/































































































































































































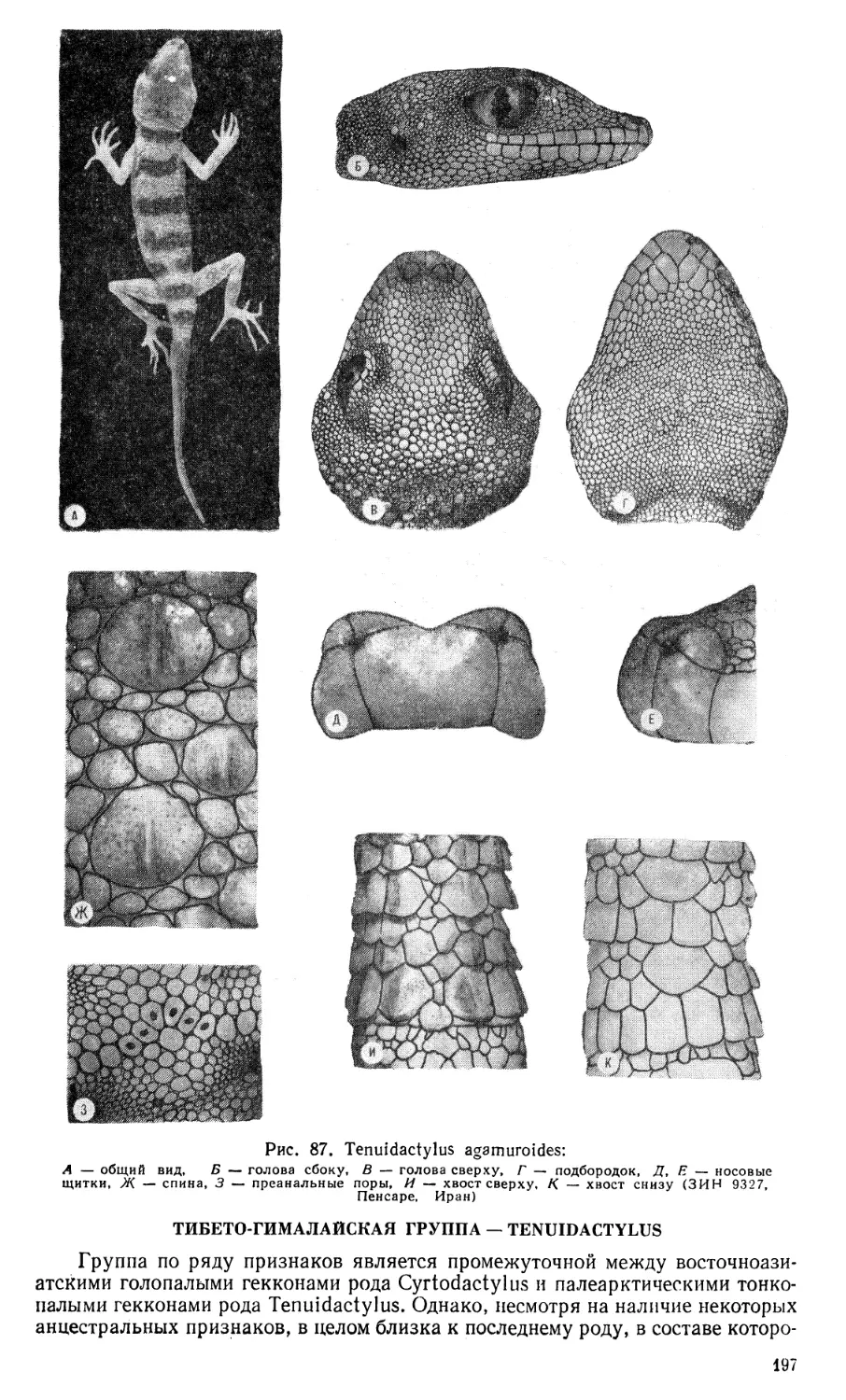






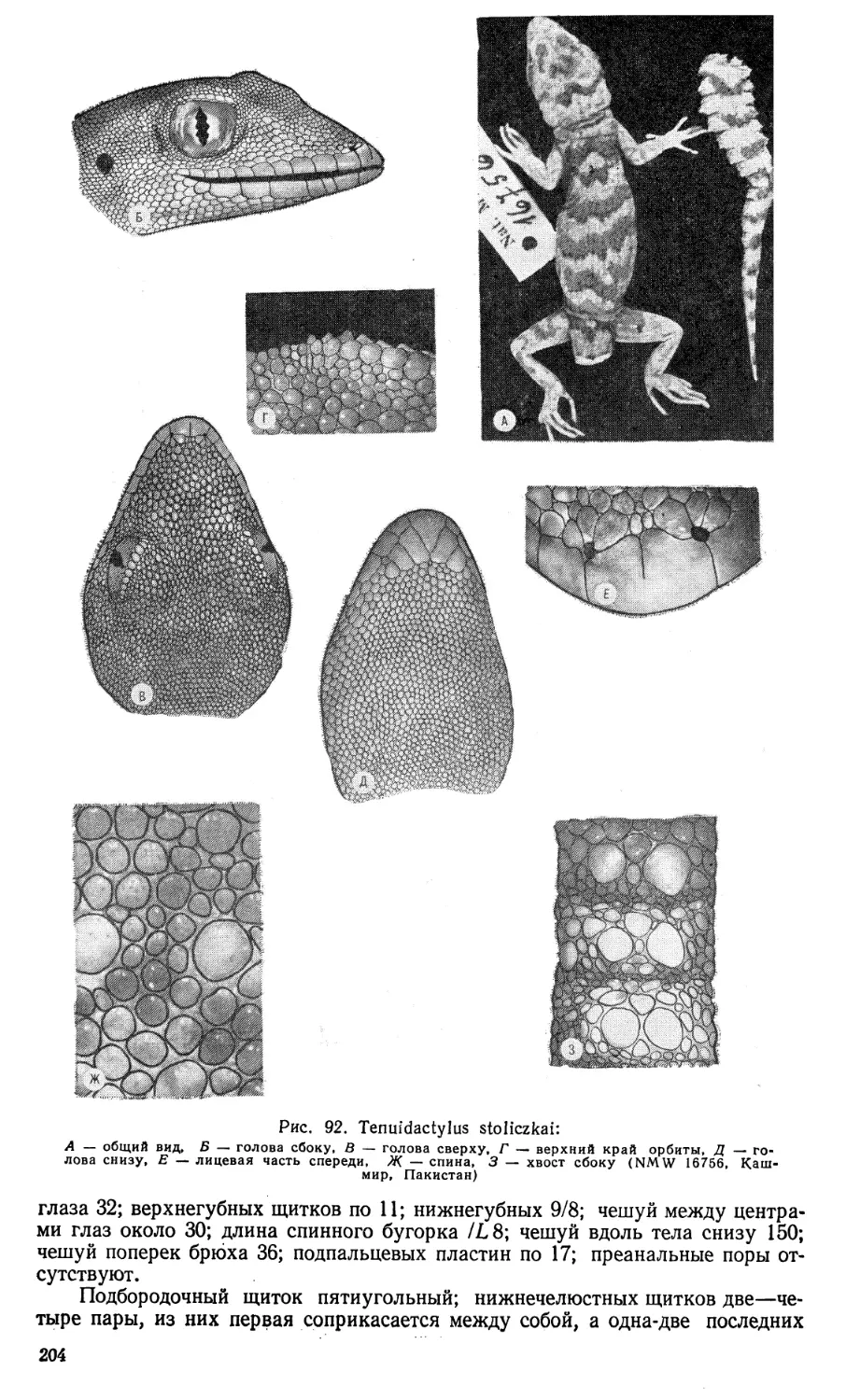




































Author: Щербак Н.Н. Голубев М.Л.
Tags: reptilia рептилии пресмыкающиеся герпетология фауна фауна ссср
Year: 1986




Text
ЩЕРБАК. M.JI ГОЛУБЕВ
ГЕККОНЫ
ФАУНЫ
СССР
И СОПРЕДЕЛЬНЫХ
СТРАН
АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР
ИНСТИТУТ ЗООЛОГИИ им. И. И. ШМАЛЬГАУЗЕНА
Н. Н. ЩЕРБАК, М. Л. ГОЛУБЕВ
ГЕККОНЫ
ФАУНЫ
СССР
И СОПРЕДЕЛЬНЫХ
СТРАН
Определитель
КИЕВ НАУКОВА ДУМКА 1986
УДК 598.112.1 (4—013)
Гекконы фауны СССР и сопредельных стран / Щербак Н. Н., Голубев М. Л.— Киев :
Наук, думка, 1986.— 232 с.
В определителе обобщены сведения об одной из наиболее слабо изученных групп ящериц,
ведущих сумеречно-ночной и скрытый образ жизни, среди которых много редких и исчезающих
видов. Приведены детальные карты распространения с кадастром мест находок, материалы о
филогенетических отношениях гекконов, а также оригинальные определительные таблицы.
Изучена географическая изменчивость гекконов.
Для зоологов, герпетологов, специалистов в области охраны природы, работников музеев
и заповедников, преподавателей и студентов вузов.
Табл. 7. Ил. 122. Библиогр.: 217—226 с.
Ответственный редактор
М. А. ВОИНСТВЕНСКИЙ
Рецензенты
И. С. ДАРЕВСКИЙ, в. А. КОТЛЯРЕВСКАЯ
Редакция общей биологии
НИКОЛАЙ НИКОЛАЕВИЧ ЩЕРБАК
МИХАИЛ ЛЕОНИДОВИЧ ГОЛУБЕВ
ГЕККОНЫ ФАУНЫ СССР
И СОПРЕДЕЛЬНЫХ СТРАН
Определитель
Утверждено к печати ученым советом
Института зоологии им. И. И. Шмальгаузена АН УССР
Редактор С. В. Вечерская
Оформление художника В. Л4. Флокса
Художественный редактор Р. И. Кал ыш
Технический редактор А. М. Капустина
Корректоры П. С. Бородянская, И. В. Кривошеина
И Б. 7465
Сдано в набор 17.06.85. Подо, в печ. 29.01.86. БФ 04513. Формат 70x108/16.
Бум. тип. № 1. Лит. гарн. Выс. печ. Усл. печ. л. 21,0. Усл. кр.-отт. 23,1.
Уч.-изд. л. 22,94. Тиране 1200 экз. Заказ 5—1760. Цена 2 р.
Издательство «Наукова думка»». 252601, Киев 4, ул. Репина, 3.
Головное предприятие республиканского производственного объединения
«Полиграфкнига». 25205? Киев, ул. Довженко, 3.
2005000000-045_
М221(04)-86
© Издательство «Наукова думка», 1986
ПРЕДИСЛОВИЕ
Гекконовые ящерицы — группа пресмыкающихся, замечательная во многих отношениях.
Во-первых, это одно из древнейших семейств рептилий, их предки прослеживаются до мезозоя
[238] и несут ряд примитивных черт [111]. Во-вторых, гекконы — животные южного происхож-
дения, подавляющее большинство их обитает в тропических областях [307]. В Палеарктику
проникло мало видов этих ящериц. В числе объектов нашего исследования роды Eublepharis,
Tefatoscincus, Crossobamon, Alsophylax, Bunopus, Tropiocolotes, Carinatogecko, Tenuidactylus,
Agamura, представители которых встречаются на территории СССР и сопредельных стран, т. е.
подавляющее большинство палеарктических форм.
Гекконы резко отличаются от всех ящериц многими особенностями: у них характерный
вертикальный зрачок немигающих глаз, прикрытый, как у змей, неподвижной прозрачной обо-
лочкой; они способны издавать звонкие звуки, ведут преимущественно ночной образ жизни;
в отличие от iбольшинства рептилий — откладывают яйца в твердой известковой оболочке.
Основная адаптивная радиация у гекконов идет по пути видоизменения пальцев, многие виды
способны передвигаться по вертикальным поверхностям.
До недавнего времени систематика палеарктических гекконов была изучена недостаточ-
но, хотя представлялась более или менее благополучной. Было известно, что взгляды специа-
листов на отдельные таксоны не всегда одинаковы. Так, голопалые гекконы в работах европей-
ских герпетологов [307, и др.] были известны под родовым названием Gymnodactylus. амери-
канские зоологи, а в последние годы и многие западногерманские исследователи относят их к
роду Cyrtodactylus [191, 205, и др.]. Особенно много спорных моментов существовало в системе
мелких и редких видов, относимых в разное время к родам Alsophylax, Bunopus, Tropiocolotes
и Microgecko. При ближайшем рассмотрении выяснилось, что существуют расхождения во
взглядах при определении родовой принадлежности ряда видов гекконов; в основу их системы
положены признаки, имеющие сомнительную таксономическую ценность; не изучена географи-
ческая изменчивость отдельных видов; описание ряда форм нуждается в ревизии [149]. Цель
нашей работы — изучение систематического положения и филогенетических отношений гекко-
нов фауны СССР и сопредельных стран, проведение ревизии их таксономии, составление на
большом серийном материале достаточно обоснованных описаний и изучение географической
изменчивости видов, создание достоверных определительных таблиц, изучение распространения
и экологии, выявление редких и исчезающих видов, составление рекомендаций по их охране.
Наши материалы по редким видам, кроме того, вошли в первое (1977) и второе (1984) издания
Красной книги СССР.
Детальное изучение и описание видов из сопредельных территорий обусловлено тем, что,
во-первых, нельзя провести ревизию наших видов, понять их таксономию без исследований ро-
довых группировок в целом (в ряде случаев и близких родов), во-вторых, до сих пор в отечест-
венной литературе отсутствуют описания и определительные таблицы видов гекконов из сосед-
них стран, хотя они часто поступают в коллекции музеев СССР и их обработка представляет
определенные трудности. Многолетний опыт нашей работы показал, что на территории ряда со-
юзных республик еще возможны находки видов, проникающих с юга, а их констатация воз-
можна только при наличии соответствующей справочной литературы.
Ревизия систематического положения гекконов, проведенная нами, привела к сущест-
венным и неожиданным результатам. Обнаружено своеобразие группы голопалых палеаркти-
ческих видов, она выделена в новый род Tenuidactylus [153]. Новый род (Carinatogecko) описан
из Ирана [151]. Усовершенствование таксономической структуры привело к выделению трех
новых подродов: Mediodactylus [150], Asiocolotes [40], Altiphylax [56]. В процессе работы
были открыты новые виды — Gymnodactylus turcmenicus [144], Tropiocolotes levitoni
[43], G. mintoni [44], Alsophylax tadjikiensis [38], A. tokobajevi [56] и новые подви-
ды— G. caspius insularis [14], Teratoscincus scincus rustamovi [146], A. loricatus szczerbaki
[42], а также перемещены в другой род A. spinicauda и A. tibetanus, выявлены новые виды для
фауны СССР (Bunopus tuberculatus и G. longipes microlepis) [48], значительно уточнены ареалы
изученных видов, получены новые данные по экологии, размножению и образу жизни.
Настоящая работа выполнялась в лабораториях Зоологического музея Института зооло-
гии и ЦНПМ АН УССР в 1972—1984 гг. За эти годы авторы провели широкие полевые экспе-
диционные исследования на территории Крыма, Кавказа и Закавказья, Средней и Центральной
Азии, имели возможность ознакомиться с коллекционными материалами крупнейших музеев
3
нашей страны, Западной Европы и США. В общей сложности было исследовано свыше 5 тыс.
экземпляров гекконовых ящериц, хранящихся в фондах Зоологического музея Института зооло-
гии АН УССР (Киев — ИЗАНУ), а также любезно представленных нам И. С. Даревским,
Л. Н. Лебединской и Н. Л. Орловым (Зоологический институт АН СССР, Ленинград — ЗИН),
В. Ф. Орловой и Т. И. Александровской (Зоологический музей Московского университета —
ММГУ), Я. Д. Давлятовым и Т. Я. Ядгаровым (Институт зоологии и паразитологии АН УзССР,
Ташкент — ИЗПАНУ), С. А. Саид-Алиевым (Институт зоологии и паразитологии АН ТаджССР,
Душанбе — ИЗПАНТ), С. М. Шаммаковым и Ч. А*. Атаевым (Институт зоологии АН ТССР,
Ашхабад — ИЗАНТ), 3. К. Брушко и Р. А. Кубыкиным (Институт зоологии АН КазССР,
Алма-Ата — ИЗАНК), В. К. Еремченко (Институт биологии АН КиргССР, Фрунзе — ИБК),
В. Н. Кудашевой (Биолого-почвенный институт ДВНЦ АН СССР — ДВНЦ), Л. М. Писаревой
(Зоологический музей Киевского университета — МКГУ), В. И. Ведмедерей (Зоологический
музей Харьковского университета —МХГУ), В. В. Неручевым (Горьковский пединститут —
ГПИ), Т. С. Сатторовым (Душанбинский пединститут — ДПИ), В. П. Великановым (Красно-
водский заповедник — КЗ), А. П. Маркевичем (сборы из АРЕ), а также много материалов мы
получили из зарубежных хранилищ — от И. Айзельта, Ф. Тидеманна и М. Хепля (Венский
естественно-исторический музей, Австрия — NMW), Е. Арнольда, А. Грандиссон и Б. Кларке
(Британский естественно-исторический музей, Лондон — BMNH), В. Беме (Зоологический
институт и Музей А. Кенига, Бонн, ФРГ — ZFMK), М. И. Бунни (Естественно-исторический
музей, Багдад, Ирак — NHMB), Г. де Витте (Королевский институт естественных наук и зоо-
музей, Брюссель, Бельгия — BZ), О. Дели (Венгерский национальный музей, Будапешт —
UNM), Р. Мертенса и Клеммера (Зенкенбергский музей, Франкфурт-на-Майне, ФРГ — SMF),
Г. Маркса и А. Резетара (Чикагский музей естественной истории, США —CMNH), Л. Капокач-
чиа и М. А. Черзи (Институт зоологии MIZG и зоомузей, Генуя, Италия — MCZN), А. Г. Клю-
ге (Зоологический музей Мичиганского университета, Энн Арбор, США — UMMZ), А. Е. Леви-
тона (Калифорнийская АН, Сан-Франциско, США—CAS), Минтона (частная коллекция, Индиа-
наполис, США—SAM), Г. Петерса (Зоологический музей Берлинского университета им. А. Гум-
больта, ГДР—ZMB), Поггези (Зоологический музей «Ла-Спеккола», Флоренция, Италия —
MZSF), П. Розадо (Музей сравнительной зоологии Гарвардского университета, Кембридж,
Массачусетс, США — MCZ), 3. Рочека (Пражский Национальный Музей, ЧССР — PNM),
Р. Ру-Эстев (Национальный музей естественной истории, Париж, Франция — MNHP),
Г. Свинни (Королевский Шотландский музей, Эдинбург, Англия — RSM), У. Р. Хейер,
Р. И. Кромби, Ф. Маккулог (Смитсоновский институт, Вашингтон, США — USNM), М. С. Хуг-
муда (Королевский Естественно-исторический музей, Лейден, Нидерланды — RMNH),
Р. Г. Цвайфеля (Американский музей естественной истории, Нью-Йорк, США — AMNH),
Цедерхолма (Зоологический музей Лунда, Швеция — ZML), Б. Шатти (Зоологический музей
университета в Цюрихе, Швейцария — ZMZ), И. Шмидтлера (частная коллекция, Мюнхен,
ФРГ — ZSM) и К. Эдельстама (Шведский музей естественной истории, Стокгольм — ZMS),
Б. К. Тикадера (директора Зоологической службы, Калькутта).
Кариопрепараты гекконов подготовлены В. В. Жуковой-Манило, беспозвоночные, най-
денные в пищеварительном тракте, определены В. М. Ермоленко.
Ценные замечания при подготовке рукописи к печати были получены от И. С. Даревского.
Большую помощь в проведении экспериментов и обработке коллекционных материалов
оказали нам Н. П. Бондарева, Ю. Н. Исаев и А. А. Токарь, а в сборе материала в полевых
условиях — многие сотрудники Зоологического музея Института зоологии АН УССР. В изго-
товлении иллюстраций принимал участие В. Ю. Раевский.
Всем перечисленным выше лицам авторы выражают искреннюю благодарность.
СПИСОК СОКРАЩЕНИЙ
Водохр.— водохранилище
Возв.— возвышенность
В.— восточный
В. д.— восточная долгота
Г.— город
Дор.— дорога
3.— западный
Зал.— залив
Кишл.— кишлак
Кол.— колодец
Колл.— коллекция
Креп.— крепость
Н. у. м.— над уровнем моря
О.— остров
Оз.— озеро
Окр.— окрестности
Обл.— область
Пер.— перевал
Письм. сообщ.— письменное
сообщение
П-ов — полуостров
Пос.— поселок
Пров.— провинция
Пуст.— пустыня
Разв.— развалины
Р.— река
С.— селение
Сев.— северный
С. ш.— северная широта
Ст.— станция
Стр.— страна
Уроч.— урочище
Устн. сообщ.— устное со-
общение
Ущ.— ущелье
Хр.— хребет
Ц.— центральный
Ю.— южный
ОБЩАЯ ЧАСТЬ
КРАТКИЙ ОБЗОР ИССЛЕДОВАНИЙ ГЕККОНОВ
ФАУНЫ СССР И СОПРЕДЕЛЬНЫХ СТРАН
Гекконы настолько своеобразная группа ящериц, что были выделены в от-
дельный таксон еще на заре развития герпетологии (Laurenti, 1768; Latrei,
1825 [цит. по 111]; 1111, 215], и др.). Наиболее устойчивой оказалась система
на уровне семейств [199, 200]. Ее пересмотр стал возможен лишь в последнее
время, после накопления соответствующих коллекционных материалов и ис-
пользования новых признаков. Первый, кто исследовал данную группу под
нестандартным углом зрения, был Г. Андервуд [3051, который, используя при-
знак «форма зрачка», различал три традиционных семейства — Eublepharidae,
Sphaerodactylidae и Gekkonidae, причем последнее было разделено на два
подсемейства: Cekkoninae и Diplodactilinae. А. Г. Клюге [237] установил, что
форма зрачка варьирует и не может быть основанием для выделения подсе-
мейств. Указанный автор в фундаментальной работе, посвященной высшим так-
сономическим категориям гекконов и их эволюции, сгруппировал 82 рода миро-
вой фауны в 4 подсемейства (Eublepharinae, Diplodactylinae, Gekkoninae
и Sphaerodactylinae) семейства Gekkonidae. Эта точка зрения безоговорочно
была принята большинством авторов современных сводок [17, 191, и др.].
Однако, внимательно знакомясь с работой А. Г. Клюге, нетрудно заметить,
что автор, объективно показав различия перечисленных выше подсемейств,
проявил субъективизм в оценке их уровня, в решении вопроса «семейство —
подсемейство». Это привело к тому, что такая важная таксономическая катего-
рия, как семейство, в его понимании имеет весьма нечеткий диагноз, а разли-
чия между подсемействами превышают уровень различий ряда семейств других
ящериц. Особенно это касается различий между эублефарами и гекконами (14
признаков) [237], а это больше, чем между такими стабильными семействами,
как Agamidae и Iquanidae. Эублефары и гекконы различаются прежде всего по
характеру позвонков (у первых они процельные, у вторых — амфицельные).
В систематике лягушек характер позвонков является основанием даже для вы-
деления подотрядов (Opistocoela, Procoela). По-видимому, структуру подотря-
да ящерицы (Sauria) следует ревизовать комплексно, с применением соот-
ветствующих критериев. Мы рассматриваем эублефаров и собственно гекконов
в качестве отдельных семейств, однако признаем близость указанных групп
и разделяем точку зрения других исследователей [111, и др.1, объединявших
их в надсемейство (или подотряд) Gekkota.
* * *
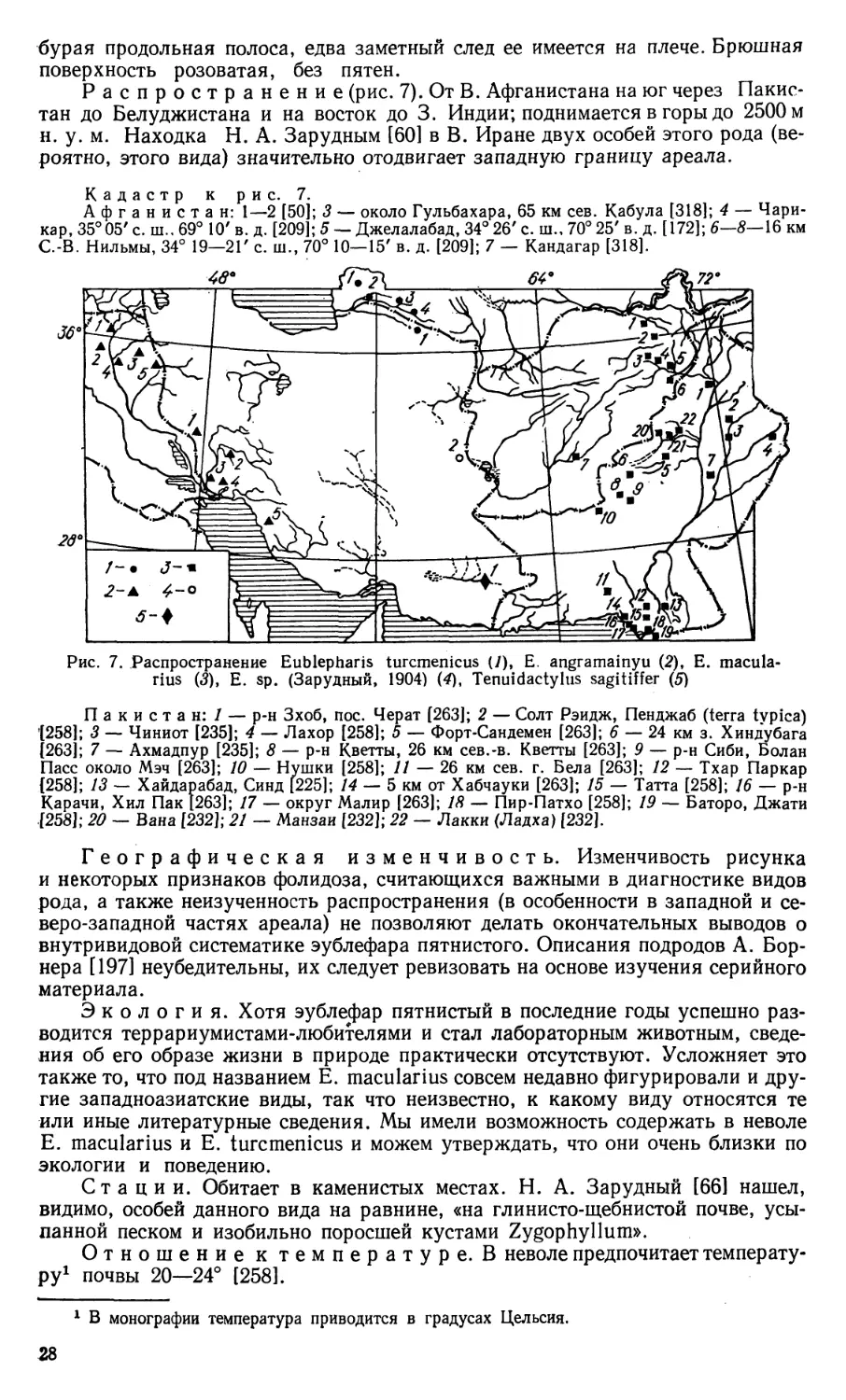
Представители рода Eublepharis (установлен Греем в 1827 г., типовой вид
Е. hardwickii), обитающие в Юго-Западной Азии, до недавнего времени не прив-
лекали внимания исследователей. Описанный в середине прошлого века вид
Cyrtodactylus macularius [190] был переведен в указанный род спустя тридцать
лет [200], это название существует до сих пор. Только однажды А. М. Николь-
ский [96] заметил, что у экземпляра из Туркмении имеется ряд отличий от осо-
бей из Индии и высказал сожаление по поводу отсутствия у него сравнительного
материала. Г. Буланже [202] и М. К. Михайловский [76], впервые исследовав-
5
шие эублефара из Копетдага, отнесли его к индийскому виду Е. macularius;
эта точка зрения была принята последующими герпетологами [23, 112, 121,
и др.].
Видовая самостоятельность ранее описанного из Хайдарабада (3. Пакис-
тан) Е. fasciolatus Gunther, 1864 подвергалась сомнению еще в прошлом веке
[202], впоследствии этот вид был сведен М. Смитом [291] в синоним Е. macu-
larius. Недавно С. Андерсон и А. Левитон [171] установили, что в Ю.-З. Иране
обитает отличный от Е. macularius вид — Е. angramainyu. Все это послужило
стимулом для дальнейших попыток ревизии ящериц, правда, не всегда удач-
ных. Именно таким, на наш взгляд, является выделение нового вида —
Е. gracilis, автором которого является А. Р. Борнер [195]. Он описал ящерицу
по одному живому экземпляру неизвестного происхождения, экспонируемому
в Кельнском зоопарке. Описание основано на мелких и недиагностических при-
знаках, в частности очень изменчивых окраске и рисунке.
Изменение окраски у эублефаров, как мы установили, зависит от физиоло-
гического состояния животного, особенно гормональных факторов. Кстати,
различия Е. fasciolatus и Е. macularius также заключались в окраске; хотя
С. Андерсон и А. Левитон [171] высказывали неуверенность по поводу их иден-
тификации (по нашему мнению, без достаточных оснований), мы относим пер-
вую форму в число младших синонимов последней. Впоследствии А. Борнер
[196, 1971, пользуясь уже коллекционным материалом, но с прежним методи-
ческим подходом, описал новый вид Е. afghanicus из р-на Кабула — Джела-
лабада и три новые формы Е. macularius fuscus из окр. Бомбея, Е. m. smithi
из окр. Дели (Индия) и Е. т. montanus из окр. Карачи (Пакистан), а также
различает, кроме номинативной формы (окр. Лахора, Сев.-В. Пакистан) еще
два неописанных подвида из г. Нушки и округа Зхоб (3. Пакистан), а также
Е. m. fasciolatus [225].
Такое «дробительство» только усложнило и запутало тексономию эубле-
фаров (ареалы отдельных форм Борнера находятся в 150 км друг от друга),
мы считаем его необоснованным и несерьезным. До недавнего времени эубле-
фары фауны СССР также относились к виду Е. macularius. И. С. Даревский,
несколько лет назад побывавший в Иране и наблюдавший там живых эубле-
фаров, заподозрил неладное в их систематике. Проанализировав имеющиеся
материалы, он убедился, что в Туркмении встречается особый вид, которому
в 1978 г. присвоил название Е. turcmenicus. Мы имели возможность детально
ознакомиться с живыми и фиксированными особями этого вида и Е. macularius
и убедились, что они действительно различаются по ряду признаков фолидоза,
особенно окраске и рисунку. Эти различия более значительны, чем между дру-
гими формами. Все сказанное свидетельствует, что назрела необходимость в спе-
циальной ревизии таксономии эублефаров. Не исключено, что в результате
этого в Ю.-З. Азии будет насчитываться лишь один-два вида с несколькими
подвидовыми формами. До проведения подобного исследования нам представ-
ляется возможным различить в пределах этой группы три вида: Е. macularius,
Е. angramainyu и Е. turcmenicus. Наиболее полные сведения по экологии и по-
ведению Е. turcmenicus содержатся в статье Н. Н. Щербака [145].
* * *
Сцинковый геккон (под названием Stenodactylus scincus) известен с 1858 г.
благодаря описанию Г. Шлегеля [283] по материалам из Прибалхашья. В 1863 г.
А. Штраух выделил род Teratoscincus, типом которого был Т. keyzerlingii —
вид, описанный им по сборам из Ирана. Впоследствии Г. Буланже [200] устано-
вил идентичность названных выше видов. Однако С. А. Чернов [125] полагал,
что гекконы из среднеазиатских республик относятся к номинативной форме,
а в Иране распространен Т. scincus keyzerlingii, отличающийся большим ко-
личеством чешуй вокруг туловища (31—36 вместо 28—32) и многочисленными
(42—50 вместо 30—48) зернистыми чешуями между глазами. Описанный
А. М. Никольским [87] Т. zarudnyi из В. Ирана рассматривался С. А. Черновым
как младший синоним Т. s. keyzerlingii [112, 125], им же был также изучен го-
6
лотип Т. roborowskii Bedriaga, 1907 из оазиса Сачжоу (КНР), отличающийся
от особей из Средней Азии лишь особенностями окраски и признан подвидом
сцинкового геккона. Таким образом, в соответствии с современными представ-
лениями [17, 3071 сцинковый геккон из пустынь Средней Азии и Прибалхашья
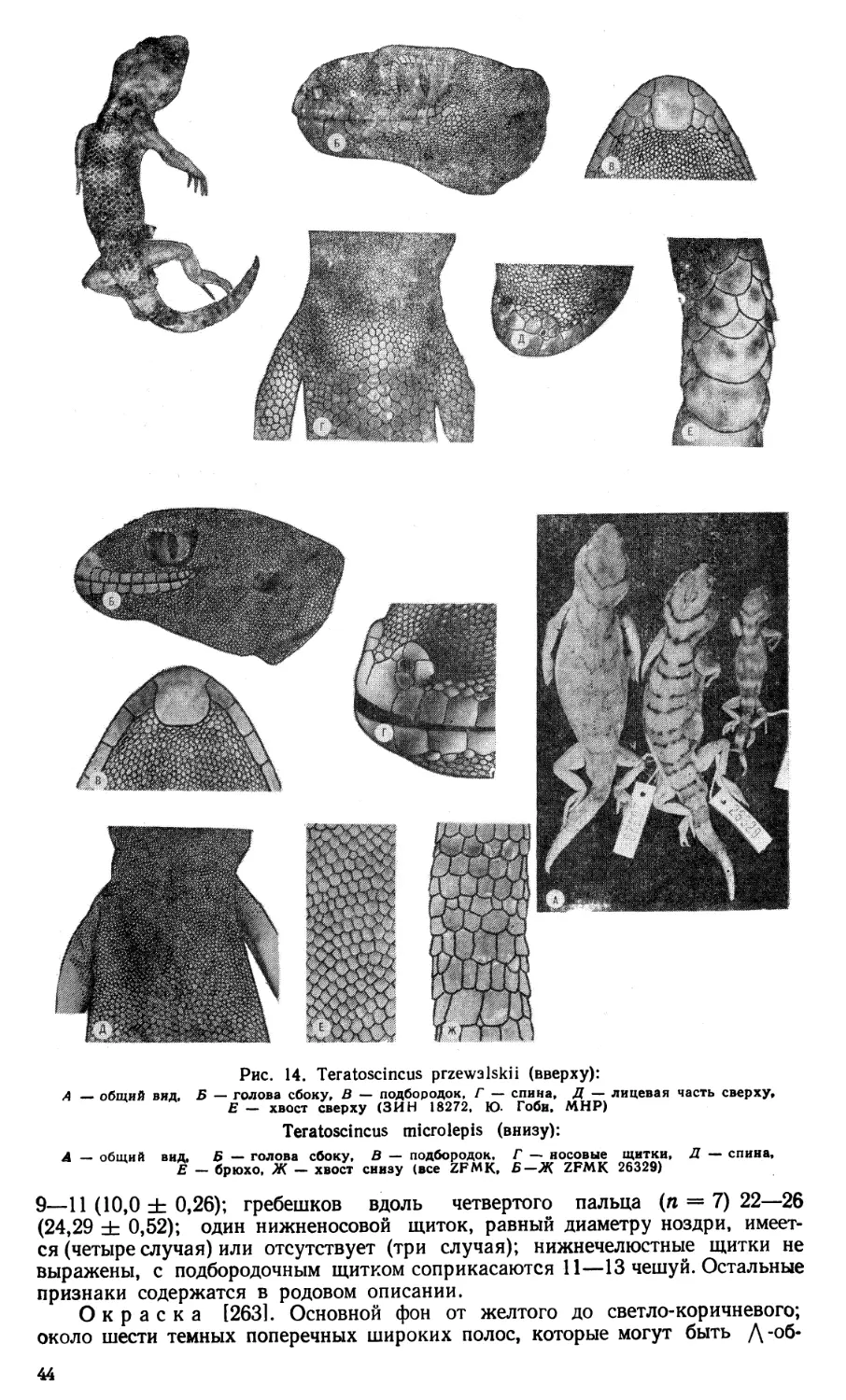
относится к номинативной форме, а всего род объединяет четыре вида: в Цент-
ральной Азии обитает Т. przewalskii Strauch, 1887, в Иране — Т. bedriagai
Nik., 1899 и Т. microlepis Nik., 1899. Существующие ныне описания данных
видов и выводы таксономического характера основаны на изучении небольшого
количества экземпляров, географическую изменчивость сцинковых гекконов
не изучали. Проведенное нами в 1979 г. исследование выборок популяций дан-
ного вида из территории СССР [145] показало их неоднородность, а наиболее
уклоняющаяся (изолированная) популяция из Ферганской долины описана
в качестве подвида Т. scincus rustamovi Szczerbak.
* * *
История изучения гребнепалых гекконов начинается с описания его типо-
вого вида под названием Gymnodactylus eversmanni Wiegmann, 1834 (был вна-
чале отнесен Вигманном к роду голопалых гекконов). Это несоответствие
обнаружил А. Штраух [222], который в 1887 г. выделил особый род Ptenodacty-
lus, но ошибочно применил название, уже использованное для обозначения рода
игуанид. Годом позже указанная ошибка была исправлена О. Беттгером [1931
и этот род получил название Crossobamon. Долгие годы род гребнепалых гек-
конов считался монотипическим, однако в 1967 г. А. Г. Клюге [237] отнес к дан-
ному роду еще три южноазиатских вида (Stenodactylus orientalis Blanford,
1876, S. lumsdeni Boulenger, 1877 и S maynardi Smith, 1933). С тех пор ревизия
рассматриваемой группы не проводилась, но автор сводки по гекконам рода
Stenodactylus [176] тем не менее подтвердил правомерность перевода указанных
видов в род гребнепалых гекконов.
Материалы по экологии С. eversmanni широко представлены в трудах со-
ветских зоологов [22, 23, 98. 107, 125, 130, и др.].
* * *
Начало исследований гекконов рода Alsophylax связано с именем П. С. Пал-
ласа [258], описавшего по материалу, собранному им у горы Большой Богдо
в районе оз. Баскунчак (ныне Астраханская обл.), новый вид ящерицы «Lacerta
pipiens». В дальнейшем этот вид был отнесен к роду Ascalabotes [247]. К. Эйх-
вальд [213] переместил его в род Gymnodactylus, а Л. Фитцингер [215] выделил
в самостоятельный монотипический род Alsophylax. Г. Буланже [200] объеди-
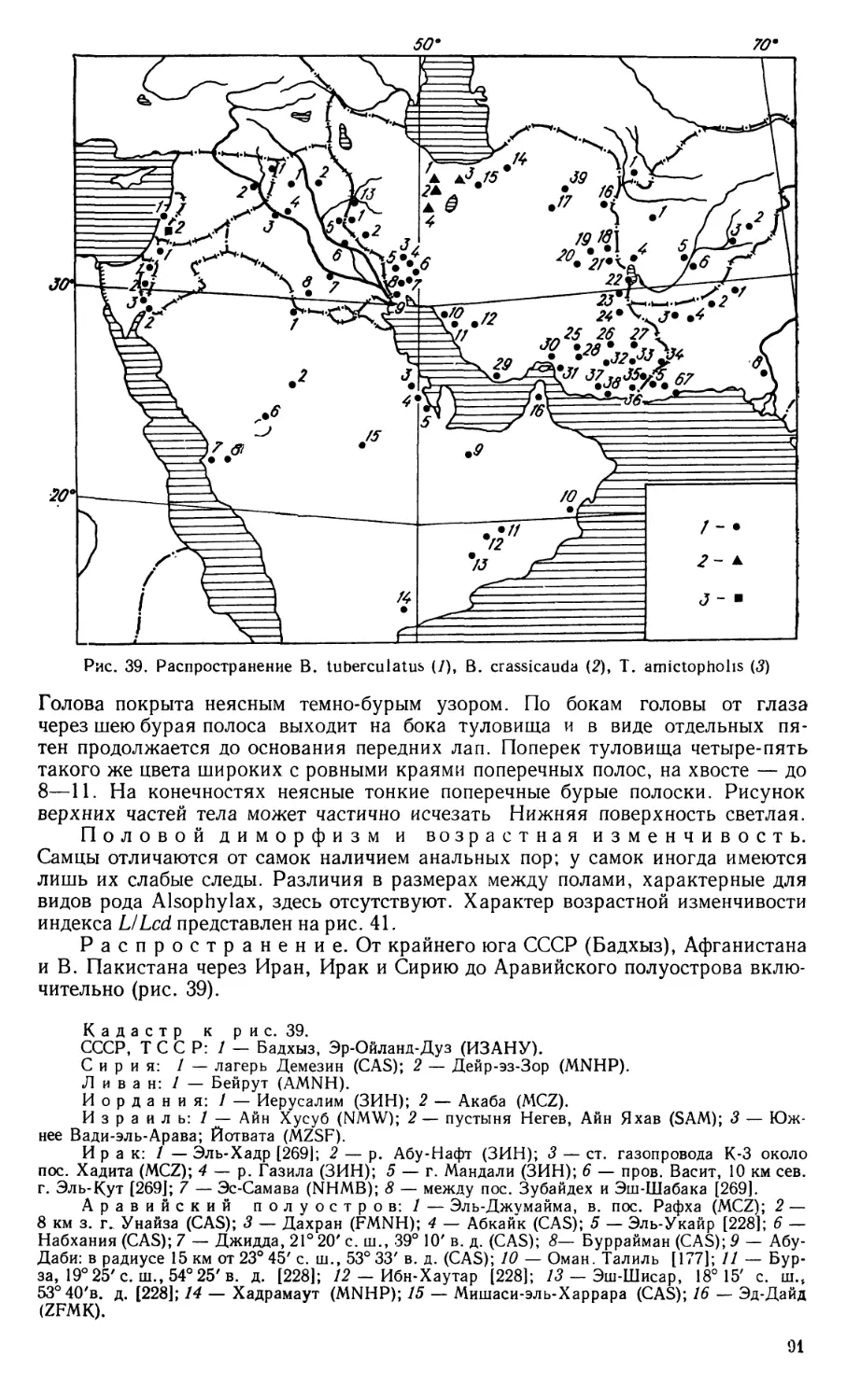
нил его с родом Bunopus Blanford, 1874 (типовой вид В. tuberculatus; Белуджис-
тан). Это привело к существенному изменению первичных родовых диагнозов,
так как они характеризовались взаимоисключающими признаками: гладкими
(Alsophylax) или бугристыми (Bunopus) подпальцевыми пластинками. Кроме
того, Г. Буланже объединил с видом П. С. Палласа другой вид — Gymnodac-
tylus microtis Blanford, не обосновав, однако, свою точку зрения сравнительным
анализом. Возникшая в результате этого путаница сохранилась до настоящего
времени, поскольку подавляющее большинство дальнейших исследователей
приняло точку зрения Г. Буланже.
Основатель фондовых коллекций Зоологического музея в Петербурге
А. Штраух [300] описал сразу три новых вида рода Alsophylax: A. przewalski
(3. Китай), A. loricatus (Ферганская долина) и A. spinicauda (Сев. Иран).
Очень точные и подробные видовые описания А. Штрауха не были, к сожалению,
сопровождены обобщающей родовой характеристикой. В этой же работе был
описан и Bunopus blanfordi (Египет).
Все виды, включенные в то время в род Alsophylax, происходили из Сред-
ней и Юго-Западной Азии и сопредельных территорий. Тем не менее Г. Буланже
[202] отнес к этому роду свой новый вид из Тибета A. tibetanus.
7/
Дж. Андерсон [165], базируясь на коллекции рептилий из Южной Аравии,
описал вид В. spatalurus, также характеризующийся ключевым диагностиче-
ским признаком рода Bunopus — бугристыми подпальцевыми пластинками.
Почему-то этот вид выпал из внимания специалистов и более 60 лет не упоми-
нался в герпетологической литературе.
Первую попытку разделения рода Alsophylax предпринял А. М. Николь-
ский [93], описавший из Сев. Ирана В. crassicauda. Он обратил внимание на один
из признаков первичного диагноза рода Bunopus — шиповатость боковой паль-
цевой чешуи. Однако его мнение о гетерогенности этого рода не нашло под-
держки у герпетологов того времени. В 1907 г. А. М. Никольский описал
A. laevis из Ю. Туркмении, близкий к A. pipiens. С. А. Чернов [121] свел это на-
звание в синоним с пискливым геккончиком без критического анализа (что в
дальнейшем привело к путанице в некоторых вопросах систематики рода Тго-
piocolotes). Гладкий геккончик был восстановлен в ранге вида О. П. Богда-
новым [19, 20].
Следующая попытка восстановить род Bunopus была сделана А. Левито-
ном и С. Андерсоном [243], которые предложили отделить его от рода Also-
phylax на основании наличия спинных бугров. Р. Мертенс [257] обратил внима-
ние на необоснованность такого подхода. Учтя это, названные авторы верну-
лись к признаку «шиповатые свободные края пластинок снизу пальцев», но
прибавили еще один: «шиповатые некоторые или все чешуи брюха» [244, с. 166]
и включили в род Bunopus виды В. tuberculatus, В. blanfordi, В. crassicauda,
В. abudhabi (описанный в этой же работе и происходящий из В. Аравии).
Однако в таком виде диагноз вызвал возражения Ю. К. Горелова и др.
[48], обнаруживших отдельные зазубренные брюшные чешуйки у типового вида
рода Alsophylax — A. pipiens. Кроме того, эти авторы отметили, что А. Леви-
тон и С. Андерсон не указали, к какому роду они относят бугорчатый вид А. 1о-
ricatus. Такая недоработка позволила присоединиться к мнению Р. Мертенса
[258] и рассматривать Bunopus (с видами tuberculatus и loricatus в фауне СССР)
только как подрод Alsophylax (с видами A. pipiens, A. spinicauda и A. laevis
в фауне СССР).
Критерием для разделения подродов должно было бы служить наличие
четких продольных и поперечных спинных бугров (Bunopus) либо их
отсутствие или неупорядоченное расположение (Alsophylax). На ос-
новании этих данных А. Банников и др. [17] ограничили объем рода 13 видами,
не очертив конкретно объем каждого подрода. Однако, используя предложен-
ные диагностические признаки, в каждый подрод следовало бы ввести виды
с гладкими бугристыми подпальцевыми пластинками, что по-прежнему не со-
ответствовало их первичным диагнозам. Мы опубликовали разъяснение по дан-
ному вопросу [150] и предложили, исходя из данных обработки серийного мате-
риала с учетом первоописаний, свой вариант диагнозов родов Alsophylax и
Bunopus. Однако недавняя находка на Тянь-Шане нового вида A. tokobajevi
[56], отличающегося рядом морфологических признаков от других видов
рода, привела к выделению нового подрода Altyphylax и уточнению родового
диагноза
Включение в род Bunopus нового вида В. aspratilis [170] из Ю.-З. Ирана не
вызвало возражений специалистов. Е. Арнольд [175] восстановил вид В. spa-
talurus, ошибочно описанный как Trachydactylus jolensis [227] и сведенный
в синоним с Т. spatalurus [244], и подчеркнул, что это оправданно по крайней
мере до тех пор, пока не будет проведена ревизия рода в целом, а в последующей
работе [177] он свел в качестве младшего синонима В. tuberculatus виды
В. blanfordi и В. abudhabi.
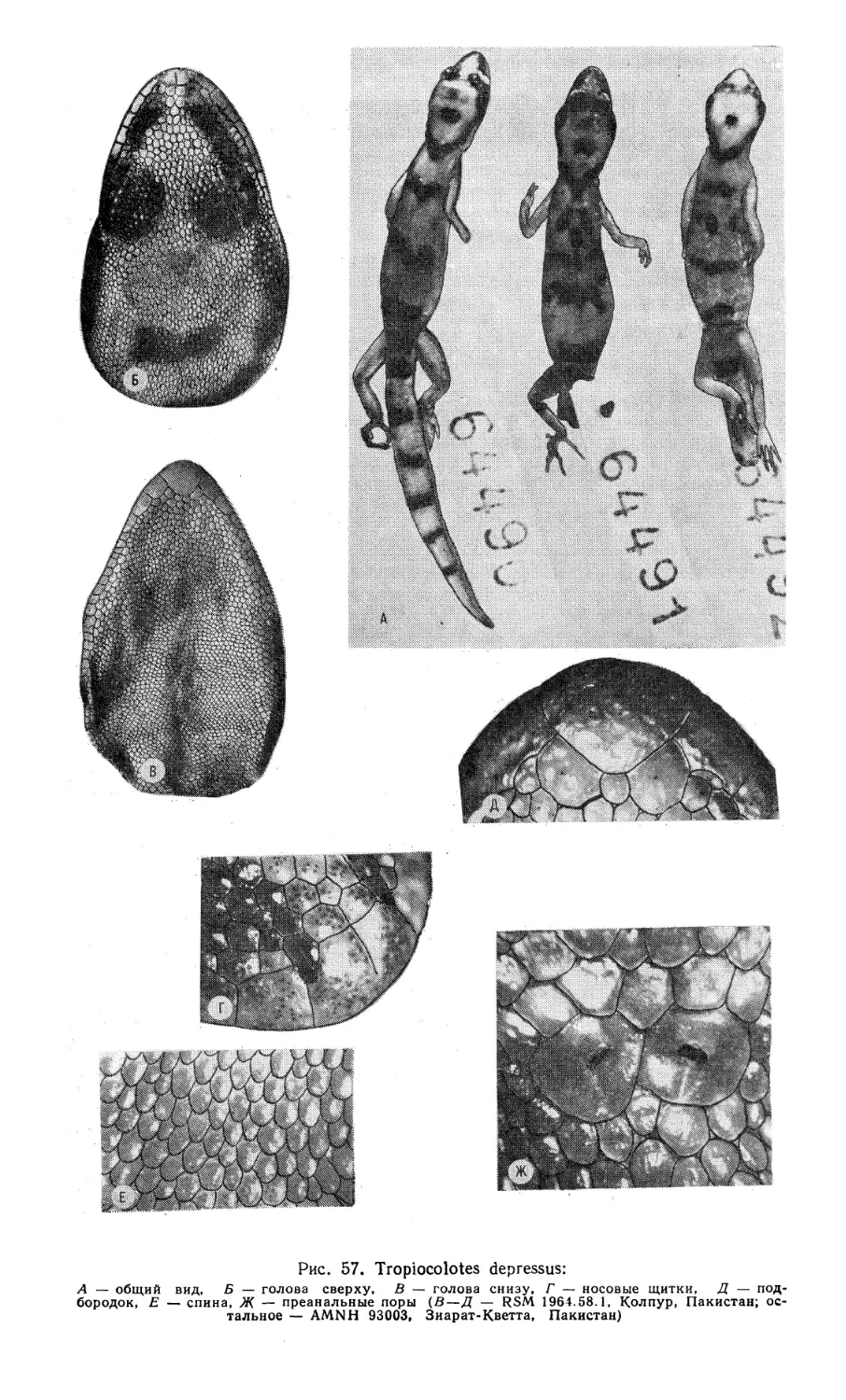
Таксономия рода Tropiocolotes не менее сложна и запутана. Так, описан-
ный В. Петерсом [276] род Tropiocolotes первоначально включал лишь один
вид — Т. tripolitanus, известный из Ливии. Г. Буланже [204] включил в этот
род Gymnodactylus steudneri, описанный В. Петерсом [275] из Сев. Судана.
Ф. Штейндахнер [294] описал Т. nattereri из Синая, который впоследствии был
признан младшим синонимом предыдущего вида. К 1947 г. [249] выяснилось,
что ареалы видов этого рода охватывают Сев. Африку к северу от 10 парал-
8
лели, причем типовой вид обитает здесь повсеместно и образует четыре подви-
да, второй вид встречается только в северо-восточной части континента. Об-
щими признаками этих видов были ребристость подпальцевых пластин, а также
некоторых или всех чешуй туловища, отсутствие спинных бугров, малые разме-
ры и ряд других, менее весомых признаков. В этот же род был включен вскоре
еще один вид, обнаруженный на юге Аравийского полуострова,— Т. scortecci
[208]. Диагноз его не противоречил родовому диагнозу.
Однако с конца 50-х до начала 70-х годов возник вопрос: включать или не
включать единственный вид рода Microgecko — М. helenae — в род Tropioco-
lotes? Этот вид был описан А. М. Никольским [94] по добытым Н. Зарудным
в Ю.-З. Иране нескольким экземплярам. Ряд черт — гладкие подпальцевые
пластинки и гладкая чешуя туловища — противоречили диагнозу рода Tropio-
colotes, другие — определенное расположение некоторых щитков головы,
отсутствие спинных бугров, мелкие размеры — соответствовали ему. В пользу
включения Microgecko в род карликовых гекконов (Tropiocolotes) высказались
ряд исследователей [167, 256, 258, 262, 263, 307]. Другие отстаивали самостоя-
тельность рода Microgecko [168, 223, 302]. Как выяснилось впоследствии [265],
почти все авторы базировались на неверно определенном материале и было по-
казано, что за Т. helenae ошибочно принимали очень близкий вид — Т. persi-
cus, первоначально описанный А. М. Никольским [94] как Alsophylax и оши-
бочно идентифицированный [168] как Bunopus. Ш. Минтон и др. [265] доказали
принадлежность Microgecko к роду Tropiocolotes и выделили три подвидовые
формы Т. persicus. Одновременно они описали Т. heteropholis из Ирака, один из
основных признаков которого — разнородность спинной чешуи — противоре-
чил диагнозу рода.
За это время род Tropiocolotes пополнился еще двумя южноазиатскими
видами с гладкими подпальцевыми пластинками и однородной спинной че-
шуей — Т. depressus из Пакистана [264] и Т. latifi из Ц. Ирана [246]. Пос-
ледние авторы отметили, что, по их мнению, род Tropiocolotes состоит из двух
групп. К южноазиатской группе видов («helenae» — complex) они отнесли Т. he-
lenae, Т. persicus, Т. depressus и Т. latifi, остальные виды (Т. tripolitanus,
Т. steudneri, Т. nattereri, Т. scortecci) — к другой, североафриканской группе.
Однако какой-либо таксономический статус этим группам придан не был.
Список подвидовых форм видов рода Tropiocolotes также пополнился дву-
мя названиями: Т. Папенфусс [271] описал Т. t. apoklomax из Мали, Й. Шмидт-
лер и др. [286] — Т. helenae fasiatus из 3. Ирана. Последние авторы не сравни-
вали свои экземпляры с типами вида, считая их утерянными.
Таким образом, к настоящему времени стало известно, что ареал рода Tro-
piocolotes включает Сев. Африку и Ю.-З. Азию, но внутриродовые, подвидо-
вые связи и ряд других вопросов систематики рода остались нерешенными.
Сведения, касающиеся видов обсуждаемых здесь родов, содержатся во
многих работах [3—7, 15, 19—25, 59—61, 106—108, 123—125, 130, 131, 216,
250—252, и др.]. Характерной их особенностью является то, что в отечествен-
ных работах в той или иной мере освещаются различные стороны экологии ука-
занных гекконов, в зарубежных источниках такие сведения почти полностью
отсутствуют. Следовательно, существующая точка зрения, согласно которой
Bunopus является подродом рода Alsophylax, не базируется на результатах
специального исследования и нуждается в пересмотре. Внутриродовая структу-
ра родов Alsophylax, Bunopus и Tropiocolotes, их внутри- и межвидовые отно-
шения, а также географическая изменчивость включенных в эти роды видов
почти не изучены. Вследствие слабой изученности видов указанных групп от-
сутствует единый взгляд на таксономический вес диагностических признаков
различных уровней.
При изучении систематического положения гекконов родов Bunopus
Blanf. и Tropiocolotes Peters выяснилось, что в составе этих таксонов ошибоч-
но числятся виды, имеющие с остальными их представителями лишь конвер-
гентное сходство. Анализ характера, степени и границ изменчивости фолидоза
на серийном материале позволил уточнить состав перечисленных родов и пос-
ле соответствующих таксономических перестановок очертить их границы более
9
четко. В роде Tropiocolotes, как указывалось выше, числится вид Т. hetero-
pholis[169].
Неоднородный фолидоз — один из основных признаков этого вида, что,
кстати, отражено в видовом названии. В данном случае неоднородность фоли-
доза обусловлена наличием на спине 12 продольных рядов крупных чешуй.
Этот признак, а также очень выраженная ребристость подпальцевых пластин
и спинной чешуи склонили авторов описания указанного вида включить его
именно в род Tropiocolotes. Однако от остальных видов рода его отличает раз-
нородность спинных чешуй. В результате изучения типа этого вида мы пришли
к выводу, что его пребывание в роде Tropiocolotes необоснованно. Впоследст-
вии [1611 описали новый вид из Ирана — Bunopus aspratilis, также харак-
теризующийся сильной ребристостью всех чешуй, исключая крупные щитки
головы. Кроме того, данный вид имел отчетливые спинные и хвостовые бугор-
ки в сочетании с ребристыми подпальцевыми пластинками, что побудило ав-
тора отнести новый вид к роду Bunopus. Он отметил, что этот вид близок к
Т. heteropholis, но не придал этому особого значения. После изучения типов
указанных видов мы пришли к заключению, что они имеют ряд особенностей,
позволяющих выделить их в самостоятельный род — Carinatogecko Golubev
et Szczerbak, 1981.
* * *
Наибольшее количество видов гекконов до недавнего времени объединяли
в род голопалых гекконов — Gymnodactylus Spix, 1825, описанный по типово-
му виду G. geckoides из Нового Света. Двумя годами позже Грэй опубликовал
диагноз рода Cyrtodactylus (типовой вид С. pulchellus) по материалам из Ю.-В.
Азии (Сингапура), который отличался от предыдущего порядком располо-
жения преанальных пор. Ненадежность указанного признака дала право ав-
торам каталогов [96, 200, 212, 215, 300, и др.] при том или ином содержании
объема рода сохранить первое название в качестве валидного.
В современной литературе отсутствует общепринятое мнение относительно
объема рода голопалых гекконов (Gymnodactylus — Cyrtodactylus). Европей-
ские герпетологи [257, 307, и др.], а эту точку зрения разделяют и советские
специалисты [17], принимают род голопалых гекконов (Gymnodactylus), ох-
ватывающий виды, распространенные космополитически. Г. Андервуд [305]
разделил род Gymnodactylus (типовой — южноамериканский вид G. geckoides)
на основании строения зрачка глаза и некоторых других признаков на два рода.
Для видов из Старого Света он предложил родовое название Gyrtodactylus
(типовой — южноазиатский вид С. pulchellus), которое было принято амери-
канскими герпетологами [170, 245, 263, и др.]; оно в последнее время фигури-
рует в работах зарубежных европейских авторов 1194].
Мы [153] присоединились к точке зрения Г. Вермута, автора каталога гек-
конов мировой фауны [305], и рассматривали группу евроазиатских голопалых
гекконов в качестве подрода Cyrtodactylus. Одновременно были уточнены опи-
сания родов и подродов гекконов фауны СССР Такое решение, казалось,
было вполне удовлетворительным, однако оно не выдержало испытания време-
нем. В 1983 г. обнаружен новый вид геккона в Ц. Тянь-Шане [56], который име-
ет основные признаки родов Gymnodactylus и Alsophylax. Диагноз первого
рода к этому времени содержал следующие основные признаки: 1) пальцы тон-
кие, кривые, так как две или три последние фаланги сжаты с боков и соединены
друг с другом под углом; 2) пальцы снизу покрыты одним продольным рядом
подпальцевых пластинок; 3) туловище покрыто однородной зернистой чешуей,
между которой часто расположены более крупные ребристые чешуи (бугор-
ки, образующие более или менее правильные продольные ряды; чешуи брюха
крупнее боковых; 4) у самцов имеются обычно преанальные и бедренные поры
или только преанальные.
Проведенная нами комплексная ревизия гекконов фауны СССР обнаружи-
ла ряд неожиданностей. Беглое знакомство с представителями рода Alsophylax
фауны СССР показало, что все признаки, за исключением первого (искривление
10
пальцев), свойственны и представителям данного рода. У видов A. spinicauda
и A. tibetanus искривлены пальцы так же, как и у голопалых гекконов. Изуче-
ние евроазиатских голопалых гекконов показало их неоднородность. Здесь
заметно выделяются две группы видов: средиземноморских и среднеазиатских.
К первой группе следует отнести G. kotschyi, G. russowi и некоторые другие,
ко второй относятся G. caspius, G. fedschenkoi, G. longipes. Группа средизем-
номорских видов характеризуется более слабо искривленными пальцами лап,
что сближает их с некоторыми геккончиками, выделяемыми в группу Bunopus.
Изучение голопалых гекконов выявило различия указанных групп и по
другим признакам. Так, средиземноморские гекконы отличаются от средне-
азиатских (в нашем понимании) отсутствием бедренных пор (имеются только
преанальные) и рядом других признаков фолидоза. Эти данные позволили нам
выделить группу средиземноморских гекконов в подрод Mediodactylus Szczer-
bak et Golubev, 19771. Проведенная нами ревизия рода Alsophylax показала
также, что вид A. spinicauda более близок к представителям подрода Medio-
dactylus и должен быть перенесен в род голопалых гекконов, что стабилизиру-
ет диагнозы указанного рода и геккончиков. По совокупности признаков A. ti-
betanus мы также перенесли в род Gymnodactylus, а его диагноз дополнили
признаком «на нижней поверхности хвоста имеется ряд увеличенных щитков
или чешуи».
Возвращаясь к находке 1983 г. на Тянь-Шане, необходимо подчеркнуть,
что задние лапы нового вида геккона имеют такие же искривленные пальцы,
как и у некоторых голопалых гекконов. По большинству признаков он был
включен на правах особого подрода Altiphylax в род Alsophylax, но одновремен-
но стала очевидной несостоятельность традиционного описания рода Gymno-
dactylus (диагноз рода Alsophylax требует также серьезного дополнения).
Знакомство с типовым видом Cyrtodactylus pulchellus из Сингапура показало,
что между ним и группой палеарктических видов голопалых гекконов существу-
ет ряд весьма важных различий (у первого базальные фаланги значительно
расширены и покрыты снизу рядом широких пластин, дистальные фаланги рез-
ко утончены; у вторых пальцы одинаково тонкие на всем протяжении).
Строение пальцев, особенно расположение и форма подпальцевых пласти-
нок, настолько постоянны для разных видов, что имеют решающее значение
в систематике гекконов. Именно по этим признакам таксономисты различают
группы голопалых, полупалых, листопалых, вееропалых и других гекконов.
Адаптация к передвижению по вертикальным поверхностям у разных родов
решается совершенно различным способом. До сих пор никто не обратил
внимания на такие различия в строении палеарктических и южноазиатских
видов, которые не позволяют объединять их в одном роде. Еще Г. Андервуд
[305] заметил, что в рамки его диагноза рода Cyrtodactylus не укладывались
бывшие в его распоряжении каспийский и туркестанский гекконы.
А. Клюге [239] убедительно показал несостоятельность указанного выше
рода применительно к некоторым тропическим группам гекконов. Он пишет:
«Набор андервудовских признаков оказался не в состоянии диагностировать
и четверти видов Cyrtodactylus из-за их индивидуальной и межвидовой изменчи-
вости или из-за обычной плезиоморфности. Таким образом, имеющиеся доказа-
тельства показывают, что «Cyrtodactylus... (учитывая изменчивость в размерах,
форме увеличенных спинных бугров и их килеватости, расположении преаналь-
ных пор) является, возможно, полифилетической группой» (перевод наш).
Валидность указанной группы гекконов Старого Света была взята под
сомнение в результате нашей работы на севере ее ареала и А. Клюге — на юге.
Это привело нас к мысли о выделении евроазиатских тонкопалых гекконов
в особый род Tenuidactylus (типовой вид Т. caspius) [153]. В соответствии с этой
работой и принят объем рода в настоящей монографии. Каспийский геккон вы-
бран в качестве типового не случайно. Во-первых, он первым описан среди
1 В новом каталоге рептилий Европы и Ю.-З. Азии, изданном в США [322], этот подрод
без каких-либо оснований возведен в ранг рода, признается наличие в данном регионе как
рода Gymnodactylus, так и Cyrtodactylus, содержатся другие ошибки.
11
наших тонкопалых гекконов; во-вторых, вид содержит весь комплекс очень
хорошо выраженных, характерных для группы признаков. По предваритель-
ным данным, род Tenuidactylus объединяет 18 видов палеарктических гекко-
нов, относящихся к трем подродам — номинативному и описанным нами
недавно Mediodactylus, включающему четыре средиземноморских вида, а так-
же Cyrtopodion — из шести видов, обитающих в Ю.-З. и Ц. Азии.
Выделение группы видов тибетско-гималайского региона в особый под-
род, видимо, дело недалекого будущего. Род Cyrtodactylus, по нашим представ-
лениям, включает большую и разнородную группу тропических, преимущест-
венно южноазиатских видов (около 40). Наличие группировок внутри рода го-
лопалых гекконов отмечал еще М. Смит [281], хотя его исследование касалось
преимущественно индийских видов. Тем не менее даже краткое знакомство
с палеарктическими формами убедило этого герпетолога в их своеобразии.
Первая группа указанного автора (типа «scaber») по составу и по некоторым ха-
рактеризующим ее признакам во многом соответствует подроду Cyrtopo-
dion. Последняя группа не столь однородна и состоит из двух групп: одна
занимает промежуточное положение по многим показателям между Te-
nuidactylus и Mediodactylus и состоит из четырех видов (Т. kachensisT. scaber,
Т. watsoni и Т. montiumsalsorum). Вторая (Т. agamuroides и Т. elongatus)
небольшая группа «проагамуроидных» гекконов (по некоторым признакам
напоминают представителей рода Agamura). Тем не менее их нельзя объеди-
нять с агамуроидными гекконами, поскольку они являются носителями основ-
ных черт палеарктических тонкопалых гекконов.
Процесс описания всех видов этого рода, видимо, еще не закончен. Сви-
детельством тому служит описание вида Т. turcmenicus Szczerbak, 1977, который
обнаружен на такой хорошо исследованной зоологами территории, как Бад-
хыз. Следует ожидать новых находок на территории Ирана, Афганистана,
особенно в Тибетско-Гималайском регионе.
Как показали работы в области кариологии [73, 154, и др.], наши номенк-
латурные перестройки и отнесение отдельных видов к родам Alsophylax и Te-
nuidactylus убедительно подтверждаются особенностями строения хромосом-
ного аппарата.
* * *
Небольшая родовая группировка так называемых крысохвостых гекконов
(Agamura), описанная в 1874 г. известным английским натуралистом У. Блан-
фордом [181] из Иранского Белуджистана, объединяет два вида — A. persica —
типовой и A. femoralis [211, 290], обитающих в пустынных местностях В. Ира-
на, Афганистана, Пакистана и Сев.-З. Индии [49]. В 1874 г. была описана форма
A. cruralis Blanf. (под этим названием она числилась и в каталоге Буланже
[200]), но М. Смит [291] доказал ее идентичность A. persica. Вид A. femoralis,
известный только из типового местонахождения, считается валидным [307],
но, видимо, требует более пристального внимания со стороны систематиков.
Первоначально A. persica был отнесен к роду Gymnodactylus, однако свое-
образный внешний вид и ряд признаков чешуйчатого покрова настолько четко
его отличают от представителей предыдущего рода, что новое родовое описание
после его публикации никем не подвергалось сомнению. Тем не менее род Aga-
mura, на наш взгляд, близок к роду Tenuidactylus.
Как мы установили, описанная из Ирана агамуроидная ящерица Gymno-
dactylus gastropholis Werner, 1917, впоследствии сведенная по чисто внешнему
сходству в синоним G. agamuroides Wettstein, 1951, в действительности отве-
чает диагнозу рода Agamura и рассматривается нами в его составе.
Бельгийский зоолог де Витте описал из Ирана новый род Rhinogekko
(типовой вид—R. misonei) [320], который характеризуется очень вздутыми
носовыми щитками, образующими канал ноздри, заметно приподнятый над по-
верхностью морды. По всем остальным признакам указанный вид относится
к роду Agamura. Приподнятые носовые чешуйки известны (хотя развиты сла-
бее) у представителя рода Tenuidactylus (Т. elongatus), поэтому родовая при-
надлежность вида «misonei» требует уточнения.
12
* * *
Недавно [323] немецкий герпетолог Ю. Фрицше во время пребывания в ту-
ристской поездке в Ашхабаде якобы на стене здания добыл экземпляр нового
вида для фауны СССР — Hemidactylus turcicus. Мы специально искали данно-
го геккона в указанном районе, но безрезультатно. Очевидно, особь турецкого
полупалого геккона была случайно завезена кем-то, а затем отловлена или дан-
ный факт относится к категории музейных ошибок и вносить этот вид в список
фауны СССР нецелесообразно. Поэтому мы и не рассматриваем здесь род Hemi-
dactylus.
МАТЕРИАЛ И МЕТОДИКА
Основой для работы послужили прежде всего сборы и наблюдения авторов,
проведенные во время полевых экспедиционных исследований. Материалы по
гекконам мы собирали до начала плановой работы по данной группе и исполь-
зовали при выполнении региональных сводок [ 139] попутно со сбором других
пресмыкающихся [143]. В этот период были изучены систематика и экология
крымского геккона [138, 139], сделан ряд новых находок [147], причем полевые
экспедиционные работы в то время регулярно проводились в Крыму (1956—
1964, 1975, 1980, 1981 и 1982 гг.), а также Азербайджане (1958), Грузии и Ар-
мении (1963), Туркмении (1963, 1964, 1966, 1970 и 1971), Узбекистане (1964,
1968), Таджикистане (1964), Киргизии (1957, 1960, 1962, 1972), Казахстане
(1973 г.). С 1974 г. мы проводили обширные полевые работы, основной задачей
которых был сбор материала по гекконам.
Экспедиционные исследования охватывали территорию Кавказа и Закав-
казья (1974, 1975), Туркмении (1975—1984), Узбекистана (1976), Таджикистана
(1976, 1980, 1981), Казахстана (1976) и Киргизии (1984). Особенно обширные
исследования в Средней Азии были проведены в 1976 г.— автомобильный про-
бег по маршруту Киев — Баку — Красноводск — Кушка — Керки — Тер-
мез — Ферганская долина — пустыня Кызылкумы — Устюрт — Гурьев —
Астрахань — Волгоград — Воронеж — Киев охватывал более 20 тыс. км.
За эти годы только пустыню Каракумы мы пересекли восемь раз в направлении
с юга на север и обратно, а также с запада на восток. Животных мы добывали
разными способами. В дневное время их чаще всего извлекали из убежищ
при переворачивании камней, раскопках нор насекомых и грызунов, термит-
ников, оснований отмерших кустарников. Активный поиск позволил собрать
много видов ящериц и змей, которые до тех пор считались очень редкими, при-
сутствовали в коллекциях единичными экземплярами. В сумерках и ночью
с помощью фонарей осматривали скальные и лессовые обрывы или барханы.
На равнинах (в степных и пустынных районах) использовали автомобиль
Газ-66 с установленными дополнительными фарами, а также несколько
наблюдателей. С медленно двигающейся машины хорошо просматривалась
полоса шириной до 5 м (различались даже муравьи). Этот способ был незаменим
при сборе мелких наземных гекконов, проведении количественного учета (ско-
рость 4—5 км/ч, расстояние учитывалось по спидометру) Часть добытых яще-
риц фиксировали в спирте на месте поимки, других пересаживали в садки для
последующего изучения в лабораторных условиях.
Собранные обширные коллекции и наблюдения по экологии, а также коллек-
ционные материалы, полученные из многих отечественных и зарубежных музе-
ев, позволили нам с принятых позиций концепции полйтипического вида изу-
чить серийный материал, состоящий преимущественно из выборок популяций
различных районов видового ареала. Это дало возможность составить доста-
точно обоснованные описания таксонов (мы могли ознакомиться с большинст-
вом сохранившихся типовых материалов), изучить индивидуальную, возраст-
ную и (что особенно важно) географическую изменчивость, провести ревизию
систематического положения изученных таксонов. Всего мы исследовали
13
свыше 5 тыс. экземпляров, при составлении описаний и изучении географиче-
ской изменчивости использовали около 20 признаков.
Кроме традиционных, мы наметили и использовали ряд новых признаков.
По мере обработки материала стали отчетливо проявляться границы их измен-
чивости, что позволило установить признаки и их комбинации большого так-
сономического веса с малой степенью изменчивости от родового до подвидового
уровней и впервые применить их в качестве диагностических. При этом родовые
диагнозы потеряли одно достоинство — легкость и краткость, но приобрели
другое — более высокую степень надежности. Мы использовали такие признаки:
1. Длина туловища (L) (от конца морды до анальной щели).
2. Длина нерегенерированного хвоста (Led) (от анальной щели до конца
хвоста). Указанные выше измерения проводили штангенциркулем, признаки
3—9 измеряли микрометром бинокуляра МБС-2.
3. Длина головы (от переднего края ноздри до переднего края уха).
4. Длина морды (от переднего края ноздри до переднего края орбиты).
5. Поперечный диаметр глаза.
6. Максимальный (вертикальный) диаметр ушного отверстия.
7. Ширина головы (за глазами).
8. Высота головы (за глазами).
9. Длина спинного бугорка (в одном из двух центральных рядов посередине
спины).
Часть этих промеров была использована для получения индексов:
1. ЫLed.
2. Отношение высоты головы к ее ширине (Х100).
3. Отношение ушного отверстия к диаметру глаза (X 100).
4. Отношение длины спинного бугорка к длине тела (X 1000).
Было подсчитано количество чешуй и щитков:
1. Чешуй поперек головы (между центрами глаз; чешуйки на надглазнич-
ных козырьках не учитывались).
2. Чешуй вдоль головы (только для сцинковых гекконов; от носовых
щитков до укрупненной спинной чешуи).
3. Верхнегубных щитков отдельно слева и справа (учитывалось также,
через какой из этих щитков проходит проекция переднего края орбиты).
4. Носовых щитков.
5. Чешуй, разделяющих первые носовые щитки.
6. Нижнегубных щитков.
7. Пар нижнечелюстных щитков слева и справа или чешуй, их заменяющих
(для Bunopus, некоторых Teratoscincus и др.).
8. Чешуй, окружающих спинной бугорок (в одном из двух центральных
рядов посередине спины).
9. Продольных и поперечных рядов спинных бугорков.
10. Чешуй вдоль тела снизу от первой пары нижнечелюстных щитков до
анальной щели (если граница между анальными щитками и зернышками между
ними и анальной щелью была резкой, то эти зернышки не учитывали).
11. Чешуй поперек брюха (посередине туловища).
12. Чешуй вокруг середины туловища (только для видов, не имеющих
спинных бугорков и резкой границы между брюшной и боковой чешуей).
13. Подпальцевых пластинок на IV пальце задних конечностей слева
и справа (для сцинковых гекконов — количество боковых гребешков на тех же
пальцах).
14. Преанальных (бедренных) пор (для самок подрода Tenuidactylus учи-
тывалось количество чешуй, могущих нести на себе эти поры).
Кроме того, учитывались форма, соотношение размеров, состояние поверх-
ности и взаиморасположение всех чешуй и щитков и нижней стороны тела гек-
конов, описывались окраска и рисунок.
Для размерных признаков (1, 2), признаков фолидоза (1, 6, 8, 10—14),
а также индексов (1—4) вычисляли показатели Min—max, М ± m, CV. Выбор-
ки сравнивали на достоверность различия (/). Статистическую обработку при-
знаков и сравнение на достоверность различия (t) проводили по общепринятым
14
методикам [100, 101] на микрокалькуляторе «Электроника БЗ-21» по програм-
мам, разработанным для этой цели Л. И. Францевичем [115].
Каждому видовому очерку предшествуют русское и латинское название ви-
да. Для части видов, подродов и родов русские названия применены нами впер-
вые. Далее указано типовое местообитание, в некоторых случаях оно уточня-
лось при изучении типовых экземпляров. В соответствии с процедурными
правилами Международного кодекса зоологической номенклатуры [74] опреде-
ляли валидность названия и выделяли, когда это было необходимо, лектотипы.
Приведены данные по кариотипу, большая часть их — оригинальные, полу-
ченные в нашей лаборатории. В списке синонимов указаны в хронологическом
порядке работы, в которых название применялось впервые. Описанию пред-
шествуют диагнозы (часто новые, составленные авторами. При изучении расп-
ространения мы уточнили современные ареалы видов и сделали первую попыт-
ку дать карту распространения с учетом всех известных находок; перечисление
их содержится в кадастре. При изучении биотопов приводили описание почвы,
рельефа, растительности, учитывали высоту над уровнем моря (н. у. м.),
экспозицию склона и его угол наклона. Количественные данные получены либо
общепринятым маршрутным способом (при пешеходном или автомобильном уче-
те), либо проводили абсолютный учет переворачиванием камней и раскопкой
убежищ. И в том и в другом случае возможен пересчет численности на 1 га.
Отношение к температуре изучали, измеряя температуру субстрата и воз-
духа при находке на поверхности активной ящерицы, а также в лабораторных
условиях при исследовании в термоградиентприборе (модифицированный при-
бор Гертера), что позволяло определить температурный преферендум (пред-
почитаемую температуру) для исследуемой выборки гекконов.
Данные по суточному циклу активности были получены при полевых ис-
следованиях (появление и уход в убежища), пик активности — при проведении
количественного учета и изучении отношения к температуре. Сезонный цикл
активности выясняли при непосредственных поисках в природе и при сопостав-
лении дат находок коллекционных материалов.
Линьку регистрировали при сборе материала в природе, а также при со-
держании ящериц в неволе.
Сведения по питанию получены при анализе содержимого желудков, а так-
же наблюдений в природе и террариуме. Поскольку питание изучали особенно
активно в последние годы, мы вскрывали желудки только тогда, если литератур-
ные сведения были ограничены или отсутствовали.
Материалы по размножению, описание их кладок и инкубирования яиц мы
предпочитали получать при содержании гекконов в террариуме. Точные све-
дения по этим вопросам в природных условиях получить невозможно. Различие
в сроках инкубации отдельных кладок зависит от темпов получения необходи-
мой суммы температур. В последние годы также стало известно, что температура
инкубации яиц может влиять на соотношение полов и даже на особенности ри-
сунка. Мы считаем изучение репродуктивных особенностей пресмыкающихся
в лабораторных условиях весьма перспективным и актуальным.
Темп роста изучали преимущественно статистическим методом при сопо-
ставлении размеров туловища особей, добытых в разное время.
Данные о врагах гекконов мы получили (наряду с литературными источни-
ками), добывая хищных животных в биотопах ящериц, а также при осмотре
гнезд хищных птиц в период выкармливания птенцов. Причины колебания
численности интересующих нас ящериц выяснялись при сопоставлении антро-
погенных факторов, изучении их влияния на биотоп.
Большое внимание мы уделяли также исследованию особенностей поведе-
ния. Наряду с наблюдениями в природе особое место здесь занимают данные,
полученные при содержании в неволе. При этом мы описывали способ передви-
жения, нападения и защиты, поведение в брачный период и особенности звуко-
вой коммуникации. Материалы о летних и зимовочных убежищах получены
главным образом при изучении в природе.
Практическое значение определяли прежде всего с точки зрения сохране-
ния генофонда редких видов и в необходимых случаях мы даем рекомендации
15
по охране. Исходя из характера питания все гекконы в целом насекомоядные
животные, поедают много вредных насекомых, занимают определенное место
в биоценозах (некоторые виды — фоновые), полезны.
Сведения по экологии многих видов, обитающих за пределами СССР,
крайне скудны (хотя мы использовали всю доступную литературу), к сожале-
нию, многие зарубежные герпетологи не уделяют этим вопросам должного
внимания. Отсюда определенная неравномерность материала в видовых
очерках.
Подавляющее количество рисунков и фотографий оригинальны. Слайды
многих видов мы приводим впервые.
ФИЛОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ
И ЗООГЕОГРАФИЧЕСКИЙ АНАЛИЗ
Представление о древности гекконов (а иногда и их примитивности, что
в конечном итоге часто не одно и то же) стало хрестоматийным. Однако изуче-
нию филогении данных ящериц до сих пор уделяется недостаточное внимание.
Частично это оправдано тем обстоятельством, что нежные кости ископаемых
гекконов не сохраняются и палеонтологические материалы, как правило,
отсутствуют. Пользуясь сравнительно-морфологическим методом, мы можем
в значительной степени приблизиться к решению данной проблемы.
Известный советский герпетолог П. В. Терентьев [111] подвел итог работы
по филогении ящериц и предложил схему их родственных отношений. У ос-
нования ветви гекконообразных он поставил представителей юрских ящериц
Ardeosauridae (ископаемые остатки представителей двух родов известны из
Баварии, ФРГ и Маньчжурии). Подобная точка зрения принята и в настоящее
время. Специальное исследование эволюции гекконовых ящериц (и, пожалуй,
пока единственное) принадлежит А. Клюге [237]. Он рассматривает филогене-
тические связи гекконид на обширном материале на уровне подсемейства при
сравнении внутренней и внешней морфологии (18 диагностических показателей),
устанавливает их эволюционные уровни, примитивные и прогрессивные приз-
наки, обсуждает возможные параллелизмы.
По его данным, примитивными признаками являются наличие настоящих
век (их отсутствие считается прогрессивным признаком; далее в скобках при-
водим прогрессивное состояние признака); развитие премаксиллярных костей
из двух центров окостенения (одного центра); посткраниальный эндолимфати-
ческий аппарат без кальцинированной сумки (сумка присутствует); участки
с неравномерным щиткованием отсутствуют (присутствуют); способны издавать
звуки (неспособны); откладывают два яйца (одно); есть надвисочная кость (от-
сутствует); количество склеральных косточек в глазу больше 14 (равно или мень-
ше); клоакальные сумки и кости имеются (отсутствуют); отмечена угловая кость
(отсутствует); есть пластинчатая кость (spleniale) (отсутствует); лобная кость од-
на (парная); носовые кости парные (слиты в одну); теменная кость одна (парная);
позвонки в поясничной области процельные (амфицельные); гиоидная дуга
очень ороговела (слабо или отсутствует ороговение); во второй висцеральной
дуге цератобранхиально-эпибранхиальное соединение продолжительное (сло-
мано, но соединены в разных вариантах или не соединены); чешуйчатая кость
имеется (отсутствует).
Большинство указанных признаков можно принять как примитивные,
значение таких, как «амфицельные-опистоцельные позвонки» или «в кладке два
или одно яйцо» — спорное. С помощью статистики для каждой из исследован-
ных групп автор вычисляет количество тех или иных признаков и в зависимо-
сти от этого помещает их на определенный эволюционный уровень. И хотя в
оценке признаков имеется доля субъективизма, в целом выводы представляют
интерес, соответствуют действительности. Сравнительно-морфологический ана-
лиз фолидоза, проведенный нами, позволил выявить следующие альтернатив-
ные значения признаков (примитивный — прогрессивный): чешуя головы и ту-
ловища мелкая (крупная), фолидоз слабо дифференцирован или чешуя од-
16
нородная (хорошо дифференцирован), наличие только преанальных пор (име-
ются и бедренные поры), базальные фаланги пальцев расширены (не расширены),
сегментация хвоста не выражена (четко выражена), на затылке имеется
опоясывающая голову темная полоса (полоса отсутствует); ареал вида распо-
ложен южнее (севернее; здесь нередки исключения).
П. В. Терентьев [111] ближе к ардеозаврам помещает семейство чешуеноги
(Pygopodidae), затем семейство ночные ящерицы (Xantusiidae), плоскохвостые
гекконы (Uroplatidae) и, наконец, эублефары (Eublepharidae) и семейство гек-
Рис. I.» Филогенетические отношения родов гекконов ’фауны СССР и сопредель-
ных стран
коны (Gekkonidae). По нашему мнению, эублефаров более правильно рассмат-
ривать в качестве семейства, но семейства весьма примитивного и его место
должно быть расположено на филогенетической ветви ниже, чем хорошо спе-
циализированное семейство чешуеноги. Ночные ящерицы занимают как бы
промежуточное положение между гекконами и сцинками [49], видимо, ближе к
последним; окончательное решение данного вопроса требует специальных ис-
следований. И, наконец, плоскохвостые гекконы сейчас рассматриваются [49]
в качестве рода семейства гекконы. Таким образом, филогения гекконообраз-
ных должна выглядеть в виде ветви от ардеозавров к эублефарам, от которой
ответвляются гекконы, и в начале ветви последних — чешуеноги.
А. Клюге [237] сгруппировал всех гекконов в четыре подсемейства. У ос-
нования этой ветви он помещает подсемейство эублефар (в нашем понимании
семейство), затем от основного ствола ответвляется ветвь подсемейства австра-
лийские шипохвостые гекконы (Diplodactylinae) и в конце его — разветвление
к настоящим гекконам и подсемейству круглопалые гекконы (Sphaerodactyli-
пае); последняя группа расположена на более высоком эволюционном уровне
как наиболее прогрессивная ветвь. Таким образом, мы принимаем (с небольшой
поправкой) филогению на уровне подсемейств А. Клюге. Его взгляды хорошо
согласуются с зоогеографией: Ю.-В. Азия — центр происхождения циркум-
глобального семейства эублефар (отдельные роды обитают, кроме того, в Афри-
ке и Америке — проникли туда, очевидно, через Берингийскую сушу в палео-
цене, что еще раз свидетельствует о древности указанной группы).
С достаточной долей уверенности можно предположить, что все гекконы
произошли на протяжении верхней юры и нижнего мела в Ю.-В. Азии. Отно-
сительно примитивное подсемейство Diplodactylinae привязано к Австралий-
скому региону и Новой Зеландии (автохтон австралийского региона). Их пре-
док произошел в Ю.-В. Азии от эублефаридных предков где-то в позднем
мезозое и проник в Австралию через Индо-Австралийские острова. Видимо,
произошли от эублефаридных предков и гекконы, мигрировавшие на запад
и в Африку. В настоящее время Gekkoninae являются доминантной группой,
в ее пределах наблюдается до трех уровней эволюционной радиации: два
17
ранних уровня привели их к проникновению в Африку, на Мадагаскар и в Ю.-З*
Азию. Гекконы Нового Света возникли от палеотропических форм, предки ко*
торых обитали в Африке и Средиземноморье (не учитывается более поздний за-
воз человеком). Так, А. Клюге высказывает мысль о том, что Sphaerodactylinae
Нового Света произошли от африканских предков, которые воспользовались
трансатлантическим путем.
Филогенетические схемы родов и видов палеарктических гекконов в извест-
предпринята попытка представить род-
ственные отношения в этой группе пу-
тем построения дендрограмм, которые
получены в результате морфологиче-
ских исследований (рис. 1,2) Несомнен-
но, что род Tenuidactylus связан в про-
исхождении с палеотропическим родом
Cyrtodactylus и занимает центральное
положение. Его проникновение в пале-
арктическую Азию через Гималаи бес-
спорно. Правые ветви (Bunopus— Ste-
nod act у 1 us—Crossob a mon) объединяют
роды, адаптированные к псаммофиль-
ному образу жизни, характеризуются
развитием ребристости подпальцевых
пластинок, образованием на пальцах
гребешков и общей мелкочешуйчато-
стью, но с участками крупной чешуи.
Видимо, это прогрессивная ветвь, так
как песчаные пустыни — образование
более молодое, чем тропические леса и
скалы. Родсцинковых гекконов и роды
геккончиков и микрогекконов разви-
вались независимо и пока трудно
представить их предковые группы.
Не исключено, что род микрогекконов
связан с африканским родом Sauroda-
ctylus (их сближают мелкие размеры
и щиткование головы).
Мы доказали самостоятельность
ной нам литературе отсутствуют. Нами
Рис. 2. Филогенетические отношения видов
родов:
а — Bunopus, б — Carinatogecko, в — Teratosci-
ncus, г — Agamura, д — Crossobamon
родов Alsophylax и Bunopus, которых
ранее объединяли. Указанное положение хорошо подтверждается и данными зо-
огеографии. Род Alsophylax эндемичен для туранской равнинно-пустынной,
центральноазиатской пустынной и горноазиатской провинций [148, 301]. Род
Bunopus характерен для пустынной сахаро-аравийской, средиземноморской
и переходной ирано-афганской провинций. Сказанное позволяет сделать вывод
не только о самостоятельности этих родов, но и об отсутствии близкого родства
между ними (отсутствует общность ареалов). Филогению рода Alsophylax
можно представить следующим образом (рис. 3): род делится на две неравные
части — реликтовый подрод Altiphylax с видом A. tokobajevi и номинативный
подрод. В центре последнего располагаются близкие виды A. laevis и A. pipiens.
Их сближает наименьшая дифференциация фолидоза в области ноздри, и, хотя
последний вид достигает северных пределов видового ареала (зоны степи), его,
видимо, следует рассматривать в качестве более древнего вида, так как биотоп
A. laevis — такыры, образованные вследствие аллювиальных процессов уже
в четвертичное время,— более молодое ландшафтное образование. Следует
здесь учитывать и обширный ареал A. pipiens: от берегов Волги до Монголии.
От вида A. laevis в недавнее время отделился второй «гладкий» вид — A. ta-
djikiensis. Центральноазиатская ветвь — вид A. przewalskii— тяготеет к под-
роду Altyphylax (ареал которого лежит в пределах центральноазиатского гор-
ного округа), a A. loricatus по степени развития спинных бугорков занимает
несколько обособленное положение, но все же близок к A. przewalskii.
18
Как свидетельствуют морфологические данные, род Tropiocolotes близок
к роду Alsophylax, особенно ветвь подрода Asiocolotes, которая, видимо, и яв-
ляется предковой для рода Alsophylax (рис. 4). Р. Мертенс 1256] считал род
Tropiocolotes североафриканским, который проник глубоко в 3. Азию (она слу-
жила мостом для проникновения многих североафриканских рептилий на ази-
атский континент). Это мнение согласуется с данными А. Клюге [237], который
на обширном остеологическом материале показал, что распространение подсе-
мейства Gekkoninae, сопровождавшееся видообразованием, на всех континен-
Рис. 3. Филогенетические отношения видов рода Alsophylax
тах происходило в направлении от тропиков к полюсам. Таким образом, рас-
пространение рептилий из Сев. Африки через Ю.-З. в Среднюю Азию представ-
ляется достаточно обычным явлением. С этим направлением совпадает направ-
ление изменчивости некоторых признаков фолидоза внутри рода Tropiocolotes.
В ветви Т. tripolitanus — Т. steudneri — Т. depressus — Т. levitoni при
переходе от вида к виду постепенно увеличиваются количество анальных пор
(соответственно 0 — иногда 2 — 2—5 — 6—7), подпальцевых пластинок (11—
17 — 15—18 — 16—18 — 17—23), размеров тела (до 26 мм, на востоке ареала —
до 27—32—45 мм), постепенно формируется ряд подхвостовых щитков (нет —
нет — только в последней трети — вдоль всего хвоста) и др. У части особей
Т. levitoni появляется третья носовая чешуйка и, что особенно важно, бугорки
в области крестца. У этого вида рисунок тела близок к таковому у Alsophylax
(среди видов последнего рода к Asiocolotes ближе всех стоит A. laevis). Совре-
менные ареалы указанных видов размещаются в той же последовательности:
Т. tripolitanus — Сев. Африка; Т. steudneri — восток Сев. Африки и Ю.-З.
Азия; Т. depressus — Сев.-В. Белуджистан (хр. Ц. Брагуй); Т. levitoni —
Ю.-З. Гиндукуш; A. laevis — юг Средней Азии. Все это позволяет допустить,
что между предками Т. levitoni и предками A. laevis имелось несколько проме-
жуточных форм.
Исключительно сходное в некоторых случаях щиткование носовой области
у Microgecko и Alsophylax (A. loricatus) объясняется, на наш взгляд, одним из
положений закона гомологических рядов Н. И. Вавилова, согласно которому
«не только генетически близкие виды, но и роды проявляют сходство в рядах
наследственной изменчивости» [32, с. 24]. В данном случае путь становления
признаков (определенное сочетание носовых щитков и чешуек) в ряду Tropioco-
lotes — Asiocolotes — Alsophylax значительно длиннее по количеству рецепт-
ных «промежуточных» форм, чем в ряду Tropiocolotes — Microgecko, что, ве-
роятно, вызвано различными темпами эволюции этих групп гекконов. Мнение
Р. Мертенса о североафриканском происхождении рода Tropiocolotes в еще
большей степени относится к номинативному подроду, включающему три вида.
19
Их общие особенности: ребристые подпальцевые пластинки, в основном не диф-
ференцированные носовые чешуи, довольно крупная спинная чешуя, отсутствие
подхвостовых щитков и четко выраженных, присутствующих у всех самцов
анальных пор.
Имеющиеся отклонения от диагноза позволяют установить связи с другими
подродами. Так, у отдельных самцов Т. steudneri есть две слабо выраженные по-
ры, что позволяет предположить его большую, чем у остальных видов подрода,
близость к Asiocolotes (другие соображения обсуждались ранее), у Т. scortecci
интерназальные щитки с ноздрями не соприкасаются, но они уже несколько
увеличены и сходны по форме с таковымы у Microgecko. Вероятно, три вида но-
минативного подрода произошли от одного общего предка (сходные размеры
уплощенной чешуи и колебания в степени ее ребристости). В отличие от Т. scor-
tecci, у всех микрогекконов интерназальные щитки не только увеличены, но
и соприкасаются с ноздрями. Две дополнительные носовые чешуйки и одна-
две пары крупных постинтерназальных щитков — обязательный подродовой
признак (очень редко здесь наблюдается отклонение от нормы: первая допол-
нительная чешуйка оттесняется от ноздри второй и интерназальным щитком).
Сочетание этих признаков с гладкими подпальцевыми пластинками (не более
15) и отсутствием подхвостовых щитков полностью отграничивает виды этого
подрода от остальных. Если обратить внимание на длинный список синонимов
Т. persicus, возникший за последние 25 лет, то близость этих видов станет еще
более очевидной.
У Т. persicus и Т. helenae по многим признакам фолидоза, в том числе и ди-
агностическим, есть некоторое перекрывание, что также свидетельствует об их
близости. Эти виды, по нашим данным, надежно различаются лишь по количест-
ву верхнегубных щитков и по цвету конца регенерированного хвоста (у первого
он белый, у второго — черный). Отсутствие репрезентативных коллекций
Т. latifi и ненадежность его основного диагностического признака (отсутствие
нижнечелюстных щитков, как показывают наши наблюдения, очень часто яв-
ляется признаком ниже видового и даже подвидового ранга, как, например,
у A. laevis, Т. depressus, В. spatalurus и др.) не позволяют с уверенностью ут-
верждать о его видовой самостоятельности и о большей близости к какому-
либо из видов подрода. Но по малому количеству нижнегубных щитков он более
сходен с Т. helenae.
По двум направлениям, ведущим от Т. tripolitanus (Т. steudneri — Т. dep-
ressus — Т. levitoni и Т. scortecci — Т. helenae), наблюдается параллельная
изменчивость количественных признаков: уменьшение спинной чешуи (увели-
чение чешуй вокруг середины тела), увеличение числа чешуй вдоль тела снизу,
чешуй поперек головы. Центральное положение (рис. 4) видов номинативного
подрода определяется менее дифференцированным фолидозом, слабым разви-
тием или отсутствием анальных пор, общностью их распространения. Внутри
подрода Microgecko четко прослеживается тенденция к потере нижнечелюст-
ных щитков: у Т. persicus они почти всегда в контакте, у Т. helenae — почти
всегда разделены, у Т. latifi — отсутствуют совсем. У видов подрода Asioco-
lotes наблюдаются увеличение количества пор и выраженность подхвостовых
щитков, а также географическая близость их ареалов.
Ареалы родов Tropiocolotes и Bunopus частично совпадают, но разобщены
вертикальным распределением по биотопам (переходная ирано-африканская про-
винция). Для средиземноморской провинции эндемичен один вид — В. tuber-
culatus, но представители рода Tropiocolotes здесь отсутствуют; аравийский
округ пустынной сахаро-аравийской провинции характеризуется обилием
форм рода Bunopus (два вида, три подвида), эта территория явилась, вероятно,
центром образования рода. Лишь на крайнем юге округа (Гадрамаут) ареалы
родов совпадают (В. spatalurus и Т. scortecci). Следовательно, род Bunopus,
с одной стороны, близок к роду Tenuidactylus (вид В. crassicauda сближается
с группой Cyrtopodion отсутствием или слабым развитием ребристости фоли-
доза и щиткованием верхней стороны хвоста), с другой — к роду Stenodactylus
(В. spatalurus характеризуется увеличением количества чешуй туловища — ее
измельчением, уменьшением выраженности анальных пор) (рис. 2).
20
Филогенетическая ветвь Carinatogecko отходит от основного ствола, дав-
шего группу Mediodactylus, по-видимому, вблизи Т. heterocercus [45]. Связь
между С. aspratilis и Т. heterocercus выражается в большей ребристости многих
чешуй тела по сравнению с другими видами подрода.
Филогения рода Teratoscincus (рис. 2), на наш взгляд, характеризуется
более или менее одинаковым развитием ветвей, однако внутри рода прослежи-
вается тенденция к увеличению туловищной чешуи и ее продвижению к затылку
у Т. microlepis и Т. scincus, в то время как ветви Т. bedriagai и Т. przewalskii
Рис. 4. Филогенетические отношения видов рода Tropiocolotes
близки в этом плане. В соответствии с данными зоогеографии, наиболее древним
очагом пустынной фауны в нашем регионе является центральноазиатский, более
молодым — среднеазиатский и его производный ирано-афганский горностепной
очаг [142]. Следовательно, более древняя ветвь Т. przewalskii, самая молодая—
Т. microlepis и Т. bedriagai. Наиболее специализированной псаммофильной
формой, а следовательно, и прогрессивной является ветвь Т. scincus.
Проста филогения и рода Crossobamon (рис. 2).
Наиболее сложной нам представляется филогения рода Tenuidactylus
(рис. 5). Было установлено, что эта ветвь отходит от рода Cyrtodactylus, а к
последней наиболее близка группа тибетско-гималайских видов (не случайно
они соседствуют и территориально). Расположение видов в этой исходной группе
сделано с учетом последовательного приобретения наиболее характерных черт
рода Tenuidactylus. В первую очередь это касается появления сегментирован-
ного хвоста, увеличения размеров спинных и хвостовых бугорков, удлинения
конечностей (chitralensis — stoliczkai — kirmanensis). Виды Т. fasciolatus,
Т. lawderanus и Т. himalayanus, вероятно, должны располагаться на ветви
Т. chitralensis, ближе к Т. tibetanus. Разрыв между Т. chitralensis и Т. stoliczkai
довольно велик, не исключено, что позже будут найдены «промежуточные
звенья», в этом регионе, несомненно, имеются еще не описанные виды.
Ближе всех к исходной тибетско-гималайской группе стоят средиземномор-
ские гекконы (территориально связь осуществлялась через ирано-афганскую
провинцию). Они сближаются щиткованием хвоста, его некоторой вздутостью,
относительно мелкими овальными или овально-треугольными спинными бу-
горками. Внутри подрода центральное положение занимает «среднестатисти-
ческий» Т. kotschyi. Линия kotschyi — sagittifer — heterocercus — Carina-
togecko (aspratilis — heteropholis) характеризуется увеличением ребристости
с последующим уменьшением размеров тела. Т. russowi — единственный сред-
неазиатский вид, обитающий, как правило, на равнине; он отклоняется от дру-
гих представителей подрода и по числу хромосом (44 вместо 42) [73]. Т. amicto-
21
pholis и T. spinicauda — горные виды, с чем связаны уменьшение их общих
размеров и гомогенизация спинного фолидоза.
Представители подрода Mesodactylus больше отклонились от исходных,
чем виды группы Mediodactylus. У них более крупные спинные бугорки, уве-
личиваются чешуи головы, но отдельные представители (Т. agamuroides
и Т. elongatus) сближают указанные подроды. Последних два вида внутри под-
рода образуют несколько обособленную группу, которая по ряду признаков
близка и к роду Agamura. У тех и других отмечен утонченный хвост и удли-
Рис. 5. Филогенетические отношения видов рода Tenuidactylus
ненные конечности, имеются вздутие носовых щитков, гомогенизация фолидо-
за хвоста, появление на нижней поверхности трех колец чешуи, увеличение
чешуй с анальными порами. В группе Т. watsoni, Т. kachensis, Т. scaber и
Т. montiumsalsorum наблюдается увеличение брюшных чешуй и спинных
бугорков. Характерный для этой группы фолидоз верхней стороны хвоста
позволяет высказать предположение о близости ее к роду Bunopus.
Номинативный подрод, несомненно, наиболее близок к подроду Mesodacty-
lus через вид Т. montiumsalsorum. Об их родственных отношениях свидетель-
ствуют появление зачаточных бедренных пор, продвижение на север. Централь-
ным видом подрода Tenuidactylus, по-видимому, является Т. fedtschenkoi.
Отсутствие серийного материала по Т. montiumsalsorum не позволяет нам сей-
час выяснить, какой из видов рассматриваемой группы наиболее близок к нему.
Удлиненные конечности и некоторые другие признаки (в том числе наиболее
юго-западный ареал) позволяют усмотреть сходство Т. longipes с представите-
лями рода Mesodactylus.
Учитывая, что Т. caspius наиболее пластичный вид, переходит к обитанию
на равнине, продвинулся далее всех других на запад и север, т. е, от исходных
форм, его следует считать самым прогрессивным видом группы. Если сравни-
вать на спине бугорки (их величину, ребристость, продвинутость к головному
отделу), то мы получим ряд, состоящий из caspius — turcmenicus — fedtschen-
koi — longipes, у которого в направлении справа налево возрастает бугрис-
тость спины, увеличиваются брюшные чешуи и становится более широким
контакт первой пары нижнечелюстных щитков. Несомненная близость сущест-
вует и между Т. turcmenicus и Т. caspius, о чем свидетельствуют многие призна-
ки (до недавнего времени эти виды даже не умели различать). В целом в
подроде наблюдается весьма компактная группа видов, среди них наиболее
уклонившаяся форма — Т. longipes.
И наконец, небольшой, но своеобразный род Agamura. Много признаков
22
свидетельствует о том, что он близок к роду Tenuidactylus. В его подроде (Cyr-
topodion) имеется группа видов (Т. agamuroides и Т. elongatus) — носителей
«агамуроидных» признаков. Внутри рода Agamura от A. persica к A. misonnei
заметно усиление «агамуроидности»: становятся более выпуклыми носовые
щитки, удлиняются конечности, укрупняются нижнебедренные щитки, умень-
шается выраженность пор. Внутри рода наблюдается некоторое сближение ви-
дов A. persica и A. gastropholis, с одной стороны, и видов A. femoralis и A. mi-
sonnei — с другой (рис. 4). В результате можно сделать следующее заключение:
1. Нашими материалами подтверждается вывод А. Клюге [237] о проникнове-
нии гекконов в Азии с юга на север и на запад (более примитивная, исходная
группа Cyrtodactylus, более прогрессивная и специализированная — род
Tenuidactylus). 2. Установлено, что переход гекконов из одной зоогеографиче-
ской категории в другую сопровождается таксономической перестройкой
(в Индомалайской области — род Cyrtodactylus, в Палеарктике — род Tenu-
idactylus, в ирано-афганской провинции — род Bunopus, в туранской и горно-
азиатской провинциях — род Alsophylax, в пустынной сахаро-аравийской
провинции — роды Stenodactylus и Tropiocolotes, в туранской пустынной про-
винции — роды Crossobamon, Alsophylax и Teratoscincus, хотя ряд из них —
Stenodactylus, Tropiocolotes, Crossobamon и Teratoscincus — встречаются в
ирано-афганской переходной провинции; в среднеазиатском горном округе —
A. tadjikiensis, в центральноазиатском горном округе — A. tokobajevi и др.).
Таблица для определения родов гекконов
1 (2). Раздельные веки имеются ............... Eublepharis (семейство Eublepharidae)
2 (1). Раздельных век нет, глаза покрыты прозрачной оболочкой.......................
....................... ................................(семейство Gekkonidae)
3 (4). Пальцы резко расширены (кроме конечных фаланг)..................Hemidactylus
4 (3). Пальцы плавно утончаются от основания к концу
5 (8). Пальцы снизу покрыты многочисленными мелкими чешуйками.
6 (7). Хвост сверху покрыт рядом крупных ногтевидных щитков; чешуя брюха крупная,
плоская — «сцинкоидная» ...............................................Teratoscincus
7 (6). Чешуя хвоста однородная, мелкая; чешуя брюха мелкая, обычно полукониче-
ская ..................................................................Stenodactylus
8 (5). Пальцы снизу покрыты одним рядом поперечно расширенных пластинок.
9 (16). Подпальцевые пластинки гладкие.
10 (13). Три (изредка два) более или менее равных друг другу мелких выпуклых носовых щитка;
обычно имеются спинные бугорки; бугорки также выражены почти на всем протяже-
нии хвоста; пальцы обычно резко искривлены.
11 (12). Конечности очень тонкие и длинные: передняя лапа, вытянутая вперед вдоль тела,
достает кистевым суставом до конца морды; длина стопы с пальцами заметно меньше
длины голени; хвост резко утончается у основания, не ломкий, как правило, короче
тела ..................................................................... Agamura
12(11). Конечности обычно умеренной длины: передняя лапа, вытянутая вперед вдоль тела,
достает или незначительно заходит концами пальцев за конец морды; длин^ стопы с
пальцами почти равна длине голени; хвост ломкий, как правило, длиннее тела (если ра-
вен или короче, то он толстый, мясистый) ..............................Tenuidactylus
13 (10). Носовые щитки плоские, первый значительно крупнее остальных одного-двух, которые
могут отсутствовать вовсе (если имеются два-три более или менее равных друг другу но-
совых щитка, то спинные бугорки отсутствуют); бугорки на хвосте почти всегда отсутст-
вуют.
14 (15). Спинных бугорков нет; самки (часто и самцы) лишены анальных пор; верхнегубные
щитки заворачиваются на верхнюю поверхность морды так, что каждый расположен
в разной плоскости ..................................Tropiocolotes
15 (14). Спинные бугорки почти всегда имеются; самки обычно с анальными порами (если спин-
ных бугорков нет, то самцы и самки с анальными порами, если у самок нет анальных
пор, то спинные бугорки выражены хорошо); верхнегубные щитки находятся в одной
плоскости и отвесно опускаются к краю рта ................................Alsophylax
16 (9). Подпальцевые пластинки с шипиками или ребрышками.
17 (20). Хорошо выраженных спинных бугорков нет (если они есть, то едва отличимы от мелкой
спинной чешуи и разбросаны в беспорядке).
18 (19). Спинная чешуя крупная, плоская; с ноздрей соприкасаются два плоских носовых
щитка, не отличающихся размерами от остальных чешуй морды . . . Tropiocolotes
19 (18). Спинная чешуя мелкая, выпуклая; с ноздрей соприкасаются три заметно вздутых но-
совых щитка, значительно крупнее остальных чешуй морды.................Stenodactylus
20 (17). Спинные бугорки выражены более или менее четко и упорядочены в ряды.
23
21 (22). Боковая чешуя пальцев образует хорошо заметную бахрому .... Crossobamon
22 (21). Боковая чешуя пальцев с шипиками или ребрышками.
23 (24). Хвостовые бугорки в полукольцах одного сегмента не соприкасаются друг с другом;
первый носовой щиток крупнее остальных; высота первого верхнегубного щитка от но-
здри до края рта заметно меньше его ширины вдоль края рта .... Carinatogecko
24 (23). Хвостовые бугорки в полукольцах одного сегмента широко соприкасаются между со-
бой боковыми гранями; носовые щитки равны друг другу; высота первого верхнегуб-
ного щитка от ноздри до края рта заметно больше (если меньше, то незначительно)
его ширины вдоль края рта ................................................... Bunopus
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО ЭУБЛЕФАРОВЫЕ — EUBLEPHARIDAE
Позвонки передневогнутые (процельные). Непарная теменная кость.
Предчелюстная кость образуется из двух центров окостенения. Надвисочная
кость часто имеется, угловая — отсутствует редко. Кальцинированные эндо-
лимфатические мешки в затылочной области отсутствуют. Настоящие подвиж-
ные веки имеются. Яйца откладываются в мягкой оболочке.
Известно два рода в Азии, два в Африке и один на юге Сев. и в Ц. Америке.
Род эублефары — Eublepharis Gray, 1827
Типовой вид: Е. hardwickii Gray.
1827 — Eublepharis Gray, Philos. Mag., London, (2), Vol. 2, p. 56.
Диагноз. Пальцы короткие, прямые, цилиндрические, без зубчиков
по краям, снизу с одним продольным рядом пластинок. Тело покрыто мелкой
чешуей, среди которой расположены отдельные более крупные конусовидные
чешуи. Подбородочные щитки большие. У большинства видов подмышечные впа-
дины передних лап преобразованы в глубокие кожные карманы. Самцы с хорошо
выраженными преанальными порами. Хвост значительно короче туловища
с головой (в 1,5—2 раза), вздут посередине, оканчивается остроконечно.
Известно восемь видов, из которых шесть распространены в Ю.-З. Азии
и Сев. Индии, два на о-ве Хайнань, о-вах Норвей в Тонкинском заливе и о-вах
Рюкю в Японии. В СССР встречается один вид.
Группа палеарктических, юго-западноазиатских видов объединяет формы,
различающиеся лишь деталями рисунка, окраски и небольшим количеством
стабильных признаков фолидоза. Все это позволяет нам привести описание этой
группы, выделить признаки, общие для рассматриваемых здесь видов: Е. ma-
cularius, Е. angramainyu и Е. turcmenicus.
ГРУППА ЮГО-ЗАПАДНОАЗИАТСКИХ ЭУБЛЕФАРОВ
Описание. Крупные ящерицы с длиной туловища до 160 мм; голова
массивная, яйцеобразная, слегка приплюснутая, резко отграничена от шеи;
все чешуи тела, кроме подпальцевых пластинок, гладкие. Ноздря прорезана
в задней части носового щитка, окружена широким межчелюстным, первым
верхнегубным, крупным, плоским межносовым щитками и шестью—восемью
мелкими носовыми чешуйками. Межчелюстной щиток четырехугольный, с не-
ровным верхним краем. Межносовые щитки разделены одной — тремя круп-
ными чешуями. Чешуя морды крупная, плоская, многоугольная, неоднород-
ная, резко уменьшается в предглазничной области, где встречаются округлые
бугорки, которые по мере приближения к затылку становятся все более выпук-
лыми. В задней части верхнего века имеется несколько ресницеобразных вы-
ростов.
Верхнегубные щитки мелкие, плавно уменьшающиеся назад; высота пер-
вого верхнегубного щитка от ноздри до края рта равна его ширине вдоль края
25
pi а ушное отверстие крупное, овальное. Подбородочный щиток пятиугольный,
его ширина вдоль края рта в два раза превосходит его высоту; первая пара
крупных нижнечелюстных щитков обычно соприкасается между собой или раз-
делена одной-двумя чешуйками; за ней может следовать одна-две пары более
мелких щитков, ниже которых располагаются несколько сильно увеличенных
верхнегорловых чешуй; нижние горловые чешуйки мелкие, слабо выпуклые.
Нижнегубные щитки широкие, плавно уменьшающиеся назад.
Спинная чешуя мелкая, округло-многоугольная, несколько более увели-
ченная на боках, где граница с брюшной чешуей определяется не вполне четко;
длина бугорков в двух центральных рядах посередине спины меньше проме-
жутков между ними, по направлению к бокам туловища размеры бугорков
увеличиваются, их диаметр становится равным или больше промежутков. Спин-
ные бугорки конические или сферические, образуют около 20 продольных и 40
поперечных не очень четких рядов. Чешуя брюха крупная, округло-много-
угольная, слабо черепитчатая; ряд анальных пор расположен углообразно,
иногда бывает прерван неперфорированными чешуйками (чаще посередине).
Сегменты хвоста сверху покрыты рядами мелкой прямоугольно-округлой че-
шуи, среди которой кольцами расположены конические или сферические бугор-
ки (до 10), как правило, не соприкасающиеся между собой; снизу на одном сег-
менте три-четыре ряда плоских многоугольных чешуй, увеличивающихся от
первого ряда к последнему и от боков к центру, но не достигающих размеров
щитков. Регенерированный хвост покрыт прямоугольно-округлой чешуей,
упорядоченной в более или менее правильные ряды только сверху, хвостовых
бугорков нет.
Конечности умеренной длины: передняя лапа, вытянутая вперед вдоль
тела, достает концами пальцев до середины промежутка между глазом и нозд-
рей, а задняя — до локтя отведенной назад передней лапы или до лопатки;
передняя конечность покрыта однородной плоской чешуей, более мелкой на
нижних поверхностях; на предплечье имеются отдельные более крупные вы-
пуклые чешуи, аналогичные спинным бугоркам; на верхних поверхностях зад-
них конечностей разбросаны многочисленные четко выраженные конические
бугорки; несколько мелких бугорков расположено на внутренней поверхности
бедер. Чехол когтя состоит из верхней, нижней и двух боковых пластинок.
Таблица для определения видов рода Eublepharis Ю.-З. Азии
1 (2). Подпальцевые пластинки гладкие.......................Е. angramainyu
2 (1). Подпальцевые пластинки бугристые.
3 (4). Бугорки на подпальцевых пластинках выражены слабо; анальных пор восемь или мень-
ше ......................................................... Е. turcmenicus
4(3). Бугорки на подпальцевых пластинках выражены резко; анальных пор восемь или боль-
ше ............................................................Е. macularius
ЭУБЛЕФАР ПЯТНИСТЫЙ — EUBLEPHARIS MACULARIUS (Blyth, 1854)
Типовая территория: Соляной хребет, Пенджаб, С.-З. Индия.
Кариотип: 2п = 38, все акроцентрики, NF = 38.
1854 — Cyrtodactylus macularius Blyth, Jour. Asiat. Soc. Bengal, vol. 23, p. 737.
1864 — Eublepharius fasciolatus Giinther, Ann. Mag. nat. Hist., London (3), 14, p. 429 (terra
t у p i с а: Синд, Хайдарабад).
1871 — Eublepharis macularius, John Anderson, proc. Zool. Soc., London, p. 163.
Диагноз. Подпальцевые пластинки отчетливо бугристые; 8—11 аналь-
ных пор.
Голотип. Хранится в Индийском музее, Калькутта [245].
Описание (рис. 6) (по 5 экз. из колл. ИЗАНУ).
L взрослых 92,3—128 мм (по литературным данным — до 150 мм); самцы круп-
нее самок; L/L cd (п = 3) 1,35—1,53; индекс высоты/ширины головы (п = 5)
54—68 (61,6 ± 2,56); диаметр уха/глаза (п = 5) 0,78—1,0 (0,91 ± 0,04); верх-
негубных щитков (п = 10) 8—12 (9,6 ± 0,37); нижнегубных (гг = 10) 8—10
(9,0 ± 0,21); чешуй между центрами глаз (и = 5) 24—33 (28,0 ± 1,87); длина
спинного бугорка/L (п = 5) 9,0—11,0 (9,80 ± 0,37); чешуй, окружающих
26
спинной бугорок (п = 5) 12—13 (12,8 ± 0,2); чешуй вдоль тела снизу (п = 4)
153—165 (158,5 ± 2,5); чешуй поперек брюха (п = 4) 24—26 (24,75 ± 0,48);
преанальных пор 8—11 (по литературным данным — 11—17); подпальцевых
пластинок (п = 9) 14—21 (17,44 ± 0,69); добавочных носовых чешуй 5 (один слу-
чай), 6 — не встречалось, 7 (один случай), 8 (три случая); межносовые щитки
разделены одной (два случая), двумя (один случай) или тремя (два случая) че-
шуйками. Первая пара нижнечелюстных щитков соприкасается друг с другом
(четыре случая) или разделена одной чешуйкой (один случай); в двух случаях
эта пара щитков отделена от нижнегубных рядом мелких зернышек; за нею сле-
Рис. 6. Eublepharis macularius:
А — голова сбоку, Б — голова сверху, В — подбородок, Г — спина, Д — хвост свер-
ху, Е — хвост снизу, Ж — пальцы задней лапы снизу (ИЗАНУ SR 2763, род. в тер-
рариуме)
дуют одна-две пары значительно менее выраженных щитков; до уровня передне-
го края орбиты доходит пять-шесть верхнегубных щитков.
Прижизненная окраска. Основной тон верхней стороны те-
ла — лимонно-желтый, более активный на туловище. Сочетание многочислен-
ных темно-коричневых пятен и светлых тонких промежутков между ними со-
здает на голове сетчатый рисунок; переходя на туловище, эти пятна увеличива-
ются в размерах и располагаются в продольном направлении, иногда сливаясь
друг с другом, однако более или менее узкая полоса вдоль позвоночника от
затылка до крестца остается свободной от пятен; на хвосте пятна могут сли-
ваться в поперечные полосы. Конечности сверху также покрыты пятнами.
Нижняя часть головы и хвоста в менее четких, размытых пятнышках; ос-
тальные нижние поверхности белые. У особи из Пакистана (окр. Татта) тем-
ные спинные пятна вытягиваются продольно таким образом, что между ними
образуются три продольные светлые полосы. Кроме того, у этой особи четко
выражены три светлые поперечные широкие полосы: лопаточная, среднетуло-
вищная и поясничная; снизу рисунок имеется только на регенерированной
части хвоста.
Половой диморфизм и возрастная изменчивость.
Самцы крупнее самок. У молодых гекконов основной фон верхней стороны те-
лесный; верхняя часть головы от уровня ушей до ноздрей покрыта темно-бу-
рой «шапочкой», в теменной области которой, между глазами, есть светлый узор,
напоминающий птичий след, направленный тремя концами к затылку; на за-
тылке от уха до уха — сплошная темно-бурая полоса, между «шапочкой»
и этой полосой промежуток белый; на туловище три четкие широкие попереч-
ные бурые полосы; на хвосте четыре-пять таких же полос; на бедрах сверху
27
бурая продольная полоса, едва заметный след ее имеется на плече. Брюшная
поверхность розоватая, без пятен.
Распространение (рис. 7). От В. Афганистана на юг через Пакис-
тан до Белуджистана и на восток до 3. Индии; поднимается в горы до 2500 м
н. у. м. Находка Н. А. Зарудным [60] в В. Иране двух особей этого рода (ве-
роятно, этого вида) значительно отодвигает западную границу ареала.
Кадастр к рис. 7.
Афганистан: 1—2 [50]; 3 — около Гульбахара, 65 км сев. Кабула [318]; 4 — Чари-
кар, 35° 05' с. ш.. 69° 10' в. д. [209]; 5 — Джелалабад, 34° 26' с. ш., 70° 25' в. д. [172]; 6—8—16 км
С.-В. Нильмы, 34° 19—21' с. ш., 70° 10—15' в. д. [209]; 7 — Кандагар [318].
Рис. 7. Распространение Eublepharis turcmenicus (7), Е. angramainyu (2), Е. macula-
rius (5), Е. sp. (Зарудный, 1904) (4}, Tenuidactylus sagitiffer (5)
Пакистан: 1 — р-н Зхоб, пос. Черат [263]; 2 — Солт Рэидж, Пенджаб (terra typica)
[258]; 3 — Чиниот [235]; 4 — Лахор [258]; 5 — Форт-Сандемен [263]; 6 — 24 км з. Хиндубага
[263]; 7 — Ахмадпур [235]; 8 — р-н Кветты, 26 км сев.-в. Кветты [263]; 9 — р-н Сиби, Болан
Пасс около Мэч [263]; 10 — Нушки [258]; 11 — 26 км сев. г. Бела [263]; 12 — Тхар Паркар
[258]; 13 — Хайдарабад, Синд [225]; 14 — 5 км от Хабчауки [263]; 15 — Татта [258]; 16 — р-н
Карачи, Хил Пак [263]; 17 — округ Малир [263]; 18 — Пир-Патхо [258]; 19 — Баторо, Джати
[258]; 20 — Вана [232]; 21 — Манзаи [232]; 22 — Лакки (Ладха) [232].
Географическая изменчивость. Изменчивость рисунка
и некоторых признаков фолидоза, считающихся важными в диагностике видов
рода, а также неизученность распространения (в особенности в западной и се-
веро-западной частях ареала) не позволяют делать окончательных выводов о
внутривидовой систематике эублефара пятнистого. Описания подродов А. Бор-
нера [197] неубедительны, их следует ревизовать на основе изучения серийного
материала.
Экология. Хотя эублефар пятнистый в последние годы успешно раз-
водится террариумистами-любителями и стал лабораторным животным, сведе-
ния об его образе жизни в природе практически отсутствуют. Усложняет это
также то, что под названием Е. macularius совсем недавно фигурировали и дру-
гие западноазиатские виды, так что неизвестно, к какому виду относятся те
или иные литературные сведения. Мы имели возможность содержать в неволе
Е. macularius и Е. turcmenicus и можем утверждать, что они очень близки по
экологии и поведению.
Стации. Обитает в каменистых местах. Н. А. Зарудный [66] нашел,
видимо, особей данного вида на равнине, «на глинисто-щебнистой почве, усы-
панной песком и изобильно поросшей кустами Zygophyllum».
Отношение к температуре. В неволе предпочитает температу-
ру1 почвы 20—24° [258].
1 В монографии температура приводится в градусах Цельсия.
28
Суточный цикл активности. Ведет сумеречно-ночной образ
жизни. В природе [60] был найден на поверхности «на солнечном закате».
Сезонный цикл активности. По коллекционным материалам
[197] известна самая ранняя находка из 3. Пакистана — 15.II 1976 г., самая
поздняя 19.XII 1960 г. В лабораторных условиях обходится без спячки.
Линька. В неволе наблюдалась в марте и апреле.
Питание. В террариуме хорошо поедает сверчков, тараканов и даже
мышат.
Размножение. Отмечены кладки Е. macularius в неволе 7. IV
и 8.IV 1984 г., 1 .VIII 1984 г., размеры яиц (п=6): 24,4—30,5 X 14,5—15,5 мм,
масса 2—2,1 г. Откладывает по два яйца. В нашем террариуме инкубация
длилась 61—63 дня (температура 26—29°, влажность 75—100 %). Первая яще-
рица вывелась 27.IX 1984 г., вторая 29.IX 1984 г., их размеры: L = 49 мм,
Led = 31—32,5 мм. По сведениям Г. Эмста [214], за год одна самка отложила
четыре кладки с интервалом в четыре недели, их инкубация длилась 42—56
дней. Яйца закапывались во влажный песок.
Темп роста. По нашим наблюдениям в неволе, за первые два месяца
(X, XI) прирост составлял 10 мм (L — 5,8—5,9), за следующий (XII) месяц
еще 10 мм (L = 70—71 мм), а далее темп ускорился, за месяц (1) они увеличи-
лись почти на 20 мм (L = 90—92 мм).
Враги. В природе их могут поедать лисицы [277].
Особенности поведения. Менее пуглив, чем туркменский эуб-
лефар, берет корм с пинцета, отсутствует пауза в питании летом. При совмест-
ном обитании нескольких особей агрессивны только самцы к особям своего по-
ла, при драках они могут оторвать и съесть хвост, вырвать кусок кожи на боку.
Отмечен случай спаривания Е. macularius (</) и Е. turcmenicus ($>), кладка
яиц, к сожалению, погибла.
ЭУБЛЕФАР ИРАНСКИЙ — EUBLEPHARIS ANGRAMAINYU 1
ANDERSON ЕТ LEVITON, 1966
Типовая территория: между Мешед-и-Сулейман и Бетвендом, Хузестан,
Иран.
Кариотип: неизвестен.
1963 — Eublepharis macularius Anderson S. C., Proc. Calif. Acad. Sci., ser. 4, vol. 31, p. 435.
1966 — Eublepharis angramainyu Anderson S. C., Leviton, Occas Papers Calif. Acad. Sci.,
N 53, p. 1.
Диагноз. Подпальцевые пластинки гладкие; 11 —13 анальных пор в од-
ном ряду.
Голотип [171] CAS 86384. ad. Добыт вдоль старой дороги между
Мешед-и-Сулейман и Бетвендом, пров. Хузестан, Иран, сборщик С. Ц. Андер-
сон; 20.V 1958 г.
L = 147 мм; Led = 100 мм. Межносовые щитки разделены единственной почти
пятиугольной межносовой чешуйкой, ширина которой превышает длину;
5—7 мелких дополнительных носовых чешуек; по 10 верхне- и нижнегубных
щитков; ухо большое, его длина в 2,5 раза превосходит ширину; за пятиуголь-
ным подбородочным щитком следует четыре ряда увеличенных чешуй; спинные
бугры на боках туловища почти соприкасаются между собой; поперек брюха
24 чешуйки; 13 анальных пор; 24 гладких подпальцевых пластинок.
Окраска и рисунок (в спирте). Вдоль хребта светлая непрерывная
полоса от затылка до основания хвоста; с обеих сторон она граничит с темными
прерывистыми полосами, которые могут сливаться между собой в продольном
направлении, а также с поперечными метками; на голове черно-белый сетчатый
рисунок; подковообразной затылочной темной или светлой полосы нет; на ко-
нечностях многочисленные пятнышки; хвост покрыт многочисленными нерегу-
лярными темными поперечными метками, которые шире светлых промежутков
между ними. Брюхо светлое, желтовато-коричневое.
1 angramainyu — «дух ночи».
29
Паратипы по сравнению с голотипом имеют отличия: у самок И—13 слабо
выраженных пор, а у № 86333 — ниже ряда из 13 имеется дополнительный ряд
из четырех анальных пор. У молодых три темные поперечные спинные полосы:
вокруг задней части шеи и лопаток, посередине и в области поясницы. Суммар-
ные размеры: L = 142—154; Led = 97—100; L $ = 126—137; Led = 86—
90; Ljuv = 90, хвост регенерирован.
Распространение. Западные предгорья хр. Загрос и север Месо-
потамской равнины в Иране и Ираке (рис. 7). Отмечен между 300 — 900 м
н. у. м. [200].
Кадастр к рис. 7.
Ирак [269]: / — пров. Ниневех, развалины Ниневии (300 м н. у. м.) около Эль-Мав-
зил; 2 — пров. Ниневех, Эль-Хадр; 3 — пров. Киркук, пос. Чалдаю; Чамчамаль; 4 — слияние
рек Малый Заб и Тигрис; 5 — пров. Сулеймания, пос. Мера-ди и окр. дороги из Дарбанди-
Хан к Сулеймании.
Иран: 1 — Хузистан, Дехлоран, Али Хош (32° 41' с. ш., 47° 16' в. д.) [302]; 2 — ста-
рая дорога между Месджеде — Сулейман и Бетвендом, пров. Хузистан [168]; 3—5 — [50].
Экология. Не изучена. Известно, что эти ящерицы обитают в камени-
стых предгорьях и руинах Ниневии на Месопотамской равнине, ведут ночной об-
раз жизни. В Иране откладка яиц происходит в конце мая — начале июня [59].
В иранском Хузистане чаще встречаются за час до полуночи, среди камней ло-
вят сверчков, также поедают скорпионов, сольпуг, крупных пауков и жуков,
мелких гекконов своего и другого вида [277].
ЭУБЛЕФАР ТУРКМЕНСКИЙ — EUBLEPHARIS TURCMENICUS
DAREVSKY, 1978
Типовая территория: окр. Бахардена (Копетдаг), Туркмения.
Кариотип: не изучен.
1980 — Eublepharis macularius Boulenger, Ann. Mag. Nat. Hist., v. 6, p. 352.
1978 — Eublepharis turcmenicus Darevsky, Tp. Зоол. ин-та, Ленинград, т. 61, с. 204.
Диагноз. Подпальцевые пластинки слабо бугристые; пять — восемь
анальных пор.
Голотип [50]. ЗИН АН СССР № 10103, </ad; окр. Бахардена (Копет-
даг), 17.IV 1903 г., колл. И. В. Васильева.
L = 125 мм; Led = 80 мм. Межносовые щитки разделены двумя чешуйка-
ми; верхнегубных и нижнегубных щитков по 10; пара крупных нижнечелюст-
ных щитков соприкасается друг с другом позади подбородочного и за ними сле-
дует девять увеличенных многоугольных верхнегорловых чешуй; каждый ниж-
нечелюстной щиток отделен от первого нижнегубного зернышками; поперек
брюха 32 чешуйки; пять анальных пор (в двух местах ряд прерван несколькими
лишенными пор чешуйками). Бугорки на подпальцевых пластинках развиты
слабо.
Описание (по 4 экз. из колл. ИЗАНУ) (рис. 8). L взрослых (п = 4)
111,9—129,3 мм; Led = 64,0; самцы крупнее самок; LILcd(n=2) 1,94—2,61; ин-
декс высоты/ширины головы (п = 3) 0,59—0,68; диаметр уха/глаза (п = 3)
0,75—0,86; верхнегубных щитков (п = 8) 9—11 (9,75± 0,25); нижнегубных
(п = 5) 10—11 (10,6 ± 0,24); чешуй между центрами глаз (п = 3) 26—30; дли-
на спинного бугорка/L (п = 4) 8,06—10,05 (9,16 ± 0,43); чешуй, окружающих
спинной бугорок (п = 4), 10—13 (11,75±0,63); чешуй вдоль тела снизу (п = 4)
148—156 (152,25 ± 1,93); чешуй поперек брюха (п = 4)20—22 (21,25 ± 0,48);
анальных пор (п = 2) 8; подпальцевых пластинок (и = 7) 20—23 (21,43 ± 0,37);
добавочных носовых чешуек 7 (два случая), 6 или 8 (по одному случаю); межно-
совые щитки разделены 1 (два случая), 2 или 3 (по одному случаю) чешуйками;
первая пара нижнечелюстных щитков соприкасается между собой широким (два
случая) или точечным (один случай) швом или разделена чешуйкой (один слу-
чай); эта пара щитков отделена от нижнегубных рядом зернышек; за ней сле-
дуют одна — две пары значительно менее выраженных щитков; до уровня пе-
реднего края орбиты доходит шесть верхнегубных щитков. Масса до 62 г.
П ри жизненная окраска. Основной фон верхней стороны тела
охристый, по которому расположены сиренево-коричневые пятна и точки;
30
на голове эти пятна расположены густо, почти без промежутков; на туловище
они вытянуты вдоль светлой полосы, проходящей по хребту от затылка до крест-
ца; между ними имеются отдельные пятнышки, образующие иногда тонкие по-
перечные линии; между лопатками, по середине тела ив поясничной области —
три широкие светлые поперечные полосы; хвост и верхние поверхности лап бес-
порядочно покрыты темными пятнышками и крапинками; нижние поверхности
белые, лишь на регенерированной части хвоста узор из темных крапинок.
снизу, Д — брюхо,
ущ.)
Рис. 6. Eublepharis turcmenicus:
А — голова сбоку, Б — голова сверху, В — подбородок, Г — хвост
Е — преанальные поры (Re 8/1 Копетдаг, Чандырское
Половой диморфизм и возрастная изменчивость.
Lmax d 143 мм, Lmax $ 135 мм [329]. У молодых особей рисунок такой же,
как у сеголеток эублефаров пятнистых.
Распространение. Копетдаг и Туркмено-Хорасанские горы
(рис. 7).
Кадастр к рис. 7.
Туркмения:
1 — 3. Туркмения, хр. Кюрендаг, родник Кемал [23]; Даната [100]; 2 — Чандырское ущелье
в Каракалинском р-не [145]; Междуречье Сумбара и Чандыря, уроч. Дамган, уроч. Шихиндере
[109]; 3 — долина р. Арваз, окр. Бахардена [23]; 4 — Чули, Фирюза [13].
Иран: 1 — Туркмено-Хорасанские горы к сев.-з. от Мешхеда [50]; 2 — Чах-и-Зи-
РУ [60].
Систематика и географическая изменчивость.
На изученном материале географическая изменчивость не выражена.
Стации и количественные данные (рис. 9). В Копетдаге
встречается в каменистых предгорьях, занимает склоны холмов из крошки
шиферного сланца с редкими кустами полыни, эфедры, отдельными деревьями
держидерева, обломками скал. По находкам четырех особей выяснилось, что
они предпочитают северную и северо-восточную экспозиции, уклон около 30°,
высота от подножия холма 100—120 м, абсолютная высота до 812 м н. у. м. Как
правило, по соседству с местом находок этой ящерицы располагались колонии
афганских полевок или норы персидских песчанок и рыжеватых охотон. На
участке в 2000 м2 найдены две особи на расстоянии 35—37 м одна от другой. Вид
очень редкий, встречается спорадически, на территории Советского Союза из-
вестно всего 12 находок.
Отношение к температуре. Наблюдалась активная ящери»
ца весной при температуре 18°, в террариуме предпочитали 30°.
31
Рис. 9. Биотоп Tenuidactylus spinicauda и Eublepharis turcmenicus (щебнисто-каме-
нистые осыпные склоны в окр. с. Даната, Зап. Копетдаг, ТССР)
Суточный цикл активности. Ведет сумеречно-ночной образ
жизни. Одна особь была добыта в окр. Чули около 24 ч (сообщение С. Шамма-
кова), в террариуме выходит из убежища при наступлении сумерек.
Сезонный цикл активности. Самая ранняя находка, сделан-
ная нами 22.IV 1977 г., не совсем отражает сроки появления этого вида после
зимовки. Видимо, это происходит в начале апреля. Не исключено, что эубле-
фары исчезают с поверхности еще летом (последняя известная находка 22.VII
1973 г. [12]), так как по наблюдениям в неволе в это время они резко сокращают
потребление корма. Впрочем, в террариуме они обходились и без зимней
спячки.
Линька. Самец, добытый 12.V 1984 г., линял 13.VII 1984 г. В неволе
наблюдалась пятикратная линька за год у другого экземпляра с наибольшим
интервалом между IV иVIII месяцами. Линька проходит в течение двух дней,
шкурку съедает.
Питание. В экскрементах добытой нами в Туркмении в мае 1977 г.
ящерицы обнаружены остатки клопа, перепончатокрылого, жука и ящурки,
видимо, Eremias strauchi. В желудке были найдены остатки многоножки, мок-
рицы и туркестанского термита [127]. Возможно, поедает мелких грызунов.
Наши экземпляры в террариуме охотно поедали сверчков, тараканов и личинок
мучного хруща.
Размножение. Соотношение полов по известным нам сборам (и =
= 10) 1 : 1. Откладывает два овальных белых яйца в мягкой кожистой оболочке.
По устному сообщению Ч. Атаева, самка, добытая 16.VII 1972 г., отложила в
неволе яйца размером 38 X 19 и 40 X 19,5 мм (масса 7 и 7,5 г). В нашем тер-
рариуме самка 20.VII 1984 г. отложила одно яйцо размером 32 X 19,4 мм, а
23.VII 1984 г.— второе (30 X 19 мм). Спаривание в последнем случае происхо-
дило не позже начала апреля (был отсажен самец), а явные признаки беремен-
ности отмечены 20.V 1984 г. По сообщению Н. Л. Орлова, самка, добытая в мае
1984 г. под Фирюзой, в июле того же года в течение двух недель отложила 3,
а затем 2 яйца.
Темп роста не изучен. Известная продолжительность жизни в неволе
взрослой особи — 7 лет (сообщение С. Шаммакова). Молодая особь в годичном
32
возрасте имела размеры: L = 71 мм, Led = 32 мм, масса 5,1 г (сообщение
Ч. Атаева).
Враги. Одна особь была отрыгнута коброй, добытой в окр. пос. Кара-
Кала [109].
Особенности поведения. Передвигается эублефар туркмен-
ский осторожно, на вытянутых лапах, бегает редко. На каменистых осыпях
перед каждым шагом ящерица раскачивается взад и вперед. Добычу хватает
цепко, ощутимо и для человека, старается убить ее крепкими челюстями (их
сжимает так, что опускаются глаза в орбитах и голова заметно уплощается).
Если добычу что-то удерживает — вращается вокруг своей оси, как желтопу-
зик. Пьет воду, как кошка, зачерпывая ее широким языком. В мягком грунте
может рыть норы (по наблюдениям в террариуме). В природе, видимо, занимает
норы грызунов, так как все ящерицы найдены под камнями или плитами (до
1 м длиной) после дождей и ливней, очень увлажняющих землю. Здесь же они
и зимуют. В природе и террариуме у них имеется определенное место — «туа-
лет», где скапливаются экскременты. Такой «туалет» был найден нами вблизи
Кара-Калы под плитой известняка рядом с убежищем ящерицы. Эублефар —
очень пугливое, нервное животное, плохо привыкает к неволе. Оно издает два
вида звуков — лающие или (попав в руки, при беспокойстве) тонкие и звеня-
щие, которые можно передать как «я-я-ззь». Состояние хвоста ящерицы служит
показателем ее упитанности: у истощенного животного (такие тоже встречают-
ся в природе) он морщинистый, у упитанного — гладкий, выпуклый. В природе
встречаются чаще одиночные особи. Две самки, посаженные в террариум,
вскоре подрались и одна другой поломала нижнюю челюсть. Самец при спарива-
нии хватал самку за шкуру, начиная от основания хвоста до шеи, и, удерживая
в таком положении, подводил хемипенисы к ее клоаке сбоку. Весь процесс спа-
ривания продолжался пару минут.
Ослепленный светом эублефар не останавливается, а старается скрыться.
Практическое значение и охрана. Эублефар туркменский
в Копетдаге обитает на северной границе ареала; холодные зимы, видимо, резко
снижают численность южного вида. Эта своеобразная ящерица представляет боль-
шой научный интерес. До сих пор она не найдена на территории туркменских
заповедников (встречается у их границ). Включена в Красную книгу СССР. Це-
лесообразно провести работы по разведению в неволе данного вида с последую-
щим выпуском приплода в соответствующие биотопы заповедных угодий.
СЕМЕЙСТВО ГЕККОНОВЫЕ — GEKKONIDAE
Позвонки двояковогнутые (амфицельные), сохраняются остатки хорды.
Теменная кость парная. Предчелюстная кость образуется из одного центра окос-
тенения. Надвисочная дуга и угловая кость почти всегда отсутствуют. Каль-
цинированные эндолимфатические мешки в затылочной области имеются. Под-
вижных век нет (глаза покрыты сросшимися прозрачными веками). Яйца от-
кладываются в жесткой обызвествленной оболочке.
Ареал семейства охватывает поясом тропическую, субтропическую и
(отчасти) умеренные зоны земного шара между 50° с. ш. и 40° ю. ш. (в Новом
свете между 35° с. ш. и 48° ю. ш.). Известно 82 рода.
Род сцинковый геккон — Teratoscincus Str., 1863
Типовой вид: Teratoscincus keyzerlingii Str.
1863 — Teratoscincus Strauch, Bull. Acad. Sci. St. Petersbourg, T. 4, S. 397.
Диагноз. He расширенные и не сжатые с боков прямые пальцы покрыты
снизу многочисленными мелкими шиповатыми чешуйками, по бокам — ба-
хромой из длинных и плоских чешуек; туловище покрыто однородной, как пра-
вило, очень крупной ромбовидно-округлой сцинкоидной чешуей, а голова —
33
Рис. 10. Teiatoscincus bedriagai (вверху):
A — общий вид, Б — голова сбоку, В — лицевая часть сбоку, Г — голова сверху, Д — го-
лова снизу, Е — спина (ЗИН 9161. Ходжи-и-ду-Чаги, Иран)
Teratoscincus scincus (внизу):
А — общий вид, Б — спина, В — надглазничный козырек, Г, Д — носовые щитки, Е —
брюхо, Ж — пален снизу (А, В, Ж — Т s. keyzerlingi — MZSF 24152, 30° 10' с. ш., 60° 50'
в д., Иран)
мелкой, слабо выпуклой чешуей, более крупной на морде; на хвосте сверху
в последних 2/8 — ряд крупных ногтевидных пластинок, снизу — мелкая,
однородная чешуя; анальные и бедренные поры отсутствуют.
Дополнительные признаки. Голова крупная, массивная, те-
ло брусковидное, хвост короче тела. Зрачок вертикальный с ровными краями.
Все чешуи тела гладкие; носовые щитки слабо выпуклые (три крупных и час-
то — один-два мелких нижненосовых), кроме них с ноздрей соприкасается
34
крупный четырехугольный межчелюстной щиток; высота первого верхнегуб-
ного щитка по линии от ноздри до края рта больше или равна его ширине
вдоль края рта; ушное отверстие крупное, овальное; хорошо развит «надглаз-
ничный козырек» (рис. 10). Межчелюстной щиток крупный, четырехуголь-
но-трапециевидный; по бокам от него, примыкая к межчелюстным щиткам, мо-
гут располагаться одна — три пары мелких нижнечелюстных щитков, в первой
из них щитки никогда не соприкасаются между собой; горловая чешуя мелкая,
слабо выпуклая; спинные бугорки отсутствуют; хвостовые сегменты не выраже-
ны. Передняя конечность, вытянутая вперед и вдоль тела, достигает концами
пальцев конца морды, задняя — почти подмышечной впадины; за исключени-
ем задних поверхностей бедер, покрытых мелкой выпуклой чешуей, конеч-
ности покрыты аналогичной туловищной сцинкоидной чешуей.
Род включает четыре вида, распространенных в основном в центральной,
средней и восточной частях 3. Азии.
Таблица для определения видов рода Тeratoscincus
1 (2). Вокруг середины туловища более 80 чешуй........................... Т. mi его! ер is
2 (1). Вокруг середины туловища не более 50 чешуй.
3 (4). Крупная спинная чешуя оканчивается на уровне затылка..................Т. scincus
4 (3). Крупная спинная чешуя оканчивается на уровне лопаток.
5(6). Вокруг середины туловища более 40 чешуй..............................Т. bedriagai
6(5). Вокруг середины туловища менее 40 чешуй..............................Т. przewalski
СЦИНКОВЫЙ ГЕККОН ОБЫКНОВЕННЫЙ — TERATOSCINCUS SCINCUS (SCHLEGEL., 1858)
Типовая территория: р. Или в «Туркестане».
Кариотип: 2п = 36, метацентриков 12, акроцентриков 28, субтелоцентриков 6,
NF = 44.
1858 — Stenodactylus scincus Schlegel, Handl. Beoefen. Dierk., vol. 2, p. 16.
1863 — Teratoscincus keyserlingii Strauch, Bull. Acad. Sci. St. Petersbourg, T. 4, S. 397.
Terra typica: Seri Tschah, Persia.
1885 — Teratoscincus scincus, Boulenger, Cat. Liz. Brit. Mus., vol. 1, p. 12.
1896 — Teratoscincus zarudnyi Nikolsky, Ежег. Зоол. Муз., т. 1, с. 370. Terra typica
restricta: «Rume in Persia orientali».
1905 — Teratoscincus roborowskii, Bedriaga, Ежег. Зоол. Муз., т. 10, № 314 (terra t у р i -
c а: Оазис Сачжоу).
Диагноз. Крупная спинная чешуя оканчивается на уровне затылка;
вокруг середины туловища 26—36 чешуй.
Лектотип. Хранится в Лейденском музее [234].
Описание (рис. 10) (по 216 экз. из колл. ИЗАНУ, ЗИН, MZSF).
L ad(n = 66) 50,9—92,8 (75,24 ± 1,34) мм; L $ ad (п = 67) 50,6—116,2
(77,15 ± 1,57) мм; L/Lcd (п = 122) 1,15—2,13 (1,49 ± 0,02); высота/ширина
головы (п = 30) 57—72 (65,27 ± 0,69); диаметр уха/глаза (п = 25) 60—ПО
(84,52 ± 2,60); верхнегубных щитков (и = 416) 8—13 (10,57 ± 0,07); нижне-
губных (и = 409) 8—12 (9,95 ± 0,05); чешуй между центрами глаз (п = 154)
29—45 (36,07 ± 0,26); чешуй вдоль головы (п = 142) 50—80 (62,42 ± 0,47);
чешуй вдоль тела снизу (п = 176) 84—113 (95,85 ± 0,44); чешуй вокруг сере-
дины туловища (п = 111) 26—36 [30,62 ± 0,21); надхвостовых пластинок (п =
= 140) 9—19 (15,36 ± 0,02); гребешков вдоль четвертого пальца (п = 34) 20—
30 (24,0 ± 0,35). Нижненосовых щитков 1 (85,4 %) или 2 (8,3 %), у 6 % осо-
бей они отсутствуют, в 1 случае (0,3 %) их 3 (соотношение размеров нижне-
носовых щитков и диаметра ноздри имеет значение при определении одного из
подвидов); нижнечелюстных щитков 1 (30,2 %), 2 (62,7 %) или 3 (7,1 %).
Остальные признаки содержатся в родовом описании. Масса до 30 г.
Вариации окраски даны при описании подвидов.
Половой диморфизм и возрастная изменчивость.
Самки не крупнее самцов (различие недостоверно: t = 0,95). Самца от самки
в сезон размножения можно отличить по припухлости в заклоакальной области.
Молодые имеют более четкий рисунок.
Распространение. Средняя Азия, восточная часть Западной Азии
(включая восточную часть Аравийского полуострова); известна находка в за-
падной части Центральной Азии (оазис Сачжоу).
35
Кадастр к рис. 12,
Туркмения: 1 — возвыш. Дузкыр [130]; 2 — Сарыкамыш (ИЗАНУ); 3 — крепосгь
Измыкшир [41]; 4 — кол. Дикче, кол. Ербурун [130]; 5 — Кумсебшен [130]; 6 — 3. Узббй
[102]; 7 — пески Учтаган [130]; 8 — Дояхатын (ИЗАНУ); 9 — сев.-в. отроги Чиль-Мамед-Ку-
мов (ЗИН); 10 — Чаарли, 70 км з. Дарвазы (ИЗАНУ); 11 — Дарваза [130]; окр. Серного за-
вода; 40 км ю. Серного завода; 30 км ю. Дарвазы; 12 — Ясха (ИЗАНУ); кол. Капланлы [130];
13 — Узун-Ада (Узын-Ада) [98]; 14 — Молла-Кара (ИЗАНУ); Джебел [80]; 15 — Ахча-Куй-
ма [135]; 16 — Челекен [130]; 17 — кол. Кирпили [130]; 18 — Мессерианская равнина [130];
19 — уроч. Кара-Бсгаз, 40 км сев. Кизыл-Арвата (ЗИН); 20 — 50 км сев.-в. Кизыл-Арвата;
22 км сев.-в. Кизыл-Арвата; 35 км в. Кизыл-Арвата; 21—250 км сев. Мары (ИЗАНУ); 22 — кол.
Рис. 11. Свежевылупившийся сцинковый геккон и скорлупа от яйца (фото А. Д. Ба-
утина)
Яраджи [130]; 23 — с. Бугдайлы [130]; 24 — Фараб (ЗИН); 25 — Мешхед (ИЗАНУ); Мадау
[133]; 26 — оз. Малое Делили [130]; 27 — окр. Бахардена (ИЗАНУ); кол. Синекли [130]; 28 —
кол. Порсыкую; Каррыкуль [130]; 29 — ст. Ербент [130]; Бахардок (ИЗАНУ); 30 — окр. Чар-
джоу (ИЗАНУ); 31 — сев. Геок-Тепе (ИЗАНУ); 32 — окр. Ашхабада; ст. Аннау; оз. Куртлин-
ское [130]; 33 — Репетек; 20 км сев. Репетека; 37 км з. Репетека (ИЗАНУ): 34 -— 100 км
ю. Чарджоу [23]; 35 — Гяуре [130]; 36 — Карабата [130]; 37 — Уч-Аджи (ИЗАНУ); 38 — Кы-
зылча-Баба [23]; 39 — с. Чекич [133]; 40 — ст. Керкичи [23]; 41 — пос. Достлук [133]; 42 —
20 км ю. пос. Иолотань (ИЗАНУ); 43 — окр. Нички (ИЗАНУ); 44 — Келиф [96]; 45 — Пул-и-
Хиатум [96]; 46 — окр. Ташкепри (ИЗАНУ); 47 — уроч. Кагазлы [130]; 48 — окр. Тахта-База-
ра (ИЗАНУ); 49 — Мангышлак, пески Ак-Тюбе [98].
Узбекистан: / — оз. Судочье [113]; 2 — ю.-в. Нукуса (МГУ); 3 — 150 км в. Нукуса
по трассе на Учкудук, местность Барыкбай-шин (ИЗАНУ); 4 — Кара-Катта (ИЗАНУ); 5 —
впадина Аякагитме (ИЗАНУ); 6 — низовья Зеравшана, ст. Алат и Каракуль [22]; 7 — Самар-
канд [96]; 8 — окр. Коканда; ст. Бувайды [22]; 9 — Кара-су, 20 км от Ширабада [22]; 10 —
г. Джаркурган (МХГУ); 11 — Термез [96]; 12 — 25—30 км сев. Чабанказгана (Шабанказган)
(ИЗАНУ).
Таджикистан: / — Шаартузский р-н, Бешкентская долина (ИЗАНУ); Шаартуз-
ский р-н, пос. Чирик (ИЗАНУ); 3 — ю. граница заповедника Тигровая Балка (ИЗАНУ).
Казахстан: / — пески Малые Барсуки близ Койлибай (ЗИН); 2 — 100 км сев.-в.
Аральска (ИЗАНУ); 3 — Аральская противочумная ст. (ММГУ); 4 — п-ов Кара-Тюп [98];
5 — Устюрт, пески Сам [84]; 6 — устье р. Или (МГУ); 7 — между реками Или и Караталом [98];
8 — оз. Алаколь (ИЗАНУ); 9 — Куван-Дарья (Куван-Джарма) [96]; 10 — ст. Соль-Тобе (Со-
лотобе) (ЗИН); // — окр. Баканаса (ИЗАНУ); 12 — ст. Байгакум [96]; 13 — Дубунская пере-
права на р. Или (ИЗАНУ); 14 — Муюнкум, ст. Акыр-Тюбе (ЗИН); 15 — пески Арасан [98];
16 — пески Арыскум [98]; 17 — Асматай-Матай [98].
Китай: / — Сачжоу [300] (вставка «б»); 2 — Янгигиссар [189]; 3 — Яркенд [189].
Иран: / — пров. Тегеран, окр. Меренджаба [216]; 2 — Чехардех [194]; 3 — Ахангерун
в стр. Зиркух [88]; 4 — Руме [96]; 5 — Сарр-Чахская котловина [59]; 6 — с. Хуник [59]; 7 —
р. Бемпур около с. Махмудаббад, 12 км ю.-з. пос. Ираншехр [256]; 8 — Минаб [175]; 9 —
Зия-и-Болох и Зия-и-Лагун, 30° 10' с. ш., 60° 50' в. д. (MZSF).
Катар (вставка «а»): 1 — Доха [175].
Объединенные Арабские Эмираты (вставка «а»): / — Шарджа [175];
2 — Эд-Дайд [175]; 3 — Джебел ь-Фая, 25° 08' с. ш.» 55° 50' в. д. [175].
36
Афганистан:/ — р. Герируд около г. Герат [193]; 2 — 48 км з. г. Диларам, 32° 15*
с. ш., 62650' в. д. [194]; округ Герат и Кандагар, дорога Фаях-Диларам [172]; 3 — ю. пос. Кала-
и-Канг, 31° 05' с. ш., 61° 52' в. д. [172].
Систематика и географическая изменчивость.
Внутри этого широко распространенного вида следует различать три формы.
В отличие от других исследователей [17, 112], мы считаем целесообразным ис-
пользовать для диагностики южного подвида не количество чешуй вокруг сере-
дины туловища и поперек головы, где имеется значительное перекрывание,
а по рисунку взрослых особей. При определении юго-восточного подвида необ-
ходимо учитывать, что между ним и номинативной формой в Южном Таджики-
стане существует зона интеградации. Описанные ранее Т. zarudnyi [86] и Т. го-
borowskii [182] не имеют таксономического значения.
Таблица для определения подвидов Teratoscincus scincus
1 (2). Если нижненосовой щиток есть, то его диаметр равен или меньше половины диаметра
ноздри................................................................Т. s. rustamowi
2 (1). Диаметр одного-двух нижненосовых щитков составляет больше половины диаметра
ноздри.
3 (4). Спина взрослых особей с поперечными пятнами...................................
........................................................................ Т. s. scincus
4 (3). Спина взрослых особей с продольными полосами..................................
....................................................................Т. s. keyserlingi
СЦИНКОВЫЙ ГЕККОН ОБЫКНОВЕННЫЙ — TERATOSCINCUS SCINCUS SCINCUS (SCHLEGEL,
1858)
Типовая территория: как у вида.
1858 — Stenodactylus scincus Schlegel, Hand. Beoefen. Dierk., Vol. 2, p. 16.
1905— Teratoscincus roborowskii Bedriaga, Ann. Mus. zool. Acad. Sci. St. Petersbourg, T. 10,
p. 159.
Диагноз. Между первым верхнегубным щитком и ноздрей имеется
один или более дополнительных нижненосовых щитков; их диаметр всегда бо-
лее половины диаметра ноздри; спина взрослых особей в поперечных пятнах.
Дополнительные признаки (по 122 экз. из коллекций
ИЗАНУ и ЗИН). L ad (п = 58) 52,4—92,8 (76,72 ± 1,49) мм; L j ad (п =
= 54) 50,6—93,1 (75,12 ± 1,48) мм; чешуй вдоль тела снизу (п = 122) 81—113
(97,38 ± 0,51); чешуй вокруг середины туловища (п = 80) 26—33 (29,71 ±
± 0,16); чешуй поперек головы (п = 106) 29—45 (35,94 ± 0,34). Остальные
признаки содержатся в видовом описании. Тип видовой.
Прижизненная окраска. Основной фон серовато-лимонно-жел-
тый. На голове серо-коричневые пятна (иногда с фиолетовым оттенком) и поло-
сы, образующие грубопятнистый рисунок, частично переходящий на горло. От
затылка до крестца семь-восемь широких темных поперечных полос; по бокам
туловища с каждой стороны от лопаток до основания задних конечностей тянет-
ся тонкая темная полоса; чешуи туловища, особенно в зонах пятен и полос,
по свободному (заднему) краю окаймлены тонкой белой окантовкой; некоторые
из этих чешуй темные в центре, а к периферии окрашены в красновато-лиловые
или охристые тона; хвост сверху сажисто-черный, каждая ногтевидная чешуйка
с тонким белым кантом вдоль свободного края; на конечностях рисунок отсутст-
вует. Горло белое со слабым лимонным оттенком; брюхо белое; низ хвоста тем-
ный; внутренние поверхности лап розовые.
Распространение. Северная часть видового ареала от Каспий-
ского моря до Семиречья; южная граница проходит по восточным отрогам Турк-
мено-Хорасанских гор и по западным отрогам Паропамиза. В Ферганскую доли-
ну не проникает, в Ю. Таджикистане образует зону интерградации со сцинко-
вым гекконом Рустамова. Подвидовая принадлежность гекконов из Ц. Азии
(оазис Сачжоу) нуждается в уточнении.
37
СЦИНКОВЫЙ ГЕККОН КАЙЗЕРЛИНГА — TERATOSCINCUS SCINCUS KEYZERLINGII STR.
Типовая территория: Сери-Чах, В. Иран.
1863 — Teratoscincus keyzerlingii Strauch, Bull. Acad. Sci. St. Petersbourg, T. 4, S. 397.
Terra typica: Seri Tschah, Persia.
1896 — Teratoscincus zarudnyi Nikolsky, Ежег. Зоол. Муз., т. 1, с. 370. Terra typica
restrict a: «Rume in Persia oriental!».
1949 — Teratoscincus scincus keyzerlingii, Терентьев, Чернов, Определитель пресмыкающихся и
земноводных, М., Сов. Наука, с. 128.
Диагноз. Между первым верхнегубным щитком и ноздрей имеется
один или больше дополнительных нижненосовых щитков; их диаметр всегда
больше половины диаметра ноздри; спина взрослых особей в продольных поло-
сах (рис. 10).
Дополнительные признаки (по 8 самкам из коллекций ЗИН
и MZSF). L § ad (п = 5) 62,3—116,2 (98,14 ± 9,43) мм; чешуй вдоль тела сни-
зу (и = 6) 100—118 (108,83 zb 2,43); чешуй вокруг середины тела (п = 4)
31—35 (32,5 zb 0,96); чешуй поперек головы (л = 5) 39—46 (43,20 ± 1,24).
Остальные признаки содержатся в видовом диагнозе.
Лектотип. ЗИН 2396. $> ad «Seri Tschah, Кайзерлинг, 1862»: L =
= 104,1 мм; хвост оторван; чешуй поперек головы 42; верхнегубных щитков
13/12; нижнегубных — 13/12; нижнечелюстных щитков одна пара; чешуй вдоль
тела снизу 118; чешуй вокруг середины тела 35; надхвостовых пластинок 18.
Окраска [263]. Цветовой фон спины подвержен изменениям в пределах
желтого, оранжевого и различных оттенков коричневого, перемежающихся
с участками светло-серого. Обычно есть две широкие продольные темно-корич-
невые полосы, однако на хвост они не заходят; последний лишен определенного
рисунка. Бока и брюхо от светло-розового до белого. Молодые особи от темно-
желтых до светло-оранжевых; есть по четыре-пять сажисто-черных попереч-
ных полосок на теле и хвосте.
Распространение. Видовой ареал к югу от Паропамиза:
Ц. и Ю.-В. Иран, 3. Афганистан, Сев.-З. Пакистан, восток Аравий-
ского п-ова.
СЦИНКОВЫЙ ГЕККОН РУСТАМОВА — TERATOSCINCUS SCINCUS RUSTAMOWI SZCZERBAK
Типовая территория: пески в окр. Коканда и Кайраккума, Ферганская
долина.
1979 — Teratoscincus scincusrustatnowi, Щербак, в кн.: Охрана природы Туркменистана, вып. 5,
Ашхабад, с. 129.
Диагноз. Между первым верхнегубным щитком и ноздрей дополнитель-
ный нижненосовой щиток отсутствует; если он имеется, то его диаметр меньше
или равен половине диаметра ноздри; спина взрослых особей в поперечных
пятнах.
Дополнительные признаки. L ad (n = 13) 61,7—77,5
(68,64 zb 1,50) мм; L $ ad (n = 15) 61,9—81,8 (72,93 zb 1,94) мм; чешуй вдоль
тела снизу (и = 37) 84—97 (91,0 zb 0,60); чешуй вокруг середины туловища
(и = 37) 28—33 (30,60 zb 0,35); чешуй поперек головы (п = 37) 30—40 (35,40 ±
zb 0,60). Остальные признаки содержатся в видовом описании. Окраска совпа-
дает с окраской номинативного подвида.
Голотип. Re № 9 ИЗАНУ, 8.V 1976 г., окр. Коканда. rfadL =
= 84,0 мм; Led = 43,0 мм; чешуй поперек головы 38; верхнегубных щитков
И—10; нижнегубных — по 11; нижнечелюстных щитков 3 пары; чешуй вдоль
тела снизу 88; чешуй вокруг середины туловища 31; надхвостовых пластинок
15; нижненосовой щиток слева отсутствует.
Распространение. Ферганская долина; в Ю. Таджикистане об-
разует зону интерградации с номинативным подвидом.
Стации и количественные данные. Сведения о местах
обитания сцинкового геккона имеются во многих работах [22, 23, 98, 107,
125, 130, 133, и др.] (рис. 11, 12). По единодушному мнению всех зоологов, это
38
5(Г 60*
80*
Рис. 12. Распространение Т. scincus
типичный псаммофильный вид. Как нам удалось установить, он населяет раз-
нообразные места в песчаных пустынях, однако в ряде районов обитает на
твердых глинистых и солончаковых такыровидных почвах. В разных местах
отменена и различная их численность.
В центральных районах ареала (Каракумы) сцинковый геккон предпочита-
ет полузакрепленные и развеянные пески, здесь встречается чаще, до 40 осо-
бей на 1 км, а на севере ареала (Кызылкум) и востоке (Ю.-З. Таджикистан) оби-
тает на твердых и солончаковых почвах, местами достигая плотности до 20 осо-
бой на 1 км маршрута.
Отношение к температуре. По наблюдениям в Туркмении, са-
мая низкая температура почвы (14°), при которой встречались активные сцин-
ковые гекконы, отмечена нами 29. IV 1979 г. у пос. Чаирлы, западнее Дарва-
за. Обычно они встречаются на поверхности при температуре почвы 19—27°,
воздуха 15,5—30°.
По сообщению А. Д. Баутина, сцинковые гекконы откладывали в терра-
риуме яйца в место с температурой песка 34—36°. Инкубация яиц, по наблю-
дениям в неволе, происходила при температуре 28—30° [117].
Суточный цикл активности. Выходят из убежищ, как пра-
вило, около 21 ч, после наступления темноты. Наибольшая численность яще-
риц наблюдалась в период с 23 до 1 ч, а затем их количество на поверхности
уменьшается, хотя отдельные особи были активными до рассвета (5 ч). Осенью,
в октябре, их наблюдали с 19 ч 40 мин [133].
Сезонный цикл активности. Сцинковый геккон в Туркмении
появляется из убежищ рано — встречен уже 15.1 II 1966 г. сев. Безмеина [133];
массовый выход этих ящериц наблюдается в апреле. В Таджикистане гекконы
данного вида пробуждаются после зимней спячки в конце марта (18, 20 и
31.III 1959 г.) [107]. В Казахстане они появляются в Кызылкуме во второй
половине марта, в Муюнкумах — начале апреля, в Ю. Прибалхашье — в пер-
вой половине апреля [98].
39
На зимовку в Туркмении, очевидно, уходит в конце октября — начале но-
ября (последние наши находки датируются 17. X 1982 г., ст. Репетек). В Аф-
ганистане активные гекконы добыты в серэдине ноября [172], в Иране (Хора-
сан) Н. А. Зарудный добывал их 12.XI 1900 г. (ст. стиль) [96]. В Ю. Прибал-
хашье сцинковые гекконы уходят на зимовку в первых числах сентября [98].
Л и н ь к а. За сезон сцинковый геккон линяет не менее трех раз. Линька
наблюдалась в середине апреля — конце мая — начале июня и во второй по-
ловине августа [23, 96, 128]. Проследить в природе линьку трудно, так как гек-
коны линяют в норах. По наблюдениям в неволе, линька длится пять-шесть
дней. Линочную шкурку животное снимает лапами и съедает.
Питание изучено достаточно хорошо [22, 23, 98, 107, 130]. В Кара-
кумах сцинковый геккон в основном питается жуками [130]. Весной
(п = 50) встречаемость жуков составляет 86,0 %, летом (п = 87)—82,7
осенью (п = 13) 46,1 %. Среди жесткокрылых доминируют чернотелки и хру-
щи, которых особенно много геккон поедает в апреле и мае. Видное место в пи-
тании гекконов занимают перепончатокрылые, преимущественно муравьи.
Другими видами беспозвоночных (таракановые, термиты, саранчовые, мокрицы,
фаланги) гекконы питаются изредка. Состав пищи ящериц, добытых в Узбеки-
стане и Таджикистане, принципиально не отличался. В желудках гекконов,
собранных в Муюнкумах [98], преобладали саранчовые. В неволе сцинковые
гекконы нормально размножаются при питании личинками мучного хруща,
сверчками и тараканами, однако один-два раза в неделю с кормом давали
глицерофосфат кальция, а раз в месяц — «тетравит» из расчета одна капля на
взрослую ящерицу [117].
Размножение. Соотношение полов по коллекционным материалам
близкое 1:1. Половозрелость наступает при достижении размеров туловища
68—70 мм и больше. Спаривание происходит в апреле. Как было эксперимен-
тально установлено В. Е. Фроловым, достаточно одного спаривания, чтобы сам-
ка отложила две оплодотворенные кладки. Одно-два яйца откладываются в
июне — июле, реже — начале августа [22, 125]. А. Д. Баутин получил кладку
яиц у самок, отловленных в Ю. Таджикистане, 20.VII 1976 г. В лабораторных
условиях гекконы у него откладывали яйца в январе, апреле, июне и августе.
Самка, содержащаяся в неволе [126], отложила яйца 16—22.III и 22.1; по дан-
ным В. Е. Фролова (в печати), отмечено две кладки в период с 20. III по 26.VI
с интервалом приблизительно в три месяца. Родители происходили из Турк-
мении. Яйца белые, почти шарообразные, с неглубокой вмятиной, их размеры
17—21 X 16—19 мм, масса 2340—3050 мг [116]. По данным А. Д. Баутина,
самки, добытые в Ю. Таджикистане, откладывали яйца (л = 12) со средними
размерами 18,7 X 17,1 мм. Инкубация яиц длится 60 дней (условия неизвест-
ны) [13], при температуре 28—30° — 72—93 дня [116], а при температуре 34°
всего 54 дня (сообщение А. Д. Баутина). Сеголетки отмечены в Туркмении с
13.IX по 18.X [133], в Казахстане — 15—18.VIII 1969 г. [98].
Темп роста. Только вышедшие из яиц гекконы имеют следующие
размеры: L = 37—38 мм, Led = 22—25 мм, масса 1770 = 1890 мг [116]. При
содержании в неволе спустя год они принесли потомство. После появления на
свет в июле 1976 г. молодая самка через шесть месяцев в январе 1977 г. (без спяч-
ки) стала половозрелой и отложила яйца (наблюдения А. Д. Баутина). Эти
сведения показывают, что гекконы первых генераций могут участвовать в раз-
множении уже в конце следующего сезона, но чаще это происходит, видимо, в
возрасте двух лет. Максимальных размеров они достигают не ранее чем в пяти-
летием возрасте.
Враги. Сцинковых гекконов поедают пегий путорак, малая белозубка,
саксаульная сойка, песчаный и восточный удавчики, поперечнополосатый по-
лоз, гребнепалый геккон [22, 23, 130, 133, 107, 98]. 28.IV 1979 г. у кол. Кызыл-
Гаты (185 км сев. Бахардена) мы нашли убитого геккона в гнезде с птенцами
домового сыча.
Особености поведения. Распространенное мнение о том, что
сцинковый геккон — неуклюжее животное, не соответствует действительности.
Освещенная ярким светом, ящерица прижимается к грунту и замирает или мед-
40
ленно старается уйти. При наблюдениях в лунные ночи мы видели, как быстро
передвигается эта ящерица. После выхода из норки первое время (15—20 мин)
геккон не удаляется от убежища и при беспокойстве мгновенно уходит в нору.
В случае опасности, если нет поблизости нор, он скрывается в кусты. Сцинко-
вый геккон передвигается на вытянутых лапах, не касаясь туловищем субстра-
та, хвост обычно приподнят кверху. Во время охоты эта ящерица осторожно
приближается к насекомому, приподнимается на лапах й сверху, резким дви-
жением хватает жертву, разжевывает ее и проглатывает.
Летними и зимними убежищами этого вида являются норы. Геккон выбира-
ет небольшое углубление, как правило, под склоном песчаного бугра или под
кустом, начинает его копать передними лапками, откидывая йочву под брюшко
и по бокам, затем задними лапками отталкивает дальше. Потолок норки уплот-
няется головой, выход, как правило, забивается песчаной пробкой (наблюде-
ние А. Д. Баутина). Норы бывают длиной 20—150 см, глубиной 10—60 см,
диаметром 4—5 см. Наиболее содержательные сведения о норах сцинковых гек-
конов принадлежат К. П. Параскиву Р 8]. По наблюдениям автора, в мягких
почвах гекконы сами роют норы, помогут пользоваться и норами других жи-
вотных (большая жужелица (Scarites sp.), жук-навозник, мохноногий и малый
тушканчики, полуденная песчанка). Геккон лишь проникает в ход норы хозяи-
на и от хода уже роет себе отнорок в слое увлажненного песка, в котором и
укрывается Геккон закрывает отверстие главного хода пробкой из песка или
лесса. Иногда пробка закрывает вход в камеру (гнездо) и располагается в ходе
перед ней.
Сцинковому геккону присущ голос — писк его похож на звучание метал-
лической пластинки, зажатой с одной стороны; он обладает хорошим слухом.
Живут гекконы поодиночке. Очень редко встречаются в одной системе хо-
дов две самки; самцы и молодь обитают по одному в норе. В течение ночи самки
и молодь не уходят далеко от постоянной норы. Самцы, напротив, уходят от но-
ры на значительное расстояние (до 300 м) и часто оставляют нору, в которой
провели день. Каждое животное имеет определенную территорию, на которой
ловит насекомых, не заходя на участок другой особи. Исключение составляют
самцы, передвигающиеся в брачный период и, вероятно, расселяющаяся молодь.
По сведениям В. Е. Фролова, драка самцов из-за укрытия наблюдалась при со-
держании в неволе только однажды. Беременные самки довольно часто травми-
руют приблизившихся самцов. Таким образом, появление травмированных сам-
цов свидетельствует о беременности самок и требует отсаживания их во из-
бежание дальнейшей травматизации. У сцинкового геккона, в отличие от всех
гекконовых ящериц нашей фауны, не только очень ломкий хвост, но легко по-
вреждаются и чешуйчатые покровы, которые затем регенерируют.
При спаривании самец прижимает самку мордой к почве, затем изгибается,
подводит ногу под самку и копулирует (наблюдение в неволе А. Д. Баутина).
Он так описывает процесс кладки яиц. Самка за 30—40 мин до кладки начина-
ет волноваться, искать подходящее место, находит углубление в почве и, более
его углубив, становится задними лапами под ямкой, очень опустив туда заднюю
часть. Откладывается мягкое яйцо, после этого самка часто начинает его обка-
тывать задними лапками. Через 5—10 мин яйцо затвердевает и самка закапыва-
ет его или оставляет на поверхности. Известен случай коллективной откладки
(5 яиц) в одной норе [22]. После появления на свет новорожденный геккон ве-
дет себя активно, приступает к постройке норки и берет корм.
Кроме обычного голоса, сцинковый геккон способен издавать характерный
треск, получающийся при быстрых движениях хвоста от трения расположенных
на его верхней поверхности ногтевидных щитков. Известно [981, что треск, из-
даваемый отброшенным извивающимся хвостом, воспринимается другими осо-
бями как сигнал опасности.
Практическое значение и охрана. Сцинковый геккон,
несомненно, полезное животное, важный фоновый вид песчаных пустынь Сред-
ней Азии и Казахстана, его следует охранять от бесцельного уничтожения.
Обширный ареал и довольно высокая численность этого вида свидетельствуют
о том, что необходимость в проведении специальных охранных мероприятий
41
отсутствует. Он охраняется на территории многих заповедников (Репетекский
в Туркмении, Кызылкумский в Узбекистане, Тигровая балка в Таджикистане
и др.).
СЦИНКОВЫЙ ГЕККОН ПРЖЕВАЛЬСКОГО — TERATOSCINCUS PRZEWALSKII STRAUCH, 1887
Типовая территория: оазис Хами (3. Китай).
Кариотип: не изучен.
1887 — Teratoscincus przewalskii Strauch, Mem. Acad. Sci. St. Petersbourg, Ser. 7, Vol. 35, N 2,
S. 71.
Диагноз. Крупная спинная чешуя оканчивается на уровне лопаток;
вокруг середины туловища 31—38 чешуй.
Лектотип. ЗИН 6564. g ad «Хами, 1879, Н. М. Пржевальский». L =
= 89,7 мм; хвост оторван; чешуй поперек головы 42; верхнегубных щитков по
Рис. 13. Биотоп Crossobamon eversmanni, Teratoscincus scincus и Tenuidactylus russowi
(полузакрепленные пески у оз. Ясха, ТССР)
11; нижнегубных — 9—10; нижнечелюстных щитков одна пара; чешуй вдоль
тела снизу 108; чешуй вокруг середины туловища 35; надхвостовых пласти-
нок 14.
Описание (рис. 14) (по 66 экз. из колл. ЗИН). L ad (п = 29)
51,7—93,4 (72,8 ± 2,4) мм; L $ ad (п = 34) 52,6 —93,0 (76,54 ± 2,17) мм;
L/Lcd (п = 48) 1,06—2,12 (1,58 ± 0,035); высота/ширина головы (п = 31)
57—71 (61,97 ± 0,61); диаметр уха/глаза (и = 93) 72—92 (82,79 ± 1,12); верх-
негубных щитков (и = 132) 9—13 (10,42 ± 0,07); нижнегубных (п = 132)
7—12 (9,74 ± 0,09); чешуй между центрами глаз (и = 56) 38—50 (42,71 zb
± 0,35); чешуй вдоль головы (л = 45) 97—112 (104,24 zb 0,61); чешуй вдоль
тела снизу (п = 22) 102—115 (108,82 ± 0,73); чешуй вокруг середины ту-
ловища (и = 56) 31—38 (34,71 zb 0,20) (по [300] — 37—39); надхвостовых
пластинок (п = 62) 8—16 (13,0 ± 0,22); гребешков вдоль IV пальца (п =
= 59) 19—25 (21,97 ± 0,18). Нижненосовой щиток всегда один, его диаметр
больше или равен половине диаметра ноздри (95,5 %); нижнечелюстных щит-
ков 1 (1,5 %), 2 (59,1 %) или 3 (39,4 %) пары. Остальные признаки содержатся
в родовом описании.
Прижизненная окраска. Основной фон взрослых животных
сверху охристо-серый, молодых — охристо-желтоватый, более яркий в передней
части туловища. На туловище широкие поперечные бурые полосы (едва замет-
ные у взрослых и четкие, почти черные у молодых особей). На спине их пять-
42
шесть, на хвосте пять, на боках они размыты. Более заметна поперечная полоса
между плечами; на шее имеется дугообразно изогнутая темно-бурая полоса.
На затылке несколько темных пятен, такие же пятна выражены на губных щит-
ках. Бедра и голени охристо-буроватые с неясным пятнистым рисунком. Ниж-
няя сторона беловатая или желтоватая, кроме буроватого хвоста.
Половой диморфизм и возрастная изменчи-
вость. Самки не крупнее самцов (/ = 0,38). Рисунок у молодых особей очень
четкий, темно-бурые широкие полосы с узкими желтоватыми просветами.
Распространение. Центральная и западная часть пустыни Гоби.
Западная граница проходит по линии Джунгарская Гоби — оазисы Хами и Сач-
жоу, восточная — по западным районам Восточно-Гобийского аймака (МНР);
с севера и юга распространение ограничено отрогами Монгольского Алтая и
Наньшаня (рис. 14).
Кадастр к рис. 15.
МНР [78]: / — Алтай-Сомон; 2 — Шарахулс; 3 — Эхингол; 4 — Ногонцав; 5 — Ин-
гени-Хубрых-Булак; 6 — Дзулганай; 7 — Борузон-Гоби; 8 — Галбын-Гоби.
Китай: 1 — оазис Хами [300]; 2 — оазис Куфи [300]; 3 — р. Эдзин-Гол за Нарыном;
Цаган-Тохой; Бейли [96]; 4 — от Сачжоу до Бугаса (ЗИН); 5 — Черчен [96]; 6 — Ния [300];
7 — Чарлык (Чахарлык) [300].
Систематика и географическая изменчивость.
В пределах изученных выборок изменчивость не обнаружена.
Экология почти не изучена. По наблюдениям в Заалтайской Гоби
[311 обычно держится на песчаных участках (здесь их количество 15—20 осо-
бей на 1 га), но заходит и на каменистую равнину — гаммаду, где насчитыва-
ли 1—2 особи на 1 га. Сцинковые гекконы Пржевальского сами копают норы,
хотя используют и норы грызунов. При преследовании они могут взбираться
на кусты высотой 80 см, имеют стабильный индивидуальный участок. Питаются
в основном жуками-чернотелками, хрущами, долгоносиками, поедают также
крупных паукообразных — скорпионов, фаланг, в террариуме нападают на
мелких ящериц.
Откладывают два белых шаровидных яйца (16—17 X 15—16 мм) в песке
у основания куста саксаула, на глубине 30 см. 26.VIII 1982 г. уже были сфор-
мированные эмбрионы (устное сообщение Н. Л. Орлова). Среди вероятных вра-
гов — ушастый еж. Отброшенный ящерицей извивающийся хвост издает гром-
кий шуршащий звук.
Вид обычный, широко распространен в пустынях Ц. Азии, охраняется на
территории Гобийского биосферного заповедника (МНР).
СЦИНКОВЫЙ ГЕККОН МЕЛКОЧЕШУЙНЫЙ — TERATOSCINCUS MIGROLEPIS NIKOLSKY, 1889
Типовая местность: Дуз-аб, В. Керман, Иран.
Кариотип: не изучен.
1899 — Teratoscincus microlepis Nikolsky, Ежег. Зоол. Муз., № 2, с. 145.
Диагноз. Спинная чешуя мелкая, на уровне лопаток плавно переходит
в затылочную; вокруг середины туловища около 100 чешуй.
Голотип. ЗИН 9164, ad «Дуз-аб в Восточном Кирмане, 15.VI 1898,
Н. Зарудный». L = 53,2 мм; L cd = 34,0 мм; L/L cd = 1,56; верхне- и нижне-
губных щитков по 11; нижнечелюстные щитки отсутствуют; чешуй между цент-
рами глаз около 50 [88]; чешуя на затылке плавно переходит в спинную;
чешуй вдоль тела снизу 160; чешуй вокруг середины туловища около 100; над-
хвостовых пластинок около 10.
Описание (рис. 14) (по 7 экз. из колл. ЗИН и ZFMK). L общ. (п = 6)
53,2—73,0 мм (64,42 ± 3,07); L/L cd (п = 5) 1,56—1,95 (1,78 ± 0,07;
высота/ширина головы (п = 5) 63—66 (64,0 ± 1,0); диаметр уха/глаза (л =
= 4) 53—89 (71,0 ± 9,90); верхнегубных щитков (п= 14) 10—12 (11,07 0,13);
нижнегубных (п = 14) 9—11 (9,78 ± 0,24); чешуй между центрами глаз
(л = 7) 48—58 (50,57 ± 1,29); чешуй вдоль головы (/г = 3) 135—154; чешуй
вдоль тела снизу (л = 7) 150—188 (169,57 ± 4,67); чешуй вокруг середины
туловища (л = 7) 85—110 (96,14 ± 3,39); надхвостовых пластинок (л = 6)
43
Рис. 14. Teratoscincus przewalskii (вверху):
A — общий вид, Б — голова сбоку, В — подбородок, Г — спина, Д — лицевая часть сверху,
Е — хвост сверху (ЗИН 18272, Ю- Гоби, МНР)
Teratoscincus microlepis (внизу):
А — общий вид, Б — голова сбоку, В — подбородок, Г — носовые щитки, Д — спина,
Е - брюхо, Ж - хвост снизу (все ZFMK, Б—Ж ZFMK 26329)
9—11 (10,0 ± 0,26); гребешков вдоль четвертого пальца (п = 7) 22—26
(24,29 ± 0,52); один нижненосовой щиток, равный диаметру ноздри, имеет-
ся (четыре случая) или отсутствует (три случая); нижнечелюстные щитки не
выражены, с подбородочным щитком соприкасаются 11—13 чешуй. Остальные
признаки содержатся в родовом описании.
Окраска [2631. Основной фон от желтого до светло-коричневого;
около шести темных поперечных широких полос, которые могут быть Д-об-
44
разными, темно-коричневыми или сероватыми у взрослых, красноватыми у мо-
лодых на хвосте пять-шесть темных поперечных полос.
Распространение. Котловины Харан, Бемпур, Деште-лут в
Ю.-В. Иране и 3. Пакистане; возможны находки в Ю. Афганистане (рис. 15);
Кадастр к рис. 15.
Иран: / — Деште-лут близ Кермана, Иран [2631; 2 — Дуз-аб (Добаз-Захедан) (ЗИН)
[88]; 3 — Бампур, р. Руд-и-Бампур (ЗИН).
Пакистан: 1—округ Чагаи, 3 км з. пос. Нушки; 14—17 км сев.-з. пос. Нушки (AMNH);
2 — ю. пос. Далбандин (28°, 53' с. ш., 64° 26' в. д.) [258]; 3 — Танк-и-Граваг (27° 13' с. ш.,
63° 25' в. д.) (2581.
Рис. 15. Распространение Teratoscincus przewalskii (/), Tenuidactylus elongatus (2)
Географическая изменчивость не обнаружена.
Экология не изучена. Н. А. Зарудный [60] сообщал, что добытый им
экземпляр «держался на сильно солонцеватой почве, вспученной и покрытой
хрустевшею под ногами соленою корою». По наблюдениям Ш. Минтона [2631,
он «привычками напоминает сцинкового геккона, но не столь раздражителен,
как последний, кожа его не так легко обрывается. Один экземпляр, содержав-
шийся в неволе несколько недель, отказывался от любой пищи, за исключени-
ем точно не определенных личинок крупных песчаных жуков» (перевод наш).
СЦИНКОВЫЙ ГЕККОН ВЕДРЯГИ — TERATOSCINCUS BEDRIAGAI NIKOLSKY, 1899
Типовая местность: Ходжи-и-ду-Чахи, В. Иран.
Кариотип: не изучен.
1899 — Teratoscincus bedriagai Nikolsky, Ежег. Зоол. Муз., № 2, с. 147.
Диагноз. Крупная спинная чешуя оканчивается на уровне лопаток;
вокруг середины туловища 44—48 чешуй.
Лектотип. ЗИН 9161. dad «Ходжи-и-ду-Чахи, 5.V 1898, Н. Заруд-
ный». L = 63,0 мм; хвост отсутствует; чешуй поперек головы 32; верхнегуб-
ных щитков 11/10; нижнегубных 10/9; нижнечелюстные щитки отсутствуют;
чешуй вдоль тела снизу 103; чешуй вокруг середины туловища 48; надхвосто-
вых пластинок 12.
Описание (рис. 10) (по 5 экз. из колл. ЗИН). L d ad (п = 4) 61,8—
63,0 мм; L g ad (п = 1) 73,4 мм; хвосты оторваны; высота/ширина головы (п =
= 5) 63—71 (67,40 ± 1,47); диаметр уха/глаза (п = 5) 86—93 (89,75 ± 1,49);
верхнегубных щитков (п = 10) 10—11 (10,70 ± 0,15); нижнегубных (л == 10)
(9,40 ± 0,16); чешуй между центрами глаз (л = 5) 31—33 (32,0 ± 0,45); чешуй
вдоль головы (п = 5) 69—90 (76,80 ± 3,62); чешуй вдоль тела снизу (л = 5)
93—103 (98,50 ± 2,10); чешуй вокруг середины туловища (л = 5) 44—48
45
(46,0 ± 0,71) 188); надхвостовых пластинок (п = 5) 10—12 (11,0 ±0,45);
гребешков вдоль четвертого пальца (п = 5) 17—21 (19,0 ± 0,84). Нижнехвос-
товой щиток, равный по размерам ноздре, всегда имеется; нижнечелюстные
щитки не выражены. Остальные признаки содержатся в родовом описании.
Окраска [881. Тело сверху белесоватое, на затылке имеются косые ли-
нии, на спине четыре темные широкие поперечные полосы; снизу тело белое.
Распространение. Обитает в Иране вдоль ирано-афганской гра-
ницы, а также в Ю. Афганистане (рис. 16).
Кадастр к рис. 16.
Иран: / — Ниаз-Абад, страна Хаф [60]; 2 — Бамруд (Бемруд), стр. Зиркух [601; 3 —
кол. Чах-и-Гюшие; Ходжи-и-Ду-Чаги (Ходжи-до-чахи) [60]; 4 — Доруне [303].
Афга н иста н: / - 16 км ю. пос. Кала-и-Канг, 30° 58' с. ш., 61° 54' в. д. [172]; 2 —
Кандагар, 31° 36' с. ш., 65° 47' в. д. [172].
Экология не изучена. По наблюдениям Н. А. Зарудного [60], «найден
на хрящеватой почве, усыпанной редким мелким щебнем и местами покрытой тон-
ким слоем песка. Несколько экземпляров наблюдалось... на сильно солонцева-
той почве, рыхлой и покрытой тонкой соляной корой. Очень обыкновенен в
Сейстанской котловине, где попадался как по невозделанным участкам куль-
турной части этой страны, так и, в особенности, вне ее, далее к югу, в пустыне,
поросшей тамарисковыми кустами. Здесь он держался на гладких, плотных
глинах и суглинках, местами покрытых невысокими песчаными буграми».
Известно [245], что этот вид доходит как минимум до высоты 4700 футов (1400 м)
н. у. м.
РОД ГРЕБНЕПАЛЫЙ ГЕККОН — CROSSOBAMON BOETTGER, 1888
Типовой вид: Gymnodactylus eversmanni Wiegman.
1887 — Ptenodactylus Strauch, Mem. Acad. Sci. St. Petersbourg, VII Ser., Vol. 35, N 2, S. 64.
1888 — Crossobamon Boettger (nomen novum), Zool. Anz., Vol. 11, S. 260.
Диагноз. Пальцы когтистые, прямые, плавно утончаются к концам,
оторочены по бокам чешуйками с роговыми зубчиками, а снизу покрыты одним
рядом поперечно расширенных, слабо ребристых или пильчатых вдоль свобод-
ного края пластинок; чешуя (за исключением крупных щитков головы и иног-
да чешуй брюха) с более или менее выраженными ребрышками; зрачок верти-
кальный с зазубренными краями; у самцов почти прямой ряд анальных пор,
очень часто прерванный посередине; на спине выражены мелкие бугорки; ниж-
нечелюстных и подхвостовых щитков, хвостовых бугров и сегментов нет; хвост
тонкий, стройный, не ломкий.
Дополнительные признаки. Тело и голова несколько при-
плюснутые. Межчелюстной щиток четырехугольный; три слабо вздутых носовых
щитка, равных друг другу; первый может быть оттеснен от ноздри вторым и
межчелюстным; первые носовые соприкасаются между собой или разделены од-
ной — тремя чешуйками; первый верхнегубной щиток выше остальных, его
высота больше ширины; чешуя морды неправильно-многоугольная, неоднород-
ная, уменьшается к затылку; отдельные увеличенные чешуи в предглазничной
области и над глазами; ухо — вертикальный овал. Подбородочный щиток тра-
пециевидный, с округлым и неровным задним краем; размеры нижнегубных
щитков плавно убывают от первого к последнему; горловая чешуя полукону-
совидная, верхние чешуйки несколько крупнее остальных.
Спинная чешуя плоская, округло-четырехугольная или ромбовидная,
границы с брюшной чешуей нет; уплощенные мелкие округлые спинные бугор-
ки располжены среди этой чешуи почти беспорядочно или слабо выраженными
продольными и поперечными рядами. Брюшная чешуя плоская, ромбическая
или треугольная, слабо черепитчатая; анальные поры расположены на более
крупных чешуйках, у самок они не перфорированы; между рядом пор и аналь-
ной шелью около 15 рядов мелких зернышек. Хвост покрыт поперечными коль-
цами чешуй, которые несколько крупнее на его нижней поверхности. Передняя
конечность, вытянутая вперед и вдоль тела, достает концами пальцев до кон-
ца морды, задняя — до подмышечной впадины. На голени и бедре имеется
46
несколько бугорков, аналогичных спинным. Сверху пальцев между рядами бо-
ковой бахромы три-четыре ряда однородных чешуй, к чехлу когтя подходит
одна широкая чешуйка, снизу между этими рядами и рядом подпальцевых
пластинок по одному-два ряда мелких зернышек.
Род содержит два вида, распространенных в Средней Азии, Казахстане,
Иране, Афганистане и Пакистане.
Таблица для определения видов рода Crossobamon
1 (2). Хвост длиннее тела; у самцов пять и более анальных пор..................С. eversmanni
2 (1). Хвост короче тела; у самцов пять и менее анальных пор....................С. orientals
ГРЕБНЕПАЛЫЙ ГЕККОН ЭВЕРСМАННА — CROSSOBAMON EVERSMANNI (WIEGMANN, 1834)
Типовая территория: Агитма (Кызылкум).
Кариотип: 2л = 42; акроцентриков 14, субтелоцентриков 4, субметацентриков 2,
метацентриков 2, NF =70.
1834 — Gytnnodactуlus eversmanni Wiegmann, Herpetol. mexic., Vol. 1, p. 19.
1887 — Stenodactylus lumsdenii Boulenger, Cat. Liz. Brit. Mus., London, Vol. 3, p. 479.
1888 — Crossobamon eversmanni, Boettger, Zool. Hb. Syst., Jena, vol. 3, S. 880.
Диагноз. Хвост длиннее тела; у самцов пять и более анальных пор.
Типовой экземпляр. Место нахождения не известно.
Описание (рис. 17) (по 308 экз. из колл. ИЗАНУ, ЗИН, МГУ, UMMZ,
AMNH, SAM). L <? ad (п = 144) 34,0—52,2 (42,02 ± 0,36) мм; L £ (п = 129)
34,0—59,0 (45,92 ± 0,66) мм; LILcd (п = 276) 0,57—0,82 (0,66 ± 0,003);
индекс высотьГширины головы (и = 301) 52—57 (64,37 ± 0,27); диаметр уха/
глаза (п = 257) 0,27—0,71 (0,47 ± 0,0040); верхнегубных щитков (и = 616)
10—16 (12,70 ± 0,05); нижнегубных (и = 616) 9—14(10,82 + 0,14); чешуй
между центрами глаз (п = 306) 22—37 (29,16 ± 0,28); длина спинного бугра/L
(п = 2£4) 5,45—14,1 (9,01 ± 0,089); чешуй, окружающих спинной бугор
(п = 300), 8—12 (9,37 ± 0,04); чешуй вдоль тела снизу (п = 258) 165—291
(230,06 ± 1,63); анальных пор (п = 130) 5—12 (8,29 ± 0,15); подпальцевых,
пластинок (п = 611) 25—38 (32,08 ± 0,12).
Рис. 16. Распространение Teratoscincus microlepis (/), Т. bedriagai (2), Carinatogecko hetero-
pholis (5), C. aspratilis (4)
Прижизненная окраска. Основной фон сверху песочно-розо-
ватый. От ноздри через глаз над ухом и над передней лапой вдоль обоих
боков туловища тянется по коричневой полосе, которая в последней трети может
разбиваться на отдельные метки; на голове и конечностях есть неясный
узор из червеобразных темных полос. Нижние поверхности белые. Остальные
детали рисунка имеют значение при определении форм.
47
Рис. 17. Crossobamon eversmanni;
Л — голова сбоку, Б — носовые щитки, В — голова сверху, Г — подбородок, Д — спи-
на, Е — брюхо, Ж — преанальные поры, 3 — хвост сверху (ИЗАНУ SR 550. ТССР. Ба-
хардок)
Половой диморфизм и возрастная изменчивость.
Самки крупнее самцов, у них отсутствуют анальные поры. У сеголеток нижняя
сторона хвоста оранжево-лимонная.
Систематика и географическая изменчивость.
Как показало исследование 10 экземпляров формы maynardi, она вполне укла-
48
дывается в диагноз данного вида. Различия в рисунке между белуджистанскими
гребнепалыми гекконами и более северными особями позволяют рассматривать
форму maynardi в качестве самостоятельного подвида. Не исключено, однако,
что в дальнейшем будут обнаружены переходные формы, поскольку особи из
Сев. Ирана деталями рисунка отличаются от номинативной формы и имеют
незначительное сходство с южным подвидом.
Таблица для определения подвидов
Crossobamon eversmanni
1 (2). Вдоль хребта и хвоста расположены тонкие поперечные червеобразные полоски; с подбо-
родочным щитком соприкасается 5—11 (М — 7,77) горловых чешуек ..
................................................................................................ С. е. eversmanni
2 (1). Вдоль хребта и хвоста расположены две продольные полосы, пространство между кото-
рыми лишено поперечных полос; с подбородочным щитком соприкасается 10—13 (М =
= 10,80) горловых чешуек........................ С. е. lumsdenii
ГРЕБНЕПАЛЫЙ ГЕККОН ЭВЕРСМАННА ОБЫКНОВЕННЫЙ —
CROSSOBAMON EVERSMANNI EVERSMANNI (WIEGMANN, 1834)
Типовая территория: видовая.
Описание (по 298 экз.). Вдоль хребта располагается 10—12 тонких
коротких темных червеобразных поперечных полосок; темные полоски, тяну-
щиеся от ноздрей через глаз и над ухом, оканчиваются, как правило, на боках
в первой трети туловища; на хвосте до 30 поперечных полосок, которые на боках
хвоста могут сливаться в продольные полосы; 5—И (7,77 ± 0,07) горловых
чешуек, соприкасающихся с подбородочным щитком. Все остальные признаки
соответствуют видовому описанию.
Географическая изменчивость. Исследование 17 выборок
номинативного подвида показало, что у восточных и южных популяций (Беш-
кентская долина; правый берег р. Мургаб; Эр-Ойлан-Дуз; Сеистан) меньше че-
шуй вдоль нижней стороны тела и вокруг середины туловища; границей между
ними и остальными является Бадхызское холмогорье и долина р. Мургаб. Од-
нако севернее, при исчезновении водной преграды, изменчивость этих призна-
ков носит клинальный характер и, возможно, маркирует пути расселения греб-
непалого геккона. Сходно изменяется и количество горловых чешуй, соприка-
сающихся с подбородочным щитком, хотя для восточных выборок характерны
более высокие, максимальные значения признака, которые уменьшаются к за-
паду и югу. Последнее обстоятельство должно облегчить диагностику подвидо-
вых форм.
Распространение. Средняя Азия, Казахстан, на юге доходит до
Систана (Иран) (рис. 18).
Кадастр к рис. 18.
СССР, Казахстан: 1 — Иргиз (ЗИН); 2 — пески около г. Аральск [98]; 3 — пе-
ски Ак-Тюбе [98]; 4 — сухое русло Кувандарья [98]; 5 — ст. Байгакум, около Жулека (ЗИН);
6 — зал. Кендирли [98]; 7 — кол. Карай-кудук и близ. г. Туркестан [95]; 8 — пески Муюн-
кум; правый берег р. Чу, кол. Чигил-Тма [98]; 9 — 70 км сев. г. Джамбул, берег р. Телас;
оз.Каргалы-Куль (ЗИН); 10 — ст. Акыртобе [98]; 11—Приаральские Каракумы, кол. Илля-
кудук [98]; 12 — Устюрт, родник Кин-Тычке [98]; 13 — пески Арыскум [98]; 14—16 — юг
песков Кызылкум [98].
Туркмения: / — Ю.-В. Сарыкамыш (ИЗАНУ); 2 — пески Уч-Таган [130]; 3 — Шах-
сенем; Мере (ЗИН); 4 — з. берег Кара-Богаз-Гола [130]; 5 — Тахтинский р-н, 40 км ю. колхо-
за «Коммунар» (ИЗАНУ); 6 — Чильмамедкум [130]; 7 — Кумсебшен [130]; 8 — возв. Дузкыр,
120 км ю.-в. Куня-Ургенча [130]; 9 — 70 км з. пос. Дарваза (наши данные); 10 — 100—150 км
ю. колхоза «Коммунар» (наши данные); 11 — Красноводск (ЗИН); 12— п-ов Дарджа [110];
13 — 185 км сев. пос. Бахарден, кол. Кызыл-Каты (наши данные); 14 — 30 км ю. пос. Дарваза
(ИЗАНУ); 15 — пос. Дарганата (наши данные); 16 — Челекен (ИЗАНУ); 17 — ст. Джебел,
пески на юг; Моллакара (ЗИН); 18— Ясхан (ИЗАНУ); 19—Куртышбаба [130]; 20 — сев.
Ак-Молла, Унгуз, 136 км в. Серного завода (наши данные); 21 — ст. Перевальная [130]; 22 —
Кирпили [130]; 23 — Дояхатын (ИЗАНУ); 24 — Бугдайлы [130]; 25 — ст. Ахча-Куйма [128];
26 — 50 км сев.-в. Кизыл-Арвата (ЗИН); 27 — кол. Култакыр [130]; 28 — 250 км сев. г. Мары
(ИЗАНУ); 29 — Мадау [130]; 30 — Бахардок (ИЗАНУ); 31 — 180 км сев. г. Байрам-Али (наши
данные); 32 — левый берег р. Амударьи по дор. на Фараб (наши данные); 33 — ст. Фараб (ЗИН);
34 — 100 км сев.-з. Репетека (наши данные); 35 — Репетек (ИЗАНУ); 36 — Каракала (ЗИН);
49
37 — Бахарден [23]; 38 — сев. пос. Геок-Тепе [130]; 39 — 37 км з. Репетека (наши данные);
40 — ст. Пески [23]; 41 — Уч-Аджи (ИЗАНУ); 42 — 9 км сев.-з. Гасан-Кули [130]; 43 — оз.
Малое Делили [130]; 44 — ст. Гяуре; пос. Анау; пос. Ниса (ИЗАНУ); 45 — Дорткую [23];
46 — Карабата (МГУ); 47 — Шахсенем (ЗИН); 48 — кол. Джапар [130]; 49 — окр. Керкичи
[130]; 50 — Достлук [130]; 51 — Келиф (ЗИН); 52 — пос. Иолотань (ИЗАНУ); 53— пос. Имам-
баба (ИЗАНУ); 54 — Эр-Ойлан-Дуз (ИЗАНУ); 55 — Кепели (ЗИН); 56 — окр. Тахта-Базара,
правый берег р. Мургаб (ХГУ); 57 — сев. пос. Кушка [23]; 58 — пос. Серахс (МГУ).
Узбекистан: / — 25—30 км сев. Чабанказгана (ИЗАНУ); 2 — р. Караколь (ЗИН);
3 — Чабанказган (ИЗАНУ); 4 — кол. Тамды (ЗИН); 5 — 100 км от Тартикуль (ХГУ); 6 — Хи-
Рис. 18. Распространение С. eversmanni
ва (ЗИН); 7 — Аякагитма и Ташкудук (ИЗАНУ); 8 — 100 км сев. пос. Шафрикан, Кызыл-
кумская биостанция (МГУ); 9 — Шафрикан [22]; 10 — Самарканд (ЗИН); 11 — ст. Каракуль
[22]; 12 — Бухара; Каган; Кызыл-Кудук [22]; 13 — пески Учкум, в. Кагана [22); 14 — пески
около пос. Джаркурган (ХГУ); пойма р. Сурхандарьи и остров Арал-Пайгамбар [158]; 15 —
15 км сев. пос. Термез (ИЗАНУ); 16 — Урганчи, 18 км сев.-з. Коканда [88]; 17— Кемпер-
Тюбе (МГУ); 18 — Кизылрабат (МГУ); 19 — 20 км сев.-з. Газли (МГУ).
Таджикистан: 1 — Шаартузский р-н, Бешкентская долина (ИЗАНУ); 2 — южная
граница ТаджССР и УзССР (ИЗАНУ); 3 — 8 км от пос. Чирик (ИЗАНУ); 4 — Тигровая Бал-
ка [88]; 5 — колхоз Абад, пески Карадум (ЗИН).
Иран: 1 — с. Мондехи; с. Фейзабад (в.— в Сарр-Чахской котловине) [59]; 2—Можнабад
и Фендукт [60]; 3 — Хоуз в стр. Зиркух (ЗИН); 4 — равнина Таг-и-Дорох [60]; 5 — кол. Чах-
Иси, Сейстан (ЗИН); 6 — Тайабаг 30° 47' с. ш., 60° 47' в. д., пров. Хорасан (CAS).
Афганистан: 1 — 20кмю. Андхой (CAS); 2 — 30 км сев.-з. Шибирган (CAS).
ГРЕБНЕПАЛЫЙ ГЕККОН ЮЖНЫЙ —
CROSSOBAMON EVERSMANNI LUMSDENII (BLGR., 1877) STAT. NOV.
Типовая территория: Белуджистан возле афганской границы.
1877 — Stenodactylus lumsdenii Boulenger, Cat. Liz. Brit. Mus., Vol. Ill, p. 479 (между Нушки
и Гильмендом).
1933 — Stenodactylus maynardi Smith, Rec. ind Mus., Calcutta, Vol. 35, p. 18.
1967 — Crossobamon maynardi, Kluge, Bull. Amer. Mus. Nat. Hist., N 4, Vol. 135, art. 1, p. 23.
Диагноз. Вдоль хребта и хвоста расположены две продольные полосы,
пространство между которыми лишено поперечных полос; с подбородочным
щитком соприкасается 10—13 (М = 10, 80) горловых чешуек. Все остальные
признаки соответствуют видовому описанию (рис. 20).
50
Типы хранятся в Британском музее естественной истории (Лондон) и в Ин-
дийском музее (Калькутта) [245].
Распространение. Белуджистан из района афгано-пакистанской
и афгано-ирано-пакистанской границ (рис. 21).
Кадастр к рис. 21.
Иран: / — Хормек [88].
Афганистан: 1 — между Нушки (3. Пакистан) и р. Гильменд — terra typica [245].
Пакистан: 1 — Нушки; 13—16 км сев.-з. Нушки (AMNH); 2 — Харан [258].
Систематика и географическая изменчивость.
Описание формы maynardi [290] позволяет почти безошибочно идентифициро-
вать ее с южным подвидом гребнепалого геккона Эверсманна, несмотря на имею-
Рис. 19. Три яйца туркестанского геккона и одно (меньшее) гребнепалого геккона из
Таджикистана (фото А. Д. Баутина)
щиеся небольшие различия. Два носовых щитка отмечены нами у незначитель-
ной части особей обоих видов рода (первый носовой щиток оттеснен от ноздри
вторым и межчелюстным); кроме того, А. М. Никольский [89], определив гек-
кона из Гурмука как lumsdenii, отметил наличие трех носовых щитков и более
ребристой подхвостовой чешуи. Колебания этого признака, таким образом,
следует отнести к индивидуальной изменчивости.
Стации и количественные данные (рис. 13). Псаммофиль-
ный вид. Обитает в закрепленных и полузакрепленных песках. Как правило,
встречается вместе с другим псаммофилом — сцинковым гекконом, однако
соотношение их различное в разных биотопах. Численность ящериц может слу-
жить показателем их отношения к данной стации. По нашим наблюдениям
(проводились в апреле — мае, экскурсия после сумерек в течение трех часов),
явное преобладание гребнепалых гекконов над сцинковыми отмечено на край-
нем западе Туркмении в окр. пос. Челекен. Здесь они обитают на буграх из ра-
кушки и осолоненного серого песка на берегу моря с растительностью из соле-
росов. На маршруте в 1 км встречено 2 геккона, а всего — 13 гребнепалых на
20 сцинковых. В окр. оз. Ясхан, на чистых развеваемых песках с кандымом,
саксаулом, песчаной акацией и селином, на 4 гребнепалых был добыт 31 сцинко-
вый геккон. На левом берегу Амударьи в песчаной пустыне южнее с. Дояхатын
на 8 гребнепалых отмечено 25 сцинковых.
В подобном пустынном биотопе в окр. с. Уч-Аджи (В. Туркмения) на 4
гребнепалых геккона приходился 21 сцинковый. В центре пустыни Каракумы
(250 км севернее г. Мары) встречен 1 гребнепалый на 12 сцинковых, а севернее
51
Ашхабада на 2 гребнепалых — 20 сцинковых гекконов. В пустыне Кызылкум
в окр. с. Аяк-Агитме, среди бугристых песков с тамариксом и верблюжьей
колючкой 1 гребнепалый геккон приходился на 20 сцинковых. На закрепленных
пылевых песках в окр. с. Имам-Баба на правом берегу р. Мургаб (Ю. Туркме-
ния) было добыто 9 гребнепалых гекконов и ни одного сцинкового. На закреп-
ленных песках с кустами солянок, прилегающих к шору во впадине Эр-Ойлан-
Дуз и Намаксар (Бадхыз), обитают только гребнепалые гекконы.
В окр. Гяурса на слабо закрепленных и развеянных песках с примесью
лесса (окраина песков в предгорьях Копетдага) на площади 600 х 200 м было
добыто 15 гекконов интересующего нас вида, т. е. 1 ящерица встречена на 0,8 га.
Следовательно, хотя гребнепалый и сцинковый гекконы обитают симпатри-
чески, у них различные требования к биотопам: первый предпочитает более или
менее закрепленные пески с примесью лесса или засоленные, а второй — чис-
тые, слабо закрепленные или развеянные пески. В Казахстане С. eversmanni
обитает на закрепленных и развеваемых песках с редкими кустиками джузгуна,
терескена и верблюжьей колючки [981. С середины июля по сентябрь наблю-
дается резкое сокращение численности гекконов с 15—25 особей на 1 км до
5—15 [3261.
Отношение к температуре. С. Шаммаков [130] наблюдал
активных гекконов в интервале температуры воздуха 10—24°. В Таджикистане
(южнее Бабатага, 9.IV 1980 г.) при температуре песка 17° гекконы не появля-
лись на поверхности (выкапывались из почвы). 14.IV 1977 г. в окр. Иолотани
гекконы были активны при температуре воздуха 22°, а при падении ее до 18°
они исчезли. 18.IV 1978 г. вполне активные гекконы наблюдались в песках
южнее с. Чирик Шаартузского р-на ТаджССР при температуре воздуха и поч-
вы 19°. 27.V 1982 г. в окр. Гяурса при температуре воздуха 27° задул ветер и
стал накрапывать дождь, гекконы встречались с подветренной стороны, за
буграми и кустиками; после падения температуры до 18° они исчезли. В терра-
риуме гребнепалые гекконы предпочитали температуру почвы около 30°.
Суточный цикл активности. Ведут сумеречно-ночной образ
жизни. Весной изредка встречаются днем (8.IV 1977 г. добыт во впадине На-
максар, Бадхыз). Обычно добывали мы их после наступления сумерек. В весен-
нее время гекконы, как правило, исчезали после похолодания к 1 ч 30 мин.
С апреля по сентябрь активны всю ночь, в середине октября наблюдались толь-
ко вечером [3261. В пасмурную погоду эти ящерицы появляются несколько
раньше, чем в безоблачную. В лунную ночь их активность падает.
Сезонный цикл активности. Первая находка этого геккона
после зимовки известна в Туркмении 15.III, массовый выход с начала апреля
[130], что согласуется с нашими наблюдениями. В Таджикистане [107] первая
ящерица была добыта 3.IV, в Казахстане [981 гекконы активны в конце апреля.
На зимовку они уходят в Туркмении во второй половине октября, последняя
активная особь была добыта 25.X 1967 г. [130]. 10.X 1981 г. под Султанбентом
на левом берегу Мургаба активные ящерицы уже не наблюдались, а одна особь
была выкопана из зимовочной норы. В Казахстане, песках Муюнкум, гекконы
уже не встречались в конце сентября [981.
Линька. Отмечена у особи, добытой 14.IV 1977 г. под Иолотанью (Турк-
мения).
П и т а н и е. По сведениям из Туркмении [130], гекконы поедают (особен-
но весной) хрущей (32,5 % по встречаемости), муравьев (30,2 %) и долгоно-
сиков (16,3 %), а также гусениц бабочек, пауков, термитов и сверчков. Дважды
в их желудках были обнаружены молодые особи сцинкового геккона. В Узбе-
кистане [23] в желудках этих ящериц найдены преимущественно жуки (60 %
встречаемости — хрущи, долгоносики), а также пауки, равнокрылые хобот-
ные, гусеницы бабочек. В Таджикистане [107] гребнепалые гекконы поедают
главным образом жуков (чернотелок, хрущей, долгоносиков, листоедов, щел-
кунов, златок, жужелиц), а также бабочек и их гусениц, муравьев, мух, кло-
пов, пауков, богомолов.
Размножение. Соотношение полов, по коллекционным материалам,
близкое 1 : 1. Самцы достигают половой зрелости при достижении размеров ту-
52
ловища более 35 мм, самки — 42 мм. Спаривание, судя по оживленным крикам
гекконов, отмечено 14.IV 1977 г. в окр. Иолотани и 13.IV 1979 г. в окр. ст.
Уч-Аджи, О. П. Богданов [23] наблюдал спаривание 14.V 1959 г., которое про-
должалось до начала июня [326]. По наблюдениям в Таджикистане [107], спа-
ривание происходит в апреле, а кладка яиц растянута и длится около двух ме-
сяцев. Предполагается в Туркмении две-три кладки за сезон [3261. Кладка из
одного-двух яиц в Туркмении происходит с конца мая до конца июля [23, 130]у
в Узбекистане — с начала июня по 19.VII [22], в Таджикистане отмечена в се-
редине — последней декаде июня [107]. К. П. Параскив [98] пишет, что в пес-
ках Ак-Тюбе на Мангышлаке нашел свежую кладку яиц 28.VII 1947 г., сформи-
рованные яйца в яйцеводах самок, добытых на южной окраине песков Муюн-
кум, найдены 20. V 1949 г. В яичниках этих самок имелись пятна от вышедших
фолликул и по два увеличенных фолликула, что свидетельствует о возможности
повторной кладки. Яйца белые, овальные, при откладке могут склеиваться.
Размеры яиц, отложенных гекконами из Туркмении (п=7), 11, 15—12,30x8,0—
9,85 мм, масса 0,458—0,848 г [281]. По сведениям из Узбекистана, величина яиц
И—12 X 8—9 мм [62], наибольшие размеры яиц, добытых из яйцевода самок
в Таджикистане, близки указанным выше — 11,5 X 8 мм [107]. Как установ-
лено в террариуме, инкубация яиц продолжается 45—53, в среднем 48 дней,
а вышедшие из яиц детеныши достигают длины туловища 21—26 мм, хвоста
33—41 мм, массы 0,365—0,501 г. [281]. Сеголетки в Туркмении встречены в окр.
Иолотани в конце июля — начале августа (первая генерация), а севернее Гяур-
са— 13—15.IX (повторная генерация) [22, 130]. Там же 26.VII 1947 г. было
выкопано яйцо, внутри которого обнаружен готовый к вылуплению гек-
кон [22].
Темп роста. По данным С. Шаммакова [130], гекконы после выхода из
яиц за 9—11 месяцев вырастают на 10—18 мм, а через два-три года достигают
максимальных размеров (L & в среднем 42, L g = 52 мм). Половая зрелость
у ящериц первой генерации наступает на следующий после выхода из яиц
год, а у особей второй генерации — в двухлетнем возрасте.
Враги. В Туркмении гребнепалых гекконов поедают поперечнополоса-
тый и краснополосый полозы (Coluber karelini, С. rhodorhachis), а также песча-
ный удавчик (Eryx miliaris) [130, 326]. 22.VI 1976 г. в желудках обыкновенно-
го сыча, добытых нами в руинах крепости Дарган-Ата, обнаружены гекконы
(в одном шесть, в другом — один). В Узбекистане на них также нападает по-
перечнополосатый полоз [22], в Таджикистане геккон был найден в желудке
пятнистого полоза (Spalerosophis diadema) [107].
Особенности поведения. Обычно передвигаются медленно,
потревоженные, могут быстро бегать; отбежав 5—6 м, затаиваются. По свиде-
тельству ряда авторов [23, 98, 136], гребнепалые гекконы могут лазать по вет-
вям кустов и там добывать себе пищу, перепрыгивать с ветки на ветку на рас-
стояние до 10 см. В поисках корма гребнепалый геккон удаляется от норы на
значительное расстояние, до 100—150 м, возвращаясь под утро в оставленную
накануне норку. Летом часто закапывается в песок на глубину до 5 см, исполь-
зует норы грызунов, ящериц, может сам выкапывать норы. О. П. Богданов
[23] сообщал о том, что отложенное яйцо было выкопано с глубины 15 см.
Зимующую особь мы выкопали 10.X 1981 г. под кустом кандыма в бугре
с глубины 60 см. Длина зимовочных нор [130] на южных склонах закреплен-
ных песков достигала 40—42 см, глубина — 27—30 см; одна особь зимовала
в норе мохноногого тушканчика. Длина летних нор достигала ПО см, глубина
не превышала 20 см, диаметр 1,5 см [23]. По наблюдениям в Казахстане [98],
норы гребнепалых гекконов просты по устройству и представляют прямой ход,
в конце которого имеется расположенное в зоне увлажненного песка незначи-
тельное расширение, в 30—40 см от поверхности; длина хода не более 70 см,
обычно 40—50 см.
К. П. Параскив [98] неоднократно находил этих гекконов в совместных но-
рах с жуком-навозником и жужелицей. Пользуясь открытым ходом этих жуков,
геккон самостоятельно рыл отнорок от основного хода, длина отнорка не пре-
вышала 20—30 см. На день ход норы или отнорок геккон забивает пробкой из
53
песка длиной до 10 см. Норки могут располагаться недалеко одна от другой:
три жилые норы были обнаружены на площади 1 м2. По наблюдениям А. Д. Бау-
тина (письменное сообщение), гребнепалый геккон при закапывании работает
хвостом: высовывает хвост из норы наружу и начинает с краев сгребать песок
до тех пор, пока норка не будет засыпана, затем хвост втягивается в нору.
В этот момент наиболее рабочей частью хвоста является его кончик. Хвост у
этой ящерицы не ломкий, как у других видов, однако утраченный кончик ре-
генерирует.
В брачный период, как мы установили, геккон издает звуки, голос более
низкий, чем у тонкопалых гекконов, его можно передать как «ек...ек...ек».
При спаривании самец хватает самку за бок или в области передней лапы
(груди) и придерживает при копуляции хвостом. Для кладки яиц самка выка-
пывает ямку и откладывает туда пару яиц с интервалом 5—6 мин, во время ко-
торого отдыхает лежа. Яйца откладываются мягкими, затем быстро твердеют.
Практическое значение, охрана. Судя по характеру пита-
ния, гребнепалый геккон является полезным животным, он занимает определен-
ное место в биоцинозе песчаной пустыни. Поскольку территория пустынь,
где обитают эти ящерицы, довольно обширна, а они там встречаются часто, необ-
ходимость в проведении охранных мероприятий этого вида отсутствует. Греб-
непалый геккон в числе других животных охраняется на территории девяти
заповедников (главнейшие из них Репетекский и Красноводский в Туркмении;
Бадай-Тугай, Каракульский и Кызылкумский в Узбекистане; Тигровая Бал-
ка в Таджикистане).
ГРЕБНЕПАЛЫЙ ГЕККОН ВОСТОЧНЫЙ —
CROSSOBAMON ORIENTALIS (BLANFORD, 1876)
Типовая территория: Рохри и Шикарпур, Верхний Синд (Пакистан).
Кариотип: не изучен.
1876 — Stenodactylus orientalis Blanford, J. asiat. Soc. Bengal, Calcutta (2), vol. 45, p. 21.
1884 — Stenodactylus dunstervillei Murray, Vertebr. Zool. Sind, p. 363.
1967 — Crossobamon orientalis, Kluge, Bull. Amer. Mus. Nat. Hist., Vol. 135, art. l,p. 23.
Диагноз. Хвост короче тела; у самцов пять и менее анальных пор.
Типовой экземпляр хранится в Индийском музее, Калькутта
Описание (рис. 20) (по 15 экз. из колл. UMMZ, AMNH и SAM).
L d ad (п = 9) 29,5—47,8 (41,04 ± 1,67) мм; L $ ad (п = 5) 27,0—56,5
(43,04 ± 5,33) мм; LIL cd (п = 6) 1,18—1,55 (1,31 ± 0,05); индекс высоты/ши-
рины головы (п = 15) 54—64 (53,73 ± 0,81); диаметр уха/глаза (п = 15) 0,45—
0,59 (0,53 ± 0,012); верхнегубных щитков (п = 30) 9—14 (11,97 ± 0,23); ниж-
негубных (п = 30) 9—14 (11,40 ± 0,26); чешуй между центрами глаз (п = 15)
20—26 (22,47 ± 0,48); длина спинного бугорка/L (п = 15) 6,80—12,70 (9,91 ±
± 0,42); чешуй, окружающих спинной бугорок (п = 15), 7—9 (8,47 ± 0,16);
чешуй вдоль тела снизу (п = 14) 185—225 (204,43 ± 3,17); анальных пор (п =
= 7) 2—5 (2,71 ± 0,42); подпальцевых пластинок (п = 29) 24—29 (26,55 ±
± 0,24).
Прижизненная окраска [263]. Основной фон верхней части
тела — от коричневого до палево-серого, с тремя — пятью неясными темно-се-
рыми поперечными полосами; хвост желтоватый, с ясными темными поперечны-
ми широкими полосами. Брюхо белое. У особи из Лас Белы (Пакистан) отмече-
на оригинальная вариация рисунка: темные спинные бугорки расположены
правильными рядами на почти совершенно однородном черном фоне.
Половой диморфизм и возрастная изменчивость.
Самки крупнее самцов. Возрастная изменчивость не изучена.
Распространение. Синд (Пакистан) (рис. 21).
Кадастр к рис. 21.
Пакистан:/ — Меруи, Гаурбенд 28° 57' с. ш., 63° 48' в. д. [2581:2 — Рохри, terra-
typica [258]; 3 — Шилра, ю. Шахдаткота, 27° 45' с. ш., 67° 55' в. д. [258J; 4 — Халла [2581;
5 — 4 км ю. Сехвана (AMNH); 6 — р-н Санхгар, 18 км ю.-з. от Джемрао, песчаные склоны око-
54
ло Джемрао Хэд (AMNH); 7 — р-н Санхгар, 1 км з. Бурра; 8 км ю. Санхгара (AMNH); 8 —Ор-
мара [258]; 9 — р-н Лас Бела, зал. Миянихор (AMNH); 10 — Сонмияни (там же); 11 — Накак-
харари (там же); 12 — р-н Дабу, Тханобулахан (там же); 13 — р-н Татта, 3 км в. Гхаро (там
же); 14 — Татта; Джунгшахи, 16 км сев.-з. от Татты [258]; 15 — Дипло [258].
Экология не изучена.
Род североазиатский геккончик—Alsophylax Fitz, 1843
Типовой вид: Lacerta pipiens Pallas.
1843 — Alsophylax Fitzinger, Syst. Reptilium, Fasc. 1, p. 90.
Диагноз. Ноздря расположена между пятиугольным межчелюстным,
первым верхнегубным, крупным плоским межносовым щитком и часто одним-
двумя мелкими добавочными носовыми чешуйками; верхнегубные щитки нахо-
дятся в одной плоскости и опускаются отвесно к краю рта; высота первого верх-
негубного от ноздри до края рта заметно меньше его ширины вдоль края рта.
Зрачок вертикальный с зазубренными краями. Реснитчатых выростов в верхней
части орбиты нет. Все мелкие чешуи тела гладкие; на спине кроме чешуи могут
располагаться бугорки (иногда ребристые); они могут заходить за основание
хвоста (не далее V3). Хвостовые сегменты сверху неразличимы или различимы
слабо; снизу хвост покрыт продольным рядом пластинок (обычно по две на сег-
менте). Анальных пор не более 13, у § они могут отсутствовать. Пальцы пря-
мые или слабо искривленные, цилиндрические, когтистые, снизу покрыты
рядом гладких пластин; боковая чешуя пальцев бахромы не образует. Мелкие
гекконы, L — до 46 мм.
Дополнительные признаки. Чешуя морды крупная, выпук-
лая, округлая, уменьшается к затылку. До переднего края орбиты доходит
2,5—4 верхнегубных щитка. Ушное отверстие мелкое, округлое. Верхнегорло-
вые чешуйки увеличены; горловая чешуя округлая, слабо выпуклая. Спина
покрыта выпуклой чешуей, увеличивающейся к бокам тела, где плавно перехо-
дит в брюшную. Среди спины может находиться до 12 продольных рядов бугор-
ков. Брюхо покрыто крупной плоской черепитчатой чешуей. Анальные поры
обычно расположены Д-образно. Хвост толстый, цилиндрический, в последней
четверти резко утончается; покрыт кольцами крупных плоских чешуй, увеличи-
вающихся к бокам, регенерированный хвост начинается вздутием и беспорядоч-
но покрыт гладкой чешуей.
Верхняя поверхность конечностей покрыта чешуей, сходной со спинной,
нижняя — чешуей, сходной с брюшной; кроме того, на верхней поверхности
голени имеются бугорки, обычно превосходящие размерами спинные. Четвер-
тый палец задней конечности длиннее остальных, сверху покрыт одним, с бо-
ков — двумя рядами чешуй; основание когтя покрыто чехлом, состоящим из
верхней и нижней пластинок; с чехлом соприкасается по одной чешуйке сверху
и с боков. Подпальцевые пластинки почти одного размера.
Род включает шесть видов, разделяемых на два подрода, распространенных
в Казахстане, Средней и Центральной Азии.
Таблица для определения видов и подвидов рода Alsophylax
1 (4). Увеличенный носовой щиток один, добавочные носовые чешуйки отсутствуют.
2 (3). Спинные бугорки отсутствуют.....................................A. laevis Nik.
3 (2). Спинные бугорки имеются......................................A. pipiens (Pall.)
4 (1). Кроме крупного носового щитка к ноздре примыкают более мелкие дополнительные
носовые чешуйки.
5 (10). Мелкая дополнительная носовая чешуйка одна.
6 (7). Анальных пор более семи....................................А. 1. loricatus Str.
7 (6). Анальных пор менее семи.
8 (9). Спинные бугорки отсутствуют............................ A. tadjikiensis Golubev
9 (8). Спинные бугорки имеются ..................................... A. przewalskii Str.
10 (5). Мелких дополнительных носовых чешуек две.
11 (12). Анальных пор менее семи ..............A. tokobajevi Jeriomtschenko et Szczerbak
12 (11). Анальных пор более семи......................А. 1. szczerbaki Golubev et Sattorov
55
Подрод равнинный геккончик — Alsophylax Fitz., 1843
Типовой вид: A. pipiens
Диагноз: Пальцы прямые; анальные поры имеются у особей обоих
полов; длина передней конечности составляет около 25 % длины тела, достига-
ет концами пальцев до переднего края орбиты (кроме A. przewalski), задняя ко-
нечность, вытянутая вперед, достигает локтевого сустава направленной назад
передней конечности; хвостовые сегменты не различимы; хвостовые бугорки
не опускаются на боковые поверхности хвоста. Подрод включает пять видов.
СЕВЕРОАЗИАТСКИЙ ГЕККОНЧИК ПИСКЛИВЫЙ —
ALSOPHULAX PIPIENS (PALL., 1811)
Типовая территория: гора Большой Бог до (Астраханская область, РСФСР).
Кариотип: 2и=36; акроцентриков—30, субтел оцентриков—4, субметацентриков—2,
NF = 42.
1811 — Lacerta pipiens Pallas, Zoogr. Rosso-Asiatica, Vol. 3, S. 27 (t e г г a t у p ic a: «Berg.
Boedo, Kirgizensteppe»).
1823 — Ascalabotes pipiens, Lichtenstein (in Eversmann), Reise von Orenburg nach Bukhara,
Berlin, S. 145.
1831—Gymnodactylus pipiens, Eichwald, Zoologia specialis, Vilno, Vol. 3, p. 181.
1843 — Alsophylax pipiens, Fitzinger, Syst. Rept., p. 90.
1856 — Gymnodactylus atropunctatus Lichtenstein, Nomenkl. Rept. Amph. Mus. Zool. Berlin,
S. 6 (t e г r a t у p i c a: «Tartarei»).
1885 — Alsophylax microtis, Boulenger, Cat. Lis. Brit. Mus., London, Vol. 1, p. 19 (err. syn.).
Диагноз. Мелкие округлые спинные бугорки (иногда со следами реб-
ристости) расположены более или менее часто, беспорядочно; 8—13 анальных
пор; имеется один крупный межносовой щиток, мелкие добавочные носовые че-
шуйки отсутствуют; поперек туловища четыре — семь темных широких полос;
хвост снизу белый.
Н е о т и п. ЗИН 3600. d ad «Mt. Bogdo magnus, 1872, leg Becker.» L =
= 34,3 мм; хвост регенерирован; из носовых имеются только крупные меж-
носовые, соприкасающиеся между собой; замежносовые щитки выражены и раз-
делены чешуйкой; верхнегубных щитков 9/10, нижнегубных — по шесть;
чешуй поперек головы 13; анальных пор 11; чешуй вдоль средней линии тела
снизу 103; подпальцевых пластинок 18—19. Спинные бугорки расположены
более или менее правильными рядами; каждый такой бугорок в центре спины
окружен восемью-девятью чешуйками.
Описание (рис. 22) (по 690 экз. из колл. ИЗАНУ, ЗИН, МГУ, ИЗАНК,
ИЗАНУЗ, ВПВ, NMW, MNHP, FMNH, USNM и UMMZ). L d ad (п = 230)
25,0—38,0 (31,88 ± 0,21) мм; L g ad (n=269) 25,0—41,6 (33,71 ±0,26) мм; LIL cd
(n = 188) 0,80—0,99 (0,880 ± 0,004); индекс ширины/высоты головы (n =
- 420) 53—74 (62,27 ± 0,18); диаметр уха/глаза (n = 360) 6—29 (18,57 ±
± 0,20); верхнегубных щитков (n = 1305) 5—9 (7,03 ± 0,02); нижнегубных
{n = 1249) 4—7 (5,49 ± 0,02); чешуй между центрами глаз (п = 636) 11—20
(14,76 ± 0,06); длина спинного бугорка/Л (и = 571) 9—16 (12,38 ± 0,07);
чешуй, окружающих спинной бугорок (п = 286), 7—И (8,59 ± 0,05); чешуй
вдоль тела снизу (п = 515) 84—112 (96,54 ± 0,23); анальных пор (и = 646)
8—13 (9,44 ± 0,04); подпальцевых пластин (п = 1148) 13—22 (17,35 ± 0,04).
Масса до 1,5 г. Ноздря находится между межчелюстным, первым верхнегуб-
ным и крупным межносовым щитком; в восточной части ареала у части особей
(до 18 %) может появляться также одна мелкая добавочная носовая чешуйка;
межносовые шитки выражены четко (27,7 %), неясно (29,7 %) или совсем не вы-
ражены (42,6 %). Подбородочный щиток четырех-пятиугольный (74 %), шес-
тиугольный (16,5 %) или трапециевидный (9,4 %). Нижнечелюстных щитков
нет или две пары. Спинные бугорки округлые, гладкие или со следами ребрис-
тости, иногда образуют слабо выраженные ряды.
Прижизненная окраска. Сверху песочно-охристого цвета. От
первого верхнегубного шитка через глаз и значительно выше ушного отверстия
с каждой стороны головы тянется темно-коричневая полоса. Эти полосы могут
сливаться друг с другом на затылке, образуя подковообразный рисунок. Между
56
рКлО'З
Рис. 20. Crossobamon е. lumsdenii:
А — общий вид
С. orientalis:
Б — общий вид, В — голова сверху, Г — спина, Д — палец снизу, Е — палец сверху (А —
ИЗАНУ SR 1450, окр. Нушки, Пакистан; остальное — ИЗАНУ SR 1449, окр. Бурра.
Пакистан)
носовым отверстием и глазом полосы сверху окантованы светлым кантом, а в
промежутке между светлыми кантами на верхней стороне морды от верхней час-
ти межчелюстного щитка до пространства между перед! ими краями орбит или
далее имеется неясный темно-коричневый узор. От затылка до поясницы попе-
рек тела есть четыре—семь темно-коричневых полос, которые значительно шире
57
Рис. 21. Распространение Crossobamon е. lumsdenii (/), С. orientalis (2)
промежутков между ними. В центре спины они могут быть разорванными и сме-
щенными друг относительно друга вдоль продольной оси тела. На хвосте свер-
ху до 11 такого же цвета широких поперечных полос. Конечности сверху также
покрыты неясными поперечными полосами. Брюшная сторона белая.
Половой диморфизм и возрастная изменчивость.
У пискливого геккончика, по нашим данным, половой диморфизм проявляется
в более крупных максимальных и средних размерах самок по сравнению с
самцами; анальные поры у самок развиты слабее, чем у самцов и с возрастом
могут почти полностью исчезать. Возрастной изменчивости подвержены некото-
рые пропорции тела. Молодые особи этого вида более короткохвостые (у
новорожденных длина хвоста меньше длины тела) (рис. 23). Кроме того, у
молодых больше диаметр глаза относительно длины тела. Значения этих
индексов становятся более однородными у половозрелых геккончиков. В связи
с этим мы учитывали значения этих индексов только для взрослых особей.
Распространение (рис. 27, 28). Низовья Волги, Устюрт, Казах-
ский мелкосопочник, Сев. Кызылкум, Бетпак-Дала, Семиречье, Джунгария,
Ю. Гоби и прилежащие районы Китая; граница в юго-восточной части ареала
точнее не установлена.
Кадастр к р и с. 27, 28.
Казахстан и Средняя Азия: / — Астраханская обл., г. Богдо (ЗИН);
2 — Киргизские степи Букеевской Орды [96]; 3 — Оренбургский край, г. Алмас-тау [58, 94];
4 — 4 км з. пос. Эмба [151]; 5 — г. Каражар вблизи Эмбы [90]; 6 — хр. Жильтау, Гурьевская
обл. [65]; 7 — Дангуз-тау (ЗИН); 8 — сев.-з. берег Арала, уроч. Каска-Джол [98]; 9 — о-в
Барсакельмес [16]; 10 — Жаслык [29]; 11 — Кырк-кыз [29]; 12 — Сор-Барсакельмес (ИЗАНУ);
13 — чинки у оз. Судочье (ИЗАНУ); 14 — коса Кыпыллар-кыр, сев.-в. берег оз. Сарыкамыш
[34]; 15 — Куня-Дарья, 15 км з. от Бутентау [23]; 16 — 3. Кызылкум, кол. Чабанказган
(ММГУ); 17 — Сев. Кызылкум, 80 км к сев.-в. от Чабанказгана (ММГУ); 18—Кум-Кала
(ММГУ); 19 — между пос. Ерембет и Тупбугут (ММГУ); 20 — 21 км к з. от Тупбугута (ММГУ);
21 — 50—70 км к ю.-з. от ст. Джусалы (ММГУ); 22 — между Сырдарьей и Жанадарьей (ММГУ);
23 — ст. Джусалы [98]; 24 — низовья р. Сарысу, Кок-Аладжар [98]; 25 — холмы Уч-чоку [61];
26 — р. Иргиз [96]; 27 — кол. Торт-мола, верховья р. Тургай [98]; 28 — Тургай, правый бе-
рег р. Джулыгалы, Джиланчик (Р. Кубыкин, уст. сообщ.); 29 — г. Улытау (ЗИН); 30 — пос.
Джезды (ИЗАНУ); 31 — г. Актау, ю. ст. Бассага (ЗИН); 32 — г. Акчатау (ИЗАНУ); 33 — Бет-
пак-Дала (ЗИН); 34 — Бурубайтал [123]; 35 — уроч. Мын-Арал [96]; 36 — мыс Коражун-Тю-
бек [96]; 37 — о-в Тас-арал [96]; 38 — между реками Или и Каратал [137]; 39 — близ п-ова Шау-
кар [85]; 40 — в. берег оз. Балхаш (ММГУ); 41 — кол. Тумарча [137]; 42 — Голова
58
Рис. 22. Alsophylax tadjikiensis (вверху):
A _ голова сверху, Б — голова сбоку, В — подбородок, Г — брюхо, Д — преанальные поры
(ИЗАНУ SR 942, окр. Сумбула, ТаджССР)
Alsophylax pipiens (внизу):
А — лицевая часть сверху, Б — подбородок. В, Г - спина, Д — преанальные поры (А —
В — ЗИН 3599, Богдо, РСФСР, Г, Д — BNM MG/8, Байн—Дзон, МНР)
Баканаса (ЗИН); 43 — Бартогой у Турайгыра (ИЗАНУ); 44 — Турайгыр у р. Чи-
лик (ИЗАНУ); 45 — правый берег р. Чарын к з. от с. Чунджа (ИЗАНУ); 46 — предгорья
Кетменя, среднее течение р. Пияздык (Р. Кубыкин, устн. сообщ.); 47 — г. Кату-тау, предгорья
Джунгарского Алатау (Р. Кубыкин, устн. сообщ.); 48 — Панфилов (Джаркент) [96]; 49 — Ки-
тай, Харк-Укюр [3]; 50 — среднее течение р. Тахта (Р. Кубыкин, устн. сообщ.); 51 —
Ргайты (он же); 52 — о-в Средний [70]; 53 — о-в Кишкене-Аралтобе [70]; 54 — между р. Эмель-
су и Ргайты [137]; 55 — 60 км к з. от г. Зайсан [151]; 56 — ст. Жангизтобе (Р. Кубыкин, устн.
сообщ.); 57 — окр. Семипалатинска, у моста через р. Мукур (ИЗАНУ); 58 — г. Иман-Кара
(письм. сообщ. В. В. Неручева); 59 — г. Бесбай (письм. сообщ. В. В. Неручева); 60 — плато
59
Ак-Толагай (он же); 61 — плато Ак-Керегеше (колл. Е. Ю. Кудакина); 62 — Кара-Калпакская
АССР, Тахта* Купы реки й р-н, 30 км к сев. от пос. Чабанказган (ИЗАНУ); 63 — КазССР,
Кзыл-Ординская обл., пос. Кекрели; 35 км к сев. от пос. Кекрели (ИЗАНУ); 64 — 40 км к
сев.-з. от пос. Кекрели, развилины крепости Чирик-Робат (ИЗАНУ).
Центральная Азия: МНР: 1 — 5 км сев. г. Уэнч; 2 — В. Джунгария
(18]; 3 — Бур-Нур (BNM); 4 — МНР, Хатан-Хайрхан-ула [78]; 5 — Сомон-Хайрхан-ула
I78J; 6 — Дзун-Мод (BNM); 7 — Улдзийт-гол [78]; 8 — Ногон-Цав [78]; 9 — Яман-ус [78];
10 — хр. Цаган-Богдо (ИЗАНУ); Шара-Хулс [78); 11 — Эхин-Гол [78]; 12 — Нарын-Булак
(ММГУ); 13 — Торын-Худук (BNM); 14 — Хайчин-Ула [78]; 15 — кол. ДжирГаланту-Худук
[78]; 16 — хр. Нэмэгэту [15]; 17 — Мухор (BNM); 18 — Обот-Хурал (BNM); 19 — Ноен-Бог-
до (ММГУ); 20 — 25 км сев. Булган-Сомон (BNM); 21 — Шовон-Хувэр [78?; 22 — Тэвш-Хайр-
хан [78]; 23 — 58 км з.-ю.-з. Баян-Далай (BNM); 24 — Сухайтын-Журэн-Тологой, ю. склон
Золон-Ула [77]; 25 — Дзурамтай [78]; 26 — хр. Хухэ [15].
К и т а й: 27 — округ Нинся [278]; 28 — пров. Ганьсу [118]; 29 — оазис Сачжеу [18];
30— 50 км сев. Севрей-Ула, р-н Дзурган-Худук (ИЗАНУ); не найдены на картах МНР, око-
ло хр. Барун-Баян (ММГУ);МНР, уроч. Ушин-Хундей (ММГУ); Werow (Nerow?) — Bulag
(BNM); Bajn-Dzon (Dzak?) (BNM).
Систематика и географическая изменчивость.
13 выборок, охватывающих весь ареал вида, сравнивались по 21 признаку фо-
лидоза. Можно констатировать наличие очень слабых различий между ящери-
цами, обитающими к западу и востоку от Аральского моря. Западные особи
несколько более мелкие, у них слабее выражены спинные бугорки и упорядо-
ченность их в ряды; существуют незначительные отличия и в средних значениях
количественных признаков. Однако в южной части ареала (оз. Судочье, Сев.
Кызылкум, оз. Балхаш) эти различия нивелируются. Лишь на крайнем востоке
ареала (на территории Монголии) возрастает частота встречаемости таких приз-
наков, как наличие мелкой добавочной носовой чешуйки и слабой ребристости
спинных бугров. Как известно, эти признаки свойственны, в частности, близ-
кому виду — A. przewalski, который обитает в непосредственной близости
с монгольскими" популяциями данного вида.
Стации .й количественные данные. Наблюдения, прове-
денные нами з окр. пос. Эмба [143], в горных массивах Улытау и Акчатау,
в окр. Семипалатинска (Ц. и Сев.-В. Казахстан) и данные ряда авторов [3—5,
15, 22, 23, 29, 34, 35, 55, 70, 77, 98, 112, 118, 125, Неручев, Чхиквадзе —
письм. сообщ.] позволяют дать обобщенную характеристику стаций пискливого
геккончика. Этот вид является типичным обитателем полупустынь, глубоко
проникающим местами в зону степей, где он все же придерживается участков
полупустынного типа. Встречается преимущественно по склонам невысоких
поднятий (до 150—350 м в Средней Азии и Казахстане и 600—1550 м в Джунга-
рии и Гоби), покрытых крупно- и мелкообломочным материалом, в зонах вывет-
ривания с плитовидными обломками, реже — на глинисто-щебнистых равни-
нах. Известны единичные случаи проникновения вида на окраины песков
(в пустыне Гоби; Головушкин, уст. сообщ.). Для всех стаций характерна очень
скудная травянистая растительность, состоящая из боялыча, тасбиюргуна,
терескена, полыни, солянок и низкорослых злаков. Указания ряда авторов на
находки этого вида на стволах саксаула [64, 67, 79, 80, 121] относятся, несом-
ненно, к серому голопалому геккону.
Наши наблюдения, проведенные в 1978 г. в нескольких пунктах Казахско-
го мелкосопочника, показывают, что пискливый геккончик всюду является фо-
новым видом и может достигать весьма высокой численности. У моста через
р. Му кур (В. Казахстан) на участках, наиболее богатых убежищами (разруша-
ющиеся склоны сланцевых сопок), плотность пискливого геккончика достигает
10 особей на 1 м2. Несколько меньшая плотность отмечена в горах Улытау
и Акчатау: соответственно 7 и 3 особи на 1 м2.
Отношение к температуре. Активные геккончики отмечены
при температуре 21—25°.
Суточный цикл активности. В конце июля — первых чис-
лах августа 1979 г. в Казахском мелкосопочнике мы наблюдали выход этого гек-
кончика на поверхность вскоре после захода солнца, около 20 ч 30 мин — 21 ч.
Максимальная численность наблюдалась в течение 1,5—2 ч после выхода. Яще-
рицы были активными до восхода солнца. В светлое время суток они скрываются
в убежищах. Отмечены на поверхности в пасмурную погоду [98].
60
Сезонный никл активности. В Ю. Прибалхашье появляется
на поверхности после зимовки в конце марта — начале апреля. На Ю.-В. Ус-
тюрте геккончики ушли в зимние убежища 3. XI [98].
Линька. В горном массиве Акчатау,, видимо, повторная линька взрос-
лых, а также сеголеток наблюдалась 3—17.VIII 1938 г. [41. В Сарыкамышской
котловине линяющих особей добывали в июне и сентябре [34]. По нашим наблю-
дениям, новорожденные ящерицы линяют через несколько часов после выхода
из яйца.
Питание. Основным кормом пискливого геккончика являются насеко-
мые (прямокрылые, жуки, перепончатокрылые, мухи и их личинки), кроме того,
l/Lccf
^F *
Atoxotejeirt - •
Aprzeuratoa - +
он поедает паукообразных, в том числе
мелких скорпионов [4, 17, 22, 98,
1321. По нашим данным (проанализи-
ровано 25 желудков ящериц, собран-
#?L_
20
-I—4--------1---------1
JO 40 LMM
Рис. 23. Возрастная изменчивость индекса Рис. 24. Размерно-возрастные группы А.
L/Lcd у A. przewalski и A. tokobajevi pipiens
ных в окр. Семипалатинска 10. VIII 1978 г.), по встречаемости преобладали бабоч-
ки (22 %), мухи (14,3 %), перепончатокрылые и муравьи (соответственно 9,1
и 7,8 %), а также пауки (6,5 %). В желудках геккончиков из хр. Ахча-Тау
(п = 25), добытых 19.VIII 1978 г., чаще встречались клопы (29,1 %), муравьи
(20 %), мухи (12,8 %) и пауки (12,8 %). Всего в указанных желудках обнаруже-
но не менее 20 видов беспозвоночных.
Размножение. Половой зрелости геккончики достигают при длине
туловища 25—28 мм. Соотношение полов по коллекционным материалам близ-
ко 1 : 1. Спаривание, судя по наиболее увеличенным гонадам, происходит в кон-
це апреля — мае [701. Готовые к откладке яйца были обнаружены в мае — июне
[15, 34, 123, 137]. Самка откладывает одно-два яйца. Известны следующие даты
откладки яиц: 29.V 1983 г. из окр. Чабанказгана (первая кладка) и 10.VIII
1978 г. из окр. Семипалатинска (вторая кладка; наши данные). 15.VIII 1969 г. в
Гоби найдены яйца с развивающимися эмбрионами длиной 13 мм [77], а 26.VIII
1982 г. найдены20 яиц размером 9—10,5x6—7(9,65 ± 0,52 х 6,82 ± 0,1) мм
(письм. сообщ. Н. Л. Орлова). Размеры яиц из Казахстана (п = 30), 8,0—
11,0Х 5,0—7,0(9,72 ±0,14 X 6,81 ± 0,84) мм. Выход молоди из кладок, най-
денных в окр. Семипалатинска, отмечен нами 16—28.VIII 1978 г. По литера-
турным данным [17], молодые геккончики начинают появляться с июля.
Темп роста. Сеголетки, вышедщие из яиц, достигают длины туловища
16,0—18,0 мм (в этих пределах колебались размеры особей, добытых нами в при-
роде и выведенных из яиц в лабораторных условиях). К концу сезона активности
они достигают размеров (без хвоста) 24 мм, следовательно, особи первой генера-
ции могут принимать участие в размножении уже на следующий год. В популя-
ции данного вида характерно наличие 3—4 возрастных групп (рис. 24), причем
старшая группа включает в себя особей трехлетнего и более возраста.
Враги, причины сокращения численности. Среди
врагов пискливого геккончика отмечена сорока [3]. О. П. Богданов [27] устано-
вил сокращение численности A. pipiens в 600—700 раз за 20 лет — с 1956
по 1976 г., вызванное усыханием Аральского моря и отступлением береговой
61
линии от скал на 5—6 км, которое вызвало (в свою очередь) резкое сокращение
численности беспозвоночных. Эти ящерицы вытесняются из своих биотопов
также при распашке целины.
Особенности поведения. В период активности передвигается
по почве короткими перебежками, высоко приподнимая над субстратом и из-
гибая туловище. Может свободно передвигаться и по вертикальным поверхно-
стям, которых, однако, избегает. Далеко от убежищ не уходит. Охотятся за на-
секомыми у входа щелей и нор, в пространстве под камнями, гоняются друг за
другом. Возбужденный геккончик издает отдельные «металлические» цикающие
звуки, которые можно слышать не только ночью, но и днем, почему их нередко
принимают за птичьи. В гортани пискливого геккончика наиболее хорошо по
сравнению с другими гекконами фауны СССР развит голосовой аппарат. На
небольшом участке щели гортани стенки ее утолщены и выпуклы [98].
Летними убежищами геккончиков данного вида служат пространства под
камнями и плитами, трещины в почве, пустоты прикорневой части саксаула,
норы позвоночных, щели искусственных каменных сооружений [98] (наши на-
блюдения). Зимуют в норах, щелях, дуплах, известен случай зимовки в но-
ре в каменной стене загона для скота [98]. Брачное поведение геккончиков
из-за их скрытого образа жизни до сих пор не изучено.
На выходах сланцев в окр. р. Мукур в конце июля — начале августа, по
нашим данным, пискливые геккончики откладывают яйца в трещинах и пусто-
тах на склонах юго-восточной экспозиции на глубине 5—20 см от поверхности.
Яйца откладываются одиночно и группами, по 5—10 штук от разных самок
в одном убежище. Эти убежища используются для откладки постоянно в тече-
ние нескольких лет. Так, в разрушающемся склоне сопки нам удалось обнару-
жить полость на глубине 13 см площадью 30 X 50 см, в которой скопилось не
менее чем 250—300 остатков скорлупы и неразвившихся яиц, отложенных
в разные годы. Здесь же были и свежие кладки. Н. Л. Орлов (письм. сообщ.)
в Ю. Гоби, в песчаном бугре у основания саксаула, на южной и восточной экс-
позии склона на глубине 30 см обнаружил 20 яиц (коллективная кладка).
Практическое значение и охрана. Пискливый геккончик
является полезным видом, занимающим, как правило, участки, не используе-
мые человеком. Однако здесь имеются исключения: так, в районе типового мес-
тонахождения — на горе Большой Богдо (Астраханская обл.) его местообита-
ния уничтожаются вследствие выработки коренной породы для получения гра-
вия. На этой территории пискливый геккончик нуждается в срочных охранных
мероприятиях: необходимо включить его в Красную книгу РСФСР, поскольку
г. Богдо — единственное местообитание этого вида в Европе. Учитывая неболь-
шой ареал и сокращение местообитаний геккончика в Туркмении, этот вид вклю-
чен в Красную книгу ТССР [104].
СЕВЕРОАЗИАТСКИЙ ГЕККОНЧИК ПРЖЕВАЛЬСКОГО —
ALSOPHYLAX PRZEWALSKU 'STR ., 1887
Типовая территория: Низовье р. Тарим, оазисы Хами и Черчен, р. Черчен
Лобнор.
Кариотип: не изучен.
1875 — Gymnodactylus microtis Blanford, J. Asiat. Soc. Bengal., Calcutta, Vol. 44, part. 2,
p. 193 («homen oblitum», Голубев, 1984).
1885 — «Alsophylax microtis (Blanf.)», Boulenger, Cat. Liz. Brit. Mus., London, Vol. 1, p. 19
(nomen oblitum).
1887 — Alsophylax przewalski Strauch, Mem. Acad. Sci. St. Petersburg, (7), N 2, S. 55.
Диагноз. Крупные овальные слабо ребристые спинные бугорки обра-
зуют продольные и поперечные ряды; пять-шесть анальных пор; кроме круп-
ного межносового щитка имеется одна мелкая добавочная носовая чешуйка;
широкие поперечные полосы на туловище отсутствуют; хвост снизу белый.
Лектотип. ЗИН 5144. d'ad р. Тарим (2500), 1878, Н. М. Пржевальский.
L = 29,2 мм; хвост регенерирован; из носовых щитков имеется крупный меж-
носовой и одна мелкая добавочная носовая чешуйка; межносовые соприкаса-
ются между собой, за ними расположена пара крупных плоских замежносовых
62
щитков, разделенных чешуйкой. Между центрами глаз 11 чешуй. Верхнегуб-
ных щитков по восемь, нижнегубных — по шесть. Вдоль тела снизу 92 чешуй-
ки; 6 анальных пор; подпальцевых пластинок 22—21.
Описание (рис. 26) (по 31 экз. из колл. ЗИН, NMW, MNHP, UMMZ,
USNM и MCZ).
L <? ad (п = 12) 25,4—31,0 (28,85 ± 0,49) мм; L j ad (п = 17) 25,5—33,8
(30,89 ± 0,59) мм; LIL cd (общая для и g) (п = 7) 0,66—0,85 (0,743 ± 0,028);
индекс ширины/высоты головы (п = 18) 63—71 (67,89 ± 0,66); диаметр уха/гла-
за (п = 23) 19—29 (24,83 ± 0,60); длина спинного бугорка/L (п = 29) 17—22
Рис. 25. Возрастная изменчивость индекса L/Lcd у A. pipiens
(18,59 ± 0,28); верхнегубных щитков (п = 60) 7—10 (8,40 ± 0,08); нижнегуб-
ных щитков (п = 60) 5—7 (5,83 ± 0,07); чешуй между центрами глаз (и = 30)
9—13 (11,10 ± 0,15); чешуй, окружающих спинной бугорок (п = 30), 8—11
(9,53 ± 0,15); чешуй вдоль тела снизу (п = 26) 84—98 (92,69 ± 0,74); анальных
пор (п = 30) 5—6 (5,30 ± 0,09); подпальцевых пластинок (и = 54)
16—22 (19,63 ± 0,20).
Ноздря находится между межчелюстным, первым верхнегубным, крупным
межносовым щитками и мелкой добавочной носовой чешуйкой; межносовые
щитки соприкасаются между собой (93,3 %) или разделены чешуйкой; замеж-
носовые щитки выражены четко (19,4 %), слабо (38,7 %) или не выражены
(41,9 %); если они выражены, то соприкасаются между собой (44,4 %) или раз-
делены одной-двумя (по 27,8 %) чешуйками. Подбородочный щиток пятиуголь-
ный, нижнечелюстных две-три пары; первая соприкасается между собой. Круп-
ные овальные слабо ребристые спинные бугорки расположены правильными про-
дольными и поперечными рядами. Щиткование хвоста несколько отличается от
такового у остальных представителей рода; на основании хвоста расположены
крупные овальные слабо ребристые бугорки, плавно исчезающие в первой трети
хвоста и не опускающиеся ниже его верхне-боковой поверхности. Снизу хвос-
та продольный ряд крупных щитков либо отсутствует, либо слабо выражен и
образует сочетания: на одном сегменте ближе к основанию хвоста — 1 = 1 = 1
или 2 = 2 = 2, в середине — 1=2 или 1 = 1, а в конце — только 1 = 1.
Рисунок и окраска (по спиртованным экземплярам). Сверху тело
песочно-охристое. От середины первого — третьего верхнегубного щитков через
глаз, над ухом и затем вдоль всего тела с обеих сторон тянутся темно-бурые,
шириной три — пять чешуек, полосы, которые на основании хвоста переходят
на его боковые поверхности. Каждая такая полоса с верхней стороны на всем
протяжении окантована светлой полоской, в полтора—два раза уже первой.
Эта светлая полоса на участках между межчелюстным щитком и передне-верх-
ним краем орбиты и в последней трети тела вновь окантована более тонкой
63
Рис. 26. Alsophylax przewalskii:
A — общий вид, Б — голова сбоку, В — морда спереди, Г — голова сверху, Д — голова снизу, Е —
спина, Ж — преанальные поры, 3 — хвост снизу, И — лапа сверху (Б — Д — ЗИН 15690, от Бугаса
до Люкчуна, Китай; остальные — ЗИН 7030, р. Черчен, Китай)
темно-бурой полосой, которая, сливаясь у основания хвоста с противополож-
ной, тянется вдоль его середины до конца. На теле и хвосте отсутствуют какие-
либо поперечные полосы. Нижние поверхности белые.
Половой диморфизм и возрастная изменчивость.
По нашим данным у самок по сравнению с самцами анальные поры выражены
слабее; самки несколько крупнее самцов. Для изучения возрастной изменчивости
в нашей выборке количество особей было недостаточным (рис. 23),
Распространение. Кашгария; на востоке доходит до оазиса Хами
(рис. 28).
64
Кадастр к рис. 28.
1 — Чилан-су (ЗИН); 2 — Кашгар [189]; 3 — Яркенд (MCZ); 4 «— Янгигисар [189]; 5 —
Хотан (ЗИН); 6 — Ниядарья (ЗИН); 7 — Кара-Сай (ЗИН); 8 — оазис Черчен (ЗИН); 9 — ни-
зовья р. Черчен (ЗИН); 10 — Низовья Тарима (ЗИН); 11 — оз. Лобнор (ЗИН); 12 — Люк-
чун (NMW); 13 — от Бугаса до Люкчуна (ЗИН); 14 — оазис Хами (ЗИН).
Систематика и географическая изменчивость.
На изученном материале географическая изменчивость не отмечена.
Экология не изучена. Из работ У. Бланфорда [187, 189] следует,
что в Западной Кашгарии геккончик Пржевальского встречается в жилых и за-
брошенных постройках, высота мест обитания данного вида 700—1400 м н. у. м.
Судя по описанию маршрутов коллекторов, ящерицы активны с апреля по ок-
тябрь.
СЕВЕРОАЗИАТСКИЙ ГЕККОНЧИК ПАНЦИРНЫЙ —
ALSOPHYLAX LORICATUS STR., 1887
Типовая территория: Мурзарабат, Моголтау (Сев. Таджикистан, Ферган-
ская долина).
Кариотип: 2п = 36; акроцентриков 30, субтелоцентриков 4, субметацентриков 2,
NF = 42.
1887 — Alsophylax loricatus Strauch, Mem. Acad. Sci. St. Petersb., Ser.~7, Vol. 35, N 2, p. 59.
Диагноз. Крупные овальные или треугольные спинные бугорки с чет-
кими ребрами расположены строгими продольными и поперечными рядами;
8—12 анальных пор; кроме крупного межносового щитка имеется 1—2 мелкие
добавочные носовые чешуйки; широкие поперечные полосы на туловище от-
сутствуют; низ хвоста белый.
Лектотип. ЗИН 4197. <? ad кишл. Мурза-Рабат, 1870, leg. Кушаке-
вич. L = 28,8 мм; L cd = 41,8 мм; L/L cd = 0,69; из носовых щитков есть круп-
ный межносовой и один-два мелкие добавочные носовые чешуйки; позади сопри-
касающихся между собой межносовых щитков расположена пара также сопри-
касающихся замежносовых; между центрами глаз десять чешуек; верхнегуб-
ных щитков семь-восемь; нижнегубных — по пять с каждой стороны; подбо-
родочный щиток пятиугольный; позади него две пары нижнечелюстных щитков,
первая соприкасается между собой; в центре спины вокруг спинного бугорка
13 чешуй; вдоль продольной оси тела снизу 91 чешуйка; анальных пор 10; под-
пальцевых пластинок 18—19.
Описание (рис. 29) (по 229 экз. из колл. ИЗАНУ, ЗИН, ДПИ,
ИЗАНТадж, ИЗАНТ, ИЗАНУз, CAS, UMMZ, USNM и NMW).
Рис. 27. Распространение Alsophylax pipiens в Средней Азии и Казахстане
L ad (п = 70) 22,0—30,8 (26,43 ± 0,28) мм; L g ad (п = 83) 22,3—
32,8 (28,38 ± 0,29) мм; LIL cd (общая для и g ad) (п = 66) 0,64—0,86
(0,755 ± 0,007); индекс ширины/высоты головы (п = 164) 57—75 (65,04 ±
± 0,33); диаметр уха/глаза (п = 108) 17—33 (23,01 ± 0,39); длина спинного
бугра/L (п = ПО) 18—28 (22,77 ± 0,19); верхнегубных щитков (п = 458)
6—9 (7,07 ± 0,03); нижнегубных (п = 458) 3—6 (4,96 ± 0,02); чешуй между
центрами глаз (п = 227) 8—15 (11,22 ± 0,09); чешуй вдоль тела снизу (п =
= 183) 82—101 (90,80 ± 0,30); анальных пор (п = 192) 8—12 (9,52 ± 0,06);
65
подпальпевых пластинок (и = 403) 16—22 (18,46 ± 0,06): чешуй вокруг спин-
ного бугорка (п = 227) 10—15 (12,25 ± 0,08).
Ноздря находится между межчелюстным, первым верхнегубным, крупным
межносовым щитком и одним-двумя мелкими добавочными носовыми чешуйка-
ми; позади соприкасающихся между собой (99,1 %) межносовых щитков
почти всегда (96,6 %) имеется пара соприкасающихся между собой (94,6 %)
замежносовых щитков. Подбородочный щиток пятиугольный; нижнечелюстных
щитков одна-две пары, первая соприкасается между собой. Крупные овальные
или треугольные ребристые спинные бугорки расположены четкими продоль-
ными и поперечными рядами.
Рис. 28. Распространение в Центральной Азии Alsophylax pipiens (/) и A. przewalskii (2)
Прижизненная окраска. Сверху от темно-бурого до светло-
кофейного цвета. От заднего края носовых щитков до передне-верхнего края
орбиты с каждой стороны морды тянется светлая полоса, часто окаймленная
сверху темным; надглазничные чешуйки темные; от заднего края глаза длиной,
равной его диаметру или несколько больше, тянется темная полоса; спина
без рисунка или с семью-восемью неясными узкими темными поперечными по-
лосками; на основании хвоста сверху может быть неясное темное пятно; вдоль
хвоста до 12 неясных темных тонких поперечных полоток; эти полоски посереди-
не хвоста могут сливаться друг с другом, образуя X-образный рисунок, который
на хвосте и теле может частично или полностью исчезать. Нижняя поверхность
тела белая.
Половой диморфизм и возрастная изменчивость.
У самок панцирного геккончика в сравнении с самцами (и с самками других
видов рода) анальные поры выражены в меньшей степени, а с возрастом могут
почти полностью исчезать. Кроме того, максимальные размеры самок превос-
ходят таковые самцов, а в западной части ареала между средними значениями
размеров самцов и самок выявлено достоверное различие (t =3,72). Возраст-
ная изменчивость проявляется в некоторых пропорциях тела: молодые особи
более короткохвостые. Значения этого индекса становятся более однородными
у половозрелых геккончиков, в связи с чем мы учитывали его значения только
для взрослых особей.
Распространение. По современным данным, ареал панцирного
геккончика разорван на две части, отстоящие друг от друга не менее чем на
600 км. На западе ареала он обитает в В. Туркмении вдоль левого берега Аму-
дарьи, на востоке — в Ферганской долине (рис. 30).
€6
Кадастр к рис. 30.
А. 1. szczerbaki (1—7): 1— Куня-Ургенч (ИЗАНУ); 2 — 27 км по дор. Ташауз —
Куня-Ургенч, крепость Гульдумсаз [41]; 3 — Тахтинский р-н, крепость Измыкшир [41]; 4 —
крепость Бедеркенд (ИЗАНУ); 23 км ю.-в. Ташауза [129]; 5 — пос. Тахта (ИЗАНУ); 6 —
г. Хива [41]; 7 — пос. Дарганата (ИЗАНУ).
А. I. 1 о г i с a t u s: 8 — водораздел р. Ангрен и Сырдарьи (ЗИН); 9 — Ленинабад
(ИЗАНУ); 10 — Кок-Курак (ЗИН); 11 - Канибадам (ИЗАНУ); 12 — Кирово [160]; 13 —
Наукат [160]; 14 — Моголтау (ЗИН); 15 — Мирзорабад (ЗИН); 16 — кишл. Курганча [147].
Систематика и географическая изменчивость.
По данным М. Голубева и Т. Саттарова [42], при сравнении западных и восточ-
ных популяций выяснилось, что, несмотря на частичное или полное совпа-
дение крайних значений почти всех признаков, средние их величины с запада на
восток либо уменьшаются (число окружающих спинной бугорок чешуй; отно-
шение длины спинного бугорка к длине тела; число чешуй вдоль тела снизу;
число верхнегубных щитков), либо, наоборот, увеличиваются (LIL cd, число
анальных пор и подпальцевых пластинок). Ряд признаков (L сГ и $>; диаметр
уха/глаза; число нижнегубных щитков) стабилен и географической изменчи-
вости не подвержен. Изменение размеров чешуй и щитков верхней поверхности
тела выражается в уменьшении с востока на запад размеров скуловых и замеж-
носовых щитков, всех чешуй морды, увеличении количества чешуй между цент-
рами глаз, уменьшении размеров спинных бугорков и, как следствие этого,
числа окружающих их чешуй. Однако появление у особей на западе ареала вто-
рой добавочной носовой чешуйки образует хиатус между западными и восточ-
ными популяциями, что дало основание к выделению туркменских экземпля-
ров в особую форму.
Таблица для определения подвидов Alsophylax loricatus
1(2). Ноздря расположена между межчелюстным, первым верхнегубным, крупным межносо-
вым щитком и одной (100 %) мелкой носовой чешуйкой .... А. 1. loricatus Str.
2(1). Ноздря расположена между межчелюстным, первым верхнегубным, крупным межносо-
вым щитком и двумя (94 %, очень редко одной — 6 %) мелкими носовыми чешуйками*..
.................................А. 1. szczerbaki Golubev et Sattorov
ГЕККОНЧИК ПАНЦИРНЫЙ ОБЫКНОВЕННЫЙ
alsophylax loricatus loricatus str.
1979 — Alsophylax loricatus loricatus Голубев, Саттаров, Вести, зоол., Киев, № 5, с. 9 (типо-
вая территория: видовая)..
Диагноз. Ноздря расположена между межчелюстным, первым верхне-
губным, крупным межносовым щитком и одной мелкой добавочной носовой че-
шуйкой; замежносовые щитки четко выражены и всегда соприкасаются между
собой; два — четыре крупных скуловых щитка, равных по размерам межно-
совым.
Описание (по 125 экз. из Ферганской долины). Тип видовой.
L <? ad (п = 25) 22,0—30,08 (27,28 ± 0,52) мм; L % ad (п = 42) 22,5—
32,8 (28,87 ± 0,34); L/L cd (п = 27) 0,64—0,82 (0,718 ± 0,008); диаметр
уха/глаза (п = 34) 13—33 (22,94 ± 0,82); чешуй вокруг спинного бугорка (п =
= 122) 10—15 (13,22 ± 0,09); длина спинного бугорка/L (и = 90) 19—28
(23,18 ± 0,19); чешуй вдоль туловища снизу (п = 87) 84—101 (92,05 ± 0,41);
чешуй между центрами глаз (п = 123) 8—13 (10,44 ± 0,09); верхнегубных щит-
ков (п = 250) 6—9 (7,23 ± 0,04); нижнегубных (п = 250) 3—6 (4,98 ± 0,03);
анальных пор (п = 123) 8—И (9,29 ± 0,06); подпальцевых пластинок (п =
= 243) 16—22 (18,15 ± 0,08). Кроме крупного межносового, имеется только
одна мелкая добавочная носовая чешуйка (98,4 %), межносовые щитки сопри-
касаются между собой (99,2 %), за ними следует пара замежносовых щитков,
также соприкасающихся между собой (98,4 %); между первым верхнегубным
щитком и передне-верхним краем орбиты два — четыре крупных скуловых
щитка, вытянутых в ряд, которые незначительно меньше или равны межносо-
вым щиткам.
Распространение. Ферганская долина.
67
ГЕККОНЧИК ПАНЦИРНЫЙ ЩЕРБАКА —
ALSOPHYLAX LORICATUS SZCZERBAKI GOLUBEV ET SATTAROV, 1979
Типовая территория: В. Туркмения, Куня-Ургенч.
1979 — Alsophylax loricatus szczerbaki Голубев, Саттаров, Вести, зоол., № 9, с. 22.
Диагноз. Ноздря расположена между межчелюстным, первым верхне-
губным, крупным межносовым щитком и двумя мелкими добавочными носовы-
ми чешуйками; замежносовые щитки выражены в различной степени и разделе-
ны между собой одной (89 %) или двумя (11 %) чешуйками; три—шесть мел-
ких скуловых щитков заметно меньше межносовых.
Голотип. № 13. ИЗАНУ. ad Туркмения, Ташаузская обл. г. Куня-
Ургенч, 26.VI 1976 г., leg. Н. Щербак и М. Голубев. L = 25,0 мм; L cd =
= 32,0 мм; LIL cd = 0,78. Кроме крупного межносового щитка, есть две мел-
кие добавочные носовые чешуйки; за соприкасающимися между собой межносо-
выми щитками расположена пара разделенных одной чешуйкой слабо выражен-
ных замежносовых; между центрами глаз 13 чешуй; верхнегубных щитков по 7;
нижнегубных — по 6; две пары нижнечелюстных щитков; вокруг спинного бу-
горка 11 чешуй; вдоль тела снизу 89 чешуй; анальных пор 9; подпальцевых
пластинок по 18.
Описание (по 104 экз. из В. Туркмении).
L d ad (п = 40) 22,0—29,3 (25,89 ± 0,49) мм; L $ ad (п = 37) 22,3—32,4
(27,89 ± 0,31) мм; LIL cd (п = 32) 0,70—0,95 (0,801 ± 0,009) (различие с вос-
точным подвидом недостоверно — t = 2,50); диаметр уха/глаза (и = 30) 17—33
(23,21 ± 0,43; t = 0,18); чешуй вокруг спинного бугорка (п = 98) 10—13
(26,00 ± 0,075; t = 10,6); длина спинного бугорка/L (п = 50) 14—30 (19,68 ±
± 0,34; t = 8,99); чешуй вдоль тела снизу (п = 78) 82—100 (89,52 ± 0,42;
t = 3,83); чешуй между центрами глаз (п = 99) 10—15 (12,14 ± 0,12; t =
= 7,38); верхнегубных щитков (п = 208) 6—8 (6,78 ± 0,04; t = 4,84); нижне-
губных (и = 208) 3—6 (4,95 ± 0,03; t = 0,20); анальных пор (п = 101) 8—12
(9,96 ± 0,09; t = 4,20); подпальцевых пластинок (п = 203) 16—21 (18,67 ±
ч- 0,08; t = 1,45). Кроме крупных межносовых, с ноздрей соприкасаются
2 (94,2 %), редко — одна (5,8 %), мелкие добавочные носовые чешуйки; меж-
носовые щитки соприкасаются между собой (99,5 %); за ними следует пара за-
межносовых щитков, выраженных четко (52 %) или неясно (42,2 %), иногда
они отсутствуют (5,8 %); между первым верхнегубным щитком и передне-
верхним краем орбиты три — шесть мелких скуловых щитков, заметно меньше
межносовых.
Распространение. Левобережье низовьев Амударьи, Сев.-В.
Туркмения.
Стации и количественные данные (рис. 31). В Ферган-
ской долине добыт в трещинах и руинах старых построек [105, 108]. В В. Турк-
мении найден на небольшом глинисто-солончаковом участке на берегах старого
арыка между полями хлопчатника [129]. Все известные места обитания этого
вида связаны с деятельностью человека, а располагаются они в непосредствен-
ной близости от глинисто-солончаковых такыров с зарослями тамарикса, верб-
люжьей колючки, дерезы и древовидной солянки. Многочисленные поиски этого
вида в естественных биотопах, проведенные нами, не увенчались успехом.
Видимо, они там исчезли в связи с многолетним освоением земель. Все находки
панцирных геккончиков известны из древних оазисов, местообитания их рас-
положены в пределах 200—400 м н. у. м. По нашим наблюдениям, в соответст-
вующих биотопах геккончики данного вида обычны и даже многочисленны.
26—27. VI 1976 г. у подножия стены старой крепости из необожженного лессо-
вого кирпича западной экспозиции (самая высокая численность, на северной
стороне они отсутствовали) длиной 80 м добыто 70 особей. Отдельные геккончи-
ки встречались здесь на расстоянии 30—40 см друг от друга. Здесь же 6.VI
1979 г. отмечено 52 особи. Судя по обилию звуков, отмеченных 18—20. VI 1979 г.
в окр. Канибадама, эта ящерица здесь также весьма многочисленна. С. Шам-
маков [129] на берегу арыка под с. Дарган-Ата длиной 150 м добыл 34 геккон-
чика.
68
Рис. 29. Alsophylax laevis (вверху):
A — голова сбоку, Б — носовые щитки, В, Г — подбородок, Д, Е — спина, Ж. —
преанальные поры (Г — ИЗАНУ SR 1438, Кызылкум; остальные — ИЗАНУ SR 2618,
Мешхед-и-Мисриан, ТССР)
Alsophylax loricatus (внизу):
А — голова сбоку, Б — лицевая часть сверху, В — подбородок, Г, Д — лицевая часть
сбоку, Е, Ж — спина, 3 — преанальные поры (В, Д, Ж — ИЗАНУ SR 2725, Хива,
УзССР —А. 1. szczerbaki; остальное — ИЗАНУ SR 940, Канибадам, ТаджССР— А. 1.
loricatus)
Широко распространенное раннее мнение [23, 112, 121, 122] о том, что пан-
цирный геккончик является редким, следует отнести за счет скрытного образа
жизни и небольшого, разорванного ареала.
Отношение к температуре. По наблюдениям С. Шаммакова
[129, 130], в мае геккончики были активными при температуре воздуха 21—24°.
69
В Куня-Ургенче в конце июня 1976 г. и начале июня 1979 г. активные ящерицы
этого вида отмечены в 23 ч при температуре воздуха 21—26°, почвы 23—29°.
Понижение температуры воздуха до 17° (Канибадам, конец июня 1978 г.) не
влияло на активность ящериц. По наблюдениям Т. Саттарова (устн. сообщение),
в Ферганской долине впервые после зимовки панцирные геккончики появляются
при температуре воздуха 18—20°, летом они активны при 30—35°, а при выше
37° уходят в убежища.
Таблица 1. Содержимое желудков панцирного геккончика
(Куня-Ургенч, 3. Туркмения, 7.VI 1979 г., п — 40)
Объект питания Стадия Абсолютное чис- ло встреч Число встреч, % Общее число экземпляров
Isopoda 2 6,7 4
Arachnida
Pseudoscorpionidae Imago 3 10,0 4
Fam. gen sp. » 1 3,3 1
Phalangidae 9 30,0 12
Arachne idae » 1 3,3 2
Ixodidae > 1 3,3 3
Insecta
Orthoptera » 3 10,0 7
Copeognatha » 1 3,3 1
Homoptera
Cicadellidae » 3 10,0 6
Psyllidae » 2 6,7 6
Homoptera » 2 6,7 2
Hemiptera » 4 13,3 6
Coleoptera
Histeridae I 3,3 1
Elatheridae » 1 3,3 1
Cocci nel J idae » 1 3,3 1
Ch г у some 1 idae » 1 3,3 1
Curculionidae » 1 23,3 51
Coleoptera Imago 3 10,0 3
Coleoptera Larvae 1 3,3 1
Lepidoptera Imago 5 16,7 7
Lepidoptera Larvae 1 3,3 1
Hymenoptera
Formicidae Imago 17 56,7 103
Hymenoptera 6 20,0 6
Diptera 1 3,3 2
Reptilia, линная шкурка 1 3,3 1
A. loricatus
Всего 78 233
Суточный цикл активности. Мы отметили первых особей
в Куня-Ургенче 26.VI 1976 г. и 6.VI 1979 г. в 21 ч. Здесь они были активны
всю ночь до рассвета. Наибольшее количество геккончиков отмечено около
23 ч. По данным Т. Саттарова (письм. сообщ.), в Сев. Таджикистане эти яще-
рицы в мае— июне и сентябре—октябре переходят на двувершинный пик
активности: они отмечены в первую половину дня (до 12—14 ч.) и во вторую
(с 18—21 ч). Эти наблюдения проведены в жилых постройках г. Канибадам,
которые изобилуют укромными затененными участками, где создаются днем
условия, сходные с сумеречными.
Сезонный цикл активности. Т. Саттаров (письм. сообщ.
отметил выход геккончиков из зимовочных убежищ в Канибадаме 10—
11.111 1977 г., массовый выход — 7—12.IV 1978 г. По нашим данным, массо-
вый выход этих ящериц в окр. Ленинабада происходил 25.III 1979 г., в это
время преобладают молодые особи. На зимовку в Ферганской долине панцир-
ные геккончики уходят в конце октября..
Линька. Среди 49 геккончиков, добытых в В. Туркмении в конце
1972 г., две особи линяли [1291. Т. Саттаров встречал линных геккончиков
70
в Сев. Таджикистане в мае и августе 1977—1978 г. У особей из Куня-Ургенча
отмечена линька 3.V и 27.V 1983 г.)
Питание. По данным С. Шаммакова [129, 130], панцирные геккончик и
питаются мелкими пауками (58,3 % встречаемости) и муравьями (33,3 %),
редко встречаются щелкуны, нарывники, чернотелки, листоеды, двукрылые
и долгоносики (анализировано 53 желудка). По нашим данным из Куня-Ургенча
(вскрыто 40 желудков геккончиков, добытых 7.VI 1979 г.), первое место по
встречаемости занимают муравьи (56,7 %), мелкие фаланги (30 %), долгоносики
Таблица 2. Содержимое желудков панцирного геккончика
(ТаджССР, окр. Канибадама; Т. Саттаров, п = 25)
Объект питания Стадия Абсолютное чис- ло встреч Число встреч, % Общее число ’ экземпляров
Crustacea isopoda Imago 3 12 10
Arachnoidea Aranei » 2 8 5
Insecta Orthoptera » 1 4 1
Acrid idae » 1 4 1
Aphidodea » 3 12 10
Coleoptera Elateridae » 2 8 6
Carabidae » 1 4 3
Curcu lion idae » 1 4 3
Neuroptera Chrvsopidae » 1 4 3
Hymenoptera Formicidae » 19 76 152
Hvmenoptera Imago 2 8 4
Hymenoptera Larvae 1 4 3
Psocoptera Imago I 4 4
Гастролиты 2 8 2
Всего 40 207
(23,3 %), перепончатокрылые (20,0 %), бабочки (16,7 %), пауки, прямокрылые,
цикады, клопы встречаются реже (табл. 1). Анализ желудков геккончиков из
Ферганской долины (Саттаров, письм. сообщ.) показал, что и здесь в их питании
преобладают муравьи (76 %), мокрицы (12 %) и тли (12 %), однако рацион
менее разнообразен (табл. 2). В террариуме геккончики охотно поедали тлей.
Размножение. Соотношение самок и самцов у данного вида, по све-
дениям из В. Туркмении [130], 1,0 : 1,4. Половозрелость отмечена при мини-
мальной длине туловища 22 мм. В Сев. Таджикистане, по наблюдениям Т. Сат-
тарова, геккончики приступают к размножению в апреле — начале мая. В кон-
це июня, июле и первой половине августа они откладывают одну-две кладки по
одному-два яйца размером 8,6—8,8 X 5,5—6,1 мм (п = 5). В середине августа
начинается вылупление из яиц молоди. Инкубация яиц, по нашим наблюдени-
ям, длится 42 дня. Длина туловища геккончиков, только что вышедших из яиц,
15—17 мм.
Панцирные геккончики (и = 20), просмотренные в Куня-Ургенче 14.IV
1983 г., еще не спаривались. У трех самок, добытых здесь в конце июня 1979 г.,
найдено по одному сформированному яйцу, спустя три года здесь же добытая
самка 28.VI отложила два яйца.
Темп роста. Сведения в литературе отсутствуют. Как указывалось,
геккончики, появившиеся на свет, достигают длины туловища 15—17 мм.
Т. Саттаров сообщил нам, что в октябре встречаются сеголетки с размером ту-
ловища 18—19 мм, после зимовки (в апреле) 20—21 мм, в мае — июне — 23—
25 мм, а в июле — августе — 27—29 мм. Предполагают, что особи, рано вышед-
шие из яиц, уже весной следующего года участвуют в размножении. При этом
бывают исключения: в выборке из окр. Ленинабада, добытой в конце марта
71
1979 г. (n = 42), встречены молодые особи размером 15—19 мм, видимо, из
поздних генераций.
Враги. В Ферганской долине этих ящериц поедают узорчатый полоз,
сорока и домашняя кошка (Саттаров, письм. сообщ.). В желудках двух попереч-
нополосатых полозов и шести домовых сычей, добытых в местах обитания гек-
кончиков в Куня-Ургенче, ящериц данного вида не обнаружено.
Резкое сокращение численности панцирных геккончиков происходит из-за
хозяйственного освоения их биотопов. Так, в тех местах, где С. Шаммаков [129]
отметил высокую численность геккончиков, в 1976 г. мы их уже не нашли, так
как берега арыков недавно были перекопаны и подсыпаны за счет грунта, вы-
нутого со дна.
Особенности поведения. Перемещается короткими быстрыми
перебежками, предпочитает горизонтальные поверхности, но может лазать по
вертикальным стенкам; по кирпичной кладке поднимается на высоту до 0,5 м.
Может вылезать в поисках пищи на травянистые растения (наблюдения в тер-
рариуме). Летними убежищами (по наблюдениям в Куня-Ургенче) служат про-
странства под грудами глины или щели в стене между лессовыми кирпичами
на глубине 5—20 см. В Сев. Таджикистане они найдены в трещинах почвы у ос-
нования дувалов и других старых глинобитных построек (в таких условиях об-
наружена и старая скорлупа яйца), в кучах бытового мусора и старых стеблей
хлопчатника. В одном убежище встречалось до трех особей. Зимовочные убе-
жища расположены в тех же местах, но более глубже (20—70 см).
Весной (в брачный период) особенно часто можно слышать характерный
голос этой ящерицы. Его можно передать словами «чек...ниг». Возбужденный
геккончик может издавать и другие звуки, передаваемые как «цок...цок...цок»,
напоминающие постукивание металлических палочек.
Практическое значение и охрана. Существование пан-
цирного геккончика как вида целиком зависит от человека. Эта ящерица пред-
ставляет большой интерес для науки как своеобразный компонент — реликт
исчезнувших теперь глинисто-солончаковых биотопов. Ареал данного вида,
и без того малый (он известен всего из нескольких точек), постоянно сокращает-
ся, поскольку старые постройки в местах обитания геккончика, имеющие ис-
торическую ценность, активно реставрируются, а старые глинобитные бытовые
помещения заменяются кирпичными и бетонными. Вид включен во второе из-
дание Красной книги СССР и это дает основание объявить заповедным хотя бы
по одному участку его угодий величиной до 1 га (сделать микрозаповедник)
в Куня-Ургенче и под Ленинабадом или Канибадамом, поскольку панцир-
ный геккончик не обитает ни в одном из существующих заповедников.
СЕВЕРОАЗИАТСКИЙ ГЕККОНЧИК ГЛАДКИЙ —
ALSOPHYLAX LAEVIS NIK., 1907
Типовая территория: кишлак Карры-Бент (Ю. Туркмения).
Кариотип: 2п = 36; акроцентриков 28, телоцентриков 2, субтелоцентриков 4,
метацентриков 2, субметацентриков 2, NF = 44.
1907 (1905) — Alsophylax laevis Никольский, Ежег. Зоол. Муз., т. 10, № 3—4, с. 333.
1934 — Alsophylax microtis Чернов, Тр. совета по изуч. производит, сил. Л., вып. 6, с. 257
(part., err. syn.).
1949 — Alsophylax pipiens Терентьев,Чернов, Определитель пресмыкающихся и земноводных.
М., Сов. наука, с. 130 (part.).
1964 — Alsophylax kashkarovi Андрушко, Миккау. Вести. ЛГУ, № 9, сер. биол., вып. 2, с. 14.
Диагноз. Спинные бугорки отсутствуют; 9—11 анальных пор; имеется
единственный крупный межносовой щиток, мелкие добавочные носовые чешуй-
ки отсутствуют; поперек туловища четыре — семь темных широких полос; хвост
снизу белый.
Голотип утерян, типовая территория залита водами Тедженского во-
дохранилища.
Н е о т и п. SR N 314 ИЗАНУ, gad, Туркмения, Тедженский р-н, ст. Та-
кыр, 10.V 1976 г. Колл. М. Л. Голубев, Н. Н. Щербак, L = 33,6 мм; L cd =
= 35,0 мм; верхнегубных щитков по 8; нижнегубных 5—6; чешуй поперек голо-
72
вы 14; вдоль брюха снизу 93; анальных пор 9; подпальцевых пластинок 16—15;
межносовые разделены одной чешуйкой; замежносовые отсутствуют; нижне-
челюстных нет, к трапециевидному подбородочному примыкает 5 горловых
чешуек, из которых угловые несколько крупнее остальных.
Неотип происходит из ближайшего ныне к типовому известного местооби-
тания этого вида.
Описание (рис. 29) (по 290 экз. из колл. ИЗАНУ, ЗИН, ИЗАНТ).
L ad (п = 98) 25,0—35,4 (31,07 zb 0,24) мм; L 2 ad (п = 114) 25,0—37,8
55' 70'
Рис. 30. Распространение Alsophylax laevis (/), A. tadjikiensis (2), A. loricatus (3)
(33,12 ± 0,33) мм; L/L cd (n = 66) 0,82—1,00 (0,900 ± 0,001); индекс ширины/
высоты головы (п = 231) 58—72 (62,73 zb 0,22); диаметр уха/глаза (и = 169)
13—26 (19,54 ± 0,23); верхнегубных щитков (п = 578) 5—9 (7,05 ± 0,02);
нижнегубных (п = 579) 4—7 (5,76 zb 0,03); чешуй между центрами глаз
(п = 290) 12—19 (15,08 ± 0,08); чешуй вдоль тела снизу (п — 90) 87—105
(95,07 ± 0,42); анальных пор (и = 158) 8—13 (10,37 ± 0,08); подпальцевых
пластинок (и = 556) 12—19 (15,46 zb 0,05). Ноздря находится между меж-
челюстным, первым верхнегубным и крупным межносовым щитками (до 7 %
особей могут иметь одну мелкую дополнительную носовую чешуйку); межносо-
вые щитки соприкасаются между собой или разделены одной — тремя чешуйка-
ми; позади межносовых могут находиться выраженные в различной степени за-
межносовые щитки; подбородочный щиток от пяти- или треугольной формы до
трапециевидной; нижнечелюстных щитков 0—2 пары; спинные бугорки отсут-
ствуют, однако у некоторых особей из разных выборок в области крестца
имеются слабо выраженные бугорки, иногда заходящие в поясничную область
и очень редко — на спину.
Прижизненная окраска. Сверху песочно-охристого цвета.
От первого верхнегубного щитка с каждой стороны головы через глаз и значи-
тельно выше ушного отверстия тянется широкая (в две-три чешуйки шириной)
темно-коричневая полоса. Эти полосы на затылке могут сливаться друг с дру-
гом, образуя рисунок в виде подковы. Между носовым отверстием и глазом по-
лосы сверху окантованы более светлым, а в промежутке между светлыми кан-
тами, на верхней поверхности морды, от верхней части межчелюстного щитка
до пространства между передними краями орбит, выражен неясный темно-ко-
ричневый узор. От затылка до поясницы поперек тела идут четыре — семь тем-
73
но-коричневые полосы разной ширины (как правило, шире промежутков между
ними). Эти полосы в центре спины могут быть разорванными и смещенными друг
относительно друга вдоль продольной оси тела. На верхней поверхности хвоста
имеется до 11 такого же цвета широких поперечных полос. Верхние поверхности
конечностей также покрыты неясными поперечными полосками. Тело снизу
белое.
Половой диморфизм и возрастная изменчивость.
По нашим данным, половой диморфизм у гладкого геккончика проявляется
в более крупных максимальных и средних размерах самок и более слабом,
чем у самцов, проявлении преанальных пор, которые с возрастом могут почти
полностью исчезать.
Возрастной изменчивости подвержены некоторые пропорции тела. Как
видно из рис. 33, молодые особи более короткохвостые (у новорожденных длина
хвоста меньше длины тела). Кроме того, для молодых особей характерен боль-
ший диаметр глаза по сравнению с длиной тела. Значения этих индексов ста-
новятся более однородными у половозрелых геккончиков, в связи с чем мы учи-
тывали их значения только для половозрелых особей.
Распространение (рис. 30). 3. и Ю. Туркмения, Ц. и Ю. Узбе-
кистан. Вероятно, может быть обнаружен в Сев. Иране и Сев. Афганистане.
Кадастр к рис. 30 (по нашим данным, [36]).
1 — Мешхед-и-Мисриан; 2 — такыр между хр. Кюрен-Даг и Малый Балхан; 3— 4 км ю.-з.
Кизыл-Арвата; 4 — 4 км сев.-з. Кизыл-Арвата; 5 — 10 км сев.-з. Кизыл-Арвата; 6 — 30 км
сев. Кизыл-Арвата; 7 — 40 км сев. Кизыл-Арвата; 8 — 8 км. сев.-з. ст. Вами; 9 — окр. ст.
Бахарден; 10 — ci. Такыр; 11 — 33 км з. Теджена, /2—24 км от Теджена на Серахс;
13 — долина р. Теджен; 14 — 22 км от Серахса на Теджен; 15 — пос. Шурук, ю. шлейф Куль-
джуктау; 16 — 30 км з. Аякагытмы; 17 — Аякагытма; 18 — впадина Каракатта; 19 — впадина
Мынбулак; 20 — Карасу у Термеза.
Систематика и географическая изменчивость.
Гладкий геккончик долгое время идентифицировался с близкородственным
пискливым геккончиком [98, 112, 121, и др.]. После того как валидность дан-
ного вида была подтверждена [20], из него выделили самостоятельный вид —
A. kashkarowi [6, 9]. С. Шаммаков и Ч. Атаев [131] отрицали существование
этого вида, не приводя, однако, доводов в пользу своей точки зрения. Нами [361
изучено около 300 особей гладкого геккончика из различных частей ареала
и сделан вывод о невалидности формы A. kashkarowi. Как выяснилось, ряд ме-
ристических и пластических признаков этого вида подвержен клинальной из-
менчивости. Так, с юго-востока на северо-запад заметны «огрубление» и увели-
чение размеров чешуй морды, увеличение размеров интерназальных и постин-
терназальных щитков, повышение частоты встречаемости их соприкасания
между собой (линия Мешхед — ст. Такыр является изофеной клины последнего
признака), некоторое увеличение абсолютных размеров тела и количества
анальных пор. В этом же направлении на каждые 100 км около 15 % геккончи-
ков теряют трапециевидную форму подбородочного щитка и заметно выражен-
ную пару нижнечелюстных щитков.
Несмотря на то что кызылкумские особи имеют ряд особенностей, в част-
ности более низкое среднее значение числа подпальцевых пластинок и охристо-
коричневые (вместо темно-коричневых) полосы на верхней стороне тела, хвоста
и конечностей, выделение их в качестве самостоятельной формы в настоящее
время нецелесообразно.
Стации и количественные данные (рис. 32). Гладкий гек-
кончик — типичный обитатель такыров, т. ё. голых, почти без растительности,
плоских глинистых поверхностей в зоне пустынь. На территории Туркмении мы
добывали этот вид на гладком и растрескавшемся такыре с редкой травянистой
растительностью из эфемерных злаков и сухих солянок, реже на такыровидных
осолоненных участках, среди кочек от отмерших кустарников саксаула и соля-
нок на границе глинистой и песчаной пустынь (на Мессерианском плато, в окр.
хр. Малый Балхан, у г. Кизыл-Арват и ст. Вами, ст. Такыр). В Узбекистане мы
наблюдали геккончиков данного вида на осолоненном такыре с редкими кусти-
ками древовидной солянки, участках с растительностью и лишенных ее (окр.
74
Рис. 31. Биотоя A. loricatus и Tenuidactylus caspius (руины крепостного вала, г. Ку-
ня-Ургенч, ТССР)
с. Аякагытма, юго-западнее хр. Кульджуктау). Осолоненность такыра в Кызыл-
куме не влияет на численность этих ящериц. Иное дело в 3. Туркмении. На
Мессерианском плато 5.V 1984 г. мы раскапывали геккончиков из убежищ вско-
ре после дождя. Осолоненный такыр приобрел темный цвет, а чистоглинистые
участки, которые не впитывают влагу, более светлые. Их пятна были беспоря-
дочно вкраплены ’в массив осолоненного такыра. Результаты работы быстро
подсказали нам, что вести раскопки необходимо только на глинистых участках,
так как на осолоненных, рядом, ящерицы отсутствовали. Это объясняется, ви-
димо, тем, что западнотуркменские такыры весной часто заливаются дождевой
водой, а она не проникает сквозь слой глины даже толщиной 5 см. Осолоненные
почвы легко проницаемы для воды и долго ее удерживают. Места обитания глад-
ких геккончиков расположены на абсолютных высотах 200—250 м н. у. м.
По данным количественного учета с помощью автомобильных фар на Мес-
серианском плато 2.V 1975 г. учтено на 16 км маршрута 13 особей, максималь-
ная численность — до 3 особей — отмечена на участке в 10 м2; в этом же районе
12.V 1975 г. на 12 км маршрута добыто 14 особей, a 7.V 1983 г. на 17 км маршру-
та и полосе осмотра 5 м наблюдалось 25 особей. В эту же ночь обнаружена самая
высокая численность на участке площадью 100 X 25 м — 5 особей, т. е. 1 особь
на 500 м2. В окр. руин Мешхеда-и-Мессериана 5.V 1984 г. встречались участки,
на которых были найдены ящерицы на расстоянии 5—6 м друг от друга. Здесь
зарегистрирована самая высокая в Туркмении численность гладкого геккончи-
ка. В других местах нам попадались только единичные особи, хотя, по лите-
ратурным данным [25, 131], ящерицы данного вида недавно были многочислен-
ны в предгорьях Копетдага и Малого Балхана, в долине Теджена, у ст. Такыр.
Об упорных поисках гладких геккончиков может свидетельствовать то, что нами
за 12 экскурсий в последние годы тщательно осмотрена территория биотопов
этой ящерицы площадью в 930 000 м2. Они не были обнаружены на такырах
долины р. Теджен и окр. Хаусхана, г. Теджена, в междуречьеМеаны и Чаачи,
ст. Такыр, между Компрессорной и Уч-Депе, между Арчманом и Бахарденом,
25 км в. Казанджика, между Кизыл-Арватом и Казанджиком, у Бутентау
(Заунгузские Каракумы), у ст. Бами (последний раз здесь был добыт геккон-
чик 8.V 1975 г.), на многих участках в Ю. и Ц. Каракумах.
75
В Узбекистане в окр. с. Аякагытмы 14.VI 1976 г. отмечена самая высокая
численность — пять особей на 3 км маршрута, т. е. одна особь на 600 м. 25—
29.VI 1979 г. большая плотность популяции этого вида обнаружена в окр. хр.
Кульджуктау: на участке 500 X 200 м отмечались ящерицы через каждые 5—
8 м, т. е. здесь обитало их не менее чем 1600 особей. 18.VI 1976 г. гладких гек-
кончиков не обнаружили на такырах Мынбулакской впадины.
Все сказанное выше показывает, что в настоящее время отмечены прогрес-
сирующее снижение численности и сокращение ареала данного вида.
Таблица 3. Содержимое желудков гладкого геккончика
(3. Туркмения, Мессерианское плато, май 1975 г., п = 30)
Объект питания Стадия Абсолютное число встреч Число встреч, % Общее число экземпляров
Crustacea
Isopoda Imago 8 22,2 12
Arachnida — — —
Pseudoscorpionidae — — —
Chelifer sp. » 3 8,3
Arachnida > 4 11,1 3
Insect a 6
Mantoidea » 1 2,8 1
Homoptera » 1 2,8 1
Hemiptera » 2 5,6 3
Coleoptera
Staph ilinidae 1 2,8 1
Elatendae I 2,8 1
Scarabaeidae » 1 2,8 1
Hymenoptera
Formicidae » 5 13,9 10
Lepidoptera Larvae 1 2,8 1
Diptera Imago 1 2,8 1
Insecta sp 5 13,9 5
Всего 36 48
Отношение к температуре. ВЗ. Туркмении активные геккон-
чики в мае (1975, 1983, 1984) наблюдались на поверхности при температуре воз-
духа 21°, почвы 18°, при температуре воздуха 18° они отсутствовали. 26.VI
1979 г. в окр. хр. Кульджуктау температура почвы к 22—23 ч упала до 15° и ак-
тивность геккончиков резко снизилась, хотя отдельные редкие попискивания
их еще продолжались до 2—3 ч ночи.
Суточный цикл активности. По нашим наблюдениям в
3. Туркмении 2.V 1975 г., геккончики появились около 22 ч, наибольшее их
количество встречено в 23 ч, а исчезли они в 24 ч. Там же 12.V 1975 г. первая
ящерица встречена в 23 ч 30 мин; особи были активны до 2 ч 30 мин. 7.V 1983 г.
они обнаружены с 22 ч 30 мин, самая высокая численность отмечена в 1 ч 45 мин,
а активные геккончики (экскурсия была закончена в 2 ч 10 мин), судя по голо-
сам, были до рассвета.
В Кызылкуме 14. VI 1976 г был добыт первый геккончик в 21 ч; 26.VI 1979 г.
они были отмечены вскоре после захода солнца (первых можно было наблюдать
без фонаря) в 20 ч 30 мин, а массовый выход — в течение получаса после этого.
В течение 2—3 ч после выхода у геккончиков наблюдался максимальный пик
активности, который спадал к 23—1 ч. При благоприятных условиях отдельные
особи находились на поверхности в неактивном состоянии до рассвета — 4—
4 ч 30 мин. Суммируя эти наблюдения, можно заключить, что геккончики появ-
ляются из убежищ в сумерках с 20 ч 30 мин — 22 ч, пик их активности отме-
чен спустя 2—3 ч после выхода на поверхность, исчезают они в массе после 24—
1 ч, хотя отдельные особи могут оставаться на поверхности до рассвета. В ве-
сеннее время период активности этих ящериц несколько сокращен и длит-
ся 2—3 ч.
Сезонный цикл активности. Известна первая находка гек-
кончика с наполненным желудком — 10.III 1968 г. [25]. Как правило, они по-
76
являются позже, в начале апреля. С. Шаммаков [1301 встречал их впервые 6
и 7.IV 1970 г., в это время в 1971 г. они еще были на зимовке. По его данным,
последних активных геккончиков добывали 21.X 1967 г. и 20.X 1968 г., а пол-
ный уход их на зимнюю спячку предполагается в начале ноября. 1.Х 1981 г. на
Мессерианском плато, по нашим наблюдениям, геккончики уже не встре-
чались.
Л и н ъ к а. С. Шаммаков [130] линных особей отмечал в мае, сентябре и
октябре, а в желудках геккончиков обнаружены шкурки в мае, июле и сентяб-
ре. Мы отмечали массовую линьку геккончиков в Ю. Кызылкуме в конце
июня 1979 г., в Туркмении — 10.VIII 1983 г. Свежая линная шкурка этого ви-
да найдена в термитнике у Мешхед-и-Мессериана 28.IX 1981 г. Новорожденные
геккончики линяют через несколько часов после выхода из яйца. По-видимому,
они линяют не менее четырех раз за сезон.
Питание. Наиболее полные сведения о питании данного вида приведе-
ны в работе С. Шаммакова [1301. Автор проанализировал 184 желудка ящериц,
добытых с апреля по октябрь в 1967—1970 гг. Как выяснилось, главное место
в рационе гладких геккончиков занимают пауки (50,5 % встречаемости), а так-
же термиты (20,6 %) и жуки (10,9 %), остальные группы беспозвоночных
встречаются редко. В апреле — мае геккончики питаются преимущественно
термитами (40,2 %) и мелкими жуками (31,2 %), а летом и осенью — пауками
(до 80 % встречаемости). Результаты анализа желудков гладких геккончиков
из 3. Туркмении, проведенного нами (п = 30), показали, что они чаще поедают
мокриц (22,2 % встречаемости), муравьев и мух (по 13,9 %), пауков (11,1 %)
(табл. 3). Как правило, это мелкие беспозвоночные с мягким хитиновым пок-
ровом.
Размножение. Соотношение полов в наших сборах близкое 1:1.
Половозрелыми становятся геккончики по достижении длины туловища 25—
29 мм. Спаривание, судя по наибольшей массе семенников [130], происходит
в начале апреля. У восьми самок, добытых нами 16.V 1975 г. у руин Мешхед-
и-Мессериана, в яйцеводах обнаружено по одному яйцу, почти готовому к от-
кладке (8—9 х 5 мм). У вскрытых семи ящериц размеры крупного яйца были
несколько меньшими (6—8 мм), но имелось и второе, менее развитое яйцо
(2—4 мм), что свидетельствует о наличии порционной кладки. Самка геккончи-
ка, добытая одновременно с указанными выше и там же, отложила одно, другая
отложила два яйца 22.V 1975 г. С. Шаммаков [130] готовое к откладке яйцо
обнаружил 14.VII 1972 г. Размеры яиц, по нашим материалам (п = 14), 8,5—
10,0 X 5,9—7,2 (9,27 ± 0,13 X 6,51 ±0,1) мм. Из яйца, найденного 25.VI
1979 г. в Ю. Кызылкуме, 5.VIII 1979 г. вывелся геккончик с длиной туловища
18,5 мм, хвоста 16,5 мм, 10.VIII 1979 г. из яйца, добытого там же, вышел ново-
рожденный с размерами соответственно 17,3 и 17,0 мм. Судя по представленным
выше данным, инкубация яиц продолжалась не менее 42—47 дней.
Поздние находки новорожденных могут свидетельствовать о второй
кладке.
Темп роста. По материалам С. Шаммакова [130], длина туловища
геккончиков за первые 9—10 месяцев увеличивается на 6—10 мм. Предполага-
ется, что в июне и июле некоторые особи входят в группу половозрелых и вклю-
чаются в репродуктивный цикл. В последующие годы различия между двух-
и трехлетними особями сглаживаются.
Враги. Гладкий геккончик был обнаружен в желудке средней ящурки
(Eremias intermedia), поперечнополосатого полоза (Coluber karelini) и эфы
(Echis carinatus). Возможно, их поедают фаланги, многочисленные в биотопах
этого вида. Как указывалось, в последние годы отмечены прогрессивное сни-
жение численности и сокращение ареала гладкого геккончика на территории
Туркмении. Основная причина этого — преобразование его биотопов в резуль-
тате хозяйственной деятельности человека. Туркменский канал, прошедший
по полосе предгорных такыров Копетдага, дал мощный стимул к их распашке.
Так, в окр. ст. Такыр, где эти ящерицы были обычны, после распашки такыра
исчезли. Сейчас канал доходит до Кизыл-Арвата и далее пойдет по Мессериан-
скому плато, что угрожает полному уничтожению этого вида.
77
Уже исчезли геккончики и в долине р. Теджен.
Типовое местообитание, уроч. Каррыбент, было особенно богато геккончи-
ками. Так, О. П. Богданов [251 сообщал, что здесь на протяжении 200 м он вы-
копал из убежищ 62 особи. Здесь же, по сообщению С. Шаммакова, в 1971 г.
в течение часовой экскурсии он собрал до 10 ящериц. В последние годы упорные
и многочисленные поиски не увенчались успехом. После наводнения 1977 г.,
вызванного прорывом дамбы водохранилища, гладкие геккончики здесь пол-
ностью исчезли. 5.V 1984 г. на Мессерианском плато мы выкапывали геккончи-
ков из трещин и нор насекомых, причем они находились так близко от поверх-
ности (пасмурный день), что даже простой пробег автомобиля по такыру, не-
сомненно, многих убивает.
Особенности поведения. Эта ящерица передвигается в типич-
ной для равнинных гекконов манере: держит изогнутые дугой туловище и
хвост приподнятыми над землей. Потревоженный геккончик быстро пробегает
2—5 м и затаивается. По наблюдениям в террариуме, они могут передвигаться
и по вертикальным поверхностям, подниматься на высоту 50 см.
Летними убежищами геккончикам служат трещины в почве, ходы в термит-
никах, старые норы жуков, пустоты в саксауловых пнях и древовидной солян-
ки, норы других ящериц. Они были обнаружены в норах с почти полностью
закрытым или замытым дождем входным отверстием. Как мы установили, гек-
кончики во влажном субстрате сами могут рыть норы диаметром 0,5—0,6 см.
В пасмурные дни они держатся на глубине 3—5 см, в жаркие — глубже.
Здесь же геккончики и откладывают яйца. Так, мы находили кладки в 3. Турк-
мении в старом термитнике на глубине 15—20 см (шесть яиц — групповая клад-
ка), под старым кустом солянки на глубине 5 см. В термитнике диаметром 2 м
и высотой 20 см на границе сухого и влажного субстрата были обнаружены
фрагменты 16 яиц геккончиков, сконцентрированных в северном секторе купо-
ла. Зимовочные убежища отличаются от летних только глубиной залегания;
С. Шаммаков [130] находил геккончиков в термитниках на глубине 5—11 см,
в норах такырной круглоголовки (глубина 12 см, длина норы 17—30 см), в кор-
нях и стволах сухого саксаула, в норе крапчатой месалины. Обычно зимует
одна-две особи, однажды нашли пять ящериц вместе.
Гладкие геккончики издают отдельные отрывистые звуки, передаваемые
как «иик...иик». При обнаружении добычи или при опасности частота звуков
увеличивается. Особенно часто можно их слышать в брачный период.
Практическое значение и охрана. Как вид с уменьшаю-
щейся численностью и ареалом гладкий геккончик по нашему предложению
был включен во второе издание Красной книги СССР. К сожалению, он не из-
вестен на территории существующих заповедников. Целесообразно выделить
для его охраны участок засоленного такыра севернее ст. Вами площадью 6—
10 км2 (в качестве филиала Копетдагского заповедника), где, кроме гладкого
геккончика, обитает уникальный вид ящерицы — пятнистая круглоголовка
(Phrynocephalus maculatus).
СЕВЕРОАЗИАТСКИЙ ГЕККОНЧИК ТАДЖИКСКИЙ —
ALSOPHYLAX TADJIKIENSIS GOLUBEV 1979
Типовая территория: долина р. Вахш, Ю. Таджикистан.
Кариотип: 2 л==36, акроцентриков 30, субтелоцентриков 4, субмета центриков 2,
NF=42.
1962 — Alsophylax pipiens Саид-Алиев, Тр. ИЗиП АН ТаджССР, т. 22, с. 103.
1977 — Alsophylax laevis Муратов Н., Муратов Р., в кн.: Вопр. герпетол., авгореф. докладов,
Л., Наука, с. 150.
1979 — Alsophylax laevis tadjikiensis, Голубев, Тр. ЗИН АН СССР, т. 89. Экол. и сист., амф.
и репт., Л., изд. АН СССР, с. 62.
1984 — Alsophylax tadjikiensis, Голубев, Вести, зоол., № 2, с. 73.
Диагноз. Спинные бугорки отсутствуют; пять — семь анальных пор;
кроме крупного межносового щитка, есть одна (около 93 %), очень редко две
(около 7 %), мелкие добавочные носовые чешуйки; широкие спинные полосы от-
сутствуют; хвост у живых особей охристо-оранжевый.
78
Голотип. ЗИН 18 929, с? ad Таджикистан, Кургантюбинский р-н,
7 км от пос. Кызылкала вниз по реке Вахш, 25.IV 1965 г., коллектор неизвестен.
L — 28,2 мм; Lcd= 34,5 мм; L/Lcd = 0,82; из носовых имеются только крупный
межносовой щиток и одна мелкая добавочная носовая чешуйка; позади сопри-
касающихся между собой межносовых щитков расположена пара слабо выра-
женных замежносовых щитков, разделенных чешуйкой; верхнегубных и ниж-
негубных щитков по шесть; между центрами глаз 15 чешуй; анальных пор шесть,
вдоль тела снизу 98 чешуек; подпальцевых пластин 16—17.
Описание (рис. 22) (по 20 экз. из колл. ИЗАНУ и ЗИН). L сГ ad
(п = 8) 27,4—30,2 (28,30 ± 0,34) мм; L £ ad (п = 6) 25,0—31,0 (28,18 ± 1,05);
L/L cd (п = 3) 0,80—0,88 (0,833 ± 0,024); индекс ширины/высоты головы (п =
= 11) 60—70 (64,82 ± 0,91); диаметр уха/глаза (и = 16) 13—26 (19,50 ±
± 0,94); верхнегубных щитков (п = 40) 5—7 (6,10 ± 0,08); нижнегубных (п =
= 40) 5—6 (5,38 ± 0,08); чешуй между центрами глаз (п =13) 12—15 (13,58 ±
± 0,23); чешуй вдоль тела снизу (п = 14) 89—102 (96,50 ± 1,08); анальных
пор (п = 18) 5—7 (6,22 ± 0,15); подпальцевых пластин (п — 34) 14—18
(15,88 ± 0,18). Ноздря находится между межчелюстным, первым верхнегуб-
ным, крупным межносовым щитком и одной (около 93 %), очень редко двумя
(около 7 %), мелкими добавочными носовыми чешуйками; межносовые щитки
соприкасаются между собой (90 %) или разделены чешуйкой; позади них в 30 %
случаев имеются замежносовые щитки, выраженные в разной степени и разде-
ленные между собой чешуйкой (90 %) или соприкасающиеся друг с другом.
Подбородочный щиток трапециевидный (60 %), шестиугольный (25 %), тре-
или пятиугольный (15 %); у 70,5 % имеется одна пара нижнечелюстных щит-
ков, у 27,3 % — две пары, у 2,2 % эти щитки отсутствуют. Спинных бугорков
нет, но, как и у предыдущего вида, изредка они могут проявляться в крестцовой
и поясничной областях.
Прижизненная окраска и рисунок. Цвет верхней поверх-
ности тела охристый с оливковым оттенком (хаки). На туловище могут быть от
передних конечностей до основания задних пять более светлых, чем основной
тон, поперечных полос. На границе с хвостом бурая поперечная волнистая по-
лоса, хвост однообразно ярко-охристый с оранжевым оттенком, на нем могут
быть беспорядочно разбросаны бурые точки или полоски (до восьми). От кон-
чика морды через глаз до височной области и боков шеи идет темно-бурая поло-
са, которая может в виде отдельных бурых точек продолжаться по границе спи-
ны и брюха до основания задних конечностей. От ноздри над этой полосой такой
же ширины до надглазничных щитков идет желтоватая полоса. Светлые над-
глазничные полосы сходятся на конце морды, промежуток между ноздрями и
глазами более темный, чем основной фон. Регенерированный хвост окрашен так
же, как и неповрежденный. Нижняя сторона туловища (брюхо, бедра, лапы)
светло-сероватые, низ хвоста охристо-оранжевый.
Половой диморфизм и возрастная изменчивость.
У самок таджикского геккончика по сравнению с самцами анальные поры
выражены меньше; самки несколько крупнее самцов. Несмотря на относительно
небольшое количество исследованных особей, замечена тенденция к изменению
некоторых пропорций тела, характерная для всех видов этого рода: увеличение
с возрастом длины хвоста и уменьшение диаметра глаза относительно длины
тела.
Распространение (рис. 30). Ю. Таджикистан.
Кадастр к рис. 30.
/ — 7 км от пос. Кызыл-Кала вниз по р. Вахш [81]; 2 — пос. Сумбулак (ИЗАНУ); 3 —
Колхозабад (ЗИН); 4 — правый берег р. Вахш против Джиликуля у подножья гор Аруктау
[107]; 5 — 53—56 км дороги на Шаартуз [81]; 6 — 18 км от переправы через Кызылсу в пред-
горьях Каратау ([107]; нуждается в подтверждении).
Географическая изменчивость не обнаружена. Близок к
гладкому геккончику, с которым объединялся до недавнего времени.
Стации и количественные д а н н ы е (рис. 34). Мы добыва-
ли этих ящериц в основании разрушенных лессовых построек, но основным мес-
тообитанием данного вида являются глинистые склоны с россыпями камней
79
Рис. 32. Биотоп A. laevis (такыр на Мессерианском плато, ТССР)
на возвышениях и норами грызунов, отдельными кустиками эфедры и травя-
нистых растений (наблюдение у с. Сумбулак, 13 км южнее пос. Кызыл-Кала).
Встречались они также и в сухих галечниковых руслах, оставшихся после селей
(из конгломератов вымыта глина, в нагромождениях камней множество щелей
и полостей). Местообитания этого вида находятся на высоте 450 м н. у. м. На
участке 500 X 500 м, по нашим подсчетам по голосам, обитает около 20 особей,
отдельные особи встречались на расстоянии 25 м друг от друга.
Отношение к температуре. Активные ящерицы 27—
28. IV 1980 г отмечены при температуре почвы 20—27°, температуре воздуха 17—
18°. При температуре воздуха 12° 22.IV 1978 г. геккончики не появлялись.
Суточный цикл активности. По наблюдениям в конце апре-
ля под с. Сумбулак, крики геккончиков стали раздаваться еще до наступления*
темноты, в 20 ч 45 мин, первая ящерица была добыта в 21 ч, к 22 ч 30 мин ак-
тивность резко сократилась, почти перестали раздаваться их голоса, послед-
няя особь была добыта около 1 ч.
Сезонный цикл активности. Из убежищ после зимовки
выходит в марте [811, в 1984 г. после холодной и затяжной весны в начале ап-
реля ящерицы на поверхности еще отсутствовали. Уход на зимовку не отмечен.
Линька наблюдалась 29. IV 1975 г. [811.
Питание. По нашим исследованиям (вскрыто шесть желудков геккон-
чиков, добытых в апреле), преобладают пауки (37,5 % встречаемости), осталь-
ные группы беспозвоночных представлены одинаково редко (псевдоскорпионы,
цикады, тли, жуки и бабочки по 12,5 %).
Размножение не изучено. При раскопках в биотопах геккончика
найдена скорлупа, по которой нами установлены следующие размеры яиц —
8,3—8,7 х 5,1—6,1 мм (п = 6).
Темп роста не изучен. Судя по аналогии с близким видом (гладким
геккончиком), возможно наступление половой зрелости к началу следующего
сезона.
Враги не обнаружены. В стациях этой ящерицы обитают змеи-стрелы,
фаланги и скорпионы, которые при случае могут нападать и на геккончиков.
Особенности поведения. В отличие от гладкого геккончика,
таджикский часто передвигается в природе по вертикальным поверхностям,
несколько раз мы находили их на камнях высотой до 0,5 м, а однажды он сидел
80
на боку большого валуна размером 1,5 м вниз головой на вертикальной поверх-
ности с отрицательным углом.
Убежищами этому виду служат щели в нагромождениях камней, простран-
ства под камнями, норы грызунов и щели в лессовых руинах. Здесь они могут
и зимовать (на глубине до 40 см). Кладки яиц мы находили при раскопке лес-
совых развалин построек, у их основания на глубине 5—20 см от поверхности.
Судя по их количеству, можно предположить, что в одно место могут отклады-
вать яйца две-три самки.Остатки кладки геккончика мы также обнаружили
в ходах грызуна (песчанки) на глубине 40 см и расстоянии от вхо-
да 1 м.
Весной часто после наступления сумерек слышен голос таджикского гек-
кончика, который можно передать словом «пець» с металлическим оттенком.
Эти крики раздаются сериями по три-четыре звука, а затем замолкают, чтобы
повториться через интервал 0,5—1 мин и более.
Практическое значение и охрана. Таджикский геккон-
чик — редкий узкоареальный вид, обитающий в зоне активной хозяйственной
деятельности человека. Его нет на заповедных территориях Таджикистана.
Данный вид должен быть включен в Красную книгу Таджикской ССР, а учас-
ток с высокой плотностью его (13 км южнее с. Кызыл-Кала) между шоссе и
р. Вахш площадью 500 X 1000 м следует объявить микрозаповедником.
Подрод высокогорный североазиатский геккончик —
Altiphylax Jeriomtschenko et Szczerbak, 1984
Типовой вид: A. tokobajevi
1984 — Altiphylax Еремченко, Щербак, Вести, зоол., № 2, с. 46.
Диагноз. Пальцы слабо искривлены; анальные поры имеются только
у самцов; длина передней конечности составляет около 30 % длины тела, дости-
гает концами пальцев конца морды; задняя конечность, вытянутая вперед, до-
стает концами пальцев середины шеи; хвостовые сегменты слабо различимы (до
9—10 сегмента); до 6—8 сегмента хвостовые бугорки опускаются на боковые по-
верхности хвоста. Подрод включает один вид.
ТЯНЬ-ШАНЬСКИЙ ГЕККОНЧИК —
ALSOPHYLAX TOKOBAJEVI JERIOiVlTSCHENKO ET SZCZERBAK, 1984
Типовая территория: окрестности с. Байгончек, Нарынская обл., Киргизия.
Кариотип: 2л = 34; акроцентриков — 26, субтел оцентр и ков — 4, субмета центр и-
ков — 4, NF = 44.
1984 — Alsophylax tokobajevi, Еремченко В. К., Щербак Н. Н., Вести, зоол., № 2, с. 46—50.
Диагноз. Мелкие округлые гладкие спинные бугорки расположены более
или менее беспорядочно; шесть-семь анальных пор; имеются крупный межносо-
вой щиток и две мелкие добавочные носовые чешуйки; верхнегубных щитков
8—13; поперек туловища четыре — семь широких полос; хвост охристый.
Г о л о т и п. RE. № 16 (ИЗАНУ) dad КиргССР, Нарынская обл., отроги
хр. Байбиче-Тоо, окр. с. Байгончек, 1800 м н. у. м., 11.V 1983 г., колл.
В. К. Еремченко. L = 34,2 мм; L cd (регенерирован) = 30 мм; чешуй поперек го-
ловы 18, верхнегубных щитков 10—12, нижнегубных по 7; нижнечелюстных
1—3; чешуй вдоль тела снизу 100; анальных пор 6; подпальцевых пласти-
нок 26.
Описание (рис. 35) (по 61 экз. из колл. ИЗАНУ). L d ad (п = 22)
33,6—45,4 (40,58 ± 0,60) мм; L g ad (п = 27) 30,8—49,5 (40,80 ± 1,08) мм;
LILcd (п = 18) 0,68—0,87 (0,77 ± 0,01); индекс высоты/ширины головы (п =
= 51) 47—64 (55,33 ± 0,53); диаметр уха/глаза (п = 33) 18—28 (23,64 ± 0,44);
чешуй вокруг спинного бугорка (п = 61) 7—10 (8,34 ± 0,09); длина спинного
бугорка/L (п = 49) 7—12 (9,57 ± 0,16); верхнегубных щитков (п = 119) 8—13
(10,44 ± 0,09); нижнегубных (и = 122) 5—8 (6,72 ± 0,07); чешуй между цент-
рами глаз (п = 60) 13—21 (16,37 ± 0,21); чешуй вдоль тела снизу (п = 56)
91—109 (102,04 ± 0,53); анальных пор (п = 21) 5—7 (6,33 ± 0,16); подпальце-
вых пластин (п = 121) 20—27 (23,5 ± 0,12).
81
Кроме крупных межносовых щитков, с ноздрей соприкасаются две (100 %)
мелкие д. бавочные носовые чешуйки; межносовые щитки соприкасаются между
собой (19,7 %) или разделены одной (75,4 %) или двумя (4,9 %) мелкими доба-
вочными носовыми чешуйками; замежносовые выражены очень слабо (14,7 %)
или не выражены вовсе (85,3 %); подбородочный щиток шестиугольный (42,6 %)
или трапециевидный (57,4 %); нижнечелюстных щитков одна (4,2 %), две
(59,7 %) или 3 (36,1 %) пары; первая пара нижнечелюстных щитков разделена
одной (42,6 %), двумя (54,1 %) или тремя (3,3 %) горловыми чешуйками; мел-
кие округлые гладкие спинные бугорки расположены не вполне четкими про-
дольными и поперечными рядами; размеры их заметно возрастают от середины
спины к бокам туловища.
Прижизненная окраска и рисунок. Основной фон верх-
ней стороны туловища сероватый с фиолетовым оттенком, может темнеть до бу-
рого, хвост серовато-охристый. От межчелюстного щитка до верхней части ор-
бит проходит пара охристо-бурых полос. Такие же полосы идут от ноздри через
глаз и височную область к затылку, где они смыкаются или разделены основ-
ным тоном. На голове сверху неясный буроватый рисунок. От шеи до основания
хвоста поперек туловища проходит шесть-семь угловатых полос буроватого
цвета, задние края которых окантованы черным. Подобные полосы проходят
и по верхней стороне хвоста (до 13), где черные линии преобразованы в неболь-
шие черные пятна треугольной формы Верхняя сторона конечностей также
имеет бурый поперечный рисунок, но менее яркий и без черной окантовки. Ниж-
няя сторона беловатая.
Половой диморфизм и возрастная изменчивость.
Как и у других видов рода, самки крупнее самцов. У самок анальные поры не
выражены (однако перфорированные чешуйки с порами обнаружены у 7,4 %
изученных самок). Пропорции тела с возрастом изменяются аналогично дру-
гим видам рода (рис. 23). У неполовозрелых особей хвост оранжево-желтый.
Распространение. Киргизия, Ц. Тянь-Шань, Нарынская обл.,
адыры правобережья р. Алабуги от с. Угют на сев.-в. до с. Кош-Дебе на ю.-з.
на высотах от 1800 до 2500 м н. у. м. (рис. 89).
Кадастр к рис. 89 [56, 57|.
1 — с. Байгончек; 2 — с. Конгорчок; 3 — с. Джергетал; 4 — с. Кош-Дебе.
Экология [57].
Стации и количественные данные (рис. 36). Типичное мес-
тообитание этого вида — глинистые денудированные адыры, почти лишенные
растительности или покрытые редкими кустами солянки. Геккончики встреча-
ются здесь под плитами рыхлого сланца и в промоинах на склонах. Крутизна
склонов достигает 45°. Ящерицы обнаружены на южных, западных и восточных
склонах. Относительная высота холмов до 250 м, геккончики встречаются от
подножья до верха хребта. В окр. с. Конгорчок тянь-шаньский геккончик обна-
82
Рис. 34. Биотоп A. tadjikiensis (каменистая полупустыня вдоль подножья хр. Актау
в окр. пос. Сумбула, ТаджССР)
ружен нами также на лессовых обрывах сезонных потоков и ручьев — притоков
р. Алабуги, которые соединяют адыры с поймой реки. В этих оврагах встречает-
ся древесная растительность из ивы и кустарников (шиповник, боярышник).
На обрывах южной экспозиции (маршрут 500 м) высотой до 2 м отмечено две
особи. Пользуясь обрывами оврагов, тянь-шаньские геккончики могут засе-
лять и постройки человека, особенно руины средневековых крепостей. Так, на
остатках лессовых стен крепости Чолок-Коргон (высота 1950 м н. у. м.), на ее
территории площадью 30 X 50 м, где остались бугры от построек, обнаружена
значительная плотность геккончиков. Учитывая периметр стен и остатки пост-
роек внутри двора, что составляет маршрут длиной 170 м, мы обнаружили здесь
30 геккончиков, в среднем одну особь на 5,6 м. При этом ящерицы встречались
в подавляющем большинстве на стене южной экспозиции, реже западной и вос-
точной; на северной они отсутствовали. Наибольшая плотность геккончиков
здесь (три особи) обнаружена на куче лессовых кирпичей с норами грызунов
площадью 2 х 2 м, высотой до 40 см. Рядом, на расстоянии 1 км от крепости,
на склонах глинисто-щебнистых холмов за 2 ч экскурсии под камнями было до-
быто пять особей.
Западнее этого места, в окрестностях с. Кош-Дебе количество обломков
сланцев и щебнистость склонов возрастает, чаще встречаются и тянь-шаньские
геккончики. Так, 6.VI 1984 г. за 1ч экскурсии (обследована территория 200 X
X 20 м) найдено шесть особей (наименьшее расстояние между ними — Юм).
Учитывая отсутствие здесь серых гекконов и очень низкую численность ящу-
рок Никольского, следует рассматривать тянь-шаньского геккончика в качест-
ве фонового вида пустынных адыров, характеризующихся крайней скудностью
фауны и флоры.
Отношение к температуре. Территория, населенная тянь-
шаньскими геккончиками, характеризуется континентальностью климата,
резкими перепадами температуры, летними осадками. По измерениям темпера-
туры субстрата в местах находки активных геккончиков на поверхности в ноч-
ное время (п = 10) установлено 4.VI 1984 г., что средняя температура появле-
ния их из убежища 18°. Минимальная температура активности отмечена 5.VI
1984 г. в окр. с. Конгорчок — 14,5°. При исследовании в лаборатории пяти осо-
83
бей в термоградиентприборе 3.VII 1984 г. получены данные по 182 отметкам
температуры: ящерицы занимали места с температурой 6—40° (М ± т —
= 25,95 ± 0,84). Предпочитаемая температура этого вида значительно ниже
таковой других видов гекконов фауны СССР (у Т. russowi — 33,3 ± 0,83°.
У Т. caspius — 32,7 ± Г).
Таблица 4. Содержимое желудков A. tokobajevi
(п = 25, 16 и 9 J), добытых 4. VI 1984 г. в окр. с. Конгорчок
Объект питания Встречаемость Общее количество
экз | экз. %
Arthropoda
Arachnoidea'
Araneida 10 40 14 18,4
Acarina 1 4 1 1,3
Trombicidae 2 8 2 2,6
Chilopoda 1 4 1 1,3
Isopoda Insecta 1 4 1 1,3
Homoptera 1 4 1 1,3
Cicadidae 6 24 12 16,3
Coleoptera 2 8 2 2,6
Carabidae 1 4 1 1,3
Curculionidae 11 44 20 26,3
Elateridae 1 4 1 1,3
Scarabeidae 1 4 1 1,3
Chrysomelidae 2 8 2 2,6
Hemiptera 4 16 4 5,*2
Pentatomidae (Eungaster) 1 4 1 1,3
Tysanura Hymenoptera 2 8 2 2,6
Formicidae 3 12 3 3,9
Lepidoptera (1.) 5 20 6 7,8
Lepidoptera (i.) 1 4 1 1,3
Суточный цикл активности. Геккончики поедают насеко-
мых, которые попадают к ним в убежища в дневное время. 4.VI 1984 г. отмечен
их выход из убежищ в 22 ч 20 мин местного времени, сразу после наступления
сумерек. Наибольшее их число отмечено с 23 до 24 ч, а после 24 ч резкое похо-
лодание не позволило им находиться на поверхности. Можно предполагать,
что их питание и спаривание обеспечиваются в основном достаточно высокими
(до 30°) дневными температурами, на поверхности они появляются в сумерки
и начале ночного времени.
Сезонный цикл активности не изучен. Исходя из климатиче-
ских условий исследуемой территории и состояния гонад можно предполагать,
что тянь-шаньские геккончики активны с начала мая до середины сен-
тября.
Линька отмечена нами 5.VI 1984 г., 8.VI 1984 г. она наблюдалась у
большинства особей, отловленных для лабораторных исследований. Пара лин-
ных ящериц отмечена 16.VI 1984 г. и 24.VI 1984 г., еще одна особь линяла
15.VII 1984 г., две — 30.VII 1984 г.
Питание. Проанализировано содержимое 25 желудков геккончиков,
собранных в окр. с. Конгорчок 4—5.VI 1984 г. (табл. 4) и 45 образцов экскре-
ментов, полученных от свежеотловленных ящериц. В рационе тянь-шаньских
геккончиков преобладают по встречаемости и от общего количества долгоноси-
ки (44 и 26,3 % соответственно), цикадовые (24 и 16,3 %), гусеницы бабочек
(20 и 7,8 %), а также пауки (40 и 18,4 %). Приведенные данные преимуществен-
но соответствуют материалам, полученным в результате анализа экскре-
ментов.
В 45 пробах экскрементов обнаружены остатки 44 насекомых (в одной
найдены песок и неопределенные фрагменты хитина). По встречаемости най-
84
A — голова сбоку, Б — голова сверху, В — подбородок, Г — спина, Д — преанальные
поры, £ — лапа сверху, Ж — палец снизу, 3 — хвост сверху, И — хвост снизу (ИЗАНУ
RF 16/3, Байгончек. КиргССР)
денные здесь беспозвоночные распределяются следующим образом: долгоноси-
ки — 22,7 % (10 экз.), клопы — 20,4 (9 экз.), равнокрылые — 18,2 (8 экз.)г
гусеницы бабочек — 13,8 (6 экз.), муравьи и жуки по 6,8 (по 3 экз.), мухи —
4,5 (2 экз.), пауки — 2,3 % (1 экз.). Таким образом, результаты анализа желуд-
ков и экскрементов свидетельствуют о том, что геккончик предпочитает поедать
жуков-долгоносиков, полужесткокрылых, цикад и гусениц бабочек. При сравне-
нии с данными по анализу питания серого геккона из Киргизии [162] хорошо
заметны отличия в рационах этих видов: серый геккон предпочитает поедать
прямокрылых (84,2 %), полужесткокрылых (36,8 %) и жуков (15,7 %).
Размножение. Тянь-шаньские геккончики становятся половозре-
лыми при достижении размеров туловища самок 38 мм, самцов — 37 мм. Спа-
ривание происходит, вероятно, вскоре после появления из зимовочных
85
Рис. 36. Биотоп A. tokobajevi (руины крепости и адыры в пойме р. Алабуга, Ц. Тянь-
Шань, КиргССР)
убежищ, в начале мая, так как уже в начале июня встречалось большинство взрос-
лых самок с явными признаками беременности (фолликулы особи, добытой 4.VI
1984 г., достигали размеров 8,2 X 6 мм). 5.VI 1984 г. в трещинах стены средне-
вековой крепости Чолок-Коргон была обнаружена свежая кладка из двух яиц.
Отловленные здесь геккончики откладывали яйца 13, 20, 27.VI и 2.VII 1984 г.
(в этот день было получено три кладки). Как правило, взрослые самки откла-
дывали по два яйца, молодые — по одному (последние встречаются в два раза
реже). Размеры яиц (и — 28) 10,5—12,5 X 6,8—9,0 мм (соответственно М ±
±т = 11,56 ± 0,11 X 8,48 ± 0,09). Яйца откладываются в ночное время (по
наблюдениям в лаборатории). Инкубация яиц длилась 94 дня. Яйца, получен-
ные от оплодотворенных в природе самок, инкубировались в лаборатории в тер-
мостате при температуре 26—30° и детеныши вылупились из яиц З.Х 1984 г.
(рис. 37). Длина туловища свежевылупившихся геккончиков 21,4 мм, хвос-
та — 19,2 мм.
Темп роста. Самые мелкие экземпляры с ювенильными признаками
(яркая окраска низа хвоста), добытые в начале мая, достигали размеров тулови-
ща 20—22 мм, в начале июня — 27—34 мм. Сравнивая их с размерами свеже-
вылупившихся геккончиков, видно, что за период с зимовки по июнь (три ме-
сяца активности) их длина туловища увеличилась в среднем на 10 мм. Учитывая
последующее замедление темпа роста, можно предположить, что геккончики
достигают половозрелости на третий календарный год после рождения, а мак-
симальных размеров — на четвертый-пятый год жизни 45,5 мм, $ 47,0 мм).
Враги. Данные о численности геккончиков в разных биотопах могут
свидетельствовать о возможных конкурентных взаимоотношениях с серым гек-
коном. Нами установлено, что там, где эти виды обитают вместе, A. tokobajevi
редок, в местах, где отсутствует серый геккон, он достигает большей численно-
сти. В биотопе тянь-шаньского геккончика мы находили узорчатого полоза
(Elaphe dione). Молодой экземпляр этой змеи, найденный под камнем на склоне,
где встречались и геккончики, отрыгнул молодую ящурку Никольского. При
случае полоз, несомненно, нападает и на геккончиков. Как выяснилось, боль-
шинство особей A. tokobajevi с регенерированными хвостами (59,5 %). По на-
блюдениям в террариуме, они могут терять хвост в драках с особями своего
86
вида. В местах обитания геккончиков несколько раз была отмечена пустель-
га — потенциальный враг ящериц.
Особенности поведения. Как следует из приведенных выше
данных, тянь-шаньский геккончик — самый высокогорный вид геккона фауны
СССР и самый холодостойкий. Обитание в таких суровых условиях горной пус-
Рис. 37. A. tokobajevi, вылупление из яйца с одновременной линькой
тыни привело к возникновению у него ряда особенностей поведения. Он мо-
жет свободно передвигаться по вертикальным поверхностям, что позволяет,
в сущности, занимать экологическую нишу, характерную для тонкопалых
гекконов. Как правило, геккончики обитают у основания стен, но встреча-
ются на них до высоты 1 м, а внутри найдены на высоте до 2 м. Об этой способ-
ности свидетельствует и совершенно нетипичное для представителей рода Al-
sophylax строение задних конечностей, очень близкое к видам рода Tenuidac-
tylus. Во время охоты за насекомыми тянь-шаньские геккончики могут даже со-
вершать прыжки на расстояние, превышающее длину их туловища, хотя они не
такие быстрые, как серые гекконы.
Судя по остаткам насекомых в желудках и экскрементах, среди которых
много дневных форм, геккончики часто питаются днем. Эта особенность
характерна и для других сумеречно-ночных высокогорных животных, в част-
ности для полиплоидных жаб. В ночные часы здесь резко снижается темпера-
тура, что ограничивает возможности пойкилотермных животных.
Убежищами тянь-шаньским геккончикам служат пространства под облом-
ками скальных пород (как правило, сланцев, но под гипсовыми корочками и
кусками галечниковых конгломератов они отсутствуют), щели в лессовых об-
рывах, промоины на глинистых склонах, пространства между необоженными
кирпичами древних руин, иногда норы грызунов. Здесь же они откладывают
яйца, но поскольку удобных для инкубации прогреваемых мест мало, чаще яйца
откладывают коллективно — несколько самок в одно место. Нами кладки были
найдены под плитами сланца (шесть яиц кладки прошлых лет у с. Конгорчок),
в щелях лессовых обрывов (одно прошлогоднее яйцо на высоте 1,6 м) и в пус-
тотах стены старой постройки из необожженных лессовых кирпичей. В руинах
крепости Чолок-Коргон обнаружен своеобразный «инкубатор», куда геккончи-
ки коллективно откладывали яйца на протяжении многих лет. Так, в выступе
стены, расположенной в направлении с севера на юг, в его южной оконечности
(он обогревался на протяжении всего дня солнцем с востока, юга и запада),
на высоте 1,2 м от уровня почвы и на глубине 10 см (площадь этой территории
50 X 50 см) найдены старые высохшие яйца, кладка этого года и скорлупа,
оставшаяся после вылупления ящериц (не менее 60 яиц). Группы яиц достигали
десятка, они были приклеены к стенкам щелей или лежали на горизонтальных
87
поверхностях (рис. 37). Здесь же были добыты и две беременные самки, а в од-
ном месте найдено скопление помета, свидетельствующее о частом нахождении
здесь геккончиков. Также ограничено здесь количество и зимовочных убе-
жищ — глубоких промоин и нор грызунов.
Индивидуальная территория геккончика ограничивается укрытием и его
ближайшими окрестностями. В целом они весьма терпимы друг к другу, о чем
свидетельствуют находки двух однополых особей под одним камнем, коллектив-
ная откладка яиц в одно место и наблюдения при содержании их в террариуме.
Находки тянь-шаньских геккончиков в руинах построек, расположенных на
расстоянии до 1 км от их естественных мест обитания, соединенных лессовыми
оврагами, которые не являются типичными местообитаниями, свидетельствуют
о способности к миграциям. По-видимому, подобные миграции наиболее часты
к месту откладки яиц и зимовальным убежищам.
При спаривании, судя по следам на туловище самок, самцы удерживают
их укусами за бока брюха выше бедер.
При содержании в террариуме геккончики хорошо приспосабливаются
к новым условиям (необходимо делать укрытия), не взыскательны к корму
(поедают мелких личинок мучных червей, тлей, других мелких насекомых, на-
кошенных сачком), спариваются в неволе и откладывают яйца (беременная
самка случайно погибла 16.11.1984 г., ее два яйца в оболочке достигали разме-
ров 7,5—7,8 мм X 6,2—6,5 мм).
Практическое значение и охрана.
Полезный вид; как редкий узкоареальный эндемик Советского Тянь-Шаня
подлежит охране и включению в Красную книгу КиргССР. Учитывая то, что
тянь-шаньский геккончик обитает на территории, не включенной в активную
хозяйственную деятельность человека,численность его по наблюдениям 1983—
1984 гг. стабильна, проводить какие-либо особые мероприятия по охране
данного вида нецелесообразно.
Род южноазиатский геккончик — Bunopus Blanf., 1874
Типовой вид: Bunopus tuberculatus Blanf.
1874 — Bunopus Blanford, Ann. Mag. Nat. Hist., London, Vol. 13, p. 454.
1885 — Alsophylax, Boulenger, Cat. Liz. Brit. Mus., Vol. 1, p. 20 (partim).
Диагноз. Пальцы слабо искривлены, когтистые, цилиндрические, не
расширенные и не сжатые с боков, снизу покрыты одним рядом поперечно рас-
ширенных подпальцевых пластинок, бугристых или шиповатых вдоль свобод-
ного края. Боковая пальцевая чешуя с более или менее выраженными шипика-
ми, бахромы (как у Crossobamon или у части Stenodactylus) не образует. Зрачок
вертикальный, с зазубренными краями. Увеличенные спинные бугорки, рас-
положенные более или менее четкими продольными или поперечными рядами,
всегда имеются. Анальные поры (3—16) хорошо выражены только у самцов.
Два — пять выпуклых носовых щитков почти одинаковы по размеру. Хвост
сегментирован. Первый сегмент ближе к проксимальной части хвоста покрыт
двумя-тремя кольцами чешуй и кольцом, состоящим из хвостовых бугорков.
В центре сегмента они могут быть разделены одной-двумя чешуйками на два
полукольца. В полукольцах бугорки плотно прилегают друг к другу боковыми
краями. Снизу хвоста на тех же сегментах есть три кольца чешуй, они в середи-
не каждого кольца могут иногда заменяться крупным щитком. Высота первого
верхнегубного щитка от ноздри до края рта больше, равна или незначительно
меньше его ширины вдоль края рта. Каждый верхнегубной щиток находится
в одной плоскости и опускается отвесно к краю рта. Реснитчатые выросты в
верхней части орбиты есть.
Дополнительные признаки. Ноздря расположена между
четырехугольным межчелюстным, первым верхнегубным и тремя — пятью почти
равными друг другу мелкими выпуклыми носовыми щитками; первые носовые
щитки разделены между собой одной — тремя чешуйками, замежносовые
щитки отсутствуют; чешуя морды гладкая или слабо ребристая, выпуклая,
88
неоднородная, неправильно-многоугольная, уменьшается к затылку, где сме-
шивается с бугорками; ушное отверстие овальное. Горловая чешуя коническая,
с порой на вершине или плоская. Спина покрыта гладкой или ребристой
плоской многоугольной чешуей, увеличивающейся к бокам туловища так, что
границу ее с боковой чешуей провести невозможно; брюшная чешуя ромбо-
видная или субтреугольная, черепитчатая, гладкая или ребристая; анальные
поры расположены почти прямой линией на более крупных, чем брюшные,
чешуйках; между рядом анальных пор и анальной щелью имеется несколько
крупных, сходных с брюшными, чешуй, плавно уменьшающихся к анальной
щели. Передняя конечность, вытянутая вперед вдоль тела, достигает концами
пальцев конца морды, задняя — подмышечной впадины; сверху и снизу покры-
ты плоской, слабо ребристой или гладкой чешуей, более или менее однородной
и сходной с чешуей туловища, среди которой на верхних поверхностях могут
быть разбросаны бугорки, сходные со спинными. Пальцы сверху и с боков по-
крыты двумя рядами чешуй; с чехлом когтя соприкасаются по две чешуйки свер-
ху и с боков; чехол состоит из мелкой верхней и крупной нижней пластинок,
которые могут иметь много продольных ребрышек.
Распространены на Аравийском полуострове и в Ю.-З. Азии до Инда.
Северная граница ареала проходит около 35° с. ш. Род включает три вида,
ошибочно объединяемых ранее с видами рода Alsophylax.
Таблица для определения видов рода Bunopus
1(4). Нижнечелюстные щитки есть; анальных пор у самцов три — шесть.
2(3). Увеличенные подхвостовые щитки есть; чешуй вдоль продольной оси тела снизу менее
130; горловая чешуя плоская ...............................В. crassicauda Nik.
3(2). Увеличенных подхвостовых щитков нет; чешуй вдоль продольной оси тела снизу более
130; горловая чешуя полуконическая........................В. spatalurus J. Anderson
4(1). Нижнечелюстных щитков нет; анальных пор у самцов 5—16......................
. ....................................................В tuberculatus Blanf.
ЮЖНОАЗИАТСКИЙ ГЕККОНЧИК БУГОРЧАТЫЙ — BUNOPUS TUBERCULATUS BLANF.. 1874
Типовая территория: «Gedrosia in Persique meridionali» (terra typica
restrict a: Smith, 1935, Белуджистан).
Кариотип: 2n = 42 (более детальные сведения отсутствуют) [312].
1874 — Bunopus tuberculatus Blanford, Ann. Mag. Nat. Hist London, Vol. 13, p. 454.
1885 — Alsophylax tuberculatus, Boulenger. Cat. Liz. Brit Mus., Vol. 1, p. 20.
1887 — Bunopus blanfordii Strauch, Mem. Acad. Sci. St. Petersbourg, 35 (7), 2, p. 61.
1938 — Bunopus biporus Werner, Zool. Anz., Leipzig, vol. 121, S. 267.
1967 — Bunopus abudhabi Leviton et Anderson, Proc. Calif. Acad. Sci., 35, 9, p. 164.
Диагноз. Нижнечелюстных щитков нет; у самцов. 5—16 аналь-
ных пор.
Лектотип. BMNH 74.11.23.85 RR 1946. 8.22.84 «near Bampur, Baluc-
histan, coll. W. T. Blanford» $> ad L = 50,7 мм; хвост регенерирован. Ноздря
расположена между межчелюстным, первым верхнегубным и тремя мелкими вы-
пуклыми носовыми щитками, средний из них несколько крупнее остальных.
Между центрами глаз 14 чешуй. Верхнегубных щитков 13—14, нижнегубных —
по 10. Подбородочный щиток трапециевидный, с неровным задним краем, к кото-
рому примыкают четыре мелкие горловые чешуйки. От подбородочного щитка
до анальной щели снизу 153 чешуйки, поперек середины брюха — около 30.
Снизу четвертого пальца задних конечностей по 18 пластинок.
Описание (по 132 экз. из колл. ИЗАНУ, ЗИН, AMNH, BMNH, CAS,
FMNH, MCZ, MNHP, MZSF, MNMB, NMW, SAM, UMMZ, USNM рис. 38).
(L <?ad(n = 54) 24,6—53,8 мм (41,98 ± 0,72); L $ ad (n = 59) 31,9 —55,7 мм
(43,58 ± 0,78); L/Lcd(pjin и $ ad) (n = 54)0,69—1,09 (0,83 ± 0,030), индекс
высоты/ширины головы (n = 69) 50 — 68 (60,78 ± 0,41); диаметр глаза
(n = 75) 37—61 (48,20 ± 0,74); верхнегубных щитков (n = 220) 10—15
(12,42dz0,09); нижнегубных щитков (и = 220) 7—14 (9,97 ± 0,12); чешуй
между центрами глаз (п = 115) 10—19 (14,72 ± 0,23); отношение длины
спинного бугорка к длине тела (и = 96) 11—39 (11,88 ± 6,12); чешуй, ок-
ружающих спинной бугорок (п = 103), 11—18 (12,82 ± 0,20); чешуй вдоль
продольной оси тела снизу (п = 105) 122—170 (145,31—1,28); чешуй поперек
89
Рис. 38. Bunopus tuberculatus:
A — голова сбоку, Б — лицевая часть спереди, В — голова сверху, Г — подбородок,
Д — носовые щитки, Е — спина, Ж — брюхо, 3 — палец снизу, И — преанальные по-
ры, К — хвост снизу, Л — хвост сверху (Л—В — CAS 97847, Абу Даби; остальное —
ЗИН 11488, Абинефт, Месопотамия)
брюха (п = 76) 25—34 (30,17 ± 0,26); анальных пор d (п = 50) 6—16 (8,26 ±
± 0,36), 2 (п = 6) 6—13 (9,33 ± 1,05); подпальцевых пластинок (п = 236)
16—24 (20,15 ± 0,10). Первый носовой щиток может быть оттеснен от ноздри
двумя (21,01 %), тремя (78,99 %) другими чешуйками и с ней не соприкасаться,
они соприкасаются (0,8 %) или разделены между собой одной (69,9 %), двумя
(15,5 %) или тремя (13,8 %) чешуйками. Ухо овальное. Подбородочный щиток
трапециевидный (100 %), с неровным задним краем. Сзади к нему примыкают
три (6,67 %), четыре (24,4 %), пять (50,54 %), шесть (11,01 %) или семь
(7,34 %) горловых чешуек. Горловая чешуя субконическая, с порой на вершине.
Спина покрыта гладкой или слабо ребристой плоской чешуей и 12—16 продоль-
ными рядами крупных ребристых трехгранных бугорков. Чешуя брюха глад-
кая или ребристая.
Прижизненная окраска сверху светло-коричневых тонов.
90
50* 70*
Рис. 39. Распространение В. tuberculatus (/), В. crassicauda (2), Т. amictopholis (3)
Голова покрыта неясным темно-бурым узором. По бокам головы от глаза
через шею бурая полоса выходит на бока туловища и в виде отдельных пя-
тен продолжается до основания передних лап. Поперек туловища четыре-пять
такого же цвета широких с ровными краями поперечных полос, на хвосте — до
8—11. На конечностях неясные тонкие поперечные бурые полоски. Рисунок
верхних частей тела может частично исчезать Нижняя поверхность светлая.
Половой диморфизм и возрастная изменчивость.
Самцы отличаются от самок наличием анальных пор; у самок иногда имеются
лишь их слабые следы. Различия в размерах между полами, характерные для
видов рода Alsophylax, здесь отсутствуют. Характер возрастной изменчивости
индекса L/Lcd представлен на рис. 41.
Распространение. От крайнего юга СССР (Бадхыз), Афганистана
и В. Пакистана через Иран, Ирак и Сирию до Аравийского полуострова вклю-
чительно (рис. 39).
Кадастр к рис. 39.
СССР, Т С С Р: 1 — Бадхыз, Эр-Ойланд-Дуз (ИЗАНУ).
Сирия: / — лагерь Демезин (CAS); 2 — Дейр-эз-Зор (MNHP).
Ливан: 1 — Бейрут (AMNH).
Иордания: 1 — Иерусалим (ЗИН); 2 — Акаба (MCZ).
Израиль: 1 — Айн Хусуб (NMW); 2 — пустыня Негев, Айн Яхав (SAM); 3 — Юж-
нее Вади-эль-Арава; Йотвата (MZSF).
Ирак: 1 — Эль-Хадр [269]; 2 — р. Абу-Нафт (ЗИН); 3 — ст. газопровода К-3 около
пос. Хадита (MCZ); 4 — р. Газила (ЗИН); 5 — г. Мандали (ЗИН); 6 — пров. Васит, 10 км сев.
г. Эль-Кут [269]; 7 — Эс-Самава (NHMB); 8 — между пос. Зубайдех и Эш-Шабака [269].
Аравийский полуостров: 1 — Эль-Джумайма, в. пос. Рафха (MCZ); 2 —
8 км з. г. Унайза (CAS); 3 — Дахран (FMNH); 4 — Абкайк (CAS); 5 — Эль-Укайр [228]; 6 —
Набхания (CAS); 7 — Джидда, 21° 20' с. ш., 39° 10' в. д. (CAS); 8— Буррайман (CAS); 9 — Абу-
Даби: в радиусе 15 км от 23° 45' с. ш., 53° 33' в. д. (CAS); 10 — Оман. Талиль [177]; 11 — Бур-
за, 19° 25'с. ш., 54° 25'в. д. [228]; 12 — Ибн-Хаутар [228]; 13 — Эш-Шисар, 18° 15' с. ш.,
53°40'в. д. [228]; 14 — Хадрамаут (MNHP); 15 — Мишаси-эль-Харрара (CAS); 16 — Эд-Дайд
(ZFMK).
91
Иран: 1 — р. Генгир (ЗИН); 2 — г. Дехлоран (UMMZ); 3 — Дизфуль (ЗИН); 4 —
г. Шустер (ЗИН); 6 км з. пос. Гетвенд (MCZ); 5 — Махур Биринджи (MCZ); 6 — пос. Гольгир
(Гулып или Гуржир) (ЗИН); 7 — дор. Ахваз — Хефтгель (CAS); 8 — Ахваз (Насриэ-и-Ахва-
зе) (ЗИН); Р— р. Карун от места Харма, Зиарет Макрам-Эмин-эль-Мумунад (ЗИН); 10 — 52—
56 км сев.-з. пос. Генаве по дор. на Бехбехан (CAS); 11 — 10—13 км ю. пос. Бораздан по дор.
из Бушира (CAS); 12 — Ахрам, пров. Фарс (CAS); 13 — Касре-Ширин (ЗИН); 14 — Дамган
(ЗИН); 15 — 65 км в. г. Тегеран, г. Семнан (MNHP); 16 — пос. Ниазабад в стр. Хаф (ЗИН);
17 — Хельван (NMW); 18 — кол. Чах-и-Бена [59]; 19 — Сарр-Чах [59]; 20 — между Фай-
забадом и Бесираном [92]; 21 — кол. Ча-и-Гюише (ЗИН); между кол. Хаджи-и-Ду-Чаги и
Гюише [59]; 22 — Нейзар в Сейстане (ЗИН); 10 км ю.-з. р. Гильменд, 31° 03' с. ш., 61° 38' в. д.
(CAS); Хоуздар; Чах-и-Новар в Сейстане; Гумбез-и-Новар в Сейстане (ЗИН); 23 — Хормек
(ЗИН); 24 — ю. г. Захедан, 29° 08' с. ш., 61° 20' в. д. (CAS); 25 — Себзеваран [315]; стр. Сар-
гад, уроч. Андо (ЗИН); 26 — уроч. Дуз-аб на равнине Тагаб между пос. Шураб и Пенсаре
[60]; 27 — Калган (ЗИН); 28 — пос. Хану, 100 км ю. пос. Себзеваран (NMW); 29 — Бендер-
Ленге [223]; 30 — 13 и 40 км сев. г. Бендер-Аббас (NMW); 31 — сев. г. Минаб (CAS); 32 —
32 км з. г. Бемпур (27° 10' с. ш., 60° 10' в. д.) (CAS); окр. Бемпура (BMNH); Ираншехр (ЗИН);
33 — Шураб в Мегасе (ЗИН), Хаш [303]; 34 — пос. Дехек (ЗИН); гора Раган [172]; 35 — пос.
Пишин [172]; 36 — Бахукелат [172]; 37 — Зиарет (NMW); Макран, Багу (ЗИН); 38 — Чах-
бехар [223]; 39 — Дору не [303].
Афганистан: / — Адраскан (Герат) (NMW); 2 — 30 км ю. г. Мукур [209]; 3 — Кан-
дагар [172]; 4 — 35 км ю. г. Фарах (CAS); 5 — Чах-и-Ангир (CAS); 6 — 10 км в. с. Дарвешан
(CAS).
Пакистан: / — округ Сиби; Кач [263]; 2 — окр. Нушки [263]; 3 — Далбандин
[258]; 4 — Харан [258]; 5 — с. Манд [172]; 6 — р. Дашт [172]; 7 — ю.-в. с. Пасни [258]; 8 —
г. Хайдарабад [263].
Не найдены на картах: а) Зиарет Сеид-Ханат, Месопотамия (ЗИН);
б) И р а н: Isfandak, Tumb Island; Пакистан: Baluchistan, Saman[ 1 721; в) Leb-e-Kal [310].
Систематика и географическая изменчивость.
К. Шмидт [283] выразил сомнение в том, что бугорчатый геккончик, обитающий
на обширной территории, не проявляет географической изменчивости. Ю. Го-
релов и др. [48] предположили, что на востоке ареала (в частности, в Бадхызе)
этот вид представлен особым подвидом, более мелким, с меньшим количеством
анальных пор и чешуй позади подбородочного щитка. В то же время Е. Ар-
нольд [177] объединил с этим видом два других — В. blanfordi Str. и В. abud-
habi Leviton et Anderson — на основании отсутствия у этих форм четких раз-
личительных признаков.
Результаты изучения бугорчатых геккончиков из разных частей ареала
склоняют нас принять последнюю точку зрения. Однако заметна слабо выражен-
ная географическая изменчивость по некоторым признакам. Так, от Бадхыза
и Белуджистана на востоке ареала к Месопотамии постепенно возрастает число
анальных пор, но затем оно вновь убывает в направлении к югу Аравийского
полуострова. С востока на запад и далее на юг возрастает выраженность про-
дольной ребристости чешуй морды, брюха и нижней поверхности хвоста. На
крайнем юге ареала (от Объединенных Арабских Эмиратов до султаната Оман)
ребристость чешуй брюха вновь падает, уменьшается шиповатость подпальце-
вых пластин и появляется ряд подхво?товых щитков в конце хвоста. Однако
последние признаки в той или иной степени характерны для всех популяций,
населяющих Аравийский полуостров, что и послужило основанием для объеди-
нения указанных видов [1771. Таким образом, слабая дифференциация призна-
ков и наличие переходов не позволяют пока выделить внутри этого вида какие-
либо формы, имеющие таксономическое значение.
Стации и количественные данные. Н. Зарудный [601
сообщал, что этот вид широко распространен в В. Иране, где во многих котлови-
нах весьма обычен и по соответствующим почвам поднимается в горы. Автор
отмечает, что... «она по возможности избегает слишком каменистых почв и, не
особенно долюбливая слишком рыхлые и глубокие пески, с наибольшей охотой
показывается на глинах, суглинках и супесках, частью обнаженных, частью по-
крытых тонким слоем песка и иногда солонцеватых» (с. 8—9) В Объединенных
Арабских Эмиратах бугорчатый геккончик встречается на такырах, сцементи-
ровавшихся песках, и на песках рыхлых, эоловых [177]. Ш. Минтон [263] счи-
тает, что эта ящерица в окр. г. Нушки (Пакистан) обычна только в песчаных
местностях. Однако в горах Белуджистана геккончик был добыт на высоте около
700 м н. у. м. в пойме небольшой горной речки среди валунов и низкорослой рас-
92
Рис. 40. Биотоп Bunopus tuberculatus и Tenuidactylus caspius (гипсово-солончако-
вая впадина Эр-Ойлан-Дуз, Бадхыз, ТССР)
тительности. По левому притоку Гильменда — р. Торнак (Афганистан) — он
поднимается до высоты 1425 м, а у Гаоми-Фаринги до 2100 м н. у. м. [172; 245].
По письм. сообщ. Ш. Минтона, в Палестине геккончик обитает в весьма
аридной местности на песчаной почве с разреженной кустарниковой раститель-
ностью. Возле заброшенного здания он наблюдал три ящерицы данного вида.
Впервые на территории СССР две особи бугорчатого геккончика были добыты
в апреле 1972 г. в котловине Эр-Ойлан-Дуз (Бадхыз на высоте 315—460 м н. у. м.)
(рис. 40). Ящерицы обнаружены под камнем на эффузивном останце и у вхо-
да в нору песчанки. Здесь же мы 15.V 1976 г. добыли геккончика у подножия
невысокого глинистого холма, полностью лишенного растительности, в непосред-
ственной близости от влажного шора (солончака). 10.IV 1977 г. еще один эк-
земпляр геккончика был добыт нами в 2—3 м от шора на довольно крутом гли-
нистом растрескавшемся склоне без растительности, а выше, через 10 м, начина-
лось основание останца с кустами солянок и щебнистыми склонами. В последу-
ющие годы (май 1982 г., апрель 1984 г.) тщательные поиски этой ящерицы во
впадине Эр-Ойлан-Дуз не увенчались успехом, что позволяет считать ее одним
из самых редких видов фауны СССР.
Отношение к температуре. Мы добыли активного геккончика
на поверхности при температуре почвы 17°. По литературным данным [172],
встречены активные ящерицы данного вида с температурой тела 26—34°, а у
особей, выкопанных из нор, температура тела была выше — 33,4—37°.
Суточный цикл активности. Н. Зарудный [60] относил бу-
горчатого геккончика к «настоящим ночным ящерицам», другие авторы [288]
подтверждают это или относят данный вид к сумеречным животным [313].
Первый экземпляр был добыт нами в 21 ч, второй — в 21 ч 30 мин. Более де-
тальные сведения до сих пор отсутствуют.
Сезонный цикл активности. Первая активная особь в
Ю. Туркмении добыта нами 10.IV 1977 г. Весна в том году была прохладная,
так что в данном случае указанную находку на территории СССР можно считать
первой. Коллекционные материалы из Ирака собраны с января по октябрь.
В Иране, по сборам Н. Зарудного,— от 13.11 1901, 10.1, 18.11 и 25.11 1904 г.—
находки весной и 8.Х 1903 г. (ст. стиль) — последняя ящерица встреченная
осенью.
93
Линька. Данные о ней отсутствуют.
Питание. Бугорчатый геккончик в неволе поедал термитов [48]. В двух
желудках этих ящериц, вскрытых нами, обнаружены остатки паука (1), прямо-
крылого (1), муравья (1) и перепончатокрылого (ближе не определенного, 1).
Размножение. Самки, добытые 14 и 21.V в Сев. Белуджистане,
имели одно-два яйца, почти полностью готовые к откладке [263]. Сведения о
размножении этого вида на территории СССР отсутствуют.
В р а г и. А. Левитон [242] обнаружил бугорчатого геккончика в желудке
афганского литоринха (Lytorhynchus ridgewayi).
Особенности поведения. Еще Н. Зарудный [60] отмечал, что
эта ящерица «в противоположность большинству... гекконов не стенная, а зем-
ная... так как сравнительно редко ее приходится видеть на вертикальных по-
верхностях» (с. 8—9). Этот вид, бегающий по земле и случайно лазающий по
вертикальным поверхностям, отмечали и другие авторы [313]. Убежищами гек-
кончику, видимо, служат норы грызунов, возможно, соляные корки на берегу
шора, пространства под камнями. По крайней мере геккончик в Эр-Ойлан-Дузе
однажды был добыт у норы песчанки [48], на Аравийском полуострове их добы-
вали у входа в норы, в норах грызунов и под кусками дерева 1177].
Практическое значение и охрана. Как чрезвычайно ред-
кий вид включен во второе издание Красной книги СССР (1984). Обитает в на-
шей стране на территории Бадхызского заповедника, в дополнительных ох-
ранных мероприятиях пока нет необходимости.
ЮЖНОАЗИАТСКИЙ ГЕККОНЧИК АРАВИЙСКИЙ —
BUNOPUS SPATALURUS J. ANDERSON, 1901
Типовая территория: Вади Джамал вблизи Адена, НДРЙ.
Кариотип: не изучен.
1901 — Bunopus spatalurus J. Anderson, Proc. Zool. Soc. London, N 3/4, p. 137.
1959 — Trachydactylus jolensis Haas and Battersby, Copeia, no. 3, p. 200.
1967 — Trachydactylus spatalurus, Leviton and Anderson, Proc. Calif. Acad. Sci., 4th Ser., Vol.
35, no. 9, p. 166.
Диагноз. Нижнечелюстные щитки есть; у самца три — шесть анальных
пор; горловая чешуя коническая; вдоль продольной оси тела снизу более 130
чешуй; подхвостовых щитков нет.
Голотип. BMNH 99.12.13.40RR 1946.8.23.59 «Jimil Valley, Southern
Arabia, leg. A. B. Percival». <? juv. L = 34,6 мм, хвост регенерирован и на
конце слабо вздут. Ноздря расположена между четырехугольным межчелюст-
94
Рис. 42. Bunopus spataluius:
A — общий вид, Б, В — спина, Г, Д — брюхо, Ж — палец
поры (Л, В, Д - BMNH 1978. 1095, Дофар, Оман — В. s.
BMNH 1971 1207 - В с hajarensis)
снизу, Е — преанальные
spatalurus; остальное —
ным, первым верхнегубным и тремя выпуклыми мелкими носовыми щитками,
более или менее равными друг другу. Первые носовые щитки разделены между
собой одной чешуйкой. Между центрами глаз 14 чешуй. Верхнегубных щитков
по 15, нижнегубных — 11/10; четыре анальные поры; подпальцевых пластинок
по 16.
О п и с а н и е (по 16 экз. из колл. BMNH и MNHP, рис. 42). L ad(n = 12)
42,6—66,5 мм; (50,88 + 1,79); половой диморфизм не выражен. L/L cd общ.
(п = 8) 1,20—1,45 (1,29 ± 0,03); индекс ширины/высоты головы (п = 12)
95
30*
40*
50* 60*
Рис. 43. Распространение Bunopus spatalurus (/), Tropiocolotes scortecci (2),
T. tripolitanus (5), T. steudneri (4)
57—67 (63,08 ± 0,84); диаметр уха/глаза (и = 11) 38—53 (45,50 ± 1,49); верх-
негубных щитков (n = 32) 13—17 (15,10 ± 0,20); нижнегубных — (п = 32)
10—13 (11,50 ± 0,16); чешуй между центрами глаз (п = 16) 12—16 (13,81 —
0,32); длина спинного бугорка/L (п = 16) 7—10 (8,13 ± 0,18); чешуй вдоль тела
снизу (п = 15) 133—160 (143,4 ± 1,96); чешуй поперек брюха (п = 12) 30—50
(36,3 ± 1,54); анальных пор (п = 2) 4—5; подпальцевых пластинок (п = 32)
14—17 (16,09 ± 0,16).
Три — пять носовых щитков; первые носовые разделены между собой
одной (14,2 %), двумя (73,7 %) или тремя (7,1 %) чешуйками; высота первого
верхнегубного щитка от ноздри до края рта больше его ширины вдоль края рта;
до орбиты доходит 7—7,5 верхнегубных щитков. Чешуя морды гладкая
или слабо ребристая. Подбородочный щиток пяти- (50 %), шести- (35.8 %),
семиугольный или трапециевидный с неровным задним краем (по 7,1 %); пара
нижнечелюстных щитков, иногда почти не выраженных (около 30 % просмот-
ренных экземпляров имеет очень слабо выраженную вторую пару), соединена
между собой швом (46,5 %), разделена одной (40 %) или двумя (13,5 %) чешуй-
ками; горловая чешуя коническая. Чешуя брюха гладкая или ребристая. Глад-
кие или ребристые спинные бугорки выражены в различной степени. Конеч-
ности покрыты однородной чешуей, среди которой только на верхней поверх-
ности бедра можно различить отдельные крупные чешуи, напоминающие спин-
ные бугорки. Подпальцевые пластинки несут на себе вдоль свободного края ряд
из нескольких бугорков, заканчивающихся шипиком. Каждый бугорок окру-
жен сзади полукольцом из значительно более мелких шиповатых бугорков.
Анальные поры выражены значительно хуже, чем у остальных представи-
телей рода.
Окраска и рисунок (после фиксации в спирте). Основной фон
верхних поверхностей коричневатый. Голова лишена рисунка. От шеи до пояс-
ницы проходит четыре-пять темно-шоколадно-коричневых широких поперечных
полос. Каждая полоса со слабо изогнутым передним и ровным задним краем.
На хвосте до 9—10 такого же цвета широких ровных полос. Конечности покры-
ты несколькими тонкими такого же цвета поперечными полосками. Нижние
поверхности белые.
96
Распространение. Южное и юго-восточное побережье Аравий-
ского полуострова (рис. 43).
Кадастр к рис. 43.
1 — Вади-Джимил (BMNH) [175]; 2 — с. от Замах, Хадрамаут (BMNH); 3— Оман;
16 км ю. Марказ-Тамарид [177]; 4 — о. Масира [177]; 5 — Султанат Оман; Аваби [177]; 6 —
Вади-Сехтан [177]; 7 — Эр-Рустак [177]; 8 — Вади-Кунд, Джебел Асвад [177]; 9 — Маскат
(MNHP); 10 — ю. Аква [177]; 11 — Идхн [177]; 12 — Манама [177]; 13—Сиджи [177]; 14— Вади-
Асима [177]; 15 — Мазафи [177]; 16— равнина Тайябад [177]; 17 — Джебел Фаях [177];
18 — Хаур-эль-Факкан [177]; 19 — Рас-эль-Дибба [177]; 20 — Дхая, с. пос. Рас-эль-Хайма
[177]; 21 — ю. пос. Эш-Шаам [177].
Половой диморфизм и возрастная изменчивость.
Отличие самцов от самок этого вида заключается только в присутствии аналь-
ных пор у первых, при этом у самцов аравийских геккончиков анальные поры
выражены значительно слабее, чем у самцов остальных видов рода. Недостаточ-
ное количество молодых особей в нашей выборке не позволило изучить возраст-
ную изменчивость вида. Можно предположить, что у данного геккончика значе-
ние индекса L/Lcd (в противоположность видам Alsophylax) с возрастом увели-
чивается.
Систематика и географическая изменчивость.
Аравийский геккончик — вид, восстановленный Е. Арнольдом [1751 в роде
Bunopus. В настоящее время он представлен северной (побережье Персидского
залива) и южной (побережье Аденского залива) группами популяций, которым
придан статус подвидов [177].
Таблица для определения подвидов В. spatalurus
1 (2). Чешуя спины гладкая .......................В. s. spatalurus J.Anderson
2 (1). Чешуя спины ребристая....................... В. s. hajarensis Arnold
ГЕККОНЧИК АРАВИЙСКИЙ ОБЫКНОВЕННЫЙ —
BUNOPUS SPATALURUS SPATALURUS J. ANDERSON
1980 — Bunopus spatalurus spatalurus Arnold, J. Oman. Stud. Spec. Rep. N 2, p. 273—332.
Диагноз. Чешуя головы и спины (включая слабо выраженные бугорки)
гладкая; носовых щитков три, изредка четыре; Lmax = 50—55 мм.
Тип видовой. Описание подвида, основанное на типовых экземплярах вида
и Trachydactylus jolensis (BMNH 1953,1.6.96), а также экземпляре из Омана
(BMNH 1978.1095) по остальным признакам существенно не отличается от ви-
дового.
Обитает на юге Аравийского полуострова от Адена до Дофара [177].
ГЕККОНЧИК АРАВИЙСКИЙ ХАДЖАРСКИЙ —
BUNOPUS SPATALURUS HAJARENSIS ARNOLD, 1980
Типовая территория: 25° 18,5' с. ш., 56° 10' з. д., Объединенные Арабские
Эмираты.
1980 — Bunopus spatalurus hajarensis Arnold, J. Oman Stud. Spec. Rep. N 2, p. 277.
Диагноз. Чешуя головы и спины (включая крупные, но сильно упло-
щенные спинные бугорки) отчетливо ребристая; носовых щитков четыре, редко
ПЯТЬ, Lmax 65—70 ММ.
Голотип [1771. $ ad Wadi Hamm Masafi (25° 18,5'N; 56° 10'E) United
Arab Emirates: BM 1973. 1801, (E. N. Arnold, 27.5. 1973).
L — не указано. Межносовые щитки разделены двумя чешуйками; ноздри
расположены между межчелюстным, первым верхнегубным, межносовым и тре-
мя мелкими заносовыми щитками; 15—14 верхнегубных щитков (12—11 до сере-
дины глаза); по 11 нижнегубных (по 9 до середины глаза). Позади подбородоч-
ного щитка пара крупных чешуй (слабо выраженных нижнечелюстных щитков),
разделенных между собой двумя горловыми чешуйками; по 18 подпальцевых
пластинок.
Описание подвида составлено по 10 паратипам из Британского
музея и трем ювенильным особям из Парижского музея; по остальным призна-
97
кам, кроме указанных в диагнозе, существенно не отличается от видового.
Обитает в В. Арабских Эмиратах, Сев. Омане на юг до о. Масира [177].
Экология почти не изучена. Некоторые сведения по данному вопросу
имеются только в работе Е. Арнольда и М. Галлагера [178], которые сообщают,
•что Bunopus spatalurus — обычный сумеречный земляной геккон, широко рас-
пространенный в разнообразных участках каменистых равнин, а также в вади
(сухих руслах) и каменистых холмогорьях, примыкающих к скалам. Именно
в таких местах были добыты все 59 экз. коллекции, указанные авторами. Все
местообитания расположены на высоте 270—500 м н. у. м. Соотношение полов:
41,6%. $50,0 %, juv. 8,4 %.
Судя по коллекционным материалам, ящериц добывали с февраля по де-
кабрь.
Активные животные часто приподнимаются на прямых ногах в манере, ти-
пичной для многих открыто-наземных пустынных гекконов, медленно и часто
перемещаются в поисках пищи. Температура тела двух кормящихся особей сра-
зу же после поимки была 35°.
Дневными убежищами этим ящерицам часто служат совсем мелкие камни.
В песчаных участках по краям возвышенностей их замещают В. tubercula-
tus и виды рода Stenodactylus, в частности S. leptocosymbotus Leviton et S. An-
derson.
ЮЖНОАЗИАТСКИЙ ГЕККОНЧИК ТОЛСТОХВОСТЫЙ — BUNOPUS CRASSICAUDA NIK., 1907
Типовая территория: Кум, Малят-Абад и Хара-Магомед-Абад в пров. Ирак*
Аджеми (Сев. -3. Иран).
Кариотип: не изучен.
1907 — Bunopus crassicauda Никольский, Ежег. Зоол. Муз., т. 10, № 3—4, с. 261.
1963 — Alsophylaxcrassicauda, S. Anderson, Proc. Calif. Acad. Sci., 4thSer., Vol. 31, p. 474.
Диагноз. Нижнечелюстные щитки имеются; у самцов три — шесть
анальных пор; горловая чешуя плоская, гладкая; чешуй вдоль продольной оси
тела снизу менее 130; подхвостовые щитки есть.
Лектотип. Вследствие того что вид был описан по серии синтипов, мы
выделили из них наиболее сохранившийся экземпляр в качестве лектотипа:
ЗИН 10233 сГ ad Хара-Магомед-Абад, 10.V 1904 г., leg. Н. Зарудный.
L = 40 мм; хвост регенерирован. Ноздря расположена между четырех-
угольным межчелюстным, первым верхнегубным и тремя выпуклыми мелкими
носовыми щитками, из которых средний незначительно крупнее остальных;
первые носовые щитки разделены между собой одной чешуйкой. Между центра-
ми глаз 14 чешуй. Верхнегубных щитков девять-десять, нижнегубных слева
девять, справа ряд поврежден. Чешуй вдоль продольной оси тела снизу около
120. Анальных пор пять, они расположены почти по прямой линии; подпальце-
вых пластинок 19—20.
Описание (по 21 экз. из колл. ЗИН, AMNH, CAS, SAM и MNHP)
(рис. 44).
L d ad (n = 12) 39,9—45,3 (44,35 ± 0,82) мм; L $ ad (n = 9) 37,3—54,5
(46,30 ± 1,70) mm; L/Lcd общ. (n = 4) 0,97—1,13 (1,06 ± 0,03); индекс шири-
ны/высоты головы (n = 18) 57—71 (61,61 ± 0,83); диаметр уха/глаза (п = 18)
27—46 (38,0 ±1,17); верхнегубных щитков (п = 41) 9—12 (10,40 ± 0,12);
нижнегубных (п = 41) 8—11 (9,07 ± 0,12); чешуй между центрами глаз (п =
= 21) 13—17 (15,10 ± 0,24); длина спинного бугорка/L (п = 20) 11—16
(13,2 ± 0,31); чешуй, окружающих спинной бугорок (и = 20), 8—10 (9,40 ±
±0,15); чешуй вдоль тела снизу (п = 21) 101—127 (112,3 ± 1,46); чешуй попе-
рек брюха (п = 19) 26—31 (27,60 ± 0,34); анальных пор (п = 12) 3—6 (5,16 ±
± 0,27); подпальцевых пластинок (п = 40) 16—20 (17,45 ± 0,19).
Три выпуклых носовых щитка, из которых средний часто несколько круп-
нее остальных или равен им; первые носовые щитки разделены одной (81%)
или двумя чешуйками; высота первого верхнегубного от ноздри до края рта не-
значительно меньше его ширины вдоль края рта; до орбиты достигают 4,5—
6 верхнегубных щитков; подбородочный щиток пятиугольный; две (16 %), три
98
Рис. 44. Bunopus crassicauda:
A — общий вид, Б — голова сбоку, В — голова сверху, Д — спина, Е — брюхо, Ж — па-
лец снизу, 3 — хвост снизу, И — хвост сверху (Л, Д, 3, И — CAS 140599, Газвин, Иран;
остальное —AMNH 99663, пров. Тегеран, Иран)
(63 %) или четыре (21 %) пары нижнечелюстных щитков, первая значительно
крупнее остальных и соединена между собой широким швом. Горловая чешуя
крупная и гладкая. Среди гладкой спинной чешуи в 9—12 продольных рядов
расположены округлые в основании полуконические гладкие, со слабой гранью
в конце бугорки. Брюшная чешуя крупная, плоская, гладкая, округло-шести-
угольная, значительно крупнее спинной. Чешуя хвоста сверху гладкая, на
99
хвостовых бугорках следы ребристости; снизу хвоста на базальных сегментах
три ряда чешуй, каждая центральная чешуйка увеличивается от первого коль-
ца к последнему (1 > 1 > 1; иногда 1 > 1 > 2 или 1 > 2 > 2); через два-три
сегмента появляются щитки; на протяжении х/4 длины хвоста на первом сегмен-
те их три (1 = 1 = 1), дальше — по два (1 = 1). Боковая чешуя пальцев, в от-
личие от других видов рода, гладкая или с очень слабыми следами ребристости.
Окраска и рисунок (после фиксации в спирте). Основной фон
верхних поверхностей от светло-шоколадного до песочно-желтого. От затылка
к пояснице проходят четыре — шесть темно-коричневых широких поперечных
полосы с почти прямыми краями. На хвосте восемь — десять такого же цвета
полос. На голове имеется неясный темный узор. Конечности покрыты попереч-
ными тонкими темно-коричневыми полосами. Нижние поверхности белые.
Распространение. Обладает небольшим ареалом к югу и западу
от Тегерана (Иран) (рис. 39).
Кадастр к рис. 39.
И р а н: 1 — г. Казвин (CAS); 2 — Хара-Магометабад; Чалджатабад [92]; 3 —- 8 км ю.
Тегерана (SAM); 4 — г. Кум [92].
Половой диморфизм и возрастная изменчи-
вость. Самцы отличаются от самок этого вида наличием анальных пор; у са-
мок иногда имеются лишь их следы. У исследованных нами экземпляров самки
несколько крупнее самцов, однако это различие не достоверно. Вследствие от-
сутствия молодых особей в нашей выборке детально изучить возрастную из-
менчивость не представилось возможным.
Систематика и географическая изменчивость.
Данный узкоареальный вид не проявляет географической изменчивости, однако
он представляет интерес в связи с тем, что по некоторым признакам отличается
от других представителей рода: у него полностью или частично отсутствует
ребристость боковой чешуи пальцев, спинных бугорков, горловых чешуй
и чешуй хвоста; горловая и брюшная чешуя крупная и плоская — следствием
этого является меньшее число чешуй вдоль продольной оси тела снизу; меньше
нижне- и верхнегубных щитков; имеются очень крупные подхвостовые щитки
(у «В. t. abudhabi» они также есть, однако выражены значительно меньше).
Ф. Вернер [309] отмечал, что с его точки зрения этот вид является переходным
между видами родов Bunopus и Gymnodactylus. Вид, в отличие от остальных
представителей рода, обитает в более умеренном климате (на высоте 1000—
2000 м), что, вероятно, явилось причиной уменьшения ребристости чешуй и щит-
ков, а также других различий, связанных, видимо, с водообменом.
Экология не изучена. Две особи этого вида были добыты в полдень
5.VIII 1965 г. [265] на краю окультуренной аллювиальной равнины [265].
Температура в тени была 53°. Одна ящерица обнаружена под комком земли,
другая, вероятно вспугнутая из убежища, передвигалась среди невысокого кус-
тарника.
Род карликовый геккон — Tropiocolotes Peters, 1880
Типовой вид: Tropiocolotes tripolitanus Peters, 1880.
1880 — Tropiocolotes Peters, Mon. Berlin. Acad., S. 306.
1885 — Stenodactylus, Boulenger, Cat. Liz. Brit. Mus., Vol. 1, p. 16 (part.).
Диагноз. Носовые щитки и чешуйки плоские; свободные (верхние)
края верхнегубных щитков загибаются за верхнюю поверхность морды так, что
каждый щиток находится не в одной плоскости; высота первого верхнегубного
щитка от ноздри до края рта заметно меньше его ширины вдоль края рта; спин-
ная чешуя однородная, плоская или несколько выпуклая, ребристая или глад-
кая; спинных и хвостовых бугорков нет; хвостовые сегменты сверху неразличи-
мы; снизу хвост покрыт однородной чешуей или одним рядом увеличенных
щитков (по два на сегменте); анальные поры отсутствуют или их не более семи
(только у самцов); пальцы слабо искривлены, не расширены и не сдавлены с бо-
100
ков, оснащены когтями, их боковая чешуя бахромы не образует (может нести
ши пики), снизу покрыты одним рядом гладких или ребристых подпальцевых
пластинок. Зрачок вертикальный, с зазубренными краями. Реснитчатые вы-
росты в верхней части орбиты есть.
Распространены в Сев. Африке (сев. 10° с. ш.), на Аравийском полуострове
и в Ю.-З. Азии до р. Инд. Северная граница ареала рода проходит около
35° с. ш. Род включает восемь видов, которые мы разделяем на три подрода.
Таблица для определения подродов рода Tropiocolotes
1 (2). Подпальцевые пластинки ребристые...................... Tropiocolotes Peters
2(1). Подпальцевые пластинки гладкие.
3(4). С ноздрей соприкасаются три носовых щитка, из них первый (интерназальный) значи-
тельно крупнее остальных ........................................Microgecko Nik.
4 (3). С ноздрей соприкасается два-три носовых щитка, почти равных друг другу по размерам
.............................................................Asiocolotes Golubev
Подрод африканский карликовый геккон — Tropiocolotes Peters, 1880
Типовой в и д: Т. tripolitanus (s. str.).
Диагноз. Подпальцевые пластинки ребристые; пара плоских межносо-
вых щитков с ноздрями не соприкасается; замежносовые щитки не выражены;
кроме межчелюстного и первого носового к ноздре примыкают две чешуйки,
существенно не отличающиеся по размерам ни от интерназальных, ни от осталь-
ных чешуй морды (кроме Т. scortecci, у которого межносовые щитки значительно
крупнее остальных чешуй, однако они также с ноздрями не соприкасаются);
верхние края верхнегубных щитков загибаются на верхнюю поверхность мор-
ды так, что каждый щиток находится не в одной плоскости; чешуя тела плоская,
ромбическая, ребристая или гладкая; спинная иногда может быть несколько
мельче брюшной (обычно крупнее или равна); подхвостовые щитки отсутству-
ют; анальные поры, как правило, отсутствуют (у отдельных самцов Т. steudneri
и Т. scortecci иногда бывают два слабо выраженных пятна на месте пор).
Дополнительные признаки. Чешуя морды плоская, непра-
вильно-многоугольная, незначительно уменьшается к затылку. Ухо мелкое,
округлое. Подбородочный щиток тре- или пятиугольный, иногда со скошенным
задним краем. Нижнечелюстные щитки могут отсутствовать, но обычно имеется
первая пара, соприкасающаяся между собой или разделенная горловой чешуей;
за ней следует вторая пара, более мелкая; часто за нею вдоль нижнегубных
щитков следуют одна-две пары убывающих в размерах щитков. Количество
и взаиморасположение нижнечелюстных щитков подвержены у разных видов
изменчивости. Горловая чешуя плоская, неправильно-многоугольная; несколь-
ко верхних горловых чешуй могут быть крупнее остальных. Спинная чешуя
плоская, однородная, ромбическая, черепитчатая; брюхо также покрыто пло-
ской однородной ромбической чешуей. Хвост цилиндрический, несколько утол-
щенный вначале и утончающийся в последней четверти, покрыт сверху и снизу
плоской черепитчатой чешуей. Вытянутая вперед передняя конечность доста-
ет концами пальцев до середины промежутка между ноздрей и передним краем
орбиты или до конца морды, задняя — почти до лопатки. Конечности сверху
и снизу покрыты чешуей, сходной с чешуей туловища. Пальцы покрыты сверху
одним, с боков двумя рядами чешуй.
Распространены в Сев. Африке (сев. 10° с. ш.), на Синайском и Аравийском
полуостровах; известна одна находка в Ю. Иране. Подрод включает три вида.
Таблица для определения видов подрода Tropiocolotes
1 (4). Все чешуи морды, в том числе и соприкасающиеся с ноздрей, почти равны друг другу
по размерам.
2(3). Спинная чешуя заметно крупнее брюшной; все чешуи туловища с четкими продольными
ребрами ................................................... Т. tripolitanus Peters
3 (2). Спинная чешуя равна или незначительно крупнее брюшной; чешуя туловища гладкая
............................................................Т. steudneri (Peters)
4(1). Межносовые щитки заметно крупнее остальных чешуй морды, но с ноздрями не сопри-
касаются ...........................................Т. scortecci Cherchi et Spano
101
АФРИКАНСКИЙ КАРЛИКОВЫЙ ГЕККОН ТРИПОЛИТАНСКИЙ —
TROPIOCOLOTES TRIPOLITANUS PETERS, 1880
Типовая территория: «Wadi M’belem» (Триполитания, Ливия).
Кариотип: не изучен.
1880 — Tropiocolotes tripolitanus Peters, Mon. Berlin. Akad., S. 306.
1942 — Tropiocolotes sotnalicus Parker, Bull. Mus. Comp. Zool., Vol. 91, N 1, p. 46.
1942 — Tropiocolotes occidentalis Parker, Bull. Mus. Comp. Zool., Vol. 91, N 1, p. 47.
Диагноз. Чешуя туловища очень ребристая; чешуя спины заметно
крупнее брюшной; интерназальные щитки не крупнее остальных, с ноздрями
не соприкасаются.
Описание (по 106 экз. из колл. AMNH, BMNH, USNM, CAS, MNHP,
MZSF) (рис. 45). Типовой экземпляр вида утерян (ZMB 9668, д-р Г. Петерс,
письм. сообщ.). L общ. = 11,7—39,8 (26,31 ± 0,59) мм; индекс ширины/вы-
соты головы 56—67 (62,47 ± 0,46); чешуй между центрами глаз 8—13 (10,52 ±
± 0,10); верхнегубных щитков 7—10 (8,81 ± 0,05); нижнегубных щитков 5—8
(7,05 ± 0,04); чешуй вдоль тела снизу 77—115 (100,43 ± 1,20); чешуй попе-
рек брюха 34—48 (43,48 ± 0,28); подпальцевых пластинок 11—17 (14,10 + 0,11).
До орбиты достает 2,7—3 верхнегубных щитка. Нижнечелюстные щитки,
как правило, выражены хорошо, первая пара соприкасается между собой ши-
роким швом. Ребра на чешуйках туловища, совмещаясь друг с другом, образу-
ют характерные продольные ряды килей. У самцов пор нет.
Окраска и рисунок (после фиксации в спирте). Основной фон
тела светло-желтоватый. От ноздри через глаз к уху с обеих сторон тянутся ши-
рокие темно-коричневые полосы, которые исчезают, не достигая плеча (у особей
из Сомали эти полосы сливаются на затылке, образуя подковообразный рису-
нок). Голова и верхняя часть туловища обычно густо покрыта множеством тем-
но-коричневых точек и мелких пятен; иногда на теле есть до семи неясных попе-
речных полос; такие же полосы (до 10) могут быть на хвосте. Нижние поверх-
ности светлые.
Распространение. Сев. Африка, от 10° с. ш. на востоке до ни-
зовьев долины Нила и Аденского залива (Джибути и Сомали) (рис. 43, 46).
Кадастр к рис. 43.
Сомали: / — 10° 20' с. ш., 42° 50' в. д. [272]; 2 — окр. Борама: 10° 45' с. ш., 43° в. д.
[272]; 3 — Биджи [272]; 4 — 11° 25' с. ш., 43° 15' в. д. [272]; 5 — Джибути (MNHP); 6 — Обок
(MNHP).
Кадастр к рис. 46.
АРЕ:/- Гебель Мокаттам, в. Каира [249]; 2 — пров. Гиза, Абу-Раваш (USNM); 3 —
Эль-Гиза [249]; 4 — пров. Эль-Файюм, окр. оз. Карун (USNM).
Л и в и я: 5 — оазис Джало [249J; 6 — Эль-Агейла (MCZN); 7 — Сирт [249]; 8 — Джа-
афер [249];
Тунис: 9 — Фум-Татахуин [249]; 10 — Габес [249]; 11 — Гафса [249]; 12 — Оум
(Окум) Али, окр. пос. Гафса [249]; 9 км сев.-в. пос. Гафса (CAS); 13 — Тунис (ZFMK); 14 —
Таферма, между пос. Гафса и Тозер [249].
Алжир: 15 — Бон (SMF); 16 — Бискра [2491, 17 — Гардая (MZSF); 18 — Джеб ел -
Джара, Айн-Сефра (MNHP); 19 — Фигиг [249]; 20 — Бени-Униф (MNHP); 21 — Бени Аббез
(MNHP); 22 — Коломб-Бешар (CAS); 23 — Кенадза [249].
Марокко: 24 — Бу-Хайра [198]; 25 — Бу-Дениб, Бу-Анан [198]; 26—Хамада-
Гир [198]; 27 — Мески (Тафилалит) [198]; 28 — 40 км сев.-в. Тиндуф (CAS); 29 — 7 км с.
г. Танальт (RMNH); 30 — 10 км ю. г. Танальт (RMNH); 31 — Гулимин (MNHP); 32 — Эль-Хас-
сан [198]; 33 — Авинет-Торкоз (CAS); 34 — 15 км ю.-з. г. Авинет-Торкоз (CAS); 35 — 10 км ю.
г. Авинет-Торкоз (CAS); 36 — Асса [198]; 37 — Авинет-Айт-Оусса [198]; 38 — Таскала [198];
39 — Бу-Гуджоф [198]; 40 — Уэд-Дра (AMNH).
Западная Сахара: 41 — Сегует-Эль-Хамра: Амисерат [198]; 42 — 10 км сев. г.
Смара (CAS); 43 — 20 км з. г. Эль-Аюн (CAS); 44 — бухта Рио-де-Оро [249].
Мавритания: 45 — Порт Этьен [249]; 46 — 15 км сев. аэропорта Нуакшот (CAS).
М а л и: 47 — Санга [271]; 48 — Бандиагара [271]; 49 — Хомбори [271].
Н и г е р: 50 — 18 км сев. г. Ниамей (CAS); 51 — Вади Аззель [271]; 52 — Дабага [271];
15 км ю. г. Арлит (ZFMK); 8—10 км сев. г. Агадес (ZFMK); 53 — Теуар [271]; 54 — 80 км ю.
г. Агельхок (CAS); 55 — Тесалит (CAS).
А л ж и р: 56 — горы Ахаггар [249]; Куэд Амра (ZFMK); Тин-сиг (ZFMK).
Не найдены на картах: Uadi M’Belem, Tripolitania [249]; Tunisia: Djebel Bu
Hedma [249]; Govrnm. de Gabes: Gabes, 21 km. W., 19 km N. Busack (USNM); Gol. de Tafana
(MNHP); Algerie Carie (MNHP): Sahara et Souss. Jusqu’a [274]; Mauritania N’Keder (MNHP);
Rio-de-Oro Audebdinet [271]; Марокко: zw. Tazzarine und Alnif: О El Arba, Trummerfeld
(ZFMK); 16 km W. Bouanane sandiges Trocken oued (ZFMK); 5 km SW Bou-Rbia Fuss des I. Bani
(ZFMK); Touantia (ZFMK).
102
Рис. 45. Tropiocolotes tripolitanus:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — голова снизу, Д — лицевая
часть сверху, Е — подбородок, Ж — спина, 3 — брюхо, И — палец снизу, К — верхний
край орбиты, Л — хвост сверху, М — хвост снизу (USNM 195376 Тунис, Габес)
Половой диморфизм и возрастная изменчивость.
Самцов этого вида можно отличить от самок лишь по незначительным вздутиям
у основания хвоста снизу. Самки несколько крупнее самцов, однако незначи-
тельные выборки, имевшиеся в нашем распоряжении, не позволили выявить это
различие достоверно. Возрастная изменчивость пропорций тела сходна с тако-
вой у видов рода Alsophylax (рис. 47).
103
Систематика и географическая изменчивость.
Г. Паркер [272] описал два новых вида, близких к триполитанскому карликово-
му геккону. Т. somalicus, по его мнению, отличался от последнего наличием
задних нижнечелюстных щитков и мелкими щитками надглазничной области.
Рис. 46. Распространение Т. tripolitanus
Для Т. occidentalis, напротив, характерны очень неясные, слаборазличимые
задние нижнечелюстные щитки и крупные щитки надглазничной области. Оба
новых вида также отличались от типового 35—41 рядом чешуй вокруг середи-
ны туловища (у Т. tripolitanus их 44—48).
Проводя ревизию африканских гекконов, А. Лоуридж [249] рассматри-
вал оба вида, описанных Г. Паркером, уже в качестве подвидовых форм Т. tri-
L/Lccf.
0,90-
Q05-
Q<30~
Q75 -
------1----1-----1-----1----1--.--I____I_____I_____l____I_____i____i
/7 /J Zf 77 19 21 2J 25 27 29 JI JJ Lhm
Рис. 47. Возрастная изменчивость индекса L/Lcd у Т. tripolitanus
politanus. Кроме того, он описал подвид Т. t. algericus (его признаки — корот-
кий шов между первой парой нижнечелюстных и вторыми нижнегубными щит-
ками; мелкие чешуи надглазничной области; 44 ряда чешуй вокруг середины ту-
ловища; обитают в Алжирской Сахаре) и обозначил номинативный подвид
(широкий шов между первой парой нижнечелюстных щитков и вторыми нижне-
губными щитками; отсутствие соприкасания между второй парой нижнечелюст-
ных и первыми нижнегубными щитками, 42—48 рядов чешуй вокруг середины
104
туловища; обитают от Египта через Ливию до Туниса). Он уточнил, что у Т. t.
somalicus вторая пара нижнечелюстных щитков соединяется широким швом
с первыми нижнегубными, а у Т. t. occidentalis нижнечелюстные щитки вообще
слабо выражены и первая пара не может соприкасаться с нижнегубными. А. Лоу-
ридж не указал, какое количество особей он использовал в своей работе, од-
нако по косвенным данным можно заключить, что в его распоряжении были еди-
ничные экземпляры.
Т. Папенфусс [271] описал из нескольких пунктов Мали подвид Т. t. apoklo-
max, у которого обнаружил 46—54 ряда чешуй вокруг середины туловища и от-
сутствие соприкасания между второй парой нижнечелюстных и первыми ниж-
негубными щитками. Однако он отметил, что первый признак изменчив: особи из
Комбори имеют 46—52 чешуйки вокруг середины тела, а из Сангхи и Банди-
агары (120 миль к юго-западу) — 52—54.
Таким образом, в настоящее время известны пять подвидов этого вида,
различающихся преимущественно количеством чешуй вокруг середины тулови-
ща, различной степенью выраженности нижнечелюстных щитков и наличием
соприкасания с нижнегубными щитками.
Сравнение небольших выборок и отдельных экземпляров из Марокко,
Испанской Сахары, Рио-де-Оро, Мавритании, Мали (севернее ареала Т. t.
apoklomax), Нигера, Алжира, Туниса, Египта и Сомали показало, что признаки,
использующиеся для диагностики этих форм, возможно, проявляют клиналь-
ную изменчивость. Так, с юго-запада на северо-восток увеличиваются размеры
первой и второй пар нижнечелюстных, появляются третья и четвертая пары
щитков, поэтому ширина швов между первой парой нижнечелюстных, а также
между этими и вторыми нижнегубными щитками заметно увеличивается. Кро-
ме того, по линии, соединяющей западное и северное побережья Африки (Ал-
жир), наблюдается относительно однородное (изофена) количество чешуй во-
круг середины туловища и вдоль продольной оси тела снизу, а от этой линии
в глубь материка количественные показатели этих признаков возрастают.
Исключение составляют лишь особи из Сомали. Учитывая полную неизучен-
ность распространения триполитанских карликовых гекконов на столь обшир-
ной территории, мы считаем выделение у них подвидовых форм преждевремен-
ным. Можно предположить, что реально существует лишь сомалийская фор-
ма, имеющая дискретные показатели указанных признаков и изолированная от
остальных Абиссинским нагорьем, и, возможно, малийская. Однако оконча-
тельное решение этого вопроса должно базироваться на более обширном кол-
лекционном материале.
Экология не изучена. А. Лоуридж [249], основываясь на своих наблю-
дениях, сообщает, что номинативная форма в Алжире и Египте (рядом с пирами-
дами в Гизе) обитает под камнями среди галечников и на развеянных песках
(высота 100—300 м н. у. м.). Питается кровососущими насекомыми, мелкими
пауками, очень мелкими личинками жуков (мучными червями) и сверчками.
По его данным, сомалийский подвид ограничен в распространении арид-
ной береговой зоной. Семь из восьми экземпляров, составляющих типовую се-
рию, были добыты на грунте в песчаной местности, покрытой камнями, почти
полностью лишенной растительности, на высоте около 960 м н. у. м.
Алжирский подвид на восточных склонах гор в окр. Фермы Дюфоур был
найден под камнями (высота мест его обитания — 100—500 м н. у. м.). Коллек-
ционные материалы собраны с февраля по декабрь. Известно также [219],
что он питается муравьями и термитами, возможно, мелкими бабочками. Счита-
ют (по наблюдениям в неволе), что это ночной вид. Дважды его добывали днем
в глубине норы и под камнем. Весной, летом и осенью карликовый геккон вы-
ходит на охоту с заходом солнца на всю ночь. Вероятно, он активен также
и зимой, особенно на джебелах, где температура всегда выше. По данным
В. Клингельгоффера [236], Т. tripolitanus встречается под камнями в камени-
стой, перемещающейся с полосами песка, местности. Активен в вечернее время.
В террариуме питается мелкими пауками, дрозофилами, восковой молью.
105
АФРИКАНСКИЙ КАРЛИКОВЫЙ ГЕККОН ШТЕЙДНЕРА —
TROPIOCOLOTES STEUDNERI (PETERS, 1869)
Типовая территория: Сеннар, Судан.
Кариотип: 2л = 38 (детальнее не описан [312]).
1869 — Gymnodactylus steudneri Peters, Monats. Acad. Wiss., Berlin, S. 788.
1891 — Stenodactylus petersii Boulenger, Transact. Zool. Soc. London, Vol. 13, p. 108.
1900— Tropiocolotes nattereri Steindachner, Denkschr. Acad. Wiss. math.-naturw. KI., Bd 69,
no. 1, S. 326.
1900—Tropiocolotes steudneri Steindachner, Denkschr. Akad. Wiss. math.-naturw. KI, Bd 69,
no. 1, S. 326.
Диагноз. Чешуя туловища гладкая или со слабыми следами ребристо-
сти; спинная чешуя равна или незначительно больше брюшной; интерназальные
щитки не крупнее остальных, с ноздрей не соприкасаются.
Описание (рис. 48) (по 38 экз. из колл. ИЗАНУ, FMNH, USNM, CAS,
MNHP, MZSF). Типовой экземпляр утерян (д-р М. Хепль, NMW, письм. со-
общ.).
Аобщ. = 15,3—34,8(25,24 ± 0,87) мм; индекс ширины/высоты головы 55—
68 (60,17 ± 0,82); чешуй между центрами глаз 12—17 (14,10 ± 0,21); верхне-
губных щитков 8—10 (9,13 ± 0,08); нижнегубных щитков 6—8 (6,87 ± 0,07);
чешуй вдоль тела снизу 95—118 (107,90 ± 0,98); чешуй вокруг середины туло-
вища 46—55 (50,68 zb 0,45); подпальцевых пластинок 15—18 (16,70 ± 0,12).
До орбиты достает 3,5—3,8 верхнегубных щитков. Нижнечелюстных щит-
ков две (28,6 %), три (31,4 %) или четыре (40 %) пары; первая пара самая круп-
ная, щитки соединены широким швом. Иногда у самцов имеется пара слабо вы-
раженных анальных пор, часто разделенных одной-двумя чешуйками.
Рисунок и окраска (после фиксации в спирте). Основной фон тела
песочно-желтый. От ноздри через глаз к уху тянутся с обеих сторон головы две
широкие темно-бурые полосы, не достигающие лопатки. Рисунок спины варьи-
рует: для особей из Египта характерно большое количество темных пятен и то-
чек или слабо выраженные широкие поперечные полосы. Особям из Палестины
свойственны четыре-пять тонких извилистых темных поперечных полос, каждая
из которых окантована сзади светлой полоской. На хвосте имеется до 13 темных
поперечных полос, выраженных в разной степени.
Распространение. Сев.-В. Африка от Алжира на западе до Сев.
Судана на юге и р. Иордан на востоке; известна изолированная находка в Ю.-В.
Иране (рис. 43, 49).
Кадастр к р и с. 43, 49.
Иран:/ — Бендер-Ленге [223].
А л ж н р: 1 — Уэд-Игаргар, Ц. Сахара [249]; 2 —Амгид (MNHP); 3 — 5 км сев.-в.
г. Таманрассет (CAS); 4 — 3 км в. г. Таманрассет (CAS); 5 — Тассилин-Адджер (MNHP).
Л и в и я: 6 — 25° 30' с. ш., 16° в. д. [273]; 7 — оазис Куфра, Эт-Тадж [249]; 8 — оазис
Куфра, Эль-Джауф (MCZN); 9 — оазис Куфра, Эль-Тебиб (MCZN); 10 — оазис Куфра, Эз-
Зуг [306].
А Р Е: // —Эль-Гиза [249]; 12 — Эль-Харрания (USNM); 13 — Абу-Раваш [249]; 14 —
р-н Эмбаба; 15 — 24 км з. г. Каир [251]; 16 — Вади-Гувейбба [251]; 17 — 60 км от Каира в
Суэц (ИЗАНУ); 18 — Гебель-эль-Мукаттам, в. г. Каир [249]; 19 — Гебель-эль-Анквабия [249];
20 — Вади-Гиндали [251]; 2/— Вади-Маади [251]; 22 — Вади-Гельван [251]; 23 — Вади-
Хоф [249]; 24—Вади-Асьютин [251]; 25 — Рас-Гариб [251]; 26— Каламшах, Фаюм [249];
27 — Хургада [249]; 28 — Луксор [249); 29 — пуст, в окр Эль-Карнак (ZFMK); 30 — пуст, в
г. Иена [249]; пуст. в. г. Асфун-эль-Матаана [216]; 31 — Вади-Аббад [251]; 32 — Филе [249];
33 — Бир Абрак (USNM).
С у д а н: 34 — Халаиб [249] (на картах — Египет); 35 — Нубия [249]; 36 — Кошех
[249]; 37 — Сеннар [249]; 38 — Кутум (ZFMK); АРЕ: 38 — Синайский п-ов, Нувейба, побере-
жье зал. Акаба [295]; 39 — Синайский п-ов, Бир-аль-Маши (Машия) [295].
Израиль: 40 — Вади-Роман, пуст. Негев [249]; 41 — Айн-Джеди (SAM).
Не найдены на картах: Алжир, 15 km Terhenanet Raud lines Wadi m. Fel-
sen; Ливия: El Airenat [249]; Израиль: пуст. Negev, Hamakhtesh Haqatan (MZSF).
Половой диморфизм и возрастная изменчивость.
У самцов этого вида часто имется пара слабо выраженных анальных пор. Кро-
ме того, с нижней стороны основания хвоста слабо выражены вздутия. Вероят-
но, самки несколько крупнее самцов, однако незначительные выборки, имев-
шиеся в нашем распоряжении, не позволили выявить различие достоверно. По
этой же причине не представилось возможным изучить возрастную изменчи-
вость.
S06
Рис. 48. Tropiocolotes steudneri:
A — голова сверху, Б — лицевая часть сбоку, В — голова снизу, Г — спина, Е — брюхо,
Д — преанальные поры (Г—Е — USNM 136355, окр. Каира, АРЕ; остальное — CNHM 78393,
берег Красного моря, АРЕ)
Систематика и географическая изменчивость.
Ш. Минтон и др. [263], изучая систематическое положение некоторых видов это-
го рода, обратили внимание на различия между особями из Египта и Палести-
ны. Они отметили, что у западных экземпляров несколько большее количество
верхне- и нижнегубных щитков (6—10 и 6—8 против 7—8 и 5—7), чешуй во-
круг середины туловища (38—50 против 41—43) и подпальцевых пластинок (14—
19 против 14—16). Отмечены различия в окраске.
107
Нами предпринято сравнение по некоторым признакам небольших выборок
из ^л? ира, Египта, Палестины и единственного экземпляра из Ирана; оно по-
казало в северных частях ареала действительно наблюдается уменьшение ко-
личества чешуй вдоль продольной оси тела снизу и вокруг середины туловища
в направлении с запада на восток. Однако мы не отметили изменчивости коли-
чества верхне- и нижнегубных щитков и подпальцевых пластинок.
Если учесть, что распространение этого вида внутри ареала изучено в на-
стоящий момент недостаточно хорошо, отсутствует сравнительный материал из
10'
JO9
Рис. 49. Распространение Т. steudneri
типового местонахождения (Сев. Судан), а выборки, имеющиеся в коллекциях,
недостаточно велики, то вопрос о внутривидовой систематике Т. steudneri пока
еще не решен.
Стации. Встречается в каменистых и песчаных пустынях, под камнями,
а в Сахаре живет на стволах деревьев. Все местообитания расположены на бес-
плодных, продуваемых ветром и сильно прогреваемых солнцем сухих камени-
стых холмах. Высота мест обитания 100—300 м н. у. м. [249].
Особенности поведения. В террариумах эти ящерицы нор не
роют, а проводят все время на вершинах крупных камней. Очень осторожны
и подвижны, однако быстро приручаются. Коллекционные материалы собраны
в период с февраля по октябрь [249].
Питание. Дважды было замечено, что геккончики Штейднера поедали
щетинохвосток (Lepisma sakharina). В неволе ежедневно питались различными
летающими насекомыми [249].
АРАВИЙСКИЙ КАРЛИКОВЫЙ ГЕККОН СКОРТЕЧЧИ —
TROPIOCOLOTES SCORTECCI CHERCHI ET SPANO, 1963
Типовая территория: Эль-Сафа, Хадрамаут, НДРЙ.
Кариотип: не изучен.
1963 — Tropiocolotes scortecci Cherchi et Spano, Boll. Mus. 1st. Biol, dell Univ. Genova, Vol.
32, no. 188, p. 29.
Ди а г н о з. Чешуя туловища слабо ребристая; спинная чешуя несколько
больше брюшной; интерназальные щитки заметно крупнее остальных чешуй
морды (с ноздрями не соприкасаются).
Голотип. MIZG — I. «Е1 Safa, Hadramaut 2.V 1962, G. scortecci leg.»
d* ad L = 21,5 mm; L cd — 20,2 mm, LIL cd = 1,06. Ноздря расположена меж-
ду четырехугольным межчелюстным, первым верхнегубным и двумя мелкими
плоскими носовыми щитками (первый несколько крупнее второго), не отлича-
ющимися от остальных чешуй морды; интерназальные щитки крупнее окружаю-
щих чешуй, но с ноздрями не соприкасаются. Чешуй между центрами глаз 11.
108
Рис. 50. Tropiocolotes scortecci:
А, Б общий вид, В — голова сбоку, Г — голова сверху, Д — лицевая часть сверху, Е,
Ж, — подбородок, 3, И — спина, К — брюхо, Л — преанальные поры (В, Г, Е — MIZG 1,
Хадрамаут, Йемен; А — MIZG 2, там же; остальное — BMNH 1978. 1098, Дофар. Оман)
Верхнегубных щитков по 9 (согласно первоописанию — по 10); нижнегубных —
по 7 (согласно первоописанию — по 8). Подбородочный щиток шестиугольный;
две пары нижнечелюстных щитков (согласно первоописанию — одна пара),
щитки в первой паре разделены одной горловой чешуйкой; чешуй вдоль тела
снизу 106; вокруг середины туловища 50. Подпальцевых пластинок 15—16
(согласно первоописанию — 17).
Описание. Составлено по всем известным в настоящее время экземпля-
рам (3 типовых MIZG и 11 — BMNH), рис. 50.
Лобщ. = 17,2—29,8(23,99 ± 1,07); UL cd общ. (и = 6)0,87—1,31 (1,14 ±
± 0,06); индекс ширины/высоты головы (п = 10) 58—70 (62,20 ± 1,07); чешуй
109
между центрами глаз (п = 14) 10—16 (12,36 ± 0,45); верхнегубных щитков
(п = 28) 9—11 (9,61 ± 0,13); нижнегубных щитков (п = 28) 6—9 (7,36 ± 0,14);
чешуй вдоль тела снизу (п = 13) 85—107 (94,54 ± 2,50); чешуй вокруг середи-
ны туловища {п = 13) 36—50 (43,54 ± 1,43); подпальцевых пластинок (п =
= 28) 12—19 (14.75 ± 0,81).
Подборедочный щиток тре- или пятиугольный (78,7 %), шестиугольный
(14,2 %) или трапециевидный (7,1 %); одна (28,6 %), две (57,1 %) или три
(14,3 %) пары нижнечелюстных щитков, первая из которых соприкасается меж-
ду собой (78,7 %) или разделена одной (14,2 %) или двумя (7,1 %) чешуйками.
Иногда у самцов есть пара слабо различимых анальных пор, часто разделенных
одной-двумя чешуйками За второй парой нижнечелюстных щитков вдоль ниж-
негубных может следовать одна (34,6 %), две (23,1 %), три (7,7 %) пары еще бо-
лее мелких, постепенно уменьшающихся в размерах щитков; у 34,6 % особей
дополнительные нижнечелюстные щитки отсутствуют
Окраска и рисунок Основной фон тела — светло-желтый (по
другим данным [1781, прижизненная окраска розово-желтая). От ноздри через
глаз к уху с обеих сторон тянутся широкие темно-коричневые полосы, оканчи-
вающиеся за лопаткой. Кроме того, над лопаткой от каждой стороны отходят
ответвления, которые на шее соединяются между собой, окантовывая голову на-
подобие ошейника. Спина лишена полос; каждая спинная чешуйка в середине
окрашена в более темный, чем основной фон, цвет. На хвосте от его основания до
конца имеется 10—12 темно-коричневых широких полос. Верхние поверхности
конечностей окрашены так же, как спина; нижние поверхности беловатые.
Распространение. Юг Аравийского п-ова (рис. 43).
Кадастр к рис. 43.
1 - Эль-Сафа, Хадрамаут, Южный Йемен (MIZG); 2 — Вади-Райкут [167]; 3—Вади-
Хаиз [167]; 4 — Дюн [1671; 5 — Марказ-Тамарид [167], 6 — 16 км ю. Марказ-Тамарид [1671;
7—Бирба Шуайтан.Оман [167].
Половой диморфизм и возрастная изменчивость.
Как у других видов подрода, у геккончиков Скортеччи половой диморфизм вы-
ражен слабо. У самцов в основании хвоста снизу иногда заметны вздутия.
Самки, вероятно, несколько крупнее самцов, однако незначительная выборка,
имевшаяся в нашем распоряжении, не позволила подтвердить достоверность
этого различия. По этой же причине осталась неизученной и возрастная измен-
чивость.
Систематика и географическая изменчивость.
Уже четвертый экземпляр, добытый вслед за тремя типовыми, показал изменчи-
вость некоторых признаков фолидоза. Прежде всего это выразилось в наличии
соприкасания между первой парой нижнечелюстных щитков (у всех типовых
экземпляров эти щитки разделены одной-двумя чешуйками). В сравнении с ти-
пами у этого экземпляра больше чешуй между центрами глаз, верхнегубных
щитков и подпальцевых пластинок и меньше чешуй вокруг середины туловища
и вдоль продольной оси тела снизу. Если учесть, что этот экземпляр был добыт
в 500—600 км восточнее типовой территории [1851, можно предположить, что
по указанным признакам наблюдается географическая изменчивость.
Экология не изучена. На типовой территории эти гекконы были най-
дены под камнями в местности, изобиловавшей кустарниковой растительностью,
а также на песчано-шебнистых и щебнистых участках. Одна из особей была заме-
чена днем: скрываясь от преследования, спряталась под камень, где находился
другой геккон — Pristurus karterii [2081. Места обитания этого вида (типовая
территория) расположены на востоке 400 м н. у. м.
Подрод микрогеккон — Microgecko Nik, 1907
Типовой вид: Microgecko helепае Nikolsky, 1907.
Диагноз. Подпальцевые пластинки гладкие; межносовые щитки зна-
чительно крупнее остальных носовых чешуек и всегда соприкасаются с нозд-
рями; одна-две пары замежносовых щитков хорошо выражены; чешуя тела
(включая подпальцевые пластинки) гладкая; чешуя спины заметно мельче
110
брюшной; увеличенных подхвостовых щитков и анальных пор нет. Распро-
странен на юге Ю.-З. Азии. Подрод включает три вида.
Дополнительные признаки. Морда покрыта выпуклой
разноразмерной округло-многоугольной гладкой чешуей, заметно уменьша-
ющейся к затылку; ушное отверстие очень мелкое, округлое. Горловая чешуя
округло-многоугольная, выпуклая, гладкая, однородная. Спина покрыта сла-
бо выпуклой гладкой овальной мелкой чешуей, увеличивающейся к бокам
таким образом, что границу между ней и брюшной чешуей определить невоз-
можно. Брюшная чешуя ромбовидно-округлая, крупная, плоская, гладкая,
черепитчатая, заметно крупнее спинной. Хвост цилиндрический, несколько
утолщенный у основания и довольно резко сужающийся в последней трети;
покрыт округлой плоской гладкой чешуей; подхвостовых щитков нет. Вытяну-
тая передняя конечность достает концами пальцев до переднего края орбиты или
чуть дальше, задняя — до локтя вытянутой назад передней конечности или по-
чти до лопатки. Сверху конечности покрыты чешуей, сходной со спинной, сни-
зу — сходной с брюшной. Пальцы покрыты сверху одним, с боков двумя ряда-
ми чешуй; основание когтя скрыто чехлом, состоящим из верхней и нижней
гладких пластинок; к чехлу примыкают сверху одна, с боков— по две чешуйки.
Таблица для определения видов подрода Microgecko
1 (4). Нижнечелюстные щитки есть.
2 (3). Верхнегубных щитков не более семи..................Т. helenae (Nik.)
3 (2). Верхнегубных щитков не более восьми........ Т. persicus (Nik.)
4 (1). Нижнечелюстных щитков нет.................Т. latifi Leviton et Anderson
МИКРОГЕККОН ХУЗЕСТАНСКИЙ —TROPIOCOLOTES HELENAE (NIK., 1907)
Типовая территория: Альхоршир в Хузестане (хр. Загрос, 3. Иран).
Кариотип: не изучен.
19071 — Microgecko helenae Никольский, Ежег. Зоол., Муз., 1905, т. 10, № 3—4, с. 265.
1970 — Tropiocolotes helenae, Minton, S. Anderson and Jr. Anderson, Proc. Calif. Acad. Sci.,
4th Ser., Vol. 37, no. 9, p. 345.
Диагноз. Верхнегубных щитков шесть-семь; нижнечелюстные щитки
имеются (в первой паре они обычно не соприкасаются).
Лектотип. ЗИН 10242. j ad Персия, с. Альхоршир, 30.XII 1903 г.,
Н. Зарудный. L = 27,5 мм; L cd = 30,5 мм; LIL cd = 0,90. Ноздря расположе-
на между пятиугольным межчелюстным, первым верхнегубным, крупным пло-
ским межносовым щитком и двумя мелкими носовыми чешуйками; межносовые
щитки разделены между собой одной чешуйкой; за межносовыми щитками рас-
положена пара крупных замежносовых щитков, разделенных между собой од-
ним рядом, состоящим из двух чешуек. Чешуй между центрами глаз 19. Верх-
негубных щитков по шесть, нижнегубных — пять-шесть; первая пара нижне-
челюстных щитков разделена двумя горловыми чешуйками; чешуй вдоль тела
снизу 121; вокруг середины туловища — 68. Подпальцевых пластинок по 14.
Описание (по 60 экз. из колл. ИЗАНУ, ЗИН, USNM и MCZ), рис. 51.
L общ.= 15,5—28,5 (22,03 zb 0,46) мм; индекс высоты/ширины головы
56—66 (60,41—0,48); чешуй между центрами глаз 15—23 (18,57 ± 0,23); верх-
негубных щитков 6—7 (6,68 ± 0,04); нижнегубных — 5—7 (5,66 zb 0,05);
чешуй вдоль тела снизу 101 —122 (113,08 zb 0,71), чещуй вокруг сере-
дины туловища 60—75 (66,41 zb 0,51); подпальцевых пластинок 11—15
(12,90 ± 0,07). Межносовые щитки соприкасаются между собой (55 %) или раз-
делены рядом чешуек (45 %); пара крупных замежносовых щитков соприкаса-
ется между собой (12 %), разделена одной (78 %) или 2 (10 %) чешуйками; до
переднего края орбиты доходят 2,3—3 верхнегубных щитка. Подбородочный
щиток трапециевидный (87 %), изредка пяти- (8 %) или треугольный (5 %);
одна (95,8 %), очень редко — две (4,2 %) пары нижнечелюстных щитков (вто-
рая пара очень мелкая); первая пара соприкасается между собой (11,7 %) или
разделена одной (25 %), двумя (55 %) или тремя (8,3 %) горловыми чешуями.
1 X том Ежегодника Зоологического музея (за 1905 г.) вышел в свет в 1907 г. Таким об-
разом, датой опубликования следует считать 1907 г., а не 1905 г.
111
Рис. 51. Tropiocolotes helenae:
А, Б — общий вид, В — лицевая часть сверху, Г, Д — подбородок, Е — спина, Ж —
брюхо (В. Д — ЗИН 10242. Альхоршир, Иран; Б — ZSM 500/68 Мекух, Иран: остальное —
ИЗАНУ SR 569)
Окраска и рисунок. Основной фон верхней поверхности тела
желтовато-серый; от ноздри через глаз и над ушным отверстием проходит тем-
но-коричневая полоса, оканчивающаяся на уровне затылка или лопатки.
Такого же цвета тонкие поперечные полосы имеются на спине (пять — семь)
и хвосте (до десяти). Все полосы в задней части окантованы светлым. Спинные
полосы прямые или зигзагообразные; иногда они могут быть разорванными
112
по середине и смещенными вдоль продольной оси тела таким образом, что две
половины одной полосы не соприкасаются друг с другом. Регенерированный
хвост темно-коричневый, почти черный. Нижние поверхности светлые. Ко-
нечности лишены рисунка. Рисунок спины у отдельных особей может частич-
но или полностью исчезать [286].
Распространение. Найден во многих пунктах вдоль хр. Загрос
(3. Иран); вероятно, обитает в Сев. Ираке (рис. 52).
Кадастр к рис. 5 2.
Иран:/ — Диз, 125 км з. г. Керманмах по дор. на Багдад (ZSM): 2 — 25 км з. г. Хор-
рембад по дор. на Ахваз, Лурестан (ZSM); 3 — г. Месджеде-Солейман (ИЗАНУ); 4 — пров.
Хузестан, 16 км ю. г. Месджеде-Солейман (USNM); 5 — пров. Хузестан : Бидезар (ЗИН);
Рис. 52. Распространение Tropiocolotes helenae (/), Т. depressus (2), Т. levitoni (5), Т. latifi
(4)
6 — Агуляшкер (ЗИН); 7 — Альхоршир (ЗИН); 8 — г. Исфахан [92]; 9 — пров. Хузе-
стан, 35 км в. пос. Гечсаран, по дор. из Гечсарана в Казерун (USNM); 10 — 80 км з. г. Шираз
(ZSM); 11 — пров. Мекух, 80 км ю. г. Шираз и Мианкоталь (ZSM).
Половой диморфизм и возрастная изменчи-
вость. Половой диморфизм не выражен. Возрастная изменчивость проявля-
ется в более крупных размерах диаметра глаза в сравнении с длиной тела у моло-
дых особей.
Систематика и географическая изменчивость.
Гекконы из западной части ареала (провинции Лурестан и Керманшах) были
выделены недавно в подвид Т. h. f asci at us [286]. Единственным отличием от но-
минативной формы (типовая местность, по мнению авторов,— «Бидезар»,
Бид Зард) явились прямые полосы поперек спины. Однако у вновь найденных
нами синтипов рисунок отсутствует (обозначенный лектотип происходит не
из Бид Зарда, а из Альхоршира, что вызвано плохой сохранностью остальных
типов). Ознакомившись с коллекцией этого вида из разных частей ареала, а
также с литературными источниками, мы обратили внимание на то, что в рабо-
тах А. М. Никольского [92] и Р. Така [302] у изображенных гекконов спинные
полосы слабо извилистые. Такие же полосы и у имеющихся у нас гекконов из
Месджеде-Солейман (рис. 51). Однако у геккончиков из юго-восточной части
ареала (пров. Фарс) спинные полосы резко извилисты и часто разорваны по-
середине (рис. 51). Таким образом, довольно четко заметна клинальная измен-
чивость по этому признаку, в связи с чем разделение Т. helenae на подвиды не
представляется целесообразным. Изучение изменчивости фолидоза подтвер-
ждает этот вывод.
Экология. Краткие сведения о биотопах хузестанского микрогекко-
на содержатся в статье И. и Ф. Шмидтлеров [286], из которой следует, что
особи этого вида были добыты у г. Мекух на склонах гор, покрытых разрежен-
ными зарослями фисташки и одиночными дубами, среди которых встречаются
выходы коренных пород. Геккончики укрывались под небольшими — величи-
на
Рис. 53. Tropiocolotes persicus (вверху):
А, Б — общий вид, В — спина, Г — лицевая часть сверху, Д, Ж — подбородок, 3 — хвост
сверху, И — хвост снизу, К — палец снизу (Б, Г, Ж — MNHP 1966 17 — Т. р. persicus,
Ираншехр, Иран; В, Е — CAS 86408, Месджере Солейман-Сари Гах, Иран, — Т р. bakhtiari;
остальное — USNM 166178, Хаб Чаукн, Пакистан — Т. р. euphorbiacola)
Tropiocolotes latifi (внизу):
А — общий вид, Б — лицевая часть сверху, В — подбородок (CAS 134365 Кирман, Иран)
ной с человеческую ладонь — плоскими камнями в затененных местах, на высо-
те около 1200 м н. у. м.
Р. Так [3021 сообщает, что у дороги Гечсаран — Казерун в сухом русле
на з. склоне хр. Загрос 8.II 1964 г. были добыты несколько гекконов этого вида.
Местность — межгорная равнина с крутыми склонами из гипса и известняка,
почва усыпана мелкими плоскими камнями. Немногочисленная раститель-
114
ность — редкие злаки, лишайники, колючие кустарники, молочаи — эуфорбии и
реликтовый дуб. Дневная температура 19—42°, относительная влажность89 %;
дождь, высокая облачность. Гекконы были извлечены из-под камней посреди
вади. К югу от Месджеде-Солейман, также на западном склоне хр. Загрос
26.11 1964 г. добыт молодой микрогеккон. Местность пересечена сухими русла-
ми, покрыта очень скудной растительностью, имеются выходы известняков.
Температура воздуха была 28—44° Автор отмечает, что среди восьми самок
не было ни одной беременной.
По словам И. С. Даревского, он добывал этих гекконов в окрестностях
населенного пункта в щелях лессовых дувалов и развалин.
Коллекционные материалы, известные нам, добыты с января по декабрь.
МИКРОГЕККОН ПЕРСИДСКИЙ—TROPIOCOLOTES PERSICUS (NIK., 1903)
Типовая территория: «Vikus Degak» Дехек (Дизек) Иранский Белуджистан.
Кариотип: не изучен.
1903 — Alsophylax persicus, Никольский А. М., Ежег. Зоол. Муз. АН, т. 8, с. 95.
1970—Tropiocolotes persicus, Minton, S. Anderson and Jr. Anderson, Proc. Calif. Acad. Sci.,
4th Ser., Vol. 37, no. 9, p. 348.
Диагноз. Верхнегубных щитков 8—10; нижнечелюстные щитки име-
ются (в первой паре они обычно соприкасаются).
Голотип. ЗИН 10 005, «Vikus Degak in terra Dizak, Persia Orientalis,
9.II. 1901, leg. N. Zarudny».
L = 25,3 мм; хвост регенерирован; ноздря расположена между пятиуголь-
ным межчелюстным, первым верхнегубным, крупным, плоским межносовым
щитком и двумя мелкими дополнительными носовыми чешуйками; межносовые
щитки соприкасаются; за межносовыми щитками расположены две пары замеж-
носовых щитков, следующих друг за другом и соприкасающихся Чешуй между
центрами глаз 19. Верхнегубных щитков по 9, нижнегубных — по 7; 2 пары
нижнечелюстных щитков, из них первая самая крупная, щитки в ней разделе-
ны одной горловой чешуйкой.
Описание (по 61 экз. из колл. ЗИН, ИЗАНУ, AMNH, USNM, UMMZ,
CAS, MNHP), рис. 53.
L общ = 17,8—35,9 (26,18 ± 0,52) мм; индекс высоты /ширины головы
56—70 (53,31 ± 0,75); чешуй между центрами глаз 15—20 (17,64 ± 0,18);
верхнегубных щитков 8—10 (8,81 ± 0,06); нижнегубных щитков 6—9 (7,18 ±
±0,04); чешуй вдоль тела снизу 113—130(121,03 ±0,64); чешуй вокруг середи-
ны туловища 66—78 (71,21 ± 0,42); подпальцевых пластинок 11—15
(13,50 ± 0,08).
115
Дополнительные признаки. Межносовые щитки соприкаса-
ются (47,5 %) или разделены чешуйкой (52,5 %); за ними расположена одна-
две пары замежносовых щитков, в первой паре они соприкасаются (95 %);
до переднего края орбиты доходит 3,4—3,8 верхнегубных щитка. Подборочный
щиток тре- или пятиугольный с острым (очень редко со скошенным) задним кра-
ем; количество нижнечелюстных колеблется от трех до одной слабо выражен-
ной пары; первая пара, как правило, соприкасается или очень редко разделена
горловой чешуйкой.
Окраска и рисунок (послефиксации в спирте). Основной фон
верхней поверхности тела светло-желтоватый. От ноздри через глаз и ушное
Рис. 55. Возрастная изменчивость индекса LlLcd у Т. helenae (/), Т. persicus (2), Т. latifi (5)
отверстие до лопатки (иногда вдоль боков туловища до основания задних
конечностей) тянется темно- или шоколадно-коричневая широкая полоса. От
шеи до крестца на спине идут пять — семь таких же полос, иногда окантованных
сзади белым. У некоторых особей интенсивность окраски нарастает к дисталь-
ной части каждой полосы (признак подвержен географической изменчивости);
соотношение ширины полосы и ширины промежутка между полосами имеет
важное значение при определении подвидов. На хвосте 6—10 полос. Голова и
конечности лишены рисунка. Нижние поверхности белые. Регенерированный
хвост белый.
Распространение. Обитает в Ю.-З. иЮ. Иране и Ю. -В. Пакиста-
не (рис. 54).
Кадастр к рис. 5 4.
Т. р. bakhtiari:/ — Иран, между г. Месджеде-Солейман и Сар-и-Гач, 31° 57*
с. ш., 45° 2Г в. д. (CAS).
Т. р. persicus: 1— округ Чагай, пос. Мирджаве (в Иране, окр. Чагай — Пакистан)
(SAM); 2 — 100 км с. г. Ираншехр (MNHP); 3 — 20 км ю.-з. пос. Пиб [256]; 4 — Дегак в стр.
Дизак (ЗИН).
Т. р. euphorbiacola: Пакистан: 1 — 36 км сев. Бела (AMNH); 2 — верховья
р. Хаб, провинция Лас-Бела (MNHP); 3 — 10 км сев. пос. Дивана (AMNH); 4 — 10 км сев. пос.
Утхал (AMNH); 5 — округ Лас-Бела, хр. Паб (CAS); 6 — 10 км сев.-з. г. Карачи (UMMZ);
7—8 км в пос. Ландхи (AMNH); 8—35 км сев.-з. г. Бандмурадхан [263]; 9 — 9 км ю. г. Сехван
(AMNH); 10 — 8 км в. г. Тханобулахан [263]; 11 — окр. Татта, оз. Халеджи (AMNH); 12 —
г. Набисар (AMNH).
Половой диморфизм и возрастная изменчивость.
Самки несколько крупнее самцов. Возрастная изменчивость проявляется в
большем значении признака отношения диаметра глаза к длине тела у молодых
особей. Молодые более короткохвосты в сравнении со взрослыми (рис. 55).
Систематика и географическая изменчивость.
Долгое время этот вид, описанный как Alsophylax и близкородственный
11G
T. helenae, не различался специалистами, следствием чего явился большой спи-
сок синонимов Т. persicus. Ш. Минтон и др. [265], разобравшись в его запутан-
ной систематике, доказали принадлежность данного вида к роду Tropiocolotes
и описали три подвидовые формы. Различия между ними состоят в различном
соотношении ширины спинных полос и промежутков между ними.
Изучение небольших выборок и отдельных экземпляров персидского мик-
рогеккона из разных мест показало, что внутри ареала восточного подвида
(наиболее полно представленного в наших выборках) изменчивость некоторых
признаков варьирует в довольно больших пределах, однако диагностические
признаки преимущественно образуют хиатус между подвидами. Не исключено,
однако, что между восточным и номинативным подвидами будут обнаружены
переходные формы. В настоящее время можно различить три подвида персид-
ского микрогеккона.
Таблица для определения подвидов Т. persicus
1(2). Первая пара нижнечелюстных щитков разделена одной горловой чешуйкой или соеди-
нена между собой точечным швом ..................... . Т. р. persicus (Nik.)
2 (1). Первая пара нижнечелюстных щитков соединена между собой широким швом.
3 (4). Ширина спинных полос превышает ширину промежутков между ними ..........
..............................................Т. р. bakhtiari Minton et al.
4 (3). Ширина спинных полос равна или незначительно уже промежутков между ними (но
всегда более чем половина ширины промежутков)..................................
........................................Т. р. euphorbiacola Minton et al.
ПЕРСИДСКИЙ МИКРОГЕККОН ОБЫКНОВЕННЫЙ —
TROPIOCOLOTES PERSICUS PERSICUS (NIK., 1903)
Типовая территория: видовая.
1903 — Alsophylax persicus Никольский, Ежег. Зоол. Муз., т. 8, с. 95.
1956 — Tropiocolotes helenae, Mertens, Jeh. Wer. Naturk. Wiirtemberg, Vol. 3, S. 95.
1963 — Bunopus persicus, S. Anderson, Proc. Calif. Acad. Sci., 4th Ser., Vol. 31, p. 474.
1966 — Microgecko helenae, Guibe, Bull. Mus. Nat. Hist. Natur., Ser. 2, Vol. 38, p. 98.
1970 — Tropiocolotes persicus persicus Minton, S. Anderson and Jer. Anderson, Proc. Calif. Acad.
Sci., 4th Ser., Vol. 37, no. 9, p. 348.
Диагноз. Ширина спинных полос уже половины ширины промежут-
ков между ними; каждая полоса с четкой светлой окантовкой сзади; ширина
хвостовых полос уже промежутков; первая пара нижнечелюстных щитков раз-
делена одной горловой чешуйкой или соединена между собой точечным швом.
Описание (по 3 экз. из колл. ЗИН, MNHP и SAM).
L общ = 25,3 — 31,8 мм; чешуй между центрами глаз 16—19; верхнегуб-
ных щитков 9 — 10; нижнегубных щитков 7—9; чешуй вдоль тела снизу 130; че-
шуй вокруг середины туловища 78; подпальцевых пластинок 13/14. Межносовые
щитки соприкасаются между собой (один случай) или разделены чешуйкой (два
случая); замежносовые щитки соприкасаются между собой; подбородочный
щиток пяти- или треугольный со скошенным задним краем (два случая)
или трапециевидный (один случай); две пары нижнечелюстных щитков;
самая крупная (первая) пара разделена чешуйкой (два случая) или соединена
между собой точечным швом (один случай).
Распространение. Белуджистан вдоль Ирано-Пакистанской гра-
ницы.
ПЕРСИДСКИЙ МИКРОГЕККОН БАХТИЯРСКИЙ —
TROPIOCOLOTES PERSICUS BAKHTIARI MINTON ET AL., 1970
Типовая территория: Месджеде-Солейман, Хузестан, Иран.
1961 — Tropiocolotes helenae (not Microgecko helenae Nik., 1907), S. Anderson, Washmann Journ.
Biol., Vol. 19, p. 287.
1963 — Microgecko helenae (not Nikolsky, 1907), S. Anderson, Proc. Calif. Acad. Sci., 4th Ser.,
Vol. 31, p. 440.
1970 — Tropiocolotes persicus bakhtiari Minton, S. Anderson and Jer. Anderson, Proc. Calif. Acad.
Sci., 4th Ser., Vol. 37, no. 9, p. 351.
117
Диагноз. Ширина спинных и хвостовых полос превышает ширину
промежутков между ними; белой окантовки позади этих полос нет. Первая пара
нижнечелюстных щитков соединена между собой широким швом.
Описание. Составлено по голотипу: CAS 86408, 6я ad (?) «между
Месджеде-Солейман (Masjed-Soleyman) и Сар-и-Гач, пров. Хузестан, Иран
13.V1958 S. С. Anderson».
L = 20,5 мм; Led = 29,8 мм; LILcd = 0,96; чешуй между центрами глаз
18 (согласно первоописанию — 22); верхнегубных щитков 9—10; нижнегуб-
ных щитков по 7. чешуй вдоль тела снизу ИЗ; чешуй вокруг середины тулови-
ща 74; подпальцевых пластинок по 12.
Межносовые щитки разделены между собой чешуйкой, а замежносовые —
двумя Подбородочный щиток пятиугольный, за ним следуют две пары нижне-
челюстных щитков (первая значительно крупнее второй), соединенных между
собой широким швом. В первоописании указано, что у этого экземпляра под-
пальцевые пластинки слабо ребристые. Ознакомившись с голотипом, мы пришли
к выводу, что образование структур, несколько напоминающих ребристость
подпальцевых пластин, возникло за счет высыхания экземпляра, в естествен-
ном состоянии они должны быть гладкими.
Распространение. Добыт только в типовом местонахождении.
ПЕРСИДСКИЙ МИКРОГЕККОН МОЛОЧАЙНЫЙ —
TROPIOCOLOTES PERSICUS EUPHORB1 ACO<LA MINTON ET AL, 1970
Типовая территория: Провинция Лас-Бела, Окр. Карачи, Пакистан.
1962 — Tropiocolotes helenae (not Nikolsky, 1907), Minton, Amer. Mus. Nat. Hist., Vol. 134, p. 81.
1970 — Tropiocolotes persicus euphorbiacola Minton, S. Anderson and J er. Anderson, Proc. Calif.
Acad. Sci., 4th Ser., Vol. 37, no. 9, p. 354.
Диагноз. Ширина спинных полос равна или незначительно уже про-
межутков между ними (но всегда более чем половина ширины промежутков);
регенерированный хвост желтый, темных поперечный полос нет.
Голотип. CAS 93939. «3. Пакистан, округ Лас-Бела, Хабчауки, около
150 м н. у. м.— хр. Паб; 27.XII 1962. Jer. A. Anderson». $> ad. L = 28,5 mm;
L cd = 32,5 mm; L/L cd = 0,87; межносовые щитки соприкасаются между со-
бой; за ними следует пара крупных замежносовых щитков, также соприкасаю-
щихся друг с другом; верхнегубных щитков 8/9. Нижнегубных — восемь;
две пары нижнечелюстных щитков; первая значительно больше второй и соеди-
нена между собой широким швом. Подпальцевых пластинок по 14.
Описание (по 58 экз. из колл. ИЗАНУ, CAS, SAM, AMNH, MNHP,
SMF, USNM).
L общ. = 17,8—35,9 (26,01 ±0,53) мм; чешуй между центрами глаз 15—
20 (17,67 ±0,19); верхнегубных щитков 8—10 (8,78 ±0,06); нижнегубных 6 —
8 (7,18 ± 0,04); чешуй вдоль тела снизу 111 —130 (120,89 ± 0,60); чешуй во-
круг середины туловища 66—77 (71,00 ±0,41); подпальцевых пластинок 11—
15 (13,55 ± 0,09).
Межносовые щитки соприкасаются между собой (45,6 %) или разделены
одной чешуйкой (54,4 %); замежносовые щитки соприкасаются между собой
{96,5 %), очень редко (3,5 %) разделены чешуйкой. Подбородочный щиток
пяти- или треугольный; за ним следуют одна (18,4 %), две (65,8 %) или три
(14,0 %) пары нижнечелюстных щитков, из которых первая значительно круп-
нее остальных (у единственного экземпляра, что составляет 1,8 % всех просмот-
ренных, имеется одна пара слабо выраженных нижнечелюстных щитков);
в первой паре нижнечелюстные щитки соединены широким швом (лишь у
указанного экземпляра нижнечелюстные щитки разделены горловой чешуйкой).
Распространение. Данный подвид занимает восточную часть
видового ареала: Ю. Пакистан до р. Инд на востоке.
Экология. Некоторые сведения по экологии содержатся в единствен-
ной работе Ш. Минтона [2631 и Ш. Минтона и др. [265]. Наблюдения сделаны
в Пакистане и касаются подвида Т. р. euphorbiacola.
Стации и количественные данные. Этот микрогеккон
широко распространен от г. Бела до Халеджи (вблизи г. Татта), здесь плотность
118
его весьма высока. Гекконы придерживаются участков, где скапливаются от-
мершие части молочаев Euphorbia caudiciphola и находят убежища в полых
стволах растений Вероятно, гекконы также обитают в зарослях молочаев и в
период их вегетации, однако в это же время поиск гекконов здесь затруднен.
Локальные популяции встречены авторами в аналогичных условиях в окр.
Лакки (ю. г. Сехван), Дивана (долина в верховьях р. Хаб) и Набисар (пусты-
ня Тар к в. от р. Инд). В окр. Лакки персидский микрогеккон добыт на высоте
около 225 м н. у. м., номинативная форма — на высоте 1880 м н. у. м. [223].
Суточный и сезонный цикл активности. Много-
численен с середины ноября до конца февраля; бывает обилен после редких
зимних ливней и туманов. Их можно наблюдать влажными теплыми ночами
в сентябре и октябре на поверхности бегающими среди камней. По коллекци-
онным материалам этот вид добывали с января по декабрь.
Питание. В желудке вскрытой нами ящерицы, добытой 28. VII 1958 г.
в Ю. Пакистане, обнаружены остатки клопа и муравья. Содержащиеся в терра-
риуме персидские микрогекконы пили воду, слизывая ее с часового стекла.
Питались ящерицы неразборчиво, поедая разнообразных беспозвоночных под-
ходящего размера, в частности молодых пауков и термитов. Подсчитано, что
один геккончик съедает за раз более 20 экз. термитов. У только что отловленных
гекконов хвост более толстый, чем у длительно содержавшихся в неволе и пло-
хо питавшихся.
Размножение. Взрослые особи обычно живут парами. Самка откла-
дывает единственное яйцо размером 8 х 5,7 мм в разрушенных полых корнях
молочаев, в расщелинах или под небольшими корнями других растений. Период
кладки длится с апреля по август. Сразу после откладки яйцо мягкое, но через
несколько часов оболочка твердеет и становится хрупкой. Оно удлиненное
на концах, глянцево-белое, с продольной полосатостью, исчезающей через три-
четыре недели после откладки. За это время яйцо несколько увеличивается
в размерах Продолжительность инкубации составляет пять—восемь недель
(35—60 суток) Длина тела новорожденных геккончиков составляет 13—15 мм,
молодые окрашены сходно со взрослыми.
Особенности поведения. Потревоженные ящерицы стремятся
скрыться в убежище. Двигаются при этом змеевидно, сильно изгибая тело
и хвост и почти не используя конечности. Кроме того, они могут спасаться
бегством, ловко прыгая с ветки на ветку мелких кустарников. Такое поведе-
ние особенно характерно ночью, вовремя поиска пищи. Исследователи обраща-
ют внимание на то, что в террариуме персидские микрогекконы охотно влеза-
ют на стволы деревьев, однако в природе на деревьях они ни разу замечены не
были.
МИКРОГЕККОН ЛЯТИФИ —
TROPIOCOLOTES LATlFl LEVITON ЕТ S. ANDERSON. 1972
Типовая территория: Керман, Иран.
Кариотип: не изучен.
1972 — Tropiocolotes latifi Leviton and S. Anderson, Occas. Papers Calif. Acad. Sci., no. 96, p. 37.
Диагноз. Нижнечелюстных щитков нет.
Описание (рис. 53) (по голотипу, единственному известному экзем-
пляру)САБ 134365. «Керман, пров Керман, Иран 1965»? сГ juv. L = 17,1 mm;
L cd = 15,7 mm; LIL cd = 1,09; отношение диаметра уха/глаза 0,38.
Ноздря расположена между пятиугольным межчелюстным, первым верх-
негубным, крупным плоским щитком, одной (слева) или двумя (справа) допол-
нительными носовыми чешуйками (по первоописанию, с обеих сторон имеется
только по одной дополнительной чешуйке); межносовые щитки соприкасаются
между собой. За ними располагается пара крупных замежносовых щитков, ко-
торые отделены друг от друга чешуйкой; чешуй между центрами глаз 16; верх-
негубных щитков шесть-семь (согласно первоописанию — по семь); нижнегуб-
ных — по шесть (согласно первоописанию — по пять); чешуй вдоль тела снизу
120; чешуй вокруг середины туловища около 72; подпальцевых пластинок 14/13
119
(согласно первоописанию — по 14). Подбородочный щиток трапециевидный
с неровным задним краем, где к нему примыкает восемь горловых чешуек.
Окраска и рисунок (после фиксации в спирте). Основной фон
верхней поверхности — желтовато-белый.От заднего края глаза через теменную
область и над лопаткой с каждой стороны тянется темная полоса, оканчиваю-
щаяся на уровне локтевого сустава вытянутой назад передней конечности.
В центре между лопатками имеется небольшое темное пятно; такого же цвета
и размера два пятна имеются на спине перед основанием задних конечностей.
На хвосте заметны девять нешироких (уже промежутков) темных ровных
поперечных полос. Голова и конечности лишены рисунка. Нижние поверхности
белые.
Распространение. Известен только из типового местонахожде-
ния в Ц. Иране, на границе южной оконечности хр. Загрос и крупного пустын-
ного массива.
Кадастр к рис. 5 2.
Иран: 1 — г. Керман (CAS).
Систематика. Отсутствие у этого экземпляра нижнечелюстных
щитков позволило А. Левитону и С. Андерсену выделить его в самостоятельный
вид. Однако у наиболее близких к нему видов — Т. persicus (номинативный
подвид) и Т. helenae — частота проявления признака «отсутствие контакта
между первой парой нижнечелюстных щитков» весьма высока (особенно у
Т. helenae), что свидетельствует о наличии тенденции к исчезновению нижнече-
люстных щитков. Кроме того, изменчивость этого признака у других видов
(Т. tripolitanus, Т. depressus, Т. scortecci, A. laevis и др.) показывает, что ис-
пользование его для дифференциации видов вряд ли целесообразно. Однако
в связи со слабой изученностью данной группы видов мы рассматриваем Т. la-
tifi в качестве самостоятельного вида условно.
Экология — не изучена. Добыт на высоте 1710 м н. у. м.
ПОДРОД АЗИАТСКИЙ КАРЛИКОВЫЙ ГЕККОН —
ASIOCOLOTES GOLUBEV, 1984
Типовой вид: Tropiocolotes levitoni Golubev et Szczerbak.
Диагноз. Подпальцевые пластинки гладкие; пара плоских межносо-
вых щитков с ноздрями не соприкасается; замежносовые щитки не выражены;
кроме межчелюстного и первого верхнегубного щитков к ноздре примыкают
еще две (очень редко три) чешуйки, существенно не отличающиеся размерами
ни от межносовых, ни от остальных чешуй морды; чешуя тела гладкая, спины —
мелкая, заметно мельче брюшной; по крайней мере в последней х/3 нерегенери-
рованного хвоста имеются различные сочетания увеличенных подхвостовых
щитков; у самцов всегда есть две—восемь хорошо выраженных анальных пор.
Распространен в восточной части Ю.-З. Азии (Пакистан и Афганистан). Под-
род включает два вида.
Дополнительные признаки. Морда покрыта слабо выпук-
лой гладкой многоугольной чешуей, заметно уменьшающейся к затылку. Гор-
ловые чешуи плоские, гладкие, округло-многоугольные; верхнегорловые
чешуи несколько крупнее остальных. Спина покрыта плоской, гладкой, округ-
ло-четырехугольной мелкой чешуей, увеличивающейся к бокам туловища так,
что границу между ней и брюшной чешуей провести невозможно. Брюшная че-
шуя плоская, гладкая, округло-многоугольная, черепитчатая, заметно круп-
нее чешуи спины. Анальные поры расположены «Л-образно под небольшим уг-
лом на более крупных, чем брюшные, чешуйках; между ними и анальной щелью
около пяти рядов чешуй Хвост несколько уплощен, покрыт кольцами гладких,
овально-прямоугольных, плоских, увеличивающихся к бокам чешуй; снизу
покрыт различными сочетаниями крупных щитков. Передняя конечность, вы-
тянутая вперед, достает концами пальцев до конца морды, задняя — почти
до лопатки. Пальцы слегка сдавлены с боков, покрыты сверху одним, с боков —
120
двумя рядами чешуй; основание когтя скрыто чехлом, состоящим из верхней
и нижней гладких пластинок; сверху и с обеих сторон с ним соприкасается по
одной чешуйке.
Таблица для определения видов подрода Asiocolotes
1(2). Анальных пор у самцов две — пять; подхвостовые щитки имеются только в последней
трети хвоста ...................Т. depressus Minton, S. Anderson et Jer. Anderson
2 (1). Анальных пор шесть-семь; подхвостовые щитки имеются на всем протяжении хвоста
............................................Т. levitoni Golubev et Szczerbak
АЗИАТСКИЙ КАРЛИКОВЫЙ ГЕККОН ЛЕВИТОНА —
TROPIOCOLOTES LEVITONI GOLUBEV ET SZCZERBAK, 1979
Типовая территория: Афганистан; Кабул, р. Пагман; 10 км к ю.-з. от Кабу-
ла; пров. Погар, Бараки Барак.
Кариотип: не изучен.
1960 — Cyrtodactylus(~ Gymnodactylus) persicus (= Alsophylax persicus), Wettstein, Zool. Anz.,
Bd 165, Haft 5/6, S. 190.
1963 — Alsophylax pipiens, Leviton and S. Anderson, Proc. Calif. Acad. Sci., 4th Ser., Vol. 31,
no. 12, p. 334.
1979 — Tropiocolotes levitoni Голубев, Щербак. Докл. АН УССР, № 4, сер. «Б», с. 309.
Диагноз. У самцов шесть-семь анальных пор; подхвостовые щитки
есть на всем протяжении хвоста.
Голотип. CAS 120283 d ad. Афганистан, Кабул, 6.V 1968 г., Р. и
Э. Кларк, L = 39, 5 мм; L cd= 46,2 мм; L/L cd — 0,85. Ноздря расположена
между четырехугольным межчелюстным, первым верхнегубным и тремя плос-
кими носовыми щитками, не отличающимися размерами от остальных чешуй
морды. Межносовые щитки плоские, с ноздрями не соприкасаются, существен-
но не отличаются от остальных чешуй морды, замежносовые отсутствуют.
Чешуй между центрами глаз 18. Верхнегубных щитков по 10, нижнегубных —
7/8; две пары нижнечелюстных щитков, щитки внутренней пары соприкасаются
между собой и с треугольным подбородочным щитком. Чешуй вдоль тела снизу
133, чешуй вокруг середины туловища 80; анальных пор 6 ; подпальцевых плас-
тинок по 23.
Описание (по 24 экз. из колл. CAS , USNM, NMW и ZML) (рис. 56).
L <? ad = 27,4—44,9 (37,30 ± 1,27) мм; L $ ad = 22,0—44,8 (37,91 ±
±0,98) мм; ULcd общ. = 0,80—0,85; индекс высоты/ширины головы 43—
55 (48,00 ± 0,70); чешуй между центрами глаз 16—21 (18,23 ± 0,29); верхне-
губных щитков 9—11 (9,80 ± 0,09); нижнегубных 6—9 (7,63 ± 0,10); чешуй
вдоль тела 123—146 (129,84 ± 1,02); вокруг середины туловища 71—82
(77,93 ± 0,84); подпальцевых пластинок 17—23 (20,02 ± 0,23); анальных
пор у самцов 6 (11 экз.) или 7 (2 экз.).
До орбиты доходит четыре-пять верхнегубных щитков; ушное отверстие
овальное, над ним расположены 0—2 увеличенные чешуйки. Подбородочный
щиток пяти- или треугольный, со скошенным (60,9 %) или острым (39,1 %) кон-
цом; за ним следуют две пары нижнечелюстных щитков, из которых первая со-
единена между собой швом (21,8 %) или разделена одной (60,9 %) или двумя
(17,3 %) чешуйками. Иногда в области крестца и на верхнебоковых поверх-
ностях у основания хвоста имеются увеличенные чешуи, напоминающие спин-
ные бугорки, на некоторых из них есть слабо различимые продольные ребра.
Каждому хвостовому сегменту снизу соответствуют на протяжении 2/3 длины
от основания хвоста чередующиеся один и два крупных щитка, далее до его
конца — по одному. На внешней поверхности голени (и единично на бедре)
есть крупные ребристые бугорки, перемешанные в беспорядке с более мелкими
гладкими чешуями.
Окраска и рисунок. Основной фон спины при жизни [209]
палево-желтовато-коричневый; задние края темных поперечных полос окан-
тованы белым. Сквозь просвечивающееся белое или сероватое брюхо видны внут-
ренние органы. Губные щитки в серых точках.
От ноздри через глаз до ушного отверстия тянется коричнево-бурая поло-
са; такого же цвета полоски могут быть в теменной и затылочной областях,
121
Рис. 56. Tropiocolotes levitoni:
Д — общий вид, Б — голова сбоку, В — голова сверху, Г — голова снизу, Д — носовые щитки,
Е — преанальные поры, Ж — хвост сдизу, 3 —палец сбоку (CAS 96217, Кабул, Афганистан)
причем в последней — М-образные. Семь-восемь аналогичных поперечных по-
лос расположены от шеи до крестца, до 10—11 полос — от основания до кончика
хвоста; при этом полосы в два—четыре раза уже промежутков между ними; эти
полосы могут быть разорванными посередине и смещенными на месте разрыва
вдоль продольной оси тела. На конечностях есть неясный темно-бурый узор.
Распространение. Узкоареальный вид: известен из нескольких
пунктов в р-не Кабула и несколько южнее (рис. 52).
Кадастр к рис. 5 2.
Афганистан: 1 — Окуак [257]; 2 — Чарикар [244]; 3 — окр. р. Пагман (CAS);
4 — Кабул (CAS); 5 — Баракибарак (USNM).
Половой диморфизм и возрастная изменчивость.
Самцов можно отличить от самок по наличию анальных пор. В отличие от более
мелких гекконов этого рода, у Т. levitoni нет различий в размерах между пола-
ми. Индекс Аи/О подвержен возрастной изменчивости, как и у видов рода
Alsophylax.
Систематика и географическая изменчивость.
В результате ошибочной идентификации трех видов североазиатских геккончи-
ков (A. pipiens, A. laevis и A. microtis [121]) для большинства аридных районов
Средней и Центральной Азии обычно указывался пискливый геккончик [112],
который в таком объеме характеризовался широким диапазоном изменчивости
многих признаков. Это обстоятельство привело к тому, что зарубежные авторы
[209, 243, 245, 253, 257, 263, 289, и др.], не представляя реально ни ареала,
ни границ изменчивости вида A. pipiens, отнесли к нему особей из Афганистана.
Ознакомившись с коллекционными материалами, мы пришли к заключению
[43], что эти особи относятся к новому виду, принадлежащему к роду Tropio-
colotes.
Вид с узким ареалом не проявляет географической изменчивости по изу-
ченным признакам. Однако границы изменчивости некоторых из признаков
(наличие подхвостовых щитков и бугров в области крестца, количество аналь-
ных пор и др ) позволяют предположить, что между карликовым гекконом Леви-
тона и видами рода Alsophylax (в частности, A. laevis) были общие предковые
формы.
Немногочисленные сведения, касающиеся экологии этого вида, имеются
почти исключительно в работе Г. Сойфера [289], который наблюдал этих яще-
риц в 1971 —1973 гг. в естественных условиях, а в 1977 г.— в террариуме.
Стации и количественные данные. В Афганистане этот
вид встречается только в зоне деятельности человека, где часто является
фоновым. В Кабуле на глиняных стенах домов он иногда образует большие
скопления. Обитает до высоты 2300 м н. у. м. [2091.
От ношение к температуре. Г. Сойфер [289] отмечает, что
в Кабуле, расположенном на высоте 1800 м н. у. м., зимы холодные, снежный
покров достигает метровой высоты. Средняя температура января здесь — 1,3°.
По ночам температура может иногда опускаться до —25°. Тем не менее геккончик
Левитона хорошо адаптировался к таким условиям. Содержавшиеся этим авто-
ром в неволе ящерицы активно перемещались по террариуму при весьма низкой
для рептилий температуре (+8—10°)^
Несколько экземпляров этих геккончиков были добыты указанным авто-
ром майским вечером при температуре воздуха 20° на стене глинобитного дома на
высоте 1 м от земли.
Суточный и сезонный цикл активности. Покидают
убежища в ранних сумерках, активны ночью. В естественных условиях зимов-
ка этих геккончиков продолжается с ноября до конца февраля; в некоторые хо-
лодные годы она затягивается до конца апреля.
Питание. Излюбленные места охоты — наружные поверхности окон-
ных противомоскитных сеток, где скапливается много насекомых. Охотно по-
едают комаров (Anopheles), что обеспечивает этому виду хорошую кормовую
базу до глубокой осени. Кроме того, в состав пищи входят жуки, пауки, ночные
бабочки, гусеницы, клопы, муравьи и тли.
12&
Размножение. Спаривание геккончиков наблюдать указанному
автору не удалось. Из приведенных в работе фотографий видно, что у самок
бывает одно или два яйца одновременно. Откладка яиц в неволе происходила
с февраля по декабрь; размеры их 13 X 9 мм. Самки, как правило, закапыва-
ли яйца в почву или оставляли их открытыми на коре деревьев, имевшейся в
террариуме. Яйца содержались в инкубаторе при температуре 20—28° Молодые
выходили через 44—93 дня (большинство — через 53—58 дней) после начала
инкубации. Молодые геккончики питались гусеницами вощинной моли, мелки-
ми медведками и сверчками; достигали размеров взрослых через 1,5 года.
Особенности поведения. Р. Кларк и др. [209] отмечают, что
эти гекконы имеют постоянные охотничьи участки, где их можно наблюдать
каждую ночь, часто сидят на участках стен, освещаемых уличными фонарями.
Очень осторожны; потревоженные, быстро скрываются в трещинах или в полу-
сгнивших досках дверей и фрамуг.
АЗИАТСКИЙ КАРЛИКОВЫЙ ГЕККОН ПЛОСКИЙ —
TROPIOCOLOTES DEPRESSUS MINTON ET JER. ANDERSON, 1965
Типовая территория: Пров. Кветта, Пакистан.
Кариотип: не изучен.
1965 — Tropiocolotes depressor Minton et Jer. Anderson, Gerpetologica, Vol. 21, по. 1, p. 59.
Диагноз. У самцов — две—пять анальных пор; подхвостовые щитки
есть только в последней трети хвоста.
Голотип. AMNH 93003 «Кач (округ Кветта), 3. Пакистан, покинутая
железная дор. Зиарат — Кветта, 2200 м н. у. м. 18.V 1962. S. A. Minton».
L 9 juv = 26,8 мм; L cd — около 24,5 мм; L/L cd = 1,09. Ноздря расположе-
на между четырехугольным межчелюстным, одним верхнегубным и двумя
плоскими мелкими носовыми щитками (первый незначительно крупнее второ-
го), не отличающимися размерами от остальных чешуй морды. Межносовые
щитки плоские, с ноздрями не соприкасаются, не отличаются размерами от ок-
ружающих чешуй; замежносовые щитки отсутствуют. Чешуй между центрами
глаз 17; верхнегубных щитков по девять, нижнегубных щитков 6/7. Позади
трапециевидного подбородочного щитка есть пара мелких нижнечелюстных щит-
ков, разделенных двумя чешуйками, почти равными нижнечелюстным щиткам.
Чешуй вдоль тела снизу 133, вокруг середины туловища — 76. Анальных пор
нет. Подпальцевых пластинок 16/17.
Описание. Составлено по всем известным в настоящее время 5 экз.
(из колл. AMNH, RSM и SMF), рис. 57.
L общ. = 26,8—32,2 (29,82 ± 0,89) мм; LIL cd = 0,94—1,09; индекс вы-
соты/ширины головы 47—53 (49,33 ± 1,86); чешуй между центрами глаз 16—
20 (17.80 ± 0,73); верхнегубных щитков 8—10 (8,80 ± 0,25), нижнегубных —
6—8 (7,00 ± 0,15); чешуй вдоль тела снизу 129—139 (134,00 ± 1,79); вокруг
середины туловища 74—76 (75,00 ± 0,32); подпальцевых пластинок 16—
18 (17,30 ± 0,26); анальных пор у самцов 2—5 (3,67 ± 0,80).
До орбиты доходит 3,7—4 верхнегубных щитка; ушное отверстие округлое,
над ним увеличенных чешуй нет. Подбородочный щиток трапециевидный с неров-
ным задним краем; за ним идут одна-две пары мелких нижнечелюстных, из ко-
торых первая между собой не соприкасается, разделена одной (20,0 %) или
двумя (80 %) горловыми чешуйками. Бугорки, напоминающие спинные, в об-
ласти крестца отсутствуют. Хвост снизу покрыт кольцами плоских гладких
чешуй, которые в последней трети замещаются крупными щитками, сочета-
ющимися на одном сегменте сначала как 2 = 1, а затем — 1=1. На верхних
поверхностях конечностей увеличенных бугорков нет.
Прижизненная окраска [264; 2631: основной фон спины шаф-
ранно-желтый; три— пять темно-коричневых поперечных полос на спине и шесть
на хвосте; все полосы четко ограничены, густо окрашены, не окантованы белым,
уже промежутков между ними. Темно-коричневая короткая полоса есть на те-
мени. Такого же цвета полосы тянутся от ноздри через глаз и ушное отверстие
с обеих сторон головы и, соединяясь на затылке, образуют подковообразный
124
Рис. 57. Tropiocolotes depressus:
A — общий вид, Б — голова сверху, В — голова снизу, Г — носовые щитки, Д — под-
бородок, Е — спина, Ж — преанальные поры (В—Д — RSM 1964.58.1, Колпур, Пакистан; ос-
тальное — AMNH 93003, Зиарат-Кветта, Пакистан)
рисунок. Конечности лишены рисунка. Нижние поверхности палево-розово-бе-
лые.
Половой диморфизм и возрастная изменчивость.
Самцы отличаются от самок наличием хорошо выраженных анальных пор. По
известным пяти экземплярам этого вида судить о возрастной изменчивости
не представляется возможным.
Распространение. Известен из трех пунктов вблизи Кветты
(Пакистан) (рис. 52).
Кадастр к рис. 52.
Пакистан: / — р-н Кветты, Кач на заброшенной железнодорожной линии из Зиарата
в Кветту (AMNH); 2 — в. Чилтан-Бергес, 27 км ю. г. Кветта и 22 км сев. г. Мастунг [259];
3 — Колпур (RSM).
Систематика и географическая изменчивость.
Т. depressus имеет локальный ареал (известен из нескольких пунктов пров.
Кветта в Пакистане), что позволяет предположить отсутствие у этого вида
каких-либо форм. Вероятно, здесь может идти речь лишь об индивидуальной
изменчивости: у вновь добытых особей [265] в сравнении с типами оказалось
больше анальных пор, подпальцевых пластинок и чешуй между центрами глаз.
Экология. Сведения о ней имеются в немногих работах [263, 264,
265]. Геккончики этого вида были добыты ранними сумерками между двумя
небольшими гранитными поднятиями на высоте около 2700 м н. у. м. на каме-
нистом склоне холма, расположенном под углом 45° к линии горизонта и покры-
том скудной кустарниковой и травянистой растительностью. Они обитают в ста-
рых норах рыжей пищухи (Ochotona г. rufescens) и под камнями. Самец и самка
были добыты под одним камнем. Довольно быстро перемещаются по грунту
и обломкам скал, на которые часто залазят. В спокойном состоянии движения
замедлены: геккончики стоят, змееобразно изогнув тело, затем, сильно изгиба-
ясь, пробегают некоторое расстояние и вновь занимают исходную позицию.
Потенциальные враги: скорпионы (Androchtonus astral is,
Ruthus occitanus), сольпуга (Galeodes caspius), многоножки (Skolopendra sp.),
молодые змеи (C. rhodorachis, C. Karelini, V. lebetina), ящерица (E. schneideri
blitanus), белуджистанские землеройки и еж (Hemiechinus megalotus).
Дневные температуры этого региона в октябре достигают 15° днем, ночью
падают до —4°. Летом исследователи здесь геккончиков не обнаруживали.
Род ребристый геккон — Carinatogecko Golubev et
Szczerbak, 1981
Типовой вид: Bunopus aspratilis S. C. Anderson, 1973.
1981 — Carinatogecko Голубев, Щербак, Вести, зоол., № 5, с. 34.
Диагноз. Все чешуи тела, кроме крупных щитков головы, очень реб-
ристые; с ноздрей соприкасаются три носовых щитка, из них первый несколько
выпуклый, в 1,5—2 раза больше второго, который незначительно больше тре-
тьего; межносовые и замежносовые щитки отсутствуют; высота первого верх-
негубного щитка от ноздри до края рта заметно меньше его ширины вдоль края
рта; верхнегубные щитки верхним краем загибаются на верхнюю поверхность
морды. До орбиты доходят 3,5—4 верхнегубных щитка, всего их 8—10. Зрачок
вертикальный, с зазубренными краями. Спина покрыта плоской чешуей и бу-
горками. Преанальные поры у самок отсутствуют (самцы пока неизвестны).
Хвост сегментирован (сегменты слабо различимы); с обеих сторон одного сегмен-
та имеется два полукольца из трех-четырех бугорков, расположенных основа-
ниями в середине сегмента, не соприкасающихся между собой и отделенных
от заднего края сегмента кольцом чешуй; серединные чешуи снизу сегмента
могут быть несколько увеличены, но не достигают размеров подхвостовых щит-
ков, характерных для многих гекконов. Пальцы слабо искривлены, когтистые,
не расширены, снизу покрыты одним продольным рядом поперечно расширенных
пластинок, боковая бахрома отсутствует.
126
Дополнительные признаки. Мелкие гекконы (L = 20—
30 мм). Межчелюстной щиток пятиугольный; первые носовые щитки разделены
между собой чешуйкой; заносовые впадины выражены, лобная — нет; чешуя
морды неоднородная, многоугольная, на затылке чередуется с бугорками;
реснитчатые выросты выражены слабо; ушное отверстие округлое. Подборо-
дочный щиток пятиугольный; за ним следуют четыре пары нижнечелюстных
щитков, плавно уменьшающихся назад так, что последняя пара слабо отличима
от горловых чешуй; первая пара соединена между собой широким швом, по-
следние две пары с нижнегубными щитками не соприкасаются; верхнегорловые
чешуи несколько крупнее остальных. Среди плоской мелкой спинной чешуи
правильными продольными (12—14) и поперечными (22—24) рядами располо-
жены спинные бугорки; брюшная чешуя плоская, ромбическая, черепитчатая;
граница ее с боковой не выражена. Бугорки прослеживаются почти до конца
хвоста. Основание когтя скрыто чехлом, состоящим из верхней и нижней плас-
тинок с несколькими невысокими продольными килями; сверху к чехлу примы-
кают одна, с боков — по две чешуйки.
Распространены вдоль хр. Загрос в В. и 3. Иране. Род включает два вида.
Таблица для определения видов рода Carinatogecko
1(2). Чешуя середины спины заметно меньше брюшной, хвостовые бугорки остроконечные,
высок е, с увеличенной задней гранью; аналогичные спинным бугорки имеются на пред-
плечье; подпальцевых пластинок на IV пальцах задних конечностей 17—18 . .
...................... . Carinatogecko aspratilis (S. Anderson)
2(1). Чешуя середины спины почти равна брюшной; хвостовые бугорки неостроконечные, уп-
лощенные, их задняя грань невысокая; аналогичные спинным бугорки на предплечье
отсутствуют; подпальцевых пластинок на IV пальцах задних конечностей 15.
............Carinatogecko heteropholis (Minton, S. Anderson et J. Anderson)
РЕБРИСТЫЙ ГЕККОН ИРАНСКИЙ —
CARINATOGECKO ASPRATiLlS (S. C. ANDERSON, 1973)
Типовая территория: 35 км в. пос. Гечсаран (30° 20' с. ш., 50° 48' в. д.,
пров. Фарс, Иран).
Кариотип: не изучен.
1973 — Bunopus aspratilis S. С. Anderson, Herpetologica, Vol. 29, N 4, p. 35.
1981 — Carinatogecko aspratilis, Голубев, Щербак, Вести, зоол., Киев, № 5, с. 35.
Диагноз. Чешуя середины спины заметно меньше брюшной; хвосто-
вые бугорки очень шиповатые, остроконечные, высокие; боковая пальцевая
чешуя нижнего ряда, ближе к концу пальца, плоская, с невысоким продольным
килем, не оканчивающимся шипом; подпальцевых пластинок 17—18; чешуя
верхней стороны предплечья разнородная.
Голотип. USNM 193961, g ad Иран, Хузестан, 35 км в. от Гечсаран,
колл. Дж. В. Нилл, 10.11 1964. L = 27,4 мм; L cd = 29,6 мм; LIL cd = 0,93;
чешуй поперек головы 14; верхнегубных щитков девять, нижнегубных семь;
нижнечелюстных щитков четыре пары; чешуй вдоль тела снизу НО, поперек
брюха 28; подпальцевых пластинок 18; преанальные поры отсутствуют.
Описание (по двум известным самкам из колл. USNM) (рис. 58).
L g = 26,2—27,4 мм; L cd = 29,6 мм; LIL cd = 0,93; индекс высоты/ширины
головы 55—56; диаметр уха/глаза 29—30; верхнегубных щитков 9—10; нижне-
губных 6—8; чешуй между центрами глаз 14—16; длина спинного бугорка /
L 23—25; чешуй, окружающих спинной бугорок, 10—11; чешуй вдоль тела сни-
зу НО—111; чешуй поперек брюха около 28; подпальцевых пластинок 17—18.
Над глазами на голове два — четыре укрупненных надбровных щитка; за
последним верхнегубным щитком через три-четыре чешуйки следует ряд из
трех крупных бугорков; спинные бугорки треугольно-овальные, на боках туло-
вища соприкасаются между собой в поперечных рядах. Вдоль хвостового сег-
мента в центре четыре-пять чешуй; хвостовых бугорков в полукольце с каж-
дой стороны сегмента три-четыре, по крайней мере первый и второй отделены
от заднего края сегмента рядом чешуй; каждый бугорок крупный, остроконеч-
но-шиповатый, с очень крупной задней гранью, расположенной под отрица-
127
Рис. 58. Carinatogecko aspratilis:
A — общий вид, Б — голова сверху, В —- носовые щитки, Г — спина, Д — брюхо, Е — пред-
плечье сверху, Ж — хвост сверху, 3 — хвост снизу, И — подпальцевые пластины (USNM 193961,
окр. Гечсаран, Иран)
Рис. 59. Carinatogecko heteropholis:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — голова снизу, Д — предплечье
сверху, Е — спина, Ж — пален снизу, 3 — хвост сверху, И — хвост снизу (CNHM 74549,
Салахеддин, Ирак)
тельным углом к поверхности сегмента; снизу хвоста на сегменте две централь-
ные пары чешуй, несколько крупнее остальных. Вытянутая вперед и вдоль
тела передняя лапа достает концами пальцев почти до ноздри, задняя — до
подмышки; среди однородной чешуи на предплечье, бедре и голени разбросаны
бугорки, аналогичные спинным (на предплечье они менее выражены). Нижний
ряд боковой чешуи ближе к основанию пальца состоит из утолщенных, с высо-
ким килем чешуй, которые в дальнейшем уплощаются и несут на себе только
невысокий продольный киль без шипа; каждая подпальцевая пластинка имеет
129
два-три продольных киля, не оканчивающихся шипом и менеэ выраженных
ближе к концу пальца.
Окраска и рисунок. Основной фон верхней стороны тела серо-
голубой; на голове заметен неясный узор, образованный тонкими изогнутыми
короткими темными линиями; от затылка до поясницы пять очень изогнутых
М-образных темных полос; местами они сливаются выступающими углами, об-
разуя продольный узор; на хвосте 13 поперечных полос такого же цвета; на ко-
нечностях неясный рисунок, состоящий из слабо выраженных поперечных тем-
ных полос. Нижние поверхности светлые, со множеством пигментных пятен.
Распространение. Известны только из типовой местности.
Экология не изучена. Судя по месту добычи, обитает в зоне предгорий
на высоте 200—500 м н. у. м. Экземпляр был добыт под галькой на берегу су-
хого русла среди редкой травянистой и древесной растительности [169].
РЕБРИСТЫЙ ГЕККЦН ИРАКСКИЙ —
CARINATOGECKO HETEROPHOLIS (MINTON, S. ANDERSON ЕТ JEB. ANDERSON, 1970)
Типовая территория: Ирак, пров. Эрбиль, Салах-эд-Дин.
Кариотип: не изучен.
1959 — Alsophylax persicus Nik., Reed. Marx, Transact. Kansas, Acad. Sci., Vol. 62, N 1, p. 97
(err. det).
1970 — Tropiocolotes heteropholis Minton, S. C. Anderson, J. A. Anderson, Proc. Calif. Acad.
Sci., 4th ser., Vol. 37, N 9, p. 357.
1981 — Carinatogecko heteropholis. Голубев, Щербак, Вести, зоол., № 5, с. 37.
Диагноз. Чешуя середины спины незначительно больше или равна
брюшной; хвостовые бугорки отчетливые, но уплощенные, неостроконечные,
их задняя грань невысокая; боковые пальцевые чешуи нижнего ряда с высоким
продольным килем, оканчивающимся шипиком; подпальцевых пластинок 15;
чешуя верхней стороны предплечья однородная.
Описание (по голотипу FMNH 74549) (рис. 59). g ad? L = 20,4 мм;
Led = 19 мм; индекс ширины/высоты головы 56; диаметр уха/глаза 19; верхне-
губных щитков по 8; нижнегубных по 6; чешуй между центрами глаз 15; длина
спинного бугорка / L 19; чешуй, окружающих спинной бугорок, 8—9; чешуй
вдоль тела снизу около 100; чешуй поперек брюха 28—30; подпальцевых плас-
тин по 15.
Надбровные щитки не выражены; за последним верхнегубным щитком че-
рез одну чешуйку следует ряд из трех крупных бугорков; спинные бугорки тре-
угольные, друг с другом не соприкасаются. Индикаторами хвостовых сегмен-
тов служат невысокие бугорки. Хвостовые бугорки располагаются в цент
ре сегмента и отделены отего заднего края рядом чешуй. Вдоль сегмента три —
пять чешуй, хвостовые бугорки четких полуколец не образуют и не соприкаса-
ются. Вытянутая вперед и вдоль тела передняя лапа достает концами пальцев
до переднего края орбиты, задняя — до середины плеча вытянутой назад
передней лапы; на верхней стороне бедра и голени имеются бугорки, аналогич-
ные спинным; на плече и предплечье чешуя однородная. Боковая пальцевая че-
шуя плоская, с невысоким, но отчетливым килем, не оканчивающимся шипом.
Окраска и рисунок. Из-за плохой фиксации рисунок верхней
стороны тела не сохранился. Чешуя нижних поверхностей в многочисленных
темных пигментных пятнах.
Распространение. Известен только из типовой местности.
Экология. Не изучена. Экземпляр вида [280] добыт под куском ко-
ры на полу гостиницы. Автор полагает, что он был завезен с дровами.
Род тонкопалый геккон — Tenuidactylus Szczerbak
et Golubev, 1984
Типовой вид: Gymnodactylus caspius Eichwald, 1831.
1825 — Gymnodactylus Spix (part.) Spec. nov. Lacert. Brasil. S. 17.
1827 — Cyrtodactylus Gray (part.) Philos. Mag. (2), 3, p. 55.
1843 — Cyrtopodion Fitzinger, Syst. Rept, s. 18, 93*.
* Последнее название нельзя использовать как родовое, так как его диагноз противоречит
диагнозу Tenuidactylus.
130
Диагноз. Пальцы длинные, когтистые, базальные фаланги почти не
отличаются по толщине, плавно переходят в дистальные (некоторые утолщения
могут быть на суставах), снизу покрыты одним рядом поперечно расширенных
гладких пластин; вторые дистальные фаланги образуют угол с проксимальной
частью пальца; концы пальцев слабо или совершенно не сжаты с боков; бахро-
мы и иных выростов на боковой чешуе пальцев нет. Зрачок вертикальный с за-
зубренными краями. В лобной области головы продольная вогнутость, как
правило, отсутствует или выражена слабо; между центрами глаз обычно не
более 30 чешуек; преанальные и (реже) бедренные поры имеются только у сам-
цов. Сегментация хвоста выражена хорошо.
Дополнительные признаки. Мелкие и средние гекконы
(L = 60—80 мм). Спинные бугорки, как правило, крупные, от треугольных
или трехгранных до округлых, обычно ребристые, образуют 8—14 продоль-
ных и 20—28 поперечных (от затылка до крестца) рядов; три (очень редко два)
заметно увеличенных носовых щитка, из которых первый иногда несколько
крупнее остальных; выросты в задневерхней части орбиты часто хорошо выра-
жены; поперек брюха до 35 чешуй; вдоль тела снизу до 150 чешуй (обычно до
130); преанальные поры не отделены промежутком от бедренных и расположе-
ны почти прямой линией; крупные бугорки, достигающие нижнебоковых по-
верхностей хвоста (по три—пять с каждой стороны), почти доходят до его кон-
ца; как правило, на его нижней поверхности есть ряд увеличенных щитков (два
или две пары на сегменте). Рисунок неяркий, состоит из четырех— семи попереч-
ных туловищных и 7—12 хвостовых полос.
Включает 16 видов, обитающих к з. от р. Инд, в Гималаях и к сев. от них;
состоит из четырех подродов.
Таблица для определения подродов рода Tenuidactylus
1(2). Шипообразно выступающие хвостовые бугорки в полукольцах на сегменте не соприкаса-
ются между собой; они окружены однородной чешуей; высота первого верхнегубного
щитка от ноздри до края рта заметно меньше его ширины вдоль края рта; основной фон
и рисунок выдержаны в светло- и темно-серых тонах...................
........................................Mediodactylus Szczerbak et Golubev
2 (1). Невысокие хвостовые бугорки широко соприкасаются между собой в полукольцах од-
ного сегмента, окружены одним-двумя более мелкими бугорками и еще более мелкой хвос-
товой чешуей; высота первого верхнегубного щитка заметно больше его ширины или не-
значительно меньше ее; основной фон и рисунок выдержаны в светло- и темно-охристо-
коричневых тонах.
3 (4). У самцов не менее 20 бедренных пор; ряд подхвостовых щитков занимает почти всю ши-
рину хвоста; если шов между первой парой нижнечелюстных щитков имеется, то его дли-
на обычно не превышает половины длины подбородочного щитка..........
........................................Tenuidactylus Szczerbak et Golubev
4 (3). У самцов не более 10 преанальных пор; ряд подхвостовых щитков заметно уже ширины
хвоста; длина шва между первой парой нижнечелюстных щитков превосходит половину
длины подбородочного щитка .............Cyrtopodion Fitz.
Подрод тонкопалый геккон —
Tenuidactylus Szczerbak et Golubev, 1984
1984 — Tenuidactylus Щербак, Голубев, Вест, зоол., Киев, № 2, с. 53 (т и п о в о й вид:
G. caspius Eichw).
Диагноз. У самцов имеются преанальные и бедренные поры (не менее
20), вытянутые в почти прямую линию; если шов между первой парой нижне-
челюстных щитков имеется, то его длина обычно не превышает половины длины
подбородочного щитка; форма спинных бугорков варьирует от трехгранной до
округло-треугольной; хвостовые бугорки широко соприкасаются между собой
боковыми гранями; спереди у основания такого бугорка есть один-два более
мелких бугорка, превышающих по размерам окружающую чешую; ряд подхвос-
товых щитков занимает почти всю ширину хвоста (высота щитка более чем
в двое меньше его ширины).
Дополнительные признаки. Высота первого верхнегубного
щитка от ноздри до края рта заметно больше или незначительно меньше его
131
ширины вдоль края рта; уплощенно-треугольные реснитчатые выросты в задне-
верхней части орбиты слабо выражены; среди мелкой чешуи задней поверхнос-
ти бедра обычно имеется два—десять мелких трехгранных или конических
бугорков. Хвост тонкий. Основной фон и рисунок в светло- и темно-коричне-
вых тонах.
Включает четыре вида, распространенных в Средней Азии; частично про-
никает в Закавказье и центральную часть Ю.-З. Азии.
Таблица для определения видов номинативного подрода
рода Tenuidactylus
1 (2). Бедренных пор не более 30; чешуй вдоль тела снизу не более 115; чешуй поперек головы
не более 11...............................................Т. caspius (Eichw.)
2 (1). Бедренных пор обычно не менее 30; чешуй вдоль тела снизу обычно не менее 120; чешуй
поперек головы обычно больше 11.
3 (4). Первая пара нижнечелюстных щитков разделена горловыми чешуями или соприкасается
между собой узким швом (у 90 % особей его длина не превышает 10 % длины подбородоч-
ного щитка) ..............................................Т. longipes (Nik.)
4 (3). Первая пара нижнечелюстных щитков соприкасается между собой широким швом (у 85 %
особей его длина колеблется в пределах 10—40 % длины подбородочного щитка).
5 (6). Спинные бугорки очень ребристые, заходят вперед дальше области передних ног;
обычно три нижнечелюстных щитка; между рядом бедренных пор (или чешуй, соответст-
вующих им у самок) и анальной щелью обычно два ряда чешуй................
Т. turcmenicus (Szczerbak)
6 (5). Спинные ребристые бугорки обычно не заходят вперед дальше области передних конеч-
ностей; обычно два нижнечелюстных щитка; между рядом бедренных пор и анальной
щелью обычно три ряда чешуй..........................Т. fedtschenkoi (Strauch)
КАСПИЙСКИЙ ГЕККОН — TENUIDACTYLUS CASPIUS (EICHW., 1831)
Типовая территория: Баку.
Кариотип: 2п == 42, субметацентриков 2, акроцентриков 38, субтелоцентриков 2,
NF = 46.
1831 —Gymnodactylus caspius Eichwald, Zool. spec. Ross. Polon., Vol. 3, p. 181.
1954 — Cyrtodactylus caspius, Underwood, Proc. zool. Soc. London, p. 475.
1984 — Tenuidactylus caspius, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 53.
Диагноз. У самцов не более 32 преанальных и бедренных пор; не более
115 чешуй вдоль тела снизу и 31 чешуя поперек брюха; чешуй поперек головы
не более 11; спинные бугорки крупные, трехгранные, почти всегда с ними
соприкасаются один—четыре более мелких дополнительных бугорка; две-три
пары нижнечелюстных щитков; в первой нижнечелюстной паре щитки соприка-
саются или разделены одной-двумя чешуйками; обычно два ряда чешуй между
рядом бедренных пор и анальной щелью; вытянутая вперед вдоль тела перед-
няя лапа достает концами пальцев до конца морды, общая длина предплечья
и плеча составляет 29—37 % (31,8 ± 0,1) длины туловища.
Лектотип. 3182 ЗИН. ad «Баку, 1830, Менетрие». L = 61,0 мм;
Led = 81,9 мм; LILcd = 0,74; чешуй поперек головы 9; верхнегубных щит-
ков 11, нижнегубных 8; чешуй вдоль тела снизу 96, поперек брюха 28, окружа-
ющих спинной бугорок, 13 + 1 дополнительный бугорок; бедренных пор 24;
подпальцевых пластин 26/24; нижнечелюстных щитков 2/3.
Описание (рис. 60) (по 565 экз. из колл. ИЗАНУ, ЗИН, МЛГУ,МХГУ,
NMW, MNHP).
Ldad (п= 249) 40,6—72,00 (56,06 ± 0,48) мм; L $ ad (п = 293) 40,2—
68,5 (55,33 ± 0,49) мм; LIL cd 0,62—0,95 (0,76 + 0,004); индекс высоты / ши-
рины головы (п = 100) 55—67 (61,09 + 0,26); диаметр уха / глаза (п = 60) 41—68
(55,88 ± 0,95); верхнегубных щитков (и = 1121) 9—14 (10,43 ± 0,02); нижне-
губных (п = 1117) 6—10 (8,04 ± 0,02); чешуй между центрами глаз (и =
= 559) 7—11 (8,68 + 0,04); длина спинного бугорка / L (п = 439) 23—40
(32,11 ± 0,13); чешуй, окружающих спинной бугорок (п = 549), 13—22
(17,39 ± 0,06); чешуй вдоль тела снизу (п = 501) 89—113 (103,44 ± 0,20),
поперек брюха (п = 464) 22—31 (26,60 ± 0,10); бедренных пор (п = 298) 22—
32 (26,65 ± 0,11); бедренных чешуй у $> (п = 185) 25—33 (30,86 ± 0,14);
подпальцевых пластинок (и = 1095) 20—29 (23,85 ± 0,05).
132
Рис. 60. Tenuidactylus caspius:
A — голова сбоку, Б — лицевая часть спереди, В — голова сверху, Г — голова снизу, Д —•
подбородок, Е — спина, Ж. — бедренные поры (Д — ИЗАНУ Re 7,0. Вульф АзССР; осталь-
ное — ИЗАНУ SR 378, Мешхед-и-Мисриан, ТССР)
Подбородочный щиток всегда тре- или пятиугольный; две (56,7 %)
или три (43,2 %) пары нижнечелюстных щитков, щитки первой могут разде-
ляться одной-двумя горловыми чешуйками; первые носовые щитки разделе-
ны чешуйкой (37,7 %) или соприкасаются (62,3 %); с центральными спинными
бугорками соприкасаются один (41,3%), два (32,5 %), три (13,3 %) или четыре
(2,5 %) дополнительных бугорка, у 10,4 % такой контакт отсутствует.
133
Рис. 61. Распространение Т. caspius
Окраска и рисунок. Более светлый, чем остальные виды подро-
да: основной фон бледно-песочно-сероватый; темные поперечные М-образные
полосы на туловище (пять-шесть) и хвосте (8—12); на конечностях неясные по-
перечные полоски; на голове имеется узор из тонких темных полос: затылочная
соединяет вершины ушных отверстий, параллельно ей от заднего края каждого
глаза отходят полоски, прерывающиеся на темени; между свободными краями
их располагается темное пятно, над которым почти между глазами находятся
одно—три пятнышка; от передне-верхнего края орбиты к ноздре, от переднего
края орбиты к первому верхнегубному щитку и от передне-нижнего края орби-
ты к пятому-шестому верхнегубному щитку тянутся три параллельные полоски,
заходящие и на нижнегубные щитки; они у каспийского геккона выражены
значительно более четко, чем у других видов рода, и могут служить одним из
его полевых определительных признаков. Нижние поверхности белые. Масса
до 6,5 г.
Распространение. Северной границей ареала каспийского гек-
кона является линия, проходящая от сев.-в. части Каспийского моря (зал. Ком-
сомолец) до сев. побережья Аральского моря и Сырдарьи. Найден вдоль запад-
ного берега Каспийского моря до Калмыкии (с. Нарын-Худук), в Закавказье, на
юго-западе крайними пунктами являются р. Акера (естественное местообитание)
и г. Джульфа в Азербайджане, г. Тбилиси (синантропно). На востоке найдена
нами в 500 км от основного ареала изолированная популяция в Ферганской
долине. В последние годы наблюдается расширение ареала за счет использова-
ния водных (Каспийское море) и железнодорожных коммуникаций (Кавказ,
Средняя Азия), распространяется с перевозимыми товарами. Достаточ-
но уверенно можно утверждать, что своим появлением в европейской части
ареала они обязаны человеку. В статье [147] относительно новых нахо-
док в Кызылкумах вкралась опечатка: фраза «мы находили его и севернее
Бухары у Мынбулака и Учкудука» относится не к туркестанскому, а к кас-
пийскому геккону (с. 69).
134
Южная граница в Иране и Афганистане четко не определена. Находки в
Центральном и Западном Иране и Сирии нуждаются в подтверждении, возмож-
на путаница здесь с близкими видами (в частности, до недавнего времени кас-
пийского геккона не отличали от туркменского) (рис. 61).
Кадастр к рис. 61.
СССР, РСФСР: 1— Калмыцкая АССР, Лиманский р-н, пос. Нарын-Худук (МХГУ);
2 — Дагестанская АССР, окр. г. Махачкала; г. Буйнакск (ИЗАНУ); 3 — г. Дербент (ИЗАНУ).
Грузия: 1 — Тбилиси [82].
Азербайджан: 1 — Нахичевань, Джульфа (ИЗАНУ); 2 — Шемаха (ЗИН); 3 —
р. Сумгаит-Чай, 35 км сев. г. Баку (ИЗАНУ); 14 км сев. г. Сумгаит (МХГУ); 4 — Апшеронский
п-ов, Баку, Мардакьяны, Загульба, Вишневка, Бильга (ИЗАНУ); 5 — окр. г. Приморск; Ко-
быстан (ИЗАНУ); пос. Сангачалы (ЗИН); 6 — о. Вульф (ИЗАНУ); 7 — о. Обливной (ИЗАНУ);
8 — Сальяны (ЗИН); 9 — Физулинский р-н, р. Акера, с. Керимбейли (ИЗАНУ).
Т С С Р: / — Куня-Ургенч (ИЗАНУ); 2 — Деу-Кескен-Кала (ИЗАНУ); 3 — креп. До-
ярбекир; Шахсенем [1301; 4 — оз. Сарыкамыш, р. Дарьялык (ИЗАНУ); сев. берег оз. Сарыка-
мыш [130]; 5 — Кунядарьинская равнина [130]; Кунядарья, 3 км ю. Чапрак-Кала [23]; 6 —
27 км от г. Ташауз на Куня-Ургенч, креп. Гульдумсаз (ИЗАНУ); 7 — з. граница впадины Са-
рыкамыш [23]; 8 — ю.-в. чинк впадины Сарыкамыш (ИЗАНУ); коса Капылларкыр [130]; 9 —
Тахтинский р-н, креп. Измыкшир [130]; 10 — 25 км ю. пос. Тахта, креп.Бедеркенд (ИЗАНУ);
11 — 90 км в. южной границы впадины Сарыкамыша (ИЗАНУ); 12 — ю.-в. берег зал. Кара-
Богаз-Гол, кол. Чайги-Бей-Булак [23]; чинк Кулон-даг (ЗИН); 13 — Заунгузские Каракумы,
кол. Даудыр-Енедже (ЗИН); 14 — Кумсебшен [130]; 15 — кол. Дахлы [130]; 16 — с. чинк Ка-
планкыра [130]; 17 — пос. Сульмен (МХГУ); 18 — 25 км сев. пос. Серный завод (ИЗАНУ);
19 — Красноводск (ИЗАНУ); 20 — с. Нарген; о. Святой [130]; 21 — ст. Джебел [130]; 22 —
хр. Большой Бал хан [130]; 23 — Западный Узбой; кол. Йылгынлы; кол. Тоголак [130]; 24 —
пос. Серный завод; 10—15 км з. Серного завода (ИЗАНУ); 25 — Дорджа (ЗИН); 26 — ле-
вый берег Узбоя, 4 км з. пос. Молла-Кара (ЗИН); 27 — Ахча-Куйма [130]; 28 — Ясхан (ЗИН);
29 — Киркили [130]; 30 — 180 км сев. г. Бахарден, Кизыл-Гаты (ИЗАНУ); 31 — Малый Бал-
хан [130]; 32 — Казанджик, 20 км ю. ст. Искандер (ИЗАНУ); ст. Искандер [130]; 33 — 44 км
сев.-в. Кизыл-Арвата, окр. аула Демерджан (ЗИН); 34—Култакыр [130]; 35 — Даната
(ИЗАНУ); 36 — Кизыл-Арват (ИЗАНУ); 15 км ю. Кизыл-Арвата (ЗИН); 14 км сев. Кизыл-Арва-
та у Такырабада [23]; 37 — Челекен [130]; 38 — Бахарденский р-н, Шехр-Ислам (ИЗАНУ); 39—
Мешхед (ИЗАНУ); Мессерианская равнина (ИЗАНУ); 40 — г. Казанжик (наши данные);
Кюрендаг [130]; 41 — Карагез [130]; 42 — 15—20 км сев. Бами (ИЗАНУ); Вами [130]; 43 —
53 км сев. Бахардена [130]; 44 — 40 км ю. Мадау [130]; 45 — Кара-Кала; междуКара-Калой
и Чандырем; 20 км ю. Кара-Калы (ИЗАНУ); Кара-Калинский р-н, совхоз им. Ленина
(МХГУ); Чандырская долина (ЗИН); ущелье Елдере [130]; 46 — Айдере (ИЗАНУ);47 — Арч-
ман [130]; 48 — Синекли [130]; 49 — 50 км с. Кара-Имам (ИЗАНУ); 50 — с. Шарлоук
(ИЗАНУ); 51 — долина р. Чандырь, 20 км ю. пос. Кизылимам (ИЗАНУ); 52 — пос. Бахар-
ден; 15 км ю. пос. Бахарден, р. Арваз (ИЗАНУ); между Бахарденом и Дуруном [23]; пос.
Дурун, Караган, Яраджи [130]; 53 — Ашхабад, Берзенги; Куры-Хоудан; 20 км ю. Ашхаба-
да, дор. на Гаудан (ИЗАНУ); гора Нах-Дуйн (Гоудан) [23]; 54 — 57 км от Ашхабада на
Теджен (ИЗАНУ); 55 — Чули (ИЗАНУ); 15 км з. г. Ашхабад, Багир (ММГУ); пос. Верхне-
Скобелевский [23]; пос. Гермаб; Келенджар; Фирюза; сев. Геок-Тепе; сев. Безмеина; кол.
Челтек, кол. Порсыкую [130]; 56 — пос. Калининский (ИЗАНУ); 57 — окр. Большое и Малое
Делили [130]; 58 — уроч. Баят-Ходжа (ЗИН); 59— пос. Гасан-Кули (ИЗАНУ); бугор Ак-Пат-
лаук [23]; 60 — пос. Дейнау [80]; 61 — Чарджоу [130]; 62 — 20 км сев. Репетека (ИЗАНУ);
ст. Репетек [130]; 63 — Марыйская обл., ст. Равнина (ИЗАНУ); 64 — г. Керки [130]; 65 —
Керкичи; Достлук [130]; Талимарджан (ММГУ); 66 — Ходжепиль (ИЗАНУ); 67 — древнее го-
родище Каахка (ИЗАНУ); 68 — Байрам-Али (ИЗАНУ); 69 — бывшее село Базардепе, нижний
пояс гор Кугитанг [130]; 70—Каахкинский р-н, пос. Меана (ИЗАНУ); сев. пос. Меана
[130]; 71 — Карабата [130]; 72 — Султанбент; Иолотань [130]; 73 — Копетдаг, ю. с. Чаача
(ИЗАНУ); 74 — ст. Такыр [130]; окр. Теджена [130]; 20 км от ст. Теджен вверх по течению
реки [23]; 75 — Имамбаба (ИЗАНУ); 76 — Серахс (ИЗАНУ); 77 — Ташкепри [130]; 78 — Ка-
рабиль, пос. Хумлы (ИЗАНУ); 79 — Пул-и-Хачум; уроч. Науруз-Абад; Ширдепе (ИЗАНУ);
р. Теджен, близ ирано-афганской границы [23]; 80 — Серахский р-н, уроч. Адамулен
(ИЗАНУ); 81 — сев. станции Кала-и-Мор (ИЗАНУ); 52 — 40 км от Тахта-Базара; Орна-Дав-
лет; р. Рават-Качан, уроч. Тарашек (ИЗАНУ), Кашан, к сев. от Тарашека; оазис Пенде [23];
83 — уроч. Пеленговели (ИЗАНУ); 84 — Бадхыз, хр. Гез-Гядык (ИЗАНУ); 85 — Кушка;
Агашлы (ИЗАНУ); 5 км сев. пос. Моргуновский (ЗИН); Чемен-и-Бит [130]; 86 — Бадхыз,
Эр-Ойлан-Дуз (ИЗАНУ); 87 — Бадхыз, Акар-Чешме; Пенхан-Чешма; Шакмакли (ИЗАНУ);
Акрабат; Кепели; Кызыл-Джар [130]; 88 — Ничка; 65 км ю.-з. пос. Ничка [130]; 89 — Чарджоу-
ская обл., Дарганата (ИЗАНУ); 90 — Ак-Молла, 150 км в. кишл. Серный завод (наши данные).
Узбекистан: / — ю.-з. побережье Арала (Урга) [22]; 2 — Каракалпакия, 30—40 км
сев.-в. Тахта-Купыр, подножье Бельтау (ЗИН); 3 — Каракалпакия, 25—30 км сев. Чабанказ-
гана (ИЗАНУ); 4 — Каракалпакия, 45 км от Нукуса на Турткуль (ИЗАНУ); 5 — Каракалпа-
кия, з. Бодай-Тугай, креп. Джампык-Кала (ИЗАНУ); 6— Каракалпакия, 25 км сев.-в. Бируни,
креп. Кызыл-Кала (ИЗАНУ); 7 — Хорезмская обл., г. Хива; 30 км сев.-в. Бируни, креп.
Топрак-Кала; 8 — Турткуль [22]; 9 — Бухарская обл., 100 км к сев. от Шафрикана, Мынбу-
лак, Учкудук (ИЗАНУ); 10 — кишл. Аякагитме (ИЗАНУ); 11 — окр. Термеза (ИЗАНУ); 12 —
Бельтау [22]; 13 — хр. Нуратау, кол. Темиркаук (ЗИН); 14 — Бухара [22]; 40 км ю.-в.
г. Бухара (ММГУ); 15 — Каракульский заповедник, 20 км ю. ст. Алат (ММГУ); 16 — сев.-в.
135
Бухарской обл., останцевые возв. (ММГУ); 17 — Бухарская обл., Джингильды (ММГУ);
18 — 60 км сев. г. Карши, уроч. Уч-тепе (ММГУ).
Таджикистан: 1 — Ленинабадская обл.; 2 — Куляб, окр. кишл. Калимтай [107];
3 — долина р. Вахш, окр. пос. Джиликуль и Кумсангир [107]; 4 — Бешкентская долина, окр.
кол. Тулхар, Кабадиан [107].
Казахстан: 1 — о. Кокарал [2]; 2 — г. Шевченко (бывший форт Александровск)
(ЗИН); 3 — Мангышлак (ЗИН); Таучик [98]; Ново-Александровск [98]; 4 — Кзыл-Ординская
обл., 40 км сев.-з. пос. Кекрели, развалины креп. Чирик-Рабат (ИЗАНУ); 5 — котловина Шах-
Пахты и западный край впадины Ассаке-Аудан [22]; 6 — сев. чинк Устюрта [98]; 7 — сор Ас-
матай-Матай [98]; 8 — сев.-з. берег Аральского моря [98].
Иран: 1 — Горган, Сев.-В. Иран (MNHP, NMW); 2 — Астрабад (Дамган), Ак-Кала,
в 15 км от Астрабада; Насиабад (ЗИН); 3— Шахруд (МЛГУ); 4— 40 км сев. г. Мешхед (ЗИН);
5 — Боз-Хоуз-Паин (ЗИН); 6 — окр. старого Бахарса, с. Кериз-и-Ноу (ЗИН); 7 — стр. Хошта-
дан, Керат (ЗИН); 8 — Керманшах (ЗИН), точка нуждается в подтверждении; 9 — Мириндуз
(ЗИН); 10 — Хоуз в стр. Зиркух (ЗИН); 11 — Хорасан, кол. Чах-Иси (ЗИН); 12 — Сейстан,
Нейзар (ЗИН); 13 — Шираз (NMW); место нахождения требует подтверждения; 14 — Бирджан
(ЗИН); 15 — Серахс, Сев.-В. Иран (MHNP); 16 — Зюрабад [80]; Gymnodactylus sp., находка
нуждается в подтверждении; 17 — Солеабад (Сольтанабад) [80]; находка нуждается в под-
тверждении; 18 — Мешхед [80], находка нуждается в подтверждении.
Афганистан: 1 — около 100 км в. г. Файзабад (CAS); 2 — 25 км ю.-з. пос. Акча
(CAS); 3 — Мазари-Шариф, 36° 43' с. ш., 67° 05' в. д. [172]; 4 — 10 км з. г. Ташкакурган
(Хульм) (CAS); 5 — окр. с. Сари-Пуль, Вагиех (ZML); 6 — дор. Герат — Исламкала (CAS);
7 — Герат (МЛГУ); 8 — Пагман, 34° 36' с. ш., 68° 56' в. д. [172].
С и р и я: (на вставке): 1— Хеме, ю. Халеба (ММГУ—находка нуждается в подтвержде-
нии).
п оловои диморфизм и возрастная изменчивость.
Самцы, в отличие от самок, имеют бедренные поры; у них несколько более мас-
сивная голова. Самцы чуть крупнее самок по максимальным размерам (см.
выше), у них более длинный хвост (L cd <2 = 95 мм, L cd g = 89,1 мм). В брач-
ный период самцы окрашены более
Рис. 62. Возрастная изменчивость индекса
Li Led у Т. caspius
ярко и контрастно, чем самки, у них
основной фон желтоватый, у самок —
сероватый. Это мы установили при
инъекциях гонадотропного гормона:
через сутки самцы резко посветлели
и стали хорошо отличаться по окрас-
ке от самок.
У молодых особей голова и конеч-
ности относительно более крупные,
хвост — более короткий (рис. 62), чем
у взрослых гекконов.
Различий в окраске и рисунке в
зависимости от места обитания не от-
мечено, но неоднократно наблюдалось,
как в связи с физиологическим состо-
янием окраска ящериц то темнела (до
полного исчезновения рисунка), то
светлела.
Систематика и геогра-
фическая изменчивое ть.
15 материковых выборок, охватывающих весь ареал, при сравнении по основ-
ным признакам фолидоза не показали существенных различий (даже такая
удаленная, как популяция из Ферганской долины). Наиболее изменчивыми
признаками оказались количество подпальцевых пластинок, бедренных пор,
вер хне- и нижнегубных щитков, чешуй поперек брюха, вдоль тела снизу и по-
перек головы, а также соотношение длин спинного бугорка и тела. По всем
этим признакам существует выраженная в различной степени клинальная из-
менчивость по направлению запад — восток. Однако наиболее крупные осо-
би обнаружены почти в центре ареала — под г. Кушка (Ю. Туркмения).
От всех указанных выборок существенно отличаются гекконы с о. Вульф
в Каспийском море, описанные как отдельный подвид [14]. Для этой формы
характерно почти полное или полное отсутствие соприкосновения первой пары
нижнечелюстных щитков (признак, общий с одним из близких видов — Т. Ion-
136
gipes). Характерно и то, что на материковой части — в Азербайджане, Кара-
калпакии, Иране, Туркмении — этот признак отсутствует у 93—100 % особей,
но на Апшеронском п-ове, на ближайшем к острову берегу, его нет только
у 70 %.Вероятно, причины своеобразия данной популяции следует искать не в
особенностях формирования острова (он тектонического происхождения),
а в действии генетических механизмов.
Представленные данные позволяют сделать заключение о том что данный
вид на Кавказ проник из Ю. Туркмении через С. Иран, о чем свидетельствует
минимальное количество отличий у соответствующих выборок.
Таблица для определения подвидов Т. caspius
1(2). Первая пара нижнечелюстных щитков соединена между собой широким швом.
..............Т. с. caspius (Eichwald)
2 (1). Первая пара нижнечелюстных щитков не соприкасается или соединена между собой
точечным швом .......................Т. с. insularis (Akhmedow et Szczerbak
КАСПИЙСКИЙ ГЕККОН ОБЫКНОВЕННЫЙ —
TENUIDACTYLUS CASPIUS CASPIUS (EICHWALD, 1831)
Типовая территория: видовая.
Диагноз. Первая пара нижнечелюстных щитков соприкасается между
собой широким швом.
Остальные признаки содержатся в видовом описании.
Тип видовой.
Распространение. Видовой ареал за исключением о. Вульф в
Каспийском море в 12 км от Апшеронского п-ова.
КАСПИЙСКИЙ ГЕККОН ОСТРОВНОЙ —
TENUIDACTYLUS CASPIUS INSULARIS (AKHMEDOW ET SZCZERBAK, 1978)
Типовая территория: о. Вульф (Бакинский зал. Каспийского моря).
1978 — Gymnodactylus caspius insular is, Ахмедов, Щербак, Вест, зоол., Киев, № 2, с. 80.
Диагноз. Первая пара нижнечелюстных щитков разделена горловыми
чешуйками (65,5 %) или соприкасается в одной точке (34,5 %).
Голотип. Re № 7 ИЗАНУ, $ ad, о. Вульф, 28.III 1976 г., М. И. Ах-
медов. L = 55,7 мм; L cd = 69,0; верхнегубных 10/9; нижнегубных щитков
9/9. От подбородочного щитка до анальной щели в продольном ряду 105
чешуй, чешуй поперек брюха 28, между центрами глаз восемь, первая пара
нижнечелюстных щитков не соприкасаются.
Описание (п = 30 экз.). Приведены только те признаки, по которым
имеются достоверные отличия от других (4—13) выборок. L/L cd =
= 0,81—0,88 (0,83 ± 0,01). Верхнегубных щитков 9—13 (11,0 ± 0,1); нижне-
губных щитков 7—10 (8,45 ± 0,11). Длина спинного бугорка /£ 26—32
(28,66 ± 0,29) (этот и последующий наиболее уклоняющиеся признаки); ко-
личество подпальцевых пластинок 24—29 (26,3 ± 0,2).
Остальные признаки соответствуют видовому описанию.
Распространение. Известен только из типовой местности.
Стации и количественные данные (рис. 31, 40). Каспий-
ский геккон в кавказской части ареала обитает, как правило, на вертикальных
поверхностях скал, обрывов, стенах построек. Стены построек, скальные обры-
вы чинка Узбоя, Унгуза и Устюрта также являются обычными биотопами этого
вида в 3. Туркмении. Однако в равнинной части Каракумов они встречаются
в норах больших, краснохвостых и полуденных песчанок, других норников. Эта
особенность более выражена в направлении на юг и восток (Эр-Ойлан-Дуз,
Ферганская долина), исключая лессовые стены древних построек, например
Мервского оазиса (Байрам-Али). Поскольку на востоке ареала каспийский
геккон встречается вместе с близкими видами (туркменским, Федченко, длин-
ноногим), то возникает подозрение, что причиной этого являются конкурент-
ные взаимоотношения с ними [24].
137
В Бадхызе скальные останцы и обрывы хребта Гез-Гядык заняты длинноно-
гим гекконом, скальные обнажения в Кушко-Кашанском междуречье — турк-
менским, а в среднем течении Амударьи — гекконом Федченко. Однако в тех
местах, где нет указанных «конкурентных» видов (Ц. и В. Копетдаг), каспий-
ский геккон, как правило, также избегает скальных обнажений и встречается
в зоне предгорий в кяризах, норах, расселинах и глинистых промоинах. Да и
трудно представить себе, что эвритопный вид — каспийский геккон, может
быть вытеснен таким стенотопным видом, как длинноногий. Причина здесь, оче-
видно, в другом. На территории ареала каспийского геккона аридизапия кли-
мата нарастает с запада на восток.
Как свидетельствуют географы [30, и др.], относительная влажность воз-
духа в мае в закавказской части ареала 50—70 %, в 3. Туркмении, приблизи-
тельно до Казанджика,— 40—50 %, а далее за линией Ашхабад — Нукус —
35 %. Это подтверждает и схема распределения осадков за теплый период года;
в Закавказье не менее 200 мм, 3. Туркмении от 100 до 50 мм, в Ц. Каракумах
и Кызылкуме — менее 50 мм. Годовая сумма осадков в Закавказье не ниже
300 мм (речь идет о биотопах каспийского геккона), в 3. Туркмении — 100—
200 мм, восточнее — менее 100 мм. Следовательно, каспийский геккон занимает
скальные местообитания, где относительная влажность воздуха в мае не ни-
же 40 %, около 100 мм осадков в теплый период года, годовая сумма осадков пре-
вышает 100 мм. Установлено также, что относительная влажность более высо-
кая в песчаном и глинистом субстрате по сравнению со скальным (более бога-
тая растительность в песчаных пустынях, самая скудная — в щебнистых).
Поэтому мы полагаем, что биотопы каспийского геккона лимитируются влаж-
ностью. Близкие к нему ирано-афганские виды, имеющие более южное проис-
хождение,— длиннопалый и туркменский гекконы, являются более устойчивы-
ми к низкой влажности, что подтверждается и при наблюдении в лабораторных
условиях.
В горах каспийские гекконы найдены до высоты 900 мн. у. м. (Копетдаг),
С. Андерсон и А. Левитон [172] сообщали о находках этого вида в Афганистане
до высоты 1653 м н. у. м.
На Апшеронском п-ове встречается до 12—15 экз. на 10 м2 выходов извест-
ковых скал. В Туркмении на руинах кирпичных построек средневекового горо-
да Мессериана одна особь встречена на 1—2 м2. На лессовых стенах древнего
Мерва (окр. г. Байрам-Али) на площади стен южной экспозиции 2000 м2 учте-
но 35 гекконов, или 185 экз. на 1 га (15.IV 1977 г.; здесь же в сумерки 22.V 1982 г.
в пасмурную погоду с дождем отмечено до 125 особей на 1 га). В Бадхызе на
незаповедной части шора Эр-Ойлан-Дуза на маршруте в 1 км при ширине по-
лосы 5 м встречены единичные особи, а на песчаной арене с кустиками солянок
у нор песчанок отмечено до 18 ящериц на 1 га (16.V 1976 г., 10.IV 1977 г.,
8.Х 1981 г., 24.IV 1984 г.). В закрепленных песках, перемежающихся с такы-
рами, между Серахсом и Хаусханом 28.IV 1984 г. встречен в среднем 1 гек-
кон на 1 км маршрута. Такая же численность отмечена в Кызылкуме на осоло-
ненном такыре в окр. с. Аяк-Агытма (14.VI 1976 г.).
Единичные особи здесь встречены в зимниках, на туранговых деревьях;
в Каракумах — в основаниях и руинах лессовых построек, скальных останцах
по Унгузу, стенах колодцев среди песчаных барханов, буграх на такыре с ос-
татками кустов солянок и саксаулов, глинистых промоинах, лессовых обрывах,
осыпях камней под обрывом чинка ю.-з. Сарыкамыша и Устюрта; в Бадхызе —
на лессовом плато с кустиками полыни, в трещинах телефонных столбов; на
Карабиле (кроме нор песчанок) на лессовых обрывах; в Беш.кентской долине
(Таджикистан) — в норах грызунов в эфемеровой пустыне. В окр. Репетека,
в черных саксаульниках на 1 га приходится в среднем три, а в поселке — 6,5
особей [120]. Единичные особи встречаются также на крайнем востоке ареала —
в Ферганской долине. Таким образом, наибольшей численности каспийские
гекконы достигают в западных частях ареала и в постройках (особенно в руи-
нах).
Отношение к температуре. Активные гекконы отмечены на-
ми при температуре воздуха 15—30°, в среднем (п = 14) 2Г. О. П. Богданов
138
[23] активных ящериц этого вида отмечал при температуре 17—36°. Одновремен-
но мы определяли температуру субстрата мест, где они встречались; оказалось,
что на поверхности гекконы наблюдались при температуре 16—22°, в среднем
(и = 14) 19,6°. Большая часть промеров сделана весной, осенью (8.Х 1981 г.)
при температуре воздуха 20° ящерицы не наблюдались. Прямая зависимость
между температурой воздуха и почвы при континентальном климате Сред-
ней Азии часто отсутствует (почва может быть нагрета больше воздуха и на-
оборот). Так, гекконы не выходили из убежищ при температуре воздуха 17°,
почвы — 21,5°(30.IV 1979 г.) или воздуха и почвы 18° (16.IV 1977 г., 24.IV
1977 г.), однако были на поверхности в пасмурную погоду 3.V 1984 г. при тем-
пературе воздуха и почвы 16°. В мае наблюдались ящерицы на поверхности
даже во время мелкого дождя. При исследовании каспийских гекконов в
термоградиентприборе (и = 42) они занимали места с температурой 13—4Г,
в среднем 32,7 ±1,1.
Температура тела гекконов в зимовочных убежищах при температуре воз-
духа 6,5° 6.II 1971 г. достигала 8—13° [12].
Суточный цикл активности. Весной гекконы часто появля-
ются днем (в феврале в 13—17 ч [23]), сначала они греются на солнце, затем охо-
тятся в тени. Часто в Ю. Туркмении мы наблюдали, как гекконы, сидящие
днем у входа в нору, хватали скрывающихся туда от жаркого солнца насекомых.
Летом активны в сумерках и ночью, осенью, в прохладные дни, снова часто
наблюдаются утром и днем. Днем они активны также при рассеянном свете
в постройках и пещерах. В сумерки и ночью охотятся с 19 ч до рассвета — 5—
6 ч, однако пик активности приходится на 19—20 ч [130]. Первое появление
весной (апрель) отмечено нами с 20 до 21 ч, осенью (октябрь) в 19 ч 20 мин.
Сезонный цикл активности. В Ю. Туркмении каспийские
гекконы в отдельные годы появляются в теплые дни даже зимой (25.1 1964 г.,
7.II 1971 г. [11]; 25.11 1977 г., 27.11 1964 г. [130]). Однако массовое появление
здесь отмечено в конце марта, начале апреля (по нашим наблюдениям,
31.III 1977 г. у Серахса, 6.IV 1978 г. у пос. Берзенги, 7.IV 1979 г. в уроч.
Куры-Хоудан в окр. Ашхабада, 13.IV 1983 г. у пос. Берзенги и 14.IV 1963 г.
у г. Казанджик). Близкие даты весеннего появления гекконов известны из
Азербайджана (26.III 1975 г. из Сумгаита и 28.III 1976 г. с о. Вульф) и Калмы-
кии (3.IV 1951 г.). Поздние находки каспийских гекконов по нашим коллекци-
ям из Туркмении: 26.X 1966 г. у оз. Ясха, 28.X 1966 г. в окр. ст. Казанджик,
4.Х 1981 г. ю. г. Кара-Кала, 8.Х 1981 г. из Эр-Ойлан-Дуза, 18.X 1981 г.
в окр. ст. Уч-Аджи. Суммируя приведенные данные и сведения из литературы
[1, 23, 98, 107, 130, и др.], можно заключить, что на обширной территории ареа-
ла единичные каспийские гекконы появляются после зимовки во второй поло-
вине марта, массовый выход происходит в первой половине апреля, уход на
зимовку — в конце октября, отдельные особи могут встречаться еще в ноябре,
в теплые зимы, на юге ареала, они могут также кратковременно покидать зимо-
вочные убежища.
Линька. По данным литературных источников [1, 8, 107, 130], линяю-
щие особи встречаются с апреля по июль, изредка в начале августа. Мы отме-
чали линьку 8.Х 1981 г. в Эр-Ойлан-Дузе. По наблюдениям в неволе линяет
трижды: весной, летом и осенью; эта картина может смазываться за счет нездо-
ровых и истощенных особей, линяющих чаще. Линька начинается, как обыч-
но, с помутнения роговицы глаз и приобретения белесого оттенка основного
цвета туловища. Затем роговой слой лопается в подчелюстной области, с брюш-
ной стороны он отходит почти полностью, большие лоскуты сходят и со спинной
стороны. Ящерица укусами снимает в себя старую кожу, как правило, съедает
линкую шкурку.
Питание. При вскрытии 27 желудков каспийских гекконов из Азер-
байджана [1] были обнаружены остатки саранчевых, долгоносиков, жужелиц,
сверчков, клопов, муравьев, пчел, богомолов, гусениц, а также пауков.
Анализ содержимого 154 желудков из Туркмении, добытых в разное время
[130], показал, что первое место по встречаемости в рационе этих ящериц за-
нимают жуки (31,2 %), затем термиты (28 %), прямокрылые (24,6 %), сверчки
139
(17,5 %), пауки (13 %), бабочки и их гусеницы, а также мокрицы (по 7,8 %),
В апреле и мае (и == 20) встречаемость жуков составляет 50 %,бабочек 25 %,
прямокрылых и пауков — по 20 %. В июне (п = 120) в желудках гекконов,
добытых в старых глинобитных домах, термиты составляли 32,5 %, жуки —
30 и прямокрылые, преимущественно сверчки, — 25,8 %. В сентябре и октябре
(п = 14) чаще встречаются пауки (35,7 %), прямокрылые, термиты (по 28,6 %)
и мокрицы (7,1 %). В Туркмении [23] отмечен случай поедания гекконом
мелкой ящурки (Eremias velox).
По сведениям из Узбекистана (п = 15 [22]), они чаще поедают жуков
(80 % встреч, преобладали долгоносики — 33,3 %, чернотелки и пластинчато-
усые — по 12—6 %), двукрылых (26,3 %), мокриц (20 %). В неволе каспий-
ские гекконы хорошо поедают кузнечиков, мелких тараканов, личинок мучных
червей, молодые особи — дрозофил и гусениц восковой огневки.
Размножение. Каспийские гекконы становятся половозрелыми
при достижении длины туловища более 40 мм. В популяциях обычно преобла-
дают самки (от 1,4 до 2,1 на 1 самца [130]). Вскоре после выхода из зимовочных
убежищ гекконы приступают к спариванию. Об этом свидетельствуют ранние
находки беременных самок — в конце мая в Азербайджане [1], в Туркмении
даже в конце апреля (26.IV 1979 г., Каракумы, 50 км сев. Бахардена, наши дан-
ные) и мае (15.V 1979 г., уроч. Хумлы, Карабиль, наши данные [130]), в Узбе-
кистане — также в конце апреля [22], в Казахстане — в конце мая — начале
июня [98]. Кладка яиц двукратная, в редких случаях, возможно, и трехкратная,
продолжается до августа [130] и даже сентября [1]. По наблюдениям в неволе
отмечены кладки осенью и в феврале. Известны достоверные даты кладок:
30.IV 1979 г., Ц. Каракумы, кол. Джарли и 5.V 1975 г. ю. Кара-Калы,
наши данные; 5.V 1952 г., долина Мургаба, Туркмения [23]; 15.V 1979 г.,
уроч. Хамли, Карабиль, наши данные; 6.VI 1931 г., окр. ст. Искандер [8];
13.VI 1967 г., сев. Геок-Тепе [130]; 14.VI и 19.VI 1979 г., окр. Берзенги, наши
данные; 16.VI 1947 г., п-ов Мангышлак [98]; 26—28.VI 1979 г., Кызылкум, Пус-
тынная ст.; 26.VI 1983 г., Каракалпакия, окр. с. Чабанказган; 6.VII 1979 г.,.
Кызылкум, Пустынная ст., наши данные; 27—30 VIII 1946 г. и 7—16.IX 1947 г.,
Апшеронский п-ов [1]. В литературе [1, 22, 130| приводятся несколько умень-
шенные размеры яиц каспийского геккона, так как они, видимо, извлекались
из яйцевода не совсем зрелыми. Мы получили от пойманных ящериц и нашли
в природных условиях нормально отложенных 16 яиц. Их размеры 10,9—12,5 X
X 9,0—10,2 мм, в среднем 11,1 X 9,1 мм. Обычно откладывается два яйца,
реже одно (40 % [130]). Яйца белые, овальные, со слабым розоватым оттенком,
их масса 380—670 мг. В одно место могут откладывать яйца несколько самок
(находили скопления до шести—восьми яиц). При содержании в неволе инкуба-
ционный период длится 68—81 дней. Известны даты находок свежевыл у пив-
шихся гекконов в Туркмении 8.VII 1963 г., 24.VII 1931 г., 8.VIII 1962 г.,
20—27.IX 1968 г., 13.Х 1967 г., 26.Х 1969 г. [130], в Азербайджане — 5—
26.IX 1952 и 17.Х 1951г.; 14—15.IX 1948 г. найдены яйца со сформирован-
ным зародышем [1].
Темп роста. Свежевылупившиеся из яиц гекконы имеют размеры
L = 17—23 мм; L cd = 22—28 мм. До ухода на зимовку сеголетки увеличива-
ются в размерах минимум на 7—8 мм, к середине следующего лета они достига-
ют 40—45 мм ИЗО] и становятся половозрелыми. По наблюдениям в неволе
половозрелость наступает ранее двух лет. Через два года размеры всех ящериц
превышают 46—48 мм ИЗО]. Максимальных размеров туловища (68—72 мм)
гекконы достигают не ранее чем в пятилетием возрасте.
Враги. В Закавказье на каспийских гекконов нападают ящеричная
змея (Malpolon monspessulanus) и гюрза (Vipera lebetina) [1]. В Туркмении
их поедают поперечнополосатый полоз (Coluber karelini), краснополосый полоз
(С. rhodorhachis), стрела-змея (Psammophis lineo latum), эфа (Echis carinatus),
желтопузик (Ophisaurus apodus), домовой сыч (Athene noctua), черный коршун
(Milvus korschun), лисица (Vulpes vulpes) [8, 23, 52, 53, 130; наши наблюдения].
Сведения о врагах каспийского геккона из других мест отсутствуют.
Как выяснено нами, много гекконов данного вида гибнет в зимовочных
£40
убежищах и после появления на поверхности при заморозках. Так, 27.VI 1976 г.
мы обнаружили в полости лессовой стены под Куня-Ургенчем скопление 14
скелетов каспийского геккона, в этом же районе в подобной ситуации в щели
6.VI 1979 г. было обнаружено 18 скелетов, а рядом еще остатки двух ящериц;
И.IV 1984 г. в норе у пос. Берзенги были обнаружены два мумифицированных
геккона, что, несомненно, явилось результатом сильных и внезапных морозов
(до —13°) в конце февраля. О находках 5—17 скелетов этих ящериц в одном мес-
те сообщал и А. М. Алекперов [1].
Особенности поведения. Каспийские гекконы хорошо пе-
редвигаются по вертикально и потолочно расположенным поверхностям; бы-
стро бегают по скалам, лессовым обрывам, вертикальным норам, вырытым
в рыхлом грунте. Они способны передвигаться во время охоты у подножия скал
в траве, мигрировать на весьма длительные расстояния, встречаются в степи
и на такырах. Отдельные взрослые и неполовозрелые особи постоянно мигри-
руют в поисках подходящих мест. На равнине под Ашхабадом в бетонированной
яме объемом 2 м3, расположенной в полынной степи, были выловлены все яще-
рицы и при прохладной погоде с частыми дождями (апрель 1978 г.) за 10 дней
здесь появилось два геккона, за следующих 10 дней — три. Живут гекконы по
одиночке или в подходящих местах группами по 10—15 особей [1] (наблюдали
их скопление до 50). Заметив добычу, геккон медленно приближается к ней, а
затем хватает резким броском и поедает. Особи этих ящериц привязаны к сво-
им убежищам: выпущенные на расстояние до 20—30 м от него, возвращались
обратно (981. Убежищами им служат трещины обрывов и скал, пустоты между
камнями в осыпях и постройках, норы грызунов. Здесь же они и зимуют.
Ч. Атаев [12] на холмистых участках под Ашхабадом выкапывал каспийских
гекконов из нор на глубине 8—10 см (было обнаружено вместе 16 самок и 12
самцов), а в 80 см в стороне — еще пять особей. В брачный период гекконы час-
то дерутся, издают металлические звуки (писк более тихий и более низкого
тона, чем у серого геккона). При дневном свете видят на расстоянии не менее
двух метров. Кладки каспийских гекконов находили в пустотах старых лессо-
вых построек, яйца были слегка погружены в лесс или песок.
Практическое значение и охрана.
Особенно полезны каспийские гекконы при синантропном образе жизни.
Они охотно поедают закаспийских термитов, которые разрушают здания, а так-
же долгоносиков, мух и других вредных насекомых. Широкий ареал, высокая
и стабильная численность в настоящее время обеспечивают виду нормальное
существование, поэтому отсутствует необходимость в проведении каких-либо
природоохранных мероприятий.
ТУРКМЕНСКИЙ ГЕККОН —
TENUIDACTYLUS TURCMENICUS (SZCZERBAK, 1978)
Типовая территория: ТССР, Бадхыз, окр. Кушки, уроч. Агашлы.
Кариотип: 2п= 42 субтелоцентрика 4, акропентриков 38, NF = 46.
1978 — Gymnodactylus turcmenicus, Щербак, Вест, зоол., Киев, № 3, с. 39.
1984 — Tenuidactylus turcmenicus, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 53.
Диагноз. У самцов не менее 30 бедренных пор; чешуй вдоль тела сни-
зу обычно более 115 (до 130); поперек брюха 25—32; поперек головы 8 — 12;
спинные бугорки треугольно-округлые или трехгранные, обычно крупные,
дополнительных бугорков обычно один—три (около 94 %); нижнечелюстных
щитков две-три пары (обычно три), первая соприкасается между собой широким
швом (92 %); обычно два ряда чешуй между рядом бедренных пор и анальной
щелью; вытянутая вперед вдоль тела передняя лапа достает концами пальцев
до конца морды (общая длина предплечья и плеча составляет 33—38 % (35,4 ±
± 0,2) длины туловища).
Голотип. Re № 10 ИЗАНУ, в' ad «Туркмения, окр. Кушки, уроч.
Агашлы, 21.IX 1977 г., L = 78,3 мм. L cd (половина регенерирована) 97,7 мм;
чешуй между центрами глаз 10; верхнегубных щитков по 11; нижнегубных
8/9; чешуй вдоль тела снизу 128; чешуй поперек брюха 30; чешуй, окружающих
141
Рис. 63. Tenuidactylus turcmenicus:
A — голова сбоку, Б — голова сверху, В — голова снизу, Г — спина, Д — брюхо, Е —
бедренные поры, Ж — лапа (ИЗАНУ SR 561)
спинной бугорок, 12 + 2 дополнительных бугорка; бедренных пор 35; подпаль-
цевых пластинок 25/27; нижнечелюстных щитков две пары, первая соприкаса-
ется между собой широким швом.
Описание (рис. 63) (по 132 экз. из колл. ИЗАНУ, ЗИН, NMWи
FMNH).
L <f ad (п — 51) 41,4—80,0 (70,76 ± 1,09) мм; L ? ad (п = 51) 40,6—
71,0 (60,76 ± 0,79) мм; масса 16,3 г. L/L cd (п = 31) 0,67—0,89 (0,73 ± 0,008);
индекс высоты/ширины головы (п = 21) 50—66 (58,10 ± 1,00); диаметр уха/
глаза (п = 24)40—62(50,54 ± 1,20); верхнегубных щитков (п = 235) 10— 14
142
60' 66* 76*
Рис. 64. Распространение Tenuidactylus fedtschenkoi (/), Т. turcmenicus (2),
Т. longipes microlepis (5), Т. 1. longipes (#), Т. 1. voraginosus (5)
(11,43 ± 0,05); нижнегубных (и = 238) 6—10 (8,32 ± 0,05); чешуй между цен-
трами глаз (и = 120) 8—12 (9,81 ± 0,07); длина спинного бугорка IL (п =
= 118) 21—32 (27,79 ± 0,22); чешуй, окружающих спинной бугорок (п =
= 132), 14—21 (16,76 ± 0,13); вдоль тела снизу (п = 108) 106—133 (119,31 ±
± 0,65); поперек брюха (и = 102) 25—32 (28,20 ± 0,18); бедренных пор
(и = 55) 31—39 (35,04 ± 0,23); бедренных чешуй у $ (п = 49) 35—41 (37,63 ±
± 0,23); подпальцевых пластин (п = 222) 23—30 (25,75 ± 0,10).
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
две (28,4 %) или три (71,2 %) пары (в одном случае — 0,4 % — зарегистриро-
вано четыре); в первой паре щитки соприкасаются широким швом (92,2 %),
у 5,2 % этот шов точечный, а у 2,6 % отсутствует; первые носовые щитки сопри-
касаются или разделены одной чешуйкой (по 50 %); с центральными спинными
бугорками соприкасаются один (30,3 %), два (45,5 %), три (18,1 %), четыре
(1,5 %) дополнительных бугорка, у 4,6 % такого контакта нет; у самцов ряд
бедренных пор посередине не прерван.
Окраска и рисунок. Цвет спинной стороны охристый, имеется
пять размытых нечетких буроватых поперечных полос на туловище и около
десятка таких же темных полос на хвосте. Брюшная сторона светлая, без пятен.
Распространение (рис. 64). Крайний юг СССР (Туркмения) и
Афганистан: от Кушко-Мургабского междуречья и южных окраин возвышен-
ности Карабиль на восток и юг через Паропамиз до западных отрогов Гиндуку-
ша. Возможны находки в Пакистане. Особи из Афганистана были определены
как Cyrtodactylus fedtschenkoi и С. caspius (материалы зарубежных музеев).
Кадастр к рис. 64.
СССР, Туркмения: / — возв. Карабиль, ущ. Пеленговели (ИЗАНУ); 2 — Чемен-
и-Бит (ИЗАНУ); 3 — ущ. Агашлы (ИЗАНУ).
Афганистан: / — 60 км сев. пос. Талукан (CAS); 2 — Мазари-Шариф (FMNH);
3 — 24 км в. пос. Ханабад (CAS); 4 — в. г. Файзабад, 36° 32' с. ш.> 71° 2Г в. д. (FMNH); 5 —
143
Рис. 65. Биотоп Т. turcmenicus (песчаниковые плиты вдоль русла р. Кушка, ст. Че-
мен-и-Бит, ТССР)
пос. Вагиех, 10 км от Сари-Пуль 1317]; 6 — пос. Чешмех-Шер, 17 км от Пули-Хумри (NMW);
7 — пос. Пагман (FMNH); 8 — Шибирган (ZFMK); 9 — Кундуз (ZFMK).
Половой диморфизм и возрастная изменчив ость.
У этого вида самцы крупнее самок. Остальные показатели сходны с другими
видами (см. Т. caspius).
Систематика и географическая изменчивость. Четыре
выборки из разных частей ареала сравнивали для выявления географической
изменчивости. Результат сравнения показал, что у гекконов из Чемен-и-Бита,
по сравнению с близко расположенной популяцией из Агашлы, более низкое
число брюшных щитков, чешуй вдоль нижней стороны тела и между центрами
глаз; средние величины признаков: количество бедренных пор и верхнегубных
щитков также плавно возрастает к востоку (Мазари-Шариф, Афганистан).
Количество чешуй, окружающих спинной бугорок, наиболее велико у особей
из Карабиля. Признаков, резко выделяющих какую-либо из исследованных
выборок, не обнаружено.
Экология до настоящего исследования не изучалась, все данные при-
водятся впервые.
г тации и количественные данные. Впервые этот вид был
найден в уроч. Агашлы под Моргуновкой (Бадхыз) в холмогорье с деревьями
фисюшки и хорошо выраженным травяным покровом на обрывах из песчани-
ка. Здесь на обрыве длиной 400 м, высотой 3 м отмечено 27 гекконов, или 216 экз.
на 1 га.
В уроч. Пеленговели (Карабиль) туркменские гекконы встречались только
на вертикальных скальных (известняк) обрывах; на осыпях и отдельных
скальных глыбах они отсутствовали. Обрывы здесь без лишайников, со щелями,
склон южной экспозиции. На склоне северной экспозиции их встречалось в
10 раз меньше. Любопытно, что каспийские гекконы здесь встречены только
у подножия склонов, в лессовых промоинах, а на скалах не отмечено даже еди-
ничных особей. Здесь на расстоянии 500 м, высоте 2,5 м учтено 20 гекконов,
или 160 экз. на 1 га площади скал.
Обнаружены нами эти гекконы и на обрывах скал из конгломератов на ле-
вом берегу р. Кушки и у места слияния ее с р. Ислим (рис. 65). Здесь узкая
полоса скальных обрывов (около 2 м) на песчаном склоне холма тянется на
33 км от г. Кушки (с перерывами). Отдельные особи гекконов встречаются на
расстоянии 1—10 м друг от друга, на расстоянии в 1 км и высоте полосы учета
144
в 2 м нами встречено 60 особей, или 300 экз. на 1 га. Высота мест обитания турк-
менских гекконов 570—720 м н. у. м.
Отношение к температуре. Весной гекконы появились при
температуре скалы 17° (9.V 1979 г., уроч. Агашлы, 12.V 1979 г., уроч. Пеленго-
вели), наибольшее их количество отмечено при температуре субстрата 25°.
Осенью эти ящерицы появлялись при температуре субстрата и тела 22°
(9.Х 1981 г., Чемен-и-Бит), при температуре скалы 21° они исчезали; затем
Таблица 5. Анализ содержимого желудков туркменского геккона
(Т. turcmenicus, п = 50) (п = 30, уроч. Пеленговели, 11.V 1979 г.; п == 20, уроч. Агащлы,
14.V 1978 г.)
Объект питания Встречаемость Число съеденных экз
абсолютная % абсолютное | %
Crustacea
Isopoda Arachnoidea 2 4 2 1,6
Phalangidae 4 8 6 4,8
Araneidae 2 4 2 1,6
Hylopoda Insecta 1 2 1 0,8
Orthoptera 2 4 3 2,4
Tettigoniidae 1 2 1 0,8
Acrididae 2 4 2 1,6
Blattoptera 2 4 2 1,6
Homoptera 4 8 5 4,0
Hemiptera 6 12 9 7,2
Reduviidae 2 4 3 2,4
Coleoptera 1 2 1 0,8
Carabidae 1 2 1 0,8
Chrysomelidae 2 4 4 3,2
Curculionidae 14 28 42 30,2
Hymenoptera 1 2 1 0,8
Formicidae 5 10 21 21,6
Diptera 2 4 2 1,6
Tipulidae 2 4 2 1,6
Lepidoptera 11 22 13 10,6
через 45 мин наблюдалось падение температуры воздуха до 15°, скалы до 18°.
При сильном ветре гекконы не выходили из щелей. При изучении туркменских
гекконов в термоградиентприборе (п = 46) ящерицы отмечены при температу-
ре 9—44° (29,5 ± 1,5).
Суточный цикл активности. Ведут сумеречно-ночной об-
раз жизни. 13.V 1978 г. в уроч. Агашлы они появились, спустя час после на-
ступления сумерек — в 21 ч 30 мин, там же 9.V 1979 г. они вышли из убежищ
в 22 ч, в уроч. Пеленговели 11.V 1979 г. первая ящерица отмечена в 21 ч, а мас-
совый выход — в 23 ч. Последние гекконы были на поверхности до 2 ч. Осенью
(9.Х 1981 г., окр. ст. Чемен-и-Бит) ящерицы данного вида появились в
20 ч 45 мин и в 21 ч 15 мин исчезли в убежищах. Таким образом, туркменские
гекконы, в отличие от каспийских, выходят из убежищ позже.
Сезонный цикл активности. Самая ранняя дата добычи
этого вида — И.IV 1977г., поздняя — 9.Х 1981 г., видимо, правильно отражает
период его активности (возможны сдвиги в ту или иную сторону на 10—15 дней
в зависимости от погодных условий).
Линька. По наблюдениям в террариуме ящерицы линяли в период
17—27.IV, 13—20.V и 1—10.IX. Сеголетки линяют вскоре после вылупления.
9.Х 1981 г. в окр. Чемен-и-Бита отмечена массовая линька.
Питание. Сведения о питании туркменского геккона нами приводятся
впервые. Вскрыто для анализа питания 50 желудков туркменских гекконов
(остатки пищи обнаружены только в 17) (табл. 5). Эти гекконы поедают долго-
носиков (28 % встречаемости), бабочек (22 %); в щелях скал в это время было
много совок (мы наблюдали 11.V 1979 г., как ящерица схватила одну из них),
полужесткокрылых (12 %), муравьев (10 %), цикад и фаланг (по 8 %). Во
145
время охоты гекконы спускаются к подножию скал, мы наблюдали их даже в
траве.
Размножение. Соотношение полов в популяции 1:1. Период спари-
вания не наблюдался. 13.V 1978 г. у самок из уроч. Агашлы отмечены хоро-
шо развитые яйца. Геккон, добытый под Чемен-и-Битом 18.V 1982 г., отло-
жил пару яиц размером 12,4—13,0 х 8,5—9,0 мм. 9.Х 1981 г. здесь добы-
вали сеголеток; в щели скалы, занятой гнездом воробья, обнаружено яйцо
туркменского геккона размером 13,7 х 9,5 мм. Оно было неосторожно раздав-
лено и внутри оказался живой сформированный геккон.
Темп роста не изучен.
Враги. В биотопе туркменского геккона под Чемен-и-Битом нами добыта
бойга (Boiga trigonatum), которая при случае может нападать и на гекконов.
Особенности поведения. По характеру передвижения не от-
личается от каспийского и туркестанского гекконов, хотя это более крупная
ящерица и бегает несколько быстрее. Летними и зимними убежищами являются
щели в скалах из песчаника, конгломератов или известняка. При ветреной по-
годе охотятся на насекомых, которые скрываются в щелях, в тихую погоду
спускаются к подножию скал и ловят добычу в траве. Судя по находкам старой
скорлупы яиц, их откладывают в щелях скал. Пойманные гекконы издают
тихий писк.
Практическое значение и охрана. На территории нашей
страны туркменский геккон обитает только в трех пунктах, причем общая
численность их здесь невелика, не превышает порядка 500 экз. Для сохранения
генофонда этого редкого вида по нашей рекомендации он включен в Красную
книгу СССР и ТССР. К счастью, биотопы туркменских гекконов не вовлечены
в сферу хозяйственной деятельности человека, поэтому им пока ничего не угро-
жает. Однако природоохранным службам Туркмении необходимо в перспекти-
ве хотя бы одно из мест, где обитают гекконы данного вида (например, уроч.
Агашлы), объявить заповедным.
ТУРКЕСТАНСКИЙ ГЕККОН — TENUIDACTYLUS FEDTSCHENKOI (STRAUCH, 1887)
Типовая территория: Самарканд, Узбекистан.
Кариотип: 2п = 42, субметацентриков 2, акроцентриков 38, субтелоцентриков 2,
NF = 46.
146
Рис. 67. Tenuidactylus iedtschenkoi:
A — голова сбоку, Б — голова сверху, В — носовые щитки, Г — голова снизу, Д — спи-
на, Е — бедренные поры, Ж — бедро сзади (ИЗАНУ SR 1081, Каратау, ТаджССР)
1870 — Gymnodactylus scaber Федченко, Изв. общ. любит, естеств., антропол., этногр., VIII,
1,с. 165.
1887 — Gymnodactylus fedtschenkoi Strauch, Mem. Acad. Sci. St. Petersbourg, ser. 7, Vol. 35,
N 2, S. 46.
1954 — Cyrfodactylus fedtschenkoi Underwood, Proc. zool. Soc. London, p. 475.
1984 — Tenuidactylus fedtschenkoi Щербак, Голубев, Вести, зоол., Киев, № 2, с. 53.
Диагноз. У самцов обычно не менее 30 бедренных пор (26—29 бедрен-
ных пор встречено менее чем у 5 % обследованных самцов). Чешуй вдоль тела
снизу, как правило, 105—130 (только у выборки из Нурека число брюшных
чешуй доходило до 144, у остальных выборок (около 3 % случаев) — более чем
130 чешуй, в пределах 131—136). Поперек брюха 24—37, поперек головы 9—18
чешуй; спинные бугорки треугольно-округлые, обычно средних размеров;
147
как правило, дополнительные бугорки с ними не соприкасаются. Два — четыре
(обычно два) нижнечелюстных щитка, первая пара их соприкасается между со-
бой широким швом. Обычно три ряда чешуй между рядом бедренных пор и
анальной щелью. Вытянутая вперед вдоль туловища передняя лапа достает
концами пальцев до конца морды (общая длина предплечья и плеча составляет
31—38 (35,1 ±0,1) % длины туловища).
«Лектотип. ЗИН 6354, d'ad «Самарканд, 1874, В. Руссов». L = 60, 7 мм;
хвост регенерирован; чешуй поперек головы 13; верхнегубных щитков 12/10;
нижнегубных 8/9; чешуй вдоль тела снизу 127; чешуй поперек брюха 28;
чешуй, окружающих спинной бугорок, 13 и один несколько увеличенный до-
полнительный бугорок; бедренных пор 33; подпальцевых пластинок по 26;
нижнечелюстных щитков по два, первая пара соприкасается между собой ши-
роким швом.
Описание (рис. 67) (по 461 экз. из колл. ИЗАНУ, ЗИН, ММГУ,
ИЗТ адж)
L ad (п = 162) 48,2—76,7 (59,66 ± 0,53) мм; L ? ad (и = 166) 48,0—
70,0 (58,79 ±0,42) мм; LIL cd (п = 100) 0,68—0,91 (0,76 ± 0,004); индекс
высоты/ширины головы (п = 100) 48—60 (54,35 ± 0,29); диаметр уха/глаза
(и = 100) 33—67 (45,53 ± 0,57); верхнегубных щитков (п = 919) 9—14
(11,51 ± 0,03); нижнегубных (гг = 921) 7—11 (8,28 ± 0,02); чешуй между
центрами глаза (и = 461) 9—18 (12,36 ± 0,07); длина спинного бугорка/L
(п = 442) 15—32 (23,39 ± 0,14); чешуй, окружающих спинной бугорок (п =
= 434), 10—18 (13,92 ± 0,09); чешуй вдоль тела снизу (п = 428) 105—144
(120,73 ± 0,31); чешуй поперек брюха (п = 403) 24—37 (29,47 ± 0,10); бед-
ренных пор (и = 217) 26—44 (33,86 ± 0,21); бедренных чешуй у самок (и —
= 204) 28—48 (37,11 ± 0,27); подпальцевых пластинок (и = 893) 19—31
(25,02 ± 0,08). Подбородочный щиток тре- или пятиугольный, нижнечелюст-
ных щитков две (27,1 %), три (69,1 %) или четыре (3,8 %) пары, первая из
которых соприкасается между собой широким (88,8 %) или точечным (8,5 %)
швом или разделена горловой чешуйкой (2,7 %); первые носовые щитки разде-
лены между собой одной (71,0 %) или двумя (0,7 %) чешуйками или соприка-
саются (28,3 %); с центральными спинными бугорками соприкасаются один
(32,4 %), два (10,6 %) или три (0,7 %) дополнительных бугорка, у 56,3 % осо-
бей такого контакта не наблюдается, у 95 % самцов ряд бедренных пор посе-
редине не прерван. Масса до 9,0 г.
Рисунок и окраска. В отличие от каспийского геккона основной
фон спины светло- или кофейно-коричневый, а поперечных спинных и хвосто-
вых полос — темно-коричневый; еще одним отличием от предыдущего вида
является почти полное исчезновение рисунка головы. Особи, добытые в окр.
Нурека на темных скалах, имели редкий темно-коричневый цвет с лиловым
или красноватым оттенком.
Распространение (рис. 64). Широко распространен в горных
системах 3. Памира и примыкающих к нему равнинах; проникает в Ю. Кызыл-
кум и В. Каракумы, куда, вероятно, был занесен человеком; южной границей
этого вида, по-видимому, являются реки Амударья и Пяндж.
Кадастр к рис. 64.
СССР, Туркмения:/ — пос. Фараб (ММГУ); 2 — Чарджоуская креп. (ИЗАНУ); 3 —
Репетек (ЗИН); 4 — 85 км ю.-в. Чарджоу, Карабек-аул (ИЗАНУ); 5—80 км сев.-в. Керкичи,
Талимарджан (ИЗАНУ); 6—Керкичи (ИЗАНУ); Достлук [130]; 7 — хр. Кугитанг, окр. Ше*
рабада; пос. Карлюк; Ходжепиль; пос. Кугитанг (ИЗАНУ); Базар-Тепе [23]; Кызыл-Алма:
Шалкан, Зарабак, Майдан (кишлаки в предгорьях хр. Кугитанг) [22]; Свинцовый рудник,
возвышенность Юрек-тепе, пос. Кайнарата [130]; Ак-Булак (ЗИН); 8 — 80 км ю.-в. Керки-
чи (ИЗАНУ); 9 — пос. Келиф (ЗИН).
Узбекистан: / — 3. Актау, пос. Кокча [161]; 2 — пос. Илланды; ст. Кермине [22];
3 — г. Пистелитау (сев.-в. предгорья хр. Нуратау [98]); окр. Джизака (ЗИН); 4 — Хавает;
Джулянгар (ЗИН); 5 — Бухара (ИЗАНУ); г. Каган (МХГУ); 6 — пос. Акташ (ИЗАНУ); 7 —
Ката-Курганское водохр. [22]; 8 — г. Самарканд (ИЗАНУ); Кара-Тюбе (ЗИН); 9 — р. Сангар-
дакдарья (ЗИН); 10—окр. кишлака Падахан, р. Тулаланга; пос. Чинары; хр. Бабатаг (МХГУ);
11 — Денау на Сурхандарье (ЗИН); 12 — Сурхандарьинская обл., пос. Дербент (ЗИН); 13 —
Кашкадарьинская обл., Ак-Рабат (ИЗАНУ); 14 — пос. Шурчи (ИЗАНУ); 15 — Лейлякан
(МХГУ); 16—Кугитангтау, пос. Вандоб (МХГУ); /7—пос. Карасу (между Термезом и Шираба-
дом) [22]; 18 — пос. Кокайты (ЗИН); 19 — з. пос. Джаркурган; 11 км по дор, из Джаркургана
148
в Ангор (МХГУ); Ангор (ММГУ); 20 — пос. Сурхан; 8 км з. ст. Сурхан (МХГУ); 21 — окр.
Термеза, креп. Султан-Саадат (ИЗАНУ).
Казахстан: / — Ц. Кызылкум, кол. Аяккудук [161]; 2 — оз. Тузкан, 100 км сев. хр.
Нуратау [161].
Таджикистан: 1 — пос. Ура-Тюбе [161]; 2 — пос. Шинг (ИЗАНУ); 3 — Гармский
р-н, р. Сарбач, уроч. Шинглич (ИЗАНУ); 4 — ю. склон Гиссарского хр., кишл. Сары-Пул и
Шурхок (ИЗИП ТаджССР); 5 — пос. Варзоб [125]; 6 — Рамитское ущ. Чуянгарон (ИЗАНУ);
7 — Регар (ЗИН); 8 — Душанбе (ИЗАНУ); 20 км в. Душанбе (ЗИН); 9 — 90 км ю.-з. Душанбе,
пос. Исамбай (ИЗАНУ); окр. г. Гиссар (ЗИН); 10 — г. Иван (ЗИН); 11 — пер. Чермакзак
(1670 м н. у. м.); Нурек (ИЗАНУ); 12— ю. Нурека, дор. в Турткаул [125]; 13— Каратау,
860 м н. у. м. сев. г. Пяндж (ИЗАНУ); 14 — Сурхоб (ИЗАНУ); 15 — Куляб, дор. на Шур-
абад (ИЗАНУ); 16—Даштиджум (ИЗИПТадж.); 17—Дарвазский хр. [96]; 18—12—20 км ю.
пос. Кызыл-Кала, по дор. на Шаартуз (ИЗАНУ); 19 — г. Курган-Тюбе; кишл. Уялы по дор.
Душанбе — Курган-Тюбе (ЗИН); 20 — окр. Шурабада (ИЗАНУ); 21 — Вахшстрой [125];
22 — Ходжа-Мумин (ИЗАНУ); 23 — Шаартузский р-н, Бешкентская креп. (ИЗАНУ); кишл.
Чиль-Чор-Чашма [125]; 24 — Пархар, между Сараем и Чубеком [119]; 25 — окр. Иола между
Чубеком и Тогмаем [119]; 26 — хр. Туюн-Тау (ИЗАНУ); 27 — 12 км ю. пос.Джиликуль (ЗИН);
28 — 26 км от Пянджа на Пархар (ИЗАНУ); 29 — Кировабад (ИЗИПТадж.); 30 — Шаар-
туз (ИЗАНУ); 31 — 18 км от пос. Чирик (ИЗАНУ); 32 — кишл. Айвадж [125].
Киргизия: 1 — Фрунзе (ИЗАНУ).
Половой диморфизм и возра стная изменчивость
соответствуют таковым каспийского геккона (см. выше).
Систематика и географическая изменчивость.
Для изучения географической изменчивости проведено сравнение по основным
признакам фолидоза между 16 выборками из всего ареала. Оно показало некото-
рое свсеобразие западных равнинных популяций: у гекконов из Чарджоу, Кара-
бекаула, Термеза, пос. Шурчи, Керкичи, в 80 км к востоку от последнего пункта,
наблюдаются более низкие средние значения признаков: количества бедренных
пор и заменяющих их чешуй у самок, количества дополнительных бугорков,
соприкасающихся с центральными спинными бугорками, относительной длины
последних и количества окружающих их чешуй. У этих выборок наблюдается
также увеличение средних показателей признаков: количество подпальцевых
пластин и чешуй поперек головы. Для равнинных популяций Ю. Таджикиста-
на и высокогорных гекконов характерны противоположные значения этих
признаков. Такие выборки, как самаркандская или карлюкская (т. е. рас-
положенные в предгорьях), по многим признакам обладают промежуточными:
показателями. У этого вида можно констатировать наличие вертикальной зо-
нальности в изменчивости некоторых признаков. Из особенностей популя-
ций обращает внимание резкое увеличение количества чешуй поперек брю-
ха и вдоль тела снизу у гекконов из окр. Нурекского водохранилища, чего,
однако, на наш взгляд, недостаточно для выделения их в особый подвид.
Стации и количественные данные. По свидетельству
литературных источников [22, 23, 98, 107, 125, 130, и др.], а также по нашии
наблюдениям, туркестанские гекконы обитают исключительно на вертикальных,
поверхностях: обрывах скал, лессовых оврагов, в развалинах, на дувалах,
в жилых и хозяйственных постройках, изредка встречаются на стволах деревьев.
Наибольшее количество этих ящериц мы находили на лессовых обрывах в пред-
горьях. Так, в окр. с. Карлюк (Кугитанг) на участке выходов сланца длиной
20 м и высотой 2 м было добыто 30 ящериц (из щелей выглядывали часто по
два и более геккона), недалеко отсюда в заброшенной цементированной яме
размером 2 X 1 X 1 м (общая плошадь 6 м2) было отмечено 11 гекконов; в окр..
с. Талимарджан (80 км сев.-в. Керкичи) на лессовых обрывах вдоль соленого-
ручья учтена самая высокая плотность: на 1 м2 шесть — восемь особей.
У хр. Каратау на обрыве пористой гипсовой скалы (100 X 2 м) добыто
шесть ящериц, в окр. с. Сурхоб (Таджикистан) на глинисто-осолоненном обрыве
площадью 10 м2 добыто 12 гекконов; в окр. Нурека на обрыве из желези-
стого песчаника встречался один геккон на 10 м маршрута, на лессовых обрывах
близ Душанбе на площади 10 м2 учтено 7—13 особей. Единичные особи отмече-
ны на скалах в окр. ст. Акташ, в карстовых воронках и на выходах каменной
соли и гипса в окр. пос. Московский (г. Ходжа-Мумин), между с. Шинг и Коста-
раш. Многочисленны эти ящерицы (1 экз. через 1—5 м маршрута) на древних
лессовых постройках, стенах крепостей (Чарджоу, Карабекаул, Туркмения),
149.
их руинах (Бешкентская креп, у с. Насир Хисров, Таджикистан; Кырк-Кыз
и Султан-Садат, Узбекистан). Очень редко туркестанские гекконы встречаются
на каменистых склонах в фисташковой саванне (хр. Каратау, Таджикистан):
здесь был проведен абсолютный учет на полосе 900 м шириной 6 м и обнаружен
под камнем лишь один геккон. Литературные данные о численности туркестан-
ских гекконов [107, 130] часто нельзя сравнить с нашими, так как ящерицы
учитывались в течение 1 ч (при неизвестной скорости, необозначенном биотопе)
или на неопределенном точно расстоянии (6—8 км, 7—9 км). Однако максималь-
ное количество гекконов в Таджикистане [107] указано в окр. с. Ганджина:
на маршруте 8—10 км было зарегистрировано 50—60 особей, или шесть особей
на 1 км, что является очень заниженным результатом. По другому источнику
[327] средняя плотность 11,3 особей на 100 м2 (в зоне Нурекского водохр.).
Встречается на высоте до 2300 м [125].
Отношение к температуре. К. П. Параскив [98] отметил
появление гекконов на поверхности после повышения температуры камней
до 23—25°. 23.IV 1980 г. в окр. Шураба (Таджикистан) отмечен днем при тем-
пературе воздуха 13° (высота 1520 м, в оврагах еще лежал снег). При темпера-
туре воздуха 19° в сумерках отмечены активные гекконы 8.IV 1980 г. на высоте
580 м н. у. м. в окр. с. Сумбула, а при температуре воздуха 16° в сумерках в окр.
с. Сурхоб. А. Д. Баутин (письм. сообщ.) инкубировал яйца туркестанского
геккона при температуре 36°, влажности 60—80 %.
Суточный цикл активности. Ранней весной и поздней осе-
нью гекконы данного вида активны днем, летом часто можно их также наблю-
дать в нежаркие часы или в затененных местах днем, но наиболее деятельны
они в сумерки [22, 23, 98, 125, наши наблюдения]. Из укрытий они выходят
не раньше 8—9 ч, в апреле — мае активные ящерицы встречены ночью в 22—1 ч
(8.IV 1979 г., окр. с. Сумбула; 10.IV 1980 г., Ю. Туюн-Тау; 20.V 1979 г., окр.
г. Чарджоу).
Сезонный цикл активности. В предгорьях Кугитанга
в 1954 г. первые особи после зимовки появились в середине февраля, а на Зе-
равшанском хр. они отмечены в начале апреля [22]. После зимовки под Душан-
бе гекконы появляются во второй половине, а в долине среднего течения Вахша
даже в первой половине марта [125]. С. А. Саид-Алиев [107] наблюдал еди-
ничные особи 7.II 1963 г. в г. Душанбе и в пос. Чор-Теппа, 23.11 1963 г.—
в окр. с. Зигар, но массовое появление отмечено в марте. На юге Казахстана
туркестанские гекконы появляются в первой половине марта [98]. Конкретные
сроки появления этих ящериц зависят от погодных условий года и вертикаль-
ной зональности. Добытые гекконы в окр. с. Сумбула 8.IV 1980 г. уже питались
(прохладная весна, высота 580 м н. у. м.), а 23.IV 1980 г. в окр. Шурабада (вы-
сота 1520 м) только оставили зимовочные убежища. Несомненно, ящерицы,
обитающие в высокогорье (3. Памир), могут оставлять зимовку в конце апре-
ля — начале мая. Уход на зимовку в Таджикистане (под Душанбе) отмечен
в декабре [125], в конце ноября и начале декабря 1963 г. они еще встреча-
лись в окр. Гульбиста, 15—18 и 29—30.XI 1964—1968 г. в долине р. Ка-
фирниган [107]. В Ю. Казахстане эти ящерицы исчезают в октябре — нояб-
ре [98].
Линька. Линные особи в Туркмении встречались в апреле — мае,
массовая линька в Таджикистане также отмечена в апреле — мае [125, 130].
При наблюдении в террариуме линька отмечена у разных особей 8—14.IV 1982 г.
и 13—25.V 1982 г. Несомненно, существует и осенняя линька.
Питание. В желудках 35 гекконов, добытых в августе в Кугитанге
[23], преобладали термиты (74,4 % встречаемости) и жуки (34,3 %, среди них —
жужелицы, чернотелки, пластинчатоусые, долгоносики), кроме того, равно-
крылые хоботные (14,3 %), клопы-щитники, перепончатокрылые и саранчевые
(по 11,4 %), а также бабочки (5,7 %). В незначительном количестве они пита-
ются уховертками, сетчатокрылыми, гусеницами бабочек, пауками, сколо-
пендрами, мокрицами и фалангами. В пище 10 гекконов, добытых в августе в
долине Сурхандарьи [22], преобладали жесткокрылые (70 %), перепончатокры-
лые (50 %); второстепенные компоненты пищи — полужесткокрылые, двукры-
150
лые, кричащие прямокрылые, чешуекрылые, термиты, равнокрылые хоботные
и пауки.
Состав пищи 12 гекконов, добытых в том же месяце в Бабатаге, состоял
из равнокрылых хоботных и чешуекрылых (33,3 % встречаемости) и полужест-
кокрылых (25 %); второстепенные объекты: жесткокрылые, перепончатокрылые,
прямокрылые, двукрылые, тараканы, мокрицы, скорпионы, пауки, фа-
ланги. По сведениям М. В. Калужиной [63], желудки 50 гекконов, добытых
в районе рисовых полей в Узбекистане, были набиты комарами: за одну ночь,
как установила автор, каждый геккон съедает до 73 комаров.
Анализ 35 желудков гекконов, собранных в окр. г. Душанбе в апреле —
мае [125], показал, что основной их корм в это время — паукообразные (100 %
встречаемости), все остальные беспозвоночные — многоножки, насекомые и их
личинки — имеют второстепенное значение (встречаемость около 40 %). Летом
пауки исчезают из пищевого рациона, а насекомые и их личинки становятся
основной пищей. Наиболее частой добычей гекконов являются жуки, мелкие
саранчевые, двукрылые, перепончатокрылые, в частности муравьи и пчелиные.
Размножение. Соотношение полов в наших выборках близкое 1:1.
Половозрелость наступает при достижении размеров туловища 48 мм и выше.
По сведениям из Узбекистана [22], наибольшие семенники обнаружены
в первой декаде апреля. Видимо, в это время гекконы спариваются. Готовые
к откладке яйца в окр. Термеза указанный автор находил в середине мая (он
полагает, что с начала развития яиц до их откладки проходит примерно 1,5 ме-
сяца). В это же время готовые к откладке яйца находили и в Туркмении [130].
Период размножения в окр. Термеза заканчивается к середине июля (вторая
кладка проходит через две-три недели). В указанный период в предгорьях и го-
рах, а также более северных районах размножение продолжается: последние
самки с яйцами были добыты в Бабатаге у Кашка-Булака 6.VIII, в Иллисае —
12.VIII. Чаше самка откладывает два яйца (108 случаев), реже одно (32 случая)
и совсем редко три (один случай) [22]. Последнее обстоятельство указано
только для туркестанского геккона. По наблюдениям в Таджикистане [125],
самка чаще откладывает два яйца.
Начало откладки в окр. Душанбе отмечалось во второй половине мая
(в особо холодные годы задерживается до середины июня). Последующие клад-
ки происходят, видимо, в конце июня или даже в июле. Во второй половине
июля еще встречались самки с готовыми к откладке яйцами; предполагается
возможность третьей кладки. Размер яиц варьирует в пределах 11 X 12,6 х
X 8,8 X 10,1 мм (п = 20) Часто несколько самок вместе откладывают яйца,
находили от 10 до 14 яиц в конце июля — начале августа, причем эмбрионы в
них находились на разных стадиях развития: от готовых к вылуплению до
едва сформировавшихся. Предполагаемый срок инкубационного развития —
60—65 дней.
По нашим наблюдениям, первая кладка найдена 17.V 1979 г. в окр. Кер-
кичи (здесь же обнаружено около 20 старых яиц — коллективная кладка),
пара свежих яиц геккона данного вида найдена также 18.V 1979 г. в щели лес-
сового обрыва в 80 км сев.-в. пос. Керкичи. Массовая кладка отмечена нами
в лессовом обрыве в окр. ст. Шурчи 24.V 1964 г. 30.V 1976 г. геккон, добытый
в окр. с. Карлюк, отложил одно яйцо. Коллективная кладка (яйца прошлогод-
ние, около 10) обнаружена нами также в лессовой постройке в окр. с. Самбу л а
8.IV 1980 г. Размеры яиц 10,5—12 X 8,6—9,6 мм (п = 5). По данным из Ле-
нинского р-на Таджикистана, в 1976 г. яйца туркестанских гекконов были най-
дены с конца мая по конец сентября. В неволе эти ящерицы приступили к спа-
риванию в конце ноября, в конце декабря было отложено первое яйцо, через
два дня — второе, его размер 13,3 X 10,2 мм, масса 0,69 г. Средние размеры
яиц 12,8 X 9,9 мм, средняя масса 0,6 г (п = 5). Инкубация длилась 62 дня,
масса сеголетка 0,45 г (письм. сообщ. А. Д. Баутина).
Первая молодая ящерица добыта в Бухаре 2.VII 1955 г. [22]. Новорожден-
ные в окр. Душанбе начинают появляться в конце июля — начале августа [125L
Темп роста. Размеры туловища новорожденных ящериц достигают
20—24 мм [107, 125]. По наблюдениям С. А. Чернова в Таджикистане [125],
151
через 1,5 месяца после выхода из яйца гекконы вырастают приблизительно
на 5 мм, а к концу мая следующего года, т. е. через четыре-пять месяцев актив-
ной жизни (не считая зимнего периода покоя), на 12—13 мм и в среднем дости-
гают длины 42—45 мм. К концу июля они вырастают еще на 9,5—10 мм, а к зи-
ме становятся вполне взрослыми. Таким образом, полнорослость достигается
к середине второго года жизни. С удлинением тела до 65—68 мм дальнейший
рост прекращается, хотя, очень редко, единичные особи вырастают до 70—
71 мм. Самки, вышедшие из первой по времени кладки, к следующей весне
достигают половозрелости. Особи из последующих кладок достигают полово-
зрелое™ позже.
Автор отмечает темп роста хвоста гекконов в первый месяц жизни: у только
что вышедших из яиц гекконов хвост лишь немного длиннее туловища с голо-
вой (L/L cd в среднем 0,93), но уже через месяц он становится значительно длин-
нее и отношение LIL cd в среднем равно 0,75—0,78. При дальнейшем росте это
отношение не изменяется.
По наблюдениям в террариуме, туркестанские гекконы становились по-
ловозрелыми и приступали к размножению через 15—16 месяцев с момента
появления на свет (сообщение А. Д. Баутина).
Враги. В Узбекистане этих ящериц поедают поперечнополосатый по-
лоз (Coluber karelini) и эфа (Echis carinatus) [221. Туркестанский геккон най-
ден нами в желудке поперечнополосатого полоза, добытого 24.V 1964 г. в окр.
ст. Шурчи. 21.V 1979 г. в руинах креп. Карабекаул (100 км южнее Чарджоу)
мы наблюдали, как сизоворонка склевывала утром гекконов, появившихся
из убежищ. А. Д. Баутин фотографировал в Бешкентской долине нападение
на гекконов крупной фаланги (рис. 66) и поперечнополосатого полоза.
Особенности поведения. По характеру передвижения и охоты
туркестанские гекконы не отличаются от каспийских. В одной трещине может
обитать до десятка и более гекконов. Драки между ними возникают постоянно,
причем, как правило, дерутся только половозрелые самцы. Достаточно одному
геккону двинуться с места, как начинают двигаться и другие; то там, то здесь
между ними вспыхивают ссоры. Если на стенку трещины садится насекомое,
то к нему устремляются две-три особи. Писк потревоженного геккона хорошо
слышен [98].
По наблюдениям в Узбекистане [22] и по нашим наблюдениям в Туркмении,
местом их укрытия служат различные щели и трещины в скалах, лессовых об-
рывах и между кирпичами построек.
Зимует геккон в тех же щелях, забираясь на глубину 20—30 см. По на-
блюдениям М. В. Калужиной [63], в окр. Каттакургана в одном месте зимует
по 5—11 особей.
По свидетельству С. А. Чернова [125], в Таджикистане туркестанский гек-
кон особенно многочислен на лессовых обрывах. Здесь в трещинах, рассели-
нах, норах птиц и насекомых гекконы находят убежища и места для откладки
яиц и зимовки. От этих мест гекконы далеко не отходят, что подтверждается
специально проведенным мечением.
В окр. Душанбе яйца встречаются в тех же норках в лессовых обрывах,
которые служат убежищами этим животным. Как правило, яйца встречаются
главным образом только в тех из них, которые имеют входное отверстие, при-
ближающееся по размерам к таковому норки сизоворонки. Яйца лежат в 12—
20 см от входа, присыпаны тонким слоем лесса. Днем в этом месте норки темпе-
ратура около 33—34°.
Спаривание может происходить на вертикальной стене. По наблюдениям
в террариуме, к неподвижно лежащей самке подошел самец, приподнимаясь
высоко на лапах, слегка покачиваясь из стороны в сторону и подняв волнооб-
разно извивающийся хвост, так что это походило на своеобразный танец. Он
схватил самку за основание задней конечности (в тазовой области), припод-
нял ее, слегка опрокинул и приступил к копуляции (письм. сообщ. А. Д. Бау-
тина, Душанбе). Он же наблюдал и откладку яиц: самка, выбрав укрытие
в террариуме (расщелина коряги), успокоилась, через 20 мин слегка приподня-
лась, повернула голову, как бы наблюдая появление яйца. Яйцо появилось
152
жидким и, выливаясь каплеобразно, скатилось на дно расщелины, его цвет
был розовый. Спустя короткое время, прямо на глазах его оболочка стала твер-
деть и побелела. В конце инкубации новорожденный пробил маленькое, почти
круглое отверстие диаметром примерно 1,5 мм, отдохнув 15 мин, стал расши-
рять его резкими импульсивными движениями. Это продолжалось до тех пор,
пока геккон полностью не освободился от скорлупы. Вылупившись, он тотчас
исчез в укрытии; спустя 4 ч после появления на свет стал принимать пищу.
Практическое значение и охрана. Туркестанский гек-
кон по характеру питания полезное животное. Этот вид имеет широкий ареал
и высокую численность, достаточно легко переходит к синантропному образу
жизни, встречается на территории многих заповедников (Кугитангский в ТССР,
Кизылсуйский, Миракинский и Нуратинский в УзССР, Рамитский в ТаджССР
ид р.). Поэтому он не нуждается в специальных природоохранных мероприятиях.
ДЛИННОНОГИЙ ГЕККОН — TENUIDACTYLUS LONGIPES (NIK., 1896)
Типовая территория: Hex, В. Иран.
Кариотип: 2л = 42, акроцентриков 30, NF = 42
1896 — Gymnodactylus longipes Никольский, Ежег. Зоол. Муз., т. 1, № 4, с. 369.
1918 — Gymnodactylus microlepis Lantz, Proc. Zool. Soc. London, N 11—17, p. 11.
1963 — Cyrtodactylus longipes S. Anderson, Proc. Calif. Acad. Sci. 4th ser., Vol.
31, N 16, p. 474, San-Francisco.
1984 — Cyrtodactylus voraginosus Leviton, Anderson, I. Herpetol., Vol. 18, №3, p. 270.
1984 — Tenuidactylus longipes, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 53.
Диагноз. У самцов не менее 30 бедренных пор; чешуй вдоль тела снизу
обычно более 130; поперек брюха 30—40 чешуй; поперек головы 12—19 чешуй;.
спинные бугорки треугольно-округлые, обычно мелкие, дополнительные бугор-
ки чаще не соприкасаются с центральными спинными бугорками; нижнечелюст-
ных щитков две-три пары; первая соприкасается между собой; обычно два-три
ряда чешуй между рядом бедренных пор и анальной щелью; вытянутая вперед
вдоль тела передняя лапа заходит концами пальцев за конец морды.
Лектотип. ЗИН 8810. 1 g ad «В. Персия, Hex, 1896, Н. Зарудный».
L = 62,7 мм; хвост отсутствует; чешуй поперек головы 16; верхнегубных
щитков по 13; нижнегубных 9/8; чешуй вдоль тела снизу 153; чешуй поперек
брюха 33; чешуй, окружающих спинной бугорок, 12, дополнительные спинные
бугорки с ним не соприкасаются; бедренных чешуек 40; подпальцевых пластин
22/23; нижнечелюстных щитков две пары, первая разделена двумя чешуйками.
Описание, (рис. 68) (по 117 экз. из коллекций ИЗАНУ, ЗИН, ММГУ,
NMW, FMNH, ZML).
L <7 ad (п = 40) 45,3—68,8 (60,41 ± 1,48) мм; L g ad (п = 35) 45,0—66,4
(59,05 ± 1,91) мм; LIL cd (п = 27) 0,71—0,82 (0,78 ± 0,02); масса 3,75 г; ин-
декс высоты/ширины головы (п = 95) 39—58 (49,89 ± 0,57); диаметр уха/гла-
за (п = 55) 31—56 (44,68 + 0,78); верхнегубных щитков (п = 234) 11—16
(13,27 ± 0,08); нижнегубных (п — 234) 7—11 (8,84 ± 0,05); чешуй между
центрами глаз (п = 115) 12—19 (15,95 ± 0,27); длина спинного бугорка/L
(п = 80) 15—22 (18,82 ± 0,20); чешуй, окружающих спинной бугорок (п =
= 117), 10—15 (11,92 zb 0,13); чешуй вдоль тела снизу (и = 98) 119—169
(141,67 zb 1,27); чешуй поперек брюха (и = 106) 30—40 (34,05 ± 0,39); бед-
ренных пор (п = 58) 31—44 (35,86 zb 0,29); бедренных чешуй у g (п = 21)
35—44 (39,45 zb 0,77); подпальцевых пластин (п = 220)20—30 (25,17 zb 0,19).
Подбородочный щиток тре- и пятиугольный; нижнечелюстных щитков две
(58,9 %) или три (40,2 %) (четыре щитка отмечены в одном случае, что состав-
ляет 0,9 %) пары; первая из них соприкасается широким или точечным швом
(по 19,7 %) или разделена одной (57,3 %) или двумя (3,3 %) чешуйками; первые
носовые щитки разделены чешуйкой (96,6 %), этих чешуек бывает две или они
отсутствуют совсем (по 1,7 %); с центральными спинными бугорками соприкаса-
ется один (30,2 %) или два (8,6 %) дополнительных бугорка, но чаще (61,2 %)
они не соприкасаются. У самцов ряд бедренных пор посередине не прерван.
Окраска и рисунок. Основной фон охристо-буроватый, иногда
с оливковым или сиреневатым оттенком. Рисунок более темный, чем основной
153
Рис. 68. Tenuidactylus longipes:
A — голова сбоку, Б — голова сверху, В — лицевая часть спереди, Г — голова снизу,
Д, Е — подбородок, Ж — бедренные поры, 3 — хвост сверху, И — хвост снизу (е —
ЗИН 8810, Hex, Иран: остальное — ИЗАНУ SR 307, Акарчешме, ТССР)
фон, но слегка размытый, нечеткий. Спинные бугорки чуть светлее, все это
создает впечатление некоторой бархатистости. В целом этот геккон более свет-
лый, чем туркменский и туркестанский, и не такой яркий, как каспийский.
На спине пять — семь поперечных полос, на хвосте 8—12, на лапах шесть —
девять. Ширина полос равна промежутку между ними или шире их. Полосы
часто не заходят на бока туловища. Нижняя поверхность белая.
Половой диморфизм. Самцы отличаются от самок наличием
бедренных пор.
Возрастная изменчивость соответствует таковой каспий-
ского геккона (см. выше).
Распространение (рис. 64). Крайний юг СССР (Туркмения),
В. Иран, Афганистан от западной части плоскогорья Бадхыз на юг через Кайен-
ские горы до хр. Пеленган и к востоку от последнего по южным предгорьям
Гиндукуша до Кандагара. Возможны находки в Пакистане.
154
Кадастр к рис. 6 4.
Т. longipes microlepis. СССР, Туркмения: / — уроч. Пули-Хатум
(ИЗАНУ); 2 — Бадхыз, кордон Акар-Чешме (ИЗАНУ); 3 — Бадхыз, уроч. Керелек, источник
«Каменный» (МХГУ); 4 — уроч. Зульфагар (ИЗАНУ).
Т. 1. longipes. Иран: / — русло Бааза в стр. Зиркух [89]; 2 — Бирдженд [80];
3— Чахи-Зиру в стр. Зиркух (ЗИН); 4 — Чах-и-Гюише в стр. Зиркух [89]; 5 — пос. Нэх
(ЗИН); 6 — пос. Алиабад [59]; 7 — Нейзар в Сейстане [89]; 10 км ю.-в. Руде-Гирменд, дор.
Заболь — Мохаммедхан [325].
Афганистан (в зарубежных музеях был определен как G. fedtschenkoi): / —
г. Сиахкох, р-н Нейзар (ZML); 2 — пос. Хуссейнабад [59].
Т. 1. voraginosus: Афганистан: / — г. Фарах (NMW); 2 — пров. Канда-
гар, 48 км з. г. Диларам (FMNH); 3 — 55 и 64 км з. г. Гиришк (CAS); 4 — 32 км з. г. Кандагар-
(CAS); 5 — Панджваи (ZML); Кандагар 31°36' с. ш., 65°47' в. д. [172].
Систематика и географическая изменчивость.
Недавно установлено [48], что Т. microlepis (Lantz) является подвидом Т. lon-
gipes Nik. Первая из названных форм обитает на юге Туркмении (Бадхыз),
вторая — на большей части ареала, в В. Иране и Ю. Афганистане. Совершенным
диссонансом сказанному выше прозвучала публикация [97], в которой автор
утверждал, что отыскал в фондах ЗИН четыре геккона номинативной формы,
якобы добытые В. Я. Лаздиным 15.V 1915 г. в окр. Термеза «на пути к Гиссар-
скому хребту». Результаты специального исследования, проведенного нами,
опровергают это мнение:
1. Пятнадцатого мая 1915 г. В. Я. Лаздин находился не «на пути к Гиссар-
скому хребту», как считает Н. Л. Орлов, а в Патта-Гиссаре (к югу от Термеза),,
что отмечено на первичной этикетке и в дневнике исследователя [71].
2. Серия ЗИН 19215 состоит из трех взрослых гекконов (а, в, d) Т. longi-
pes разной сохранности и молодого геккона 19215 с, переопределенного нами
как Т. fedtschenkoi. Это свидетельствует о неодновременности помещения эк-
земпляров в одну емкость: несомненно, три длинноногих геккона были случай-
но помещены туда не в Патта-Гиссаре в 1915 г., а позже, в лаборатории. Сохра-
нившаяся этикетка В. Я. Лаздина относится к туркестанскому геккону 19215 с.
3. Всего в коллекцию ЗИН поступил 21 экз. длинноногого геккона (со-
гласно каталогу), но в настоящее время из них сохранилось только 11 (кстати,
серия из четырех гекконов ЗИН 9877, ранее определенных как G. longipes,
оказалась принадлежащей к виду Т. fedtschenkoi; судя по месту ее наход-
ки — «Хорасан, стр. Бэхарс, развалины Пэш-Рабат» — это тоже явная ошиб-
ка). Пока известна судьба лишь одного из 10 (NMW 17393). Видимо, три
геккона 19215 а, в и d происходят именно из девяти недостающих особей,
добытых в В. Персии Н. А. “Зарудным.
Кроме того, в указанной работе имеются еще две досадных оплошности.
Селение Hex в В. Персии здесь названо «terra typica restricts» необоснованно,
поскольку ограничение типовой территории было произведено еще за 80 лет
до этой публикации А. М. Никольским [87]. Ограничение типовой серии тремя
самками привело к тому, что очень важный в диагностике всей группы при-
знак — количество бедренных пор — в ней не представлен, хотя существовала
реальная возможность подобрать подходящий экземпляр среди остатков,
серии 8809 (теперь — бывшей типовой).
Таким образом, мнение об обитании длинноногого геккона в Ю. Узбекиста-
не базируется на музейной ошибке. В заключение следует заметить, что
Н. Н. Щербак в 1980 г. детально обследовал территорию Ю. Таджикистана
и Ю. Узбекистана вдоль советско-афганской границы, но, как и следовало ожи-
дать, Т. longipes здесь не обнаружен.
Запутанная ситуация сложилась также в связи с описанием нового вида
из Афганистана [325]. Имевшиеся в нашем распоряжении типы этой формы
из Чикагского музея убедили нас в том, что под этим названием можно разли-
чать гекконов из юго-западных предгорий Гиндукуша, придавая им, однако,
лишь статус подвида. Но сами авторы описания включили в число дополни-
тельных материалов также особь из В. Ирана (пров. Систан), с чем нельзя со-
гласиться: в системе предгорий восточноиранских хребтов Кайен и Пеленган
обитает номинативная форма. Кроме того, как следует из приведенной в пер-
воописании таблицы, именно этот экземпляр обладает признаками, не позво-
155.
ляющими однозначно отнести его к новому подвиду. Возможно, отсутствие су-
щественных физических преград могло привести к образованию в настоящее
время широкой зоны интерградации между этими формами в равнинной части
Ю.-З. Афганистана и примыкающих районах В. Ирана.
Таблица для определения подвидов Т. longipes
1(2). Длина передней лапы (предплечья и плеча) составляет 36—40 % длины туловища с голо-
вой; первая пара нижнечелюстных щитков обычно (65 %) разделена горловыми чешуйка-
ми ..........................................Т. longipes microlepis (Lantz)
2(1). Длина передней лапы (предплечья и плеча) составляет 38—45 % длины туловища с голо-
вой; первая пара нижнечелюстных’щитков обычно (75 %) соприкасается между собой.
3(4). Чешуй вдоль тела снизу 135—158; чешуй поперек головы 13—17.....
.......................................... Т. longipes longipes (Nik.)
4(3). Чешуй вдоль тела снизу 156—169; чешуй поперек головы 17—19.....
...............................Т. longipes voraginosus (Leviton et Anderson)
ДЛИННОНОГИЙ ГЕККОН ОБЫКНОВЕННЫЙ —
TENUIDACTYLUS LONGIPES LONGIPES (NIK ,1896)
Типовая территория: Hex, В. Иран, 31° 30' с. ш., 60° в. д.
1974 — Gymnodactylus longipes longipes, Горелов, Даревский, Щербак, Вести, зоол., Киев,
№ 4, с. 35.
Диагноз (по 16 экз. из колл. ЗИН, NMW и ZML). Длина передней
лапы (предплечья и плеча) составляет 38—45 % длины туловища с головой;
первая пара нижнечелюстных щитков соприкасается между собой относитель-
но широким (50 %) или точечным (37,6 %) швом или разделена одной-двумя
(12,4 %) чешуйками; чешуй вдоль тела снизу (и = 16) 135—158 (146,64 ± 1,64);
чешуй поперек головы (п = 16) 13—17 (15,93 ± 0,32); чешуй поперек брюха
(и = 16) 30—36 (32,79 ± 0,47); бедренных пор (п = 9) 32—38 (34,56 ± 0,67).
Остальные признаки содержатся в видовом описании.
Тип видовой.
Распространение. В. Иран, Ю.-З. Афганистан: Кайенские горы,
хр. Пеленган.
ДЛИННОНОГИЙ ГЕККОН МЕЛКОЧЕШУЙНЫЙ —
TENUIDACTYLUS LONGIPES MICROLEPIS (LANTZ, 1918)
Типовая территория: долина р. Теджен близ границы с Ираном.
1918 — Gymnodactylus microlepis Lantz, Proc. Zool. Soc. London, p. 11.
1975 Gymnodactylus (Cyrtodactylus) longipes microlepis, Горелов, Даревский, Щербак, Вест,
зоол., Киев, № 4, с. 35.
Диагноз (по 96 экз. из колл. ИЗАНУ, ЗИН и ММГУ). Длина перед-
ней лапы (предплечья и плеча) составляет 34—40 % длины туловища с голо-
вой; первая пара нижнечелюстных щитков разделена одной (65,1 % ) или дву-
мя (1,8 %) чешуйками или соприкасается между собой точечным (16,1 %) или
относительно широким (17 %) швом; чешуй вдоль тела снизу (п = 78) 119—
150 (137,61 ± 0,83); чешуй поперек головы (п = 95) 12—16 (13,86 ± 0,09);
чешуй поперек брюха (п = 87) 30—40 (34,15 ± 0,22); бедренных пор (п = 28)
32—39 (35,79 ± 0,35). Остальные признаки содержатся в видовом описании.
Лектотип. ММГУ 151а, d ad «долина р. Теджен близ Персидской гра-
ницы, IV—IX 1914, Н. В. Мариакри». L = 60,2 мм; L а/= 78,6 мм; LIL cd =
= 0,77; чешуй поперек головы 12; верхнегубных щитков 12/13; нижнегубных
по 9; чешуй вдоль тела снизу 136; чешуй поперек брюха 32; чешуй, окружаю-
щих спинной бугорок, 10 и два дополнительных бугорка; бедренных пор 34;
подпальцевых пластин по 23; нижнечелюстных щитков три пары; первая разде-
лена чешуйкой.
Распространение. 3. часть плоскогорья Бадхыз, по-видимому,
сопредельные районы Ирана и Афганистана.
156
ДЛИННОНОГИЙ ГЕККОН ЮГО-ВОСТОЧНЫЙ —
TENUIDACTYLUS LONGIPES VORAGINOSUS (LEVITON
ET ANDERSON, 1984) COMB ET STAT. N.
Типовая территория: Афганистан, 55 км западнее г. Гиришк.
1969 — Cyrtodactylus fedtschenkoi, Clark, Clark, Anderson, Leviton, Proc. Calif. Acad. Sci., Ser.
4, Vol. 36, p. 300.
1970 — Cyrtodactylus sp., Leviton, Anderson, Proc. Calif. Acad. Sci., Ser. 4, Vol. 38, p. 185.
1984 — Cyrtodactylus voraginosus Leviton, Anderson, J. Herpetol., Vol. 18, no. 3, p. 270.
Диагноз (no 5 экз. из колл. FMNH, NMW и ZML). Длина передней
конечности (предплечья и плеча) составляет 38—45 % длины туловища с голо-
вой; первая пара нижнечелюстных щитков соприкасается между собой точеч-
ным (40 %) или относительно широким (20 %) швом, часто (40 %) она разделе-
на горловыми чешуйками; чешуй вдоль тела снизу (п = 5) 156—169 (161,40 ±
± 2,32); чешуй поперек головы (п = 5) 17—19 (17,60 ± 0,40) (по 13251 — 16—
19); чешуй поперек брюха (п = 5) 34—39 (37,0 ± 1,05) (по [325] — 36—40);
бедренных пор (и = 3) 35—44 (по [325] — 35—46). Остальные признаки содер-
жатся в видовом описании.
Голотип [325]. CAS 120332, 2 ad «55 км западнее г. Гиришк, Афгани-
стан, 10 мая 1968, Р. и Э. Кларк». L = 60 мм; L cd = 85 мм; LIL cd = 0,71;
верхнегубных щитков 17; нижнегубных 11/9; чешуй поперек брюха 36;
бедренные поры отсутствуют; подпальцевых пластинок 25; нижнечелюстных
щитков одна-две пары, первая соприкасается между собой точечным швом.
Распространение. Юго-западные предгорья Гиндукуша в Аф-
ганистане.
Сведения по экологии в литературе отсутствуют. В разделе Определителя
земноводных и пресмыкающихся СССР [17], написанном Н. Н. Щербаком, со-
общалось, что данный вид в Бадхызе живет на выходах скал, встречается на
каменных и глинобитных заборах и стенах зданий.
Стадии и количественные данные. Как мы установи-
ли, длинноногий геккон проникает на территорию СССР из Ирана и Афганиста-
на по хр. Гез-Гядыку и Зульфагарскому, расположенным в меридиональном
направлении. У первого участки скальных обнажений имеются в долине р. Тед-
жен, они состоят из известняков, здесь гекконы встречаются единичными осо-
бями. Севернее, в холмогорье Бадхыза с фисташковой саванной, имеется груп-
па останцев и скальных обрывов в окр. кордона заповедника Акар-Чешме
(рис. 69). Скалы здесь из песчаника и конгломератов вулканических пород,
сцементированных песчанником. И хотя здесь убежищ меньше, чем в долине
р. Теджен, численность гекконов выше. Так, на скале «Зуб», на высоте 1,5 м
от основания в полосе длиной 20 м 13.V 1976 г. отмечено 13 особей. Здесь же
22.IV 1984 г. в полосе 2 X 50 м учтено 7 гекконов. Все встречены на склонах
юго-западной экспозиции.
В районе Зульфагарского хребта они также встречаются (единичные)
на обрывах к Теджену, далее — севернее в промоинах плато с гипсовыми вы-
ходами (на площади 2 X 300 м было добыто три геккона 18.V 1982 г.) и, нако-
нец, далее на новой гряде возвышенностей юго-западной экспозиции, на обры-
вах плато и останцах темных вулканических пород и конгломератов, лишенных
растительности. Здесь они встречаются сравнительно редко, через 7—10 м
маршрута (18.V 1982 г.; 25.IV 1983 г.). Высота биотопов 760—900 м н. у. м.
На останцах Акар-Чешме на 50 длинноногих гекконов встречено 2 каспийских.
Отношение к температуре. Вялые гекконы впервые наблю-
дались 4—7.IV 1977 г. в Акар-Чешме при минимальной температуре воз-
духа 14° и субстрата 17°. Самая высокая температура скалы, отмеченная нами
в сумерки в период активности гекконов,— 25°. Осенью, 7.Х 1981 г. они были
на поверхности при температуре воздуха 18,5°, скалы 25,0°, температура тела
ящериц была 25—26°. Длинноногие гекконы, вероятно, более теплолюбивы,
чем каспийские; при фотографировании на солнце первый был спокоен, вто-
рой пытался скрыться от перегрева (перед этим они содержались в одинаковых
условиях).
Суточный цикл активности. Весной и осенью длинноно-
гие гекконы наблюдаются греющимися на солнце» 6. IV 1977 г. ночью было
157
Рис. 69. Биотоп Т. longipes (песчаниковые останцы в окр. Акар-Чешме, ТССР)
8—11°, но утром в щелях они появились на местах, освещенных солнцем.
Осенью, когда солнце нагрело скалу, после 14 ч гекконы стали выходить на
поверхность. В целом это сумеречное животное, весной отмечены активные
ящерицы с 22 до 1 ч, осенью с 19 до 23 ч. Ночью при луне скрываются в зате-
ненные места.
Сезонный цикл активности. Первые особи у зимовальных
убежищ встречены нами 4.IV 1977 г., последние — 7.Х 1981 г. (Акар-Чешме).
По-видимому, они активны с конца марта — начала апреля до середины ок-
тября, в отдельные годы, возможно, позже, до ноября.
Линька. В природных условиях линные особи встречены 13.V 1976 г.
и 24.IV 1984 г. В террариуме эти ящерицы линяли 8—14.IV и 13—23.V.
Питание. Анализ содержимого желудочно-кишечного тракта 25 яще-
риц, добытых в апреле в Ю. Туркмении, показал (табл. 6), что в их рационе
преобладают по встречаемости насекомые: бабочки и их гусеницы (100 %),
двукрылые (24 %), жужелицы и долгоносики (по 20 %), а также пауки (16 %).
Среди других групп беспозвоночных в незначительных количествах (4—8 %
встреч) поедаются сольпуги, скорпионы и иксодовые клещи. У длинноногого
геккона в рационе преобладают чешуекрылые, что отличает его от других
видов.
Размножение. Соотношение полов в нашей выборке близкое 1 :
: 1. Половозрелыми эти гекконы становятся при достижении длины туловища
45 мм и выше. Спаривание, видимо, происходит вскоре после выхода из зимо-
вочных убежищ, в апреле, так как 12.V 1978 г. у большинства самок уже были
видны хорошо развитые яйца. Откладывают одно-два (чаще) яйца в конце мая.
Судя по скорлупе старых яиц, найденных в щелях скал в ур. Зульфагар, их
диаметр 10 мм. Сроки появления сеголеток не выяснены.
Темп роста не изучен. Наименьшие размеры сеголеток, бывших
в нашем распоряжении, L = 27,1 мм; L cd = 28,4 мм. По аналогии с близкими
видами можно предполагать, что половой зрелости они достигают в возрасте
15—16 месяцев, т. е. к середине второго года жизни, а максимальных разме-
ров — к четырем-пяти годам.
Враги. Конкретные сведения отсутствуют. Не исключено, что за ними
охотятся обыкновенные пустельги, гнездящиеся на тех же останцах, где оби-
тают гекконы.
Особенности поведения. По наблюдениям в природе и терра-
риуме, длинноногие гекконы передвигаются и охотятся так же, как и каспий-
ские, хотя длинные конечности дают им некоторый шанс уберечься от перегре-
158
ва: эти ящерицы выше приподнимают свое тело при движении и остановке по
раскаленной солнцем скале. Они встречаются только на вертикальных поверх-
ностях. Летние и зимовальные убежища их расположены в щелях скал, тут
же гекконы и откладывают яйца. Попав в руки, издают тихий писк. Брачное
поведение не наблюдалось.
Практическое значение и охрана. Как редкий вид
фауны СССР, имеющий ограниченное распространение, взят под охрану зако-
Таблица 6. Анализ содержимого желудков I. longipes microlepis из Ю. Туркмении
(апрель — май (п = 25)
Объект питания Встречаемость Число съеденных экз
абсолютная % абсолютное %
Crustacea Isopoda 1 4 1 0,9
Arachnoidea Solpugidae 1 4 1 0,9
Scorpionidae 2 8 2 1,8
Araneinae 4 16 5 4,5
Acarina Ixodidae 1 4 1 0,9
Hylopoda 1 4 1 0,9
Insecta Orthoptera 2 8 2 1,8
Coleoptera 4 16 10 9
Carabidae 5 20 6 5,4
Staphylinidae 1 4 2 1,8
Dermestidae 1 4 1 0,9
Curculionidae 5 20 6 5,4
Neuroptera Mirmeleonidae 1 4 1 0,9
Plasmodea 1 4 1 0,9
Diptera 6 24 18 17,5
Lepidoptera Гусеница 15 60 24 26,1
Имаго 10 40 21 20,4
на и включен в Красные книги ТССР и СССР. Охраняется на территории
Бадхызского заповедника. Возможно искусственное расселение его в под-
ходящих местах на территории указанного заповедника.
ПОДРОД СРЕДИЗЕМНОМОРСКИЙ ТОНКОПАЛЫЙ ГЕККОН-
MEDIODACTYLUS SZCZERBAK ET GOLUBEV. 1977
Типовой вид: Gymnodactylus kotschyi Steindachner, 1870.
1977 — Mediodactylus Щербак, Голубев, Герпетол. сб. Тр. ЗИН АН СССР, т. 74, Ленинград,
с. 130.
Диагноз. У самцов имеется не более 10 преанальных пор; если шов
между первой парой нижнечелюстных щитков имеется, то его длина обычно
не превышает половины длины подбородочного щитка; спинные бугорки оваль-
ные или округлые; шипообразные хвостовые бугорки, расположенные среди
однородной чешуи, друг с другом в полукольцах не соприкасаются; ряд под-
хвостовых щитков занимает не всю нижнюю поверхность хвоста (высота одного
щитка менее чем вдвое меньше его ширины).
Дополнительные признаки. Высота первого верхнегуб-
ного щитка от ноздри до края рта заметно меньше его ширины вдоль края рта;
в задне-верхней части орбиты часто выражены четкие конические выросты;
чешуя сверху хвоста однородная, в основании крупных основных бугорков
нет дополнительных. Хвост слабо вздутый, ряд подхвостовых щитков может
замещаться двумя рядами или обычной чешуей. Основной фон и рисунок вы-
держаны чаще в серых тонах.
Подрод включает шесть-семь видов, распространенных в Средиземноморье;
проникают в Среднюю Азию.
159
Таблица для определения видов подрода Mediodactylus
1(2). Спинных бугорков нет ..............................Т. amictopholis (Hoofien)
2(1). Спинные бугорки есть.
3(4). Спинные бугорки округлые, гладкие ................. Т. spinicauda (Strauch)
4(3). Спинные бугорки овальные или овально-треугольные, ребристые.
5(6). У взрослых диаметр ушного отверстия превышает половину продольного диаметра гла-
за ..........................................................Т. russowi (Strauch)
6(5). Диаметр ушного отверстия меньше половины диаметра глаза.
7(8). Все чешуи и щитки хвоста гладкие (не считая бугорков); передне-верхние поверхности
конечностей гладкие или с едва заметными следами ребристости.....................
.................................................Т. kotschyi danilewski (Steindachner)
8(7). Все чешуи и щитки хвоста, а также передне-верхние поверхности конечностей с от-
четливыми ребрышками.
9(10). Вдоль нижней стороны тела не более 90 чешуй; вдоль середины нерегенерированного
хвоста снизу увеличенные чешуи (по две пары на сегменте) . . . Т. sagittifer (Nik.)
10(9). Вдоль нижней стороны тела не менее 90 чешуй; снизу хвоста нет заметно увеличенных
чешуй ...............................................Т. heterocercus (Blanford)
СРЕДИЗЕМНОМОРСКОЙ ГЕККОН —
TENUIDACTYLUS KOTSCHYI (STEINDACHNER, 1870)
Типовая территория (Mertens, L. Muller, 1928): о. Сирое, Киклады.
Кариотип: 2п = 42, акроцентриков 42, NF = 42.
1870 — Gymnodactylus kotschyi Steindachner, Sber. Akad. Wiss. Wien, math.-naturwiss. KI., Abt*
1, 62, S. 329.
1981 — Cyrtodactylus kotschyi Beutler, Handb. Amph. Rept. Europ. Bd 1, 1, S. 53.
1984 — Tenuidactylus kotschyi Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Форма и размеры спинных бугорков очень вариабельны:
ребристые или гладкие, овальные, сердцевидные или удлиненные. Диаметр
ушного отверстия меньше половины (0,3) диаметра глаза. Вдоль нижней сторо-’
ны туловища не менее 90 чешуй. Передне-верхние поверхности конечностей
гладкие или ребристые.
Лектотип. с? № 10868, Insel Syra (= Syros) и 7 паратипов с этикет-
кой «Syra, 1866, CD 8а, don. Steind». Хранятся в NMW. Первоначальное мес-
то добычи в описании [294] значилось как «Goree, Senegal», там же упоминается
и о. Сирое. Впоследствии типовая территория была уточнена и ограничена
о. Сирое [260, 314].
Описание (по 231 экз. [183]). L сГ шах = 51 мм (А4 = 42,6 мм);
L $ шах = 56 мм (М = 44,4 мм). L/L cd = 0,71—1,0. Между носовыми от-
верстиями обычно расположены три-четыре, редко два или более четырех меж-
носовых; верхнегубных восемь — десять, нижнегубных шесть — восемь. Голова
сверху покрыта зернистой чешуей, между которой встречаются отдельные или
группы увеличенных, несколько ребристых чешуй. Спина покрыта выпуклой
чешуей, среди которой расположены 8—14 продольных рядов увеличенных бу-
горков. Между спинными бугорками чаще одна — три зернистых чешуйки в
продольном и две — четыре в поперечном рядах. Брюшная чешуя крупная, плос-
кая с пильчатым или зубчатым задним краем, расположена ,в 23- 40 продольных
рядов. Самцы с двумя —девятью преанальными порами или без них. За клоакой
обычно один —три постанальных бугорка с каждой стороны. Нерегенерирован-
ный хвост на верхней стороне покрыт мелкой чешуей, сегментирован, с четырь-
мя — шестью увеличенными шиповатыми бугорками на каждом сегменте, они
расположены полукругом. На хвосте 17—24 поперечных кольца. Чешуйча-
тый покров хвоста снизу очень вариабельный. Регенерированный хвост по-
крыт гомогенной или циклоидной чешуей, у некоторых форм нерегулярными
щитками. Вдоль боков туловища часто заметна боковая складка кожи.
Окраска и рисунок. Основной фон от желтоватого, оранжева-
того или красноватого (Т. k. stepaneki, Т. к. adelphiensis), серого до темно-
коричневого и почти черного цвета. На спине обычно шесть-семь темных М-
образных поперечных полос, угол которых направлен в сторону хвоста. От
конца морды вдоль боков головы к барабанной перепонке тянется обычная
темная полоска. Интенсивность окраски и рисунка у одной и той же особи
может изменяться, зависит от физиологического состояния. Брюшная сторона
160
белая или сероватая, слегка желтоватая или оранжевая. В преанальной об-
ласти часто заметна пятнистость.
Распространение. Ю. Италия, Балканский п-ов, о-ва Эгейско-
го моря, о-ва Крит и Кипр, Ю. Крым, Малая Азия, Ю.-З. Закавказье, Сирия
и Палестина (приводится карта с кадастром только для формы фауны СССР —
Т. k. danilewskii, см. ниже).
Систематика и географическая изменчивость.
Исключительно вариабельный вид. По данным А. Бойтлера 1183], максималь-
ные размеры туловища (56) мм наблюдаются у особей из Ю. Киклад, в то вре-
мя как многие южноэгейские формы едва достигают 40 мм. Регенерированный
хвост с беспорядочно расположенными снизу пластинами найден у гекконов
из Греции, Италии и Македонии, а мелкими чешуйками — на Крите, в Болга-
рии и Крыму, Нижняя поверхность нерегенерированного хвоста также значи-
тельно варьирует: наряду с формами с одним рядом увеличенных щитков встре-
чаются такие, у которых подхвостовые щитки делятся или представляют
собой циклоидную чешую (Крит, Болгария, Крым). У особей из Крыма, Болга-
рии и Ю.-В. Эгейских островов наблюдается за анальной щелью один пост-
анальный бугорок, в то время как их бывает два-три. На Сев. Спорадах, Крите
и Сиросе у взрослых самцов вообще отсутствуют преанальные поры. На Ю.-В.
Эгеях у самцов только две преанальные поры, в Крыму и Болгарии —шесть —
девять, на остальной территории ареала — три—пять. У большинства форм
брюшная чешуя расположена в 26—32 продольных ряда; на Крите и
Ю.-В. Эгеях, в Югославии и материковой Греции их число значительно меньше,
у популяций из Сев. Спорад имеется 34—38 продольных рядов.
Спинные бугорки обычно расположены в 12 продольных рядов, однако
у ящериц из Ю.-В. Эгеев, В. Крита, Югославии и о. Сирое, как правило, их
только 8—10. Спинные бугорки у форм из материка и Сев. Киклад имеют дли-
ну четырех-пяти окружающих их зернистых чешуек, особи из Сев. Спорад,
Ю. Киклад и Крита имеют значительно меньшие бугорки; у форм из Ю.-В. Эге-
ев и 3. Крита они достигают величины 1,5—2,5 чешуек, гладкие и трудно раз-
личимы. Подобная картина отмечена с особенностями щиткования верхних
и нижних поверхностей бедер; бугорки у особей Т. k. solerii, Т. к. bartoni
и у форм из о. Гавдос и Ю.-В. Эгеев полностью отсутствуют. Особенно хорошо
развиты бугорки сверху на бедрах у Т. к. fuchsi, Т. к. schultzewestrumi и
Т. к. tinensis. Обычно сверху основания хвоста шесть продольных рядов бу-
горков; у Т. к. schultzewestrumi и особей из Ю.-В. Эгеев их чаще семь—девять
[165, 184, 296].
Географической изменчивости подвержена и окраска тела. Сочетание ука-
занных признаков позволяет сгруппировать отдельные популяции в формы
и группы форм. В разное время было описано много подвидов средиземномор-
ского геккона, но в настоящее время валидными признаются только 25 [183].
Ниже приведены их диагнозы, которые можно использовать для определения
отдельных особей и выборок. В силу большой изменчивости создание надеж-
ных определительных таблиц пока не представляется возможным. Выделение
отдельных форм часто приведено на основании незначительных признаков и
ограниченного числа особей.
I группа ’’kotschyi”
Т. k. kotschyi (Steindachner, 1870): самцы без преанальных пор, продоль-
ных рядов брюшных чешуй до 33, спинных бугорков 12, хвостовых — 6; на
верхней поверхности бедер по 3 свободно стоящих бугорка (архипелаг Сирое).
Т. k. fuchsi (Beutler et Gruber, 1977) (=C. k.gruberi): близок к T. k. kotschyi,
но брюшных чешуй 34—40 продольных рядов, на бедрах сверху по 6 свободно
стоящих бугорка (сев. цепь островов Сев. Спорады).
Т. k. schultzewestrumi (Beutler et Gruber, 1977): близок к Т. k. kotschyi,
но хвостовые бугорки собраны в 7—8 продольных рядов, на бедрах сверху по
6 свободно стоящих бугорков (о. Валакса, на других островах архипелага
Скирос переходит в Т. к. tinensis).
И61
Т. к. tinensis (Beutler et Fror, 1980): самцы с 3—5 преанальными порами;
верхнебедренные бугорки в 4 раза, спинные в 4—5 раз шире зернистых чешуек
спины; по 6 свободных бугорков сверху на бедрах, продольных рядов спинных
бугорков 12, брюшных — 30 (Сев. Киклады; на о. Серифос, Миконос, Наксос
и Икария переходит в Т. k. saronicus).
Т. к. saronicus (Werner, 1939): близок к Т. k. tinensis, но с 3 свобод-
ными верхнебедренными бугорками (3. и Ц. Киклады, Саронские о-ва).
Т. k. buchholzi (Beutler et Gruber, 1977): близок к Т. k. saronicus, но с
10 продольными рядами спинных бугорков (архипелаг Сифнос).
Т. k. solerii (Wettstein, 1937): близок к Т. k. saronicus, но со спинными
и верхнебедренными бугорками, в 3 раза длиннее зернистых чешуек спины
(Ю. Киклады, Астипалая; на Ю.-З. Кикладах переходит в Т. k. saronicus).
К зоне интерградации относится Т. к. christianae (Buchholz, 1955, nomen nu-
dum).
T. к. rumelicus (L. Miiller, 1939): близок к T. k. saronicus, но подхвостовые
щитки на регенерированном хвосте разделены (Ю.-З. Болгария).
Т. k. bibroni (Beutler et Gruber, 1977): близок к Т. saronicus, но брюшные
расположены в 24 продольных ряда (Апулия, юг Балканского п-ова, о. Китира,
о. Китнос).
Т. k. skopjensis (Karaman, 1965): как Т. k. saronicus, но спинные чешуи
расположены в 10 продольных рядов (Сев. Македония).
II группа ’’danilewskii”
Т. k. danilewskii (Strauch, 1887) (= С. k, bureschi (Stepanek, 1937) (Крым,
Болгария, Фракия). Ниже будет приведено подробное описание. Единственная
форма, представляющая данный вид в фауне СССР.
Подвиды Т. k. steindachneri (Stepanek, 1937) из Ю.-З. Анатолии и Т. к.
lycaonicus (Mertens, 1952) из Анатолии недавно [1811 сведены в синоним Т. к.
danilewskii.
Т. к. colchicus (Nik., 1902): на туловище до 12 продольных рядов бугорков;
26—29 продольных рядов брюшных чешуй; снизу хвоста 3—4 продольных ря-
да чешуй, преданальных пор 2—4. Брюхо желтоватое, низ хвоста оранжевый
(видимо, у неполовозрелых) (Сев.-В. Анатолия. Возможны находки в Аджарии
на территории СССР).
Т. k. syriacus (Stepanek, 1937): крупные ребристые спинные бугорки рас-
положены в 10—14, чаще 12 продольных рядов; преанальных пор 4—5 (в сред-
нем 4,5), поперек брюха 24—26 чешуй, нижняя сторона нерегенерированного
хвоста покрыта у клоаки 2 чешуйками, остальные 3/4 — одним увеличенным
рядом чешуй (Сев. Сирия).
Т. k. orientalis (Stepanek, 1937): спинные бугры расположены в 10 несколь-
ко нерегулярных продольных рядов, ребристые, сердцевидные. Брюшных
чешуй в поперечном ряду 26—30 (Сирия, Ливан, Палестина).
Т. k. fitzingeri (Stepanek, 1937): спинные бугорки очень ребристые и про-
долговатые, расположены в 10—12 продольных рядов. Брюшных чешуй попе-
рек 20—25 рядов. Преанальные поры отсутствуют или их 2—3. Постанальных
бугорков 1—1. Нерегенерированный хвост покрыт 2 рядами циклоидных че-
шуй, регенерированный — мелкой чешуей (Кипр).
Т. k. karabagi (Baran et Gruber, 1982): спинных бугорков 12 и более про-
дольных рядов, брюшных чешуй — 30—34 (в среднем 32) продольных рядов,
преанальных пор 3—4, постанальных бугорков 2—2 (о. Фенер в Мраморном
море).
Т. k. beutleri (Baran et Gruber, 1981): продольных рядов спинных бугров
10—12, брюшных чешуй 25—33, в среднем 30, преанальных пор 1—6
(в среднем 3,8), постанальных бугорков 2—2 (Ю.-З. Турция и прилежащие
острова в Эгейском море).
Т. k. ciliciensis (Baran et Gruber, 1982): на голове ребристая чешуя (у
Т. к. danilewskii — гладкая), продольных рядов спинных бугорков 10—12,
брюшных 23—26 (в среднем 23,8), преанальных пор 2, постанальных бугорков
162
1—1, подпальцевых пластинок 14—18 (в среднем 15,5), у Т. k. danilewskii —
18—24 (ю. побережье Средиземного моря в Турции).
Т. к. ponticus (Baran et Gruber, 1982): продольных рядов спинных чешуй
10—14, чаще 12, брюшных—26—31 (в среднем 29,2); преанальных пор 2—5
(в среднем 3,5), постанальных бугорков 1—2, подпальцевых пластинок 17—
21 (в среднем 19,9) (известен из окр. г. Токат, Амасия и Сивас на сев. Ц. Тур-
ции).
III группа ’’bartoni”
Т. k. bartoni (Stepanek, 1934): спинные бугорки в 2 раза длиннее спинных
зерновидных чешуек (3. Крит).
Т. k. wettsteini (Stepanek, 1937) (=G. k. rarus Wettstein, 1952, == G. k. stub-
bei Wettstein, 1952): спинные бугорки в 3 раза длиннее спинных зерновидных
чешуек (побережье В. Крита).
IV группа ’’oertzeni”
Т. k. kalypsae (Stepanek, 1939): 6 продольных рядов бугорков у основания
хвоста, 26—29 продольных рядов брюшных чешуй (о. Гавдос около Крита).
Т. k. oertzeni (Boettger, 1888) (= G. k. unicolor Wettstein, 1937): продоль-
ных рядов хвостовых бугорков 7—8, брюшных чешуй 23—27, основной цвет
серый (архипелаг Карпатос).
Т. k. stepaneki (Wettstein, 1937): близок к Т. k. oertzeni, но основной цвет
красный (о. Мегали Софраноу у Сирина).
Т. k. adelphiensis (Beutler et Gruber, 1978): близок к Т. k. stepaneki, но
брюшные чешуи расположены более чем в 28 продольных рядов (побережье
о. Сирина).
КРЫМСКИЙ ГЕККОН — TENUIDACTYLUS KOTSCHYI DANILEWSKII (STRAUCH, 1887)
Типовая территория: Ялта, Крым.
1887 — Gymnodactylus danilewskii Strauch, Mem. Acad. Sci. St. Petersbourg, ser. 7, vol. 35, N 2,
S. 48.
1940 — Gymnodactylus kotschyi danilewskii, Mertens u. Muller, Abh. Senckenberg. naturf. Ges.,
Frankfurt am Main, Abh. 451, S. 23.
1966 — Gymnodactylus kotschyi bureschi, Щербак, Herpetologia taurica, Киев, c. 80.
1981 —Cyrtodactylus kotschyi danilewskii, Beutler, Handb. Amph. Rept. Europ. Bd 1, 1, S. 63.
1982 — Cyrtodactylus kotschyi steindachneri, C. kotschyi lycaonicus, Baran u. Gruber, Spixiana,
Munchen, 5, 2, S. 132.
1983 — Tenuidactylus kotschyi danilewskii, Щербак и Голубев, Вести, зоол.,Киев, № 2, с. 54.
Диагноз. Спинные бугорки овальные, ребристые, расположены в
10—13, чаще 12 продольных рядов, чешуй поперек брюха 22—30; преанальных
пор 2—15, постанальных бугорков по 1 с каждой стороны; все чешуи и щитки
хвоста (не считая спинных бугорков) гладкие; передне-верхние поверхности
конечностей гладкие или с едва заметными следами ребристости.
Лектотип. Хранится в Зоологическом институте АН СССР, Ленинград
ЗИН 3688. «Крым, Ялта, 1868, Данилевский».
Описание (рис. 70) (по 75 экз. из колл. ИЗАНУ из Крыма). L $ ad
(п = 25) 32,9—43,0 (38,68 ± 0,64) мм; L $ ad (п = 30) 31,5—45,0 (39,79 ±
± 0,78) мм; LIL cd (п = 20) 0,86—0,98 (0,91 ± 0,0018); индекс высоты/ширины
головы (и = 29) 52—74 (54,41 ± 0,75); диаметр уха/глаза (п = 32) 23—48
(34,20 ± 1,00); верхнегубных щитков (/? = 146) 5—8 (6,15 ± 0,06); нижнегуб-
ных (п = 148) 7—10 (7,87 ± 0,08); чешуй между центрами глаз (м = 75)
12—19 (14,41 ± 0,17); длина спинного бугорка/L (п = 70) 14—25 (18,67 ±
± 0,26); чешуй, окружающих спинной бугорок (п = 73), 9—15 (10,85 ± 0,14);
чешуй вдоль тела снизу (п = 48) 89—112 (100,75 ± 0,81); чешуй поперек
брюха (п = 54) 22—30 (25,67 ± 0,27); подпальцевых пластин (п = 138) 14—
21 (18,93 ± 0,13), в Малой Азии — до 24 [1811; преанальных пор (п = 27)
2—9 (6,44 ± 0,31) (в Малой Азии и на прилежащих островах 6—15, в
среднем 8) [181]. Подбородочный щиток тре- (73,6 %) или пятиуголь-
163
Рис. 70. Tenuidactylus amictopholis (вверху):
A — общий вид, Б — голова сверху, В — подбородок, Г — брюхо, Д — хвост сверху,
Е — хвост снизу, М — преанальные поры (TAUR 10351 г. Хермон, Ливан)
Tenuidactylus kotschyi danilewskii (внизу):
А — голова сверху, Б — подбородок, В — верхний край орбиты, Г — преанальные поры,
Д — хвост сверху, Е — хвост снизу, Ж — лапа снизу (ИЗАНУ SR 794, Херсонес, Крым)
ный (26,4 %); обычно три пары (96,4 %) нижнечелюстных щитков, очень редко
их бывает две (1,2 %) или четыре (2,4 %) пары; первая пара соприкасается
между собой (96,2 %) или разделена чешуйкой (3,6 %); три носовых щитка;
первые носовые щитки соприкасаются между собой (34,3 %) или разделены
одной (60, 0 %) или двумя (5,7 %) чешуйками; дополнительные спинные бугор-
ки почти всегда отсутствуют, выражены у 6,8 % и соприкасаются с основными
164
бугорками. Подхвостовые щитки хо-
рошо выражены, гладкие, на одном
сегменте образуют сочетание 1—1 или
1—2; остальная чешуя хвоста, кроме
хвостовых бугорков, гладкая; поверх-
ность регенерированного хвоста покры-
та мелкой циклоидной чешуей; на пе-
редне-верхних сторонах передних ко-
нечностей могут быть чешуи со слабыми
ребрышками, несколько больше вы-
раженными на бедрах.
Окраска и р и с у н о к. Ос-
новной фон сверху пепельно- или песо-
чно-серый. На туловище сверху имеет-
ся пять—восемь, чаще шесть темных М-
образных полос, имеющих на середине
Рис. 71. Распространение Т. kotschyi dani
lewski i
спины вид угла, направленного верши-
ной назад; на хвосте 10—12 таких полос, причем у молодых особей они окан-
тованы охристым или светло-оранжевым. Брюшная сторона тела окрашена в
светлый желтовато-зеленоватый или грязновато-белый цвет, низ хвоста крас-
новато-охристый или оранжеватый, более яркий у молодых особей. Регенери-
рованный хвост окрашен в серо-синюшный или серо-коричневый цвет, только
начинающий регенерировать (до 0,5 см) почти черный.
Распространение (рис. 71). Южное побережье Крыма (не под-
нимается выше 200 м н. у. м.) от Севастополя до Алушты, черноморское побере-
жье Болгарии южнее Варны, 3. и Ю.-З. Турция.
Кадастр к рис. 71.
СССР: / — Крым, Севастополь, разв. Херсонеса (ИЗАНУ); 2 — Батилиман; 3 — Си-
меиз; 4 — Ялта; Гурзуф; 5 — Карабах; Алушта; (2—5 [141]).
Болгария: 1 — Русе [317]; 2 — Варна [66]; 3 — Камчия [66]; 4 — Несебыр :[66]*
5 — Бургас [66]; 6 — Созопол; о. Святого Ивана [207]; 7 — окр. Приморско, Мыс Маслен-
Нос [181].
Турция: 1 — Кыркларели [181]; 2 — Энез [181]; 3 — Текирдаг [181]; 4 — Стамбул
[181]; 5 — о. Эзек Адаси [181]; 6 — о-ва Пашалиманы [179]; 7 — о. Имралы [179]; 8 — Прин-
цевы о-ва [179]; 9 — о. Хаирс [179]; 10 — о. Кефкен [179]; 11 — г. Бурса [181]; 12 — г. Балы-
кесир: 50 км ю.-в. г. Бурса [181]; 13 — Анкара [181]; 14 — Саваштепе [181]; 15 —- Эгридир
[181]; 16 — Конья [181]; 17 — Аксеки [181].
Экология — впервые изучена одним из авторов [139, 141].
Стации и количественные данные. Населяет руины
древних построек, современные жилые и хозяйственные каменные и деревян-
ные постройки, каменные кладки, укрепляющие дороги, сложенные из дикого
камня заборы, скалы, изредка встречается под корой старых можжевеловых
деревьев и пней, находящихся вблизи руин и построек (рис. 72). По нашим
наблюдениям, там, где не проводятся какие-либо хозяйственные работы
(реставрация, ремонты — штукатурка стен и их окраска и др.), на протяжении
последних 25 лет численность гекконов стабильна. В местах обитания они рас-
пространены неравномерно: максимальная плотность наблюдается в Херсоне-
се у фонарей на стенах домов — на 1 м2 до трех-четырех особей, а среди антич-
ных руин Херсонесского музея в 1958—1981 гг. одна особь приходилась на
100—1600 м2.
Отношение к температуре. В летнее время гекконы в Крыму
не встречались при температуре воздуха ниже 18°. В апреле—мае они отмечены
при температуре воздуха 14,5°, 13° и даже 12°. У двигающейся ящерицы отме-
чено повышение температуры тела на 1° по сравнению с температурой окружа-
ющей среды. Наибольшее число гекконов наблюдалось при температуре возду-
ха 26°. По исследованию в термоградиентприборе, температурный оптимум дан-
ного вида лежит в пределах 34—40°. Яйца инкубировались при температуре
18—35°.
Суточный цикл активности. Сумеречно-ночной тип актив-
ности, вид активен вечером и в первую половину ночи. Первые особи выходят
165
Нис. 72. Биотоп I. kotscfiyi danilewskii (руины г. Херсонес, Крым, УССР)
из убежищ еще до захода солнца (в затененных местах) в 19 ч 10 мин, массовый
выход около 21 ч, наибольшая численность отмечена к 22 ч. К 24 ч они уже
исчезают, кроме особей, обитающих у уличных фонарей; последние уходят
в убежища на рассвете, когда выключают свет. В ранневесеннее время и позд-
ней осенью греющиеся гекконы наблюдались и в дневные часы.
Сезонный цикл активности. Наиболее ранние сроки появ-
ления после зимовки — 24.IV 1959 г. (Батилиман) и 25.III 1960 г. (Херсонес).
В Херсонесе последний раз ящерицы отмечены 3.XI 1958 г., в 1959 г. в середи*
не октября, а в 1960 г. они исчезли уже в конце сентября.
Линька. Массовая линька не наблюдалась, отдельные линные особи
встречены 27.V 1958 г. и 8.VII 1960 г. В неволе гекконы линяли 24.VII и
15.VIII 1961 г.
Питание. Было проанализировано содержимое 75 желудков [141].
В числе предпочитаемых объектов питания по встречаемости пауки (38,8 %),
бабочки (45,3 %), двукрылые (36,9 %), перепончатокрылые (32,7 %), жуки
(28,0 %). Кроме того, в незначительном количестве обнаружены прямокрылые,
равнокрылые, сетчатокрылые насекомые и равноногие ракообразные.
Размножение. Соотношение полов, по нашим выборкам, близкое
к 1 : 1. Максимальные размеры туловища половозрелого самца 36,5 мм, бере-
менной самки — 41 мм. Спаривание происходит в мае. Прослежены кладки
яиц 1 и 7.VI 1961 г., 17 и 20.VII 1960 г., 25, 26 и 27.VII 1963 г. (Херсонес).
В яйцеводах сформированные яйца найдены 27.V 1958 г., 7 и 8.VII 1960 г.
и 15.V 1961 г. (Херсонес). Откладывается одно-два яйца размером 5,8—8,1 X
X 7,5—9,7 мм. Форма яиц овальная, цвет белый с розоватым оттенком. Клад-
ка одна, но сроки ее растянуты. Сеголетки появляются в июле — августе (толь-
ко что вылупившиеся добыты 2, 4 и 22.VIII 1958 г.). В условиях лаборатории
инкубация яип продолжалась 50 дней, а выход молоди из яиц отмечен 11 и
12.IX 1963 г. Откладываются яйца и появляется из них молодь в ночное время.
Темп роста. Размеры свежевылупившихся гекконов: L = 17,8—
18,5 мм; Lcd= 16—16,1 мм. Мечение их показало, что наиболее интенсивно рас-
тут они в первый год жизни. С момента вылупления за 8,5 месяцев (5 приходи-
лось на зимнюю спячку) длина туловища этих ящериц увеличилась с 18 до
23,5 мм. Через два года геккон достигает размера взрослой ящерицы — 39 мм,
еще через год — 41,8 мм. Максимальных размеров (L $ = 50,5 мм, L d* =
= 43,5 мм) они достигают в пятилетием возрасте.
Враги. По наблюдению в Крыму, на них нападают домашние кошки,
среди возможных врагов этой ящерицы следует назвать домового сыча, обык-
новенного ежа и каменную куницу, обитающих в тех же биотопах.
166
Особенности поведения. Хорошо передвигаются по верти-
кальным и потолочным поверхностям, вертикальное положение для ящериц
данного вида даже в состоянии покоя является обычным. Двигаются перебеж-
ками с короткими остановками. В местах, часто посещаемых человеком, гек-
коны становятся пугливыми и исчезают, услышав шаги. Обычно охотятся, не-
подвижно поджидая добычу, у фонарей преследуют насекомых, которые, ослеп-
ленные светом, ударяются несколько раз о стену. Если одна ящерица хватает
добычу, то остальные возбужденно попискивают, почесывают задней лапой
морду, облизывают языком глаза. Иногда возникают в местах их скоплений
ссоры: у возбужденной ящерицы змеевидно изгибается хвост при неподвижном
туловище, она попискивает, угрожающе бросается на противника и прогоняет
его. Часто им оказывается другой половозрелый самец. При опасности и пресле-
довании гекконы прыгают со стены. Если они падают в траву, то затаиваются,
а затем снова вылезают на стену. Если геккон падает на открытый грунт, то
быстро убегает, пока не доберется до подходящей щели.
Летом дневными убежищами гекконам служат щели, кучи камней, про-
странство под крышами домов, прикрепленные к стенам домов металлические
номерные знаки и др. Обычно они находятся на высоте 2—Зм. Зимовочные убе-
жища — глубокие щели в стенах руин и жилых зданий, их находили даже в жи-
лых квартирах, сараях.
Спаривание не наблюдалось. Возбужденные гекконы издают характерный
звук, который передается как «пик...пик...пик...к...к...к...». В тех местах,
где они обычны, ночью часто можно слышать эти тихие, но четкие звуки. Писк
гекконов из убежищ иногда слышен и днем.
Крымские гекконы существуют сейчас в основном как синантропы. В по-
стройках они находят убежища, приспосабливаясь даже к свету и теплу улич-
ных фонарей, охотясь за насекомыми и согреваясь.
Практическое значение и охрана. Крымский геккон
является интересным видом с точки зрения науки, своеобразным памятником
природы Крыма, редок и его следует всемерно охранять. В настоящее время
охраняется в Херсонесском археологическом заповеднике. Включен в Крас-
ную книгу СССР и «Червону книгу УкрашськоТ РСР». Нами был проведен опыт
по привлечению гекконов в искусственные убежища [141]. Это особенно следует
применять там, где проводятся широкие реставрационные и ремонтные работы.
Для увеличения численности и лучшей охраны крымских гекконов желательно
их поселить в Крыму на территории Карадагского заповедника.
СЕРЫЙ ГЕККОН — TENUIDACTYLUS RUSSOWI (STRAUCH, 1887)
Типовая территория: развалины старого укрепления Ново-Александровское \
Кариотип: 2л =44, метацентриков, 14 акроцентриков 30, NF = 58.
1887 — Gymnodactylus russowi Strauch, Mem. Acad. Sci. Petersbourg, ser. 7, vol. 35, N 2, S. 49.
1899 — Gymnodactylus zarudnyi Никольский, Ежег. Зоол. Муз., т. 4, с. 385.
1954 — Cyrtodactylus russowi, Underwood, Proc. Zool. Soc. London, vol. 124, N 3, p. 475.
1984 — Tenuidactylus russowi, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Спинные бугорки овальные или овально-треугольные,
ребристые; у взрослых диаметр ушного отверстия превышает половину продоль-
ного диаметра глаза; вдоль нижней стороны тела не менее 90 чешуй; чешуя
хвоста (не считая бугорков) и конечностей гладкая.
Лектотип. ЗИН 3658. <? ad «Ново-Александровск, 1842, Леманн».
L = 44,3 мм; L cd = 52,9 мм; L/L cd = 0,84; чешуй между центрами глаз
19; верхнегубных щитков по 9; нижнегубных по 7; чешуй вдоль тела снизу
ПО; чешуй поперек брюха 29; три пары нижнечелюстных щитков, первая со-
прикасается между собой; преанальных пор четыре; подпальцевых пластин
по 15.
1 В литературе [17] утверждалось ошибочное мнение, что типовой территорией является
Форт Шевченко (б. Александровск). В действительности серый геккон описан из развалин ста-
рого укрепления Ново-Александровское, расположенного на 30 км восточнее.
167
Рис. 73. Tenuidactylus russowi:
A — голова сбоку, Б — носовые щитки, В — гълъьа сверху, Г, Д — подбородок, Е, Ж —
спина, 3 _ хвост сверху, И, К — хвост снизу (Г, Е, К — ЗИН 9334, Сейстан, Иран; осталь-
ное — ИЗАНУ SR 971, КиргССР)
Описание (рис. 73) (по 285 экз. из колл. ИЗАНУ, ЗИН, ММГУ)-
L <? ad (п — 104) 25,1—49,6 (39,73 ± 0,54) мм; L % ad (п = 112) 25,5—53,2
(42,0 ± 0,61) мм; масса до 3 г; L/L cd (п = 126) 0,69—1,02 (0,869 ± 0,01);
индекс высоты/ширины головы (л = 100) 53—66 (57,65 ± 0,27); диаметр уха/
глаза (л = 79) 36—81 (56,60 ± 0,50); верхнегубных щитков (п = 568) 8—11
(9,78 ± 0,03); нижнегубных (л = 570) 6—10 (7,89 ± 0,03); чешуй между цен-
трами глаз (л = 200) 15—24 (18,36 ± 0,10); длина спинного бугорка /L (п =
= 100) 14—28 (19,93 ± 0,22); чешуй, окружающих спинной бугорок (л =
= 190), 11—18 (13,92 ± 0,08); чешуй вдоль тела снизу (л = 204) 94—130
168
(113,33 ± 0,47); чешуй поперек брюха (п = 198) 23—34 (28,81 ± 0,16); под-
пальцевых пластин (п = 554) 15—24 (19,67 ± 0,07); преанальных пор (п =
= 119) 2—6 (3,16 ± 0,09).
Подбородочный щиток тре-или пятиугольный, нижнечелюстных щитков од-
на (10 %), две (55,7 %) или три (34,3 %) пары; первая пара соприкасается между
собой (71,7 %) или разделена одним-двумя щитками (28,3 %); первые носовые
щитки разделены одной (22,3 %), двумя (65,0 %) или тремя (12,7 %) чешуйка-
ми; носовых щитков три; дополнительные спинные бугорки имеются (71,0 %)
или отсутствуют (29 %); более или менее выраженные гладкие подхвостовые
щитки образуют на одном сегменте сочетание 1—1 (очень редко 1—2); остальная
хвостовая чешуя (не считая бугорков) и чешуя конечностей гладкая (на отдель-
ных чешуйках предплечья иногда могут быть едва заметны следы ребрышек).
Окраска и рисунок. Сверху пепельно-серого или буровато-серо-
го цвета с темными узкими М-образными или поперечными полосами; верхняя
поверхность головы в мелких темных пятнах, по бокам головы темная продоль-
ная полоса, часто окантованная светлым, переходящая на бока шеи, а иногда
и туловище. Серые гекконы изменяют окраску и рисунок: в оптимальных ус-
ловиях последний у них четкий, контрастный, основной фон — более светлый.
При неблагоприятных условиях их окраска становится темной, рисунок исче-
зает. Замечено также, что популяции, обитающие на стволах саксаулов, при-
обретают очень сложный рисунок (рис. 22, цв. табл. 3) из продольных элемен-
тов, который делает их на фоне древесины почти незаметными. Особи, обитаю-
щие в постройках и на лессовых обрывах, часто вообще теряют рисунок или
имеют только поперечные полосы на туловище и хвосте.
Распространение. Основной ареал расположен восточнее Кас-
пия в Средней Азии и Казахстане, на восток до Сев.-З. Китая, на юге найден
в Сев.-В. и В. Иране. Известна одна находка в В. Предкавказье (ст. Староглад-
ковская, Чечено-Ингушетия) (рис. 74).
Кадастр к р ис. 74.
Казахстан: / — зал. Паскевича близ мыса Турангли (ЗИН); 2 — кишл. Джарбу^
лак [98]; 3—уроч. Кос-Казах [98]; 4—р. Чеган (ЗИН); 5 — п-ов Тюп-Караган [98]; 6 — зал.
Комсомолец [98]; 7 — Устюрт у сора Асматай-Матай [98]; 8 — о. Барсакельмес [98]; 9 — Форт
Шевченко; у Таушика; сев. хр. Каратау [98]; 10 — Ново-Александровск [98]; уроч. Беке
(ЗИН); 11 — между Кунградом и Бейнеу, Устюрт (наши данные); 12 — Кугарал [98]; 13 — пес-
ки Арыскумы [98]; 14 — Кзыл-Орда (ЗИН); 15 — ст. Байгакум [98]; 16 — левый берег р. Чу
[98]; 17 — хр. Каратау [98]; 18 — Чимкент [98]; 19 — Хумсан, Бричмула [23]; 20 — разв.
креп. Чирик-Рабат (ИЗАНУ); 21 — ю.-з. берег Балхаша, окр. Буру-Байтала (ЗИН); 22 —
Ю.|Прибалхашье, между реками Или и Каратал [137]; 23 — Междуречье Или и Каратал [98];
24 — пос. Баканас (ИЗАНУ); 25 — предгорья Джунгарского Алатау [98]; 26 — пос. Панфилов
[98]; 27 — оз. Алаколь (ИЗАНУ); 28 — Джунгарские ворота, окр. оз. Джаланаш-куль [98];
29 — междуречье Чарын и Чилик [98]; 30 — госграница, долина р. Или [98]; 31 — с. Тасарал
[17]; 32 — Желтау [83, 328]; 33 — Кой-Кара, Иман-Кара, Сарынияз, Ак-Толагай [328].
Узбекистан: / — чинки Устюрта у оз. Судочье (ИЗАНУ); 2 — Чабан казган (ИЗА-
НУ); 3 — Ургенч [23]; 4 — Хазарасп [23]; 5 — р. Ангрен, Той-Тюбе (ЗИН); 6 — Ташкент
(ЗИН); 7 — хр. Чаткальский [17]; 8 — Чиназ (ЗИН); 9 — Голодная Степь, окр. ст. Урсатьев-
ская (ЗИН); 10 — кишл. Джуленгар (ЗИН); 11 — Хавает [106]; 12 — Заамин (МХГУ); 13 —
пос. Кассансай [33]; 14 — Балыкчи (ЗИН); 15 — Учкурган близ Нарына (ЗИН); 16 — ст. Бу-
вайда [33]; 17 — Андижан (ЗИН); 18 — Коканд (ИЗАНУ); 19 — долина Кувеч (ММГУ); 20 —
Фергана (ММГУ); 21—23 — Ю. Кызылкум [17].
Туркмения1: /— разв. Деу-Кескен-Кала (ИЗАНУ); 2—Куня-Ургенч (ИЗАНУ); 3 —
Сарыкамыш (ИЗАНУ); 4 — 10 км з. Буджуную-Даудака [130]; 5 — кол. Чарышми [130]; 6 —
100 км сев.-з. пос. Дарваза (ЗИН); 7 — Унгуз, 2,5 км сев. Серного завода (ИЗАНУ); 8 — п-ов
Дарджа (ЗИН); 9 — ст. Джебел; ст. Молла-Кара [130]; 10 — оз. Ясха [23]; //— Тогалак
[23]; 12 — верховья Узбоя, кол. Чарышлы [23]; 13 — 180 км сев. пос. Бахарден (ИЗАНУ);
14 — кол. Ата-Кую, 40 км з. пос. Дарваза (наши данные); 15 — 20 км в. пос. Дарваза, кол.
Джарлы (ИЗАНУ); 16 — 20 км в. Серного завода, колхоз Окуз (ИЗАНУ); 17 — Киркили
[130]; 18 — Дояхатын (ИЗАНУ); 19 — Култакыр [130]; 20 — ст. Ахча-Куйма [130]; 21 — уроч.
Кара-Богаз, 40 км сев. г. Кизыл-Арвата (ЗИН); 22 — ю.-в. Малого Балхана, родник Чал-Су
[130]; 23 — сев. г. Кизыл-Арвата (ИЗАНУ); 24 — окр. Данаты (ИЗАНУ); 25 — подножье Кю-
рендага [130]; 26 — ст. Искандер [8]; 27 — Кизыл-Арват [130]; 28 — 50—100 км сев. пос. Ба-
харден (ЗИН); 29 — 100 км сев.-з. Репетека, кол. Ер-Аджи (ИЗАНУ); 30 — 30 км ю. пос. Ка-
ра-Кала [130]; 31 — ущ. Айдере, пос. Сапарбакар [130]; 32 — 37 км з. Репетека (ИЗАНУ);
33 — Репетек (ИЗАНУ); 34 — Байрам-Али [130]; 35 — Карабата [130]; 36 — 20 км з. пос. Нич-
1 Указания на находки этого вида в Айдере [23, 24, 109] ошибочны и относятся к колюче-
хвостому геккону.
169
Рис. 74, Распространение Tenuidactylus russowi (/) и Т. spinicauda (2)
ка [130]; 95 км ю.-в. ст. Уч-Аджи (ИЗАНУ); 37 — Керки [23]; 38 — ст. Дорткую [130]; 39 —
Иолотань (ИЗАНУ); 40 — Келиф [128]; 41 — Ташкепри (ИЗАНУ).
Киргизия: 1 — Орто-Токойское водохр. [22]; 2 — сев. р. Нарын по дороге на То-
гуз-Тороу [162]; 3 — пос. Сарыг-Булун, не доезжая 15 км до Кавака [162]; 4 — среднее течение
р. Нарын, от Ала-Буги до г. Нарын [162]; 5 — Джалал-Абад (ИЗАНУ); 6 — г. Ош (ЗИН);
7 — Ирка-Кашка, между Араваном и Андижаном (ЗИН); 8 — пос. Гульча (ИЗАНУ);
9 — Фрунзе (ИЗАНУ).
Таджикистан: 1 — ущ. Оби-Ашт (Бобоиоб) [106]; 2 — Аштинский р-н, обрывы
старого русла р. Сырдарья [106]; 3 — Могол-Тау; Мурза-Рабат (ЗИН); 4 — ю. склон Курамин-
ского хребта [106]; 5 — Кырккудук [106]; 6 — пос. Курганча (ИЗАНУ); 7 — Кок-Курак
[106]; 8 — окр. Кайраккума (ИЗАНУ); 9 — Канибадам (ИЗАНУ); 10— окр. г. Ура-Тюбе (ЗИН);
11 — Каттаксай у кишл. Джойлангар [106]; 12 — Яван [106]; 13 — заповедник «Тигровая Бал-
ка» (ЗИН).
Кавказ, Чечено-Ингушетия (вставка «я»): станица Старогладковская (ЗИН).
Иран (вставка «б»): 1 — стр. Бехарс (ЗИН); 2 — Сейстанская котловина (ЗИН).
Половой диморфизм и возрастная изменчивость.
Самки крупнее самцов (Ad" = 49 мм; L g = 53,2 мм), у них отсутствуют
анальные поры.
Молодые особи отличаются от взрослых относительно более крупной го-
ловой, тонким туловищем и коротким хвостом (L/L cd juv=0,8— 1,0) (рис. 75).
Систематика и географическая изменчивость.
Исследование девяти выборок (п = 285) по 14 признакам показало, что значе-
ния некоторых из них увеличиваются от центра ареала к периферии (L/L cd).
Возрастают (контакт первого нижнечелюстного и второго нижнегубного щит-
ков) или убывают (количество подпальцевых пластинок; чешуй поперек брю-
ха и вдоль тела снизу) к северу и югу от линии Небит-Даг— Фергана — Джа-
лал-Абад. С юга на север возрастают количество преанальных пор, частота
встречаемости дополнительных спинных бугорков и максимальные размеры
тела. Признаки отношение диаметра глаза к длине морды и головы, длины мор-
ды к длине четвертого пальца задней ноги, а также длины задних конечностей
к длине тела, которые были использованы для описания подвида из Семире-
чья [1371, не свойственны какой-либо одной популяции, что не позволяет счи-
тать форму G. г. copalensis самостоятельной. Напротив, гекконы из Ирана
(Систан), описанные как отдельный вид [89], но впоследствии сведенные в си-
ноним [125], характеризуются некоторыми оригинальными чертами, на основа-
нии чего эта форма восстановлена в ранге подвида [152, 282].
Таблица для определения подвидов Т. russowi
1 (2). Чешуй в продольном ряду на брюшной стороне туловища (от нижнечелюстных до аналь-
ной щели) 100—130 (114,99 ± 0.44) ................Т. russowi russowi (Str.)
2 (1). Чешуй в продольном ряду на брюшной стороне туловища (от нижнечелюстных до аналь-
ной щели) 94—105 (99,2 ± 0,99) ................Т. russowi zarudnyi (Nik.)
СЕРЫЙ ГЕККОН ОБЫКНОВЕННЫЙ —
TENUIDACTYLUS RUSSOWI RUSSOWI (STRAUCH, 1887)
Типовая территория; видовая.
1887 — Gymnodactylus russowi Strauch, Mem. Acad. Sci. St. Petersbourg, ser. 7, vol. 35, N 2,
S. 49.
1928 — Gymnodactylus russowicopalensis, Шнитников, Тр. о-ва изуч. Казахстана, Кзыл-Орда,
т. 8, в. 3, с. 24.
Диагноз. Первая пара нижнечелюстных щитков позади подбородоч-
ного обычно (72 %) в контакте между собой. Чешуй вдоль брюха от нижнече-
люстных до анальной щели 100—130 (114,99 ± 0,44); чешуй поперек середины
брюха 25—34 (29,09 ± 0,14).
Тип — видовой.
Описание. Остальные признаки соответствуют видовым.
Распространение. От Сев.-В. Предкавказья по всей Средней
Азии и Казахстану до Джунгарии и Сев. Ирана.
171
СЕРЫЙ ГЕККОН ЗАРУДНОГО — TENUIDACTYLUS RUSSOWI ZARUDNYI (NIKOLSKY, 1900)
Типовая территория: Нейзар в Сейстане, В. Иран.
1900 — Gymnodactylus zarudnyi Никольский, Ежег. зоол. муз., т. 4, с. 385.
1981 — Gymnodactylus russowi zarudnyi, Scerbak, Handbuch Rept. Amph. Europ. Bd 1, 1, S. 79.
Диагноз. Первая пара нижнечелюстных щитков позади подбородоч-
ного всегда разделена одной-двумя чешуйками; чешуй вдоль брюха от нижне-
челюстных до анальной щели 94—105 (99,2 ± 0,99; t = 13,93); чешуй поперек
середины брюха 23—26 (24,3 ± 0,35; t = 12,71). Дополнительные спинные бу-
горки всегда отсутствуют.
Лектотип. ЗИН 9334 a, d ad «Нейзар в Сейстане, 21—24.V 1899,
Н. Зарудный». L = 31,4 мм; L cd = 38,9 мм; L/L cd = 0,81; чешуй между цент-
рами глаз 17; верхнегубных щитков по 10; нижнегубных 9/8; чешуй вдоль
тела снизу 102; чешуй поперек брюха около 25; чешуй, окружающих спинной
бугорок, 11; нижнечелюстных щитков две пары, первая разделена между собой
чешуйкой; преанальных пор две; подпальцевых пластин 18/17; подхвостовых
щитков на базальном сегменте нет, далее — по два в один ряд (1—1).
Описание. Остальные признаки соответствуют видовым.
Распространение. Обитает в Сейстанской котловине (Ю.-В. Иран).
Стации и количественные данные. Северная граница аре-
ала G. russowi не выходит за пределы северной границы пустынной зоны.
В Предкавказье также его находили на участке песчаной пустыни. Хотя этот
геккон и обитает на вертикальных поверхностях, далеко в горы не идет. Осо-
бенно это касается западных районов ареала. Так, в Туркмении данный вид
избегает даже предгорий Копетдага, хотя в приферганских районах Киргизии
встречается на высоте 900—1200 м и по Нарынской котловине поднимается до
2000 м н. у. м. Населяя песчаные пустыни и полупустыни (Туркмения, Ю. Тад-
жикистан, Прибалхашье), G. russowi обитает на стволах саксаула и других
деревьев. В глинистых пустынях и лессовых предгорьях (Узбекистан,
Ю. Казахстан, Сев. Таджикистан) он обычен на обрывах в долинах рек и овра-
гов, под камнями и (особенно часто) на стенах жилых и заброшенных домов
и других построек из глины и камня. Некоторые ящерицы приспособились
к обитанию у ламп уличного света и питаются насекомыми, летящими на свет.
В Туркмении, Ц. Каракумах, окр. кол. Кирпили на каждом четвертом —
шестом стволе саксаула встречено по одному геккону [130]. На западе указан-
ной пустыни, в окр. с. Моллакара 7.V 1983 г. отмечено на каждом старом сак-
сауле высотой 5—8 м по одной-две особи. В заповеднике «Тигровая Балка»
(Ю. Таджикистан) в роще старых саксаулов найдено 12.IV 1980 г. на каждом
дереве по одному — три геккона. В Узбекистане и Сев. Таджикистане в посел-
ках на лессовых дувалах (заборах) наблюдается 10—15 особей на 10 погонных
метров [22, 125]. Значительно реже здесь (единичные экземпляры) встречается
серый геккон по лессовым и конгломератовым обрывам, дуплам деревьев.
21.VIII 1972 г. мы обнаружили на окраине г. Джалал-Абада (Киргизия) на
дувалах общей длиной 1 км 16 гекконов. 14.IV 1980 г. в сарае из досок в «Тиг-
ровой Балке» (Ю. Таджикистан) с площадью стен 64 м2 наблюдалось 15 гек-
конов.
На северной границе ареала в Приэмбье (письм. сообщ. В. В. Неручева1)
серые гекконы не выходят за нулевую горизонталь — границу низменности —
и обитают на останцах с крутыми склонами и выходами скал — песчаных кон-
креций; здесь их находили до восьми особей за вечернюю экскурсию.
На крайнем востоке ареала, в Ц. Тянь-Шане, 2.VI 1984 г. мы на адырах
в окр. с. Байгончек наблюдали на 5 км маршрута два серых геккона; 3.VI 1984 г.
в 15 км выше устья р. Алабуга, на верхней террасе правого берега, на скаль-
ных обрывах и среди глыб — обломков скал — на 500 м маршрута отмечено
10 гекконов. В окр. с. Конорчок на щебнистых и каменистых склонах холмов
1 На г. Жельтау, как установил В. В. Неручев, обитает серый геккон, а не каспийский,
как это утверждал В. А. Киреев [65, с. 64],
172
гекконы занимают низ склонов, а геккончики Токобаева обитают здесь у вер-
шин холмов.
Отношение к температуре. Известно, что в Узбекистане
серые гекконы наблюдались весной весьма активными при температуре воздуха
17°, поверхности обрывов 37°, охотящуюся ящерицу летом — при температу-
ре воздуха 37°, осенью — на поверхности обрыва при температуре воздуха
10°, почвы — 24° [22]. По нашим наблюдениям, голоса активных гекконов
13.IV 1977 г. в зарослях саксаула под Ташкепри (Туркмения) были слышны
при ветреной погоде и температуре воздуха 20°, а в 25 км восточнее Дарваза
при температуре воздуха 17°, почвы 21,5° геккон был добыт на скальном ос-
танце 30.IV 1979 г. При температуре субстрата ниже 17° эти ящерицы не появ-
лялись из убежищ. Активных гекконов в долине р. Алабуга (Ц. Тянь-Шань)
мы наблюдали в июне 1984 г. в сумерках при температуре воздуха 18°.
При исследовании серых гекконов в термоградиентприборе они занимали
места с температурой субстрата 19—41°, предпочитаемая температура — 33,3 ±
± 0,83° (п = 62).
Суточный цикл активности. Ранней весной гекконы встре-
чаются днем, они греются у убежищ и охотятся. 27.11 1955 г. их активность от-
мечена с 14 до 18 ч, пик активности — с 16 до 17 ч [22]. Летом эти ящерицы мо-
гут греться на солнце только в утренние часы, а охотятся в сумерки и ночью,
часто до рассвета. В апреле, по наблюдениям в Туркмении, серые гекконы, как
правило, были активны после 21 ч. 12.IV 1980 г. в Ю. Таджикистане активность
этих ящериц, обитающих на деревьях, была хорошо выражена днем, а с на-
ступлением сумерек, после 21 ч, прекращалась. В прохладные дни сентября—
октября серые гекконы снова чаще встречаются днем.
Сезонный цикл активности. Наиболее ранняя находка се-
рого геккона в Туркмении, по нашим сборам,— 27.III 1977 г. Выход их после
зимовки в Узбекистане, долине Ангрена, отмечен 16.11 1955 г., а 27.11 1955 г.
там же наблюдался их массовый выход [22]. В окр. ст. Урсатьевской они отме-
чены несколько позже — 15—20.III [54]. В Таджикистане после зимовки гек-
коны появляются в марте [107], на юге Казахстана — в начале марта, Прибал-
хашье — начале апреля [98].
На зимовку в Туркмении они уходят, видимо, в начале ноября, так как
еще 15.Х 1981 г., по нашим наблюдениям под Репетеком, эти ящерицы были
вполне активны и обычны. В Узбекистане они исчезают к середине ноября
[22]. В 1950 г. в Ю. Прибалхашье уход серых гекконов на зимовку отмечен
в начале сентября [96].
Линька. Массовая линька в Узбекистане отмечена в конце апреля —
мае [54], в Киргизии находили линных особей в конце июля [162].
Питание. В составе кормов серого геккона обнаружены представите-
ли 34 семейств насекомых и 2 семейств других членистоногих. В Киргизии (ис-
следовано 39 желудков гекконов, добытых в июле) он поедает преимуществен-
но прямокрылых (84,2 % встречаемости), клопов (36,8 %), жуков (15,7 %),
а также перепончатокрылых и бабочек (по 5,3 %) [162]. В Узбекистане, в окр.
ст. Урсатьевской (вскрыто 24 желудка гекконов, добытых в сентябре) его пищу
составляют жуки (50 %), клопы (29 %), перепончатокрылые, в том числе му-
равьи (25 %), бабочки (25 %), и пауки (16 %) [22]. В Ташкентской обл., в до-
лине Ангрена, исследовали содержимое 35 желудков гекконов, добытых
2.VIII 1955 г., и 36 желудков, собранных 25.VIII 1955 г. [28]. В первом случае
по встречаемости преобладали жуки (46,8 %), среди которых чаще были най-
дены представители жуков быстрянок (28,13 %) и чернотелки (15,6 %), а за-
тем перепончатокрылые (25,2 %, в том числе муравьи составляли 12,5 %) и
пауки (15,6 %). Во втором случае первое место по встречаемости занимали пе-
репончатокрылые (40,6 %, в том числе муравьи 28,1 %), затем следовали кло-
пы (18,75 %), жуки (15,6 %) и двукрылые (15,6 %). В Туркмении вскрыли
32 желудка гекконов, добытых в разное время [130]. Здесь первое место по
встречаемости занимали перепончатокрылые (46,8 %), в том числе муравьи
(43,7 %), бабочки и их гусеницы (34,4 %), и жуки (15,6 %). Сравнивая мате-
риалы по питанию серого геккона, собранные в разное время, можно заключить,
173
что с приближением осени интенсивность питания геккона снижается (увели-
чивается число особей с пустыми желудками), несколько изменяется состав
кормов. Так, уменьшается число жуков и прямокрылых, увеличивается коли-
чество перепончатокрылых (особенно муравьев) клопов, мух.
Размножение. Соотношение полов в наших выборках равно 1:1,
в Нарынской котловине (Киргизия) самцов и самок 1 : 1,9 [162]. Половозрелы-
ми серые гекконы становятся при достижении размеров туловища самцов
37 мм, самок — 40 мм и более. Брачные игры и спаривание мы наблюдали
12.IV 1980 г. в окр. пос. Нижний Пяндж (Ю. Таджикистан). По ряду данных,
между выходом из зимовки и размноже-
нием у серого геккона проходит некото-
рое время. Так, в Узбекистане [22]
спаривание происходит с середины мая
до конца июня, т. е. спустя два месяца,
а откладка яиц — с середины июня до
конца июля. Сеголетки появляются здесь
с конца июля — начала августа по ко-
нец октября. На западе ареала (Ман-
гышлак) самки откладывают яйца с на-
чала июня до середины июля — около 45
дней, а на востоке (Прибалхашье) рань-
ше — с конца мая; сеголетки здесь начи-
нают встречаться с начала августа [98].
Яйца эллиптические, белые. Их
размеры: из Узбекистана — 7,5—9 X
х 9—11 мм [22], Казахстана — 8 X
х 10—11,5 мм [98] и Киргизии — 8—9 X
X 10—12 мм [162]. Размеры кладок,
/О 20 JO 40 50 L'MM
Рис. 75. Возрастная изменчивость индекса
ыLed у Т. russowi
собранных нами, следующие: г. Джалал-
Абад в августе 1972 г.— 7,4—7,9 X 10,0—10,8 мм (п = 7); из Таджикистана
(с. Курганча) 26.VI 1979 г.— 7,5 X 10,0—10,5 мм (и = 2); из Ц. Тянь-Шаня
(Киргизия, с. Конорчок) 29.VI 1984 г.—7,7—7,8 X 11,2—11,8 мм (п = 2).
Как следует из приведенных данных, самые крупные размеры яиц у гекконов
из Ц. Тянь-Шаня. Яиц в кладке одно-два, возможны две кладки за сезон. Не-
сколько самок часто откладывают яйца в одно место, что позволило некоторым
авторам [54, 107] допустить большее количество их в кладке. Опровергается
это исследованиями О. П. Богданова [22] и нашими материалами. Инкубация
яиц продолжается 50—54 дня [28], 45—55 дней [17]. Первый сеголеток был
добыт нами в г. Джалал-Абаде 21.VIII 1972 г. Из яиц, собранных здесь, в лабо-
ратории вывелся детеныш 21 IX 1972 г.
Темп роста. Размеры сеголетка, который вывелся в нашей лабора-
тории из кладки, найденной в г. Джалал-Абаде: А = 16,6 мм, L cd = 16,9 мм.
Перезимовавшие молодые гекконы, добытые в марте — апреле в Таджикистане,
имели размеры 20,1—22,2 мм [107], в Казахстане — 21—25 мм [98], в июле
в Ц. Тянь-Шане — 27—34 мм [1621. Представленные данные свидетельствуют,
что половая зрелость у этих ящериц наступает в возрасте года, но приступают
они к размножению в возрасте 20 месяцев, после второй зимовки. Максималь-
ных размеров серые гекконы достигают к четырем годам (рис. 75).
Враги. По нашим наблюдениям, на гекконов данного вида нападают
поперечнополосатый полоз (Coluber karelini) и стрелка (Psammophis lineola-
tum). Известна находка геккона в желудке волкозуба (Lycodon striatus) [54].
Серые гекконы найдены в желудках чайконосой крачки (встречаемость 2,2 %)
[155]. Возможны в числе врагов и некоторые другие виды птиц (сорокопуты,
домовые сычи), змеи,~а также беспозвоночные (яйца иногда поедают муравьи,
на гекконов нападают сольпуги). Значительно больше на популяции гекконов
влияют другие факторы. Численность данного вида (по многолетним наблюде-
ниям в Средней Азии) более или менее стабильна. Возможна гибель в зимовоч-
ных убежищах в особенно суровые зимы [22]. Резко снижается численность
ящериц после ремонта глинобитных построек (особенно заборов), часто они ис-
174
чезают совсем. По свидетельству того же автора [241, освоение целинных и
залежных земель приводит к резкому сокращению числа пресмыкающихся: уже
через год на освоенных землях серый геккон отсутствует.
Особенности поведения. Серый геккон — наиболее быстрый
и ловкий. Он превосходно лазит по стволам и веткам деревьев, потревоженный
бежит по стволу винтом, может быстро бежать и вниз головой по вертикальной
поверхности, но при опасности — немедленно прыгает. Он бегает и по потолоч-
ным поверхностям камней и веток. О. П. Богданов [22] наблюдал, как серый
геккон во время охоты прыгнул на расстояние 1 м от обрыва вниз и поймал ли-
чинку саранчи.
Убежищами гекконам служат трещины стволов саксаулов, лессовых обры-
вов и построек, пустоты в нагромождениях камней и между кирпичами зда-
ний, реже ходы грызунов, пустующих муравейников и термитников, полости
у сгнивших стволов кустарников. В песчаной пустыне под Казанджиком в ап-
реле 1977 г. мы выкопали из норы грызуна в бархане, на глубине 40 см, яще-
рицу. Здесь же была найдена и скорлупа ее яиц. В норах в песчаной пустыне,
трещинах лессовых обрывов и остатках лессовых построек эти гекконы зимуют.
Оцепеневшие ящерицы были найдены на глубине 8—25 см от поверхности [22].
В одном укрытии живут три-четыре неполовозрелых особи, половозрелые яще-
рицы встречаются одиночно [98], от своего убежища далеко не уходят. По на-
блюдениям в Узбекистане [22], гекконы летом с наступлением сумерек спуска-
ются с обрывов, в кишлаках — с заборов на дороги, где бегают в поисках пи-
щи. В ущельях они иногда уходят от своих убежищ на 100—200 м, перебегая
от одного камня к другому и вылавливая под ними спрятавшихся насекомых.
К утру гекконы вновь поднимаются к своим норкам в обрывах.
В брачный период самцы ожесточенно дерутся, часто, сцепившись челюс-
тями, сваливаются со стены. Возбужденные ящерицы извивают хвост и издают
громкие звуки. Звуки гекконов можно передать как быстро следующее друг
за другом «эк...эк...эк». В пустыне Каракумы весной эти голоса четко слыш-
ны со многих деревьев саксаула. Схваченные гекконы также издают слабый
писк. При дневном рассеянном свете видит на расстоянии до 2 м [282].
Практическое значение и охрана. Обычный многочис-
ленный вид. Полезен. В природоохранных мероприятиях не нуждается. Боль-
шие популяции охраняются в заповедниках Репетекском (Туркмения) и «Тиг-
ровая Балка» (Таджикистан).
КОЛЮЧЕХВОСТЫЙ ГЕККОН — TENUIDACTYLUS SPINICAUDA (STRAUCH, 1887)
Типовая территория: Шахруд, Иран.
Кариотип: 2п = 42, метацентриков 2, акроцентриков 38, субтелоцентриков 2, NF = 46.
1887 — Alsophylax spinicauda Strauch, Mem. Acad. Sci. St. Petersbourg, ser. 7, Vol. 35, N 2,
S. 58.
1977 — Gymnodactylus spinicauda, Щербак, Голубев, Tp. зоол. Ин-та, т. 74, Ленинград, с. 130.
1984 — Tenuidactylus spinicauda, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Спинные бугорки округлые, гладкие; диаметр ушного от-
верстия меньше половины продольного диаметра глаза; вдоль нижней стороны
тела не менее 90 чешуй; чешуя хвоста (не считая бугорков) и конечностей
гладкая.
Голотип. ЗИН 4047, 2 ad «Шахруд, 1875, Христоф». L = 37,2 мм;
Led = 30,5 мм; LILcd = 1,21; чешуй между центрами глаз 17; верхнегубных
щитков 8/9; нижнегубных по 7; чешуй вдоль тела снизу 107; чешуй поперек
брюха 27; одна пара нижнечелюстных щитков разделена между собой чешуйкой;
преанальные поры отсутствуют; подпальцевых пластин 14/15. Носовых щит-
ков слева — один, справа — два.
Описание (рис. 76) (по 41 экз. из колл. ИЗАНУ, ЗИН, ИЗАНТуркм,
МКГУ, МХГУ).
L<? ad (п = 10) 26,2—43,1 (34,22 ± 1,95) мм; L g ad (п = 19) 28,7—
48,2 (39,46 ± 0,95) мм; масса до 2,5 г; L/L cd (п = 16) 1,02—1,21 (1,09 ±
± 0,01); индекс высоты/ширины головы (/г = 31) 51—68 (59,87 ± 0,86); диа-
метр уха/глаза (п = 31) 14—33 (25,16 ± 0,71); верхнегубных щитков (п = 80)
175
Рис. 76. Tenuidactylus spinicauda:
A — голова сбоку, Б — голова сверху, В, Г — носовые щитки, Д — подбородок, Е — спина,
Ж — преанальные поры, 3 — лапа снизу, И — хвост сверху, К — хвост снизу (Г — ЗИН
4047 Шахруд, Иран; остальное — ИЗАНУ SR 364, Даната, ТССР)
8—11 (9,50 ± 0,09); нижнегубных (л = 80) 6—9 (7,81 ± 0,08); чешуй между
центрами глаз (п = 41) 16—21 (17,90 ± 0,20); длина спинного бугорка/L
(л = 41) 10—16 (13,27 ± 0,25); чешуй, окружающих спинной бугорок (л =
= 41), 8—10 (8,80 ± 0,09); чешуй вдоль тела снизу (л = 32) 103—127
(113,41 ± 1,34); чешуй поперек брюха (и = 23) 26—32 (28,22 ± 0,33); под-
пальцевых пластин (л = 80) 13—18 (15,45 ± 0,14); преанальных пор
(л = 12) 2—3 (2,17 ± 0,11).
176
Подбородочный щиток пяти- (2,4 %), шестиугольный (65,9 %) или трапе-
циевидный с изогнутым задним краем (31,7 %); дополнительных спинных
бугорков нет (100 %); иногда (около 5 %) могут быть слабо выраженные под-
хвостовые щитки (1—2, 1—1); остальная чешуя хвоста и конечностей гладкая.
Первые носовые щитки обычно разделены одной (66,4 %) или двумя (21,6 %)
чешуйками, редко их бывает три (4,8 %) или не бывает вовсе (7,2 %). Носовых
щитков обычна по три с каждой стороны; нижнечелюстных: 0 (1,7 %), один
(49,0 %), два (34,0 %), три (15,3 %). В отличие от других видов подрода, на
шиповатых хвостовых бугорках очень слабо выражена продольная ребристость.
Регенерированный хвост в средней части заметно утолщен.
Окраска и рисунок. Основной фон спинной поверхности пепель-
но-серый, редко с охристым или буроватым оттенком. На боках головы от ноз-
дри через глаз до уха тянется бурая полоска, пилеус и верхнегубные испещре-
ны мелкими бурыми точками. На спине от шеи до основания хвоста есть семь-
восемь волнистых, часто очерченных черным, бурых поперечных полосок. По-
перечные полоски и пятна имеются на хвосте и конечностях. Низ серовато-
белый. У молодых особей хвост сверху и снизу (как и основной фон тела) розо-
вато-буроватый, но светлее (охристее), чем туловище.
Распространение (рис. 74). В СССР встречается в горах Копет-
даг и Бадхызе (Туркмения), в Иране — в типовом местонахождении и, вероят-
но, во всем массиве Туркмено-Хорасанских гор.
Кадастр к рис. 7 4.
СССР, Туркмения: 1 — пос. Даната; Казанджик; кол. Эйшем; Обой (ИЗАНУ);
2 — уроч. Монджуклы (ИЗАНУ); 3 — 20 км ю. пос. Кара-Кала (ИЗАНУ): Кара-Кала, хр.
Монджуклы [107]; 4 — Копетдаг, Тузлы-Тепе (ИЗАНУ); Айдере [107]; 5 — уроч. Каранки
(ИЗАНУ); 6 — пер. Гаудан (ИЗАНУ); 7 — окр. пос. Калининский (ИЗАНУ); родник Мей-
мили [13]; 8— 3. Бадхыз [13]; 9 — уроч. Чаача (ИЗАНУ).
Иран: 1 — Шахруд (ЗИН).
Половой диморфизм и возрастная изменчи-
вость. Молодые особи более короткохвостые. Самки лишены преаналь-
ных пор.
Систематика и географическая изменчивость.
Первоначально был отнесен к роду Alsophylax, однако наличие у него искрив-
ленных пальцев и ряда других признаков позволило нам перевести этот вид
в группу тонкопалых гекконов [150], правильность чего впоследствии была
подтверждена кариологическим анализом [73]. Отличие от голотипа по коли-
честву носовых щитков от остальных 40 исследованных экземпляров связано,
видимо, с индивидуальной изменчивостью.
Стации и количественные данные. Обитает в предгорьях
на глинистых холмах с редкой сухолюбивой растительностью (полынь, типчак)
в окр. Казанджика, ст. Искандер или на склонах холмов, покрытых шиферной
крошкой с камнями, редкими кустиками эфедры, полыни, злаков, губоцветных
(проективное покрытие 10—100 %) или без них. Экспозиция склонов чаще се-
верная, реже восточная или западная, наклон до 45°, хотя встречаются эти
ящерицы также и в каменисто-щебнистых долинах (окр. с. Даната, Айдере,
Монжуклы), а также на скалистых склонах с выходами плит известняка или
сланцев (верховья р. Чулинка, ущ. Большие Каранки в Ц. Копетдаге, уроч.
Чаача в В. Копетдаге). Высота биотопов колючехвостого геккона 500—
1800 м н. у. м. Встречается единичными особями. Численность этих гекконов не
подвержена колебаниям из-за неблагоприятных условий года: с 1963 по 1982 г.
они отмечались в окр. пос. Даната стабильно, хотя многие обычные фоновые
виды резко сокращали численность после суровых зим. В соответствующих
местообитаниях площадью 300 X 200 м (южнее ст. Искандер, 4.V 1983 г.)
было добыто 3 экз., т. е. 1 особь на 2 га; в отдельных случаях они встречались
чаще: в окр. пос. Даната 24.IV 1982 г. на площади 150 м2 было найдено две яще-
рицы, а в 6.IV 1982 г. ю. Кара-Калы на участке 2000 м2 обнаружен один гек-
кон. Обычно, чтобы добыть колючехвостого геккона, одному сборщику при-
ходилось переворачивать камни от 5 до 10 ч.
177
Рис. 77. Новорожденный Т. spinicauda
Отношение к температуре. Активных ящериц добывали при
температуре воздуха 22—24°. Под камнем 20.IV 1977 г. найден геккон, актив-
ный при температуре воздуха 8°; потревоженная ящерица пыталась уйти. Яй-
ца инкубировались при температуре 20—27°.
Суточный цикл активности. Ведет сумеречный и ночной
образ жизни, первую активную ящерицу мы наблюдали 27.IV 1975 г. около
21 ч. В дневное время встречаются только в убежищах.
Сезонный цикл активности. Весной первые экземпляры
встречаются в середине марта [17]. В наших сборах самая ранняя находка
датируется 15.IV 1963 г. (окр. пос. Даната). Последние особи были добыты
там же 29.IX 1981 г., а 30.IX 1981 г. в сумерках был слышен их писк.
Линька. Линька взрослых особей не наблюдалась. Сеголетки, вышед-
шие из яиц, линяли спустя шесть дней после появления на свет, 15—16. VIII
1983 г. и во время выхода из яиц (рис. 77).
Питание. Из десяти желудков ящериц, добытых весной, остатки пи-
щи (цикада) содержались в одном. По литературным данным [171, колюче-
хвостые гекконы поедают мелких членистоногих, преимущественно пауков и
клопов. В неволе эти ящерицы поедали тлей, мелких личинок мучного хруща
и тараканов.
Размножение. Соотношение полов в нашей выборке: самок почти
в два раза больше самцов (на 10 d приходится 19 $>). Становятся половозре-
лыми эти ящерицы при достижении размеров туловища 26—28 мм. Самки с яй-
цами размером 5 X 7мм добыты в 3. Копетдаге 1 и 19.IV. Мы добывали бере-
менных самок 27.IV 1975 г., 20.IV 1977 г. Кладки (одно, чаще два яйца) полу-
чены нами от гекконов 10.V 1983 г. (30 км ю. ст. Искандер), 14.V 1975 г. (из
пос. Данаты), 30.V 1983 г. (ущ. Большие Каранки, Ц. Копетдаг), 10.VI 1983 г.
(добыты там же). Размеры яиц (п — 8) 6,7—8,0 X 8,2—10 мм, в среднем
7,1 X 9 мм. Яйца овальные, после откладки белые, спустя два дня становятся
розовыми (если развиваются), за месяц до окончания развития они бледнеют,
затем появляются внутри темные пятна. Инкубация продолжается, по нашим
наблюдениям, 71—80 дней. Яйца из одной кладки, инкубирующиеся в одина-
ковых условиях, могут развиваться с разницей в сутки. Первые сеголетки поя-
вились у нас 8.VIII 1983 г.; в природе были добыты 29.IX 1981 г.
178
Темп роста. Гекконы, появившиеся на свет, достигают размеров ту-
ловища 17,5—18,0 мм, хвоста — 14 мм. Половой зрелости, видимо, достигают
после второй зимовки.
Враги не известны. Однажды под камнем вместе с колючехвостым гек-
коном был обнаружен волкозуб (Lycodon striatus), который при случае может
нападать и на этих ящериц.
Особенности поведения. Гекконы данного вида, внешне на-
поминают серых, однако менее ловки и быстры, чем последние. Они не прыга-
ют, медлительны, и, хотя обитают преимущественно на горизонтальных скло-
нах, по наблюдениям в неволе, способны передвигаться по вертикальным и по-
толочным поверхностям. Дневные убежища их — это пространства под камня-
ми, у корней кустов, щели в почве. Зимуют в щелях полузасыпанных почвой
камней, норах грызунов. Скорлупу их яиц мы наблюдали под корнями куста
полыни, в старой осыпи на глубине 50 см, под камнем. Брачное поведение не
наблюдалось. Издают голос, похожий на писк серого геккона, но более тихий.
Практическое значение и охрана. С целью сохранения
генофонда этого редкого вида, имеющего ограниченное распространение в на-
шей стране, колючехвостый геккон включен в Красную книгу СССР и ТССР.
Охраняется на территории Копетдагского заповедника и его Меано-Чаачин-
ского заказника.
ЛИВАНСКИЙ ГЕККОН -
TENUIDACTYLUS AMICTOPHOLIS (HOOFIEN, 1967)
Типовая территория: гора Хермон в окр. пос. Мегделъ Шаме, Ливан, около
2000 м н. у. м.
Кариотип: не изучен.
1967 — Cyrtodactylus amictopholis Hoofien, Isr. J. Zool., Tel Aviv, Vol. 16, p. 205.
1981—Gymnodactylus (Mediodactylus) amictopholis, Щербак, Голубев, Вести, зоол., Киев,
№ 5, с. 40.
1984 — Tenuidactylus amictopholis, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Спинные бугорки отсутствуют; диаметр ушного отверстия
меньше половины продольного диаметра глаза; вдоль нижней стороны тела
не менее 90 чешуй; чешуя хвоста (не считая бугорков) и конечностей гладкая.
Голотип [230] TAU 7072 ad «Mt. Hermon above Megdel Shams
about 2000 м elevation, 20.VIII 1967, Hoofien et all». L = 36 мм; хвост регенери-
рован; два носовых щитка; первые носовые разделены между собой тремя че-
шуйками; восемь верхнегубных щитков; подбородочный щиток треугольный,
первая пара нижнечелюстных щитков соприкасается между собой, за ней
следует еще три-четыре пары плавно уменьшающихся щитков; чешуя брюха
крупная: вокруг середины тела около 70 чешуй, из них только 20 приходится
на брюшную; преанальные поры отсутствуют; подпальцевых пластинок по 13.
Описание (рис. 70) (по 2 экз. из коллекции TAU). L = 34,3—35,4 мм;
хвосты регенерированы; индекс высоты/ширины головы 49—53; диаметр
уха/глаза 29—30; верхнегубных щитков 8—9; нижнегубных 6—7; чешуй
между центрами глаз 15—17; чешуй вдоль тела снизу 95—102; чешуй поперек
брюха 18—20; подпальцевых пластин 17—18; преанальных пор 2.
Подбородочный щиток тре- или пятиугольный; три-четыре пары нижнече-
люстных щитков, первая соприкасается между собой; два носовых щитка, пер-
вые разделены двумя чешуйками; подхвостовые щитки на нерегенерированной
части хорошо выражены, по два на сегмент (1—1); остальная чешуя хвоста и
конечностей без ребрышек.
Окраска и рисунок. Этот вид отличается по цвету и рисунку
от всех видов подрода. При жизни [2301 его тело отливает золотисто-бронзовым
оттенком; на спине около 10 темных тонких поперечных червеобразных полосок,
перемешанных с отдельными пятнышками; на основании хвоста две заметные
желтые поперечные полоски, граничащие с черной окантовкой нерегенериро-
ванного основания хвоста; после поимки или фиксации яркая окраска теряется,
остается лишь коричневато-серый фон.
Распространение (рис. 39). Известен только из типовой мест-
ности.
179
Кадастр к рис. 39.
Ливан: ю.-з. подножье хр. Хермон, с. Мегделъ Шаме [230].
Экология почти не известна. Все сведения принадлежат автору опи-
сания [230L
Стации. Обитает на каменистых участках со скудной растительностью,
видимо, горного плато на высоте 2000 м н. у. м.
Отношение к температуре. Обитает в суровых условиях
высокогорья, где снег на отдельных участках лежит все лето. Температура воз-
духа днем не превышает 22°, ночью — заморозки.
Суточный цикл активности. Дневной вид (1), активен в са-
мое теплое время года после полудня, когда субстрат и воздух достаточно про-
греваются.
Сезонный цикл активности, линька, питание
и другие вопросы экологии — не изучены.
Особенности поведения. Очень осторожен и проворен, автор
заметил, что ловить этих гекконов тяжело.
БЕМПУРСКИП ГЕККОН —
TENUIDACTYLUS SA.G1TTJFER (NIKOLSKY, 1899)
Типовая территория: Бемпур в В. Кирмане, В. Иран.
1899 — Gymnodactylus sagittifer Никольский, Ежег. зоол. Муз., т. 4, № 4, с. 379.
1974 — Cyrtodactylussagittifer (ex errore), S. Anderson, Field : Zool., Chicago, Vol. 65, N 4, p. 33.
1984 — Tenuidactylus sagittifer, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Спинные бугорки овальные, ребристые; диаметр ушного
отверстия меньше половины продольного диаметра глаза; вдоль нижней сто-
роны тела не более 90 чешуй; чешуя и щитки хвоста, а также чешуя передне-
верхних поверхностей конечностей с заметными ребрышками.
Лектотип. ЗИН 9331, d” ad «р. Бампур в В. Кирмане, 30.VI 1898,
Н. Зарудный». L = 29,7 мм, хвост поврежден; чешуй между центрами глаз
14; верхнегубных щитков по 10; нижнегубных по 7; чешуй вдоль тела снизу
85; чешуй поперек брюха 17; две пары нижнечелюстных щитков,первая сопри-
касается между собой; четыре преанальные поры; подпальцевых пластин по 18;
носовых щитков по три.
Описание (рис. 78) (по 5 экз. из колл. ЗИН). £общ. (п = 5) 24,1—
31,9 (29,4 ± 1,37) мм; LILcd 0,95; индекс высоты/ширины головы (и = 5)
64—68 (66,40 ± 0,98); диаметр уха/глаза (п = 2) 36—38; верхнегубных щит-
ков (п = 10) 9—10 (9,70 ± 0,15); нижнегубных (и = 10) 7—8 (7,40 ± 0,16);
чешуй между центрами глаз (и = 5) 14—15 (14,40 ± 0,24); длина спинного
бугорка/L (и = 5) 19—26 (23,40 ± 1,21); чешуй, окружающих спинной буго-
рок (п = 5), 12—13 (12,21 ± 0,20); чешуй вдоль тела снизу (и = 5) 83—88
(85,60 ± 0,87); чешуй поперек брюха (п = 5) 16—19 (16,97 ± 0,99); подпальце-
вых пластинок (п = 10) 18—19 (18,55 ± 0,16); преанальных пор (п = 3) 4.
Подбородочный щиток пятиугольный; дополнительных спинных бугорков нет;
подхвостовых щитков по две пары на сегменте (2—2), они ребристые, как и ос-
тальная чешуя хвоста и передне-верхних поверхностей лап; нижнечелюстных
щитков одна-две пары, иногда они не выражены; носовых щитков три, первые
носовые обычно разделены чешуйкой, но могут и соприкасаться между собой.
Окраска и рисунок (после фиксации в спирте). Основной цвет
светло-серый. По бокам головы от глаза в области уха и по границе теменной
области и височной две бурые полоски, пилеус с беспорядочно разбросанными
редкими бурыми пятнами. На туловище сверху пять-шесть поперечных М-об-
разных полос, которые могут распадаться на фрагменты, боковые части слива-
ются, как у серого геккона, в продольные линии по бокам спины. Хвост с попе-
речными бурыми пятнами, также сливающимися в продольные полосы по бокам
хвоста. На передних и задних конечностях три-четыре неясных поперечных
полосы. Брюхо светлое.
Распространение. Известен только из типовой местности
(рис. 7). Иран: Бемпур; Фарра (Пурра) (ЗИН); 30 км з. пос. Бемпур,
27° 10' с. ш., 60° 10' в. д. (CAS).
180
Рис. 78. Tenuidactylus sagittifer:
4 — общий вид, Б — голова сверху, В — подбородок, Р — спина, Д — плечо спереди,
Е — хвост снизу, Ж преанальные поры (ЗИН 9331, Бемпур, Иран)
Половой диморфизм и возрастная изменчивость.
Преанальные поры у самок отсутствуют. Отсутствие молодых особей не позво-
ляет судить о возрастной изменчивости.
Систематика и географическая изменчивость.
О. Веттштейн [315] ошибочно синонимизировал этот вид с Т. heterocercus
[3071. На изученном материале изменчивость не выявлена.
Экология не изучена. Н. А. Зарудный [601 сообщал, что этот вид
найден «в Бампурской котловине и по окружающим ее высотам». Из трех эк-
земпляров два пойманы на древесных стволах в акациевом лесу, а один — на
стене подземного жилья. Днем малоподвижен и ловится без всякого труда.
181
Рис. 79. Tenuidactylus heterocercus:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — носовые щитки, Д — голова
снизу, Е — передняя лапа сверху, Ж. — спина, 3 — боковая чешуя брюха, И — хвост свер-
ху, А — хвост снизу (Г, 3, Е — NMW 7286, Персеполис, Иран; остальное — TZM 2532,
Хамадан, Иран)
ПЕРЕДНЕАЗИАТСКИЙ ГЕККОН —
TENUIDACTYLUS HETEROCERCUS BLANFORD, 1874)
Типовая территория: Хамадан, 3. Иран.
Кариотип: не изучен.
1874 — Gymnodactylus heterocercus Blanford, Ann. Mag. Nat. Hist., ser. 4, vol. 13—14, p. 453.
1974 — Cyrtodactylus heterocercus, S. Anderson, Field. : Zool. Chicago, vol. 65, N 4, p. 33.
1984 — Tenuidactylus heterocercus, Щербак, Голубев, Вести, зоол., Киев, №> 2, с. 54.
182
Диагноз. Спинные бугорки овальные, ребристые; диаметр ушного
отверстия меньше половины продольного диаметра глаза; вдоль нижней сто-
роны тела не менее 90 чешуй; чешуя и щитки хвоста, а также чешуя передне-
верхних поверхностей конечностей с заметными ребрышками.
Лектотип. TZM № R2532 (рис. 79) g ad «Persia dono Doria». L =
— 47,3 мм; хвост поврежден; чешуй между центрами глаз 14; верхнегубных
щитков по 9, нижнегубных по 8; чешуй вдоль тела снизу 118; чешуй поперек
брюха 30; продольных рядов спинных бугорков 12; три пары нижнечелюстных
щитков, первая соприкасается между собой; преанальные поры отсутствуют;
подпальцевых пластин по 21; первые носовые оттеснены от ноздри вторым и
третьим носовыми щитками; брюшная чешуя ребристая.
—О-п-и х^а н и е (рис, 79) (по 17 экз. из колл. TZM, SMF, NMW, FMNH).
L общ. (п = 17) 36,2—49,9 (43,92 ± 1,06) мм; LlLcd В,86; индекс высо-
ты/ширины головы (л — 12) 48—59(53,42 ± 0,91); диаметр уха/глаза (и = 10)
28—48 (38,50 ± 1,97); верхнегубных щитков (п = 34) 9—12 (9,41 ± 0,17);
нижнегубных (и = 34) 7—9 (7,50 ± 0,09); чешуй между центрами глаз (л =
= 15) 12—16 (13,80 ± 0,28); длина спинного бугорка/L (и = 26) 15—24
(20,25 ± 0,54); чешуй, окружающих спинной бугорок (л = 17), 10—13 (11,41 ±
±0,19); чешуй вдоль тела снизу (л = 15) 98—122 (109,07 ± 1,63); чешуй
поперек брюха (л = 16) 24—30 (26,00 ± 0,41); подпальцевых пластин (л —
— 34) 18—23 (20,56 ± 0,23); преанальных пор (л ~ 4) 2.
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
три (82,3 %) или четыре (17,7 %) пары; первая обычно (94,1 %) соприкасается
между собой; дополнительных спинных бугорков нет; носовых щитков два-три,
обычно они разделены одной чешуйкой (82,3 %), иногда этих чешуек две-три
или они отсутствуют (по 5,9 %); чешуя нижней стороны хвоста однородная, она,
как и чешуя передне-верхних поверхностей конечностей, отчетливо ребристая;
у этого вида шипообразные хвостовые бугорки настолько велики, что в отличие
от остальных видов подрода могут соприкасаться между собой на одном сег-
менте.
Окраска и рисунок (после фиксации в спирте). Основной фон
серый, рисунок сверху из многих неясных бурых поперечных полос, образую-
щих мраморный рисунок. На голове и хвосте рисунок неясен, так как изученные
экземпляры длительное время находились в фиксаторе.
Распространение (рис. 80). Анатолия (Ю. Турция) и Хамадан
(3. Иран); находки этого вида в Персеполисе (пров. Фарс, 3. Иран — колл.
NMW и FMNH) требуют подтверждения. Также требует подтверждения наход-
ка в Эль-Басре [269]; видимо, здесь авторы приняли за этот вид Т. scaber.
Кадастр к рис. 80.
Иран: 1 — Хамадан (NMW); 2 — разв. Персеполис (на вставке).
Половой диморфизм и возрастная изменчивость.
У самок отсутствуют преанальные поры. Возрастная изменчивость не изучена.
Систематика и географическая изменчивость.
Р. Мертенс [254] выделил анатолийских гекконов в самостоятельный подвид.
Впоследствии [255] он выразил сомнение в том, что эта форма принадлежит
к G. heterocercus, а не к G. kotschyi. Действительно, оба вида очень близки
друг к другу, однако переходные формы между ними не обнаружены. Более
сомнительным кажется существование двух популяций, оторванных друг от
друга не менее чем на 600 км. Вероятно, это связано со слабой изученностью
герпетофауны региона. Нет сомнения в том, что данный вид будет найден в се-
верных районах Сирии и Ирака; вероятно, в 3. Иране он распространен также
более широко.
Таблица для определения подвидов Т. heterocercus
1(2/. 12—14 продольных рядов спинных бугорков . . . Т. heterocercus heterocercus (Blanford)
2(1). 10 продольных рядов спинных бугорков. .... Т. heterocercus mardinensis (Mertens)
183
ПЕРЕДПЕАЗИАТСКИЙ ГЕККОН ИРАНСКИЙ -
TENUIDACTYLUS HETEROCERCUS HETEROCERCUS (BLANFORD, 1874)
Типовая территория: Хамадан, 3. Иран.
1924.— Gymnodactylus heterocercus heterocercus, Mertens, Senckenbergiana, Frankfurt am Main,
vol. 6, S. 84.
Диагноз. 12—14 продольных рядов спинных бугорков; чешуя брюха,
особенно на боках, часто с заметными ребрышками; первый носовой щиток час-
то оттеснен от ноздри вторым и третьим.
Остальные признаки содержатся в видовом описании.
Тип видовой.
Рис. 80. Распространение Т. heterocercus mardinensis (/), Т. h. heterocercus (2)
Распространение. Известен из типового местонахождения —
Хамадана (3. Иран); находки этого геккона в Персеполисе нуждаются в про-
верке.
ПЕРЕДНЕАЗИАТСКИЙ ГЕККОН ТУРЕЦКИЙ —
TENUIDACTYLUS HETEROCERCUS MARDINENSIS (MERTENS, 1924)
Типовая территория: Мардин, Ю. Турция.
1924 — Gymnodactylus heterocercus mardinensis Mertens, Senckenbergiana, Frankfurt am Main,
vol. 6, S. 84 (Terra typica designata (Mertens, 1952); Mardin, Kurdistan, Tiirkei).
Диагноз: 10 продольных рядов спинных бугорков; чешуя брюха глад-
кая; с ноздрей соприкасаются три носовых щитка.
Остальные признаки содержатся в видовом описании.
Н е о т и п. SMF 8198 с? ad «Mardin, SO-Anatolien. О. Wolter, 1918».
Экземпляр высох, размеры не определяются. Чешуй между центрами глаз 15,
верхнегубных щитков по 9; нижнегубных по 7; чешуй вдоль тела снизу 102;
чешуй поперек брюха около 25; продольных рядов спинных бугорков 10; три
пары нижнечелюстных щитков, первая соприкасается между собой; две преа-
нальные поры; подпальцевых пластин 21/20; носовых щитков три; брюшная
чешуя гладкая.
Распространение. Анатолия; возможны находки в северных
районах Сирии и Ирака (рис. 80).
Кадастр к рис. 80.
Турция: 1—Малатья [181]; 2 — Битлие [181]; 3— Сиирт [181]; 4 — Диярбакыр
[181]; 5 — Газиантеп [181]; 6 — Низип (SMF); 7 — Дерик [181]; 8 — Мардин [181].
Ирак: 1 — пров. Ниневия, Аль-Наваран, 30 км сев. Аль-Мавзила [269].
Экология не изучена. По данным М. Бажоглу и И. Барана [1801,
обитает на стенах жилых помещений, в трещинах. Издает звуки, передаваемые
как «чек—чек», которые слышны после захода солнца. Ловят насекомых у элек-
троламп, очень подвижны, наблюдались с конца весны до конца июля.
184
Подрод циртоподион — Cyrtopodion Fitzinger, 1843, stat. n.
T и по вий вид: Stenodactylus scaber Hey den, 1827.
1843 — Cyrtopodion Fitzinger, Syst. Rept. S. 18, p. 93.
1984 — Mesodactylus Szczerbak et Golubev, Вести, зоол., 2, с. 54.
Диагноз: У самцов имеется не более 10 преанальных пор; длина шва
между первой парой нижнечелюстных щитков превышает половину длины под-
бородочного щитка; форма спинных бугорков от трехгранной до округло-тре-
угольной; хвостовые бугорки широко соприкасаются между собой боковыми
гранями; спереди, у основания такого бугорка есть один-два более мелких бу-
горка, превышающих размерами окружающую чешую; ряд подхвостовых щит-
ков уже ширины хвоста; высота одного щитка менее чем вдвое меньше его ши-
рины.
Дополнительные признаки. Высота первого верхнегуб-
ного щитка от ноздри до края рта заметно больше или немного меньше его
ширины вдоль края рта; в задне-верхней части орбиты могут находиться слабо
выраженные уплощенно-треугольные выросты; среди мелкой чешуи задней по-
верхности бедра обычно имеется 2—10 мелких трехгранных или конических бу-
горка; хвост плавно утоньшается к концу; ряд подхвостовых щитков может за-
мещаться двумя их рядами или обычной чешуей. Основной фон и рисунок вы-
держаны в светло- и темно-коричневых тонах.
Включает шесть видов. Известен в Ю.-З. Азии, один вид проникает в
Ц. Азию.
Таблица для определения видов подрода Cyrtopodion
1 (2). Есть два носовых щитка...............................Т. elongatus (Blanford)
2 (1). Есть три носовых щитка.
3 (4). Чешуи, на которых расположены две преанальные поры, значительно крупнее окру-
жающих; хвостовой сегмент покрыт снизу тремя кольцами однородной чешуи ....
................................................Т. agamuroides (Nikolsky)
4 (3). Чешуи, на которых расположены четыре — девять преанальных пор, не отличаются
по размерам от окружающих; хвостовой сегмент покрыт снизу одним-двумя продольными
рядами крупных щитков.
5 (8). Чешуй поперек брюха не более 25.
6 (7). Есть не более девяти четко выраженных преанальных пор . . . . Т. scaber (Heyden)
7 (6). К восьми четко выраженным преанальным порам с обеих сторон примыкает по 10—12
очень слабо выраженных пор, заходящих на бедра....................................
..............................................Т. montiumsalsorum (Annandale)
8 (5). Чешуй поперек брюха не менее 25.
9(10). Чешуй вдоль тела снизу 100—128 ....................................Т. kachensis (Stoliczka)
10 (9). Чешуй вдоль тела снизу 124—142 ........................ ............Т. watsoni (Murray^
ГЕККОН КАЧСКИЙ —
TENUIDACTYLUS KACHHENSIS (STOLICZKA, 1872)
Типовая территория: штат Кач, Ю.-З. Индия.
1872 — Gymnodactylus kachhensis Stoliczka, Proc. Asiat. Soc. Bengal, Calcutta, (1), p. 79.
1884 — Gymnodactylus petrensis Murray, Vertebr. Zool. Sind., p. 362.
1935 — Gymnodactylus fcacft/iens/s kachhensis, M. A. Smith, Fauna brit. Ind., Rept., Amph., vol. 2,
p. 43.
1954 — Cyrtodactylus kachensis, Underwood Proc. zool. Soc. London, 1954, vol. 124,
N 3, p. 475.
1984 — Tenuidactylus kachhensis, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Три носовых щитка; чешуи, на которых расположены пре-
анальные поры, равны окружающим; снизу хвостового сегмента имеются круп-
ные щитки; чешуй вдоль нижней стороны тела 100—128; чешуй поперек брю-
ха 28—35; четыре — восемь четко выраженных преанальных пор.
Судя по фотографиям типа G. brevipes, присланным из Калькутты Б. К. Тикадером,
небольшое количество преанальных пор (4) и некрупные спинные бугорки сближают этот эк-
земпляр, принадлежащий к данному подроду, с Т. kachhensis, а наличие крупных брюшных
чешуй (20—22) — с Т. scaber. Отсутствие материала не позволяет нам решить вопрос о его
таксономическом положении.
185
Рис. 81. Tenuidactylus kachhensis:
A — общий вид, Б — голова сверху, В — голова снизу, Г — верхний край орбиты, Д — спи-
на, Е — передняя лапа сверху, Ж — хвост сверху, 3 — хвост снизу (ИЗАНУ SR 1446, Ха-
леджи, Пакистан)
Лектотип. NMW 17383:5, <f ad «Katch, Stoliczka, 1874; 11.98»
L = 42,3 mm; Led = 50,3 mm; LILCd = 0,84; чешуй между центрами глаз 13;
верхнегубных щитков по 10; нижнегубных по 8; чешуй вдоль тела снизу
100; чешуй поперек брюха 27; преанальных пор семь; подпальцевых пла-
стин 20/21.
Описание (рис. 81) (по 86 экз. из колл. ИЗАНУ, NMW, FMNH,
MNHP, UMMZ, AMNH).
186
64*
Рис. 82. Распространение Tenuidactylus watsoni (/), Т. kachhensis (2), Т. montium-
salsorum (5), T. agamuroides (4)
L<? ad (n = 29) 35,3—45,8 (38,91 ± 0,58) мм; L $ ad (n = 24) 35,3—43,8
(39,24 ± 0,51) mm; LILcd (n = 15) 0,76—0,98 (0,83 ± 0,01); индекс высо-
ты/ширины головы (п = 32)55—65(59,31 ± 0,56); диаметр уха/глаза (п = 11)
38—48 (42,01 ± 0,01); верхнегубных щитков (п = 172) 10—12 (10,85 ± 0,05);
нижнегубных (п = 172) 7—10 (8,30 ± 0,05); чешуй между центрами глаз
(п = 85) 13—18 (14,92 ± 0,12); длина спинного бугорка/Л (п = 62) 17—25
(21,40 ± 0,83); чешуй, окружающих спинной бугорок (и = 83), 11 —15
(12,69 + 0,11); чешуй вдоль тела снизу (п = 79) 100—128 (113,04 ± 0,62);
чешуй поперек брюха (п = 76) 28—35 (30,47 ± 0,21); подпальцевых пластин
(п = 162) 18—23 (21,42 ± 0,10); преанальных пор (п = 37) 4—8 (5,57 ±
± 0,19).
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
две (23,4 %) или три (76,6 %) пары, первая пара соприкасается между собой
у 98,8 %; дополнительных спинных бугорков нет (90,6 %), иногда один (7,8 %)
187
или два (1,6%); носовых щитков всегда три; первые носовые разделены одной
(86,1 %) или двумя (12,3 %) чешуйками, у 1,6 % они соприкасаются; подхвосто-
вые щитки образуют на одном сегменте сочетания 1—2—2, 2—2—2, 0—0—2,
1—1—2, а в конце хвоста обычно 1—1 или 2—2.
Рисунок и окраска очень похожи с таковой близкородственного
Т. scaber: от затылка до поясницы следы в виде пятен от четырех-пяти попереч-
ных полос, каждой полосе соответствуют три — пять пятнышек, соединенных
иногда между собой тонкими линиями; семь—десять поперечных полос вдоль
хвоста; на конечностях неясный поперечный узор; снизу белый.
Распространение (рис. 82). Ю. Пакистан, крайний ю.-з. Индии:
п-ов Кач, Синд, береговая зона Лас-Белы; на север проникает по руслам неко-
торых рек, однако выше 29° с. ш. достоверно не известен; указания Р. Мертен-
са [258] и М. Хана [2351 на находки этого вида в северных районах Пакистана
(Сват и северо-западная пограничная провинция) сомнительны и требуют уточ-
нения.
Кадастр к рис. 82.
Пакистан: 1 — Мирпур (MNHP); 2 — округ Даду, Унапур (AMNH); 3 — 26' км
сев. г. Бела [263]; 4 — округ Даду, 8 км ю. г. Сехван (AMNH); 5 — Лас-Бела, 13 км ю. пос.
Дивана (AMNH); 6 — 67 км ю.-з. пос. Лияри [263]; 7 — округ Татта, Джеррук (AMNH); 8 —
Хабчауки (UMMZ), Бхавани [259]; 5 км ю.-з. Бандмурадхан [263]; 10 — округ Татта,
оз. Халеджи (AMNH); 11 — 12 км с.-з. Карачи (UMMZ); Карачи (AMNH); Ландхи; Хоранги
[258]; 12 — 5 км в. Ладхи; Булейджи [263]; 13 — 1,6 км ю. пос. Гуджо [263]; 14 — 3 км з. пос.
Гхаро [263]; 15 — округ Татта, окр. г. Пир-Патхо (AMNH); 16 — п-ов Кач, Индия (NMW);
17 — Сонда [258]; 18 — Аттркк Равалпинди ([258] — нуждается в переопределении); 19 — Зи-
арат ([258] — нуждается в переопределении); 20 — горы Карана, около Лалиена ([235] — ну-
ждается в переопределении).
Не найдены: Rabwah [235]; Ladha [279].
Половой диморфизм и возрастная изменчи-
вость. Самцы отличаются от самок наличием преанальных пор, возрастная
изменчивость не изучена.
Географическая изменчивость. На доступном нам мате-
риале не выражена.
Экология практически не изучена; имеющиеся скудные сведения
приведены по данным Ш. Минтона [263].
Стации и количественные данные. Обитает в сухих
каменистых местах, днем прячется под камни и кусты молочаев. В жилых до-
мах встречается редко. Многочисленный вид.
Сезонный цикл активности. Судя по коллекционному ма-
териалу (крайние даты находок 24—25.1 1959; 20.11 1960 и 6.XII 1959), вид
активен круглогодично.
Размножение. Откладывает два яйца размером 7 X 9,5 мм.
ГЕККОН ПАКИСТАНСКИЙ —
TENUIDACTYLUS WATSONI (MURRAY, 1892)
Типовая территория: Кветта, 3. Пакистан.
Кариотип: не изучен.
1892 — Gymnodactylus watsoni Murray, Zool. Baloochist. S. Afghanis!., p. 68.
1923 — Gymnodactylus ingoldbyi Procter, J. Bombey nat. Hist. Soc., vol. 29, p. 121.
1935 — Gymnodactylus kachhensis watsoni, M. A. Smith, Fauna brit. Ind., Rept. Amph., Vol. 2,
p. 44.
1966 — Cyrtodactyluswatsoni, Minton. Bui. amer. Mus. Nat. Hist., (N. Y.), Vol. 134, art. 2, p . 79.
1984 — Tenuidactylus watsoni, Щербак, Голубев, Вестн. зоол., Киев, № 2, с. 84.
Диагноз. Три носовых щитка; чешуи, на которых расположены пре-
анальные поры, равны окружающим; снизу хвостового сегмента имеются круп-
ные щитки; чешуй вдоль нижней стороны тела 124—142, чешуй поперек брюха
28—35; пять—девять четко выраженных преанальных пор.
Тип. Местонахождение не известно.
Описание (рис. 83) (по 20 экз. из колл. ИЗАНУ, AMNH, SAM, SMF).
L <? ad (п= 14) 42,0—53,1 (48,09 ± 1,03) мм; ad (n = 5) 35,0—54,0 (48,58 ±
± 3,48) mm; LI Led (n = 5) 0,75—0,87 (0,805 ± 0,019); индекс высоты/ширины
головы (n = 18) 48—60 (56,4 ± 0,66); диаметр уха/глаза (п = 18) 37—52
(44,2 ± 0,011); верхнегубных щитков (п = 40) 10—13 (11,40 ± 0,14); нижне-
188
Рис. 83. Tenuidactylus scaber (вверху):
A — общий вид, Б — голова снизу, В — голова сбоку, Г — голова сверху, Д — спина, Е —
палец снизу (ИЗАНУ SR 1451, окр. Сангара, Пакистан)
Tenuidactylus watsoni (внизу):
А — общий вид, Б — голова сбоку, В — лицевая часть спереди, Г — голова снизу, Д — голова
сверху, Е — преанальные поры, Ж — хвост снизу (ИЗАНУ SR 1447, Вад, Пакистан)
губных (п = 40) 8—10 (8,57 ± 0,11); чешуй между центрами глаз (п = 20)
12—16 (13,80 ± 0,29); длина спинного бугорка/Z, (п = 19) 17—26 (22,16 ±
± 0,42); чешуй, окружающих спинной бугорок (га = 19), 10—15 (12,15 ±
± 0,29); чешуй вдоль тела снизу (п = 17) 124—142 (132,11 ± 1,39); чешуй
поперек брюха (п = 18) 30—40 (35,55 ± 0,57); подпальцевых пластин (п =
189
= 38) 19—27 (22,76 ± 0,31); преанальных пор (п = 14) 5—9 (7,36 ± 0,29).
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
две (10 %) или три (90 %) пары, первая всегда соприкасается между собой;
дополнительных спинных бугорков нет у 90 %, по 5 % особей имеют один-два
таких бугорка; носовых щитков всегда три; первые носовые разделены одной
(85 %) или двумя (15 %) чешуйками; обычное сочетание подхвостовых щитков
на одном сегменте — 1—1, на первом 2—2, но иногда встречаются и 1—2—1
и 2—2—2. Спинные бугорки трехгранные.
Рисунок и окраска. Общий фон серый, есть пять—восемь попе-
речных М-образных спинных и 10—12 хвостовых темно-бурых полос; по пять-
шесть неясных полосок на конечностях, на голове — неясный узор; нижние
поверхности белые.
Распространение (рис. 82) Сев.-В. Афганистан, Сев. и Ц. Паки-
стан; почти не опускаются ниже 28° с. ш.
К а д а с т р к р и с. 82.
Афганистан: 1 — Джелалабад [245].
Пакистан: 1 — северо-западная пограничная провинция, склоны над пос. Удиграм
(AMNH); 2 — Мансехра [258]; 3 — 13 км ю. г. Кемпбелпур (AMNH); 4 — округ Сиби, пос. Кач
(AMNH); пос. Урак [258]; 5 — г. Кветта; 16 км ю. г. Кветта (AMNH); 6 — округ Кветта,
5 и 11 кмю. пос. Колпур (AMNH); 7 — округСиби, окр. пос. Мач, р. Болан (AMNH); 8 — 13 км
ю. г. Калат (AMNH); 9 — 3 км ю. пос. Вад (AMNH), 10 — 53 км ю. пос. Вад (AMNH); 11 — Лас-
Бела, 32 и 37 км сев. г. Бела (AMNH).
Не найден: Ladha [279].
Половой диморфизм и возрастная изменчивость.
У самок отсутствуют преанальные поры. Возрастная изменчивость не изучена.
Географическая изменчивость не обнаружена.
Экология не изучена. Известны находки этого вида на высоте 1500—
6000 футов (соответственно 461—1884 м [263]). Самые ранние и поздние наход-
ки по коллекционным материалам: 18.III 1961 г. и 24.IX 1961 г.
ГЕККОН ШЕРШАВЫЙ—TENUIDACTYLUS SCABER (HEYDEN, 1827)
Типовая территория: окр. с. Тор, Синай.
Кариотип: не изучен.
1827 — Stenodactylus scaber Heyden in Riippel, Alt. Reisenordl. Afr. Rept., S. 15.
1836 — Gymnodactylus scaber, Dumeril et Bibron (part.) Erpetol. gen., vol. 3, S. 421.
1954 — Cyrtodactylus scaber, Underwood, Proc. zool. Soc. London, vol. 124, N 3, p. 475.
1982 — Cyrtodactylus basoglui Baran, Gruber, Spixiana, 5, 2, Mtinchen, S. 121.
1984 — Tenuidactylus scaber, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Три носовых щитка; чешуи, на которых расположены пре-
анальные поры, равны окружающим; снизу хвостового сегмента имеются круп-
ные щитки; чешуй вдоль нижней стороны тела 85—106; чешуй поперек брюха
16—23; четыре — девять четко выраженных преанальных пор.
Лектотип. Хранится в Зенкенбергском музее, Франкфурт-на-Майне,
ФРГ [245].
Описание (рис. 83) (по 137 экз. из колл. ИЗАНУ, ЗИН, ММГУ,
UMMZ, SAM, AMNH, NMW).
L <? ad (n = 62) 34,4—50,0(43,65 ± 0,49) мм; L $ ad (n = 52) 34,3—53,2
(45,32 ± 0,73) mm; LIL cd (n = 45) 0,65—0,97 (0,76 ± 0,059); индекс высоты/
ширины головы (п = 47) 45—62 (53,00 ± 0,53); диаметр уха/глаза (п = 55)
30—60 (49,03 ± 0,009); верхнегубных щитков (п = 206) 10—14 (11,38 ±
± 0,06); нижнегубных (п = 264) 7—10 (8,31 ± 0,04); чешуй между центрами
глаз (п = 137) 10—16 (12,72 ± 0,11); длина спинного бугорка/Z, (п = 117)
20—31 (24,74 ± 0,21); чешуй, окружающих спинной бугорок (и = 132), 12—
18 (15,05 ± 0,11); чешуй вдоль тела снизу (п = 119) 85—106 (90,87 + 0,36);
чешуй поперек брюха (и = 120) 16—23 (19,47 ± 0,14); подпальцевых пласти-
нок (п = 260) 19—27 (22,82 ± 0,09); преанальных пор (и = 68) 4—9 (5,78 ±
± 0,09).
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
две (21,1 %), три (76,9 %) или четыре (2,0 %) пары, первая пара всегда сопри-
касается между собой; дополнительных спинных бугорков нет у 93,7 % особей,
иногда их бывает один (4,7 %) или два (1,6 %); носовых щитка три, вытеснен-
ный первый носовой обнаружен у одного геккона с одной стороны; первые носо-
190
Рис. 84. Распространение Tenuidactylus scaber
вые разделены одной (85,5 %) или двумя (15,3 %) чешуйками, у 2,2 % эти щит-
ки соприкасаются; обычное сочетание подхвостовых щитков на одном сегменте
1—1. Спинные бугорки трехгранные, крупные, в продольных рядах почти ка-
саются друг друга.
Рисунок и окраска (рис. 83). Похожи на Т. kachhensis
(см. выше).
Распространение (рис. 84). Встречается от африканского побе-
режья Красного моря через Аравийский п-ов и Ю.-З. Азию до В. Сирии, Ю. Тур-
ции, Ирака, Ирана, Афганистана, Пакистана до западных границ пустыни
Тар; вероятны находки на западе Индии.
Кадастр к рис. 84.
Турция: / — Сюроч [46]; 2 — Урфа [46]; 3 — Кызыл тепе [46]; 4 — Мардин [46].
Египет:/ — окр. Каира [249]; 2 — Эт-Тур (Tor, Sinai — terra typica restricta [307]);
3 — Эль-Кусейр [249]:
Судан: / — Порт-Судан [249].
Эфиопия: / — Эритрея [249].
Сирия: Абу-Кемаль [173].
Ирак: / — Акра [269]; 2 — Талль-Кайф; Мосул [269]; 3 — Эрбиль; Кей-Санджак
[269]; 4 — Хальва [269]; 5 — Рава [269]; 6 — Ханакин (ЗИН); 7 — Ана [269]; 8 — Эль-Хадита
[269]; 9 — Эт-Тартар [269]; 10 — Балад [269]; // — Дияла [284]; Эль-Халис [269]; 12 — Баг-
дад [269]; 13 — Рамади Эль-Хабания [269]; 14 — Хилла [269); 15 — Эд-Дивана [269]; 16 — Эль-
Куфа; Эн-Наджар [269]; 17 — Кербела; Хан-эль-Хаммад [269]; 18 — Али-эль-Гарби [269];
19 — Амара [269]; 20 — Эн-Насирия [269]; 21 — Басра [269]; 22 — Сафван [269].
Иран: / — Тегеран (ЗИН); 2 — Бошруйе [285]; 3 — Шуш, 32° 1Г с. ш.,48° 15' в. д.
(CAS); 4 — пров. Хузестан, 31° 17' с. ш., 48° 49' в. д. [168]; 5 — Месджеде-Солейман (CAS);
6 — Агаджари (CAS); 7 — окр. Бирдижанда (ЗИН); 8 — Иезд (NMW); 9 — Казерун (CAS)
10 — Шираз (CAS); 11 — 1 км з. пос. Бендер-Ленге (CAS); Бендер-Ленге [223]; 12 — Бендер-
Аббас [223]; 13 — о. Хормоз (NMW); 14 — Чахбехар [303]; 15 — Эхрем (CAS).
Аравийский п-ов:/ — Эль-Хаба (CAS); 2 — Дахран; Рас-Танура; Эль-Хубар
[226]; 3 — Абкаик (CAS); 4 — о. Хавар, Бахрейн (ММГУ); 5 — Эт-Тариф (CAS); 6 — Эр-Ри-
яд (CAS).
Афганистан: / — Джелалабад [240]; 2 — между Гератом и Исламкалой, 34°22'
с. ш.» 62° 10' в. д. [209]; 3 — Хост (Матун) (CAS); 4 — 32 км ю. пос. Мукур, 32° 38' с. ш.,
67° 30' в. д. [209]; 5 — Шархи-Сафа, 60 км сев.-в. г. Кандагар (CAS); 6 — Кандагар,
31° 30' с. ш., 65° 47' в» д. [172].
191
Пакистан: 1 — Лакки [232]; 2 — Кхирги; Танк [232]; 3 — Вана [232]; 4 — Дери-
смаилхан [232]; 5 — Ноккунди [258]; 6 — Далбандин [258]; 7 — Джамалдини [258]; 8 —
Шахдадкой [258]; 9 — Суккур [202]; 10 — разв. Мохенджодаро [258]; Ларкана (AMNH); 11 —
с. Бубак, 15 км з. г. Сехван [258]; 12 — окр. Дас-Бела, 65 км ю.-з. пос. Лиари (AMNH);
13 — горы Зараин [258]; 14 — пещеры Гхандран (AMNH); 15 — Мирпурсакро (MNHP); 16 —
Хала [258]; 17 — 3 км сев.-з. пос. Сангхар; Джемрао (AMNH) 8 км ю. пос. Сангхар [263]; 18—
9 км ю. г. Сехван (AMNH); 4 км сев.-з. Тирс Лакхми [263], Тирс (SAM); 19 — окр. Хайда-
рабада, 1,6 км з. пос. Котри [263]; 20 — Бхавани [235]; 21 — Чиниот [235].
Не найдены на карте: Abu Guraib, Аг Rashdiyah, Al Kadhaniyah, Al Adha-
miyax, Babylon Prov. all from Khalaf, Monsaib [269]; Shah Baran, Mansuabad [284]; Rabwah,
Labian [235]; Kaur Bridge, Sarwekai [216]; Khasawiayah (CAS).
Половой диморфизм и географическая изменчи-
вость. Как и у остальных представителей рода, у самок этого вида отсут-
ствуют преанальные поры. Молодые особи более короткохвосты, чем взрослые.
Систематика и географическая изменчивость.
Недавно [1811 был описан из Турции новый вид Cyrtodactylus basoglui. Как
нам удалось установить [46], по всем признакам он соответствует виду Т. sca-
ber. Таким образом, И. Баран и У. Грубер обнаружили не новый для науки вид,
а новый для фауны Турции. Мы сравнивали семь выборок с территории всего
ареала. Оказалось, что у шершавого геккона отсутствует географическая из-
менчивость в фолидозе, однако ей подвержены размеры туловища: наиболее
мелкие гекконы данного вида обитают в Пакистане, на востоке ареала, наиболее
крупные — в Египте и Иране — на западе.
Экология изучена недостаточно.
Стапии и количественные данные. Обитает на камени-
стых склонах холмов с сухим травянистым покрытием или на песках с ксеро-
фитной растительностью [263]. Наиболее обычный вид в домах у подножия
гор [168].
Отношение к температуре. Найден 5 декабря при довольно
холодной погоде [263].
Суточный цикл активности. Активен в сумерки и
ночью [168].
Сезонный цикл активности. Встречается с конца июня до
ноября [168]. По коллекционным материалам, первая особь добыта 17.11 1962 г.
в окр. Лиари, Пакистан, последняя — 5.XII 1959 г. в окр. с. Даду, Пакистан.
Размножение. Откладка яиц с середины августа, их размер 7 X
X 10,5 мм [168].
Особенности поведения. Часто ловят летящих на свет насе-
комых [168]. Видимо, ведут синантропный образ жизни.
ГЕККОН ПЕНДЖАБСКИЙ —
TENUIDACTYLUS MONTIUMSALSORUM (ANNANDALE, 1913)
Типовая территория: Соляной хр., Зап. Пенджаб, Пакистан.
Кариотип: не изучен.
1913 — Gymnodactylus montiumsalsorum Annandale, Rec. ind. Mus., Calcutta, vol. 9, part. 5, p. 313.
1966 — Cyrtodactylus montiumsalsorum, Minton, Bull. Amer. Mus. Nat. Hist., N. Y., Vol. 134,
art. 2, p. 78.
1984 — Tenuidactylus montiumsalsorum, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Три носовых щитка; чешуи, на которых расположены пре-
анальные поры, равны окружающим; снизу хвостового сегмента имеются круп-
ные щитки; чешуй вдоль тела снизу 86; чешуй поперек брюха 19; к 8 четко вы-
раженным порам с обеих сторон примыкают по 10—12 очень слабо выраженных
пор, заходящих на бедра.
Описание* (рис. 85) (по голотипу BMNH 1904. 11.19.1. о” ad Salt
Range, Punjab.) L = 37,2 мм; хвост оборван; индекс высоты/ширины головы 58;
диаметр уха/глаза 44; верхнегубных щитков по 10; нижнегубных 7/8; чешуй
между центрами глаз 11; длина спинного бугорка/L 30; чешуй, окружающих
спинной бугорок, 15; чешуй вдоль тела снизу 86; чешуй поперек брюха 19;
* Использовалась также фотография паралектотипа, хранящаяся в Зоологической
службе Индии, Калькутта.
192
Рис. 85. Tenuidactylus montiumsalsorum:
A —• общий вид, Б — голова сбоку, В — лицевая часть спереди, Г — голова сверху, Д —
голова снизу, Е — спина, Ж — преанальные поры (BMNH 1904.11.19. i, Пенджаб Пакистан)
подпальцевых пластин 21/20; преанальных пор 8, к ним с обеих сторон примы-
кает по 10—12 очень слабо выраженных пор, заходящих на бедра.
Подбородочный щиток пятиугольный; две пары нижнечелюстных щитков,
первая соприкасается между собой; дополнительные спинные бугорки с основ-
ными не соприкасаются; носовых щитка три, первые разделены чешуйкой;
спинные бугорки трехгранные.
Рисунок и окраска. Экземпляр обесцвечен.
Распространение. Известен только из типовой местности
(рис. 82).
193
Систематика. Как отмечалось ранее 1153], наличие у обоих экземп-
ляров дополнительных слабо выраженных пор, по-видимому, аномалия; по
всем остальным признакам они укладываются в рамки диагноза Т. scaber. Одна-
ко окончательно идентифицировать Т. montiumsalsorum и Т. scaber будет воз-
можно только после получения дополнительного сравнительного материала.
Экология не изучена.
ГЕККОН КАШГАРСКИЙ — TENUIDACTYLUS ELONGATUS
(BLANFORD, 1875)
Типовая территория: Янгигиссар, 3. Китай.
Кариотип: 2п = 42, акроцентриков 19, субтелоцентриков 1, телоцентриков 1,
NF = 42.
1875 — Gymnodactyluselongatus Blanford, J. Asiat. Soc. Bengal, Calcutta, vol. 44, part. 2, p. 193.
1954 — Cyrtodactylus elongatus, Underwood, Proc. zool. Soc. London, vol. 124, N 3, p. 475.
1984 — Tenuidactylus elongatus, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54;
Диагноз. Два заметно вздутых носовых щитка; чешуи, на которых
расположены преанальные поры, более или менее равны окружающим; снизу
хвостового сегмента имеются крупные щитки; чешуй вдоль нижней стороны
тела 102—132; чешуй поперек брюха 26—30; 4—7 четко выраженных пре-
анальных пор.
Типы хранятся в Индийском музее, Калькутта.
Описание (рис. 86) (по 33 экз. из колл. ИЗАНУ, ЗИН и ММГУ).
L<?ad(n = 10) 43,9—55,6 (49,27 ± 1,05) мм; L g ad (п = 18) 42,2—56,8
(50,03 ± 0,94) мм; L/L cd (п = 12) 0,7—0,8 (0,76 ± 0,01); индекс высоты/ши-
рины головы (п — 29) 54—67 (59,03 ± 0,006); диаметр уха/глаза (п = 12)
39—44 (41,25 ± 0,46); верхнегубных щитков (п = 66) 10—13 (11,80 ± 0,10);
нижнегубных (п = 65) 8—11 (9,72 ± 0,11); чешуй между центрами глаз (и =
— 33) 15—19 (16,71 ± 0,18); длина спинного бугорка/L (п = 27) 14—19
(16,41 ± 0,26); чешуй, окружающих спинной бугорок (п = 33), 8—12 (10,45 ±
± 0,16); чешуй вдоль тела снизу (п = 32) 102—132 (119,37 ± 1,13); чешуй
поперек брюха (п = 29) 26—30 (27,41 ± 0,23); подпальцевых пластин (и = 64)
19—23 (21,23 ± 0,13); преанальных пор (п = 14) 4—7 (5,80 ± 0,24).
Подбородочный щиток пятиугольный; нижнечелюстных щитков одна (3,7%),
две (87 %) или три (9,3 %) пары, первая соприкасается между собой; дополни-
тельные спинные бугорки соприкасаются с основными только у 14,7 %, у
остальных лишены контакта или отсутствуют; носовых щитка всегда два, они
довольно значительно вздуты, первые носовые разделены двумя (41,2 %), тремя
(53 %) или четырьмя (5,8 %) чешуйками; подхвостовые щитки на одном сегмен-
те образуют сочетания: на первых сегментах (до II—VIII) — 1—1—1, далее
до конца — 1—1, реже—1—2. Спинные бугорки округло-треугольные.
Рисунок и окраска. Общий фон сверху светло-серый; на боках
головы слабо выражена полоска от глаза до затылка и в углу рта; пять-шесть
бурых поперечных полос на туловище от затылка до поясницы, на хвосте —
7—10 полос такого же цвета, промежутки между ними равны толщине полос;
нижняя сторона белая.
Распространение (рис. 15; КНР, МНР: Кашгария и пустыня
Гоби до разв. г. Хара-Хото и прилегающих с сев.-з. районов Заалтайской Гоби).
Кадастр к рис. 15.
КНР:/- Янгигиссар [187]; 2 — разв. Хара-Хото, Ц. Гоби (ЗИН).
МНР: 1 — Ногонцав [78]; 2 — Шара-Хулсны-Булак [78]; 3 — Цугалбар-Ус [78]; 4 —
Цувра-Хара-Ула [78]; 5 — Хуцын-Хара-Ула [78].
Половой диморфизм и возрастная изменчи-
вость. У самок отсутствуют бедренные поры. Возрастная изменчивость не
изучена.
Систематика и географическая изменчивость.
В нашем распоряжении были выборки из Гоби. Здесь географическая изменчи-
вость не обнаружена. Следует учесть, однако, что этот вид описан из Кашгарии,
расположенной около 2000 км к западу. Не исключено, что западные выборки
будут отличаться от восточных.
Экология изучена недостаточно.
194
Рис. 86. Tenuidactylus elongatus:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — голова снизу, Д — лицевая
часть спереди, Е — спина, Ж — брюхо, 3 — преанальные поры, И — хвост сверху, К —
хвост снизу (ИЗАНУ SR 2642, Юж. Гоби, МНР)
Стации и количественные данные. Обитает в скалистой
местности, в ущельях и останцах. Редкий вид [31].
Отношение к температуре. Геккон данного вида, бывший
в нашем распоряжении, исследовался в термоградиентприборе (п = 42).
Он отмечен в диапазоне температур 28,5—40°, предпочитаемая температура
34,1 ± 0,5°. Видимо, это теплолюбивый вид.
Суточный цикл активности. По наблюдениям в террариу-
ме — сумеречно-ночной вид.
Сезонный цикл активности не изучен. Известна поздняя
находка из окр. Хара-Хото 10—12.XII 1926 г. (по коллекционным материалам,
ЗИН).
Питание не изучено. В неволе охотно поедал личинок мучных хрущей.
Размножение. В нашей выборке почти в два раза больше самок,
195
чем самцов (на 10—18). Детальные сведения по размножению отсутствуют.
В р д г и. Известен случай нападения на этого геккона стрелы-змеи
(Psammophis lineolatum) [31].
Особенности поведения. По нашим наблюдениям в террари-
уме, кашгарские гекконы, в отличие от других видов рода, хорошо роют норы
в песке и ведут себя не как обитатели вертикальных поверхностей, а как псам-
мофилы. В то же время в Гоби их находили в трещинах скал, пещерах и под
камнями [31].
Практическое значение и охрана. Вид редкий и неизу-
ченный. Включен в Красную книгу МНР, охраняется на территории Гобий-
ского биосферного заповедника.
ГЕККОН АГАМУРОВИДНЫЙ — TENUIDACTYLUS AGAMUROIDES
(NIKOLSKY, 1900)
Типовая территория: Пенсаре (Пенджсара), В. Керман, Иран.
Кариотип: не изучен.
1900 — Gymnodactylus agamuroides Nikolsky, Ежег. зоол. Муз. за 1895 г., Спб, т. 4, № 4, с. 384.
1963 — Cyrtodactylus agamuroides, S. Anderson, Proc. Calif. Acad. Sci. San Francisco, ser. 4,
vol. 31, p. 438 (err. det.).
1966 — Agamura agamuroides, Minton, Bull. Amer. Mus. Nat. Hist., N. Y., Vol. 134, Art. 2,
p. 80 (err. det.).
1984 — Tenuidactylus agamuroides, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 54.
Диагноз. Три носовых щитка; чешуй, на которых расположены пре-
анальные поры, значительно больше окружающих; снизу хвостового сегмента
имеются три кольца чешуй; чешуй вдоль нижней стороны тела 120; чешуй по-
перек брюха около 28; 2 четко выраженные преанальные поры.
Описание (рис. 87) (по лектотипу ЗИН 9327, ad «Пендж-Сара,
В. Кирман, 10.VIII 1898, Н. Зарудный»). L = 38,7 мм; L cd = 49,1 мм; L/
Led = 0,79; индексвысоты/ширины головы 63; диаметр уха/глаза 40; верхнегуб-
ных щитков 15/14; нижнегубных по 10; чешуй между центрами глаз 16; длина
спинного бугорка/И8; чешуй, окружающих спинной бугорок, 10; чешуй вдоль
тела снизу 120; чешуй поперек брюха около 28; подпальцевых пластин 20/21;
преанальных пор 2; дополнительные поры отсутствуют.
Подбородочный щиток пятиугольный; две пары нижнечелюстных щитков,
первая соприкасается между собой; дополнительные спинные бугорки отсут-
ствуют; первые носовые щитки разделены чешуйкой; спинные бугорки округло-
треугольные с довольно слабой ребристостью.
Рисунок и окраска (после фиксации в спирте). Напоминает
Т. kachhensis или Т. kirmanensis: между затылком и поясницей семь попереч-
ных полос, каждая из которых образована тремя—пятью пятнами; на хвосте
13 поперечных сплошных полос; на конечностях четыре—шесть поперечных по-
лосок; на голове неясный узор; нижняя поверхность белая.
Распространение (рис. 82). Достоверно известен только из ти-
повой местности.
Кадастр к рис. 82.
Иран:/ — Пенсаре [89]; 2 — Нейзар в Сейстане ([89] — находка нуждается в
подтверждении).
Систематика. Как показано [47], С. Андерсон [168] и Ш. Минтон
[263] ошибочно определили свой материал, приняв этот вид за агамур (cf.
persica). Это объясняется не только тем, что диагноз агамуровидного геккона со-
ставлялся по экземплярам, принадлежащим к видам из разных родов, но и их
близостью.
Экология не изучена. По данным Н. А. Зарудного [60], встречается
в изобилии на руинах глинобитных построек, в тенистых местах глинистых
и скальных обрывов. Рано утром и по вечерам встречается в нескольких сотнях
шагов от дневных убежищ на ровных местах. Активен и днем, но только.в тени.
196
Рис. 87. Tenuidactylus agamuroides:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — подбородок, Д, Е — носовые
щитки, Ж — спина, 3 — преанальные поры, И — хвост сверху, К — хвост снизу (ЗИН 9327,
Пенсаре, Иран)
ТИБЕТО-ГИМАЛАЙСКАЯ ГРУППА — TENUIDACTYLUS
Группа по ряду признаков является промежуточной между восточноази-
атскими голопалыми гекконами рода Cyrtodactylus и палеарктическими тонко-
палыми гекконами рода Tenuidactylus. Однако, несмотря на наличие некоторых
анцестральных признаков, в целом близка к последнему роду, в составе которо-
197
го и рассматривается. Почти все виды в музейных коллекциях представлены
единственными или единичными экземплярами, включая Т. fasciolatus, который
также, возможно, принадлежит к этой группе. Типовые экземпляры четырех
видов—Т. lawderanus1, Т. fasciolatus1 2, Т. himalayanus3и Т. dattensis4— оказа-
лись недоступными для нас, а опубликованные описания не учитывают мно-
гих основных признаков, использованных в настоящей работе. Это вынудило
нас не приводить здесь диагнозы указанных видов. Исходя из неполно-
го описания очень сложно установить не только подродовую принадлеж-
ность, но и его видовую самостоятельность. С другой стороны, необходимо
указать на довольно высокую степень изменчивости группы в целом, что отмеча-
лось нами ранее [153]. Это явление уже повлекло за собой появление многочис-
ленных синонимов: иногда их число приближается к числу известных экземп-
ляров (Т. lawderanus, Т. stoliczkai). Знакомство с типами видов, обитающих
непосредственно в Гималаях, вероятно, увеличит их число.
Следовательно, приведенная ниже таблица для определения и краткая ха-
рактеристика группы носят предварительный характер.
Краткая характеристика группы тибето-гималайские
тонкопалые гекконы. Хвост заметно приплюснут у основания, тол-
стый, довольно резко утоньшается в последней трети, равен длине тела, не-
значительно короче или длиннее; у части видов сегменты не выражены, а
хвостовые бугорки и подхвостовые щитки выражены очень слабо; три-четыре
носовых щитка, из которых первый крупнее остальных; высота первого верхне-
губного щитка обычно заметно меньше его ширины; чешуя головы обыкновенно
мелкая, однородная; лобная впадина часто выражена лучше, чем у остальных
видов рода (но не так сильно, как у Cyrtodactylus); нижнечелюстные щитки
хорошо выражены, первая пара всегда соприкасается между собой; спинные
бугорки мелкие, обычно округлые, иногда овально-треугольные со слабо вы-
раженной ребристостью, дополнительные бугорки отсутствуют; остальная че-
шуя всегда гладкая; вдоль нижней стороны тела 120—160 чешуй; до четырех
—шести преанальных пор; конечности иногда немного укорочены; базальные
фаланги пальцев у некоторых видов несколько расширены. Рисунок и окраска
значительно варьируют: у части видов на голове сохраняется мраморный
узор, на затылке — остатки затылочной полосы, а на теле — поперечные
полосы, разорванные посередине; у части видов этот рисунок преобразуется
в обычные для палеарктических гекконов М-образные полосы, иногда со-
храняющиеся лишь фрагментарно в виде пятен; эти полосы могут быть выра-
жены также в виде тонких пбперечных червеобразных линий.
Таблица для определения видов группы
тибето-гималацские тонкопалые гекконы
1 (6). Хвостовые сегменты не выражены.
2 (3). Вдоль нижней стороны тела не более 130 чешуй.........Т. tibetanus (Boulenger)
3 (2). Вдоль нижней стороны тела не менее 130 чешуй.
4 (5). Спинные поперечные полосы очень тонкие, червеобразные......................
......................................................Т. mintoni (Colubev, Szczerbak)
5 (4). Спинные поперечные полосы широкие, часто прерванные посередине.............
..................................................... Т. chitralensis (М. A. Smith)
6 (1). Хвостовые сегменты выражены отчетливо.
7 (8). На спине сплошные широкие поперечные полосы; L/L cd не менее 0,8; подхвостовые
щитки отсутствуют .......................................Т. stoliczkai (Steindachner)
8 (7). Фрагменты поперечных спинных полос в виде отдельных пятен; L/Lcd не более 0,8; под-
хвостовые щитки выражены не менее, чем в последней трети хвоста...................
......................................................Т. kirmanensis (Nikolsky)
1 Распространен [по 291] в Индии (рис. 89): / — округ Симла, Гималаи; 2 — Амбала;
3 — Техри-Гархвал; 4 — Алмора; 1—3. Непал.
2 Распространен (рис. 89): 1 — Индия, Джамму и Кашмир, г. Дода, г. Киштвар [210].
3 Распространен (рис. 89): 2 — Индия, округ Симла; 3 — Кумаон, Субату, Алмора, 1500 м
н. у. м. ]291].
4 Распространен (рис. 89) в Пакистане: / — Сев.-Зап. Пограничная пров., окр. пос. Дат-
та [330] (возможно, экз. SMF 63548 на рис. 90 относится к этой форме).
198
Рис. 88. Tenuidactylus tibetanus:
A — общий вид, Б — голова сбоку, В — лицевая часть спереди, Г — голова сверху, Д —
голова снизу (BMNH 1946.8.23.41, переправа Чангпо, Тибет)
ГЕККОН ТИБЕТСКИЙ — TENUIDACTYLUS TIBETANUS
(BOULENGER, 1905)
Типовая территория: переправа Чаксам, долина р. Чангпо (Брахмапутра),
Тибет, КНР.
Кариотип: не изучен.
1905 — Alsophylax tibetanus Boulenger, Ann. Mag. nat. Hist., London, ser. 7, Vol. 15, p. 378.
1.981 — Gymnodactylus tibetanus, Голубев, Вопр. герпетологии, Ленинград, с. 39.
1984 — Tenuidactylus tibetanus, Щербак, Голубев, Вестн. зоол., Киев, № 2, с. 55.
199
Рис. 89. Распространение Tenuidactylus lawderanus (/), Т. himalayanus (2)t T. chitralensis
(JJ, T. tibetanus (4), T. mintoni (5), T. stoliczkai (6), A. tokobajevi (7), T. fasciolatus (5), T. kir-
manensis (9), T. dattensis (10)
Диагноз. Чешуй вдоль нижней стороны тела не более 130; спинные по-
перечные полосы широкие, разорванные посередине; хвостовые сегменты не вы-
ражены; 2/3 нижней стороны хвоста покрыты щитками; L/L cd больше 1; под-
хвостовые щитки выражены в последних 2/3 хвоста; чешуй поперек головы \25.
Описание (рис. 88) (по голотипу BMNH 1905.2.8.5 RR 1946 8.23.41,
$ ad «Tsangpo Valley, Tibet. Pres: Lt Col. L. A. Waddel») L = 52,3 мм; L cd =
— 45 mm; L/L cd = 1,16; индекс высоты/ширины головы 57; диаметр
уха/глаза 60; верхнегубных щитков 9/10; нижнегубных 7/8; чешуй между цен-
трами глаз 25; длина спинного бугорка/L 26; чешуй, окружающих спинной бу-
горок, 8—9; чешуй вдоль тела снизу 126; чешуй поперек брюха около 30; под-
пальцевых пластин по 21; преанальные поры отсутствуют.
200
Подбородочный щиток треугольный; три-четыре пары нижнечелюстных
щитков, первая соприкасается между собой; первые носовые щитки также
соприкасаются между собой; три носовых щитка; подхвостовые щитки выражены
в последних 2/3 (в сочетании 1—1 на сегменте); хвостовые бугорки выражены
только на основании хвоста; спинные бугорки гладкие, округлые, расположе-
ны относительно регулярными рядами; конечности несколько укороченные;
передняя лапа, вытянутая вперед вдоль тела, немного заходит концами паль-
цев на передний край орбиты; базальные фаланги пальцев не расширены.
Рисунок и окраска. Основной фон кофейно-коричневый; на го-
лове мраморный узор, образованный темными пятнышками и червеобразными
полосками; на затылке заметны фрагменты затылочной полосы, характерной
для видов рода Cyrtodactylus; между лопатками и поясницей расположены че-
тыре более темных, чем основной фон, широких, с ровными краями поперечных
полос; каждая полоса окантована спереди и сзади темно-коричневым и пре-
рвана посередине; на хвосте и конечностях рисунок отсутствует; нижняя сторо-
на белая.
Распространение (рис. 89). Известен только из типовой мест-
ности.
Систематика. Переведен в род тонкопалых гекконов из рода Also-
phylax [37]. Очень близок к Т. chitralensis и, вероятно, к Т. lawderanus, Т. hi-
malayanus и Т. fasciolatus.
Экология не изучена. Известно, что типовой экземпляр был добыт
на высоте 12 000 футов (3648 м н. у. м. [205]).
ГЕККОН ЧИТРАЛЬСКИЙ — TENUIDACTYLUS CHITRALENSIS
(М. A. SMITH, 1935)
Типовая территория: Каракал в Бамхотской долине, Читрал, Сев.Пакистан.
Кариотип: не изучен.
1935 — Gymnodactylus chitralensis М. A. Smith, Fauna brit. Ind., Rept. Amph., Vol. 2, p. 46.
1969 — Gymnodactylus (Cyrtodactylus) chitralensis, Mertens, Stutt. Beitr. Naturk., Stuttgart,
N 197, S. 24.
1984 — Tenuidactylus chitralensis, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 55.
Диагноз. Чешуй вдоль нижней стороны тела не менее 130; спинные по-
перечные полосы широкие, разорванные посередине; хвостовые сегменты не вы-
ражены; в последней трети хвоста снизу может быть ряд крупных щитков;
L/L cd = 0,98; чешуй поперек головы 26—32.
Типы хранятся в Индийском (Калькутта) и Британском (Лондон) му-
зеях.
Описание (рис. 90) (по 3 экз. из колл. SMF и MNHP). L = 36,3—
59,5 мм; L/L cd = 0,80; индекс высоты/ширины головы 49—56; диаметр уха/
глаза 63; верхнегубных щитков (п = 6) 10—12 (11,31 ± 0,28); нижнегубных
(п = 6) 8—9 (8,89 ± 0,01); чешуй между центрами глаз 26—32; длина спинного
бугорка/L 8—9; чешуй вдоль тела снизу 142—163; чешуй поперек брюха 30—
34; подпальцевых пластин (п = 6) 15—22 (17,80 ± 1,31); преанальных пор 8.
Подбородочный щиток тре- или пятиугольный; две-три пары нижнечелюст-
ных щитков, первая соприкасается между собой; первые носовые щитки раз-
делены чешуйкой, три-четыре носовых щитка; подхвостовые щитки могут быть
выражены в последней трети хвоста (в сочетании 1—2, 1—1 на сегменте); хвос-
товые бугорки выражены только на основании хвоста; спинные бугорки округ-
лые или слабо овальные, гладкие, расположены относительно регулярными
рядами; конечности несколько укороченные: передняя лапа, вытянутая вперед
вдоль тела, не доходит концами пальцев до конца морды; базальные фаланги
пальцев не расширены.
Окраска. Сходна с окраской предыдущего вида или отличается более
узкими поперечными слегка зигзагообразными полосами.
Распространение (рис. 89). Обитает в княжестве Читрал в
Сев. Пакистане.
Экология не изучена. Ранняя находка в Абботабаде — 12.V 1958 г.
(по коллекционным экземплярам).
201
Рис. 90. Tenuidactylus chitralensis:
А, Б — общий вид, В — лицевая часть спереди, Г — подобородок, Д — спина, Е —
брюхо, Ж — преанальные поры, 3 — хвост сверху, И — хвост снизу (Б, Д — MNHP
1916.63, верховья Инда, Пакистан; остальное — SMF 63548, Абботабад, Пакистан)
ГЕККОН МИНТОНА — TENUIDACTYLUS MINTON]
(GOLUBEV, SZCZERBAK, 1981)
Типовая территория: Удиграм, Сват, Пакистан.
Кариотип: не изучен.
1981 — Gymnodactylus mintoni Голубев, Щербак, Вести, зоол., Киев, № 3, с. 40.
1984 — Tenuidactylus mintoni, Щербак, Голубев, Вестн. зоол., Киев, № 2, с. 55.
202
Рис. 91. Tenuidactylus mintoni:
Л общий вид, Б — голова сбоку, В — носовые щитки, Г — голова сверху, Д — голова
снизу, Е— спина, Ж — хвост сверху, 3 — палец снизу (SAM 683, Сват, Пакистан)
Диагноз. Чешуй вдоль нижней стороны тела не менее 130; спинные
поперечные полосы тонкие, червеобразные; хвостовые сегменты не выражены;
подхвостовых щитков нет на всем протяжении хвоста; L/L cd = 1,18; чешуй
поперек головы около 30.
Описание (рис. 91) (по голотипу SAM 683, $> ad. (?) «Udigram in
Swat, W. Pakistan, 30 Murch.S. Minton»).
L = 38,4 mm; LIL cd =; 1,18; индекс высоты/ширины головы 60; диаметр уха/
203
Рис. 92. Tenuidactylus stoliczkai:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — верхний край орбиты, Д — го-
лова снизу, Е — лицевая часть спереди, Ж — спина, 3 — хвост сбоку (NMW 16756, Каш-
мир, Пакистан)
глаза 32; верхнегубных щитков по 11; нижнегубных 9/8; чешуй между центра-
ми глаз около 30; длина спинного бугорка ZL8; чешуй вдоль тела снизу 150;
чешуй поперек брюха 36; подпальцевых пластин по 17; преанальные поры от-
сутствуют.
Подбородочный щиток пятиугольный; нижнечелюстных щитков две—че-
тыре пары, из них первая соприкасается между собой, а одна-две последних
204
слабо отличимы от горловой чешуи; первые носовые щитки разделены двумя
чешуйками; три-четыре носовых щитка, последний слабо различим; подхвосто-
вые щитки отсутствуют; спинные и хвостовые бугорки почти не выражены;
конечности несколько укороченные: передняя лапа, вытянутая вперед вдоль
тела, достигает концами пальцев переднего края орбиты; базальные фаланги
пальцев несколько расширены.
Прижизненная окраска и рисунок [263]. Основной фон
спинной стороны янтарный, переходящий на хвосте в лимонный; на спине серия
из восьми неправильных прерывистых темных очень тонких поперечных по-
лос, большинство из которых окантованы сзади белым; на боках и дорсальных
поверхностях головы, хвоста и конечностей разбросаны небольшие черные пят-
на и точки; брюхо палево-желтое; на губных щитках чередующиеся черные и бе-
лые полоски; от ноздри через глаз до уха проходит коричневая полоса, также
окантованная сверху белым.
Распространение (рис. 89). Известен только из типовой местности.
Экология не изучена.
ГЕККОН СТОЛИЧКИ — TENUIDACTYLUS STOLICZKAI
(STEINDACHNER, 1868)
Типовая территория: Кароо, сев. Драса, Кашмир.
Кариотип: не изучен.
1868 — Gymnodactylus stoliczkai Steindachner, Reise Novara, Zool., 1, Rept., s. 15.
1872 — Cyrtodactylus уarkandensis J. Anderson, Proc. zool. Soc. London, p. 381.
1922 — Gymnodactylus walli Ingaldby, J. Bombay nat. Hist. Soc., Vol. 28, p. 1051.
1954 — Cyrtodactylus stoliczkai, Underwood, Proc. Zool. Soc., London, Vol. 124, N 3, p. 475.
1984 — Tenuidactylus stoliczkai, Щербак, Голубев, Вестн. зоол., Киев, № 2, с. 55.
Диагноз. Чешуй вдоль нижней стороны тела менее 150; спинные по-
перечные полосы широкие, М-образные; хвостовые сегменты хорошо выраже-
ны; увеличенных подхвостовых щитков нет на всем протяжении хвоста;
L/L cd = 0,85—0,90; чешуй поперек головы 17—20.
Описание (рис. 92) (по голотипу NMW 16 756, $ ad «Karoo bei Dras,
Kashmir, 1866, Stoliczka», типам Gymnodactylus walli BMNH 1910. 7.12.1.
и Cyrtodactylus yarkandensis BMNH 72.3.22. 4). L = 27—48 мм; L/L cd =
= 0,85—0,98; индекс высоты/ширины головы 52—59; диаметр уха/глаза
28—36; верхнегубных щитков (п = 6) 10—11 (10,87 ± 0,02); нижнегубных
(п = 6) 7—8 (7,88 ± 0,02);чешуй между центрами глаз 17—20; длина спинного
бугорка/L 8—10; чешуй вдоль тела снизу 120—149; чешуй поперек брюха
27—32; подпальцевых пластин (п = 6) 23—24 (23,5 ± 0,01); преанальные поры
отсутствуют (все $>).
Подбородочный щиток тре- или пятиугольный; три-четыре пары нижне-
челюстных щитков, первая пара соприкасается между собой; первые носовые
щитки разделены одной-двумя чешуйками; три носовых щитка; конечности
умеренной длины; передняя лапа, вытянутая вперед вдоль тела, достигает
концами пальцев конца морды; базальные фаланги пальцев не расширены;
уплощенность хвоста и хвостовые сегменты выражены очень четко (8—10 сег-
ментов); три-четыре конических бугорка расположены по обеим сторонам сег-
мента в его средней части и друг с другом не соприкасаются; подхвостовые щит-
ки не выражены. Спинные бугорки округлые, гладкие, расположены почти
беспорядочно; мелкая чешуя хвоста гладкая.
Рисунок и окраска. На светлом фоне от затылка до крестца
семь-восемь широких поперечных М-образных темных полос; затылочная по-
лоса подходит с обеих сторон головы через глаза к ноздрям; каждая попереч-
ная полоса от передней (ближе к голове) к задней части темнеет и оканчивается
тонкой буро-коричневой окантовывающей линией; на хвосте около 12 такого
же цвета поперечных полос; на голове сверху неясный узор; конечности в по-
перечных тонких полосках; нижние поверхности белые.
Распространение (рис. 89). Известен из Читраля ^Сев. Паки-
стан) и 3. Кашгарии (Китай); обитает между этими областями в горных системах
Сев. Индии.
205
Рис. 93. Tenuidactylus kirmanensis:
A — общий вид, Б — голова сбоку, В — голова сверху, Г — голова снизу, Д — преанальные
поры, Е — хвост сверху, Ж — хвост снизу, 3 — палец снизу (ЗИН 9330, Вост. Кирман<
Иран)
Кадастр к рис. 89.
К и т а й:/ —Яркенд [193].
Пакистан: 2 — Читрал [258].
И н д и я: 3 — Кашмир, окр. Кароо, сев. пос. Драс (NMW); 4 — Кашмир, Каргил [263];
5 — Ладак; Саспул-Гомпа; Геми [324].
Систематика и географическая изменчивость
не изучены.
Экология не изучена. Ранняя находка сделана в Читрале —
26.IV 1962 г. (по коллекционным экземплярам).
206
ГЕККОН КЕРМАНСКИЙ — TENUIDACTYLUS KIRMANENSIS
(NIKOLSKY, 1900)
Типовая территория: гора Кухе-туфтан, В. Кирман, Иран.
Кариотип: не изучен.
1900 — Gymnodactylus kirmanensis Nikolsky, Ежегод. зоол., Муз. за 1889 г., т. 4, с. 381.
1963 — Cyrtodactylus kirmanensis, S. Anderson, Proc. Calif. Acad. Sci., San Francisco, Ser. 4,
vol. 31, N 16, p. 474.
1984 — Tenuidactylus kirmanensis, Щербак, Голубев, Вести, зоол., Киев, № 2, с. 55.
Диагноз. Чешуй вдоль нижней стороны тела менее 140; поперечные
полосы спины выражены фрагментами в виде пятен; хвостовые сегменты хоро-
шо заметны; подхвостовые сегменты выражены на всем протяжении хвоста;
L/L cd = 0,71—0,78; чешуй поперек головы 17—20.
Лектотип. ЗИН 9330, d* ad «В. Кирман, 17.VI 1898, Н. Зарудный».
L = 50,4; хвост поврежден; чешуй между центрами глаз 17; верхнегубных
щитков по 11; нижнегубных по 8; чешуй вдоль тела снизу 131; чешуй поперек
брюха около 30; подпальцевых пластин по 22; преанальных пор 4.
Описание (рис. 93) (по 8 экз. из колл. ЗИН и NMW). LO6m (ft = 8)
37,4—50,7 мм; LIL cd (п = 2) 0,71—0,78; индекс высоты/ширины головы
(n = 1) 53; диаметр уха/глаза (п = 7) 32—42 (39,14 ± 1,69); верхнегубных
щитков (и = 15) 11—12 (11,47 ± 0,13); нижнегубных (п = 16) 7—9 (8,0 ±
±0,16); чешуй между центрами глаз (п = 7) 15—20 (17,43 ± 0,65); дли-
на спинного бугорка/L (и = 8) 15—16 (15,62 ± 0,18); чешуй вдоль тела
снизу (п = 7) 120—132 (128,0 ± 1,59); чешуй поперек брюха (п = 3) 26—
30; подпальцевых пластинок (и = 14) 20—24 (22,51 ± 0,34); преанальных
пор 4.
Подбородочный щиток тре- или пятиугольный; нижнечелюстных щитков
три-четыре пары, первая соприкасается между собой; первые носовые щитки
разделены чешуйкой; три носовых щитка; конечности умеренной длины: перед-
няя лапа, вытянутая вперед вдоль тела, достигает концами пальцев до конца
морды; базальные фаланги пальцев несколько расширены; хвостовые сегменты
выражены четко, бугорки расположены так же, как у предыдущего вида;
подхвостовые щитки образуют на одном сегменте сочетания 1—1—2, 1—1—1,
2—1—1, 2—2—2; спинные бугорки на затылке и лопатках округлые и гладкие,
к задней части и бокам туловища вытягиваются в овальные и овально-треу-
гольные, на них появляются слабые ребрышки; мелкая чешуя хвоста слабо реб-
ристая.
Рисунок и окраска (после фиксации в спирте). На светлом фоне
спины около восьми темных поперечных полос, образованных отдельными пят-
нами, местами образующими шахматный узор; на хвосте восемь — десять такого
же цвета полос; на голове мелкие темные пятна, на конечностях — поперечные
полоски; нижние поверхности белые.
Распространение. Известен только из типовой местности
(рис. 89).
Половой диморфизм и возрастная изменчивость.
Самки лишены преанальных пор; среди изученных гекконов молодые отсутство-
вали.
Систематика. Ранее [46] этот геккон был помещен в подрод Medio-
dactylus, однако затем [153] в группу тибето-гималайских видов. Причина
этого — близость указанного вида к обеим группам и, кроме того (в меньшей
степени), к видам подрода Cyrtopodion. С тибето-гималайской группой его
связывает большее количество основных признаков фолидоза.
Экология не изучена. Приводятся отдельные сведения по наблюде-
ниям Н. А. Зарудного [60].
Стации и количественные данные. Встречается в изоби-
лии в горной местности между Сейстанской и Бампурской котловинами. Най-
ден в долине и в горах, где обитает на отвесных каменистых выступах (обрывах)
в горах, в берегах речек и сухих русел, среди тенистых карнизов, трещин, ниш,
иногда среди осыпей скальных глыб. Чаще встречается на гранитах, реже на
конгломератах и других горных породах.
207
Суточный цикл активности. Чаще встречается утром и ве-
чером, чем ночью.
Особенности поведения. Общителен по отношению к другим
особям. Днем отлично видит, поэтому добывать данный вид трудно. Каждый
экземпляр имеет свой наблюдательный пункт, на который возвращается после
того, как был спугнут и загнан куда-нибудь в трещину. Автор добывал их
в мае.
Род крысохвостый геккон, или агамура —
Agamura Blanford, 1874
Типовой вид: Agamura crural is Blanf.
1874 — Agamura Blanford, Ann. Mag. nat. Hist., London, ser. L, Vol. 13, p. 455.
1973 — Rhirtogekko de Witte, Bull. Inst. roy. Sci. nat. Belg., Vol. 49, N 1, p. 1.
1984 — Agamura Щербак, Голубев, Вести, зоол. Киев, № 2, с. 55.
Диагноз. Конечности очень тонкие и длинные: передняя лапа, вытя-
нутая вперед и вдоль тела, достает кистевым суставом до конца морды1;
длина стопы с пальцами заметно меньше длины голени; хвост резко утоньшается
у основания, не ломкий, короче или незначительно длиннее тела; преанальные
поры выражены только у самцов; пальцы слабо искривлены, не расширены,
иногда слабо сжаты с боков, когтистые, покрыты снизу продольным рядом по-
перечно расширенных гладких пластинок; зрачок вертикальный с зазубренны-
ми краями; все чешуи и щитки тела гладкие (кроме хвостовых бугорков, на ко-
торых могут проступать слабые продольные ребрышки).
Дополнительные признаки. Межчелюстной щиток четырех-
или (слабо выражено) пятиугольный; три носовых щитка; чешуя морды округло-
многоугольная, незначительно уменьшается к затылку, где перемешивается с
коническими бугорками; высота первого верхнегубного щитка равна или не-
значительно меньше его ширины вдоль края рта; ухо мелкое, овальное; заносо-
вые впадины выражены, лобная — нет. Подбородочный щиток пятиугольный
или трапециевидный; одна — три пары нижнечелюстных щитков, из которых
первая соприкасается между собой, может замещаться рядом из более крупных,
чем остальные, горловых чешуй. На спине, покрытой мелкой округлой, плавно
увеличивающейся к бокам чешуей, располагаются слабо выраженными про-
дольными (около 26) и поперечными (8—10) рядами мелкие конические спинные
бугорки; брюшная чешуя шестиугольно-округлая, часто с пильчатым свобод-
ным (задним) краем; по три- четыре соприкасающихся друг с другом боковыми
гранями слабо выраженных бугорка по обеим сторонам хвостового сегмента;
снизу хвоста расположен продольный ряд из крупных пластинок или несколь-
ко увеличенных чешуй, по две-три на сегменте. Основание когтя скрыто чех-
лом, состоящим из верхней и нижней пластинок, с которыми соприкасается
сверху один, с боков по два ряда чешуй. Среди мелкой чешуи верхней стороны
задних лап расположены округлые плоские бугорки, отсутствующие на перед-
ней лапе.
Половой диморфизм и возрастная изменчивость
проявляются, по-видимому, у всех видов рода сходно: у самок отсутствуют
преанальные поры; у молодых особей агамуроидный габитус (длинные субтиль-
ные конечности, относительно крупная голова) выражены менее четко, чем
у взрослых (что неоднократно приводило к ошибочному определению их как
голопалых гекконов).
Род включает четыре вида, распространенных в Ю.-З. Азии (Афганистан,
Иран, Пакистан).
Таблица для определения видов рода Agamura
14). На нижней поверхности бедер чешуя однородная.
2(3). Поперек живота больше 30 чешуй..................A. persica (Dumeril)
3(2). Поперек живота меньше 20 чешуй................A. gastropholis (Werner)
4(1). На нижней поверхности бедер ряд из увеличенных чешуй.
1 Кроме A? femoral is, у которой конечности более короткие [290],
208
Рис. 94. Agamura persica (восточная популяция):
А — общий вид, Б — голова сбоку, В, Г — подбородок. Д — спина, Е — преанальные по-
ры, Ж — палец снизу, 3 — хвост сверху, И — хвост снизу (А, Б, Ж — ZML 57/3791. Хас-
сан-Гуйлан, Пакистан; остальные — CAS 120282, Гиришк, Афганистан)
5(6). Носовые щитки заметно вздуты, поперек живота 17—22 чешуйки . . A. femoralis (Smith)
6(5). Носовые щитки резко вздуты и вытянуты в трубочки; поперек живота 26—28 чешуй . .
.................................................................A. misonnei (de Witte)
ПЕРСИДСКАЯ АГ АМУ РА — AGAMURA PERSICA
(DUMERIL, 1856)
Типовая территория: «Персия».
Кариотип: не изучен.
1856 — Gymnodactylus persicus Dumeril, Arch. Mus. Hist. Nat. Paris, vol. 8, p. 481.
209
Рис. 95. Распространение Agamura persica (7), A. temoralis (2), A. gastropholis (5), A. mison-
nei (4)
1874 — Agamura cruralis Blanford, Ann. Mag. Nat. Hist., London, Ser. 4, vol. 13, N 78, p. 455.
1876 — Agamura persica, Blanford, East. Persia, Zool., vol. 2, p. 358.
Диагноз. Поперек живота 30—50 чешуй; носовые щитки слабо взду-
ты; на нижних поверхностях бедер нет ряда из увеличенных чешуй; 2—4 слабо
выраженных преанальных поры; хвост короче туловища.
Типы. Хранятся в Парижском музее [245].
Описание (рис. 94, 96) (по 10 экз. из колл. ЗИН, CASh ZML); Ьобщ.
(и = 9) 41,1—72,0 (52,21 ± 3,77) мм; L/L cd (п = 8) 1,05—1,42 (1,22 ± 0,05);
индекс высоты/ширины головы (л = 7) 53—60 (56,86 ± 0,96); диаметр
уха/глаза (п = 7) 20—40 (30,57 ± 2,84); количество верхнегубных щитков
(п = 20) 9—12 (11,19 ± 0,21); нижнегубных (п = 20) 12—16 (14,62 ± 0,30);
чешуй между центрами глаз (л = 8) 21—27 (23,62 ± 0,75); длина спинного
бугорка/L (л = 7) 11—12 (11,57 ±0,20); чешуй, окружающих спинной буго-
рок (п = 8), 8—11 (10,25 ± 0,37); чешуй вдоль тела снизу (л = 8) 130—
158 (148,86 ± 3,67); чешуй поперек брюха (л = 9) 37—50 (40,86 ± 1,62);
преанальных пор (л = 4) 2—4; подпальцевых пластинок (л = 14) 16—24
(20,57 ± 0,71).
Подбородочный щиток трапециевидный (80 %) или пятиугольный (тре-
угольный) (20 %); нижнечелюстные щитки имеются (20 %) или отсутствуют
(80 %), в этом случае с подбородочным соприкасаются три-четыре (по 25 %),
пять (37,5 %) или шесть (12,5 %) горловых чешуек; первые носовые щитки раз-
делены двумя (30 %), тремя (50 %), четырьмя или пятью (по 10 %) чешуй-
ками.
Рисунок и окраска (после фиксации в спирте). Основной фон
тела охристый. На голове неясный бурый узор; от затылка до поясницы попе-
рек тела располагаются пять — семь темных М-образных широких полосы, из
которых затылочная и следующая за ней могут быть выражены значительно
лучше остальных; на хвосте и конечностях по несколько поперечных полосок
такого же цвета. Нижние поверхности тела светлые.
Распространение (рис. 95). Ц. и В. Иран, Ю. и Ю.-В. Афга-
нистан, Ц. и 3. Пакистан.
Кадастр к рис. 95.
Иран:/ — окр. Шах-Аббас, пров. Тегеран, 34° 44' с. ш., 52° 10' в. д. (CAS); 2 — Пир-
бекран, около Исфахана [315]; 3 — Галлатаппех, пров. Исфахан [315]; 4 — г. Сиахкух, пров.
Тегеран (на картах — пров. Исфахан) [216]; 5 — между Фейзабадом и Чахар-Фарсангом в
Сарр-Чахской котловине [59]; 6 — Бесиран [87]; 7 — пров. Керман, хр. Джебальбарез [315];
8 — 32 км з. пос. Захедан, 29° 28' с. ш., 60° 41' в. д. [216]; 9 — В. Кирман, Дуз-Аб (ЗИН;
определен как G. agamuroides); 10 — стр. Сиб (ЗИН); 11 — Эскан, Белуджистан [291]; 12 —
Бахукелат [181]; 13 — Торбете-Хейдерие [303]; 14 — 6 км з. Аюби, 34° 18' с. ш.,
60° 35' в. д. [303].
210
Афганистан:/ — Патман, 34° 36' с. ш., 68е 56' в. д. [172]; 2 — Чахардех (Мазан-
деран) [96]; 3 — Гассан-Гуилан (Джайлан), ю.-з. пос. Гиришк [317]; 4— 50 км з. пос.
Гиришк (CAS).
Пакистан:/ — Вана [232]; 2 — Калат [258]; 3 — сев. пос. Ноккунди [258]; 4 — Ме-
руи [258]; 5 — 30—37 км сев. пос. Бела [263]; 6 — Дживани [258]; 7 — Ормара [258]; 8 —
в. пос. Монз, около Карачи, Вазиристан [245].
Не найдены: Zaudi Achmed-Wiiste [310]; Kidsh, Mekrankuste (ZFMK).
Половой диморфизм и возрастная изменчивость.
Кроме отмеченного уже наличия преанальных пор у самцов, в области клоаки
у них наблюдается вздутие, отсутствующее у самок. Возрастную изменчивость
см. выше.
Систематика и географическая изменчивость.
М. Смит [2911 синонимизировал A. persica и A. cruralis Blanf., и это мнение
в настоящее время стало общепринятым. Однако в пределах Иранского Белуд-
жистана (типовая территория вида Блэнфорда) и к северу до Сейстанской кот-
ловины и пустыни Деште-Лут у особей этого вида отмечено появление крупных
нижнечелюстных щитков. Эти особи имеют отклонения и по некоторым другим
показателям. Недостаток сравнительного материала не позволяет пока делать
окончательные таксономические выводы. Однако, учитывая то, что в указанном
регионе наблюдается высокая степень эндемизма внутри рода агамур (A. fe-
moralis, A. misonnei), можно допустить наличие здесь особого подвида —
A. persica cruralis Blanf. (рис. 96).
Экология изучена недостаточно.
Стации. По наблюдениям И. С. Даревского в Иране (письм. сообщ.),
персидская агамура обитает в каменистой полупустыне, покрытой черной
щебенкой, на стыке с песчаной полупустыней, на склонах холмов. Н. А. Заруд-
ный [601 добывал этих ящериц среди крупных обломков скал «у подошвы ка-
менной розсыпи» и на твердой очень щебнистой возвышенной степи, скудно по-
росшей низкорослой полынью. Ш. Минтон [263] встречал их на скальных тер-
расах на высоте 100—3000 футов (30—942 м н. у. м.)
Суточный цикл активности. Ведет ночной образ жизни
[49].
Сезонный цикл активности. Известна по коллекционным
материалам находка 22.XII 1962 г. в Иранской пров. Исфаган и 9.II 1901 г.
в стр. Сиб (Белуджистан).
Размножение. Яйца откладывает в июне, молодые появляются
в конце сентября [263].
Особенности поведения. Ходит на приподнятых лапах, дви-
жения напоминают собачьи (по наблюдениям в неволе). Хвост почти не лом-
кий [49].
ФАРСИЙСКАЯ АГАМУРА — AGAMURA GASTROPHOLIS
(WERNER. 1917) COMB. N.
Типовая территория: Иран, пров. Фарс.
Кариотип: не изучен.
1917 — Gymnodactylus gastropholis Werner, Verh. zool.-botan. Ges. Wien, Vol. 67, S. 194.
1951 — Gymnodactylus agamuroides, Wettstein, S. ber. Osterr. Acad. Wiss. Wien, math.-natur-
wiss. KI., Abt. 1, Vol. 160, p. 432.
Диагноз. Поперек живота 14—18 чешуй; носовые щитки слабо взду-
ты; на нижних поверхностях бедер чешуя однородная; 4 четко выраженные
преанальные поры; хвост не длиннее тела.
Лектотип. ZFMK 27095, d ad Пров. Фарс, Иран [3081. L = 49,4 мм,
хвост поврежден; чешуй между центрами глаз 20; верхнегубных щитков 12—
13; нижнегубных 9—10; чешуй вдоль тела снизу 90; чешуй поперек брюха 16;
чешуй вокруг спинного бугорка 8; преанальных пор 4; подпальцевых пласти-
нок по 22.
Описание (рис. 96) (по 4 экз. типовой серии из колл. ZFMK).
Ьобщ.. (и = 3) 44,6—50,0 (48,0 ± 1,71) мм; хвосты повреждены у всех особей;
индекс высоты/ширины головы (и = 3) 58—63 (60,0 ± 1,53); диаметр уха/гла-
211
Рис. 96. Agamura persica (вверху) (западная популяция «cruralis»):
А — общий вид, Б — носовые щитки, В — подбородок, Г — брюхо, Д — спина, Е — хвост снизу
(ЗИН 9328, страна Сиб. Иран)
Agamura gastropholis (внизу):
А — общий вид, Б — голова сбоку, В — лицевая часть сверху, Г — голова снизу,
Д — брюхо, Е — хвост сверху, Ж — преанальные поры, 3 —- бедро (ZFMK 27095, пров. Фарс,
Иран)
за (п = 3) 20—37 (31,0 ± 5,51); количество верхнегубных щитков (п — 8)
12—14 (12,87 ± 0,23); нижнегубных (п = 8) 9—10 (9,25 ± 0,16); чешуй меж-
ду центрами глаз (п = 4) 18—20 (18,75 ± 0,48); длина спинного бугорка/L
(п = 3) 11—14 (12,33 ± 0,88); чешуй, окружающих спинной бугорок (п = 3),
8—9 (8,25 ± 0,25); чешуй вдоль тела снизу (п = 4) 84—94 (91,0 ± 1,08); че-
шуй поперек брюха (п = 4) 14—18 (16,0 ± 0,82); преанальных пор (п = 4) 4;
подпальцевых пластинок (п = 8) 22—24 (23,0 ± 0,33).
212
Подбородочный щиток пяти- или треугольный (100 %); нижнечелюстных
щитков три пары (100 %), первая соприкасается между собой широким швом
(100 %); первые носовые щитки разделены между собой тремя чешуйками
(100 %).
Рисунок и окраска (после фиксации в спирте). Гекконы в зна-
чительной степени утратили окраску и рисунок, но по сохранившимся ее эле-
ментам можно заключить, что они не отличаются от таковой у близких видов:
по более светлому фону проходит пять — семь спинных и пять — семь хвостовых
поперечных темных полос; неясные темные полосы имеются на конечностях;
нижняя поверхность белая.
Распространение. Известны только из типового местонахожде-
ния (рис. 95).
Систематика. Этот вид, описанный первоначально как голопалый
геккон, был идентифицирован О. Веттштейном [315] с G. agamuroides. Однако
при сравнении этих и других близких видов отчетливо видно, что форма «ga-
stropholis» является не тонкопалым, а типичным агамуроидным гекконом, кото-
рый среди видов последнего рода выделяется очень крупной брюшной чешуей
и может рассматриваться как самостоятельный вид. Вероятнее всего, что оба
молодых экземпляра «G. agamuroides» из Кермана, использованные О. Ветт-
штейном при сравнении с G. gastropholis, относятся именно к последнему виду.
Экология не изучена.
АГАМУРА-НОСОРОГ — AGAMURA MISONNEI
(DE WITTE, 1973), COMB. N.
Типовая территория: Иран, пуст. Деште-Лут, ЗГ 13' с. ш., 58° 47' в. д.
Кариотип: не изучен.
1973 — Rhinogekko misonnei de Witte, Bull. Inst. r. Sci. nat. Belg., Bruxelles, vol. 49, N 1, p.l.
Диагноз. Поперек брюха 26—28 чешуй; носовые щитки резко вздуты
и вытянуты в трубочки; на нижних поверхностях бедер ряд из 9—12 укруп-
ненных чешуй; 4—8 очень слабо выраженных пор; хвост несколько длиннее
тела.
Голотип хранится в Королевском бельгийском институте естествен-
ных наук (Брюссель), добыт «desert du Dast-I-Lut, Iran, 30° 13' N et 58° 47' Е»;
паратип — «Kuh-I-Bakhtu, desert du Lut, Iran Oriental, 31°40'N, 58° 2Г E,
12.IV 1972 leg. X. Misonne». .
Описание (рис. 97) (по паратипу из той же коллекции и [320, 321].
L = 56,9—61,0 мм; L cd = 58,0—73,0 мм; UL cd = 0,84—0,96; индекс
высоты/ширины головы 56; диаметр уха/глаза 53; количество верхнегубных
щитков 9—12; нижнегубных 8—11; чешуй между центрами глаз 16; длина спин-
ного бугорка/Л 13; чешуй, окружающих спинной бугорок, 8—9; чешуй вдоль
тела снизу 120; чешуй поперек брюха 26—28; преанальных пор 4—8.
Подбородочный щиток пятиугольный, пара крупных нижнечелюстных
щитков соединена широким швом; снизу хвоста на одном сегменте вначале
продольный ряд из трех более крупных, чем остальные, чешуй, а в дальней-
шем — из крупных четырехугольных пластинок (по две на сегменте).
Рисунок и окраска. Основной фон верхней стороны тела ко-
фейно-серый; широкие поперечные темно-коричневые полосы на туловище (пять)
и хвосте (семь), на конечностях также имеются темные полоски, которые свет-
лее остальных; губы и межчелюстной щиток покрыты темно-коричневыми пят-
нами; нижние поверхности беловатые.
Распространение (рис. 95). В настоящее время известен только
из пуст. Деште-Лут и ю. предгорий хр. Чагайи (Иран и Пакистан).
Кадастр к рис. 95.
Иран: 1.— Кух-и-Бахту, 31° 40' с. ш., 58° 21' в. д. (BZM); 2 — Деште-Лут, 30® ЗГ с.
ш., 58° 47' в. д. [320].
Пакистан: 1 — сев. Ноккунди [320].
Систематика. Родовая самостоятельность этого оригинального гек-
кона повторно защищалась автором его описания [321]. Однако именно из
213
Рис, 97. Agamura misonnei:
A — голова сбоку, Б >-» лицевая часть спереди, В —> голова сверху, Р — голова снизу,
Д — спина, Е — брюхо, X — бедро снизу, 3 — преанальные поры, И — хвост сверху,
К — хвост снизу (BZ 24.703 Reg.25/6, Кухе-Бахту, Иран)
этой работы, как и из других источников [290, 263, 172], вытекает, что форма
«misonnei» настолько близка к A. femoralis, что выделение ее в самостоятельный
род не представляется целесообразным.
Экология не изучена. Автор описания [320] отмечает, что голотип
был добыт в пустыне без всякой растительности. Паратип — в гравийной пу-
стыне с очень скудной растительностью.
214
АГ АМУРА ХАР АНСКАЯ — AGAMURA FEMORALIS SMITH, 1933
Типовая территория: Харан, Белуджистан в Пакистане.
К а р и от и п: не изучен.
1933 — Agamura femoralls М. A. Smith, Rec. ind. Mus., Calcutta, vol. 35, p. 17.
Диагноз. Поперек живота 17—21 чешуя; носовые щитки заметно взду-
ты; на нижних поверхностях бедер ряд из увеличенных чешуй; пять-шесть
четко выраженных пор; хвост незначительно длиннее тела.
Тип. Хранится в Британском музее.
Описание [290 и 2631 (рис. 98). Lad = 52—60 мм; ULcd = 0,91; диаметр
уха менее половины диаметра глаза; верхнегубных щитков 12; нижнегубных —
11; пара нижнечелюстных щитков, за которыми следует еще одна пара более
мелких щитков; поперек брюха 17—21 чешуя, брюшная чешуя увеличивается
в задней части тела; у d 6 (реже — 5) преанальных пор; передняя конечность,
вытянутая вперед и вдоль тела, достает только до затылка; вдоль нижней
поверхности каждого бедра тянется ряд из 9—12 укрупненных чешуй; вдоль
Рис. 98. Agamura femoral is:
А общий вид сверху, Б — общий вид снизу, В — лицевая чисть сбоку, Г — анальные поры
(голотип BMNH 1912.3.26.12; фото Е. N. Arnold).
215
нижней стороны каждого сегмента несколько более крупных, чем остальные,
чешуй, по две в трех продольных рядах.
Окраска и рисунок. На более светлом фоне пять темных бурых
поперечных полос на туловище и 8—10 — на хвосте. Нижняя сторона белая.
Распространение (рис. 95). Известна только из пуст. Харан и
прилегающих к ней с севера гор Чагай на Афгано-Пакистанской границе.
Кадастр к рис. 95.
Иран: / — г. Кухе-Тефтан [263]; окр. Мирджаве [321].
Пакистан:/ — сев. пос. Ноккунди [258]; 2 — г. Харан [258].
Экология не изучена. По сообщению Ш. Минтона [263], этот вид
обитает на выходах скал среди песков; 7 июня у самок отмечены крупные
яйца.
СПИСОК ЛИТЕРАТУРЫ
1. Алекперов А. М. Земноводные и пресмыкающиеся Азербайджана.— Баку : Элм, 1978.—
264 с.
2. Аленицын В, Гады островов и берегов Аральского моря.— Тр. Арало-Касп. экспедиции,
• 1876, вып. 3, с. 1—64. Прил. к Тр. Санкт-Петербург, о-ва естествоиспытателей.
3. Алфераки С. [Alpheraki]. Кульджа и Тянь-Шань : Путевые заметки. — Спб/, ИРГО,
1891.— 193 с.
'4. Андрушко А. М. Материалы по биологии Alsophylax pipiens (Pallas).— Науч. бюл. Ле-
нингр. ун-та, 1949, № 23, с. 25—39.
5. Андрушко А. М. Пресмыкающиеся Казахского Нагорья и их хозяйственное значение.—
Учен. зап. Ленингр. ун-та. Сер. биол. наук, 1955, № 181, вып. 38, с. 19—43.
6. Андрушко А. М. Новый вид геккончика Alsophylax kashkarovi sp. n. (Reptilia, Sauria,
Gekkonidae) из Средней Азии.— Зоол. журн., 1968, 47, вып. 2, с. 308—311.
7. Адрушко А. М. Современное понимание статуса и структуры родов Alsophylax (Fitzinger,
1843) и Bunopus (Blanford, 1874) (Reptilia, Sauria, Gekkonidae) — В кн.: Вопросы герпето-
логии : Автореф. докл. IV Всесоюз. герпетол. конф. Л. : Наука, 1977, с. 10—12.
J. Андрушко А. М., Ланге Н- О., Емельянова Е. Н. Экологические наблюдения над репти-
лиями в районе Кызыл-Арват, станции Искандер и в районе г. Красноводска. Туркмения.—
Вопр. экологии и биоценологии, 1939, вып. 4, с. 207—252.
9. Андрушко А. М., Миккау И. Е. Распространение и образ жизни афганского литоринха
(Lytorhynchus ridgewayi Boulenger, 1887) с эколого-географическим обзором рода Lytorhyn-
chus Peters, 1862.— Вести. Ленингр. ун-та, 1964, № 9, вып. 2, Сер. биол., с. 15—19.
10. Атаев Ч. Проникновение пресмыкающихся гор в речные долины Туркмении.— Изв.
АН ТССР. Сер. биол. наук, 1974, № 3, с. 38—43.
11. Атаев Ч. О зимней активности пресмыкающихся на Копетдаге и в Бадхызе.— Там же,
1975, № 4, с. 63—67.
12. Атаев Ч. К распространению и экологии некоторых видов пресмыкающихся Туркмениста-
на.— Там же, 1977, № 1, с. 80—82.
13. Атаев Ч., Горелов Ю. К., Шаммаков С. Материалы по редким и исчезающим видам пресмы-
кающихся фауны Туркменистана.— Там же, 1978, № 4, с. 81—8?.
14. Ахмедов М. И., Щербак И. И. Gymnodactylus caspius insularis ssp. n. (Reptilia, Sauria) —
новый подвид каспийского геккона с острова Вульф в Каспийском море.— Вести, зоологии,
1978, № 2, с. 80—82.
15. Банников А. Г. Материалы по фауне амфибий и рептилий Монголии.— Бюл. Моск, о-ва
испытателей природы, Отд. биол., 1958, 58, № 2, с. 71—91.
16. Банников А. Г. По заповедникам Советского Союза.— М. : Мысль, 1974.— 235 с.
17. Банников А. Г., Даревский И. С,, Ищенко В. Г. и др. Определитель земноводных и пресмы-
кающихся фауны СССР.— М. : Просвещение, 1977.— 414 с.
18. Бедряга Д. В. Научн. результаты путешествия Н. М. Пржевальского по Центр. Азии.
Отд. зоол. т. 3, ч. 1. Земноводные и пресмыкающиеся, вып. 3, Lacertilia. Спб., 1909, с. 73—
502.
19. Богданов О. П. О видовой самостоятельности Alsophylax laevis и его распространении в Узбе-
кистане.— Докл. АН СССР, 1955, 101, № 5, с. 959—960.
20. Богданов О. П. О распространении Alsophylax laevis Nik. в Узбекистане.— Докл. АН
Уз ССР, 1955, № 6, с. 43—45.
21. Богданов О. П. Несколько поправок к статье В. П. Костина о земноводных и пресмыкаю-
щихся древней дельты Аму-Дарьи и Устюрта.— Тр. Ин-та зоологии и паразитологии АН
УзССР, 1956, вып. 8, с. 194—195.
22. Богданов О. П. Фауна Узбекской ССР. Ч. 1. Земноводные и пресмыкающиеся.— Ташкент :
Изд-во АН УзССР, I960.— 254 с.
23. Богданов О. П. Пресмыкающиеся Туркмении.— Ашхабад : Изд-во АН ТССР, 1962.— 235 с.
24. Богданов О. П. Экология пресмыкающихся Средней Азии.— Ташкент : Наука, 1965.—
259 с.
25. Богданов О. П. Экология гладкого геккончика в весенний период.— Экология, 1971, № 1,
’ с. 105—107.
26. Богданов О. П. Новые данные по распространению панцирного геккончика в Туркмении.—
Изв. АН ТССР. Сер. биол. наук, 1972, № 1, с. 91,
217
27. Богданов О. П. О сокращении численности некоторых видов беспозвоночных и рептилий
Устюрта в связи с усыханием Арала.— Вести. Каракалп. фил. АН УзССР, 1977, № 3, с. 57.
28. Богданов О. П., Петрусенко О. А. Про живлення гекона ciporo — Gymnodactylus russowi
Strauch, 1887.— 36. праць зоомузею, 1973, № 35, с. 66—67.
29. Бондаренко Д. А., Антонова Г. С. Ландшафтное распределение рептилий на плато
Устюрт.— В кн.: Вопросы герпетологии : Автореф. докл. IV Всесоюз. герпетол. конф.
Л. : Наука, 1977, с. 41—42.
30. Борисов А. А. Климат СССР.—М. : Просвещение, 1967.— 296 с.
31. Боркин Л. Д., Мунхбаяр Х-> Семенов Д. В. Амфибии и рептилии Заалтайской Гоби.—
Природа, 1983, № 10, с. 68—75.
32. Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Линнеевский
вид как система.— Л. : Наука, 1967.— 92 с.
33. Вашетко Э. В., Камалова 3. Д. Черепахи, ящерицы.— В кн.: Позвоночные животные фер-
ганской долины. Ташкент : ФАН, 1974, с. 60—75.
34. Великанов В. П. О новых находках пискливого геккончика и обыкновенного щитомордника
в Туркмении.— Изв. АН ТССР. Сер. биол. наук., 1977, № 5, с. 81—82.
35. Великанов В. П. О герпетофауне Сарыкамышской котловины.— В кн.. Вопросы герпето-
логии : Автореф. докл. IV Всесоюз. герпетол. конф., Л. : Наука, 1977, с. 56—57.
36. Голубев М.. Л. О географической изменчивости и таксономии гладкого геккончика Alsophy-
lax laevis Nikolsky, 1905 (Sauria, Gekkonidae).— В кн.: Экология и систематика амфибий
и рептилий. Л. : 1979, Зоол. ин-т АН СССР, с. 55—64 (Тр. Зоол. Ин-та АН СССР; Т 89).
37. Голубев М. Л. Об объеме рода Alsophylax Fitzinger, 1843 (Reptilia, Gekkonidae).— В кн.:
Вопросы герпетологии, 5-ая Всесоюз. герпетол. конф., Ашхабад, 1981. Л., 1981, с. 39.
38. Голубев М.Л. Alsophvlax tadjikiensis Golubev, stat. n. (Alsophylax laevis tadjikiensis Golu-
bev, 1979 : 62).— Вести, зоологии, 1984, № 2, c. 73.
39. Голубев M. Л. Alsophylax przewalski (Reptilia, Gekkonidae); некоторые номенклатурные
трудности.— Там же, № 4, с. 18—22.
40. Голубев М. Л. Структура рода Tropiocolotes (Reptilia, Gekkonidae).— Там же, 1984, № 6,
с. 12.
41. Голубев М. Л. Новые точки находок Alsophylax loricatus szczerbaki Golubev et Sattorov
(Reptilia, Gekkonidae).— Там же, 1985, № 2, с. 87.
42. Голубев М. Л., Саттаров Т. С. О подвидах у панцирного геккончика Alsophylax loricatus
Strauch, 1887 (Reptilia, Sauria, Gekkonidae). — Там же, 1979, № 5, с. 18—24.
43. Голубев М. Л., Щербак Н. Н. Новый вид рода Tropiocolotes Peters 1880 (Reptilia, Sauria,
Gekkonidae) из Афганистана.— Докл. АН УССР. Сер. Б, 1979, № 4, с. 309—312.
44. Голубев М. Л., Щербак Н. Н. Новый вид рода Gymnodactylus (Reptilia, Gekkonidae) из Па-
кистана.— Вестн. зоологии, 1981, № 3, с. 40—45.
45. Голубев М.. Л., Щербак Н. Н. Carinatogecko gen. n. (Reptilia, Gekkonidae) — новый род
гекконовых ящериц из Юго-Западной Азии.— Там же, № 5, с. 34—41.
46. Голубев М. Л., Щербак Н. Н. Gymnodactylus scaber (Heyden), 1827 = Cyrtodactylus basog-
lui Baran et Gruber, 1982, syn. n.— Там же, 1983, № 2, с. 54.
47. Голубев М. Л., Щербак Н. Н. О взаимоотношениях двух родов палеарктических гекконов:
Tenuidactylus и Agamura (Reptilia, Gekkonidae).— В кн.: Вопросы герпетологии : Автореф.
докл. 6-й Всесоюз. герпетол. конф., Ташкент, 1985, с.
48. Горелов Ю. К.» Даревский И С., Щербак Н. Н. Два новых для фауны СССР вида ящериц
из семейства гекконов.— Вестн. зоологии, 1974, №4, с. 33—39.
49. Даревский И. С. Ящерицы.— В кн.: Жизнь животных, Т. 4, ч. 2, М. : Просвещение, 1969,
с. 202—316.
50. Даревский И. С. Какой вид эублефара (Sauria, Gekkonidae) встречается в Средней Азии.—
Тр. Зоол. ин-та АН СССР. Новые виды животных, 1978, 61, с. 204—209.
51. Даревский И. С. Изучение функциональных особенностей признаков чешуйчатого покрова
у пресмыкающихся.— В кн.: Состояние и перспективы развития морфологии : (Материалы
к Всесоюз. совещ.). М. : Наука, 1979, с. 182—185.
52. Дементьев Г. П., Карташев Н. Н., Солдатова А. Н. Питание и практическое значение не-
которых хищных птиц в Юго-Западной Туркмении.— Зоол. журн., 1953, 32, вып. 3, с. 361—
375.
53. Дементьев Г. П.» Рустамов А. К-, Спангепберг Е. П. Материалы по фауне наземных
позвоночных Юго-Восточной Туркмении.— Тр. Туркм. с.-х. ин-та, 1955, т. 7, с. 125—183.
54. Дубинин В. Б. Эколого-фаунистический очерк земноводных и пресмыкающихся Хаваст-
ского района Ташкентской области УзССР.— Тр. Ин-та зоологии и паразитологии АН
УзССР, 1954, т. 3, с. 159—170.
55. Дубровский Ю. А. Новые находки рептилий в степях Казахстана.— Бюл.Моск. о-ва испы-
тателей природы, 1967, 72, № 1, с. 146—147.
56. Еремченко В. К., Щербак И. И. Новый вид геккона — Alsophylax tokobajevi sp. nov. из
Тянь-Шаня (Reptilia, Sauria, Gekkonidae).— Вестн. зоологии, 1984, № 2, с. 46—50.
57. Еремченко В. К-> Щербак Н. И. Распространение и экология геккончика Токобаева Alsophy-
lax tokobajevi Jeriomtschenko et Szczerbak 1984 (Reptilia, Gekkonidae).— Изв. AH
КиргССР, 1985, № 3, c. 60—63.
58. Зарудный H- Материалы для фауны амфибий и рептилий Оренбургского края.— Бюл.
Моск, о-ва испытателей природы, 1896, № 3, с. 361—370.
59. Зарудный Н. Заметка о чешуйчатых и голых гадах из северо-восточной Персии.— Ежегод-
ник Зоол. музея АН, 1897, 2, № 3, с. 349—361.
60. Зарудный И. О гадах и рыбах Восточной Персии,— Зап. ИРГО, 1904, 36, № 3, с, 1—42.
218
61. Зарудный Н* А. Поездка по Аральскому морю летом 1914 года. It. Гады Арала.— Изв.
Туркест. отд. РГО, 1915, 11, вып. 1, с. 113—125.
62. Захидов Т. Биология рептилий южных Кзыл-Кумов и хребта Нура-Тау.— Ташкент,
1938.— 52 с. (Тр. Среднеаз. ун-та. Сер. 8, Зоология, вып. 54).
63. Калужина М. В. Морфология и биология отряда ящериц Зеравшанской долины.— Тр.
биол.-почвенного фак. Узб. ун-та. Зоология. Н. С. 1951, вып. 46, с. 75.
64. Карташев Н. Н. Материалы по амфибиям и рептилиям Ю.-З. Туркмении.— Учен. зап.
Моск, ун-та, 1955, вып. 171, с. 173—202.
65. Киреев В. А. Земноводные и пресмыкающиеся хребта Жельтау.— В кн.: Вопросы герпе-
тологии : Автореф. докл. V Всесоюз. герпетол. конф. Л. : Наука, 1981, с. 64—65.
66. Ковачев В. Принос за изучване земноводнитъ и влечучитв въ Болгария.— Сб. за нар. умо-
творения, 1905, кн. 21, с. 1—13.
67. Костин В. П. Заметки по распространению и экологии земноводных и пресмыкающихся
древней дельты Аму-Дарьи и Кара-Калпакского Устюрта.— Тр. Ин-та зоологии и парази-
тологии АН УзССР, 1956, вып. 8, с. 47—65.
68. Красная книга СССР / Гл. редкол. : А. М. Бородин и др.— М. : Лесн. пром-сть, 1978.—
460 с.
69. Красная книга СССР / Гл. редкол.: А. М. Бородин и др.— М. : Лесн. пром-сть, 1984.—
390 с.
70. Кубыкин Р. А. Эколого-фаунистический обзор рептилий островов оз. Алаколь (Вост. Ка-
захстан).— Изв. АН КазССР. Сер. биол., 1975, № 3, с. 10—16.
71. Лаздин В. В. Маршрут поездок с зоологической целью в Восточную Бухару и на западный
Памир летом 1915 года.— Ежегодник Зоол. музея АН, 1915, т. 20, с. LIV — LVIII.
72. Майр Э. Принципы зоологической систематики.— М. : Мир, 1971.— 454 с.
73. Манило В, В., Щербак Н. Н. Кариотипы гекконов подрода Mediodactylus (Reptilia, Gekko-
nidae) фауны СССР.— Вести. зоологии, 1984, № 3, с. 81—83.
74. Международный кодекс зоологической номенклатуры / Под ред. А. Н. Световидова,
Я. И. Старобогатова. М. ; Л. : Наука, 1966.— 100 с.
75. Медников Б. М. Современное состояние и развитие закона гомологических рядов в наследст-
венной изменчивости.— В кн.: Проблемы новейшей истории эволюционного учения. Л. :
Наука, 1981, с. 127—135.
76. Михайловский М. К- К герпетофауне Закаспийской области.— Ежегодник Зоол. музея
АН, 1904, т. 9, с. 39—44.
77. Монхбаяр Хорлоогийн. Земноводные и пресмыкающиеся Монгольской Народной Республи-
ки : Автореф. дис. ... канд. биол. наук.— 1978.— 38 с.
78. Монхбаяр Хорлоогийн. Новые данные о распространении некоторых амфибий и рептилий
Монгольской Народной Республики.— В кн.: Фауна и экология амфибий и рептилий пале-
арктической Азии. Л. : Зоол. ин-т АН СССР, 1981, с. 52—56 (Тр. Зоол. ин-та АН СССР;
Т. 101).
79. Мориц Л. Д. Список пресмыкающихся, собранных в 1921 г. в Закаспийской области, Турк-
менистане и Северной Персии.— Тр. Ставроп. с.-х. ин-та, 1922, 1, № 11, с. 35—47.
80. Мориц Л. Д. Пресмыкающиеся Туркменистана и сопредельной Персии.— Туркменоведе-
ние, 1929, 4, № 6/7, с. 17—19.
81. Муратов Ш. X.» Муратов Р. Ш. О гладком геккончике Южного Таджикистана.— В сб.:
Вопросы герпетологии : Автореф. докл. IV Всесоюз. герпетол. конф. Л. : Наука, 1977,
с. 150.
82. Мусхелишвили Т. А. О фауне ящериц (Sauria, Reptilia) окрестностей Тбилиси.— Сообщ.
АН ГССР, 1964, 35, № 1, с. 199—205.
83. Киреев В. А. О северной границе ареала серого голопалого геккона.— В кн.: Экология и
фаунистика амфибий и рептилий СССР и сопредельных стран. Л., 1984, Зоол. Ин-т АН СССР,
с. 141—142.
84. Неручев В. В.» Васильев Н- Ф. Фауна рептилий Северо-Восточного Прикаспия.— Вести,
зоологии, 1978, № 6, с. 36—41.
85. Никольский А. М. О фауне позвоночных животных дна Балхашской котловины.— Тр.
Сиб. о-ва естествоиспытателей, Отд. зоол., 1887, 18, с. 59—190.
86. Никольский А. М. Diagnoses Reptilium et Amphibiorum novorum in Persia orientali a N. Za-
rudny collectorum.— Ежегодник Зоол. музея АН, 1896, 1, № 4, с. 369—372.
87. Никольский А. М. Пресмыкающиеся, амфибии и рыбы, собранные Н. А. Зарудным в Во-
сточной Персии.— Там же, 1897, 2, № 3, с. 306—348.
88. Никольский А. М. Два новых вида Teratoscincus из Восточной Персии.— Там же, 1899,
№ 2, с. 145—147.
89. Никольский А. М. Пресмыкающиеся, амфибии и рыбы второго путешествия Н. А. Заруд-
ного в Персию в 1898 г.— Там же, 1900, 4, с. 375—417.
90. Никольский А. М. Gymnodactylus danilewskii Str. et Gymnodactylus colchicus n. sp.
(Lacertilia, Gekkonidae).— Там же, 1902, 7, с. 3—6.
91. Никольский А. М. Новые виды гадов из Восточной Персии, привезенные Н. А. Зарудным
в 1901 г.— Там же, 1903, 8, с. 95—98.
92. Никольский А. М. Пресмыкающиеся и земноводные, собранные Н. А. Зарудным в Персии
в 1903—1904 гг,— Там же, 1907, 10, № 3/4, с. 260—301.
93. Никольский А. М. Alsophylax laevis sp. nov. (Geckonidarum).— Там же, с. 333—335.
94. Никольский А. М. Определитель пресмыкающихся и земноводных Российской Империи.—
Харьков : Русс, тип. и литография, 1907,— 182 с.
219
95. Никольский Л. М. Материалы по герпетологии Русского Туркестана.— Ежегодник Зоол.
музея АН, 1908, 13, № 3, с. 336—344.
96. Никольский А. Л4. Фауна России и сопредельных стран.— Пресмыкающиеся (Reptilia).
Т. 2. Chelonia и Sauria.— Пг.; Изд-во Имп. ак. наук, 1915.— 532 с.
97. Орлов Н. Л. О восточной границе длинноногого голопалого геккона Gymnodactylus longi-
pes Nikolsky, 1896.— В кн.: Фауна и экология амфибий и рептилий палеарктической
Азии. Я. : Зоол. ин-т АН СССР, 1981, с. 89—91 (Тр. Зоол. ин-та АН СССР ; т. 101).
98. Параскив К- П. Пресмыкающиеся Казахстана.— Алма-Ата : Изд-во АН КазССР, 1956. —
228 с.
99. Пащенко Ю. М> Новые данные о распространении некоторых пресмыкающихся в Туркме-
нии.— В кн.: Вопросы герпетологии : (Материалы герпетол. конф., 12—14 окт. 1964 г.)
Л. : Изд-во Ленингр. ун-та, 1964, с. 52—53.
100. Плохинсхий Н. А. Биометрия.— Новосибирск : СО АН СССР, 1961.— 366 с.
101. Рокицкий П. Ф. Основы вариационной статистики для биологов.— Минск : Изд-во
Белорус, ун-та, 1961.— 221 с.
102. Рустамов А. А. Птицы пустыни Кара-Кум.— Ашхабад : Изд-во АН ТССР, 1954.— 344 с.
103. Рустамов А. К., Атаев Ч. Новые данные по гернетофауне Туркменистана.— Изв.
АН ТССР, 1976, № 5, с. 47—53.
104. Рустаков А. А., Шаммаков С, Редкие и исчезающие виды рептилий Туркменистана.—
Охрана природы Турменистана, 1979, вып. 5, с. 139—146.
105. Саид-А лиев С. А. О новых находках панцирного и пискливого геккончиков из рода Al-
sophylax в Таджикистане.— Тр. Ин-та зоологии и паразитологии ТаджССР, 1962, вып. 22,
с. 103—105.
106. Саид-Алиев С. А. Материалы к фауне пресмыкающихся и земноводных Сев. Таджикиста-
на.— Изв. АН ТаджССР, Отд. биол., 1963, № 3, с. 81—94.
107. Саид-Алиев С. А. Земноводные и пресмыкающиеся Таджикистана.— Душанбе : Дониш,
1979.— 145 с.
108. Саттаров Т. О новых находках некоторых пресмыкающихся в Северном Таджикистане.—
Докл. АН ТаджССР, 1976, 19, № 12, с. 49—51.
109. Скалой Н. В. Земноводные и пресмыкающиеся Юго-Западного Копетдага.— В кн.: При-
рода Западного Копетдага. Ашхабад : Ылым, 1982, с. 146—157.
НО. Соловкин Н* Отчет о сборе фаунистических коллекций по берегам Астрабадского залива,
а также в районе Красноводского залива и на о. Челекене в 1913 г.— Ежегодник Зоол.
музея АН, 1915, 20, № 4, с. I—XVIII.
111. Терентьев П- В. Герпетология.— М. : Высш, шк., 1961.— 335 с.
112. Терентьев /7. В., Чернов С. А. Определитель пресмыкающихся и земноводных.— М. ; Л. :
Сов. наука, 1949.— 340 с.
113. Утемисов О. Ящерицы культурного ландшафта Каракалпакии.— В кн.: Вопросы герпе-
тологии : Автореф. докл. III Всесоюз. герпетол. конф. Л. : Наука, 1973, с. 184—185.
114. Федченко А. /7. Отчет Туркестанской ученой экспедиции Общества с 16 апреля 1869 г.
по 15 апреля 1870 г.— Изв. о-ва любителей естествознания, антропологии и этнографии,
1870, 8, № 1, с. 135—189.
115. Францевич Л. И. Обработка результатов биологических экспериментов на микро-ЭВМ
«Электроника БЗ-21».— Киев : Наук, думка, 1979.— 91 с.
116. Фролов В. Е. Размножение четырех видов гекконов в Московском зоопарке.— В кн.:
Вопросы герпетологии : Автореф. докл. V Всесоюз. герпетол. конф. Л. : Наука, 1981,
с. 138-139.
117. Фролов В. Е. Неоднократное размножение сцинкового геккона (Teratoscincus s. scincus)
в Московском зоопарке.— Вести, зоологии, 1985.
118. Ху Шу-цинь, Ху Бу-цин, Дин Хань-no, Хуан Чжу-цзянъ. Описание животного мира Китая
(пресмыкающиеся).— Пекин : Наука, 1962.— 67 с.
119. Царевский С, Ф. Пресмыкающиеся и земноводные, собранные В. Я- Лаздиным в Южной
и Восточной Бухаре и в с.-в. части Закаспийской области летом 1916 года.— Ежегодник
Зоол. музея АН, 1922. 22, № 1/3, с. 79—90.
120. Целлариус А. Ю. Фауна, биотопическое размещение и численность пресмыкающихся
Репетекского заповедника.— Изв. АН ТССР. Сер. биол. наук, 1975, № 6, с. 42—47.
121. Чернов С. А. Пресмыкающиеся Туркмении.— Тр. совета по изуч. произвол, сил, 1934,
вып. 6, с. 255—290.
122. Чернов С. А. Ящерицы (Sauria) Таджикистана.— Тр. Тадж. базы АН СССР, 1935, № 5,
с. 469—475.
123. Чернов С. А. Материалы к герпетофауне Казахского нагорья, северного побережья Бал-
хаша и гор. Кин-Тау.— Изв. АН КазССР. Сер. зоол., 1947, вып. 6, с. 120—124.
124. Чернов С. А. Эколого-фаунистический обзор пресмыкающихся юга междуречья Волга —
Урал.— Тр. Зоол. ин-та АН СССР, 1954, т. 16, с. 137—158.
125. Чернов С. А. Фауна Таджикской ССР. Т. 18. Пресмыкающиеся.— Сталинабад : Изд-во
АН ТаджССР, 1959.— 202 с.
126. Чегодаев А. Е. Сцинковый геккон.— Природа, 1975, № 9, с. 98—100.
127. Шаммаков С. О некоторых редких видах пресмыкающихся Туркмении.— Изв. АН ТССР.
Сер. биол. наук, 1964, № 6, с. 86—87.
128. Шаммаков С. Фаунистические материалы о пресмыкающихся малых хребтов (Малый Бал-
хан, Кюрен-Даг, Кара-Гоз) : Зап. Туркмении.— В кн.: Герпетология Средней Азии. Таш-
кент : Ин-т зоологии и паразитологии АН УзССР, 1968, с. 10—15.
129. Шаммаков С. К экологии панцирного геккончика (Alsophylax loricatus Str.) в Туркмении.—
Изв. АН ТССР. Сер. биол. наук, 1974, № 2, с, 76—77.
220
130. Шаммаков С. Пресмыкающиеся равнинного Туркменистана.— Ашхабад : Ылым, 1981.—
311 с.
131. Шаммаков С., Атаев Ч. О гладком геккончике (Alsophylax laevis Nikolsky) в Туркмении.—
Изв. АН ТССР. Сер. биол. наук, 1971, №3, с. 65—69.
132. Шаммаков С., Великанов В. П. О распространении и экологии пискливого геккончика*
в Туркменистане.— Там же, 1982, № 2, с. 72—73.
133. Шаммаков С., Сопыев О. С., Федорова Н- М. Экология сцинкового геккона в Каракумах.—
Изв. АН ТССР. Сер. биол. наук, 1982, № 3, с. 36—42.
134. Шарпило В. ГЕ Паразитические черви пресмыкающихся фауны СССР.— Киев : Наук,
думка, 1976.— 287 с.
135. Шестоперов Е. Л. Предварительное обследование в зоологическом отношении Ахча-
Куйминского заповедника.— Изв. Туркм. между ведомств, ком. по охране природы и раз-
витию природ, богатств. Сер. биол., 1935, № 2, с. 161—192.
136. Шибанов И. В. Ящерицы.— В кн.: Жизнь животных по А. Э. Брему. М. : Учпедгиз,
1939, т. 3, с. 596—698.
137. Шнитников В. Пресмыкающиеся Семиречья.— Тр. о-ва изуч. Казахстана. Кзыл-Орда,
1928, 8, вып. 3, с. 24—28.
138. Щербак М. М. До вивчення кримського гекона (Gymnodactylus kotschyi danilewskii
Strauch).— Доп. АН УРСР, 1960, 7, № 7, с. 970—973.
139. Щербак Н. Н. Новые данные о крымском гекконе (Gymnodactylys kotschyi danilewskii
Strauch).— Зоол. журн., 1960, 39, вып. 9, с. 1390—1397.
140. Щербак Н. Н. Новые данные о размножении крымского геккона (Gymnodactylus kotschyi
danilewskii Str.).— Там же, 1965, 44, вып. 9, с. 1421.
141. Щербак Н. И. Земноводные и пресмыкающиеся Крыма.— Киев : Наук, думка, 1966.—
240 с.
142. Щербак Н. Н. Систематика рода ящурка— Eremias (Sauria, Reptilia) в связи с очагами раз-
вития пустынно-степной фауны Палеарктики.— Вестн. зоологии, 1971, № 2, с. 48—55.
143. Щербак Н. Н. Ящурки Палеарктики.— Киев : Наук, думка, 1974.— 294 с.
144. Щербак Н. Н. Gymnodactylus turcmenicus sp. n. (Reptilia, Sauria) новый вид геккона иа
Южной Туркмении.— Вестн. зоологии, 1978, № 3, с. 39—44.
145. Щербак Н. Н. Эублефар туркменский.— Природа, 1979, № 6, с. 83—85.
146. Щербак Н. Н. Новый подвид сцинкового геккона (Teratoscincus scincus rustamowi ssp.
n., Sauria, Reptilia) из Узбекистана и систематика вида.— Охрана природы Туркменистана,.
1979, вып. 5, с. 129—138.
147. Щербак И. Н. Новые находки ящериц и змей на территории Средней Азии.— Вестн.
зоологии, 1979, № 1, с. 68—70.
148. Щербак И. И. Основы герпетогеографического районирования территории СССР.— В кн.:
Вопросы герпетологии : Автореф. докл. V Всесоюзн. герпетол. конф. Л. : Наука, 1981,
с. 157—158.
149. Щербак Н. Н.» Голубев М. J1. Взаимоотношения родов Gymnodactylus и Alsophylax и их
внутриродовая структура.— В кн.: Вопросы герпетологии : Автореф. докл. IV Всесоюзн.
герпетол. конф. Л. : Наука, 1977, с. 237—238.
150. Щербак ЕЕ.Н-, Голубев М. Л. Материалы к систематике палеарктических гекконов (роды
Gymnodactylus, Bunopus, Alsophylax).— В кн.: Герпетологический сборник. Л. : Зоол.
ин-т АН СССР, 1977, с. 120—133 (Тр. Зоол. ин-та АН СССР ; Т. 74).
151. Щербак Н. И., Голубев М. Л. Новые находки земноводных и пресмыкающихся в Средней
Азии и Казахстане.— Вестн. зоологии, 1981, № 1, с. 70—72.
152. Щербак Н. Н., Голубев М. Л. Географическая изменчивость и таксономия серого голопа-
лого геккона (Gymnodactylus russowi Str., Gekkonidae, Reptilia).— В кн.: Физиологиче-
ская и популяционная экология. Саратов : Изд-во СГУ, 1983. с. 160—162.
153. Щербак Н. Н., Голубев М. Л. О родовой принадлежности и внутриродовой структуре па-
леарктических голопалых гекконов (Reptilia, Gekkonidae, Tenuidactylus gen. n.).—
Вестн. зоологии, 1984, № 2, с. 50—56.
154. Щербак Н. Н., Жукова В. В., Писанец Е. М. Кариотипы гекконов подрода Cyrtodactylus
(Gymnodactylus, Gekkonidae, Sauria, Reptilia) фауны СССР. — Докл. АН УССР. Сер. Б,
1981, № 8, с. 85—87.
155. Эминов А. Экология чайконосой крачки в Южной Туркмении.— В кн.: Фауна и экология
птиц Туркмении. Ашхабад : Ылым, 1974, с. 149—161.
156. Яблоков А. В. Прыткая ящерица.— М.: Наука, 1976.— 374 с.
157. Яблоков А. В. Проблема признака в морфологии.— В кн.: Современные проблемы эволю-
ционной морфологии животных : Тез. докл. Междунар. симпоз. М. : Наука, 1981, с. 121—
122.
158. Ядгаров Т. Состав, распределение и численность пресмыкающихся о. Арал-Пайгамбар-
(бассейн р. Сурхандарьи).— В кн.: Вопросы герпетологии : Автореф. докл. III Всесоюз.
герпетол. конф. Л. : Наука, 1973, с. 229—230.
159. Ядгаров Т. Гладкий геккончик (Alsophylax laevis) в пустыне Кызылкум.— Зоол. журн.*
1975, 54, вып. 9, с. 1412—1413.
160. Ядгаров Т. Новые данные о распространении и экологии панцирного геккончика (Also-
phylax loricatus) в Узбекистане.— В кн.: Вопросы герпетологии: Автореф. докл. 5-й Все-
союз. конф., Ашхабад, 1981, Л., 1981, с. 159.
161. Ядгаров Т. Близкие виды голопалых гекконов рода Gymnodactylus на стыке их ареалов,
в Средней Азии.— Узб. биол. журн., 1982, № 3, с. 41—43.
162. Яковлева И. Д. Пресмыкающиеся Киргизии.— Фрунзе : Ылым, 1964.— 272 с.
221
163. Anderson J. On some Indian Reptiles.— Proc. Zool. Soc., London, 1871, 9, p.'149—211.
164. Anderson J, On some Persun, Himalayan and other Reptiles.— Ibid., 1872, 2, p. 371—404.
165. Anderson J. A list of reptiles and batrachians obtained by Mr. A. Blayney Percival in southern
Arabia.— Ibid., 1901, 2, N 3/4, p. 135—152.
166. Anderson L. G. Catalogue of Linnean Type — Specimens of Linnaeus’s Reptilia in the Royal
Museum in Stockholm. Bihang tile K- Sven. Vet.— Akad. Handl., 1900, 26, N 1, p. 1—29.
167. Anderson S. C. A note on the synonymy of Microgecko Nikolsky with Tropiocolotes Peters.—
Wasmann J. Biol., 1961, 19, N 2, p. 287—289.
168. Anderson S. C. Amphibians and Reptiles from Iran.— Proc. Calif. Acad, Sci., 1963, 31,
N 16, p. 417—498.
169. Anderson S. C. A new species of Bunopus from Iran and key to lizards of the genus
Bunopus.— Herpetologica, 1973, 29, N 4, p. 355—358.
170. Anderson S. C. Preliminary key to the Turtles, Lizards and Amphisbaenians of Iran.—
Fieldiana : Zool., 1974, 65, N 4, p. 27—44.
171. Anderson S. C., Leviton A. E. A new species of Eublepharis from Southwestern Iran (Reptilia:
Gekkonidae).— Occas. Pap. Calif. Acad. Sci., 1966, N 53, p. 1—5.
172. Anderson S. C., Leviton A. E. Amphibians and Reptiles collected by the street expedition to
Afganistan, 1965.— Proc. Calif. Acad. Sci., 1969, 37, N 2, p. 25—56.
173. Angel F. Reptiles et Batrachiens de Syrie et de Mesopotamie recoltes par M. P.Pallary.—
Bull. Inst. Egypt, 1936, 18, p. 107—116.
174. Annandale N. The Indian Geckos of the Genus Gymnodactylus sp. n.— Rec. Ind. Mus.,
1913, 9, pt 5, p. 313.
175. Arnold E. N. Little-known geckoes (Reptilia : Gekkonidae) from Arabia with descriptions
of two new species from the Sultanate of Oman.— In: The Scientific Results of the Oman
Flora and Fauna Survey 1975, [J. Oman Stud. Spec. Rep. (N 1)] 1977, p. 81 —110.
176. Arnold E. N. A review of the lizard genus Stenodactylus (Reptilia : Gekkonidae).— In:
Fauna of Saudi Arabia. Basle : Pro Entomol. and Ci ba Geigy, 1980a, vol. 2, p. 368—404.
177. Arnold E. N. The Reptiles and Amphibians of Dhofar, Southern Arabia.— J. Oman Stud.
Spec. Rep., 1980b, N 3, p. 273—332.
178. Arnold E. N., Gallagher M. D. Reptiles and Amphibians from the mountains of Oman with
special reference to the Jebel Akhdar region.— In: The scientific Results of the Oman Flora
and Fauna Survey 1975, [J. Oman Stud. Spec. Rep. (N 1)], 1977, p. 59—80.
179. Baran Gruber U. Taxonomische Untersuchungen an tiirkischen Inselformen von Cyrtodac-
tylus kotschyi (Steindachner, 1980). Teil. I. Die Populationen der nordlischen Agais, des
Marmarmeeres und des Schwarzen Meers (Reptilia : Gekkonidae).— Spixiana, 1981, 4, N 3,
S. 255—270.
180. Baran l.t Gruber U. Taxonomische Untersuchungen an tiirkischen Gekkoniden.— Ibid.,
1982, 5, N 2, S. 109—138.
181. Basoglu M., Baran I. Tiirkije siirungenleri.— Bornova; Izmir : Ilker matbaasi, 1977'.— К- 1.
182. Bedriaga von. Verzeichnis der von der Central — Asiatischen Expedition unter Stabs-
Kapitan W. Roborowski in den Jahren 1893—1895 gesammelten Reptilien.— Ежегодник
Зоол. музея АН, 1905, 10, № 3/4, с. 159—200.
183. Beutler A. Cyrtodactylus kotschyi (Steindachner, 1870) — Agaischer Bogenfingergecko.—
In: Handbuch der Reptilien und Amphibien Europas / Hrsg. W. Bohme. Bd. 1. Echsen.
Wiesbaden : Akad. Verlagsges, 1981, S. 53—74.
184. Beutler A., Gruber U. Intraspezifische Untersuchungen an Cyrtodactylus kotschyi (Stein-
dachner, 1870); Reptilia : gekkonidae. Beitragzu einer mathematischen Definition desBegriffs
Unterart.— Spixiana, 1977, 1, N 2, S. 165—202.
185. Beutler A., Fror E. Die Amphibien und Reptilien der Nordkykladen (Griechenland).— Mitt.
Zool. Ges. Braunau, 1980, 8, N 10/12, S. 255—290.
186. Blanford U7. T. Descriptions of new Reptilia and Amphibia from Persia and Baluchistan.—
Ann. Mag. Natur. Hist. Ser. 4, 1874, 13, N 78, p. 453—455.
187. Blanford W. T. List of Reptilia and Amphibia collected by the late Dr. Stoliczka in Kashmir,
Ladak, Eastern Turkestan, and Wakhan, with descriptions of new Species.— J. Asiat. Soc.
Bengal, 1875, 44, p. 191—196.
188. Blanford W. T. On some new lizards from Sind with descriptions of new species of Ptyodac-
tylus, Stenodactylus and Trapelus.— Ibid., 1876, 45, pt 2, p. 18—26.
189. Blanford W. T. Scientific results of the Second Yarkand Mission based upon the collections
and notes of the late Ferdinand Stoliczka, Ph. D. Reptilia and Amphibia.— Calcutta : Office
of the Superintendent of Government Printing, 1878.— p. [1—11] + 1—26.
190. Blyth E. Proceedings of the Society. Report of the curator, zoological department.— J. Asiat.
Soc. Bengal, 1854, 23, p. 737—740.
191. Bohme W. Handbuch der Reptilien und Amphibien Europas.— Wiesbaden : Akad. Verlags-
ges., 1981.—Bd 1.
192. Boettger O. Die Reptilien und Batrachier Transkaspiens.— Zool. Jahrb., Abt. Syst., 1888,
3, s. 871—1014.
193. Boettger O. Materialen zur herpetologischen Fauna von China II.— Ber. Offenbah. Ver.
Naturk., 1888, N 26/28, S. 53—191.
194. Boettger O. Katalog der Reptilien-Sammlung in Museum der Senckenbergischen Naturfor-
schenden geseltschaft in Frankfurt am Main. Teil 1.— Frankfurt am Main : Knauer, 1893.—
140 S.
195. Borner A. R. Ein neuer Lidgecko der Gattung Eublepharis Gray, 1827.—Misc. Art. SauroL,
1974, N 4, p. 5—14.
222
196. Borner A. R. Second contribution to the systematics of the southwest Asian lizard of the ge-
ckonid genus Eublepharis Gray, 1827 : Materials from the Indian subcontinent.— Saurologi-
ca, 1976, N 2, p. 1—15.
197. Borner A. R. Third contribution to the systematics of the southwest Asian lizards of the
geckonid genus Eublepharis Gray, 1827 : Further materials from the Indian Subcontinent.—
Ibid., 1981, N 3, p. 1—7.
198. Bons J. Les Lacertiliens du Sud-Ouest Marocain.— Trav. Inst. Sci., cherif. Ser. ZooL, 1959,
N 18. 1—130 p.
199. Boulenger G. A. Description of a new Genus of Geckos.— Ann. Mag. Natur. Hist., 1883, 5,
N 11, p. 174—176.
200. Bouenger G. A. Catalogue of the lizards in the British Museum.— London : Taylor et Francis,
1885.— Vol. 1. 436 p.
201. Bouenger G. A. Catalogue of the lizards in the British Museum.— London : Taylor et
Francis, 1887,— Vol. 3. 575 p.
202. Bouenger G. A. The Fauna of british India, including Ceylon and Burma. Reptilia and
Batrachia.— London : Taylor et Francis, 1890.— 541 p.
203. Boulenger G. A. The Zoology of the Afghan Delimination Comission (Reptiles and Batrachi-
ans).— Trans. Linn. Soc. Ser. 2, ZooL, 1891, 5, pt 3, p. 94—106.
204. Boulenger G. A. Catalogue of the Reptiles and Batrachians of Barbary (Morocco, Algeria,.
Tunisia).— Trans. Zool. Soc., 1891, 13, pt 3, p. 93—164.
205. Boulenger G. A. On some Batrachians and Reptiles from Tibet.— Ann. Mag. Natur. Hist.,
1905, 15, p. 378—379.
206. Brongersma L. D. Contributions to Indo-Australian Herpetology.— Zool. meded., Rijksmus.
natuurl. Hist., 1934,.17, Afl. 3/4, p. 161—251.
207. Bures I., Zonkow J. Untersuchungen fiber die Verbreitung der Reptilien und Amphibien in
Bulgarien und auf der Balkanhalbinsel.— Mitt, aus den Konig. Naturwiss. Instituten, Sofia.
1. Schildkroten und Echsen, 1933, S. 150—207.
208. Cherchi M. A., Spano S. Una nuova specie di Tropiocolotes del Sud Arabia Spedicione Scor-
tecci nell’Hadramut (1962).— Boll. Mus. e 1st. Biol. Univ. Genova Ser. biol. anim., 1963,
32, N 188, p. 29—34.
209. Clark JClark E.D., AndersonS. C., Levi ton A. E. Report on a collections of Amphibians and
Reptiles from Afghanistan.— Proc. Calif. Acad. Sci. Ser. 4, 1969, 36, N 10, p. 279—315.
210. Duda P. L., Sahl D. N. Cyrtodactylus himalayanus: a new gekkonid species from Jammu,
India.—J. Herpetol., 1978, 12, N 3, p. 351—354.
211. Dumeril A. M. Description des Reptiles nouveaux ou imparfaitment connus de la Collection
du Museum d’Histoire naturelie.— Arch. Mus. nat. hist, nat., 1856, 8, p. 437—588.
212. Dumeril A. M., Bibron G. Erpetologie generale geckos et les varans.— Paris, 1836.— Vol. 3.
517 p.
213. Eichwald E. Zoologia specialis quam expositis animalibus turn vivis...— Vilnae, 1831.—
404 S.
214. Ernst G. Eublepharis macularius.— Leopardgecko.— Aquaria, 1983, 30, N 10, S. 169—171.
215. Fitzinger L. Systema reptilium. Fasc. 1. Amblyglossae.— Vienna, 1843.— 106 p.
216. Forcart L. Amphibien und Reptilien von Iran.— Verh. Naturforsch. Ges., 1950, 61, S. 141—
156.
217. Flower S. Notes on the recent reptiles and amphibians of Egypt with a list of the species re-
corded from that Kingdom.— Proc. Zool. Soc., 1933, pt 3, p. 735—851.
218. Gasperetti J. Survey of the Reptiles of the Sheikhdom of Abu Dhabi, Arabian Peninsula. Pt 1.
A geographical sketch of the Sheikhdom of Abu Dhabi.— Proc. Calif. Acad. Sci. Ser. 4, 1967,
35, N 8, p. 141—156.
219. Gauthier R. Ecologie et Ethologie des Reptiles du Sahara Nord-Occidental (Region de Beni-
Abbes).— Ann. Mus. Afr. centr., 1967, ser. in.— 8, N 155, p. 1—83.
220. Golubev M. L. The content of the genus Bunopus (Reptilia: Gekkonidae).— In: I. conf,
herp. resp. soc. Budapest, 1981, p. 17.
221. Gray J. E. A synopsis of the Genera of the Saurian Reptiles in which some new Genera are
indicated, and the other rewieved by actual Examination.— Phil. Mag. Ser. 2, 1827, 3,
p. 53—56.
222. Gray J. E. Catalogue of the Specimens of Lizards in the Collection of the British Museum.—
London, 1845.— 289 p.
223. Guibe J. Reptiles et Amphibiens recoltes par la mission Franco-Iranienne.— Bull. Mus. nat.
hist, natur. Ser. 2, 1966a, 38, N 2, p. 97—98.
224. Guibe J. Contribution a 1’Etude des genres Microgecko Nikolsky et Tropiocolotes Peters
(Lacertilia, Gekkonidae).— Ibid., 1966b, 38, N 4, p. 337—346.
225. Gunther A. Description of a new species of Eublepharis.— Ann. Mag. Natur. Hist. Ser. 3,
1964, 14, p. 429—430.
226. Haas G. Amphibians and reptiles from Arabia.— Proc. Calif. Acad. Sci. Ser. 4, 1957, 29,
N 3, p. 47—86.
227. Haas G., Battersby J. C. Amphibians and Reptiles from Arabia. — Copeia, 1959, N 3, p.196—
202.
228. Haas G.> Werner У. L. Lizards and snakes from Southwestern Asia, collected by Henry Field.—
Bull. Mus. comp. zool. harv., 1969, 138, N 6, p. 327—406.
229. Heyden С. H. G. Reptilien.— In: Ruppel, E.— Atlas zu der Reise im nordlichen Africa.
Frankfurt-am-Main, 1827.— 24 S.
230. Hoofien J. H. Contributions to the Herpetofauna of Mount Hermon No 1. Cyrtodactylus
amictopholis n. sp. (Sauria, Gekkonidae).— Isr, J. Zool., 1967, 16, p. 205—210.
223
231. Ingoldby С, M. A new Stone-Gecko from the Himalaya.—J. Bombay Natur. Hist. Soc.,
1923, 28, N 4, p. 1051.
232. Ingoldby R., Procter B. Notes on a collection of reptilia from Waziristan and the djoining
portion of the N. W. Frontier Province.— Ibid., 1924, 29, pt 1/2, p. 117—130.
233. Karaman M. S. Eine neue Unterart der Eideche Gymnodactylus kotschyi aus Mazedonien,
Gymnodactylus kotschyi scopjensis n. ssp.— Zool. Anz., 1965, 174, N 4—5, S. 348—351.
234. Kasimir M. J. Zur Herpetofauna der Provinz Badghis (N. W. Afghanistan).— Dtsch. Aquar.-
Terr. Z., 1971, 24, S. 244—246.
235. Khan M. S. Checklist and key to the lizards of Jhangh District, West Pakistan.— Herpetolo-
gica, 1972, 28, N 2, p. 94—98.
236. KUngelhofter W. Terrarienkunde. Bd. 3. Echsen.— Stuttgart : Kernen, 1957.— 264 S.
237. Kluge A. G. Higher taxonomic categories of gekkonid lizards and their evolution.— Bull.
Amer. Mus. Natur. Hist., 1967, 135, art. 1, p. 1—60.
238. Kluge A. G. An interpretation of the status of Gymnodactylus tenius Hallowell (Sauria,
Gekkonidae).— Copeia, 1969, N 3, p. 623—624.
239. Kluge A. G. Cladistic relationships among Gekkonid Lizards.— Ibid., 1983, N 2, p. 465—
475.
240. Kral B. Notes on the Herpetofauna of Certain Provinces of Afghanistan.— Zool. listy, 1969,
18, N 1, p. 55—66.
241. Lantz L. A. Reptiles from the River Tajan (Transcaspia).— Proc. Zool. Soc., 1918, 7, p. 11—
13.
242. Leviton A. E. Report on collection of reptiles from Afghanistan.— Proc. Calif. Acad. Sci.
Ser. 4, 1959, 29, N 12, p. 445—463.
243. Leviton A. E., Anderson S. C. Third contribution to the herpetology of Afganistan.— Ibid.,
1963, 31, N 12, p. 329—339.
244. Leviton A. Е.» Anderson S. C. Survey of the reptiles of the Sheikhdom of Abu Dhabi, Arabian
Peninsula. Pt 2. Systematic account in the Sheikhdom of Abu Dhabi by John Casperetti.—
Ibid., 1967, 35, N 9, p. 157—192.
245. Leviton A. E., Anderson S. C. The amphibians and reptiles of Afghanistan, a checklist and
key to the herpetofauna.— Ibid., 1970, 38, N 10, p. 163—206.
246. Leviton A. E., Anderson S. G. Description of a new species of Tropiocolotes (Reptilia : Gekko-
nidae) with a revised key to the genus.— Occas. Pap. Calif. Acad. Sci., 1972, N 96, p. 1—7.
247. Lichtenstein H. Naturhistorische Anhang.— In: Eversmann E. Reise von Orenburg nach
Buchara. Berlin, 1823, S. 1—141.
248. Lichtenstein H. Nomenclator Reptilium et Amphibiorum Musei Zoologici Berolinensis.—
Berlin : Buchdruck. Konigl. Akad. Wiss., 1856.— 48 S.
249. Loveridge A. Revision of the African lizards of the family Gekkonidae.— Bull. Mus. Comp.
Zool. Harvard College, 1947, 98, N 1, p. 1—469.
250. Marx H. Keys to the lizards and snakes of Egypt.— NAMRU-3, Cairo, Egypt, 1956.— 8 p.
251. Marx H. Checklist of the Reptiles and Amphibians of Egypt.—Cairo, 1968.— 91 p.—
(Spec. publ. U. S. NAMRU-3).
252. Marx H. Supplementary Catalogue of Type Specimes of Reptiles and Amphibians in Field
Museum of Natural History.— Fieldiana : Zool., 1976, 69, N 2, p. 33—94.
253. Meeuwen H. M., van. Enkele herpetologische aantekeiningen van een reis naar Afghanistan.—
Lacerta, 1977, 35, N 8, S. 115—124.
254. Mertens R. Ein neuer Gecko aus Mesopotamien.— Senckenbergiana, 1924, 6, S. 84—85.
255. Mertens R. Amphibien und Reptilien aus der Turkei.— Istambul univ. fen fak. mecm. B,
1952, cilt XVII, Say. I, S. 41—75.
256. Mertens R. Amphibien und Reptilien aus SO—Iran 1954.— Jahrb. Ver. vaterland. Naturk.
Wurtt., 1956, 111, S. 90—97.
257. Mertens R. Bemerkungen fiber einige Eidechsen aus Afghanistan.— Senckenberg. biol.,
1965, 46, H. 1, S. 1—4.
258. Mertens R. Die Amphibien und Reptilien West-Pakistans.— Stuttgart. Beitr. Naturk.,
1969, 197, S. 1—96.
259. Mertens R. Die Amphibien und Reptilien West-Pakistans.— Senckenberg. biol., 1971, 52,
N 1/2, S. 7—15.
260. Mertens R., Muller L. Liste der Amphibien und Reptilien Europas.— Abh. Senckerberg.
naturforsch. Ges., 1928, 41, Lief. 1, S. 1—62.
261. Mertens R., Muller L. Die Amphibien und Reptilien Europas (Zweite Liste, nach dem Stand
vom 1. Jannar 1940).— Abh. Senckenberg. naturforsch. Ges., 1940, 451,— 56 S.
262. Minton S. A• An annotated key of the amphibians and reptiles of Sind and Las Bela, West
Pakistan.— Amer. Mus. Novit., 1962, 2081, p. 1—60.
263. Minton S. A. A contribution to the herpetology of West-Pakistan — Bull. Amer. Mus. Natur.
Hist., 1966, 134, art. 2, p. 1—184.
264. Minton S. A., Anderson Jer. A new dwarf gecko (Tropiocolotes) from Baluchistan.—
Herpetologica, 1965, N 1, p. 59—61.
265. Minton S. A., Anderson S. C., Anderson Jer. Remarks on some geckos from Southwest Asia,
with descriptions of three new forms and a key to the genus Tropiocolotes.— Proc. Calif.
Acad. Sci. Ser. 4, 1970, 37, N 9, p. 333—362.
266. Miiller L. Ober die von den Herren Dr. v. Jordans und Dr. Wolf im Jahre 1938 in Bulgarien
gesammelten Amphibien und Reptilien.— Mitt. Konigl. Nat. Inst., Sofia, 1939, 13, S. 1—17.
267. Murray J. A- The vertebrate zoology of Sind.— Bombay, 1884.— XIV + 424 p.
268. Murray J. A. The Zoology of Beloochistan and Southern Afghanistan.— Bombay, 1892.—
83 p.
224
269. Nader J. AJawdat S. Z. Taxonomic Study of the Geckos of Iraq (Reptilia : Gekkonidae).—
Bull. Biol. Res. Centre. Publ., 1976, N §, p. 1—41.
270. Pallas P.-S. Zoographia Rosso-Asiatica.— Petropolis, 1811, 3.— 428 p.
271. Papenfuss T. J. Preliminary analysis of the Reptiles of arid central west Africa.— Wasmann J,
Biol., 1969, 27, p. 249—325.
272. Parker H. W. The Lizards of British Somaliland. — Bull. Mus. Comp. Zool., 1942, 41, p. 1—
101.
273. Pasteur G. Redecouverte et validite probable du Gekkonide Tropiocolotes nattereri Steind.—
C. r. Soc. sci. natur. phys. Maroc., 1960, 26, N 8, p. 143—145.
274. Pasteur G., Boris J. Catalogue des Reptiles actuels du Maroc.— Trav. Inst. sci. cherif., Ser.
Zool. Rabat, 1960, N 21, p. 1—132.
275. Peters W. С. H. Ober neue Saurier.— Monatsber. Dtsch. Acad. Wiss. Berlin, 1869, S. 786—
790.
276. Peters W. С. H. Ober die von Hern Gerhard Rohlfs und Dr. A. Stecker auf der Reise nach
der Oase Kufra gesammelten Amphibien.— Ibid., 1880, S. 305—309.
277. PetersG. In Urania Tierreich. Bd 4. Reptilia.— Leipzig etc.: Urania VerL, 1973. S. 508.
278. Pope С. H- The reptiles of China.— Nat. Hist. Centr. Asia, 1935, 10, 1—604 p.
279. Procter J. В. Further Lizards and Snakes from Persia and Mesopotamia.— J. Bombay Na-
tur. Hist. Soc., 1923, 28, N 1, p. 251—253.
280. Reed C. A., Marx H. A Herpetological Collection from Northeastern Iraq.— Trans. Kans.
Acad. Sci., 1959, 62, N 1, p. 91—122.
281. Rossler H. Fang, Haltung und Eiblage des Kammzehengeckos Crossobamon eversmanni
[(Wiegmann, 1834).— Aquarien Terrarien, 1983, 31, H. 1, S. 33—36.
282. Scerbak N. N. Cyrtodactylus russowii (Str., 1887) — Transkaspischer Bogenfingergecko.—
In: Handbuch der Reptilien und Amphibien Europas/ Hrsg. W. Bohme. Bd. 1. Echsen Wies-
baden : Akad. VerL, 1981, S. 75—83.
283. Schlegel H. Handleiding tot de Beoefening der Dierkunde.— Breda, 1858, II, S. 16.
284. Schmidt К. P, Reptiles and amphibians from southwestern Asia.— Publ. Field Mus. Natur.
Hist. Ser. zool., 1939, 24, N 7, p. 49—92.
285. Schmidt К. P. Amphibians and Reptiles from Iran.— Vid. Medd. Dan. naturhist. foren.
Kbenhaven, 1955, 117, N 3, S. 193—207.
286. Schmidtler J. J., Schmidtler J. F. Zwerggeckos aus dem Zagros-Gebirge (Iran). — Sala-
mandra, 1972, 8, N 2, S. 59—66.
287. Shcherbak N. N. Grundzuge einer herpetogeographischen Gliederung der Palaarktis.— Ver-
tebr. Hung., 1982, 21, S. 227—239.
288. Shockley С. H. Herpetological notes for Ras Jiunri, Baluchistan.— Herpetologica, 1949,.
5, pt 6, p. 121—123.
289. Seufer H. Der Kaspische Geradfinger— Gecko (Alsophylax pipiens Pallas).— Herpetofauna,
1979, 1 Jahr., S. 10—15.
290. Smith M. A. Remarks on some Old World geckos.— Rec. Indian Mus., 1933, 35, p. 9—19.
291. Smith M. A. Fauna of British India, including Ceylon and Burma. Reptilia and Amphibia*
Vol. 2. Sauria.— London : [Taylor, Francis] 1935.— XIII + 440 p.
292. Spix J. B. Animalia nova sive Species novae lacertarum, quas in itinere per Brasiliam Annis
1817—20 collegit et descripsit.— Monacho, J925.— 26 p.
293. Steindachner F. Reptilien.— In: Reise der Osterreichischen Fregate Novara, Wien, 1868.—
98 S.
294. Steindachner F. Herpetologische Notizen IL Reptilien gesammelt wahrend einer Reise in«
Senegambien.— Sitzungsber. Akad. Wiss. Wien, 1870, 62, Abt. 1, S. 326—335.
295. Steindachner F. Expedition S. M. Schiff «Pola» in das Rothe Meer. 17. Bericht uber die Her-
petologischen Aufsamm mlungen.— Denkschr. Acad. Wiss. Math.-naturwiss. KL 1901, 69,
N 1, S. 325—335.
296. Stepanek O. Sur le Gymnodactylus danilewskii en Bulgarie.— Sb. zool. odd. narod. Mus*
Praze, 1934, N 1, p. 31.
297. Stepanek O. Gymnodactylus kotschyi und sein Rassenkreis.— Arch. Natur. Leipzig. N. F.»
1937, N 6, S. 258—280.
298. Stoliczka F. Notes on the reptilian and amphibian fauna of Kachh.— Proc. Asiat. Soc. Ben-
gal, 1872, 5, p. 71—85.
299. Strauch A. Characteristik zweier neuen Eidechen aus Persien.—Bull. Acad. Sci. St.-Petersb.,.
1863, 4, S. 393—398.
300. Strauch A. Bemerkungen uber die Geckoniden-Sammlung im Zoologischen Museum der
Kaiserlichen Academie der Wissenschaften zu St.— Petersburg.— Mem. Acad. Sci. St.-
Petersb. Ser. 7, 1887, 35, N 2, S. 1—74.
301. Szczerbak N. N. Grundzuge der herpetogeographischen Gliederung des palaarktischen Gebie-
tes.— In: Auszuge der Vortage der ersten herpetologischen Konferenz sozialischer Lander.
Budapest, 1981, S. 42.
302. Tuck R. G. Rediscovery and redescription of the Khuzistan dwarf Gecko, Microgecko helenae
Nikolsky (Sauria : Gekkonidae).— Proc. biol. Soc. Wash., 1971, 83, N 42, p. 477—482.
303. Tuck R. G. Amphibians and reptiles from Iran in the United States National Museum col-
lection.— Bull. Md. Herpetol. Soc., 1971b, 7, N 3, p. 48—86.
304. Tuck R. G. Additional Notes on Iranian Reptiles in the United States National Museum col*
lection.— Ibid., 1973, 9, N 1, p. 13.
305. Underwood G. On the classification and evolution of geckos.— Proc. Zool. Soc., London,
1954, 124, N 3, p. 469—492.
225.
306. Vincigguerra D. Spedizione scientifica all Oasi di Kurfa — Rettili.— Ann. Mus. civ. stor.
natur. Genova, 1931, 55, p. 248—258.
307. Wermuth H. Liste der recenten Amphibien und Reptilien Gekkonidae, Pygopodidae, Xantu-
siidae.— Tierreich, 1965, Lief. 80, S. 1—246.
308. Werner F. Reptilen aus Persien (Provinz Fars).— Verh. Zool.-bot. Ges. Wien, 1917, 67,
S. 191—220.
309. Werner F. Reptilien und Gliedetiere aus Persien.— Festschrift fur Prof. Dr. Embrik Strand,
1937, vol. 2, S. 193—204.
310. Werner F. Reptilien aus Iran und Belutschistan.— Zool. Anz., 1938, 121, N 9/10, S. 265—271.
311. Werner F. Die Amphibien und Reptilien von Syrien.— Abh. Berl. Mus. Nat.—u. Heimatr
(Naturk. Vorgesch.) Magdeburg, 1939, 7, H- 1, S. 211—223.
312. Werner Y. L. Chromosome numbers of some male geckos (Reptilia: Gekkonidae).— Bull.
Res. Counc. Isr., 1956, SB, N 3/4, p. 319.
313. Werner Y. L. Eye size in geckos of various ecological types (Reptilia : Gekkonidae and
Sphaerodactylidae).— Isr. J. Zool., 1969, 18, p. 291—316.
314. Wettstein 0. Vierzehn neue Reptilien — Rassen von den agaischen Inseln.— Zool. Anz.,
1937, 118, S. 79—90.
315. Wettstein O. Ergebnisse der . Osterreichischen Iran — Expedition 1949/50.— Amphibien
und Reptilien.— Sitzungsber. Osterr. Akad. Wiss. Math.-naturw. KL Abt. 1, 1951, 5, H. 9,
S. 427—448.
316. Wettstein O. Dreizehn neue Reptilienrasse von den Agaischen Inseln.— Anz. Osterr. Akad.
Wiss, Math.-naturwiss., 1952, 15, S. 251—256.
317. Wettstein O. Lacertilia aus Afghanistan. Contribution a 1’etude de la faune d’Afghanistan.
3.— Zool. Anz., 1960, 165, H. 1/2, S. 58—63.
318. Wettstein— Westersheimb O. Drei seltene Echsen aus Siidwest— Asien.— Zool. Anz., 1960,
165, H. 5/6, S. 190—193.
319. Wiegmann A. F. 4. Herpetologia mexicana, seu Descriptio Amphibiorum Novae Hispania.
Pars prima, Saurorum species.— Berlin, 1834, p. 54.
320. Witte G. F., de. Description d’un Gekkonidae nouveau de LHran (Reptilia, Sauria).— Bull.
Inst. roy. sci. natur. Belg. Biol., 1973, 49, N 1, p. 1—6.
321. Witte G. F., de Note ralative a Rhinogekko misonnei de Witte et Agamura femoral is M.
Smith (Reptilia, Saurin).— Ibid., 1980, 52, N 8, p. 1—3.
322. Welch R. R. G. Herpetology of Europe and southwest Asia: A Checklist and Bibliography
of the Orders Amphisbaena, Sauria and Serpentes.— Malabar, Florida : Krieger, 1983.—
135 p.
323. Обет Ф. Ю. Находка турецкого полупалого геккона Hemidactylus turcicus (L., 1758) в
Туркмении.— В кн.: Экология и фаунистика амфибий и рептилий СССР Л. : Зоол. ин-т
АН СССР, 1984, с. 142—143.
324. Gruber (J. Notes on the Herpetofauna of Kashmir and Ladakh.— Brit. J. Herpetol., 1981,
6, N5,p. 145—150.
325. Leviton A. E., Anderson S. C. Description of a new species of cyrtodactylus from Afghanistan
with remarks on the status of C. longipes and C. fedtschenkoi.— J. Herpetol., 1984, 18,
N 3, p. 270—276.
326. Резник E. П. К экологии гребнепалого геккона [Crossobamon eversmanni) в среднем тече-
нии р. Мургаб.— В кн.: Вопросы герпетологии, 6. Л. : Наука, 1985, с. 175.
327. Ржепаковский В. Т. К изучению экологии туркестанского голопалого геккона в Таджи-
кистане.— В кн.: Вопросы герпетологии. 6, Л. : Наука, 1985, с. 176.
.328. Смирнов С. И., Шкунов В. Ф., Кудакина Е. Ю. Гекконы [Gekkonidae] Северного Прикас-
пия.— В кн.: Вопросы герпетологии, 6. Л. : Наука, 1985, с. 195.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Агамура 12, 196, 207,211
— носорог 213
— персидская 208, 211
— фарсийская 211
— харанская 213
Ардеозавры 17
Белозубка малая 40
Бойга 146
Волкозуб 174, 179
Геккон австралийский шипохвостый 17
— агамуровидный 196, 212
— анатолийский 183
— бемпурский 180
— вееропалый 11
— голопалый 3, 7, 10, 11, 208, 212, 220
----восточноазиатский 196
----длинноногий 219
----серый 60, 218, 220
— гребнепалый 7, 40, 46, 49, 51, 52, 53, 54
----восточный 54
----Эверсманна 47, 51
----Эверсманна обыкновенный 49
— — южный 50
— длинноногий 137, 138, 153, 155, 157, 158
—. — мелкочешуйный 156
Геккон длинноногий обыкновенный 156
----юго-восточный 157
— карликовый 100, 105
---- азиатский 120
--------Левитона 121, 123
-------- плоский 124
----аравийский Скортеччи 108, ПО
----африканский 101
--------триполитанский 102, 104, 105
--------Штейднера 106, 108
— каспийский 11, 132, 134, 135, 137, 138,
139, 140, 141, 144, 145, 146, 148, 149, 152,
154, 157, 158, 216
----обыкновенный 137
---- островной 137
— качский 185
— кашгарский 193, 195
— керманский 205
— колючехвостый 175, 177, 178, 179
— круглопалый 17
— крымский 163, 167, 220
— крысохвостый (см. Агамура)
— ливанский 179
— листопалый 11, 17
— Минтона 201
— пакистанский 188
— пенджабский 192
— переднеазиатский 182
иранский 184
турецкий 184
— полупалый 11
турецкий 13
— ребристый 126
---- иракский 130
---- иранский 127
— серый 83, 85, 86, 87, 167, 169, 173, 175,
179, 180, 217
----обыкновенный 171
----Зарудного 172
— средиземноморский 160, 161
— Столички 205
— сцинковый 6, 7, 14, 18, 33, 37, 38, 39, 40,
41, 45, 51, 52, 219, 220
----Бедряги 45
----Кайзерлинга 38
----мелкочешуйный 43
----обыкновенный 35, 37
----Пржевальского 42, 43
----Рустамова 38
— тибетский 198
— тонкопалый 12, 87, 131, 177, 196, 201,
212
---- евроазиатский 11
----средиземноморский 159
— туркестанский (Федченко) 11, 134, 137,
138, 146, 149, 150, 151, 152, 154, 155
— туркменский 135, 137, 138, 141, 144, 145,
146, 154
— читральский 201
— шершавый 190, 192
Гекконы тонкопалый тибето-гималайские
196, 198
Гекконовые 3, 4, 16, 17, 33, 41
Геккончик высокогорный североазиатский 81
Геккончик равнинный 56
— североазиатский 55, 123
Геккончик североазиатский гладкий 72, 74,
75, 77, 78, 79, 80, 216, 217, 218, 220
----панцирный 65, 66, 68, 69, 70, 71, 72,
216, 217, 219, 220
--------обыкновенный 67
--------Щербака 68
----пискливый 8, 56, 58, 60, 62, 72, 123,
217, 219, 220
----Пржевальского 62, 65
----таджикский 78, 79, 80, 81
— Токобаева 173, 217
— тяныпанский 81, 82, 83, 84, 85, 86, 87,
88
— южноазиатский 88
----аравийский 94, 97
--------обыкновенный 97
--------хаджарский 97
227
-----бугорчатый 89, 92, 93, 94
—- — толстохвостый 98
Гюрза 140
Еж обыкновенный 166
— ушастый 43
Желтопузик 140
Змея ящеричная 140
Кобра 33
Коршун черный 140
Крачка чайконосая 174, 220
Круглоголовка пятнистая 78
— такырная 78
Куница каменная 166
Лисица обыкновенная 29, 140
Литоринх афганский 94, 216
Месалина крапчатая 78
Микрогеккон 18, 20, 110
— лятифи 119
—персидский 115, 117, 119
----- бахтиярский 117
----- молочайный 118
-----обыкновенный 117
— хузестанский 111, 113
Пещанка краснохвостая 137
— полуденная 41, 137
Пищуха рыжая 126
Полоз краснополосный 53, 140
— поперечнополосатый 40, 53, 72, 77, 140,
152, 174
— пятнистый 53
— узорчатый 72, 86
Пустельга 87
Путорак пегий 40
Сизоворонка 152
Сорока 61, 72
Сорокопут 174
Стрела-змея 80, 140, 174, 195
Сцинки 17
Сыч домовой 40, 72, 140, 166, 174
— обыкновенный 53
Тушканчик малый 41
— мохноногий 41, 53
Удавчик восточный 40
— песчаный 40, 53
Циртоподион 185
Щитомордник обыкновенный 217
Эублефар 5, 6, 33, 217
— иранский 29
— пятнистый 26, 28, 31
— туркменский 29, 33, 220
Эублефаровые 17, 25
Эфа 77, 140, 152
Ящерица прыткая 220
Ящерицы ночные 17
Я щурка Никольского 83
— средняя 77
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Agamidae 5
Agamura 3, 12, 17, 18, 22, 23, 207, 208
agamuroides 196
cruralis 12, 208, 210
femoralis 12, 18, 23, 208, 210, 211, 213, 225
gastropholis 18, 23, 208, 210, 211, 212
misonnei 18, 23, 208, 210, 211, 213, 214
persica 12, 18, 23, 208, 209, 210, 212
persica cruralis 211, 212
Alsophylax 3, 7, 8, 9, 10, 11, 12, 17, 18, 19,
21, 23, 55, 56, 87, 88, 89, 91, 97, 116, 123,
201, 216, 217, 219, 220
crassicauda 98
kashkarovi 72, 216
laevis 8, 18, 19, 20, 55, 69, 72, 73, 78, 80,
82, 120, 123, 216, 217, 218, 220, 228
laevis tadjikiensis 78
loricatus 7, 8, 18, 19, 65, 69, 70, 73, 75,
217, 219, 220
loricatus loricatus 55, 67, 69
loricatus szczerbaki 3, 55, 67, 68, 69, 217,
228
microtis 56, 62, 72, 123
persicus 115, 117, 121, 130
pipiens 8, 18, 19, 55, 56, 59, 61, 63, 65, 66,
72, 78, 121, 123, 216, 224, 228
przewalski 7, 18, 19, 55, 61,62,64,66,217
spinicauda 3, 8, 11, 175
tadjikiensis 3, 18, 19, 23, 55, 59, 73, 78,
83, 217, 228
tibetanus 3, 7, 11, 198 J
tokobajevi 3, 8, 18, 19, 23, 55, 61, 81, 84,
85, 200, 217, 228
tuberculatus 89
Altiphylax 3, 8, 11, 18, 19, 81
Ardeosauridae 16
Asca la botes 7
pipiens 56
Asiocolotes 3, 19, 20, 21, 101, 120, 121
Athene noctua 140
Boiga trigonatum 146
Bunopus 3, 7, 8, 9, 10, 11, 14, 17, 20, 24,
88, 100, 216, 220, 221, 222
abudhabi 8, 89, 92, 100
aspratilis 8, 10, 126, 127
biporus 89
blanfordi 7, 8, 89, 92
crassicauda 8, 20, 89, 91, 94, 98, 99
persicus 117
spatalurus 8, 20, 89, 94, 95, 96, 97
spatalurus hajarensis 95, 97
spatalurus spatalurus 95, 97
tuberculatus 3, 7, 8, 20, 88. 89, 90, 91, 93,
94, 228
Carinatogecko 3, 10, 17, 18, 21, 24, 126, 127, 217
aspratilis 18, 21, 47, 127, 128
heteropholis 18, 21, 47, 127, 129, 130
Coluber karelini 53, 77, 126, 140, 152, 174
rhodorhachis 53, 126, 140
Crossobamon 3, 7, 17, 18, 23, 24, 46, 47, 48,
49, 50, 88
eversmanni 7, 18, 42, 47, 49, 224, 225, 228
eversmanni eversmanni 49
eversmanni lumsdenii 49, 50, 57, 58
lumsdenii 51
maynardi 50, 51
orientalis 18, 47, 54, 58
Cyrtodactylus 3, 10, 11, 12, 18, 21, 22, 23,
130, 196, 198, 220
agamuroides 196
amictopholis 179, 222
basoglui 190, 192, 217
caspius 132, 143
elongatus 194
fedtschenkoi 143, 147, 157, 225
heterocercus 182
himalayanus 222
kachhensis 185
kirmanensis 206
kotschyi 160, 221
kotschyi danilewskii 163
longipes 153, 225
macularius 5, 26
montiumsalsorum 192
persicus 121
pulchellus 10, 11
russowi 167, 224
sagittifer 180
scaber 190
stoliczkai 205
voragi nosus 153, 157
watsoni 188
yarkandensis 205
Cyrtopodion 12, 20, 22, 23, 130, 185, 207
Diplodactylinae 5, 17
Echis carinatus 77, 140, 152
Elaphe dione 86
Eremias 220
intermedia 77
velox 140
Eryx miliaris 53
Eublepharidae 5, 17, 23, 25
Eublepharis 3, 5, 23, 25, 26, 221, 222
afghanicus 6
angramainyu 6, 25, 26, 28, 29
fasciolatus 6, 26
gracilis 6
hardwickii 5, 25
229
macularius 6, 25, 26, 27, 28, 29, 30, 228
macularius fasciolatus 6
macularius fuscus 6
macularius montanus 6
macularius smithi 6
turcmenicus 6, 25, 26, 28, 30, 31, 32, 228
Eumeces schneideri blithanus 126
Gekkonidae 5, 17, 23, 33
Gekkoninae 5, 17, 19
Gymnodactylus 3, 7, 10, 11, 12, 100, 130,
217, 220, 221
agamuroides 12, 196, 210, 211, 213
amictopholis 179
atropunctatus 56
brevipes 185, 197
caspius 11, 130, 131, 132
caspius insularis 3, 137, 216
chitralensis 201
colchicus 218
danilewskii 163, 218, 224
elongatus 194
eversmanni 7, 46, 47
fedtschenkoi 147, 155
gastropholis 12, 211, 213
geckoides 10
heterocercus 183, 184
heterocercus mardinensis 184
ingoldbyi 188
kachhensis 185
kachhensis kachhensis 185
kachhensis watsoni 188
kirmanensis 205
kotschyi 11, 159, 160, 183, 223, 224
kotschyi danilewskii 163, 220
longipes 11, 153, 155, 219
longipes longipes 156
longipes microlepis 3, 156
microlepis 153
microtis 7, 62
mintoni 3, 201
montiumsalsorum 192
persicus 208
petrensis 185
pipiens 56
russowi 11, 167, 171, 217, 220
russowi copalensis 171
russowi zarudnyi 172
sagittifer 180
scaber 147, 190, 217
spinicauda 175
steydneri 8, 106
stoliczkai 205
tenius 223
tibetanus 198
turcmenicus 3, 141, 220
walli 205
watsoni 188
zarudnyi 167, 172
Hemidactylus 13, 23
turcicus 13, 225
Hemiechinus megalotus 126
Iguanidae 5
Lacerta pipiens 7, 55, 56
Lycodon striatus 174, 179
Lytorhynchus ridgewayi 94, 216
Malpolon monspessulanus 140
Mediodactylus 3, 11, 12, 21, 22, 130, 159,
160, 207, 218
Mesodactylus 185
Microgecko 3, 9, 19, 20, 21, 101, 110, 221,
222, 224
helenae 9, 110, 111, 117
Milvus korshun 140
Ochotona rufescens rufescens 126
Ophisaurus apodus 140
Pristurus karterii 110
Psammophis lineolatum 140, 174, 195
Ptenodactylus 7, 46
Ptyodactylus 221
Pygopodidae 17
Rhinogekko 12, 207
misonnei 12, 213, 225
Saurodactylus 18
Sphaerodactylidae 5
Sphaerodactylinae 5, 17, 18
Stenodactylus 7, 17, 18, 20, 23, 88, 100, 221
dunstervillei 54
lumsdenii 7, 47, 50
maynardii 7, 50
orientalis 7, 54
petersii 106
scaber 185, 190
scincus 6, 35, 37 ,
sthenodactylus 228
Tenuidactylus 3, 11, 12, 14, 17, 18, 20, 21,
22, 23, 87, 130, 131, 132, 196, 220
agamuroides 12, 22, 23, 185, 196, 197
amictopholis 21, 22, 91, 160, 164, 179
caspius 11, 22, 75, 84, 93, 132, 133, 134,
136, 137, 228
caspius caspius 137
caspius insularis 137
chitralensis 21, 22, 198, 200, 201, 202
dattensis 196
elongatus 12, 22, 23, 45, 185, 193, 194, 228
fasciolatus 21, 196, 200, 201
fedtschenkoi 22, 132, 143, 146, 147, 155, 228
heterocercus 21, 22, 160, 182, 183
heterocercus heterocercus 183, 184.
heterocercus mardinensis 183, 184
himalayanus 21, 196, 200, 201
kachhensis 12, 22, 185, 186, 187, 190
kirmanensis 21, 22, 198, 200, 205, 206
kotschyi 21, 22, 160
kotshyi danilewskii 160, 161, 162, 163,
164, 165, 166, 228
lawderanus 21, 196, 198, 200, 201
longipes 22, 132, 153, 154, 155, 156, 158,
228
longipes longipes 143, 155, 156
longipes microlepis 143, 155, 156, 159
longipes voraginosus 155, 156, 157
microlepis 155
mintoni 22, 198, 200, 201, 203
montiumsalsorum 12, 22, 185, 187, 192, 193
russowi 21, 22, 42, 84, 160, 167, 168, 170,
171, 174
russowi russowi 171
russowi zarudnyi 171, 172, 228
sagittifer 21, 22, 28, 160, 180, 181
scaber 12, 22, 185, 189, 190, 191, 192, 193,
194
spinicauda 22, 32, 160, 170, 175, 176, 178,
228
stoliczkai 21, 22, 198, 200, 204
tibetanus 21, 22, 198, 200
turcmenicus 12, 22, 132, 141, 142, 143, 144,
145, 228
watsoni 12, 22, 185, 187, 188, 189
Teratoscincus 3, 6, 14, 17, 18, 21, 23, 33, 35, 218
bedriagai 7, 18, 21, 34, 35, 47
keyzerlingii 6, 33, 35, 38
microlepis 7, 18, 21, 35, 43, 44, 47
przewalskii 7, 18, 21, 35, 42, 44, 45, 228
scincus 18, 21, 34, 35, 39, 228
scincus keyzerlingii 6, 33, 37, 38
230
scincus scincus 37, 219
scincus rustamowi 3, 7, 37, 38, 220
roborowskii 7, 35, 37
zarudnyi 6, 35, 38
Trachydactyl us 8
jolensis 8, 94
spatalurus 94
Trapelus 221
Tropiocolotes 3, 8, 9, 10, 17, 19, 20, 21, 23,
100, 101, 117, 217, 221, 222, 223
depressus 9, 19 , 20, 21, 113, 120, 121, 124,
125
helenae 9, 20, 21, 111, 112, 113, 117, 118,
120
helenae fasciatus 9
heterop hoi is 9, 10, 130
latifi 9, 20, 21, 111, 113, 114, 119, 120
levitoni 3, 19, 20, 21, 113, 120, 121, 122,
123
nattereri 8, 9, 106, 224
occidentalis 102, 104
persicus 9, 20, 21, 111, 114, 115, 117, 120
persicus bakhtiari 115, 116, 117
persicus euphorbiacola 115, 116, 117, 118
persicus persicus 115, 116, 117
scortecci 9, 20, 21, 96, 101, 108, 109, 120
somalicus 102, 104
steudneri 9, 19, 20, 21, 96, 101, 106, 107,
108
tripolitanus 8, 9, 19, 20, 21, 96, 100, 101,
102, 103, 105, 120, 228
tripolitanus algericus 104
tripolitanus apoklomax 9, 105
tripolitanus occidentalis 105
Uroplatidae 17
Vipera lebetina 126, 140
Vulpes vulpes 140
Xantusiidae 17
ОГЛАВЛЕНИЕ
Предисловие................................................................... . 3
ОБЩАЯ ЧАСТЬ
Краткий обзор исследований гекконов фауны СССР и сопредельных стран................ 5
Материал и методика............................................................... 13
Филогенетические отношения и зоогеографический анализ ............................ 16
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Семейство эублефаровые — Eublepharidae ........................................... 25
Род эублефары — Eublepharis Gray. 1827 ..................................... 25
Группа юго-западноазиатских эублефаров ....................................... 25
Семейство Гекконовые — Gekkonidae ................................................ 33
Род сцинковый геккон — Teratoscincus Str., 1863 .............................. 33
Род гребнепалый геккон — Crossobamon Boettg., 1888 ............................. 46
Род североазиатский геккончик — Alsophylax ..................................... 55
Подрод равнинный геккончик — Alsophylax Fitz., 1843 ........................ 56
Подрод высокогорный североазиатский геккончик — Altiphylax Jeriomtschenko
et Szczerbak, 1984 ......................................................... 81
Род южноазиатский геккончик — Bunopus Blanf., 1874 ............................. 88
Род карликовый геккон — Tropiocolotes Peters, 1880 ............................ 100
Подрод африканский карликовый геккон — Tropiocolotes Peters, 1880 ......... 101
Подрод микрогеккон — Microgecko Nik., 1907 .............................. 110
Подрод азиатский карликовый геккон — Asiocolotes Golubev, 1984 ............ 120
Род ребристый геккон — Carinatogecko Golubev et Szczerbak, 1981.................126
Род тонкопалый геккон — Tenuidactylus Szczerbak et Golubev, 1984 ........ • • 130
Подрод тонкопалый геккон — Tenuidactylus Szczerbak et Golubev, 1984 ....... 131
Подрод средиземноморский тонкопалый геккон — Mediodactylus Szczerbak et
Golubev, 1977 ............................................................. 159
Подрод циртоподион — Cyrtopodion Fitzinger, 1843 . ........................ 185
Тибето-гималайская группа — Tenuidactylus ..................................197
Род крысохвостый геккон, или агамура — Agamura Blanford, 1874 ................. 208
Список литературы.................................................................217
Алфавитный указатель русских названий животных................................. 227
Алфавитный указатель латинских названий животных 229
ТАБЛИЦА 1:
/ — Eublepharis macularius, ad (фото А. Д. Баутина), 2 — Eublepharis macularius, juv. (фото
В. Миропольского), 3— Eublepharis turcmenicus (фото H. И. Щербака),
4 — Teratoscincus scincus (фото H. H. Щербака), 5 — Teratoscincus przewalskii (фото
Н. Л. Орлова), 6 — Crossobamon eversmanni (фото Н. Л. Орлова),
ТАБЛИЦА 2:
7—Stenodactylus stenodactylus (фото П. Кодыма), о — Alsophylax tokobajevi (фото
Н. Н. Щербака)
1 — Alsophylax pipiens (фото Н. Н. Щербака),
2 — Alsophylax laevis (фото H. H. Щербака), 3 — Alsophylax tadjikiensis (фото Н. Н. Щер-
бака), 4 — Alsophylax tadjikiensis (фото Н. Н. Щербака),
5 — Alsophylax loricatus szczerbaki (фото И. Л. Орлова), 6 — Bunopus tuberculatus (Турк-
мения) (фото H. H. Щербака), 7— Bunopus tuberculatus (Иран) (фото П. Кодыма),
ТАБЛИЦА 3:
8— Tropiocolotes tripolitanus (фото П. Кодыма)
1—Tenuidactylus caspius (фото Н. Н. Щербака), 2— Tenuidactylus turcmenicus (фото
Н. Н. Щербака),
3— Tenuidactylus fedtschenkoi (фото H. H. Щербака), 4 — Tenuidactylus longipes (фото
Н. Н. Щербака), 5 — Tenuidactylus elongatus (фото Н. Л. Орлова),
6 — Tenuidactylus kotschyi danilewskii (фото H. H. Щербака), 7 — Tenuidactylus russowi
(фото H. Л. Орлова), 8 — Tenuidactylus spinicauda (фото Н. Н. Щербака)