность в количестве и качестве добываемых
кормов, что и определяет благополучный исход зимовки хищников. У других
представителей семейства - обыкновенной ласки, горностая и лесной куницы,
для которых характерна интенсивность двигательных реакций и энергообмена,
необходимость в высококалорийном рационе ощущается практически во все
сезоны. Для них зимний период оказывается особенно напряженным, что
усугубляется еще и уменьшением в эти месяцы калорийности добычи. И, наконец,
крайний случай трофической адаптации представляет собой барсук, который в
связи с интенсивными сезонными изменениями уровня обмена веществ и
образованием мощного слоя жировых резервов может проводить зиму в состоянии
сна, не испытывая потребности в пище.
Взаимосвязь показателей кормовой специализации с морфологическими
особенностями пищеварительного тракта куньих находит своё подтверждение в
специальной литературе (Боголюбский, 1941; Соколов, 1941; Халилов, 1955;
Михеичев, 1955,1960). Согласно нашим данным, у куньих в целом прослеживается
тенденция к росту относительной длины кишечника с увеличением размеров тела
животных (табл. 8-9). Так, если у ласки индекс этого органа равен в среднем
275%, то у лесной куницы 459, а у росомахи 651%. В то же время данная связь
не пропорциональна и не абсолютна. Например, в ряду таких крупных
хищников, как барсук - росомаха - выдра, с увеличением длины тела размеры
кишечника закономерно снижаются. Это можно объяснить особенностями
питания видов - переходом от эврифагии (барсук) к некрофагии (росомаха) и
плотоядности (выдра).
Вообще повышение в рационе изучаемых хищников доли высококалорий-
ных и легкоусвояемых кормов сопряжено с уменьшением индекса кишечника.
Это можно видеть при сопоставлении его размеров у лесной куницы и барсука,
обитающих на Северо-Западе России и на Кавказе, где значительную долю в
питании южных форм составляют корма растительного происхождения. По
нашим данным, относительная длина кишечника у отловленных в горах самцов
лесной куницы была равна 514% (и=6), а барсука - 980% (и=8), т.е. заметно
больше таковой у особей из северного региона (табл. 10-11).
Обращают на себя внимание и значительные размеры кишечника росомахи.
Это соответствует положению, согласно которому хищники, питающиеся такими
трудно перевариваемыми кормами, как падаль, кости, разнообразные отбросы,
имеют более длинный со сложной конфигурацией кишечник, чем узко-
специализированные плотоядные виды (Юргенсон, 1956; Жеденов, 1965).
Половой диморфизм в размерах пищеварительного тракта прослеживался
у исследуемых куньих достаточно отчётливо. В целом у самцов индекс
кишечника больше, чем у самок. Достоверность различий по этому признаку
составляла у обыкновенной ласки, горностая и европейской норки 98-99%, у
39
Таблица 1 О/Table 10
Относительная длина кишечника и его отделов у самцов куньих
из северо-западного региона России (%) (по: Туманов, Смелов, 1980)
Relative length (in %) of intestine and its parts in males of mustelids in the North-West
region of Russia
Виды/ Species n Длина тела, см/ Length of body, cm Кишечник/ Intestine Тонкий отдел/ Small intestine Отношение тонкого отдела к кишечнику/ The ratio of small intestine to total intestine Толстый отдел/ Large intestine Ободочная кишка/ Colon
Ласка/
Mustela nivalis Горно- 12 18*1 294*8.2 267*18.2 91*0.5 27*1.8 16*1.6
стай/ Mustela erminea Лесной 9 26*0.4 363+12.2 326+11.4 90*0.4 37*1.5 24*1.1
хорёк/ Mustela putorius Европ. 24 35*0.5 500*12.4 464*12.3 93*0.2 36*0.8 22*0.7
норка/ Mustela lutreola Америк. 8 36*0.5 590*20.3 551+20.3 93*0.2 39*0.6 24*0.7
норка/ Mustela vison Лесная 10 37*0.5 483*14.3 445*13.1 92*0.3 38*1.4 24*0.8
куница/ Martes martes Барсук/ 33 43*0.5 474+7.8 436*8.5 92*0.2 38*1 24*0.8
Meles meles Росомаха 9 72*3.5 772*31.2 724+23.3 94*0.6 48*0.9 29*0.6
/Gulo gulo Выдра/ 5 79*1.5 680*45.1 - - - -
Lutra lutra 14 82*3.7 556*22.5 - - - -
лесной куницы и американской норки - 90-95%, а у остальных видов надежность
вывода оказалась невысокой. Анализ материалов показал, что выявленные
половые различия определяются прежде всего разной длиной у самцов и самок
тонкого отдела кишечника, где происходит переваривание основной массы
белковой пищи.
Особенности питания оказывают заметное влияние не только на общую длину
кишечника, но и на развитие его частей. Так, степень плотоядности хищников
в значительной мере определяет размеры тонкого отдела, отношение которого
40
Таблица 11/Table 11
Относительная длина кишечника и его отделов у самок куньих
из северо-западного региона России (%) (по: Туманов, Смелов, 1980)
Relative length (%) of intestine and its parts in females of mustelids in the North-West
region of Russia
Виды/ Species n Длина тела, см/ Length of body, cm Кишечник/ Intestine Тонкий отдел/ Small intestine Отношение тонкого отд. к кишечнику/ The ratio of small intestine to total intestine T олстый отдел/ Large intestine Ободоч- ная кишка/ Colon
Ласка/ Mustela 10 16±0.9 256±9.2 231+12.4 90±l 25±1.2 14±1.5
nivalis Горно- стай/ Mustela erminea Лесной 5 22+0.4 313±5.3 278±5.3 89±0.7 35±2 23+2.5
хорёк/ Mustela 12 32±0.7 486±10.3 451+10.1 93±0.4 35±2.5 23±1.8
putorius Европ. норка/ Mustela lutreola 9 33±0.4 529+8.1 489+8.4 92+0.3 40±l.l 24±1.4
Америк, норка/ Mustela 9 33+0.5 449±11.4 414+10.3 92±0.2 35±1.6 22±1.3
vison Лесная куница/ Martes martes 19 39+0.3 444±8.8 405±8.2 91+0.3 39±1.4 25±0.9
Барсук/ Meles 7 68±2.9 734±28.1 685+26.1 93±0.7 49+1.3 29±0.8
meles Росомаха/ Gulo gulo 3 74+1.3 622+67.8 - - - -
Выдра/ Lutra lutra 12 76±3.9 515±32.6 - - -
к кишечнику у всех куньих составляет 89-94%. В нём происходит основной
процесс ферментативного расщепления легко перевариваемых продуктов
питания, поэтому у мелких плотоядных видов (ласка, горностай, солонгой) он
оказался практически таким же, как у более крупных (куниц, соболя и барсука)
с их широким спектром питания.
Известно, что значительное содержание клетчатки и других трудно
перевариваемых компонентов пищи находит своё наибольшее отображение у
млекопитающих в морфологии толстого отдела и его частей (Воронцов, 1967;
Башенина, 1969; Большаков, 1972; Ивантер и др., 1985; Наумов, 1981; Rensh,
1948; Myrcha, 1964). В полной мере это относится и к представителям семемейства
куньих. Исследования показали, что виды, рацион которых в большой степени
41
состоит из низкокалорийных, грубых (в том числе и растительных) кормов,
характеризуются относительно более длинным толстым отделом, прежде всего
его ободочной кишкой, где происходит бактериальное брожение химуса и основное
всасывание минеральных солей и воды (табл. 10-11).
Адаптивная особенность пищеварительного тракта хищников просматри-
вается не только в оптимальном соотношении различных составляющих
кишечника, но и в специфической архитектонике его слизистой оболочки. Так,
абсорбционная поверхность тонкого отдела заметно возрастает за счёт
образования различных микроструктур, увеличивающих площадь всасывания,
в частности развития кишечных ворсинок.
У куньих, как и у других хищных млекопитающих (Халилов, 1955; Саблина,
1973; Саблина, Жарова, 1985), ворсинки достигают значительной высоты, которая
уменьшается в направлении от двенадцатиперстной кишки к толстому отделу. У
представителей разных видов их величина оказалась различной. Так, в средней
части двенадцатиперстной кишки высота ворсинок составляет у лесного хорька
930 мкм, у лесной куницы 855, у барсука 793, а у отловленной на Кавказе
каменной куницы 761 мкм. Заметно меньше их размеры у американской норки
(625 мкм) и ласки (540 мкм) (Смелов, 1985).
Следует также отметить, что у лесного хорька, ласки и американской норки
ворсинки имеют сравнительно простую пальцевидную форму, а у барсука, соболя,
лесной и каменной куниц они обладают поперечной складчатостью и фестонча-
тостью. Отмеченные складки иногда бывают очень глубокими, что придаёт
ворсинкам раскидистый или ветвистый вид. Всё это усложняет форму рельефа,
увеличивая у животных с широким спектром питания абсолютную площадь
поверхности ворсинок и кишечника в целом.
Изучению печени как самой крупной железы пищеварительной системы в
эколого-морфологических исследованиях отводится одна из ведущих ролей (Rensch,
1948; Яблоков, 1966; Шварцидр., 1968). Это обусловливается мультифункцио-
нальностью данного органа и в частности участием в процессах пищеварения
и обмена веществ. Большая изменчивость величины печени в семействе куньих
является результатом адаптивной радиации видов, которая привела их< к
расхождению по размерам и массе тела, определяя тем самым интенсивность их
обменных процессов. Видимо поэтому очень высокий индекс печени (5.5%)
как углеводного и жирового депо организма характерен для самого
маленького хищника - обыкновенной ласки, обладающей значительным
уровнем обмена веществ.
Зависимость размеров печени от массы тела прослеживается внутри
семейства куньих достаточно отчётливо (табл. 12), однако и здесь наблюдаются
некоторые особенности, связанные с пищевой специализацией изучаемых
хищников. Эту зависимость в разных отрядах класса млекопитающих
отмечали и другие исследователи (Юргенсон, 1956; Жеденов, 1965; Ивантер
и др., 1985).
Анализ собранных материалов по представителям семейства куньих
показал, что относительная масса печени оказалась выше у более плотоядных
42
Таблица 12/Table 12
Индекс печени у взрослых особей куньих северо-западного региона России (%)
The index of liver (%) in adult individuals of mustelids in the North-West region of Russia
Виды/Species Количество/ Number Индекс печени/ Index of liver
<?<? 1 25 <?<? 1 59
Ласка/ Mustela nivalis 64 27 5.4±0.6 5.7±1.1
Горностай/ Mustela erminea 54 35 4.6±0.8 4.7±1.1
Лесной хорёк/ Mustela putorius 84 25 4.4±0.6 5.0±1.2
Европ. норка/ Mustela lutreola 45 30 4.4+0.6 4.5+1.1
Америк, норка/ Mustela vison 96 57 4.3±0.6 4.4±0.9
Лесная куница/ Martes martes 48 37 3.4±0.6 3.5±0.7
Барсук/ Meles meles 16 11 3.3±1.2 3.4±1.8
Росомаха/ Gulo gulo 7 3 3.3±1.5 3.4±1.8
Выдра/ Lutra lutra 15 12 4.4±1.6 4.7+1.1
видов по сравнению с хищниками, потребляющими смешанные и мало-
калорийные корма. Например, среди крупных представителей семейства
прослеживается тенденция к увеличению данного показателя у выдры
(4.4-4.7) по сравнению с росомахой и барсуком (3.3-3.4%).
По нашим данным, у взрослых самцов соболя из Восточных Саян
относительная масса печени (3.3±О.8%) практически идентична таковой лесной
и каменной куниц (в среднем 3.4±0.6%), но ниже, чем у лесного хорька или
норок (в среднем 4.5±0.5%). Близкие к этим материалы по индексу печени у
соболя Зауралья получены В.Г. Монаховым (2001) - у самцов ее относительный
вес был равен 3.3%, а у самок - 3.7%. По имеющим сведениям (Барабаш-
Никифоров и др., 1968), самым высоким показателем относительной мас-
сы печени обладает такой стенофаг, как калан (в среднем 6.7±0.3%).
М.И. Урманов (1971) также подтвердил наличие крупной печени у самцов
(4.0%, п=9) и самок (5.2%, п= 10) этого вида. Понятно, что несколько меньший
индекс печени и других органов у самцов куньих по сравнению с самками
определяется большими размерами (массой) их тела.
Статистический анализ степени различий сравниваемых мелких видов
куньих, обитающих на Северо-Западе России, по индексу печени с помощью
критерия Стьюдента показал, что наименьший её размер характерен для
эврифагов и прежде всего для лесной куницы. Так, если в группе взрослых
особей достоверность различий между нею и лесным хорьком составляла
только 80%, то между нею и обыкновенной лаской 98%.
43
Зависимость размеров печени от особенностей питания отчётливо
прослеживается и на животных одного вида, но обитающих в разных
географических зонах. Так, если в северо-западном регионе России индекс
этого органа у взрослых самцов лесной куницы и барсука был равен в среднем
соответственно 3.4±0.6 и 3.3±1.2%, то в Закавказье, где эврифагийность зверей
заметно выше, только 2.7±0.2 и 2.8±0.6%. В Южном Приаралье относительная
масса печени у взрослых самцов барсука оказалась ещё более низкой -
2.3±1.2% (Реймов, 1972).
Таким образом, в процессе эволюции у куньих установилась прочная связь
между кормовой специализацией видов и строением их желудочно-кишечного
тракта. Степень плотоядности во многом определяет строение, а также общую
длину кишечника и его отделов. Количество и калорийность потребляемой
белковой пищи оказывают влияние прежде всего на размеры тонкого отдела и
его частей, а сочетание животных и растительных кормов - на длину ободочной
кишки и всего толстого отдела. Кроме того, хищникам, склонным к эврифагии,
свойственно увеличение активной всасывающей поверхности ворсинок в тонком
отделе кишечника за счёт усложнения их конфигурации. В то же время плотоядное
питание обусловливает значительные размеры печени при менее сложном рельефе
абсорбционной поверхности ворсинок слизистой оболочки кишечника.
Семейство собачьих (Canidae)
Среди представителей данного семейства енотовидная собака (Nyclerenles
procyonoides Gray) обладает наибольшей эврифагийностью. В южных областях
Северо-Запада России, где она весьма обычна, рацион зверей составляют корма
животного происхождения, хотя остатки растительной пищи встречаются в
биопробах регулярно (табл. 13).
Мышевидные грызуны представлены в питании енотовидной собаки серыми
и рыжими полёвками. Заметно реже она потребляет полевых и лесных мышей.
Остатки зайцев, ондатры и насекомоядных встречаются сравнительно редко. <
Значение птиц в рационе хищников также невелико. Летом они в небольшом
количестве ловят птенцов, а осенью подбирают подранков утиных и тетеревиных
птиц, не найденных охотниками. Во время жировок на берегах водоёмов поедают
моллюсков, мелкую рыбу, лягушек, иногда яйца куликов, чаек и других птиц,
отыскивают водных и почвенных насекомых. Лягушек они потребляют с
большой охотой. В отдельные годы в рационе увеличивается доля рыбы, которую
звери собирают в поймах рек и по отмелям крупных водоёмов. Заметное место
в их питании в осенне-зимний период имеет падаль, прежде всего остатки
копытных, отстреливаемых охотниками или добытых волками.
Из растительных кормов енотовидная собака предпочитает зёрна злаков (овса,
ржи), ягоды кустарничков (черники, голубики, брусники), плоды диких фруктовых
деревьев (прежде всего яблони) и в значительно меньшей степени поедает
корневища, луковицы, вегетативные части растений (осок, злаков и т.п.).
44
Таблица 13/Table 13
Состав пищи енотовидной собаки (//=154) в октябре-декабре
The food composition in the racoon-dog (Nyctereutes procyonoides) (//=154)
in October-December
Вид корма/ Встречаемость/
Food component Percentage, %
Млекопитающие/Mammals 20.1
Птицы/Birds 12.3
Яйца птиц/Eggs of birds 7.8
Падаль/Carrion 20.8
Амфибии (лягушки)/Атр1пЫап5 13.5
Рептилии (ящерицы)/ЯерЦ1е5 1.9
Рыбы/Fish 5.2
Н асекомые/Insects 6.5
Моллюски/Molluscs 1.9
Дождевые черви/Earthworms 2.6
Растительные корма/Vegetarian food 9.1
Сезонная смена пищевых компонентов выражена достаточно ярко и в
значительной мере определяется различной доступностью кормов в течение
года. Так, в ноябре-декабре (в дни бодрствования) в питании хищников
преобладают мышевидные грызуны (30.4%) и зёрна злаков (26.0%), которые
они собирают на полях после уборки урожая. Из-за недостатка полноценной
пищи в это трудное время звери используют также падаль (6.7%), выкопанные
из-под снега желуди и ягоды.
Весной и летом енотовидные собаки благодаря своей всеядности хорошо
обеспечены кормом. Потребление мышевидных грызунов и рептилий несколько
уменьшается (21.8%), зато возрастает кормовая роль насекомых (29.5%), ягод
(27.3%), амфибий (10.3%) и птиц (6.1%). Чаще они едят рыбу (2.4%), моллюсков
(2%), червей и корневища растений (0.6%). Из насекомых енотовидная собака
больше всего добывает жуков: хрущей, жужелиц, долгоносиков, навозников.
Ранней осенью заметно увеличивается потребление лягушек, мышевидных
грызунов и растительных кормов (Морозов, 1953). Поданным М.Х. Геллера
(1959), осенью встречаемость мышевидных грызунов в желудках и экскре-
ментах хищников составляла 48.3%, амфибий 19.1, растительных остат-
ков 68.3%. Из последних наиболее часто они потребляют ягоды брусники,
клюквы, крушины, черники, зерна овса, ржи, ячменя, а также семена льна и
желуди.
Следует отметить, что вред, причиняемый енотовидной собакой спортивно-
охотничьему хозяйству, не так велик, как принято думать. Действительно, летом
она может поедать молодых зайчат, яйца и птенцов тетеревиных птиц и уток,
45
однако эти корма не являются для неё основными. Гораздо проще ей добыть
имеющихся в этот период в большом количестве лягушек, насекомых и грызунов,
а в истреблении промысловых животных енотовидная собака повинна не более,
чем любой другой хищник, например лисица (Данилов и др., 1979).
Количество потребляемой хищником пищи заметно меняется по сезонам
года. У добытой в августе молодой самки (Псковская обл.) весом 1.8 кг в
желудке находились остатки лягушек весом 100 г и 3 навозных жука. У другой
молодой самки с массой тела 2.5 кг, добытой там же в октябре, желудок был
наполнен остатками яблок и лягушек и весил 280 г. В сентябре-октябре, в
период интенсивного нагула, потребность в кормах как у молодых, так и у
взрослых особей наибольшая. Это хорошо видно из анализа содержимого
наполненных желудков взрослых зверей (и=12), добытых на одной территории
(Локнянский район Псковской обл.) в разные месяцы. В августе вес
содержимого желудков составлял 160-200 г, в сентябре-октябре 140-500, а в
ноябре 95-110 г.
Интересные данные по сезонным изменениям потребности в кормах были
получены в Санкт-Петербургском (Ленинградском) зоопарке. По этим сведениям
в среднем одна особь в вольерах съедает в январе (в период бодрствования) 340 г,
в марте 465, а осенью (сентябрь) 735 г мясо-рыбного корма.
В тесной связи с динамикой веса потребляемой пищи находятся и колебания
массы тела зверей (рис. 15). Её изменения в течение года весьма показательны для
характеристик периодических сдвигов в уровне теплообмена. Весной и летом, когда
показатели энергообмена наибольшие, животные имеют минимальную массу тела.
С конца лета отмечается постепенное её нарастание, продолжающееся до
октября-ноября, когда вес зверей достигает максимума. Осенью енотовидные
собаки интенсивно питаются и жиреют, масса жировой ткани в этот период
нередко превышает у них 30% от общего веса.
В декабре-январе вес хищников уменьшается, однако расходование
энергетических резервов в начале зимы происходит очень экономно из-за
сравнительно низкого обмена веществ в организме. Во второй половине зимы
жировые запасы расходуются более интенсивно, и показатели массы тела зверей
резко падают. Эти изменения находятся в тесной связи с весенним подъёмом
уровня энергообмена и активизацией деятельности половых желёз. Следует
также отметить, что у енотовидной собаки, как и у большинства куньих, имеет
место небольшой весенний скачок веса, который обычно отмечался в мае-
июне, реже в апреле.
По свидетельству Ю.С. Заболотских (1983), в условиях неволи енотовид-
ные собаки способны переносить длительное зимнее голодание, экономно
расходуя свои жировые резервы. При отсутствии пищи в течение 130 дней
случаев падежа или патологических явлений в их организме не наблюдалось.
По наблюдениям автора, лисицы оказались наименее приспособленными к
отсутствию корма: они способны зимой прожить без него в среднем около
месяца. Продолжительность же жизни песцов при тех же условиях была почти
в 2 раза больше. Однако максимальной устойчивостью к голоданию все-таки
46
i п ш iv v м vn vm ix х xi хп
Рис. 15. Сезонные изменения массы тела четырех взрослых енотовидных собак
в условиях неволи (средние многолетние данные).
1 - самки; 2 - самцы.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, кг.
Fig. 15. Seasonal changes of the body mass in four adult racoon-dogs
(Nyctereutes procyonoides) under conditions of captivity (average multiyear data).
1 - females; 2 - males. Abscissa - months; ordinate - body mass, kg.
отличались енотовидные собаки, которые почти в 5 раз превосходили по этому
показателю лисиц и при этом не теряли способности размножаться весной.
Песец (Alopex lagopus L.) - мало разборчивый в пище, в основном
плотоядный хищник. Растительные корма существенного значения в его жизни
не имеют (табл. 14). В континентальных тундрах Кольского полуострова он
питается преимущественно мелкими грызунами: рыжими и серыми полёвками
или норвежским леммингом. Обилие грызунов в большой степени и определяет
численность этого хищника. К основным его кормам следует отнести также
белую куропатку, мелких воробьиных и водоплавающих птиц, их яйца,
остатки погибших оленей и снулую рыбу, которую он собирает по берегам
водоёмов. Наблюдения, проведенные нами на Северном Таймыре и в
Большеземельской тундре, показали, что основу пищи песца здесь составляют
лемминги, серая и красная полёвки, молодняк птиц и зайца-беляка. Он
регулярно посещает колонии гусей, места гнездования уток, куликов и чаек,
где ловит птиц и разоряет их гнёзда. На побережье Баренцева моря и озера
Таймыр песцы в основном питаются их дарами - выброшенными волной снулой
рыбой, ракообразными, трупами птиц, а на морских отмелях - тюленей и
других водных млекопитающих. На северном берегу озера Таймыр нам
неоднократно приходилось наблюдать «мышкующих» или скрадывающих
песцов у нор сибирских и копытных леммингов. Во время рыбного промысла
в годы с низкой численностью леммингов они летом, не боясь человека, бегали
около вынутых из воды сетей и выхватывали оставленную мелкую рыбу.
47
Таблица 14/ТаЫе 14
Встречаемость основных кормов в летнем рационе песца ("/и)
Occurrence of main foods (%) in the summer ration of arctic fox (Alopex lagopus)
Вид корма/Food component Континентальные тундры Кольского п-ова, //-51 (Золотов, 1940)/ Continental tundra of the Kola Peninsula Большеземельские тундры, побережье Баренцева моря, //=55 (наши данные)/ Bolshezemel’skaya tundra, the coast of the Barents Sea
Мелкие грызуны/ Small rodents 80 7 6.3
Падаль (северный олень, нерпа)/ Carrion (Rangifer tarandus, Phoca 18 9.1
hispida) Птицы/ Birds 16 10.9
Ракообразные/ Crustaceans 2 3.6
Моллюски/ Molluscs 2 0
Растительная пища (ягоды)/Вегпез 4 5.5
Зимой песец очень осторожен. В это время года он наиболее активен:
выкапывает из-под снега мышевидных грызунов, ловит зайцев и белых
куропаток. Следуя за росомахой или белым медведем, подбирает остатки их
добычи. В период сильных морозов, в январе-феврале, он чаще выходит на
припай морского побережья и питается выбросами моря - павшей водопла-
вающей птицей, рыбой и останками морских млекопитающих, около которых
могут подолгу жить даже несколько зверьков.
В биопробах по питанию песцов, обитающих в тундрах Кольского
полуострова (Золотов, 1940), практически во все месяцы года обнаруживались
части грызунов, павших оленей и белой куропатки. Кости и перья водо-
плавающих птиц встречались в летнее время (май-сентябрь), а их яйца-только
в июне. В трудное время года (декабрь-март) большое значение в рационе
животных имеет падаль. В конце августа-сентябре недалеко от нор попадались
грибы (сыроежки), частично объеденные песцом, и экскременты с остатками
ягод. Однако даже в годы низкой численности полёвок и леммингов эти грызуны
служат основным кормом хищника.
На европейском Севере (Шиляева, 1972) песец выступает как типичный
миофаг. Свойственная ему полифагия больше проявляется в условиях зимней
бескормицы. Основу его питания здесь составляет сибирский лемминг
(встречаемость 37.2%) и узкочерепная полёвка (17.4%), от обилия которых
зависит состояние популяции песца. Из прочих кормовых компонентов обычны
птицы, особенно белая куропатка (16.8%), их яйца (3.7%), рыба (6.9%), заяц-
беляк (2.6%) и другие.
Потребность в пище у песцов заметно изменяется по сезонам года.
Например, в Санкт-Петербургском (Ленинградском) зоопарке наибольшее
количество корма (в сутки) им требовалось осенью, а минимальное - зимой. В
среднем суточная норма пищи на одного зверька была равна в сентябре 500 г,
январе 340, мае 390 и в июне 360 г мясо-рыбного корма. Эти сезонные измене-
48
Рис. 16. Сезонная динамика массы тела трех взрослых песцов в условиях неволи
(средние многолетние данные).
1 - самки; 2 - самец.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, кг.
Fig. 16. Seasonal dynamics of the body mass in three adult arctic foxes (Alopex lagopus)
under conditions of captivity (average multiyear data).
1 - females; 2 - males. Abscissa - months; ordinate - body mass, kg.
ния потребности в пище способствуют пониманию особенностей кормо-
добывающей деятельности песцов в разное время года. Анализ имеющихся
материалов свидетельствует о том, что сезонные колебания их массы тела
полностью согласуются с изменениями показателей суточного потребления
кормов. Осенью и весной вес у песцов бывает наибольшим, а летом снижается
до минимального уровня. Так, по наблюдениям за животными, содержащимися
в вольерах, и самцы, и самки песца имели максимальную массу тела весной и
особенно в осенний (предзимний) период (рис. 16). При длительном голодании
у пушных зверей (Заболотских, 1996, 2000) наиболее заметно снижались
весовые индексы брыжейки, печени и особенно внутреннего и подкожного жира.
В процессе зимнего голодания индекс одного только внутреннего жира
уменьшается у взрослых песцов с 4.5 до 2%, а у енотовидной собаки - с 2.5
до 1.3%.
Лисица (Vulpes vulpes L.) в европейской части России, как и песец,
растительные корма потребляет в небольшом количестве. Основу её рациона
во все сезоны составляют мелкие млекопитающие и птицы (рис. 17). В
Мурманской области в пробах по питанию представителей вида встречаемость
мышевидных грызунов достигает 93%, а тетеревиных птиц - до 40%
(Насимович, 1948). В Карелии в их рационе большое значение имеют
мышевидные грызуны (60-80%), заяц-беляк (12-24%), боровая дичь (8-30%)),
насекомые (15-17%>), снулая рыба и другая падаль (6-30%), реже ягоды
(Ивантер, 1969).
В южных областях зоны, поданным О.С. Русакова (Данилов и др., 1979),
основными жертвами лисицы становятся мышевидные грызуны (табл. 15). Среди
них по встречаемости преобладает обыкновенная полёвка (21.2%). Мышей и
49
Рис. 17. Годовой спектр питания лисицы на Северо-Западе России (по: Данилов,
Русаков, Туманов, 1979).
1 - мышевидные грызуны; 2 - зайцы; 3 - падаль; 4 - птицы; 5 - прочие корма.
По оси абсцисс: 7 - в Мурманской обл.; II- в Карелии; III- в Ленинградской обл.;
IV-в Новгородской обл.; К- в Псковской обл.; по оси ординат - встречаемость,
% от общего количества проб.
Fig. 17. Year spectrum of red fox feeding (Vulpes vulpes) in the North-West of Russia.
1 - small rodents; 2- hares; 3 - offal; 4 - birds; 5 - other food.
Abscissa: I - in the Murmansk Prov., II - in Karelia; III - in the Leningrad Prov., IV -
in Novgorod Prov., V- in Pskov Prov.; ordinate - occurrence, % from total number of
samplings.
насекомоядных хищник поедает заметно реже (0.3-0.6% встреч). Насекомые
представлены преимущественно жуками (18.2%). Среди них преобладают
навозники (5.8%) и листоеды (3.7%). Растительные корма встречаются в
весеннем рационе (8.6%) нечасто. Обычно они представлены различными
семенами, ягодами или вегетативными частями злаков.
Роль насекомых, гнездящихся на земле тетеревиных и воробьиных птиц,
амфибий, рептилий и растительных остатков заметно возрастает в летнем
рационе лисицы. В осенне-зимней ее добыче преобладают мышевидные
грызуны, ондатра, молодняк зайцев, а также падаль (Новиков и др., 1970). По
материалам С.А. Корытина (1968), к весне отмечается заметное уменьшение
роли мышевидных грызунов. Так, в экскрементах лисицы в декабре-феврале
их остатки были отмечены в 65% проб, марте - в 62 и в апреле - в 40% проб. В
целом, по свидетельству автора, в Кировской области среди основных ее
кормов встречались мышевидные грызуны (58.8%), заяц-беляк (39.2%), птицы
(35.3%) и падаль (29.4%). Семена злаков, плоды и ягоды кустарничков в
просмотренных пробах также попадались регулярно. Спрятанные ранее остатки
еды наиболее часто посещались в феврале и марте, т.е. в месяцы наибольших
трудностей с кормом (глубокий снег, наст и пр.). В декабре на 10 км следов
раскопанных лисицами кладовых приходилось в среднем 0.6, в январе 0.3,
феврале 1.6, марте 1.1 и в апреле 0.3.
50
Таблица 15/Table 15
Встречаемость основных кормов в весеннем рационе лисиц (/>=325)
из Ленинградской и Псковской областей (по: Данилов и др., 1979)
Occurrence of main foods in the spring ration of foxes (Vulpes vulpes) (n=325) in the
Leningad and Pskov Provinces
Вид корма/ Food component Встречаемость/ Percentage, %
Мелкие грызуны/Small rodents 98.9
Насекомоядные/Insectivores 0.3
Птицы/Birds 2.8
Насекомые/Insects 18.2
Растительные остатки/Vegetarian food 8.6
Таблица 16/Table 16
Состав зимней и ранневесенней пищи волка (л=780) в Псковской области
в 1968-1977 гг. (по: Данилов и др., 1979)
The composition of winter and early spring food of wolf (Cams lupus) (n=7Sff) in the Pskov
Province in 1968-1977
Вид корма/ Food component Встречаемость/ Percentage, %
Мелкие грызуны/Small rodents 7.9
Ондатра/Muskrat 0.5
Заяц-беляк/Lepus timidus 25
Мелкие и средние хищные звери/ 3
Small and average predators species
Домашние животные/Livestock 9.2
Лось/Moose 79.5
Кабан/Wild boar 3.1
Птицы/Birds 0.6
Рыба/Fish 0.1
Растительные остатки/Vegetarian food 12.8
В Карелии численность лисицы определяется не столько обеспеченностью
каким-либо одним кормом, сколько запасами и доступностью всех основных
зимних кормов - мышевидных грызунов, зайца-беляка и, в меньшей степени,
тетеревиных птиц. При этом связь между обилием мышевидных грызунов и
численностью лисицы бывает менее чёткой, чем между нею и зайцем (Ивантер,
1969).
51
Рис. 18. Сезонные изменения массы тела взрослых лисиц (5^ и 4$>)
в неволе (средние многолетние данные).
1 - самцы; 2 - самки.
По оси абсцисс - время, месяцы; по оси ординат - масса тела, кг.
Fig. 18. Seasonal changes in the body mass in the adult red foxes (5 g and 4$>) under
conditions of captivity (average multiyear data).
1 - males; 2 - females. Abscissa - months; ordinate - body mass, kg.
У лисиц обоего пола, содержащихся в вольерах, наибольшая потребность
в пище наблюдалась в сентябре - октябре, когда они за сутки съедали до 450-
500 г мясного корма. Зимой им требовалось меньше еды - в среднем 250-280 г,
а летом - до 330-350 г. Сезонная изменчивость массы тела у подопытных
животных также прослеживалась достаточно отчетливо (рис. 18). В июне они
были наименее упитанными (в среднем вес самцов 6.45 кг, самок - 4.15 кг).
Затем масса их тела постепенно увеличивалась, достигая максимума к
декабрю. В это время самцы уже весили 7.35, а самки - 4.91 кг. В январе -
апреле, когда у лисиц активно протекает репродуктивный процесс, масса их
тела заметно снижалась, а в мае, в период бурной линьки, вновь несколько
возрастала.
Волк (Canis lupus L.). Как и в других регионах, основу пищи волков на
Северо-Западе России составляют копытные. Если в Мурманской области в
зимне-весенний период они добывают преимущественно диких и домашних
оленей, реже лося, то в Карелии и других территориях региона последний
преобладает в кормовом рационе хищников (Троицкий, 1974; Новиков и др.,
1970; Данилов и др., 1985). Так, по материалам О.С. Русакова (Данилов и
др., 1979), в Псковской области доля лосей в исследованных экскрементах
волков составляла 79.5% по встречаемости (табл. 16) и 47% по массе.
Существенную роль в зимнем и ранневесеннем питании этих хищников имели
52
Рис. 19. Сезонные изменения питания волка в Карелии (по: Данилов, Русаков,
Туманов, 1979).
1 - дикие копытные (лось, северный олень); 2 - домашние животные,
преимущественно овцы; 3 - заяц-беляк; 4 - птицы.
По оси абсцисс: I - весна; II -лето; III - осень; IY - зима;
по оси ординат - встречаемость, % от общего количества проб.
Fig. 19. Seasonal changes in the feeding of wolf (Canis lupus) in Karelia.
1 - wild ungulates (moose, reindeer); 2 - livestock, mostly sheep;
3 - alpine hare (Lepus timidus); 4 - birds.
Abscissa: I - spring, II - summer, III - autumn, IY - winter; ordinate - occurrence,
% from total number of samplings.
зайцы (встречаемость 25%, масса 15.4%), мышевидные грызуны (соответ-
ственно 7.9 и 4.7%), а также домашние животные - собаки, овцы и козы (9.2 и
6%). Незначительное место в их питании занимали кабан, ондатра, тетеревиные
птицы, рыба, мелкие и средние по размеру хищные звери. Довольно часто
(встречаемость 12.8%) в остатках пищи волков попадалась сухая трава,
преимущественно осока, потребление которой, вероятно, способствует
очищению желудочно-кишечного тракта от гельминтов. Однако общая ее доля
невелика и составляла всего 0.6% от всего объема экскрементов.
Сезонная динамика основных компонентов пищи просматривается у волка
достаточно отчётливо. В Карелии (Данилов и др., 1979,1985) летом и осенью
частота потребления волками лосей не превышала 10-14.3%, а весной и зимой
колебалась в пределах от 42 до 59.3% (рис. 19). Однако летом и осенью заметно
чаще, чем зимой и весной, в пище волков встречались домашние животные,
преимущественно овцы и собаки (летом 80, осенью 71.4%). Зимой и весной их
доля составляла только 16.3 и 33.3%. Потребление зайцев и птиц менялось по
сезонам в меньшей степени.
Следует также заметить, что летнее питание волка на Северо-Западе России
изучено еще недостаточно. Судя по отдельным наблюдениям, в частности по
нахождению остатков пищи у логова, его состав разнообразнее, чем зимой.
53
Рис. 20. Сезонные изменения массы тела взрослых волков (2^ и 2§>) в условиях
неволи (средние многолетние данные).
7 - самцы; 2 - самки.
По оси абсцисс - месяцы; по оси ординат - масса тела, кг.
Fig. 20. Seasonal changes in body mass in adult wolves (Cants lupus) (2^ и 2<J>) in the
conditions of captivity (average multiyear data).
7 - males; 2 - females. Abscissa - months; ordinate - body mass, kg.
Более существенное кормовое значение имеют птицы. На севере региона, в
тундре, волк в массе уничтожает леммингов, лесных полёвок, гнездящихся на
земле птиц. В южных областях изучаемой территории в его летнем питании
увеличивается доля птиц, грызунов и растительных кормов (черника, брус-
ника). Однако наибольшее значение приобретает молодняк копытных и зайцев,
численность которых в этот период бывает высокой. Взрослые копытные в
летнее время редко становятся добычей хищников.
В летне-осеннее время волк употребляет как полноценный корм плоды
ряда растений: ягоды черники, брусники, ежевики, калины, боярышника,
рябины и шиповника, а также плоды диких фруктовых деревьев. В северных
регионах встречаемость растительной пищи (прежде всего ягод кустарничков)
в рационе волков не превышает 5-6%. По мере продвижения с севера на юг
ареала вида значение растительной пищи в жизни хищников возрастает
(Руковский, Куприянов, 1972; Калецкая, 1973; Руковский, 1985).
Как и у других представителей семейства, потребность в пище у волков во
многом определяется сезоном года (Мертц, 1953). В Санкт-Петербургском
(Ленинградском) зоопарке осенью суточный рацион взрослых особей состав-
лял 2.5-3 кг мяса с костями, а зимой и летом - не более 2 кг. По данным
И.С. Козловского (1986), в среднем объем желудка волка составляет 6.9 л, а
масса его содержимого - 474 г. Максимальная величина разовой массы
перевари-ваемой пищи достигает 3.3 кг. По свидетельству автора, зимой волки
54
Таблица 17/Table 17
Сравнительная характеристика массы печени и длины кишечника у некоторых
взрослых представителей семейства Canidae со сходной массой тела
Comparative characteristics of liver mass and intestine length in some adult representatives
of the family Canidae with a similar body mass
Виды/ Species Пол/ Sex Масса тела, кг/ Body mass, g Масса печени/ Mass of liver Длина тела, см/ Length of body, cm Длина кишечника/ Length of intestine
г/g % к массе тела/ % of body mass см/ cm % к длине тела/ % of body length
Енотовидная собака/ 3 6.6 155 2.2 69 230 333
Nyctereutes 3 7 230 3.3 67 226 337
procyonoides $ 4.8 170 3.5 73 252 345
Песец / 3 4.5 165 3.7 63 202 321
Alopex lagopus 5 3.7 140 3.8 65 210 323
Лисица 3 5 200 4 70 212 303
обыкновен./ Vulpes vulpes 5 4.5 190 4.2 68 196 288
часто голодают, и пережить этот трудный период им помогают жировые
отложения. Зимой они имелись практически у всех осмотренных взрослых
особей, и лишь в 5.3% случаев упитанность зверей была низкой. Общая масса
подкожного и внутреннего жира достигала у добытых животных 15.5% от массы
туши.
В сезонных изменениях веса волков имеются некоторые общие черты и вместе
с тем определённые половые различия (рис. 20). Как у самцов, так и у самок
резкое падение массы тела наблюдалось в январе, во время гона, когда
энергозатраты организма резко возрастают. Последующее увеличение этого
показателя в феврале и некоторое снижение его к июню имели место у зверей
обоего пола. Однако если у самцов отмечался скачкообразный характер
изменения массы тела, то у самок кривая веса менялась сравнительно плавно. К
декабрю по сравнению с июнем масса тела зверей за счёт лучшей упитанности
увеличивалась в среднем на 10-15%.
Особенности питания волков отражают и специфику строения их пище-
варительного тракта. У взрослых особей среднерусского волка из Смоленской обл.
(Саблина, Жаркова, 1985) общая длина кишечника была равна 374-520 см, что сос-
тавляло 304-386%от дпинытела животных. Тонкий отдел кишечника, где происходит
основное всасывание белка у этих хищников, также имел значительные размеры -
317-340 см. Его слизистая образует по всей поверхности крупные ланцетовидные
55
Таблица 18/Table 18
Масса тела и размерно-весовые показатели некоторых внутренних органов
енота-полоскуна в Дагестане (по: Гинеев, 1972)
The body mass and size-weight parameters of some inner organs in the racoon
(Procyon lotor) in the Daghestan
Показатели/ Parameters Единица измерен./ Абсолютная величина/ Absolute value Относительная величина, “/</ Relative value
Unit п 33 п 9? 33 99
Масса тела/ Body mass кг/kg 29 6.1±0.3 33 4.7+О.З
Длина/Length of: тела/body ем/cm 29 56.3±1 33 51.2+0.8
кишечника/ intestine см/cm 25 4О3.6±2 22 352.0+1 716.9±24.9 687.5±16.8
Macca/Mass of:
ПОДКОЖНОГО жира/ кг/kg 24 1.4+0.4 21 1.2+0.4 21.1±1.7 22.2+1.6
subcutaneous fat
большого сальника/ г/g 21 81.0+6.8 19 75.6+8.2 1.6+0.2 1.6+0.1
caul печени/ liver г/g 25 169.5±11.6 23 107.7+4.4 2.8±1.4 2.3±1.4
поджелудочной железы/ г/g 24 14.6+0.7 24 11.4+0.6 0.2+0.1 0.2+0.1
pancreas
желудка без содержимого/ stomach without г/g 21 37.1+1.9 22 27.6+1.4 5.8+O.7 5.6+0.5
content
ворсинки, высота которых увеличивается в дистальном направлении. Если в начале
двенадцатиперстной кишки они не превышают 300 мкм, то ближе к тощей высота
ворсинок достигает уже 600-800 мкм, обеспечивая интенсивное всасывание
питательных веществ. Анализ толстого отдела кишечника показал его большую длину
у волков по сравнению с собаками сходных размеров, что авторы связывают с
постоянным потреблением дикими хищниками растительной пищи. Возможно, этот
же фактор определяет у лесных волков из северных регионов, где в летнее время в их
рационе растительные корма играют важную роль, и большую общую длину
кишечника. У хищников, добытых на европейском Северо-Востоке, этот показатель
был равен 550-675 см, что составляет 478-542% длины их тела (Полежаев, 1998).
Различия в индексе печени, способной депонировать питательные вещества (в
основном гликоген) на случай изменения состояния кормовой базы и ухудшения
56
условий существования, прослеживаются у животных из разных регионов
недостаточно отчётливо. Так, у волков в возрасте 1-3 лет и более из северных
территорий России (л= 18) индекс печени был равен у самцов 1.2-2.1%, а у самок
2.5-2.6% (Полежаев, 1998). У хищников из Беларуси (/г=76) данный показатель
составлял для самцов 2.4 ± 0.82 и для самок 2.5 ± 0.86% (Козло и др., 1985). У
добытых в Псковской области двух взрослых самцов (масса тела 38 и 40 кг, длина
120 и 126 см) индекс кишечника был равен 418 и 406%, печени 2.7 и 2.8%.
Известно, что у животных, питающихся разнообразными кормами
(эврифагов), печень менее массивна, а кишечник заметно длиннее, чем у
плотоядных видов (Юргенсон, 1951; Жеденов, 1965, и др.). В полной мере эта
закономерность прослеживается и на примере представителей семейства Canidae
со сходной массой тела (енотовидная собака, лисица, песец). Среди них
енотовидную собаку можно отнести к типичным эврифагам, что обусловливает
наличие у неё сравнительно низкого индекса печени и большую относительную
длину кишечника (табл. 17), однако эти тенденции проявляются не столь отчётливо,
как предполагалось ранее (Гептнер и др., 1967; Морозов, 1970). Между тем
С.Н. Боголюбский (1939) отмечает, что енотовидные собаки как эврифаги по
размеру кишечника заметно отличаются от песцов и лисиц. По данным автора,
длина тонкого отдела кишечника превышает размеры тела у енотовидной собаки
в 5.62 ± 0.03, у лисицы 2.95 ± 0.07 и у песца в 3.17 ± 0.04 раза. Кроме того, у
енотовидной собаки в строении зубной системы прослеживается менее выраженная
плотоядность по сравнению с другими представителями семейства.
Семейство енотовых (Procyonidae)
Акклиматизация енота-полоскуна (Procyon lotor L.) на юге СССР началась
в 1936 г. В настоящее время он широко распространен в Дагестане и
Азербайджане. Зверьки питаются смешанной пищей, отдавая предпочтение
более обильным и доступным в данный период ее компонентам. По характеру
питания енот-полоскун близок к барсуку. На Кавказе нам приходилось
добывать зверей (и=22), в желудочно-кишечном тракте которых находились
остатки яблок, мушмулы, орехов, алычи, ежевики, насекомых, лягушек,
ящериц, мышевидных грызунов, воробьиных птиц и фазана.
По имеющимся сведениям (Руковский, 1953, 1963; Г. Новиков, 1956), на
территории Кавказа встречаемость насекомых в рационе енота состаляет 82%,
растительных остатков 64, амфибий 53, рептилий и моллюсков - по 13, птиц
12, млекопитающих 5.5, рыбы и раков 5%. Из позвоночных животных хищники
чаще потребляют полевок, мышей, мелких воробьиных птиц, птенцов уток,
черепах, ящериц и лягушек, из насекомых - жуков, чешуекрылых, прямо-
крылых, стрекоз и ручейников. С большой охотой еноты употребляют в пищу
яйца птиц и орехи, плоды дикого винограда, яблони, груши, алычи, терна,
ягоды шиповника и боярышника. Обитая вблизи селений, они могут добывать
ночующую здесь на низких деревьях сельскохозяйственную птицу, которую
обычно поедают в убежище - дупле или густой кроне древесных пород. Добыв
57
жертву у водоема, зверек может прополоскать ее в воде, прежде чем съесть,
что и послужило основанием для его видового названия.
Весной и в зимние дни, когда еноты бывают активны, они больше
потребляют корма животного происхождения, а летом и осенью - раститель-
ного. Взрослые самцы (6 особей), отловленные нами в августе-октябре, весили
4.5-5.6 кг. К началу зимнего сезона за счет увеличения жировых запасов масса
тела хищников заметно возрастает, и вес добытых 4-х енотов того же пола
составлял уже 6-6.8 кг. Истощенные или не набравшие осенью должного
количества жировых отложений, чаще всего молодые звери долгое время не
впадают в зимний сон, занимаются бродяжничеством и довольно легко
попадают в капканы местных охотников.
Как сообщает А.М. Гинеев (1972), отловленные для расселения в августе-
сентябре в Дагестане еноты весили: самцы (и=8) - 4.3±0.6 кг, а самки (и=7) 3.3±0.6 кг.
Звери, добытые в осенне-зимний сезон, были заметно крупнее, имели
существенные жировые резервы (табл. 18). Как и другие хищники-эврифаги, они
обладали сравнительно небольшим индексом печени, поджелудочной железы и
длинным кишечником, близким по размеру к таковому барсука.
Семейство медвежьих (Ursidae)
Среди представителей семейства медвежьих, обитающих на европейском Севере,
бурый медведь (Ursus arctos L.) отличается большей эврифагийностью по
сравнению с белым (Ursus maritimus Phipps), рацион которого почти целиком
состоит из кормов животного происхождения. В период бодрствования, в теплые
сезоны года, бурый медведь предпочитает питаться зелеными частями и
корневищами растений, ягодами, плодами диких яблонь и лещины, желудями,
насекомыми и их личинками. В конце лета медведей можно встретить на полях
поспевающего овса. Его остатки постоянно доминируют среди набора кормов,
обнаруженных в желудках зверей, добытых вблизи сельскохозяйственных угодий
в летне-осенний и предзимний периоды (табл. 19). Доля ягод кусгарничкорых
пород (черника, брусника, клюква, голубика) и рябины в осеннем рационе бурого
медведя также достаточно велика, особенно в урожайные на них годы. Средняя
масса содержимого желудков в августе-ноябре была равна у самцов 3.8 и у
самок 3.6 кг. В декабре у еще не залегших в берлоги зверей она снизилась
соответственно до 3.0 и 1.8 кг. Максимальное количество корма, обнаруженное
в желудках самцов и самок, 10-12 кг. Животные с таким содержанием
растительной пищи в желудочно-кишечном тракте были добыты в основном в
августе-сентябре, в период интенсивного накопления жировых резервов. При
содержании в условиях неволи (в Санкт-Петербургском зоопарке) в августе-
октябре среднесуточный рацион бурых медведей состоял из 5.8 кг смешанного
корма (мясо, рыба, хлеб, яблоки, овощи), тогда как весной они съедали его не
более 4 кг в день.
Мясной корм звери также потребляют с большой охотой. По наблюдениям
П.И. Данилова (Данилов и др., 1979), количество съедаемой за сутки пищи,
58
Таблица 19/Table 19
Динамика средних размеров тела и массы содержимого желудков у бурых медведей
по месяцам (Новгородская обл., 1979-1984 гг.)
Dynamics of average body sizes and weight of stomach content in brown bears
(Ursus arctos) by month (Novgorod Province, 1979-1984)
Показатели/ Parameters Август/ August Сентябрь/ September Октябрь/ October Ноябрь/ November Декабрь/ December
Масса тела, кг/ Самцы/Males (n=105) 115 120 126 131 102
Body mass, kg
Длина тела, см/ 154 152 152 147 143
Length of body, cm
Высота в холке, см/ 83 91 89 97 80
Height in withers, cm
Масса содержимого желудков, кг/ 3.7 3.7 4.3 3.2 3
Weight of stomach’s content, kg
Состав кормов/ Food component (%):
овес/oats 100 89 71 100 83
ягоды/berries 46 27 29 - 17
рябина/rowan 7 7 14 17 -
яблоки/apples - 1 29 - 33
зонтичные и корешки/ 2 29
Uinbelliferae and roots
желуди/acoms - 5 14 17 -
насекомые и падаль/ 6
insects and carrion
Масса тела, кг/ Самки/Females (n=38) 86 107 122 135 118
Body mass, kg
Длина тела, см/ 156 157 185 160 145
Length of body, cm
Высота в холке, см/ 69 79 88 82 80
Height in withers, cm
Масса содержимого желудка, кг/ 5 3.7 3.5 2 1.8
Mass of stomach, kg
Состав кормов/ Food component (%):
овес/oats 100 92 100 100 33
ягоды/berries 33 23 33 - 67
рябина/rowan - 15 10 15 -
яблоки/apples - 23 - - 33
зонтичные и корешки/ 17 8 33
Uinbelliferae and roots
насекомые и падаль/ 8 16
insects and carrion
Примечание. Масса обескровленной туши животного дается без головы, шкуры,
внутренностей н нижней части ног, так называемый «убойный вес».
59
состоящей только из компонентов животного происхождения (главным образом
мяса копытных), значительно превосходит таковое растительной. Так, средний
вес мясной пищи, обнаруженной в желудках медведя, 8.2 кг (;г=7), ягод и зерна
5.2 кг (zz=73). Вес смешанных кормов - трава, ягоды, овес, мясо, насекомые
(/г= 17) - также был больше, чем только растительных, и составлял 6.6 кг.
Хищнический инстинкт у бурых медведей наиболее отчетливо проявляется
ранней весной, при выходе из берлог, когда в природе еще наблюдается дефицит
растительных кормов. В это время года учащаются случаи нападений на
копытных и домашних животных, разорения кладок птиц, поедания птенцов и
падали. С появлением травянистой растительности, ягод и насекомых
хищничество бурых медведей уменьшается. В летне-осенний период в кормовом
рационе зверей, обитающих на юге Северо-Запада и в центральных областях
России, основное место занимает овес, а затем ягоды и яблоки. Такие компоненты
пищи, как насекомые, листья зонтичных, стебли и корешки других травянистых
растений, имеют второстепенное значение. В северных регионах Российской
Федерации в июле -сентябре они потребляют прежде всего ягоды брусники,
морошки, клюквы; при обильном урожае охотно кормятся смородиной,
малиной, черникой, голубикой и шикшей. Важную роль в питании медведей
играют рыжие муравьи, их личинки, осы, шмели и пчелы, и все же основным
кормом зверей в этот период служат ягоды (Руковский, 1991; Вайсфельд, 1993).
Замечено, что в последние 20-30 лет участились случаи проникновения бурых
медведей вдоль облесенных речных долин глубоко в тундру. Данный факт можно
объяснить увеличением численности животных, хорошим состоянием в тундре
их кормовой базы, а также снижением уровня фактора беспокойства за счет
интенсивной миграции местного населения из арктического бассейна. Нам
неоднократно приходилось встречать следы этих хищников в Болыиеземельской
тундре, севернее 68° с.ш., где некоторые из них порой достигают прибрежной
зоны Баренцева моря и в поисках ягодных мест или выброшенной волнами пищи
кочуют в направлении от Печорской губы до Хайпудырской. Медведи заходят
сюда в конце июня-июле, а уже во второй половине августа откочевывают на
юг, в сторону тундрового редколесья.
Появляющиеся в тундре звери на 70-80% питаются растительным кормом,
преимущественно ягодами. Кроме того, они добывают леммингов, птиц, их яйца,
подбирают на берегу разную падаль. Зимуют они в южных кустарниковых тундрах
и лесотундре, устраивая берлоги по долинам рек - в естественных пустотах, под
выворотнями, корнями упавших деревьев и в каменистых осыпях. Интересно, что
уже в конце сентября-начале ноября, т.е. после ледостава, нередки случаи появления
на побережье Печорского моря белых медведей, которые также питаются
выбросами моря, ловят нерпу и рыбу, а некоторые из них, удаляясь в глубь
тундры на 3-5 км, кормятся разной падалью или выкапывают и поедают
леммингов и других грызунов. Подобные формы поведения зверей просле-
живаются и в других регионах. Так, на Западном Таймыре известны случаи
встреч белых медведей в тундре на расстоянии 10-12 км от побережья в северной
части Енисейской губы (Рыбкин, 1994).
60
На северо-востоке Сибири в бассейне р. Колымы и в приохотской части
ареала бурые медведи также преимущественно растительноядны, хотя рыба (в
основном лосось) и некоторые млекопитающие в их рационе встречаются
регулярно. Из растительных кормов особенно важны ягоды (брусника,
голубика, шикша), орешки кедрового стланика, хвощи. На Анадырском
плоскогорье встречаемость животных кормов особенно велика; из них к
преобладающим видам относятся дикий северный олень, длиннохвостый
суслик и яйца птиц (Чернявский и др., 1993).
Основная пища белых медведей - это морские млекопитающие, прежде всего
арктические тюлени и рыба. По материалам В.И. Цалкина (1936), который в
летнее время исследовал их кормовой рацион, среди 145 обследованных
желудков 44.2% оказались пустыми, 37.9% содержали остатки кольчатой нерпы,
15.1%- моржа и морского зайца, 2.1%-водорослей и 0.7% - птиц. Находясь на
суше, они при случае могут напасть на северного оленя, поймать лемминга или
рыжую полевку, добыть линяющего гуся или сидящую на гнезде птицу.
Существенное значение в питании хищников, которые в районах Арктического
бассейна нередко испытывают чувство голода, имеют выбросы моря, отходы
зверобойного промысла и другая падаль (Попов, 1939; Lono, 1970; Russel, 1975).
В весеннее время в остатках пищи зверей (особенно самок) встречаются части
травянистых растений, мхи и лишайники, водоросли и некоторые злаки (Russel,
1971; Беликов и др., 1977), однако наиболее важными кормовыми объектами
белых медведей служат кольчатая нерпа и морской заяц (лахтак), обеспеченность
которыми во многом определяет численность их популяций. По мнению
С.Е. Беликова (1993), растительная пища не служит для этих хищников
энергетическим источником: за ее счет они пополняют в организме недостаток
минеральных солей, витаминов и микроэлементов. Особенно необходима
растительная пища лактирующим самкам, которым для успешного выращивания
потомства требуется хорошо сбалансированный и полноценный кормовой рацион.
Расчеты показали, что голодный белый медведь может съесть кольчатую
нерпу за два приема, хотя в среднем в желудках добытых зверей находили от
15 до 20 кг пищи (Беликов, 1993). Известны случаи, когда в желудках этих
хищников обнаруживали до 41 кг и более моржового мяса (Цалкин, 1936;
Успенский, 1989). По-видимому, столь значительную вместимость данного
органа следует рассматривать как адаптивную особенность вида, позволя-
ющую животным благополучно переживать голодные дни при недостатке
традиционных кормов в природе.
Различия в пищевой специализации рассматриваемых хищников
обуславливают и заметную разницу в строении их пищеварительной
системы, которая у белых медведей по сравнению с бурыми имеет черты
большей плотоядности. В частности, первые характеризуются укоро-
ченной длиной кишечника, размеры которого превышают длину тела
только в 6 раз (Успенский, 1989). По нашим данным, у бурого медведя
это превышение достигало 7-10 раз. Вообще питание преимущественно
растительной пищей обусловило развитие у бурых медведей длинного
61
кишечника и сравнительно небольшой печени. Так, относительная длина
кишечника у медвежат в возрасте 4-6 мес была равна у самцов 740-762% и
у самок 802-835%. У взрослых животных этот показатель равнялся
соответственно 733-785 и 862-907%. Индекс печени у молодых и взрослых
животных колебался в пределах 2.8-3.5%. Белый медведь обладает заметно
более крупной печенью с необычайно высоким содержанием витамина А (до 9-
10 тыс. мг/г), что связано с питанием жирной пищей, необходимой для
существования организма в условиях постоянного холода. В жире этого органа
находится большое количество жирных кислот и мало фосфолипидов и
холестерина, что свидетельствует о высоком уровне плотоядности вида
(Успенский, 1989).
В отличие от бурых медведей желудочно-кишечный тракт и ферментативная
система белых, питающихся преимущественно мясной пищей, приспособлена к
расщеплению прежде всего белков, жиров и в меньшей степени крахмала. Как
сообщает Т.Б. Саблина (1973), организм белого медведя способен переваривать
крахмал только на 64-74%. Во время зимнего сна в клетках его желудочно-
кишечного тракта происходит интенсивная утилизация запасенных организмом
жировых резервов. Строение слизистой желудка и кишечника зверей
свидетельствует о высокой степени приспособленности всей системы хищников
к расщеплению такого легко усвояемого корма, как жир. Обнаруженные в
двенадцатиперстной кишке белых медведей ворсинки большого размера, по-
видимому, значительно увеличивают ферментативную и всасывающую
деятельность данного участка кишечника, куда из желудка поступает часть
еще не полностью переваренной пищи. Вероятно, этим же можно объяснить
наличие у них и мощного слизистого слоя в тонком отделе кишечника, за
пределами двенадцатиперстной кишки, а также крупных кровеносных сосудов,
обеспечивающих интенсивную воскуляризацию тканей органов пищеварения.
Многолетние материалы показывают, что упитанность бурых медведей,
добытых в Новгородской области (табл. 19), начинает уменьшаться уже в
декабре. Судя по размерам животных и весу сдаваемых на мясозаготовки
туш, наиболее крупные и упитанные звери встречаются в октябре-ноябре. В
данной выборке в ноябре средний убойный вес самцов составлял 131 кг, а
самок -135 кг. Уже в декабре он снизился соответственно до 102 и 118 кг, что
свидетельствует о начавшемся процессе расходования жировых резервов в
организме бурых медведей, залегших в берлоги.
Семейство кошачьих (Felidae)
Семейство кошачьих на Северо-Западе России представлено обыкновенной
рысью (Lynx lynx L.). Если на севере Швеции и в финской Лапландии в зимнее
время рысь добывает преимущественно одомашненных оленей, а летом - мелких
животных и тетеревиных птиц (Haglund, 1965,1968; Pulliainen, Siivonen, 1972),
то на рассматриваемой территории основу её питания составляют заяц-беляк,
62
Таблица 20/Table 20
Состав пищи рыси (»=268) иа Северо-Западе России (по: Данилов и др., 1979)
The composition of food in lynx (Lynx lynx) (»=268) in the North-West of Russia
Вид корма/ Food component Встречаемость/ Percentage, %
Млекопитающие/Mammals: 74
дикие копытиые/wild ungulates 8
заяц-беляк/Lep/w timidus 37.6
заяр-русак/Lepus europaeus 2.2
бобр/Castor fiber 3.7
мышевидные грызуны/small rodents 5.9
домашние животные/livestock 6.2
Птицы/Birds: 26
глухарь/ГеГгао urogallus 2.2
тетерев/Lyrurus tetrix 9.7
рябчик/Tetrastes bonasia 10
тетеревиные птицы и мелкие млекопитающие (Данилов и др., 1979). Из-за
сравнительно низкой численности копытных, доступных для рыси, их доля в
рационе хищника невелика. В летне-осеннее время в остатках её добычи
встречаются и домашние животные (табл. 20). В зимние сезоны в Ленинград-
ской и Псковской областях при троплении зверей нам приходилось фиксировать
нападение хищника из засады на косулю и бобров. Последних он подстерегал у
постоянных вылазов из воды.
В южных областях региона рацион рыси более разнообразен, чем на севере,
где она добывает в основном мелких млекопитающих и боровую дичь. В Карелии
абсолютное большинство нападений на добычу, предпринимаемых хищником,
приходится на зайца-беляка (80%), немногим более 16%-на тетеревиных птиц
и лишь 3.7% - на прочих животных. Результативность охоты на основных жертв
(зайцев и тетеревиных птиц) довольно схожа: 28.7 и 27.8% (Данилов и др., 1979).
В Центрально-Лесном заповеднике (Желтухин, 1979,1987) основу питания
рыси также составляет заяц-беляк, доля которого в экскрементах (67.6%) и
остатках добычи (64.8%) примерно одинакова. Встречаемость белки в добыче
зверей была равна 2-2.2%, мышевидных грызунов - 2.2-4.4, рябчика - 8.8-9.3
и глухаря - 2.2-3.4%. Сезонные различия в питании проявляются прежде всего
в уменьшении в рационе (в бесснежный период года) доли зайца-беляка (с 73.2
до 41.7%) и увеличении значения тетеревиных и других птиц (с 11.9 до 38.8%).
Успешность охоты рыси в среднем составляет на зайца-беляка 22.7%, на
рябчика 30.4, на глухаря 16.7 и на белку 13.3%.
63
В других регионах кормовой спектр рыси более разнообразен (Матюшкин,
1974; Зырянов, 1980; Малафеев и др., 1986). Так, в верховьях Енисея (Смирнов,
1997) основными кормовыми объектами рыси являются заяц-беляк, косуля,
кабарга, рябчик. К второстепенным кормам можно отнести марала, лося,
кабана, сибирского горного козла, мышевидных грызунов, тетерева, глухаря
и других. В редких случаях она нападает на домашних животных, в основном
на овец. По сообщению А.Н. Зырянова (1997), зимой в пределах Западно-
Сибирской низменности заяц-беляк, несомненно, служит главным компонентом
рациона рыси. В то же время в заповеднике «Столбы» места ее обитания
тяготеют к районам концентрации косули, кабарги и марала (встречаемость
среди остатков жертв 69%), затем зайца-беляка (10.6%) и тетеревиных птиц
(7.7%).
В различных регионах Восточной Сибири важная роль в рационе рыси отводится
копытным (Лавов, 1972). В Прибайкалье и приленских районах основу ее питания
составляет косуля, а на Витимском плоскогорье - кабарга, запасы которой
достаточно велики. Заяц-беляк (как и тетеревиные птицы) играет небольшую
роль в питании рыси, что связано со сравнительно малой его численностью в
угодьях. Заметно чаще он встречается в питании хищников на Витимском
Таблица 21/ТаЫе 21
Остатки обнаруженной зимой добычи рыси (экз.) и доля отдельных компонентов
(%) в питании хищника с учетом веса жертвы (по: Лавов, 1972)
Remains of lynx’s prey, discovered in winter and the portion of individual components (%)
in the food of predator with the account of weight of the prey
Вид добычи/ Kind of prey Прибайкалье/ Baikal area Приленские р-ны/ The Lena River area Ветимское плоскогорье/ Vitim Plateau Всего/ Total
« % n | % n % %
Косуля/ 23 66 14 73 5 32 42 60
Capreolus capreolus Кабарга/ 10 14 5 13 16 51 31 21
Moschus moschiferus Изюбр/ 1 11 - - 1 6
Cervus elaphus Заяц-беляк/ 24 7 20 10 23 15 67 10
Lepus timidus Рябчик/ 5 1 6 1 4 1 15 1
Tetrastes bonasia Глухарь/ 3 1 3 2 1 1 7 1
Tetrao urogallus Тетерев/ - - 3 I - - 3 1
Lyrurus tetrix Дятел-желна/ Dryocopus martius 1 - - - - - 1 -
Итого/Total 67 100 51 100 49 100 167 100
64
Таблица 22/Table 22
Изменение массы тела самки обыкновенной рыси и питательная ценность её
суточного рациона по месяцам (по: Обухова, Шахназаров, 1949)
Changes of body mass in female of lynx (Lynx lynx) and nutritional value of its daily
ration by month
Показатели/ Parameters Месяцы/Months
апрель/ April май/ Мау июнь/ June июль/ July август/ August сентябрь/ September октябрь/ October ноябрь/ November
Масса тела, кг/ 11.1 10.9 11.3 - 11.2 11.6 12.1 12.9
Body mass, kg
Калорийность
суточного
рациона, кал/
987 1039 1025 1083 1157 1283 1370 1289
The calorific
value of daily
ration, cal
Содержание
переваримого
белка в корме, г/
146 151 149 158 168 186 196 188
Content of
digestible protein
in food, g
плоскогорье, где плотность населения зверьков выше (табл. 21). На лося,
изюбра и северного оленя рысь нападает редко, выбирая в качестве жертвы в
основном больных и ослабленных животных.
Судя по наблюдениям в природе, рысь за сутки потребляет не более 1.5 кг
корма (Haglund, 1966; Новиков и др., 1970; Pulliainen, Siivonen, 1972). В
Тверской области (Желтухин, 1984) рысь съест зайца-беляка в два приема за
1.5-2 суток. За это время, если хищника не беспокоить, он съедает пойманную
добычу почти полностью, оставляя лишь желудочно-кишечный тракт, а иногда
и задние ступни жертвы. Рысь, потревоженная при потреблении пищи, может
возвращаться к добыче вновь и доедать ее остатки даже спустя длительное
время.
В Санкт-Петербургском (Ленинградском) зоопарке взрослый хищник в
среднем съедал осенью до 1.5, а зимой и летом-до 1.2 кг мясного корма. В
Рижском зоопарке (Стоянова, Сваринская, 1964) при норме мясного корма 2 кг
на одну рысь часть пищи оставалась несъеденной, поэтому суточный рацион
был сокращен до 1.3 кг. В опытах А. Д. Обуховой и Г.М. Шахназарова (1949),
проведенных в Московском зоопарке, самки рыси в течение года потребляли
до 1.3-1.4 кг мяса в сутки, однако в весенне-летний период они съедали корма
меньше, чем осенью. Так, в мае живой вес самки был равен в среднем 10.9 кг,
а средний уровень ее суточного рациона составлял 1039 кал и 151 г
3 Зак. № 4309
65
Таблица 23/Table 23
Некоторые весовые н размерные показатели камышового и лесного котов
(по: Насибов, 1970)
Some weight and size parameters in the chaus (Felis chaus) and the wild cat
(Felis silvestris)
Показатели/ Parameters Камышовый кот/ Felis chaus (n=26) Лесной кот/ Felis silvestris (n=39)
<5<S 22 <33 22
% « 1 % n % « 1 %
Масса, г/Mass of, g; тела/body 7000 - 5400 - 3900 - 3300 -
печени/liver 216 3.1 143.2 2.7 109.9 2.8 83.7 2.5
Длина, см/Length of, cm: тела/body 75.7 - 71.5 - 60 - 54.4 -
хвоста/tail 29.3 38.7 25 35 32.4 54 31.4 57.7
кишечника/intestine 242.6 320.5 212.2 296.8 184.8 308 179.1 329.2
Таблица 24/Table 24
Встречаемость отдельных компонентов пищи в рационе камышового и лесного
котов на территории Кавказа (по: Насибов, 1970)
The occurrence of individual components of food in the ration of the chaus (Felis chaus)
and the wild cat (Felis silvestris) in the Caucasus
Вид пищи/ Food component Камышовый кот/ Felis chaus (252 пробы) Лесной кот/ Felis silvestris (124 пробы)
fl | % n | %
Млекопитающие/ Mammals: 126 50 105 84.6
мелкие грызуны/ 88 34.9 99 79.8
small rodents
землеройки/ 4 1.6 2 1.6
Soricidae
заяц-русак/ 1 0.4 4 3.2
Lepus europaeus
нутрия/ 33 13.1
Myocastor coypus
Птицы/ Birds 106 42 36 29
Пресмыкающиеся/ Reptiles 7 2.8 - -
Рыбы/ Fish 10 4 1 0.8
Насекомые/ Insects 20 7.9 3 2.4
Растительные остатки/ 22 8.7 18 14.5
Vegetative remains
66
переваримого белка (табл. 22). С конца сентября показатели поедаемости корма
и массы тела увеличивались. В октябре живой вес зверя составлял в среднем
12.1 кг при суточной норме питания 1370 кал и 196 г переваримого белка. У
взрослого самца, потребляющего мясную пищу, калорийность суточного
рациона составляла весной 998 кал, в начале лета 1258, в конце лета 1360 и
осенью 1565 кал.
Рысь, обитающая в Восточной Сибири, более прожорлива, чем звери из
европейских регионов России, что, видимо, объясняется разницей в их размерах.
Так, в желудках 2 отловленных самцов находилось по 3-3.5 кг мяса и шерсти
(в первом - кабарги, а во втором - взрослой косули). Кроме того, известен
случай, когда самка рыси с 2 детенышами съели 8-10 кг мяса - примерно 3/4
туши только что добытой ими сибирской косули (Лавов, 1972).
Как и другие плотоядные животные, рысь обладает сравнительно высоким
индексом печени и небольшим - кишечника. У добытых в Новгородской области
взрослого самца (масса тела 15 кг, длина- 86 см) и молодой самки (масса тела 7.8 кг,
длина - 75 см) индекс печени был равен соответственно 2.8 и 3.5%, а относительная
длина кишечника-320 и335%. Эти данные совпадают с материалами, полученными
в Республике Коми (Пыстин, 1998). Здесь у 4 отловленных самок разного возраста
индекс печени составлял 2.7-3.8%, а длина кишечника - 319-386%.
У другого вида семейства кошачьих - лесного кота {Felis silvestris Schreb.),
которого нам приходилось добывать в Дагестане и горных районах
Азербайджана (п=8), показатели относительной массы печени (2.8-3.3%) и
длины кишечника (320-342%) мало отличались от подобных у рыси, что,
видимо, объясняется сходством их кормового рациона.
По свидетельству С.Б. Насибова (1970), несмотря на значительные различия по
массе тела и абсолютному весу внутренних органов, у обитающих на Кавказе
лесного и камышового (Felis chaus Gueld.) котов их индексы печени и длины
пищеварительного тракта оказались практически одинаковыми (табл. 23). Размеры
последнего у представителей обоих видов в 3 раза превышают длину их тела, а по
относительному его показателю в среднем в 2 раза уступают таковому у склонных
к эврифагии енота-полоскуна или барсука. Примерно 2/3 длины пищеварительного
тракта котов приходится на долю тонкого отдела кишечника, где осуществляется
основной процесс всасывания белковой фракции пищи. У самцов камышового кота
он составляет 61.8%, лесного - 62.1%, а у самок соответственно 62.8 и 63.2% от
размеров кишечника. Общая длина пищеварительного тракта этих зверей оказалась
значительно короче, чем у шакала (Canis aureus L.). У последнего, питающегося
разнообразной (часто грубой) пищей, кишечник в среднем в 5 раз превышает длину
тела, что, видимо, связано с меньшей плотоядностью шакала по сравнению с
представителям семейства кошачьих.
В просмотренных нами 8 желудках лесных котов растительные корма
отсутствовали, зато постоянно встречались остатки мелких грызунов и птиц, реже
насекомых и рыбы. В то же время имеются сведения (Г. Новиков, 1956), что в
небольшом количестве они иногда потребляют зеленые части злаков и осок,
иногда фрукты. Обитая в районах горных селений, звери могут нападать на
67
домашнюю птицу, а около сельскохозяйственных угодий - на молодых зайчат
и фазанов. У лесных котов, добытых в районе Кавказского заповедника, доля
отдельных пищевых компонентов по встречаемости такова: мышевидные
грызуны (в основном кустарниковая полевка и лесная мышь) - 94%, соня-
полчок, заяц-русак и насекомые - по 2%, крот и землеройки - по 4%, мелкие
птицы - 9% и растительные остатки - 11% (Теплов, 1938).
В Дагестане и Азербайджане основу питания камышового и лесного котов
составляют млекопитающие и птицы (табл. 24). По сообщениям С.Б. Насибова
(1967, 1970), в рационе первого хищника из грызунов преобладают водяная
(50%) и общественная (29.5%) полевки, а у второго - кустарниковая полевка
(43.4%) и лесная мышь (30.3%). В составе пищи камышового кота обнаружены
17 видов птиц, относящихся к 3 отрядам. Среди них преобладают кряква
(18.9%), пастушковые (19.8%) и голенастые (8.5%). В питании лесного кота
зарегистрированы лишь 7 видов орнитофауны, относящихся к 3 отрядам. Из
них ведущее место принадлежит мелким воробьиным птицам - 52.8%. Вообще
кормовой рацион камышового кота разнообразнее такового лесного, зато
последний активнее добывает мышевидных грызунов, молодых зайчат, чаще
потребляет побеги злаков, тамариска и других растений.
Сезонная смена кормов более отчетливо просматривается у камышового кота.
По наблюдениям того же автора, зимой мыши и полевки составляют в его питании
19.2%. В это время года он заметно больше, чем в другие сезоны, добывает нутрию,
которая из-за ледостава на водоемах вынуждена выходить на сушу, где и становится
добычей хищника. Кроме того, в зимний период берега заливов, озер и каналов
заполняются прилетающей на зимовку птицей, что и определяет резкое увеличение
ее доли (до 57.7%) среди кормов камышового кота. Весной и летом в его питании
заметно нарастает встречаемость мышевидных грызунов (43.4-44.9%),
пресмыкающихся (3.8-4.4%), насекомых (5.8-21.7%) и рыбы (до 5.8%), но падает
потребление птиц (2.7-32%) из-за отлета в места гнездовий представи-телей
зимующих здесь видов, а также нутрий, зайца-русака и насекомоядных.
В отличие от камышового у лесного кота такой резкой смены пищевого
рациона не наблюдается. У него во все сезоны года основную роль среди кормов
играют мышевидные грызуны. Большое значение в зимнем питании
принадлежит дендрофильным видам птиц (40%), доля которых в отдельные
сезоны не превышает 24-25%. В весенне-летнее время кормовой спектр
хищника несколько увеличивается за счет частей растений (12.5-18.8%),
насекомых (9.4%) и рыбы (3.1%). Из-за хорошей обеспеченности пищей
численность обоих видов котов на Кавказе не подвергается резким
колебаниям, однако на состояние популяции камышового кота, который
относится к хищникам более южного происхождения, суровые зимы могут
оказывать заметное негативное влияние (Насибов, 1970).
Осенью потребность к калорийных кормах у представителей семейства
кошачьих резко возрастает; они стараются больше добывать пищи; масса тела и
упитанность зверей увеличиваются, что и обеспечивает им благополучный
исход зимовки.
68
Таким образом, анализ питания разных видов хищных млекопитающих
позволил выявить у них четко выраженные особенности кормовой специализа-
ции, изменчивость объёма и компонентов пищевого рациона по сезонам года.
В основном у них прослеживается единая адаптивная реакция, направленная
на подготовку к неблагоприятному зимнему периоду. У всех рассмотренных
видов потребность в высококалорийной пище возникает в конце лета-осенью
и сопровождается увеличением массы тела за счёт образования внутренних
жировых резервов. В зимнее время им требуется меньше корма, чем в тёплые
сезоны. Происходит экономное расходование созданных в организме
энергозапасов, что в итоге и определяет благополучный исход зимовки
хищников.
Резюме
Специфика питания, сезонная динамика массы тела и потребность в
пище во многом определяют физиологическое состояние, готовность к гону,
воспроизводственный потенциал, а в конечном итоге численность хищных
млекопитающих в природе. Сезонные изменения энергетики рациона
проявляются у них достаточно отчетливо и представляют собой стойкую
адаптивную реакцию, направленную на экономное расходование создаваемых
в организме энергозапасов. При зимнем падении уровня обмена веществ
потребность хищников в пище резко снижается, что и определяет благополуч-
ный исход зимовки. В этот период они бывают малоактивны и потребляют
кормов немного, постепенно расходуя свои внутренние резервы. Перед сезоном
размножения и весенней линькой интенсивность питания и масса тела особей
вновь несколько увеличиваются. Когда животные находятся в гонном или
предгонном состоянии, они едят сравнительно мало. В этот период запасы
питательных веществ в их организме вновь быстро расходуются, а масса тела
уменьшается.
Летом, в жару, хищники потребляют мало пищи. Существенное
уменьшение суточной нормы у большинства видов становится заметным уже
в июне и продолжается до середины-конца августа, после чего уровень этого
показателя начинает возрастать. В летнее время они бывают наименее
упитанны, а их вес заметно меньше, чем весной и осенью. В сентябре-октябре
потребность животных в кормах увеличивается. К середине осени они
достигают наибольшей упитанности, после чего их потребности в пище
снижаются. Осенний нагул имеет большое значение, так как обеспеченность
жировыми запасами способствует выживаемости хищников в неблагоприятные
периоды года. В конце осени они становятся менее активными, масса тела у
самцов и у самок снижается, и цикл повторяется. Следовательно, осенний
период, когда интенсивность обменных процессов в организме падает, а
жировой резерв, или энергетический потенциал, возрастает, является очень
важным в жизни зверей. По-видимому, их выживаемость зимой во многом
69
определяется наличием и доступностью высококалорийных кормов в осеннее
время, когда потребность в пище особенно велика.
Экологическая радиация рассматриваемых видов по степени кормовой
специализации прослеживается достаточно отчетливо. В целом хищные
млекопитающие потребляют разнообразные корма. В то же время по
значимости основных компонентов пищи среди них можно выделить миофагов
{Mustela nivalis, М. erminea, М. altaica), ихтиофагов {Lutra lutra), стенофагов
{Enhydra lutris) и некрофагов {Gulo gulo). К видам с широким кормовым
спектром (полифаги) относятся М. putorius, М. eversmanni, М. vison, М. lutreola,
а также представители семейств Canidae и Felidae. Наибольшая эврифагий-
ность свойственна Martes martes, М. foina, М. zibellina, Meles meles, Procyon
lotor и Ursus arctos. Потребность в высококолорийной добыче тесно связана с
массой тела и уровнем теплообмена животных. У самых мелких хищников -
Mustela nivalis и М. erminea - удельная величина энергии, потребляемая с
пищей, оказалась в 4-6 раз больше, чем у Lutra lutra и Meles meles, а у Martes
martes выше, чем у Mustela vison и М. lutreola. Установлена прочная связь
между кормовой специализацией и строением пищеварительного тракта
представителей рассматриваемых видов. Ббльшая калорийность белковой
пищи у миофагов определяет размеры тонкого отдела кишечника и его частей,
а сочетание животных и растительных кормов у эврифагов - длину ободочной
кишки и всего толстого отдела. Кроме того, видам с широким спектром питания
свойственно увеличение активной всасывающей поверхности ворсинок в
тонком отделе за счет усложнения их конфигурации. В то же время плотоядное
питание обусловливает значительные размеры печени как углеводного и
жирового депо организма при упрощенной форме абсорбционной поверхности
ворсинок слизистой оболочки кишечника. Таким образом, строение
пищеварительного тракта хищных млекопитающих в полной мере отражает
пищевую специализацию видов, которая проявляется не только в характерных
размерах печени и кишечника, но и в специфической архитектонике слизистой
оболочки его отделов.
Summary
Specificity of feeding, seasonal dynamics of the body mass and food requirement
to a great extent determine the physiological state, readiness to rut, reproductive
potential and, finally, the number of carnivore mammals in nature. The seasonal
changes of ration energetics are displayed in them rather distinctly and represent a
stable adaptive reaction directed towards economic expenditure of energy resources
produced in the organism. With a winter decrease of the metabolic rate, the
requirement of predators in food is decreasing dramatically, which defines favorable
result of wintering. During that period they are not active and eat a little amount of
food, gradually spending their inner resources. Before the season of breeding and
spring molting the intensity of feeding and body mass of individuals is again
somewhat increasing. When animals are in rut or pre-rut state they eat comparatively
70
little. In that period resources of nutritional substances in their organism again are
used quickly and body mass decreases.
In the summer, during hot days, carnivores consume a little amount of food.
Essential decrease of circadian norm in most species is noticeable already in June
and it continues up to the middle or end of August, after that the level of this
parameter starts to increase. In the summer time carnivores are least fat and their
weight is considerably less than in spring and autumn.
In September through October the requirement of animals in food is increasing.
To the middle of autumn they achieve the maximum of fatness afterwards their
requirements in food are decreasing. The autumn fattening is of great importance
as fat resources promote survival of carnivores in the unfavorable periods of the
year. In the end of the autumn they become less active, body mass in males and
females decreasing and the cycle repeats.
Hence, the autumn period when the intensity of metabolic processes in the
organism falls and fat reserve or energetic potential increases is of great importance
in the life of animals. Apparently, their survivorship in winter to a great extent is
determined by the occurrence and availability of highly caloric food in the autumn
time, when requirements in food are particular high.
The ecological radiation of the species considered by the degree of food
specialization is defined quite clearly. Carnivorous mammals on the whole consume
various food. At the same time the following groups could be distinguished among
them by the significance of the main components: myophagous {Mustela nivalis,
M.erminea, M.altaica), ichtyophagous {Lutra lutra), stenophagous {Enhydra lutris)
and necrophagous {Gulo gulo). Among Mustelidae M. putorius, M. eversmanni,
M. vison, M. lutreola and representatives of the families Canidae and Felidae are
referred to the species with a wide food spectrum (polyphagous). The greatest
euryphagy is typical of Martes martes, M.foina, M. zibellina, Meles meles, Procyon
lot or and Ursus arctos. The need for the highly caloric prey is closely related to the
body mass and heat exchange of the animals. In most small carnivores - Mustela
nivalis and M. erminea the specific energy value consumed with food appeared to
be 4-6 times greater than in Lutra lutra and Meles meles, and in Martes martes is
higher than in Mustela vison and M. lutreola. The firm relation between the food
specialization and the structure of alimentary tract in representatives of the species
considered has been established. The high calorific value of protein food in the
myophagous determines the sizes of the small intestine and its parts and combination
of animals and vegetable food in the euryphagous - the length of the colon and
large intestine in total. In addition, the increase of active absorbing surface of villus
in the small intestine owing to the complication of their configuration is typical of
species with a wide spectrum of food. At the same time carnivorous food stipulates
considerable sizes of the liver as a carbohydrate and fat depot of the organism with
a simplified form of absorbing surface of villus in the mucous membrane of the
intestine. Thus, the structure of the alimentary tract of the carnivores in full measure
reflects the food specialization of species, which is displayed both in optimal sizes of
the liver and intestine and in the specific architectonics of mucous membrane of its
parts.
Глава 2. ДВИГАТЕЛЬНАЯ АКТИВНОСТЬ И ЕЁ
СВЯЗЬ С КОРМОВОЙ СПЕЦИАЛИЗАЦИЕЙ
ВИДОВ
Активность хищных млекопитающих - один из показателей их жизне-
деятельности. Она во многом определяется условиями внешней среды и,
сопровождается значительной тратой внутренней энергии, поэтому периоды
бодрствования и покоя животных в течение суток закономерно чередуются.
По сезонам года их подвижность также очень заметно меняется: у большинства
видов она возрастает весной, к началу гона и осенью, в связи с повышенной
потребностью в кормах и расселением молодняка. Благодаря суточной смене
фаз активности и покоя поддерживается оптимальный уровень физиоло-
гических процессов в организме. Распределение интенсивности двигательных
реакций в течение суток и связь этой ритмики с сезонными изменениями
физиологического состояния имеют большое значение в жизни хищников. Из
многих факторов, оказывающих влияние на их активность, пищевая
специализация и доступность корма определяют приспособительные
возможности животных, которые реализуются в виде различных ритмов
жизнедеятельности (Lockie, 1961,1966; Erlinge, 1975,1981; Шилов, 1977,1982;
Соколов, Кузнецов, 1978; Dunstone, Birks, 1987; Бигон и др., 1989).
Достаточно отчётливо данная зависимость прослеживается на предста-
вителях семейства куньих (Mustelidae) с их широким спектром экологической
радиации. Все они в холодное время года затрачивают много энергии на поиск
и добычу пищи, что находится в соответствии с размерами животных, уровнем
теплообмена, количеством и качеством потребляемого корма. У хищников,
близких по пищевой специализации, прослеживается тенденция к прямой
зависимости величины суточного хода от массы тела зверьков. В то же время
представители мелких видов для удовлетворения потребности в пище в
количестве, необходимом для поддержания интенсивного энергообмейа,
вынуждены кормиться чаще крупных.
Исследуемые животные заметно различаются между собой по манере
отыскания добычи. Если росомаха, барсук и лесная куница имеют сравнительно
прямолинейный охотничий поиск, то у мелких видов он более сложный и
извилистый. В среднем на одинаковом отрезке суточного хода интенсивнее
обыскивают территорию такие хищники, как ласка и горностай (см. табл. 14).
Это объясняется сравнительно высокими уровнями их обмена веществ и удельной
величины энергии, потребляемой с пищей. Значительная напряженность охот
(на единицу пути) характерна также для норок, которые в отличие от лесной
куницы, близкой к ним по массе тела, больше потребляют малокалорийные корма
(земноводных, рыбу), что вызывает необходимость частых поисков добычи.
Охотничий поиск хищников-амфибионтов различен. У выдры он проходит
вдоль береговой линии и изобилует выходами на сушу и спусками к воде. В
72
Карелии и Архангельской области она зимой может по льду пересекать
небольшие озёра, направляясь из устья одной реки в другое. Во время таких
кочевок по суше зверек срезает излучины, заходит в прибрежные леса, но
кормится, как правило, у воды. Подобные формы поведения выдры отмечены
в разных частях её ареала (Данилов, 1969; Вшивцев, 1972; Mason, Macdonald,
1986; Sidorovich, 1991; Ozolins, Rantins, 1992; Reuther, 1993; Ruis-Olmo,
Delibes, 1998).
Европейская норка в отличие от выдры не совершает длительных пере-
ходов по суше, а, придерживаясь узкой полосы долины рек, ведет относительно
оседлый образ жизни. Американская норка хотя и близка по пищевой
специализации к европейской, однако в поисках пищи часто отходит от реки в
лесные заросли, останавливаясь на дневку в удобных местах на краю болота
или около мелкого ручья. Во время таких кочёвок она охотится на суше,
нередко залезает в человеческие постройки, а порой её след петляет по лесу в
направлении соседнего водоёма на протяжении 2-3 км. Таким образом,
американская норка не имеет такой строгой приуроченности к водной среде
обитания, как европейская. Ее природные популяции в основном представлены
потомками зверьков, бежавших со звероферм, поэтому клеточное содержание
животных в течение многих генераций могло привести к ослаблению
физиологически сложившихся реакций организма, связанных с добычей пищи
в воде.
Снеговой покров оказывает заметное влияние на возможность передви-
жения и охоты хищников. Животные, которые обитают в глубокоснежных
таёжных районах страны (например, лесная куница, соболь, колонок) или
в тундрово-таёжной зоне (ласка, горностай, росомаха), испытывают
зимой дефицит корма. У них, как правило, опорная площадь лап больше,
чему представителей других видов (Туманов, 1976а). Это уменьшает
весовую нагрузку на единицу площади опоры и облегчает передвижение зверей
по рыхлому снегу. Наши исследования показали, что весовая нагрузка
на площадь лап составляет зимой у ласки и горностая 7.2-7.4 г/см2, у
лесной куницы, соболя и колонка 11.3-13.2, у норок 19-20.8, а у выдры 59.8-
60.3 г/см2.
У лесного хорька, имеющего зимой склонность к синантропному образу
жизни, весовая нагрузка (20.7-22.3 г/см2) на опорную поверхность лап
оказалась больше таковой у лесной куницы, близкой по массе тела. По-
видимому, эта особенность лесного хорька служит одной из причин его
отсутствия в северных регионах Европейской России. У барсука, впадающего
в зимний сон, обсуждаемый показатель был в 2-3 раза выше (88.7-124.4 г/см2),
чем у свободно передвигающейся по снегу росомахи (41.2-42.6 г/см2).
На Кавказе в малоснежные годы, т.е. при глубине снега в горах менее 6-7 см,
в хорошую погоду следы барсука можно встретить даже в зимние месяцы.
Интересно, что за счёт слабой опушенности лап каменная куница, или
белодушка, по сравнению с лесной имеет большую весовую нагрузку на
единицу опорной поверхности. У зверьков, отловленных в Дагестане, она
73
составляла 22.5-25.4 г/см2, что свидетельствует о меньшей её способности к
обитанию зимой на снежных склонах высокогорий (Туманов. 1982). Каменная
куница заметно проваливается в рыхлом снегу и вынуждена избегать глу-
боко-снежных участков (Динник, 1914; Г. Новиков, 1956; Соков, 1972а).
Возможно, поэтому, а также в связи с большей, чем у лесной куницы,
термофильностью белодушка в зимние месяцы нередко встречается в селах,
устраивая свои гнезда на чердаках, в сараях или различных брошенных
строениях. Вертикальное распределение видов во многом соответствует их
широтному распространению. В обоих случаях лесная куница обитает в
местах с более холодным климатом и с глубоким снеговым покровом, куда
каменная не заходит.
В Предкавказье (Гинеев, 1991) расселение каменной куницы идет широким
фронтом от Каспия до Азовского и Черного морей. Все чаще поступают
сообщения и о появлении ее в поселках. На пойменных участках она держится в
густых захламленных лесонасаждениях, в горных районах - по опушкам леса и
в каменистых осыпях, в плавнях и степных районах - по балкам и крутым берегам
рек. Величина участка обитания (3-5 га) и суточный ход зверьков (5-8 км) во
многом зависят от кормности угодий. Белодушка - сумеречно-ночной зверек,
хотя в голодные годы ее можно встретить и в дневные часы. Чаще всего она
проявляет активность вечером, от 20 до 23 ч, и утром - от 4 до 7 ч. В Белгородской
области (Г. Новиков, 1962) каменная куница, жившая в вольере, была активна
с 17 до 20 ч и (с небольшими перерывами) до 6-7 ч утра. В естественных
местообитаниях она добывает пищу в основном на поверхности почвы.
Значительную часть пути зверек идет ходом, не задерживаясь, останавливаясь
только в зарослях кустарников, где он обычно охотится на полевок и мышей.
При троплении суточных следов разных особей этого вида на протяжении 17 км
белодушки только 129 м прошли в кронах деревьев. На протяжении наземного
пути они осмотрели 107 различных «ухоронок» грызунов, 34 раза копались в
снегу на открытых местах и 23 - около деревьев. Таким образом, каменной
кунице, как и другим мелким хищникам, свойствен высокий уровень охотничьей
деятельности, что связано с особенностями ее энергообмена.
Лесная куница - зверёк с сумеречным и ночным типом суточной
активности. Добывает пищу она чаще на земле. Её передвижения в кроне
деревьев непродолжительны и связаны либо с охотой на белку и птиц, либо с
выбором места на днёвку. В Карелии (Данилов, Туманов, 1976а) при
троплении куниц общей протяженностью 412 км удалось установить, что
«верхом» она прошла лишь 8 км (2%). При этом количество заходов на деревья
колебалось от 2 до 22 на 10 км маршрута. По сравнению с самкой самец менее
разборчив в выборе гнездового убежища. Обычно, поднявшись на дерево (чаще -
дуплистую старую осину), он остаётся здесь на днёвку или, перепрыгнув на
ветви близко расположенных елей, устраивается на отдых в беличьем или
сорочьем гнезде. Самка же, выбирая место для днёвки, может до 2-3 раз
подниматься на деревья и спускаться вниз, пройти 100-200 м по ветвям деревьев
и только после этого, облюбовав убежище, устроится в нём на отдых.
74
Отмечается некоторая связь в характере поведения хищника с обилием или
недостатком того или иного корма. Так, в годы высокой численности полёвок
куница ведёт в основном наземный образ жизни, а в годы, отличающиеся
обилием белок или рябчика, чаще поднимается на деревья, где и охотится.
Активность хищника во многом определяется наличием корма и погодными
условиями. При обилии пищи суточный наслед его невелик (1.5-3 км), в
малокормные годы он заметно возрастает и достигает 7-8 км. Насытившись,
зверёк прекращает охотничью деятельность и уходит на днёвку. В снегопады
и последующие дни куница может по 2-3 суток не выходить из своего укрытия,
довольствуясь сделанными запасами и внутренними резервами организма. В
сильные морозы хищник, как правило, малоактивен и охотится сравнительно
недолго.
В ноябре-январе куница обычно остается на дневку в верхнем ярусе леса -
в беличьем гайне, дуплистой осине или березе. В период глубокоснежья (в
феврале-марте) она чаще устраивает гнездо в комлевой части старых пней,
под ветровалом и стволами деревьев, занесенных снегом. В этих надежных
ухоронках зверек находится в условиях более мягкого микроклимата и
свободно передвигается в подснежных пустотах, добывая концентрирующихся
здесь мышевидных грызунов.
У соболя размеры участка обитания и длина суточного наследа близки к
таковым у куниц. По нашим наблюдениям, в Восточных Саянах при хорошем
урожае кормов (кедровый орех, сеноставки, полевки) его участок обитания
был равен 4-6 км2, а жировочный ход не превышал 5-7 км у самцов и 3.5-4.5 км -
у самок. Однако в пределах всего ареала вида размеры участков зверей
могут колебаться от 4 до 30 км2, а суточный наслед - до 12-15 км (Гептнер и
др., 1967). Обычно они определяются состоянием кормовой базы, доступностью
пищи, метеоусловиями, полом зверьков и некоторыми другими факторами.
Глубокий рыхлый снег несколько ограничивает передвижение соболя, особенно
самок, которые заметно мельче и слабее самцов.
В Восточной Сибири (Комаров, 1972) старые особи занимают в 2-3 раза
большие площади, чем сеголетки. Индивидуальные участки состоят из
нескольких «суточных». У старых зверьков их бывает до 7, у сеголеток - не
более 3. Суточные участки охватывают 1-2 стации, на индивидуальных
разнообразие местообитаний гораздо шире - гари, ветровалы, россыпи,
стланик, кедровник и ягодник. На каждом суточном участке зверьки имеют
постоянные гайна, однако нередко соболь отдыхает и во временных убежищах.
Наибольшая его активность отмечается после выпадения пороши, когда
особенно подвижны полевки и пищухи - основной объект охоты хищника во
многих регионах Сибири.
В годы с неблагоприятными кормовыми условиями сеголетки первыми
покидают семейные участки. В такие годы они представляют первую волну
миграции, которая постепенно захватывает и более старшие возрастные
группы зверьков. Отмечено, что самые старые особи даже в бескормные годы
оставались на своих постоянных участках, занимая при этом и освободившиеся
75
соседние. В различных популяциях соболя встречаются особи, которые зимой
постоянно перемещаются и не имеют своей территории. Часто мигрантами
становятся половозрелые особи и почти всегда-самцы. Так, в январе 1971 г.
взрослый самец с характерным отпечатком лап за 2 суток проделал 21 км по
относительно прямолинейному маршруту. Вновь в старые поселения он больше
не возвратился, хотя урожай кормов в тех местах был хорошим. Другой
помеченный соболь (самец), местами используя лыжню, прошел за сутки 15 км.
Через день его следы встречались уже за 28-30 км от места, где впервые видели
след этого хищника (Комаров, 1972). В Байкальском заповеднике при
повторном отлове маркированных в разные годы соболей (42 особи) 6 зверьков
были отловлены на расстоянии от 6 до 25 км от места их мечения (Сутула,
Попов, 2001).
Данные А.И. Хлебникова (1977) свидетельствуют о том, что с увеличением
мощности снежного покрова существенно меняется характер отыскания
соболем корма - продолжительность суточного наследа снижается, он начинает
тщательно и систематически осматривать все доступные укрытия, где может
находиться добыча. Обычно во второй половине зимы количество его жировок
в россыпях снижается из-за меньшей их доступности и в то же время значительно
возрастает посещаемость менее засыпанного снегом валежника (табл. 25). В
глубокоснежный период подснежные пустоты, образованные валежником, с
их более «мягким» микроклиматом создают благоприятные условия как для
концентрации здесь мышевидных грызунов, так и для успешного преследо-
вания жертв соболем. Так, в Западном Саяне на маршруте длиной 30 км
автором был проведен учет валежника с регистрацией посещения его
Таблица 25/Table 25
Характер поиска пищи соболем в тайге центральной части Западного Саяна,
32 км троплений (по: Хлебников, 1977)
The character of food search in sable (Martes zibellina) in the taiga of the central part of
the Zapadnyi Sayan, according to 32 km of counts
Действия соболя/ The actions of sable Общее кол-во жировок/ The total number of fattening sites Кол-во жировок на 1 Kkt троплений/ The number of fattening sites per 1 km of transect count
n | %
Забирался: под снег и копался в снегу/ Went under snow and was digging in snow в россыпи, под камни/ went into gravel, under stones под валежины, вывороты корней/ under fallen wood, inverted roots на деревья/climbed trees Обследовал прикомлевую часть деревьев и кустарников/Examined the butt part of trees and bushes 21 5.7 0.6 14 3.8 0.4 267 73 8.3 15 4.1 0.4 49 13.4 1.5
76
хищником. Из 121 валежины, выделявшейся над поверхностью снега частично
или полностью, 109 (90.8%) имели следы обследования зверьками. Важная
роль валежника в биологии вида, а также четкая зависимость между
количеством характерных для соболя жировок и мощностью снежного покрова
прослежена и в Баргузинском заповеднике (Гусев, 1966).
Росомаха относится к стопоходящим животным, поэтому имеет большую
площадь опоры и легко перемещается по рыхлому снегу, осваивая глубинные
заснеженные участки тайги. За счёт широкой стопы и незначительной массы
тела её весовая нагрузка на снег в 8-10 раз меньше, чем у копытных. Это
позволяет зверю зимой преодолевать большие (до 50-70 км) расстояния в
поисках корма, будь то олени, заяц-беляк или привада, поставленная у капкана.
Известен случай, когда в Туруханской тайге одна росомаха за сутки обошла
участки трёх охотников. Постоянно чувствуя заметённую лыжню, она
безошибочно следовала по путикам, срывая приманку и расстораживая
ловушки. Оседло живущие звери, обладая хорошей памятью, могут «спрямлять
ход» и точно выходить на уловистый капкан, в который часто попадаются
белка или соболь. Обнаружив в нем еще живого зверька, росомаха давит его,
вырывает из ловушки и либо съедает, либо относит в сторону и прячет. Обычно
её «тайники» размещаются в радиусе до 60-100 м, а иногда и до 500 м. Свою
добычу она прячет как на открытых местах (полянах, тундровых участках),
так и под кронами старых деревьев. Раскопав в снегу ямку глубиной до 30-35 см,
сытая росомаха заталкивает в нее тушку и забрасывает снегом. Затем,
крутясь на месте, она утрамбовывает снег, образуя площадку диаметром 50-
70 см, на которой оставляет мочевую точку. Иногда как бы обеспечивая
надёжность захоронений, зверь делает 2-3 «ложные» ямки, утаптывает снег и
метит его, а добычу прячет где-нибудь в другом, более потаённом месте. В
течение долгой зимы содержимым подобных кладовых, по-видимому,
пользуются не только оседло живущие хищники, но и росомахи-мигранты,
которые, попадая в новые места, часто следуют маршрутом своих собратьев-
резидентов. Острый слух, природный инстинкт и тонкое чутьё помогают
хищникам избегать на путиках встреч с человеком, несмотря на то что в
голодные зимы они регулярно проверяют выставленные охотником капканы
(Туманов и др., 1999).
В условиях сибирской тайги зима является трудным периодом в жизни
зверей. Среднемесячные температуры достигают здесь -30- -36°С, а в отдельные
дни понижаются до -55- -60°С. Благодаря длинному остевому волосу, густой
подпушке и наличию жировой прослойки росомаху можно считать видом,
хорошо адаптированным к холоду. Однако в такое время года расход энергии
на поддержание температуры тела животных повышается, что заставляет их
увеличивать суточные передвижения в поисках корма. Замечено, что в
морозные дни (при -30 и -40°С) они активнее охотятся и чаще проверяют
настороженные охотниками самоловы. На маршруте росомаха передвигается
прыжками (до 1.15-1.20 м), а почуяв падаль или приваду - переходит на шаг,
долго топчется и принюхивается (Туманов и др., 1999).
77
В Красноярском крае (Кожечкин, 1995, 1999) большая выносливость и
площадь опоры, позволяющие легко передвигаться в зимнее время,
обеспечивают росомахе успешность охоты в верхней, глубокоснежной части
горно-таёжного комплекса, где ею добывается около 64% оленей. Чаще всего
хищник нападает на них во второй половине зимы, когда высота снежного
покрова достигает максимума. При этом протяженность преследования,
нападения и умерщвления росомахой самцов марала составляла 150-5000 м,
а самок - 50-400 м. В отдельных случаях сеголетки копытных, зимующие при
глубокоснежье, погибали практически на месте. Судя по пяти троплениям,
хищник стремится согнать оленей со склона в заснеженную пойму, где они
быстро выбиваются из сил. По мнению автора, прослеживается определённая
зависимость между результативностью нападения росомах на маралов и
глубиной снежного покрова (табл. 26).
По материалам П.И. Данилова (Данилов, Туманов, 1976а), полученным
при троплениях зверей в Карелии (п-10), средняя величина суточного наследа
хищника - 19.5 км (2.5-35 км). В шведской и финской Лапландии испуганная
росомаха может убежать на расстояние до 40 км, а наибольшая протяженность
охотничьего хода самца достигала 70 км (Krott, 1959,1960). По наблюдениям,
проводимым в Швеции (Bjarvall et al., 1996), за сутки росомаха наиболее
активна в период от 18 до 6 ч утра. При этом известны случаи, когда она
отдыхала в ночные часы или, наоборот, интенсивно охотилась в промежуток
Таблица 26/ТаЫе 26
Успешность охот росомахи при разной высоте снежного покрова
(по: Кожечкин, 1999)
Success of wolverine’s (Gulo gulo) hunting with different height of the snow cover
Месяц/ Month Длина учетных маршрутов, км/ The length of count routes, km Высота снежного покрова, см/ The height of the snow cover, cm Встречаемость следов на 10 км маршрута/ Occurrence of tracks per 10 km of the route Обнаружено маралов, задавленных хищником, особей/ The number of discovered red deer (individuals), killed by predator
Октябрь/ October 160 8.4 ±0.39 0.06 -
Ноябрь/ November 378 22.4 ± 1.34 0.15 -
Декабрь/ December 132 33 ± 1.95 0.07 -
Январь/ January 350 60 ±0.52 0.31 1
Февраль/ February 178 64 ± 0.75 0.84 4
Март/ March 297 77.2 ±0.67 0.97 6
Апрель/ April 18 81.3 ±3.77 1.10 1
78
времени от 1 до 3 ч ночи. П. Кротт (Krott, 1960) вообще считает, что этот
хищник может попеременно бодрствовать и спать через 3-4 ч. Автор делает
вывод, что голод, оттепель, полнолуние и северное сияние значительно
повышают её активность, а затяжные дожди, снегопад или жаркая погода
увеличивают продолжительность периода покоя.
По наблюдениям А.В. Кречмара (устное сообщение), в Магаданской
области росомаха, передвигаясь постоянно со скоростью 40 км/ч, способна
упорно (измором) добирать зайца-беляка, не отвлекаясь при этом на другие
жертвы, встречающиеся на ее пути.
Ухудшение условий добывания пищи, особенно при высоком снеговом
покрове, может быть причиной перемещения хищных млекопитающих на новые
места, подобно тому как это происходит при неурожае кормов. Такие сезонные
кочевки регулярно отмечаются даже у видов, плохо приспособленных к
передвижению по глубокому и рыхлому снегу, т.е. имеющих большую весовую
нагрузку на площадь опоры. Так, норок и хорьков вынуждают совершать
подобные перекочевки трудности, связанные с добычей корма в связи с
промерзанием нор мышевидных грызунов и мелких водоёмов, где они
отлавливают земноводных. Обитающие в этих местах хищники смещаются к
устьевым участкам ручьев и речек, а часто и к населенным пунктам. Помимо
Рис. 21. Суточная активность горностая в разные месяцы
(по наблюдениям в неволе).
7 - октябрь; 2 - февраль.
По оси абсцисс - время суток, ч; по оси ординат - активность, мин.
Fig. 21. Circadian activity of the ermine (Mustela erminea) in various months
(according to observation in captivity).
1 - October; 2 - February.
Abscissa - hours of the day; ordinate - circadian activity, min.
79
названных факторов на изменение активности куньих заметное влияние
оказывают физиологическое состояние животных и погодные условия, такие
как наст, ветер, осадки и температура воздуха: в европейской части России,
когда она опускается ниже -20- -25°С, зверьки покидают свои убежища редко
и ненадолго. Вообще изменения двигательной активности в зависимости от
сезона года и температурного фактора у всех куньих проявляются очень
отчётливо (Волков, 1969; Вшивцев, 1972; Вайсфельд, 1977; Войлочников,
1977; Сидорович, 1990, 1991; Gerell, 1967, 1968; Baskirk et al., 1988; Kruuk,
1989, 1995; Jedrzejewski et al., 1989, 1993). Ниже приводятся материалы,
собранные нами совместно с П.И. Даниловым в различных областях Северо-
Запада европейской части России (Данилов, Туманов, 1976а, б).
Зависимость активности ласки и горностая от погодных условий особенно
отчётливо проявляется в зимнее время. В сильные морозы (ниже -25°С) они
редко оставляют своё убежище. Так, во время наблюдений зимой за тремя
горностаями удалось установить, что они не выходили на охоту по 2-3 дня,
когда температура воздуха опускалась до -27- -32°С. Во время поиска добычи
горностай тщательно обследует места возможного пребывания грызунов,
пробегая за сутки в среднем 2.1 км. В малокормные годы его охотничий нарыск
увеличивается очень заметно. Зимой зверёк нередко охотится под снегом,
проходя под ним до 10-40 м. В случае преследования горностай может забраться
на куст или дерево, иногда на высоту до 4-5 м. Горностай хорошо плавает.
Так, 6 ноября 1970 г. в Псковской области зверёк, спасаясь от преследования
собак, переплыл канаву шириной 2.5-3 м и забрался на молодую сосну.
Ласка, так же как и горностай, тщательно обследует все укрытия
мышевидных грызунов, вследствие чего наслед зверька чрезвычайно извилист.
Проходы её под снегом невелики и обычно не превышают 15-17 м. Охотится
ласка преимущественно в сумеречное и предрассветное время, однако при
недостатке корма её можно встретить и в дневные часы. Особенности
фотопериода северных широт заметно влияют на цикличность суточной
активности хищника. В Мурманской области и Карелии в весенне-летний
период чёткой периодичности в активности ласки не прослеживается: зверьки
довольно равномерно встречаются в любое время суток. В более южных
областях зоны ласка, как и горностай, во все сезоны года активна
преимущественно в сумеречно-ночное время, однако при недостатке корма в
холодную погоду ее можно встретить и в дневные часы. Зимой миофаги менее
подвижны и реже выходят из гнездовой камеры (рис. 21). В зависимости от
уровня обилия мышевидных грызунов длина суточного хода ласки достигает
0.5-0.8 км.
По данным Дж.Д. Локки (Lockie, 1966), дневная потребность в пище и
размеры тела горностаев и ласок находятся в отношении 2:1, а размеры
охотничьих территорий - приблизительно 8:1. Эта разница объясняется тем,
что ласки могут эксплуатировать популяцию полёвок эффективнее горно-
стаев, так как они много охотятся в норах, под землёй. В то же время горностай,
чтобы добыть соответствующее его потребности количество полёвок, вынужден
охватывать заметно большую территорию.
80
Суточная активность лесных хорьков на Северо-Западе России в
значительной мере определяется погодными условиями и наличием пищи. В
метели и сильные морозы они могут в течение нескольких суток не покидать
свои убежища. Содержащиеся в вольерах зверьки при температуре -25°С и
ниже не выходили из гнёзд и не поедали корм. В такие дни они в основном
крепко спали, зарывшись в сено и свернувшись клубком.
Характер охотничьего поиска лесного хорька во многом определяется типом
биотопа. На полях и в лесу его охотничий нарыск имеет резко выраженные изломы
и нередко пересекается. У зверьков, живущих по берегам рек, суточный ход
относительно прямолинеен, хотя и следует изгибам русла. В среднем длина его
охотничьего маршрута, по нашим наблюдениям, составляла 4 км, но при дефиците
корма увеличивалась примерно в 2 раза. Хорьки хорошо плавают и ныряют:
зимой 1969 г., отличавшейся низкой численностью грызунов в Псковской области,
основными их жертвами были земноводные. В январе нам удалось установить по
следам на снегу, что один зверек, многократно ныряя в полынью, вытаскивал на
лёд лягушек и относил их по тропе в сарай, который находился в 30 м от ручья.
Судя по всему, хорёк жил под сараем давно и в большие морозы отсиживался в
своём убежище, питаясь имеющимися запасами.
Длительные перемещения на большие расстояния лесному хорьку, по-
видимому, не свойственны. Так, в 80-х годах в верховьях р. Добрицы (бассейн
р. Псковы, Псковская область) нами было выпущено 50 зверьков, преиму-
щественно молодых особей, родившихся в неволе от пойманных в природе
родителей. Все они были маркированы ушными метками. Через 2-3 недели
следы их жизнедеятельности и самих хорьков обнаруживали на расстоянии 2-
3 км от места выпуска. Несмотря на режим охраны, в осеннее время, спустя
2.5-3 мес после начала опыта, 4 особи (1 самка и 3 самца) были отловлены
охотничьими собаками: самка - на расстоянии 2 км, самцы - 7 и 13 км от
исходной точки. Судьба остальных зверьков осталась неизвестной. Учеты
численности местной популяции лесного хорька в последующие 2-3 года
существенного роста их поголовья не выявили.
Европейская норка, как и другие мелкие куньи, в основном активна в
вечернее и предрассветное время. Весной и летом суточная активность
увеличивается, и её можно встретить практически в любое время, хотя
наибольшее число встреч приходится на утренние и вечерние часы. Осенью
зверьки часто кочуют, что связано с интенсивным расселением молодняка и с
увеличенной потребностью в кормах. В связи с тем что рацион представителей
данного вида сравнительно разнообразен (полёвки, лягушки, рыба и т.д.),
суточные передвижения зверьков невелики и обычно не превышают 2.5-3 км.
В отличие от американской норки европейская зимой редко совершает
большие переходы и даже в случае опасности не забирается на деревья. В
декабре-феврале она ведет скрытный образ жизни, сосредоточиваясь вблизи
кормных мест у незамерзающих участков рек. Лишь однажды, в декабре
1963 г., в Ленинградской области (Тосненский р-н) нами была добыта
молодая самка в старом лесном завале, примерно в 500-600 м от реки.
81
При начавшемся промерзании верховий ручьев и небольших речушек
европейские норки, которые обычно селятся на мелких и нешироких водотоках,
начинают «скатываться к большой воде» - к устьевым участкам, где в холодное
время года в омутах стоит рыба и концентрируются на зиму лягушки. Так, в
ноябре 1975 г. на 5 км участке по р. Чернушка (Псковская обл., бассейн р.
Локня) нами были выставлены 15 капканов на норку. Из них «работал» только
один, установленный на берегу в комлевой части старого пня в 100 м от устья,
где за неделю попались 4 зверька (2 взрослых самца и 2 сеголетка). Судя по
следам, все особи оказались здесь в результате миграции с верховий реки.
У американской норки на Северо-Западе России суточный цикл активности
поздней осенью и зимой можно разбить на 2 периода: утренний (с 5-6 до 9-10 ч)
и вечерний (с 16-17 до 21-22 ч). В весенне-летний сезон, особенно в северных
областях региона, строго выраженной циклики в суточной активности зверьков
не наблюдается, и они часто выходят на жировку даже в дневные часы. При
сильном морозе активность американской норки заметно снижается.
Наблюдения за самкой, проведенные в Карелии в феврале при температуре
воздуха -30- -40°С и глубине снегового покрова до 80-90 см, показали
следующее. За 4 суток зверёк 2 дня (10 и 13 февраля) не показывался на
поверхности снега, а 2 других (11 и 12 февраля) выходил из убежища, пересекал
несколько раз ручей и вновь скрывался под снегом. Суточный его наслед в эти
дни не превышал 50-60 м, хотя в хорошую погоду при слабой обеспеченности
кормом он может составлять 5-8 км.
В период глубокоснежья, особенно на водоёмах с подмытыми каменистыми
берегами, норка много ходит пустолёдкой. При этом максимальная длина
непрерывного подснежного хода достигает 100-150 м. Продолжительность её
пребывания под водой около минуты. Обычно, проплыв 5-8 м, зверёк
выныривает или, достигнув берега, уходит от преследователей пустолёдкой,
бобровыми ходами и естественными пустотами под корнями деревьев. В случае
опасности довольно свободно забирается на деревья на высоту до 4-7 м, чего
нельзя сказать о европейской норке. По результатам радиотроплений,
проведенных в Швеции (Gerell, 1970), холодная погода в зимнее время является
причиной общей низкой активности американских норок и проявления чёткой
тенденции к увеличению их дневных перемещений. В период беременности
(апрель-май) самки демонстрировали очень низкую активность. В декабре
зверьки могли в пургу и после неё по 2 суток не выходить из гнезда. В обычные
дни их суточный наслед вдоль берега составлял 1-2 км.
Как и лесному хорьку, американской норке свойственно делать запасы
пищи, которые можно обнаружить в любой сезон. Например, в мае 1969 г. в
Ленинградской области в гнезде щенной самки нами были найдены 4 налима
длиной 17-19 см и водяная полёвка. В конце июня в другой выводковой норе
был обнаружен налим длиной 20 см, а в июле, уже в Лапландском заповеднике,
около убежища самца были найдены 26 тушек норвежского лемминга.
Очевидно, инстинкт запасания пищи выражен сильнее у самок, что помогает
им и благополучно выращивать молодняк, и пережидать периоды с
82
неблагоприятными погодными условиями. Так, в ноябре 1973 г. в Суоярвском
районе Карелии в норах, принадлежащих 2 самкам, были обнаружены 9 и 11,
а в декабре 1999 г. в Волховском районе Ленинградской области, в убежище
еще одного зверька - 26 обездвиженных лягушек. Предварительные наблюдения
за ними показали, что все 3 самки большую часть времени проводили в норах,
расположенных в крутом берегу реки, и редко отлучались от них. В оттепели
они еще оставляли след, но даже в эти дни отбегали от своего постоянного
гнезда не более чем на 50-100 м. В морозные дни (-20- -23°С) зверьки по 3 дня
не выходили из нор. Наблюдения за поведением самки в Волховском районе
показали, что уже на следующий день около её убежища отсутствовали запасы
земноводных, которые накануне были нами вынуты из гнезда и положены на
снег. За ночь зверёк перетаскал всех лягушек в свою новую, более сложно
устроенную нору. В те же морозные дни некоторые самцы предпринимали
значительные переходы вдоль берегов рек к новым, более кормным местам.
Интересные данные о перемещениях американских норок получены
А.Я. Васеневой (19726) в Хабаровском крае. Здесь на р. Хунгари (один из
правых притоков р. Амур) в августе 1966-1970 гг. было поймано и помечено
ушными «сережками» с дополнительной ампутацией фаланги пальца 272
зверька, из которых 67 особей выпустили в места их поимки, а 205 перевезли
на новые, близлежащие территории. В первом случае повторно удалось
отловить 21 норку. Из них 16 животных попались в местах исходного обитания
или на расстоянии от него до 1 км, а 3 - до 2-3 км и только одна молодая самка
за 6 дней после первого мечения ушла дальше 17 км.
2 4 6 8 10 12 14 16 18 20 22 24
Рис. 22. Суточная активность барсука в летнее (7) и осеннее (2) время
(по: Данилов, Туманов, 1976).
По оси абсцисс - часы суток; по оси ординат - относительное количество встреч
животных, % от общего числа встреч в сутки.
Fig. 22. Circadian activity of badger (Meles meles) in summer (7) and autumn (2).
Abscissa - hours of the day; ordinate - relative number of animals’ occurrence, % of
total number of occurrence during the day.
83
Из 205 норок, перевезенных после маркировки на новые территории, было
отловлено 68 особей, в том числе 32 (47%) возвратились в места их
первоначальной поимки. Среди последних взрослые самцы составляли 31.3%,
взрослые самки - 46.9%, молодые самцы - 21.8%, а из самок-сеголеток ни
одна, даже перемещенная всего на 200-300 м, в свои прежние угодья не
вернулась. Максимальное расстояние, которое преодолевали норки,
возвратившиеся назад, оказалось равным у самцов взрослых 3 км, у молодых
8.5 и у взрослых самок 6 км.
Сводные данные по миграции зверьков на реке за 5 лет наблюдений
показали, что большинство из них (64.7%) перемещались на небольшие
расстояния и были выловлены повторно в радиусе 2 км от места первоначальной
поимки. На 3-10 км ушли 14.8% американских норок, а 9.2% особей удалились
от него на 10-20 км. Наибольшее расстояние в 60-80 км оказалось пройденным
5 зверьками (4 самца и 1самка), при этом все они в год мечения были
сеголетками.
Европейский барсук поисками пищи занят преимущественно в утренние
и вечерние часы суток (рис. 22), однако осенью его можно встретить и во
второй половине дня, после 16 ч. Так, в сентябре 1982 г. в Ленинградской
области на поле нескошенного овса наши собаки (лайки) дважды
обнаруживали жирующих зверей в дневные часы. Весной барсуки активны
также бывают днём. В апреле-мае уже в Псковской области встречи с ними
отмечались в 11 ч и после полудня. По-видимому, в это время на жировку
чаще выходит молодняк (годовалые звери), не успевший набрать с осени
достаточное количество жировых запасов, которые интенсивно расходуются
в зимние месяцы.
Наблюдения охотоведа П.Д. Иванова (устное сообщение) за актив-
ностью барсуков вне нор в Ленинградской области показали, что их кормо-
Рис. 23. Суточная активность выдры (по: Данилов, 1969).
Условные обозначения как на рис. 22.
Fig. 23. Circadian activity of the otter {Lutra lutra).
Abscissa - hours of the day; ordinate - relative number of animals’ occurrence,
% of total number of occurrence during the day.
84
добывающая деятельность летом начинается около 19 ч, а прекращается к
6-7 ч утра. За это время звери 2-3 раза возвращаются к норам, от которых
не отходят более чем на 1.5-2.5 км. В Карелии (Ивантер, 1973) барсук
обычно дважды за сутки покидает нору. Первый раз он уходит на кормежку
в 20-22 ч и возвращается в 1-2 ч ночи. Во второй раз, покинув нору в 3-4 ч,
возвращается около 7 ч утра. Однако в период гона барсука удается
наблюдать и в дневные часы.
Сходные данные были получены и во Франции (Bonnin-Laffargue, Canivenc,
1961), где животные были активны только в сумеречно-ночное время,
отправляясь на охоту после захода солнца и заканчивая её с восходом. За этот
период они могли несколько раз возвращаться в жилище и снова покидать его.
При этом наибольшее их удаление от нор составляло 1-2 км.
Активная жизнь барсука продолжается до конца октября. Последние
известные нам случаи добычи зверей вне нор датируются 14 и 25 октября, а
также 7 ноября. С продвижением на север сезонная смена образа жизни этих
хищников проявляется более отчетливо. Если в Псковской и Новгородской
областях выходы барсуков из нор в зимне-весенние оттепели нередки, то в
Карелии они практически не наблюдаются. Во время таких выходов звери
обычно не удаляются от своих жилищ далее 50-70 м. В южной части ареала
вида зимний сон барсука непродолжителен, а в Крыму отсутствует вовсе
(Алгульян, 1940; Корчмарь, 1962; Гептнер и др., 1967). В горных районах
Дагестана и Азербайджана нам также не раз приходилось встречать молодых
зверей на жировке в декабре и январе. Первые выходы зверей из нор после
зимнего сна отмечены в Псковской и Новгородской областях 15 и 20 марта, а
массовые - в начале апреля. Таким образом, этот период продолжается у них
около 5 месяцев. В Карелии и на юге Архангельской области начало выхода
барсуков из нор наблюдали в середине апреля, а массовый - в первой декаде
мая. Соответственно продолжительность зимнего сна здесь дольше, чем на
юге региона (Данилов, Туманов, 1970).
Подвижность выдры заметно увеличивается в конце зимы-начале весны
(февраль-март). В это время её следы (даже по рыхлому снегу) тянутся на
большие расстояния вдоль реки по направлению к незамерзающим, богатым
кормом участкам. Так, на юго-востоке Карелии (Пудожский р-н) нам
неоднократно приходилось наблюдать следы одиночных зверей или самки с
молодыми, которые по береговой кромке леса и запорошенному снегом льду
пересекали часть Водлозера, направляясь к устью р. Водла. Длина этих
маршрутов была не менее 1.5-2 км. В Шотландии (Kruuk, 1995) расстояние
между основными убежищами выдры составляло около 1 км, а длительность
пребывания под водой во время охоты не превышала 49 с, причем только
19.2% её ныряний в поисках пищи оказывались успешными. В течение суток
выдра активна в вечерние и утренние часы, при этом утренний период более
продолжителен (рис. 23). В северных регионах (Кольский п-ов, Карелия и
Архангельская обл.) в летний сезон строгой цикличности в её активности нет
(Владимирская и др., 1953; Данилов, 1969; Вайсфельд, 1977).
85
Собранные материалы по охотничьей деятельности изучаемых хищников
показывают, что они с разной степенью интенсивности осваивают участки
обитания. Миофаги на добычу пищи затрачивают больше времени и заметно
тщательнее по сравнению с другими видами обыскивают кормовой участок.
В Западной Сибири (Терновский, 1977), в местах, где скапливались грызуны,
суточный маршрут ласки не превышал 1 км, а горностая и колонка -1.5-2 км,
однако в местах, «бедных» полёвками, они совершали переходы по 5-15 км,
делая в поисках пищи до 60 «нырков» в снег. В годы высокой численности
водяной полёвки суточный наброд колонка в поисках пищи составлял от 50 до
2500 м или в среднем 450 м (по 32 троплениям). В следующем сезоне, при
обедненной кормовой базе, охотничий маршрут колонка в течение суток
варьировал по протяженности от 400 до 4500 м, составляя в среднем 1700 м
(по 25 троплениям). По свидетельству автора, в Западной Сибири светлый
хорёк на участках с различной степенью кормности проходил за сутки от 1 до
11 км. За неделю охоты маршрут зверька составил 26 км. За время переходов
он сделал 24 «нырка» в снег в поисках жертвы, которую при удачной охоте
поедал в её же убежище.
Таким образом, для всех исследованных видов характерна тесная
зависимость между длиной суточного хода животных и состоянием кормовой
базы. В этом плане хищники-стенофаги наиболее заметно реагируют на
уменьшение количества пищи в природе. Например, на Северо-Западе России
при низкой численности полёвок (1-2 особи на 100 ловушко-суток - л.с.)
суточный ход обыкновенной ласки и горностая увеличивается примерно в 2
раза по сравнению с таковым в годы обилия грызунов (7-9 особей на 100 л.с.).
В то же время у эврифага (лесной куницы) тесной связи между величиной
охотничьего пути и наличием одного из видов кормов не отмечается. Она
прослеживается только при изменении численности одновременно 2-3 основных
компонентов пищи (Данилов, Туманов, 1976а). Всем хищникам в холодное
время года приходится затрачивать много энергии на поиск жертвы, что
находится в соответствии с количеством и качеством потребляемого корма,
массой тела и уровнем их теплообмена. В целом представители крупных видов
имеют большую протяженность суточного хода и площадь охотничьего участка
по сравнению с мелкими.
Многие куньи характеризуются одинаковым набором кормов и ритмом
суточной активности, что, казалось бы, должно обусловливать конкурентные
отношения между ними, однако нам не приходилось наблюдать ни прямого
уничтожения, ни даже преследования мелких хищников крупными. Нет ни
одного достоверного случая, подтверждающего конкурентную борьбу
лесного хорька с норками или разных видов норок между собой. Чаще
наблюдается другая картина - использование чужой добычи хищниками
малых размеров, т.е. явление комменсализма. Сведения об отсутствии острой
конкуренции между представителями семейства куньих можно найти в
работах П.И. Данилова (1968), Д.В. Терновского (1972), Н.Н. Гракова
(1981), В.Е. Сидоровича (1991), а также В.В. Рожнова с соавторами (Соколов,
86
К rt Я Й 1* П Ж ЯГ Ч <№ l> 11 13 31 t I 3 13 3 7 3 3
лети жшгтпгш!
0 Я Й й Иг Ч <3 0 ft li № г> ti 33 &г 1 t 3 M 4 # 7 J S
•fmejwwowwiiirj
Время суток, ч
Рис. 24. Актограммы суточной активности мышевидных грызунов.
1 - желтогорлая мышь; 2 - полевая мышь; 3 - рыжая полевка;
4 - обыкновенная полевка.
Fig. 24. Actograms of circadian activity of myomorph rodents.
1 - yellow-necked mouse (Apodemus flavicollis)-, 2 - stripped field mouse (Apodemus
agrarius)'. 3 - bank vole (Glethrionomys glareolus):, 4 - common vole (Microtus arvalis).
Рожнов, 1979, 1989; Рожнов, Петрин, 1998), который изучал поведение
мелких хищников в полигонах-вольерах. Кормовая специализация куньих
связана не только с длиной охотничьего поиска, но и с периодичностью
изменений активности в течение суток. Так, для удовлетворения потребности
в пище, необходимой для поддержания интенсивного энергообмена, особи
мелких видов выходят на кормёжку заметно чаще крупных, что объясняется
более высоким уровнем их обменных процессов.
Активность животного представляет собой ответную реакцию организма
на ряд эндогенных и экзогенных факторов, что делает её показатели весьма
лабильными не только для мелких, но и крупных хищников семейства (Krott,
1960; Magoun, 1978, 1982; Б. Новиков, 1933; Bjarvall et al., 1996). Тем не менее
в природных условиях большинство куньих демонстрируют два пика активности:
утренний и вечерний (Терновский, 1958; Данилов, Ивантер, 1967; King, 1983,
1989; Сидорович, Яковец, 1989). При содержании в вольерах у мелких хищников
четко прослеживались два основных периода сумеречной активности (дифазиый
тип), длительность которых редко превышала 2-2.5 ч. В то же время у самого
мелкого представителя семейства (обыкновенной ласки) интервалы между
периодами бодрствования настолько коротки, что ее следует относить к видам
с полифазиым типом активности (Туманов, 19766; Ивантер и др., 1985).
С увеличением размеров животных общая продолжительность их
двигательной активности сокращается, а период времени, когда они отдыхают,
заметно возрастает. Существенные различия в продолжительности суточной
87
Таблица 27/Table 27
Некоторые показатели двигательной активности мышей и полёвок в эксперименте
Some parameters of moving activity of mice (Murinae) and voles (Microtinae)
in experiment
Виды/ Species п Скорость передвижения, м/мин/ The velocity of movement, m/min Расстояние, пробегаемое за сутки в актографе, км/ The distance covered per day in the aktograph, km
n <3<3 n ?$
Желто гордая мышь/ Apodemus flavicollis Полевая мышь/ Apodemus agrarius Рыжая полёвка/ Clethrionomys glareolus Обыкновенная полёвка/ Microtus arvalis 14 58.6 ±2.2 15 12.3 ±0.7 14 11.6 ±0.6 18 53.3 ±0.9 11 10.1 ±0.8 10 9.5 ±0.9 16 49.3 ±1 12 8.9 ±0.7 13 8.5 ±0.6 14 36.2 ±0.7 9 8.2 ±0.7 9 7.5 ± 0.7
активности и её распределении по часам достаточно отчетливо просле-
живаются в пределах всего отряда хищных млекопитающих, в том числе и
куньих (Калабухов, 1950; Соколов, Кузнецов, 1978). Например, у отловленных
в природе животных, не успевших свыкнуться с режимом кормления в неволе,
средняя продолжительность суточных передвижений в октябре была равна у
ласки 365 мин, у горностая 337, у американской норки 258 и у выдры 212 мин.
После утреннего периода активности дневная фаза отдыха длилась у ласки с
10 до 14 ч, у горностая до 16 ч, у американской норки до 18 ч и у выдры с 8 до
18 ч (Туманов, 19766). Среди семейства куньих представители рода Martes
заметно выделяются своей подвижностью. У животных, разводимых в вольерах,
она достигала у соболя 601-647 мин, у лесной куницы 411-568, а у более крупной
харзы 396-433 мин в сутки (Пономарёв, 1944). У хищников амфибионтов,
близких к ним по таксономическому рангу и массе тела, этот показатель в
летний период имел меньшую величину: у американской норки 332-379 мин, а
у выдры, близкой по весу к харзе, 101-110 мин (Ефремов, 1968; Шило,
Немшилова, 1989).
Таким образом, собранные материалы демонстрируют тесную связь
поведенческих реакций куньих с их размерами и кормовой специализацией.
Увеличение спонтанной активности у мелких видов можно рассматривать как
приспособительную реакцию организма к поддержанию высокого уровня
энергообмена в результате интенсивного использования кормовых ресурсов.
Основу рациона большинства представителей семейства куньих
составляют мышевидные грызуны. В этой связи интересно сопоставить
распределение периодов бодрствования и покоя у хищников и их жертв - мышей
и полёвок. Результаты инструментальной записи суточной активности грызунов
свидетельствуют о том, что трудности поиска концентрированных кормов
(семян), которыми питаются мыши, обусловливают и более сложные формы
их поведения по сравнению с полёвками, потребляющими сочные, зелёные части
88
Таблица 28/ Table 28
Сравнительная характеристика индекса мозга некоторых видов куньих,
разводимых на зверофермах (по: Боголюбский, 1941)
Comparative characteristics of brain index in some species of mustelids, bred in fur-farm
Виды/ Species Средняя масса тела/ Average body mass Индекс мозга/ Index of brain
п 1 г/g п | %о
Соболь/ Martes zibellina 19 1057 7 19.08 ± 1.83
Куница лесная/ Martes martes 33 1214 11 14.10 ± 1.1
Норка американская/ Mustela vison 8 725 12 10.17 ±0.4
Скунс/ Mephitis mephitis 10 1965 5 5.47
растений. Для первых характерны высокая скорость передвижения и
продолжительные периоды покоя (рис. 24).
Самая короткая в течение суток активность (в среднем с 19 до 3 ч утра)
наблюдается у лесной и желтогорлой мышей (монофазный тип). Полевая
мышь, которая наряду с семенами потребляет еще и зеленые мало-
калорийные корма, подвижна в основном в сумеречные часы - с 16 до 22 и
с 3 до 7 ч (дифазный тип). Среди полёвок рыжая и красная проявляют
активность примерно через равные промежутки времени (2.5-3 ч), усиливая
её в сумерки. У серых полёвок наблюдаются частые, но короткие периоды
бодрствования. Они следуют через каждые 1.5-2 ч отдыха, что, вероятно,
объясняется питанием зверьков сочными, но малокалорийными вегета-
тивными частями растений (полифазиый тип). Кроме того, общая
продолжительность суточной активности у грызунов возрастала с
увеличением доли сочных зелёных кормов в их рационе. Так, средняя
продолжительность спонтанной активности, записанной с помощью
актографа, была равна у желтогорлой мыши 183 ± 14.7 мин, а у зеленоядной
обыкновенной полёвки 375 ± 28 мин. Полевая мышь и рыжая полёвка,
потребляющие смешанные корма, занимают по этому показателю про-
межуточное положение: фазы активности составляют у них соответственно
266 ± 27 и 225 ± 27.7 мин (Туманов, 1969, 19766).
По нашим исследованиям (Ивантер и др., 1985), мыши по сравнению с
полёвками передвигаются быстрее, а их кормодобывающая деятельность
занимает сравнительно короткий промежуток времени, в течение которого
они способны преодолевать значительные расстояния. По устному сообщению
акад. П.А. Свириденко, суточный пробег, установленный по следам на снегу
в природе, у желтогорлой мыши составляет 500-600 м, у полевой - до 200 м, у
рыжей полёвки - до 100-200 м, а у обыкновенной - 45-60 м. Характер
передвижения мышей и полёвок неодинаков: полёвки передвигаются бегом, а
мыши - бегом и прыжками. Эти особенности явились причиной морфо-
89
Таблица 29/ Table 29
Характеристика осенне-зимней охотничьей деятельности куньих и относительные
размеры их головного мозга (Данилов, Туманов, 1976а, с добавлениями)
Characteristics of autumn-winter activity of mustelids and relative sizes of their brain
Виды/ Species n Средняя длина суточного хода, км/ Average length of circadian way, km Количество попыток добыть корм/ The number of food- obtaining attempts n Индекс/ Index, %c
за сутки/ during a day на 1 км пути/ on I km of way головного мозга/ brain мозжечка/ little brain
Ласка обыкн./ Mustela II 0.7±0.1 18.2±1.2 26 20 35.8±1.1 4.2±0.4
nivalis Горностай/ Mustela 31 2.1+0.1 16±0.8 7.6 13 24.8+1.6 2.9±0.1
erminea Хорёк лесной/ 24 4+0.4 10.2+0.8 2.6 13 14.8+1.5 1.6+0.3
Mustela putorius Куница лесная/ 32 5.5+0.4 9.7+O.5 1.9 12 18.4+0.8 2.1+0.1
Martes martes Норка европ./ Mustela 20 1.8±0.2 11.0+1.1 6 10 11.9±0.8 1.6±0.3
lutreola Норка америк./ Mustela 23 2.6±0.4 11.6±1 4.3 15 ll.l±0.6 1.6±0.2
vison Выдра/ 15 8+0.6 9 10.6+0.4 1.2+0.1 '
Lutra lutra Росомаха/ Gulo gulo 10 19.5±3.5 - - 5 7.7±1 0.9±0.1
Барсук/ 6 1.6+0.4 5 6.1±1.3 0.8+0.2
Meles meles
логических различий в строении костей их таза и задних конечностей (Becker,
1954).
Результаты наших экспериментальных работ показали, что скорость
передвижения мышей и полёвок различна (табл. 27). Наибольшей она оказалась
у желтогорлой мыши, которая прекрасно ориентируется в пространстве и
обладает точной и быстрой координацией движений, а наименьшей - у
малоактивной обыкновенной полёвки. Близкие данные по рыжей полевке
получены К. Вельтеном (Velten, 1954), по материалам которого скорость
пробежек зверьков этого вида составляла 42-62 м/мин.
90
Таблица 30/ Table 30
Размеры головного мозга и мозжечка у мелких видов куньих разного пола (ноябрь)
The sizes of brain and cerebellum of mustelid small species of different sex (November)
Вицы/ Species Пол/ Sex п Масса, г/Mass of, g
тела/body головного мозга/ brain мозжечка/ little brain
Ласка обыкновенная/ Mustela nivalis <3<3 6 78.4 ± 3.2 2.4 ±0.1 (31.7) 0.30 ±0.01 (3.8)
$? 5 41.3 ±3.9 1.6 ±0.1 (39.2) 0.18 ±0.01 (4.3)
Г орностай/ <s<s 5 223.5 ± 10.7 4.5 ±0.3 (20.3) 0.63 ±0.02 (2.8)
Mustela erminea $? 6 108.3 ± 6 3.1 ±0.1 (28.3) 0.33 ± 0.02 (3)
Солонгой/ <s<s 3 312.3 ±8.3 5.5 ±0.3 (17.7) 0.66 ±0.03 (2.1)
Mustela altaica 3 235.4 ± 5.6 4.8 ± 0.4 (20.4) 0.52 ± 0.02 (2.2)
Хорек лесной/ 33 5 765.4 ± 15.3 9.8 ±0.4 (12.8) 1.03 ±0.02 (1.3)
Mustela putorius ?? 5 354.2 ± 12.9 6.1 ±0.4 (17.2) 0.62 ±0.01 (1.7)
Примечание. Цифры в скобках обозначают весовой индекс головного мозга и
мозжечка, %о к массе тела.
Кормовая специализация мышей и полёвок находится во взаимосвязи не
только с общей продолжительностью их двигательной активности, но и с
изменением её в течение суток. Для удовлетворения высокой потребности в
пище, необходимой для поддержания интенсивного обмена веществ,
зеленоядные полёвки вынуждены кормиться чаще. У мышей, питающихся
калорийными кормами, в процессе эволюции выработалась способность
быстрее передвигаться и за меньший период активности пробегать в поисках
пищи большие расстояния. Данные обстоятельства, а также часто повторя-
ющиеся выходы полёвок из нор могут служить причиной их большего отлова
даже при равной численности с мышами, что и подтверждается анализом
пищевого рациона всех куньих. Однако необходимо отметить, что у хищников-
миофагов периоды суточной активности по времени в основном совпадают с
таковыми у мышей, свидетельствуя о высокой степени их специализации к
питанию разными видами грызунов.
Способ добывания корма и сложность локомоторных функций определяют
ряд поведенческих и физиологических особенностей животных, которые в
значительной мере сказываются на проявлении видовых различий в величине
и стоении центральной нервной системы. Двигательная активность является
хорошим показателем деятельности мозгового центра и служит одним из
факторов, которые определяют его величину в процессе цефализации
(Боголюбский, 1941; Стрельников, 1953, 1970; Полякова, 1956, 1960;
Никитенко, 1963,1966; Яскин, 1980,1981). Ещё в 1868 г. А. Брандт указал на
то, что чем мельче организм, тем деятельнее в нем физиологические процессы
и при прочих равных условиях больше относительные размеры головного
мозга. Его величина, а также мозжечка как центра координации движения,
91
связана с экологическими условиями жизни вида, в частности с уровнем его
двигательной активности, которая во многом определяется его кормо-
добывающей деятельностью и физиологическим состоянием организма. У
хищников прослеживается обратная зависимость относительной величины
центральной нервной системы от массы тела, возраста, пола и уровня
энергообмена. Среди куньих, близких по массе тела, большим индексом
головного мозга обладают представители видов со сложными и разнообраз-
ными локомоциями. Например, у лесной куницы он был достоверно выше,
чем у норок (/=6.5; />=99.9%) и лесного хорька (/—4.5; />=99.9%), примерно
равных по массе тела, а у выдры - по сравнению с барсуком (/=3.5; />=95%).
По свидетельству С.Н. Боголюбского (1941), среди хищников, разводимых
на зверофермах, представители рода Martes обладали большей относительной
массой мозга. Индекс этого органа у соболя был заметно выше такового у
скунса и американской норки, несмотря на её меньший живой вес (табл. 28).
Сложность локомоторных функций обусловливает и размеры мозжечка как
центра регуляции и координации движения животного, од нако здесь эти тенденции
не проступают достаточно отчётливо (табл. 29). Последнее, вероятно, объясняется
тем, что степень развития отделов головного мозга определяется не только
величиной, но и особенностями микростроения этого органа. Все исследования
по цитоархитектонике центральной нервной системы разных видов млекопита-
ющих из природных популяций свидетельствуют о тесной взаимосвязи строения
двигательных зон мозга с характером мышечной деятельности животных (Rensch,
1954; Астанин, Шевченко, 1954; Стрельников, 1970).
На основании ряда исследований по грызунам (Полякова, 1956, 1960;
Яскин, 1980,1981) известно, что гистологическое строение мозга разных видов
также четко различается по ширине слоёв, густоте клеток коры и проводящих
путей. При этом, несмотря на мелкоклеточность и нечеткую горизонтальную
стратификацию двигательного поля головного мозга, клеточные слои лучше
выражены у животных, которые быстро передвигаются и обладают сложными
локомоторными реакциями. Ганглионарный слой с характерными для
двигательной области коры мозга крупными пирамидальными клетками
выражен у разных видов неодинаково. Он наиболее широк и содержит крупные
пирамидальные клетки у активных видов - мышей. У менее подвижных форм
(полёвок) он выражен нечетко, а пирамидальные клетки имеют у них
сравнительно малые размеры.
Поскольку сравниваемые хищники обладают неодинаковой степенью
развития локомоторных функций, то, видимо, и двигательные отделы головного
мозга как рефлекторно-автоматического центра локомоций отражают в своей
структуре эти различия.
Взаимосвязь размеров головного мозга и мозжечка с уровнем подвижности
и кормодобывающей активностью куньих исследовалась методами множе-
ственного регрессионного, корреляционного и дисперсионного анализов.
Установлена высокая степень достоверности и надежности зависимости
индекса головного мозга и мозжечка изучаемых видов от специфики добывания
92
пищи и эффективности охот (табл. 29). В целом у представителей семейства
куньих коэффициент корреляции между этими показателями составлял для
головного мозга 0.998 и для мозжечка 0.997. Коэффициент детерминации был
равен соответственно 99.6 и 99.8%.
Известно, что при наличии полового диморфизма по массе тела у
млекопитающих прослеживается существенная разница между самцами и
самками по относительной величине внутренних органов, в том числе и
центральной нервной системы (Pucek, 1964; Шварц и др., 1968; Большаков,
1970, 1972; Ивантер, 1975). У мелких видов куньих взрослые особи разного
пола достаточно отчётливо различаются по массе тела, меньшей у самок, что
согласуется и с большей относительной массой их головного мозга и его
отделов (табл. 30). Кроме того, при сравнении представителей одного
семейства, близких между собой не только в филогенетическом отношении, но
и по особенностям образа жизни и кормовой специализации, наблюдается
закономерное снижение индекса головного мозга и мозжечка с увеличением
массы тела хищников.
Интенсивность охотничьей деятельности других представителей отряда
хищных млекопитающих, так же как и куньих, определяется в первую очередь
состоянием кормовой базы, временем года, погодными условиями и
индивидуальными особенностями животных разного пола.
Представители семейства кошачьих (Felidae) в поисках пищи мо-
гут осваивать значительные расстояния, однако эти перемещения чаще все-
го совершаются в пределах определенного участка обитания. Длитель-
ные кочевки им не свойственны. Тропления охотничьих маршрутов
обыкновенной рыси в разных частях ареала показали, что суточные ее
перемещения невелики. В Псковской и Новгородской областях в январе-
феврале в разные по обилию корма годы (л=8) они составляли 4-7 км.
Наблюдения за поведением зверей свидетельствуют о том, что при глубоком
снеге часть пути они проделывали по лесным дорожкам или по лыжне.
Нередко, взбираясь на поваленные и лежащие наклонно деревья, хищники
некоторое время на них задерживались, как бы прослушивая темноту леса,
а затем продолжали свой путь. Передвижение рысей по валежнику
наблюдалось не только в очень снежные зимы, но и при высоте снега 3-5 см.
По-видимому, это позволяет им бесшумно и неожиданно подкрадываться к
жертве и, обнаружив, добыть ее в прыжке, длина которого может достигать
2.5-3 м.
По материалам П.И. Данилова (Данилов и др., 1979), в южной части
Карелии за 8 сезонов средняя длина охотничьего маршрута рыси равна 5.7 км
(/г=57). При этом отдельные кросс-ходы зверей, не живущих постоянно на одной
территории, достигали 11-12.5 км. Такие перемещения отличались прямолиней-
ностью и малым количеством охот. Во время длительных переходов хищники
не караулили и не скрадывали добычу, а нападали на зайца-беляка или рябчика,
только наткнувшись на них. На Северо-Западе России, где в питании рыси
ведущая роль принадлежит зайцу-беляку, протяженность ее охотничьего хода
93
Таблица 31/Table 31
Средняя длина суточного хода рыси в разные месяцы зимнего периода
(по: Желтухин, 1984)
The average length of circadian way of lynx (Lynx lynx) in various months of winter period
Месяц/ Month Кол-во троплений/ The number of tracks Средняя длина суточного хода, км/ Average length of circadian way, km
Ноябрь/ November 7 8.9
Декабрь/ December 14 10.6
Январь/ January 10 11.2
Февраль/ February 14 10.2
Март/ March 8 8.9
находится в тесной связи с обилием жертвы (Данилов, Русаков, 1974). При
увеличении плотности населения зайца-беляка в 1.5-2 раза длина суточного
хода хищника резко сокращалась. В годы депрессии численности зайца она
составляла 8.4 км, а при его обилии - 3.2-3.9 км.
В Красноярском крае, в заповеднике «Столбы» (Зырянов, 1997), размеры
индивидуальных участков и охотничьих маршрутов рыси зависят от времени
года, мощности снежного покрова, обилия жертв и физиологического состояния
особей. По данным тропления 9 суточных наследов средняя длина пути зверей
составляла 8 км, хотя в отдельные дни они перемещались вдоль реки на 10-15
км и более. При остром дефиците пищи и неблагоприятных климатических
условиях охотничий поиск хищников заметно увеличивается. Оптимальными
для них следует считать сезоны со средним количеством зимних осадков и
умеренным температурным режимом.
Основные приемы добывания рысью пищи - это скрадывание и
подкарауливание жертвы из засады. Первый способ приводит к успеху чаще
других, второй также служит обычным элементом ее охотничьего поведения.
Обнаружив жировку зайца-беляка или след кабарги, рысь обычно ложится и
поджидает подхода жертвы, а затем выскакивает на нее из засады. В случае
неудачи рысь продолжает поиск новой добычи (Матюшкин, 1974 в; Зырянов,-
1980, 1997).
Близкие к этим данные получены М.Н. Смирновым (1997) в верховьях
Енисея, где автору удалось протропить зверей в феврале 1977 г. и ноябре 1985 г.
В первом случае (при обилии корма) длина пройденного хищником пути была
равна 3 км. На этом маршруте он сделал 9 мочевых меток, оставляя их на
тропе, поваленных деревьях, пнях и других предметах. Высота снежного
покрова достигала 50 см, но поскольку рысь шла старыми тропами, то
проваливалась только на глубину свежевыпавшего снега (3 см). Во втором
случае (при ухудшении кормовой базы и глубине снега 27-35 см) протяженность
охотничьего хода хищника составляла 8 км. Здесь рысь шла по рыхлому снегу,
94
после пороши, оставив 17 мочевых меток в основном в местах пересечения ее
маршрута с тропами копытных. Хищник явно предпочитал идти берегом озера,
опушкой леса, промерзшим болотом, редколесьем, избегая при этом густых
зарослей карликовых березок и участков, где не было дичи. Суточные
перемещения рысь обычно совершает в пределах своего охотничьего участка,
размеры которого варьируют в зависимости от состояния кормовой базы.
В европейской части России осенью и зимой уровень активности зверя
возрастает в утренние и вечерние часы, т.е. в период наибольшей подвижности
его основной жертвы - зайца-беляка. Весной и летом активность рыси, видимо,
не имеет строгой цикличности, что также объясняется особенностями поведения
добываемых животных и широким ассортиментом кормов в природе.
Массовых миграций хищников в одном направлении не наблюдается. Однако
известно, что в малокормные годы они более подвижны, их охотничьи
маршруты увеличиваются, учащаются случаи появления зверей вблизи
населенных пунктов, где они осмеливаются нападать на домашних животных
(Данилов и др., 1979).
Интересные наблюдения за суточной активностью и охотничьим
поведением рыси в районе южной тайги Верхневолжья проведены
А.С. Желтухиным (1984, 1987). По этим данным максимум двигательной
активности зверей приходится на сумеречно-ночные часы. Ее нарастание в
дневное время наблюдается в период гона. Наиболее часто они активны
днем в годы низкой численности зайца-беляка - основного компонента их
рациона питания. Более 70% встреч с рысями, приходящихся на дневные
часы, относится к бесснежному периоду года (май-октябрь), когда в светлое
время суток наблюдается и наибольшая активность их жертв. Протя-
женность суточного хода колеблется от 0.3 до 28 км, составляя в среднем
10.1 ±0.8 км. У самцов (?г=28) этот показатель равен 11.1 ±1.3 км, у самок
(л=19) - 9.3± 1.2 км. Длина суточного пути рыси заметно меняется по годам,
однако четкой ее зависимости от изменений численности зайца-беляка не
обнаружено. От начала зимы и до конца января величина суточного хода
зверей имеет тенденцию к росту, а затем, в феврале-марте, - к снижению
(табл. 31).
В первую половину зимы суточные перемещения зверей увеличиваются
параллельно нарастанию высоты снежного покрова и понижению температуры
среды. В феврале-марте, когда глубина снега достигает максимальной
величины (в среднем 40-50 см), а температура воздуха повышается,
наблюдается противоположная тенденция. В эти дни тонкая ледяная корка,
образующаяся после непродолжительных оттепелей, с заметным шумом
проваливается под тяжестью зверя, что затрудняет его охоту.
Пик двигательной активности хищников приходится на самый холодный
месяц. При слабых морозах, в мягкую погоду, наблюдаемую в начале и конце
зимы, суточный ход заметно уменьшается, что может объясняться успешностью
их охот. С увеличением мощности снежного покрова рысь отдыхает чаще. В
ненастную погоду (чередование обильных снегопадов с дождями при резкой
95
смене температуры) суточная активность подавляется. В эти дни звери могут
не выходить из убежищ иногда до 3 суток. Участок обитания самок составляет
в среднем около 70 км2, у самцов - 130-250 км2. Семейные участки (самка с
котятами) достигают 160-180 км2. Основными способами охоты является
скрадывание в процессе активного поиска жертвы. Из 59 зайцев-беляков,
добытых рысью, 75% были пойманы во время жировок, 22% - на переходах и
лишь 3% - на лежках (Желтухин, 1984).
При средней массе тела 15-17 кг рысь имеет сравнительно большую
площадь опоры. Ее весовая нагрузка на субстрат составляет 40-42 г/см2. Это
в 2 раза меньше, чем у енотовидной собаки, и позволяет ей даже в период
глубокого снега легко передвигаться в поисках добычи на расстояние в
несколько километров.
Другой представитель семейства кошачьих - лесной кот, за которым мы
наблюдали в Дагестане и ряде районов Азербайджана, явно избегает участки
леса с рыхлым снеговым покровом высотой более 15-20 см. При массе тела 4-
5 кг его весовая нагрузка на снег значительна - 100-125 г/см2. Это
обстоятельство обусловливает возможность обитания хищника только на
равнинной части региона, в предгорьях и по склонам гор с широколиственным
лесом, т.е. на территории с мягким климатом и неглубоким снежным покровом.
Обычно следы лесного кота встречаются на склонах, поросших кустарником,
буком или грабом, до высоты 1300-1500 м. Выше, в пояс хвойных лесов и
субальпийской зоны, он практически не поднимается.
Основу кормового рациона хищника составляют мышевидные грызуны,
поэтому его охотничьи маршруты пролегают по местам их концентрации -
долинам горных рек и ручьев, захламленным ветровалом распадкам, берегам
болот и малых озер или по краям каменистых россыпей. Посещая свои
постоянные участки у водотоков лесной кот кроме полевок-гигрофилов
добывает здесь уток, куликов и воробьинообразных птиц, разоряет их кладки,
ловит на мелководье рыбу (форель) и насекомых. Передвигается он бесшумно,
обычно шагом, реже легкой рысью, но потревоженный уходит прыжками
длиной более 1.5 м. В связи с высокой кормовой емкостью участков обитания
его суточные передвижения невелики и обычно не превышают 1.5-2 км. Иногда
он селится около поселков и горных селений, где добывает синантропных
грызунов и нападает на домашнюю птицу. Лесной кот - преимущественно
ночной хищник, поэтому большую часть дневного времени проводит в трещинах
и расселинах скал, в сухих убежищах под обрывами у воды и в дуплах старых
деревьев. Кроме того, нам приходилось обнаруживать его в заброшенных норах
барсуков и лисиц, а также во временных убежищах - укромных сухих местах,
в зарослях ежевики.
Обычно мягкий и рыхлый снег не позволяет лесному коту делать далекие
переходы (Диниик, 1914). В такие дни он подолгу отдыхает в теплых гнездах.
Осенью, во время нагула, зверь часто охотится не только ночью, но и днем. Об
этом свидетельствуют факты обнаружения в желудках добытых перед заходом
солнца хищников почти свежих остатков лесных мышей. В особо суровые,
96
многоснежные зимы лесной кот утрачивает обычную осторожность. Порой
даже в дневные часы он забегает на фермы в поисках тепла и пищи. Эти
наблюдения еще раз свидетельствуют о недостаточной его приспособленности
к морозам и глубокоснежью. В обычные годы состояние кормовой базы
определяет размеры индивидуального участка животного. Так, в богатых
пищей долинных кустарниках участок обитания лесного кота занимает 1-2 га,
а в горах, где запасы корма ограничены, - 50-60 га.
Камышовый кот, или хаус, на территории Кавказского региона в
основном селится в густых зарослях тростника и осоки по низким берегам
морских заливов, рек и озер. Сухие места среди кустарников он
предпочитает открытым стациям и лесным массивам по склонам гор, куда
выше 700-800 м над ур. м. не поднимается. Активен чаще ночью, но в
малокормные годы добывает себе пищу в дневные часы, выслеживая
водоплавающую и водно-болотную птицу, фазанов, молодых зайчат или
мелких грызунов. Хищник хорошо плавает и может забираться на деревья,
но охотится обычно в местах, покрытых камышами, ежевикой, осокой или
колючим кустарником. Пробирается по ним он очень осторожно, часто
прислушивается и, обнаружив жертву, в 2-3 прыжка настигает и хватает ее
(Динник, 1914).
Интересно, что длина хвоста камышового кота (в среднем у самцов 29.3
см, у самок 25) заметно меньше таковой у лесного (в среднем у самцов 32.4 см,
у самок 31.4). Отмеченная разница составляет у самцов 9.6, а у самок 20.4%.
Данные видовые различия, вероятно, объясняются особенностями образа жизни
зверей. Если камышовый кот обитает в зарослях кустарников, то лесной
представляет собой ярко выраженного обитателя лесных биотопов. Он хорошо
лазает по деревьям, совершая порой большие прыжки, во время которых его
хвост выполняет роль балансира (Насибов, 1970).
Обитающие на Кавказе еноты-полоскуны (семейство енотовых -
Procyonidae) встречаются в лесах по склонам гор и поймам рек, где устраивают
гнезда в дуплах широколиственных древесных пород (ясень, дуб, граб, бук,
белолистный тополь) или в старых норах лисиц и барсуков. В теплое время
года они часто ищут добычу вокруг водоемов, нередко вдали от леса. На зиму
откочевывают в лесные массивы, где в удобных дуплах впадают в зимний
сон, который в регионах с продолжительными морозными днями может длиться
у них с декабря и по конец февраля. В местах с непродолжительной и
сравнительно теплой зимой еноты также прекращают свою активность в
снежный период, но в солнечные дни, при таянии снега, они вновь начинают
бродяжничать в поисках добычи.
Как и другие впадающие в зимний сон хищники, еноты-полоскуны осенью
интенсивно кормятся. В их организме образуются значительные жировые
резервы, что позволяет благополучно пережить холодное время года. Звери
без жировых отложений или с их малым количеством в зимний сон не впадают,
бывают достаточно активными в холодное время года и до весны, видимо, не
доживают.
4 Зак. № 4309
97
Рис. 25. Время выхода и жировки медведей на овсах в сентябре на Северо-Западе
России (по: Данилов, Русаков, Туманов, 1979).
По оси абсцисс - время суток, ч; по оси ординат - встречаемость, % от общего числа
регистраций (п =144).
Fig. 25. The time of emergence and fattening in the brown bears (Ursus arctos) on the
oat grasses in September in the North-West of Russia
Abscissa - time of the day; ordinate - occurrence, % from total number of records.
На территории Кавказа (Гептнер и др., 1967) размеры участка обитания
взрослых животных связаны с обилием в их пределах пищи и с удаленностью
кормовых угодий от убежищ. Площадь такого участка обычно равна 4-9 км2
при радиусе 1-1.5 км от гнезда. Кормящая самка не уходит от выводка далее
чем на 600 м, при этом площадь ее обитания не превышает 144 га. Енот-
полоскун - сумеречно-ночной зверь: из дупла выходит на закате солнца, а
возвращается на рассвете. Он хорошо лазает по деревьям, но добывает пищу
в основном на земле - по берегам рек, ручьев и на болотах. В поисках пищи
нередко бродит по мелководью и береговым отмелям, где можно добыть
полевок-гигрофилов, рыбу, насекомых, птенца или лягушку.
Как свидетельствует А.М. Гинеев (1987), одной из специфических
особенностей вида считается чрезвычайно тонкая способность зверей
ощупывать, распознавать и схватывать пальцами передних лап мелкие
предметы. Благодаря этому они тщательнее, продуктивнее и на сравнительно
небольшом участке ведут поиск кормов, с успехом добывая их в воде, на
земле и деревьях. Енот-полоскун неприхотлив к пище и довольно быстро
переключается с одного вида корма на другой, который чаще встречается в
текущем сезоне. Ему присуща способность к активному знакомству с новой
обстановкой и предметами. На кормежку эти хищники выходят поодиночке,
парами или семьями. Их кормовые участки налегают друг на друга и иногда
на обсыхающих водоемах, где их не трогают, можно одновременно наблюдать
до десятка жирующих животных.
Представители семейства медвежьих (Ursidae) заметно различаются
между собой по формам поведения и характеру двигательной активности.
Как белый, так и бурый медведи - довольно медлительные животные,
которые передвигаются обычно шагом. При необходимости они могут
переходить на галоп или неспешную рысь, т.е. двигаться некоторое время с
достаточно большой скоростью. Напуганные выстрелом или преследуя
98
120
Рис. 26. Суточная активность бурого медведя на юге Камчатки,
1989-1991 гг.; по материалам 920 наблюдений (по: Ревенко, 1993).
1 - апрель; 2 - май; 3 - июнь; 4 - июль; 5 - август; 6 - сентябрь.
По оси абсцисс - время суток, ч; по оси ординат - количество наблюдений.
Fig. 26. Circadian activity of the brown bear (Ursus arctos) in the south of Kamchatka,
1989-1991; based on 920 observations.
1 - April; 2 - May; 3 - June; 4 - July; 5 - August; 6 - September.
Kbscissa - time of the day; ordinate - quantity of occurrence.
добычу, первый может пробежать некоторое расстояние галопом со ско-
ростью до 20-30, а второй - до 40 км/ч (Павлинов, 1999).
Бурый медведь, в отличие от белого, живет оседло. Расстояние, которое
он проходит за сутки во время летней жировки, обычно не превышает 10 км.
При кажущейся неуклюжести этот хищник достаточно ловок, а во время охот
на копытных движения его расчетливы и стремительны. Он хорошо плавает,
легко преодолевает любые водные преграды в лесу и способен на мелководье
догнать и задавить молодого лося или оленя. На Кавказе в урожайные годы
медведи взбираются на деревья и, обламывая ветки, питаются плодами диких
яблонь, груш и тутовника. В альпийской зоне они могут часами пастись на
зеленых лугах или, переворачивая дерн и большие камни, добывать все
съестное - насекомых, их личинок, грызунов и корешки травянистых растений.
В лесах Псковской и Ленинградской областей мы неоднократно находили
старые осины, на которые залезали медведи. Обнаружив в них гнезда пчел,
они разламывали дупла и поедали соты с медом и личинками насекомых.
В центральной части России (Пажетнов, 1990, 1993) после гона медведи
интенсивно кормятся травянистой растительностью и сравнительно равномерно
99
размещаются по территории. Их индивидуальные участки составляют 400-
600 га, а перемещения на значительные расстояния редки. В нажировочный
летне-осенний период они концентрируются в местах с обильной и доступной
пищей - в рябинниках, около ягодников и овсяных полей. Обычно в этот период
их индивидуальные участки не выражены, а одними и теми же тропами могут
пользоваться разные особи. Выделы леса, часто посещаемые людьми с целью
сбора грибов и ягод, медведи избегают. При этом они достаточно терпимы к
работе лесозаготовителей, и регистрация следов зверей вблизи лесосеки, а то
и на ней - явление обычное. Перед залеганием в берлогу в стациях зимнего
переживания кормовой участок, посещаемый медведем, может быть меньше
летнего иредко превышает 100-150 га. Таким образом, при добывании корма
этот всеядный хищник демонстрирует разные формы пищевого поведения,
свидетельствующие о его сообразительности, сметливости и высокой степени
экологической адаптации к условиям существования.
Как сообщает И.Я. Павлинов (1999), оседлый образ жизни бурых медведей
не мешает им совершать сезонные кочевки в места, богатые пищей и где она им
наиболее доступна. В горных районах такие переселения регулярно совершаются
на 50-100 км. При неурожае кормов звери способны пройти в поисках более
благоприятных кормовых угодий расстояние до 200-350 км. Так, в горных
районах Сибири ранней осенью они жируют в зоне гольцов, где находят изобилие
орехов кедрового стланика, брусники и смородины. С началом снегопадов в
высокогорьях животные покидают эти места и перебираются в нижний пояс гор,
в так называемые «берложистые» угодья. В лесах Соединенных Штатов сезонные
перемещения самцов медведей, помеченных радиоошейниками, обычно
составляли 96-128 км, в то время как самки с медвежатами редко проходили за
лето более 16-24 км (Hugie, 1977).
Охотничьи маршруты бурых медведей, как правило, индивидуальны. В
Карелии (Данилов и др., 1979, 1993) в весеннее время у одних они очень
извилисты, прерываются разорением муравейников, копанием снега в поисках
падали, кормежкой клюквой на болотах и корешками растений на суходолах.
Другие звери следуют прямо, почти не затрачивая времени на поиск пищи. В
конце лета-осенью, когда начинает поспевать овес, медведи, идя на жировку,
проходят значительные расстояния. Их экскременты, содержащие овес,
встречаются иногда в 4-8 км от ближайшего овсяного поля. Особенно большие
переходы они совершают, направляясь к местам зимовки - к берлогам.
Например, только часть такого маршрута, установленная по трем троплениям,
равнялась 8,10 и 12 км. Приуроченность зимнего жилья к постоянному участку
обитания медведя весьма сомнительна. Этому противоречат большие
переходы зверей как перед залеганием в спячку, так и после их выхода из
берлог, а также известная концентрация зимних убежищ в пределах
ограниченной территории.
В летне-осенние месяцы в течение суток медведи активны длительное
время. Данный факт объясняется тем, что они питаются в основном
малокалорийным кормом, и для поддержания нормального жизнеобеспечения
100
Рис. 27. Суточная активность самца белого медведя по результатам 4-дневных
наблюдений на мысе Блоссом, апрель 1976 г. (по: Беликов, 1993).
1 - кормежка; 2 - «прогулка»; 3 - отдых; 4 - плохая видимость; 5 - вне поля зрения;
6 - смена характера активности.
Fig. 27. Circadian activity of the polar bear male (Ursus maritimus) according to the
results of 4 daily observations at the Cape Blosso, 1976.
1 - feeding; 2 - «promenade»; 3 - rest; 4 - poor visibility; 5 - outside of visual field;
6 - change in the character of activity.
организма его требуется большое количество. В Новгородской и Псковской
областях нам приходилось встречать медведей весной в 3, 7 и 10 ч утра, а
также в вечернее время - в 19, 20 и 22 ч. Вообще следует отметить, что их
активность в основном приурочена к сумеречному и ночному периодам,
поэтому осенью, с уменьшением светлого времени суток и увеличением
потребности в кормах, встречи с жирующими медведями происходили и в
101
более ранние сроки - после 16 или 17 ч. Из сообщений многих охот-
корреспондентов северо-западных областей России следует, что в конце
августа-сентябре звери наиболее часто появляются на засеянных овсом полях
в период от 18-19 идо 22-23 ч (рис. 25).
На Северо-Востоке России (Чернявский и др., 1993) у зверей также
преобладает сумеречная активность, что в наибольшей степени свойственно
крупным особям (64.3% встреч). В меньшей степени к сумеречному образу
жизни тяготеют звери небольших размеров и самки с молодняком. Возможно,
данный факт объясняется необходимостью (у этой части популяции)
затрачивать больше времени на поиск пищи, поскольку кормные угодья
оказываются занятыми более сильными и агрессивными особями. В осенний
период в данном регионе, когда кормовая база медведей заметно обогащается
за счет орешков кедрового стланика, ягод и проходных лососей, практически
все встречи со зверями приходятся на вечернее время.
На Камчатке в суточной активности медведей четко наблюдаются два пика
(Ревенко, 1993): первый - с 11 до 15 ч местного времени, второй - с 17 до 20 ч (рис.
26). Наиболее заметно повышение активности в эти часы с мая по июль (в период
питания растительной пищей) и в сентябре-октябре (при питании ягодами и
орехами). В течение первого месяца после выхода из берлог большинство зверей
активны только вечером, с 15 до 20 ч. В рыбный же период (с июля по сентябрь)
подъемы активности сглажены, и охотничья деятельность распределена
равномерно в течение сумеречного и всего светлого времени суток.
Белые медведи ведут преимущественно странствующий и одиночный образ
жизни. Их скопление в местах, богатых кормом, - явление редкое.
Многолетние наблюдения, проведенные в российской Арктике С.Е. Беликовым
(1992, 1993), показали, что, вследствие высокой динамичности ледовых
условий, эти хищники вынуждены время от времени менять свои участки
обитания, подыскивая районы, где их основной корм (тюлени) более
многочислен, а среди ледяных полей имеются разводья, облегчающие им
добычу жертвы. Такие места обычно приурочены к припайной зоне, поэтому
белые медведи часто концентрируются здесь в зимнее время. В поисках более
подходящих мест для охоты они проходят порой не одну сотню километров,
поэтому их участок обитания значительно меняется даже в течение одного
сезона. При отсутствии у этих хищников территориализма отдельные особи
или семейные группы на какое-то время осваивают сравнительно небольшой
кормовой участок, но в случае изменения обстановки они перекочевывают в
другие места. В этом плане интересны наблюдения биолога В.Б. Погребова,
сообщившего нам, что в 1994 г. при подходе судна к о-ву Виктория (Баренцево
море) в брошенном поселке была замечена группа белых медведей в количестве
15-16 особей. Судя по всему они поселились здесь довольно давно и питались,
видимо, старыми пищевыми отходами на помойках и оставленными для
следующей экспедиции запасами мясных консервов и сгущенного молока.
Белый медведь ведет постоянно кочующий образ жизни и практически не
привязан к какой-либо конкретной территории. Во время весенних и осенних
102
Рис. 28. Изменение суточной активности енотовидных собак в течение года при
содержании в неволе (по: Заболотских, 1999а).
I - молодые; 2 - взрослые.
По оси абсцисс - месяцы года; по оси ординат - двигательная активность, ч/сут.
Fig. 28. Change in circadian activity of the racoon dogs (Nyctereutes procyonoides)
during the year in the conditions of captivity.
1 - young; 2 - adult.
Abscissa - months of the year; ordinate - moving activity, hours/day.
миграций звери проходят за сутки 40-80 км. В условиях малоподвижных
морских льдов размах их кочевок может составлять до 750, а иногда и до
1000 км от основных местообитаний (Павлинов, 1999).
Представители этого вида хорошо плавают и ныряют, проявляя способность
находиться под водой до 2 мин. Известны случаи встреч с ними в открытом
море, на большом расстоянии от ближайшей кромки припайного льда или суши.
Так, в августе 1994 г. сотрудник ААНИИ В.Б. Погребов (устное сообщение) с
борта судна наблюдал в Баренцевом море белого медведя, поедающего нерпу
на льдине размером 2.5 на 2 м. При подходе парохода хищник бросил добычу и
поплыл в сторону айсберга, который был удален от этого места на расстояние
20-25 км. В августе 1995 г. сотрудница того же института М.В. Таврило (устное
сообщение) с борта судна видела в воде белого медведя в 3-4 км от ближайшего
из островов архипелага Земля Франца Иосифа.
Четырехсуточные наблюдения за крупным самцом, проведенные
С.Е. Беликовым (1993), выявили два периода его активности: ночной и
вечерний. Хронометражные исследования показали, что 21 и 23 апреля зверь
дважды выходил на длительную кормежку: в первом случае - в период с 19 до
21чис2доЗчЗО мин, а во втором-с 15 ч 20 мин до 19 ч не 3 ч40 мин до 6 ч
утра. 22 и 24 апреля пищевая активность медведя носила монофазный характер.
Он кормился только вечером, соответственно с 19 ч 30 мин до 24 ч и с 17 ч 20
мин до 22 ч (рис. 27).
Белый медведь прекрасно ориентируется во льдах, может быстро уходить
от преследования, ловко и легко преодолевать ледяные торосы. Однако,
постоянно пребывая в сходных условиях, не требующих разнообразных
навыков для добывания пищи, он часто не испытывает страха перед другими
103
Периоды наблюдений/periods
Подопытные
животные/
species
до голодания/
berfore the
hunger
при голодании в
течение 20 ди/
hunger during 20
days
после голодания/
after the hunger
Енотовидная
собака/
Nyctereutes
procyonoides
Песец/
A lopex
lagopus
- 1
-2
-3
Лисица/
Vulpes vulpes
Рис. 29. Соотношение периодов активности и покоя у некоторых хищников
зимой при наличии и отсутствии корма (по: Заболотских, 1999 б).
1 - интенсивная активность; 2 - локальная активность; 3 - сон.
Fig. 29. Correlation of periods of activity and rest in some carnivores in winter with
the availability and absence of food.
1 - intensive activity; 2 - local activity; 3 - sleep.
хищниками и человеком. Это не способствует развитию сложных поведенческих*
реакций на возникающие ситуации, поэтому по сравнению с бурым медведем
он кажется менее осторожным, контактным и сообразительным. Возможно, что
разные черты поведения этих животных обусловили и особенности строения
их центральной нервной системы. Установлено (Авксентьева, Богусловская,
1973), что у бурого медведя при весе в 200 кг масса головного мозга составляла
296 г, а его индекс 1.48%». У белого медведя весом в 450 кг эти показатели
были равны соответственно 509 г и 1.13%». Таким образом, по размерам индекса
головного мозга белый медведь заметно уступает бурому. В то же время авторы
отмечают, что у белых медведей этот орган заметно отличается от такового
других хищников своими сложными очертаниями и топографией мозговых
извилин больших полушарий. Если у бурого медведя головной мозг имеет
сравнительно простой рисунок расположения борозд, то у белого он
усложняется за счет сети мелких вторичных извилин.
104
Среди представителей семейства собачьих (Canidae) енотовидная собака
в основном активна в сумеречное и ночное время, однако во второй половине
лета и особенно осенью, когда звери накапливают жир, их можно встретить и
в дневные часы. Весной и в начале лета они бывают очень осторожны, проявляя
основную активность в сумерки. Днем их охотничья деятельность прекра-
щается, и хищники скрываются в норах. В это время года они хорошо
обеспечены пищей и кормятся недалеко от логова. В августе суточный ход и
продолжительность активности животных увеличиваются. Они начинают
интенсивно питаться, причём не только в темное, но и в светлое время суток. В
середине-конце августа нам приходилось встречать как одиночных молодых
особей, так и самок с подросшими детенышами в утренние и вечерние часы и
даже в дневное время. В сентябре наиболее активными бывают сеголетки,
которые в ночное и в дневное время бродят по окраинам болот, лесным низинам
и берегам водоемов в поисках лягушек, полевок и насекомых. В это же время
интенсивно жируют и взрослые особи, которые, в отличие от молодняка, к
концу сентября бывают уже хорошо упитанными.
В октябре жирующие енотовидные собаки также попадаются как в сумерки,
так и в дневное время, особенно в интервале от 12 до 15 ч. По-видимому, осенью
периоды отдыха и бодрствования чередуются у них сравнительно равномерно.
С конца октября хищники начинают встречаться парами (самец и самка) и до
весны обычно живут вместе. Следует отметить, что осенью енотовидные собаки
постоянно посещают брошенные хутора, где в большом количестве потребляют
опавшие с кустов ягоды, плоды яблонь и других культур.
В ноябре активность зверей заметно падает. Они нерегулярно выходят из
нор на кормежку и подолгу отлёживаются в местах, защищенных от холода, -
в старых землянках и осыпавшихся бобровых норах, под стогами, в торфяных
кучах, заломах тростника и т.п. Микроклимат таких убежищ способствует
уменьшению теплоотдачи, что обеспечивает минимум расхода резервных
жировых отложений. В теплые дни енотовидные собаки вновь начинают
бродяжничать в поисках корма, но охотятся большей частью ночью или в
сумерках. В таком относительно малоактивном состоянии они, в зависимости
от погодных условий, могут пребывать до второй половины ноября-начала
декабря. С наступлением постоянных низких температур и выпадением снега
глубиной 10-15 см звери забираются в свои убежища и только в оттепели, в
тихие ночи выходят на кормёжку. Осенью 1971 г. в Локнянском районе
Псковской области следы енотовидной собаки регулярно встречались на
постоянных маршрутах до 20 ноября, когда температура воздуха упала до
-20- -24°С и снежный покров достиг 15 см. Морозная погода простояла до
конца месяца; следы енотовидной собаки за это время ни разу не отмечались.
2-3 декабря температура повысилась до -2-0°С, днем - до 0- +5°С, снег заметно
осел и стал таять. Сразу же появились следы зверей, которые ходили парами.
8 декабря опять начались сильные морозы, температура воздуха днем упала
до -15°С, участились снегопады, и следы енотовидных собак нам больше не
встречались.
105
В теплую малоснежную осень эти хищники кормятся и в дневное время,
посещая берега водоемов и окрайки болот. В солнечную погоду они могут и не
забираться на отдых в норы, а оставаться на дневку в сухих возвышенных
местах - на кочках в зарослях тростника, в густом кустарнике с толстой
подстилкой из опавших листьев, в скирдах и стогах сена. В связи с
разнообразием потребляемых кормов их жировочный след невелик и имеет
протяженность до 3-5 км, а максимальные перемещения - до 10 км. В январе-
феврале в центральных и северных областях европейской части России обычно
устанавливаются постоянные низкие температуры, а глубина снежного
покрова превышает 20 см. Несмотря на это, в оттепели хищники иногда выходят
из нор и удаляются от них на 1-2 км. Очевидно, это характерно для зверьков с
небольшими запасами жировых резервов.
Енотовидная собака плохо приспособлена к передвижению по глубокому
рыхлому снегу. Обладая сравнительно короткими конечностями и отно-
сительно небольшой поверхностью лап, она вязнет в снегу, оставляя глубокие
борозды. Весовая нагрузка на площадь опоры довольно значительна: в
декабре-январе 75-80 г/см2, а в марте, когда запасы жира в основном
израсходованы, 33-37 г/см2 (Попов, 1956). Для сравнения укажем, что у
росомахи весовая нагрузка на снег составляет зимой 41.2-42.6 г/см2. Зимний
сон енотовидных собак в северных областях России продолжается до середины
марта. С наступлением теплых солнечных дней и появлением наста звери
выходят из нор, их активность резко возрастает, и они начинают широко
перемещаться по угодьям в поисках пищи.
По материалам Ю.С. Заболотских (1999а, б), долгое время изучавшего
поведение енотовидных собак, содержащихся в неволе, уровень суточной
активности молодых зверей во все месяцы года был заметно выше, чем у
взрослых особей. Так, у сеголеток среднегодовая двигательная активность за
сутки составляла 11 ч 45 мин, а у половозрелых зверей лишь 6 Ч (рис. 28).
Соответственно коэффициент активности (суточное отношение периодов
активности к покою) в среднем за год был равен для молодых животных 0.97,
а для взрослых - 0.36. Это связано с инстинктом молодняка к расселению и
большой потребностью в кормах, необходимых для благополучной зимовки.
Наибольшую активность звери проявляли в вечерние часы, а её спад
регистрировался в период с 4 до 7 ч утра. Минимальный уровень активности
прослеживался в конце января-марте, максимальный - в апреле-сентябре.
По данным автора, в зимний период у енотовидных собак наблюдался
однофазный тип суточной активности с пиком в вечерние часы, а летом -
двухфазный, с увеличением подвижности в дневное и ночное время. Весной и
осенью активность зверей в течение суток распределялась относительно
равномерно, без четко выраженных ее пиков и спадов. Изучение влияния
голодания на различные формы поведения животных в течение суток зимой
дало следующие результаты. Если у енотовидных собак при голодании
отмечалось некоторое увеличение периода покоя и сна, то у лисиц возрастала
доля интенсивной двигательной активности, а у песцов - сна и локальной
106
подвижности без резких перемещений по клетке (рис. 29). По-видимому,
видоспецифичность жизнедеятельности хищников является приспособительной
формой выживаемости и снижения напряженности в конкурентной борьбе за
пищевые ресурсы (Заболотских, 19996).
Песец в зимнее время, при коротком световом периоде, кормится
преимущественно в сумеречное и ночное время суток. Днем большинство
зверьков отдыхает в норе, вырытой в сугробе, или в своих старых жилищах, в
защищенных от ветра местах - за камнем, холмиком, торосами и проч. В
сильные бураны и метели песцы малоактивны, а в ясные морозные ночи, даже
с небольшим ветром, их подвижность увеличивается. Весной и летом
активность зверьков снижается в дождливую и ветреную погоду. Наоборот, в
тихие или пасмурные, но безветренные дни они выходят на жировку.
В июле-августе на Таймыре и в Большеземельских тундрах нам
неоднократно приходилось наблюдать охотящихся на леммингов песцов в
утренние и дневные часы. В эти месяцы их суточный ход невелик, поскольку
даже в годы с низкой численностью грызунов добычей хищников становятся
птенцы гнездящихся в тундре многочисленных птиц. В то же время необходимо
отметить, что при необходимости песец легко и свободно преодолевает
значительные расстояния. В предгорьях Бырранга (Северный Таймыр)
мышкующий днем зверек, заметив направляющегося к нему человека, убегал
на расстояние до 0.8-1 км и только потом успокаивался. В бинокль отчетливо
было видно, как, пробежав 200-300 м, песец останавливался, поворачивался
в сторону опасности, садился или даже привставал на задние лапы, пытаясь
разглядеть напугавший его объект, а затем снова с периодическими
остановками продолжал движение в направлении гор.
Об увеличении дневной активности песцов в летнее время свидетельствуют
и экспериментальные исследования Н.И. Калабухова (1950) с использованием
суточных актографов. Общая активность белых песцов при высокой
температуре обычно ниже, чем при отрицательной. Так, активность зверьков
при температуре среды -10- -18°С почти в 2.5 раза выше, чем при температуре
+12-+20°С. В естественных условиях зимой при ограниченной кормовой базе
песцы обеспечивают себе необходимое количество пищи, увеличивая
активность, не носящую циклического характера. Установлено, что у белых
песцов осенью, особенно при недостатке корма, двигательная активность
сравнительно равномерно распределяется в течение суток, т.е. носит
полифазный характер. В зимнее время, при снижении уровня энергообмена,
животным требуется меньше пищи и их охотничья деятельность сдвигается в
основном на сумеречно-ночной период. В эти дни в поисках пищи они
пробегают 5-8 км и более.
Леммингов и других грызунов песцы отлавливают, раскапывая их норы,
либо подолгу карауля появление жертв из норы. В последнем случае хищник
мгновенно прыгает и давит грызуна. По наблюдениям А.Ф. Чирковой (Гептнер
и др., 1967), прислушиваясь к писку или шороху лемминга, песец в несколько
секунд разрывает снег глубиной до 0.5 м и более. Сотрудник Мурманского
107
морского биологического института В.В. Семёнов (устное сообщение)
наблюдал, как зимой в районе пос. Дальние Зеленцы зверек, заслышав писк
полевок, выкопал жировочный ход в сугробе на глубину не менее 2 м.
Сезонные перекочевки песцов - явление весьма обычное. Причиной их
служит недостаток пищи в зимнее время. Осенью они начинают откочевывать
из мест норения в более кормные угодья, преодолевая 200-300, иногда до 1 000 км
(Сдобников, 1940; Г. Новиков, 1956; Якушкин, 1967). Путь их до ледостава
проходит преимущественно по побережью рек вдоль кромки воды, а после
ледостава - по льду. Передвижение зверей происходит как бы в несколько
этапов - отдельные группы появляются в местах наблюдения в разное время.
Такое явление имеет место в районе пос. Дальние Зеленцы (Мурманская обл.),
где первые подкочевки песца отмечаются в конце октября, а наиболее
многочисленным он становится в морозную погоду, в январе-феврале. По
наблюдениям охотников-корреспондентов Западного отделения ВНИИОЗ,
направление зимних кочевок в этих местах постоянно западное - зверьки
проходят из Иоканга-Понойской тундры, откуда они также двигаются к
Терскому берегу и на юг полуострова, встречаясь даже около пос. Кузомень.
Кочуют преимущественно молодые особи и самцы. Самки более привязаны к
своим выводковым стациям.
На севере Кольского полуострова песцы зимой перемещаются из приморской
тундры во льды моря и к островам, а также в южном направлении - в глубь
материка. Особенно много зверей кочует по долинам рек Каменки и Кочковки.
Весной перезимовавшие песцы двигаются в обратном направлении - к местам
вывода молодняка. По материалам Л.М. Шиляевой (1972), в разных районах
севера России преобладающее направление миграций песца имеет отличия,
обусловленные причинами орографического и климатического характера.
Дальние миграции песцов в западном и восточном направлениях характерны для
хищников обского севера, в южном - енисейского и якутского. По-видимому,
интенсивность и направление миграций представляют собой наследственно
закрепленную особенность популяций, поэтому различие между территори-
альными группировками автор расценивает как следствие их генетической
гетерогенности. Процент мигрантов из ареала норения, потенциально имеющих
возможность осесть на новых территориях, невелик и вряд ли превышает 25-26%.
Канадский исследователь А.Х. Макферсон (Macpherson, 1969) считает,
что песцы обладают хорошо развитыми навигационными способностями и
чувством «инстинкта дома». Свои выводы он основывает на факте возврата
(за 1100 км) одного зверька на родительский участок (к месту отлова) после
того, как песец 2 года прожил в условиях неволи.
Перемещение песцов на новые места в поисках пищи прослеживаются во
многих северных регионах (Pulliainen, 1965; Якушкин, 1967, 1977; Шиляева,
1974,1982; Крылов, 1986; Линейцев, 1988; Полежаев, 1998, и др.). Наблюдение
за состоянием ресурсов вида на полуострове Таймыр показывают, что при
ухудшении кормовых условий в ареале его норения и, следовательно, снижении
интенсивности размножения маточного поголовья отмечается сокращение
108
I П Ш IV
Рис. 30. Суточная активность обыкновенной лисицы в
Лапландском заповеднике (по: Насимович, 1948).
/ - утро; // - день; /// - вечер; IV - ночь.
По оси абсцисс - время суток; по оси ординат - количество встреч.
Fig. 30. Circadian activity of the red fox (Vulpes vulpes)
in the Lapland Natural Reserve.
I - morning; II - day; III - evening; IV - night.
Abscissa - time of the day; ordinate - quantity of occurrence,
численности популяции за счет главным образом активной миграции сеголеток.
При этом они уходят далеко от границ ареала, и их возврат крайне ограничен.
Более ценная в репродуктивном аспекте взрослая часть популяции, переживая
сезонные трудности в добывании пищи внутри ареала, перемещается на
ближайшие от границы зоны норения участки лесотундры и северной тайги.
Здесь динамика численности мышевидных грызунов и показатели их
биоразнообразия заметно отличаются от таковых в тундре, а кормовая база
может быть намного богаче. В дальнейшем из этих районов и идет возврат
мигрантов в весенний период (Линейцев, 1988). По-видимому, данный
механизм возникновения миграционных потоков песца в поисках корма
характерен для всего ареала, где происходит норение зверей.
Обыкновенная лисица в отличие от песца способна более эффективно
использовать кормовые ресурсы района обитания. Эффективность её охоты
на мелких грызунов в годы их низкой численности заметно выше. При
недостатке полёвок или мышей хищник переключается на добычу птиц, зайца-
беляка, потребляет насекомых, ягоды и разную падаль (Полежаев, 1998),
поэтому в фазу депрессии грызунов он испытывает меньшее напряжение, чем
песец. Длительные перемещения прослеживаются редко и имеют слабую
выраженность. По свидетельству А.А. Насимовича (1948), в Лапландском
заповеднике (Мурманская обл.) сезонные кочевки лисицы при обилии грызунов
составляют 10-20 км, а в годы бескормицы увеличиваются до 30-50 км от
места летнего обитания. Автор отмечает, что в обычные годы зимой лисица
выходит на охоту 1-4 раза в течение суток и ее маршрут составляет по лесу до
8, а в горной тундре до 12-15 км. В поисках грызунов она за один период
кормления делает от 5 до 20 покопок, из которых только в 40% случаев охота
109
25
20
15
10
до 8
до 11 до 14
до 17 до 20
0
4
Рис. 31. Активность волка в Беловежской Пуще в светлое время суток
(по: Буневич и др., 1999).
По оси абсцисс - время суток; по оси ординат - количество встреч.
Fig. 31. Activity of the wolf (Canis lupus) in the Bielowieza in the light time
of the day.
Abscissa - time of the day; ordinate - quantity of occurrence.
бывает успешной. Близкие данные были получены в Ленинградской области,
где для добычи одной полевки лисице приходится делать до 15 покопок и
пробегать как минимум 1.6 км (Новиков идр., 1970).
В Карелии длина суточного хода лисицы (Ивантер, 1969) составляет в
среднем 7.4 км (4.3-11.6 км, н=7).Этот показатель, как и размер участка
обитания зверя, меняется в зависимости от наличия кормов и сезона года. В
обильные грызунами периоды размеры суточного пробега и индивидуального
участка зверя сравнительно небольшие. Там, где есть падаль, лисица вообще
может жить долгое время на участке в 2-5 км2. В голодные годы жировочный
ход и территория местообитания хищника увеличиваются. Размеры последних
составляют от 10 до 35 км2. Зависимость размеров участка обитания и длины
суточного хода от состояния кормовой базы прослеживается в различных
частях ареала вида (Гептнер идр., 1967; Macdonald, 1978).
Интересные наблюдения по расселению молодых лисиц, снабженных
кнопочными ушными метками, были проведены в Воронежском заповеднике
(Сапельников, 1999). Всего были помечены 123 лисенка и получено 14
сообщений об их расселении (11.4%). Дальность перемещения самок была
минимальной и не превышала 6 км. Самец-сеголеток был добыт осенью
примерно в 50 км по прямой от места мечения, что подтверждает общепринятое
мнение исследователей о большей активности и дальних миграциях самцов
хищников в первый год жизни.
В годы высокой численности мышевидных грызунов лисица в основном
концентрируется в местах их обитания: на полях зерновых культур, по берегам
водоемов, в смешанных лесах с богатой кустарниковой растительностью. В
такие годы в Псковской и Новгородской областях (1996-1997 гг.) за утренние
ПО
и вечерние осенние сумерки можно было встретить по 2-3 мышкующих зверя. В
дневное время некоторые из них отдыхали прямо на поле, на стогах или
небольших копнах сена. При низкой численности полевок (1998-1999 гг.)
хищники шире осваивали угодья и чаще встречались в лесах разного типа,
где кроме грызунов старались добыть зайца-беляка, тетеревиных или
воробьиных птиц. Нередко они приходили кормиться на скотомогильники, к
скотным дворам или на свалки. В малокормные годы на севере европейской
части России лисица много времени проводит на морском побережье, где
подбирает выбросы моря и отходы рыбного промысла.
Летом хищник чаще охотится в ночное и сумеречное время, зимой - в
основном в утренние часы (рис. 30). При низкой численности грызунов его
жировочный след в осенне-зимний сезон можно встретить практически в любое
время суток (Насимович, 1948). По нашим наблюдениям (Псковская обл.), в
районе свалок и скотомогильников в ноябре-декабре лисицы были наиболее
активны в вечернее и ночное время. Способность добывать корм, изменяя
формы своего охотничьего поведения в соответствии с местными условиями и
видом добычи, свидетельствует о высокой сообразительности и экологической
пластичности лисицы, что обеспечивает ей успешное существование в
различных наземных экосистемах.
Волк относится к видам, обладающим широким спектром питания. Каки
у других неспециализированных к определенным объектам добычи хищников,
его трофические связи и характеристики охотничьего поведения весьма
разнообразны. По материалам О.С. Русакова (Данилов и др., 1979), в северных
регионах европейской части России, отличающихся длинным полярным днем,
в летнее время ночной образ жизни волков не столь ярко выражен, как в более
южных областях, где эти хищники в любой сезон активны преимущественно
ночью (с вечерних сумерек и до раннего рассвета). Летом они заняты поисками
пищи обычно не более 5-6 ч в сутки, а зимой 12-15 ч. Однако в конце теплого
периода года, когда волки-родители обучают своих подросших щенков охоте,
они нередко нападают на пасущихся овец и днем. Осенью агрессивность
хищников по отношению к домашнему скоту заметно увеличивается: даже в
дневное время отмечаются случаи их нападения на овец, коз, телят и коров.
Пользуясь паникой стада, волки отделяют от него одно или несколько
животных, которых настигают и загрызают.
Волк, хоть и считается ночным хищником, но часто бывает активен и в
дневные часы. В Белорусском Полесье период дневной активности увеличи-
вается у него в период гона и вскармливания потомства (Воронецкий и др.,
1999). Наибольшая подвижность зверей прослеживается на вечерних и
утренних зорях, однако преследование лосей, косуль или кабанов волчьей
семьей в 5-7 особей авторы наблюдали и в светлый период, в основном с 10 до
16 ч.
В Беловежской Пуще (Буневич и др., 1999), по среднемноголетним данным
(/7=390), наибольшее количество встреч с волками приходится на утренние
часы (24.6%). К полудню их двигательная активность несколько снижается, а
111
Таблица 32/ Table 32
Характеристика двигательной активности и относительные размеры головного
мозга у представителей семейства собачьих
Characteristics of moving activity and relative sizes of brain in representatives of canids
family
Виды/ Species Длина суточного хода, км/ The length of circadian way, km Дальность кочевок, км/ The amplitude of wandering, km Индекс головного мозга/ Index of brain, %о
Енотовидная собака/ Nyctereules procyonoid.es 3-5 до 10 5.8 ±0.12
Песец белый/ Alopex lagopus 5-8 200-300 и более 8.7 ±0.31
Лисица обыкновенная/ Vulpes vulpes 7.4(4.3-11.6), до 8-15 до 6-50, 10-50 9.9 ± 0.23
Волк/ Canis lupus 18.8(15-26), 5-40 100-200, 150-300 5.2 ±0.35
Примечание. В таблице кроме своих данных использованы материалы следующих
авторов: Боголюбский, 1939; Сдобников, 1940; Насимович, 1948; Слудский, 1962; Якушкин,
1963; Ивантер, 1969; Филимонов, 1980; Бибиков и др., 1985; Сапельников, 1999.
наименьшее количество встреч приходится на период с 14 до 17 ч. В это время
большинство волков залегает на дневку и только к вечеру вновь уходит на
поиск добычи, в результате чего после 17 ч наблюдается увеличение количества
встреч хищников (рис. 31). Прослеживается некоторая сезонная изменчивость
в распределении активности волков. Так, зимой дневной период их охотничьей
деятельности более продолжителен - с 8 и до 17 ч, что можно объяснить
трудностями поиска и добывания пищи в холодный снежный сезон, а также
увеличением подвижности хищников во время гона. Весной и летом они в
основном активны в утренние (22% встреч) и вечерние часы (24% встреч), а
осенью, когда светлая часть суток уменьшается, - с утра и до 13-14 ч (Буневич
и др., 1999).
В тундрах Кольского полуострова, где основная жертва волка- северный
олень, его сезонные миграции нередко вызывают соответствующие, иногда
довольно длительные кочевки хищников и постоянную мобильность их
популяции. В то же время в южной части Северо-Запада России при обилии и
многообразии кормов прослеживается сокращение подвижности волков и
уменьшение площади их обитания.
Перемещение волков связано с поисками добычи. Следуя по своим
охотничьим маршрутам, они посещают знакомые места, где возможны их
встречи с жертвами, - вдоль русла рек и ручьев, по опушкам или молодым
посадкам леса. Передвижение зверей по территории направлено на
эффективную и равномерную эксплуатацию популяции жертвы. Так, на
112
Чукотке стая из 6 волков контролировала 5-6 стад домашних северных оленей
на площади около 7.5 тыс. км2. При этом хищники постоянно посещали каждое
стадо через 12-15 дней (Железнов, 1990). В Кавказском заповеднике стая (также
из 6 зверей) регулярно обходила свой охотничий участок площадью 100 км2 за
5-7 дней, задерживаясь у крупной добычи на несколько дней (Кудактин, 1979,
1980).
Размеры участков волчьей стаи во многом определяются особенностями
ландшафта: на открытых пространствах они больше; в лесной зоне или в
горах, где много надежных укрытий, а копытные размещены локально,
заметно меньше. Например, в лесной равнинной части Северной Америки
они составляют 175-260 км2, в горах Аляски - 585-788 км2, а в тундровой
зоне Канады-до 1250 км2 (Corbyn, 1980). Плотность населения хищника и
кормность участков обитания также имеют большое значение. Например, в
Дарвинском заповеднике (центральная часть России) при снижении плотности
населения волков их участки обитания увеличивались в 4 раза (Калецкая,
1973). В Якутии в годы пика численности зайца-беляка (до 170 особей/км2)
волчья семья имела кормовой участок размером 100-120 км2, а в фазы
депрессии численности жертвы охотничья территория хищников увеличива-
лась в несколько раз (Лабутин, 1977).
Молодые, еще не размножающиеся одинокие звери много кочуют в поисках
добычи. Они составляют резервное ядро популяции. Добыть крупное животное
им сложно, и это может служить причиной объединения молодых зверей в
группы. Их доля в популяциях составляет 30-40%, а дальность кочевок
достигает 200-300 км (Stenlund, 1955; Kuyt, 1962; Meeh, 1970, 1977). По
имеющимся данным (Семенов, 1954; Слудский, 1962; Макридин, 1978;
Филимонов, 1980; Бибиков и др., 1985), дальность передвижения волков вслед
за стадами северного оленя в тундровой зоне нередко составляет 150-300 км, а
в южных регионах (за табунами сайгака) - 100-200 км.
Обеспеченность кормом играет важную роль в жизни хищников и заметно
влияет на протяженность охотничьего маршрута. При высокой численности
копытных суточный ход волков бывает более коротким и прямолинейным по
сравнению с годами, когда запасы корма ограничены. При локальном скоплении
копытных на ограниченных участках продолжительные кочевки хищников
прекращаются и следы их жизнедеятельности отмечаются в одних и тех же местах.
Протяженность суточного хода волков - показатель изменчивый. По многолетним
материалам С.Г. Приклонского (Бибиков и др., 1985), в среднем она была равна
18.8 км. При обитании зверей на открытой местности суточные передвижения
увеличивались до 26 км, а в лесных угодьях составляли 15 км.
Сходные данные получены в Белорусском Полесье (Воронецкий и др., 1999).
По наблюдениям специалистов, расстояние, пройденное за сутки семьей волков
в поисках пищи, определяется, с одной стороны, кормностью угодий, а с другой -
наличием в них надежных укрытий. При высокой плотности поголовья диких
копытных (лось 5-7, кабан 20-25, косуля 15-20 особей/1000 га) и полном
отсутствии фактора беспокойства хищники проходили 5-10 км в сутки, а при
ИЗ
низкой (лось 0.5, кабан 5-7, косуля 3-5 особей/1000 га) - 35-40 км и более.
Отмечено, что на переходах волки часто пользовались лесными дорогами и
тропами. Их ход был неторопливым, длина шага составляла 50 см, а средняя
скорость передвижения - 10-15 км/ч. Отдельные особи в поисках жертвы
периодически уходили в сторону от основного маршрута семьи на 0.5-0.7 км.
Способы охоты хищников на диких копытных и зайцев весьма
разнообразны и в каждом отдельном случае очень рациональны, что
свидетельствует о совершенстве высшей нервной деятельности этих зверей.
Обычно волки скрадывают или подкарауливают добычу, а затем накоротке
(10-30 м) резким броском настигают ее. Длительное преследование жертвы
для них нехарактерно, однако в отдельных случаях стаи волков идут по
следу лосей (особенно подранков) на протяжении нескольких километров.
Стайно-семейный образ жизни выработал у них целый ряд интересных
поведенческих особенностей, направленных на рациональное исполь-
зование усилий всех зверей при их совместной охоте. Нападая на стадо
овец или оленей, хищники, как правило, упорно преследуют определенных
животных, как бы не замечая других. Так же они ведут себя и при охоте на
лосей, где отдают явное предпочтение молодым животным и взрослым
самкам, на которых обычно охотятся коллективно и достаточно успешно
(Данилов и др., 1979).
Сезонные особенности охотничьего поведения волков проявляются
прежде всего в том, что летом они добывают животных, подкарауливая
добычу в одиночку. В это время года взрослые хищники держатся парами,
их добыча многочисленнее и разнообразнее. Зимой волки чаще практикуют
охоту загоном или преследуют крупных животных стаями, в которые они
сбиваются в этот сезон. В Ленинградской области в годы низкой численности
волков показатель стайности равнялся летом 1.75, а зимой 2.49 (Новиков и
др., 1970). Через несколько сезонов, в конце 70-х годов, когда численность
волка в Ленинградской и Псковской областях увеличилась, летний
показатель стайности изменился и (по встречам 128 волков) составлял уже
2.8, а зимний 3.9 (Данилов и др., 1979; 1985). Таким образом, степень
стайности изменяется не только по сезонам, но и в зависимости от уров-
ня численности волка: в годы депрессии она минимальна, а на пиках наи-
большая.
Глубокий снег при наличии наста, который проваливается под ногами
копытных и держит волков, способствует успеху охот последних. Весовая
нагрузка на снег у сайгака и лося больше в 2.3-3.0, у джейрана - в 3.1 и у
дзерена - в 3.7 раза по сравнению с этими хищниками (Слудский, 1962;
Peterson, Allen, 1974; Корытин, Бибиков, 1985), поэтому зимние охоты волков
достаточно эффективны. По наблюдениям А.В. Кречмара (устное сообщение),
дикие северные олени более выносливы, чем лоси, и при неглубоком снежном
покрове способны длительное время уходить от преследования со скоростью
до 50 км/ч. Лоси, спасаясь от волков, бегут со скоростью до 30 км/ч, при этом
быстро устают и в зимнее время часто становятся добычей хищников.
114
Наметив жертву, волки преследуют ее на протяжении 2-4 км, но при
обнаружении физически ослабленных особей гонят до тех пор, пока она не
обессилит окончательно. В Карелии и Мурманской областях в ноябре-декабре
по следам на снегу нам неоднократно приходилось восстанавливать картину
нападения волчьих стай (по 5-6 голов) на лосей, которых хищники выгоняли на
запорошенный снегом лед озера, где жертвы скользили и часто падали. Обычно
волки загрызали лося при его попытках выбраться на высокий берег. Наличие
подледных пустот («пустоледок») заметно осложняло положение жертвы, мешая
быстро преодолеть заснеженный или скользкий склон. Настигнув лося, волки
перегрызали ему сухожилия задних ног, а затем хватали за горло. В некоторых
случаях копытные проваливались в полынью недалеко от берега и, ломая
непрочный лед, стремились достичь мелководья. Здесь, обессилев, они все равно
становились добычей хищников. Судя по оставленным на снегу лежкам, волки
кормились на месте удачной охоты не менее 3 суток, устраиваясь здесь же и на
дневку. Потревоженные звери бросали добычу и больше не возвращались к ней,
а около остатков лося появлялись следы горностая, куницы или росомахи.
Вышеизложенный материал свидетельствует о том, что рассматриваемые
виды собачьих, как и представители семейства куньих, характеризуются разным
уровнем двигательной активности, обусловленным особенностями их питания.
Среди хищников со сходной массой тела песец и лисица по сравнению с
енотовидной собакой обладают большими длиной охотничьего хода, периодом
общей активности, дальностью и продолжительностью кочевок (табл. 32). Эти
черты поведения животных находятся в тесной взаимосвязи с морфофункци-
ональными особенностями видов, в частности со степенью развития центральной
нервной системы. Так, например, сравнительно малоактивная енотовидная собака
заметно отличается от песца и лисицы своим более низким показателем
относительной массы головного мозга.
Близкие к этим материалы получены Ю.С. Заболотских (1999а, б) при
содержании хищников в условиях неволи. По свидетельству автора, у хорошо
упитанных взрослых зверей индекс мозга был у енотовидной собаки 0.48%, а
у песца 0.8%. После длительного голодания, расхода жировых резервов и
падения массы тела данный показатель увеличивался соответственно до 0.7 и
0.9%. Молодые енотовидные собаки, добытые в природе, были менее упитанны,
чем взрослые, что соответствовало и разнице в относительном весе их
головного мозга: если у молодых животных из природных популяций индекс
этого органа был равен 0.75%, то у взрослых - 0.6%.
Таким образом, пищевая специализация изучаемых хищников определяет не
только продолжительность двигательных реакций и интенсивность охотничьей
деятельности, но и размеры головного мозга животных как рефлекторно-
автоматического центра локомоторных функций. Взаимосвязь величины и
строения центральной нервной системы с массой тела, активностью и сложностью
поведения является общебиологической закономерностью, которая наиболее
отчетливо проявляется среди близких в таксономическом отношении видов.
115
Резюме
Двигательную активность хищных млекопитающих следует рассматри-
вать как ответную реакцию организма прежде всего на ряд внешних факторов,
что делает ее показатели весьма лабильными у видов различного веса и
линейных размеров. При наличии кормов все они обычно демонстрируют 2
основных пика активности - утренний и вечерний (дифазный тип). Однако у
ласки периоды отдыха настолько непродолжительны, что ее можно
охарактеризовать как хищника с полифазным типом активности.
У близких в систематическом плане видов с увеличением веса животных
общая продолжительность их двигательных реакций сокращается, а время,
когда они отдыхают, заметно возрастает. Собранные данные демонстрируют
тесную связь поведенческих особенностей с размерами и кормовой
специализацией животных. Продолжительность их суточной активности и ее
распределение по часам во многом определяются состоянием кормовой базы.
У представителей видов малого размера увеличение периода суточной
активности можно рассматривать как адаптивную реакцию организма к
под держанию высокого уровня энергообмена за счет интенсивного освоения
пищевых ресурсов.
Количество выпавшего зимнего снега и наличие наста оказывают
заметное влияние на возможности передвижения хищников в поисках добычи.
Животные, обитающие в глубокоснежных лесных районах или в тундрово-
таежной зоне, в зимний сезон испытывают определенный дефицит корма и
трудности при его отыскании. Обычно они имеют большую площадь опоры
конечностей, что уменьшает весовую нагрузку и облегчает передвижение по
рыхлому снегу. Так, весовая нагрузка на площадь опоры лап зимой составляет
у Mustela nivalis и М. erminea 7.2-7.4 г/см2, у Martes martes, М. zibellina и
Mustela sibirica 11.3-13.2, а у мало приспособленных к передвижению по снегу
Mustela vison, М. lutreola - 19-20.8, Lutra lutra -59.8-60.3 и Meles meles-
88.7-124.4 г/см2.
Каменная куница по сравнению с лесной за счет слабой опушенности лап
имеет большую весовую нагрузку (22.5-25.4 г/см2) на единицу их опорной
поверхности. Она заметно проваливается в глубоком снегу и вынуждена избегать
заснеженные биотопы. Возможно поэтому (а также за счет большей, чем у Martes
martes, термофильности) каменная куница зимой нередко встречается в селах,
устраивая гнезда на чердаках домов и в брошенных строениях. Вертикальное
распределение этих видов во многом соответствует их широтному распреде-
лению. В обоих случаях лесная куница может обитать в местах с более холодным
климатом и глубоким снежным покровом, куда каменная не заходит.
Рассматриваемые виды хищных млекопитающих обладают разным уровнем
двигательных реакций, связанным с особенностями пищевой специализации.
Среди них наиболее длительные перемещения в поисках богатых в кормовом
отношении мест могут совершать песец, волк и белый медведь. Последний
постоянно ведет кочующий образ жизни, задерживаясь в местах, богатых кормом.
116
В период весенних и осенних миграций белые медведи способны за сутки
преодолевать до 40-80 км. При недостатке пищи дальность их кочевок достигает
750, а иногда и 1000 км от основных местообитаний. В осенне-зимнее время в
поисках хорошей кормовой базы на расстояния до 200-300 км и более может
откочевывать и песец.
Среди хищников из семейства Canidae примерно равного веса песец и
лисица по сравнению с енотовидной собакой обладают большими длиной
суточного хода, периодом кормодобывающей деятельности, дальностью и
продолжительностью кочевок. Подобное поведение животных находится в
тесной взаимозависимости с морфофункциональными особенностями видов. В
частности, малоактивная в зимний период енотовидная собака заметно
отличается от песца и лисицы своим сравнительно низким показателем
относительной массы головного мозга и возможностью организма создавать на
зиму большие жировые резервы. Подобные зависимости прослеживаются и во
всех других рассматриваемых семействах хищных млекопитающих.
Summary
The moving activity of carnivorous mammals is a response of the organism to a
number of endogenous and exogenous factors, which makes its parameters relatively
labile in species of different weight and sizes. They usually demonstrate two main
peaks of activity with availability of food - morning and evening (biphase type).
However, in Mustela nivalis the intervals between the periods of wakening are so
short that it may be characterized as a predator with a polyphase type of activity.
The total duration of moving reactions decreases with the increase of the body
weight and the period of time when they are having rest noticeably increases. The
data obtained demonstrate the close relation of behavioral features with the sizes
and food specialization of animals. The duration of their circadian activity and its
distribution by hours to a great extent are determined by the state of food supply.
The increase of spontaneous activity in representatives of small species may be
regarded as an adaptive reaction of the organism to the maintenance of high level
of energy exchange as a result of intensive using of food resources.
The snow cover has a great influence on the possibilities of moving of carnivores in
the search of prey. The representatives of species inhabiting forest regions with a deep
snow cover or tundra-taiga zone experience a certain shortage of food and difficulties in
obtaining it in the winter. Usually they have a large surface of limb support, which
decreases the weight load and facilitates moving in the loose snow. Thus, the weight
load on the surface of paw support comprises in Mustela nivalis and M. erminea 7.2-
7.4 g/cm2, Martes martes, M. zibellina and Mustela sibirica 11.3-13.2, and in poorly
adapted to the moving in loose snow Mustela vison, M. lutreola - 19.0-20.8, Lutra
lutra - 59.8-60.3 and Meles meles - 88.7-124.4 g/cm2.
Martes foina owing to the thin down of paws, as compared to Martes martes,
has greater weight load (22.5-25.4 g/cm2) per unit of their support surface. It
117
noticeably falls in loose snow and is enforced to avoid the areas with deep snow.
Probably, as result, Martesfoina also being more thermophilous than Martes martes,
not infrequently occurs in winter in villages, making nests in attics and abandoned
buildings. The vertical distribution of these species largely corresponds to their
latitude distribution. In both cases M. martes can inhabit places with colder climate
and deep snow cover where M. foina is not occurring.
Carnivores of middle and large sizes as representatives of small species of the
family Mustelidae are characterized by different levels of moving activity, stipulated
by characteristic features of their feeding behavior. Alopex lagopus, Cams lupus
and Ursus maritimus may accomplish the longest movements in the search of places
with abundant food. The latter constantly leads nomadic life and is practically not
confined to any particular territory. During spring and autumn migrations of animals
are able to cover 40-80 km per day. Under conditions of slowly moving sea ice the
range of their wandering is up to 750 and sometimes 1000 km from the main habitats.
The arctic fox Alopex lagopus may roam up to 200-300 and more km in the search
of good food supply in the autumn-winter time.
Among the carnivores of the family Canidae with a similar body mass Alopex
lagopus and Vulpes vulpes have a longer circadian rhythm, period of total activity,
length and duration of wandering as compared to Nyctereutes procyonoides. These
features of animals’ behavior are closely related to morphofunctional characteristics of
species, in particular, to the degree of development of the central nervous system. It has
been established that low-active in the winter period Nyctereutesprocyonoides noticeably
differs from Alopex lagopus and Vulpes vulpes by lower value of relative brain weight.
Similar relations are followed also among representatives of the family Mustelidae.
Thus, the food specialization of carnivores determines not only duration of
moving reactions and intensity of hunting activity, but also the sizes of animal
brain as a reflective-automatic center of locomotory functions. The relation of
values and structure of the central nervous system with body weight, activity and
complexity of behavior is a general biological pattern, which most distinctly is
displayed in taxonomically close species.
Глава 3. УРОВЕНЬ ТЕПЛООБМЕНА И ПУТИ
ПОДДЕРЖАНИЯ ЭНЕРГЕТИЧЕСКОГО
БАЛАНСА
Большой расход энергии на терморегуляцию и воспроизводство определяет
у мелких и средних по размерам млекопитающих соответствующую
потребность в пище и высокий уровень двигательной активности, связанный с
кормодобывающей деятельностью. Известно, что при малых размерах тела
поверхность теплоотдачи увеличивается, поэтому мелким животным в
сравнении с крупными требуется вырабатывать больше тепла на единицу массы
для поддержания энергетического баланса на необходимом уровне (Слоним,
1962, 1976; Шилов, 1982, 1985; Башенина, 1969, 1977; Rauch, Hayword, 1969;
Wolk, 1969; Большаков, 1972; Gebczynski et al., 1972; Шмидт-Ниельсен, 1982;
Бигон и др., 1989). В свете сказанного тот факт, что среди хищных
млекопитающих виды, связанные с водной средой обитания (норки, выдра,
калан, белый медведь), имеют достаточно крупные размеры тела, следует
рассматривать как возникшую в процессе эволюции адаптацию, направленную
на сохранение тепла в холодных условиях водоемов. Подобная закономерность
прослеживается и среди землероек и полевок, где наибольшие размеры имеют
кутора и водяная полевка.
Соотношение скорости теплопродукции и выделения тепла создает условия
для поддержания термического гомеостаза. В данной связи следует отметить,
что у большинства мелких хищных млекопитающих некоторые свойства
физической терморегуляции проявляются неотчетливо. Так, за исключением
очень быстрых и ловких в движениях представителей рода Martes, у всех куньих
слабо развиты такие механизмы теплоотдачи, как потоотделение и тепловая
одышка - полипноэ. Это позволяет сохранить тепло в условиях низких
температур, но делает их мало устойчивыми к перегреву. Как справедливо
отмечает И.А. Шилов (1977,1985), у млекопитающих усиление потоотделения
не прямо зависит от температуры окружающей среды, а происходит при
определенном пороге ее повышения, сопровождающегося началом перегрева
организма. Обычно интенсивность потоотделения возрастает пропорционально
повышению температуры окружающей среды и увеличению эндогенного
теплообразования при мышечной работе. При этом не менее эффективен и
охлаждающий механизм, включающий испарение влаги с поверхности
слизистых оболочек ротовой полости и верхних дыхательных путей.
Наблюдения показывают, что у куниц, лисиц и волка в теплую погоду
после продолжительного бега (как и у собак) при тепловой одышке
выдыхаемый воздух проходит через ротовую полость, обусловливая испарение
влаги с ее слизистой и поверхности высунутого языка. Кроме того, у таких
быстро передвигающихся и резких в движениях видов, как лесная и каменная
куницы, при длительном их преследовании с собаками в теплые месяцы года
119
подушечки лап всегда оказывались влажными, порой даже покрытыми
пенообразной жидкостью. При быстром беге, который повышает температуру
тела, ни у норок, ни у хорьков нами не отмечалось включения подобного
механизма активной испарительной теплоотдачи.
Известно, что в жизни млекопитающих сосудистые реакции и сезонные
различия в уровне теплопроводности мехового покрова имеют существенное
значение, определяя, в конечном счете, соответствующие изменения
теплоотдачи и теплоизоляции при изменении внешних условий (Irving, Krog,
1955; Hart, 1956; Когтева, 1963; Яблоков, 1966, 1987; Hill, 1976; Давыдов,
1982; Ивантер и др., 1985; Одум, 1986; Гайдук и др., 1999). В работах
отмеченных авторов показано, что фотопериод является пусковым фактором
линьки зверей, а температурный уровень корректирует сроки наступления и
скорость протекания этого процесса.
По данным В.Е. Гайдука (1977, 1999), весенняя и осенняя линьки у
млекопитающих с ярким сезонным диморфизмом окраски меха (ласка,
горностай, песец) приходятся соответственно на период разрушения или
образования устойчивого снежного покрова. Их линька протекает при сходном
термическом режиме в различных частях ареала видов. Тем не менее диапазон
температур, при которых звери меняют наряд, видоспецифичен. Здесь на сроки
и характер линьки оказывают влияние особенности их экологической и
этологической терморегуляции. По мнению автора, сроки начала и окончания
весенней и осенней смены меха и ее продолжительность во многом определяются
размерами животных: чем больше масса тела, тем продолжительнее линька.
Начинается этот процесс весной и осенью у мелких зверей (ласка, горностай)
на 5-10 и более дней позже, а заканчивается на столько же дней раньше, чем у
сравнительно крупных животных (песец, лисица). Индексы линьки, или
отношение продолжительности популяционной смены меха к индивидуальной,
у зверей с различными образом жизни и формами поведения прослеживаются
достаточно четко. Так, у млекопитающих, которые в своей жизнедеятельности
используют постоянные утепленные гнезда и норы, индекс линьки оказался в
2-3 раза больше, чем у зверей-логовиков, имеющих просто устроенные,
открытого типа убежища, которые используются на короткий срок.
Наряду с меховым покровом важную термоизоляционную роль в жизни
млекопитающих играют и подкожные жировые отложения. По нашим данным,
масса подкожного жира, расположенного в одной только паховой области
(генитальное депо), составляет осенью у обыкновенной ласки 3.5-11.8%о и у
горностая 2.5-11.6%о, тогда как летом она у них практически отсутствует. У
горностая, соболя и куниц жировые резервы, периоды их накопления и
расходования заметно меньше, чем у норок и хорьков, что, видимо, объясняется
существенной разницей в формах поведения, показателях суточной активности
и энергообмена этих видов. Осенью, когда интенсивность обменных процессов
в организме уменьшается, жировой запас (или энергетический потенциал)
снижающих свою активность зверьков резко возрастает. Например, у
сравнительно малоактивных хорьков и норок в июне-июле вес генитального
120
жирового депо как одного из индикаторов упитанности не превышал 3-4%» от
массы их тела. В то же время в ноябре-декабре он составлял у самцов 14-28%»,
а у самок 12-15%». Явно выраженный половой диморфизм по рассматри-
ваемому показателю может служить одной из причин большей элиминации
самок в природе в условиях трудной зимовки.
Динамика массы тела, а следовательно, и упитанности у большинства видов
куньих характеризуется двухвершинной кривой с заметным увеличением
уровня значимости этих величин осенью и весной. В то же время у барсука
сезонные изменения веса характеризуются одной четко выраженной вершиной,
свидетельствующей о подъеме упитанности животных от летних месяцев к
осени. Так, в конце августа-сентябре, в период наибольшей упитанности,
количество подкожного жира у добытых барсуков достигало у самок 3.2, а у
самцов 3.8 кг, что составляло соответственно 29 и 23% от их живого веса.
Подкожные и внутренние жировые резервы имеют важное энергетическое
и термоизоляционное значение, позволяя животным благополучно пережить
экстремальные условия зимнего периода. В осеннее время они достигают
максимальной величины практически у всех хищных млекопитающих. Так, к
началу зимы масса жировой ткани составляла у енотовидной собаки 30-32%,
у волков 15.5 (Козловский, 1986)иурыси 12-13%. Большое количество жира
в осенне-зимние месяцы содержится также в организме бурого и белого
медведей. У представителей первого вида (весом 200-250 кг) жировой запас
может достигать 40-50 кг, что составляет 20% от живой массы тела. Обычно за
зиму жировой резерв в организме этих зверей расходуется очень интенсивно,
однако в теплые зимы часть его может сохраняться и до лета. Например, в
Прибайкалье отстрелянный 19 мая бурый медведь имел более 30 кг, а добытый
23 июня - 2-3 кг подкожного жира (Устинов, 1993).
У обитающего в холодной Арктике белого медведя (Успенский, 1989) общая
масса подкожной жировой ткани осенью часто достигает 100 кг, причем
интересно, что значительные ее отложения у зверей наблюдаются на подошвах
лап. Даже у самок, уже покидающих весной берлоги, толщина жира здесь равна
3-5 см. Кроме того, эти хищники имеют густо опушенную поверхность нижней
части ступни, благодаря чему площадь голых участков на ней оказывается
меньше, чем у бурого медведя, что также снижает уровень теплоотдачи.
Исследование особенностей энергообмена белого медведя показало, что
подкожный жир и густой шерстный покров играют важную роль в процессе
терморегуляции вида, хотя и не обеспечивают ему полную защиту от холода.
Недостаточное совершенство морфологических механизмов регулирования
теплового баланса эти животные компенсируют специфическими формами
поведения. Так, в морозные дни с сильным ветром они отыскивают надежные
убежища, укрываются за торосами, вырывают яму или временную берлогу в
снегу, где могут подолгу пережидать непогоду (Oritsland, 1970; Frisch et al.,
1974; Беликов, 1993).
Кроме того, сохранению тепла в организме многих видов млекопитающих
способствуют такие поведенческие реакции, как окучивание детенышей,
121
утепление самкой гнезда с новорожденными, зарывание зверьков в снег, уход в
нору или свертывание в клубок, когда поверхность тела значительно
уменьшается (Понугаева, 1953, 1960; Проссер, Браун, 1967; Башенина, 1977;
Терновский, 1977; Пантелеев, 1983). Все это особенно важно, так как у мелких
животных (в частности, у куньих) новорожденные не способны поддерживать
собственную температуру тела на постоянном уровне, и данную функцию берет
на себя самка. Наши наблюдения за семьей горностая показали, что в первые
дни после щенения самка редко отлучается из гнезда, обогревая детенышей
своим телом. Выходя на кормежку, она закрывала входное отверстие подстилкой
из гнезда или тушкой несъеденной полевки. В жаркие дни самка выносила
щенков из гнезда и укладывала их открыто на подстилку из травы и листьев
(Туманов и др., 1970). Подобное же поведение мы отмечали у приносивших
приплод самок ласки, солонгоя, норок и лесной куницы. Учитывая огромную
роль поведения в поддержании температуры тела, И. Д. Стрельников (1955) даже
предлагал ввести термин «экологическая терморегуляция» для характеристики
уменьшающейся амплитуды колебания температуры среды обитания мелких
млекопитающих под влиянием поведенческих реакций.
Используя теплые убежища (норы, дупла, берлоги) в своей жизнедеятельности,
хищные звери в значительной мере нивелируют воздействия неблагоприятных
факторов и сохраняют в организме тепло. Гнездовые убежища позволяют
животным в холодное время года поддерживать свой термический гомеостаз с
минимальными энергозатратами. Их способность находить или создавать
микроклиматические условия, необходимые для рождения и выкармливания
детенышей, обеспечивает возможность осуществлять разные жизненные процессы
в широком диапазоне температур и других абиотических факторов внешней
среды. Исследования показали, что в обнаруженных гнездовых камерах, где
находились детеныши, температура была выше таковой наружного воздуха в
разные дни у ласки на 8-18°С, а у горностая-на 5-19°С (Гайдук, 1983,1998).
Подобная разница между температурой наружного воздуха и гнезда
прослеживается и у других млекопитающих (Harington, 1968; Стрельников,
1970; Baskirk, Harlow, 1989). Об условиях обитания белых медведей в берлогах
можно составить представление на основе наблюдений С.Е. Беликова (1976,
1993). По показаниям датчиков в 1974 г. температура у края основной камеры
берлоги была на 14-17°С, а в 60 см от входа в нее - на 5-8°С выше таковой
наружного воздуха. В 1975 г. во время проведения на одной из берлог
аналогичных наблюдений температура наружного воздуха в разные дни
колебалась от -14 до -27°С, а в убежище - от-5.6 до +5°С и от-0.2 до +6.5°С.
По заключению автора уровень температур в занятой берлоге определяется в
первую очередь толщиной потолка, которая зависит от глубины снега и
ветрового режима. В то же время у берлоги с тонким потолком (меньше 20-
25 см) потери тепла бывают столь велики, что самка вынуждена либо рыть
новую камеру, либо покидать родовую берлогу и устраивать временную. В
удобном убежище, периодически меняя позу, звери обеспечивают себе более
комфортные условия для сохранения тепла. Замечено, что чем холоднее погода,
122
тем плотнее сворачивается в клубок лежащий в убежище медведь, уменьшая
тем самым поверхность отдачи тепла в окружающую среду.
Одним из показателей интенсивности обменных процессов, протекающих
в живых организмах, служит ректальная температура. В известной мере у
взрослых животных она определяется величиной (массой) их тела. Как правило,
самые маленькие зверьки имеют более высокие температурные параметры
(Калабухов, 1950; Слоним, 1952; Ивантер идр., 1985). Вместе с тем у близких
видов естественный диапазон температуры тела сравнительно небольшой, и
уровень средних величин определяется прежде всего не размером животных, а
их экологическими особенностями. Например, среди представителей семейства
Mustelidae самую высокую температуру тела имели горностай, лесная куница
и соболь, т.е. очень энергичные, быстрые и активные виды (Туманов, Левин,
1974; Туманов идр., 1982). Следует отметить, что у мелких куньих разница в
температуре тела между обыкновенной лаской и горностаем была лишь близка
к статистически достоверной (р=85%), а между горностаем и солонгоем вполне
достоверна (р>99%) и в группе самцов, и в группе самок (табл. 33).
Среди изучаемых видов куньих при температуре среды 20-25°С самую
низкую полостную температуру (35-35.5°С) регистрировали у зверьков
небольших размеров - пустынной ласки и перевязки, т.е. представителей
аридной зоны. Данную особенность названных видов, вероятно, можно
Таблица 33/ Table 33
Ректальная температура у мелких видов куньих с разной массой тела
(май-июнь 1975-1976)
The rectal temperature of mustelid small species with different body mass
(May-June 1975-1976)
Вид, подвид/ Species, subspecies Пол/ Sex п Масса тела, г/ Body mass, g Температура тела/ Temperature of body, °C
Ласка обыкновенная/ <3<3 9 91 ±4.2 37.4 ± 0.3
Mustela nivalis 22 6 63 ± 2.5 37.6 ±0.3
Ласка закавказская/ M. n. boccamela <3<3 3 149 37.2 ±0.6
Ласка пустынная/ M. п. heptneri 33 2 234 35.3
Горностай/ 33 7 241 ±13.2 38 ± 0.2
М. erminea 22 13 140 ±6.2 38.2 ±0.3
Солонгой/ 33 6 497 ± 12.1 37.1 ±0.1
М. altaica 22 6 360 ±21.7 37.2 ±0.1
123
Таблица 34/ Table 34
Температура тела (°C) и частота дыхания (в 1 мин) у взрослых самок некоторых
видов семейства куньих (июль-август 1979)
Body temperature (degrees) and breath frequency (per 1 min) of adult females of some
mustelid species (July-August 1979)
Объект исследования/ Objects of research Масса тела, г/ Body mass, g Температура тела/ Temperature of body Частота дыхания/ Respiration rate
« 1 M ± m n M (lim)
Колонок/ Mustela sibirica 504 ± 42.7 7 38.5 ±0.5 7 60 (52-68)
Хорек степной/ Mustela eversmanni 691 ±37.3 6 37.0 ± 0.7 12 30 (26-36)
Хорек белый - фуро/ Mustela putorius furo 885 ± 23.4 5 36.7 ± 0.5 8 55 (42-64)
Хорек лесной/ Mustela putorius 797 ± 22 4 37.6 ± 0.6 8 57 (44-66)
Норка европейская/ Mustela lutreola 762 ± 11.5 14 36.2 ±0.3 14 41 (36-48)
Норка американская/ Mustela vison 891 ±47.3 5 37.3 ±0.3 7 48 (38-56)
Куница лесная/ Martes martes 1000±5.6 3 39.8 ±0.5 8 31 (28-36)
Куница каменная/ Martes foina 1197 ± 2.9 3 38.4 ±0.3 7 30 (28-32)
Соболь/ Martes zibellina 1234 ±44.1 5 39.9 ± 1 5 53 (48-62)
Росомаха/ Gulo gulo 9000 2 37.5 5 43 (32-60)
Примечание, л- количество замеров.
рассматривать как наследственно закрепленную и поэтому достаточно
консервативную реакцию организма на существование в условиях жаркого,
климата (Терновский и др., 1981). Вообще видовые различия мелких
млекопитающих по сопротивляемости к перегреву или переохлаждению очень
велики и во многом определяются условиями той среды, в которой животные
развивались.
Норки и хорьки характеризуются невысокими показателями температуры
тела (табл.34). У европейской норки она была достоверно ниже, чем у
американской (рис. 32). У самцов эта разница составляла 0.6°С (р=90%), а у
самок 1.1°С (р=95%). Сравнительно низкая ректальная температура постоянно
отмечалась также у степного хорька (М. eversmanni) и белого - фуро (альбиносная
форма лесного хорька). Причем разница между особями черного хорька и
малоактивного фуро была статистически достоверной (р=95%), что согласуется
с данными о разном уровне их химической терморегуляции (Калабухов, 1950).
У лесной куницы ректальная температура была постоянно выше таковой у норок
и хорьков (рис. 33).
124
Таблица 35/ Table 35
Ректальная температура и некоторые показатели дыхательной и выделительной
систем у взрослых самцов семейства куньих (многолетние данные)
The rectal temperature and some parameters of respiratory and excretory systems of
mustelid adult males (multiyear data)
Объект исследования/ Objects of research n Ректальная температура, ?С/ Temperature of body Частота дыхания/ Respiration rate n Индекс/ Index, %c
в 1 мин/ in 1 min в 1 мин/кг веса/ in 1 min/kg of body mass легких/ lunds почек/ kidneys
Ласка обыкновенная/ 20 37.8 ± 0.2 84±0.9 923 64 16.8+0.4 13.9±0.3
Mustela nivalis Ласка закавказская/ Mustela nivalis boccainela Ласка 3 37.2 ±0.6 77+0.8 516 4 18.5+0.5 11.4+O.G
пустынная/ Mustela nivalis 2 35.3 72 307 7 15.9±2.6 11.4+0.1
heptneri Г орностай/ 18 38.2 + 0.3 60±2.4 249 52 15.2+0.3 10.3+0.2
Mustela erminea Солонгой/ Mustela altaica 16 37.3 + 0.2 111 5 15.2+0.8 10.2±1.5
55±1.4
Колонок/ Mustela sibirica Перевязка/ 10 37.6 ± 0.4 61±4.8 83 2 15 10.2
Vonnela 4 35.2 ±0.4 41±3.4 54 2 15.8 10.2
peregusna Хорек лесной/ Mustela putorius 26 37.8 ± 0.3 55±3.8 40 86 15.6+0.3 8.4+O.2
Хорек степной/ Mustela 8 37.3 ± 0.4 32±6.7 24 2 15.7 8.2
eversmanni Хорек белый - фуро/ Mustela putorius furo 8 36.4 ±0.4 56±3.5 35 2 13.5 7.5
Норка европейская/ Mustela lutreola 18 37.0 ± 0.2 48±2.8 41 45 17.8+0.4 10. 1+0.2
Норка американская/ Mustela vison 25 37.6 ± 0.3 51 ±3.9 43 98 16.6±0.3 9+0.1
Куница лесная/ Martes martes 15 39.2 ±0.3 59±3.5 48 49 15.5+0.4 7. l±0.1
Куница каменная/ 3 38.6 ± 0.3 55±3.8 43 5 13. 1+0.5 6.8+0.2
Martes foina Соболь/ Martes zibellina 9 39.4 ±0.4 68±7.8 32 2 15.1 7.3
Росомаха/ Gulo t>ulo 3 38.0 ± 0.2 33±2.5 3 8 16.2±1.2 7.4+O.7
Барсук/ Meles meles 10 37.3 ± 0.2 28±5.3 4 14 12.6+1 6.4+О.З
Выдра/ Ultra lutra 3 37.7 ± 0.5 27±2.5 4 15 21.4+0.7 9.8+O.2
Примечание. Данные по индексу легких и почек пустынной ласки взяты из работы
Р. Реймова, 1972.
125
Таблица 36/ Table 36
Изменение температуры тела (ректальной) и бедра
у некоторых грызунов (масса тела 20-28 г) при температуре среды 15 и 10 °C
Changes of body (rectal) and femur temperature of some rodents (body mass 20-28 g)
with ambient temperature 15 and 10 °C
Виды/ Species Пол/ Sex п 15°С п 10°С
тело/ body бедро/ thigh тело/ body бедро/ thigh
Мышь жслтогорлая/ Apodemus flavicollis 33 22 6 7 34.4±6.1 33.8±4.5 33.1±7.1 32.5±4.9 7 9 32.7±4.9 31.7±3.8 28.2±5.6 27.5±2.7
Мышь полевая/ 33 10 33.5±3.5 32.5±7.8 14 30.3±2.5 27.8±2.3
Apodemus agrarius 22 5 33.3±8.4 32.3±8.5 7 29.1±4.4 25.3±3.8
Полевка рыжая/ 33 11 33.4±3.3 31.6±3.8 12 30.4±2.8 27.8±2.6
Glethrionomys glareolus 22 7 32.6±4.9 31.5±8.2 13 29.5±2.6 26.5±2.9
Полевка 33 5 33.6±7.2 31.7±9.3 4 30.2±8.5 26.6±7.5
обыкновенная/ Microlus arvalis 22 11 33.3±3.4 31.5±5.9 4 29.2±8.2 25.7±6.9
Среди представителей рода Martes сравнительно низкими термо-
метрическими, как и морфометрическими, показателями выделялась каменная
куница (табл. 35). Отмеченные видовые различия не только по термофильности,
но и по величине терморегуляции могут обусловливать и соответствующую
разницу в обменных процессах видов. По-видимому, лесная куница обладает
большим уровнем теплообмена, что и позволяет ей иметь широкий ареал,
продвигаясь далеко на север. Подобные эколого-физиологические различия
обнаруживаются и среди хорьков (Калабухов, 1950), объясняя особенности их
территориального распределения.
Измерение контактным способом кожно-мышечной температуры тканей
двигательного аппарата свидетельствует о том, что в покое соболь и лесная
куница обладают большей величиной этого показателя по сравнению с другими
куньими сходного размера. Так, кожно-мышечная температура задне-
бедренной группы мышц у них оказалась в среднем на 0.3-0.5°С выше, чем у
хорьков и норок, хотя в целом и у тех, и у других она была на 1-1.2°С ниже
ректальной. Таким образом, собранные материалы свидетельствуют о высокой
степени васкуляризации тканей (следовательно, и обменных процессов в них)
локомоторного аппарата у очень активных и быстрых животных. Необходимо
также отметить, что у мелких хищников кожно-мышечная температура
поверхностей тела мало изменяется при некоторых естественных перепадах
температуры наружного воздуха. В отличие от них у грызунов даже при ее
небольшом падении уменьшались и ректальная, и кожно-мышечная
температура (табл. 36).
126
38
1
37 -=-----/
36 --
35 --
34 Ч--1---1--1---1--1---1--1---1--1--1---1
i nmivvvivnvmixxxixn
Рис. 32. Сезонная динамика ректальной температуры самок американской (7) и
европейской (2) норок.
По оси абсцисс - месяцы года; по оси ординат - температура тела, °C.
Fig. 32. Seasonal dynamics of the rectal temperature in females of the American
(Mustela visori) (7) and European (Mustela lutreola) (2) minks.
Abscissa - months of the year; ordinate - temperature of body, °C.
Рис. 33. Сезонная динамика ректальной температуры самок лесной куницы (7) и
лесного хорька (2).
Условные обозначения как на рис. 32.
Fig. 33. Seasonal dynamics of the rectal temperature in females of the common marten
(Martes martes) (7) and polecat (Mustela putorius) (2).
Abscissa - months of the year; ordinate - temperature of body, °C.
Рис. 34. Сезонная динамика ректальной температуры самцов американской норки
(7) и ласки (2).
Условные обозначения как на рис. 32.
Fig. 34. Seasonal dynamics of the rectal temperature in males of the American (Mustela
vison) (7) and European (Mustela lutreola) (2) minks.
Abscissa - months of the year; ordinate - temperature of body, °C.
127
Рис. 35. Сезонная динамика ректальной температуры самцов лесного хорька (7) и
европейской норки (2).
По оси абсцисс - месяцы года; по оси ординат - температура тела, °C.
Fig. 35. Seasonal dynamics of the rectal temperature in males of polecat {Mustela
putorius) (1) and European mink {Mustela lutreola) {2).
Abscissa - months of the year; ordinate - temperature of body, °C.
Вполне возможно, что у грызунов реакция сосудов кожи и распределение в
ней крови имеют заметно большее значение, чем у хищных млекопитающих.
По-видимому, высокая и сравнительно постоянная температура тела
последних сопровождается и малым диапазоном таковой на разных участках
кожи. Интересно также и то, что пустынная ласка, для которой характерна
сравнительно низкая температура тела, в одинаковых условиях с «северной»
формой - обыкновенной лаской - имела кожно-мышечную температуру в
среднем на 0.5°С выше. Вероятно, высокий уровень температуры кожных
покровов вообще характерен для представителей аридной зоны. Интенсивная
Рис. 36. Многолетняя динамика ректальной температуры самца европейской
норки.
7- 1986; 2- 1987; 3- 1988; 4- 1989.
По оси абсцисс - месяцы года; по оси ординат - температура тела, °C.
Fig. 36. Multiyear dynamics of the rectal temperature in the male of the European
mink {Mustela lutreola).
1 - 1986; 2- 1987; 3- 1988; 4- 1989.
Abscissa - months of the year; ordinate - temperature of body, °C.
128
теплоотдача с поверхностных тканей у них может иметь существенное значение
как адаптивная реакция к обитанию в условиях жаркого климата.
Сезонная изменчивость температурных реакций у представителей
семейства куньих прослеживается вполне отчетливо (рис. 34,35). Наблюдение
за содержащимися в условиях неволи животными (опыты 1986-1989 гг.)
показали, что в целом наиболее высокая температура у них отмечалась весной
и летом, а низкая - зимой (рис. 36-39). Если в апреле температура тела самок
горностая колебалась в пределах 38.6-40°С, а в июле - 38.4-39.2°С, то к
декабрю-январю она снижалась до 37.7-38.4°С. У обыкновенной ласки и
солонгоя полостная температура летом в среднем на 0.8-1.2°С выше, чем
зимой. Зимнее ее падение можно, вероятно, рассматривать в качестве одной
из форм термоадаптации, так как уменьшение амплитуды температур между
телом животного и окружающей средой (воздух, почва, вода) способствует
уменьшению интенсивности теплоотдачи (Шилов, 1982, 1985; Buskirk et al.,
1988).
Заметная разница летней и зимней температур прослеживается и у барсука,
который среди крупных хищников семемейства куньих имеет наиболее низкие ее
показатели. У самцов ректальная температура изменялась в среднем от 37.3°С
летом до 35.5°С зимой, а живой вес за этот период уменьшался на 35-44%. Известно,
что у барсуков в холодный период года основной обмен снижается на 30-50%
и более по сравнению с летними месяцами (Слоним, 1952; Harlow, 1981а, б;
Harlow, Miller, 1985). Низкий уровень температуры тела и обменных процессов
позволяет им создавать в организме значительные энергорезервы, которые
экономно расходуются в период зимнего сна. Интенсивное жиронакопление
барсука следует рассматривать как видовое приспособление терморегуляции,
благодаря которому он снижает зимой теплоотдачу и расход энергии.
Температура тела енотовидной собаки менее постоянна, чем, например, у
лисиц и песцов, однако эти изменения обычно незначительны. По нашим данным,
в июле при температуре воздуха +22°С ректальная температура взрослых особей
(самца и самки) колебалась в пределах 37.6-38.2°С. Зимой при температуре
воздуха -10°С она снижалась до 37-37.5°С. Следует отметить, что у енотовидных
собак вообще температура тела ниже, чем у других представителей семейства.
Так, при температуре среды+15- +18°С у обыкновенной лисицы она составляла
осенью 38.6-39°С, у белого песца 38.8-39.3°С, а у енотовидной собаки 37.4-
37.6°С. У песцов сезонные колебания температуры тела, уровня теплообмена и
потребностей в корме прослеживаются достаточно четко. Зимой, когда в природе
наблюдается определенный дефицит кормов и затрудняется их добывание,
потребность в пище и ректальная температура как индикатор процесса
энергообмена находятся на минимальном уровне. Отдача тепла в окружающую
среду ограничена благодаря густому меховому покрову и значительным
подкожным жировым отложениям. Весной в связи с линькой и гоном
энергозатраты, а следовательно, и потребность в пище возрастают. К лету песцы
освобождаются от густого мехового покров и жировой прослойки, в результате
чего теплоотдача увеличивается. Летний песец выглядит тощим, мелким и «бедно
5 Зак. № 4309
129
Рис. 37. Многолетняя динамика ректальной температуры самца американской
норки.
Условные обозначения как на рис. 36.
Fig. 37. Multiyear dynamics of the rectal temperature in the male of the American mink
{Mustela visori).
Look fig. 36.
Рис. 38. Многолетняя динамика ректальной температуры самки лесного хорька.
Условные обозначения как на рис. 36.
Fig. 38. Multiyear dynamics of the rectal temperature in the female of the polecat
{Mustela putorius).
Look fig. 36.
Рис. 39. Многолетняя динамика ректальной температуры самки европейской
норки.
Условные обозначения как на рис. 36.
Fig.39. Multiyear dynamics of the rectal temperature in the female of the European
mink {Mustela lutreola).
Look fig. 36.
130
Таблица 37/ Table 37
Изменение температуры тела и уровня потребления кислорода у хищных
млекопитающих при разной температуре среды (по: Слоиим, 1952)
Changes of body temperature and level of oxygen consumption of carnivores with various
ambient temperature
Температура среды, °C / Temperature of Лисица обыкновенная/ Vulpes vulpes Псссц белый/ Alopex lagopus Собака енотовидная/ Nyctereutes procyonoides Барсук/ Meles meles Медведь бурый/ Ursus arctos
°C 1 2 1 2 1 2 1 2 1 2
25 738 38.7 897 39.5 470 38.2 468 38.4 229 36.9
20 714 38.9 819 38.8 479 38.1 421 37.5 243 36.8
15 790 39.1 685 39.3 467 37.8 556 37.5 357 36.9
10 713 38.4 1036 38.6 468 37.6 647 37.5 487 35.7
5 1025 38 1418 39.3 583 37.4 - - - -
Примечание. 1- потребление кислорода, см’ на кг/ч; 2 - температура тела, °C
одетым». Вместе с тем при сравнительно высокой температуре окружающей среды
животным для поддержания оптимального теплового баланса не требуется
большого количества пищи, а обилие кормов в летнее время освобождает
организм от лишних затрат энергии, связанных с их добычей.
В конце августа-сентябре, когда в природе имеется большое количество
кормов, а энергозатраты на их добывание незначительны, показатели
температуры тела и уровня обменных процессов падают. Это способствует
накоплению в организме жировых запасов, которые интенсивно расходуются
зимой.
У взрослых бурых медведей ректальная температура в летние месяцы равна
в среднем 37°С. Зимой во время сна обмен веществ у зверей понижается на 50-
70%, что ведет к большой экономии энергии, однако температура тела в это
время снижается незначительно - на 1.5-2°С (Слоним, 1962).
Температура тела взрослого белого медведя близка к таковой или несколько
выше, чем у бурого и колеблется в пределах 36.8-38.8°С. Реакция сосудистого
русла кожных покровов на изменение условий среды выражена у него очень
отчетливо. Снижение внешней температуры вызывает у этого хищника
соответственное падение и кожно-мышечной температуры. Если в тихую теплую
погоду она равна 30-36°С, то на ветру опускается до 27°С, а при плавании зверей
падает даже до 10-14°С (Успенский, 1989). По-видимому, при низкой температуре
среды у них происходит резкое сужение просветов капилярного русла кожных
покровов, что способствует снижению теплоотдачи. По мнению С.М. Успен-
ского (1989), уровень метаболизма у белого медведя, очевидно, выше, чем
у бурого. У него обнаруживается необычайная стойкость к воздействию низких
температур не только за счет более совершенной терморегуляции, но и по причине
низкой «критической температурной точки», при которой достигается тепловой
131
Таблица 38/ Table 38
Сезонные изменения потребления кислорода и величины теплопродукции у
енотовидных собак (по: Соколов, 1949а)
Season changes in oxygen consumption and value of heat production
of racoon dogs (Nyctereutes procyonoides)
Месяц/ Month Живой вес, кг/ Body mass, kg Потребление кислорода/ Oxygen consumption Теплопродукция/ Heat production
см3/(кг-ч) cm3/kg in 1 hour % к январской норме/ % to January norm кал/(кг-ч) cal/kg in 1 hour % к январской норме/ % to January norm
Март/ March 6 353.6 126.7 1.69 127
Апрель-май/ April-May 6.1 407.1 145.9 1.92 144
Июнь-июль/ 6 514.8 184.5 2.39 180
June- July Сентябрь/ September 6.8 124.3 119
346.4 1.59
Октябрь/ October 7.2 298 106.8 1.42 107
Январь/ January 7.2 279.1 100 1.33 100
баланс co средой. Даже при -50°C у белого медведя не наблюдается заметного
повышения уровня газообмена, т.е. еще не возникает необходимости в
максимальном использовании организмом механизма химической термо-
регуляции, связанной с большим расходом энергии.
Вообще следует отметить, что хищники, активные в течение всего года,
обладают более высокими показателями обменных процессов и термо-
регуляции по сравнению с животными, впадающими в зимний сон (табл. 37).
Например, если у обыкновенной лисицы и белого песца при падении
температуры среды от 25 до 10°С они изменяются в незначительных пределах,
то у бурого медведя проявления химической терморегуляции выражены очень
резко, составляя 6-10% на 1°С (Слоним, 1952).
По сообщению Е.А. Соколова (1949а), который содержал енотовидных
собак в условиях эксперимента на одинаковом рационе (суточная дача корма
составляла 343 г при калорийности 1 г, равной 8.195 ккал) в течение 3 лет,
наиболее интенсивный газообмен наблюдался у них в апреле-августе, наи-
меньший - осенью и зимой (сентябрь-январь), а в марте он снова усиливался
(табл. 38).
В соответствии с газообменом изменяется и теплопродукция животных.
Осенью и зимой она характеризуется минимальными показателями. В этот
период сбережению тепла способствуют густой мех и отложенные под кожей
значительные жировые резервы. Такая физическая терморегуляция дает
возможность зверям поддерживать температуру тела на необходимом уровне
при ослаблении распада веществ в организме и уменьшении теплопродукции.
132
Таблица 39/ Table 39
Изменение температуры тела (°C) и частоты дыхания (в мнн) у некоторых видов
млекопитающих при понижении температуры и длительном голодании
(по: Заболотских, 2000)
Changes of body temperature (degrees) and breath frequency (per 1 min) of mammal some
species with temperature fall and long starvation
Показатели/ Parameters Сурок степной/ Marmota bobac Сурок черношапочный/ Marmota camtschatica Собака енотовидная/ Nyctereutes procyonoides Песец белый/ Alopex lagopus
Температура при/ Temperature during: активном состоянии/activity 36-38 37-38 39 40.2
длительном голодании/ hunger-strike 1.5-3 0.9-2 38.1 39.6
Частота дыхания при/ Respiration rate during: активном состоянии/activity 25-30 40-55 30-35 25-35
длительном голодании/ hunger-strike 0.2-0.3 0.2-0.3 20-25 25-38
Весной и летом процессы линьки и гона (а у самок еще беременности и
лактации) сопровождаются увеличением энергетических затрат. Это приводит к
потреблению и переработке большого количества пищи. Повышение тепло-
продукции в организме животных в теплый период прослеживается достаточно
четко (табл. 38), что хорошо согласуются и с имеющимися данными по теплообмену
песцов. Так, максимальная теплопродукция у них наблюдается в летние месяцы,
когда в сутки продуцируется в среднем 106.2 кал на 1 кг веса, а минимальная -
зимой (66.9 кал). При этом суточная теплопродукция на 1 кг живого веса
повышается у песцов весной на 23.5%, летом - на 58.6%, а осенью - на 14.7% по
сравнению с зимним сезоном (Поздняков, 1953). Подобные изменения характерны
и д ля обыкновенной лисицы. Наибольшая суточная потребность в корме (в среднем
450 г) у особей обоего пола отмечается осенью, когда уровень энергообмена
снижается у них на 15-20% по сравнению с зимними месяцами (Ильина, 1952).
Зимой при дефиците кормов теплообмен и потребность в пище находятся на
минимальном уровне (до 270 г).
Известно, что у зимоспящих животных в период глубокого оцепенения
уровень физиологических процессов в организме заметно понижен
(Калабухов, 1946). По материалам Ю.С. Заболотских (1999а, б, 2000), у
степного и черношапочного сурков во время зимней спячки наблюдалось
резкое падение температуры тела и частоты дыхания (табл. 39). Из хищных
млекопитающих семейства собачьих некоторое снижение физиологических
параметров при низких температурах и длительном голодании отмечалось
только у енотовидной собаки. По мнению автора, данные особенности
имеют явно приспособительное значение, направленное на экономное
расходование внутренних энергоресурсов и повышающее выживаемость
этих зверей в период длительной зимней бескормицы. Таким образом,
133
отмеченная сезонная динамика потребности в пище, температуры тела и
уровня теплообмена способствует пониманию характера кормодобывающей
активности и адаптивных поведенческих особенностей исследуемых хищ-
ников в разные сезоны года.
Как известно, наиболее наглядным и важным показателем энергетических
реакций в организме животных служит уровень основного обмена, который
обычно определяется методом непрямой калориметрии, т.е. по потреблению
кислорода в единицу времени. Согласно полученным нами данным, у куньих
наблюдается тесная взаимосвязь уровня основного обмена и размеров животных.
При прочих равных условиях он тем больше, чем меньше масса тела зверьков.
Данная зависимость хорошо прослеживается в семейства куньих при сравнении
видов с разной массой тела (Туманов, 1972, 1974а, б). У обыкновенной ласки
на единицу веса он оказался выше, чем у горностая и других хищников, что
прежде всего является следствием ее мелких размеров. Так, в летние месяцы
уровень потребления кислорода был равен у молодых самцов ласки 4 ± 0.69,
горностая 3.1 ± 0.08 и солонгоя 2.3 ± 0.05 мл/(г-ч). У самок данный показатель
составлял соответственно 4.2 ± 0.66, 3.3 ± 0.13 и 2.6 ± 0.43 мл/(г-ч).
Собранные материалы свидетельствуют о том, что тенденция к половым
различиям по интенсивности теплообмена прослеживается у рассматриваемых
видов вполне отчетливо. Она выражается в большем продуцировании тепла
на единицу веса у самок (по сравнению с самцами) и объясняется характерным
для зверьков половым диморфизмом по массе тела, играющим основную роль
в величине теплоотдачи (Сегаль, 1975а, б). При расчете расхода энергии не на
массу, а на единицу поверхности тела разница между самцами и самками по
интенсивности метаболизма нивелируется (Мелькина, 1966).
Сезонная динамика обменных процессов приобретена животными в
результате естественного отбора под влиянием периодически изменяющихся в
течение года условий существования. В целом у куньих она характеризовалась
увеличением газообмена (соответственно и теплопродукции) в весенне-летние
месяцы и падением его к декабрю-январю. В соответствии с этим менялись по
сезонам года масса тела и потребность зверьков в кормах: летом они имели
сравнительно низкий живой вес и жировой запас, а к началу зимы становились
более упитанными. Разница в весе за отмеченный период составляла у самцов
лесной куницы и соболя 7-10%, американской норки 22-30 и лесного хорька
28-35%. Среди хищников примерно равного веса зимнее понижение энерге-
тического обмена лучше выражено у представителей видов, способных
создавать в организме жировые резервы.
По имеющимся сведениям, в условиях клеточного содержания тепло-
продукция в декабре была ниже, чем в июле, у норок на 30-33, а у соболей - на
17-18% (Титова, 1950; Афанасьев, Перельдик, 1966). По нашим данным, уровень
потребления кислорода за этот период понизился в среднем у лесной куницы на
16-20%, у американской норки - на 31-35 и у лесного хорька - на 35-38%. Так,
у самок американской норки, отловленных в природе, уровень потребления
кислорода достигал в декабре 0.77 ± 0.6, а в июле 1.13 ± 0.3 мл/(г ч).
134
Подобные сезонные изменения показателей газообмена наблюдались и у лесных
хорьков того же пола, у которых он составлял зимой 0.78 ± 0.3 и летом 1.25 ±
± 0.5 мл/(г ч). Необходимо отметить, что снижение величины энергообмена в
зимнее время представляет собой приспособительную реакцию организма,
направ-ленную на создание дополнительного терморезерва и свидетельствую-
щую о широких возможностях химической терморегуляции, а также о высокой
степени экологической пластичности видов (Шилов, 1977; Пантелеев, 1983;
Buskirk et al., 1988).
Интересные данные по основным показателям энергетического баланса
мелких куньих в Якутии получены В.М. Сафроновым (2002). Они свиде-
тельствуют о том, что у ласки термонейтральная зона не выражена. При
отрицательных температурах (-15--17°С) интенсивность метаболизма
возрастала на 200%. Относительная масса печени, дающая 12-24% общей
теплопродукции организма, у ласки в 1.3 раза больше, чем у колонка и соболя.
При высоких энергетических требованиях (потребление пищи до 70% от массы
тела) она удовлетворяется 1-2 полевками в сутки. Суточное потребление корма
у соболя около 20% от массы тела, что требует добычи 7-8 полевок и
длительного охотничьего поиска.
У колонка минимальное потребление кислорода в Якутии наблюдалось
при 20 °C (1.48±0.24 л/кг в 1 ч), зона термонейтральности располагалась в
интервале 15-20 °C. Химическая терморегуляция развивалась до -15- -30 °C
(134-147% от минимального уровня). При -35 °C ее интенсивность снижалась,
свидетельствуя об ограниченности реакций теплообразования. Теплоизоляция
улучшалась хорошо выраженным слоем жира. Большие затраты на
химическую терморегуляцию повышают требования этого вида к корму как
источнику энергии.
По данным автора (Сафронов, 2002), соболь характеризуется широкой
термонейтральной зоной (2-22 °C). При понижении температуры воздуха
интенсивность потребления кислорода повышалась на 39-68% и стаби-
лизировалась при -5- -8 °C. Эта умеренность химической терморегуляции в
состоянии покоя объясняет тесную привязанность якутского соболя к утепленным
норовым убежищам, а также быструю его гибель от переохлаждения в ловушках,
ограничивающих свободу движения. У зверьков в зимнем меху предпочитаемая
температура находится в большом интервале положительных температур 7.5-
17.5 °C (Пономарев, 1944). У клеточных соболей при беге теплопродукция
возрастала в 3.3-4 раза (Титова, 1950). Очевидно, мышечная деятельность,
связанная со значительным теплообразованием, выполняет важную роль в
поддержании термического гомеостаза у соболя в зимних условиях. В свою
очередь минимизация обмена веществ в пределах широкой термонейтральной
зоны во время отдыха в гнезде способствует сбережению энергии, обеспечивая
устойчивость энергообмена на холоде.
У горностая (Сегаль, 1975а), как и у ласки, термонейтральная зона не
выражена. Интенсивность химической терморегуляции (4.65 на 1 °C) заметно
выше, чем у американской норки. По сравнению с последней у горностая на
135
единицу массы тела расход энергии и потребность в пище оказались более
значительными. В среднем за сутки зверек потребляет количество пищи, равное
60% массы тела, в то время как у американской норки этот показатель не
превышает 30%.
Интенсивный обмен веществ у куньих сопровождается и высоким
уровнем легочного дыхания. Если у грызунов в нормальных условиях темп
дыхания - относительно постоянная величина, то у мелких хищников в
бодрствующем состоянии частота дыхания отличается вариабельностью: у
зверьков спонтанно может происходить задержка дыхания или его учащение.
Во время сна наблюдается стабилизация темпа дыхания, что даже дает
возможность произвести его визуальную регистрацию. В целом сезонные
изменения частоты дыхания у всех изучаемых хищников выражены довольно
отчетливо: летом темп их дыхания увеличивается, зимой несколько
понижается.
У ласки, горностая, солонгоя и хорьков наиболее высокий уровень
внешнего дыхания и потребления кислорода отмечался в июне-июле. Вполне
вероятно, что снижение интенсивности дыхания и газообмена в зимние месяцы
в значительной степени связано с изменениями теплоизоляционных свойств
покровов - увеличением слоя подкожного жира и улучшением термозащитных
качеств меха. Правда, уровень основного обмена снижался зимой и у тех
особей ласки и горностая, которые в связи с содержанием в теплом помещении
вылиняли только частично и не сменили полностью летний коричневый мех на
зимний белый. Следовательно, эти специфические реакции не просто связаны
с физическими свойствами кожных покровов, а являются сложными видовыми
особенностями нервного и гуморального механизмов терморегуляции (Слоним,
1952). Весной и в начале осени частота дыхания и количество потребляемого
кислорода у ласки, горностая и других куньих возрастали, однако этот подъем
по отношению к зимним месяцам был на 40-60% ниже летнего.
Регистрация внешнего дыхания показала, что уровень легочной вентиляции
находится в обратной зависимости от массы тела животных. Эта закономерность
особенно наглядно проявляется при расчете удельной величины легочного
дыхания, которая, как и теплопродукция, оказалась наиболее высокой у самых
маленьких представителей семейства - ласки и горностая (см. табл. 35).
Необходимо также отметить, что у куньих, обладающих высокой степенью
двигательных реакций, энергичная работа легких может обеспечиваться не
только большой частотой, но и глубиной дыхательного акта. Например, у
степного хорька по сравнению с одомашненным белым дыхание оказалось
относительно редким, но более глубоким, что значительно увеличивает
вентиляцию легких. Последняя, вероятно, усиливается еще и за счет сравнительно
больших размеров его респираторных органов (табл. 35).
Одним из важнейших факторов, определяющих возможности пребывания
мелких млекопитающих в глубоких норах, является соотношение количества
кислорода и углекислоты в почве. По данным И.Д. Стрельникова (1955), в
связи с большой воздухопроницаемостью песчаных почв количество
136
углекислоты в воздухе нор, вырытых в них, невелико и достигает 0.1%. В
суглинистых почвах содержание повышается, а в солончаковых достигает
уже 3-4%. Воздухопроницаемость почв уменьшается при повышении их
влажности, в результате чего могут создаваться неблагоприятные условия для
дыхания мелких животных.
Н.В. Башенина (1956) пишет, что в воздухе подснежных ходов серой
полевки количество углекислоты может достигать 4%. В зимних гнездах
слепыша содержание углекислого газа составляет 2.5-4.8%, в то время как в
летних свежевырытых его ходах оно заметно меньше - 0.6-2.8% (Arieli, 1979).
Высокое содержание углекислого газа отмечается в ходах и европейского
крота (Olzewski, Skoczen, 1965). Поданным В.И. Висковой (1983), в глубинных
убежищах этого зверька содержание углекислоты примерно в 1.2 раза выше,
чем в поверхностных. Автор сообщает, что если на поверхности земли ее
количество колебалось в пределах 0.03-0.04%, то в нежилых норах оно уже
достигало 1.10±0.21% в основном за счет жизнедеятельности почвенных
микроорганизмов. В жилых ходах крота, вырытых на низинных участках,
содержание углекислоты в весенне-летний период составляло 2.35-2.40%.
По материалам Н.В. Некипелова (1958), в конце зимы в закрытых ходах
полевок Брандта количество углекислого газа составляло 1%, а в отдельных
норах на глинистых почвах-даже 5-6%. В убежищах длиннохвостого суслика,
по данным того же автора, оно достигало 1.8%.
По нашим наблюдениям, проведенным в августе-сентябре в окрестностях
Киева, в нежилых поверхностных ходах мышевидных грызунов количество
углекислоты и кислорода не отличалось от их содержания в атмосферном
воздухе (соответственно 0.03 и 20.93%). По-видимому, поверхностные ходы
хорошо аэрируются за счет приточно-вытяжной вентиляции. В жилых
поверхностных норах, имеющих 2-3 и более выходных отверстий, содержание
углекислоты также не превышало нормы, что можно объяснить их хорошей
вентиляцией. В то же время в заглубленных убежищах, в гнездовых камерах с 1
или 2 зверьками, количество углекислого газа увеличивалось до 0.8-1.2%
(Туманов, 1966а, б, 1968). Таким образом, во время пребывания в норах мелкие
млекопитающие (в том числе и куньи) могут находиться в условиях повышенного
содержания углекислого газа. В качестве убежищ они часто используют ходы и
гнезда грызунов или глубокие естественные пустоты под корнями и стволами
упавших деревьев. В осенне-зимний сезон ласок нередко отлавливают в кротовых
ходах, где они активно добывают их обитателей и, следовательно, должны быть
хорошо адаптированы к условиям обитания под землей.
Известно, что у животных, не приспособленных к пребыванию в воздушной
среде с высоким содержанием углекислого газа, его концентрация порядка
2-3% вызывает функциональные сдвиги, которые выражаются в первую очередь
в изменении темпа дыхания и сердечной деятельности (Голодов, 1946;
Сергиевский, 1950).
Наши опыты, выполненные в лабораторных условиях на ласках и грызунах,
показали, что придыхании зверьков газовой смесью с 1-1.2% углекислоты у
137
Таблица 40/ Table 40
Относительная масса легких у взрослых куньих с разной массой тела
в октябре-ноябре
Relative mass of lungs of mustelid adults with different body mass in October-November
Виды/ Species Пол/ Sex n Масса тела, г/ Body mass, g Индекс легких/ Index of lungs, %o
Ласка/ 3 <3 10 81 ±2.5 15.6 ±0.7
Mustela nivalis $ $ 8 52± 1.8 17.5 ±0.3
Горностай/ 3 3 19 209 ± 5.8 13.8 ±0.4
Mustela erminea $ $ 14 114 ± 3.1 13.5 ±0.4
Солонгой/ 3 3 3 282 ± 16.4 13.2 ±2.3
Mustela altaica $ $ 3 205 ± 12.5 13.1 ±2.5
них наблюдалось лишь незначительное урежение и углубление дыхательных
движений. Так, в эксперименте частота дыхания уменьшилась у желтогорлых
мышей на 12-14%, а у ласок - на 13-17%, т.е. была в пределах допустимой
нормы. Таким образом, у мелких хищников, как и у грызунов-норников, в
отличие от наземных животных воздействие на дыхательный центр повы-
шенных концентраций углекислого газа не оказывает существенного влияния
на вентиляцию легких. По-видимому, у тех видов куньих, жизнь которых
постоянно связана с норой, снижение чувствительности дыхательного центра
к накоплению углекислоты во вдыхаемом воздухе представляет собой
выработанную в процессе эволюции важную адаптивную реакцию, способ-
ствующую обитанию в подземных убежищах, где они могут успешно охотиться
и укрываться в непогоду.
У крупных хищников, как и у мелких, вентиляция легких заметно
увеличивается в летнее время за счет большей частоты и глубины дыхания.
Падение уровня энергообмена в зимний период обусловливает у них и
снижение интенсивности дыхательного акта. У белых медведей при
температуре среды +20- +25°С темп дыхания достигает 30 движений в мин.
При падении температуры воздуха до -10- -20°С он становится заметно реже
в среднем до 5.3 в мин. На о-ве Врангеля у спокойно лежащих в берлоге белых
медведиц частота дыхания снижалась до 2.5-3.5 в мин (Беликов, 1993).
Подобные изменения вентиляции легких при зимнем падении уровня
энергообмена прослеживаются и у бурых медведей. Так, по нашим
наблюдениям в Санкт-Петербургском (Ленинградском) зоопарке в июне-июле
частота дыхания у отдыхающих зверей колебалась в пределах 25-30 в мин, а
зимой у залегших в берлогу особей темп дыхания был более редким - 3-5 в
мин. По данным многих исследователей, за зимний период бурый медведь
интенсивно потребляет накопленные летом питательные вещества. Объем его
жирового слоя уменьшается за это время в среднем на две трети всей массы, а
138
сжигание жиров в клетках (грамм жира дает грамм воды) полностью
обеспечивает зверю зимнюю потребность в воде (Hesse et al., 1951; Слоним,
1952; Kayser, 1965).
Интенсивность легочной вентиляции у мелких млекопитающих по
сравнению с крупными определяет и большие относительные размеры
респираторных органов. В целом можно сказать, что индекс легких зависит от
массы тела животного (см. табл. 35). С увеличением этого показателя
абсолютная масса легких возрастает, а относительная падает. Так, упитанные
мелкие хищники в осеннее время имели несколько меньший индекс этого
органа по сравнению с особями, добытыми в весенне-летний сезон, когда их
жировые резервы были незначительными. Среди взрослых представителей
семейства куньих, относящихся к видам, близким и в систематическом плане, и
по образу жизни, прослеживается закономерное падение индекса легких с
увеличением массы тела зверьков, причем эта зависимость наблюдается как в
группе самцов, так и самок (табл. 40).
Следует отметить, что размеры респираторных органов определяются не
только величиной животных, но и их экологическими особенностями. Четкой
взаимосвязи между частотой дыхания и индексом легких у куньих не
регистрируется. Последний возрастает у них только по мере увеличения
специализации видов к водной среде обитания. Если у самцов американской
норки он равен 16.6 ± 0.3%о, то у выдры того же пола 21.4 ± 0.7, а у калана
(Барабаш-Никифоров и др., 1968) даже 48.4 ± 3%о.
Известно, что из всех внутренних органов почки по уровню своего развития
четко реагируют не только на частный водно-солевой обмен, но и на любые
изменения метаболизма (уменьшение общих размеров тела, усиление
двигательных реакций, увеличение теплоотдачи с продвижением в высокие
широты или в высокогорье и т.д). Это позволяет использовать данный показатель
в качестве индикатора популяционной напряженности метаболических
процессов и масштаба обмена веществ (Rensch, 1948; Warburton, 1955; Шварц,
1958,1960; Межжерин, Мельникова, 1966; Шварц и др., 1968; Большаков, 1972;
Ивантер, 1976; Ивантер и др., 1985).
В целом прослеживается обратная зависимость индекса почек от массы
тела изучаемых хищников. Достаточно наглядно она проявляется у куньих
разного веса, но близких по образу жизни и кормовой специализации (см.
табл. 35). Среди хищников сходного размера относительная масса почек
больше у амфибионтов. Например, у выдры она была достоверно выше,
чем у росомахи и барсука (д>99%). Вообще адаптация к водному образу
жизни сопровождается структурными преобразованиями этого органа: если
у самцов американской норки его индекс равен в среднем 9 ± 0.1 %о, то у
выдры 9.8 ± 0.2, а у калана (Барабаш-Никифоров и др., 1968) достигает
21.6 ± 1.6%». Причем в отличие от других видов, у выдры и калана почки
дольчатые, что, вероятно, связано с их усиленной функциональной деятель-
ностью, в том числе и с потреблением кормов, содержащих высокий процент
воды.
139
Таблица 41/ Table 41
Сравнительная характеристика массы легких и почек у некоторых взрослых
представителей хищных млекопитающих
Comparative characteristics of lungs and kidney mass of some representatives of carnivore
adults
Виды/ Species II Масса тела, кг/ Body mass, kg Легкие/Lungs Почки/Kidneys
г/g % от массы тела/ % of body mass г/g % от массы тела/ % of body mass
Самцы/Males
Песец/ Alopex lagopus 5 4.5 50 1.11 32 0.71
Лисица обыкновенная/ 8 5 54 1.08 43 0.86
Vulpes vulpes Собака
енотовидная/ Nyctereutes procyonoides Шакал/ 8 6.8 54 0.79 38 0.56
4 9.5 80 0.84 68 0.72
Cants aureus Волк/
6 38 440 1.16 110 0.29
Cams lupus
Кот лесной/ 0.86
Felis silvestris Рысь/ DU 118
5 15 0.79 91 0.61
Lynx lynx Гепард/ Acinonyx jubatus 1 21 244 99
1.16 0.47
Тигр/ Panthem tigris 1 160 1024 0.64 688 0.43
Самки/Females
Песец/ Alopex lagopus Лисица 4 3.7 42 1.14 28 0.76
обыкновенная/ Vulpes vulpes Собака 5 4.5 49 1.09 42 0.93
енотовидная/ Nyctereutes procyonoides Шакал/ 6 4.8 39 70 0.81 30 0.63
0.88
4 8 29 60 0.75
Canis aureus
Волк/ 370
Canis lupus Кот лесной/ 5 1.28 1.08 89 35 0.31 0.88
3 4 7.8 43
Felis silvestris
Рысь/ 0.83
Lynx lynx 3 65 62 0.79
Лев/ Panthem lea 1 125 750 0.60 663 0.53
Примечание. Данные по гепарду, льву и тигру амурскому получены в Московском
зоопарке.
140
Таблица 42/ Table 42
Характеристика массы легких и почек у некоторых животных, добытых
в Дагестане и Азербайджане (по: Насибов, 1970; Гинеев, 1972)
Characteristics of lungs and kidney mass of some animals obtained in Daghestan and
Azerbaijan
Виды/ Species Пол/ Sex Средняя масса тела, кг/ Average body mass, kg Легкие/Lungs Почки/Kindneys Селезенка/Lien
г/g % от массы тела/ % ofbody mass г/g % от массы тела/ % ofbody mass г/g % от массы тела/ % ofbody mass
Кот лесной/ <?<? 3.9 48 1.23 31 0.79 7.7 0.20
Felis silvestris (л=39) 3.3 41 1.24 26 0.79 6.4 0.19
Кот камышовый/ <?<? 7 92 1.31 83 1.19 26.5 0.38
Felis chans («=26) $$ 5.4 64 1.19 43 0.80 17.8 0.33
Енот-полоскун/ 6.1 51 0.84 28 0.46 9.8 0.16
Procyon lotor (п=4Ъ $$ 4.7 34 0.72 20 0.43 6.6 0.14
В семействах собачьих и кошачьих, так же как и у куньих, в целом
прослеживается тенденция к уменьшению индекса легких и почек с увеличением
размеров представителей видов, что объясняется разницей в массе тела животных
(табл. 41). Эта зависимость наблюдается в разных отрядах млекопитающих, что
подтверждает связь между относительными размерами рассматриваемых органов
и уровнем метаболизма. Увеличение массы тела влечет за собой уменьшение
теплоотдачи на единицу веса, что сопровождается снижением интенсивности
обмена веществ и сокращением размеров легких и почек (Ивантер и др., 1985).
Самки хищных млекопитающих имеют либо больший, либо равный с самцами
относительный вес этих органов, что соответствует хорошо выраженному
половому диморфизму по массе тела. Например, по данным В.Г. Монахова (2001),
у соболя индекс легких и почек (по отношению к массе тушки без шкурки) был
равен соответственно у самок 2.2 и 0.99%, а у самцов 2.1 и 0.86%.
При исследовании размеров легких у хищников из семейства собачьих и
кошачьих (табл. 41) можно отметить довольно низкий их индекс у енотовидной
собаки по сравнению с песцом и лисицей, а также высокий у таких видов,
обладающих большой массой тела, как волк и гепард, что обусловлено
особенностями их экологии. Представители двух последних видов способны к
длительному и быстрому бегу. Причем гепард, добывая копытных (прежде
всего джейрана) путем преследования, может развить на коротком расстоянии
скорость до 72, а по некоторым данным и до 112 км/ч (Г. Новиков, 1956; Гептнер
и др., 1967).
От других представителей семейства собачьих примерно равного веса
енотовидная собака отличается также сравнительно низким показателем индекса
почек, что совпадает с наблюдениями С.Н. Боголюбского (1939). Подобная
141
Таблица 43/ Table 43
Содержание эритроцитов и их диаметр у самок грызунов с разной массой тела
(июль-август)
The content of erythrocytes and their diameter of rodent females with different body mass
(July-August)
Виды/ Species n Масса тела, г/ Body mass, g Кол-во эритроцитов, млн/мм3/ Erythrocytes number, million/mm3 Диаметр эритроцитов, мк/ Erythrocyte size, /rk Концентрация гемоглобина, г%/ Hemoglobin concentration, g%
Полевка рыжая/ Glethrionomys 9 21 ± 0.2 9.3 ± 0.4 6.3 ± 0.07 12.7 ±0.5
glareolus Мышь полевая/ Apodemus agrarius Полевка 9 23 ± 0.2 9.2 ±0.5' 6.2 ± 0.08 12.5 ±0.3
обыкновен./ Microtus arvalis Мышь 8 28 ± 0.2 9.3 ± 0.4 6.4 ±0.08 13.3 ±0.3
желтогорлая/ Apodemus flavicollis 8 30 ±0.2 9.8 ± 0.7 6.3 ± 0.06 14.6 ±0.4
Крыса серая/ 172 +
Rattus 5 6.1 7.9± 1.1 6.6 ± 0.07 13.5 ±0.8
norvegicus Суслик крапчатый/ Cilellus 5 255 ± 8.2 7.5 ± 1.2 7 ±0.09 12.5 ±0.5
suslieus Хомяк обыкновен./ 4 280 ± 7.2± 1.2 7.1 ±0.08 14.9 ±0.5
Cricetus cri cetus 12.3
Сурок степной/ 5 3653 ± 23.1 5.4 ± 1.5 7.4 ±0.06 11.8 ±0.4
Marmota bobac
Дикобраз/ Hystrix leucura 1 8600 4.8 7.5 12.1
картина характерна и для барсука, который, как и енотовидная собака, в
холодный период года впадает в длительный зимний сон. Эти звери ведут сходный
образ жизни, при котором они часть года проводят в малоактивном состоянии,
сопровождающемся снижением уровня метаболических процессов и размеров
почек. Их индекс в декабре-феврале, в период зимней депрессии метаболизма,
наиболее низкий. В весенне-летнее время в связи с расходованием жировых
резервов, падением веса и большой напряженностью энергетического баланса,
обусловленного состоянием линьки и половой активностью, относительная масса
почек у взрослых животных увеличивается.
Отмеченные зависимости хорошо прослеживаются и среди некоторых
представителей фауны Кавказа (табл. 42). Так, по имеющимся данным
142
Таблица 44/ Table 44
Сравнительная характеристика дыхательной функции крови и относительного
веса селезенки у взрослых самцов куньих (июль-декабрь)
Comparative characteristics of respiratory 'function of blood and relative spleen weight
of mustelid adult males (July^December)
Виды/ Species n . Содержание/Number n Индекс селезенки/ Index of lien, %o
эритроцитов, млн/мм3/ erythrocytes, million/mm3 гемоглобина, г%/ hemoglobin, g%
Ласка/ Mustela nivalis 6 7.4 ± 0.2 14.8 ± 0.4 60 4.9 ± 0.2
Горностай/ Mustela erminea 8 6.9 ±0.1 14.6 ± 0.2 51 3.3 ±0.1
Хорек лесной/ Mustela putorius 7 6.5 ± 0.2 14.1 ±0.2 85 3.1 ±0.1
Норка европ./ Mustela lutreola 6 7.5 ± 0.3 17.6 ± 0.5 41 2.8 ± 0.2
Норка америк./ Mustela vison 8 7.8 ± 0.4 17.3 ± 0.3 95 2.5 ±0.1
Куница лесная/ Martes martes 1 - 16.5 47 2.2 ±0.1
Куница каменная/ Martes foina - - - 5 2.8 ±0.4
Барсук/ Meles meles 6 7.3 ±0.8 16.7 ±0.3 16 3.4 ± 0.2
Выдра/ Lutra lutra 1 7i6 15.8 15 3.5 ±0.1
Росомаха/ Gulo gulo - - - 8 5.4 ± 0.5
Калан/ 6 3.5 ±0.3 15.1 ±0.5 8 3.9 ±0.4
Enhydra lutris 1 5.1 17.9 - -
Примечание. Данные по калану взяты из следующих работ - Барабаш-Никифоров и
др., 1968; Маминов, 1971.
(Насибов, 1970; Гинеев, 1972), высокий относительный показатель внутренних
органов прослеживается у более активной формы - камышового кота, а у
впадающего в зимний сон енота-полоскуна индекс морфологических
показателей дыхательной и выделительной систем оказался заметно ниже
такового у обоих видов котов. Это еще раз подтверждает тесную
функциональную связь образа жизни и строения внутренних органов
исследованных животных.
Интенсивность газообмена между внешней средой и тканями организма
во многом определяется количеством и размерами эритроцитов крови. Связь
функции газообмена с массой тела и экологическими особенностями видов по
уровню эритроцитоза прослеживается в рядах различных систематических
групп животных. Например, у мышевидных грызунов концентрация
эритроцитов в крови оказалась в среднем в 2 раза выше, чем у таких крупных
143
видов, как сурок, нутрия, бобр (Галанцев, Туманов, 1970), но особенно наглядно
закономерное снижение числа эритроцитов с увеличением массы тела
проявляется у грызунов, сходных по образу жизни (табл. 43).
Данные особенности наблюдаются и у полуводных животных. Например,
содержание эритроцитов в крови обыкновенного бобра составляет 2.40-
5.69 млн/мм3, нутрии 4.95-6.63, ондатры 4.44-8.50, а водяной полевки 9.24-
13.29 млн/мм3 (Кацнельсон, Орлова, 1956; Салганская, 1962; Галанцев, 1977).
Следовательно, специализация к водному образу жизни и обитанию в глубоких
норах в условиях избыточного содержания углекислого газа не оказывает
столь существенного влияния на концентрацию эритроцитов в крови, как
величина тела.
В известной мере выявленная закономерность прослеживается и среди
мелких хищников. Так, содержание эритроцитов в крови самцов ласки со
средней массой тела 85 г равно 7.4 млн, горностая (185 г) - 6.5, а лесного
хорька (896 г) - 6.1 млн/мм3 (Туманов, 1974а).
Следует также отметить, что у видов, представители которых резко
различаются между собой по показателям двигательной активности,
наблюдаются и отличия в количестве содержащихся в крови эритроцитов
(Булатова, 1953; Коржуев, 1955, 1960; Шварц, 1960, 1968; Большаков, 1972),
причем у более активных форм эта величина бывает заметно выше. По нашим
данным, у хорьков, норок, выдры и барсука, различающихся между собой по
образу жизни, содержание эритроцитов в крови колеблется в незначительных
пределах, т.е. они близки по уровню эритроцитоза (табл. 44). В их крови имеется
в среднем 6-7 млн/мм3 эритроцитов размером 6.6-6.8 мкм. Несколько иные
показатели (эритроциты -11.3-11.4 млн/мм3, размер 5.5-5.7 мкм) обнаружены
у соболя и лесной куницы (Уткин, 1956,1963). При большом количестве мелких
эритроцитов увеличивается общая поверхность соприкосновения гемоглобина
с кислородом воздуха, а следовательно, и кислородная емкость крови, что
согласуется с интенсивным энергообменом этих активных и быстрых
хищников.
Анализ собранных материалов свидетельствует о том, что у куньих, так
же как и у грызунов, адаптация к обитанию в водной среде не сопровождалась
увеличением концентрации эритроцитов в крови. По уровню эритроцитоза
они заметно уступают высокогорным животным, в крови которых насчи-
тывается до 15-25 млн/мм3 мелких эритроцитов (Булатова, 1953; Коржуев,
1955,1964; Большаков, 1967,1972). Сравнительно малое количество красных
кровяных клеток (3.5-4.9 млн/мм3), не превышающее таковое у калана, и
лейкоцитов (58-83 тыс/мм3) обнаружено также и у молодых белых медведей
(Переверзева, Веретенникова, 1973). По-видимому, специализация хищных
млекопитающих к водному образу жизни, обитанию в берлогах или глубоких
норах, где они могут находиться в условиях избыточного содержания
углекислого газа, не оказывает столь существенного влияния на концентрацию
эритроцитов в крови, как это прослеживается у животных, адаптированных к
высокогорной гипоксии.
144
В результате исследований форменных элементов крови у ласок было
обнаружено, что зверьки, отловленные летом в Ленинградской области, имели
более высокие показатели содержания эритроцитов в крови (в среднем
7.4 млн/мм3) по сравнению с таковыми у особей, пойманных в июле-августе
под Киевом (6.8 млн/мм3). Подобные различия по уровню эритроцитоза
выявлены нами и в популяциях желтогорлых мышей из названных регионов
(Ивантер и др., 1985). Данный факт еще раз свидетельствует о том, что у
мелких млекопитающих северные формы по сравнению с южными хара-
ктеризуются более высокими энергетическими показателями, в том числе и
дыхательной функции крови.
Необходимо отметить, что у теплокровных животных существенное
значение для жизнеобеспечения имеет не только количество эритроцитов в
крови, но и их размеры, которые во многом определяют интенсивность
газообмена между внешней средой и тканями органов. Например, в отряде
грызунов прослеживается прямая зависимость между диаметром эритроцитов
и массой тела представителей видов, а также обратная - между размерами и
количеством красных кровяных телец (табл. 43). Обращает на себя внимание
и тот факт, что у обитающих в холодных условиях Арктики белых медведей
все форменные элементы крови имеют крупные размеры и их диаметр оказался
заметно больше такового у человека (Переверзева, Веретенникова, 1973).
Среди хищников из семейства куньих данные тенденции не прослеживаются, и
разные по массе тела животные, за исключением видов рода Martes (Уткин,
1963), имеют примерно одинаковый диаметр эритроцитов. Кроме того,
исследования показали, что их размер у амфибионтов - европейской и
американской норок практически не отличается от такового у наземных видов
(лесного хорька или колонка), близких к ним по массе тела. Такого же диаметра
эритроциты оказались и у выдры. Следовательно, у полуводных хищников
адаптация дыхательной функции крови к условиям гипоксии не сопро-
вождалась увеличением ни количества эритроцитов, ни их диаметра, как это
наблюдается у специализированных водных животных (Галанцев, 1977,1980).
Как известно, большое значение в транспорте кислорода от легких к
тканям органов и углекислоты в обратном направлении принадлежит
содержащемуся в эритроцитах гемоглобину. Его количество в крови во многом
обусловливается особенностями поведения и экологии животных. Сезонная и
географическая изменчивость этого признака у представителей разных отрядов
млекопитающих также прослеживается очень отчетливо (Калабухов, 1950,
1969; Коржуев, 1952,1964; Верболович, 1961; Берестов, 1971,1981; Башенина,
1977, 1981; Покровский, Большаков, 1979 идр.).
Собранные материалы свидетельствуют о том, что, как и у грызунов, у
мелких хищников количество гемоглобина в эритроцитах, по-видимому,
определяется не размерами тела, а характерными чертами их экологии -
особенностями питания, двигательной активности и др. Так, концентрация
гемоглобина в крови ласки (1.5-16.1 г %) и горностая (13.8-15.5 г %) летом
оказалась такой же, как и у более крупного миофага - лесного хорька (13.7-
145
Таблица 45/ Table 45
Содержание гемоглобина, показатели относительной массы крови и костного
мозга у взрослых представителей семейства собачьих (по: Коржуев, Спасская, 1985)
The content of hemoglobin, relative blood and bone marrow mass parameters of adult
representatives of canid family
Виды/ Species Пол/ Sex п Масса тела, кг/ Body mass, kg Гемоглобин, г%/ Hemoglobin, g% Отношение (%) к массе тела массы/ Precentage of body mass, %
крови/ blood костного мозга/ medulla
Волк/ Cams lupus 3 в 3 41 18 9.2 4.6
9 $ 1 32.7 19 9 4.3
Шакал/ 3 3 3 7.2 14.6 8.2 3.5
Canis aureus 9 9 1 8.1 13.8 7.7 3.8
Собака енотовидная/ 3 3 3 6.6 16.8 6.6 3
Nyctereutes procyonoides 9 $ 4 5.3 15.2 5.6 3.4
Лисица обыкн./ 3 3 5 4.6 18 8.3 4.3
Vulpes vulpes 9 9 3 5.8 16.6 8 4.4
Корсак/ 3 3 2 3.9 16.2 7.3 4
Vulpes corsac 9 9 2 4.1 15.4 8 4.2
15.3 г %). Примерно в тех же пределах она колеблется и у других наземных
хищников из данного семейства (Уткин, 1956, 1963; Лобачев, 1976). В то же
время содержание дыхательного пигмента у полуводных видов куньих -
европейской и американской норок - было более высоким (15.1-18.6 г %),
подтверждая тесную взаимосвязь дыхательной функции крови животных с их
экологической спецификой (табл. 44).
Известно, что типично наземные виды грызунов по сравнению с амфи-
бионтами независимо от размеров особей имеют меньшую насыщенность
эритроцитов гемоглобином (Irving, 1939; Кацнельсон, Орлова, 1956;
Салганская, 1962). Например, средний уровень содержания дыхательного
пигмента в крови мышей, полевок, сусликов и хомяков (табл. 43) оказался
заметно ниже, чем у водяной полевки, -17.5 ± 1.6 г%, ондатры - 20 ± 0.8 или
нутрии - 16 ± 1.8 г% (Галанцев, 1967; Галанцев, Туманов, 1969). Еще выше
этот показатель у такого специализированного гидробионта, как обыкновенный
бобр - 21.3 г% (Никитенко, 1966).
Высокая степень насыщения крови гемоглобином отмечается и у полуводных
насекомоядных. Так, общая обеспеченность организма гемоглобином (в
пересчете на 1 кг массы тела) составляет у куторы 19.3, а у обыкновенной
землеройки 10.5 г% (Коржуев, Корецкая, 1962). У выхухоли среднее содержание
гемоглобина в крови равно 16.9 ± 0.8 г% (Галанцев, 1977), что заметно
превышает количество дыхательного пигмента в эритроцитах ушастого ежа -
146
12.5 г%(Филатова, 1958). Между тем у хищников-амфибионтов специализация
к обитанию в воде не сопровождается достоверным повышением гемоглобина в
крови, как это наблюдается, например, у животных, адаптированных к
высокогорью (Булатова, 1953; Коржуев, 1964; Большаков, 1967, 1972). У
молодых белых медведей (Переверзева, Веретенникова, 1973) этот показатель
был равен 66-84% по Сали (11-14 г%). По данным разных авторов, содержание
дыхательного пигмента в крови американских норок колебалось в пределах
15.3-18 г%, выдры -13.7-17.4 и калана-12.4-17.9 г% (Маминов, 1971; Данилов,
Туманов, 1976а, б; Галанцев, 1977). Как показали результаты исследований,
адаптация многих видов млекопитающих (в том числе и хищников) к водной
среде сопровождается увеличением прежде всего мышечного гемоглобина
(миоглобина), общего объема крови и концентрации гемоглобина в одном
эритроците, в результате чего его поверхность становится более эффективной в
функциональном отношении (Scholander, 1940; Клейненберг, 1956; Галанцев,
1967, 1980).
Следует также отметить, что обитающий в многоярусных норах барсук по
насыщенности крови гемоглобином практически не отличается от активного
наземного представителя того же семейства - лесной куницы (табл. 44). По-
видимому, как у грызунов, так и у куньих зависимости уровня концентрации
гемоглобина от размеров тела или степени приспособления животных к
обитанию в условиях измененной газовой среды не прослеживается.
У другого впадающего в зимний сон представителя хищных млеко-
питающих - енотовидной собаки показатели морфологического состава крови
заметно отличались от таковых у белого песца. Как сообщает Ю.С. Заболотских
(2000), у енотовидных собак количество эритроцитов в конце ноября составляло
в среднем 6.9 млн/мкл при насыщенности гемоглобином 14.5 г%. У песцов
уровень данных показателей был выше - соответственно 8 млн/мкл и 17.7 г%,
что можно рассматривать как характерную особенность этого северного вида.
По наблюдениям автора, на отсутствие пищи рассматриваемые животные
реагируют спокойно, как на обычную ситуацию. При дефиците кормов их
организм переходит на эндогенное питание за счет использования резервных
жировых запасов, как подкожных, так и внутренних органов. В периоды
частичного голодания зверей показатели температуры тела, частоты дыхания и
гематологические параметры не выходили за пределы физиологической нормы.
Кроме того, не были отмечены нарушения кислотно-щелочного равновесия
внутренней среды организма, не наблюдалось ацидоза и необратимых
дистрофических явлений. При длительном отсутствии корма содержание в крови
общего белка и глюкозы заметно снижалось, но быстро восстанавливалось до
исходного уровня при наличии пищи.
Исследования, выполненные на животных, отловленных на юге России
(Коржуев, Спасская 1985), показали, что волк по сравнению с енотовидной
собакой и другими представителями семейства Canidae обладает более
высокими показателями гемоглобина и индекса массы крови (табл. 45).
Данный факт, вероятно, объясняется его очень активным, подвижным образом
147
жизни и интенсивным расходом энергии на добывание пищи. Так, у самок
енотовидной собаки, корсака и шакала содержание гемоглобина в крови
оказалось на 19-27% ниже, чем у волка. По свидетельству авторов, у всех
представителей данного семейства костномозговая фракция скелета щенков
была заметно больше, чем у их родителей, что обусловлено интенсивным
расходом энергии молодого организма на рост и развитие. Если у взрослых
волков масса костного мозга составляет 4.3-4.6% от веса тела, то у детенышей
5.4%. Волки старших возрастов характеризуются не только высоким
уровнем основных компонентов крови, но и костного мозга как очага
гемопоэза. Показатели дыхательной функции крови взаимосвязаны с
размерами кроветворных органов, в том числе и селезенки. Она выступает
не только как регулятор кровотока и энергетическое депо крови, связанное с
обменными процессами в организме, но и как индикатор кроветворения, в
частности эритропоэза. У исследуемых хищников явной зависимости
размеров селезенки от массы тела животных не прослеживается. Например,
у сравнительно крупного камышового кота индекс селезенки был выше, чем
у животных с меньшей массой тела - лесного кота и енота-полоскуна (см.
табл. 42). Отсутствие такой зависимости можно отнести за счет высокой
изменчивости величины данного органа под влиянием различных эколо-
гических воздействий - сезонных и возрастных изменений метаболитических
процессов (Ивантер и др., 1985).
Среди куньих наиболее низкий индекс селезенки отмечался у куниц и норок
- видов с высоким содержанием гемоглобина в крови (табл. 44). По-видимому,
его количество находится в обратной зависимости от относительного веса
селезенки, снижение которого сопровождается увеличением гематологиче-
ских показателей. Исследования, проведенные методом множествен-
ного регрессионного анализа, показали, что у куньих величина индекса
селезенки определяется взаимодействием таких факторов, как количество
эритроцитов и гемоглобина в крови, с увеличением концентрации которых
данный показатель снижался. При этом коэффициент детерминации был
равен 81%.
Сходные зависимости прослеживаются и среди других млекопитающих.
Так, у енотовидных собак, для которых характерен низкий уровень
метаболизма, показателей концентрации эритроцитов и гемоглобина в крови,
индекс селезенки оказался сравнительно высоким - 2.24-3.57%о. У волка, рыси
и лисицы, добытых в Ленинградской области, он был одинаков и составлял у
самцов 2.52-2.67%о, а у самок 2.56-2.69%о. Данные по волку из северо-
западного региона России почти полностью совпадают с материалами
П.Г. Козло с соавторами (1985) по Белоруссии и Алтайскому краю (самцы 2.2-
2.4%о, самки 2.5-2.6%о). В то же время у хищных зверей из северных популяций,
обладающих высокими значениями гематологических показателей, относитель-
ный вес селезенки был ниже. У песцов, поступивших к нам из Ненецкого АО, и
у волков, отстрелянных в Республике Коми (Полежаев, 1998), он составлял в
среднем у самцов 1,73%о и у самок 1,76%о.
148
Таким образом, высокие показатели дыхательной функции крови и уровня
энергообмена отражают, вероятно, одну из приспособительных реакций
организма, призванную обеспечить представителям наиболее активных и
подвижных видов полноценную работу мышц путем создания лучших условий
для связывания и переноса кислорода кровью. Выявленная взаимосвязь
дыхательной функции крови с интенсивностью потребления кислорода,
размерами органов дыхания, выделения и кроветворения дает представление
об уровне окислительно-восстановительных процессов и энергетическом
обмене в клетках тканей хищных млекопитающих, близких по таксо-
номическому положению, но обладающих различной экологической
валентностью. Однако эти сведения будут недостаточно полными без учета
особенностей работы сердечно-сосудистой системы животных, обусловленных
их образом жизни.
Резюме
Интенсивный расход энергии на терморегуляцию и процессы размножения
определяет у мелких и средних по размерам хищников большую потребность в
пище, высокий уровень поведенческих реакций и двигательной активности,
связанных с кормодобывающей деятельностью. Поскольку при малых
размерах животного поверхность теплоотдачи увеличивается, то предста-
вителям мелких видов (в сравнении с крупными) требуется вырабатывать
больше тепла на единицу массы тела для поддержания на необходимом уровне
энергетического баланса. Жировые запасы имеют важное (не только
энергетическое, но и теплоизоляционное) значение, позволяя хищникам
благополучно пережить трудный зимний период. Осенью и у самцов, и у самок
они достигают максимальной величины. Исследования показали, что к началу
зимы масса жировой ткани достигает у енотовидной собаки 30-32%, у волка
15-16, у рыси 12-13 и у бурого медведя до 20% от массы тела.
Ректальная температура как один из индикаторов интенсивности обменных
процессов во многом определяется массой тела и экологическими особен-
ностями хищников. Среди представителей семейства Mustelidae наиболее
высокая температура тела прослеживалась у горностая, соболя и лесной
куницы, т.е. очень активных, быстрых и подвижных видов.
Сезонная динамика температурных реакций и уровня основного обмена
прослеживается у хищников достаточно отчетливо. Она находится в тесной
взаимозависимости с изменениями массы тела и характеризуется подъемом
ректальной температуры и газообмена в летний период, когда вес животных
наименьший. К началу зимы уровень обменных процессов заметно снижается,
что согласуется с уменьшением в этот период потребности живитных в кормах.
Среди мелких куньих примерно равного веса зимнее понижение энергети-
ческого обмена лучше выражено у представителей тех видов, которые
способны с осени создавать в организме жировые резервы (норки, хорьки). У
представителей семейства Canidae заметное снижение физиологических
149
параметров при падении температуры внешней среды отчетливо просле-
живалось только у енотовидной собаки, впадающей в зимний сон. Для нее
данный факт имеет важное адаптивное значение, направленное на экономное
расходование внутренних энергоресурсов и повышающее выживаемость
животных в неблагоприятные периоды года. Выявленная сезонная динамика
потребности в пище, температуры тела и уровня теплообмена способствует
пониманию кормодобывающей активности и адаптивных поведенческих
особенностей хищных млекопитающих в разные сезоны года.
Интенсивность обменных процессов у наиболее активных и подвижных
видов сопровождается высоким уровнем легочного дыхания и гематоло-
гических показателей. Связь основных параметров, характеризующих
энергообмен с массой тела, строением органов дыхания и выделения, а также
с поведенческими особенностями прослеживается у хищных млекопитающих
вполне отчетливо. Большое значение в транспорте кислорода от легких к
тканям органов и углекислоты в обратном направлении принадлежит
содержащемуся в эритроцитах гемоглобину. Его количество в крови во многом
определяется особенностями поведения и экологии животных. В целом
содержание гемоглобина у хищников-амфибионтов было выше, чем у
наземных видов. Вместе с тем их специализация к обитанию в воде не
сопровождается достоверным повышением уровня эритроцитоза и содержания
гемоглобина в крови, как, например, у животных, адаптированных к жизни
высоко в горах.
Собранные сведения показывают, что среди представителей семейства
Canidae волки обладают более высокими показателями гемоглобина и
индексом массы крови, по сравнению с енотовидной собакой и другими
хищниками. Это, вероятно, объясняется их очень активным образом жизни и
интенсивным расходом энергии на добывание пищи. Взаимосвязь дыхательной
функции крови с интенсивностью легочной вентиляции и рядом морфо-
метрических показателей дает представление об уровне энергетического
обмена в клетках тканей рассматриваемых животных, близких по таксо-
номическому положению, но различающихся по особенностям поведения й
образу жизни.
Summary
The intensive energy expenditures for thermoregulation and reproduction in
small and middle-sized carnivores determine high food requirements and high level
of moving activity related to food procurement. As with a small body size the
surface of the heat loss increases, the small animals need to develop more heat per
unit of weight for the maintenance of energetic balance on the needed level as
compared to large animals.
The fat reserves have both important energetic and heat insulation significance,
permitting animals to survive successfully the hard winter period. In autumn fat
reserves in all carnivores attain maximum value. Thus, in the beginning of winter
150
the mass of the fat tissue in Nyctereutes procyonoides is 30-32%, in Canis lupus 15-
16, in Lynx lynx 12-13 and in Ursus arctos up to 20% of the body mass.
The rectal temperature an indicator of intensity of metabolic processes is largely
determined by the body mass and ecological characteristics of carnivores. Among
representatives of the family Mustelidae the highest body temperature was observed
in Mustela erminea, Martes martes, M. zibellina, i.e. very energetic and active
species.
The seasonal dynamics of temperature reactions and basal metabolic rate is
relatively distinct in carnivores. It is closely related to body mass changes and is
characterized by a dramatic increase of the rectal temperature and gas exchange in
the summer period. To the beginning of winter the values of these parameters
decrease notably; in this connection, the weight of animals and food requirements
are changing by season of the year. Among the carnivores of approximately equal
size the winter decrease of energetic exchange is more pronounced in representatives
of those species, which are able to accumulate fat reserves in the organism. In the
representatives of the family Canidae a distinct decrease of physiological parameters
with the fall of the ambient temperature was observed only in Nyctereutes
procyonoides. The given fact may have an adaptive significance directed toward
economic expenditure of inner energy resources and increasing survival ability of
individuals of this species in the unfavorable periods of the year. The above-
mentioned seasonal dynamics of food demand, body temperature and heat exchange
level enhances the understanding of food obtaining activity and adaptive behavioral
characteristics of carnivores in different seasons of the year.
Haemoglobin, contained in erythrocytes, plays an important role in the transport
of oxygen from the lungs to the tissues of organs and carbon dioxide in back direction.
Its quantity in the blood is largely determined by characteristic features of behavior
and ecology of animals. On the whole, the content of haemoglobin in amphibiont
carnivores was higher than in terrestrial species. At the same time, their specialization
to the aquatic mode of life is not followed by significant increase of erythrocytosis
level and content of haemoglobin in the blood, as it is observed in animals adapted
to life at high altitudes in mountains.
The data obtained show that among the representatives of the family Canidae
Canis lupus have higher values of haemoglobin and blood mass index as compared
to Nyctereutesprocyonoides. This, probably, may be explained by their very active
mode of life and intensive energy expenditures on food procurement.
The relation of blood respiratory function and intensity of oxygen consumption,
sizes of respiratory organs, excretory and haemopoiesis organs gives an idea of the
level of energetic exchange in the tissue cells of carnivorous mammals close in
taxonomic position, but differing in characteristic features of behavior and life
history.
Глава 4. ФУНКЦИОНАЛЬНЫЕ
ОСОБЕННОСТИ СЕРДЕЧНО-СОСУДИСТОЙ
СИСТЕМЫ
В оценке функционального состояния организма большое значение имеют
показатели сердечного ритма, который не только определяет его компенсатор-
ные возможности, но и может служить индикатором энергетического
метаболизма (Соколов, Задальский и др., 1984; lohnson, Gessaman, 1973; Pauls,
1980; Соколов, 1990).
В норме у рассматриваемых хищников регистрируются характерные для
видов темп сердечной деятельности и форма электрокардиограммы (ЭКГ). У
куньих в отличие от грызунов и насекомоядных сходного веса частота пульса -
величина вариабельная. Необычная окружающая среда и различные внешние
раздражители могут вызывать усиление функциональной деятельности
сердечной мышцы, однако эти изменения носят кратковременный характер и
укладываются в ту норму, которая свойственна каждому виду. При записи
пульса у представителей семейства куньих в начале опыта регистрируется
аритмия, которая исчезает, когда зверьки успокаиваются. Такая «стартовая
реакция» длится у мелких видов от 1 до 4 мин, а у более крупных она
продолжается до 5-10 мин. Отмеченные «сбои» сердечного ритма имеют, по-
видимому, неврогенный характер, т.е. не наблюдаются на ЭКГ, снятых у
детенышей на ранних стадиях постнатального онтогенеза и у взрослых
животных, находящихся под наркозом (Туманов и др., 1981,1982).
Установлено, что внешние раздражители не вызывают столь резкого
изменения сердечного ритма, как физическая нагрузка (бег, прыжки и т.п.).
Реакция на такие раздражители, как световые вспышки, свист, незнакомые
предметы, проявляются обычно в кратковременном увеличении скорости
сердечных сокращений в среднем на 20-25%, тогда как даже после
непродолжительного бега пульс учащался в 1.6-2 раза. Эти изменения
сопровождались подъемом вольтажа основных зубцов ЭКГ и наиболее четко
были выражены у представителей таких быстрых и активных видов, как
горностай, соболь и куницы.
В целом среди изучаемых видов куньих прослеживается обратная
зависимость частоты пульса и индекса сердца от массы тела животных. В то же
время на фоне общего падения сердечного ритма по мере увеличения веса
хищников (особенно в пересчете на единицу их массы) наблюдается тесная
зависимость уровня этого показателя и размеров сердца от экологической
специфики видов (табл. 46). Например, среди всех представителей семейства
закавказская ласка (Mustela nivalis boccamela) обладает наибольшей
относительной массой этого органа, которая у нее оказалась достоверно выше,
чем у мелкой обыкновенной ласки (р>0.99). Высокий индекс сердца, уровень
его сокращений и вольтаж основных положительных зубцов желудочкового
152
Таблица 46/ Table 46
Сердечный ритм и индекс сердца у взрослых самцов куньих
Cordial rhythm and heart index of mustelid adult males
Вид, форма/ Objects of research Средняя масса тела, кг/ Average body mass, kg n Сердечный ритм/Heart rate ll Индекс сердца/ Index of heart, %0
в 1 мин/ per 1 min в 1 мин на кг веса/ per 1 min per 1 kg of body mass
Ласка обыкновенная/ Mustela nivalis Ласка закавказская/ 0.09 12 364+ 13.5 4044 64 10.7 ±0.2
Mustela nivalis boccamela 0.15 3 345 2300 4 12.7 ±0.3
Ласка пустынная/ Mustela nivalis 0.23 2 300 1282 - -
heptneri Горностай/ 0.25 12 288 ±5.2 1152 52 11.1 + 0.2
Mustela erminea Солонгой/ Mustela altaica 10 567 5 10.3 ±0.4
0.48 272 ±11.4
Колонок/ 0.68 8 285 ± 8.3 419 5 8.8 + 0.5
Mustela sibirica Перевязка/ Vormela peregusna 4 409
0.76 312 ± 9.9
Хорек лесной/ Mustela putorius Хорек белый 1.03 15 283 + 7.5 275 86 8.6 ±0.2
(фуро)/ Mustela putorius furo 1.63 8 290 ± 19.4 178 - -
Хорек степной/ Mustela eversmaimi 1.68 3 255 ±18.3 151 - -
Норка европейская/ Mustela lutreola 1.08 12 218 ± 4.3 202 45 9.5 + 0.2
Норка америк./ Mustela vison 1.15 18 235 ± 6.4 204 98 9.3 ±0.1
Куница лесная/ Martes martes 1.28 8 325 ±8 254 49 9.3 ± 0.2
Куница каменная/ Martes foina 1.37 6 359 ± 10.5 257 5 6.5 ± 0.2
Соболь/ Mattes zibellina 1.52 6 328 ±2.3 216 4 9.1+ 0.3
Выдра/ Lutra lutra 6.44 3 150+ 13.4 23 15 10.0 ±0.3
Барсук/ Meles meles 7.83 8 188 ± 15.6 24 14 6.8 ± 0.3
Росомаха/ Gulo gulo 9.68 2 193 20 8 8.5 ± 0.3
комплекса ЭКГ могут отражать адекватную реакцию сердечной мышцы на
жизнь закавказской ласки в горных условиях.
Известно, что у млекопитающих, населяющих горные местности или
обладающих большой скоростью передвижений, одно из важных адаптивных
преобразований сердечно-сосудистой системы заключается в гипертрофии
153
миокарда при снижении частоты пульса (Машковцев, 1935; Аршавский, 1958,
1967; Слоним, 1962, 1976; Большаков, 1972; Покровский, Большаков, 1979).
Они обусловливают соответствующие изменения в работе сердечной мышцы,
которые сводятся к увеличению общего объема крови, выбрасываемой в аорту
в единицу времени (Барбашова, 1956; Walker, 1964; Morrison, 1964). Подобная
зависимость четко прослеживается также среди куниц, из которых каменная
имеет достоверно более низкий индекс сердца (д>0.99). По сравнению с лесной
куницей она менее приспособлена к обитанию в высокогорье и вообще к
условиям холодного климата (Туманов, 1982).
Особенно наглядно связь размеров сердца и темпа его деятельности с
экологическими особенностями животных выступает у видов, близких по массе
тела, но отличающихся по образу жизни. Так, у европейских норок индекс сердца
был достоверно выше, чем у лесных хорьков (р>0.99), что можно объяснить разной
степенью связи зверьков с водной средой обитания. В то же время частота пульса
у норок оказалась на 60-100 уд./мин меньше, чем у хорьков, свидетельствуя о
разной направленности адаптивного процесса. Достаточно большим по размеру
сердцем обладают также и другие полуводные хищники, такие как калан и выдра.
Например, у самцов выдры средний индекс сердца равен 10%о (табл. 46), а у
такого крупного стенофага, как калан, - 8.3%о (Барабаш-Никифоров и др., 1968).
По-видимому, необходимый для жизнедеятельности организма минутный объем
крови обеспечивается у амфибионтов за счет систолического, или ударного,
объема, а у наземных видов, обладающих меньшей массой сердечной мышцы,
путем повышения частоты пульса.
Прослеживается тенденция к увеличению индекса сердца у самок по
сравнению с самцами, что, видимо, связано с половым диморфизмом по массе
тела животных (табл. 47). Кроме того, у них отмечается и большая частота
пульса, которая после прозревания детенышей заметна во всех возрастных
группах. Сравнительный анализ достоверности этого положения по /-критерию
Стьюдента для сопряженных пар наблюдений показал, что самки взрослых
особей характеризуются очень высоким ритмом сердца. Доверительная
вероятность вывода в целом для представителей всех изучаемых видов была
высокой (р-0.99), однако изменчивость данного показателя у них велика. Если
у норок и куниц наблюдается только тенденция к повышению частоты пульса
у самок по сравнению с самцами, то у обыкновенной ласки (р=0.85), лесного
хорька (р=0.95) и горностая (д=0.99) различия были высокодостоверными.
При расчетах удельного (на единицу массы тела) темпа сердцебиений, так же
как энергообмена и уровня дыхания, половая специфика отчетливо проявляется
у всех видов.
У крупных хищных млекопитающих имеет место тенденция к снижению
индекса сердца с увеличением массы тела животных (табл. 48). Вместе с тем
следует отметить сравнительно высокий (соотносительно большого веса тела)
абсолютный и относительный показатель массы сердца у волков, что, видимо,
связано с активным и подвижным образом их жизни. Полученные нами
материалы по относительному весу этого органа у волков и шакалов близки к
154
Таблица 47/ Table 47
Сердечный ритм и индекс сердца у взрослых самок куньих
The cordial rhythm and heart index of mustelid adult females
Вид, форма/ Objects of research Средняя масса тела, кг/ Average body mass, kg n Сердечный ритм/ Heart rate n Индекс сердца/ Index of heart, %o
в 1 мин/ per 1 min в 1 мин на кг веса/ per 1 min per 1 kg of body mass
Ласка обыкновенная/ Mustela nivalis Ласка закавказская/ 0.06 4 388 ± 10.3 6467 24 11.3 + 0.4
Mustela nivalis boccamela 0.07 - - - 1 12.9
Горностай/ Mustela erminea 0.14 13 384 ± 12.3 2743 35 11.8 + 0.3
Солонгой/ Mustela altaica 0.36 6 300 ± 12.5 833 3 10.5 + 0.6
Колонок/ Mustela sibirica 0.50 7 291 ± 14.2 577 - -
Хорек лесной/ Mustela putorius 0.68 6 323 ± 14.3 475 27 8.8 ± 0.3
Хорек степной/ Mustela eversmanni 0.69 6 260 ± 10.0 376 - -
Хорек белый (фуро)/ Mustela putorius furo 0.93 3 373 ±31.8 401 - -
Норка европейская/ Mustela lutreola 0.76 14 223 ± 14.2 292 32 9.6 + 0.2
Норка американская/ Mustela vison 0.78 7 264 ± 16.5 338 58 9.3 ± 0.3
Куница лесная/ Martes martes 0.98 3 345 + 17.8 352 38 9.5 ±0.2
Куница каменная/ Martes foina 1.15 3 370 ± 16.4 322 3 6.6 ± 0.3
Соболь/ Martes zibellina 1.23 5 332 ± 23.2 269 2 9.2
Выдра/ Lutra lutra 4.81 3 152 ± 18.3 32 12 10.1 ±0.3
Барсук/ Lutra lutra 7.25 4 192 ± 16.7 26 10 6.8 ±0.3
Росомаха/ Gulo gulo 9 2 225 25 2 8.7
таковым других авторов. Так, по сообщению М.Ф. Ковтуна и Л.П. Осинского
(1985), индекс сердца у самок шакала (л=25) был равен 0.90%, а у самцов
(л=29) - 0.83%. У добытых в Белоруссии волков он составлял соответственно
1 и 0.94% (Козло и др., 1985). Собранные сведения демонстрируют также
достаточно высокие показатели относительной массы рассматриваемого
органа у более мелких по сравнению с самцами самок. Данная особенность
характерна практически для всех видов хищных зверей. Скорее всего, она не
155
Таблица 48/ Table 48
Сравнительная характеристика массы сердца у некоторых взрослых
представителей хищных млекопитающих
Comparative characteristics of heart mass of some adult representatives of carnivores
Масса тела, кг/ Body mass, kg Ce рдце/Heart
Виды/Species n масса, г/ mass, g % от массы тела/ % to body mass
Самцы/Males
Песец / Alopex lagopus 5 4.5 46 1.02
Лисица обыкновенная/ 8 1.12
Vulpes vulpes J
Собака енотовидная/ 6.8 0.99
Nyctereutes procyonoides
Шакал/ Canis aureus 4 9.5 81 0.85
Волк/ 6 350 0.92
Canis lupus 3o
Кот лесной/ Felis silvestris 3 5 40 0.80
Рысь/ Lynx lynx 5 15 100 0.67
Гепард/ Acinonyx jubatus 1 21 127 0.61
Тигр/ Panthera tigris 1 160 432 0.27
Самки/Females
Песец / Alopex lagopus 4 3.7 38 1.03
Лисица обыкновенная/ Vulpes vulpes 5 4.5 53 1.18
Собака енотовидная/ 6 4.8 1.08
Nyctereutes procyonoides Шакал/ 4
0.91
Canis aureus Волк/ 5 8 73 300
1.03
Canis lupus Кот лесной/ 3 29
0.83
Felis silvestris Рысь/ 3 4 7.8 33
0.77
Lynx lynx Лев/ 1 60
Panthera leo 125 675 0.54
Примечание. Данные по гепарду, льву и тигру амурскому получены в Московском
зоопарке.
156
отражает каких-либо функциональных преобразований в организме животных,
а является следствием существенной разницы в их массе тела.
По имеющимся данным (Насибов, 1970; Гинеев, 1972), индекс сердца у
отловленных осенью на Кавказе средних по размеру хищников - енота-
полоскуна (0.41%), лесного (0.55%) и камышового (0.64%) котов оказался
заметно ниже, чем у представителей семейства собачьих - лисицы (1.13%) и
шакала (1.06%), что объясняется высоким уровнем двигательной активности
у последних и разными формами охотничьего поведения животных.
Анализ ЭКГ куньих показал, что продолжительность сердечного цикла
(А-/?) находится в обратной зависимости от частоты пульса, с увеличением
которого она уменьшается. Эти изменения происходят в основном за счет
вариабельности диастолы (Т-Р), а продолжительность предсердно-желудочко-
вой проводимости (P-Q) и активной систолы желудочков (QRST) меняется
незначительно. Таким образом, учащенный пульс при меньшей фазе отдыха
сердца свидетельствует о повышении биоэлектрических процессов в миокарде
и более интенсивной его деятельности у самок по сравнению с самцами.
У изучаемых мелких хищников сердечный ритм соответствует определен-
ной норме, которая может быть сходной с таковой у других видов, близких по
размеру, образу жизни и поведению. Например, темп сердечной деятельности
был сравнительно низким у амфибионтов - норок и учащенным - у активных,
обладающих быстрой двигательной реакцией представителей рода Martes
(Туманов и др., 1981, 1982). Характерные различия в скорости сокращений
сердца между лесной и каменной куницами статистически не достоверны
(/2=0.75), в то же время у европейской и американской норок надежность
данного признака была более высокой и у самцов (р=0.85), и у самок (р=0.90).
Хорьки (лесной и альбинос - фуро) имеют примерно одинаковый сердечный
ритм. У степного хорька по сравнению с ними прослеживается замедленный
пульс. Известно, что степной хорек имеет высокий индекс сердца и хорошо
адаптирован к раскапыванию нор своих основных жертв - обыкновенного
хомяка, сусликов или водяной полевки, которых добывает прямо в убежищах
(Терновский и др., 1981). По-видимому, более редкий пульс степного хорька
можно рассматривать как компенсаторную реакцию организма, направленную
на увеличение «разрешающей способности» сердца во время повышения
физической нагрузки, при копании.
Различия в частоте пульса между близкими видами семейства куньих часто
оказываются статистически недостоверными. В то же время образ жизни и
поведение животных во многом определяют основные показатели
биоэлектрической активности сердца. Например, соболь и куницы заметно
отличаются от других видов по форме кривой его биотоков. Для ЭКГ этих
зверьков характерно отсутствие аритмии, высокие темп сокращений сердца и
вольтаж основных положительных (при слабой выраженности отрицательного -
У) зубцов желудочкового комплекса. Данные особенности, вероятно, имеют
свою экологическую обусловленность, поскольку состояние положительных
зубцов комплекса Q-T служит индикатором функционального состояния
157
/ /
5
Рис. 40. Нормальная ЭКГ мелких куньих (ориг.).
1 - показатель усиления, I мВ = 10 мм; 2 - обыкновенная ласка; 3 - пустынная
ласка; 4 - закавказская ласка; 5 - горностай; 6 - солонгой.
Fig. 40. Normal ECG of small mustelids.
1 - coefficient of intensification, 1 mV = 10 mm; 2 - weasel (Mustela nivalis); 3 - desert
weasel (Mustela nivalis heptneri)', 4 - Transcaucasia weasel (Mustela nivalis boccamela);
5 - ermine (Mustela erminea); 6 - Alpine weasel (Mustela altaica).
миокарда и организма в целом. Их увеличенный вольтаж отражает
интенсивность работы сердечной мышцы при нагрузках - усиление
биохимических процессов в ней и повышение биоэлектрических явлений в
желудочках (Рощевский, 1972, 1978; Меерсон, 1986; Pauls, 1980; Ploucha,
Aulerich, 1982).
158
4
о
Рис. 41. Нормальная ЭКГ колонка, хорьков и перевязки (ориг.).
1 - показатель усиления, 1 мВ=10 мм; 2 - колонок; 3 - степной хорек; 4 - фуро
(белый хорек); 5 - лесной хорек; 6 - перевязка.
Fig. 41. Normal ECG of Siberia weasel, polecats and marbled polecat.
1 - coefficient of intensification, 1 mV = 10 mm; 2 - Siberia weasel {Mustela sibirica)',
3 - Siberia polecat {Mustela eversmanni}\ 4 - Furo {Mustela putorius furo)',
5 - European polecat {Mustelaputorius)', 6 - marbled polecat {Vormela peregusna).
Ласка и горностай также имеют много общих черт в характере рисунка
ЭКГ (рис. 40). У зверьков наблюдается малая продолжительностью сердечного
цикла {R-R), а также довольно высокий вольтаж положительных {Р, R,T)vt
отрицательного зубца S. Близко к ним стоит солонгой, однако от ласок и
горностая он отличается растянутостью интервалов сердечного цикла и
сравнительно низким вольтажом всех зубцов ЭКГ.
По величине положительных зубцов желудочкового комплекса QRST
закавказская ласка скорее напоминает горностая, чем представителей
номинального подвида. Так, зубец R, характеризующий начало распростра-
нения волны возбуждения по миокарду желудочков, и зубец Т, который
обозначает окончание фазы систолы и восстановление покоя в сердечной
мышце, были разными по высоте. У самцов закавказской ласки они составляли
соответственно 23 и 6 мм, а у обыкновенной - 16.7 и 4.7 мм. Обычно
увеличенный вольтаж зубца R специфичен прежде всего для активных,
подвижных животных, и не случайно высокий зубец R наблюдается на ЭКГ
такого быстрого и резкого в движениях зверька, как горностай (табл. 49).
Второй положительный зубец {Т) желудочкового комплекса Q-Tявляется
одним из важнейших индикаторов функционального состояния миокарда и
организма в целом. Как правило, его большие размеры связаны с повышенным
обменом веществ, значительными физическими нагрузками или воздействием
высотной гипоксии на сердечно-сосудистую и дыхательную системы (Никулин,
1956; Фогельсон, 1957; Черногоров, 1962; Долабчян, 1968). Следовательно,
159
s 1
JiaJaJ^aJaMaJaI
z
5
Рис. 42. Нормальная ЭКГ норок, куниц и росомахи (ориг.).
7 - европейская норка; 2 - американская норка; 3 - каменная куница; 4 - лесная
куница; 5 - соболь; 6 - росомаха.
Fig. 42. Normal ECG of minks, martens, and wolverine.
1 - European mink {Mustela lutreola); 2 - American mink {Mustela vison); 3 - stone
marten {Martes foina); 4 - pine marten {Martes martes); 5 - sable {Martes zibellina);
6 - wolverine {Gulo gulo).
характерный для закавказской ласки большой индекс сердца и специфика
распространения волны возбуждения по миокарду могут отражать адаптивную
реакцию сердечной мышцы на особенности обитания зверьков в горных
условиях (пониженное парциальное давление кислорода, повышенная
инсоляция, физические нагрузки и т.д.).
У хорьков (степного, лесного и белого - фуро) различий в размерах
основных зубцов кривой биотоков сердца не прослеживается (рис. 41). В то же
время по большей продолжительности сердечного цикла {R-R), общей систолы
{PQRS Т) и особенно диастолы (Т-Р) степной хорек заметно отличается от двух
других. Отметим и то, что кариотипы лесного хорька и фуро также морфологи-
чески идентичны, но несколько различны с хромосомным набором степного
хорька (Волобуев и др., 1974). От ЭКГ ласок и горностая кривая биотоков
сердца хорьков отличается не только длительностью интервалов, но и меньшей
высотой зубца Т и особенно отрицательного S (из комплекса ORST),
соответствующего полному охвату возбуждением желудочков сердца. По этим
показателям к хорькам ближе всего стоит перевязка (рис. 41), а рисунок ЭКГ
колонка больше совпадает с таковым ласки, горностая и солонгоя (Туманов и
ДР-, 1982).
Для ЭКГ норок характерен сравнительно низкий вольтаж основных зубцов
(рис. 42). Анализ ее у исследованных видов позволил выявить существенные
расхождения между ними. Так, на ЭКГ американской норки отчетливо проступает
высокий зубец Т, по вольтажу которого она сходна с куницами и соболем. Размер
160
зубца Т, отражающего уровень обменных процессов в сердце, а следовательно,
и в целом организме, у американской норки, соболя и куниц оказался в 1.4-2 раза
больше, чем у европейской норки. В то же время у последней отрицательный
зубец S выражен более отчетливо, чем у представителей американского вида
(табл. 49). По форме кривой биотоков сердца к соболю и куницам близка
росомаха, на ЭКГ которой низкий зубец S и положительный Р, отражающий волну
возбуждения в обоих предсердиях, имеют сходные размеры с представителями
рода тИаг/ел’(Терновскийидр., 1981).
Таблица 49/ Table 49
Характеристика некоторых показателей сердечного ритм взрослых самок куньих
Characteristics of some parameters of cordial rhythm of mustelid adult females
Объекты исследования/ Objects of research Кол-во снятых ЭКГ/ Number of EKG Вольтаж основных зубцов желудочкового комплекса, мм/ Voltage of dents, mm Систолический показатель/ Systolic index, % Отношение систолы к диастоле/ Tenseness index, %
R 5 T
Ласка обыкновенная/ Mustela nivalis 4 16.2±0.5 6.3±0.7 4+0.5 50 306
Горностай/ Mustela 13 18.8+0.8 5.2+0.7 6.5±0.6 50 312
erminea Солонгой/ Mustela altaica 6 14.6±1.8 3.0±0.6 3.2±0.3 45 235
Колонок/ Mustela sibirica Хорек степной/ 7 19.1±0.4 4.9±0.8 5.1+0.7 49 253
Mustela eversmanni 6 17.2±1.6 2±0.5 4+0.4 43 204
Хорек белый/ Mustela 3 17.9±3.6 3±0.4 27+0.9 46 276
putorius furo Хорек лесной/ Mustela 6 17.6±1.8 2.5±0.5 3.3+О.8 47 283
putorius Норка европ./ Mustela lutreola 14 16+1.7 4.2±0.3 3.6+0.2 42 166
Норка америк./ Mustela vison 7 15.6±1.6 21+0.3 5.8±0.3 44 172
Куница лесная/ Martes martes 3 19.5±2.3 2±0.2 7.5+0.4 58 424
Куница каменная/ 3 17+1.5 1.5+0.2 5+0.6 53 462
Martes foina Соболь/ Martes zibellina 6 17.2±1.3 2.6±0.4 5.8±0.9 55 502
Барсук/ Meles meles 4 14.6±1.4 1.2±0.3 7.2±0.7 47 167
Росомаха/ Gulo gulo 2 17 2 4 49 282
6 Зак. № 4309
161
Функциональная деятельность сердечной мышцы барсуков также
находится в тесном соответствии с их образом жизни. Сравнительно небольшая
двигательнаяактивность зверей, рытье нор, обитание в глубоких убежищах в
условиях возможного дефицита кислорода нашли свое отражение и в
характеристике основных элементов ЭКГ. На них отчетливо просматриваются
сравнительно редкий пульс, большая продолжительность сердечного цикла
(R-R) и особенно диастолы (Т-Р). Электрокардиограммы барсуков
демонстрируют низкий вольтаж зубцов R и S при достаточно высоких Т
(компонент желудочкового комплекса) и Р (предсердный комплекс). Подобное
состояние элементов ЭКГ может свидетельствовать об адаптированности
сердечно-сосудистой системы организма к физическим нагрузкам и условиям
гипоксии (Никулин, 1956; Рощевский, 1972, 1978; Меерсон, 1986).
При оценке функционального состояния миокарда используют такие
индикаторы, как процентное отношение размеров всей систолы к диастоле
(Р-ТТГ-РхЮО %), а также продолжительности систолы желудочков к
сердечному циклу (Q-T/R-Rx.100 %), или систолический показатель (Никулин,
1956; Фогельсон, 1957; Долабчян, 1968). Их уровень определяет напряженность
работы сердца. У крупных представителей семейство куньих примерно равного
веса они оказались высокими у росомахи (соответственно 282 и 49%) и низкими
у барсука (167 и 47%). Из других видов по комплексной оценке величины
данных тестов наименьшую напряженность работы сердечной мышцы
демонстрируют европейская и американская норки (в среднем 169 и 43%), а
Таблица 50/ Table 50
Комплексная оценка напряженности работы сердца у самок куньих
Comprehensive estimate of intensity of cordial muscle work of mustelid females
Номер группы,
объект
исследований/
Groups, object of
_______research
1 группа (норк^
хорьки)/
1 group (minks,
polecats)
2 группа (ласка,
горностай,
солонгой)/
2 group (Mustela
nivalis, M. erminea,
M. Altaic a)
3 группа (соболь,
куницы)/
3 group (Martes sp.)
Отношение
систолы к
диастоле/
Tenseness
index, %
Систолический
показатель/
Systolic index,
%
33 206 ±4.9
14.7 99.9
44 ±0.2
48 ± 0.3
41.6 99.9
12 463 ± 7.2 55 ± 0.5
11.1 99.9
12 99.9
162
наиболее интенсивную - соболь, лесная и каменная куницы (463 и 55%). У
последних она определяется значительной частотой пульса й связанной с ней
малой продолжительностью сердечного цикла (R-R) и диастолической фазы
(Т-Р). Предсердно-желудочковая, или атриовентрикулярная, проводимость
(P-Q) с увеличением сердечного цикла животных удлиняется, поэтому у
представителей рода Martes она была меньше, чем у хорьков и норок.
Статистический анализ изучаемых величин у мелких хищников,
объединенных в группы по степени активности, особенностям питания и
поведения, показал достоверность различий между ними. В этих группах
напряженность работы сердечной мышцы у животных возрастает в
соответствии с увеличением скорости передвижений и быстроты двигательных
реакций (табл. 50).
Найденные различия между группами, установленные по f-критерию
Стьюдента, оказались статистически значимы. Достоверность различий была
очень высокой и превышала 99%.
Анализ материалов по влиянию низких температур на организм мелких
хищников позволил сделать вывод, что характер изменений частоты пульса и
контура ЭКГ зависит от массы тела животных, а также степени и длительности
воздействия на них холода: обычно чем они крупнее, тем дольше могут
противостоять действию отрицательных температур, и чем ниже температура
среды, тем быстрее наступает гибель зверьков, находящихся вне теплого
укрытия. Эти наблюдения совпадают с данными Д.В. Терновского (1977) по
влиянию охлаждающего действия воды на организм некоторых видов куньих.
В воде, отличающейся высокой теплопроводностью, у млекопитающих идет
интенсивная и некомпенсированная потеря тепла, приводящая их к гибели. По
данным автора, при пребывании в холодной воде (+8°С) переохлаждение
зверьков наступило у американской норки при массе тела 660 г через 118 мин,
у степного хорька (442 г) через 28 и у горностая (284 г) через 26 мин. В целом
американская норка плавала в холодной воде в 2.7-5.4 раза дольше горностая
и светлого хорька, что говорит о ее более высоких возможностях переносить
охлаждение.
Результаты наших опытов свидетельствуют о значительной индиви-
дуальной изменчивости сердечного ритма и рисунка ЭКГ при действии
гипотермии на организм мелких хищников, однако в целом у них, как и у
грызунов (Ивантер и др., 1985), можно отметить два периода, характеризу-
ющихся различной реакцией сердечной мышцы на охлаждение организма.
Вначале при понижении температуры окружающей среды частота сердечных
сокращений несколько возрастает. При этом вольтаж зубцов либо не меняется,
либо уменьшается незначительно. Данный период у ласок и горностаев,
находящихся вне гнезда, при температуре среды от -15 до -18°С продолжается
примерно 30-45 мин, после чего наблюдается падение пульса. По-видимому,
эту реакцию следует рассматривать как компенсаторное приспособление
организма мелких млекопитающих к существованию в условиях холода.
Затраты энергии на поддержание оптимальной температуры тела в таком
163
случае возрастают, в результате чего сердечно-сосудистой системой
осуществляется энергичная доставка кислорода и питательных веществ к
органам. Поскольку интенсивная работа сердечной мышцы долго продол-
жаться не может, то при расходовании организмом резервных возможностей и
с углублением гипотермии наблюдается второй период - постепенное
ослабление сердечной деятельности.
Действие холода может вызывать изменение всех зубцов и интервалов
ЭКГ, но в разной степени. С понижением температуры тела зверьков зубец Р
(предсердный комплекс) заметно увеличивался по вольтажу и приобретал более
острую форму. При умеренно низких температурах среды он был положи-
тельным, но в фазу глубокой гипотермии нередко становится отрицательным.
Зубцы желудочкового комплекса - A, ThS (если он был выражен на ЭКГ)-в
этот период резко возрастали по вольтажу. Длительность их в фазу глубокой
гипотермии и у мелких хищников, и у грызунов заметно увеличивалась, а
зубец Гк тому же становился зазубренным или двухвершинным. Одновременно
с урежением пульса наблюдалось увеличение продолжительности интервала
P-Q, комплекса QRS и отрезка Q-T. У ласок и молодых горностаев в 30-40%
случаев при длительном влиянии холода на организм отмечалось смещение
вверх сегмента RS-Т, возникновение аритмии и экстрасистолии. Последние
нарушения обычно являлись предвестниками остановки сердца. При
умеренной гипотермии аритмия и экстрасистолия у мелких хищников
возникала редко.
Материалы по динамике физиологических показателей, в частности по
изменению сердечного ритма при действии на организм представителей
семейства куньих низких температур или избыточного содержания углекисло-
ты, позволяют глубже понять их адаптивные возможности в разных условиях
существования. Повышенное содержание углекислого газа в воздухе жилых
нор мелких млекопитающих является вполне обычным явлением (Некипелов,
1958; Olzewski, Skoczen, 1965; Туманов, 1967, 1968; Arieli, 1979; Kochonen,
1980; Вискова, 1983, и др.). В то же время большая его концентрация во
вдыхаемом воздухе вызывает у животных определенные функциональный
изменения в работе сердца и дыхательного аппарата (Голодов, 1946;
Желудкова и др., 1964).
Регистрация ЭКГ у обыкновенных ласок, помещенных в герметическую
камеру с высоким содержанием углекислоты, показала следующее. Как и в
наших опытах с мышевидными грызунами, перевод ласок с дыхания
атмосферным воздухом на смесь, содержащую 1-1.5% углекислого газа и
нормальное количество кислорода, приводило к устойчивому замедлению
частоты пульса (брадикардии) в среднем на 13-20%. При замене этой газовой
смеси атмосферным воздухом сердечная деятельность зверьков возвращалась
к исходному уровню. Пребывание их в камерах с высокой концентрацией
углекислого газа сопровождалось увеличением вольтажа зубцов R и Гна ЭКГ.
Последнее, видимо, указывает на усиление процессов метаболизма в сердечной
мышце (Логунова, 1954; Райскина, 1962). При переключении мелких
164
млекопитающих (ласки, желтогорлые и полевые мыши, серые и рыжие полевки)
на дыхание атмосферным воздухом вольтаж зубцов А и Ту них возвращался
к первоначальной величине. Наблюдения показали также, что при переходе
животных с дыхания атмосферным воздухом на смесь газов с небольшим
недостатком кислорода (19.5-20%) и нормальным содержанием углекислоты
существенных преобразований в работе сердца не происходит. Подобный
характер изменения темпа сердечной деятельности наблюдался и у грызунов
во время их пребывания в естественных норах (Туманов, 1966а, б, 1967),
поэтому все вышеизложенное может свидетельствовать об устойчивости
организма животных-норников к гипоксии.
Отчетливые изменения функционального состояния сердечно-сосудистой
системы мелких млекопитающих следует, вероятно, рассматривать в основном
как проявление компенсаторных реакций их организма, однако у представи-
телей всех исследуемых «наземных» видов замедление частоты пульса во время
пребывания в норах не выражено так четко, как у полуводных животных в
момент погружения (Галанцев, 1977, 1980). У мелких млекопитающих
брадикардия в гнездах под землей наступает, по-видимому, как ответная
реакция на действие повышенного содержания углекислоты в убежищах. У
зверьков, роющих норы, в условиях среды с увеличенной концентрацией
углекислого газа брадикардия возникает вследствие изменения тонуса
блуждающих нервов и служит одним из важных проявлений физиологической
адаптации организма.
По материалам В.П. Галанцева (1983), у хищников, связанных с водной
средой обитания, функциональные адаптации сердечно-сосудистой системы
характеризуются приспособительным изменением рабочего состояния сердечной
мышцы и развитием ряда циркуляторных реакций при погружении животных в
воду. При нырянии у них сразу же развивается торможение автоматизма работы
сердца и пульс становится более редким (табл. 51). У амфибионтных хищников
и ластоногих темп сердечной деятельности при погружении в воду резко
замедляется, при этом латентный период составляет не более 1 с. Как сообщает
автор, особенности проявления ряда структурных или функциональных адап-
тивных признаков сердечно-сосудистой системы во многом зависят от уровня
экологической специализации видов к водной среде обитания. Приспособитель-
ное изменение сердечного ритма при нырянии млекопитающих связано с
преобладающим влиянием на сердце в этот период определенных регуляторных
механизмов. Под водой у них резко возрастает концентрация в крови ацетилхолина
и повышается активность холинэстеразы. Атропинизация или двухсторонняя
ваготомия животных не только не снимает эффекта брадикардии при погружении
в воду, но и не сокращает сроки ныряния. Проявление сердечного компонента
приспособительных реакций, наблюдаемое у хищных млекопитающих при
нырянии, выражается в развитии под водой рефлекторной брадикардии, которая
характерна для всех амфи- и гидробионтных видов.
Как и у куньих, у более крупных представителей отряда хищных
млекопитающих скорость сердечных сокращений определяется размером
165
Таблица 51/ Table 51
Характер изменения уровня сердечного ритма при нырянии хищных
млекопитающих и ластоногих (по: Галанцев, 1983)
The character of changes in level of cordial rhythm with diving of carnivores (Carnivora)
and pinnipeds (Pinnipedia)
Виды/ Species Частота пульса, в 1 мин/ Heart rate, per 1 min
до погружения/ before diving под водой/ under water
Норка европейская/ 267 ± 13 3 57 + 9 i
Mustela lutreola
Норка американская/ 235 ± 15.8 45 + 7.3
Mustela vison
Выдра/ Lutra lutra 172 ± 9.7 51 ± 8.5
Калан/ Enhydra lutris 139 ±5.8 48 ± 4.8
Нерпа каспийская/ Phoca caspica 75 ± 9.3 21 ± 0.6
Нерпа кольчатая/ Phoca hispida 71 ± 18.7 19 ±5.8
животных, их экологическими особенностями и формами поведения. Так, если у
лисицы и песца в спокойном состоянии частота пульса равна 150-210 уд./мин, то у
медведей - 50-80 уд./мин. При непродолжительном беге и активных перемещениях
сердечный ритм этих зверей увеличивался в 1.6-2.4 раза (Слоним, 1976; Успенский,
1989). Известно, что при переходе животных к пассивному состоянию энергозатраты
организма заметно снижаются. Наблюдения за спящими бурыми медведями
(Лобачев, 1951) показали, что интенсивность зимнего оцепенения возрастает ко
второй половине зимы, когда их сон становится более глубоким. Возможность
перехода ко сну обусловли-вается наличием у зверей жировых резервов,
достаточных для поддержания жизненных функций в неактивный период.
Сердечный ритм и частота дыхания заметно снижаются, а температура тела может
упасть до 29-37°С. Резко снижаются уровни потребления кислорода и выделения
углекислоты. Дыхательные движения становятся редкими и аритмичными. Обычно
после каждых 5-10 циклов дыхания наступает пауза, которая может длиться от 2 до
4 мин. Во время зимнего сна в естественных берлогах скорость сокращений сердца
уменьшается у белого медведя с 60 до 27, а у бурого - до 8 уд./мин (Folk et al., 1970).
Собранные материалы свидетельствуют о том, что у пребывающих в норах
или зимних берлогах хищных млекопитающих адаптивные реакции Сердечно-
сосудистой системы способствуют наиболее полной и рациональной утилизации
имеющегося в их организме резерва кислорода и питательных веществ,
обеспечивая тем самым животным высокую степень резистентности к гипоксии
и уменьшение энергетических затрат в неблагоприятные периоды года.
166
Резюме
Показатели сердечного ритма не только определяют компенсаторные
возможности организма, но и могут служить индикатором энергетического
метаболизма. Прослеживается обратная зависимость частоты пульса и
индекса сердца от массы тела рассматриваемых хищных млекопитающих. В
то же время на фоне общего падения частоты пульса по мере увеличения веса
животных (особенно в пересчете на единицу их массы) наблюдается тесная
зависимость уровня этого показателя от экологической специфики видов.
Анализ функционального состояния миокарда по основным показателям,
характеризующим напряженность работы сердца хищников, объединенных
по степени активности и особенностям питания, показал высокий уровень
достоверности различий. В этих группах интенсивность сердечной деятельности
животных возрастала в соответствии с увеличением их двигательных реакций.
Следует отметить, что сердечный ритм и характер рисунка ЭКГ у
представителей одного вида соответствуют определенной норме, которая
может быть сходной с таковой у другого животного, близкого по массе тела,
образу жизни и поведению. Например, у обоих видов норок темп работы сердца
был сравнительно низким, а у активных, обладающих быстротой двигательных
реакций куниц и соболя - учащенным. Различия в частоте пульса между Martes
foina и М. martes оказались статистически недостоверными (р=0.75), в то же
время между Mustela lutreola и М. vison надежность данного признака была
более высокой и у самцов (р=0.85), и у самок (р=0.9).
Материалы о влиянии низких температур на организм мелких хищников
показали, что характер изменений скорости их сердечных сокращений и контура
ЭКГ определяется прежде всего массой тела животных, степенью и длительно-
стью воздействия холода. Обычно чем крупнее особи, тем дольше они могут
противостоять действию отрицательных температур, и чем ниже температура
среды, тем быстрее наступает гибель животных, находящихся вне теплого укрытия.
Собранные материалы по динамике основных физиологических
показателей (и в частности, по изменению сердечного ритма при действии на
организм низких температур или избыточного содержания углекислого газа)
позволяют глубже понять адаптивные возможности хищных млекопитающих,
находящихся в разных условиях существования. Повышенное содержание
углекислого газа в жилых норах животных является вполне обычным явлением.
В то же время большая его концентрация во вдыхаемом воздухе вызывает у них
определенные функциональные изменения в работе сердца и органов дыхания.
Summary
The parameters of cordial rhythm, which determine compensatory abilities of
the organism and may serve as indicators of its energetic metabolism, play an
important role in the assessment of the functional state of the organism.
167
In carnivorous mammals an inverse negative relationship of pulse frequency,
heart index and body mass is recorded. At the same time, on the background of
general pulse frequency decrease with the increase of the animal body weight
(especially if calculated per unit of body mass), close interrelationship of this
parameter with the ecological specificity of species is observed.
Analysis of functional state of the myocardium by the main parameters
characterizing intensity of heart work in carnivores united by the degree of activity
and food specialization, showed a high level of significance of the differences. In
these groups the intensity of heart activity of animals increased in accordance with
the increase of their moving reactions.
It should be noted that heart rhythm and the character of ECG pattern in
representatives of one species correspond to a certain norm, which may be similar
to such in the other, close in body mass, life history and behavior. For example, the
rate of heart work was comparatively low in Mustela lutreola and M. vison and
rapid in the active, possessing quick moving reaction representatives of the genus
Martes. The differences in the pulse frequency between Martes foina and M. martes
appeared to be statistically insignificant (/>=0.75). At the same time the reliability
of this parameter between Mustela lutreola and M. vison was higher in both males
(p=0.85) and females (p=0.9).
Materials on the influence of low temperatures on the organism of small
carnivores have shown that the character of changes of their pulse frequency and
ECG contour is determined first of all by the body mass of animals, degree and
duration of cold effect. Commonly, the larger the animals the longer they can resist
the impact of negative temperatures, and the lower the ambient temperature, the
faster is the death of animals that are outside the warm shelter.
The data on the dynamics of the main physiological parameters considered in
the current work, particularly, on changes of the heart rhythm with the low
temperatures impact or excessive content of carbon dioxide in the organism of
carnivores, allow us a deeper insight into adaptive possibilities in various habitat
conditions. The increased content of carbon dioxide in the living holes of animals is
a quite common phenomenon. At the same time its high concentration in the air
causes in them certain functional changes in the work of the heart and respiratory
organs.
The adaptive reactions of cardiovascular system in mammals occurring in
deep holes or winter refuges promote the most complete utilization of oxygen reserves
and nutritional substances available in the organism. This provides them
minimization of energetic expenses in the unfavorable periods of the year.
Щенок ласки в возрасте 15 дней. (Фото автора).
A weasel cub at the of 15 days. (Photo by the author).
В прохладном гнезде во время отдыха детеныши ласки собираются вместе и
обогревают друг друга своим теплом. (Фото автора).
In a chilly nest of weasel the cubs warming each other. (Photo by the author).
Щенок горностая в возрасте 4 дней. (Фото автора).
An ermine cub at the age of 4 days. (Photo by the author).
Горностай в зимнем наряде. (Фото А. Кречмара).
Ermine in winter coloration. (Photo by A. Krechmar).
У европейской норки всегда хорошо видны белые пятна на верхней
и нижней губе. (Фото автора).
European mink has distinct white spots on the upper and lower lips.
(Photo by the author).
Европейская норка в настороженной позе. (Фото автора).
European mink in alarmed position. (Photo by the author).
Американская норка в поиске добычи. (Фото автора).
American mink in search for food. (Photo by the author).
В летнее время лесной хорек часто встречается по берегам водоемов.
(Фото автора).
In summer polecat often occurs along rivers and streams. (Photo by the author).
Соболь — «жемчужина» Сибири. (Фото Г. Монахова).
Sable — «gem» of Siberia. (Photo by G. Monakhov).
Зимой при глубоком снежном покрове добычей соболя может стать кабарга.
(Фото Г. Москова).
In winter with deep snow cover musk deer may become sabel’s prey.
(Photo by G. Moskov).
На кормежку барсук выходит из норы обычно в сумеречно-ночное время.
(Фото Ю. Пукинского).
Badger usually leaves his barrow for feeding in twilight and night time.
(Photo by J. Pukinsky).
Барсук бывает активен только в теплый период года. (Фото П. Иванова).
Badger is active in the warm period of the year. (Photo by P. Ivanov).
Росомаха — типичный представитель млекопитающих северных регионов России.
(Фото А. Макарова).
Wolverine — a typical representative of mammals from northern regions of Russia.
(Photo by A. Makarov).
В поисках пищи, особенно в зимний период, росомаха способна преодолевать
значительные расстояния. (Фото А. Кречмара).
Wolveme, hunting. (Photo by A. Krechmar).
Песец в летнем наряде. (Фото А. Кречмара).
Arctic fox with summer fur. (Photo by A. Krechmar).
Акклиматизированный в нашей стране енот-полоскун широко расселился
на Кавказе. (Фото А. Гинеева).
Racoon is widely spread over the Caucasian Region. (Photo by A. Gincyev).
Глава 5. АДАПТИВНЫЕ ЧЕРТЫ СТРОЕНИЯ
СОСУДИСТОГО РУСЛА
Выявленные приспособительные реакции сердечной мышцы хищных
млекопитающих находятся в тесном соответствии и с особенностями строения
кровеносной системы, которая обеспечивает доставку кислорода и
питательных веществ к различным органам. В ее преобразованиях
прослеживается строгая взаимосвязь морфофункциональных признаков,
отражающих уровень приспособления видов к определенным условиям
существования. Для них характерны хорошо выраженные различия в
строении прежде всего артериальной сети, которые обусловлены
особенностями экологии и поведения животных. Эти адаптивные изменения
проявляются по-разному, но имеют одну направленность - на интенсивное
кровоснабжение тканей работающих органов - и наиболее отчетливо
прослеживаются на примере семейства куньих, обладающих широким
спектром экологической радиации.
Сердце рассматриваемых зверей почковидной формы. Оно располагается
между 6-м и 10-м ребрами. Его верхушка направлена каудолатерально.
Правая (большая) часть основания заходит за среднюю линию грудной полости
и имеет краниодорсальное положение.
У куньих аорта представляет собой мощный сосуд эластического типа,
линейные размеры которого по отношению к длине тела у разных видов
заметно колеблются. Если у норок, выдры и калана ее относительная длина
равна 40-42%, то у остальных видов колеблется в пределах 35-38%.
Пропорции отделов аорты ко всей ее длине также неодинаковы: у мелких
хищников относительно самый короткий грудной отдел этого сосуда
характерен для куниц и соболя (53.5 ± 0.3%), а наиболее длинный - для норок
(56.8 ± 0.4%). Среди крупных представителей семейства по размерам
относительной длины аорты также выделяются полуводные животные и
барсук, у которых данный показатель был больше (58.8 ± 0.2%), чем у
росомахи (54.8 ± 0.3%). Выявленные различия, по-видимому, объясняются
характерной вытянутостью грудного отдела позвоночника, а следовательно,
и грудной аорты у полуводных и роющих животных по сравнению с
наземными видами (И. Соколов, А. Соколов, 1970, 1971).
Диаметр аорты у куньих (рис. 43) уменьшается каудальнее почечных
артерий (a.a. renales). При этом отмеченная диспропорция согласуется с
уровнем их специализации к водной среде обитания. Так, если у калана и
выдры калибр грудного отдела аорты превосходит брюшной в 2.8-3.1 раза, то
у норок уже в 2.3-2.6, а у остальных видов только в 1.7-2 раза. Резкое
изменение такого магистрального ствола, как аорта, при сравнительно
больших ее линейных размерах прослеживается также у грызунов (рис. 44-
45), которые ведут полуводный образ жизни (водяная полевка, ондатра) или
обитают в глубоких порах (суслики, сурки), что заметно отличает их от
7 Зак. № 4309
169
Рис. 43. Артериальная система горностая (а) и ласки (б).
1 - общая сонная артерия; 2 - плечеголовной ствол; 3 - левая подключичная
артерия; 4 - грудной отдел аорты; 5 - брюшной отдел аорты.
Fig. 43. Arterial system of ermine (Mustela erminea) (a) and weasel (M. nivalis) (6).
1 - common carotid; 2 - arm-cephalic stem; 3 - left clavicular artery; 4 - pectoral part
of aorta; 5 - abdominal part of aorta.
Рис. 44. Изменение диаметра аорты у водяной полевки.
1 - общие сонные артерии; 2 - грудной отдел аорты; 3 - брюшной отдел аорты.
Fig. 44. Change in aorta diameter of water vole (Arvicola terrestris).
1 - common carotids; 2 - pectoral part of aorta; 3 - abdominal part of aorta.
170
Рис. 45. Артериальная система сурка-байбака.
7 - грудной отдел аорты; 2 - межреберные артерии; 3 - почечные артерии;
4 - брюшной отдел аорты; 5 - общая подвздошная артерия; 6 - внутренняя
подвздошная артерия; 7 - наружная подвздошная артерия; 8 - бедренная артерия.
Fig. 45. Arterial system of steppe marmot (Marmota bobac).
1 - pectoral part of aorta; 2 - intercostal artery; 3 - renal arteries; 4 - abdominal part of
aorta; 5 - common iliac artery; 6 - internal iliac artery ; 7 - external iliac artery;
8 - femoral artery.
Рис. 46. Артериальная система обыкновенной (а) и рыжей (б) полевки.
1 ~ общая сонная артерия; 2 - подключичная артерия; 3 - дуга аорты; 4 - грудной
отдел аорты; 5 - брюшной отдел аорты; 6 - общая подвздошная артерия.
Fig. 46. Arterial system of the common (Microtus arvalis) (a)
and bank (Glethrionomys glareolus) (o) voles.
1 - common carotids; 2 - clavicular artery; 3 - arch of aorta; 4 - pectoral part of aorta;
5 - abdominal part of aorta; 6 - common iliac artery.
171
Рис. 47. Артериальные магистрали енотовидной собаки.
1 - грудной отдел аорты; 2 - брюшной отдел аорты; 3 - наружные подвздошные
артерии.
Fig. 47. Arterial system of raccon dog (Nyctereutesprocyonoides).
1 - pectoral part of aorta; 2 - abdominal part of aorta; 3 - external iliac arteries.
Рис. 48. Артериальные магистрали обыкновенной лисицы.
1 - грудной отдел аорты; 2 - брюшной отдел аорты; 3 - наружная подвздошная
артерия; 4 - внутренняя подвздошная артерия.
Fig. 48. Arterial system of red fox (Vulpes vulpes).
1 and 2 - pectoral and abdominal parts of aorta; 3 and 4 - external and internal iliac
arteries.
172
Рис. 49. Магистральные сосуды переднего отдела туловища лесного хорька.
1 - общие сонные артерии; 2 - подключичные артерии; 3 - грудной отдел аорты.
Fig.49. Mainstream vessels of anterior part of body polecat (Mustela putorius).
1 - common carotids; 2 - clavicular arteries; 3 - pectoral part of aorta.
наземных видов (рис. 46). Данная особенность сосудистого русла может иметь
немаловажное значение для животных-амфибионтов и норников. Оно улучшает
кровоснабжение органов, удаленных от сердца, так как согласно законам
гемодинамики при уменьшении диаметра сосуда должно происходить
увеличение в нем давления и скорости кровотока (Лесгафт, 1905). Как и у
наземных видов куньих, у собачьих (лисица, енотовидная собака) резкого
уменьшения диаметра аорты каудальнее почечных артерий не наблюдается
(рис. 47-48).
Наружный диаметр аорты около сердца у соболя, куниц, норок и хорьков
примерно одинаков (табл. 52). В то же время среди крупных представителей
семейства росомаха характеризуется меньшим диаметром аорты в грудной
полости по сравнению с барсуком, каланом и выдрой, т.е. животными,
приспособленными к жизни в норах или водной среде.
Высота дуги аорты у куньих также оказалась различной (табл. 52). В целом
в ряду видов прослеживается увеличение ее размеров с нарастанием массы
тела зверьков. Среди крупных хищников семейства примерно равного веса
наиболее высокую дугу имеют выдра и калан (22-23 мм), т.е. виды с большими
размерами грудной клетки.
173
Таблица 52/ Table 52
Некоторые показатели артериальной системы представителей семейства куньих
Some parameters of arterial system of mustelid representatives
Виды/ Species n Отношение длины/ Relation of Наружный диаметр аорты, мм/ External diameter of aorta, mm Высота дуги аорты, мм/ Height of aorta arch, mm Относит, длина общей сонной артерии, %/ Relative length of common carotid, %
аорты к длине тела, %/ aorta length to body length, % грудного отдела аорты ко всей ее длине, %/ pectoral part of aorta to its total length, % У сердца/ at heart У трифур- кации/ at threefur- cation
Ласка/ Mustela 6 37.6 55.2 1.8 1 1.5 17.8
nivalis Горно- стай/ 10 37 54.1 2.7 1.4 2.6 19.4
Mustela erminea Хорек лесной/ 12 36.2 55.8 4.9 2.6 10.2 20.2
Mustela putorius Норка америк./ 14 40.2 56.9 5 2.2 9.3 22.3
Mustela vison Куница лесная/ 7 36.5 53.2 4.8 2.5 15.5 19.3
Martes martes Соболь/ Martes 6 36.8 53.8 5.0 2.5 15.3 19.1
zibellina Барсук/ Meles 8 35.3 57.7 12.5 5.7 20.5 22.7
meles Выдра/ Lutra 8 40.8 58.9 13.8 4.4 22.1 21.5
lutra Калан/ Enhydra 2 41.3 59.4 14.6 5.1 22.8 15.7
lutris Росомаха/ Gulo gulo 2 36.9 54.8 11.3 5.5 16.5 20.1
Существенная разница по высоте дуги аорты и ветвлению артериальных
магистралей прослеживается и между такими мелкими хищниками, как
обыкновенная ласка и горностай. У последнего дуга этого сосуда хорошо
очерчена и по высоте достигает 2.5-3 мм. У ласки она почти в 2 раза меньше
(1.4-1.6 мм) и не всегда выражена отчетливо. Выйдя из левого желудочка сердца,
174
грудная аорта ласки без заметного подъема сразу же направляется в дорсально-
каудальном направлении к позвоночному столбу, а затем вдоль него - к
брюшной полости. В среднем дуга аорты у рассматриваемого хищника в 2.5-
3 раза ниже, чем у мелких грызунов - мышей и полевок. Отмеченные
морфологические особенности ласки, по-видимому, являются одной из
адаптивных черт ее строения. Они уменьшают потери напора кровотока в
сосудистом русле и могут быть связаны с высоким уровнем метаболических
процессов в организме данного вида.
Как и у других хищных млекопитающих, у куньих от вершины дуги аорты
в краниальном направлении отходят два сосуда, которые снабжают кровью
голову, шею и передние конечности (рис. 49). Это плечеголовной ствол - truncus
brachiocephalicus communis - и левая подключичная артерия - a. subclavia
sinistra. Диаметр последней у всех изучаемых видов в 1.2-1.6 раза меньше
диаметра плечеголовного ствола. У ласки в связи со слабой выраженностью
дуги плечеголовной ствол отходит непосредственно от основания аорты, что
обеспечивает лучшее кровоснабжение головы и передних отделов туловища.
От каудальной части дуги аорты как у наземных, так и у полуводных видов
куньих отходят левая и правая передние межреберные артерии (а.а. intercostales
suprema sinistra et dextra). Интересно, что у собачьих (лисица, песец) они
ответвляются от подключичных артерий, т.е. имеют совершенно другие места
отхождений.
Плечеголовной ствол последовательно дает начало левой и правой общим
сонным артериям (а.а. carotides communes sinistra et dextra) и далее переходит
в правую подключичную артерию (a. subclavia dextra). У ласки в связи со
слабой выраженностью дуги плечеголовной ствол отходит не от ее вершины, а
непосредственно от основания аорты. В результате этого, вероятно, резко
снижаются потери напора кровотока и обеспечивается лучшее кровоснабжение
всей краниальной части туловища, однако в ветвлении плечеголовного ствола
изучаемых видов наблюдается большое разнообразие. Так, у некоторых
особей норок, росомахи, барсука левая и правая общие сонные артерии отходят
от этого сосуда последовательно на расстоянии 4-7 мм друг от друга. С
дорсальной стороны между ними отделяется правая внутренняя грудная
артерия, чего не наблюдается у других хищников (кошачьих и собачьих).
У горностая, хорька, соболя и лесной куницы общие сонные артерии
ответвляются в начале единым общим стволом, а затем делятся. Правая
внутренняя грудная артерия отходит у них непосредственно от правой
подключичной. У ласки, выдры и калана общие сонные артерии отделяются
от основного ствола одновременно, образуя бифуркацию. При этом правая
внутренняя грудная артерия может отходить как от плечеголовной, так и от
подключичной артерии.
Как показали исследования, ветвление плечеголовного сосуда у куньих
отличается значительной вариабельностью и внутри каждого вида можно
встретить особей с отхождением сосудов по типу, позволяющему замерить
углы, образованные общими сонными артериями.
175
Известно, что интенсивность питания органов во многом зависит от угла
отхождения сосудов от основного ствола, поскольку острые углы и более
прямолинейный ход артерий дают меньшую потерю напора кровотока
(Ковешникова, 1936; Мажуга, 1966). У активных видов куньих, характеризу-
ющихся высоким уровнем энергообмена, угол между общими сонными
артериями, обеспечивающими кровоснабжение головы и всего кранильного
отдела туловища, оказался более острым, чем у сравнительно мало подвижных
форм сходного веса: например, у куниц и соболя он был равен 22-25°, а у
норок и хорьков 31-33°. Данная зависимость прослеживалась и среди других
хищников этого семейства: если у горностая и росомахи угол общих сонных
артерий составлял 28-32°, то у барсука, калана и выдры - 35-40°. У ласки этот
показатель колебался в пределах 18-22°. Более острыми у нее оказались и
углы ветвления наружных и внутренних сонных артерий. Отмеченные
морфологические особенности демонстрируют важные адаптивные черты
строения сосудистого русла представителей семейства куньих. Высокий
уровень обменных процессов и интенсивное кровообращение, характерные
для активных видов, стали возможными только при соответствующей
приспособительной перестройке всего организма, в том числе и артериальной
системы.
Еще в 1878 г. В. Ру (Roux, 1878) установил, что на сосудистое русло
животного наряду с фактором наследственности влияет величина его
энергетической деятельности. В связи с этим становится понятным, что у ласки
(при относительно малых ее размерах и сравнительно высоком уровне обменных
процессов) усиление интенсивности кровообращения вызывает и соответству-
ющие изменения в строении артериальной системы.
Общие сонные артерии, следуя по латеральным поверхностям трахеи,
дают ветви к мышцам шеи, околоушной и щитовидной железам. Наружный
калибр их равен у горностая 1-1.3 мм, у норки и соболя 1.7-2 мм, у барсука
2.6-2.8 мм, а у росомахи и выдры 4.0-4.4 мм. Линейные размеры общих
сонных артерий у изучаемых видов тесно связаны с величиной их шейного
отдела позвоночника. Так, у куньих (И. Соколов, А. Соколов, 1970, 1971)
при равном количестве шейных позвонков относительная длина этого отдела
изменяется более чем в 1.5 раза, что связано с размерами каждого
отдельного сегмента, неодинаковыми у разных видов. Наиболее длинный
шейный отдел позвоночника характерен для барсука. Близко к нему по
этому признаку стоит выдра, а у калана относительная длина шеи в 1.5
раза меньше соответствующего индекса у предыдущих видов.
Как показали наши исследования, линейные размеры левой общей сонной
артерии (по отношению к длине тела) оказались наименьшими у калана и ласки,
а наибольшими - у барсука, выдры и норки (табл. 52), что вполне согласуется
с данными указанных выше авторов. Дальнейшее ветвление общих сонных
артерий принципиально не отличается от такового у хищников с различными
формами поведения и уровнем энергообмена и поэтому специально нами не
рассматривается.
176
За исключением тех случаев, когда правая внутренняя грудная артерия
отходит от плечеголовного ствола, строение правой и левой подключичных
артерий одинаково. От каждой из них отделяются следующие артерии:
поперечная шейная, позвоночная, внутренняя и наружная грудные, а также
плечешейный ствол. От грудного отдела аорты берут начало межреберные и
задние диафрагмальные артерии.
Передним конечностям всех хищных млекопитающих свойствен широкий
диапазон функциональных адаптаций. Характер ветвления сосудов и
соотносительные размеры кровеносных магистралей грудных конечностей
подробно изучались П.М. Мажугой (1966). Он показал их прямую зависимость
от особенностей строения и функции локомоторного аппарата. По данным
автора, у куньих a. brachialis отдает мощную глубокую артерию, плеча,
которая в отличие от таковой у представителей семейства кошачьих отходит
не двумя, а одним стволом. Далее от нее ответвляются артерии: плечевой кости -
a. nutritia humeri, двуглавого мускула- a. bicipit is и предплечья -a. antibrachii
superficialis. Последняя у выдры и барсука имеет самостоятельное начало, а у
лесного хорька и куниц совмещена в этой части с артерией двуглавого
мускула.
В области плечевого сустава плечевая артерия у представителей
семейства куньих и кошачьих проходит вместе со срединным нервом через
подмыщелковое отверстие, в то время как у собачьих она располагается
свободно. Деление плечевой артерии на конечные ветви - срединную,
межкостную и локтевую - происходит по-разному. У выдры, хорька и лесной
куницы эти сосуды отходят самостоятельно, а лучевая артерия отделяется от
серединной. У барсука плечевая артерия делится на 2 ствола, из которых
один является общим для срединной и лучевой, а другой - для межкостной и
локтевой артерий. Лучевая артерия у этого вида имеет больший калибр по
сравнению со срединной, в то время как у хорька и лесной куницы она
уступает ей по мощности. Замечено, что у животных со сходной функцией
конечностей по особенностям строения их магистрального русла можно
обнаружить более четко выраженное сходство, чем у представителей одной
систематической группы, но с функционально различными конечностями
(Мажуга, 1966).
Кровоснабжение органов краниальной части брюшной полости осущест-
вляется чревной артерией - a. coeliaca, начинающейся от вентральной стенки
брюшной аорты на уровне 1-го поясничного позвонка. Как и у других хищных
млекопитающих, у куньих она делится на печеночную, желудочную и
селезеночную артерии, которые снабжают кровью одноименные органы и,
кроме того, отдают по одной ветви к стенкам желудка.
Таким образом, к желудку у всех изучаемых видов подходят по 3 артерии.
Многократно ветвясь, они образуют густую сеть кровеносных сосудов,
соединенных между собой анастомозами. Большее их количество отмечается
у барсука и видов-амфибионтов, что, вероятно, связано с экологическими
особенностями этих животных.
177
1
3
Рис. 50. Ветвление брюшного отдела аорты лесного хорька.
7- брюшная аорта; 2 - наружная подвздошная артерия; 3 - внутренняя
подвздошная артерия.
Fig. 50. Diversification of abdominal part of aorta of polecat {Mustela putorius).
1 - abdominal aorta; 2 - external iliac artery; 3 - internal iliac artery.
Краниальная брыжеечная артерия {a. mesenterica cranialis) отделяется от
вентральной стенки брюшной аорты в области второго поясничного позвонка,
т.е. несколько каудальнее чревной артерии. Она васкуляризирует почти весь
тонкий и толстый отделы кишечника.
На уровне 2-го поясничного позвонка от аорты отходят парные брюшные
артерии, которые делятся на ветви II—III порядков и несут кровь к мышцам
поясницы и брюшной стенки.
В области 3-го поясничного позвонка брюшная аорта отдает почечные
артерии - a. a. renales, левая из которых почти вдвое короче правой и отходит
несколько каудальнее ее. Калибр почечных артерий равен у ласки и горностая
0.5-0.7 мм, у куницы, норки и хорька 1-1.5 мм, а у выдры, росомахи и барсука
4-6 мм. У американских норок нередко встречаются сдвоенные а.а. renales,
которые идут двумя стволами к почке (Тихонов, 1966). Подобное раздвоение
почечной артерии отмечено нами и у одного из исследованных экземпляров
речной выдры.
Васкуляризация надпочечников осуществляется сосудами, которые
отходят либо от аорты, либо от соответствующей почечной артерии.
Каудальная брыжеечная {a. mesenterica caudalis) и 6 (у барсука 5) поясничных
артерий (a.a. lumbales) у куньих не отличаются по характеру ветвления от
одноименных сосудов у других видов хищных млекопитающих - песца, лисицы
178
или волка. Парные пояснично-подвздошные артерии отходят от аорты на
уровне 5-го поясничного позвонка. Они снабжают кровью поясничные и
ягодичные мышцы.
Тазовая область и задние конечности изучаемых животных васкуляризуются
конечными ветвями брюшной аорты (рис. 48,50). Ее деление происходит у
куницы, соболя, хорька и росомахи на уровне краниальной, а у остальных
видов - каудальной части 6-го (у барсука 5-го) поясничного позвонка. У
представителей семейств куньих и собачьих аорта отдает мощные наружные
подвздошные артерии (a.a. iliacae externae) и переходит в короткий ствол
внутренних подвздошных. Его длина в ряду видов возрастает от 1-1.5 мм у
ласки и горностая до 4-5 мм у куницы и соболя и до 6-8 мм у барсука и росомахи.
От этого сосуда у куньих отходят парные внутренние подвздошные (a. a. iliacae
internae) и хвостовая (a. coccyged) артерии. Угол, образованный наружными
подвздошными артериями, васкуляризующими задние конечности, оказался
различным у хищников, обладающих неодинаковым уровнем активности и
локомоторных функций. Так, у горностая он был более острым (60-65°), чем у
обыкновенной ласки (70-76°), а соболь и лесная куница по величине данного
показателя (50-58°) превышали европейскую и американскую норок (62-68°),
близких к ним по массе тела. Эти особенности строения кровеносной системы
характерны и для крупных представителей семейства. Например, у барсука и
росомахи угол, образованный рассматриваемыми сосудами, был заметно острее
(60-68°), чем у выдры и калана (70-78°).
Наружная подвздошная артерия (которая в дальнейшем переходит в
бедренную) отдает 2 сосуда: глубокую артерию бедра (a. profunda femoris) и общий
ствол каудальной надчревной и срамной артерий (truncu.s pudendoepigastricus),
дальнейшее ветвление которых схоже у куньих и собачьих. Относительная длина
наружной подвздошной и бедренной артерий (в сравнении с длиной бедренной
кости) у наземных видов оказалась больше таковой у хищников-амфибионтов,
что, видимо, обусловлено разницей в пропорциях частей локомоторного аппарата
и испытываемых им функциональных нагрузках. Внутренняя подвздошная
артерия, делясь на париетальную и висцеральную ветви, снабжает кровью органы,
расположенные в тазовой полости, и мышцы таза.
Интересно, что у выдры и калана ветвление кровеносных сосудов бедра I
и II порядка носит более рассыпной характер, чем у наземных форм (таких,
например, как лесная куница, росомаха или соболь). При этом между
отдельными ветвями имеются как артериальные, так и артерио-венозные
анастомозы.
Полученные материалы показывают, что хищные млекопитающие,
близкие в таксономическом и филогенетическом отношении, имеют ряд
существенных различий в характере ветвления основных артериальных
магистралей. Эти различия тесно связаны с образом жизни изучаемых видов и
особенностями их локомоции. Сравнительно небольшая величина суточных
передвижений ласки и горностая обусловливает и особенности строения
сосудистого русла. Они характеризуются прежде всего слабо выраженной
179
адаптацией артериальной системы локомоторного аппарата к интенсивному
бегу. Между тем ласка по сравнению с другими видами имеет наибольшую
величину поверхности теплоотдачи, а следовательно, обладает высоким
уровнем энергообмена. Строение ее артериальной системы - низкая дуга
аорты, острые углы отхождения сосудов краниальной части туловища,
большое количество анастомозов и др. - демонстрирует направленность
магистрального русла к быстрому кровоснабжению всех тканей организма
этого мелкого и очень активного зверька.
Особенности экологии лесной куницы, соболя, хорька и росомахи также
находят свое отражение в строении кровеносной системы. Обладая
разнообразными формами локомоций, высокими показателями активности и
суточных перемещений, они имеют и некоторые своеобразные черты строения
артериальной системы по сравнению, например, с лаской или норкой. Сущность
этих различий заключается в более интенсивном снабжении питательными
веществами и кислородом тканей локомоторного аппарата. Так, в отличие от
других хищников семейства куньих у перечисленных видов отмечаются
относительно короткая аорта, ответвление от нее подвздошных артерий под
более острым углом и прямолинейный ход этих сосудов, что обеспечивает
наибольшую скорость тока крови к мышцам задних конечностей.
У барсука в связи с роющим образом жизни выявляется ряд особенностей
в строении артериальных сосудов передних конечностей, а также такие
отличительные черты, как увеличение относительной длины общих сонных
артерий и грудного отдела аорты. Кроме того, для этого зверя характерны
относительно короткая аорта, большой диаметр магистральных сосудов и
увеличенные углы расхождения основных артерий. Например, ветвление
наружных подвздошных артерий у него происходит под бблыпим углом по
сравнению с росомахой, что может быть связано со значительными
статическими нагрузками, испытываемыми задними конечностями при рытье
нор.
У видов-амфибионтов (американской норки, выдры и калана) полуводный
образ жизни также обусловил ряд преобразований, которые произошли в их
артериальной системе. Эти адаптивные изменения находятся в тесной связи с
особенностями экологии видов и заключаются в увеличении диаметра и
линейных размеров грудного отдела аорты, в значительной диспропорции
калибра ее основных частей, усиленной васкуляризации внутренних органов
и больших углах отхождения магистральных сосудов. Следует отметить и
наличие у американской норки относительно низкой дуги аорты по сравнению
с наземными видами, а у выдры и калана - артериальных и артерио-венозных
анастомозов между отдельными ветвями сосудов бедра, деление которых часто
происходит по рассыпному типу.
Артериальная система ныряющих млекопитающих в процессе адапта-
циогенеза не претерпела таких значительных структурных изменений, как
венозная, однако и в ней, особенно у наиболее высокоспециализированных
форм, можно отметить некоторые приспособительные перестройки. По
180
материалам В. Галанцева (1983), у каланов наблюдается развитие бульбо-
образного расширения в восходящей части дуги аорты. В этом месте диаметр
просвета аорты увеличивается почти в 2 раза. С обнаруженной структурной
особенностью аорты, видимо, связано под держание энергии диастолического
давления крови в период сложных гемодинамических перестроек во время
ныряния. У высокоспециализированных к водной среде обитания видов
значительного развития достигает и правый желудочек сердца, стенки которого
у калана по толщине лишь немногим уступают левому. Развитие этой
адаптивной особенности сердца, видимо, было связано с изменяющимися
гемодинамическими условиями под водой, что приводит к сжатию легких и
повышению давления в легочной артерии.
Исследование артериальных магистралей различных представителей
семейства Canidae показало, что характер их ветвления не несет четко
выраженных видоспецифических черт и в целом соответствует таковому у
наземных куньих. Это совпадает с мнением М.Ф. Ковтуна и Л.П. Осинского
(1985) о том, что у волка не наблюдается анатомических особенностей в
ветвлении артериального русла, которые были бы характерны только для этого
вида. В то же время, по сообщению авторов, в микростроении сосудов
некоторые различия между хищниками проявляются отчетливо. Так, из 3
оболочек, составляющих основу артерий и дуги аорты собачьих, большая ее
часть представлена средней оболочкой (tunica media), которая занимает 66-
93% всей толщины стенки. Гладкомышечные клетки в дуге аорты имеют
преимущественно спиралевидную направленность. Количество их рядов у
волков (75-145) было достоверно большим, чем у енотовидной собаки, лисицы
или белого песца (26-44).
По данным указанных специалистов, эластические элементы средней
оболочки, или медии, представлены мембранами и волокнами разного раз-
мера. При этом внутренняя эластическая мембрана всюду выражена очень
отчетливо. В стенках дуги аорты ее величина достигала у волков, добытых в
тундровых регионах, 2 мкм, а на Кавказе - 3.5 мкм. В области передних
конечностей внутренняя эластическая мембрана хорошо контурирована. Ее
толщина в подмышечной артерии волков равна 1.6-2.1 мкм, а у других
представителей семейства - 0.7-1.1 мкм. Наибольшее количество эластических
мембран в медии подмышечной артерии волков с Кавказа - 23-28. Меньше их
у животных из тундры (18-23) и Украинского Полесья (15-19). У енотовидной
собаки и обыкновенной лисицы в данной артерии содержится 6-9 рядов
эластических мембран, а у песца их в 2 раза больше. В медии плечевой артерии
всех видов собачьих количество этих элементов уменьшается до 5-6.
В артериальных магистралях задних конечностей также наиболее развита
средняя оболочка. Количество эластических мембран в толще медии наружной
подвздошной и бедренной артерий составляет у волков до 4-8, а у лисицы,
песца и енотовидной собаки-до 2-3.
Необходимо отметить, что у волков, добытых в горных условиях Кавказа,
калибр всех сосудов и толщина их стенок больше, чем у зверей из других
181
Таблица 53/ Table 53
Основные параметры дуги аорты разных видов куньих
(по: Коршунова, Туманов, 1979, с дополнениями)
The main parameters of aorta arch of mustelid various species
Виды/ Species Отношение к просвету сосуда/ Relation to the vessel opening Эластические мембраны/ Elastic membranes
его стенки/ its wall толщины медии/ thickness of medial layer кол-во/ number толщина, мкм/ thickness, pm
Ласка, горностай, солонгой, хорек лесной, перевязка/ Mustela nivalis, М. erminea, М. altaica, М. putorius, Vormela 1 : 5-6 1 : 6-8 14-20 5-6
peregusna Норки европейская и американская/ Mustela lutreola, М. vison Соболь, куницы 1 :4-5 1 : 6-8 22-35 4-6
лесная и каменная/ Martes zibellina, М. martes, М. foina Росомаха/ i :7-8 1 : 6-7 1 :9-12 1 :9-10 20-30 58-60 5-10
5-10
Gulo gulo Барсук/ Meles те les 6-7
1 : 7-8 1 :9-10 60-62
Выдра/ Lutra lutra Калан/ 1 : 12-15 1 :19 37-39 6-8
10-12
Enhydra lutris 1 : 12-15 1 : 17 21-23
регионов. Например, диаметр просвета дуги аорты и толщина ее стенки были
равны соответственно у волков кавказской популяции 22 500 и 2808 мкм,
тундровой - 12 500 и 1690 мкм, а из Украинского Полесья - 19 800 и 2648 мкм.
У волков из зоопарка эти показатели составляли 10 500 и 2811мкм. Анализ
собранных материалов показал, что у всех собачьих дуга аорты представляет
собой относительно тонкостенный артериальный сосуд. По мере его ветвления
артерии утончаются, их просвет становится более узким. Звери, добытые в
природе, имеют более тонкие, эластичные и с большим внутренним диаметром
артерии по сравнению с животными, выращенными в неволе. У волков
кавказской популяции основные артериальные магистрали имели мощный
просвет, а стенки сосудов - большее содержание эластических элементов по
сравнению с особями из других регионов (Ковтун, Осинский, 1985).
Среди представителей семейства куньих существенные видовые различия
в строении магистрального артериального русла прослеживаются очень
отчетливо (Алексеева, 1968; Алексеева и др., 1968; Туманов, 1974а). При
182
Рис. 51. Гистологическое строение стенки дуги аорты
(орсеин, об. 9, ок. 12.5).
1 - ласка; 2 - солонгой; 3 - лесной хорек; 4 - лесная куница; 5 - росомаха; 6 -
барсук; /-американская норка; 8- речная выдра.
Fig. 51. Histological structure of arch of aorta (orsein, 9x12.5).
1 - weasel {Mustela nivalis)-, 2 - Alpine weasel (M. altaica)-, 3 - polecat (M. putorius)-,
4 - pine marten (Martes martes)', 5 - wolverine (Gulo gulo)', 6 - badger (Meles meles)-,
7 - American mink (Mustela vison); 8 - otter (Lutra lutra).
183
этом адаптивная перестройка их организма сопровождалась преобразованиями
основных артериальных сосудов не только в макро-, но и микроструктуре. В
целом можно отметить, что у наиболее активных (куницы, соболь) и
специализированных к обитанию в глубоких норах или водной среде животных
(барсук, выдра, калан) высокая дуга аорты имеет широкий просвет. Тонкие
стенки ее медии состоят в основном из крупных мембран эластического типа и
узкого слоя мышечных элементов, который заметно увеличен у барсука.
Диаметр просвета дуги аорты превосходит толщину всей ее стенки у калана и
выдры в 12-15 раз, у соболя, куниц и барсука - в 7-8, а у остальных видов - в
4-6 раз (табл. 53).
Анализ гистосрезов, проведенный нами совместно с Л.А. Коршуновой
(Коршунова, Туманов, 1979), показал, что дуга аорты у куньих, как и у других
хищных млекопитающих, представляет собой сосуд эластического типа,
состоящий из 3 оболочек. Внутренняя - интима - наименее широкая. Она
занимает 1-4% толщины стенки и у разных видов равна 2.5-22 мкм. В ней
хорошо развит слой эндотелиальных клеток, уплощенных и плотно
соединенных между собой, с ярко окрашенными ядрами. Подэндотелиальный
слой выражен слабо.
Наружная оболочка - адвентиция - у всех представителей семейства куньих
сходна. Она состоит из рыхлой соединительной ткани с большим количеством
коллагеновых и эластических волокон, имеющих спиральную или продольную
направленность. Если последние сосредоточены главным образом в глубоких
слоях адвентиции, ближе к эластической мембране средней оболочки, то
коллагеновых волокон заметно больше в периферической ее части. Среди них
равномерно распределены фибробласты и гистиоциты. В адвентиции
расположены кровеносные сосуды (vasa vasorum), питающие стенку аорты, и
содержится много жировой ткани. Толщина адвентиции больше интимы, но в
2-3 раза меньше таковой средней оболочки.
Существенные различия по размерам и структуре основных элементов
наблюдаются в средней оболочке, или в медии. У сходных по образу жизни
видов - ласки (Mustela nivalis), горностая (М. erminea), солонгоя (М. altaica),
перевязки (Vormela peregusna), лесного и степного хорьков (М. putorius, М.
eversmanni) - средняя оболочка развита хорошо и занимает от 62 до 81%
толщины стенки аорты (рис. 51). Отношение ширины медии к просвету у них
равно 1 : 6-8, а отношение толщины всей стенки сосуда к просвету - 1 : 5-6.
Вся средняя оболочка у этих видов представлена окончатыми эластическими
мембранами в количестве от 14 до 20 размером 5-6 мкм. Они слабо
фрагментированы, почти прямые, неразветвленные. Между ними имеются
гладкомышечные волокна, расположенные циркулярно, а ближе к адвентиции -
продольно. Количество их также небольшое - от 16 до 30. Продольно
расположенные мышечные волокна единичны, а циркулярные преобладают.
У более активных и энергичных зверьков - соболя (Martes zibellina), куниц
лесной (М. martes) и каменной (М. foina) - увеличивается как толщина средней
оболочки, так и диаметр просвета сосуда. Отношение толщины медии к
184
просвету аорты заметно возрастает и составляет 1 : 9-12. Эластические
мембраны средней оболочки более извилисты и фрагментарны. Толщина
эластических мембран 5-10 мкм, количество их 20-30. Между мембранами
имеется сеть тонких эластических волокон, образующих подобие войлока, в
котором находятся веретенообразные гладкомышечные клетки и коллагеновые
волокна.
У крупного представителя семейства - росомахи (Gulo gulo) - средняя
оболочка достаточно развита и занимает до 90% толщины стенки. Отношение
ее к просвету дуги аорты близко к таковому у соболя и куниц (1 : 9-10). Вся
медия состоит из эластических мембран, плотно прилегающих друг к другу и
слабо фрагментарных (рис. 51). Ширина эластических мембран 5-10 мкм, но
количество их больше, чем у предыдущих видов, - 58-60. Между ними
циркулярно расположены гладкомышечные клетки. На границе с адвентицией
эластические мембраны медии истончаются, и в этих местах небольшими
пучками лежат продольно идущие гладкомышечные волокна. Они делают
стенки сосуда более прочными.
У сравнительно малоактивного, постоянно живущего в глубоких норах
барсука (Meles meles) в отличие от росомахи интима отделена от средней
оболочки присутствующим в ней внутренним эластическим слоем толщиной
10-12 мкм. Вся медия плотно заполнена тонкими эластическими мембранами
размером до 6-7 мкм, количество которых больше, чем у других видов, - 60-
62. В некоторых местах они плотно соприкасаются. Ближе к адвентиции, как и
у росомахи, гладкомышечные клетки образуют пучки продольно распо-
ложенных волокон размером 300x80 мкм, отдаленных друг от друга на 400-
700 мкм. Здесь эластические мембраны заметно истончаются. По количеству
гладкомышечных волокон в медии (25-40) барсук явно превосходит остальных
представителей семейства. Отношение к просвету аорты толщины стенки этого
сосуда у него равно 1 : 7-8, а медии - 1 : 9-10.
Микростроение дуги аорты мелких полуводных видов - европейской и
американской норок (М. lutreola и М. vison) - мало чем отличается от такового
наземных видов. Отношение к просвету толщины стенки дуги аорты и медии у
них оказалось практически одинаковым. Оно составляло соответственно 1 : 4-
5 и 1 : 6-8, т.е. было таким же, как и у других представителей рода Mustela.
Толщина эластических мембран колебалась от 3.5-5 мкм у европейской норки
до 4-6 мкм у американской. Их количество у рассматриваемых видов заметно
разнилось и составляло соответственно 33-35 и 22-28.
У специализированных амфибионтов - речной выдры (Lutra lutra) и
калана (Enhydra lutris) - средняя оболочка дуги аорты занимает 70-82% ее
стенки. Отношение размеров медии к просвету было равно соответственно
1:19 и 1:17, а всей стенки сосуда к внутреннему диаметру - 1:12-15. Размер
эластических мембран у речной выдры 6-8, а у калана 10-12 мкм. У обоих
видов они почти прямые, усилены коллагеновыми волокнами, особенно в
периферическом слое. Как и в случае с норками, с увеличением толщины
эластических мембран их количество уменьшалось. Так, у рассматриваемых
185
амфибионтов количество данных элементов в медии выдры было равно 37-
39, а у калана 21-23.
Таким образом, сравнительно-гистологический анализ показал, что у
подавляющего большинства видов семейства куньих тканевое строение
стенок дуги аорты довольно однотипно. Она представляет собой сосуд
эластического типа с хорошо выраженной средней оболочкой, состоящей в
основном из эластических мембран. Более тонкими (4-6 мкм) они оказались
у представителей рода Mustela, а самыми мощными (10-12 мкм) - у калана.
Наибольшее количество эластических мембран (58-62) содержалось в медии
росомахи и обыкновенного барсука, а наименьшее (14-16) -у ласки и солонгоя.
Интима и адвентиция построены однообразно. Небольшие различия касаются
структуры средней оболочки: расположения, количества гладкомышечных
волокон и эластических мембран, а также степени разветвленности
последних.
Необходимо отметить, что у активных представителей семейства куньих
дуга аорты имеет широкий просвет и относительно тонкую стенку, состоящую
из большого количества эластических элементов и незначительного -
мышечных. В то же время у барсука, отличающегося меньшей по сравнению
с другими видами активностью и постоянными физическими нагрузками
при копании, в средней оболочке дуги аорты увеличивается масса мышечных
элементов (Коршунова, Туманов, 1979).
Венозная система хищных млекопитающих характеризуется густой сетью
сосудов, наличием в них тонкой стенки и большого просвета. По данным
М.Ф. Ковтуна и Л.П. Осинского (1985), по мере приближения к сердцу вены
утончаются еще больше. В их стенке хорошо развита наружная оболочка,
толщина которой становится заметно меньше вблизи сердца. В медии
рассматриваемых сосудов имеется небольшое количество гладкомышечных
волокон, но основной материал, из которого построена венозная стенка, - это
волокнистая соединительная ткань и прежде всего коллагеновые элементы.
Как сообщают авторы, при сопоставлении относительной толщины стенки
основных венозных магистралей передних и задних конечностей у предста-
вителей семейства собачьих существенных различий не наблюдается. Например,
у волка диаметр плечевой вены больше толщины ее стенки в 31.4, а бедренной -
в 33.3 раза. Особенно тонкостенным сосудом передней части туловища зверя
следует признать краниальную полую вену. Толщина ее стенки меньше калибра
сосуда в 48 раз. У других видов собачьих данная венозная магистраль имеет
более массивную стенку. У енотовидной собаки отношение размеров
краниальной полой вены к ее просвету равно 1: 41, а у обыкновенной лисицы -
1 : 32. Из трех оболочек, образующих стенку вены, лучше всего развита
наружная. У хищников она составляет от 53 до 73% толщины ткани сосуда.
Средняя оболочка развита меньше (27-44%), а внутренняя не превышает 1.5-2%
ширины стенки вены. По мере следования крови от периферии к центру отмечается
некоторое уменьшение относительной толщины адвентиции и увеличение медии.
Однако размеры интимы остаются практически неизменными.
186
Рис. 52. Венозные магистрали обыкновенной полевки (а) и желтогорлой мыши (б).
1 - наружная яремная вена; 2 - подключичная вена; 3 - передняя полая вена;
4 - сердце; 5 - задняя полая вена в грудной полости; 6 - печеночная вена;
7 - задняя полая вена в брюшной полости; 8 - почечная вена; 9 - пояснично-
подвздошная вена.
Fig. 52. Venous main ways of common vole (Microtus arvalis) (a)
and yellow-necked mouse (Apodemus flavicollis) (6).
1 - external jugular vein; 2 - clavicular vein; 3 - anterior hollow vein; 4 - heart;
5 - posterior hollow vein in pectoral cavity; 6 - hepatic vein; 7 - posterior azygous vein
in abdominal cavity; 8 - nephric vein; 9 - lumbar-iliac vein.
Внутренняя оболочка состоит из эндотелиальных клеток и волокон
соединительной ткани, а средняя - из спиралевидно расположенных гладко-
мышечных клеток. Между их рядами в небольшом количестве встречаются
элементы волокнистой соединительной ткани. Внутренняя эластическая
мембрана очень тонкая и не во всех венах четко выражена. Среди
представителей семейства собачьих она в основном прослеживается в
краниальной полой вене, а у обыкновенной лисицы - еще и в плечевом сосуде.
В большинстве других вен ее заменяет густая сеть эластических волокон. В
толще средней оболочки они имеют продольную направленность, а отдельные
короткие нити - спиралевидную. Таким образом, основные отличия в строении
сосудов центрального и периферического кровообращения собачьих состоят
в различных структурных характеристиках артерий и вен, обусловленных
особенностями поведения и уровнем двигательной активности животных
(Ковтун, Осинский, 1985).
187
Таблица 54/ Table 54
Наружный диаметр венозных сосудов у грызунов с массой тела 40-70 г
(средние данные)
External diameter of venous vessels of rodents with body mass 40-70 g (average data)
Виды/ Species Масса тела, г / Body mass, g Вена, мм/Уеиа, mm
наружная яремная/ jugularis externa Подклю- чичная/ subclavia задняя полая/ cava caudalis
у входа в печень/ near entry to liver в грудной полости/ in pectoral cavity
Наземные/Terrestrial animals
Соня лесная/ Dryomus nitedula Летяга/ 43 1.6 1.3 2 1.7
Pteromys volans Полевка Прометеева/ 70 2.5 1.4 2.2 Полуподземные/Semi-fossorial animals 1.6
Prometheomys schaposchnikovi 70 4.5 3.3 3.4 1.4
Полевка Брандта/ Microtus brandti Хомячок 58' 4.2 3 3.3 1.5
джунгарский/ Phodopus sungorus Слепушонка 41 3.8 2.9 Подземные/Fossorial animals 3.7 1.3
обыкновен./ Ellobius talpinus 65 2.8 1.2 2.4 2
Схема ветвления основных венозных магистралей хищных млеко-питающих
в общих чертах сходна с таковой у грызунов. У последних приспособительная
перестройка вен наиболее основательно изучена на полуводных видах (Mosch,
1951; Koch, 1953; Андреева, 1954, 1956; Галанцев, 1967, 1977). Согласно
проведенным исследованиям у этих животных в связи с нырянием, во время
которого, как известно, выключается внешнее дыхание и происходит резкое
замедление сердечной деятельности, строение венозной системы приобретает
адаптивные черты, не свойственные наземным формам. Венозные сосуды
полуводных грызунов превращаются в объемистые коллекторы, депонирующие
кровь на период нахождения под водой.
По материалам В.П. Галанцева (1977), у грызунов-амфибионтов (от
водяной полевки до бобра) топография венозных магистралей, характер
развития венозных коллекторов и их локализация имеют много общего. В
плечешейной области зверей самыми крупными коллекторами являются
наружные яремные и подключичные вены, между которыми устанавливаются
анастомозы. Их каудальная полая вена имеет мощный просвет на всем ее
протяжении в пределах брюшной полости. Особенно заметно увеличивается
188
Таблица 55/ Table 55
Наружный диаметр венозных сосудов у грызунов с массой тела 250-280 г
(средние данные)
External diameter of venous vessels of rodents with body mass 250-280 g
(average data)
Виды/ Species Масса тела, г/ Body mass, g Вена, мм/ Vena, mm
наружная яремная/ jugularis externa подклю- чичная/ subclavia задняя полая/ cava caudalis
у входа в печень/ near entry to liver в грудной полости/ in pectoral cavity
Белка обыкновенная/ Sciurus vulgaris Наземные/Terrestrial animals 265 2.5 1.5 3.5 3.2
Полуподземные/Semi-fossorial animals
Суслик крапчатый/ Citellus suslicus 250 5.2 3.0 3.9 1.4
Хомяк обыкновен./ Cricetus cricetus 255 3.8 3.0 3.7 1.3
Тушканчик большой/ Allactaga major 276 4.1 3.1 4.3 1.5
Подземные/Fossorial animals
Слепыш обыкновен./ Spalax microphthalmus 257 2.4 1.2 3.2 2.4
Слепыш белозубый/ Spalax leucodon 266 2.1 1.3 2.7 2.0
Цокор алтайский/ Myospalax myospalax 278 2.2 1.3 3.2 3.0
калибр этого сосуда краниальнее устья почечных вен, а максимально он
расширяется в области печени.
Значительные венозные образования обнаружены нами и у роющих
грызунов, ведущих полуподземный образ жизни (Туманов, 1966а, б, 1967).
При изучении их кровеносной системы обращает на себя внимание значи-
тельная диспропорция артериального и венозного русла. Так, задняя полая
вена обыкновенной полевки и желтогорлой мыши у печени превосходит по
своему диаметру брюшную аорту в том же месте в 2.3-2.5 раза. Особенно
большая разница (в 5-6 раз) наблюдается у этих зверьков в диаметре сонной
артерии и наружной яремной вены, а также подключичных артерии и вены.
Анализ строения венозных магистральных сосудов у различных
представителей отряда грызунов (полевок, мышей, сусликов, сурков) показал,
что наиболее развитые коллекторы обнаруживаются только у животных,
ведущих полуподземный образ жизни и использующих норы как временное
убежище (рис. 52). Например, у прометеевой полевки диаметр наружных
яремных вен равен 4.3-4.6, а подключичных - 3.2-3.5 мм, в то время как у
189
Рис. 53. Основные венозные магистрали крота.
1- передняя полая вена; 2 - задняя полая вена в грудной полости; 3 - задняя полая
вена в брюшной полости; 4 - почечная вена.
Fig. 53. The main venous ways of mole (Talpa europaea).
1- anterior hollow vein; 2 - posterior hollow vein in pectoral cavity; 3 - posterior
hollow vein in abdominal cavity; 4 - nephric vein.
видов, сходных по массе тела, но не связанных с обитанием в глубоких норах,
он был заметно меньше (табл. 54).
Следует также отметить, что и у мелких, и у крупных по размеру грызунов,
использующих норы как временное убежище, в грудной полости за диафрагмой
задняя полая вена резко сужается и ее диаметр в среднем в 2.5-3 раза меньше,
чем перед входом в печень. У наземных видов, не использующих норы
(обыкновенная белка, летяга и др.), диаметр задней полой вены в грудной
полости только в 1.2-1.4 раза меньше, чем перед печенью. Интересно также,
что для всех норников характерна хорошо развитая система непарных вен
(у. у. azygos), которые осуществляют связь каудальной и передних полых вен.
В отличие от видов, использующих норы в качестве временных убежищ, у
постоянно подземноживущих грызунов (слепыши, цокоры, слепушонки)
венозная система не имеет заметных расширений и по своему строению
напоминает таковую наземных видов (табл. 55). Так, у обыкновенного слепыша
задняя полая вена на всем протяжении меняется очень незначительно, и если ее
диаметр перед слиянием общих подвздошных вен равен в среднем 2.7-3 мм, то
перед входом в печень он не превышает 3.2-3.3 мм. В грудной полости задняя
190
Рис. 54. Венозные магистрали американской норки.
1 - передняя полая вена; 2 - задняя полая вена в грудной полости; 3 - задняя полая
вена в брюшной полости.
Fig. 54. The main venous ways of American mink {Mustela vison).
1 - anterior hollow vein; 2 - posterior hollow vein in pectoral cavity; 3 - posterior
hollow vein in abdominal cavity.
полая вена сужается у него очень незначительно - до 2.3-2.5 мм. Малый калибр
венозных сосудов характерен также для горного слепыша, алтайского цокора
и обыкновенной слепушенки, т.е. для тех видов, у которых подземная среда
является постоянной и единственной средой обитания.
Существенные изменения венозной системы, выражающиеся в основном в
образовании кровяного депо в различных отделах магистрального русла,
наблюдаются также у насекомоядных - куторы и выхухоли - как приспо-
собительная особенность этих млекопитающих, связанная с водной средой
обитания. Если у куторы в плечешейной области наибольшего развития
достигает наружная яремная вена, то у выхухоли емкость кровеносного русла
плечевого пояса увеличивается главным образом в результате развития
обширного венозного сплетения на дорсальной поверхности грудной клетки и
в межлопаточной области. В системе каудальной полой вены у куторы
коллектор формируется лишь в области печени за счет увеличения просвета
этого сосуда и открывающихся в него крупных печеночных вен, а у выхухоли
мощный синус обнаруживается только в преренальном отделе этой вены
(Иванова, 1964, 1975; Черняев, 1964; Галанцев, 1983). В то же время у крота,
191
который постоянно живет под землей, где и добывает себе пищу, четко
выраженные специальные депо крови не отмечаются (рис. 53), хотя задняя полая
вена в его брюшной полости имеет значительный просвет.
Таким образом, сравнительный анализ строения венозных магистралей у
грызунов и насекомоядных, ведущих полуподземный или полуводный образ
жизни, позволил выявить у них специфические расширения сосудов
(коллекторы), расположенные в плечешейной, брюшной и пояснично-тазовой
областях. Данные образования следует, по-видимому, рассматривать как
адаптивное свойство организма, связанное с периодическим пребыванием
зверьков в норах или под водой. Они могут иметь особое значение, которое
заключается в способности (благодаря разности диаметра сосудов)
регулировать давление и скорость кровотока в период замедления темпа
сердечной деятельности в измененных условиях. При этом предотвращается
переполнение кровью правой половины сердца, так как узкий просвет полых
вен в грудной полости, вероятно, способствует равномерной регуляции
поступления крови в правое предсердие.
Функциональные сдвиги в работе сердца и дыхательного аппарата, по-
видимому, явились главной причиной адаптивной перестройки кровеносной
системы животных. Одной из таких приспособительных особенностей
магистрального русла можно считать увеличение его емкости за счет
образования коллекторов - заметного расширения некоторых участков
венозных сосудов.
Газовый состав воздуха в гнездовых камерах мелких млекопитающих
может резко изменяться за счет повышенного содержания углекислоты в норах.
В результате этого у зверьков, находящихся в маловентилируемой части
убежища, происходит замедление скорости кровотока подобно тому, которое
наблюдается у амфибионтов. У животных, которые ведут полуподземный образ
жизни, обитая в двух средах (наземной и подземной), быстрая и частая смена
экологических условий сопровождается четкими изменениями со стороны
дыхания и кровообращения. Это обстоятельство, в свою очередь, в процессе
эволюционного развития могло привести к формированию венозных
коллекторов перед входом сосудов в грудную клетку. Резкое увеличение их
диаметра согласно законам гемодинамики вызывает уменьшение давления
и скорости кровотока в венах, что препятствует переполнению кровью
предсердия в момент замедления темпа сердечной деятельности у особей,
находящихся в норах. В то же время небольшой просвет полых вен в грудной
полости, по-видимому, способствует лучшей регуляции крови, поступающей
к правой половине сердца.
У представителей наземно- и подземноживущих видов, которые все время
находятся в постоянных условиях газовой среды, венозные образования не
обнаруживаются. Вполне вероятно, что отсутствие коллекторов у них связано
с деятельностью органов дыхания и кровообращения в одном режиме, при
котором равномерное течение крови не вызывает изменения диаметра сосудов.
Вместе с тем необходимо отметить, что у всех полуподземноживущих,
192
Таблица 56/ Table 56
Наружный диаметр некоторых венозных сосудов в грудной и плечешейной
областях у взрослых представителей семейства куньих (средние данные)
External diameter of some venous vessels iu pectoral and arm-cervical areas of mustelid
adult representatives (average data)
Виды/ Species Вены, vMlVena, mm
передняя полая/ cava cranialis наружная яремная/ jugularis externa внутренняя яремная/ jugularis interna подключичная/ subclavia
Ласка/ Mustela nivalis 3.1 2.3 1.3 0.8
Г орностай/ Mustela erminea 3.1 2.7 1.3 1.3
Хорек лесной/ Mustela putorius 5.5 3.4 2.2 3.2
Норка америк./ Mustela vison 6 3.6 2.1 2.8
Куница лесная/ Martes martes 5.4 3.5 2 2.8
Соболь/ Martes zibellina 5.2 3.2 1.9 3.3
Барсук/ Meles meles 13.5 5.8 3.5 4.6
Выдра/ Lutra lutra 10.2 3.5 2.6 4.4
Калан/ Enhydra lutris 12.8 4 2.8 4.2
Росомаха/ Gulo gulo 12 4.8 3 4.2
питающихся на поверхности почвы и использующих норы в качестве надежных
убежищ, морфологические преобразования кровеносной системы не достигли
такого совершенства, как у амфибионтов.
Исследование венозной системы животных показало (Туманов, 1974а),
что у хищных млекопитающих в отличие от роющих и полуводных видов
из отрядов грызунов и насекомоядных отчетливо выраженные коллек-
торы в их плечешейной области отсутствуют (рис. 54). Венозная кровь из
краниального отдела туловища по передней полой вене собирается в правое
предсердие. Если у грызунов передние полые вены (v. v. cava craniales) парные,
то у Carnivora это один ствол, диаметр которого у животных бывает различным.
Например, в ряду видов из семейства куньих он увеличивается от 2.5-3.5 мм
(ласка, горностай) до 10-14.6 мм (росомаха, калан). Передняя полая вена
хищников образуется от слияния наружной и внутренней яремных (y.v.jugulares
externae et internae), а также подключичных вен (v. v. subclaviae). С дорсальной
стороны в нее впадают сосуды, собирающие кровь от грудной клетки.
Наружные яремные вены принимают кровь от челюстных вен - v.v.
maxillares externa et interna. Они лежат поверхностно под кожей, представляя
193
Рис. 55. Венозные магистральные сосуды барсука.
1 - наружные яремные вены; 2 - подключичные вены; 3 - передняя полая вена.
Fig. 55. The main venous ways of badger {Meles meles).
1 - external jugular veins; 2 - clavicular veins; 3 - anterior hollow vein.
собой узкие, длинные сосуды, диаметр которых почти не изменяется на всем
протяжении. У рассматриваемых зверей преобладание поверхностных вен над
глубокими заметно не только по относительно большему диаметру, но и по
более широким зонам их ветвления (Мажуга, 1966). У представителей
семейства куньих калибр наружных яремных вен в среднем в 1.5-2 раза больше
внутренних, которые проходят магистрально по шее вдоль общих сонных
артерий. Интересно, что для калана и выдры характерно наличие многих
анастомозов между яремными и челюстными венами, образующими общую
густую сеть кровеносных сосудов.
На уровне первого грудного позвонка в наружные яремные вены впадают
надключичные. Изменение их диаметра в ряду видов незначительно - от 2 до
4 мм. От плечевого пояса и передних конечностей кровь собирается в
подмышечные, а затем в подключичные вены (v. v. subclaviae), наружный диаметр
которых у хищников-амфибионтов в отличие от грызунов не превышает этот
показатель у наземных куньих со сходной массой тела (табл. 56).
По нашим наблюдениям, в плечевом поясе полуводных видов каких-либо
функциональных расширений или сплетений венозных сосудов не наблюдается
и этот отдел их венозного русла практически не отличается от такового других
представителей данного семейства, ведущих наземный, лазающий или норный
образ жизни.
194
l’iic- 56. Задняя полая вена барсука.
1 - задняя полая вена в брюшной полости; 2 - почечные вены
Fig. 56. Posterior hollow vein of badger (Meles meles).
1 - posterior hollow vein in abdominal cavity; 2 - nephric veins.
С каудальной стороны туловища в правое предсердие впадает задняя полая
вена - V. cava caudalis. Ее начало образуют общие подвздошные вены (v.v.
iliacae communes) и непарная средняя крестцовая вена (v. caudalis), собирающие
кровь от задних конечностей, хвоста и органов тазовой области.
В брюшной полости задняя полая вена представляет собой сосуд с
большой площадью поперечного сечения. У выдры и калана на участке до
почечных вен он может быть представлен двумя стволами, после слияния
которых диаметр данной магистрали заметно увеличивается. Задняя полая
вена принимает в себя кровь из сосудов, идущих от брюшной стенки,
половых органов, почек, печени и диафрагмы. На уровне пятого
поясничного позвонка в нее впадают парные пояснично-подвздошные
вены - v.v. circumflexae ilium profunda. Однако у барсука они обычно
подходят непосредственно к общим подвздошным венам, хотя встречаются
особи и с характерным для других куньих ветвлением названных сосудов.
Пояснично-подвздошные и общие подвздошные вены у этого вида пред-
ставляют собой значительное кровяное русло. Следует отметить, что у
барсука в связи с особенностями его экологии как задняя, так и передняя
полые вены представляют собой крупные сосуды с большой площадью
поперечного сечения (рис. 55-56).
Диаметр v. cava caudalis у изучаемых животных несколько увеличивается
краниальнее почечных вен до диафрагмы. Особенно это характерно для калана,
195
Таблица 57/ Table 57
Наружный диаметр (мм) некоторых венозных сосудов в брюшной и пояснично-
тазовой областях у взрослых представителей семейства куньих (средние данные)
External diameter (mm) of some venous vessels in abdominal and lumbar-sacral areas of
mustelid adult representatives (average data)
Виды/ Species Вена, мм/Vena, mm
почечная/ renalis непарная/ azygos воротная/ portae задняя попая/cava caudalis
в грудной полости/ in pectoral cavity в брюшной полости/ in abdominal area
У печени/ at liver У бифуркации/ at bifurcation
Ласка/ Mustela nivalis 1.4 2.3 1.7 2.5 3.2 1.9
Горно- стай/ Mustela ertninea 1.5 2.6 1.7 2.1 3.5 2.4
Хорек лесной/ Mustela 3.3 4.7 3.1 5.3 5.8 4.5
putorius Норка америк./ Mustela vison 4.1 5.1 4.6 5.2 6.6 4.3
Куница лесная/ 3.5 4.3 3.3 5.5
Martes 4.3 4.5
mattes Соболь/ Martes 3.4 4.2 3.1 4.3 5.4 4.4
zibellina Барсук/ Meles meles Выдра/ 6.5 5.2 5.1 11.6 16.3 10.8
Lutra lutra Калан/ 8.5 1.5 5.7 11.4 17.2 10.3
Enhydra lutris 9.2 1.2 6 11.5 18.6 10.7
Росомаха/ Gulo gulo 6.3 5.1 5 11.2 13.5 9.6
выдры и барсука (табл. 57). У них наружный диаметр данной магистрали
заметно возрастает на отрезке между почечными венами и печенью. Если у
выдры калибр задней полой вены в месте впадения в нее общих подвздошных
вен равен 7.3-12.7 мм, то перед входом в печень-уже 14.2-20.5 мм.
196
Рис. 57.Венозные магистрали горностая.
1 - передняя полая вена; 2 - задняя полая вена в грудной полости; 3 - задняя полая
вена в брюшной полости.
Fig. 57. The main venous ways of ermine {Mustela erminea).
1 - anterior hollow vein; 2 - posterior hollow vein in pectoral cavity;
3 - posterior hollow vein in abdominal cavity.
Следует отметить, что у барсука и полуводных видов прослеживается
значительная диспропорция в диаметре артериального и венозного русла.
Например, их задняя полая вена у печени превосходит по своему диаметру
брюшную аорту в том же месте в 2.8-3.6 раза, в то время как у наземных форм -
только в 2-2.4 раза.
Для выдры и калана характерна также большая площадь поперечного
сечения почечных и печеночных вен - v. v. renales et hepaticae, впадающих в v.
cava caudalis, которая вместе с этими сосудами представляет собой объемистый
резервуар венозной крови. По наблюдениям М.И. Урманова (1971), у калана,
как и у ластоногих, в области дорсального края печени имеется расширенный
отрезок каудальной полой вены (печеночный синус), обусловленный
адаптацией сосудистого русла этих животных к полуводному образу жизни. У
речной выдры и норок обоих видов подобные образования нами не обнаружены.
В грудной полости задняя полая вена куньих в отличие от грызунов и
насекомоядных сужается очень незначительно (рис. 57). Более четко этот
перепад обозначен только у таких специализированных к водной среде обитания
видов, как калан и выдра. У них просвет задней полой вены в грудной полости
в среднем в 1.5-1.6 раза меньше по сравнению с той частью сосуда, которая
197
лежит за ее пределами, образуя синусообразное расширение. Обращает на себя
внимание и относительно большой калибр непарных вен (v. v. azygos) у этих
видов. Как и почечные вены, они могут служить вместительным депо венозной
крови при нырянии животных.
Размеры воротной вены (v. portae) у куньих невелики, и ее диаметр перед
впадением в печень у барсука, росомахи, выдры и калана не превышает
таковой брюшного отдела аорты. У американской норки она имеет больший
диаметр по сравнению с воротной веной хорька и куницы, что, видимо, связано
с экологическими особенностями этого вида.
Как свидетельствует В.П. Галанцев (1977, 1983), у калана в начале
грудного участка каудальной полой вены за счет диафрагмальных мышц
формируется сфинктер, который неполным кольцом охватывает ее наружную
поверхность. Функциональная роль этого мышечного образования
заключается, по-видимому, в регуляции кровотока во время ныряния и после
него. Весьма значительное синусообразное расширение каудальной полой
вены в области печени вместе с мощными впадающими в нее печеночными
венами у калана и выдры можно рассматривать как наиболее выражен-
ный венозный коллектор. При этом система непарных вен обеспечивает
у них тесную связь каудальной и краниальной полых вен с внутрипозво-
ночными.
По мнению автора, единая задача депонирования венозной крови при
нырянии и соответствующее увеличение емкости венозной системы многими
видами решалась по-своему. Даже относительно близкие в систематическом
отношении животные по-разному реагировали в процессе филогенетического
освоения водной среды свойственными им формообразовательными
процессами в сосудистой системе. В то же время у видов из отрядов грызунов и
хищных млекопитающих прослеживается определенное сходство в развитии
морфологических преобразований гомологичных структур сосудистого русла
(т.е. параллелизм), интенсивность которых обусловлена уровнем эколо-
гической специализации животных к нырянию и добыванию пищи в воде.
Вполне понятно, что стратегия природных адаптаций организма хищных
млекопитающих к водной среде обитания и жизни в глубоких норах не
ограничивается структурным преобразованием только сосудистой системы.
Вместе с тем она является одним из основных ее элементов. Выявленная
адаптивная перестройка кровеносных магистралей имеет основную
направленность на быструю доставку к тканям работающих органов запасов
кислорода и рациональное их использование.
Таким образом, рассматриваемые виды, близкие в таксономическом и
филогенетическом отношении, имеют как сходные черты, так и существенные
различия в характере ветвления и строении сосудистого русла. При этом
отмеченные преобразования определяются прежде всего образом жизни и
поведением животных, в том числе и особенностями их локомоторной
деятельности.
198
Резюме
Адаптивные реакции сердечной мышцы хищных млекопитающих тесно
связаны с особенностями строения кровеносной системы, которая обеспечивает
доставку кислорода и питательных веществ к различным органам. В строении
сосудистого русла наблюдается строгая взаимосвязь морфофункциональных
признаков, отражающих уровень приспособления видов к условиям
существования. У хищников выявлены четко выраженные различия в строении
прежде всего артериальной сети, которые обусловлены особенностями
экологии и поведения животных. Эти адаптивные изменения проявляются по-
разному, но имеют одну направленность - на интенсивное кровоснабжение
тканей работающих органов. Они хорошо прослеживаются на примере
семейств Mustelidae и Canidae, т.е. зверей, обладающих широким спектром
экологической адаптаций.
Особенности экологии хищных млекопитающих, представители
которых ведут активный образ жизни, находят наиболее яркое отображение в
строении их кровеносной системы. Обладая разнообразными формами
локомоций и высокими показателями суточных перемещений, они имеют
некоторые специфические черты в строении артериального русла. Эти
особенности позволяют улучшить кровоснабжение тканей локомоторного
аппарата быстро передвигающихся животных. Обычно у представителей
активных видов отмечаются относительно короткая аорта, ответвление от нее
артерий под более острым углом и прямолинейный ход этих сосудов, что
обеспечивает наибольшую скорость тока крови к мышцам конечностей.
Сравнительно-гистологический анализ показал, что у подавляющего
большинства видов из семейства Mustelidae тканевое строение стенок аорты
однотипно. Она представляет собой сосуд эластического типа с хорошо
выраженной средней оболочкой, состоящей в основном из эластических
мембран. Более тонкими (4-6 мкм) они оказались у представителей рода
Mustela, а самыми мощными (10-12 мкм) - у калана. Наибольшее количество
эластических мембран (58-62) содержалось в средней оболочке у росомахи и
барсука, а наименьшее (14-16)-у ласки и солонгоя.
Функциональные сдвиги в работе сердца и дыхательного аппарата
являются важной причиной адаптивной перестройки и венозной системы
животных. Одной из таких приспособительных особенностей магистрального
русла можно считать наличие у роющих и полуводных видов крупных
венозных сосудов с большой площадью их поперечного сечения перед входом
в грудную полость. Данные вместительные образования (коллекторы) способны
депонировать венозную кровь во время ныряния полуводных хищников или
нахождения в глубоких норах роющих форм, обеспечивая им высокую
резистентность к гипоксии.
Понятно, что стратегия природных адаптаций организма хищных
млекопитающих к водной среде обитания и жизни в глубоких норах не
ограничивается структурными изменениями только в сосудистой системе. В
199
комплексе различных преобразований последняя является одним из основных
ее элементов. Выявленная адаптивная перестройка артериальных магистралей
имеет основную направленность на быструю доставку к тканям работающих
органов запасов кислорода, а венозных сосудов - к регуляции венозного
кровотока. При этом отмеченные морфофункциональные преобразования
определяются прежде всего образом жизни и особенностями поведения
животных.
Summary
The adaptive reactions of heart muscle of carnivores are in close
correspondence with characteristic features of circulatory system structure, which
provides delivering of the oxygen and nutritional substances to various organs. A
strict interrelation of morphofunctional characters, reflecting the level of species
adaptation to certain environmental conditions is noticeable in its transformations.
Pronounced differences in the structure of arterial net in the first turn, are typical
of them. These differences are stipulated by ecological and behavioral
characteristics of animals. These adaptive changes are variously displayed, but
are aimed at intensive blood supply of tissues of working organs and most distinct
in the families of Mustelidae and Canidae, i.e. animals with a wide spectrum of
ecological radiation.
Characteristic features of ecology of carnivorous mammals with an active
mode of life are reflected in the structure of the circulatory system. They posses
various forms of locomotion and high parameters of daily movements and, hence,
some characteristic features in the structure of arterial flow. The essence of these
features is in the more intensive oxygen and nutritional substances supply of
locomotory organs’ tissues as compared to species with a less active mode of life.
A relatively short aorta with arteries branching at a more acute angle and
rectilinear way of these vessels are commonly recorded in the active animals. All
these provide the greatest velocity of blood flow to the limb muscles.
Comparative histologic analysis shows that in the overwhelming majority of
species of the family Mustelidae the tissue structure of the walls of aorta is of one
type. It is a vessel of elastic type with a well-pronounced middle membrane,
mainly composed of elastic membranes. They appeared to be thinner (4-6 mm) in
representatives of the genus Mustela, and most powerful (10-12 mm) in Enhydra
lutris. The greatest number of elastic membranes (58-62) was found in the middle
membrane of Gulo gulo and Meles meles, the least (14-16) in Mustela nivalis and
M. altaica.
The functional shifts in the work of the heart and respiratory apparatus are
the important cause of adaptive transformation of venous system of animals. The
formation of collectors — notable extension of some areas of venous vessels may
be considered to be one of such adaptive features of the mainstream flow. These
capacious structures (collectors) are able to deposit venous blood during diving in
semiaquatic carnivores, providing them high resistance to hypoxia.
200
The strategy of natural adaptations of carnivorous mammals organism to the
water environment and life in deep holes is not limited to the structural changes
only in the blood-vessel system. At the same time it is one of its elements. The found
adaptive transformation of circulatory ways is mainly directed toward the fast
delivery of oxygen resources to the tissues of working organs and their rational use.
Therewith, the morphofunctional transformations recorded in the first turn are
determined by the mode of life and behavior of animals.
8 Зак. № 4309
Глава 6. ЭКОЛОГИЧЕСКАЯ
ОБУСЛОВЛЕННОСТЬ РЕПРОДУКТИВНЫХ
ФУНКЦИЙ
Многие виды хищных млекопитающих заметно различаются между собой
по времени наступления половой зрелости, срокам гона, плодовитости и
продолжительности процесса беременности. В ходе эволюционного развития
у них сложились определенные биологические ритмы, которые позволяют
приносить приплод и воспитывать молодняк в благоприятные по клима-
тическим и пищевым факторам сроки. Представители одних видов
спариваются весной; другие, у которых беременность протекает с длительной
задержкой имплантации зародышей, проявляют готовность к гону в более
поздние сроки - в начале или в течение всего лета. Однако за счет разной
продолжительности беременности рождение детенышей в естественных
условиях происходит у большинства животных в весенне-летнее время.
Необходимо, правда, отметить, что самки обоих видов медведей и росомахи
приносят малышей в теплых берлогах и логовах в зимнее время, но покидают
их звери только весной - в конце марта или апреле. Следовательно, самки
хищных млекопитающих имеют возможность, благополучно, с малыми
энергозатратами пережив трудные зимние месяцы, приурочить выращивание
молодняка к богатому кормами весенне-летнему сезону. По окончании
репродуктивного периода, заметно потеряв в весе, они успевают до наступления
холодов перелинять и создать необходимое количество энергозапасов, что
снижает зимой теплоотдачу организма в окружающую среду.
Данные биоритмы сохраняются и у таких специализированных к водной
среде обитания хищников, как калан и речная выдра. Они характеризуются
существенной разницей в сроках гона и продолжительности беременности,
равной у калана 8-9 мес, а у речной выдры 2-2.5 мес, но рождение молодых
представителей обоих видов наблюдается в основном в теплое время года
(Барабаш-Никифоров и др., 1968; Шило, Немшилова, 1989). Например, судя
по материалам гистологической обработки гонад, взрослые особи речной
выдры на Северо-Западе России и в Белоруссии могут спариваться в разные
сезоны, однако более 90% известных здесь случаев обнаружения в природе
гнезд со слепыми щенками приходится на апрель-начало октября (Туманов,
Сидорович, 1994). Сезонные особенности биологии и, в частности, вос-
производственных функций данного вида прослеживаются и в других частях
его ареала (Erlinge, 1967, 1968; Fairley, 1972; Вшивцев, 1972; Вайсфельд,
1977; Robin, 1989; Weber, 1990; Kruuk, 1995, и др.).
Большинство мелких хищников из семейства Mustelidae (от ласки до
хорьков и норок включительно) достигают половой зрелости и принимают
участие в размножении в конце первого года жизни, примерно в возрасте 9-12
мес. Обычно к весне молодые и взрослые особи практически не различаются
202
по размерам половых желез и массе тела. В то же время у горностая, как и у
американской длиннохвостой ласки {Mustela frenata), половая зрелость
молодых самок наступает уже в 2-3 мес и даже раньше (Hamilton, 1958; Fog,
1969; Muller, 1970; Терновский, 1972а, 1974). По-видимому, эта биологическая
особенность и позволяет горностаю иметь более широкий по сравнению с
другими видами ареал, достаточно высокую численность популяций, а также
быструю ее восстанавливаемость после периодов депрессии.
Для остальных более крупных представителей семейства характерно
относительно позднее становление половой зрелости зверей, и основная часть
особей в популяции впервые начинает участвовать в размножении только в
конце второго или на третьем году жизни. В целом прослеживается
определенная зависимость между размерами тела, энергообменом и скоростью
полового созревания животных: чем экономичнее в энергетическом отношении
представители изучаемых видов, тем позже они вступают в период полового
созревания и, следовательно, популяция в целом может медленнее восста-
навливать свою численность после депрессии.
Морфологические изменения, происходящие в репродуктивных органах
куньих и характеризующие физиологическое состояние животных в течение
года, происходят по одной схеме, хотя и сдвинуты во времени (Данилов,
Туманов, 1972, 1975а, б). Процесс созревания половых продуктов в гонадах
взрослых самцов характеризуется тремя основными периодами, приуро-
ченными к определенным сезонам: пассивный сперматогенез, активный и
стадия редукции сперматогенного эпителия. Половой цикл самок синхронен
таковому самцов тех же видов, однако время готовности их к продуктивному
спариванию довольно ограничено по сравнению с длительным периодом
активного сперматогенеза самцов. Эта особенность позволяет последним
обеспечивать максимальное покрытие взрослых самок, готовых принять
участие в размножении.
Мелкие куньи, обладающие высоким уровнем энергообмена, испы-
тывают на себе сильное воздействие неблагоприятных факторов. Видимо,
данное обстоятельство и определило в процессе их филогенеза отбор на
повышенную плодовитость, раннее половое созревание, короткую по
продолжительности беременность у большинства видов и готовность самцов
к продуктивному спариванию в течение сравнительно длительного периода.
Сказанное относится прежде всего к ласке, солонгою, колонку, хорькам и
норкам. Наблюдения за мелкими хищниками при их содержании в вольерах
(Ленинградская обл.) показали, что в период с сентября по январь ни самцы,
ни самки не демонстрируют своей готовности к спариванию. Заметное
увеличение размеров репродуктивных органов и способность самцов к
размножению прослеживали у американской норки в феврале-марте, лесного
хорька и европейской норки - в марте-июле, а горностая - в апреле-июле
(табл. 58). В отдельные годы самцы ласки могли находиться в состоянии
половой активности с марта по август, а лесной куницы - с конца мая и до
середины августа.
203
Таблица 58/ Table 58
Продолжительность репродуктивного периода у самцов мелких хищников,
содержащихся в неволе
Duration of reproductive period of small carnivore males, kept in captivity
Виды/ Species Годы/ Years Готовность к спариванию (+) по месяцам/ Readiness to copulation (+) by month
11 | III IY Y YI Yll | YIII
Ласка 1986 4- 4- 4- 4- + +
обыкновен./ 1987 + 4- 4- 4- 4-
Mustela 1988 4- 4- 4-
nivalis 1989 4- 4- 4-
Горностай/ 1986 4- 4- 4-
Mustela 1987 4- 4- 4- 4-
erminea 1988 4- 4- 4-
1989 4- 4- 4- 4-
Хорек 1986 4- 4-
лесной/ 1987 4- 4- 4- 4-
Mustela 1988 + 4- 4- 4- 4-
putorius 1989 4- 4- 4-
Норка 1986 + +
америк./ 1987 + +
Mustela vison 1988 4-
1989 + +
Норка 1986 4- 4- 4-
европ./ 1987 + 4- 4-
Mustela 1988 4- 4- 4-
lutreola 1989 4- 4- 4- 4-
Куница 1986 4- 4- 4-
лесная/ 1987 4- 4- 4-
Martes 1988 4- 4- 4-
martes 1989 + 4- 4-
Интенсивный гон в условиях неволи наблюдался у американских норок во
второй половине февраля и в двух первых декадах марта, у европейских - с
марта по май, а у лесных хорьков - с марта и до начала июня (Туманов,
Сорина, 1997а, б). Довольно поздно вступали в стадию готовности к гону
взрослые самки горностая. У них состояние проэструса регистрировали обычно
в марте-апреле, а способность к спариванию - во второй половине апреля-
мае (табл. 59).
Наибольшая продолжительность репродуктивного периода прослеживалась
у ласок. При вольерном содержании зверьков фазу проэструса отмечали у самок
в разные годы в конце марта-мае, а эструса - в апреле-начале августа. Позднее
всех приходили в состояние репродуктивной активности лесные куницы, у
которых, судя по анализу вагинальных мазков, проэструс наступал в мае-июне,
а эструс - в июне-июле и до начала августа в разные годы.
204
Таблица 59/ Table 59
Продолжительность репродуктивного периода у самок мелких куньих,
содержащихся в неволе
Duration of reproductive period of small mustelid females, kept in captivity
Готовность к спариванию (+) по месяцам/
о ИДЫ/ i иды/ Years Readiness to copulation (+) by month
II | III | IY | Y | YI | YII | YIII
Ласка 1986 + + + +
обыкновен./ 1987 + +
Mustela nivalis 1988 + +
1989 + + +
Горностай/ 1986 + +
Mustela erminea 1987 + +
1988 + +
1989 + +
Хорек лесной/ 1986 + +
Mustela putorius 1087 + + + +
1988 + +
1989 + + +
Норка америк./ 1986 + +
Mustela vison 1987 +
1988 +
1989 +
Норка европ./ 1986 + + +
Mustela lutreola 1987 + + +
1988 + + +
1989 + + +
Куница лесная/ 1986 + +
Martes martes 1987 + +
1988 + +
1989 + +
Беременность у европейской норки протекает без задержки имплантации.
Впервые ее продолжительность точно установили в Московском зоопарке
(Рубецкая и др., 1933). По этим данным самка, покрытая 22-25 апреля,
принесла щенков 6 июня, т.е. через 42-45 дней. В наших опытах роды у
европейской норки проходили в конце мая-июне, а длительность беременности
(по 5 выводкам) составляла 40-43 дня. Следует отметить, что у самок этого
вида, как и у лесного хорька, в период половой охоты влагалище увели-
чивается в диаметре в несколько раз по сравнению с состоянием покоя, что
может служить косвенным показателем их готовности к спариванию. У
американских норок подобные морфологические изменения выражены крайне
слабо.
Самки американской норки приносят щенков обычно в конце апреля-мае.
Диапазон продолжительности беременности у них заметно больше такового у
аборигенного вида. В опытах Р.В. Клера (1941) она составляла 40-65 дней,
205
Д.В. Терновского (1977) - 45-56, М.Д. Абрамова (1974) - от 40 до 73 дней. В
Ленинградском (Санкт-Петербургском) зоопарке известны 3 случая, когда
самки, спаривавшиеся в период с 15 по 19 февраля, приносили щенков два
раза 1-го и один раз 7-го мая. Продолжительность беременности у них
составляла 71-75 дней. Максимальная длительность вынашивания эмбрионов
выявлена у горностая. По свидетельству Д.В. и Ю.Г. Терновских (1994), у
этого вида она колеблется от 224 до 393 дней (и=96).
Как показал гистологический анализ репродуктивных органов у мелких
куньих, добытых на Северо-Западе России, гон в естественных условиях также
проходит в обозначенные выше сроки (Данилов, Туманов, 1972, 1976а, б).
Таким образом, морфологические изменения, наблюдаемые в половых органах
изучаемых хищников и характеризующие их физиологическое состояние в
течение года, имеют циклический характер, следуя по единой схеме, хотя и
сдвинуты во времени.
Необходимо отметить и тесную взаимосвязь периодов половой активности
с энергетикой питания представителей видов. Так, у всех рассматриваемых
животных начало гона сопровождалось повышением уровня показателей
ректальной температуры, массы тела и суточной потребности в пище.
Последняя у беременных и воспитывающих молодняк самок продолжала
оставаться высокой в течение всего летнего сезона.
В монографии Г.А. Новикова (1939) имеются ссылки на работы, в которых
сообщается о возможностях повторного гона и щенения у европейских норок.
По данным В.Я. Генерозова (1922), подобное явление имеет место и у скунса
{Mephitis mephitis), самки которого приносили молодых обычно один раз в
год, «однако бывали случаи и вторичного помета». О повторном спаривании
и щенении степных и лесных хорьков {Mustela eversmanni, М. putorius) после
гибели первого выводка сообщали многие исследователи (Афанасьев и др.,
1953; Rempe, 1957; Строганов, 1962).
По свидетельству Н.М. Курис (1972), самки колонка {М. sibirica) при
содержании в условиях неволи способны приходить в эстральное состояние
по 2-3 раза за сезон размножения. Повторность эструса наступала после
гибели щенков первого помета или при несостоявшемся оплодотворении во
время предыдущего покрытия. Автор сообщает о случаях, когда у 4 самок
на 4-10-е сутки после гибели приплода вновь наблюдались характерные
признаки течки. Повторно покрытые, все они благополучно щенились и
вырастили потомство. Кроме того, еще 2 самки из опытного поголовья,
которые уже нормально щенились и некоторое время кормили молодых
детенышей, после их отсадки спаривались вновь, но щенков не принесли. В
том же году Д.В. Терновский (19726) опубликовал материалы о том, что в
1971 г. от одной самки колонка в вольерах Института биологии СО РАН
были получены 2 помета, однако детеныши сразу же погибли из-за отсутствия
молока у матери. Все это свидетельствует о большой лабильности
репродуктивной цикличности самок данного вида и о значительном диапазоне
изменчивости сроков их гона.
206
В Ленинградском (Санкт-Петербургском) зоопарке в 1983-1988 гг. гон
колонков регулярно наблюдался в конце апреля-мае. Если после первого
спаривания беременность не наступала, то самки через 14-15 дней вновь
приходили в состояние половой охоты. В 1987 г. продуктивное спаривание
зверьков отмечалось 3-5 мая, а в 1988 г. - 24-28 апреля. В первом случае щенки
родились 5 июня (5<5 и 2$), во втором - 3 мая (2 <5 и 3 $). Несмотря на ранние
сроки родов и короткую по продолжительности беременность (33 и 36 дней по
первому покрытию), т.е. наличие условий для полноценной реализации их
репродуктивного потенциала, самки, вырастив молодняк, повторно в
астральное состояние не приходили.
Широкий диапазон репродуктивных возможностей демонстрируют и
другие мелкие хищники. Так, 8 апреля 1968 г. пойманная в Казахстане
беременная самка степного хорька (Л/, eversmanni) родила в неволе 9 щенков
(4 <5 и 5 $?). Семью доставили в Ленинградский (Санкт-Петербургский) зоопарк,
но в дороге начался падеж молодняка, и к 18 апреля все детеныши погибли.
Через неделю самка повторно пришла в состояние эструса и была готова к
спариванию. Известен также случай, когда в Каунасском зоопарке у пары
степных хорьков коитус наблюдался 9 и 10 марта 1973 г., а через 38 суток (16
апреля) в гнезде самки были обнаружены 2 (1 <5 и 1 §) мертворожденных
детеныша. Спустя 1-2 дня ее вновь соединили с самцом, и 29 мая от них был
получен повторный приплод - 5 <5 и 5 $ (Туманов, 1977).
В 1981-1983 гг. у содержащихся в Ленинградском (Санкт-Петербургском)
зоопарке степных хорьков гон проходил в одни и те же сроки - в марте-апреле.
В 1983 г. они спаривались 4-6 апреля, all мая (длительность беременности 37
суток) самка щенилась. В гнезде находились 8 детенышей (3 <5 и 5 $?). Как и в
случае с колонками (1987-1988 гг.), благополучно вырастив потомство, она
повторно в процесс размножения не включалась.
Близкий родственник колонка и степного хорька - солонгой (М altaica)
представляет собой сравнительно редкий для фауны России вид, который
хорошо размножается в неволе, обладает короткой по продолжительности
беременностью и высокой плодовитостью. У содержащейся в нашей вольере
пары солонгоев в конце марта 1972 г. были отмечены признаки гона и
спаривание. Вскоре вес самки заметно увеличился. Она редко выходила из
гнезда и, судя по внешнему виду и поведению, была беременна. Однако по
невыясненным причинам, видимо, произошла резорбция эмбрионов, и роды
не наступили. В первых числах мая того же года самка вновь пришла в
астральное состояние. 3 мая она была покрыта и 10 июня (через 38 дней)
принесла приплод - 5 самцов и 5 самок. Выращенный молодняк был передан
нами в Ленинградский (Санкт-Петербургский) зоопарк, где 23 мая 1973 г. два
уже годовалых солонгоя спарились. Через 40 дней (2 мая) молодая самка
родила 13 детенышей (5 <5 и 8 $), которые по разным причинам погибли в
возрасте 20-25 дней. Примерно через неделю после смерти щенков она вновь
стала очень беспокойной, а взятые вагинальные мазки подтвердили ее
повторную готовность к размножению.
207
В 1974 г. от той же пары вновь удалось получить помет (4 и 3$). Коитус
зверьков наблюдался 28 и 29 марта, а рождение щенков - 8 мая, т.е.
беременность длилась 40-41 день. Вскоре (24 мая) весь приплод погиб, а уже
30-31 мая самка повторно спаривалась с тем же самцом. Вторично щенки (3
и 4 ?) родились у нее 11 июля и в дальнейшем развивались вполне нормально.
Подобный же случай имел место и в Каунасском зоопарке, где двухлетняя
самка солонгоя, покрытая 28 февраля 1973 г., принесла первый помет 2 апреля.
На другой день щенков в гнезде нашли мертвыми, а через неделю она
спаривалась вторично. Беременность развивалась нормально, но 17 мая по
чистой случайности самка погибла. При вскрытии в рогах матки было
обнаружено 7 крупных эмбрионов на последней стадии развития.
По мнению Ю.А. Баевского (1968) и С.А. Корытина (1974), многие виды
потенциально способны размножаться более интенсивно, чем обычно.
Доказательством наличия скрытых периодов полового возбуждения у хищных
млекопитающих, как справедливо отмечают авторы, могут служить факты
получения 2 пометов в год и различные отклонения в сроках гона у лисиц,
песцов, рысей, скунсов, енотовидных собак и енотов-полоскунов. Некоторые
животные (с короткой по продолжительности беременностью) обладают
подобными репродуктивными особенностями, которые необходимо исполь-
зовать в хозяйственных целях.
В этом плане интересные материалы по увеличению биологической
продуктивности самок ряда видов мелких куньих (с продолжительностью
беременности до 1.5 мес) приводятся в работах Д.В. Терновского (19726,
1977). В них автор сообщает о фактах получения у самок искусственным путем
(методом ранней отсадки щенков) по 2 приплода за один сезон размножения.
Среди куньих повторное состояние готовности к спариванию (в случае гибели
или отсадки молодняка) прослеживалось у хорьков, колонка, европейской
норки, ласки и солонгоя.
Приведенные данные позволяют считать, что у многих хищных млеко-
питающих с коротким сроком беременности могут иметь место повторно^
спаривание и рождение молодняка в течение одного сезона размножения, т. е.,
по нашему определению (Туманов, 1977а, б, в), они обладают потенциальной
полиэстричностью. Получение второго помета возможно только от самок тех
видов, в эмбриональном развитии которых отсутствует длительная диапауза.
По-видимому, и в природных условиях в случае рассасывания эмбрионов или
гибели молодняка на ранних этапах постнатального онтогенеза часть самок
вновь приходит в состояние половой охоты, спаривается и дает второй приплод.
Этим можно объяснить факты добычи в природе поздней осенью щенков
некоторых видов хищных млекопитающих (хорьков, енотовидных собак,
лисиц и песцов) в возрасте 2-3 мес. Например, на Северо-Западе России гон у
лесного хорька (М. putorius) протекает в марте-апреле, а массовое щенение
наблюдается в апреле-мае (Данилов, Русаков, 1969; Новиков и др., 1970;
Данилов, Туманов, 1972, 1975а, б). Беременность у хорька, по нашим
наблюдениям при вольерном содержании (6 выводков), продолжается 40-43
208
дня. Вместе с тем известны случаи отлова в сентябре-конце октября молодых
хорьков весом 250-300 г, т.е. не старше 3 мес. Так, в 1971 г. 10 сентября к нам
поступили пойманные охотниками в природе 2 детеныша (<$ и $) этого вида.
Они весили 270 и 250 г и имели мягкий темно-серый ювенильный волос. У
хорьков только заканчивалась смена временных зубов на постоянные, что
соответствовало возрасту 2-2.5 мес. В следующем году егерь Западного
отделения ВНИИ охотничьего хозяйства и звероводства также поймал в конце
октября в Псковской области 2 молодых самцов лесного хорька весом по 300 г,
т.е. опять же в возрасте не более 2.5-3 мес.
Выявленные случаи повторного участия самок некоторых видов хищных
млекопитающих в процессе размножения (причем даже особей, благополучно
вырастивших потомство) свидетельствуют о значительных их репродуктивных
возможностях. Такая лабильность физиологического состояния позволяет
зверькам быстро реагировать на действия факторов внешней среды и,
приспосабливаясь к новым условиям существования, изменять сроки и
интенсивность размножения. Все это дает основание считать данные виды
близкими к полиэстричным животным, а отмеченный феномен получения
естественными путем двух приплодов за сезон размножения следует
рассматривать как один из механизмов регуляции численности популяции и
возможность увеличения уровня биологической продуктивности самок в
зверохозяйствах.
К интересной особенности репродуктивной цикличности представителей
рода Martes следует отнести явление так называемого «ложного гона». В
настоящее время не вызывает сомнения, что течка и спаривание у соболей и
куниц происходит в июне-первой половине августа, тем не менее природу
ложного гона исследователи трактуют по-разному. Согласно Р.В. Клеру (1941,
1948), его начало совпадает с началом роста фолликулов и имплантацией
зародышей у беременных самок после латентной стадии. Другие авторы
совместное появление весной следов самцов и самок соболей или куниц
связывают с образованием семейных пар, которое следует сразу же после
имплантации зародышей (Донауров, 1949; Павлюченко, 1962; Граков, 1964).
В процессе многолетних наблюдений за поведением лесных и каменных
куниц в Ленинградском (Санкт-Петербургском) зоопарке нам удалось
выявить, что в отдельные годы некоторые особи начинают испытывать весной
сильное возбуждение. Поведение их в этот период в точности соответствует
играм во время гона. Самец бегает за самкой, делает садки, но коитуса не
происходит. Судя по собранным данным, половое возбуждение зверьков имеет
место только в летний или весенний сезоны. В первом случае спаривание
происходило в обычные для куниц и соболей сроки - в июне-начале августа. В
это время нам неоднократно приходилось наблюдать у зверьков коитус
продолжительностью от 15-20 до 40 мин, а в некоторых случаях и до 1.5 ч. В
период же весеннего возбуждения куниц (конец февраля-март) отмечались
только кратковременные садки самцов на самок без коитуса, т.е. без
фактического спаривания. Часто весной после проявления взаимного интереса
209
зверьки начинали драться, и их приходилось рассаживать в разные клетки. В
монографии В.В. Тимофеева и В.Н. Надеева (1955) сказано, что ложный гон
у соболей бывает не ежегодно. Это подтверждается и нашими наблюдениями
за поведением куниц в неволе.
Анализируя собранные материалы, мы пришли к выводу, что предположения
большинства авторов о природе ложного гона ошибочны. В конце зимы-ранней
весной его признаки можно отметить и у куниц, никогда ранее не спаривавшихся.
В эти сроки сексуальное поведение наблюдалось как у молодых (около 2 лет),
впервые готовящихся к гону зверьков, так и у старых особей, которые в
предшествующие годы по разным причинам не приходили в охоту и не
спаривались. Следовательно, происхождение ложного гона не зависит от
имплантации зародышей у беременных самок или от образования семейных пар.
В этой связи заслуживает внимания работа Т.В. Шварц (1974), в которой
показаны изменения активности яичников и щитовидной железы у молодых
самок соболей в возрасте до 15-16 мес. Функциональная активность
генеративных органов прослеживалась на гистологических срезах, которые
демонстрировали изменения диаметра зрелых полостных фолликулов и
размера клеток teca interna, обусловленные специфической гормональной
деятельностью гонад. По материалам автора, прослеживаются 2 пика
активности яичников - в марте и в июне-июле. По данным Р.В. Клера (1941),
в конце февраля-начале марта у некоторых самок соболей и куниц происходят
энергичный рост фолликулов, набухание слизистой оболочки матки и
ороговение эпителия влагалища, т.е. наблюдается состояние, характерное
для периода проэструса.
По-видимому, ложный гон представляет собой атавистическое явление,
близкое по природе к осеннему токованию тетеревиных птиц. Идея измене-
ния интенсивности размножения под влиянием оледенения обсуждалась
в литературе неоднократно, о чем подробно сообщалось в работе
Ю.А. Баевского (1968). При этом автор считает вполне вероятной не только
возможность редукции числа репродуктивных циклов, но и вторичное их
восстановление в областях наибольшего потепления климата (например, в
Западной Европе).
С.А. Корытин (1974) предполагает наличие двух сроков гона и двух
пометов в год у предков соболей и куниц, обитавших в условиях мягкого
климата третичного периода. В дальнейшем, в четвертичном периоде, вслед
за оледенением произошло похолодание климата и ухудшение кормовой базы.
По мнению автора, это могло привести к трансформации половой циклики
животных и угасанию одного из периодов размножения, в результате чего гон
стал проходить в наиболее благоприятные сроки. В развитии зародыша
появилась длительная эмбриональная диапауза, способствующая повышению
выживаемости молодняка, рождаемого весной. Понятно, что подобного рода
адаптация, направленная на сохранение вида, вместе с тем вела к снижению
интенсивности размножения за счет уменьшения числа репродуктивных циклов
во время воспроизводительной активности особей. Таким образом,
210
потенциальная полиэстричность животных, по-видимому, обусловлена
особенностями их исторического развития.
Интересно, что мелкие виды куньих имеют довольно продолжительный
репродуктивный период, который у многих видов длится с весны и до конца
лета. Обладая малыми размерами и относительно очень высоким уровнем
метаболизма, они в большей степени испытывают на себе воздействие
неблагоприятных факторов внешней среды. Это, вероятно, в значительной
степени и определило в процессе филогенеза большую их плодовитость в
сравнении с крупными хищниками, а также способность повторно продук-
тивно спариваться в случае гибели приплода.
За время гона и выращивания молодняка самки представителей семейства
куньих расходуют значительную часть энергозапасов. Их масса тела за период
от начала гона и до отделения молодняка снижается в среднем на 20-25%, т.е.
становится ниже веса, при котором нормально протекает овуляция (Курис,
Бакеев, 1974). Скорее всего, в природных условиях второй помет самки
мелких куньих могут приносить в основном в случае рассасывания эмбрионов
или гибели щенков на ранних этапах постнатального онтогенеза. Однако это
не исключает возможности получения двух жизнеспособных пометов в год от
одной самки при клеточном разведении не только мелких, но и других хищных
млекопитающих.
Рассматриваемые виды заметно различаются между собой по срокам гона
и родов, продолжительности беременности самок и плодовитости. Биология
размножения куньих изучена с разной степенью подробности, причем для
отдельных представителей семейства, обитающих в России, до сих пор
сохраняется необходимость в освещении фактического материала о мало-
известных, интимных сторонах их жизни. Это относится, прежде всего, к
обыкновенной ласке, перевязке, харзе, росомахе, выдре, барсуку и куницам.
Обыкновенная ласка (AT. nivalis). В условиях вольерного содержания у
нас под наблюдением находилось в общей сложности 15 самцов и 15 самок,
из последних только 12 участвовали в размножении. Все они отлавливались в
Пол/
Таблица 60/ Table 60
Динамика массы репродуктивных органов у сеголеток ласок (мг, М±т)
Dynamics of reproductive organs mass of weasel (Mustela nivalis) youngs
Месяцы/Months
Sex
YII YIII IX X XI XII
Females (3) 65±8'4 (6) 68±8.8 (5) 52±4.2 (3) 65±4.5 (3) 63±9.6 (1)70
Самцы/ (3) 74±4.2 (16)76±5.8 (5)46±4.I (3) 25±4.2 (3)28±2.1 (3) 32±2.4
Males
Примечание. Здесь и далее цифры в скобках обозначают количество просмотренного
материала.
211
Таблица 61/ Table 61
Динамика массы репродуктивных органов у взрослых ласок (мг, М±т)
Dynamics of reproductive organs mass of adult weasels {Mustela nivalis)
Пол/ Месяцы/Months
Sex I-II III-IY Y-YI YII-YIII IX-X X1-XII
Самки/ Females (3) (2) (7) (4) (3)
80±5.4 280±8.6 138±8.4 102±9.1 77 ±4.2
Самцы/ (4) (4) (7) (17) (14) (7)
Males 135±6.8 211±9.4 232±8.8 130±6.9 88±10.6 85±5.1
Ленинградской и Псковской областях. За период с 1978 по 1987 г. в условиях
неволи было получено 13 приплодов с 56 щенками (25<? и 31$).
Анализ материала, поступившего от охотников и кротоловов, в ловушки
которых нередко попадались ласки, показал, что молодые самки и самцы
начинают принимать участие в размножении только в конце первого года
жизни. У самок сеголеток, добытых в июле-сентябре (возраст 2- 4 мес) матка
и яичники несут в себе черты инфантильности и весят в среднем в 2 раза меньше,
чем у взрослых зверьков. Осенью они практически не меняются в размерах, но
уже к началу зимы вес репродуктивных органов взрослых и молодых самок
довольно близок (Туманов, 1988в).
Самцы сеголетки, отловленные в июле-августе, имели несколько увеличенные
семенники по сравнению с осенне-зимним периодом (табл. 60), однако в это время
их вес был в среднем в 1.5-2 раза меньше, чем у взрослых животных. В связи с
растянутостью сроков гона и рождения детенышей в конце августа в природе
могут встречаться молодые ласки с семенниками разного веса, но чаще всего он
колеблется в пределах от 40-50 до 70-80 мг. В то же время известны случаи добычи
молодых самцов, у которых гениталии достигали 100-10 мг, т.е. по массе они
практически мало отличались от таковых у половозрелых особей. Интересно, чтб
в августе 1985 г. в Ленинградской области был отловлен молодой самец, который
оказался крипторхом. Его единственный семенник весил 70 мг.
Наблюдения за содержащимися в вольерах ласками показали, что молодые
самцы из поздних выводков имеют в августе-сентябре мелкие недоразвитые
семенники. У зверьков из ранних пометов в это время они хорошо видны, а в
их поведении проявляются элементы сексуального характера (преследование
самок, садки, характерная вокализация), хотя к продуктивному спариванию
сеголетки еще не готовы. К концу февраля-марту молодые и взрослые самцы
практически не различаются по размерам тела и половых желез.
У взрослых особей заметное увеличение массы семенников наблюдается
уже в январе-феврале. В этот период у добытых животных они весят 100-160 мг,
т.е. бывают примерно в 1.6-1.8 раз больше, чем в декабре. Однако готовые
половые продукты в гонадах самцов впервые обнаруживаются только во II
212
Таблица 62/ Table 62
Время спаривания, родов и величина выводков у обыкновенной ласки
в условиях неволи
The time of copulation, labour and litter size of weasel (Mustela nivalis)
in conditions of captivity
№ самки/ N of female Возрастная группа, возраст, лет/ Age groups, age, years Дата/Date Продолжит, беременности, сут./ Gestation period, day Детеныши/Cubs
спаривания/ copulation родов/ birth всего/ total
1 Взрослая/ Adult 12-14.05.78 18.06.78 37-35 6 3:3
1 Взрослая/ Adult 1-3.08.78 5.09.78 35-33 4 2:2
2 1 12-15.05.79' - - -
2 1 27-30.06.79 - - - -
2 2 15-18.05.80 - - - -
2 3 23-26.04.81 30.05.81 37-34 5 1:4
2 4 29.04.82 15.06.82 37 3 2:1
2 4 19.06.82 23.07.82 34 5 2:3
3 Взрослая/ Adult - 7.08.80 - 8 4:4
4 1 7-8.06.82 13.07.82 36-35 5 3:2
5 2 29.04-1.05.83 4.06.83 36-34 4 2:2
5 2 11-12.06.83 - - - -
5 3 29-31.05.84 - - - -
6 2 11-13.05.87 17.06.83 37-35 1 0:1
6 2 29-30.06.87 - - - -
7 1 19-20.05.87 24.06.87 36-35 4 2:2
8 1 6-7.06.87 11.07.87 35-34 4 1:3
9 1 9-12.06.87 - - - -
10 1 29.06-1.07.87 - - - -
11 1 23-24.06.87 26.06.87 33-32 1 0:1
Взрослая/
12 Adult 23-24.06.87 29.06.87 37-36 6 3:3
половине марта. В апреле-июне генеративные органы наиболее крупные, а в
их канальцах хорошо заметны сперматозоиды. В июле размеры семенников
заметно снижаются, но еще до конца августа в биопробах встречаются особи
с весом гениталий в 170-190 мг, т.е. таким же, как в марте.
У добытых в природе взрослых самок наибольший вес органов размно-
жения отмечался в мае-июле (табл. 61). Матка зверьков в эти месяцы была
213
заметно увеличена и гиперемировала, а в яичниках имелись хорошо
выраженные желтые тела, что свидетельствовало о прошедшей овуляции.
Беременные самки попадались чаще всего в июле. Их матки с эмбрионами и
яичниками в этом месяце весили 1250-1520 мг. Судя по всему, для ласок
характерен высокий коэффициент эмбриональной смертности. Так, при
вскрытии 3 отловленных в июле беременных самок было обнаружено 10,7 и 3
эмбриона, из которых соответственно 4,3 и 1 находились в состоянии резорбции.
Таким образом, средняя доля рассасывающихся эмбрионов в этом случае
составила 40%. К середине августа органы размножения самок заметно
уменьшались в размерах и в дальнейшем до конца зимы находились в состоянии
функционального покоя.
Содержащиеся в вольерах ласки размножались и приносили приплод
регулярно, однако в 3 случаях молодые самки в возрасте около года весной
участия в размножении не принимали и вскоре погибли по разным причинам.
Спаривание зверьков всегда проходило в одни сроки - в конце апреля-начале
августа, но больше всего в мае-июне (табл. 62). Обычно взрослые самки
демонстрировали состояние половой готовности раньше молодых. Во время
гона коитус можно было наблюдать в любое время суток, но чаще ранним
утром и вечером. При совместном обитании ласки спаривались регулярно в
течение 2-4 дней, потом либо переставали интересоваться друг другом, либо
самка отказывалась подпускать самца. В этот период коитус регистрировался
от 1 до 3 раз в сутки. Какой-либо закономерности в частоте спаривания и его
продолжительности не отмечалось. Коитус мог длиться от 5-15 мин до 1 ч 45 мин
и даже до 3 ч. В среднем по 33 точно зафиксированным наблюдениям его
продолжительность составляла у ласок 47 ± 7.3 мин.
Новозеландский исследователь Л. Хартман (Hartman, 1964) и американские
зоологи Дж. Хейт с соавторами (Heidt et al., 1968) смогли получить по 2-3
приплода в год от одних и тех же самок ласок, впервые установив, что период
беременности у этого вида не превышает 35-37 дней. Ласки относятся к видам с
короткой и сравнительно консервативной беременностью. По нашим наблю-
дениям, она равна 33-37 дням. Чаще всего роды у зверьков отмечали в утреннйе
(с 5 до 10) и реже в вечерние (с 10до23)часы. При этом их длительность составляла
от 40 до 60 мин. В случае резорбции эмбрионов или гибели молодняка самки
через 1-2 недели снова приходили в охоту и спаривались повторно. Интересно,
что в 1978 г. одна взрослая самка принесла второй приплод после уже
благополучно выращенных щенков. Первые роды у нее были 18 июня. Самка
родила 3 самцов и 3 самок, которых выкормила, и, когда детенышам было
42-43 дня, она снова пришла в состояние охоты. Второй раз она спаривалась
1-3 августа, а 5 сентября у нее родились 4 щенка. Вполне вероятно, что и в природе,
вблагоприятные по условиям годы, рано покрытые взрослые самки способны
вырастить молодняк и вновь в том же сезоне принять участие в размножении.
Отмеченные репро-дуктивные особенности ласок, по-видимому, позволяют виду
чутко реагировать на изменения внешних факторов, при наступлении благо-
приятных в кормовом и климатическом отношении годов быстро увеличивать
214
Таблица 63/ Table 63
Время спаривания, сроки родов и величина выводков у лесной куницы и харзы,
содержащихся в неволе
The time of copulation, labour and litter size of pine marten (Martes martes) and Indian
marten (Martes flavigula), kept in captivity
Виды/ Species Дата/Date Продолжит, беременности, сут./ Gestation period, day Детеныши/Cubs
спаривания/ copulation родов/ birth всего/ total
Лесная куница/ 20-23.07.85 19.03.86 242 3 1:2
Martes martes 7-8.07.86 28.03.87 264 1 0:1
26-27.07.87 23.03.88 240 3 3:0
24-26.07.88 26.03.89 245 3 2:1
Харза/ 18-21.08.65 19.04.66 244 2 1:1
Martes flavigula 25-29.10.86 27.03.87 153 1 1:0
численность популяции и широко расселяться по природным территориям. По
устному сообщению Д.В. Терновского, попытка скрестить закавказскую ласку с
обыкновенной закончилась неудачей: крупный самец загрыз мелкую самку, хотя та и
находилась в состоянии эструса.
Содержащиеся в вольерах две взрослые самки таксономически близкого к ласке
вида - перевязки (Vormelaperegusnd) - приходили в состояние половой охоты в
весеннее время. Просмотренные нами вагинальные мазки зверьков
свидетельствовали о готовности их к размножению в апреле-начале мая. В
то же время в Новосибирском зоопарке гон у перевязок отмечался в июле,
роды - в марте-апреле следующего года, а эмбриональный период равнялся
250-270 дням. Половозрелость у самок наступала в возрасте одного года
(Шило, Батенева, 1989).
Все представители рода Martes в большинстве своем достигают половой
зрелости на третьем году жизни, хотя некоторые самки демонстрируют
готовность к размножению в возрасте 1.5 лет. Так, в популяциях лесной куницы
их доля составляет 10-15% и более (Старков, 1947; Данилов, 1967).
Гистологический анализ гонад взрослых самцов лесной куницы (Martes
martes) показал, что зрелые сперматозоиды появляются в семенниках в конце
мая, а атрезия эпителия извитых семенных канальцев начинается в конце
августа. Таким образом, сперматозоиды постоянно присутствуют в
гениталиях самцов в течение июня-августа, когда они обычно и способны к
продуктивному спариванию.
Половая циклика самок синхронна таковой самцов. Макро- и
микроизменения половой сферы зверьков, готовящихся к размножению,
становятся заметны в конце февраля-начале марта. У молодых, впервые
215
вступающих в фазу размножения куниц наблюдается активизация поло-
вой сферы, выражающаяся в утолщении слизистой влагалища до 4-6 слоев
эпителиальных клеток. Яичники содержат уже довольно крупные расту-
щие фолликулы, размеры которых увеличиваются с 330-450 мкм в январе-
феврале до 550-580 мкм в марте. У беременных самок существенные
изменения половых органов отмечаются в феврале, когда начинается
подготовка к имплантации зародышей. Уже в середине февраля слизистая
матки на поперечном разрезе имеет древовидный характер (типичная
реакция на прогестрон), маточные железы активно секретируют, заметна
общая гиперемия тканей. Функционирующие желтые тела беременности
достигают 1300-1500 мкм в диаметре. Подготовка к имплантации и сам
этот процесс, очевидно, продолжаются до середины-конца марта, переходя
затем в собственно беременность. В это время слизистая матки очень
извилиста, желтые тела достигают наибольшего размера (1700-1800 мкм),
а их лютеиновые клетки крупные и четкие. Беременность после имплантации
зародыша продолжается около полутора месяцев (Данилов, Туманов, 1972,
1976а).
В Ленинградском (Санкт-Петербургском) зоопарке спаривание лесных куниц
в возрасте 2 лет и более (1972-1987 гг.) наблюдалось в интервале с 20 июня по 4-7
августа. Из 10 известных нам случаев 3 раза гон проходил в конце июня, 5 - в
июле и 2 раза - в начале августа. В эти же сроки готовность к размножению
демонстрировали самки данного вида и на биостанции ВНИИОЗ (г. Киров), где
удалось получить кидусов - гибрид лесной куницы и соболя (Граков, 1969 а, 1970).
Во время спаривания коитус зверьков продолжется от 2-3 до 40 мин и даже до
1.5 ч. В среднем (п=9) он составлял 26 мин. Беременность самок длилась 240-
264 дня (табл. 63), при этом роды наблюдались всегда утром - не позднее 11ч.
Сезон размножения у каменной куницы (Martesfoina) несколько превышает
таковой у лесной и может продолжаться до середины сентября. Так, 2 самки,
родившиеся в неволе, поступили в Ленинградский (Санкт-Петербургский)
зоопарк летом 1984 г. В состояние половой охоты они пришли в возрасте 1.5
лет. Одна самка была готова к размножению в период с 7 по 20 августа, вторая -
со 2 по 10 сентября. При подсадке к самцам спаривание зверьков состоялось,
но по неясным причинам обе самки остались не покрытыми. Судя по
вагинальным мазкам, в 70-80-х годах XX в. другие самки каменных куниц в
зоопарке приходили в состояние половой активности в конце июня-июле. По
наблюдениям в Воронежской области (Рябов, Бойко, 1982) гон у каменных
куниц в вольере отмечался в июне-начале августа, а продолжительность
беременности составляла 234-279 дней. Наши (совместные с сотрудником
зоопарка И.В. Корнеевым) попытки скрестить их с лесными куницами остались
безуспешными. Зверьки разных видов просто не реагировали друг на друга. В
Новосибирске на экспериментальной базе Института биологии СО РАН опыты
по соединению в период гона самца соболя и самки каменной куницы
закончились дракой зверьков и гибелью последней (Терновский, Терновская,
1994).
216
У дальневосточного представителя рода Martes - харзы (Martes flavigula)
гон происходит в несколько иные сроки по сравнению с лесной и каменной
куницами. Так, в Новосибирском зоопарке продуктивное спаривание зверей
отмечалось с 19 октября по 25 апреля, а половозрелость самок наступала в
2.5 года (Шило, Батенева, 1989). Из 9 известных нам случаев гона харз в
Ленинградском (Санкт-Петербургском) зоопарке за период 1965-1987 гг. по
3 раза он приходился на август и октябрь, а по 1 - на ноябрь, декабрь и
февраль. За это время самки дважды приносили детенышей, при этом
продолжительность беременности составляла 153 и 244 дня (см. табл. 61).
Росомаха (Gulo gulo) также относится к видам, у которых в развитии
эмбрионов прослеживается длительная эмбриональная диапауза. Собранный
в природе и проанализированный нами материал (Данилов, Туманов, 1975а,
б) показывает, что половой зрелости самцы и самки достигают на третьем
году жизни. У молодых самцов (1+), добытых в феврале-марте, признаков
активизации половой системы отмечено не было. Гистологический анализ
гениталий молодых самок, добытых в эти же сроки, также не обнаружил в них
каких-либо заметных изменений в сторону активизации овогенеза.
Специалисты, изучающие размножение росомахи на Аляске, показали,
что все взрослые самцы, добытые в ноябре-январе, не были готовы к гону,
зато в мае-июне они демонстрировали наибольшую активность сперматогенных
процессов (Wright, Rausch, 1955; Rausch, Pearson, 1972).
Материалы по половому циклу взрослых самцов росомахи (6 особей) из
северных регионов Европейской России свидетельствуют о том, что в январе-
феврале гонады зверей находились в фазе пассивного сперматогенеза. Зрелых
половых продуктов в них не было. Средний диаметр извитых семенных
канальцев не превышал 90-100 мкм. Однако уже в середине марта-апреле их
диаметр увеличивался до 170-187 мкм, а в канальцах придатка семенников
наблюдалось большое количество сперматозоидов.
Взрослая самка, пойманная охотниками в середине февраля, была
беременной. Ее половая система характеризовалась следующими признаками:
матка гиперемирована, ее мышечные стенки утолщены, в яичниках имелись
большие (до 1500 мкм) функционирующие желтые тела беременности и крупные
атрезирующие полостные фолликулы. Судя по состоянию половой сферы, в
это время у нее могла происходить имплантация зародышей. В то же время две
другие взрослые самки, добытые в сереДине-конце марта, находились в
состоянии, близком к проэструсу: общий вес репродуктивных органов достигал
13-15 г, а длина рогов матки - 93-108 мм. В яичниках обеих самок наблюдалось
много фолликулов на разных стадиях развития - от примордиальных (50-
60 мкм) до близких к граафовым пузырькам (700-1000 мкм), а ткань этих органов
была пронизана заметно увеличенными в диаметре кровеносными сосудами
(Данилов, Туманов, 1976 а).
Зоолог А.В. Кречмар (устное сообщение) на Северном и Центральном
Таймыре неоднократно отмечал наличие парных следов росомахи в
середине мая, а добытая им в конце июня взрослая самка имела на рогах
217
матки три отчетливых плацентарных пятна, свидетельствующих о
прошедших родах. Кроме того, проводимый нами анализ вагинальных
мазков двух взрослых росомах, содержащихся в неволе, показал их
готовность к спариванию в июле. На основании всего вышеизложенного
можно сделать заключение, что гон у представителей этого вида на Севере
России, как и в других частях ареала (Krott, 1959, 1960; Raush, Pearson,
1972), приходится на конец весны-лето.
По устному сообщению (1987 г.) финских сотрудников из Хельсинкского
зоопарка, у приобретенных ими двух самок росомахи, отловленных в
Мурманской и Архангельской областях, половозрел ость наступила на третьем
году жизни. Всего здесь было получено 6 приплодов с 12 щенками (6<?и 6$>).
Спаривание животных наблюдалось в конце мая-начале июня, а роды - в
последней декаде февраля-марте, т.е. беременность длилась 8-9 мес. Звери
содержались в больших вольерах, где самки в снегу делали родовые берлоги.
По мнению финских специалистов, самцы, вероятно, приносят пищу самкам,
которые после родов по 1.5-2 мес не выходят из своих убежищ.
По-видимому, в те же сроки самки приносят детенышей и в естественных
местах обитания в России, о чем можно судить по срокам обнаружения зрячих
щенков (т.е. в возрасте старше 1 мес) в убежищах, вырытых в снегу, чаще в
корнях или под стволами поваленных деревьев. Так, в конце апреля 1970 г. в
Мончегорском районе Мурманской области было найдено гнездо росомахи с
2 самцами и 2 самками, а 5 мая 1973 г. в Архангельской области - с 1 самцом.
Кроме того, известен случай, когда в конце марта 1981 г. зоолог А.Н. Зырянов
(устное сообщение) в одном из урочищ заповедника «Столбы» (Красноярский
край) нашел выводок с 3 (1«? и 2$) зрячими детенышами этого вида.
Как и у других видов крупных представителей семейства Mustelidae,
большинство самцов и самок барсука (Meles meles) на Европейском Севере и
Северо-Западе России становятся половозрелыми в двухлетнем возрасте, о
чем свидетельствует состояние гонад зверей, добытых в природе. В то же время
в Центральной Европе, Англии и Швеции основная часть молодых зверей
участвует в размножении уже на втором году жизни (Notini, 1948; Harrisqn,
Neal, 1956; Canivenc, 1966; Conivenc et al., 1968; Stubbe, 1970; Kruuk, 1989).
Известно также, что и в Московском зоопарке молодые самцы покрывали
самок в возрасте полутора лет (Осмоловская, 1948).
Анализ собранного в полевых условиях материала показал, что взрослые
самцы после выхода из нор (конец марта-апрель) имеют увеличенные
гениталии весом до 16.7 г. В их семенных канальцах прослеживаются все
клеточные элементы, включая зрелые спермин. В последующие месяцы (май-
июль) наблюдается наиболее активная стадия цикла - массовое появление
зрелых половых продуктов. Вес семенников достигает максимума - 20-22 г.
Активный сперматогенез у барсуков, обитающих в северных и северо-
западных регионах европейской части России, продолжается до окончания
первой декады августа, а затем затухает. Уже в сентябре состояние семенных
канальцев гонад самцов свидетельствует о глубокой атрезии герминативного
218
Таблица 64/Table 64
Показатели размножения барсуков в Московском зоопарке в 1932 - 1933 гг.
Parameters of reproduction of badgers {Meles meles) in Moscow Zoo in 1932 - 1933
Дата/Date Продолжительность беременности, сут./ Gestation period, day Детеныши/Cubs
спаривания/ copulation родов/ birth всего/ total
- 29.03.32 - 2 1:1
4.05.32 13.04.33 335 5 4:1
3-13.07.32 21.04.33 292 2 0:2
»
эпителия и о функциональном покое. Аналогичное состояние гениталий
обнаружено у них и в октябре-ноябре.
Быстрая редукция сперматогенного эпителия в семенных канальцах
барсуков имеет, по-видимому, адаптивное значение. За сравнительно короткий
промежуток времени (конец лета-осень) животным необходимо накопить
достаточное количество жировых запасов для зимовки, а физиологические
процессы, связанные с функционированием гонад, требуют увеличения
дополнительного расхода энергии (Данилов, Туманов, 1971,1972).
Анализ вагинальных мазков взрослых самок, содержащихся в разные годы
в условиях неволи, показал их готовность к спариванию в период с 20-23 мая
по 5-7 августа. При этом в 8 из 10 известных нам случаев гон зверей проходил
в июне. Пойманная в Киришском районе Ленинградской области и переданная
в зоопарк беременная самка барсука 23 марта 1983 г. родила 1 детеныша ($>),
а уже 28 июня она пришла в половую охоту и была готова к спариванию. У
другой самки, которую отловили в Чудовском районе Новгородской области
и содержали в вольерах, роды наступили 19 марта 1986 г. В выводке
находились 2 щенка (l^1 и 1$>), которых затем передали в зоопарк.
Обнаруженные в разных районах Ленинградской области в конце апреля-
первых числах мая 3 помета этого вида состояли из детенышей месячного
возраста: у них только начинали открываться глаза. В двух гнездах находилось
по 2 самца и 1 самке, а в третьем - 1 самец и 1 самка.
Интересные первичные материалы о размножении барсуков в Москов-
ском зоопарке в 1932-1933 гг. были переданы нам известным зоологом
В.И. Осмоловской. По этим данным, роды у 2 самок, содержавшихся в неволе,
проходили в марте-апреле. Беременность по первому покрытию длилась 292
и 335 дней, а средняя величина выводка составляла 3 детеныша (табл. 64).
Осмотренные нами гонады выдры {Lutra lutra) и наблюдения за
животными, содержащимися в неволе, убеждают в том, что большинство особей
включаются в процесс размножения в двухлетнем возрасте или на третьем
году жизни (Sidorovich, Tumanov, 1994). В Новосибирском зоопарке гон у
зверей зарегистрирован в период с февраля по август (табл. 65). Спаривания
проходили в воде и на суше с длительностью коитуса от 2 до 30 мин. В период
219
Таблица 65/ Table 65
Показатели размножения выдр, содержащихся в Новосибирском зоопарке
(по: Шило, Немшнлова, 1989)
Parameters of reproduction of otters {Lutra lutra), kept in Novosibirsk Zoo
Дата/Date Продолжительность беременности, сут./ Gestation period, day Детеныши/Cubs
спаривания/ copulation родов/ birth всего/total
28.02-6.03.79 7.05 68 3 3:0
29.04-2.05.79 - - - -
27.02-12.03.80 7.05 69 2 2:0
16.05-27.05.80 23.07 68 2 1:1
2.04-24.04.80 8.06 67 2 1:1
27.02-7.03.84 9.05 71 1 1:0
1984 8.06 - 2 1:1
22.07-29.07.84 23.09 63 2 1:1
27.06-5.07.85 30.08 64 1 0:1
14.05-1.06.87 26.07 73 3 2:1
9.07-4.08.87 21.09 74 2 0:2
Итого/Total 7.05-23.09 63-74 20 12:8
гона самец был способен покрыть 2-3 самок. Роды у них (4 особи) наступали в
мае-сентябре, а продолжительность беременности равнялась (по первому
покрытию) 63-74 дням (Шило, Немшилова, 1989; Шило, Батенева, 1989). На
ферме Института цитологии и генетики СО РАН (г. Новосибирск) гон у зверей
наблюдался в апреле-октябре. Длительность коитуса составляла от 20-30 мин
до 1.5 ч, а беременность в среднем - 60 дней, причем у некоторых самок
половрзрелость наступала в возрасте 21 мес (Трапезов, 1992). Интересно, что
у другого вида рода Lutra - канадской выдры {Lutra canadensis) беременность
самок колеблется от 9.5 до 12.5 мес (Liers, 1951, 1958).
Репродуктивную циклику выдр из природных популяций мы изучали
методом гистологической обработки гонад 39 самцов и 43 самок в возрасте
старше года и 20 сеголеток (10 <3 и 10$>), добытых на Северо-Западе России и
в сопредельных районах Белоруссии (Туманов, Сидорович, 1994). Семенники
молодых особей (и=19) в возрасте 0+ и 1+ неразвиты. Их масса вместе с
придатками не превышала 1.8 г, в то время как у самцов старше 2 лет их
средняя масса осенью и зимой составляла 8.2 г. В январе-мае все отловленные
взрослые самцы {п= 16) старше 2 лет находились в функционально активном
состоянии. У особи, добытой в июне, в семенных канальцах происходил
процесс, который можно охарактеризовать как начало активного спермато-
генеза. В то же время у самца, отловленного в июле, общее состояние гонад
220
было близким к половому покою. В сентябре-декабре исследованные взрослые
выдры (12 особей) имели в канальцах семенников все клеточные элементы от
сперматогониев до сперматозоидов, что свидетельствует об их готовности к
спариванию. Однако необходимо отметить неоднородность данной выборки
по репродуктивным показателям. Например, в ноябре одни самцы находились
в функционально очень активном состоянии, другие - на начальной фазе
активного сперматогенеза, а у некоторых особей наблюдался спад этого
процесса. У последних семенные канальцы в гонадах располагались
сравнительно далеко друг от друга, интерстициальная ткань была достаточно
велика, а зрелые половые продукты единичны и сохранились не во всех
канальцах.
Взрослые самки, готовые к размножению, как и самцы, находились в
возрасте старше 2 лет. Матки зверей 1-го года жизни (0+, 10 особей) были
тонкими и инфантильными. Сходная картина наблюдалась и у годовалых
самок (1+, 13 особей), которые также оказались неполовозрелыми. Их
влагалищный эпителий имел всего 2-3 слоя, а в яичниках преобладали мелкие,
реже средней величины, растущие и атрезирующие фолликулы диаметром до
400-500 мкм. Взрослая самка, поступившая к нам в январе, находилась в
состоянии проэструса, другая, добытая в начале февраля, оказалась
беременной и имела по 2 небольших эмбриона в каждом роге матки.
Среди 8 выдр, отловленных в марте-апреле, 4 оказались беременными.
Три из них имели на рогах матки по 2 крупных околоплодных вздутия, а одна -
3 достаточно развитых эмбриона. Остальные самки находились в состоянии
либо проэструса, либо эструса. В последнем случае в их яичниках
наблюдались граафовы пузырьки диаметром 1200-1500 мкм с готовыми к
овуляции яйцеклетками.
Из 6 самок, пойманных в мае-июле, 2 находились в состоянии эструса и
имели в яичниках крупные, близкие к разрыву фолликулы диаметром до
1650 мкм. Среди других зверей одна самка готовилась к родам и имела
3 крупных эмбриона, а остальные находились в фазе проэструса.
Примерно такое же состояние половой сферы демонстрировали животные
(5 особей) в августе-сентябре. В этот период встречались звери, прошедшие
овуляцию и готовящиеся к ней. К первой группе можно отнести 2 самок, в
яичниках которых были обнаружены соответственно 3 и 4 крупных желтых
тела размером 1100-1200 мкм; ко второй - остальных особей, причем у 2 из
них отмечалась только ранняя фаза активизации половой сферы: видимо, самое
начало проэструса.
Заметная половая активность прослеживалась и у 4 самок, отловленных
в октябре. В этом месяце одна выдра оказалась беременной и имела 2
околоплодных вздутия в рогах матки, а состояние гонад еще 3 животных
свидетельствовало о недавно прошедшем у них периоде половой охоты.
Из 7 самок, отловленных в ноябре-декабре, 4 имели крупную, сильно
гиперемированную матку, а в яичниках - готовые к овуляции фолликулы
размером 1500-1600 мкм. Три другие самки были беременными. У одной из
221
них в яичниках просматривались три желтых тела, а также много мелких и
средних по размеру растущих фолликулов. Она имела крупную матку массой
23 г с 3 околоплодными вздутиями на ее утолщенных рогах. Внутри плодовых
тел находились изогнутые зародыши размером до 10 мм. В половой системе
еще двух других беременных выдр были обнаружены соответственно 2 и 3
мелких эмбриона.
Судя по результатам обследования 10 упомянутых выше беременных выдр,
плодовитость самок в регионе равна в среднем 2.6 (от 2 до 4) щенка на одну
особь. При этом в природных популяциях вида выводки с 2 щенками, по-
видимому, составляют не менее 50%.
Наши материалы, многочисленные сведения, полученные от охотников-
корреспондентов и специалистов, у которых выдры разводятся в условиях
неволи, показывают, что роды у самок этого вида не имеют четкой
приуроченности к одному сроку. Однако чаще всего они происходят во второй
половине весны-начале осени (апрель-начало октября).
Собранные данные свидетельствуют о существенной разнородности
популяции выдры в репродуктивном отношении и об отсутствии у нее четко
выраженного брачного периода. Известно, что у большинства видов хищных
млекопитающих отправными моментами активизации половой сферы являются
реакции организма и прежде всего гипоталамо-гипофизарной системы на
изменения продолжительности и спектрального состава светового дня. Однако
у самок выдры готовность к размножению может наступить в разные месяцы,
что, видимо, регулируется не только отмеченными причинами, но и такими
факторами, как физиологическое состояние особей и сроки наступления их
физиологической зрелости. При этом в популяции в любой сезон года имеются
самцы, способные оплодотворять самок, которые пришли в состояние половой
активности.
Половозрелость морской выдры, или калана (Enhydra lutris), наступает на
третьем году жизни. Определенных сроков гона и рождения детенышей, по-
видимому, нет. Однако чаще брачные игры наблюдаются в период с марта по
август. Продолжительность беременности 8-9 мес, спаривание происходит в
воде и длится 3-9 мин, но роды бывают на берегу или на залитых водой камнях.
Самка приносит одного или (очень редко) двух детенышей (Ильина, 1950,1952;
Гентнер и др., 1967).
По продолжительности беременности у куньих в России достаточно
отчетливо выделяются 3 группы (Данилов, Туманов, 1976). К первой относятся
виды с длительной эмбриональной диапаузой в развитии зародыша (горностай,
перевязка, соболь, лесная и каменная куницы, харза, росомаха, барсук и калан);
ко второй - животные с относительно небольшой, но лабильной беременностью
(американская норка, возможно, речная выдра) и к третьей - хищники с
коротким, четко фиксированным периодом вынашивания щенков (ласка,
солонгой, европейская норка, хорьки, колонок). Если у самок I группы
беременность имеет продолжительность более 9-10 мес, то у американской
норки (II группа) - только 40-75 дней, а у III группы не превышает 40-45 дней.
222
Таблица 66/ Table 66
Влияние сроков спаривания на продолжительность беременности самок
некоторых видов семейства куньих (по: Териовский, 1977; Туманов, 1984)
The inflnence of time of copulation on the duration of pregnancy in some species of mustelids
Ввды/Species n Дата/Date Продолж. беремен., сут/ Gestation period, day t p
спаривания/ copulation родов/ birth
Соболь/Martes zibellina: самки 3 лет/ females 3 years old самки 5 лет/ females 5 years old 35 1-30.06 2-24.04 293.9 ±1.8 5.1 99 12 1-15.07 2-21.04 278.5 ±2.4 4.6 99 2 2-4.08 14-18.04 255.0 11 15-20.06 5-19.04 301.2 ±1.2 4.8 99 68 21-30.06 5-29.04 294.3 ±0.8 8.0 99 26 1-15.07 2-22.04 279.8 ±1.6
Горностай/ Mustela erminea 3 1.04-9.05 25-29.04 368 ± 16.8 29 gg 7 1-28.06 24.04-13.05 318.4 ±3.5 17.5 99 4 1-25.07 14.03-14.05 275.2 ±1.4 0 1 6.08 6.05 273.0
Норка америк./ Mustela vison 3 15-19.02 1-7.05 73.7 ±1.1 13.8 99 6 2-18.03 25.04-13.05 54 ±0.9 5.2 99 13 21.03-2.04 9-23.05 49.1 ±0.3
Солонгой/ Mustela altaica 10 6-31.03 17.04-10.05 39.6 ±0.4 1.8 90 8 1-25.04 10.05-2.06 38.7 ±0.3 0 4 3.05-3.06 10.06-11.07 39 ±0.6
Колонок/ Mustela sibirica 5 1-14.04 9-20.05 36.4 ±0.8 2.1 90 3 17-24.04 24-27.05 34.7 ±1.7 0 6 10.05-12.06 16.06-16.07 34 ±0.4
Примечание. Материалы по соболю получены в зверохозяйстве «Заря» Ленинградской
области.
Изменчивость ее длительности определяется размерами подвижной эмбрио-
нальной диапаузы (Баевский, 1964,1968). Установлено, что срок собственно
беременности, т.е. периода от имплантации до родов, составляет у американ-
ской норки около 30, а у соболя - 35-40 суток (Клер, 1941; Залекер, 1950;
Р. Enders, A. Enders, 1963; Абрамов, 1974). Время задержки имплантации у
животных III группы составляет всего несколько суток (Колповский, 1971;
Цыцорина, Терновский, 1974).
223
Таблица 67/ Table 67
Продолжительность беременности и плодовитость самок у некоторых видов
хищных млекопитающих (по: Терновский, 1977; Туманов, 1984)
Duration of pregnancy and fertility of some species of carnivores
Виды/Species п Продолж. беременности, сут/ Gestation period, day Плодовитость на 1 самку, особей/ Fertility of 1 female, ind.
Соболь/Martes zibellina: 291-325 2.7 ±0.15
самки 3 лет/ 24
females 3 years old 16 281-290 3±0.21
9 254-280 2.9 ± 0.35
самки 5 лет/ 63 291-311 2.8 ±0.10
females 5 years old 31 281-290 2.7 ±0.13
И 266-280 3.1 ±0.29
Г орностай/ 4 333-393 5.2 ± 2.24
Mustela erminea 6 310-325 8 ± 1.92
5 240-295 8.4 ± 1.93
Американская норка/ 3 71-75 3.6 ± 0.73
Mustela vison 9 51-56 3.4 ±0.59
10 45-50 5.1 ±0.81
У куньих I группы и у американской норки период спаривания самок очень
растянут, а родов - ограничен. Длительность беременности у них во многом
определяется сроками спаривания; чем раньше гон, тем она длиннее (табл.
66). При этом достоверность различий достигала 98-99%. Понятно, что у
хищников III группы (например, солонгоя или колонка) влияние сроков
спаривания на продолжительность беременности отчетливо не прослеживается
(Туманов, 1984).
Интересные данные по размножению норки в неволи приводит М.Д. Абра-
мов (1974). На большом фактическом материале (3800 самок) автор показал,
что у зверьков, покрытых в феврале, беременность продолжается 63-73 дня, а
позже 8 марта - 40-58 дней. При этом разница в средней ее продолжительности
у самок, спарившихся в эти сроки, составляла 21 день. Значительная
изменчивость длительности беременности у хищных зверей обусловлена тем,
что после прохождения через яйцеводы бластулы вступают в стадию
относительного покоя разной продолжительности, а сокращение или удлинение
беременности определяется прежде всего рядом эндокринных факторов (Беляев,
1950; Беляев и др., 1963).
224
Таблица 68/ Table 68
Показатели воспроизводственного процесса у разных представителей семейства
куньих (по: Данилов, Туманов, 1976а)
Parameters of reproductive process of various representatives of mustelids
Виды/ Species Средний вес взрослых самок, г/ Average body mass of adult female, g Максим, продолжит, жизни, лет/ Max. lenght of life, years Количество детенышей/ Cubs number Выживае- МОСТЕ молодняка, %/ Survivorship of young
в гнездах/ in litters перед распадом выводка/ before disintegration of litter
Ласка обыкн./ Mustela nivalis 51.5±1.5 5-6 5.6 4 71
Горностай/ Mustela erminea 114.1±3.2 6-7 7.1 4.7 65
Хорек лесной/ Mustela putorius 448.0±24.2 7-8 6.4 4.6 72
Норка европ./ Mustela lutreola 540.3+15.8 7-8 4.7 3.5 74
Норка америк./ Mustela vison 604.6+26.6 7-9 4.5 3.4 75
Куница лесная/ Martes martes 878.3±18.3 12-15 3.4 2.6 76
Выдра/ Lutra lutra 5121±268.4 15-16 2.6 2 77
Барсук/ Meles tneles 8214±334.2 15-16 3 2.3 77
Росомаха/ Gulo gulo 9615±308.8 16-17 2.5 2 80
Анализ собранных материалов показал, что при благоприятных условиях,
когда гон хищных зверей проходит в нормальные для вида сроки, наблюдается
обратная зависимость между продолжительностью беременности и факти-
ческой плодовитостью. Естественное сокращение срока беременности в
пределах, характерных для вида, сопровождается некоторым увеличением
размера выводка, и наоборот (табл. 67). Эта тенденция прослеживается у самок
разных хищных зверей, у которых развитие зародышей протекает с длительной
задержкой имплантации, - от горностая и соболя до белого и бурого медведей.
По-видимому, удлинение срока беременности определяет большую вероят-
ность резорбции эмбрионов от действия на организм неблагоприятных
факторов внешней среды, т.е. приводит к снижению размера выводка. Вместе
с тем очень короткая беременность у предельно поздно покрытых самок также
может сопровождаться уменьшением количества новорожденных в помете,
что связано с затуханием фолликулярной деятельности яичников. Отмеченные
репродуктивные особенности хищников, способствующие увеличению их
плодовитости, следует учитывать при разведении редких и ценных пушных
зверей.
225
Таблица 69/ Table 69
Показатели массы тела самок и величины выводков у куньих
при вольерном содержании (по: Териовский, 1977)
Parameters of body mass of females and litter size of mustelids with cage keeping
Виды/ Species Средний вес самок, г/ Average body mass, g Кол-во выводков/ Litter number Средняя плодовитость на 1 самку, особей/ Average fertility per 1 female
Ласка/ Mustela nivalis 46.5 ±5.7 3 6
Горностай/ Mustela erminea 121.5 + 5.4 16 7.4
Солонгой/ Mustela altaica 220 ± 10.9 20 8.6
Колонок/ 309.9 ± 11.6 4.9
Mustela sibirica
Хорек лесной/ Mustela putorius 646.8 ±86.1 5 5
Хорек степной/ Mustela eversmanni 653.6 ± 26 13 7.6
Норка европ./ Mustela lutreola 686.4 ± 44 17 3.5
Норка америк./ Mustela vison 706 ± 22.2 13 5.6
Соболь/ Martes zibellina 832.1 ±60.2 3 2.7
Поскольку ранние или очень поздние покрытия самок куньих, для которых
характерна длительная задержка имплантации в развитии зародышей, могут
сопровождаться уменьшением размеров приплодов, то гон животных в условиях
неволи следует проводить в оптимальные для конкретного вида сроки, смещая
его к середине брачного периода. Опыты на соболях и американских норках,
разводимых в зверохозяйствах, показали, что пропуск раннего покрытия
оказывает положительное влияние на биологическую продуктивность самок -
уменьшает количество оставшихся без приплода и не снижает их плодовитости
(Докукина, Куличкова, 1980; Вернадский, Маштак, 1983).
Известно, что животные разных размеров неодинаково реагируют на
воздействие факторов внешней среды: чем меньше их вес, тем интенсивнее
протекает обмен веществ в организме, теснее взаимодействие с условиями
существования и сильнее влияние среды на отдельных особей в популяции. В
процессе исторического развития видов это могло определить большую
плодовитость мелких хищников по сравнению с крупными.
У представителей видов небольшого размера, характеризующихся
высоким уровнем энергообмена, интенсивное размножение не только
компенсирует малую продолжительность жизни, но и способствует быстрому
росту численности популяции после депрессии (Jversen, 1972; Erlinge, 1983;
Стрельников, 1980; Вигони др., 1989).
226
Собственные материалы, а также многочисленные сведения, полученные
от охотников-корреспондентов, позволили отметить у куньих прямую
положительную связь между продолжительностью жизни и массой тела
хищников (г = 0.94), а также резко выраженную линейную отрицательную
зависимость (г = - 0.86) между последней и плодовитостью самок (см. табл. 68).
Из этой схемы несколько выпадает обыкновенная ласка, плодовитость которой
оказалась ниже, чем у более крупных зверьков - горностая, солонгоя и хорьков.
Установленный факт, вероятно, связан с характерной для вида высокой
эмбриональной смертностью, достигающей 40%. Однако известная
способность ласки приносить и благополучно выращивать щенков из двух
приплодов в течение года (Hartman, 1964; Heidt et al., 1968; Туманов, 1988в)
может компенсировать сравнительно низкую ее плодовитость.
Следует также отметить тесную линейную положительную связь (г=0.85)
массы тела самок с выживаемостью молодняка куньих в выводковый
период. В то же время у ласки она оказалась выше, чем у более крупного и
многоплодного горностая, что практически нивелирует существенную
разницу в их плодовитости (табл. 68). В среднем у куньих выживаемость
детенышей за выводковый период увеличивалась от 65-71% у мелких
(ласка, горностай) до 77-80% у более крупных (выдра, барсук) хищников.
При этом надежность выводов о различиях по этому показателю у ласки
или горностая с норками составляла 90%, а с барсуком и росомахой -
99.8%. У норок и лесного хорька достоверность различий с крупными
хищниками семейства по сохранности молодняка также оказалась очень
высокой (р=99.8%).
Характерной особенностью всех представителей изучаемых видов
может служить положительная линейная связь продолжительности жизни
животных и выживаемости щенков (г=0.81). Кроме того, наблюдается
отрицательная зависимость между плодовитостью самок и выживаемостью
детенышей (г= - 0.93), а также между первым показателем и продолжи-
тельностью жизни родителей (г=-0.90).
Тенденция к снижению плодовитости зверьков с увеличением их размеров
хорошо прослеживается в группе мелких представителей семейства со средней
массой тела самок от 47 до 832 г (табл. 69). По материалам Д.В. Терновского
(1977), высокая плодовитость, наблюдаемая у специализированного в питании
хищника-миофага степного хорька, имеет адаптивную направленность на
быстрое изменение численности популяции при благоприятной ситуации. При
этом она сочетается у данного вида с малыми размерами новорожденных.
Так, у степного хорька они весят примерно в 108 раз меньше своей матери, в
то время как у горностая - в 70 раз, у солонгоя - в 65 и у соболя - в 40 раз. Если
у степного хорька вес родившихся щенков равен 4.5-7.6 г, то у близкого к нему
лесного - 7.6-11.1 г.
Таким образом, важной адаптивной репродуктивной особенностью
представителей семейства куньих, во многом определяющей численность
популяции, является для мелких видов значительный воспроизводственный
227
Таблица 70/ Table 70
Показатели репродуктивного процесса у волков и лисиц, содержащихся в неволе
(по: Туманов, 1984)
Parameters of reproductive process of wolves (Canis lupus) and foxes (Vulpes vulpes), kept
in captivity
Виды/ Species /1 Дата/Date Продолжигель- ность беременности, сут/ Gestation period, day Величина выводка, особей/ Number of cubs in litter, ind.
спаривания/ copulation родов/ birth
Волк/ Canis lupus 6 6-30.01 15.03-16.04 69.5 ± 1.94 4.3 ±0.71
7 10-21.02 16.04-2.05 68.7 ± 1.91 3 ±0.31
10 2-14.03 3-27.05 67.7 ± 1.51 4.2 ± 0.64
3 21-24.03 25-26.05 64 ± 1.26 6 ±0.84
Лисица обыкновен./ 3 21-22.01 1-19.02 20.03 29.03-17.04 57.3 ± 0.42 4.6 ± 0.83 4.6 + 0.51
6 56 ±0.53
Vulpes vulpes 2 1-15.03 18.04-3.05 48.5 5
потенциал, а для крупных - высокий уровень выживаемости молодняка и
большая продолжительность жизни особей.
В процессе изучения ожидаемой плодовитости самок соболя, куниц,
горностая и других животных, у которых зародышевый период протекает с
длительной задержкой имплантации, нами был разработан и применен экспресс-
метод, основанный на вымывании бластоцист из их гениталий. Он менее
трудоемок, чем способ гистологической обработки яичников, и повышает
точность исследований в среднем на 16.5% (Туманов, 19886, в; Туманов,
Девяткин, 1992). В отличие от традиционного этот метод позвол'яет
устанавливать в промысловых пробах долю прохолоставших самок и
плодовитость беременных даже в полевых условиях. Кроме того, его
применение дало возможность выявить в отдельные годы более растянутые
сроки до имплантационного периода у лесной куницы по сравнению с
литературными сведениями (Старков, 1947; Граков, 1981). Случаи позднего
обнаружения свободноплавающих бластоцист у трехлетних самок, добытых
в период с 18 по 23 марта 1984 г., подтверждают высокую пластичность
организма куниц и широкий диапазон их репродуктивных возможностей.
Использование данного метода позволяет вполне обоснованно планировать
годовой прирост популяций животных и нормы их промыслового изъятия
задолго до предстоящего охотничьего сезона.
У самок хищных млекопитающих из семейства собачьих (Canidae)
беременность протекает без длительной эмбриональной диапаузы. Разница по
228
Таблица 71/Table 71
Продолжительность беременности и плодовитость самок волка
и лисицы, содержащихся в неволе (по: Туманов, 1984)
Duration of pregnancy and fertility of wolf (Canis lupus) and red fox ( Vulpes vulpes), kept
in captivity
Виды/ Species п Продолжит, беременности, сут/ Gestation period, day Плодовитость на 1 самку, особей/ Fertility of 1 female, ind.
Волк/ Canis lupus 7 71-76 2.7 ± 0.45
9 66-70 4.5 ±0.47
10 62-65 4.7 ± 0.65
Лисица обыкновен./ 8 56-58 4.5 ±0.39
Vulpes vulpes 3 48-55 4.8 ±0.84
срокам ее продолжительности у большинства видов, обитающих в европейской
части России, составляет 10-14 дней. Например, данные, полученные нами из
Ленинградского (Санкт-Петербургского), Московского и Таллинского
зоопарков, свидетельствуют о том, что беременность самок волка (п=26) равна
62-76 суток, обыкновенной лисицы (и=11) - 48-58, а песца (и=6) - 46-58
суток. Как и у представителей семейства куньих с лабильной беременностью
(II группа), у них наблюдается тенденция к ее сокращению при поздних
покрытиях. Однако относительно короткая амплитуда длительности периода
вынашивания щенков у самок этих видов и малое количество собранного
материала не позволяют отчетливо проследить связь сроков спаривания с
продолжительностью беременности и величиной выводков (табл. 70-71).
У обыкновенной лисицы (Vulpes vulpes) большинство особей становится
половозрелыми в конце первого года жизни, в 9-10 мес. В природных условиях,
судя по сведениям охотников-корреспондентов (п-77), гон у нее протекает в
основном в феврале-марте (83.1% от общего количества сообщений), реже - в
январе (10.4%) и апреле (6.5%). Щенки появляются в мае (60% случаев), а также
в апреле (25%) и июне (15%). В 16 полученных в неволе выводках (в 5 случаях
сроки спаривания неизвестны) содержалось от 2 до 6 детенышей на самку.
В естественных местообитаниях на Северо-Западе России средняя
плодовитость лисицы составляла 4.6 ± 0.23 щенка на одну размножающуюся
самку. В 45 выводках из известных 56 (80.2% встреч) находилось по 3-6
детенышей. Малоплодные пометы с 1-2 щенками и многоплодные (7-9 особей)
сравнительно редки: на их долю приходится всего 19.8% встреч от общего
количества обнаруженных выводков (Данилов и др., 1979). Отмечено, что
плодовитость лисицы резко увеличивается в годы обилия мышевидных
грызунов и, наоборот, снижается в «неурожайные» для них годы (Ивантер,
1969; Когтева, Морозов, 1972).
229
Таблица 72/ Table 72
Продолжительность беременности и плодовитость у енотовидных собак
в Ленинградском (Санкт-Петербургском) зоопарке в 1960-1980 гг.
Duration of pregnancy and fertility of racoon dogs (Nyctereutes procyonoides)
in Leningrad (St. Petersburg) Zoo in 1960-1980
Дата спаривания/ Copulation time Сроки родов/ Birth time Продолжительность беременности, сут/ Gestation period, day Детеныши/Cubs
всего/ total
15-25.01 24.03 68 6 4:2
21.01-5.02 10.04 73 9 5:4
2-6.02 11.04 68 6 3:3
3-7.02 4.04 60 И 5:6
2-9.02 6.04 63 8 4:4
12.02 30.04 77 3 1:2
16.02 4.05 77 6 -
21.02 20.04 58 8 -
24.02 16.04 51 10 -
24.02 18.04 53 6 3:3
5.03 5.05 61 11 6:5
17.03 16.05 60 8 4:4
Примечание. Прочерк означает отсутствие данных.
У енотовидной собаки {Nyctereutes procyonoides) период гона заметно
растянут. По-видимому, это объясняется разными сроками размножения
молодых и взрослых самок, более поздним созреванием гонад у истощенных
за зиму зверей и погодными условиями в весенний период.
По данным анкетного опроса охотников-корреспондентов (30 сообщений),
гон у енотовидной собаки в северо-западных областях России происходит с
конца февраля по апрель, но преимущественно в марте. Вес пары семенников
с придатками у 5 взрослых самцов, добытых в конце февраля, был равен 5.5-
7.5 г, а в середине марта (6 особей) - 8.2-10.8 г. В это время гонады зверей
находятся в состоянии наибольшей половой активности, которая затем
начинает снижаться. В конце июня-июле зрелых спермиев в семенных канальцах
зверей, разводимых в вольерах, не отмечали (Соколов, 19496). Осмотр
репродуктивной системы 5 самок, отловленных в конце февраля-марте, так
же как и у самцов, свидетельствовал об их полной готовности к спариванию.
В условиях вольерного содержания гон у енотовидных собак в разные годы
проходил в одно время - со второй половины января и до середины марта.
Роды у самок (л=23) отмечали в период с 20 марта по 24 мая: в марте родили
230
3 самки (13%), в апреле 15 (65%) и в мае 5 самок (22%). Судя по интервалу
между датами спаривания и родов, в тех случаях, когда они были точно
известны (н—12), продолжительность беременности у енотовидной собаки имеет
большую амплитуду, чем у других видов, и колеблется в пределах от 51 до 77
дней (табл. 72). Половозрелыми звери становятся к весне следующего года,
т.е. в возрасте примерно 9-11 мес.
В природных условиях енотовидные собаки, по-видимому, моногамы.
Формирование пар происходит у них осенью, однако в период гона самку
могут преследовать 2 и даже 3 самца. В естественных местообитаниях сроки
щенения более растянуты, чем при клеточном разведении зверей. Так,
В.Ф. Морозов (1953) отмечает, что в 1950 г. в Новгородской области бере-
менная самка была поймана в середине сентября. А.Г. Банников (Гептнер
и др., 1967) сообщает об известных ему фактах, когда большинство самок
приносило детенышей до июня, а иногда и до сентября. В.Т. Шималов (1977)
указывает, что в Полесье новорожденных щенков этого вида находили даже в
сентябре. Столь поздние сроки рождения детенышей объясняются, вероятно,
не задержкой гона, а способностью самок в случае резорбции эмбрионов,
рождения мертвых щенков или их ранней гибели вновь спариваться и приносить
второй помет, что мы неоднократно наблюдали у куньих.
По материалам охотников-корреспондентов (и=87), плодовитость
енотовидных собак в природе сравнительно высока - 6.3 (2-11) особей на
рожавшую самку. При этом выводки с 6-8 детенышами составляют примерно
70% от общего их количества.
Молодые песцы (Alopex lagopus) становятся готовыми к участию в
размножении в возрасте 9-11 мес. По материалам Н.М. Полежаева (1998), на
европейском Севере процессы сперматогенеза у молодых самцов идут менее
интенсивно, а зрелая сперма появляется в придатках на месяц позже, чем у
взрослых особей. Активный сперматогенез наблюдается в семенниках,
достигших массы 2.5-2.6 г. Половая активность молодых самок также
проявляется позже, чем у взрослых, что вызывает значительную растянутость
сроков гона и рождения молодняка.
В. А. Золотов (1940), ссылаясь на наблюдения охотников-промысловиков,
сообщает, что гон материкового песца на Кольском полуострове происходит
во второй половине марта. По его же данным, на о-ве Кильдин спаривание
зверей приходится на период со второй пятидневки марта и до последних чисел
апреля, однако массовое покрытие самок - в конце марта-начале апреля. По-
видимому, и на материке период гона у песца растянут, что обусловлено разной
готовностью к размножению молодых и взрослых особей, упитанных и
отощавших. По устному сообщению сотрудника Мурманского морского
биологического института В.В. Семенова, неоднократно тропившего зверьков
по следам на снегу, спаривание песца в зависимости от погодных условий
происходит в конце марта-апреле, но может быть и в мае. Однажды в середине
мая он был свидетелем брачного поведения двух зверьков, которые играли и
возились в утренние часы на холме у небольшого озера. В этом году снег в
231
Таблица 73/ Table 73
Показатели размножения песца в Ненецком автономном округе за 1957-1966 гг.
(по: Шиляева, 1972)
Parameters of reproduction of arctic fox (Alopex lagopus) in Nenets Autonomous Center
in 1957-1966
Численность Вскрыто взрослых Кол-во плаце нтарных пятен на самку, шт./ The number of placental spots per 1 female Обследовано нор/ The number of examined dens Кол-во щенков в выводке,
грызунов/ Rodents number самок, особей/ The number of dissected adult females, ind. всего, шт./ total, ind. занято выводками, %/ occupied by litters особей/ Cubs number in litter
Высокая /High Низкая/ 24 14.2+1.3 709 68.3 ± 1.7 7.8 ±0.3
121 13.5 ±0.8 1086 16.6 ± 1.1 6.2 ±0.3
Средняя/ Average 16 3+1 730 24.1 + 1.5 7.2 ± 0.4
тундре сошел только в конце июня. В другом случае тропление парных следов
песца проводилось в конце марта. При приближении человека зверьки (видимо,
брачная пара) выскочили из 2 снежных нор, вырытых на расстоянии 15-20 м
друг от друга.
В Ленинградском (Санкт-Петербургском) зоопарке половая активность
белых песцов в течение ряда лет также наблюдалась в марте и апреле, чаще во
время оттепелей. В это время самцы (иногда по двое) бегают за самкой, много
кричат и мочатся, обнюхивают друг у друга половые органы. Коитус обычно
отмечался в утренние часы, продолжительность его была от 5 до 20 мин. При
спаривании зверей 11 апреля самка принесла детенышей (1 3 и 1 §) 27 мая, т.е.
через 46 дней. В двух других случаях (сроки спаривания не зафиксированы)
самки родили щенков 14 мая (3 3 и 2 §) и 20 мая (1 и 1 $>).
По результатам разведения песцов в зверохозяйствах продолжительность
беременности самок составляет 46-58 дней (Ильина, 1952; Полежаев, 1998).
Как сообщает А.Ф. Чиркова (Гептнер и др., 1967), в неволе и у песцов, и у
лисиц при резорбции эмбрионов или гибели новорожденных наблюдаются
случаи повторной течки и спаривания. В некоторые годы до 5-8% самок этих
видов приходит в состояние эструса в тот же репродуктивный сезон второй раз.
По-видимому, и в природных популяциях многие самцы сохраняют способность
покрывать самок, которые по тем или иным причинам поздно пришли в
состояние эструса.
В условиях Болыпеземельской тундры массовое щенение самок песца
наблюдается в конце мая-начале июня. Основная часть детенышей рождается
в норе. В их выкармливании принимают участие оба родителя. В первые дни
самка постоянно находится с выводком, не выходит из норы, а пищу получает
от самца. При хорошей обеспеченности кормами она не отлучается от щенков
232
весь период лактации (примерно 1.5 мес) и большую часть времени проводит в
убежище или около него (Полежаев, 1998).
При вольерном содержании белые песцы приносили от 2 до 5 щенков.
Примерно такое же количество молодых (2-6) находил в жилых норах этого
вида на Кольском полуострове А.Н. Дубровский (1939).
Средняя величина выводка на о-ве Врангеля в 80-х годах XX в. была заметно
выше - 8.6 щенков. В отдельные годы этот показатель изменялся в пределах от
4 до 11.6 особей на размножавшуюся самку (Овсяников и др., 1991). На
Таймыре на разных фазах динамики численности в выводках находилось в среднем
от4.9до6.7щенков(Куксов, 1989). Близкие к этим данные за 70-80-е годы по средней
плодовитости песца (3.7-6.6) в северной и южной подзонах тундры Таймырского
полуострова приводит М.К. Крылов (1989).
В «урожайные» на грызунов годы в Большеземельской тундре (Полежаев,
1998), когда популяция песца вступает в фазу пика численности, среднее
количество плацентарных пятен на рогах матки, свидетельствующих об
интенсивности размножения, достигает 14.4 ± 1.4, а в годы депрессии - 8.2 ±
± 0.7 на рожавшую самку. Количество желтых тел в яичниках этих зверей
превышает таковое у обыкновенных лисиц в среднем в 2-3 раза. Подобная
высокая плодовитость вида, по-видимому, представляет собой важное
средство поддержания его популяционного гомеостаза в условиях тундры. В
то же время, как и у других многоплодных хищников, у песца отмечается
высокий уровень гибели молодняка на разных этапах постнатального
онтогенеза. Так, в природных условиях в норный период смертность щенков
составляет 45-67%, или в среднем 57%. В дальнейшем величина гибели
детенышей определяется кормовыми условиями сезона. Наиболее ощутима
она в годы депрессии численности леммингов и других мелких грызунов.
Эти материалы полностью совпадают с данными Л.М. Шиляевой (1971,
1972), свидетельствующими о существенной разнице между потенциальной и
реальной плодовитостью вида. По свидетельству автора, в годы высокой
численности мышевидных грызунов в Большеземельской тундре и на
Югорском полуострове в основных очагах норения песца бывает занято
выводками около 70% нор и размножается 40-41% самок. В этот период
наиболее высока потенциальная плодовитость вида и крупны выводки (табл.
73). Весной среди самок разного возраста могут численно преобладать (до
46%) молодые половозрелые особи рождения прошлого года, которые часто
остаются бесплодными, поэтому чем выше в популяции процент молодых
(годовалых) самок, тем больше доля животных, оставшихся без выводка,
особенно при неблагоприятных условиях обитания. В неблагоприятные сезоны
гибель молодняка в популяциях песца с учетом эмбриональной смертности
может достигать 70%.
Молодые волки (Canis lupus) в естественных условиях становятся
половозрелыми в конце второго года жизни, однако при содержании в неволе
они в редких случаях могут демонстрировать готовность к продуктивному
спариванию и заметно раньше (Medjo, Meeh, 1976; Seal et al., 1979)
233
9 Зак. № 4309
В Ленинградском (Саект-Петербургском) зоопарке самец, родившийся
весной 1971 г., принял участие в гоне в декабре 1972-январе 1973 г., т.е. в
возрасте менее 2 лет. Покрытая им волчица (1968 г. р.) в марте 1973 г. принесла
нормальный помет. Подобные случаи наблюдались в зоопарке и раньше. В
природе молодые самцы (до 2 лет), очевидно, от пришедших в состояние
половой охоты самок отгоняются взрослыми и могут принимать участие в
размножении только в «разреженных» популяциях, в которых были
истреблены особи-доминанты.
Спаривание волков в лесной зоне Северо-Запада и европейского севера России
происходит в основном в январе-начале марта. По материалам анализа сведений,
полученных от охотников-корреспондентов (Данилов и др., 1979), из 70
зарегистрированных случаев гона на январь приходилось 27 (38.6%), а на февраль
34 (48.6%). Заметно реже он наблюдался в конце декабря (1.4% случаев) или в
марте (11.4%).
Проведенные нами гистологические исследования гонад от 5 волков-
переярков (возраст около 2 лет), добытых в Ленинградской области в конце
февраля-марте, позволили установить их полную готовность к спариванию.
Гениталии зверей были увеличены, а канальцы семенных придатков заполнены
спермой. Подобная же картина прослеживалась и на срезах репродуктивных
органов взрослых животных. Вместе с тем имеются сведения (Данилов и др.,
1985а) о том, что в природе встречаются переярки, которые не достигают
половозрелости в указанном возрасте. У этих животных даже в конце марта
зародышевый эпителий развивается только до сперматид, и дальнейшая их
дифференциация не происходит.
Просмотренные половые органы самок (8 особей) в возрасте старше
1 года свидетельствуют о том, что в конце февраля-начале марта взрослые
волчицы были либо беременны, либо в фазе эструса. Гонады молодых зверей,
впервые готовящихся к размножению, также находились в функционально
активном состоянии - просвет матки имел звездообразную форму, ее мышечная
стенка утолщена и гиперемирована, а в яичниках содержались крупные
фолликулы с четко обозначенными яйцеклетками (фаза проэструса).
Все эти материалы подтверждают высказывания П.И. Данилова (1994)
о том, что данные факты следует рассматривать как популяционные
механизмы, способствующие восстановлению численности волка в
нарушенных группировках хищника. По мнению автора, для молодых
волчиц, впервые вступающих в размножение, поздние сроки течки весьма
характерны. Данная адаптивная особенность вида обеспечивает в
популяции так называемый возрастной кросс - большую вероятность
покрытия молодой самки средневозрастным самцом (субдоминантом), не
имеющим пары. Поздняя течка у самок-переярок может также частично
компенсировать потери популяции волков, вызванные интенсивным
преследованием их со стороны человека, особенно в конце зимы. Если в
результате истребления семья теряет матерую самку, находящуюся в течке
или в начале беременности, то ее место занимает самка-субдоминант из
234
переярок. Приходя в течку позже первой, она образует пару с оставшимся
самцом. Если семья лишается обоих матерых, их место занимают переярки,
образующие пару. И в первом, и во втором случае воспроизводительный
потенциал зверей практически не сокращается, а вновь сформировавшаяся
семья сохраняет участок обитания.
Кроме того, необходимо отметить и большую продолжительность половой
активности самцов по сравнению с самками за репродуктивный сезон. Даже
волки-переярки, только что достигшие половой зрелости, бывают готовы к
спариванию (в гонадах зрелые сперматозоиды) до начала течки взрослых
волчиц. Способность самцов к продуктивному спариванию сохраняется и после
появления потомства. В случае его ранней гибели гон может повториться.
Отмеченное явление, характерное для многих млекопитающих, можно
рассматривать в качестве компенсаторного механизма, призванного
поддерживать оптимальную численность вида. В случае депрессии или
нарушения половой структуры популяции этим путем обеспечивается
максимально возможная репродуктивная потенция за счет участия в
размножении большого числа самок (Данилов идр., 1985а).
В обозначенные выше сроки проходит гон у волков и на севере европейской
части России. Так, по имеющимся данным (Полежаев, 1998), одна из молодых
самок (возраст около 2 лет), добытая 8 марта, находилась в состоянии
проэструса, а другая, отловленная в начале апреля, была на ранней стадии
беременности. В рогах матки у нее находились 5 зародышей. Две трети из
осмотренных автором взрослых самок в первой декаде марта также были уже
беременными. Наибольшее количество эмбрионов (10 шт.) имела матерая
волчица, а самое малое (5 шт.) - самка в возрасте около 2 лет. При этом
среднее их количество у зверей из северных территорий составляло 6.3 эмбриона
на размножавшуюся самку.
На Северо-Западе России роды у волков большей частью наблюдаются в
апреле-мае. Поданным охотников-корреспондентов, из 49 зарегистрированных
случаев щенения на апрель пришлось 21 (42.8%) и на май - 24 (48.9%). Реже
роды наблюдались в марте - 3 раза (6.1%) и в июне - 1 раз (2.2%). В выводках
чаще встречается по 3-7 щенков. Пометы с меньшим (1-2) и большим (8-9)
количеством детенышей относительно редки и не превышают 4.5-7.5% от общего
количества обнаруженных выводков. Средняя плодовитость самок в регионе
(по 66 случаям) - 5 щенков. Поскольку смертность волчат велика и может
достигать 60-80% (Наумов, 1967), то зимой семья-стая волков состоит обычно
из 2 взрослых животных, 1-2 переярков и 2-3 прибылых зверей. Таким образом,
в популяции волка в целом преобладают неполовозрелые звери, и потенциал его
размножения не столь высок, как у более мелких плодовитых и скороспелых
видов того же семейства (Данилов и др., 1979).
Подтверждением этому наблюдению служит состав 2 волчьих стай,
представители которых были отстреляны с вертолета в Ленинградской области
20-25 марта 1987 г. В состав первой брачной стаи (5 особей), добытой в торосах
Ладожского озера, входили 2 матерых зверя (^ и £), самец переярок и двое
235
прибылых Qj и §). Вторая группа волков (7 особей), отстрелянных в лесном
массиве Тихвинского района, состояла из 3 самок и 4 самцов. Среди последних
находились матерый зверь, переярок и 2 прибылых. Одна из самок оказалась
сеголеткой (0+). Она имела анемичную матку (весом до 3 г), тонкие рога, которые
в длину не превышали 10-12 см, и мелкие яичники размером 4x7 мм. В них были
отчетливо видны растущие фолликулы до 400-500 мкм в поперечнике. Другая
самка - переярок (1+) - находилась в состоянии окончания эструса. Ее матка
весом 8.5 г имела звездообразный просвет, мышечные стенки гиперемированы, а
утолщенные рога достигали в диаметре 8-9 мм и в длину до 19-20 см. В яичниках
размером 8x14 мм имелись развивающиеся желтые тела и крупные полостные
фолликулы. Третья самка в возрасте 5-6 лет была беременной. Ее матка весила
31 г, а рога толщиной до 10 мм имели длину 24-25 см. В яичниках размером 11x18
мм находились четко отграниченные от окружающей ткани желтые тела - 4 на
одном и 5 на втором. Однако околоплодных вздутий в матке было меньше, чем
желтых тел, соответственно 4 и 3 на каждом ее роге.
По имеющимся сведениям (Данилов и др., 1985а, б), сроки гона у волков
заметно варьируют в зависимости от широты местности, фенологии сезонов
года и продолжительности светового дня. Раньше, чем в других местах, этот
процесс отмечается у зверей, обитающих на юге России, в частности на Кавказе -
в начале декабря. Позднее (конец этого месяца) спаривание хищников
происходит в лесостепной и степной зонах. Затем с продвижением в северном
и восточном направлениях время гона сдвигается на более поздние сроки. Так,
по материалам И.С. Козловского (1996), на северо-востоке европейской части
России взрослые особи спариваются в феврале-марте, иногда в апреле.
Щенение происходит во второй половине апреля и до 10-12 мая (около 60%
случаев). Молодые самки приносят детенышей в конце мая-июне. В Кировской
области средний уровень рождаемости составлял (по 20 выводкам) 7.1 щенка
на самку. При этом волчицы чаще приносили 7 (25%) и 9 (20%), реже 6 или 4
(по 15%) детенышей.
Как у песцов, лисиц и волков, у представителей семейства кошачьих
(Felidae), таких как европейская рысь, лесной и камышовый коты,
беременность протекает без длительного периода эмбриональной диапаузы и
продолжается примерно 2-2.5 мес. По имеющимся данным, гон у лесного кота
(Felix silvestris) на Кавказе происходит в феврале-марте (Динник, 1914;
Г. Новиков, 1956). По нашим наблюдениям, готовность зверей к спариванию
отмечалась в конце января-марте, реже - до середины апреля. В это время в
сумеречно-ночные часы самцы и самки много бегают по территории своего
участка, постоянно метят его мочевыми метками, громко и часто заунывно
кричат. Просмотренные нами животные были добыты местными охотниками в
горных районах Дагестана и Азербайджана в период с 25 января по 15 марта
1976 и 1977 гг. В выборке находились 7 самцов в возрасте от 2 до 4 лет, которые
весили 3.4-5.2 кг и имели длину тела 54-66 см, а хвоста - 30-35 см. Их
репродуктивная система находилась в функционально активном состоянии.
Средний диаметр семенных канальцев достигал 85-93 мкм, вес пары семенников
236
с придатками - 3.5-4.7 г. Канальцы последних были заполнены готовыми
половыми продуктами - сперматозоидами.
Отловленные в эти же сроки 5 взрослых самок (возраст 2-3 года) весом
2.8-4.2 кг характеризовались следующими размерами: длина тела 52-58 см,
хвоста - 30-33 см. Два зверя, пойманные в последнюю неделю января, имели
крупные генеративные органы с утолщенной гиперемированной маткой,
просвет которой был звездообразной формы. В яичниках наблюдались большие
фолликулы с обособленными яйцеклетками, готовыми к овуляции. Три другие
самки, поступившие к нам на исследование в конце февраля, оказались
беременными. На рогах матки у них были отчетливо видны соответственно 4
и 5 околоплодных вздутий.
Весовые и линейные размеры органов размножения самок лесного кота в
разные сезоны года заметно изменяются. У трех взрослых самок, отловленных
в Дагестане в ноябре (состояние покоя), вес яичников был равен 150-300 мг, а
длина рогов матки 90-140 мм. Две другие взрослые особи, добытые в тех же
местах в первой декаде февраля, имели уже увеличенные яичники весом 240-
480 мг и рога матки длиной 150-190 мм.
На основании опроса местных охотников из кавказского региона можно
сделать вывод о том, что самки лесного кота приносят приплод в апреле-мае,
иногда даже в конце июня. В помете бывает от 3 до 6 (чаще 3-4) котят, забота
о которых лежит только на матери. Самец участия в воспитании молодняка не
принимает.
У камышового кота (Fells chaus) гон обычно протекает в период с конца
января и до начала апреля. В это время между самцами происходят
ожесточенные драки, сопровождающиеся громкими криками (Г. Новиков,
1956; Павлинов, 1999). Примерно через 2-2.5 мес, в апреле-июне, самка
приносит от 3 до 6 (обычно 4-5) котят, которых воспитывает в одиночку.
Как сообщает С.Б. Насибов (1970), заметное увеличение веса и линейных
размеров семенников, а следовательно, и готовность к спариванию у
взрослых особей камышового кота на Кавказе зарегистрированы в конце
декабря-начале марта. У некоторых зверей, по разным причинам поздно
вступающих в размножение, рост этих показателей прослеживался даже в
летнее время. Длина семенников от периода покоя и до гона увеличивается с
2.2 до 3.1 см, а их масса-от 4.1 до 7.7 г. Вес двух яичников у добытых самок
в период покоя был равен в среднем 300 мг, а во время готовности к спарива-
нию - 490 мг. За это время размеры матки увеличились от 15 до 25 см, а длина
ее рогов - от 10 до 16 см. У самок обоих видов в постоянных убежищах, где
происходят роды, гнездовая камера выстилается сухими листьями, травой,
иногда перьями птиц. Диаметр логовища колеблется у камышового кота в
пределах 71-98 см, а у более мелкого лесного - 43-69 см.
У близкого родственника лесного и камышового котов - каракала (Fells
caracal), который на территории бывшего Советского Союза встречался в
Туркмении, при содержании в неволе половозрелость наступала у самцов в
возрасте 24-25 мес, аусамок-в 14-16мес. Продолжительность беременности
237
Таблица 74/ Table 74
Некоторые репродуктивные показатели европейских рысей, содержащихся
в условиях Ленинградского (Санкт-Петербургского) зоопарка
Some reproductive parameters of European lynx {Lynx lynx), kept
in Leningrad (St. Petersburg) Zoo
Год/ Year Сроки/Date Продолж. беременности, сут/ Gestation period, day Величина выводка, особей/ Number ind. in litter Соотношение полов/ Sex ratio,
первого спаривания/ first copulation родов/ birth
1959 25.03 27.05 63 2 1:1
1960 30.03 29.05 60 2 1:1
1963 27.03 23.05 57 2 0:2
1964 23.03 30.05 66 1 1:0
1965 24.03 26.05 63 2 0:2
1979 10.04 11.06 62 3 1:2
1980 12.04 24.06 73 3 1:2
1981 20.03 01.06 73 2 1:1
1985 16.03 09.05 54 1 0:1
1986 10.03 21.05 72 3 2:1
1987 17.03 26.05 70 1 0:1
1988 13.03 24.05 72 3 2:1
В Среднем/Average 65 2.1 1:1.5
no 7 выводкам составляла 77 (74-79) дней. В этих случаях спаривание зверей
наблюдалось в январе-июне, а рождение молодняка - в апреле-начале
сентября. Причем одна самка дважды (1993 и 1997 гг.) приносила по2приплода
за один репродуктивный период. Плодовитость у каракала (л=22) составляла
1-7, в среднем 2.6 детенышей, а смертность молодняка 56.1% (Stehlik, 2001).
Рысь {Lynx lynx) относится к видам с консервативными сроками
репродуктивных процессов (Tumanov, 1996). Наблюдения показали, что
молодые животные достигают половой зрелости в возрасте 1 года 10 мес и
старше, а половозрелые самки способны спариваться и приносить выводок
каждый год. Анализ многолетних материалов свидетельствует о том, что в
Ленинградском (Санкт-Петербургском) зоопарке спаривание европейских
рысей происходит в основном в марте (табл. 74). Здесь за период с 1959 по
1988 г. самки принесли 12 выводков, причем только в 2 случаях (16.7%) гон
зверей проходил не в марте, а в первой половине апреля. В эти годы роды были
зафиксированы в середине и конце июня, хотя обычно самки приносили котят
238
в мае. Продолжительность беременности самок имела небольшую амплитуду
колебаний и изменялась в интервале 54-73, или в среднем 65 суток. В 1985 г.,
когда беременность самки составила 54 дня, детеныш родился мертвым и,
видимо, недоношенным.
Близкие к нашим данные получены в Чешской Республике, в зоопарке
г. Острава (Stehlik, 1984,2000). Здесь готовность к продуктивному спариванию
у молодых самцов и самок обыкновенной рыси (Lynx lynx) наблюдалась в
конце второго года жизни (в возрасте 22 мес). В период с 1967 по 1976 г. гон
у зверей (6cJ и 6$) регистрировался в марте и только один раз - в апреле. Из
19 нормальных родов 13 (68.4%) проходили в мае, а 6 (31.6%) - в июне. За
время гона звери могли спариваться по 5-15 раз в сутки с длительностью коитуса
от 1 до 3 мин. Средняя продолжительность беременности была 73 (70-75) дня,
плодовитость - 2.3 (1-3) детеныша, а отход молодняка составлял 51% в группе
из 49 родившихся котят.
Гистологические исследования репродуктивных органов от 6 взрослых
самцов, добытых в Карелии, свидетельствуют о том, что и в природных
условиях гон у рысей проходит примерно в те же сроки - в конце февраля-
марте (Данилов и др., 1979). В этом убеждают не только результаты
гистологического анализа состояния гонад, но и особенности поведения зверей:
встречи следов 1-2 самцов, следующих за самкой, стычек между ними и т.д.
Период течки, а следовательно, и спаривания растянут, о чем свидетельствуют
близкие по срокам встречи котят из разных выводков, сильно отличающихся
по развитию. Однако продолжительное пребывание самцов в состоянии
половой активности значительно сокращает возможность прохолостания
самок. На готовность рыси к продуктивному спариванию в марте указывают
и другие специалисты. Так, по наблюдениям Е.К. Рымаревой (1933), в
Московском зоопарке спаривание животных отмечалось в марте, а рождение
молодых - в конце мая-июне. Продолжительность беременности была равна
67-74 сут. Время готовности самок к оплодотворению длилось примерно 2
недели. В эти сроки коитус партнеров (7-15 мин) происходил многократно, в
основном в сумеречные и ночные часы.
По материалам В.К. Стояновой и В.Г. Сваринской (1964), в Рижском
зоопарке в течение ряда лет рыси регулярно спаривались во второй половине
марта. Роды у самок наступали в конце мая-первых числах июня, а
продолжительность беременности составляла 60-70 сут. Авторы отмечают,
что в 1959 г. одна из самок, потревоженная посетителями, сразу же после
родов (4 июня) съела новорожденных, а через месяц вновь была покрыта
самцом и 9 сентября родила 1 котенка. В том же 1959 г. подобная картина,
т.е. необычное повторение гона (конец апреля-май) после ранней гибели
первого приплода, наблюдалось и у одной из самок (с рождением мертвого
детеныша), содержащейся в Ленинградском (Санкт-Петербургском) зоопар-
ке (Андреевская, 1964). По-видимому, самки европейской рыси, лесного и
камышового котов, как и некоторых других видов млекопитающих с
коротким, четко фиксированным сроком беременности, в случае
239
рассасывания эмбрионов или гибели молодняка в раннем возрасте
способны вновь спариваться и давать повторный приплод. Таким образом,
они обладают потенциальной полиэстричностью, которая способствует
выживанию популяций этих видов в неблагоприятные для них годы
(Туманов, 1977а, б, в).
Интересные данные по размножению европейских рысей были получены
также в 1988-1997 гг. на экспериментальной базе «Черноголовка» под
Москвой, где вели наблюдения за взрослыми особями и детенышами из 19
выводков (Найденко, 1997). Здесь коитус у животных отмечался с 5 по 27
марта, а роды - в период с 24 мая по 6 июня. Продолжительность беременности
самок была сходна с таковой в зоопарках и составляла 67-74 сут.
Рысь относится к малоплодным животным. В Ленинградском (Санкт-
Петербургском) зоопарке за все годы наблюдений у них рождалось от 1 до 3
котят при средней плодовитости 2.1 особи на одну самку. Среди размно-
жавшихся зверей пометы с одним детенышем составляли 25%, с двумя - 42%
и с тремя - 33% от общего их количества. Сходная плодовитость у этого вида
отмечалась и в других местах, где занимались разведением европейской рыси.
Так, в Московском зоопарке молодые самки обычно приносили весной по 1
котенку (Рымарева, 1933). В Рижском зоопарке в 6 из 12 просмотренных
выводков преобладали двойни (50%). Пометы с 1 и 3 котятами были
представлены равной долей - по 25%. При этом средняя плодовитость
рожавших самок составляла 2 особи, а отход молодняка в первый же месяц
после родов - 54.2% (Стоянова, Сваринская, 1964). По материалам С.В. Най-
денко (1997), на экспериментальной базе «Черноголовка» (Московская обл.)
средняя плодовитость европейской рыси (по 19 выводкам) была равна
2.1 детеныша. Причем в восьми выводках было по 2 котенка (42%), в шести -
по 3 (32%) и в пяти - по 1 (26%). Смертность молодняка в первые месяцы
достигала 33.3%.
Судя по результатам собственных наблюдений, при троплений самок рыси
с котятами (8 случаев) и анкетным данным (71 сообщение), полученным в 70-х
годах XX в.от егерей и охотоведов из Карелии, Ленинградской и Псковской
областей (Данилов и др., 1979), в естественных условиях в ее выводках также
бывает от 1 до 3 детенышей. В среднем к концу осени-началу зимы в природе
на одну рожавшую самку приходится 1.7-1.8 молодых.
У принадлежащего к семейству енотовых (Procyonidae) енота-полоскуна
(Procyon lotor), по данным Н.Н. Руковского (1953, 1963), самки становя-
тся половозрелыми в 10-11 мес, самцы - несколько позже. Гон у взрос-
лых зверьков происходит в феврале-марте, у молодых - нередко в апреле.
Щенение наблюдается в апреле-мае, иногда в июне. Самка приносит от 3 до
8 детенышей.
Наиболее подробно особенности репродуктивного процесса у этого вида
изучены А.М. Гинеевым (1973) на Северном Кавказе. По материалам автора,
в зимний период у самцов-сеголеток относительный вес семенников не
превышает 0.25%о, в то время как у взрослых особей он был больше - 0.6%о.
240
Увеличение размеров гениталий у самцов начинается в декабре - начале января
и становится максимальным в марте. Половой зрелости они достигают в 2-
летнем возрасте.
У самок, добытых в феврале-марте, рога матки увеличены больше, чем у
зверьков, отстрелянных в ноябре-январе. Судя по материалам гистологической
обработки репродуктивных органов и полевым наблюдениям, спаривание у
них происходит в следующие месяцы: в феврале в гоне участвует 22.5% самок,
в марте 27.5, в апреле 17.5, в мае 10, в июне 12.5, в июле и августе - по 5%.
Таким образом, гон у енота-полоскуна на Северном Кавказе может протекать
в период с февраля и до начала августа. Молодые самки в возрасте 10-12 мес
приходят в охоту позднее взрослых особей (чаще в апреле-июне). В апреле-
мае рождается до 50% детенышей, однако в целом в популяции вида роды
наблюдаются в период с апреля и до первой декады октября. Угасание половой
активности самок прослеживается в 8-9-летнем возрасте.
Количество щенков в помете - от 1 до 7, в среднем 4 (стандартная ошибка
0.27). В обнаруженных 24 гнездах этого вида находилось следующее
количество детенышей: в одном - 1, в шести - по 3, в девяти - по 4, в четырех
- по 5, в трех - по 6 и в одном - 7 особей.
У самок из семейства медвежьих (Ursidae) беременность протекает с
длительной задержкой имплантации. Период эмбриональной диапаузы и
развития зародыша у них продолжается в среднем 220-240 дней.
Репродуктивные особенности белого медведя (Ursus maritimus) до
настоящего времени остаются малоизученными. Вместе с тем вопросы,
связанные с воспроизводственным потенциалом вида, представляют интерес
для практических целей - разведения в неволе и разработки стратегии охраны
его природных популяций.
Известно, что гон у белых медведей происходит главным образом весной.
Данные о сроках их спаривания и родов, а также репродуктивном поведении
крайне отрывочны и разрозненны. По свидетельству редких наблюдателей, в
период гона самцы неотступно ходят за самками и между ними часто случаются
драки (Lono, 1970; Ramsay, Stirling, 1986, 1988; Wiig et al., 1992).
В Канадской Арктике и Восточно-Сибирском море спаривание белого
медведя отмечается в марте-апреле (Harington, 1968; Беликов, 1993), а на
Аляске - в марте-мае (Lentfer et al., 1980). На Земле Франца-Иосифа и
Шпицбергене готовность к гону приходится на конец апреля-начало июля
(Паровщиков, 1964; Lono, 1970). Большинство этих сведений основано на
встречах парных следов животных и на гистологическом исследовании органов
размножения самцов.
За последнее десятилетие появились новые интересные сообщения о
репродуктивных особенностях белых медведей, обитающих в некоторых
арктических регионах (Derocher, Stirling, 1994; Derocher, 1999, и др.), однако
многие интимные стороны жизни и биологические особенности диких животных
можно проследить только при постоянном наблюдении за ними, что
обеспечивается содержанием зверей в вольерах.
241
Таблица 75/ Table 75
Зависимость продолжительности беременности самок белого медведя от сроков
их спаривания в условиях неволи
Dependence of pregnancy duration of polar bear (Ursus maritimus) on the time
of copulation under conditions of captivity
Сроки/Date Количество выводков/ Litter number Средняя продолж. беременности, сут/ Average gestation period, day
спаривания/ copulation родов/ birth
11-28.02 19.11-6.12 9 281 ± 2.74
2-31.03 10.11-22.12 15 254 ±2.93
1-30.04 10.11-30.12 20 237+ 1.60
8-16.05 2-7.12 4 207 ± 3.09
5-13.06 21-24.12 2 166
Нами проанализирован архивный материал по разведению белого медведя
в условиях Ленинградского (Санкт-Петербургского) зоопарка. Здесь за период
с 1932 по 1988 г. было получено 50 выводков с 88 медвежатами. Многолетние
наблюдения показали, что основной период спаривания животных (70%)
приходится на март-апрель, хотя были случаи, когда гон проходил в феврале,
мае и даже июне (табл. 75). Продолжительность беременности самок
колебалась в пределах 164-294 дней. В среднем она составляла 224 дня. Коитус
длился от 10 до 40 мин.
Анализ собранных материалов позволил выявить интересную биоло-
гическую особенность. В целом для вида характерна большая лабильность
периода гона, в то время как сроки родов - показатель строго консервативный.
Так, в связи с возрастными и физиологическими особенностями самки белого
медведя могут спариваться в отдельные годы в конце зимы, весной или в начале
лета, однако независимо от сроков гона или температуры окружающей среды
в осенне-зимний сезон роды проходят только в ноябре-декабре. Таким образом,
продолжительность беременности самок в основном определяется временем
спаривания: при поздних сроках она сокращается, а при ранних - удлиняется
(табл. 75).
По имеющимся сведениям в российской Арктике (о-в Врангеля) средняя
величина выводка в конце берложного периода (п=310) равнялась 1.8 особи.
Выводки с 2 молодыми составляли 71.3%, с 1 - 23.9% и с 3 - 4.8% (Беликов,
1993).
В Ленинградском (Санкт-Петербургском) зоопарке средняя величина
выводка (н=50) также была равна 1.8 особи. Обычно самки рожали 2 или 1
детеныша, и только в 2 случаях (4%) выводок состоял из 3 особей (1962 и
1987 гг.). Прослеживается определенная тенденция к увеличению размеров
выводков у взрослых самок (табл. 76), которые спаривались в наиболее
благоприятные для вида сроки - в марте-мае (за июнь материалов для анализа
242
Таблица 76/ Table 76
Плодовитость самок и соотношение полов в выводках белого медведя
в условиях вольерного содержания
Female’s fertility and sex ratio in the litters of polar bear (Ursus maritimus)
under conditions of captivity
Время спаривания/ Copulation time Кол-во выводков/ Litter number Кол-во детенышей, особей/ Cubs number, ind. Процент самцов/ % of males
всего/ total на 1 самку/ per 1 female
Февраль/ February 9 14 1.55 ±0.10 50
Март/ March 15 27 1.80 ±0.08 51.9
Апрель/ April Май/ May Июнь/ June 20 35 1.75 ±0.12 54.3
4 9 2.25 ± 0.28 55.6
2 3 1.50 33.3
недостаточно). При очень раннем (февраль) спаривании наблюдалось снижение
уровня плодовитости самок.
Выявленные особенности подтверждаются наличием некоторой связи между
продолжительностью беременности и фактической плодовитостью самок (табл.
77). Как показывают собранные материалы, сокращение периода беременности
в пределах, характерных для вида, может сопровождаться увеличением
среднего размера выводка, и наоборот. По-видимому, удлинение срока
беременности, которое наблюдается при спаривании животных в феврале,
определяет большую вероыятность резорбции эмбрионов от действия внешних,
неблагоприятных факторов, т.е. приводит к снижению размеров выводков. В
условиях вольерного содержания, находясь во время родов в своих зимних
Таблица 77/ Table 77
Продолжительность беременности и плодовитость самок белого медведя,
содержащихся в неволе
Pregnancy duration and fertility of polar bear (Ursus maritimus), kept in captivity
Продолжительность беременности, сут/ Gestation period, day Кол-во выводков/ Litter number Кол-во детенышей на 1 самку, особей/ Cubs number per 1 female, ind.
260-294 17 1.64 ± 0.07
239-256 17 1.76 ±0.07
164-233 16 1.88 ±0.13
243
Таблица 78/ Table 78
Зависимость продолжительности беременности самок бурого медведя от сроков их
спаривания при вольерном содержании
Dependence of pregnancy duration of females of brown bears (Ursus arctos) on the time
of their copulation under conditions of captivity
Сроки/Date Кол-во выводков/ Litter number Средняя продолжительность беременности, сут/ Average gestation period, day
спаривания/ copulation родов/ birth
5-31.05 2-25.01 7 240 ± 1.05
10-23.06 18-22.01 3 218 ±3.78
3-7.07 25.12-31.01 5 195 ±6.12
убежищах («берлогах»), самки белого медведя приносили приплод преимуще-
ственно в ночное и утреннее время.
Молодые самцы бурого медведя (Ursus arctos) достигают половой зрелости
на четвертом, иногда пятом году жизни, что подтверждается гистологическим
анализом их генеративных органов. Однако в этом возрасте они еще заметно
уступают взрослым особям по массе тела и в ненарушенных по демографической
структуре популяциях, видимо, отгоняются ими от самок и не принимают участия
в размножении (Данилов и др., 1979). По наблюдениям В.С. Пажетнова (1993а)
реакция страха у молодых зверей поддерживается в течение нескольких лет и
является важным барьером к преждевременному спариванию. Только на 4-5-м
году жизни, в период, предшествующий гону, медведи начинают обнаруживать
стремление к контактам с другими особями и перемещаются на новые места.
Такие одиночные животные представляют популяционный резерв на период гона
и естественного расселения. Перемещаясь на свободные территории, они уже
способны к продуктивному спариванию. По данным автора, в центральной
части, как и на Северо-Западе России (Данилов, Туманов, 1991), гон у медведей
проходит в мае-июне, а рождение молодых - в декабре-январе. Супружеские
пары находятся вместе от 4-6 до 12-15дней. Они живут на небольшой территории.
В это время самка интенсивно питается, а самец почти ничего не ест, неотступно
следуя за ней, сидит или лежит невдалеке. Иногда в местах спаривания зверей
образуются «токовища», которые характеризуются множеством коротких троп
с четко очерченными вытоптанными площадками.
Самки становятся половозрелыми в 3.5, иногда 4.5 года. Звери (4 особи),
добытые осенью в возрасте двух лет, судя по всему, в гоне не участвовали. Их
репродуктивные органы были анемичными и инфантильными, а на срезах
яичников наблюдались лишь примордиальные и мелкие полостные фолликулы.
Возрастные изменения веса и размеров генеративных органов у самок,
отстрелянных в январе 1988 г. в Лодейнопольском районе Ленинградской
области, прослеживались отчетливо. У двухлетней особи вес матки с рогами и
яичниками был равен 2.1 г, масса каждого яичника 210 мг, длина тела
244
Таблица 79/ Table 79
Плодовитость самок и соотношение полов в выводках бурых медведей,
спаривающихся в разные месяцы
Fertility of females and sex ratio in litters of brown bears (Ursus arctos) copulating
in various months
Время спаривания/ Copulation time Кол-во выводков/ Litter number Кол-во детенышей, особей/ Cubs number, ind. % самцов/ % of males
всего/total на 1 самку/ per 1 female
Май/Мау 7 19 2.7 ± 0.2 57.9
Июнь/June 3 9 3 55.6
Июль/July 5 11 2.2 ±0.2 18.2
матки 50 мм, а рогов - 5.8 и 6.3 см. У трехлетней самки половые органы
были заметно крупнее: их вес составлял 9.8 г, а каждого из яичников - по 1.1 г. Тело
матки имело в длину 56 мм, а ее рога -10.2 и 12.5 см. Судя по всему, летом она
могла принять участие в размножении.
Взрослая самка (возраст 4-5 лет), добытая в том же районе в середине
февраля, была с 3 медвежатами весом по 19-20 кг каждый. Она имела
толстостенную матку с крупными рогами и яичниками общим весом 18.6 г.
Масса яичников достигала 1.8 и 2.1 г. Длина тела матки 85 мм, а ее рогов -
16.5 и 20.5 см. В яичниках отчетливо были видны некрупные полостные
фолликулы, свидетельствующие об отсутствии активизации в половой сфере
зверя. Это подтверждает данные о том, что в норме взрослая медведица
приносит потомство не ежегодно - через год, а на севере нередко и через два
(Данилов, 1994). Вместе с тем самки, «потерявшие» медвежат в раннем
возрасте, могут участвовать в размножении каждый год. По свидетельству
А.М. Коровиной (1964), в Московском зоопарке, в случае отъема от матери
медвежат в возрасте 4-5 мес, она способна приносить приплод ежегодно.
Данную репродуктивную особенность вида можно использовать в
практических целях - для увеличения биологической продуктивности
разводимых животных.
Известно, что у бурых медведей беременность протекает с длительной
задержкой имплантации бластоцист. По результатам анализа материалов (15
случаев), полученных в Ленинградском (Санкт-Петербургском) и Московском
зоопарках, с точно установленными сроками первого дня спаривания и родов,
средняя ее продолжительность у самок равнялась 221 (174-257) дню.
Всего мы располагаем данными по 52 случаям рождения медвежат в неволе
за период 1952-1994 гг. Исследования показали, что для бурого медведя, как
и для белого, характерны достаточная растянутость периода, когда они могут
спариваться, и строгая консервативность сроков родов. Так, в условиях
вольерного содержания в разные годы гон у зверей мог проходить в мае, июне
или начале июля. Между тем, независимо от сроков спаривания или
245
Таблица 80/ Table 80
Плодовитость бурого медведя и смертность медвежат в Карелии
(по: Данилов и др., 1979)
Fertility of brown bear (Ursus arctos) and mortality of young bears in Karelia
Возраст/ Age Сезон/ Season Встреч самок с кол-вом медвежат/ The number of occurrence of females with cubs Среднее кол-во медвежат на 1 самку/ Average cubs number per 1 female Смертност %/ Mortality, %
1 2 3 4 5
Новорожден./ Newborn Сеголетки/ Youngs, Оч- Лончаки/ Yearlings, 14- ЗИМа/ 2 8 10 1 1 2.58 winter лето- осень/ ю 49 18 2 . 2.15 16.6 summer- autumn лето- осень/ 21 23 8 - - 1.75 18.6 summer- autumn
температуры окружающей среды в летне-осенний сезон, роды всегда проходили
в январе и только в 4 случаях (7.7%) - во второй половине декабря. Таким
образом, длительность беременности определяется временем спаривания
самок: при поздних сроках она сокращается, а при ранних - удлиняется.
Особенно наглядно это прослеживается, когда имеется точная датировка
сроков спаривания и родов (табл. 78).
Наблюдается также тенденция к снижению размеров выводка при спа-
ривании медведя в конце сезона размножения - в июле (табл. 79). В данном
случае плодовитость зверей не определялась возрастом самок, поскольку все
они уже были взрослыми и ранее приносили приплод. По-видимому, в июле
происходит затухание фолликулярной деятельности яичников, и спаривание в
этом месяце может приводить к уменьшению размера выводка.
В естественных условиях медведицы чаще приносят 2-3 медвежат, пометы
с 4-5 детенышами встречаются редко (Мельницкий, 1915; Hensel et al., 1969;
Данилов и др., 1979; Пажетнов, 1990, и др.). По наблюдениям в Эстонии,
молодые медведицы чаще рожают 1-2 детенышей, старые - 2-3. Из 154
выводков 18.9% содержали по одному, 61.6 - по два, 17.5 - по три, 1.3 - по
четыре и 0.7% - по пять детенышей (Каал, 1972). Вероятно, потенциальная
плодовитость вида весьма значительна, но она либо вообще не реализуется,
либо нивелируется довольно высокой эмбриональной смертностью.
Как сообщает П.И. Данилов (Данилов и др., 1979, 1994), средняя пло-
довитость медведя в Карелии, рассчитанная по встречам новорожденных
в берлогах, - 2.58 особей на 1 рожавшую самку. При этом гибель детенышей
в первые месяцы жизни относительно невелика, как, впрочем, и смертность
медвежат в течение второго года жизни (табл. 80)
246
По результатам анализа 52 случаев рождения медвежат в Ленинградском
(Санкт-Петербургском) и Московском зоопарках, в выводках в разные годы
также преобладали пометы с 2-3 детенышами. Случаи рождения одного или
четырех медвежат встречались сравнительно редко. В рассматриваемой
выборке только у одной самки выводок состоял из 4 детенышей и у трех - из 1
(5.8%). Средняя плодовитость медведиц, содержащихся в условии неволи,
оказалась достаточно высокой и составляла 2.5 ± 0.09 особи на самку.
Интересно, что и у бурого, и у белого медведей при поздних покрытиях (по
сравнению с ранними) рождалось меньшее количество медвежат и соотношение
полов в выводках часто сдвигалось в пользу самок.
Таким образом, представленные в разделе материалы демонстрируют
многообразие биологических особенностей репродуктивного процесса хищных
млекопитающих, что следует рассматривать как единую адаптивную
стратегию, способную обеспечить видам поддержание численности на
оптимальном уровне.
Резюме
Прослеживается определенная зависимость между массой тела, уровнем
энергообмена и скоростью полового созревания хищных млекопитающих. Чем
экономичнее в энергетическом отношении представители видов, тем позже
они вступают в период полового созревания и, следовательно, популяция в
целом медленнее восстанавливает численность после депрессии.
Особи мелких видов семейства Mustelidae по сравнению с крупными
обладают высоким уровнем энергообмена и могут испытывать заметное
воздействие на организм неблагоприятных факторов. Это обстоятельство,
видимо, и определило в процессе филогенеза отбор на большую их
плодовитость, раннее половое созревание, короткую (у большинства видов)
продолжительность беременности. Для более крупных представителей отряда
Carnivora характерно позднее становление половой зрелости, малая
плодовитость и длительный период развития молодняка. В случаях ранней его
гибели у самок с короткой по продолжительности беременностью выявлены
случаи их повторного спаривания и рождения детенышей в течение одного
репродуктивного периода. Отмеченная лабильность физиологического
состояния позволяет хищникам четко реагировать на действия факторов
внешней среды, что проявляетя в изменении сроков гона и интенсивности
размножения. Возможность получения в природе двух приплодов за сезон
размножения следует рассматривать как один из механизмов регуляции
численности популяций. Это дает основание считать данные виды близкими к
полиэстричным животным. При их содержании в неволе, применяя метод
ранней отсадки молодняка от матери, можно имитировать его гибель и вызывать
повторное спаривание родителей, что позволяет увеличивать уровень
биологической продуктивности самок.
Животные разного веса и размеров неодинаково реагируют на воздействие
факторов внешней среды. Интенсивный обмен веществ в организме мелких
247
хищников в процессе исторического развития определил большую их
плодовитость по сравнению с крупными. У особей мелких видов, характе-
ризующихся высоким уровнем энергообмена, интенсивное размно-жение не
только компенсирует малую продолжительность жизни, но и способствует
быстрому росту численности популяций после депрессий.
Среди куньих прослеживается прямая положительная связь (г = 0.94)
между продолжительностью жизни и массой тела животных, а также
отрицательная (г = - 0.86) - между их весом и плодовитостью самок. Отмечается
положительная линейная связь продолжительности жизни родителей и
выживаемости щенков (г - 0.81). Кроме того, наблюдается отрицательная
зависимость между плодовитостью самок и выживаемостью детенышей
(г = - 0.93), а также между первым показателем и продолжительностью жизни
зверьков (г - - 0.9). Необходимо отметить и большую выживаемость детенышей
у крупных представителей семейства куньих (70-80%) по сравнению с мелкими
(65-71%). Таким образом, важной адаптивной репродуктивной особенностью
хищников, во многом определяющей численность популяций, является для
мелких видов - значительный воспроизводственный потенциал, а для круп-
ных - высокий уровень выживаемости молодняка и большая продол-
жительность жизни особей.
Интересные черты воспроизводственного процесса прослеживаются и
среди представителей других семейств хищных млекопитающих. Например,
для медведей характерна большая лабильность периода гона, в то время как
сроки родов - показатель строго консервативный. В неволе, в связи с
возрастными и физиологическими особенностями, самки белого медведя могут
спариваться в отдельные годы в конце зимы, весной или в начале лета. Однако
независимо от сроков гона и температуры окружающей среды в осенне-зимний
сезон роды у них наблюдаются только в ноябре-декабре, а беременность
колеблется в пределах 164-294 дней. Продолжительность ее у обоих видов
медведей в основном определяется временем спаривания животных: при
поздних сроках она сокращается, а при ранних - удлиняется.
Анализ репродуктивных особенностей белого и бурого медведей позволил
также выявить наличие связи между продолжительностью беременности и
фактической плодовитостью. Обычно сокращение периода беременности в
пределах, характерных для вида, сопровождается увеличением размера
выводков, и наоборот. Удлинение периода вынашивания детенышей
определяет большую вероятность гибели части эмбрионов от действия на
организм неблагоприятных факторов внешней среды, что обусловливает
уменьшение размера выводка. Подобная зависимость прослеживается и у
других видов хищных млекопитающих {Mustela erminea, Martes martes, M.
zibellina') с длительным периодом эмбрионального развития щенков. Для
рассмотренных представителей семейства собачьих и кошачьих характерен
короткий период задержки имплантации при развитии зародышей, или
эмбриональной диапаузы, что обусловливает малую продолжительность
беременности у этих видов.
248
Summary
A certain dependence between the body sizes, energy exchange and rate of
sexual maturing of animals is observed. The more economic in the energetic
relation the representatives of species the later they enter the period of sexual
maturation and, hence, the population on the whole slowly recover its number
after depression.
The small species of the family Mustelidae with a high level of energy exchange
may experience a strong impact of unfavorable factors on the organism. This
circumstance, apparently, determined selection for high fertility, early sexual
maturation, short pregnancy duration in most species and readiness of males to
productive copulation during a long period in the process of their phylogenesis.
Late sexual maturation, low fertility and long period of young development are
typical of larger representatives of the order Carnivora. The cases of repeated
copulation and second breed during one reproductive season were revealed in females
with a short duration of pregnancy in case of early death of the young. Such flexibility
of physiological state allows animals quickly to react on impacts of environmental
factors and to adapt to new conditions, to change time and intensity of reproduction.
All this makes it possible to consider the given species to be close to polyestrus
animals. The phenomenon of two breeds during one reproductive season recorded
in nature should be considered as a mechanism of population number control. It is
possible to imitate the death of the young and to induce repeated copulation of
parents applying the method of early withdrawal of young from a mother, when
keeping animals in captivity. This allows one to increase the level of biological
productivity of females.
Carnivores of different weight and size react to the impact of environmental
factors in a different way. The lower their weight, the more intensive is the metabolism
of the organism, the closer the interaction with the habitat conditions and the stronger
the influence of environment on the individuals in population. This could have
determined the greater fertility of small carnivores as compared to large ones in the
process of historic development of species. In small species, which are characterized
by a high level of energy exchange, the intensive breeding not only compensates the
low lifetime of individuals, but promotes fast increase of population number after
depressions.
The direct positive correlation (r = 0.94) between the lifetime and body mass of
predators and dramatically pronounced negative relationship (/• = - 0.86) between
their weight and female fertility were found among the species of the family
Mustelidae. The positive linear correlation between the lifetime of animals and
survivorship of pups (r - 0.81) is the characteristic feature of species of the given
family. In addition, the negative relationship between the fertility of females and
survivorship of young (r = - 0.93) and between fertility of females and lifetime of
parents (r - - 0.9) is observed.
An important adaptive reproductive feature of carnivores largely determining
the population number for the small species is considerable reproductive potential,
249
and, for the large species, high level of survivorship of the young and long lifetime
of individuals.
Interesting reproductive characteristics are observed in large carnivores, such
as Ursus maritimus and U. arctos. More flexible period of the rut is typical of them,
whereas time of labour is very conservative parameter. For example, in captivity,
females of Ursus maritimus may copulate, in some years, in the end of winter, in
spring or in the beginning of summer depending on the age and physiological
characteristics. However, independent of the time of rut and ambient temperature
the labour in this species in autumn-winter season is observed only in November-
December and pregnancy duration fluctuates within 164-294 days. The duration of
pregnancy in females of both species of bears, is mainly determined by the time of
copulation: with late timings it decreases and with early - elongates.
The analysis of reproductive characteristics of Ursus maritimus и U. arctos
showed the occurrence of relation between the duration of pregnancy and actual
fertility of females. Commonly the decrease of period of pregnancy within the limits
typical for the species is followed by the increase of the size of the litter and vice
versa. Seemingly, the elongation of pregnancy time determines high probability of
embryos resorption owing to the impact of unfavorable environmental factors on
the organism, i.e. leads to a decrease of the litter size. Similar dependence is observed
also in other species of carnivorous mammals {Mustela erminea, Martes martes, M.
zibellina) with a long period of embryonic development.
All recorded diversity of biological features of reproductive process in
carnivorous mammals should be regarded as a single adaptive strategy directed on
the survivability of species and maintenance of local population number at the
optimal level.
Глава 7. НЕКОТОРЫЕ ЗАКОНОМЕРНОСТИ
РОСТА И РАЗВИТИЯ МОЛОДНЯКА
Известно, что общий рост млекопитающего можно характеризовать по
динамике весовых изменений, которые служат надежным индикатором его
онтогенетического развития. Наиболее наглядное представление о данном
процессе дает такой показатель, как привес массы тела за единицу времени.
При изучении возрастной динамики массы тела животных она рассчитыва-
лась по формуле: A =(y2-v,)/t, где А - суточный привес, г; v2~v‘ - разница в
массе тела за определенный отрезок времени, г; t - продолжительность
периода, дни.
Среди представителей семейства куньих по мере увеличения массы тела
взрослых особей весовые и линейные размеры новорожденных также
возрастают. По нашим наблюдениям, проводимым в виварии Западного
отделения ВНИИОЗ и в Ленинградском (Санкт-Петербургском) зоопарке,
детеныши, измеряемые в день своего рождения, имели следующий вес: у ласки
1.2-1.7 г, солонгоя 2.5-4.3, колонка 6.7-8.1, лесного хорька 7.5-9.6, степного
хорька 5.8-8.3, европейской норки 7.9-10.5, американской норки 7.8-10.8,
лесной куницы 22.7-32.5, харзы 50.2 и у барсука 75-77 г. Этаже зависимость
прослеживается и при сравнении веса эмбрионов на последней стадии
развития, обнаруженных у самок некоторых видов с различной массой тела.
Так, у добытых в природе особей они весили: у лесной куницы 28-30 г, соболя
27-31, выдры 69-73, барсука 70-74 (Лобачев, 1976), росомахи 73-83 (Теплов,
1955) и у калана 1450 г (Барабаш-Никифоров и др., 1968). По данным
финских исследователей, рожденные в вольерах детеныши лесной куницы (8
выводков) весили 28-35 г (Nyholm, 1980).
Обычно у самок близких видов со сходной массой тела вес ново-
рожденных практически одинаков. В то же время у таких многоплодных
видов, как горностай, хорек степной и перевязка, детеныши рождаются
сравнительно мелкими. Так, у степного хорька, близкого по массе тела к
лесному, новорожденные весят 4.5-7.6 г, а у горностая, который заметно
крупнее ласки, - 0.8-2.6 г (Терновский, 1977). Пойманная в Ленинградской
области самка горностая, содержавшаяся в домашних условиях у биолога
В.А. Смелова, 22 апреля 1981 г. родила мертвыми 7 щенков (3<? и 4(J). Вес
самцов был равен 1.1-1.2 г, а самок - 0.6-1.1 г. Еще одна беременная самка
горностая, отловленная нами в Ленинградской области, родила щенков (6<?и 5$>)
26 апреля 1968 г. Измеренные на другие сутки, 5 особей имели длину тела в
среднем 43 мм, а вес - 3.2-4.2 г (Туманов и др., 1970). У взрослых самок
перевязки, размерные показатели которых сходны с таковыми лесного хорька,
масса тела новорожденных детенышей колебалась в пределах 3.2-4.7 г
(Флинт, 1962; Шило, Батенева, 1989). По-видимому, данную особенность
узкоспециализированных видов-миофагов следует рассматривать как одну
из черт своеобразной адаптации, позволяющей этим многоплодным животным
251
Таблица 81/ Table 81
Возрастные изменения массы тела (М ± /и) горностая и колонка
в первый месяц жизни, г (по: Туманов и др., 1982)
Age changes of body mass (M ± in) of ermine (Mustela erminea) and Siberia weasel
(Mustela sibirica) in the first month of life, g
Воз- раст, дни/ Age, days Г орностай/Л/му/^/« erminea Колонок/Л/ил7е/а sibirica
(/7=9-19) $$ (//=8-19) 33 £$ (/7=3-8)
масса тела/ body mass прнвсс в сутки/ weight growth per day масса тела/ body mass привес в сутки/ weight growth perday масса тела/ body mass привес в сутки/ weight growth per day масса тела/ body mass привес в сутки/ weight growth per day
1 2.110.1 - 2.2+0.1 - 7.9+O.2 7.510.3 -
5 4.9±0.3 0.7 510.2 0.7 16.6+0.3 2.2 16+0.3 2.1
10 9.2±0.5 0.8 9.310.3 0.8 32.210.9 3.1 34.1+1.8 3.6
15 14.710.5 1.1 14.110.4 0.9 50.8+0.9 3.7 51.912 3.5
20 22.711.4 1.6 22.6+1.1 1.7 68.510.8 3.5 6712.4 3
25 32.2+1.8 1.9 33+0.9 2.1 105+0.5 7.3 8913 4.4
29 43+2.1 2.7 4111.5 2 136.412.3 7.8 125.5+7.3 9.1
35 55.4+3 2.1 50.812.2 1.6 177.415.4 6.8 170+5.2 7.4
при возникновении благоприятных условий среды быстро реагировать на
них путем полной реализации своего репродуктивного потенциала.
Интересно, что у новорожденных детенышей горностая на дорсальной
стороне шеи, в выйной области, нами впервые обнаружено крупное железистое
образование (Туманов и др., 1970). Оно имеет вид кожной складки, которая
вскоре темнеет и покрывается длинным густым волосом, напоминая львиную
гриву. Примерно к 20-му дню это образование становится малозаметным и в
дальнейшем инвалюирует. Возможно, что на ранних этапах постнатального
онтогенеза горностая оно имеет важное приспособительное значение,
предохраняя мягкие ткани кожных покровов молодняка от повреждения.
Обычно многоплодная самка переносит расползающихся из гнезда щенков
или (в случае опасности) перетаскивает их на новое место, хватая детенышей
преимущественно за загривок. По своей природе это образование, видимо,
близко к мощно развитой надвыйной железе новорожденных у американской
норки, функции которой до конца не ясны (Азбукина, 1970, 1973; Петской,
Колповский, 1970).
Скорость весового и линейного роста куньих изучена достаточно полно
(Осмоловская, 1948; Абрамов, 1961; Мелькина, 1966; Heidt, 1970; Туманов,
1971, 1972; Терновский, 1974, 1977; Сегаль, 1975а; Данилов, Туманов, 1976а,
б; Курис, 1980; Heggberget, 1993; Reuther, 1993, 1999, и др.).
Для молодняка мелких представителей семейства характерно постепенное
нарастание абсолютного суточного привеса (табл. 81) за период от рождения
до прозревания. После открытия глаз он резко возрастает, достигая максимума'
252
Таблица 82/ Table 82
Динамика массы тела {М ± /л) ласки и солонгоя в первые 2 месяца
постнатального развития, г (по: Туманов и др., 1982)
Dynamics of body mass {M ± m) of weasel {Mustela nivalis) and Alpine weasel {Mustela
altaica) in the first 2 months of postnatal development, g
Воз- раст, дни/ Age, days Ласка/Mustela nivalis Сопонгой/Mustela altaica
&3 (//=3-8) $$ (/1=3-8) (»=3-ll) $£ (//=2-11)
масса тела/ body mass привес всут/ weight growth per day масса тела/ body mass привес в сут/ weight growth per day масса тела/ body mass привес в сут/ weight growth per day масса тела/ body mass привес в сут/ weight growth per day
1 1.5+0.1 - 1.4±0.1 - 4.1±0.1 - 4±0.1 -
5 4.7+0.4 0.8 4.6+О.З 0.9 11.9+0.3 1.9 10.5±0.4 1.6
10 7.1±0.7 0.5 7.6+0.7 0.6 24.6+0.4 2.5 21±0.3 2.1
15 11.3+0.4 0.8 ll.l±0.4 0.7 28.8±0.8 0.8 25.6+0.6 0.8
20 16.5+1.1 1.4 17.1+1.3 1.2 39.5±0.9 2.1 33±0.8 1.5
25 20.9+1.6 0.9 21.2+1.4 0.8 55+1.3 3.1 45.7±0.9 2.5
30 35.4+3.4 2.9 30.8±0.7 1.9 74.8+8.3 3.9 63.5 3.5
35 42.9+6.1 1.5 37.6±1.2 1.3 110.6+8.8 7.1 89.5±13.5 5.2
40 47.1+5.6 0.8 40.4±0.8 0.6 146.3+10.2 7.1 102.5+13.3 2.6
45 49.3+7 0.4 42.4±1.2 0.4 178+11.7 6.3 145.7+16.7 8.6
50 53.6+8.5 0.8 46.8±0.9 0.9 208±5.2 6 162+9.5 3.3
55 60.3±5 1.3 49.3+1 0.5 217.7±6 1.9 180+9.7 3.6
60 67.2+8.3 1.4 53.6+0.9 0.9 248.3+9.7 6.1 201.5+13 4.3
к моменту, когда щенки начинают выходить из гнезда и потреблять много
дополнительной пищи, а затем вновь снижается (табл. 82).
Вслед за открытием глаз абсолютный суточный привес самцов ста-
новится больше, чем у самок, хотя скорость общего роста последних
(как и мелких видов по сравнению с крупными) заметно выше. Например,
в условиях неволи масса тела у молодых самок ласки в 60-дневном возрасте
составляла 84.4%, у горностая 83.1 и у колонка 66.2% от массы тела роди-
телей, а у молодых самцов соответственно 73.6, 61.6 и 54.5%. Величина
относительного привеса наиболее высока в первые 10 дней жизни щенков,
а затем заметно уменьшается в связи со снижением их общей скорости роста.
Интенсивное изменение массы тела у детенышей прослеживается только в
первые два-три месяца, после чего оно замедляется. В отмеченный период
их линейные размеры составляют уже 90-95% по сравнению со взрослыми
особями, хотя по весу они достигают дефи-нитивного уровня несколько
позже - у представителей мелких видов примерно к 4-6, а у крупных - к 8-
10 мес.
253
Таблица 83/ Table 83
Возрастные изменения массы тела (М ± /и) степного и лесного хорьков в первые
месяцы жизни, г
Age changes of body mass (M ± m) of Siberia {Mustela eversmanni) and European
{Mustela putorius) polecats in the first months of life, g
Возраст, дни/ Age, days Хорек степной/Л/tf.tfe/fl eversmanni Хорек лесной/Л/м5/е/л putorius
<3<3 ?9(n=3-13) $$(n=3-6)
масса тела/ body mass привес в сут/ weight growth, per day масса тела/ body mass привес в сут/ weight growth, perday масса тела/ body miss привес в сут/ weight growth, perday масса тела/ body mass привес в сут/ weight growth, perday
1 9.5±О.З - 8±0.8 - 93*0.6 - 8.4+O.4 -
5 18.8*0.5 2.3 16.2*1.1 2.1 20.1*1.8 2.7 19.4*1.1 2.7
10 41.5*2.1 4.5 37.5*1.5 4.3 45.1*2.1 5 37.8*2.1 3.7
15 69.2*3.2 5.5 61.6*2.6 4.8 80.2*3.8 7 64.8*2.2 5.4
20 90*4.7 4.2 81.3*3.5 3.9 109.7*4.2 5.9 89.5*7.4 4.9
25 114.2±3.7 4.8 106.3*2.9 5 147*1.3 7.5 124.5*5.3 7
30 150*5.9 7.2 127.2*4.3 4.2 214.3*1.7 13.5 169.8*10.3 9.1
35 195.3*12.6 9.1 167.6*9 8.1 300.3*12.1 17.2 239.5*11.1 13.9
40 309*6.7 22.7 249*8.1 16.3 360*8.4 11.9 283*8.5 8.7
45 414±7.9 21 333*7.6 16.8 450.7*30.2 18.1 350*9.3 13.4
50 473.3*26.4 11.9 396.5*8.5 12.7 521.6*17.3 14.2 408.4*21.3 11.7
55 570*46.2 19.3 468.5*7.6 14.4 - - - -
Взрослые/ Adults 1285±21.5 - 691*37.3 - 1232*24.9 - 797*28.3 -
В отличие от наиболее мелких представителей семейства куньих (ласка,
горностай, солонгой), у которых однодневные детеныши разного пола
практически не различаются по массе тела, у хорьков и норок в том же возрасте
самцы уже заметно крупнее самок, хотя разница эта не всегда существенна.
Достоверные различия по данному тесту прослеживаются у них очень рано - в
возрасте 10-12 дней, в то время как у мелких куньих - в конце первого месяца
жизни, когда происходит их прозревание.
В раннем возрасте у рассматриваемых хищников изменение весовых и
линейных размеров происходит очень быстро. Так, на пятые сутки масса тела
увеличивается у хорьков и норок в 2-2.5, а у соболя и куницы - в 2.7-2.9 раза.
К концу второй недели жизни у европейской и американской норок, лесного и
степного хорьков она превышает таковую однодневных детенышей в 7.3-8.6
раза, а у белого хорька - фуро, лесной куницы и соболя - в 6.3-6.7 раза. В
целом для молодняка хорьков и норок характерно ровное и достаточно плавное
нарастание суточного привеса в период от рождения и до прозревания (рис.
58). Во время процесса открытия глаз (30-35-й день) и в последующие 3-5 суток
он существенно возрастает, что, видимо, связано с участившимися выходами
детенышей из гнезда (исследовательские реакции), увеличением двигательной
активности и потреблением дополнительной пищи. Если до этого суточный
254
Рис. 58. Возрастные изменения массы тела и суточного привеса лесного (д)
и степного (б) хорьков.
Масса тела: /-самцы, II-самки; суточный привес: 1 - самцы, 2 - самки.
По оси абсцисс - дни; по оси ординат: слева - масса тела, г; справа - суточный
привес, г.
Fig. 58. Age changes of body mass and circadian weight growth rate of European
(Mustela putorius) (a) and Siberia (Mustela eversmanni) (6) polecats.
Body mass: I - males, II - females; daily weight growth: 1 - males, 2 - females.
Abscissa - days; ordinate: to the left - body mass, g; to the right - circadian weight
growth rate, g.
привес щенков разного пола примерно одинаков (табл. 83-84), то вслед за
открытием глаз он в целом больше увеличивается у самцов (рис. 59). Однако
масса тела самок по отношению к дефинитивному уровню уже к двухмесяч-
ному возрасту была выше, чем у самцов, и составляла у степного хорька 67.8%,
европейской норки и лесного хорька - по 51.2 и у фуро 46.4%. У самцов этот
показатель равнялся соответственно 44.4, 35.8, 42.3 и 43.2%.
Сходная картина изменений скорости весового роста прослеживается
также у молодых соболей и лесных куниц. В первый месяц жизни их абсолютный
суточный привес колебался в пределах 8.2-10.6 г. Позднее, после прозревания,
у 1.5-3-месячных детенышей, как и у других видов, он возрастал и составлял
уже 10.8-22.7 г в сутки.
255
Рис. 59. Возрастные изменения массы тела и суточного привеса европейской
норки (а) и фуро (б).
Масса тела: / -самцы, II- самки; суточный привес: 1 - самцы, 2 - самки.
По оси абсцисс: дни; по оси ординат: слева - масса тела, г; справа - суточный
привес, г.
Fig. 59. Age changes of body mass and circadian weight growth rate
of European mink {Mustela lutreola) (a) and Furo {Mustela putorius furo) {6).
Body mass: / -males, II- females; daily weight growth: 1 - males, 2 - females.
Abscissa - days; ordinate: to the left - body mass, g; to the right - circadian weight
growth rate, g.
Показатели относительного прироста хорьков и норок сравнительно высоки
в первые 2 недели поснатального онтогенеза, после чего начинают плавно
снижаться. Заметный рост молодых животных наблюдается в возрасте 3 мес, а
затем он замедляется. В это время линейные размеры детенышей составляют
уже 90% от размеров родителей, хотя по массе тела они достигают уровня
последних только к 6-7 мес.
В исследованных нами 3 выводках барсуков на ранних этапах пост-
натального развития масса тела у самок была несколько выше таковой у
самцов. После открытия глаз (36-40-й день) эта разница стала особенно
очевидной (табл. 85). Когда животным исполнилось 2.5 мес, их вес практически
сравнялся, а затем самки начали отставать в росте от самцов. Наиболее
интенсивное нарастание среднесуточного привеса наблюдалось вслед за
прозреванием, а затем после того, как детеныши стали пробовать мясную пищу.
256
Таблица 84/ Table 84
Возрастные изменения массы тела {М ± т) европейской норки и фуро в первые
месяцы жизни, г
Age changes of body mass {M ± m) of European mink {Mustela lutreola) and furo
{Mustela putorius furo) in the first months of life, g
Возраст, дни/ Age, days Европейская норка/Л/млгеМ lutreola &vvolMustela putorius furo
33 0>=3-9) 22 01=з-б) 33 01=3-10) 22 («=3-9)
масса тела/ body mass привес в сут/ weight growth, per day масса тела/ body mass привес в сут/ weight growth, per day масса тела/ body miss привес в сут/ weight growth, per day масса тела/ body mass привес в сут/ weight growth, per day
1 10.5±0.4 9.05±0.7 11.8±0.3 I0.2±0.8
5 24.3+1.3 3.5 22.7±2.4 3.4 25.2±1 3.4 21±0.7 2.7
10 54.4+4 6 42.3+2.2 3.9 51.9+0.6 5.3 42.9+1 4.4
15 77.9±3 4.7 66.4±2.7 4.8 79.4±1.4 5.5 64+1.1 4.2
20 125.5±7 9.5 93±11.8 5.3 104.7±5.9 5.1 78.6±2.3 2.9
25 139.5±8.6 2.8 110+5.6 3.4 182.5±4.8 15.6 95.2±6.9 3.3
30 180.7±3.9 8.2 173.1±4.7 12.6 267.5±6.3 17 134±10.9 7.8
35 230.5±7.3 9.9 212.5±7.3 7.9 301±4.5 6-7 189+8.8 11
40 282±3.8 10.3 238±5.4 5.1 333.7±4.2 6.5 227.3±18.1 7.7
45 319±6.7 7.4 253±8.1 3 376±5.1 8.4 287±17.6 11.9
50 349±7.5 6 279±6.3 5.2 443.7±29.8 13.5 334.7±18.1 9.5
55 381±5.6 6.4 329±6.5 10 501.5±17.9 11.6 384.7±12.3 10
60 422.8±4 8.4 390±5 12.2 574±21.4 14.5 429+16 8.9
65 485±9.1 12.4 435±6.2 9 641.5+18.3 13.5 485.5±15.6 11.3
70 548.3±23.1 12.7 503±18.1 13.6 759.5±15.7 23.6 564±16.4 15.7
Взрослые/ Adults 1180+41.3 - 762±11.5 - 1329±45.2 - 925+29.1
Ходить барсуки начинают поздно - в возрасте полутора месяцев. Взятые
нами из гнезда щенки (37-38 дней) едва ползали и большую часть суток спали.
Проснувшись, они пили из соски теплое молоко и после этого снова засыпали.
Перед едой им приходилось подолгу массажировать животы, так как
самостоятельно оправляться они начинают не ранее 46-47 дней. В обычных
условиях самки всегда тщательно вылизывают щенков, массажируя их и таким
образом помогая освободить кишечник. В полтора месяца барсуки уже много
играли друг с другом, напоминая по манере поведения щенков собаки.
Интересно, что если детенышей брали за загривок даже в самый разгар игры,
то они сразу же складывали хвосты и задние лапы на живот, передние опускали
вниз и в таком «транспортабельном» виде затихали. Вероятно, эта адаптивная
реакция выработалась у животных в связи с тем, что при избыточном
увлажнении гнезда весной или заселении его эктопаразитами самка сразу же
переносит щенков по узким проходам в другую гнездовую камеру или более
сухой отнорок.
Долгое время молодые барсуки, за которыми велось наблюдение, сами не
ели, и их приходилось кормить из соски (Туманов, 1971, 1972). В возрасте
257
Таблица 85/ Table 85
Динамика размеров (мм) и массы тела (г) у барсуков
на ранних этапах постнатального онтогенеза
Dynamics of sizes (mm) and body mass (g) of badgers (Meles meles) at the early steps
of postnatal development
Возраст, дни/ Age, days Самец/Male Самка/Female
длина/length масса тела/ body mass привес в сут/ weight growth, per day длина/length масса тела/ body mass привес в сут/ weight growth, per day
тела/ body хвоста/ tail тела/ body хвоста/ tail
1-2 120 32 75 - 120 32 77 -
10-11 185 40 131 6.2 186 40 135 6.4
20-21 230 48 304 17.3 235 48 312 17.7
25-26 262 55 412 21.6 264 55 423 22.2
37-38 295 62 900 40.7 305 64 970 45.6
41-42 305 70 1000 25 315 65 1050 20
46-47 310 73 1045 9 320 70 1065 7.5
48-49 315 75 1080 17.5 325 73 1100 17.5
51-52 335 80 1280 66.7 345 75 1380 93.3
58-59 390 93 1800 74.2 390 90 1870 70
64-65 435 95 2140 56.7 435 93 2280 68.3
70-71 450 105 2450 51.7 460 105 2600 53.3
74-75 460 105 2900 112.5 470 105 3000 100
79-80 480 106 3370 94 480 106 3350 70
89-90 520 110 4300 93 500 108 3900 55
2.5 мес они перешли на самостоятельное питание, и у них наблюдалась
интенсивная смена зубов, которая закончилась к 3 мес. Видимо, в этом возрасте
в природе они начинают выходить из нор и приучаются разыскивать пищу
сами. Так, 1 июля 1968 г. в Ленинградский (Санкт-Петербургский) зоопарк
поступил молодой барсучонок весом 3.1 кг, который был обнаружен утром в
лесу один. В естественных условиях молодняк остается при матери до конца
лета, а осенью мы неоднократно находили сеголеток, которые жили отдельно
от взрослых животных в неглубоких, просто устроенных норах.
У другого крупного представителя семейства - речной выдры - в отличие
от обитателя морских побережий калана молодые рождаются слепыми, с
закрытыми слуховыми проходами. По наблюдениям О.В. Трапезова (1999),
проведенным на ферме Института цитологии и генетики СО РАН (г. Но-
восибирск), масса щенков при рождении колебалось от 70 до 120 г, а длина
258
Таблица 86/ Table 86
Динамика размеров (мм) н массы (г) тела молодых выдр в первые месяцы жизни
Dynamics of body sizes (mm) and mass (g) of young otters {Lutra lutra)
in the first months of life
Дата/ Date Самец/Male Самка/Female
длина/length масса тела/ body mass привес в сут/ weight growth, per day длина/length масса тела/ body mass привес в сут/ weight growth, per day
тела/ body хвоста/ tail тела/ body хвоста/ tail
30.08 350 157 950 - 310 142 870 -
5.09 360 160 1020 11.6 320 145 950 13.3
12.09 375 172 ИЗО 15.7 335 150 1050 14.3
18.09 381 179 1200 11.6 348 151 1150 16.6
25.09 393 193 1780 82.8 362 153 1580 61.4
5.10 400 200 2150 37 370 155 1860 28
25.10 415 208 2580 28.6 383 160 2190 22
тела - от 120 до 200 мм. В возрасте 20 дней выдрята начинают впервые поедать
мясной корм, у них открываются слуховые походы, а в 4-5 недель - глаза. В
50-60 дней у самок масса тела достигает 1-1.3, а у самцов - 1.3-1.5 кг.
Как сообщает К. Ройтер (Reuther, 1999), в Германии отчетливые половые
различия у детенышей выдры прослеживаются по массе тела в 50-60, а по длине -
в 60-70 дней. По его данным, в первые 2 мес жизни за каждые 10 суток их
размерные показатели увеличиваются на 3-5 см, а весовые - на 200-300 г.
В двух гнездах выдры, обнаруженных в природе (Ленинградская область)
14 мая и 20 сентября, находилось по 2 щенка (в каждом по Id1 и 1$). К периоду
прозревания они мало различались по размерным показателям. В это время в
первом выводке самец весил 690, самка 600 г, во втором соответственно 705 и
620 г. К 2-месячному возрасту самцы стали заметно обгонять самок по скорости
весового роста, и к 3 месяцам различия по массе тела были уже отчетливо
выражены.
Еще одно гнездо выдры было найдено в той же области 28 октября. В нем
находились еще слепые 2 самки весом 570 и 460 г при длине тела 260 и 220 мм, а
хвоста-145 и 140 мм (Туманов, 1971). Глазау них открылись через 7-8днейпосле
отлова. К этому времени, т.е. в месячном возрасте, они весили 680 и 650 г.
Изменения размеров тела выдрят из одного помета, найденного в природе
30 августа, представлены в табл. 86. Судя по опубликованным данным о
динамике размерных показателей молодняка этого вида, родившегося в
вольерах (Reuther, 1999), щенкам было 40-45 дней. Малыши только начали
ходить, пили молоко из соски, а от мясо-рыбного фарша отказывались. В первое
259
время они росли довольно медленно, прибавляя в весе за 10 суток в среднем на
130-150 г. К 25 сентября, т.е. в возрасте 9-10 недель, активность выдрят
повысилась, они стали более подвижными и игривыми. Темпы их весового и
размерного роста увеличились, а суточный привес достигал 60-80 г, что по
времени совпадало с переходом животных на дополнительное питание пищей
животного происхождения. В дальнейшем отмечалось некоторое снижение
интенсивности роста детенышей. Самостоятельно есть рыбу и кусочки мяса
они стали только 3-4 октября, а уже к 18-19-му (возраст 3 мес) полностью
отказались пить предлагаемое им молоко. Несколько раньше, при массе тела
около 2 кг, выдрята стали плавать, вначале робко и неуверенно, но затем
свободно и с большой охотой.
По-видимому, и в природных условиях при среднем весе в 2 кг молодые
животные способны хорошо плавать. Так, 16 октября 1968 г. в Ленинградской
области мы встретили взрослую самку с 2 детенышами, которые плавали, ныряли
и неотступно следовали за матерью. Отловленные выдрята оказались самцом и
самкой, у которых проходила интенсивная замена молочных клыков на постоянные.
Вес самца был равен 2.2 кг, самки - 1.8 кг. Длина тела и хвоста у первого щенка
составляли соответственно 430 и 215 мм, у второго - 390 и 155 мм.
У близкого родственника выдры - калана новорожденные детеныши имеют
в длину 50-56 см и весят 1.4-1.6 кг. Они рождаются зрячими, в плотном
ювенильном меху, имея все 26 молочных зуба. В первые дни после рождения
молодняк малоактивен и много лежит на берегу или в прибрежных водах на
спине. Активно плавать начинает в 2.5-3 недели, а нырять - заметно позднее.
Детский мех меняется на постоянный в возрасте около 6-7 мес. К этому же
времени заканчивается смена молочных резцов и первых ложнокоренных на
постоянные. В возрасте 10-12 мес вес зверей достигает 17-19 кг при длине тела
1.25-1.3 5 м, а временный зубной ряд уже полностью заменен на постоянный,
состоящий из 32 зубов (Ильина, 1950; Гептнер и др., 1967).
Следует отметить, что у молодых околоводных хищников, за исключением
калана, тесная связь с водной средой обитания устанавливается сравнительно
поздно. Например, детеныши европейской и американской норок начинают
периодически покидать гнездо практически сразу же после открытия глаз, т.е. в
месячном возрасте, а входить в воду и плавать - только в 2-2.5 мес. По нашим
наблюдениям, щенки, родившиеся в вольерах 1 мая от пойманной в природе
беременной самки американской норки, стали плавать и робко нырять 28-29
июня. Известен также случай, когда 25 июня в Псковской области было найдено
гнездо с детенышами европейской норки в возрасте 40-45 дней. Одну молодую
самку взяли из помета и вырастили в неволе. Во время прогулки 27-28 июля она
начала самостоятельно подходить к канаве с водой и пробовать ее лапой, а
затем плавать. Во время полевых работ 24 июня 1967 г. на Карельском перешейке
(Ленинградская обл.) нами было обнаружено гнездо с 4 молодыми амери-
канскими норками в возрасте около 2 мес. Они выскочили из норы и подбежали
к кромке воды, но лишь одна из них рискнула переплыть ручей шириной 2 м,
остальные же повернули назад и скрылись в прикорневых пустотах.
260
Таблица 87/ Table 87
Возрастные изменения массы и длины тела самца росомахи
Age changes of body length and mass of male of wolverine (Gulo gulo)
Показатели/ Parameters Время измерений/Censusing time
4 апреля/ 14 апреля/ 24 апреля/ 1 мая/ 9 мая/ 3 июля/
4 April 14 April 24 April 1 May 9 May 3July
Возраст, дни/ Age, days 35-40 45-50 55-60 62-67 70-75 125-130
Масса тела, кг/ Body mass, kg 1.8 2.5 3.6 4.4 5 7.8
Длина тела с хвостом, см/ Length of body and tail, cm 45 53 56 64 70 97
Примечание. Данные получены E.A. Крутовской в заповеднике «Столбы».
Сведения по онтогенезу росомахи крайне ограничены. Интересные
наблюдения за ростом и развитием молодого самца этого вида провела
сотрудница заповедника «Столбы» (Красноярский край) зоолог Е.А. Крутов-
ская (архив заповедника). Детеныш (возраст 30-35 дней) был передан сюда 29 мар-
та 1981 г. биологом-охотоведом А.Н. Зыряновым, который обнаружил в одном
из таежных урочищ гнездо самки росомахи с 3 щенками (2<? и 1§). Зверек был
покрыт серым ювенильным пухом, зубы не просматривались. У него, видимо,
только что открылись глаза. При содержании в неволе он первое время
потреблял только молоко и жидкую кашу, но уже через 10 дней стал охотно
пробовать сырые яйца, есть мясной фарш и кусочки мяса, т.е. перешел на
смешанное питание. Потребность в высококалорийном свежем мясном корме
появилась у него только осенью. В 2.5 мес детеныш начал активно прятать
излишки корма в домик, а позднее - в выкопанные в земле ямки, прикрывая их
сверху разными предметами.
В 45-50-дневном возрасте общая окраска меха самца заметно побурела,
на спине появилась темная ость, а на боку - светлые волосы, образующие его
«мантию». В 4 мес он был уже хорошо одет, имел густую темно-бурую шерсть
с длинной желтовато-белой полосой на боку. В весенне-летний период рост
молодого самца росомахи протекал очень интенсивно, и за 3 мес (апрель-июнь)
его размеры удвоились (табл. 87). К 3 июля (возраст 125-130 дней) детеныш
уже достиг размеров небольшой взрослой самки и имел длину тела 77 см, хвос-
та - 20, задней ступни - 18 и высоту в холке 35 см.
Изучение основных процессов роста и развития молодняка куньих
показало, что в целом у этих хищников формирование морфофизиологических
систем в процессе онтогенеза происходит раньше у представителей тех видов,
взрослые особи которых имеют меньшие весовые и линейные размеры и,
следовательно, обладают высоким уровнем энергообмена. По-видимому, это
обстоятельство обеспечивает лучшую выживаемость их щенков, смертность
которых в детский период очень велика.
261
Из молочных зубов у мелких куньих (от ласки до хорьков и норок
включительно) обычно первыми прорезаются верхние, а затем нижние клыки
и верхние крайние резцы (Z ;). Затем, через 1-2 дня следуют второй и третий (по
счету от клыков) предкоренные зубы на обеих челюстях, а еще через 3-4 дня -
очень мелкие первые верхние и нижние премоляры. Таким образом, общая
формула молочных зубов для этой группы животных -1 ‘/п C!/t Рт3/3 =18.
Выявленная последовательность в прорезании молочных зубов иногда
может нарушаться. Так, даже в одном выводке солонгоя, горностая и ласки
встречаются отдельные особи, у которых первыми появлялись не верхние, а
нижние клыки. Причем крайние верхние резцы (/•’) у них могут следовать уже
после прорезания средних предкоренных зубов на обеих челюстях. Необходимо
также отметить, что у американской норки (а нередко и у перевязки) верхние
крайние резцы вырастают раньше клыков. На черепах молодых американских
норок в возрасте около 1 мес хорошо видны все 3 пары молочных верхних
резцов, из которых над поверхностью десен выступают только крайние резцы
(Р), поэтому их можно считать единственно функционирующими временными
резцами (Aulerich, Swindler, 1968; Данилов, Туманов, 1976а).
По нашим наблюдениям, первые зубы молочной группы прорезаются у
ласки на 12-14-е сутки, солонгоя-на 13-14-е, горностая-на 15-16-е, хорьков
и европейской норки - на 16-17-е, а у американской - на 18-21-е сутки от
рождения, что в целом совпадает с данными Д.В. Терновского (1977). У более
крупных представителей семейства этот процесс происходит несколько позднее:
у лесной куницы - на 20-25-й день (Старков, 1947), соболя - на 23-25-й (Тернов-
ский, 1977), барсука - на 28-33-й (Осмоловская, 1948), а у росомахи - на 30-
37-й день (Krott, 1959, 1960). У выдрят первые молочные зубы появляются,
видимо, в те же сроки, что и у лесной куницы (Туманов, 1971; Данилов,
Туманов, 1976).
У крупных хищников семейства также отмечается заметная редукция
зубов молочной группы. Например, формула временного зубного ряда
барсука и выдры выглядит следующим образом: I С'/, РпР/2 = 14. У них,
как и у щенков мелких видов, все нижние и две средние пары молочных резцов
(7л2/;,7) отсутствуют. В то же время из характерных для взрослых осо-
бей постоянных предкоренных зубов (барсук - Рт4/4, выдра - Рт4/3) у
детенышей над поверхностью десен выступают только 2 последние пары их
предшественников (соответственно Рт3’4/3 4 и Рт34/гз). Заканчивается
формирование зубной системы временного ряда у молодняка этих видов к 65-
70-му дню.
По материалам Е.А. Крутовской, хранящимся в фондах заповедника
«Столбы», у росомахи формула зубов молочной группы несколько иная -
1°/в С'/1 Рт3/3-16. Первыми (в возрасте 32-37 дней) появляются верхние, а еще
через 3 дня - нижние клыки. Затем прорезаются первые от клыка предкоренные
зубы на нижней челюсти, а также первый и второй на верхней {Рт' 2/^. Далее
следуют 2 нижних премоляра, и заканчивается рост молочных зубов
появлением последнего верхнего предкоренного в возрасте 83-88 дней от
262
рождения. По свидетельству автора, у молодого самца росомахи формиро-
вание постоянного зубного ряда закончилось в 5 мес. В возрасте 125-130
дней у него уже сменились основные предкоренные зубы, а рядом с мелкими
саблевидными молочными клыками были видны толстые и менее изогнутые
постоянные. К этому периоду детеныш уже перелинял в зимний мех.
У мелких хищников временные зубы заменяются постоянными в течение
второго месяца жизни, и весь процесс их смены завершается к 65-70-му дню. У
крупных представителей семейства он более растянут. Как сообщают Д.В. и
Ю.Г. Терновские (1994), у соболей окончательное формирование постоянного
зубного ряда завершалось к 120-136-му дню. У содержащихся в наших вольерах
детенышей барсука только на 70-71-й день появлялась первая пара средних
постоянных резцов на верхней челюсти (/'), а двумя днями позже - вторая пара
на нижней (/,). С этого дня у них происходил интенсивный рост зубов, который
завершился к 4-4.5 мес. Смена клыков отмечалась на 90-96-й день от рождения.
В этот период можно было наблюдать прорезание мощных постоянных клыков,
появляющихся рядом с еще не выпавшими тонкими молочными.
Пробовать мясную пищу, которую приносит в гнездо самка, щенки
куньих начинают до открытия глаз, в лактационный период, и некоторое
время их питание носит смешанный характер. В этом возрасте они еще не
могут самостоятельно разорвать шкурку грызуна, хотя с жадностью сосут
кровь и выедают слабыми молочными зубами мягкие ткани частично
объеденной самкой мелкой жертвы. Впервые пробовать вскрытую полевку
содержащиеся у нас в вольерах щенки ласки стали на 16-22-й день, солонгоя -
на 17-22-й, а хорьков и норок - на 20-28-й день. Самостоятельно забивать
грызунов молодняк рассматриваемых хищников начинает заметно позднее.
По материалам Д.В. Терновского (1977), интенсивная реакция преследования
и добыча жертвы были зарегистрированы у мелких представителей семейства
в следующем возрасте: ласка - 37-50 дней, степной хорек - 51-60, европейская
норка - 52-62 и горностая - 58-72 дней. Последним в этом ряду стоит соболь
(75-85-й день), что, видимо, обусловлено большей продолжительностью
периода формирования у него зубного ряда.
Таким образом, у детенышей куньих, кроме калана, щенки которого
рождаются зрячими и со всеми молочными зубами, отмечается существенная
редукция последних. Особенно это относится к временным резцам, из которых
над поверхностью десен обычно выступают только крайние верхние (/ *), а
остальные отсутствуют. Данная особенность, видимо, имеет адаптивное
значение, поскольку молодые животные поздно переключаются на самостоя-
тельное питание. Продолжительность периода лактации у мелких хищников
составляет 1.5-2, а у крупных - до 2.5 мес. Частичная редукция временных
резцов может объясняться тем, что щенки куньих долгое время кормятся
молоком матери и сравнительно поздно начинают самостоятельно охотиться
за добычей. По-видимому, монофиодонтизм 10 резцов является одним из
характерных признаков представителей семейства Mustelidae (Терновский,
1977).
263
Формирование молочной зубной системы у молодняка куньих
происходит очень рано, до открытия глаз и выхода их из гнезда, что
позволяет самкам активно подкармливать еще слепых щенков мясной пищей
задолго до окончания лактации и обеспечивает потомству лучшую
выживаемость. Следует также отметить, что среди всех видов у молодых
самок по сравнению с самцами прослеживается тенденция к более раннему
созреванию организма, о чем можно судить не только по скорости весового
роста, но и по времени открытия глаз и ушных проходов, формированию
зубной и половой систем.
Процесс открытия глаз у мелких хищников протекает в определенные,
характерные для вида сроки и может длиться у отдельной особи от 1 до 3-5
дней. В известной мере они обусловлены уровнем энергообмена, весовыми и
линейными размерами взрослых животных. В просмотренных нами выводках
глаза открывались у детенышей в следующие дни: ласка - 22-28-й, горностай
- 29-35-й, солонгой - 26-32-й, колонок - 32-35-й, хорек лесной - 27-33-й,
хорек степной, норка американская и европейская - 30-34-й. У крупных
представителей семейства глаза открываются обычно на ЗО-й день и несколько
позднее. Так, по имеющимся сведениям, детеныши прозревают у соболя,
каменной и лесной куниц на 30-36-й день (Старков, 1947), росомахи - на 32-
35-й (Krott, 1960), канадской и речной выдр-на 30-35-й (Liers, 1951; Шило,
Батенева, 1989) и у барсука-на 32-42-йдень (Осмоловская, 1948). Слуховые
проходы у всех куньих открываются на несколько дней раньше, чем глаза.
Продолжительность этого процесса варьирует от 2 до 6 суток (Терновский,
Терновская, 1994).
Возрастные изменения морфометрических показателей выражены у куньих
достаточно отчетливо. Они характеризуются прежде всего закономерным
нарастанием абсолютного и снижением относительного веса основных
внутренних органов с увеличением возраста животных. Резкое падение индекса
органов наблюдается лишь на ранних этапах постнатального онтогенеза и
становится малозаметным, когда молодые достигают линейных размеров
взрослых особей. Для новорожденных показателен довольно высокий
относительный вес печени. В первый месяц он еще достаточно велик, но после
прозревания животных быстро снижается. Некоторое его увеличение у мелких
видов хищников отмечается только в возрасте 5-6 мес, когда по весу и
линейным размерам они практически не отличаются от родителей, хотя
организм зверьков еще не закончил свое окончательное формирование. Эти
колебания, по-видимому, отражают ход гликогенообразовательных процессов,
происходящих в печени. Они могут объясняться усиленным распадом в ней
гликогена в период наиболее интенсивного роста щенков, а также изменением
режима питания в процессе их развития.
В течение первых 4-6 недель жизни у детенышей куньих обнаруживается
сравнительно большой индекс почек и селезенки, уровень которого затем с
возрастом плавно уменьшается. Закономерное снижение индекса почек,
вероятно, определяется усовершенствованием в ходе онтогенеза абсорб-
264
Таблица 88/ Table 88
Возрастные изменения величины сердца и легких у самцов
некоторых видов куньих
Age changes of heart and lungs size of males of some mustelid species
Виды/ Species Возраст, мес/ Age, month n Сердце/Heart Легкие/Lungs
%v t P %o t P
Хорек лесной/ Mustela putorius 1-1.5 5 10.3±0.3 21.5±0.4 2.6 98 6.7 99 5-6 17 9.2±0.3 16.7±0.6 1.7 88 6.6 88 взрослые/adults 86 8.6+0.2 15.2+0.3
Норка америк./ Mustela vison 1-1.5 7 6.7+0.1 21.3+1.2 7.3 99 4.4 99 5-6 15 9+0.3 16+0.2 0.9 70 1.7 90 взрослые/adults 98 9.3±0.1 16.6±0.3
Росомаха/ Gulo gulo 6-7 3 9.1±0.3 21.5±2.2 1.2 75 2.1 92 взрослые/adults 8 8.5+0.3 16.2+1.2
Барсук/ Meles meles 1-1.5 3 5.7+0.2 26.1±1.2 2.1 92 6.2 99 5-7 6 6.3±0.2 18.6±O.l 1.4 80 6 99 взрослые/adults 14 6.8+О.З 12.6±1
Выдра/ Lutra lutra 1-1.5 3 7.5±0.2 21.4±1.6 3.5 99 3.7 99 5-7 6 9.4±0.5 15.5+0.2 1 50 8.1 99 взрослые/adults 15 10±0.3 21.4+0.7
ционных механизмов, а усиленный рост селезенки в течение первого месяца
жизни зверьков - их длительной физиологической незрелостью. При этом
интенсивное развитие селезенки как кроветворного органа и биологического
фильтра способствует увеличению жизнестойкости организма за счет большого
образования лимфоцитов.
Если у наземных видов куньих в период роста щенков наблюдается
постепенное падение относительного веса сердца, то у полуводных и роющих
форм (норки, выдра, барсук) с возрастом он увеличивается (табл. 88).
Некоторое его снижение прослеживается только на ранних этапах пост-
эмбрионального развития. Начиная с 4-5 мес индекс сердца заметно нарастает,
что, видимо, объясняется значительной функциональной нагрузкой, испытываемой
организмом полуводных и роющих животных при переходе к самостоятельному
образу жизни.
Относительный вес легких у всех видов куньих, кроме полуводных, с
возрастом уменьшается. У норок и выдры это падение прослеживается
примерно до 4 мес, после чего индекс легких вновь увеличивается, что связано
с адаптивной перестройкой организма амфибионтных животных.
Характер возрастной изменчивости веса внутренних органов у хищни-
ков обоих полов принципиально одинаков. Раннее достижение самками
10 Зак. №4309
265
Таблица 89/Table 89
Возрастная динамика температуры тела (°C) у самок ласки и солонгоя
(по: Туманов и др., 1982)
Age dynamics of body temperature (degrees) of females of weasel (Mustela nivalis)
and Alpine weasel (Mustela altaica)
Возраст, дни/ Age, days Ласка/ Mustela nivalis Солонгой/ Mustela altaica
5 (7) 30.2+0.8 (10) 31.4*0.6
10 (7) 31.8+0.9 (10) 31.8±0.5
15 (8) 32.5+0.3 (10) 32.8+0.4
20 (8) 33.1+0.2 (И) 33.8±0.4
25 (4) 33.7±0.4 (4) 34.2+0.3
30' (7) 34.8+0.2 (2) 35.3
35 (7) 34.2+0.2 (3) 34.9±0.4
40 (7) 34.6+0.2 (3)35.2±0.4
45 (4) 35.6+0.2 (3) 35±0.8
50 (3) 35.8±0.3 (5)36.8+0.4
55 (3) 36.2±0.2 (4) 37.8±0.4
60 (4) 36.7±0.3 (4) 37.1+0.3
70 (4) 37.2+0.4 (И) 37.3+0.3
.,<80 - (4) 37'.6±О.2
Взрослые/Adults (6) 37.6±0.3 (6)37.2+0.1
Примечание. Здесь и далее в скобках указано количество исследованных животных,
дефинитивных размеров согласуется с высокой скоростью роста всего
организма и различных его систем. На начальных этапах постнатального
развития детенышей половая специфика проявляется неотчетливо, однако уже
к 3 мес молодые самцы, как и взрослые особи, имеют более крупные внутренние
органы, что определяется бблыпими их весовыми и линейными размерами. В
то же время для самок характерно наличие высоких показателей индексов
сердца, легких, печени и почек, обусловленных не только меньшей массой
тела, но и интенсивностью их энергообмена (Сегаль, 1975а; Ивантер и др.,
1985).
В процессе роста детенышей мелких видов куньих наблюдаются и
существенные изменения физиологических параметров у животных, которые
до месячного возраста не способны длительное время самостоятельно
поддерживать постоянную температуру тела. Вместе с тем прослеживается
заметная устойчивость их организма к действию низких температур.
266
Таблица 90/ Table 90
Изменение температуры тела (°C) горностая и колонка на ранних этапах
постнатального онтогенеза (по: Туманов и др., 1982)
Changes of body temperature (degrees) of ermine {Mustela erminea) and Siberian weasel
{Mustela sibirica) at the early steps of postnatal ontogenesis
Возраст, дни/ Age, days Горностай/МизГе/а erminea Колонок/Mustela sibirica
S3 99 99
15 (4) 30.2±0.3 (4) 29.2+0.3 (2) 34.9 (3) 35.3+0.1
20 (9) 31.1+0.3 (8) 29.6+0.1 (2)35 (3) 34.9+0.2
25 (9) 30+0.1 (8) 30.2+0.6 (2) 34.8 (3) 35.6±0.4
30 (9) 32.1±0.3 (8) 32.4+0.4 (5) 35.2+0.2 (8) 36.1+0.3
35 (9) 34+0.5 (8) 33.9+0.3 (3) 34.5+0.2 (5) 35.2+0.3
90 (3) 37±0.5 (3) 37.5+0.4 - -
Взрослые/ Adults (7) 38+0.2 (13) 38.2±0.3 (10) 37.6+0.4 (7) 38.5±0.5
Наибольшую выносливость к длительной и глубокой гипотермии молодняк
куньих обнаруживает именно в первый месяц жизни. Некоторые легко
возбудимые самки хорьков и норок при частом беспокойстве могут
выбрасывать из гнезда 1-2 еще слепых детенышей, которые, пролежав на полу
без подстилки по 10-12 ч при температуре 15-17 °C, способны после искус-
ственного обогрева полностью восстанавливать все жизненные функции.
Нам известен случай, когда в мае из звероводческой клетки, где содержа-
лась самка дикой американской норки с пометом, один щенок в возрасте 8
дней при уборке жилища выпал из гнезда. Он провалился сквозь сетку клетки
и оставался незамеченным на цементном полу до следующего утра (около 17 ч).
Утром его подобрали, отогрели под настольной лампой и подложили под самку.
Щенок выжил и в дальнейшем ничем не отличался от других особей из того же
выводка.
Постоянная температура в гнезде мелких хищников поддерживается в
основном матерью, которая обладает хорошо развитыми механизмами
терморегуляции. Содержавшиеся в неволе лактирующие самки в холодную
погоду закрывали вход в свое убежище подстилкой гнезда, а в жаркое время,
наоборот, выбрасывали ее, оставляя щенков на прохладном полу клетки.
Кроме того, в случае отсутствия матери совместное пребывание детенышей в
группе (эффект скучивания) способствует сохранению тепла в гнезде и
поддержанию температуры их тела на должном уровне. Следует отметить, что
лесные хорьки, американские и европейские норки в возрасте 3-4 мес тяжело
переносят высокие температуры и, оставленные на 1-2 ч в жаркую погоду
закрытыми в плохо вентилируемом гнездовом домике, погибают от перегрева.
Закономерности весового роста достаточно четко согласуются с
изменениями интенсивности физиологических процессов в организме, которые
в полной мере прослеживаются по таким параметрам, как энергообмен,
температура тела и уровень сердечной деятельности. Возрастная динамика
величины потребления кислорода и ректальной температуры выражена у
куньих очень отчетливо. У детенышей ласки, солонгоя, горностая, хорька и
норок потребление кислорода в пересчете на единицу массы с возрастом
снижается, а температура тела увеличивается.
Мелкие хищники относятся к незрелорождающимся животным, поэтому
ректальная температура у новорожденных во многом определяется
температурой окружающей среды. Извлеченные из гнезда в пятидневном
возрасте, самки ласки имели среднюю температуру тела 30.2 ± 0.8, а солонгоя -
31.4 ± 0.6°С (табл. 89). За короткий период пребывания в лаборатории при
температуре воздуха 20-22°С она падала почти до температуры окружающей
среды - 25-26°С. До такого уровня температура тела снижалась у зверьков,
изолированных от братьев и сестер, через 8-12 мин, а при групповом содержа-
нии -через 15-20 мин.
С увеличением массы и размеров животных, развитием волосяного покрова
и становлением химической терморегуляции температура тела детенышей
повышалась, достигая в среднем к месячному возрасту у горностая 32.3°С, а
у колонка 35.6°С (табл.90). Как показывают материалы исследований, в
раннем периоде постнатального онтогенеза ректальная температура более
крупных хищников - колонка и солонгоя - в среднем на 2-3°С выше, чем у
ласки и горностая того же возраста. Интересно, что в течение первого месяца
жизни самая низкая ректальная температура наблюдалась у детенышей
горностая, хотя по своим размерам они больше, чем ласки. В то же время
полостная температура взрослых горностаев довольно высокая (табл. 90).
Причем разница между взрослыми особями горностая и ласки по этому тесту
близка к статистически достоверной (р>95%), а между горностаем и солон-
гоем - достоверна и в группе самцов, и в группе самок (р>99%).
Следует отметить, что на ранних этапах постнатального онтогенеза ректальная
температура обычно выше у детенышей тех видов, которые имеют большую массу
тела. Заметно раньше она достигает у них и дефинитивного уровня (табл. 91),
хотя среднесуточные темпы ее роста выше у щенков мелких видов.
У исследованных хищников интенсивность терморегуляции возрастает к
концу первого месяца жизни зверьков, когда они прозревают и становятся
более активными и подвижными. В период открытия глаз и усиления
локомоторных функций увеличиваются затраты энергии на окислительный
метаболизм, изменяются уровень потребления кислорода и ректальная
температура животных. Обычно в конце первого месяца жизни у особей разного
пола последняя увеличивалась на 0.4-1.1°С (табл. 89-90). В дальнейшем она
опять повышалась довольно плавно. Некоторый ее скачок отмечался только в
полутора-двухмесячном возрасте. По времени этот период совпадал с
окончанием лактации и переходом на мясную пищу. Например, у ласки
268
Таблица 91/ Table 91
Возрастная динамика температуры тела (°C) у самцов некоторых видов куньих
Age dynamics of body temperature (degrees) of males of some mustelid species
Возраст, дни/ Age, days Ласка обыкновенная/ Mustela nivalis Солонгой/ Mustela altaica Норка европейская/ Mustela lutreola
температ./ temp. % к температ. взрослых/ % to adult temp. температ./ temp. % к температ. взрослых/ % to adult temp. температ./ temp. % к температ. взрослых/ % to adult temp.
5 (4) 29.7+0.9 79.4 (Н) 31±0.6 83.6 (2) 32.1 86.8
10 (4) 32.2±0.1 85.1 (Н) 32.4±0.3 87.3 (2) 32.2 87
15 (8) 32.4+0.3 86.6 (П) 33+0.3 88.9 (2) 33.1 89.5
20 (8) 32.9+0.3 88 (Н) 34.4±0.4 92.7 (2) 34.4 93
25 (6) 33.1+0.4 88.5 (5) 34.6±0.3 93.3 (2) 34.7 93.8
30 (5) 34+0.2 90.9 (5) 35.7±0.3 96.2 (3) 35.7±0.3 96.5
35 (4) 3.6+0.3 89.8 (8) 35+0Л 94.3 (3) 35.2±0.2 95.1
40 (4) 34.1±0.4 91.2 (8) 34.2+0.4 92.2 (5) 35.8±0.2 96.8
45 (3) 35.2±0.3 94.1 (8) 35.8±0.3 96.5 (5) 36.3±0.3 98.1
50 (3) 35.5±0.2 94.9 (8) 36.6+0.2 98.7 (5) 36.7+0.3 99.2
55 (3) 36±0.3 96.3 (4) 36.9+0.4 99.5 (6) 36.8+0.4 99.5
60 (3) 36.3+0.2 97.1 (4) 37+0.3 99.7 (5) 37+0.3 100
Взрослые/ Adults (9) 37.4+0.3 100 (6) 37.1 ±0.1 100 (18) 37±0.2 100
заметный подъем температуры тела наблюдался на 45-48-й, а у солонгоя - на
54-55-й день от рождения. Интересно, что по данному показателю детеныши
достигают уровня взрослых особей у европейской норки примерно на 50-й,
солонгоя - на 55-й, ласки - на 70-й, а у горностая - на 90-й день от рождения.
Новорожденные хорьки, куницы и норки, как и детеныши самых мелких
представителей семейства (ласки, горностая, солонгоя), отнятые от матери,
быстро приобретают температуру, близкую к таковой окружающей среды. У
однодневного молодняка этих видов она была практически одинаковой - 26-
28.3°С. У сравнительно крупных по размеру щенков альбиносной формы хорь-
ков - фуро в том же возрасте полостная температура оказалась заметно выше
(табл. 92). В первую неделю жизни детеныши рассматриваемых видов растут
быстро, и повышение температуры тела прослеживается у них очень отчетливо.
269
Таблица 92/ Table 92
Возрастная динамика температуры тела хорьков (°C)
Age dynamics of body temperature of polecats (degrees)
Возраст, дни/ Age, days Хорек лесной/ Mustela putorius Хорек степной/ Mustela eversmanni Хорек белый (фуро)/ Mustela putorius furo
33 2$ 33 <₽? 33 1 ??
1 (6) (4) (6) (2) (5) (2)
28±0.9 26.8±1.9 26.3±0,1 26 29.2±0.6 29.5
5 (3) (5) (3) (3) (Ю) (9)
32.8±0.3 31.8±0.6 31.3±0,4 32±0.3 32.5±0.4 32±0.7
10 (3) (5) (Ю) (12) (Ю) (9)
33.4±0.3 32.7±0.5 33.1±0,3 32.7±0.5 33.2±0.2 33.5+0.3
15 (3) (4) (10) (13) (Ю) (9)
34.1±0.3 34±0.3 34.2±0,3 34.4±0.2 33.9±0.5 32.7±0.3
20 (3) (4) (Ю) (13) (7) (7)
34.5±0.3 34.4±0.2 34.6±0,2 34.3±0.3 34.9±0.3 35.1+0.2
25 (3) (4) (Ю) (13) (2) (3)
34.5±0.2 34.6±0.4 34.8±0,2 35±0.2 34.5 35.2+0.3
30 (3) (4) (4) (6) (2) (3)
35.3±0.04 35.4±0.2 35.4±0,4 35.5±0.5 35.4 35.5+0.2
35 (4) (4) (3) (5) (2) (3)
36.3±1.1 36.5±0.7 35±0,3 35.1±0.3 35 35.2+0.4
40 (3) (2) (2) (1) - -
35.5±0.3 36.3 35.4 35.2
45 (3) (2) (2) (1) (2) (2)
36.2±0.7 36.7 5.7 35.4 35.4 35.6
50 - - (3) (2) (2) (2)
36±0.4 36.3 35.8 36.0
55 (2) (2) (3) (2) - -
36.8 37 36,1±0.8 35.8
60 - - (2) (2) (3) (2)
36.4 36.5 36.2±0.3 36.2
65 (2) (2) - - (3) -
37.2 37.4 36.3±0.4
70 - - (2) (2) (3) (2)
37.2 36.9 36.5±0.2 36.6
75 - - - - (8) -
36.4±0.3
Взрослые/ (26) (4) (8) (6) (8) (5)
Adults 37.8±0.3 37.6+0.6 37.3±0.4 37±0.7 36.4±0.4 36.7+0.5
К пятидневному возрасту она достигала уже 31.3-32.8°С. В связи с постоян-
ным нарастанием массы и размеров тела, развитием волосяного покрова и
включением механизмов химической терморегуляции прослеживается даль-
нейший рост этого показателя, достигающего 35-36.7°С у хорьков и норок
к 1 -1.5 мес. Интересно, что в этом же возрасте молодые соболя и лесные куницы
имели ректальную температуру, равную 36.1-37.8°С, что, видимо, объясняется
большими их весовыми размерами (следовательно, меньшей теплоотдачей) и
уровнем теплообмена. Сходные различия наблю-даются и у взрослых животных
этих видов. Так, если у 2-3 летних самцов хорьков и норок полостная температура
составляет в среднем 37.4°С, то у соболей и куниц - 39.1°С.
270
Таблица 93/ Table 93
Возрастные изменения уровня потребления кислорода у ласки и горностая (мл/г в 1 ч)
Age changes of oxygen consumption level of weasel (Mustela nivalis)
and ermine (Mustela erminea), ml/g per h
Возраст, мес/ Age, months Ласка/Mustela nivalis (n=l 1) Горностай/Л/u.vZe/a erminea (n=9)
9? <s<s 9?
1 6.2±0.16 7.5±0.18 3.9+0.09 4.1+0.11
1.5 6.6+0.15 8.3±0.09 4.2+0.12 4.5±0.13
2 5.7±0.08 6.8±0.11 3.6+0.07 3.8±0.09
3 5.2±0.12 5.8±0.68 3.4±0.05 3.6+0.05
4 4.8±0.67 5.2±0.75 3.2±0.08 3.4+0.12
5 4.5±0.48 4.6+0.28 3.3±0.06 3.5±0.07
6 4±0.69 4.2±0.66 3.1+0.08 3.3±0.13
В целом в первые месяцы постнатального онтогенеза температура тела
молодых хорьков и куниц увеличивается сравнительно плавно, как и у мелких
миофагов. Заметный скачок, выражающийся в повышении ее в среднем на
0.7-0.8°С, прослеживается только во время открытия глаз. В этот период,
сопровождающийся у детенышей усилением локомоторных функций и
увеличением затрат энергии на окислительный метаболизм, резко изменяются
как уровень потребления кислорода, так и ректальная температура животных
(Tumanov, 1993а).
Близкую к взрослым особям температуру тела молодые хорьки и норки
имеют уже в 2-2.5, а куницы и соболя - в 3-4 мес. Следует также отметить, что
существенных половых различий по данному показателю, дыхательному и
сердечному ритмам у мелких хищников в первые 3-4 мес жизни не
прослеживается.
Известно, что наиболее наглядным и важным индикатором энергетических
реакций в организме животных служит уровень основного обмена, который
обычно определяется методом непрямой калориметрии, т.е. по потреблению
кислорода в единицу времени. Согласно полученным материалам, его уровень
во многом зависит от размеров и возраста хищных млекопитающих и при
прочих равных условиях тем больше, чем они меньше. Зависимость
интенсивности обмена от веса животного хорошо прослеживается как при
сравнении отдельных видов с различной массой тела, так и среди особей одного
вида, но разной величины (Слоним, 1952,1961; Grodzinski, 1961; Gebczynski,
1971, 1975; Туманов, 1974а, б; Ивантер и др., 1985).
Возрастные изменения уровня потребления кислорода (на единицу массы)
в процессе роста и развития молодняка рассматриваемых хищников
271
Таблица 94/ Table 94
Возрастные изменения уровня потребления кислорода
у американской норки (мл/г в 1 ч)
Age changes of oxygen consumption of American mink (Mustela vison), ml/g per h
Возраст, дни/ Age, days Самцы/Males (п = 9) Самки/Females (п = 13)
масса тела, г/ body mass, g потребление кислорода/ oxygen consumption масса тела, г/ body mass, g потребление кислорода/ oxygen consumption
7 26±0.1 2.03±0.08 25±0.11 2.11 ±0.07
25 112±0.9 1.52±0.09 108±0.08 1.21±0.08
30 155±2 1.66±0.06 131±1 1.86±0.08
32 165±2 1.61±0.06 138±1 1.47±0.06
35 198±2.3 1.53±0.05 185±2.1 1.38±0.10
38 251±2.8 0.87+0.12 238±2.4 1.25±0.05
40 272±2.4 0.96±0.15 255±2.5 1.61±0.06
45 346+3.5 0.84±0.13 314±3.2 1.37±0.07
50 421 ±3.8 0.79±0.18 344±3.4 1.17±0.05
демонстрируют некоторый подъем их энергообмена к месячному возрасту, т.е.
к периоду прозревания (Мелькина, 1966; Сегаль, 19756). По-видимому,
усиление локомоторных функций, следующее за процессом открытия глаз,
значительно увеличивает затраты энергии организма на окислительный
метаболизм и тем самым изменяет величину теплообмена. В целом же падение
интенсивности энергообмена в процессе постнатального онтогенеза у куньих
выражено достаточно четко. Например, у самок горностая в месячном возрасте
уровень потребления кислорода достигал 4.1 ±0.11 мл/(г ч). К 6 мес он снизился
до 3.3±0.13 мл/(г ч) и в дальнейшем практически не изменялся (Туманов,
Левин, 1974).
В возрасте старше месяца у детенышей ласки и горностая некоторое
увеличение интенсивности энергообмена наблюдается только на 40-45-й день
от рождения (табл. 93). Это может быть связано с участившимися выходами
зверьков из гнезда, резким изменением их поведения и пищевого рациона. При
этом во все периоды роста рассматриваемый показатель обменных процессов
у молодняка ласки был выше, чем у других куньих того же возраста, что
является следствием ее малых размеров.
Как и у других видов, у лесного хорька общее потребление кислорода с
возрастом увеличивается, однако относительная величина поглощенного
кислорода как один из индикаторов интенсивности окислительного метаболизма
уменьшается. Так, у двухмесячных хорьков потребление кислорода на единицу
272
Таблица 95/ Table 95
Потребление кислорода детенышами солонгоя (9 особей) при групповом
содержании (мг/г в 1 ч)
The oxygen consumption of young (9 ind.) of Alpine weasel (Mustela altaica) kept
in groups (ml/g per h)
Возраст, дни/ Age, days Средняя масса тела, г/ Average body mass, g Потребление кислорода группой/ Oxygen consumption by the group Возраст, дни/ Age, days Средняя масса тела, г/ Average body mass, g Потребление кислорода группой/ Oxygen consumption by the group
4-5 9.1+0.4 2.34±0.08 20-21 39.3±1.4 1.12±0.27
9-10 18.4+0.5 2.18+0.07 25-26 57.5±1.6 1.44+0.23
14-15 26.5±0.8 1.64±0.18 29-30 74.8±2.2 1.55±0.31
массы тела равно 1.92-2.17, у пятимесячных - 0.88-1.72 и у годовалых (в весенний
период) - 0.83-1.42 мл/(г ч). Подобная динамика уровня основного обмена на
ранних этапах постнатального онтогенеза характерна и для американской норки,
у которой его падение с возрастом проявлялось достаточно четко (табл. 94).
Поглощение кислорода (в пересчете на единицу массы тела) несколько
увеличивалось у щенков только в возрасте 38-40 дней, т.е. после прозревания и
участившихся выходов из гнезда в поисках добычи.
Рассмотренные изменения показателей энергообмена представляют собой
общебиологическую закономерность и наблюдаются среди представителей
разных отрядов и видов млекопитающих. Например, в отряде насекомоядных,
по материалам М.Гебжинского (Gebczynski, 1965, 1971), у взрослых особей
обыкновенной бурозубки при средней массе тела 11:6г потребление кислорода
составляло 82.8, а у молодых весом 7.8 г - 99.1 см3 на 10 г массы в час. У
самого мелкого вида из этого отряда - крошечной бурозубки - взрослые особи
(средняя масса 5 г) потребляли кислорода 119.3, а молодые (3 г) - до 150 см3 на
10 г массы в час.
Известно, что у многих животных явление групповой терморегуляции
способствует экономии вырабатываемого организмом тепла. Собираясь вместе
и взаимно обогревая друг друга, они уменьшают поверхность теплоотдачи, а
следовательно, и интенсивность обменных процессов. Групповой эффект
прослеживается прежде всего у всех мелких млекопитающих, снижая
потребление кислорода примерно на 10-30% (Понугаева, 1960; Gebczynski,
1975; Башенина, 1977). Совместное пребывание в гнезде детенышей горностая
или ласки приводило к сокращению у них уровня этого поазателя в 1.5-2 раза.
Интенсивность потребления кислорода у отдельно лежащих на подстилке
детенышей солонгоя в 10 дней составляла 2.86±0.09, в 20 - 2.31±0.35 и в 30
дней, когда у зверьков открываются глаза, -2.52±0.27 мл/(г ч). В группе из 9
273
Таблица 96/ Table 96
Возрастные изменения частоты дыхания (в 1 мин) у самок ласки и горностая
при температуре 22-24°С
Age changes in the breath frequency (per 1 min) of females of weasel (Mustela nivalis)
and ermine (Mustela erminea) with the temperature 22-24°C
Возраст, мес/ Age, months Ласка/Mustela nivalis (п=6) Горностай/Л/и^е/а erminea (n=l)
масса тела, г/ body mass, g частота дыхания/ respiration rate масса тела, г/ body mass, g частота дыхания/ respiration rate
1 28±0.3 113±3.8 36+1.2 106±5.3
2 52±0.2 88±3.5 85±1.4 93+6.5
3 55±0.9 102±3.4 105±0.7 101±4.7
4 59±0.8 96+2.6 112+1.3 98+3.5
5 62±0.8 94+3.7 118+1.5 91±2.8
6 64±0.9 98±3.5 121±1.2 82+3.1
10 65+1.3 93±3.4 126+0.9 85±2.5
Взрослые/ Adults 66+2.3 91±2.1 14O±3.4 87±2.2
зверьков эти величины были заметно меньшими (табл. 95). Интересно, что у
молодых солонгоев половые различия по уровню энергообмена проявляются
сразу же после того, как они начинают активно потреблять мясную пищу.
Например, если у самок (и=10) уровень потребления кислорода на единицу
массы тела в среднем равен в возрасте 40 дней 2.14, а в 50- 1.95 мл/(гч), то у
самцов (и=12) он был соответственно ниже - 1.77 и 1.59 мл/(г-ч).
Интенсивный обмен веществу куньих сопровождается постоянно высоким
уровнем легочного дыхания. В первый месяц жизни дыхательные движения
совершаются очень часто и поверхностно. Плоскостное расположение ребер
молодого организма ограничивает грудное дыхание, так что изменение объема
легких происходит главным образом за счет движения диафрагмы. Однако
диафрагменный компонент дыхательного акта в этот период лимитирован
большой печенью, поскольку ее размеры у новорожденных достаточно велики.
С возрастом происходит морфологическая перестройка организма, а вместе с
ней - замедление и углубление дыхательных движений, в результате чего
респираторный объем легких заметно нарастает (Меерсон, 1986).
В целом в пределах вида с увеличением возраста и массы тела зверьков
темп дыхания замедляется. У самых маленьких представителей семейства -
ласки и горностая - интенсивный энергообмен возможен только при большой
частоте дыхания. Экспериментальные исследования показали, что с
изменением возраста от 1 мес до взрослого состояния темп дыхания снижался
у самок обыкновенной ласки со 113±3.8 до 91±2.1 в минуту, а горностая - со
109±5.8 до 87±2.2 в минуту (табл. 96).
274
Таблица 97/ Table 97
Возрастные изменения ритма дыхания (в 1 мин)
у самцов хорьков и европейской норки
Age changes of the breathing rhythm (per 1 min) of males of polecats
and European mink (Mustela lutreola)
Возраст, дни/ Age, days Хорек лесной/ Mustela putorius Хорек степной/ Mustela eversmanni Норка европейская/ Mustela lutreola
5 (2)84 (2)72 (2)66
10 (5) 64±3.9 (5)70+12.4 (3) 77±12.6
15 (6) 63±8.5 (8) 50±5.3 (3) 90±6.3
20 (6) 60+4.3 (8) 57+6.4 (3) 90±8.3
25 (6) 71±8.1 (8) 61±6.4 (2)92
30 (6) 60±5 (7) 49±4.8 (3) 90+7.6
35 (6) 63±6.4 - (2) 105
40 (2)72 - (2)86
45 (5) 64±7.7 (3) 51±8.7 -
Взрослые/ Adults (26) 55±3.8 (8) 32±6.7 (18)48±2.8
Регистрация внешнего дыхания лесного и степного хорьков также
демонстрирует обратную зависимость его темпа от массы тела и возраста
животных (табл. 97). Так, по сравнению с пятым днем от рождения в конце
первого месяца жизни частота дыхания снизилась у лесного хорька на 28-29%, а
у степного - на 30-32%. Дефинитивного уровня по данному тесту они достигали
в 2.5-3.5 мес. Обращает на себя внимание замедленный темп дыхания у взрослых
степных хорьков в сравнении с лесными, что согласуется с большей абсолют-
ной и относительной массой их легких (Терновский, 1977). Отмеченный факт,
вероятно, объясняется интенсивной роющей деятельностью и постоянным
пребыванием степных хорьков в условиях затрудненного дыхания - в глубоких
норах, куда они проникают в поисках основного корма - грызунов.
В отличие от хорьков у норок на ранних этапах постнатального онтогенеза
происходит усиление темпа дыхания, продолжающееся до 1.5-2-месячного
возраста. Затем, в период энергичного развития организма, связанного с
изменением режима питания и форм пищевого поведения, которое у
полуводных животных сопровождается увеличением массы легких, а
следовательно, и их объема, дыхание молодых норок становится более редким
и глубоким. Например, у европейской норки за первый месяц жизни
дыхательный ритм детенышей возрастал на 36-59%, а в дальнейшем, за период
от 1-1.5 мес до года, он уже снижался на 47-54% (табл. 97). Отмеченные
возрастные изменения дыхательного ритма молодых животных, начинающих
вести активную самостоятельную жизнь, связанную с водной средой обитания,
275
Таблица 98/ Table 98
Характеристика ЭКГ барсуков разного пола н возраста
Characteristics of ECG of badgers (Meles meles) of different age and sex
Пол/Sex п Частота пульса, уд./мин/ Heart rate, heart beats/min Ср. продолжительность, мин/ Average lenght, min Вольтаж зубцов, мм/ Voltage of dents, mm
R-R P-Q QRST T-P R2 T2
Самцы/ Двухмесячные/2 months
males Самки/ 3 180-240 0.24 0.04 0.11 0.09 8 3
females Самцы/ 3 210-270 0.23 0.04 0.10 0.09 Годовалые/12-15 months 7 3
males Самки/ 4 150-180 0.35 0.05 0.17 0.13 15 7
females 3 160-180 0.34 0.05 0.16 0.13 14 7
могут иметь направленность на усиленную вентиляцию их респираторных
органов.
Наблюдения показывают, что у амфибионтов возрастная амплитуда
параметров внешнего дыхания имеет большую величину по сравнению с другими
видами сходного размера. У детенышей американской норки за период от одного
месяца и до года частота дыхательного акта уменьшается в 2 раза и более, что связано
с увеличением объема легких. У взятых из гнезда месячных щенков этот
показатель был равен 84-120 в минуту. К 4-5 мес уровень внешнего дыхания
снизился у нихдо60-90, акгодоваломувозрасту-до40-60движенийвминуту.
В процессе индивидуального роста выдрят также прослеживаются
закономерные сдвиги в энергии их физиологических процессов, о чем можно
судить по изменениям легочного дыхания, полостной температуры, нарастанию
индекса легких и сердца. При увеличении возраста детенышей от 2-3 мес до
года и массы тела от 2.5-3 до 5.5-6.7 кг частота дыхания уменьшалась очень
заметно - с 40-45 до 24-30 движений в минуту. За этот же период ректальная
температура снизилась у молодых выдр в среднем на 1 °C (с 38.6 до 37.7°С), что
может обусловливать их меньшую теплоотдачу в водную среду при плавании
и нырянии. Возрастное увеличение у представителей данного вида
относительной массы легочно-сердечного комплекса и специфическое строение
почек, по-видимому, объясняются большой функциональной нагрузкой на эти
органы в связи с амфибионтностью животных. Строение у выдры таких
276
Таблица 99/ Table 99
Возрастные изменения содержания эритроцитов (млн/мм3) и гемоглобина (г %)
в крови некоторых видов куньих (июнь-октябрь)
Age changes of the erythrocyte (mln/mm1) and hemoglobin (g%) content in the blood
of some mustelids species (June-October)
Виды/ Species Возраст, мес/ Age, months n Содержание/Concentration
эритроцитов/ гемоглобина/ erythrocytes hemoglobin
Ласка обыкн./ Mustela nivalis 3 5 6.5-7.3 13.2-14.8 6 4 6.8-7.6 13.6-15.3 Старше года/ 5 68-7.9 13.5-16.1 1+ and older
Горностай/ Mustela erminea 3 8 6-7.3 13-15.1 6 6 6.5-7.5 13.5-15.6 Старше года/ 5 6.5-7.6 13.8-15.5 1+ and older
Хорек лесной/ Mustela putorius 3 6 6-6.8 13.0-13.8 6 5 5.6-7.2 13.3-15.1 Старше года/ 5 s.8-7.2 13.5-15.3 1+ and older
Норка европ./ Mustela lutreola 3 4 6.3-7.8 14.3-15.5 Старше года/ 5 6 g g 7 15.1-18.6 1+ and older
Норка америк./ Mustela vison 3 6 6.4-7.5 13.6-14.8 Старше года/ д 69.89 15 3.18 1+ and older
Барсук/ Meles meles 3 3 5.5-8.6 14.9-15.7 Старше года/ 3 6.3-8.9 16.1-17.3 1+ and older
жизненно важных систем, как дыхательная, сердечно-сосудистая и выдели-
тельная, носит черты явно выраженной специализации к водной среде обитания.
У другого крупного представителя семейства - росомахи - изменение
температуры тела, частоты дыхания и пульса в разные периоды жизни
свидетельствует о сравнительно высоком уровне ее энергообмена. По
имеющимся сведениям (Iversen, 1972), большая, чем у других хищных
млекопитающих, близких по массе тела, интенсивность основного обмена
у щенков росомахи в первые месяцы жизни, по-видимому, связана с
энергичным ростом тканей внутренних органов, характеризующихся
высокой теплопродукцией (печень, почки, сердце, мозг). Кроме того, резкое
изменение интенсивности метаболизма у животных весом около 3 кг
совпадает по времени с окончанием лактационного периода и переходом
на самостоятельное добывание кормов.
277
В возрасте 6-7 мес росомахи уже мало отличаются по своим размерам и
физиологическим параметрам от взрослых особей. Так, у содержащихся в неволе
двух молодых самцов в августе-сентябре ректальная температура была равна
37.6-38.3°С, а через год в те же месяцы- 37.8-38.5°С. За этот срок частоты дыха-
ния (31-35 в 1 мин) и пульса (130-150 уд./мин) у них также практически не изменились.
У барсука возрастная динамика темпа дыхания выражена незначительно. У
детенышей в возрасте 1.5-2мес онбыл равен 32-40, ав 10-15 мес-25-32дыхательных
движений в минуту. Падение скорости сердечных сокращений прослеживалось
более отчетливо. За названный период частота пульса снизилась в среднем с 222до
165 уд./мин, что сопровождалось изменением продолжительности и вольтажа
основных элементов ЭКГ (табл. 98).
Возрастная изменчивость дыхательной функции крови у рассматри-
ваемых видов прослеживается достаточно отчетливо (табл. 99). Повышение
количества эритроцитов и насыщения их гемоглобином интенсивно про-
ходило у щенков только в возрасте 3-4 мес и в дальнейшем эти величины
существенно не менялись. Относительная стабилизация показателей красной
крови наблюдалась у них осенью (в полугодовалом возрасте) и совпадала
с началом распада выводков. Таким образом, отмеченные возрастные
изменения показателей дыхательной функции крови имеют направленность
на увеличение ее кислородной емкости к периоду активной самостоятельной
жизни молодых животных, повышая тем самым устойчивость организма к
возможным экстремальным условиям.
Работы с разными видами пушных зверей, разводимых в звероводческих
хозяйствах, показали, что у взрослых самок кровь отличается от таковой
самцов более высоким насыщением эритроцитов гемоглобином при
сравнительно равном соотношении красных кровяных клеток (Берестов, 1971,
1981). Это обусловливает высокий общий уровень гемоглобина в крови и ее
дыхательной функции у самок по сравнению с самцами (Иржак, 1964).
У молодых американских норок, отловленных в октябре в Карелии,
половые различия по показателям красной крови также проявляются
достаточно отчетливо. Они выражаются в заметном повышении количества
эритроцитов и концентрации гемоглобина у самок по сравнению с самцами.
При этом содержание лейкоцитов и общего белка сыворотки крови у норок
обоих полов остается либо одинаковым, либо с небольшим преобладанием в
группе самцов (табл. 100).
Существенные возрастные изменения наблюдаются у куньих и в работе
сердечной мышцы. Необходимый для жизнедеятельности организма минутный
объем сердца обеспечивается у молодых особей в основном за счет увеличения
частоты пульса, а у взрослых - силы сердечных сокращений (систолический
объем). Это способствует экономии энергии при функционировании взрослого
организма (Аршавский, 1967). Изложенное в полной мере относится и к
рассматриваемым хищникам, у которых в процессе индивидуального развития
отмечается зависимость частоты сердцебиения от возраста, массы тела и
интенсивности энергообмена животных.
278
Таблица 100/Table 100
Состав крови у молодых американских норок, отловленных в Карелии (октябрь)
Blood composition of young American minks (Mustela vison), trapped in Karelia (October)
Вес тела, г/ Body mass, g Кол-во в 1 мл крови/ Number in 1 ml of blood Содержание, г %/ Concentration, g %
эритроцитов, млн/ erythrocytes, mln лейкоцитов, тыс/ leucocytes, thousands гемоглобина/ hemoglobin ; общего белка сыворотки крови/ common protein of the blood serum
Самцы/Males
664 6.15 5.75 .14.8 9.91
707 8.59 6.95 12.5 8.28
750 6.27 5.85 12 8.70
793 5.72 10.90 12.2 7.01
В среднем/ Average 6.68 7.36 12.9 8.48
Самки/Females
430 8.88 7.75 12.5 8.74
457 6.29 5.25 14 8.15
470 6.75 6.84 14.6 7.83
В среднем/ Average 7.31 6.61 13.7 8.24
Примечание. Все измерения проводились зоологом В.Я. Каньшиевым.
Для детенышей куньих характерен достаточно частый пульс с относительно
постоянной для каждого возраста скоростью сокращений сердца. Динамика
этих преобразований наиболее полно прослежена нами на мелких предста-
вителях семейства. С первых же дней после рождения темп сердечной
деятельности у щенков регулярно нарастает. Особенно резкий его подъем
наблюдается в период открытия глаз, когда у них регистрируется и повышение
уровня энергообмена. Увеличение частоты пульса заканчивается у зверьков к
1.5-2 мес, после чего происходит ее спад и появляется присущая взрослым
особям дыхательная аритмия.
У молодняка обыкновенной ласки она возникает в возрасте около 2 мес,
когда детеныши по массе и размерам тела близки к родителям. У
10-15-дневных ласок частота пульса колебалась в пределах 210-255 ударов
в минуту. Затем темп сердечной деятельности увеличивается, достигая к
месячному возрасту 450-480, а к полутора месяцам - 420-540 уд./мин. В
дальнейшем пульс становится реже. У взрослых самцов (при средней массе
91 г) он был равен 364±13.5, а у самок (масса 63 г) - 388±103 уд./мин.
Длительность сердечного цикла (R-R) находится в обратной зависимости от
279
Таблица 101/Table 101
Изменение частоты пульса и некоторых элементов ЭКГ у колонков
на ранних этапах постнатального онтогенеза
Changes in the pulse rate and some elements of ECG of Siberian weasel (Mustela sibirica)
at the early stages of postnatal development
Возраст, дни/ Age, days Самцы/Males Самки/Females
сердечный ритм, уд./мин/ heart rate, heart beats/min сердечный цикл, с/ heart cycle, second вольтаж зубца /?, мм/ voltage of dent R, mm сердечный ритм, уд./мин/ heart rate, heart beats/min сердечный цикл,с/ heart cycle, second вольтаж зубца /?, мм/ voltage of dent R, mm
15 (2) 375 0.16 5 (3) 415+16 0.15±0.007 5±0.6
20 (2) 390 0.15 6.5 (3) 420±14.5 0.14±0.007 6±0.6
25 (2) 420 0.14 10 (3) 412+9 0.15±0.007 8.7+1.2
30 (5) 457±7.5 0.13±0.02 12.8+0.8 (8) 455±9.2 0.13±0.006 9+0.9
35 (3) 450±17.3 0.13±0.03 13.5±0.7 (5) 450±9.5 0.13±0.02 13.3±0.8
Взрослые/ Adults (8) 285±8.3 0.19±0.03 20+1.2 (7) 291±14.2 0.21±0.02 19. l±0.4
Примечание. Здесь и далее цифры в скобках - количество исследованных зверьков.
частоты пульса. Ее снижение сопровождалось увеличением сердечного цикла,
изменение которого происходит в основном за счет вариабельности
диастолической фазы сердца (Т-Р) и активой систолы жедудочков (QRST).
Продолжительность предсердно-желудочковой проводимости (P-Q) при этом
практически не менялась. Сходным образом прослеживались изменения темпа
сердечной деятельности и у других мелких куньих.
На ЭКГ детенышей колонка в возрасте до 30-35 дней дыхательная аритмия
отсутствовала. По-видимому, у них она начинает проявляться в более позднем
возрасте, когда масса зверьков достигает 70-80% дефинитивного веса. В
первый месяц жизни темп сердечной деятельности молодняка постоянно растет
и ко времени открытия глаз оказывается в среднем на 150-160 ударов в минуту
больше, чем у родителей (табл. 101).
В дальнейшем сердечный ритм постепенно снижался до уровня,
свойственного животным старших возрастных групп (280-300 уд./мин).
С изменением частоты пульса менялись и размеры сердечного цикла (интервал
R-R). На ранних стадиях постнатального онтогенеза колонков он сокращался
в среднем с 0.16 до 0.13 с, а потом вновь увеличивался до 0.19-0.21 с у взрослых
особей (табл. 101). Вольтаж основных зубцов ЭКГ молодых колонков с
возрастом изменялся мало, за исключением зубца R, который характеризует
распространение волны возбуждения по миокарду желудочков. В процессе
роста и развития зверьков его вольтаж постоянно нарастал. Так, если размеры
280
Таблица 102/Table 102
Возрастная динамика сердечного ритма (уд./мин) и вольтажа зубца R (мм)
у горностая и солонгоя (по: Туманов и др., 1982)
Age dynamics of the heart rate (strike/min) and voltage of the spike R (mm)
of ermine {Mustela sibirica) and Alpine weasel {Mustela altaica)
Возраст, дни/ Age, days Г орностай/Л/м5/е/л erminea Солонгой/Mustela altaica
33 <"=3-12) («=з-1 з) 33 («=3'11) («=3-ii)
сердечн. ритм/ heart rate, heart вольтаж зубца RJ voltage of dent R сердечн. ритм/ heart rate, heart вольтаж зубца /?/ voltage of dent R сердечн. ритм/ heart rate, heart вольтаж зубца Rl voltage of dent R Сердечн. ритм/ heart rate, heart вольтаж зубца RJ voltage of dent R
5 - - - - 276+12.7 5.5±0.4 3OO±23.3 6.4+О.8
10 - - - - 315+10.5 5.2+0.3 313±1O 5.3+03
15 346±17.3 8.5±1.3 345±12.2 8.3+0.6 332±6.5 6.6+0.4 334+11.2 5.5±0.4
20 356±14.4 10±1.2 357±8.7 9.9+0.8 347±10.4 6.2±0.6 344±18.3 6.2+0.6
25 348±26.8 11.2±1 405±13.8 11.1+0.4 350+12.3 7.3±0.5 355+10.5 6.8+0.4
30 420+6.1 12.4+0.8 412±17 12.9+0.7 354±28.2 9.8±1.7 39O±l 1.6 7+0.5
35 432±1О 14.2±0.9 500+10 15.1+0.5 374±15.8 6.9+0.7 383±41.1 8.3±1.8
40 416±12.3 15.3+0.6 470±8.6 15.3±0.4 390±17.9 8.6±0.8 427±26 8.1+0.8
45 430±10.5 15.3+0.8 463±7.5 15.1+0.5 399±12.5 7.5+0.5 428+33 7+0.6
50 402±7.3 15.5+0.4 480±10 15.5+0.7 421±11 8.4±0.5 441±21.2 9+0.5
55 385±6.3 16.2+0.5 436±8.6 16.1+0.4 436±22.7 11.7±1.9 387±13.8 12.1+0.7
60 - - - - 382±25.7 12±1.2 421±18 12±1.6
70 - - - + 375±19.2 11.5±1.4 413±23.4 11+0.8
80-90 350±10 17.7±1.3 39О±13.5 17.5±1.1 354±10.2 11.8±1 362±13.7 11±1.2
Взросл./ Adults 288±5.2 21.4±1.2 384±12.3 18.8+0.8 272±11.4 15±1.5 300+12.5 14±1.8
предсердного комплекса (зубец P) в первый месяц жизни колонков увели-
чивались в среднем с 2 до 2.5 мм, то вольтаж зубца R - с 5 до 13.5 мм.
Динамика возрастных изменений сердечного ритма горностая и солонгоя
также прослеживалась очень отчетливо (табл. 102). В первые дни после
рождения пульс зверьков сравнительно редкий и по частоте близок к таковому
у взрослых животных. С возрастом скорость сердечных сокращений заметно
увеличивалась. У большинства детенышей во время открытия глаз наблюдалось
учащение пульса, которое ранее отмечал А.Н. Сегаль (1975а). По времени этот
подъем сердечной деятельности совпадал с нарастанием массы, уровня
энергообмена и температуры тела у щенков. Таким образом, имеет место четкая
взаимосвязь массы тела с основными показателями физиологического состояния
организма.
Увеличение темпа сердечных сокращений заканчивается у горностая в
возрасте 45-50 дней, а у солонгоя - 50-55 дней, после чего происходит спад
частоты пульса. У самок падение скорости сердечных сокращений наблюда-
281
Таблица 103/Table 103
Возрастная динамика сердечного ритма (уд./мин) у самцов норки европейской
и хорьков
Age dynamics of the heart rate (strike/min) of males of European mink (Mustela lutreola)
and polecats
Возраст, дни/ Age, days Норка европейская/ Mustela lutreola Фуро/ Mustela putorius furo Хорек степной/ Mustela eversmanni
1 (5) 260±8.3 (7) 251 ±4.5 (4) 240±16.9
5 (5) 300±25.7 (17) 279±6.3 (15) 306±7.8
10 (4) 420±3.5 (17) 299±4.2 (15) 331±6.2
15 (7) 372±18.2 (19) 322±3.8 (15) 364±8.3
20 (3) 350±17.8 (14) 365+9.8 (15)393±4.6
25 (3) 420±7.6 (2) 450 (15) 404±6.9
30 (2) 453 (3) 465±8.6 (8) 425±6.2
35 (2)413 (2) 440 (2) 375
40 (2) 450 (2) 440 (3) 363±12.6
45 (2) 450 (3) 430±12.7 (2) 358
50 (3) 443±8.6 (3) 422±7.6 (3) 349±9.7
55 (3) 440±15.8 (3) 390±8.6 (3) 345±8.6
60 (3) 355±25.2 (3) 376±13.1 (2) 340
65 (3) 420+7.5 (2) 330 -
70 (8) 409±19.9 (3) 375±9.3 -
75 (8) 386±12.5 (2) 345 -
80 (3) 353±63 (2) 330 -
Взрослые/ Adults (12) 218±4.3 (8) 290±19.4 (3) 255±18.3
лось на 5 дней раньше, чем у самцов. Обращает на себя внимание тенденция к
повышению частоты пульса и величины ректальной температуры у самок по
сравнению с самцами на всех стадиях роста (от прозревания до полово-
зрелое™), однако эти различия статистически недостоверны. При расчетах
удельного (на единицу массы) ритма сердца и дыхания, так же как и уровня
энергообмена, половая специфика выступает очень отчетливо.
Аритмия, характерная для ЭКГ взрослых особей, у детенышей горностая
начинала прослеживаться только в 55-дневном возрасте. У 3-месячных
зверьков в начале записи биотоков сердца она регистрировалась уже постоянно.
282
Расшифровка их рисунка у солонгоя показала, что у молодняка этого вида
аритмия отсутствует до 50-дневного возраста. На ЭКГ 55-60-дневных щенков
она встречалась в 28%, а 65-дневных - в 35% случаев. У 70-дневных солонгоев
аритмия в начальном периоде записи биотоков сердца отсутствовала только у
небольшого количества зверьков (23%).
В возрасте 80-90 дней сердечный ритм горностаев и солонгоев (особенно
самок) близок к таковому взрослых особей (табл. 102). Это сходство прослеживается
и по вольтажу основных зубцов ЭКГ, прежде всего комплекса QRS (зубец R). Однако
окончательное совпадение ЭКГ молодых и полово-зрелых зверьков по
продолжительности сердечного цикла, основных интервалов ЭКГ и вольтажу ее
зубцов происходит уже в более поздние сроки.
Пульс однодневных детенышей хорьков и норок близок по частоте к
взрослым особям (табл. 103). С первых же дней после рождения он неуклонно
увеличивается. Если у новорожденных зверьков сердечный ритм был равен
223-268 уд./мин, то уже к 5-му дню возрастал до 273-326, а к 20-му - до 332-
398 уд./мин. Во время прозревания частота пульса у детенышей хорьков и
норок оказалась в среднем на 130-220 уд./мин больше, чем у новорожденных
или взрослых животных. Нарастание темпа сердцебиений заканчивается у
разных особей к 1.5-2-месячному возрасту, после чего наблюдается его спад,
и на записях ЭКГ появляется присущая взрослым животным аритмия.
Сходная динамика сердечного ритма прослеживается также у молодых
соболей и куниц. Например, у 10-дневных самцов соболей он был равен 260-
270 уд./мин. К месячному возрасту частота пульса достигала в среднем 375, а
к 45 дням - 405 уд./мин. Затем темп сердцебиений замедлялся, и в возрасте 4-
5 мес частота пульса детенышей (330-350 уд./мин) уже не отличалась от таковой
у взрослых особей.
Как уже отмечалось выше (см. гл. 4), при оценке функционального
состояния миокарда используют такие индикаторы, как процентное отношение
систолы желудочков к сердечному циклу, или систолический показатель
(Q- T)/(R-R)xl 00%, и отношение размеров всей систолы к диастоле, или индекс
напряженности (/-’-7)/(Г-/’)х100%. Их уровень определяет интенсивность
работы сердечной мышцы (Никулин, 1956; Фогельсон, 1957). Анализ
изучаемых величин показал существенную разницу по этому тесту между
молодыми и взрослыми животными. Интенсивность сердечной деятельности у
первых оказалась достоверно более высокой, чем у взрослых особей.
Например, у детенышей европейской норки, степного и лесного хорьков в
возрасте от 1 до 6 мес систолический показатель в среднем был равен 52-63%,
а индекс напряженности - 250-460%. У взрослых особей эти признаки имели
меньшую величину - соответственно 41-49% и 170-280%.
В процессе постнатального онтогенеза вольтаж основных зубцов ЭКГ
куньих меняется незначительно, за исключением положительных зубцов RvlT
желудочкового комплекса QRST. Во время интенсивного роста и увеличения
массы тела детенышей особенно заметно нарастал вольтаж зубца R, который
характеризует распространение волны возбуждения по миокарду и служит
283
Таблица 104/Table 104
Изменение массы (г) и размеров (см) тела у самок (л=4) песцов
на ранних этапах постнатального онтогенеза (средние данные)
Changes of body mass (g) and sizes (cm) of females (и=4)
of arctic foxes (Alopex lagopus') at the early stages of postnatal development (average data)
Показатели/ Parameters Возраст, дни/Age, days
1 10 15 | 20 25 30 35 40
Масса тела/ 63.5 171.3 321.5 453.4 586.2 693.6 875.2 1120.3
Body mass Длина/Length of: 18.4 21.3 22.5 23.6 28.5 32.3 34.6 37.2
тела/body головы/head 5.6 6.8 7 8.4 10.6 12 11.6 12.4
хвоста/tail 5.5 6.6 7.8 8.6 10.3 12.6 15.8 17.6
индикатором возрастных изменений сердечной мышцы и функционального
состояния организма в целом (Меерсон, 1986). По нашим данным, у самцов
хорьков, норок, соболей и куниц за период от 5 дней до месяца вольтаж зубца
R увеличивался в среднем с 5.5 до 8.7 мм. У полуторамесячных детенышей
этот показатель был равен уже 11-13 мм, а у половозрелых особей он колебался
в пределах 16-20 мм.
В возрасте 3-4 мес молодняк хорьков, норок и куниц (особенно самки) по
показателям сердечного ритма, продолжительности интервалов и вольтажу
основных зубцов ЭКГ близок к таковым родителей, однако окончательное
совпадение элементов ЭКГ молодых и взрослых особей у разных видов
происходит в более поздние сроки.
Отмеченные возрастные изменения терморегуляции, сердечного и дыха-
тельного ритмов в процессе посгнатального онтогенеза куньих демонстрируют
определенную взаимосвязь физиологических функций с морфологическими
особенностями весового роста животных. Характер возрастных изменений ЭКГ
прослеживается у них очень отчетливо и имеет направленность на экономное
обеспечение гомеостаза организма у взрослых животных. Учащенный пульс
при меньшей фазе отдыха сердца, высокий уровень индекса напряженности
его работы и систолического показателя свидетельствуют о большей
интенсивности биоэлектрических процессов в миокарде молодых особей по
сравнению с родителями. Следовательно, для них характерен менее
экономичный режим работы сердечной мышцы и больший, чем у взрослых
особей, расход энергии организма. По-видимому, при неблагоприятных
условиях это может стать важным фактором, который определяет высокую
долю естественной элиминации молодняка куньих в природной среде.
Сведения о росте и развитии средних и крупных по размеру хищных
млекопитающих России довольно скудны, что связано с трудностями
проведения наблюдений за ними в природе, содержания и разведения в неволе.
284
1
Рис. 60. Изменение массы тела щенков (2с? и 2<J) песца (средние данные).
Масса тела: 7 - самцы, 2 - самки.
По оси абсцисс - возраст, мес; по оси ординат - масса тела, кг.
Fig. 60. Changes of body mass of arctic fox (Alopex lagopus) cubs (2c? и 2^)
Body mass: 1 - males, 2 - females.
Abscissa - age, months; ordinate - body mass, kg.
Виды из семейства собачьих характеризуются сравнительно быстрыми
темпами роста - изменением весовых и размерных показателей, а также
ранними сроками становления основных физиологических функций у
животных. Период открытия глаз, слуховых проходов и появления зубов
молочного ряда наступает у них намного раньше, чем у крупных предста-
вителей семейства куньих (выдры, барсука, росомахи).
Осмотренные в день своего появления детеныши песца из двух приплодов
(в каждом по 3 с? и 2 $) весили 60-76 г, имели длину тела 18-18.8 см, хвоста -
5-5.7 см и были опушены коротким темно-серым мехом. Росли они быстро, и к
10-му дню их вес увеличился в среднем в 3 раза, к 20-му - в 7, к 30-му - в 11 раз
(табл. 104). Глаза у щенков открылись на 12-18-й день, а на 2-3 дня раньше
стали заметными и наружные слуховые проходы.
У зверьков, за которыми велись наблюдения, молочные зубы появились в
2-недельном возрасте. К 16-17-му дню у них прорезались временные клыки и
резцы, а двумя сутками позже - верхние и нижние последние предкоренные
зубы. К 25-27-му дню животные имели все молочные зубы и были покрыты
густым шелковистым темно-бурым мехом с редкими остевыми волосами.
Детеныши песца рано переходят на смешанные корма и к 1.5 мес практически
прекращают питаться молоком матери. На 40-42-й день они уже способны
самостоятельно забивать предлагаемых им белых мышей и поедать их целиком,
а на 60-е сутки - с жадностью потреблять ту же пищу, что и родители (рыбу,
тушки голубей, куски мяса и т.д.). Смена молочных зубов на постоянные
началась в возрасте 3 мес и закончилась через 1-2 мес, когда молодые животные
по окраске меха, размерным и весовым показателям практически не
отличались от взрослых особей (рис. 60). В этом возрасте в природных условиях
они пытаются уже самостоятельно охотиться, распределяются по временным
убежищам, а к осени покидают родительский участок.
285
Таблица 105/Table 105
Возрастная динамика массы тела (г) у молодых енотовидных собак (3 и 3 <j>)
в первые месяцы жизни (средние данные)
Age dynamics of body mass (g) of young racoon dogs (Nyctereutesprocyonoides)
(3 с? и 3 $>) in the first months of life (average data).
Пол/ Май/ Июнь/ Июль/ Август/ Сентябрь/ Октябрь/ Ноябрь/ Декабрь/
Sex Мау June July August September October November December
Самцы/ Males 72 546 1320 2780 5240 6320 7210 7730
Самки/ Females 80 573 1150 2320 4370 5870 6430 6740
В ходе многолетних работ Ю.С. Заболотских (1997) удалось установить
положительное влияние пищевых разгрузок на организм белых песцов и других
хищных млекопитающих. Молодняк, полученный от взрослых особей,
прошедших период голодания, отличался от такового из контрольной группы
жизнестойкостью, скоростью роста и физиологическими параметрами. Он
своевременно достигал половой зрелости, активно размножался и имел высокие
репродуктивные показатели. По данным автора, щенки песца, родившиеся от
самок, которые некоторое время содержались при дефиците корма, в возрасте
30 сут весили в среднем 603 г, а в контрольной группе - 534 г, т.е. почти на 12%
меньше (р<0.01). В процессе дальнейшего их роста эта разница увеличивалась.
Так, масса тела 4-месячных детенышей от подопытных самок составляла в
среднем 4.49 кг, а в контрольной группе - на 410 г меньше (р<0.01).
У енотовидной собаки молодые также рождаются покрытыми короткой
и мягкой черной шерстью без остевых волос, с закрытыми глазами и слуховыми
проходами. Вес новорожденных 60-110 г. Глаза открываются на 9-12-й день,
ушные проходы - на 2-3 дня раньше. На 14-16-е сутки у них появляются
временные зубы - вначале верхние и нижние клыки, затем резцы и пред-
коренные. Лактация продолжается 40-60 дней, но уже с 3-недельного возраста
детеныши начинают есть пищу, которую приносит самка. Остевые волосы
интенсивно отрастают у них в 1.5-2 мес. В это время преобладающий в окраске
меха черный цвет меняется на более светлый темно-серый.
Молодняк енотовидной собаки характеризуется высокой скоростью
весового роста. Например, новорожденные самцы, родившиеся в начале мая,
имели средний вес 72 г. К месячному возрасту масса тела зверьков увеличилась
в среднем до 546 г, к 2 мес-до 1320 г и к 6 мес-до 6.3 кг (табл. 105). Обычно
осенью сеголетки по данному показателю близки к взрослым особям.
Распад выводков происходит в конце августа-сентябре. Так, 12 августа
1972 г. в Ленинградской области мы встретили недалеко от брошенного
барсучника еще не распавшуюся семью этого вида на жировке. Она состояла
из взрослой самки и 6 молодых. В то же время в другие годы в Псковской и
286
Таблица 106/Table 106
Снижение массы тела и выживаемость некоторых хищных млекопитающих
разного возраста в опытах по искусственному голоданию (по: Заболотских, 1997)
Decrease of body mass and survivorship of some carnivores of different age
in the experiments on artificial starvation
Виды/ Species Взрослые особи/Adults Молодые особи/Youngs
снижение массы тела, усл. ед./ decrease of body mass выживаемость, сут/ survivorship, days снижение массы тела, усл. ед./ decrease of body mass выживаемость, сут/ survivorship, days
Енотовидная собака/ Nyctereutes procyonoides 1 115-150 1.2 94-110
Песец/ Alopex lagopus 1.4 40-56 1.5 38-42
Лисица обыкновен./ Vulpes vulpes 1.3 28-34 1.4 19-29
Норка америк./ Mustela vison 2.8 9-25 2.9 10-22
Примечание. За 1 уел. ед. принято процентное снижение массы тела животных при
голодании.
Новгородской областях наши собаки-лайки неоднократно находили
кормящихся уже в одиночку молодых зверьков в конце августа и первой декаде
сентября. В октябре прибылые соединяются в пары и часто вместе уходят на
зимовку.
По материалам Ю.С. Заболотских (1997), в опытах по влиянию искусственного
голодания на организм енотовидных собак интенсивность падения массы тела у
взрослых особей была достоверно ниже, чем у молодых. Если у последних индекс
упитанности за 50 сут голодания уменьшался на 37%, то у животных старше года -
на 31% от исходного уровня. Интересно, что наибольшее снижение массы тела у
особей разных возрастов отмечалось в первые 10 сут. Молодые и взрослые
енотовидные собаки, у которых при голодании резко снижались температура тела
и частота дыхания, отличались от других видов хищников медленным снижением
массы тела и заметно большей продолжительностью жизни в условиях отсутствия
корма(табл. 106).
По данным автора, несмотря на существенную вариабельность пороговых
границ выживаемости у разных видов хищных зверей при голодании,
287
Таблица 107/Table 107
Масса (г) и размеры (см) тела новорожденных детенышей лисицы
Body mass (g) and sizes (cm) of red fox (Vulpes vulpes) newborns
Показатель/ Parameters Самцы/ Males (/;=6) Самки/ Females (n=4)
Масса тела/Body mass 118(105-125) 105(105-110)
Длина/Length of: тела/body 17.6(17-18) 17.5(16-19)
головы/ head 5.3 (4-6) 5.6 (5.5-6.S)
хвоста/tail 7.9 (7.5-9) 8.2 (7.3-9.5)
выявляется взаимосвязь между этими показателями и интенсивностью
уменьшения их веса: чем выше уровень снижения массы тела, тем меньше
продолжительность жизни животных при голодании, и наоборот. Некоторое
отступление от этой зависимости наблюдалось только между песцами и
лисицами (табл. 106), однако последнее объясняется тем, что исходная
упитанность первых перед голоданием была существенно выше, чем у лисиц.
Индекс снижения массы тела у песцов в период голодания составлял 100-110
Рис. 61. Изменение массы тела щенков лисицы (3<? и 3§>) на ранних
этапах постнатального онтогенеза.
Масса тела: 1 - самцы, 2 - самки.
По оси абсцисс - возраст, дни; по оси ординат - масса тела, г.
Fig. 61. Changes of body mass of red fox (Vulpes vulpes) cubs (3<S и 3<J) at the early
stages of postnatal ontogenesis.
Body mass: 1 - males, 2 - females.
Abscissa - age, days; ordinate - body mass, g.
288
6
5
4
3
2
1
1 2 3 4 5
Рис. 62. Возрастные изменения массы тела молодых самцов лисицы (п = 5)
в первые месяцы жизни.
По оси абсцисс - возраст, месяцы; по оси ординат - масса тела, кг.
Fig. 62. Age changes of body mass of red fox (Vulpes vulpes) young males (n = 5)
in the first months of life.
Abscissa - age, months; ordinate - body mass, kg.
усл. ед., а у лисиц был в среднем на 30 усл. ед. меньше. Высокая исходная
упитанность песцов могла положительно сказаться на их выносливости к
отсутствию пищи.
У обыкновенной лисицы новорожденные щенки несколько крупнее, чем у
белого песца. По наблюдениям в неволе масса их тела у самцов составляет
105-125 г, у самок - 105-110 г (табл. 107). В первые дни после родов самка не
выходит из норы, и корм ей приносит самец. В дальнейшем, по мере
подрастания молодняка, он все реже навещает семейное гнездо и живет
одиночно, но по соседству.
Детеныши лисицы рождаются слепыми, с закрытыми слуховыми
проходами и имеют темно-серый окрас шерстного покрова. Прозревают они
на 13-15-й день от рождения, несколько раньше открываются слуховые
проходы. Временные клыки появляются у них на 14-15-е сутки. Смена молочных
зубов на постоянные заканчивается в конце третьего-начале четвертого месяца
жизни. Лактационный период длится 6-7 недель. Детский мех начинает
выпадать у щенков уже на втором месяце жизни, примерно с 35-37-го дня от
рождения. В августе отмечается интенсивная смена ювенильного волоса на
дефинитивный наряд. Как и у взрослых особей, у молодняка этот процесс
длится до конца осени.
Темпы абсолютного прироста массы тела у особей в возрасте 20-25 дней
уступают таковым в последующий период активного роста и развития зверьков
(рис. 61). Вероятно, это связано с тем, что в возрасте 3-4 недель они начинают
выходить из норы, много двигаться и питаться не только материнским молоком,
но и мясной пищей, которую приносят им родители. В 1.5-2 мес молодняк все
больше осваивает территорию вблизи гнезда, пытается самостоятельно
289
Рис. 63. Изменение массы (7), длины тела (2), обхвата груди (3) и длины хвоста (4) у
щенков волка в условиях неволи
(по: Данилов, Русаков, Туманов, 1979).
1 - самцы (п-6); II - самки (л=5).
По оси абсцисс - возраст, дни; по оси ординат: А - масса тела, кг; В - промеры, см.
Fig. 63. Changes of mass (I), body length (2), diameter of thorax (3) and tail length (4)
of wolf (Canis lupus) cubs in conditions of captivity.
1 - males (n=6); II- females (л=5).
Abscissa - age, days; ordinate; A - body mass, kg; В - measurements, cm.
охотиться на мышей и полевок, а в 3-3.5 мес полностью оставляет свое
гнездовое убежище.
В процессе постнатального онтогенеза динамика весового роста
обыкновенной лисицы особенно отчетливо прослеживается в течение первых
4-5 мес. В среднем в месячном возрасте самцы весят уже около 1 кг, в 2.5 меС -
3, в 3.5 - 4.3 и в 4.5 мес - 5.2 кг (рис. 62). Сходным образом изменяется масса
тела и у самок. Все это свидетельствует о том, что у лисицы, как и у песца,
скорость весового (соответственно и размерного) роста очень высока. К 6-7
мес по данным показателям детеныши мало отличаются от родителей,
приобретая размеры и пропорции взрослых животных.
У волков детеныши при рождении имеют массу тела 250-400 г. В
Воронежской области (Рябов, 1993) два выводка волков были обнаружены
5 и 9 мая. В гнездах находились слепые щенки весом 500-600 г. Еще в
одном логове, найденном несколько позже - 20 мая, масса уже зрячих
детенышей колебалась от 1.5 до 2 кг. В природе, пока самка кормит
молодых, самец приносит ей пищу. В дальнейшем он вместе с нею под-
кармливает подрастающих щенков, отрыгивая куски частично переварен-
ного мяса.
290
30
Рис. 64. Изменение массы тела самки волка в неволе в течение первого года жизни.
По оси абсцисс - возраст, месяцы; по оси ординат - масса тела, кг.
Fig. 64. Changes of body mass of wolf (Canis lupus) female in captivity during the first
year of life.
Abscissa - age, months; ordinate - body mass, kg.
В условиях неволи открытие глаз у волчат отмечалось на 11-14-й день от
рождения, а появление первых временных зубов (клыков)-на 14-16-й день. В
3-недельном возрасте они уже выходят из гнезда, много играют и охотно
пробуют предлагаемую мясную пищу. В 30 дней у них появляются элементы
охотничьего поведения, такие как затаивание, подкрадывание и нападение на
партнера. Однако самостоятельно забивать мышей и полевок щенки начинают
в возрасте 2.5-3 мес. Период лактации у самок волка, как и у лисицы,
продолжается до 6-7 недель.
В естественных местообитаниях 2-3-месячные волчата переходят на
интенсивное потребление мясной пищи, которую приносят им родители, или сами
отыскивают ее вблизи логова (грызунов, насекомых, мелкую птицу). Осенью
молодняк вместе с родителями принимает участие в групповых охотах на диких, а
порой и на домашних копытных. В конце авгусга-сентябре они покидают логово,
бродят в его окрестностях, меняя места дневок и осваивая новые близлежащие
участки леса.
Материалы наблюдений за 11 щенками из разных выводков, динамика
роста которых была прослежена в условиях неволи, показывают, что темп
физического развития волчат очень высок. Уже к 5-му дню от рождения масса
тела самцов составляла в среднем 760, а самок 726 г. В возрасте 30 дней
детеныши весили соответственно 3.1 и 2.9 кг, в 60 дней-7.4 и 7.7 кгив70дней-
9.1 и 9.6 кг. Длина тела у самцов увеличивалась от 32 см в 5-дневном возрасте
до 53 в месячном и 71 см - в 2-месячном. У самок соответствующие показатели
были равны 32,52 и 72 см. (Данилов и др., 1979). Судя по полученным данным
(рис. 63), линейные изменения размеров щенков (длина тела, обхват груди и
длина хвоста) заметно уступают темпам прироста их массы тела.
291
Интенсивный рост молодых волков, продолжающийся в течение первых 6
мес жизни, затем резко приостанавливается и в дальнейшем изменяется мало
(рис. 64). Например, волчица Ракша в Ленинградском (Санкт-Петербургском)
зоопарке весила в 6-месячном возрасте 23.6 кг, а к концу первого года жизни
-в среднем 26.3 кг. Вес взрослой самки Кичи (1968 г. р.) в разные сезоны 1973
г. также колебался незначительно - от 28 до 31.9 кг. Следовательно, уже к
концу первого года жизни размеры молодых особей приближаются к
таковым взрослых животных, однако полного физического развития самки
достигают не раньше чем в 2-3 года, а самцы, вероятно, еще позже (Данилов и
др., 1979).
Как и среди представителей семейства куньих, у собачьих в целом
отмечается существенное возрастное увеличение показателей абсолютной
массы внутренних органов и падение относительных. У взрослых волков
белорусской популяции, по данным П.Г. Козло с соавторами (1985), масса
сердца по сравнению с прибылыми увеличена у самцов в 1.8, а у самок в 1.4
раза. В то же время индекс сердца за период жизни детенышей в логове
снижается практически в 2 раза. С переходом зверей к более активному
самостоятельному образу жизни он начинает нарастать. Анализ возрастных
изменений абсолютной массы легких показал, что они достигают дефинитивных
размеров только на втором году жизни. В процессе постнатального онтогенеза
волчат индекс легких с возрастом резко снижается. Если у самок эта
зависимость прослеживается в процессе всего активного периода роста и
развития животных, то у взрослых самцов индекс легких (14.3±0.76%о) оказался
достоверно больше по сравнению с прибылыми и переярками (11,9±0.46%о),
но меньше, чем у зверей в первые месяцы жизни.
Из материалов этих же авторов (Козло и др., 1985) следует, что во время
обитания в логове щенки волка имеют также очень высокий индекс печени,
который по мере их роста закономерно снижается. Это может свидетельствовать
о повышенной функциональной значимости печени в первые месяцы жизни
молодняка и депонировании в ней большого количества гликогена.
Интенсивное поступление из нее в организм щенков питательных веществ
подстраховывает волчат от гибели при недостаточном количестве корма,
добываемого родителями. Данный факт следует рассматривать как важную
адаптивную особенность, повышающую выживаемость щенков.
Возрастная динамика массы селезенки характеризуется стабильным
увеличением ее абсолютной величины и падением относительной до того
периода, когда вес молодых и взрослых особей выравнивается. В дальнейшем
эти показатели изменяются мало. Абсолютная масса почек в процессе роста и
развития волчат постоянно увеличивается. В среднем у хищников из белорусской
популяции в возрасте 2 мес они весили 74 г, в 3 - 85 и в 4 мес - 135 г. Даже у
подрастающих зверей, уже ведущих более активный образ жизни, масса почек
также продолжала нарастать примерно до 2-летнего возраста. У прибылых она
достигала 182±9.57 г, у переярок - 228±11.64 и у взрослых животных -
28 5± 14.47. Интересно, что за время пребывания волчат в логове индекс почек
292
Таблица 108/Table 108
Возрастная изменчивость гематологических показателей и массы костного мозга
у представителей семейства собачьих из Дагестана (по: Коржуев, Спасская, 1985)
Age variation of hematological parameters and marrow mass in representatives
of canids family from Daghestan
Виды/ Species Возрастная группа/ Age group n Масса тела, кг/ Body mass, kg Гемоглобин, г%/ Hemoglobin, g% Отношение к массе тела, %/ Relation to the body mass, %
массы крови/ blood mass массы костного мозга/ medulla mass
Волк/ Adults 4 36.9 18.6 9.1 4.5
Canis lupus Youngs 4 1.4 12.6 7.2 5.4
Шакал/ Adults 4 7.7 14.2 8 3.7
Canis aureus Youngs 3 1.1 11.3 6 4
Лисица Adults 8 5.2 17.3 8.2 4.4
обыкновен./
Vulpes vulpes Youngs 8 0.5 13:4 9.1 4.8
Собака Adults 7 6 16 6.1 3.2
енотовидн./
Nyctereutes Youngs 3 0.9 11 4.3 3.7
procyonoides
закономерно снижался, а затем, с переходом щенков на самостоятельное
питание, постепенно увеличивался до уровня взрослых животных.
Таким образом, в постнатальном развитии волка можно выделить 2
периода, характеризующихся различными темпами развития основных
внутренних органов. В первом, охватывающем малоподвижный логовищный
период жизни волчат, наблюдается строгая обратная зависимость между
массой тела и относительным весом органов. По-видимому, это связано с
малой подвижностью щенков во время интенсивного роста и увеличения массы
тела животных. Во втором периоде, когда звери ведут типично хищнический
образ жизни, индексы органов уже не обнаруживают обратной корреляции с
массой тела и, следовательно, их развитие не подчиняется «правилу рядов»
Гесса. Из этого можно заключить, что у взрослых волков интенсивность
обменных процессов не связана с величиной поверхности тела, а определяется
спецификой экологии вида, требующей повышенной двигательной активности,
мобилизации физической силы и необычайной выносливости (Козло, 1981;
Козло и др., 1985).
Сравнение ряда эколого-физиологических показателей у волка и других
представителей семейства собачьих в процессе онтогенеза представляет
293
Таблица 109/ Table 109
Динамика массы тела и суточного прироста у молодняка (7<? и 5$)
европейской рыси (средние данные)
Dynamics of body mass and circadian growth rate of young (7^ н 5^)
of European lynx (Lynx lynx) (average data)
Возраст, мес/ Age, months Масса тела, г/ Body mass, g Среднесуточный прирост, г/ Weight growth per day, g Относительный прирост (% к массе тела однодневных особей)/ Relative growth rate (% to the body mass of one day individuals)
Однодневные/
One day 250 - 100
individuals
0.5 420 11.5 168
1 860 29.3 344
1.5 1200 22.7 480
2 1500 20 600
3 3100 53.3 1240
4 5000 63.3 2000
определенный интерес для понимания адаптивных морфофункциональных
особенностей этих хищников, обладающих разными скоростью роста,
активностью и формами поведения. Данные П.А. Коржуева и Т.Х. Спасской
(1985) свидетельствуют о том, что щенки собачьих в первый месяц жизни
обладают меньшими по сравнению с взрослыми особями показателями
концентрации гемоглобина в эритроцитах и массы крови (табл. 108). Вместе с
тем основной очаг гемопоэза позвоночных животных (скелет) у всех
рассматриваемых видов имеет больший относительный весу новорожденных по
сравнению с особями старших возрастных групп. То же можно сказать и о
костномозговой фракции скелета. Все это обусловливает быстрое нарастание
в организме уровня гематологических показателей, который заметно увели-
чивается после перехода хищников на самостоятельное добывание пищи,
сопровождающееся резким повышением их двигательной активности.
У детенышей обыкновенной рыси (семейство кошачьих), взятых из разных гнезд,
скорости роста и увеличения массы тела могут иметь некоторые различия,
определяемые размерами выводков и условиями содержания животных в неволе,
однако в целом в первые месяцы жизни они растут и развиваются наиболее
быстро. Молодняк рыси рождается слепым, с закрытыми слуховыми проходами,
светло-желтого цвета, со слабо выраженными темными пятнами на спине и лапах.
Вес однодневных детенышей 230-280 г при длине тела 22-25 и хвоста - 4-6 см.
Глаза открываются на 12-13-й, а ушные проходы - на 10-11-йдень. В 1.5-2 не-
дели они весят уже 400-450 г. К месячному возрасту масса их тела вновь
удваивается, а к 4 мес котята достигают 5 кг и более (табл. 109). До этого периода
достоверных половых различий по массе тела не прослеживается.
294
12
Рис. 65. Изменение массы тела сеголеток рыси (/г=20) в течение осени-весны
(по:‘Данилов, Русаков, Туманов, 1979).
По оси абсцисс - месяцы; по оси ординат - масса тела, кг.
Fig. 65. Changes of body mass of lynx (Lynx lynx) youngs (n=20) during
the autumnr-spring.
Abscissa - months; ordinate - body mass, kg.
В возрасте 3 недель молодые особи часто выглядывают из убежища и пробуют
выходить из него на прогулки, однако регулярно покидать родительское гнездо и
лазать начинают только к 1.5месяцам. Вэтотпериод они двигаются еще медленно,и
неуверенно, а оборонительный рефлекс явно превалирует над ориентировочно-
исследовательским. Детеныши очень пугливы,, ходят осторожно, а любой звук или
незнакомый предмет вызывает активно- оборонительную реакцию и желание
спрятаться в материнском убежище.
Когда животным исполняется 2 мес, они большую часть времени проводят
вне логова и заходят в него только для сна, кормежки или.в момент опасности..
В этом возрасте котята весят в среднем 1.5 кг, бывают очень подвижны и
игривы, хотя их движения еще не столь координированны, как у взрослых особей.
И ериод лактации продолжается 3-3.5 мес. В течение суток молодые звери питаются
молоком матери 10-12 раз, но уже в 45-50-дневном возрасте начинают пробовать
мясную пищу, которую приносит в гнездо самка.
В 2.5 мес детеныши рыси много играют, хорошо бегают, самостоятельно
находят и в небольшом количестве едят мясо. Их постоянные игровые контакты
являются не только основной тренировкой организма, но и имеют большое
значение для формирования охотничьих навыков и разнообразных форм
поведения.
В возрасте около 3 мес котята активно лазают, с жадностью отрывают
мясо от целого куска, но еще периодически (иногда до 4-6 раз в сутки) кормятся
молоком матери, потребность в котором вскоре пропадает. Если раньше самка
забивала и приносила для них добычу, которую молодые хищники активно
«терзали» и ели, то теперь мать учит их самостоятельно ловить мелких
животных. К 4-м месяцам детеныши начинают сами умерщвлять жертву и
охотно потребляют свежую добычу. В этом возрасте они весят уже 5-5.5 кг.
Переход на регулярное питание мясной пищей обусловливает и заметное
нарастание массы тела, которая у 4-месячных рысей составляет в среднем
63.3 г в сутки, что в 2.7-3 раза больше, чем у 1.5-2-ме.сячных животных,
295
питающихся молоком матери (табл. 109). О высоких темпах скорости роста
молодняка в данный период свидетельствует и такой показатель, как уровень
прироста его массы тела по отношению к таковой у новорожденных.
Например, по нашим наблюдениям, его величина у 4-месячных котят в среднем
в 4-4.5 раза больше, чем у 1.5-месячных особей, которые только начинают
пробовать мясную пищу.
Молодые рыси в отличие от большинства представителей семейства
Canidae поздно приступают к самостоятельной жизни. Обычно они покидают
логово в 3 мес, и мать начинает водить их за собой. Выводок распадается к
середине зимы или перед началом течки у самки (Гептнер, Слудский, 1972).
Это подтверждается и нашими наблюдениями, поскольку в ноябре-декабре
неоднократно приходилось тропить след самки с детенышами в различных регионах
России. Ко времени распада выводков масса их тела чаще всего составляет 50-70%
от таковой у взрослых зверей.
По имеющимся сведениям (Данилов и др., 1979), наибольшего веса
подросший молодняк достигает в декабре, затем он несколько снижается
(рис. 65). Падение массы тела, вероятно, происходит потому, что детеныши,
отделяясь от матери, вынуждены голодать. На данный период приходится и
наибольшая их гибель. Известен случай, когда 25 января 1972 г. в Карелии
был обнаружен мертвый рысенок, который весил всего 4.6 кг и погиб, видимо,
от голода, так как следов травм или признаков болезни при вскрытии у него не
наблюдалось. По сообщениям охотников-корреспондентов, за 1975-1976 гг.
в январе-марте в Карелии и Ленинградской области были найдены 3 мерт-
вых молодых зверя, которые весили не более 5 кг и были сильно истощены.
В обычные годы самцы-сеголетки рыси в первую зиму достигают максималь-
ного веса 9 кг (промеры по 7 особям) при длине тела до 80 см, кондилобазаль-
ной длине черепа 116 и скуловой ширине 90 мм. Однако известны и
более крупные экземпляры сеголеток, весившие 10.8 и 11.2 кг (Новиков и
др., 1970).
Интересные данные по постнатальному онтогенезу обыкновенной рыси
получены в Чешском зоопарке г. Острава (Stehlik, 1983). Здесь под
наблюдением находились 36 особей (16 и 20 £) из 16 пометов. Открытие
глаз у них отмечалось в интервале 7-15 дней, координированная ходьба - в
возрасте 24-30, а бег - 32 дней. Смена ювенильного шерстного покрова на
дефинитивный прослеживалась в 10-недельном возрасте. Продолжительность
этого процесса 4-5 недель. Временные клыки прорезались на 6-12-й день, рез-
цы - на 9-29-й и премоляры - на 24-63-й день от рождения. Средний вес
новорожденных (н=14) составлял 289 г при длине тела 252 мм. Половые
различия по этим показателям были незначительными. Самцы (л=8) имели массу
260-360 г и длину тела 245-261 мм, самки (п-6) соответственно 260-300 г
и 242-260 мм. В годовалом возрасте звери в среднем весили 18.4 кг (13.1-22).
По данным Стехлика (Stehlik, 1979), молодая рысь впервые самостоятельно
смогла добыть в вольере морскую свинку в возрасте 78 дней, курицу - 83 и
кролика - 86 дней. Элементы коллективной охоты впервые проявились у
296
хищников в возрасте 100-101 дня. Индивидуальные различия в развитии
охотничьего поведения у детенышей рыси выражены существенно. Основные
его формы - скрадывание и добывание пищи - врожденные, однако при-
обретаются они с опытом, в процессе обучения, поэтому потеря матери на
первом году жизни молодняка ставит под угрозу возможности выживания всего
выводка. Как правило, в первые 6-8 мес молодые рыси еще не способны
самостоятельно обеспечивать себя необходимым для нормальной жизнедея-
тельности количеством пищи.
Как и у рыси, детеныши лесного и камышового котов рождаются слепыми, с
закрытыми слуховыми проходами. Они покрыты густым серовато-охристым
пухом, местами имеющим темный оттенок. Прозревают довольно быстро - на
9-12-й день. По материалам С.Б. Насибова (1970), лактационный период у
самок продолжается около 2 мес. В кормлении, охране и воспитании молодняка
самец не участвует. Темпы роста детенышей достаточно высокие. В 3-ме-
сячном возрасте котята камышового кота весят до 2 кг, а лесного - до 1.5 кг. Масса
тела молодняка, родившегося ранней весной, к началу промыслового сезона
достигает у первого вида 3750-4250 г, а у второго -1200-2200 г, т. е. примерно
половины веса родителей. В годовалом возрасте молодые и взрослые особи
практически не различаются. Вместе с матерью выводок держится до осени. В
октябре-начале ноября он распадается, и котята приступают к самостоятельной
жизни.
У енота-полоскуна (семейство енотовых) новорожденные весят около 80 г.
Они рождаются слепыми и прозревают в среднем на 20-й день. К этому времени
у щенков прорезаются временные резцы и клыки. Растут они быстро и в
возрасте 2-2.5 мес уже переходят на самостоятельное питание. В сентябре-
ноябре молодые еноты по размерам не уступают взрослым, но еще заметно
отстают от них по массе тела (Г. Новиков, 1957; Гептнер и др., 1967).
Среди сеголеток, добытых осенью на Северном Кавказе, вес самцов (л=9)
составлял в среднем 4 кг (2.4-4.8), а самок (?z=21) - 3.7 кг (1.9-4.9). Взрослые
животные, отловленные в это же время, имели следующую массу тела: самцы
(/г=12) 5.7 кг, самки (п=12) 5.2 кг. Максимальный вес хищников достигал
соответственно 8.8 и 7.9 кг (Гинеев, 1972, 1973).
Самки бурого медведя (семейство медвежьих) рожают детенышей
зимой, в берлогах. На Севере и Северо-Западе России они остаются с
матерью в убежищах до начала-середины апреля. Медвежата рождаются
слепыми, с закрытыми слуховыми проходами и бывают покрыты редкой
белесой шерстью длиной 2-3 мм. Подошвы лап мягкие, голые, с мало пигмен-
тированными когтями. Вес 2 детенышей, родившихся у самки бурого
медведя 13 января 1952 г. в Ленинградском (Санкт-Петербургском)
зоопарке, был равен 438 г у самца и 397 г у самки при длине тела 25 и 23 см.
Близкие к этим данные получены в Московском зоопарке, где вес
однодневных медвежат составлял 501 и 510 г, а размеры - 23.5 и 23 см
(Гуляева, 1934). Глаза у молодняка открываются на 30-35-й день, а ушные
проходы - несколько раньше. На 14-15-е сутки у медвежат заметна
11 Зак. № 4309
297
Таблица 110/Table ПО
Возрастные изменения массы тела (кг) молодняка бурого медведя-
Age changes of body mass (kg) of young of brown bears (Ursus arctos)
Возраст, мес/ Age, months Самец/Male Самка/Female
масса тела/ body mass среднесуточный привес, г/ weight growth per day, g масса тела/ body mass среднесуточный привес, г/ weight growth per day, g
1.5 2.5 - 2.1 -
2 3.4 60 3 60
2.5 5 106 4.1 73
3 7 133 5.8 113
3.5 9.5 166 8.1 153
4 11.8 153 10.3 146
5 14.1 77 12.8 83
пигментация когтей, а шерстный покров становится густым, мягким,
буровато-коричневого цвета. В месячном возрасте они весят 1.4-1.8 кг и
достигают в длину 32-38 см (хвост 3-3.5 см). Примерно в том же возрасте
были медвежата (1<? и 2$), обнаруженные мертвыми 23 января 1978 г. в
зимнем убежище зверя (Волховский район Ленинградской области)
биологом В.А. Смеловым. Признаков гибели матери не прослеживалось.
Видимо, кем-то потревоженная, она была вынуждена покинуть гнездо
открытого типа, и детеныши замерзли. Они были слепыми, без зубов, с
закрытыми слуховыми проходами и хорошо развитым шерстным покро-
вом. Самки весили 1625 и 1785 г, самец - 1855 г. Длина тела и хвоста
равнялись соответственно у самок 36.5 и 38.5, 3 и 3.5 см, у самца - 39 и
3.7 см. Длина задней ступни медвежат колебалась в пределах 6.4-6.7 см.
У месячного молодняка бурого медведя обычно хорошо видно белое горлбвое
пятно, которое после 5-6 мес пропадает. В этот период они бывают очень цепкими, но
еще плохо стоят на ногах. В возрасте 1.5 мес детеныши довольно быстро передвигаются
и уже освобождают кишечник без предварительного массажа живота. В берлоге в
первые недели жизни детенышей медведица постоянно вылизывает их каудальную
часть тела, способствуя тем самым дефекации.
Поступившие от охотников 2 февраля 1957 г. в Ленинградский (Санкт-
Петербургский) зоопарк медвежата (2с? и 2$) в возрасте примерно 3 недель имели
средний вес 1.2 кг. Через 2 недели самцы весили 2.8 и 3 кг, самки - 2.1 и 2.2 кг.
Более четкий половой диморфизм по весу проявляется у молодняка заметно позд-
нее -в возрасте 2.5-3 мес. Двухмесячные детеныши очень игривы, неплохо бегают
и лазают, копают землю. В 3 мес они весят примерно 6-7 кг, ав4мес-10-12 кг.
В природе подросший молодняк выходит из берлоги вместе с матерью,
когда ему исполняется 3-4 мес. Детеныши всюду следуют за ней и уже
298
самостоятельно разыскивают пищу - ягоды, корешки, зеленую травку,
муравьев и других насекомых. Тем не менее они еще долго сосут мать: лактация
продолжается 5-6 мес. Однако и потом, в течение всего лета, самка находится
с медвежатами и обычно залегает с ними осенью в одну берлогу. В естественных
местообитаниях медведица, видимо, приходит в течку не чаще 1 раза в 2 года
(Данилов и др., 1979; Данилов, Туманов, 1991). По данным В.С. Пажетного
(1981), молодняк, зимующий без матери, за зиму теряет до 40% своего веса.
Высокая скорость роста детенышей прослеживается в течение первых 3-4
мес, а затем она постепенно снижается. Так, 2 медвежонка (самец и самка),
родившиеся в неволе в январе 1979 г., в возрасте 3 мес весили в среднем 6.5 кг,
а еще через месяц вес самца достиг 8.9 кг, а самки -7.1 кг. Длина тела была
равна у самца 75 см, хвоста - 7 см, а у самки - 66 и 5 см. Обхват груди
достигал соответственно 43 и 42 см. В 5-месячном возрасте самка (данные по
самцу отсутствуют) весила уже 12.9 кг при длине тела 82 см, хвоста - 6 и обхвате
груди - 51 см. Затем скорость ее весового роста снизилась и к 7 мес масса тела
составила всего 15-16 кг.
Наблюдения за изменением весового роста у медвежат, родившихся в
Ленинградском (Санкт-Петербургском) зоопарке в 1961 г., показали, что в
первые месяцы жизни (табл. 110) наибольший среднесуточный привес отмечался
у них в возрасте 3-4 мес, что совпадает и с данными Т.А. Адольф (1949). По ее
материалам, 1.5-месячные медвежата имеют клыки и по одному (четвертому)
предкоренному зубу в каждой челюсти. Затем, в возрасте 50 дней и старше,
происходит прорезание 3-го предкоренного зуба и дружное подрастание резцов:
сначала на верхней, а потом и на нижней челюсти (в возрасте 2-2.5 мес). К 2.5 мес
у зверей начинают появляться слабозаметные верхние и нижние 1-й и 2-й
предкоренные зубы. Молочный зубной ряд функционирует у них в апреле-мае,
а позже отмечается смена временных резцов на постоянные (в возрасте 3-4 мес).
В начале мая происходит набухание и потемнение десен позади последних
предкоренных и к июню-июлю прорезается первая, а затем вторая пара коренных
зубов, сначала на верхней, а потом и на нижней челюсти (в возрасте 5-5.5 мес).
У 6-8-месячных детенышей наблюдается смена предкоренных зубов, затем у 8-
10-месячных меняются 3-й и 4-й резцы, а у годовалых - клыки. К ноябрю-январю
у молодых зверей (в возрасте 10-12 мес) прорезается 3-й коренной зуб на нижней
челюсти. По свидетельству Т.А. Адольф (1949), окончательная смена зубов у
животных заканчивается уже на втором году жизни. При этом индивидуальная
изменчивость последовательности замены молочных зубов на постоянные в
выводках бывает выражена отчетливо.
Температура тела 1.5-2-месячных медвежат (при средней температуре
гнезда 20°С) колеблется от 36.9 до 37.2°С. У взрослых бурых медведей,
содержащихся в тех же условиях, она была равна летом 37-38°С. Зимой, во
время сна, обмен веществ (как у молодых, так и у взрослых зверей) понижается
примерно на 50-70%, что ведет к уменьшению энергорасходов. При этом
температура тела животных в зимний период снижается только на 1.5-2%
(Слоним, 1952, 1961).
299
По материалам В.С. Пажетнова (19936), в природных условиях у большин-
ства медведиц лактация прекращается к середине лета и в их организ-
ме начинается интенсивный процесс жиронакопления для успешной зимов-
ки. К этому периоду у медвежат уже сформирована основа пищевого по-
ведения, завершается формирование оборонительного и проявляется
способность к ориентации на местности. Материнский инстинкт, направленный
на охрану детенышей, значительно ослабевает. Внутрисемейная связь
под держивается в основном со стороны медвежат через реакцию следования,
а также с помощью «социального фактора», выражающегося в потребности
общения членов семьи друг с другом. Большое значение в их жизни приобретают
игровое и комфортное поведение, связанное прежде всего с устройством
временных лежек.
В берлогу семья ложится вместе. Весной, после выхода из нее, связь
медведицы с медвежатами под держивается с помощью сохранившейся на этот
год реакции следования, звуковых и запаховых сигналов. В период гона самка
объединяется с самцом, а отошедшие от нее лончаки нередко остаются жить
вместе до осени и могут даже лечь в одну берлогу. Весной такие пары
распадаются. Иногда в семьях медведей появляются пестуны. Обычно это -
медвежата третьего года жизни, которые вернулись к матери после ее
спаривания с самцом. Какого-либо участия пестуна в воспитании медвежат-
сеголеток не наблюдалось (Пажетнов, 1985).
Самки белого медведя в условиях неволи приносят детенышей либо в зимних
берлогах, либо в открытых, просто устроенных убежищах. По сообщению Т.Т.
Воккен (1933), в Ленинградском (Санкт-Петербургском) зоопарке 17 декабря
1932 г. медведица родила медвежат (2 самок) в небольшом углублении на снегу.
Один из них появился на свет в 11 ч, второй - в 15 ч. Они весили 708и712г при
длине тела 26.5 см и хвоста - 2 см. Детеныши были слепыми, с закрытыми
слуховыми проходами, покрытые светло-серой шерстью длиной 6-7 мм.
Масса тела медвежат (1<? и 1$), родившихся в Московском зоопарке
(Афонская, Крумина, 1958), составляла 560 и 690 г при длине тела 28 и 30 см.
Самка родила их 24 ноября при температуре среды 6-7°С ниже нуля. Несмотря
на холодную погоду, потреблять пищу она начала только на 3-й день после
родов, а полностью съедать обычную норму суточного рациона - через 2.5 мес.
В отмеченный период взрослые звери уже интенсивно расходуют свои
внутренние энергетические резервы. Глаза у малышей открылись на 30-35-й
день, слуховые проходы - на несколько дней раньше. В 2 мес они весили в
среднем 8.1 кг, в 2.5 - 10.5 и в 3 мес - 12.8 кг. К этому времени детеныши уже
охотно потребляли мясную пищу и некоторые растительные корма, однако
питаться молоком матери они продолжали регулярно до 10-11 мес, т.е. намного
дольше, чем молодняк бурого медведя.
У рожденных в пятидесятых годах в Ленинградском (Санкт-Петербург-
ском) зоопарке детенышей белого медведя (Г. Новиков, 1956) молочные зубы
(вначале резцы и клыки, а затем и остальные) появлялись в 1.5-2-месячном
возрасте, а их смена на постоянные начиналась в 5.5 мес и заканчивалась к 10-
300
11 мес. В 45 дней молодняк активно играл и ползал в логове, а вскоре стал
выбираться наружу и некоторое время проводить вне гнезда. В возрасте 2 мес
медвежата весили в среднем 6.6 кг, в 4 мес - 22.5, в 6 - 41 кг и в 8 мес - 60 кг.
В дальнейшем темпы их роста замедлялись. В природных условиях 6-месячные
детеныши достигают в длину около ПО см и способны съедать за время
кормежки до 2.5 кг сала.
В теплое время года, появляясь с матерью в прибрежной тундре, часто у
береговой кромки моря, сеголетки подбирают его выбросы, едят яйца птиц,
ловят леммингов. В их экскрементах в небольшом количестве встречаются
остатки побегов ивы, злаков и мхов, морошки и других ягод. К 5-5.5 мес
медвежата уже не боятся холодной воды, хорошо и охотно плавают. К началу
зимы они достаточно подготовлены к существованию в условиях высоких
широт. Густой мех, практически не намокающий в воде, а также мощный
слой подкожного жира надежно защищают их от низких температур и
увеличивают плавучесть. Белая окраска меха делает зверей не только
малозаметными во льдах и снежных торосах, но и играет важную роль в
снижении уровня теплоотдачи.
Многолетние наблюдения С.Е. Беликова (1993) за поведением разных
семейных групп белого медведя в Арктике показали, что с момента залегания в
берлогу и до периода, когда детеныши могут самостоятельно передвигаться, самки
редко покидают убежище. После того как малышам исполняется 2-2.5 мес,
медведица в тихие дни, когда нет ни сильного мороза, ни ветра, выводит их па
прогулку продолжительностью от 10-15 мин до 1.5-2 ч. Во время прогулки
медвежата затевают игры, в которых проявляются элементы охотничьего
поведения, -скрадывание, затаивание, прыжки, завершающиеся ударом лапой,
хватанием зубами и когтями различных предметов. Окончательно покинув
берлогу (обычно в последней декаде марта-начале апреля), семья отправляется к
морю. По пути следования самка делает остановки, чтобы покормить медвежат.
Кормит она их, чаще лежа на снегу. При этом в ненастную погоду медведица
ложится спиной к ветру, а при наличии глубокого снега выкапывает и забирается
в небольшую яму или временную берлогу, что способствует сохранению тепла в
организме животных. По свидетельству автора, в конце апреля-первой половине
мая на весь период короткого лета семьи белых медведей уходят во льды. После
того как они оставляют сушу и переходят на морской лед, самка начинает охотиться
на тюленей. Чаще всего ее добычей становятся кольчатая нерпа и ее детеныши,
находящиеся в подснежных убежищах на припайном льду. Медвежата остаются
при матери до конца третьей зимы, т.е. примерно до 2.5 лет.
Анализируя собранные материалы по особенностям постнатального онтогене-
за рассматриваемых хищных млекопитающих, можно отметить, что возрастные
изменения скорости весового и размерного роста, а также структурно-
функциональные преобразования различных систем органов проявляются у них
очень отчетливо. Они характеризуются видоспецифичностью и имеют направ-
ленность на более экономное обеспечение гомеостаза организма взрослых
животных.
301
Резюме
В ряду видов хищных млекопитающих из семейства куньих прослежива-
ется закономерное увеличение размерно-весовых показателей новорожденных
детенышей с повышением массы тела взрослых особей. У видов, представители
которых близки по этому признаку, вес новорожденных практически одинаков.
В то же время у горностая, степного хорька и перевязки детеныши рождаются
сравнительно мелкими. Эту особенность узкоспециализированных видов
(миофагов) можно рассматривать как одну из черт экологической адаптации,
позволяющей многоплодным животным при благоприятных условиях среды
быстро реагировать на них путем полной реализации своего репродуктивного
потенциала.
Динамики весового и линейного роста молодняка свидетельствуют о том,
что у куньих формирование морфофизиологических систем происходит
раньше у представителей тех видов, взрослые особи которых имеют меньшие
размеры, а следовательно, высокий уровень энергообмена. Это, вероятно,
обеспечивает лучшую выживаемость их щенков, смертность которых в
выводковый период весьма значительна. В целом выживаемость детенышей у
мелких хищников во многом обеспечивается скоростью их роста, развития и
формирования морфофизиологических систем в процессе онтогенеза. Если
молодняк этих видов по массе тела достигает дефинитивного уровня к
4-6 мес, то у крупных хищников семейства Mustelidae - к 8-10 мес. Важное
значение для первых имеет также раннее формирование временной зубной
системы (до открытия глаз), а для вторых - длительный период частичной
редукции молочных резцов у щенков. Эта особенность постнатального
онтогенеза позволяет самкам мелких многоплодных видов начинать рано
подкармливать малышей мясной пищей, а детенышам крупных позднеспелых
животных - долгое время кормиться молоком матери. Возрастные измене-
ния морфометрических показателей выражены у хищных млекопитающих
достаточно отчетливо. С увеличением возраста детенышей прослеживается
закономерное нарастание абсолютного и снижение относительного веса
основных внутренних органов. Интенсивное падение величины последнего
показателя (соотношение массы органов и тела) наблюдается лишь на ранних
этапах развития молодого организма и становится малозаметным ко времени
распада выводков.
Виды из семейств Canidae и Felidae характеризуются сравнительно
быстрыми темпами роста - изменением весовых и размерных показателей,
а также ранними сроками становления у хищников основных физио-
логических функций. Период открытия глаз, слуховых проходов и появления
временных зубов наступает у них заметно раньше, чем у крупных
представителей семейства Mustelidae - барсука, выдры и росомахи. В
отличие от большинства видов семейства собачьих у обыкновенной рыси
молодняк довольно поздно приступает к самостоятельной жизни. Котята
долго питаются молоком матери, родительское логово покидают только в
302
трехмесячном возрасте, но при этом еще до середины зимы остаются с
самкой. Основные формы их охотничьего поведения - врожденные, но
приобретаются они в процессе обучения, поэтому потеря матери в
выводковый период ставит под угрозу возможности выживания сеголеток
данного вида.
Молодняк бурого и белого медведей демонстрирует высокую скорость
линейно-весового роста в первые 3-4 мес жизни, когда у них отмечается и
наибольший показатель среднесуточного увеличения массы тела. В
дальнейшем темпы роста замедляются. Детеныши обоих видов характеризу-
ются поздним формированием морфологических систем и физиологических
функций, длительными сроками пребывания в родительской берлоге и
питания молоком матери. Они до 2-2.5 лет продолжают поддерживать
внутрисемейные отношения.
Функциональные изменения основных систем органов в процессе
постнатального онтогенеза молодняка разных видов отряда хищных
млекопитающих прослеживаются достаточно отчетливо. Если в первый
месяц жизни дыхание детенышей очень поверхностное, а пульс - частый, то с
возрастом происходят углубление дыхания, снижение темпа работы сердца и
увеличение кислородной емкости крови. Возрастная брадикардия
проявляется у них за счет удлинения фазы отдыха сердца (диастолы), при
этом продолжительность активной систолы практически не меняется.
Следовательно, молодым особям хищных млекопитающих свойствен менее
экономный темп функциональной деятельности сердца и органов дыхания,
что обусловливает большие энергозатраты и их высокую смертность при
неблагоприятных условиях среды.
Summary
Among the representatives of the family Mustelidae with the increasing of
body mass in adult individuals the weight and linear sizes of newborns also increase.
In females of closely related species with a similar body mass the weight of newborns
is practically equal. At the same time, in such fecund carnivores as Mustela erminea,
M. evesmanni, Vormelaperegusna, the young are born comparatively very small.
Apparently, the given peculiarity of myophagous specialized on feeding on rodents
should be regarded a feature of peculiar adaptation allowing these fecund animals
in favorable environmental conditions quickly to react by the full realization of
their reproductive potential.
The study of the main parameters of growth dynamics and development of the
young evidence that in Mustelidae the formation of morphophysiological systems
occurs earlier than in representatives of those species whose adult individuals have
smaller weight and linear sizes and, hence, high level of energy exchange. This,
probably, provides better survivorship of their pups, mortality of which in the child
period is rather high.
303
On the whole, survivorship of the young in small carnivores is largely provided
by the rate of weight growth and formation of morphophysiological systems in the
process of ontogenesis. If the pups of these species achieve by body mass the definite
level at 4-6 months, in large carnivores of the family Mustelidae at 8-10 months.
Highly significant for the former is the early formation of dentition (before eyes
opening), whereas for the latter - partial reduction of milk incisors in pups. This
characteristic of ontogenesis allows females of small fecund species to start early
feeding of the young with meat, whereas the young of large, late maturing animals
feed on mother’s milk for a long time.
Age changes of morphometric parameters are pronounced in all carnivores rather
distinctly. They are characterized by regular enlargement of absolute and decrease of
relative weight of the main inner organs with the increase of the age of an individual.
Dramatic decrease of organs mass relative to the body weight of animals is observed
only on early stages of ontogenesis and becomes less noticeable when the young
achieve the size of the parents.
The species of the family Canidae, Felidae are characterized by comparatively
fast rate of growth - change of weight and size parameters and early timings of
formation of main physiological functions. The period of opening of eyes, auditory
passages and appearance of deciduous teeth begins in them earlier than in the large
representatives of the family Mustelidae - Meles meles, Lutra lutra and Gulo gulo.
The high growth rate of the young of Ursus maritimus and U. arctos is observed
only during the first 3-4 months and then it gradually decreases. As in bears, the
young of Lynx lynx, unlike most of representatives of the family Canidae, start the
independent life late. They commonly leave the den at the age of 3 months, but up
to the middle of winter they could be met together with female. Up to the time of
disintegration of breeds the body mass of the young comprises not more than 50-
70% of the body weight of the adult individual.
The functional changes of the main systems of organs in the process of
ontogenesis in various representatives of the order Carnivora are followed rather
explicitly. In the first month of life the breath of the young is very superficial and
pulse frequent, with age the deepening of the breath and slowing of the heart rate
take place. The age bradycardia is displayed in them owing to the elongation of the
rest phase of the heart (diastole) and the duration of the active systole practically
not changing. Consequently, less economic rate of heart and respiratory organs
activity is typical of young individuals of carnivores. It may determine high energy
expenses and their high mortality with unfavorable environmental conditions.
Глава 8. ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ
СООТНОШЕНИЯ ПОЛОВ И ИХ
БИОЛОГИЧЕСКАЯ НАПРАВЛЕННОСТЬ
Одним из основных показателей демографической структуры популяции
служит половой состав, изучение которого позволяет иметь дополнительные
сведения о сложных механизмах саморегуляции видов и динамике их
численности. Животные разных возрастов различаются по формам поведения,
прежде всего по кормодобывающей деятельности, показателям уровня
энергообмена и репродуктивным возможностям. Для каждой из старших
возрастных групп свойственна определенная интенсивность размножения. При
этом возрастная динамика соотношения полов имеет важное значение, так
как определяет энергетический баланс популяции (Кубанцев, 1961, 1964;
Шварц, 1968,1980; Шилов, 1977; Ostby et al., 1978; Bucher et al., 1983;
Большаков, Кубанцев, 1984; Novak et al., 1987).
Как справедливо отмечает П.И. Данилов (1994), связь данного показателя
с уровнем численности животных можно считать одним из проявлений
механизма авторегуляции, направленного на максимальную компенсацию
потерь в популяции посредством включения в размножение возможно большего
количества самок. Под воздействием факторов внешней среды самцы и самки
разного возраста, в силу характерного для них уровня физиологических
процессов, элиминируют с разной степенью интенсивности. В связи с этим
рассматриваемый показатель у молодых и взрослых особей необходимо
учитывать, поскольку он определяет темпы воспроизводства природных
группировок вида.
В качестве первичного соотношения полов у млекопитающих принимается
количество самцов и самок, складывающееся при оплодотворении. Оно
определяется сочетанием половых хромосом в процессе мейоза и обычно
бывает близко 1 : 1 (Хатт, 1969; Яблоков, 1987).
В практике изучения динамики численности животных принято выделять и
такие ступени половой структуры, как вторичное соотношение (среди зрелых
эмбрионов и новорожденных), ювенильное (в группе молодняка перед его
расселением), субтретичное (среди молодняка, еще не участвовавшего в
размножении) и третичное, которое наблюдается у взрослых животных.
Изменение соотношения полов в пределах каждой из этих возрастных групп
имеет важное значение для жизнеобеспечения популяции, влияя на темпы ее
воспроизводства, использование энергетических ресурсов, поведение и
выживание особей (Ивантер,1962; Граков, 1980; Большаков, Кубанцев, 1984).
Показатель соотношения полов у мелких хищников семейства Mustelidae
характеризуется относительно высоким уровнем динамической стабильности.
В общих чертах у них прослеживается сравнительно выровненный половой
состав в выводках и заметное преобладание самцов в старших возрастных
305
группах. При этом субтретичное соотношение полов демонстрирует
наибольшую лабильность. Если в годы резкого подъема численности в
популяции заметно преобладают молодые особи (и среди них - самцы), то при
депрессии происходит перестройка половой структуры в сторону увеличения
доли взрослых животных, а в группе сеголеток-самок (Кукарцев, 1970,1972;
Белов, 1972, 1980; Бакеев, 1973, 1976; Монахов, 1974, 1975; Граков, 19696,
1976). Считается, что высокий уровень рождаемости самцов характеризует
благополучие популяции хищников (Теплов, 1954; Монахов, 1968; Корытин,
1971; Глушков и др., 1999). Регулярное отклонение от равного вторичного
соотношения полов в ту или иную сторону при благоприятных условиях
обитания животных (например, при вольерном содержании), вероятно,
свидетельствует о видоспецифичности данного признака.
Собранные нами материалы (гнезда со щенками, обнаруженными в
природе, и детенышами, родившимися в неволе), а также данные по
размножению животных, опубликованные Д.В. Терновским (1977), пока-
зывают, что у мелких представителей семейства куньих (от ласки до хорьков
и норок включительно) в целом соотношение полов в пометах при рождении
либо примерно равное, либо имеет некоторую направленность на увеличение
доли самок (табл. 111). Большинство литературных источников по данному
вопросу подтверждает это положение. Так, в выводках ласки самки составляли
50 и 52% (Heidt et al., 1968; Blomquist et al., 1981), горностая - 53.3 и 50.2%
(Копеин, 1977;Терновский, 1983), колонка-52.3 (Курис, 1972), хорька лесного
- 50% (Данилов, Туманов, 1976а), норки европейской - 59.1% (Мошонкин,
1981), а американской - 60 и 62.8% (Терновский, 1972в; Данилов, Туманов,
1976а, б).
Следует также отметить, что статистический анализ доли самцов в целом
среди всех выводков из группы мелких видов куньих (ласка - норки) и более
крупных (соболь - выдра) хищников семейства по /-критерию Стьюдента
показал наличие достоверных различий между ними (р=95%). У последних
доля самцов в пометах составляла 56.5±3.5%, а у мелких видов -48.2± 1.2%.
О половом составе населения хищных млекопитающих вне гнездового
периода обычно судят по данным их отлова или отстрела в сезон охоты.
Промысловая выборка включает такие материалы, как заполненные лицензии
на отлов животных, их тушки и шкурки. Однако следует учитывать, что
большинство современных способов добычи зверей имеет явную направ-
ленность на вылов самцов и не всегда отражает истинное соотношение полов
в природных популяциях (Русанов, 1973; Большаков, Кубанцев, 1984;
Глушков и др., 1999). При охоте на крупных представителей фауны выбивают
преимущественно взрослых полноценных самцов. При отлове пушных зверей
их также добывают чаще, чем самок, но уже по причине большей подвижности,
меньшей осторожности и тщательности в подборе мест для дневок.
Некоторые способы охоты, позволяющие в значительной степени
исключить избирательность промысла, показывают, что преобладание
самцов в выборках бывает и не столь существенным. Об этом же
306
Таблица 111/Table 111
Соотношение полов в гнездовой период у мелких видов куньих
Sex ratio in the den period of small species of mustelids
Виды/ Species Данные автора/Our data Данные Терновского, 1977/Temovsky, 1977
кол-во/number % самок/ % of females кол-во/number % самок/ % of females
выводков/ litter щенков/ cubs выводков/ litter щенков/ cubs
Ласка обыкновен./ Mustela nivalis 17 74 52.7 3 18 50
Горностай/ Mustela 9 73 52.1 16 118 50
erminea Солонгой/ Mustela 8 74 54.1 20 171 50.3
altaica Колонок/ Mustela sibirica 6 32 46.9 10 49 46.9
Хорек лесной/ 52.8
Mustela putorius Хорек степной/ 12 72 37 5 25 52
54.1
Mustela eversmanni Норка 5 13 99 50.5
европ./ Mustela 8 28 46.4 17 60 46.7
lutreola Норка америк./ Mustela vison 23 102 55.9 13 73 58.9
Примечание. В табл. Ill и 114 приводятся сведения автора по соотношению полов
среди детенышей, родившихся в неволе, и обнаруженных в гнездах, в природе.
свидетельствуют полевые наблюдения, когда половую принадлежность
животных устанавливают либо визуально, либо методом тропления - по
размерам следов и расположению мочевых точек (Meeh, 1970,1977; Туманов,
1972, 1980; Пажетнов, 1985, 1993а; Сидорович, 1990; Данилов, 1988, 1994;
Железнов, 1990; Bjarvall et al., 1996, и др.). Данные о структуре промысловых
выборок необходимы для общей охотхозяйственной оценки поголовья
природных группировок, его изменчивости в разные фазы популяционных
циклов и сезоны года. Они позволяют разрабатывать нормативы рацио-
нального использования ресурсного потенциала видов в интересах как
человека, так и сохранения самих животных.
Промысловые выборки из популяций мелких хищников в целом за ряд
лет характеризуются стабильным преобладанием в них доли самцов (табл. 112),
причем среди взрослых особей эта диспропорция имеет наибольшую величину
(Lockie, 1966; Данилов, 1968,1969; Fog, 1969; Васенева, 1972а, 1974; Данилов
307
Таблица 112/Table 112
Соотношение полов в выборках из популяций представителей семейства куньих
в 1972-1986 гг. (северо-западный регион России)
Sex ratio in the samples from populations of representatives of mustelids in 1972-1986
(North-West region of Russia)
Виды/ Species Шкурки зверьков/ Skins Тушки сеголеток/ Carcasses of youngs Тушки взрослых/ Carcasses of adults
всего/ total доля самцов, %/ percentage of males всего/ total доля самцов, %/ percentage of males всего/ total доля самцов, %/ percentage of males
Ласка/ Mustela nivalis 364 66.5 310 54.2 350 67.4
Горностай/ Mustela 1420 65.1 420 55.2 380 66.6
erminea Хорек лесной/ Mustela 4314 60.6 520 57.3 475 67.2
putorius Норка европ./ Mustela lutreola 1755 52.7 318 50.3 310 52.6
Норка америк./ Mustela 11540 53.4 426 47.9 408 57.6
vison Куница лесная/ Martes martes 4145 53.1 316 48.1 188 57.4
Росомаха/ Gulo gulo 68 52.9 9 55.6 15 60
Барсук/ Meles meles 82 47.6 42 52.4 58 55.2
Выдра/ Lutra lutra 618 52.4 64 54.7 83 57.8
и др., 1977; Войлочников, 1977; Синицын, Кукарцев, 1982; Erlinge, 1983,1986;
King, 1983, 1989; Синицын, 1985, 1988).
В основных местообитаниях мелких видов куньих, судя по материалам их
троплений, в большинстве случаев самцы также встречаются чаще. Эта разница
становится наиболее очевидной на фазе роста и пика численности животных.
По наблюдениям В.Е. Сидоровича с соавторами (1997), зимой 1991/92 и
1992/93 гг. в стабильной по численности малоопромышляемой группировке
европейской норки (Локнянский р-н, Псковская обл., данные троплений)
соотношение полов было примерно одинаковым - 13 самцов и 11 самок
308
Таблица 113/ Table 113
Смертность молодняка и соотношение полов в выводках американской норки
стандартного окраса разного возраста (количество особей на 1 рожавшую самку)
The mortality' of young and sex ratio in the litters of American mink (Mustela vison)
of standard color and different age (individuals per 1 delivering female)
Годы/ Years Возраст самок, лет/ Age of females Обследованный молодняк/Youngs Соотношение полов при рождении/ Sex ratio of cubs
всего выводков/ number of itters смертность/ mortality всего выводков/ number of litters <?<?
1975 1 455 0.8±0.05 273 3+0.06 2.9±0.06
1975 2 629 0.6±0.09 438 3.4±0.06 3.3±0.06
1975 3 365 0.5±0.03 235 3.1±0.08 3±0.07
1976 1 86 0.6±0.10 48 3.1±0.15 3.2±0.17
1977 2 86 0.5±0.11 59 3.7+0.16 3±0.15
1978 3 86 0.4±0.09 54 3.2±0.16 3.3±0.18
1979 1 88 0.9±0.07 43 2.8±0.10 3.2+0.08
1979 2 55 0.6±0.16 30 2.6±0.09 3.5±0.13
1979 3 24 0.4±0.15 15 3.3±0.12 2.7+0.18
1979 4 18 0.3±0.12 13 3±0.34 2.8±0.34
1980 1 122 0.9±0.10 60 3.1+0.10 3.2±0.09
1980 2 118 0.6±0.16 76 3.2±0.08 3.3±0.06
1980 3 55 0.5±0.14 31 2.6+0.08 3.2+0.09
1980 4 14 0.5±0.18 7 2.9±0.32 2.8+0.36
(%2 = 0.1,/>>0.1). При этом в осмотренной промысловой выборке количество
отловленных в районе самцов и самок (27 с? и 23 $>) также не имело достоверных
различий (%2 = 0.2;р>0.1). В других угодьях, занятых американской норкой,
когда численность ее постоянно росла, чаще преобладали самцы, но при резком
снижении плотности населения зверьков доля самок в естественных
местообитаниях увеличивалась с 43% (л=58) до 58% (л=298).
Следует отметить, что природные группировки американской норки во
многих регионах России имеют звероводческое происхождение, поскольку при
целенаправленном ее расселении животных брали из зверохозяйств. Кроме
того, нельзя не учитывать и процесс «естественной интродукции», когда норки
со звероферм убегали в природу, где и происходила их натурализация.
Учитывая эти обстоятельства, в 1975-1980 гг. в хозяйстве «Заря»
(Ленинградская область) были проанализированы сведения по племенному
разведению американской норки стандартного окраса, что позволило выявить
309
величину смертности и соотношение полов в пометах зверьков, постоянно
находящихся в стабильных и благоприятных условиях содержания.
Данные учета смертности молодняка за период от его рождения и до отсадки
свидетельствует о том, что в выводках американской норки естественный отход
щенков снижается с увеличением возраста самок. Эта разница между молодыми
и взрослыми особями по величине элиминации детенышей прослеживается очень
отчетливо (табл. 113). Она объясняется особенностями физиологии рожавших и
не приносивших приплод самок, а также приобретенным опытом выращивания
детенышей у норок старших возрастов.
Для проверки установленного факта и устранения возможных искажений
результатов за счет целенаправленного влияния селекции на показатели
продуктивности норок, нами выборочно были просмотрены регистрационные
ведомости по размножению одной и той же группы самок (86 особей). Они
приносили приплод в 1976-1978 гг. Полученные материалы подтвердили
наблюдения о меньшей естественной смертности молодняка в группе самок,
которые уже выращивали потомство (р <0.005). При этом во всех случаях доля
погибших до регистрации молодых самцов была несколько выше таковой самок.
Сходные данные о большей детской смертности среди самцов получены и
другими исследователями. Например, по материалам Н.В. Граковой и Н.Н.
Гракова (1977), среди просмотренных мертворожденных детенышей разводимой
в хозяйстве американской норки соотношение полов было практически равным
(248 самцов и 251 самка). В то же время в группе павших до регистрации щенков
самцы явно преобладали (55±2.21%). Авторы считают, что в пометах норки
постоянно проявляется тенденция к преимущественной послеродовой гибели
самцов. За длительный период полевых работ, который включал разные фазы
популяционного цикла американской норки, нами было обнаружено 15 гнезд с
57 щенками разного возраста. Среди них самки составляли 56.1% (32 особи).
Судя по всем этим фактам, к концу выводкового периода в популяциях
американской норки несколько преобладают молодые самки, что способствует
увеличению ее репродуктивного потенциала, а следовательно, и общей
численности зверьков. Наблюдаемая у европейской норки в последние годы
сравнительно низкая плодовитость самок и примерно равное соотношение полов
в выводках могут свидетельствовать о низком уровне воспроизводственных
возможностей природных группировок данного вида.
Определенный интерес представляет изменчивость соотношения полов в
пометах самок разного возраста и в целом в группировках в отдельные годы.
Рассматривая величину этого показателя у американской норки, содержащейся
в зверохозяйсгвах (табл. 113), можно отметить, что вне зависимости от возраста
самок соотношение полов в выводках было близко к 1:1. Всего же за период
с 1975 по 1980 г. среди 8730 просмотренных щенков самцы составляли 50.2%
(4390 особей). Вместе с тем годы, когда в выводках американской норки чаще
рождались самки, явно преобладали. Так, доля самцов (%) от всего
родившегося молодняка составляла: в 1975 г. - 50.7, 1976 - 49.1, 1977 - 55.1,
1978-49.2,1979-47 и в 1980 г. -48.1 (Туманов, 1983). Близкое соотношение
310
полов у разводимой в неволе американской норки прослежено и в других
регионах страны (Ильина, 1952; Абрамов, 1961; Большаков, Кубанцев, 1984).
В группе молодых американских норок (и=426), добытых в 70-80-х годах
в угодьях южных областей Северо-Запада России, доля самцов-сеголеток
также была ниже таковой самок (табл. 112). В то же время среди взрослых
особей во все промысловые сезоны чаще добывались самцы, доля которых в
среднем за ряд лет была равна 57.6±3.8%. При этом надежность вывода о
возрастных различиях составляла 95%. Отмеченное общее преобладание
самцов в промысловых выборках и природных популяциях американской
норки определяется прежде всего их большей долей в старших возрастных
группах. По-видимому, при интенсивном расселении животных ресурсный
потенциал вида увеличивается не только за счет изменения плодовитости
самок старших возрастов, но и большей рождаемости и выживаемости
молодых самок в помете.
Взрослые самки за время щенения, лактации и осенней линьки интенсивно
расходуют свои внутренние энергетические резервы и за короткий период
осеннего нагула не успевают создать достаточного количества жировых
запасов. В результате они оказываются хуже подготовленными к зимовке, и
их естественная смертность по сравнению с самцами может быть заметно
большей, что подтверждается средней продолжительностью жизни взрослых
зверьков. По нашим данным, она составляла в рассматриваемых выборках
из природных группировок американской норки у самцов 3.7 и у самок 2.6
года, а европейской соответственно 3.3 и 2.3 года.
В биопробах из популяции американской норки среди старых особей
диспропорция по полу прослеживалась вполне отчетливо. Из 255 добытых
на Северо-Западе России взрослых зверьков, возраст которых определяли
по слоистым структурам клыков (Клевезаль, Клейненберг. 1967), только 12
(4.7%) принадлежали к особям 5-6 лет. В этой группе 9 зверьков (75%)
оказались самцами и 3 (25%) - самками.
В угодьях, занятых европейской норкой (до вселения в них особей
американского вида), в слабоопромышленных популяциях соотношение
полов у молодых и взрослых животных оказалось в среднем одинаковым.
Так, в конце 60-х-начале 70-х годов XX в. в ряде районов Псковской и
Новгородской областей плотность населения аборигенной норки на многих
водоемах достигала 5-12 особей на 10 км побережья. В промысловой
выборке, состоящей из тушек и шкурок добытых зверьков (п=486),
соотношение полов было следующим: 255 самцов и 231 самок (%2= 1.2;
р>0.50). Примерно равный половой состав среди отловленных сеголеток и
взрослых особей прослеживался и позднее (табл. 112). В обнаруженных нами
в природе 4 выводках было 8 самцов и 8 самок. В пометах, рожденных в
условиях неволи (табл. 111), этот показатель также был близок к 1.
Очевидно, что для устойчивых популяций этого вида вообще характерно
равное соотношение полов в выводках и биопробах среди сеголеток, а
увеличение доли самцов в группе взрослых животных выражено не так
311
отчетливо, как у американской норки. В выборке из 58 взрослых
европейских норок до 4-7-летнего возраста дожили только 10 зверьков
(17.2%), среди которых оказалось 6 самцов и 4 самки. Из них один самец
был семилетним, а два других самца и две самки - шестилетними. Возраст
остальных 5 норок определен в 4-5 лет.
В 4 гнездах лесного хорька, обнаруженных в природе, было 10 самцов и
10 самок (4с? и 3<J; 3<? и 2<ф; 2с? и 2<j>; 1<? и 3$). В 12 выводках зверьков,
рожденных в неволе, это соотношение было несколько сдвинуто в сторону
увеличения доли самок (табл. 111). В просмотренных в 60-х-начале 70-х годов
промысловых пробах (Данилов, Туманов. 1976а) из популяций этого вида без
разделения по возрастным группам достоверно преобладали самцы. В Карелии
их оказалось 56% (и=47), в Псковской области - 66% (и=530), а в целом по
северо-западному региону России - 61.5% (х2=29; р>0.99). Среди сеголеток
соотношение полов было близким к 1:1. Следовательно, большая доля самцов
в промысловой пробе объясняется их значительным преобладанием среди особей
старших возрастов. В 70-80-х годах в биопробах из популяций этого вида
(н=4314) доля самцов всех возрастов была равна 60.6% (табл. 112). В пробе из
38 взрослых особей, добытых в Ленинградской области, 4-6-летние зверьки
составляли 21% от общего их количества, причем до 5-6 лет дожили только 5
самцов и 1 самку. В Псковской области за один промысловый сезон охотниками
Локнянского района было отловлено 82 взрослых лесных хорька, из которых
5-6-летние (11 особей) составляли только 13.4%, причем до указанного возраста
дожили 8 самцов (72.7%) и 3 самки (27.3%), что также может свидетельствовать
о большей элиминации последних в природных группировках.
Для изучения возрастного состава рассмотренных популяций в группе
американской норки были проанализированы 238 тушек зверьков, добытых
преимущественно в Ленинградской области в 70-80-е годы. Установлено, что
доля сеголеток в пробе была равна 51.3%, годовиков - 19.7, двухлеток - 16,
трехлеток - 9.7, а особей 4 лет и старше - 3.3%. Выборка из популяции
европейской норки, отловленной в тот же период в Псковской и Новгородской
областях, состояла из 106 тушек животных. Сеголетки в ней составляли 41.5%,
год овики-21.7, двухлетки- 16.1, трехлетки- 11.3 и зверьки более старшего
возрата - 9.4%. Возрастной состав лесного хорька из тех же областей оказался
следующим: сеголетки - 59.8%, годовалые особи - 17.1, двухлетние - 13.7,
трехлетние - 6, а животные в возрасте 4 лет и старше составляли 3.4%. Большая
доля сеголеток в выборках из популяций американской норки и лесного хорька,
а также равномерное уменьшение количества взрослых особей разного возраста
свидетельствуют о высокой численности и благополучном состоянии
группировок этих видов в обозначенный временной период.
Изменчивость демографической структуры популяции колонка изучена
недостаточно. В просмотренных нами выводках этого вида, полученных при
вольерном разведении зверьков, соотношение полов было практически равным
(табл. 111), что совпадает с материалами других авторов (Курис, 1972;
Войлочников, 1977; Терновский, 1977). В промысловых выборках из популяции
312
вида данный показатель заметно менялся по годам, однако все исследователи
отмечают тот факт, что в целом в угодьях самцов отлавливают в 2-3 раза
больше, чем самок (Бромлей, 1956; Корытин, 1963; Войлочников, 1977; Глуш-
ков идр., 1999). Особенно велика эта разница в начале охотсезона.
Следует иметь в виду, что у всех мелких куньих (включая колонка) на
соотношение полов в биопробах существенное влияние оказывают такие
факторы, как способы и сроки отлова зверьков. Самцы крупнее, активнее и
смелее самок. Они отличаются большей длиной суточного (жировочного) хода,
меньшей осторожностью и используют для дневок сравнительно просто
устроенные убежища. Все это влияет на структуру выборки из популяций вида
и соотношение полов в ней. Обычно в начале охотсезона (прежде всего при
использовании самоловов) сеголеток и самцов всех возрастов добывают
заметно больше, чем самок. Доля последних увеличивается только в конце
зимы, когда многие самцы уже выловлены.
По материалам В. М. Сапаева (1969), в биопробах из популяции колонка
(//=534), собранных в течение 3 сезонов на юге Хабаровского края, среди
отловленных сеголеток оказалось 19,34 и 39% самок, а среди взрослых особей
соответственно 24, 38 и 35%. По этим данным в первый год наблюдений,
характеризующийся благоприятными экологическими условиями и высокой
численностью зверьков, доля самок в добыче охотников была наименьшей. В
последующие сезоны, при низкой плотности населения колонков в угодьях и
ухудшении состояния кормовой базы, количество отловленных самок по
сравнению с первым годом заметно возросло. Отмеченный факт можно
объяснить увеличением их доли в выводках (а следовательно, в популяции) и
суточной активности особей, в том числе обычно «осторожных» самок, при
дефиците кормов в природе.
Собранные сведения позволяют предположить, что половая структура
колонка очень динамична. В то же время необходимо учитывать и то
обстоятельство, что его добывают преимущественно в первую половину
охотсезона, когда из популяции изымаются прежде всего «малоопытные»
сеголетки и более активные самцы, что, видимо, и находит свое отображение
в характеристике биопроб.
В выводках мелких хищников-миофагов (ласка, горностай, солонгой) доля
самцов среди новорожденных редко превышает таковую самок. В дальнейшем
(субтретичное и третичное соотношение полов) эта разница проявляется более
отчетливо, особенно в группе особей старше года (табл. 111,112).
В этом плане интересны наблюдения К. И. Копеина (1977), проведенные в
Западной Сибири за популяциями горностая. В 7 обнаруженных им выводках
из северных районов Тюменской области было 45 щенков (21<? и 24$>). Среди
добытых летом 65 молодых особей на долю самцов пришлось 52.3%. В
обследованных в конце зимы выборках из популяций этого вида самцов (прежде
всего взрослых) оказалось заметно больше, чем самок. Так, на севере
Тюменской области (//=4109) их доля была равна 71.8%, на Урале (л=287) -
67%, а в Северном Казахстане (//=500) - 60.2%.
313
Преобладание самцов горностая в зимних сборах наблюдалось также на'
Сахалине и Камчатке. При этом установлено, что по мере снижения численности
вида в этом регионе доля самцов и разнополых сеголеток в промысловых
выборках достоверно уменьшалась (Воронов, 1977; Илюшкин, 1988).
В разных областях России в начале охотсезона среди мелких хищников
самцов отлавливается заметно больше, чем самок. По сообщению К.И. Ко-
пеина (1977), в Омской области в охотничий сезон 1963/64 г. из всех добытых
горностаев (л=1018) на долю самцов приходилось в ноябре 72.5%, в декабре
61.7, в январе 43.2 и в феврале 34.8%. В октябре 1964 г. в промысловой
выборке из 127 зверьков они составляли 87%, причем пойманные в первой
половине месяца особи (п=36) все, кроме одной, оказались самцами. Таким
образом, по мере изъятия из популяции самцов их относительное количество
неуклонно снижается, а доля самок в конце промыслового сезона заметно
возрастает.
Влияние промысла на половой состав местных группировок горностая
достаточно подробно отражено в работах В.А. Кукарцева (1970, 1974),
выполненных в Ханты-Мансийском АО. Им установлено, что в выводках этого
вида соотношение самцов и самок соответствует 1:1.6. В то же время по
материалам анализа биопроб в ноябре на одну самку приходится 2.17-3.82
самца, а в конце охотсезона (в феврале) - 1.09-1.62. Плавное увеличе-
ние количества самок в выборках к концу промысла свиде-тельствует о
том, что на протяжении всего охотсезона сохраняется непреднамеренная
избирательность, направленная на отлов самцов. Следовательно, одна из
главных предпосылок рационального исполь-зования ресурсов горностая
заключается в оперативном слежении за динамикой добычи особей разного
пола.
Многолетние тропления мелких хищников, проведенные нами на
стационарных участках, свидетельствуют о большей встречаемости следов
самцов по сравнению с самками, что подтверждается наблюдениями и других
исследователей (Терновский, 1958, 1977; Сидорович и др., 1997). В целом
можно отметить, что значительная доля самцов в биопробах из популяций
рассматриваемых видов куньих определяется их заметным преобладанием
среди особей старших возрастов. Так, при поголовном отлове лесных хорьков
на типичном участке площадью до 80 км2 (Новгородская область) в сезон
1967/68 г. были добыты 18 зверьков. Среди них мы обнаружили 7 взрослых особей
(5<? и 2<j>) и 11 сеголеток (6<? и 5$>).
Необходимо отметить и частую встречаемость самцов в выборках из
популяций ласки, специальной охоты на которую нигде не проводится. Обычно
она попадается в ловушки, установленные в подземных ходах обыкновенного
крота без какой-либо пищевой приманки. Поэтому высокий процент отлова
взрослых самцов ласки может быть обусловлен как их большей подвижностью,
так и численностью в угодьях. Если соотношение полов в выборках из
популяций мелких хищников и не полностью адекватно таковому в природе,
то все равно преобладание взрослых самцов над самками вполне очевидно.
314
Таблица 114/ Table 114
Соотношение полов в гнездовой период у видов из рода Martes
и некоторых крупных представителей семейства куиьих
Sex ratio in the species of genus Martes and some large representatives
of mustelids family during den period
Виды/ Species Количество/Number
выводков/ litters щенков/ cubs самок/ females % самок/ % of females
Соболь/ Martes zibellina 6 15 6 40
Куница лесная/ Martes martes 10 33 15 45.5
Харза/ Martes flavigula 3 5 2 40
Росомаха/ Gulo gulo 8 18 9 50
Барсук/ Meles meles 11 27 13 48.1
Выдра/ Lutra lutra 15 30 14 46.7
Оно, вероятно, связано с их лучшей подготовленностью к зимовке, а следовательно,
и меньшей элиминацией в холодный период года.
Анализ небольшой промысловой выборки из популяции горностая (л=32)
показал наличие в ней (из долгожителей) шести 4-летних зверьков (4<? и 2<j>) и
одного самца, возраст которого был определен в 6 лет. Среди 30 ласок,
отловленных в Ленинградской области, самыми старыми особями оказались 5
самцов и 1 самка в возрасте 3 лет, а также один пятилетний самец. По-
видимому, это - предельный возраст для мелких миофагов, обитающих в
природе.
Данные о вторичном соотношении полов у представителей рода Martes
немногочисленны, однако в своем большинстве они свидетельствуют либо о
равном количестве новорожденных самок и самцов, либо о незначительном
преобладании последних в выводках (Юргенсон, 1950, 1956; Гептнер и др.,
1967; Лобачев, 1973; Терновский, Терновская, 1994, и др.). Собранные нами
материалы (табл. 114) также подтверждают эти зависимости.
Из практики разведения соболей в зверохозяйствах известно, что в
некоторые годы в пометах могут преобладать и самки (Залекер, 1962; Г. Мо-
нахов, 1975), но чаще всего доля самцов в них составляет 52-57% (Нуме-
ров, 1969; Монахов, Бакеев, 1981). Сходное вторичное соотношение полов
наблюдалось и среди соболей, разводимых в зверохозяйстве «Заря»
Ленинградской области (1976-1980 гг.). В первой группе среди 140 щенков,
полученных от 49 самок в возрасте 3 лет, самцы составляли 52.1%. Во второй
имелось 295 детенышей, которые родились у 105 самок в возрасте 5 лет. Среди
них также преобладали самцы (51.2%). Судя по имеющимся материалам,
возраст самок, по всей видимости, не определяет соотношение полов в пометах.
315
Таблица 115/ Table. 115
Соотношение полов в выборках из популяции лесной куницы в Карелии •
(по: Данилов, Туманов, 1976а)
Sex ratio in the samples from populations of pine marten (Martes martes) in Karelia
Возраст/ Species Количество исследованных куниц/Number
всего/total доля самцов, %/ percentage of total
Молодые/Youngs:
До 2 мес/1-2 months old 23 52.2
6-9 мес/6-9 months old 51 45.1
Взрослые/Adults:
1.5-2 года/1.5-2 years old 41 43.9
2.5-3 года/2.5-3 years old 30 60
3.5 года и старше/3.5 years old and older 25 80
Анализ данных по половой структуре 1057 выводков соболей из
Салтыковского зверохозяйства (Монахов, Бакеев, 1981) за 1959-1964 гг.
показал, что среди новорожденных щенков (л=3622) из разных генераций
процент самцов в целом неизменно оказывался близким к среднемноголетнему
значению, равному 52.7%. За 6 лет наблюдений только в один год самки
несколько преобладали (50.5±1.8%) среди детенышей (и=757), причем это
отклонение было статистически несущественным. По-видимому, и в
природных условиях каждая новая генерация соболей и куниц включается в
популяционные процессы с выявленными показателями соотношения полов.
По сообщению Н.Н. Гракова (19696, 1973), в обнаруженных на севере
европейской части России 5 гнездах лесной куницы находилось в общей
сложности 18 щенков с соотношением полов 1:1. В Карелии и Ленинградской
области (Данилов, Туманов, 1976а) в пометах этого вида (5 гнезд с 19
детенышами и тушка самки с 4 крупными эмбрионами) количество самцов и
самок также оказалось практически равным (12<? и 11 $>).
Материалы, характеризующие субтретичное и третичное соотношение
полов у лесной куницы и соболя по промысловым выборкам из популяций,
свидетельствуют о том, что тенденция к общему преобладанию самцов
сохраняется во многих частях Их ареалов (Тимофеев, Надеев, 1955; Данилов,
Ивантер, 1967; Когтева, Морозов, 1972; Бакеев, 1973, 1976; Г. Монахов,
1974,1975; Соколов, 1978,1992а, б; Белов, 1972,1980; Граков, 1981; Глушков
и др., 1999; В. Монахов, 1983а; Зырянов, 1990, 1998; Валенцев, 1992;
Седалищев, 1998, и др.). Эти виды имеют относительно невысокие показатели
плодовитости, потенциала размножения локальных группировок и плотности
населения зверьков в угодьях, поэтому большее количество самцов в
природных комплексах, по-видимому, обеспечивает полное включение в
процесс размножения всех готовых к гону самок, а следовательно, и
увеличение численности животных в популяциях, разреженных промыслом.
316
Таблица 116/Table 116
Половозрастной состав биопроб (%) из популяции лесной куницы
в разные периоды осенне-зимнего сезона (по: Треничев, 1985)
Sex-age composition of biological samplings (%) from populations of pine marten
{Martes martes) in various periods of autumn-winter season
Возраст/Age II Период добычи/Censusing period
1 сентября-10 декабря/ 1 Septemb.-10 Decemb. 11 декабря- 20 января/ 11 Decemb.-20 Jan. 21 января— 1 марта/ 21 Jan.-l March
Самцы/Males
Взрослые/ Adults 75 41.3±5.7 38.7±5.6 20±4.6
Сеголетки/ Youngs, 0+ 139 52.5±4.2 27.3+3.8 20.2±3.4
Самки/Females
Взрослые/ Adults 79 44.3±5.6 34.2±5.3 21.5±4.6
Сеголетки/ Youngs, 0+ 104 65.4±4.7 22.1 ±4.1 12.5±3.2
Среди молодых и взрослых особей лесной куницы показатель соотношения
полов в выборках заметно различается. Если в среднем за ряд лет у сеголеток из
Северо-Западных областей доля самцов и самок была примерно одинаковой, то у
взрослых куниц первые постоянно преобладали (см. табл. 112). Материалы,
собранные в Карелии (Данилов, Туманов, 1976а), также наглядно демонстрируют
возрастную изменчивость полового состава биопроб из популяции этого вида
(табл. 115). В гнездовой период у щенков он был близок к 1 : 1, среди зверьков
первого (0+) и второго (1+) годов несколько преобладали самки, асреди куниц
старших возрастов - самцы. Вполне вероятно, что бблыпая доля молодых самок
по сравнению с самцами объясняется их лучшей выживаемостью в ювенильный
период и повышенной активностью при расселении.
В то же время в некоторые промысловые сезоны при высокой численности
лесной куницы в биопробах среди сеголеток могут преобладать самцы. Так, в
ноябре-декабре 2001 г. в северных районах Псковской области плотность
населения зверьков была достаточно высокой и достигала 1.5-1.8 особей на
1 000 га. На одном из участков, где куницу промышляли активным способом -
с собаками, среди добытого молодняка оказалось 6 самцов и только 2 самки.
Сходная картина отмечалась и на других участках охотников. Отмеченный
факт можно объяснить действительно большим количеством молодых самцов
в природе в данном году - в период пика численности куницы. В этом случае
рабочая собака скорее обнаружит след самца, суточная активность которого
и по времени, и по длине пройденного пути выше, чем у самки.
Изменчивость половой структуры промысловых выборок из локальных
группировок лесной куницы европейского Севера прослежена Н.Н. Граковым
(19696, 1976). Им выявлено, что в период нарастания численности вида в
317
биопробах преобладали самцы всех возрастов (52.9±0.8%) и сеголетки
(57±0.5%). Среди последних самки составляли 51.2%, а в группе половозрелых
животных - 39.5%.
Интересные сведения по поведению и составу промысловых выборок лесной
куницы из Уральского региона приводятся в работе Е.М. Треничева (1985). По
его наблюдениям, в начале зимы угодья, освободившиеся от части зверьков,
добытых зимой прошлого года, интенсивно осваиваются сеголетками обоего
пола и взрослыми самцами. Молодые самцы (0+) до начала весны нередко
продолжают жить на прежнем материнском участке. В то же время самок-
сеголеток добывали обычно в новых местах, где следов других куниц либо не
было, либо они появлялись здесь лишь в конце сезона охоты. Повышение следовой
активности в позднезимний период характерно для всех половозрастных групп
куницы. Осенью самок-первогодок добывают достоверно больше, чем особей
других половозрастных групп (табл. 116). В предзимний период, когда
происходит расселение зверьков, молодым самкам свойственна наибольшая
следовая активность. У самцов-сеголеток она оказалась несколько ниже, чем у
самок. К середине зимы количество прибылых куниц в пробах существенно
уменьшается. В просмотренных выборках, собранных в это время, доля самок-
первогодок составляла лишь 22.1% от общей их добычи за сезон охоты и
уменьшилась по сравнению с первым периодом на 43.3%. Для молодых самцов
аналогичные показатели составляли соответственно 27.3 и 25.2%. Наряду с
этим процент добычи взрослых особей продолжал сохраняться на достаточно
высоком уровне. В конце зимы основная масса зверьков оказывается уже
отловленной, а удельный вес каждой половозрастной группы, изымаемой
промыслом на последней его стадии, бывает минимальным по отношению к их
доле в начале и середине охотсезона (табл. 116).
Материалы, характеризующие демографическую структуру природной
популяции лесной куницы, были получены в январе-марте 1972 г. в
Выборгском районе Ленинградской области. Здесь сотрудниками Госохот-
инспекции проводился опытный поголовный отлов зверьков этого вида на
одном из участков, прилегающих к заказнику «Дымово». Всего удалось добыть
17 особей (9с? и 8$). Среди них молодых самцов и самок (0+) оказалось равное
количество (З^1 и 3$). Они составляли 35.3% от общего количества
отловленных животных. В возрасте 1.5-2.5 лет были 3 самца и 4 самки, а среди
особей старше 3 лет - 3 самца и 1 самка.
По-видимому, у лесной куницы, так же как и у мелких куньих, в природных
условиях среди взрослых особей наблюдается большая смертность самок как
менее подготовленных к холодному зимнему периоду, о чем сообщают и
некоторые другие исследователи (Теплов, 1954; Граков, 19696). Кроме того,
взрослые самцы зимой бывают упитаны заметно лучше сеголеток, что в
малокормные годы может служить причиной высокого уровня естественной
элиминации последних.
В целом на Северо-Западе России в различных группировках лесной
куницы первогодки (0+) составляют 39.9%, годовики (1+) - 30.8, двухлетки
318
(2+) - 18.3, а особи в возрасте 3 лет и более - 11% (Когтева, Морозов,
1972). На ее возрастной состав, очевидно, заметное влияние оказывает
состояние кормовой базы в зимний период, предшествующий промыслу.
Например, в охотничьи сезоны 1960/61 и 1962/63 гг., следовавшие за
зимами с обилием различных пищевых компонентов, сеголеток добывали
больше (соответственно 44 и 45%), чем в 1961/62 и 1964/65 гг. (38 и 22%),
которым предшествовали годы, неблагоприятные в кормовом отношении
(Данилов, Туманов, 1976а). Вероятно, наличие и доступность пищи в зимние
месяцы сказываются прежде всего на сохранности эмбрионов, т.е. на
плодовитости самок, в то время как фазы популяционного цикла обусло-
вливают соотношение полов в выводках.
Возрастная изменчивость полового состава в группировках соболя сходна с
таковой у лесной куницы. Практически всегда среди взрослых особей в пробах
оказывается больше самцов, чем самок, а среди сеголеток, наоборот, последние
нередко преобладают. Сложилось мнение (Глушков и др., 1999), что подобные факты
не являются следствием непреднамеренной избиратель-ности добычи, а
представляют собой нормальную структурно-популяционную характеристику
вида.
В работе Г.И. Монахова и Н.Н. Бакеева (1981) показана сравнительно
слабая корреляция между плотностью населения соболя и соотношением
полов в выборках (г =0.46), однако они констатируют тенденцию к увеличе-
нию доли самцов в них в годы высокой численности вида. Отмеченная
стабильность в соотношении полов у рассматриваемых животных на разных
фазах популяционного цикла неслучайна и является одним из параметров,
который определяет динамику их численности. Из материалов В.Г. Монахова
(1985), собранных в Приобье, следует, что территориальное регулирование
промысла соболя может являться эффективным методом охраны и под-
держания на высоком уровне его численности. При этом различные воз-
растные группы имеют и разную территориальную локализацию. В
припойменных стациях большую часть населения вида составляют самки-
сеголетки (50%), несколько меньшую - молодые самцы (45%) и только 5% -
взрослые животные. В материковых стациях и на водоразделах группы
молодых самок и самцов не так многочисленны - 24.6 и 20.3% соответ-
ственно. Среди взрослых зверьков во всех возрастных группах преобладают
самцы. В материково-водораздельных стациях возрастная структура
оказалась более сложной, поскольку в пробах встречались особи почти всех
старших возрастов, в то время как в припойменных лесах обнаружены соболя
лишь из средневозрастных групп. По данным автора, основная масса взрослых
зверьков, дающая более 90% годового прироста поголовья, сосредоточена
здесь, в материково-водораздельных лесах, которые занимают около 30%
лесопокрытых площадей. Таким образом, в Приобье для территориального
регулирования промысла соболя следует учитывать биологические
особенности расселения молодняка и дифференцированно подходить к
использованию запасов вида.
319
Судя по результатам анализа биопроб, у соболя в различных частях ар’еала,
как и у лесной куницы, промыслом из природных группировок изымаются
прежде всего сеголетки и взрослые самцы. В Красноярском крае преобладание
самок в выборках прослеживалось лишь в группе сеголеток (68.6%) и
годовиков (60.5%). Среди половозрелых особей во все промысловые сезоны
самцов было больше, что, вероятно, представляет своеобразный «механизм
выравнивания» возрастной структуры популяции вида. Свойственная взрослым
самкам осторожность обусловливает их меньший вылов. Так, если при отлове
зверьков капканами доля взрослых самок в биопробах редко превышала 20-
25%, то на охотах с собакой, которые менее избирательны, этот показатель
возрастал до 48.5% (Зырянов, 1998).
Возможно, в слабоопромышленных популяциях соболей половое соотношение
зверьков по сравнению с таковым в биопробах более выровнено. Необходимо
учитывать тот факт, что зимой суточная активность самок заметно меньше, чем у
самцов. Это увеличивает вероятность встреч следов последних собакой, а
следовательно, и долю их добычи промысловиками. Например, в Магаданской
области при дневной температуре воздуха 30-35°С ниже нуля суточный наслед
самцов составлял в среднем около 3, а самок-не более 2 км (п-10). С повышением
температуры воздуха до 10-14°С мороза охотничий ход хищника увеличивался до
5-12 км в зависимости от обилия корма, но все равно активность самок всегда была
меньше, чем у самцов (Девяткин, 1998).
Анализ многолетних данных, характеризующих воспроизводственный
потенциал популяций соболя в Красноярском крае (Зырянов, Туманов,
2000), показал, что доля взрослых самок в биопробах (л=10 180 особей)
колеблется от 10.9-14.4% (северная тайга и Западно-Сибирская равнина)
до 17.8-18.2% (Саяны и средняя тайга). По материалам А.И. Хлебникова
(1977), в малоосвоенных угодьях красноярской тайги самцов-сеголеток
оказывалось в 3 раза меньше, чем самок того же возраста. В зоне
интенсивного промысла это соотношение было близко к 1 : 1. Дальнейшее
возрастное изменение структуры популяции в сторону увеличения доли
самцов прослеживается очень отчетливо. На не осваиваемых ранее
промыслом территориях, судя по размерам встреченных следов и
количеству выловленных самцов, установлено заметное их преобладание
в популяции. Так, в пробе из 12 зверьков, добытых на ходовых охотах с
сабакой, оказалось 9 половозрелых самцов и только 3 молодые самки
первого и второго года жизни.
Сравнительный анализ демографической структуры различных группи-
ровок соболя свидетельствует об увеличении доли молодых зверьков на
интенсивно опромышляемых территориях, как и в случаях с лесной куницей;
заполнение свободных участков происходит в основном за счет молодняка
(преимущественно самок), мигрирующего с неопромышляемых угодий, что
было подтверждено методом мечения животных (Валенцев, 1980,1981).
Выявленные возрастные особенности соотношения полов в выборках из
популяций рассматриваемого вида прослеживаются в разных регионах России
320
(Тимофеев, Надеев, 1955; Нумеров, 1973; Мельников, 1975). Например, в
биопробах из Магаданской области (сезон 1991/92 гг.) преобладали сеголетки
(/г=247), среди которых доля самок достигала 60%. Если низкая выживаемость
молодых соболей (группы 0+ и 1 +) обусловлена здесь в основном влиянием
промысла, то для особей старших возрастов (особенно самок) наибольшее
значение имеет фактор естественной смертности (Девяткин, 1992, 1998). В
Колымских районах Якутии за период с 1983 по 1991 г. была обследована
выборка из 1477 тушек соболей (Седалищев и др., 1992). В целом среди
сеголеток в ней преобладали самки, а у взрослых особей - самцы. Во все годы
работы доля самок (49-70%) в группе молодых зверьков постоянно
превалировала над таковой у особей старших возрастов (26-60%). При этом
количество сеголеток в биопробах было достаточно большим (42-75%),
свидетельствуя о стабильном состоянии популяции, и близким по уровню
(47.9%) к таковому в западносибирских группировках (В. Монахов, 1983а).
На Камчатке, как и в других частях ареала вида, в начале охотсезона
молодых соболей отлавливали чаще, чем в конце промысла. Средний возраст
самцов, добытых в ноябре-декабре, составлял 3.8, а в январе-феврале - 5.1
года. Продолжительность их жизни в природе, видимо, больше, чем у самок, у
которых этот показатель был равен соответственно 3.2 и 4.2 годам (Валенцев.
1981, 1992).
В зоопарках некоторые соболя и лесные куницы разного пола живут до 13-
15 лет. И.Д. Старков (1940) считает, что в неволе продолжительность их жизни
может достигать 18 лет. По-видимому, это - предельный возраст, до которого
в условиях вольерного содержания доживают соболь и куница. Среди
просмотренных нами в Восточных Саянах 35 черепов соболей из старших
возрастных групп самыми старыми особями оказались самцы 7-9 лет. В то же
время В.Г. Монахов (1985) отмечает, что в биопробах из Приобья находились
зверьки «всех возрастов до 14 лет включительно». В выборке из 48 черепов
лесных куниц, добытых на Северо-Западе России, самцы старше 7-8 и самки
4-5 лет встречены не были (Данилов, Туманов, 1976 а).
Таким образом, собранные материалы свидетельствуют о том, что
возрастная структура популяций куницы и соболя представляет собой один из
важных факторов, определяющих общее соотношение полов, а следовательно,
и численности этих видов. Очевидно, что морфологические и поведенческие
различия самцов и самок (по размерам тела, возможности использовать
встречающиеся убежища, уровню осторожности, продолжительности
суточного хода и т.д.) в значительной степени определяют соотношение полов
в промысловых выборках. Ежегодный достаточно высокий уровень изъятия
взрослых самцов из природных группировок, казалось бы, должен привести к
увеличению доли самок в биопробах последующих сезонов, однако этого не
наблюдается. Сказанное может свидетельствовать о постоянном численном
превышении половозрелых самцов в угодьях, что, видимо, объясняется
естественной элиминацией рожавших самок, которые, как правило, хуже
подготовлены к трудным условиям зимовки. Постоянное преобладание
321
взрослых самцов в популяциях рассматриваемых видов представляется нам
одним из важных факторов, обусловливающих характерное соотношение
полов во времени и пространстве.
По мнению Г.А. Соколова (1992, 2001), величина репродуктивного
потенциала в группировках соболя трансформируется через половую и
возрастную структуры, поэтому в одних случаях промысловое изъятие зверей
из популяций должно основываться на адекватности той структуры, которая
свойственна совокупности особей на момент их ухода в состояние покоя перед
новым репродуктивным циклом, в других экологически вполне оправдан
усиленный пресс охоты на сеголеток - при высоком приросте их численности -
и на самцов, когда они существенно преобладают над самками. Исследования
автором демографии группировок саянского и тунгусского подвидов соболя
позволили выявить некоторые различия в колебаниях их третичного соотноше-
ния полов. Так, у саянской популяции, отличающейся высокой численностью
животных в угодьях (до 25 особей на 1 тыс. га), увеличение доли самцов над
самками (/>2) достигает ббльших величин, чем у тунгусской, характеризую-
щейся низкой плотностью населения зверьков (4-5 особей на 1 тыс. га).
Превышение в подавляющем большинстве случаев количества самцов над
самками у саянского соболя можно рассматривать как один из механизмов
регуляции численности популяции. Воспроизведя бопьшесамцовпривысокойплотности
населения зверьков, популяция тормозит процесс дальнейшего увеличения ее размеров,
а следовательно, обеспечивает свое гомеостатическое состояние.
В пометах росомахи, выдры и барсука наблюдается довольно устойчивое,
примерно равное соотношение полов (табл. 114). В промысловых выборках из
популяций этих зверей на Северо-Западе России в целом доля самцов
составляла у барсука 47.6%, а у выдры и росомахи соответственно 52.4 и
52.9%. Анализ материалов по субтретичному и третичному половому составу
этих видов показал, что если среди сеголеток соотношение полов близко к
1 : 1, то с возрастом, как и у других куньих, количество добываемых самцов
несколько увеличивается (табл. 112). Отмеченная зависимость прослеживается
в разных частях ареала крупных хищников семейства Mustelidae и характерна
для популяций барсука (Stubbe, 1969,1970; Лобачев, 1976), выдры (Данилов,
1969; Вшивцев, 1972; Green et al., 1984; Сидорович, 1990, 1991), росомахи
(Паровщиков, 1960, 1964; Rausch, Pearson, 1972; Hornocker, Hash, 1981;
Б. Новиков, 1993; Кожечкин, 1999) и калана (Ильина, 1950; Барабаш-Ники-
форов и др., 1968).
В известных нам 11 выводках барсука, родившихся в неволе и обнаруженных
в природе, находилось 27 щенков, среди которых самки (13 особей) составляли
48.1%. В 3 пометах этого вида, полученных в Московском зоопарке, соотношение
полов также оказалось практически равным - 5 самцов и 4 самки. Среди 31
барсука разного возраста, отловленного в 60-х-начале 70-х годов в Южной
Карелии, Псковской и Ленинградской областях, превышение доли самцов
(51.6%, %2 = 0.13, р>0.75) было незначительным (Данилов, Туманов, 1976а).
Примерно таким же оно оставалось и в последующие годы.
322
Таблица 117/ Table 117
Величина выводка и соотношение полов в пометах росомах из разных
регионов России (по: Туманов, Кожечкии, 2000)
The size of the litter and sex ratio in the litters of wolverines (Gulo gulo)
from different regions of Russia
Кол-во щенков Соотнош.
Регион/ в выводке/ полов/ Источник/
Region Number of cubs Sex ratio Authors
in litter (<?<?/£?)
Левобережье р. Енисея (Красноярский 2 0/2 Туманов,
край)/ Кожсчкин,
The left bank of the Yenisey (Krasnoyarsk Territory) 2 0/2 2000
Бассейн p. Оби (Новосибирская обл.)/ The Ob’ River drainage (Novosibirsk Prov.) 4 2/2 Тсрновскнй, Тсрновская, 1994
Восточный Саян (заповедник «Столбы»)/ -5 1/7 Зырянов,
Eastern Sayan (Natural Reserve «Stolby») J 1989
Северо-Запад европейской части России/ 4 2/2 Данилов,
North-west of European Russia 3 1/2 Туманов,
1 1/0 1976а
Архангельская область/ 2 0/2 Паров щиков,
Arkhangelsk Prov. 2 0/2 1960
2 1/1
2 0/2
1 0/1
2 2/0 Паров щиков,
1964
Bccro/Total 30 10/20
В Ленинградском (Санкт-Петербургском) зоопарке барсуки-долгожители
(2 самки и самец) прожили по 10 лет, а еще 1 самка и 1 самец - соответственно
11 и 12 лет. Очевидно, и в природе максимальная продолжительность жизни
особей этого вида вряд ли превышает 12-15 лет, что подтверждается
наблюдениями и других исследователей (Гептнер и др., 1967; Stubbe, 1970).
Судя по результатам анализа слоистой структуры зубов, максимальный
возраст самок, отловленных в Псковской области, составлял 8-9, а самцов -
10-11 лет. Поскольку все эти звери были хорошо упитаны, а самки еще и
беременны, то можно предположить, что и в природных условиях данный
возраст не является предельным для барсуков. При раскопках старых,
заброшенных городков мы дважды находили кости и черепа зверей, гибель
которых, вероятно, была естественной. Возраст одного из них определен в 7-
8, а второго в 13-14 лет. Зубы последнего оказались стертыми до корней, а
их режущая и жевательная поверхности - гладко отшлифованными.
323
В пометах выдры, судя по наблюдениям в условиях вольерного
содержания, смертность щенков разного пола примерно одинакова, а
соотношение полов близко к 1:1 (табл. 114). В Новосибирском зоопарке среди
20 детенышей (10 выводков) находились 12 самцов и 8 самок (Шило,
Немшилова, 1989). Существенных половых различий в продолжительности
охотничьего поиска у подросшего молодняка из естественных местообитаний
не отмечается, что, видимо, и определяет примерно равное соотношение
самцов и самок среди добытых сеголеток. К периоду становления полово-
зрелости у выдры, как и у барсука, количество самцов в биопробах
увеличивалось незначительно - в среднем на 3% (табл. 112).
В Ленинградской, Псковской и Новгородской областях в выборках из
популяций выдры (tt-618) доля самцов разного возраста в целом составляла
52.4%. В Карелии этот показатель был равен 53% (Данилов, 1969), а на
Сахалине - 56.2% (Вшивцев, 1972). Поскольку выдру добывают преиму-
щественно капканом, то, вероятно, из-за меньшей осторожности самцов
отлавливают чаще, чем самок, поэтому в биопробах их доля может быть
больше, чем в естественных местообитаниях. Исследования, проведенные в
Белоруссии (Сидорович, 1992), показали, что для охраняемых природных
территорий характерен практически равный половой состав в популяциях
выдры с небольшим отклонением в сторону увеличения доли самцов, однако
статистически достоверным оно оказалось только для взрослых особей из
Березинского заповедника (%2=4.6,/?=0.03).
Разведение выдры в условиях неволи свидетельствует о том, что ее предельная
продолжительность жизни может составлять 13-15 лет. В промысловой выборке
из 35 взрослых особей, добытых в Ленинградской, Псковской и Новгородской
областях, самки-долгожители были в возрасте 4-5, а самцы - 5-6 лет. В
Белоруссии (Сидорович и др., 1997) средний возраст зверей, отловленных на
территории охотпользования, составлял 3.6 года, а максимальный - 9-10 лет
(//=73). На охраняемых природных территориях средняя и наибольшая
продолжительность жизни выдры, вероятно, выше. Так, в Березинрком
заповеднике в период его временной ликвидации (50-е годы XX в.) кроме
нескольких молодых животных были добыты 2 выдры в возрасте 7 и 12 лет.
В известных нам 8 пометах росомахи (18 щенков) количество самцов и
самок было равным (см. табл. 114). Таким же оно оказалось и в 6 выводках (6с?
и 6<£), полученных от 2 самок в Финляндии, в Хельсинкском зоопарке (устное
сообщение его сотрудников). Среди детенышей в одном помете находился
1 щенок, во втором - 3, а в остальных - по 2.
Сведения о плодовитости росомах, а также количестве самцов и самок в
выводках, обнаруженных в различных регионах России, представлены в табл.
117. По этим данным, соотношение полов часто оказывалось сдвинутым в
сторону увеличения в пометах доли самок. О некотором их преобладании в
выводках рассматриваемого хищника сообщают также и норвежские зоологи
(Myrberget, Sorumgard, 1979). Так, в осмотренных ими гнездах с 83 щенками,
самки составляли 58% от общего количества молодняка.
324
Таблица 118/Table 118
Доля самцов росомахи в промысловых выборках из некоторых регионов
Приеиисейской Сибири (по: Туманов, Кожечкин, 1999)
The portion of males wolwerine in the game samples from some regions of Siberia
Регион/ Region Сезон охоты/ Season of hunting Добыто зверей/Number
всего/ total в том числе самцов/ including males
n | %
Северная тайга/ Northern taiga 1980/81 4 3 75
1981/82 3 . 2 67
Средняя тайга/ Middle taiga 1988/89 4 2 50
1989/90 5 4 80
1992/93 3 2 67
1993/94 5 3 60
Саяны/Sayans 1989/90 4 3 75
Танну-Ола/ Tannu-Ola 1985/86 3 2 67
Итого/Total 31 21 67.7
Если в гнездовой период среди детенышей росомахи наблюдалось либо
равное соотношение полов, либо явное увеличение в них доли самок, то уже
среди первогодок, добытых в природе в период промысла, самцы обычно
несколько преобладали (Rausch, Pearson, 1972; Данилов, Туманов, 1976а;
Данилов идр., 1978; Myrberget, Sorumgard, 1979; Б. Новиков, 1981). Однако
в штате Монтана (США) из 24 пойманных росомах среди взрослых животных
было 9<? и 8$>, а в группе молодых - 2<? и 5$> (Hornocker, Howard, 1981).
В целом среди осмотренных нами на пушно-меховом холодильнике
(г. Санкт-Петербург) 68 шкур добытых зверей доля самцов составляла 52.9%
(см. табл. 112). В группе взрослых особей эти различия оказываются наиболее
выраженными. Например, из 15 половозрелых зверей, отловленных в Карелии
и Мурманской области, 9 оказались самцами. В Республике Коми во взрослой
части популяции вида они преобладают постоянно. В Печоро-Илычском
заповеднике из 147 зверей, пол которых был определен по следам на снегу,
самцы составляли 66%, а среди зверей, добытых охотниками, - 61 %. При этом
капканный лов увеличивает долю самцов. Из 8 росомах, пойманных на севере
Республики Коми, только 2 особи оказались самками (Полежаев, 1998).
325
Самцы росомахи представляют собой наиболее активную и подвижную
часть популяции, возможно поэтому в период промысла их добывают чаще
самок. Собранные В.В. Кожечкиным (Туманов, Кожечкин, 2000) сведения об
отлове росомахи в угодьях Приенисейской Сибири свидетельствуют о постоян-
ном преобладании в биопробах самцов, доля которых здесь достигает 67.7%
(и=31) от общего количества отловленных животных (табл. 118).
О большей встречаемости взрослых самцов в популяциях росомахи из
разных регионов Голарктики сообщает также Б. В. Новиков (1993). По его
данным, в Британской Колумбии соотношение самцов к самкам составляет
1 : 0.9, на Аляске и Чукотке - 1 : 0.8, в скандинавских странах и на Камчатке -
1 : 0.7, а в Приуралье -1 : 0.6. При этом демографическая структура популяций
росомахи под воздействием различных факторов может изменяться в
относительно короткий срок, равный 2-3 годам. В связи с тем что репродуктивный
период у представителей вида очень растянут, летом часть зверей проявляет
большую подвижность по сравнению с теми особями, которые заняты
воспитанием молодняка, родившегося ранней весной. По данным автора, в
летний сезон группировки росомах состоят из животных, участвующих в гоне,
воспи-тывающих потомство, и неполовозрелого молодняка. Основное ядро
производителей в популяциях вида составляют особи старших возрастов, среди
которых количественно преобладают взрослые самцы. Эта особенность,
вероятно, обусловлена поздними сроками становления половозрелости у
молодых самцов по сравнению с самками и жизненно необходима для популяции
вида с разреженным распределением половых партнеров по территории.
Увеличение доли новорожденных самок в пометах росомахи, по-видимому,
следует рассматривать как один из возможных компенсаторных механизмов
адаптации вида, восполняющих низкую плодовитость зверей, сравнительно
малое количество в популяции взрослых самок и позднеспелость молодняка.
В то же время преобладание самцов в группах старших возрастов является
биологически оправданным, поскольку они обеспечивают максимальное
включение в процесс размножения всех половозрелых самок даже в годы с
низкой численностью животных (Туманов, Кожечкин, 2000).
Максимальный возраст 2 самцов, добытых в природе, был равен 8 и 9
годам, однако в неволе продолжительность жизни зверей бывает заметно
большей. Так, годовалая самка, поступившая в Ленинградский (Санкт-
Петербургский) зоопарк в июне 1952 г. из Кандалакши (Мурманская обл.),
прожила в нем 15 лет.
Анализ всего собранного материала показал, что у представителей семей-
ства Mustelidae наблюдается обратная зависимость между уровнем диспропорции
по полу и размерами животных. Возрастные изменения полового состава в
промысловых выборках из популяций мелких хищников прослеживаются очень
отчетливо. Например, у ласки, горностая и лесного хорька разница в
соотношении полов между группами сеголеток и взрослых особей составляла в
среднем 11.5% (/>=99%). В пробах из популяций американской норки и лесной
куницы данные различия были выражены в меньшей степени (7.4%), а надежность
326
Таблица 119/Table 119
Масса надпочечников у взрослых особей куньих, отловленных капканами
в осеине-зимиий (1) и весеиие-летиий (2) сезоны
The mass of adrenal glands of adult individuals of musteiids, caught with traps in the
autumn-winter (1) and spring-summer (2) seasons
Виды/ Species Сезон/ Season n Масса надпочечников/ Paranephros mass мг/mg | %o t p
Самки/Females
Ласка обыкновен./ 1 4 17.5+1.4 0.4Q+0.08 1.2 73
Mustela nivalis 2 4 22±4.5 0.61±0.16
Горностай/ 1 22 22.9±1.4 0.20+0.02 0.5 50
Mustela enninea 2 6 26.4±6.3 0.25±0.11
Хорек лесной/ 1 35 1O3.3±3.2 0.28±0.01 3.1 99
Mustela putorius 2 6 131.5+8.6 0.31±0.02
Норка свроп./ 1 14 70.6±3.3 0.15+0.01
Mustela lutreola 2 3 110±12.6 0.24±0.01
Норка амсрик./ 1 14 70.6+3.6 0.13±0.01 2.1 95
Mustela vison 2 4 98.8±9.8 0.16±0.02
Куница лесная/ 1 27 85.6±4.7 0.12±0.01 1.8 93
Martes martes 2 5 96.3+3.5 0.15±0.02
Выдра/ 1 8 792.5±82.5 0.20+0.01 2.9 95
Lutra lutra 2 4 1093.5±64.8 O.23±O.O2
Самцы/Males
Ласка обыкн./ 1 12 16+1.4 0.21+0.01 3.1 99
Mustela nivalis 2 6 23.3±3.6 0.31 ±0.03
Горностай/ 1 37 28.2±1.1 0.15±0.01 2.9 99
Mustela erminea 2 11 44.5±3.5 0.19±0.01
Хорек лесной/ 1 42 147.8±5.3 0.21±0.01 1.4 85
Mustela putorius 2 8 162.6±18.6 0.22+0.02
Норка свроп./ 1 14 90.8±4.1 0.14±0.01 6.4 99
Mustela lutreola 2 3 140+21 0.23+0.01
Норка амсрик./ 1 45 103.4±2.3 0.12±0.01 2.2 97
Mustela vison 2 14 142.5±4.9 0.15±0.01
Куница лесная/ 1 26 112.2±5.4 0.11±0.01 0.7 65
Maries martes 2 5 149.3±22.3 0.14±0.04
Выдра/ 1 9 1165.6±70.2 0.21 ±0.02 3.3 99
Lutra lutra 2 5 1528±61.3 0.22±0.02
327
вывода не превышала 95%. У выдры, барсука и росомахи, добытых в северо-
западных областях России, достоверных возрастных изменений в размерах этого
показателя обнаружено не было. Учитывая все вышесказанное, можно
предположить, что естественная элиминация самцов и самок в популяциях
крупных хищников семейства более выровнена. По сравнению с мелкими
видами они меньше расходуют тепловой энергии на единицу массы тела. При
этом неблагоприятные воздействия факторов внешней среды сказываются на
организме взрослых самок, которые обычно хуже самцов подготовлены к
зимовке, не столь губительно, как, например, в популяциях ласки или горностая.
Кроме того, небольшая плодовитость, а, возможно, и неежегодное рождение
молодняка у части самок крупных представителей семейства Mustelidae также
способствуют лучшей их выживаемости в неблагоприятные годы. У самок
мелких, узкоспециализированных в кормовом отношении видов высокие
показатели плодовитости и энергообмепа обусловливают большую их естест-
венную элиминацию по сравнению с самцами, что и увеличивает диспропорцию
по полу.
Демография природных группировок рассматриваемых хищников заметно
меняется по сезонам года на участках с разными степенью интенсивности
охотпользования и плотностью населения животных (Большаков, Кубанцев, 1984;
Сидорович, 1992; Б. Новиков, 1993). Количество и доступность основных кормов
сказываются на физиологическом состоянии особей, воспроизводственном
потенциале и демографической структуре популяций видов. При этом адаптация
животных к изменяющимся условиям среды сопровождается гормональной
перестройкой всего организма. Одним из показателей его физиологической
напряженности, четко реагирующих на любые преобразования, могут служить
изменения в строении надпочечной железы у особей разного пола. Оживление
функциональной деятельности коры надпочечников влечет за собой гипертрофию
желез, в результате чего прослеживается и их адаптивная перестройка (Selye, 1946;
Christian, 1957; Добринский, 1967; Ардашев, Горбачев, 1980).
Результаты анализа биопроб из популяций куньих показали, что величина
и строение надпочечников у самцов и самок во многом зависят от способа
добычи зверьков (Туманов, Коровник, 1981). Так, долгое их пребывание в
ловушках усиливает стрессовую ситуацию в организме и активизацию
гормональной деятельности коркового слоя железы по сравнению с теми
особями, которые изымались из естественных местообитаний путем отстрела.
Данный факт был учтен при использовании рассматриваемого теста в качестве
индикатора состояния популяций животных. Взрослые особи характеризуются
большей абсолютной массой железы у самцов и отчетливо выраженной
тенденцией к увеличению относительного ее веса у самок, причем эта разница
наблюдалась у них во все сезоны года.
Сравнение размеров надпочечников у куньих в целом позволяет
проследить, согласно правилу величины Гесса, закономерное увеличение
абсолютного и падение относительного их веса в порядке нарастания массы
тела хищников. Например, при отлове зверьков капканами в осенне-зимний
328
Таблица 120/Table 120
Масса надпочечников у взрослых особей куньих, добытых с ружьем и собакой
в осенне-зимний (1) и весенне-летний (2) сезоны
The mass of adrenal glands of adult individuals of mustelids, obtained with a gun and dog
in the autumn-winter (1) and spring-summer (2) seasons
Виды/ Species Сезон/ Season n Масса надпочечников/ Paranephros mass t P
мг/mg | %0
Самки/Females
Хорек лесной/ 1 17 101.2+4.9 0.22+0.01 5.2 95
Mustela putorius 2 5 136.3+3.5 0.32±0.02
Норка америк. 1 16 60.5+4.4 0.11+0.01 5.7 99
Mustela vison 2 8 93.7±4 0.19±0.01
Куница лесная/ 1 28 89.1±5.5 0.11±0.01 0.5 38
Martes martes 2 7 94+13.2 0.12±0.02
Барсук/ 1 5 566+94.1 0.09+0.02 1.1 75
Meles meles 2 6 761.7+63.8 0.12+0.02
Самцы/Males
Горностай/ 1 6 34+1.8 0.18±0.01
Mustela erminea 2 2 36.2 0.24
Хорек лесной/ 1 29 153.1±7.4 0.20±0.01 1.8 92
Mustela putorius 2 8 201.3±21.9 0.24±0.02
Норка европ./ 1 21 101.8±3.7 0.13+0.01
Mustela lutreola 2 2 162.5 0.21
Норка америк./ 1 23 93.4±5 0.10±0.01 1.4 83
Mustela vison 2 5 115±12.9 0.13±0.02
Куница лесная/ 1 35 96.5+6.3 0.08+0.01 0.5 42
Martes martes 2 7 118.6+10.2 0.10+0.02
Барсук/ 1 3 6O3.3±89.1 0.06±0.02 1.4 75
Meles meles 2 6 675±55 0.10±0.02
сезон средний индекс железы был равен у самок ласки 0.40±0.08%о, у норки
европейской 0.15±0.01 и у лесной куницы 0.12±0.1%о. У самцов тех же видов
эти показатели составляли соответственно 0.21±0.01%о, 0.14±0.01 и
0.11 ±0.01 %о (табл. 119). Подобная зависимость прослеживается и при добыче
животных активным способом, т.е. с помощью собаки и ружья (табл. 120).
Следовательно, у хищников, как и у представителей других отрядов
12 Зак. № 4309
329
млекопитающих (Шварц и др., 1968; Ивантер, 1974), мелкие виды,
обладающие сравнительно высоким уровнем энергообмена по сравнению с
крупными, имеют и наибольший индекс надпочечников как показатель
напряженности энергетического баланса в организме. В то же время у выдры
их абсолютный вес оказался в 1.5-2 раза больше, чем у барсука, а по величине
индекса этого органа она близка даже к мелким миофагам. Отмеченный факт
следует рассматривать как видоспецифическую особенность, присущую этому
околоводному животному. По-видимому, данное исключение из правила рядов
Гессе может свидетельствовать об энергетических тратах, не обусловленных
величиной поверхности тела.
Среди самцов разного возраста существенных различий в индексе
надпочечников не обнаруживалось. В то же время у молодых самок он был
меньше, чем у взрослых, что свидетельствует о постоянной физиологической
напряженности организма последних.
В процессе сезонных изменений условий внешней среды отмечается'
гормональная перестройка организма животных и, в частности, физиоло-
гической активности коры надпочечников. Эти периодические сдвиги находят
свое выражение в закономерных колебаниях их размеров. Увеличение
абсолютной массы надпочечников в весенне-летнее время прослеживается у
взрослых особей всех видов куньих. Согласно полученным данным, половой
диморфизм в реакциях адреналовой железы на календарные сроки проявляется
у них по-разному (Туманов, Коровник, 1986). Среди норок, лесного хорька,
выдры и барсука ее весенняя гипертрофия была более заметна-у самок, чем у
самцов. При добыче зверьков на ходовых охотах (с ружьем и собакой) весенний
рост массы надпочечников (по отношению к зимнему периоду) составлял у
самок норки американской 55% и барсука 35%, а у самцов соответственно
23 и 12%. В то же время у самых активных видов, характеризующихся быстрой
двигательной реакцией (ласки, горностая и лесной куницы), сезонные различия
у самок оказались существенно меньшими, чем у самцов. Например, при
отстреле лесной куницы эта разница в размерах адреналовой железы составляла
в среднем у самок 6%, а у самцов 23%. Отмеченный факт, видимо, объясняет-
ся тем, что самки рассматриваемых видов приходят в состояние готовности
к гону, сопровождающееся высокой энергетической напряженностью
организма и гиперфункцией надпочечников, позднее самцов - уже в летний
период, а основная масса собранного нами материала относилась к началу
весны.
Сезонная ритмика размеров надпочечников особенно наглядно проявляется
при анализе их относительного веса (табл. 119-120); У большинства видов
индекс рассматриваемого органа весной и летом был выше, чем в осенне-
зимнее время, причем во многих группах различия были статистически
достоверны или величина этого показателя стремилась к достоверности.
Наибольшее оживление гормональной деятельности эндокринных желез
происходит в репродуктивный период - во время гона, беременности и
лактации, что обусловливает своеобразие хода сезонных изменений размера
330
Таблица 121/Table 121
Изменение размеров коркового слоя надпочечников у взрослых особей куньих
в разные сезоны года
Changes in the cortical layer of adrenal glands of adult individuals of mustelids
in the different seasons of the year
Виды/ Species n Осенне-зимний/ Autumn-winter n Весенне-летний/ Spring-summer
корковый слой, мм/ corticate, mm отношение коркового слоя к мозговому/ Relation of cortical layer to medullar корковый слой, мм/ corticate, mm отношение коркового слоя к мозговому/ Relation of cortical layer to medullar
Самки/Females
Ласка обыкн./ Mustela nivalis 4 0.67±0.11 0.97+0.08 3 0.82+0.12 0.85+0.09
Горностай/ Mustela erminea 12 0.73+0.04 0.61+0.06 5 0.78±0.13 0.86+0.03
Хорек лесной/ Mustela putorius 15 1.30+0.09 0.90±0.07 3 1.44±0.15 0.92±0.12
Норка европ./ Mustela lutreola 7 0.95±0.07 0.59±0.06 3 1.24±0.13 0.71 ±0.09
Норка америк./ Mustela vison 5 0.92+0.11 0.59+0.14 8 1.21±0.09 0.63+0.11
Куница лесная/ Martes martes 7 0.90±0.05 0.53±0.06 6 1.05±0.18 0.58±0.23
Выдра/ Lutra lutra 3 2.37±0.25 0.70±0.05 3 2.47±0.23 0.72±0.13
Барсук/ Meles meles 4 1.78±0.54 0.78+0.21 5 2.32+0.37 0.82±0.05
Самцы/Males
Ласка обыкн./ Mustela nivalis 6 0.71+0.05 0.92+0.19 3 0.85+0.08 0.87±0.12
Горностай/ Mustela erminea 17 0.84+0.06 0.75+0.05 6 0.87±0.09 0.83+0.03
Хорек лесной/ Mustela putorius 20 1.73±0.10 0.95±0.05 5 1.83±0.75 O.98±O.3O
Норка европ./ Mustela lutreola 9 1.02+0.17 0.58±0.09 3 1.28+0.35 0.73+0.09
Норка америк./ Mustela vison 11 0.96+0.07 0.56±0.03 10 1.26+0.09 0.59±0.06
Куница лесная/ Martes martes 3 1.11+0.15 0.49±0.03 5 1.18+0.32 0.61 ±0.15
Выдра/ Lutra lutra 4 2.44±0.12 0.67±0.04 3 2.53±0.21 0.76±0.08
Барсук/ Meles meles 3 2.22+0.13 0.85+0.15 4 2.82+0.52 0.93±0.22
331
надпочечников у разных видов животных. Подготовка к гону и размножение
протекают у них на фоне значительного физиологического напряжения
эндокринной системы и повышения энергетических расходов, в результате чего
достигает своего максимума функциональная активность коркового слоя
надпочечников (Christian, 1957; Myers, 1967; Шварц и др., 1968; Краснощеков,
1975; Чернявский, Ткачев, 1982). Данное положение наглядно иллюстрируется
материалами гистологической обработки этого органа у куньих (табл. 121).
Исследования показали, что у самцов, обладающих большей абсолютной
массой рассматриваемых эндокринных желез, корковый их слой оказался
шире, чем у самок. У большинства видов в весенне-летний период по
сравнению с осенне-зимним происходит заметное увеличение объема коркового
слоя, прежде всего за счет гипертрофии пучково-сетчатой зоны. При этом
мозговой слой надпочечников изменяется незначительно, о чем можно судить
по его абсолютным показателям и на основании стабильного нарастания в
весенне-летнее время соотношения размеров ткани коркового и мозгового
слоев (табл. 121).
Анализ материалов, характеризующих функциональную активность коры
надпочечников, свидетельствует о том, что уровень гормональной деятельности
у хищных млекопитающих существенно изменяется по сезонам года. В осенне-
зимнее время морфологические проявления функциональной активности
коркового слоя отмечаются у них заметно слабее, чем весной и летом. На
основании парных сравнений по /-критерию Стьюдента для сопряженных
величин оценивалась достоверность различий весеннего увеличения размеров
коркового слоя надпочечников (по отношению к осеннему сезону). Оцен-
ка проводилась в целом для группы самцов и самок. В обоих случаях
достоверность различий прослеживалась отчетливо и превышала 95%. При
этом у всех видов весенняя гипертрофия коркового слоя (в процентном
отношении к осенним размерам) была более заметна у самок, чем у самцов,
свидетельствуя о высоком уровне напряженности их энергетического баланса.
Половозрелые многоплодные самки за время гона, беременности, родов и
выращивания молодняка резко теряют в весе. Данный факт особенно четко
заметен у тех видов, которые, обладая короткой эмбриональной диапаузой в
развитии детенышей, в случае их гибели повторно спариваются и приносят
второй приплод. Например, у самок колонка масса тела после выращивания
молодняка снижалась на 20-25%, а после двукратных щенений - на 38-44% от
таковой перед беременностью (Курис, Бакеев, 1974).
В конце лета-начале осени, когда интенсивность обменных процессов в
организме животных снижается, их жировой резерв, или энергетический
потенциал, увеличивается. Существенные жировые отложения появляются
прежде всего у самцов, поскольку самки долгое время заняты выращиванием
и воспитанием молодняка. Естественный отход взрослых самок зимой может
быть следствием их более интенсивных энергозатрат во время репродуктивного
периода, недостаточным состоянием упитанности и теплозащитных покровов
организма. Особенно это характерно для мелких и высокопродуктивных видов,
332
Таблица 122/Table 122
Изменение соотношения полов по возрастам у некоторых представителей
семейства собачьих (по: Данилов и др., 1979, с дополнениями)
Changes of sex ratio by age of some representatives of canids family
Виды/ Species Щенки в гнездах/ Cubs in litter Сеголетки/ Youngs Взрослые/ Adults
п % самцов/ % of males п % самцов/ % of males п % самцов/ % of males
Песец/ Alopex lagopus Собака 17 52.9 62 54.8 84 56
енотовидная/ Nyctereutes procyonoides 68 51.5 97 53.6 200 56.5
Лисица обыкн./ Vulpes vulpes 27 55.6 136 46.3 148 52
Волк/ Canis lupus 107 53.3 48 54.2 1661 56
таких как ласка, горностай, солонгой, хорьки, чем и объясняется резкая
диспропорция соотношения полов в группах старших возрастов. В то же время
более сильные самцы, не занятые воспитанием потомства, испытывают меньше
трудностей при добывании корма и бывают лучше подготовлены к зимовке. В
этом плане показательно изменение жировых отложений у взрослых лесных
хорьков и американских норок, добытых в северо-западном регионе России:
если в июне все подкожные запасы зверьков не превышают 2-4 г, то в декабре
масса только одного пахового жира достигает у самцов 13-25 г, а у самок - 8-
9 г. По-видимому, в холодную малокормную осень последним трудно набрать
необходимое для организма количество энергетических запасов, и доля
элиминирующих взрослых самок в популяции может увеличиваться. У
представителей рода Martes (соболя и куниц) даже в благоприятные в кормовом
отношении годы упитанность самок заметно ниже, чем самцов, что и опре-
деляет разную степень воздействия на них экстремальных условий (Залекер,
1962; Нумеров, 1966; Г. Монахов, 1968).
Неодинаковая продолжительность жизни самцов и самок куньих в
природных условиях вполне очевидна. Среди старых особей мелких хищников
диспропорция между самцами и самками по их соотношению в популяциях
выражена в большей степени, чем у крупных, обладающих меньшим уровнем
теплоотдачи на единицу массы тела. Судя по промысловым выборкам из
группировок куньих, самки ласки и горностая до 5-6-летнего возраста, видимо,
вообще не доживают, а у лесного хорька и норок обоих видов они встречаются
в 3-4 раза реже, чем самцы. Сходные особенности половой структуры
рассматриваемых животных, характеризующиеся разным уровнем естествен-
ной смертности разнополых особей, прослеживаются в различных частях
ареала видов (Erlinge, 1968, 1983; Г. Монахов, 1974; Соколов, 1978; Граков,
1969 б, 1981; Синицын, 1985; Б. Новиков, 1993; Глушков и др., 1999).
333
Таблица 123/Table 123
Половозрастная структура промысловой пробы (л=508) из таймырской популяции
песца в сезоне 1986/87 г. (по: Куксов, 1989)
Sex-age structure of the game sampling (л=508) from Taimyr population of arctic fox
(Alopex lagopus) in season 1986/87
Показатели/ Parameters Возраст особей, лет/ Age, years
0+ 1+ 2+ 3+ 4+ 5+ и старше/ 5+ and older
Всего особей/ Total 5 2 78 170 129 124
Самок/ Females 2 0 30 93 80 62
Самцов/ Males 3 2 48 77 49 62
Процент самцов/ % of males 60 100 62 45 38 50
Описанные выше зависимости можно проследить и на примере предста-
вителей других семейств отрада хищных млекопитающих. Особенно наглядно они
проявляются у собачьих (табл. 122). В среднем в пометах этих зверей наблюдается
примерно равное соотношение полов с незначительным преобладанием самцов,
которое с возрастом обычно увеличивается.
В известных нам 5 выводках песца, полученных в Ленинградском (Санкт-
Петербургском) и Московском зоопарках, находилось 17 детенышей, среди
которых было 9 самцов и 8 самок. Видимо, и в природе половое соотношение
щенков в гнездах в среднем близко к Г. 1. В промысловых пробах из
Архангельской области и Ненецкого АО (1993-1997 гг.) среди сеголеток и
взрослых особей самцов оказалось несколько больше (табл. 122).
Следует отметить, что половозрастная структура природных группировок
песца изменчива и во многом определяется фазами популяционного цикла и
связанными с ними изменяющимися формами поведения. По многолетним
наблюдениям В.А. Куксова (1989), проведенным на Таймыре, популяция
данного вида очень динамична. Анализ половозрастной структуры биопроб
из гнездового ареала животных и с его периферии показал, что с наименьшей
ошибкой поголовье песцов и половозрастную структуру популяции можно опре-
делить при минимальной их численности, когда почти все они половозрелы и
участвуют в размножении. На фазах подъема и пика популяционной динамики
большинство молодых особей в течение первого года жизни обитает вне
гнездовых участков. В эти периоды отлавливают преимущественно молодых
особей (на одного взрослого до 10 сеголеток) примерно в равном соотношении
или с небольшим преобладанием самцов. Ресурсы популяции в данной ситуации
позволяют вести промысел практически без ограничения объемов добычи,
поскольку из-за хорошей обеспеченности зверей кормом максимально
разредить плотность группировки не удается. На фазе спада численности песца
334
Таблица 124/Table 124
Половозрастной состав (%) промысловых проб из популяции песца,
обитающего в районе плато Путорана (по: Линейцев, 1988)
Sex-age structure of the game samplings (%) from the population of arctic fox
(Alopex lagopus), inhabiting the region of Putoran Plateau
Возраст 1978/79 («=15 8) 1979/80 („=106) 1980/81 (л=128)
животных/ Age 6# $>$> <S<3 $>$> <?<? $5
0+ 67.7 30.5 54.8 35.8 7 3.9
1+ 0.6 0.6 0.9 3.9 9.3 5.5
2+ - - - - 13.3 7.8
3+ - - 2.8 - 10.2 10.1
4+ - 0.6 0.9 - 5.5 10.1
5+ - - - - 4.7 5.5
6ь - - 0.9 - 2.3 2.4
7+ - - - - - 0.8
8+ - - - - 0.8 -
9+ - - - - 0.8 -
Bcero/Total 68.3 31.7 60.3 39.7 53.8 46.2
в центре ареала размножения сеголеток в биопробах очень мало (табл. 123).
Особей прошлого года рождения практически не встречается, а свыше 80%
поголовья составляют животные 3 лет и старше, среди которых самки
составляют 55.6%. Миграции хорошо выражены. Время и причины их
наступления обычно связаны с резким ухудшением состояния кормовой базы.
Соотношение полов в центре гнездового ареала в разных возрастных группах
неоднозначно, а среди престарелых особей оно часто оказывается выров-
ненным. В целом за все фазы популяционного цикла промыслом на Таймыре
изымаются в основном престарелые и молодые особи.
Материалы С.Н. Линейцева (1988) показали, что в районе плато Путорана
(Долгано-Ненецкий АО) половозрастной состав песцов-мигрантов цели-
ком зависит от состояния кормовых условий в ареале норения. При хоро-
ших и средних условиях миграционный поток формируют сеголетки,
преимущественно самцы. По мере ухудшения кормовой базы возрастает доля
молодых самок и взрослых животных. В начале сезона массовых перемещений
зверей на осваиваемых территориях чаще встречаются самцы. Количество
самок здесь заметно увеличивается уже во второй его половине. Так, в 1978-
1984 гг. во время миграционного потока песцов (отловлено 474 особи) доля
самцов, добытых в ноябре-декабре, составляла 68.3-73.5%, а в январе-марте
снижалась до 26.7-57.8%.
335
По свидетельству автора, при резком ухудшении кормовых условий (сезон
1980/81 г.) в ареале норения песца молодняка рождается мало (табл.. 124).
Значительная часть взрослых особей мигрирует в лесную зону, а популяция
переживает глубокую депрессию. В миграционном потоке уже за гнездовой
территорией появляются звери старших возрастных групп, которые в обычные
годы здесь отсутствуют. Интересно, что среди зверей старше 4 лет соотношение
полов было примерно равным, хотя в целом во все сезоны самцов разных возрастов
отлавливали чаще самок (табл. 124).
В биопробах из популяции песца полуострова Ямал постоянно преобла-
дают молодые особи с равным соотношением полов. Вероятность добычи
сеголеток здесь в среднем в 4 раза выше, чем взрослых зверей. При допустимой
интенсивности промысловой нагрузки это обстоятельство может играть
положительную роль, так как позволяет сохранять к периоду размножения основное
репродуктивное ядро популяции, в том числе и взрослых самок (Смирнов, Корытин,
1979).
В известных нам промысловых выборках особи старше 8-9 лет не
встречались. В Ленинградском (Санкт-Петербургском) зоопарке три белых
песца прожили 9 лет, один - 10 и еще двое - 11 лет, причем некоторые из них
были выбракованы без видимых признаков старости.
В пометах енотовидной собаки соотношение полов среди новорожденных
детенышей близко к 1:1 с незначительным преобладанием самцов. При
разведении зверей в неволе их доля в выводках, по сообщению Л.А. Пав-
линского (1937), составляла 52%, а М.Х. Геллера (1959) - 50.9%. Сходные с
этими данные (51.5%) по 9 выводкам получены и в Ленинградском (Санкт-
Петербургском) зоопарке (табл. 122).
Опрос охотников-корреспондентов из северо-западных областей России
показал, что в обнаруженных в природе 13 выводках (87 щенков) енотовидной
собаки соотношение полов было в пользу самцов (55.2%), что статистически
оставалось не доказанным (х2=0.9,р<0.75). Примерно таким же оно оказалось
в промысловых выборках в группах сеголеток (53.6%) и взрослых особей (56.5%).
При содержании в условиях неволи многие енотовидные собаки жили до
9-11 лет. В естественных местообитаниях в таком возрасте они практически не
встречаются.
У обыкновенной лисицы при рождении в большинстве случаев наблю-
дается некоторое увеличение количества самцов в пометах (Данилов и др.,
1979). Среди 27 щенков (6 выводков), обнаруженных в лисьих норах, на их
долю приходилось 55.6% (15 особей). Еще заметнее эта разница оказа-
лась в пометах, полученных при разведении зверей в вольерах Ленинград-
ского (Санкт-Петербургского) зоопарка. Здесь в 10 выводках (46 щенков)
соотношение полов было следующим - 31 самец и 15 самок (соответственно
67.2 и 32.8%), причем в данном случае диспропорция в количестве детенышей
разного пола подтверждается статистически (х2=5.7,р>0.97). Среди зверей,
добытых в период промысла в северо-западном регионе России (табл. 122),
существенных различий по соотношению полов в выборках из популяции вида
336
Таблица 125/Table 125
Соотношение полов среди щенков волка, отловленных на логовах
в северо-восточной части европейской России (по: Козловский, 1996)
Sex ratio among of wolfs cubs (Canis lupus), trapped in the dens
in north-eastern part of European Russia
Место отлова/ Region Кол-во щенков/ Cubs number Самцы/Males Самки/Females t
п % п %
Кировская обл./ Kirov Prov. Удмуртия/ Udmurtiya Пермская обл./ Perm Prov. 1095 449 41 646 59 4.09 120 58 48 62 52 0.43 22 8 36 14 64 1.31
Всего/ Total 1237 515 42 722 58 5.65
не прослеживалось. В группе сеголеток несколько преобладали самки, а у
взрослых особей-самцы. По материалам Э.В. Ивантера(1969), в 1958-1968 гг.
в Карелии среди взрослых лисиц самцы составляли 60% от их общего
количества. Увеличение доли самцов среди сеголеток у этого вида было
выражено в меньшей степени.
В условиях неволи обыкновенная лисица живет до 10-12 лет, однако в
естественных местообитаниях особи старше 7-8 лет встречаются крайне редко.
Известная максимальная продолжительность жизни лисицы в природе -11 лет
(Н. Корытин, 1983).
В популяции волка из северо-западных областей России (Данилов и др.,
1979) среди 107 щенков, обнаруженных в логовах, самцы составляли 53.3%
(Х2=0.46, р>0.50), а среди добытых взрослых зверей - 56% (х2=24.1, р>0.99).
Примерно таким же соотношение полов оказалось и среди сеголеток (табл.
122). Выявленный уровень этих показателей, видимо, вообще характерен для
биопроб из популяций волка данного региона. Так, в Ленинградской области
в 1956-1961 гг. было отравлено фторацетатом бария 662 зверя, из которых
367 (55.4%) оказались самцами (Иванов, 1963). В просмотренных нами в 1975-
1980 гг. на пушно-меховом холодильнике (г. Санкт-Петербург) партиях шкур
от добытых волков (н=225) без разделения по возрастам самцы (54.7%) также
несколько преобладали.
На северо-востоке европейской части России (Козловский, 1996) соот-
ношение самцов и самок среди щенков, взятых из логова, в разные годы
заметно варьирует, однако в целом в период с 1962 по 1986 г. доля самок
была здесь достоверно выше (табл. 125). Позднее, в осенне-зимний сезон, среди
добытого молодняка самцов всегда оказывалось больше, иногда эта разница
достигала 25%. Основная масса зверей в выборке (п-382) - сеголетки (50.3%)
и переярки (18.3%). Наиболее продуктивное ядро популяции (звери от 2 до 5
лет) составляло 26.6%, доля волков старше 8 лет не превышала 0.8%.
337
Рекордный возраст добытого самца в рассмотренной группировке был равен
17 годам. Зимой встречались стаи с 4-9 особями и разным соотношением полов.
Показатель стайности менялся по годам: в периоды высокой численности вида
он был наибольшим.
В разные годы даже в одном регионе соотношение полов в выводках и
группах зверей старших возрастов может быть различным. Это определяется
многими факторами, в том числе численностью животных в популяции, ее
биологической продуктивностью, состоянием кормовой базы и другими. В
Воронежской области на рубеже 40-50-х годов, по данным П.А. Мертц (1953),
в обнаруженных выводках (л=19) волков доля самцов была равна 42.8%. В
более поздние годы их количество в природных группировках оказалось более
высоким и составляло среди молодняка в гнездах 54%, а добытых взрослых
животных - 53.1%. Количество пометов, в которых было больше самцов,
достигало 53.3%, самок - 30%, а с равным соотношением полов -16.7% (Рябов,
1993).
Как и у других хищных млекопитающих, у волков прослеживается характерное
изменение соотношения полов на разных фазах их популяционного цикла. В Карелии
(Данилов, 1994) в годы низкой численности животных среди добытых особей (л=627)
самцы составляли только 51.9% (х2=1-53). Вгоды ее подъема доля самцов (57%)
заметно превышала таковую самок (?i=643,
X2=12.9). Та же картина наблюдалось и в годы падения численности зверей. В
выборке из 1167 особей доля самцов была равна 53.1% (%2 =4.57). Связь
соотношения полов с плотностью населения в популяции волка отмечали и
другие исследователи (Meeh, 1975; Бондарев, 1979). Поскольку основные
способы добычи этого хищника избирательны в отношении самцов,
предполагается, что, по крайней мере в годы пессимума, в популяции реально
могут преобладать самки.
При отстреле зверей с помощью авиации непреднамеренная избира-
тельность сказывается меньше, чем при иных способах охоты. В Республике
Коми из 50 особей, добытых с вертолета, самцы составляли 52%. В выборке
было больше прибьшых и переярков, а среди них-самок. В группе взрослых
животных самцов старше 4 лет не отмечалось, а возраст самых старых 3 самок
определен в 8-9 лет (Полежаев, 1998).
Обобщив сведения, содержащие в совокупности данные о 23 797 взрослых
волках и 2168 щенках, В.С. Смирнов и Н.С. Корытин (1985) пришли к выводу,
что в целом в первой группе доля самцов равна 56.21±0.32%, а во второй -
53.05± 1.07%, причем эти различия оказались вполне достоверными. По мере
увеличения возраста зверей соотношение полов все больше меняется в пользу
самцов, достигая максимума в 6-7 лет и более. В группе из 13 хищников старше
9 лет было 9 самцов и 4 самки, из которых один самец оказался 15-летним. В
естественных условиях эти хищники в возрасте 8-9 лет встречаются очень редко.
В неволе, при условии хорошего содержания, волки живут до 12-16 лет, причем
звери в возрасте 10-12 лет даже не обнаруживали каких-либо признаков
старения и тем более дряхлости.
338
Материалы о соотношении полов среди представителей семейства
кошачьих крайне ограничены. В 2 известных нам выводках лесного кота,
найденных местными охотниками в горных районах Дагестана, находилось
равное количество самцов и самок (3 : 3 и 2 : 2). Таким же оно оказалось и в
группе первогодков (9<? и 8$). Среди 32 шкурок взрослых зверей,
просмотренных нами на пункте приемки пушнины в г. Махачкале, самцы
составляли 56.3% (18 особей). Анализ данных, собранных С.Б. Насибовым
(1970) на Кавказе, показал, что в промысловой выборке из популяции лесного
кота (л=41) самцы составляли 48.8% (20 особей), а камышового кота (и=28) -
60.7% (17 особей). Среди отловленных хищников, возраст которых удалось
установить, находилось у камышового кота 10 (50%) молодых особей, 8 (40%)
двухлеток и 2 (10%) зверя были в возрасте 3-5 лет. Для лесного кота отмеченные
показатели распределялись следующим образом: 25 (67.6%), 11 (29.7%) и
1 (2.7%). Судя по этим сведениям, доля долгожителей в популяциях обоих ви-
дов невелика, а продолжительность их жизни в природе, вероятно, не превы-
шает 6-7 лет.
При содержании в условиях неволи продолжительность жизни и физиоло-
гическая активность зверей значительно больше. Например, у близкого к ним
еще одного представителя семейство Felidae - каракала - максимальная продол-
жительность жизни в зоопарке составляла у самцов 9-12 лет, а у самок 9-13.7
лет. При этом 1 самец активно участвовал в размножении до 12 лет, а самка -
до 9 лет. Что касается соотношения полов у каракала, то среди родившихся 54
котят было 34 самца и 20 самок (Stehlik, 2001).
В 12 выводках рыси, полученных в Ленинградском (Санкт-Петербургском)
зоопарке, соотношение полов у котят (10<?и 15$) было несколько смещено в
сторону самок и составляло 60% от общего количества новорожденных. На
экспериментальной базе «Черноголовка» под Москвой в 1988-1997 гг. удалось
получить 19 приплодов рыси (39 особей) с практически равным количеством
самцов и самок (Найденко, 1997). Близкие ко всем этим данные были получены
в Чешском зоопарке г. Острава. Здесь в период с 1967 по 1976 г. был получен
21 выводок с 24<J и 25$, а в 1977-1996 гг. - 15 выводков, в которых было 10<?
и 17$ (Stehlik, 1984, 2000).
В обнаруженных охотниками-корреспондентами (1976-1983 гг.) из
Новгородской и Псковской областей 4 выводках рыси было 3 самца и 5 са-
мок. Отловленная в 1993 г. в окрестностях г. Сыктывкара семья этого ви-
да состояла из взрослой самки и 2 котят-самочек (Пыстин, 1998). Можно
предположить, что сдвиг соотношения полов в сторону последних в пометах
рыси случается не так уж редко. Среди сеголеток, добытых в северо-западных
областях России, как и у новорожденных, несколько преобладали самки.
Так, в пробе из 51 особи оказалось 22 самца, что составляло 43.1% (х2=0.9)
от общего количества зверей этого возраста. В группе взрослых животных
(п=61) самцов было заметно больше (60.7%; х2=2.8).
Некоторый тренд соотношения полов в сторону самок у молодых рысей
прослеживается и в других регионах (Малафеев и др., 1986; Козловский, 1999).
339
Например, в Кировской области (1977-1997 гг.) среди отловленных сеголеток
(л=76) доля самок равнялась 61.9%, а у годовалых особей- 53.8%. При этом
соотношение полов среди взрослых животных было близко к 1 :1 (Козловский,
1999). Многолетнее изучение состояния популяции рыси на Среднем Урале и
других территориях России позволило не только выявить соотношение полов
и выживаемость зверей в разных возрастных группах, но и оценить их роль в
динамике численности вида (Матюшкин, 19746, в, 1986; Малафеев, Кря-
жимский, 1988).
На основании исследований биопроб, полученных на Северо-Западе
России, максимальная продолжительность жизни взрослых рысей (л=25 особей)
в природе, установленная по структуре костной ткани зубов, не превышает 5-
7 лет. В то же время в Ленинградском (Санкт-Петербургском) зоопарке
некоторые особи рыси жили до 19-20 лет, что свидетельствует о высоких
потенциальных возможностях организма этого вида, а в Чешской Республике,
в зоопарке г. Острава (Stehlik, 2000), одна самка прожила 21 год, причем
потомство она приносила до 12 лет.
Опрос местных охотников, проведенный нами в горных районах Кавказа,
показал, что соотношение полов при рождении у енота-полоскуна (семейство
енотовых) близко к 1 : 1. В 5 известных выводках находилось 9 самцов и 11
самок (1 : 1, 2 : 1, 2 : 2, 2 : 3 и 2 : 4). Примерно таким же оно оказалось и в
промысловой выборке, состоящей из молодых животных (23<? и 20$). По
сообщению А.М. Гинеева (1973), в охотсезонах 1968-1971 гг. в Дагестане им
было проведено обследование 88 добытых взрослых зверей, среди которых
находилось 38 самцов и 50 самок. В группе сеголеток, состоящей из 30 особей
(9с£ и 21$), доля самок достигала 70%. Судя по материалам автора,
максимальная продолжительность жизни зверей в природе составляет не менее
9-10 лет.
У представителей семейства медвежьих, как и у большинства видов
хищных млекопитающих, в целом в репрезентативных выборках в гнездах с
новорожденными количество самок и самцов обычно одинаково, с небольшим
преобладанием в них последних. В выводках белого медведя (л=50) среди 88
детенышей, родившихся в разные годы в Ленинградском (Санкт-Петер-
бургском) зоопарке, самцы составляли 52.3% (46 особей). Подобное (примерно
равное) соотношение полов вообще характерно для многих представителей
отряда Carnivora, когда звери находятся в благоприятных условиях и не
испытывают недостатка в кормах.
Интересные сведения о количестве разнополых особей в выводках белого
медведя и их смертности на о-ве Врангеля приводятся в работе С.Е. Беликова
(1993). На всей обследованной территории в берложный период соотношение
полов было несколько отклонено в сторону увеличения доли самцов (95^ и
81$), которая по отношению к общему количеству обнаруженных медвежат
была равна 54%. В пометах зверей, обитающих на западных и северо-
западных участках острова, отношение самцов и самок соответствовало 71
: 51 (за 7 сезонов наблюдений), а на восточных -24:30 (за 5 сезонов).
340
Таблица 126/Table 126
Доля самцов среди медведей, добытых разными способами в Новгородской области
(1969-1974 гг.)
The portion of males among brown bears (Ursus arctos), obtained by different methods in
Novgorod Province (1969-1974)
Годы/ Years Лончаки, добытые/ Yearlings at Взрослые особи, добытые/ Adults at
на овсах/ oat tields в берлогах/ dens на овсах/ oat tields в берлогах/ dens
всего/ total % самцов/ % males всего/ total % самцов/ % males всего/ total % самцов/ % males всего/ total % самцов/ % males
1969 3 67 1 0 13 69 1 100
1970 11 64 1 0 16 75 2 50
1971 2 100 2 100 34 85 5 60
1972 1 100 - - - - - -
1973 3 100 - - 14 86 3 67
1974 7 71 - - 12 83 1 100
Итого/ Total 27 74 4 50 90 82 12 67
По уровню смертности разнополый молодняк белого медведя практически
не различается. У сеголеток она достигает 20%, а всего за первые 2 года жизни
хищников их погибает около 50%. В последующие периоды естественная
элиминация молодых животных снижается до 5% за каждый год. Обычно их
гибель связана с неблагоприятными ледовыми и кормовыми условиями, а также
каннибализмом крупных самцов (Lono, 1970; Jonkel, 1970; Успенский, 1989).
При чрезмерной плотности населения зверей на ограниченной территории
между ними могут возникать драки, от которых особенно страдают более
слабые молодые особи, в том числе самки. Скопление белых медведей около
крупной добычи и на лежбищах моржей способствует встрече половых партне-
ров и тем самым поддерживает репродуктивный потенциал популяции (De-
rocher, Stirling, 1989; Беликов, 1993).
Соотношение полов у взрослых особей близко к 1:1. Продолжительность
жизни зверей в различных частях ареала примерно одинакова, но обычно не
превышает 30 лет (Природа Ямала, 1995). В зоопарках белые медведи живут
25-30, а некоторые даже до 33-40 лет (Потелов, 1998).
В пометах бурого медведя при содержании животных в вольерах
соотношение полов примерно равное. В Киевском зоопарке в 12 выводках
оказалось 16 самцов и 18 самок (Пажетнов, 1990). В 33 пометах, полученных
в Ленинградском (Санкт-Петербургском) и Московском зоопарках, сре-
ди новорожденных (и=83) находились 43 самца и 40 самок. Кроме того, в
341
Московском зоопарке, по сообщению Т.А. Адольф (1949), в 40-х годах в
2 полученных выводках было по 1 самцу и 2 самки, а в 3 других, поступивших
от охотников, по 1 самцу и 1 самке. Материалы специалистов Лейпцигского
зоосада, которые приводит в своей работе Е. Пуллиайнен (Pulliainen, 1972),
свидетельствуют о том, что среди родившихся в нем за длительный период
наблюдений 332 медвежат самцы составляли 50.6% (163 особи).
Близкое к этому соотношение полов наблюдается и в пометах медведиц,
обитающих в природе. Например, в среднесибирской тайге в 1970-1975 гг.
при добыче 25 самок в их убежищах были обнаружены медвежата, в их числе
33 самца и 28 самок (Завадский, 1993). Изучение полового состава в разных
возрастных группах зверей в Карелии (Данилов и др., 1979) показало, что
среди детенышей, взятых из берлог или отловленных сразу после выхода
молодняка из убежищ, количество самцов и самок было либо равным, либо
первые несколько преобладали. С возрастом количество самцов в популяции
заметно увеличивалось: если среди лончаков (1+) они составляли 52% (и=48),
то среди половозрелых медведей, добытых в Псковской, Новгородской и
Ленинградской областях (н=58), - 56.8%, а в Карелии (л=42) - 60.9%.
Интересно, что в Финляндии (Pulliainen, 1972) в 1960-1971 гг. были отстрелены
40 молодых медведей (0+) с равным соотношением полов (20<? и 20$).
Следует отметить, что разные способы добывания медведей в той или
иной степени избирательны (табл. 126), поэтому они, видимо, не отражают
истинного соотношения полов в популяции. Крупных зверей, т.е. самцов,
отстреливают чаще. Судя по имеющимся сведениям (Данилов, 1993, 1994),
в дореволюционный период, когда не было ограничений в отстреле зверей ни
по полу, ни по возрасту, в южных губерниях Северо-Запада России доля
самцов в выборках (54.7-58.7%) лишь немногим превышала таковую самок.
В Карелии (1955-1959 гг.), когда за уничтожение медведей выплачивались
премии и добыча медведиц, у которых были детеныши, разрешалась,
существенного преобладания самцов среди трофеев не наблюдалось. Доля
последних в группе зверей, изъятых из популяции в периоды до и после запрета
охоты на самок с сеголетками, составляла соответственно 57.7% (/1=322) и
73.1% (и=354).
Примерно такая же картина прослеживается и в других регионах России
(Варнаков, 1979; Лоскутов и др., 1993). Так, в Кировской области в прошлом,
по данным анкетного опроса лесников и охотников, среди добытых медве-
дей самки и самцы встречались поровну. При этом свыше 70% зверей
отстреливали в 1-м и 4-м кварталах года, т.е. в основном на берлогах. В
сезоны 1981-1986 гг. в просмотренных выборках среди животных, пол
которых был установлен (л=701), постоянно преобладали самцы, доля
которых в разные годы составляла 61.7-69.8% (Лоскутов и др., 1993). По-
видимому, большее их изъятие в этом регионе из популяции в последнее время
является следствием того, что теперь медведей чаще добывают способами,
избирательными на самцов, - подкарауливанием и отстрелом крупных зверей
на овсах или у падали, а также во время объезда полей на автомашине.
342
В то же время, по устному сообщению биолога А.М. Величко, в 1988-
1998 гг. среди 22 осмотренных им зверей, добытых охотниками в конце
января-феврале на берлогах в Ленинградской области, подавляющее
большинство оказалось самками с медвежатами. Все убежища были
обнаружены случайно при охоте на пушнину с собаками, а последним,
видимо, легче найти логово лактирующующей медведицы с активно
сосущими ее детенышами.
Данные, опубликованные В.П. Тепловым (1960), свидетельствуют о том,
что из 58 хищников, взятых на берлогах в 1937-1949 гг. в окрестностях Печоро-
Илычского заповедника (Республика Коми), взрослые особи составляли 52%,
сеголетки 31, а годовалые 17%. В группе половозрелых животных самцы и
самки встречались поровну, среди молодых преобладали самцы, доля которых
у сеголеток была равна 67%, а у годовалых зверей 57%. В 1972-1990 гг. в
этом регионе среди добытых и найденных погибшими взрослых животных
(л=21) самцов оказалось 66.6% (Полежаев, Нейфельд, 1998).
По материалам Б.В. Завадского (1993), в промысловой выборке (л=93) из
Средней Сибири в целом самцы также составляли 66.6%. С увеличением
возраста животных этот показатель заметно увеличивался: если у сеголеток
он был равен 59.5%, то у размножающейся части популяции - 68.6%, а у старых
особей - 87%.
Замечено, что и в природных группировках вида взрослые самцы нередко
преобладают над самками. Например, на Северо-Востоке Сибири, судя по
анализу следов и визуальным наблюдениям (л=176) медведей в бассейне
р. Анадырь, среди крупных особей (31.8% от общего количества) чаще
встречались самцы. В Северном Приохотье они составляли 39.8%, а взрослые
самки - 26.8% от общего количества (л=123) обнаруженных в разные годы
зверей (Чернявский и др., 1993).
В отряде хищных млекопитающих бурого медведя, как и белого, с
полным правом можно отнести к долгожителям. В Ленинградском (Санкт-
Петербургском) зоопарке 2 самца дожили до 26 и 29 лет, а затем были
выбракованы по старости. Известно также, что в неволе предельный возраст
медведей, умерших своей смертью, был равен 30-34 годам (Мантейфель,
1948). В то же время в природе (в частности, на северо-западе европейской
части России) звери старше 20 лет встречаются редко. В выборке из
сибирской тайги (Завадский, 1981,1993) средний возраст 25 самцов старше
5 лет составлял 15.4, а 13 самок старше 4 лет - 10.2 года. Как показали
результаты определения возраста более чем у 100 добытых особей, самый
старый самец оказался 39, а самка - 29 лет. Оба зверя не имели признаков
дряхлости и находились в состоянии хорошей упитанности. Среди
долгожителей наиболее старая самка, которая имела приплод, была в
возрасте 18 лет.
343
Резюме
У всех видов хищных млекопитающих, несмотря на имеющиеся различия
по уровню энергообмена и потенциалу размножения, возрастная динамика
соотношения полов прослеживается отчетливо. В большинстве случаев для
них характерно примерно равное количество самцов и самок в выводках,
иногда с небольшим отклонением в ту или иную сторону. На разных фазах
динамики численности это соотношение может изменяться. С увеличением
возраста животных диспропорция по количеству самцов и самок в популяции
становится вполне очевидной. Выявленная разница в третичном соотношении
полов резко увеличивается в направлении от крупных хищников к мелким, у
которых рожавшие многоплодные самки бывают хуже самцов подготовлены
к зимовке, а следовательно, в большей степени подвержены естественной
элиминации. Если в популяции видов из семейства Canidae на Северо-Западе
России среди взрослых животных доля самцов в среднем составляет 52-56%,
то у миофагов {Mustela nivalis, М. erminea, М. putorius') - 66-68%. В годы
подъема и пика численности животных доля самцов разных возрастов в
промысловых пробах бывает особенно велика.
Большая доля взрослых самцов в популяции обеспечивает включение в
репродуктивный процесс максимального количества готовых к размножению
самок, которые оплодотворяются наиболее активными и сильными самцами.
Причем у многих видов молодые самцы проявляют готовность к гону заметно
позднее самок того же возраста, что создает условие для разновозрастного
спаривания и является одним из важных механизмов популяционного
гомеостаза. Все это способствует сохранению жизнеспособности популяции и
регуляции полового состава при рождении в соответствии с экологической
ситуацией.
Summary
In all species of carnivorous mammals despite of the differences in the level of
energy exchange and reproductive potential, the age dynamics of the sex ratio is
observed very distinctly. In most cases approximately equal number of males and
females in breeds, sometimes with a small deviation in this or that side, is typical of
them. At the different phases of abundance dynamics this ratio may change. With
the increase of animals’ age the disproportion in the number of males and females
in the population became quite evident. The difference in the tertiary sex ratio
dramatically increases in direction from large to small carnivores. In the latter the
fecund females that already delivered the young appeared to be less prepared for
wintering and, hence, to a greater extent are subjected to natural elimination. In the
population of species from the family Canidae in the North-West of Russia the
portion of males among the adult animals comprises on the average 52-56%, in
myophagous {Mustela nivalis, M. erminea, M. putorius) 66-68%. In the years of
344
increase and peak of abundance the portion of males of various ages in game samples
is particularly high.
Unequal rate of sexual maturation and lifetime of individuals of different sexes
is typical of most carnivorous mammals. The same is true for the time of readiness of the
young to rut, starting the reproduction for the first time, and adult animals. These
features have an important biological implication. Great portion of adult males in the
population provides the inclusion into the reproduction process of the maximal number
of females ready for breeding, which are inseminated by most active and strong males.
Therewith, in many species the young males manifest readiness to rut noticeably later
than females of the same age, which creates the premise for the copulation between
animals of different age and is one of important mechanisms of population homeostasis.
All this promotes maintenance of population viability and control of sex ratio at birth in
accordance with the ecological situation.
Глава 9. ГЕЛЬМИНТОЗЫ И ИХ ВЛИЯНИЕ
НА ПОПУЛЯЦИИ ХИЩНЫХ
МЛЕКОПИТАЮЩИХ
Среди болезней хищных млекопитающих гельминтозы играют важную
роль, и их значение необходимо учитывать при оценке состояния популяций
рассматриваемых животных. При разведении зверей в неволе в случае
отсутствия правильно организованных мер борьбы гельминтозы могут
поражать значительную часть имеющегося поголовья в хозяйствах и нередко
приводить к падежу ослабленных особей, прежде всего молодняка. Болезни,
вызываемые гельминтозами, часто протекают в очень важный и ответ-ственный
период роста и развития детенышей, ослабляя молодой организм и способствуя
его восприимчивости к разного рода иным заболеваниям. У взрослых животных
паразиты служат причиной замедления линьки, ухудшения состояния мехового
покрова, истощения и уменьшения плодовитости самок.
В природных группировках промысловых млекопитающих паразити-
ческие черви могут.выполнять функцию регуляторов численности хозяев,
снижая их репродуктивный потенциал и биологическую продуктивность
(Граков, 19626; Контримавичус, 1963, 1969; Шиляева, 1968; Савельев,
1975; Анисимова, Сидорович, 1996а, б). Все это свидетельствует о том, что
вопросы, связанные с изучением влияния гельминтов на состояние зверей и
в целом на их популяции, имеют не только теоретическое, но и практическое
значение.
Гельминтозы и экологию доминирующих видов патогенных червей у
хищных млекопитающих исследовали многие зоологи (Петров, 1941;
Шихобалова, 1950; Fahmy, 1954; Шахматова, 1964, 1966; Hansson, 1967,
1968, 1970; Карасев, 1972, 1975; Казлаускас, Прусайте, 1976; Козлов, 1977;
Schierhorn et al., 1981; Feliu et al., 1992; Miguel et al., 1994, и др.). По
материалам А.М. Петрова (1941), их видовой состав у зверей, содержащихся
в зверохозяйствах, очень разнообразен и представлен у лисицы 61 видом,
у песца 28, у енотовидной собаки 19,усоболя 14, у лесной и каменной куниц
19, у американской и европейской норки 27 видами гельминтов.
У хищников из семейства Mustelidae наиболее полно видовой состав
гельминтофауны, пути циркуляции паразитических червей и циклы их развития
представлены в работах В.Л. Контримавичуса (1963,1969). По этим данным,
мировая фауна гельминтов лесной куницы насчитывает 37 видов, лесного
хорька - 38, европейской норки - 20, американской - 64 и выдры - 26 видов.
В Восточной Европе у куньих насчитывается 55 видов паразитиче-
ских червей, относящихся к 4 классам (Trematoda, Cestoda, Nematoda и
Acanthocephala). У выдры обнаружены 17 видов гельминтов, у горностая 18,
у ласки 19, у норки европейской 21, у куницы лесной 33 иу хорька лесного 34
вида (Анисимова, 1997).
346
Следует отметить, что в разных частях ареала любого хищника
гельминтофауна неодинакова, что объясняется рядом причин (климатические
условия, состояние кормовой базы, плотность населения хозяина и др.).
Например, гельминтоценоз популяции выдры в Белорусском Полесье включает
15 видов (Shimalov et al., 2000), на севере республики -11 видов (Сидорович и
др., 1997), в Карелии - 3 (Шахматова, 1966), Хабаровском крае - 5
(Контримавичус, 1963)инаСахалине-2вида(Беньковскийидр., 1973). Судя
по этим данным, состав паразитических червей у выдры из разных мест
обитания оказался небогатым. В полной мере это относится и к другим крупным
представителям семейство Mustelidae - росомахе и барсуку.
Зараженность выдры гельминтами на северо-западе европейской части
России незначительна. В Карелии у нее обнаружено лишь 3 вида нематод при
очень слабой интенсивности и экстенсивности инвазии (Шихматова, 1964). В
группе из 36 тушек зверей, добытых в Псковской и Ленинградской областях,
ни у одного из них нами не были отмечены наиболее часто встречаемые у
куньих региона нематоды родов Skrjabingylus и Filaroides. В Белоруссии
зараженность выдры (л=74) нематодой S. nasicola за длительный период
времени (1960-1995 гг.) оказалась минимальной и составляла 4.0-8.2%
(Шималов, 1963). Среди 25 особей этого вида из Белорусского Полесья
широко распространенная у мелких куньих нематода F. mortis не была отмечена ни
разу (Shimalov, 2000).
Среди полуводных хищников выдра характеризуется наименьшим разно-
образием видов гельминтофауны, экстенсивностью и интенсивностью
заражения, что может объясняться особенностями ее питания и малой
восприимчивостью к паразитическим червям. В Белоруссии (Сидоро-
вич и др., 1997) гельминтоценозы выдры активно изучали в 1960-1980 и 1987-
1995 гг. В первый период исследований у нее было заре-гистрировано 11 ви-
дов гельминтов, а общая зараженность популяции составляла 72%. При этом
32% особей оказались инвазированными одним, а 40% - двумя и более
видами паразитических червей. Из класса нематод часто встречались
С. putorii (Aonchoteca putorii) - 28% и С. mucronata (Pearsonema mucronata) -
20%, co средней интенсивностью инвазии 3 и 2 экз. соответственно.
Во второй период работы в гельминтоценозе популяции выдры отмечали
только 6 видов гельминтов, хотя совокупная зараженность зверей изменялась
мало, при этом у 36.7% особей был обнаружен 1 вид паразитических червей
(Х2=0.2, р>0.1), у 31% - 2 (х2=0.6, р>0.1) и только у 7.3% выдр регистри-
ровалиь по 3 вида гельминтов (х2=П.8, /><0.01). В среднем на одну
инвазированную особь их приходилось по 1.6±0.7 экз. Зараженность выдр
S. erinaceha. просматриваемый период выросла с 8 до 57.1% (х2=14.2,/?<0.01),
а С. putorii (A. putorii) во второй период исследований не регистрировали ни
разу.
Мировая фауна гельминтов, паразитирующих на росомахе, насчитывает
21 вид, относящийся к 18 родам и 15 семействам. Из них на европейской части
ареала хищника отмечены в России только 12 видов (Контримавичус, 1969).
347
На севере этой территории популяция вида часто оказывается свободной от
паразитических червей (Данилов, Туманов, 1976а; Макарова, 1982). Среди
просмотренных нами 15 тушек росомахи из Мурманской области и Карелии
носители скрябингилезной или филяроидозной инвазии не найдены. Все
животные были здоровыми и хорошо упитанными.
В Предуралье зараженность зверей нематодами оказалась незначительной
(Теплое, 1955). Их инвазированность паразитическими червями в тундровых
биоценозах полуострова Таймыр также невелика. Здесь росомаха оказалась
хозяином всего 2 видов гельминтов - Taenia martis и Trichinella spiralis
(Савельев, 1975). В то же время на севере Дальнего Востока этот хищник
практически не встречается свободным от эндопаразитов. Так, в Анадырском
районе Чукотки в 1971-1978 гг. зараженность популяции росомахи три-
хинеллезом составляла 70-100% (Овсюкова, Новиков, 1982). Замечено, что
у зверей, обитающих вблизи морских побережий, интенсивность заражения
трихинеллезом увеличивается. В материковой же части региона (бассейн р.
Омолон) до 50% осмотренных особей было инвазировано другой нематодой -
Aelurostrongulus pridhami (Домнич, Обушенков, 1983; Б. Новиков, 1993). По
наблюдениям Б.В. Новикова (1993), какой-либо связи между упитанностью
росомах и экстенсивностью их заражения трихинеллами не отмечено. Даже
сильно зараженные ими звери оставались хорошо упитанными, а в годы
высокой интенсивностью инвазии их добывали даже больше, чем обычно.
Барсук на северо-западе европейской части России инвазирован 8 видами
гельминтов, среди которых преобладают нематоды (Шахматова, 1964,1966).
Из них наиболее часто встречаются нематоды Uncinaria stenocephala и
Molineus patens, а также цестода Mesocestoides lineatus. Гельминты, обна-
руженные у барсука, известны и как паразиты других видов куньих.
В отношении патогенности гельминтозных инвазий для барсука
существуют лишь отдельные предположения. Например, Д.Н. Рухлядев (1948)
считает наиболее опасными элюростронгилюс и кренозоматоз. Интересно,
что у барсуков не обнаружены паразиты лобных пазух - скрябингилюсы,
характерные для большинства видов куньих (Контримавичус, 1969), хотя
легочные нематоды (филяроидесы) встречаются в популяции вида сравни-
тельно регулярно. Среди 24 тушек зверей, поступивших из Псковской и
Ленинградской областей, очаги филяроидоза (1-3 экз.) были обнаружены нами
у 8 особей (33.3%). Даже у годовалого самца весом 3.9 кг, добытого в апреле
1974 г., в трахеях имелись 2 очага нематод этого вида. Необходимо также
отметить, что среди всех просмотренных животных 2 особи были поражены
нематодой, локализующейся в подкожной соединительной ткани (Filaria
martis) в количестве от 300 до 500 экз. Невосприимчивость барсука к
скрябингилезной инвазии, весьма распространенной среди куньих, и
сравнительно слабая интенсивность заражения другими видами гельминтов
(при активном потреблении в пищу их промежуточных хозяев) свиде-
тельствует об устойчивости организма данного хищника к воздействию
паразитических червей.
348
В Республике Татарстан у барсука отмечены 16 видов гельминтов, в том
числе по 1 виду трематод и скребней, а также 4 вида цестод и 10 нематод.
Экстенсивность и интенсивность заражения невелики. Эпидемиологическое и
эпизоотологическое значение могут иметь только 4 вида: цестоды Diphyllo-
bothrium latum и Mesocestoides lineatus, нематоды Thominx aerophilus
(Eucoleus aerophilus) и Trichinella sp. (Троицкая, Горшков, 1972; Горшков,
1974). По материалам П.К. Горшкова (1997), в Татарстане на участках, где
барсуки могут добывать рыбу, среди зверей увеличивается степень их
зараженности дифиллоботриями, а при питании грызунами - такими
нематодами, как Molineus patens и Aelurosirongylus spiralis. Потребление
падали способствует большей инвазированности животных Trichinella sp.
Последней в Мордовии заражено 6.3% обследованных барсуков, в Башкирии
- 80%, а в Нижегородской области - 20% (Третьякова, 1956; Романов, 1964;
Мачинский, Семов, 1971).
В Белорусском Полесье у рассматриваемого хищника обнаружены 17 видов
паразитических червей, среди которых доминировали нематоды семейства
Capillariidae(B.B Шималов, В.Т. Шималов, 1999а).
У барсуков Джунгарского Алатау были зафиксированы 5 видов
гельминтов из 17, известных для него в Казахстане (Лобачев, 1976). Нередко
у одного зверя находили до 4 видов гельминтов. Начиная с последней декады
мая и до середины лета при вскрытии желудочно-кишечных трактов барсуков
постоянно регистрировали единичные экземпляры Ascaris columnaris
(Baylisascaris columnaris). У 8 животных, отловленных в сентябре-первой
декаде октября, данных паразитических червей также обнаруживали у всех
особей, но уже в количестве от 10 до 250 экз. (в среднем 105 экз.). Из 6 зверей,
добытых и вскрытых в конце октября, только у одной особи в кишечнике был
выявлен 1 экз. Л. columnaris (В. columnaris). По-видимому, перед зимним сном
барсуки способны очищать организм от этого паразита, потребляя большое
количество кормов растительного происхождения. При обследовании 20
животных на трихинеллез нематода Trichinella sp. ни в один из сезонов
обнаружена не была.
Гельминтоценоз соболя и воздействие патогенных червей на его популяции
исследовали многие специалисты (Петров, 1941,1948; Мальчульский, Тощев,
1953; Романов, 1959; Губанов, 1964; Контримавичус, 1969; Ковальчук, 1979;
В. Монахов, 1981, и др.). По материалам А.М. Петрова (1941), у этого хищника
обнаружены 14 видов паразитических червей. Среди них 2 вида относятся к
трематодам, 2 - к цестодам и 10 - к нематодам, локализующимся в лобных
пазухах, органах дыхания, пищеварения и выделения.
На востоке России у соболя найдены 13 видов гельминтов (Контримавичус,
1969), из которых к наиболее массовым и часто встречаемым следует отнести
нематод и цестод, паразитирующих в желудке, кишечнике, трахее и в легочной
ткани. Остальные обнаруживаются редко, и интенсивность заражения ими
животных невелика. Однако длительность наблюдений и массовость
исследуемого материала позволили Н.А. Трабенковой (2001) констатировать
349
Таблица 127/Table 127
Инвазироваиность соболей (%) разного пола н возраста (по: Трабенкова, 1992)
Invasibility (%) of sables (Martes zibellina) of different sex and age
Инвазия/ Invasion Пол/ Sex Возрастная группа (roflbi)/Age group (years)
0+ 1+-2+ 3+ и старше/ 3+ and older
Соболифнмы/ 62.7+7.1 (94) 58.8±8.2 (160) 46.6±5.9 (231)
Soboliphyme 44.5+7 (115) 53.1 ±7.1 (198) 74.4±12.3 (136)
Аскариды/ 11.8+5.8 (92) 17.3±5.6 (228) 7.2±3.2 (220)
Ascaris 55 19.3±6.1 (118) 11.1±3.1 (199) 12.8±5.7 (143)
Цестоды/ 20.4±7.8 (95) 13.1±5.7 (223) 4.4±3.3 (227)
Cestoda 55 11±4 (112) 11.8±5.3 (193) 3.2±1.9 (142)
Кренозомы/ 40.8±6.6 (95) 34.8±7.4 (223) 14.5±3.6 (227)
Csenosoma 55 22.4±9.8 (112) 23.1±10.3 (193) 19.9±5.6 (142)
Томинксы/ 13.3±5.8 (95) 20.2±6.8 (220) 11.6±3.6 (227)
Thominx 55 5.4±2(114) 13.3+3.6(196) 6.7±2.9 (147)
Примечание. В табч. 127-129 в скобках указано количество просмотренных особей.
наличие в Камчатской области 19 видов гельминтов. Среди них выделяются
паразиты, имеющие сплошной ареал либо встречающиеся спородически. Они
относятся к трем классам - цестод, скребней и нематод. Все зарегис-
трированные представители гельминтофауны соболя Камчатки локализуются
в органах дыхания, желудочно-кишечном тракте и мышечной ткани. В пе-
риоды депрессии популяции хищника зараженность некоторыми паразитами
снижается настолько, что их могут не регистрировать. Этим объясняется
отсутствие некоторых детальных характеристик даже для фоновых инвазий,
не говоря уже о редких.
По мере продвижения на север Камчатской области (Трабенкова, 1992)
прослеживаются обеднение видового состава и снижение напряженности
воздействия инвазий на популяцию соболя. Так, выше 60-й параллели в его
организме отсутствуют соболифимы, а томинксы встречаются крайне редко.
Самцы соболя в возрасте до 3 лет чаще и интенсивнее самок инвазированы
различными видами гельминтов. Исключение составляет зараженность
сеголеток аскаридами (табл. 127). В старшей возрастной группе уровень
экстенсивности гельминтизации самок соболифимами, аскаридами и
кренозомами оказался заметно выше такового у самцов. Негативное влияние
паразитов на хозяина выражается в снижении массы тела и плодовитости
зараженных зверьков (табл. 128). Весовые различия между здоровыми и
инвазированными самками 3 лет и старше по соболифимам составляли 11.5%,
а по аскаридами 10.7%, при этом потенциальная плодовитость зараженных
350
Таблица 128/Table 128
Влияние некоторых инвазий на массу тела (г) и плодовитость самок (экз.) соболей
3 лет и старше (по: Трабенкова, 1992)
The influence of some invasions on the body mass (g) and fertility of sable (Martes zibellind)
females (ind.) of 3 years and older
Инвазия/ Invasion Масса тела/ Body mass Достоверность различий/ Significance of differences Кол-во желтых тел на беременную самку/ Number of corpus luteum per 1 gestation female Достоверность различий/ Significance of differences
Без инвазии/ Healthy 817.2±15.8 (52) - 2.6±0.4 (46) -
Соболифимы/ Soboliphyme 722.8+84.4(19) 1.85 1.7±0.2(35) 2.34
Аскариды/ Ascaris 729.6+52(12) 2.18 1.7±0.6 (10) 2.54
особей падала на 34.6%. Для центральных районов Камчатки, где автором
исследовано более 3 тыс. тушек, выявлена положительная достоверная
корреляция численности хищника и экстенсивности инвазии желудочно-
кишечными паразитами. При заражении аскаридами и цестодами коэффициент
корреляции составил 0.43 и 0.41 (за 25 и 23 года наблюдений соответственно),
а соболифимами - 0.39 (за 27-летний период). Уровень значимости первых
двух был выше 95%, а последнего - близок к этому. Анализ материалов по
инвазированности соболей, содержащихся в зверохозяйствах, свидетельствует
и о заметном ухудшении качества меха у больных животных - позднее
Таблица 129/Table 129
Влияние филяроидоза иа показатели размножения самок соболей старше 2 лет
(по: Трабенкова, Валеицев, 1992)
The influence of Filaroides on the parameters of reproduction of sable (Martes zibellina)
females over 2 years
Наличие инвазии/ Invasion Кол-во самок/ Females number Среднее кол-во желтых тел/ Number of corpus luteum Потенциалы прирост поголовья, °Х Potential grow rate of population, %
всего/ total из них беременных, абс. (%)/ including gestation на каждую самку/ per 1 female на беременную самку/ per 1 gestation female
Незараженные/ Healthy 23 11 (47.8) 1.65 3.45 74.5
Зараженные/ Infected 14 1 (7.1) 0.29 4 12.9
351
Таблица 130/Table 130
Влияние интенсивности филяроидозной инвазии (%) на некоторые показатели
размножения соболя (р. Жупанова, Камчатская обл.)
(по: Трабенкова, Валенцев, 1992)
The influence of Filaroides invasion intensity (%) on some parameters of reproduction
of sable {Martes zibellina), River Zhupanova, Kamchatka Peninsula
Участок реки/ Area of the river Экстенсивность заражения/ Percentage of total infestation Исследовано самок/ Research females total Из них беременных/ Including gestation Потенциальный прирост, %/ Potential growth rate of population, %
n %
Верхний/ 30-44 46 2 4.3 2.6
Upper Средний/ 7.1-21.1 28 6 24.4 33.9
Middle Нижний/ Under 0 14 4 28.6 45
созревание, отсутствие должной густоты, блеска и прочие дефекты
(Любашенко, Петров, 1962).
Одним из наиболее массовых и патогенных паразитов легочной ткани
соболей на Камчатке является нематода Filaroides martis. По материалам
Н.А. Трабенковой и А.С. Валенцева (1992), ею чаще бывают заражены особи
в возрасте старше 2 лет. Среди 1-2-летних зверьков инвазированность самцов
(37.3%) оказалась несколько выше, чем самок (33.3%), однако с увеличением
возраста последние бывают заражены даже больше. Так, из 39 обследованных
самцов старше 2 лет филяроидесы были найдены у 18 особей (46.2%), а из 9
самок того же возраста - у 5 (55.6%). Если учесть, что напряженность
рассматриваемой инвазии усиливается к периоду самого продуктивного
возраста зверьков, то трудно переоценить ее негативное влияние на популяцию
соболя. По расчетным данным, наматода Filaroides martis может снижать в
популяции уровень размножения хозяина в 6-7 раз (табл. 129). На Камчатке
она зарегистрирована только у соболя, поэтому любое воздействие на*
численность хищника так или иначе сказывается и на паразите. Отлов во время
промысла зараженных особей снижает и напряженность гельминтозной
нагрузки на популяцию соболя. Поток инвазии, количество оплодотворенных
яиц, выделяемых в окружающую среду, ослабевает. Происходит некоторое
оздоровление местообитаний и поголовья вида. Проведенные исследования
свидетельствуют о том, что количество зверьков, зараженных филяроидозом,
на отдельных участках, где велись наблюдения, доходило до 30-44.4%.
Показатели воспроизводства здесь оказались заметно ниже, чем на периферии
очага или за его пределами (табл. 130), при этом на группировки вида да
данной территории воздействия другого инвазионного начала не было.
Зараженность соболей филяроидозом обычно увеличивается в годы
высокой численности мышевидных грызунов, что, вероятно, связано с участием
последних в передаче этим хищникам гельминтов. Среди животных, добытых
352
Таблица 131/Table 131
Заболеваемость филяроидозом соболей Северного Зауралья и Западного Саяна
(по: В. Монахов, 19836)
Disease incidence of Filaroides of sables (Martes zibellina) in Northern Trans Urals and
Zapadnyi Sayan
Интенсивн. инвазии/ Intensity of infestation Север Зауралья/ North of Transurals (/i=170) Хакасия/ Khakassiya (/1=94) Бассейн p. Кизир/ Drainage of the River Kizir (//=609)
кол-во инвазирован. особей/ number of infestation ind. среди них доля самцов, %/ percentage of males кол-во инвазирован. особей/ number of infestation ind. среди них доля самцов, %/ percentage of males кол-во инвазирован. особей/ number of infestation ind. среди них доля самцов, %/ percentage of males
Кол-во цист/ Number of cysts:
1-3 34 67.6 16 75 175 59
4-6 11 45.5 1 100 78 62.8
>7 9 33.3 - - 23 43.5
Заражено(%)/ Infestation:
самцов/males 57.4+3.78 76.5+ 10.3 58.8±1.94
самок/females 42.6+3.78 23.5± 10.3 41.2± 1.94
Достоверн.
различий/ Significance of 2.77 3.64 6.41
differences
в Северном Зауралье и Западном Саяне, наиболее зараженными были особи в
бассейне р. Кизир, а наименее-в Хакасии (В. Монахов, 1981,19836). Влегких
больных соболей чаще встречались 1-3 очага (цисты) филяроидоза, причем в
пробе из Хакасии животные с такой интенсивностью инвазии резко
преобладали (табл. 131).
Как отмечает В.Г. Монахов (1983), экстенсивность инвазии у самцов во
всех выборках была достоверно выше, чем у самок, однако интенсивность их
заражения оказалась ниже таковой у последних. Большая инвазированность
самцов рассматриваемым видом и другими паразитическими червями
прослеживалась в популяции соболя и ранее (Романов, 1959; Губанов, 1964).
Заболеванию филяроидозом в процентном отношении подвержены в основном
зверьки-первогодки, а среди них самцы (табл. 132). Показатель интен-
сивности заражения у подавляющего большинства сеголеток, как и у особей
других возрастных групп, составляет 1-3 цисты на всю массу легких. С
возрастом напряженность инвазии в популяции соболя снижается, и доля
больных животных в возрасте 3-10 лет становится минимальной. Это
согласуется с данными других авторов, которые работали с представителями
рода Martes (Романов, 1959; Грибов, 1959; Граков, 19626; Губанов, 1964).
Некоторые исследователи (Петров, 1948; Мачульский, Тощев, 1953)
объясняют данный факт способностью организма хозяина освобождаться от
гельминтов через 1-1.5 года после их внедрения в легкие.
353
Таблица 132/Table 132
Заболеваемость филяроидозом соболей разного возраста (по: В. Монахов, 19836)
Disease incidence of Filaroides of sables {Martes zibellina) of different age
Возрастная группа (лет)/ Age group (years) Доля каждой группы, %/ Percentage of groups Доля самцов, %/ Percentage of males
в пробе/ at probe среди зараженных/ among infestation в пробе/ at probe среди зараженных/ among infestation
До 1 года/ 0+ Бассейн р. Кизир/Drainage of the River Kizir 74.8 84.6 69.9 83.5
1 + 6.3 6.8 6.8 6.7
2+ 6.2 6.1 6.5 6.7
3+ и больше/ 3+and older 12.7 2.5 16.8 3.1
Северное Зауралье/North of Transurals
До 1 года/ 0+ 38.9 61.1 37.1 61.3
1 + 10.8 14.8 12 12.9
2+ 9.9 13 12 12.9
3+ и больше/ 3+ and older 40.4 11.1 38.9 12.9
Хакасия/Khakassiya
До 1 года/ 0+ 37.2 47.1 37.5 38.4
1+ 51.1 41.2 47.9 46.2
2+ и больше/ 2+ and older 11.7 11.8 14.6 15.4
Среди взрослых соболей преобладают переболевшие особи. Хищники, не
подвергшиеся инвазии в первые годы жизни, могут, по-видимому, заболеть позднее,
хотя в целом восприимчивость зверьков к филяроидозу с возрастом, судя по
имеющимся данным, снижается (В. Монахов, 1981,19836).
По сообщению А.А. Вершинина и Г.А. Белова (1973), в популяции соболя
наиболее плодовиты крупные, хорошо упитанные и свободные от парази-
тических червей самки. У инвазированных кишечными гельминтами зверьков
плодовитость в среднем ниже на 38%, чем у здоровых. Минимальная
плодовитость наблюдалась при одновременном недостатке основных кормов
и массовом заражении соболей гельминтами. Такая картина отмечалась,
например, в промысловом сезоне 1959/60 г., когда в популяции прохолостало
85 % самок, а прирост поголовья к началу сезона составлял 15% против 50-
81% в хорошие по кормовым условиям годы.
В таежной зоне Зауралья, по свидетельству В.Г. Монахова и С.П. Тру-
шина (2001), в целом без разделения по полу средняя упитанность больных
филяроидозом соболей ниже (/=4.16, р=0.0043), чем неинвазированных,
однако внутри половозрастных групп значимые связи зараженности с
упитанностью характерны лишь для взрослых самок. Среди здоровых самцов
354
различия в упитанности между сеголетками и взрослыми особями не выяв-
лены, в то же время в группе больных животных они оказались значи-
мыми (/=1.83, /2=0.03). У самок различия по этому показателю выражены
сильнее: у взрослых особей жировые запасы были выше, чем у молодняка
(р<0.01). У них также прослеживается снижение упитанности у зараженных
животных разного возраста по сравнению со здоровыми (/=2.44-2.50,
р=0.0068-0.0072).
По материалам этих авторов, практически во всех возрастных группах
потенциальная плодовитость самок была выше у не зараженных филярои-
дозом хищников. Особенно четкие различия проявляются с 3-летнего воз-
раста. Зараженных самок старше 6 лет, участвующих в размножении, не
отмечено. В среднем на каждую взрослую самку среди здоровых соболей
приходилось 4.01 ±0.12 желтых тел беременности, у инвазированных -
3.52±0.24 (/=1.84, р=0.028). Многолетние исследования показали, что
интенсивность филяроидозной инвазии соболей Зауралья оказалась наивысшей
у 1-летних самок и 1-4-летних самцов. У последних она увеличивалась (д<0.05)
от 3.97 у годовалых особей до 5.2 у 3-4-летних зверьков. Основными
носителями инвазии следует считать прибылых особей, возможно, потому, что
в популяции они численно преобладают. В то же время интенсивность поражения
филяроидесом выше у хищников старших возрастов. Анализ собранных
данных показал, что в популяции соболя смертность инвазированных особей
больше таковой у здоровых в группе самцов в среднем в 1.6, самок - в 2.8
раза. Следовательно, патогенное влияние филяроидоза на популяции соболя
существенно и особенно сильно сказывается на самках.
В обширной сводке по болезням пушных зверей А.М. Петровым (1941)
отмечено наличие у каменной куницы 19 видов паразитических червей. Два из
них относятся к трематодам, 3 - цестодам и 14 - к классу нематод. В
Заилийском Алатау у этого хищника (Тазиева, Лобачев, 1970) обнаружены
10 видов гельминтов, из которых наиболее обычны паразиты тонкого отдела
кишечника - цестоды Mesocestoides lineatus и Taenia martis, а также нематода
Molineuspatens. В Джунгарском Алатау у каменной куницы выявлены только
9 видов гельминтов, среди которых преобладали кишечные нематоды Ascaris
colunmaris (В. columnaris) и Molineus patens. Все зараженные зверьки были
хорошо упитанными, а у одной самки, несмотря на обилие в ней трихинелл,
наблюдались еще и 3 нормально развитых эмбриона (Лобачев, 1973).
Гельминтоценоз мелких хищников от ласки до лесной куницы вклю-
чительно, обитающих на европейском Севере и Северо-Западе России,
характеризуется сравнительно небольшим видовым разнообразием
(Шахматова, 1964, 1966; Контримавичус, 1969; Юшков, 1995). К общим
для них и наиболее часто встречаемым в популяциях следует отнести широко
распространенные здесь нематоды родов Skrjabingylus и Filaroides.
Считается, что они оказывают сильное патогенное воздействие на организм
хозяев (Петров, 1948; Мачульский, Тощев, 1953; Грибова, 1959; Граков,
19626; Когтева, Морозов, 1970а, б).
355
Скрибингилюсы представляют собой сравнительно крупных, нитевидных,
ярко-красного цвета гельминтов, паразитирующих в лобных и гайморовых
пазухах. Филяроидесы - тонкие, светло-серые нематоды, которые плотно
переплетают бронхи и легочную ткань, образуя округлой формы очаги или
цисты. Степень их инвазии учитывается по наличию данных очагов в легких.
Циклы развития возбудителей филяроидоза легких и скрябингилеза лобных
пазух куньих подробно рассмотрены в работах отечественных гельминто-
логов (Петров, Гагарин, 1938а, б; Петров, 1941; Контримавичус, 1969).
Заболеваемость хищников патогенными червями происходит при попадании
в их организм моллюсков (промежуточные хозяева), инвазированных
личинками паразитов. С куньими паразитирующие черви также связаны в
основном по линии трофических цепей. К ним относится около 82% видов
гельминтов от общего их количества - это трематоды, цестоды, скребни и до
67% нематод (Контримавичус, 1969). Следует отметить, что представители
большинства видов рассматриваемого семейства сравнительно редко поедают
моллюсков, рачков-циклопов и других беспозвоночных, которые служат
промежуточными хозяевами паразитов. В организм хищников они обычно
попадают при потреблении традиционных кормов - мелких позвоночных
животных (мышевидные грызуны, земноводные, рыбы), являющихся
одновременно резервуарными и дополнительными хозяевами гельминтов,
поэтому в замыкании циклов развития паразитических червей у куньих они
имеют огромное значение (Любашенко, Петров, 1962; Sidorovich, Anisimova,
1996).
Негативное влияние гельминтов в результате прямого воздействия на ткани,
а также продуктов их жизнедеятельности на организм хищников несомненно.
Филяроидесы вызывают специфическое изменение ткани, в результате чего
отдельные участки легких теряют способность к интенсивному газообмену:
известны случаи гибели соболей и хорьков в зверохозяйствах при поражении
ими органов дыхания. При этом у зверьков отмечались очаговые изменения в
легких близ мест локализации паразитов. Кроме того, у них наблюдалась
лейкоцитарная инфильтрация в районе филяроидесных капсул, а также
очаговая эмфизема и даже гепатизация около мест расположения паразитов.
Их бронхи обычно бывают расширены и находятся в стадии воспаления
(Петров, 1941; Любашенко, Петров, 1962).
Локализующиеся в лобных пазухах мелких хищников скрябингилюсы
вызывают утончение и даже перфорацию костей черепа. Вследствие
интоксикации данным видом гельминтов в популяциях горностая и лесной
куницы регистрировалось снижение плодовитости и сокращение численности
животных (Лавров, 1944; Граков, 1962 б; Когтева, Морозов, 1970а, б).
На европейском северо-востоке России состав паразитических червей у
горностая представлен 13 видами, принадлежащими к 10 семействам
(Юшков, 1995). В северной части региона гельминтофауна этого вида
заметно беднее, чем в южной (Козлов, 1969). Зараженность горностая
паразитами составляет в среднем 70.1 ±3.5%. Из них только 3 вида
356
(Mesocestoides lineatus, Hepaticola hepatica, Trichinella nativa) свойственны
широкому кругу хозяев (главным образом хищным млекопитающим), остальные
10 видов являются специфичными для куньих. Доминируют нематоды -
Skrjabingylus nasicola и Ascaris devosi (Baylisascaris devosi). Среди 164
просмотренных тушек зверьков только в 2 случаях отмечена перфорация у них
лобных костей при поражении скрябингилезом (Юшков, 1995).
В Карелии инвазированность горностая гельминтами достигала 70%
(Шахматова, 1964, 1966). Здесь зарегистрированы всего 7 видов этих
паразитов, из которых 6 относятся к нематодам. Наиболее часто встречаются
представители родов Skrjabingylus и Filaroides. Большинство паразитирующих
у горностая гельминтов оказываются общими с таковыми у лесного хорька,
европейской норки, ласки и лесной куницы, что, видимо, объясняется сходным
характером питания и выбором местообитаний.
Отловленные в Ленинградской области в конце 60-х-начале 70-х годов
горностаи (л=53) имели слабую инвазированность филяроидесами, экстен-
сивность которой составляла 11.3% при незначительной (1 -2 очага) интенсивности
заражения. Скрябингилезная инвазия была выше и достигала 58.5% при средней
встречаемости паразитов в лобных пазухах, равной 7.2 экз. Доля пораженных
самцов в этот период здесь составляла 77, а самок - 43%.
В составе гельминтофауны горностая из Татарстана (Троицкая, 1960,1967)
обнаружены 9 видов, из которых 3 относятся к сосальщикам и 6 - к нематодам.
В. А. Попов (1943,1947) особенно большое значение в динамике численности
горностая этого региона придает нематоде Skrjabingylus nasicola. По его
данным, зараженность зверьков этим паразитом за 11 промысловых сезонов
колебалась от 20.5 до 45.2% при средней многолетней 27.9%.
В обследованных в Башкирии (Лавров, 1944,1956) 99 тушках горностая
обнаружены только 5 видов паразитов- Capillariaputorii (Aonchoteca putorii),
Molineus patens, Skrjabingylus nasicola, Strongyloides martis и Taenia tenuicollis.
К наиболее патогенным из них автор относит скрябингилюсы, отмечая
обратную зависимость между величиной сезонных заготовок шкурок зверька
и степенью зараженности популяции этими гельминтами. По свидетельству
Ю.С. Лобачева (1973), в горах Юго-Восточного Казахстана у горностая най-
дены 8 видов гельминтов, из которых общими для Заилийского и Джунгарского
Алатау являются Insinuarotaenia apasskii и Molineus patens. Экстенсивность
заражения гельминтами составляла 37.6%, причем в выборке из 109 особей у
11.9% животных скрибингилюсами была повреждена лобная кость.
На Камчатке популяция горностая инвазироваиа гельминтами в
достаточно сильной степени. Среди 184 зверьков 3 5.4% особей имели паразитов
кишечного тракта, а 7.7% - органов дыхания. Общая экстенсивность инвазии
в сезоне 1959/60 г. составляла 74%, при этом доля зараженных самцов оказалась
выше таковой у самок (Контримавичус, Скрябина, 1963).
По материалам А.А. Вершинина (1972, 1977), на Камчатке у горностая
выявлено всего 6 видов гельминтов, т.е. в 2 раза меньше, чем на северо-востоке
европейской части России. Наиболее широко распространены скрябингилюсы,
357
обнаруженные у 69% исследованных особей. Интенсивность инвазии
колебалась от 1 до 32 экз., составляя в среднем 11.3 экз. Мелкие нематоды
Capillariaputorii (A onchotecaputorii) и Molineuspatens выявлены в кишечниках
20% исследованных зверьков, а распространенные у соболей паразиты
(соболифимы) встречены в 5.4% случаев.
В Республике Коми гельминтологическому исследованию ласки были
подвергнуты 7 зверьков, из которых у 2 особей обнаружили только нематод
Ascaris devosi (Baylisascaris devosi) co средней интенсивностью поражения 2.5
экз. (Юшков, 1995).
В Ленинградской и Псковской областях в 1968-1974 гг. зараженность ласок
филяроидесами в небольшой биопробе (п=30) составляла 10%, а скрябин-
гилюсами - 53.3%. Интенсивность инвазии паразитами лобных пазух оказалась
равной в среднем 7.2 экз. при максимуме 15 экз. Существенных половых
различий в экстенсивности заражения ими зверьков не отмечалось. Среди
самцов инвазированные особи составляли 51.7%, среди самок-54.8%.
Лесной хорек в Карелии заражен всего 5 видами гельминтов (Шахматова,
1966). По экстенсивности инвазии здесь преобладают Skrjabingylus nasicola -
80%, Capillaria putorii (Aonchoteca putorii) - 55%, Molineus patens - 30% и
Filaroides martis - 30%. Специфичным для хищника паразитом оказалась
цестода Taenia tennicolis. В целом экстенсивность инвазированности хорька
здесь достигала 90%. В южной части северо-западного региона России
(Данилов, Русаков, 1969) в период 1960-1965 гг. зараженность зверьков (и=35)
филяроидесами была заметно выше (91.5%), а скрябингилюсами - несколько
меньше (70.3%), чем в Карелии. Средняя интенсивность инвазии составляла
соответственно 3.3 и 14.1 экз. При этом даже в случаях сильного заражения
паразитами заметного истощения хорьков не наблюдалось.
Следует отметить, что южные популяции лесного хорька заражены в
большей степени, чем северные. Так, в Белоруссии гельминтоценоз его
природных группировок состоит из 22 видов, относящихся к 3 классам: 6 -
трематод, 2 - цестод и 14 - нематод (Сидорович и др., 1997). Зараженность
зверьков паразитами достигает 92.5%. Среди нематод максимальная
встречаемость отмечена для S. nasicola, F. martis, С. putorii (Aonchoteca
putorii) co средней интенсивностью инвазии 7 экз., а также для С. mucronata
(Pearsonema mucronata) - 3 экз. и М. patens - 4 экз. Видовой состав
гельминтоценоза лесного хорька в Белоруссии оказался богаче такового у
полуводных куньих и лесной куницы. Во многом это, видимо, объясняется
обитанием значительной части особей по берегам водоемов вместе с норками и
выдрой, где циркулирует обильное инвазионное начало разных видов
паразитов. Кроме того, лесной хорек заселяет не только многие околоводные
биотопы, но также лесные, болотные экосистемы, постройки человека и
агроценозы, т.е. может иметь больший контакт с разнообразными про-
межуточными хозяевами паразитов.
Гельминтофауна колонка насчитывает 22 вида (Контримавичус, 1977).
По материалам анализа опубликованных работ А.Т. Войлочников (1977)
358
приходит к выводу, что зараженность гельминтами популяций колонка как в
видовом, так и в количественном отношении возрастает с запада на восток и с
севера на юг. Если в Предуралье и Западной Сибири гельминтоценоз хищника
насчитывает 9-12 видов паразитических червей, то в Средней Сибири и
Забайкалье -14-16. В суровой по климатическим условиям Якутии у колонка
обнаружены всего 6 видов гельминтов, а общая зараженность ими популяции
достигает 18% при сравнительно невысокой инвазированноста особей (Губанов,
1964). В то же время гельминтоценоз этого хищника из Приморья и Приамурья
насчитывает 19 видов, причем практически все осмотренные здесь особи
оказывались зараженными. Экстенсивность инвазии такими распро-
страненными паразитами, как соболифимы и скрябингилюсы, достигала.39 и
85%, а интенсивность - 80-90 экз. (Контримавичус, 1963).
По мнению А.Т. Войлочникова (1977), массовые виды нематод (например,
скрябингилюсы) способны снижать численность колонков лишь в те годы, когда
они из-за недостатка пищи оказываются очень истощенными. При благо-
приятных кормовых условиях случаев уменьшения запасов вида не
наблюдалось даже в те сезоны, когда популяция, хищника была сильно
заражена гельминтами.
Изучение инвазированности европейской норки (л=33) из Ленинградской
и Псковской областей нематодами в период с 1968 по 1974 г. показало, что
экстенсивность заражения ее популяции здесь достигает 77.1%. Большая часть
исследованных зверьков (72.7%) оказалась инвазирована скрябингилюсами,
и только у 11 особей (33.4%) были отмечены филяроидесы. В среднем на 1
зверька приходилось 2.3' (от. 1 до 8) очага филяроидеса и 10.2 (от 2 до 23)
паразитов лобных пазух. По экстенсивности инвазии самцы (72.2%) и самки
(73.4%) практически не различались.
У европейских норок, обитающих в Белоруссии, обнаружены 18 видов
гельминтов. Общая зараженность ими популяции достигает 88%. За
тридцатилетний период ее инвазированность возрасла с 50% в 60-е годы до
95-100% в 90-е, причем интенсивность инвазии увеличилась заэто время
почти в 20 раз. Интересно, что если в 60-е годы у европейских норок в данном
регионе были выявлены всего 5 видов паразитов, то в 80-е - уже 15, из них 5
- трематод, 2 - цестод, 7 - нематод и 1 - акантоцефал. В последнее десятилетие
исследований список гельминтов пополнился еще тремя высокопатогенными
видами: Opisthorchis relineus, Trichinella spiralis и Crenasoma taiga (B.E. Ши-
малов, B.B. Шималов, 1999a).
Американская норка в Карелии (Шахматова, 1964, 1966) инвазирована
10 видами гельминтов. Наиболее часто встречаются нематоды (7 видов), реже -
трематоды (2 вида) и акантоцефалы (1 вид). Преобладают следующие
паразиты: Capillaria putorii (Aonchoteca putorii) - 52.4%, Skrjabingylus nasico-
la - 21.7%, Capillaria mucronata (Pearsonema mucronata) - 20.8% и Filaroides
martis - 6.4%.
В Ленинградской области в сезоны 1968-1974 гг. зараженность зверьков
гельминтами была невысокой: из 81 просмотренной тушки только у 8 (9.8%)
359
были отмечены очаги филяроидесов и у 27 (33.4%) обнаружены скрябингилюсы.
Интенсивность инвазии также незначительна. Обычно на одного пораженного
зверька приходилось 1-2 очага филяроидесов (в среднем 1.5) и 1-6 экз. (в среднем
2.8) скрябингилюсов. Экстенсивность инвазии самцов (39%) и самок (37.5%) в
данной выборке оказалась практически одинаковой.
В Белоруссии гельминтоценоз рассматриваемого хищника оказался заметно
разнообразнее (Sidorovich, Anisimova, 1996). Например, в выборке из популяции
американской норки, обитающей на р. Дрисса, выявлены 17 видов паразитов, из
которых 5 относились к трематодам, 2 - цестодам, 9 - нематодам и 1 - к
акантоцефалам. Преобладающими среди них оказались следующие гельминты:
Spirometra erinacei, larvae - 54.2%, Euparyphium metis {Icthmiophora metis) - 36%,
Capillaria putorii {Aonchoteca putorii) - 22.7%, C. mucronata {Pearsonema
mucronata) - 20% и Skrjabingylus nasicola- 16%.
В Камско-Вятском междуречье 62.1% популяции американской норки
были зараженными гельминтами (Стерлягов, 1991). Здесь обнаружены 6 видов,
из которых 1 относился к классу трематод и 5 - нематод. Наиболее широко
распространены Е. metis {Icthmiophora metis) и С. mucronata {Pearsonema
mucronata), зараженность которыми хищников составляла по 36.3%.
Экстенсивность инвазии Capillaria putorii (Aonchoteca putorii) достигала 55-
60% при интенсивности до 69-219 экз. на одного зверька. Трихинелез среди
околоводных животных распространен повсеместно и имеет тенденцию к росту.
По данным автора, в 1 г мышечной ткани передних конечностей и в диафрагме
при анализе под компрессорием обнаружено в среднем по 47 личинок
трихинелл. Интересно, что ранее на данной территории такие виды паразитов,
как Е. metis {Icthmiophora metis), С. mucronata {Pearsonema mucronata) и F.
martis, у куньих не были отмечены (Контримавичус, 1969).
В равнинной части Западной Сибири американская норка слабо инва-
зирована гельминтами (Синицын, 1988). В промысловых выборках филяроидесы
не обнаружены, а случаи поражения соболифимами и капилляриями единичны.
Экстенсивность инвазии норки скрябингилюсами составила 3.6% при средней
интенсивности 1.5 экз. на одну пораженную особь.
У норок из Приморского края выявлены 6 видов гельминтов: Euparyphium
metis {Icthmiophora melis), Soboliphyme baturini, Ascaris columnaris, Molineus
patens, Skrjabingylus nasicola и Capillaria putorii {Aonchoteca putorii). Здесь
ими бывает заражено до 96% всех обследованных в промысловых выборках
животных. При этом максимальная интенсивность инвазии в отдельные годы
достигала 150-162 экз. паразитических червей на одного больного зверька
(Васенева, 1967, 1969).
Таким образом, анализируя материалы по гельминтофауне американской
норки, можно отметить, что видовой состав ее паразитических червей и
интенсивность заражения особей в популяции заметно снижаются, прежде всего
в направлении с севера на юг, и подобная географическая изменчивость
состояния гельминтоценоза отчетливо прослеживается в пределах рассма-
триваемого ареала большинства представителей семейства куньих.
360
Популяция лесной куницы из Архангельской области и Республики Коми
в сезонах 1953-1955 гг. была поражена нематодами на 100% (Граков, 19626,
1978). В просмотренных выборках у 70% особей обнаруживались скрябинги-
люсы, а у 54% - филяроидесы. Инвазированность ими хищников в западной
части Архангельской области заметно сильнее таковой в ее восточных райо-
нах и на севере Республики Коми.
В последней в бассейнах рек Печоры, Мезень, Вычегды и Сысолы в 60-80-х
годах зараженность лесной куницы достигала 77.9% (Юшков, 1995). Всего у
нее были выявлены 12 видов паразитических червей, относящихся к нематодам
(10 видов) и цестодам (2 вида). В целом у хищника здесь доминируют нематоды
F. martis, S. petrovi и С.putorii {Aonchotecaputorii). Промежуточными хозяевами
для них служат наземные моллюски и олигохеты, а резервуарными - мелкие
грызуны. Редко встречающиеся паразиты (71 mustelae, Т. nativd) попадают в
организм хищника при потреблении грызунов и падали. На Северном Урале
пик инвазированности хищника нематодой F. martis приходится на декабрь
(66.6%). В последующие месяцы наблюдается постепенное снижение в
популяции доли больных особей, а в марте ею бывают поражены только 25%
животных. За исключением A. devosi (Baylisascaris devosi) и Т. aerophilus
{Eucoleus aerophilus), экстенсивность инвазии паразитическими червями на
севере региона ниже, чем в южных районах. Так, в приполярном Урале
зараженность куницы F. martis и S', petrovi была равна соответственно 20.4±6
и 2.6±2.2%, а на Северном Урале - 53.5±5.9 и 8.4±3.2% (Юшков, 1995).
В Карелии популяция рассматриваемого хищника заражена гельминтами
на 89.1%. Здесь лесная куница инвазирована в основном нематодами (87.7%),
реже цестодами (9.6%) и трематодами (6.1%). Экстенсивность ее заражения S.
petrovi достаточно высока (71%), a F. martis (56.3%) - заметно ниже (Шахмато-
ва, 1964, 1966).
Инвазированность куниц скрябингилюсами и филяроидесами в Ленин-
градской области в 60-х годах составляла соответственно 85.3 и 50.4%, в
Новгородской - 62.5 и 45.1, а в Псковской области - 63.6 и 43.5% (Когтева,
Морозов, 1970 а). Интенсивность заражения зверьков филяроидесами в регионе
невысокая -1-7 очагов, скрябингилюсами - значительна: в среднем на 1 особь
приходится 11.6, а максимально - до 53 паразитов. В случае смешанного
заболевания, т.е. когда куницы кроме паразитов лобных пазух инвазированы
еще и легочными нематодами, зараженность первыми бывает почти в 2 раза
выше (15.1 экз.), чем при наличии только одних скрябингилюсов (8.6 экз.).
В Тверской области, по свидетельству В.А. Волкова (1971), влияние
основных паразитов лесной куницы (возбудителей филяроидоза и скрябингиле-
за) на состояние популяции хищника в условиях южной тайги незначительно.
Экстенсивность и интенсивность инвазий не превышает таковую в Псковской
и Новгородской областях, а случаев массовой гибели животных при сильном
заражении паразитами не отмечалось.
В Белоруссии (Сидорович, Анисимова, 1997) гельминтоценоз популяции
лесной куницы включает 11 видов, из них 1 - трематод, 1 - цестод и 9 - нематод.
361
Таблица 133/ Table 133
Зараженность (%) нематодами норок разного возраста и пола
на Северо-Западе России
Infection (%) by nematodes of minks of different age and sex in North-West of Russia
D Сеголетки/Youngs Взрослые/Adults
Инвазия/ D среднем / Average В в среднем/
Invasion среднем/ average 99 average 99
Норка европейская/Л/uwM lutreola
(101) (48) (27) (21) (53) (23) (30)
Общая/Total 63.4 56.3 55.6 57.1 69.8 69.6 70
Скрябингилюсами/ Skrjabingylus 61.4 56.3 55.6 57.1 66 65.2 66.7
Филяроидесами/ Filaroides 40.6 41.7 40.7 42.9 39.6 52.2 30
Норка американская/Ми.«е/а vison
(235) (ИЗ) (66) (47) (122) (70) (52)
Общая/Total 21.7 16.8 16.7 17 26.2 25.7 26.9
Скрябингилюсами/ Skrjabingylus 18.7 15 13.6 17 22.2 22.9 21.2
Филяроидесами/ Filaroides 14.5 14.2 12.1 17 14.8 12.9 17.3
Примечание. В скобках указано количество просмотренных животных.
В среднем на 1 зараженную особь приходится 1.6±0.9 экз. паразитов. Общая
же инвазированность ими хищника составляла 65.7%. Встречаемость С. putorii
(Aonchotecaputorii) здесь достигала 34.8%, Spirometra erinacei-europaei - 34.3,
Skrjabingylus petrovi - 26.8, F. martis - 21.7%. В сравнении с полуводными
животными зараженность популяции лесной куницы паразитическими червями в
Белоруссии оказалась заметно ниже, а их видовой состав - менее
разнообразен.
Работы по инвазированности природных группировок куньих нематодами
родов Filaroides и Skrjabingylus обычно содержат данные о гельминтозах
зверьков, экстенсивности и интенсивности воздействия паразитов на их
локальные группировки. В то же время сведения о влиянии гельминтов на
основные элементы демографической структуры популяции, ее репродук-
тивные возможности и морфологические параметры, характеризующие
физиологическое состояние животных, крайне отрывочны и редки, что и
побудило нас заняться этим вопросом.
С целью выяснения взаимосвязи уровня зараженности мелких куньих с
их плотностью населения в Псковской области (1969-1988 гг.) проводились
учетные работы и сбор биологического материала по европейской норке и
горностаю, а в Ленинградской (1969-1990 гг.) - по американской норке и
лесному хорьку. За данный период было собрано от охотников и
362
Таблица 134/Table 134
Интенсивность заражения нематодами (экз.) норок разного возраста и пола
Intensity of infection by nematodes (ind.) of minks of different age and sex
Инвазия/ Invasion В среднем/ Average Сеголстки/Y oungs Взрослые/Adults
М Нт в среднем/ average 99 в среднем/ average 33 99
Норка свропсйская/Ми.це/а lutreola
Скрябингилюс./ (62) (27) (15) (12) (35) (15) (20)
Skrjabingylus 7 1-21 6.2 5.7 6.8 7.7 7.5 7.9
Филяроидсс./ (41) (20) (И) (9) (21) (12) (9)
Filaroides 4.2 1-20 5.2 7.8 2 3.2 3.2 3.3
Норка амсриканская/Мш/е/а vison
Скрябингилюс./ (44) (17) (9) (8) (27) (16) (И)
Skrjabingylus 4 1-13 4.3 4.3 4.4 3.7 4.3 2.8
Филяроидсс./ (34) (16) (8) (8) (18) (9) (9)
Filaroides 2 1-4 2.4’ 2.8 2 1.6 1.6 1.6
Примечание. В скобках указано количество зараженных животных.
проанализировано по каждому виду от 550 до 680 тушек зверьков или их
фрагментов (головы, легкие), что позволило проследить динамику заражения
нематодами особей в популяциях.
Влияние зараженности на различные структурные элементы популяций
зверьков, их вес, упитанность, размеры и морфологические параметры
устанавливали только при обработке свежего материала, количество которого
указано в соответствующих таблицах. При этом индексы внутренних органов
рассчитывали по отношению к живому весу зверьков.
По уровню зараженности паразитическими червями европейская и
американская норки различаются очень существенно. Показатель
инвазированности зверьков первого вида в рассматриваемой выборке был
равен 63.4%, а второго-21.7%. Выявленные различия отчетливо сохранялись
у них в группах и молодых, и взрослых особей (табл. 133), причем филяроидесы
встречались в популяциях обоих видов в среднем в 1.5 раза реже
скрябингилюсов.
Известно, что изначально все поступающие в природу американские норки
имели звероводческое происхождение и в хозяйствах постоянно подвергались
дегельминтизации. Вероятно, слабую их восприимчивость к паразитам следует
рассматривать либо как достаточно консервативную реакцию, выработанную
в процессе строгого ветнадзора при клеточном содержании многих поколений
животных, либо как изначально высокую степень устойчивости их организма
к заражению гельминтами.
В популяциях обоих видов взрослые особи были инвазированы скрябин-
гилюсами заметно больше сеголеток, хотя по уровню филяроидозного
363
Таблица 135/Table 135
Длина (мм) и масса тела (г) у здоровых и инвазированных нематодами норок
Body sizes (mm) and mass (g) of healthy minks and infected by nematodes
Возрастная группа/ Age group Пол/ Sex 3 доро вые/Healthy Инвазированные/Infected
п длина тела/ length of body масса тела/ body mass п длина тела/ length of body масса тела/ body mass
Норка европейская/МиЛela lutreola
Сеголетки/ Youngs 8 356.9+2 762±22.6 19 326.8±3.2 706.7±20.8
4 342±2.8 557.5±13.2 17 322.6±4.2 523.8+12.3
Взрослые/ 9 370.2±4.4 854.4±33.3 14 356.8±4.5 732.5±31
Adults 12 346.8±2.3 579.2±15.7 18 329.2±2.3 455.6±15.3
Норка американская/Л/илТе/я vison
Сеголетки/ Youngs 55 385.5±2.3 916.6±17.5 11 366.6±3.2 825.5±37.9
39 328.1±2.2 549.6±9.5 8 316.9±3.3 500.6±10.9
Взрослые/ 52 387.3±2.9 997.7±17.2 18 370.6±4.7 863.9±26.7
Adults 38 340.3±2.3 588.3±16.3 14 330.4±2.9 510.7±20.4
поражения легочной ткани они практически не различались (табл. 133).
Существенной разницы в инвазированности нематодами разнополых
особей не прослеживалось. Исключение составляет только группа
взрослых особей у европейской норки, среди которых самцы были
поражены филяроидесами больше самок в среднем на 22%. Интересно,
что в Белоруссии в популяциях американской норки и лесного хорька
зараженность взрослых особей разными паразитическими нематодами
(65.2-88%) была выше, чем у сеголеток (61.5-76.8%), а обнаруженные
различия оказались вполне достоверными (/=2.58, р=0.05). Инвазиро-
ванность самцов и самок у этих видов в республике одинакова и
составляет в среднем 73-74% (Шималов и др., 1993). Экстенсивность
заражения разнополых особей европейской норки также сходна. У самцов
она достигала 96.3%, у самок - 93.7%. Достоверных различий по уровню
инвазированности молодых (96.5%) и старых (93.3%) зверьков не выявлено
(Бычкова, Сидорович, 1994).
В наших сборах интенсивность заражения (табл. 134) европейской норки
скрябингилюсами (7 экз.) и филяроидесами (4.2 очага) была достоверно выше
таковой у американской (соответственно 4 и 2). По интенсивности распростра-
нения паразитических нематод американские норки разного возраста
практически не различалась. В популяции аборигенного вида сеголетки были
364
Таблица 136/Table 136
Упитанность (г) здоровых и инвазированных нематодами норок
разного возраста
The fatness (g) of minks of different age, healthy and infected by nematodes
Пол/ Sex Наличие инвазии/ Invasion 0+ 1 + 2+ 3+ и более/ and older
п масса пахового жира/ Mass of inguinal fat n масса пахового жира/ Mass of inguinal fat n масса пахового жира/ Mass of inguinal fat n масса пахового жира/ Mass of inguinal fat
Норка европейская/Л/ил/е/л luireola
Самцы/ Males Здоровые/ healthy Иивазнрован./ infected 8 19 2.18 2.07 3 5 3.20 1.13 3 4 2.66 5.63 3 5 4.13 2.61
Самки/ Здоровые/ healthy 4 1.83 5 1.72 3 5 4 2.52
Females Инвазировап./ infected 17 1.56 6 0.71 7 1.19 5 1.04
Норка американская/Л/w.v/e/n vison
Здоровые/ 55 8.74 22 6.72 15 8,69 15 6.92
Самцы/ healthy
Males Инвазировап./ infected 11 9.43 8 10.59 7 7.18 3 9.33
Самки/ Здоровые/ healthy 39 3.92 17 4.50 10 3.26 11 5.52
Females Ипвазирован./ infected 8 3.29 5 2.40 4 3.16 5 2.50
больше, чем половозрелые особи, инвазированы филяроидесами, но меньше -
паразитами лобных пазух. В целом самцы и самки обоих видов заражены
паразитическими червями примерно в равной степени, и только у сеголеток
европейской норки самцы оказались инвазированы больше самок в 3.3-4 раза.
Определенный интерес представляет вопрос о возможном влиянии
паразитических червей на основные морфологические параметры куньих.
Собранные материалы свидетельствуют о том, что у обоих околоводных видов
зараженные зверьки имеют заметно меньшие линейные размеры и массу тела
по сравнению со здоровыми (табл. 135). Эти различия прослеживались у них в
группах как сеголеток, так и взрослых животных. В среднем разница в массе
тела между здоровыми и зараженными самцами европейской норки составляла
у сеголеток 55 г, а у взрослых особей 122 г. У самцов американского вида она
достигала соответственно 91 и 134 г.
Результаты однофакторного дисперсионного анализа показали, что у норок
обоих видов фактор зараженности не влияет на вес сеголеток. Однако у
взрослых разнополых особей масса тела здоровых зверьков была достоверно
выше, чем у инвазированных (доверительная вероятностьр>95%).
Одним из основных показателей упитанности куньих может служить масса
подкожных жировых отложений, расположенных в паховой области, или их
365
Таблица 137/Table 137
Относительный вес внутренних органов (%о) здоровых и
инвазированных нематодами норок
Relative weight of inner organs (%<>) in healthy and infected by nematodes minks
Пол/ Sex Наличие инвазии/ Invasion п Сердце/ Heart Легкие/ Lungs Печень/ Liver Почки/ Kidneys Селезенка/ Lien
Норка европейская/Л/илГе/а lutreola
Самцы/ Здоровые/ healthy 10 10.3±0.5 17.8+0.8 44.2±1.8 10.1 ±0.4 3.3±0.4
Males Инвазирован./ invasion 23 9.7±0.3 16.5±0.6 41.3±1.2 10.5±0.4 3±0.2
Самки/ Здоровые/ healthy 8 10.8±0.7 17.8±0.8 45.8±1 10.7±0.5 3.7±0.4
Females Инвазирован./ invasion 20 10.3±0.5 16.3±0.7 43.2±1.2 10.8±0.3 3.3±0.2
Норка американская/Миз/е/а vison
Самцы/ Здоровые/ healthy 77 9.6±0.2 17.1 ±0.3 43.8±0.7 9 ±0.1 2.6±0.1
Males Инвазирован./ invasion 13 9±0.3 16±0.9 37.5±1.6 7.6±0.4 2.3±0.2
Самки/ Здоровые/ healthy 50 9.8±0.5 17.3±0.4 44.6±0.7 9.4±0.2 2.8±0.1
Females Инвазирован./ invasion 12 9.5±0.5 1б.5±1.7 42.1±1.3 8.9±0.5 2.7±0.2
энергетическое депо (его размеры чрезвычайно лабильны). Они определяются
рядом факторов и служат индикатором физиологического состояния животных.
Проведенные исследования свидетельствуют о том, что в разных
возрастных группах здоровые самки норок бывают более упитанными, чем
зараженные. В то же время у самцов в большинстве вариантов данная
зависимость не прослеживается (табл. 136). Дисперсионный анализ,
проведенный по фактору инвазированности норок, подтвердил достоверность
влияния зараженности на упитанность только взрослых самок (доверительная
вероятность р>99%).
Сеголетки и взрослые особи мелких хищников в осенне-зимний период
различаются между собой по массе тела ненамного: самцы в среднем на 12-
14%, самки - на 8-10%. Это дает возможность объединять в пределах вида
животных разного возраста, разделяя их только по половому признаку.
Анализ состояния основных внутренних органов здоровых и зараженных
особей показал, что рассматриваемые паразиты не вызывают у норок заметного
уменьшения веса жизненно важных органов (табл. 137). Вместе с тем
белорусские гельминтологи В. Т. и В. В. Шималовы (1999а) неоднократно
отмечали случаи интенсивного патогенного воздействия на европейскую норку
некоторых видов нематод, проявляющегося в дегенерации внутренних органов
и тканей.
366
Таблица 138/Table 138
Масса тела и упитанность норок при разной интенсивности
скрябингилезной инвазии
Body mass and fatness of minks with various intensity of Skrjabingylus invasion
Виды/ Species Интенсивн. инвазии, экз./ Intensity of invasion, ind. Самцы/Males Самки/Females
п масса, г/mass, g п масса, г/mass, g
тела/ body пахового жира/ inguinal fat тела/ body пахового жира/ inguinal fat
Норка европ./ 1-5 6 и более/ 9 721±24.7 1.8±0.98 7 491.4±33.2 1.1+0.66
Mustela lutreola 6 and older 10 777.1±45.7 2.8±1.12 8 513.8±22.6 1.2+0.39
Норка америк. 1-5 6 и более/ 16 872.5±32.9 8.8±1.88 11 513.6±20.3 4±1.07
1 Mustela vison 6 and older 8 805±45.1 9.3±3.32 8 492.5±38.6 6.6+2.88
По данным этих авторов, трематоды О. felineus, Р. truncation и М. albidus
(М. bills) приводят к механическому раздражению слизистых оболочек желчных
протоков, печень покрывается множеством бугорков различной формы и
величины с общей желтушностью тканей. В просвете желчных протоков
содержится мутная, слегка тягучая масса со значительным количеством
гельминтов (до 40-60 экз.), а в брюшной полости зверьков отмечается скопление
желтоватого транссудата с нитями фибрина. Нематоды С. taiga вызывают
припухлость слизистых оболочек легких, бронхов и трахеи с многочисленными
точечными кровоизлияниями, а нематоды F. martis - очаговые изменения
легочной ткани. Установлена также отрицательная корреляционная
зависимость между зараженностью и весом европейских норок. Она
свидетельствует о том, что инвазия паразитами общим количеством 800-1200
экз. резко уменьшает массу тела хозяина.
Исследование материалов о влиянии скрябингилезной инвазии разного
уровня на вес и упитанность здоровых и больных норок (табл. 138) не выявило
существенных различий между ними. Более того, во многих случаях сильно
зараженные зверьки были даже упитаннее слабо инвазированных особей.
Хотя однофакторный дисперсионный анализ и не выявил достоверных
различий между этими группами животных, паразитические черви, видимо,
все-таки способны активно развиваться и достигать высокой численности,
прежде всего на хорошо упитанных хозяевах с большим содержанием
жировых резервов. Нельзя исключать также и возможность быстрой
естественной элиминации в малокормные годы ослабленных и истощенных
гельминтозами особей. Например, при росте интенсивности заражения лесной
куницы скрябингилезом от 1-10 до 51-60 экз. доля сильно инвазированных
367
Рис. 66. Плотность населения (а), экстенсивность (б) и интенсивность (в)
заражения нематодами европейской норки в Псковской области.
1 - скрябингилюсы; 2 - филяроидесы.
По оси абсцисс - годы; по оси ординат: а - особей на 10 тыс. га; б-зараженность, %;
в - зараженность, экз.
Fig. 66. Population density (a), extensity (б) and intensity (в) of infection by
nematodes of European mink (Mustela lutreola) in Pskov Province.
1 - Skrjabingylus., 2 - Filaroides.
Abscissa -years; ordinate: a-ind./10000 ga; 6 - infectiousness, %;
e - infectiousness, ind.
368
Рис. 67. Плотность населения (а), экстенсивность (б) и интенсивность (в)
заражения нематодами американской норки в Ленинградской области.
I - скрябингилюсы; 2 - филяроидесы.
По оси абсцисс - годы; по оси ординат: а - особей на 10 тыс. га; б - зараженность, %;
в - зараженность, экз.
Fig. 67. Population density (a), extensity (б) and intensity (e) of infection by
nematodes of American mink (Mustela vison) in Leningrad Province.
1 - Skrjabingylus: 2 - Filaroides.
Abscissa - years; ordinate: a - ind./10000 ga; 6 - infectiousness, %;
в - infectiousness, ind.
369
Таблица 139/Table 139
особей в популяции постепенно уменьшалась от 57.4 до 0.8% (Когтева,
Морозов, 1970а).
Интересно, что, как и у норок, у копытных присутствие в организме
паразитических червей не всегда сопровождается снижением веса и
упитанности зверей. По материалам Н.К. Железнова (1990), некоторые старые
самцы лосей, добытые на Чукотке, были в большой степени заражены гельминтами,
но имели притом мощные жировые отложения на спине, груди и внутренних
органах. У лосей Северной Америки (Samuel, 1976; McNeill, Rausch, 1987) степень
инвазированности увеличивается с возрастом животных и достигает максимальной
величины у крупных, хорошо упитанных особей старше года.
Сходная картина прослеживается и в популяциях других видов копытных.
Например, наблюдения Н.К. Железнова (1990) за снежными баранами в Якутии
и на Чукотке показали, что максимальное разнообразие гельминтофауны и
высокий уровень интенсивности поражения тканей часто обнаруживается у
крепких, нагулявших жировые запасы зверей среднего возраста, а молодые и
старые слабо упитанные особи были инвазированы заметно ниже. У ягнят этого
вида при незначительной интенсивности заражения цестодами порой
отмечалось сильное истощение организма, а стенки их кишечника при вскрытии
оказывались гиперемированы и покрыты мелкими точечными изъязвлениями.
У взрослых упитанных животных даже при наличии паразитов патологических
изменений в системе внутренних органов не обнаруживалось. Автор считает,
что только при определенных условиях, когда происходит наложение одних
факторов на другие, отягощающий существование животных паразитарный
фактор может быть элиминирующим и активно влиять на структуру и состояние
популяции в целом. Так, имеются сообщения о массовой гибели зверей разного
возраста от гельминтозной пневмонии при высокой степени их заражения
Protostrongulus stillesii (Woodgard et al., 1974). Подобное совпадение у
копытных гельминтозных заболеваний с повышением бактериальной флоры в
организме может сокращать размеры популяции на 80-90% (Stelfox, 1971;
Железнов, 1990).
Сведения об изменении зараженности норок приразнойплотности их населения
в угодьях практически отсутствуют. По нашим наблюдениям, за весь период работы
пики численности следовали у них с интервалом 3-5 лет. Обычно в эти годы, а часто
и в последующем сезоне в пробах из популяций норок обоих видов прослеживалось
увеличение доли инвазированных особей и интенсивности их поражения (рис. 66- I
67).
Необходимо отметить, что на Северо-Западе России, в связи со слабо
развитым сельскохозяйственным производством, поймы рек и берега водоемов
служат основными местообитаниями не только норок, но и выдры, хорька,
горностая. Данные биотопы богаты необходимой для них пищей - полевками
и мышами, рыбой и земноводными, гнездящейся птицей и водными
насекомыми, поэтому при высокой плотности населения куньих на береговых
Зараженность (%) нематодами мелких хищников разного возраста и пола
Infection (%) by nematodes of small carnivores of different age and sex
Инвазия/ Invasion В среднем/ Average Сеголетки/Youngs Взрослые/Adults
в среднем/ average 99 В среднем/ average 99
Ласка!Mustela nivalis
Общая/ (126) (51) (34) (17) (75) (56) (19)
Total 38.1 39.2 38.2 41.2 37.3 37.5 36.8
Скрябингилюс./ Skrjabingylus 37.3 37.3 35.3 41.2 37.3 37.5 36.8
Филяроидес./ Filaroides 10.3 14.3 11.8 17.6 10.6 10.7 10.5
Горностай/ЛЛмге/а erminea
(79) (18) (11) (7) (61) (47) (14)
Общая/ Total 64.5 44.5 45.5 42.9 70.4 70.2 71.4
Скрябингилюс./ Skrjabingylus 63.3 44.4 45.5 42.9 68.9 68.1 71.4
Филяроидес./ Filaroides 25.3 27.8 27.3 28.6 24.6 25.5 21.4
Примечание. В табл. 139-140 в скобках указано количество просмотренных животных.
Таблица 140/Table 140
Возрастные изменения экстенсивности заражения нематодами ласки,
горностая и норок (%)
Age changes of extensity of infection by nematodes of weasel (Mustela nivalis),
ermine (Mustela erminea), and minks (%)
Виды/ Species Инвазия/ Invasion Возраст животных/Age
0+ 1+ 2+ 3+ и более/ 3+ and older
(51) (48) (18) (9)
Ласка/ Скрябингилюс/ Skrjabingylus 37.3 33.3 38.9 55.6
Mustela nivalis Филяроидес/ 14.3 10.4 11.1 11.1
Filaroides (18) (30) (15) (16)
Горностай/ Скрябингилюс/ Skrjabingylus 44.4 63.3 66.7 81.3
Mustela erminea Филяроидес/ 27.8 26.7 26.7 18.8
Filaroides (48) (15) (17) (21)
Норка европ./ Скрябингилюс/ Skrjabingylus 56.3 60 58.8 76.2
Mustela lutreola Филяроидес./ 41.7 46.7 47.1 28.6
Filaroides (113) (53) (40) (29)
Норка америк./ Mustela vison Скрябингилюс/ Skrjabingylus Филяроидес./ Filaroides 15 14.2 22.6 11.3 25 12.5 17.2 13.8
370
371
Таблица 141/Table 141
Интенсивность заражения нематодомн (экз.) ласки и горностая
Intensity of infection by nematodes (specimens) of weasel {Mustela nivalis) and
ermine {Mustela erminea)
Виды/ Species Инвазия/ Invasion В среднем/ Average Сеголетки/Youngs Взрослые/Adults
М Нт В среднем/ average 99 В среднем/ average 99
Ласка/ (47) (19) (12) (7) (28) (21) (7)
Mustela Скрябингилюс./ Skrjabingylus 7.5 1-25 6.6 7.8 4.6 8.1 7.8 9
nivalis Филяроидес./ (15) (7) (4) (3) (8) (6) (2)
Filamides 1.8 1-4 2 2.3 1.5 1.6 1.7 1.5
Горно- (50) (8) (5) (3) (42) (32) (Ю)
стай/ Скрябингилюс./ Skrjabingylus 9 1-33 6.6 7.7 4.7 9.5 9.3 10.2
Mustela Филяроидес./ (20) (5) (3) (2) (15) (12) (3)
erminea Filaroides 2.1 1-9 2.2 2.7 1.5 2.1 1.9 2.7
Примечание. В табл. 141-142 в скобках указано количество зараженных животных.
магистралях контакты между ними усиливаются, что и создает условия для
большей инвазированности зверьков.
Для околоводных хищников, постоянно обитающих у водоемов,
характерны сезонные их концентрации в данных местах. Они собираются
на участках, где зимуют и нерестятся лягушки, концентрируются в омутах
раки и рыба. Как справедливо отмечает В.Е. Сидорович (1990, 1991), при
высокой численности куньих, а также лисицы и енотовидной собаки,
привлекаемых сюда богатой добычей, здесь можно обнаружить большое
количество их экскрементов. В результате в почве и на других субстратах
концентрируются яйца гельминтов, обильно циркулируют инвазионные
агенты, способные внедряться в организм хищников, в том числе и норок.
При этом в организме резидентов, имеющих столь богатый в кормовом
отношении участок обитания, всегда содержится достаточное количество
жировых резервов. Подобные взаимосвязи, по-видимому, и определяют
хорошую упитанность даже интенсивно зараженных гельминтами норок,
заселяющих прибрежные биотопы.
Среди самых мелких представителей семейства куньих высокая общая
зараженность нематодами наблюдалась в популяции горностая (табл. 139).
Особенно четко эта разница прослеживалась в группе взрослых особей, где
его инвазированность скрябингилюсами оказалась сильнее таковой у ласки в
среднем в 1.9, а филяроидесами - в 2.3 раза. По экстенсивности заражения
нематодами разнополые особи обоих видов, а также молодые и взрослые
зверьки в популяции ласки практически не различались.
У горностая зараженность взрослых особей скрябингилюсами оказалась
выше таковой у сеголеток в среднем на 25%. Обращает на себя внимание и тот
372
Таблица 142/ Table 142
Возрастные изменения интенсивности заражения нематодами (экз.)
ласки, горностая и норок
Age changes in the intensity of infection by nematodes (specimens)
of weasel (Mustela nivalis), ermine (Mustela erminea) and minks
Виды/ Species Инвазия/ Invasion Возраст животных/Age
(И 1+ 2+ 3+ и более/ 3+ and older
Ласка/ Скрябингилюсами/ (19) (16) (7) (5)
Mustela nivalis Skrjabingylus 6.6 7.3 6.7 12.8
Филяроидесами/ (7) (5) (2) (1)
Filaroides 2 1.8 1.5 1
Горностай/ Скрябингилюсами/ (8) (19) (Ю) (13)
Mustela erminea Skrjabingylus 6.6 9.8 8.5 10.2
Филяроидесами/ (5) (8) (4) (3)
Filaroides 2.2 2.5 1.8 1.3
Норка Скрябингилюсами/ (27) (9) (Ю) (16)
европейская/ Skrjabingylus 6.2 7.6 6.8 8.3
Mustela lutreola Филяроидесами/ (20) (7) (8) (6)
Filaroides 5.2 3.8 3 2.8
Норка американская/ Скрябингилюсами/ (17) (12) (Ю) (5)
Skrjabingylus 4.3 5 2.5 3.3
Mustela vison Филяроидесами/ (16) (6) (8) (4)
Filaroides 2.4 2 1.5 1
факт, что в целом в группе миофагов и околоводных хищников особи-
долгожители (возраст 3 года и старше) по сравнению с молодняком были
больше заражены скрябингилюсами, но меньше-филяроидесами (табл. 140).
У американской норки эти изменения отчетливо не прослеживались.
По интенсивности заражения сеголетки ласки и горностая практически не
различаются. В этой группе зверьков в целом самцы оказались инвазированы
нематодами сильнее самок (табл. 141). Среди взрослых особей меньшая степень
поражения характерна для ласок. Следует отметить, что у миофагов взрослые самки
обычно инвазированы скрябингилюсами сильнее самцов и особенно (в среднем в
2 раза) самок-сеголеток. Молодые зверьки по сравнению с взрослыми в меньшей
степени заражены скрябингилюсами, хотя по уровню интенсивности
филяроидозной инвазии они очень близки.
Анализ собранных материалов свидетельствует также о том, что в
популяциях ласки, горностая и норок обоих видов прослеживается возрастное
увеличение не только экстенсивности, но и интенсивности поражения зверьков
скрябингилюсами и падение уровня филяроидозной инвазии (табл. 142). В то
Таблица 143/Table 143
Длина (мм) и масса тела (г) здоровых и инвазированных нематодами
мелких хищников
Sizes (mm) and body mass (g) of healthy and infected by nematodes small carnivores
Возрастная группа/ Age group Пол/ Sex Здоровые/Healthy Инвазированные/Infected
n длина тела/ length of body масса тела/ body mass n длина тела/ length of body масса тела/ body mass
Сеголетки/ Youngs 33 21 Ласка/Mustela nivalis 165.4+1.8 57.8+1.7 10 163+1.6 61.5+1.9
99 10 150.6+2.4 37.7±1.4 7 154±3.8 39.2+2
Взрослые/ Adults <?<? 34 175.4±1.7 75.3+1.5 18 178.3±2 76.2+2
12 154.7±1.8 47.8±2.6 7 152.8±2.3 43±2.7
Сеголетки/ 7 Горностай/Мн.йе/а erminea 250.4±3 138.3±6.6 4 247.5±5.6 121±14
Youngs 9? 5 214±2.1 109.2±7.7 2 212 98
Взрослые/ Adults 33 14 254+2 202.6+7.9 34 250.2+2.7 191.5+4.1
99 4 218±9.8 129.5±9.3 10 221.5±2.4 137.5±11.1
Таблица 144/Table 144
Упитанность (г) здоровых и инвазированных нематодами ласок и горностаев
разного возраста
The fatness (g) of weasels (Mustela nivalis) and ermines (Mustela erminea)
of different ages, healthy and infected by nematodes
Виды/ Species Пол/ Sex Наличие инвазии/ Invasion 0+ 1+ 2+ и более/ 2+ and older
n масса пахового жира/ mass of inguinal fat n масса пахового жира/ mass of inguinal fat n масса пахового жира/ mass of inguinal fat
33 Здоровые/ healthy 21 0.32 24 0.47 10 0.56
Ласка/ Mustela nivalis 99 Инвазирован./in fected Здоровые/ healthy 10 10 0.10 0.31 10 6 0.23 0.39 8 6 0.18 0.34
Инвазирован./ infected 7 0.10 4 0.32 3 0.25
33 Здоровые/ healthy 7 0.67 9 0.61 5 0.73
Горно- стай/ Инвазирован./ infected 4 0.18 15 0.57 19 0.49
Mustela erminea 99 Здоровые/ healthy 5 0.62 3 0.57 1 1.22
Инвазирован./ infected 2 0.31 3 0.21 7 0.62
374
1969 1971 1973 1975 1977 1979 1981 1983 1985 1987
1969 1971 1973 1975 1977 1979 1981 1983 1985 1987
Рис. 68. Плотность населения (а), экстенсивность (б) и интенсивность (в)
заражения нематодами горностая в Псковской области.
1 - скрябингилюсы; 2 - филяроидесы.
По оси абсцисс - годы; по оси ординат: а - особей на 10 тыс. га; б - зараженность, %;
в - зараженность, экз.
Fig. 68. Population density {a), extensity (б) and intensity (e) of infection by
nematodes of ermine {Mustela erminea) in Pskov Province.
7 - Skrjabingylus', 2 - Filaroides.
Abscissa - years; ordinate: a - ind./ЮООО ga; 6 - infectiousness, %;
в - infectiousness, ind.
375
Таблица 145/Table 145
Относительный вес внутренних органов (%о) здоровых
и инвазированных мелких хищников
Relative weight of inner organs (%o) of healthy and infected small carnivores
Пол/ Sex Наличие инвазии/ Invasion п Сердце/ Heart Легкие/ Lungs Печень/ Liver Почки/ Kidneys Селезенка/ Lien
Ласка/Mustela nivalis
Самцы/ Здоровые/ healthy 51 10.2±0.2 16.5±0.4 54.9±0.9 13.9±0.3 4.2±0.1
Males Инвазирован./ infected 23 10.6±0.3 . 16.1 ±0.4 50.4±0.9 12.1±0.4 4.9±0.2
Самки/ Здоровые/ healthy 18 12.1±0.6 17.2±0.9 56.9±1.2 15.1±0.5 5.2+0.2
Females Инвазирован./ infected 10 9.4±0.6 17.8±1.5 52.3±2.5 12.6±0.9 5.8±0.2
Горностай/Л/изГе/а erminea
Самцы/ Здоровые/ healthy 13 11.3+0.5 15.8±1.2 45.5±1.5 10.3±0.8 3.1 ±0.4
Males Инвазирован./ infected 21 10±0.5 15.1±0.9 40.5±1.3 10.6+0.5 3.5±0.2
Самки/ Females Здоровые/ healthy 8 11.1+0.5 16.7±0.9 47.5±2.3 12±0.6 3.9±0.5
Инвазирован./ infected 9 11.8±0.6 15.3±0.8 49.4±2.7 10.5±0.5 2.5±0.4
Таблица 146/Table 146
Зараженность (%) нематодами лесного хорька и лесной куницы
на Северо-Западе России
Infection (%) by nematodes of polecat (Mustela putorius) and pine marten (Martes martes)
in North-West of Russia
Виды/ Species Инвазия/ Invasion в среднем/ Average Сеголетки/Y ounps Взрослые/Adults
В среднем/ average <?<? ?? в среднем/ average Зв ??
(238) (140) (105) (35) (98) (72) (26)
Хорек Общая/Total 68.9 66.5 70.5 54.3 72.4 72.2 73.1
лесной/ Mustela Скрябингилюс./ Skrjabingylus 66 65.7 70.5 51.4 66.3 65.3 69.2
putorius Филяроидес./ Filaroides 37.8 37.9 41.9 25.7 37.8 37.5 38.5
Куница Общая/Total (231) 40.7 (160) 41.9 (90) 43.3 (70) 40 (71) 38 (41) 36.6 (30) 40
лесная/ Martes Скрябингилюс./ Skrjabingylus 32 31.9 31.1 32.9 32.4 31.7 33.3
martes Филяроидес./ Filaroides 25.1 26.9 30 22.9 21.1 22 20
Примечание. В табл. 146 и 147 в скобках указано количество просмотренных
животных.
376
Таблица 147/Table 147
Возрастные изменения экстенсивности заражения нематодами
лесного хорька и лесной куницы (%)
Age changes of extensity of infection by nematodes of polecat (Mustela putorius)
and pine marten (Martes martes) (%)
Виды/ Species Инвазия/ Invasion Возраст животных/Age
0+ 1+ 2+ 3+ и более/ 3+ and older
Хорек (140) (39) (32) (27)
Скрябингилюсами/ 65.7 64.1 68.8 66.7
Mustela Skrjabingylus
putorius Филяроидесами/ Filaroides 37.9 38.5 43.8 29.6
Куница лесная/ Martes Скрябингилюсами/ Skrjabingylus (160) 31.9 (37) 29.7 (19) 36.8 (15) 33.3
martes Филяроидесами/ Filaroides 26.9 21.6 21.1 20
Таблица 148/ Table 148
Интенсивность заражения нематодами (экз.) лесного хорька и лесной куницы
на Северо-Западе России
Intensity of infection by nematodes (specimens) of polecat (Mustela putorius)
and pine marten (Martes martes) in North-West of Russia
Инвазия/ Invasion В среднем/ Average Сеголетки/Y oungs Взрослые/Adults
М Нт В среднем/ average В среднем/ average <?<? 99
Скрябингилюс./ Skrjabingylus Филяроидес./ Filaroides (157) 9.5 (90) 2.7 Хорек лесым/Mustela putorius (92) (74) (18) 1-70 9.6 9.5 9.8 (53) (44) (9) 1-11 2.9 3 2.1 (65) 9.3 (37) 2.5 (47) 8.8 (26) 2.3 (18) 10.6 (П) 2.8
Куница лесная/Л/аг/ел' martes
Скрябингилюс/ Skrjabingylus (74) 6.8 1-34 (51) 6.7 (28) 7.6 (23) 5.5 (23) 7 (12) 7.3 (И) 6.6
Филяроидес./ Filaroides (58) 3.5 1-12 (43) 3.6 (27) 3.9 (16) 3 (15) 3.2 (9) 3.2 (6) 3.2
Примечание. В табл. 148 и 149 в скобках указано кол-во зараженных животных.
377
Таблица 1497 Table 149
Возрастная изменчивость интенсивности заражения нематодами (экз.)
лесного хорька и лесной куницы
Age variation of intensity of infection by nematodes (specimens)
of polecat (Mustela putorius) and pine marten (Martes martes)
Виды/ Species Инвазия/ Invasion Возраст животных/Age
0+ 1+ 2+ 3+ и более/ 3+ and older
Хорек лесной/ Mustela putorius Скрябингилюсами/ (92) (25) (22) (18)
Skrjabingylus 9.6 6.4 10 12.5
Филяроидесами/ (53) (15) (14) (8)
Filaroides 2.9 2.6 2.4 2.3
Куница лесная/ Скрябингилюсами/ (51) (И) (7) (5)
Skrjabingylus 6.7 6 7.2 9
Martes martes Филяроидесами/ (43) (8) (4) (3)
Filaroides 3.6 3.5 3 2.7
Таблица 150/Table 150
Длина (мм) и масса тела (г) здоровых и инвазированных нематодами особей
лесного хорька и лесной куницы
Sizes (mm) and body mass (g) of polecat (Mustelaputorius)
and pine marten (Martes martes), healthy and infected by nematodes
Возраст- ная группа/ Age group Пол/ Sex Здоровые/Healthy Инвазированные/Infected
п длина тела/ length of body масса тела/ body mass п длина тела/ length of body масса тела/ body mass
Сеголетки/ <3<3 32 Хорек лесной/Mustela putorius 366.7+3.6 775+24.8 73 355.1±1.7 689.9±10.8
youngs 16 318.3±5.3 446.5±12.4 19 310.3±3.2 403.4±10.9
Взрослые/ <3<3 20 377.7±3.7 806±17.8 52 360.9+2.6 746.2+10.7
adults 7 322.5±6.1 510±27.2 19 312.5±2.9 413.4±10.2
Сеголетки/ <3<3 48 Куница лесная/Martes martes 420.7±2.1 1041±13.9 39 416.3±4.2 1007.4± 19.7
youngs 99 42 385.5±3.2 769.8±12.7 28 382.6±3.1 756.9±14.3
Взрослые/ <3<3 25 423.5±4.6 1089±21.4 15 421.2+3.2 1051.7±38.5
adults 99 18 387.5 ±4 774.2+22 13 385.3±5.5 812.1±24
378
Таблица 151/Table 151
Упитанность (г) и инвазированность нематодами лесного хорька и лесной куницы
разного возраста
Fatness (g) and infection by nematodes of polecat {Mustela putorius)
and pine marten {Martes martes) of different ages
Пол/ Sex Наличие инвазии/ Invasion 0+ 1+ 2+ 3+ и более/ 3+ and older
п масса пахового жира/ mass of inguinal fat n масса пахового жира/ mass of inguinal fat n масса пахового жира/ mass of inguinal fat n масса пахового жира/ mass of inguinal fat
Хорек лесной/Mustela putorius
Самцы/ Здоровые/ healthy 32 6.03 9 9.80 4 8.15 7 7.12
Males Инвазирован./ infected 73 4.53 19 4.48 19 5.22 14 4.54
Самки/ Здоровые/ healthy 16 3.29 3 4.10 1 3.82 3 3.53
Females Инвазирован./ infected 19 2.56 8 2.13 8 3.05 3 1.47
Куница пссная/Martes martes
Самцы/ Здоровые/ healthy 48 1.09 14 1.33 4 1.62 7 1.34
Males Инвазирован./ infected 39 1.02 7 0.57 3 0.51 5 0.89
Самки/ Здоровые/ healthy 42 0.68 6 0.82 9 0.67 3 0.53
Females Инвазирован./ infected 28 0.27 8 0.69 4 0.48 1 0.25
же время у американской норки закономерного увеличения количества
паразитов в лобных пазухах у зверьков старших возрастов отмечено не было.
Влияние нематодной инвазии на размеры и массу тела миофагов
несущественно (табл. 143), что подтверждается данными однофакторного
дисперсионного анализа (при уровне значимости ос=0.05, ^расчетное не превышало
Fкритическое). Вместе с тем свободные от паразитических червей ласки и горностаи
в целом оказались более упитанными (табл. 144). Причем по результатам
однофакторного дисперсионного анализа у этих видов зараженность достоверно
негативно влияет на упитанность разнополых сеголеток (р>95%) и взрослых самцов
ласок (р>99%).
Как показали материалы морфологических исследований, существенной
разницы между здоровыми и инвазированными зверьками в величине основных
внутренних органов не прослеживается. В обеих группах хищников их
относительный вес у здоровых и зараженных самцов или самок оказался
практически одинаковым (табл. 145).
Собранные сведения о движении численности ласки и горностая в угодьях
показали, что ее изменения во многом определяются фактором обилия
379
мышевидных грызунов. При этом у горностая подъемы и спады численности
закономерно следуют за соответствующими изменениями таковой у мелких
млекопитающих с запозданием на один год, что связано с особенностями
размножения этого вида (Данилов, Туманов, 1976а). Колебания численности
миофагов в северных территориях Северо-Запада России выражены заметно
сильнее, чем в южных, где состав кормов разнообразнее и зависимость
хищников от обилия мышевидных грызунов менее отчетлива. Однако и здесь
некоторое увеличение плотности их населения в характерных местообитаниях
прослеживается в интервале 2-3 лет. В год пика численности (а нередко и в
последующем осенне-зимнем сезоне) наблюдается высокий уровень инвази-
рованности миофагов (и в частности, горностая) паразитическими червями
(рис. 68).
Лесной хорек и лесная куница по экстенсивности заражения нематодами
различаются очень заметно. Инвазированность зверьков первого вида в
популяции достигает 68.9% и близка к таковой у горностая и европейской
норки. Зараженность лесной куницы заметно меньше (40.7%): она сходна с ее
уровнем у ласки.
Молодые и взрослые зверьки у обоих видов по экстенсивности их
поражения скрябингилюсами и филяроидесами практически не различались.
Инвазированность ими разнополых особей в популяциях лесной куницы (а у
хорька - в группе взрослых особей) примерно одинакова. Среди молодых
лесных хорьков доля пораженных нематодами самцов была выше, чем у самок
(табл. 146).
По свидетельству Е.З. Когтевой и В.Ф. Морозова (1970 а, б), на Северо-Западе
России в популяциях лесной куницы разница в экстенсивности филяроидозной
инвазии у разнополых особей не превышала 1.1 -6.7%, т.е. была близка к таковой
в наших сборах. В Марийской, Татарской и Башкирской республиках она равна
1.5% (Троицкая, 1965), а в Архангельской области - 10-15% (Граков, 19626). В
большинстве регионов России степень зараженности лесной куницы филяро-
идесами и скрябингилюсами незначительна и патогенная их роль для популяции
вида невелика. Однако Н.Н. Граков (19626,1964) отмечает, что в годы высокой
численности зверьков взрослые самки бывают поражены нематодами интенсивнее,
чем самцы и сеголетки обоих полов, и заметно чаще гибнут от инвазий. К такому
же выводу приходит В.Ф. Юшков (1982, 1985). По его данным, в Республике
Коми в бассейне р. Вычегды общая зараженность лесной куницы гельминтами
составляет для самок 838±65, для самцов 83.6±4.7%, а на водосборе р. Илыч
соответственно 80±6.7 и 86.1±5.8%. В целом зараженность ими разнополых особей
несущественна (/<2), однако в годы высокой плотности населения вида
самки бывают поражены сильнее самцов. Так, в промысловый сезон 1976/77 г.
при плотности населения хищника 2 особи на 1000 га интенсивность инвазии F.
martis составляла у самок 5.3, у самцов 2.1 цисты, a S', petrovi соответственно 9.6
и 7.1 экз. Автор считает, что при высокой плотности населения куницы в
угодьях интенсивность заражения гельминтами самок заметно усиливается, что
может оказывать влияние на состояние ресурсов вида.
380
Возрастные изменения скрябингилезной инвазии у хорька и лесной куницы
выражены неотчетливо, доля же зверьков, зараженных филяроидесами, в группе
долгожителей оказалась ниже, чем у сеголеток (табл. 147). Многие
исследователи данный факт объясняют возрастным усилением у куньих
приобретенного иммунитета к гельминтам, физиологической устойчивостью
и способностью организма хозяина самостоятельно освобождаться от паразитов
(Романов, 1959; Грибова, 1959; Граков, 19626; В. Монахов, 1983; Юшков,
1995, и др.). В то же время нельзя исключать и возможность естественной
элиминации старых ослабленных особей с высокой степенью заражения, что
также позволяет объяснить малое количество инвазированных филяроидесами
зверьков-долгожителей.
Следует отметить, что рассматриваемые виды мало различаются между
собой по интенсивности филяроидозной инвазии, хотя уровень заражения
скрябингилюсами среди молодняка и взрослых особей был заметно выше у
лесного хорька (табл. 148). Количество нематод, паразитирующих у особей
разного пола, в группах сеголеток и взрослых животных старшего возраста
оказалось практически одинаковым, за исключением первогодок лесной
куницы, у которых, как и у миофагов, зараженность самок скрябингилюсами
была меньше, чем у самцов, на 28%.
В целом молодые и взрослые особи обоих видов инвазированы рассматри-
ваемыми паразитическими червями примерно в равной степени. В то же время,
как и у других видов куньих, в популяциях хорька и лесной куницы старые
особи по сравнению с сеголетками имеют тенденцию к большей зараженности
скрябингилюсами, но меньшей - филяроидесами (табл. 149).
Размерно-весовые показатели у здоровых и больных особей лесной
куницы оказались примерно одинаковыми (табл. 150). У лесного хорька
зараженные нематодами особи были заметно мельче неинвазированных, а
по массе тела они достоверно (р>95%) уступали животным, свободным от
паразитов. По уровню упитанности в популяции лесной куницы больные и
здоровые самцы-сеголетки достоверно не различались (табл. 151), в то время
как неинвазированные взрослые особи и молодые самки были упитаны
больше зараженных (р>95%).
Анализируя состаяние популяций лесной куницы европейского Севера и
Северо-Востока России, Н.Н.Граков (19626, 1981) отметил определенную
зависимость между упитанностью и интенсивностью заражения лесной куницы
скрябингилюсами и филяроидесами. По его данным, сильно инвазированные
особи всегда имеют меньше жировых отложений, чем здоровые. При этом
упитанность зверьков, пораженных нематодами обоих видов, в большей
степени зависит от наличия цист F. martis, чем от количества 5. petrovi.
По сообщению Е.З. Когтевой и В.Ф. Морозова (1970а, б), в 60-х годах
XX в. в Ленинградской и Псковской областях среди плохо упитанных разнополых
особей лесной куницы филяроидоз и скрябингилез были распространены сильнее
(93.3%), чем у зверьков с хорошим и средним уровнем содержания жировых
резервов (88.6%). Интенсивность инвазии скрябингилюсами у слабо упитанных
381
Таблица 152/Table 152
Относительный вес внутренних органов (%») здоровых и инвазированных
нематодами лесных хорьков и лесных куниц
Relative weight of inner organs (%o) of healthy and infected by nematodes polecats
(Mustela putorius) and pine martens (Martes martes)
Пол/ Sex Наличие инвазии/ Invasion п Сердце/ Heart Легкие/ Lungs Печень/ Liver Почки/ Kidneys Селезенка/ Lien
Хорек лесной/МихГе/а putorius
Самцы/ Здоровые/ healthy 38 8.8±0.3 15.6±0.3 44.1 ±0.9 8.7±0.2 3.1±0.1
Males Инвазирован./ infected 90 8.4+0.2 15+0.3 44+0.6 9+0.1 2.8±0.1
Самки/ Здоровые/ healthy 17 9.2+0.3 16.1+0.5 50.4±1.6 10.3+0.3 3.3±0.2
Females Инвазирован./ infected 25 8.8±О.З 15.3±0.5 48.1±1.5 9.3±0.2 3±0.3
Куница лесная/Martes martes
Самцы/ Здоровые/ healthy 57 9.4+0.1 15.5±0.4 33.8±0.7 7.1±0.1 2.2±0.1
Males Инвазирован./ infected 36 8.7±0.2 13.6±0.3 33±0.8 7+0.2 2.2±0.1
Самки/ Здоровые/ healthy 50 9.6±0.2 15.8±0.4 34.5+0.6 7.7±0.1 2.4±0.1
Females Инвазирован./ infected 30 9.1±0.2 15.3±0.5 33.1±0.8 7.3±0.2 2.2±0.1
особей также оказалась выше и составляла 19.7 против Ю.бэкз. Обычнокконцу
зимы встречаемость истощенных куниц с признаками интенсивного заражения
заметно возрастала. По данным В.А. Волкова (1971), в условиях южной тайги в
разные промысловые сезоны упитанность лесной куницы была ниже среднего
уровня только при высокой степени экстенсивности и интенсивности фидяроидозной
и скрябингилезной инвазии. Здоровые особи всегда имели жировые отложения1 в
паховой области и брюшной полости.
В отличие от куницы масса тела лесного хорька за счет интенсивного
жиронакопления к осенне-зимнему сезону увеличивается очень заметно, причем
хорошо упитанные хищники часто встречаются среди как здоровых, так и
пораженных нематодами зверьков. Однако в целом в промысловых пробах
свободные от гельминтов особи самцов и самок имели бблыпую массу жировой
ткани. По данным однофакторного дисперсионнного анализа, различия между
ними по уровню упитанности были вполне достоверны (р>95%).
Интенсивная инвазированность мелких куньих (в том числе и лесного
хорька), обитающих по берегам водоемов, объясняется большей плотностью
их населения на участках, богатых кормом, и концентрацией здесь яиц
гельминтов в процессе маркировки зверьками территории. Кроме того,
значительное потребление ими пораженных гельминтами жертв и высокий
382
уровень поглощения яиц паразитических червей связаны с близким
расположением маркировочных пунктов куньих к местам отдыха, потребления
пищи и ухода за волосяным покровом (Сидорович и др., 1997). Для лесной
куницы эти зависимости менее характерны прежде всего из-за диффузного ее
распределения по территории и меньшего скопления зверьков на охотничьих
участках.
Как и у норок, у рассматриваемых видов существенной разницы в индексе
основных внутренних органов пораженных и здоровых животных не наблюдается.
Например, у самцов лесной куницы свободные от нематод особи (и=57) имели в
среднем относительный вес печени, равный 33.8±0.7, а почек-7.1±0.1%о. У
инвазированных гельминтами хищников (п=36) данные показатели составляли
соответственно 33±0.8 и 7±0.2%о (табл. 152).
Отрицательное влияние гельминтов на состояние ресурсов лесной куницы
в северо-западных областях России прослежено Е.З. Когтевой и В.Ф. Мо-
розовым (1970а, б). Ими отмечено, что зараженность популяции вида
филяроидесами в сезоны 1957-1967 гг. изменялась от 27 до 66.6%, составляя в
среднем 50.3%. Наименьшая инвазированность зверьков наблюдалась в
периоды низкой численности куницы. На следующий сезон после года с высокой
степенью поражения популяции запасы хищника заметно сокращались. Резкое
снижение филяроидозной инвазии отмечалось во время обильных урожаев
плодов рябины, когда их значение в питании куниц возрастало до 63% против
12.5% в обычные годы. В эти промысловые сезоны уменьшалась роль
мышевидных грызунов в рационе хищников (с 50 до 30%), благодаря чему
возможность заражения их паразитами сокращалась.
На четко выраженную зависимость степени инвазированности группи-
ровок лесной куницы от урожая плодов рябины указывает также и А.А. Тро-
ицкая (1962, 1963). По-видимому, они оказывают дегельминтизирующее
действие не только на кишечных паразитов, но и на паразитических червей,
локализующихся в органах дыхания и лобных пазухах.
Снижение численности лесной куницы после годов с высокой степенью
экстенсивности и интенсивности заражения зверьков гельминтами наблю-
далось также в Республике Коми, Архангельской и других областях. По
наблюдениям Н.Н. Гракова (1978), инвазированность популяции вида
фляроидозом и скрябингилезом в известной степени влияет на потенциальную
плодовитость этих хищников. Так, в Кировской области здоровые самки лесной
куницы имели 3.23±0.37 желтых тела беременности, а инвазированные
скрябингилюсами - 2.90±0.46 на одну рожавшую особь. При заражении
филяроидесами данный показатель был равен в среднем 3, а обоими
паразитами одновременно - 2.7 на половозрелую самку. Автор считает, что,
несмотря на статистическую недостоверность этих различий, тенденция к
снижению их плодовитости при инвазиях прослеживается здесь достаточно
отчетливо и вряд ли случайна.
На севере европейской части России плотность популяции лесной куницы
тесно связана с видовым составом гельминтофауны и степенью обилия
383
Рис. 69. Плотность населения (а), экстенсивность (б) и интенсивность (в)
заражения нематодами лесного хорька в Ленинградской области.
1 - скрябингилюсы; 2 - филяроидесы.
По оси абсцисс - годы; по оси ординат: а - особей на 10 тыс. га; б - зараженность, %;
в - зараженность, экз.
Fig. 69. Population density («), extensity (6) and intensity (e) of infection by
nematodes of polecat (Mustela putorius) in Leningrad Province.
1 - Skrjabingylus., 2 - Filaroides.
Abscissa - years; ordinate: a - ind./ЮООО ga; 6 - infectiousness, %; в - infectiousness,
ind.
384
Таблица 153/Table 153
Масса тела и упитанность некоторых видов куньих при разной интенсивности
скрябингилезной инвазии
The body mass and fatness of some mustelids species with various intensity
of Skrjabingylus invasion
Виды/ Species Интенсивн. инвазии, экз./ Intensity of infestation, cyst Самцы/Males Самки/Females
п масса, г/mass, g n масса, г/mass, g
тела/ body пахового жира/ inguinal fat тела/ body пахового жира/ inguinal fat
Ласка/ 1-5 12 68.4±2.8 0.2+0.04 10 40.6±2.3 0.2+0.11
Mustela nivalis 6 и более/ 6 and > 10 73.5±2.4 0.3+0.05 9 41.6+1.7 0.3±0.06
Горно- 1-5 15 188.5±9.7 0.5±0.18 10 128±63.5 0.4±0.18
стай/ 6 и более/
Mustela erminea 6 and > 11 191.7±14.9 0.6+0.21 8 129-4±11.4 0.7±0.26
Хорек 1-5 28 734.6±18.2 4.1+0.79 10 410+12.9 1.8+0.61
лесной/ 6 и более/
Mustela putorius 6 and > 26 736.7±22.2 7+1.41 10 409.4±18.6 2.5±0.98
Куница 1-5 13 1110.4+41.6 0.5±0.19 15 768.1±23.4 0.4±0.11
лесная/ 6 и более/
Martes 6 and > 10 1096±77.3 1.3±0.58 18 816±45.8 0.8±0.23
martes
паразитов у зверьков. Как сообщает В.Ф. Юшков (1995), в фазе роста
численности хищника общая его зараженность паразитами изменилась с
63.6±8.4 до 83.6±5.1%. Наряду с этим их состав становился разнообразнее, а
степень заражения особей разными видами гельминтов увеличивалось. Рост
инвазированности был наиболее характерен для тканевых и полостных
паразитов (F. martis, S. petrovi) и менее свойствен кишечным формам. При
нарастании плотности населения куниц в междуречье Вычегды и Мезени с
0.85 до 2 особей на 1000 га поражение зверьков нематодами F. martis
повысилась от 30.3±7.9 до 52.7±6.7%, а 5. petrovi - от 27.3±7.8 до 50.9+6.6.
Таким образом, по мнению автора, основной фон паразитических червей
лесной куницы определяется филяроидозной и скрябингилезной инвазиями. К
наиболее важным экологическим факторам, обусловливающим ее зара-
женность, относятся трофические связи, возраст и плотность населения
зверьков. Кроме того, большое значение имеют также годовые и сезонные
изменения погодных условий, влияние которых проявляется опосредованно,
через промежуточных и резервуарных хозяев.
Численность лесного хорька в различных областях северо-западной зоны
России не остается неизменной, хотя масштабы ее колебаний выражены нерезко
(рис. 69). Плотность его населения в угодьях до 60-х годов XX в. была заметно
385
Таблица 154/Table 154
Масса тела и упитанность самцов лесного хорька и лесной куницы
при разной интенсивности филяроидозной инвазии
Body mass and fatness of males of polecat (Mustela putorius)
and pine marten (Martes martes) with various intensity of Filaroides invasion
Виды/ Species Интенсивность инвазии, цист/ Intensity of infestation, cyst п Масса, г/Mass, g
тела/ body пахового жира/ inguinal fat
Хорек лесной/ 1-2 11 736+27.4 5.7±1.82
Mustela putorius 3 и более/ 3 and > 8 820.2±86.1 8.2±3.08
Куница лесная/ 1-2 12 982.1±32.3 1±0.57
Martes martes 3 и более/ 3 and > 10 1018.7+42.4 1.2±0.39
выше современной. Причины данного явления до конца не ясны, но, вероятнее
всего, связаны с изменением интенсивности сельскохозяйственного произ-
водства, что сопровождалось ухудшением кормовой базы хищника. Если в
довоенный период основу его питания составляли мышевидные грызуны
(Новиков и др., 1970), то теперь - представители фауны земноводных. В
Ленинградской области за время исследований периоды подъема и спада
численности лесного хорька наблюдались в интервале 3-4 лет. Как и у других
мелких куньих, в годы высокой численности вида экстенсивность и
интенсивность поражения зверьков увеличивалась, причем нередко отмеченная
тенденция прослеживалась и в следующем сезоне.
Анализ собранных материалов свидетельствует о том, что скрябингилезная и
филяроидозная инвазии разного уровня интенсивности существенного влияния
на массу тела или упитанность рассматриваемых хищников не оказывают (табл.
153 и 154), что под тверждается данными однофакторного дисперсионного анализа.
Здесь во всех вариантах Fрасчетное не превышало Fкритическое при уровне
значимости а =0.05. Во многих случаях у разных видов интенсивно зараженные
гельминтами особи имели даже большую массу тела и упитанность, чем слабо
инвазированные зверьки.
Следует отметить, что сложившаяся в историческом развитии система
«паразит-хозяин» обладает определенной устойчивостью, которую можно
рассматривать как адаптивную реакцию организмов к существованию
в природной среде. Видимо поэтому собранные многолетние данные
не позволяют проследить резкого снижения морфофункциональных показате-
лей даже у сильно зараженных животных. Тем ни менее нельзя исклю-
чать негативное влияние паразитических червей на иммунитет хищных
млекопитающих к другим видам заболеваний, таким как вирусные или
бактериальные. При разведении пушных зверей в хозяйствах хронические
386
гельминтозы в случае отсутствия правильной организации борьбы с ними
нередко принимают характер эпизоотий, увеличивают эмбриональную
смертность и вызывают падеж животных. В период роста молодняка
нематодные инвазии ведут к его истощению, недоразвитию и сильной
восприимчивости к разного рода заболеваниям (Петров, Гагарин, 1938а, б;
Петров, 1941).
Замечено, что интенсивное поражение представителей некоторых видов
куньих скрябингилюсами может приводить не только к утончению, но и к
перфорации костной ткани. Инвазированные зверьки при наличии 5-20
скрябингилюсов в лобных пазухах постоянно встречаются среди мелких
хищников. Так, в 1972 и 1973 гг. из 12 ласок, попавшихся в Ленинградской
области в ловушки кротоловов, у 6 особей наблюдалась перфорация костной
ткани. В их лобных полостях находилось от 5 до 9 срябингилюсов. В 1986 г. в
Псковской области среди добытых самок лесного хорька (6 особей) и
европейской норки (4 особи) по 1 зверьку каждого вида имели четко
выраженные признаки деструкции лобных костей. Интенсивность инвазии у
них составляла соответственно 17 и 15 паразитов на самку. Вообще перфорация
черепа от действия скрябингилюсов чаще проявляется у особей мелких видов,
т.е. с более тонкой костной тканью, - ласок и горностаев, а также у самок
европейской норки, лесного хорька и куницы. Американские норки с
подобными признаками нами встречены не были, что, возможно, объясняется
низким уровнем интенсивности их скрябингилезного заражения.
По мнению В.Ф. Юшкова (1995), это заболевание при значительном
поражении животного может вызывать анемичное состояние центральной
нервной системы, катар лобных пазух и воспаление головного мозга.
Постоянное присутствие паразитов в лобных полостях приводит к механи-
ческому сдавливанию окончаний обонятельного и глазничного нервов. Все
это, вероятно, влияет на формы поведения зверьков и уровень их естественной
элиминации. У особей, пораженных филяроидесами, бронхи бывают
расширены и находятся в стадии воспаления. Для них характерна очаговая
эмфизема, а нередко и гепатизация легочной ткани в местах образования
цист. Судя по всему, поражение куньих паразитическими червями оказывает
негативное воздействие на физиологическое состояние, репродуктивный
потенциал и численность особей в популяции. Рассматриваемые виды
гельминтов играют не последнюю роль в характере изменения численности
хищных млекопитающих, в популяциях которых показатель их инвази-
рованности во многом определяется состоянием кормовой базы. Максималь-
ный уровень заражения наблюдается в малокормные годы и при большой
численности зверьков в угодьях. В такие периоды экстенсивность и
интенсивность инвазий усиливаются, что ухудшает общее состояние
популяций, а иногда вызывает случаи падежа животных.
Результаты наших работ показали, что уровень потенциальной пло-
довитости на фазе бластоцист и развивающихся эмбрионов в среднем у
здоровых куниц в возрасте 2-3 лет заметно выше, чем у зараженных (табл.
387
Таблица 155/Table 155
Некоторые показатели потенциальной плодовитости у здоровых и зараженных
нематодами лесных куниц в Ленинградской области
Some parameters of potential fertility of healthy and infected by nematodes pine martens
(Martes martes) in Leningrad Province
Кол-во бластоцист у самок/ Number of female blastocysts Кол-во эмбрионов у самок/ Number of female embryos
здоровых/ healthy инвазированных/ infected здоровых/ healthy инвазированных/ infected
дата/ date экз./ ind. дата/ date экз./ ind. дата/ date экз./ ind. дата/ date экз./ ind.
28.01.84 2 25.01.84 2 19.03.84 5 19.03.84 3(1)
30.01.84 4 28.01.84 3 21.03.84 4(1) 27.03.84 3(1)
2.02.84 3 28.01.84 2 25.03.84 3 28.03.84 5(2)
6.02.84 5 5.02.84 5 18.03.85 4(1) 18.03.85 3(1)
20.03.84 3 8.02.84 4 23.03.85 2 20.03.85 4
14.02.85 4 10.02.84 2 - - 21.03.85 4(1)
15.02.85 4 15.03.84 4 - - 24.03.85 3
15.02.85 2 23.01.85 3 - - - -
18.02.85 4 25.01.85 2 - - - -
20.02.85 5 12.02.85 3 - - - -
26.02.85 3 15.02.85 4 - - - -
- - 18.02.85 2 - - - -
В среднем по бласто- цистам/ Average blastocysts 3.55 - 3 В среднем по развивающ. эмбрионам/ Average embryos 3.2 - 2.71
Примечание. В скобках указано количество резорбирующих эмбрионов.
155). Подобная картина прослеживалась и в популяции европейской норки в
Псковской области, где у 5 здоровых самок, добытых в конце апреля-мае
1970 и 1971 гг., в рогах матки было обнаружено в среднем 4.6 (4-6) крупных
нерезорбирующих эмбриона, а у инвазированных нематодами шести особей -
только 3.7 (3-4).
Таким образом, негативное влияние гельминтов на популяцию куньих
вполне очевидно. Вероятно, у мелких хищников основная роль паразитических
червей сводится к ослаблению иммунитета, снижению функциональной
деятельности и общей сопротивляемости организма. Все это способствует
проникновению в него бактериальных и вирусных агентов, обусловливая
388
развитие болезней, снижение плодовитости и повышение уровня смертности
животных.
Гельминтофауна собачьих в основном состоит из широко распро-
страненных видов, которые в своем большинстве свойственны разным
представителям семейства. Как сообщает В.Ф. Юшков (1989, 1995), на
европейском северо-востоке России у песца выявлено 22 вида паразитических
червей, принадлежащих к 4 классам: цестоды - 9 видов, трематоды - 2,
Скрябины - 1 и нематоды - 10 видов. Как и у куньих, основные виды
паразитических червей попадают в организм этого хищника через
промежуточных или резервуарных хозяев. Ими обычно служат грызуны,
насекомоядные, зайцеобразные, копытные, рыбы, насекомые, олигохеты,
моллюски и амфибии. Основные виды гельминтов могут проникать в организм
песца путем потребления им разных пищевых объектов. Например, для
цестоды М. lineatus резервуарными хозяевами, помимо грызунов, могут быть
птицы, рептилии и амфибии; для нематод С. skrjabini и S. arctica, кроме насе-
комых, также и птицы. По данным автора, молодые и взрослые особи обитают
в сходных экологических условиях и по видовому составу паразитов
практически не различаются. Общая зараженность хищника в обеих
возрастных группах одинакова и равна 100%. Различия в экстенсивности
инвазии сеголеток и взрослых особей доминирующими видами, за
исключением Т. leonina, несущественны, однако показатели интенсивности
заболевания молодых зверей выше, чем взрослых. Самки и самцы
инвазированы в равной степени (100%), и набор паразитических червей у
них также оказался сходным. Достоверных различий в уровне зараженности
разнополых особей основными видами гельминтов не выявлено. Совпадение
фазы роста численности 2-3 представителей отряда грызунов приводит к
увеличению интенсивности размножения и зараженности паразитами этого
хищника.
При изучении гельминтоценозов песца из тундровой зоны Ненецкого
АО паразитические черви были обнаружены Л.М. Шиляевой (1968, 1970)
только в желудочно-кишечном тракте, и лишь Dioctophyme renale выявлен
на брыжейке. Всего здесь зарегистрированы 11 видов паразитов, из
которых токсаскаридоз и унцинариоз относятся к геогельминтозам, т.е.
заражение ими происходит по преимуществу с субстрата. Другие виды
паразитов попадают в организм вместе с объектами питания (прежде всего
грызунами и птицами) - промежуточными или резервуарными хозяевами
гельминтов.
Судя по результатам обследования автором 443 песцов, существенной
разницы в зараженности самцов и самок не наблюдалось. Известно, что
гельминтотоксины оказывают отрицательное влияние на центральную нервную
систему, функции пищеварения и размножения, при этом молодые особи
бывают более подвержены паразитарным заболеваниям (Шихобалова, 1950;
Догель, 1962). В Ненецком АО сеголетки также оказались заражены сильнее
взрослых особей, что указывает на наличие возрастного иммунитета и,
389
Таблица 156/ Table 156
Возрастная изменчивость заражения молодых (л=256) и взрослых (л=156) песцов
основными видами гельминтов (по: Шиляева, 1968)
Age variation of infection by main species of helminthes of young (л=256)
and adult (л=156) arctic foxes {Alopex lagopus)
Виды гельминтов/ Helminth species Экстенсивность инвазии, %/ Percentage of total infestation Интенсивность инвазии, экз. Intensity of infestation, ind.
сеголетки/ youngs взрослые/ adults t сеголетки/ youngs взрослые/ adults t
/'. leonina 99.5 74.3 6.1 22.6 11.7 24
S. lupi 11.7 30.7 2.1 40.3 21.1 15
A. multilocularis 18.7 10.8 0.6 16.6 10.4 7
T. crassiceps 29.6 15.3 1.6 20.5 11 10
T. polyacantha 29.2 13.4 1.8 15.4 7.7 11
Прочие/Other 33.5 18.5 1.7 5.1 6.3 2.4
Примечание. Интенсивность инвазии A. multilocularis дана в тыс. экз.
Таблица 157/ Table 157
Инвазированность размножавшихся и оставшихся без приплода взрослых самок
песца в Ненецком АО (по: Шиляева, 1970)
Invasiveness of adult females of arctic foxes {Alopex lagopus) that participated
in reproduction and those who have no litters in Nenets Autonomous Territory
Группы самок/ Female groups n Экстенсивность инвазии, %/ Percentage of total infestation Интенсивность инвазии, экз./ Intensity of infestation, ind.
Размножавшиеся/ Breeding 38 89.4±4.9 11.9±0.7
Неразмножавшиеся/ Non breeding 42 78.5±6.2 22±5.1
возможно, является одной из причин высокого уровня элиминации животных в
первый год жизни (табл. 156).
Сравнение инвазированности размножавшихся и оставшихся без приплода
взрослых самок позволило установить с необходимой долей достоверности
(/ >2), что последние менее часто, но вдвое более интенсивно заражены всеми
видами гельминтов (табл. 157).
Анализ собранных данных позволил Л.М. Шиляевой (1970) выявить
определенную зависимость между величиной помета (учеты по относительному
показателю - наличию плацентарных пятен) и количеством гельминтов у
390
размножавшихся самок. Отмечено, что у малоплодных особей наблюдается в
среднем до 10 экз. паразитов, в то время как у мно го плодных их количество обычно
превышает 30 экз. Поскольку самки с большими выводками инвазированы
сильнее малоплодных, то, видимо, это обусловливает более интенсивное
заражение щенков гельминтами и меньшую жизнестойкость молодняка из
крупных пометов. В Ненецком АО среднеупитанные песцы оказались сильнее
поражены гельминтами, чем звери с низким содержанием жировых резервов в
организме. Данный факт находит объяснение в большей сопротивляемости
организма особей, имеющих жировые запасы, и в отсутствии нормальных
условий существования для паразитов в истощенных животных, вследствие
чего происходит отмирание гельминтов. Вероятно поэтому в природе случаи
непосредственной гибели хищников от паразитов сравнительно редки, однако
их паразитирование приводит к истощению организма хозяина и ослаблению
сопротивляемости неблагоприятным условиям среды и инфекциям, причем в
первую очередь у малоприспособленного и неиммуиного молодняка.
Инвазированность популяции песца по годам претерпевает заметные
изменения в связи с динамикой численности и его, и мышевидных грызунов -
основных кормов, а также источников заражения хищника. В годы «урожая»
песца, когда одновременно наблюдается высокая численность леммингов и
других грызунов, происходит увеличение инвазии зверей гельминтами, так
как популяция в основном состоит из сеголеток, которые сильнее подвержены
паразитарным заболеваниям. В эти промысловые сезоны животные бывают в
достаточном количестве обеспечены кормом и хорошо упитаны, тем не менее
они оказываются интенсивно зараженными паразитами, которые в дальнейшем
негативно воздействуют на популяцию хозяина. При ухудшении кормовой
базы и сокращении численности песца выживают в основном иммунные особи,
не ослабленные болезнями. В эти сезоны происходит снижение интенсивности
размножения зверей и в популяции преобладают взрослые особи. Ухудшение
условий питания и приобретенный иммунитет избавляют на некоторое время
хищников от высокого уровня заражения гельминтами. В годы с нарастанием
численности мышевидных грызунов и песца вновь улучшаются показатели
размножения хищника, повышается роль молодняка в популяции, что в свою
очередь повторно приводит к некоторому ухудшению санитарного состояния
всей группировки вида (Шиляева, 1968, 1970).
В тундровых биоценозах полуострова Таймыр у песца обнаружено уже 10
видов паразитических червей (Савельев, 1974,1975), т.е. с продвижением на
восток прослеживается уменьшение разнообразия его гельминтофауны.
Экстенсивность заражения животных (и=293) составляет 98.3%. Чаще всего
здесь отмечали смешанную цестодно-нематодную инвазию, которая в
популяции достигала 71.5%. Ленточные черви были обнаружены у 14.4%,
круглые - у 12.4% осмотренных особей. Сравнение гельминтофаун двух
представителей семейства Canidae (песца и полярного волка) показывает
существенное и закономерное их различие. У последнего в кишечном
паразитоценозе преобладают цестоды Taenia krabbei и Т parenchymatosa,
391
личиночные фазы которых паразитируют на северном олене. У песцов же из
ленточных червей чаще встречаются Т. crassiceps, Tetratirotaeniapolyacantha и
Alveococcus multicularis, промежуточными хозяевами которых служат
мышевидные грызуны. Отмеченные различия находятся в полном соответствии
со спецификой питания этих хищников.
Заражение песцов ленточными червями, личинки которых развиваются в
грызунах, обнаруживает прямую зависимость от состояния запасов последних
в тундре. Резкие изменения численности леммингов (основных объектов
питания хищников) приводят к заметному снижению инвазированности песцов
в годы, когда популяция грызунов находится в депрессионном состоянии, и
наоборот, к увеличению - в годы их обилия.
Важным фактором, влияющим на процессы циркуляции гельминтов в
биоценозах Таймырского полуострова, следует признать климатический и, в
первую очередь, температурный режим и снежный покров. Снег большую часть
года ограничивает доступ инвазированных элементов к основным хозяевам, а
мороз приводит к гибели яиц многих паразитических червей. Именно поэто-
му отличительной чертой ряда гельминтов, обнаруженных на Таймыре,
оказывается известная исключительная устойчивость яиц и личинок к низким
температурам. Суровый климат северных территорий косвенно воздействует
и на половозрелые фазы гельминтов. В зимний период (вследствие коли-
чественных и качественных изменений питания хозяев) происходит дестро-
биляция некоторых цестод, яйцепродукция которых к концу зимнего периода
заметно снижается. Низкие температуры зимы, вызывая быстрое промерзание
трупов животных и гибель личинок паразитов (в том числе Trichinella spiralis),
уменьшают их численность и уровень зараженности хищников в природе
(Савельев, 1974).
По сравнению с песцом гельминтофауна обыкновенной лисицы на
европейском Севере заметно беднее. В тундровой зоне она в основном включает
альвеококки и трихинеллы, интенсивность инвазированности которыми
составляет соответственно 16.7 и 21.6% (Сороченко, 1971). У зверей со Средней
и Нижней Печоры были выявлены 7 видов гельминтов, преимущественно
нематод (Козлов, 1969). У лисиц таежной зоны европейского Северо-Востока
России (н=21) удалось обнаружить уже 14 видов паразитов, в том числе цестод
4, трематод 1 и нематод 9 видов (Юшков, 1995). Фауна паразитических червей
состояла из широко распространенных форм и была близка по составу к таковой
у песца и волка.
Заражение лисицы происходит главным образом при поедании мелких
грызунов, амфибий, олигохет, моллюсков. Реже встречаются виды гельминтов,
связанные своим развитием с насекомыми (S. arctica, С. skrjabini), копытными
(Г krabbei) и зайцеобразными (Г. pisifirmis). Видовой состав паразитических
червей обычно отражает особенности трофических и топических связей
рассматриваемого хищника. Чаще у него преобладают гельминты, разви-
вающиеся у позвоночных (57.1%), реже у беспозвоночных (21.4%) жи-
вотных. При этом инвазия происходит через трофические связи основного
392
Мышевидные грызуны — основная пища самого маленького хищника, ласки.
(Фото В. Сидоровича).
Small rodents — the major food of weasel, the smallest predator.
(Photo by V. Sidorovich),
В марте у горностая начинается весенняя линька: его мех тускнеет, а на голове
появляются бурые волосы. (Фото В. Сидоровича).
Spring molt of ermine begins in March: its fur dims, brown on the head.
(Photo by V. Sidorovich).
У этой лесной куницы гнездо находилось в старом штабеле не вывезенных из леса
дров. (Фото В. Сидоровича).
Nest of this marten is situated in an old woodpile in a forest. (Photo by V. Sidorovich).
Даже в летнее время лисица выглядит очень нарядной. (Фото Г. Смирнова).
Even is summer time the fox has bright colouration. (Photo by G. Smirnov).
Выдра — типичный представитель околоводной фауны. (Фото В. Сидоровича).
Otter — a typical representative of fauna inhabiting near water bodies.
(Photo by V. Sidorovich)
Мелкая речная рыба — основной корм выдры. (Фото В. Сидоровича).
Small river fish — major food of otter. (Photo by V. Sidorovich).
Енотовидная собака в настоящее время широко расселена по многим областям
России. (Фото В. Сидоровича).
Racoon dog is now widely spread in many regions of Russia. (Photo by V. Sidorovich).
Леса таежного типа — основные местообитания волков. (Фото автора).
Forests of taiga type — major habitats of wolves. (Photo by the author).
Волк на территории России распространен очень широко, (Фото В. Бологова).
Wolf is widely spread in Russia. (Photo by V. Bologov).
Зимой волки часто объединяются в стаи. (Фото В. Бологова).
In the winter wolves often form prides. (Photo by V. Bologov).
Рысь в поисках добычи. (Фото автора).
Lynx, hunting. (Photo by the author).
Рысь — редкий и осторожный хищник. (Фото автора).
Lynx is a rare and cautious predator. (Photo by the author).
Молодняк бурого медведя на овсяном поле. (Фото С. Пажетного).
Juveniles of brown bear in oat field. (Photo by S. Pazhetnov).
Осенью бурый медведь не прочь полакомиться ягодами рябины.
(Фото С. Пажетнова).
In the autumn brown bear sometimes enjoys ashberries. (Photo by S. Pazhetnov).
Подрастающий молодняк белого медведя постоянно следует за матерью, без
которой он становится беспомощным и часто не может обеспечить себя пищей.
(Фото М. Гаврило).
Growing juveniles of polar bear follows their mother, otherwise they become helpless
and often cannot procure food. (Photo by M. Gavrilo).
На этот раз охота белого медведя была удачной. (Фото М. Таврило).
The hunt of a polar bear for seal was a success. (Photo by M. Gavrilo).
хозяина. Другая часть паразитических червей (21.5%) попадает в организм
лисицы уже по линии топических связей. В Республике Коми и Ненецком АО в
просмотренной биопробе звери были заражены в основном Т leonina (66.6%),
A. alata (52.4%), а также С. vulpis и U. stenocephala (по 47.6%). Интенсивность
инвазии животных первыми двумя видами паразитов оказалась максимальной
и достигала соответственно 137 и 144 экз. (Юшков, 1995).
Структура тундровых биоценозов отличается сравнительной неслож-
ностью. Обедненность видового состава животного мира, простые и короткие
пищевые связи -характерные черты арктических сообществ. В соответствии
с этим гельминтофауна лисицы из тундровой зоны полуострова Таймыр
оказалась инвазированной только 4 видами паразитических червей, характерных
также и для песца. Три из них относились к классу цестод и один (Т leonina) - к
нематодам (Савельев, 1975).
С продвижением на юг гельминтоценоз обыкновенной лисицы, как и у
других хищников, заметно нарастает. В Татарстане (Горшков, 1997) у
добытых зверей обнаружены 12 видов гельминтов, среди которых
превалировали нематоды (6 видов) и цестоды (4 вида). Кроме того, в
этом регионе у них встречено также по одному представителю скребней
(Macro canthorhynchus catalinus) и сосальщиков (Euparyphium melts). Таким
образом, как справедливо отмечает А.Ф. Чиркова (Гептнер и др., 1967), в
целом у лисицы из природных популяций различных территорий России
может насчитываться не менее 20 видов гельминтов, которые, безусловно,
оказывают негативное влияние на состояние локальных группировок
данного хищника.
В Белорусском Полесье обыкновенная лисица является хозяином и
распространителем уже 24 видов гельминтов (В.В. Шималов, В.Т. Шималов,
19996). Наибольшая интенсивность инвазии характерна для Alaria alata и
Uncinaria stenocephala (по 40%), а также Taenia crassiceps (28.9%) и Тахосага
canis (24.4%). Интенсивность заражения зверей первыми двумя видами
достигала соответственно 600 и 50 экз., вторыми -100 и 40 экз.
Копроскопические исследования также указывают на высокую заражен-
ность паразитами не только лисицы, но и других диких видов собачьих. Яйца
и личинки гельминтов обнаружены здесь в экскрементах волков в 100%
просмотренных образцов, енотовидной собаки - в 91.1 и лисицы - в 90.2%.
Встречаемость яиц многих паразитических червей значительна. Например,
яйца A. alata найдены в 51.8% экскрементов представителей рассматриваемого
семейства (количество яиц колебалось в пределах 25-15 600), U. stenocephala
- в 12.5 % (25-1980), a Toxocara sp. - в 23.2% (25-3900).
Мелиоративные каналы Полесья представляют собой очаги, где проис-
ходит концентрация гельминтов. Установлена высокая степень загрязнения
их берегов пометом хищников с инвазионными элементами. В 90-х годах
XX в. на 100 м береговой полосы каналов здесь приходилось 11.5 г
экскрементов, содержащих до 3870 яиц и личинок паразитов. Авторы считают,
что при высокой численности особи диких видов собачьих, посещая в поисках
393
14 Зак. № 4309
пищи водоемы и (прежде всего) мелиоративные каналы, активно маркируют
территорию и приводят ее к биологическому (экскременты) и паразитарному (яйца
и личинки гельминтов) загрязнению. При наличии благоприятных климатических
условий в этих местах формируются и укореняются устойчивые очаги
гельминтозов, имеющие эпидемиологическое значение. В дальнейшем инвазия
может распространяться на прилегающие к каналам территории, приводя к
паразитарному загрязнению экосистемы.
Енотовидная собака во многих регионах бывает меньше других собачьих
заражена гельминтами. В Новгородской области в годы ее высокой чис-
ленности встречаемость в популяции инвазированных зверей оказалась
незначительной. У 29 добытых и обследованных животных отмечалось от 1 до 86
экз. паразитических червей, в основном из класса нематод (Морозов, 1953,1977).
Гельминты (общей численностью от 12 до 75 экз. на одну зараженную особь)
обнаружены и нами у зверей (и= 12), отловленных в 80-х годах в Псковской и
Новгородской областях, причем все инвазированные животные были хорошо
упитаны.
Количество видов паразитических червей, которыми заражена еното-
видная собака в Татарстане, в среднем на 33% меньше, чем у лисицы, и на
50% - по сравнению с барсуком. Ее гельминтофауна включает 4 вида нематод,
3 - цестод и 1 вид сосальщиков. Все они найдены также и у широко рас-
пространенных в данном регионе уже отмеченных норников - барсука и
обыкновенной лисицы (Горшков, 1980, 1997). Всего у енотовидной собаки
известны 32 вида гельминтов. Из них у зверей, обитающих на Дальнем Востоке,
обнаружены 18 видов, а остальные - у особей из районов акклиматизации
(Гептнер и др., 1967). Для северо-западного региона европейской части России
отмечена большая зараженность животных (до 96%) трематодами A laria alata
и несколько меньшая (33.7%) - нематодой Uncinaria stenocephala (Геллер,
1959). Для Татарстана также выявлено значительное количество зверей,
инвазированных A laria alata, но экстенсивность заражения его популяции здесь
не превышала 76.2% (Гептнер и др., 1967).
В соседних с территорией России районах Белоруссии у енотовидной собаки
встречается до 17 видов гельминтов, экстенсивность инвазии которыми
колеблется от 1.3-2.6 (Р. truncatum, М. albidus, S. erinacei, М. lineatus, А.
caninum) до 20.5-52.6% (J. melis, A. alata, U. stenocephala, T. canis).
Интенсивность заражения зверей A. alata и U. stenocephala оказалась
максимальной и соответственно колебалась в пределах 4-800 и 1-40 экз. В
целом популяция рассматриваемого хищника в Полесье заражена гельмин-
тами на 83.3% (В.В. Шималов, В.Т. Шималов, 1999в). Необходимо также
отметить наличие здесь очагов описторхидозной инвазии (Opisthorchidae,
Trematoda), дефинитивными хозяевами которых могут быть некоторые дикие
животные, в том числе и енотовидная собака. Роль промежуточных хозяев в
циркуляции основных видов описторхид в природной среде выполняют
моллюски семейство Bithyniidae и карповые рыбы - елец, язь, плотва, лещ и
др. Медицинское значение имеют О. felineus, Р. truncatum и М. albidus
394
(Metorchis bills'), которые могут вызывать сложнейшее заболевание человека
описторхоз с локализацией паразитов в желчном пузыре, протоках печени и
поджелудочной железе. В местных локальных популяциях описторхидами были
заражены енотовидные собаки на 7.7±3%, лисицы - на 9.1 ±3.1 и волки - на
3.9±2.7%. Интенсивность инвазии колебалась соответственно в пределах 1-4,
1-6 и 3-7 экз. Увеличение в республике площадей сельскохозяйственных угодий
с сетью мелиоративных каналов, берега которых активно посещают
хищники, повысило вероятность контакта с ними человека, в результате чего возрос
и риск его заражения паразитами диких животных (В.В. Шималов, В.Т. Шималов,
1998).
Гельминтофауна волка представлена широко распространенными видами,
среди которых отсутствуют характерные только для него узкоспецифичные
формы. В Большеземельской тундре его популяция оказалась на 33.3%
инвазирована эхинококками и на 65.4% - трихинеллами (Сороченко, 1971).
На обширной территории северо-востока европейской части России (Юшков,
1995), в бассейнах рек Нижняя Печора, Вычегда, Мезень и Сысола из 46
добытых и обследованных животных 86.9% особей оказались зараженными 16
видами паразитов: цестоды - 8, трематоды - 2 и нематоды - 6 видов. К числу
характерных основных гельминтов волка относятся цестоды Е. granulosus
(уровень заражения популяции 32.6±6.8%), Т. hidatigena (50±7.5), нематода
Т. nativa (45.8±7.3) и трематода A. alata (26.1±6.8%). Эти виды составляют
основной фон гельминтофауны этого хищника в природных экосистемах.
Интенсивность инвазии отмеченными цестодами здесь составляла соответ-
ственно 1.5-12 тыс. и 1-6 экз., а двумя другими паразитами - 2-34 и 4-87 экз. на
одну особь.
Анализ биоценотических связей гельминтов, проведенный В.Ф. Юшковым
(1977,1995), показывает, что большинство из них волк приобретает вместе с
добытой пищей и только 12.5% форм могут попасть в организм хищника
топическим путем. Таким образом, состав кормов представляет собой один из
основных экологических факторов в становлении фауны гельминтов этого
зверя. Зараженность самок и самцов в целом в регионе оказалась практически
одинаковой - соответственно 81.6 и 91.7%. По составу паразитов, за
исключением некоторых редко встречающихся форм, они также не разли-
чались. Несколько более высокая инвазированность самцов по сравнению с
самками трематодой Л. alata и нематодой Т. nativa объясняется, скорее всего,
значительным охватом первыми территории при отыскании добычи и большей
потребностью в пище.
По данным автора, прибылые и взрослые особи по показателям общей
зараженности гельминтами не различались (соответственно 90.9 % и 92.3 %).
Их видовой состав (табл. 158) у молодняка был менее разнообразен (9 видов),
чем у взрослых животных (12 видов). При этом только у прибылых обнаружены
гельминты Т. aerophilus (Eucoleus aerophilus) и С. vulpis, промежуточными
хозяевами которых служат беспозвоночные. По частоте встречаемости и
обилию паразитов, связанных жизненными циклами с копытными, молодые и
395
Таблица 158/ Table 158
Зараженность гельминтами (%) волков разного пола и возраста
на северо-востоке европейской части России (по: Юшков, 1995)
Infection by helminthes (%) of wolves (Canis lupus) of different sex and age
in the North-Eastern part of Russia
Виды гельминтов/ Helminth species Самцы/ Males (n=24) Самки/ Females (n=22) Прибылые/ Youngs (n=ll) Переярки и взрослые/ Yearlings and adults («=13)
Т. hydatidena 50+10.2 45.5±10.6 54.5+1.5 53.8±13.8
Т. parenchimatosa 4.2+4.2 9.1±6.3 9.1±8.6 7.7±7.5
Т. pisiformis 4.2+4.2 9.1±6.3 - 7.7±7.5
Т. eras sleeps 4.2+4.2 9.1±6.3 - 7.7±7.5
Т. krabbei 12.5±6.7 9.1±6.3 - 15.4±9.9
A. multilocularis 4.2±4.2 9.1±6.3 18.2±11.5 -
Е. granulosis 33.3+9.4 31.8+9.9 36.4± 14.4 38.5+13.5
M.lineatus - 4.5±4.5 - 7.7+7.5
A. alata 33.3+9.4 18.2±8.4 18.2±11.5 7.7+7.5
P. elegans 4.2±4.2 - - -
T. nativa 50±10.2 40.9±10.3 45.4+1.5 69.2+12.8
T. aerophilus 4.2±4.2 9.1±6.3 9.1±8.6 -
C. vulpis 4.2±4.2 13.6±7.5 9.1 ±8.6 -
U. stenocephala 12.5+6.7 31.8+9.9 18.2±11.5 15.4±9.9
T. leonina 25±8.7 13.6±7.4 - 7.7+7.5
S. lupi 4.2±4.2 - - 7.7+7.5
взрослые животные довольно схожи. Очевидно, половые и возрастные отличия
в инвазированности особей определяются прежде всего особенностями набора
кормовых объектов - их обилием, составом и доступностью в разные годы. У
зараженных зверей также неоднократно отмечались очаговые и расплывчатые
воспалительные участки слизистой оболочки кишечника, чаще всего при
паразитировании цестод, особенно эхинококков. Несмотря на это все
исследованные особи, в том числе и инвазированные разными видами
гельминтов, были средней или высокой упитанности.
Из добытых в Кировской области 216 волков 61.1% животных были
заражены трихинеллами (Козловский, 1996). Интенсивность инвазии составля-
ла в среднем 44 личинки. Высокая их морозоустойчивость обеспечивает
длительное сохранение паразитов в трупах павших зверей, которые часто
396
потребляют хищники. В указанной биопробе самки были заражены интенсивнее
самцов, причем достоверность различий составляла 0.95%.
На полуострове Таймыр у полярного волка обнаружены только 7 видов
гельминтов, которые встречены у 95.1% просмотренных хищников (Савельев,
1975). Средняя интенсивность инвазии составляла 53.7 экз. Более половины
обследованных зверей (и=41) были заражены цестодами (70.7%). В остальных
случаях наблюдалась смешанная цестодно-нематодная инвазия. В целом
гельминтофауну таймырского волка можно охарактеризовать как цестодную
с постоянными компонентами Taenia krabbei и Т. parenchymatosa, заражение
которыми происходит при потреблении дикого северного оленя. Уровень
инвазированности популяции хищника в течение многих лет не обнаруживал
заметных изменений, что связано с отсутствием резких перепадов в численности
копытных, а следовательно, и зараженности волков.
Как и в случаях с другими видами хищных млекопитающих, с продви-
жением на юг гельминтофауна популяции волков возрастает. Так, в
Белорусском Полесье зараженные особи составляли 80.8%; они были
инвазированы 18 видами гельминтов, причем экстенсивность заражения
Trichinella spiralis достигала 19.2%, Toxocara canis-2\.2 м Taenia hydatidena-
26.9%. В целом в выборке из 52 зверей инвазированные особи составляли 80.8%.
Считается, что в Полесье волк и другие крупные хищные млекопитающие
являются хозяевами и распространителями 25 видов гельминтов, способных
заражать домашних животных и представляющих опасность для здоровья человека
(В.В. Шималов, В.Т. Шималов, 1999в).
Гельминтоценоз представителей семейства кошачьих, в частности рыси,
изучен сравнительно слабо. По материалам В.Ф. Юшкова (1995), этот хищник
представляет собой один из резерватов возбудителя трихинеллеза. У зверя,
добытого в Республике Коми, были выявлены 4 вида нематод: Thominx
aerophilus (Eucoleus aerophilus), Ancylostoma caninum, Trichinella nativa и
Toxocara mustax. Из них только последняя специфична для данного
представителя семейства кошачьих, а остальные свойственны и другим
хищным млекопитающим.
Полное гельминтологическое вскрытие тушек 30 рысей (1631 и 14£) из
Белоруссии показало 100%-ную зараженность зверей паразитическими чер-
вями (В.Т. Шималов, В.В. Шималов, 1999а). У них были обнаружены 19 видов
гельминтов, в том числе 8 цестод и 11 нематод. У 93.3% исследованных
животных встречалось от 2 до 8 видов паразитов со значительной степенью
интенсивности инвазии (в среднем 475 экз. на зараженную особь). Цестоды
выявлены у 70% хищников, нематоды-у 80%. Среди паразитов преобладали
нематоды - токсокары (Г. canis и Т. cat Г), количество которых у рысей
колебалось в пределах 1-36 экз. Установлены случаи гибели молодых слабо
упитанных особей (возраст до 1 мес) от токсокароза и дирофиляриоза, что не
исключает интраутеринный путь передачи инвазии.
По результатам копрологического анализа обыкновенных рысей (200
проб от 54 особей) из Чешского зоопарка в г. Острава, встречаемость
397
паразитических червей составляла 61% от просмотренных проб. Среди них
были 6 видов нематод и 11 - цестод. Экстенсивность поражения разными
видами была следующая: Toxocara mystax-33.5%, Toxoscaris leonina-26.5,
Toxocara canis - 5 и Trichocephalus vulpis - 4%. Встречаемость в пробах
таких паразитов, как Capillaria aerophila, Thominx aerophilus и Taenia
pisiformis, не превышала 1-2%. Инвазированность молодых рысей первой
из перечисленных нематод оказывала большее негативное воздействие на их
рост и формирование организма. У 22 взрослых зверей, отловленных в
словацкой части Карпат, экстенсивность заражения гельминтами достигала
86%. В основном рыси были инвазированы Toxocara mystax (63.6%),
Trichocephalus vulpis (22.7%) и Toxoscaris leonina (9.1%). Встречаемость
Capillaria aerophila, Toxocara canis и Taenia pisiformis достигала 4.5% от
общего объема выборки (Stehlik, 1985).
Зараженность лесного и камышового котов невысокая. У первого
обнаружено 2 вида гельминтов, у второго - 8 (4 вида нематод и 4 цестод).
Существенного их влияния на численность популяций этих хищников не
отмечено (Насибов, 1970).
Гельминтофауна енота-полоскуна (семейство енотовых) более разно-
образна. На Кавказе у него выявлено 6 видов паразитических червей: 2 вида
трематод, 3 - нематод и 1 - акантоцефал (Садыхов, 1962). В работе А.М. Ги-
неева и В. Л. Гаркави (1974) приводятся данные об инвазированности енота-
полоскуна в Дагестане личинками трихинеллеза. В промысловой выборке из
61 зверька 5 особей (8.2%) оказались зараженными данным паразитом, при
этом интенсивность инвазии составляла от 4 до 6 личинок на 1 г мышечной
ткани. Среди больных животных находились 4 самца в возрасте от 1 до 6 лет и
самка-сеголетка. По весу и упитанности все они не отличались от здоровых
особей.
У крупного хищника высоких широт - белого медведя - многие исследо-
ватели также отмечают наличие в тканях личинок трихинелл. Инвазированные
ими хищники на о-ве Врангеля встречаются достаточно часто, а на Земле
Франца-Иосифа в некоторые годы их зараженность достигает 100%. Известны
случаи, когда мясо таких зверей служило причиной групповой вспышки
трихинеллеза среди местного населения (Озерецковская и др., 1957, 1969;
Сороченко, 1971; Переверзева, Веретенников, 1973, и др.).
Среди гельминтов, паразитирующих у бурого медведя на Северо-Западе
России, наибольшее распространение имеют личинки трихинелл. Зверей,
сильно зараженных ими, добывали в 60-70-х годах XX в. в Калевальском,
Кандопожском и Беломорском районах Карелии. Они регулярно приходили
кормиться на свалки отходов со звероферм, где, вероятно, и происходило
заражение животных (Данилов и др., 1979).
Имеются сообщения об обнаружении личинок трихинелл у медведей,
добытых в лесной и тундровой зонах европейского Северо-Востока России.
Их мясо послужило источником заражения других хищных млекопитающих и
человека (Сороченко, Колесов, 1962; Сороченко, 1971). В подзоне средней
398
тайги Республики Коми, кроме трихинелл, у бурых медведей найдены
нематоды Dirofilaria ursi в количестве 20-30 экз. на одну особь (Юдин, 1995),
а восточнее - в зоне тундрового редколесья Таймыра - лишь один вид
паразитических червей, Toxoscaris transfuga (Савельев, 1975).
В более южных частях ареала вида гельминтоценоз хищника заметно
богаче. Обследование 63 образцов экскрементов медведей, собранных в
Башкирском заповеднике, показало наличие в них яиц Ascaridae - 25%
встреч, Strongyloides - 24, Trichospongilata - 5 и Dicrocoelim - 6, а личинок
Strongilata - 15% встреч. В большинстве случаев зараженность зверей была
слабой, а в 40% просмотренных экскрементов наличие гельминтов не
выявлено. Анализ образцов тканей добытых животных на трихинеллез дал
отрицательный результат, хотя у других хищных млекопитающих Башкирии это
опасное для человека заболевание зарегистрировано в 12 районах (Лоскутов и
др., 1993).
В Сибири и Казахстане гельминтофауна и воздействие паразитов на
локальные популяции бурого медведя изучены недостаточно, однако многие
исследователи не считают существенным их влияние на состояние запасов
вида в регионах, особенно в благоприятные годы. Подтверждением этому
служит его устойчивая численность в заповедниках и трудно посещаемых
человеком местах, где действуют только естественные факторы смертности
(Грачев, 1981; Жиряков, Грачев, 1993, и др.). Экстенсивность заражения
медведей на этих территориях невелика. На Южном Алтае личинки трихинелл
обнаружены у 1 из 12 обследованных зверей, на Западном Тянь-Шане - у 2 из
5, а в Казахстане (и=21) инвазированность популяции вида данным паразитом
составляла 19% (Боев и др., 1969). В Центральном Тянь-Шане ткани внутренних
органов добытого хищника оказались поражены только нематодой Toxoscaris
transfuga и цестодой Mesocectoides lineatus (Янушевич и др., 1972).
Обследование 21 медведя из Туруханского района средней Сибири
(Завадский, 1993) показало, что 7 из них (33.3%) были инвазированы
гельминтами. У 5 особей удалось обнаружить нематоды Dirofilaria ursi в
количестве 6-9 экз. на 100 см2, а у двух выявлены аскариды и легочные черви,
по 2-3 экз. каждого вида. Проверка мяса зверей на зараженность личинками
трихинелл не дала положительных результатов, хотя на юге региона их наличие
отмечается регулярно.
По наблюдениям С.К. Устинова (1987, 1993), в Прибайкалье в голодные
годы у шатунов (а в обычные - только у старых или ослабленных животных)
количество гельминтов в организме заметно увеличивается. Мышечная ткань
и внутренние органы таких зверей бывают сплошь поражены разными видами
нематод, цестод и личинками трихинелл. Паразитические черви у добытых в
Прибайкалье медведей обнаруживаются практически постоянно, но в массе
они встречаются только при общей ослабленное™ организма хозяина.
Собранные материалы показывают, что обеспеченность пищей и ее состав
служат основным экологическим фактором, формирующим гельминтофауну
и уровень инвазированности хищных млекопитающих. В природных
399
биоценозах на процессы циркуляции паразитических червей существенное влияние
оказывают климатические факторы (низкие температуры, высота снежного
покрова, осадки и т.п.), поэтому их вцдовой состав и степень зараженности животных
снижаются по мере продвижения вида к северным и северо-восточным границам
ареала. В целом у хищников проявляется негативное воздействие гельминтов
на размерно-весовые показатели и упитанность особей.
Резюме
В благоприятные в кормовом отношении годы, при обильном потреблении
прежде всего мышевидных грызунов, которые являются промежуточными
хозяевами паразитов, плотность населения животных и содержание у них
жировых резервов увеличиваются. При этом возрастает и их зараженность
гельминтами, которые в дальнейшем негативно воздействуют на популяцию
хозяина. При ухудшении кормовой базы, а следовательно, и меньшем
потреблении пищи инвазированность хищников заметно падает. Возможно,
поэтому даже в такие малокормные годы зараженные особи разных видов
нередко оказываются хорошо упитанными.
В период максимального жиросодержания (осень-зима) в организме мелких
хищников неинвазированные самки всех возрастов имеют большую массу
пахового жира по сравнению с инвазированными. Среди самцов данные
различия отчетливо не прослеживаются. Следует также отметить, что у
изучаемых видов нематодные инвазии не вызывают заметного уменьшения
веса жизненно важных внутренних органов. По-видимому, наличие жировых
резервов способствует повышению иммунитета и общей сопротивляемости
организма паразитарному воздействию. Вероятно, в популяциях мелких и
средних по размеру хищных млекопитающих роль гельминтов сводится к
снижению иммунной деятельности организма, что обусловливает развитие
болезней, уменьшение плодовитости самок и выживаемости молодняка.
Влияние гельминтов на локальные популяции крупных хищников при обилии
в природе кормов несущественно и только при их дефиците старые,
ослабленные и больные особи заражаются гельминтами в сильной степени.
Summary
Food supply and its composition are the main ecological factors forming
helminthofauna and the level ofinvasibility of carnivores. In natural biocenoses the
climatic factors (low temperatures, the level of snow cover, precipitation etc.) have
an essential influence on the processes of circulation of parasitic worms, therefore,
their species composition and degree of infection of animals are decreasing as the
species advances the north and north-eastern boundaries of the range. On the whole
400
the helminthes exert a negative impact on the size-weight parameters and fertility
of carnivorous animals. In the years favorable in relation to food’with abundant
consumption, in the first turn, of rodents, which are the intermediate hosts of
parasites, the density of animal population and their fat resources are increasing.
Therewith the infection by helminthes, which further on exert a negative impact on
the population, also increases. With deterioration of food supply, and hence, lower
food consumption, the invasion of carnivores decreases notably. Probably, as a
result, infected individuals of various species often appear to be fat.
Non-infected females of all ages had greater mass of groin’s fat as compared to
non-infected in the period of maximum content of fat (autumn-winter) in the
organism of small carnivores. It should be also noted that nematode invasions in
the species studied did not cause pronounced decrease in weight of vitally important
inner organs.
Apparently the occurrence of fat resources promotes the increase of immunity
and general resistance of organism to parasite’s impact. Probably, the role of
helminthes in the populations of carnivores of small and average size reduces to the
decrease of immune activity of organism, which stipulates the development of
diseases, decrease of fertility of females and survivorship of the young. The influence
of helminthes on the local populations of large carnivores with, the abundance of
food is insignificant, and only with their shortage old, weak and sick individuals are
infected by helminthes to a high degree.
Заключение
В ходе эволюционного развития реализация структурно-функциональных
адаптаций важнейших систем органов у представителей отряда хищных
млекопитающих осуществлялась по-разному. В основе этого процесса лежат
приспособительные морфофизиологические преобразования, обусловленные
образом жизни и уровнем энергообмена животных. Особенности их экологии
в значительной степени определяют характер физиологической активности и
прогрессивной перестройки организма. В связи с различными показателями
кормовой специализации и суточного ритма жизнедеятельности хищников
важное значение приобретают разнообразные адаптивные изменения,
затрагивающие основные системы органов.
Специфика питания, наличие и доступность кормов влияют на
физиологическое состояние особей, воспроизводственный потенциал,
демографическую структуру и в конечном итоге на численность популяций.
У видов, стабильно освоивших прибрежные биотопы, основу пищевого
рациона составляют амфибии, мышевидные грызуны и рыба. Регулярное
питание этими группами кормов следует рассматривать как важную
трофическую адаптацию, которая позволяет хищникам широко расселяться
по угодьям. В холодное время года, особенно при недостатке пищи, сходство
рациона может обусловливать конкурентные отношения между ними, однако
большое разнообразие и количество добычи, которая встречается у водоемов,
смягчают трофическую конкуренцию, и она практически не оказывает
влияния на численность популяций.
Энергетическая емкость суточного рациона зверей существенно возрастает
осенью, а затем с разной степенью интенсивности снижается в зимнее время.
На большей части территории России сезонные изменения кормового спектра
и энергетики питания отчетливо проявляются у обитателей прибрежных
биотопов (норки - Mustela lutreola, М. vison, лесной хорек - М. putorius, коло-
нок - М. sibirica, выдра - Lutra lutra), представляя собой стойкую адаптивную
реакцию, направленную на экономное расходование создаваемых в организме
энергетических запасов. У них при зимнем падении уровня обмена веществ
потребность в пище резко снижается, что и определяет благополучный исход
зимовки. Для других представителей отряда хищных млекопитающих, прежде
всего для горностая (Mustela erminea), солонгоя (М. altaica), куниц (Martes
martes, М. foina), лисы (Vulpes vulpes) и песца (Alopex lagopus), характерна
высокая интенсивность двигательных реакций и энерго обмена, в связи с чем
необходимость в калорийном рационе отмечается практически во все сезоны.
Для этих видов снежный период оказывается очень напряженным, что
усугубляется слабым развитием жировой ткани, а также уменьшением в зимние
месяцы калорийности добычи. Наконец, крайний случай трофической
адаптации представляют виды, впадающие в зимний сон (барсук - Meles meles,
енотовидная собака - Nyctereutesprocyonoides, медведи - Ursus arctos, U. mari-
timus), которые в связи с интенсивным сезонным изменением уровня обмена
402
веществ и образованием мощного слоя жировых резервов в зимнее время
прекращают активность, не испытывая потребности в пище. Таким образом,
в основе приспособительных реакций, направленных на успешную пере-
зимовку, лежат процессы минимизации энергообмена, снижения энергозатрат,
а у некоторых видов и общего торможения жизнедеятельности.
Пищевая специализация оказывает заметное влияние на адаптивные
возможности рассматриваемых животных, которые реализуются в виде
различных по интенсивности процессов отыскания корма и ритмов
жизнедеятельности. В холодное время года активные формы затрачивают
много энергии на поиск добычи, что находится в соответствии с их массой
тела, уровнем теплообмена, количеством и качеством потребляемой пищи. В
целом среди систематически близких видов с увеличением размеров хищников
общая продолжительность двигательных реакций сокращается, а фазы отдыха
возрастают. При наличии кормов для них характерны два основных
сумеречных периода охотничьей деятельности (дифазный тип) и только у ласки
{Mustela nivalis) отмечается частая смена состояний бодрствования и покоя
(полифазный тип). Повышенная продолжительность суточной активности у
мелких видов при поиске корма представляет собой приспособительную
реакцию организма зверьков к поддержанию высокого уровня энергообмена.
Многообразие пищевых связей, заметная биологическая активность и
высокий уровень адаптированности к меняющимся условиям среды
характерны также для хищных млекопитающих среднего и крупного размеров.
Их значение как важных элементов биоценозов не ограничивается только
регулированием численности и воздействием на демографическую структуру
популяций добываемых животных. При высокой плотности населения в угодьях
(прежде всего представителей семейства собачьих) наблюдается интенсивное
использование запасов копытных, зайца-беляка, тетеревиных и водопла-
вающих птиц, что несомненно создает сложную проблему, решение которой
возможно только при ограничении численности хищников. Необходимо
учитывать и их негативную роль как распространителей возбудителей
трансмиссивных заболеваний. Однако, с другой стороны, сокращая
численность многих носителей инфекционных и инвазионных агентов (прежде
всего грызунов), они способствуют оздоровлению ландшафтов и препятствуют
экспансии природных очагов болезней. Важное значение имеет деятельность
крупных хищников, способствующая обеспечению пищей видов-коммен-
салов, в основном птиц и мелких хищных зверьков, нередко питающихся
остатками чужой добычи. Следует также отметить, что большинство хищных
млекопитающих относится не только к промысловым, но и к спортивно-
охотничьим животным, ценность которых как охотничьих трофеев постоянно
растет (Данилов и др., 1979).
Различия в образе жизни, величине и ритме суточной активности
обусловливают многообразие адаптивных изменений, затрагивающих основные
системы организма представителей изучаемых видов. Прослеживается тесная
связь относительных размеров головного мозга и мозжечка как рефлекторно-
403
автоматического центра локомоторных функций с показателями пищевого
поведения - интенсивностью охот хищников. У амфибионтов специализация к
водному образу жизни сопровождается увеличением индекса легких, сердца и
почек, а у обитателя высоких широт - росомахи (Gulo gulo) - кроветворного
органа - селезенки. Однако не только размеры основных внутренних органов, но и
структурные преобразования мышечной, артериальной и венозной систем
характеризуются у хищных млекопитающих видоспецифичносгью и взаимосвязаны
с образом жизни и двигательной активностью животных.
Необходимо также отметить, что большая относительная величина
поверхности теплоотдачи у особей мелких видов сопряжена с ростом уровня
обменных процессов, респираторной и сердечной деятельности. Обладая
повышенным энергообменом, они постоянно испытывают сильное воздействие
неблагоприятных факторов среды. В процессе филогенеза это обстоятельство
и определило их большую плодовитость по сравнению с крупными предста-
вителями отряда, а также способность самок некоторых хищников приносить
повторный приплод в случае ранней гибели молодняка. К другим репро-
дуктивным особенностям, регулирующим численность популяций, следует
отнести у мелких видов - высокий воспроизводственный потенциал и темп
роста щенков, а у крупных - низкий уровень смертности молодняка и большую
продолжительность жизни взрослых особей.
Возрастные изменения морфологических признаков у всех представителей
отряда Carnivora прослеживаются вполне отчетливо и в целом характеризуются
закономерным снижением индекса основных внутренних органов, однако
особенности образа жизни вносят заметные изменения в направленность этого
процесса. Например, у полуводных животных падение относительного веса
легких и сердца отмечается только на ранних этапах постэмбриогенеза. При
переходе их к самостоятельной жизни, с повышением функциональной
нагрузки на организм, эти показатели возрастают.
Отмеченная за период постнатального онтогенеза особей динамика
уровня терморегуляции, сердечного и дыхательного ритмов наглядно
демонстрирует взаимосвязь функциональных преобразований с морфо-
логическими особенностями роста и развития животных. Выявленные
закономерности в скорости весового роста, изменениях терморегуляции,
легочной вентиляции и работы сердечной мышцы прослеживаются очень
отчетливо и имеют направленность на более экономное обеспечение
гомеостаза организма особей изучаемых видов.
У всех представителей отряда хищных млекопитающих возрастная
динамика соотношения полов четко выражена. В основном для них характерно
примерно равное количество самок и самцов в выводках и постоянное
увеличение доли последних в группах старших возрастов. Выявленная разница
в третичном соотношении полов резко увеличивается в направлении от
крупных хищников к мелким. Высокий уровень плодовитости и энергозатрат
на выращивание потомства у многоплодных видов, характеризующихся
малыми размерно-весовыми показателями, служит причиной большей по
404
сравнению с самцами естественной элиминации, определяющей диспропорцию
по полу. Наблюдаемое увеличение в их выводках доли новорожденных самок
наряду с высоким потенциалом размножения половозрелых особей следует
рассматривать как один из компенсаторных механизмов адаптации мелких
видов к факторам неблагоприятного воздействия внешней среды. В то же время
избыток самцов в группах старших возрастов у всех исследуемых хищников
является биологически целесообразным, поскольку они обеспечивают
максимальное включение в процесс размножения половозрелых самок в годы
с низкой численностью животных.
Обеспеченность пищей и ее состав служат основным экологическим
фактором, формирующим гельминтофауну и уровень инвазированности
хищных млекопитающих. Негативное воздействие паразитов на их размерно-
весовые показатели и плодовитость вполне очевидно. При обильном
потреблении прежде всего мышевидных грызунов и земноводных, которые
являются промежуточными хозяевами гельминтов, жировые резервы в
организме хищников увеличиваются, однако при этом возрастает и их
инвазированность паразитическими червями, которые в дальнейшем снижают
репродуктивный потенциал популяций.
Таким образом, использованный комплексный подход к выяснению
механизмов адаптации позволил установить многообразие путей приспособ-
ления хищных млекопитающих к среде обитания. Совокупность экологических,
поведенческих и структурно-функциональных преобразований у видов,
различающихся по кормовой специализации, размерам тела и уровню
энергообмена, определяет их общую адаптивную стратегию, направленную
на успешное выживание в изменяющихся условиях существования.
Полученные данные о кормовой специализации и энергетике питания,
репродуктивном потенциале и особенностях онтогенеза хищных млеко-
питающих могут служить основой при разработке путей управления их
популяциями в хозяйственных целях, а также мер по сохранению ценных и
редких видов. Выявленные в процессе работы признаки экологической
адаптации необходимо учитывать при организации природных резерватов для
охраны исчезающих элементов биоты и мероприятий по рациональному
использованию ресурсов промысловых животных.
Conclusion
Structural-functional adaptations of crucial systems of organs in representatives
of the order of carnivores were realized in the course of evolutionary development
in different ways. Underlying this process were morpho-physiological
transformations stipulated by the life history and metabolic rate of animals.
Characteristic features of their ecology to a large extent define the character of
physiological activity and progressive reconstruction of the organism. Various
adaptive changes involving the main systems of organs acquire important meaning
405
related to different parameters of food specialization and circadian activity of
carnivores.
The specificity of feeding, occurrence and availability of food influence the
physiological state of individuals, their reproductive potential, demographic
structure and, finally, the population number. In species that inhabit coastal
biotopes, amphibians, myomorph rodents and fish form the basis of the diet. The
regular feeding on this group of food should be regarded as an important trophic
adaptation, which allows carnivores to disperse widely along home ranges. In the
cold period of the year, especially with a shortage of food, the similarity of the diet
may stipulate the competitive relations between them. However, high diversity and
number of prey occurring near the water bodies facilitates the trophic competition
and practically has no influence on the population number.
The energetic capacity of daily ratio of animals considerably increases in
autumn and then with a different degree of intensity declines in winter. The seasonal
changes of the food spectrum and energy are clearly pronounced in inhabitants of
coastal biotopes {Mustela lutreola, M. vison, M. putorius, M. sibirica, Lutra lutra)
in the most part of the territory of Russia. These changes are a stable adaptive
reaction directed to the economic expenditure of energetic resources developed in
the organism. With a winter decrease of metabolism the level of food requirements
dramatically decreases, which determines the favorable completion of wintering.
High intensity of moving reactions and energy exchange are typical of other
representatives of the order of carnivores {Mustela erminea, M. altaica, Martes
martes, M.foina, Vulpes vulpes, Alopex lagopus) and other active forms. In this
relation necessity in caloric ration is recorded in them practically during all seasons.
The snowy period appears to be very tense for these species, which deepens by the
poor development of fat tissue in males and, particularly, in females and decreases
calorific value of prey in the winter months. Finally, the extreme case of the trophic
adaptation represent the hibernating species (badger - Meles meles, racoon dog -
Nyctereutesprocyonoides, bears - Ursus arctos, ll. maritimus), which in relation to
the intensive seasonal change of metabolic level and development of powerful layer
of fat resources cease activity in the winter time and do not experience any necessity
in food. Thus, the processes of minimization of energy exchange, decrease of energy
expenditures and in some species general inhibition of vital activity are the basis of
adaptive reactions directed to the successful wintering.
The food specialization has a notable influence on the adaptive possibilities of
the species under study. These possibilities are realized in a form of differing in
intensity processes of food search and rhythms of vital functioning. In the cold
period of time the active forms spend a lot of energy on the search of prey, which
conforms to their body mass, level of heat exchange, quantity and quality of food
consumed. On the whole among the systematically close species with the increase of
the body size in predators the general duration of moving reactions decrease and
phases of rest increase. Two main nocturnal periods of hunting activity (diphase
type) are typical of them with the availability of food and only in the weasel {Mustela
nivalis) the frequent change of the states of waking and rest (polyphase type) is
recorded. The increasing duration of circadian activity in the small species during
406
the food search is an adaptive reaction of the organism to the maintenance of high
energy exchange level.
The diversity of food connections, the notable biological activity and high
level of adaptation to the changing environmental conditions are also typical of
carnivorous mammals of middle and large sizes. Their significance as important
elements of biocenosis is not limited only to the number control and impact on the
demographic structure of prey populations. When the population number in habitats
is high, first of all if this concerns representatives of the Canidae family, an intensive
use of reserves of ungulates, hares {Lepus timidus), game (Tetraonidae) and
waterfowl birds is observed. This, in its turn, definitely, creates a problem, the
solution of which is possible only with the limitation of the predators’ number. The
negative role of carnivores as distributors of causative agents of transmissive
diseases should be taken in account. However, on the other hand, decreasing the
number of many carriers of infection and invasion causative agents (first of all
rodents) they promote recovering of landscapes and prevent expansion of natural
foci of diseases. The activity of large carnivores promoting food provision for
commensal species, first of all birds and small predators often feeding on remnants
of prey, plays an important role. It should be also noted that most of carnivores are
referred not only to game, but to the sport-hunting animals too, which value as a
hunting trophy is constantly increasing.
The differences in the mode of life, character and rhythm of circadian activity
stipulate the diversity of adaptive changes concerning main systems of organs of
species under study. The close connection between the relative sizes of the brain and
cerebellum, as reflective-automatic center of locomotion functions and parameters
of food behavior-intensity of hunting of predators is recorded. In amphibionts the
specialization to the aquatic mode of life is followed by the increase of index of
lung, heart and kidney, whereas in the inhabitant of high latitudes, the wolverine
(Gulo gulo) - of the blood generating organ, spleen. Both the sizes of the main inner
organs and structural transformations of muscle, arterial and venous systems in
carnivores are distinguished by species specificity and are interrelated with the
mode of life and moving activity of animals.
It should be also emphasized that the greater the relative size of heat loss surface
in individuals of small species is coupled with the increase of the level of metabolic
processes of respiratory and cordial activity. Having high energy exchange they
constantly experience a strong impact of unfavorable environmental factors. In the
process of evolution this circumstance determined their greater fertility as compared
to large representatives of the order and ability of females of some carnivores to
deliver the second breed in the case of early death of young. Other characteristic
reproductive features that control population number are: in small species - high
reproductive potential and growth rate of cubs, in large - the low level of young
mortality and greater lifespan of adult individuals.
The age changes of morphological characters in all representatives of the order
Carnivora are quite distinct and on the whole are characterized by determinate
decrease of the main inner organs index. However, characteristic features of the life
history introduce notable changes into the direction of this process. For example, in
semi-aquatic animals the decrease of the lung and heart weight is recorded only at
407
the early stages of postembryogenesis. These parameters increase with the transition
to the independent life, with the increase of functional load on the organism.
The dynamics of thermoregulation level, heart and respiratory rhythms
recorded in the postnatal ontogenesis clearly demonstrate the interrelation of
functional transformations with morphological characteristics of animal growth
and development. The regularities revealed in the rate of weight growth, in changes
of thermoregulation, lung ventilation and heart muscle activity are followed very
distinctly and are directed to more economic provision of organism homeostasis in
individuals of the species studied.
In all representatives of the order carnivores the age dynamics of sex ratio was
very distinct. Approximately equal number of females and males in litters and
gradually increase of the portion of males in the groups of older age are, generally,
typical of them. The difference revealed in the tertiary sex ratio is dramatically
increasing from large to small carnivores. The high level of fertility and energy
expenditures on the nursing of offspring in repeatedly reproducing species
characterized by the small size-weight parameters cause greater, as compared to
males natural elimination, which determines disproportion in sex ratio. The
increased proportion of newborn females in litters of such species alongside with a
high reproductive potential of mature individuals should be regarded as one of
compensatory mechanisms of adaptations of small species to unfavorable impact
of environmental factors. At the same time an excess of males in the groups of older
ages in all studied carnivores is biologically justified as far as they provide maximum
inclusion of sexually mature females in years with a low population number into the
process of reproduction.
The availability of food and its composition are the main ecological factors
that form helminth fauna and the level of invasiveness in carnivorous mammals.
The negative impact of parasites on their size-weight parameters and fertility is
quite evident. With a plentiful consumption, first of all myomorph rodents and
amphibians, which are the intermediate hosts of helminths, the fat resources in the
organism of predators are increasing. However, therewith their invasiveness with
parasitic worms, which later on decrease the population reproductive potential, is
increasing.
Thus, the integrated approach to the revealing of mechanisms of adaptations
allowed us to demonstrate the diversity of ways of adaptation of carnivorous
mammals to the environment. The combination of ecological, behavioral and
structural-functional transformations of species differing in food specialization,
body sizes and energy exchange level determine their common adaptive strategy,
directed to the successive survival in changing habitat conditions.
The data obtained on food specialization and energetics of feeding, reproductive
potential and characteristic features of ontogenesis in carnivores may serve as the
basis in the development of ways of control over their populations for economic
purposes and measures on conservation of valuable and rare species. The characters
of ecological adaptation revealed should be taken into account when organizing
natural reserves for the conservation of threatened species and measures on rational
using of game animal resources.
Литература
Абеленцев В.И. Куницевк Фауна Украши. Ссавщ. - Кшв, 1968. Т. 1, вып. 3. 279 с.
Абрамов М.Д. Разведение норок. - М., 1961. 176 с.
Абрамов М.Д. Норководство. - М., 1974. 205 с.
Авксентьева Л.И, Богословская Л.С. Анатомия головного мозга белого медведя//Экология
и морфология белого медведя. - М., 1973. С. 107-113.
Адольф Т.А. Рост и развитие медвежат по наблюдениям в Московском зоопарке в 1940 г. //
Тр. Моск, зоопарка. - М., 1949. Т. 4. С. 78-91.
Азбукина М.Д. Специфические кожные железы щенков американской норки (Mustela vison) //
Сб. науч.-техн, информ. ВНИИОЗ. - Киров, 1970. Вып. 30. С. 60-67.
Азбукина МД. Специфические кожные железы американской норки (Mustela vison)'. Автореф.
дис. ... канд. биол. наук. - М., 1973. 27 с.
Алексеева Т.Г. К вопросу вариабельности дуги аорты, плечеголовной и подключичной
артерий соболя // Науч. тр. Омского вет. ин-та. 1968. Т. 25, вып. 2. С. 80-86.
Алексеева Т.Г, Шульц БД. Некоторые особенности ветвления дуги аорты, плечеголовной
и подключичной артерий норки // Науч. тр. Омского вет. ин-та. 1968. Т. 25, вып. 2. С. 22-38.
Алгульян С.Г. Питание и сезонные особенности образа жизни крымского барсука (Meles
meles tauricus Ogn.) // Зоол. журн. 1940. Т. 19, вып. 3. С. 120-123.
Андреева Е.Г. Кровеносная система речного бобра И Тр. Воронеж, заповеди. 1954. Вып. 5.
С. 30-50.
АндрееваЕ.Г. Кровеносная система речного бобра//Тр. Воронеж, заповеди. 1956. Вып. 6.
С. 47-58.
Андреевская В.С. Некоторые данные по размножению и постэмбриональному раз-
витию кошачьих в Ленинградском зоопарке // Материалы Прибалтийской конф. - Рига, 1964.
С. 27-64.
Анисимова Е.И. Определитель гельминтов куньих Европы // Куньи в Беларуси. - Минск,
1997. С. 21-29.
Анисимова Е.И., Сидорович В.Е. Анализ формирования гельминтоценозов в популяции
американской норки Белорусского Поозерья // Сохранение биол. разнообразия Белорусского
Поозерья: Тез. докл. конф. - Витебск, 1996а. С. 55-56.
Анисимова Е. И., Сидорович В. Е. Сравнительная характеристика гельминтофауны
европейской норки и лесной куницы в Беларуси // Систематика, таксономия и фауна
паразитов: Материалы конф. - М., 19966. С. 9-10.
Ардашев А.А., Горбачев АЛ. Функциональная активность надпочечников копытного
лемминга в зависимости от сезона и плотности популяции // Механизмы регуляции численности
леммингов и полевок на Крайнем Севере. - Владивосток, 1980. С. 112-145.
Аршавский И.А. К сравнительно-онтогенетической характеристике некоторых черт
физиологии кроликов и зайцев в связи с особенностями их экологии // Совещ. по экол., физиол.:
Тез. докл. - М.; Л., 1958. Вып. 1. С. 3-15.
Аршавский И.А. Очерки по возрастной физиологии. - М., 1967. 475 с.
Астанин Л.П., Шевченко Ю.Г Экологическая морфология головного мозга некоторых
млекопитающих // Науч, сессия по вопр. морфологии: Тез. докл. - Л., 1954. С. 86-90.
Афанасьев А.В., Бажанов В.С., Коречов М.Н. и др. Звери Казахстана. - Алма-Ата, 1953.
435 с.
Афанасьев В.А., Перельдик Н.Ш. Клеточное пушное звероводство. - М., 1966. 359 с.
Афонская Р.И., Крумина М.К. Наблюдения за белыми медведями // Моск, зоопарк. 1958.
Вып. 2. С. 56-63.
Баевский Ю.А. Эмбриональная диапауза млекопитающих и ее эволюционно-биологическое
значение//Темп индивидуального развития животных и его изменения в ходе эволюции. -М.,
1968. С. 129-175.
15 Зак. № 4309
409
Бакеев Н.Н. Лесная и каменная куницы (Кавказ) // Соболь, куницы, харза. - М., 1973.
С. 193-200; С. 213-219.
Бакеев Н.Н. Северные популяции соболя в СССР, их продуктивность и рациональ-
ное использование // Биол. проблемы Севера. - Петрозаводск, 1976. С. 56-61.
Барабаш-Никифоров И.И., Мараков С.В., Николаев А.М. Калан (Морская выдра). - Л.,
1968. 160 с.
Барбашова З.И. Тканевые процессы при акклиматизации к кислородному голоданию //
Материалы по эволюционной физиологии. -М.; Л., 1956. Т. 1. С. 140-152.
БашенинаН.В. Влияние особенностей подснежного воздуха на расположение зимних гнезд
полевок // Зоол. жури. 1956. Т. 35, вып. 6. С. 940-943.
Башенина Н.В. Интерьерные показатели мелких грызунов и их связь с уровнем
энергетического обмена И Учен. зап. Перм. пед. ин-та. 1969. Т. 79. С. 75-116.
Башенина Н.В. Энергетический обмен и терморегуляция И Европейская рыжая полевка. -
М., 1981. С. 118-133.
Беликов С.Е. Телеметрические наблюдения за температурным режимом в берлоге белого
медведя // Научные основы охраны природы. - М., 1976. Вып. 4. С. 220-226.
Беликов С.Е. Численность, распределение и миграции белого медведя в Советской Аркти-
ке // Крупные хищники. - М., 1992. С. 74-84.
Беликов С.Е. Белый медведь // Медведи. - М., 1993. С. 420-478.
Беликов С.Е., Богданов НЕ., ВеховВ.Н. Состав растительных кормов лактирующих самок
белого медведя на о-ве Врангеля // Белый медведь и его охрана в Советской Арктике. - М., 1977.
С. 55-65.
Белов ГА. О некоторых механизмах саморегуляции численности в популяциях камчатского
соболя И VIII Всесоюз. конф, по природной очаговости болезней животных и охране их
численности. - Киров, 1972. Ч. 2. С. 18-21.
Белов Г.А. Изменение биологической структуры и репродуктивных особенностей популяций
камчатского соболя под воздействием интенсивного опромышлеиия ресурсов // Влияние хоз.
деятельности человека на популяции: Материалы конф. - Киров, 1980. Т. 2. С. 8-9.
Беляев Д.К. Роль света в управлении биологическими ритмами млекопитающих // Общая
биология. 1950. Т. 11. С. 39-50.
Беляев Д.К., Клочков Д.В., Железова А.И. Влияние световых условий на воспроиз-
водительную функцию и плодовитость норок (Mustela vison Schreb.) //Бюл. МОИП. 1963. Т. 63,
вып. 2. С. 107-125.
Беньковский Л.Н., Головина Т.Н., Шербина Р.Д. Изучение болезней и паразитов выдры на о-ве
Сахалин // Вести, зоол. 1973. № 6. С. 21-24.
Берестов В.А. Биохимия и морфология крови.пушных зверей. - Петрозаводск, 1971. 291 с.
Берестов В.А. Лабораторные методы оценки состояния пушных зверей. - Петрозаводск,
1981. 150 с.
Бернацкий В.Г, Маштак З.А. Воспроизводительность норок в зависимости от времени
спаривания // Кролиководство и звероводство. 1983. № 1. С. 10-11.
Бибиков Д.И., Кудактин А.Н., Филимонов А.Н. Использование территории, перемещение //
Волк. -М., 1985. С. 415-430.
Бигон М., Харпер Дж., Таунсенд К. Экология // Особи, популяции и сообщества. - М.,
1989. Ч. 1 и 2. 1114 с.
Боголюбский С.Н. Опыт анализа комплекции разводимых зверей сем. Canidae // Тр. Ин-та
эвол. морфол. АН СССР. 1939. Т. 2, вып. 5. С. 48-51.
Боголюбский С.Н. Соотношение массы органов и размеров тела у разводимых Mustelidae //
Тр. Моск, зоотехн. ин-та. 1941. Вып. 1. С. 143-165.
Боев С.Н, Бондарева В.И., Соколова И.Б., Газиева З.Х. Роль диких и домашних животных
в эпидемиологии трихинеллеза в Казахстане // Работы по гельминтологии в Казахстане. - Алма-
Ата, 1969. С. 58-66.
410
Большаков В.Н. О путях приспособления мелких млекопитающих к горным условиям //
Бюл. МОИП. Отд. биол. 1967. Т. 22, вып. 5. С. 151-152.
Большаков В.Н. Экологическая обусловленность некоторых интерьерных признаков мелких
млекопитающих гор в связи с характером питания // Экология. 1970. № 6. С. 70-79.
Большаков В.Н., Кубанцев Б.С. Половая структура популяций млекопитающих и ее
динамика. - М., 1984. 232 с.
Бондарев А.Я. Особенности размножения волка Алтайского края И Экологические основы
охраны и рац. использования хищных млекопитающих. - М., 1979. С. 89-90.
Борзых П.Л. Осеннее питание и миграционные перемещения енисейского соболя в юго-
восточной части ареала // Рациональное использование ресурсов соболя в России. - Красноярск,
2001. С. 89-92.
Брандт А. Об отношении количества мозга к массе тела у различных животных // Сб. тр. 1 -го
съезда русских естествоиспытателей в Петербурге. - СПб., 1868. С. 57-68.
Бромлей ГФ. Материалы по биологии соболя и харзы, распространенных в Приморском
крае // Материалы по результатам изучения млекопитающих в государственных заповедниках. -
М., 1956. С. 5-19.
БромлейГ.Ф. Харза (южная часть Дальнего Востока) // Соболь, куницы, харза. - М., 1973.
С. 223-228.
Булатова Н.Н. Некоторые особенности дыхательной функции крови высокогорных
животных: Автореф. дне. ... канд. биол. наук. - М., 1953. 23 с.
Буневич Л.А., Буневич А.Н., Гайдук В.Е. Дневная активность волка Беловежской пущи в
различные сезоны года // Биологические ритмы: Материалы междунар. науч.-практ. конф. -
Брест, 1999. С. 54-56.
Бычкова Е.И., Сидорович В.Е. Заражение гельминтами вымирающей популяции европейской
норки в Беларуси И Вести. АН Беларуси. 1994. № 3. С. 105-109.
Вайсфельд М.А. Выдра // Колонок, горностай, выдра. - М., 1977. С. 175-205.
Вайсфельд М.А. Северо-Восток Европейской территории России. Медведи. - М., 1993.
С. 37-50.
ВаленцевА.С. К вопросу о хоминге соболей// Сельское хоз-во Крайнего Севера: Тез. докл.
IV Всесоюз. совещ. Биол. ресурсы и охрана природы. - Магадан, 1980. С. 206-208.
Валенцев А. С. Миграции соболей на Камчатке и их влияние на промысел // Охрана и рац.
использования ресурсов соболя в РСФСР: Тез. докл. науч.-произв. совещ. - Красноярск, 1981.
С. 39-41.
Валенцев А.С. Мониторинг, прогнозирование и управление структурой попу-
ляции камчатского соболя // Материалы III Всерос. науч-произв. совещ, - Красноярск, 1992.
С. 10-13.
Варнаков А.П. Добыча и численность медведя в Вологодской области // Экологические
основы охраны и рационального использования хищных млекопитающих. - М., 1979. С. 213-
214.
Васенева А.Я. Материалы по гельминтофауне норки, акклиматизированной в Приморском
крае // Тр. ВНИИЖП: Охота, пушнина и дичь. - М., 1967. Вып. 15. С. 41 -45.
Васенева А.Я. Американская норка (Mustela vison Schreber, 1777) Приморья и Приамурья:
Автореф. дисс. ... канд. биол. наук. - Владивосток, 1969. 27 с.
Васенева А.Я. Изменение структуры популяций американской норки под воздействием
промысла // Тез. докл. VIII Всесоюз. конф, по природной очаговости болезней животных и
охрана их численности. - Киров, 1972а. Т. 2. С. 23-25.
Васенева А.Я. Изучение перемещений американской норки с помощью мечения //
Материалы к науч, конф., посвящ. 50-летию ин-та. - Киров, 19726. Ч. 2. С. 100-103.
Васенева А.Я. Рациональное использование американской норки на юге Дальнего Востока //
Современное состояние и пути развития охотоведческой науки в СССР: Тез. докл. 1-й Всесоюз.
конф. - Киров, 1974. С. 72-74.
411
Верболович П.А. Миоглобин и его роль в физиологии и патологии животных. - М., 1961.
214 с.
Вершинин А.А. Биология и промысел камчатского горностая // Бюл. МОИП. Отд. биол.
1972. Т. 77, вып. 2. С. 132-141.
Вершинин А.А. Горностай (Камчатка) // Колонок, горностай, выдра. - М., 1977. С. 146-153.
Вершинин А.А., Белов Г.А. Соболь: Камчатка и о-в Карагинский//Соболь, куницы, харза. -
М., 1973. С. 118-131.
Вискова В.И. Содержание углекислого газа в ходах крота (Talpa europaea) И Зоол. журн.
1983. Т. 62, вып. 6. С. 966-967.
Владимирская М.И., Лебедев В.Д., Насимович А.А. Новые данные по экологии выдры //
Бюл. МОИП. Отд. биол. 1953. Т. 58, вып. 3. С. 146-154.
Войлочников А. Т. Колонок (Предбайкалье и Забайкалье) // Колонок, горностай, выдра. - М.,
1977. С. 44-50.
Воккен ГГ. Наблюдения за размножением белого медведя в Лензоосаде // Бюл. зоопарков и
зоосадов. 1933. Вып. 3. С. 6-9.
Волков В.А. Значение куницы и животных потребителей семян ели в насаждениях южной
тайги: Автореф. дис. ... канд. биол. наук. - Л., 1971. 20 с.
Волобуев В.Т, Терновский Д.В., Графодарский А.С. Таксономический статус белого
африканского хоря, или фуро, в свете кариологических данных// Зоол. журн. 1974. Т. 53,
вып. 11. С. 1738-1739.
Воронецкий Н.Н., Дунин В.Ф., Пискунов В. С., Киреенко К.М. Суточная активность и питание
волка в Полесском государственном радиационно-экологическом заповеднике // Биологические
ритмы: Материалы Междунар. науч.-практ. конф. - Брест, 1999. С. 65-66.
Воронцов Н.Н. Эволюция пищеварительной системы грызунов (мышеобразные). -
Новосибирск, 1957. 239 с.
Вшивцев В.П. Выдра Сахалина. - Новосибирск, 1972. 107 с.
Гайдук В.Е. Факторы, обусловливающие линьку и пигментацию волосяного покрова
горностая (Mustela erminea) II Зоол. журн. 1977. Т. 56, вып. 8. С. 1226-1231.
Гайдук В.Е. Ритмы линьки зверей с сезонным диморфизмом окраски шерстного покрова И
Журн. общей биол. 1983. Т. 44, № 2. С.269-279.
Гайдук В.Е. Сравнительный анализ биоритмов доминирующих видов млекопитающих
Беларуси: Автореф. дис. ... д-ра биол. наук. - Петрозаводск, 1998. 43 с.
Гайдук В.Е. Экология ритмов млекопитающих в Палеарктике И Биологические ритмы:
Материалы Междунар. науч.-практ. конф. - Брест, 1999. С. 16-19.
Гайдук В.Е., Баранов В.М., Жук В.А., Абрамова И.В. Биология промыслово-охотничьих
наземных позвоночных Брестской области. - Брест, 1999. 134 с.
Галанцев В.П. Анатомо-физиологические адаптации ондатры и водяной полевки к
полуводному образу жизни и нырянию // Тр. НИИ с.-х. Крайнего Севера. - Красноярск, 1967.
Т. 14. С. 97-106.
Галанцев В.П. Эволюция адаптаций ныряющих животных // Эколого-морфологические
аспекты. - Л., 1977. 191 с.
Галанцев В.П. О направлениях и путях морфофункциональных адаптаций полуводных
пушных зверей // Адаптационные реакции пушных зверей. - Петрозаводск, 1980. С. 15-22.
Галанцев В.П. Структурно-функциональные адаптации сердечно- сосудистой системы к
нырянию у млекопитающих: Автореф. дис. ... д-ра биол, наук. - Л., 1983. 34 с.
Галанцев В.П., Туманов И.Л. Эколого-физнологическая характеристика сердечной
деятельности грызунов // Зоол. журн. 1969. Т. 48, вып. 7. С. 1066-1073.
Галанцев В.П., Туманов И.Л. Эколого-физиологическая характеристика дыхательной
функции крови и легочного дыхания грызунов//Вести, зоологии. 1970. №4. С. 47-52.
Геллер М.Х. Биология уссурийского енота, акклиматизированного на северо-западе евро-
пейской части СССР И Тр. НИИ с.-х. Крайнего Севера. - Красноярск, 1959. Вып. 9. С. 51-56.
412
Генерозов В.Я. Промышленное разведение скунсов. - Петроград, 1922. 40 с.
Гентнер В.Г, Наумов Н.П., Юргенсон П.Б и др. Млекопитающие Советского Союза. Т. 2.
Морские коровы, хищные. - М., 1967. 1003 с.
Гептнер В.Г, Слудский А.А. Млекопитающие Советского Союза. - М., 1972. Т. 2, ч. 2.
С. 134-139.
Гинеев А.М. Описание и морфологические особенности енота-полоскуна Дагестана //
Материалы к науч, конф., посвящ. 50-летию ин-та: Тез. докл. - Киров, 1972. Ч. 2. С. 91-94.
Гинеев А.М. Размножение енота-полоскуна на Северном Кавказе // Вопр. экологии
позвоночных животных. - Краснодар, 1973. С. 94-98.
Гинеев А.М. Особенности экологии енота-полоскуна, способствующие его успешной
акклиматизации в антропогенных ландшафтах // Влияние антропоген. трансформ, ландшафта
на население назем, позвоноч. животных: Тез. Всесоюз. совещ. - М., 1987. Ч. 1. С. 241-242.
Гинеев А.М. К экологии каменной куницы Северо-Кавказского региона // Экология, охрана
и воспроизводство животных Ставропольского края и сопредельных территорий: Материалы
науч.-практ. конф. - Ставрополь, 1991. С. 69-72.
Гинеев А.М., Гаркави В.Л. О трихинеллезе у енота-полоскуна, каменной куницы и лесной
кошки пойменных лесов Дагестанской АССР // Сб. НТИ ВНИИОЗ. - Киров, 1974. Вып. 40-41.
С. 125-127.
Глушков В.М., Граков Н.Н. и др. Управление популяциями охотничьих животных. - Киров,
1999. 195 с.
Голодов И.И. Влияние высоких концентраций углекислоты на организм. - Л., 1946. 235 с.
Горшков П.К. К экологии енотовидной собаки в Волжско-Камском заповеднике // Волжско-
Камский заповедник: Тез. докл. - Казань, 1980. С. 52-53.
Горшков П.К. Барсук в биоценозах Республики Татарстан. - Казань, 1997. 176 с.
ГраковН.Н. Роль белки в питании куницы на Европейском Севере//Тр. ВНИИЖП. 1962а.
С. 154-162.
Граков Н.Н. Филяроидоз и скрябингилез лесной куницы (Martes martes L.) и их влияние на
состояние популяций этого вида // Тр. ВНИИЖП. 19626. С. 298-314.
Граков Н.Н. К вопросу о ложном гоне лесной куницы И Сб. науч.-техн. информ. (ВНИИЖП). -
Киров, 1964. Вып. 10. 68-72 с.
Граков Н.Н. Запасы пищи и их роль в жизни лесной куницы // Сб. науч.-техн. информ.
«Охота, пушнина, дичь». - Киров, 1965. Вып. 13. С.10-14.
Граков Н.Н. Поедание некоторых естественных кормов лесными куницами, содержащимися
в вольерах // Сб. науч.-техн. информ. «Охота, пушнина, дичь». - Киров, 1966. Вып. 16. С. 15-21.
ГраковН.Н. О гибридизации соболя и лесной куницы//Сб. науч.-техн. информ. (ВНИИОЗ). -
Киров, 1969а. Вып. 26. С. 8-16.
Граков Н.Н. Половая, возрастная структура и прирост популяций лесной куницы // Тр.
ВНИИЖП. - Киров, 19696. Т. 22. С. 3-18.
Граков Н.Н. Кидусы на биостанции ВНИИОЗ И Сб. науч.-техн. информ. (ВНИИОЗ). -
Киров, 1970. Т. 33. С. 48-53.
Граков Н.Н. Лесная куница (Европейский Север) // Соболь, куницы, харза. - М., 1973.
С. 143-154.
Граков Н.Н. Возрастная структура популяций и ее влияние на динамику численности
лесной куницы И Сб. науч.-техн. информ. ВНИИОЗ. - Киров, 1976. Вып. 54-55. С. 3-8.
Граков Н.Н. Экологические основы рационального использования ресурсов лесной куницы
в СССР: Автореф. дис.... д-ра биол. наук. - Свердловск, 1978. 36 с.
Граков Н.Н. Лесная куница. - М., 1981. 108 с.
Гракова Н.В., Граков Н.Н. Об элиминации самцов и самок в потомстве у американской
норки // Биология и патология клеточ. пушных зверей: Тез. докл. - Киров, 1977. С. 53-54.
Грачев Ю.А. Семейство медвежьи - Ursidae // Млекопитающие Казахстана. - Алма-Ата,
1981. Т. 3, ч. 1. С. 148-191.
413
Грибова З.А. О зараженности лесной куницы в Вологодской области филяроидозом и
скрябингилезом // Тр. ВНИИ животного сырья и пушнины. 1959. Вып. 18. С. 51-56.
Губанов Н.М. Гельминтофауна промысловых млекопитающих Якутии. - М., 1964. 115 с.
Гуляев С.В. К биологии размножения бурого медведя // Бюл. зоопарков и зоосадов. 1934.
№ 2 (3). С. 13-21.
Гусев О.К. Экология соболя. - М., 1966. 124 с.
Давыдов А.Ф. Гомойотермные организмы бореальной климатической зоны//Экологическая
физиология животных. - Л., 1982. Ч. 3. С. 110-130.
Данилов П.И. Сравнительно-экологическая характеристика семейства куньих Карелии //
Конф, молод, биологов Карелии. - Петрозаводск, 1968. С. 48-51.
Данилов П.И. Биология выдры в Карелии // Вопросы экологии животных. - Петрозаводск,
1969. С. 35-38.
Данилов П.И. Демография бурого медведя в СССР // Фауна и экология наземных
позвоночных. - Петрозаводск, 1988. С. 138-139.
Данилов П.И. Бурый медведь Северо-Запада России и охота на него // Медведи России и
прилегающих стран. - М., 1993. Ч. 1. С. 75-76.
Данилов П.И. Экологические основы охраны и рационального использования крупных
хищников Северо-Запада России: Автореф. дне. ... д-ра биол. наук. - М., 1994. 69 с.
Данилов П.И., Ивантер Э. В. Лесная куница в Карелии//Учен. зап. Петрозавод. гос. ун-та.
1967. Т. 15, вып. 4. С 112-115.
Данилов П.И., Ивантер Э. В. и др. Изменение численности охотничьих зверей в Карелии
по данным зимиих маршрутных учетов // Фауна и экология птиц и млекопитающих таежного
Северо-Запада СССР. - Петрозаводск, 1978. С. 128-159.
Данилов П.И., Русаков О.С. Особенности экологии черного хоря в северо-западных
областях европейской части СССР // Зоол. журн. 1969. Т.48, вып. 9. С. 1112-1115.
Данилов П.И., Русаков О.С. К вопросу о роли оценки кормности угодий при относительных
учетах охотиичье-промысловых животных // Вопросы экологии животных. - Петрозаводск,
1974. С. 174-179.
ДаниловПИ.РусаковО.С., ТумановИ.Л. Горностай. Региональные особенности динамики
запасов, экологии и хозяйственного использования. Северо-Запад И Колонок, горностай, выдра. -
М., 1977. С. 79-92.
ДаниловП.И., Русаков О.С., Туманов И.Л. Хищные звери Северо-Запада СССР. - Л., 1979.
160 с.
Данилов П.И., Русаков О.С., ТумановИ.Л. Северо-Запад европейской части СССР //Волк. -
М., 1985. С. 479-482.
Данилов П.И., Туманов И.Л. Барсук северо-западных областей//Охота и охот, хозяйство.
1970. № 3. С. 16-17.
Данилов П.И., Туманов И.Л. Материалы по биологии барсука на Северо-Западе РСФСР //
Науч.-техн. информ. ВНИИЖП. - Киров, 1971. Вып. 34. С. 25-31.
Данилов П.И., Туманов И.Л. Репродуктивные циклы самцов некоторых куньих (Musteli-
dae) // Зоол. журн. 1972. Т. 51, вып. 6. С. 871-880.
Данилов П.И., Туманов И.Л. Репродуктивные циклы самок некоторых куньих. Мелкие
хищники//Бюл. МОИП. 1975а. № 1. С. 137-146.
ДаниловП.И, ТумановИ.Л. Репродуктивные циклы самок некоторых куньих. Выдра, барсук,
росомаха // Бюл. МОИП. 19756. № 5. С. 35-47.
Данилов П.И., Туманов И.Л. Куньи Северо-Запада СССР. - Л., 1976а. 256 с.
Данилов П.И., Туманов И.Л. Экология европейской и американской норок на Северо-
Западе СССР // Экология птиц и млекопитающих Северо-Запада СССР. - Петрозаводск, 19766.
С. 118-146.
Данилов П.И., Туманов И.Л. Рост, развитие и особенности размножения бурого медведя //
Медведи СССР - состояние популяций. - Ржев, 1991. С. 71-77.
414
Данилов П.И., Туманов И.Л., Русаков О. С. Северо-Запад европейской территории России.
Медведи. - М., 1993. С. 21-36.
Девяткин ГВ. Половая и возрастная структуры популяции соболя в Магаданской области //
Материалы III Всерос. науч.-произв. совещ. - Красноярск, 1992. С. 22-25.
Девяткин ГВ. Соболь Магаданской области и Чукотки: реакклиматизация, особенности
экологии и поведения // Соболь. Состояние ресурсов и перспективы пушного промысла. -
Киров, 1998. С. 23-29.
Динник Н.Я. Звери Кавказа. Хищные. - Тифлис, 1914. Т. 2. 536 с.
Добринский Л.Н. Опыт использования относительного веса надпочечников при изучении
экологии птиц // Экологические основы адаптации животных. - М., 1967. 232 с.
Догель В.А. Общая паразитология. - Л., 1962. 461 с.
Докукина Ю.М., Куличков А.Б. Новое в технике гона соболей // Кролиководство и
звероводство. 1980. № 3. С. 24.
Долабчян З.Л. Синтетическая электрокардиология. - Ереван, 1963. 257 с.
Домнич И.Ф., Обушенков И.Н. Гельминтофауна хищных млекопитающих северо-восточной
Азии. - Магадан, 1983. С. 5-18.
Донауров С.С. Некоторые данные по биологии лесной куницы в Кавказском заповеднике //
Тр. Кавказ, заповедника. - М., 1949. Т. 3. С. 46-72
Дубровский А.Н. Песец Кольского полуострова // Тр. НИИ поляр, земледелия, жи-
вотноводства и промысл, хоз-ва. Сер. промысл, хоз-ва. - Л., 1939. Вып. 6. С. 43-52.
Ефремов Ю.М. Некоторые данные по адаптивности соболей и иорок // Материалы XLIII
иауч. конф, аспирантов и студентов Моск. вет. акад. - М., 1968. С. 45-51.
Жеденов В.Н. Анатомия домашних животных. - М., 1965. 320 с.
Железнов Н.К Дикие копытные Северо-Востока СССР. - Владивосток, 1990. 478 с.
Желтухин А.С. Зимнее питание и охотничье поведение рыси в Цеитрально-Лесном
заповеднике // Экологнч. основы охраны и рац. использования хищных млекопитающих. - М.,
1979. С. 266-267.
Желтухин А. С. О суточной активности и размерах участков обитания рыси в южной
тайге Верхневолжья // Бюл. Моск, о-ва испытателей природы: Отд. биол. 1984. Т. 89, вып. 1.
С 54-62.
Желтухин А.С. Рысь южной тайги Верхневолжья (экология, поведение, вопросы
использования и охраны): Автореф. дис. ... канд. биол. наук. - М., 1987. 15 с.
Желудкова Т.Н., Загрядский В.П., Сулимо-Самуйло З.К. Влияние на организм длительного
пребывания в газовой среде с повышенным содержанием углекислоты // Функции организма в
условиях измененной газовой среды. - М.; Л., 1964. С. 187-192.
Жиряков В.А., Грачев Ю.А. Бурый медведь. Центральная Азия и Казахстан // Медведи. -
М., 1993. С. 170-205.
Заболотских Ю. С. Экология поведения некоторых видов зверей сем. Canidae // Экология и
промысел охотничьих животных (Сб. науч. тр.). - М., 1983. С. 63-71.
Заболотских Ю.С. Этология, гомеостаз и воспроизводнтельность пушных зверей на фоне
алиментарного голодания // Сельхоз. биология. 1997. № 2. С. 48-58.
ЗаболотскихЮ.С. Динамика сезонной и суточной ритмики енотовидной собаки в неволе //
Биологические ритмы: Материалы Междуиар. науч.-практич. конф. - Брест, 1999а. С. 88-90.
Заболотских Ю.С. Влияние пищевого фактора на суточный ритм у пушных зверей И
Биологические ритмы: Материалы Междуиар. научно-практич. конф. - Брест, 19996. С. 90-92.
Заболотских Ю.С. Научные основы и практика пищевых разгрузок в пушном звероводстве:
Автореф. дис. ... д-ра биол. наук. - М., 2000. 39 с.
Заварзин А.А., Румянцев А.В. Курс гистологии. - М., 1946. 354 с.
Завацкий Б.П. Индивидуальная изменчивость и половой диморфизм бурого медведя средней
Енисейской тайги // Экология, морфология и охрана медведей в СССР. - М., 1981. С. 22-24.
Завацкий Б.П. Бурый медведь. Средняя Сибирь // Медведи. - М., 1993. С. 249-275.
415
Залекер В.Л. Материалы по половому циклу соболя (Marteszibellina L.) // Тр. ВНИО. - М.,
1950. Вып. 9. С. 135-151.
Залекер ВЛ. Изменение возрастного, полового состава популяции и плодовитости соболя //
Тр. ВНИИ животного сырья и пушнины. - М., 1962. Вып. 19. С. 54-58.
Злобин БД. Питание солонгоя в Южном Прибалхашье // Материалы к науч, конф., посвящ.
50-летию ин-та. - Киров, 1972. Ч. 2. С. 82-84.
Золотов В.А. Материалы по распределению и биологии песца Кольского полуострова //
Изв. географ, о-ва. 1940. Т. 72, вып. 1. С. 47-64.
Зырянов А.И. К экологии рыси и росомахи в Красноярском крае И Вопр. экологии: Тр.
заповедника «Столбы». - Красноярск, 1980. Вып. 12. С. 3-28.
Зырянов А.Н. Ресурсы соболя в Красноярском крае и перспективы улучшения их
использования // Экология диких животных и растений и их использование. - Красноярск,
1990. С. 14-24.
Зырянов А.Н. Рысь в Красноярском крае//Фауна и экология наземных позвоночных Сиби-
ри. - Красноярск, 1997. С. 177-195.
Зырянов А.Н. Популяционный гомеостаз и промысел соболя // Соболь. Состояние ресурсов
и перспективы пушного промысла. - Киров, 1998. С. 39-46.
Зырянов А.И., Туманов И.Л. Региональный кадастр соболя Красноярского края // Тр.
ВНИИОЗ. - Киров, 2000. № 1 (51). С. 162-166.
Зырянов А.Н., Туманов И.Л., Кожечкин В.В. Соболь и кабарга // Сб. науч, исследований в
енисейских заповедниках по проблеме «Хищник-жертва». - Шушенское, 1997. С. 15-18.
Зырянов А.Н., Туманов И.Л., Кожечкин В.В. Об особенностях охоты соболя на кабаргу //
Сб. материалов IV Всерос. конф. «Рациональное использование ресурсов соболя в России». -
Красноярск, 2001. С. 101-103.
Иванов П.Д. Истребление волков в Ленинградской области // Промысловая фауна и охот,
хоз-во Северо-Запада РСФСР. - Л., 1963. Вып. 2. С. 277-282.
Иванова Е.И. Адаптивные особенности морфологии кровеносной и дыхательной сис-
тем русской выхухоли // Морфологические особенности водных млекопитающих. - М., 1964.
С. 82-127.
Иванова Е.И. Функциональная морфология сосудистой системы млекопитающих. - М.,
1975. 56 с.
Ивантер Э.В. К вопросу о соотношении полов у голубых песцов // Вопросы экологии. -
М., 1962. Т. 6. С. 187-205.
Ивантер Э.В. Питание лесной куницы в Карелии // Науч. конф. Ин-та биологии КФ АН
СССР. - Петрозаводск, 1963. С. 68-73.
Ивантер Э.В. К вопросу о взаимоотношениях лесной куницы и белки // Материалы III
зоол. конф. пед. ин-тов РСФСР. - Волгоград, 1967. С. 120-123.
Ивантер Э.В. Материалы по биологии и численности лисицы в Карельской АССР // Учен,
зап. Петрозавод. ун-та. 1969. Т. 17, вып. 4. С. 196-211.
Ивантер Э.В. К изучению барсука на северном пределе ареала // Тр. гос. заповедника
«Кивач». - Петрозаводск, 1973. Вып. 2. С. 164-173.
Ивантер Э.В. Морфологические особенности обыкновенной бурозубки (Sorex araneus
L.) в свете ее сезонной и возрастной экологии // Вопросы экологии животных. - Петрозаводск,
’1974а. е. 36-94.
Ивантер Э.В. Млекопитающие. - Петрозаводск, 19746. 214 с.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада
СССР. - Л., 1975. 247 с.
Ивантер Э.В. Питание и некоторые морфологические особенности мышевидных грызунов
Карелии // Экология птиц и млекопитающих Северо-Запада СССР. - Петрозаводск, 1976. С. 68-95.
Ивантер Э.В., Ивантер Т.В., Туманов И.Л. Адаптивные особенности мелких млекопитаю-
щих. - Л., 1985. 317 с.
416
Ильина Е.Д. Островное звероводство. - М., 1950. 256 с.
Ильина Е.Д. Звероводство. - М., 1952. 335 с.
ИржакЛ.И. Дыхательная функция крови в индивидуальном развитии млекопитающих. -
М.; Л., 1964. 183 с.
Каал М.И Бурый медведь в Эстонии // Экология, морфология, охрана и использование
медведей: Материалы совещ. - М., 1972. С. 41-44.
Казлаускас Ю., Прусайте Я. Гельминты животных отряда хищных (Carnivora) в Литве //
Acta parasitologica Lituanica. 1976. № 14. С. 33-44.
Калабухов Н.И. Физиологические особенности близких видов животных // Успехи соврем,
биол. 1940. Т.13, вып. 3. С. 403-429.
Калабухов Н.И. Эколого-физиологические особенности животных и условия среды. -
Харьков, 1950. 267 с.
Калабухов Н.И. Методика экспериментальных исследований по экологии наземных
позвоночных. - М., 1951. 177 с.
Калабухов Н.И. Периодические (сезонные и годичные) изменения в организме грызунов,
их причины и последствия. - Л., 1969. 248 с.
Калецкая М.Л. Волк и его роль как хищника в Дарвинском заповеднике // Тр. Дарв.
госзаповед. 1973. Вып. 11. С. 41-58.
Карасев Н.Ф. Экологический анализ гельминтофауны млекопитающих Березинского
заповедника. Березинский заповедник. - Минск, 1972. Вып. 2. С. 181-185.
Карасев Н.Ф. Цестоды хищных млекопитающих//Березинский заповедник. - Минск, 1975.
Вьш. 4. С. 221-223.
Кацнельсон З.С., Орлова И.И. Морфология крови речного бобра //Тр. Воронеж, заповед. -
Воронеж, 1956. Вып. 6. С. 61-65.
Кацнельсон З.С., Рихтер А.Л. Практические занятия по гистологии и эмбриологии. - М.;
Л., 1959. 241 с.
Клевезаль Г.А., Клейненберг С.Е. Определение возраста млекопитающих.-М., 1967. 142с.
Клер Р.В. Течка и беременность у некоторых Mustelidae // Тр. Моск, зоотехн. ин-та. 1941.
Т. 1. С. 20-60.
Клер Р.В. Ложный гон и плацентация у соболей // Каракулеводство и звероводство. 1948.
№ 1. С. 15-18.
Кляйненберг С.Е. Млекопитающие Черного и Азовского морей. - М., 1956. 350 с.
Ковальчук Е.И. Гельминты диких промысловых млекопитающих Тюменской области и
некоторые вопросы их экологического анализа // Экология и морфология гельминтов Западной
Сибири. - Новосибирск, 1979. С. 56-61.
Ковешникова А.К. О кровоснабжении мышц с различной функцией // Изв. ин-та им. П.Ф. Лес-
гафта. 1936. Т. 20. С. 118-124.
Ковтун М.Ф., ОсинскийЛ.П. Сердечно-сосудистая система// Волк.-М., 1985. С. 226-239.
Когтева Е.З. Сезонная изменчивость и возрастные особенности строения кожи и волосяного
покрова крота, зайца-беляка и енотовидной собаки // Промысловая фауна и охотничье хозяйство
Северо-Запада РСФСР. - Л., 1963. С. 213-271.
Когтева Е.З., Морозов В.Ф. О зараженности лесной куницы филяроидозом в северо-
западных областях РСФСР // Промысловая фауна и охот, хоз-во Северо-Запада РСФСР. - Л.,
1963. С. 204-206.
Когтева Е.З., Морозов В. Ф. О зараженности лесной куницы (Martes martes) нематодами родов
Filaroides и Skrjabindylus на Северо-Западе РСФСР // Зоол. журн. 1970а. Т. 49, вып. 1. С. 131-136.
Когтева Е.З., Морозов В. Ф. О региональных особенностях некоторых гельминтозов лесной
куницы (Martes martes L.) на Северо-Западе СССР И Тр. IX междунар. конгр. биологов-
охотоведов. - М., 19706. С. 634-639.
Когтева Е.З., Морозов В.Ф. Состояние популяции лисицы и ее промысловое использование
на Северо-Западе РСФСР//Тр. НИИ охот, хоз-ва и звероводства. - М., 1972. Вып. 24. С. 170-181.
417
Кожечкин В.В. Охота росомахи на марала // Бюл. МОИП. 1995. Т. 10, вып. 6. С. 14-20.
Кожечкин В.В. Росомаха (Gulo gulo L.) в лесных экосистемах северо-западной части
Восточного Саяна: Автореф. дис. ... канд. биол. наук. - Красноярск, 1999. 24 с.
Козло П.Г. Особенности развития внутренних органов волка // Экология. 1981. №3. С. 89-92.
КозлоП.Г, Филимонов А.Н., Бондарев А.Я. Морфометрия внутренних органов // Волк. -
М„ 1985. С. 254-258.
КозловД.П. К изучению гельминтофауны бассейна р. Печоры // Тр. гельмиитол. лаб. АН
СССР. 1969. Т. 20. С. 71-78.
КозловД.П. Определитель гельминтов хищных млекопитающих СССР. - М., 1977. 275 с.
Козловский И.С. Зимнее питание волка Северо-Востока европейской части СССР // Сб.
науч. тр. КСХИ. - Пермь, 1986. С. 26-29.
Козловский И.С. Волк Северо-Востока европейской части России: Автореф. дис. ... канд.
биол. наук. - Киров, 1996. 24 с.
Кстовский В.М. Определение возраста эмбрионов американской норки (Mustela vison Вг.) //
Сб. науч.-техн. информ. (ВНИИОЗ). - Киров, 1971. Вып. 33. С. 54-62.
Комаров А.В. К этологии соболя // Материалы науч, конф., посвящ. 50-летию ин-та. -
Киров, 1972. Ч. 2. С. 46-48.
Контримавичус В.Л. Гельмиитофауна куньих Дальнего Востока // Тр. Гельмиитол. лаб.
АН СССР. 1963. Т. 13. С. 26-47.
Контримавичус В.Л. Гельмиитофауна куньих (Mustelidae) СССР // Тр. Гельмиитол. лаб.
АН СССР. 1966. Т. 17. С. 76-79.
Контримавичус В.Л. Гельмиитофауна куньих и пути ее формирования. - М., 1969. 132 с.
Контримавичус В.Л., Скрябина Е.С. Гельминты соболя и горностая Камчатки // Тр.
Гельмиитол. лаб. АН СССР. 1963. Т. 13. С. 63-68.
Копеин КИ. Горностай. Урал и прилегающая часть Западной Сибири // Горностай, колонок,
выдра.-М„ 1977. С. 127-136.
Коржуев ПА. О формах локализации и количестве гемоглобина в крови различных живот-
ных//Успехи соврем, биол. 1952. Т. 33, вып. 3. С. 391-398.
Коржуев ПА. Дыхательная функция крови и скелета позвоночных // Успехи совр. биол.
1955. Т. 39, вып. 2. С. 163-182.
Коржуев П.А. Различия в количестве крови и весе скелета в эмбриональ-ный период у
млекопитающих, птиц и их биологические причины // Тр. Ин-та морфол. животных АН СССР. -
М„ 1960. Вып. 31. С. 51-54.
Коржуев П.А. Гемоглобин. - М., 1964. 287 с.
Коржуев П.А., Корецкая Т.И. Эколого-физиологические особенности крови землероек и
кротов // Тр. Ин-та морфол. животных АН СССР. - М., 1962. Вып. 41. С. 129-136.
Коржуев П.А, Спасская Т.Х. Весовые особенности крови, скелета, костного мозга и
мускулатуры // Волк. - М., 1985. С. 259-260.
Коровина А.М. Размножение животных Московского зоопарка // Материалы Прибалт, конф. -
Рига, 1964. С. 173-178.
Коршунова Л.А., Туманов И.Л. Морфологические особенности дуги аорты некоторых
представителей семейства куньих // Вести, зоологии. 1979. № 3. С. 35-40.
Корчмарь Н.Д. Некоторые данные о распространении, биологии и хозяйственном значении
барсука в Молдавии // Вопросы экологии и практического значения птиц и млекопитающих
Молдавии. - Кишинев, 1962. С. 56-63.
Корытин Н.С. Демографический анализ популяций обыкновенной лисицы в условиях
роста и снижения численности: Автореф. дис. ... каид. биол. наук. - Свердловск, 1983. 24 с.
Корытин С.А. Зимняя экология лисицы в Кировской области // Бюл. МОИП. 1968. Т. 73,
вып. 5. С. 33-44.
Корытин С.А. Об избирательности промысла по полу и соотношение самцов и самок у
хищных млекопитающих // Тр. ВНИИОЗ. - М., 1971. Вып. 23. С. 73-84.
418
Корытин С.А. Сезонные изменения обонятельного поведения песцов и лисиц // Биология
и патология пушных зверей. - Петрозаводск, 1974. С. 83-87.
Корытин С.А., Бибиков Д.И. Охотничье поведение // Волк. - М., 1985. С. 311-324.
Краснощеков Г.П. Гипотеза эндокринной регуляции численности популяции И Материалы
по экологии мелких млекопитающих Субарктики. - Новосибирск, 1975. С. 112-124.
Крылов М.К К вопросу о миграции песцов на Таймыре И Науч.-техн. бюл. ВАСХНИЛ.
Сиб. отд. - Новосибирск, 1986. Вып. 33. С. 12-17.
Крылов М.К. Биологическая продуктивность песцовых угодий Таймыра // Млекопитающие
и птицы севера Средней Сибири. - Новосибирск, 1989. С. 113-119.
Кубанцев Б.С. Соотношение полов в потомстве млекопитающих и его зависимость от
условий жизни // Учен. зап. Сталинград, пед. ин-та. 1961. Вып. 13. С. 59-68.
Кубанцев Б.С. Условия существования и пол у млекопитающих // Учен. зап. Сталинград,
пед. ин-та. 1964. Вып. 16. С. 26-84.
Кудактин А.Н. Территориальное размещение и структура популяции волка в Кавказском
заповеднике // Бюл. МОИП. 1979. Т. 84, вып. 2. С. -56-65.
Кудактин А.Н. О поведении волков в условиях заповедной экосистемы И Поведение волка. -
М., 1980. С. 90-102.
Кукарцев В.А. К вопросу о влиянии промысла на половую структуру популяций горностая //
Сб. НТИ ВНИИОЗ: Охота, пушнина и дичь. - Киров, 1970. Вып. 29. С. 27-29.
Кукарцев В.А. Методика регулирования численности горностая промыслом // Материалы
к науч, конф., посвящ. 50-летию ин-та: Вопросы экологии. - Киров, 1972. Ч. 2. С. 64.
Кукарцев В.А. Экология горностая и рациональное использование его ресурсов в пойме
нижней Оби: Автореф. дис. ... канд. биол. наук. - Иркутск, 1974. 24 с.
Куксов В.А. Интенсивность размножения и планирование заготовок песца на Таймыре //
Млекопитающие и птицы севера Средней Сибири: Тр. НИИСХ Крайнего Севера. - Новосибирск,
1989. С. 103-113.
Курис Н.М. Биология размножения колонков в условиях клеточного содержания//
Тр. Всесоюз. НИИ охот, хоз-ва и звероводства. 1972. Вып. 24. С. 350-356.
Курис Н.М. Особенности биологии, размножения, развития и морфологии колонка: Автореф.
дис. ... канд. биол. наук. - Казань, 1980. 20 с.
Курис Н.М., Бакеев Н.Н. Об особенностях размножения некоторых видов семейства
Mustelidae // Бюл. МОИП. Отд. биол. 1974. Т. 79. С. 22-30.
Лабутин Ю.В. Волк // Млекопитающие Якутии. - М., 1971. С. 364-377.
Лавов М.А. Влияние волка и рыси на диких копытных в Восточной Сибири // Материалы
науч, конф., посвящ. 50-летию ин-та. - Киров, 1972. Ч. 2. С. 112-115.
Лавров Н.П. Биология размножения горностая И Тр. науч.-исслед. лаб. биологии, охотн.
промысла и тавароведения животного сырья. - М., 1944. Вып. 6. С. 112-118.
Лавров Н.П. Особенности и причины длительной депрессии численности горностая в лесо-
степной и степной зонах СССР // Вопр. биологии пушных зверей. - М., 1956. Вып. 16. С. 89-107.
Лесгафт П.Ф. Основы теоретической анатомии. - СПб., 1905. Ч. 1. 348 с.
Линейцев С.Н. Миграции песцов // Животный мир плато Путорана, его рациональное
использование и охрана. - Новосибирск, 1988. С. 63-71.
Лобачев С.В. Охота на медведя. - М., 1951. 125 с.
Лобачев Ю.С. Экология некоторых куньих в горах Юго-Восточного Казахстана: Автореф.
дис. ... канд. биол. наук. - Алма-Ата, 1973. 24 с.
Лобачев Ю.С. Экология барсука в горах Юго-Востока Казахстана // Бюл. МОИП. 1976.
Т. 81, вып. 5. С. 7-22.
Логунова К. С. О зависимости электрических потенциалов от обмена веществ сердца // Изв.
АН СССР. 1954. Вып. 6. С. 52-61.
Лоскутов А.В., Павлов М.Н., Пучковский С.В. Бурый медведь. Волжско-Камский край //
Медведи. - М., 1993. С. 91-135.
419
Любашенко С.Я., Петров А.М. Болезни пушных зверей. - М., 1962. 215 с.
Макарова О.А. Исследование на трихинеллез лис и росомах Кольского полуострова.
Млекопитающие // III съезд Всесоюз. териол. о-ва: Тез. докл. - М., 1982. Т. 2. С. 280.
Макридин В.П. Волк // Крупные хищники и копытные звери. - М., 1978. С. 8-50.
Малафеев Ю.М., Кряжимский Ф.В., Добринский Л.Н. Анализ популяции рыси Среднего
Урала. - Свердловск, 1986. 116 с.
Малафеев Ю.М., Кряжимский Ф.В. Значение демографического анализа населения
промысловых животных для оптимизации промысла (на примере рыси) // Научно-технич.
прогресс - в практику перестройки охотничьего хоз-ва: Науч. конф. - М., 1988. С. 53-54.
Мамедов А.Л. Экологические особенности некоторых представителей семейства куньих и
нх хозяйственное значение в Азербайджане: Автореф. дис. ... канд. биол. наук. - Баку, 1972.
26 с.
МаминовМ.К. Некоторые результаты исследования эколого-морфологических особенностей
крови калана (Enhydra lutris) И Изв. тихоокеан. НИИ рыбн. хоз-ва и океанографии. 1971. Т. 80.
С. 240-246.
Мантейфель П.А. Жизнь пушных зверей. - М., 1948. 96 с.
Матюшкин Е.Н. Крупные хищники и падальщики среднего Сихотэ-Алиня И Бюл. МОИП.
Отд. биол. 1974а. Т. 79, вып. 1. С. 5-21.
Матюшкин Е.Н. Рысь, ее настоящее и будущее И Охота и охот, хоз-во. 19746. № 11. С. 20-22.
Матюшкин Е.Н. Рысь. - М., 1974в. 62 с.
Матюшкин Е.Н. География населения рыси // Охота и охот, хоз-во. 1986. № 3. С. 18-20.
МачинскийА.П., СемовВ.Н. Трихинеллез диких животных в Мордовии//Мед. паразитол.
и паразитарн. болезни. 1971. Вып. 5. С. 35-39.
Мачульский С.Н., Тощее А.П. Гельминтозные заболевания соболей в Иркутской области и
Бурят-Монгольской АССР // Тр. Всесоюз. НИИ охоты. - Киров, 1953. Вып. 12. С. 32-35.
Машковцев А.А. Влияние горного климата на конституцию млеко-питающих // Тр. лаб.
эвол. морфологии. - М., 1935. Т. 2, вып. 3. С. 5-30.
Меерсон Ф. 3. Основные закономерности индивидуальной адаптации // Физиология
адаптационных процессов. - М., 1986. С. 10-69.
Межжерин В.А., Мельников ГЛ. Адаптивное значение сезонных изменений некото-
рых морфологических показателей землероек-бурозубок // Acta theriol. 1966. Vol. 11(25).
Р. 503-521.
Мелькина А.Н. Закономерности роста и энергетического метаболизма в постнатальном
онтогенезе американской норки: Автореф. дис. ... канд. биол. наук. - М., 1966. 25 с.
Мельников В.К. Избирательность промысла и определение фактической структуры населения
соболя // Проблемы охотоведения и охраны природы. - Иркутск, 1975. С. 111-115. '
Мельницкий НА. Медведь и охота на него. - СПб., 1915. 223 с.
Мертц П.А. Волк в Воронежской области. Преобразование фауны позво-ночных нашей
страны (Биотехнические мероприятия). - М., 1953. С. 117-135.
Михеичев В.Я. Сравнительно анатомические исследования кишечника разводимых пушных
зверей: Автореф. дис. ... канд. биол. наук. - М., 1955. 10 с.
Михеичев В.Я. О границах отделов кишечника у некоторых млеко-питающих // Тр.
Башкирского с.-х. ин-та. - Уфа, 1960. Т. 9, вып. 2. С. 65-70.
Монахов В.Г. О заболевании филяроидозом соболей разного пола и возраста // Биология и
патология пуш. зверей: Тез. докл. - Петрозаводск, 1981. Ч. 2. С. 51-53.
Монахов В.Г. Избирательность промысла соболя в Северном Зауралье И Количеств, методы
в экологии позвоночных: Урал. науч, центр. - Свердловск, 1983а. С. 29-37.
Монахов В.Г. Заболеваемость фидяроидозом соболей разного пола и возраста // Бюл. МОИП.
Отд. биол. 19836. Т. 88, вып. 4. С. 67-69.
Монахов В.Г. Размещение и рациональное использование ресурсов соболя иа Урале. -
Свердловск, 1985. С. 22-24.
420
Монахов В.Г. Эколого-морфологический очерк соболя Юганского заповедника // Рац.
использование соболя в России. - Красноярск, 2001. С. 118-124.
МонаховВ.Г., Трушин С.П. О влиянии филяроидоза на репродукцию и упитанность соболя
Зауралья // Рац. использование ресурсов соболя в России. - Красноярск, 2001. С. 131-134.
Монахов ГИ. Численность, структура и воспроизводство в популяциях соболя Предбайкалья
и Забайкалья // Оптимальная плотность и оптимальная структура популяций животных. -
Свердловск, 1968. С. 19-21.
Монахов Г.И. Стабильность и изменчивость возрастной структуры популяций соболя //
Зоол. журн. 1974. Т. 53, вып. 9. С. 1654-1662.
Монахов Г.И. Изменчивость и стабильность соотношения полов в популяциях соболя //
Бюл. МОИП. 1975. Т. 80, вып. 4. С. 113-114.
Монахов Г.И., Бакеев Н.Н. Соболь. - М., 1981. 240 с.
Морозов В.Ф. Акклиматизация уссурийского енота как пример успешного преобразова-
ния фауны пушных зверей европейской территории СССР // Зоол. журн. 1953. Т. 32, вып. 3.
С. 120-125.
Морозов В.Ф. Материалы по питанию лесной куницы на Северо-Западе РСФСР //
Промысловая фауна и охотничье хоз-во Северо-Запада РСФСР. - Л., 1963. С. 124-132.
Морозов В.Ф. Особенности питания лесной куницы (Martes martes) в различных областях
Северо-Запада СССР // Зоол. журн. 1976. Т. 55, вып. 12. С. 1886-1892.
Морозов В.Ф. Енотовидная собака И Охота на пушных. - М., 1977. С. 98-140.
Мошонкин Н.Н. Потенциальная полиэстричность европейской норки // Зоол. журн. 1981.
Т. 60, вып. 11. С. 1731-1732.
Найденко С.В. Социальное поведение рыси (Lynx lynx L.) и особенности его формирования
в онтогенезе: Автореф. дис. ... канд. биол. наук. - М., 1997. 24 с.
Насибов С.Б. К экологии камышового кота в Азербайджане // Изв. АН АзССР. 1967. № 3.
С. 31-35.
Насибов С.Б. Эколого-морфологические особенности и хозяйственное значение камышо-
вого и лесного котов в Азербайджане: Автореф. дис. ... канд. биол. наук. - Баку, 1970. 21 с.
Насимович А.А. Экология лесной куницы // Тр. Лапландского заповедника. - М., 1948.
Вып. 3. С. 81-106.
Насимович А.А. Биология ласки на Кольском полуострове в связи с конкурентными
отношениями с горностаем // Зоол. журн. 1949. Т. 38, вып. 2. С. 334-339.
Наумов Н.П. Биология волка // Млекопитающие СССР. - М., 1967. Т. 2, ч. 1. С. 146-193.
Некипелов Н.В. Количество углекислого газа и кислорода в зимних норах грызунов // Изв.
Иркутск, противочум. ин-та Сибири и Дальнего Востока. 1958. Т. 19. С. 138-139.
Никитенко М.Ф. Головной мозг и пути его эволюции у позвоночных // Успехи соврем,
биол. 1963. Т. 56, вып. 3 (6). С. 142-145.
Никитенко М.Ф. Головной мозг, эволюция и систематика млеко-питающих И Внутривидовая
изменчивость наземных позвоночных животных и микроэволюция.-Свердловск, 1966. С. 119-136.
Николаев А.М. Материалы по биологии калана острова Урупа // Сообщ. Сахалин, комплексн.
НИИ. Вост.-Сиб. отд. АН СССР - Южно-Сахалинск, 1958. Вып. 6. С. 48-64.
Никулин НГ. Основы и техника электрокардиографии. - М., 1956. 204 с.
Новиков Б.В. Материалы по морфологии и биологии северо-восточной росомахи // Хищные
млекопитающие: Тр. ЦНИИЛ Главохоты РСФСР. - М., 1981. С. 57-63.
Новиков Б.В. Росомаха. - М., 1993. 135 с.
Новиков ГА. Европейская норка. - Л., 1939. 173 с.
Новиков ГА. Хищные млекопитающие фауны СССР. - М.; Л., 1956. 398 с.
Новиков ГА. К экологии каменной куницы в лесостепных дубравах // Бюл. МОИП. Отд.
биол. 1962. Т. 47, вьш. 6. С. 87-91.
Новиков ГА. О русской научной номенклатуре хищных млекопитающих фауны СССР //
Бюл. МОИП. 1971. Т. 76, вып. 6. С. 115-121.
421
Новиков Г.А., Айрапетьянц А.Э. и др. Звери Ленинградской области // Фауна, экология и
практическое значение. - Л., 1970. 359 с.
Нумеров К.Д. К истории разведения соболя в неволе//Тр. ВНИИЖП. - М., 1969. Вып. 22.
С. 274-292.
Нумеров К.Д. Собопъ. Средняя Сибирь // Соболь, куницы, харза. - М., 1973. С. 70-77.
Обухова А.Д., Шахназаров ГМ. О нормах кормления животных семейства кошек // Тр.
Моск, зоопарка. - М., 1949. Т. 4. С. 180-219.
Овсюкова НИ., Новиков Б.В. Географическая изменчивость некоторых паразитарных заболе-
ваний животных севера Дальнего Востока // Промысловые звери РСФСР. - М., 1982. С. 206-215.
Овсянников НГ,Марюхнич П. В. идр. Структура поселений и динамика популяций песца на
острове Врангеля И Популяция и сообщества животных о-ва Врангеля. - М., 1991. С. 5-23.
Одум Ю. Экология. - М., 1986. Т. 1. 325 с.
Озерецковская Н.Н., Романова В.И., Бронштейн А.М. Трихинеллез в Советской Арктике от
мяса белого медведя // Белый медведь и его охрана в Советской Арктике. - Л., 1969. С. 157-168.
Озерецковская Н.Н., Успенский С.М. Групповое заражение трихинеллезом от мяса белого
медведя в Советской Арктике//Мед. паразитология и паразитарн. болезни. 1957. №2. С. 152-159.
Осмоловская В.И. К биологии барсуков по материалам Московского зоопарка // Бюл.
МОИП. 1948. Т. 53, вып. 2. С. 37-49.
Павлинов И.Я. Природа России: жизнь животных. Млекопитающие. ~М., 1999. Ч. 1.608 с.
ПавлинскийЛ.А. Материалы по размножению и содержанию енотовидной собаки в неволе И
Тр. Новосибирск, зоосада. 1937. Т. 1. С. 37-45.
Павлюченко В.М. Некоторые особенности размножения баргузинского соболя // Тр. Моск,
вет. акад. 1962. Т. 41. С. 41-43
Пажетнов В.С. Мои друзья медведи. - М., 1985. 141 с.
Пажетнов В.С. Бурый медведь. - М., 1990. 215 с.
Пажетнов В.С. Центр Европейской территории России//Медведи. -М., 1993а. С. 51-59.
Пажетнов В. С. Экологические основы охраны и управления популяциями бурого медведя
центральной части Европейской России: Автореф. дис. ... д-ра биол. наук. - М., 19936. 48 с.
Пантелеев П.А. Биоэнергетика мелких млекопитающих. - М., 1983. 271 с.
ПаровщиковВ.Я. К биологии росомахи и выдры //Зоол. журн. 1960. Т. 39, вып. 7. С. 1111.
ПаровщиковВ.Я. Размножение белого медведя архипелага Франца-Иосифа //Бюл. МОИП.
Отд. биол. 1964. Т. 69, вып. 1. С. 127-129.
Переверзева Э.В., Веретенникова Н.Л. К изучению гельминтозных заболеваний у белых
медведей о-ва Врангеля // Экология и морфология белого медведя. - М.,1973. С. 43-54.
Петров А.М. Глистные болезни пушных зверей. - М., 1941. 219 с.
Петров А. М. Опыт полного оздоровления соболей зверосовхоза от легочноглистной болезни
(филяроидоза) и от глистного поражения лобных пазух (скрябингилеза) // Каракулеводство и
звероводство. 1948. № 1. С. 12-13.
Петров А.М., Гагарин В.Г. Изучение цикла развития возбудителей филяроидоза {Filaroides
bronchialis) легких и скрябингилеза лобных пазух {Skrjabingylus petrowi) пушных зверей // Тр.
ВИГИС. 1938а. Т. 3. С. 127-133.
Петров А.М., Гагарин В.Г. Прижизненная дифереициальная диагностика легочных
гельминтозов пушиых зверей // Тр. Всесоюз. ин-та гельминтологии им. акад. К.И. Скрябина.
19386. Т. 3. С. 33-45.
Петров В.В. К вопросу о систематическом положении американских норок // Учен. зап.
Ленингр. пед. ин-та им. А.И. Герцена. 1958. Вып. 179. С. 61-65.
Пианка Э. Эволюционная экология. - М., 1981. 399 с.
Пикулик М. М., Сидорович В. Е. Оценка структурно-функциональных отношений популяций
полуводных хищников и амфибий в Белоруссии // Экология. 1991. № 6. С. 28-36.
Поздняков Е.В. Сезонные изменения в обмене веществ у голубых песцов // Тр. Моск,
пушно-мехового ин-та. 1953. Т. 4. С. 72-76.
422
Покровский А.В., Большаков В.Н. Экспериментальная экология полевок. - М., 1979. 147 с.
Полежаев НМ. Отряд хищные: Енотовидная собака, волк, песец, лесная куница // Фауна евро-
пейского северо-востока России. Млекопитающие. - СПб., 1998. Т. 2, ч. 2. С. 22-61; С. 104-122.
Полежаев Н.М., Нейфелъд Н.Д. Бурый медведь // Фауна европейского северо-востока
России. Млекопитающие. - СПб., 1998. Т. 2, ч. 2. С. 66-79.
Полякова Р.С. О взаимосвязи величины и строения мозга некоторых млекопитающих
(грызунов) с их образом жизни и двигательной активностью // Докл. АН СССР. 1956. Т. 108,
№ 1. С. 733-758.
Полякова Р.С. Межвидовые различия в величине мозга и строении коркового конца
двигательного анализатора некоторых грызунов в связи с их двигательной активностью И Арх.
анатом., гистол. и эмбриол. 1960. Т. 39, № 8. С. 58-64.
Пономарев А.Л. Реакция некоторых куньих (Mustelidae) на градиент температуры // Зоол.
журн. 1944. Т. 23, вып. 1. С. 51-55.
Понугаева А.Г. Стадные реакции обмена веществ у млекопитающих // Опыт изучения
регуляции физиол. функций. - М.; Л., 1953. Т. 2. С. 81-93.
Понугаева А.Г. Физиологические исследования инстинктов млеко-питающих. - М.; Л.,
1960. 180 с.
Попов В.А. Влияние скрябингилезной инвазии горностая на изменение его численности //
ДАН СССР. 1943. Т. 39, № 4. С. 170-172.
Попов В.А. Возрастной состав, кормовая база и гельминтозы горностая как индикаторы
колебаний численности этого вида // Тр. о-ва естествоиспытателей при Казанск. ун-те. 1947.
Т. 57, вып. 3-4. С. 122-125.
Попов Л.Н. Промысловые млекопитающие восточного побережья Таймырского полуострова //
Тр. НИИ полярн. земледелия, животноводства и промыслов, хоз-ва. 1939. Вып. 8. С. 87-121.
Потелов В.А. Белый медведь // Фауна европейского северо-востока России. Млекопитаю-
щие. - СПб., 1998. Т. 2, ч. 2. С. 80-88.
Проссер Л., Браун Ф. Сравнительная физиология животных. - М., 1967. 766 с.
ПыстинА.Н. Рысь //Фауна европейского северо-востока России: Млекопитающие. - СПб.,
1998. С. 182-185.
Райскина М.В. Биохимия нервной регуляции сердца. - М., 1962. 326 с.
Ревенко И.А. Бурый медведь. Камчатка // Медведи. - М., 1993. С. 380-403.
Рожнов В.В., Петрин А.А. Материалы по межвидовым отношениям европейской иорки
(Mustela lutreola L.) с близкородственными видами в период покоя половой системы животных //
Поведение, коммуникация и экология млекопитающих. - М., 1998. С. 67-105.
Романов И.В. Зависимость гельминтофауны соболей Красноярского края от окружающей
ее среды // Зоол. журн. 1959. Т. 38, вып. 9. С. 195-201.
Романов И.В. Изучение очагов дифиллоботриоза в Горьковской обл И Мед. паразитол. и
паразитарн. болезни. 1964. Т. 33, № 3. С. 112-115.
Рощевский М.П. Эволюционная электрокардиология. - Л., 1972. 250 с.
Рощевский М.П. Электрокардиология копытных животных. - Л., 1978. 167 с.
Рубецкая, Щербаков, Зейдель. К биологии европейской норки (Mustela lutreola L.) в
Московском зоопарке // Бюл. зоопарков и зоосадов. - М., 1933. № 10. С. 55-58.
Руковский Н.Н. Взаимоотношение енота в Восточном Закавказье с представителями местной
фауны // Тр. ВНИИ охот, промысла. - М., 1953. № 13. С. 125-128.
Руковский Н.Н. Енот-полоскуи в СССР // Акклиматизация животных в СССР. - Алма-Ата,
1963. С. 230-236.
Руковский Н.Н. Значение птиц в Питании лесной куницы и их влияние на некоторые
особенности этого вида // Тр. Моск. пед. ин-та. 1974. Вып. 5. С. 125-132.
Руковский Н.Н. Питание // Волк. - М., 1985. С. 325-335.
Руковский Н.Н. Бурый медведь Вологодской области // Медведи СССР - состояние
популяций. - Ржев, 1991. С. 220-227.
423
Руковский Н.Н., Куприянов А.Г. Некоторые особенности распространения волка на
Онежском полуострове // Зоол. журн. 1972. Т. 51, вып. 10. С. 1593-1596.
Русаков О. С. Некоторые данные по питанию черного хоря в Ленинградской, Псковской и
Новгородской областях // Промысловая фауна н охот, хоз-во Северо-Запада РСФСР. - Л., 1963.
С. 195-199.
Русанов Я.С. Охота и охрана фауны (влияние охоты на структуру популяций охот,
животных). - М., 1973. 144 с.
Рухлядев Д.Н. Паразиты и паразитозы диких копытных и хищных животных горно-лесного
Крыма // Паразитофауна и заболевания диких животных. М., 1948. С. 41-48.
Рыбкин А.В. Материалы по фауне окрестностей бухты Медузы (Биологическая станция
«Виллем Беренц», Северо-Западный Таймыр) // Арктические тундры Таймыра и островов
Карского моря: Природа, животный мир и проблемы их охраны. - М., 1994. Т. 2. С. 11-21.
Рымарева Е.К К биологии рыси И Бюл. зоопарков и зоосадов. - М., 1933. №3. С. 21-27.
РябовЛ.С. Волки Черноземья. - Воронеж, 1993. 167 с.
Рябов Л. С., Бойко ГМ. Каменная куница в природе и неволе // Охота и охот, хоз-во. 1982.
№ 3. С. 18-20.
Саблина Т.Б. Строение пищеварительного тракта белого медведя // Экология и морфология
белого медведя. - М., 1973. С. 107-113.
Саблина Т.Б., Жарова ГК. Пищеварительная система // Волк. - М., 1985. С. 240-253.
Савельев В.Д. Ленточные черви песцов полуострова Таймыр // Бюл. науч.-техн. информ.
НИИСХ Крайнего Севера. 1974. № 3-4. С. 40-42.
СавельевВ.Д. Паразитические черви промысловых млекопитающих и пути их циркуляции
в тундровых биоценозах полуострова Таймыр: Автореф. дис. ... канд. биол. наук. - Л., 1975.
23 с.
Садыхов И.А. Гельмиитофауна нутрии и енота в условиях их акклиматизации в Азербайджа-
не// Вопросы экологии. - Киев, 1962. Т. 8. С. 120-125.
СалганскаяЛ.О. Про морфолопю кров! ондатри // Докл. АН УРСР. 1962. № 6. С. 809-811.
Сапаев В.М. Половой и возрастной состав промысловых проб колонка // Сб. НТИ ВНИИОЗ:
Охота, пушнина и дичь. - Киров, 1969. Вып. 27. С. 3-5.
Сапельников С.Ф. Методика отлова и результаты мечения лисят в Воронежском заповедни-
ке // Тез. докл. VI съезда териол. о-ва. - М., 1999. С. 225.
Сафронов В.М. Зимняя экология млекопитающих Якутии: Автореф. дис. ... д-ра биол.
наук. - Петрозаводск, 2002. 47 с.
СдобниковВ.М. Опыт массового мечения песцов//Пробл. Арктики. 1940. № 12. С. 21-24.
Сегаль А.Н. Рост, метаболизм и терморегуляция в постнатальном онтогенезе горностая //
Экология, 1975а. № 1. С. 38-44. '
Сегаль А.Н. Очерки экологии и физиологии американской иорки. - Новосибирск, 19756.
260 с.
Седалищев В.Т. К экологии соболя Северо-Восточной Якутии // Соболь: Состояние ресурсов
и перспективы пушного промысла. - Киров, 1998. С. 125-132.
Седалищев В.Т., Аникин Р.К., Плеснивцев В.В. Реакклиматизация соболя в Колымских
районах Якутии // Рац. использ. ресурсов соболя. - Красноярск, 1992. С. 51-56.
Семенов Б.Т. Волки Архангельской области и нх истребление. - Архангельск, 1954. 56 с.
Сергеевский М.В. Дыхательный центр млекопитающих животных и регуляция его
деятельности. - М., 1950. 395 с.
Сидорович В.Е. Особенности экологии полуводных хищников на мелио-ративных каналах
Белоруссии // Интенсификация воспроизводства ресурсов охотничьих животных. - Киров,
1990. С. 90-100.
Сидорович В.Е. Питание и взаимоотношения выдры (Lutra lutra) и американской норки
(Mustela visori) в Белоруссии // Вести. АН Беларуси. - Минск, 1991. 98 с. (Деп. в ВИНИТИ
02.04, № 1431-В91).
424
Сидорович В.Е. Структура популяции выдры (Lutra lutra) в Беларуси // Бюл. МОИП. 1992.
Т. 97, вып. 6. С 43-51.
Сидорович В.Е. и др. Куньи в Беларуси. - Минск, 1997а. 263 с.
Сидорович В.Е., Яковец Н.Н. Суточная активность полуводиых хищников в Белоруссии //
Весщ АН БССР. Сер. б!ял. Навук. 1989. № 1. С. 106-107.
Синицын А.А. Использование запасов колонка и американской норки в Западной Сибири //
Ресурсы охотничье-промысл. хоз-ва и прогноз их использования. - М., 1985. С. 59-72.
Синицын А.А. Особенности экологии и хозяйственное использование американской норки,
акклиматизированной в равнинной части Западной Сибири: Автореф. дис.... канд. биол. наук. -
М„ 1988. 23 с.
Синицын А.А., Кукарцев В.А. Динамика структуры популяций мелких куньих под
воздействием промысла // Обогащение фауны и разведение охотничьих животных: Материалы
Всесоюз. науч.-произв. конф., посвящ. 100-летию со дня рождения П.А. Мантейфеля. - Киров,
1982. С. 158.
Слоним А.Д. Животная теплота и ее регуляция в организме млеко-питающих. - М.; Л.,
1952. 327 с.
Слоним А.Д. Основы общей экологической физиологии млекопитающих. - М.; Л., 1961.
432 с.
Слоним А.Д. Частная экологическая физиология млекопитающих. - М.; Л., 1962. 498 с.
Слоним А.Д. Среда и поведение: Формирование адаптивного поведения. - Л., 1976. 209 с.
Слудский А.А. Взаимоотношения хищников и добычи (на примере антилоп и других
животных и их врагов) // Тр. Ин-та зоол. АН КазССР. - Алма-Ата, 1962. Т. 17. С. 24-143.
Смирнов В.С., Корытин Н.С. Избирательность отлова животных и возможности ее
использования в экологических исследованиях. - Свердловск, 1979. С. 1-78.
Смирнов М.Н. Особенности распространения и экологии рыси (Felis lynx L.) в верховьях
Енисея // Фауна и экология наземных позвоночных Сибири. - Красноярск, 1997. С. 196-209.
Соков А.И. Материалы по биологии каменной куницы (Martes foina Severtzov) в
Таджикистане // Изв. АН ТаджССР. - Душанбе, 1972а. Сер. 3, вып. 48. С. 91-97.
Соков А.И. Новые данные о солонгое (Mustela altaica Pall.) из Таджикистана // Докл. АН
ТаджССР. - Душанбе, 19726. Т. 15, № 5. С. 70-72.
Соколов А.С., Соколов Н.Н. Некоторые особенности органов движения лесной куницы (Martes
martes L.) в связи с образом жизни // Бюл. МОИП. Отд. биол. 1971. Т. 76, вып. 6. С. 40-51.
Соколов А.Я. Частота пульса у северного оленя (Rangifer tarandus) в условиях свободного
поведения //Журн. эвол. биохим. и физиол. 1990. Т. 26, № 2. С. 182-184.
Соколов А.Я., Задалъский С.В. и др. Энергетическая стоимость различных форм поведения
у северного оленя в летнее и зимнее время И Биоэнергетика и термодинамика живых систем. -
Новосибирск, 1984. С. 32-34.
Соколов В.Е., Кузнецов ГВ. Суточный ритм активности млекопитающих: Цитологические
и экологические аспекты. - М., 1978. 263 с.
Соколов В.Е., Рожнов В.В. Территориальность, агрессивность и маркировка у куньих
(Mustelidae) И Млекопитающие. Исследования по фауне Советского Союза: Сб. тр. Зоол. музея
МГУ. - М„ 1979. С. 163-214.
Соколов В.Е., Рожнов В.В. Изменчивость реакции черных хорей на запаховые сигналы
особей разного пола и вида // Изв. АН СССР. Сер. Биол. 1989. № 1. С. 5-10.
Соколов ГА. Экологическая структура популяции соболя и пути рационального
использования его численности И Пути и методы рациональной эксплуатации и повышения
продуктивности охот, угодий. - М., 1978. С. 215-217.
Соколов ГА. Экологические основы управления репродуктивным потенциалом соболя И
Рац. использование ресурсов соболя. - М., 1992. С. 62-67.
Соколов ГА. Экологические основы управления популяциями соболя // Рац. использование
ресурсов соболя в России. - Красноярск, 2001. С. 82-88.
425
Соколов Е.А. Емкость пищеварительного тракта и показатели перевариваемости у пушных
зверей // Тр. Моск, зоотехн. ин-та. - М., 1941. Т. 1. С. 72-83.
СоколовЕ.А. Охотничьи животные // Корма и питание промысловых зверей и птиц. - М.,
1949а. Вып. 1. 259 с.
Соколов Е.А. Сезонные изменения в основном обмене у енотовидной собаки И Тр. Моск,
пушио-мехового ин-та. 19496. Вып. 2. С. 3-27.
Соколов И.И, Соколов А.С. Некоторые особенности органов движения речной выдры и
калана в связи с образом жизни // Бюл. МОИП. Отд. биол. 1970. Т. 75, вып. 5. С. 5-17.
Сороченко Е.В. Распространение и особенности эпидемиологии альвео-коккоза и
трихинеллеза в Ненецком национальном округе: Автореф. дис. ... д-ра мед. наук. - Л., 1971.
24 с.
Сороченко Е.В., Колесов Н.В. Заражение трихинеллезом голубых песцов и серебристо-
черных лисиц мясом медведя в Ненецком национальном округе // Мед. паразитология и
паразитари. болезни. 1962. № 3. С. 366-367.
Смелов В.А. Питание и некоторые морфологические особенности пищеварительной системы
куиьих (Mustelidae): Автореф. дис. ... канд. биол. наук. - Л., 1985. 18 с.
Старков И.Д. Биология и разведение соболей и куниц. - М., 1947. 132 с.
Стерлигов А.В. Особенности экологии, охраны и использования американской и
европейской норок в Камско-Вятском междуречье: Автореф. дис. ... канд. биол. наук. - М.,
1991. 20 с.
Стоянова В.К, Сваринская В.Г. Наблюдения за рысятами в Рижском зоосаде//Материалы
Прибалт, конф. - Рига, 1964. С. 21-25.
Стрельников И.Д. Особенности величины мозга и теплопродукции у грызунов // Докл. АН
СССР. 1953. Т. 88, № 2. С. 377-380.
Стрельников ИД. Микроклимат нор и гнезд грызунов//Зап. ЛСХИ. 1955. Вып. 9. С. 140-166.
Стрельников И.Д. Анатомо-физиологические основы видообразования позвоночных. -
Л., 1970. 367 с.
Строганов СУ. Звери Сибири. Хищные. - М., 1962. 458 с.
Сутула В.И., Попов В.Н. Соболь в Байкальском заповеднике // Рац. использование ресурсов
соболя в России. - Красноярск, 2001. С. 147-152.
Тазнева З.Х., Лобачев Ю.С. Гельминты каменной куницы (Martes foina Erxl.) и горно-
стая (Mustela erminea L.) Джунгарского и Заилийского Алатау // Изв. АН КазССР. 1970. № 4.
С. 56-63.
Гейлов В.П. Материалы по биологии дикого кота (Felis silvestris caucasicus Satunin) в условиях
Кавказского гос. заповедника//Тр. Кавказского гос. заповедника. 1938. Вып. 1. С. 331-341.
Теплое В.П. К вопросу о соотношении полов у диких млекопитающих // Зоол. журн. 1954.
Т. 33, № 1. С. 837-839.
Теплое В.П. К зимней экологии росомахи в районе Печоро-Илычского заповедника // Бюл.
МОИП. 1955. Т. 9, вып. 1. С. 3-11.
ТеплоеВ.П. Динамика численности и годовые изменения в экологии промысловых животных
Печерской тайги // Тр. Печеро-Илыч. заповедника. 1960. № 8. С. 63-71.
Терновский Д.В. Биология и акклиматизация американской норки (Lutreola vison Brisson)
на Алтае. - Новосибирск, 1958. 138 с.
Терновский Д.В. Исследования репродуктивного цикла у горностая // Оптимальная плотность
и оптимальная структура популяций животных. - Свердловск, 1972а. Вып. 3. С. 31-33.
ТерновскийД.В. Материалы по размножению куницеобразных//Оптимальная плотность и
оптимальная структура популяций животных. - Свердловск, 19726. Вып. 3. С. 35-36.
Терновский Д.В. Особенности размножения ласки и горностая II Охота и охот, хоз-во.
1974. № 6. С. 16-18.
Терновский Д.В. Биология куницеобразных (Mustelidae). - Новосибирск, 1977. 280 с.
ТерновскийД.В., Терновская Ю.Г Экология куницеобразных. - Новосибирск, 1994. 200 с.
426
ТерновскийД.В., ТерновскаяЮ.Г, ТумановИ.Л. Экологическая обусловленность сердечной
деятельности у Mustelidae // Вести, зоол. 1981. № 1. С. 62-69.
Тимофеев В.В., Надеев В.Н. Соболь. - М., 1955. 402 с.
ТитоваМ.И. Нормы поддерживающего кормления взрослых соболей//Тр. ЦНИЛ пушного
звероводства. 1950. Вып. 5. С. 124-127.
Тихонов И.Ф. Кровоснабжение органов брюшной полости норок // Вопр. растениеводства
и животноводства Карелии: Учен. зап. Петрозаводского ун-та. 1966. Т. 14, вьгп. 3. С. 64-68.
Трабенкова Н.А. Мониторинг и регуляция пресса глистных инвазий соболя И Материалы
III Всерос. науч.-произв. совещ. - Красноярск, 1992. С. 78-82.
Трабенкова Н.А. Особенности территориального размещения гельмии-тозных инвазий у
соболей Камчатской области // Рац. использование ресурсов соболя в России. - Красноярск,
2001. С. 153-155.
Трабенкова НА., Валенцев А.С. Регулирование плодовитости соболей // Материалы III
Всерос. науч.-произв. совещ. - Красноярск, 1992. С. 82-86.
Трапезов О.В. Доместикация как один из возможных путей сохранения биоразнообразия
(на примере речной выдры Lutra lutra L.) // Генетика. 1997. Т. 33, вып. 8. С. 1162-1167.
Треничев Е.М. Особенности поведения и освоения запасов куниц разного пола и возраста //
Млекопитающие в системе природопользования на Урале. - Свердловск, 1985. С. 38-40.
Третьякова О.Н. О трихинеллезе диких животных Башкирии // Тр. Башкирского с.-х. ин-та.
1956. Т. 7. С. 58-63.
Троицкая А.А. Гельминтофауна диких пушных зверей Татарской АССР //Уч. зап. Казанского
ун-та. 1960. Т. 120, вып. 6. С. 335-357.
Троицкая А.А. К экологической характеристике гельминтофауны лесной куницы Волжско-
Камского края // Материалы науч. конф. Всесоюз. о-ва гельминтологов. - М., 1962. Ч. 2. С. 202-
204.
Троицкая А.А. Зависимость скрябингилезной и филяроидозной инвазий лесной куницы от
урожая ягод рябины // Материалы науч. конф. Всесоюз. о-ва гельминтологов. - М., 1963. Ч. 2.
С. 136-137.
Троицкая А.А. К изучению гельминтофауны диких пушных зверей Среднего Поволжья и
Башкирской АССР //Тр. Всесоюз. НИИ животного сырья и пушнины. - М., 1967. Вып. 21. С. 266-277.
Троицкая А.А., Горшков П.К. К изучению фауны гельминтов барсука и его роли в
поддержании природных очагов некоторых гельминтов в Волжско-Камском крае // Восьмая
Всесоюз. конф, по природным очаговым болезням животных и охране их численности. - Киров,
1972. С. 128-129.
Троицкий ГА. Численность и промысел лося в Карелии // Вопр. экологии животн. -
Петрозаводск, 1974. С. 161-173.
Туманов И.Л. Деяк! функцюнальи! особливост! будови серцево-судинно! системи
мышевидных грызушв //Доповщ АН УРСР. 1966а. № 9. С. 263-266.
Туманов И.Л. Деяк1 морфо-ф!з!олопчш особливост! серцево-судинно! системи грызушв,
зумовлеш умовами кнування // Биология корисних та шкщливих тварин Украши. - Кшв, 19666.
С. 134-138.
Туманов И.Л. Еколого-морфолопчна адаптация венизних мапстралей грызушв у зв’язку з
перебуванням в норах И Доповщ! АН УРСР. 1967. № 4. С. 361-364.
Туманов И.Л. Взаимосвязь дыхательной функции крови с особенностями образа жизни
грызунов // Биологические науки. 1968. №8. С. 17-20.
Туманов И.Л. Межвидовые различия в показателях двигательной активности мышевидных
грызунов // Вести, зоологии. 1969. № 2. С. 15-19.
Туманов И.Л. О размножении и постэмбриональном развитии барсука и выдры // Биол.
науки. 1971. № 9. С. 20-26.
Туманов И.Л. Возрастные изменения морфо-физиологических пока-зателей некоторых видов
куньих (Mustelidae) И Зоол. журн. 1972. Т. 51, вып. 5. С. 694-703.
427
Туманов И.Л. Распространение куньих и промысловое использование их запасов на северо-
западе РСФСР // Науч.-техн, информ. ВНИИОЗ. - Киров, 1973. Вып. 40-41. С. 23-28.
Туманов И.Л. Морфофункциональные особенности кровеносной системы куньих II Тр.
Зоол. ин-та АН СССР. - Л., 1974а. Т. 54. С. 123-147.
Туманов И.Л. Возрастная изменчивость морфологических показателей у американской
норки и солонгоя // Биология и патология пушных зверей. - Петрозаводск, 19746. С. 76-78.
Туманов И.Л. Сезонные изменения веса и суточной нормы корма у черного хоря и
американской норки // Материалы конф, по фенологии млекопитающих и птиц. - М., 1976а.
С. 34-36.
Туманов И.Л. Некоторые черты поведения куньих И Науч.-техн, информ. ВНИИОЗ:
Поведение охотничьих животных. - Киров, 19766. Вып. 51-52. С. 20-27.
Туманов И.Л. О потенциальной полиэстричности куньих // Зоол. журн. 1977а. Т. 56, вып. 4.
С. 619-625.
Туманов И.Л. Уссурийский енот-перспективный вид для клеточного разведения //Биология
и патология клеточных пушных зверей: Тез. докл. - Киров, 19776. С. 110.
Туманов И.Л. О потенциальной полиэстричности норок и других мелких куньих // Биология
и патология клеточных пушных зверей: Тез. докл. - Киров, 1977в. С. 110-111.
Туманов И.Л. Влияние различных способов и сроков добычи иорок на структуру
промысловых проб // Влияние хоз. деятельности человека на популяции охот, животных и среду
их обитания: Тез. докл. - Киров, 1980. Т. 2. С. 56.
Туманов И.Л. Особенности вертикального распределения лесной и каменной куниц в горах
Азербайджана И Сезонная ритмика природы горных областей: Тез. докл. 1-го Всесоюз. совещ.
по горной фенологии. - Л., 1982. С. 162-163.
Туманов И.Л. Возрастные изменения соотношения полов у американской норки // Фауна и
экология птиц и млекопит. северо-запада СССР. - Петрозаводск, 1983. С. 126-132.
Туманов И.Л. О некоторых репродуктивных особенностях хищных млекопитающих // Вести,
зоол. 1984. № 6. С. 55-60.
Туманов И.Л. Определение плодовитости куниц и соболей: экспресс-метод И Охота и
охот, хоз-во. 1988а. № 10. С. 15.
Туманов И.Л. Простой и надежный способ определения плодовитости некоторых
промысловых животных И Рационализация методов изучения охотничьих животных. - Киров,
19886. С. 127-136.
Туманов И.Л. Некоторые черты экологии ласки (Mustela nivalis) И Фауна и экология наземных
позвоночных. - Петрозаводск, 1988 в. С 130-138.
Туманов И.Л., Девяткин Г.В. Методика определения плодовитости соболей // Рац.
использование ресурсов соболя. - Красноярск, 1992. С. 86-87. '
Туманов И., Кожечкин В., Буянов Ю. Росомаха - «гиена севера» // Охота и охот, хоз-во.
1999. № 9. С. 6-8.
Туманов И.Л., Кожечкин В.В. Величина выводков и соотношение полов в пометах росомахи
(Gulo gulo L. 1758) И Мониторинговые исследования в заповедниках Южной Сибири. - Кемерово,
2000. Вып. 1. С. 140-144.
Туманов И.Л., Коровники.Л. О реакциях надпочечной железы куньих при разных способах
их добычи // Экология наземных позвоночных северо-запада СССР. - Петрозаводск, 1981.
С. 136-142.
Туманов И.Л., Коровник И.Л. Сезонные ритмы функциональной активности надпочечной
железы и ее связь с плотностью населения у куньих // Экология наземных позвоночных северо-
запада СССР. - Петрозаводск, 1986. С. 95-103.
Туманов И.Л., Левин В.Г. Возрастные и сезонные изменения некоторых физиологических
показателей у ласки и горностая // Вести, зоол. 1974. № 2. С. 25-30.
Туманов И.Л., Левин В.Г., Лысов Е.С. К биологии горностая в Ленинградской области //
Науч.-техн, информ. ВНИИЖП. - Киров, 1970. Вып. 31. С. 22-26.
428
Туманов И.Л., Сидорович В.Е. Репродуктивные особенности выдры (Lutra lutra) И Зоол.
журн. 1994. Т. 73, вып. 9. С. 167-171.
Туманов И.Л., Смелов В.А. Материалы по питанию некоторых куньих на Северо-Западе
СССР // Фауна и экология птиц и млекопитающих таежного северо-запада СССР. - Петрозаводск,
1978. С. 167-174.
ТумановИ.Л., СмеловВ.А. Кормовые связи куньих на Северо-Западе РСФСР//Зоол. журн.
1980. Т. 59, вып. 10. С. 1536-1544.
Туманов И.Л., Сорина Е.А. Динамика энергетики питания самок куньих (Mustelidae) //
Вести. СПб. ун-та. 1997а. Сер. 3, вып. 1. С. 16-25.
Туманов И.Л., Сорина Е.А. Сезонная изменчивость энергетики питания самцов мелких
видов куньих (Mustelidae) // Вести. СПб. ун-та. 19976. Сер. 3, вып. 4. С. 38-44.
Туманов И.Л., Сорина Е.А. Особенности энергетики питания и репро-дуктивной циклики
некоторых видов куньих (Mustelidae) // Экология. 1999. № 1. С. 60-66.
Туманов И.Л., Терновская Ю.Г, Терновский Д.В. Некоторые физиологические показатели
и особенности поведения куницеобразных (Mustelidae) // Поведение охотн. животных. - Киров,
1981. С. 31-37.
Туманов И.Л., Терновский Д.В., Терновская Ю.Г. Постнатальные изменения массы,
температуры тела и сердечного ритма куньих // Физиол. состояние пушных зверей и пути его
регуляции. - Петрозаводск, 1982. С. 162-173.
Урманов М.И. Сравнительно морфологическая характеристика печени ластоногих и калана//
Тр. ВНИИ мор. и рыб. хоз-ва и океанографии. 1971. Т. 82. С. 289-300.
Успенский С.М. Белый медведь. - М., 1989. 189 с.
Устинов С.К. Бурый медведь Прибайкалья // Человек и природа. - М., 1987. Вып. 1. С. 22-77.
Устинов С.К. Прибайкалье. Медведи. - М., 1993. С. 275-300.
УткинЛ.Г. Морфологические и физиологические свойства крови пушных зверей: Автореф.
дис. ... канд. биол. наук. -М., 1956. 25 с.
Уткин Л.Г. Картина кровн у пушных зверей в зависимости от возраста и пола // Тр.
НИИ пушного звероводства и кролиководства. 1963. Т. 4. С. 83-112.
Фетисов А. С. Зимние кормовые объекты куницеобразных Забайкалья // Изв. биол.-геогр.
НИИ при Вост.-Сиб. гос. ун-те. - Иркутск, 1942. Т. 9, вып. 3-4. С. 118-124.
Филатова Л.Г. Реакция на гипоксемию у млекопитающих с различной экологической
специализацией // Совещ. по экол. физиологии: Тез. докл. - М.; Л., 1958. Вып. 1. С. 75.
Филимонов А.Н. К вопросу о внутрипопуляционной численности волка // Сб. науч. тр.
ВНИИ охраны природы и заповедного дела МСХ СССР. 1980. С. 37-43.
Флинт В.Е. К биологии размножения перевязки // Бюл. МОИП. Отд. биол. 1962. Т. 67,
вып. 4. С. 143.
Фогельсон Л.И. Клиническая электрокардиография. - М., 1957. 158 с.
Халилов Ф.К К сравнительной морфологии кишечника млекопитающих в связи с характером
питания // Зоол. журн. 1955. Т. 34, вып. 2. С. 415-426.
Хатт Ф. Генетика животных. - М., 1969. 345 с.
Хлебников А.И. Экология соболя Западного Саяна. - Новосибирск, 1977. 117 с.
Цалкин В.И. К биологии белого медведя архипелага Франца-Иосифа // Бюл. МОИП. 1936.
Т. 45, вып. 5. С. 335-363.
Цицорина Т.Н., Терновский Д.В. Морфологические параметры некоторых Mustelidae в
доимплантационный и ранний имплантационный периоды И Первый междуиар. териологический
конгресс. - М., 1974. Т. 2 С. 315.
Черногоров И.А. Нарушение ритма сердца. - М., 1962. 371 с.
Чернявский Ф.Б., Кречмар А.В., Кречмар М.А. Бурый медведь. Север Дальнего Востока //
Медведи. - М„ 1993. С. 318-348.
Чернявский Ф.Б., Ткачев А.В. Популяционные циклы леммингов в Арктике. - М., 1982.
352 с.
429
Черняев Э.Г. Особенности строения задней полой вены некоторых насекомоядных // Докл.
АН СССР. 1964. Т. 156, № 6. С. 1483-1485.
Шахматова В.И. Гельминты куньих Карелии и цикл развития Taenia intermedia Rudolphi,
1809: Автореф. дис. ... канд. биол. наук. - М., 1964. 24 с.
Шахматова В.И. Гельминты куньих Карелии // Тр. гельмиитол. лаб. АН СССР. 1966.
Т. 17. С. 277-289.
Шварц С.С. Метод морфофизиологических индикаторов в экологии наземных позвоночных
животных // Зоол. журн. 1958. Т. 37, вып. 2. С. 161-173.
Шварц С.С. Некоторые закономерности экологической обусловленности интерьерных
особенностей наземных позвоночных животных // Проблемы флоры и фауны Урала. - Свердловск,
1960. С. 113-177.
Шварц С.С. Эволюционная экология животных. - Свердловск, 1968. 198 с.
Шварц С.С. Экологические закономерности эволюции. - М., 1980. 278 с.
Шварц С.С., Смирнов В.С.,ДобринскийЛ.Н. Метод морфофизиологических индикаторов
в экологии наземных позвоночных // Тр. Ин-та экологии растений и животных. - Свердловск,
1968. Вып. 58. 386 с.
Шварц ТВ. Функциональная активность щитовидной железы у самок соболей в различ-
ные стадии полового цикла // Биология и патология пушных зверей. - Петрозаводск, 1974.
С. 91-94.
Шило Р.А., Батенева Н.А. Опыт разведения некоторых куницеобразных (Mustelidae) в
Новосибирском зоопарке // Достижения зоопарков в изучении биол. разведения редких и
исчезающих животных: Информ, материал междунар. науч. конф. - М., 1989. С. 7-8.
Шило Р.А., Немшилова И.В. Суточная активность речной выдры в условиях зоопар-
ка // Зоопарки и их роль в сохранении диких животных: Сб. науч. тр. - Алма-Ата,
1989. С. 136-143.
Шилов И.А. Эколого-физиологические основы популяционных отношений у животных. -
М„ 1977. 263 с.
Шилов И.А. Популяционный гомеостаз у животных // Бюл. МОИП. 1982. Т. 87, № 4. С. 23-32.
Шилов И.А. Физиологическая экология животных. - М., 1985. 328 с.
Шиляева Л.М. Гельминты песца и их воздействие на популяцию хозяина в Ненецком
национальном округе // Зоол. журн. 1968. Т. 47, вып. 3. С. 331-336.
Шиляева Л.М. Гельминты как фактор динамики численности песца европейского Севера //
Тр. IX междунар. конгр. биологов-охотоведов. - М., 1970. С. 616-620.
ШиляеваЛ.М. Структура популяции песца (Alopex lagopus L.) и роль различных генераций
в динамике его численности // Зоол. журн. 1971. Т. 50, вып. 12. С. 1112-1125.
Шиляева Л.М. Экология и основы прогнозирования численности песца на примере
североевропейской популяции: Автореф. дис. ... канд. биол. наук. - М., 1972. 29 с.
Шиляева Л.М. К вопросу о популяционной структуре материкового песца // Экология,
1974. № 1. С. 54-61.
Шиляева Л.М. Смертность и выживаемость песца Енисейского Севера на разных фазах
популяционного цикла // Промысловая териология. - М.,1982. С. 116-126.
ШималовВ.Т. Зависимость поведения животных в Белоруссии от различных факторов //
Управление поведением животных. - М., 1977. С. 329-331.
Шималов В.Т., Сидорович В.Е., Шималов В.В. Гельминты куньих, которые живут около
водоемов Беларуси // Вести. АН Беларуси. 1993. № 4. С. 96-101.
Шималов В.В., Шималов В.Т. Черви семейства Opisthorchidae (Trematoda, Fasciolida) у
диких животных Белорусского Полесья и их роль в медицине // Весщ нацыян. Акадэмп навук
Беларусг Серыя б!ялапчных навук. 1998. № 2. С. 102-105.
ШималовВ.Т., ШималовВ.В. К изучению гельминтофауны редких видов млекопитающих
(рысь, европейская норка) Беларуси // Материалы междунар. науч.-практ. конф. «Биол. ритмы». -
Брест, 1999а. С. 183-185.
430
Шималов В.В., Шималов В Т. Хищные млекопитающие семейства Canidae и Mustelidae
(Mammalia, Carnivora) Белорусского Полесья - хозяева гельминтов, паразитирующих у человека //
Весшк Брэсцкага ушверспэта. 19996. № 2. С. 77-80.
Шималов В.В., Шималов В.Т. Дикие псовые Белорусского Полесья - источники
распространения гельминтов, паразитирующих у человека // Современная паразитология:
проблемы и перспективы: Тр. науч. конф. - Витебск, 1999в. С. 137-141.
Шихобалова Н.И. Вопросы иммунитета при гельминтоценозах. - М.; Л., 1950. 134 с.
Шмидт-Ниелъсен К. Физиология животных: Приспособление и среда. - М., 1982. Т. 1-2. 785 с.
Юргенсон П.Б. Опыт сравнительной экологии рода Martes'. Автореф. дис. ... канд. биол.
наук. - М., 1950. 10 с.
Юргенсон П.Б. Эколого-географические аспекты в питании лесной куницы и географическая
изменчивость эколого-географических адаптаций ее жевательного аппарата // Зоол. журн. 1951.
Т. 30, вып. 2. С. 172-185.
Юргенсон П.Б. Очерки по сравнительному изучению соболя и куниц // Сб. материалов по
результатам изучения млекопитающих в гос. заповедниках. - М., 1956. С. 52-55.
ЮшковВ.Ф. Материалы по гельминтофауне диких млекопитающих Приполярного Урала //
Тр. Коми филиала АН СССР. - Сыктывкар, 1977. № 34. С. 102-124.
Юшков В. Ф. Гельминты малоизученных видов млекопитающих Коми АССР//Тр. Комн
НЦ УрО СССР. - Сыктывкар, 1989. № 100. С. 102-111.
Юшков В.Ф. Фауна европейского северо-востока России. - СПб., 1995. Т. 3. 202 с.
Яблоков А.В. Изменчивость млекопитающих. - М., 1966. 364 с.
Яблоков А.В. Популяционная биология. - М., 1987. 303 с.
ЯзанЮ.П. Повинна ли куница в сокращении численности белки И Зоол. журн. 1962. Т. 41,
вып. 4. С. 633-635.
ЯкушкинГ.Д. Песец Таймыра и рациональное использование его запасов: Автореф. дис....
канд биол. наук. - М., 1967. 20 с.
Якушкин ГД. Песец // Охот, хоз-во Енисейского севера. - Красноярск, 1977. С. 29-37.
Янушевич А.И., Айзин В.М. и др. Млекопитающие Киргизии. - Фрунзе, 1972. 462 с.
Яскин В.А. Изменение веса головного мозга и веса гиппокампа рыжих полевок в связи с
динамикой численности популяции //Проблемы экологии, рац. использования и охраны природн.
ресурсов иа Урале. - Свердловск, 1980. С. 112-115.
Яскин В.А. Сезонная и годичная изменчивость некоторых особенностей морфологии
головного мозга полевок: Автореф. дис. ... канд. биол. наук. - Свердловск, 1981. 23 с.
Arieli R. The atmospheric environment of the fossorial mole rat (Spalax ehrenbereji)'. Effects
of season, soil texture, rain, temperature and activity // Comp. Biochem. Physiol. 1979. Vol. A63,
N 4. P. 569-575.
Aulerich R.J., Swindler D.R. The dentition of the mink (Mustela vison) И J. Mammal. 1968.
Vol. 49(3). P. 488-494.
Baskirk S. W., Harlow HL Body-fat dynamics of the American marten (Martes americana) in
winter// J. Mammal. 1989. Vol. 70. P. 191-193.
Becker K. Geschlechtsunterschiede am Becken von Miusen (Murinae) und Wihlmiusen
(Microtinae) // Zool. Jahrb. Abt. 3. 1954. Bd 82, N 5. S. 143-156.
Bjarvall A., Danielsson S. et al. Experiences with the first radio - collared wolverins in Sweden //
J. Wild Res. 1996. Vol. 1. P. 3-6.
Bonnin-Laffarque M., Canivenc R. Etude de 1’activite du blaireau Europeen badgers (Meles
meles L.) // Mammalia. - Paris, 1961. T. 25, N 4. S. 476-484.
Brandford P.R.S. Biology of the Polecat Mustela putorius'. a literature review // Mammal Rev.
1987. Vol. 17, N 4. P. 155-198.
Brzezinski M.,Jedrzejewski W.,JedrzejewskaB. Diet of otter (Lutra lutra) inhabiting small rivers
in the Bialowieza National Park, eastern Poland // J. Zool. - London, 1993. Vol. 230. P. 495-501.
431
Bucher P, Warner R., Jenkins S. Population density, colony composition, and local movements
in two Sierra Nevada beaver populations // J. Mammal. 1983. Vol. 64, N 2. P. 314-318.
Canivenc R. A study of progestation in the european badger (Meles meles L.) // Comparative
biology of reproduction in Mammals. - London, 1966. P. 76-81.
Canivenc R., Bonnin-Laffargue M., Relexans M.-C. Cycles genitaux de quelques mustellides euro-
peens // Cycles genit. Saisonn. Mammifer sauvages: Ser. physiol. - Paris, 1968. N 1. P. 134-138.
Christian J. J. A review of endocrine, response in rats and mice to increasing population size
including delayed effects on offspring // Naval. Med. Pres. Inst. Leet. Rev. 1957. N 57. P. 443-462.
Derocher A.E. Latitudinal variation in litter size of polar bears: ecology or methodology// Polar
Biol. 1999. Vol. 22. P. 350-356.
Derocher A.E., Stirling J. Age - specific reproductive performance of female polar bears //
J. Zool. - London, 1994. Vol. 234. P. 527-534.
Drescher H.E. Die Homologisierung der Darmabschuitte bei Musteliden // Zool. Anz. - Jena,
1976. Vol. 196, N 3-4. S. 279-288.
Dunstone N., Birks J.D.S. The feeding ecology of mink (Mustela vison) in a coastal habitat //
J. Zool. - London, 1987. Vol. 212. P. 69-83.
Dunstone N. The Mink. - London, 1993. 232 p.
Enders R.K., Enders A.C. Morphology of the female reproductive traet during delayed
implantation in the mink // Delayed Implantation / Univ. Chicago. Chicago, 1963. P. 129-140.
Erlinge S. Food habits of the fish-otter, Lutra lutra L., in south Swedish habitats // Viltrevy.
1967. Vol. 4. N 6.
Erlinge S. Territoriality of the otter Lutra lutra L. // Oikos. 1968. Vol. 19. P. 186-209
Erlinge S. Feeding habits of the weasel Mustela nivalis in relation to prey abundance // Oikos.
1975. Vol. 26. P. 378-384.
Erlinge S. Food preference, optimal diet and reproductive output in stoats Mustela erminea in
Sweden// Oikos. 1981. Vol. 36. P. 303-315.
Erlinge S. Demography and dynamics of a stoat Mustela erminea population in a diverse
community of vertebrates // J. Anim. Ecol. 1983. Vol. 52. P. 705-726.
Erlinge S. Specialists and generalists among the mustelids // Lutra. 1986. Vol. 29. P. 5-11.
Fahmy M.A.M. Studies on same Mammals (Carnivores and Rodents) // Ztschr. fur Parasitenk.
1954. N 25. S. 135-147.
Fairley J. Food of otter (Lutra lutra) from Co Galway, Ireland and notes on other aspects of
their biology // J. Zool. - London, 1972. N 166. P. 469-474.
Feliu C.,Tores J. et al. On the helminthfaunas of semiaquatic carnivores in the Iberian Peninsula.
Semiaquatische Saugetiere // Wiss. Beitr. Univ. Hall. 1992. P. 353-356.
Fog M. Studies on the weasel (Mustela nivalis) and the stoat (Mustela erminea) in Denmark //
Dan. Rev. Game Biol. 1969. Vol. 6, N 2. P. 14.
Folk G.E., Brewer M.D., Sanders D. Cardiac physiology of polar bear in winter dens // Arctic.
1970. Vol. 23, N 2. P. 130.
Frisch J., Qristsland N.F., KrogJ. Insulation of furs in water// Comp. Biochem. and Physiol.
1974. Vol. 47. P. 402-410.
Gebczynski M. Seasonal and age changes in the metabolism and activity of Sorex araneus // Acta
theriol. 1965. Vol. 10, N 22. P.303-331.
Gebczynski M. The rate of metabolism of the lesser shrew (Sorex minutus) // Acta theriol. 1971.
Vol. 16, N 20. P. 329-339.
Gebczynski M. Heat economy and the energy cost of growth in the bank vole during the first
month of postnatal life // Acta theriol. 1975. Vol. 20, N 29. P. 379-434.
Gebczynski M., Gorecki A., DrozdzA. Metabolism, food assimilation and bioenergetics of three
species of dormice (Ciliridae) // Acta theriol. 1972. Vol. 17, N 21. P. 271-294.
Gerell R. Dispersal and Acclimatization of the Mink (Mustela vison Schreb.) in Sweden //
Viltrevy. 1967. Vol. 5, N 1. P. 3-38.
432
Gerell R. Population studies on the mink (Mustela vison Schreb.) in Sweden // Viltrevy. 1968.
Vol. 8. P. 83-114.
Gittleman J.L. Carnivore behaviour, ecology and evolution. - New York, 1996. 235 p.
Gorecki A. Energy of body in small mammals//Acta theriol. 1965. Vol. 10, N 23. P. 333-352.
Green J., Green R., Jefferies D.J. A radio-tracking survey of otters Lutra lutra on a Perthshire
river system // Lutra. 1984. Vol. 27. P.85-145.
Grodzinski W. Metabolism rate and bioenergetics of small rodents from the deciduons forest //
Bull. Acad. Polon. Sci. Ser. sci. biol. 1961. N 12. P. 67-84.
Haglund B. Winter food habits of the lynx (Lynx lynx) and the wolverine (Gulo gulo) as
revealed by tracking in the snow // Trans, int. Un. Game Biol. 1965. Vol. 7. P. 51-55.
HaglundB. De store rovdjareus vintervanor // J. Viltrevy. 1966. Vol. 4, N 3. P. 79-310.
Haglund B. De fura stora rovdjuren // Kgl. skogs-och lantbruksakad. tidskr. 1968. Vol. 107.
P. 134-147.
Hamilton W.J. Early sexual maturity in the female start-tailed weasel // Science. 1958. Vol. 127,
N 3305. P. 1057-1062.
Hansson J. Transmission of the parasitic nematode Skrjabingylus nasicola (Leuckart, 1842) to
species of Mustela (Mammalia) // Oikos. 1967. N 18. P. 247-252.
Hansson J. Cranial helminth parasites in species of Mustelidae: 1. Frequency and damage in
fresh mustelids from Sweden // Oikos. 1968. N 19. P. 217-233.
Hansson J. Cranial helminth parasites in species of Mustelidae: II Regional frequencies of
damage in preserved crania from Denmark, Finland, Sweden, Greenland and the N.E. of Canada
compared with the helminthinvasion in fresh mustelid skull from Sweden // Ark. Zool. 1970. N 22.
P. 571-594.
Harington C.R. Denning habits of the polar bear (Ursus maritimus Phipps) // Canad. Wildlife
Serv. Rep. Ser. 1968. N 5. P. 1-30.
Harlow H.J. Torpor and other physiological adaptation of the badger Taxidea taxus to cold
environments // Physiol. Zool. 1981a. Vol. 53. P. 267-273.
Harlow H.J. Metabolic adaptations to prolonged food deprivation by the American badger
Taxidea taxus // Physiol. Zool. 1981b. Vol. 54. P. 276-284.
Harlow H.J., Miller B. Non-shivering thermogenesis in the American badger // Comp. Biochem.
Physiol. 1985. Vol. 80A. P. 159-161.
Harrison R.J., Neal E. G. Ovulation during delayed implantation and other reproductive phenomena
in the badger (Meles meles L.) // Nature. 1956. Vol. 177, N 4517. P. 122-128.
Hart J.S. Seasonal changes in insulation of the fur // Canad. J. Zool. 1956. Vol. 34,
N 1. P. 53-57.
Hartman L. The behavior and breeding of captive weasels (Mustela nivalis L.) // New Zealand J.
Science. 1964. Vol. 7(2). P. 147-156.
Heidt G.A. The least weasel, Mustela nivalis Linnaeus. Developmental biology in comparison
with North American Mustela H Biological series. 1970. Vol. 4, N 7. P. 227-282.
Heidt G.A., Petersen M.K., Kirkland G.L. Mating behavior and development of least weasels
(Mustela nivalis) in captivity // J. Mammal. 1968. Vol. 49(3). P. 413-419.
Heggberget Th. M. Age determination of Eurasian otter (Lutra lutra) cubs // Reproductive
strategy and feeding ecology of the Eurasian otter Lutra lutra'. Dr. scient. thesis in terrestrial ecology /
Univ. Trondheim. Depart, of Zool. 1993.
Hensel R.J., Troyer W.A., Erickson A. W. Reproduction in the female brown bear // J. Wildlife
Manag. 1969. Vol. 33, N 2. P. 357-365.
Hesse R., Allee W., Schmidt K. Ecological animal geography. - New York, 1951. 415 p.
Hill R. W. Comparative Physiology of animals. An environmental approach. - New York, 1976.
656 p.
Hornoker M.G., Hash H.S. Ecology of the Wolverine in north-western Montana// Canad. J.
Zool. 1981. Vol. 59. P. 1286-1301.
433
Hugie R. How to bug a bear// Maine fish wildlife. 1977. Vol. 19, N 4. P. 6-9.
Irving L. The respiration in diving mammals // Amer. J. Physiol. 1939. Vol. 19, N 1. P. 72-94.
Irving L., Krog J. Temperature of skin in the arctic as a regulator of heat // J. Appl. Physiol.
1955. Vol. 7. P. 355-364.
Iversen J.A. Basal energy metabolism of mustelids // J. Physiol. 1972. Vol. 81. P. 341-344.
Johnson S.F., Gessaman J.A. An evaluation of heart as an indirect monitor of free-
living energy metabolism // Ecological Energetics of Homeotherm. - Logan, 1973. P. 44-54.
Jonkel Ch. J. Some comments on polar bear management //Biol. Conserv. 1970. Vol. 2, N 2.
P. 94-96.
JKdrzejewski W., JKdrzejewska B. Seasonal Surplus Killing as Hunting Strategy of the Weasel
Mustela nivalis - Test of a Hypothesis // Acta theriol. 1989. Vol. 34, N 25. P. 347-359.
JKdrzejewski W., JKdrzejewska B., Brzezicski M. Winter habitat selection and feeding habits of
polecats (Mustela putorius) in Bialowieza National Perk, Poland // Z. Saugetierkunde. 1993. Vol. 58.
P. 75-83.
Kayser C. Hibernation // Physiological Mammalogy. - New York, 1965. Vol. 2. P. 175-185.
King C.M. Factors regulation mustelid populations // Acat. Zool. Fennica. 1983. Vol. 174.
P. 217-220.
King C.M. The Natural History of Weasels and Stoats. - London, 1989. 253 p.
Koch T. Beitrnge zur Anatomie des Sumpfbibers. - Leipzig, 1953. Bd 8. 168 S.
Kochonen K. Ventilation in the subnivean tunnels of the voles Microtus agrestis and M. economusH
Ann. zool. fenn. 1980. Vol. 17, N 1. P. 1-4.
KrottP. Der Vielfrass: Monographik der Wildsaugetiere. -lena, 1959. Bd 13. 159 S.
Krott P. Der vielfrass oderjirv. - Wittenberg; Lutherstadt, 1960. 56 S.
Kruuk H. The Social Badger. - Oxford, 1989. 155 p.
Kruuk H. Wild Otter. Predation and Populations. - Oxford, 1995. 90 p.
Kuyt E. Movements of young wolves in the Northwest territories of Canada // J. Mammal.
1961. Vol. 43, N 2. P. 270-271.
Lentfer J. W. Alaskan polar bear denning // Bears - their biology and management. - Kalispell
(Mont.), 1980. P. 101-108.
Liers Emil E. Notes on the river otter (Lutra canadensis) // J. Mammal. 1951. Vol. 32, N 2. P. 1-9.
Liers Emil E. Early breeding in the river otter // J. Mammal. 1958. Vol. 39, N 3. P.438.
LockieJ.D. The food of the pine marten Martes martes in west Ross-Shire, Scotland //Proc. zool.
Soc. London. 1961. Vol. 136. P. 187-195.
LockieJ.D. Territory in small Carnivores // Proc. Zool. Soc. London. 1966. N 18. P. 143-165.
Lono O. The polar bear (Ursus maritimus Phipps) in the Svalbard area // Norsk Polarinst. skr.
1970. N 149. P. 1-115.
Macdonald D.W. The sociable fox // Wildlife. 1978. Vol. 20, N 6. P. 272-277.
Macdonald D.W. European Mammals. Evolution and Behaviour. London, 1995. 352 p.
Macpherson A.H. The dynamics of Canadian arctic fox populations // Canad. Wildl. Serv.
Reports Series. 1969. N 8. P. 51-67.
Magoun A. J. Ecology of wolverines on the North Slope of the Brooks Range, Alaska // Alaska
Coop. Wildlife Res. Unit. 1978. Vol. 30, N 1. P. 16-18.
Magoun A.J. Ecology of wolverines in an arctic ecosystem / Alaska Coop. Wildlife Res. Unit.
Prog. Rep.; Univ. Alaska. - Fairbank, 1982. 27 p.
Mason C.F., Macdonald S.M. Otters. Ecology and conservation. - Cambridge, 1986. 236 p.
McNeil M.A., Ran M.E. Echinococcus granulosus (Cestoda: Taeniidae) infections in moose
(Alces alces) from Southwestern Quebec // Wildlife Disease. 1987. Vol. 23, N 3. P. 418-421.
Meeh D. The wolf: The ecology and behavior of an endangered species / The Natur. Hist. Press
Garden City. - New York, 1970. 834 p.
Meeh D. Disproportionate sex rations of wolf pups // J. Wildlife Manag. 1975. Vol. 39, N 4.
P. 737-740.
434
Meeh D. Productivity, mortality and population trends of wolves in Northeastern Minnesota //
J. Mammal. 1977. Vol. 58, N 4. P. 559-574.
Miguel J., Torres J. et al. Helmints parasits de camivors silvestres a Catalunya. Museu de
Granollers. - Granollers, 1994. 231 p.
Moors P.J. The food of weasels (Mustela nivalis) of farmland in northeast Scotland //
J. Zool. Lond. 1975. Vol. 177. P. 455-461.
Morrison P. Adaptation of small mammals to the arctic // Fed. Proc. 1964. Vol. 23, N 6.
P. 1202-1206.
Mosch K. Iber die Anatomic der Blutgeif system des Suntpfbibers (Nutria) // Diss. - Berlin, 1951.
Bd 28. S. 112-125.
Miller Heinrich. Beitrige zur Biologie des Hermelins, Mustela erminea Linne // Siugetierk. Mitt.
1970. Vol. 18, N 4. S. 293-380.
Myers K. Morphological changes in the adrenal glands of wild rabbits //Nature. 1967. Vol. 213,
N 2. P. 147-150.
Myrberget S., Srumgard R. Fodselstidspunkt og Kullstrrelse hosjjerv // Fauna. 1979. Vol. 32,
N 1. P. 9-13.
Myrcha A. Variations in the length and weight of the alimentary tract of Clethrionomys glareolus
(Schreber, 1780) // Acta theriol. 1964. Vol. 9, N 10. P. 1-10.
Notini G. Biologiska Undersoknigar over gravlingen // Uppsala. 1948. S. 48-54.
Novak M., Baker J., Obbard V, Malloch B. Wild furbearer management conservation in North
America. - Ontario, 1987. 1150 p.
Nyholm E.S. Niidin (Martes martes L.) tarhakas- vatuskokeilusta Kuusamossa // Turkistalous. -
Turku, 1980. N 4. S. 1-7.
Olszewski J., Skoczen T. The airing of burrows of the mole Tapia europaea Linn., 1758 // Acta
theriol. 1965. Vol. 10, N 12. P. 10-17.
Oritsland N.A. Temperature regulation of the polar bear (Thalarctos maritimus) // Comp.
Biochem. and Physiol. 1970. Vol. 37, N 2. P.225-235.
OstbyE., SkarH.-J., SvalastogD. etal. Arctic fox (Alopex lagopus) and red fox (Vulpes vulpes)
on Hardangeridda: den ecology, distribution and population status // Medd. Norsk viltforsk. 1978.
Ser. 3, N 4. P. 66-82.
OzolinA J., Rantin/ъ M. Some preconditions for the present development of otter Lutra lutra (L.),
number and distribution in Latvia // Semiaquatische. Wiss. Beitr. Univ. Halle. 1992. Bd 37. S. 365-384.
Palazon S., Ruiz-Olmo J. Estatus actual del vison Europeo (Mustela lutreola) у del vison
Americano (Mustela vison) en Espano. El Vison Europeo. - Madrid, 1992. 172 p.
Pauls R. IV. Heart rate as an index of energy expenditure in red squirrels (Taniiasciurus hudsonicus) //
Comp. Biochem. and Physiol. 1980. Vol. 67, N 3. P. 525-548.
Peterson R., Allen D. Snow conditions as a parameter in moose wolf relation-ships // Canad.
Natur. 1974. Vol. 101, N 7. P. 481-492.
Pucek Z. Morphological changes in shrews kept in captivity // Acta theriol. 1964. Vol. 8, N 9.
P. 137-166.
Pulliainen E. On the distribution and migrations of the Arctic for (Alopex lagopus L.) in
Finland // Aguilo. Ser. Zoologica. 1965. Vol. 2. P. 25-40.
Pulliainen E. Distribution and population structure of the bear (Ursus arctos L.) in Finland //
Ann. Zool. Fennici. 1972. N 9. P. 199-207.
Pulliainen E. Pohjooissuomtn mikkien ravinnon koostumuksesta runsaan pikkujyrsijakanna
oloissa // Suomen riista. 1984. N 31. P. 43-46.
Pulliainen E., Siivonen L. Ilves // Suomen nisikkiit. - Helsinki, 1972. T. 2. S. 286-306.
Ramsay M.A., Stirling I. On the mating system of polar bears // Canad. J. Zool. 1986. Vol. 64.
P. 2142-2151.
Ramsay M.A., Stirling I. Reproductive biology and ecology of female polar bear (Ursus
maritimus) // J. Zool. 1988. Vol. 214. P. 601-634.
435
Rauch J.C., Hayward J.S. Topography and vascularization of brown fat in small nonhibemator
mammals (deer mouse, Peromyscus maniculatus) // Canad. J. Zool. 1969. Vol. 47, N 6. P. 1301-1314.
Rausch R.A., Pearson A.M. Notes on the Wolverine in Alaska and the Yukon territory//
J. Wildlife Manag. 1972. Vol. 36, N 2. P. 249-268.
Rensch B. Organproportionen und Kerpergrosse bei Vegeln und Siugetieren // Zool. Jahrb.
Abt. 1. 1948. Bd 61, N 4. S. 337-412.
Rensch B. The relation between the evolution of central nervous functions and body size of
animals. Evolution as a Process. - London, 1954. 215 p.
Rempe U. Beobachtungen ьЬег Brunst, Paarung, Tragzeit, Geburt und Rreuzungen bei Mitgliedem
der Untergattung Putorius // Saugetierkundl Miit. 1957. Vol. 5 (3). P. 215-246.
Reuther C. Development of weight and length of Eurasian otter (Lutra lutra) cubs // IUCN.
Otter specialist group bulletin. - Vienna, 1999. Vol. 16, N 1. P. 11-25.
Robin K. Haltung und Zucht des Fischotters Lutra lutra in zoologischen Garten: Ein Oberblick //
Populations okologie marderartider Saugetiere. - Halle, 1989. P. 181-192.
Roux И Uber die Virschweigungen der Blutgefabs des Menschen//Ztschr. Naturwiss. 1878. Bd 12.
S. 44-57.
Ruiz-Olmo J., Delibes M. La nutria en Espana. Barcelona; Sevilla; Malaga, 1998. 300 p.
Russel R.H. The food habits of mainland and island populations of polar bear: M. Sci. thesis. -
Edmonton, 1971. 84 p.
Russel R.H. The food habits of polar bear of James Bay and southwest Hudson Boy in summer
and autumn // Arctic. 1975. Vol. 28, N 2. P. 117-129.
Samuel W.M. Helminths in moose of Alberta // Canad. J. Zool. 1976. Vol. 54, N 3. P. 57-81.
Schierhorn K., Stubbe M. et al. Helminthofauna of the otter (Lutra lutra L., 1758) in East
Germany // V Intern. Otter Colloquium. 1981. N 6. P. 133-142.
Scholander P.F. Experimental investigations on the respiration in diving mammals and birds //
Norse. Vid. Acad. - Oslo, 1940. P. 1-31.
Selye H. The general adaptation syndrome and the diseases of adaptation // J. Clin. Endocrinol.
1946. Vol. 6 P. 117-230.
Shimalov V.V. et al. Helminths of the Eurasian otter (Lutra lutra L., 1758) in Belorussian
Polesie // Otter specialist group Bulletin. - Vienna, 2000. Vol. 17(2). P. 89-90.
Sidorovich V. E. Structure, reproductive status and dynamics of the Otter population in Byelorussia //
Acta theriol. 1991. Vol. 36, N 1-2. P. 153-161.
Sidorovich V. E. Seasonal variation in the feeding habits of riparian mustelids in river valleys of
NE Belarus // Acta theriol. 2000. Vol. 45, N 2. P. 233-242.
Sidorovich V. E., Anisimova E.I. Peculiarities of helminthocenosis in the American mink
population inhabiting a severely polluted river ecosystem (the Svisloch River, Belarus) // Helmintologia.
1997. Vol. 34, N 1. P. 45-52.
Sidorovich V. E., Tumanov I.L. Reproduction in otters in Belarus and north-western Russia //
Acta theriol. - Warsaw, 1994. Vol. 39, N 1. P. 59-66.
Stehlik J. Ontogenesis of behaviour of lynx (Lynx lynx L.) toward prey // Folia Venatoria. 1979.
N 9. P. 323-328.
Stehlik J. Postnatal development or the lynx (Lynx lynx L.) // Folia Venatoria. 1983. N 13.
P. 147-161.
Stehlik J. Reproduction of the lynx (Lynx lynx L.) in captivity // Folia Venatoria. 1984. N 14.
P. 163-179.
Stehlik J. Results of coprological investigations in lynx (Lynx lynx L.) in the Zoological garden
in Ostrava // Folia Venatoria. 1985. N 15. P. 269-273.
Stehlik J. Reproductive biology of the European lynx (Lynx lynx L.) at Ostrava Zoo // Zool.
Garten N. F. 2000. Vol. 70, N 6. S. 351-360.
Stehlik J. Haltung und Zucht von Karakalen (Caracal caracal) im Zoo Ostrava // Zool. Garten
N. F. 2001. Vol. 71, N 5. S. 310-315.
436
StelfoxJ.G. Sheep in the Canadian Rockies: A History 1800-1970 // Canad. Field-Natur. 1971.
Vol. 85, N 2. P. 101-122.
Stenlund M. A. field study of the timber wolf (Canis lupus) on the superior national forest,
Minnesota // Techn. Bull. Dep. Conserv. 1955. N 4. P. 55.
Stubbe M. On population biology of the badger (M. meles) // Trans, the IX intern. Congress of
game biologists. - Moscow, 1970. P. 74-76.
Tumanov I.L. Ecological determination of the level of energy metabolism in the some mustelids //
J. Lutreola. - Moscow, 1993a. N 1. P. 17-21.
Tumanov I.L. Some eco-physiological features of the weasel (Mustela nivalis L.)//J. Lutreola. -
Moscow, 1993. N 2. P. 1-6.
Tumanov I.L. Information on the breeding and development of the young of the lynx (Lynx
lynx lynx L.) in captivity // J. Lutreola. - Moscow, 1996. N 7. P. 10-12.
Velten C. Aites und Neuls uber die Lebensweisen der Rotelmaus // Aus Heimat. - Berlin, 1954.
Bd 62, H. 5-6. S. 75-86.
Walker E.P. Mammals of the World. - Baltimore, 1964. Vol. 1. 646 p.
Warburton F.E. Feed-back in development and its evolutionary significance // Amer. Natur.
1955. Vol. 89. P. 846-858.
Weber J.-M. Seasonal exploitation of amphibians by otters (Lutra lutra) in north-east Scotland //
J. Zool. - London, 1990. Vol. 220. P. 641-651.
Wiig O., Gjertz J., Hansson R., Thomassen I. Breeding behaviour of polar bears in Homsund,
Svalbard//Polar Record. 1992. Vol. 28. P. 157-159.
WolkE. Body weight and daily food in take in captive shrews//Acta theriol. 1969. Vol. 14,N4.
P. 35-47.
Woodard TN., Gutierrez R.J., Rutherford W.H. Bighorn lamb production, survival, and mortality
in South-Central Colorado // J. Wildlife Manag. 1974. Vol. 38, N 4. P. 771-774.
Wright Ph.L., Rausch R. Reproduction in the wolverine, Gulo gulo // J. Mammal. 1955. Vol. 36,
N 3. P. 346-355.
Оглавление
Введение............................................................5
Материал и методики.................................................8
Глава 1. Сравнительно-экологическая характеристика питания и особенности
строения пищеварительного тракта................................12
Глава 2. Двигательная активность и её связь с кормовой специализацией видов 72
Глава 3. Уровень теплообмена и пути поддержания энергетического баланса 119
Глава 4. Функциональные особенности сердечно-сосудистой системы...152
Глава 5. Адаптивные черты строения сосудистого русла............. 169
Глава 6. Экологическая обусловленность репродуктивных функций.....202
Глава 7. Некоторые закономерности роста и развития молодняка......251
Глава 8. Возрастные изменения соотношения полов и их биологическая
направленность.................................................305
Глава 9. Гельминтозы и их влияние на популяции хищных
млекопитающих..................................................346
Заключение........................................................402
Литература........................................................409
Contents
Introduction...................................................................5
Material and methods...........................................................8
Chapter 1. Comparative- ecological characteristics of feeding and characteristic
features in the structure of alimentary tract.................................12
Chapter 2. Moving activity and its relation to food specialization of species.72
Chapter 3. The level of heat exchange and means of maintenance
of energetic balance........................................................ 119
Chapter 4. Functional characteristics of the cardio-vascular system.........152
Chapter 5. Adaptive patterns in the structure of the vascular system....... 169
Chapter 6. Ecological stipulation of reproductive functions...................202
Chapter 7. Some regularities of growth and development of young...............251
Chapter 8. Age changes in the sex ratio and their biological directivity......305
Chapter 9. Helminthosis and its influence on the populations of carnivores..346
Conclusion...................................................................402
References...................................................................409
Научное издание
Игорь Леонидович Туманов
БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ХИЩНЫХ МЛЕКОПИТАЮЩИХ РОССИИ
Редактор Т. А. Асанович
Художник Е. В. Кудина
Компьютерная верстка В. А. Решетова
Оригинал-макет предоставлен И. Л. Тумановым
Лицензия ИД №02980 от 06 октября 2000 г. Подписано к печати 20.06.03.
Формат 70 х 100 !/i6. Бумага офсетная. Печать офсетная.
Усл. печ. л. 35.7. Уч.-изд. л. 27.7. Тираж 1000 экз. Тип. зак. №4309. С 154
Санкт-Петербургская издательская фирма «Наука» РАН
199034, Санкт-Петербург, Менделеевская лин., 1
main@nauka.nw.ru
Санкт-Петербургская типография «Наука» РАН
199034, Санкт-Петербург, 9 лин., 12