



















































































































Author: Ананьевой Н.Б. Боркина Л.Я.
Tags: reptilia рептилии пресмыкающиеся герпетология экология зоология фауна амфибии академия наук ссср палеарктическая азия
Year: 1981




। 4
АКАДЕМИЯ НАУК СССР
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
ФАУНА И ЭКОЛОГИЯ
АМФИБИЙ И РЕПТИЛИЙ
ПАЛЕАРКТИЧЕСКОЙ АЗИИ
АКАДЕМИЯ НАУК СССР
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА
Том 101
ФАУНА И ЭКОЛОГИЯ
АМФИБИЙ И РЕПТИЛИЙ
ПАЛЕАРКТИЧЕСКОЙ АЗИИ
Под редакцией Н. Б. Ананьевой и Л. Я. Боркина
ЛЕНИНГРАД
1981
ACADEMY OF SCIENCES OF THE USSR
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
Vol. 101
THE FAUNA AND ECOLOGY OF AMPHIBIANS
AND REPTILES OF THE PALAEARCTIC ASIA
Edited by N. B. Ananjeva and L. J. Borkin
Главный редактор
директор Зоологического института АН СССР
О. А. С КАР Л АТ О
Редакционная коллегия:
А. Н. Световидов (редактор серии), И. С. Даревский, В. А. Заславский,
И. М. Кержнер, Я. И. Старобогатов (заместитель редактора), В. А. Тряпицын.
| К. А. Юдин |
Рисунок на обложке — амурский полоз, Elaphe schrenckii (Strauch, 1873) —
художника Н. Н. Кондакова
2005000000—002
Ф---------------- Без объявления © Зоологический институт АН СССР, 1981
055(02)3—81
ОСОБЕННОСТИ СТРОЕНИЯ ЧЕРЕПА, ЗУБНОЙ СИСТЕМЫ
И ГИОИДА ЯЩЕРИЦ РОДА AGAMA ФАУНЫ СССР
Н. Б. Ананьева
Зоологический институт АН СССР (Ленинград)
Род Agama насчитывает около 60 видов, распространенных в юж-
ной Европе, Азии и Африке (Терентьев, 1961; Wermuth, 1967). На тер-
ритории СССР до настоящего времени были отмечены 6 видов: Agama
sanguinolenta (Pallas, 1814), Agama ruderata Olivier, 1804, Agama cau-
casia (Eichwald, 1831), Agama lehmanni (Nikolsky, 1896), Agama ery-
throgastra (Nikolsky, 1896), Agama himalayana (Steindachner, 1869).
Известная по одному экземпляру Agama pawlowskii Cernov et Dubinin,
1946 не представляет собой самостоятельного вида и должна быть
исключена из списка агамовых ящериц фауны мира и СССР (Ананьева
и Петерс, 1981). Agama chernovi Ananjeva, Peters et Rzepakovsky, 1981,
описание которой приводится в этом сборнике (с. 23) — седьмой вид
агам нашей фауны.
Некоторые агамы (A. sanguinolenta, A. caucasia) занимают обшир-
ный ареал в пределах нашей страны и являются одними из наиболее
массовых видов рептилий степных и горных районов. К изучению эко-
логии агам герпетологи обращались довольно часто (Богданов, 1958;
Хонякина, 1965; Хонякина и Кузеева, 1967; Атаев, 1974; Камалова,
1975; Соколовский, 1976), в то время как исследования по таксономии
до сих пор остаются значительно менее популярными.
Некоторые аспекты систематики и эволюции ящериц семейства
агамовых фауны СССР (роды Agama и Phrynocephalus) изучались
с использованием метода сравнительной кариологии (Соколовский,
1974, 1975, 1977). Попытки таксономической интерпретации данных ка-
риологии успешно практикуются и на других агамовых ящерицах,
в частности, видах рода Agama (Gorman a. Shochat, 1972; Moody
a. Hutterer, 1978). Однако имеющие сами по себе большую ценность
результаты кариологического исследования пока не имеют аналогов
в других подходах к систематике этой группы. Как это ни странно,
виды агамовых ящериц слабо изучены классическими, стандартными
методами зоологии. В частности, почти не обращалось внимание на
таксономические различия в строении черепа, зубной системы и подъ-
язычного аппарата, хотя отдельные работы, посвященные различным
сторонам этой проблемы, показали плодотворность такого подхода к си-
стематике агамид (Siebenrock, 1895; Царевский, 1929). В связи с этим
в настоящей работе сделана попытка проанализировать различия
в строении черепа, зубной системы и подъязычного аппарата агам
фауны СССР и оценить их таксономическое и функциональное зна-
чение.
3
Материал и методика
При изучении строения черепа и зубной системы агам фауны СССР
были использованы отпрепарированные черепа, отдельные кости черепа
и подъязычные аппараты следующих видов: A. sanguinolenta— 17 пол-
ных и расчлененных на фрагменты черепов, азиатская часть ареала;
26 полных черепов, Ногайская степь, Восточное Предкавказье;
A. caucasia — 5 полных черепов и около 50 отдельных фрагментов,
Закавказье, Туркмения; A. lehmanni — 3 полных и 3 расчлененных
на фрагменты черепа, Таджикистан; A. himalayana— 2 полных
и 1 расчлененный на фрагменты черепа, Таджикистан; A. erythro-
gastra—1 полный и 2 расчлененных черепа, Бадхыз; A. ruderata —
1 полный и 1 расчлененный черепа, Азербайджан. Строение зубной
системы дополнительно изучалось на ящерицах из коллекции отделения
герпетологии Зоологического института АН СССР, при этом исполь-
зовались экземпляры без повреждений зубов. Отдельные детали
строения черепа и подъязычного аппарата были уточнены на специально
изготовленных препаратах агам с глубокой окраской костных элемен-
тов ализарином и последующим просветлением в глицерине (Якубов-
ски, 1970). Строение подъязычного аппарата изучено по 2—5 отпрепа-
рированным гиоидам каждого вида. В данной работе я пользуюсь на-
званиями элементов гиоидной дуги, принятыми в работах К. Кэмпа
(Camp, 1923), Д. Эвери и В. Таннера (Avery a. Tanner, 1971) и П. Дуда
(Duda, 1966), установив их синонимию с более редко встречающимися
названиями (Соре, 1892; Beddard, 1905, 1907).
Я считаю своим приятным долгом выразить глубокую признатель-
ность В. П. Природиной, оказавшей мне большую методическую по-
мощь в проведении этой работы.
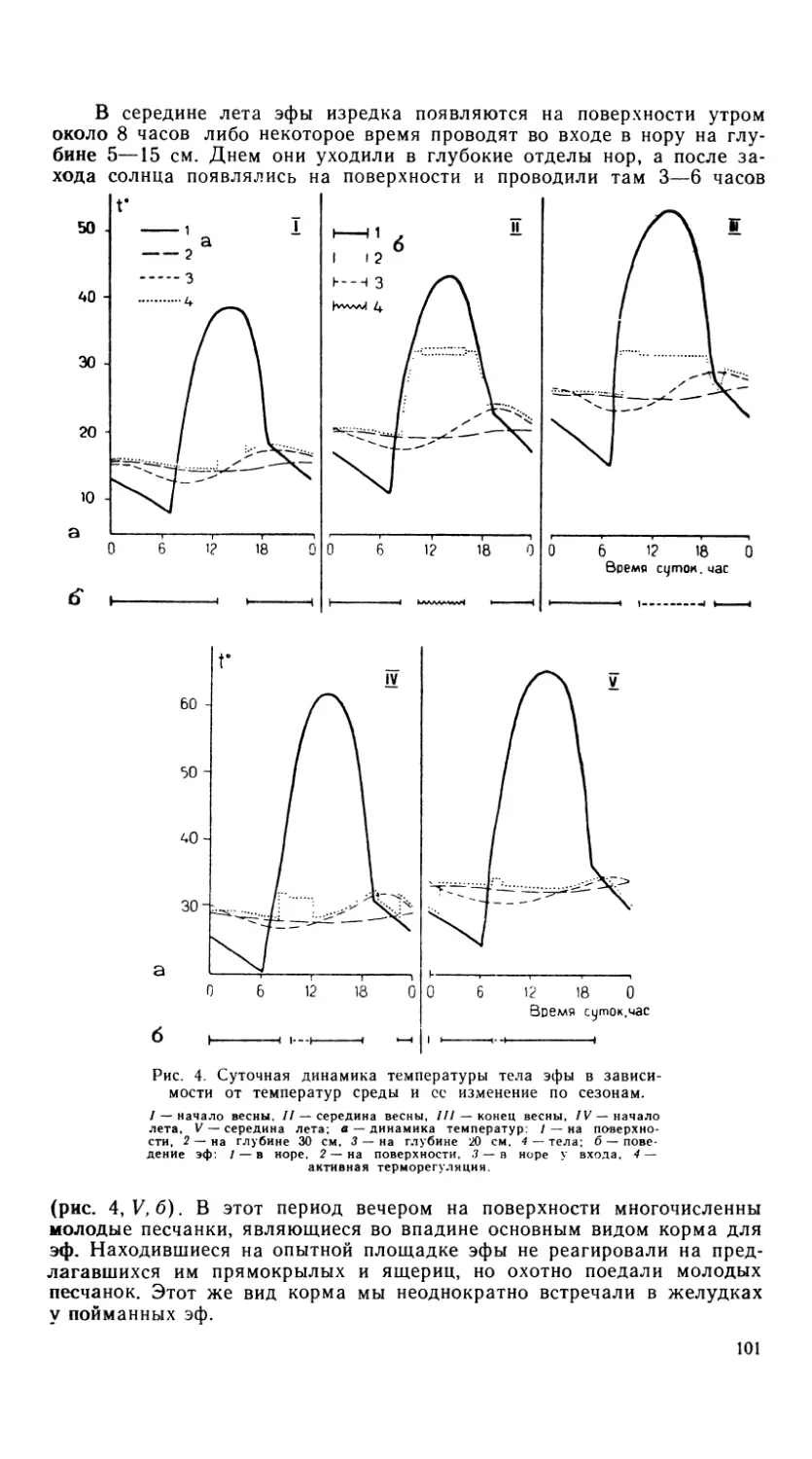
Результаты
Череп ящериц рода Agama, как и других агамид, широкий, с ко-
роткой лицевой частью. Форма черепа пирамидальная. Дорзальная,
вентральная и латеральная поверхности по своим очертаниям напоми-
нают треугольники. Основание черепа имеет форму равнобедренного
треугольника. Затылочное сочленение находится на одной оси с зад-
ними углами черепа, образованными соединением чешуйчатых костей
(os squamosum) с супратемпоральными отростками теменных костей
(os parietale) в основании этого треугольника. По сравнению с чере-
пом агам у ящериц рода Calotes височная область более узкая и глу-
бокая (Duda, 1965). Краниологические отличия от ящериц рода Uro-
mastix (Duda, 1965) и Phrynocephalus (Ананьева, 1977) во многом опре-
деляются различиями в пропорциях полушарий мозга и глаз.
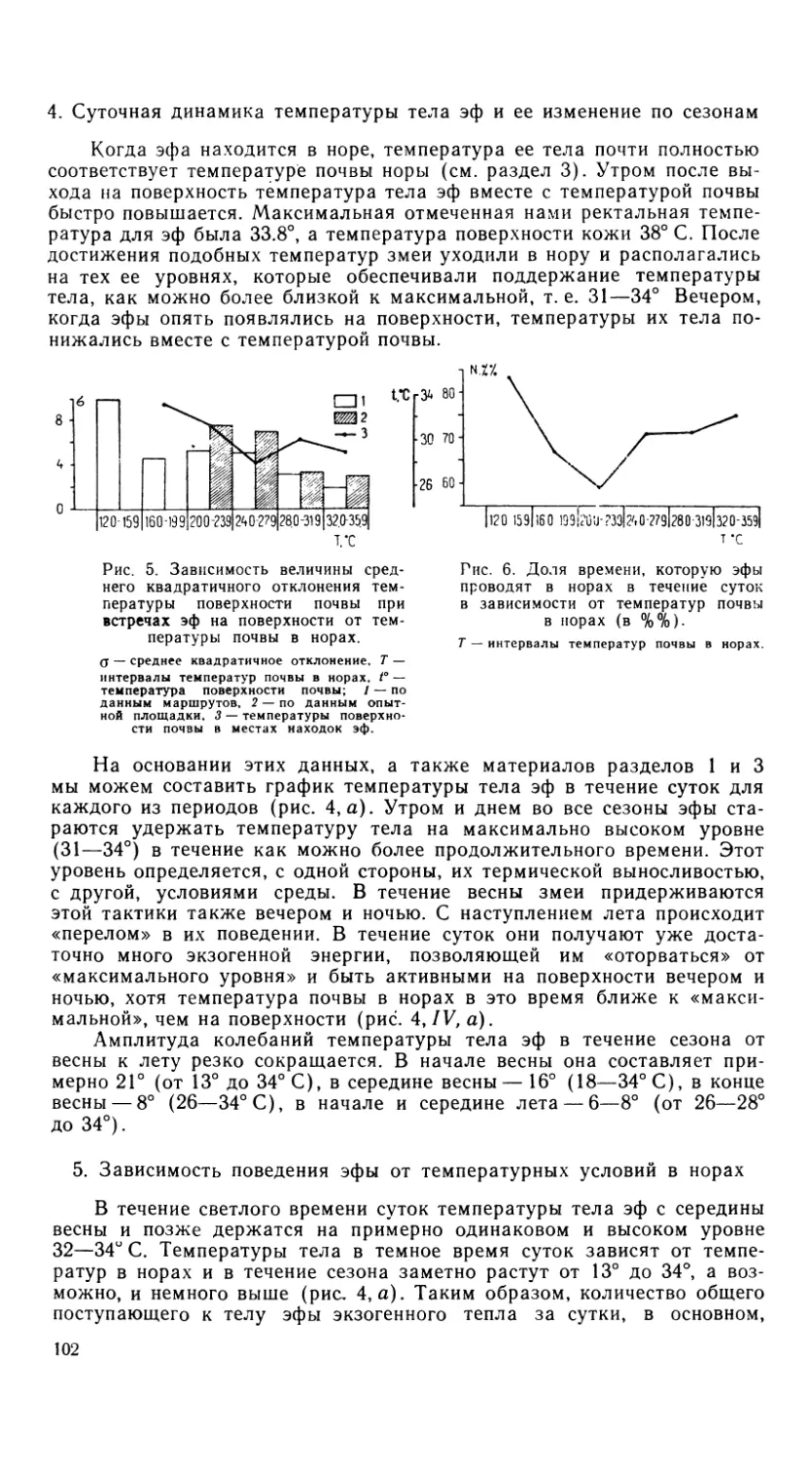
Зубы агам (за исключением передних) акродонтные (Wagler, 1830;
Siebenrock, 1895), т. е. прикреплены к верхнему краю челюсти, а зубная
система гетеродонтна. В нее входят зубы различного размера и формы,
выполняющие разные функции. Зубы располагаются только на челюст-
ной, предчелюстной и зубной костях, в отличие от некоторых других
ящериц (игуаниды, лацертиды), обладающих, кроме того, еще и неб-
ными зубами.
Предчелюстная кость (os praemaxillare) имеет шилообразную
форму и несет 2—3 острых конических зуба, как правило, не выступаю-
щих относительно высоты всего зубного ряда на верхней челюсти.
Верхнечелюстная кость (os maxillare) по форме напоминает вытя-
нутый треугольник (рис. 5), ее основание образует crista dentalis, на
котором укреплены зубы. Ветви верхней челюсти расположены под
острым углом, а зубные ряды наклонены несколько внутрь, лингуально.
Самый передний зуб, соседний с предчелюстными, сходен с ними по
4
форме и по размеру. По внешней аналогии с гетеродонтной зубной си-
стемой млекопитающих эти зубы часто называют «резцами» (dentes
incisivi). Второй зуб также конической формы, но значительно выше
и мощнее; его принято называть «клыком» (dens canini). Эти зубы,
«резцы» и «клыки», располагаются по одному в каждой половине верх-
ней челюсти, прикрепляясь к ней плевродонтно. Далее следуют более
широкие, типично акродонтные зубы с треугольными коронками, раз-
мер которых увеличивается, а форма усложняется в направлении спе-
реди назад.
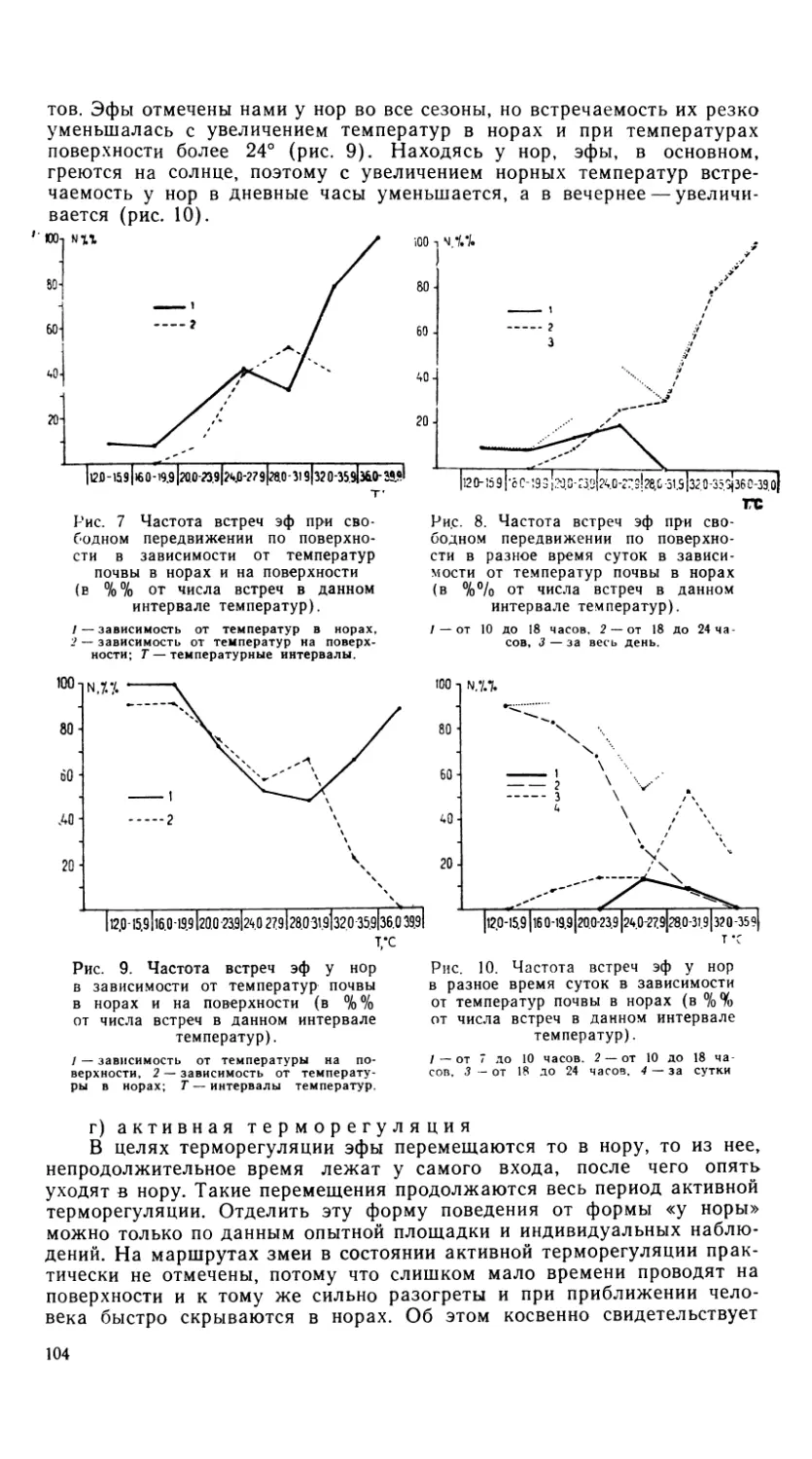
Нижняя челюсть образована двумя асимметричными ветвями, со-
единенными связками. Каждая половина состоит из следующих костей:
зубная (os dentale), сочленовная (os articulare), предсочленовная (os
praearticulare), угловая (os angulare), надугловая (os supraangulare),
венечная (os согопаге) и пластинчатая (os spleniale) Зубная кость
несет передние плевродонтные и задние акродонтные зубы. Они представ-
ляют собой по форме и расположению зеркальное отражение верхне-
челюстных зубов. Премаксиллярные зубы при окклюзии входят в про-
межуток между резцами нижней челюсти в области симфиза. Сочле-
новная кость образует вытянутый ретроартикулярный отросток для
сочленения с квадратной костью.
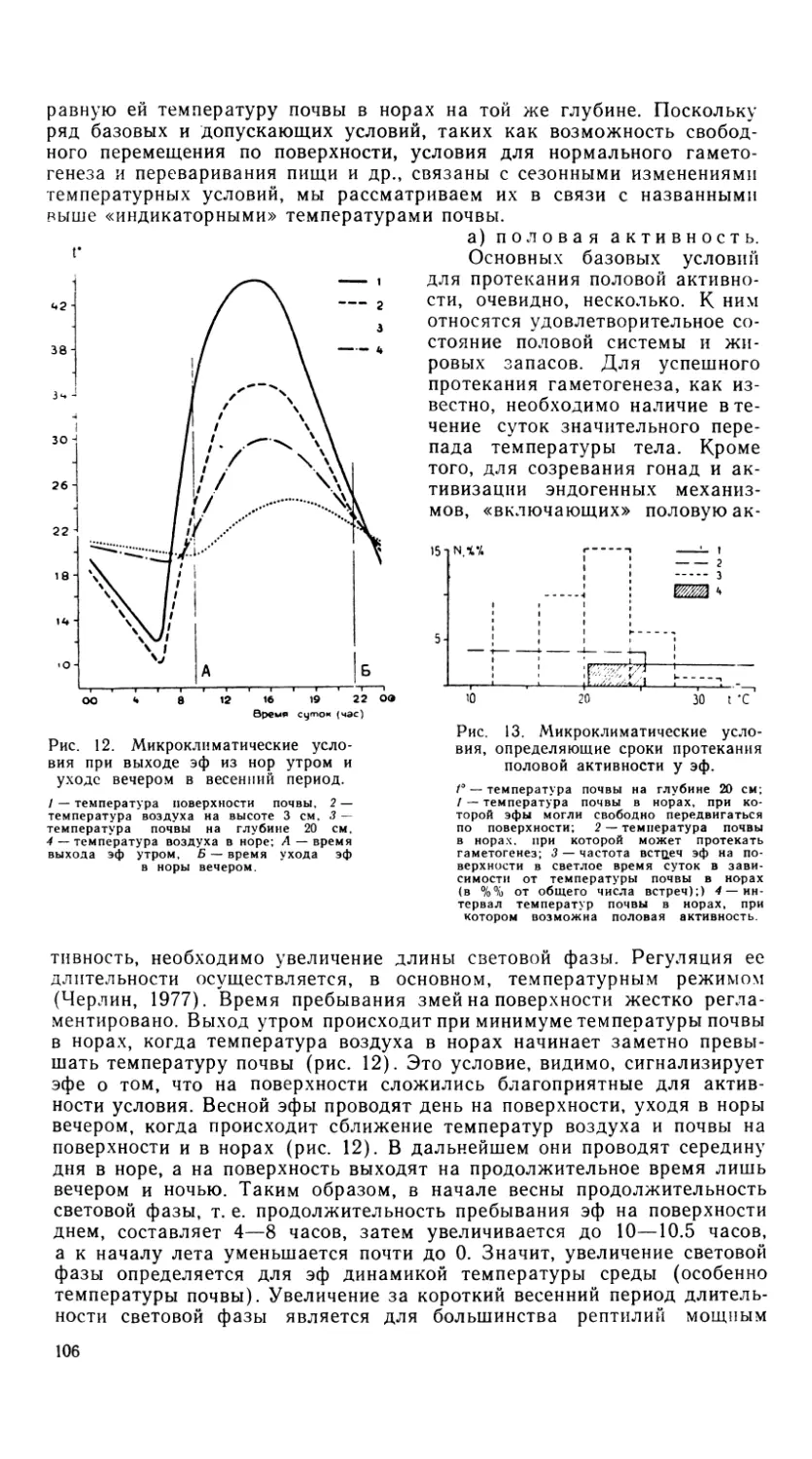
Своеобразна смена зубов у агамид, отличающаяся от типично реп-
тильной. У других ящериц зубы заменяются в течение всей жизни и
зубная система характеризуется неопределенным числом их генераций.
У взрослых же агамовых ящериц большая часть зубного ряда состоит
из акродонтных зубов, которые, однажды развившись, никогда более
не заменяются. Сравнение формы, размера и способа прикрепления пе-
редних зубов у молодых и половозрелых ящериц рода Agama показы-
вает, что с возрастом происходят их замещения (до четырех), в резуль-
тате которых акродонтное состояние заменяется плевродонтным, а го-
модонтная зубная система становится гетеродонтной (Cooper et al.,
1970). Это явление послужило основой для еще одной аналогии между
агамовыми ящерицами и млекопитающими, имеющими задние постоян-
ные зубы. И у тех, и у других животных замещение происходит только
в ряду передних зубов. Акродонтные зубы с возрастом стираются до
такой степени, что остается одна зубная полоска. Стирание зубов про-
исходит в большей степени на 4—5 передних акродонтных зубах, зад-
ние же сохраняются значительно дольше.
Видовые различия агам фауны СССР касаются целого ряда мор-
фологических признаков и будут рассмотрены ниже.
Степная агама, Agama sanguinolenta (Pallas, 1814)
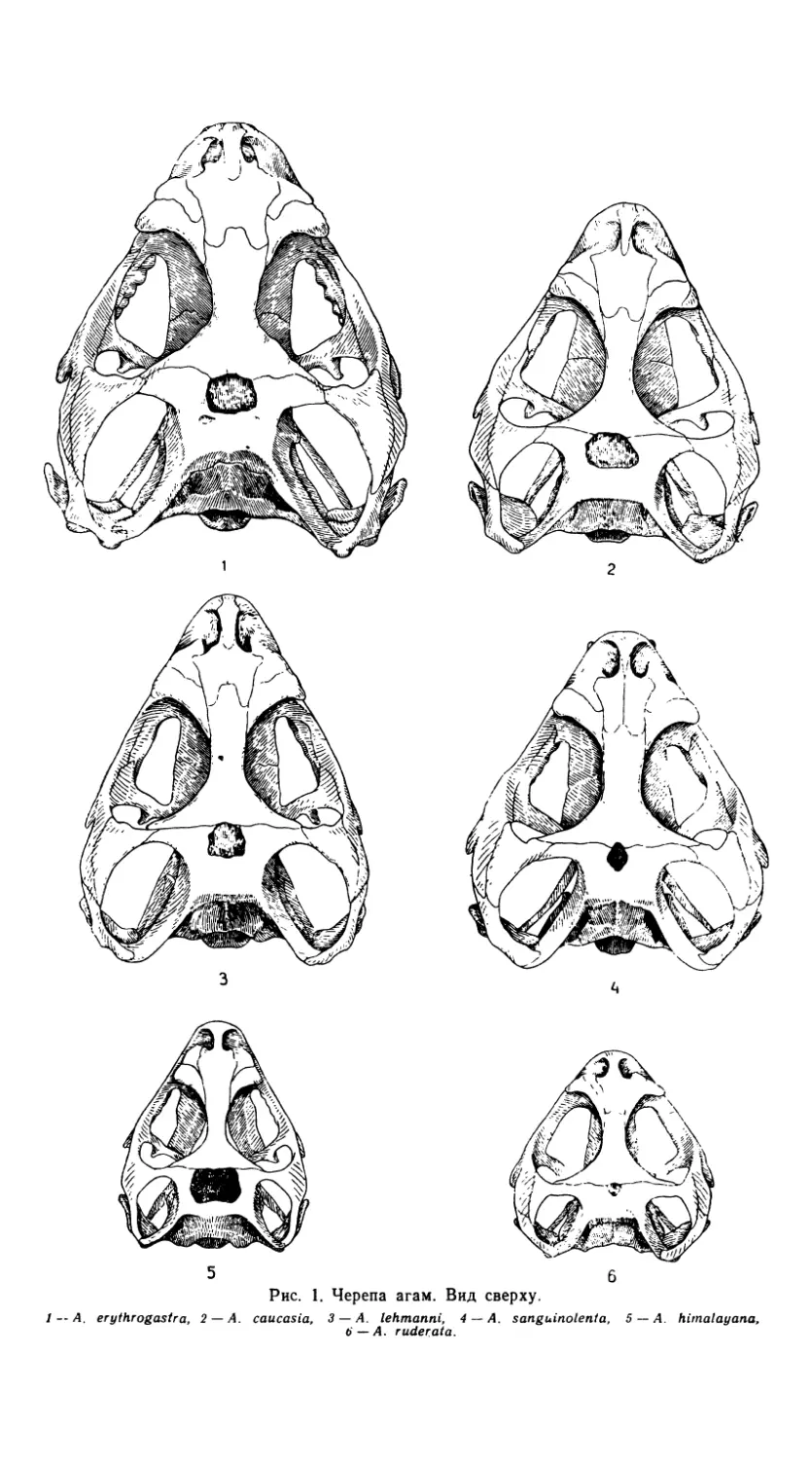
Череп A. sanguinolenta (рис. 1 и 2) относительно широкий и вы-
сокий, треугольной формы. Крыша черепа, слагающаяся из теменной и
лобной костей (os parietale и os frontale), выпуклая. Теменное отвер-
стие, расположенное между этими костями, очень мало. Поверхность
теменной и особенно лобной костей скульптурировано небольшими бу-
горками и бороздами. Задние выступы лобной кости соединяются с те-
менной костью в области теменного отверстия, а передние входят в кон-
такт с носовыми костями (рис. 1 и 4). Небно-птеригоидный шов распо-
ложен под острым углом. Сдвиг затылочного сочленения по отношению
к задним углам черепа весьма незначителен. Два последних признака
характерны и для других видов рода Agama, отличая их, в частности,
от видов рода Phrynocephalus (Ананьева, 1977).
Верхнечелюстная кость (os maxillare) имеет два отростка: pro-
cessus praefrontalis и processus praemaxillaris. Определенная ориента-
ция (табл. 2) к оси верхнечелюстной кости обусловила своеобразную
конфигурацию передней части черепа A. sanguinolenta (рис. 3 и 5)
5
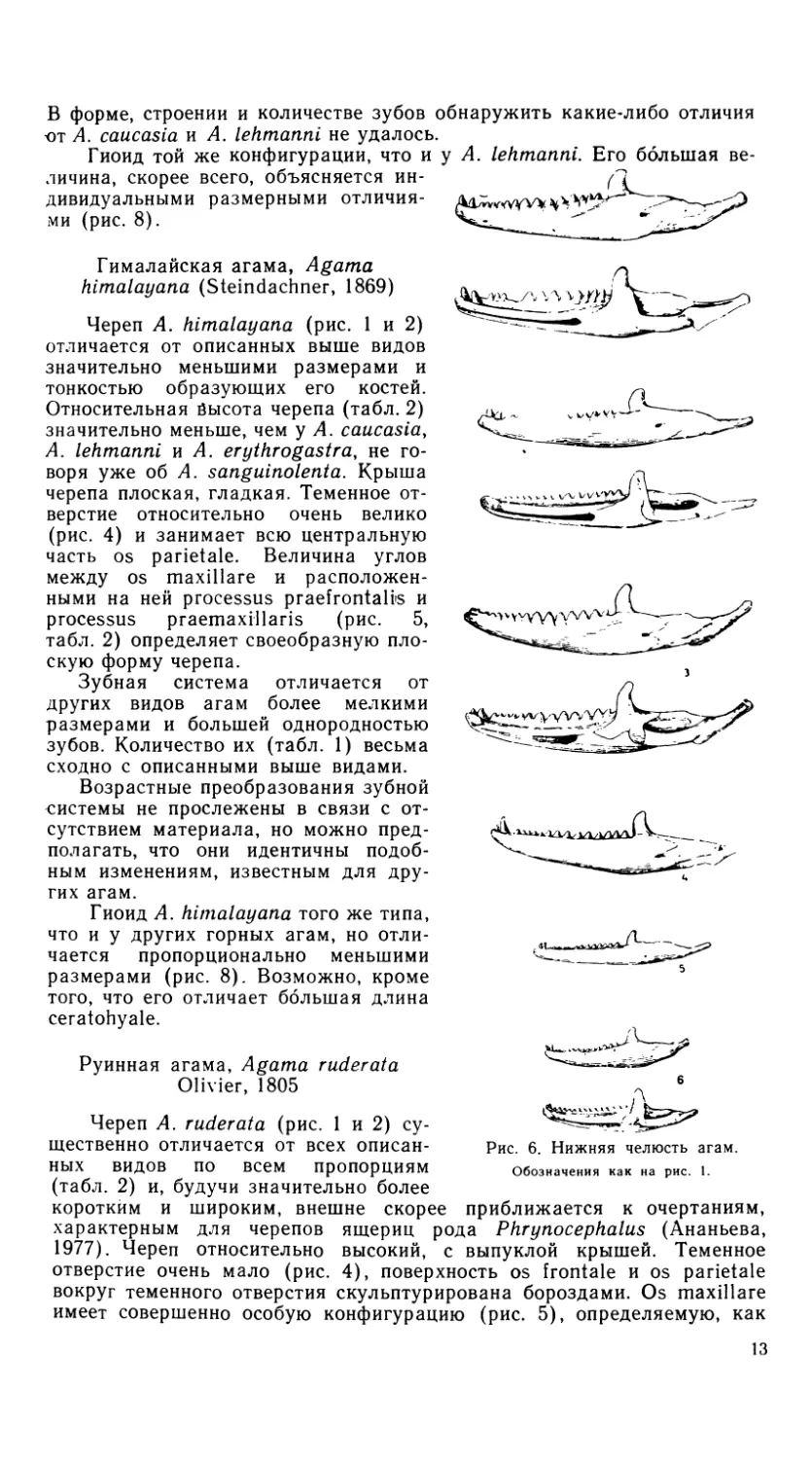
5 6
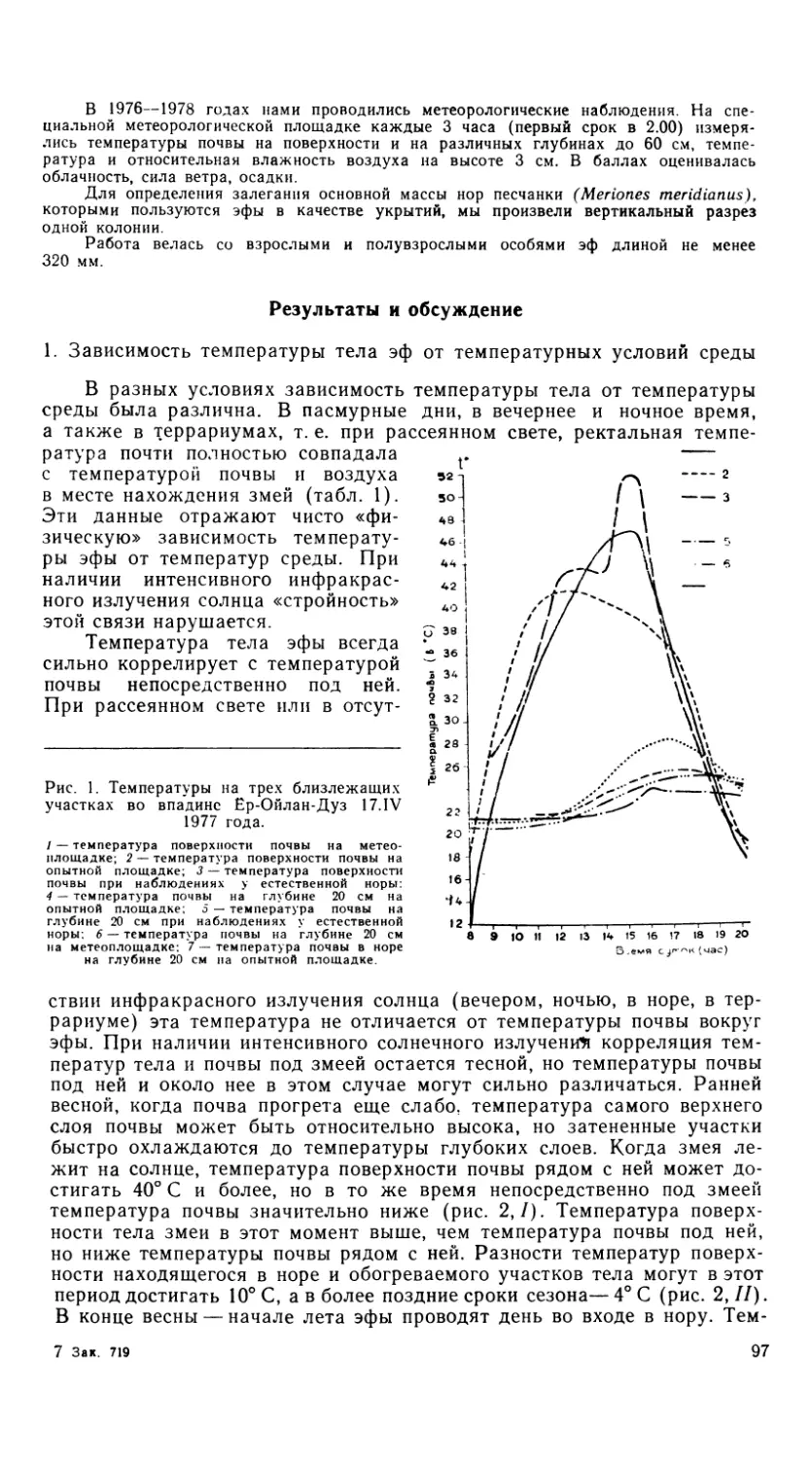
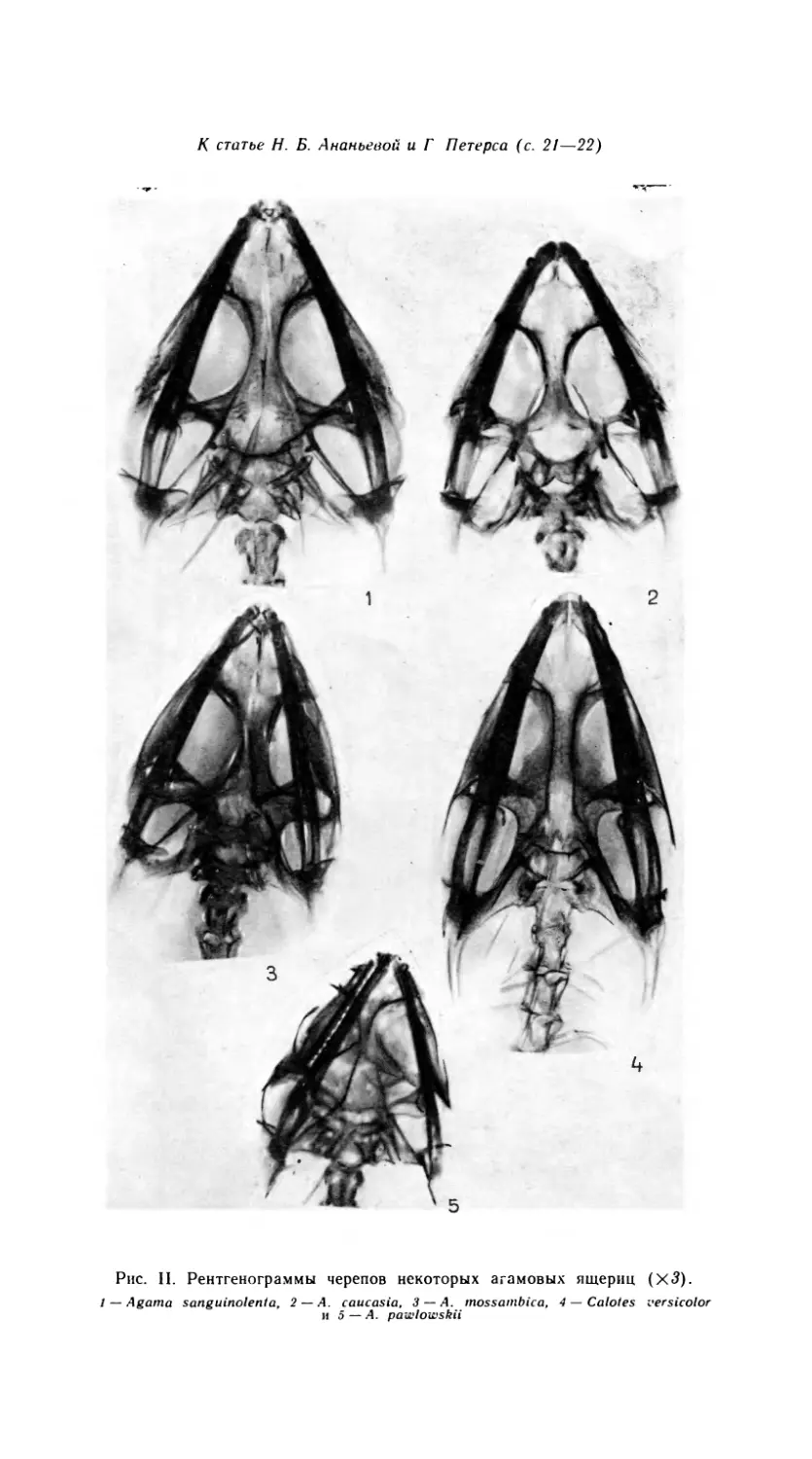
Рис. 1. Черепа агам. Вид сверху.
1 -- A. erythrogastra, 2 — A. caucasia, 3 — A. lehtnanni, 4 — A. sanguinolent а, 5 — A. hirnalayana,
6 — A. ruderata.
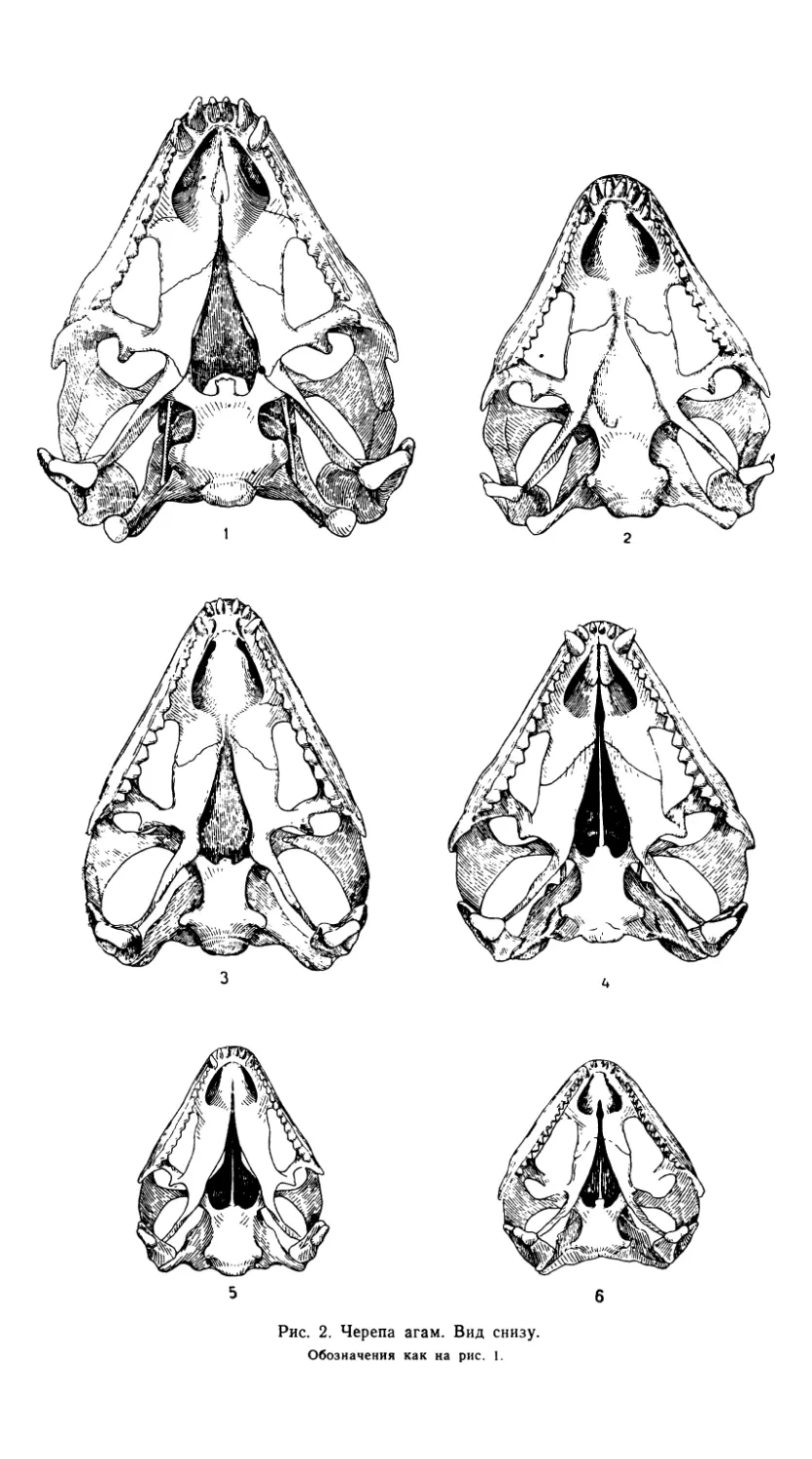
Рис. 2. Черепа агам. Вид снизу.
Обозначения как на рис. 1.
с выпуклой носовой частью и носовыми отверстиями, направленными
вперед. Нижняя челюсть с типичным для агам вытянутым ретроартику-
лярным отростком (рис. 6).
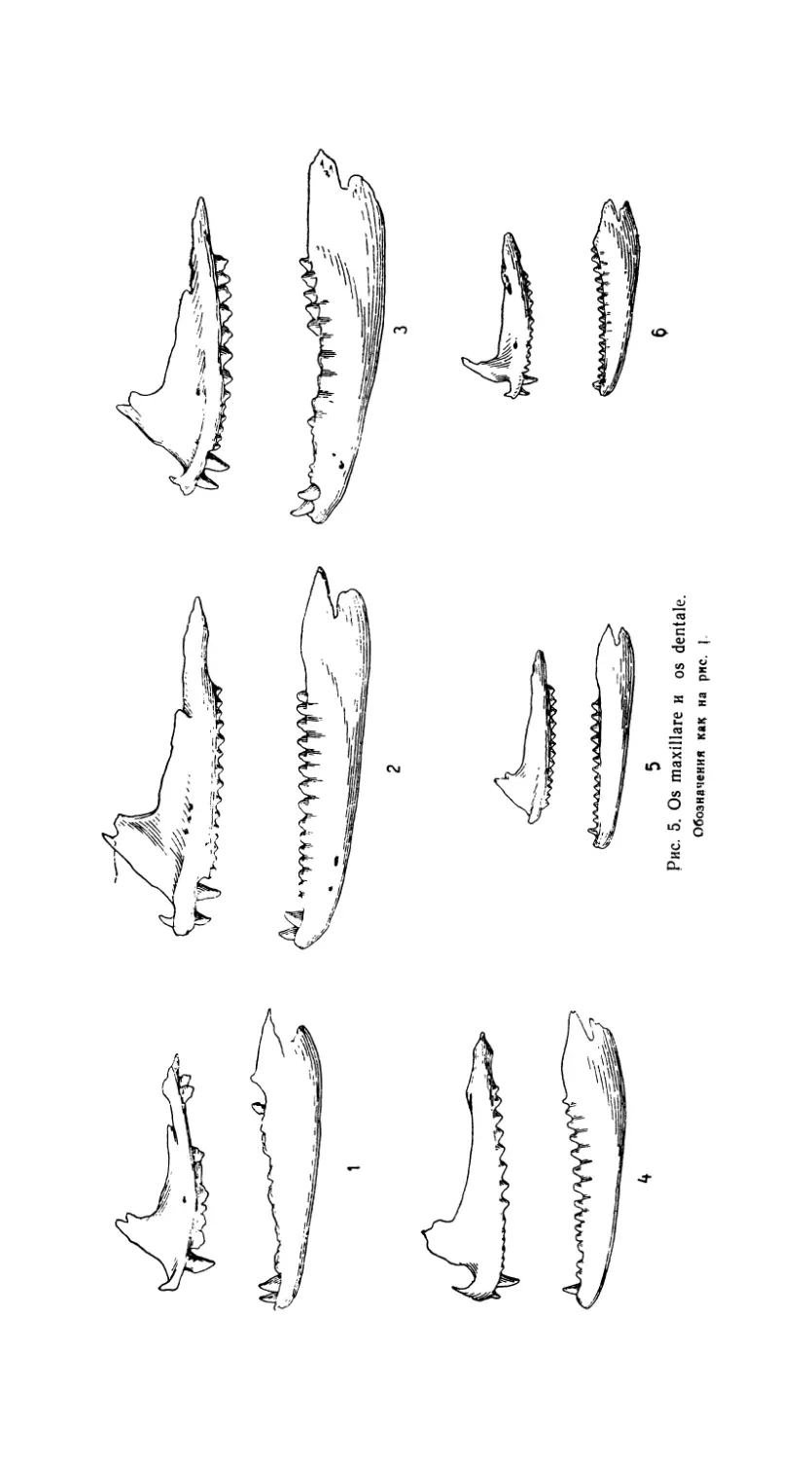
Зубная система хорошо дифференцирована (табл. 1, рис. 3 и 5).
Исследование серии черепов степной агамы (и = 26, Ногайская степь,
Восточное Предкавказье) показало, что в 8 случаях на предчелюстной
кости располагаются 3 зуба, в то время как на просмотренных до этого
черепах степной агамы из азиатской части ареала к этой кости по-
стоянно прикрепляются 2 зуба (Ананьева, 1977). Отметим также, что
серийный материал позволяет обнаружить наличие двух находящихся
очень близко друг к другу резцов вместо одного, разную величину и
ориентацию резцов и клыков в левой и правой половине челюстей
и т. д. Такая вариабильность состояния и количества передних зубов
может, вероятно, объясняться тем, что изученные черепа демонстрируют
различные стадии замещения зубов агам разного возраста. Возможно,
кроме того, что большее количество зубов на предчелюстной кости (3)
чаще встречается у предкавказских степных агам, чем у азиатских,
однако недостаток фактов, свидетельствующих о географической из-
менчивости в строении зубной системы, не позволяет рассмотреть этот
вопрос более подробно.
Таблица 1
Зубные формулы ящериц рода Agama
Виды Количество зубов на костях челюстей Автор
praema- xillare maxillare dentale
A. sanguinolenta 2 I1C1M12-15 I1C1M12-15 Siebenrock, 1895
2 (3) 11С1М1з_ц I1C1M13-15 данные автора
A. ruder ata 2 11С1М12-Ц I1C1M13-14 »
A. caucasia 2 (3) I1C1M.12-14 I1C1M12-14 »
A. lehmanni 2 (3) I1C1M12-14 I1C1M12-14 »
A. erythrogastra 2 (3) I1C1M12-15 I1C1M12^16 »
A. himalayana 2 I1C1M12 I1C1M12 Siebenrock, 1895
2 (3) I1C1M12-14 I>C1M12-14 данные автора
A. pallida 2 IiCiMu Siebenrock, 1895
A. hispida 2 I1C1M13 IlC!M13 »
A. atra 1 IlCiMi4_tf IiCiM^ »
A. colonorum 1 I1C1M18-20 I1C1M17 »
A. tuberculata 3 I1C1M15-16 I1C1M15-13 »
3 IiCiMie IiCiMu Duda, 1965
A. stellio 2 IiCiMe-13 IiCiMio-i Siebenrock, 1895
Примечание. I — «резцы», С — «клыки», М — задние акродонтные зубы, «ко-
ренные».
Таблица 2
Пропорции черепа ящериц рода Agama фауны СССР
Виды Lt. cr./Lg. cr. Lt. сг./At. cr. I П
A. sanguinolenta 11 0.85 1.75 4 72 61
A. ruderata 1 0.93 2.00 1 80 71
A. caucasia 4 0.79 2.12 8 55 45
A. lehmanni 2 0.78 2.02 3 59 51
A. himalayana 2 0.80 2.38 2 — 40
A. erythrogastra 1 0.80 2.01 1 — 52
Примечание, п — число изученных черепов, Lt. сг. — ширина черепа, Lg. сг. —
длина черепа, At. сг. — высота черепа, I—величина угла между maxillare и processus
praemaxillaris, II — величина угла между maxillare и processus praefrontalis. Индексы
вычислялись на основании промеров черепов без нижней челюсти, что объясняет
отклонения от указанных в работе П. В. Терентьева и С. А. Чернова (1949) пропор-
ций головы.
8
На верхнечелюстной кости размещаются все три категории зубов.
Первый, dens incisivi, находится рядом с предчелюстными зубами. Вто-
рой, dens canini, значительно выше и мощнее, сильно выдается относи-
тельно высоты всего зубного ряда. Далее следуют более широкие ти-
пично акродонтные зубы с треугольными коронками. В конце зубного
ряда эти зубы (dentes molares) приобретают триконодонтную форму,
их число обычно варьирует от 13 до 15 (табл. 1).
Возрастные изменения структуры черепа выражаются в его боль-
шем окостенении у взрослых особей. На примере степной агамы можно
наблюдать возрастную смену зубов, типичную для агамовых ящериц.
Гетеродонтная и постоянная в количестве зубов зубная система форми-
руется к моменту наступления половозрелости. До этого в передней
части челюсти происходит несколько смен зубов, причем каждая по-
следующая генерация представлена более крупными зубами, чем пре-
дыдущая (Cooper et al., 1970). Число акродонтных зубов увеличивается
с возрастом, причем новые зубы отрастают в задней части челюстной
и зубной костей.
Гиоид A. sanguinolenta, состоящий из обызвествленного хряща,
представлен элементами первой и второй висцеральных дуг и находится
между ветвями нижней челюсти. Третья дуга, как и у большинства
ящериц, у степной агамы отсутствует. Редукции подвергся также эле-
мент второй дуги—epibranchiale II (Camp, 1923). По своей конфигу-
рации гиоид этого вида напоминает подъязычный аппарат некоторых
древесных игуановых ящериц (Camp, 1923; Gnanamuthu, 1937; Avery
a. Tanner, 1971) и калота Calotes versicolor (Camp, 1923). Однако у ка-
лота и степной агамы, в отличие от игуанид, относительную редукцию
претерпевает ceratohyale (рис. 8). Гиоид A. sanguinolenta характери-
зуется очень тонким processus entoglossus, образующим скелет языка.
Особенно обращает на себя внимание расположение ceratobranchiale II.
Эти элементы подъязычного аппарата у степной агамы очень тонки и
расположены очень близко друг к другу, почти сливаясь зрительно
в один непарный элемент (рис. 8).
Кавказская агама, Agama caucasia (Eichwald, 1831)
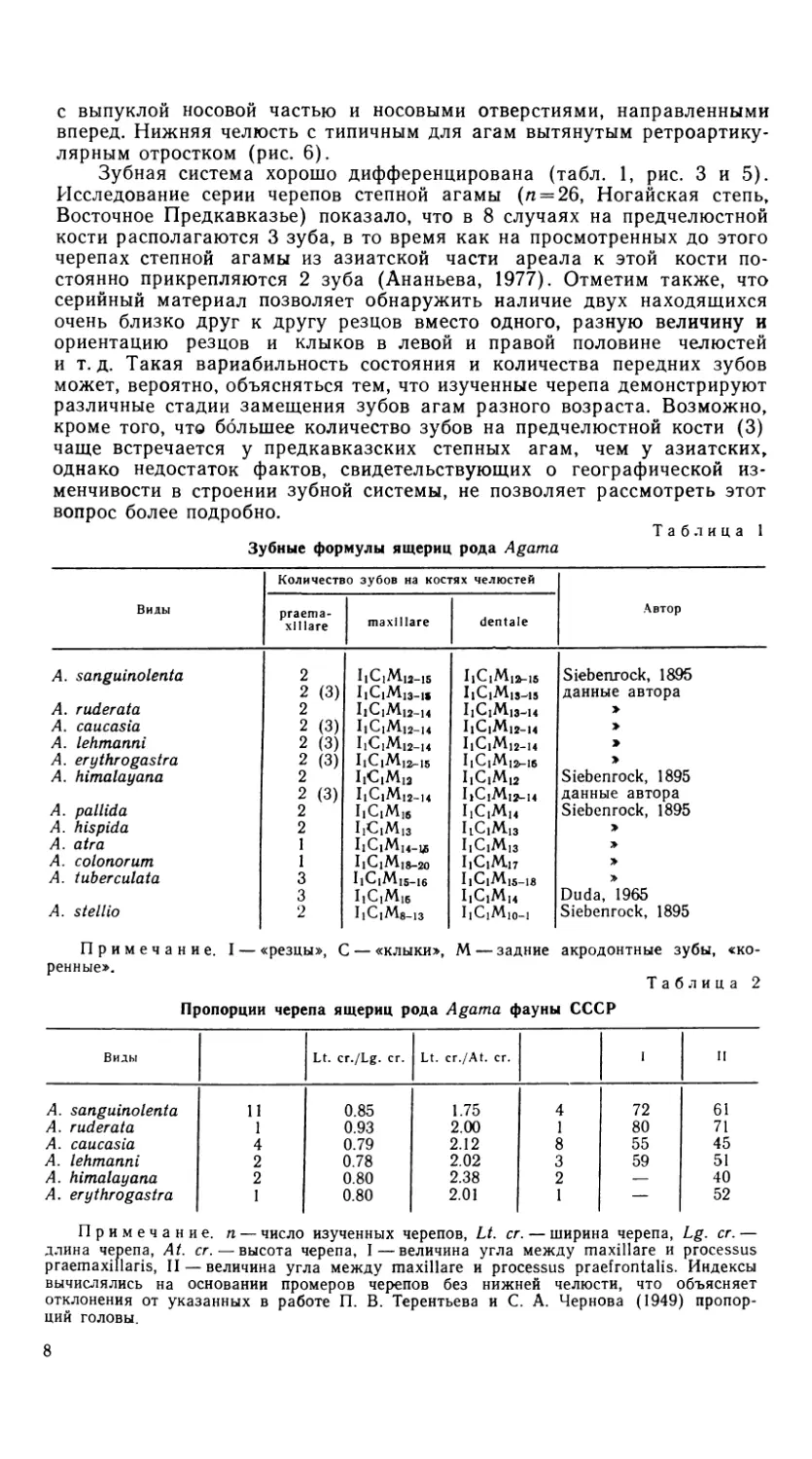
Череп (рис. 1 и 2) относительно широкий, треугольной формы, как
и у степной агамы. От последней A. caucasia значительно отличается
по индексу относительной высоты черепа (рис. 3, табл. 2). Полное опи-
сание скелета этого вида было впервые сделано в середине прошлого
века (Eichwald, 1841), а также недавно, наряду с анатомией других
система органов (Гуртовой и др., 1978).
Крыша черепа очень плоская, скульптурировка лобной и теменной
костей почти не выражена, их поверхность гладкая. Теменное отверстие
(foramen parietale) намного крупнее, чем у степной агамы. На темен-
ной кости оно имеет округлую форму (рис. 4). A. caucasia заметно
отличается от степной агамы формой верхнечелюстной кости (os maxil-
lare) и, в первую очередь, расположением отростков processus praefron-
talis и processus praemaxillaris (рис. 5, табл. 2). Это определяет типич-
ную для кавказской агамы плоскую форму черепа. Следует отметить
в перечне видовых отличий также иную по сравнению с A. sanguino-
lenta форму os maxillare в области processus praefrontalis (рис. 5) и
значительно более удлиненный, вытянутый и заостренный задний отро-
сток этой кости. Ретроартикулярный отросток нижней челюсти также
значительно более длинный, острый и вытянутый, чем у степной агамы
(рис. 6).
Зубная система хорошо дифференцирована (табл. 1, рис. 3 и 5).
В отличие от A. sanguinolenta длина клыков и резцов на нижней че-
люсти бывает примерно одинаковой. В остальном зубные формулы
9
этих двух видов очень сходны. На предчелюстной кости обычно укреп-
лены 2 или 3 зуба. Возрастные изменения в строении зубной системы
типичны для агамовых ящериц и аналогичны выше описанным. Как
на проявление полового диморфизма следует обратить внимание на,
относительно большую ширину черепа у самцов.
Гиоид A. caucasia имеет более широкий и более короткий по срав-
нению с A. sanguinolenta processus entoglossus с небольшой заметной
щелевидной фонтанелью по средней вертикальной линии и более ко-
роткие ceratohyale. Наиболее существенные отличия от A. sanguinolenta
состоят во взаимном расположении парных элементов ceratobran-
chiale II, которые расходятся от basihyale таким образом, что между
ними образуется значительное расстояние (рис. 8).
Рис. 3. Черепа агам. Вид сбоку.
Обозначения как на рис. 1
Туркестанская агама, Agama lehmanni (Nikolsky, 1896)
Череп A. lehmanni (рис. 1 и 2), как и у описанных выше видов,
относительно широкий (табл. 2), треугольной формы. По сравнению
с A. sanguinolenta череп более плоский, отличия особенно выражены
в той части черепа, которая слагается из os parietale и os frontale.
По сравнению с A. caucasia, однако, череп туркестанской агамы выгля-
дит более высоким (рис. 3, табл. 2). Скульптурировка костей крыши
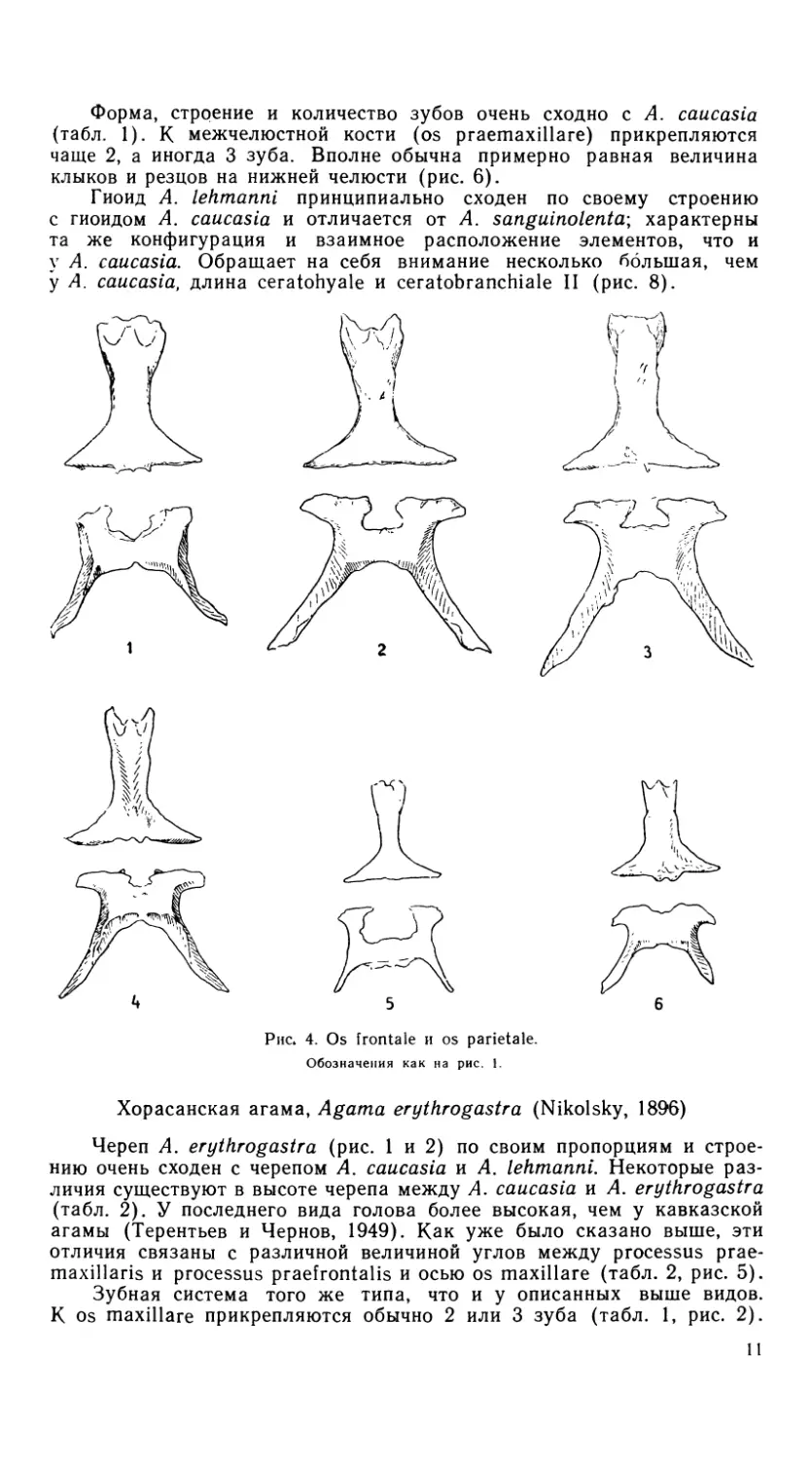
черепа выражена довольно слабо. Теменное отверстие значительной
величины, оно отличается по форме от описанных выше видов (рис. 4);
на os parietale оно имеет приблизительно треугольную форму. Наибо-
лее четкие видовые различия заключаются в форме os maxillare. Более
конкретно их можно определить по величине углов между os maxillare
и расположенными на ней processus praemaxillaris и processus ргае-
frontalis (рис. 5, табл. 2).
10
Форма, строение и количество зубов очень сходно с A. caucasia
(табл. 1). К межчелюстной кости (os praemaxillare) прикрепляются
чаще 2, а иногда 3 зуба. Вполне обычна примерно равная величина
клыков и резцов на нижней челюсти (рис. 6).
Гиоид A. lehmanni принципиально сходен по своему строению
с гиоидом A. caucasia и отличается от A. sanguinolenta; характерны
та же конфигурация и взаимное расположение элементов, что и
у A. caucasia. Обращает на себя внимание несколько большая, чем
у A. caucasia, длина ceratohyale и ceratobranchiale II (рис. 8).
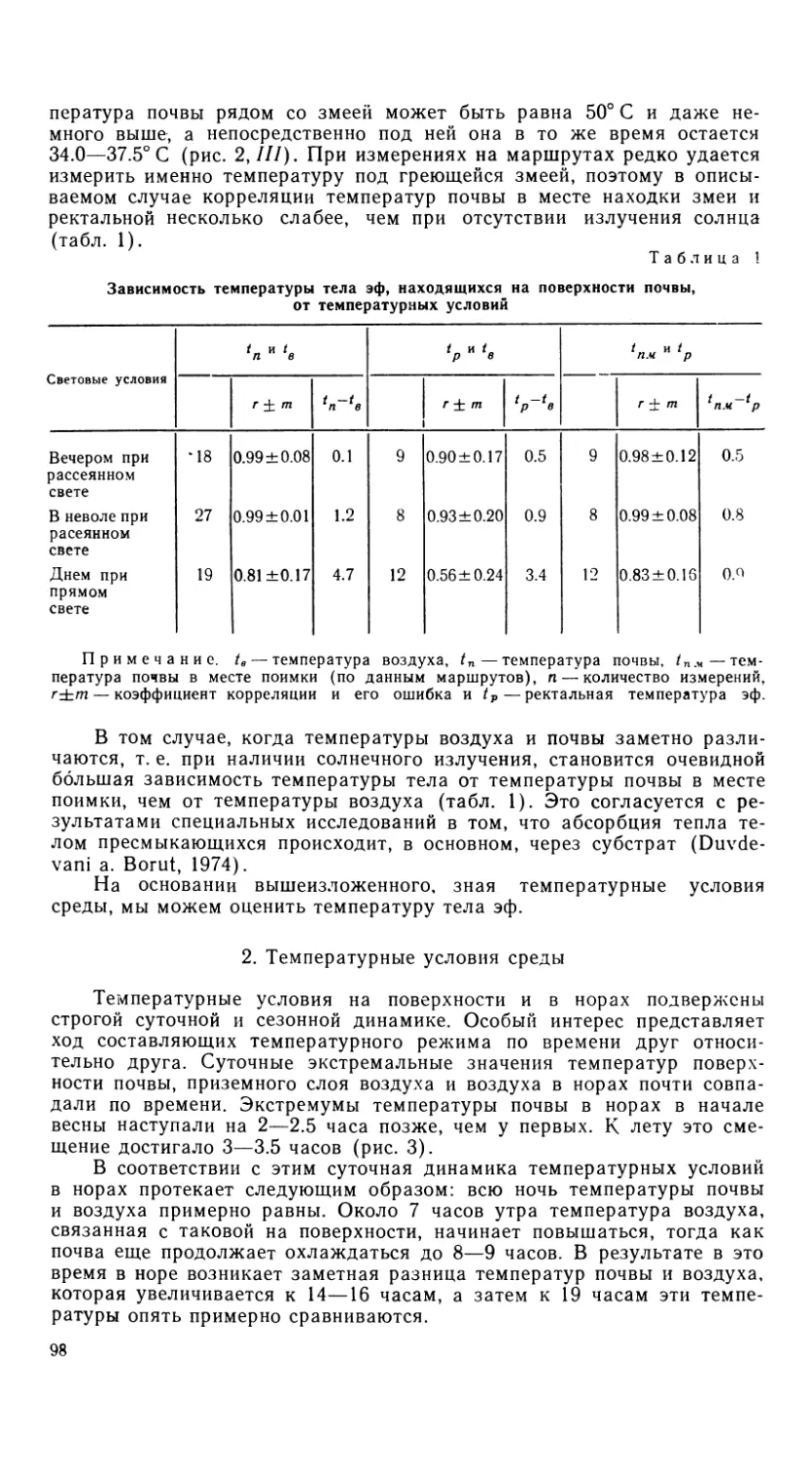
Рис. 4. Os frontale и os parietale.
Обозначения как на рис. 1.
Хорасанская агама, Agama erythrogastra (Nikolsky, 1896)
Череп A. erythrogastra (рис. 1 и 2) по своим пропорциям и строе-
нию очень сходен с черепом A. caucasia и A. lehmanni. Некоторые раз-
личия существуют в высоте черепа между A. caucasia и A. erythrogastra
(табл. 2). У последнего вида голова более высокая, чем у кавказской
агамы (Терентьев и Чернов, 1949). Как уже было сказано выше, эти
отличия связаны с различной величиной углов между processus ргае-
maxillaris и processus praefrontalis и осью os maxillare (табл. 2, рис. 5).
Зубная система того же типа, что и у описанных выше видов.
К os maxillare прикрепляются обычно 2 или 3 зуба (табл. 1, рис. 2).
И
2
5
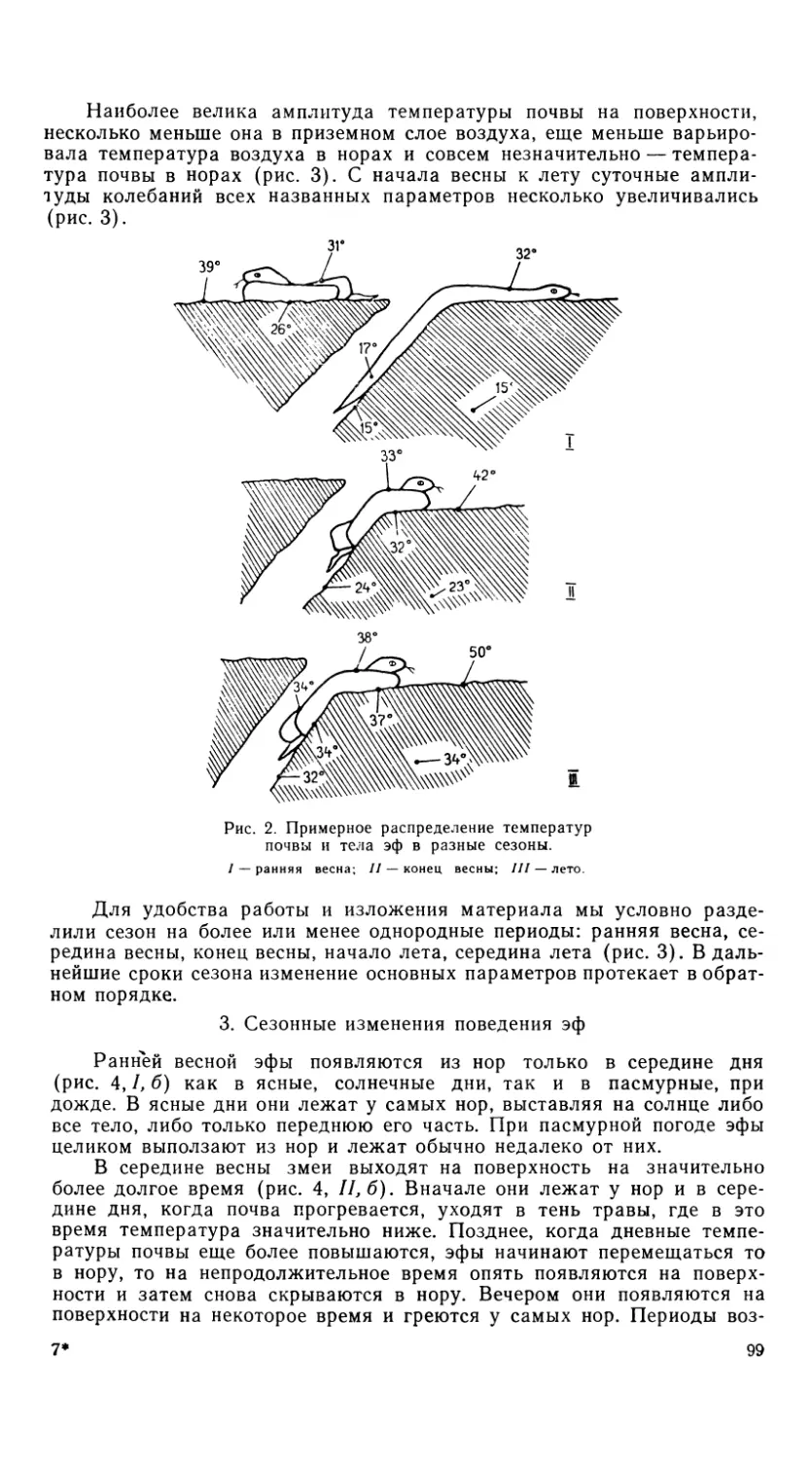
Рис. 5. Os maxillare и os dentale.
Обозначения как на рис.
б
В форме, строении и количестве зубов обнаружить какие-либо отличия
ют A. caucasia и A. lehmanni не удалось.
Гиоид той же конфигурации, что и у Л. lehmanni. Его большая ве-
личина, скорее всего, объясняется ин-
дивидуальными размерными отличия-
ми (рис. 8).
Гималайская агама, Agama
himalayana (Steindachner, 1869)
Череп A. himalayana (рис. 1 и 2)
отличается от описанных выше видов
значительно меньшими размерами и
тонкостью образующих его костей.
Относительная йысота черепа (табл. 2)
значительно меньше, чем у A. caucasia,
A. lehmanni и Л. erythrogastra, не го-
воря уже об Л. sanguinolenta. Крыша
черепа плоская, гладкая. Теменное от-
верстие относительно очень велико
(рис. 4) и занимает всю центральную
часть os parietale. Величина углов
между os maxillare и расположен-
ными на ней processus praefrontalis и
processus praemaxillaris (рис. 5,
табл. 2) определяет своеобразную пло-
скую форму черепа.
Зубная система отличается от
других видов агам более мелкими
размерами и большей однородностью
зубов. Количество их (табл. 1) весьма
сходно с описанными выше видами.
Возрастные преобразования зубной
системы не прослежены в связи с от-
сутствием материала, но можно пред-
полагать, что они идентичны подоб-
ным изменениям, известным для дру-
гих агам.
Гиоид Л. himalayana того же типа,
что и у других горных агам, но отли-
чается пропорционально меньшими
размерами (рис. 8). Возможно, кроме
того, что его отличает большая длина
ceratohyale.
Руинная агама, Agama ruderata
Olivier, 1805
Череп Л. ruderata (рис. 1 и 2) су-
щественно отличается от всех описан-
ных ВИДОВ ПО ВСеМ пропорциям Обозначения как на рис. 1.
(табл. 2) и, будучи значительно более
коротким и широким, внешне скорее приближается к очертаниям,
характерным для черепов ящериц рода Phrynocephalus (Ананьева,
1977). Череп относительно высокий, с выпуклой крышей. Теменное
отверстие очень мало (рис. 4), поверхность os frontale и os parietale
вокруг теменного отверстия скульптурирована бороздами. Os maxillare
имеет совершенно особую конфигурацию (рис. 5), определяемую, как
13
и у других видов агам, расположением отростков (табл. 2). Ретроарти-
кулярный отросток нижней челюсти заметно укорочен по сравнению
с другими изученными видами (рис. 6).
Зубная система, как и у других мелких форм (например, A. hima-
layana), дифференцирована менее заметно, чем у крупных агам. Зубы
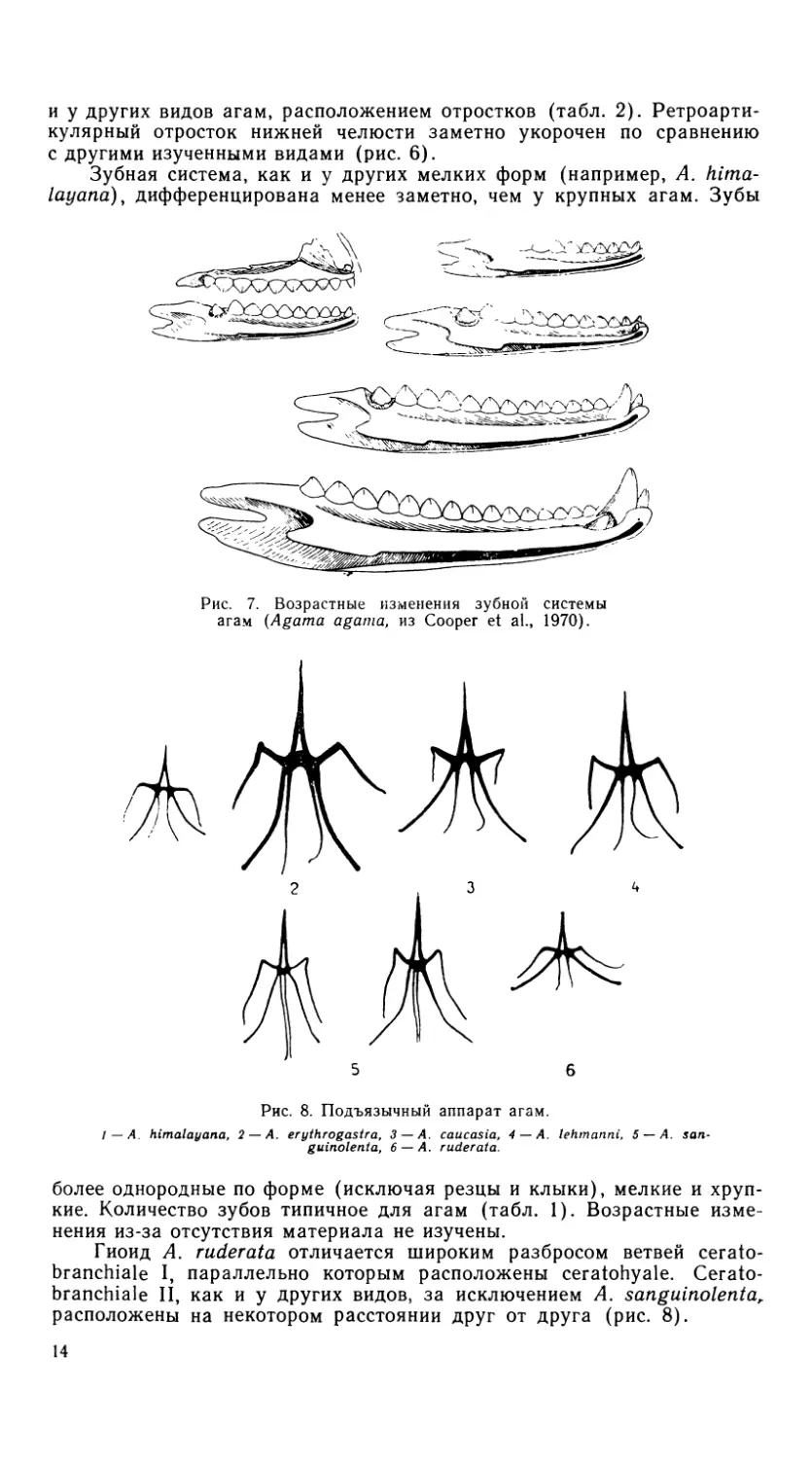
Рис. 7. Возрастные изменения зубной системы
агам (Agama agama, из Cooper et al., 1970).
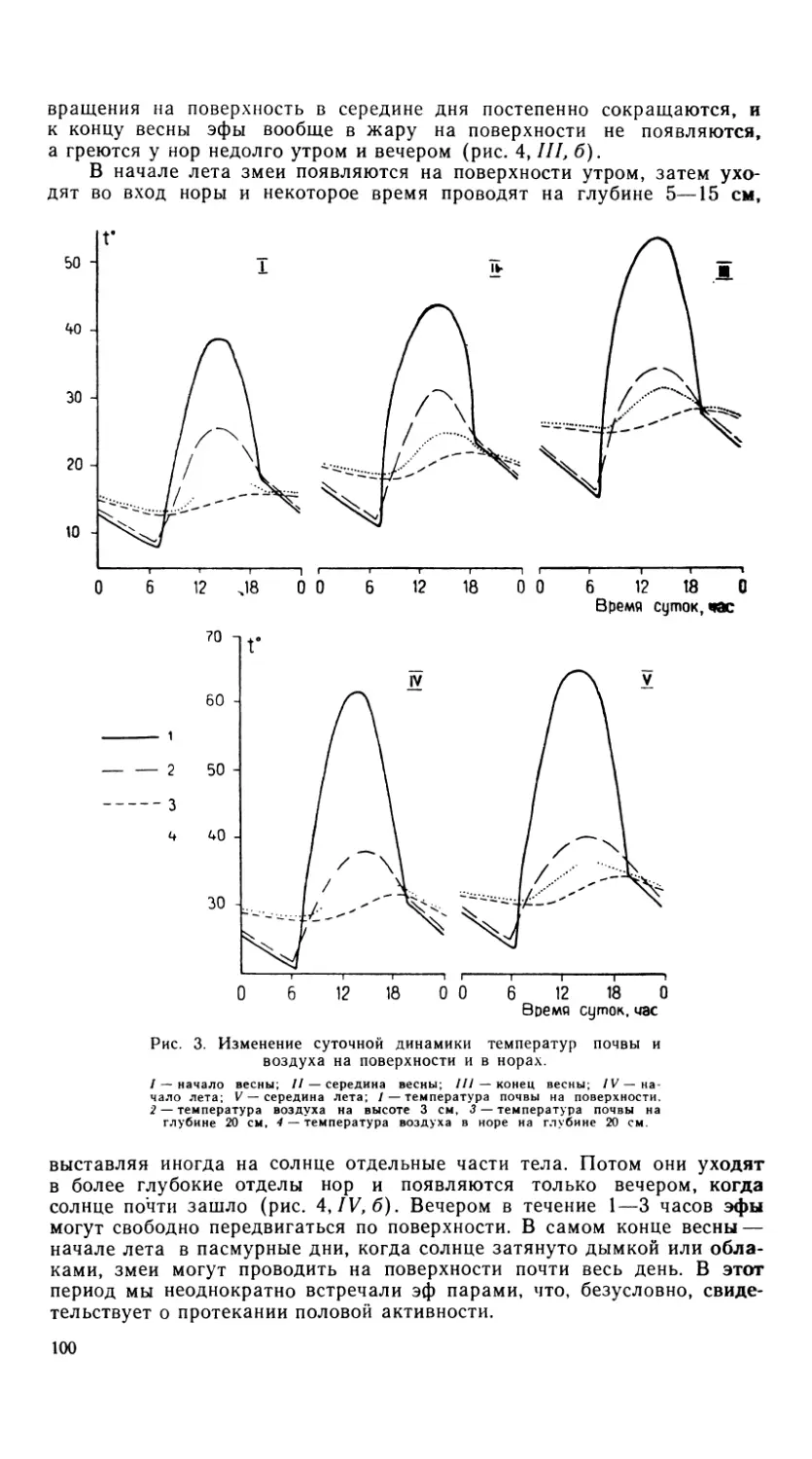
Рис. 8. Подъязычный аппарат агам.
/ — A. himalayana, 2 — A. erythrogastra, 3 — A. caucasia, 4 — A. lehmanni, 5 — A. san-
guinolenta, 6 — A. ruder at а.
более однородные по форме (исключая резцы и клыки), мелкие и хруп-
кие. Количество зубов типичное для агам (табл. 1). Возрастные изме-
нения из-за отсутствия материала не изучены.
Гиоид A. ruderata отличается широким разбросом ветвей cerato-
branchiale I, параллельно которым расположены ceratohyale. Cerato-
branchiale II, как и у других видов, за исключением A, sanguinolenta,
расположены на некотором расстоянии друг от друга (рис. 8).
14
Обсуждение результатов
Таксономическое и адаптивное значение изученных признаков
морфологии
Анализ строения черепа, зубной системы и подъязычного аппарата,
в первую очередь, свидетельствует о морфологической неоднородности
видов агам, входящих в герпетофауну СССР.
По своим пропорциям черепа и форме отдельных костей можно
отнести их к трем группам видов: 1. Череп плоский и длинный (A cau-
casia,А- lehmanni,A. himalayana, A. erythrogastra). 2. Череп высокий
и длинный (A. sanguinolenta). 3. Череп высокий и короткий (A. rude-
rata) (табл. 2). Это распределение по морфологическим типам отчасти
совпадает с принятым ранее таксономическим делением рода Agama
на подроды Agama sensu stricto (A. sanguinolenta, A. ruderata) и Stellio
(A. caucasia, A. lehmanni, A. erythrogastra, A. himalayana), подтверж-
денным, кроме того, кариологическими данными (Gorman a. Shochat,
1972; Соколовский, 1974, 1977; Moody a. Hutterer, 1978). Однако данные
о строении черепа и подъязычного аппарата позволяют отнести A. san-
guinolenta и A. ruderata к разным комплексам видов внутри рода
Agama. Пропорции черепа и строение гиоида сближают A. sanguino-
lenta с древесными агамами, a A. ruderata — с ящерицами рода Phryno-
cephalus (Ананьева, 1977). Сходство A. ruderata с круглоголовками
подтверждается также анализом кариотипов и особенностей экологии
(Соколовский, 1976).
В то же самое время сближение A. sanguinolenta и ящериц рода
Phrynocephalus, основанное на данных кариологии (Соколовский, 1977),
не вполне согласуется с полученными мною морфологическими резуль-
татами. Наиболее серьезные отличия заключаются в строении подъ-
язычного аппарата (рис. 8) A. sanguinolenta. По этому признаку степ-
ная агама противопоставлена всем изученным видам агам фауны СССР.
Наличие в гиоиде степной агамы длинных, расположенных очень близко
друг к другу ceratobranchiale II весьма напоминает состояние подъязыч-
ного аппарата, описанное для агамовых ящериц родо*в Draco, Catotes
и Sitana (Соре, 1892; Camp, 1923; Gnanamuthu, 1937) и игуановых
ящериц родов Anolis, Brachylophus, Ctenosaura, Iguana, Dipsosaurus
(Cope, 1892; Camp, 1923; Gnanamuthu, 1937; Oelrich, 1956; Avery a. Tan-
ner, 1971).
Адаптивное значение гиоида было установлено еще Э. Копом
(Соре, 1892), который отмечал, что у агамид и игуанид близко распо-
ложенные ceratobranchiale II характерны для древесных форм, тогда
как у наземных они находятся на некотором расстоянии друг от друга.
В дальнейшем при рассмотрении строения подъязычного аппарата раз-
личных ящериц морфологи обычно были единодушны в том, что эта
часть гиоида играет важную роль в поддержании и растяжении горло-
вого мешка (Beddard, 1905; Camp, 1923; Gnanamuthu, 1923). Более того,
было показано, что мускулатура гиоида и языка участвует в движе-
ниях, обеспечивающих демонстрации окрашенных участков горлового
мешка. У ящериц родов Calotes, Anolis и Sitana гиоидный аппарат
служит рычагом первого рода, позволяющим растягивать горловой
мешок. Благодаря прикреплению соединенных задних концов cerato-
branchiale II к коже горла она оттягивается как складка при действии
musculus mylohyoideus posterior и m. constrictor colli. M. omohyoideus
возвращает ceratobranchiale II в прежнее состояние.
Отмеченное морфологами адаптивное значение гиоида для демон-
страций горлового мешка, имеющих важное значение в территориаль-
ном и брачном поведении, подтверждено недавно экспериментальными
исследованиями. При изучении действия различных факторов на ре-
15
продуктивное поведение анолисов (Anolis carolinensis) было показано;
что решающую роль в цепи брачных демонстраций играет именно спо-
собность самцов к растяжению горлового мешка. Половое поведение
самок, находящихся вместе с самцами, подвергшимися эксперименталь-
ному удалению гиоида, было нарушено, в то время как искусственное
изменение цвета (перекрашивание) демонстрируемого нормального
горлового мешка для них не имело значения (Crews, 1975). Столь важ-
ное и явное функциональное значение такой структуры гиоида под-
тверждается еще и тем, что он независимо развился у агамид и игуанид
для одной и той же цели. Ч. Кэмп (Camp, 1923) полагает, что гиоид-
ный механизм Calotes и Sitana представляет собой определенное от-
клонение от общего типа внутри семейства агамид. На вторичность
такого типа подъязычного аппарата, вероятно, указывает и его строе-
Рис. 9. Подъязычный ап-
парат 8-миллиметрового
эмбриона Calotes versi-
color (из Ramaswami,
1946).
ние у 8-миллиметрового эмбриона Calotes versi-
color (Ramaswami, 1946), у которого ветви сега-
tobranchiale II разведены друг от друга (рис. 9).
Можно на этом основании предполагать, что рас-
положение ceratobranchiale II у взрослых кало-
тов (и, вероятно, других ящериц с таким же ти-
пом гиоида) является производным по сравне-
нию с тем, что обнаружено на эмбриональной
стадии.
Изложенные соображения о значении гиоида
позволяют утверждать, что его строение имеет
важное таксономическое значение при изучении
близких видов. Его использование при изучении
систематики игуановых ящериц (Avery a. Tan-
ner, 1971) и агамовых ящериц рода Amphibolu-
rus (Badham, 1976) уже доказало свою эффек-
тивность.
Особое строение гиоида степной агамы
можно расценивать как одно из морфологиче-
ских свидетельств того, что разные виды рода
Agama формировались в различной экологической обстановке. Для гор-
ных агам характерен более скрытный образ жизни, они очень осто-
рожны и при любом сигнале опасности скрываются в узких щелях
между скалами, служащих им убежищами. Наблюдаемое у них брач-
ное и апосематическое поведение менее демонстративно и, вероятно,
его зрительный эффект рассчитан на относительно небольшие расстоя-
ния. Изучение их экологии делает понятными и отсутствие горлового
мешка, и своеобразную очень плоскую форму головы, и существование
своеобразного механизма автотомии хвоста. Все эти особенности харак-
терны для целостной группы агам, куда, кроме видов фауны СССР,
входят A. atricollis, A. stellio, A. tuberculata, обладающих 36-хромосом-
ным кариотипом. Эта группа ящериц, очевидно, сформировалась
именно в условиях гор и, как предполагает В. В. Соколовский (1977),
независимо от агам подрода Agama sensu stricto.
A. sanguinolenta относится к группе видов с кариотипом 2п = 46—48,
в которую входят из изученных в этом отношении видов также A. ru-
derata, A. pallida, A. sinaita, A. planiceps (Gorman a. Shochat, 1972).
Эта кариологически единая группа, однако, представляется мне неодно-
родной. В частности, A. sanguinolenta обладает рядом экологических
и поведенческих особенностей, отличающих ее от A. ruderata. Ее де-
монстративное поведение (Полынова, 1979) включает в себя не только
обычные для большинства агамид приседания и кивки головой
(«bobbing»), но и раздувание горлового мешка и изменение окраски.
Последние компоненты часто используются в брачных ритуалах и
в территориальном поведении, действуя дистантно на особей с других
16
индивидуальных участков. Зрительный эффект усиливается, когда
агама демонстрирует свой ярко окрашенный горловой мешок, сидя на
возвышении (ветви кустарников). Поведение же руинной агамы, ви-
димо, значительно ближе к типичному для ящериц рода Phrynocephalus
(Соколовский, 1976). Эти отличия, связанные с описанными выше осо-
бенностями морфологии, вероятно, говорят о более сложной, чем это
предполагалось до сих пор, структуре рода Agama. Очевидно, что ее
нельзя ограничить всего лишь разделением на группировки Agama и
Stellio. Предложение вернуть этим группам видов самостоятельный
статус отдельных родов кажется мне хотя и весьма убедительным, но
преждевременным до проведения полной кариологической и морфоло-
гической ревизии всего рода. Замечу также, что представления о путях
формирования этих групп видов, основанные на данных о возможных
эволюционных преобразованиях кариотипа, также в силу своей проти-
воречивостиДСоколовский, 1977; Moody a. Hutterer, 1978) пока не могут
считаться убедительными.
Строение зубной системы, весьма существенное для установления
различий между отдельными родами семейства агамид (Cooper a. Poole,
1973: Ананьева, 1977), при изучении систематики рода Agama оказа-
лось менее эффективным. Приведенная таблица (табл. 1) показывает,
что зубные формулы разных видов очень сходны, а отличия, касаю-
щиеся количества предчелюстных зубов, также не могут считаться до-
статочно стабильным признаком. Заметные отличия обнаружены только
между горными агамами (равная величина клыков и резцов) и A. san-
guinolenta (более выраженная триконодонтность зубов в конце ряда).
Более существенное значение при определении фрагментов черепа имеет
конфигурация костей, несущих зубы (os maxillare и os dentale).
Отдельные кости черепа могут иметь значение как для определения
современных агамид, так и для сопоставления их с вымершими фор-
мами. Наряду с os maxillare и os dentale, видовые различия в форме
которых были описаны выше, следует более подробно остановиться на
os frontale и os parietale. Кроме видоспецифических различий в форме
этих костей (рис. 4), сравнительное изучение черепов агам обнаружило
также разнообразие размера и формы теменного отверстия, foramen
parietalq.
Теменное отверстие, имеющее у агамовых ящериц различное рас-
положение (Siebenrock, 1895; Trost, 1956), у представителей рода Agama
размещается на os parietale, ограничиваясь лобно-теменным швом. Его
размер заметно отличается у A. sanguinolenta и A. ruderata, с одной
стороны, и 4-х видов горных агам, с другой стороны. Значительно
более крупные размеры foramen parietale горных агам имеют у разных
видов различные очертания выемок в передней части os parietale
(рис. 4). По сравнению с ними теменное отверстие очень мало у A. ru-
derata и A. sanguinolenta, отчасти благодаря выступанию в его полость
отростка на заднем крае os frontale.
Видовые различия в размере foramen parietale мне кажется умест-
ным обсудить в связи с вопросом о функции теменного глаза. Теменной
(париетальный) орган расположен в теменном отверстии, развивается
вместе с эпифизом из крыши промежуточного мозга и имеет глазопо-
добное строение (имеет сетчатку, склеру, хрусталик, стекловидное тело)
(Schmidt, 1909; Новиков, 1910; Чугунов, 1966). Светочувствительные
клетки сетчатки по своему строению близки фоторецепторам боковых
глаз. Сложный для методического разрешения вопрос о существовании
связи теменного глаза с мозгом (наличие nervus parietalis) дискути-
руется до сих пор и, вероятно, представление о функционально слабо-
развитом теменном глазе игуановых и агамовых ящериц (Schmidt,
1909; Camp, 1923) нельзя считать окончательным. В настоящее время
доказана способность париетального глаза^мшишнимать свет, а также
2 Зак. 719
17
становится все более вероятным его влияние (под действием света/
на функцию эпифиза и других эндокринных органов, обеспечивающих
регуляцию репродуктивных циклов (Bethea a. Walker, 1978). Светочув-
ствительность делает теменной глаз дозиметром радиации, в связи
с этим вероятна его роль в сложной системе поведенческой терморегу-
ляции. Обе эти функции (контроль терморегуляции и сезонных циклов
размножения), возможно, имеют более существенное значение в усло-
виях резких климатических изменений, и именно поэтому теменной
глаз, вероятно, значительно чаще развит у ящериц, распространенных
относительно далеко от тропиков (семейства Agamidae, Iguanidae, La-
certidae, Scincidae) (Gundy et al., 1975).
Сравнение изученных мною агам показало, что размер теменного
отверстия значительно больше у горных форм (A. caucasia, A. lehmanni,
A. erythrogastra, A. himalayana), чем у равнинных A. ruderata и A. san-
guinolenta. Изложенные выше соображения о вероятных функциях
теменного глаза представляют особый интерес в связи с различиями
в температурном и световом режиме районов обитания горных и рав-
нинных видов. Хотя до сих пор не существует определенной точки зре-
ния в вопросе о корреляциях размера и развития теменного глаза и
теменного отверстия (Camp, 1923; Trost, 1956), можно все же предпо-
ложить, что различный размер foramen parietale имеет определенное
адаптивное значение, вероятнее всего, связанное с фоточувствительной
функцией теменного органа ящериц.
В заключение мне хочется отметить, что изучение строения черепа
и подъязычного аппарата ящериц рода Agama еще раз показало зна-
чение морфологических признаков как материала не только для уста-
новления сходства и различия, но и для анализа истории формирования
видов.
ЛИТЕРАТУРА
Ананьева Н. Б. Таксономические различия в строении черепа и зубной системы
агамовых ящериц (Sauria, Agamidae) фауны СССР. — Зоол. ж., 1977, т. 56,
вып. 7, с. 1062—1070.
Ананьева Н. Б. и Петерс Г О видовой самостоятельности Agama pawlowskii
Cernov et Dubinin, 1946. — В кн.: Фауна и экология амфибий и рептилий пале-
арктической Азии (Тр. Зоол. ин-та АН СССР, т. 101), 1981, с. 21—22.
Ананьева Н. Б., Петерс Г. и Ржепаковский В. Т. Новый вид горных
агам из Таджикистана, Agama chernovi sp. nov. — В кн.: Фауна и экология
амфибий и рептилий палеарктической Азии (Тр. Зоол. ин-та АН СССР, т. 101),
1981, с. 23—26.
Атаев Ч. Особенности зимовки кавказской агамы на Копет-Даге. — Экология, 1974,
№ 2, с. 76—78.
Богданов О. П. Материалы к экологии хоросанской агамы (Agama erythrogastra
Nik.)—Тр. Ин-та зоол. и паразитол. АН Туркм. ССР, Ашхабад, 1958, т. 2,
с. 145—147.
Гуртовой Н. Н., Матвеев Б. С. и Дзержинский Ф. Я. Практическая
зоотомия позвоночных. Земноводные, пресмыкающиеся. М., «Высшая школа»,
1978, 407 с.
Камалова 3. Я. К экологии хоросанской агамы (Agama erythrogastra Nik.)
в юго-восточной Туркмении. — В кн.: Экология и биология животных Узбеки-
стана. Часть 2 (Позвоночные). Ташкент, 1975, с. 63—66.
Новиков М. М. Исследования о теменном глазе ящериц. — Учен. зап. Имп.
Московск. ун-та, отд. естеств.-истор., 1910, вып. 27, с. 1 — ПО.
Полынова Г В. Поведение и популяционная структура степной агамы. — Вестник
Московск. гос. ун-та, 1979, биол., № 1, с. 50—54.
Соколовский В. В. Сравнительно-кариологическое изучение ящериц сем. Agami-
dae. I. Хромосомные наборы восьми видов рода Phrynocephalus. — Цитология,
1974, т. 16, № 7, с. 920—925.
Соколовский В. В. Сравнительно-кариологическое изучение ящериц сем. Agami-
dae. II. Кариотипы пяти видов рода Agama. — Цитология, 1975, т. 17, № 1,
с. 91—93.
Соколовский В. В. К биологии руинной агамы в юго-западном Азербайджане.—
Экология, 1976, № 5, с. 102—103.
18
Соколовский В. В. Систематические взаимоотношения в семействе Agamidae
по наркологическим данным. — В кн.: Вопросы герпетологии. Автореф. докл.
IV Всесоюзн. герпетол. конф. Л., «Наука», 1977, с. 195.
Терентьев П. В. Герпетология. М., «Высшая школа», 1961, 335 с.
Терентьев П. В. и Чернов С. А. Определитель пресмыкающихся и земновод-
ных. М., «Советская наука», 3-е изд., 1949, 340 с.
Хонякина 3. П. Биология кавказской агамы Дагестана. — В кн.: Вопр. физио-
логии, биохимии, зоологии и паразитологии. Махачкала, 1965, с. 126—134.
Хонякина 3. П. и Кузеева Ю. А. Материалы по питанию степной агамы
в Дагестане. — В кн.: Вопр. физиологии, биохимии, зоологии и паразитологии.
Махачкала, 1967, вып. 2, с. 97—99.
Царевский С. Ф. Contribution to the classification and distribution of the lizards
of the genus Phrynocephalus. — Докл. АН СССР, 1929, c. 415—419.
Чернов С. А. Пресмыкающиеся. — В серии: Фауна Таджикской ССР, т. 18 (Тр.
Ин-та зоол. и паразитол. АН Тадж. ССР, т. 48), Сталинабад, 1959, 203 с.
Чугунов Ю. Д. Строение и возможные функции теменных органов позвоночных
животных. — Усп. совр. биол., 1966, т. 62, вып. 2 (5), с. 265—273.
Якубовски М. Методы выявления и окраски системы каналов боковой линии и
костных образований у рыб in toto. — Зоол. ж., 1970, т. 49, вып. 9, с. 1398—
1402.
Avery D. F. a. Tanner W. W. Evolution of the iguanine lizards (Sauria, Iguanidae)
as determined by osteological and myological characters. — Brigham Young Univ.
Sci. Bull., 1971, biol. ser., vol. 12, n. 3, p. 1—79.
Badham J. A. The Amphibolous barbatus species-group (Lacertilia: Agamidae).—
Austral. J. Zool., 1976, vol. 24, p. 423—443.
Beddard F. E. Some notes on the cranial osteology of the mastigure lizard, Uro-
mastix. — Proc. Zool. Soc. London, 1905, p. 2—9.
Beddard F. E. Contribution to the knowledge of the systematic arrangement and ana-
tomy of certain genera and species of Squamata. — Proc. Zool. Soc. London, 1907,
p. 35—68.
Bethea C. L. a. W a 1 k e r R. F. Parietal eye-pineal gland interactions in the lizard
Sceloporus occidentalis (Reptilia, Lacertilia, Iguanidae).—J. Herpetol., 1978,
vol. 12, n. 1, p. 83—87.
Camp C. L. Classification of the lizards. — Bull. Amer. Mus. Nat. Hist., 1923, vol. 48,
art. 11, p. 289—481.
Capel-Williams G. a. PrattenD. The diet of adult and juvenile Agama bibroni
(Reptilia : Lacertae) and a study of the jaw mechanisms in the two age groups. —
J. Zool., 1978, vol. 185, pt. 2, p. 309—318.
Cooper I. S. a. P о о 1 e D. F. G. The dentition and the dental tissues of the agamid
lizard Uromastix.—J. Zool., 1973, vol. 169, pt. 1, p. 85—100.
Cooper I. S., Poole D. F. G. a. Lawson R. The dentition of agamid lizards with
special reference to tooth replacement. — J. Zool., 1970, vol. 162, pt. 1, p. 85—88.
Cope E. D. The osteology of the Lacertilia. — Proc. Amer. Philos. Soc., 1892,
p. 185—221.
Crews D. Psychobiology of reptilian reproduction. — Science, 1975, vol. 189, n. 4208,
p. 1059—1665.
Duda P. L. On the cranial osteology of Agama tuberculata with a note on the com-
parative study of skull in the family Agamidae. — Vestn. Ceskosl. Spolecn. Zool.,
1965, t. 29, n. 1, p. 156—174.
Duda P. L. Hyoid arch skeleton of Agama tuberculata. — Proc. 2nd All-India Con-
gress Zool., 1966, pt. 2, p. 179—184.
E i c h w a 1 d E. Fauna Caspio-Caucasia nonnulis observationibus novis. Petropoli,
1841, IV+233 p.
Gnanamuthu С. P. Comparative study of the hyoid and tongue of some typical
genera of reptiles. — Proc. Zool. Soc. London, 1937, vol. 107, ser. B. Systematic
a. Morphology, p. 1—64.
Gorman G. C. a. Shochat D. A. A taxonomic interpretation of chromosomal and elec-
trophoretic data on the agamid lizards of Israel with notes on some East African
species. — Herpetologica, 1972, vol. 28, n. 2, p. 106—112.
G u n d у С. C., R a 1 p h C. L. a. Wur s t G. Z. Parietal eyes in lizards — zoogeo-
graphical correlates. — Science, 1975, vol. 190, n. 4215, p. 671—673.
MoodyS. a. Hutterer R. Karyotype of the agamid lizard Lyriocephalus scutatus
(L., 1758) with a brief review of chromosomes of the lizard family Agamidae. —
Bonn. Zool. Beitr., 1978, Bd. 29, H. 1—3, S. 165—170.
Oelrich T. M. The anatomy of the head of Ctenosaura pectinata (Iguanidae). — Misc.
Publ. Mus. Zool., Univ. Michigan, 1956, n. 94, p. 1 —122.
Ramaswami L. S. The chondrocranium of Calotes versicolor (Daud.) with a descrip-
tion of the osteocranium of a just-hatched young. — Quart. J. micr. Sci., 1946,
vol. 87, pt. 3, p. 237—297.
Schmidt W. J. Beitrage zur Kenntnis der Parietalorgane der Saurier. — Z. wiss.
Zool. 1909, Bd. 92, H. 3, S. 359—426.
2*
19
Siebenrock F. Das Skelet der Agamiden.— Sitz.-Ber. Akad. Wiss. Wien, 1895,
Bd. 104, S. 1089—1196.
Trost E. Uber die Lage des Foramen parietale bei rezenten Reptilien und Labyrintho-
dontia. — Acta anat., 1956, vol. 26, n. 4, S. 318—339.
Wagler J. Natiirliches System der Amphibien mit vorangehender Klassification der
Saugethiere und Vogel. Ein Beitrag zur vergleichenden Zoologie. Miinchen —
Stuttgart — Tubingen, 1830, 354 S.
Wermuth H. Agamidae. — In: Das Tierreich, Liste der recenten Amphibien und Rep-
tilien, Berlin, 1967, Lief. 86, XIV+127 S.
Structural characteristics of skull, dentition and hyoid of lizards of the genus Agama
from the fauna of the USSR
N. B. Ananjeva
Zoological Institute, Academy of Science, U.S.S.R. (Leningrad)
Skulls, dentitions and the structure of hyoid of 6 species of Agama genus in the
USSR fauna are described. Specific differences in the shape and proportions of the
skull, in shape of single bones, in particular of os maxillare, os dentale, os frontale
and os parietale have been noted. Dental formulae of A. sanguinolenta, A. caucasia,
A. lehmanni, A. erytrogastra, A. himalayana and A. ruderata are given. A. sanguinolenta
is sharply different in its hyoid structure from the other species studied and resembles
in this respect some tree agamid and iguanid lizards. Adaptive importance of hyoid for
demonstrative movements of dewlap is discussed, specific differences in the size of fora-
men parietale are considered. Analysis of the studied characteristics shows morphological
heterogeneity of Agama species in the USSR fauna. They can be considered as belon-
ging to three groups: 1) mountain species A. caucasia, A. lehmanni, A. himalayana,
A. erythrogastra, 2) A. sanguinolenta and 3) A. ruderata.
О ВИДОВОЙ САМОСТОЯТЕЛЬНОСТИ AGAMA PAWLOWSKI!
CERNOV ET DUBININ, 1946
H. Б. Ананьева и Г. Петерс
Зоологический институт АН СССР (Ленинград) и Музей природоведения
университета имени Гумбольта (Берлин)
Агама Павловского (Agama pawlowskii) была описана С. А. Чер-
новым и В. Б. Дубининым (1946) по единственному экземпляру, пой-
манному В. Б. Дубининым 29 мая 1944 года в Хавастском районе
Узбекской ССР, граничащем с Ура-Тюбинским районом Таджикской
ССР, в 15 км к югу от станции Урсатьевская. Ящерица была найдена
на склоне каменистого ущелья среди северных предгорных холмов
Туркестанского хребта, на высоте около 1300 м над уровнем моря.
За прошедшие 35 лет делались неоднократные попытки получить новые
данные по этому виду, однако они не увенчались успехом.
Уникальная ящерица все это время хранилась в коллекции отде-
ления герпетологии Зоологического института АН СССР (ЗИН.15637),
не вызывая сомнений в идентичности данного экземпляра с описанием.
Этот вид неоднократно упоминался в списках мировой фауны и фауны
СССР (Терентьев и Чернов, 1949; Чернов, 1959; Богданов, 1960; Wer-
muth, 1967; Банников и др., 1977; Саид-Алиев, 1979). Однако отсут-
ствие новых находок A. pawlowskii не могло не вызывать сомнений
в ее видовой самостоятельности. Высказывалось предположение, что
эта ящерица является случайно завезенным в этот район Средней Азии
экземпляром одного из видов рода Calotes. Предполагалось также, что
агама Павловского представляет собой некое отклонение от нормаль-
ного типа A. sanguinolenta, с которой ее, на первый взгляд, сближает
относительно высокая голова и характер фолидоза хвоста, неразделен-
ного на кольца. Уже один из авторов описания вида (С. А. Чернов)
отмечал существенные отличия агамы Павловского от остальных из-
вестных видов палеарктических агам. Изложенные выше причины по-
будили нас переопределить данный экземпляр.
Результат переопределения по определительным таблицам каталога
рептилий Британского музея (Boulenger, 1885) оказался довольно не-
ожиданным. Экземпляр агамы Павловского без всяких трудностей
определяется как A. mossambica Peters, 1854 (вклейка, рис. I),
распространенная в юго-восточной Африке. Предварительное опреде-
ление по таблицам было подтверждено более детальным сравнением
изучаемого экземпляра с типовой серией A. mossambica, хранящейся
в ЗИН (№ 5306) и в коллекциях Берлинского музея природоведения
(БМП). У данной особи были обнаружены все признаки, приводящие
к такому результату определения: увеличенный затылочный щиток,
равная величина спинных чешуй, наличие небольшого гребня вдоль
затылка и спины, диагональное расположение хвостовых чешуй. Внутри
группы видов с увеличенным теменным щитком эта ящерица выделяется
по столь редкому для агам признаку, как существование у нее гребня
из увеличенных чешуй. Этот признак характеризует не только A. mos-
sambica, но и A. kirkii. Однако идентификация с последним видом не-
возможна хотя бы из-за наличия у него спинных и брюшных чешуй
равной величины, тогда как у A. mossambica спинные значительно
крупнее брюшных. Кроме того, A. kirkii (сравнение проводилось по
экземплярам БМП) отличается иными пропорциями и телосложением,
чем A. mossambica и экземпляр A. pawlowskii.
Следует отметить также, что упоминаемая в оригинальном описа-
нии (Чернов и Дубинин, 1946) близость к африканской A. agama
исключается по той причине, что у этого вида значительно более
плоская голова и гребень расположен только вдоль затылка, а не про-
21
должается вдоль спины. Необходимо уточнить описание такого при-
знака A. pawlowskii, как наличие двойного ряда анальных пор. У изу-
ченного экземпляра передний ряд состоит из недоразвитых пор, как это
имеет место и у Л. mossambica. Единственное отмеченное нами отли-
чие экземпляра агамы Павловского от изученных особей A. mossam-
bica состоит в наличии более килеватых чешуй в области горла между
челюстями. Проведенный анализ рентгенограмм черепов агамы Пав-
ловского, некоторых других видов агам и калота, позволяющий полу-
чить косвенные данные об особенностях краниологии типового экземп-
ляра, подтверждает выводы, сделанные на основании признаков фоли-
доза (вклейка, рис. II).
Идентификация экземпляра A. pawlowskii с A. mossambica не ре-
шает, однако, вопроса о происхождении загадочной находки этого
южноафриканского вида на границе Таджикистана и Узбекистана.
История его описания не дает оснований считать его всего лишь так
называемой «музейной находкой», остается только предположить слу-
чайный завоз единичного экземпляра A. mossambica в этот район Сред-
ней Азии.
Таким образом, A. pawlowskii впредь должна считаться синонимом
A. mossambica и как вид должна быть изъята из списка фауны СССР
и мировой фауны.
ЛИТЕРАТУРА
Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К. и
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны
СССР. М., «Просвещение», 1977, 414 с.
Богданов О. П. Земноводные и пресмыкающиеся. — В серии: Фауна Узбекской
ССР, Ташкент, 1960, т. 1, 260 с.
Саид-Алиев С. А. Земноводные и пресмыкающиеся Таджикистана. Душанбе,
«Дониш», 1979, 145 с.
Терентьев П. В. и Чернов С. А. Определитель пресмыкающихся и земновод-
ных. М., «Советская наука», 1949, 340 с.
Чернов С. А. Пресмыкающиеся. — В серии: Фауна Таджикской ССР, т. 18 (Тр.
Ин-та зоол. и паразитол. АН Тадж. ССР, т. 48), Сталинабад, 1959, 203 с.
Чернов С. А. и Дубинин В. Б. Новый эндемик гор Средней Азии — Agama
pawlowskii sp. nov. (Reptilia, Sauria).—Докл. АН СССР, 1946, т. 52, № 8,
с. 747—749.
Boulenger G. A. Catalogue of the lizards in the British Museum (Natural History).
Vol. 1. Gekkonidae, Eublepharidae, Uroplatidae, Pygopodidae, Agamidae. London,
Taylor, a. Francis, 1885, XII4-436 p.
Wermuth H. Agamidae. — In: Das Tierreich, Liste der recenten Amphibien und Rep-
tilien, Berlin, 1967, Lief. 86, XIV 4-127 S.
On the validity of Agama pawlowskii Cernov et Dubinin, 1946
N. B. Ananjeva and G. Peters
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad)
and Museum fiir Naturkunde der Humboldt-Universitat (Berlin)
The history of description of the Agama pawlowskii is considered. Comparison
of descriptions and museum specimens of A. pawlowskii and A. mossambica, distributed
in south-eastern Africa, has shown their identity. A. pawlowskii should be considered as
synonym of A. mossambica and excluded from the lists of the USSR and world fauna.
X-ray pictures of skulls of A. mossambica, A. pawlowskii and some other agamid lizards
are given.
НОВЫЙ ВИД ГОРНЫХ АГАМ ИЗ ТАДЖИКИСТАНА,
AGAMA CHERNOVI SP. NOV.
Н. Б. Ананьева, Г. Петерс и В. Т. Ржепаковский
Зоологический институт АН СССР (Ленинград), Музей природоведения
университета имени Гумбольта (Берлин)
и Отдел охраны и рационального использования природных ресурсов
АН Таджикской ССР (Душанбе)
Летом и осенью 1976 и 1977 годов в окрестностях города Нурек
(юго-западный Таджикистан) на хребте Сурхку было поймано не-
сколько своеобразных агам. Эти ящерицы (6 самцов) внешне напоми-
нают гималайскую агаму (Agama himalayana), но в то же время за-
метно отличаются от этого вида комплексом признаков. Заслуживает
внимания и то обстоятельство, что все они найдены на высоте 900—
1400 м над' уровнем моря, где следует ожидать и где действительно
обитает туркестанская агама (Agama lehmanni), а не высокогорная
гималайская агама.
По ходу изучения заинтересовавших нас агам, которые хранятся
в фондах отделения герпетологии Зоологического института АН СССР
(ЗИН) в Ленинграде, в коллекции Дрезденского музея зоологии (ДМ3)
нами были обнаружены еще 4 подобные агамы (3 самца и 1 самка),
собранные в 1970, 1974 и 1977 годах также в юго-западном Таджики-
стане.
Сравнивая эти 10 агам не только с экземплярами A. himalayana
из разных точек ареала, но и с особями A. badakhshana, A. stoliczkana,
A. tuberculata, A. nuristanica, A. agrorensis, A. lehmanni, мы пришли
к выводу, что они, по всей видимости, свидетельствуют о существова-
нии на юге Средней Азии неизвестного до сих пор вида агамовых
ящериц, для которого мы предлагаем русское название «агама Чернова».
Agama chernovi Ananjeva, Peters et Rzepakovsky, sp. nov.
Материал: всего 10 экземпляров (8 взрослых самцов, 1 полу-
взрослый самец, 1 взрослая самка).
Голотип хранится в ЗИН, № 19385, взрослый самец, пойман
на хребте Сурхку вблизи города Нурек, юго-западный Таджикистан,
окрестности кишлака Дашти-Хонако, 1050 м над уровнем моря, 25.X.
1977, коллектор В. Т. Ржепаковский (вклейка, рис. III).
Паратипы: а) в коллекциях ЗИН (коллектор В. Т. Ржепаков-
ский): № 19386, взрослый самец, хребет Сурхку, окрестности кишлака
Чинор, 950 м над уровнем моря, 4.VI.1977; № 19387, взрослый самец,
хребет Сурхку, окрестности кишлака Дашти-Хонако, 900 м над уров-
нем моря, 26.Х. 1977; № 19388, взрослый самец, хребет Сурхку, окрест-
ности кишлака Санги-Джиозак, 1400 м над уровнем моря, 15.VI.1976;
№ 19389, взрослый самец, хребет Сурхку, окрестности кишлака Дашти-
Хонако, 970 м над уровнем моря, 16.X.1977. Еще один экземпляр, полу-
взрослый самец, хребет Сурхку, окрестности кишлака Дашти-Хонако,
левый берег ручья Нолинг, 950 м над уровнем моря, 18.X.1977, передан
в герпетологическую коллекцию Берлинского музея природоведения
(БМП № 46508).
б) в коллекциях ДМ3: № 8513, взрослый самец, река Кштут, Гис-
сарский хребет, около 90 км к северо-западу от города Душанбе,
2900—3500 м над уровнем моря, VIII.1970, коллектор Г Шене
(G. Schone); № 11421, половозрелый самец, река Матча (верхнее тече-
ние реки Зеравшан), окрестности кишлака Худгиф, VIII.1974, коллек-
тор Г Шене; № 14313, половозрелая самка и № 14314, половозрелый
23
самец, на месте Нурекского водохранилища, 1020 м над уровнем моря,
15.Х. 1977, коллектор В. Т. Ржепаковский.
Вид назван в честь известного советского герпетолога профессора
С. А. Чернова.
Диагноз: Горная агама с уплощенной головой и туловищем;
хвост с круглым сечением. Все чешуи спины и боков туловища ребри-
стые; вдоль хребта четкая полоса из 8 рядов увеличенных, ребристых,
шиповатых чешуй; чешуи двух внутренних и двух крайних рядов по
величине уступают чешуям в остальных рядах, на спиннобоковых склад-
ках мелкие, сильно заостренные чешуи. Среди очень мелких чешуй
между спинной полосой и спиннобоковыми складками расположены
отдельные более крупные чешуи. У половозрелых самцов большое
овальное пятно мозолистых чешуй на брюхе; в каждом хвостовом сег-
менте по три кольца чешуй; спинная сторона тела однотонно-светлая
с темными пятнышками (вклейка, рис. III).
Сравнительное описание. Агамы средних размеров
(в среднем крупнее A. himalayana), длина тела половозрелых особей
8—12 см, длина хвоста 15—25 см. Телосложение стройное (вклейка,
рис. III), хвост в 1.75—2.2 раза длиннее тела; задние ноги достигают
73—85% длины тела, на длину головы приходится в среднем 27.5—28%
(от 26.2 до 28.4%) всей длины тела (у половозрелой самки относитель-
ная длина головы на 1.5% меньше). У взрослых самцов A. himalayana
и A. badakhshana хвост в 1.7—2.0 раза длиннее тела, задние ноги до-
стигают всего 65—79% длины тела, а голова 25.9—27.3% длины тела
(х = 26.5%) у самцов и 24.0—27.0% (х = 25.5%) у самок.
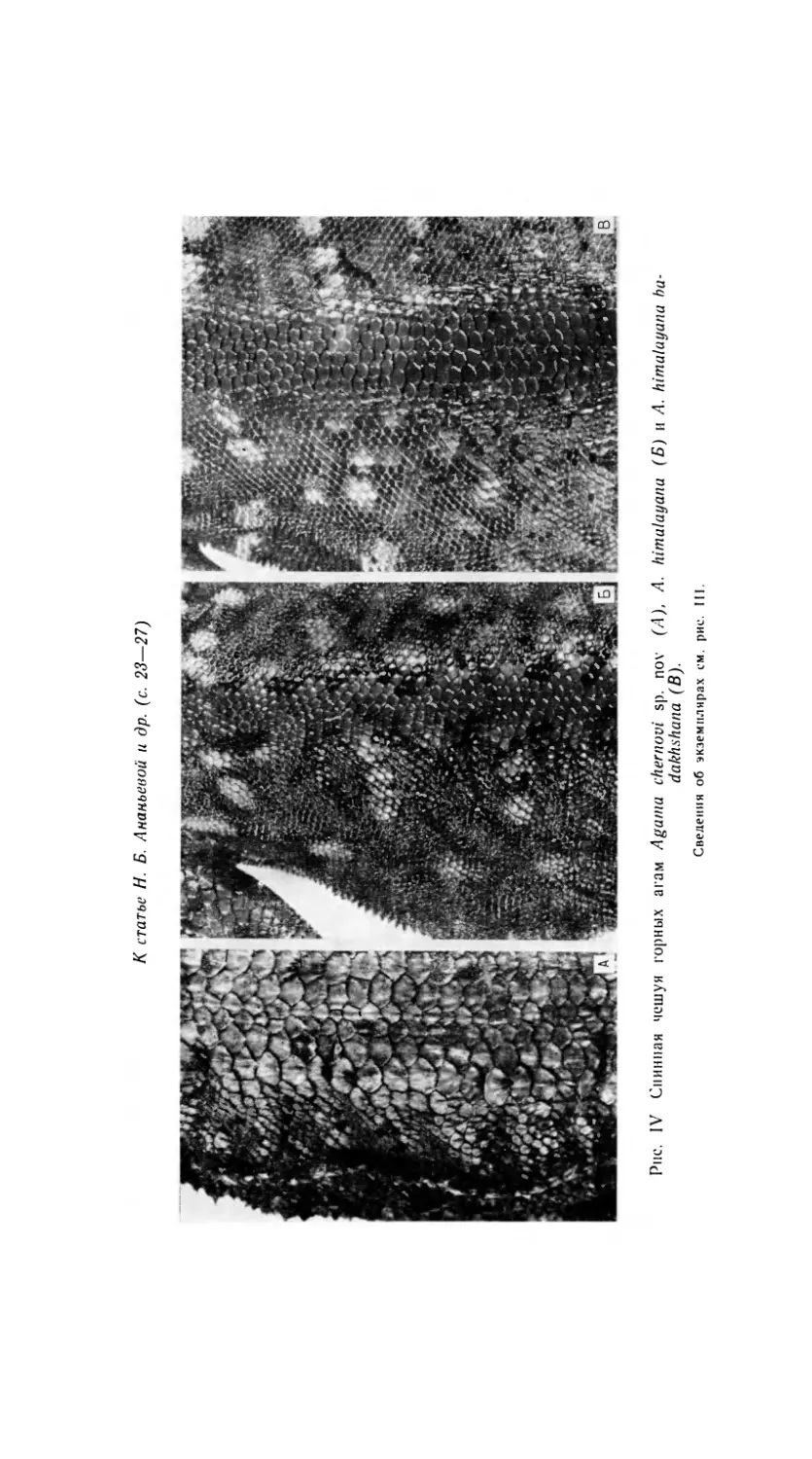
По характеру фолидоза A. chernovi ближе всего стоит к A. hima-
layana и A. badakhshana (вклейка, рис. IV). От этих горных агам но-
вый вид отличается целым рядом признаков.
В височной области, перед ухом расположено пятно очень мелких
и плоских чешуй, окруженное крупными килеватыми чешуями; чешуи
спинной полосы заметно крупнее спиннобоковых. Как и спиннобоковые
чешуи, они не гладкие или едва заметно ребристые (как у A. badakh-
shana и в отдельных случаях у A. himalayana), а килеватые с шипи-
ками. Сама спинная полоса состоит довольно строго из 8 рядов чешуй;
внутри рядов чешуи одинаковой величины (отличие от A. lehmanni),
однако чешуи внутренней пары рядов и двух внешних рядов меньше
чешуй в остальных рядах (вклейка, рис. IV). На затылке у большин-
ства особей выступает слабый, иногда едва заметный короткий про-
дольный ряд чуть увеличенных чешуй; спиннобоковые и боковые чешуи
еще меньше, чем у гималайской агамы; но в то же время всегда ребри-
стые; среди спиннобоковых чешуй по обеим сторонам спинной полосы
выступают несколько отдельных, заметно увеличенных чешуй, четко
выступающие спиннобоковые складки заняты по всей длине слабо уве-
личенными шиповатыми чешуями, хорошо заметно боковое поле уве-
личенных шиповатых чешуй, особенно вдоль боковой складки. Кроме
преанальных пор в 2—4 ряда, у самцов всегда имеется овальное пятно
мозолистых чешуй на животе, которое есть и у самцов A. badakhshana,
но в большинстве случаев отсутствует у A. himalayana.
Рисунок своеобразен. У изученных агам на спинной стороне туло-
вища отсутствуют светлые «глазчатые» пятна в темном окружении
(обычно более или менее заметно собранные в поперечные ряды), ха-
рактерные для A. himalayana, A. badakhshana и A. stoliczkana. Верх
головы, спина и хвост светлого тона с узором из мелких темных пятен
(вклейка, рис. III). Бока тела и грудь темные; на подбородке, горле
и отчасти на груди светлые пятна, иногда брюхо, грудь и бока светло
окрашены, с темными пятнышками (в этих случаях бока лишь немного
24
темнее спины). Окраска живых агам неизвестна. Судя по фиксирован-
ным особям, спина или коричневая (у более молодых ящериц), или
зеленоватая. Участки шиповатых чешуи на шее синие или зеленые.
В окраске горла и боков туловища, вероятно, имеются также синие
тона.
Распространение. Ареал, судя по имеющимся данным, отно-
сительно небольшой: южные склоны и боковые хребты Гиссарских гор
к западу, северу, к востоку от города Душанбе и прилегающие районы
Зеравшанского хребта (река Матча). По склонам названной реки встре-
чается также гималайская агама (4 экз., ДМ3.10330-41).
Согласно точкам находок, нанесенным на схему ареала A. hima-
layana (Банников и др., 1977, карта 43), памиро-алайские популяции
этого вида отделены пространственным разрывом от западно-таджик-
ских, не говоря уже о популяциях на хребте Кугитангтау. В связи
с находкой в западном Таджикистане нового вида агам, близкого к ги-
малайской, коллекционные материалы из западной части ареала
A. himalayana в будущем следует изучить более внимательно. Не исклю-
чено, что среди них могут оказаться экземпляры A. chernovi. Логично
предположить также существование A. chernovi в пограничных с Тад-
жикской ССР районах Афганистана.
Изменчивость. Все агамы в коллекциях ЗИН и ДМ3, кроме
двух экземпляров с Нурекского водохранилища, имеют темноокрашен-
ные бока, на которых (особенно на шее) могут выступать более свет-
лые пятна; у нурекских же особей (ДМ3. 14313-14) бока туловища
в спирту имеют светлокоричневую окраску с неотчетливым узором из
более светлых и темных пятнышек. По характеру фолидоза агамы,
пойманные у речек Матча и Кштут, отличаются от остальных 8 экземп-
ляров тем, что у них полностью отсутствуют выступающие чешуи
в области затылка, а также слабой ребристостью спинных чешуй и от-
сутствием чередования рядов с мелкими и крупными чешуями внутри
спинной полосы. Кроме того, у этих экземпляров низ тела целиком
темный. По относительной длине хвоста и задних ног эти агамы
в меньшей степени, чем остальные известные пока особи A. chernovi,
отличаются от A. himalayana.
Более обширный материал в будущем позволит выяснить законо-
мерности изменчивости признаков и возможную обособленность высоко-
горных популяций A. chernovi на Гиссарском и Зеравшанском хребтах.
Обсуждение. Из-за отмеченного сходства с A. himalayana и
A. badakhshana описанная выше агама сравнивалась, в первую оче-
редь, именно с этими видами. A. lehmanni в среднем заметно крупнее
A. chernovi и имеет более плотное телосложение и более короткие ко-
нечности. По щиткованию спины туркестанская агама сильно отли-
чается от гималайской, бадахшанской и агамы Чернова: среди круп-
ных килеватых чешуй внутри спинной полосы расположено много
мелких чешуй. A. tuberculata, A. nuristanica и A. agrorensis по многим
признакам фолидоза отличаются от только что названных форм.
В частности, у этих видов отдельные сегменты хвоста состоят не из 3,
а из 4 и даже 5 колец чешуй (Smith, 1935; Anderson a. Leviton, 1969)
4 кольца чешуй в одном сегменте имеется также у A. stoliczkana, у ко-
торой к тому же спинная полоса из более крупных чешуй слабо выра-
жена (Peters, 1971).
Описанная здесь агама на основании морфологических данных
должна быть выделена как вид, в то время как отличия A. badakhshana
от A. himalayana таковы, что в первую форму следует считать лишь
подвидом последней, распространенным в горах Гиндукуша, в чем не-
трудно убедиться, сравнивая описание бадахшанской (Anderson a. Le-
viton, 1969) и гималайской агам (Чернов, 1959). A. himalayana sacra
25
из Тибета, с другой стороны, очевидно, в будущем должна быть при-
нята за самостоятельный вид (Peters, в печати).
Постепенно складывается впечатление о том, что в горной системе
Гиндукуша, Памира и Гималаев произошло и еще происходит видо-
образование внутри азиатских горных агам группы Stellio. Сильное рас-
членение рельефа, очевидно, способствует изоляции популяций и в тоже
время препятствует экологической дифференциации близкородственных
форм. В редких случаях существования экологической дифференциа-
ции она прежде всего выступает в виде приуроченности к разным вы-
сотным поясам. Поэтому топографическая и географическая обособлен-
ность затрудняют проверку таксономического статуса морфологически
различающихся агам без проведения полевых и экспериментальных
исследований. Вероятно, именно в такой ситуации находится A. cher-
novi по отношению к A. himalayana. 8 из 10 экземпляров нового вида
были пойманы в нижнем поясе гор. Однако, как нам любезно сообщил
дрезденский герпетолог Ф. Ю. Обет (Obst, in litt.), на реке Матча
в одном и том же месте с тремя гималайскими агамами был пойман
самец A. chernovi (ДМ3.11421). Этот факт свидетельствует о хотя бы
частичной симпатрии обоих видов. Альпинист Г Шене сообщил, что
он видел на этом участке среди гималайских агам несколько более
крупных ящериц «сине-серой или зеленоватой» окраски, которых
он принимал за A. lehmanni; одного самца этой формы он
поймал.
То обстоятельство, что ареал агамы Чернова, по всей вероятности,
весьма невелик, не делает ее исключением среди агамовых ящериц.
У агамид (как и у игуанид) нередки случаи, когда в одном и том же
роде наряду с видами, имеющими широкое распространение, есть и
виды с почти «точечными» ареалами (Agama zonura в юго-восточной
Эфиопии, A. nuristanica в восточном Афганистане, несколько видов
лесных калотов в горах южной Индии, часть видов Amphibolurus в раз-
личных районах Австралии, немало видов Anolis в Вест-Индии и т. д.).
В этих случаях трудно утверждать, что виды с узкими ареалами, не-
сомненно, являются реликтовыми или «угасающими». Помимо ограни-
ченных размеров территорий, пригодных к заселению определен-
ными видами, причиной малых размеров ареалов нередко является
экологическая конкуренция между соседними близкородственными
видами.
Родственные, т. е. филогенетические отношения азиатских горных
агам, как и вообще всех видов рода Agama еще не вполне ясны. На
данном уровне знаний все же допустимо считать, что A. chernovi
вместе с A. himalayana (включая badakhshana как ее подвид), A. sto-
liczkana и A. sacra составляют одну близкородственную группу. Анализ
этой и других группировок внутри рода Agama — задача будущего.
ЛИТЕРАТУРА
Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К. и
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР
М., «Просвещение», 1977, 414 с.
Чернов С. А. Пресмыкающиеся. — В серии: Фауна Таджикской ССР, т. 18 (Тр.
Ин-та зоол. и паразитол. АН Тадж. ССР, т. 48), Сталинабад, 1959, 203 с.
Anderson S. С. a. Leviton А. Е. Amphibians and reptiles collected by the Street
Expedition to Afghanistan. — Proc. California Acad. Sci.. 1969, vol. 37, n. 2,
p. 26—56.
Peters G. Die Wirtelschwanze Zentralasiens (Agamidae: Agama). — Mitt. Zool. Mus.
Berlin, 1971, Bd. 47, H. 2, S. 357—381.
Smith M. A. Reptilia and Amphibia. Vol. II, Sauria — In: The Fauna of British
India, including Ceylon and Burma. London, Taylor a. Francis, 1935, XIII4-440 p.
26
New species of the mountain agamas from Tadjikistan, Agama chernovi sp. nov.
N. B. Ananjeva, G. Peters and К T. Rzepakovsky
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad),
Museum fiir Naturkunde der Humboldt-Universitat (Berlin) and
Department of Wildlife Protection and Management, Academy of Sciences
of the Tadjik SSR (Dushanbe)
From 10 specimens, collected in different sites in western Tadjikistan, Agama
chernovi is described as a new species. Type-locality is Dashti-Khonaco in the Surkhku
mountain ridge near Nurek. By its morphological features the new species is most simi-
lar to A. himalayana, which prefers higher altitudes. Only in one locality of the Zerav-
shan mountains specimens of both species were observed in the same habitat (at about
^200 m). In order to elucidate the relationships between A. chernovi and A. himalayana
further field study will be necessary. Comparison of the new species with material
of other Agama species in the Pamir, Hindukusch and Himalayan mountains has shown,
that A. badakhshana, described by Anderson and Leviton in 1969, can be considered only
as a subspecies of A. himalayana. The holotype of the new species is deposited in the
herpetological collection of the Zoological Institute, Academy of Sciences, Leningrad,
USSR.
ДВЕ НОВЫЕ НАХОДКИ ПОЛОСАТОГО ПОЛОЗА,
COLUBER SPINALIS (PETERS, 1866)
НА ДАЛЬНЕМ ВОСТОКЕ СССР И В ВОСТОЧНОМ КАЗАХСТАНЕ
Ж. Ш. Бердыбаева, В. Ф. Орлова и В. Е. Фролов
Усть-Каменогорский государственный педагогический институт,
Зоологический музей Московского государственного университета
и Московский зоологический парк
Полосатый полоз, Coluber spinalis (Peters, 1866) широко распространен в север-
ной части Китая, Монголии и Корее, а в пределах Советского Союза несколько раз
был найден в Зайсанской котловине (Банников и др., 1977). Первая находка этого
полоза в СССР принадлежит К. М. Параскиву (1956), а 20 лет спустя он был добыт
в Маркакольском районе Восточно-Казахстанской области (Орлова и Баранов, 1977).
Настоящее сообщение касается двух новых местонахождений этого вида, первое
из которых является единственной достоверной находкой полосатого полоза на Даль-
нем Востоке СССР. Единственный экземпляр, по-видимому, неполовозрелый самец,,
был пойман О. Шубравым 25 августа 1979 года на краю болота в бухте Пензовая,
залив Посьета, южная часть Приморского края и хранится сейчас в отделе герпето-
логии Зоологического музея Московского государственного университета (№ 4336).
Длина тела самца 363, длина хвоста 112 мм, L/Lcd = 3.24. Брюшных щитков 198, под-
хвостовых 87, вокруг середины туловища 17 чешуй; чешуи туловища гладкие, без
ребрышек. С каждой стороны головы по 2 предглазничных и заглазничных щитка,
верхнегубных щитков 8, из них четвертый и пятый касаются глаза. Окраска из-за
фиксации в формалине темная, но дорзальная белая полоса четко выражена.
Предположение о нахождении полосатого полоза в Уссурийском крае было
высказано еще А. М. Никольским (1916). Позднее А. А. Емельянов (1929) описал
один экземпляр этого вида, который был найден учениками на левом берегу Амура
против города Хабаровск. Еще один экземпляр был получен им от преподавателя
местной школы Эмме с указанием на то, что эта змея была подобрана мертвой в ки-
тайском квартале Хабаровска. В связи с этим А. А. Емельянов высказал предположе-
ние о том, что оба эти экземпляра были вывезены фокусниками-китайцами из Китая.
Следует отметить, что более поздние зоологические и специальные герпетологические
обследования территории советского Дальнего Востока, в том числе и юга Приморья,
не привели к положительным результатам. Вполне вероятно, что полосатый полоз,
заходящий из северной части Кореи на юг Приморья, не является массовым видом,
но может быть обнаружен там и в дальнейшем.
Как уже было указано выше, первые достоверные местонахождения полосатого
полоза в пределах СССР известны из южной части котловины озера Зайсан. В ок-
тябре 1979 года взрослый экземпляр этого вида (самец) был добыт на севере Зайсан-
ской котловины в Курчумском районе близ села Аменгелыб и характеризуется сле-
дующими признаками: длина тела 755 мм, длина хвоста 270 мм, брюшных щитков 209,
подхвостовых 99 пар. Данный экземпляр хранится в коллекциях Зоологического инсти-
тута АН СССР, Ленинград (№ 19338).
Указанное местонахождение лежит значительно севернее двух первых находок
рассматриваемого вида и может свидетельствовать о его расселении в пределах Зай-
санской котловины к северу.
ЛИТЕРАТУРА
Банников А. Г., Д а р е в с к и й И. С., Ищенко В. Г., Рустамов А. К. и
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР.
М., «Просвещение», 1977, 414 с.
Емельянов А. А. Змеи Дальнего Востока. — Зап. Владивостокск. отд. гос. Русск.
геогр. о-ва, 1929, т. 3(20), вып. 1, с. 1—208.
Никольский А. М. Пресмыкающиеся (Reptilia), том II. Ophidia. — В серии:
Фауна России и сопредельных стран. Петроград, 1916, III4-356 с.
Орлова В. Ф. и Баранов А. С. Новое местонахождение полосатого полоза
в СССР. — В кн.: Вопросы герпетологии. Автореф. докл. IV Всесоюзн. герпетол.
конф. Л., «Наука», 1977, с. 164—165.
Параскив К. П. Пресмыкающиеся Казахстана. Алма-Ата, изд. АН Каз. ССР„
1956, 228 с.
Two new records of the Coluber spinalis (Peters, 1866)
in the Far East of the USSR and eastern Kazakhstan
Zh. Sh. Berdybaeva, V. F. Orlova and V E. Frolov
Ustj-Kamenogorsk State Pedagogical Institute, Zoological Museum, Moscow State
University and Moscow Zoological Park
28
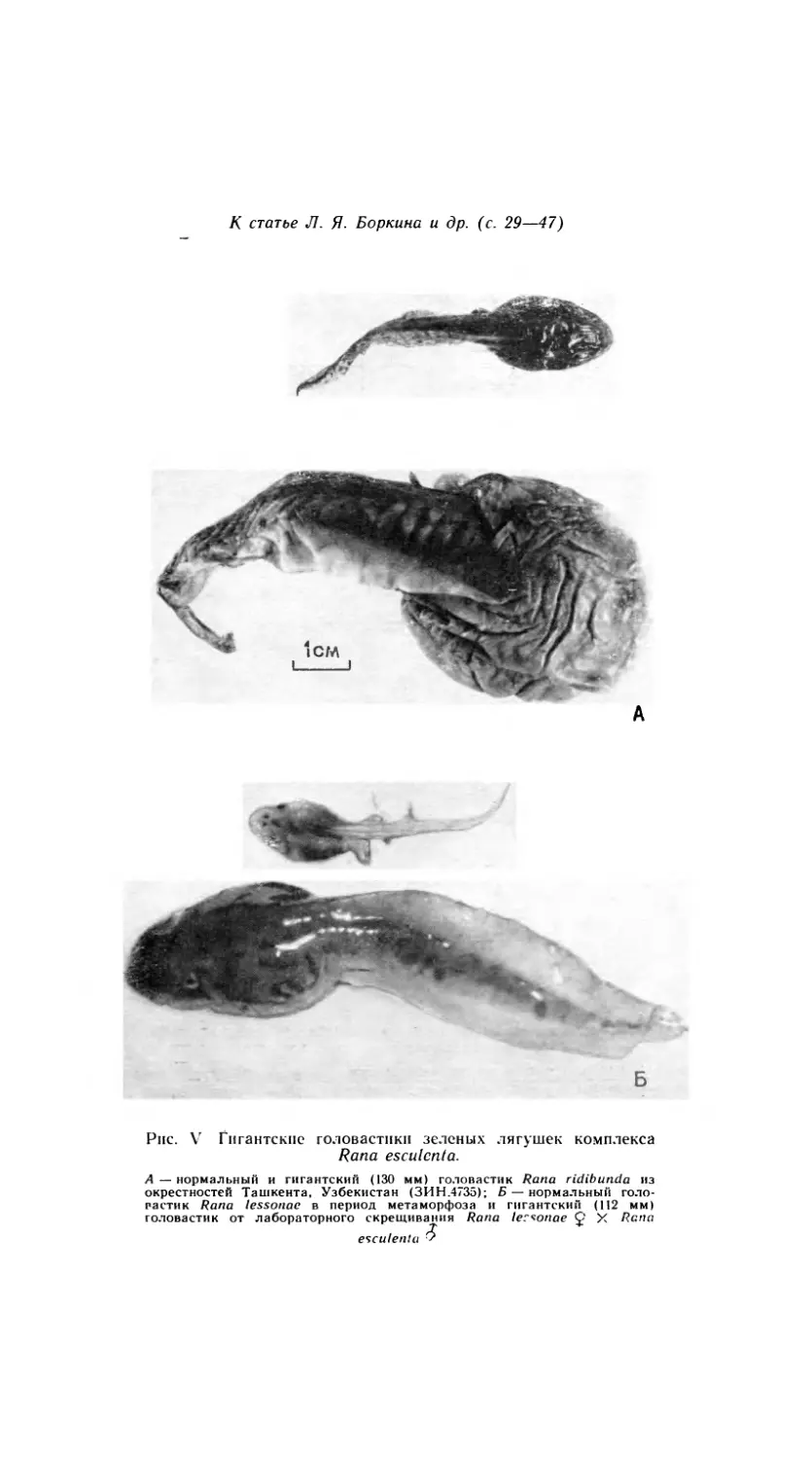
О ГИГАНТСКИХ ГОЛОВАСТИКАХ ЗЕЛЕНЫХ ЛЯГУШЕК
КОМПЛЕКСА RANA ESCULENTA
Л. Я. Боркин, Л. Бергер и Р. Гюнтер
Зоологический институт АН СССР (Ленинград),
Отдел агробиологии Польской Академии наук (Познань) и Музей природоведения
университета имени Гумбольта (Берлин)
Время от времени в литературе появляются сведения о нахожде-
нии в природе гигантских головастиков европейских зеленых лягушек
комплекса Rana esculenta. Эти головастики своей колоссальной вели-
чиной (до 18.5 см) внешне напоминают личинок чесночницы Pelobates
fuscus, наиболее крупных среди европейских бесхвостых амфибий, и
их иногда ошибочно относили к этому виду (Pfliiger, 1883; Kollmann,
1884; Wesenberg-Lund, 1922; Gislen a. Kauri, 1959; Боркин, 1979).
В данной работе сообщается о некоторых новых находках и обсуж-
даются возможные причины личиночного гигантизма у зеленых ля-
гушек.
Прежде всего необходимо уточнить, каких головастиков следует
считать «гигантскими». В табл. 1 приведены данные об обычных раз-
мерах головастиков зеленых лягушек в разных географических районах.
Как видно, они всюду меньше 100 мм. Поэтому под «гигантскими» мы
будем понимать головастиков с длиной тела (туловище + хвост) более
100 мм.
Таблица 1
Размеры (в мм) головастиков зеленых лягушек комплекса Rana esculenta
в разных частях ареала перед метаморфозом
Район I R. esculenta + R. lessonae i R ridibunda Автор
Швеция до 81.6 Gislen a. Kauri (1959)
Дания 60—70 — Wesenberg-Lund (1922)
Польша до 79 не больше 100 Berger a. Michalowski (1963)
Чехословакия — до 90 Boulenger (1898)
Великобритания до 74 — Boulenger (1898)
ФРГ — 70—75 Pfliiger (1883)
Румыния — 43—88, обычно 77 Fuhn (1960)
Московская область 60—73 70—90 Банников (1957)
Дельта Волги — до 68.5 Белова (1964)
Украина 40—50 40—50 Таращук (1959)
Крым — до 62 Щербак (1966)
Армения — до 50 Папанян (1952)
Казахстан — 40—70 Искакова (1959)
Киргизия — меньше 100, обычно 50—60 Янушевич (1976)
Туркмения — 50—52 । Денисова (1969)
В старой герпетологической литературе примерно до 20—30-х годов
нашего столетия европейские зеленые лягушки Rana lessonae и Rana
ridibunda обычно считались внутривидовыми формами вида «Rana
esculenta». Мы знаем теперь (см. обзоры Бергер, 1976; Berger, 1977; Du-
bois, 1977; Боркин и Тихенко, 1979), что зеленые лягушки, живущие
в центральной Европе, принадлежат к трем морфологическим формам,
две из которых (Rana lessonae и Rana ridibunda) являются обычными
биологическими видами, а третья (Rana esculenta) — формой, образо-
вавшейся в результате их гибридизации и существующей в природе
благодаря особому механизму размножения — «гибридогенезу» (Tunner,
1974). Сейчас во многих случаях трудно выяснить, к какой именно
29
Таблица
Размеры (в мм) крупных головастиков и сеголетков с хвостами зеленых лягушек
комплекса Rana esculenta в окрестностях города Познань (Польша)
1 Фенотип Головастики Лягушата с хвостами
N длина тела длина тела (без хвоста)
min—max х min—max х
Водоем I (Rana lessonae — Rana esculenta)
Rana esculenta 54 85—112 95.9 36 30.8—34.5 32.2
Rana lessonae 2 77—95 86.0 4 28.2—29.4 28.9
Вод о e м II (только Rana esculenta)
Rana esculenta 4 86—98 91.5 12 24.7—35.5 32.2
Rana ridibunda — — — 1 — 32.0
Водоем III (только Rana ridibunda)
Rana ridibunda 5 87—105 i 93.8 i 6 29.5—35.0 32.8
Примечание. В скобках указана структура популяции зеленых лягушек
водоема. Особь фенотипа ridibunda в водоеме II произошла, по-видимому, от скре-
щивания Rana esculentay^Rana esculenta (см. текст на с. 32).
форме этих лягушек в современном таксономическом понимании сле-
дует отнести гигантских головастиков, упоминаемых тем или иным
автором, почему мы и цитируем их как «Rana esculenta».
Новые находки гигантских головастиков
1. Два головастика длиной в 130 и 114 мм были пойманы Б. В. Ве-
ригиным 12 октября 1967 года в окрестностях Ташкента, Узбекистан,
устье реки Чирчик, рыбное хозяйство Чирчик (Боркин, 1979). Один
из них хранится в коллекции отделения герпетологии Зоологического
института АН СССР, Ленинград (ЗИН. 4735, вклейка, рис. V, А),
а другой — в Зоологическом музее Московского государственного уни-
верситета (№ 1041). Некоторые промеры этих экземпляров приводятся
в табл. 3. Спиракулюм, расположенный на левой стороне туловища и
направленный назад и вверх; отверстие клоаки, открывающееся вправо
от нижнего гребня хвоста; расположение губных зубчиков (2 ряда
сверху и 3 снизу); характерные сосочки, окаймляющие ротовой диск
с боков и снизу — все это явно указывает на принадлежность этих голо-
вастиков к Rana ridibunda, обитающей в Узбекистане.
2. Головастик длиной в 109 мм (другие промеры см. табл. 3) из
окрестностей города Казалинск, восточное побережье Аральского моря,
относимый рядом авторов (Елпатьевский, 1903; Никольский, 1918;
Искакова, 1959) к Pelobates fuscus, на самом деле, как показало его
переисследование, является личинкой Rana ridibunda, обычного в этом
районе вида (Боркин, 1979).
3. Головастик длиной около 150 мм был пойман осенью в Чуйской
долине, Киргизия. А. И. Янушевич (1976) определил его как Pelobates
fuscus. Однако эта находка очень сильно удалена от известных границ
ареала чесночницы и в то же время лежит в пределах ареала Rana ri-
dibunda. Кроме того, несмотря на поиски, больше не удалось найти
ни других головастиков, ни взрослых особей чесночницы. Л. Я. Боркин
30
Таблица 3
Размеры некоторых гигантских головастиков зеленых лягушек,
найденных в природе
Место находки №№ Длина, мм Автор
туловище
Ташкент, Узбеки- стан 1 1 130 56 74 9.2 наши данные
Казалинск, восточ- ное побережье Аральского мо- ря, Казахстан 1 Лиепайский район, Латвия 2 2 1 . 1 1145 109 — 120 51 46 635 63 10.0 4.5 нет А. С. Северцов
Познань, Польша 3 2 3 4 5 1 122 100 164.6 115 163 58 44 50 106.6 71 113 нет 5 4.1 5.2 9.0 (in litt.) » » » » наши данные
Ла-Рошфуко, Тар- дуаза, Шаранта, Франция 4 1 125 41 84 6 Angel (1937)
Хэм, Суррей, Анг- лия 4 1 122 42 80 — Smith (1954)
Примечание. 1 — Rana ridibunda (определение наше); 2 — Rana ridibunda
(определение Я. Я. Лусиса); 3 — этого головастика определить не удалось, но он про-
исходит от скрещивания Rana esculentay^Rana exculenta; 4 — «Rana esculenta» (Angel,
1937; Smith, 1954); 5 — кончик хвоста оборван.
(1979) предположил, что скорее всего это — гигантский головастик
озерной лягушки. К сожалению, личинка не сохранилась, и поэтому
установить ее истинную видовую принадлежность невозможно.
Эти три находки являются первым указанием гигантских голова-
стиков зеленых лягушек в Азии.
4. Несколько других находок гигантских головастиков известно
из Латвии. Я. Я. Лусис (in litt.) любезно сообщил нам о 3 экземпля-
рах, пойманных им в местечке Априки севернее города Айзпуте Лиепай-
ского района, и о 8 головастиках из местечка Лигуты, расположенного
на берегу озера Дурбес, также Лиепайского района. Находки были
сделаны им в 1952—1954 годах и определены как Rana ridibunda.
Кроме того, в районе города Айзпуте близ местечка Вецпили в 1975 году
были пойманы еще 2 гигантских головастика этого вида, фотография
которых была опубликована в одном из местных журналов (И. А. Цауне,
in lift.). А. С. Северцов (in litt.), просмотревший 5 головастиков из
сборов Я. Я. Лусиса, любезно прислал нам их описания (размеры см.
табл. 3, нумерация особей одинаковая): № 1—с хорошо развитой
икрой, задних конечностей нет, кожа сильно пигментирована; № 2 —
с нормально развитыми семенниками, задних конечностей нет, пигмен-
тация пятнистая (пегая), челюсти, кишечник и подъязычно-жаберный
аппарат типично личиночные, без всяких признаков метаморфоза;
№ 3 — частичный альбинос (окрашены глаза и перитонеум), гонады
не развиты, имеется конический зачаток задних конечностей; № 4 —
нормально пигментирован, гонады не развиты, челюстной и подъязычно-
жаберный аппараты типично личиночные; кишечник длинный, извитой;
№ 5 — нормально пигментирован, судя по зачаткам передних конечно-
стей (длиной 2.2 мм) и изменениям челюстного и подъязычно-жабер-
31
ного аппаратов, особь явно вступила в метаморфоз; пойманная 12.X.
1953, она погибла в аквариуме 4.IV.1954.
5. В Польше нам известны гигантские личинки, многие из которых
были пойманы в природе, а одна выращена в лаборатории.
В окрестностях города Познань живут все три формы зеленых
лягушек (Berger, 1966). Небольшие водоемы заселены смешанной по-
пуляцией Rana lessonae — Rana esculenta, а крупные — Rana ridi-
bunda — Rana esculenta. В искусственных прудах на реке Варта оби-
тает очень большая по численности популяция Rana ridibunda. В 1963—
1970 годах во многих водоемах были произведены сборы материала,
включая головастиков и лягушат. Выяснилось, что длина тела личинок
в разных водоемах была очень различна. В табл. 2 приведены размеры
головастиков и лягушат с хвостами из тех водоемов, где были обнару-
жены личинки крупнее 100 мм. В водоеме I наряду с многочисленными
особями Rana esculenta встречаются также единичные Rana lessonae.
В нем 18.VIII.1968 были собраны 90 особей (73 самки и 17 самцов)
Rana esculenta, из них 54 головастика и 36 лягушат, еще имеющих
хвост, а также 6 особей Rana lessonae, из них 2 головастика и 4 лягу-
шонка с хвостами (табл. 2). Вес тела головастиков составлял 6.0—
13.1 г (в среднем 9.88 г). Среди 54 личинок Rana esculenta длина тела
16 особей была равна 101 —112 мм. В водоеме II, где обитают только
Rana esculenta, мы собрали 30.VIII.1967 60 особей (45 самок и 15 сам-
цов) Rana esculenta, из них 4 головастика, 12 лягушат с хвостами (их
размеры см. табл. 2), остальные — сеголетки без хвостов, а также
одного лягушонка (самка) фенотипа ridibunda с хвостом (табл. 2),
который, несомненно, произошел от скрещивания Rana esculentaXRana
esculenta. В начале августа в этом же водоеме были собраны десятки
личинок; длина тела у некоторых была равна 100—115 мм, а 29 сен-
тября была найдена гигантская личинка длиною в 163 мм (вклейка,
рис. VI, Б) и весом 31.5 г. Кишечник достигал длины более 73 мм
(другие промеры см. табл. 3). Этот головастик оказался мужского пола
и имел довольно крупные семенники: левый 3, правый 2 мм длиной.
В водоеме III, заселенном только Rana ridibunda, 17 VIII.1963 было
отловлено 55 особей (26 самок, 26 самцов и 3 особи с гонадами про-
межуточного типа). Среди них было 5 личинок, в том числе одна длин-
нее 100 мм, и 6 лягушат с хвостами (табл. 2). Личинки esculenta в во-
доеме I могли появиться только вследствие скрещивания особей Rana
lessonae и Rana esculenta (Berger, 1968a) путем «гибридогенеза»
(Tunner, 1974), в пользу чего говорит также преобладание самок над
самцами (Berger, 1971). В водоеме II, как мы теперь знаем (Berger
a. Roguski, 1978), особи Rana esculenta были, вероятно, триплоидами
типа LRR, т. е. с одним геномом lessonae и двумя геномами ridibunda,
что видно по их морфологии (D. р./С. int. 1. = 2.45—2.90, х = 2.74). Круп-
ные размеры личинок из этих прудов связаны однако не с гибридиза-
цией, а скорее с условиями обитания, так как в других водоемах ли-
чинки очень редко достигали длины в 100 мм. Это же подтверждает
и пример с личинками Rana ridibunda из водоема III (табл. 2).
В мае 1963 года были проделаны многочисленные опыты по скре-
щиванию особей всех трех форм познаньских зеленых лягушек (Berger,
1967). Все головастики в опытах находились под наблюдением до их
метаморфоза или гибели. В некоторых скрещиваниях отмечались еди-
ничные случаи задержки развития, однако эти особи проходили мета-
морфоз или погибали на следующий год. Особого внимания заслужи-
вают три головастика, личиночное развитие которых с мая 1963 года
продолжалось около 2 лет. Их задние конечности достигали лишь
3—4 мм длины. Наиболее крупная личинка, появившаяся в результате
скрещивания самки Rana lessonae и самца Rana esculenta 18 мая,
к концу 1963 года достигла длины в 125 мм (вклейка, рис. VI, Д).
32
В это время у нее уже было заметно искривление позвоночника, кото-
рое в 1964 году усилилось, а 1.II.1965 эта личинка погибла. Спираку-
люм этой личинки не развился, яичников также почти не было. Второй
головастик, также женского пола, на этот раз от скрещивания самки
Rana lessonae и самца Rana ridibunda, погиб 15.V.1965, достигнув
в длину лишь 60 мм. И, наконец, третий головастик, самец-альбинос
с небольшими семенниками, оба родителя которого были Rana lessonae,
ко дню смерти (31.III.1965) имел длину в 58 мм.
6. 30.Х.1970 Р. Гюнтер скрестил в лаборатории самца и самку
Rana ridibunda, пойманных близ Ашхабада, Туркмения. Из 13 икринок
вывелись 4 личинки, 3 из которых погибли в первые же недели своей
жизни, а четвертая дожила на личиночной стадии до 28.VII.1971, до-
стигнув в длину 100 мм и превратившись в лягушку. 29.XII.1971 она
была умерщвлена; исследование показало, что семенники ее были раз-
виты нормально и содержали много сперматозоидов.
30.X.1970 эта же самка Rana ridibunda из Ашхабада была скре-
щена с самцом Rana ridibunda из старицы Альте Одер близ города
Лебуз (ГДР) недалеко от польской границы. Из 64 икринок вывелись
только 6 нормально развитых личинок, 5 из которых погибли в первые
месяцы своей жизни, не дойдя до метаморфоза. Шестая достигла
длины 104 мм, прожив на стадии личинки 18 месяцев, после чего она
погибла 3.IV.1972. В ноябре 1971 года ее длина составляла 102 мм,
а к апрелю 1972 года она увеличилась лишь на 2 мм. Яичники этого
головастика были крупнее и содержали значительно более развитые
яйцеклетки, чем у нормальных личинок. Однако яйцеводы и даже за-
чатки задних конечностей обнаружить не удалось.
Большое количество погибших икринок и головастиков указывает
на нарушение фертильности самки Rana ridibunda из Ашхабада, и,
возможно, что появление гигантских головастиков связано с наруше-
нием физиологических процессов формирования яйцеклеток. Несмотря
на очевидные нарушения в гормональной системе, которые будут рас-
смотрены нами ниже, гигантские головастики, вероятно, обладают не-
кими свойствами, предоставляющими им селективные преимущества,
ибо в обоих выше описанных скрещиваниях выживали только они.
17.Х.1973 были скрещены диплоидный самец Rana esculenta из ста-
рицы Альте Одер и самка Rana lessonae из окрестностей Берлина. Из
91 икринки вывелись 63 головастика. Они хорошо развивались, и из
15 личинок, оставленных на выращивание, 12 в январе 1974 года про-
шли метаморфоз, один погиб, а два продолжали расти, оставаясь ли-
чинками. Исследование кариотипа одного из них в июле 1974 года по-
казало, что он имел диплоидное (2м = 26) число хромосом. В этом
опыте данный головастик погиб, достигнув в длину 106 мм. Второй го-
ловастик погиб 28.IX.1974. Длина тела составляла 112 мм (вклейка,
рис. V, Б), половые органы были развиты очень слабо, и точно опреде-
лить его пол не удалось. Длина туловища была равна 35, хвоста — 77,
задних конечностей — 3 мм. Из 12 головастиков, превратившихся в ля-
гушат, 9 принадлежали к форме Rana lessonae, а 3 — к Rana esculenta.
Исходя из этого, отцовская особь (Rana esculenta), взятая из смешан-
ной популяции Rana ridibunda — Rana esculenta, дала разные типы
гамет (см. Uzzell et al., 1977). Так как личинки Rana lessonae с трудом
отличимы от Rana esculenta (Giinther, 1978), нельзя с достоверностью
утверждать, принадлежат ли упомянутые гигантские головастики
к форме Rana esculenta или же Rana lessonae.
7. Р. Грегер (Groger, 1978) неоднократно наблюдал в 1977 году
гигантских головастиков «Rana esculenta» в пруду, расположенном
на Райнгарцевом поле около городка Бад Шмидеберг, округ Биттер-
фельд, северо-восточнее города Лейпциг, ГДР. Пять пойманных личи-
нок имели размеры 175—183 мм.
3 Зак. 719
33
8. Ш. Феннер (Ch. Fenner, in litt.) обнаружила в 1979 году в окрест-
ностях города Киль, ФРГ, большое количество очень крупных личинок
зеленых лягушек, плававших в пруду на участке, поросшем элодеей.
15 личинок, отловленных 5 августа, имели следующие размеры:
3 особи — 7.5, 7—8.5—9, 2 — 9.5, 1 — 10 и 2—И см; обычная же ве-
личина головастиков до 7 см. Часть из этих 15 особей уже имела зад-
ние конечности, часть нет. В отличие от обычных головастиков, описы-
ваемые личинки выделялись более массивным туловищем и более
широкой каймой хвостового плавника. Отловленные 15 особей были
помещены в акватеррариум. 18 августа появился первый лягушонок
с нередуцированным до конца хвостом, 7 сентября — последний. Мета-
Рис. 1. Распространение гигантских головастиков зеленых ля-
гушек комплекса Rana esculenta.
1 — Ла-Рошфуко, долина реки Тардуаза, Шаранта, Франция (125 мм,
Angel, 1937, 1946); 2 — Хэм, Суррей, Англия (122 мм, Smith, 1954);
3 — Нойдорф близ Базеля, Швейцария (до 105 мм, Fatio, 1872;
Kollmann, 1884); 4 — Прага, Чехословакия (111 мм, Pfliiger, 1883),
5 — Бад Шмидеберг, округ Биттерфельд, северо-восточнее города
Лейпциг, ГДР (до 183 мм, Grdger, 1978); 6 — Киль, ФРГ (до ПО мм,
Ch. Fenner, in litt.); 7 — Хиллеред и Эмдруп, Фреденсборг, остров
Зеландия, Дания (до 145 мм, Wesenberg-Lund, 1922); 8 — Анебода,
Смоланд, Швеция (Gislen a. Kauri, 1959); 9 — Познань, Польша (до
163 мм, наши данные); 10 — Априки, Лигуты и Вецпили, Лиепайский
район, Латвия (до 165 мм, Я. Я. Лусис и И. А. Цауне, in litt.);
11 — Казалинск, Казахстан (109 мм, Боркин, 1979); /2 — Чирчик близ
города Ташкент, Узбекистан (до 130 мм, Боркин, 1979) и 13 — Чуйская
долина, Киргизия (150 мм, «Pelobates fuscus»?, Янушевич, 1976).
морфоз личинок с задними конечностями протекал 8—10 дней; тех жег
которые их еще не имели, 16—23 дня. Все головастики кормились до-
зированным кормом для собак. Молодые лягушата достигали в длину
2.5—2.8 см. В природе многочисленные лягушата с остатками хвоста
были отмечены 18 августа и имели размеры тела около 2.5 см.
9. Кроме уже отмеченных ранее (Angel, 1937, 1946) двух гигант-
ских головастиков длиной ПО и 125 мм, пойманных 8.IX.1936 во фран-
цузском департаменте Шаранта (табл. 3, рис. 1), нам (Р. Гюнтер)
удалось обнаружить в коллекциях Национального музея естественной
истории в Париже еще 3 гигантских личинки, хранящиеся вместе
с 4 нормальными под общим номером и этикеткой (127416, «зверинец
в Париже»). Эти 3 личинки достигали в длину 100, 102 и 103 мм, их
задние конечности, в отличие от особей из Шаранты, также хранящихся
в этом музее, были хорошо развиты, а сами они были близки к мета-
морфозу. Длина 4 обычных личинок составляла около 65 мм, их задние
конечности были развиты немного лучше.
Помимо описанных выше, на рис. 1 показаны также все известные
нам прежние находки гигантских головастиков зеленых лягушек.
Следует отметить, что изредка встречающиеся в природе гигант-
ские головастики известны не только для зеленых лягушек, но и для
других бесхвостых амфибий Европы и западной Азии. Они были также
описаны для травяной лягушки, Rana temporaria в Финляндии (пойман
в октябре, длина 63.5 мм — Luter, 1917) и Швеции (до 70 мм, обычный
размер 45 мм — Gislen a. Kauri, 1959), квакши Hyla arborea в Израиле
(до 56 мм, норма около 30 мм — Boschwitz, 1961), чесночниц Pelobates
fuscus в Германии и Чехословакии (до 150 мм — Pfliiger, 1883), Польше
(Grube, 1873), Швейцарии (Kollmann, 1884), Дании (до 180 мм — We-
senberg-Lund, 1922), Швеции (Gislen a. Kauri, 1959), Казахстане (до
150 мм — Искакова, 1959) и Pelobates syriacus в Израиле (до 205 мм,
норма до 107 мм — Boschwitz, 1957).
Причины гигантизма
Каковы же причины встречающегося в природе гигантизма голова-
стиков зелёных лягушек? Ниже мы обсуждаем значимость ряда фак-
торов, упоминаемых в литературе.
1. Географическая изменчивость. «Головастики на се-
вере достигают 40—50 мм общей длины, а на юге (Франция) —до 125».
(Терентьев и Чернов, 1949, с. 90). Однако гигантские головастики от-
мечены в самых разных частях ареала зеленых лягушек (рис. 1). Кроме
того, размеры нормальных головастиков на севере не меньше, чем на
юге (табл. 1 и 2). Поэтому это предположение приходится отклонить
(см. также Gunther, 1978).
2. Доступность и качество пищи, плотность скопле-
ний. Как установлено большой серией работ (Berger, 1968b; Heusser
u. Blankenhorn, 1973; Шварц и др., 1976), высокая плотность скопле-
ний головастиков может вызывать как торможение, так и ускорение
метаморфоза. В любом случае, однако, появление гигантизма у голо-
вастиков, вызванного «метаболитами скоплений», не отмечается. Можно
было бы предположить, что при низкой или оптимальной плотности
скоплений и обилии хорошей пищи головастики будут достигать боль-
ших, чем обычно, размеров.
Так, например, Э. Юнг (Yung, 1905) упоминает очень крупных го-
ловастиков «Rana esculenta» из окрестностей Женевы: личинки с двумя
задними конечностями в среднем имели длину туловища около 29.5 мм,
а лягушата после метаморфоза — 28.2 мм, тогда как размеры туловища
у соответствующих контрольных личинок и лягушат были равны 16
и 18 мм. Этот автор отмечает обилие пищи в водоеме, где были пой-
маны крупные головастики, а также значительно более длинный кишеч-
ник у крупных личинок, заполненный органическими веществами.
Исходя из этого, Э. Юнг считает обилие хороших кормов главной при-
чиной гигантизма. Следует, однако, отметить, что судя по приводимым
им размерам туловища личинок и лягушат головастики из-под Женевы
были крупными, до 90—100 мм (сравни с нашими данными в табл. 2),
но не «гигантскими» в нашем понимании. Л. Бергер (Berger, 1957)
описывает лягушат с хвостами, длина туловища которых достигала
35—40 мм. Следовательно, на личиночной стадии перед метаморфозом
они, несомненно, были значительно длиннее 100 мм, как это видно из
таблицы 2.
Хотя при хороших условиях питания головастики явно достигают
более крупных размеров, по-видимому, в некоторых случаях прибли-
жаясь к «гигантским», однако вряд ли обилие хорошей пищи может
быть причиной рассматриваемого нами гигантизма. Во-первых, гигант-
ские головастики, как правило, немногочисленны, тогда как при пред-
полагаемом действии трофического фактора достижение гигантских
размеров должно быть более или менее массовым явлением. Во-вторых,
Зф
35
при лабораторном выращивании, когда условия примерно одинаковы,
все же время от времени спонтанно появляются редкие гигантские
особи, как это отмечено для «Rana esculenta» в старом понимании
(Hahn, 1912), Rana ridibunda (наши данные) и Xenopus laevis (Toivo-
nen, 1952; Jurand, 1955) Голодание и некоторые особенности диеты
(например, нехватка иода) могут замедлять развитие и препятствовать
метаморфозу (см. обзоры Lynn a. Wachowski, 1951; Toivonen, 1952),
но вряд ли будут причиной гигантизма.
3. Полиплоидия, межвидовая гибридизация и гете-
розис. Как известно, при гибридизации разных видов животных и
растений получаемые гибридные особи часто значительно крупнее роди-
телей. Полиплоиды (авто- и аллополиплоиды) по размерам могут пре-
восходить диплоидных особей. Несколько гигантских головастиков-аль-
биносов были получены при скрещивании жаб Bufo bufo vxBufo viri-
dis <3 (Hertwig u. a., 1959) (см. однако c. 40). Гигантские личинки были
получены нами при скрещивании Rana lessonae и Rana esculenta (см.
с. 32 и 33) Познаньская личинка длиной в 163 мм, по-видимому, также
произошла от скрещивания особей Rana esculenta (см. с. 32). Кажется
очень соблазнительным связать происхождение гигантских головастиков
зеленых лягушек с их межвидовой гибридизацией, особенно если учесть
открытие гибридной природы Rana esculenta (Berger, 1967; Giinther,
1969; Tunner, 1970) и частую встречаемость в природе триплоидных
особей этой формы (Giinther, 1970, 1975; Uzzell a. Berger, 1975),
а также тот факт, что некоторые типы триплоидных личинок растут
быстрее и, как правило, достигают больших размеров, чем диплоидные
(Бергер, 1976; Berger a. Uzzell, 1977).
Однако целый ряд возражений не позволяет принимать эту гипо-
тезу. Во-первых, наши данные не говорят в пользу полиплоидности по-
лученных нами гигантских головастиков. Во-вторых, находки гигант-
ских головастиков Rana ridibunda в Средней Азии, где этот вид алло-
патричен, а также гигантизм личинок из скрещиваний в пределах этого
вида (см. с. 33) не связаны с их гибридным происхождением. Во мно-
гих работах показано, что как алло-, так и автополиплоидные (Зи и
4п) личинки амфибий по величине тела от диплоидных особей сильно
не отличаются. Хотя объем ядер и клеток у полиплоидов больше, чем
у диплоидов, однако размеры органов и тела примерно те же. Отсут-
ствие гигантизма у полиплоидов является следствием компенсаторного
уменьшения количества клеток (Fankhauser a. Watson, 1949; Fischberg,
1958; Gurdon, 1959; Brandom, 1960; Volpe a. Dasgupta, 1962). В то же
время, несмотря на сильные различия в размерах тела, величина клеток
эпителия пищевода и желчного пузыря, а также печени и желез же-
лудка у гигантских и нормальных головастиков Rana ridibunda одина-
кова (Hahn, 1912) Более крупные у «гигантов», рано дифференцирую-
щиеся клетки ганглиев не показательны, так как для них характерна
корреляция с размерами тела (Levi, 1906 — цит. по Hahn, 1912). Таким
образом, вся совокупность данных позволяет отклонить гипотезу о гиб-
ридизации как главной причине гигантизма головастиков.
4. Влияние освещенности. В опытах по содержанию личи-
нок Xenopus laevis в условиях 1.5 часов света в сутки удалось вызвать
задержку развития по сравнению с контрольными особями, содержа-
щимися в условиях круглосуточного света. Гистологические особенности
щитовидной железы указывали на ее гипотрофию. Автор (Toivonen,
1952) полагает, что световой фактор может быть причиной «частичной
неотении» (sensu Kollmann, 1884). Ранее на это указывал П. Каммерер
(Kammerer, 1906), работавший с Alytes obstetricans. Однако можно вы-
двинуть и ряд возражений. Во-первых, сам автор (Toivonen, 1952) ука-
зывает, что различия в строении щитовидной железы между подопыт-
ными и контрольными головастиками не столь значительны по срав-
36
нению с «неотеническими» личинками, спонтанно появляющимися
в культуре. Опыты и наблюдения других авторов (Ирихимович, 1947;
Riihmekorf, 1958; Топоркова и Клименко, 1977) не подтверждают силь-
ного влияния светового фактора на рост и развитие бесхвостых амфи-
бий. Кроме того, экспериментальное укорочение светового дня, что,
собственно говоря, и наблюдается осенью в природе, вызывает слабое
ускорение метаморфоза (Ирихимович, 1947). Хотя фотопериодическая
реакция лягушек разных широт различна, гигантизм головастиков
в опытах не отмечается (Топоркова и Клименко, 1977). В опытах
Т. М. Ивановой (1952) путем разрушения зрительных путей при уда-
лении промежуточного мозга, а также удаления глазных яблок у личи-
нок Rana ridibunda не удалось вызвать задержки развития. Рост, раз-
витие и метаморфоз как слепых, так и контрольных головастиков про-
ходили примерно одинаково. Таким образом, по крайней мере, зритель-
ная рецепция не имеет сколько-нибудь существенного значения для
развития гигантских головастиков зеленых лягушек. Кроме того, в при-
родных условиях головастики зеленых лягушек практически никогда
не подвергаются действию только света или только темноты в ходе
своего развития, как это бывает в опытах.
5. Влияние температуры. Повышение температуры воды,
как известно, ускоряет развитие. Понижение ее замедляет темпы раз-
вития и отодвигает наступление метаморфоза, понижая, по-видимому,
реактивность тканей головастиков на гормон щитовидной железы
(Huxley, 1929; Белкин, 1934; Войткевич, 1938; Lynn a. Wachowski, 1951)
или активность гипофизарно-тиреоидной системы (Saxen et al., 1957).
Хотя низкие температуры «консервируют» личиночное состояние, они
также отрицательно сказываются и на росте головастиков. Даже если
рост головастиков продолжается, то вряд ли они успевают достичь ги-
гантских размеров за одно лето. Так, например, головастики Rana ridi-
bunda, обнаруженные в Крыму близ Симферополя 28 января — 9 фев-
раля (т. е. 10—22 февраля по сегодняшнему календарю), достигали
длины лишь 40—60 мм (Kessler, 1879). Кроме того, влиянию темпера-
туры в качестве причины гигантизма противоречит также и тот факт,
что единичные гигантские головастики встречаются как на севере
ареала вида, так и на юге (рис. 1).
6. Перезимовка. Обычные сроки развития головастиков зеле-
ных лягушек охватывают 10—16 недель, хотя в зависимости от благо-
приятных или плохих условий метаморфоз может наступать раньше
или позже, через 8—40 недель (наши данные, см. также Giinther, 1978).
Перезимовка головастиков приводит к сохранению личиночной стадии
особи в течение значительно большего срока, чем обычно. Для обозна-
чения такого явления был предложен термин «неотения» в двух моди-
фикациях: «тотальная неотения», если личинки достигали половой зре-
лости (например, аксолотль Ambystoma mexicanum), и «частичная нео-
тения», когда метаморфоз не наступает, но и половозрелость личинками
не достигается (Kollman, 1884)* Многие авторы, описывавшие гигант-
ских головастиков зеленых лягушек, указывали, что они или были пой-
маны после зимовки, или же способны к длительному в течение года
и более существованию в аквариуме без метаморфоза. Так, например,
в культуре они жили 10—12 месяцев и достигли длины 11.6—12.0 см
(Hahn, 1912; Smith, 1954). Согласно И. Кольманну (1884), обнаружен-
ные им гигантские головастики «Rana esculenta» пережили две зимы.
В наших опытах две гигантских личинки, полученные при скрещивании
Rana lessonae и Rana esculenta, жили 11.5 (см. с. 33) и 20.5 месяцев
у
* Исторически любопытно, что термин «неотения», который можно перевести
с древнегреческого как «сохранение юности», был предложен И. Кольманном (1884)
как раз в связи с обсуждением найденных им в окрестностях Базеля гигантских голо-
вастиков Rana esculenta.
37
(см. с. 32—33). Третья личинка от скрещивания особей Rana ridibunda
жила 18 месяцев (см. с. 33). Личинки в возрасте до 2—2.5 лет и 4 лет
2 месяцев описаны также для жабы-повитухи, Alytes obstetricans (Wie-
dersheim, 1878; Brunk, 1882; Kammerer, 1906); предполагается даже,
что в высокогорных озерах они доживают до 20 лет (Angelier et Ange-
lier, 1964). Головастики обыкновенной жабы, Bufo bufo, жили в аква-
риуме почти до 3 лет, а травяной лягушки, Rana temporaria— 17 меся-
цев (Knauer, 1878). Давно известным примером перезимовывающих
личинок являются головастики Pelobates fuscus (Pfliiger, 1883). Инте-
ресно, что среди североамериканских видов лягушек рода Rana наибо-
лее крупные головастики (до 100 мм и выше) известны для видов, зи-
мующих одну или даже две зимы в личиночном состоянии (рис. 2).
которых завершается после 1—2 зимовок (Л) или за одно лето (Б).
1 — R. catesbeiana, 2 — R grylio, 3 — R. septentrionalis, 4 — R. heckscheri, 5 — R. virgatipes,
6 — R. tarahumarae, 7 — R. muscosa, 8 — R. pustulosa, 9 — R. capita, 10 — R. pipiens pipiens,
11 — R. aurora draytoni, 12 — R. palustris, 13 — R. pipiens sphenocephala, 14 — R. cascadae,
15 — R. pretiosa, 16 — R. clamitans, 17 — R. boylii, 18 — R. areolata circulosa, 19 — R. sylvatica.
(no Wright a. Wright, 1949 and Zweifel, 1955).
Следует однако отметить, что в ряде достоверных случаев перези-
мовки или задержки развития в аквариуме головастики Rana ridibunda
не достигали гигантских размеров (Kolazy, 1871; Fischer-Sigwart, 1885;
Wesenberg-Lund, 1922). Так, наряду с гигантами в 10.5 см И. Кольманн
(1884) отмечает и головастиков только в 5—6 см, которые, по его мне-
нию, моложе первых на год, так как пережили только одну зиму. Среди
5 «неотенических» личинок Rana ridibunda в возрасте 1 год четыре были
гигантскими (11.6—12.0 см), а один головастик был всего лишь 6 см
в длину (Hahn, 1912). В наших опытах среди трех личинок, живших
в аквариуме 20—24 месяца, только одна стала гигантом в 12.5 см,
тогда как две остальные достигли только 6 см (см. с. 32—33). А. А. Войт-
кевич (1952) в течение 4 лет изучал массовые перезимовки головасти-
ков (несколько тысяч!) Rana ridibunda в окрестностях города Алма-
Ата, Казахстан. Длина этих головастиков, у которых обычно в середине
следующего лета наступал метаморфоз, колебалась в пределах 7.6—
8.3 см. Таким образом, хотя между гигантизмом головастиков зеленых
лягушек и перезимовками существует положительная связь, однако
сама по себе зимовка не гарантирует достижения «сверхкрупных» раз-
меров. Иначе говоря, личинки могут быть «частично неотеническими»,
но не обязательно гигантскими.
7. Гормональный контроль. Теория регуляции метамор-
фоза амфибий гормональной системой, главными членами которой
38
являются щитовидная железа и гипофиз, а также гипоталамус, была
блестяще доказана в большой серии экспериментальных работ еще
в первой четверти нашего столетия и сейчас общепринята. Как хорошо
известно, гормон щитовидной железы (скорее всего, трииодотиронин —
Leloup et Buscaglia, 1977) индуцирует метаморфоз. В свою очередь,
функция щитовидной железы контролируется гормоном (тиреотропи-
ном), выделяемым передней долей гипофиза — аденогипофизом, дея-
тельность которого регулируется гипоталамусом (Lynn a. Wachowski,
1951; Etkin, 1963, 1964). Такие особенности гигантских головастиков
зеленых лягушек и некоторых других бесхвостых, как спорадическая
встречаемость, длительная задержка развития и их сосуществование
с нормальными головастиками, отмеченные как в природных, так и
в лабораторных условиях, несомненно, указывают на внутренние при-
чины этого явления, как это справедливо предполагал еще К. Везен-
берг-Лунд (Wesenberg-Lund, 1922).
Имеющиеся данные подтверждают это. Анатомические и гистоло-
гические исследования показали полное отсутствие щитовидной железы
у «неотенических» гигантских головастиков шпорцевой лягушки, Xeno-
pus laevis, спонтанно появляющихся в польской (Jurand, 1955, 1959)
и финской линиях разведения (Saxen, 1957; Saxen et al., 1957). Отсут-
ствие щитовидной железы обнаружено также у гигантских головасти-
ков Pelobates syriacus (Boschwitz, 1957). Искусственное удаление щито-
видной железы (тиреоидэктомия) вызвало появление гигантских голо-
вастиков Rana pipiens (Allen, 1918; Etkin a. Lehrer, 1960) и Rana sylva-
tica (Hoskins a. Hoskins, 1919). В то же время кормление личинок трех
североамериканских видов Rana порошком из щитовидной железы вы-
звало метаморфоз при карликовых размерах личинок по сравнению
с контрольными особями (Swingle, 1918).
А. С. Северцов (in litt.) отпрепарировал под бинокуляром двух ги-
гантских головастиков Rana ridibunda из Латвии и не нашел у них
щитовидной железы, в том числе у самого крупного экземпляра
в 16.5 см. Удаление щитовидной железы резко замедляет процесс око-
стенения, что было показано на гигантских тиреоидэктомированных
личинках Rana pipiens (Terry, 1918). Недоразвитие щитовидной железы
и слабое окостенение скелета обнаружены также у гигантской личинки
Hyla arborea (Boschwitz, 1961). Слабое развитие костного скелета от-
мечается у гигантских «Rana esculenta» Дании (Wesenberg-Lund, 1922),
а также обнаружено нами при рентгенографическом изучении гигант-
ского головастика Rana ridibunda из Ташкента. Косвенным указанием
на нарушение гормональной системы является, кроме того, пегая
окраска гигантских личинок.
Влияние гипофиза на функцию щитовидной железы было открыто
уже в ранних работах по эндокринологии метаморфоза амфибий. Так,
разрушение гипофиза вызвало атрофию щитовидной железы и появле-
ние трех гигантских головастиков Rana temporaria (Adler, 1914). В то
же время пересадка ткани передней доли гипофиза взрослых лягушек
«частично неотеническим» личинкам Rana clamitans повлекла за собой
усиление активности щитовидной железы и метаморфоз (Ingram, 1929).
Тиреотропный гормон, стимулирующий активность щитовидной железы,
вырабатывается базофильными клетками pars distalis переднего отдела
гипофиза (Saxen et al., 1957). Предполагается, что тиреотропная актив-
ность аденогипофиза, в свою очередь, может регулироваться механиз-
мом обратной связи путем ингибирующего влияния повышенной кон-
центрации гормона щитовидной железы (Etkin, 1964). Исходя из всего
этого, следует ожидать, что появление гигантизма головастиков может
быть вызвано нарушениями не только непосредственно щитовидной же-
лезы, как это отмечено выше, но и отклонениями в деятельности адено-
гипофиза.
3S
В отношении гигантских головастиков, полученных при гибридиза-
ции жаб Bufo bufo sXBufo viridis d, упоминается без каких-либо под-
робностей о наличии дефектов гипофиза, щитовидная железа была раз-
вита нормально (Hertwig и. а., 1959). В других случаях сообщается
об увеличенных размерах гипофиза (гигантские головастики «Rana
esculenta» в старом понимании—Hahn, 1912; Wesenberg-Lund, 1922;
Rana sylvatica — Hoskins a. Hoskins, 1919; Xenopus laevis— Jurand,
1955: Hyla arborea—Boschwitz, 1961) и гиперактивном состоянии ба-
зофильных клеток аденогипофиза у гигантских головастиков Xenopus
laevis (Saxen, 1957; Saxen et al., 1957). Такая гипертрофия может быть
вызвана тиреоидной недостаточностью.
Влияние гипоталамуса на рост и метаморфоз головастиков также
довольно хорошо изучено. Установлено, что секреция тиреотропного
гормона, выделяемого дистальной частью гипофиза, контролируется
гипоталамусом. Этот контроль осуществляется через нейросекреторные
клетки и портальную систему гипофиза (Etkin, 1963, 1964). Удаление
гипоталамуса вызывает задержку метаморфоза и приводит к крупным,
превышающим нормальные, размерам головастиков (Иванова, 1952;
Remy et Bounhiol, 1966b). Так, личинки леопардовой лягушки, Rana
pipiens, искусственно лишенные гипоталамуса на ранней стадии раз-
вития, не смогли пройти метаморфоз в течение 375 дней. Щитовидная
железа у них была явно меньше, а высота ее фолликулярных эпите-
лиальных клеток значительно ниже, чем у контрольных особей. Эта
атрофия щитовидной железы была обусловлена понижением уровня
секреции тиреотропного гормона в гипофизе. По своей величине опери-
рованные головастики были вдвое крупнее нормальных личинок, кото-
рые метаморфизировали на 135-й день после оплодотворения. Голова-
стики с удаленным гипофизом также остались на личиночной стадии
даже на 375-й день, но росли очень медленно и по размерам тела лишь
незначительно превышали контроль (Hanaoka, 1967).
Таким образом, выше приведенные факты недвусмысленно указы-
вают на нарушения эндокринной системы у спорадически появляющихся
гигантских головастиков бесхвостых амфибий и, в частности, Rana ri-
dibunda и хорошо согласуются с общей теорией эндокринных факторов
метаморфоза амфибий.
Почему же все-таки эти головастики достигают гигантских разме-
ров, хотя другие личинки также с задержкой метаморфоза, иногда из
той же культуры, остаются значительно более мелкими (см. с. 38)?
По-видимому, основной особенностью таких гигантских головастиков
являются их неспособность к метаморфозу * из-за врожденных глубо-
ких и необратимых нарушений щитовидной железы (аплазия) или ги-
пофиза, а также усиленное действие гормона роста, в отличие от других
случаев «частичной неотении» зеленых лягушек и других бесхвостых.
Иначе говоря, эндокринные нарушения у этих гигантских головастиков
вызваны сугубо внутренними, генетическими причинами, а не являются
следствием обратимого понижения тиреоидной активности из-за влия-
ния факторов внешней среды (похолодание, особенности пищи и т. д.).
Изменения эндокринных желез в последнем случае не столь суще-
ственны, как показывает изучение головастиков Xenopus laevis с «экс-
периментальной неотенией», вызванной влиянием темноты или холода
(Toivonen, 1952; Saxen et al., 1957).
Необычные размеры гигантских головастиков зеленых лягушек, не-
сомненно, связаны с эффективным действием гормона роста. Процессы
* А. С. Северцов (in litt.) указывает,- что один из гигантских головастиков Rana
ridibunda из Латвии длиной в 11.5 см явно начал метаморфоз после перенесен!'/: его
в аквариум (треугольная голова, зачатки передних конечностей длиной в 2.2 мм,
изменения челюстного и подъязычно-жаберного аппаратов), но погиб 4.IV. 1954 через
6 месяцев после его поимки в природе. См. также с. 42.
40
роста и метаморфоза являются, как хорошо известно, антагонистиче-
скими. В норме скорость роста личинок замедляется по мере увеличе-
ния их размеров, и в период метаморфоза головастики не растут,
а, напротив, сокращаются в размерах из-за резорбции хвоста. В то же
время ацидофильные клетки аденогипофиза, вырабатывающие гормон
роста, нормально функционируют у необратимо «неотенических» личи-
нок, как это было обнаружено у гигантских головастиков Xenopus laevis
(Saxen, 1957), т. е. рост головастиков продолжается.
Относительно недавно было выяснено, что регуляция роста у амфи-
бий осуществляется двумя разными гормонами: пролактин стимулирует
рост у головастиков, а так называемый ростовой гормон, или сомато-
тропин, — рост у лягушек после метаморфоза (Berman et al., 1964 Etkin
a. Gona, 1967; Brown a. Frye, 1969a, b; Snyder a. Frye, 1972, 1974). Хотя
в опытах были использованы гормоны млекопитающих, соответствую-
щие гормоны у амфибий должны быть, по-видимому, примерно та-
кими же. Усиленный рост, приведший к гигантизму головастиков Rana
pipiens, был отмечен после трансплантации аденогипофиза (Etkin а.
Lehrer, 1960), в pars distalis которого, как известно, вырабатывается
пролактин. Удаление гипофиза вызывает задержку метаморфоза и за-
медленный рост, размеры головастиков не превышают нормы * (Etkin
a. Lehrer, I960; Remy et Bounhiol, 1966b). В результате инъекции про-
лактина ** головастикам Alytes obstetricans с удаленным гипофизом
были получены гигантские личинки (Remy et Bounhiol, 1966а, b). Из-
быточный рост был отмечен также при обработке пролактином личинок
Rana pipiens (Brown a. Frye, 1969а). Все эти и многие другие опыты
подтверждают ростовое действие пролактина у гигантских головасти-
ков (см. также Guardabassi et al., 1969, 1971).
В этом плане интересны также карликовые головастики Telmato-
bius hauthali schreiteri (семейство Leptodactylidae), обнаруженные в вы-
сокогорных популяциях в Андах. В гипофизе этих личинок была выяв-
лена явная недостаточность или отсутствие ацидофильных клеток,
вырабатывающих гормон роста (Pisano, 1958) Показано также, что
пролактин и гормон щитовидной железы являются антагонистами
(Etkin a. Gona, 1967; Hsti et al., 1976; Clemons a. Nicoll, 1977; Wright
et al., 1979). Если учесть, что у головастиков, лишенных щитовидной
железы, ингибирующее влияние на рост отсутствует, а стимуляция со
стороны пролактина продолжается, то становится понятным их гиган-
тизм. При отсутствии контроля над дистальной частью гипофиза со
стороны гипоталамуса, что, в частности, наблюдается при тиреоидэкто-
мии, происходит повышенная секреция пролактина, результатом чего
является усиленный рост и, в конечном счете, личиночный гигантизм
(Guardabassi et al., 1971). При достижении размеров тела, при которых
у нормальных личинок наступает метаморфоз, гипофиз тиреоидэктоми-
рованных головастиков выделяет гормон роста еще более эффективно
(Etkin a. Lehrer, 1960).
Генетика гигантизма
Известные нам данные по наследованию способности к неотении
довольно скудны и получены, главным образом, при гибридизации раз-
ных видов хвостатых амфибий рода Ambystoma. Хотя эти результаты
не всегда четки, все же в ряде случаев можно с определенной долей
* Появление 3 гигантских головастиков Rana temporaria в опытах Л. Адлера
(Adler, 1914), по-видимому, вызвано неполным удалением гипофиза.
** Гигантизм был вызван также инъекцией соматотропина (Remy et Bounhiol,
1966°), хотя в работах американских авторов это не подтвердилось (Etkin a. Gona,
1967; Snyder a. Frye, 1972, 1974); возможно, что он был обусловлен примесью пролак-
тина в препаратах соматотропина (Brown a. Frye, 1969а).
41
уверенности предполагать, что неспособность к метаморфозу насле-
дуется как простой рецессивный признак (Freytag, 1947; Humphrey,
1967; Brandon, 1976). Следовательно, неотенические особи должны быть
рецессивными гомозиготами по данному признаку. Спорадическое по-
явление гигантских головастиков зеленых лягушек наиболее логично
также объяснить выщеплением рецессивной мутации, непосредственно
или в качестве плейотропного эффекта нарушающей гормональный кон-
троль метаморфоза. В пользу этого может свидетельствовать возник-
новение единичных гигантских личинок в культуре, когда условия со-
держания примерно одинаковы для всех головастиков. Например,
сверхкрупные личинки Xenopus laevis спорадически появляются с часто-
той 0.1% в польской (Jurand, 1955) и 1—4% в финской линиях разве-
дения (Saxen, 1957; Saxen et al., 1957).
Тем не менее, не исключено, что гигантские «неотенические» голо-
вастики в разных случаях могут иметь разную (в том числе и полиген-
ную) генетическую природу. Возможно также, что при массовом появ-
лении гигантских головастиков зеленых лягушек (Wesenberg-Lund,
1922; наши данные см. с. 32) важную роль играют негенетические фак-
торы, нарушающие взаимодействие системы гипоталамус—гипофиз—
щитовидная железа. Известны случаи, когда гигантские (или прибли-
жающиеся к ним по размерам) личинки в природе благополучно прохо-
дят метаморфоз и дают лягушат, более крупных, чем обычно (см. наши
данные на с. 32 и данные Ш. Феннер на с. 34). Не исключено, что в таких
случаях имеет место или гиперстимуляция роста особей по неизвестным
нам причинам, или же нарушения (генетические и/или средовые) гор-
мональной системы гипоталамус—гипофиз—щитовидная железа не
столь глубоки, как в выше обсуждавшихся примерах, и обратимы или
же компенсируются каким-либо фактором. Любопытно, что в обоих
достоверных случаях гигантизм головастиков, прошедших метаморфоз,
имел массовый характер.
Можно также предположить, что в некоторых случаях гигантизм
головастиков обусловлен развитием в гипофизе так называемых гор-
монально активных опухолей (ацидофильные аденомы), для которых
характерно выделение повышенного количества гормона роста. Именно
эти опухоли являются причиной патологического гигантизма (акроме-
галии) у человека (Schurr, 1966), причем рост некоторых больных пре-
вышал 250 см.
Судьба гигантских головастиков
В отличие от неотенических хвостатых амфибий, случаи размноже-
ния гигантских головастиков зеленых лягушек и, по-видимому, бес-
хвостых вообще не известны. Развитие гонад так же, как и функцио-
нирование щитовидной железы или рост головастиков, находится под
гормональным контролем передней доли гипофиза (аденогипофиза).
В ряде случаев сообщалось о значительно более мощном развитии по-
ловой системы у гигантских головастиков по сравнению с нормальными,
находящимися на сходной стадии развития. Так, например, хорошо
развитые яичники, заполненные крупными, но еще не зрелыми яйце-
клетками, богатыми желтком, описаны у гигантских головастиков «Rana
esculenta» (Hahn, 1912). У гигантских головастиков Rana sylvatica от-
мечаются крупные, но незрелые яичники с ооцитами, яйцеводы не раз-
виты; однако у личинок мужского пола семенники достигали полной
зрелости (Hoskins a. Hoskins, 1919). А. С. Северцов (in litt.) сообщает
о двух гигантских личинках Rana ridibunda из Латвии с хорошо разви-
той икрой и семенниками (см. также наши данные на с. 32 и 33). У ги-
гантских головастиков Xenopus laevis, достигающих возраста 9—17 ме-
сяцев, гонады, как яичники, так и семенники, были хорошо развиты и
42
соответствовали по уровню развития гонадам взрослых особей такого же
возраста (Jurand, 1955). Таким образом, теоретически не исключена
возможность достижения полной половозрелости «гигантов» и их раз-
множения. В пользу нормального развития половой системы может
говорить также тот факт, что гигантские головастики Xenopus laevis
с врожденным отсутствием щитовидной железы после трансплантации
ее от нормальных особей или добавления в виде порошка в воду
успешно проходили метаморфоз и достигали взрослой половозрелой
стадии с явными вторичнополовыми признаками (Jurand, 1959).
Однако даже если теоретически представить себе, что гигантские
личинки достигают стадии половой зрелости, то и в этом случае, на наш
взгляд, они не будут способны к размножению из-за нарушения брач-
ного поведения, вызванного отсутствием конечностей (это же обстоя-
тельство отмечает А. С. Северцов, in litt.). Таким образом, гигантские
«частично неотенические» личинки зеленых лягушек и других бесхво-
стых амфибий, потерявшие способность к метаморфозу, эволюционно
бесперспективны, в отличие от хвостатых амфибий, для многих форм
которых характерны или же факультативная способность к успешному
существованию на личиночной стадии развития, или же постоянно вод-
ный личиночный образ жизни.
В заключение мы хотели бы сердечно поблагодарить И. С. Дарев-
ского (Ленинград), Я. Я. Лусиса и Й. А. Цауне (Рига), А. С. Северцова
(Москва) и Ш. Феннер (Киль) за любезное предоставление нам не-
опубликованных материалов и/или обсуждение рукописи статьи.
ЛИТЕРАТУРА
Банников А. Г Об особенностях в сроках размножения и развития бесхвостых
земноводных различных биологических групп. — Учен. зап. Московск. гор. пед.
ин-та, каф. зоол., 1957, т. 65, с. 103—107.
Белкин Р. И. Взаимодействие внешних и внутренних факторов в процессе онтогенеза
амфибий. 1. Влияние температуры на тироидный метаморфоз головастиков Rana
temporaria. — Докл. АН СССР, 1934, т. 4, № 8—9, с. 462—465.
Белова 3. В. Питание головастиков Rana ridibunda Pall, в разных зонах дельты
Волги. — Бюлл. Московск. о-ва испыт. прир., 1964, отд. биол., т. 69, № 5,
с. 40—46.
Бергер Л. Является ли прудовая лягушка Rana esculenta L. обыкновенным гибри-
дом. — Экология, 1976, № 2, с> 37—43.
Боркин Л. Я. О некоторых находках обыкновенной чесночницы, Pelobates fuscus
(Laurenti) на юго-востоке ареала. — В кн.: Экология и систематика амфибий
и рептилий (Тр. Зоол. ин-та АН СССР, т. 89), 1979, с. 118—120.
Боркин Л. Я. и Тихенко Н. Д. Некоторые аспекты морфологической изменчи-
вости, полиморфизма окраски, роста, структуры популяции и суточной актив-
ности Rana lessonae на северной границе ареала. — В кн.: Экология и система-
тика амфибий и рептилий (Тр. Зоол. ин-та АН СССР, т. 89). 1979, с. 18—54.
Войткевич А. А. О роли температурного фактора в период метаморфоза голова-
стиков лягушки. — Биол. ж., 1938, т. 7, № 4, с. 749—762.
Войткевич А. А. О температурной стадии в развитии бесхвостых амфибий.—
Ж. общ. биол., 1952, т. 13, № 4, с. 286—297.
Денисова М. Н. Отряд Бесхвостые земноводные (Anura). — В кн.: Жизнь живот-
ных. Т. 4, ч. 2. Земноводные, пресмыкающиеся. М., «Просвещение», 1969,
с. 63—134.
Елпатьевский В. Гады Арала (Амфибии и рептилии берегов и островов Араль-
ского моря). — Научн. результаты Аральской экспедиции, Ташкент, 1903, вып. 4,
с. 1—32.
Иванова Т. М. Роль нервной системы в развитии амфибий. — Ж. общ. биол., 1952,
т. 13, № 3, с. 182—210. ‘
Ирихимович А. И. Влияние света на скорость развития головастиков Rana tem-
poraria. — Докл. АН СССР, 1947, т. 55, № 2, с. 177—180.
Искакова К. И. Земноводные Казахстана. Алма-Ата, изд. АН Каз. ССР, 1959,
92 с.
Никольский А. М. Земноводные (Amphibia). В серии: Фауна России и сопре-
дельных стран. Петроград, 1918, 311 с.
Папанян С. Б. Данные по экологии и хозяйственное значение озерной лягушки
в Армянской ССР.—Изв. АН Арм. ССР, Ереван, 1952, биол. и сель.-хоз. н.,
т. 5, № 11, с. 39—48.
43
Та ращу к В. I. Земповодш нлазуни. В серии Фауна УкраТнп. Kills. 1959,
т. 7, 246 с.
Терентьев П. В. и Чернов С. А. Определитель пресмыкающихся и земновод-
ных. М., «Советская наука», 3-е изд., 1949, 340 с.
Топоркова Л. Я. и Клименко И. А. Влияние фотопериода на рост и разви-
тие личинок рода Rana. — Экология, 1977, № 3, с. 101 —103.
Шварц С. С., П я сто лов а О. А., Добринская Л. А. и Рункова Г Г
Эффект группы в популяциях водных животных и химическая экология. М.,
«Наука», 1976, 152 с.
Щербак Н. Н. Земноводные и пресмыкающиеся Крыма. Herpetologica Taurica.
Киев, «Наукова думка», 1966, 240 с.
Янушевич А. И. Материалы по земноводным Киргизии. — Изв. АН Кирг. ССР,
Фрунзе, 1976, № 3, с. 47—50.
Adler L. Metamorphosestudien an Batrachierlarven. I. Exstirpation endokriner Drusen.
A. Exstirpation der Hypophyse. — Archiv f. Entwicklungsmechanik d. Organismen,
1914, Bd. 39, S. 21—45.
Allen В. M. The results of thyroid removal in the larvae of Rana pipiens. — J. Exp.
Zool., 1918, vol. 24, n. 3. p. 499—519.
Angel F Sur deux tetards geants de Rana esculenta L. — Bull. Mus. Nation. Hist.
Nr.tur.. 193~, 2 ser.. i. 9. n. 1. p. 54—55.
Angel F Reptiles et Amphibiens. In: Faune de France, Paris, Paul Lechevalier, 1946,
vpi. 45. 0 i p.
Angel ier E. e t Ange Her M.-L. Etude d’une population de crapauds accoucheurs
dans un hac de haute montagne. — C. R. Acad. Sci. Paris, 1964, t. 258, n. 2,
p. 701—7' o.
Berger L. Trudno ci orzy oznaczaniu zab krajowych (Ranidae) i ich kiianek. — Przegl.
Zool.. 1957, t. 1, n. 1, s. 31—38.
Berger L. Biometrical studies on the population of green frogs from the environs
of Poznan. — Annal. Zool., Warszawa, 1966, t. 23, n. 11, p. 303—324.
Berger L. Embrional and larval development of FI generation of green frogs diffe-
rent combinations. — Acta zool. cracov., 1967, t. 12, n. 7, p. 123—160.
Berger L. Morphology of the F( generation of various crosses within Rana esculenta-
complex. — Acta zool. cracov., 1968a, t. 13, n. 13, p. 301—324.
Berger L. The effect of inhibitory agents in the development of green-frog tadpo-
les. — Zool. Polon., 1968b, vol. 18, fasc. 3, p. 381—390.
Berger L. Sex ratio in the Fi progeny within forms of Rana esculenta complex.—
Genet. Polon, 1971, t. 12, n. 1—2, p. 87—101.
Berger L. Systematics and hybridization in the Rana esculenta complex. — In:
D. H. Taylor a. S. I. Guttman (eds.), The reproductive biology of amphibians.
New York — London, Plenum Press, 1977, p. 367—388.
Berger L. a. Michalowski L. Amphibia (Plazy). In: Klucze do oznaczania
krpgowcdw Polski, 1971, pt. 2, 78 p. (Amer. ed.).
Berger L. a. Roguski H. Ploidy of progeny from different egg size-classes of Rana
esculenta L. — Folia Biol., Krakow, 1978, t. 26, n. 4, p. 231—248.
Berger L. a. Uzzell T. Vitality and growth of progeny from different egg size
classes of Rana esculenta L. (Amphibia, Salientia). — Zool. Polon. 1977, t. 26.
fasc. 3—4, p. 291—317.
Berman R., Bern H. A., N i с о 11 C h. S. a. S t г о h m a n R. C. Growth-promoting
effects of mammalian prolactin and growth hormone in tadpole of Rana cates-
beiana.— J. Exp. Zool., 1964, vol. 156, n. 3, p. 353—360.
Boschwitz D. Thyroidless tadpoles of Pelobates syriacus Boettgcr. — Copeia, 1957,
n. 4, p. 310—311.
Boschwitz D. A giant tadpole of Hyla arborea L. — Bull. Res. Council Israel, Jeru-
salem, 1961, sect. B, zool., vol. 10 B, n. 4, p. 173—174.
Bon longer G. A. The tailless batrachians of Europe. Part II, London, Printed for
the Ray Society, 1898, p. 211—376.
Brandom’W. F. Gene-dosage studies in polyploid hybrids of California newt. —
J. Exp. Zool., 1960, vol. 143, n. 3, p. 323—345.
Brandon R. A. Spontaneous and induced metamorphosis of Ambystoma dumerilii
(Duges), a paedogenetic Mexican salamander, under laboratory conditions. — Her-
petologica, 1976, vol. 32, n. 4, p. 429—438.
Brown P. S. a. F г у e В. E. Effects of prolactin and growth hormone on growth and
metamorphosis of tadpoles of the frog, Rana pipiens. — Gen. a. Comp. Endocrinol.,
1969a, vol. 13, n. 1, p. 126—138.
Brown P. S. a. F г у e В. E. Effects of hypophysectomy, prolactin, and growth hor-
mone on growth of postmetamorphic frogs. — Gen. a. Comp. Endocrinol., 1969b,
vol. 13, n. 1, p. 139—145.
Brunk A. Ein neuer Fall von Entwickelungshemmung bei der Geburtshelferkrote. —
Zool. Anzeiger, 1882, 5. Jahrg., N 104, S. 92—94.
Clemons G. K. a. Nico 11 C. S. Effects of antisera to bullfrog prolactin and growth
hormone on metamorphosis of Rana catesbeiana tadpoles. — Gen. a. Comp. Endo-
crinol., 1977, vol. 31, p. 495—497
4
Dubois A. Les problemes de 1’espece chez les Amphibiens Anoures. — In: Ch. Bocquet,
J. Genermont et M. Lamotte (eds.), Les problemes de I'espece dans le regne
animal. Paris, 1977, t. 2, p. 161—284.
Etkin W. Metamorphosis-activating system of the frog. — Science, 1963, vol. 139,
n. 3557, p. 810—814.
Etkin W. Metamorphosis. — In: J. A. Moore (ed.), Physiology of the Amphibia.
New York — London, Acad. Press, 1964, p. 427—468.
Etkin W. a. G о n a A. G. Antagonism between prolactin and thyroid hormone
in amphibian development. — J. Exp. Zool., 1967, vol. 165, n. 2, p. 249—258.
Etkin W. a. Lehrer R. Excess growth in tadpoles after transplantation of the
adenohypophysis. — Endocrinology, 1960, vol. 67, n. 4, p. 457—466.
Fankhauser G. a. Watson R. C. The effects of pituitary implantation on diploid
and triploid larvae of the newt, Triturus viridescens, with special reference to the
• gonads. — J. Exp. Zool., 1949, vol. Ill, n. 3, p. 349—391.
Fat io V Faune des Vertebres de la Suisse. Volume III. Histoire naturelle des Rep-
tiles et des Batraciens. Geneve et Bale, H. Georg, Libraire-ed., 1872, 602+XII p.
Fischberg M. Experimental tetraploidy in newts. — J. Embryol. exp. Morphol.,
1958, vol. 6, pt. 3, p. 393—402.
Fischer-Sigwart H. Das Uberwintern der Lurche im Larvenzustande. — Zool.
Garten, 1885, 26. Jahrg., N 10, S. 299—312.
Freytag G. E. Die Neotenie der Urodelen.— Mitt. Mus. Naturkunde u. Vorgeschichte
u. naturwiss. Arbeitskreis, Magdeburg, 1947, Bd. 1, H. 1, S. 5—11.
Fuhn I. E. Amphibia. In: Fauna Rep. Pop. Romine, 1960, vol. 14, fasc. 1, 288 p.
Gislen T. а. К a u r i H. Zoogeography of the Swedish amphibians and reptiles with
notes on their growth and ecology. — Acta Vertebratica, 1959, vol. 1, n. 3,
p. 195—397
Groger R. Beitrage zur Erfassung der Amphibienfauna des Kreises Bitterfeld und der
Diibener Heide. Diplom-Arbeit, Martin-Luther-Universitat, Halle—Wittenberg,
1978.
Grube. [Ein ungewohnlich grosses Exemplar einer Kaulquappe, der Larve der Was-
ser- oder Knoblauchkrote (Pelobates juscus)]. — 50. Jahres-Bericht Schlesisch. Ges.
vaterland. Cultur, Breslau, 1873, S. 55.
Guardabassi A., Campantico E., Ferraro G. a. Olivero M. Effects of pro-
tection on thyroidectomized Bufo bufo larvae. — Monitore zool. ital., N. S., 1971,
vol. 5, n. 2, p. 99—115.
Guardabassi A., Cocito A. M. a. Ponzetti G. Ectopic pituitary homotrans-
plantations from larvae and adult donors into Xenopus laevis Daud. intact
larvae. — Monitore zool. ital., N. S., 1969, vol. 3, n. 4, p. 213—224.
G й n t h e r R. Untersuchungen zum Artproblem an europaischen Anuren der Gattung
Rana (Amphibia). Diss. Humboldt-Univ. Berlin, 1969.
G й n t h e r R. Der Karyotyp von Rana ridibunda Pall, und das Vorkommen von Trip-
loidie bei Rana esculenta L. (Anura, Amphibia). — Biol. Zentralbl., 1970, Bd. 89,
H. 3, S. 327—342.
G ii n t her R. Zum natiirlichen Vorkommen und zur Morphologic triploider Teichfro-
sche, «Rana esculenta», L., in der DDR (Anura, Ranidae).—Mitt. Zool. Mus.
Berlin, 1975, Bd. 51, H. 1, S. 145—158.
Giinther R. Zur Larvenmorphologie von Rana ridibunda Pall., R. lessonae Cam.
und deren Bastard R. «esculenta» L. (Anura, Ranidae). — Mitt. Zool. Mus. Berlin,
1978, Bd. 54, H. 1, S. 161 — 179.
Gurdon J. B. Tetrapioid frogs. — J. Exp. Zool., 1959, vol. 141, n. 3, p. 519—543.
Hahn A. Einige Beobachtungen an Riesenlarven von Rana esculenta. — Archiv f.
mikroskop. Anat. I. Abt. f. vergleich. u. exp. Histol. u. Entwicklungsgeschichte,
1912, Bd. 80, H. 1, S. 1—38.
Hanaoka Y. The effects of posterior hypothalectomy upon the growth and meta-
morphosis of the tadpole of Rana pipiens. — Gen. a. Comp. Endocrinol., 1967,
vol. 8, n. 3, p. 417—431.
Hertwig G., Weiss I. u. Ziemann C. Unterschiedliche Ergebnisse reziproker
Kreuzungen der drei europaischen Krotenarten unter besonderer Beriicksichtigung
ihrer Kern- und ZellgroBen. — Biol. Zentralbl., 1959, Bd. 78, H. 5, S. 675—702.
Heusser H. u. Blankenhorn H. J. Crowding-Experimente mit Kaulquappen aus
homo- und heterotypischen Kreuzungen der Phanotypen esculenta, lessonae und
ridibunda (Rana escw/en/a-Komplex, Anura, Amphibia). — Rev. Suisse Zool., 1973,
t. 80, fasc. 2, S. 543—569.
Hoskins E. R. a. H о s к i n s M. M. Growth and development of Amphibia as affec-
ted by thyroidectomy. — J. Exp. Zool., 1919, vol. 29, n. 1, p. 1—69.
Hsii Chih-yiin, Yii Nai-wen a. Liang Hsii-mu. The changes of growth
activities in pituitaries of tadpoles at different metamorphic stages. — Gen. a.
Comp. Endocrinol., 1976, vol. 30, n. 4, p. 424—431.
Humphrey R. R. Albino axolotls from an albino tiger salamander through hybridi-
zation.— J. Heredity, 1967, vol. 58, n. 3, p. 94—101.
Huxley J. S. Thyroid and temperature in cold-blooded vertebrates. — Nature, London,
1929, vol. 123, n. 3106, p. 712.
45
Ingram W. R. Studies of amphibian neoteny. II. The interrelation of thyroid and
pituitary in the metamorphosis of neotenic anurans. — J. Exp. Zool., 1929, vol. 53,
n. 3, p. 387—409.
Jurand A. Zjawisko neotenii u «Xenopus laevis» Daud. — Folia Biol., Krakow, 1955,
t. 3, n. 4, s. 315—330.
Jurand A. Futher investigations on the partial neoteny of Xenopus laevis Daud. —
Folia Biol., Krakow, 1959, vol. 7, n. 1, p. 17—22.
Kammerer P. Experimentelle Veranderung der Fortpflanzungstatigkeit bei Geburts-
helferkrote (Alytes obstetricans) und Laubfrosch (Hyla arborea).—Archiv f. Ent-
wicklungsmechan. d. Organismen, 1906, Bd. 22, H. 1—2, S. 48—140.
Kessler K. Uber einen Fall der Uberwinterung von Kaulquappen der Rana esculen-
ta L. — Bull. Soc. Imp. Natur. Moscou, 1879, t. 54, n. 2, p. 207—217.
Knau er F. K. Hemmungsbildungen bei Caudaten und Batrachiern. — Zool. Anzeiger,
1878, 1 Jahrg., N 8, S. 173—174.
Kolazy J. Ueber Kaulquappen der Batrachier. — Verh. Zool.-Botan. Ges. Wien, 1871,
Bd. 21, S. 38—40.
К о 11 m a n n J. Das Uberwintern von europaischen Frosch- und Tritonlarven und die
Umwandlung des mexikanischen Axolotl. — Verh. Naturf. Ges. Basel, 1884,
Bd. 7, S. 387—398.
Leloup J. et Buscaglia M. La triiodothyronine, hormone de la metamorphose
des Amphibiens. — C. r. Acad. Sci., Paris, 1977, sect. D, t. 284, n. 22, p. 2261—2263.
Luther A. Jattelik grodlarv.—Medd. Soc. Fauna Flora Fenn. (1916—1917), Helsing-
fors, 1917, p. 37—39, 286.
Lynn W. G. a. W a chowski H. E. The thyroid gland and its functions in cold-bloo-
ded vertebrates. — Quart. Rev. Biol., 1951, vol. 26, n. 2, p. 123—168.
P f 1 u g e г E. Das Ueberwintern der Kaulquappen der Knoblauchkrote. (Ein Beitrag
zur Lehre von der Anpassung der Organismen an die ausseren Lebensbedingungen
und zur Diagnose der Batrachierlarven). — Archiv f. gesammte Physiol., 1883,
Bd. 31, S. 134—145.
Pisano A. Variaciones somaticas en larvas de Telmatobius hauthali schreiteri de la
Cordillera Andina. — Acta Zool. Lilloana (Revista de Zool.), Tucuman, 1958,
t. 16, p. 5—12.
Remy C. et Bounhiol J. J. Croissance exageree des tetards de Crapaud accou-
cheur, entiers ou prives de leur hypophyse, et subissant un traitement par la
prolactine. — C. r. Soc. BioL, Paris, 1966a, t. 159, n. 7 (1965), p. 1532—1535.
Remy C. et Buonhiol J. J. Gigantisme experimental obtenu par intervention
chirurgicale et traitements hormonaux chez le tetard du Crapaud accoucheur. —
Annal. Endocrinol., Paris, 1966b, t. 27, n. 3 bis (suppl.), p. 377—382.
Riihmekorf E. Beitrage zur Okologie mitteleuropaischer Salientia. III. Wirkungen
des Lichtes auf Laich und Larven. — Z. Morph, u. Okol. Tiere, 1958, Bd. 47,
H. 1, S. 37—53.
Saxen L. Schilddriisenaplasie als Ursache von partieller Neotenie. — Acta Endocri-
nol., 1957, Bd. 24, S. 271—281.
Saxen L., Saxen E., Toivonen S. a. SalimakiK. The anterior pituitary and
the thyroid function during normal and abnormal development of the frog. —
Annal. Zool. Soc. Zool. Botan. Fenn. «Vanamo», Helsinki, 1957, t. 18, n. 4,
p. 1—44.
Schur r P. H. Pituitary tumours in man. — In: G. W. Harris а. В. T. Donovan (eds.),
The pituitary gland, 1966, vol. 2, London, Butterworths, p. 498—518.
Smith M. The British amphibians and reptiles. London, Collins, 1954, 2nd ed., XIV+
+ 322 p.
Snyder B. a. F г у e В. E. Physiological responses of larval and postmetamorphic
Rana pipiens to growth hormone and prolactin. — J. Exp. Zool., 1972, vol. 179,
n. 3, p. 299—313.
Snyder B. a. F г у e В. E. Effect of prolactin and growth hormone on proteins syn-
thesis in larval and adult frogs (Rana pipiens).—J. Exp. Zool., 1974, vol. 190,
n. 2, p. 167—174.
Swingle W. W. The acceleration of metamorphosis in frog larvae by thyroid feeding,
and the effects upon the alimentary tract and sex glands.—J. Exp. Zool., 1918,.
vol. 24, n. 3, p. 521—543.
Terry G. S. Effects of the extirpation of the thyroid gland upon ossification in Rana
pipiens. — J. Exp. Zool., 1918, vol. 24, n. 3, p. 567—587.
Toivonen S. Ein Fall von partieller Neotenie bei Xenopus laevis Daudin und expe-
rimentelle Untersuchungen zu seiner kausalen Erklarung. — Archiv Soc. Zool.
Botan. Fenn. «Vanamo», 1952, vol. 6, n. 2, S. 107—123.
Tunner H. G. Das SerumeiweiBbild einheimischer Wasserfrosche und der Hybridcha-
rakter von Rana esculenta. — Verh. Deutsch. Zool. Ges., 1970, 64. Jahrversamml.,
S. 352—358.
Tunner H. G. Die klonale Struktur einer Wasserfroschpopulation. — Z. zool. Syst. u.
Evolutionsforsch., 1974, Bd. 12, H. 4, S. 309—314.
Uzzell T. a. Berger L. Electrophoretic phenotypes of Rana ridibunda, Rana les-
46
sonae, and their hybridogenetic associate, Rana esculenta. — Proc. Acad. Natur..
Sci. Philadelphia, 1975, vol. 127, n. 2, p. 13—24.
Uzzell T., Gunther R. a. Berger L. Rana ridibunda and Rana esculenta:
a leaky hybridogenetic systems (Amphibia Salientia). — Proc. Acad. Natur. Sci.
Philadelphia, 1977, vol. 128, n. 9, p. 147—171.
Volpe E. P. a. Dasgupta S. Effects of different doses and combinations of spot-
ting genes in the leopard frog, Rana pipiens. — Develop. Biol., 1962, vol. 5,
n. 2, p. 264—295.
Wesenberg-Lund C. Contributions to the biology of Danish Amphibia. II. Rana
esculenta. — Int. Rev. gesamt. Hydrobiol. u. Hydrograph., 1922, Bd. 10, N 3,
p. 209—232.
Wiedersheim R. Zweijahrige Alytes-Larven. — Zool. Anzeiger, 1878, 1. Jahrg., N 5,
S. 104—105.
Wright A. H. a. W r i g h t A. A. Handbook of frogs and toads of the United States
and Canada. Ithaca, Comstock Publ. Co., 1949, 3rd ed., XIV + 640 p.
W r i g h t S. M. L., M a j e г о w s к i M. A., L u к a s S. M. a. Pike P. A. Effect
of prolactin on growth, development and epidermal cell proliferation in the hind-
limb of the Rana pipiens tadpole. Gen. a. Comp. Endocrinol., 1979, vol. 39,
n. 1, p. 53—62.
Yung E. Les larves geantes de Rana esculenta.—Archiv. Sci. phys. et natur., Geneve,
1905, 4eme per., t. 20, p. 595—597.
Z w e i f e 1 R. G. Ecology, distribution, and systematics of frogs of the Rana boylei
group. — Univ. California Publ. Zool., 1955, vol. 54, n. 4, p. 207—292.
On giant tadpoles of the green frogs of the Rana esculenta complex
L. J. В or kin, L. Berger and R. Giinther
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad), Department
of Agrobiology, Polish Academy of Sciences (Poznan) and Museum fCir Naturkunde
der Humboldt-Universitat (Berlin)
Occasional giant tadpoles (100 millimeters and more) of the green frogs of the Rana
esculenta complex both collected in various regions of the USSR (Latvia, Kazakhstan,
Uzbekistan) and in Poland, and obtained in crossing experiments are recorded. All
known records of natural giant tadpoles are mapped. Various factors, probably, caused
such a gigantism are analysed (geographic variability, quality of food and crowding
effect, polyploidy, hybridization and heterosis, light, temperature, wintering, hormonal
control). The main cause of occasional occurence of giant tadpoles of the green frogs
seems to be a disturbance of hormonal system (hypothalamus — pituitary — thyroid axis}
evoked by genetical factors (mutations etc.).
О СЕВЕРНОЙ ГРАНИЦЕ АРЕАЛА ОБЫКНОВЕННОЙ ГАДЮКИ,
VIPERA BERUS (L.) В ЯКУТИИ
Л. Я. Боркин и Ф. Н. Кириллов
Зоологический институт АН СССР (Ленинград)
и Институт биологии Якутского филиала Сибирского отделения АН СССР (Якутск)
Северная граница ареала обыкновенной гадюки, Vipera berus (L.), заходя в евро-
пейской части СССР за Полярный круг, к востоку резко снижается в более южные
широты. Считается (Ларионов, 1958, 1961), что в Якутии этот вид по долине реки
Лена доходит примерно до 61° северной широты (поселок Кытыл-Дюра на левом
берегу и примерно 30-й километр южнее поселка Тит-Ары на правом берегу Лены).
Эти же пункты указаны в последней сводке по герпетофауне СССР (Банников и др.,
1977, карта 135 на с. 389). П. Д. Ларионов (1958, 1961, 1977) справедливо полагает,
что распространение гадюки к северу определяется наличием благоприятных мест
зимовок в виде талых участков среди вечномерзлого грунта.
Мы располагаем данными о находке обыкновенной гадюки в Якутии, лежащей
в значительном удалении от выше указанных точек. Один экземпляр (самец) гадюки
был пойман 15.IX.1976 около северной оконечности Вилюйского водохранилища, не-
далеко от границы с Красноярским краем. Эта наиболее северная точка находок дан-
ного вида в Якутии имеет примерные координаты 63° 30' северной широты и 110°
восточной долготы и удалена от ближайшей находки (район Ленска — Банников и др.,
1977, карта 135 на с. 389) примерно на 400 км к северо-западу. Таким образом,
сообщаемая нами находка, переданная в коллекции Зоологического института АН
СССР, Ленинград (ЗИН. 19274), значительно изменяет представление о северной гра-
нице ареала обыкновенной гадюки в Восточной Сибири.
Следует также отметить, что находка этого вида в районе поселка Северо-
Енисейск Красноярского края (несколько севернее 60°), сопровождаемая на карте
135 знаком вопроса (Банников и др., 1977, с. 389), является вполне достоверной и
подтверждается экземплярами, собранными здесь А. А. Костиным (ЗИН.18310, 18311
и 19390).
ЛИТЕРАТУРА
Банников А. Г Даревский И. С., Ищенко В. Г., Рустамов А. К. и
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР
М., «Просвещение», 1977, 414 с.
Ларионов П. Д. Некоторые данные о северной границе распространения змей
по долине реки Лены. — Учен. зап. Якутск, гос. ун-та, 1958, вып. 4, с. 157—159.
Ларионов П. Д. Об условиях зимовки гадюк на северной границе их распро-
странения в долине Лены. — Зоол. ж., 1961, т. 40, вып. 2, с. 289—290.
Ларионов П. Д. К экологии обыкновенной гадюки (Vipera berus) в Якутии.—
Зоол. ж., 1977, т. 56, вып. 6, с. 919—923.
On northern range limit of the adder, Vipera berus (L.) in Yakutia
L. J. Borkin and F N. Kirillov
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad)
and Institute of Biology, Yakutian Department of Siberian Branch
of Academy of Sciences, U.S.S.R. (Yakutsk)
Northernmost record (63° 30' N and 110°E) of the adder, Vipera berus (L.) in
Eastern Siberia is reported.
НОВЫЙ ДЛЯ ФАУНЫ СССР РОД
и вид сцинковои ЯЩЕРИЦЫ
CHALCIDES OCELLATUS (FORSKAL)
И. С. Даревский
Зоологический институт АН СССР (Ленинград)
Глазчатый халцид, Chalcides ocellatus (Forskal) —один из широко
распространенных палеарктических видов сцинковых ящериц, ареал
которого простирается от южной Европы и северной и северо-восточ-
ной Африки на западе до южного Ирана и Пакистана на востоке.
В восточной части этого ареала немногочисленные известные местона-
хождения халцида, судя по литературным данным, расположены
в узкой прибрежной полосе на юге Ирана и Пакистана (Werner, 1917;
Schmidt, 1955;'Minton, 1966; Mertens, 1969; Tuck, 1971; Anderson, 1974),
причем в Ираке (Khalaf, 1959) и Афганистане (Leviton a. Anderson,
1970) этот вид до настоящего времени не обнаружен. Достаточно не-
ожиданным выглядит поэтому нахождение халцида в пределах СССР
на юге Туркмении в значительном отрыве от установленной ранее се-
верной границы его ареала. Происходящий отсюда взрослый экземпляр
был добыт с целью изучения хромосом В. В. Соколовским 22 мая
1971 года в окрестностях селения Акар-Чешме на юго-западе Бадхыза
и провизорно отнесен к виду Mabuya aurata. Под этим названием яще-
рица некоторое время хранилась в коллекции Биолого-почвенного ин-
ститута Дальневосточного научного центра АН СССР во Владивостоке,
а позднее вместе с другими герпетологическими коллекциями посту-
пила в герпетологическое отделение Зоологического института АН
СССР, где числится сейчас под инвентарным № 19332 (вклейка,
рис. VII).
Некоторое габитуальное сходство глазчатого халцида с предста-
вителями близкого рода Mabuya позволяло предполагать, что в кол-
лекционных материалах из южной Туркмении могут находиться и дру-
гие ошибочно определенные особи этого вида. Действительно, в сборах
Н. А. Зарудного нами обнаружен молодой экземпляр глазчатого хал-
цида, добытый в 1892 году близ русско-иранской границы в долине
реки Мерген-Улья. Маршрут этой поездки опубликован не был, однако
судя по некоторым географическим привязкам, Мерген-Улья протекает
в южных предгорьях Копет-Дага севернее советско-иранской границы,
примерно на долготе поселка Гермаб в Туркмении. Таким образом, это
второе местонахождение халцида также находится в пределах СССР.
В литературе последний экземпляр (ЗИН № 8472) ошибочно приводится
А. М. Никольским (1897, 1915) и самим Н. А. Зарудным (1903) под
названием Mabuya septemtaeniata (=Mabuya aurata septemtaeniata
Reuss). Отметим, что указываемый этими авторами другой экземпляр
(ЗИН № 8471), добытый Н. А. Зарудным в 1892 году также в долине реки
Мерген-Улья, действительно, относится к виду Mabuya aurata. Оба при-
веденные выше местонахождения халцидов в Туркмении расположены
на расстоянии около 400 км по прямой друг от друга и лежат более
чем на 1100 км севернее ближайших находок на юге Ирана (см. рис.).
По устному сообщению Ю. К. Горелова, по всей видимости, взрослые
особи глазчатого халцида наблюдались им также близ Разнатурского
перевала на хребте Гяз-Гядык и у родника Нардеванлы в северо-за-
падной части Бадхыза.
Вопрос о подвидовой принадлежности наших экземпляров остается
открытым. Согласно Р. Мертенсу (Mertens, 1969), халциды из южных
районов Пакистана и Ирана относятся к номинативному подвиду Chai-
4 Зак. 719
49
cides ocellatus ocellatus (Forskal), распространенному также в северной
Африке, Греции, некоторых островах Эгейского и Средиземного морей,,
южной Турции и юго-западной Азии. При этом Р. Мертенс особо обра-
щает внимание на тот факт, что если на западе ареала данный вид
образует несколько подвидовых форм, то на востоке он не обнаружи-
Основные известные местонахождения Chalcides ocellatus в юго-западной Азии и на
юге Туркмении.
1 — Акар-Чешме; 2 — Мерген-Улья.
вает сколько-нибудь выраженной географической изменчивости. Как
показывает сделанное нами сравнение, особи из Туркмении характери-
зуются несколько большим числом чешуй вокруг середины туловища,
и не исключено, что они образуют здесь самостоятельную подвидовую
форму (см. табл.). Окончательно этот вопрос может быть решен лишь
при получении нового коллекционного материала из Туркмении и при-
лежащих районов Ирана.
Характеристика некоторых морфологических признаков Chalcides ocellatus
из различных районов восточной части ареала
Район ареала Длина туловища с головой Число чешуй вокруг середины тела Источники
Южная Туркмения 120 31 ЗИН № 19332
(окрестности селения Акар-Чешме) Южная Туркмения (реч- 58 (молодой) 32 ЗИН № 8472
ка Мерген-Улья) Южный Иран (Хордж) 71—86 28 Зоомузей МГУ, № 2200
Южный Пакистан 108 28—29 Minton, 1966
Южная Турция 26—31 Baran, 1977
Оман 90—97 28 Haas a. Werner, 1969
Оман 112 28—30 Werner, 1911
50
Нужно подчеркнуть, что районы южной Туркмении, включая пло-
скогорье Бадхыз, в герпетогеографическом отношении представляют
особый интерес, так как здесь проходит крайняя северная граница рас-
пространения ряда видов пресмыкающихся, основные ареалы которых
лежат значительно южнее в пределах юго-западной Азии. Местонахож-
дения некоторых из этих видов носят в Туркмении ярко выраженный
реликтовый характер и в настоящее время далеко оторваны от ближай-
ших находок на юге Ирана. К числу их относятся, например, иранская
кошачья змея (Telescopus rhynopotna), а из ящериц — туркменский
эублефар (Eublepharis turkmenicus), дивергировавший здесь до уровня
самостоятельного вида. Яркий пример такого реликтового разрыва
ареала, как было показано выше, демонстрирует своим распростране-
нием и глазчатый халцид. Ранее мною было высказано предположение,
что участившиеся в последние годы случаи обнаружения на юге Турк-
мении новых для нашей фауны видов ящериц и змей являются след-
ствием колебания численности или, что более вероятно, расширением
их ареалов к северу (Даревский, 1977). Если эта точка зрения верна,
то в ближайшее время мы можем ожидать на юге Туркмении новых
находок также и глазчатого халцида.
ЛИТЕРАТУРА
Даревский И. С. Расширение ареалов или колебание численности (О нахожде-
нии на юге Средней Азии ряда новых для фауны СССР видов пресмыкаю-
щихся).— В кн.: Вопросы герпетологии. Автореф. докл. IV Всесоюзн. герпетол.
конф., Л., «Наука», 1977, с. 77—79.
Зарудный Н. А. О чешуйчатых и голых гадах восточной Персии. — Зап. Русск.
геогр. о-ва, 1903, т. 36, № 3, с. 1—38.
Никольский А. М. Пресмыкающиеся, амфибии и рыбы, собранные Н. А* Заруд-
ным в восточной Персии. — Ежегодн. Зоол. муз. Акад. Наук, 1897, т. 2,
с. 306—348.
Никольский А. М. Пресмыкающиеся, т. 1. — В серии: Фауна России и сопре-
дельных стран, Петроград, 1915, 352 с.
Anderson S. С. Preliminary key to the turtles, lizards, and amphisbaenians of
Iran. — Fieldiana: Zoology, 1974, vol. 65, n. 4, p. 27—44.
Baran I. Turkiye’ de Scincidae Ramilyasi Turlerinin taksonomisi. — Doga, 1977,
n. 1, p. 217—223.
Haas G. a. Werner Y. L. Lizards and snakes from southwestern Asia, collected
by Henry Field. — Bull. Mus. Comp. Zool., 1969, vol. 138, n. 17, p. 327—405.
К h a 1 a f К. T. Reptiles of Iraq with some notes on the amphibians. Baghdad, Ar-Ra-
bitta, 1959, 96 p.
Leviton A. E. a. Anderson S. C. The amphibians and reptiles of Afghanistan,
a checklist and key to the herpetofauna. — Proc. California Acad. Sci., 1970,
vol. 38, n. 10, p. 163—206.
Mertens R. Die Amphibien und Reptilien West-Pakistans. — Stuttgart. Beitr. Naturk.r
1969, N. 197, 96 S.
Minton S. A. A contribution to the herpetology of West-Pakistan. — Bull. Amer. Mus.
Nat. Hist, 1966, vol. 134, p. 27—184.
Schmidt К. P. Amphibians and reptiles from Iran. — Videskabelige Meddel. Dansl
Naturhist. Forening, 1955, vol. 117, p. 193—207.
Tuck R. G. Amphibians and reptiles from Iran in the United States National Museum
collection. — Bull. Maryland Herpetol. Soc., 1971, vol. 7, n. 3, p. 48—86.
Werner F. Reptilien aus Persien (Provinz Fars). — Verh. Zool. Botan. Ges. Wien,
1917, Bd. 67, S. 191—200.
A first record of the ocellated skink, Chalcides ocellatus (Forskal) in the fauna
of the USSR
7. S. Darevsky
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad)
The ocellated skink (Chalcides ocellatus) was for the first time recorded in the
USSR in southern Turkmenistan, in two localities separated by more than 1000 km from
the earlier known northern range limit in south Iran and Pakistan. In their morpholo-
gical features specimens from Turkmenistan are somewhat different from ones from
other range regions. The recognition of subspecies identity is need of additional mate-
rials.
4*
НОВЫЕ ДАННЫЕ О РАСПРОСТРАНЕНИИ
НЕКОТОРЫХ АМФИБИЙ И РЕПТИЛИЙ
МОНГОЛЬСКОЙ НАРОДНОЙ РЕСПУБЛИКИ
X. Монхбаяр
Монгольский государственный педагогический институт (Улан-Батор)
За 20 лет, прошедших после опубликования первой сводки по гер-
петофауне Монгольской Народной Республики (Банников, 1958), нако-
пились новые сведения по географическому распространению амфибий
и рептилий этой страны (Эрэгдэндагва, 1958, 1961; Obst, 1962, 1963;
Дементьев и др., 1966; Монхбаяр, 1967, 1973, 1976а, б, 1977; Шагдар-
сурэн и Монхбаяр, 1968; Peters, 1971а, b; Хотолхуу, 1971). В данной
работе приводятся материалы, уточняющие распространение, в основ-
ном, редких и мало изученных видов герпетофауны МНР, собранные
автором в последнее десятилетие и частично опубликованные на мон-
гольском языке.
Сибирский углозуб, Hynobius keyserlingii (Dybowski)
В пределах МНР известен с восточного берега озера Хубсугул и
на севере страны. В последние годы был найден (рис. 1) также в окрест-
ностях поселка Баян-Дзурх около Улан-Батора (Obst, 1962), в Дархат-
ской котловине (Монхбаяр, 1967) и на реке Богдын-Гол в Хэнтее (Хо-
толхуу, 1971).
Зеленая жаба, Bufo viridis Laurenti
Г. Петерс (Peters, 1971а) обнаружил этот вид в урочище Улястай
и на реке Уенч-Гол. Кроме того, Э. Хухэндэй и О. Доржра в 1967—
1971 годах поймали особей зеленой жабы в ряде других пунктов за-
падной Монголии: Ушигин-Ус, Ихэр-Толь, Шара-Булак, Увдэк, Хуйтэн-
Булак, Баян-Мод (рис. 1). Эти сборы хранятся в коллекциях Монголь-
ского государственного педагогического института (МГПИ) в Улан-
Баторе.
Дальневосточная лягушка, Rana chensinensis David
Этот новый для фауны Монголии вид амфибий был найден нами
в восточной части страны и описан в качестве Rana sp. (Монхбаяр,
1973). Определение вида впоследствии было уточнено сотрудником
Зоологического института АН СССР (ЗИН АН СССР) Л. Я. Борки-
ным. Более подробно данные о распространении и некоторых особен-
ностях морфологии и экологии этой лягушки будут опубликованы нами
отдельно совместно с Л. Я. Боркиным.
Сцинковый геккон Пржевальского,
Teratoscincus przewalskii Strauch
Первое сведение о нахождении этого вида на территории МНР
опубликовал Д. Эрэгдэндагва (1961) под названием Teratoscincus sp.
Ф. Ю. Обет (Obst, 1962, 1963) в сопровождении автора совершил
поездку по северной и южной Монголии и собрал несколько особей
сцинкового геккона Пржевальского в Галбын-Гоби. Кроме того, в 1976—
1979 годах этот вид был найден нами в ряде мест Заалтайской Гоби
(оазис Дзулганай, Ногон-Цав, Эхин-Гол, Шара-Хулс, Ингэни-Хуврын-
Булак) и в Алтай-сомоне Гоби-Алтайского аймака (рис. 2).
52
Рис. 1. Точки находок амфибий и рептилий в Монголии.
Hynobius keyserlingii (А): 1 — Баян-Дзурх 2 — Богдын-Гол, 3 — Дархатская котловина; Bufo viri-
dis (Б): 1 — Ушигин-Ус, 2—6 — Ихэр-Толь, Шара-Булак, Увдэг, Хуйтэн-Булак, Баян-Мод; Alsophylax
pipiens (В): 1 — Тэвш-Хайрхан, 2 — Шовон-Хувэр, 3 — Дзолон-Ула, 4 — Дзурамтай, 5 — Хайчин-Ула,
6 — Эхин-Гол, 7 — Шара-Хулс. 8 — Яман-Ус, 9— Ногон-Цав, 10 — Улдзийт-Гол, 11 — Дзун-Мод,
12 — Сомон-Хайрхан-Ула, 13 — Хатан-Хайрхан-Уда
Рис. 2. Точки находок рептилий в Монголии.
Teratoscincus przewalskii (А): 1 — Галбын-Гоби, 2 — Бордзон-Гоби, 3 — Дзулганай, 4 — Эхин-Гол,
5 — Ногон-Цав, 6 — Ингэни-Хубрын-Булак; 7 — Шара-Хулс, 8 — Алтай-сомон; Eremias argus (Б):
1 — Джаргалант, 2 — Хишиг-Ундэр, 3 — Барун-Сайхан, 4 — Атас-Ула; Eryx tataricus (В): / — Борд-
зон-Гоби, 2 — Толь-Булак, 3 — Эхин-Гол, 4 — Ногон-Цав.
Пискливый геккончик, Alsophylax pipiens (Pallas)
По литературным данным (Банников, 1958), этот вид был известен
в ряде пунктов Северной Гоби: Барун-Баян, Арц-Богд, Хурх, Нэмэгт.
В 1969—1978 годах найден нами в Северной и Заалтайской Гоби (Тэвш-
Хайрхан, Шовон-Хувэр, Дзолон-Ула, Ногон-Цав, Яман-Ус, Улдзийт-Гол,
Дзун-Мод, Шара-Хулс, Хатан-Хайрхан-Ула, Сомон-Хайрхан-Ула, Эхин-
гол, Дзурамтай (рис. 1).
Гобийский голопалый геккон, Gymnodactylus elongatus Blanford
Впервые в пределах МНР этот вид был обнаружен нами (Монх-
баяр, 19766, 1977) в Заалтайской Гоби в Ногон-Цав. Кроме того,
в 1977—1978 годах найден нами в Цувра-Хара-Ула, Хуцын-Хара-Ула,
Шара-Хулсны-Булак и Цугалбар-Ус (рис. 3). Экземпляры, собранные
в этих местах, переданы в коллекции МГПИ (20 особей) и ЗИН АН
СССР (3 экземпляра).
Монгольская ящурка, Eremias argus Peters
Распространена в МНР в степной, полупустынной и лесостепной
зонах. Самые западные и юго-западные точки находок образуют озеро
Орок-Нур, среднее течение реки Туйн-Гол и Ноен-Богд (Щербак, 1974).
В последние годы этот вид найден также в следующих пунктах страны:
Джаргалант, Барун-Сайхан, Хишиг-Ундэр, местность севернее Атас-Ула
(рис. 2). Последняя находка (2 экземпляра, МГПИ, сборы О. Доржра)
оказалась очень интересной, так как раньше этот вид не был найден
так далеко в пустыне.
Восточная разноцветная ящурка, Eremias arguta potanini (Bedriaga)
Этот вид впервые был найден в пределах МНР в 1966 году в Хонин-
Усны-Гоби (Шагдарсурэн и Монхбаяр, 1968). В ЗИН АН СССР и Зоо-
логическом музее Берлинского университета хранятся особи, пойман-
ные близ озера Хиргис-Нур и Уенч-сомона в Джунгарской Гоби (Щер-
бак, 1974).
Центральноазиатская, или пестрая ящурка,
Eremias vermiculata Blanford
Этот до недавних пор считавшийся редким вид раньше был изве-
стен в МНР только из двух мест: около границы с Алашанем и в 150—
160 км к югу от хребта Дэлгэр-Хангай (Банников, 1958). В 1976—1978
годах он был найден нами в ряде новых пунктов Заалтайской Гоби:
севернее горы Атас-Ула, Хайчин-Ула, Хэрмэн-Цав, Дзулганай, Ногон-
Цав, Толь-Булак, Эхин-Гол, Улдзийт-Гол, Хуцын-Шанд (рис. 4). Цен-
тральноазиатская ящурка обычно встречается в закрепленных мало-
мощных песках около оазисов.
Живородящая ящерица, Lacerta vivipara Jacquin
Это — единственный вид ящериц рода Lacerta в Монголии. По ли-
тературным данным (Банников, 1958), живородящая ящерица обитает
только в самых северных частях страны: в районе озера Хубсугул и
в Хэнтэе. В 1962 году нам удалось найти этот вид в урочище Бага-
Бдян, Хандгайт, примерно в 15—25 км к северу от Улан-Батора. Кроме
того, С. Дуламцэрэн любезно передал нам 3 экземпляра живородящей
ящерицы, пойманные им в июле 1978 года в Монгольском Алтае. Они
были обнаружены около верхней границы леса на высоте 2900 м над
уровнем моря южнее озера Хотон-Нур (рис. 4).
54
Рис. 3. Точки находок рептилий в Монголии.
Gymnodactylus elongatus (А): 1 — Ногон-Цав, 2 — Цувра-Хара-Ула, 3 — Хуцын-Хара-Ула, 4 — Шара-
Хулсны-Булак, 5 — Цугалбар-Ус; Eremias ar gut а (Б): Хонин-Усны-Гоби: Coluber spinalis (В):
1 — местность южнее Увэр-Хангайского аймака, 2 — Сулан-Хэрэ.
Рис. 4. Точки находок рептилий в Монголии.
Eremias vermiculaia (А): / — местность севернее Атас-Ула, 2 — Хайчин-Ула, 3— Хэрмэн-Цав,
4 — Дзулганай, 5 — Ногон-Цав, 6 — Толь-Булак, 7 —Эхин-Гол, 8 — Улдзийт-Гол, 9 — Хуцын-Шанд;
Lacerta vivipara (Б): / — местность южнее Хотон-Нур, 2 — Хандгайт, 3 — Бага-Баян.
Восточный удавчик, Eryx tataricus Lichthenstein
Редкая змея в Монголии. Нами найдена в ряде пунктов Заалтай-
ской Гоби: Бордзон-Гоби, Толь-Булак, Ногон-Цав, Эхин-Гол (рис. 2).
Полосатый полоз, Coluber spinalis (Peters)
Широко распространенная, но очень редкая змея Монголии. Нами
найдена в местности южнее Увэр-Хангайского аймака и в Сулан-Хэрэ
в Восточной Гоби (рис. 3).
В заключение я хочу выразить глубокую благодарность профес-
сору И. С. Даревскому, предоставившему мне возможность ознако-
миться с центральноазиатскими коллекциями амфибий и рептилий, хра-
нящимися в ЗИН АН СССР, а также любезно просмотревшему пра-
вильность моего определения ящериц МНР. Я также благодарен
Л. Я. Боркину за редактирование текста статьи.
ЛИТЕРАТУРА
Банников А. Г. Материалы по фауне и биологии амфибий и рептилий Монго-
лии. — Бюлл. Московск. о-ва испыт. прир., 1958, отд. биол., т. 63, вып. 2,
с. 71—91.
Дементьев Г. А., Шагдарсурэн О. и Болд А. Зоогеографический обзор
монгольской пустыни Гоби. — В кн.: Материалы к фауне Монголии. Учен. зап.
Монгольск. гос. ун-та, Улан-Батор, 1966, т. 9, № 2 (18), с. 16—26.
Монхбаяр X. Сибирский углозуб Hynobius keyserlingi (Dybowsky et God lewsky
1870). — Известия АН МНР, Улан-Батор, 1967, № 2, с. 26—30 (на моиг. языке).
Монхбаяр X. Земноводные и пресмыкающиеся Монгольской Народной Республики.
Автореф. канд. дисс., Ташкент, 1973, 38 с.
Монхбаяр X. Земноводные и пресмыкающиеся МНР. Улан-Батор, 1976а, 167 с.
(на монг. языке).
Монхбаяр X. Ящерицы пустыни Гоби. — Наука и жизнь, Улан-Батор, 19766,
№ 6, с. 88—92 (на монг. языке).
Монхбаяр X. Гобийский голопалый геккон, Gymnodactylus elongatus Blanford —
новый для Монголии вид ящериц.— В кн.: Герпетологический сборник (Тр. Зоол.
ин-та АН СССР, т. 74), Л., 1977, с. 73—74.
Хотолхуу Н. Новое местонахождение сибирского углозуба. — Известия АН МНР,
Улан-Батор, 1969, № 2, с. 99—102 (на монг. языке).
Шагдарсурэн О. и Монхбаяр X. Eremias arguta Pall. — новый для фауны
Монголии вид ящериц. — Studia museologia. Улан-Батор, 1968, № 1, с. 125—129
(на монг. языке).
Щербак Н. Н. Яшурки Палеарктики. Киев, «Наукова думка», 1974, 296 с.
Эрэгдэндагва Д. Распространение агамы в Монголии. — Наука и техника, Улан-
Батор, 1958, № 1(53), с. 25 (на монг. языке).
Эрэгдэндагва Д. Находка нового вида ящериц в нашей стране. — Известия
АН МНР, Улан-Батор, 1961, № 3, с. 66 (на монг. языке).
Obst F. J. Eine herpetologische Sammelreise nach der Mongolei. — Aquarien Terrarien,
1962, 9. Jhrg., H. 11, S. 333—342.
Obst F. J. Amphibien und Reptilien aus der Mongolei. — Mitt. Zool. Mus. Berlin,
1963, Bd. 39, H. 2, S. 361—370.
Peters G. Materialien zur Okologie und Verbreitung der Amphibien in der Mongo-
lei. — Mitt. Zool. Mus. Berlin, 1971a, Bd. 47, H. 2, S. 315—348.
Peters G. Die Wirtelschwanze Zentralasiens (Agamidae: Agama). — Mitt. ZooL
Mus. Berlin, 1971b, Bd. 47, H. 2, S. 357—381.
New data on the distribution of some amphibians and reptiles in Mongolia
Kh. Monkhbayar
Mongolian State Pedagogical Institute (Ulan-Bator)
New records of 3 species of amphibians (Hynobius keyserlingii, Bufo viridis, Rana
chensinensis) and 9 species of reptiles (Teratoscincus przewalskii, Alsophylax pipiens,
Gymnodactylus elongatus, Eremias argus, Eremias arguta potanini, Eremias vermiculata,
Lacerta vivipara, Eryx tataricus, Coluber spinalis) in Mongolia over the last 20 years
have been reported.
ХВОСТАТЫЕ И БЕСХВОСТЫЕ ЗЕМНОВОДНЫЕ
МЕЛА КЫЗЫЛКУМОВ
Л. А. Несов
Ленинградский государственный университет
Остатки наземных позвоночных мела нашей страны происходят
более чем из 60 крупных и мелких местонахождений, расположенных,
в основном, на территории Узбекистана, Таджикистана, Киргизии, юж-
ного Казахстана и северной Туркмении. Подавляющее большинство
остатков тетрапод из этих местонахождений принадлежит обычно трем
группам рептилий — черепахам, динозаврам, крокодилам. Кости назем-
ных ящериц мела, лишь недавно обнаруженные в нашей стране (Несов,
1978, 1979, 1980), очень редки, а птицы (Шилин, 1974, 1977) и млеко-
питающие мелового периода известны (Бажанов, 1972; Несов, 1979;
Несов и Трофимов, 1979) только по единичным находкам. Остатки зем-
новодных мела на территории СССР ранее не были известны.
Осенью 1975 и в летние сезоны 1977—80 годов в Кызылкумах
автору при визуальных поисках костного материала на развеваемых
поверхностях коренных пород позднемелового возраста удавалось нахо-
дить локальные скопления остатков позвоночных, содержащие позвонки,
кости черепов и конечностей амфибий. Основная масса материала была
получена в результате последующего просеивания значительных объе-
мов костеносных пород на ситах с отверстиями 2 мм. Сортировке под-
верглись более 100 000 извлеченных мелких костей, зубов и их фрагмен-
тов, принадлежавших различным позвоночным. Это позволило обнару-
жить тысячи костей различных хвостатых земноводных (Caudata), ко-
торые являются первыми находками остатков меловых представителей
этого отряда в нашей стране. Более редкими в материале оказались
кости бесхвостых амфибий (Salientia), впервые обнаруженные в мезо-
зое СССР и поэтому являющиеся древнейшими известными у нас на-
ходками бесхвостых. Краткие предварительные сведения о результатах
поисков были сообщены ранее (Несов, 1979). Все изображения остат-
ков, упоминаемых в работе, приведены на вклейках (рис. VIII—XII).
Выраженная латеральная уплощенность многих найденных туло-
вищных позвонков хвостатых земноводных, их амфицельность, относи-
тельно сильнбе развитие узкого медиального «киля» снизу, некоторое
смещение углубления для хорды в верхнюю часть тела, а также ряд
особенностей строения нижних челюстей и зубов позволили определить
(Несов, 1979) принадлежность большей части остатков (рис. VIII,
2—6, 8, 12—15} IX, 1—20} X, 1—7} XI, 20—26} XII, 1—7) двум видам,
отмечаемым для 27 точек сбора костей и относящимся к семейству
Scapherpetontidae Auffenberg et Goin, 1959. Представители скаферпе-
тонтид, ранее известные только для конца позднего мела (кампан,
Маастрихт) и палеогена Северной Америки, как было показано (Estes,
1969е; Edwards, 1976), обнаруживают близость к некоторым современ-
ным североамериканским уклоняющимся Ambystomatoidea Noble, 1931
и причисляются к указанному надсемейству, в частности, по особенно-
стям взаиморасположения спинномозговых нервов и дуг позвонков и
в связи с развитием gonio-angulare в нижней челюсти. Атласы, встре-
чающиеся совместно с туловищными позвонками скаферпетонтид и от-
несенные по этой причине к тем же видам, имеют необычно большой
нижний отросток, или гипапофиз (рис. VIII, 5, 8} IX, 16, 20} X, 3).
Сильное развитие этого отростка на атласе однако не отмечалось
среди Scapherpetontidae. Такая ранняя черта уклонения от состояния,
обычного для Caudata (см. Teege, 1958) и остальных скаферпетонтид,
т. е. апоморфная черта, у очень ранних представителей семейства, сви-
57
детельствует, что животные принадлежали особой филогенетической
ветви, которую следует противопоставлять уже известным, существо-
вавшим в более позднее время членам семейства Scapherpetontidae.
Оба рода этой ветви в строении неба имеют черты определенного
сходства с саламандроидеями. В Кызылкумах найдены остатки и более
мелкого хвостатого земноводного, имевшего протяженную меккелеву
борозду (рис. XI, 10, И, 14—19). Наиболее высокая часть зубной кости
располагается сразу позади зубного ряда. Позвонки этой амфибии
амфицельные с округлыми сочленовными поверхностями тел, относи-
тельно уплощенной сверху невральной дугой, тонкими носителями ребер
(рис. XI, 15—18), иногда с передними базапофизами. Передние фасетки
атласа округлые, межсуставной бугорок широкий, слабо выступающий
вперед (рис. XI, 14). Все эти особенности позволяют сближать амфи-
бию с Palaeoproteus Herre, 1935 и относить ее (Несов, 1979), следуя
Б. Нейлору (Naylor, 1978) и Р. Эстесу (Estes, 1975) к семейству Batra-
chosauroididae Auffenberg, 1958 и подотряду Proteida Соре, 1866. Неко-
торые из найденных зубных костей Caudata (рис. VIII, 1) имеют зубы,
увеличивающиеся в размерах на участке, лежащем несколько позади
от симфиза, и очень короткую борозду для меккелева хряща, что отме-
чается (Estes, 1964) для другого семейства — Prosirenidae Estes, 1969,
ранее известного из мела Северной Америки и Ближнего Востока и
миоцена Европы.
В материале представлены скульптированные кости черепов бес-
хвостых амфибий (рис. X, /9; XII, 10—12, 14). Рельеф и форма покров-
ных костей черепа, в том числе лобнотеменной кости, а также особен-
ности положения заднего края окна в эндокрании позволяют отнести
эти и некоторые сопутствующие остатки к Pelobatidae Stannius, 1856 и
сопоставить их с известными формами рода Eopelobates Parker, 1929.
В ряде локальных верхнетуронских и верхнетурон-сантонских скопле-
ний костей вместе с остатками Eopelobates найдены амфицельные туло-
вищные позвонки (рис. XII, 18, 19), прободенные каналом для хорды,
их невральная дуга налегает на таковую последующего позвонка. Об-
наружены атласы (близкие к типу II по Trueb, 1973, fig. 2—10, но
с более узкими передними сочленовными поверхностями) с небольшим,
расщепленным посредине межсуставным бугорком (ЦГМ № 205—207/
/11657). Найдены и крестцовые позвонки со слабо расширенными по-
перечными отростками, спереди вогнутые, а сзади с выпуклой двойной
сочленовной поверхностью (рис. XII, 20), а также длинные, тонкие
уростили с парой скошенных назад поперечных отростков и с двойной
передней сочленовной поверхностью (рис. XII, 22; ЦГМ № 67—69/
/11657). Сходные уростили найдены и в сеномане Шейх-Джейли (см.
ниже). По формальным признакам эти остатки могут быть отнесены
к распространенному в Австралийской области семейству Myobatrachi-
dae Schlegel, 1850 (Lynch, 1973; Heyer a. Liem, 1976), возможно, пред-
ставленному (Indobatrachus) и в палеогене Индии (см. Noble, 1930;
Savage, 1973). Таким членом миобатрахид может быть одна из бес-
хвостых амфибий пока неясного систематического положения (рис. X,
20—24, 26; XI, 1—3; XII, 21). Однако учитывая случаи неприрастания
межпозвоночных дисков к позвонкам даже у некоторых современных
Pelobatidae (Trueb, 1973) (обычно имеющих, однако, одинарную сочле-
новную поверхность уростиля и не сохраняющих во взрослом состоянии
канал для хорды в позвонках), можно ожидать принадлежности ука-
занных остатков и архаичному представителю Pelobatidae, например,
отмечаемому в материале Eopelobates (рис. X, 19, 25; XII, 9—15, 23).
Систематическую принадлежность ряда костей хвостатых и бесхвостых
амфибий уточнить не удается.
Находки остатков позднемезозойских земноводных редки (см. при-
ложение) и относительно мало известны. Однако, видимо, именно в до-
58
кайнозойское время происходила дивергенция как хвостатых, так и
бесхвостых амфибий на основные семейства, часть которых представ-
лена и ныне (Estes, 1965b, 1970b; Romer, 1966; Savage, 1973). В то же
время выяснение былого хода эволюции (точнее, конкретных этапов
на ее пути) в пределах указанных отрядов на основании только лишь
данных о современных формах бывает затруднено, в частности из-за
достаточно широкого распространения параллельных и конвергентных
преобразований в разных филогенетических ветвях (см. Noble, 1922;
Griffiths, 1963; Trueb, 1973). Важность находок, а также разнообразие
форм, представленных в коллекции, требуют описания многих из най-
денных костей, в то время как своеобразие изучаемых амфибий, рас-
члененность костных остатков и не всегда хорошая их сохранность
-создают трудности в их изучении.
В Юго-Западных Кызылкумах кости как хвостатых, так и бесхво-
стых амфибий были найдены в верхней части разреза ходжакульской
свиты (сеноман) близ возвышенности Шейх-Джейли. Незначительный
материал по хвостатым земноводным сеномана был собран нами и
в верхней части разреза той же свиты в Ходжакульсае. Остатки Cau-
data и Salientia обнаружены и в верхней части обрыва, расположен-
ного на северо-восток от озера Ходжакуль, а также на юго-западном
склоне горы Челпык. Отложения, вмещающие здесь остатки, относятся
уже к нижней части бештюбинской свиты и датируются ранним туро-
ном (Шульц, 1972).
В Центральных Кызылкумах кости хвостатых и бесхвостых земно-
водных найдены на уровне нижней части тайкаршинской пачки, прибли-
зительно на середине линии северо-западного склона системы обрывов
Джара-Кудука ( = Биссекты, =Кульбеке), в крупном овраге, называемом
нами Айдахарсай, в верхней части упомянутой пачки на востоке
той же системы (близ оврага Темирагачсай с остатками найденных
нами древних лесов), а также в средней и верхней частях тайкаршин-
ской пачки Джара-Кудука севернее колодца Биссекты (овраг Тасба-
лыксай и другие). Указанная пачка относится к верхнему турону (Пят-
ков и др., 1967), коньяк — сантону (Сочава, 1968) или верхнему
турону — сантону (Мартинсон, 1969). Отмеченное выше совместное
•обнаружение в локальных местонахождениях остатков мезозойских
представителей как хвостатых, так и бесхвостых амфибий является
очень редким (Hecht a. Estes, 1960; Estes, 1964; Nevo, 1968, Nevo a.
Estes, 1969; Broin et al., 1974).
Азиатские хвостатые земноводные из Scapherpetontidae древнее
других известных, их черты сходства с криптобранхоидеями симплезио-
морфны, а с саламандроидеями — появились, возможно, за счет упроч-
нения неба при склерофагии. Представители Batrachosauroididae
и Pelobatidae из мела Кызылкумов, видимо, являются древнейшими
и, в то же время, достаточно примитивными членами соответствующих
семейств. Можно считать, что ранние этапы развития указанных групп
проходили на территории Азии. Следует отметить, что все эти семей-
ства, как и Prosirenidae, впервые отмечаются для мезозоя нашей страны.
На основании всего комплекса находок устанавливаются черты
сходства древней западноазиатской фауны земноводных с западно-севе-
роамериканской и, видимо, в несколько меньшей степени — с евро-
пейской.
Датировки для североамериканских геологических формаций даны
в соответствии с обычно принимаемыми исследователями позвоночных
мела Северной Америки.
Голотипы установленных форм амфибий и экземпляры с № 11 по
№ 252 хранятся в отделении герпетологии Зоологического института
АН СССР (коллекция ZIN, РИА № К77); дополнительные найденные
59
и изученные 302 экземпляра переданы в Центральный геологоразведоч-
ный музей имени Ф. Н. Чернышева (коллекция № 11657).
Трудности экспедиционных работ в Кызылкумах разделили с авто-
ром Н. П. Коломийцев, Т. Б. Красовская, С. А. Бурсиан, Л. Ф. Хари-
тонова, Л. Б. Головнева, Г О. Черепанов, М. Н. Казнышкин,
А. О. Иванов. Автор признателен Л. П. Татаринову за прочтение ру-
кописи и ряд ценных замечаний, Н. С. Лебедкиной, В. Г. Борхвардту
и Л. Я. Боркину за консультации, И. Ф. Ортянову, Л. Б. Головневой
и Г О. Черепанову за помощь в разборке части материала, получен-
ного в результате просеивания породы, М. Н. Зубину за предоставление
разновозрастных экземпляров Ambystoma.
Отряд Caudata Oppel, 1801
Подотряд Ambystomatoidea Noble, 1931
Семейство Scapherpetontidae Auffenberg et Goin, 1959
Подсемейство Scapherpetontinae Auffenberg et Goin, 1959
Типовой род — Scapherpeton Cope, 1876, конец позднего мела
(кампан — Маастрихт) —палеоген Северной Америки.
Диагноз. Верхнечелюстная кость без направленного вниз кост-
ного гребня, лежащего снаружи от зубного ряда. Атлас без гипапо-
физа или следы его присутствия лишь намечаются. Передние сочленов-
ные поверхности шейного позвонка вытянуты в горизонтальном направ-
лении, отверстие канала для спинного мозга на этом позвонке относи-
тельно широкое.
Состав. Роды Scapherpeton Соре, 1876; Lisserpeton Estes, 1965*
и, видимо, Piceoerpeton Meszoely, 1967.
Подсемейство Eoscapherpetontinae Nessov, subfam. nov.
Типовой род — Eoscapherpeton gen. nov., поздний мел (поздний
турон — сантон) Кызылкумов.
Диагноз. Верхнечелюстная кость имеет направленный вниз кост-
ный гребень, лежащий кнаружи от зубного ряда. Атлас с мощным, ла-
терально сильно уплощенным гипапофизом, округлыми или вытянутыми
в вертикальном направлении передними сочленовными поверхностями.
Отверстие канала для спинного мозга на атласе во взрослом состоянии
узкое. На задней стороне боковых выростов шейного позвонка имеются
хорошо развитые гребни.
Состав. Кроме типового рода, еще Horeztnia gen. nov. из сено-
мана и раннего турона Юго-Западных Кызылкумов.
Сравнение. Новое подсемейство отлично от Scapherpetontinae
Auffenberg et Goin, 1959 по чертам, указанным в диагнозе.
Род Eoscapherpeton Nessov, gen. nov
Название от ео (лат.)—до того (как указание на древность) и
рода Scapherpeton Соре, 1876.
Типовой вид — Е. asiaticum sp. nov., поздний турон — сантон
Кызылкумов.
Диагноз. Относительно мелкие земноводные. Верхнечелюстная
кость длинная, довольно резко уменьшающаяся в высоту спереди. Вос-
ходящий отросток этой кости относительно невысокий, значительно вы-
тянутый спереди назад. Латеральнее от зубного ряда на верхнечелюст-
ной и предчелюстной костях располагается длинное, выступающее на-
ружу костное разрастание, имеющее хорошо развитый, выступающий
вниз продольный гребень, почти параллельный зубному ряду. Этот гре-
бень особенно сильно выражен спереди и отделен от зубного ряда
продольной бороздой. Лобная кость относительно широкая, контакти-
60
рующая с носовой и предлобной костями. Теменная кость длинная,
с мощным, косо расположенным, дугообразным гребнем снизу. На на-
ружной переднебоковой стороне этой кости нет резкого углубления
или вырезки. Зубная кость позади зубного ряда относительно резко,
но незначительно уменьшается в высоту. Меккелева борозда длинная,
но не достигающая симфиза, спереди она значительно изгибается в на-
правлении к нижней стороне зубной кости. Зубы с высоким основанием,
тонкие, сходные по величине, округлые в поперечном сечении, располо-
женные в один неровный ряд, их около 12—19 в 5 мм. Под длинным,
несколько наклоненным вниз межсуставным бугорком атласа находится
небольшая треугольная вогнутая площадка, обращенная своей пло-
скостью, в основном, вперед и несколько вниз и ограниченная по сто-
ронам двумя невысокими гребнями. Последние, сливаясь, и дают вы-
ступающий вниз очень сильно развитый гипапофиз. Подвздошная кость
высокая, поверхность на ней, являющаяся частью вертлужной впадины,
имеет округлую форму. Седалищная кость мощная, распространяю-
щаяся вперед за середину вертлужной ямы. Конечности хорошо раз-
витые.
Замечания. Новый род обнаруживает значительное сходство
со Scapherpeton Соре, 1876 прежде всего по строению позвонков, но
отличается их меньшими размерами, формой теменной кости, а от Lis-
serpeton Estes, 1965 — отсутствием выраженного углубления для мышц
на передне-боковой стороне теменной кости снаружи, от обоих родов —
широким в основании восходящим отростком верхнечелюстной кости,
а от Piceoerpeton (Meszoely, 1967; Estes, 1969е), иногда сближаемого
с батрахозауроидидами (Estes a. Hutchison, 1978) —существенно мень-
шей величиной тела. Род Eoscapherpeion gen. nov. имеет некоторые
черты сходства с Bairachosauroides Taylor et Hesse, 1943 (Batracho-
sauroididae), в частности, по форме и относительным размерам спинно-
мозгового канала атласа, по присутствию двух вытянутых углублений,
располагающихся вдоль остистого отростка атласа почти на боковых
его сторонах. Род Eoscapherpeion gen. nov. обнаруживает также сход-
ство с позднемеловым и современным представителями семейства
Amphiumidae Gray, 1825 (Teege, 1958; Auffenberg a. Goin, 1959; Estes,
1969f), в частности, по укороченности атласа, а также по некоторому
смещению углубления для хорды в верхнюю часть позвонка, с Sala-
mandroidea — по форме сошника.
Следует отметить, что части мелких зубных костей, определяемые
нами как Eoscapherpeion sp. найдены в верхнем туроне—сантоне не-
посредственно близ кровли тайкаршинской пачки Джара-Кудука (ЦГМ
№ 81—90/11657) в 2 км севернее оврага Айдахарсай.
Видовой состав. Один вид — Е. asiaticum sp. nov. поздний
турон—сантон Кызылкумов.
Eoscapherpeton asiaticum Nessov, sp. nov.
Голотип — ZIN, PHA № K77-1, левая верхнечелюстная кость;
Узбекская ССР, Центральные Кызылкумы, Джара-Кудук; верхний ту-
рон, нижняя часть тайкаршинской пачки (обнажение ЦДЖ-17а).
Описание. Правая и левая предчелюстные кости не слиты друг
с другом (ЦГМ № 208/11657). На них, как и на верхнечелюстных ко-
стях, снаружи имеется гребень, параллельный зубному ряду. Верхний
медиальный отросток узкий, не длинный, расположен у средней линии.
Латеральный выступ предчелюстной кости входил в карманообразную
щель на внутренней стороне переднего отростка верхнечелюстной кости
(рис. IX, 16). Гребень на последней, расположенный латеральнее от
зубного ряда, у крупных экземпляров (рис. IX, /), в отличие от мелких
(рис. IX, 2, 3), сильнее выступает вниз. Спереди и снизу на этом гребне
61
(рис. IX, 16, в) могут быть мелкие выросты. На небном отростке
(рис. IX, 16) имется вырезка, к которой, возможно, прилегал сошник.
Сзади, на небольшом протяжении эта вырезка могла входить в край
хоан. Задний (птеригоидный) отросток кости (ЦГМ № 91/11657) был
длинный (обычно он оказывается обломанным), с довольно узкой пло-
щадкой сверху. На конце этого отростка, несколько отогнутого к ме-
диальной линии, имеются продольные гребни, отмечающие, вероятно,
место прикрепления ligamentum quadrato-maxillare (см. Лебедкина,
1964). Несколько сзади от вершины восходящего отростка с внутрен-
ней стороны обычно бывает хорошо выражено углубление с неровно-
стями на дне, открытое назад и несколько к медиальной линии (IX, 16)
и служившее, видимо, местом прикрепления мощной связки ligamentum
rostro-pterigoideum. Зубы на верхнечелюстной кости многочисленные.
Наружная поверхность черепа спереди неровная, особенно вблизи ниж-
него края верхней челюсти (рис. IX, 1а, 4, 5), где имеются извилистые
гребешки и канавки. Шов quadratum — squamosum длинный, прочный.
Задний конец лобной кости (рис. XI, 20) заостряется, вытягиваясь
вдоль медиальной линии. Спереди на этой кости находятся две шовные
поверхности, на том же уровне сбоку еще одна, поменьше, а ниже ее —
узкий длинный шов. К медиальной из указанных передних поверхно-
стей сверху прилегала носовая кость, к другой — вершина восходящего
отростка верхнечелюстной кости. К боковой сочленовной поверхности
прилегала предлобная кость, а к узкому шву — глазничноклиновидная.
Снизу лобная кость вогнута, своей задней частью она несколько нале-
гала на теменную. На нижней стороне последней (рис. IX, 10\ ЦГМ
№ 92, 93/11657) от передне-боковой стороны к ее задне-внутреннему
углу тянется мощный гребень. Затылочный мыщелок боковой затылоч-
ной кости (рис. XI, 21\ ЦГМ № 94—96/11657) слабо вытянут в верти-
кальном направлении (он был почти округлым). Затылочное отверстие
невысокое, яремное отверстие некрупное. Парасфеноид (рис. XI, 22)
сзади со значительным, в сравнении с Ambystoma, углублением на вну-
тренней стороне и двумя небольшими каротидными отверстиями, ле-
жащими довольно далеко от латеральных краев кости (плезиоморфный
признак). Сошник (ЦГМ № 171, 172, 209, 210/11657) с длинным, сужи-
вающимся назад отростком, протягивающимся вдоль латерального
края парасфеноида (характерно уже для Salamandroidea, а не Amby-
stomatoidea). Передняя кромка сошника имеет вид широкой дуги, при-
острена, значительно выступает вниз. Следов прирастания зубов к кости
нет, в углублении позади передней кромки, на нижней поверхности
сошника, хорошо заметны отверстия многочисленных каналов.
Борозда для меккелева хряща (рис. IX, 66, 7, 9\ X, /; ЦГМ № 97—
105/11657) на зубной кости спереди значительно суживается (рис. IX,
11). Поверхность симфиза ровная, без складок. Зубная кость снизу
снаружи обычно более или менее гладкая. Однако на крупных экземп-
лярах, лишь условно относимых к Е. asiaticum sp. nov., наружная поверх-
ность испещрена многочисленными грубыми продольными бороздами.
Такие кости могли принадлежать старым животным указанного вида,
но не исключена их принадлежность и особой форме Scapherpetontidae.
В материале имеются и задние части нижних челюстей (рис. XI, 23:
ЦГМ № 106/11657), которые позволяют говорить, что угловое и ге-
ниальное окостенения были обособленными (это есть черта симплезио-
морфного сходства с Hynobiidae и Cryptobranchidae). Гониальная кость
(рис. XI, 13) с характерным косым гребнем на медиальной стороне.
В подъязычном аппарате окостенения были (рис. XI, 24).
Все позвонки позади атласа амфицельные, углубления передних
и задних сочленовных поверхностей тел выстланы тонкой пленкой от-
ложений солей. Остистый отросток атласа высокий, его межсуставной
бугорок (рис. IX, 16- X, 3; XI, 25; ЦГМ № 107—115/11657) имеет только,
62
очень слабое сужение в основании. На нижней стороне межсуставного
бугорка с каждой стороны находится по узкой сочленовной поверх-
ности, почти достигающей средней линии и переходящей сбоку непо-
средственно в указанные выше передние сочленовные поверхности шей-
ного позвонка. Атлас и туловищные позвонки, особенно у взрослых
экземпляров, на задней стороне остистого отростка над постзигапофи-
зами имеют обширную площадку, покрытую гребнями (рис. XI, 25б\
XII, /б), к которой, очевидно, причленялись мощные связки коротких
межостистых мышц. Позвонки, интерпретируемые нами как передние
туловищные (рис. IX, 77; ЦГМ № 116—130/11657) имеют, как и атлас,
очень длинные гипапофизы, которые могут быть равны по длине вер-
тикальному диаметру сочленовной поверхности тела позвонка. Гипапо-
физы наиболее сильно выступают в передней и задней частях каждого
туловищного позвонка или же спереди. Невральная дуга с ясным греб-
нем по верхней кромке, длинная, простирается за задний край тела
позвонка, относительно круто поднимаясь вверх. Невральный канал на
туловищных позвонках бывает более или менее уплощен дорсовен-
трально (рис. IX, 77), сзади он обычно более высокий и треугольный
в сечении. Носители ребер прочные, не очень длинные, имеют вид вер-
тикально вытянутых пластин. Позвонки из задней части туловищного
отдела (рис. IX, 18, 19\ XII, 7; ЦГМ № 131 —140/11657) имеют в срав-
нении с передними туловищными гипапофизы меньшего размера и менее
вытянутые в вертикальном направлении сочленовные поверхности тел.
Длина тел большинства найденных туловищных позвонков 3.5—4.5 мм.
Крестцовый позвонок (рис. XI, 26\ ЦГМ № 141 —150/11657) обычно
не имеет гипапофиза. Тело его округлое в сечении или слегка уплощено
дорсовентрально за счет того, что снизу оно бывает почти плоским,
здесь на позвонке может быть два продольных гребня, разделенных
углублением, и небольшие базапофизы, обычно несколько лучше выра-
женные спереди. Отверстия межсегментарных артерий на крестцовом
позвонке обычно сильно увеличены, носители ребер необыкновенно
мощные, расширенные в направлении места прикрепления крестцового
ребра. Последнее (рис. X, 7; XII, 4) длинное, хорошо развитое, его
верхняя сочленовная поверхность, однако, несколько более узкая и вы-
сокая, чем, например, у Ambystoma.
Тела хвостовых позвонков (рис. X, 4\ XII, 2, 3; ЦГМ № 151 —155/
11657) снизу могут нести два продольных гребня, часто неравные по
высоте, разделенные глубокой бороздой. Некоторые из найденных хво-
стовых позвонков (очевидно, не передние в ряду) имеют очень длинные
гемальные дуги (рис. XII, 2). На хвостовых позвонках остистый отро-
сток тонкий, незначительно суживающийся к вершине. В отличие от
Ambystoma он обычно субтреугольный в сечении. Постзигапофизы на
указанных позвонках тесно сближены, сочленовные поверхности тел по
форме близки к округлым.
Отверстие спинномозгового нерва на атласе крупное (рис. IX, 166),
расположено вполне типично для Caudata в передней части позвонка.
На крестцовом позвонке подобное отверстие тоже имеет довольно боль-
шие размеры, располагаясь в задней части дуги. Степень развития та-
ких отверстий на других позвонках сильно варьирует, обычно они очень
узкие, порой оказываются выраженными только с одной стороны или же
отсутствуют. Тем не менее удается найти по одному очень узкому от-
верстию в задней части дуг у ряда обследованных экземпляров туло-
вищных и хвостовых позвонков, что можно рассматривать, в частности,
как промежуточное состояние между отмечаемым (Edwards, 1976)
у Ambystomatidae и известных ранее Scapherpetontidae.
В средней части тайкаршинской пачки Джара-Кудука найдены до-
вольно крупные позвонки (ЦГМ № 156—159/11657): два туловищных,
хвостовой и атлас. Длина тел первых трех из них равна соответственно
5, 7.3 и 5.8 мм. Эти позвонки имеют неровную, порой эродированную
поверхность и, видимо, принадлежали старым особям.
Лопатка (?скапулокоракоид) высокая, с большим по площади, за-
кругленным по заднему краю пластинчатым участком (ЦГМ № 192,
193/11657), обломанным на изображенном экземпляре (рис. XII, 16).
Верхний край вертлужной впадины на подвздошной кости (рис. IX,
12, 13) значительно выступает. Контактная поверхность седалищной
кости (рис. XII, 7) и лобкового хряща толстая.
Плечевая кость (рис. IX, 14; X, 5; ЦГМ № 160—162/11657) в обла-
сти диафиза относительно тонкая, довольно резко расширенная на
дистальном конце. В сравнении с плечевой костью Ambystoma у нее
более сильно отставлен дельтопекторальный гребень. Бедренная кость
(рис. X, 6; ЦГМ № 163—167/11657) относительно короткая. По срав-
нению с Ambystoma она имеет значительно расширенные эпифизарные
области (особенно прочна ее проксимальная часть). Гребень трохантера
мощный, значительно выступающий, он простирается в дистальном на-
правлении существенно дальше середины длины кости. Лучевая (ЦГМ
№ 211/11657), локтевая (рис. XII, 5), большая берцовая (рис. XII, 6)
и малая берцовая кости (ЦГМ № 168/11657) с более расширенными
эпифизарными частями, но относительно тонкими диафизами, в срав-
нении с тем, какие они бывают у половозрелых аксолотлей. В мате-
риале имеются отдельные фаланги и другие элементы аутоподия (ЦГМ
№ 169, 170/11657).
Замечания. Новое земноводное по наличию шовного контакта
лобных, предлобных и носовых костей, значительной длине верхнече-
люстных костей, степени развития конечностей не имело столь замет-
ных неотенических черт, как американские Scapherpetontidae и Муп-
bulakia surgayi gen. et sp. nov. (см. ниже). Форма предчелюстной и
верхнечелюстной костей и, особенно, малый угол между плоскостями
восходящего и небного отростков на последних (около 35°) указывают
на определенную уплощенность черепа Е. asiaticum sp. nov. в дорсовен-
тральном направлении. Мощный, порой даже зазубренный нижний
гребень на указанных костях, вероятно, содействовал увеличению
прочности челюстей, что могло иметь значение при захвате и удержа-
нии твердой добычи, например, мелких ракообразных, остатки которых
нередко встречаются совместно с костями Е. asiaticum sp. nov. Вместе
с тем тонкие, плотно сидящие зубы явно не могли обеспечивать эффек-
тивное разрушение таких объектов в ротовой полости. Очертания те-
менных костей вполне типичны для мелких хвостатых (см. Parker, 1882:
Bolkay, 1928). Мощные гребни на нижней стороне этих костей, вероятно,
указывают на существование значительных усилий, передаваемых от
передней части черепа назад. В связи с этим можно думать, что раз-
витие всех указанных гребней могло определяться также защитными
функциями при рытье. Значительные размеры остистого отростка и ги-
папофиза на атласе у £. asiaticum sp. nov. позволяют предполагать, что
к ним могли прикрепляться сильные мышцы, обеспечивавшие усилия
при движении передней части тела в вертикальной плоскости, что мо-
жет быть важным, в частности, для зарывающегося животного. Вытяну-
тость передних фасеток атласа в вертикальном направлении, которая
могла быть связана со значительной амплитудой движения головы
вверх й вниз, кажется, согласуется с данным предположением. Однако
надо иметь в виду, что мышцы и связки, участвующие в приведении
нижней челюсти, у хвостатых земноводных крепятся не только на че-
репе, но и на остистых отростках 1-го и 2-го позвонков (Hinderstein,
1971; Саломатина, 1975). Поэтому не исключена связь степени разви-
тия остистого отростка атласа и с характером движения челюстей жи-
вотного.
64
Как указывалось, у Caudata обычно нет выраженных гребней на
нижней стороне первого позвонка. Лишь иногда на атласе может при-
сутствовать едва заметный гребень снизу, как, например, у Lisserpeton,
Opisthotriion и Prodesmodon (Estes, 1965b; Naylor, 1979). Этот гребень
у Lisserpeton спереди, кажется, расширяется или разделяется на два,
что в более резкой форме можно видеть у Eoscapherpeton gen. nov. и
Horezmia gen. nov.
' Судя по значительной высоте подвздошных костей и крестцовых
ребер, задняя часть туловища у Е. asiaticum sp. nov. была довольно
высокой. Значительная длина остистых и гемальных отростков хвосто-
вых позвонков свидетельствует о выраженной сжатости с боков и зна-
чительной высоте хвостовой лопасти. Последнее могло обеспечивать
хорошие возможности для плавания. Конечности у животных, в сравне-
нии с половозрелыми аксолотлями, были довольно короткими, но креп-
кими.
Исходя из*строения позвонков нового земноводного, можно пред-
полагать, что костный элемент, отнесенный к Hemitrypus jordanianus
(Соре, 1876b; Auffenberg a. Goin, 1959, fig. 3; Estes, 1964), может пред-
ставлять не второй позвонок, а один из задних туловищных.
Распространение. Поздний турон—сантон Центральных Кы-
зылкумов.
Материал. Не менее 3000 костей (среди которых более 160 зуб-
ных. 180 атласов) из нижней, средней и верхней частей тайкаршинской
пачки.
Род Horezmia Nessov, gen. nov.
Название от Хорезмского оазиса.
Типовой вид — Н. gracile sp. nov. ранний турон Юго-Западных
Кызылкумов.
Диагноз. Относительно мелкие земноводные. Строение предче-
люстных, верхнечелюстных и зубных костей в основных чертах сходно
с таковым у Eoscapherpeton. Передние сочленовные поверхности атласа
округлые, разделены под межсуставным бугорком большой треугольной
вогнутой площадкой, ограниченной по сторонам двумя невысокими,
сливающимися сзади гребнями. Указанная площадка обращена своей
плоскостью в большей степени вниз, чем вперед. Гипапофиз развит уме-
ренно, направлен скорее назад, чем вниз. Латеральные выросты атласа,
несущие передние фасетки, тонкие, гребни на их задней стороне раз-
виты хорошо. Остистый отросток атласа высокий, несколько наклонен-
ный назад, нижняя часть его невральной дуги немного выступает
вперед за края передних сочленовных поверхностей.
Сравнение. Род Horezmia gen. nov. обнаруживает сходство
с Eoscapherpeton gen. nov., но отличается от последнего округлыми
очертаниями передних сочленовных поверхностей атласа, малой толщи-
ной латеральных выростов этого позвонка, меньшей длиной основания
гипапофиза, а также, видимо, несколько менее выступающим межсу-
ставным бугорком, более высоким гребнем на гениальной кости.
Видовой состав. Типовой вид.
Horezmia gracile Nessov, sp. nov.
Название от gracilis (лат.) — стройный.
Голотип — ZIN, PHA № К77-2, атлас; Каракалпакская АССР,
Юго-Западные Кызылкумы, Ходжакуль, нижний турон, нижняя часть
бештюбинской свиты (обнажение СХ-20).
Описание. Сечение остистого отростка атласа (рис. VIII, 5, 8)
близко к треугольному, углубление для хорды на этом позвонке прак-
5 Зак. 719
65
тически не смещено в верхнюю часть тела. Остальные элементы скелета
(рис. VIII, 2—4, 6, 9, 12—15) сходны с костями представителя преды-
дущего рода (но см. гениальные кости, ЦГМ № 226-232/11657).
Замечания. Земноводное имеет черты сходства с видом рода
Opisthotriton Auffenberg, 1961 (Batrachosauroididae). Это касается
формы передних сочленовных поверхностей атласа, значительной вы-
соты и формы остистого отростка, несущего спереди резкий гребень.
Атласы из Ходжакуля, судя по их сохранности, не принадлежат
к тем остаткам амфибий, которые могли быть перемытыми из более
древних отложений (см. раздел по Mynbulakia nongratis sp. nov.).
Распространение. Сеноман и ранний турон Юго-Западных
Кызылкумов (Шейх-Джейли, Ходжакульсай, Челпык, Ходжакуль).
Материал. Более 12 атласов, другие элементы скелета.
Подотряд Proteida Соре, 1866
Семейство'Batrachosauroididae Auffenberg, 1958
Род Mynbulakia Nessov, gen. nov.
Название от бессточной центральнокызылкумской впадины Мын
булак.
Типовой вид — М. surgayi sp. nov., поздний турон—сантон Цен-
тральных Кызылкумов.
Диагноз. Мелкие земноводные. Верхнечелюстная кость довольно
короткая, резко уменьшающаяся в высоту спереди. Восходящий отро-
сток этой кости относительно высокий, не очень сильно вытянутый
спереди назад. Задний (птеригоидный) отросток относительно корот-
кий, близ конца значительно изогнутый внутрь. Наибольшую высоту
зубная кость имеет непосредственно позади зубного ряда. Меккелева
борозда длинная, спереди она, сильно суживаясь, достигает симфиза.
Зубы относительно крупные, располагающиеся не очень плотно друг
к другу с частотой около 12 штук в 5 мм, расширенные в поперечном
направлении, сильно притупленные к вершине. Передние сочленовные
поверхности атласа округлые, межсуставной бугорок относительно
большой по сечению, но слабо выступающий вперед. Спинномозговой
канал атласа шире межсуставного бугорка и задней сочленовной по-
верхности тела позвонка. Туловищные позвонки относительно длинные,
сочленовные поверхности их тел округлые, медиальный гребень снизу
выражен слабо или отсутствует, многие позвонки имеют слабые перед-
ние базапофизы. Остистый отросток невысокий, носители ребер тонкие,
длинные, с двумя довольно близко расположенными сочленовными
участками на контакте с каждым ребром. Зигапофизы обычно значи-
тельно вытянуты в передне-заднем направлении. Бедро с относительно
толстым диафизом, мощным, пластинчатым, очень сильно выступающим
гребнем трохантера, короткое.
Сравнение. Род Mynbulakia gen. nov. сходен с Palaeoproteus
Негге, 1935 из палеогена Франции и Германии (Негге, 1935; Estes et al.,
1967) по небольшим размерам элементов скелета, по укороченности
верхнечелюстной и зубной костей, по наличию возвышения зубной
кости позади окончания зубного ряда. Для обоих родов отмечается
амфицельность позвонков, значительная длина теменных костей, отно-
сительно крупные размеры зубов, которые несколько расширены в по-
перечном направлении и имеют по относительно крупному отверстию
в основании с внутренней стороны. Новый род отличается от Palaeopro-
teus существенно более выступающим гребнем трохантера, меньшей
относительной высотой зубной кости позади зубного ряда, более про-
тяженной меккелевой бороздой, достигающей области симфиза, округ-
лыми передними сочленовными поверхностями атласа, межсуставной
бугорок которого имеет более широкое основание, существенно более
66
крупным отверстием для спинного мозга на первом шейном позвонке.
Род Mynbulakia gen. nov. сходен с Opisthotriton Auffenberg, 1961 по
значительной высоте зубной кости позади конца зубного ряда, по форме
передних сочленовных поверхностей атласа, но отличается более мощ-
ным основанием межсуставного бугорка атласа, амфицельностью по-
звонков (имеющих, однако, в глубине сочленовных поверхностей тел
отложения солей), отсутствием задних базапофизов, поперечно расши-
ренными зубами.
Замечания. Указанный выше палеогеновый род Palaeoproteus,
имеющий ряд черт сходства с Mynbulakia gen. nov., сближался ранее
с Proteidae (Негге. 1935), относился к Salamandridae (Estes a. Hecht,
1967; Edwards, 1976) или предположительно к Batrachosauroididae
(Naylor, 1978, 1979). Устанавливаемый здесь род сходен с Prodesmodon
(Prosirenidae) относительно шпрот им основанием межсуставного бу-
горка и формой спинномозгового канала атласа, но отличен иным
строением симфиза нижних челюстей, меккелевой борозды, зубов.
По степени удлинения туловищных позвонков и по наличию, по край-
ней мере, на некоторых из них передних базапофизов новый род
обнаруживает сходство с Proamphiuma Estes, 1969 (Amphiumidae). На-
конец, можно отметить наличие отдельных черт сходства позвонков
Mynbulakia gen. nov. с таковыми у различных Proteidae (Estes a. Darev-
sky, 1977; Naylor, 1978).
Род Mynbulakia gen. nov. с меккелевой бороздой, достигающей сим-
физа) вместе с Palaeoproteus, также имеющим амфицельные позвонки,
относится нами к особой примитивной группе внутри Batrachosauroidi-
dae, видимо, сравнительно мало изменившейся в сравнение с формами,
которые могли дать, с одной стороны, начало ветвям Proteidae, продви-
нутым на путях педоморфоза, а с другой, — менее педоморфным Opi-
sthotriton и Batrachosauroides (Batrachosauroididae), получившим осо-
бый (примитивный) тип опистоцельности позвонков (Estes, 1969е).
Можно думать, что две указанные группировки Batrachosauroididae
вместе с Proteidae и, возможно, Prosirenidae образуют группу относи-
тельно близко родственных стволов.
Следует отметить, что некоторая поперечная расширенность зубов
ранее отмечалась и среди Scapherpetontidae (Edwards, 1976). Это яв-
ление можно рассматривать как приспособление, содействующее уве-
личению их прочности при поперечных нагрузках. Это, а также значи-
тельная притупленность коронок у Mynbulakia gen. nov. говорит о воз-
можной склерофагии животных; выраженность педоморфных черт ука-
зывает на водный образ жизни этих длиннотелых земноводных.
Mynbulakia surgayi Nessov, sp. nov.
Название в честь коллектора В. Т. Сургая, осуществлявшего в 1932
году специальные поиски остатков позвоночных мела в Джара-Кудуке
до приезда А. Ф. Соседко (1937).
Голотип — ZIN, РНА № К77-3, левая верхнечелюстная кость;
Узбекская ССР, Центральные Кызылкумы, Джара-Кудук, верхний ту-
рон—сантон, средняя часть тайкаршинской пачки (обнажение ЦБИ-4в).
Описание. Заостренный задний (птеригоидный) отросток верх-
нечелюстной кости (рис. XI, 10) сверху имеет относительно широкую,
постепенно суживающуюся назад площадку, несущую на медиальном
крае выступающий вверх гребень. Снаружи верхнечелюстная кость не-
ровная, с рядом отверстий, самые крупные из которых находятся
в области передней части восходящего отростка. Здесь относительно
высоко над нижним краем челюсти находится небольшой гребень,
ориентированный не вниз, а в сторону. На голотипе на участке в 4.5 мм
находилось 11 педицеллярных зубов (рис. XI, 10), плевродонтность ко-
5*
67
торых, как и на зубных костях, выражена не очень резко. Теменная
кость (ЦГМ № 173, 174/11657) длинная. На уровне медиального шва
кость имеет ориентированный вниз гребень, увеличивающий площадь
соприкосновения двух теменных костей. На боковой кромке спереди
имеется узкая бороздка. Зубные кости (рис. XI, П\ ЦГМ № 175—
177/11657) относительно длинные, несут обычно по 13—15 зубов. Сна-
ружи кость относительно гладкая, лишь в ее верхней части есть ряд
небольших отверстий.
Сравнение. См. сопоставление для Mynbulakia nongratis sp. nov.
Замечание. Остатки представителей рода Mynbulakia gen. nov.
встречаются в местонахождениях существенно реже, чем Scapherpeton-
tidae.
Распространение. Поздний турон—сантон Центральных Кы-
зылкумов.
Материал. 5 верхнечелюстных костей, 20 зубных, 4 теменных,
3 атласа и около 100 других позвонков, а также бедренные кости из
средней и верхней частей тайкаршинской пачки Джара-Кудука.
Mynbulakia nongratis Nessov, sp. nov.
Название в связи с условиями поисков осенью 1975 года от поп
(лат.) —не и gratis (лат.)—даром.
Голотип — ZIN, РНА № К77-8, часть зубной кости; Каракалпак-
ская АССР, Юго-Западные Кызылкумы, Ходжакуль, нижний турон
(?альб), нижняя часть бештюбинской свиты (обнажение СХ-20).
Описание. На фрагменте зубной кости длиной 6.2 мм (рис. VIII,
И) помещается 12 зубов, вершины которых обломаны. Зубы овальные
в сечении, длинный радиус овала расположен поперек продольной оси
тела и может быть вдвое больше короткого. Пространство между зу-
бами заполнено прочной массой, видимо, фосфатизированной породой.
Сравнение. Отличается от М. surgayi sp. nov. из позднего ту-
рона—сантона Центральных Кызылкумов меньшей величиной и зубами,
видимо, сильнее расширенными в поперечном направлении.
Замечания. Фрагмент зубной кости М. nongratis sp. nov. из
Ходжакуля (рис. VIII, //), найденный вместе с позвонками и другими
остатками ящериц (Несов, 1977а, 1978), первоначально из-за попереч-
ной расширенности зубов был причислен (Несов, 1977г) к ящерицам
из группы Iguania, рассматриваемой в объеме, принятым в издании
«Основы палеонтологии» (Рождественский, Татаринов, 1964). У неко-
торых ящериц, относимых ранее к этой группе, например, у Polyglypha-
nodontidae (Gilmore, 1942, 1943), теперь причисляемых к примитивным
Scincomorpha (Sulimski, 1975) или порой непосредственно к Teiidae
(МйПег, 1966; Estes, 1969с), имеются поперечно расширенные зубы.
Однако у этих ящериц зубы акродонтные, а не плевродонтные, как
у Mynbulakia gen. nov.
Возраст нижней части бештюбинской свиты, из которой происхо-
дят остатки М. nongratis sp. nov., определен по аммонитам (Шульц,
1972) как раннетуронский. Однако в локальном местонахождении
с остатками земноводных и ящериц встречались целые и окатанные
зубы хрящевых рыб, часть которых — Paraisurus macrorhiza (Pict, et
Camp.) имеет альбский возраст (определения Л. С. Гликмана). Учиты-
вая это, а также наличие на поверхности голотипа породы иного со-
става в сравнении с вмещающей и окатанность кости, нельзя исклю-
чать возможности перемыва данных остатков М. nongratis sp. nov.
из альбских отложений.
68
Распространение. Ранний турон (?альб) Юго-Западных Кы-
зылкумов.
Материал. Часть зубной кости.
Семейство Prosircnidae Estes, 1969
Род Nukusurus Nessov, gen. nov.
Название от города Нукус.
Типовой вид — V insuetus sp. nov ранний турон Юго-Запад-
ных Кызылкумов.
Диагноз. Очень мелкие земноводные с относительно невысокой
зубной костью. Борозда для меккелева хряща замыкается в канал
лишь немного далее вперед от заднего конца зубного ряда. Задние зубы
нижней челюсти мелкие, остальные заметно увеличиваются в размерах
по направлению к участку, лежащему несколько вперед от середины
зубного ряда. Плевродонтность зубов выражена нерезко, особенно в зад-
ней части зубного ряда. На месте соединения двух ветвей нижней че-
люсти имеется своеобразный «замок», формирующийся за счет системы
противолежащих выступов и углублений, самое заметное из последних
лежит на нижней стороне симфиза.
Сравнение. Зубная кость Nukusurus gen. nov. сходна с одно-
именными костями, приписываемыми Prodesmodon Estes, 1964 (по по-
воду последних см. возражения у Naylor, 1979) по наличию некоторой
«разнозубости», по укороченности меккелевой борозды, наличию «замка»
в симфизе. Однако новый род отличается меньшими размерами, еще
меньшей протяженностью меккелевой борозды, не столь заметным воз-
вышением края нижней челюсти в области, где зубы несколько увели-
чены, более слабым развитием выростов и углублений в области сим-
физа. От Prosiren Goin et Auffenberg, 1958 (Estes, 1969b) новый род
отличен строением меккелевой борозды, относительно более редким
расположением зубов, от Albanerpeton Estes et Hoffstetter, 1976 из позд-
него миоцена Франции (Estes, Hoffstetter, 1976) — более короткой мек-
келевой бороздой (апоморфный признак), меньшей относительной вы-
сотой зубной кости, ее меньшей величиной и относительно гладкой
наружной поверхностью.
Замечания. Челюсть Nukusurus gen. nov. по сравнению с че-
люстями, приписываемыми Prodesmodon по ряду признаков (кроме сте-
пени замыкания канала для меккелева хряща) демонстрирует состоя-
ние, которое можно рассматривать как примитивное. Зубная кость
(ЦГМ № 203/11657) очень мелкого представителя Prosirenidae найдена
в верхней части разреза ходжакульской свиты близ Шейх-Джейли (се-
номан).
Видовой состав. Типовой вид.
Nukusurus insuetus Nessov, sp. nov.
Название от insuetus (лат.) —необычный.
Голотип — ZIN, PHA № К77-4, левая зубная кость; Каракалпак-
ская АССР, Юго-Западные Кызылкумы, Челпык, нижний турон, ниж-
няя часть бештюбинской свиты (обнажение СЧ-1).
Описание. Длина зубной кости (рис. VIII, /), видимо, лишь
немного обломанной сзади, 4.8 мм. В области симфиза кость довольно
толстая. На кости могло находиться 17—23 зуба.
Замечания. Повышение прочности соединения двух ветвей
нижней челюсти у Nukusurus gen. nov., отмеченное увеличение разме-
ров некоторых зубов, а также превращение зубной кости на одном из
69
участков в трубчатую конструкцию, видимо, содействовало упрочнению
аппарата схватывания добычи и могло быть связано с питанием отно-
сительно твердыми, хотя и очень мелкими объектами.
Распространение. Ранний турон Юго-Западных Кызылкумов.
Материал. Зубная кость.
Caudata fam. indet.
Род Bissektia Nessov, gen. nov.
Название от родника (колодца) Биссекты в Центральных Кызыл-
кумах.
Типовой вид — В. папа sp. nov., поздний турон—сантон Цен-
тральных Кызылкумов.
Диагноз. Мелкие земноводные с гладкой, слабо выпуклой на-
ружной поверхностью невысокой верхнечелюстной кости, которая лишь
постепенно и незначительно увеличивается в высоту кпереди. Небный
отросток гребневидный, длинный, несколько увеличивающийся в ши-
рину сзади, где на его верхней кромке находится длинный гребнистый
след, возможно, для прикрепления ligamentum quadrato-maxillare.
Углубление, где, видимо, крепилась другая связка, ligamentum rostro-
pterigoideum, хорошо выраженное, крупное. Зубы мелкие, округлые
в поперечном сечении, поверхность прикрепления их к кости невысокая.
Сравнение. Отличается от Eoscapherpeion gen., nov. и Mynbula-
kia gen. nov. строением зубов, небного гребня, гладкой наружной по-
верхностью верхнечелюстной кости.
В и до в о й с о с т а в. Типовой вид.
Bissektia папа Nessov, sp. nov.
Название от nanus (лат.) —маленький.
Голотип— ZIN, РНА № К77-9, левая верхнечелюстная кость;
Узбекская ССР, Центральные Кызылкумы, Джара-Кудук; верхний ту-
рон- сантон, средняя часть тайкаршинской пачки (обнажение ЦБИ-4в).
Описание. Восходящий отросток кости обломан (рис. XII, 24),
на голотипе зубы располагаются с частотой около 20 штук в 5 мм.
Замечание. Представителю этого же рода может принадлежать
и задняя часть верхнечелюстной кости (рис. XII, 26), найденная в верх-
ней части тайкаршинской пачки Джара-Кудука; две очень небольшие,
невысокие зубные кости (ЦГМ № 204, 205/11657, обнаружены в сред-
ней и верхней частях пачки) с мелкими зубами. На одной из указан-
ных зубных костей хорошо заметно, что меккелева борозда на значи-
тельном протяжении (видимо, большем, чем у Nukusurus из Prosireni-
dae) была замкнута.
Распространение. Поздний турон—сантон Центральных Кы-
зылкумов.
Материал. Верхнечелюстная кость.
Caudata fam., gen. et sp. indet.
Из нижней части тайкаршинской пачки Джара-Кудука (верхний турон) проис-
ходит ряд своеобразных остатков хвостатых земноводных.
1. Атлас, сходный с одноименными позвонками Eoscapherpeton gen. nov., имеет,
однако, несколько повернутую вверх заднюю сочленовную поверхность (рис. X, 9).
2. На небольшой по размеру теменной кости (рис. X, 10) изнутри не имеется
резкого гребня, отмеченного у Eoscapherpeton gen. nov. В то же время снаружи
кость усеяна мелкими бугорками неправильных очертаний, не известными для Myn-
bulakia gen. nov. На наружной передне-боковой части кости имеется хорошо выра-
женное продольное узкое углубление.
70
3. На редко встречающихся небольших, тонких бедренных костях, имеющих
слабые расширения диафизов (рис. X, 5), и, возможно, не принадлежащих Е. asiati-
сит sp. nov., гребень трохантера выступает относительно слабо. Он остается доста-
точно явственным приблизительно лишь до середины длины кости. В этом, а также
в строении проксимального окончания кости проявляется внешнее сходство с Ambysto-
matidae (Hecht a. Estes, 1960).
4. Плечевая кость (рис. XI, 28) с относительно хорошо окостеневшей областью
диафиза может принадлежать животному из Prosirenidae или близкому к этому се-
мейству.
5. Верхнечелюстная кость (без наружного гребня) еще одной, новой амфибии
(ЦГМ № 233/11657) найдена выше середины указанной пачки.
6. В верхней части тайкаршинской пачки отмечается появление остатков до-
вольно крупных хвостатых земноводных, похожих на Eoscapherpeton (высота зубной
кости более 8.5 мм, длина не менее 40 мм), имеющих грубую, гребнисто-ямчатую
скульптуру наружной поверхности черепа (ЦГМ № 212—219/11657).
Отряд Salientia Laurenti, 1768
Семейство Pelobatidae Stannius, 1856
Род Eopelobates Parker, 1929
Типовой вид — Е. anthracinus Parker, 1929, средний олигоцен
Германии.
Eopelobates sosedkoi Nessov, sp. nov.
Название в честь геолога А. Ф. Соседко, руководившего геологи-
ческими работами и осуществлявшего поиски остатков позвоночных
в Джара-Кудуке в 1932 году.
Голотип — ZIN, РНА № К77-5, задняя часть правой лобнотемен-
ной кости; Узбекская ССР, Центральные Кызылкумы, Джара-Кудук,
верхний турон—сантон, средняя часть тайкаршинской пачки (обнаже-
ние ЦБИ-4в).
Описание. Некоторые покровные кости черепа (рис. X, 19\ XII,
10—12, 14) снаружи несут хорошо развитую скульптуру, образованную
более или менее плотно расположенными ячейками, разделенными
анастомозирующими гребнями. Лобнотеменная кость (рис. XII, 14)
в задней своей части плоская или слабовогнутая сверху, с резким греб-
нем на заднебоковом отростке кости. Последний направлен относи-
тельно круто назад. Латеральная кромка скульптированной зоны лобно-
теменной кости на некотором участке впереди указанного отростка
остается примерно параллельной медиальному костному шву, а затем
кость образует очень небольшой, приостренный латеральный выступ,
за которым эта кромка относительно круто поворачивает в направлении
к средней линии черепа. Канал лобнотеменной кости (рис. XII, 14а)
открывается в углублении, лежащем медиальнее от заднебокового от-
ростка; переднее отверстие канала (рис. XII, 146) лежит в ямке, почти
точно под указанным очень небольшим латеральным выступом. Ме-
диальное ответвление этого канала, имеющееся у Pelobates Wagler,
1830, здесь, видимо, было не выражено или оно представлено очень
узким отверстием в нижнем латеральном гребне кости. Лобное окно
в хрящевом эндокрании, заметное по отпечатку на нижней стороне
лобнотеменной кости (Spinar, 1976) в заднебоковой своей части было
закруглено. Верхнечелюстная кость (рис. X, 79; XII, 10—12\ ЦГМ
№ 220—225/11657) на середине своей длины в области глазницы невы-
сокая, но увеличивающаяся в высоту спереди и сзади. Небный гребень
(рис. X, 106) выступает умеренно, он выражен сильнее в задней части
кости. Над небным гребнем (рис. X, 196) расположена суживающаяся
вперед площадка (рис. XII, 12) с продольной гребнистостью — место
прикрепления мощной крыловидной кости. Спереди от нее отделен ко-
сым гребнем участок, к которому причленялась небная кость. Над ука-
занным гребнем близ верхнего края кости расположен желобок
71
(рис. XII, 10)л в котором находился нижний край носовой кости. Под
глазницей элементы рельефа на верхнечелюстной кости выражены
сильно, а спереди кость была наиболее заглаженной. Зубы на верхне-
челюстной кости расположены с частотой около 20 штук в 5 мм; поверх-
ность, к которой осуществлялось их прикрепление, невысокая. Пред-
челюстная кость (рис. XII, 9), условно отнесенная к Е. sosedkoi sp. now,
в целом сходна с таковой у Pelobates fuscus (L.), но с относительно
высоким восходящим отростком. Снаружи эта кость гладкая. Чешуй-
чатая кость (ЦГМ № 199—202/11657) с ямчатой наружной поверх-
ностью, существенно более широкая, чем у амфибии из мела США
(Estes, 1964, fig. 31а, b), причисляемой к lEopelobates sp. (Estes, 1970а).
Боковая затылочная кость (рис. XII, 13) с сильно развитым около-
затылочным отростком, несущим два бугорка, из которых нижний круп-
ный, гребнистый. Затылочный мыщелок относительно широкий, за-
метно суживающийся книзу, большое затылочное отверстие, видимо,
было уплощенным, субтреугольным в очертаниях. Латеральный отро-
сток очень длинный, утолщающийся в дистальном направлении, не-
сколько изогнутый вверх. Передне-верхний и задне-нижний края углуб-
ления овального окна значительно выступают. Яремное отверстие
крупное, в виде вертикально вытянутого овала. Два близко располо-
женных перилимфатических отверстия разделены только тонкой кост-
ной перемычкой. Они лежат глубже от наружного края яремного от-
верстия, чем у Pelobates. На внутренней стороне кости пара отверстий
для слухового нерва располагается в общем углублении, лишь далее
вглубь эти отверстия разделяются тончайшей костной балочкой
(рис. XII, 136). Переднеушная кость (рис. XII, 23), условно относимая
к Е. sosedkoi sp. nov., является обширным, хорошо развитым окостене-
нием. Отверстие для тройничного нерва выражено на ней в виде отно-
сительно глубокой, суживающейся вглубь щели.
Крыло подвздошной кости (рис. X, 25; XII, /5; ЦГМ № 186/11657)
относительно сильно уплощено в латеральном направлении, верхний
отросток хорошо развит, в виде косого гребня он переходит на внутрен-
нюю сторону кости. Передний медиальный отросток выдается вперед
умеренно, но меньше, чем у Pelobates fuscus. Передне-нижний край
хорошо оформленной вертлужной впадины значительно выступает в сто-
рону. Седалищные кости (ЦГМ № 187/11657), условно отнесенные
к Е. sosedkoi sp. nov., с далеко выступающим назад, закругленным по
краю выступом, без гребня на его боковой плоскости, но с изломом ли-
нии задней кромки, как у современного Scaphiopus hatnmondi (Baird)
(Zweifel, 1956, fig. 15).
Сравнение. Новый вид рода Eopelobates обнаруживает наиболь-
шее сходство с Е. anthracinus Parker, 1929 (Estes, 1970) из олигоцена
Германии, в частности, по ямчатому, а не бугорчатому рельефу покров-
ных костей черепа, по небольшой высоте верхнечелюстной кости в обла-
сти глазницы, по общей форме латерального края лобнотеменной кости
сзади. Новое земноводное отличается от Е. anthracinus наличием рез-
кого сужения лобнотеменных костей близ переднего отверстия прони-
зывающего их канала, хорошо выраженным медиальным костным
швом между указанными костями, а также, видимо, более узкими и
длинными латеральными отростками боковых затылочных костей. От
Е. guthriei (Estes, 1970а) из эоцена США, имеющего резкое сужение
лобнотеменных костей близ середины их длины, отличен ямчатым
рельефом покровных костей черепа. От Е. leptocalaptus из кампана
Монголии (Borsuk-Bialynicka, 1978) отличается существенно меньшей
высотой и, видимо, более выраженной скульптировкой верхнечелюстной
кости в области глазниц. От ^Eopelobates sp. из Маастрихта США
(Estes, 1964, 1970а) новый вид отличен, в частности, иной формой зад-
72
ней части верхнечелюстной кости и вертлужной впадины, более широ-
кой чешуйчатой костью.
Замечания. Eopelobates sosedkoi sp. nov. — это древнейший из-
вестный вид рода Eopelobates (если считать последний естественным)
и всего семейства Pelobatidae, представленного и в современной фауне
в Северном полушарии. Данная находка, как и ряд других, сделанных
в Кызылкумах (рис. XI, 30; Несов, 1977в) показывают, что история
многих представленных ныне семейств позвоночных прослеживается
почти с начала позднего мела.
Вид Е. sosedkoi sp. nov. обнаруживает черты сходства с предста-
вителями североамериканского современного рода и подрода Scaphiopus
Holbrook, 1836, особенно с плиоценовым его представителем с терри-
тории США — S. alexanderi Zweifel, 1956 (Zweifel, 1956; Kluge, 1966;
Estes, 1970a), в частности, по форме и значительной длине латераль-
ного отростка боковой затылочной кости (этот отросток более корот-
кий у известных ранее Eopelobates). Новый вид отличен от различных
Scaphiopus формой лобнотеменных костей, иной конфигурацией дор-
сального окна в хрящевом эндокрании. От современных азиатских Pelo-
batidae родов Leptobrachium Tschudi, 1838 и Megophrys Kuhl et Van
Hasselt, 1822 (Megophryinae) амфибия из мела Кызылкумов отличается,
в частности, совершенно иным характером экзостоза костей черепа (см.
Trueb, 1973; Estes, 1970а); от представителей указанных родов, а также
от Eopelobates grandis Zweifel, 1956 новый вид, вероятно, отделен от-
сутствием горизонтального гребня на боковой плоскости седалищной
кости (Zweifel, 1956). От Macropelobates osborni из олигоцена Монго-
лии (Noble, 1924) новый вид отличается присутствием скульптуры на
верхнечелюстной кости, более мелкими размерами, но, видимо, сходен
с этим видом, Pelobates fuscus, а также Scaphiopus hammondi по строе-
нию седалищной кости.
У новой амфибии чешуйчатая и лобнотеменная кости были, оче-
видно, широко разделенными, а на месте заглазничной дуги, форми-
рующейся у ряда древних и современных Pelobatidae (Ba^oglu a. Za-
loglu, 1964), могла находиться лишь относительно слабая связка, ме-
диальный конец которой, видимо, крепился к небольшому латеральному
выступу лобнотеменной кости. Е. sosedkoi sp. nov. в целом демонстри-
рует ряд примитивных черт в строении скелета. Относительно образа
жизни Е. sosedkoi sp. nov. трудно говорить, однако отметим, что орна-
ментация покровных костей черепа встречается обычно у бесхвостых
амфибий аридных и полуаридных местообитаний (Trueb, 1973). Среди
Eopelobates наблюдается выраженная разнородность форм. В составе
этого рода, по некоторым признакам промежуточного между Megophryi-
nae и Pelobatinae (Zweifel, 1956; Estes, 1970а; Spinar, 1972), ныне, ви-
димо, объединяются формы, представляющие собой разные линии эво-
люции примитивных Pelobatidae.
Распространение. Поздний турон—сантон Центральных Кы-
зылкумов.
Материал. Ряд костей черепа и пояса задних конечностей.
Salientia fam. indet.
Род Kizylkuma Nessov, gen. nov.
Название от пустыни Кызылкум.
Типовой вид — К. antiqua sp. nov. поздний турон—сантон Цен-
тральных Кызылкумов.
Диагноз. Некрупные земноводные с гладкой наружной поверх-
ностью верхней челюсти. Верхнечелюстная кость имеет сильно высту-
пающий небный гребень, довольно резко обрывающийся назад и при-
обретающий утолщение по своей верхней кромке спереди. Линия верх-
73
него края кости над окончанием небного гребня выпукла при взгляде
сбоку, далее вперед она приобретает плавное понижение. Шовная пло-
щадка с крыловидной костью немного выпуклая спереди назад и рас-
положена над небным гребнем. Линия зубного ряда несколько выпукла
при взгляде сбоку. Поверхность, к которой осуществлялось прикрепле-
ние зубов, относительно высокая, позади небного гребня она постепенно
уменьшается в высоту. Зубы расположены с частотой около 20—25
штук в 5 мм, основания коронок задних зубов несколько расширены
в поперечном направлении.
Видовой состав. Типовой вид.
Сравнение. Отличается от меловых Eopelobates и Scotiophryne
Estes, 1969 гладкой наружной поверхностью верхнечелюстной кости,
отсутствием вогнутости ее верхней кромки, сильно развитым небным
гребнем. Среди известных автору бесхвостых выделяется наличием по-
перечной расширенности задних зубов верхнечелюстной кости.
Kizyikuma antiqua Nessov, sp. nov.
Название от antiquus (лат.) —древний.
Голотип — ZIN, PHA № К77-10, левая верхнечелюстная кость;
Узбекская ССР, Центральные Кызылкумы, Джара-Кудук, верхний ту-
рон—сантон, средняя часть тайкаршинской пачки (обнажение ЦБИ-4в).
Описание. Земноводное, видимо, могло достигать больших раз-
меров, чем Е. sosedkoi sp. nov. Над небным гребнем (рис. X, 20\ XII,
17) различим вытянутый спереди назад желобок, в котором находится
углубление, являвшееся, вероятно, местом причленения латерального
конца небной кости. На наружной стороне сзади располагается очень
неглубокое, вытянутое вдоль кости понижение.
Распространение. Поздний турон—сантон Центральных Кы-
зылкумов.
Материал. Три верхнечелюстных кости.
Род Araiobatrachus Nessov, gen. nov.
Название от Аральского моря.
Типовой вид — A. robustus sp. nov., поздний турон Центральных
Кызылкумов.
Диагноз. Относительно крупные земноводные с продольно струй-
чатой скульптурой на наружной поверхности верхнечелюстной кости.
Нижняя и верхняя кромки последней прямые и приблизительно парал-
лельные друг другу. Поверхность, к которой осуществлялось прикреп-
ление зубов, относительно высокая. Шовная площадка на контакте
с крыловидной костью значительно вытянута в вертикальном направле-
нии. Зубы довольно редкие, их было около 11 —12 в 5 мм.
Сравнение. Род Araiobatrachus gen. nov. выделяется струйчатой
наружной поверхностью верхнечелюстной кости, относительно крупными
размерами последней, редким расположением зубов.
Видовой состав. Типовой вид.
Araiobatrachus robustus Nessov, sp. nov.
Название от robustus (лат.) —крепкий.
Голотип — ZIN, PHA № К77-7, средняя часть верхнечелюстной
кости; Узбекская ССР, Центральные Кызылкумы, Джара-Кудук, верх-
ний турон, нижняя часть тайкаршинской пачки (обнажение ЦДЖ-17а).
Описание. Над не очень высоким небным гребнем, вплотную
прилежащим сверху к зубному ряду (рис. X, 22), для связи с крыловид-
ной костью имеется большая, относительно высокая площадка, несущая
74
резкие шовные гребни и борозды. На этом участке кость значительно
утолщена, тогда как на противоположной стороне фрагмента она пред-
ставляет собой относительно тонкую пластинку.
Распространение. Поздний турон Центральных Кызылкумов.
Материал. Верхнечелюстная кость.
Род Itemirella Nessov, gen. nov.
Название от колодцев Итемир в Центральных Кызылкумах.
Типовой вид — I. cretacea sp. nov. поздний турон Центральных
Кызылкумов.
Диагноз. Некрупные земноводные с гладкой наружной поверх-
ностью верхнечелюстной кости. Верхняя кромка последней имеет слабое
понижение приблизительно над наиболее выступающим участком неб-
ного гребня, нижняя кромка кости здесь почти прямая. Небный гребень
выступает сильно, но относительно плавно понижается кзади. Поверх-
ность, к которой осуществлялось прикрепление зубов, невысокая; по-
зади небного гребня она довольно резко обрывается. Верхнечелюстная
кость, продолжая быть довольно высокой, протягивается несколько
назад и за конец зубного ряда. Зубы расположены относительно редко
друг к другу, их около 12 штук в 5 мм.
Видовой состав. Типовой вид.
Сравнение. Род Itemirella gen. nov. отличается от других родов
бесхвостых земноводных мела Кызылкумов наличием понижения на
верхней кромке недалеко от заднего конца верхнечелюстной кости, не-
большой высотой оснований зубов, относительно редким их располо-
жением.
Itemirella cretacea Nessov, sp. nov.
Название от cretaceus (лат.) —меловой.
Голотип — ZIN, PHA № К77-6, задняя часть правой верхнече-
люстной кости; Узбекская ССР, Центральные Кызылкумы, Джара-Ку-
дук, верхний турон, нижняя часть тайкаршинской пачки (обнажение
ЦДЖ-17а).
Описание. Над небным гребнем (рис. X, 21) располагается хо-
рошо заметный желобок, к которому, видимо, причленялась небная
кость. Непосредственно позади зубного ряда верхнечелюстная кость
продолжает быть относительно толстой и высокой.
Распространение. Поздний турон Центральных Кызылкумов.
Материал. Верхнечелюстная кость.
Salientia fam., gen. et sp. indet.
Ряд неопределимых остатков бесхвостых происходит из верхнего турона Джара-
Кудука (нижняя часть тайкаршинской пачки).
1. Дистальная часть плеча (рис. X, 24) с сильно эродированной крупной голов-
кой имеет слабые латеральный и медиальный бугорки, fossa cubitalis ventralis ка-
жется несколько более слабой, чем у Rana (Gaupp, 1896).
2. Проксимальная часть кости предплечья (рис. X, 23) несет умеренно развитый
локтевой отросток. Лучевой отросток оставлен перпендикулярно тонкому, значительно
уплощенному стержню кости. На неизображенной стороне кости заметна борозда
sulcus longitudinalis.
3. Обломок подвздошной кости (рис. X, 26) относительно крупного бесхвостого
земноводного имеет медиальную поверхность соприкосновения двух подвздошных
костей размером 6.8X 2.5 мм. Передняя ость таза длинная, заостренная.
Найденные кости голени принадлежат, видимо, не менее чем двум формам.
4. Кости первого типа (рис. XI, 1, 2) относительно тонкие в проксимальной
части. В хорошо выраженной борозде sulcus longitudinalis недалеко от дистального
и проксимального концов кости можно видеть по небольшому отверстию, связанному
с особой более узкой бороздкой. В средней части кости располагается сквозное отвер-
75
стие для подколенной артерии, arteria poplitea (Gaupp, 1896; Терентьев, 1950); снаружи
на проксимальной части имеется слабый гребень.
5. Кости голени второго типа (рис. XI, 5), видимо, имеют более мощную прокси-
мальную часть, несколько более сильный изгиб. Небольшое отверстие в проксималь-
ной части кости расположено относительно ближе к концу кости. Следует отметить,
что кость голени такого типа, возможно, принадлежащая Pelobatidae, найдена и в верх-
ней части ходжакульской свиты (сеноман) близ Шейх-Джейли в Юго-Западных Кы-
зылкумах (ЦГМ № 191/11657).
6. В Ходжакулс (нижний турон, нижняя часть бештюбинской свиты) найдена
кость голени, отличающаяся от предыдущих более крупными размерами (рис. VIII, 16),
но имеющая отдельные черты сходства с костями второго типа.
Остальные находки происходят из тайкаршинской пачки (верхний турон — сантон)
Джара-Кудука.
7. Сфснэтмонд (рис. XII, 21) имеет крупные относительно широко расставлен-
ные отверстия для обонятельных нервов, как у Pelobates, но канал, расположенный
латеральнее от каждого из указанных отверстий, существенно более узкий, чем у со-
временного рода.
8. Значительный интерес представляют отмеченные выше позвонки и уростили
(рис. XII, 18—20, 22), обнаруженные в нижней, средней и верхней частях тайкар-
шичског пачки Джара-Кудука и сеномане Шейх-Джейли, которые имеют уникальный
набор черт и могут принадлежать к Myobatrachidae или же Pelobatidae. Среди миоба-
•тра?пд поперечк 1ми отростками уростиля обладает только Rheobatrachus (Heyer а.
Licm, 1976), но _ последнего имеется одна сочленовная поверхность на этой кости.
В то же время у очень многих Pelobatidae уростиль срастается с крестцовым позвон-
ком, поперечные отростки которого обычно очень сильно расширены. В тех же слу-
чаях, когда уростиль свободен (Megophryinae), он имеет одинарную сочленовную по-
верхиость (Griffiths, 1963; Zweifel, 1956; Estes, 1970а). Парный мыщелок на уростиле,
кажется, имеется у Pelodytinae Bruch, 1861 (см. Zweifel, 1956; Татаринов, 1964а; но
Lynch, 1973). однако для этой своеобразной группы Pelobatidae отмечаются процель-
ные позвонки
У известных Eopelobates, характеризующихся процельными позвонками, уростиль
порой не срастается с крестцовым позвонком, но последний несет очень широкие диа-
пофизы. В то же время у олигоценового Е. grandis (Zweifel, 1956), имеющего сво-
бодный хвостец, как и у рассматриваемых амфибий имелись короткие, скошенные
назад поперечные отростки. На позвонках из Кызылкумов (рис. XII, 18) заметен не-
который наклон поперечных отростков туловищных позвонков вперед, что нередко
бывает среди Pelobatidae (Noble, 1924; Zweifel, 1956); отмечаются при этом и отдель-
ные черты сходства с некоторыми Discoglossidae.
Хотя вопрос об указанных элементах осевого скелета не может быть пока
решен, автор полагает, что здесь мы имеем дело с состоянием чрезвычайно архаичным
именно для Pelobatidae, среди ранних представителей которых могли быть формы
с чертами, сохранившимися доныне у миобатрахид. Указанные два семейства, видимо,
не случайно стоят рядом в некоторых системах бесхвостых амфибий (Savage, 1973)
или сближаются как члены более крупных стволов (Estes a. Reig, 1973).
Любопытно, что в США в составе комплекса палеоценовых остатков, в котором
представлен, в частности, известный и для позднего мела род Scotiophryne (Estes,
1969а), был найден амфицельный позвонок бесхвостой амфибии, пронизанный кана-
лом для хорды (Estes, 1976). Указанный род Scotiophryne, которому, вероятно, может
принадлежать этот позвонок, характеризуется скульптированными костями черепа и
относится к Discoglossidae (Estes, 1969а) или сближается (видимо, справедливо)
с Pelobatidae (Vergnaud-Grazzini et Wenz, 1975). К последнему семейству может быть
близок и Indobatrachus из эоцена Индии.
Обсуждение
Достаточно важным является вопрос о среде обитания отмеченных
амфибий. Известно, что в начале позднего мела на территории Кызыл-
кумов (Пятков и др., 1967; Шульц, 1972; Пак, 1978) располагалась
неровная по очертаниям и неустойчивая по положению юго-западная
окраина азиатского массива суши. Климат на этой территории был от-
носительно близким к тропическому и обладал достаточно выраженной
аридностью (Синицын, 1966, 1967; Пак, 1978), проявлявшейся, однако,
не так сильно (Сочава, 1968), как в более восточных районах, напри-
мер, в Фергане (Верзилин, 1967, 1975). Близость к местам существова-
ния древних земноводных Узбекистана соленых вод, определенный уро-
вень аридности климата и общие экологические особенности амфибий
(Neill, 1958; Терентьев, 1961) указывают, что разнообразная фауна
этих животных могла, очевидно, существовать на указанной территории
76
только при наличии более или менее устойчивого стока пресных вод
с суши. Последний мог обеспечиваться, в частности, за счет зоны пере-
менно влажного климата, лежавшей к северу от Кызылкумов (Пак,
1978), а также вследствие увлажняющего действия моря на климат
прибрежной полосы. В пользу наличия хорошо увлажненных, хотя,
может быть, и не всегда очень больших по площади участков на рас-
сматриваемой территории говорят достаточно обильные находки отпе-
чатков листьев платанов и тополей (Несов, 1977а), окремненной древе-
сины, а также локальных скоплений железистых конкреций, возникших
вокруг разрушенных в прошлом веток, корней, стволов. Группы ука-
занных конкреций, отмечающие автохтонно захороненные участки сено-
манского леса (Шульц, 1965, 1972; Давиташвили и Захариева-Ковачева,
1975), известны в Юго-Западных Кызылкумах. Шесть прекрасно сохра-
нившихся участков захороненного очень густого леса обнаружены авто-*
ром в 1977 году и в верхнем туроне—сантоне Центральных Кызылку-
мов, в верхней части тайкаршинской пачки Джара-Кудука (Несов,
1979, 1980). Один из участков имеет некоторые внешние черты сход-
ства с манграми. Встречи на этом уровне и здесь же остатков акул
и водных черепах указывают на непосредственную близость лесов к бе-
реговой линии водоема. Засыпание прибрежных лесов песком на вы-
соту до 5—6 м без участия эоловых процессов (песок косослоистый,
с пылеватыми глинистыми частицами, слюдистый) могло осуществляться
лишь при наличии определенного подъема местности вглубь суши.
В целом в местонахождениях с остатками земноводных преобла-
дают мелко- и среднезернистые песчаники. Эти, а также другие геоло-
гические данные свидетельствуют об относительной удаленности
сколько-нибудь возвышенных областей сноса от мест осадконакопления
и о наличии достаточно обширной и выровненной приморской низмен-
ности. По крайней мере, часть этой низменности была покрыта боло-
тами. В последних шло осаждение из вод соединений железа, в обилии
представленных в некоторых слоях (Архангельский, 1916; Соседко,
1937, 1938; Измайлов и Доминик, 1938; Сочава, 1968).
В местонахождениях Челпык, Ходжакуль, Джара-Кудук (Несов,
1977а, г; 1979, 1980) в локальных скоплениях совместно найдены
остатки существ, обычно обитающих в море, а также принадлежащих
к группам, современные представители которых тяготеют к пресным
водам. По наблюдениям автора, местонахождения с подобными комп-
лексами остатков имеют очень широкое распространение в мелу Сред-
ней Азии и представлены, в частности, в Кызылкумах, Приташкентских
Чулях, в Фергане (Несов, 1980). В этих районах автору не известны
важные местонахождения остатков континентальнььх позвоночных, в ко-
торых совсем не было бы, например, зубов хрящевых рыб. Часто в та-
ких местонахождениях в одном тонком слое вместе с указанными остат-
ками обнаруживаются фрагменты панцирей раков и крабов, порой зубы
плезиозавров, а остатки древесины имеют следы работы моллюсков-
древоточцев. Вместе с тем в составе кызылкумских ориктоценозов
с костями земноводных отсутствуют непереотложенные остатки орга-
низмов нормальной морской солености. Так, встречающиеся на Челпыке
и Ходжакуле немногочисленные части раковин аммонитов имеют осо-
бый характер сохранности, располагаясь внутри и на поверхности ока-
танных фосфатизированных галечек, резко отличных по составу породы
от той, которая вмещает кости. Некоторые из обследованных локальных
скоплений формировались близ береговых линий. Так, в нижней части
тайкаршинской пачки Джара-Кудука слой с костями позвоночных без
видимых следов размыва перекрывается на одном из участков слоем
с остатками автохтонно захороненных деревьев, найденных в 1979 году
и произраставших, очевидно, на суше или при небольшой глубине воды.
На уровне ходжакульского скопления костей, неподалеку, в 1978 году
77
найдены захороненные волноприбойные ниши (Несов, 1979), в Ходжа-
кульсае—трещины усыхания.
Наблюдения показывают, что ориктоценозы позвоночных сеномана
и позднего турона—сантона Кызылкумов представляют собой, в основ-
ном, ассоциации остатков существ, обитавших совместно или в непо-
средственной близости друг от друга, вероятнее всего, в полузамкнутых
водоемах, связанных с морем, а также в прилегавших к ним болотах,
протоках. Указанные водоемы порой называют лиманами в широком
смысле этого слова (Одум, 1975). Ныне в условиях тропической и суб-
тропической зон в таких водоемах, разнообразных по солености вод
(Невесская, 1971), в соответствующих их достаточно опресненных участ-
ках может быть встречено, по крайней мере 52 вида и подвида амфи-
бий (Neill, 1958). По-видимому, именно в таких окраинных сильно
опресненных и пресных участках лиманов, в прилегавших болотах, про-
токах и жили описываемые амфибии мела. Отмеченная аридизация
климата в позднем мелу Средней Азии должна была в значительной
степени ограничивать возможности существования амфибий вдали от
этих водоемов.
Отсутствие в Кызылкумах находок скоплений остатков исключи-
тельно пресноводных организмов сеномана—сантона говорит о том, что
сток пресных вид с суши в этом районе скорее всего не был массиро-
ванным, а осуществлялся по мелким водотокам или некрупным прото-
кам дельт. В этих условиях достаточно устойчивое сохранение опрес-
ненности бассейнов или их участков могло достигаться лишь при силь-
ной изрезанности береговых линий, затруднявшей смешение относи-
тельно малых объемов пресных вод и больших объемов относительно
соленой воды. Общая неустойчивость среды в указанной переходной
зоне, которая, видимо, не была очень широкой, могла временами вызы-
вать массовую гибель животных, содействуя формированию достаточно
концентрированных местонахождений, включавших остатки амфибий.
В то же время высокая продуктивность отмеченной зоны (Одум, 1975)
должна была обеспечивать в ней обилие жизни.
Надо сказать, что крайне малое количество данных о находка?;
остатков земноводных современных отрядов в мезозойских отложениях
и некоторые особенности нынешнего распространения этих животных
позволили И. И. Шмальгаузену (1964а, б) высказать гипотезу об от-
ступании хвостатых и бесхвостых амфибий во время расцвета пресмы-
кающихся вверх по течению рек, в горные местности. Эта гипотеза
нашла отражение, в частности, и в учебных пособиях (Волчанецкий и
Ларина, 1973; Наумов и Карташев, 1979). Однако отмечаемые в данной
работе разнообразные земноводные существовали на приморских низ-
менностях, на которых в условиях климата, относительно близкого
к тропическому (Пак, 1978), обитало большое количество рептилий
(Несов, 1977г, 1978, 1980а). Среди последних были и разнообразные
мелкие формы (рис. VIII, 7, 18; X, 12—14; XI, 27); здесь же были
представлены разные по размерам млекопитающие (рис. IX, 22,
23; XI, 29, 30). В условиях приморских равнин обитали и северо-
американские позднеюрские и позднемеловые амфибии (Hecht a. Estes,
1960; Estes, 1964, 1965b; Estes a. Berberian, 1970). Таким образом, ви-
димо, нет оснований считать, что хвостатые и бесхвостые амфибии при-
морских низменностей мезозоя не могли иметь своих специфических
экологических ниш в условиях биологического прогресса архозавров
и других рептилий.
Комплексы остатков из мела Средней Азии и западной части Се-
верной Америки по ряду черт сходны (Несов, 1979). Так, родственные
формы хвостатых, Eoscapherpeton asiaticum и Scapherpeton tectum,
являются доминирующими формами среди позднемеловых земноводных
в Кызылкумах и в Северной Америке, вероятно, занимавшими важное
78
место в биоценозах. В то же время Batrachosauroididae и Prosirenidae
в Средней Азии отмечаются в меньшем количестве, чем на западе США
(Estes, 1964, 1970b). Любопытно отметить, что в туроне—сантоне Цен-
тральных Кызылкумов, как и в североамериканских формациях Ланс,
Хелл-Крик и некоторых других, вместе с костями хвостатых и бесхво-
стых земноводных присутствуют части черепов и высокие чешуи цельно-
костных рыб из отряда Aspidorhynchiformes (рис. XI, 4, 5), описто-
цельные позвонки и зубы со складчатостью дентина в основании
(рис. XI, 8, 9), принадлежавшие цельнокостным рыбам из отряда пан-
цирных щук Lepisosteiformes, зубы, челюсти и зубные пластинки раз-
личных других лучеперых рыб, в основном, склерофагов (рис. VIII, 17,
19—21; X, 15, 16; XI, 6) (Lambe, 1902; Osborn, 1902; Estes, 1964; Berbe-
rian a. Meszoely, 1969; Estes a. Berberian, 1970; Sahni, 1972), в том
числе Phyllodontidae и Albulidae. В обоих регионах присутствуют
в местонахождениях зубы, плавниковые и головные шипы акул семей-
ства Hybodontidae, в Джара-Кудуке попадаются зубы cf. Lonchidion
(Estes, 1964). Найдены ростральные зубы (рис. XII, 25; ЦГМ № 17/
11822) мелких пил-рыб Pristidae (Ischyriza), отмечаемые и в мелу США
(Estes, 1964), часто встречаются относительно мелкие элементы зуб-
ных аппаратов своеобразных скатов (рис. XI, 7; ЦГМ № 19—30/11822),
сходные с зубами Myledaphus bipariitus Соре. Последние являются
обычными в континентальных отложениях позднего мела на западе
США и Канады (Соре, 1876а; Lambe, 1902; Estes, 1964). В Средней
Азии и в Северной Америке среди черепах позднего мела отмечаются
представители Trionyx из Trionychidae, Adocus из Adocidae и Basilemys
из Nanchiungchelyidae, есть и некоторые другие семейства тестудинат,
общие для двух регионов (Estes, 1964; Estes et al., 1969; Несов, 19776,
в, г, 1978). Определенное сходство комплексов архозавров позднего
мела и Северной Америки достаточно хорошо известно (Рождествен-
ский и Татаринов, 1964; Рождественский, 1974). Обнаруживаются и
отдельные общие черты в фауне меловых наземных ящериц Средней
Азии и Северной Америки (рис. VIII, 18), о чем подробнее предпола-
гается сообщить в отдельной работе. В комплексах обоих регионов от-
мечаются насекомоядные млекопитающие (Insectivora) *, в том числе,
видимо, Leptictidae (рис. XI, 29) и Palaeoryctidae, однако в средне-
азиатских комплексах уже в сеномане и туроне, т. е. в самом начале
позднего мела, на приморских низменностях с климатом, близким
к тропическому, а не в умеренной зоне в глубине материка (Трофимов
и Решетов, 1975), насекомоядные доминировали среди Mammalia и
были весьма разнообразны (представлено несколько семейств)**.
Остатки земноводных, ящериц и млекопитающих в большинстве
(но не во всех) кызылкумских местонахождений отмечаются суще-
ственно реже, чем в Северной Америке (Estes, 1964; Estes et al., 1969;
Estes a. Berberian, 1970), тогда как остатки акул, костных рыб и вод-
* Интервал времени, когда в Азии насекомоядные и другие плацентарные уже
устойчиво доминировали в составе комплексов млекопитающих, все более точно
начинает соответствовать времени широкого распространения покрытосеменных расте-
ний и некоторых связанных с ними насекомых (см. Трофимов и Решетов, 1975;
Жерихин, 1978).
** Надо отметить, что интенсивные поиски остатков млекопитающих в орикто-
ценозах мела Кызылкумов были предприняты в связи с обнаружением сходства со-
става комплексов с североамериканскими по низшим позвоночным, прежде всего,
по земноводным (Несов, 1979). Различия между комплексами двух указанных регио-
нов к 1978 году касались в основном лишь отсутствия находок в Кызылкумах млеко-
питающих, что позволило до некоторой степени увериться в возможности нахождения
здесь их остатков и направить усилия на специальные поиски зубов и костей Mam-
malia. Само обнаружение в локальных верхнемеловых скоплениях остатков амфибий
рассматривается нами теперь как признак, благоприятствующий нахождению остатков
млекопитающих, костей птиц (Ichthyornithes и другие; тайкаршинская пачка).
79
ных черепах встречаются чаще. Это может указывать на формирование
кызылкумских местонахождений в зоне, расположенной несколько
ближе к внешней границе зоны развития лиманов и связанных с ними
прибрежных болот.
Ранее отмечалось значительное сходство комплексов рептилий и
амфибий, представленных ныне на юго-востоке США (в Луизиане, Мис-
сисипи, Флориде, Джорджии) и позднемеловых комплексов этих живот-
ных из западных районов Соединенных Штатов (Estes, 1969f, 1970b;
Estes a. Berberian, 1970). Все сказанное выше обращает внимание не
только на определенную генетическую связь между отдельными пред-
ставителями позвоночных из удаленных регионов Земли, связанную
с историческими возможностями дальних распространений, но и на
определенное сходство в систематическом составе позвоночных, входя-
щих в сообщества, представленные на современных приморских равни-
нах юго-востока США, на подобных древних равнинах в западной части
Северной Америки (палеоцен, Маастрихт, кампан), а также в Средней
Азии (сантон—турон, сеноман). Любопытно, что некоторые сред-
неазиатские комплексы позднего мела существовали, видимо, почти на
30 миллионов лет раньше известных североамериканских. С одной сто-
роны, это может указывать на длительный гомеостаз экологических си-
стем, включавших отмеченные организмы, на становление некоторых
важных особенностей структуры отдельных кайнозойских экосистем еще
в начале позднего мела, а с другой — на возможность существенного
смещения в пространстве за длительный срок комплексов организмов,
в значительной степени сохраняющих свою целостность. «Дивергенция»
среднеазиатских и богатых архаичными млекопитающими североамери-
канских комплексов могла произойти до становления разнообразной
фауны насекомоядных в Средней Азии. Приведенные данные, а также
сохранение ряда родов, а порой, как указывается, и некоторых видов
амфибий и других позвоночных мела после известных перестроек эколо-
гических систем на рубеже мезозоя и кайнозоя (см. приложение) по-
зволяют думать, что в процессе изменений в указанных экосистемах
приморских равнин чередование длительных периодов ценотического
равновесия и критических периодов смены доминирующих групп (Кра-
силов, 1977) могло быть выражено не очень резко. Даже при наиболее
крупном отмечаемом нами изменении в составе среднеазиатских позд-
немеловых комплексов позвоночных близ рубежа раннего и позднего
турона происходили, в основном, лишь частичные замены в составе не-
которых доминировавших групп, не сопровождаемые их вымиранием
(на уровне семейств) в масштабах всей мировой фауны. Среди черепах
это — исчезновение многочисленных в сеномане и раннем туроне Caret-
tochelyidae (Kizylkunietnys) и биологический прогресс с позднего турона
Dermatemydidae (Lindholmemys)—явление, отмечаемое и для Монго-
лии (Несов, 1981), замена крупных Ferganemys на Shachemys и Adocus.
Вместо мелкого редкого крокодила из Goniopholidae, сходного с Par al-
ligator ulgicus Ef., появляются доминант из гониофолид—крупный
Kansaisuchus borealis Ef.; длинномордая форма с огромным предче-
люстным клыком; мелкий вид с очень короткой, плоской dentale, дру-
гой— со сближенными альвеолами двух нижних клыков (Джара-Ку-
дук). Произошли смены среди акул, скатов, амиид, альбулид, регресс
Lepidotes, Dapedium, Anomoedus, распространение Megalopidae, ряда
других костистых. Среди динозавров к позднему турону неожиданно
сокращаются в числе, если не исчезают в регионе, архаичные Protoce-
ratopsidae и появляются древнейшие Ceratopsidae. Именно к этой, об-
наруженной нами смене в составе комплексов и приурочен биологиче-
ский регресс хвостатых амфибий рода Horezmia и распространение
Eoscapherpeton.
80
Приложение
Важнейшие находки остатков докайнозойских хвостатых и бесхвостых амфибий *,
а также животных, ранее причислявшихся к этим группам **
Caudata. Хвостатые
1. Vaughniella urodeloides Kuhn, 1964. Позвонок из копролита. Отличается от
Cryptobranchus немногими чертами (Vaughn, 1963 по МйПег, 1966). О. Кюном выделен
в семейство Vaughniellidae и отнесен к Cryptobranchoidea, однако интерпретация на-
ходки трудна (Estes a. Reig, 1973) и принадлежность позвонка хвостатому земно-
водному порой считается недоказанной (Татаринов, 1968). Ранняя пермь США.
2. Permotriturus herrei Tatarinov, 1968. Челюстная кость, отнесенная к пред-
ставителю нового семейства Permotrituridae (Татаринов, 1968), принадлежность к хво-
статым не вполне ясна (Ивахненко, 1978). Поздняя пермь Татарии.
3. Triassurus sixtelae Ivachnenko, 1978. Скелет личинки земноводного, причислен-
ного к новому семейству Triassuridae в составе Urodela incertae sedis (Ивахненко,
1978). Поздний триас (ранний — средний кейпер) Киргизии.
4. Boomgartia salamandriformis Huene, 1948. Мелкое животное с головной
частью длиной 1.4 мм, отнесенное первоначально к хвостатым (Huene, 1948), но
затем исключенное из их состава (см. Huene, 1956). Принадлежит к членистоногим
(Татаринов, 19646) Средняя юра Германии.
5. Атлас хвостатого земноводного и неописанные нижние челюсти из средней
юры Франции, которые имеют (Seifert, 1969b) черты связи с Desmognathinae (sensu
Estes, 1964; см. данные о Prodesmodon и Opisthotriton).
6. Cteniogenis antiquus Gilmore, 1928. Зубные кости, отнесенные первоначально
к ящерицам (Gilmore, 1928). Ф. Хюне (Huene, 1956) указал на возможность принад-
лежности их Salamandridae. Поздняя юра (формация Моррисон) Вайоминга.
7. Comonecturoides marshi Hecht et Estes, 1960. Бедро. Caudata incertae sedis,
есть сходство c Necturidae (Hecht a. Estes, I960). Обитал на низменностях, изобилую-
щих текущей водой. Поздняя юра (формация Моррисон) Вайоминга.
8. Karaurus sharovi Ivachnenko, 1978. Полный скелет представителя семейства
Karauridae (Воробьева, 1977; Ивахненко, 1978), отнесенного к подотряду Crypto-
branchoidei и обитавшего в спокойном озере. По строению туловищных позвонков
обнаруживает определенное сходство с хвостатыми амфибиями некоторых других
семейств, представленных в мезозое. Поздняя юра южного Казахстана.
9. Фрагмент нижней челюсти хвостатого (Hecht, 1963), изучавшийся, в част-
ности, Р. Эстесом и, вероятно, отнесенный им (Estes, 1965b) к Prodesmodon
(см. ниже). Ранний мел (альб) Техаса.
10. Hylaeobatrachus croyii Dollo, 1885. Скелет. Черты сходства с Proteidae и
Salamandridae (Dollo, 1885). Иногда выделяется в особое семейство. Обычно причис-
ляется к Proteidae (Татаринов, 19646; Romer, 1966), порой рассматривается как
форма неясного положения среди хвостатых (см. Naylor, 1978). Ранний мел, вельд
(Dollo, 1885) или апт — ? ранний альб Бельгии (Жерихин, 1978).
11. Prosiren elionorae Goin et Auffenberg, 1958. Позвонки, части черепа и конеч-
ностей (Estes, 1969b). При описании причислен к Sirenidae, затем к Prosirenidae
(Estes, 1969b) из Batrachosauroidoidea (Edwards, 1976). Ранний мел (альб) Техаса.
12. Ramonellus longispinus Nevo et Estes, 1969. Остатки водных хвостатых из
раннего мела Израиля (Nevo, 1964), предположительно из Prosirenidae (Nevo а.
Estes, 1969; Estes, 1969b).
13. Предполагаемые остатки крупного хвостатого земноводного из верхнего мела
(нижнего сенона) Нигера ^Broin et al., 1974).
14. Habrosaurus dilatus Gilmore, 1928. Позвонки, части черепов, Вид, первона-
чально причисленный к ящерицам (Gilmore, 1928), позже был отнесен к Sirenidae
(Estes, 1964, 1965b; Edwards, 1976). Поздний мел, Маастрихт (формация Ланс) Вайо-
минга. Род указан и для кампана (Estes, 1969f), вид — для формации Джудит-Ривер
Монтаны того же возраста (Sahni, 1972). Сирениды прослеживаются в Северной
Америке, возможно, с раннего мела (Estes, 1965b).
15. Adelphosiren olivae Goin et Auffenberg, 1958. Позвонок. Название рассматри-
вается как младший синоним для Habrosaurus dilatus Gilmore, 1928, (Estes, 1964;
Romer, 1966).
16. Proamphiuma cretacea Esies, 1969. Позвонки. Доказывается принадлежность к Am-
phiumidae (Estes, 1969f; Edwards, 1976. Поздний мел (формация Хелл-Крик) Монтаны.
17. Scapher peton tectum Cope, 1876. Позвонки и другие части скелета. Первона-
чально указывалась близость к Ambystomidae и Cryptobranchus (Соре, 1876b). Род
Scapherpeton был выделен в семейство Scapherpetonidae (Auffenberg a. Goin, 1959),
отнесенное к Cryptobranchoidea, как имеющее черты сходства с криптобранхоидеями
* Остатки мезозойских безногих амфибий (Apoda) пока не найдены, древнейшей
известной формой безногих является Apodops pricei (Caeciliidae) из позднего палео-
цена Бразилии (Estes a. Wake, 1972).
** Названия таксонов выше рода приведены в написаниях, данных в цитируемых
работах.
6 Зак. 719
81
и немного с амфиумидами (Estes, 1964). Род причислялся к Ambystomidae (Татари-
нов, 19646), предполагалась связь с Batrachosauroididae (Estes, 1969f). На основании
строения позвонков Scapherpeton в составе Scapherpetontidae был отнесен к Dicampto-
dontidae Tihen, 1958 (см. Edwards, 1976) и включен в Ambystomatoidea (Estes. 1969е.
1975). Другие описанные виды рода Scapherpeton, S. laticolle, S. excisum и S. favosum
(Cope, 1876b), порой (Auffenberg a. Goin, 1959) рассматриваются как младшие сино-
нимы для S. tectum. Считается водной формой (Estes, 1969f; Estes a. Berberian,
1970). Поздний мел — формации Джудит-Ривер (Osborn, 1902; Sahni, 1972), Белли-
Ривер (Lambe, 1902), Ланс и Хелл-Крик Северной Америки. Род Scapherpeton (Estes,
1969е; Naylor, 1978), а иногда даже вид S. tectum (Соре, 1876b; Estes, 1964, 1975)
отмечаются и для позднего палеоцена. Этот род предположительно указывается и для
раннего эоцена (Estes a. Hutchison, 1978). Данные о присутствии Scapherpeton в ран-
нем мелу (Romer, 1966), вероятно, ошибочны.
18. Hemitry pus jordanianus Соре, 1876. Позвонок (Соре, 1876b) и некоторые дру-
гие материалы (Auffenberg a. Goin, 1959). Причисляется к Scapherpeton tectum (Estes,
1964; Romer, 1966), но порой рассматривается как самостоятельная форма не вполне
ясного систематического положения (Auffenberg a. Goin, 1959; Muller, 1966) или же
как представитель Ambystomidae (Татаринов, 19646). Поздний мел (формация Джу-
дит-Ривер), США.
19. Hedronchus sternbergi Соре, 1876. Часть позвонка Scapherpeton tectum (Estes,
1964), описанная первоначально как зуб хрящевой рыбы (Соре, 1876а).
20. Lisserpeton bairdi Estes, 1965. Позвонки, челюсти. Отнесен к Scapherpetonidae
(Estes, 1965а). Рассматривается как водная форма (Estes a. Berberian, 1970). Позд-
ний мел, формации Джудит-Ривер (Sahni, 1972), Хелл-Крик Монтаны и Ланс Вайо-
минга. Род представлен и в палеоцене.
21. Opisthotriton kayi Auffenberg, 1961. Череп, позвонки. Предположительно отно-
сился к Plethodontidae или к вымершему семейству из Salamandroidea (Auffenberg,
1961), к «? Desmognathinae» (Estes, 1969е), к Batrachosauroididae в составе группы
Ambystomatoidea (Estes, 1969е). Принадлежность к Batrachosauroididae была под-
креплена Д. Эдвардсом (Edwards, 1976), который отнес последнее семейство вместе
с Prosirenidae к Batrachosauroidoidea. Рассматривается как водная форма (Estes,
1969f). Поздний мел — формации Джудит-Ривер Монтаны (Sahni, 1972) и Ланс Вайо-
минга, указывается и для палеоцена (Estes, 1975; Naylor, 1978).
22. Автору остается не до конца ясной систематическая принадлежность архаич-
ных хвостатых амфибий с опистоцельными позвонками из мела Вайоминга, упомя-
нутых В. Ауффенбергом и К. Гойном (Auffenberg a. Goin, 1959).
23. Prodesmodon copei Estes, 1964. Разные части скелетов. Первоначально отнесен
к Desmognathinae внутри Plethodontidae (Estes, 1964), позже был включен в Prosi-
renidae (Estes, 1969b), которое помещалось Р. Эстесом у основания расходящихся
стволов плетодонтид и амбистоматид. Однако, позвонки Р. copei отличаются от тако-
вых у Ambystomatidae и Plethodontidae рядом важных черт. Род Prodesmodon при-
числяется к Batrachosauroididae; принадлежность к Р. copei многих, ранее отнесенных
к нему остатков ставится под сомнение (Naylor, 1979). Позднемеловые формации
Ланс Вайоминга (Estes, 1964), Джудит-Ривер (Sahni, 1972) и Хелл-Крик Монтаны,
Олдмен и Аппе-Милк-Ривер Альберты (Naylor, 1979), раннемеловая формация Энт-
лерс Техаса (Estes, 1965b; Naylor, 1979). Младшим синонимом для Р. copei рас-
сматривается (Naylor, 1979) Cuttysarcus mcnallyi Estes, 1964, относившийся ранее
к ящерицам (Estes, 1964).
Salientia. Прыгающие или бесхвостые.
1. Lasalia cutlerensis Vaughn, 1965 из Lasaliidae Vaughn, 1965. Позвонок, для
которого указывается определенное сходство с одноименным элементом скелета бес-
хвостого земноводного (Vaughn, 1965), однако некоторые черты строения тела по-
звонка -не позволяют сближать животное с Salientia (Estes a. Reig, 1973). Ранняя
пермь Юты.
2. Отпечатки следов из перми (формация Иксе) Южной Африки, приписанные
земноводному из отряда Proanura (Griffiths, 1963). Р. Эстес и О. Рейг (Estes a. Reig,
1973) не интерпретируют их таким образом.
3. Triadobatrachus massinoti (Piveteau, 1937). Скелет. Рассматривается как пред-
ставитель отряда Proanura (семейство Triadobatrachidae Kuhn, 1962) со специфиче-
скими примитивными чертами (Piveteau, 1937; Татаринов, 1964а) или же, с опреде-
ленными оговорками, как личинка на поздней стадии метаморфоза (Griffiths, 1956,
1963), а также как водное животное, вероятно, молодое, принадлежавшее анцестраль-
ному стволу Proanura (Estes a. Reig, 1973). Ранний триас Мадагаскара.
4. Vieraella herbsti Reig, 1961. Скелет. Амфибия причислялась к семействам
Vieraellidae или Notobatrachidae. Ныне на основе подробного морфологического ана-
лиза рассматривается предположительно как представитель Ascaphidae с некоторыми
чертами сходства с Discoglossidae (Estes a. Reig, 1973); сходство с круглоязычными
отмечалось и ранее (Hecht, 1963). Ранняя юра Аргентины.
5. Notobatrachus degiustoi Reig, 1957. Скелет. Земноводное причислялось к No-
tobatrachidae (Reig, 1957 по Estes a. Reig, 1973; Romer, 1966; Nevo, 1968) или к Asca-
phidae (Griffiths, 1963; Estes a. Reig, 1973). Поздняя юра Аргентины.
82
6. Eobatrachus agilis Marsh, 1887. Плечевая кость. Форма неясного системати-
ческого положения (Marsh, 1887; Hecht a. Estes, 1960). Иногда относилась к Bufonidae
(Moodie, 1912, 1914), Montsechobatrachidae или Discoglossidae (см. Hecht a. Estes,
1960; Estes a. Reig, 1973), сближалась c Pipidae или Pipoidea (Hecht, 1963; Nevo,
1968). Отнесена к Pipoidea, но нет фактов (Estes a. Reig, 1973), которые позволили бы
однозначно отнести ее к Pipidae или Palaeobatrachidae. Поздняя юра Вайоминга.
7. Comobatrachus aenigmathus Hecht a. Estes, 1960. Плечевая кость. Системати-
ческое положение неясно, возможна близость к Leptodactylidae (Hecht a. Estes, 1960).
Более продвинутая форма, чем Eobatrachus agilis (Hecht, 1963) или не обяазательно
продвинутая (Estes a. Reig, 1973). Поздняя юра Вайоминга.
8. Montsechobatrachus gaudryi (Vidal, 1902). Скелет. Форма неясного системати-
ческого положения (Fejervary, 1921; Татаринов, 1964а; Romer, 1966). Плохая сохран-
ность образца (Vergnaud-Grazzini et Wenz, 1975) дает мало оснований (Hecht, 1963),
чтобы выделять особый род или вид, тем не менее амфибия порой выделяется в осо-
бое семейство Montsechobatrachidae. Есть черты сходства с Discoglossidae (Vergnaud-
Grazzini et Wenz, 1975). Возможна принадлежность к Palaeobatrachidae; поздняя юра
или ранний мел Испании (Estes a. Reig, 1973).
9. Eodiscoglossus santonjae Villalta, 1957. Скелеты. Представитель Discoglossi-
dae (Hecht, 1970; Estes a. Reig, 1973), обладавший зубами на верхней челюсти (Ver-
gnaud-Grazzini et Wenz, 1975). Остатки впервые изображены Ж- Пивто (Piveteav,
1955). Земноводное причислялось и к Leiopelmatidae (Ascaphidae) (Romer, 1966)
Поздняя юра или ранний мел Испании (Estes a. Reig, 1973).
10. Neusibatrachus wilferti Seifert, 1972. Скелет. Форма, рассматриваемая как
предковая для Palaeobatrachidae и Ranidae (Seifert, 1972). Причислена к Palaeobatra-
chidae в составе Pipoidea (Estes a. Reig, 1973). Поздняя юра или ранний мел Испании.
11. Очень мелкая кость, рассматриваемая как предгрудинник среднеюрской амфи-
бии, сближаемой с Ranidae (Seifert, 1969а). Эта кость может быть иным элементом
плечевого пояса бесхвостых других семейств; поздняя юра Франции (Estes a. Reig, 1973).
12. Stremtneia scabra Nopcsa, 1930 (laps. cal. — Stremmia scaber in Hecht, 1963;
Nevo, 1968). Кость конечности рептилии (Stremme, 1916), отнесенная (Nopcsa, 1930;
Huene, 1956) к Pelobatidae. Принадлежность к бесхвостым справедливо отвергается
(Hecht, 1963; Estes a. Reig, 1973). Эпифизы хорошо оформленные, кость принадлежит
представителю Amniota, но вряд ли может быть пряжкой (carpo-metacarpus) птицы
(см. Stremme, 1916; Hecht, 1963). Поднял юра Танзании.
13—15. Амфибии Thoraciliacus rostriceps Nevo, 1968, Cordicephalus gracilis Nevo,
1968 и C. longirostris Nevo, 1968 (Pipidae) из раннего мела Израиля (Nevo, 1956,
1968), возможно, неокомского возраста (Nevo a. Estes, 1969), иногда ошибочно при-
числявшиеся к Pelobatidae (Griffiths, 1963).
16. Shomronella jordanica Estes, Spinar et Nevo, 1978. Отпечатки личинок при-
митивных Pipidae. Ранний мел (готтерив) западного побережья реки Иордан (Estes,
Spinar a. Nevo, 1978).
17. Бесхвостые амфибии из раннего мела Техаса (Zangerl a. Denison, 1950).
Кости относятся к двум или трем формам (Hecht, 1963), условно причисляемым
к Leptodactylidae, последнее положение подвергается сомнению (Estes, 1970b; Estes
a. Reig, 1973).
18. Saltenia ibanezi Reig, 1959. Скелет. Амфибия рассматривалась как раннеме-
ловой представитель безъязычных с процельными позвонками (Reig, 1959). Порой
включалась среди последних в Eoxenopoididae (см. Nevo, 1968) или относилась
к Discoglossidae (Estes, 1964). Возраст этого представителя Pipoidea, имевшего описто-
цельные позвонки, определяется ныне как позднемеловой, точнее раннесенонский
(Estes a. Reig, 1973).
19— 21. Три формы: Xenopus и бесхвостое с промежуточными чертами между
Hymenochirus и Pipa (Pipidae), а также предположительно Ranidae из раннего сенона
Нигера (Broin et al., 1974).
22. Eopelobates leptocalaptus Borsuk-Bialynicka, 1978. Неполный скелет. Peloba-
tidae. Поздний мел, ?средний кампан, Монгольская Народная Республика (Borsuk-
Bialynicka, 1978).
23. ?Eopelobates sp. (Pelobatidae). Остатки из позднемеловой формации Ланс
Вайоминга. К ним предположительно относится (Estes, 1970а; Estes a. Reig, 1973)
и подвздошная кость, ранее определявшаяся как «?Ascaphidae», а также остатки,
отмеченные как «near Hylidae?» (Estes, 1964), происходящие из той же формации.
24. Scotiophryne pustulosa Estes, 1969. Части скелета. Амфибия отнесена к Disco-
glossidae (Estes, 1969а) или сближается с Pelobatidae (Vergnaud-Grazzini et Wenz,
1975). Поздний мел, формации Хелл Крик и Ланс, соответственно, Монтаны и Вайо-
минга, поздний палеоцен Монтаны. К этому же роду относилась (Estes, 1969а)
подвздошная кость, описанная ранее как «?Ascaphidae» (см. также данные по ?Еоре-
lobates) и чешуйчатая кость, указанная как «near Leptodactylidae?» из формации
Ланс Вайоминга (Estes, 1964).
25—31. Части скелетов, определенные как a) «?Pelobatidae»; б) «cf. Barbourula
sp. (Discoglossidae»; в) «near Hylidae?»; г) «near Leptodactylidae»; д) некоторые не-
определимые формы, все из позднемеловой формации Ланс Вайоминга (Estes, 1964);
е) неописанный род Discoglossidae; ж) некоторые неопределимые точно формы из
6*
83
фсфмации Хелл Крик Монтаны (Estes et al., 1969); з) несколько форм Discoglossidae
из позднемеловых формаций Ланс и др. (Estes, 1970а). Находки б) и в) были ука-
заны (Estes et al., 1969) как Pelobatidae?—Eopelobates sp.? Находка б) на основе
дополнительного материала из Монтаны (Estes et al., 1969) рассматривается как Ра-
laeobatrachidae (Estes a. Reig, 1973). По поводу находки в) см. также раздел об
? Eopelobates, о находке г) и, видимо, е) см. раздел по Scotiophryne pustulosa.
32. Кости, причисленные к трем формам Discoglossidae и к одной из Pelobati-
dae. Процельный позвонок бесхвостой амфибии, систематическое положение неясно.
Поздний мел, формация Джудит-Ривер Монтаны (Sahni, 1972).
33. Неописанные остатки, предположительно отнесенные к Leptodactylidae или
близкой к ним группе. Мел Бразилии (Estes a. Reig, 1973).
34. Eoxenopoides reuningi Haughton, 1931. Рассматривался как предположительно
позднемеловой вид Pipidae (Haughton, 1931). Принадлежность к этому семейству под-
тверждена Р Эстесом (Estes, 1977). Иногда относится к семейству Eoxenopoididae.
Эоцен (? олигоцен) Южной Африки.
ЛИТЕРАТУРА
Архангельский А. Д. Верхнемеловые отложения Туркестана. — Тр. Геол, ко-
митета, нов. сер., 1916, вып. 151, с. 1—98
Бажанов В. С. ‘Первое мезозойское млекопитающее (Beleutinus orlovi Bashanov)
из Советского Союза. — Териология, Новосибирск, «Наука», 1972, т. 1, с. 74—80.
Верзилин Н. Н. Меловые отложения юга Ферганской впадины и их нефтенос-
ность. — Тр. Ленинградск. о-ва естествоиспыт., отд. геол, и минерал., 1967,
т. 71, вып. 2, с. 3—139.
Верзилин Н. Н. Закономерности аридного литогенеза и методы их выявления
(на примере меловых отложений Ферганы). Л., изд. Ленинградск. ун-та, 1975,
140 с.
Волчанецкий И. Б. и Ларина Н. И. Пособие по зоологии позвоночных. Изд.
Саратовск. ун-та, 1973, 229 с.
Воробьева Э. И. Морфология и особенности эволюции кистеперых рыб. М.,
«Наука», 1977, 239 с.
Давиташвили Л. Ш. и Захариева-Ковачева К. Происхождение камен-
ных лесов. Тбилиси, «Мецниереба», 1975, 195 с.
Жерихин В. В. Развитие и смена меловых и кайнозойских фаунистических ком-
плексов (трахейные и хелицеровые). — Тр. Палеонтол. ин-та АН СССР, 1978,
т. 165, с. 1 — 198.
Ивахненко М. Ф. Хвостатые амфибии из триаса и юры Средней Азии. — Па-
леонтол. ж., 1978, № 3, с. 84—89.
Измайлов Ф. Ф. и Доменик П. Н. Местонахождение динозавров в Кара-
Калпакских Кзыл-Кумах. — Соц. наука и техника, 1938, № 5, с. 87—91.
Красилов В. А. Эволюция и биостратиграфия. М., «Наука», 1977, 256 с.
Лебедкина Н. С. Развитие покровных костей основания черепа хвостатых амфи-
бий сем. Hynobiidae. — В кн.: Морфология позвоночных животных (Тр. Зоол.
ин-та АН СССР, т. 33), М. — Л., «Наука», 1964, с. 75—172.
Мартинсон Г. Г. Биостратиграфия и фауна меловых континентальных отложений
Таджикской депрессии, Кызылкумов и Приташкентских Чулей. — В кн.: Кон-
тинентальные образования восточных районов Средней Азии и Казахстана (ли-
тология и биостратиграфия). Л., «Наука», 1969, с. 18—51.
Наумов Н. П. и Карташев Н. Н. Зоология позвоночных. Часть I. М., «Выс-
шая школа», 1979, 333 с.
Невесская Л. А. К классификации древних замкнутых и полузамкнутых водоемов
на основании характера их фаун. — В кн.: Современные проблемы палеонтоло-
гии (Тр. Палеонтол. ин-та АН СССР, т. 130), М., «Наука», 1971, с. 258—278.
Не со в Л. А. Новый род двукоготных черепах из верхнего мела Каракалпакии.—
Палеонтол. ж., 1977а, № 1, с. 103—114.
Не со в Л. А. О некоторых особенностях строения черепа двух позднемеловых чере-
пах.— Вестник Ленинградск. ун-та, 19776, № 21, сер. биол., вып. 4, с. 45—48.
Не со в Л. А. Филогения и систематика мезозойских черепах СССР и их связи
с современными группами. Автореф. канд. дисс., 1977в, Л., 19 с.
Нес о в Л. А. Черепахи и некоторые другие рептилии мела Каракалпакии. — В кн.:
Вопросы герпетологии. Автореф. докл. IV Всесоюзн. герпетол. конф. Л., «Наука»,
1977г, с. 155—156.
Не со в Л. А. Архаичная позднемеловая черепаха из Западного Узбекистана. —
Палеонтол. ж., 1978, № 4, с. 101 —105.
Не со в Л. А. Сходство некоторых комплексов позвоночных мела Средней Азии
и Северной Америки в связи с динамикой биогеоценотических преобразований. —
В кн.: Палеонтология и эволюция биосферы. Тез. докл. XXV сессии Всесоюзн.
палеонтол. о-ва, Л., 1979, с. 40—43.
Нес о в Л. А. Комплексы остатков организмов как индикаторы гидрохимического
состояния водоемов мела Средней Азии. — В кн.: Гидрология и гидрохимия во-
доемов различных климатических зон, Владивосток, 1980, с. 137—153.
84
Несов Л. А. О черепахе семейства Dermatemydidae из мела бассейна реки Амур
и некоторых других редких находках остатков древних черепах Азии. — В кн.:
Герпетологические исследования в Сибири и на Дальнем Востоке. Л., изд. Зоол.
ин-та АН СССР, 1981, с. 69—73.
Несов Л. А. и Трофимов Б. А. Древнейшее насекомоядное мела Узбекской
ССР. — Докл. АН СССР, 1979, т. 247, № 4, с. 952—955.
Одум Ю. Основы экологии. М., «Мир», 1975, 740 с.
Пак А. И. Мезозойские и кайнозойские коры выветривания Западного Узбекистана.
Ташкент, «Фан», 1978, 154 с.
Пятков К. К., Пяновская И. А., Бухарин А. К. и Быковский Ю. К.
Геологическое строение Центральных Кызылкумов. Ташкент, «Фан», 1967, 177 с.
Рождественский А. К. История динозавровых фаун Азии и других материков
и вопросы палеогеографии. — В кн.: Фауна и биостратиграфия мезозоя и кайно-
зоя Монголии (Тр. Совм. Советско-Монгольской палеонтол. эксп., вып. 1), М.,
«Наука», 1974, с. 107—131.
Рождественский А. К- и Татаринов Л. П. (ред.). Земноводные, пресмы-
кающиеся и птицы. В серии: Основы палеонтологии, М., «Наука», 1964, 722 с.
Саломатина Н. И. К вопросу о биомеханике черепа амфибий. — Докл. АН
СССР, 1975, т. 224, № 6, с. 1456—1459.
Синицын*В. М. Древние климаты Евразии. Часть 2. Мезозой. Л., изд. Ленин-
градск. ун-та, 1966, 166 с.
Синицын В. М. Введение в палеоклиматологию. Л., «Наука», 1967, 232 с.
Соседко А. Ф. Кладбище позвоночных в центре пустыни Кзыл-Кум. — Соц. наука
и техника, 1937, № 5, с. 106—111.
Соседко А. Ф. К статье Ф. Ф. Измайлова и П. Н. Доменика «Местонахождение
динозавров в Кара-Калпакских Кзыл-Кумах. — Соц. наука и техника, 1938, № 7,
с. 90—91
Сочава А. В. Красноцветы мела Средней Азии. Л., «Наука», 1968, 120 с.
Татаринов Л. П. О роли условий жизни в филогенезе земноводных. Автореф.
канд. дисс., М., 1953, 10 с.
Татаринов Л. П. Надотряд Salientia. Прыгающие или бесхвостые. — В кн.: Основы
палеонтологии. Земноводные, пресмыкающиеся и птицы. М., «Наука» 1964а,
с. 125—133.
Татаринов Л. П. Подкласс Lepospondyli. Лепоспондильные. — В кн.: Основы
палеонтологии. Земноводные, пресмыкающиеся и птицы. М., «Наука», 19646,
с. 144—164.
Татаринов Л. П. Находка примитивного хвостатого земноводного в верхней
перми Поволжья. — В кн.: Верхнепалеозойские и мезозойские земноводные и
пресмыкающиеся СССР. М., «Наука», 1968, с. 7—10.
Терентьев П. В. Лягушка. М., «Советская наука», 1950, 345 с.
Терентьев П. В. Герпетология. Учение о земноводных и пресмыкающихся. М.,
«Высшая школа», 1961, 334 с.
Трофимов Б. А. и Решетов В. Ю. Азия как центр развития млекопитаю-
щих. — Природа, 1975, № 8, с. 32—43.
Шилин П. В. Сенонская флора Восточного Приаралья и Центрального Казахстана
и история ее формирования. — В кн.: Материалы по истории фауны и флоры
Казахстана, Алма-Ата, 1974, т. 6, с. 121 —129.
Шилин П. В. Новая находка отпечатка пера меловой птицы в Казахстане.—
Тез. докл. Всесоюзн. орнитол. конф., Черкассы, 1977, часть 1, с. 33.
Шмальгаузен И. И. Положение хвостатых амфибий среди низших наземных
позвоночных. — В кн.: Морфология позвоночных животных (Тр. Зоол. ин-та
АН СССР, т. 33), М. — Л., «Наука», 1964а, с. 5—33.
Шмальгаузен И. И. Происхождение наземных позвоночных. М., «Наука», 19646,
271 с.
Шульц С. С. мл. Меловые леса в низовьях Аму-Дарьи. — Тез. докл. IX сессии
Всесоюзн. палеонтол. о-ва, Л., 1963, с. 77—78.
Шульц С. С. мл. Геологическое строение зоны сочленения Урала и Тянь-Шаня.
М., «Недра», 1972, 207 с.
Auffenberg W. A new genus of fossil salamander from North America. — Amer.
Midland Naturalist, 1961, vol. 66, n. 2, p. 456—465.
Auffenberg W. a. Goin C. J. The status of the salamander genera Scapherpeton
and Hemitrypus of Cope. — Amer. Mus. Novitates, 1959, n. 1979, p. 1 —12.
Ba$oglu M. a. Zaloglu S. Morphological and osteological studies in Pelobates
syriacus from Izmir Region, Western Anatolia (Amphibia, Pelobatidae). — Senken-
berg. biol., 1964, Bd. 45, H. 3/5, S. 233—242.
Bolkay S. J. Die Schadel der Salamandrinen mit besonders Rucksicht auf ihre sys-
tematische Bedeutung. — Z. Anat. Entwicklungsgesch., 1928, Bd. 86, H. 3/4, p. 259—319.
Borsuk-Bialynicka M. Eopelobates leptocalaptus sp. n. — the first Upper Creta-
ceous pelobatid frog from Asia. — PalaeontoL Polonica, Res. Pol.-Mong. Palaeontol.
Exp., 1978, pt. 8, n. 38, p. 57—63.
Broin F. de, Buffetaut E., Koeniguer J. C., Rage J. - C., Russel D.,
Taquet Ph., Vergnaud-Grazzini C. et Wenz S. La faune de Verte-
85
brcs contjncntau.x du gisement d’In Beceten (Senonien du Niger). —C. R. Acad.
Sci. Paris, 1974, i. 279, p. 469—472.
Cope E. D. Descriptions of some vertebrate remains from the Fort Union beds of Mon-
tana. — Proc. Acad. Nat. Sci. Philadelphia, 1876a (1877), pt. 3, p. 248—261.
Cope E. D. On some extinct reptiles and Batrachia from the Judith River an : Fox
Hills beds of Montana. — Proc. Acad. Nat. Sci. Philadelphia, 1886b (1887), pt. 3,
p. 340—359.
Do 11 о L. Note sur batracien de Bernissart. — Bull. Mus. Roy. Hist. Nat. Belg., 1885
(1884—1885), t. 3, p. 85—93.
Edwards J. L. Spinal nerves and their bearing on salamander phylogeny. — J. Morph..
1976, vol. 148, n. 3, p. 305—327.
Estes R. Fossil vertebrates from the Late Cretaceous Lance Formation, eastern Wyo-
ming.— Univ. California Publ., Geol. Sci., 1964, vol. 149, p. 1 —180.
Estes R. A new fossil salamander from Montana and Wyoming. — Copeia, 1965a,
n. 1, p. 90—95.
Estes R. Fossil salamanders and salamander origins. — Amer. Zoologist, 1965b, vol. 5,
n. 2, p. 319—334.
Estes R. A new discoglossid frog from Montana and Wyoming. — Breviora, 1969a,
n. 328, p. 1—7
Estes R. Prosijenidac, a new family of fossil salamanders. — Nature, 1969b, vol. 224,
n. 5214, p. 87—88.
Estes R. Relationships of two Cretaceous lizards (Sauria, Teiidae). — Breviora, 1969c,
n. 317, p. 1—8.
Estes R. Studies on fossil phyllodont fishes : interrelationships and evolution in the
Phyllodontidae (Albuloidei). — Copeia, 1969d, n. 2, p. 317—331.
Estes R. The Batrachosauroididae and Scapherpetontidae, Late Cretaceous and Early
Cenozoic salamanders. — Copeia, 1969e, n. 2, p. 225—234.
Estes R. The fossil record of amphiumid salamanders. — Berviora, 1969f, n. 322,
p. 1—11.
Estes R. New fossil pelobatid frogs and a review of the genus Eopelobates. — Bull.
Mus. Comp. Zool., 1970a, vol. 139, n. 6, p. 293—339.
Estes R. Origin of recent North American lower vertebrate fauna: an inquiry into
the fossil record. — Forma et Functio, 1970b, vol. 4, p. 139—163.
Estes R. Lower vertebrates from the Fort Union formation, Late Paleocene, Big Horn
Basin, Wyoming. — Herpetologica, 1975, vol. 31, n. 4, p. 365—385.
Estes R. Middle Paleocene lower vertebrates from the Tongue River formation, sou-
theastern Montana.—J. Paleontol., 1976, vol. 50, n. 3, p. 500—520.
Estes R. Relationships of the South African fossil frog Eoxenopoides reuningi (Anura,
Pipidae). — Annals South Afric. Mus., 1977, vol. 73, pt. 2, p. 49—80.
Estes R. a. Berberian P. Palaeoecology of a Late Cretaceous vertebrate commu-
nity from Montana. — Breviora, 1970, n. 343, p. 1—35.
Estes R., Berberian P. a. Meszoely С. A. M. Lower vertebrates from the Late
Cretaceous Hell Creek Formation, McCone County, Montana. — Breviora, 1969,
n. 337, p. 1—32.
Estes R. a. Darevsky I. Fossil amphibians of the Miocene of the North Caucasus,
U.S.S.R. — J. PalaeontoL Soc. India, 1977, vol. 20, p. 164—169.
Estes R., Hecht M. a. Hoffstetter R. Paleocene amphibians from Cernay,
France. — Amer. Mus. Novitates, 1967, n. 2295, p. 1—25.
Estes R. a. Hoffstetter R. Les Urodeles du Miocene de La Grive-Saint-Alban
(Isere, France). — Bull. Mus. Nat. d’Hist. Nat., 1976, ser. 3, n. 398, sci. ter. 57,
p. 297—343.
Estes R. a. Hutchison H. Eocene lower vertebrates from Ellesmire Island. —
Abstracts with programs, Geol. Soc. America 1978 Annual Meetings, Toronto,
n. 10, p. 398.
Estes R. a. R e i g O. A. The early fossil record of frogs: a review of the evidence. —
In: Vial J. L. (ed.), Evolutionary biology of the anurans. Contemporary research
on major problems. Columbia, Missouri; Univ, of Missouri Press, 1973, p. 11—63.
Estes R., S p i n a r Z. V. a. Nevo E. Early Cretaceous pipid tadpoles from Israel
(Amphibia: Anura). — Herpetologica, 1978, vol. 34, n. 4, p. 374—393.
Estes R. a. Wake M. H. The first fossil record of caecilian amphibians. — Nature,
1972, vol. 239, p. 5369, p. 228—231.
Fejervary G. J. von. Kritische Bemerkungen zur Osteologie, Phylogenie and Syste-
matik der Anuren. — Archiv Naturgesch., 1921, Jahrg. 87, Abt. A, H. 3, S. 1—30.
G a u p p E. A. Ecker’s und R. Wiedersheim’s Anatomie des Frosches auf Grund eigener
Untersuchungen durchaus neu bearbeitet. Erste Abteilung. Lehre vom Skelet und
vom Muskelsystem. Friedrich Vieweg und Sohn, Braunschweig, 1896, 234 S.
Gilmore Ch. W. Fossil lizards of North America. — Mem. Nat. Acad. Sci., 1928,
vol. 22, p. 1—201.
Gilmore C h. W. Osteology of Polyglyphanodon, an Upper Cretaceous lizard from
Utah.—Proc. U. S. Nat. Mus., 1942, vol. 92, n. 3148, p. 229—265.
Gilmore Ch. W. Osteology of Upper Cretaceous lizards from Utah, with a descrip-
tion of a new species. — Proc. U. S. Nat. Mus., 1943, vol. 93, p. 209—214.
86
Goin C. J. a. Auffenberg W. New salamanders of the family Sirenidae from
the Cretaceous of North America. — Fieldiana: Geology, 1958, vol. 10, n. 33.
p. 449—4o9.
Griffiths J. Status of Protobatrachus massinoti. — Nature, 1956, vol. 177, n. 4503,
p. 342—343.
Griffiths J. The phylogeny of the Salientia. — Biol. Rev. 1963, vol. 38, p. 241 —
292.
Haughton S. N. On a collection of fossil frogs from the Clays at Banke. — Trans.
Roy. Soc. South Africa, 1931 (1930—1931), vol. 19, n. 1—4, p. 233—249.
Hecht M. K. Reevaluation of the early history of the frogs. Part II. — Syst. Zool.
1963, vol. 12, n. 1, p. 20—35.
Hecht M. K. The morphology of Eodiscoglossus, a complete jurassic frog. — Amer.
Mus. Noxitates, 1970, n. 2424, p. 1 —17.
Hecht M. K. a. Estes R. Fossil amphibians from Quarry Nine. — Postilia, Yale
Peabody Mus. Nat. Hist., 1960, n. 46, p. 1 — 19.
Her re W. Die Schwanzlurche der mitteleocanen (oberlutetischen) Braunkohle Geisel-
tales und Phvlogenie der Urodelen unter Einschluss der fossilen Formen. — Zoolo-
gica, Stuttgart. 1935, Bd. 33, Lief. 4, H. 87, S. 1—85.
Heyer R. W. a. Liem D. S. Analysis of the intergeneric relationships of the
Australian frog family Myobatrachidae. — Smithsonian Contrib. Zool., 1976, n. 233,
p. 1—29.
Hinderstein B. The desmognathine jaw mechanism (Amphibia: Caudata: Pletho-
dontidae). — Herpetologica, 1971, vol. 27, n. 4, p. 467—476.
Huene F. von. Ein echter Urodele aus dem unteren Dogger. — Neues Jahrbuch Mi-
neral., Geol., Palaontol., Jahrg. 1945—1948, 1948, Abt. B: Geol., Palaontol.,
S. 33—39.
Huene F. von. Palaontologie und Phylogenie der niederen Tetrapoden. Jena, Gustav
Fischer Verlag, 1956, 716 S.
Kluge A. G. A new pelobatine frog from the Lower Miocene of South Dacota with
description of evolution of the Scaphiopus — Spea complex. — Contrib. Sci. Los
Angeles County Mus., 1966, n. 113, p. 1—26.
L a m b e L. M. On Vertebrata of the Mid-Cretaceous of the North West Terri-
tory. 2. New genera and species from the Belly River Series (Mid-Cretaceous).—
Contrib. Canad. Palaeontol., 1902, vol. 3, pt. 2, p. 23—81.
Lynch J. D. The transition from archaic to advanced frogs. — In: Vial J. L. (ed.),
Evolutionary biology of the anurans. Contemporary research on major problems.
Columbia, Missouri; Univ, of Missouri Press, 1973, p. 133—182.
Marsh О. C. American Jurassic mammals. — Amer. J. Sci., 1887, ser. 3, vol. 33,
p. 327—348.
Meszoely С. A. M. A new cryptobranchid salamander from the Early Eocene of Wyo-
ming. — Copeia, 1967, n. 2, p. 346—349.
Moodie R. L. An American Jurassic frog. — Amer. J. Sci., 1912, ser. 4, vol. 34,
p. 286—288.
Moodie R. L. The fossil frogs of North America. — Amer. J. Sci., 1914, ser. 4, vol. 38,
p. 531—536.
Miiller A. H. Lehrbuch der Palaozoologie. Band III. Vertebraten. Teil 1. Fische im
weiteren Sinne und Amphibien. Jena, Gustav Fischer Verlag, 1966, 638 S.
Miiller A. H. Lehrbuch der Palaozoologie. Band III. Vertebraten. Teil 2. Reptilien
und Vogel. Jena, Gustav Fischer Verlag, 1968, 657 S.
Naylor B. G. The earliest known Necturus (Amphibia, Urodela) from the Paleocene
Ravenscrag Formation of Saskatschevan. — J. HerpetoL, 1978, vol. 12, n. 4,
p. 565—569.
Naylor B. G. The Gretaceous salamander Prodesmodon (Amphibia: Caudata).—
Herpetologica, 1979, vol. 35, n. 1, p. 11—20.
Neill W. T. The occurence of amphibians and reptiles in saltwater areas, and a bib-
liography.— Bull. Mar. Sci. Gulf and Caribbean, 1958, vol. 8, n. 1, p. 1—97.
Nevo A. Fossil frogs from a Lower Cretaceous bed in southern Israel (central Ne-
gev). — Nature, 1956, vol. 178, n. 4543, p. 1191 — 1192.
Nevo E. Fossil urodeles in early Lower Cretaceous deposits of Makhtesh Ramon,
Israel. — Nature, 1964, vol. 201, n. 4917, p. 415—416.
Nevo E. Pipid frogs from the Early Cretaceous of Israel and pipid evolution. — Bull.
Mus. Comp. Zool., 1968, vol. 136, n. 8, p. 255—318.
Nevo E. a. E s t e s R. Ramonellus longispinus, an Early Cretaceous salamander from
Israel. — Copeia, 1969, n. 3, p. 540—547.
Noble G. K- Phylogeny of the Salientia. I. The osteology of the thigh musculature,
their bearing on classification and phylogeny. — Bull. Amer. Mus. Nat. Hist., 1922,
vol. 46, p. 1—87
Noble G. K. A new spadefoot toad from the Oligocene of Mongolia with a summary
of the evolution of the Pelobatidae. — Amer. Mus. Novitates, 1924, n. 132,
p. 1 — 15.
Noble G. K. The fossil frogs of the intertrapped beds of Bombay, India. — Amer. Mus.
Novitates, 1930, n. 401, p. 1 — 13.
87
Nopcsa F. von. Notes on Stegocephalia and Amphibia. — Proc. Gen. Meet. Sci.
Bus. Zool. Soc. London, 1930, pt. 4, p. 979—995.
Osborn H. F. On Vertebrata of the Mid-Cretaceous of the North West Territory. 1. Di-
stinctive characters of the Mid-Cretaceous fauna. — Contrib. Canad. Palaeontol.,
1902, vol. 3, pt. 2, p. 7—21.
Parker W. K- On the structure and development of the skull in the Urodeles.— Trans.
Zool. Soc. London, 1882, vol. 11, pt. 6, n. 11, p. 171—214.
Piveteau J. Paleontologie de Madagascar. XXIII. Un amphibien du trias inferieur.
Essai sur 1’origine et 1’evolution des amphibiens anoures. — Ann. Paleontol., 1937,
t. 26, p. 135—178.
Piveteau J. Anoura. — In: Traite de Paleontologie. Paris, Masson et Cie, 1955,
t. 5, p. 250—274.
Reig O. A. Primeros datos descriptivos sobre los anuros del eocretaceo de la provin-
cia de Salta (Rep. Argentina). — Ameghiniana, 1959, t. 1, n. 4, p. 3—8.
Romer A. S. Vertebrate Paleontology. Chicago and London, The University of Chicago
Press, 1966, 468 p.
Sahni A. The vertebrate fauna of the Judith River Formation, Montana. — Bull.
Amer. Mus. Nat. Hist., 1972, vol. 147, art. 6, p. 321—412.
Savage J. M. The geographical distribution of frogs: patterns and predictions.—
In: Vial J. L. (ed.), Evolutionary biology of the anurans. Contemporary research
on major problems. Columbia, Missouri; Univ, of Missouri Press, 1973, p. 65—132.
Seifert J. Sternalelement (omosternum) eines mitteljurassischen Anuren von SE-
Aveyron (Siidfrankreich). — Z. zool. System, u. Evolutionforsch., 1969a, Bd. 2,
S. 145—153.
Seifert J. Urodelen-Atlas aus dem obersten Bajocien von SE-Aveyron (Siidfrank-
reich). — Palaontol. Z., 1969b, Bd. 43, S. 32—36.
Seifert J. Ein Vorlaufer der Froschfamilien Palaeobatrachidae und Ranidae im Grenz-
bericht Jura — Kreide. — Neues Jahrbuch Geol. Palaontol., Monatshefte, Jahrg.
1972, H. 2, S. 120—131.
S p i n a r Z. V. Tertiary frogs from Central Europe. Prague, Academia, 1972, 286 p.
Spinar Z. V. Endolymphatic sacs and dorsal endocranial pattern: their significance
for systematics and phylogeny of frogs. — Vestnik Ustfedniho ustavu geologickeho,
1976, n. 51, p. 285—290.
Stremme H. von. Uber die durch Baudverknocherung hervorgerufene proximale
Verschmelzung zweiter Mittelhand- oder Mittelfussknochen eines Reptiles.—Archiv
f. Biontologie, 1916, Bd. 4, H. 1, S. 143—144.
Sulimski A. Macrocephalosauridae and Polyglyphanodontidae (Sauria) from the
Late Cretaceous of Mongolia. — Palaeontol. Polonica, Res. Pol.-Mong. Palaeontol.
Exp., 1975, n. 33, p. 25—102.
Taylor E. H. a. Hesse C. J. A new salamander from the Upper Miocene beds
of San Jacinto County, Texas. — Amer. J. Sci., 1943, vol. 241, n. 3, p. 185—193.
Teege M. J. Studien zur Entwicklung und Gestalt der Urodelenwirbel. — Z. wiss.
Zool., 1958, Bd. 160, H. 1—2, S. 95—163.
Trueb L. Bones, frogs, and evolution. — In: Vial J. L. (ed.), Evolutionary biology
of the anurans. Contemporary research on major problems. Columbia, Missouri;
Univ, of Missouri Press, 1973, p. 65—132.
Vaughn P. P. Frog-like vertebrae from the Lower Permian of southeastern Utah.—
Contrib. Sci. Los Angeles County Mus., 1965, n. 87, p. 1 —18.
Vergnaud-Grazzini C. et Wenz S. Les discoglossides du Jurassique supe-
rieur du Montsech (Province de Lerida, Espagne). — Ann. Paleontol. (Vertebres).
1975, t. 61, fasc. 1, p. 19—48.
Zangerl R. a. Denison R. H. Discovery of Early Cretaceous mammals and frogs
in Texas. — Science, 1950, vol. 112, n. 2398, p. 61.
Z w e i f e 1 R. G. Two pelobatid frogs from the Tertiary of North America and their
relationships to fossil and recent forms. — Amer. Mus. Novitates, 1956, n. 1762,
p. 1—45.
Cretaceous salamanders and frogs of Kizylkum Desert
L. A. Nessov
Leningrad State University
Cretaceous salamanders (Scapherpetontidae, Batrachosauroididae, Prosirenidae, fam.
indet. — 6 new species of 5 genera in all) and anurans (Pelobatidae, fam. indet.—
4 new species of 4 genera) which had existed on coastal plains are for the first time
described for the USSR from Turonian — Santonian of central and south-western parts
of Kizylkum Desert. Representatives of these orders have been reported also from Ceno-
manian of Kizylkum. All materials consist of several thousands of bones. Similarity has
been noted between Middle Asian Turonian — Santonian vertebrate faunae amphibians
included and much later ones of Campanian and Maerstrichtian age of North America
also associated with coastal plains. A world list of the main known Precenozoic uro-
delan and anuran amphibians is given.
88
О ВОСТОЧНОЙ ГРАНИЦЕ
АРЕАЛА ДЛИННОНОГОГО ГОЛОПАЛОГО ГЕККОНА
GYMNODACTYLUS LONGIPES NIKOLSKY, 1896
Н. Л. Орлов
Зоологический институт АН СССР (Ленинград)
По современным представлениям (Wermuth, 1965; Горелов и др.,
1974; Банников и др., 1977; Щербак и Голубев, 1977; Щербак, 1979)
длинноногий голопалый геккон, Gymnodactylus longipes Nikolsky, 1896
распространен в восточном Иране, северо-западном Афганистане и
южной Туркмении. Восточная граница ареала этого вида проходит по
междуречью Мургаба и Теджена (Горелов и др., 1974, рис. 1).
Недавно при разборке неопределенных материалов в коллекции
отделения герпетологии Зоологического института АН СССР (ЗИН)
были обнаружены 4 экз. (3 взрослых и 1 молодой) гекконов, опреде-
ленные мною как G. longipes (вклейка, рис. XIII). Они были собраны
В. Я. Лаздиным 15 мая 1915 года в окрестностях города Термез и хра-
нятся сейчас в коллекциях ЗИН (№ 19215a-d). Местопребывание
В. Я. Лаздина и соответственно место сбора гекконов 15 мая 1915 года
подтверждается описанием маршрута его поездки (Лаздин, 1915). Эта
точка отстоит от ближайших находок примерно на 500 км на северо-
восток и имеет координаты: 67°20' восточной долготы и 37°18' северной
широты.
В настоящее время различают два подвида длинноногого голопа-
лого геккона: G. longipes longipes Nikolsky, 1896 и G. I. microlepis
Lantz, 1918 (Горелов и др., 1974; Банников и др., 1977), однако рас-
пространение их остается неясным, что частично объясняется недоста-
точно четким пониманием объема этого вида, а также некоторыми но-
менклатурными неточностями. Так, например, в качестве типовой тер-
ритории G. longipes, а следовательно, и номинативного подвида G. I.
longipes указывают (Банников и др., 1977) долину реки Теджен, южная
Туркмения. Однако, как известно, А. М. Никольский (1896) описал
этот вид по сборам Н. А. Зарудного с территории восточного Ирана.
Из записей в каталоге отделения герпетологии ЗИН вытекает, что
экземпляры, по которым был описан этот вид, были пойманы в окрест-
ностях селения Hex в южной части провинции Хорасан, где Н. А. За-
рудный (1896) находился 19—20 мая и 8—9 июня 1896 года. Таким
образом, этот пункт, имеющий координаты 66° восточной долготы и
31°30' северной широты, следует рассматривать в качестве terra typica
restricta. Один из этих экземпляров типовой серии я предлагаю обозна-
чить в качестве лектотипа (ЗИН.8810.1), а другие экземпляры соответ-
ственно в качестве паралектотипов (ЗИН.8810.2; ЗИН.8811). Одновре-
менно указанные типовая территория и лектотип являются таковыми
и для номинативного подвида G. I. longipes. В то же время указанная
в качестве terra typica для этого вида долина реки Теджен (Банников
и др., 1977) может считаться таковой только для подвида G. I. micro-
lepis Lantz, 1918, описанного в свое время в ранге самостоятельного
вида (Lantz, 1918). Ю. К. Горелов и др. (1974) выделили один из трех
типовых экземпляров Л. А. Лантца, хранящихся в коллекциях Зооло-
гического музея Московского государственного университета, в каче-
стве голотипа (№ 151а). Однако, так как сам Л. А. Лантц типового
экземпляра не выделил, то в соответствии с Международным кодексом
зоологической номенклатуры этот экземпляр может считаться только
лектотипом, а остальные два соответственно — паралектотипами.
Долгое время этот вид смешивали с Gymnodactylus fedtschenkoi
Strauch, 1887. Так, например, С. А. Чернов (1931) свел G. microlepis
89
в синонимы G. fedtschenkoi. В свою очередь, Р. И. Кларк и др. (Clark
et al., 1969) рассматривают G. longipes также в качестве синонима
G. fedtschenkoi, хотя ранее один из авторов (Anderson, 1963, 1968)
считал G. longipes самостоятельным видом. Вновь G. longipes в каче-
стве самостоятельного вида, а также G. I. microlepis в качестве новой
комбинации рассматривается Ю. К. Гореловым и др. (1974), что при-
нято также в последнем определителе герпетофауны СССР (Банников
и др., 1977).
В описаниях коллекций, собранных В. Я. Лаздиным, С. Ф. Царев-
ский (1917) упоминает только G. fedtschenkoi. По-видимому, обнару-
женные мною 4 экз. G. longipes, пойманные также В. Я. Лаздиным,
остались незамеченными или были оставлены С. Ф. Царевским среди
неопределенных материалов в связи с возникшими трудностями в видо-
вой идентификации этих особей. Три экземпляра (ЗИН. 19215а, Ь, с),
Расположение нижнечелюстных щитков у Gymnodacty-
lus longipes Nikolsky из района города Термез, Узбеки-
стан.
Слева — типичное для вида (ЗИН.19215а), справа — более ха-
рактерное для G. fedtschenkoi (3HH.19215d). Увеличение Х2.
на мой взгляд, полностью характеризуются как G. longipes (вклейка,
рис. XIII). Четвертый же экземпляр (ЗИН. 19215d), плохо сохранив-
шийся, имеет сходящиеся друг с другом нижнечелюстные щитки, неот-
деленные горловыми чешуйками, что более типично для G. fedtschenkoi
(см. рис.). Однако шов, по которому соприкасаются нижнечелюстные
щитки, очень короткий, что может иметь место и у G. longipes (Николь-
ский, 1896; Горелов и др., 1974; Банников и др., 1977). Других отличий
в фолидозе и габитусе от G. longipes этот экземпляр не имеет. Таким
образом, и этот экземпляр следует, по-видимому, отнести к G. longipes.
До сих пор все экземпляры длинноногого голопалого геккона
с территории СССР относили к подвиду G. I. microlepis (Горелов и др.,
1974; Банников и др., 1977). Однако по таблице, предложенной для
определения подвидов G. longipes (Горелов и др., 1974), два взрослых
хорошо сохранившихся экземпляра (ЗИН. 19215 а и Ь) идентифици-
руются как представители номинативного подвида, G. I. longipes Ni-
kolsky. В частности, на это четко указывает относительная длина перед-
них и задних конечностей этих экземпляров (см. табл.). Подвидовую
принадлежность двух других экземпляров (ЗИН. 19215 с и d) трудно
определить из-за плохой сохранности одного и малого размера другого
экземпляра.
Таким образом, описанная мною находка длинноногого голопалого
геккона, сильно удаленная к северо-востоку от известной границы
Относительная длина конечностей у Gymnodactylus longipes
longipes Nikolsky, 1896 из района города Термез
ЗИН № L ^ра ~Т~ АРР L
19215 а 67 0.52 0.70
19215 b 62 0.50 0.69
Примечание. L — длина тела (туловище с головой),
Lpa и ЬрР—длина передних и задних конечностей, соот-
ветственно, в мм.
90
ареала, относится к номинативному подвиду G. I. longipes, впервые
указываемому для территории СССР.
В заключение считаю своим приятным долгом поблагодарить
И. С. Даревского и Л. Я. Боркина за консультации в работе.
ЛИТЕРАТУРА
Банников А. Г., Д а р е в с к и й И. С., И щ е н к о В. Г., Р у с т а м о в А. К. и
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР.
М., «Просвещение», 1977, 414 с.
Горелов Ю. К., Даревский И. С. и Щербак Н. Н. Два новых для фауны
СССР вида ящериц из семейства гекконов. — Вестник зоологии, 1974, № 4,
с. 33—39.
Зарудный Н. А. Маршрут Н. А. Зарудного по восточной Персии в 1896 г.—
Ежегодник Зоол. муз. Ймп. Акад. Наук, С.-Петербург, 1896, т. 1, с. XVIII—
XXI.
Л аз дин В. Я. Маршрут поездки с зоологической целью в Восточную Бухару и
на Западные Памиры летом 1915 года. — Ежегодник Зоол. муз. Имп. Акад.
Наук, Петроград. 1916, т. 20 за 1915, с. IV—VIII.
(Никольский А. М.) Nikolsky А. М. Diagnoses Reptilium et Amphibium
biorum novorum in Persia orientalia a N. Zarudny collectorum. — Ежегодник
Зоол. муз. Имп. Акад. Наук, С.-Петербург, 1896, т. 1, с. 369—372.
Даревский С. Ф. Пресмыкающиеся и земноводные, собранные В. Я. Лаздиным
в Южной и Восточной Бухаре и в С.-В. части Закаспийской области летом
1915 года. — Ежегодник Зоол. муз. Росс. Акад. Наук, Петроград, 1922, т. 22
за 1917—1921, с. 79—90.
Чернов С. А. Об идентичности Gymnodactylus microlepis Lantz и G. fedtschenkoi
Strauch. — Докл. АН СССР, 1931, № 2, с. 59—61.
Щербак Н. Н. и Голубев М. Л. Материалы к систематике палеарктических
гекконов (роды Gymnodactylus, Bunopus, Alsophylax). — В кн.: Герпетологиче-
ский сборник (Тр. Зоол. ин-та АН СССР, т. 74), 1977, с. 120—133.
Щербак Н. Н. Новые находки ящериц и змей на территории Средней Азии. —
Вестник зоологии, 1979, № 1, с. 68—70.
Anderson S. С. Amphibians and reptiles from Iran. — Proc. California Acad. Sci.,
1963, vol. 31, n. 16, p. 417—498.
Anderson S. C. Zoogeographic analysis of the lizards fauna of Iran. — In: The
Cambridge history of Iran, vol. I. The Land of Iran. London, 1968, p. 305—371.
Clark R. J., Clark E. D., A n d e г s о n S. C. a. Levi ton A. E. Report on a col-
lection of amphibians and reptiles from Afghanistan. — Proc. California Acad. Sci.,
1969, vol. 36, n. 10, p. 279—315.
Lantz L. A. Reptiles from the River Tajan (Transcaspia). — Proc. Zool. Soc. London,
1918, n. 11—17.
Wermuth H. Gekkonidae, Pygopodidae, Xantusiidae. — Das Tierreich, Liste der recen-
ten Amphibien und Reptilien, Berlin, 1965, Lief. 80, 60 S.
About the eastern range limit of the gecko Gymnodactylus longipes Nikolsky, 1896
V. L. Orlov
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad)
Four specimens of the gedko Gymnodactylus longipes Nikolsky are described. They
have been collected by V. Ya. Lazdin in 1915 in the region Termez town, 500 km north-
east of the known range limit. Lectotype is distinguished and corrected type-locality of this
species has been ascertained. Described specimens belong to the nominative subspecies,
o. /. longipes Nikolsky, 1896 for the first time indicated in the USSR fauna.
91
НОВЫЙ ВИД ЭФЫ, ECHIS MULTISQUAMATUS SP. NOV.
ИЗ ПЕРЕДНЕЙ И СРЕДНЕЙ АЗИИ
В. А. Черлин
Зоологический институт АН СССР (Ленинград)
В настоящее время известно 3 вида эф: Echis carinatus (Schneider,
1801), Е. coloratus (Giinther, 1864), E. ocellatus (Hughes, 1976). Для пер-
вого вида описано большое число подвидов: Е. с. pyramidutn (Geoffroy,
1827) из Египта, Е. с. sinhaleyus (Deranijagala, 1951) с острова Цейлон,
Е. с. leakeyi (Stemmier & Sochurek, 1969) с севера Кении, Е. с. sochu-
reki (Stemmier, 1969) из Ирана, Пакистана, СССР, Афганистана и се-
верной Индии, Е. с. astolae (Mertens, 1970) с острова Астоле в Персид-
ском заливе, Е. с. leucogaster (Roman, 1972) с юга Сахары, Е. с. alia-
borri (Drews & Saharer, 1974) с северо-востока Кении. Традиционно
для фауны СССР.указывалась Е. с. pyramidutn, а с 1969 года — Е. с. so-
chureki (Stemmier, 1969). Однако, еще С. А. Чернов (1959), отмечал, что
у эф из СССР несколько увеличено число брюшных щитков. В резуль-
тате осуществляемой нами ревизии рода Echis мы пришли к выводу,
что на территории Средней Азии и прилегающих районов Передней
Азии обитает новый для науки вид. Материалы статьи были доложены
на семинаре отделения герпетологии Зоологического института АН
СССР (Ленинград) 24 мая 1979 года.
Материал
Для удобства изложения мы в дальнейшем будем пользоваться
следующими сокращениями названий музеев: ЗИН — Зоологический
институт АН СССР, Ленинград; МГУ — Зоологический музей Москов-
ского государственного университета; AM — The American Museum of
Natural History, New York; БМ — The British Museum (Natural History),
London; ЗМБ — Zoologisches Forschungsinstitut und Museum A. Koenig,
Bonn; 3MB — Naturhistorisches Museum, Wien; ЧМ — Field Museum of
Natural History, Chicago; MC3 — Museum of Comparative Zoology, Cam-
bridge, USA.
В работе использовались коллекционные экземпляры (количество
указано в скобках) —из СССР: ЗИН (46), 6 змей, содержащихся в Ле-
нинградском зоопарке, которые в дальнейшем будут переданы в кол-
лекцию ЗИН, МГУ (25); из Ирана: ЗИН (И), ЗМВ (5), ЧМ (12); из
Афганистана: ЗМБ (1), AM (1), ЧМ (2); из Пакистана: БМ (2), ЗМБ
(1), AM (18), ЗМВ (6), ЧМ (2), МСЗ (2); из северной Индии: БМ (1),
ЗМВ (4), ЧМ (2), МСЗ (5). Всего было обработано 152 эфы. Кроме
того, мы использовали опубликованные данные о фолидозисе 35 особей
(Boulenger, 1869; Constable, 1949; Mertens, 1956, 1957; Briik, 1968;
Stemmier u. Sochurek, 1969; Biswas a. Sanyal, 1977).
При изучении щиткования, кроме общепринятых показателей, мы
ввели новый — количество «боковых» чешуй тела — lateralia (later.),
т. е. количество чешуй в поперечном ряду посередине тела от брюшного
щитка до последней косой чешуйки с одной стороны.
Echis multisquamatus Cherlin, sp. nov
Echis arenicola: Штраух, 1868, c. 22;
Echis carinatus: Никольский, 1905, c. 319; 1916, c. 261; Терентьев и Чернов, 1949,
с. 275; Чернов, 1959, с. 173; Богданов, 1962, с. 216;
Echis carinatus pyramidum: Банников и др., 1977, с. 327;
Echis carinatus sochureki: Stemmier, 1969, S. 118 (part.).
Голотип. ЗИН. 19059.1, самка, поселок Байрам-Али, Марыйская
область, Туркмения, О. П. Богданов, 1972 (вклейка, рис. XIV, А).
92
Описание голотипа. Размеры и щиткование голотипа ука-
заны в табл. 1. Брюхо белое, окраска верхней стороны тела коричне-
вато-сероватая. По бокам туловища по одной белой волнистой полосе.
Ее вершины соединены на спине узкими поперечными полосками, отде-
ленными от боковых полос черными участками. В некоторых местах
вершины боковых полос сдвинуты друг относительно друга. Тогда белые
полоски на спине либо скошены, либо «разорваны». На голове рисунок
в виде белого креста (вклейка, рис. XIV, Л).
Паратипы. ЗИН.19059.2—6, местонахождение то же, что и у го-
лотипа; пол, размеры и щиткование сведены в табл. 1.
Таблица 1
Размеры и щиткование особей типовой серии Echis multisquamatus sp. nov.
Экземпляры Пол L L. cd. Ventr. Scd Sq later inter- ocular.
Голотип ЗИН. 19059.1 $ 556 43 182 32 38 11 11
Паратип » .2 d 380 40 177 33 39 12 10
» .3 в 452 42 187 33 36 10 10
» .4 в 433 44 185 34 40 11 11
» .5 $ 462 3j 184 31 38 10 11
.6 2 518 50 189 31 37 12 11
Таблица 2
Сравнение щиткования
Е. carinatus sochureki и Е. multisquamatus sp. nov.
F.. с. sochureki F mulisquamatus
Признаки —
mln max min •'2 max
Ventr. 75 160 173.4 187 73 169 188.1 199 108.6
Scd. 70 27 31.7 37 71 27 33. i 38 104.4
Sq. 69 27 32.7 36 76 34 37.1 40 114.9
interocular. 30 8 10.4 12 70 10 11.2 12 107.7
later. 31 8 8.8 11 76 10 10.3 11 117.0
Чешуи верха морды Силы зобугор )чатые или Гл [адкие ИЛИ бу] горчать те
Надглазничные щитки С одной или обеих сто- Не увеличены
рон обычно сильно
увеличены
Диагноз. Эфы средних и крупных размеров, отличающиеся от
остальных видов увеличенным количеством брюшных щитков (в сред-
нем 188.1) и спинных чешуй (в среднем 37.1), а также отсутствием
увеличенных надглазничных чешуй, рисунком спины и головы.
Отличие от близких видов. От всех известных видов рода
Echis новый вид отличается увеличенным числом спинных (в среднем
37.1) и боковых (в среднем 10.3) чешуй; количество брюшных щитков
(в среднем 188.1) сходно лишь с Е. coloratus, у которой их в среднем
189. Окраска особей Е. multisquamatus sp. nov. красновато-коричнева-
тая, желтовато-коричневатая или почти серая. Рисунок на спине мало-
вариабилен. Его изменения связаны лишь с различным количеством
косых или «разорванных» спинных белых полосок. Брюхо пятнистое
(78.5%) или белое (21.5%). У других видов все элементы рисунка
остаются, но находятся на разной стадии редукции или слегка изме-
няются. У всех подвидов Е. carinatus боковая полоса разбита на отдель-
ные дуги, обращенные к спине, либо представлена отдельными белыми
штрихами по бокам спинных пятен. Она есть только у трех изученных
нами змей из Пакистана. Спинные пятна имеют различную форму от
93
поперечно вытянутых (Е. с. sochureki), ромбовидных (Е. с. carinatus,
Е. с. sochureki, Е. с. pyramidum, Е. с. leucogaster, Е. с. leakeyi) до округ-
лых разной величины (Е. с. sochureki, Е. ocellatus, Е. coloratus). У всех
этих эф спинные пятна контрастно окаймлены черным и соединяются
более или менее ясно выраженной темной полосой, чего практически
не бывает у Е. multisquamatus sp. nov. Внутри дуг боковой полосы
у всех эф расположен темный участок, который у описываемого вида
имеет различные очертания, а у остальных видов боковой полосой огра-
ничивается в более или менее ясное пятно, превращаясь у Е. ocellatus
Распространение и terra typica(*) Echis multisqumatus sp. nov.
(1) и Echis carinatus sochureki (2)
в «глазки». Рисунок головы у Е. multisquamatus в виде светлого креста,
у которого горизонтальные ветви иногда обращены назад. У остальных
эф рисунок на голове обычно иной (кроме нескольких экземпляров
Е. с. sochureki из Пакистана). Особый интерес представляет сравнение
Е. multisquamatus с обитающей рядом с ней Е. с. sochureki по фолидозу
(табл. 2) и по окраске (вклейка, рис. XIV).
Распространение. Е. multisquamatus sp. nov. обитает в Сред-
ней Азии по всей зоне южных пустынь умеренного пояса, на севере до
южного чинка Устюрта и Аральского моря, на северо-востоке доходит
до линии от южного берега Арала до Ташкента, отмечена в Ферганской
долине (ЗИН. 16013), на востоке обходит горы Алая, поднимаясь по
речным долинам до высоты не более 900 м, на юге до Гиндукуша, Па-
ропамиза, Копет-Дага и Эльбурса, заходит на южный берег Каспий-
ского моря (ЗМВ 23972.1—2); кроме того, встречается в восточном
Иране и южном Афганистане (см. рис.).
Обсуждение
Все изученные нами эфы из юго-восточного Ирана (ЗИН. 9274г
9989, 10010, 10011, 10286, 10309; 3MB.23974. 1—2; ЧМ. 141596—141599,
166971, 166972) относятся к Е. с. sochureki. С севера и востока Ирана
и юга Афганистана нам известны Е. multisquamatus (ЗИН. 1095, 9276,
94
9277, 9990; ЗМБ.8647; ЗМВ.23972), у которых спинных чешуй не менее
37, тогда как у Е. с. sochureki из восточного Ирана (ЗИН.17992; ЗМВ.
23997), Афганистана (AM.70593; ЧМ.161072, 161073) и прилегающих
районов Пакистана (AM.21457, 96236, 96237) их не более 36. Змеи этого
подвида с количеством спинных чешуй менее 36 отмечены в Заболи
в восточном Иране, в районе Кветты в западном Пакистане и в Синде
в нижнем течении Инда. Крайние западные точки нахождения Е. с. so-
chureki в восточном Иране, Заболь и Захедан (ЗМВ.29977), «перекры-
вают» крайнюю восточную точку нахождения Е. multisquamatus в юж-
ном Афганистане, Кандагар (ЗВМ.8647; ЧМ.161072—161073) на 400 км.
Эта зона симпатрии обеих форм свидетельствует в пользу их видовой
самостоятельности.
Выражаю свою глубокую признательность за предоставление кол-
лекций для обработки Е. Арнольду (Лондон), В. Беме (Бонн), И. С. Да-
ревскому (Ленинград), X. Марксу (Чикаго), Д. Розадо (Кембридж),
М. Хойплкх (Вена) и Р. Цвейфелю (Нью-Йорк), а также Л. Я. Боркину
(Ленинград) за консультации в работе и замечания по рукописи.
ЛИТЕРАТУРА
Банников А. Г., Даревский И. С., Ищенко В. Г., Рустамов А. К-,
Щербак Н. Н. Определитель земноводных и пресмыкающихся фауны СССР.
М., «Просвещение», 1977, 414 с.
Богданов О. П. Пресмыкающиеся Туркмении. Ашхабад, АН Туркм. ССР, 1962, 235 с.
Никольский А. М. Пресмыкающиеся и земноводные Российской Империи. — Зап.
Имп. Акад. Наук, физ.-мат. отд., СПб., 1905, т. 12, № 1, 518 с.
Никольский А. М. Пресмыкающиеся (Reptilia), т. 2. Ophidia. — В сер.: Фауна
России и сопредельных стран, Петроград, 1916, 344 с.
Терентьев П. В. и Чернов С. А. Определитель пресмыкающихся и земно-
водных. М., «Советская наука», 1949, 3-е изд., 340 с.
Чернов С. А. Пресмыкающиеся. — В сер.: Фауна Таджикской ССР (Тр. Ин-та
зоол. и паразитол. АН Тадж. ССР, т. 98), Сталинабад, 1959, 204 с.
Штраух А. О ядовитых змеях, водящихся в России. — Тр. 1 съезда русск. есте-
ственен. по отд. зоол., СПб., 1868, с. 1—27
Biswas S. a. S а п у а 1 D. Р. Fauna of Rajisthan, India. — Rec. Zool. Surv. India,
1977, vol. 73. n. 1—4, p. 247—269.
Boulenger G. A. Catalogue of snakes in the British Museum (Natural History).
Vol. 3. Colubridae (Opistoglyphae and Proteroglyphae), Amblycephalidae, and
Viperidae. London, 1896, 727 p.
Briik G. Zur Herpetofauna Afghanistans.—Vestnik Cs. spol. zool. (Acta soc. zool.
Bohemoslov.), 1968, t. 32, n. 3, S. 201—208.
Constable I. D. Reptiles from the Indian Peninsular in the Museum of Compara-
tive Zoology, Cambridge, Mass., 1949. — Bull. Mus. Comp. Zool., 1949, vol. 103,
n. 2, p. 159—160.
Mertens R. Amphibien und Reptilien aus SO-Iran 1954 (Ergebnisse der entomolo-
gischen Reisen Willi Richter, Stuttgart, im Iran 1954 und 1956 Nr. 5). — Jh. Ver.
vaterl. Naturk. Wiittemberg, 1956, Bd. Ill, S. 90—97.
Mertens R. Weitere Unterlagen zur Herpetoiauna von Iran 1956. — Jh. Ver. vaterl.
Naturk. Wiittemberg, 1957, Bd. 112, H. 1, S. 118—128.
Stemmier O. Die Sandrasselotter aus Pakistan: Echis carinatus sochureki subsp.
nov. —Aquaterra, 1969, Bd. 6, H. 10, S. 118—125.
Stemmier O. u. Sochurek E. Die Sandrasselotter von Kenya: Echis carinatus
leakeyi subsp. nov. — Aquaterra, 1969, Bd. 6, S. 89—94.
The new saw-scaled viper, Echis multisquamatus sp. nov. from South-Western
and Middle Asia
V A. Cher Un
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad)
The saw-scaled viper from the Middle Asia, northern and eastern Iran and
southern Afghanistan is described as a new species: Echis multisquamatus sp. nov. It is
separated from the all other snakes of this genus by the large number of ventral
(x= 188.1), dorsal (x = 37.1) and lateral scales (x=10.3). Moreover it lacks enlarged
scales over eyes. The light wavy stripes on the sides of the body are present and are
not devided in separate arcs as in all other species of Echis. The white stripes on the
back are transverse and narrow. There is a crosslike figure on the top of the head. In sou-
thern Afghanistan Echis multisquamatus sp. nov. and E. carinatus sochureki are sym-
patric.
95
ЗАВИСИМОСТЬ ПОВЕДЕНИЯ ПЕСЧАНОЙ ЭФЫ,
ECHIS MULTISQUAMATUS CHERLIN, 1981
ОТ ТЕМПЕРАТУРНЫХ УСЛОВИЙ В ЮЖНОЙ ТУРКМЕНИИ
В. А. Черлин и А. Ю. Целлариус
Зоологический институт АН СССР (Ленинград)
и Бадхызский государственный заповедник (Кушка)
Первоначальной целью наших исследований являлось изучение за-
висимости поведения песчаной эфы от различных микроклиматических
факторов. Литературные данные, посвященные этому вопросу (Пестин-
ский, 1936; Богданов, 1950, 1962, 1965; Персианова, 1969, 1972; Хозац-
кий и Захаров, 1970; «Содержание ядовитых змей Средней Азии в не-
воле», 1977 и др.), в большинстве своем рисуют слишком общую кар-
тину; результатом исследований, как правило, оказывается выявление
«оптимального интервала температур». Между тем проблема имеет не
только теоретический, но и несомненный практический интерес. Яд эф
обладает очень редкими и ценными биохимическими свойствами: его
компоненты действуют непосредственно на протромбин. В лаборатор-
ной практике подобными свойствами обладает очень дорогостоящий
препарат из яда редкой австралийской змеи тайпана (Oxyuranus scut-
tellatus), которым могут воспользоваться лишь немногие учреждения.
Недостаточная изученность связи экологии с микроклиматом не позво-
ляет подолгу успешно содержать эф в питомниках. Мы сочли целесо-
образным подробно изучить этот вопрос. Однако, уже в процессе поле-
вых работ было выявлено, что эфы значительную часть времени прово-
дят в норах на глубине до 50 см, где условия температуры и влаж-
ности достаточно стабильны. Выход их на поверхность, а значит, и воз-
можность подвергаться воздействию солнечного света (видимых, инфра-
красных и ультрафиолетовых лучей), резких колебаний температуры и
влажности, оказался тесно связан с температурными условиями среды
и их динамикой. Вследствие этого, в дальнейшем мы акцентировали
внимание на изучении именно температурных условий и на зависимость
поведения эф от их изменений.
Материал и методика
Работы проводились нами в весенне-летний период 1975—1978 и осенью 1977—
1978 годов во впадине Ер-Ойлан-Дуз (юго-восточная Туркмения, Бадхыз, около 70 км
к северо-западу от города Кушка).
На регулярных маршрутах мы отмечали состояние погоды, проводили общие
фенологические наблюдения. При встречах эф мы регистрировали время поимки и
размеры змей, температуру почвы и воздуха на высоте 1—3 см непосредственно
в месте поимки, те же параметры в норе, если змея находилась около нее, при воз-
можности измеряли у нее ректальную температуру. Кроме того, мы отмечали пове-
дение эф и метили их. Всего нами было встречено 112 эф, произведено 49 измерений
ректальных температур и 326 измерений параметров окружающей среды.
В разные сезоны за 5 особями велись непрерывные индивидуальные наблюде-
ния в течение 3—5 суток за каждой. В процессе наблюдений хронометрически реги-
стрировались все передвижения эф относительно норы, освещенных или затененных
участков, ее поведение; по возможности, отмечалась глубина, на которую змея ухо-
дила в нору. В течение индивидуальных наблюдений каждый час измеряли темпера-
туру почвы и воздуха на высоте 1—3 см и в норах на глубине 20 см. Продолжи-
тельность наблюдений суммарно за всеми особями составляла 21 сутки.
За 4 особями наблюдения проводились на опытной площадке размером 3X4 м,
огороженной заборчиком из полиэтиленовой пленки высотой 60 см. На площадке
были устроены искусственные норы и стационарно установлены термометры. Темпера-
турный режим искусственных и естественных нор оказался примерно одинаковым
(рис. 1). Поведение эф регистрировали по той же схеме, что при индивидуальных
наблюдениях. Кроме того, каждые 3 часа круглосуточно отмечалось положение эф
на площадке и измерялись температуры среды (как и при индивидуальных наблю-
дениях). Продолжительность наблюдений на опытной площадке составила 61 сутки.
96
В 1976—1978 годах нами проводились метеорологические наблюдения. На спе-
циальной метеорологической площадке каждые 3 часа (первый срок в 2.00) измеря-
лись температуры почвы на поверхности и на различных глубинах до 60 см, темпе-
ратура и относительная влажность воздуха на высоте 3 см. В баллах оценивалась
облачность, сила ветра, осадки.
Для определения залегания основной массы нор песчанки (Meriones meridianus),
которыми пользуются эфы в качестве укрытий, мы произвели вертикальный разрез
одной колонии.
Работа велась со взрослыми и полувзрослыми особями эф длиной не менее
320 мм.
Результаты и обсуждение
1. Зависимость температуры тела эф от температурных условий среды
В разных условиях зависимость температуры тела от температуры
среды была различна. В пасмурные дни, в вечернее и ночное время,
а также в террариумах, т. е. при рассеянном свете, ректальная темпе-
ратура почти полностью совпадала г ---
с температурой почвы и воздуха
в месте нахождения змей (табл. 1).
Эти данные отражают чисто «фи-
зическую» зависимость температу-
ры эфы от температур среды. При
наличии интенсивного инфракрас-
ного излучения солнца «стройность»
этой связи нарушается.
Температура тела эфы всегда
сильно коррелирует с температурой
почвы непосредственно под ней.
При рассеянном свете или в отсут-
Рис. 1. Температуры на трех близлежащих
участках во впадине Ер-Ойлан-Дуз 17.IV
1977 года.
1 — температура поверхности почвы на метео-
площадке; 2 — температура поверхности почвы на
опытной площадке; 3 — температура поверхности
почвы при наблюдениях у естественной норы:
4 — температура почвы на глубине 20 см на
опытной площадке; 5 — температура почвы на
глубине 20 см при наблюдениях у естественной
норы; 6 — температура почвы на глубине 20 см
на метеоплощадке: 7 — температура почвы в норе
на глубине 20 см на опытной площадке.
Б.емя cjn^K(uac)
ствии инфракрасного излучения солнца (вечером, ночью, в норе, в тер-
рариуме) эта температура не отличается от температуры почвы вокруг
эфы. При наличии интенсивного солнечного излучений корреляция тем-
ператур тела и почвы под змеей остается тесной, но температуры почвы
под ней и около нее в этом случае могут сильно различаться. Ранней
весной, когда почва прогрета еще слабо, температура самого верхнего
слоя почвы может быть относительно высока, но затененные участки
быстро охлаждаются до температуры глубоких слоев. Когда змея ле-
жит на солнце, температура поверхности почвы рядом с ней может до-
стигать 40° С и более, но в то же время непосредственно под змеей
температура почвы значительно ниже (рис. 2,/). Температура поверх-
ности тела змеи в этот момент выше, чем температура почвы под ней,
но ниже температуры почвы рядом с ней. Разности температур поверх-
ности находящегося в норе и обогреваемого участков тела могут в этот
период достигать 10° С, а в более поздние сроки сезона—4° С (рис. 2, II).
В конце весны — начале лета эфы проводят день во входе в нору. Тем-
7 Зак. 719 97
пература почвы рядом со змеей может быть равна 50° С и даже не-
много выше, а непосредственно под ней она в то же время остается
34.0—37.5° С (рис. 2,111). При измерениях на маршрутах редко удается
измерить именно температуру под греющейся змеей, поэтому в описы-
ваемом случае корреляции температур почвы в месте находки змеи и
ректальной несколько слабее, чем при отсутствии излучения солнца
(табл. 1).
Таблица 1
Зависимость температуры тела эф, находящихся на поверхности почвы,
от температурных условий
Световые условия f и t п в f и t р в ‘пм н ‘р
г ± т п в г ± т 'р-'в г ± т i пм *р
Вечером при рассеянном свете ‘18 0.99 ±0.08 0.1 9 0.90±0.17 0.5 9 0.98 ±0.12 0.5
В неволе при расеянном свете 27 0.99 ±0.01 1.2 8 0.93±0.20 0.9 8 0.99 ±0.08 0.8
Днем при прямом свете 19 0.81 ±0.17 4.7 12 0.56±0.24 3.4 12 0.83 ±0.16 ОД
Примечание. te — температура воздуха, tn—температура почвы, tnM — тем-
пература почвы в месте поимки (по данным маршрутов), п — количество измерений,
r±m — коэффициент корреляции и его ошибка и tp — ректальная температура эф.
В том случае, когда температуры воздуха и почвы заметно разли-
чаются, т. е. при наличии солнечного излучения, становится очевидной
большая зависимость температуры тела от температуры почвы в месте
поимки, чем от температуры воздуха (табл. 1). Это согласуется с ре-
зультатами специальных исследований в том, что абсорбция тепла те-
лом пресмыкающихся происходит, в основном, через субстрат (Duvde-
vani a. Borut, 1974).
На основании вышеизложенного, зная температурные условия
среды, мы можем оценить температуру тела эф.
2. Температурные условия среды
Температурные условия на поверхности и в норах подвержены
строгой суточной и сезонной динамике. Особый интерес представляет
ход составляющих температурного режима по времени друг относи-
тельно друга. Суточные экстремальные значения температур поверх-
ности почвы, приземного слоя воздуха и воздуха в норах почти совпа-
дали по времени. Экстремумы температуры почвы в норах в начале
весны наступали на 2—2.5 часа позже, чем у первых. К лету это сме-
щение достигало 3—3.5 часов (рис. 3).
В соответствии с этим суточная динамика температурных условий
в норах протекает следующим образом: всю ночь температуры почвы
и воздуха примерно равны. Около 7 часов утра температура воздуха,
связанная с таковой на поверхности, начинает повышаться, тогда как
почва еще продолжает охлаждаться до 8—9 часов. В результате в это
время в норе возникает заметная разница температур почвы и воздуха,
которая увеличивается к 14—16 часам, а затем к 19 часам эти темпе-
ратуры опять примерно сравниваются.
98
Наиболее велика амплитуда температуры почвы на поверхности,
несколько меньше она в приземном слое воздуха, еще меньше варьиро-
вала температура воздуха в норах и совсем незначительно — темпера-
тура почвы в норах (рис. 3). С начала весны к лету суточные ампли-
туды колебаний всех названных параметров несколько увеличивались
(рис. 3).
Рис. 2. Примерное распределение температур
почвы и тела эф в разные сезоны.
/ — ранняя весна; // — конец весны; /// — лето.
Для удобства работы и изложения материала мы условно разде-
лили сезон на более или менее однородные периоды: ранняя весна, се-
редина весны, конец весны, начало лета, середина лета (рис. 3). В даль-
нейшие сроки сезона изменение основных параметров протекает в обрат-
ном порядке.
3. Сезонные изменения поведения эф
Ранней весной эфы появляются из нор только в середине дня
(рис. 4, /, б) как в ясные, солнечные дни, так и в пасмурные, при
дожде. В ясные дни они лежат у самых нор, выставляя на солнце либо
все тело, либо только переднюю его часть. При пасмурной погоде эфы
целиком выползают из нор и лежат обычно недалеко от них.
В середине весны змеи выходят на поверхность на значительно
более долгое время (рис. 4, 11,6). Вначале они лежат у нор и в сере-
дине дня, когда почва прогревается, уходят в тень травы, где в это
время температура значительно ниже. Позднее, когда дневные темпе-
ратуры почвы еще более повышаются, эфы начинают перемещаться то
в нору, то на непродолжительное время опять появляются на поверх-
ности и затем снова скрываются в нору. Вечером они появляются на
поверхности на некоторое время и греются у самых нор. Периоды воз-
7*
99
вращения на поверхность в середине дня постепенно сокращаются, и
к концу весны эфы вообще в жару на поверхности не появляются,
а греются у нор недолго утром и вечером (рис. 4, III, б).
В начале лета змеи появляются на поверхности утром, затем ухо-
дят во вход норы и некоторое время проводят на глубине 5—15 см,
Рис. 3. Изменение суточной динамики температур почвы и
воздуха на поверхности и в норах.
/ — начало весны; // — середина весны; /// — конец весны; IV — на-
чало лета; V — середина лета; 1 — температура почвы на поверхности.
2 — температура воздуха на высоте 3 см, 3 — температура почвы на
глубине 20 см, 4 — температура воздуха в норе на глубине 20 см.
выставляя иногда на солнце отдельные части тела. Потом они уходят
в более глубокие отделы нор и появляются только вечером, когда
солнце почти зашло (рис. 4, IV, б). Вечером в течение 1—3 часов эфы
могут свободно передвигаться по поверхности. В самом конце весны —
начале лета в пасмурные дни, когда солнце затянуто дымкой или обла-
ками, змеи могут проводить на поверхности почти весь день. В этот
период мы неоднократно встречали эф парами, что, безусловно, свиде-
тельствует о протекании половой активности.
100
В середине лета эфы изредка появляются на поверхности утром
около 8 часов либо некоторое время проводят во входе в нору на глу-
бине 5—15 см. Днем они уходили в глубокие отделы нор, а после за-
хода солнца появлялись на поверхности и проводили там 3—6 часов
Рис. 4. Суточная динамика температуры тела эфы в зависи-
мости от температур среды и се изменение по сезонам.
/ — начало весны, // — середина весны, /// — конец весны, IV — начало
лета, V — середина лета; в — динамика температур: / — на поверхно-
сти, 2 — на глубине 30 см, 3 — на глубине 20 см, 4 — тела; б — пове-
дение эф: / — в норе, 2—на поверхности, 3 — в норе у входа, 4 —
активная терморегуляция.
(рис. 4, V, б). В этот период вечером на поверхности многочисленны
молодые песчанки, являющиеся во впадине основным видом корма для
эф. Находившиеся на опытной площадке эфы не реагировали на пред-
лагавшихся им прямокрылых и ящериц, но охотно поедали молодых
песчанок. Этот же вид корма мы неоднократно встречали в желудках
у пойманных эф.
101
4. Суточная динамика температуры тела эф и ее изменение по сезонам
Когда эфа находится в норе, температура ее тела почти полностью
соответствует температуре почвы норы (см. раздел 3). Утром после вы-
хода на поверхность температура тела эф вместе с температурой почвы
быстро повышается. Максимальная отмеченная нами ректальная темпе-
ратура для эф была 33.8°, а температура поверхности кожи 38° С. После
достижения подобных температур змеи уходили в нору и располагались
на тех ее уровнях, которые обеспечивали поддержание температуры
тела, как можно более близкой к максимальной, т. е. 31—34° Вечером,
когда эфы опять появлялись на поверхности, температуры их тела по-
нижались вместе с температурой почвы.
Рис. 6. Доля времени, которую эфы
проводят в норах в течение суток
в зависимости от температур почвы
в норах (в % %).
Т — интервалы температур почвы в норах.
Рис. 5. Зависимость величины сред-
него квадратичного отклонения тем-
пературы поверхности почвы при
встречах эф на поверхности от тем-
пературы почвы в норах.
(j — среднее квадратичное отклонение, Т —
интервалы температур почвы в норах, t° —
температура поверхности почвы; 1 — по
данным маршрутов, 2 — по данным опыт-
ной площадки, 3 — температуры поверхно-
сти почвы в местах находок эф.
На основании этих данных, а также материалов разделов 1 и 3
мы можем составить график температуры тела эф в течение суток для
каждого из периодов (рис. 4, а). Утром и днем во все сезоны эфы ста-
раются удержать температуру тела на максимально высоком уровне
(31—34°) в течение как можно более продолжительного времени. Этот
уровень определяется, с одной стороны, их термической выносливостью,
с другой, условиями среды. В течение весны змеи придерживаются
этой тактики также вечером и ночью. С наступлением лета происходит
«перелом» в их поведении. В течение суток они получают уже доста-
точно много экзогенной энергии, позволяющей им «оторваться» от
«максимального уровня» и быть активными на поверхности вечером и
ночью, хотя температура почвы в норах в это время ближе к «макси-
мальной», чем на поверхности (рис. 4, IV, а).
Амплитуда колебаний температуры тела эф в течение сезона от
весны к лету резко сокращается. В начале весны она составляет при-
мерно 21° (от 13° до 34° С), в середине весны— 16° (18—34° С), в конце
весны — 8° (26—34° С), в начале и середине лета — 6—8° (от 26—28°
до 34°).
5. Зависимость поведения эфы от температурных условий в норах
В течение светлого времени суток температуры тела эф с середины
весны и позже держатся на примерно одинаковом и высоком уровне
32—34и С. Температуры тела в темное время суток зависят от темпе-
ратур в норах и в течение сезона заметно растут от 13° до 34°, а воз-
можно, и немного выше (рис. 4, а). Таким образом, количество общего
поступающего к телу эфы экзогенного тепла за сутки, в основном,
102
определяется именно температурой норы. Значит, и свободное переме-
щение эф по поверхности тоже определяется, в основном, температурой
норы (раздел 3). От температуры норы также зависит и осуществление
активной терморегуляции (см. раздел 6).
На возможное отношение эфы к высоким температурам поверх-
ности оказывает большое влияние наличие температурного «противо-
веса»— низкой температуры почвы в норах. При низких температурах
нор (13—17° С) эфы встречаются на поверхности при 13—40° В даль-
нейшем нижняя температурная граница встречаемости возрастает вме-
сте с ростом температуры почвы в течение сезона, а верхняя — сни-
жается с 40° (при 17—20° в норах) до 34° (при 30—34° в норах), по-
скольку у змей нет более возможности охладиться. В результате «раз-
брос» температур поверхности почвы, при которых мы встречали эф,
а также абсолютные значения этих температур уменьшаются с ростом
температуры нор (рис. 5).
Кроме того, температура почвы в норах, соответствующая темпе-
ратуре на глубине почвы, является четким показателем фенологического
состояния природы данного района. Все это дает возможность исполь-
зовать температуру почвы в норах или на глубине 20 см как косвенный
показатель активности, возможного поведения и физиологического со-
стояния эф данной популяции.
6. Поведение, обусловленное температурами среды
Поведение эф в связи с необходимостью поддержания определен-
ного интервала температуры тела складывается из ряда компонентов.
Мы различаем следующие формы этого поведения: в норе, свободное
передвижение по поверхности, у норы, активная терморегуляция. Каж-
дая из них характеризуется своим уровнем двигательной активности
и протекает в определенных микроклиматических условиях.
а) в норе
Эфы более или менее продолжительное время находятся в глубине
норы. Судя по визуальным наблюдениям с поверхности и сделанному
нами разрезу колонии песчанок, основная масса нор лежит в горизонте
15—50 см, в среднем около 20—30 см. Выделить эту форму поведения
можно по данным маршрутов, индивидуальных наблюдений и опытной
площадки. Ранней весной эфы проводят в норах 83% времени, в конце
весны — начале лета 58%, в середине лета — 75% (рис. 6). Они поки-
дают норы в разные сезоны на разное время, обусловленное темпера-
турной обстановкой и физиологическими потребностями. Температура
тела у змей в норах практически равна температуре почвы на соответ-
ствующей глубине и вместе с ней медленно растет от начала весны
к середине лета с 10—11° до 34° или, возможно, немного выше, а затем
так же медленно падает к осени.
б) свободное передвижение по поверхности
Эфы в течение длительного времени активно передвигаются по по-
верхности вдали от нор при условиях, описанных в разделе 4. Выде-
лить эту форму поведения можно по массовым находкам змей на марш-
рутах вдали от нор. Встречи при этой форме поведения отмечены при
температурах поверхности не ниже 20° Чаще всего змеи встречались
при температурах поверхности 28—32° и почвы в норах (на глубине
20 см) более 32° (рис. 7). При увеличении температуры почвы в норах
эфы все чаще встречаются на поверхности, и при 36° встречаемость
при этой форме поведения достигает 100%, смещаясь с утренне-дневных
на вечерние часы (рис. 8).
в) у норы
В течение продолжительного времени эфы лежат рядом с норой.
Наиболее удобно выделить эту форму поведения по данным маршру-
103
тов. Эфы отмечены нами у нор во все сезоны, но встречаемость их резко
уменьшалась с увеличением температур в норах и при температурах
поверхности более 24° (рис. 9). Находясь у нор, эфы, в основном,
греются на солнце, поэтому с увеличением норных температур встре-
чаемость у нор в дневные часы уменьшается, а в вечернее — увеличи-
бодном передвижении по поверхно-
сти в
почвы в норах и на поверхности
(в % % от числа встреч в данном
интервале температур).
/ — зависимость от температур в норах,
2 — зависимость от температур на поверх-
ности; Т — температурные интервалы.
зависимости от температур
бодном передвижении по поверхно-
сти в разное время суток в зависи-
мости от температур почвы в норах
(в % % от числа встреч в данном
интервале температур).
/ — от 10 до 18 часов. 2 — от 18 до 24 ча-
сов, 3 — за весь день.
Рис. 10. Частота встреч эф у нор
в разное время суток в зависимости
от температур почвы в норах (в % %
от числа встреч в данном интервале
температур).
/ — от 7 до 10 часов. 2 — от 10 до 18 ча-
сов, 3 — от 18 до 24 часов. 4 — за сутки
Рис. 9. Частота встреч эф у нор
в зависимости от температур почвы
в норах и на поверхности (в % %
от числа встреч в данном интервале
температур).
/ — зависимость от температуры на по-
верхности, 2 — зависимость от температу-
ры в норах; Т — интервалы температур.
г) активная терморегуляция
В целях терморегуляции эфы перемещаются то в нору, то из нее,
непродолжительное время лежат у самого входа, после чего опять
уходят в нору. Такие перемещения продолжаются весь период активной
терморегуляции. Отделить эту форму поведения от формы «у норы»
можно только по данным опытной площадки и индивидуальных наблю-
дений. На маршрутах змеи в состоянии активной терморегуляции прак-
тически не отмечены, потому что слишком мало времени проводят на
поверхности и к тому же сильно разогреты и при приближении чело-
века быстро скрываются в норах. Об этом косвенно свидетельствует
104
Таблица 2
Температурные условия и частота встреч эф на поверхности почвы
при активной терморегуляции
/о.) х±т *тах
20.5—21.4 40.8± 1.3 ±3.79 11.25 44.5
21.5—22.4 43.3± 1.1 ±4.10 18.75 55.7
22.5—23.4 48.5 ±1.5 ±6.35 22.50 56.0
23.5—24.4 48.4 ±1.2 ± 5.45 26.25 54.2
24.5—25.4 45.5±1.4 ±4.19 11.25 49.0
25.5—26.4 42.0 ±0.9 ±2.45 10.00 48.0
Примечание. /20—температура почвы в норах на глубине 20 см, л + ш —
средняя температура почвы при встречах эф и ее ошибка, о — среднее квадратичное
отклонение температур поверхности почвы при встречах эф, п — количество встреч
в данном интервале температур почвы в норах в % % от числа встреч при активной
ТерМОрегуЛЯЦИИ, tmax абсолютный максимум температуры поверхности почвы в ме-
стах нахождения эф.
и то, что на маршрутах эфы ни разу не были встречены при высоких
температурах поверхности почвы, характерных для этой формы поведе-
ния. Активная терморегуляция наблюдается в середине весны. Темпе-
ратуры на глубине 20 см уже достаточно высоки, чтобы стимулировать
30 1 Ы.1Л
Рис. 11. Проявление активной тер-
морегуляции в зависимости от тем-
пературы почвы в норах (в % % от
общего числа встреч при активной
терморегуляции)
Т — интервалы температур.
J120 T5s[l50 iss|?co 23з|240-гго|г80 3!э|з20 359I
т ’С
змей к началу половой активности, но еще слишком низки, чтобы обес-
печить эту активность. Необходимо дополнительное тепло, но темпера-
турная ситуация для прогрева тела достаточно «неудобная»: во входе
в нору температура почвы резко падает, в то же время температура
поверхности уже слишком высока. С утра эфы еще могут лежать
у входа в нору, но позже на поверхности становится горячо. Переход
от невысокой норной температуры к высокой поверхностной происходит
на небольшом участке входа норы — всего около 10 см. Все это не по-
зволяет змее «удобно расположиться» в «комфортном» диапазоне тем-
ператур, и она вынуждена то вылезать из норы греться, то быстро ухо-
дить вглубь охлаждаться. Эти передвижения мы и называем активной
терморегуляцией. Данная форма поведения отмечена нами при темпе-
ратурах поверхности 35—56° и в норах (на глубине 20 см) 20.5—26.5°
(табл. 2). Наиболее ярко она выражена при температурах в норах
21.5—24.5° (рис. 11).
7. Зависимость половой и пищевой активности
от температурных условий
Большой интерес представляют такие сложные и важные формы
поведения, как пищевая и половая. Для успешной реализации каждой
из них должно существовать два рода условий: базовые, связанные,
в основном, с физиологическим состоянием организма, и допускающие,
которые связаны с реализацией комплекса соответствующего поведения
и обусловлены в значительной степени микроклиматом среды. В каче-
стве «индикатора фенофазы» для данного района мы рассматриваем
не календарные сроки, а температуру почвы на глубине 20 см или почти
105
равную ей температуру почвы в норах на той же глубине. Поскольку
ряд базовых и допускающих условий, таких как возможность свобод-
ного перемещения по поверхности, условия для нормального гамето-
генеза и переваривания пищи и др., связаны с сезонными изменениями
температурных условий, мы рассматриваем их в связи с названными
выше «индикаторными» температурами почвы.
а) половая активность.
Основных базовых условий
для протекания половой активно-
сти, очевидно, несколько. К ним
относятся удовлетворительное со-
стояние половой системы и жи-
ровых запасов. Для успешного
протекания гаметогенеза, как из-
вестно, необходимо наличие в те-
чение суток значительного пере-
пада температуры тела. Кроме
того, для созревания гонад и ак-
тивизации эндогенных механиз-
мов, «включающих» половую ак-
Рис. 12. Микроклиматические усло-
вия при выходе эф из нор утром и
уходе вечером в весенний период.
/ — температура поверхности почвы, 2 —
температура воздуха на высоте 3 см, 3 —
температура почвы на глубине 20 см,
4 — температура воздуха в норе; А — время
выхода эф утром, Б — время ухода эф
в норы вечером.
Рис. 13. Микроклиматические усло-
вия, определяющие сроки протекания
половой активности у эф.
/° — температура почвы на глубине 20 см;
/ — температура почвы в норах, при ко-
торой эфы могли свободно передвигаться
по поверхности; 2 — температура почвы
в норах, при которой может протекать
гаметогенез; 3 — частота встцеч эф на по-
верхности в светлое время суток в зави-
симости от температуры почвы в норах
(в % % от общего числа встреч);) 4 — ин-
тервал температур почвы в норах, при
котором возможна половая активность.
тивность, необходимо увеличение длины световой фазы. Регуляция ее
длительности осуществляется, в основном, температурным режимом
(Черлин, 1977). Время пребывания змей на поверхности жестко регла-
ментировано. Выход утром происходит при минимуме температуры почвы
в норах, когда температура воздуха в норах начинает заметно превы-
шать температуру почвы (рис. 12). Это условие, видимо, сигнализирует
эфе о том, что на поверхности сложились благоприятные для актив-
ности условия. Весной эфы проводят день на поверхности, уходя в норы
вечером, когда происходит сближение температур воздуха и почвы на
поверхности и в норах (рис. 12). В дальнейшем они проводят середину
дня в норе, а на поверхность выходят на продолжительное время лишь
вечером и ночью. Таким образом, в начале весны продолжительность
световой фазы, т. е. продолжительность пребывания эф на поверхности
днем, составляет 4—8 часов, затем увеличивается до 10—10.5 часов,
а к началу лета уменьшается почти до 0. Значит, увеличение световой
фазы определяется для эф динамикой температуры среды (особенно
температуры почвы). Увеличение за короткий весенний период длитель-
ности световой фазы является для большинства рептилий мощным
106
стимулом развития гонад и проявления полового поведения (Licht,
1973; Singh, 1974). Можно предположить, что температурные условия,
особенно температуры почвы, определяя время нахождения змеи на
поверхности, регулируют через длину световой фазы половую актив-
ность и «подстраивают» ее под самые благоприятные сроки сезона. Для
успешного протекания половой активности необходимы условия для
поиска полового партнера — возможность свободного перемещения по
поверхности. В районе исследований это условие особенно важно, по-
скольку эфы здесь зимуют диффузно, не образуя скоплений.
Температурные условия протекания половой активности отмечены
на рис. 13.
б) пищевая активность
Основными базовыми условиями для протекания пищевой актив-
ности мы считаем наличие аппетита и возможности переваривания
пищи. Наблюдения за эфами, содержавшимися в Ленинградском зоо-
парке, показали, что если ночная температура тела
змей становится ниже 15—17°, т. е. суточная ам-
плитуда колебаний температуры их тела превы-
шает примерно 15°, то эфы либо вообще не пи-
таются, либо не могут успешно переварить корм
Рис. 14. Микроклиматические условия, определяющие время
протекания пищевой активности.
Л — температура почвы на глубине 20 см; / — температура почвы
в норах, при которой возможно переваривание пищи; 2 — температура
'почвы в норах, при которой возможна охота; 3 — температура почвы
в норах после завершения половой активности; а — интервал темпе-
ратур почвы в норах, при которых возможно питание не размножав-
шихся в данный сезон особей, б — интервал температур почвы в но-
рах. при котором возможно питание размножавшихся в данный сезон
особей.
и срыгивают его. Для реализации комплекса поведения, связанного
с охотой, змеям необходима возможность для свободного передвиже-
ния по поверхности и в норах, что возможно при температурах в норах
не ниже 20—22° Кроме того, необходимо учесть, что в период половой
активности пищевая подавлена. Значит, питание у приступивших
к размножению особей может начаться только после прохождения
пика половой активности. Часть эф, которая по ряду причин в дан-
ный сезон не принимает участия в размножении, а также неполово-
зрелые особи могут начинать питаться, очевидно, несколько раньше.
Г И. Ишунин (1949) указывал, что в более северных районах Турк-
мении и Узбекистана эфы начинают питаться не сразу после выхода
из зимовок, а только с мая.
Температурные условия протекания пищевой активности отмечены
на рис. 14.
Все описанное выше показывает глубокие тонкие механизмы адап-
тации песчаной эфы к суровым условиям среды, чуткую «подстройку»
ее жизненного цикла к конкретной климатической обстановке данного
сезона. Это позволяет ей, оставаясь теплолюбивой змеей, с большой
эффективностью использовать жесткие условия пустынь умеренного
пояса и, «лавируя» между критически низкими и критически высокими
температурами, в кратчайшие сроки завершить цикл размножения и
успешно подготовиться к следующему сезону.
ЛИТЕРАТУРА
Богданов О. П. К экологии песчаной эфы. — Изв. АН Узбек. ССР, Ташкент,
1950, № 5, с. 86—97.
Богданов О. П. Пресмыкающиеся Туркмении. Ашхабад, изд. АН Туркм. ССР,
1962, 235 с.
107
Богданов О. П. Экология пресмыкающихся Средней Азии. Ташкент, «Наука»
Узбек. ССР, 1965, 259 с.
И ш у н и н Г. И. Распространение, экология и ядовитость песчаной эфы в Средней
Азии. Автореф. канд. дисс. Ташкент, Ин-т зоол. АН Узбек. ССР, 1949, 25 с.
Персианова Л. А. Суточная активность самцов и самок песчаной эфы после
выхода из зимовок в Южной Туркмении. — В кн.: Экология и биология живот-
ных Узбекистана. Ташкент, «Фан», 1969, с. 323—326.
Персианова Л. А. Экология песчаной эфы в природе и неволе. Автореф.
канд. дисс. Ташкент, Ин-т зоол. АН Узбек. ССР, 1972, 20 с.
Пестинский Б. В. Материалы по биологии ядовитых змей Средней Азии, их
ловля и содержание в неволе. — Тр. Узбек, зоол. сада, Ташкент, 1939, т. 1,
с. 4—62.
«Содержание ядовитых змей Средней Азии в неволе». Ташкент, «Фан», 1972, 177 с.
X оз а цк и й Л. И. и Захаров А. М. Реакция некоторых ядовитых змей на тем-
пературу и свет. — В кн.: Ядовитые животные Средней Азии и их яды. Таш-
кент, «Фан», 1970, с. 164—175.
Че р л и н В А. Зависимость поведения эф от микроклиматических условий. — В кн.:
Вопросы герпетологии. Автореф. докл. IV Всесоюзн. герпетол. конф. Л., «Наука»,
1977, с. 223—224.
D и v d е v a n i I. а. Borut A. Mean body temperature and heat absorption in four
species of Acantodactylus lizards (Lacertidae)—Herpetologica, 1974, vol. 30,
n. 2, p. 176—181.
Licht P. Photoperiodic and thermal influences on reproductive cycles in reptiles. —
In: Endocrinology, Amsterdam — New York, 1973, p. 185—190.
Singh D. P Analysis of environmental factors regulating the gonadal cycle in a tro-
pical pond turtle, Lissemys p. granosa (Schoepff). — Experientia, 1974, vol. 30.
n. 8, p. 967—968.
The behaviour dependence of the saw-scaled viper,
Echis multisquamatus Cherlin, 1981 on temperature in south Turkmenia
V. A. Cherlin and A. J. Tsellarius
Zoological Institute, Academy of Sciences, U.S.S.R. (Leningrad) and Badkhys State
Nature Reservation (settlement Kushka, Turkmen Republic)
Temperature conditions in the holes are considered as a factor determining time
and pattern of snake state on the surface, regulating duration of photoperiod and also
as indirect index of behaviour and physiological state of snake. Four main patterns
of behaviour are pointed out: in the hole, at the hole, free removal on the surface and
behaviour thermoregulation. Conditions of realisation of these behaviour patterns as
well as sexual and feeding activities are recorded.
АЛЬБЕРТ СЕБА И ЕГО РОЛЬ В РАЗВИТИИ ГЕРПЕТОЛОГИИ
К. Б. Юрьев
Зоологический институт АН СССР (Ленинград)
Имя Альберта Себа (1665—1736) мало что говорит ныне даже образованному
биологу, если только он не является специалистом в области истории этой науки.
Старший современник Карла Линнея и Жоржа Бюффона, он не оставил однако столь
яркого следа ни в истории биологии, ни, подобно Бюффону, в истории литературы.
Тем не менее его имя осталось навсегда связанным с историей зоологии, развитию
которой он оказал огромную услугу. Предлагаемая работа посвящается выяснению
роли А. Себа в истории исследования земноводных и пресмыкающихся.
Вначале приведем краткие биографические сведения. Альберт Себа родился
2/15 мая 1665 года в местечке Этцель в Восточной Фрисляндии (Голландия) в семье
фермера. С ранней юности он увлекался собиранием минералов, растений, раковин
и других произведений природы. Это увлечение не покидало его всю жизнь, и именно
оно и связало его имя с развитием зоологической систематики. По окончании местной
школы он поступил учеником в аптекарскую лавку Абрахама Крамера в Нейштадт-
Годене. Позднее он переехал в Амстердам, где поступил в ученики к фармацевту
Николаю Эрдвину Думбсторфу. Сменив еще несколько учителей в разных городах
Голландии, он возвратился в Амстердам. Здесь, сдав экзамены на звание аптекаря,
он основал собственную торговлю аптекарскими товарами.
Женившись, он купил дом на Гаарлемской улице (этот дом сохранился и по-
ныне), где обосновался с семьей и где располагалось его предприятие. Здесь же позд-
нее размещались и его знаменитые коллекции, ставшие вскоре одной из достоприме-
чательностей Амстердама.
На обогащение и организацию своего музея Себа не жалел средств. Широкие
торговые связи с крупнейшими торговыми компаниями того времени и, в первую
очередь, с Ост- и Вест-Индской, в значительной мере способствовали росту его музея.
Есть данные, что Себа совершил одно или несколько путешествий в Вест-Индию,
хотя сам он нигде не упоминает об этом. Себа вел оживленный обмен с другими
голландскими коллекционерами: Николаем Витсеном (1641 —1715), Левинусом Вин-
центом (1658—1727), Теодором Гюйгенсом (годы жизни не известны).
Он был лично знаком и вел активную переписку с рядом выдающихся натура-
листов и врачей того времени: Фридериком Рюйшем (1638—1731), Германом Бурга-
вом (1668—1738), Иоганном Линком (1674—1734), Якобом Теодором Клейном
(1685—1750), Гансом Слоаном (1660—1753), Якобом Шейхцером (1682—1738) и дру-
гими. 15/28 июня 1731 года музей Себа посетил великий Линней (1707—1778). Себа
хотел привлечь его к обработке и систематизации экспонатов своей коллекции для
третьего тома своих «Тезавров» (о них будет сказано ниже), но, занятый рядом
ботанических работ, Линней отказался и рекомендовал вместо себя своего друга
Петра Артеди (1705—1735), только что закончившего свой обобщающий труд по
ихтиологии, позднее изданный Линнеем. Артеди определил всех рыб из собрания
Себа и сделал их описание, но трагическая смерть (он утонул в городском канале,
оступившись при возвращении домой в ночное время) прервала его работу
Во время своего второго посещения Голландии в 1716—1717 годах музей Себа
посетил Петр Великий. К этому времени у Себа возникла мысль продать свое собра-
ние русскому царю, о чем он уже вел с ним переписку. Известно письмо Себа к царю
от 4 октября 1715 года с подробным описанием его музея (Пекарский, 1862, с. 558—
561). Ввиду большой редкости книги П.П. Пекарского и большого интереса письма
для истории первого отечественного музея, знаменитой «Кунсткамеры», в основу ко-
торой и легло собрание Себа, позволим себе привести его полностью. Перевод письма
на русский язык, сделанный для Петра I, ныне хранится в Государственном архиве
древних актов в Москве.
«Краткое ведение моим куриозам и самородным вещам, которые объявлены
ниже сего:
1) Кабинет сделан из самого доброго ост-индского кипарисного дерева; в нем
72 ящика — все предорогими и диковинными раковинами изукладены: привезены
из Ост и Вест Индии и прочих дальних земель; которые (раковины) с великим приле-
жанием и трудолюбием от наилучших диковинных штук сбираны в место, часто ящик
в 1000 и более штуках. О чем верно доложу, что от сих куриозов полнее числом
никто показати не может.
2) Кабинет же красного остиндского дерева с дверью того же дерева. Внутри
першпективой стеклами украшено и в нем более 400 склянок самого чистого стекла,
в которых неизреченные, чудественные, странные звери, в винном духе положенные
для содержания, в которых преузорочно презентуются. Да еще есть раритеты без
кабинета, которые утверждены на доске порядочно. Да еще ж с 400 штук скляниц
малых и больших с разными ж зверьми в винном духе положенные, а именно: род
некоторых жаб, изменяющихся в рыбу, и таких верно могу 8 показать, о которых
мы отнюдь не слыхивали и в книгах не упомянулось. Еще ж имею род больших аме-
риканских жаб, которых молодые из спины плодятся и израстающие. Також виргин-
109
ская тех, же рук с рогами, да еще ж с долгими ушми. Також необычайно великий
остиндский филандер, которых молодые, входящие внутрь и паки исходящие, до
возрасту питаются. Еще ж есть всяких рук крокодилы, армодилы, лигансы, касмансы,
гагедебы, саламандры, ленарцы, мирертесы, гавиянский дьявол, салво гардо, бавиане,
летучие змеи, летучие рыбы, полипусы, реноры, разных рук остиндские рыбы, морские
кошки, морские мыши, морские черви, морские чуда, тарантулы и превеликие пауки,
скорпионы, гекусы, милипеды, разные черепахи, раки, змеи, ревучие змеи, еще и
иных многих рук змеи, звери, которых всех поименно назвать невозможно.
3) Есть еще японского мастерства лаковый кабинет, в нем сделаны крепость
и замок на горе преизрядно презентуется и весь из самого красного чистого коралла
японского мастерства; да еще в нем суд царя Соломона и сверх вещей того всякими
бисерами унизана и иными вещми унизано.
4) Кабинет, в нем 32 ящика, в которых с 1000 вещей европейских папильенов,
весьма удивительны фигурами, пристегнутых, которые всем превосходительнее гос-
подина Винцентовых вещей и числом полная, которых комплетиса набрать невоз-
можно.
5) Еще ж есть подобием сему кабинет, в котором все странные азиатские, афри-
канские и американские гадины; частью же есть необычайно великия всяких разных
колеров и удивительные фигуры. Також презентуются папелионы всяка своего фигу-
ру ю, которых с 50 ящиков наберется, в том числе есть с серебрянными и золотыми
крылами; да еще ж, в дополику при сем, многие ящики всех рук.
6) Один кабинет китайского мастерства черепахою и серебром по дереву вы-
кладены; в нем без меры многих рук руды земляные из Ост Индии, Китая, Венгрии
из всей Германии собраны.
7) Еще же кабинет сему подобием: в нем всякие каменые вещи, которые от
потопа в твердый камень изменились, множество фигур рыбы, травы и ползущие
гадины.
8) Ящик, в нем всяких рук странные птицы, в том числе и райские, царек и
иные раритеты.
9) Ящик из слоновой кости предорогим китайским художеством вырезан и се-
ребром выложен — в нем всякие куриозитеты.
10) Ящик малой, слоновой же кости.
11) Японского мастерства лаковый кабинет, в нем разные ж раритеты.
12) Ящик, в нем великий и весьма удивительный корень притоманного дерева
Аилое местами сросся и китайским по оным местам предорогим мастерством вырезан.
Тако ж есть весьма искусного и любопытного мастерства распятие из красных
кралей, а крест финифтиной, и сей крест преж сего в Гишпании в Фигасе на пре-
столе стоял, а ныне за несколько лет англичане и голландцы оный завоевали и сюды
привезли.
Еще ж есть удивительная машина в Японии делана из ординарной трости,
с которыми и ныне ходим. Оную сумели так тонко и чисто сделать и плести шоко-
ладный, кофейный и чайный сервиз, а что во истину удивительно — числом оных 60,
а внутри лакованы красным лаком, подобно порселенным, и можно за порселеновые
держать, понеже горячую воду сдерживают и в походах лучше быть не можно, для
того оные не ломки, хотя и уронятся, но без меры гибки, да к ним же сделан футерал-
подобием байдоре. А снаружи оным же мастерством покрыта. Благовали мне в сем
поверить и верно доложу, что равного сему сервису нет у потентатов во всей Европе,
и много охотников к сему сервису имел, а отдать не хотел.
Також есть многих рук образы и фигуры с каменеями китайския работы добрыя
Також есть равного мастерства вырезаны из сандала желтого образы и всякие
лица.
Еще ж есть из слоновой кости всякия разный штуки, да есть всяких рук само-
родный бисер и всяких цветов.
Всякой морской мох чудных фигур, подобием древу насящему цвет и плод
имеющий, из красного африканского моря.
Ящик малой со всякими разными удивительными с зельями и с семенами из
дальних стран.
Еще еж есть всяких птиц яйца, в том числе птицы струса и казуария, и иных
многих крокодиловых, змеиных и лесных змеев, и еще множество всяких зверей,
которых всех поименно написать за кратким временем не умел.
Надеюсь, что его царскому пресветлому величеству ели мои куриозы весьма
нравны и угодны будут. Верно доложу, что сии штуки числом гораздо полнее докто-
ровых Рюйша, да Винцентова, которые его царское величество благоволил во Амстер-
даме видеть.
При вышеописанных куриозах пошлю преизрядные книги, в которых о всех сих
вещах описано, а именно автора славного Алдрованда комплектное описание, Даной-
стона описание о зверях, Римефиуса о раковинах, Мериана о всяких ползущих и
иных гадинах. Из того числа многое число красками расцвечены, а иные живописием
изъяснены и проч.
Из Амстердама, октября 4 дня 1715 года. Подписано Альбертус Себа.»
ПО
Приведенное письмо свидетельствует о связях Себа с русским правительством.
В своем труде Себа неоднократно упоминает о различных услугах, оказанных им
русскому царю в деле собирания коллекций, а также в снабжении страны медика-
ментами. В Архиве Академии наук в Ленинграде (фонд 120.оп.1.№ 164) хранятся
письма Себа к лейб-медику Петра I Роберту Арескину (умер в 1718 году), из кото-
рых видно, сколь тщательно выполнял он заказы царя на медикаменты. Личный
осмотр кабинета Себа Петром I, по-видимому, окончательно решил дело, и все собра-
ние было куплено за 15 000 голландских гульденов и перевезено в Петербург. О со-
ставе этой коллекции мы скажем несколько позднее.
Однако продажа собрания не умерила у Себа интереса к коллекционированию,
и он в течение нескольких лет собрал еще более богатый второй музей, ставший
одним из богатейших в Европе. Сохранились многочисленные отзывы современников
о многочисленности и разнообразии этого второго собрания. Имя владельца стано-
вится хорошо известным в кругу натуралистов. Свидетельством этого является из-
брание Себа в члены нескольких ученых корпораций. Так, в 1722 году он избирается
членом Болонского института, в 1727 — членом знаменитой Академии естественных
курьезов в Швейнфурте, в 1728—Королевского общества в Лондоне. По-видимому,
поняв все значение своей коллекции, окруженный почетом и вниманием, Себа решил
составить описание своего собрания и сопроводить его изображениями всех экспо-
натов. Это грандиозное сочинение «Locupletissimi rerum naturalium thesauri» («Бога-
тейший клад природного царства») увидело свет в 1734—1765 годах в виде огромных
(48.5x33 см) четырех томов с 449 великолепно исполненными таблицами. Последние
два тома вышли уже после смерти Себа.
2/15 мая 1736 года Себа скончался. Через 16 лет (14 апреля 1752 года) после
его смерти наследники устроили аукцион его собрания. С этой целью ими был издан
каталог, который был разослан по научным учреждениям Европы, а также многим
коллекционерам. В результате вся коллекция была продана за 24 440 голландских
гульденов. Как увидим в дальнейшем, часть этой второй коллекции тоже попала
в Россию.
Будучи ученым-дилетантом, Себа оставил после себя немного трудов. Основной
труд — это его знаменитые «Thesauri», навсегда вошедшие в историю систематической
зоологии. Помимо этого, им опубликовано еще три небольших статьи: о коричном
дереве и торговле корицей, об экзотических лекарственных средствах и о препаровке
растений (Себа, 1731 —1737)..
Отдавая должное заслугам Себа перед систематической зоологией, специалисты
разных ее разделов увековечили его имя во многих названиях животных, используя
его фамилию в качестве видового названия. Известный номенклатурный справочник
Ч. Д. Шерборна (1861 —1942) приводит 39 видовых названий животных разных клас-
сов, названных в честь Альберта Себа (Sherborn, 1930, с. 5838). Переходим к обзору
двух собраний Себа и их судьбы.
Источников сведений о первой коллекции Себа нам известно два. Первый — это
его письмо Петру I от 4 октября 1715 года, полностью приведенное нами выше.
Второй — опись собрания, хранящаяся в Библиотеке Академии наук СССР в Ленин-
граде (Отдел рукописей. Г. 188). Эта рукопись большого формата, содержащая 21 лист
и озаглавленная: «Альберта Себы собрание натуральных вещей». Эта опись занимает
16 листов указанной рукописи. 17 и 18 листы — чистые, 19 и 20 представляют опись
музея Христофора Готтвальда (1636—1700), купленного царем одновременно с собра-
нием Себа. Рукопись написана разными почерками и по палеографическим особен-
ностям, несомненно, должна быть отнесена к первой четверти XVIII века. Кроме за-
главия, вся остальная опись написана на голландском языке. Происхождение руко-
писи неизвестно. Можно думать, что это — копия описи, приложенной к коллекции
и составленной еще в Голландии перед ее отправкой в Петербург. Иначе останется
совершенно непонятным, почему в России составляли опись не на родном или же
на латинском языке — международном языке науки того времени. В этом же убеждает
и то обстоятельство, что опись коллекции Готтвальда сделана на немецком языке,
т. е. на родном языке владельца. Итак, рукопись является копией двух описей, со-
ставленных владельцами коллекций или их доверенными лицами.
Опись собрания Себа имеет три раздела: «Animalia in liquor» («Животные в жид-
кости»), «Insecta» («Насекомые») и «Diverse» («Разное»). Первый, самый крупный
раздел состоит из 345 номеров. В ряде случаев под одним номером числится не-
сколько экспонатов. В разделе насекомых, помимо насекомых собственно, упоми-
нается значительное число чучел различных животных (млекопитающих, птиц, пре-
смыкающихся, рыб). Раздел включает 80 номеров, причем некоторые из них вклю-
чают многие десятки видов. Наконец, раздел «Разное», помимо зоологических экспо-
натов, содержит ряд предметов по этнографии, а также произведения искусства.
Раздел включает 130 номеров. Итак, вся опись включает 550 номеров.
Тематика настоящей статьи не позволяет нам подробно охарактеризовать видо-
вой состав животных этой коллекции. Отметим, что ввиду краткости описаний точная
идентификация экспонатов с современными видами животных весьма затруднительна,
а порой и просто невозможна. Остановимся лишь на герпетологической части этого
собрания.
111
Всего в описи числится 170 экспонатов земноводных и пресмыкающихся, а именно
53 ящерицы, 76 змей, 11 крокодилов, 7 черепах, 22 бесхвостых и 4 хвостатых земно-
водных. Несмотря на значительный дубляж, здесь было представлено не менее
120 видов. Отметим, что Линней в десятом издании «Системы природы» (1758) на-
считывал 182 вида амфибий и рептилий. Таким образом, собрание Себа имело около
70% видового состава батрахо- и герпетофауны мира, известной в середине XVIII века.
В основном, экземпляры этой коллекции происходили из тропических областей Старого
и Нового Света: Вест-Индии, Суринама, Бразилии, Мыса Доброй Надежды, Гвинеи,
Индии, Цейлона, Явы, Амбойны. Если учесть, что в своей коллекционерской деятель-
ности Себа был связан, как уже говорилось, с Ост- и Вест-Индской компаниями,
то приведенный перечень географических районов сбора коллекций станет понятен.
В коллекции были представлены сцинки, вараны, летучие драконы, амфисбены, кро-
кодилы и кайманы, удавы и питоны, гремучие змеи, кобры, черепахи, ряд тропиче-
ских лягушек, суринамская жаба. Приведем несколько примеров описаний, которые
покажут наглядно невозможность идентификации экспонатов: «большая змея из
Америки»; «две красиво окрашенные суринамские змеи»; «очень красивая зеленая
ящерица и белая лягушка из Америки»; «редкая ящерица с Мыса Доброй Надежды».
Однако ряд экспонатов может быть определен довольно точно: «большая жаба из
Америки с молодыми, вылупляющимися из спины» — ясно, что дело идет о Pipa
americana. «Очень красивая очковая змея с Амбойны» — по-видимому, очковая кобра.
Многие животные могут быть определены лишь весьма приблизительно, не всегда
даже до семейства.
Какова же судьба этого первого собрания Себа? Оно исчезло почти бесследно:
время, неправильный уход, сырость, свет погубили его. Еще директор Зоологического
музея Академии наук Ф. Ф. Брандт писал о выставочной коллекции старой кунст-
камеры, что «только самая малая часть ее оказалась пригодной и достойной сохра-
нения» (Штраух, 1889 с. 59). Лишь единичные экземпляры пережили многочисленные
перемещения в музее и дошли до наших дней. Это — огромное чучело анаконды,
украшающее музей и поныне. Змея набита в полном соответствии с представлениями
того времени о движении змей, с изгибами в вертикальной плоскости. Кроме того,
у этого экспоната нет головы, поэтому тогдашний таксидермист, изготовляя чучело,
зашил шкуру, а с переднего конца сделал поперечный разрез, имитирующий ротовую
щель. В музее же выставлено весьма неумело изготовленное чучело крупного экзем-
пляра зеленой черепахи. В научных фондах Зоологического института хранится еще
несколько чучел черепах и крокодильих шкур, которые можно с большой долей
вероятности причислить к коллекции Себа. Кое-что из ящериц, в частности, амфисбены,
геррозавры, гекконы, быть может, тоже происходят оттуда. Все же остальное бес-
следно исчезло. Экспонаты, происходящие из Кунсткамеры, имеют однотипные эти-
кетки, на которых просто указано: «из Кунсткамеры», без каких-либо дальнейших
указаний на происхождение экспоната. Таким образом, выделить из этой массы экзем-
пляры из определенных коллекций не представляется возможным. Всего в отделении
герпетологии Зоологического института АН СССР хранится 231 экспонат из Кунст-
камеры.
Интересно отметить, что портрет Альберта Себа долгое время украшал Кунст-
камеру (Беляев, 1800, с. 240)—дань современников огромному вкладу Себа в раз-
витие первого отечественного музея.
Если о составе первой коллекции мы можем составить себе самое общее пред-
ставление, то вторая была описана, и экспонаты ее изображены в четырехтомном со-
чинении, вышедшем в свет уже после смерти Себа. По нашим подсчетам, сделанным
на основании изучения труда Себа, в коллекции насчитывалось: 120 млекопитающих,
91 птица, 14 птичьих гнезд, 587 пресмыкающихся, 60 земноводных, 141 рыба, 2084 на-
секомых, 92 ракообразных, 40 пауков и многоножек, 2225 моллюсков, 210 иглокожих,
7 червей, 118 кораллов, 23 губки. Помимо этого, в коллекции было 269 образцов
растений, 81 образец ископаемых животных и 172 минерала. Всего, таким образом,
коллекция насчитывала 5874 экспоната, из них 5412 — из животного царства.
Интересен аукционный каталог собрания Себа, ныне представляющий большую
редкость (Catalogus, 1752). Он состоит из четырех частей и в общей сложности со-
ставляет 129 страниц убористого текста. Для удобства пользования им после названия
каждого экспоната дается ссылка на соответствующее изображение в «Thesauri». Аук-
цион состоялся 14 апреля 1752 года. В библиотеке Амстердамского университета
сохранился экземпляр каталога с многочисленными пометками о лицах, приобретших
те или иные экспонаты, а также о цене. По этому экземпляру можно, таким образом,
выяснить судьбу собрания в целом. Кабинет был продан за общую сумму в 24 400 гуль-
денов. В работе М. Беземана (Boeseman, 1970) прослеживается судьба этого собра-
ния. Этому же вопросу посвящены и две работы X. Энгеля (Engel, 1937, 1961).
Мы не будем в деталях рассматривать все довольно сложные перипетии, испы-
танные отдельными частями коллекции Себа. Остановимся только на окончательном
их распределении в крупнейших музеях мира. Части коллекции находятся или нахо-
дились в следующих странах: России, Голландии, Англии, Франции, Швеции, Дании
и Германии. В Голландии они находились в университетском музее Гронингена (по-
гибли во время войны), в зоологической лаборатории университета в Утрехте, в Зоо-
логическом музее Амстердама и в Государственном музее естественной истории
112
в Лейдене. В Англин их владельцем является Британский музей естественной истории
в Лондоне, во Франции — Национальный музей естественной истории в Париже,
в Дании — Музей естественной истории в Копенгагене, в Швеции — Музей естествен-
ной истории в Стокгольме. Что касается Германии, то экспонаты коллекции Себа
находились в Океанологическом музее в Бремене (ФРГ) и в Зоологическом музее
в Берлине (ГДР). Что и когда из этого второго собрания попало к нам в Россию?
Составители некоторых путеводителей по Кунсткамере единодушно утверждают, что
этот музей сильно обогатился в результате покупки значительной части второго со-
брания Себа. Так, Иоганн Бакмейстер (умер в 1788), составитель первого путеводи-
теля по Кунсткамере, пишет: «В 1751 году прибыло множество животных, купленных
в Амстердаме на аукционе кабинета знаменитого Себы, умершего в 1736» (Backmeister,
1776. с. 173). Это указание грешит неточностью, так как известно, что аукцион со-
стоялся лишь в 1752 году. Бакмейстеру вторит и Осип Беляев (умер в 1807), автор
весьма полного путеводителя по кунсткамере (Беляев, 1800, ч. 8, с. 10).
В Архиве Академии наук в Ленинграде хранится дело о покупке с аукциона
предметов из коллекции Себа (фонд 3, on. 1, № 164, лл. 300—330). Дело открывается
рапортом адъюнкта академии Ивана Тауберта (1717—1771) от 15 апреля 1752 года
в канцеляцию академии. Поскольку этот рапорт никогда не был опубликован, мы
приведем его полностью, сохраняя орфографию.
Перед несколько днями прислан ко мне из Амстердама печатный на голанском
языке каталог натурального кабинета славного Амстердамского аптекаря Альберта
Себы, который ныне с публичного торгу продан быть имеет; я понеже во оном нахо-
дятся многие такие звери и при том, которые прежде всего были и в здешней кунст-
камере, но при случившемся в Академических палатах несщастливом пожаре згорели;
того ради Канцелярия Академии Наук сим представляю, не соблаговалено ль будет
для приведения Кунсткамеры ЕЯ Императорского Величества в прежнее состояние
употребить сию способную оказию и приказать против приложенного реестра оные
звери и птицы при помянутом аукционе купить а комиссию на то дать Амстердам-
скому материалисту Дитриху Шмиту, который в таких вещах довольное знание имеет.
Ассессор Иван Тауберт».
На следующих листах (лл. 302—305) приводится упомянутый Таубертом в его
письме реестр вещам, которые он предлагает к покупке. Всего этот реестр содержит
71 номер и касается только млекопитающих и птиц. По-видимому, канцелярия одо-
брила предложение Тауберта. Во всяком случае, рапортом от 14 мая 1752 года тот же
Тауберт извещает канцелярию, что упоминавшимся Шмитом куплено зверей и птиц
на «500 на 90 на 9 голандских гульденов и на 10 штиверов» (л. 308). Наконец, на
листах 329—330 в деле хранится «Перевод с фактуры, приложенной из Амстердама
от материалиста Дитриха Шмита, о купленных им для Императорской Кунсткамеры
зверях и птицах». Всего в фактуре числится 37 экспонатов. Что касается земноводных
и пресмыкающихся, то судя по фактуре, были приобретены «жаба американская
с спины щенят выпускающая», «ящерица цейлонская с белыми крапинками и полос-
ками» и «ящерица индийская легван называемая». Таким образом, как видно из
документов, поступления из второй коллекции Себа в Кунсткамеру были весьма
незначительны. Откуда же тогда свидетельства хорошо осведомленных Бакмейстера
и Беляева, много лет прослуживших смотрителями Кунсткамеры, о множестве живот-
ных, купленных с аукциона собрания Себа? Быть может, не все документы сохра-
нились? Ведь не сохранился же каталог аукциона Себа, присланный Тауберту. Не-
смотря на все старания, мы не смогли обнаружить его в Библиотеке Академии наук,
куда, казалось бы, он должен был быть передан из канцелярии. Дело, по-видимому,
не в этом. Как указывает М. Беземан (Boeseman, 1970, с. 179—181), многие экспо-
наты были приобретены коллекционером Т. Слютером. По его смерти его музей
также был распродан на аукционе, и через посредство комиссионеров Балгри и Дю-
лонга значительная часть экспонатов этого собрания была приобретена нашей Акаде-
мией наук. Это произошло в конце 80-х годов XVIII века. Точную дату Беземану
установить не удалось. В Архиве Академии наук мы также не обнаружили каких-либо
документов, относящихся к этому делу. Таким образом, установить совершенно точно,
что из второй коллекции Себа поступило в Кунсткамеру, помимо перечисленного
в указанном реестре, не представляется возможным.
Обратимся теперь к основному труду Альберта Себа, его знаменитым «Тезав-
рам». Труд Себа, представляющий описание его коллекции, издан в четырех томах
форматом «Большое фолио» с 449 таблицами, изображающими описанные экспонаты.
Известно два варианта этого издания: с параллельными латинским и французским
текстом, и другой — с латинским и голландским. Отметим, что экземпляры с голланд-
ским текстом очень редки. В библиотеках Ленинграда и Москвы наличие подобных
экземпляров нам не известно. Часть тиража каждого из этих вариантов была выпу-
щена в продажу с раскрашенными от руки таблицами. Но такие экземпляры были
даже в те времена очень редки. Так, известный библиограф того времени Иоганн
Самуил Шретер (1735—1808), хорошо знавший зоологическую литературу, писал
в одном из своих библиографических обзоров по поводу труда Себа: «Насколько
я знаю, имеются также иллюминованные экземпляры этой книги, однако я таковых
не видел и не могу даже вспомнить, чтобы я где-либо читал о них, поэтому о каче-
стве раскраски я ничего не могу сообщить» (Schroter, 1780, с. 170). В Зоологическом
8 Зак. 719
113
институте АН СССР в Ленинграде имеются два первых тома сочинения Себа с рас-
крашенными таблицами. По-видимому, эти тома были присланы в Академию наук
самим Себа.
Позднее, видимо, уже в начале XIX века часть тиража в несброшюрованном
виде была приобретена одной из французских книготорговых фирм. Таблицы были
раскрашены, книги одеты в переплеты и пущены в продажу. Такие экземпляры новой
раскраски по качеству значительно уступают экземплярам, раскрашенным при изда-
нии книги. Раскраска довольно небрежна, да и сами краски значительно менее яркие
и звонкие.
Весь труд вышел в 1734—1765 годах. Приведем даты публикации отдельных
томов и их пагинацию: I. 1734: (29)4-178 с., 111 таб.; II. 1735: (30)4-154 с., 114 таб.;
III. 1759: (25)+212 с., 116 табл.; IV. 1765: (42)+226 с., 108 таб.
Первый том (рис. 1) имеет великолепный фронтиспис, гравированный по рисунку
мастера, скрывшегося под инициалами L.F.D.B., известным гравером П. Танье (1706—
1761). Фронтиспис представляет собой аллегорическую картину, в центре которой по-
мещена богиня Истины. Свет, излучаемый ею, при поддержке всемогущего Времени,
помогает Науке в ее исследованиях произведений природы. В этом же томе имеется
и портрет Себа, гравированный Я. Хубракеном (1698—1780) по рисунку Я. М. Квинк-
гарда (1688—1772). Портрет, исполненный в 1731, представляет Себа в мантии уче-
ного среди коллекций его кабинета (рис. 2). Как гласит подпись под портретом, ему
в это время 66 лет. Затем после титульного листа следуют посвящение, предисловие
автора, неизбежные в то время стихи — славословия автору и регистр содержания.
Первый и второй тома изданы X. Янссоном ван Весбергом, И. Веттштейном и
В. Смитом, третий том — только Янссоном, том четвертый — X. Аркстейном, X. Мер-
ком и Т. Шутеном. Все иллюстрации изготовлены лучшими художниками и граверами
Голландии того времени: П. Танье, А. ван де Лааном, Ф. де Баккером, Я. Фолкема,
В. Иолгманом и другими.
Весь текст написан Себа по голландски и переведен на латинский язык про-
фессором медицины Лейденского университета Иеронимом Давидом Гаубиусом (1705—
1780) и профессором медицины, математики и философии Утрехтского университета
Петром Мушенброком (1692—1761). Следует отметить, что переводчики в ряде мест
дополнили текст Себа, особенно в третьем и четвертом томах, изданных уже после
смерти автора. Текст третьего тома, касающийся рыб, написан Петром Артеди. Пере-
вод на французский язык осуществлен Л. Жакуром и П. Массуэ (I, II и часть III то-
мов) и продолжен доктором Дюкло под наблюдением профессора зоологии Лейден-
ского университета Николая Себастьяна Алламанда (1716—1784).
Первый том имеет предисловие знаменитого Германа Бургава, том второй —
известного палеонтолога Якоба Байера (1677—1735), профессора медицины в Нюрн-
бергском университете.
Первый том посвящен ботаническим коллекциям, а также содержит обширные
материалы по млекопитающим, птицам, земноводным и пресмыкающимся. Встречаются
здесь и сведения об экспонатах по другим группам (насекомым, паукам, многонож-
кам и др.). Второй том почти исключительно посвящен змеям. Том третий — о рыбах,
моллюсках, иглокожих, кораллах и губках, том четвертый — о насекомых и минера-
логических коллекциях. По данным Энгеля (1937, с. 93; 1961, с. 120), текст III и
IV томов подвергся обработке Арно Возмаера (1720—1799), директора зверинца и
натурального кабинета принца Оранского. Возмаеру помогали Иоганн ле Франк Берк-
хей (1729—1812), профессор физики Лейденского университета, и знаменитый Петр
Симон Паллас (1741—1811). Однако, что именно они добавили к тексту Себа, не
установлено.
Труд Себа привлек внимание специалистов и послужил первоисточником зна-
комства со многими видами животных, особенно тропических стран. Высокая оценка,
даваемая современниками этому гигантскому сочинению, относится, собственно говоря,
к рисункам. Текст же обычно расценивался как посредственный. Так, известный биб-
лиограф, обладатель выдающейся коллекции естественно-исторических книг Иосиф
Пауль Кобрес (1754—1823) в описании своей библиотеки пишет о книге Себа: «Опи-
сания плохи, гравюры большей частью великолепны» (Cobres, 1781, т. I, с. 108—109).
Уже упомянутый И. С. Шретер дает подробное описание труда Себа и делает, на-
сколько нам известно, первую попытку идентифицировать изображенных Себа живот-
ных (в данном случае моллюсков) с уже известными видами (Schroter, 1780, с. 141 —
170). Интересно отношение Линнея к этой книге. Несмотря на недружелюбные личные
отношения с ее автором, вызванные крайней нетактичностью поведения последнего
в отношении научного наследства Артеди, имя которого Себа даже не упомянул,
Линней широко использовал сочинение Себа, его рисунки при описании видов раз-
личных животных. В «Системе природы» Линней так пишет о Себа и его труде:
«Себа собрал изумительное число неизвестных прекрасных видов и не только изобра-
зил, но и кратко их описал» (Linnaeus, 1758, с. 195). Такое отношение к труду Себа
прочно закрепилось среди зоологов. Великий Кювье (1769—1832) восхищался табли-
цами Себа как собранием материалов для описания животных. В первом издании
«Царства животных» Кювье пишет, что сочинение Себа «украшает большое число
прекрасных таблиц, но текст не имеет никакого значения, так как он написан без
разбора и без критики» (Cuvier, 1817, т. IV, с. 159).
114
Все гравированные медные доски к труду Себа оказались в результате окуп-
пации Голландии французскими войсками в 1795 году во Франции, в Музее есте-
ственной истории в Париже. Там их обнаружил известный зоолог Феликс Эдуард
Гирен-Меневилль (1799—1874). Именно ему и принадлежит идея переиздания таблиц
L О С U Р L Е ТI S S 1 М I
R Е R U М
N ATURALIUM
ACCURATA DESCRIPTIO,
ICONIBUS ARTIFICIOSISSIMIS
PER
f UNIVERSAM PHYSICES HISTORIAN. i
OPUS,
CUI, IK HOC RERUM GEKERE, \ULLUM PAR EXSTlTiT.
EX TOTO TERRARUM OR BE COLLEG1T,
PIGES5IT, &ESCRIPSTT . ET BLPINGENDUM CVRAUIT
F T Z E L A OO $ Tl U 5 I U S,
ACAOEML! СЖ5АЯЕ.Е LEOPOLD1NO CAROLINA NATURE CURIOSORVM
CCLLLGA XENOCRATfS DI CT US; SOCIETATIS REGIA ANGLICAN.!,
LT INSTITUTI BONONlENMS, SODaUS.
и Af X 7 £ t A £ E) A M /,
। Apu„ т A N S S О N 1 O-W XESBSRG 1 OS,
t T s T E N I и M i t GUL. SMITH.
-MOCCXXKH
Рис. 1. Титульный лист первого тома сочинения А. Себа.
Себа с прибавлением нового текста. С этой целью была организована рабочая группа
из сотрудников музея для написания нового текста. Как отмечается в опубликованном
проспекте этого труда (Ferussac, т. 8, 1826, с. 29—31), эта группа состояла из
И. Жоффруа Сент-Илера (1805—1861), Рене Лессона (1794—1849), Андре Ферюссака
(1786—1836), Гирен-Меневилля, Жака Одуэна (1787—1841), Ансельма Демаре (1784—
8*
115
1858). Редакторами всего издания были Гирен и Кювье. Издание должно было вы-
ходить отдельными выпусками по 25 таблиц с объяснительным текстом в каждом.
Согласно исследованию Л. Холтиса (Holthuis, 1969, с. 249), в августе 1827 года вышли
в свет три первых выпуска с 34 таблицами 3-го тома сочинения Себа. Следующие
9 выпусков (4—12), содержащие таблицы 35—116 этого же тома, вышли в конце
Рис. 2. Портрет А. Себа в возрасте 66 лет.
Художник Я. М. Квинкгард. гравер Я. Хубракеи.
1828 года. Выпуски 13—21, содержащие таблицы первого тома, были изданы также
в 1828 году. К 13 выпуску был приложен и текст к таблицам 1—48 третьего тома
(моллюски)-. Выпуски 22—45 вышли в июне 1831 года и содержали таблицы II и
IV томов оригинального издания, а также оставшиеся таблицы I тома. Дальнейшая
публикация текста не состоялась, вышло всего 8 страниц. Никогда не был издан
также и титульный лист. По-видимому, остановка публикации текста была связана
е огромными трудностями идентификации рисунков Себа. Так закончилось это изда-
ние. Отметим, что на книжном рынке оно встречается гораздо реже оригинального
издания. Раскрашенные экземпляры французского переиздания нам неизвестны.
116
Сочинение Себа было хорошо известно в России. Об этом свидетельствует нали-
чие довольно значительного числа его экземпляров в библиотеках страны, в частности,
Ленинграда. Здесь нам известны экземпляры Библиотеки Академии наук СССР
(Ленинград), Государственной Публичной библиотеки имени М. Е. Салтыкова-Щед-
рина.. Военно-Медицинской Академии имени С. М. Кирова, Ботанического института
АН СССР имени В. Л. Комарова и Зоологического института АН СССР. Отметим,
что все эти экземпляры представляют латинско-французский вариант и не раскра-
шены. Только в библиотеке Зоологического института имеется два первых тома с ве-
ликолепно раскрашенными таблицами. По-видимому, это подношение самого Себа
русскому двору. Тома третий и четвертый, выпущенные много лет спустя после
смерти автора, его наследниками присланы не были.
Неоднократно сочинение Себа предлагалось и книгопродавцами. Так, петербург-
ский книготорговец В. Грефф в 1823 году предлагал сочинение Себа за 450 рублей
(Graff, 1823, с. 29). Рижский антиквар Н. Киммель продавал труд Себа всего за
25 рублей (Киммель, 1886, с. 31, № 644). Крупнейший антиквар Петербурга Н. В. Со-
ловьев предлагал великолепно раскрашенный экземпляр за 200 рублей (Соловьев, 1910,
№ 550). Фирмой «Международная книга» в 1931 году экземпляр труда Себа про-
давался за 75 рублей. В 1961 году автору этих строк посчастливилось при посредстве
известного ленинградского букиниста-антиквара И. С. Наумова приобрести велико-
лепный экземпляр сочинения Себа с раскрашенными таблицами. Правда, раскраска
новая, начала XIX века.
Нам осталось сказать несколько слов еще об одном переиздании таблиц рас-
матриваемой книги. В 1783—1788 годах в Аугсбурге вышла в 4 томах зоологическая
книга под названием: «Angenehme und lehrreiche Geschenk fiir die Jugend»» («Прият-
ный и поучительный подарок юношеству»). Первый том посвящен млекопитающим,
второй — птицам, третий — рыбам, четвертый — земноводным и пресмыкающимся.
Подробное описание первых трех томов дает С. де Гроот (Groot, 1972). Четвертый же
том, вышедший в 1786 году и не описанный Гроотом, посвящен герпетологии и пред-
ставляет собой переиздание восьми таблиц труда Соба, изображающих лягушек и
ящериц. Гравюры исполнены Фридрихом Киршгером. Издание было прекращено на
четвертом томе. Видеть эту книгу нам не приходилось. Как и большинство детских
книг прошлых веков, она является большой библиографической редкостью. Даже
в Британском музее естественной истории имеются только 3 и 4 тома этого издания.
Отметим, что отдельные рисунки из труда Себа довольно часто перепечатывались
вплоть до середины XIX века.
История герпетологии богата славными именами, навсегда вошедшими в исто-
рию науки, в историю научной литературы: Конрад Геснер и Уллис Альдрованди,
Карл Линней и Якоб Клейн, Бернар Лясэпед и Франсуа Доден, Андре Дюмериль и
Габриэль Биброн, Иван Криницкий и Александр Штраух, Эдуард Коп и Джордж
Буленджер, Леонард Стейнджер и Роберт Мертенс. Мы не назвали, по-видимому,
десятой части имен из славного списка отцов современной герпетологии. Каждый
из названных оставил огромное научное наследство, каждый был крупным специа-
листом, широко образованным и эрудированным не только в герпетологии, но и в ряде
других разделов зоологии, а часто (Линней, Лясэпед, Коп) и биологии в целом.
Альберт Себа представляет противоположный случай. Оставленный им труд, напи-
санный не ученым, а всего лишь любителем-дилетантом, тем не менее прочно вошел
в науку. Он цитируется в научной литературе и по сей день, на него ссылаются
не только герпетологи, но и териологи, орнитологи, ихтиологи, малакологи и энтомо-
логи. Многие виды животных были введены в научный обиход и описаны только
на основании изображений и описаний Себа.
Создатель систематической зоологии, Линней, в 10-м издании «Системы при-
роды» (1758) ссылается на Себа 169 раз, в 12-м издании (1766) 197 раз, В последнем
издании этой бессмертной книги, вышедшем в 1788—1789 годах под редакцией и
с добавлениями И. Г Гмелина (1748—1804), — 265 раз. В 10-м издании он описал
182 вида земноводных и пресмыкающихся, из них 10 видов описано на основании
рисунков Себа, а именно Lacerta scutata, L. anguina, L. basiliscus, L. strumosa,
L. orbicularis, Boa hipnale, Coluber argus, C. niveus, Anguis meleagris, Rana marina.
В 12-м издании описано уже 214 видов амфибий и рептилий, из них 14 по рисункам
Себа. К описанным в 10-м издании добавлены еще Testudo Carolina, Lacerta maurita-
nia, L. dracaena, Draco praepos. В 13-м издании описано 375 видов. Но поскольку
к этому времени вышло в свет значительное количество литературы, содержащей
описание интересующих нас животных, только три вида остались описанными исклю-
чительно на основании изображений Себа: Draco praepos, Coluber ocellatus, C. ruber.
Для того, чтобы отчетливее понять роль сочинения Себа для Линнея при системати-
зации амфибий и рептилий, отметим количество ссылок на других авторов в 10-м из-
дании. Кроме него. Линней ссылается на Гроновиуса (32 раза), Кэтсби (7 раз),
а также по одному — два раза на ряд других авторов от Гесснера до Резеля фон
Розенгофа. В 12-м издании число ссылок на Себа увеличивается до 197, а на других
авторов остается в прежнем числе.
Первая специальная сводка по земноводным и пресмыкающимся (кроме чере-
пах) Иосифа Лауренти (годы жизни неизвестны), изданная в 1768 году, содержит
169 ссылок на сочинения Себа. Совпадение в количестве ссылок с десятым изданием
117
«Системы природы» Линнея, по-видимому, объясняется прямым заимствованием Лау-
ренти. В работе насчитывается 241 вид земноводных и пресмыкающихся (кроме чере-
пах, не включенных в обзор). Из них 118 видов имеют ссылки только на рисунки
Себа. Знаменитый Бернар Лясэпед (1756—1825), современник и сотрудник Бюффона,
Кювье, Ламарка, Этьена Жоффруа Сент-Илера, в первом издании своей сводки по
амфибиям и рептилиям, изданной в качестве продолжения «Естественной истории»
Бюффона (1788—1789), приводит 165 ссылок на сочинения Себа, а в последнем
издании (1832—1833), вышедшем посмертно, 219. В немецком переводе труда Лясэпеда
(1800), изданном под редакцией и с добавлениями И. М. Бехштейна (1757—1812),
насчитывается 360 ссылок. В первом издании своей сводки (1788—1789) он описывает
292 вида интересующих нас животных, из них 3 вида — только на основании рисунков
Себа. В дальнейших изданиях это число увеличивается. В немецком переводе работы
Ласэпсда Бехштейн добавляет еще 6 видов, описываемых по рисункам Себа.
Франсуа Доден (1776—1804) в своей сводке по герпетологии (1802—1803) ссы-
лается на Себа 286 раз. Он описывает 556 видов рептилий и амфибий, из них для
9 видов голотипами являются рисунки Себа. Шарль Соннини (1751 —1822) и Пьер
Лятрейль (1762—1833), авторы герпетологической части восьмидесятитомного издания
«Естественной истории» Бюффона, цитируют Себа 304 раза. Следующая по времени
ревизия амфибий и рептилий, изданная Б. Мерремом (1761 —1824) в 1820 году, со-
держит 407 ссылок на Себа. Меррем (1820) описывает, 580 видов, из них 10 только
по рисункам Себа.
Жорж Кювье в своем классическом труде «Царство животных» широко исполь-
зует сочинение Себа и не только в разделе герпетологии. В первом издании этой
книги (1817) он при описании интересующих нас классов ссылается на Себа 115 раз,
а в третьем издании (1836) — 138 раз. Перевод первого издания «Царства животных»
на немецкий язык, выполненный Генрихом Шинцем (1777—1861) с его дополнениями
(1821 —1825), содержит 174 ссылки на Себа, а перевод второго издания с добавле-
ниями Ф. С. Фойгта (1781 —1850), изданный в четырех томах в 1831 —1843 годах,—
155 ссылок. Отметим, что в списке рептилий, приложенном к английскому переводу
второго издания труда Кювье, выпущенному в 16 томах в Лондоне в 1824—1833 годах,
составленном Джоном Греем (1800—1875), имеется 109 ссылок на Себа.
Анри Дюмериль (1774—1860) и Габриэль Биброн (1806—1848), авторы ставшей
классической «Общей герпетологии» (1834—1854), все еще широко используют труд
Себа, ссылаясь на него 385 раз. Джон Грей обращался к труду Себа и в дальнейших
своих работах. Так, в каталоге змей Британского музея естественной истории (1849)
он цитирует его 59, а в каталоге ящериц того же музея (1845)—89 раз. Каталог
черепах и крокодилов того же автора (1844) дает 21 ссылку. Младший современник
Грея, Альберт Гюнтер (1830—1914) тоже неоднократно прибегал в своей работе
к сочинению Себа. В составленном им каталоге бесхвостых амфибий Британского
музея (1858) имеется 25 ссылок, а в каталоге змей семейства Colubridae (1858)—66.
Джордж Альберт Буленджер (1858—1937), произведший последнюю ревизию
всех групп амфибий и рептилий мира и составивший великолепные каталоги по этим
группам, также не обошел труд Себа своим вниманием. Его каталог змей Британ-
ского музея (1893—1896) дает 86 ссылок на изображения Себа. Отметим, что в ката-
логах других групп, составленных тем же автором, таких ссылок нет, кроме каталога
бесхвостых амфибий (1882), содержащего 27 ссылок. Буленджер был последним, кто
смог дать полную ревизию амфибий и рептилий.
Современное состояние герпетологии не дает возможности подобного труда си-
лами одного человека, однако интерес к труду Себа не угасает. Авторы многих гер-
петологических работ продолжают ссылаться на него как на источник сведений о том
или ином виде животных. Мы не станем продолжать наш подробный обзор авторов,
ссылавшихся на труд Себа. Остановимся только на нескольких примерах: Бронгерсма
(Brongersma, 1961), Дугласс (Douglass, 1975), Дэлман (Duellman, 1971), Харди и
Мак-Диармид (Hardy & McDiarmid, 1969), Гейер (Heyer, 1968), Гюи (Gyi, 1970),
Даммере (Lammeree, 1970), Лескюр (Lescure, 1976), Р Мертенс, (Mertens, 1968),
Стимсон (Stimson, 1969), Петерс (Peters, 1960).
Следует отметить, что во многих случаях изображения и описания Себа приво-
дятся в систематических работах в качестве синонимов. Выяснение конкретной судьбы
типов земноводных и пресмыкающихся, установленных разными авторами на основа-
нии изображений Себа, потребует особой работы, как и идентификация его описа-
ний и рисунков с ныне принятыми видами. Нам известна всего одна попытка подоб-
ной идентификации, принадлежащая перу известного зоолога Иоганна Георга Ваглера
(1800—1832). Рукопись этой незаконченной автором работы была обнаружена в его
бумагах и опубликована в журнале «Изида» (Wagler, 1833). Ваглер идентифицировал
только рисунки таблиц I и пяти первых таблиц II тома. Отметим, что многие из
изображений остались неопределенными Ваглером, а многие другие идентифицированы
только предположительно.
В экземпляре труда Себа, хранящемся в библиотеке Зоологического института
АН СССР, в книге между таблицами вложены небольшие листки бумаги, на которых
старинным подчерком написаны по правилам бинарной номенклатуры названия живот-
ных и номер рисунка соответствующей таблицы. По-видимому, это следы работы
по идентификации рисунков, проводившейся кем-то из сотрудников старой Кунстка-
118
меры, скорее всего, академиком А. Ф Севастьяновым (1771 —1824), переводившим
«Систему природы» Линнея на русский язык.
Работа по идентификации рисунков крайне сложна как ввиду их несовершен-
ства с точки зрения современных требований к научному рисунку, так и крайней
отрывочности, а часто и неверности описаний. Полная же идентификация возможна
лишь на основании изучения и рисунков, и текста. Проведение подобной работы
обещает пролить свет на многие вопросы номенклатуры земноводных и пресмы-
кающихся.
ЛИТЕРАТУРА
Беляев О. Кабинет Петра Великого. Т. 1—3, СПб., 1800, 215, 287, 278 с.
(Грефф В.) Graff W. Biicherverzeichniss 15. St.-Petersbourg, 1823, 56 S.
Киммель H. Каталог 29. Рига, 1886, 66 с.
«Международная книга». Новые приобретения, 1931, № И, М., 193+16 с.
Пекарский П. П. Наука и литература в России при Петре Великом. Т. 1. Спб.,
1862, 578 с.
Соловьев Н. В. Каталог № 100. Спб., 1910, 30 с.
Backmeister J. Essai sur la Bibliotheque et le Cabinet de curiosites et d’histoire
naturelie de 1’Academie des Sciences de Saint-Petersbourg. St.-Petersbourg, 1776,
254 p.
Boeseman M. The vicissitudes and dispersal of Albertus Seba’s zoological speci-
mens. — Zool. Mededel., 1970, D. 43, № 13, p. 177—206.
Boulenger G. A. Catalogue of the Batrachia salientia s. ecaudata in the collection
of the British Museum (Natural History). London, 1882, XVI+ 503 p.
Boulenger G. A. Catalogue of the chelonians, rhynchocephalians and crocodiles
in the British Museum (Natural History). London, 1889, X + 311 p.
Boulenger G. A. Catalogue of the lizards in the British Museum (Natural History)
V. I—III, London, 1885—1887, XIII + 436, XIII + 497, XII + 575 p.
Boulenger G. A. Catalogue of the snakes in the British Museum (Natural History).
V. I—III, London, 1893—1896, XIII + 448, XI + 382, ХП + 724 p.
C a t a 1 о g u s van de Uitmuntende Cabinetten met allerley soorten von ongemeene
schone Gepolijste, Dubletschelpen, Coraal en Zeegewasse... en verscheide andere
Rariteten. En nagelaten door wijlen der Heere Albertus Seba. (Amsterdam, 1752),
116 p.
Cob res J. P. Deliciae Cobresiana. Th. I, Augsburg, 1781, ХХШ + 470 S.
Cuvier G. Le regne animal. V. II, Paris, 1817, XVIII + 532 p.
Cuvier G. Das Thierreich. Obersetzt von H. R. Schinz. Bd. II, Stuttgart u. Tiibin-
gen, 1822, XVI+ 835 S.
Cuvier G. Le regne animal. Ed. 2, v. II, Paris, 1829, XV + 406 p.
Cuvier G. Das Thierreich. Obersetzt von F. S. Voigt. Bd. II, Leipzig, 1832, XVI +
+539 S.
Cuvier G. Le regne animal. Ed. Ill, Bruxelles, 1836, XXII+ 626 p.
Daudin F. M. Histoire naturelie generale et particuliere des Reptiles. Paris, 1802—
1803, v. I—VIII, 384, 432, 452, 397, 365, 447, 436, 439 p.
Douglass J. F. Bibliography of the North American land-tortoises (genus Gophe-
rus). Washington, 1975, 60 p.
Due 11m an W. E. The nomenclatural status of the names Hyla boans (Linnaeus) and
Hyla maxima (Laurenti). — Herpetologica, 1977, v. 27, n. 4, p. 397—405.
Dumer i 1 A. M. et В i b г о n G. Erpetologie generale ou histoire naturelle complete
des Reptiles. V I—IX, Paris, 1834—1854, XXIV + 447, IV + 680, V + 518; 11 + 572,
VIII + 854; XII + 610, XIX+1536, 11 + 792, XX + 440 p.
Engel H. The life of Albert Seba. — Svensk. Linne-Sallskap. Arsskrift., 1937, v. 20,
p. 75—100.
Engel H. The sale-catalogue of the cabinets of natural history of Albertus Seba
(1752). —Bull. Res. Council Israel, v. 10 B, n. 1—3, 1967, p. 119—131.
Ferussac A. E. (Prospectus de la «Planches de Seba»). — Bull. univ. sci. et Industrie
publ. par Ferussac, 1826, sect. 2, v. 8, p. 29.
Gray J. E. A synopsis of the species of the class Reptilia. In: Cuvier G. The animal
kingdom, London, 1831, v. 9, 481 + 110 p.
Gray J. E. Catalogue of the tortoises, crocodiles and amphisbaenians in the collection
of the British Museum. London, 1844, VIII+ 80 p.
Gray J. E .Catalogue of the specimens of lizards in the collection of the British
Museum. London, 1845, XXVIII+ 289 p.
Gray J. E. Catalogue of the specimens of snakes in the collection of the British
Museum. London, 1849, XV+125 p.
Groot S. J. d e. On the bibliography of a late eighteenth century German work on
natural history and on early record of Leuciscus meidingeri Haeckel, 1852 (Pisces,
Cyprinidae) in the river system of the Upper Danube an 6th April 1786. — Bijdr.
Dierkund., 1972, v. 42, p. 156—163.
Giinther A. Catalogue of the Batrachia salientia in the collection of the British
Museum. London, 1858, XVI+ 160 + 8 p.
119
Gunther A. Catalogue of colubrinae snakes in the collection of the British Museum.
London, 1858, XVI+ 281 p.
G у i K- A revision of colubrid snakes of the subfamily Homalopsinae. — Univ. Kansas
Publ. Mus. Nat. Hist., 1970, v. 20, p. 49—223.
Hardy L. M. a. McDiarmid R. W. The amphibians and reptiles of Sinaloe, Me-
xico.— Univ. Kansas Publ. Mus. Nat. Hist., 1969, v. 18, p. 39—252.
Heyer W. R. The proper name for the type-species of the genus Leptodactylus. —
Copeia, 1969, n. 10, p. 160—162.
Holthuis L. B. Albertus Seba’s «Locupletissimi rerum naturalium thesauri» (1734—
1765) and the «Planches de Seba» (1827—1831). — Zool. Mededel., 1969, v. 43,
p. 239—252.
Lacepede B. G. Histoire naturelie des quadrupedes ovipares et des serpens. Paris,
1788—1789, v. I—II, 651, 527 p.
Lacepede B. G. Naturgeschichte der Amphibien. Ubersetzt von J. M. Bechstein.,
Weimar, 1800—1802, Bd. I—V, XXXII+ 524, XX + 552, XXVI+454, XX + 298,
XII+ 200 S.
Lacepede B. G. Quadrupedes ovipares et des serpens. — In: Lacepede B. G. Ouvres
completes, v. II—IV, Paris, 1832—1833, II+ 389, 399, 428 p.
Lammeree L. Lizards of the genus Cnemidophorus from the Leeward group and the
adjacent Mainland” of South America. — Stud. Found Curasao a. other Carribean
Islands, 1970, n. 61, p. 46—72.
Laurenti J. Specimen medicum exhibens synopsin reptilium. Viennae, 1768, 214 p.
Lescure J. Contribution a 1’etude des Amphibiens de Guyane Franchise.— Bull. Mus.
Nat. Hist., 1976, ser. 3, n. 377, p. 475—524.
Linnaeus C. Systema naturae. Ed. X, Holmiae, 1758, 824 p.
Linnaeus C. Systema naturae. Ed. XII curavit J. F. Gmelin. Lipsiae, 1789, t. I,
part III, p. 1033—1516.
Me r rem B. Versuch eines System der Amphibien. Marburg, 1820, XV+191 S.
Mertens R. Remarks on the scientific name of the African Puff Adde. — Copeia,
1968, n. 3, p. 621—622.
Peters J. A. The snakes of the subfamily Dipsadinae. — Miscell. Publ. Mus. ZooL
Univ. Michigan, 1960, n. 114, 224 p.
Schroter J. S. Journal f. d. Liebhaber d. Steinreichs u. d. Konchyliologie. Weimar,
1780, Bd. 6. (18)+584+(12) S.
Seba A. An account of the Cinnamon tree in Ceylon and its several sorts. — Phil.
Trans. Roy. Soc. London, 1731, v. 36, p. 97—105.
Seba A. The anatomical preparation of vegetables. — Phil. Trans. Roy. Soc. London,
1731, v. 36, p. 441—444.
Seba A. Historia exoticorum quorundam medicamentorum simplicium. — Acta phys.-
med. Acad. Caes. Leop.-Car. Nat. Cur. exh. Ephemerides, 1737, v. 4, p. 226—232.
Sherborn C. D. Index animalium. Sect. II, part 3, London, 1930, p. 5706—6357
S о n n i n i C. S., L a t r e i 11 e P. A. Histoire naturelle des Reptiles. Paris, 1802,
v. I—IV, XXII + 280, 332, 335, 409 p.
Stimson A. F. Boidae. — In: Das Tierreich, Liste der rezenten Amphibien und Repti-
lien. 1969, Lfg. 89, XI+ 49 S.
W a g 1 e r J. Deutung den in Seba’s Thesaurus enthaltenen Abbildungen von Lurchen. —
Isis, 1833, S. 884—905.
Albert Seba and his contribution to the development of herpetology
К. B. Juriev
Zoological Institute, Academy of Sciences. U.S.S.R. (Leningrad)
The name of Seba has forever gone down in the history of zoological systematics,
since many of the pictures of animals performed him were used as types for describing
a number of species. The work elucidates Seba’s role in the development of herpetology.
The fate of both his zoological collections is considered in every detail. His famous work
«Locupletissimi rerum naturalium thesauri» and its second French edition have been de-
scribed minutely.
АВТОРЕФЕРАТЫ
УДК 598.1
Особенности строения черепа, зубной системы и гиоида ящериц рода Agama фауны
СССР. Ананьева Н. Б. — В кн.: Фауна и экология амфибий и рептилий пале-
арктической Азии. Л., 1981, Зоологический институт АН СССР, с. 3—20 (Труды
Зоологического института АН СССР, т. 101).
Описывается строение черепа, зубной системы и подъязычного аппарата 6 видов рода
Agama фауны СССР Отмечены видовые отличия в форме и пропорциях черепа и
в форме отдельных костей, особенно os maxillare, os dentale, os frontale и os parietale.
Приводятся зубные формулы агам фауны СССР. A. sanguinolenta резко отличается
по строению гиоида от остальных изученных видов, напоминая в этом отношении
некоторых древесных агамовых и игуановых ящериц. Обсуждается адаптивное значе-
ние гиоида для демонстративных движений горлового мешка, рассматриваются видо-
вые различия в размере foramen parietale. Анализ изученных признаков свидетель-
ствует о морфологической неоднородности видов агам фауны СССР. Их можно
отнести к трем группам: 1) горные агамы A. caucasia, A. lehmanni, A. himalayana,
A. erythrogastra, 2) степная агама A. sanguinolenta, 3) руинная агама A. ruderata.
Илл. 9, табл. 2, библ. 46.
УДК 598.1
О видовой самостоятельности Agama pawlowskii Cemov et Dubinin, 1946. Анань-
ева H. Б. и Петерс Г. — В кн.: Фауна и экология амфибий и рептилий палеарк-
тической Азии. Л., 1981, Зоологический институт АН СССР, с. 21—22 (Труды Зооло-
гического института АН СССР, т. 101).
Рассматривается история описания вида Agama pawlowskii. Сравнение описаний и
музейных экземпляров агамы Павловского и Agama mossambica, распространенной
в юго-восточной Африке, показало их идентичность. Приводятся рентгенограммы чере-
пов A. pawlowskii, A. mossambica и некоторых других агамовых ящериц. A. pawlow-
skii следует впредь считать синонимом A. mossambica и изъять из списков фауны
СССР и мировой фауны. Илл. 2, библ. 8.
УДК 598.1
Новый вид горных агам из Таджикистана, Agama chernovi sp. nov. Анань-
ева H. Б., Петерс Г. и Ржепаковский В. Т. — В кн.: Фауна и экология
амфибий и рептилий палеарктической Азии. Л. 1981, Зоологический институт АН
СССР, с. 23—27 (Труды Зоологического института АН СССР, т. 101).
По десяти экземплярам, собранным в разных местах юго-западного Таджикистана,
описывается новый для науки вид Agama chernovi. Типовая территория — кишлак
Дашти-Хонако, хребет Сурхку вблизи города Нурек. По своим морфологическим осо-
бенностям новый вид ближе всего к более высокогорному виду, Agama himalayana.
Особи обоих видов наблюдались в одном и том же биотопе (на высоте около 2200 м
над уровнем моря) только в одном месте на Гиссарском хребте. Для выяснения отно-
шений между A. chernovi и A. himalayana необходимо провести дальнейшие исследо-
вания их экологии. Сравнение нового вида с материалом по другим видам рода
Agama с Памира, Гиндукуша и Гималаев показало, что A. badakhshana, описанная
С. Андерсоном и А. Левитоном в 1969 г., может считаться лишь подвидом A. hima-
layana. Типовой экземпляр A. chernovi хранится в коллекции отделения герпетологии
Зоологического института АН СССР в Ленинграде. Илл. 2, библ. 5.
УДК 598.1
Две новые находки полосатого полоза, Coluber spinalis (Peters, 1866) на Дальнем
Востоке СССР и в восточном Казахстане. Бердыбаева Ж. Ш., Орлова В. Ф.
и Фролов В. Е. — В кн.: Фауна и экология амфибий и рептилий палеарктической
Азии. Л., 1981, Зоологический институт АН СССР, с. 28 (Труды Зоологического
института АН СССР, т. 101).
Сообщается о находке полосатого полоза, Coluber spinalis (Peters, 1866) на побережье
залива Посьета Приморского края. Это — первая достоверная находка полосатого
полоза на Дальнем Востоке СССР Другой описанный экземпляр был пойман на
севере Зайсанской котловины. Библ. 5.
УДК 597.6
О гигантских головастиках зеленых лягушек комплекса Rana esculenta. Бор-
кин Л. Я., Бергер Л. и Гюнтер Р. — В кн.: Фауна и экология амфибий и
121
рептилий палеарктической Азии. Л., 1981, Зоологический институт АН СССР, с. 29—47
(Труды Зоологического института АН СССР, т. 101).
Описываются гигантские головастики (длина тела более 100 мм), пойманные в СССР
(Латвия, Казахстан, Узбекистан) и Польше, а также полученные в результате лабо-
раторных скрещиваний. Приводится карта всех известных находок гигантских личинок
зеленых лягушек. Анализируются возможные факторы гигантизма (географическая
изменчивость, качество пищи и плотность скоплений, полиплоидия, межвидовая
гибридизация и гетерозис, освещенность, температура, перезимовка, гормональ-
ный контроль). Предполагается, что главной причиной спорадического появле-
ния гигантских головастиков являются нарушения гормональной системы гипоталамус-
гипофиз — щитовидная железа, вызванное генетическими факторами. Илл. 4, табл. 3,
библ. 100.
УДК 598.1
О северной границе ареала обыкновенной гадюки, Vipera berus (L.) в Якутии. Бор-
кин Л. Я. и Кириллов Ф. Н. — В кн.: Фауна и экология амфибий и рептилий
палеарктической Азии. Л., 1981, Зоологический институт АН СССР, с. 48 (Труды
Зоологического института АН СССР, т. 101).
Сообщается о находке обыкновенной гадюки около северной оконечности Вилюйского
водохранилища примерно в 400 км к северо-западу от ближайших ранее известных
находок. Подтверждается также существование этого вида в районе поселка Северо-
Енисейск Красноярского края. Библ. 4.
УДК 598.1
Новый для фауны СССР род и вид сцинковой ящерицы Chalcides ocellatus (Forskal).
Даревский И. С. — В кн.: Фауна и экология амфибий и рептилий палеарктиче-
ской Азии. Л., 1981, Зоологический институт АН СССР, с. 49—51 (Труды Зоологиче-
ского института АН СССР, т. 101).
Глазчатый халцид, Chalcides ocellatus, впервые обнаружен в СССР в пределах южной
Туркмении. Оба известных местонахождения более чем на 1000 км оторваны от ранее
известной границы ареала этого вида, проходящей на юге Ирана и Пакистана. По
своим морфологическим признакам экземпляры -из Туркмении несколько отличаются
от особей из других районов ареала. Для решения вопроса об их подвидовой при-
надлежности необходим дополнительный материал из южной Туркмении и прилежащих
районов Ирана. Илл. 1, табл. 1, библ. 14.
УДК 597.6 + 598.1
Новые данные о распространении некоторых амфибий и рептилий Монгольской На-
родной Республики. Монхбаяр X. — В кн.: Фауна и экология амфибий и рептилий
палеарктической Азии. Л., 1981, Зоологический институт АН СССР, с. 52—56 (Труды
Зоологического института АН СССР, т. 101).
Сообщается о новых находках 3 видов амфибий (Hynobius keyserlingii, Bufo viridis,
Капа chensinensis) и 9 видов рептилий (Teratoscincus przewalskii, Alsophylax pipiens,
Gymnodactylus elongatus, Eremias argus, Eremias arguta potanini, Eremias vermiculata,
Lacerta vivipara, Eryx tataricus, Coluber spinalis) в Монголии за последние 20 лет.
Илл. 4, библ. 16.
УДК 567.9 + 567.8(116.3 575.1)
Хвостатые и бесхвостые земноводные мела Кызылкумов. Несов Л. А. — В кн.:
Фауна и экология амфибий и рептилий палеарктической Азии. Л., 1981, Зоологический
институт АН СССР, с. 57—88 (Труды Зоологического института АН СССР, т. 101)
Из турона — сантона пустыни Кызылкум впервые для мела СССР описываются
остатки хвостатых земноводных (Scapherpetontidae, Batrachosauroididae, Prosirenidae,
fam. indet., всего 6 видов 5 родов) и бесхвостых амфибий (Pelobatidae, fam. indet. —
4 вида 4 родов), существовавших на приморских низменностях в зоне развития лима-
нов. Представители этих отрядов указываются и для сеномана Кызылкумов. Отме-
чается сходство среднеазиатских туронских комплексов позвоночных, включающих
земноводных, и существенно более поздних комплексов кампана и Маастрихта Север-
ной Америки, также приуроченных к приморским равнинам. Приводится перечень
основных находок остатков докайнозойских хвостатых и бесхвостых амфибий мира.
Илл. 5, библ. 140.
122
УДК 598.1
О восточной границе ареала длинноногого голопалого геккона, Gymnodactylus longipes
Nikolsky, 1896. Орлов Н. Л. — В кн.: Фауна и экология амфибий и рептилий пале-
арктической Азии. Л., 1981, Зоологический институт АН СССР, с. 89—91 (Труды
Зоологического института АН СССР, т. 101)
Описываются 4 экз. длинноногого голопалого геккона, Gymnodactylus longipes Nikol-
sky, собранные В. Я. Лаздиным в 1915 г. в районе города Термез в 500 км северо-
восточнее известной границы ареала. Выделяется лектотип и уточняется типовая
территория этого вида. Гекконы из района Термеза относятся к номинативному под-
виду, G. I. longipes Nikolsky, 1896, впервые указываемому для фауны СССР. Илл. 2,
табл. 1, библ. 13.
УДК 598.1
Новый вид эфы, Echis multisquamatus sp. nov. из Передней и Средней Азии. Ч е р -
лин В. А.— В кн.: Фауна и экология амфибий и рептилий палеарктической Азии.
Л., 1981, Зоологический институт АН СССР, с. 92—95 (Труды Зоологического
института АН СССР, т. 101).
Описывается новый вид змеи — песчаная эфа, Echis multisquamatus sp. nov. — с тер-
ритории СССР, северного и восточного Ирана и южного Афганистана. Этот вид от-
личается большим количеством брюшных щитков (в среднем 188.1), спинных чешуй
(в среднем 37.1 ряд) и боковых чешуй (в среднем 10.3 рядов). Надглазничные щитки
не увеличены. Светлая зигзагообразная полоса на боках тела присутствует целиком,
спинные пятна узкие и поперечно вытянутые. На голове рисунок в виде белого креста.
В восточном Иране и южном Афганистане Е. multisquamatus и Е. carinatus sochureki
обитают симпатрично на протяжении примерно 400 км. Илл. 2, табл. 2, библ. 15.
УДК 598.1
Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin, 1981 от тем-
пературных условий в южной Туркмении. Черлин В. А. иЦеллариус А. Ю. —
В кн.: Фауна и экология амфибий и рептилий палеарктической Азии. Л., 1981, Зооло-
гический институт АН СССР, с. 96—108 (Труды Зоологического института АН СССР,
т. 101).
Температурные условия в норах рассматриваются как фактор, определяющий время и
характер пребывания эф на поверхности почвы и регулирующий длительность световой
фазы, как косвенный показатель поведения и физиологического состояния змей. В по-
ведении выделяются 4 основных формы: в норе, у норы, свободное передвижение по
поверхности и активная терморегуляция. Определены условия реализации этих форм
поведения, а также половой и пищевой активностей. Илл. 14, табл. 2, библ. 13.
УДК 597.6 + 598.1 + 57.(09)
Альберт Себа и его роль в развитии герпетологии. Юрьев К. Б. — К кн.: Фауна
и экология амфибий и рептилий палеарктической Азии. Л., 1981, Зоологический инсти-
тут АН СССР, с. 109—120 (Труды Зоологического института АН СССР, т. 101).
Имя Альберта Себа навсегда вошло в историю зоологической систематики, так как
многие из изображенных им животных послужили типами для их описания. В работе
освещается его роль в истории исследования земноводных и пресмыкающихся. Под-
робно рассматривается судьба обоих его зоологических собраний. Детально описы-
вается его знаменитое сочинение «Богатейший вклад природного царства» и его пере-
издание во Франции. Основное внимание уделяется значению труда Себа для разви-
тия герпетологии и его использованию классиками этой науки: Линнеем, Кювье, Ля-
сэпедом, Доденом, Мерремом, Дюмерилем и др. Приводятся многочисленные примеры
привлечения материалов Себа современными герпетологами. Илл. 2, библ. 55.
СОДЕРЖАНИЕ
Н. Б. Ананьева. Особенности строения черепа, зубной системы и гиоида
ящериц рода Agama фауны СССР 3
Н. Б. Ананьева и Г Петерс. О видовой самостоятельности Agama paw-
lowskii Cernov et Dubinin, 1946 ... ‘21
H. Б. Ананьева, Г. Петерс и В. Т. Ржепаковский. Новый вид
горных агам из Таджикистана, Agama chernovi sp. nov. 23
Ж. LU. Бердыбаева, В. Ф. Орлова и В. Е. Фролов. Две новые на-
ходки полосатого полоза, Coluber spinalis (Peters, 1866) на Дальнем
Востоке СССР и в восточном Казахстане 28
Л. Я. Боркин, Л. Бергер и Р. Гюнтер. О гигантских головастиках
зеленых лягушек комплекса Rana esculenta 29
Л. Я. Боркин и Ф. Н. Кириллов. О северной границе ареала обыкно-
венной гадюки, Vipera berus (L.) в Якутии 48
И. С. Даревский. Новый для фауны СССР род и вид сцинковой ящерицы
Chalcides ocellatus (Forskal) 49
X. Монхбаяр. Новые данные о распространении некоторых амфибий и реп-
тилий Монгольской Народной Республики 52
Л. А. Несов. Хвостатые и бесхвостые земноводные мела Кызылкумов 57
Н. Л. Орлов. О восточной границе ареала длинноногого голопалого геккона,
Gymnodactylus longipes Nikolsky, 1896 . 89
В. А. Ч е р л и н. Новый вид эфы, Echis multisquamatus sp. nov. из Передней
и Средней Азии 92
В. А. Ч е р л и н и А. Ю. Це л л а р и у с. Зависимость поведения песчаной эфы,
Echis multisquamatus Cherlin, 1981 от температурных условий в южной
Туркмении 96
К. Б. Юрьев. Альберт Себа и его роль в развитии герпетологии 109
Авторефераты 121
CONTENTS
N. В. Ananjeva. Structural characteristics of skull, dentition and hyoid
of lizards of the genus Agama from the fauna of the USSR . 3
N. B. Ananjeva and G. Peters. On the validity of Agama pawlowskii,
Cernov et Dubinin, 1946 21
N. B. Ananjeva, G. Peters and V. T. Rzepakovsky. New species
of the mountain agamas from Tadjikistan, Agama chernovi sp. nov. 23
Zh. Sh. Berdybaeva, V. F. Orlova and V. E. Frolov. Two new records
of the Coluber spinalis (Peters, 1866) in the Far East of the USSR and
eastern Kazakhstan . . ............. 28
L. J. Borkin, L. Berger and R. Giinther. On giant tadpoles of the green
frogs of the Rana esculenta complex . . .29
L. J. Borkin and F. N. Kirillov. On northern range limit of the adder,
Vipera berus (L.) in Yakutia ... .48
I. S. Da revs ky. A first record of the ocellated skink, Chalcides ocellatus
(Forskal) in the fauna of the USSR ... .49
Kh. Monkhbayar. New data on the distribution of some amphibians and
reptiles in Mongolia .... 52
L. A. Nessov. Cretaceous salamanders and frogs of Kizylkum Desert . 57
N. L. О r 1 о v. About the eastern range limit of the gecko Gymnodactylus lon-
gipes Nikolsky, 1896 ... .89
V A. C h e r 1 i n. The new saw-scaled viper, Echis multisquamatus sp. nov. from
South-Western and Middle Asia 92
V A. Cherlin and A. J. Tsellarius. The behaviour dependence of the saw-
scaled viper, Echis multisquamatus Cherlin, 1981 on temperature in south
Turkmenia . . ................ . . 96
К. B. Juriev. Albert Seba and his contribution to the development of herpetology 109
Abstracts (in Russian) 121
Зоологический институт АН СССР выпустил следующие издания по герпетологии:
Герпетологический сборник (Труды Зоологического института АН СССР, т. 74),.
1977, 140 с., цена 1 руб.
Экология и систематика амфибий и рептилий (Труды Зоологического института:
АН СССР, т. 89), 1979, 126 с., цена 80 коп.
Герпетологические исследования в Сибири и на Дальнем Востоке. 1981, 147 с
цена 1 руб.
Книги можно заказать наложенным платежом по адресу:
Ленинград, 199164
Университетская наб., 1
Зоологический институт АН СССР
Отделение герпетологии
ИЛЛЮСТРАЦИИ
К статье И. Б. Ананьевой и Г Петерса (с. 21—22)
Рис. 1 Внешний вид Agama mossambica (А, Б) н A. pawlowskii (Б).
К статье Н. Б. Ананьевой и Г Петерса (с. 21—22)
Рис. II. Рентгенограммы черепов некоторых агамовых ящериц (ХЗ).
1 — Agama sanguinolenta, 2 — A. caucasia, 3 — A. mossambica, 4 — Calotes versicolor
и 5 — A. pawlowskii
К статье Н. Б. Ананьевой и др. (с. 23—27)
Рис. III. Внешний вид горных агам.
А — Agama chernovi sp. nov., голотип (ЗИН. 19385); Б — A. himalayana himalayana, половозрелый
(3ИН.9036.1); В — A. himalayana badakhshana, восточный Афганистан, провинция Газни, перевал
музей имени А. Кенига, Бонн, ZFMK-H.8608).
самец, Шугнан, долина реки Шах-дара
на Дашт-и-Навар, 3000 м (Зоологический
К статье Н. Б. Ананьевой и др. (с. 23—27)
Рис. IV Спинная чешуя горных агам Agama chernovi sp. nov (A), A. himalayana (Б) и A. himalayana ba-
dakhshana (B).
Сведения об экземплярах см. рис. HI.
К статье Л. Я. Боркина и др. (с. 29—47)
Рис. V Гигантские головастики зеленых лягушек комплекса
Rana esculenta.
А — нормальный и гигантский (130 мм) головастик Rana ridibunda из
окрестностей Ташкента, Узбекистан (ЗИН.4735); Б — нормальный голо-
вастик Rana lessonae в период метаморфоза и гигантский (112 мм)
головастик от лабораторного скрещивания Rana lersonae £ X Rana
esculenta $
К статье Л. Я. Боркина и др. (с. 29—47)
Рис. VI. Гигантские головастики зеленых лягушек комплекса Rana
esculenta.
А — гигантский (125 мм) головастик от лабораторного скрещивания Rana lesso-
гае QXRana esculenta 6 — головастики из окрестностей города Познань. Польша:
крупные личинки Rana esculenta (1) и Rana ridibunda (2). гигантский (163 мм) го-
ловастик. вероятно, от скрещивания Rana esculenta у ^Rana esculenta d (3) и
обычной величины головастик Rana ridibunda (4) Оба фото 3. Пневского.
К статье И. С. Даревского (с. 49—51)
Рис. VII. Глазчатый халцнд, Chalcides ocellatus (Forskal) —
экземпляр из Акар-Чешме, южная Туркмения (ЗИН.19332).
К статье Л. А. Лесова (с. 57—88)
Рис. VIII. Остатки земноводных и других позвоночных позднего мела.
1—21. Каракалпакская АССР, нижний турон, нижняя часть бештюбннской
свиты.
1—7 Че.тпык, / -- зубная кость Nukusurus insuetus sp. now. la — изнутри, 16 —
снизу, голотип ZIN, PHA № K77-4. X6.4. 2—6. Horezmia gracile sp. nov.
2 — верхнечелюстная кость изнутри, ЦГМ № 1/11657; 3 — зубная кость: За — снаружи,
Зб — изнутри, ЦГМ № 2/11657; 4 — зубная кость изнутри. ЦГМ 3/11657, Х4,
5 — атлас: 5а — спереди. 56 — снизу, ЦГМ № 4/11657; 6 — подвздошная кость: 6а —
сзади, 6б — сбоку, ЦГМ № 5/11657; 7 — зуб Protoceratopsidae снаружи, ЦГМ № 1/11822,
Х3.2. 8—21. Ходжакуль. 8 — атлас Horeztnia gracile sp. nov.: 8a — спереди. 8б — снизу,
8в — сзади, голотип ZIN, PHA № К77-2; 9 — туловищный позвонок Scapherpetontidae
сзади, ЦГМ № 191/11657. 10—11. Н. gracile и Mynbulakia? nongratis sp. now 10—
ЦГМ № 178/11657: 10a — снаружи, 106 — изнутри, Х5; //—голотип ZIN, PHA № К77-8:
//а — сверху, //б — изнутри, Х4. 12—15. Horezmia gracile sp. nov. /2—лучевая
кость сзади, ЦГМ № 6/11657; 13 — плечо; 13а— сзади и несколько снизу, 136 —
снизу, ЦГМ № 7/11657. 14—15. Подвздошные кости сбоку. 14 — ЦГМ № 14/11657;
/5 — ЦГМ № 15/11657. 16 — кость голени Salientia, ЦГМ № 188/11657; /7 — парасфеноид
Actinopterygii снаружи, ЦГМ № 9/11822, Х4.1: 18 — зубная кость Lacertilia изнутри,
ЦГМ № 1/11727. Х4.3; 19 — зубная кость Actinopterygii сверху, ЦГМ № 10/11822, Х3.2;
20 — зуб Dapedium sp. изнутри, ЦГМ № 11/11822. Х3.2; 2/— зуб костистой рыбы
с чертами сходства с Platacodon, ЦГМ № 3/11758. Х8; Увеличение 4.7, кроме /, 4. 7,
10—11, 17—18, 20—21.
Л’ статье Л. 1. Носова (с. 57—&8)
Рис. IX. Кости и зубы позднемеловых позвоночных.
1- 22. Узбекская ССР, Бухарская область, Джара-Кудук, верхний турон, нижняя
часть тайкаршинской пачки.
1—20. Eoscapherpeton asiaticum sp. no\. 1—5. Верхнечелюстные кости- / голотип
ZIX, PHz\ № К77-1: la снаружи, 16- изнутри, ]в снизу. Х2.4. 2 ЦГМ № 10 11637
изнутри; 3 — ЦГМ № 11 11657, изнутри, Х4.7. 4—3. Кости Е. asiaticum? снаружи.
4 — ЦГМ №1211657; 5 — ЦГМ №13 11657. 6 9. Зубные кости. 6 ЦГМ №14 11657:
ба — изнутри. 66 снаружи; 7 — ЦГМ № 15/11657 изнутри; 8 — ЦГМ № 16 11657 сна-
ружи; 9— ЦГМ № 17 11657. 10 — теменная кость: 10а — снаружи, 106 — изнутри, ЦГМ
№ 18 11657. ХЗ.Г. 11 -зубная кость снизу, ЦГМ № 19 11657. 12—13. Подвздошные кости
сбоку, Х4.7. 12- ЦГМ № 20 11657; 13 ЦГМ № 21 11657. 14 -плечо спереди. ЦГМ
№ 22 11657; /5 — бедро сзади, ЦГМ № 23 11657; 16 — атлас: 16а — спереди, 166 — сзади
и несколько сбоку, ЦГЛ\ № 24 11657. 17—19. Туловищные позвонки. /7 — ЦГМ
-№ 25/11657. спереди и несколько сбоку; 18— ЦГМ № 26 11657 снизу: 19—ЦГЛ\
№ 27/11657: 19а— сбоку. 196 сзади. 20— атлас. 20а спереди, 20б сбоку, ЦГМ
№ 28 11657. 2/ зуб ?Myledaphus или (М3?) Theria? сверху, ЦГМ № 4 117о8, Х8.2;
22— клык Deltaiheridiidae сбоку. ЦГМ -№ 5 11758.
23- Каракалпакская АССР, Шейх-Джейли, сеноман, верхняя часть ходжакх льской
свиты, эпистрофей Oxlestes grandis (Palaeoryctidae?) снизу. ЦГМ № 6 11758, Х2.4.
Увеличение 4.1, кроме 1, 3, 10, 12—14, 21, 23.
К статье Л. Л. Несова (с. 57—88)
Рис. X. Остатки позвоночных позднего мела. 1—26. Узбекская СССР, Бу-
харская область, Джара-Кудук, верхний турон, нижняя часть тайкаршин-
ской пачки.
1—7 Eoscapherpeton asiaticuni sp. nov. 1—2. Зубные кости, Х4.1. / — ЦГМ № 29/11657
изнутри; 2— ЦГМ № 30/11657 снаружи; 3 — атлас снизу, ЦГМ № 31/11657; 4 — хво-
стовой позвонок сзади и несколько сбоку, ЦГМ № 32/11657; 5 — плечо снизу, ЦГМ
№ 33/11657; 6 — бедро 6а — с проксимального конца, 6б — сверху и несколько сзади.
Х3.2, ЦГМ № 34/11657; 7 - крестцовое ребро сзади, Х4. ЦГМ № 35/11657 8—10. Caudata
<* — бедро. 8а — с проксимального конца, 86 — сзади, ЦГМ № 182/11657; 9 — атлас
9а — сбоку, 96 — спереди, ЦГМ № 180/11657; 10— теменная кость: 10а — изнутри
106 — снаружи, ЦГМ № 181/11657; // — кость Actinopterygii с углублениями для
зубов: //а — снаружи, 116 — со стороны одной из кромок, ЦГМ № 184/11657.
Зубы динозавров. 12 — Theropoda, ЦГМ № 2/11822. Х2.4: 14 — Pachycephalosauria?, ЦГМ
№ 3/11822; 13— Ceratopsidae, ЦГМ № 4/11822, ХЗ.Г, 15—17 Actinopterygii./5 — зубная
пластинка, ЦГМ № 12/11822, Х4.1; 16 — верхнечелюстная кость снизу, ЦГМ № 13/11822,
Х1.6; /7 — чешуя снаружи. ЦГМ № 14/11822, ХЗ. 18 — челюсть Daulestes kulbeckensis
(Palaeoryctidae): 18а — снаружи, 186 — сверху, ЦГМ № 1/11758, Х8.
19—26 Salientia 19—22. Верхнечелюстные кости. 19 — Eopelobates sosedkoi sp. nov.:
19a — снаружи, X4.1, 196 — изнутри, ЦГМ № 59/11657; 20 — Kizylkuma antiqua sp. nov.:
2ьи — изнутри, 206 — снизу, ЦГМ № 70/11657, X4.1; 21 — Itemirella cretacea sp. nov.:
21a — изнутри, 2/6 — снизу, голотип ZIN. PHA № K77-6; 22 — Aralobatrachus robustu^
sp. nov.: 22a — изнутри, 226 — снизу, голотип ZIN. PHA № K77-7. 23 — кость предплечья
Salientia сверху, ЦГМ № 71/11657, Х4.1. 24 — плечо Salientia: 24а — снизу. 246 — сзади.
ЦГМ № 72/11657; 25—26 Подвздошные кости c6okv. 25 — Eopelobates sosedkoi sp. nov
(?). ЦГМ № 73/11657. X4.1. 25 — Salientia, ЦГМ № 74/11657.
Увеличение 4.7, кроме 1—2, 56. 7, 12, 14—18, 19а, 26, 23, 25—26.
Л статье Л. А. Несова (с. 57—88)
Рис. XI. Кости и зубы земноводных и других позднемеловых позвоночных.
Узбекская ССР, Бухарская область, Джара-Кудук.
1—9 Верхний турон. нижняя часть тайкаршинской пачки. 1—3. Кости голени
Salientia, Х4.1. / — ЦГМ № 75/11657: 1а — с проксимального конца, /6 — сзади; 2 —
ЦГМ № 76/11657: 2а— спереди, 2б — с дистального конца; 3— ЦГМ № 77/11657:
За — сзади, 36 — сбоку 4—9 Pisces. 4—5 Aspidorhynchidae. 4 — чешуя снаружи, ЦГМ
№ 5/11822, ХЗ.Г, 5 — верхняя челюсть изнутри. ЦГМ № 6/11822, Х4 1. 6 — зубная пла-
стинк? Actinopterygii, ЦГМ № 31/11822, Х3.1. 7 — зуб IMyledaphus tritus (Myliobatoidea):
7g — снаружи, 76 — сбоку, ЦГМ № 18/11822, Х3.1. 8—9. Lepisosteidae. 8 — зтб. ЦГМ
№ 7/11822. Х2.4. 9 - позвонок. ЦГМ № 8/11822. Х3.1. 10—26. Верхний турон-
сантон. средняя часть тайкаршинской пачки. 16—11, 14—19 Mynbulakia surgayi
sp. no\. 10 — верхнечелюстная кость, Юа — изнутри, 106 — снизу, голотип Z1N,
PHA № К77-3, Х6.4; // — зубная кость, ЦГМ № 50/11657; /2 — parietale Е. asiaticum?,
ЦГМ № 51/11657; 13 — goniale Е asiaticum, ЦГМ № 52/11657; 14 — атлас,
14а— снизу. Х6.4, 146 — спереди. ЦГМ № 53/11657 15—18 Туловищные позвонки
/5 —ЦГМ № 5^11657 снизу; 16 — ЦГМ № 55/11657 спереди, /7 — ЦГМ № 56/11756 снизу.
18 — ЦГМ № oz/167 сверху. 19 — бедро сверху и несколько сзади, ЦГМ № 58/11657.
20—26 Eoscapherpeton asiaticum sp. nov. 20 — лобная кость снаружи. ЦГМ
№ 37/11657, Х2.4; 22 — парасфеноид снизу, ЦГМ № 38/11657, Х2.4; 23 — обособленная
угловая кость изнутри, ЦГМ № 39/11657; 24 — ceratobranchiale I, ЦГМ № 40/11657;
25 — атлас: 25а — спереди и несколько сбоку, Х3.1; 256 — сзади, ЦГМ № 41/11657;
26 — крестцовый позвонок снизу, ЦГМ № 42/11657, Х3.1.
27 — зуб Pterosauria, ЦГМ № 32/11822; 28 — плечо Prosirenidae (?) снизу, ЦГМ
№ 183/11657; 29 — зуб (М1) Sailestes quadrans (Leptictidae?) снизу и несколько сзади.
ЦГМ № 7/11758. Х8; 30 — задняя часть нижней челюсти землеройкп Cretasorex
arkhangelskyi (Soricidae) изнутри, ЦГМ № 2/11758, Х4.7.
Увеличение 4.7, кроме 1—16, 14а, 21—22, 25а, 26, 29, 36.
К статье Л. А. Несова (с. 57—88)
Рис. XII. Остатки позднемеловых амфибий и других позвоночных. 1—26.
Узбекская ССР, Бухарская область, Джара-Кудук.
1—25. Турон—сантон, средняя часть тайкаршинской пачки. 1—7. Eoscapherpeton
asiaticum sp. nov. / — атлас: /а — сбоку, /б — сзади, ЦГМ № 43/11657. 2—3. Хвосто-
вые позвонки, Х3.2. 2 — ЦГМ № 44/11657 сбоку и несколько спереди; .‘/ — ЦГМ
№ 44/11657 сбоку и несколько сзади. 4— крестцовое ребро сзади, ЦГМ № 46/11657;
5 — локтевая кость сзади и несколько сверху, ЦГМ № 47/11657; 6 — большая берцо-
вая кость сзади, ЦГМ № 48/11657; 7 — седалищная кость сверху, ЦГМ № 49/11657;
8 — базисфеноид Lindholmemys elegans (Dermatemydidae): 8а — сбоку, 86 — сверху;
ЦГМ № 33/11822, Х2.4. 9—15. Eopelobates sosedkoi sp. nov. 9 — предчелюстная кость
изнутри, ЦГМ № 78/11657, Х6.4. 10—12. Верхнечелюстные кости. 10 — ЦГМ № 60/11657;
// — ЦГМ № 61/11657; 12 — ЦГМ № 62/11657. 13 — боковая затылочная кость: 13а —
сзади, Х4.1; 136 — изнутри, ЦГМ №79/11657; 14 — лобнотеменная кость: 14а — снаружи.
146 — изнутри, голотип ZIN, РИА № К77-5, Х5.6, 15 — подвздошная кость снаружи,
ЦГМ № 185/11657. 16—23. Salientia. 16 — лопатка снаружи, ЦГМ № 190/11657; 17 —
верхнечелюстная кость Kizylkutna antiqua sp. nov. изнутри, голотип ZIN, PHA
№ K77-10. 18—19. Туловищные позвонки. 18— ЦГМ № 63/11657 сверху; 19 — ЦГМ
№ 64/11657 спереди. 20 — крестцовый позвонок: 20а — сзади, 206 — сверху, ЦГМ
№ 65/11657; 21 — сфенэтмоид сзади и несколько сверху, ЦГМ № 189/11657; 22 — уро-
стиль: 22а — спереди и несколько сбоку, 226 — сверху, ЦГМ № 66/11657, Х6.4; 23 — пе-
реднеушная кость Eopelobates sosedkoi sp. nov. (?): 23a — изнутри, 236 — сверху.
ЦГМ № 80/11657, Х4.1, 24 — верхнечелюстная кость Bissektia папа sp. nov. (Caudata?)
изнутри, голотип ZIN, PHA № К77-9: 25 — ростральный зхб Ischyrhiza serra (Pristi-
dae), ЦГМ № 16/11822.
Верхний турон — сантон, верхняя часть тайкаршинской пачки: вецхнечелюстн
кость Bissektia sp. изнутри и несколько снизу, ЦПМ № 179/11657
Увеличение 4.7, кроме 2—3, 8—9, 13, 14, 22—23, 26.
К статье Н Л. Орлова (с. 89—91)
Рис. XIII. Три экземпляра Gymnodactylus longipes longipes Nikolsky, 1896, пойманные В. Я. Лаздпным в 1915 году
в окрестностях города Термез, Узбекистан.
Слева направо — ЗИН.19215 а, b и с
К статье В. А. Черлина (с. 92—95)
Рис. XIV Внешний вид Echis multisquamatus sp. nov. (А) и E. carinatus
sochureki (Б и В).
ФАУНА И ЭКОЛОГИЯ АМФИБИЙ И РЕПТИЛИЙ ПАЛЕАРКТИЧЕСКОЙ АЗИИ
Труды Зоологического института АН СССР
Том 101
Утверждено к печати
Зоологическим институтом АН СССР
План 1981 г.
Редактор Т. А. Асанович
Художник Т. Г. Кашицкая
Сдано в набор 7.03.1980. Подписано в печать 18.05.1981. М-20033. Формат 70х 108/16-
Бумага № 1. Гарнитура литературная. Печать высокая. Печ. л. 8+1 вкл.
Усл.-печ. л. 11,20. Уч.-изд. л. 10. Тираж 1000 экз. Тип. зак. 719. Цена 1 руб.
Зоологический институт АН СССР, 199164, Ленинград, Университетская наб., 1.
Типография № 2 Ленуприздата, 191104, Ленинград, Литейный пр., 55