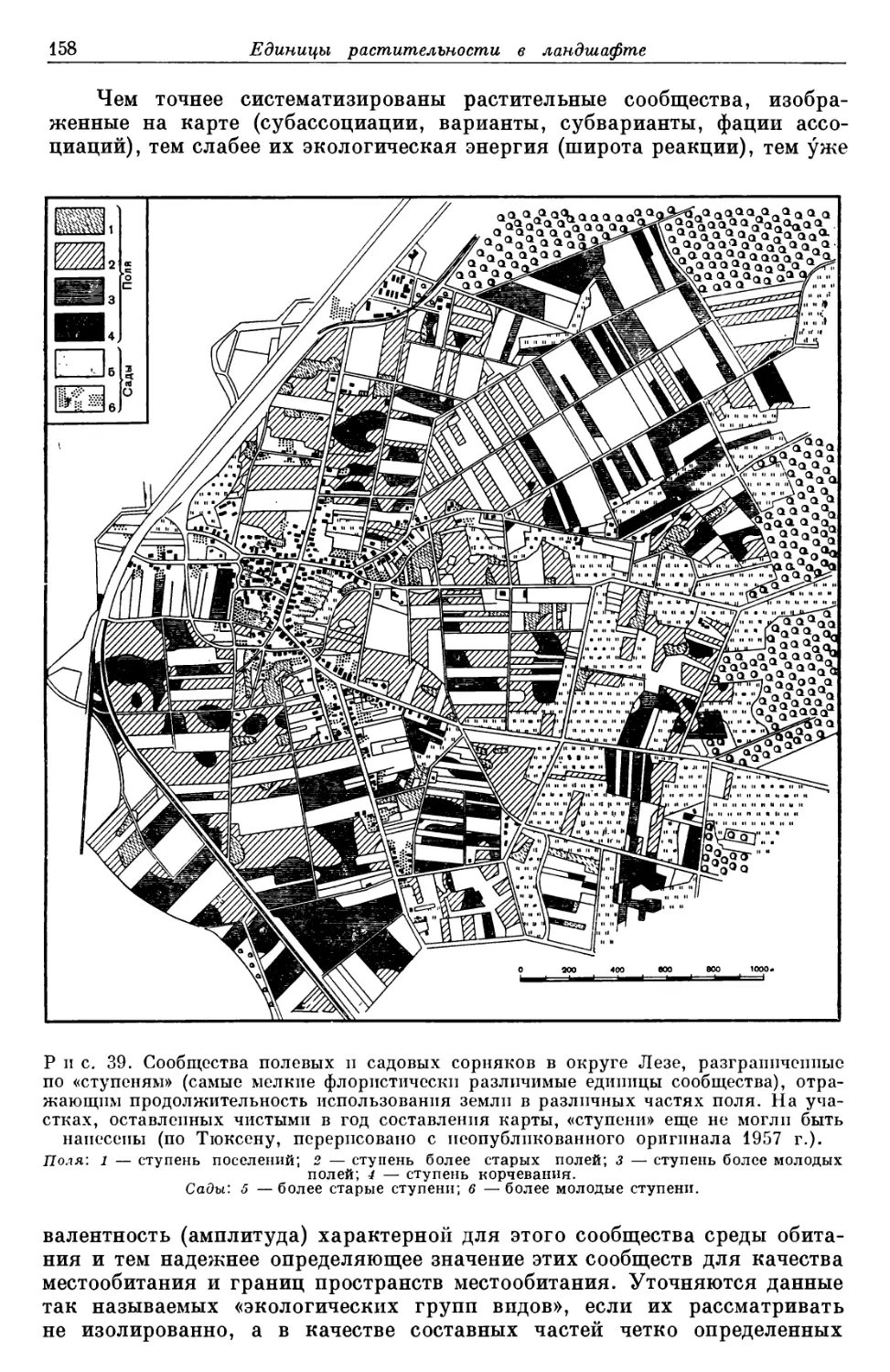
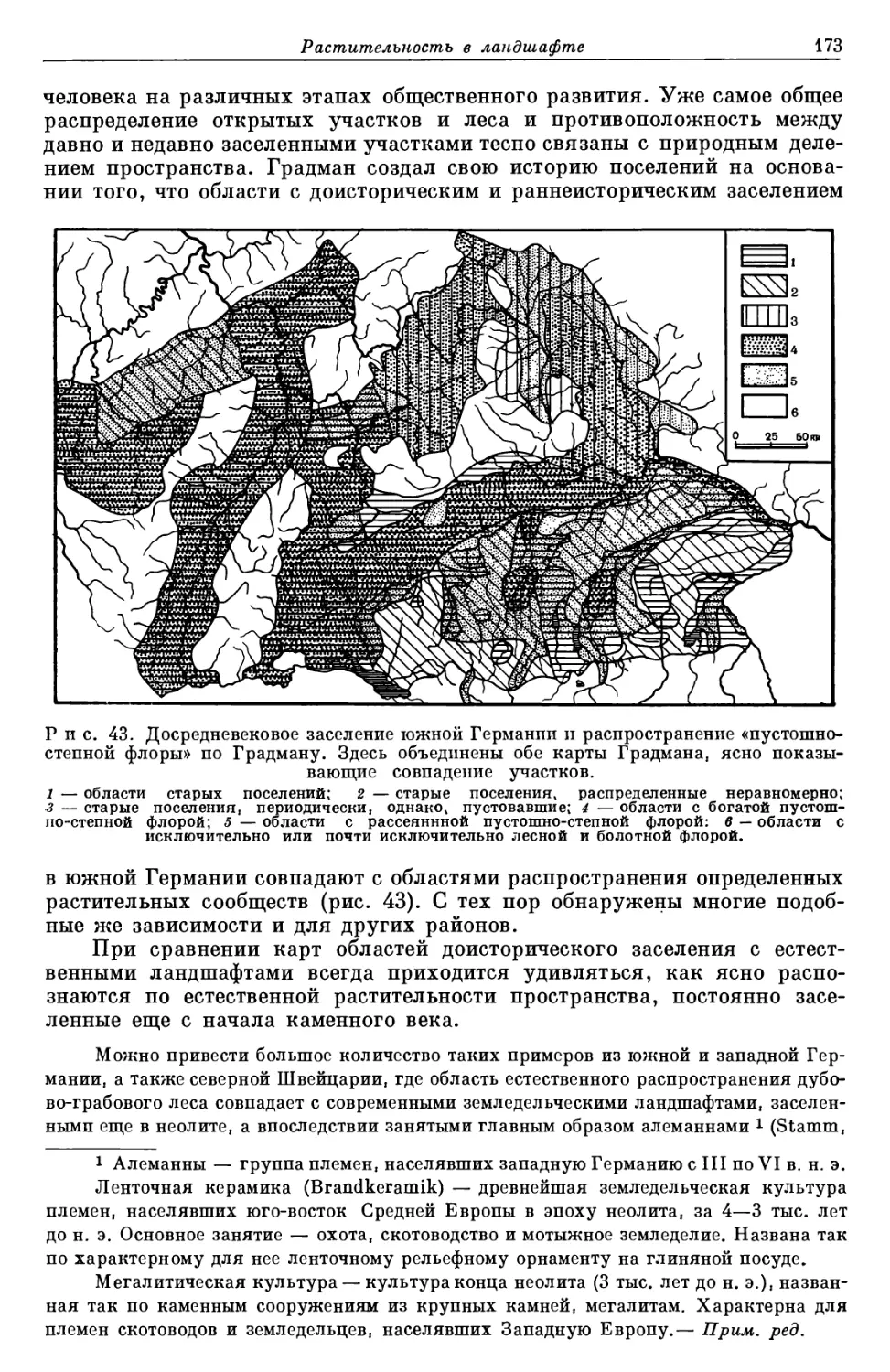
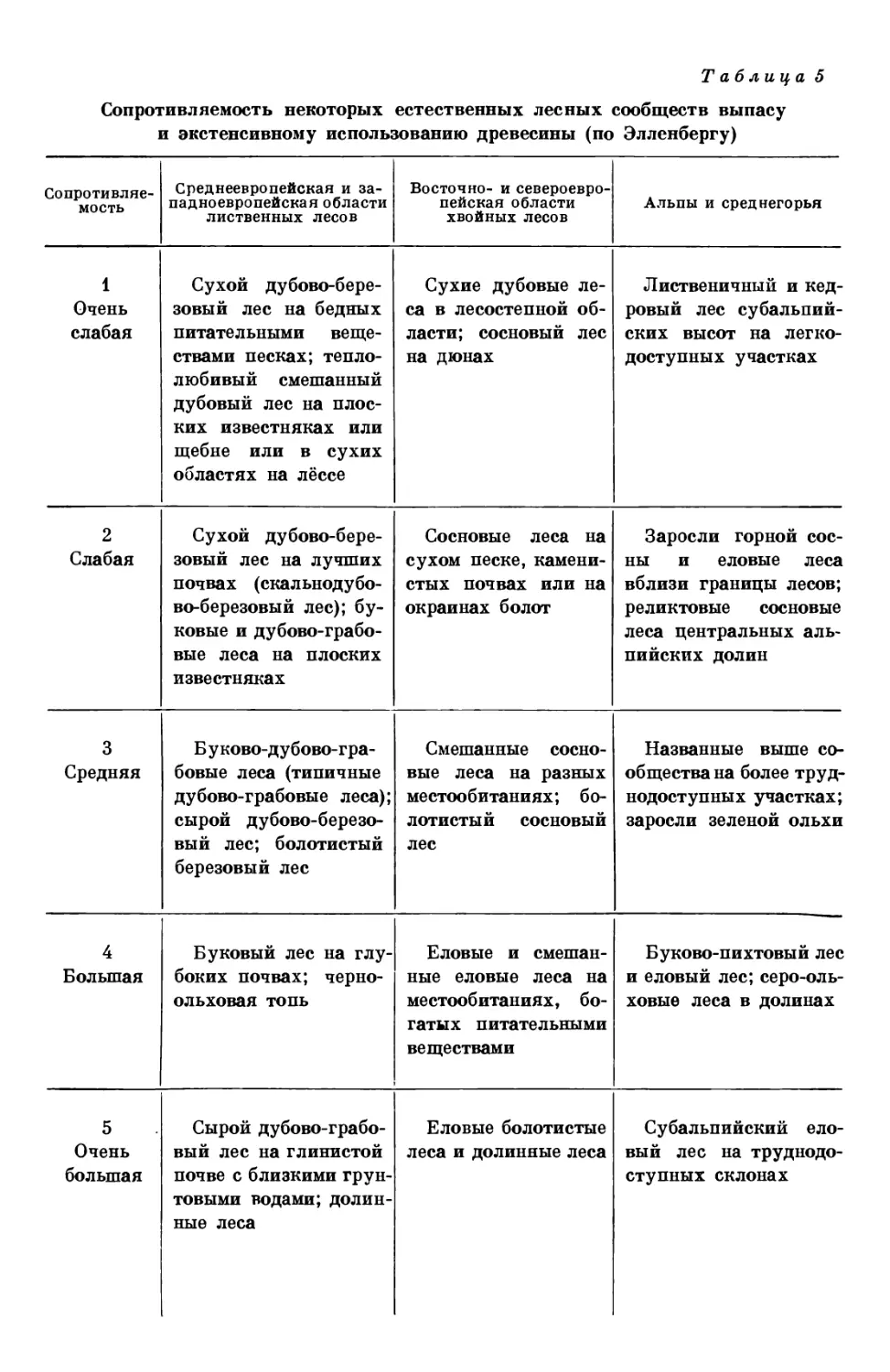
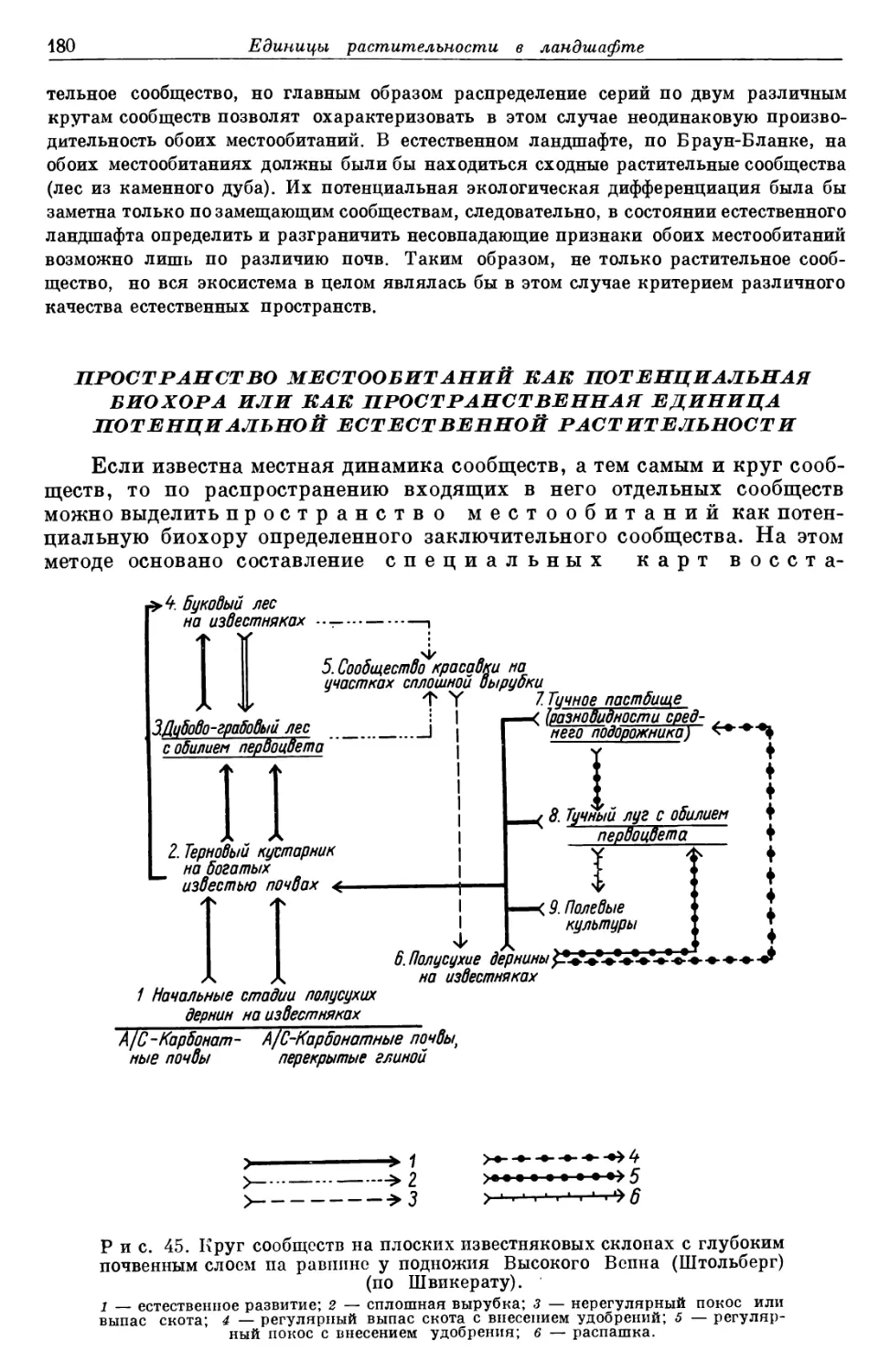
/

























































































































































































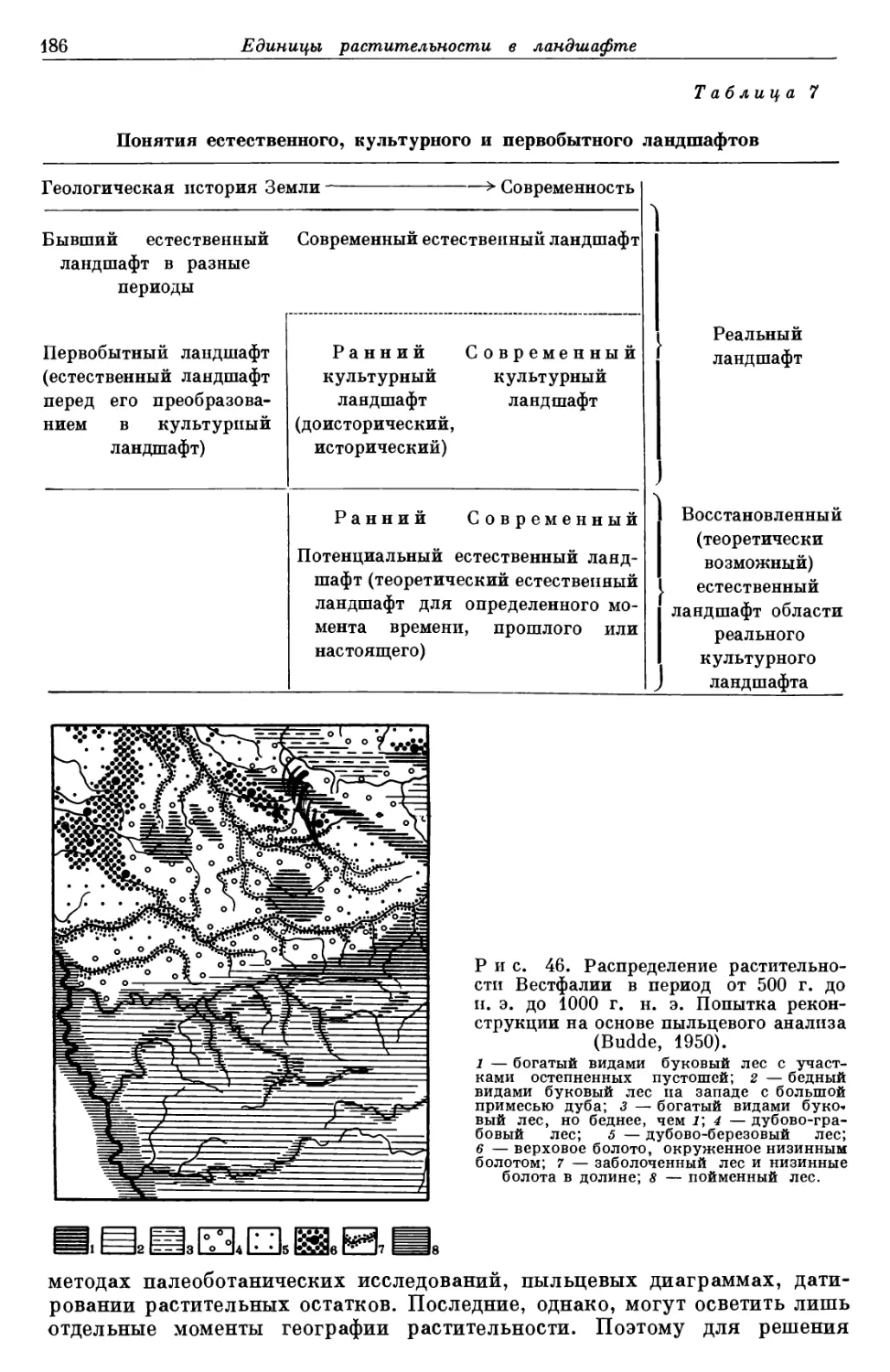









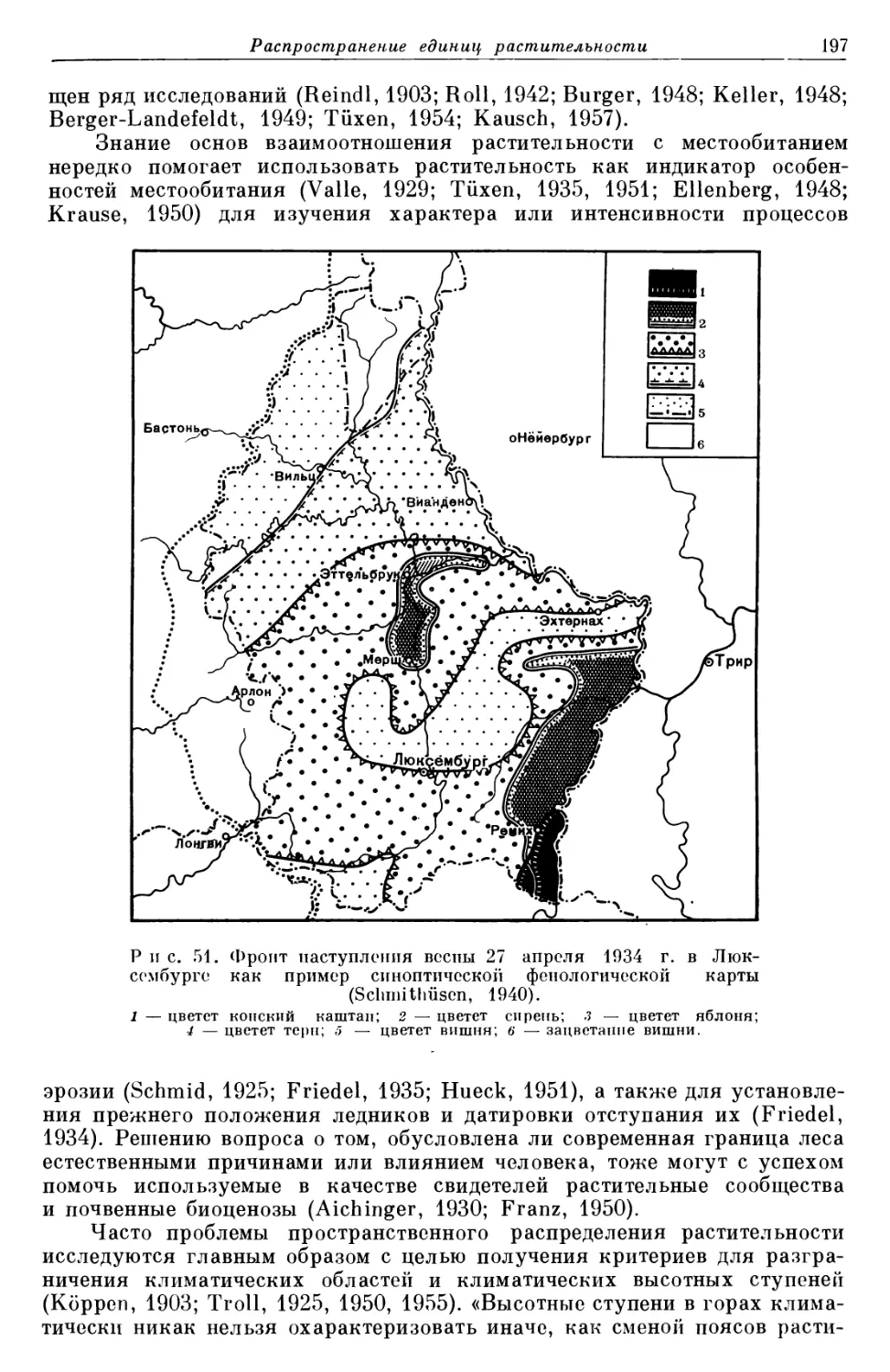







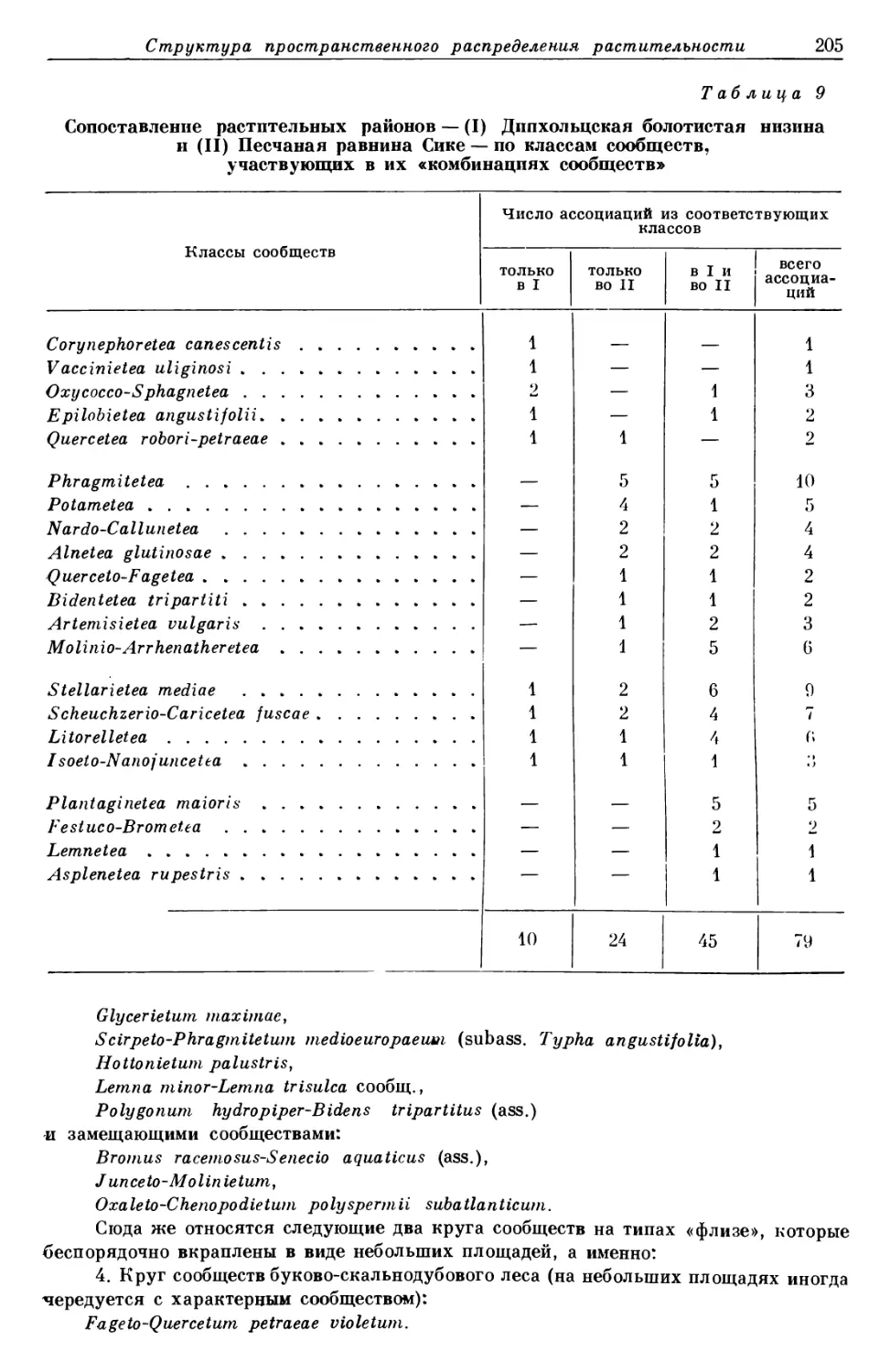



























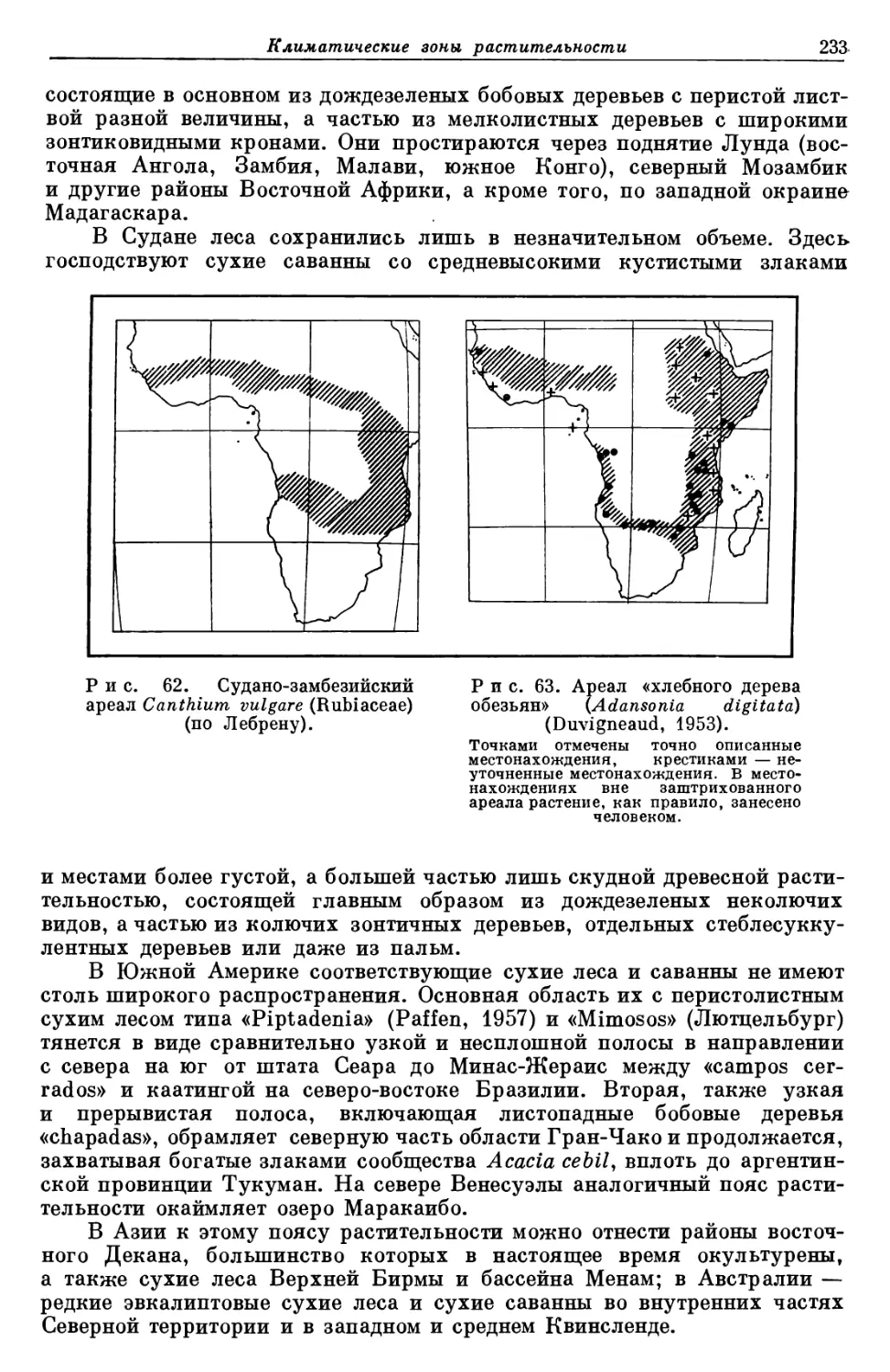









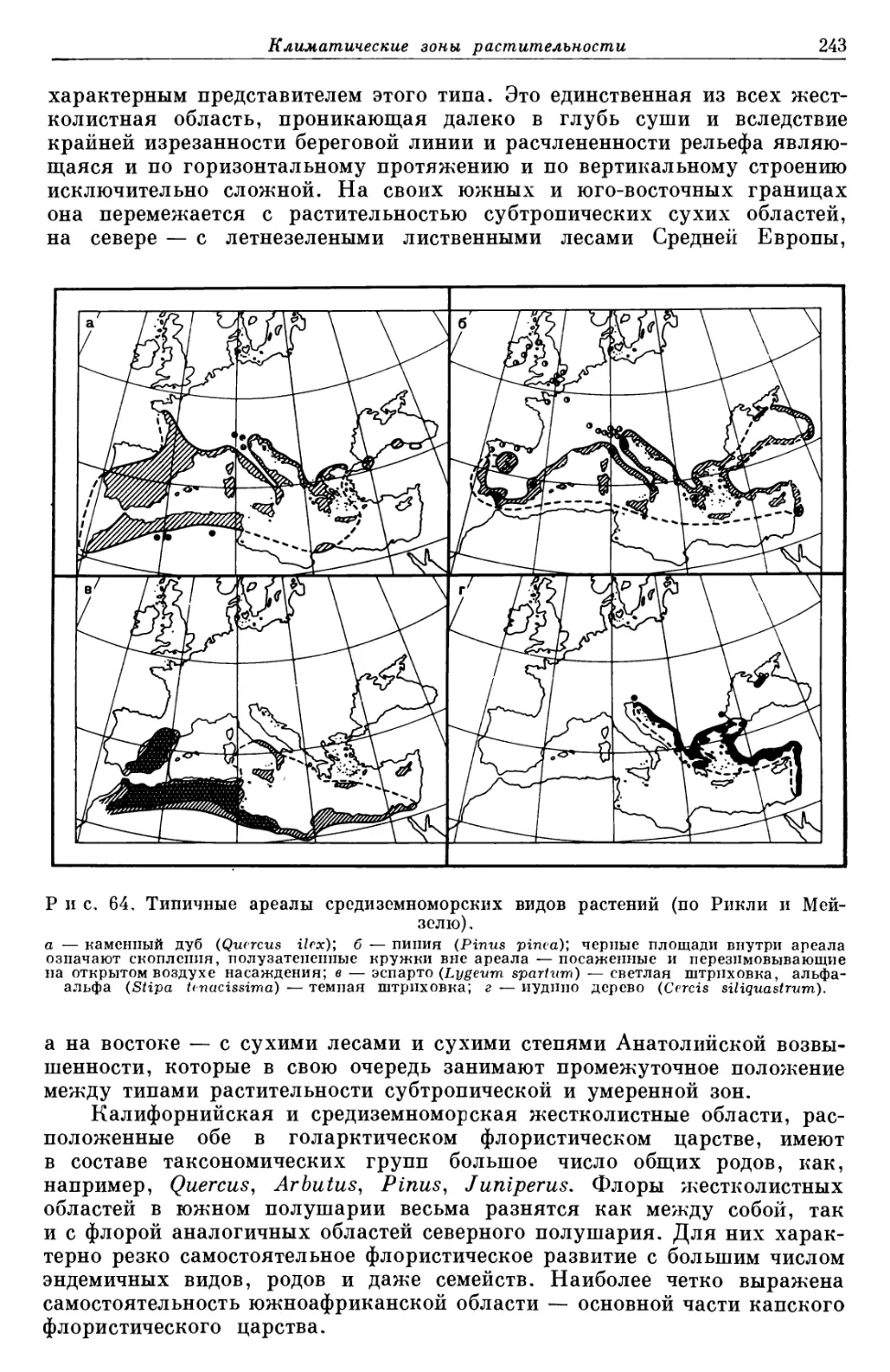
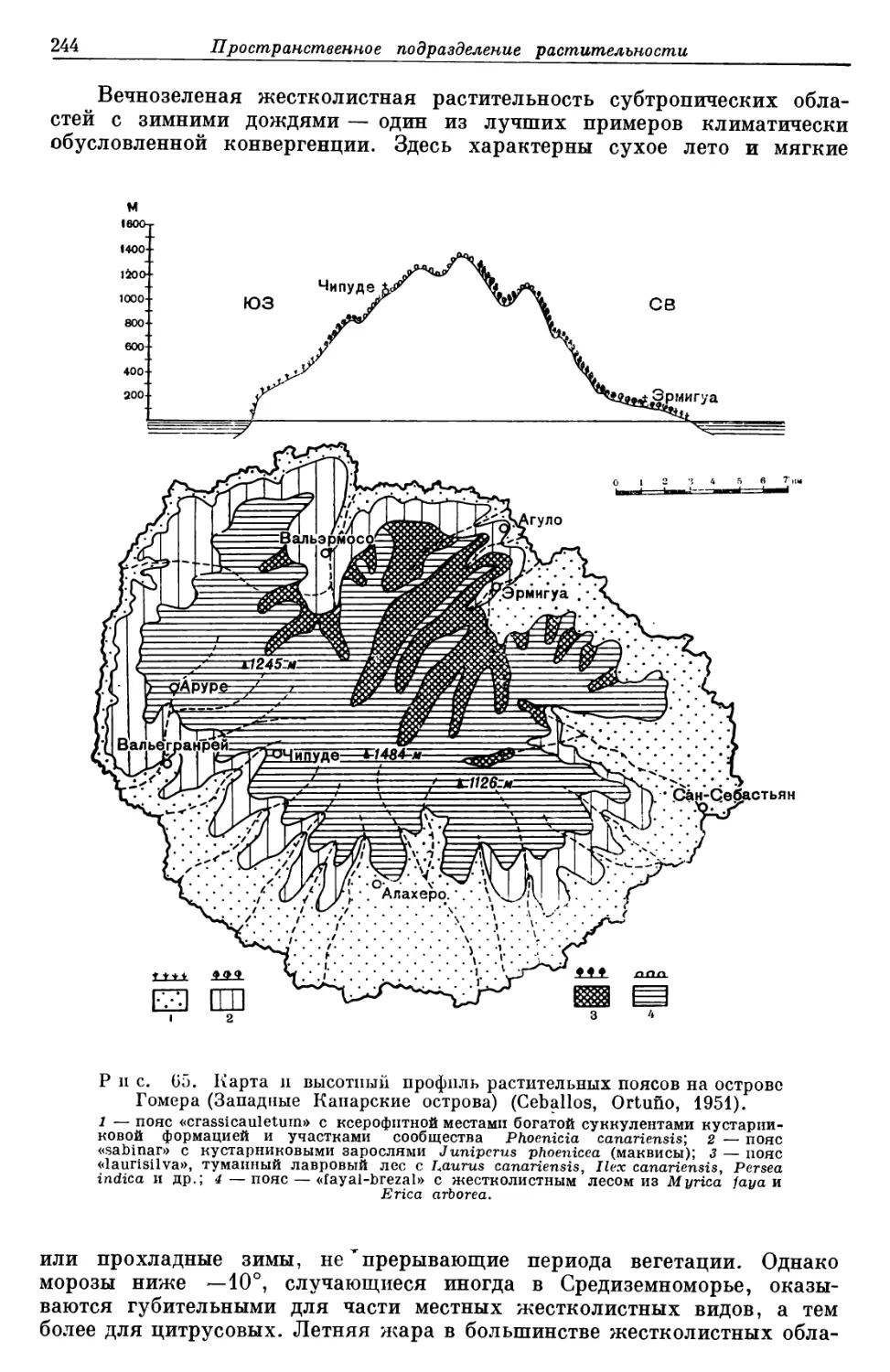







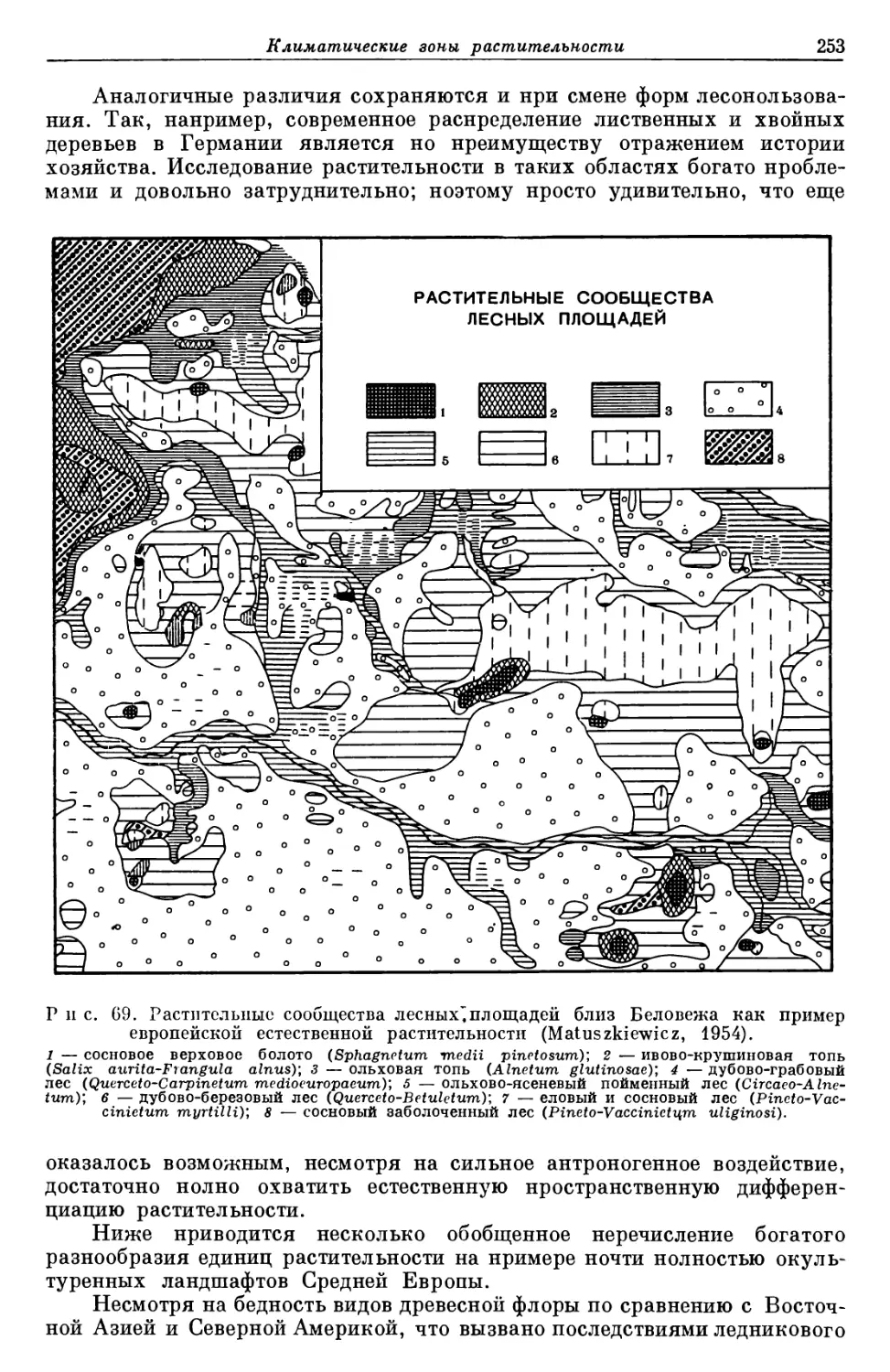






















































Text
Josef SchmithUsen,
ALLGEMEINE
VEGETATIONSGEOGRAPHIE
BERLIN 1961
И. Шмитхюзен
ОБЩАЯ
ГЕОГРАФИЯ
РАСТИТЕЛЬНОСТИ
Перевод с немецкого
В. А. ШЕРМУШЕНКО
Редакция и предисловие
Т. Б. ВЕРНАНДЕР
ИЗДАТЕЛЬСТВО «ПРОГРЕСС»
Москва 19 6 6
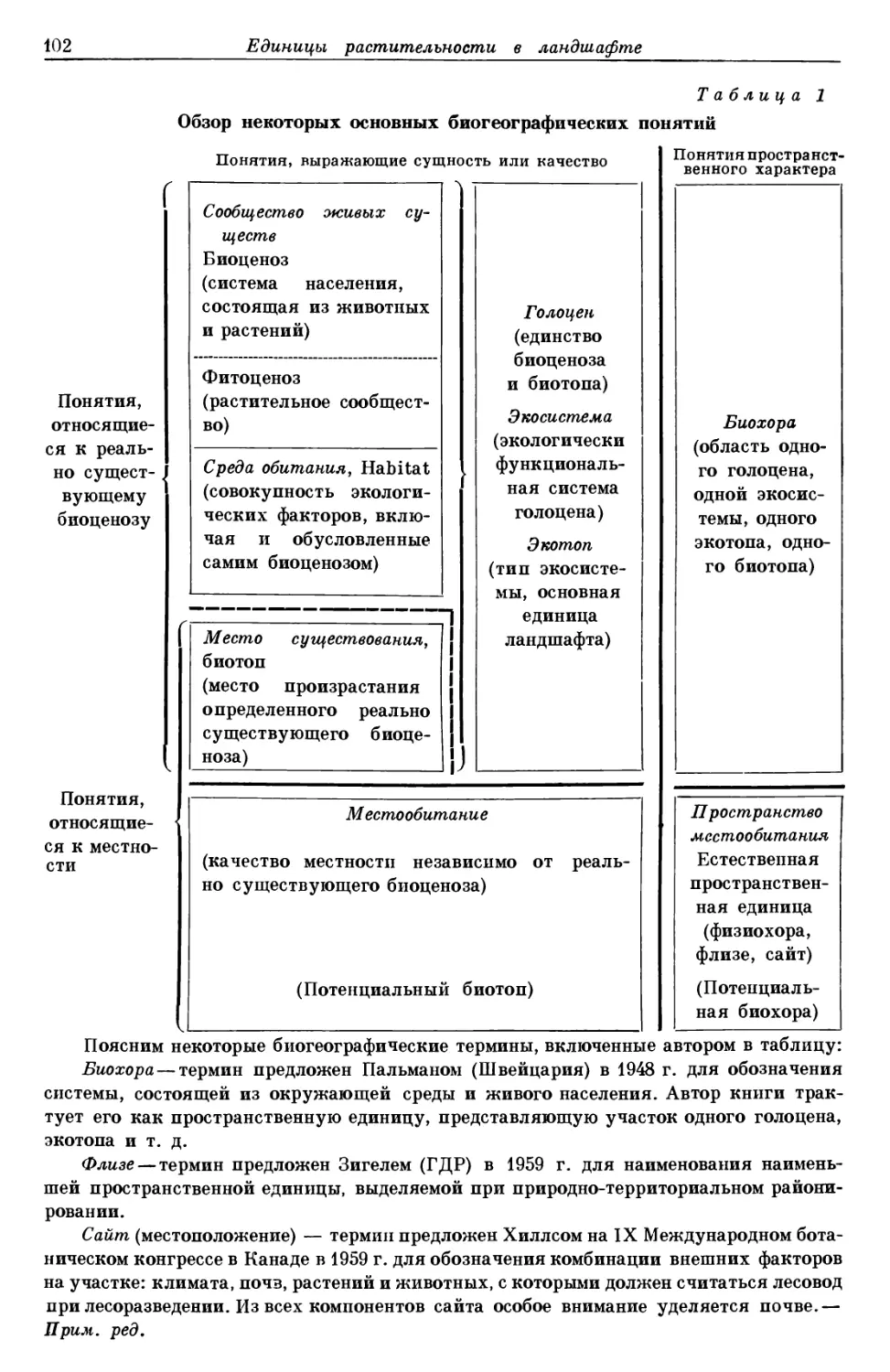
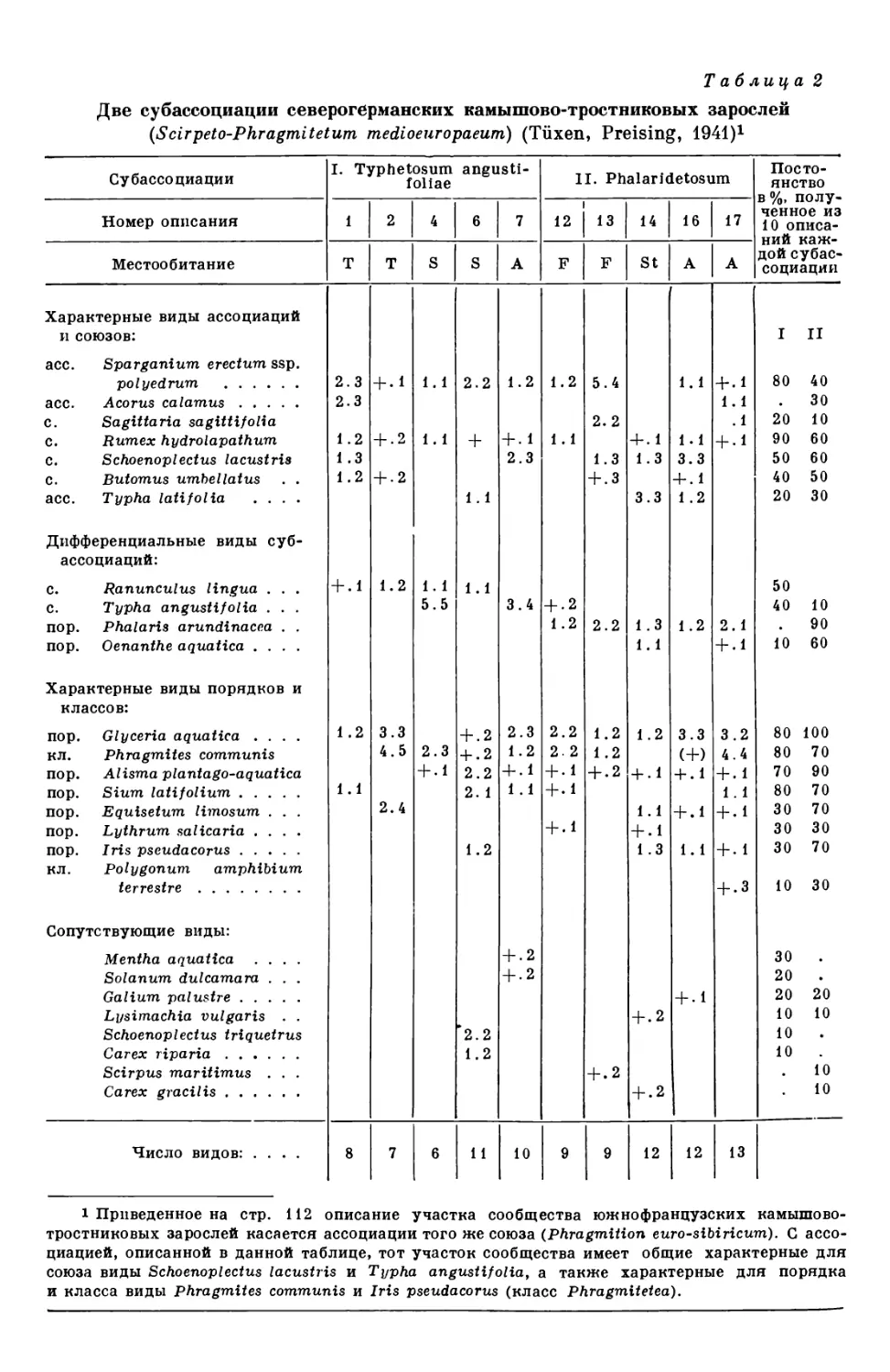
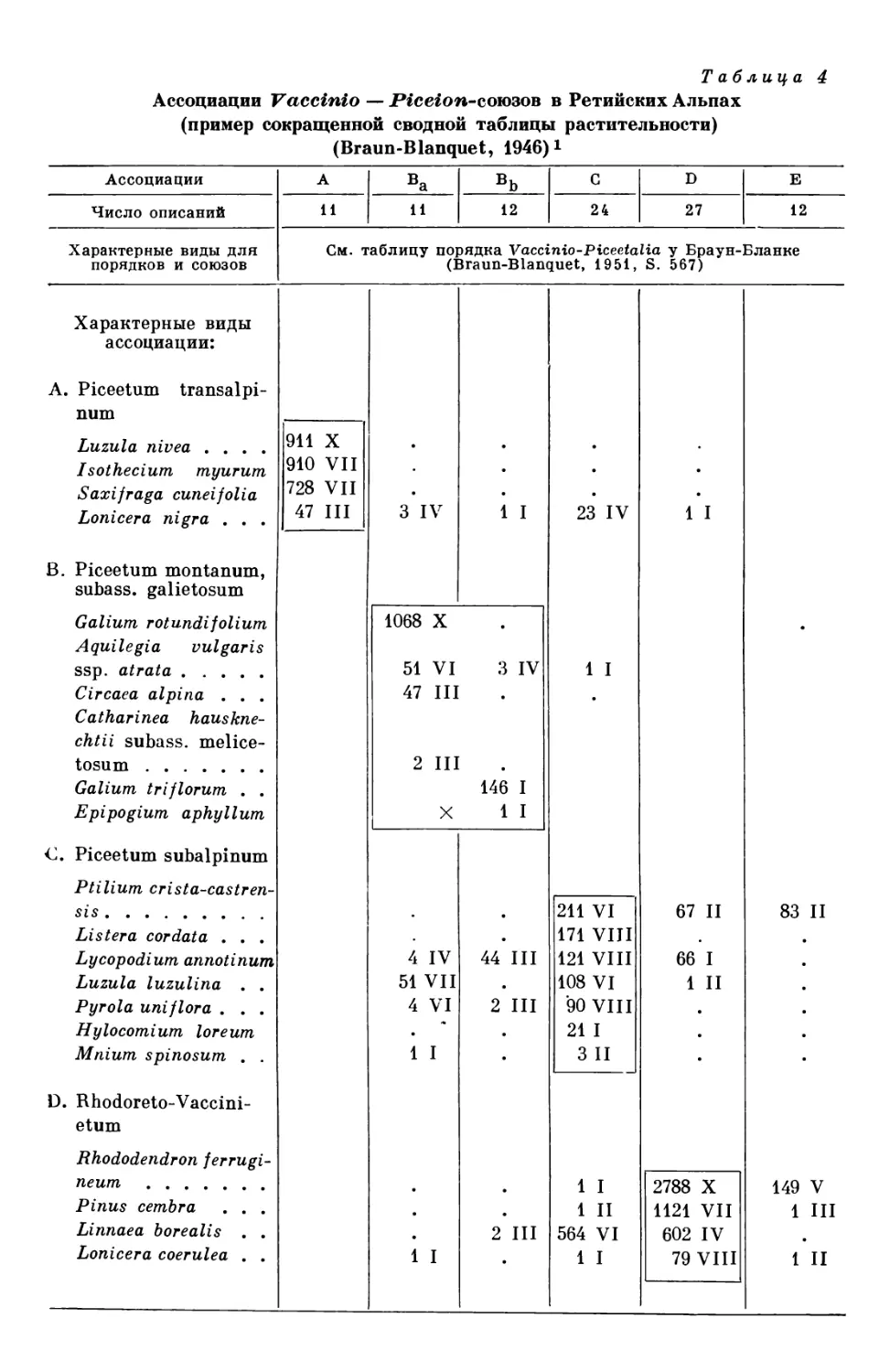
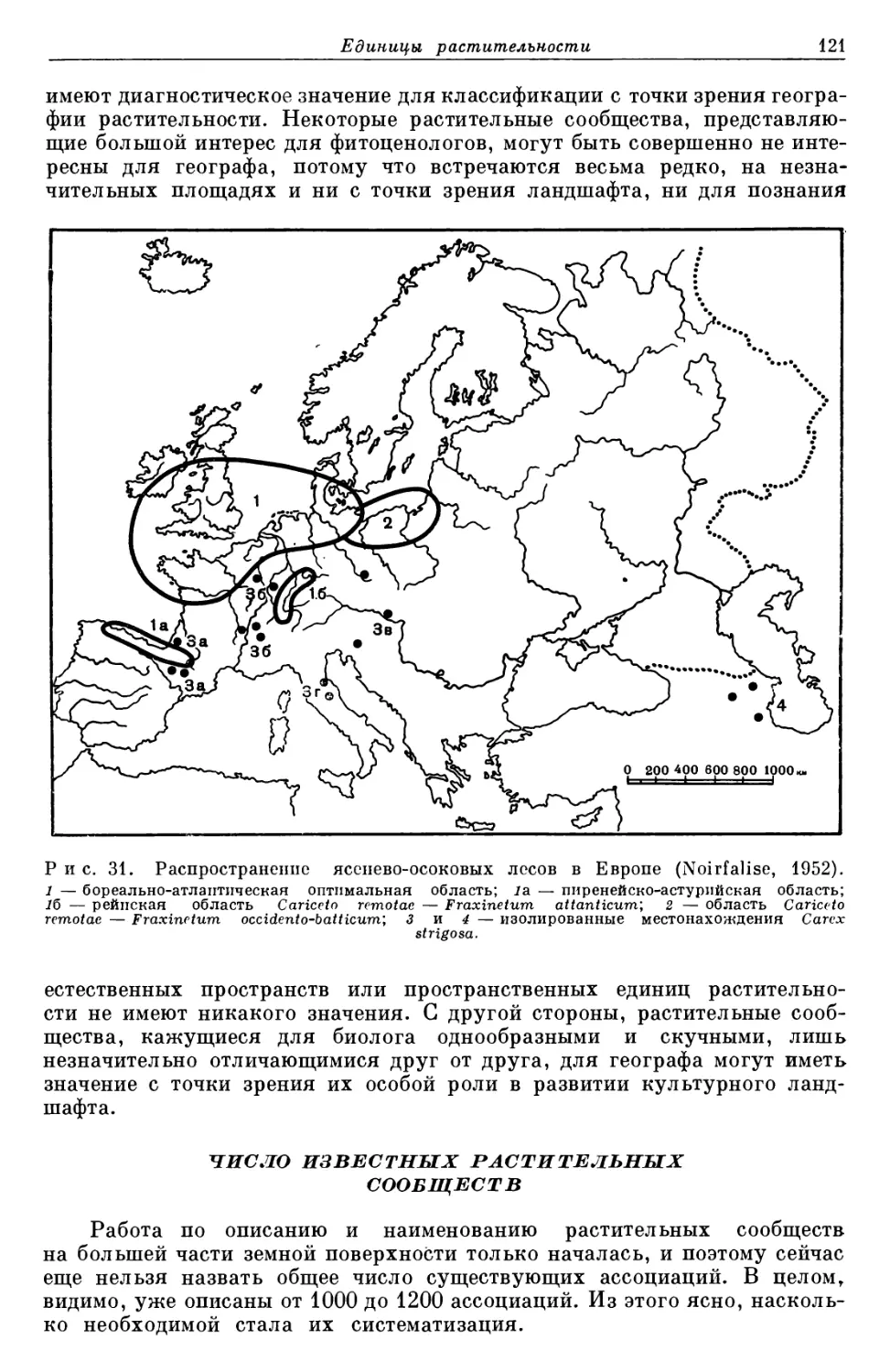
Систематические единицы
и их распространение
ЕДИНИЦЫ СИСТЕМАТИКИ РАСТЕНИИ
Элементарными составными частями растительности являются отдель-
ные индивидуумы растений, каждый из которых существует ограничен-
ное время в определенной точке Земли. Любое дерево, любая травинка
есть нечто неповторимое, обладающее своей, присущей лишь данной
особи формой и индивидуальным жизненным циклом. Однако при описа-
нии растительного покрова Земли нельзя исходить из характеристики
отдельных особей. Следует пользоваться типологическими понятиями, кото-
рые позволяют обобщать большое количество сходных индивидуумов.
Такие понятия предоставляются в наше распоряжение ботанической
системой. Она подразделяет растения по степени их родства на система-
тические единицы различного ранга. Понятие систематической единицы
вне зависимости от ее ранга в 1950 г. получило международное название
таксон.
Основной таксономической единицей является вид (Species). Один
вид растений охватывает «все растения (включая их предков и потомков),
которые обладают многими общими важными признаками» (Fitting,
Schumacher u. а., 1947). Наименование каждого вида состоит из двух частей
(например, Viola tricolor), причем первое название обозначает принад-
лежность к более высокой таксономической единице — роду. Особи,
характеризующиеся всякого рода отклонениями от обычных признаков
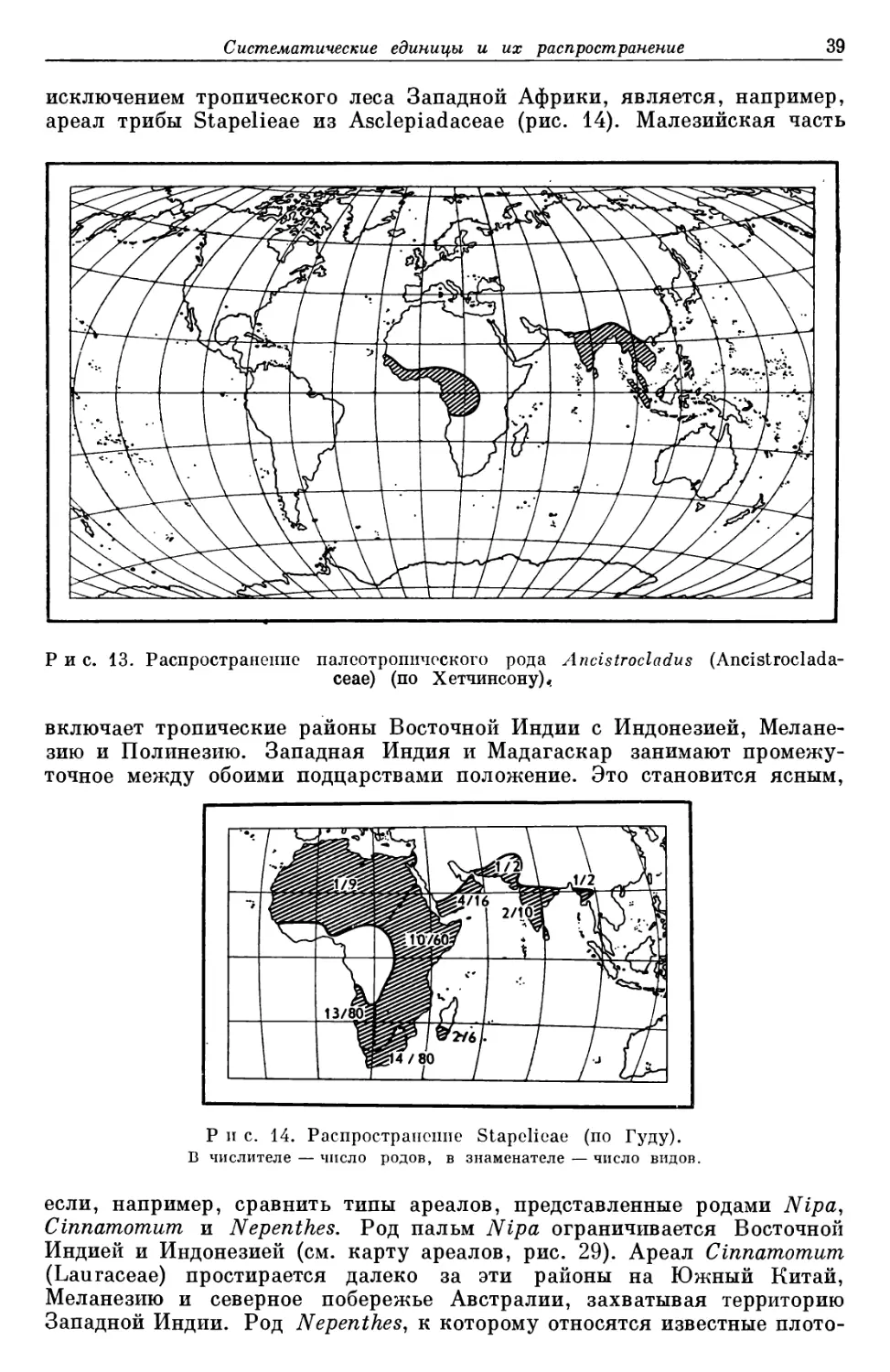
данного вида, объединяются в систематические единицы низшего порядка
(подвиды, формы, разновидности). Эти последние обозначаются с помощью
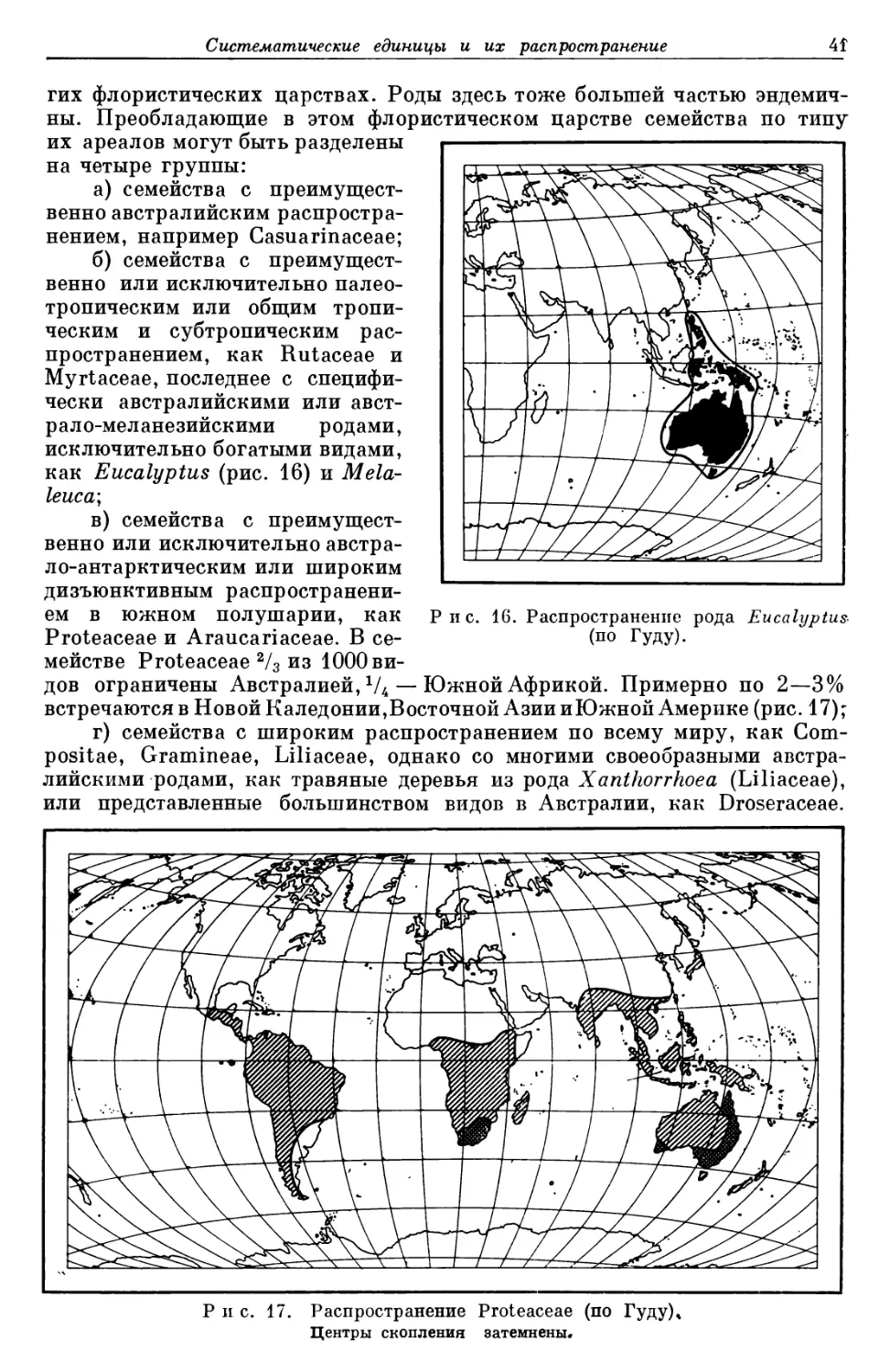
добавочных наименований к их видовым названиям (например, Viola
tricolor, подвид arvensis).
По совпадающим признакам виды объединяются в таксоны более
высокого ранга — в роды, семейства, порядки, классы и типы. При этом
могут быть и промежуточные градации. Так, например, входящие в один
род близкие группы видов объединяются в серии, подсекции, секции,
подроды. В одном семействе роды по необходимости могут группироваться
в подтрибы, трибы и подсемейства, а в высших таксономических единицах
соответственно могут различаться подпорядки, подклассы и подтипы.
Ранг таксономической единицы можно определить по форме окончания
в наименовании: подтрибы — inae; трибы — еае\ подсемейства — oideae',
семейство — асеае\ подряд или подпорядок — inales', ряд или порядок —
ales', подкласс — idae\ класс — opsida', тип — phyta.
Принцип построения системы можно пояснить примером. Сосна горная (Pinus
mugo var. mughus) — это разновидность, которая вместе с пряморастущей формой
(var. rostrata) относится к виду горных сосен (Pinus mugo). Этот вид принадлежит,
так же как и сосна лесная (Pinus Silvestris), сосна черная (Pinus nigra) и другие виды,
к секции Eupitys рода Pinus, который в двух подродах, состоящих в общем из И сек-
Предлагаемая читателю книга представляет собой руковод-
ство по географии растительности земного шара, включающее
основы флористической географии, экологии растений, фитоцено-
логии, биогеоценологии (учение об экосистеме), обзор основных
растительных формаций и их роль в ландшафте, а также описание
растительности земного шара по зонам и поясам.
Автор знакомит советского читателя со взглядами и методами
зарубежных ученых и дает подробную сводку мировой литературы
по географии растительности и связанным с ней дисциплинам.
Книга представляет большой интерес для геоботаников, лесо-
водов, географов, работников сельского хозяйства и может слу-
жить пособием для студентов старших курсов и аспирантов соот-
ветствующих специальностей.
Редакция литературы по географии
2—8—3
БЗ—18—65
Предисловие к русскому изданию
Использование естественного растительного покрова в качестве инди-
катора почвенных, геохимических и гидрогеологических условий все
больше входит в практику сельскохозяйственного районирования, поиско-
вых геологических и гидрогеологических исследований. В связи с этим
растет потребность в подготовке специалистов-геоботаников и одновре-
менно в руководствах по ботанической географии. Однако в подобного
рода работах все же ощущается недостаток и тем ценнее выход в свет
каждого нового пособия по ботанической географии. Поэтому весьма
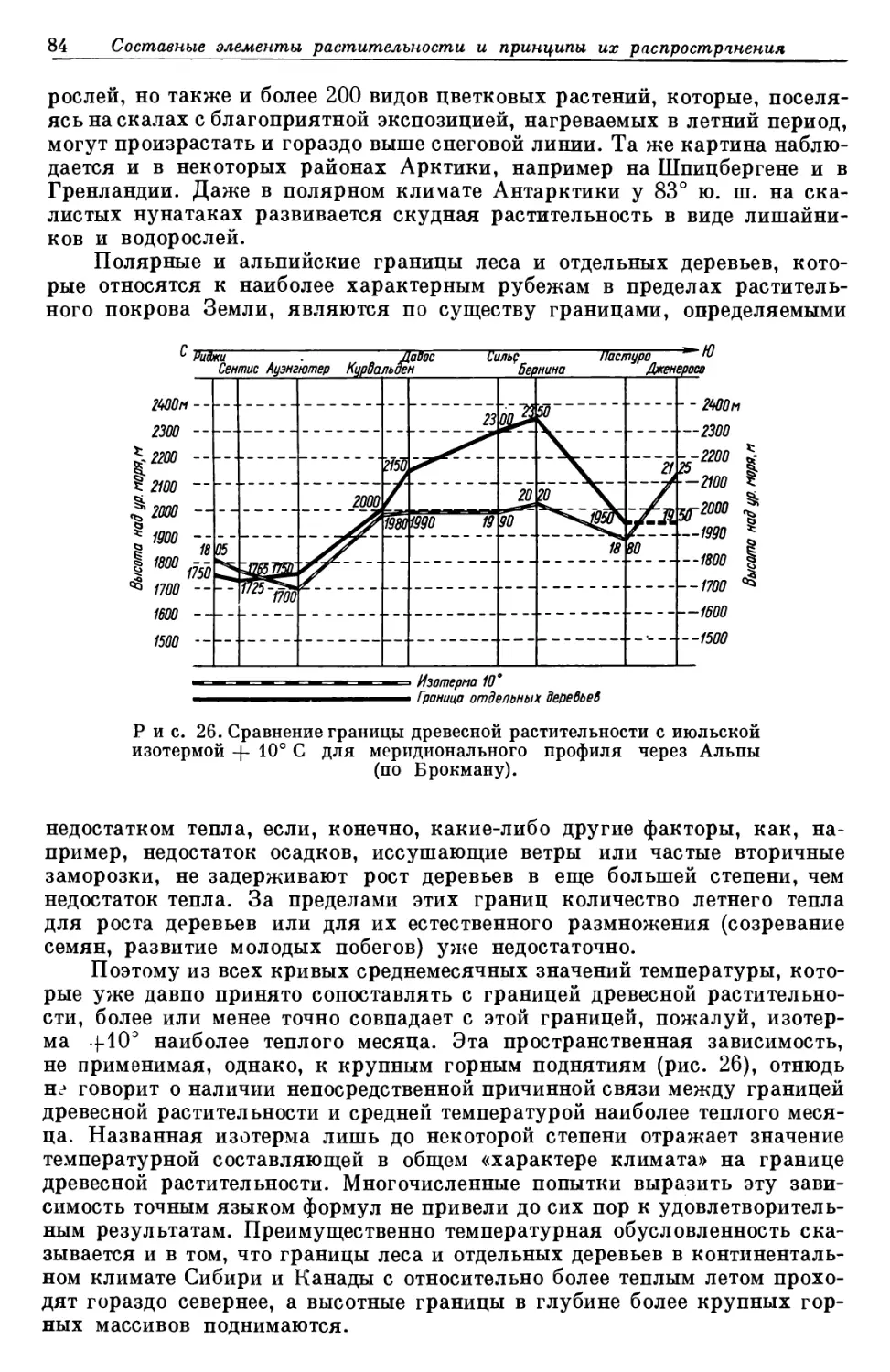
своевременным является перевод на русский язык и издание книги немец-
кого ученого проф. И. Шмитхюзена, названной им «Общая география
растительности». На наш взгляд, это заглавие гораздо вернее отражает
суть излагаемого в ней предмета, чем обычно применяемое название «Бота-
ническая география». Автор книги — по специальности географ, и это
обусловило специфику ее содержания. Растительный мир автор рассмат-
ривает с географической точки зрения, уделяя основное внимание расти-
тельным сообществам, определяющим облик ландшафта. Он подчерки-
вает, что предметом географии растительности являются «страны и ланд-
шафты и их заполнение растительностью». Однако глубокое знание бота-
ники и выяснение причинных связей между характером растительности
и условиями местообитания, по мнению автора, также обязательны при
изучении растительного покрова.
Содержание книги И. Шмитхюзена значительно шире ее названия,
ибо, помимо вопросов, связанных с распределением растительности в про-
странстве, в ней изложены и основы флористической географии, и вопро-
сы экологии растительных сообществ. Весьма интересна приведенная авто-
ром классификация форм роста (Wuchsform) и сопоставление выделен-
ных им классов форм роста с жизненными формами (Lebensformen)
Раункиера.
Излагая проблемы фитоценологии, Шмитхюзен подробно останавли-
вается на основных понятиях биогеоценологии, новой научной дисцип-
лины, получившей в последнее время широкое распространение как у нас,
так и за рубежом. Автор придерживается взглядов франко-швейцарской
школы Браун-Бланке, главенствующей сейчас не только в зарубежной
Европе (Франция, Швейцария, ФРГ, Польша, Венгрия, Болгария, Юго-
славия, Австрия: Бельгия, Италия, Испания, Португалия), но и в некото-
рых странах за ее пределами (ОАР, Израиль, Япония). Советским специа-
листам небезынтересно будет ознакомиться с некоторыми направлениями
этой школы, хотя бы они и не были согласны со многими из них. Однако
в дальнейшем при описании крупных единиц растительности автор отходит
от принципов школы Браун-Бланке и кладет в основу изложения эколо-
6
Предисловие к русскому изданию
го-физиономические признаки и доминирующие виды, что сближает взгля
ды автора со взглядами советских ученых. Значительное место в книге
отведено влиянию человека с древнейших времен по настоящее время на
распространение отдельных видов, формирование и распространение рас-
тительных сообществ, возникновение современного культурного ландшафта
и замещающих (вторичных) сообществ. Придавая большое значение хозяй-
ственной деятельности человека в происхождении современного ланд-
шафта, автор настаивает на антропогенном происхождении раститель-
ности лесостепной зоны, с чем мы не можем согласиться. Главы о воздейст-
вии человека на растительность построены в основном на примере расти-
тельности Средней Европы, однако в большинстве случаев они имеют более
широкое значение и интересны и полезны для советского читателя.
Применяемое автором разграничение растительности на горизон-
тальные зоны и пояса и высотные ступени несколько отличается от при-
нятого у нас деления, однако, стремясь сохранить стройность и самобыт-
ность авторской классификации, мы оставляем его без изменений.
Следует заметить, что не все разделы книги написаны с одинаковой
детальностью — на некоторых вопросах автор останавливается очень
подробно, в других же случаях он ограничивается лишь ссылками на
литературу, как, например, в разделе о влиянии внешних факторов на
распространение единиц растительности.
К книге приложен список литературы, содержащий около 1000 назва-
ний. К сожалению, только 16 из них принадлежат русским авторам. Сла-
бое использование работ русских ученых, которым принадлежит немало-
важная роль в развитии ботанической географии, особенно сказалось
на характеристике растительных формаций, имеющих основное распро-
странение на территории СССР. Описание их чрезвычайно кратко и гре-
шит неточностями.
Большое затруднение при редактировании вызвал перевод некото-
рых терминов, предложенных автором. Так, понятие «Gesellschaftspulk»
мы переводим как «комбинация сообществ». Выражения «Vegetations-
division» и «Divisionsgebiet» мы вынуждены оставить как «дивизионы
растительности» и «области дивизионов», однако, считая эти термины
малоудачными, мы ставим их в кавычки. Широко используемые автором
местные названия ландшафтов и единиц растительности («flats», «glades»
и т. д.) оставлены без перевода и даны латинскими буквами в тран-
скрипции автора. В целом книга издается почти без сокращений. В тех
случаях, когда взгляды автора сильно расходятся со взглядами советских
ученых или же когда автор употребляет термины и понятия, малозна-
комые советскому читателю, изложение дополнено примечаниями.
Т. Б. ВЕРНАНДЕР,
кандидат биологических наук
Памяти Лео Вайбеля посвящаю
Предисловие к немецкому изданию
Почти 25 лет назад Лео Вайбель предложил мне сотрудничать в рабо-
те над книгой о растительности Земли. Поскольку, однако, он лишился
места преподавателя в Боннском университете и вынужден был покинуть
Европу, этот план не был осуществлен.
Беседы с Вайбелем и моя попытка осуществить его заветы легли
в основу первых набросков этого труда. Я надеюсь, что мне удалось хоть
немного развить научное наследство моего уважаемого учителя.
Отдельные принципы построения этой книги я впервые высказал
еще в 1939 г. в своем конкурсном докладе на замещение должности
в Боннском университете (Schmithusen J., 1942, «Zeitschr. d. Ges. f. Erdk.
zil Berlin»). Уже тогда я думал о том, чтобы показать растительность «как
важнейшее звено в структуре ландшафтов» и одновременно сделать при-
годными для исследований именно в области географии растительности
те новые методы фитоценологии и экологического учения о местообита-
ниях, которые находились еще в стадии развития. Позднее я неоднократ-
но разными путями вновь и вновь возвращался к этой теме и поэтому
с радостью согласился на предложение Э. Обета обработать том «Геогра-
фии растений» для многотомного «Учебника общей географии». По моему
предложению заголовок был изменен на «Географию растительности».
Причины этого изложены в ранее опубликованной статье (Schmithusen,
1957).
К сожалению, я не имею возможности назвать каждого, кто помогал
мне при работе над книгой, поэтому выражаю общую благодарность мно-
гим друзьям и коллегам, облегчившим эту работу либо совместными бесе-
дами, либо предоставлением в мое распоряжение изданий своих работ.
Карлсруэ, весна 1959 г.
И. ШМИТХЮЗЕН
ВВЕДЕНИЕ
Растительность как предмет исследования
географии
БИОСФЕРА
Подобно тонкому покрывалу, сотканному из отдельных живых существ,
окутывает жизнь всю нашу Землю. Эту живую оболочку, «приют всего,
что населяет Землю» (Г. Вагнер), вместе с занимаемым ею пространством
мы и называем биосферой.
На вопрос о том, ограничено ли вообще распространение жизни только лишь
нашей планетой, мы ответить пока не в состоянии. Можно сказать одно: судя по всему,
что нам известно о сущности жизни, вовсе не следует считать, что жизнь непременно
существует и на других космических телах. Собранный палеонтологами материал позво-
ляет нам полностью представить родословное дерево развития жизни на самой Земле.
Происхождение ее, однако, пока остается неясным. Уже древнейший животный мир,
скрытый в слоях, относящихся к протерозойской эре, оказывается весьма разнообраз-
ным. Ему должен был предшествовать длительный период развития, надежных сведе-
ний о котором мы не имеем. Можно, по-видимому, полагать, что графитовые и антра-
цитовые включения в породах, насчитывающих около 1,5 млрд, лет, являются самыми
ранними свидетелями жизни на Земле. Органические остатки в этих слоях еще
не содержатся.
На рубеже протерозойской и палеозойской эр появляется сразу большое число
родов животных с тысячами различных видов. Растительная жизнь вплоть до ордовик-
ского периода ограничивается таллофитами. Только начиная с верхнего готланда
появляются сосудистые растения. «Поскольку предварительным условием развития
животных организмов является наличие кислорода, который в палеоатмосфере отсут-
ствовал и выделился в свободном виде из воды главным образом в результате жизне-
деятельности растений, можно предполагать, что возникновение растений предшест-
вовало появлению животных» (Schindewolf, 1955).
В процессе развития жизнь постепенно все более усложняется. Непре-
рывно возрастающую дифференциацию форм жизни можно рассматри-
вать как средство, с помощью которого на протяжении всей истории Зем-
ли жизнь стремилась сохранить себя в условиях окружающей среды.
Биосфера находится в неизменном, непрекращающемся движении. Не
говоря уже о становлении и исчезновении отдельных индивидуумов,
жизнь изменяется и во всей своей совокупности. Формы ее изменяются
так, что при этом родословное дерево жизни все более разветвляется;
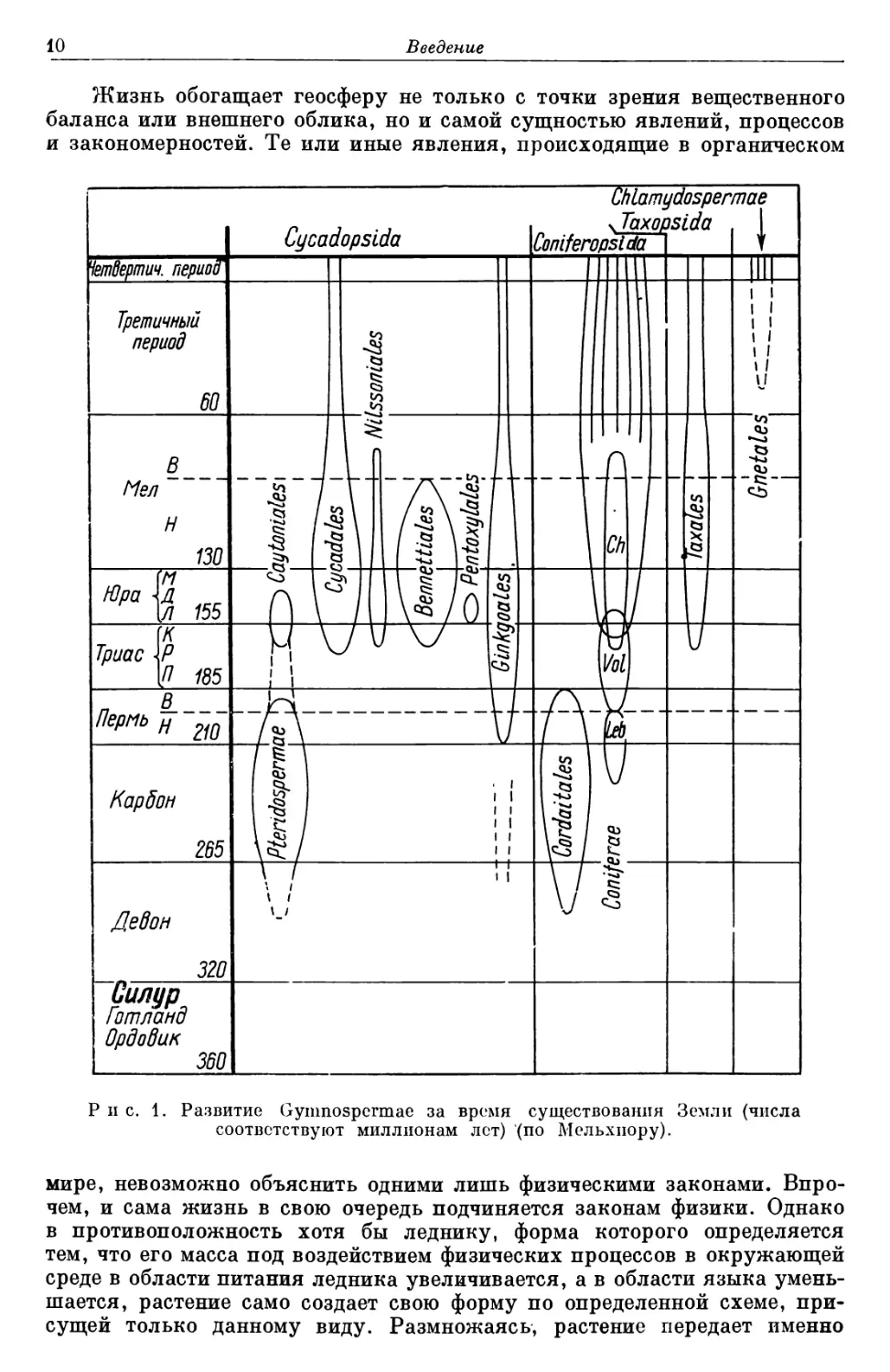
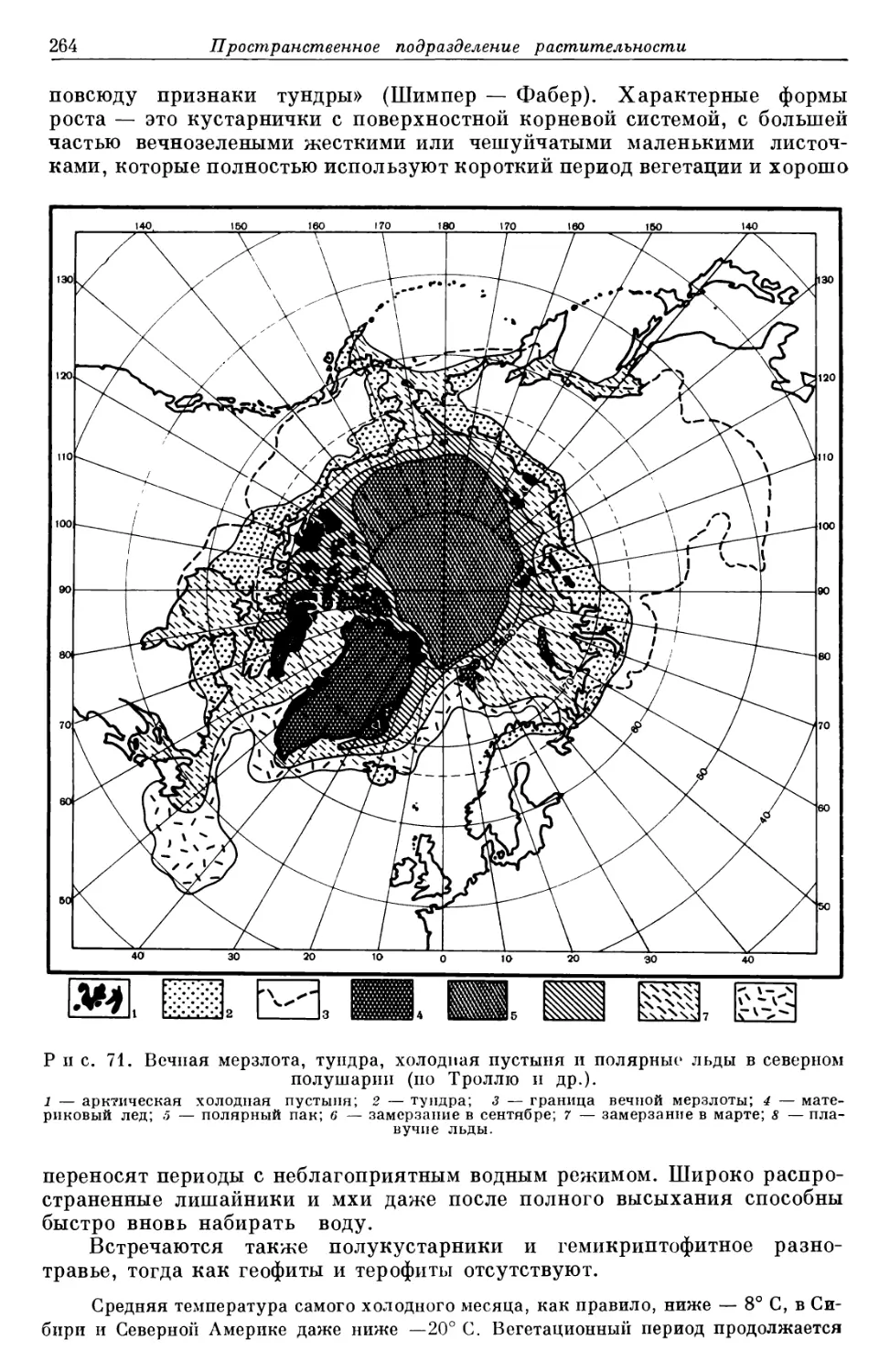
одни виды возникают, другие вымирают (рис. 1).
Процессы эволюции занимают различные по продолжительности отрезки времени.
Известно, что один из видов небольших рачков, обитающих во внутренних водоемах,
встречался еще в среднем кейпере, следовательно, данный вид существует более
180 млн. лет. С другой стороны, целый ряд видов растений и животных возник только
в современную геологическую эпоху. Таким образом, виды животных организмов, насе-
ляющих Землю, значительно разнятся по возрасту.
10
Введение
Жизнь обогащает геосферу не только с точки зрения вещественного
баланса или внешнего облика, но и самой сущностью явлений, процессов
и закономерностей. Те или иные явления, происходящие в органическом
Рис. 1. Развитие Gymnospcrmae за время существования Земли (числа
соответствуют миллионам лот) (по Мельхиору).
мире, невозможно объяснить одними лишь физическими законами. Впро-
чем, и сама жизнь в свою очередь подчиняется законам физики. Однако
в противоположность хотя бы леднику, форма которого определяется
тем, что его масса под воздействием физических процессов в окружающей
среде в области питания ледника увеличивается, а в области языка умень-
шается, растение само создает свою форму по определенной схеме, при-
сущей только данному виду. Размножаясь, растение передает именно
Введение
И
эту схему следующим поколениям. Образование новых форм тоже про-
исходит по законам, присущим только живым организмам.
Во всяком живом организме отдельные его части (например, у выс-
ших растений корни, стебли, листья и цветки) составляют единое целое.
Это целое, используя все свои возможности для добычи пищи, роста и раз-
множения, вступает в определенные отношения с окружающей средой.
При этом существование того или иного организма зависит от наличия
в среде обитания необходимых условий, соответствующего физического
состояния этой среды и присутствия в ней определенных элементов, обу-
словливающих протекание самого жизненного процесса. Поддержание
этого процесса осуществляется за счет энергии солнечного света, кото-
рая в хлоропласте растения преобразуется в энергию химическую и исполь-
зуется для создания органического вещества. Животные в свою очередь
добывают энергию для поддержания жизненных процессов, расходуя
органические вещества растений.
Родиной жизни поэтому мы можем вместе с Вагнером считать «осве-
щенную поверхность Земли» или, вернее, освещенную водную поверх-
ность Земли. Суша, представляющая собой дно воздушного океана, была
завоевана жизнью только начиная с готландского периода, уже после
того, как океан был заполнен богато дифференцированными формами
жизни. Водная оболочка и в настоящее время представляет собой боль-
шую часть биосферы. Если для Александра Гумбольдта еще не был решен-
ным вопрос, «где более широко распространена жизнь: на континентах
или в неизмеримых просторах океанов», то сейчас мы знаем, что жизнь
существует даже на величайших глубинах Мирового океана. Здесь био-
сфера является наиболее мощной, достигая толщины 10 км и более, тог-
да как распространение жизни на суше ограничено значительно более
тонким слоем.
Основная масса растительной жизни гидросферы сосредото-
чивается в верхних слоях воды до глубины около 100 м. Автотрофные,
то есть самостоятельно питающиеся, организмы возможны, по-видимому,
только на глубинах до 200 м. Гетеротрофный растительный планктон
в небольших количествах развивается даже на глубине более 3000 м.
Живые организмы наблюдаются и на значительно больших глубинах.
Таким образом, пределы, занимаемые биосферой, определяются живот-
ным миром, тогда как растения распространены, вероятно, лишь в одной
четвертой части ее объема.
Жизнь на суше отличается исключительным разнообразием
форм, что отражает многогранную способность организмов занимать под
постоянное или временное местообитание области, весьма различные по
своему характеру. Поскольку жизнь на Земле невозможна без воздуха
и воды, то ее распространение совершенно четко ограничено: например,
жизнь отсутствует внутри плотных скальных пород или на горячей лаве
действующих вулканов. В хорошо аэрируемых почвах, образующих
самый верхний покров Земли, растения и животные обитают на глубинах
до нескольких дециметров и даже метров. В пещерах жизнь может про-
никать и на большую глубину. Недостаток света не является абсолютным
ограничением при условии доступа на такие глубины органического
питания извне.
На поверхности Земли нет таких более или менее значительных про-
странств, которые, строго говоря, можно было бы назвать полностью
лишенными жизни. Где бы ни появлялся человек, его пытливый взгляд
повсюду находил какую-нибудь жизнь, пусть это были даже микроскопи-
12
Введение
ческие растения или легкие подвижные животные, которые могли быть
занесены воздушными потоками на высочайшие горные вершины или
в полярные ледяные пустыни. Конечно, следует видеть различие между
постоянным обитанием или временным, но регулярным посещением и слу-
чайным вторжением отдельных живых существ в такие области, где, как
правило, жизнь отсутствует.
Даже в самых сухих, кажущихся безжизненными пустынях зачастую
удается обнаружить жизнь в ее скрытой форме, проявляющуюся уже
после случайного дождя, когда активизируются, пусть всего лишь на
несколько часов или дней, те животные и растения, которые в обычных
условиях здесь едва заметны или находятся в состоянии длительно-
го покоя.
За исключением пекла действующих вулканов, самые крайние по
значению температуры на земной поверхности не являются препятствием
для существования растений и животных. На льду и фирне обитают глет-
черные блохи, а также некоторые водоросли зеленого или красного цвета.
Последние могут быть настолько многочисленны, что их скопление изда-
ли кажется «кровавым снегом». В горячих источниках с температурой
около + 90° обнаружены сине-зеленые водоросли и бактерии.
Что касается воздушного пространства, то лишь
нижние слои его обитаемы; кроме птиц, постоянных жителей этой среды,
здесь можно встретить и случайных обитателей — летающих животных,
растения. Активно летающие, как, например, насекомые и птицы, дер-
жатся преимущественно в нижних слоях до высоты 300—400 м. Полет птиц
на высоте до 2000 м— исключение. Обычно же на больших высотах вплоть
до 10 км находят лишь пассивно занесенные сюда воздушными течениями
микроорганизмы, пыльцу и микроскопические споры растений.
Если рассматривать вместе море, сушу и воздушную оболочку, то
максимальную протяженность биосферы по вертикали можно принять
равной немногим более 20 км.
По сравнению с размерами Земли масса имеющегося на ней органи-
ческого вещества крайне незначительна. Если бы все вещество, входящее
в состав растительных и животных организмов, равномерно распределить
по поверхности земного шара, то, по расчетам Вагнера, образовался бы
слой всего лишь в 0,5 см. Масса вещества, приходящаяся на долю живот-
ных, заняла бы только V10 такого слоя, причем в основном она сосредо-
точена в океанах, а наземные животные занимают в этой доле всего х/25о
часть. Эти данные являются отправными при дальнейшем рассмотрении
нами некоторых величин. Несмотря на кажущуюся незначительность
органической массы, жизнь оказывает огромное влияние на баланс вещест-
ва и состояние геосферы, косвенно участвуя тем самым во многих других
явлениях, изучаемых географией. Организмы состоят прежде всего из
тех же элементов, которые входят и в состав атмосферы: кислорода, водо-
рода, углерода, азота. Остальные компоненты, хотя они и весьма важны
для существования самой жизни, присутствуют в меньших количест-
вах.
Такие формы проявления жизни, как питание, рост, размножение,
а также размещение в пространстве, являются одновременно формами
вторжения жизни в круговорот веществ, составляющих геосферу. По
отношению живых существ к органическим веществам Тинеман различает
продуценты (производители), которые создают органические вещества из
неорганических (зеленые растения), консументы (потребители), погло-
щающие эти органические вещества (большей частью — животные) и реду-
Введение
13
центы (разрушители), которые их ликвидируют, упраздняют (бактерии,
грибы).
Примерно х/10 общей массы органических веществ составляет угле-
род, что соответствует приблизительно г/3 всего углерода, содержащегося
в атмосфере. В результате фотосинтеза, осуществляемого зелеными расте-
ниями, углерод воздуха поступает в состав органического вещества. Часть
связанного углерода при дыхании растений, при разложении органиче-
ского вещества или его сгорании возвращается в виде углекислоты обрат-
но в воздух. Другая часть остается в гумусных веществах (перегное)
почвы или же в виде торфа или каменного угля входит в состав твердой
земной коры. Определенная часть углерода обычно в виде карбоната каль-
ция, будучи вымыта из почвы и попав в реки и моря, участвует в созда-
нии известковой оболочки морских животных и тем самым в осадочных
и горообразовательных процессах. Баланс всех этих процессов сказы-
вается на содержании углерода в воздухе, который в свою очередь может
влиять на сохранение или изменение климата.
Все организмы в значительной степени (а некоторые в основном)
состоят из воды и в ходе своей жизнедеятельности перерабатывают воду,
участвуя тем самым в общем круговороте воды. Растения поглощают воду
из почвы и путем транспирации возвращают обратно в воздух часть выпав-
ших на почву осадков. Кроме того, растительный покров замедляет поверх-
ностный сток выпавших осадков. Изменение растительности может поэто-
му оказать влияние на водный баланс той или или иной территории.
Необходимо упомянуть и о круговороте минеральных веществ. Неко-
торые из них, извлеченные растениями из почвы в качестве пищи и вошед-
шие в состав органических тканей, возвращаются снова в почву лишь
окольным путем, пройдя через организмы животных.
Разнообразны пути воздействия органического мира на неоргани-
ческие процессы в геосфере. Живая природа создает предпосылкц для
процесса выветривания горных пород и способствует его дальнейшему
развитию; участвует в размыве земной поверхности в одном месте и созда-
нии новых морфологических форм в другом, формирует состав воды, ока-
зывает решающее влияние на образование почв и может в большой степени
обусловливать местный климат.
Если бы на Земле не было жизни, то не было бы ни «почвоведения», ни «климато-
логии». Геосфера безжизненной Земли была бы, конечно, сложной изменяющейся
системой, которая предоставила бы в распоряжение человеческой мысли достаточное
количество проблем, подлежащих исследованию. Но общая география лишенной жизни
Земли была бы несколько иной, потому что отпали бы те обусловленные жизнью кри-
терии, без которых климатология в современном понимании невозможна. Существо-
вала бы лишь физика атмосферы, потому что понятие климата прежде всего связано
с жизнью. Если мы и разграничиваем климаты по температуре, количеству осадков
ит. д., то очевидно, что это чисто физическое разграничение, потому что выбор крите-
риев, обусловливающих определенные рубежи в той или иной науке, в конце концов
производится по их значению для жизни человека. Поэтому не случайно и не произ-
вольно мы зачастую обозначаем климаты по характеру растительности в данной обла-
сти. «Климат пустынь»— это отнюдь не физическое определение, «климат тундры»—
тем менее, «средиземноморский климат»— тоже нет; в этих названиях отражен прежде
всего характер произрастающей здесь растительности. Живая природа оказывается
«определяющим элементом» для различаемых нами климатов, даже если мы и стараемся
выразить их каким-либо точным физическим «определением».
14 Введение
ЦЕЛИ И ЗАДАЧИ ГЕОГРАФИИ РАСТИТЕЛЬНОСТИ
«Les methodes sont се qui caracterise 1’etat de la science a chaque epoque et
qui determine le plus ses progres» (Alphonse De Candolle, 1855) L
Несмотря на то что обе науки, как биология, так и география, зани-
маются вопросами распространения жизни на Земле и проблемами, свя-
занными с распространением жизни (биохорологией), исходные позиции
и конечные цели у этих наук различны. Биология исследует жизнь, фор-
мы ее проявления, процессы и законы ее развития, помимо прочего, так-
же и с точки зрения их распределения в пространстве. Предметом геогра-
фии является геосфера и ее деление на страны и ландшафты, для харак-
теристики которых наряду с другими явлениями немаловажное значе-
ние имеет и их растительный и животный мир.
Под растительностью (Vegetation) мы понимаем растительный покров
Земли и его различные составные части.
При исследовании растительности геобиологическая и, строго говоря,
биогеографическая точка зрения, как правило, объединяются общим тер-
мином «география растений» (Pflanzengeographie). Кроме Альфреда Гет-
тнера (1859—1941), Роберта Градмана (1865—1950), Лео Вайбеля (1888—
1951) и некоторых других исследователей, мало кто задумывался над
тем, что цели исследований в этих двух случаях должны быть различны.
Именно поэтому в настоящее время еще довольно широко распростра-
нена неопределенность в отношении круга задач, стоящих перед геогра-
фией.
В последние десятилетия к одному из направлений в области бота-
ники привился термин «геоботаника» (Geobotanik)1 2. Однако такое поло-
жение, при котором в понятии «география растений» основной упор делал-
ся бы именно на географию, признания еще не получило. Поскольку все
же необходимо провести какое-то разграничение в терминологии, то
«география растительности» противопоставляется нами геоботанике как
отрасль географии. На практике, правда, не удается провести четкой
границы между этими областями одной науки, поскольку они в значитель-
ной степени совпадают. В каждой из этих областей, однако, постановка
вопроса осуществляется в разных плоскостях, а поэтому один и тот же
объект изучения освещается с различных точек зрения.
Геоботаника изучает систематические единицы растительного мира
(Pflanzensippen) и растительные сообщества (Pflanzengesellchaften) с точки зрения их
1 «Методы — это то, что характеризует сущность науки в каждую эпоху и чем
определяется ее прогресс».
2 Автор, следуя франко-швейцарской геоботанической школе, господствующей
в настоящее время за рубежом, понимает геоботанику очень широко, включая в нее
идиобиологпческую геоботанику, изучающую экологию и географическое распростра-
нение отдельных видов растений, и социологическую геоботанику, или фитосоциоло-
гию, объектом изучения которой являются растительные сообщества. В СССР под
«геоботаникой» понимается только учение о растительных сообществах, или фитоце-
нология (фитосоциология в терминологии зарубежных ученых). Термин «геоботаника»
был предложен в России в 1866 г. академиком Ф. И. Рупрехтом, но широкое распро-
странение в смысле учения о растительных сообществах получил лишь с 20-х годов
нашего столетия.
Идиобиологическая геоботаника, с нашей точки зрения, включает две самостоя-
тельные науки — флористическую географию растений, изучающую распростране-
ние отдельных видов растений, и экологию растений, занимающуюся изучением
взаимоотношений растений со средой. Употребляемые в дальнейшем автором термины
«социология растений» и «фитосоциология» мы заменяем принятым у нас термином
«фитоценология».— Прим. ред.
Введение
15
распространения и зависимости от условий существования. Идиобиологическая
геоботаника занимается систематическими единицами растений и охватывает как
наука основные разделы геоботаники в их классическом делении на флористическую,
историко-генетическую и экологическую геоботанику. Наряду с этим возникла ценоло-
гическая геоботаника (фитоценология), исследующая растительные сообщества.
Отдельные направления этих двух отраслей геоботаники определяются четырьмя
возможными аспектами рассмотрения вопросов: с точки зрения формы, функции, про-
странства и времени. Каждый из этих элементов является необходимой составной
частью общего направления исследований.
География растительности как одна из ветвей целого
комплекса географических наук исследует и описывает растительный
покров Земли с точки зрения его значения для характера данной местно-
сти. Предметом географии растительности являются не отдельные расте-
ния и даже не их сообщества, а страны и ландшафты и их заполнение расти-
тельностью. В своей специальной и описательной части география расти-
тельности является поэтому частью страноведения, а с точки зрения общей
постановки вопроса — частью ландшафтоведения. Такое определение дела-
ет понятным задачи, стоящие перед географией растительности, исполь-
зуемые ею методы и ее структуру. По этим же признакам определяется
и значение всех тех материалов и данных из области геоботаники, которые
предоставляются этой наукой в распоряжение географии растительности.
Мы попытаемся здесь изложить последовательно с точки зрения
географа структуру общей географии растительности. Образцов для
такой концепции мы пока не имеем, поэтому в ряде разделов будем вынуж-
дены ограничиться лишь общим изложением методики. Цель этой книги —
не компилятивное изложение отдельных фактов и точек зрения, а попыт-
ка произвести строгий анализ и систематизацию накопленного фактиче-
ского материала в определенной области науки. Автор книги не стремится
уйти от трудностей, связанных с характером самой проблемы. Геогра-
фия растительности без хорошего знания основ общей ботаники так же
мало приемлема, как и геоморфология без знания процессов, происхо-
дящих в недрах Земли, или понимания геологической истории нашей
планеты, и если кто-либо серьезно намерен заниматься этой наукой, то
он будет вынужден обратиться за помощью и к учебнику ботаники. «Кто
хочет проводить самостоятельные исследования в области географии расти-
тельного или животного мира, тот не сможет обойтись без основательно-
го изучения ботаники и зоологии» (Gradmann, 1919).
Книга построена по плану, предусматривающему последовательное и логическое
изложение всего материала. При приведении отдельных примеров автор не стремится
избежать специальных терминов и названий растений. Применение местных немецких
терминов и названий казалось нецелесообразным, и поэтому вся номенклатура
ориентирована прежде всего на общепринятые международные обозначения и правила.
Если бы автор намеренно упростил отдельные представления и понятия из области
географии растений специально «для географов» и отказался бы от научных названий
растений, то зачастую оказалось бы невозможным даже подступиться к изложению
самой проблемы. Автор стремился создать не упрощенную геоботанику для географов,
а попытался рассмотреть растительный мир с географической точки зрения, что, воз-
можно, небезынтересно и для ботаника.
При создании единой картины всего растительного покрова Земли необходимо
исходить прежде всего из рассмотрения климатических зон растительности и харак-
теризующих их растительных формаций. Тем самым большое значение придается фор-
мам роста. Необходимость сравнения поясов растительности с точки зрения климата,
16
Введение
почв и общего характера ландшафта, а также с учетом их влияния на экономику чело-
веческого общества требует привлечения многих материалов, являющихся предметом
экологической геоботаники.
При изучении особенностей отдельных зон растительности следует учитывать
различия в их флоре, то есть привлечь данные флористической геоботаники (элементы
флоры и характер флоры тех или иных местностей). Рассмотрение зон растительности
в качестве пространств обитания, обладающих индивидуальными судьбами и испыты-
вающих большей частью преобразующее влияние человека, заставляет обратить
внимание на изменение характера растительности во времени и принуждает таким
образом к историческому подходу.
При более детальном описании наш взгляд невольно обращается к единицам
растительности (Vegetationseinheiten), свойственным тому или иному ландшафту.
Ландшафт, определяющий облик данной местности, а также отдельные участки земной
поверхности, характеризуемые однородными ландшафтами, можно исследовать с точки
зрения формы (физиономия), функции (экология), пространства (хорология) и времени
(в развитии). Каждая из этих точек зрения имеет в виду изучение растительного мира
как составной части всего ландшафта.
Топографически единицы растительности различаются по растительным сооб-
ществам. Для знакомства с ними ботанико-географ использует прежде всего данные
фитоценологии, особенно если последние нашли свое отражение в геоботанических
картах. Растительные сообщества могут служить индикаторами естественного про-
странственного деления земной поверхности, поскольку они зависят от взаимодействия
всех действующих в данной местности факторов и ими, очевидно, характеризуется
окончательный эффект. Они позволяют судить о единстве, различии или подобии отдель-
ных местностей и помогают вскрыть такие пространственные зависимости, которые
каким-либо другим способом уловить трудно. Исследование причин этих зависимостей
требует, однако, предварительного знания экологии действующих факторов. Данные
геоботаники о причинных связях между характером растительности и условиями
местообитания должны поэтому пронизывать любое исследование в области географии
растительности. При изучении же экологических факторов специалист по географии
растительности часто бывает вынужден обратиться к геоботанику и в течение некото-
рого времени пройти путь совместных исследований, однако цель и направленность
этих исследований у них различна.
Специальная география растительности,
ландшафтное отображение растительного мира, имеет свои законы и пра-
во по-своему систематизировать и излагать изучаемый материал. Она
пользуется при этом понятиями и фактами, предоставляемыми в ее распо-
ряжение общей географией растительности.
Общая география растительности как наука
о заполнении земных пространств растениями строится по принципу,
позволяющему прежде всего решать стоящие перед ней задачи географи-
ческого характера. По’объему и характеру рассматриваемых вопросов
ее можно разбить на три следующие раздела: 1) основы географии расти-
тельности; 2) ландшафтно-аналитическая (топографически-ландшафтная)
география растительности; 3) хорологическая география растительности.
1. Изложение основ географии растительности носит преимуществен-
но аналитический и подготовительный характер. При этом исследуются
основные составные элементы растительного мира, их формы и причины
распространения. Эти исследования частично совпадают с исследова-
ниями, являющимися предметом идиобиологической геоботаники, дан-
ные которой должны быть отобраны и обработаны с географической точ-
ки зрения. При изложении основ географии растительности преследуется
Введение
17
не решение биологических проблем, а лишь анализ Закономерностей рас-
пределения элементов растительного мира в пространстве. Систематиче-
ские единицы растений и их формы рассматриваются как элементарные
составные части пространственной структуры биосферы и учитываются
только в той степени, в какой это необходимо для характеристики той
или иной территории. Типология растительных форм (формы роста и жиз-
ненные формы), конфигурация ареалов отдельных систематических еди-
ниц и причины их пространственного размещения — вот перечень основ-
ных проблем исследований. К ним примыкают вопросы истории и гене-
зиса флоры, имеющие, однако, подчиненное значение и привлекающиеся
лишь постольку, поскольку это необходимо для понимания современного
состояния растительного мира.
2е Ландшафтная география растительности является основой в общей
структуре географии растительности. Центральное место здесь занимают
исследования важных для данного ландшафта сообществ, их научно обос-
нованная типология и проблемы их отношений с другими составными эле-
ментами ландшафта. Географический анализ экологических связей и физио-
номических различий требует знания не только основ систематики рас-
тительных сообществ, но и знания их типологии по растительным
формациям. Не менее важной является характеристика условий место-
обитаний; одновременно следует обратить внимание на все прочие
явления, обусловливающие те или иные особенности растительного
покрова.
К указанным проблемам относятся и исследования процессов раз-
вития растительности и возможность по растительности воссоздать облик
прежних ландшафтов (палеоландшафта, палеоклимата). Историческое
рассмотрение входит, однако, в задачи географии растительности лишь
постольку, поскольку этого требует исследование современного расти-
тельного покрова. Чисто палеогеографические или биогенетические цели
в данном случае исключаются. Однако выяснение предыстории расти-
тельности зачастую оказывается решающим для понимания процессов,
происходящих в настоящее время.
Основной же целью географии растительности является анализ расти-
тельности с точки зрения характера ее воздействия на ландшафт и ее
взаимодействия с другими компонентами естественного и культурного
ландшафта (образованием почв, животным миром, возможностями посе-
ления человека, развитием и формами хозяйства). Частью этих проблем
занимается экономическая география, однако резкое разграничение здесь
едва ли возможно. Растительность на Земле в значительной степени испы-
тывает преобразующее воздействие человека и становится основной частью
культурного ландшафта.
3. Хорологическая география растительности носит более синтетиче-
ский и описательный характер. Вместе с типологией биогеографических
пространственных единиц она образует основу при построении про-
странственного деления биосферы, в которое в конце концов упирается
и общая география растительности.
ИСТОРИЯ ГЕОГРАФИИ РАСТИТЕЛЬНОСТИ
Обе науки — и география растительности и геоботаника — исто-
рически не поддаются резкому разграничению. Во всяком случае, самые
ранние сведения имеют для обеих отраслей одинаковое значение.
18
Введение
После эпохи средневековья, в которую размещение растений и животных по зем-
ной поверхности воспринималось как нечто само собой разумеющееся, конец XV в.
знаменовал собой начало новой эпохи с первыми попытками сознательного наблю-
дения природы. Результатом этого явились «Книги трав» (Brunfels, 1530; Bock, 1539;
Fuchs, 1542; Mattiolus, 1548; Dodonaeus, 1553), в которых приводились описания трав
и их зарисовки. Уже Бок указал, помимо прочих сведений, на местонахождение наблю-
давшихся им трав в Верхнерейнской области, что мы можем считать первым шагом
на пути флористической геоботаники.
Цюрихский естествоиспытатель Конрад фон Гезнер (1516—1565)
наблюдал на Гнепфштейне (Пилатус) изменение растительности с высотой.
Он различал климатические высотные пояса: зимний участок, где имеется
только одно время года (снежная область), весенний, где после долгой
зимы наступает короткая весна (альпийская область), осенний, где,
кроме зимы и весны, можно различить и осень (субальпийская область),
и наконец летний участок, где бывает теплое лето (долинная область).
Таким образом, здесь ясно виден подход с точки зрения географии расти-
тельности.
Исследования XVII в. сосредоточивались в основном на описании, наименовании
и систематизации видов. Определялся также состав растительности отдельных обла-
стей. Ознакомление с растительностью далеких стран наталкивало ученых на мысль
о возможных причинах столь неодинакового распределения растений на земле.
Рай в своей «Historia plantarum» (Ray, Rajus, 1693) посвятил целую
главу вопросу о месте произрастания. Он указал на влияние географиче-
ской широты и высоты над уровнем моря. В это же время Кристиан Мен-
дель (1622—1701) особо подчеркнул, что развитие растений находится
в зависимости от климата.
Жозеф Питтон де Турнефор (1656—1708), поднимавшийся в 1700 г.
на Арарат, в одной из своих работ охарактеризовал флористический
состав высотных поясов и сравнил его с флорой других климатических
областей и других гор. Несколько позже Альбрехт фон Галлер (1708—
1777) в своей «Альпийской поэме» (Haller, 1732) взволнованными стро-
ками, снабженными примечаниями из области ботаники, описал расти-
тельный покров гор. В этом можно видеть зародыш физиономических
представлений, которые позднее Гумбольдт довел до высокой степени
совершенства.
Галлер написал две работы о флоре Швейцарии, в которых он обратил особое
внимание на пространственное распределение растительности. В работе 1742 г. он
назвал те растения, которыми определяются границы высотных поясов в районе Грим-
зеля: каштаны, виноградные лозы, орешники, буки и дубы, ели, лиственницы, кедры
европейские, рододендроны и брусника, скудные овечьи пастбища, скалы с карлико-
выми растеньицами, глетчерные ивы и глетчерные лютики, фирн. В 1768 г. он охарак-
теризовал каждый высотный пояс сообществами произрастающей здесь растительности.
Карл Линней (1707—1778), путешествовавший по Лапландии в том
же самом году, когда вышла в свет поэма Галлера, был удивительно
многосторонним наблюдателем, обладавшим исключительной восприим-
чивостью; его интересовало все, что попадалось на пути. Многому из виден-
ного он дал впервые наименования, которые впоследствии были узаконены
в биогеографии. Места, где он находил представителей отдельных видов,
Линней характеризовал такими понятиями, как лес, болото, луг, или
какими-либо растительными сообществами. Для последних он употреблял
наименования, которыми до сих пор пользуются при обозначении расти-
Введение
19
тельных сообществ. Понятие «местообитание» Линней применял почти
в том же значении, в каком его употребляют в современной экологии.
Он считал, что различие местообитаний зависит от высоты, климата
и почвы. Он отметил также зависимость роста растений от изменения кли-
матических условий в течение года и рекомендовал регистрировать от
места к месту время распускания листьев, зацветания, созревания
плодов и листопада и на основании такого «календаря флоры» составлять
карты климатических различий.
В работах некоторых исследователей этого периода наряду с чисто
флористическими сведениями большое место занимают ботанико-геогра-
фические сопоставления. Например, Иоганн Георг Гмелин (1709—1755)
дал сравнительную характеристику флор Сибири и горных областей
Европы, Петер Симон Паллас (1741 — 1811) составил описание раститель-
ности южнорусских степей и установил зависимость ее от климатических
условий.
Накопление знаний о жизненных процессах (сокодвижение, транспирация, рост,
питание, ассимиляция углекислоты) и усовершенствование приборов, позволивших
производить сравнительные измерения температуры, привели в последней трети
XVIII в. к новому скачку в развитии географии растений, с которым связано возник-
новение экологической геоботаники.
Орас Бенедикт де Соссюр (1740—1799) и Жан Луи Жиро, более
известный под именем Сулави (1752—1813), явились первыми ботаниками,
которые путешествовали с термометром и барометром. В своем путевом
дневнике (1779—1796) Соссюр, описывая альпийскую растительность,
привел также сведения об измеренных им высотных границах отдельных
видов растений. Опубликованную в 1783 г. работу Сулави о раститель-
ности южной Франции можно рассматривать как первую попытку иссле-
довать местность с точки зрения географии растительности. Сулави
стремился установить закономерности пространственного распределения
растительности и искал их в требованиях, предъявляемых растениями
к климату, в естественном расчленении страны на отдельные ландшафты
в зависимости от климата, характера почвы и исторического развития.
Он разделил южную Францию по растительным климатам (климат апель-
синовых деревьев, маслин, виноградников, каштанов и климат альпий-
ских растений). Выделенные области он характеризовал фенологическими
и экологическими признаками и сравнивал их по существующему в них
комплексу видов.
Методы изучения растительности Александром Гумбольдтом (1769—
1859) близки к позиции Сулави. Путешествие Гумбольдта по тропическим
странам (1799—1804) явилось новым начинанием в том смысле, что впер-
вые географическая постановка вопроса была поднята до значения направ-
ляющей идеи при рассмотрении растительного мира. Гумбольдт стремился
к комплексному и всестороннему восприятию и познанию окружающего
мира. Он не руководствовался интересами какой-либо одной узкой обла-
сти науки, а подходил к изучению любого отдельного явления с точки
зрения единого целого, благодаря чему самым сильным впечатлением
от окружающего мира для Гумбольдта всегда была «полнота повсеместно
господствующей жизни». Этим взглядом ученого на природу и обуслов-
ливается тот факт, что у Гумбольдта мы встречаемся с совершенно новым
словарным составом. В центре внимания стоят у него не отдельные систе-
матические единицы (виды и роды), а весь растительный покров планеты,
разнообразно сотканный «ковер, которым богатая цветами флора покры-
20
Введение
вает обнаженное тело Земли». Растительность как явление, определяю-
щее облик каждой отдельной области земли, а тем самым и ее ландшафт,
представляет для Гумбольдта и исходный пункт, и основную цель при
исследовании растительного мира. Чтобы понять и выразить значение
растительного покрова для характера ландшафта, он предложил термин
«форма растения» (Pflanzenform). Его внимание было обращено прежде
всего на внешний облик растений, которым преимущественно опреде-
ляется физиономия растительности, а также на «географическое и кли-
матическое распространение» выделенных им 19 основных форм растений.
«Физиономика растений... должна стремиться познать законы, определяющие
физиономию природы в целом, ландшафтный характер растительности на всей земной
поверхности, то есть то живое впечатление, которое вызывает группирование кон-
трастирующих форм в различных широтных и высотных зонах».
На примере весьма обширных областей земли Гумбольдт составил
описание многих естественных ландшафтов, причем наряду с формой
растений он учитывал и образ жизни животных и человека. Кроме того,
путем широких сравнений он пытался обнаружить и понять общие черты
сходных областей Земли (саванн, степей и пустынь, пустошей), чтобы
на их основе создать пространственную классификацию растительности
и больших ландшафтных пространств Земли. Поэтому Гумбольдта можно
по праву назвать основоположником географии растительности. Прекрас-
ный популяризатор своих идей, он сумел заинтересовать этими пробле-
мами самые широкие научные круги. Благодаря Гумбольдту многие
исследователи писали свои работы по растительности выразительным
и доступным языком.
Геоботаника обязана Гумбольдту термином «ассоциация общественно живущих
видов», который впоследствии дал начало фитоценологическим исследованиям.
Одновременно с Гумбольдтом Георг Валенберг (1780—1851) вел
наряду с геоботаническими исследованиями и исследования с точки зре-
ния географии растительности. Сопоставляя растительность весьма отда-
ленных друг от друга областей, он особенно подчеркивал значение кли-
мата для распределения растений по земной поверхности. Изучая поляр-
ные и альпийские границы распространения растительности, Валенберг
нашел, что определяющим фактором здесь является не средняя годовая
температура воздуха, а сезонное распределение тепла. Кроме темпера-
туры воздуха, он учитывал также температуру почвы, прямую солнеч-
ную радиацию, осадки и влажность воздуха. В своей работе «Flora 1ар-
ponica» (1812) он различал элементы флоры по типу их ареалов. Лаплан-
дию он разделил на отдельные районы и описал их с точки зрения кли-
мата и характерных растений.
Иоахим Фридерик Скау (1787 — 1852) попытался в 1822 г. выделить области
исследований, относящиеся к географии растений, и привести их в систему. Он отделил
«историю растений» от «географии растений» в узком смысле слова. Последнюю он раз-
делил па «науку о месте растений» и на «ботаническую географию». В «ботанической
географии» Скау стремится произвести флористико-географическое расчленение зем-
ной поверхности. Своим атласом растений (Schouw, 1823) он положил основу для
создания первой ботанико-географической карты Земли. Однако работы Скау еще лише-
ны основополагающих моментов географии растительности как науки. Тем не менее
именно его работы использовали многие исследователи, путешествовавшие по далеким
странам.
Введение
21
Карл Фридрих Филипп фон Мартиус (1794—1868) описал в своем
отчете о путешествии (1823—1831) и в статье «Облик растительного мира
Бразилии» (Martius, 1824) такие сложные единицы растительного покрова
(формы растительности), как тропический дождевой лес, цдмпосы
и каатинги, по их облику, составу и условиям местообитания.
Огюст де Сент-Илер (1799—1853), который в это же время путешест-
вовал в Минас-Жераис и Гоясе, попытался провести различие между
первичной растительностью (vegetation primitive) и возникшей под влия-
нием деятельности человека вторичной растительностью, а также изучал
явление повторного заселения (сукцессии) после прекращения вмеша-
тельства человека.
Физиономические представления Гумбольдта, наполнившего жизнью
такие понятия, как «лес», «луг», «степь», «саванна», «пустыня» и т. д.,
приобрели в дальнейшем надежную поддержку в работах Августа Гри-
зебаха (1814—1879), который ввел в науку понятие «растительная фор-
мация» (Grisebach, 1838). Этот выбор физиономической единицы расти-
тельности имел для географии растительности почти столь же основное
значение, как и понятие «ландшафт» для физической географии.
География растительности получила дальнейшее развитие в XIX в.
частью в отчетах путешественников, частью в специальных исследова-
ниях о соотношениях растительности («Vegetationsverhaltnisse»), суще-
ствующих на больших или малых участках земной поверхности. Такие
работы возникали либо как «географическое введение» к большим трудам
по флоре, либо в качестве самостоятельных монографий, имеющих более
или менее ярко выраженную направленность в сторону географии расти-
тельности. Среди них труды Хеера (Heer, 1835), Унгера (Unger, 1836),
Шницлейна и Фрикингера (Schnizlein u. Frickhinger, 1848), Зендтнера
(Sendtner, 1854, 1860), Лоренца (Lorenz, 1858), Кернера фон Марилаун
(Kerner v. Marilaun, 1863 и др.), Виртгена (Wirtgen, 1864, 1865), Зенфта
(Senft, 1865), Гепперта (Goppert, 1868), Градмана (Gradmann, 1898) и др.
В более ранних работах изложение касалось в основном какой-либо
одной проблемы, как, например, влияния климата или почвы на распро-
странение растений, деления по высотным поясам и местообитаниям.
Однако внимание ученых все более привлекали проблемы, относящиеся
к географии растительности. В работах Шницлейна и Фрикингера мы
впервые находим отчетливое, характерное для географии растительности
изложение материала: условия произрастания, характер местности,
первоначальная растительность, растительность, видоизмененная чело-
веком, ландшафтные растительные формации.
Кернер уделял растительным формациям еще большее внимание.
Он проявлял большую тщательность при их физиономическом описа-
нии, а также изучал их флористический состав, экологию и изменения
во времени. Гризебах в 1872 г. дал общий обзор растительности Земли
по признаку растительных формаций; им выделены обусловленные кли-
матом пояса формаций, а также впервые обработан обширный материал,
накопленный за время путешествий, совершенных в XIX в. во все уголки
Земли (Grisebach, 1872). Работу Шимпера (Schimper, 1898) можно рас-
сматривать как продолжение трудов Гризебаха, однако с более сильным
и несколько односторонним подчеркиванием экологических проблем.
В описании «Царств растений» Геттнера (Hettner, 1935) ботанико-геогра-
фическая точка зрения становится уже главенствующей. В географиче-
ской литературе цели и задачи географии растительности получают доста-
точное освещение, в меньшей степени в теоретических трудах и все боль-
22 Введение
ше — в страноведческих работах, в отчетах о путешествиях и моногра-
фиях, посвященных отдельным проблемам. В этом отношении особенно
интересны труды Вайбеля, автора, обладавшего широким географиче-
ским кругозором
1 Излагая историю географии растительности, автор не упоминает имен русских
ученых, внесших огромный вклад в науку о растительном покрове земли и являю-
щихся в значительной степени ее основоположниками. Перечислим здесь имена неко-
торых из них: С. П. Крашенинников (1711—1755), Ф. И. Рупрехт (1814—1870),
А. Н. Бекетов (1825—1902), Д. И. Литвинов (1854—1929), И. К. Пачоский (1864—1942),
С. И. Коржинский (1861—1900), Г. И. Танфильев (1857—1928), А. Я. Гордягин (1865 —
1932), Г. Ф. Морозов (1867—1920), В. В. Алехин (1882—1946) и многие другие.—
Прим. ред.
Часть первая
СОСТАВНЫЕ ЭЛЕМЕНТЫ
РАСТИТЕЛЬНОСТИ
И ПРИНЦИПЫ ИХ РАСПРОСТРАНЕНИЯ
26
Составные элементы растительности и принципы их распространения
типов:
— бактерии.
— сине-зеленые водоросли.
ций, охватывает около 90 родов. Pinus — единственный род в подсемействе Pinoideae
семейства Pinaceae. Прочие подсемейства — это Laricoideae, к которому среди прочих
относятся роды лиственницы (Larux) и кедра (Cedrus), а также подсемейство Abietoideae,
включающее среди прочих роды пихты (Abies) и ели (Picea). С девятью другими семей-
ствами (например, Cupressaceae, Araucariaceae) семейство Pinaceae образует порядок
(ряд) хвойных Coniferae. Этот порядок вместе с известным только в ископаемом виде
порядком Cordaitales относится к классу Coniferopsida, одному из четырех классов,
входящих в тип Gymnospermae.
Система растений содержит, по Мельхиору и Вердерману (Melchior, Werder-
mann, 1954), всего 17
1. Bacteriophyta
2. Cyanophyta
3. Glaucophyta.
4. Myxophyta
5. Euglenophyta
6. Pyrrophyta
7. Chrysophyta
8. Chlorophyta
9. Charophyta
10. Phaeophyta
11. Rhodophyta
12. Fungi
13. Lichenes
14. Bryophyta
15. Pteridophyta
16. Gymnospermae
17. Angiospermae
Раньше различались только четыре типа. Типы 1—13 объединялись в один тип
таллофитов, а 16 и 17 — в тип сперматофитов (семенных растений).
— слизевики, или миксомицеты.
— эвгленовые водоросли.
— пиррофитовые водоросли.
— хризомонадовые, или золотистые, водоросли.
— зеленые водоросли.
— харовые водоросли.
— бурые водоросли.
— красные водоросли.
— грибы.
— лишайники.
— мохообразные.
— папоротникообразные.
— голосеменные.
— покрытосеменные, или цветковые.
В рамках географического рассмотрения отдельные типы имеют раз-
личное значение. Как определяющие физиономию ландшафтов и как
полезные растений наиболее важными являются последние пять типов.
Однако и типы низших растений играют в общем балансе биосферы нема-
лую роль (например, бактерии и грибы), а для жизни водных пространств
немалое значение имеют водоросли.
АРЕАЛЫ СИСТЕМАТИЧЕСКИХ ЕДИНИЦ
Распространение растений на Земле можно рассматривать как под
углом зрения заполнения земных пространств единицами растительности,
так и с точки зрения распределения отдельных систематических единиц.
Первый порядок рассмотрения принимается географами и фитоценоло-
гами, а второй — систематиками растений и флористами.
При изучении распределения отдельных систематических единиц
решающее значение имеет познание областей обитания тех или иных видов
растений. Точку, в которой обнаружено растение, называют местом
нахождения того вида, к которому относится растение. Если
все эти точки нанести на карту, то можно получить наглядное представле-
ние о распределении данного вида (рис. 2). С абсолютной полнотой это
удается, однако, сделать лишь в редких случаях. Для широко распро-
страненных видов при описании местных флор местонахождение зачастую
Систематические единицы и их распространение
27
Рис. 2. Распространение седмичника (Trientalis еигораеа) в Средней Германии.
Пример карты местонахождений (Meusel, 1953—1954). *
указывается очень приблизительно, в таких общих выражениях, как
например, «часто» или «рассеянно». Поэтому на картах в виде сплошной
площади можно представить только те области, в которых местонахожде-
ния более или менее многочисленны. При этом оказывается возможным
выделить участки, на которых данный вид не встречается.
Картирование отдельных видов в Германии начал Гофман (Hoffman, 1860, 1865,
1867—1869) для бассейна Среднего Рейна. Для других частей Германии флористиче-
ское картирование выполнялось Шульцем (Schulz, 1887), Абромейтом (Abromeit, 1892,
1900), Вангерином (Wangerin, 1915), Эйхлером — Градманом — Мейгеном (Eichler —
Gradmann — Meigen, 1905—1927). Картирование для Германии в целом в масштабе
1 * 25 000 было предпринято Маттфельдом.
Область, объединяющая все местонахождения, то есть площадь,
занимаемая всей совокупностью существующих в настоящее время инди-
видуумов, принадлежащих одному виду, называется ареалом дан-
ного вида. Точно так же можно говорить и об ареалах более мелких или
более крупных систематических единиц, например об ареале подвида
или семейства. Карты ареалов по возможности большего количества
систематических единиц (в идеальном случае — картирование ареалов
всех единиц разного ранга в пределах существующих родственных свя-
зей) являются важнейшим рабочим материалом флористической геобо-
таники и представляют большой интерес для географии растительности.
Друде в своем атласе распространения растений (Drude, 1887) впервые довольно
наглядно сопоставил большое число ареалов различных растений. Для некоторых стран
28 Составные элементы растительности и принципы их распространения
(Скандинавии, Нидерландов, Англии) существуют обзорные флористические карты,
не говоря уже о многочисленных отдельных набросках в специальных трудах или
в общих работах по геоботанике. Тщательно выполненные оригинальные карты имеют-
ся в сборном труде «Ареалы растений» (Hanning, Winkler, 1926—1938) и в «Сравни-
тельном учении об ареалах» Мёйзеля (Meusel, 1943).
В простейшем виде карты ареалов содержат только контуры области распростра-
нения какой-либо систематической единицы. Но хотя такие карты и являются лишь
схематическим изображением фактического размещения, они все же имеют большое
значение для общего представления. Для более подробного изучения ареалов требуют-
ся, однако, тщательно выполненные карты, составленные или точечным методом, или
методом комбинированного нанесения точек и площадей, позволяющих видеть разме-
щение отдельных местонахождений в пределах самого ареала.
Формы ареалов бесконечно разнообразны. Едва ли можно найти
полностью совпадающие ареалы двух разных видов. Классификация
форм ареалов может быть проведена
по разным признакам.
Замкнутым (сплошным) назы-
вают ареал, занимающий целостную
площадь. Конечно, и в этом случае
растения соответствующего вида рас-
пределены по всей площади неравно-
мерно. Каждый вид предъявляет свои
требования к местообитанию, удов-
летворить которое вряд ли может лю-
бая точка площади в пределах ареала.
Принято называть ареал сплошным,
если данный вид может преодолеть
имеющиеся пробелы с помощью своих
естественных средств распростране-
ния.
О разорванном, дизъюнктивном
ареале говорят, если отдельные его
части настолько удалены друг от дру-
га, что о природном переносе семян
через разделяющие промежутки не
может быть и речи. Многие разорван-
ные ареалы состоят из одной сплош-
Р и с. 3. Распространение вереска ной обитаемой части и большего или
(Erica arborea) как пример разорванного меньшего числа отдельных местона-
ареала (по Рикли). хождений представителей данного
вида (рис. 3). Между абсолютно
сплошными ареалами и ареалами, полностью разорванными, состоящими
лишь из отдельных местонахождений, существуют самые различные
переходы.
Как форма, так и величина ареалов весьма разнообразны. Суще-
ствуют виды, которые были найдены всего лишь в одном месте, но известны
и весьма широко распространенные.
Систематические единицы, местонахождения которых обнаружены
почти по всей земной поверхности, называются космополитами. Но и они,
строго говоря, распределены не повсеместно, поскольку их ареалы обла-
дают большим количеством пробелов. Не касаясь низших типов расти-
тельного мира, среди которых встречаются некоторые особенно быстро
Систематические единицы и их распространение
29
распространяющиеся группы, к таким космополитам относятся прежде
всего водные и болотные растения, ареалы которых охватывают всю
Землю, например рдест, ряска, жерушник и тростник. По де Кандолю,
из высших растений только у 19 видов, в том числе у однолетнего мят-
лика и обыкновенного орляка, ареал занимает более половины земной
поверхности. Примерно у 100 видов растений ареалы охватывают треть
нашей планеты.
Большинство ареалов видов ограничено небольшими участками
поверхности суши. Некоторые охватывают целые континенты, но при-
Р и с. 4. Ареал космополитного рода Drosera (росянка) (по Дпльсу, Ирмшеру, Гуду).
держиваются лишь определенных климатических поясов или специфи-
ческих местообитаний, например морских побережий. Другие располо-
жены в резко ограниченных участках земной поверхности, например
вереск крестолистный (эрика) (Erica tetralix) встречается лишь в океани-
ческих районах при атлантической Европы.
Ареалы родов, если говорить не о монотипных, то есть состоящих
только из одного вида, занимают, как правило, большее пространство,
чем ареалы видов. Космополитными родами являются, например Poly-
podium и Drosera (рис. 4). Примерно 750 родовых ареалов располагаются
в нескольких частях света одновременно, однако основная часть, как
и большинство видовых ареалов, ограничена каким-либо одним континен-
том. Генетически древние роды имеют либо очень маленькие, либо чрез-
вычайно обширные ареалы, но всегда разорванные, например ареалы
некоторых родов хвойных.
Некоторые семейства растений встречаются почти во всех флористи-
ческих областях. Широко распространены, например, семейства Grami-
пеае, Liliaceae, Compositae и др. Однако существуют семейства с очень
узким ареалом, в том числе и монотипные семейства, для которых ареал
вида и ареал семейства совпадают, например Welwitschia (Намиб) и Aexto-
xlcum (Чили). Соседние, более или менее исключающие друг друга ареалы
30 Составные элементы растительности и принципы их распространения
близких между собой таксонов называют викарирующими ареалами
(рис. 5).
С величиной ареалов связано понятие эндемизма. Оно отно-
сится всегда к одному определенному пространству, например горной
системе, острову, стране, части света или флористической области. Систе-
матическая единица называется эндемичной для данной области, если ее
ареал не выходит за пределы этой области. Так, в Альпах существуют
эндемичные виды для отдельных горных вершин (локальные эндемы).
Медовая пальма (Jubaea spectabilis) эндемична для западной части сред-
него Чили. В Бразилии существует более 12 тыс. эндемичных видов.
Из всех частей света Австралия насчитывает наибольшее число эндемич-
ных родов и семейств. Такие эндемы, относящиеся к высшим система-
тическим единицам, являются важными опорными точками для флори-
стико-географического деления земной поверхности.
Р и с. 5. Ареалы викарирующпх подвидов и разновидностей Minuartia peploides
(по Матфельду).
la — subsp. latifolia var. diffusa; 10 — subsp. latifolia var. typica; 2 — subsp. robusta;
3 — subsp. major.
Эндемизм какой-либо систематической единицы может быть обуслов-
лен тем, что данная единица возникла именно в этой области и по каким-
то причинам не смогла распространиться дальше. В этих случаях говорят
о «неоэндемах», или «прогрессивных эндемах». Острова и высокогорья
часто весьма богаты неоэндемами с резко ограниченным распростране-
нием. На Канарских островах почти половина всех видов эндемична,,
на Гавайях — более 75%, на острове Св. Елены — 85% видов эндемичны.
Многие эндемичные виды возникли отнюдь не «прогрессивным» спо-
собом, в результате новообразований в тех областях, где они встречаются
в настоящее время, а вследствие редукции их ареалов, которые и являются
остаточными ареалами часто весьма древних систематических единиц.
Их современное распространение не имеет уже никакой связи с их преж-
ней областью распространения, которая может располагаться совсем
в другой части земной поверхности. В таких случаях говорят о «регрес-
сивных», «консервативных» или «реликтовых» эндемах. Например Ginkgo'
biloba, занимающий в настоящее время очень ограниченный ареал в Ки-
тае, в раннем третичном периоде был широко распространен по всему
северному полушарию, а область возникновения этого вида так и остается
неизвестной.
Различие ареалов по форме и величине может быть в полной мере
осмыслено только с учетом географического положения конкретных ареа-
лов. Эти три критерия, вместе взятые, составляют основу понятия «тип
Систематические единицы и их распространение
31
ареала» (Wangerin, 1932; Meusel, 1943), которое позволяет систематизи-
ровать бесчисленное количество отдельных ареалов. Из большого числа
ареалов или совокупности ареалов, обладающих сходным положением,
формой и величиной, можно вывести типы ареалов, классифицируя их по
характерному пространственному принципу. К одному и тому же типу
ареалов могут относиться систематические единицы разного ранга (виды,
группы видов, роды и т. д.). Дать наименование типам ареалов довольно
трудно. Наиболее простым было бы их обозначение по (названию той
систематической единицы, ареал которой соответствует этому типу, напри-
мер тип ареала Fagus silvatica. Однако это не дает основы для создания
наглядной системы типов ареалов. Тот же недостаток обнаруживается
в обозначении типов ареалов по странам или горам (например, герцин-
ский, альпийский). Мёйзель (Meusel, 1943) попытался взять за основу
Р и с. 6. Распространение рода Larix (лиственницы). Ареал бореально-горного
пояса (по Остенфельду и Сирах-Ларсену).
номенклатуры «естественную» систему. В качестве главного принципа
классификации выступает тогда деление Земли на такие флористические
области, которые хорошо определяются границами ареалов многочис-
ленных систематических единиц высшего ранга.
Мёйзель проиллюстрировал свои предложения на примере голаркти-
ческого флористического царства. Основное деление на пояса ареалов
основано на зональном подразделении и на том факте, что многие систе-
матические единицы, распространенные в более северных поясах на рав-
нине, на юге имеют еще часть ареала, приуроченную к горным поясам.
Мёйзель различает:
1. Аркто-альпийский пояс ареалов. Сюда относятся типы ареалов,
основная часть которых находится в равнинных условиях арктической
зоны, но которые частично могут быть размещены и в горных местностях
более южных зон.
2. Бореально-горный пояс ареалов с ареалами, основная часть
которых находится в северной (бореальной и суббореальной) области уме-
ренной зоны, но которые частично простираются и до горных районов
более южных зон (рис. 6).
3. Бореально-меридиональный горный пояс ареалов с ареалами,
основная часть которых находится на равнинах южной части умеренной
зоны, но которые могут распространяться до горных районов, относя-
щихся к более южному поясу.
4. Меридионально-холмовой пояс ареалов, охватывающий те ареалы,
которые ограничены меридиональными (средиземноморскими и субсреди-
земноморскими) зонами.
32 Составные элементы растительности и принципы их распространения
Во вторую очередь Мёйзель учитывает широтное различие ареалов.
В зависимости от положения ареала — в океанических областях или во
внутренних континентальных районах и в переходных областях — он
Рис. 7. Распространение глетчерного
лютика (Ranunculus glacialis) как пример
ареала аркто-альпийско-океанического
круга типов ареалов (по Гамсу).
делит пояса ареалов на круги
типов ареалов и, наконец, на от-
дельные типы ареалов, которые
обозначаются в соответствии со
специальной флористико-геогра-
фической классификацией (рис. 7).
Круги типов ареалов образуют
систему координат из географиче-
ской широты, высоты над уровнем
моря и континентальности поло-
жения; путем соответствующей
комбинации принятых обозначе-
ний можно выразить положение
и протяженность типа ареала. Для
специальной характеристики ареа-
ла и отнесения его к тому или
иному типу ареалов, по Мёйзелю,
необходимо учитывать положение
центров происхождения и наиболь-
шей густоты скопления системати-
ческих единиц, тогда как общая
величина ареала часто оказыва-
ется не столь существенной. При
этой системе охватывается вся область распространения, на чем настаивал
еще Вангерин (Wangerin, 1932), а не только одна часть ареала, и в то же
время она дает возможность подразделять типы ареалов с любой точ-
ностью. Недостатком здесь является то, что последовательное применение
Р и с. 8. Биполярный ареал рода Empetrum (по Гуду).
Систематические единицы и их распространение
33
этой схемы приводит к слишком сложным названиям типов ареалов.
К хорологической классификации систематических единиц с более про-
стыми наименованиями можно перейти, если «элементы флоры» (Christ,
1867; Braun-Blanquet, 1919; Клеопов, 1941; Walter. 1954) называть по
ботанико-географическим регионам или флористическим областям, в кото-
рых данные систематические единицы наиболее широко распространены.
При этом, однако, охватить весь ареал целиком зачастую не удается.
Особый интерес с точки зрения истории флоры представляют собой
биполярные ареалы систематических единиц, которые встречаются в более
высоких широтах обоих полушарий, а в тропическом поясе отсутствуют.
Примерами из более чем 60 родов с таким типом распространения являются
Viola, Papaver и Empetrum (рис. 8). Примером семейства с биполярным
ареалом является семейство Fagaceae.
ФЛОРИСТИЧЕСКОЕ ДЕЛЕНИЕ ЗЕМЛИ
Если вид растения рассматривать в качестве основной систематиче-
ской единицы, то ареалы видов будут конкретными пространственными
единицами флористического деления Земли. Большое число ареалов
после их систематизации по типам ареалов становится легко обозримым,
что открывает дорогу к флористическому делению Земли. Типы ареалов
становятся при этом абстрактными элементарными единицами простран-
ственной классификации. Уже давно всю совокупность систематических
единиц, относящихся к одному и тому же типу ареалов, называют «бота-
нико-географическим элементом» (Braun-Blanquet, 1919) или «элементом
флоры».
Правда, в настоящее время точно известны ареалы лишь для части
видов растений. Поэтому их отнесение к какому-либо элементу флоры
оказывается в некоторых случаях довольно неопределенным, а подчас
оно и совсем невозможно. Тем не менее понятию «элемент флоры» мы
многим обязаны как некоторой основе при хорологической классифи-
кации систематических единиц.
Самые ранние, считающиеся исходными, представления о флористи-
ческом делении Земли были получены в ходе упорного труда многих
поколений исследователей главным образом по установлению границ
ареалов видов, родов или семейств, рассматривавшихся в качестве харак-
терных для той или иной местности.
Однако для большей части земной поверхности вышеупомянутые
представления основаны больше на методах сравнительной флористиче-
ской статистики. По флористическим спискам, которые составлялись
на протяжении трех или четырех столетий для стран, провинций или даже
произвольно ограниченных областей (например, следуя маршрутам экспе-
диций), установлено, какие части суши по составу систематических еди-
ниц более или менее сходны и где проходят основные границы между
пространствами, населенными систематически отличающимися друг от
друга единицами. Исследования «флористического контраста» (Floren-
kontrast), измеряемого количеством различных видов в двух сравнивае-
мых областях, и «флористического градиента» (Florengefalle) (число
контрастов на 100 км) позволяют установить степень флористического
различия, а также положение и точность границы между этими областями.
При определении флористических границ значительно более сущест-
венными, чем количественные, являются качественные различия в со-
U4:
составные элементы растительности и принципы их распространения
ставе систематических единиц. Одно дело, если две области различаются
только отсутствием общих видов, и другое — если в одной из областей
встречаются те роды или семейства, которые отсутствуют во второй.
Для оценки степени флористической самостоятельности большое значе-
ние имеет и то, богата или бедна область эндемами и к какому рангу
систематических единиц они относятся.
Картина распределения флоры, полученная таким путем, явилась вместе с дан-
ными систематики, генетики и палеоботаники основанием для воссоздания истории
развития растительного мира. Эта последняя в свою очередь внесла свой вклад в иссле-
дование распространения отдельных систематических единиц. В настоящее время изу-
чение отдельных ареалов происходит не изолированно, а по возможности во взаимо-
связи с ареалами более высоких по рангу систематических единиц, так, например,
рассматриваются ареалы всех разновидностей одного вида, ареалы всех видов одного
рода или всех родов одного семейства. Таким путем легче выявить причины образова-
ния ареалов.
Результаты многих попыток пространственно-флори-
стического деления Земли (Schouw, 1823; De Candolle,
1855; Engler, 1879-1882; Drude, 1884, 1887, 1890; Hemsley, 1888; Diels,
1908; Rikli, 1913; Hayek, 1926; Good, 1951 и др.) совпадают далеко не
полностью. Существует единство во взгляде на принцип иерархического
деления на флористические царства, регионы, провинции, округа, районы.
Однако уже эти названия рангов подразделений флористических областей
у разных авторов не идентичны.
Различия существуют и в самом принципе объединения низших единиц в высшие.
Они зависят прежде всего от того, преобладают ли при этом флористико-статистиче-
кий (Скау), ареал-тип ологический или историко-генетический (Энглер) методы или
же, как это имело место в ряде случаев, превалирует смешение ботанико-физиономиче-
ской и флористико-хорологической точек зрения. Пространственно-флористическое
деление не должно, однако, принимать во внимание растительные формации, а исхо-
дить только из распространения таксономических единиц (Engler, 1899; Hayek, 1926).
Только таким образом можно определить, в какой степени размещение пространст-
венно-флористических единиц совпадает с распространением растительных формаций,
обоснованных физиономическими и экологическими критериями. Можно ли совместить
оба вышеуказанных принципа при делении земной поверхности с точки зрения гео-
графии растительности — это уже другой вопрос.
Подразделение пространственно-флористических единиц, начиная
с флористически более или менее однородных районов и кончая флористи-
ческими царствами, не зависит от величины этих единиц. Оно основано
на том, различается ли флора каких-либо местностей только по система-
тическим единицам низшего ранга (разновидности, подвиды, виды) или
одновременно и по родам и по семействам. Тем самым оно соответствует
и степени самостоятельности историко-флористического развития отдель-
ных областей.
Основываясь на данных, разработанных в основном Энглером, Друде
и Дильсом, различают шесть флористических царств (рис. 9):
1. Голарктическое (северное вне тропическое) флористическое цар-
ство (Holarktis).
2. Неотропическое (тропическое флористическое царство Нового
Света) (Neotropis).
3. Палеотропическое (тропическое флористическое царство Старого
Света) (Palaeotropis).
Систематические единицы и их распространение
35
4. Австралийское флористическое царство (Australis).
5. Капское флористическое царство (Capensis).
6. Антарктическое флористическое царство (Antarktis).
Р и с. 9. Флористические царства Земли (по Дильсу).
Существуют разные точки зрения на вопрос, куда отнести Новую Зеландию,
Сахару и как разграничить пеотропическое и голарктическое царства.
«Главные разделяющие линии наземных флор» Друде (Drude, 1890) приняты почти
повсюду и сейчас, за исключением северной границы австралийского царства (Дильс).
Однако Друде применял понятие «флористическое царство» для 15 единиц низшего
ранга, которые он сводил в 6 или в 3 «группы флористических царств». Он распре-
делил 240 семейств семенных растений по бореальной (голарктической), тропической
(пеотропическая и палеотропическая) и австралийской (австралийская, капская и ан-
тарктическая) «группам флористических царств». По его данным, 38% этих семейств
встречаются во всех трех, 7 — в двух и 55% — только в одной из трех указанных
групп. Из последних 30% падают на бореальную группу, 53 — на тропическую и 17% —
на австралийскую «группу флористических царств».
ГОЛАРКТИЧЕСКОЕ ФЛОРИСТИЧЕСКОЕ ЦАРСТВО
(HOLARKTIS)
Голарктическое царство, называемое в географии животных «арктогей-
ским», охватывает большую часть северного полушария. Его южная гра-
ница с тропическими флористическими царствами проходит от северной
оконечности полуострова Нижняя Калифорния вдоль края Мексикан-
ского плоскогорья и пересекает южную часть полуострова Флорида.
Она захватывает острова Зеленого Мыса, пересекает Сахару и Аравий-
скую пустыню, прослеживается вдоль южной границы Иранского пло-
скогорья, вдоль Гималаев, пересекает Южный Китай и примерно между
северной границей провинции Фуцзянь и устьем реки Янцзы сходит
36 Составные элементы растительности и принципы их распространения
с континента. В Тихом океане эта граница проходит примерно по 30° с. ш.,
так что острова Рюкю, Огасавара и Гавайская группа остаются вне его.
Южная граница голарктического царства отделяет друг от друга
большинство ареалов семейств, частью полностью, частью по преобла-
дающему распространению их представителей. По одну сторону остаются
типично тропические семейства, которые вне тропической зоны не встре-
чаются совершенно или встречаются как исключение, а по другую —
семейства, которые, как правило, отсутствуют в тропиках, а распростра-
нены преимущественно в голарктическом царстве, например Асегасеае,
Gampanulaceae, Garyophyllaceae, Gruciferae, Cupiliferae, Ranunculaceae,
Рис. 10. Распространение морской травы Tostera marina как пример голарктиче-
ского ареала (по (Эстенфельду).
Rosaceae, Salicaceae, Saxifragaceae, Sparganiaceae, Umbelliferae. Ареал
некоторых из этих семейств биполярный; однако отдельные роды их
встречаются и во внетропической области южного полушария (рис. 8).
Единство голарктического царства проявляется в том, что не только
ареалы семейств (например, Асегасеае) и ареалы родов (например, Betula)
занимают большую часть всего пространства, но и в том, что некоторые
виды распространены как в Старом, так и в Новом Свете, например
Cardamine pratensis, Athyrium filix femina и Zostera marina (рис. 10).
Голарктика подразделяется на следующие флористические области (регионы,
Florenregionen), в каждой из которых в свою очередь, как правило, можно различить
несколько провинций:
а) Аркто-субарктическая.
б) Бореальная область с евро-сибирской и американо-бореальной подобластями
в) Восточноазиатская.
г) Тихоокеанско-североамериканская.
д) Атлантическо-североамериканская.
е) Среднеевропейско-южносибирская.
ж) Средиземноморская.
з) Центральноазиатская.
НЕОТРОПИЧЕСКОЕ ФЛОРИСТИЧЕСКОЕ ЦАРСТВО
(NEOTROPIS)
Неотропическое царство, соответствующее «неогейскому царству»
в географии животных, охватывает территорию Центральной Америки,
включая полуостров Калифорния, окраины Мексиканского плоскогорья,
юг полуострова Флорида, а также Южную Америку, за исключением
Систематические единицы и их распространение
37
южной трети Чили, небольшой части южной Аргентины и Фолклендских
островов. Большинство семейств, распространенных в неотропическом
царстве, является общим с тропиками Старого Света. Примерами таких
Рис. И. Распространение неотропического семейства Tropaeolaceae (по Гуду).
Рис. 12. Распространение пантропического рода Sapium (Euphorbiaceae) (по Паксу).
пантропических ареалов могут быть ареалы семейств Zingiberaceae,
Gesneriaceae, Ebenaceae и Hymenophyllaceae (Meusel, 1943), а также
Могасеае и Cycadaceae. Однако существуют и специфичные неотропиче-
ские семейства (рис. 11). Среди них небольшие, с малым числом родов
38
Составные элементы растительности и принципы их распространения
и видов семейства, как Marcgraviaceae и Nolanaceae, а также богатые
видами Bromeliaceae и Cactaceae. Последнее представлено несколькими
видами на Мадагаскаре и на африканском континенте. Семейство Loa-
saceae с несколькими сотнями видов ограничено Новым Светом, однако
один монотипный род (Kissenia) встречается и в Африке.
Количество неотропических родов, подродов и секций весьма велико;
40% всех встречающихся в тропиках родов характерно только для Нового
Света, 47% ограничено Старым Светом и только 13% встречаются в обеих
частях (рис. 12).
Виды, распространенные и в тропиках Старого Света, являются по
отношению к общему числу видов исключениями, поэтому выделение нео-
тропического царства в качестве самостоятельного флористического
царства вполне обоснованно. Этому соответствует и дальнейшее его деле-
ние на флористические области, некоторые из которых особенно богаты
эндемичными видами и родами.
Неотропическое флористическое царство можно разделить на следующие фло-
ристические области:
а) Нижнекалифорнийско-мексиканская.
б) Карибская.
в) Гвианская,
г) Амазонская,
д) Юго-восточнобразильская,
е) Флористическая область Ла-Платы,
ж) Флористическая область Анд (с большим числом подобластей и участков),
з) Флористическая область островов Хуан-Фернандес.
ПАЛЕОТРОПИ ЧЕСКОЕ ФЛОРИСТ И ЧЕСКОЕ ЦАРСТВО
(PALAEOTROPIS)
Палеотропис — второе по величине и наиболее богатое по количе-
ству видов флористическое царство Земли. Оно охватывает Африку, за
исключением ее северной окраины и Капской области, затем Южную
Азию до границы с голарктикой и тропические и субтропические острова
южной Атлантики, Индийского и Тихого океанов, за исключением Тас-
мании, Галапагосских островов, группы островов Хуан-Фернандес и остро-
вов у побережья Австралии и Америки. Принадлежность Новой Зелан-
дии к этому флористическому царству оспаривается. Друде (Drude, 1890)
проводил «главную разделяющую линию» через Южный остров, большую
часть которого он относил к антарктическому царству. Дильс, Хайек,
Фирбас и др. относят оба острова Новой Зеландии к палеотропическому
царству, тогда как Гуд (Good, 1951) всю эту группу включает в антаркти-
ческое флористическое царство. Многие пантропические семейства, напри-
мер уже упоминавшиеся для неотропического флористического царства
Cycadaceae, Ebenaceae, Могасеае и Zingiberaceae, представлены в Старом
Свете гораздо большим числом видов. Специфически палеотропическими
являются среди других семейства Dipterocarpaceae, Nepenthaceae, Pan-
danaceae (пример ареала палеотропического рода представлен на рис. 13).
По общему характеру флоры палеотропическое флористическое
царство делится на две основные части: индо-африканскую и малезий-
скую. Первая охватывает, помимо тропической Африки и Мадагаскара,
еще и Западную Индию. Характерным для этой части типом ареала, рас-
пространяющимся на всю индо-африканскую часть палеотрописа, за
Систематические единицы и их распространение 39
исключением тропического леса Западной Африки, является, например,
ареал трибы Stapelieae из Asclepiadaceae (рис. 14). Малезийская часть
Рис. 13. Распространение палеотропического рода Ancistrocladus (Ancistroclada-
сеае) (по Хетчинсону)<
включает тропические районы Восточной Индии с Индонезией, Мелане-
зию и Полинезию. Западная Индия и Мадагаскар занимают промежу-
точное между обоими подцарствами положение. Это становится ясным,
Р и с. 14. Распространение Stapelieae (по Гуду).
В числителе — число родов, в знаменателе — число видов.
если, например, сравнить типы ареалов, представленные родами Nipa,
Cinnamomum и Nepenthes. Род пальм Nipa ограничивается Восточной
Индией и Индонезией (см. карту ареалов, рис. 29). Ареал Cinnamomum
(Lauraceae) простирается далеко за эти районы на Южный Китай,
Меланезию и северное побережье Австралии, захватывая территорию
Западной Индии. Род Nepenthes, к которому относятся известные плото-
40
Составные элементы растительности и принципы их распространения
ядные растения, представлен большим числом видов на том же простран-
стве, однако, помимо того, один вид его встречается на Сейшельских остро-
вах и один — на Мадагаскаре (рис. 15).
Р и с. 15. Распространение рода Nepenthes (по Хстчпнсопу).
В семействах, ареал которых охватывает обе части палеотрописа,
большее число видов, как правило, ограничено одной из них. Так, напри-
мер, наибольшее распространение представители семейства Dipterocar-
расеае имеют в малезийской части.
Палеотропическое флористическое царство может быть разделено на следующие
флористические области:
Индо-африканский палеотропис
а) Североафриканско-индийская (Хайек и Фйрбас относят ее к голарктике).
б) Судано-замбезийская.
в) Конго-гвинейская.
г) Мадагаскарская.
д) Западноиндийская.
е) Флористическая область островов Св Елены — Вознесения (Асценсион).
Малезийский палеотропис
ж) Синомалайская.
з) Индонезийская.
и) Меланезийско-микронезийская.
к) Полинезийская.
л) Гавайская.
м) Североновозеландская (Гуд относит ее к антарктическому флористическому
царству).
АВСТРАЛИЙСКОЕ ФЛОРИСТИЧЕСКОЕ ЦАРСТВО
(AUSTRALIS)
Австралийское флористическое царство охватывает всю Австралию
с Тасманией. Из всех флористических царств оно наиболее своеобразно.
Примерно только 15% из встречающихся в нем видов можно найти и в дру-
Систематические единицы и их распространение
41
гих флористических царствах. Роды здесь тоже большей частью эндемич-
ны. Преобладающие в этом флористическом царстве семейства по типу
их ареалов могут быть разделены
на четыре группы:
а) семейства с преимущест-
венно австралийским распростра-
нением, например Casuarinaceae;
б) семейства с преимущест-
венно или исключительно палео-
тропическим или общим тропи-
ческим и субтропическим рас-
пространением, как Rutaceae и
Myrtaceae, последнее с специфи-
чески австралийскими или авст-
рало-меланезийскими родами,
исключительно богатыми видами,
как Eucalyptus (рис. 16) и Mela-
leuca-,
в) семейства с преимущест-
венно или исключительно австра-
ло-антарктическим или широким
дизъюнктивным распространени-
ем В ЮЖНОМ полушарии, как Рис. 16. Распространение рода Eucalyptus-
Proteaceae и Araucariaceae. В се- (по Гуду).
мействе Proteaceae 2/3 из 1000 ви-
дов ограничены Австралией, х/4 — Южной Африкой. Примерно по 2—3%
встречаются в Новой Каледонии,Восточной Азии и Южной Америке (рис. 17);
г) семейства с широким распространением по всему миру, как Сош-
positae, Gramineae, Liliaceae, однако со многими своеобразными австра-
лийскими родами, как травяные деревья из рода Xanthorrhoea (Liliaceae),
или представленные большинством видов в Австралии, как Droseraceae.
Р и с. 17. Распространение Proteaceae (по Гуду)ч
Центры скопления затемнены.
42 Составные элементы растительности и принципы их распространения
Австралийское флористическое царство может быть разделено на следующие
флористические области:
а) Юго-западноавстралийская.
б) Эремейская.
в) Юго-восточноавстралийско-тасманийская.
г) Северо-восточноавстралийская.
д) Новокаледонская (Гуд относит к палеотропическому царству).
КАПСКОЕ ФЛОРИСТИЧЕСКОЕ ЦАРСТВО
(CAPENSIS)
Самое маленькое флористическое царство — капское — насчитывает
удивительно большое число видов цветковых растений — более 6000.
В высшей степени самостоятельное развитие его систематических единиц,
с огромным числом эндемичных, позволяет выделить этот район суши
Рис. 18. Распространение Restionaceae (по Гуду).
в самостоятельное флористическое царство. При этом типы ареалов пре-
имущественно представленных здесь семейств указывают больше на
родственность с австралийской и австрало-антарктической флорой, нежели
с палеотропической африканской. Семейство Proteaceae представлено
родами Protea, Leucodendron и другими родами, присущими Южной
Африке, которые охватывают четверть всех видов, входящих в это семей-
ство. Из семейства Restionaceae 3/4 всех видов встречаются в Южной
Африке, а остальные в австралийских областях (рис. 18). Даже идентич-
ные виды Южной Африки одни и те же: с Австралией и Новой Зеландией,
например, общим является древовидный папоротник Todea barbar a (Osmun-
daceae). Род Olea связывает также капское царство с Новой Зеландией,
а с другой стороны — с европейской средиземноморской областью.
Эндемичными семействами капского флористического царства
являются, например, Репасеасеае и Geissolomaceae. Родственные им
Систематические единицы и их распространение
43
Oinaceae встречаются также в Восточной Африке; Hydrostahyaceae рас-
пространено, кроме Южной Африки, на Мадагаскаре, как и род Phylica
(Rhamnaceae), представителей которого можно найти на островах Три-
стан-да-Кунья и Новый Амстердам. Ко многим широко распространенным
семействам относятся роды, более или менее ограниченные капским фло-
ристическим царством, но порой весьма богатые видами, как, например,
Aizoaceae (Mesembryanthemum), Amaryllidaceae (Amaryllis, Clivia),
Asclepiadaceae (Stapelia), Iridaceae (Ixia, Freesia), Liliaceae (Haworthia,
Gasteria, Aloe).
Флористически капское царство подразделяется па флористические провинции
или округа, которые характеризуются многими локальными эндемами, ограниченными
в большинстве случаев весьма узкими ареалами.
АНТ АРЕТ ИЧЕСЕОЕ ФЛОРИСТ И ЧЕСЕОЕ ЦАРСТВО
(ANTARKTIS)
Кроме антарктического континента, на котором очень мало расте-
ний, к этому царству относятся острова внетропических южных морей,
самая южная часть американского континента (Огненная Земля, Магел-
ланов пролив, Западная Патагония) и небольшая часть Новой Зеландии.
Дильс включал всю Новую Зеландию в палеотропическое царство, тогда
как Гуд относит оба острова к Антарктике.
«Антарктический элемент» (Hooker, 1847) объединяет таксоны разных
рангов. Некоторые довольно широко распространены, другие, как,
например, встречающееся в южном Чили семейство Myzodendraceae,
ограничены очень небольшими ареалами. К антарктическим родам отно-
сятся, в частности, Асаепа, Aristotelia, Azorella, Gunnera, Astelia, Coprosma,
Donatia, Drapetes, Myzodendron, Nothofagus, Pernettya, Pratia, Tepualia
и эндемичные роды хвойных. Некоторые роды, как, например, четыре
первых, образуют тип ареала, который можно охарактеризовать как
антарктически-андовый, поскольку эти роды проникают и дальше к северу,
выходя за пределы антарктического флористического царства, вплоть
до Высоких Анд.
Названные 13 родов относятся к стольким же семействам, и в част-
ности, за исключением Myzodendracea и преимущественно австралий-
ского Stylidiaceae (Donatia), к семействам широко распространенным, не
связанным с определенным флористическим царством. Размещение Faga-
сеае (род Nothofagus) служит примером биполярного внетропического
типа ареала. Этот тип мы встречаем и среди родовых ареалов, например
у Empetrum (ср. рис. 8), Caltha и Veronica. Другим флористическим эле-
ментом является австрало-антарктический, представленный, например,
семействами Epacridaceae, Eucryphiaceae, Stylidiaceae, подсемейством
Luzuriagoideae (Liliaceae) и родом Metrosideros (Myrlaceae). Широко
распространены в южном полушарии семейства Proteaceae и Cunnonia-
сеае. Здесь присутствуют также элементы флоры тропических системати-
ческих единиц, которые попали в Южную Америку из неотрописа, а на
Новую Зеландию и соседние с ней острова — из палеотрописа.
Именно ими определяются некоторые различия между следующими флористи-
ческими областями:
а) Патагонская.
б) Антарктическая (островная).
в) Южноновозеландская (Дильс относит ее к палеотропису).
Формы роста растений
ПРОБЛЕМА ТИПОЛОГИИ РАСТИТЕЛЬНЫХ ФОРМ
Внешний облик растений определяет картину растительности, а тем самым в зна-
чительной степени и физиономию ландшафта. В мире растений существует столько раз-
личных типов растительных форм, сколько имеется видов, так что охватить все это
богатство с точки зрения систематики представляется едва ли возможным.
Для географии растительности прежде всего важно объединение
растительных форм по тем признакам, которые наиболее существенны
с географической точки зрения. Некоторые слова из обиходного лекси-
кона, как дерево, куст, трава, лиана, отвечают именно этим понятиям,
хотя толкование их большей частью не точно разграничено. География
растительности применяет эти понятия в смысле научных терминов. При
этом важную роль могут играть различные признаки, как, например,
величина растения, степень одревеснения, характер ветвления, форма
листьев, кора, соцветия и т. д. Продолжительность жизни растения, сме-
няющиеся во времени стадии развития, такие сезонные изменения облика,
как ежегодный листопад или отмирание надземных органов,— все эти
понятия также составляют суть внешнего облика растения; часто они
дополняются и такими признаками, как характер местожительства и спо-
соб питания (например, паразитирующие эпифиты), функции определен-
ных тканей (в частности, накопление воды), способы размножения или
распространения (например, размножение с помощью побегов) и прочие
характерные особенности образа жизни (например, растения-орнито-
филы).
При большом разнообразии признаков, по которым можно объеди-
нить все растительные формы в определенные типы, трудно создать такую
классификацию, в которой удастся учесть все точки зрения. Система таксо-
номических единиц этой задачи не решает.
Даже индивидуумы одного и того же вида, не говоря об их отличии на разных
стадиях развития, могут иметь разную форму. Так, например, можжевельник (Juni-
perus communis) встречается в пустошах северо-западной Германии в виде дерева,
на сухих задернованных средне- и южногерманских известняковых почвах это уже
невысокий пряморастущий кустарник, а на альпийских маттах он стелется по земле
или свисает по склонам в виде карликового кустарника. В составе одного рода расти-
тельные формы видов могут быть настолько различными, что пх родственность даже
трудно предположить. Достаточно вспомнить серебристую иву высотой 20 м и альпий-
Формы роста растений
45
«кие ивы с сильно разветвленной корневой системой, которые образуют над почвой
лишь низкий ковер, сотканный из круглых листочков. Во многих случаях к одному
и тому же семейству (например, Papilionaceae, Rosaceae, Euphorbiaceae, Liliaceae)
относятся как травы, так и древесные растения.
Принадлежность видов к систематическим единицам более высоких
рангов зачастую основывается на мало заметных генетических призна-
ках строения органов размножения. Однако как форма, так и строение
вегетативных органов могут быть признаками родства и особенностями
данной систематической единицы, обусловленными генетически. Это
верно не только для многих часто очень схожих по внешнему облику
низших растений, но и для некоторых представителей высших, например
для господствующей в семействе Palmae формы деревьев с пучком листьев
на вершине или формы листьев и характера ветвления у большинства
Coniferae. Это находит свое выражение в самом названии класса —
«хвойные». Некоторые особенности образа жизни, обусловливающие свое-
образие внешности растения, как, например, склонность к эпифитному
образу жизни или к паразитическому, сапрофитному способу питания,
также связаны с определенными систематическими единицами. Однако
большинство физиономических признаков — высота, степень одревесне-
ния ит. д.— могут быть в одном и том же семействе весьма различными.
С другой стороны, родственные связи проявляются в том, что древовидная
форма встречается в довольно большом, но все же ограниченном числе
семейств сосудистых растений.
Суккулентные растительные формы встречаются примерно в 40 семействах,
хотя часто только в отдельных родах их. Большинство суккулентов сконцентрировано
в 6 семействах: Aizoaceae, Asclepiadaceae, Cactaceae, Crassulaceae, Euphorbiaceae
и Liliaceae. Они не родственны и относятся к различным порядкам. Высшие рас-
тения, имеющие форму подушек, также ограничены несколькими семействами, главным
образом Caryophyllaceae, Cruciferae, Saxifragaceae, Malvaceae, Umbelliferae, Primula-
ceae nCompositae. В перечисленных семействах, существуют и совершенно другие
формы. Так, например, к Cactaceae относятся и несуккулентные облиствленные кустар-
ники, сбрасывающие листву в засушливые периоды.
КОНВЕРГЕНЦИЯ И ПРИСПОСОБЛЯЕМОСТЬ
О конвергенции и аналогии говорят, когда у каких-либо предметов,
различных по происхождению, обнаруживается сходная внешность или
одинаковые функции. Часто наблюдаемая в растительном мире конвер-
генция форм имеет место прежде всего у растений, развивающихся в одина-
ковых условиях среды. Понятие «конвергенция» относится главным обра-
зом к своеобразию внешнего вида растения, которое обусловлено отдель-
ными факторами или всей совокупностью окружающей среды и которое
обеспечивает растению возможность приспособления к жизни в данном
местообитании (например, высота растения, характер облиств ления
или защиты почек при неблагоприятных сезонных условиях, подушко-
видная форма роста, суккулентность).
Зависимость конвергенции от климата была замечена очень давно при сравни-
тельном изучении флористически различающихся растительных покровов удаленных
друг от друга горных районов. Уже Вильденов обратил внимание на сходство расти-
тельных форм Капской земли и Австралии, обладающих одинаковым климатом. Унгер
46 Составные элементы растительности и принципы их распространения
(Unger, 1836) упоминал о «модификациях», вызванных различием почв. Генетический
анализ показал, правда, что растительные формы ни в коем случае нельзя рассматри-
вать только как результат воздействия среды обитания. Тем не менее экологи на ряде
многочисленных примеров показали, насколько сильно многие растительные формы
соответствуют местным условиям существования.
Жизненные формы (биотипы)— это в значительной степени результат
приспособления растений к местам своего существования (Lebensort)
(биотопам). Форма роста отражает ту деятельность растения, которую
оно должно производить, чтобы выжить и удержаться на своем местооби-
тании. Свойства, которыми обладают отдельные систематические единицы
в соответствии с условиями среды обитания, называют «признаками при-
способления». Эти признаки генетически могут проявляться весьма раз-
лично (прогресс, редукция, изъятие мутаций, не способствующих приспо-
собляемости), что, однако, для данной работы не существенно.
Признаки родства не всегда удается отделить от признаков приспо-
собления. Некоторые присущие отдельным систематическим едини-
цам свойства, не рассматриваемые сейчас как средства приспособления,
могли первоначально быть признаками приспособления. Наоборот,
свойства, которые возникли не ради приспособления, могут стать при-
знаками приспособления в результате ограничения распространения
данных систематических единиц наличием подходящих местообитаний.
Поэтому не существует резкой границы между физиономи-
ческой классификацией растительных форм и их экологиче-
ской классификацией, основанной на признаках приспособления.
Физиономическая система оказывается всегда в какой-то степени и эко-
логической. В противном случае чисто экологическая классификация,
ограниченная экспериментально доказуемыми формами приспособле-
ния, оказалась бы явно недостаточной. Даже система Раункиера, беру-
щая за основу лишь некоторые «экологические» критерии форм, является
в значительной степени физиономической классификацией, построенной
на основе аналогии.
Если рассматривать зависимость растительных форм только от одного
из факторов окружающей среды, то и в этом случае любая система ока-
зывается ненадежной ввиду разнообразия факторов среды обитания.
С другой стороны, вся совокупность признаков внешнего и внутреннего
строения, жизненного ритма и физиологических процессов может ока-
заться приспособленной к одному решающему фактору среды обитания.
По отношению к среде обитания различают «водные растения» и «на-
земные растения», а среди наземных —«напочвенные» и «эдафофиты»,
последние обитают в самой почве. В особую группу выделены растения,
обитающие в других растениях,—«эндофиты».
Приспособленность к неодинаковым условиям существования может
проявляться в различной продолжительности жизни растений или их
частей, в величине и форме растения и отдельных его органов.
По продолжительности жизни различают:
1. Растения с малой продолжительностью жизни (терофиты, эфе-
меры).
В областях, где периодически наступают холода или засухи, теро-
фиты завершают свой жизненный цикл за ограниченный отрезок времени
и переживают неблагоприятное время года в виде семян или спор. В исклю-
чительно сухих областях с эпизодически выпадающими осадками подобные
растительные формы могут годами существовать в виде семян, с тем чтобы
Формы роста растений
47
уже после первого дождя в течение очень короткого времени развиться
и достигнуть стадии плодоношения (эфемеры).
2. Многолетние растения, которые переживают неблагоприятное
время года под землей и у которых надземные части ежегодно отмирают.
Эти растения по-иному приспособились переносить холодное или
сухое время года. Их почки возобновления перезимовывают или на извест-
ной глубине под землей (геофиты), или на уровне поверхности почвы
(гемикриптофиты).
3. Многолетние растения, переживающие неблагоприятное время года
на поверхности земли, с временной редукцией побегов.
К ним относятся виды, которые в соответствующее время года сбра-
сывают листву (тропофиты) или же пассивно теряют ее под действием
жары или холода, а также виды, которые в холодное или сухое время
года теряют части своих побегов, но восстанавливают их из почек, нахо-
дящихся на одревесневших частях побегов (полукустарники).
4. Полностью многолетние растения, непрерывно развивающиеся
на протяжении всей, продолжающейся подчас несколько лет, жизни.
Такие растения могут быть приспособлены к различным условиям
существования и иметь самый разнообразный внешний облик. Они могут
быть большими и маленькими, безлистными и вечнозелеными, древес-
ными или травянистыми.
Величина и форма растения отражают различные условия существо-
вания. Высокие, роскошные по внешнему виду экземпляры, как правило,
не могут произрастать в крайне холодных и сухих местностях. В сыром
климате по своей массе надземные части растений преобладают над корне-
вой системой, и наоборот, в сухом климате подземные части, обеспечиваю-
щие растения водой, сильнее развиты, чем надземные.
Лучше всего изучена зависимость растительных форм от воды и тепла (работы
Варминга, Шимпера, Раункиера, Фиттинга, Лундегорда, Вальтера, Иверсена, Сток-
кера).
ФОРМЫ ПРИСПОСОБЛЕНИЯ К ВОДНОМУ ФАКТОРУ
Форма и строение вегетативных органов обычно отражают приспо-
собленность растений к условиям увлажнения. Попытки подразделе-
ния с этой точки зрения предпринимались уже давно (Drude, 1890;
Schimper, 1898).
По степени приспособленности к водному фактору среди наземных
растений различают ксероморфные, мезоморфные и
гигроморфные растения, которым противостоят в качестве чет-
вертой группы водные растения — гидрофиты.
КСЕРОМОРФНЫЕ РАСТЕНИЯ
Ксероморфными мы называем растения, которые по своей форме
и строению приспособлены к условиям затрудненного поглощения воды
или ее резкой недостачи.
Не все растения сухих местообитаний ксероморфны. Надземные части
многих терофитов и геофитов также могут обладать гигроморфной струк-
турой. Далее, некоторые растения, едва ли являющиеся ксероморфными,
например многие лишайники и некоторые папоротники, могут благодаря
стойкости их плазмы к обезвоживанию высыхать при недостатке воды
почти полностью, не погибая при этом, и оживать вновь. Другие расте-
48 Составные элементы растительности и принципы их распространения
ния обеспечивают себя водой с помощью водозапасающих органов (напри-
мер, клубни, мясистые корни и т. д.) или благодаря тому, что корни их
при высыхании почвы дорастают до грунтовых вод, и таким образом эти
растения даже при наличии нексероморфных побегов могут пережить
засуху.
Совокупность различных признаков ксероморфности никогда не
встречается одновременно у одного вида растений. Поэтому существуют
весьма различные типы ксероморфных растений. Почти все они обладают
рядом приспособлений, позволяющих задерживать отдачу воды наруж-
ными покровами либо на длительное время, либо хотя бы периодически.
Такими приспособлениями являются редукция поверхности по отноше-
нию к объему растения, утолщение наружных стенок эпидермиса, покры-
тых сверху мощным слоем кутикулы, и некоторые другие защитные сред-
ства, как, например, появление воскового или лакового налета, а также
развитие волосяного покрова войлочного типа.
По прочим признакам мы можем различить пять основных
типов ксероморфных растений, у которых проблема
удовлетворительного соотношения потребления и отдачи воды, иначе
говоря, проблема стабилизации водного баланса, решена по-разному
и которые в соответствии с этим имеют разную форму.
При регулировке водного баланса основное значение имеют те струк-
турные особенности, которые задерживают водоотдачу или, наоборот,
способствуют усиленному поглощению воды:
а) если водоотдача длительное время затруднена, то при нормаль-
ном строении растения одновременно ограничивается и поглощение
воды, а тем самым и образование тканей. Растения развиваются очень
медленно и часто в течение всей, иногда продолжительной, жизни не вы-
ходят за пределы карликового роста. К ним относятся подушковидные
кустарники, которые в условиях тропического и субтропического высоко-
горья обладают необычайно медленным ростом. Этот тип называют нано-
ксероморфным*,
б) наличие исключительно разветвленной, далеко или глубоко про-
никающей корневой системы позволяет растению даже на сухих местооби-
таниях поглощать достаточное количество воды. Одновременно растение
усиленно транспирирует. Хотя при этом обильное образование питатель-
ных веществ способствует обычно развитию корневой системы, однако
оно допускает и более быстрый рост надземных побегов. Ассимилирующие
органы соответственно менее ксероморфны. Они не приспособлены также
и для периодического ограничения транспирации. Листья могут быть
нежными и большими, со слабой или умеренной ксероморфностью покров-
ной ткани. Защита от испарения усиливается часто за счет волосяного
покрова, образованного мертвыми заполненными воздухом, а потому кажу-
щимися седыми волосками. Этот покров войлочного типа препятствует
излишнему неравномерному усилению транспирации под действием пря-
мого облучения или ветра. Если транспирация становится слишком силь-
ной, то листья вянут. При длительной чрезмерной водоотдаче они отми-
рают. Подобное явление может регулярно повторяться в периодически
засушливых областях. Такой тип растений, у которых побеги оказываются
лишь слабоксероморфными (например, некоторые степные травы), мы
называем гемиксероморфнъгм.
Проблема регулирования водного баланса с ограничением водоотдачи
решается в растительном мире двумя различными путями, приводящими
к следующим основным типам, а именно: в) типу склеро-ксероморфных
Формы роста растений
49
растений, у которых интенсивность транспирации меняется во времени,
и г) типу суккулентно-ксероморфных растений с водонакапливающими
тканями, что позволяет растению в течение продолжительного времени
ограничивать расходование влаги.
Склеро-ксероморфные растения обладают в разной степени выражен-
ными приспособлениями, которые ограничивают кутикулярную транс-
пирацию. Для них часто характерна сильно редуцированная поверхность
и небольшие, слабо расчлененные, обычно блестящие листья с утолщен-
ными наружными стенками эпидермиса, мощной кутикулой и зачастую
восковым или лаковым налетом. Форма листьев разнообразна: от твердых,
овальной формы, до игольчатых, вересковой или чешуйчатой формы, очень
прочных, жестких, с густой сеткой жилок. Они могут редуцироваться,
образуя расширенные черешки (филлодии), как у некоторых акаций, или
вовсе отсутствовать. В этих случаях функции ассимиляции берут на себя
прутьевидный стебель, как у казуариновых, или расширенный стебель
(филлокладий), как у Ruscus. Листья и органы, которые их заменяют или
дополняют, имеют большое количество устьиц, а растение в целом обла-
дает мощной всасывающей корневой системой. Во влажных условиях ксеро-
морфные растения при открытых устьицах сильно транспирируют, однако
большей частью устьица либо глубоко погружены в ткань листа, либо
окружены защитными волосками. Некоторые листья имеют устьица
только на одной стороне листовой пластинки; закручиваясь или склады-
ваясь, они затрудняют свободный доступ воздуха к устьичным щелям.
Закупорка устьиц при недостатке воды снижает транспирацию (в основ-
ном за счет кутикулярной) до минимума. Однако при этом прекращается
также и поступление питательных веществ, необходимых для образова-
ния тканей.
От последствий слишком большой потери воды, увядания и возмож-
ных механических повреждений склеро-ксероморфные растения защищены
механической тканью. Они имеют мощно развитую склеренхиму, а их
побеги или листья часто превращаются в колючки.
Суккулентно-ксероморфные растения содержат в листьях (листовые
суккуленты) или в других частях своих побегов водонакопляющие ткани,
в которых скапливаются запасы воды. Сильная редукция поверхности,
ксероморфное строение наружного покрова и незначительное количество
устьиц обеспечивают исключительно бережное расходование воды. Сукку-
ленты при соответствующих обстоятельствах могут в течение месяцев
или даже нескольких лет обходиться без воды, причем они могут потерять
до двух третей всей содержащейся в них воды и при этом не погибнуть.
По окончании засухи они восполняют израсходованное количество воды
с помощью своей обычно очень разветвленной поверхностной корневой
системы.
Постоянно слабая транспирация допускает лишь медленный рост расте-
ния. Однако процессы ассимиляции продолжаются и в периоды засухи.
У безлистных стеблевых суккулентов, ксероморфное строение и физио-
логические особенности которых выражены наиболее ярко, ствол или
стебель оказывается способным к ассимиляции благодаря наличию зеле-
ных клеток под его наружным покровом. Внутренность таких растений
состоит из туго наполненной водоносной ткани, которая и придает расте-
нию прочность. Высокий колоннообразный кактус может содержать не-
сколько тысяч литров воды (до 90% своего живого веса). Кроме колонно-
образной и канделябровой формы для кактусов характерны также шишко-
образная или шаровидная формы.
50 Составные элементы растительности и принципы их распространения
Несколько менее ксероморфны, чем безлистные стеблевые суккуленты,
облиственные растения с водонакапливающими стволами (бутылочные
и бочковидные деревья).
д) и, наконец, особую группу ксероморфных растений образуют
некоторые эпифиты, У них наблюдаются выраженные в различной сте-
пени признаки, ограничивающие транспирацию, как-то: редуцированная
поверхность, ксероморфная структура наружного покрова, суккулент-
ные листья и т. д., а также способность некоторых тропических эпи-
фитов накапливать воду в воронкообразных листьях.
МЕЗОМОРФНЫЕ РАСТЕНИЯ
Мезоморфными называют растения, которые по форме и строению
не обладают ни явно выраженными ксероморфными, ни гигроморфными
признаками или у которых одновременно оба вида признаков обнаружи-
ваются либо постоянно, либо периодически. У одного и того же растения
верхние и наружные листья, более подверженные, например, облуче-
нию и действию ветра, могут иметь ксероморфное строение, в то время
как листья, находящиеся в тени, приближаются к гигроморфному типу.
Мезоморфные растения с длительно сохраняющейся листвой зачастую
оказываются вынужденными время от времени сильно сокращать водо-
отдачу путем периодического ограничения транспирации. При умень-
шении тургора многие листья теряют напряженность, однако полностью
не ослабевают, как это происходит у растений гигроморфного типа.
Тропофиты, «дождезеленые» или «летнезеленые» растения,
периодически сбрасывающие листву, приспособлены к сезонной смене
благоприятных условий транспирации сухими и холодными периодами.
Теряя листву, они сокращают свою поверхность и переживают неблаго-
приятное время с ксероморфно устроенными органами (корка, защи-
щенные почки). В облиственном состоянии тропофиты, в зависимости от
особенностей строения своих листьев, имеют мезоморфный и даже гигро-
морфный характер.
ГИГРО МОРФНЫЕ РАСТЕНИЯ
Гигроморфными мы называем наземные растения, которые приспо-
собились к обитанию в условиях избыточного увлажнения. Для них харак-
терна слабо развитая корневая система и относительно большая транспи-
рирующая поверхность. Гигроморфные листья, как правило, имеют боль-
шую поверхность, они тонки, сочны, богаты межклетниками. При умень-
шении тургора листья быстро ослабевают. Хлорофилл, содержащийся
в тонкостенных больших клетках эпидермиса, окрашивает листву в темно-
зеленый цвет. Крайне тонкая кутикула обеспечивает сильную транспи-
рацию. У ряда растений имеются некоторые специальные, усиливающие
влагоотдачу приспособления, как, например, приподнятые над поверх-
ностью листа устьица, увеличение транспирирующей поверхности за счет
живых волосков, развитие водных устьиц и особых железок, способных
выделять воду в капельно-жидком состоянии (гидатодов), если внешние
условия препятствуют транспирации. В исключительных случаях гигро-
морфные наземные растения приобретают сходство с водными расте-
ниями, как, например, нежные бескорневые папоротники (Hymenophyl-
lum) влажных тропических лесов, поглощающие влагу волосками, рас-
положенными на поверхности листа.
Формы роста растений
51
ГИДРОФИТЫ
Водные погруженные растения («submers») отличаются от наземных
растений отсутствием опорной ткани и вторичного прироста в толщину.
Они мягки и слабы, однако у них, как правило, развита воздухоносная
ткань, служащая для накопления воздуха и одновременно удерживаю-
щая листья на поверхности воды. Сосудистые пучки отсутствуют или
располагаются у растений текучих вод в центре стебля, что придает
побегам прочность на растяжение и делает их гибкими. Корни или отсут-
ствуют, или весьма слабо развиты. Листья, как правило, сильно расчле-
нены и имеют большую поверхность. Наружные стенки покровной ткани
исключительно тонки, бедны кутином и не имеют устьиц. Слабо диффе-
ренцированная ткань листьев имеет крупные клетки и богата межклет-
никами. У водных растений, листья которых плавают по поверхности
воды, строение последних сходно с листьями наземных растений, однако
без устьиц на нижней стороне. Как переходную форму к наземным расте-
ниям можно рассматривать водные растения, у которых побеги значи-
тельно возвышаются над водой. Интересны растения-амфибии, которые
могут существовать и в воде и вне воды, на земле.
ПРИСПОСОБЛЕННОСТЬ НАЗЕМНЫХ РАСТЕНИЙ
К СВЕТУ, ТЕПЛУ И ДРУГИМ ФАКТОРАМ
И СИСТЕМА ЖИЗНЕННЫХ ФОРМ ПО РАУНКИЕРУ
Климатические факторы по своему значению для формы растения
находятся во взаимной связи, а также в зависимости от водного фактора,
так что целый ряд «признаков приспособления» не удается отнести за счет
какой-либо одной особенности климата. Так, например, светолюбивые
растения часто ксероморфны, а тенелюбивые растения имеют более гигро-
морфное строение. Даже такие, казалось бы, специальные приспособле-
ния к интенсивному облучению, как волосяной покров войлочного типа,
вертикальное расположение листьев или ориентация листовых пластинок
в направлении север — юг у компасных растений, являются одновре-
менно и ксероморфными признаками.
Недостаток солнечного тепла хорошо переносят невысокие растения
с горизонтально распространенными побегами или листьями, приле-
гающими к почве в форме розеток. От холода или резкой смены темпера-
тур растения защищают покровы из плохо проводящих тепло веществ,
как-то: пробка, смола, волоски или отмершие листья (туника). Темная
окраска, например, почек, усиливает приток тепла к растению, в резуль-
тате чего температура освещенных частей растения увеличивается по срав-
нению с температурой окружающего воздуха.
Многие растения избегают воздействия холода благодаря короткому
вегетационному периоду или путем утраты надземных частей при сохра-
нении почек возобновления невысоко над поверхностью почвы или под
землей.
На основании признаков приспособления к переживанию неблаго-
приятного времени года построил свою систему Раункиер (Raunkiaer,
1905). Он выделил 30 «жизненных форм» у высших растений, которые свел
к пяти основным группам:
1. Фанерофиты. Почки возобновления находятся на значи-
тельной высоте над поверхностью почвы.
52 Составные элементы растительности и принципы их распространения
2. Хамефиты. Почки возобновления расположены на расстоя-
нии не более 0,25 м над поверхностью почвы.
3. Гемикриптофиты. Почки возобновления лежат на уровне
поверхности почвы, где они относительно защищены самой почвой и от-
дельными частями растения.
4. Криптофиты. На зиму надземные части отмирают, но почки
возобновления сохраняются в почве.
5. Терофиты. Однолетние растения, которые переживают
неблагоприятное время года в виде семян.
Впоследствии Раункиер и другие исследователи увеличили число
основных групп. Криптофиты были подразделены на геофиты, гело-
фиты (болотные растения) и гидрофиты (водные растения). Из
фанерофитов были выделены эпифиты, лианы истеблевые
суккуленты. Остальные фанерофиты по высоте роста были раз-
делены на макрофанерофиты, с почками возобновления на
высоте более 2 м над поверхностью почвы, и нанофанерофиты,
с почками на высоте от 0,25 до 2 м над поверхностью.
По Хансену (Hansen, 1956), последовательность возникновения этих жизненных
форм во времени может быть установлена, исходя из их прошлого и современного
распространения. В то время как макрофанерофиты существовали уже в меловой
период, основное развитие лиан относится к началу третичного периода. В поздний
третичный период возникли нанофанерофиты, гемикриптофиты и геофиты. Самыми
юными формами являются хамефиты и терофиты, основное развитие которых началось
только в четвертичное время.
Формы приспособления растений к тем или иным местам произ-
растания весьма многообразны. В качестве примера форм вегетативного
распространения следует упомянуть формы приспособления к подвиж-
ному субстрату (кустящиеся растения, закрепляющие склоны, растения
на развеваемых песках и др.), к периодически затапливаемым почвам
(амфибиевые формы роста, мангровые и др.), а также формы приспособ-
ления к определенным условиям питания (паразиты и полупаразиты,
сапрофиты, насекомоядные растения).
КЛАССИФИКАЦИЯ ФОРМ РОСТА
Из вышеизложенного следует, что создать пригодную на все случаи
жизни схему типизации и наглядной классификации растительных форм
едва ли удастся. В зависимости от цели такого обзора можно выдвигать
на первый план ту или инуюугочку зрения.
Гумбольдт (Humboldt, 1806), исходя из ландшафтно-физиономической точки
зрения, ввел понятие «растительная форма» и различал на основе особенно характер-
ных форм произрастания 16 (иногда 19)£основных форм, которые он именовал по назва-
ниям систематических единиц, например форма пальм, форма казуарин. В основу
своей классификации Гумбольдт положил строение «вегетативных жизненно важных
органов (питания, размножения)». Поэтому некоторые его основные формы являются
одновременно экологическими группами растений, например кактусы и тропические
лианы. И в более поздних работах, в частности Кернера (Kerner, 1863), Гризебаха
(Grisebach, 1872), физиономическая классификация содержит всегда и экологический
элемент, например подразделение на древесные растения, эпифиты, травы. Рейтер
(Reiter, 1885) сделал экологический характер растительных форм отправной точкой
Формы роста растений
53
при физиономической классификации, когда он к одной и той же форме относил рас-
тения, совпадавшие по характеру своей жизнедеятельности и по обеспечивающим
ее приспособлениям.
Друде (Drude, 1888, 1890), опираясь на труды Кернера, Гризебаха
и Рейтера, разработал принцип классификации, который явился основой
всех последующих делений, поскольку созданная им система отличается
от чисто экологических систем Варминга, Шимпера, Раункиера, бази-
рующихся на одном или нескольких критериях.
Сформулированные Друде основные положения вполне применимы
при нашей постановке вопроса с точки зрения географии растительности,
особенно его разграничение понятий «форма роста» и «класс форм роста».
Друде исходил из того, что любому виду растения в соответствии
с его наследственными свойствами и условиями существования присущи
специфическая форма и образ жизни (вегетационный период, рост, пита-
ние, размножение). Сходные формы и образ жизни многих растений
можно объединить и дать этой совокупности наименование форма
роста. Каждая форма роста в этом смысле является, следовательно,
индуктивно полученным собирательным понятием, основанным на срав-
нении. Классы форм роста и их подразделения являются,
напротив, системой дедуктивных понятий, составленных по принципу
целесообразности, что позволяет более наглядно классифицировать формы
роста. Основополагающими критериями для такой классификации Друде
(как и Рейтер) считал признаки приспособления, или, как он выражался,
«гармонию между строением и теми функциями органов, которые зависят
от внешних условий существования». Особенно важными признаками
он считал «выносливость органов и защитные средства, препятствующие
повреждению растения в период покоя». Классы форм роста Друде частич-
но соответствуют поэтому жизненным формам Раункиера (Raunkiaer,
1905), определяемым способами защиты почек или побегов возобновления
в неблагоприятное время года. Подразделение классов форм роста Друде
основывал на приспособленности их к разным условиям увлажнения,
которую Шимпер (Schimper, 1898) в своей эколого-физиономической
типологии (гигрофиты, ксерофиты, тропофиты) также считал основным
признаком классификации.
Варминг (Warming, 1895) и Варминг — Гребнер (Warming — Graeb-
ner, 1918) каждый в своей системе жизненных форм использовали при
классификации и некоторые другие критерии (способы размножения
и питания). Эти критерии, если они представляют интерес для географии
растительности, должны быть использованы и при дальнейшем выделе-
нии классов форм роста.
Все прочие многочисленные попытки классифицировать растения по их формам
роста или жизненным формам, например Гамса (Gams, 1918), Хайека (Hayek, 1926),
Браун-Бланке (Braun-Blanquet, 1928, 1951), Иверсена (Iversen, 1936), Гинцбергера
(Ginzberger, 1939), Элленберга (Ellenberg, 1956), здесь не рассматриваются.
В нижеприводимом обзоре важнейших форм роста последние под-
разделяются, по Вайбелю, на классы форм роста, которые, учитывая инте-
ресы географии растительности, созданы и расположены по принципам,
разработанным Друде.
В противоположность систематике растений, в которой за основу
классификации берется один определяющий критерий — генетическое
родство, в типологии форм роста учитываются многие весьма далекие
друг от друга признаки. Формы роста, как это подчеркнул еще Гумбольдт,
54 Составные элементы растительности и принципы их распространения
«по своей природе не поддаются строгой классификации». Это означает,
что в природе не существует стабильной «системы» форм роста, а воз-
можно любое число группировок в зависимости от того, какому крите-
рию придается наибольшее значение и какой ранг присуждается им в дан-
ной схеме классификации.
Описываемые ниже формы роста, сгруппированные с точки зрения
географии растительности, являются самостоятельными типологическими
единицами; их перечисление начинают общеизвестные характерные основ-
ные формы растений. При этом учитывается общий облик растений и прежде
всего те их признаки, которые, во-первых, определяют физиономию ланд-
шафта, а во-вторых, отражают приспособленность растения к факторам
местообитания, которые в значительной степени дифференцируют расти-
тельность Земли. Особенное значение приобретают при этом условия
существования, продолжительность жизни растения и его органов, высота
роста и положение почек возобновления относительно поверхности почвы,
одревесненность и характерные формы приспособления к условиям место-
обитания (например, суккуленты, лианы, эпифиты, растения с плаваю-
щими листьями и т. п.).
Мы объединяем все формы роста в следующие 30 классов форм
роста:
1. Кронообразующие деревья (Kronenbaiime)
2. Розеточные деревья (Schopfbaume)
3. Древовидные злаки (Baumgraser)
4. Деревья-удушители (Baumwiirger)
5. Лианы (Lianen)
6. Кустарники (Straucher)
7. Карликовые деревья (Zwergbaume) / Нанофанерофиты
8. Стеблевые суккуленты (Stammsukkulente)
(стебле-суккулентные фанерофиты)
9. Растения с травянистым стволом (Krautstammpflanzen)
(травянистые фанерофиты)
10. Эпифиты (Epiphyten)
11. Карликовые кустарники, кустарнички (Zwergstraucher)
12. Полукустарники (Halbstraucher)
13. Карликовые суккуленты (Zwerg-Sukkulenten)
14. Хамефитные травы (Chamaephytische Stauden)
15. Гемикриптофитные древесные растения (Hemikryptophytische Holz-
pflanzen)
16. Гемикриптофитные травы (Hemikryptophytische Stauden)
17. Озимые однолетники (Winterannuelle)
18. Геофитные многолетние травы (Geophyten-Stauden)
19. Терофитные однолетние травы (Therophyten-Krauter)
20. Растения с плавающими листьями (Schwimmblattpflanzen)
21. Погруженные травы (Submerse Krauter)
22. Талломные эпифиты (Thallus-Epiphyten)
23. Талломные хамефиты (Thallus-Chamaephyten)
24. Талломные гемикриптофиты (Thallus-Hemikryptophyten)
25. Талломные геофиты (Thallus-Geophyten)
26. Талломные терофиты (Thallus-Therophyten)
27. Эдафофиты (Edaphophyten)
28. Талломные гидрофиты (Thallus-Hydrophyten)
29. Планктофиты (Planktophyten)
30. Эндофиты (Endophyten)
Формы роста растений
55
Перечисленные классы, насколько это возможно, составлены так, что они могут
быть включены в основные группы жизненных форм Раункиера. Это имеет смысл, так
как последние давно уже используются для характеристики климатических поясов,
областей растительности или растительных сообществ по «спектру жизненных форм».
Этот сравнительный материал потерял бы свою ценность, если бы классы форм роста
нельзя было как-то согласовать с жизненными формами. По этой причине талломные
растения тоже подразделяются внутри основных групп Раункиера на свои собствен-
ные классы. Принятая последовательность ряда дает возможность разделения на назем-
ные и водные растения, а также на древесные, травянистые и талломные растения.
Включение классов форм роста в основные группы жизненных форм
видно из следующего обзора:
Группы жизненных форм Древес- ные рас- тения Травянис- тые рас- тения Таллом- ные рас- тения
Фанерофиты от 1 до 8 9 —
Эпифиты 10 . 22
Наземные рас- Хамефиты 11 12, 13 14 23
тения Гемикриптофиты 15 16, 17 24
Геофиты — 18 25
Терофиты — 19 26
Эдафофиты — — 27
Водные расте- ( Гидрофиты
ния [ Планктофиты
Эндофиты Эндофиты
20, 21 28
— 29
— 30
Каждый класс форм роста можно подразделить с эколого-физиономи-
ческой точки зрения. Ступени этого деления не получают специальных
наименований, чтобы не создалось впечатления жесткой системы. Прин-
ципы систематизации также не для всех классов одинаковы. Где это кажет-
ся целесообразным, классификация производится по приспособленности
к водному фактору. Затем возможна классификация по формам размно-
жения, распространения и приуроченности к месту произрастания, а также
по способам питания, если они существенны для внешнего облика расте-
ния. Помимо этого, иногда учитывают и чисто структурные физиономи-
ческие различия.
ОБЗОР ФОРМ РОСТА
КРОНООБРАЗУЮЩИЕ ДЕРЕВЬЯ
Высокие древесные растения с легко различимыми стволом и кроной, образован-
ной ветвями, растущие вертикально вверх, обладающие вторичным приростом в тол-
щину ствола, ветвей и корней. Некоторые виды, как исключение, имеют несколько
стволов или стоят на ходульных корнях. К этому классу форм роста относятся дву-
дольные покрытосеменные и хвойные, некоторые однодольные (лилейные с вторичным
приростом в толщину, например Dracaena, драконово дерево). К кронообразующим
деревьям относятся самые высокие растения с наибольшей продолжительностью
жизни (например, Sequoia, секвойя, живущая более 3000 лет, Fitzroya и Olea euro-
56
Составные элементы растительности и принципы их распространения
раса — более 2000 лет). Наибольшая измеренная высота дерева оказалась равной
111 м (Sequoia sempervirens) и 107 м (Eucalyptus regnans), наибольшая измеренная
окружность ствола — 23 м (Sequoia gigantea, обитающая в Калифорнии, и Agathis
australis — в Новой Зеландии).
а) Гигроморфные вечнозеленые кронообразующие деревья. Эта форма
деревьев является господствующей в дождевых тропических лесах; высота
таких деревьев
Р и с. 19. Ствол
Theobroma ca-
cao с плодами
(по Карстену).
весьма различна. Как правило, у них прямые, стройные,
гладкие стволы, и только в верхней части ветви обра-
зуют небольшую крону, имеющую в условиях леса узкую
веретенообразную форму. Тонкая, не защищенная коркой
кора зеленого, серебристо-серого или белого цвета. Соц-
ветия часто развиваются непосредственно на стволах
или ветвях (каулифлория) (рис. 19). Защита почек или
отсутствует, или развита совсем слабо. Листья на деревьях
большей частью средней величины, темно-зеленые, нерас-
члененные, продолговато-овальные, цельнокрайние,
иногда с так называемыми капельными остриями на кон-
цах. Такая форма листьев характерна для большинства
видов, однако встречается и другая форма, например
листья расчлененные. По сравнению со своими размерами
листья, особенно внутри кроны, тонкие, но часто с проч-
ной, более или менее кожистой поверхностью. В верх-
них наружных частях кроны листья обычно меньшей
величины и могут иметь ксероморфное строение.
Среди преобладающих низких или средней высоты
деревьев этого типа высокие «первобытные лесные вели-
каны» дождевых тропических лесов отличаются своеобра-
зием формы. Их высокие, на большом протяжении лишен-
ные ветвей колоннообразные стволы, оканчивающиеся,
как правило, небольшими шаровидными или зонтиковид-
ными кронами, удерживаются в почве с помощью высо-
ких в виде подпорок ходульных или досковидных кор-
ней. Эти боковые корни, отходящие от нижней части
ствола и напоминающие поставленные на ребро доски,
являются одновременно и дыхательными корнями.
б) Тропофитные кронообразующие деревья, периоди-
чески теряющие листву. У этих деревьев листья мезо-
морфные и гигроморфные, ежегодно обновляющиеся, опадающие в сухое
или в холодное время года, различные по величине и форме. Многолетние
части деревьев более или менее ксероморфны и имеют корку и защищен-
ные почки. Среди деревьев этого типа различают несколько форм:
1) дождезеленые лиственные деревья тропической и субтропиче-
ской зон, подразделяющиеся в свою очередь на тиковые деревья — высо-
кие, кронообразующие деревья, произрастающие в условиях тропиче-
ского и субтропического муссонного климата с влажным летним и сухим
зимним периодами, имеющие более или менее гигрофитную листву,
и дождезеленые деревья «плодового типа» сухих тропических лесов —
низкие, с широкой кроной и мощной защитной коркой, в дождливые пе-
риоды с мезоморфной до гемиксероморфной листвой разной величины и
формы;
2) летнезеленые деревья умеренной зоны. Преобладают экземпляры
средней величины, 10—40 м и выше, почти исключительно двудольные
Формы роста растений
57
покрытосеменные с ярко выраженными годовыми кольцами в древесине,
но также отдельные голосеменные, например Larix или Ginkgo. Крона
деревьев широкая (форма бука), как правило, верхушечная, лишь у неко-
торых деревьев ствол начинает ветвиться уже у самой поверхности почвы;
ствол и ветви покрыты коркой, почки возобновления защищены твердыми
плотносомкнутыми кроющими чешуйками, надежно предохраняющими
их в зимний период. Средней величины листья возобновляются каждый
вегетационный период; они распускаются весной из почек, закладывающих-
ся за год до распускания. Листья, как правило, не морозоустойчивые
и не имеют специальных приспособлений для уменьшения испарения.
При распускании они нежны и имеют свежий зеленый цвет. В глубине
кроны листья обычно больше и тоньше, на внешней части ее плотнее
и меньше, с более толстым эпидермисом. Форма листьев может быть весьма
различной (например, бук, каштаны, дуб, клен, ясень, ива).
Необходимые для растения вещества (крахмал, сахар, белок, жиры, соли)
с наступлением осени начинают поступать из листьев в ветви и ствол (зимняя и осенняя
древесина содержит больше крахмала). При этом сначала разрушаются зеленые, а затем
красные и желтые красящие вещества. Поэтому осенью окраска часто меняется с зеле-
ной на красную, потом на желтую, а в конце концов листья становятся бурыми.
Место прикрепления черешков листа к стеблю покрывается защитной тканью (кал-
люс), а сам лист отпадает.
в) Мезоморфные и ксероморфные вечнозеленые лиственные деревья,
состав которых неоднороден:
1) мезоморфные вечнозеленые деревья с прочными, кожистыми,
обычно блестящими, иногда большими (форма магнолии), иногда средней
величины и даже небольшими, иногда длинными (лавровидные) листьями;
почки защищены;
2) жестколистные деревья; вечнозеленые склеро-ксероморфные низ-
кие и средней высоты деревья с жесткими, цельнокрайними матово-зеле-
ными или гладкими блестящими, более или менее овальной формы и отно-
сительно небольшими листьями. Часто листья заменены листовыми колюч-
ками, а иногда с нижней стороны листа или с обеих сторон наблюдается
опушение. Стволы защищены мощной коркой. Описываемая форма харак-
терна для деревьев субтропических областей с дождливым зимним перио-
дом (например, вечнозеленые виды дубов и различные роды Lauraceae,
Myrtaceae и Anacardiaceae).
К этой группе относятся также некоторые древесные формы с сильно
редуцированными листьями (например, акации с филлодиями);
3) мангровые деревья; это форма'деревьев, приспособленных к суще-
ствованию на засоленных почвах тропических побережий. Крона с ровной
нижней поверхностью начинается на высоте прилива; стволы высотой
2—3 м опираются на многочисленные воздушные корни. Для этих деревьев
характерны различные формы дыхательных корней, образовавшиеся
как приспособление к недостатку кислорода в илистой почве, вечнозеле-
ная, обладающая ксероморфным строением листва, блестящие, иногда
опушенные, с мощной кутикулой листья с погруженными устьицами,
а также тканью из слизистых клеток; кроме того, интересна наблюдаемая
у этих деревьев так называемая «вивипария» плодов. Семена прорастают
еще на дереве и образуют ростки длиной до 60 см, которые после отпадания
легко укореняются в мягком иле (рис. 20).
58 Составные элементы растительности и принципы их распространения
Р п с, 20. Rhizophora mangle] справа один из проро-
сших на дереве плодов (по Эрстеду).;
Различают следующие физиономически отличные друг от друга формы корней:
Мангровые с ходульными корнями (типа Rhizophord), у которых воздушные корни
выходят из ствола на высоте прилива и по дуге устремляются к почве, прочно удержи-
вая растение в мягком иле; мангровые с коленчатыми корнями (типа Brugierd), у которых
дыхательные корни торчат из
ила наподобие колен; мангро-
вые со «спаржевыми» корнями
(типа Sonneratid) с большим
числом корней-пневматофоров,
выходящих из ила вертикаль-
но вверх на всем участке, за-
нимаемом корневой системой
(рис. 21); растения с «килевыми
корнями», которые тянутся
плоско по самой поверхности
и выступают над почвой в виде
килеобразной дыхательной
планки.
г) Мезоморфные и ксе-
роморфные вечнозеленые
хвойные деревья. Деревья
с мезоморфными и ксеро-
морфными листьями игло-
видной формы, сохраняю-
щимися на протяжении
многих лет. Преимущест-
венно хвойные, со смолистыми стволами, одетыми коркой, с защищенными
почками. Многие хвойные отличаются характерной вытянутой узкой фор-
мой кроны на прямом стволе, имеющем только верхушечную почку
(моноподий) и более или менее равномерно распределенные боковые
ветви (форма ели). Другие в позднем возрасте приобретают округлую
или зонтиковидную крону (форма сосны, форма пинии). Хвойные деревья
распространены преимущественно в областях с холодной зимой и в сухих
Рис. 21. Молодое растение Avicennia nitida (по Бёргезену).
Формы роста растений
59
внетродических областях. В тропиках они встречаются редко и ограничены
высокогорными областями. К крайне ксероморфным формам с чешуйчатой
листвой (Cupressaceae) примыкают формы тамарисков. У формы казуарин
роль листьев выполняют зеленые прутьевидные ветки.
д) Форма драконового дерева. Деревья с различно ветвящимися
стволами, с пучкоообразно расположенными на концах ветвей узкими
листьями. Эта форма встречается у представителей родов Dracaena (семей-
ство Liliaceae), достигающих высоты 18 м, и Yucca, а также у древесных
видов в семействе Euphorbiaceae.
е) Колючие деревья. Крайне ксероморфные формы сухих тропиче-
ских и субтропических областей с очень короткими влажными сезонами.
Деревья обычно низкие, с широко раскидистой кроной, с мощным, покры-
тым коркой стволом, узловатыми или причудливо искривленными сучьями
и ветвями, часто покрытыми шипами. Часть деревьев — вечнозеленые,
с простыми, маленькими, жесткими, склеро-ксероморфными листьями,
в определенных условиях редуцированными до чешуй. Другая часть —
листопадные деревья, реже с более крупными плотными кожистыми или
войлочными листьями, простыми или пальчато-расчлененными, чаще —
с маленькими дважды перистыми листьями. Много бобовых (форма зон-
тичных акаций).
ж) Стеблесуккулентные лиственные деревья. Ксероморфные деревья
периодически сухих тропиков с низкими и умеренно высокими бутылко-
образно утолщенными стволами, накапливающими влагу: «бочкоообраз-
ные стволы», «бутылочные деревья», или «толстобрюшники» («борригудос»),
главным образом из семейств Bombacaceae и Sterculiaceae. Некоторые
имеют зеленую гладкую кору, иные с развитой коркой и торчащими на
стволе шипами. Деревья преимущественно листопадные, иногда с листья-
ми довольно больших размеров.
Своеобразна выделенная Гумбольдтом «мальвообразная форма»: «колоссально
толстые стволы с шерстистыми большими сердцевидными или расчлененными листьями
и роскошными часто пурпурными цветками. К этой группе относится обезьянье хлеб-
ное дерево, которое при умеренной высоте имеет ствол до 10 м в диаметре».
РОЗЕТОЧНЫЕ ДЕРЕВЬЯ
Их называют иногда деревьями с листовой кроной в противополож-
ность деревьям, крона которых образована ветвями. Это высокие вечно-
зеленые древесные растения, лишенные ветвей, с очень большими листья-
ми, как бы смещенными на самый конец ствола, где они, выходя из одной
точки роста, образуют подобие пучка или розетки. В редких случаях
(например, пальма-дум) наблюдается слабое ветвление с большим числом
верхушечных почек и только с одним пучком листьев на конце каждой
ветви. За исключением нескольких родов семейства Liliaceae, у деревьев
этой группы нет вторичного прироста в толщину. Это древние тропиче-
ские и субтропические формы, ограниченные небольшим числом система-
тических единиц: Filices (преимущественно Cyatheaceae), Gymnospermae
(Cycadaceae), несколько семейств однодольных (Palmae, Pandanaceae,
Liliaceae) и некоторые роды двудольных, например Lobelia (Lobeliaceae)
и Senecio (Compositae).
а) Гигроморфные розеточные деревья (древовидные папоротники).
Высокие неразветвленные деревья высотой около 10 м, реже до 20 м,
с пучком мягких гигроморфных, в молодой стадии свернутых, крупных
60 Составные элементы растительности и принципы их распространения
перистых листьев. Стволы окутаны сохраняющимися нижними частями
черешков листьев, переплетенных придаточными корнями («Blattwur-
zelstamme»). Они растут в тенистых, безморозных местообитаниях, в усло-
виях повышенной влажности, чаще всего в дождевых и туманных лесах
тропических и субтропических горных районов.
б) Ксероморфные розеточные деревья. Они подразделяются на не-
сколько форм:
1) саговники и пальмы. Саговники, например Cycadaceae (Cycasr
Encephalartos, Macrozamia), высотой 10—20 м, с поникающими перистыми
листьями, напоминают общим обликом древовидные папоротники, однако
в отличие от последних несут большие шишкообразные соцветия.
Перистые пальмы объединяют многие виды пальм со стволами высо-
той от 10 до 30, реже до 50 м\ кокосовая пальма с большими твердыми
плодами, которые благодаря проводящему воздух мезокарпу не тонут
в воде, финиковая пальма с плодами в виде ягод, масличная пальма,
восковая пальма.
Веерные пальмы, например пальма-делеб (Borassus flabelliformis),
пальма-дум (Hyphaene tebaica), зонтичная пальма (Corypha umbraculi-
fera); последняя отмирает после развития расположенного на вершине
соцветия и образования плодов (гапаксантная форма) х;
2) форма травяных деревьев (форма Xanthorrhoea). Относительно
низкие стволы, вилкообразно разветвленные, несущие на вершинах
пучок длинных, узких, часто свешивающихся наружу плотных листьев.
К ним относятся Liliaceae (Xanthorroea, Kingia, Dasylirion) и Pandanaceae
(Pandanus, винтовое дерево). У последних вилкообразно разветвленные
стволы иногда поддерживаются ходульными корнями, на концах стволов
по спирали располагаются длинные со свешивающимися концами листья;
3) свечевидные розеточные деревья (форма Frailejon). Стволы высо-
той до 8 м с пучком листьев на вершине, из середины которой подни-
мается свечкообразное соцветие. Конвергентная форма роста тропических
высокогорий. Формой Frailejon обладают род Espeletia из семейства слож-
ноцветных в растительности южноамериканской парамы, древовидные
Senecio (Compositae), древовидные Lobelia (Lobeliaceae) восточноафрикан-
ских плоскогорий и виды Yucca в Мексике.
ДРЕВОВИДНЫЕ ЗЛАКИ (ФОРМА БАМБУКОВ)
Одревесневающие злаки (разные роды бамбуков), отличающиеся
исключительно быстрым ростом (до 1 м в день). Стволы толщиной до
30 см и высотой до 25 м (в исключительных случаях 40 м) несут на узлах
пучки облиственных веток. Преобладающая форма роста тропических
районов Южной Азии.
ДЕРЕВ ЬЯ-УДУШИТЕЛИ (СМОКОВНИЦЫ-УДАВЫ)
Деревья-удушители занимают промежуточное положение между эпи-
фитами, лианами и кронообразующими деревьями; их называют также
смоковницы-удавы (Wurgfeigen), потому что эта своеобразная раститель-
ная форма часто встречается в роде Ficus, Они прорастают эпифитно,
выпускают толстые воздушные корни, охватывают ими со всех сторон
дерево-опору и образуют в дальнейшем ветвистую крону, которая после
1 Гапаксантные (греч. hapaxanth; hapax — однажды, anthos — цветок) — цвету-
щие один раз.— Прим. ред.
Формы роста растений
61
гибели дерева-опоры удерживается «решетчатым стволом» из воздушных
корней. Некоторые виды Ficus представляют собой большие многостволь-
ные деревья, опирающиеся на колоннообразные воздушные корни (форма
баниан).
ЛИАНЫ (ДРЕВЕСНЫЕ ЛАЗЯЩИЕ РАСТЕНИЯ)
Рис. 22. Части концевых плетей ро-
танговой пальмы (Biinning, 1956).
Форма роста, распространенная преимущественно в тропических и суб-
тропических лесах. Это растения с длинными ветвящимися стволами.
Лианы часто напоминают канаты, брошенные на ветви деревьев. Свое-
образно строение их стволов — очень
широкие (толщиной более 0,5 см)
и очень длинные трахеи, широкие
ситовидные трубки и мощный, вхо-
дящий клинообразно в древесину
лубяной покров. У многих лиан име-
ются воздушные корни. У некоторых
видов встречается каулифлория.
Листья и цветки лиан часто можно
обнаружить только среди листьев
крон деревьев-опор. Существуют веч-
нозеленые и листопадные лианы.
Листва, как правило, ксероморфная,
однако может быть и мезоморфной и
гигроморфной. По характеру при-
способлений для лазанья различают
несколько форм:
а) Вьющиеся лианы, по спирали
обвивающие деревья-опоры. Приме-
ром в умеренной зоне является вино-
град олистный ломонос (Clematis
vitalba).
б) Корнелазящие, с воздушными хватающими корнями, как эфей
(Araliaceae) и тропические Bignoniaceae, или с усиками на воздушных
корнях, как ваниль (Orchidaceae).
в) Усиковые лианы с побегами, видоизменившимися в усики или цеп-
ляющиеся крючки (у многих тропических родов семейств Caesalpiniaceae
и Sapindaceae), или с листьями-прицепками (Bignoniaceae).
г) Растения, лазящие по подпорке, опирающиеся на жесткие боковые
побеги (например, виды бамбука) или на шипы, или, как ротанговая
пальма, имеющие цепляющиеся плети (снабженные колючками концы
перистых листьев) (рис. 22).
Длина некоторых лазящих пальм может достигать более 100 м, это
самые длинные наземные растения.
ЕУСТ АРНИКИ
Многолетние древесные растения, весьма разнообразные по форме,
у которых ветвление начинается от самой поверхности почвы и нет явно
выраженного ствола и кроны. Высота их 0,5—2 м и выше (до 8 м). По
характеру и длительности облиствленности, по степени защищенности
почек, суккулентности и т. д. среди кустарников различают большое
число разнообразных форм роста:
62 Составные элементы растительности и принципы их распространения
а) Гигроморфные вечнозеленые кустарники. Тенелюбивые растения
с большими мягкими листьями, с незащищенными почками; обитатели
влажных тропических лесов.
б) Кустарники, периодически сбрасывающие листву (летнезеленые
или дождезеленые), с защищенными почками:
1) с большими гигроморфными листьями (например, лещина, бу-
зина);
2) с мезоморфной или гемиксероморфной листвой.
в) Вечнозеленые или факультативно вечнозеленые мезоморфные
кустарники с мягкой листвой.
г) Вечнозеленые кустарники с жесткой листвой:
1) с умеренно жесткими, как правило, большими листьями;
2) с сильно склероморфными, как правило, маленькими листьями;
3) с вересковыми (эрикоидными) листьями.
д) Хвойные кустарники (например, горная сосна, можжевельник).
е) Кустарники с чешуйчатой листвой или безлистные прутьевидные
кустарники (дрок, казуарины, Ephedra).
ж) Ксероморфные колючие кустарники:
1) листопадные;
2) вечнозеленые.
з) Жесткие подушковидные кустарники. Плотные, весьма смолистые
и жесткие растения высотой до 1—2 м, как A zorella в Андах и «раститель-
ные овцы» (Raoulia и Haastia) в альпийских районах юга Новой
Зеландии.
и) Суккулентные кустарники.
КАРЛИКОВЫЕ ДЕРЕВЬЯ
Низкорослые древесные породы, имеющие ствол и крону,— редкая
форма. Представители—карликовые древовидные папоротники (Blechnum
magellanicum)} карликовая пальма (Chamaerops humilis) в ее наиболее
часто встречающейся форме (высотой до 10 м)\ виды Puya (Bromeliaceae);
Pringlea antiscorbutica («кергеленская капуста»); Welwitschia (монотип-
ное семейство кл. Gnetinae)— эндемичный обитатель пустыни Намиб, со
стержневым корнем, коротким конусообразным стволом более 1 м в диа-
метре и двумя широкими и длинными (до 3 м) листьями, растущими осно-
ваниями в течение многих лет и на концах размочаливающиеся под дей-
ствием ветра; цветки сидят на конце ствола.
СТЕБЛЕВЫЕ СУККУЛЕНТЫ
Растения с безлистными, толстыми, слабо одревесненными побегами,
с ассимилирующей зеленой корой и водонакапливающей тканью. Среди
них различают несколько форм:
а) Высокие древовидные, кронообразующие или канделябровидные
стеблевые суккуленты (до 20 м высоты).
б) Колоннообразные неветвящиеся стеблевые суккуленты (высо-
той от 0,30 до 10 м).
в) Кустарниковидные стеблевые суккуленты.
РАСТЕНИЯ С ТРАВЯНИСТЫМ СТВОЛОМ
Травянистые многолетние растения с крупными листьями и ложным
стволом высотой до 10 м, ствол образован слоями плотно охватывающих
друг друга листовых влагалищ или черешков, окружающих цветочную
Формы роста растений
63
стрелку. Распространены только во влажных тропиках, большей частью
под защитой деревьев. Эта форма роста соответствует «пизанговой, или
банановой, форме» Гумбольдта. Примером служат роды Musa, Begonia,
Impatiens и Piper.
ЭПИФИТЫ (ЭПИФИТНЫЕ СОСУДИСТЫЕ РАСТЕНИЯ)
Растения, обитающие в качестве «поверхностных» на стволах и вет-
вях других растений. Различают случайные, факультативные и облигат-
ные эпифиты. Последние с большим числом разнообразных форм роста
приспособились к обитанию в кронах древесных тропических растений.
Среди них встречаются как травянистые, так и древесные, частично
в форме кустарников, растения. Типичные эпифиты не являются пара-
зитами. Многие виды, поселившиеся в кронах деревьев, благодаря ксе-
роморфному строению своих многолетних листьев хорошо переносят
условия затрудненного водоснабжения на вершинах. Некоторые особи
(Orchidaceae, Агасеае) имеют свободно свисающие, но не достигающие
земли воздушные корни, многослойная внешняя оболочка которых,
образованная мертвыми клетками эпидермиса, всасывает дождевую влагу,
как губка. В засушливых условиях эта оболочка, заполненная воздухом,
защищает растение от сильного испарения. У некоторых видов (например,
виды Ficus и Агасеае), которые начинают свою жизнь как настоящие
эпифиты, воздушные корни достигают земли и таким образом снабжают
растение почвенной влагой и питательными веществами.
Из всего многообразия форм здесь выделены некоторые, например
орхидеи, накапливающие воду в зеленых стеблевых клубнях, виды Tilland-
sia (Bromeliaceae), лишенные корней, поглощающие воду всей поверх-
ностью чешуйчатых листьев и свисающие в виде длинной желтоватой
бороды с ветвей деревьев в южноамериканских влажных туманных лесах,
а также различные формы накопителей гумуса. Среди последних инте-
ресны воронковидные эпифиты (папоротники и Bromeliaceae), у которых
листья, расположенные на коротком стволе в виде плотной розетки,
образуют водонепроницаемый сосуд; особые клетки у основания листьев
поглощают накапливающиеся в этой воронке воду и питательные веще-
ства. У других форм перегной собирают так называемые нишевидные
листья (Nischenblatter), дальнейшим развитием которых являются
листья-урны (Urnenblatter) некоторых Asclepiadaceae; в этих естествен-
ных сосудах частично при содействии муравьев и других животных
накапливается питательный раствор и перегной; питающие корни, обра-
зующиеся на узлах на стволе, извлекают из них основную часть питатель-
ных веществ.
У гнездовых эпифитов для накапливания гумуса служит образование,
напоминающее птичье гнездо, состоящее из тесного переплетения воздуш-
ных корней.
К эпифитам относятся также полупаразитные растения (Loranthaceae), которые
хотя и ассимилируют самостоятельно, однако берут питательные вещества из сосудов,
принадлежащих растению-хозяину.
Факультативными, или случайными эпифитами, которые не приспособлены
специально к жизни на других растениях, являются в основном ризомные растения
(например, виды Polypodium), ризомы которых тянутся по покрову из мхов и лишай-
ников, окутывающему стволы деревьев. Следует упомянуть и о своеобразном типе-
волосяных папоротников, образующих в дождевых тропических лесах нежный дерни-
стый покров на стволах деревьев.
64
Составные элементы растительности и принципы их распространения
стержневыми корнями,
Рис. 23. Жесткая подуш-
ка Azorella selago (Керге-
лен) (по Шимперу).
КАРЛИКОВЫЕ КУСТАРНИКИ (КУСТАРНИЧКИ)
Эти формы роста господствуют в условиях субполярного и альпий-
ского климата с низкими температурами, частыми морозами и сильными
ветрами; они широко распространены также в аридных районах на гра-
нице растительной жизни. Это древесные растения высотой до 50 см.
часто с резко ксероморфным строением, с разветвленной корневой систе-
мой, вес которой может во много раз превышать вес надземных частей
растения, особенно в сухих местностях. Их корни либо широко развет-
влены во все стороны (в полупустыне), либо они снабжены мощными
часто достигающими нескольких метров в длину
(скальные растения). Формы кустарников
весьма различны.
а) Пряморастущие карликовые кустарники,
отличающиеся от обычных кустарников лишь
незначительной величиной; вечнозеленые (Cal-
luna) листопадные (голубика) или безлистные
{Ephedra).
б) Шпалерные карликовые кустарники,
сильно разветвленные, со шпалерного типа
коротким стволом, с горизонтально раскинув-
шимися и плотно прижатыми к земле ветвями,
иногда имеющие высоту лишь в несколько
сантиметров; растут большей частью очень
медленно, но достигают значительного возра-
ста, например у вечнозеленой Loiseleuria обна-
ружены 50-летние экземпляры со стволами
диаметром 0,5 см. Существуют также летне зе-
леные формы, например толокнянка (Ar с to s-
taphylos) и карликовые ивы.
в) Подушковидные карликовые кустар-
ники, от плосковыпуклых до полушаровидных
прижимающихся к почве подушек из тесно
сжатых разветвленных веточек, с крупным
стержневым корнем или с ползучим корнем, который оканчивается
несколькими вертикальными корнями. Растения, большей частью с
маленькими листьями, обитают в трещинах скал. Особые формы произ-
растают в сухих областях — карликовые кустарники с суккулентными
листьями или колючие подушковидные карликовые кустарники (в
Карру и Андах).
г) Жесткие подушковидные карликовые кустарники; представ-
ляют собой плоскораскинувшиеся жесткие подушки высотой до полуметра,
например род Azorella в субарктических областях (рис. 23).
ПОЛУКУСТА РНИКИ
Большей частью "не высокие растения, имеющие вид карликовых или
обычных кустарников, у которых одревесневают лишь нижние части
побегов, несущие перезимовывающие почки, тогда как травянистые верх-
ние части растения при неблагоприятных условиях отмирают. Наиболее
широко они распространены в периодически сухом, теплом и умеренно-
теплом климате. Большая часть их является дождезелеными, некоторые —
вечнозелеными. Листва преимущественно гемиксероморфная или ксеро-
морфнаяч
Формы роста растений
65
Наряду с вертикально растущими формами (например, желтофиоль)
имеется большое число низкорослых полукустарников, стелющихся в виде
ковра или со стелющимися побегами (рис. 24). В роде Ononis встречаются
Р и с. 24. Полукустарник с корневыми побегами (Dyschoriste radi-
cans) (по Лебрену).
пряморастущие и низкостелющиеся колючие виды. У многих полукустар-
ников листья имеют густой волосяной покров и богаты эфирными маслами
(Lavendula, Salvia, Artemisia).
КАРЛИКОВЫЕ СУККУЛЕНТЫ
а) Неветвящиеся, безлистные стеблевые суккуленты, шаровидные, ежеобразные,
шишковидные или в форме карликовых колонн; среди них прежде всего следует назвать
Cactaceae и Euphorbiaceae.
б) Ветвящиеся, пряморастущие (например, Asclepiadaceae) или шпалеровидные,
стелющиеся по земле безлистные суккулентные формы, часто с шарообразными или
овальными стеблями (карликовые опунции).
в) Стеблевые облиственные суккуленты; к ним принадлежат Oxalis carnosa и
виды рода Pelargonium (Geraniaceae), Haworthia (Liliaceae) и др.
г) Формы листовых суккулентов с разветвленными облиственными побегами,
например Sedum (Crassulaceae) и Nolanaceae, или с лежащими на почве розетками
листьев, как Sempervivum (Crassulaceae) и Mesembryanthemum (Aizoaceae).
ХАМЕФИТНЫЕ ТРАВЫ
Многолетние, неодревесневшие травянистые растения с почками,
сохраняющимися в период покоя на высоте до 50 см над поверхностью
почвы.
а) Гигроморфные облиственные формы, не подверженные сезонному
периодизму. Это тенелюбивые травянистые растения влажных тропиче-
ских лесов, с крупными листьями, часто окрашенными антоцианом в тем-
ный цвет, например бегонии, Агасеае.
б) Мезоморфные (тропофитные) облиственные травянистые растения,
периодически имеющие гигроморфный или мезоморфный лиственный
покров, например папоротник с розеточными листьями (мужской папорот-
ник). Распространены главным образом в умеренно-влажном климате
с мягкой зимой.
в) Кустовые высокие злаки. Тип саванновых злаков с многолетними
пучками жестких узких, часто скрученных листьев. В сезон дождей
образуются облиственные стебли высотой 1—4лг с широкими часто слегка
66 Составные элементы растительности и принципы их распространения
ксероморфными листьями. Распространены главным образом в полугу-
мидных тропиках.
г) Ксероморфные дерновинные злаки. Низкорослые дерновинные
злаки, с сильно разветвленной корневой системой и, как правило, очень
жесткими (склеро-ксероморфными)
Р и с. 25. Хамсфптпые стелющиеся расте-
ния (по Лебрену).
узкими листьями. Распространены
главным образом в полуаридных
и аридных местностях теплых зон,
например альфа-альфа (Северная
Африка), ичу (Анды).
д) Подушковидные травы.
Неодревесневшие или слабо одре-
весневшие многолетние ксероморф-
ные подушки, имеющие форму
полушарий, как проломник, или
форму плоской подушки, как Si-
lene acaults. Распространены пре-
имущественно в высокогорных
районах и на скалах.
е) Подушковидные злаки
(туссоковые злаки). Злаковые дер-
нины высотой до 1,5 м, у кото-
рых узкие щетинистые листья
образуются на толстых стеблях
или на полуметровых круглых подушках, иногда с превратившейся в
торф сердцевиной. Типичная форма для субантарктического климата
(например, виды]?Роа и Festuca, а также Carex).
ж) Хамефитные стелющиеся травянистые растения (см. рис. 25),
дерновинно-побеговые^злаки, цепляющиеся травы и др.
ГЕМИКРИПТОФИТНЫЕ ДРЕВЕСНЫЕ РАСТЕНИЯ
(ДРЕВЕСНЫЕ ДЕРНОВИНЫ)
Растения с одревесневшими подземными побегами.
Древесные растения, образующие ковер на поверхности почвы, как,
например, Salix herbacea (Европа), Adesmia subterranea (Боливия). До-
вольно редкая форма полярных и альпийских областей.
ГЕМИКРИПТОФИТНЫЕ ТРАВЫ
Многолетние травянистые растения с почками возобновления, нахо-
дящимися на уровне поверхности почвы. Встречаются преимущественно
в климатических областях с ежегодно прерывающимся вегетационным
периодом. К началу неблагоприятного времени года большей частью
уходят под почву; листья частично сохраняются на поверхности почвы,
как у многих дерновинных злаков. Побеги, отрастающие в течение веге-
тационного периода, могут достигать различной высоты (от нескольких
сантиметров до многих метров) и отличаться по форме.
Из важнейших групп форм следует назвать:
а) Лазящие растения:
1) вьющиеся, например вьюнок заборный;
2) цепляющиеся усиками, например душистый горошек;
3) цепляющиеся щетинками, например цепкий подмаренник.
б) Крупнолистные травянистые растения (ревень).
Формы роста растений
67
в) Стеблевые травянистые растения с облиственным стеблем, без розетки листьев
у основания:
1) высокотравье, например золотарник, кипрей’и многие зонтичные (Umbelli-
ferae);
2) низкие стеблевые травянистые растения.
г) Пол у розеточные растения с облиственным стеблем и прилегающей к поверх-
ности почвы прикорневой розеткой листьев, например лечебная кровохлебка (San-
guisorba officinalis).
д) Розеточные травянистые растения с прикорневой розеткой листьев и безлист-
ным (Taraxacum, Bellis, Primula) или слабо облиственным цветочным стеблем (Viola
Silvestris).
е) Побегообразующие травянистые растения с почками, перезимовывающими
на поверхности почвы (земляника), а также дернообразующие злаки, осоки, папо-
ротники.
ж) Гемикриптофитные травянистые растения, растущие гнездами, например
альпийский мак, а также кислые травы альпийских маттов.
ОЗИМЫЕ ОДНОЛЕТНИКИ И ДВУЛЕТНИКИ
Этот класс форм роста охватывает однолетние или двулетние тра-
вянистые растения. В него входят две различные формы, переходные
между многолетними и терофитными травами, жизнь которых ограни-
чивается лишь одним вегетационным периодом.
а) Озимые однолетники. Из семян, созревающих в течение того же
года, вырастают к концу вегетационного периода розетки листьев, кото-
рые переживают неблагоприятное время года как гемикриптофиты.
В последующий вегетационный период разбивается плодоносящий побег.
К озимым однолетникам принадлежат озимые зерновые злаки, пастушья
сумка, рапунцель, крупка.
б) Двулетники. Их развитие распадается на два вегетационных
периода. Растения перезимовывают в виде семян, как терофиты. В пер-
вый вегетационный период они часто образуют довольно большие розетки
листьев, которые сохраняются во второй период покоя. На следующий
год, выпустив плодоносящие побеги, они погибают. Среди представителей —
наперстянка (Digitalis), синяк (Echium) и многие чертополохи (Cirsium).
Двулетники встречаются чаще всего в периодически сухом климате уме-
ренно теплой и умеренной зон.
ГЕОФИТ НЫЕ ТРАВЫ
Растения, которые переживают холодное или сухое время года при
помощи почек возобновления, находящихся под поверхностью почвы.
Большинство из них накапливает питательные вещества, а часто и воду
в подземных органах (корневища, луковицы или клубни). Они растут
преимущественно в периодически сухом и умеренно теплом климате
холодными зимами.
Жизненный ритм геофитов может быть различным. В эпизодически влажном кли-
мате пустынь или на их окраинах надземные побеги некоторых клубневых геофитов
появляются нерегулярно, лишь после случайных дождей (Oxalis, Dioscorea). В суб-
тропическом климатес влажной зимой и в степных областях умеренной зоны некоторые
луковичные геофиты, прорастающие еще весной, заканчивают свой вегетационный
период созреванием семян уже к концу влажного времени года. У других видов (Alli-
um) плод созревает только в сухое летнее время. У некоторых клубневых геофитов
68 Составные элементы растительности и принципы их распространения
цветение и созревание плода растягивается на два вегетационных периода. Например,
шафран (Crocus albiflorus) образует плод только после летнего периода покоя. Безвре-
менник, цветущий осенью после летнего периода покоя, только на следующий год выно-
сит свой плод из почвы вместе с листвой.
Весенние цветы летнезеленого лиственного леса: лесная ветреница
(корневищное растение), хохлатка (растение со стеблевыми клубнями) и
лютичный чистяк (растениес корневыми клубнями) — имеют весьма корот-
кий надземный вегетационный период. При первом же согревании почвы,
прежде чем деревья покроются листвой, у них развиваются побеги, зало-
женные в подземных почках еще с прошлого лета, распускаются цветки,
а после созревания плодов они или полностью (как лютичный чистяк)
уходят обратно под землю, или оставляют на поверхности только часть
листьев. У некоторых видов, как у перелески, листья могут сохраняться
на почве и зимой.
Многие геофитные травы распускаются только летом, как, например,
тенелюбивые растения лиственных лесов. Это преимущественно корневищ-
ные геофиты (Convallaria, Polygonatum, Paris, Oxalis), но также и клуб-
невые растения {Arum maculatum и некоторые орхидеи).
По характеру органов вегетативного возобновления (Dduerorgane) различают
несколько форм:
а) Геофиты с корневыми почками, у которых почки возобновления в период покоя
размещаются у основания стебля под поверхностью почвы (Aconitum napellus).
б) Корневищные геофиты с более или менее горизонтально растущими под-
земными побегами, как, например, ландыши (Convallaria), некоторые папоротники
из рода Polypodium, многочисленные осоки и злаки.
в) Луковичные геофиты, особенно характерные для семейств Liliaceae (Allium,
Lilium, Scilla), Amaryllidaceae (Leuco.ium, Amaryllis, Narcissus) и Iridaceae (Gladiolus).
г) Геофиты co стеблевыми клубнями, многолетними (Cyclamen) или образующи-
мися ежегодно на боковых побегах (картофель).
д) Геофиты с корневыми клубнями, одним (Orchis) или несколькими (Asphodelus,
Ranunculus ficaria).
е) Геофиты с паразитирующими корнями; растения, берущие питательные
вещества из корней других растений и сохраняющиеся в период покоя под почвой.
Их надземные побеги не имеют зеленых листьев, например у заразихи (Orobanchaceae)
и Петрова креста (Scrophulariaceae). Обитающая в тропических лесах Южной Азии
Rafflesia arnoldi выносит на поверхность только громадный ярко-красный цветок.
К этим формам примыкает еще одна группа растений с несамостоятельным угле-
родным питанием, чьи корневища существуют в симбиозе с грибами: подъельник
(Pirolaceae), гнездовии и ряд орхидей.
ТЕРОФИТНЫЕ ТРАВЫ
Однолетние растения, которые переживают неблагоприятное время
года в виде семян и весь жизненный цикл которых от прорастания до
созревания плода завершается за один вегетационный период. Продолжи-
тельность жизни колеблется от нескольких дней или недель до года.
Основная область распространения терофитных трав — аридные и полу-
аридные местности жарких зон и районы с жарким летом в умеренной
и холодной зонах. В сухих областях, где они появляются после эпизоди-
ческих осадков или кратковременных дождливых периодов, их называют
Формы роста растений
69
эфемерными травами, или дождезелеными терофитами. В климате с холод-
ными зимами, где они относятся к летней флоре, их называют летними
однолетниками (Sommerannuele).
Среди терофитных трав различается большое количество групп:
а) высокие стеблевые травы (подсолнечник);
б) низкие стеблевые травы (Ranunculus arvensis);
в) полурозеточные травы (Centaurea cyanus)',
Г) терофитные злаки (яровые злаки);
д) розеточные травы (Draba verna, Arnoseris minima), обитающие в Акатаме виды
родов Plantago, Viola, Calandrinea, некоторые из них с суккулентными листьями;
е) стелющиеся, ползучие и побегообразующие травы (Stellaria media)',
ж) лазящие травы: вьющиеся (Polygonum convolvulus) и цепляющиеся усиками
(Vicia hirsuta);
з) паразитные терофиты (например, виды Cuscuta).
ГИДРОФИТЫ С ПЛАВАЮЩИМИ ЛИСТЬЯМИ
а) Укореняющиеся: большинство кувшинок, некоторые виды рдеста,
болотник (С alii trie he).
б) Свободноплавающие: водяной гиацинт (Eichharnia), телорез (Stra-
tiotes), сальвиния (Salvinia), ряска (Lemna).
ПОГРУЖЕННЫЕ ТРАВЫ
Водные растения с погруженными в воду побегами, например много-
численные виды рдеста (Potamogetori), уруть (Myriophyllum), водяная
чума (Helodea).
ТАЛЛОМНЫЕ ЭПИФИТЫ
Эпифитные мхи, лишайники, грибы и водоросли; растения, способ-
ные переносить длительную засуху и высыхающие при этом полностью;
в обычных условиях они поглощают влагу из атмосферы всей своей поверх-
ностью. Весьма богатый формами класс форм роста:
а) лишайники и мхи, свисающие наподобие бороды;
б) мхи, живущие на коре деревьев, образующие подобие ковра или подушек;
в) цепляющиеся за древесную кору листоватые и кустистые лишайники;
г) эпифиллы — живущие на листьях лишайники, водоросли и мхи.
ТАЛЛОМНЫЕ ХЛМЕФИТЫ
Болотные мхи, образующие кочки, рыхлые ковры или подушки.
Кустистые лишайники широко распространены в тундре и высокогорных
районах, вблизи верхней границы древесной растительности.
ТАЛЛОМНЫЕ ГЕМИКРИПТОФИТЫ
Плотно прилегающие к почве мхи (прежде всего печеночные мхи),
листоватые и корковые лишайники, прикрепленные водоросли.
ТАЛЛОМНЫЕ ГЕОФИТЫ
Губки, подпочвенные грибы с надземным плодоносящим телом.
70 Составные элементы растительности и принципы их распространения
ТАЛЛОМНЫЕ ТЕРОФИТЫ
Мхи, лишайники и грибы с малой продолжительностью жизни.
ЭДАФОФИТЫ
Микрофиты, живущие в почве, в частности — бактерии и грибы.
ТАЛЛОМНЫЕ ГИДРОФИТЫ
Нитевидные водоросли пресных вод и плавающие мхи. В морской
воде исключительное разнообразие форм, классификацию которых мы
здесь не приводим.
ПЛ АН ИТ О ФИТЫ
Обитающие во взвешенном состоянии в воде микрофиты. Сюда можно
отнести и низшие растения, обитающие на снегу и льду; скопление некото-
рых из них создает издали впечатление «кровавого снега».
ЭНДОФИТЫ
Растения, паразитирующие внутри других растений или животных,
преимущественно грибы и бактерии.
Причины пространственного
распределения систематических единиц
и форм роста
Современное распространение растений на Земле обусловлено разно-
образием растительных систематических единиц и требованиями, кото-
рые они предъявляют к окружающей среде, неодинаковостью условий
существования на поверхности Земли, распространением растений в раз-
личные геологические периоды и воздействием человека.
БИОГЕНЕТИЧЕСКИЕ ПРИЧИНЫ СОВРЕМЕННОГО
РАСПРОСТРАНЕНИЯ РАСТЕНИЙ
ОБРАЗОВАНИЕ СИСТЕМАТИЧЕСКИХ ЕДИНИЦ
Неодинаковое распределение систематических единиц растений по
поверхности Земли объясняется:
а) пространственными различиями в формировании систематических
единиц, которые обусловлены историческими особенностями развития
жизни на Земле;
б) тем, что каждая систематическая единица благодаря генетиче-
скому своеобразию предъявляет определенные требования к окружаю-
щей среде и по-разному реагирует на имеющееся в ее распоряжении про-
странство обитания.
Пространственная дифференциация, обусловленная историей разви-
тия Земли в целом, находит свое выражение во флористическом
делении земной поверхности. На процессе образова-
ния систематических единиц, который объединенными усилиями стараются
реконструировать палеонтология, историческая география, систематика
и генетика, мы не имеем возможности останавливаться.
‘ Флористическая геоботаника стремится путем сравнительного изучения ареалов
родственных систематических единиц (например, всех разновидностей и форм одного
вида пли всех видов одного рода) воссоздать историю развития систематических еди-
ниц. Исходя из положения ареалов, устанавливаются центры скопления родственных
систематических единиц, а также степень дифференциации на генетически неустойчи-
вые систематические единицы низшего ранга, что позволяет сделать заключение о цент-
рах их возникновения и развития. Надежность этих заключений должна быть под-
тверждена результатами исследований в других отраслях. Для географии раститель-
ности в общем достаточно лишь принять к сведению наличие таких центров скопления,
ajLUjncnrrioi, растительности и принципы их распространения
как особенность флористического характера флористических провинций, тем более
что относительно генетических процессов чаще имеются весьма спорные гипотезы,
нежели достоверные данные.
Основное значение для решения проблем географии растительности
имеет результат биогенеза — заполнение участков зем-
ной поверхности систематическими едини-
ц а м и, которые предъявляют различные требования к окружающей
среде.
БИОГЕНЕТИЧЕСКИ ОБУСЛОВЛЕННЫЕ
ЭКОЛОГО-СИСТЕМАТИЧЕСКИЕ ЕДИНИЦЫ
Отдельные растения, относящиеся к одному и тому же таксону низ-
шего ранга, более или менее одинаково реагируют на условия существо-
вания. Возникает понятие экологической единицы, однако
здесь возможны два варианта: или виды на основе одинаковых требова-
ний, предъявляемых ими к условиям существования, относятся к одина-
ковым экологическим единицам, или, что наблюдается столь же часто,
систематические элементы низшего ранга (разновидности, формы, линии,
или клоны, предъявляющие, как заметил еще Кернер, к окружающей среде
разные требования) следует рассматривать как экологически одинако-
вые единицы. Возможны случаи, когда отдельные растения, морфологи-
чески ничем не отличающиеся друг от друга, обладают различными эко-
логическими требованиями, закрепленными наследственно, что может
быть подтверждено экспериментально. В таких случаях говорят об эко-
типах внутри вида, подвида или разновидности. Так называемые Proveni-
enzen древесных пород, выделяемые в лесном хозяйстве, и являются
частично именно такими экотипамих. Следовательно, основные
экологические единицы могут быть разного таксономическо-
го ранга.
В обиходном лексиконе геоботаники, если нет особых причин для более точного
подразделения, основные экологические единицы называют «видами», даже если с точ-
ки зрения систематики они относятся к разновидностям, подвидам, формам и т. д.
В будущем ради упрощения мы тоже будем говорить о «видах растений» как об основ-
ных биоэкологических единицах, невзирая на их ранг в систематике, о котором доста-
точно говорит само название растения.
ЭКОЛОГИЧЕСКАЯ КОНСТИТУЦИЯ
Природные различия отдельных участков земной поверхности вли"
яют на распространение растений, так как виды растений предъявляют
различные требования к окружающей среде и при данных условиях
существования имеют неравные шансы в борьбе за жизнь.
Совокупность генетически обусловленных особенностей одного вида,
важных с точки зрения взаимоотношений с окружающей средой и, сле-
довательно, являющихся одной из причин распространения, называют
экологической конституцией вида.
Экологическая конституция вида определяет место произрастания
этого вида и возможность успешной конкуренции с другими видами при
1 Словом Provenienzen (лат. proveniens — возникающий, происходящий)
в немецком лесоводстве называют климатические расы древесных пород, возникнове-
ние которых обусловлено различиями мест их происхождения по климатическим
условиям и условиям местообитания.— Прим. ред.
Причины пространст. распределения систематических единиц и форм роста 73
менее благоприятных для него условиях. Решающими при этом являют-
ся особенности всей внешней и внутренней организации вида. Между
экологически существенными и несущественными признаками имеется
ряд переходов. Не всегда наиболее заметные черты внешнего облика
растения являются одновременно и самыми важными для жизнеспособ-
ности вида. Основные жизненные процессы, определяющие рост расте-
ния, его размножение и сохранение местообитания (питание, транспира-
ция, ассимиляция, оплодотворение, созревание семян), во многом зависят
от сил, действующих на месте произрастания, то есть от «окружающей
среды».
Поэтому среди причин распределения растений на земном шаре нема-
ловажную роль играют уже упомянутые способы приспособления форм
роста, проявляющиеся как во внешнем, так и во внутреннем устройстве
растений, к определенным экологическим факторам, и прежде всего к усло-
виям света и тепла, водному режиму, условиям питания и специфическим
условиям места произрастания. Своеобразны растения с особыми форма-
ми питания: паразиты, полупаразиты, сапрофиты и насекомоядные («пло-
тоядные») растения, а также виды, существующие в симбиозе с другими
растениями. К экологической конституции относятся также те особенно-
сти внутренней организации растения, которые регулируют функции
отдельных органов и жизненный ритм растения в целом, а также опреде-
ляют способ размножения — спорами, семенами или вегетативно с помо-
щью выводковых почек, побегов и т. д.
От характера приспособлений к распространению может зависеть
более быстрое или медленное увеличение ареала вида. Некоторые расте-
ния активно разбрасывают семена в стороны от растения-родителя (раз-
брасываемые и разбрызгиваемые плоды, например бешеный огурец).
У других наблюдается пассивный перенос семян, спор или плодов на бо-
лее далекие расстояния с помощью ветра, воды или животных. В послед-
нем случае плод должен быть съедобен, семена обладать стойкостью при
прохождении через пищеварительный тракт животного, а также быть
надежно защищенными от неблагоприятных климатических условий или
от преждевременного прорастания. Среди животных особенное значение
для распространения растений имеют прежде всего птицы, летучие мы-
ши, грызуны, обезьяны и муравьи.
Новейшими исследованиями выяснено, что величина и форма ареалов растений
не в такой степени зависит от способа распространения спор и семян, как это предпо-
лагалось ранее. Для расселения растений перенос зачатков оказывается лишь первым
предварительным условием. Решающим же моментом в конце концов является возмож-
ность растения жить в тех условиях, в которые оно попало, и возможность его победы
в борьбе за существование с другими обитателями этого места. Именно этим объяс-
няется тот факт, что низшие растения (грибы, мхи, папоротники), у которых мельчай-
шие споры, способные парить в воздухе, переносятся почти безгранично далеко,
являются, однако, немногим больше космополитными, чем цветковые растения (Christy
1910; Herzog, 1926).
СРЕДА ОБИТАНИЯ И РАСПРОСТРАНЕНИЕ
ЭКОЛОГО-СИСТЕМАТИЧЕСКИХ ЕДИНИЦ
Под средой обитания, или «местообитанием в аутэкологи-
ческом смысле» понимается весь комплекс сил, воздействующих на образ
жизни растения в месте его произрастания. От экологической конститу-
ции вида зависит возможность существования этого вида на данном месте.
74
Составные элементы растительности и принципы их распространения
Растения тесно связаны с целым рядом факторов окружающей среды, таки-
ми, как свет, тепло, вода, питательные вещества.
Причинная зависимость эколого-систематических единиц от этих
факторов выяснена лишь для весьма ограниченного числа случаев. Это
объясняется тем, что воздействия отдельных факторов в условиях естест-
венного местообитания чрезвычайно сложно переплетаются между собой
и изучить их экспериментально очень трудно. Обзор результатов подоб-
ных исследований дают Люндегорд и Вальтер.
Прежде всего еще не окончательно выяснен вопрос, в какой степени недостаточ-
ность одного фактора может быть компенсирована избытком другого. Такая взаимная
зависимость существует, например, между недостатком воды и избытком солей; недо-
статок воды в аридных почвах тем губительнее для растений, чем больше содержание
солей в почве. Линдквист (Lindquist, 1931) показал, что одни виды букового леса,
обитающие на щелочных почвах, более теневыносливы, чем экземпляры, растущие на
кислых почвах, и что другие виды при более слабой освещенности предпочитают щелоч-
ные и слабокислые почвы, тогда как при обилии света растут и на очень кислых почвах.
Во всяком случае, такая компенсация при определенном минимуме каждого отдельного
необходимого для жизни фактора имеет свои пределы. Вид прекращает свое существо-
вание, когда один из факторов достигает вышеупомянутого минимума или, наоборот,
присутствует в избытке, что также губительно для растения.
Из этого следует «закон относительного постоянства местообитания» (Walter,
1953), который еще Друде (Drude, 1876) сформулировал следующим образом: «Многие
растения, особенно вблизи границ своего ареала, путем перемены местообитания выда-
ют причину прекращения своего распространения в определенном направлении. Так,
северные растения или растения высокогорных районов ищут в теплых пограничных
районах своего ареала влажную тень лесов, что указывает на то, что увеличение инсо-
ляции и возрастающая в связи с этим транспирация препятствуют их произрастанию;
с другой стороны, можно наблюдать, как, например, растения, которые в северо-
западной Германии имеют северо-западную границу произрастания, стремятся в самых
крайних, выдвинутых к северо-западу пограничных точках найти сухие и жаркие,
открытые солнечным лучам местообитания, благодаря чему попадают в условия, близ-
кие к привычным им условиям континентального климата. Так в каждой области
в условиях локальной конфигурации местности повторяются виды климата, имеющие
широкое распространение в другой области».
Границы ареалов не всегда экологически обусловлены. Они могут
определяться и временем, например там, где область возможного обита-
ния занята распространяющимся видом еще не полностью. Чаще, однако,
границы ареалов определяются тем экологическим фактором, который
в данном участке уже не достигает минимума, необходимого для сущест-
вования соответствующего вида. В этих условиях, например, растения
вблизи границ своего ареала лишь в исключительных случаях дают зре-
лые семена или вовсе не образуют семян.
Каждый вид при определенном сочетании экологических факторов
получает оптимальные условия существования. Где эти условия выпол-
няются, там данный вид обладает максимальной жизнеспособностью
и успешно конкурирует с другими видами данного местообитания. Это
конституционно обусловленное специфическое соответствие растений опре-
деленным местообитаниям экспериментальная экология называет «при-
способляемостью». «Если мы говорим, что тенелюбивые растения как
жизненная форма приспособлены к жизни в лесной тени, то мы имеем
Причины, пространст. распределения систематических единиц и форм роста 75
в виду, что они на основании своих генотипических особенностей и своей
физиолого-морфологической организации только здесь получают опти-
мальные условия произрастания» (Lundegardh, 1949).
ЭКОЛОГИЧЕСКИЕ ГРУППЫ
Зависимость между распространением видов растений и климатом
или свойствами почвы была обнаружена еще в самом начале развития
географии растений. Давно уже делались попытки разделить растения
на экологические группы, положив в основу отношение
растений к среде в конкретной обстановке.
Сулави (Soulavie, 1783) различал виды, привязанные к климату, и виды, без-
различные к нему. «Естественным климатом» растения он называл диапазон изменений
климатических условий, в пределах которого соответствующий вид может давать
зрелые семена. По находкам ископаемых растений Сулави воссоздавал климат пред-
шествующих периодов истории Земли.
Скау (Schouw, 1822) называл следующие возможные группировки растений:
1. По среде обитания (погруженные и плавающие водные растения, растения-
амфибии, а также наземные и подземные растения суши).
2. По степени освещенности (светолюбивые, тенелюбивые и растения, обитающие
в темноте).
3. По субстрату; среди этой группы две подгруппы: по химико-геологическому
характеру (песчаные, известняковые, солончаковые растения; кремнеземные, базаль-
товые п гранитные растения) и по степени влажности почвы и плотности субстрата
(болотные, илистые, топяные и влажнолуговые растения; щебневые, галечные и скаль-
ные растения).
4. По типу растений-сожителей, преобладающих на месте произрастания.
Унгер (Unger, 1836) ввел понятия, отражающие отношение растений к почве:
«постоянные», «благосклонные» и «безразличные» растения. На этой основе в середине
XIX в. развилось учение о растениях — индикаторах почвы. Де Кандоль (De Candolle,
1874) делил растения по их отношению к теплу на мегатермы, мезотермы, микротермы
и гекистотермы, а введя такое понятие, как «ксерофилы»,— и по их требованию
к влажности. Последняя классификация позднее была уточнена Шимпером.
Варминг, Люндегорд, Вальтер, Элленберг и др. развили дальнейшее
подразделение растений на «экологические группы». В такие группы
объединяются виды, которые по своей экологической конституции более
или менее одинаково привязаны к определенному фактору местообитания.
Принцип определения локальных различий в особенностях местообитания по
«экологическим группам растений» применил еще Кернер, изучавший температурный
режим источников. При этом он использовал виды растений, для которых путем мно-
гократного изменения температуры удалось найти температурные границы, опреде-
ляющие их существование.
ПОТРЕБНОСТЬ В СВЕТЕ
Люндегорд в своей классификации растений по их потребности в све-
те, иначе говоря, по способности растения переносить сильное или уме-
ренное освещение, прибавляет к тем группам, которые выделил еще Скау,
две новые группы и различает:
76 Составные элементы растительности и принципы их распространения
а) растения, произрастающие без света: обита-
тели почвы (эдафофиты), морских глубин и некоторые паразиты (эндо-
фиты т);
б) растения сумеречного освещения: обитатели
морей, лесов (водоросли, печеночные мхи), эндолитные1 2 водоросли, расту-
щие в пещерах и на камнях, и водоросли, растущие на песках;
в) облигатно-тенелюбивые растения, например
лесные растения;
г) факультативно-тенелюбивые растенияс ши-
роким диапазоном в требованиях к свету, среди них виды, которые своп
ранний период развития проводят при слабом свете, а в дальнейшем
растут при полном освещении;
д) солнцелюбивые растения, ярким примером которых
являются высокогорные растения.
Кроме того, экологические группы могут различаться и по потреб-
ности в свете на определенных этапах развития растения, например
среди семян и спор (папоротников) имеются прорастающие на свету и про-
растающие в темноте, или группы растений, у которых раскрытие почек
ускоряется или затормаживается светом или, наоборот, темнотой. После
того как Гарнер и Аллард (Garner, Allard, 1920, 1924) установили влия-
ние периодичности света на цветение и плодоношение многих растений,
различают растения короткого дня и растения длинного дня. Обнаружено
множество вариантов зависимости роста и распространения видов от
требований их к периодичности света. Растения короткого дня — это
главным образом тропические растения. В умеренной зоне существу-
ют виды, которые по своей конституции требуют для развития сначала
несколько недель с короткими днями, а затем им необходимы длинные
дни; именно эти условия предоставляет в их распоряжение среднеевро-
пейский климат — короткие дни весной, длинные летом и, наконец, корот-
кие дни осенью (Biinning, 1955). Прекращение деятельности камбия
с наступлением осени у некоторых деревьев связано именно с укороче-
нием светового дня.
ПОТРЕБНОСТЬ В ТЕШЕ
Классификация растений по их потребности в тепле при более вни-
мательном ее рассмотрении усложняется в зависимости от особенностей
того или иного температурного фактора.
Уже давно была замечена различная степень развития растений холодных и теп-
лых зон (при достаточной влажности). Это натолкнуло на мысль сопоставить границы
распространения видов и средние значения температур. Именно эти соображения
способствовали открытию Гумбольдтом метода изотерм. Де Кандоль (De Candolle,
1874) группировал виды растений в зависимости от величины средней годовой тем-
пературы: более 20° — мегатермы, 15—20° — мезотермы, 0—14° — микротермы. Уже
Валенберг (Wahlenberg, 1812) указал на значение сезонного распределения тепла
для развития растений, а в своих исследованиях в Лапландии учитывал также тем-
пературу почвы и прямую солнечную радиацию.
1 Эдафофиты (греч. edaphophyton; edaphos — почва, phyton — растение) —
растения, живущие в почве. Эндофиты (греч. endophyton; endon — внутри) — расте-
ния, живущие внутри других растений (паразиты).— Прим. ред.
2 Эндолитные (греч. endolithos; endon — внутри, lithos — камень) — растения,
внедряющиеся в каменистый субстрат или живущие на нем.— Прим. ред.
Причины пространст. распределения систематических единиц и форм роста 77
Люндегорд (Lundegardh, 1925) группирует растения прежде всего
по их требованиям к равномерности или периодичности изменений темпе-
ратуры и уже во вторую очередь по значению температуры. При этом
необходимо различать растения, которые требуют сильной жары, рез-
ких холодов или резкой смены температуры, и растения, которые могут
выносить жару, холод и смену температур, существование которых, одна-
ко, не связано с этими условиями. Раункиер при обосновании «жизненных
форм» учитывает прежде всего способность видов переносить холодное
время года, обусловленную строением и жизненным ритмом.
Потребность видов в тепле весьма различна. Некоторых удовлетво-
ряет короткий, другим нужен длительный период ассимиляции, в тече-
ние которого температура должна превышать какое-то определенное
«пороговое» значение, а растение должно получить соответствующее
минимальное количество тепла.
Зависимость жизненных процессов в растениях от определенных количеств
получаемого тепла была известна еще в XVIII в. Розенталь (Rosenthal, 1784) говорил
о сумме тепла, в которой нуждается растение, проходя свой жизненный цикл от про-
растания до цветения и от цветения до созревания семян. В более позднее время эта
проблема подробнее исследовалась прежде всего с точки зрения сельскохозяйственной
ботаники, что привело к классификации видов по требованиям их к теплу.
Многие виды на определенных стадиях своего развития требуют
более низких температур. Другие, наоборот, требуют равномерности
тепла в течение всего года, и уже незначительное отклонение от нормы
может нарушить их жизненный процесс. Чувствительность к холоду
или жаре у одного и того же вида на разных стадиях его развития может
быть различной.
Так, например, экологическая граница распространения гигантского кактуса
(Carnegia gigantea) в северо-западной Аризоне определяется' изотермой —10° С, то есть
температурой гибели молодых растений от холода. Граница ареала европейского
бука в Северной Европе, вследствие независимой периодичности его цветения, зависит
как от поздних заморозков, так и от летней жары. Некоторые виды растений предъяв-
ляют определенные требования даже к суточному ходу температуры.
В общем же не удается коротко сформулировать температурные
пределы: ни оптимальные с точки зрения конституции вида, ни мак-
симальные или минимальные с точки зрения жизненной амплитуды
растения.
ПОТРЕБНОСТЬ В ВОДЕ
Потребность растений в воде, иначе говоря, их способность перено-
сить влажность, сухость или изменение этих факторов во времени, являет-
ся одной из важнейших причин распространения видов. Поэтому уже
давно делались попытки классифицировать виды именно с этой точки
зрения (De Candolle, 1874; Schimper, 1898) (Де Кандоль, Шимпер, Фит-
тинг, Люндегорд, Стоккер, Вальтер, Элленберг). Возможность сущест-
вования данного вида определяется тем, насколько форма роста и физио-
логическая конституция вида обеспечивают растению оптимальный вари-
ант водного баланса.
Это зависит от органов, поглощающих воду (мощность корневой
системы, всасывающая сила корней, способность набухания у кончиков
корней, а также поглощение воды талломами или побегами), и от факто-
78 Составные элементы растительности и принципы их распространения
ров регулирования водоотдачи. К таким факторам, действующим в боль-
шей или меньшей степени, непрерывно или периодически, относятся:
малая по сравнению с весом всего тела растения его общая поверхность,
ограничивающая кутикулярную транспирацию; кутинизация эпидерми-
са; восковой налет; волосяной покров; количество, величина и степень
закрывания устьиц; особые физиологические явления (изменения плазмы)
и, наконец, периодичность вегетативной жизни (тропофиты, геофиты,
терофиты).
Различное относительное потребление воды, то есть количество воды,
транспирированное растением на 1 г образовавшегося сухого вещества
(например, кукуруза требует 350 г воды, рожь — 550 г), в связи с этим
тоже следует считать признаком экологической конституции.
О важнейших формах приспособления к различным условиям увлаж-
нения, вырабатывающихся у всевозможных форм роста, было уже ска-
зано. По потребности в воде различают несколько категорий растений:
а) Растения, для которых вода является естественной средой обита-
ния (гидрофиты).
б) Наземные растения, которые требуют очень влажного воздуха
и влажной почвы (гигрофиты). Они увядают уже при малейшем недо-
статке воды.
в) Растения, которые требуют умеренной или обильной влажности,
однако могут переносить преходящую умеренную сухость (мезофиты).
г) Растения, способные переносить кратковременную (например,
в течение дня) иногда даже сильную сухость (например, древесные эпи-
фиты и растения морских побережий с суккулентными листьями).
д) Растения, которые могут переносить длительную (сезонную),
но не исключительную по величине сухость:
1) с хорошо развитой способностью регулировать транспирацию;
тропофиты (например, листопадные деревья);
2) с относительно большим сопротивлением транспирации, однако
со слабой способностью ее регулировать: гемиксероморфные, как,
например, вечнозеленые деревья с сильно опушенной умеренно-
ксероморфной листвой.
е) Растения, не погибающие даже в длительную и очень сильную
засуху:
1) со способностью регулировать транспирацию и переносить боль-
шую потерю воды без увядания — склеро-ксероморфные растения;
2) со значительной, но слабо регулируемой сопротивляемостью
к транспирации — безлистные стеблевые суккуленты;
3) растения, полностью высыхающие, как, например, таллоэпи-
фиты и скальные растения.
ж) Растения, которые переносят постоянную более или менее равно-
мерную сухость с эпизодическим увлажнением (наноксероморфные и сук-
кулентно-ксероморфные формы).
Независимо от этой общей классификации можно выделить некото-
рые специфические формы приспособления к водному фактору, напри-
мер растения, у которых семена и споры прорастают лишь при определен-
ной влажности воздуха, а также растения, живущие или погибающие
в условиях застойного увлажнения или при длительном промерзании
почвы, и т. д.
В Голландии и Германии экологические группы растений уже давно применяются
на практике для определения степени обеспеченности водой лугов и пастбищ.
Причины пространст. распределения систематических единиц и форм роста 79
ВОЗДЕЙСТВИЕ ВЕТРА
Немалое значение для растений имеет также движение воздуха,
причем растения в зависимости от экологической конституции по-разному
противостоят действию ветра. Некоторые «ветроустойчивые» растения
хорошо переносят механическое или физиологическое воздействие силь-
ных ветров (например, благодаря гибкости их побегов, образованию
почек у самого основания, подушковидной форме и т. п.). Другие,
напротив, сильно страдают от ветров. Для многих растений ветер являет-
ся положительным фактором: он способствует опылению цветков или
распространению семян.
ТРЕБОВАНИЯ К ХИМИЧЕСКИМ СВОЙСТВАМ
СУБСТРАТА
О растениях-индикаторах, указывающих на наличие определенных
свойств почвы, за полтора столетия накопилась обширная литература
(например, работы Эбермейера, Унгера, Троммера, Ратцебурга, Гофма-
на, Кернера, Линстова, Элленберга).
Растения можно классифицировать, исходя из следующих хими-
ческих особенностей субстрата, имеющих значение для распространения
видов:
Концентрация ионов водорода в субстрате
(pH) или в почвенном растворе. Эта величина зависит от веществ, вхо-
дящих в состав субстрата, и наряду с почвенным раствором может быть
подвержена сезонным изменениям. Некоторые виды требуют для опти-
мального роста щелочную реакцию, другие — кислую. Диапазон вели-
чины pH, от которого зависит жизнь растений, может быть и узким и очень
широким.
Содержание минеральных веществ и азота
в субстрате. Большинство питательных веществ, жизненно необхо-
димых для существования растений, почти всюду встречается в доста-
точном количестве. Недостаток одного вещества или вредный его избыток
определяется законом минимума, необходимого для существования вида.
Олиготрофные растения—это растения, довольствующиеся субстра-
том, бедным минеральными солями. Они поселяются на бедных квар-
цевых песчаниках, подзолистых почвах и на торфяниках. Это большей
частью медленно растущие многолетние виды, как, например, хвойные
деревья, карликовые кустарники с эрикоидными листьями, торфяные
мхи и лишайники. Исключительно на олиготрофных местообитаниях
растут некоторые насекомоядные растения и полупаразиты.
Эвтрофные растения для своего развития требуют большого коли-
чества питательных веществ. Особую группу среди них составляют виды,
которые предпочитают богатые азотом почвы, как, например, пастбищ-
ное высокотравье в районе Альмюттена.
Содержание извести в почве. При решении старого
спорного вопроса (Unger, 1836; Thurmann, 1849), почему некоторые
растения предпочитают известковые почвы, а другие их избегают или
относятся к ним безразлично, новые исследования (например, обобщения
Люндегорда и Вальтера) установили, что вопрос этот может быть объяс-
нен по крайней мере тремя различными причинами:
1) растения, которые реагируют на содержание извести в почве толь-
ко в силу физических особенностей местных условий (тепло, сухость);
30 Составные элементы растительности и принципы их распространения
2) растения, для которых решающей является величина pH из-
вестковой почвы; одним благоприятствует щелочная реакция среды,
другие, наоборот, ее не переносят, например торфяные мхи (Sphag-
num)',
3) растения, для которых содержание кальция в почве является
решающим, но в другом плане, чем с точки зрения концентрации ионов
водорода. Такие растения известны в небольшом количестве, и физио-
логия «кальцефильных» или «кальцефобных» растений представляет собой
еще не решенную проблему.
Содержание солей в почве. Известны растения, кото-
рые не нуждаются в высоком содержании солей в почве, но выносят его
и потому преобладают в таких местообитаниях из-за отсутствия конку-
рентов. Другие же при высоком содержании солей получают обусловлен-
ные их конституцией оптимальные условия для произрастания. Эти
последние называют галофитами. Большая часть их имеет суккулентные
листья. В качестве «гликофитов», не переносящих богатого солями
субстрата, им противостоит вся масса остальных растений.
Наличие определенных тяжелых металлов
в почве. Высокое содержание растворимых солей тяжелых металлов,
как, например, карбонатов свинца и цинка, вызывает отравление боль-
шей части растений. Все же имеются виды или подвиды, приспособленные
к таким местообитаниям, однако для большинства этих индивидуумов,
насколько это известно, подобные соединения вовсе не являются жизнен-
но необходимыми.
О роли микроэлементов в распространении видов известно еще очень
мало.
ЗАВИСИМОСТЬ ОТ МЕХАНИЧЕСКИХ
СВОЙСТВ СУБСТРАТА
Можно назвать целый ряд групп растений, отличающихся по своему
отношению к механическим свойствам субстрата:
а) Растения, способные жить на плотных скальных породах,— это
прежде всего водоросли, лишайники и мхи.
б) Растения, обитающие в расщелинах скал, в основном подушко-
видные кустарнички со стержневыми корнями.
в) Растения с поверхностной корневой системой, для которых глуби-
на почвенного слоя большого значения не имеет.
г) Растения с глубокими корнями, причем их можно разбить на
•большое число подгрупп в зависимости от их требований к структуре,
механическому составу почвы и другим ее свойствам.
д) Растения, способные существовать на подвижных почвах, напри-
мер на осыпях, оползающих склонах, подвижных барханах или блуждаю-
щих дюнах. Среди них имеются специально приспособившиеся формы,
например форма притупленной ивы (Salix retusa) или кустистые расте-
ния, задерживающие осыпание почвы; ползучие или стелющиеся по почве
карликовые кустарники, полукустарники или травянистые растения;
растения с побегами (укрепляющие пески или осыпи); геофиты, распола-
гающие свои клубни, луковицы или корневища в зависимости от микро-
рельефа ближе к поверхности или, наоборот, глубже; деревья на
ходульных корнях и др.
Причины пространст. распределения систематических единиц и форм роста 81
ВЛИЯНИЕ РАСТЕНИЙ ДРУГ НА ДРУГА
По характеру связи между растениями, обусловленной самой кон-
ституцией растения, можно различить несколько групп:
а) Виды, использующие другие растения в качестве места поселения
или опоры, такие, как эпифиты, лианы и другие лазящие растения.
б) Виды с особым способом питания и растения с несамостоятельным
питанием:
I) сапрофиты (обитающие на падали) и растения, живущие в сим-
биозе с корневыми или почвенными грибами и бактериями (напри-
мер, с нитрифицирующими бактериями);
2) полупаразиты — растения, которые самостоятельно ассими-
лируют углерод, но воду и минеральные вещества частью факуль-
тативно, а частью облигатно отнимают из систем снабжения других
растений, например очанка, марьянник и омела;
3) настоящие паразиты, незеленые растения, которые берут гото-
вые органические вещества из стеблей (повилика), стволов (дре-
весный гриб) (Polyporus) или корней (заразиха) (Orobanche
rapum-genistae) других растений.
РАСТЕНИЯ И ЖИВОТНЫЕ
Для некоторых видов растений животные имеют большое значение
прежде всего как их распространители. Это те растения, цветки которых
опыляются определенными видами животных или семена которых рас-
пространяются животными. Опасность быть съеденными животными для
каждого растения также различна и может оказаться решающей для
присутствия или отсутствия того или иного вида в определенном место-
обитании.
ВЛИЯНИЕ ЧЕЛОВЕКА
Растения можно группировать также по признакам, способствующим
их сосуществованию с человеком или, наоборот, их истреблению. Для
распространения растения весьма важны различия в степени его полезно-
сти для человека, которые основаны, например, на питательности его
плодов или других органов. Другая точка зрения при такой группиров-
ке — это сопротивление уничтожению при покосах, которое неодинаково
у видов растений с различными формами роста и жизненным ритмом.
Косьба, например, благоприятствует распространению многих растений,
другие же, наоборот, при регулярном скашивании лугов исчезают. Здесь
следует указать также на виды растений, произрастающих как сорняки,
вследствие их экологической конституции, или на рудеральные растения,
а с другой стороны,— на большое число растений, весьма чувствитель-
ных к сернистой кислоте промышленных дымов, естественные ареалы
которых быстро сокращаются.
ПРИЧИНЫ ПРОСТРАНСТВЕННОГО РАСПРЕДЕЛЕНИЯ
СИСТЕМАТИЧЕСКИХ ЕДИНИЦ И РАСТИТЕЛЬНЫХ ФОРМ
КЛИМАТ
Свет и тепло, минеральные питательные вещества и различные фор-
мы субстратов, на которых могут произрастать растения, распределены
на Земле неравномерно. Наличие определенных жизненных форм среди
82
Составные элементы растительности и принципы их распространения
систематических единиц зависит, не говоря об уже упоминавшихся био-
тических факторах и исторических причинах, рассматриваемых ниже,
прежде всего от характера места произрастания, кото-
рый складывается в результате совместного действия климатических
факторов и локальных особенностей данной территории.
Даже в пределах небольшого участка условия существования могут быть совер-
шенно противоположными, например на линии уреза воды, текущей среди скалистых
берегов, или там, где происходит смена видов почв, или на участке соприкосновения
склонов с различной экспозицией. Однако в общем облике Земли локальные различия
отступают на задний план перед различиями обширных областей. Разнообразие в рас-
пределении жизни, если отбросить две столь противоположные сферы, как суша и море,
обусловлено прежде всего несходством климатических условий.
Если мы под климатом вместе с Филиппсоном понимаем «харак-
терные для данного участка земной поверхности атмосферные явления»,
то это определение в зависимости от того, к пространственной величине
какого порядка мы относим «участок земной поверхности», охватывает:
а) «климат минимального пространства» (Г. Краус). Этот климат,
по Гейгеру, теперь большей частью называют «микроклиматом». По
Ансио (Ansiaux, 1948), здесь следует различать еще микроклимат 1-го
порядка (например, всего лесного сообщества), 2-го порядка (одного
из ярусов сообщества) и микроклимат 3-го порядка (непосредственное
окружение отдельного растения);
б) климат более обширного пространства, иначе «климат части мест-
ности», например склона, долины или горной вершины. Такой климат
называют «местным климатом», или по сравнению с микроклиматом —
«мезоклиматом», или, по Кноху,—«климатом местности»;
в) «региональный климат», обусловливаемый, помимо процессов
в верхних слоях атмосферы, прежде всего различиями макрорельефа.
Этот климат в отличие от предыдущих также называют «макроклиматом».
В последнее время говорят также о климате свободной атмосферы и климате
верхних слоев атмосферы. Для интересующего нас вопроса о причинах распростране-
ния растений важны лишь климаты, названные вначале. Для каждого из этих типов
следовало бы специально указать, что понимается под общим определением Филиппсо-
на «характерные атмосферные явления». Микроклимат, например, создает те условия
воздушной среды, которые образуются непосредственно под воздействием растительного
покрова, то есть затененность, защищенность от ветра, изменение содержания угле-
рода и т. д. Они могут быть, однако, несущественны для местного климата и тем более
для макроклимата. С другой стороны, некоторые решающие для местного климата
различия имеют значение и для микроклимата, поскольку они способны создать локаль-
ные отклонения в местном климате.
Среди факторов макроклиматического порядка с точки зрения причин рас-
пространения растений важны те атмосферные явления, которые определяют условия
существования, и в первую очередь свет, тепло, влажность и ветер. Как таковые
они значения не имеют. Их различия становятся ощутимыми лишь в свете осуществле-
ния оптимальной или пессимальной жизнедеятельности растений.
Оптимальные условия ассимиляции на земной поверхно-
сти возникают там, где постоянная и достаточная влажность сочетается
с более или менее равномерно высокой температурой. Поэтому в постоян-
но влажных районах жарких тропических низменностей мы находим
исключительную пышность и одновременно богатство видов и форм расти-
тельного мира. Здесь нет необходимости приспосабливаться к неблаго-
Причины пространст. распределения систематических единиц и форм роста 83
приятным условиям существования. Во всех других климатах, напротив,
могут произрастать только те растения, у которых имеются особые при-
способления, позволяющие им противостоять тому или иному климати-
ческому недостатку. Годичная периодичность растительной жизни осо-
бенно резко выражена там, где условия существования на протяжении
года сильно меняются, как, например, в периодически влажных тропи-
ках, где в сухое время года жизнь растений находится в относительном
покое и растительность кажется сухой, редкой и запыленной, или же
в умеренных и полярных зонах, где жизнедеятельность растений из-за
зимних холодов временами полностью прекращается.
Пессимальные условия, а тем самым абсолютные кли-
матические границы жизни встречаем мы там, где отсутствует одно из основ-
ных условий: всегда не хватает либо требуемого тепла, либо необходимой
влажности, либо и того и другого вместе (пустыни с недостаточным коли-
чеством тепла или сухие пустыни).
Низкая температура, вплоть до минимальной, которая была замерена
на Земле (—81,2°), не препятствует растительной жизни, если в другое
время здесь бывает достаточно тепла, а сама смена температур не отра-
жается на растениях отрицательно.
Между оптимальными и абсолютно пессимальными величинами отдель-
ных климатических факторов имеется ряд промежуточных значений
постоянного или периодического недостатка в тепле и влаге, лимитирую-
щих существование отдельных систематических единиц или определен-
ных групп жизненных форм и форм роста. Именно эти пределы и обу-
словливают в общих чертах распределение различных единиц раститель-
ности на больших пространствах.
Недостаток света как лимитирующий фактор проявляется только
в местном климате пещер, ущелий, под навесами скал и т. д., а также
в микроклимате отдельных растительных формаций, например в напоч-
венном ярусе под пологом сомкнутого леса. Макроклимат в любой части
суши предоставляет достаточное для роста растений количество света.
Изменяющиеся в зависимости от географической широты и высоты над
уровнем моря различия в интенсивности освещения, в суточном или
сезонном ходе солнечной радиации, а также в качестве освещения (напри-
мер, содержание ультрафиолетовых лучей) обусловливают сами по себе
наличие больших различий в жизненном ритме, скорости роста, способ-
ности к цветению и размножению и, наконец, во внешнем облике расте-
ний, произрастающих в приэкваториальных широтах с одинаковой про-
должительностью дня и ночи, растений более высоких широт, с длин-
ными летними днями, и растений приполярных областей, с полугодовой
сменой дня и ночи. Высокогорные области, ритм освещения в которых
не отличается от ритма равнинных областей в тех же географических
широтах, характеризуются большим обилием света и богатством ультра-
фиолетовых лучей.
Постоянный недостаток тепла является климатическим фактором,
наиболее резко лимитирующим распространение растительности. Имен-
но им обусловлено отсутствие растительности на большей части поляр-
ного пояса и на самых высоких горных вершинах. Во внутритропических
широтах с их примерно одинаковыми на протяжении всего года темпера-
турами граница растительности проходит ниже снеговой линии. В кли-
матах более высоких широт, где резче сезонная смена температур, она
располагается значительно выше. Так, в Альпах встречается большое
число растений, преимущественно лишайников, печеночных мхов и водо-
84 Составные элементы растительности и принципы их распространения
рослей, но также и более 200 видов цветковых растений, которые, поселя-
ясь на скалах с благоприятной экспозицией, нагреваемых в летний период,
могут произрастать и гораздо выше снеговой линии. Та же картина наблю-
дается и в некоторых районах Арктики, например на Шпицбергене и в
Гренландии. Даже в полярном климате Антарктики у 83° ю. ш. на ска-
листых нунатаках развивается скудная растительность в виде лишайни-
ков и водорослей.
Полярные и альпийские границы леса и отдельных деревьев, кото-
рые относятся к наиболее характерным рубежам в пределах раститель-
ного покрова Земли, являются по существу границами, определяемыми
Рис. 26. Сравнение границы древесной растительности с июльской
изотермой 10° С для меридионального профиля через Альпы
(по Брокману).
недостатком тепла, если, конечно, какие-либо другие факторы, как, на-
пример, недостаток осадков, иссушающие ветры или частые вторичные
заморозки, не задерживают рост деревьев в еще большей степени, чем
недостаток тепла. За пределами этих границ количество летнего тепла
для роста деревьев или для их естественного размножения (созревание
семян, развитие молодых побегов) уже недостаточно.
Поэтому из всех кривых среднемесячных значений температуры, кото-
рые уже давно принято сопоставлять с границей древесной растительно-
сти, более или менее точно совпадает с этой границей, пожалуй, изотер-
ма ]-10э наиболее теплого месяца. Эта пространственная зависимость,
не применимая, однако, к крупным горным поднятиям (рис. 26), отнюдь
н? говорит о наличии непосредственной причинной связи между границей
древесной растительности и средней температурой наиболее теплого меся-
ца. Названная изотерма лишь до некоторой степени отражает значение
температурной составляющей в общем «характере климата» на границе
древесной растительности. Многочисленные попытки выразить эту зави-
симость точным языком формул не привели до сих пор к удовлетворитель-
ным результатам. Преимущественно температурная обусловленность ска-
зывается и в том, что границы леса и отдельных деревьев в континенталь-
ном климате Сибири и Канады с относительно более теплым летом прохо-
дят гораздо севернее, а высотные границы в глубине более крупных гор-
ных массивов поднимаются.
Причины пространстп. распределения систематических единиц и форм роста 85
Делались попытки обосновать полярные пределы ареалов отдельных система-
тических единиц, помимо летней температуры, и другими элементами климата. Так,
по Энквисту, северная граница европейской ели должна совпадать с линией макси-
мальной суточной температуры +12,5° С, наблюдаемой по крайней мере в течение
65 дней в году. Температурная обусловленность полярной границы систематической
единицы может определяться и крайними ее значениями, проявляющимися лишь
в течение больших отрезков времени, как, например, у вида Ruscus, у которого в самых
его северных местообитаниях на юге Англии семена созревают только при условии
исключительно жаркого лета.
Большое значение для распространения растений имеет граница
морозов, линия, отделяющая безморозный климат от климата, в котором
морозы наблюдаются более или менее регулярно. В климате тропических
низменностей морозы отсутствуют. Граница между тропическим и вне-
тропическим климатом проходит в основном там, где она совпадает с гра-
ницей морозов на низменностях — весьма важным пределом в распростра-
нении многочисленных систематических единиц; кроме того, эта граница
разделяет зоны, в пределах которых правильное с точки зрения геогра-
фии растительности деление климатов основывается на разных клима-
тических факторах.
В тропиках деление растительности по климатическим поясам обу-
словливается прежде всего различием влажности, а во внетропических
широтах — недостатком тепла и зимними холодами или продолжитель-
ностью летних вегетационных периодов и обилием тепла в них.
Во внетропических климатических зонах, особенно в средних широ-
тах северного полушария, из-за различий в степени континентальности
широтное расположение теплых времен года в значительной степени
нарушено или вовсе уступило место восточно-западному их располо-
жению.
В известной мере, если различия во влажности в этих широтах не
являются решающими, смену растительности вызывают переходы от
теплых океанических климатов с мягкими, а при крайне океаническом
положении местности (умеренная зона южного полушария) и вовсе без-
морозными зимами и соответственно продолжительными вегетационными
периодами через субокеанические и субконтинентальные климаты с более
резким различием между теплыми и холодными временами года и с мень-
шей продолжительностью безморозных периодов к климатам внутрикон-
тинентальных областей с весьма холодными зимами и коротким, но очень
теплым вегетационным периодом.
Такой смене климатов в умеренной зоне в общих чертах соответству-
ет распределение форм роста деревьев: вечнозеленые лиственные деревья
в безморозном океаническом климате, летнезеленые лиственные деревья
в океаническом и умеренно-континентальном климате с продолжитель-
ным вегетационным периодом и, наконец, хвойные деревья с коротким
вегетационным периодом, способные выносить или даже требующие про-
должительных зимних морозов. Европейская ель достигает естественной
западной границы там, где суточный минимум хотя бы в течение 120 дней
бывает ниже 0° С; в климатах с более мягкими зимами она подвержена
красной гнили.
Пределы распространения растений, определяемые недостаточностью
осадков, в противоположность термическим границам не являются абсо-
лютными. Отсутствие дождей может быть компенсировано наличием рек
или грунтовых вод (оазисы). Кроме того, даже в крайне засушли-
86
Составные элементы растительности и принципы их распространения
вых областях могут выпасть случайные осадки, хотя бы и с десятилетним
перерывом. Тем не менее пустыни, почти лишенные растительности, зани-
мают значительную часть земной поверхности, располагаясь, как прави-
ло, в пределах субтропических и тропических пассатных поясов (Сахара,
Аравийская пустыня и пространственно ограниченные совместным воз-
действием рельефа и морских течений засушливые области Австралии,
Южной Африки, северного Перу, Нижней Калифорнии). Внутримате-
риковые пустыни характерны для внутренних и сухих территорий Азии
(Тарим, Гоби) и Северной Америки (Большой Бассейн, пустыни в шта-
тах Сонора и Аризона).
В пределах тропических поясов распространение расти-
тельности определяется не температурными различиями (годичная изо-
термия на экваторе и увеличение сезонных контрастов с географической
широтой), а условиями увлажнения, причем скорее имеет значение сезон-
ное распределение осадков и испарения, нежели общее количество осад-
ков. С этой точки зрения в тропиках можно выделить постоянно влаж-
ный, засушливо-влажный, влажно-засушливый и постоянно засушливый
климаты. Периодическую увлажненность создают прежде всего эквато-
риальные дожди в период равноденствия (тропические летние дожди)
или субтропические зимние дожди.
Удобной характеристикой сезонных различий влажности оказалось
число влажных или же сухих месяцев, после того как Вайбель (Waibel,
1921) на примере периодически сухих областей Африки показал возмож-
ность применения этой величины для установления границ тропических
поясов растительности. Для определения степени засушливости местно-
сти в условиях тропиков целесообразно использовать индекс аридности
Мартонна в форме, предложенной Лауэром (Lauer, 1952). Месяц считает-
ся сухим, если значение индекса меньше 20 (п— среднее количество
осадков в месяц, t — средняя месячная температура).
Во внетропических широтах в периодически сухом климате пред-
ставляет интерес взаимный ход осадков и температуры, например летняя
сухость в климатах с холодными зимами совсем не допускает роста деревь-
ев (континентальная граница леса).
Вид осадков, то есть дождь, снег, туман, роса или иней, и их аперио-
дичность также существенны для растительности, особенно в областях
с небольшим количеством осадков. Значение росы, как и ее распростра-
нение, еще не полностью изучено. Во всяком случае, в сухих областях
утренняя роса увеличивает относительную влажность воздуха, в резуль-
тате некоторые растения открывают свои устьица, которые в противном
случае оставались бы закрытыми; таким образом, роса способствует уве-
личению периода ассимиляции.
Ниже приводится перечень основных типов климата, которые обусловливают
зональность распределения растительности на больших пространствах. Вертикальные
климатические пояса при этом не выделены. В каждой климатической области основно-
му типу климата равнин соответствует ряд горных климатов.
Основные типы климата Земли:
I. Климат постоянно влажных тропиков.
II. Периодически засушливый климат влажных тропиков:
а) с коротким сухим периодом;
б) с длительным сухим периодом.
III. Периодически влажный климат сухих тропиков:
Причины пространст. распределения систематических единиц и форм роста 87
а) с длительным периодом дождей;
б) с коротким периодом дождей.
IV. Абсолютно сухой тропический или субтропический климат.
V. Периодически влажный климат сухих субтропиков:
а) с очень коротким периодом дождей;
б) с более длительным периодом дождей.
VI. Климат влажных субтропиков:
а) климат зимних дождей и сухого лета;
б) субтропический климат с влажным летом;
в) постоянно влажный субтропический климат.
VII. Абсолютно сухой климат умеренных широт.
VIII. Периодически влажный климат умеренных широт:
а) «климат полупустынь»;
б) «климат степей».
IX. Климат умеренных широт с равномерным увлажнением в течение года:
а) крайне океанический;
б) от умеренно океанического до умеренно континентального;
в) от континентального до крайне континентального;
X. Субполярный климат:
а) субполярный океанический климат;
б) субполярный континентальный климат.
XI. Полярный климат.
Пространственное распределение типов климата обусловливает в свою
очередь распространение на обширных пространствах целого ряда
физиономически важных растительных форм.
Там, где тепло или влажность минимальны, прослеживается непо-
средственная связь этих факторов с уменьшающейся интенсивностью
образования органических веществ и более медленным ростом, а тем
самым — с высотой и размерами растений. Поэтому как в субполярных
и высокогорных областях, так и на периферии крайне сухих областей
мы встречаем обычно постепенный, но иногда и скачкообразный переход
от древесных растений через кустарниковые формы к формам карликового
роста.
Помимо статистического сравнения участия разных жизненных форм во флорах
стран с различными климатами, как это сделали Раункиер и Варминг, и многочислен-
ных работ об условиях существования на границах древесной растительности, следует
указать также на широкие сравнительные исследования распространения отдельных
форм роста.
Весьма ценным материалом для подобного рода исследований являются спектры
жизненных форм (процент, занимаемый жизненными формами Раункиера), приводи-
мые во многих работах, посвященных описанию растительных сообществ.
Классификация жизненных форм Раункиера основана преимущест-
венно на тех особенностях растений, которые позволяют им переносить
неблагоприятные климатические условия, прежде всего холодные време-
на года. Доля, занимаемая высокими древесными растениями в общем
составе флоры, с увеличением географической широты, как правило,
уменьшается, тогда как доля хамефитов и гемикриптофитов в этом напра-
влении возрастает (ср. рис. 27а и 27в).
88 Составные элементы растительности и принципы их распространения
В постоянно влажных теплых климатах налицо абсолютное господ-
ство фанерофитов, и прежде всего вечнозеленых деревьев с незащищен-
ными почками; характерны также лианы, кустарники, древесные эпифи-
ты и травянистые фанерофиты. Ареал последних ограничен именно этим
климатом. В крайне сухих климатах жарких зон наиболее богато пред-
ставлены терофиты, в Центральной Сахаре, например, они составляют
больше половины всех видов. За ними следуют многочисленные хамефиты
и гемикриптофиты (рис. 276).
Рис. 27. Спектры жизненных форм.
а — Бельгия, 50—51° с. ш. (по Лебрену); б — Эль-Голеа, Сахара,
31° с. ш. (по Раункиеру); в — Янгамби, Конго, 1° с. ш. (по Луи).
Ph — фанерофиты; Ch — хамефиты; Н — гемикриптофиты;G — гео-
фиты; Th — терофиты.
В субтропических областях с зимними дождями многообразие теро-
фитов тоже велико, но особенно характерны геофиты и гемикриптофиты.
В умеренно влажных климатах гемикриптофиты составляют более
половины флоры; в большом количестве здесь встречаются также геофиты
и терофиты, тогда как доля фанерофитов уменьшается (рис. 27а).
Во флоре нивального пояса Альп (2600—3100 м) из 240 видов встре-
чающихся там цветковых растений более двух третей составляют геми-
криптофиты и одну четвертую — хамефиты. Аналогично соотношение
в полярных широтах. На Шпицбергене доля гемикриптофитов составля-
ет 60%. Их дополняют преимущественно хамефиты, тогда как терофиты
почти полностью отсутствуют.
ПОЧВА
Разнообразие почв можно рассматривать как одну из причин про-
странственного распределения растений. Отметим прежде всего зависи-
мость почвообразовательного процесса от климатических условий. Тепло
и влажность способствуют химическому распаду горных пород, тогда
как при недостатке тепла или отсутствии влаги преобладает физическое
выветривание. Поэтому направленность почвообразовательного процес-
са как в холодных, так и в сухих климатах определяется прежде всего
различиями в минералогическом составе горных пород. В остальном
в общих чертах зональному климатическому разграничению соответствует
«планетарная шкала климаксных почв» (Кубиена) от кочкарных тундр
через подзолистые почвы зоны хвойных лесов и среднеевропейские буро-
земы к субтропическим и тропическим бурым глинам, красноземам и крас-
ным глинам. В условиях умеренно влажного климата из почв прежде
Причины пространст. распределения систематических единиц и форм роста 89
всего вымываются основания, при этом глинозем, железо и кварц пассив-
но накапливаются (сиаллитное выветривание); в случае образования
гумусовых соединений (под влиянием растительности) последние, пока
из почвы не вымыты основания, оказываются связанными в верхнем гори-
зонте (бурые лесные почвы). При более сильном выщелачивании, когда
оснований для связывания гумусных кислот уже не хватает, железо
и глинозем перемещаются вниз, почвы оподзоливаются и на глубине
образуются плотные ортштейновые прослойки (подзолистые почвы). Такие
почвы бедны питательными веществами, и требовательные к ним виды
растений существовать на них не могут. На породах, богатых известью,
напротив, возникают темные рыхлые богатые перегноем и минеральными
веществами рендзины.
При аллитном выветривании в условиях жарких влажных тропиков
особенно подвижной становится кремнекислота, которая и уносится вниз.
Глинозем и железо, причем последнее в виде почти безводного соединения
интенсивно-красного цвета, накапливаются (тропические красные глины).
С образованием верхнего гумусового горизонта и в этих почвах может
начаться процесс оподзоливания и обеднения минеральными веществами.
В периодически влажном тропическом климате в дождливые периоды
происходит такой же процесс, как и во влажных тропиках, с тенденцией
в сторону обогащения аллитным материалом. В сухие периоды, напро-
тив, растворимые вещества частично снова подтягиваются к поверхности
и весь процесс в целом замедляется (почвы муссоновых лесов). Если
же выщелачивание прогрессирует, то железо и алюминий становятся
особенно подвижными и поднимаются в сухие периоды на поверхность
почвы, где этот «альферитный» материал откладывается в виде нераство-
римых соединений (латеритный краснозем), так что в конце концов обра-
зуется сплошная твердая корка (латерит).
Субтропические области с периодом зимних дождей занимают проме-
жуточное положение между тропиками и умеренной зоной. Поэтому зимой
выветривание в условиях промывания здесь ближе к сиаллитному типу,
тогда как во время сухого и жаркого лета господствуют аллитные про-
цессы. Это чередование обусловливает образование одновременно целого
комплекса почв, от бурых глинистых, богатых гумусом, через ксеросиал-
литные желтые глины до чисто аллитных почв. Последние на породах,
бедных известью, имеют большей частью желтоватую окраску, тогда
как на известняках формируются красноцветные почвы, обогащенные
в нижней части профиля железом (terra rossa). Эта красная подпочва
вследствие эрозии часто выходит на поверхность.
Широко распространены почвы сухих климатов с известковыми или
соляными корками на поверхности, на которых обитает ограниченное
число видов. Наконец, следует назвать различные формы подвижных
почв (дюны, осыпи, прибрежный ил), а также постоянно или периоди-
чески промерзающие почвы.
ИСТОРИЧЕСКИЕ ФАКТОРЫ
«Ни наличие благоприятных условий существования, ни доступность простран-
ства сами по себе не достаточны для решения вопро&а о заполнении этого пространства
органическими существами. Существенное значение имеет.исторический момент. Только
знание истории возникновения и расселения организмов помогает осмыслить проблему,
каким путем шло заселение данной территории и какие именно виды были первыми
поселенцами» (Kirchhoff, 1899).
уи Составные элементы растительности и принципы их распространения
Формирование систематических единиц растительного мира и обра-
зование их ареалов являются историческими процессами, связанными
с геологической историей Земли. Наши представления об изначальных
центрах возникновения органической жизни и эволюции систематических
единиц в значительной степени основаны лишь на гипотезах. Последние
же исходят из картины современного распределения систематических
единиц и из сравнительно бедных, базирующихся на случайных наход-
ках сведений о распределении их в прошлом.
Со всей определенностью, однако, можно сказать, что историческая
судьба континентов, бывшие связи участков суши между собой и время
возникновения морей, разделяющих эти участки, имеют в этом смысле
большое значение. Без сомнения, причинные связи распространения
систематических единиц с климатическими колебаниями, а также с изме-
няющимся на протяжении истории Земли положением поясов пустынь
и высокогорий тоже имеют место.
Многие особенности современного распределения систематических единиц
настолько ясно зависят от этих причин, что зачастую оказывается возможным по совре-
менному флористическому делению судить о тех процессах в истории геологического
развития Земли и климатов, которые установить другим путем сплошь и рядом не удает-
ся. Так, например, на основании резкого различия флоры считают, что отделение
Мадагаскара от африканского континента произошло в глубокой древности. Исходя
из родственности флоры Патагонии, Новой Зеландии и лежащих между ними островов,
заключают о существовании в раннюю геологическую эпоху антарктического моста.
Оба предположения доказать непосредственно геологическими методами невоз-
можно. Свою гипотезу перемещения материков Вегенер подкреплял наблюдаемым фло-
ристическим родством континентов. Клус, однако, считает, что строению Атлантиче-
ского океана больше соответствуют другие биогеографически оправдываемые пред-
ставления о наличии в прошлом связи между сушами, которое, кстати, высказал еще
Бюффон в 1778 г., нежели предположения о перемещении континентов в таких масшта-
бах, как это следует из гипотезы Вегенера.
Продолжительность существования сухопутной связи между конти-
нентами в прошлом и время, в которое произошло их отделение друг
от друга океанами, обусловливают степень флористического различия
или родства растительных миров, а также являются причиной противо-
положности между небольшими самостоятельными флористическими цар-
ствами на юге и охватывающим всю Землю единством голарктики. Так,
сходство флоры Британских островов и западной части Средней Европы
и отсутствие в ней эндемов объясняется сравнительно недавним (в после-
ледниковый период) отделением островов от материка.
При возникновении новых форм (неоэндемов) еще генетически
пластичных систематических единиц избирательное влияние оказывают,
по-видимому, исторические изменения условий существования, в особен-
ности изменения климата.
С другой стороны, сходные причины привели к сужению некогда
более обширных ареалов систематических единиц, не сумевших приспосо-
биться к изменившимся условиям существования (например, Ginkgo,
Sequoia, Liriodendrori). Многие дизъюнкции ареалов, повторяющиеся
в характерной форме у многих систематических единиц, как, например,
дизъюнкция Мексика — субтропическая Южная Америка или дизъюнк-
ция Восточная Азия — Северная Америка, произошли именно таким
путем. Ни в одном случае не доказано, что один и тот же вид возник само-
стоятельно в нескольких местах. Разорванные ареалы возникли либо
Причины пространст. распределения систематических единиц и форм роста 91
в результате дальнейшего расселения растений, когда вид переносится
на большие расстояния с помощью морских течений, животных или чело-
века, либо они являются следствием сокращения ареалов, сопровождав-
шегося исчезновением вида в одной из ныне разъединенных частей ареала
(рис. 28). Многие случаи реликтового обитания тех систе-
матических единиц, которые раньше имели более широкое распростране-
ние, возникли именно вследствие сокращения ареалов. Реликты разли-
чают по их отношению к пространству (например, Linnaea borealis в Гар-
це — как северный реликт), по времени их происхождения (например,
третичные реликты) или по их флористико-генетическому характеру
(например, реликты-эндемы).
Рис. 28. Схематическое изображение различных путей образования
разорванных ареалов.
а — путем исчезновения вида в части (белое поле) некогда более широ-
кого ареала; б — путем переселения или скачкообразного расширения
ареала в разных направлениях, причем в первоначальной области (штри-
ховка) вид может исчезнуть; в — то же, причем область первоначального
возникновения не является центром распространения систематической
единицы (зачернена); г — путем возникновения реликтов (крестики),
сохранившихся от некогда более широкого ареала.
В отношении небольших изолированных участков (эксклавов) какого-либо ареала
зачастую трудно бывает решить, являются ли они форпостами расширяющегося или
реликтами некогда более широкого сплошного ареала. Герцог сравнивает периодиче-
ские сужения и расширения ареалов систематических единиц с движением амебы, кото-
рая постоянно втягивает или вытягивает ложноножки то в одну, то в другую сторону
и у которой все целое может быть разделено на несколько частей, способных, однако,
снова соединяться друг с другом. Изменение границ ареалов объясняется не только
внешними условиями; при известных обстоятельствах причины генетического порядка
могут предопределить тенденцию распространения данной систематической единицы.
При благоприятных условиях существования вид распространяется
либо путем фронтального перемещения, либо единичными наступлениями
на определенные линии, либо, наконец, скачкообразно, образуя отдель-
ные форпосты. Характер границ ареалов в каждый данный момент в таких
случаях определяется только фактором времени. Подчас бывает трудно
решить, находится вид на данном участке границы своего ареала в стадии
расширения или сокращения. Для многих видов так назы-
ваемой «пустошно-степной флоры» давно уже остается спорным вопрос,
следует ли рассматривать их наличие в Средней Европе как форпосты
наступления или как реликты некогда более широкого расселения.
Результаты пыльцевого анализа показали, что некоторые местообитания,
раньше рассматривавшиеся как форпосты активного распространения,
являются признаками отступления вида.
Среди историко-климатических и исторических причин расселения
растений наиболее подробно в настоящее время исследовано воздействие
92 Составные элементы растительности и принципы их распространения
периодов оледенений на характер ареалов тех элементов растительностиу
которые составляют современную европейскую флору.
Флористическое единство голарктического царства объясняется существованием
некогда единой и исторически общей для всего царства флоры. В нее еще со времени
мелового периода входили многие роды, встречающиеся и сейчас в составе лесной фло-
ры, например Salix, Populus, Ulmus, Fagus, Quercus, Hedera. Их дополнили в ранний
третичный период Tilia, Acer, Fraxinus, Ilex и др. Флора Средней Европы, как указы-
вают ископаемые находки Liriodendron и Magnolia, была в ранний третичный период
более родственна по сравнению с настоящим временем не только североамериканской
и восточноазиатской флоре, но в основном лесным флорам тропиков и субтропиков.
В нее входили многие систематические единицы, которые сейчас ограничены тропи-
ками, например Cinnamomum, Artocarpus, Ficus, Bombax и многие роды пальм
(рис. 29). Cinnamomum является примером сокращения ареала.
Рис. 29. Современный ареал рода пальм Nipa и места, где она встречается как
реликт третичного периода (обозначены точками) (по Гуду).
В верхнем меловом периоде этот род встречался в Северной и Южной Америке,
в Гренландии, Европе, Австралии и Новой Зеландии, в эоцене — на том же простран-
стве, за исключением Южной Америки. В плиоцене Cinnamomum встречается еще
в Европе и Азии, тогда как в настоящее время его ареал ограничен лишь тропической
восточной Азией и Австралией. Часть обитающих теперь в Средней Европе родов
с листопадными видами имела здесь в ранний третичный период вечнозеленых предста-
вителей. В третичное время вплоть до плиоцена развивался основной состав мезофит-
ной флоры лиственных лесов, большая часть видов которой произрастает и в настоя-
щее время, как-то: европейский бук, дуб, граб, орешник, вяз, клен остролистный,
яблоня, крупнолистная липа, сосна, лиственница, ель. Кроме того, для позднего тре-
тичного периода характерно появление тех родов, которые сейчас встречаются в южных
районах, как Juglans и Aesculus, а также широкое развитие систематических единиц,
распространение которых в настоящее время ограничено Азией (около 30%), как
Ginkgo, и Северной Америкой (20%), как Pseudotsuga, Sequoia, Taxodium.
Причины пространст. распределения систематических единиц и форм роста 93
Разделение флоры северного полушария на океаническую и континентальную
в зависимости от удаленности от океана произошло также в позднетретичное время.
Ледниковый период углубил эту дифференциацию, однако последствия оледенения
проявились по-разному в различных областях.
Результатом ледникового периода явился разрыв ареалов, еще в тре-
тичный период охватывавших наподобие поясов все северное полушарие,
и уменьшение их до небольших остатков, зачастую реликтово-эндемично-
го характера.
Вследствие широтного простирания Альп растительность Средней
Европы испытала наибольшее воздействие периодов оледенений. В про-
тивоположность Северной Америке, где флора доледникового времени
в основном сохранилась и смогла широким фронтом вновь заселить осво-
бодившиеся ото льда территории и их окраины, в Средней Европе расти-
тельность, попавшая в зону наступания льда, была почти полностью
уничтожена.
Правда, похолодание в начале ледникового периода само по себе происходило
достаточно медленно, что допускало постепенное отступание теплолюбивой флоры
по широкому фронту, подобно тому как арктическая флора имела время вторгнуться
в Среднюю Европу до самых Альп. Однако пространство, которое могло бы быть занято
теплолюбивой флорой и на котором она могла бы удержаться, было слишком узким,
ограниченным горами и примыкающими с юга территориями с сухим летом, поэтому
оно оказалось неприспособленным для обитания всех элементов, составлявших долед-
никовую флору Средней Европы.
Между границами оледенения наступающая с севера арктическая
флора смешивалась в холодное время с альпийской третичной флорой.
Последняя, частично азиатского, частично средиземноморского происхо-
ждения, смогла удержаться в южных долинах Альп или на нуната-
ках, возвышающихся надо льдом, чтобы впоследствии вновь распростра-
ниться.
Когда после окончания ледникового периода флора холодных обла-
стей снова оказалась ограниченной северными и высокогорными района-
ми, из среднеевропейского района смешения альпийские по происхожде-
нию растения проникли в Арктику, а, с другой стороны, северные расте-
ния — в Альпы, как линнея, водяника, дриада и арктическая стелю-
щаяся ива.
Сохранившаяся в период холодов флора лиственных лесов в межлед-
никовье, хотя и в обедненном по сравнению с доледниковой флорой видо-
вом составе, смогла временно вновь частично занять первоначальные
области своего распространения. При повторном заселении Средней Евро-
пы после отступания льда, этапы которого известны на основании анали-
за ископаемой пыльцы (Firbas, 1949—1952), занять свои прежние пози-
ции смогла только часть видов, произраставших на них до ледникового
периода. Многие виды вымерли в пределах Европы, для других не наш-
лось подходящих обратных путей или же они за это время успели поте-
рять способность к распространению.
Например, два исключительно зимостойких вида хвойных Picea omorica и Pinus
рейсе, которые в межледниковую эпоху имели более широкое распространение, остались
после периода оледенения в своих убежищах (рефугиумах), ограниченных отдельными
горными вершинами Балканского полуострова, как реликтовые эндемы.
94 Составные элементы растительности и принципы их распространения
ВОЗДЕЙСТВИЕ ЧЕЛОВЕКА
НА РАСПРОСТРАНЕНИЕ РАСТЕНИЙ
ОБЩИЕ СООБРАЖЕНИЯ
Влияние человека можно в общем рассматривать как одну из исто-
рических причин распространения растений. С тех пор, как человек
появился на Земле, он во все возрастающей степени изменял характер
распространения растений. В последние тысячелетия влияние человека
было тем самым фактором, которым обусловлены наиболее значительные
изменения ареалов растений. Этому воздействию подвергалось необозри-
мое число систематических единиц. Рассматривая человека как виновни-
ка таких изменений, можно различать планомерное воздействие, при
котором распространение растений изменяется целеустремленно и созна-
тельно, и непреднамеренное воздействие.
Планомерным является культивирование полезных растений на по-
лях и лугах, в лесничествах, на плантациях, в садах и парках или вдоль
путей сообщения, а также защита определенных систематических единиц
из практических или эстетических соображений. Вполне сознательно
человек также устраняет те растения, которые мешают его сооружениям,
путем корчевания, выжигания или «планомерной борьбы с сорняками».
Непреднамеренное непосредственное воздействие на ареалы опреде-
ленных систематических единиц происходит в результате переноса семян
или хищнического хозяйничанья, как, например, при уничтожении цен-
ных древесных пород на доступных участках тропических лесов.
Степень косвенного влияния цивилизации с трудом поддается оцен-
ке. Эти воздействия в широких пределах изменяют как условия существо-
вания (почвы, грунтовые воды, климатические условия и животный мир),
так и характер конкуренции растительных единиц.
ИЗМЕНЕНИЯ АРЕАЛОВ СИСТЕМАТИЧЕСКИХ ЕДИНИЦ,
ОБУСЛОВЛЕННЫЕ ВОЗДЕЙСТВИЕМ ЧЕЛОВЕКА
Последствием влияния человека на систематические единицы расте-
ний является либо сокращение, либо увеличение их ареалов.
а) Сокращение ареалов. Неизмеримо велико число систе-
матических единиц, ареал которых был ограничен человеком то ли путем
непосредственного искоренения в одной части первоначального ареала,
то ли в результате непреднамеренных последствий вторжения в природу.
Все современные полезные площади когда-то были составными частями
естественных ареалов растений. В настоящее время в цивилизованных
странах у многих систематических единиц первобытной флоры сохрани-
лись лишь остатки их прежних ареалов. Ареалы, которые по своей при-
роде были сплошными, стали теперь разорванными или распались на
отдельные местонахождения.
Примером непреднамеренного непосредственного воздействия чело-
века является сокращение ареалов многочисленных талло-эпифитов,
которые в Европе почти совсем исчезли под влиянием бытовых и про-
мышленных дымов.
б) «Л о ж ное» увеличение ареалов. Многие виды расте-
ний, которые от природы были ограничены узкими ареалами, распростра-
нились с помощью человека по всей Земле. Здесь имеются в виду как
полезные растения, так и сорняки и прочие спутники жилых поселений,
Причины пространст. распределения систематических единиц и форм роста 95
хозяйственных сооружений, транспортных устройств. Однако поселяются
они большей частью на тех новых местообитаниях, где могут длительно
удерживаться только при постоянном содействии человека. С точки зре-
ния экологии ареалов здесь можно различать многочисленные ступени.
Случаи, когда растения находят на новых местообитаниях достаточ-
ные в соответствии со своими требованиями к климату и почве условия
существования, но без помощи человека не выдерживают конкуренции
других видов, приближаются уже к примерам настоящего увеличения
ареала. Это относится ко многим луговым и рудеральным растениям.
Р п с. 30. Пояс возделывания рода Musa (обозначен точками) и первоначальный
ареал (заштрихован) Musa paradisiaca ssp. seminifera (по Винклеру).
Ложным увеличением ареала является посадка или посев таких видов,
которые, правда, растут на соответствующих местообитаниях, однако
без помощи человека не могут размножаться, или таких, созревание
которых должно обеспечиваться специальными мерами, как, например,
защитой от морозов или искусственным орошением.
в) Настоящее увеличение ареала. Оно может про-
изойти при содействии человека
1) путем разведения полезных или декоративных растений на место-
обитаниях, которые обладают подходящими условиями существования
для данных видов, но которые не смогли бы достичь этих местообитаний
естественным путем, без помощи человека (рис. 30).
Это относится, в частности, к распространению промышленных древесных пород:
американский красный дуб и белая акация в Европе, австралийский эвкалипт во мно-
гих частях света. Ареал кокосовой пальмы своим расширением также обязан человеку;
2) путем непроизвольного занесения вида с последующим его естест-
венным размножением и спонтанным распространением.
96 Составные элементы растительности и принципы их распространения
Это касается многих луговых растений и огородных сорняков, а также элодеи
(Helodea canadense) в Европе;
3) путем самостоятельного вторжения систематических единиц на
подходящие для них местообитания, однако открытые и заново создан-
ные человеком. Примером являются многочисленные восточноевропей-
ские или азиатские по своему происхождению виды в составе среднеевро-
пейской флоры.
Часть вторая
ЕДИНИЦЫ РАСТИТЕЛЬНОСТИ
В ЛАНДШАФТЕ
Единицы растительности
СООБЩЕСТВА ЖИВЫХ СУЩЕСТВ
ВВЕДЕНИЕ
Не отдельные растения, а их сообщества на определенном про-
странстве, или участки растительных сообществ, являются мельчай-
шими единицами растительности, определяющими характер ландшафта.
Поэтому одним из важнейших предварительных условий рационального
географического отображения растительности Земли является требование
классифицировать бесконечно разнообразные и изменяющиеся раститель-
ные сообщества по их основным признакам.
Не говоря о таких общих, издавна существующих понятиях, как лес, луг, роща
или степь, значительно позже возникли определения, которые могут служить основой
для научного описания растительности.
Линней и Галлер, еще более или менее бессознательно, впервые рассматривали
растительные сообщества как единое целое. Когда Линней (Linne, 1745) охарактери-
зовал местообитания видов такими понятиями, как Pineta (сосновые сообщества)
или Ericeta (вересковые сообщества), то это было принципиально новым подходом, хотя
сейчас кажется само собой разумеющимся. Галлер различал во втором издании своей
«Флоры Швейцарии» (Haller, 1768) высотные пояса «буковых лесов», «еловых лесов»,
«коровьи пастбища с высокогорными деревьями» и «скудные каменистые овечьи паст-
бища» .
Только Гумбольдт увидел в «группировках растений», которые он
называл также «ассоциациями», особый объект для исследований. Одна-
ко потребовались дополнительные усилия многих ученых (Скау, Хеер,
Гризебах, Кернер, Мёбиус, Хульт, Градман, Флао, Шрётер и др.), пока
наконец не была найдена и признана приемлемой основа, состоящая
из трех понятий: «биоценоз»—«ассоциация»—«формация». Эти основные
понятия находятся в таком же примерно соотношении друг к другу, как
и понятия, составляющие триаду: живое существо — вид — форма роста.
ПОНЯТИЕ БИОЦЕНОЗА
«Классификация видов растений, составляющих некоторую сово-
купность... является не случайной и, несмотря на кажущуюся бессистем-
ность... вполне упорядоченной. Каждое растение имеет свое место, свое
время, свою роль и свое значение... Во всех зонах растения объединены
в определенные группы, которые предполагают наличие либо образую-
щегося, либо уже законченного, но всегда закономерного строения».
7*
100
Единицы растительности в ландшафте
В то самое время, когда Кернер (Kerner, 1863) писал эти строки, Рос-
смесслер сожалел, что в языке не хватает слова, чтобы «коротко и ясно
выразить, в какой степени лес является богатой формами совокупностью
тел и явлений... единым целым, состоящим из тысячи отдельных эле-
ментов, каждый из которых занимает свое определенное место».
Мёбиус при изучении устричных отмелей (1877) исходил из анало-
гичной точки зрения и предложил для «такой сообщности живых
существ» понятие биоценоза (греч. bios — жизнь и koinos —
объединять). Он понимал под этим «соответствующие средним внешним
условиям существования подбор и число видов и индивидуумов, которые
взаимно обусловливают друг друга и посредством размножения постоянно
удерживаются в пределах данной области».
Биоценоз является, следовательно, сообществом живых организмов
в определенном месте существования (Thienemann, 1941), или системой
населения, состоящей из растений и животных и сохраняющейся при
определенных условиях в динамическом равновесии.
СУЩНОСТЬ И СТРОЕНИЕ БИОЦЕНОЗОВ
Каждый биоценоз состоит из индивидуумов, сменяющих друг друга
на протяжении поколений и относящихся к более или менее значительно-
му числу различных видов. Подбор этих видов связан с условиями сущест-
вования, господствующими в месте произрастания. Эти условия опреде-
ляются не только характером местности (климат, почва, форма поверх-
ности, водный баланс), но и самими организмами, составляющими сооб-
щество (например, климат сообщества, водный баланс и баланс питатель-
ных веществ). Единство биоценоза и его особенность проявляются, следо-
вательно, не только в сходной приспособляемости различных видов
растений и животных к данному участку земной поверхности, но одновре-
менно и в самих жизненных процессах с их воздействием на те и другие
организмы и в совместной деятельности этих организмов, например их
участие в образовании почвы и создании микроклимата.
Взаимоотношения живых существ весьма разнообразны. Самая про-
стая форма совместного существования — это «сотрапезники» (Tischgenos-
sen). В борьбе за пространство, свет и пищу «сотрапезники» делят между
собой предоставленные им в месте произрастания жизненные условия.
Этим может быть обусловлено и пространственное строение биоценоза,
например ярусное расположение побегов и корней у различных форм
роста, причем ярусность проявляется не только над землей, но и под
землей.
Кроме того, существует много форм специальных, частично односто-
ронних, взаимозависимостей между членами биоценоза (так называемая
биоценотическая связь) («Biozdnotische Konnexe»). Жизнь в сообще-
стве — это не только борьба, но и взаимопомощь и разделение
труда.
Все животные и некоторые растения (сапрофиты, симбионты, парази-
ты) связаны по линии питания с другими компонентами биоценоза. Выше
уже отмечалась роль «цепи питания»— создающих, потребляющих и раз-
рушающих организмов в круговороте органических веществ (стр. 12).
Для многих растений другие растения необходимы в качестве поддержки
(лазящие растения) или опоры (эпифиты). Некоторые способствуют взаим-
ному росту. Так, микоризные грибы (Mycorrhiza) облегчают поглощение
Единицы растительности
101
неорганического фосфора. Связывание азота воздуха клубеньковыми
бактериями, обитающими в корнях бобовых растений, косвенно способ-
ствует развитию и других растений, входящих в биоценоз. С другой сто-
роны, соседство одного вида может непосредственно тормозить рост дру-
гих видов (аллелопатия) Примером могут служить отношения между
европейским кедром и рододендроном. Рододендрон является проме-
жуточным хозяином ржавчины, поражающей еловую хвою, и поэтому
на верхней границе лесов ослабляет конкурентную силу ели против кедра.
Таким образом последний благодаря наличию рододендрона оказывается
в выгодном положении. Следует также упомянуть о взаимоотношении
некоторых видов растений с определенными видами животных. Растения
обеспечивают пищу, защиту, предоставляют материал для постройки
гнезд, в то время как животные производят опыление цветков и распро-
страняют плоды или семена.
В строении ландшафта биоценозы являются более высокой ступенью
интеграции, чем те индивидуумы, из которых они состоят. Однако они
слабее интегрированы, чем эти последние. Дело в том, что биоценозы не
являются чем-то действительно единым, как организмы, а являются только
совокупностью таковых в частично самостоятельно существующей среде
обитания. Члены биоценоза могут меняться. Например, некоторые живот-
ные часто меняют место своего пребывания. Однако сам биоценоз при
этом не нарушается. Биоценоз только в ограниченной степени обладает
способностью к саморегулированию. Он может, например, восстанавли-
вать нарушенные части путем повторного заселения, новообразования
почвы и т. д. Однако при переселении животных или местном вымирании
видов животных или растений биоценоз не в состоянии сам по себе реге-
нерировать свой видовой состав. Происходит постепенное обеднение био-
ценоза или, наоборот, обогащение его, и подчас трудно бывает сказать,
в какой именно момент данный биоценоз превратился в другой, качест-
венно отличный от предыдущего. Следовательно, биоценоз не имеет рез-
ких границ ни в пространстве, ни во времени.
Что строение биоценоза имеет как исторические, так и случайные причины,
обусловленные историей места произрастания и его положением относительно центров
происхождения систематических единиц и преград для распространения, не требует
более подробных доказательств после того, как были изложены причины распростра-
нения растений.
БИОЦЕНОЗ И МЕСТО СУЩЕСТВОВАНИЯ
ОСНОВНЫЕ БИОГЕОГРАФИЧЕСКИЕ ПОНЯТИЯ
Прежде чем рассмотреть зависимость между биоценозами и про-
странством, на котором они обитают, уточним сначала некоторые поня-
тия. Дело в том, что в биологии, географии и прикладных науках (лесо-
водство, некоторые сельскохозяйственные науки и др.) слова «среда оби-
1 Аллелопатия — влияние одних растений на другие, одностороннее или взаим-
ное, происходящее вследствие выделения листьями или корнями растений специфи-
ческих химических веществ, действующих на другие растения. Это влияние осуще-
ствляется через почву или через приземный слой воздуха, содержащий летучие орга-
нические вещества. Влияние может осуществляться и другим путем.— Прим. ред.
102
Единицы растительности в ландшафте
Таблица 1
Обзор некоторых основных биогеографических понятий
Понятия, выражающие сущность или качество
Понятия пространст-
венного характера
Понятия,
относящие-
ся к реаль-
но сущест-
вующему
биоценозу
Сообщество живых су-
ществ
Биоценоз
(система населения,
состоящая из животных
и растений)
Фитоценоз
(растительное сообщест-
во)
Среда обитания, Habitat
(совокупность экологи-
ческих факторов, вклю-
чая и обусловленные
самим биоценозом)
Место существования,
биотоп
(место произрастания
определенного реально
существующего биоце-
ноза)
Голоцен
(единство
биоценоза
и биотопа)
Экосистема
(экологически
функциональ-
ная система
голоцена)
Экотоп
(тип экосисте-
мы, основная
единица
ландшафта)
Биохора
(область одно-
го голоцена,
одной экосис-
темы, одного
экотопа, одно-
го биотопа)
Понятия,
относящие-
ся к местно-
сти
Местообитание
(качество местности независимо от реаль-
но существующего биоценоза)
Пространство
местообитания
Естественная
пространствен-
ная единица
(физиохора,
флизе, сайт)
(Потенциаль-
ная биохора)
(Потенциальный биотоп)
Поясним некоторые биогеографические термины, включенные автором в таблицу:
Биохора — термин предложен Пальманом (Швейцария) в 1948 г. для обозначения
системы, состоящей из окружающей среды и живого населения. Автор книги трак-
тует его как пространственную единицу, представляющую участок одного голоцена,
экотопа и т. д.
Флизе — термин предложен Зигелем (ГДР) в 1959 г. для наименования наимень-
шей пространственной единицы, выделяемой при природно-территориальном райони-
ровании.
Сайт (местоположение) — термин предложен Хиллсом на IX Международном бота-
ническом конгрессе в Канаде в 1959 г. для обозначения комбинации внешних факторов
на участке: климата, почв, растений и животных, с которыми должен считаться лесовод
при лесоразведении. Из всех компонентов сайта особое внимание уделяется почве.—
Прим. ред.
Единицы, растительности
103
тания» или «местообитание» часто употребляются одновременно, но в
различных значениях. В табл. 1 приводится обзор некоторых понятий,
которые имеют немаловажное значение в географии растительности.
МЕСТООБИТАНИЕ (STANDORT)
Под «местообитанием» в его первоначальном ботанико-географическом
значении понимали место нахождения растения и те его особенности,
которые характеризуют окружающую среду. В последнее время в поня-
тии «местообитание» акцент все более делается на «качество» участка,
то есть на совокупность экологических особенностей данной местности,
обусловливающих возможность существования биоценоза. Это смещение
внесено лесоводческой и сельскохозяйственной науками. Однако оно
получило отражение и в обиходе экологии и фитоценологии. Определяют,
что производит «местообитание» или что оно могло бы «дать», судя по
характеру его природы. Планируют хозяйство в зависимости от характе-
ра местообитаний и картируют качественно равные местообитания. При
этом слово «местообитание» уже не означает только место произрастания
имеющейся единицы, а также «потенциальное место существования»,
то есть качество, которым обладает данный участок местности независи-
мо от того, произрастают на нем растения или нет.
В понятие «местообитание» влияние человека вторгается лишь настоль-
ко, насколько оно выражено в особенностях местности, возникающих,
например, в результате выторфовывания заболоченных участков, осу-
шения или разрушения почвы. Характер существующего в данный момент
растительного сообщества здесь значения не имеет, а результат хозяй-
ственного воздействия учитывается лишь как особенность местообитания.
Качество местообитания зависит от многих «факторов местности»,
из которых важнейшими являются:
а) общий климат местности, обусловленный географическим поло-
жением;
б) высота места над уровнем моря;
в) форма поверхности (экспозиция, угол наклона склонов и характер
ближайших окрестностей);
г) горные породы и их влияние на почвообразовательный процесс;
д) режим грунтовых вод;
е) возможность затопления местности;
ж) происхождение, тип, возраст почвы и ее физические, химические
и биологические особенности.
В среде обитания биоценоза влияние всех этих компонентов тесно
переплетается. Эффективное значение каждого отдельного фактора ста-
новится ясным только при анализе его взаимодействий с другими факто-
рами.
МЕСТО СУЩЕСТВОВАНИЯ (LEBENSSTАТТЕ)
Термин «место существования», или биотоп, мы применяем для обо-
значения места произрастания одного определенного, реально существу-
ющего биоценоза.
ЭКОЛОГИЧЕСКАЯ ТРАКТОВКА СРЕДЫ ОБИТАНИЯ
Под «средой обитания» (Umwelt) экология понимает совокупность
внешних условий, которые воздействуют на определенную единицу живо-
го мира в месте ее существования (Thienemann, 1941). Факторы среды
104 Единицы растительности е ландшафте
обитания — это сезонное и суточное изменение света, тепла, влажности,
содержание углекислоты в воздухе, аэрация почвы, содержание пита-
тельных веществ и концентрация ионов водорода в почвенном растворе,
а также структура почвы, возможные механические воздействия, напри-
мер ветра или со стороны человека, и, наконец, жизнь микроорганизмов
в почве и в воздухе.
Среда обитания — это совместное действие всех факторов,
непосредственно влияющих на жизнь отдельного организма или био-
ценоза в целом. Поэтому можно говорить о реальной среде обитания
отдельного растения или биоценоза или же об абстрактной типичной
среде обитания вида или ассоциации. К факторам среды обитания можно
отнести и влияние человека, например скашивание лугов. (Для обозна-
чения экологической среды обитания часто применяется также слово
«местообитание», из-за чего нередко возникают недоразумения.)
Еще Зендтнер (Sendtner, 1854) отмечал, что среда обитания, он называл ее «место-
обиталищем» (Standortlichkeit), объединяет в одно целое особенности места существо-
вания и влияние живого мира: «Растения (как живые, так и уже погибшие) сами
непосредственно участвуют в образовании местообиталища, обусловливают общий
характер растительности, изменяют существовавшие первоначальные условия. Зате-
нение и продукты распада воздействуют на микроклимат, влажность участка, содержа-
ние питательных веществ в почве и ее структуру».
Биоценоз приспособлен к месту существования не только составом
и строением. Он находится с ним в определенном взаимодействии, сам
создает свою среду обитания и развивается вместе с ней.
ГОЛОЦЕН И ЭКОСИСТЕМА
ГОЛО ЦЕН
Сообщество живых организмов и место их существования образуют
единое экологическое целое. Это единство можно рассматривать как еди-
ницу живого мира более высокого ранга, если вместе с Тинеманом (Thie-
nemann, 1941) характеризовать каждое природное образование, в которое
входят живые организмы, как некую единицу живого мира. Фридерикс
(Friederichs, 1937) обозначал единство, создаваемое сообществом живых
существ и местом их существования, понятием «голоцен» или «цен» \
У высокоразвитых сообществ голоцен предполагает также их про-
странственную и временную неоднородность, а именно вертикальную
ярусность сообщества и его горизонтальную дифференциацию, с одной
1 Термин «голоцен», или «голоценоз» (греч. holos — целый и koinos — объеди-
нять), введен Фридериксом в 1930 г. и определяется им как «комбинация экологиче-
ских факторов, как одушевленных, так и неодушевленных, объединенных их дейст-
виями и противодействиями друг с другом... «ткань жизни», в которой ни один фактор
не может быть изменен без того, чтобы не задеть все остальные» (FriederichsK., 1930,
Die Grundlagen und Gesetzmasigkeit der land- und forstwirtschaftlichen Zoologie).
«Экосистема»— термин, введенный Тзнсли в 1935 г., широко распространенный
сейчас за рубежом для обозначения естественной единицы, представляющей совокуп-
ность живых и неживых элементов в их взаимодействии. Понятие, близкое к голоцену
и биогеоценозу, но употребляемое часто в разных смыслах. Применяется к единицам
различной протяженности — от пня с его живым населением (микрозкосистема)
до океана (макрозкосистема). Биосфера является гигантской экосистемой земного
шара. Автор книги трактует экосистему как эколого-функциональную систему голо-
цена. — Прим, ред.
Единицы растительности
105-
стороны, а с другой — сезонные колебания и изменения, происходящие
в видовом составе на протяжении поколений. Тип однородных местных
голоценов обозначается также термином «экотон» (основная экологиче-
ская пространственная единица ландшафта) (Bobek, Schmithiisen, 1949);
Сукачев (1950) употребляет термин «биогеоценоз» х.
ЭКОСИСТЕМА
Существенное значение в голоцене имеет эколого-функциональное
понятие «экосистема».
В экосистеме объединяются абиотически и биотически обусловленные
процессы. Только исходя из их совместного действия можно объяснить
некоторые весьма сложные явления, как, например, микроклимат биоце-
ноза и почвы, местный водный баланс, круговорот минеральных и орга-
нических веществ, периодичность жизненных процессов, циклическая
и апериодическая смена в системе населения биоценоза и, наконец, объе-
диняющий составной элемент всей динамической системы —«почву», рых-
лый, химически и биологически активный, пронизанный корнями расте-
ний верхний покров земли с его специфическим типом профиля
и характерным составом населяющих его мелких живых организмов.
КЛИМАТ СООБЩЕСТВ И ВОДНЫЙ РЕЖИМ
Микроклимат растительного сообщества значительно отличается от
микроклимата местности, лишенной растительности. При этом влияние
оказывают не только высота и плотность растительности, но и обусловлен-
ная этой растительностью влажность и другие свойства почвы. Здесь
не место отдельно рассматривать микроклимат биоценозов, однако немно-
гие указания могут пояснить примерный характер этого влияния. Смы-
кающиеся кроны среднеевропейского летнезеленого леса ослабляют воз-
действие дневного света внутри этого сообщества и смягчают ночное излу-
чение. Микроклимат в глубине леса становится тем самым более умерен-
ным, чем климат местности. В высокоствольном буковом лесу по сравне-
нию с открытой местностью суточная амплитуда температуры может быть
на 5° меньше. Затененность, защита от ветра и перехват осадков кроной
деревьев, а также транспирация увеличивают влажность внутри сообще-
ства и уменьшают суммарное испарение на одну треть по сравнению
с открытой местностью.
Сплошной растительный покров защищает почву от прямого дей-
ствия солнечных лучей, а тем самым от воздействия крайних температур
и от высыхания. Он предохраняет почву от непосредственного ударного
воздействия осадков, корни его делают почву рыхлой и задерживают
ее смыв.
1 «Биогеоценоз»— термин, предложенный В. Н. Сукачевым в 1944 г. для обо-
значения комплекса организмов и среды их обитания: «Биогеоценоз — это совокуп-
ность на известном протяжении земной поверхности однородных природных явлений
(атмосферы, горной породы, растительности, животного мира и мира микроорганиз-
мов, почвы и гидрологических условий), имеющих свою особую специфику взаимо-
действий этих слагающих ее компонентов и определенный тип обмена веществом
и энергией их между собой и с другими явлениями природы, и представляющее собой
внутреннее противоречивое диалектическое единство, находящееся в постоянном дви-
жении, развитии» (В. Н. Сукачев, 1964, Основные понятия лесной биогеоцено-
логии, сб. «Основы лесной биогеоценологии»). Термин биогеоценоз получил широкое
распространение в СССР среди геоботаников, почвоведов, географов, агрономов, лесо-
водов и т. д.— Прим. ред.
106
Единицы растительности в ландшафте
Растительность в значительной степени регулирует и водный баланс
почвы. Часть осадков испаряется с самой поверхности растительного
покрова. Под сводом высокоствольного леса поверхностный сток наблю-
дается редко. Осадки, достигающие почвы, большей частью просачивают-
ся вглубь, тогда как на безлесных участках при тех же условиях по поверх-
ности текут обильные потоки воды. Большая часть почвенных вод погло-
щается корнями. Высокоствольный буковый лес потребляет за 6 месяцев
вегетационного периода 5 млн. л воды на 1 га и транспирирует обратно
в воздух почти две трети выпавших осадков.
ОБМЕН ВЕЩЕСТВ В ЭКОСИСТЕМЕ
Биоценозы, от скромной скальной растительности нивального поя-
са, состоящей из лишайников и водорослей, до высокоствольных лесов,
способствуют самым различным образом химическому и механическому
выветриванию пород. Корни, корневища, клубни и другие подземные
органы часто бывают расположены так же поярусно, как и надземные
части растения. Вместе с богатым животным миром почвы они образуют
как бы «лес под лесом». Корни разрыхляют, вентилируют и осушают
почву и включают ее в круговорот веществ экосистемы.
В местном обмене минеральных и органических веществ, который
со своей стороны является элементом общего для всей Земли круговорота
воды и состава воздуха, участвует вся совокупность организмов, живу-
щих над землей и под землей. Даже незаметный, невидимый невооружен-
ным глазом растительный и животный мир почвы является необходимым
участником функционального строения системы. Зеленые растения в про-
цессе фотосинтеза, используя углекислоту воздуха и минеральные почвен-
ные растворы, создают органические вещества. Бактерии и почвенные
грибки в ходе своей деятельности усваивают атмосферный азот.
Посредством целого ряда цепей питания, включающих в каче-
стве потребителей наряду с паразитными и сапрофитными высшими расте-
ниями также и низшие растения и животных, органические вещества
многократно преобразуются. Все входящие в биоценоз животные, преи-
мущественно ногохвостки, мокрицы, клещи и черви, а кроме того, грибы
и бактерии, добывают энергию для своего жизненного обихода и при
этом постепенно расходуют органические вещества. Наконец, органические
остатки минерализуются в условиях хорошо аэрируемой почвы путем
разложения, а в сырой, бедной кислородом почве — в результате
гниения, ив таком виде они снова могут быть восприняты высшими
растениями.
ГУМ УС
Совокупность органических веществ, которые в качестве промежу-
точных соединений между еще неизменившимся отмершим органическим
веществом и полностью минерализованными конечными продуктами рас-
пада временно или навсегда становятся составной частью почвы, назы-
вают «гумусом». Он состоит из большого числа разнообразных, не всегда
еще точно известных органических соединений. При распаде частично
образуются новые вещества, преимущественно гуминовые кислоты, назы-
ваемые «истинным гумусом». Содержание гумуса оказывает большое влия-
ние на структуру почвы, а также на ее водный баланс и баланс питатель-
ных веществ. Именно гумус прежде всего обусловливает обособление
горизонтов почвенного профиля, характеризующих данный экотоп.
Единицы растительности
107
Истинный гумус довольно упорно противостоит разрушаю-
щему действию микробов. Он превращается в перегной, благодаря кото-
рому в верхних горизонтах почвы могут скапливаться большие запасы
питательных веществ. Этот процесс в большом масштабе происходит путем
гумификации корней злаков в степных черноземных почвах умеренного
климата. На юге Европейской части СССР такие почвы могут содержать
до 1 млн. кг гумуса на гектар (Wittich, 1952).
Грубый гумус, образующийся в результате неполного разло-
жения из-за слабой биологической активности, содержит меньше истинно
гумусных веществ. Как кислый, недостаточно насыщенный основаниями
гумус, он может ускорить вымывание питательных веществ из верхних
горизонтов почвы (оподзоливание).
ПОЧВОО БРА 3 О В АННЕ
Почвообразование зависит от совместного действия всех факторов
местности и связано тесным взаимовлиянием с растительностью. Почва
и растительность образуют одно функциональное целое, в значительной
степени зависящее от климата. Это единство выражается в «почвенном
профиле», то есть в вертикальном расчленении почвы на горизонты и в
возникающем в почве сообществе живых организмов. Химические про-
цессы в почве, в основном обусловленные особенностями климата, но диф-
ференцирующиеся под влиянием местных горных пород, могут под влия-
нием биоценозов претерпевать существенные изменения.
ЗНАЧЕНИЕ КРУГОВОРОТА ПИТАТЕЛЬНЫХ ВЕЩЕСТВ
На местном круговороте веществ основано относительное постоян-
ство экосистем. Раньше полагали, что почвы под тропическим дождевым
лесом скрывают неисчерпаемые запасы питательных веществ. Однако, как
теперь известно, пышность этих лесов обеспечивается не столько простой
суммой питательных веществ, сколько тем фактом, что большая часть
их находится в непрерывном круговороте, то попадая в сферу деятельно-
сти живых организмов, то вновь возвращаясь в виде минеральных соедине-
ний в верхние горизонты почвы. Если лес уничтожить, плодородие почвы
может быстро иссякнуть. Естественный лес помогает удерживать в почве
питательные вещества, необходимые ему лишь в ограниченном количе-
стве; он постоянно извлекает их из почвы, чем обеспечивается возмож-
ность его роста. В среднеевропейских лесах, как показало исследование
годового баланса минеральных солей, питательные вещества, необходи-
мые растениям, в процессе круговорота также адсорбируются в верхних
слоях почвы в количестве, всегда большем, чем если бы они накаплива-
лись только в результате непосредственного выветривания пород (Klau-
sing, 1956).
БИОЦЕНОТИЧЕСКОЕ РАВНОВЕСИЕ
Мы привыкли к тому, что лес или какой-либо другой биоценоз имеет
из года в год один и тот же знакомый облик, если, конечно, он не испыты-
вает воздействия человека, и мы знаем, что таким он может сохраняться
веками. Сменяются отдельные индивидуумы, но система населения, если
она уже приспособилась к внешним условиям, остается в своем характер-
ном составе прежней. Это обстоятельство, само по себе так же мало понят-
108
Единицы растительности в ландшафте
ное, как и постоянство круговорота питательных веществ, называется
биоценотическим равновесием.
Значение биоценотического равновесия для состава биоценозов особенно заметно
по естественному равновесию размножения, и прежде всего там, где это равновесие
нарушается. У некоторых видов животных и растений один индивидуум создает тыся-
чи или миллионы зародышей. Из них в ненарушенном биоценозе выживают лишь неко-
торые. В основном их остается именно столько, сколько нужно, чтобы все сообщество
сохранило свое прежнее строение. Так, в сосновых лесах обитает сосновая совка,
являющаяся существенным элементом системы населения. Однако там, где равновесие
нарушено, например в лесохозяйственных чистых насаждениях, может наступить при
определенных условиях погоды сверхразмножение взрывного характера, приводящее
к катастрофическому увеличению числа сосновых совок, которые способны уничтожить
даже весь лес.
В областях с сезонным различием климата периодичность жизнен-
ных процессов, а тем самым и смена физиономии сообщества, является
необходимым составным элементом экологического равновесия. Фазы жиз-
ненных процессов (смена листвы, цветение, созревание плодов, прораста-
ние семян) лишь частично вызываются внешними причинами, то есть
непосредственно изменением тепла, холода, сухости или влажности, при-
чем «фенологические даты» зависят от погоды в течение года. Отчасти
же смена фаз определена генезисом видов. Процессы эти происходят авто-
номно, как, например, распускание листвы у некоторых деревьев сухих
тропических лесов еще до начала периода дождей или зимний покой сред-
неевропейских лиственных деревьев, который продолжается в течение
специфически ограниченного для каждого вида времени, даже если в неко-
торые годы зимние холода вообще не наступают.
Естественная последовательность поколений систематических еди-
ниц может, как это имеет место в североамериканских лесах, привести
к долгосрочной смене доминантов различных видов, участвующих в строе-
нии биоценоза. Поскольку при этом действительно речь идет о периоди-
ческом изменении, мы не видим в этом нарушения равновесия экосистемы,
характерной для соответствующего местообитания.
ГОЛОЦЕНОТ ИЧЕСКОЕ РАВНОВЕСИЕ
Биоценотическое равновесие проявляется как суммарный эффект
многократно переплетенных воздействий и противодействий, которые
в конечном счете связаны даже с общей динамикой планеты или космоса.
Фридерикс говорит поэтому о голоценотическом равно-
весии. Мы сможем его конкретизировать, описать и до некоторой сте-
пени «объяснить» как явление, реально существующее в природе, но
лишь при условии, что нам удастся свести единичные процессы к опре-
деленным результатам, а эти последние связать со свойствами отдельных
компонентов.
РАЗГРАНИЧЕНИЕ И КЛАССИФИКАЦИЯ БИОЦЕНОЗОВ
Наиболее целесообразная классификация биоценозов, касающаяся
их сущности, может быть произведена, если исходить из трех следующих
основных положений:
1. Биоценоз является общественно-организованным сообщест-
вом систематических единиц.
Единицы растительности
109
2. Биоценоз обладает определенной физиономией и тем самым —
сам по себе или в совокупности с другими биоценозами — определяет
облик ландшафта.
3. Биоценоз существует в голоцене, который как функционально
(в качестве экосистемы), так и генетически связан с определенным про-
странством.
Из первого положения следует: идентичность биоценоза
может быть определена лишь по образующим его систематическим едини-
цам. Поэтому только биоценология может заниматься систематическим
разграничением, классификацией и наименованием биоценозов. Эта систе-
ма должна основываться исключительно на критериях систематики так-
сонов.
Из второго положения следует: для географии растительности необ-
ходимо объединить биоценозы по их ландшафтному облику (Formations-
charakter) в типы и эти последние наглядно классифицировать.
Из третьего положения вытекает задача исследовать и описать био-
ценозы по их положению в ландшафте и в территориальном
построении растительности Земли. Систематика сооб-
ществ и типология формаций являются при этом необходимой предпосыл-
кой для понимания объектов исследования.
Чтобы полностью разобраться во всех связях, было бы желательно всегда рас-
сматривать биоценозы в целом и при разграничении отдельных единиц учитывать
как их растительный состав, так и животное население. Обычно это и делается при
изучении сообществ живых существ, обитающих в воде, поскольку в этом случае такая
необходимость особенно очевидна. Для наземных биоценозов до сих пор животный мир
привлекали к рассмотрению только в виде исключения. По сравнению с фитоценоло-
гией ценология животного мира сталкивается с большими трудностями. Совместная
деятельность фито- и зооценологов еще только начинается. Поэтому рано говорить
о систематике биоценозов, изучающей одинаково подробно и растения, и животных.
Однако этот недостаток несколько сгладился благодаря накопленным к настоящему
времени данным по зооценологии. Для наземных биоценозов доминирующее значение
при разграничении отдельных единиц имеет видовой состав растительности. Это
понятно, если учесть экологические взаимосвязи между цепями питания и условиями
обитания животных. К тому же в отношении растительных сообществ, их описания
и классификации накопился достаточно обширный материал, поэтому растительные
сообщества являются не только временно пригодной заменой, но могут рассматри-
ваться и как постоянная основа для классификации большей части наземных биоцено-
зов и их систематизации. Только более широкое исследование животного населения
растительных сообществ и разработка единиц зооценологии покажут, в какой степени
следует учитывать животный мир при разграничении и систематизации наземных
биоценозов.
ЕДИНИЦЫ РАСТИТЕЛЬНОСТИ И РАСТИТЕЛЬНЫЕ
СООБЩЕСТВА
РАСТИТЕЛЬНОЕ СООБЩЕСТВО
Выражение «растительное сообщество» применяется обычно для той
части биоценозов, которую составляют растения. Поэтому этим термином
обозначают как конкретные участки растительных сообществ, так и об-
щие понятия, имеющие в виду тип повторяющихся сочетаний видов.
110
Единицы растительности в ландшафте
Между участками, которые причисляются к одному сообществу, абсолютной иден-
тичности не существует в той же степени, в какой ее нет между индивидуумами одной
систематической единицы.
Если сознательно не замечать некоторых повсеместно существующих различий,
то только тогда можно делать общие заключения о характере растительности на боль-
ших пространствах земной поверхности. Это необходимо сознавать, иначе типовые
понятия, с которыми придется иметь дело при любом описании растительности, могут
привести к неправильным представлениям. Любое явление в природе гораздо более
дифференцированно, чем это бывает представлено в самом точном описании. Если мы
говорим об идентичности сообществ, то только в смысле их типологической оди-
наковости.
ПРИНЦИП ФЛОРИСТ И ЧЕС НО ГО ДИАГНОЗА
И ФИТОЦЕНОЛОГИЧЕСКАЯ СИСТЕМА
При проведении специальных исследований для опубликования их
результатов, проверки упорядочения и обобщения необходимо совершен-
но безошибочно установить идентичность изученного объекта. По Градма-
ну (Gradmann, 1909), при учете всего наличного состава видов следует
прежде всего дать диагностическое описание «абстрактных», по возмож-
ности одного ранга, основных единиц растительных
сообществ, в отношении важнейших «определяющих видов», и взять
их за основу при систематике растительности.
Морфология сообществ, анализирующая растительные сообщества по их флори-
стическим признакам (по форме структуры вида), является единственной объективной
основой для диагнозов, с помощью которых можно идентифицировать растительные
сообщества.
Деление на флористически однородные растительные сообщества
является для всех направлений исследования растительности предвари-
тельным условием понимания изучаемых объектов, однако подобное раз-
деление не означает предварительного знания причинных зависимостей,
а лишь стремится создать возможность для их выявления. Это заслуга
Браун-Бланке (Braun-Blanquet, 1921, 1925), что он сделал флористи-
ческий диагноз растительных сообществ, основанный на сочетаниях
характерных видов, надежным критерием законченной, ясной, однород-
но построенной идеографической системы сообществ.
Классификации единиц растительности по физиономическим или экологическим
признакам имеют, напротив, нормативный характер. В них объединения растений (или
сообщества, если таковые уже выделены) разграничиваются по общим, составленным
по дедуктивной схеме понятиям или по причинным данным. Однако такие классифи-
кации, стремясь к достаточной полноте и завершенности, заранее предполагают нали-
чие диагностического описания и наименования сообществ. Идеографическую систе-
матику сообществ не следует приравнивать или смешивать с другими видами класси-
фикации фитоценозов во избежание затушевывания ее задач и сущности.
Подобно тому как химия стремится полностью, однозначно и едино-
образно идентифицировать и классифицировать вещества только по их
атомной структуре, так и систематическая классификация растительных
сообществ может быть основана лишь на одном последовательном прин-
ципе — на флористическом строении сообществ. Как, однако, наряду
с основной классификацией имеет смысл и деление веществ, например,
Единицы растительности
Ш
по консистенции, форме кристаллов или их пригодности для человека,
так наряду с систематикой растительных сообществ возможно, а в неко-
торых случаях даже целесообразно и их группирование по физиономи-
ческим типам (растительным формациям), степени воздействия человека
на них, свойствам местообитаний и с других точек зрения.
ФИТОЦЕНОЛОГИЧЕСКИЙ МЕТОД
Этот метод, с помощью которого распознаются сообщества и произво-
дится их разграничение, заключается в исследовании отдельных участков
сообществ на местности, сравнении видового состава участков путем
составления сводных таблиц, разработке диагноза и, наконец, в присвое-
нии наименования.
Описание участков сообществ и анализ сообщества в целом
Начало планомерных исследований растительных сообществ относится к XIX в.
(Хеер, Шницлейн и Фриккингер, Турман, Лекок, Зендтнер, Лоренц, Кернер, Хульт,
Штеблер и Шрётер, Градман, Паунд и Клементс). Однако только в последние десяти-
летия были созданы единые методы, имеющие международное значение (Braun-Blan-
quet, 1928, 1951).
В основу описания участка растительного сообщества кладется пол-
ный список видов растений, встречающихся на исследуемой
пробной площади, которая в зависимости от вида сообщества должна иметь
определенную минимальную величину. Для каждого вида выявляется
мощность вида, общественность (в большинстве случаев), а по необ-
ходимости и жизненность.
Г омогенность сообщества можно проверить, определив встречаемость
видов (Frequenz), то есть выраженную в процентах величину, показывающую, насколь-
ко часто данный вид встречается на большом числе пробных площадок в одном сооб-
ществе.
Полное описание сообщества включает также характеристику его
сезонно изменяющихся аспектов. Для этого требуется произвести
неоднократное описание участков сообщества в разных климатах и в
разное время года. И наконец, все эти сведения должны быть дополнены
экологическими данными и точным описанием особенностей местообитания.
Аналитические признаки
1. Мощность вида. Под мощностью вида (Artmachtigkeit) (Schwicke-
rath, 1940) понимается введенная Браун-Бланке (Braun-Blanquet, 1928)
«суммарная оценка» числа индивидуумов (обилие — Abundanz) и степени
покрытия поверхности (доминирование — Dominanz) у отдельных видов
по следующей шкале:
г — чрезвычайно редко, с крайне незначительной площадью покрытия;
-|--редко, с крайне незначительной площадью покрытия;
1 — обильно, но с незначительной площадью покрытия или довольно редко, но
с большой площадью покрытия.
2 — весьма обильно или с площадью покрытия, равной по меньшей мере 1/2о проб-
ной площадки;
112
Единицы растительности в ландшафте
3 — площадь покрытия равна от х/4 до х/2 пробной площадки при любом числе особей;
4 — площадь покрытия равна от х/2 до 3/4 пробной площадки при любом числе особей;
5 — площадь покрытия равна более 3/4 пробной площадки при любом числе особей.
В растительных сообществах, состоящих из нескольких ярусов,
мощность вида оценивается раздельно для каждого яруса.
В особых случаях, например при исследовании пастбищ и лугов,
доля каждого вида оценивается по весу.
2. Общественность. Под общественностью понимают «характер рас-
пределения» видов в сообществе (Schouw, 1823). Общественность оцени-
вается по нижеследующей шкале (Braun-Blanquet, 1951):
1 — произрастает единично;
2 — растет группами или гнездами (кучками);
3 — растет латками (небольшие пятна или подушки);
4 — растет маленькими колониями или образует большие пятна или ковры;
5 — образует большие заросли.
Многие виды из-за их специфической формы роста имеют всегда более
или менее одинаковую способность к совместному существованию. Поэ-
тому многие авторы учитывают общественность лишь в исключительных
случаях.
3. Жизненность. Степень жизненности видов характеризуется по
следующей шкале:
1 — хорошо развитые растения, регулярно проходящие полный жизненный цикл;
2 — слабее развитые размножающиеся растения (ослабленная общественность,
скудное возобновление, низкорослость и т. д.) или пышно развитые, однако
проходящие неполный жизненный цикл;
3 — слабо вегетирующие размножающиеся растения, проходящие неполный жиз-
ненный цикл;
4 — случайно проросшие неразмножающиеся растения.
4. Пример описания участка сообщества (Tuxen, Preising, 1942).
Южнофранцузские камышово-тростниковые, Rohricht, заросли (Scirpeto —
Phragmitetum).
В Леце у Пастельно, близ Монпелье (южная Франция). Внешняя
излучина реки, глубина 1—1,5 м. Вода чистая, выше запруды почти
стоячая.
Исследование Тюксена 18 февраля 1927 г.
2.2 Typha latifolia
2.2 Typha angustifolia
1.2 S choenoplectus lacustris
4-3 Sparganium erectum^ssp. neglectum
4-1 Phragmites communis
4-2 Iris pseudacorus
4-1 Alisma plantago var. lanceolatum
4-2 Lithrum salicaria
4-1 Epilobium spec.
4-2 Polygonum amphibium terrestre.
Первая цифра означает степень мощности вида, вторая — степень обществен-
ности соответствующего вида.
Единицы растительности
113
Определяющие признаки единиц сообществ
По сведенным в таблицы спискам видов, встречающихся в однород-
ных участках сообществ, устанавливается абстрактная едини-
ца сообществ.
Определяющими признаками сообщества являются:
1. Характерное для данного сообщества сочетание видов, состоящее
из характерных видов, дифференциальных видов и видов с высоким посто-
янством.
2. Величина минимального ареала.
3. Абсолютное число видов.
Минимальным ареалом считают наименьшую величину,
которую должна иметь пробная площадка в изучаемом сообществе при
условии, что она с достаточной полнотой охватывает характеризующее
это сообщество число видов. Абсолютное число видов в ассо-
циации при достаточном числе описаний пробных площадок можно опре-
делить по кривой графического соотношения числа видов и числа опи-
сании.
Характерные сочетания видов
1. Виды с высоким постоянством. Группа постоянных видов
(steten Arten), встречающихся на 80—100% пробных площадок, выявляет
общность всех участков сообщества.
2. Характерные (определяющие) виды (Kennarten). Группа харак-
терных видов определяет флористическую особенность данного
сообщества, отличающую его от других растительных сообществ, сущест-
вующих в области его распространения. Характерные виды могут быть
выделены не по спискам видов, встреченных на участках одного сообще-
ства, а лишь на основании исчерпывающих сведений о существовании
видов во всех других сообществах.
Основанием для отнесения к характерному виду сообщества является
верность вида, или степень привязанности отдельного вида к дан-
ному сообществу. Различают следующие пять степеней верности:
5 — верный (treu), привязанный исключительно или почти исключительно к данному
сообществу;
4 — прочный (fest), предпочитающий данное сообщество, в других встречающийся
редко;
3 — благосклонный (hold), преобладающий в данном сообществе, но более или менее
широко представленный и в других; -
2 — безразличный (vag), без явно выраженной привязанности к данному сообществу;
1 — чуждый (fremd), лишь случайно встречающийся в данном сообществе, преиму-
щественно распространенный в других.
Виды со степенью верности от 3 до 5 причисляют к «характерным»
видам. Виды, безразличные к сообществу, называют «сопутствующими»
(Begleiter), а чуждые данному сообществу —«случайными» (Zufallige).
Определение верности сообществу может быть отнесено также ко все-
му ареалу вида (абсолютная верность) или к пространству, на котором
данное сообщество встречается (территориально ограниченная верность),
однако без учета распространенности вида за пределами ареала сообще-
ства. Поэтому различают абсолютно и территориально характерные виды.
114
Единицы растительности в ландшафте
Абсолютно характерные виды довольно редки. Верность сообществу относится
обычно только к части ареала вида. Поэтому один и тот же вид может быть характер-
ным для многих ассоциаций, исключающих друг друга в пространственном распро-
странении.
Часто для более точной характеристики участков сообщества привле-
каются виды, которые привязаны к соответствующему сообществу только
на определенном, строго ограниченном пространстве. В этом случае гово-
рят о локальных характерных видах.
3. Разделяющие (дифференциальные) виды (Trennarten). Дифферен-
циальными видами называют такие виды, которые, хотя и не обладают
высокой степенью верности по отношению к соответствующему сообще-
ству, однако могут быть использованы для разграничения с другими
сообществами или для подразделения ассоциации на единицы сообщества
более низких рангов.
Дифференциальный вид по отношению к характерному виду можно рассматри-
вать как вышестоящее понятие. В то время как дифференциальный вид характеризует
сообщество только с одной стороны (отличая его от другого сообщества или группы сооб-
ществ), характерный вид является «всесторонне дифференциальным видом» (Tuomi-
kowski, 1942).
СИСТЕМАТИЧЕСКАЯ КЛАССИФИКАЦИЯ РАСТИТЕЛЬНЫХ
СООБЩЕСТВ
Фитоценологическая система
Система Браун-Бланке делит растительные сообщества по их фло-
ристическому родству на единицы сообществ разного ранга.
Эта система, построенная на флористическом диагнозе сообществ, имеет также
и генетическую основу, поскольку вместе с видовым составом система учитывает и их
ареало-типологическую комбинацию, а тем самым и флористико-генетические разли-
чия сообществ.
Понятие ассоциации
Ассоциацией называют единицу сообществ совершенно определенного
ранга, как ее абстрактный тип, так и совокупность конкретных участков
сообщества, обладающих сходным видовым составом.
После Брюссельского конгресса (1910) под ассоциацией понимают «растительное
сообщество с определенным флористическим составом, однородными условиями место-
обитания и однородной физиономией». Оба последних положения не являются обяза-
тельным элементом при характеристике ассоциаций, поскольку экологические и физио-
номические признаки и без того уже определяются самим флористическим составом
сообщества.
• Ассоциация — это основная идиографическая единица в система-
тике сообществ (как вид в систематике таксономических единиц), диа-
гностически описанная и получившая наименование в соответствии с
принятыми правилами.
В пределах договоренности, что именно следует считать характерным сочетанием
видов, существует возможность более узкого или более широкого понимания ассоциа-
ции (подобно тому как понятие вида в систематике растений также может иметь и более
широкое, и более узкое значение).
Единицы растительности
115
Понятие ассоциации применяется и к той части биоценоза, которую
составляют животные, и ко всему биоценозу в целом (флористическая
и фаунистическая единицы в системе сообществ живых организмов).
Единицы сообществ более высокого ранга
Объединение ассоциаций в единицы более высокого ранга основано
на том же принципе, что и разграничение ассоциаций, а именно на верно-
сти видов.
Флористически родственные ассоциации образуют союз сооб-
ществ. Союзы отличаются друг от друга характерными для каждого
из них видами. По тому же принципу союзы объединяются в порядки
сообществ, а эти последние — в классы сообществ. Каж-
дая из этих единиц системы имеет свои характерные виды (виды, характер-
ные для порядка, и виды, характерные для класса). Число характерных
видов возрастает по мере увеличения ранга данной единицы в системе
сообществ.
Возможна ли в систематике сообществ еще более высокая ступень, чем классы
(или группы классов с соответствующими характерными родами), станет ясно лишь
в результате дальнейших исследований классов сообществ во всех частях Земли,
Единицы сообществ более низкого ранга
«Основная единица и наименьшая единица — это не обязательно
одно и то же» (Gradmann, 1909). Внутри некоторых ассоциаций удается
выделить отличающиеся друг от друга сообщества низшего ранга, у кото-
рых, правда, нет своих характерных видов, но видовой состав которых
отличается друг от друга благодаря наличию в них дифференциальных
видов. Такие единицы называют «субассоциациями», и при необходимости
они могут быть и дальше разделены на «варианты» по сочетанию видов
Для специальных целей, когда требуется очень подробное и точное разде-
ление растительных сообществ, но флористическое различие уловить уже
не удается, можно произвести и дальнейшую градацию некоторого коли-
чества видов на «ф а ц и и» 1 2.
В наиболее исследованных областях Европы растительные сообщества давно уже
используются для определения качественных различий сельскохозяйственных и лесо-
хозяйственных местообитаний. При этом оказалось неизбежным более мелкое дроб-
ление ассоциаций. Чем точнее можно подразделить ассоциации, тем вернее они будут
характеризовать особенности участков местообитаний и тем больше, соответственно,
будет их значение для крупномасштабной природной классификации ландшафтов.
Система сообществ позволяет координировать сведения, полученные по отдельным
участкам сообществ, и путем сравнения обобщать их в допустимых пределах.
Социации
«Социации» североевропейских фитоценологов, основанные не на верности видов,
а только на численном соотношении видов и однородности флористического состава,
нельзя непосредственно сравнивать с ассоциациями. По рангу они соответствуют,
скорее, низшим единицам ассоциаций (субассоциациям, вариантам, фациям).
1 В понимании большинства советских геоботаников, субассоциации предста-
вляют собой экологические варианты ассоциаций, отличающиеся или различными
со доминантами, или некоторыми сопутствующими видами.— Прим. ред.
2 Фация, в понимании франко-швейцарской геоботанической школы, выде-
ляется по одному из доминирующих дифференциальных видов.— Прим. ред.
116
Единицы растительности
в ландшафте
Наименование растительных сообществ
Названия растительных сообществ составляются таким образом,
чтобы по их окончанию был ясен ранг, занимаемый ими в системе.
Единица сообщества Класс Окончание -etea Пример Juncetea maritimi Br. Bl. 1931 (солончаковые луга)
Порядок -etalia Juncetalia maritimi Br. Bl. 1934
Союз -ion Armerion maritimi Br. Bl. et De Leuw 1936 (луга армериевые)
Ассоциация -etum Juncetum gerardi Warming (ситниковые луга)
Субассоциация -etosum Juncetum gerardi juncetosum maritimi
Вг. Bl. et Тх. 1952
Ранг вариантов, субвариантов и фаций обозначается приставкой
к названию ассоциации или субассоциации.
Примеры сводных таблиц сообществ
Способ изображения единиц сообществ в форме сводных таблиц
поясняется тремя нижеследующими примерами (табл. 2, 3 и 4).
ОБЩИЕ СВОЙСТВА И ЗНАЧЕНИЕ
ЕДИНИЦ СООБЩЕСТВ
Одновременно с изучением видового состава сообщества и его числен-
ности познается иструктура форм роста и физиономия сообще-
ства. В соответствии с входящими в него формами роста каждое сообще-
ство обладает специфическим внутренним жизненным
балансом, то есть характерным совместным действием жизненных
процессов всех участвующих в нем видов.
Полный список видов сообщества является, таким образом, одновре-
менно и отражением требований, предъявляемых этим сообществом
к местообитанию. Так как каждый вид имеет свою собственную эколо-
гическую валентность, флористический состав сообщества определяет
диапазон условий существования сообщества. За его пределами изме-
няется комбинация видов, и сообщество переходит в другое.
По комбинации видов каждое сообщество классифицируется с точки
зрения ареалов составляющих его видов и с точки зрения истории его
флоры. Оно может состоять из видов, относящихся к совершенно различ-
ным типам ареалов. Их подбор из всей флоры окружающего пространства
зависит от экологических особенностей сообщества и характера его место-
обитания.
Для сообщества, выделенного на основании флористического диагно-
за отдельных входящих в него идентичных конкретных участков, можно
установить характер его распространения и нанести его на карту ареалов
Таблица 2
Две субассоциации северогерманских камышово-тростниковых зарослей
(Scirpeto-Phragmitetum medioeuropaeum) (Tiixen, Preising, 1941)1
Субассоциации I. Typhetosum angusti- foliae 11. Phalaridetosum Посто- янство в %, полу- ченное из 10 описа- ний каж- дой субас- социации
Номер описания 1 2 4 6 7 12 13 14 16 17
Местообитание T T S S A F F St A A
Характерные виды ассоциаций и союзов: I II
асе. Sparganium erectum ssp. poly edrum acc. Acorus calamus c. Sagittaria sagittifolia c. Rumex hydrolapathum c. Schoenoplectus lacustris c. Butomus umbellatus . . acc. Typha latifolia .... 2.3 2.3 1.2 1.3 1.2 + .1 + •2 + •2 1.1 1.1 2.2 + 1.1 1.2 + .1 2.3 1.2 1.1 5.4 2.2 1.3 + .3 + .1 1.3 3.3 1.1 1.1 3.3 + .1 1.2 + .1 1.1 .1 + •1 80 40 . 30 20 10 90 60 50 60 40 50 20 30
Дифференциальные виды суб- ассоциаций:
c. Ranunculus lingua . . . c. Typha angustifolia . . . nop. Phalaris arundinacea . . nop. Oenanthe aquatica .... + .1 1.2 1.1 5.5 1.1 3.4 + •2 1.2 2.2 1.3 1.1 1.2 2.1 + .1 50 40 10 . 90 10 60
Характерные виды порядков и классов:
пор. Glyceria aquatica .... кл. Phragmites communis пор. Alisma planta go-а quatica nop. Sium latifolium nop. Equisetum limosum . . . nop. Lythrum salicaria .... nop. Iris pseudacorus кл. Polygonum amphibium terrestre 1.2 1.1 3.3 4.5 2.4 2.3 + .1 + .2 + .2 2.2 2. 1 1.2 2.3 1.2 + .1 1 .1 2.2 2 2 + -1 + .1 + .1 1.2 1.2 + .2 1.2 + .1 1.1 + .1 1.3 3.3 (+) + .1 + .1 1.1 3.2 4.4 + .1 1. 1 + .1 + .1 + .3 80 100 80 70 70 90 80 70 30 70 30 30 30 70 10 30
Сопутствующие виды:
Mentha aquatica .... Solanum dulcamara . . . Galium palustre Lysimachia vulgaris . . Schoenoplectus triquetrus Carex riparia Scirpus maritimus . . . Carex gracilis ...... 2.2 1.2 + .2 + •2 + .2 + .2 + .2 + .1 30 . 20 . 20 20 10 10 10 10 . 10 10
Число видов: .... 8 7 6 11 10 9 9 12 12 13
1 Приведенное на стр. 112 описание участка сообщества южнофранцузских камышово-
тростниковых зарослей касается ассоциации того же союза (Phragmition euro-sibiricum). G ассо-
циацией, описанной в данной таблице, тот участок сообщества имеет общие характерные для
союза виды Schoenoplectus lacustris и Typha angustifolia, а также характерные для порядка
и класса виды Phragmites communis и Iris pseudacorus (класс Phragmitetea).
Таблица 3
Piceetum subalpinum blechnetosum субальпийского елового леса
в Швейцарском Национальном парке
(Braun-Blanquet, Pallmann, Bach, 1954)
Номер описания Высота над уровнем моря, м Экспозиция Угол наклона, град Высота деревьев, м Диаметр деревьев, см Покрытие в древесном ярусе (сомкнутость крон), % Покрытие в напочвенном ярусе, % Площадь описанного участка, м% 1 1340 GCB 10 20 25-35 95 100 2 1340 C 40 30 35-60 100 80 100 3 1370 C 20 20 15-3 5 100 4 1340 GCB 20 25 30-3 5 95 90 100 5 1340 0 25 30—35 85 95 100 Посто- янство
Характерные виды ассоциаций:
Luzula luzulina (Vill.) D. T. et Sarnth . . ( + ) + + 3
Ptilium crista-castrensis (L.) + 2 • 12 • • 2
Дифференциальные виды относительно других субассоциаций:
Blechnum spicant (L.) Roth + + 2 22 + ( + ) 5
Plagiothecium undulatum (L.) 12 + 2 13 12 4
Gentiana purpurea L. (вегет.) + • • + (+) 3
Характерные виды союзов и порядков (V accini о-Piceion, Vaccinio-Piceetalia):
Picea excelsa (Lam.) Link 54 54 54 54 54 5
Vactinium myrtillus L 55 45 44 43—44 34 5
V actinium vitis-idaea L 12 12 22 + 12 5
Calamagrostis villosa (Chaix) Gmel. . . . (+) 12 + 2 3
Hieracium murorum L. em. Hudson . . . + + + 3
Melampyrum silvaticum L 11 21 2
Homogyne alpina (L.) Cass + 1
Hieracium vulgatum Fries + 1
Luzula silv. (Huds.) ssp. sieberi (T) . . . г 1
Lophozia lycopodoides (Wallr.) Cogn. . . . • + • 1
Сопутствующие виды:
Deschampsia jlexuosa (L.) Trin 21 11 12 21 21 5
Sorbus aucuparia L 11 + + + + 5
Majanthemum bijolium (L.) F. W. Schm. + + 11 3
Hylocomium proliferum LDB 33 33 33 33 22 5
Rhytidiadelphus triquetrus (L.) Warnst. 33 22 23 33 33 5
Pleurozium schreberi (Willd.) Milten . . . 33 33 33 33 23 5
Dicranum scoparium (L.) Hedwg 12 12 12 12 12 5
Sphagnum acutifolium (Ehrh.) R. et W. + + + + г 5
Polytrichum attenuatum Menzel • • • + + 2
Случайные — — 5 — 4
Таблица 4
Ассоциации Vaccinio — JPiceion-союзов в Ретийских Альпах
(пример сокращенной сводной таблицы растительности)
(Braun-Blanquet, 1946)1
Ассоциации А ва вь С D Е
Число описаний 11 И 12 24 27 12
Характерные виды для порядков и союзов См. таблицу порядка Vaccinio-Piceetalia у Браун-Бланке (Braun-Blanquet, 1951, S. 567)
Характерные виды ассоциации:
A. Piceetum transalpi- num
Luzula nivea .... Isothecium myurum Saxifraga cuneifolia Lonicera nigra . . . 911 910 728 47 X VII VII III 3 IV 1 I 23 IV 1 I
В. Piceetum montanum,
subass. galietosum
Galium rotundifolium 1068 X . .
Aquilegia vulgaris
ssp. atrata 51 VI 3 IV 1 I
Circaea alpina . . . 47 III . .
Catharinea hauskne-
chtii subass. melice-
tosum 2 III .
Galium triflorum . . 146 I
Epipogium aphyllum X 1 I
C. Piceetum subalpinum
Ptilium crista-castren-
sis ......... 211 VI 67 II 83 II
Lis ter a cor data . . . . 171 VIII
Lycopodium annotinum 4 IV 44 III 121 VIII 66 I
Luzula luzulina . . 51 VII . 108 VI 1 II
Pyrola uni flora . . . 4 VI 2 III 90 VIII .
Hylocomium loreum . . 21 I .
Mnium spinosum . . 1 I • 3 II •
D. Rhodoreto-Vaccini-
etum
Rhododendron ferrugi-
neum ....... 1 I 2788 X 149 V
Pinus cembra . . . 1 II 1121 VII 1 III
Linnaea borealis . . 2 III 564 VI 602 IV
Lonicera coerulea . . 1 I • 1 I 79 VIII 1 II
120
Единицы растительности в ландшафте
Продолжение табл. 4
Ассоциации А ва вь С D Е
Число описаний И И 12 24 27 12
Характерные виды
для порядков и союзов
См. таблицу порядка Vaccinio-Piceetalia у Браун-Бланке
(Braun-Blanquet, 1951, S. 567)
Е. Empetreto-Vaccini-
etum
Empetrum hermaphro-
diturn.............
Hieracium alpinum
ssp. halleri ....
Lycopodium alpinum
Cladonia uncialis . .
538 VIII
1 I
2980 X
232 V
43 II
42
1 Арабскими цифрами обозначается оценка покрытия, римскими — постоянство (I — мень-
ше 10%, X — больше, чем 90% описаний).
[Оценка покрытия (Deckungswert) представляет собой числовое выражение покрытия вида
в растительном сообществе. Основой для ее вычисления служит средний процент покрытия,
представляющий собой пересчет оценки мощности вида (по семибалльной шкале Браун-Блан-
ке) на среднюю оценку покрытия пробной площадки. Оценка мощности вида 5 соответствует
среднему проценту покрытия 87,5, соответственно мощность вида 4 равна среднему про-
центу покрытия 62,5 и т. д.
Для вычисления оценки покрытия сумма среднего процента покрытия вида (на одной
площадке) умножается на 100 и делится на общее число пробных площадок. Максимальная
оценка покрытия достигает 8750.— Прим, ред.}
(Knapp, 1948; Braun-Blanquet, 1951; Noirfalise, 1952; Carstensen, 1955;
Bodeux, 1955). Ареалы сообществ так же, как и ареалы систематических
единиц, можно расположить по типам ареалов. Однако до настоящего вре-
мени полностью известны ареалы только немногих сообществ.
Нуарфализ (Noirfalize, 1952) составил карту ареалов среднеевропейских ясене-
во-осоковых лесов (Cariceto remotae — Fraxinetum atlanticum) (рис. 31).
Бодё (Bodeux, 1955) на точечной карте (Punktkarte) представил все до сих пор опи-
санные случаи нахождения четырех известных в Европе сообществ черно-ольховых
болот (группа «Alnetum glutinosae» союза Alnionglutinosae), а именно
1) атлантическая ассоциация черно-ольхового болота (Cariceto laevigatae — Alne-
tum);
2) среднеевропейская ассоциация черно-ольхового болота (Cariceto elongatae —
Alnetum medioeuropaeum);
3) североевропейская ассоциация черно-ольхового болота (Cariceto elongatae —
Alnetum boreale);
4) приблизительно установленная восточноевропейская ассоциация черно-оль-
хового болота (Dryopterideto cristatae — Alnetum).
В рамках географического рассмотрения растительности разные клас-
сы сообществ неравноценны. Наибольшее значение имеют прежде всего
те сообщества, которые по их формационному признаку и распростране-
нию определяют характер ландшафта на больших пространствах или
Единицы растительности
121
имеют диагностическое значение для классификации с точки зрения геогра-
фии растительности. Некоторые растительные сообщества, представляю-
щие большой интерес для фитоценологов, могут быть совершенно не инте-
ресны для географа, потому что встречаются весьма редко, на незна-
чительных площадях и ни с точки зрения ландшафта, ни для познания
Рис. 31. Распространение ясенево-осоковых лесов в Европе (Noirfalise, 1952).
1 — бореально-атлантическая оптимальная область; 1а — пиренейско-астурийская область;
16 — рейнская область Cariceto remotae — Fraxinetum attanticum\ 2 — область Cariceto
remotae — Fraxinetum Occident o-b at ticum; 3 и 4 — изолированные местонахождения Carex
strigosa.
естественных пространств или пространственных единиц растительно-
сти не имеют никакого значения. С другой стороны, растительные сооб-
щества, кажущиеся для биолога однообразными и скучными, лишь
незначительно отличающимися друг от друга, для географа могут иметь
значение с точки зрения их особой роли в развитии культурного ланд-
шафта.
ЧИСЛО ИЗВЕСТНЫХ РАСТИТЕЛЬНЫХ
СООБЩЕСТВ
Работа по описанию и наименованию растительных сообществ
на большей части земной поверхности только началась, и поэтому сейчас
еще нельзя назвать общее число существующих ассоциаций. В целом,
видимо, уже описаны от 1000 до 1200 ассоциаций. Из этого ясно, насколь-
ко необходимой стала их систематизация.
122 Единицы растительности в ландшафте
Лишь в некоторых случаях обширные, взаимосвязанные области исследованы
настолько хорошо, что может быть построена система, доходящая до классов сооб-
ществ. Тюксен (Тйхеп, 1955) называет для северо-западной Германии 30 классов со
189 ассоциациями, входящими в 76 союзов. Для Средней Европы вместе с внесредизем-
номорской Западной Европой число классов увеличено до 34. Из них 21 встречается
в Ирландии, 20 — в Бельгии и 17 — в Голландии. На средиземноморском юге Фран-
ции встречаются, по Браун-Бланке (Braun-Blanquet, 1956), 23 класса с 58 союзами.
Для Европы, включая область Средиземного моря, до настоящего времени получили
наименование около 70 классов. Во внеевропейских областях Земли число классов
должно быть во много раз больше.
ОБЗОР КЛАССОВ РАСТИТЕЛЬНЫХ СООБЩЕСТВ,
ПРЕДСТАВЛЕННЫХ В СРЕДНЕЙ ЕВРОПЕ
(по работе Тюксена, опубликованной в 1955 г.9
и дополнениям в письмах 1957 г.)
1) Lemnetea
2) Zosteretea marinae
3) Asplenietea rupestris
4) Adiantetea
5) Thlaspetea rotundifolii
6) SaUcetea herbaceae
7) Ammophiletea
8) Corynephoretea canescentis
9) I soeto-N anojuncetea
10) Thero-Salicornietea
11) Cakiletea maritimae
12) Bidentetea tripartiti
13) Stellartetea mediae
14) Epilobietea angustifolii
15) Plantaginetea maioris
16) Artemisietea vulgaris
17) Potametea
18) Littorelletea
19) Montio-Cardaminetea
20) Phragmitetea
21) Juncetea maritimi
22) Molinio-Arrenatheretea
23) Elymo-Seslerietea
24) Caricetea curvulae
25) Festuco-Brometea
26) Scheuchzerio-Caricetea fuscae
27) Oxycocco-Sphagnetea
28) Ndrdo-Callunetea
29) Вetuleto-A denostyletea
30) Alnetea glutinosae
(сообщества ряски)
(заросли морской травы)
(сообщества в трещинах скал)
(известняково-туфовые сообщества)
(сообщества известняковых осыпей)
(сообщества снежных проталин)
(сообщества песколюба на североевро-
пейских дюнах)
(поля булавоносца)
(сообщества карликовых ситников)
(сообщества солероса)
(сообщества морской горчицы)
(сообщества череды)
(сообщества мокрицы)
(сообщества кипрея на лесосеках)
(вытаптываемый и затопляемый дерн)
(сообщества полыни на берегах и осыпях)
(сообщества плавающих листьев)
(сообщества на побережьях)
(сообщества на лугах у источников)
(тростниковые заросли и заросли круп-
ных осок)
(солончаковые луга)
(европейские сельскохозяйственные луга
и пастбища)
(сеслериевые склоны)
(естественные альпийские луга)
(сухая дернина)
(болота с низкой осокой)
(сообщества болотной клюквы и сфа-
гновых мхов и влажные пустоши)
(дерновиннозлаковые луга и кустар-
ничковые пустоши)
(субальпийские высокотравные сообще-
ства)
(болотистые ивовые кустарники и чер-
но-ольховые леса)
Единицы растительности
123
31) Vaccinietea uliginosi
32) V accinio-Piceetea
33) Quercetea robori-petraeae
34) Querceto-Fagetea
(березовые и хвойные болота)
(хвойные леса и альпийские кустар-
ничковые пустоши)
(дубово-березовые леса)
(терновые кустарники и смешанные
лиственные леса).
Из этих 34 классов 25 встречаются как в северо-западной, так и в южной Гер-
мании (Tuxen, 1955; Oberdorfer, 1957). Четыре класса, а именно Adiantetea, Salicetea
herbaceae, Elymo-Seslerietea, Caricetea curvulae в северо-западной Германии не встре-
чаются. Пять классов, а именно Zosteretea marinae, Ammophiletea, Cakiletea maritimae,
Juncetea maritimi, Vaccinietea uliginosi, отсутствуют в южной Германии.
ТИПОЛОГИЯ И КЛАССИФИКАЦИЯ ЕДИНИЦ
РАСТИТЕЛЬНОСТИ (ФИТОЦЕНОЗОВ)
ПО ИХ ВНЕШНЕМУ ОБЛИКУ
ЗНАЧЕНИЕ ФИЗИОНОМИЧЕСКОЙ ТИПОЛОГИИ
РАСТИТЕЛЬНОСТИ
Облик растительных сообществ является существенным компонен-
том в общей картине ландшафта. Поэтому для географии растительности
предметом особой заботы является физиономическая клас-
сификация фитоценозов. Подобная классификация единиц расти-
тельности важна еще и потому, что во внешнем облике растительных
сообществ в значительной степени отражается и их экологический харак-
тер. Физиономическая типология позволяет географу даже без позна-
ния растительных сообществ составить определенные представления
о классификации растительности и связях последней с другими явле-
ниями на земной поверхности. В этом географу наряду с понятием «форма
роста» помогает также понятие «растительная формация».
РАСТИТЕЛЬНАЯ ФОРМАЦИЯ
Это понятие было определено Гризебахом в 1838 г. следующим обра-
зом: «Мне хотелось бы группу растений, которая имеет замкнутый физио-
номический характер, как, например, луг, лес и т. п., назвать расти-
тельно-географической формацией». Позднее он в аналогичных случаях
употреблял более короткое название «растительная формация»; тем
самым общепринятые в языке собирательные понятия, служащие для
характеристики типичных участков биосферы, Гризебах дополнил одним
из основных научных понятий. Как видим, Гризебах выбрал в качестве
критерия для типовых понятий единицы растительности не виды расте-
ний (как Линней) и не местообитания или местности (как Унгер или
Хеер), а тип облика самой растительности, ее «замкнутый физиономи-
ческий характер».
Однако, по существу, еще в 1824 г. Мартиус под «формой растительности» уже
понимал растительную формацию. Кернер говорил о формациях как «сцеплении сооб-
ществ с одинаковыми формами роста». Исходя из известной зависимости форм роста
от местообитания, позднее в определение растительной формации была включена
и экология. Так, по Шрётеру, к формации причисляются все участки сообществ, «кото-
рые сходны по своей физиономии и в основных чертах однозначны по своей экологии».
124 Единицы растительности в ландшафте
В этом смысле понятие «тип формы биоценозов», взятое без учета систематического
состава, может быть применено как к биоценозам, охватывающим и животных, и расте-
ния (био-формации), так и к фитоценозам (растительные формации).
Даже если понятие растительной формации касается только физио-
номического момента, оно все же содержит и экологический элемент.
Дело в том, что экология и физиономия растительных форм, а тем самым
и биоценоза находятся до известной степени в причинной связи. Бла-
годаря конвергенции форм роста при аналогичных условиях существо-
вания, даже в областях, значительно друг от друга удаленных и насе-
ленных совершенно разными систематическими единицами, в одинаковых
местообитаниях встречаются физиономически сходные биоценозы, как,
например, жестколистные леса среднего Чили и средиземноморского
юга Европы.
В географии растительности мы употребляем термин «растительная
формация» только для единиц растительности высшего порядка, имею-
щих непосредственное значение для ландшафта, то есть для единиц,
являющихся самостоятельными с растительно-топографической точки
зрения. Это соответствует и взглядам Гризебаха, который свое понятие
растительной формации, данное в 1872 г., интерпретировал как «вхож-
дение растительных форм в физиономию ландшафта».
Более мелкие, топографически несамостоятельные участки растительных сооб-
ществ с одинаковым составом форм роста, как, например, моховые покровы на коре
древесных стволов, называют «синузиями». Растительная формация или содержит сину-
зии как отдельные зависимые единицы растительности, или она состоит из многих сину-
зий, мозаично расположенных на минимальном пространстве, например на кочках
и в мочажинах верхового болота.
СУЩНОСТЬ И ПРИЗНАКИ РАСТИТЕЛЬНЫХ
ФОРМАЦИЙ1
То, что было сказано выше относительно форм роста, применимо
и в данном случае: каждая отдельная растительная формация является
родовым понятием, индуктивно созданным на основе сравнения, то есть
абстрактным образом. Отнесенное к отдельному случаю, такое понятие
дает общую характеристику биоценоза, но не охватывает при этом всех
его индивидуальных особенностей. Название растительной формации
обозначает как характер формации отдельного биоценоза, так и совокуп-
ность всех биоценозов, имеющих одинаковый характер формаций. Вве-
денные в этих названиях пространственно-климатические добавления
(например, «тропический») вполне себя оправдали и прижились. Следует,
однако, подчеркнуть, что эти добавления не являются составными частями
наименования формации. Формация, названная по климатической зоне,
может в определенных случаях встречаться и вне этой зоны. Название —
только символ типа растительности, характеризуемого эколого-физионо-
мическим диагнозом. Летнезеленый лиственный лес умеренной зоны
1 Большинство советских геоботаников понимают под растительной формацией
классификационную единицу растительности, объединяющую растительные ассоциа-
ции (группы ассоциаций), в которых господствующий ярус сложен одним и тем же
доминирующим видом. Автор книги трактует растительную формацию с точки зрения
географа как ландшафтное эколого-физиономическое понятие, применяемое только
для единиц растительности высшего порядка, имеющих непосредственное значение
.для ландшафта.— Прим. ред.
Единицы растительности
125
в пределах Восточной Азии или на востоке США содержит одинаковые
со Средней Европой формы роста, сохраняющие его физиономию и отра-
жающие экологический характер: тропофитные кронообразующие
деревья и кустарники, крупное разнотравье, рано цветущие травянистые
геофиты, напочвенные мхи, образующие дерн и т. д. В обоих случаях
формация, кроме того, характеризуется ярусным строением и одинако-
вым жизненным ритмом, с осенним листопадом, зимним периодом покоя,
более ранним в сравнении с листвой на деревьях распусканием напоч-
венных растений и т. д. Как показывает этот пример, каждая формация,
кроме признаков, отражающих внешний облик (высота, плотность,
ярусность) и комбинации форм роста, может быть охарактеризована
также свойственной ей периодичностью жизни растений. Количество
сезонно изменяющихся аспектов может быть различно. Так, в летнезеле-
ном лиственном лесу Средней Европы насчитывают 6—7, в южнорусских
степях по крайней мере 10 вполне различимых фаз.
Термином «формация растительности» часто называют крупные широко распро-
страненные эколого-физиономические единицы растительности, состоящие из большого
числа различных растительных формаций. Будет более правильным называть такие
единицы «комплексами формаций» (ср. стр. 201).
ОБЗОР РАСТИТЕЛЬНЫХ ФОРМАЦИЙ
В то время как для систематики сообществ определяющим является
только один критерий, формации могут быть подразделены с нескольких
точек зрения. При этом ни в коем случае не вступают в свои права все
категории признаков одновременно.
Выделяются следующие классы растительных формаций:
I. Леса
II. Кустарниковые формации
III. Саванны и степи
IV. Луга
V. Формации кустарничков (карликовых кустарников) и полукустарников
VI. Терофитные формации
VII. Пустыни
VIII. Растительные формации внутренних водоемов
IX. Растительные формации морей.
Каждый из этих классов включает еще различные типы формаций. Лишь очень
небольшая часть растительного мира не находит в этой системе своего отражения.
Растительные формации
I. Леса
Как о понятиях, которые стремятся выразить в одном слове неко-
торый сложный элемент действительности, так и о понятии, выраженном
словом «лес», можно написать целую книгу. Лес, если он только не начисто
вырублен (что с точки зрения лесовода не исключает сохранения за ним
этого термина), состоит прежде всего из деревьев. Искусственные древес-
ные насаждения также могут предртавлять собой лес. Однако не каждый
набор деревьев является лесом. Разбросанные на большом пространстве
126 Единицы растительности в ландшафте
одиночно или группами деревья, так же как и «рощи» плодовых деревьевг
мы не называем лесом. Лес — это сложный растительный комплекс, в кото-
ром деревья расположены более или менее тесно, кроны их соприка-
саются, образуя полог и до известной степени затеняя почву; раститель-
ный покров в лесу состоит из нескольких ярусов. Ниже древесного полога,
состоящего часто из нескольких ярусов, располагается подлесок из кустар-
ников и кустарниковидного подроста молодых деревьев, а в естествен-
ном лесу всегда имеется еще ярус напочвенных растений с разнообразными
формами роста, как, например, кустарнички, многолетнее разнотравье,
злаки, подушки мхов и лишайников. Следует упомянуть и лианы,
достигающие вершин деревьев, а также более или менее многочисленные
синузии растений, живущих на стволах, ветвях или листьях. Наконец,
лес немыслим без обитающих в нем животных. Каждый ярус характери-
зуется определенными условиями существования, и произрастающие
в них растения могут весьма существенно отличаться по внешнему облику
и образу жизни.
Понятие «растительные формации лесов» при изучении их на боль-
ших пространствах расширяется. Оно охватывает не только лесные
биоценозы в узком смысле, но включает и прогалины, образующиеся при
падении стволов, и безлесные склоны, растительность скал, луга у источ-
ников или болота, лесные опушки по берегам рек и ручьев и более мел-
кие, созданные человеком открытые участки — однако все это при усло-
вии, что пространственное единство собственно лесного покрова при этом
не нарушается.
При характеристике формаций прежде всего принимаются во вни-
мание естественные участки растительности. Однако во многих частях
земной поверхности с трудом уже удается отделить естественные леса
от лесов, испытавших преобразующее воздействие человека.
По экологическому признаку, который выражается в общей физио-
номии и прежде всего в характере листвы деревьев, мы разделяем леса
на четыре основные группы: вечнозеленые дождевые леса; вечнозеленые
жестколистные и хвойные леса; листопадные леса; колючие и сукку-
лентные леса.
ВЕЧНОЗЕЛЕНЫЕ ДОЖДЕВЫЕ ЛЕСА
Тропические вечнозеленые дождевые леса низменностей. Вечнозеле-
ные дождевые леса постоянно влажных равномерно теплых тропических
низменностей являются самыми пышными растительными формациями
на Земле. При непрерывном росте их внешний вид остается неизменным
в течение всего года. Большое число видов и форм располагается многими
ярусами (рис. 32). Леса имеют три, а иногда два или четыре яруса, обра-
зуемых древесными кронами, а кроме того, ярус кустарников, содержа-
щий в основном подрост деревьев, и ярус напочвенной растительности.
Поверхность древесного полога, достигающего высоты 30—50 ле, пред-
ставляется неровной вследствие того, что отдельные виды деревьев имеют
различную высоту. Над общей поверхностью крон часто выступают вер-
шины отдельных деревьев-великанов высотой более 60 ле, а в исключи-
тельных случаях и 90 ле. Древесные растения (деревья, лианы, древовид-
ные злаки, древесные эпифиты) составляют в тропических дождевых
лесах большую часть видового состава. На одном гектаре редко встре-
чается меньше сорока видов деревьев, а порой их число доходит до ста
(рис. 33).
Единицы растительности
127
О 20 4/7 60 80 100м
Рис. 32. Профильная диаграмма ярусов тропического дождевого леса в Британской
Гвиане. Буквами обозначены различные виды деревьев (Richards, 1952).
Преобладающими формами роста являются здесь гигроморфные
вечнозеленые кронообразующие деревья. Деревья с листовой кроной,
преимущественно пальмы, часто образуют приметную, однако не опре-
деляющую общего характера этих лесов примесь. Многие возвышающиеся
над общим пологом леса де-
ревья-великаны опираются на
мощные, отходящие у основа-
ния стволов корни-подпор-
ки или досковидные корни
Рис. 34. Формы оснований стволов
трех видов деревьев тропического дож-
девого леса в Британской Гвиане
(Richards, 1952).
а — Mora excelsa', б — Mora gonggriipii",
в — Eschweilera sagotana.
Рис. 33. Число видов, при-
ходящихся на единицу пло-
щади для древесных ярусов
тропического дождевого леса
в Нигерии (Richards, 1952).
Сплошная линия относится к
деревьям с диаметром ствола бо-
лее 10 cat, пунктирная—к деревьям
с диаметром ствола более 30 см.
(рис. 34). У многих деревьев имеется поверхностная корневая система
(Wurzelteller), которая при падении стволов принимает вертикальное
128
Единицы растительности в ландшафте
положение. Распространены деревья с ходульными корнями. Светло-
зеленые или белые не защищенные коркой гладкие стволы, как правило,
бывают стройными и прямыми; у многих ветвление начинается только
в верхних ярусах, у самых крон. Стволы с диаметром более одного метра
встречаются редко. Древесный полог пропускает внутрь леса только зеле-
новатый сумеречный свет. Его яркость у поверхности почвы составляет
менее одной сотой, а иногда и менее одной тысячной доли яркости наруж-
ного дневного света; прямой солнечный луч проникает в глубь леса
только в исключительных случаях.
Внутри леса листва у поверхности почвы преимущественно крупная
и тонкая. На кронах деревьев она средней величины и более грубая,
здесь преобладают простые продолговато-овальные листья (рис. 35).
Некоторые листья имеют ка-
Р и с. 35. Типичная форма листьев у деревьев
тропического дождевого леса на острове Бор-
нео (Richards, 1952).
Показаны листья разных видов деревьев: а — бо-
лее 26 м высотой; б — от 13 до 26 At; в — менее
1 3 At.
пельные острия, ускоряющие
стекание воды. Смена листьев,
так же как цветение и созре-
вание плодов, распределена на
протяжении всего года. У мно-
гих видов деревьев этот процесс
не связан с определенным сезо-
ном. Листва меняется в харак-
терном для каждого вида ритме,
периоды которого могут быть
длиннее и короче года. У неко-
торых видов наблюдается ко-
роткий период покоя, в течение
которого растение лишено ли-
стьев. То же можно сказать и о
вторичном росте стволов и вет-
вей в толщину; поэтому на
древесине, как правило, нет
годичных колец. Цветки у де-
ревьев большей частью неяркие,
зеленоватые или белые, но
иногда встречаются большие по величине и заметные по окраске.
Лес очень богат лазящими растениями. Большинство из них —
деревянистые растения, лианы самых различных форм, которые, стре-
мясь к свету, оплетают всю поверхность древесного полога.
Обладая довольно толстыми стволами (до 20 см в диаметре) и бесчис-
ленными свисающими вниз воздушными корнями, они сужают видимое
пространство в лесу и поэтому проникнуть в глубь такого леса чрезвы-
чайно трудно. Листья и цветки распускаются в основном высоко в кро-
нах деревьев. Многие лианы украшают просеки или лесные опушки
по берегам рек и водоемов свисающими занавесями ярких цветков.
Древовидные злаки, растущие, как правило, густыми скоплениями,
делают некоторые участки тропических дождевых лесов почти непро-
ходимыми. Древесные и травянистые эпифиты, представленные боль-
шим числом своеобразных растительных форм, поселяются в основном
на ветвях и развилках стволов высоких деревьев. Многие растения,
например орхидеи и бромелиевые, имеют яркие цветки, частично опыляе-
мые птицами. Однако для наблюдателя, находящегося внизу, они про-
падают в общей массе зелени. Многочисленные эпифиты или выпускают
воздушные корни, или образуют свисающие ковры из цветков. Своеобраз-
<1> ото 1. Тропический дож-
девой лес па тихоокеанских
склонах Аид в Колумбии
(долина реки Апчнкауа,
500 м над уровнем моря).
Фото 2. Воропковпдиые
папоротниками в Западных
Кордильерах Колумбии
(близ Битако, 2700 м над
уровнем моря).
Фото 4. Субтропический
дождевой лес па острове
Окинава (фото Миявакн).
Фото 5. Умеренный
дождевой лес со свежей
гарью близ Пунтры па
Ч илоэ.
Фото 6. Умеренный дож-
девой лес близ Пунтры на
Чилоэ с бамбуковым под-
дитдииим jivvy у Jiai’U-
Тепуэйко на Чилоэ.
Фото 8. Листья чилийского
жестколистного дерева
(Peumus boldo).
Фото 9. Жестколистный лес (Quercus ,sp. и Olea cuspid ata) в северо-западных Гпма
(долина верхнего Свата, 1600 м над уровнем моря) (фото Фаутца).
Фото 10. Сухой жестколистный лес с пальмами (Jubaea spectabilis) и стеблевыми суккулен
на прогалинах (Кокалапталь).
Фото 11. Жестколистный
туманный лес с бородами
Tillandsia usiieoides (Brome-
Jiaceac) в северной части
среднего Чили.
Фото 12. Лавровый лес
Т» TATHJ Q ТТТТГЧ»» ~ —
Кетрупильяне (14UU м над
уровнем моря).
Фото 14. Мангровые
заросли из Brugiera conju-
gata па острове Ирпомоте
(фото Мияваки).
Фото 15. Лстпезелепып
лиственный лес (дубово-гра-
бовый) в Хунсрюке (долина
Вашбаха).
Ф ото 16. Дерево с сукку-
лентным стволом («бутылоч-
ППР ТТРПРИГИЙ R CVYOM TTPP.V
склоне долины Таити (фото
Фаутца).
Фото 18. Колючее дерево
(Proso pis juliflora), остаток
уничтоженного пожаром
колючего сухого леса в
северном Чили.
Ф одо 19. Жестколистный кустарник с Lucuina paradisea в северной части среднего Чили
(Лос-Внлос).
Фото 20. Формация колючих кустарников и суккулентов в северном Чили.
Фото 21. Суккулснтпая формация в туманном поясе прибрежной пустыни (Токопиль
1000 м над уровнем моря).
Ф ото 22. Вечнозеленые низкорослые заросли Nothofagus antarctica на Кетрупилья
(граница леса и снега близ Лаго-де-лос-Патос, 1650 м над уровнем моря).
Фото 23. Влажная саванна с галерейным лесом в долине Каука*
Фото 24. Колючая саванна в долине Магдалены близ Хирардота.
ф о т о 25. Высокотравье (Seiiecio smilJiii) на берегу Магелланова пролива.
ф о т о 26. Формация карликовых кустарников у Гопдолнако севернее Ла-Серены.
Фот о 27. Формация дерповинных злаков (сухая пупа) па вулканической почве (Кетру-
пильян, 1800 м над уровнем моря)..
Фото 28. Формация колючих подушек выше границы леса у Антуко.
Фото 29. Пастбище влажной пуны в поясе парамос, Эквадор (3600 л над уровнем мор
Фото 30. Парамос с «монашками» frailejones (Espeletia graitdijlora) выше Боготы (3700 м г
уровнем моря).
Фото 31. Сфагновое болото с обилием лишайников близ Далькауэ на Чнлоэ.
Фото 32. Цветущая пустыня у Копьяпо.
Единицы растительности
129
ной формой являются деревья-удушители, начинающие свою жизнь как
эпифиты (ср. стр. 60). Стволы деревьев часто покрыты споровыми эпифи-
тами: волосяными папоротниками, мхами и лишайниками. В виде спе-
циализированных форм роста листостебельные и печеночные мхи, водо-
росли и лишайники поселяются как эпифиллы на многолетней листве
деревьев или подлеска.
Растения с травянистыми стволами (например, бананы) — это форма
роста, существование которой ограничено почти исключительно тропи-
ческими дождевыми лесами; на высоких расширенных черешках, обра-
зующих ложный стебель, у них вырастают гигантские листья. Подлесок
в этих лесах может быть или очень густым, или отсутствовать вовсе.
У девственного леса подлесок часто развит слабее, чем у вторичного.
Р и с. 36. Спектры жизненных форм
(по Ричардсу и Тюксепу).
а — тропический дождевой лес в Бри-
танской Гвиане; б — летнезеленый ли-
ственный лес умеренной зоны (Герма-
ния). Е — эпифиты; Ph — фанерофиты;
Ch — хамефиты; Н — гемикриптофиты;
G — геофиты; Th — терофиты.
Кустарники с вечнозелеными большими мягкими листьями представ-
лены гораздо более скудно, чем молодая поросль деревьев и лиан. Как
и у некоторых деревьев, у многих кустарников наблюдается каулифло-
рия.
Низкорослые растения на поверхности лесных почв развиты, как
правило, слабо из-за недостатка света и большой конкуренции со сто-
роны корней. Видов в напочвенном ярусе значительно меньше, чем число
эпифитных видов. Хамефитное разнотравье наряду с молодой порослью
фанерофитов составляет основную массу растительности напочвенного
яруса. Некоторые экземпляры образуют огромные розетки из крупных
сочных листьев высотой с человеческий рост. Геофиты, гемикриптофиты
и терофиты, то есть жизненные формы, приспособленные к холодным
и сухим периодам, здесь почти полностью отсутствуют. Некоторые харак-
терные геофиты живут как паразиты на корнях или как сапрофиты
в перегное почвы и выносят на поверхность только свои большие блед-
ные или мясо-красные цветки.
Наряду с господствующими лесами, в которых ни один из видов не доминирует,
существуют леса с отдельными преобладающими видами деревьев, но, как правило,
чистые сообщества из одного доминирующего вида деревьев в тропических дождевых
лесах не встречаются.
С изменением локальных условий местообитания может меняться
и эколого-физиономический характер леса. Поэтому различают целый
ряд растительных формаций, например: леса на незатопляемых и затоп-
ляемых почвах, леса на болотистой почве, леса по берегам рек и т. д.
С высотой местности характер леса тоже меняется. В горах средней
высоты, примерно от 500 до 1000 м, лес становится двухъярусным (пред-
горный дождевой лес, фото 1).
«Сезонными дождевыми лесами» (evergreen seasonal forest) называют формации
окраинного пояса экваториальных дождевых лесов, в которых листопад приурочен
к относительно сухому времени года.
130
Единицы растительности в ландшафте
Тропические вечнозеленые горные леса (горные дождевые леса).
Вечнозеленые горные леса в постоянно влажных тропиках и туманных
районах тропических гор (преимущественно на высотах 1000—3500 м)
обладают, как леса тропических низменностей и субальпийского пояса,
непрерывной на протяжении всего года вегетативной деятельностью.
Однако рост горных лесов происходит медленнее, поскольку с высотой
количество тепла уменьшается. По сравнению с лесами тропических
низменностей они значительно беднее видами, особенно ярус деревьев.
Тем не менее и здесь этот ярус почти всегда смешанный и не замечается
никакой тенденции к образованию чистых древостоев. Высота деревьев
меньше и достигает в среднем 15—20 м. Леса, расположенные более
высоко и особенно на открытых ветру участках, не превышают 5—10 м.
Образуемый древесными кронами полог состоит из двух ярусов, между
которыми часто нет резкой границы; встречаются и одноярусные пологи.
Деревья шире и суковатее, стволы при их относительно малой высоте
много толще, чем в лесах низменностей. Досковидные и опорные корни,
а также каулифлория отсутствуют.
Листья цельнокрайние, большей частью мелкие, иногда средней
величины. Капельные острия на концах листьев отсутствуют. Распро-
странены также эрикоидные и игольчатые листья. Может встречаться
примесь хвойных деревьев, не характерных для лесов низменностей.
Лазящие растения представлены скудно. Крупные лианы, как пра-
вило, отсутствуют. С другой стороны, бамбук может играть в тропиче-
ских горных лесах большую роль. В травянистом ярусе часто преобла-
дают папоротники. Исключительно многочисленны эпифиты. Однако
среди них мало цветковых растений, тогда как листостебельные и пече-
ночные мхи, папоротники и лишайники, особенно в туманных лесах,
расположенных среди постоянной сырости и облаков, почти полностью
окутывают стволы и ветви низких, суковатых деревьев. Для тропических
горно-туманных лесов характерны деревья с листовыми кронами —
древовидные папоротники с тонкорасчлененными весьма гигроморф-
ными листьями. Встречаются и низкорослые пальмы (фото 2 и 3).
Субтропические вечнозеленые дождевые леса. Вечнозеленые субтро-
пические дождевые леса постепенно переходят в тропические и имеют
с последними много общего. По высоте и густоте они мало уступают тро-
пическим лесам. Древесный полог имеет обычно два или три яруса. Число
лиан и эпифитов в некоторых субтропических дождевых лесах почти
такое же, как и в тропических дождевых лесах, иногда значительно
меньше. Нередко здесь встречаются аналогичные формы эпифитов с круп-
ными листьями. Капельные острия чаще имеют листья средней величины.
Стройные стволы деревьев опираются на опорные или досковидные корни.
Неровная поверхность основного яруса крон достигает в среднем высоты
20—35 м, а над ней возвышаются отдельные вершины 40—50 м высотой
(фото 4).
В спектре жизненных форм субтропического леса (рис. 37), как
и тропического, характерно обилие фанерофитов и эпифитов. Однако
в субтропических дождевых лесах велика доля гемикриптофитов, которые
почти отсутствуют в дождевых лесах тропиков. Существует и ряд других
отличий: отсутствие каулифлории, появление хвойных деревьев, а глав-
ное, меньшее богатство видов в древесном ярусе и в некоторых лесах —
тенденция к явному преобладанию небольшого числа древесных видов.
Умеренные вечнозеленые дождевые леса. Эти леса частично еще
обнаруживают сходство с субтропическими дождевыми лесами. Средняя
Единицы, растительности
131
высота деревьев в отдельных древостоях 20—30 м, но встречаются экзем-
пляры и до 50 м высотой. Деревья, преимущественно с узкой кроной,
представлены большим числом видов. Иногда наблюдаются досковидные
корни, отходящие от стволов. Многочисленны древесные лианы. Среди
эпифитов встречаются также некоторые цветковые растения. В более
высоких широтах умеренные дождевые леса становятся низкорослыми
(5—20 м) с двухъярусным или одноярусным пологом. Количество видов
деревьев может уменьшаться, вплоть до древостоев, состоящих из неболь-
шого числа видов или даже из деревьев только одного вида. Многие
деревья имеют сучковатую широкую крону с очень маленькими жесткими
листьями. Рост деревьев медленный, с более или менее выраженной
сезонной периодичностью. Такие признаки тропических дождевых лесов,
как наличие каулифлории, травянистых стволов и древовидных злаков,
Р и с. 37. Спектры жизненных форм (по Рпчардсу).
а — субтропический и б — субантарктический дождевой лес. Е — эпи-
фиты; Ch — хамефиты; Me — мега- и мезофанерофиты; Н —ггеми-
криптофиты; Mi — микрофанерофиты; G —геофиты; Na — нанофа-
нерофиты; Th — терофиты.
отсутствуют; не встречаются и характерные еще для субтропических
лесов капельные острия на листьях, опорные корни; нет обилия эпифит-
ных цветковых растений. Фанерофиты не являются абсолютно домини-
рующими, как это наблюдается в тропических и субтропических дожде-
вых лесах, однако по занимаемой ими доле в спектре жизненных форм
могут стоять на первом месте (рис. 37). Крупные древесные лианы встре-
чаются в небольшом количестве, а иногда и совсем отсутствуют. Карлико-
вые кустарники (хамефиты) занимают видное место в спектре жизнен-
ных форм. Кустарники могут быть представлены большим числом видов,
чем деревья. Значительная доля принадлежит здесь гемикриптофитам.
Геофиты, как и во всех дождевых лесах, почти полностью отсутствуют.
Общим с тропическими горными дождевыми лесами является обилие
эпифитных мхов, лишайников и папоротников. Характерно для уме-
ренных дождевых лесов значительное участие в древостое хвойных
деревьев (фото 5, 6, 7).
ВЕЧНОЗЕЛЕНЫЕ ЖЕСТКОЛИСТНЫЕ И ХВОЙНЫЕ ЛЕСА
Жестколистные леса. Вечнозеленые жестколистные леса, тип расти-
тельных формаций субтропических областей с зимним периодом дождей,
являются частью сомкнутыми высокоствольными лесами, а частью пред-
ставляют собой светлые насаждения, состоящие из разреженных древо-
стоев с густым кустарниковым подлеском. Последний может переходить
в кустарниковые формации типа маквисов.
132
Единицы растительности в ландшафте
Леса достигают умеренной высоты, в среднем 15—20 м, в исключи-
тельных случаях, как лес «карри» в юго-западной Австралии, до 50 м.
Древесный полог, как правило, образует один ярус, реже два, при-
чем ксерофитные деревья с листовой кроной (например, чилийская пальма)
могут находиться в верхнем ярусе. Форма роста жестколистных деревьев,
не говоря об особых формах эвкалиптов и казуариновых, — характерный
пример конвергенции, обусловленной окружающей средой. Мощные
защищенные толстой коркой или пробкой стволы начинают ветвиться
на незначительной высоте, образуя широкие кроны. Листья, сохраняю-
щиеся в течение нескольких лет, — жесткие, кожистые, средней вели-
чины и мелкие, большей частью цельнокрайние, овальной или эллиптиче-
ской формы, которая может переходить в листовую колючку. Нередко
листья снизу опушены и имеют матовый серо-зеленый цвет. Многие
покрыты блестящим восковым или лаковым налетом и пахнут эфирными
маслами. Изредка встречаются перистые листья или крупные листья
типа лавровых. Хвойные деревья, преимущественно с широкими кронами
и пиниевидной хвоей, могут встречаться в виде примеси или, преобла-
дая, образовывать верхний ярус над жестколистным кустарником.
За исключением древесных паразитов типа омелы, эпифитные цвет-
ковые растения отсутствуют. Тонкоствольные древесные лианы встре-
чаются редко, преобладают травянистые лазящие растения. Густой под-
лесок содержит наряду с гемиксероморфными формами также и жестко-
листные кустарники с листвой разной формы и величины, вплоть до эри-
коидной и ксероморфных филлокладиев. Напочвенная растительность
в тени замкнутых лесов изреженная и бедна видами. Она состоит из кар-
ликовых кустарников, разнотравья, а также мезофитных трав и мхов;
на освещенных местах часто встречаются луковичные и клубневые рас-
тения.
Жестколистный лес не имеет настоящего сезонного вегетационного
покоя, а лишь относительные периоды покоя, наблюдающиеся холодной
зимой и сухим летом. Основной период ассимиляции падает на весну
и осень. Время цветения распространяется и на зиму (фото 8, 9, 10, 11).
Лавровые леса. Лавровым лесом называют тип растительных фор-
маций, распространенный преимущественно в туманных областях субтро-
пических нагорий и занимающий по строению и облику промежуточное
положение между жестколистными лесами и дождевыми лесами умерен-
ной зоны.
От субтропического дождевого леса лавровые леса отличаются, как
правило, меньшей высотой (10—30 м). Кроны образуют не более двух
ярусов; количество видов деревьев умеренно и даже незначительно.
Характерно отсутствие опорных корней, капельного острия на листьях
и фанерогамных эпифитов, а также крайне скудное количество древес-
ных лиан. G умеренным, вечнозеленым дождевым лесом лавровый лес
объединяет обилие папоротников в подлеске и часто довольно пышное
развитие эпифитной растительности в виде мхов, лишайников и нередко
также папоротников. Мхи могут произрастать и как эпифиллы (фото 12).
С жестколистным лесом лавровый лес имеет много общих форм роста,
особенно в кустарниковом ярусе, а также экологически сходный тип
листвы, устойчивой в сухие периоды, вечнозеленой, но не морозостой-
кой. Форма лавровых листьев, давшая название лесу из-за преобладания
этого вида деревьев, лишь несколько отличается от типа листьев, встре-
чающихся в жестколистных лесах, своим менее ксероморфным строением.
Единицы растительности
133
Относительно крупные, прочные, продолговатые или овальные листья,
как правило, не имеют волосяного покрова; они блестящи, темно-зеленой
окраски и нередко по краям слегка волнисты.
К лавровым лесам относится и часть вечнозеленых горных лесов
тропических летнедождливых областей, например Мадагаскара, Восточ-
ной Африки, Венесуэлы и Британской Гвианы. Площадь этих сравни-
тельно легко загорающихся лесов сокращается, и во многих районах их
место уже занимают горные саванны.
Деревья низкие (12—15 м), с широкой кроной и кривым стволом,
листва кожистая. Кустарниковый подлесок также вечнозеленый, но
большей частью мелколистный, частично с эрикоидной листвой.
Бореальные хвойные леса. Эти леса состоят целиком или по преиму-
ществу из вечнозеленых хвойных деревьев и имеют, как правило, одно-
ярусное строение древесного полога. Высота деревьев может быть весьма
различной. Листовой покров образуют не плоские листья, а ксероморфная
игольчатая или чешуйчатая листва, сохраняющаяся на протяжении
нескольких лет. Благодаря восковому налету и погруженным устьицам
эти листья могут периодически сильно ограничивать транспирацию.
При закрытых устьицах многие хвойные деревья при прочих равных условиях
транспирируют в пересчете на живой вес меньше, чем лиственные деревья того же раз-
мера в безлистном состоянии. Они хорошо переносят сильные зимние холода и способны
ассимилировать даже в холодные или прохладные сезоны. Поэтому хвойные деревья
могут, хотя и при медленном росте, обходиться короткими или менее теплыми, но соот-
ветственно более продолжительными вегетационными периодами и при необходимости
переживать также сухое время года.
Древесный полог часто сомкнут, внутри эти леса темны, подлесок
развит слабо. При неблагоприятных климатических условиях более высо-
ких широт эти леса разрежены. Деревья опираются на широкие поверх-
ностные корневые системы, по мере приближения к полюсам кроны их
сужаются, рост уменьшается, даже в пределах одного вида. Напочвен-
ный покров состоит в основном из кустарничков, мхов и лишайников.
Бореальные хвойные леса весьма бедны видами. Видовой состав дре-
весного яруса также ограничен. Широко распространены сообщества
с преобладанием лишь одной древесной породы. Нередко, обычно в под-
чиненной роли, в древесном ярусе в виде примеси встречаются летнезе-
леные лиственные деревья, чаще, однако, в виде кустарников в подлеске.
В остальном в спектре жизненных форм большая доля приходится
на кустарнички, полукустарники и гемикриптофитные, часто дающие
побеги травянистые растения х. Это преимущественно летнезеленые виды,
а также и зимнезеленые или полузимнезеленые, листья которых могут
сохраняться под снегом до образования новой листвы весной. В зависи-
мости от условий местообитания в напочвенном ярусе играют роль злаки,
папоротники, напочвенные лишайники, болотные мхи и сапрофитные
растения. Весенние геофиты отсутствуют.
Горные хвойные леса. В различных климатических зонах эти леса
могут быть представлены весьма различно, иметь неодинаковый состав
нижних ярусов и обнаруживать большое число вариантов экологиче-
ского характера (фото 13).
1 Полукустарники для хвойных лесов не характерны. Эта форма роста распро-
странена преимущественно в аридных областях.— Прим. ред.
134 Единицы растительности в ландшафте
Мангровые заросли. На соленых илистых грунтах тропических
морских побережий распространены вечнозеленые «земноводные леса»,
образуемые мангровыми деревьями. Почти непроходимые чащи их могут
достигать высоты 5—20 м, а в исключительных случаях, например на побе-
режье Колумбии, и более. Во время прилива кроны деревьев едва подни-
маются над водой. При отливе обнажаются нижние части стволов с опор-
ными или дыхательными корнями. Разлагающиеся органические вещества
окрашивают почву. Листва деревьев ксероморфна. Характерны, кроме
того, «вивипария» плодов (ср. рис. 20) и большое содержание дубильных
веществ в древесине, которая вследствие этого не подвергается действию
термитов и древоточцев. Кроме деревьев, эта бедная видами и на протя-
жении больших пространств довольно однородная формация содержит
также кустарники (фото 14).
ЛИСТОПАДНЫЕ ЛЕСА
Летнезеленые мезофитные лиственные леса. Летнезеленые листвен-
ные леса областей с холодной зимой и длительным влажным и теплым
вегетационным периодом умеренно высоки и в древесном ярусе не очень
богаты видами. В их составе насчитывается один-два десятка видов
деревьев. Однако чаще преобладает лишь несколько отдельных видов,
а другие являются примесью. Наряду с господствующей формой оголен-
ных зимой летнезеленых лиственных деревьев в них вкраплены — осо-
бенно у границ распространения этой формации — хвойные деревья
в большем или меньшем количестве (смешанные лиственно-хвойные леса).
Древесный полог образует один ярус, редко — два. Ярус кустарни-
ков может быть или густым, или изреженным, а может и вовсе отсутство-
вать. Травянистый ярус состоит преимущественно из многолетнего раз-
нотравья с большей или меньшей примесью кустарничков. Кроме того,
непосредственно на почве часто можно обнаружить еще ярус мхов,
а в некоторых случаях и лишайников Ч Поднимающиеся до самых крон
деревьев деревянистые лазящие или вьющиеся растения, как Clematis,
плющ, виноград, встречаются редко. Из эпифитов распространены только
мхи и лишайники, обитающие преимущественно на коре деревьев.
Строение нижних ярусов соответствует степени сомкнутости крон,
господствующей в этом сообществе и создающей благоприятные условия
для влажности и защиту от ветра. И кустарники, и деревья, за редким
исключением (например, Ilex aquifolium), сбрасывают листву. Различ-
ные виды трав имеют нежные тонкие листья со слаборазвитой колленхи-
мой и обладают способностью ассимилировать при недостатке света.
У них низкое осмотическое давление (по сравнению с травянистым яру-
сом смешанного лиственного леса сухая дернина потребляет в три раза
больше влаги).
Травянистый ярус состоит в основном из гемикриптофитного раз-
нотравья и злаков, а также геофитов. Характерны цветущие весной
растения, которые заканчивают свое надземное существование вскоре
после распускания листвы деревьев. К ним относятся и летнезеленые
теплолюбивые растения. Кроме благоприятного с точки зрения света
весеннего периода, во время которого они начинают свое развитие, они
1 В летнезеленых мезофитных лиственных лесах мхи, а также лишайники обычно
не образуют сплошного напочвенного яруса, а встречаются отдельными куртин-
ками.— Прим. ред.
Единицы растительности
135
могут использовать и осень, тогда как в промежуточное время эти расте-
ния находятся в тени и не дают ассимилирующего прироста. В странах
с мягкой зимой и своевременно появляющимся снежным покровом, защи-
щающим растения от холодов, существует группа зимнезеленых расте-
ний с зелеными листьями, перезимовывающими над почвой. Однолетние
травы отсутствуют.
Именно по сезонной периодичности летнезеленых лиственных лесов
создается наше представление обо всех четырех временах года.
После зимнего покоя весной начинается развитие растительности, происходящее
в несколько этапов, в зависимости от притока тепла и изменения света. Ранней весной
почти весь поток света достигает почвы, а после распускания листьев — только 1/5о
или 1/100 часть и даже меньше. Весной цветут многие травянистые растения, побеги
которых закладываются под почвой или непосредственно на ее поверхности еще в пред-
весенний период, а в начале лета они уже заканчивают свои жизненные функции над
поверхностью почвы (фото 15).
Лиственные деревья, из которых большинство опыляется ветром, также цветут,
как правило, до полного распускания листвы. Их основной период ассимиляции —
позднее лето. Нежная и богатая оттенками ярко-зеленая вначале листва деревьев ста-
новится позднее более грубой, однородного темпо-зеленого цвета. Осенью после изме-
нения окраски листьев большинство деревьев сбрасывает листву. У некоторых она
остается на ветвях в сухом состоянии. Листопад уменьшает площадь транспирации.
Только те части растения, которые защищены от холода и сухости, переносят при общем
снижении жизнедеятельности зимний период, во время которого поглощение воды
затрудняется, а иногда и вовсе прекращается — явление, которое при наступлении
продолжительных и сильных морозов может привести к повреждению деревьев из-за
недостатка влаги. Отсутствие листьев зимой уменьшает опасность излома ветвей под
тяжестью снега.
Тропические дождезеленые муссоновые леса. Эта группа формаций
впервые была описана на территории муссонной Азии, там, где она имеет
самое широкое распространение. Однако теперь она известна и во многих
других тропических областях, в которых осадки обильны (от 1000 до
2500 мм) и где бывает от трех до пяти сухих зимних месяцев. Эти леса
получили в литературе различные наименования (например, semi-everg-
reen и deciduous seasonal forest, foret claire tropicale, тропофитный влаж-
нотропический лес), но все же они не смогли вытеснить названия «муссо-
новый лес», впервые употребленного Шимпером. Это преимущественно
листопадный лес, который, однако, может содержать известное количе-
ство вечнозеленых деревьев. Он постепенно переходит, с одной стороны,
в тропический «сезонно-дождевой лес» (evergreen-seasonal forest), а с дру-
гой — в тропический сухой лес, однако противостоит и тому и другому
как самостоятельная формация.
Муссоновый лес в период дождей производит впечатление такой же
пышности, как тропический или субтропический дождевой лес; однако
у него есть и существенное отличие. Его древесный полог достигает
высоты от 25 до 35 (реже 40) м и состоит из двух ярусов. В верхнем ярусе
преобладают листопадные виды со сравнительно крупными мягкими мезо-
и гигроморфными листьями. Нижний ярус, напротив, может состоять
из вечнозеленых деревьев с мелкой листвой, часто более ксероморфного
характера. Ежегодно этот лес в течение 2—4 месяцев (в северном полуша-
рии с февраля по май, в южном — с июля по октябрь) остается безлистным.
При этом в зависимости от погоды в данном году наблюдаются раз-
личия в степени опадания листвы, так как многие дождезеленые древо-
136
Единицы растительности в ландшафте
стой бывают факультативно листопадными и могут в некоторые годы
сохранять свою листву в течение значительной части сухого периода.
По количеству вечнозеленых и облигатно или факультативно листопад-
ных видов в обоих ярусах древесного полога различают несколько типов
этой формации.
Сезонной периодичности соответствует образование годичных колец
и зачастую мощный покров корки на стволах. Некоторые древесные
формы напоминают по внешнему облику летнезеленые лиственные деревья
умеренной зоны. Некоторые виды, как тиковое дерево, цветут в период
дождей, но большинство — в сухое время. Новая листва распускается
обычно до начала дождей.
Древесный полог муссонового леса менее богат видами, чем в тропи-
ческом дождевом лесу, чаще в нем преобладают лишь немногие виды.
Крупные деревья-великаны с досковидными корнями, как правило, отсут-
ствуют, не наблюдается и каулифлория. Лианы, частично тоже листо-
падные, или весьма редки, или богато развиты, однако не настолько,
чтобы эти леса потеряли свой более светлый по сравнению с вечнозеле-
ным дождевым лесом характер. Среди скудно представленных эпифитных
цветковых растений деревянистые уступают место травянистым; много-
численны паразитные эпифиты (Loranthaceae).
Ярус кустарников развит слабо. Однако в подлеске большую роль
может играть бамбук. Низкие напочвенные злаки представлены или
скудно, или довольно обильно. Папоротники занимают в этом ярусе
скромное место.
Наряду с типичным муссоновым лесом различают влажный муссо-
новый лес, более богатый лианами и эпифитами и образующий благо-
даря наличию большого числа форм, характерных для дождевого леса,
переход к тропическому сезонному дождевому лесу, а также сухой муссо-
новый лес, примыкающий уже к сухому лесу. Сухой муссоновый лес
сильнее дифференцирован с точки зрения различных свойств почвы,
чем типичный или влажный.
Церрадо («cerrado») в Центральной Бразилии, высотой 10—15 м,
рассматривается как вторичная лесная формация полувечнозеленого
тропического муссонового леса.
Тропические листопадные сухие леса. Как более сухие формации
(в климате с 4—7 сухими месяцами и суммой осадков 700—1300 мм в год)
к муссоновым лесам примыкают — через ряд переходных формаций —
листопадные сухие леса. Это светлые дождезеленые леса с одним дре-
весным ярусом. Древесный полог не богат видами и в сухой период пол-
ностью или в большей своей части теряет листву. Высота деревьев около
8, чаще 10—20 м, в исключительных случаях (в Австралии) значительно
выше. Для типичного сухого леса характерна преобладающая форма
роста —«дождезеленых плодовых деревьев». У них короткий и толстый
по сравнению с их высотой ствол, с грубой коркой и широко раскинутые
сучковатые, лишенные шипов ветви, образующие плоскую разрежен-
ную крону. Листва на деревьях грубая, крепкая, часто перистая и в сухой
период образует на почве слой вялой листовой подстилки. В незначи-
тельном числе встречаются здесь и многообразные низкорослые вечно-
зеленые, частично колючие виды деревьев с более ксероморфной мелкой
листвой, в некоторых областях присутствуют также ксероморфные
высокорослые и карликовые деревья с листовой кроной (пальмы
и саговники).
Единицы растительности
137
Подлесок или отсутствует, или богато представлен вечнозелеными
мелколистными колючими кустарниками. Поверхность почвы слабо
затенена и часто имеет разреженный покров из жестких низких или
высотой до 1 м дерновинных злаков, к которым в период дождей присоеди-
няется разнотравье. В сухое время травянистый покров высыхает. Лианы
и эпифиты в лесах этого типа обычно отсутствуют. Однако и лианы, так
же, как и бамбуки в отдельных случаях, могут быть весьма многочис-
ленны. В очень редких случаях сухие леса могут иметь двухъярусный
древесный полог: верхний листопадный и нижний, большей частью веч-
нозеленый. Подобные леса, например лес «mato seco» высотой 15—20 м,
на Бразильском нагорье переходят в сухие муссоновые леса. С другой
стороны, сообщества, в которых преобладают колючие деревья с зон-
тиковидными кронами или деревья с суккулентными стволами, обра-
зуют переход к лесам колючих суккулентов (фото 16).
Как особый тип формации тропических сухих лесов можно рассматри-
вать «campos cerrados» в Бразилии. Варминг рассматривал его как «вид
открытого леса» (до 8 м высотой) и причислял «лесостепи» западного
Судана тоже к этому типу. Вайбель характеризует «campos cerrados»
как естественное «светлое насаждение» высотой 4—8 м, которое со своими
покрытыми толстой коркой стволами и крупной кожистой дождезеленой
листвой занимает промежуточное положение между лесными и травя-
нистыми сообществами. Он считает его широко распространенным в цен-
тральной Бразилии типом климаксной растительности, место которой
в значительной степени заняли возникшие после пожаров саванны (Brand-
Savannen) («сатро sujo» и «сатро limpo») (фото 17).
КРАЙНЕ КСЕРОМОРФНЫЕ ЛЕСА
Колючие и суккулентные леса. Характерным для крайне ксероморф-
ных лесов является обилие колючих растений и суккулентов. Они встре-
чаются в тропических или субтропических областях, которые на протя-
жении 8 или 9 месяцев в году остаются сухими, а в дождливые периоды
получают лишь незначительное количество осадков (от 250 до 800 мм).
Это почти всегда светлые, но труднопроходимые леса с большим разнооб-
разием форм роста. Древесный полог от 5 до 12 м высотой состоит из незна-
чительного числа видов.
Среди древесных форм господствуют преимущественно колючие
деревья с мелкой вечнозеленой, склероморфной или имеющей волосяной
покров листвой, а также деревья с зонтиковидными кронами, очень
часто с перистой листвой, опадающей в сухие периоды. Может быть отме-
чено также небольшое участие неколючих деревьев типа плодовых,
встречающихся в сухих листопадных лесах. Кроме того, встречаются
лиственные деревья с суккулентными стволами, частично со стволами,
покрытыми грубой коркой, а иногда и шипами, частично же имеющие
гладкие бутылкообразно утолщенные стволы с крупной дождезеленой
листвой. Наконец, здесь растут безлистные стеблевые суккуленты в форме
колонн или канделябров, высотой до 10 м\ иногда эти последние в зна-
чительной степени определяют общий характер формации.
Многие колючие леса, чаще всего сильно разреженные, имеют густой,
богатый видами подлесок из колючих или безлистных прутьевидных
кустарников и суккулентных растений. Нередки и ксероморфные кар-
ликовые деревья. Тонкоствольные древесные лианы и лазящие растения
в противоположность листопадному сухому лесу встречаются довольно’
198 Единицы растительности в ландшафте
часто. Кроме того, в дождливые периоды развиваются травянистые вью-
щиеся растения. Эпифиты могут отсутствовать, но в отдельных случаях
они бывают представлены весьма широко. Это крайне ксероморфные,
частично суккулентные формы, как, например, свисающие наподобие
бороды тилландсии или лишайники и паразитные Loranthaceae.
Напочвенная растительность может быть представлена весьма раз-
нообразно, в частности кустарниковыми зарослями из преимущественно
колючих и суккулентных растений. Однако чаще она развита очень
слабо. Дерновинные злаки, как правило, не играют большой роли или
отсутствуют вовсе. Колючие и суккулентные леса, как, например, вос-
точнобразильскую каатингу, с трудом удается отграничить от кустарни-
ковых формаций, имеющих сходный характер. Между ними существуют
различные переходы (фото 18).
II. Кустарниковые формации
ВЕЧНОЗЕЛЕНЫЕ КУСТАРНИКОВЫЕ ФОРМАЦИИ
Гигро- и мезоморфные вечнозеленые кустарниковые формации. Они
представляют собой главным образом временные стадии при развитии
вторичного леса в постоянно влажных тропиках.
Хвойное криволесье и эрикоидные кустарники. К этому типу фор-
маций относятся сообщества хвойных кустарников на верхней границе
лесов. Для них характерны низкостелющиеся или свисающие с горных
уступов ветви, как у стелющейся горной сосны в Альпах. Аналогичные
формации можно встретить на засушливой границе лесов, например
сообщества Juniperus в Большом Бассейне. Сюда относятся и низкорос-
лые сообщества хвойных деревьев, развивающиеся на некоторых верхо-
вых болотах.
Жестколистные кустарниковые формации. Эти формации распро-
странены в основном в субтропических областях с зимними дождливыми
периодами. Они весьма разнообразны, состоят преимущественно из веч-
нозеленых жестколистных кустарников и в различных областях носят
разные названия: средиземноморские маквисы, калифорнийский чаппа-
раль, австралийский жестколистный скрэб. Кустарниковые заросли, как
правило, густы, более или менее непроходимы и достигают 2—5 м в высо-
ту. Формации жестколистных кустарников состоят из большого числа
видов кустарников различной степени ксероморфности: от безлистных
прутьевидных кустарников и кустарников с мельчайшими чешуевидными
листьями до кустарников с лавровой или гемиксероморфной листвой.
Голубоватый восковой налет, войлочно опушенные листья, колючие или
пахнущие эфирными маслами растения определяют физиономию этих
формаций. Встречаются также вьющиеся растения. В более сомкнутых
сообществах напочвенная растительность часто совсем отсутствует, в дру-
гих же случаях она состоит главным образом из кустарников и полу-
кустарников, а на освещенных местах — из луковичных и клубневых
геофитов (фото 19). К жестколистным кустарникам близки лавровые
парамосы, которые встречаются выше границы лесов на влажных тропи-
ческих нагорьях.
Единицы растительности
139
КРАЙНЕ КСЕРОМОРФНЫЕ КУСТАРНИКОВЫЕ ФОРМАЦИИ
Колючие и суккулентные кустарники. Эта группа тоже объединяет
различные формации. Господствуют колючие склероксероморфные кустар-
ники, частью вечнозеленые, частью безлистные или с мелкой или пери-
стой листвой, опадающей в сухое время года, Суккулентные кустарники
или встречаются в небольших количествах как примесь, или могут быть
так обильно представлены, что определяют собой облик формации. Здесь
приходится отказаться от выделения отдельных типов формации. Суще-
ствуют переходы как к колючим и суккулентным лесам, колючим саван-
нам или жестколистным кустарникам, так и к формациям из суккулент-
ных карликовых кустарников (фото 20 и 21),
ЛИСТОПАДНЫЕ КУСТАРНИКОВЫЕ ФОРМАЦИИ
Формации летнезеленых лиственных кустарников умеренных и суб-
полярных широт. К этой группе относятся оголенные зимой кустарнико-
вые сообщества, развивающиеся, например, на торфяниках и на поляр-
ной и альпийской границе лесов (олыпанники и ивняки, субарктическое
березовое криволесье и субантарктические кустарниковые формации)
(фото 22),
III. Саванны и степи
(ксерофитные и мезоморфно-тропофитные травянистые пространства и открытые
богатые злаками древесные формации)
В наименовании этих групп формаций существует в литературе
большая путаница. Она произошла прежде всего потому, что слово
«степь» применяется для всех более или менее покрытых ксероморфными
травами пространств и других открытых формаций. Поскольку название
«степь» нежелательно сохранять в качестве общего заголовка для этих
групп формаций, что было бы целесообразно в отношении только таких
общепринятых производных понятий, как «степные животные», «степ
ные культуры», то предлагается вовсе исключить слово «степь» из науч-
ной терминологии или ограничить его применение только внетропиче-
с^ими «степями»,
В применении к английскому языку можно распространить предло-
женный Егером (Jaeger, 1945) термин «саванна» на тропические «колю-
чекустарниковые степи» (low grass savanna), упоминаемые более ранними
авторами. Для «степей» в первоначальном самом узком смысле можно
употребить американское выражение «прерия» или соединить его со сло-
вом «степь» («прерия-степь»). Сухие формации дерновинных злаков, кар-
ликовых кустарников и полукустарников едва ли можно называть «полу-
пустынями», так что для них неизбежен термин «сухие степи». Таким
образом, мы приходим к классификации по нижеследующей схеме, в кото-
рой для сравнения в скобках приводятся старые обозначения:
Саванны («тропические степи»)
Влажные саванны |
Сухие саванны [ высокотравные саванны («саванны»)
Колючие саванны — низкотравные саванны («колючекустар-
никовые степи»)
140 Единицы растительности в ландшафте
Прерии-степи («степи» в первоначальном смысле слова)
Полупустыни и сухие степи («пустынные степи», «кустар-
ничковые степи» и «дерновиннозлако-
вые степи»).
Дерновиннозлаковые степи относятся к основной группе «полуку-
старниковых и кустарничковых формаций».
В последнее время название «саванна» употребляют уже не для характеристики
определенного типа растительности, а для обозначения всей растительности климатов
тропических низменностей с 2 до 9!/2 влажными месяцами, без учета характера фор-
мации. Саванна при таком понимании, к которому мы не присоединяемся, охватывает
весь диапазон растительности — от муссоповых лесов до полупустынь.
САВАННЫ (ТРОПИЧЕСКИЕ ТРАВЯНИСТЫЕ ПРОСТРАНСТВА)
Под саваннами мы понимаем все покрытые злаками пространства
во влажных и периодически сухих тропиках, без учета причин их про-
исхождения и независимо от того, в какой степени в травянистое сообще-
ство вкраплены деревья и кустарники. Общим является наличие более
или менее сомкнутого травяного покрова из злаков. Поэтому саванны,
с одной стороны, противоположны лесам и кустарниковым формациям,
в которых сообщества сложены в основном древесными растениями,
а с другой — противостоят полупустыням, с сильно разреженной напоч-
венной растительностью, которую мы, даже если она во многих случаях
состоит из разнотравья и злаков (дерновиннозлаковые полупустыни),
не можем отнести к саваннам.
Наряду с широко распространенными антропогенными вторичными формациями
имеются, вне всякого сомнения, и естественные саванны, которые образовались,
например, вследствие периодических затоплений или специфических почвенных
условий.
Независимо от причин возникновения саванны можно различать
прежде всего по условиям увлажнения, обусловленным климатическими
причинами. Последние особенно сказываются на высоте травостоя и раз-
личии форм роста встречающейся здесь древесной растительности. Мы
выделяем следующие основные типы формаций саванн.
Затопляемые саванны. Это большей частью естественные, обычно
лишенные древесной растительности, покрытые злаками пространства,
с высотой травостоя до 3 м, размещающиеся в долинах тропических рек
и ежегодно один или два раза затапливающиеся на длительное время,
иногда более чем на полгода. Встречаются, как правило, на тяжелых
наносных почвах, которые после спада воды сильно высыхают. Форма-
ция может принимать характер периодически пересыхающих тростни-
ковых болот. На окаймляющих русла рек береговых валах поселяется
обычно древесная растительность, образующая так называемые «леса
береговых валов», или леса «banco» (Troll, 1936), вечнозеленые или
частично листопадные древостои, часто имеющие в древесном пологе
пальмы; в Южной Америке это преимущественно Mauri tia vinifera и Сорег-
nicia cerifera, в Африке — виды Hyphaene и пальм а-делеб (Borassus).
Местами пальмы заходят далеко в пределы травянистых пространств.
Особой формой затопляемых саванн являются «саванны с термитными
островами» (Troll, 1936), в которые вкраплены небольшие островки дре-
весной растительности, использующей термитные постройки.
Единицы растительности
141
Затопляемые саванны не связаны с растительностью определенного
климатического пояса. Они встречаются, например, в тропиках Вест-
Индии, в льяносах Венесуэлы, в дельте реки Ориноко, на прибрежных
низменностях Гвианы; занимают затапливаемые террасы в низовьях
Амазонки («campos de varzea»); приурочены к болотистой зоне «пантанал»
области Маморэ и в верхнем Парагвае; известны как «dambos» в Малави,
на берегах Конго и других рек Центральной Африки, а также в верхнем
течении Нила.
Блажные саваны (Jaeger, 1945) («savannes herbeuses» бельгийских
ботаников, «savannes subforestieres avec galeries» Шевалье, «high grass-low
tree savanna» Шанца). Периодически зеленые тропические пространства,
покрытые высокотравьем. Иногда эти пространства совершенно лишены
древесной растительности, однако чаще по травянистому покрову раз-
бросаны одиночные или рассеянными группами листопадные деревья;
в долинах, влажных ущельях и близ источников встречаются вечнозе-
леные галерейные леса.
Травянистые сообщества влажной саванны состоят из двух ярусов.
Верхний ярус образован разреженными гемикриптофитными и хамефит-
ными кустистыми злаками, стебли которых высотой от 1,5 до 3—4 м
с длинными и широкими листьями в дождливые периоды смыкаются
почти на 100% и пропускают к почве не более 5% дневного света. Число
видов этого яруса невелико, они образуют основную часть всей массы
растительности саванны. В спектре жизненных форм, относящемся
ко всему комплексу видов, они уступают геофитам, фанерофитам и теро-
фитам, которые, однако, составляют лишь малую долю всей массы рас-
тительности и появляются только на непродолжительное время. В верх-
нем ярусе, кроме злаков, участвуют и другие высокие травянистые
растения — либо самостоятельно пряморастущие, либо лазящие теро-
фиты и разнотравье. Нижний ярус высотой 0,5—1 м развивается иногда
после пожара еще до дождей и достигает оптимального развития к началу
влажного периода, до смыкания высоких злаков. Он покрывает, как
правило, около половины поверхности почвы, располагаясь между
кустами злаков, и состоит в основном из хамефитов и геофитов, значитель-
ная часть которых начинает цвести еще до начала дождей.
Большинство деревьев травянистых пространств, а изредка и раз-
бросанные по ним кустарники представляют собой пожаростойкие виды
(пирофиты). Это в основном низкие, 6—12 (реже 20) м дождезеленые
деревья со сравнительно крупными мягкими листьями и стволами, оде-
тыми толстой коркой. Большинство их сохраняет свою листву в первую
половину дождливого периода, и некоторые теряют листья на непродол-
жительное время. Встречаются также вечнозеленые виды, преимуще-
ственно пальмы.
В бездождный период растительность высыхает и большая часть
площади почти ежегодно выгорает.
Пожары не затрагивают галерейных лесов, которые пересекают
травянистые пространства сеткой вечнозеленых влажных лесов, богатых
видами сообществ типа тропического или субтропического дождевого
леса. Это пышные, густые, с обилием лиан древостои, однако значительно
менее высокие, чем дождевой лес в областях своего сплошного распро-
странения. Галерейные леса занимают не только днища долин и берега
рек и ручьев, но и крутые откосы и балки, увлажненные выходами
грунтовых вод (фото 23).
142
Единицы растительности в ландшафте
Термитные саванны. Особой формой являются термитные саванны.
Обитающие здесь термиты или (в тропиках Нового Света) муравьи-
листорезы с их громадными гнездовыми постройками обусловливают
регулярное появление гигрофильных древесных групп на пространствах,
покрытых мезофитными травами. Возникает ярко выраженный парковый
ландшафт с лесистыми островками среди безлесных травянистых или
покрытых изреженной древесной растительностью пространств. Островки
термитных лесов состоят не из обычных саванновых деревьев, а, так же
как и галерейные леса, из более требовательных к богатству и влажности
почв типов деревьев. Поэтому они не относятся собственно к саванновой
формации и не подвержены, как эта последняя, периодическому выгора-
нию трав (Troll, 1936). На участках, занятых термитными постройками,
почва перемешивается на глубину от 7 до 8 м и обогащается органическими
веществами; разрыхленность и хорошая аэрация глинистой, обычно вязкой
Р и с. 38. Формы роста травяного покрова в саванне Themeda
triandra — Heteropogon contortus Национального парка Альберт
(Lebrun, 1947).
почвы, пронизанной ходами и пустотами, способствует просачиванию
воды и препятствует испарению. Термитные постройки на пологих скло-
нах и равнинах могут поэтому создавать условия, аналогичные тем,
какие обычно существуют в балках, и тем самым способствовать поселе-
нию здесь деревьев, требующих больше влажности, чем та, которая
обусловливается климатом.
Сухие саванны (Jaeger, 1945) (Acacia-tall grass-savanna Шантца).
Сухими саваннами называют травянистые пространства, встречающиеся
главным образом в равнинных областях тропической зоны, с 5—7 влаж-
ными месяцами и с годовой суммой осадков 500—1000 мм. Их основной
тип с более или менее закономерно разреженным древостоем соответ-
ствует «садовым степям» (Obstgartensteppen) старых авторов. Злаковый
покров достигает здесь высоты 1—2 м и состоит из жестколистных
кустистых злаков, растущих разреженно и не образующих сплошного
дерна (рис. 38). Наряду с ними может встречаться дождезеленое или
ксероморфное разнотравье. Деревья, которые могут и отсутствовать,
в большинстве своем принадлежат к формам роста листопадных сухих
лесов. Они достигают 5—10 м высоты, имеют короткие одетые толстой
коркой стволы с твердой древесиной. Широкие, образованные большей
частью не имеющими шипов кривыми ветвями кроны зеленеют лишь
Единицы растительности
143
в период дождей, покрываясь сравнительно мягкими листьями средней
величины. Встречаются также и деревья с зонтиковидными кронами,
с мелкой вечнозеленой листвой, а также пальмы и отдельные лиственные
деревья с суккулентными стволами. Лианы отсутствуют. Эпифиты встре-
чаются редко. Сухие саванны бедны видами, они гораздо беднее, чем
листопадный сухой лес. Если встречаются галерейные леса, то вечнозе-
леными они бывают лишь непосредственно на берегах рек или на участ-
ках с близкими грунтовыми водами. В противном случае они состоят
из листопадных деревьев сухого леса или сухого муссонового леса.
Принятые в Бразилии обозначения «сашро limpo» и «сашро sujo»—
это физиономические понятия, которые, правда, говорят кое-что об отсут-
ствии или наличии деревьев и кустарников на травянистых простран-
ствах, однако их можно толковать также как экологически различные
типы саванн (влажные и сухие саванны).
Колючие саванны (колючекустарниковые степи). Колючие саванны
расположены в областях с 8—10 сухими месяцами и непостоянным коли-
чеством осадков от 200 до 700 мм. Эти формации, «колючекустарниковые
степи» Вайбеля, в старой литературе об Африке и зоогеографических
работах называются обычно «тропическими степями». Они гораздо более
ксероморфны, чем сухие саванны. Травяной покров повсюду имеет высоту
30—50 см, в исключительных случаях до 1 м. Злаки жестче. Кусты или
дерновины злаков расположены на большом расстоянии друг от друга.
Они зеленеют в период дождей и высыхают в сухое время; встречаются
и вечнозеленые дерновинные злаки. Большие пространства, покрытые
травами и лишенные деревьев (в Бразилии «сашро limpo» с низкими дер-
новинными злаками), являются особой формой колючих саванн. Колючая
саванна обычно содержит и древесные растения, приуроченные чаще
всего к понижениям и имеющие преимущественно характер кустарников
высотой 1—3 м. Однако встречаются и отдельно растущие низкорослые
деревья (3—8 м), а также группы деревьев, зонтиковидные колючие
деревья, лиственные деревья с суккулентными стволами и безлистные
стеблевые суккуленты. Лианы и эпифиты отсутствуют. Среди кустарни-
ков преобладают колючие кустарники, преимущественно мелколистные,
дождезеленые, реже вечнозеленые. Вместе с ними встречаются безлистные
прутьевидные кустарники и суккуленты. Разнообразные формы послед-
них принимают участие и в образовании напочвенного яруса. Злаковый
покров в некоторых случаях разнообразят также кустарнички, сукку-
лентные полукустарники, а в период дождей — мезоморфные травяни-
стые растения. Однако напочвенная растительность может состоять
и исключительно из злаков. Прибрежные заросли вдоль эпизодически
текущих рек обычно представлены ксероморфными древесными сообще-
ствами типа листопадного сухого леса или колючего леса (фото 24).
СТЕПИ УМЕРЕННОЙ ЗОНЫ
Формации этой группы обозначаются русским словом «степь» в его
первоначальном значении. Чтобы подчеркнуть существующую в терми-
нологии путаницу, заметим, что некоторые современные авторы обозна-
чают именно эту формацию как «псевдостепь».
Прерии или черноземные степи. Прерии-степи были широко распро-
странены в умеренно влажных областях с сухим летним и холодным
зимним периодами. В настоящее время из-за большой сельскохозяйствен-
144
Единицы растительности в ландшафте
ной ценности их местообитаний они постепенно исчезают. Прерии-степи
представляют собой весьма богатые видами злаковые и разнотравные
сообщества, произрастающие на черноземных или родственных им поч-
вах с большим разнообразием форм роста с высотой травостоя 0,5—2 м.
Древесные растения либо отсутствуют, что наблюдается в большинстве
случаев, либо представлены отдельными деревьями или кустарниками.
Основной состав, который, однако, не составляет большинства
встречающихся в степях растений, представлен кустистыми злаками
с глубокими корнями и узкими жесткими более или менее ксероморфными
листьями. Встречаются также низкие дерновинные злаки с мягкими
листьями. Значительное участие принимает пряморастущее и частично
вьющееся разнотравье различной высоты, а также геофиты, перезимовы-
вающие в почве, особенно луковичные и клубневые растения. Сюда же
относятся однолетние травы. Некоторые виды прорастают осенью, сохра-
няют в зимний период розетку листьев, цветут весной и к концу сухого
лета завершают свой жизненный цикл. Другие являются яровыми одно-
летниками. Они заполняют летом свободные пространства между много-
летними растениями и переживают зиму, а зачастую и сухое лето в виде
семян.
Наконец, следует упомянуть о терофитных водорослях и мхах,
появляющихся ранней весной, прежде чем разовьются высшие растения.
Характерной растительной формой являются «перекати-поле»1. Это
обычно однолетние растения, однако встречаются и многолетние геми-
криптофиты, у которых хрупкие надземные части после созревания пло-
дов обламываются у самой поверхности почвы и, гонимые ветром, пере-
катываются по открытым пространствам.
Характерной чертой этих формаций является сменяющаяся в тече-
ние года последовательность цветения с изменением окраски 1 2. Окраска
достигает особенной яркости весной, когда цветут геофиты. В конце
цветут только некоторые виды разнотравья. К началу осени большинство
растений высыхает и прерия становится желтой или коричневой.
К основным типам этой группы примыкают формации, представляю-
щие собой переходы к полупустыням, с одной стороны, и к открытым
древостоям — с другой.
В переходной к осветленным древостоям формации на деградирован-
ном черноземе резко выделяются широколистные злаки, образующие
рыхлую дерновину и высокое разнотравье 3.
Сухие степи умеренной зоны (низкозлаковые и полукустарниковые
степи). В условиях перехода от прерии к полупустыне на каштановых
почвах увеличивается количество жестких дерновинных злаков. В боль-
шом количестве появляются полукустарники. Сообщества становятся
ниже, разреженнее. Период массового цветения смещается на более
раннее время. После продолжительного сухого лета с приходом осени
эти степи часто зеленеют еще раз из-за прорастающих озимых однолет-
них растений или заново образующейся листвы злаков.
1 Форма «перекати-поле» является приспособлением для распространения плодов
и семян. Перекатываясь при помощи ветра по открытым степным пространствам,
растение постепенно рассеивает семена или плодики, для чего у них бывают развиты
особые приспособления, предотвращающие одновременное высыпание семян или
плодиков.— Прим. ред.
2 Периодичность цветения и обусловленная ею смепа окраски степи носит назва-
ние смены аспектов.— Прим. ред.
3 Автор говорит о лесостепи и луговых степях.— Прим. ред.
Единицы растительности
145
IV. Луга
(мезоморфно-тропофитные и гигроморфные злаковые и разнотравные формации)
ЛУГА И РОДСТВЕННЫЕ ИМ ФОРМАЦИИ
Лугами в широком смысле называют сомкнутую дернину из преиму-
щественно гигроморфных или слабоксероморфных травянистых расте-
ний с большим участием мягких злаков или злаковидных растений (напри-
мер, Сурегасеае).
Луга встречаются преимущественно в более или менее влажном океа-
ническом климате умеренной зоны с несуровыми зимами и без резко
выраженных сухих периодов.
В большинстве своем луга являются хозяйственными угодьями
(под посевами и пастбищами), возникшими при участии человека и сохра-
няющимися в современном состоянии благодаря постоянному хозяйствен-
ному воздействию (покос, внесение удобрений, выпас скота).
К природным луговым формациям относятся субполярные луга,
альпийские матты, некоторые солончаковые и болотистые луга, а глав-
ным образом пойменные луга, обусловленные ежегодным ледоходом
(фото 25).
Не вдаваясь в подробные объяснение, назовем лишь некоторые основные луго-
вые формации:
Разнотравные матты (альпийские матты и субполярные луга («jadar» в Исландии),
Солончаковые луга (морское побережье, ватты и марши).
Заливные луга (затопляемые и попадающие в зону ледохода луга).
Влажные п переменно влажные луга (кислые скашиваемые на подстилку луга
и луга с низкой осокой, влажные луга высокого уровня и низинные болота).
Тучные луга п тучные пастбища.
Стравливаемые луга.
Тощие луга и выгоны (пустошные луга, пастбища, тощие горные луга, дер-
новиннозлаковые луга).
Сухая дернина.
ТРОСТНИКОВЫЕ ЗАРОСЛИ И РАЗНОТРАВНЫЕ ЛУГА
Тростниковые заросли на прудах.
Тростниковые заросли у ручьев.
Крупноосоково-камышовые заросли.
Высокотравные луга.
Луга у источников.
V. Формации кустарничков и полукустарников
Встречаются главным образом на границе древесной растительно-
сти, обусловливаемой сухостью или недостатком тепла.
ПОЛУПУСТЫНИ
Формации полуаридных и аридных областей, где количество осадков,
выпадающих в течение короткого времени, не превышает 250 мм\ обычно
же их бывает значительно меньше.
Это ксероморфные открытые растительные формации с разреженным
растительным покровом, причем растения часто удалены на несколько
146
Единицы растительности в ландшафте
метров друг от друга. Их надземные части малы и низкорослы, но зато
эти растения обладают глубокой и широко разветвленной корневой
системой. Горизонтально распростертые корни смыкаются под землей,
чем определяется расстояние между отдельными растениями.
Господствующими формами роста являются здесь кустарнички,
полукустарники, низкорослые жесткие дерновинные злаки и суккулент-
ные растения. В некоторых областях как исключение в отдельные годы
или регулярно после дождей на короткое время появляются гигроморф-
ные травы. Кустарнички большей частью склероморфные, вечнозеленые,
с крайне редуцированной поверхностью листьев. Однако встречаются
и сбрасывающие листья кустарнички с более крупной листвой. Дерно-
винные злаки встречаются, как правило, редко, но в некоторых сообще-
ствах они могут играть значительную роль и занимать большие про-
странства. Полукустарники развиваются чаще всего на солончаковых
почвах. Листья галофитов обычно имеют обильный волосяной покров.
Характерны суккулентные растения самой различной формы, например
шарообразные стеблесуккулентные кустарнички, розеточные растения
с суккулентными листьями. Они могут встречаться либо как отдельные
вкрапления, либо даже как доминирующие виды (фото 26 и 27).
Различают следующие формации в зависимости от господствующих в них форм
роста:
Кустарничковые полупустыни.
Дерновиннозлаковые полупустыни.
Суккулентные полупустыни.
Галофитные полупустыни.
Полупустыни или пустыни с карликовыми деревьями.
СКАЛЬНЫЕ ФОРМАЦИИ И ФОРМАЦИИ ЖЕСТКИХ ПОДУШЕК
Растительные формации скал и трещин. Первые поселенцы на голых
скалах — это корковые лишайники и мхи. Пронизывая породу своими
ризоидами, они усиливают процесс химического выветривания. В тре-
щинах скал накапливается гумус и мелкозем, и в них могут найти приют
высшие растения, относящиеся к «хазмофитным формациям»1 высоко-
горий и скалистых обрывов. Разреженный растительный покров их пред-
ставлен в зависимости от климатических условий и горной породы следую-
щими формами роста: подушковидными кустарничками, подушковидным
и розеточным разнотравьем, а также некоторыми гемикриптофитами,
полукустарниками, ксерофитными дерновинными злаками и изредка
фанерофитами. Часто встречаются мелкие листовые суккуленты (типа
очитков).
В засушливых областях растительные формации скал могут переходить в полу-
пустыни. Обычно же они имеют самостоятельный флористический характер с только
им присущими классами сообществ (в голарктическом царстве, например Asplenietea
rupestris, сообщества скальных трещин).
Альпийские формации каменистых осыпей. В состав формаций
подвижных каменистых осыпей входят многочисленные формы роста, при-
способленные к существованию на более или менее подвижных крупно-
1 Хазмофитные формации (греч. chasmophyton chasmo — открытое, phyton —
растение) — растительные формации с несомкнутым растительным покровом.—
Прим. ред.
Единицы растительности
147
глыбистых осыпях, как, например, погруженные в осыпь (Schuttstauer),
покрывающие осыпь (Schuttdecker), ползущие по осыпи (Schuttuber-
kriecher).
Каменистые тундры или арктические луга из подушковидных расте-
ний. Они состоят из сильно разреженной растительности скал и осыпей
с низким подушковидным разнотравьем, подушковидными злаками, кор-
ковыми лишайниками и единичными кустарничками (подобные формации
можно обнаружить в Исландии вблизи границы снегов, на Шпицбергене,
Земле Франца-Иосифа и др.).
Формация жестких подушек. Частично разреженные, частично сомк-
нутые растительные формации из жестких растений-подушек, напри-
мер формация «Llareta» из видов Azorella и др. в Андах северного Чили
(около 4000 м над уровнем моря). С ними сходны формации «раститель-
ных овец» высотой до 2 м 1 в альпийском поясе Новой Зеландии (1000—
2000 м) и «bolax» (Azorella) на субантарктических островах (фото 28).
ГАРИГИ, ПУСТОШИ И ТУНДРЫ
Гариги. Средиземноморские «скальные пустоши» (фр>— гариги,
исп.— томилляры, греч.— фриганы), представляющие, как правило,
антропогенные кустарничковые формации на месте естественных лесов,
часто на каменистой смытой почве. Состав их определяется интенсивным
стравливанием. Преобладают растения колючие, ароматичные или с млеч-
ным соком, охотно поедаемые скотом. В низкорослых разреженных сооб-
ществах наряду с совсем низкими (8—10 см) или полуметровыми кустар-
ничками и полукустарниками часто встречаются луковичные или клубневые
геофиты. Более высокорослые сообщества, переходные к кустарни-
ковым, содержат до 50% хамефитов, большое количество светолюбивых
мелких кустарников (30%), а также гемикриптофиты (15%) и геофиты
(5%). Характерно полное отсутствие терофитов. Особой формой гариги
является «степь из карликовых пальм» юго-восточной Испании и Север-
ной Африки 1 2.
Альпийские горные пустоши. Низкие, часто ковровые формации
кустарничков, например альпийская рододендроновая пустошь, вклю-
чающая 60% хамефитов (ковровые кустарнички и лишайники) и при-
мерно 40% гемикриптофитов (разнотравных или злаковидных). На навет-
ренных местообитаниях они часто переходят в лишайниковые сообще-
ства (аналогично растительности «melar» в Исландии).
Парамосы. Вечнозеленые растительные формации влажного прохлад-
ного пояса тропических высокогорий (на сырых, частично заболоченных
почвах). Физиономически характерными являются хамефитное розеточное
разнотравье с длинными опушенными листьями и так называемые «монаш-
ки» («frailejones») (карликовые деревья с листовой кроной), а также дер-
новинные злаки, мелколистные кустарнички и низкое разнотравье типа
альпийских маттов (фото 29 и 30).
1 «Растительными овцами» назвали английские поселенцы в Исландии огром-
ные желтые подушки сложноцветных (Haastta ри 11 inaris, Raoulia eximia и др.), дости-
гающие 2—3 м в диаметре и имеющие свыше 60 см в высоту.— Прим. ред.
2 Пальмовые гариги других авторов.— Прим. ред.
148 Единицы растительности в ландшафте
Океанические пустоши умеренной зоны. Наиболее известным приме-
ром являются «атлантические пустоши» Северо-Западной Европы. Пре-
обладают кустарнички, частично с вечнозеленой «эрикоидной» листвой,
как Calluna vulgaris, а частично листопадные или факультативно листо-
падные. Характерны также гемикриптофитное разнотравье, низкорослые
дерновинные злаки, лишайники, мхи, а также кустарники, хвойные
деревья и летнезеленые лиственные деревья. Пустоши на сухой и влажной
почве, верховые болота и другие экологические варианты различаются
между собой и составом форм роста.
Субполярные пустоши, кустарничковые тундры. Формации кустар-
ничков и полукустарников арктической зоны и субантарктики с ковро-
выми, шпалерными и подушковидными кустарничками, как ассоциация
Асаепа adscendens на субантарктических островах и богатые мхами
и лишайниками подушковые пустоши на открытой ветрам приморской
границе леса юго-западной Патагонии.
Лишайниковые тундры. Чрезвычайно низкорослые сообщества с пре-
обладанием кустистых лишайников типа оленьего мха с крайне мед-
ленным рдстом (годовой прирост составляет примерно 0,5 см). Частично
сомкнутые лишайниковые формации, а частично открытые формации
с большими участками обнаженной почвы.
Моховые тундры. Сухие формации напочвенных мхов (например,
тундры Polytrichum и Racomitrium) или влажные моховые сообщества
с вкраплениями мелких злаков и осок, с переходами к низинным или
верховым болотам, однако обычно со скудным отложением торфа.
ВЕРХОВЫЕ БОЛОТА
Сфагновые верховые болота. Комплекс небольших участков форма-
ций на торфе, образованном мхами рода Sphagnum. В областях с боль-
шой влажностью воздуха такие болота, имеющие выпуклую поверх-
ность, способны, как губка, удерживать воду. Поверхность болота
делится на более сухие, поросшие кустарничками кочки и на влажные
углубления или окна с открытой водой (мочажины). Торф как субстрат
беден питательными веществами. Среди обитателей особенно своеобразны
насекомоядные растения (Drosera). В условиях более континентального
климата торфяные болота часто бывают облесены (сосновые болота)
(фото 31).
Тундровые болота, частично со скудным сфагновым торфообразова-
нием, частично образующие особую форму верховых болот —«торфяно-
бугристую тундру», распространенную в субарктических районах Сибири,
с круглыми торфяными буграми выше человеческого роста.
Болота с жесткими подушками. Образуются в сырых лощинах боли-
вийских Высоких Анд и характеризуются подушками чуть ли не камен-
ной твердости, образованными торфообразующим видом Distichia muscoi-
des (Juncaceae).
Единицы растительности
149
VI. Формации терофитов
Эфемерные разнотравные луга «цветущей пустыни». Формации из
однолетних трав с очень непродолжительным периодом существования
в крайних условиях произрастания, например в пустынях после эпи-
зодических дождей (фото 32).
Разнотравные луга на илистой почве. У самого уреза воды на илистых
или глинистых грунтах растут сообщества карликовых ситников (класс
Isoeto-Nanojunceted), образующие дернину высотой 5—10 см, состоящую
на 95% из терофитов. Их распространению обычно способствует ветер,
а иногда вода или птицы. Здесь же встречаются и геофиты (например,
в средиземноморском климате) или гемикриптофиты (в среднеевропей-
ском климате).
Сообщества полевых сорняков. Эти терофитные формации распро-
странены по всему миру как антропогенные сообщества полевых сорняков.
Выделяют в связи с различной обработкой почвы сообщества пропашных
и колосовых культур.
VII. Пустыни
Настоящими пустынями, или «полными пустынями», называют участки
земной поверхности, которые либо совершенно лишены растительности,
либо несут очень разреженный растительный покров. Четкую границу
с полупустынями провести здесь трудно, и она рассматривается различ-
ными авторами по-разному. В большинстве пустынь, даже на первый
взгляд совершенно безжизненных, все же встречаются одиночные, крайне-
ксероморфные растения. В строгом смысле лишенными растительности
являются некоторые участки северочилийской пустыни, пустыня Намиб
и блуждающие дюны Сахары. По субстрату пустыни очень разнообразны:
каменистые, галечниковые и песчаные, кроме того, в сухих жарких обла-
стях даже за пределами собственно пустынь могут существовать весьма
неблагоприятные для роста растений эдафические условия: почвы с
соляной или известковой коркой (северное Чили, Иран).
Сухим пустыням противостоят холодные пустыни областей оледене-
ния и высокогорий, а также лишенные растительности скалы в сходных
областях. Растительная жизнь отсутствует в холодных пустынях не пол-
ностью, но с точки зрения физиономии ландшафта она никакого значе-
ния не имеет.
В теплых сухих пустынях характерно развитие растительности
в местах выхода ключей —«оазисы»: оазисы на берегах рек, на выходах
грунтовых вод, оазисы у источников или колодцев. Растительность оази-
сов может принадлежать к различным формациям от эфемерных теро-
фитных формаций (растительность вади) до лесов (колючекустарниково-
суккулентные леса, сухие леса, пальмовые рощи).
VIII. Растительные формации внутренним водоемов
Растительность озер, прудов, рек, ручьев и источников может быть
весьма разнообразной. Классификация их здесь не приводится.
IX. Растительные формации морей
Эти формации мы здесь также не рассматриваем.
Растительность в ландшафте
ЕСТЕСТВЕННЫЙ И КУЛЬТУРНЫЙ ЛАНДШАФТ
Ландшафт, «на который природа может беспрепятственно воздействовать» (Wil-
Idenow, 1792), мы называем естественным ландшафтом. Ему противо-
поставляется «культурный ландшафт», на который оказал свое влияние человек. В куль-
турных ландшафтах лишь в ограниченной степени господствует естественно установив-
шийся порядок распределения растительности. На размещение растительности здесь
влияют другие причины, обусловленные жизненными потребностями человека. Поэтому
вопрос о том, как вписывается в эколого-функциональном и пространственном смысле
в ландшафт растительность, должен решаться отдельно для естественного и для куль-
турного ландшафтов.
Естественные ландшафты на больших пространствах встречаются
только там, где природные условия местности неудобны или трудны для
поселения и длительного пребывания человека. Это относится к ледяным
и некоторым сухим пустыням, высокогорным районам, к отдельным
участкам тропических дождевых и горных лесов, а также частично
к бореальным лесам и тундрам.
Все остальные территории земной поверхности, в особенности травя-
нистые пространства в тропиках, значительно больше преобразованы
человеком, чем это предполагалось даже несколько десятилетий тому
назад. Зачастую не удается с уверенностью разграничить культурный
и естественный ландшафты. Они часто находятся в непосредственном
соседстве друг с другом и могут перемежаться в ограниченном районе,
как, например, оазисы в пустынях или долины высокогорий. Да и сами
пространства, занятые культурным ландшафтом, иногда сохраняют
участки естественного ландшафта, например болота. Во многих куль-
турных ландшафтах наблюдается различная степень воздействия чело-
века: от исключительно сильного до крайне слабого. Если влияние чело-
века прекращается, то созданные им объекты и использованные площади
и даже целые ландшафты снова подпадают под воздействие природы.
При этом проявляется различная «склонность к одичанию», и возвраще-
ние этих участков в естественный ландшафт совершается с различной
скоростью.
Границу между естественным и культурным ландшафтом провести трудно.
Так, местное население, обитающее в тропическом дождевом лесу Суматры, практикует
«культуру подсечного земледелия». Они выкорчевывают участок девственного леса,
Растительность в ландшафте
151
сжигают деревья и возделывают рис. Так как этот рис дает новый урожай на том же поле
только через 8 лет, то корчевание приходится производить ежегодно; возникают
«кочующие поселения». У реки закладывается деревня и год за годом в ее окрестностях
выкорчевывается один участок за другим. Через 8—10 лет новые поля оказались бы на
слишком большом расстоянии от поселения, к тому же деревянные строения за это
время тоже сгнивают, поэтому деревня перемещается ближе к посевам, а старый участок
начинает зарастать лесом. Возобновление леса настолько мощно, что все неочищенное
пространство немедленно покрывается древесными всходами. Спустя десятилетие здесь
возникает молодой лес, через 50 лет это уже высокоствольный лес. Вторичность этого
леса может заметить только ботаник.
Понятие, которое мы обозначаем прилагательным «культурный», имеет в виду
«ландшафт, подвергшийся воздействию человека». Только рассматривая «культурный
ландшафт» в этом самом широком смысле, можно получить логически четкое разграни-
чение его с естественным ландшафтом, не подвергшимся влиянию человека.
При воздействии человека на растительный покров нет никаких существенных
различий между уничтожением отдельных его объектов (например, хищническая рубка
леса) и другими формами интенсивного вмешательства. Каждое .паровое поле в тот
период, когда человек его не касается, ничем не отличается от вторичного леса на Сумат-
ре. Только здесь возобновление леса, которое и в Средней Европе также наступает
немедленно, через короткое время вновь будет прервано человеком.
Пример Суматры проясняет вопрос о границах применения понятия «ландшафта».
К «ландшафту» относятся как существующие поселения с их рисовыми полями, пусты-
нями и вторичными лесами, так и первичный лес, если он связан со всеми прочими
объектами на узком пространстве. Если рассмотреть весь этот ландшафт в целом, то это
«культурный ландшафт», причем не исключается возможность в его составе различать
такие ландшафты, которые еще мало подвергались воздействию человека, а также
«отживающие», или заброшенные, предоставленные природе культурные ландшафты
заброшенные культурные поля). Границы здесь, однако, установить трудно.
БИОХОРА И ПРОСТРАНСТВО МЕСТООБИТАНИЯ
ИНДИВИДУАЛЬНОСТЬ МЕСТА ПРОИЗРАСТАНИИ
Каждое сообщество растений обусловлено и историей своего разви-
тия, и экологией местообитания. «Все исходит из местонахождения» (Send-
tner, 1854). Под «местонахождением» понимается не только качество место-
обитания, но п его географическое положение. От положения областей
развития п путей распространения систематических единиц зависит, какие
именно организмы вступают в борьбу за существование на данном место-
обитании. В этих пределах, обусловленных историей флоры, от которых
зависит видовой состав, способный заселить данный участок земной поверх-
ности, и распределяются в естественном ландшафте систематические
единицы по сообществам в соответствии с качественными различиями
отдельных местообитаний.
Б ИО ХОРА
«Биохорой» мы называем область одной единицы голоцена. Био-
хоры — это пространственные единицы ландшафта, пространственные
единицы экосистем. Это участки местности, занятые определенным биоце-
нозом и имеющие один тип почвы, что вместе создает единую экосистему.
152
Единицы растительности в ландшафте
ПРОСТРАНСТВО МЕСТООБИТАНИЯ
Это конкретное пространство, обладающее определенным качеством
местообитания. Граница проходит там, где комплекс местных факторов
резко изменяется с точки зрения возможностей существования, обуслов-
ливаемых этим комплексом. Само собой разумеется, понятие «однород-
ности» пространства местообитания предполагает неизбежно какой-то
отказ от мелких местных различий, которые по сравнению с тем, что
является общим, оказываются несущественными. Основные единицы
естественно-пространственной классифика-
ции (флизе, сайт, физиохоры) являются в этом смысле пространствами
местообитания. Это топографические участки с определенным экологиче-
ским потенциалом. Их разграничение не зависит от временного расти-
тельного покрова, который может обусловливаться характером земле-
пользования или же естественными биоценозами.
Как местообитание является качественной основой возможной среды
обитания, так и пространство местообитания — потенциальной биохорой.
При сельскохозяйственном и лесохозяйственном картировании место-
обитаний выделяют отдельные пространства местообитаний, с тем чтобы
сопоставить искусственные биохоры с хозяйственно полезными площадями
в соответствии с естественными местообитаниями. Отдельные простран-
ства местообитания могут быть одновременно биохорами, что, однако,
наблюдается не столь часто. В культурном ландшафте одно пространство
местообитания иногда делится на несколько биохор вследствие воздей-
ствия человека (например, лес и луг).
В естественном ландшафте, как правило, границы биохор совпадают
с границами отдельных пространств местообитания. Встречаются исклю-
чения, например у местообитаний, которые под влиянием исторических
условий возникли заново или изменились и у которых первичное или
повторное заселение на разных участках пространства местообитания
прогрессировало в различной степени.
УСТОЙЧИВОЕ СООБЩЕСТВО
В стационарной экосистеме, в которой растительность находится
в равновесии со всеми господствующими на занимаемом ею пространстве
условиями существования — включая антропогенные, если таковые
имеются,— одно и то же растительное сообщество может сохраняться
бесконечно долго, пока не изменится само местообитание или пока чело-
век не преобразует его своей хозяйственной деятельностью. Раститель-
ное сообщество в такой стационарной экосистеме называют «устойчивым
(длительным) сообществом» (Dauergesellschaft). «В нем каждому виду
отводится то место и пространство, которые этому виду необходимы
в соответствии с его жизнеспособностью (если мы под этим несколько
неопределенным термином объединяем рост, размножение, способность
к конкуренции и т. д.)» (Lundegardh, 1930).
Является ли устойчивое сообщество одновременно и заключи-
тельным сообществом (Schlussgesellschaft), последним зве-
ном в цепи развития растительности на данном местообитании, зачастую
решить не удается. Во многих случаях, по-видимому, это не подтвер-
ждается, особенно для многих антропогенно обусловленных устойчивых
сообществ.
Растительность в ландшафте
153
Простейшим примером являются устойчивые сообщества среднеевропейских скаши-
ваемых лугов, остающиеся весьма стабильными при сохраняющихся одинаковых усло-
виях. Если прекращается регулярное скашивание, решающий фактор в данной эко-
системе, и площадь остается неиспользуемой, то вследствие существующей в устойчивом
сообществе тенденции развития наступает сукцессия, которая приводит к образова-
нию заключительного сообщества в виде леса, присущего данному местообитанию.
ЕСТЕСТВЕННАЯ СУКЦЕССИЯ
Процессы, в результате которых в ходе естественного развития раз-
личные растительные сообщества на одном и том же месте сменяют друг
друга, называют естественными сукцессиями. Каждая
такая сукцессия развивается в определенном направлении, зависящем
от характера местообитания, экологических требований и конкурентной
способности тех систематических единиц, которые могут здесь поселиться.
Процесс сукцессии заканчивается образованием растительного сообще-
ства, образующего вместе с общим комплексом факторов местности устой-
чивую экосистему. При этом надо различать стадии первичного и повтор-
ного заселения. Там, где материнская порода заселяется в первый раз,
вместе со сменой биоценозов формируется первичный присущий данному
местообитанию тип почвы. При этом отдельные стадии сукцессии могут
следовать одна за другой довольно медленно. При повторном заселении
на уже возникшей ранее почве этот процесс происходит быстрее.
Сукцессия первичного заселения обнаженных скал была известна еще Гумбольд-
ту. Унгер (Unger, 1836) уже составлял списки видов в серо-ольховых и ивовых кустар-
никах, покрывших участки близ Китцбюгеля, занятые камнями и песком после селе-
вого потока. Аналогичные наблюдения проведены и Дарвином. Кернер описал раз-
витие степной растительности на подвижных песках в Венгрии и сукцессию близ
альпийской границы леса, от первых ее поселенцев до зарослей рододендрона, как
«конечного звена в цепи превращений» (Kerner, 1863).
Первично заселенные участки могут возникнуть в естественном
ландшафте разным путем, прежде всего в результате таких процессов,
как оползень или обвал, отложение вулканического материала, образо-
вание щебнистых осыпей или дюнных песков, а также под воздействием
водных потоков или глетчеров, как, например, при изменении русла
рек, морских прорывах или отступании ледников и т. д.
При первичном заселении сукцессия зависит от того, располагается
ли данный участок в области, уже занятой растительными видами или
сообществами, или же на голом субстрате, на котором формирование
флоры еще только должно произойти.в результате нового вселения.
В последнем случае, имевшем место, например, после ледникового
периода при новом заселении пространств Европы, освободившихся
ото льда, что наблюдается и в наше время при возникновении новых остро-
вов, сукцессия может в большой степени зависеть от «факторов мигра-
ции», то есть от исторического хода переселения систематических единиц.
Если же новое заселение происходит на площадях, окруженных уже
существующим постоянным комплексом растительных сообществ, то,
как правило, каждому типу местообитаний присуща своя определенная
последовательность стадий развития. В этом случае для подлежащей
новому заселению площади заранее можно предсказать ход сукцессии,
если известна динамика растительности на площадях с подобным же
типом «флизе» (Fliesentypus).
154
Единицы растительности в ландшафте
Повторное заселение площадей, на которых уже имелись сходные
по местообитанию устойчивые сообщества, в условиях естественного
ландшафта может быть вызвано пожаром, возникшим в результате грозы.
Однако уничтоженная пожаром растительность, соответствующая дан-
ному типу почвы, может быстро восстановиться. Например, в лесных
областях Британской Колумбии с сухим летним периодом пожары так
часты и, возникнув в одном месте, настолько беспрепятственно распро-
страняются дальше, что почти каждое поколение деревьев преждевре-
менно погибает от пожара и заключительное сообщество на некоторых
местообитаниях в этих условиях вряд ли вообще может возникнуть.
Американцы в таких случаях устойчивые сообщества, обусловленные
пожарами, называют «fire-climax» (пирогенный климакс).
Исследование смены сообществ, которое под названием «синдинамики», или «уче-
ния о сукцессиях», входит в фитоценологию, было предпринято впервые американскими
исследователями (Cowles, 1899; Clements, 1916). Клементс (Clements, 1904) ввел также
и экспериментальные методы, при которых он освобождал пробные площади от расти-
тельности, чтобы изучить процесс повторного заселения. Сукцессии можно установить
также методами пространственных сравнений, например путем сопоставления расти-
тельности долинных висячих террас разного возраста.
При сукцессиях развитие растительности и почвы тесно взаимосвя-
зано, поэтому при исследовании следует брать всю экосистему в целом.
По Тюксену (Тйхеп, 1931), каждое растительное сообщество имеет
свой специфический, биологически обусловленный и часто физиономически
распознаваемый «почвенный профиль». Это положение встретило возра-
жения со стороны авторов, которые классифицировали почвы аналити-
чески, по отдельным признакам. Однако оно остается в силе, если под
«почвенным профилем» понимать не только бросающееся в глаза морфо-
логическое различие горизонтов, но и общее вертикальное строение,
в понимании Кубиены, то есть тип развития почвы. Все, что было ска-
зано о заместимости факторов действительно и в отношении почвы. На раз-
личных первичных горных породах под влиянием климатических факто-
ров и при воздействии растительности могут образоваться сходные типы
почв. В почвенных профилях при этом сохраняются физиономические
различия, обусловленные исходным материалом. Одинаковые раститель-
ные сообщества могут встречаться фактически на различных, но функ-
ционально равноценных почвенных профилях (Pallmann, 1947; Braun-
Blanquet, Pallmann, Bach, 1954).
Только когда и растительность, и почва достигли равновесия и между
собой, и с климатом, можно говорить в строгом смысле о естественной
конечной стадии сукцессии. Замыкающим звеном этого развития является
климаксное состояние экосистемы местности.
ЕСТЕСТВЕННОЕ ЗАКЛЮЧИТЕЛЬНОЕ СООБЩЕСТВО
Заключительным сообществом данного местообитания называют еди-
ницу растительности, представляющую собой конечное звено естествен-
ной сукцессии, которая, «раз возникнув, сохраняется очень долго, если
исключить, конечно, воздействие человека и катастрофические явления»
(Braun-Blanquet, 1951). Условия для этого создает постоянство климата
и самого местообитания, если последнее не претерпевает никаких ката-
строф (извержение вулканов, горные обвалы, изменение русла рек и т. д.).
Растительность в ландшафте
155
Специфическое заключительное сообщество данного местообитания
называют его климаксной растительностью (Klimax-
vegetation).
ПОТЕНЦИАЛЬНАЯ ЕСТЕСТВЕННАЯ РАСТИТЕЛЬНОСТЬ
Там, где заключительное сообщество еще не образовалось или уже
не может возникнуть в результате воздействия человека, о нем говорят
как о потенциальной естественной растительности местообитания (Тйхеп,
1956).
СТРУКТУРА «ФЛИЗЕ» И ЕСТЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ
РАСТИТЕЛЬНОСТИ ПО УСЛОВИЯМ МЕСТООБИТАНИЯ
ТИП «ФЛИЗЕ»
«Флизе» (элементы территории), обладающие одинаковыми каче-
ствами местообитания, являются основными единицами пространствен-
ной структуры ландшафта. В пределах пространства с определенным
составом систематических единиц участки с одинаковым типом «флизе»
в естественном ландшафте несут, как правило, одинаковые биоценозы.
Различия возможны, если развитие растительности еще не достигло есте-
ственного равновесия, то есть когда характерное для данного местооби-
тания устойчивое сообщество еще не успело установиться, как это
наблюдается, например, при первичном заселении новых местообитаний
или при новом образовании растительности на участках «флизе», кото-
рые под воздействием природных явлений изменились или лишились
своей предыдущей растительности.
Если исследована целая серия сукцессий, то можно сделать заклю-
чение для участков с «флизе» одинакового типа об их потенциаль-
ной естественной растительности. Это имеет не только
теоретический интерес, но дает возможность говорить об экологическом
потенциале данного типа местообитания. Правда, такие выводы могут
быть сделаны только индуктивно для определенного типа «флизе».
На других естественно-пространственных единицах той же области иные
растительные сообщества с другими типами почвы могут образовать
последнее звено в цепи развития, то есть климакс экосистемы.
Встречающееся уже у Кернера, а позже развитое Клементсом и Браун-Бланке
положение, что только региональный климат является определяющим и что совпадение
конечных звеньев может встречаться и на первоначально различных типах местообита-
ния (теория мопоклимакса), оказалось впоследствии несостоятельным (Nichols, 1917;
Tuxen, Diemont, 1937; Schmithusen, 1950; Whittaker, 1953, 1957).
СТРУКТУРА «ФЛИЗЕ»
Пространственная структура ландшафта обусловлена в значитель-
ной степени тем, что несколько типов местообитания составляют в ней
характерный мозаичный рисунок («pattern»), который мы называем
мозаичной структурой ландшафта. Она создается в результате наложе-
ния и объединения воздействий различных факторов местности.
Структуре «флизе» (Fliesengefiige) соответствует пространственное рас-
пределение потенциальной естественной растительности. Для характера
156 Единицы растительности в ландшафте
локального заключительного сообщества решающим подчас оказывается
какой-либо один фактор местности. Поэтому «флизе» разного типа могут
под доминирующим влиянием одного фактора местности иметь одинаковую
или сходную потенциальную естественную растительность. Подобно тому
как в многоцветной печати на некоторых участках один цвет больше опре-
деляет общую картину, чем все остальные, так и среди факторов место-
обитания главным является один. Для потенциальной естественной рас-
тительности при «флизе» одного типа решающим может быть региональ-
ный климат, при других — горная порода, грунтовые воды, экспозиция
и т. д. Совпадающее качество местообитания у «флизе» одного типа не долж-
но основываться на одинаковой комбинации особенностей местности.
При определенных условиях влияние одного фактора может заменяться
влиянием другого.
ФАКТОРЫ МЕСТООБИТАНИЯ КАК ПРИЧИНЫ
ПРОСТРАНСТВЕННОГО РАЗМЕЩЕНИЯ ЕДИНИЦ
РАСТ ИТЕЛЬНОСТ И
В пространственном размещении нескольких близких растительных
сообществ можно отметить корреляционную зависимость в постепенном
или скачкообразном пространственном изменении определенного эколо-
гического фактора.
Сгруппированные в такой зависимости единицы растительности
называют «рядом местообитаний», или, используя термин
почвоведов (Вагелер), «катеной», а при более скачкообразном измене-
нии —«группой» Ч Подобные ряды местообитаний существуют как на об-
ширном, так и на небольшом пространстве. Их отдельные элементы могут
быть расположены параллельными или концентрическими поясами или,
при разнообразии экспозиций, в виде ряда соприкасающихся секторов.
Установление таких закономерностей в распределении растительных
сообществ позволяет выделить или проанализировать характерные черты
размещения растительности и ее зависимости от условий местообитания.
Не претендуя на полную законченность системы, приводим некоторые примеры
рядов или групп местообитаний:
а) ряды по световым поясам: растительные сообщества, распреде-
ленные по степени освещенности, зависящей от глубины водоема, как, например,
ясно выраженные на морских побережьях ступенчатые ряды сообществ водорослей;
б) ряды по уровню грунтовых вод и степени влажно-
сти почвы, например пояса лесных сообществ на долинных почвах;
в) ряды по степени затопления: ступени растительности в речных
долинах, разнящиеся длительностью, частотой, регулярностью и временем затопления
этих долин речными разливами;
г) солончаковые ряды: пояса растительности маршей, различающиеся
степенью засоления почвы;
д) р я д ы по степени крутизны склонов: пояса лесных расти-
тельных сообществ на склонах разной крутизны, но одинаковой экспозиции,
е) р я д ы по экспозиции: последовательность растительных сообществ,
соответствующая микроклиматическим особенностям местообитаний и изменяющаяся
в зависимости от экспозиции горных склонов (Tuxen, Diemont, 1937; Tuxen, 1954).
1 Катена (лат, catena — цепь) — последовательная смена почвенных типов,
соответствующая последовательной смене топографических условий.— Прим, ред.
Растительность в ландшафте
157
ж) группы по типам почв: группы единиц местоообитаний, обусловлен-
ные различными, более или менее закономерно взаимосвязанными горными породами
с соответствующими им различными растительными сообществами.
Следует иметь в виду, что при изображении пространственной после-
довательности сообществ в виде ряда или группы местообитаний в каждом
случае подчеркивается только одна из характерных связей. В действи-
тельности же многие связи могут многократно и разнообразно пересе-
каться. Одно и то же растительное сообщество может быть одновременно
членом ряда по степени крутизны склонов, ряда по экспозиции, и группы
по типам почвы, а кроме того, занимать определенное место в ряду сооб-
ществ, отличающихся по влажности или какому-либо другому экологиче-
скому фактору. Это и является причиной, почему на экологическом
делении биоценозов нельзя основывать системы их идентификации.
РАСТИТЕЛЬНЫЕ СООБЩЕСТВА
КАК ИНДИКАТОРЫ МЕСТООБИТАНИЙ
Картину пространственного распределения растительности в ланд-
шафте можно получить путем крупномасштабного картирования всех
растительных сообществ. Полученные карты, отражая характер подраз-
деления местообитаний, хорошо дополняют аналитические исследования
привязанности единиц сообществ к экологическим условиям, а также
исследования рядов и групп местообитаний. Таким образом, раститель-
ность служит индикатором распределения по площадям определенных
свойств местности, которые другими средствами установить либо трудно,
либо невозможно. Так, по характеру растительности можно судить
об уровне грунтовых вод, путях их передвижения или их химических
особенностях. Если в область с грунтовыми водами, бедными питатель-
ными веществами, приходит поток богатых питательными веществами
грунтовых вод, то это отражается в переходе растительных сообществ от
заболоченных или пойменных лесов к лугам и зарослям тростника.
Сообщества у источников, узко ограниченные единицы растительности, состоящие
из невзрачных видов, не могут иметь значения для физиономии ландшафта, но позво-
ляют судить о химических и термических особенностях воды источников или о пери-
одичности излияния воды на поверхность.
По распространению растительных сообществ зачастую удается
судить также о климатических различиях местности, часто точнее
и быстрее, чем по результатам метеорологических наблюдений. Расти-
тельность позволяет при известных условиях делать выводы о таких
климатических факторах, которые непосредственно определить не удается
ввиду отсутствия в этом районе метеостанций. По растительным сообще-
ствам можно легко определить и нанести на карту оттепели, «морозные
озера» и момент наступления поворота температур в долинах или глубо-
ких лощинах. Зависимость распределения единиц растительности от экспо-
зиции помогает установить среднее направление ветра (Tuxen, 1931 —
1932). В сообществах горных карликовых сосен на склонах хребта Кара-
ванке трехступенпо изменяющийся состав растительных сообществ сви-
детельствует о средней продолжительности снегового покрова (Aichinger,
1933). И островки с местным климатом, отличающимся более континен-
тальным или более океаническим характером, часто можно выделить
по особым растительным сообществам (Schulte, 1937).
158
Единицы растительности в ландшафте
Чем точнее систематизированы растительные сообщества, изобра-
женные на карте (субассоциации, варианты, субварианты, фации ассо-
циаций), тем слабее их экологическая энергия (широта реакции), тем уже
Р и с. 39. Сообщества полевых п садовых сорняков в округе Лезе, разграниченные
по «ступеням» (самые мелкие флористически различимые единицы сообщества), отра-
жающим продолжительность использования земли в различных частях поля. На уча-
стках, оставленных чистыми в год составления карты, «ступени» еще не могли быть
нанесены (по Тюксену, перерисовано с неопубликованного оригинала 1957 г.).
Поля: 1 — ступень поселений; 2 — ступень более старых полей; 3 — ступень более молодых
полей; 4 — ступень корчевания.
Сады: 5 —более старые ступени; в —более молодые ступени.
валентность (амплитуда) характерной для этого сообщества среды обита-
ния и тем надежнее определяющее значение этих сообществ для качества
местообитания и границ пространств местообитания. Уточняются данные
так называемых «экологических групп видов», если их рассматривать
не изолированно, а в качестве составных частей четко определенных
Растительность в ландшафте
159
единиц сообществ. Они могут в таком случае использоваться как индика-
торы градации отдельных факторов местообитания.
Тонкие флористические различия между участками одного и того же
сообщества можно уловить с помощью «метода совмещения» по их значе-
нию как индикаторов постепенного изменения экологического фактора
(Tuxen, 1954). Растительные сообщества позволяют, таким образом,
лучше судить об особенностях местообитаний, чем измерительные приборы.
На основе флористически распознаваемых экологических ступеней
растительных сообществ можно нанести на карту, например, глубины
залегания грунтовых вод или различное содержание гумуса в почве.
Тюксен смог уловить флористические различия сообществ полевых сор-
няков по возрасту выкорчеванных площадей. Картирование таких «сту-
пеней» сообществ позволяет по сообществам полевых сорняков опреде-
лить возраст выкорчеванных площадей (рис. 39).
НЕЗАВИСИМАЯ ПРОСТРАНСТВЕННАЯ ДИФФЕРЕНЦИАЦИЯ
РАСТИТЕЛЬНОСТИ И ВЗАИМОСВЯЗИ БИОЦЕНОЗОВ
ВНЕ МЕСТООБИТАНИЙ
НЕЗАВИСИМАЯ ПРОСТРАНСТВЕННАЯ ДИФФЕРЕНЦИАЦИЯ
РАСТИТЕЛЬНОСТИ
Границы распространения единиц растительности не всегда связаны
с классификацией местообитаний, обусловленной абиотическими факто-
рами. В особых случаях биоценозы могут сами определить свои биотопы
Р п с. 40. Схематический профиль среднеевропейского болота, возникшего
частью па месте высохшего озера, частью вследствие заболачивания леса (по
Фпрбасу).
1 — ил; 2 — камышовый торф; з — осоковый торф; 4 — лесной торф; 5 — более старый
сфагновый торф; 6 — более молодой сфагновый торф.
В центре поверхности болота заполненное водой «окно» («болотный глазок»). Болото
окаймляет заболоченный и болотистый лес.
и их границы. Примером являются верховые болота и пояса береговых
наносов на озерах, прудах и морских побережьях. и*
Предпосылкой к возникновению верхового болота является
климат. Однако субстрат растительности, отложение торфа над мине-
ральным грунтом, образуется уже самими растениями. Нарастание тор-
фяной толщи кверху, а часто и ее распространение вширь являются функ-
цией времени. Границы верхового болота могут проходить независимо
от минерального грунта, а их положение может быть обусловлено про-
должительностью развития болота. Биогенный торфяной слой уже сво-
дит на нет влияние различия горных пород или рельефа грунта на харак-
160
Единицы растительности в ландшафте
тер местообитания. Болото дифференцируется независимо в концентри-
ческом порядке; в нем выделяется внутренняя часть, состоящая из кочек
и мочажин, и наружный пояс — обычно из лесных сообществ. Единицы
местообитаний и их границы создаются здесь самой растительностью
(рис. 40).
По берегам внутренних водоемов границы раститель-
ности и местообитаний могут быть очень непостоянными. Субстрат, посте-
пенно повышающийся, возникает и здесь биогенно, преимущественно
из останков живых существ. Несмотря на непрерывность процесса, при
Р п с. 41. Последовательный ряд прибрежных сообществ па эвтрофном озере
(с границей высокого н низкого уровня воды) (по Фнрбасу).
1 — подводный дерн из Characeae; 2 — пояс рдестов; з —пояс кувшинок; 4 — тро-
стник; 5 — пояс высокой осоки; 6 — черно-ольховая топь.
этом образуются довольно резкие границы между растительными сообще-
ствами различных поясов (рис. 41). Скачкообразно сменяются доминирую-
щие в сообществах виды, однако самое положение границы соответствует
кратковременному состоянию равновесия. Пояса растительности могут
из года в год наступать все дальше в озеро. Они образуются независимо
от окружающей растительности и представляют собой характерное есте-
ственное пространственное деление в миниатюре. Положение границ обыч-
но при этом весьма эфемерно.
ВЗАИМОСВЯЗИ ВНЕ МЕСТООБИТ АНИЯ
Пространственная структура, образуемая в каждом ландшафте опре-
деленным количеством типов местообитания и растительности, не является
простой совокупностью разнородных экосистем. Между элементами
этой структуры существуют взаимные влияния и связи и вне местооби-
тания. Они наблюдаются всегда, в любом случае, потому что все про-
цессы местного жизненного баланса — это часть общего круговорота
веществ геосферы. Кроме того, соседствующие экосистемы подвержены
воздействию одного и того же регионального климата.
Имеются, однако, и другие причины, которые сближают взаимно
граничащие экосистемы функционально или по совпадающим внешним
факторам. Примером являются ряды местообитаний, находящиеся в зоне
влияния грунтовых вод пли затопления речными водами. Свойство воды
(кислотность, содержание питательных веществ) может привести к тому,
что у соседних, но разнородных единиц экосистемы возникнут общие черты
и в то же время они будут отличаться от экосистем соседних областей,
имеющих другой состав воды.
Растительность в ландшафте
161
Аналогичное явление наблюдается там, где на участки местности
с различным составом растительности наносится или навевается одина-
ковый материал (подвижные пески, лёсс, щебень осыпей).
По сходству результатов совместные воздействия отдельных факто-
ров соответствуют влиянию одного регионального климата.
Нечто другое представляют собой функциональные взаимосвязи,
которые основаны на самой жизни биоценозов. На внутренних водоемах
биоценозы открытой воды и береговой полосы — это самостоятельные,
приспособленные к определенным местным условиям единицы. Однако
экологически они зависят друг от друга. Обмен веществ между продуцен-
тами, консументами и редуцентами происходит здесь не по вертикали,
как в лесу, но в значительной мере и даже преимущественно по гори-
зонтали — между биоценозами, связанными водной средой и существую-
щими рядом друг с другом. С точки зрения биологической продуктив-
ности озера в целом являются единицей высокого ранга.
На этом основана экологическая типология озер (Thienemann, 1920, 1931), в кото-
рой для умеренных широт различают три основных типа: 1) олиготрофные (с бедным
питанием) озера со скудной береговой растительностью, выравненным обменом веществ
и с глубинными слоями воды, богатыми живыми существами и насыщенными кислоро-
дом; 2) эвтрофные (с богатым питанием) озера с высокой продуктивностью органиче-
ских веществ в береговой зоне и верхних слоях воды, с избытком неразложившегося
гнилостного ила и поэтому, несмотря на избыток питания, с недостаточностью кисло-
рода и бедным животным миром в глубинной зоне; 3) дистрофине (богатые гумусом)
озера со слабой растительной продукцией, но почти отсутствующим разложением
и поэтому с недостатком кислорода и питательных веществ и бедным животным миром
глубин.
Сходные связи вне местообитаний можно заметить и у наземных
биоценозов, особенно если не оставлять без внимания животный мир,
участвующий в их структуре. Многие виды животных, мигрируя, выхо-
дят за границы отдельных растительных сообществ, вовлекая тем самым
соседние биоценозы в общий круговорот питательных веществ; общность
создается также участием животных в опылении цветков или распростра-
нении семян и в сохранении биоценотического равновесия.
ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА КАК ФОРМИРУЮЩИЙ ФАКТОР
ОБЩИЕ ЗАМЕЧАНИЯ
«Густое население уже само по себе вызывает многочисленные изменения» (Will-
denow, 1792). Человек «является одним из звеньев природы и одновременно ее преоб-
разователем» (Thienemann, 1941). Его активное отношение к растительности прояв-
ляется различно. Он может ее переделывать, ухаживать за ней или уничтожать. Он
только не может оставить ее нетронутой, что вполне естественно. Без некоторых
веществ, производимых растениями, немыслимо существование человека. Используя
опыт многих поколений, человек преобразовывает растительность в соответствии со
своими потребностями. На месте естественных лесов, степей, болот и т. д. возникают
поля, луга, пастбища, лесные посадки и другие угодья.
На большой части земной поверхности растительность либо уничто-
жена человеком, либо настолько сильно преобразована, что от перво-
начального растительного покрова уже ничего не осталось. Поэтому
162
Единицы растительности в ландшафте
во многих странах изучение растительности является одновременно иссле-
дованием культурного ландшафта. «Мы прежде всего должны отличать
естественную растительность от растительности, образовавшейся под
влиянием человека» (Sendtner, 1854). Степень воздействия человека
на окружающую природу весьма различна, поэтому часто возникают
проблемы, которые одними естественнонаучными методами решить
не удается, а требуется провести также и исторические исследования.
Влияние человека отличается от других факторов целым рядом
особенностей, определяющих растительную жизнь и ее пространственное
распределение. Человек стремится целесообразно переделать раститель-
ность или устранить ее, если она мешает достижению его целей. Он изме-
няет состав систематических единиц, частью непосредственно, а частью
косвенно вторгаясь в жизнь ландшафта и тем самым в условия существо-
вания растительности. Таким образом изменяются условия конкуренции
между отдельными систематическими единицами по сравнению с суще-
ствовавшими в первоначальном естественном ландшафте.
Для географии растительности не всегда обязательно различать
мотивы, которыми руководствуется человек, преобразовывая расти-
тельность. Возник ли пожар из хозяйственных соображений для осво-
бождения земли под пастбище, или при борьбе с мухой цеце, или случайно
по неосторожности — последствия такого пожара для растительности оди-
наковы. Для растительного сообщества с точки зрения результатов
столь же безразлично и то, пойдут ли изъятые из него растения или
части их для нужд самого человека, на корм скоту, как горючий или
строительный материал, промышленное сырье или на украшения.
Для географии растительности мотив воздействия важен лишь с точки зрения
места, времени и пространственной протяженности этого влияния. Сельскохозяйствен-
ные воздействия большей частью распространяются на большие площади, тогда как
мероприятия, связанные со средствами сообщения, оказывают влияние на узких про-
странствах (линейно). Мотивами воздействия человека на растительность являются
также военные сооружения, идея защиты природы, научные эксперименты с раститель-
ностью в целях получения новых данных, а также эстетические соображения.
Процессы вторжения человека в природу и их воздействие на рас-
тительность иногда проявляются тотчас же, иногда протекают скрытно,
и без тщательных исследований обнаружить их бывает очень трудно.
Косвенные действия могут иметь большее значение, чем непосредственные.
С пространственной точки зрения следует различать местные, региональ-
ные и общие влияния. Самым радикальным местным воздействием является
устранение всякой растительности, если площадь, на которой она произ-
растает, нужна под строительство городов, промышленных или тран-
спортных устройств. Подобные площади, составляющие во многих стра-
нах большой процент всей поверхности, практически исключаются из рас-
смотрения географии растительности.
ФОРМЫ ВОЗДЕЙСТВИЯ ЧЕЛОВЕКА
НА РАСТИТЕЛЬНОСТЬ
Корчевание и выжигание
Из площадей, занятых растительностью, наибольшие изменения
претерпели, как правило, те, на которых человек «огнем и мечом» уни-
чтожил первоначальную растительность, чтобы освободить место для
Растительность в ландшафте
163
нужных ему растительных сообществ. Корчевание, которое в леси-
стых странах часто предшествует дальнейшему заселению этой площади
людьми, не является однородным процессом. Цель — освободить почву
от ее первоначальной растительности — может быть достигнута одно-
кратной вырубкой или выкорчевыванием тех пней, которые способны
дать побеги.
Однако лес можно уничтожить не только путем корчевания, а и мно-
гократным выжиганием или интенсивным использованием данного уча-
стка под пастбище. На землях древних культур часто лишь путем иссле-
дования истории поселений удается установить, каким именно образом
эта площадь превратилась в «выкорчеванную землю». Предшествующее
корчевание не может поэтому служить признаком различия тех форм,
которые принимает растительность в результате воздействия человека.
Результаты корчевания в значительной степени зависят и от усло-
вий местообитания. В тропическом дождевом лесу искусственно создан-
ная корчеванием свободная площадь может через небольшой проме-
жуток времени вновь подвергнуться вторжению леса. В других местно-
стях, как, например, в европейском Средиземноморье, наоборот, на
однажды обезлесенных площадях лес не возобновляется в течение
столетий.
Для изменения облика растительного покрова, подвергшегося воз-
действию человека, повторяющиеся, но кажущиеся в отдельности незна-
чительными влияния могут оказаться более ощутимыми, чем однократное
корчевание, как бы велики ни были затраченные на него усилия.
Наряду с сельскохозяйственным возделыванием и использованием
наиболее действенной формой влияния на растительность являются
пожары. Последствия пожаров выходят далеко за пределы сельскохозяй-
ственных угодий в особенности в периодически сухих тропических и суб-
тропических областях, а также в бореальных хвойных лесах. Однако
при лесных и степных пожарах, которые во многих областях земли опре-
деляют характер растительности, не всегда бывает просто установить,
какая доля участия падает на человека, потому что пожары могут воз-
никать и по естественным причинам.
В Британской Колумбии, где ежегодно в среднем выгорает около
50 тыс. га леса, большей частью виновником пожаров является человек,
но в 10% причиной их оказываются грозы. Если в настоящее время службе
охраны лесов удается ограничивать распространение пожаров, то в дев-
ственных лесах пожары, вызванные грозой, могли распространяться
беспрепятственно. Таким образом, участие человека в возникновении
пожаров обусловливает скорее количественную, чем качественную раз-
ницу условий, существующих в естественном ландшафте.
Другое дело в Средней Европе, где загорание от молнии случается редко,
и большая часть легковоспламеняющихся и горючих древостоев образовалась в резуль-
тате лесохозяйственной деятельности человека. Природный высокоствольный листвен-
ный лес нелегко поддается выжиганию. В Баварии, хотя сосна и занимает лишь незна-
чительную часть лесов, 81% площади, охваченной лесными пожарами, падает на сосно-
вые леса, 12% — на смешанные сосново-еловые леса, 2% — на еловые леса, 4% —
на лесосеки и только 1% — на лиственные леса. В Пруссии, где в среднем на 87%
площади леса уничтожены пожарами, в основном это были чистые сосновые насаж-
дения. Если при этом учесть, что в условиях среднеевропейского климата загорание
от молнии может возникнуть чаще всего при летних послеполуденных грозах и что,
за исключением болот и высокогорных склонов, лесные пожары большей частью
164
Единицы растительности в ландшафте
ночью сами затухают под воздействием ими же вызванной местной циркуляции возду-
ха, то можно предположить, что в Средней Европе выгорание растительности без
содействия человека вряд ли могло бы иметь какое-либо значение.
В травянистых пространствах периодически сухих тропиков редко
наблюдаются естественные пожары. Возможность их вообще долго оспа-
ривалась. Здесь причиной пожаров всегда является человек, который
частично ради улучшения качества пастбищ, а частью непроизвольно
вызывает выгорание травостоя, ежегодно происходящее на огромных
площадях и обусловливающее характер растительности на этих участках.
За исключением затопляемых саванн, все прочие саванны, которые
еще Шимпер из-за их зонального расположения рассматривал как кли-
матически обусловленные формации, возникли при непосредственном
воздействии человека.
Обусловленное пожарами превращение первоначальной лесной растительности
в травянистые пространства наблюдал еще Сент-Илер (Saint-Hilaire, 1831). С тех пор
многие исследователи неоднократно указывали на их антропогенный характер (Jun-
ghuhn, 1852; Schinz, 1893; Warburg, 1893; Busse, 1908 и др.). Их резкая граница с дож-
девыми лесами являлась здесь зачастую аргументом в пользу того, что травянистые
пространства не обусловлены климатом.
Перрье де ля Бати (Perrier de la Bathie, 1921) на основании деталь-
ного исследования растительности Мадагаскара по соотношению расти-
тельных сообществ убедительно доказал, что мадагаскарские саванны
являются не климаксной растительностью, а устойчивыми сообществами,
возникшими в результате пожаров.
Для доказательства пирофитного характера саванн Перрье де ля Бати нашел сле-
дующие основания:
1. Состав форм роста. Спектр жизненных форм здесь совсем иной, чем
у растительности сходных климатических поясов, не тронутой пожарами. Основные
компоненты напочвенной растительности — это растения, переживающие неблаго-
приятное время под почвой и ежегодно вырастающие заново (однолетние злаки). Сюда
же относятся растения с многолетними побегами, которые переносят уничтожение своих
надземных частей и не приспособлены к семенному распространению.
Некоторые виды, растущие в лесах, как многолетние древесные лианы и кустар-
ники в травянистых сообществах, ежегодно обновляют надземные побеги или являются
однолетниками, имеющими семенное возобновление. Например Smilax kraussiana,
лиана, добирающаяся до крон деревьев, растет в саваннах как геофит с образующимися
в период дождей побегами, достигающими всего лишь полметра в высоту.
Древесный ярус содержит пожароустойчивые деревья и кустарники с толстой
коркой, из которых более низкорослые являются вечнозелеными, как пожароустойчи-
вая пальма (Hyphaene). В открытой саванне частично господствуют виды растений,
играющие в лесу подчиненную роль, но способные благодаря своим особым свойствам
(образование семян, способность к прорастанию) к распространению после уничтоже-
ния конкурентов. Другие сохраняются лишь в виде остатков, пощаженных огнем,
и постепенно исчезают, потому что в новых условиях размножаться они уже не могут.
2. Флористический состав. Для саванн характерно малое количество видов. По
сравнению с 5000 видов, насчитывающихся в природных формациях Мадагаскара, тра-
вянистые пространства содержат всего 125 видов на в семь раз большей площади. Боль-
шая часть из них — это широко распространенные более или менее космополитные
или пантропические виды. Из 19 доминирующих видов злаков только 4 являются
автохтонными видами центрального Мадагаскара.
Растительность в ландшафте
165
Картирование растительности бывшего Бельгийского Конго также
показало, что почти во всех тропических областях распространение
саванн обусловлено прежде всего пожарами и что климатические условия
достаточно благоприятны для образования на их месте лесов или близких
к ним формаций типа бразильского «сатро cerrado» (Myers, 1933, 1936;
Germain, 1945; Lebrun, 1947; Waibel, 1948; Duvigneaud, 1955; Devred,
1956)*. Вопрос о том, следует ли рассматривать саванны как почти исклю-
чительно антропогенные формации, можно будет решить лишь после
того, как выяснится, в какой степени в периодически сухих тропиках
возможно возникновение пожаров без содействия человека.
Возделывание
«Возделываемыми» мы называем полезные площади, на которых
человек выращивает определенные растения. Сюда относятся все поля
и «посадки» в широком смысле, включая сады и искусственные древес-
ные насаждения любого вида.
С возделыванием почти всегда связана деятельность человека, способ-
ствующая росту полезных растений и помогающая им в соревновании
с дикими растеними или защищающая их от вредителей.
Предпосевная обработка почвы включает такие мероприятия, как
корчевание, выжигание, снятие дерна и т. д. В зависимости от характера
намечаемой к посеву культуры почву периодически вскапывают, пашут,
боронуют, удобряют и т. д. К этим приемам могут присоединяться меро-
приятия, изменяющие водный режим и микроклимат, как, например,
ирригация или мелиорация, устррйство полезащитных лесонасаждений,
защита от морозов методом искусственного дождевания, обогревание
посевов или устройство парников, а также защита посевов от вредите-
лей — от обычной прополки сорняков и уничтожения мелких вредителей
до применения специальных защитных средств или поселения опреде-
ленных животных для «биологического уничтожения вредителей».
Культивируемые растения также в свою очередь влияют на место-
обитания, оказывая либо механическое воздействие, либо извлекая
органические вещества с содержащимися в них почвенными солями.
Рассматриваемые в качестве растительных формаций возделываемые
растительные группировки отличаются прежде всего тем, культивируются
ли многолетние или однолетние растения и через какие промежутки
времени подвергаются обработке занятые этими растениями площади.
На ежегодно обрабатываемых полях возникают иные «сообщества сорня-
ков», нежели в виноградниках или в лесных посадках.
Часть лесов также носит характер возделываемых участков. Многие
леса возникли в результате посадки на полностью оголенных площадях
древесных пород, чуждых данной местности. Нередко в лесах почва
обрабатывается плугом или ее химический состав изменяют вносимые
удобрения. Распыляемые или разбрызгиваемые яды вызывают наруше-
ния и в составе животного мира в данном биоценозе.
Использование естественной
растительности
Эта форма хозяйственной деятельности человека широко распростра-
нена и включает столь многообразные действия, как случайный сбор
отдельных частей растений, вырубку ценных древесных пород в девствен-
166
Единицы растительности в ландшафте
ных лесах, лесохозяйственные системы вырубки с последующим естест-
венным восстановлением состава, использование участков растительно-
сти под пастбища, а также регулярное скашивание удобренных тучных
лугов. Различной степени оказываемых при этом воздействий соответст-
вует плавный ряд от почти нетронутых и едва преобразованных «квази-
природных» биоценозов до чисто антропогенных единиц растительности.
Общим является, однако, то, что видовой состав, при существующих
условиях среды, подвергающихся воздействию, представлен спонтанными
видами. Большая часть лугов, а отчасти также и пастбища занимают особое
положение, если их образованию предшествовало корчевание. На лугах
часто проводится ирригация или мелиорация, а в отдельных случаях
и искусственный подсев. Такие луга в крайних случаях примыкают к регу-
лярно возделываемым полям.
Скашиваемые луга — растительная формация, облик кото-
рой обусловлен ее хозяйственным использованием и сохраняется лишь
при его регулярном повторении. Скашивание препятствует поселению
древесной растительности на лугах. Косьба как определяющий фактор
обусловливает состав сообщества и содержащиеся в нем жизненные формы:
растения, переносящие однократное или многократное скашивание, или
растения, которые благодаря своему годичному ритму не подвергаются
скашиванию, поскольку их развитие завершается в промежутке между
покосами (Wettstein, 1904; Gradmann, 1932; Bertsch, 1947). Видовой состав
поэтому зависит от частоты и времени косьбы. Только луга, скашиваемые
осенью на подстилку, выделяются большим многообразием видов и жизнен-
ных форм, которые искоренены на лугах, скашиваемых летом на сено.
Чем интенсивнее участки местообитаний удобряются или орошаются,
тем однороднее становится их флористический состав. Тощие луга, исполь-
зование которых ограничивается скашиванием, часто в зависимости от
условий местообитания имеют значительные флористические различия.
Тучные луга, напротив, имеют удивительно сходный флористический
состав на всем пространстве Средней Европы.
Пастбищное хозяйство определяет на больших пло-
щадях характер растительности культурного ландшафта. Происходящие
при этом существенные изменения в основе своей не отличаются от подоб-
ных в естественных ландшафтах при пастьбе диких животных. Однако
в то время как в естественном ландшафте место, время и интенсивность
влияния пастьбы в конце концов регулируются естественным биоценоти-
ческим равновесием, в условиях культурного ландшафта человек может
сознательно определять распространение, длительность и интенсивность,
а путем подбора видов животных также и качественный характер воздей-
ствия этого зообиотического фактора.
Если скашивание действует одновременно и одинаково на все расте-
ния, то выпас — только выборочно. При умеренном выпасе в лесу лес
как таковой может сохраниться. Периодический выгон скота влияет
совершенно так же, как и пастьба диких животных в девственном лесу.
То же происходит при умеренном выпасе в открытых или полуоткрытых
естественных растительных формациях.
Даже в наиболее преобразованных человеком ландшафтах Средней
Европы на пастбищах и выгонах сохраняются остатки леса, а при нерегу-
лярной эксплуатации этих площадей леса могут вновь возобновиться. Чем
продолжительнее, регулярнее и интенсивнее выпас скота, тем сильнее
последовательно изменяется растительное сообщество. На месте смешан-
ного дубового леса таким образом может образоваться сухой дерновин-
Растительность в ландшафте
167
ный луг с отдельно растущими кустами терна и шиповника, а на месте
дубово-березового леса — пустошь с кустами можжевельника и отдель-
ными березами.
Вересковая пустошь на северо-западе Средней Европы, которая раньше рассмат-
ривалась как естественная формация на бедных питательными веществами почвах
в областях с влажным океаническим климатом, по современным воззрениям, возникла
в результате выпаса, сочетавшегося с вырубкой, снятием дерна и пожарами. Если луг
не подвергается хозяйственным воздействиям, особенно выпасу овец, то начинается
естественная сукцессия, которая может вновь привести к возобновлению леса.
Свободный выгон скота в Средней Европе стал редким явлением,
однако в некоторых других странах обусловленные им преобразования
растительности все еще значительны. В районе Магелланова пролива на
участках, освоенных только в последнее столетие, пастбища расширяются
за счет вытеснения субантарктического летнезеленого лиственного леса.
Во многих частях земной поверхности окончательные стадии преобра-
зования определяют уже в течение столетий и даже тысячелетий общую
картину растительности. «Играющие выдающуюся роль в ландшафтах
типы растительности старых культурных стран Средиземноморья и Ближ-
него Востока — испанские томиллары, южнофранцузские гариги, гре-
ческие фриганы — это нечто иное, как конвергентные пастбищные типы.
Они придают Средиземноморью его своеобразную прелесть и отли-
чаются особенным богатством ароматических губоцветных, ладаннико-
вых, рутовых, полыней, каучуконосных молочаев, зарослями чертополоха
и колючих кустарников {Bupleurum spinosum, Poterium spinosum, виды
Astragalus, Paliurus и т. д.) с растениями, до которых овцы и козы едва
дотрагиваются. Эти зообиотически обусловленные сообщества, к сожа-
лению, все еще распространяются и сейчас за счет жалких лесных остатков»
(Braun-Blanquet, 1951).
Избирательное воздействие пастьбы зависит от вида выпасаемых
животных и видового состава растительности. Различные виды растений
с неодинаковой охотой поедаются животными и неодинаково пригодны
для них. Некоторые растения путем особых приспособлений защищаются
против скусывания. Чувствительность растений к вытаптыванию скотом
и к связанному с пастьбой обогащению пастбища азотистыми веществами
тоже различна у разных видов. На стойбищах скота возникают особые
растительные сообщества из нитрофильных травянистых растений.
Степень воздействия пастьбы на растительность определяется прежде
всего методом ведения хозяйства. Одним из средств для регулирования
выгона является сооружение ограды (постоянные и оборотные пастбища).
Перегонная система позволяет создать лучшее, с точки зрения хозяина,
пастбище.
Формы лесопользования весьма разнообразны. Человек, срывая нуж-
ные ему части растений, представленных в естественных растительных
сообществах, старается не повредить при этом самих растений и не изме-
нить растительности в целом.
Однако, снимая плоды с диких плодовых деревьев или собирая грибы,
дикие овощи, ягоды и т. д., человек невольно способствует переносу
семян и тем самым широкому распространению отдельных видов. Более
сильное влияние на видовой состав оказывают такие формы лесопользова-
ния, при которых растения полностью изымаются, как, например, при
сборе тропических орхидей для продажи, или же совсем уничтожаются,
как чилийская пальма при добыче пальмового меда. Практика лесных
168
Единицы растительности в ландшафте
разработок, при которой в естественных лесах вырубаются стволы нужных
древесных пород (твердые древесины, цветные древесины ит. д.), часто при-
водит к обеднению состава леса и даже полному искоренению отдельных
видов. Подобные процессы происходят в различных вариантах почти повсе-
местно там, где многолетние растения изымаются из невозделываемых
растительных сообществ, поскольку такое изъятие не регулируется сточки
зрения постоянства пользования.
Принцип ведения лесного хозяйства постоянного пользования был
впервые разработан в Германии, но в течение последних десятилетий практикуется
уже во многих странах. Мероприятия по обновлению состава и уходу (прореживание
ит. д.) за лесом способствуют росту полезных видов и могут привести к образованию
чистых насаждений.
Лесохозяйственные системы пользования в широком
понимании включают использование отдельных деревьев девственного леса, создание
«непрерывно производительного леса» («Dauerwald») (Мюллер), культивируемого
в направлении сохранения естественного строения леса, улучшение качества древостоев
путем выборочной рубки (Plenterwald) и, наконец, применение самых различных спосо-
бов рубки.
Последние частично организуются так, что происходит естественное возобновле-
ние устойчивых на данных почвах деревьев (выборочные лесосеки, рубка с оставле-
нием защитных деревьев для возобновления, рубка полосами). Как и при сплошной
рубке на больших площадях, такие системы частично требуют искусственного возоб-
новления путем посадок или посева.
Системы пользования придают лесу физиономию, отличающуюся
от его естественного состояния. Наряду с упомянутыми формами высоко-
ствольного леса это особенно касается низкоствольного леса (порослевые
леса от пней с коротким периодом оборота рубки) и среднествольного
(порослевые от пней сообщества с сохранением перестойных семенных
деревьев различных периодов рубки).
В северной Швейцарии в результате введения начиная с XII в. среднествольного
лесного хозяйства ареал дубово-грабового леса был огражден от вторжения бука и бе-
лой ели. Среднествольное лесное хозяйство и дубово-грабовый лес находятся здесь
в тесном взаимодействии и одинаково распространены. Насколько с этим связано
расширение области дубово-грабового леса, обусловленное воздействием человека,
до сих пор не известно (Stamm, 1938).
Всякое регулярное лесопользование надолго изменяет видовой состав,
а зачастую также и почвенные условия. Это относится не только к непо-
средственному изъятию древесины, но и к так называемому побочному
использованию, например к сбору листвы для подстилки, срезке листвы
на корм, сбору ягод, а также охоте или к использованию леса в качестве
места отдыха населения больших городов.
В проекте классификации древесных сообществ по стадиям развития и типам
структуры Век (Week, 1956) попытался определить девственный и окультуренный
лес как взаимно исключающие друг друга понятия. «Девственным лесом» (Wald) он
называет древесные сообщества, в которых на местообитаниях, не затронутых челове-
ком, господствуют устойчивые на данных почвах виды деревьев. «Окультуренным
лесом» (Forst) он, напротив, считает сообщества, флористический состав или место-
обитание которых подверглись значительному влиянию человека и сильно изменили
свой облик.
Растительность в ландшафте
169
Однако слова «Wald» и «Forst» нельзя применять только в узком смысле, так
как это привело бы к противоречиям со значением этих слов в общем разговорном
употреблении.
Хорнштейн (Hornstein, 1951) исследовал историю хозяйствования
человека в лесах альпийских предгорий. Оказалось, что большая часть
лесов растет на почвах, которые раньше использовались под пашни или
в сменном лесо-полевом хозяйстве. Путем изучения географии поселений
во многих частях Германии обнаружено наличие лесов на месте «опусты-
ненных» лугов.
Изменение местообитаний
Корчевание, выжигание, возделывание и другие виды использования
влияют частью непосредственно, а частью косвенно через растительность
и на ее местообитания, и прежде всего на различные факторы местности.
Эти влияния, зачастую заранее предвиденные, иногда бывают локально
ограниченными, но могут изменять условия местообитания, периодически
или постоянно, и на больших пространствах.
Общее влияние на растительность, на которое мало обра-
щают внимания, оказывают бытовые и промышленные дымы. Особенно
вреден сернистый газ, попадающий в воздух повсюду, где сжигают
уголь. Уже при содержании в воздухе вблизи промышленных объектов
0,01 % SO2 происходит резкое повреждение хлорофилла, ведущее к нару-
шению дыхания и транспирации; губительно для многих растений длитель-
ное воздействие даже 0,0001-процентного содержания SO2. Из европейских
видов деревьев весьма чувствительны к дыму ясень, ель и пихта, менее
опасен дым для дуба и клена. Обитающие на коре деревьев лишайники
почти исчезли в промышленных странах с большим числом городов
именно из-за действия дыма. Соли, попадающие в почву вместе с осадками
из промышленных дымов (в некоторых промышленных районах от 30
до 80 г SO3 на 1 кв, м в год), губительны для микронаселения почвы. Осо-
бенно сильный дым может сократить число почвенных бактерий до х/10—
Vioo нормального количества.
Человек способен изменить также микроклимат, климат
почвы и водный режим местообитаний, например,
в результате дренажных работ, изменения естественного стока воды,
искусственного осушения участков по берегам озер или морей (польдеры),
понижения или повышения уровня грунтовых вод и т. д.
Во многих случаях человек изменяет форму местности,
например создавая искусственные насыпи, террасируя склоны, изменяя
наклон или экспозицию склонов. В виноградниках на Мозеле и Рейне
зачастую проводится большая работа по изменению ориентации отдель-
ных участков склонов хотя бы еще на несколько градусов к юго-западу.
Человек прокладывает каналы, роет водохранилища, искусственно
создает совершенно новые местообитания (железнодорожные насыпи, тер-
риконы). \
Изменения почвы на культивируемых участках происходят в резуль-
тате совместного воздействия всех факторов: обработки почвы и внесения
удобрения (эвтрофия), изъятия питательных веществ при жатве, извле-
чения материала (разработка торфа, снятие дерна) или добавления его
(известкование, искусственное перемешивание почв), а также намыва
или отсоса грунта человеком и, наконец, эрозии. Последняя становится
особенно заметной у границы лесов на лёссовой глине, где почва на полях
170
Единицы растительности в ландшафте
по сравнению с прилегающим профилем лесной почвы может быть снесена
вплоть до материнской породы.
Почвенный профиль изменяется в результате самых раз-
личных воздействий и частично под влиянием преобразованной челове-
ком растительности. Причинные зависимости в данном случае обоюдны.
Деградация (регрессивная сукцессия) растительности связана с «изме-
нением типа почвы» (Мюккенгаузен). Поэтому часто почвенный профиль
позволяет судить о характере хозяйственного использования данной
почвы, причем в большинстве случаев под образующимся в данный момент
почвенным профилем можно обнаружить исходный.
Классическим примером этого являются пустошные почвы северо-западной Гер-
мании. Пустошь с характерным для нее почвенным профилем возникла на месте быв-
шего леса в результате выпаса скота, снятия дерна и других воздействий. Ниже пес-
чанистого подзола с ортштейновым горизонтом, возникшего под сухой пустошью
(Calluneto-Genistetum typicum), часто еще можно видеть бурую полосу горизонта В почвы
дубово-березового леса.
В верхних горизонтах первоначальные бурые полосы превратились под воздей-
ствием пустошных гумусных растворов в черно-бурые «двойные полосы». Вкрапления
ортштейна иногда имеют форму некогда пролегавших здесь корней деревьев.
Там, где образование ортштейна зашло уже далеко, лес возникает лишь в редких
случаях. Из этого раньше делали вывод о естественном освобождении пустошных
участков от леса. Однако исследование связей между растительными сообществами
и почвенными профилями показало, что ортштейновый горизонт — это следствие
возникновения пустоши, а не первопричина ее (Тйхеп, 1939, 1954, 1955).
Изменение систематического
состава растительности
На общее обеднение систематического состава растительности куль-
турного ландшафта уже указывалось при описании воздействий промыш-
ленных и бытовых дымов. Почти все формы хозяйственного освоения
почв действуют в том же направлении. Они сильно обедняют первоначаль-
ную флору, или непосредственно уничтожая ее, или чаще, в результате
того, что отнимают у видов растений подходящие для последних
места произрастания. Вследствие того, что местообитания изменяются
и становятся непригодными для существования ранее произраставших
здесь видов, во многих культурных ландшафтах исчезают не только
многие систематические единицы, но и целые биоценозы (Тйхеп, 1955).
Примером непреднамеренного косвенного влияния человека является поражение
аппалачских каштановых лесов грибком, занесенным из Китая через Нью-Й орский
ботанический сад. Castanea dentata, некогда характернейшее наряду с дубом дерево для
Аппалачей на высотах 400—1200 м между 35 и 45° с. ш., было почти полностью унич-
тожено болезнью, упорно распространявшейся в течение трех десятилетий на юг и на
запад (Шенк).
Человек часто способствует поселению новых видов
растений, первоначально чуждых данной
области. Некоторые полезные или декоративные виды растений,
^иивезенные из чужих стран, «дичают» и в дальнейшем сохраняются и раз-
множаются без участия человека. Вместе с другими, случайно занесен-
ными и спонтанно вторгшимися видами они находят себе место среди
растительных сообществ, природному равновесию которых помешал
Растительность в ландшафте
171
человек, или же заселяют новые местообитания, созданные человеком.
Первоначально чуждые данной местности пришельцы часто образуют
здесь свои единицы растительности (рудеральные сообщества). Хотя
большинство пришельцев («п ришлая, или адвентивная
флор а») появляются без прямой помощи человека, однако в дальней-
шем они используют открытые деятельностью человека пути распростра-
нения на полях, обочинах дорог, дорожных насыпях и дамбах, укореняясь
в местных растительных сообществах, иногда навсегда. Интересно, что
менее измененные человеком местные растительные сообщества, как
правило, закрыты для внедрения новых видов. Только в исключитель-
ных случаях новому пришлому виду удается навсегда поселиться в изна-
чально обитающей здесь единице растительности. Поля, сады и другие
возделываемые участки, так же как и имеющиеся в любом культурном
ландшафте необрабатываемые, но лишенные естественной растительности
площади вдоль путей сообщения, на берегах рек и озер, окраинах селений,
каменоломнях, песчаных карьерах и т. п., могут дать приют огромному
числу переселенцев более раннего и позднего времени. Многие «древние
пришельцы», как куколь в Средней Европе, укоренились еще в доистори-
ческое или раннее историческое время, но дальнейшее их распростра-
нение ограничено «сообществами сорняков», возникшими под влиянием
человека.
Из видов, укоренившихся уже в историческое время, многие, в осо-
бенности занесенные из других частей света, заселили новые участки
в удивительно короткий срок. Молодыми пришельцами в составе средне-
европейской флоры, которые распространились только в историческое
время, являются: на полях — Senecio vernalis, Veronica tournefortii,
в садах — Mercurialis annua, Oxalis stricta и corniculata, на дорожных
насыпях — Oenothera biennis, на откосах — Datura stramonium и Lepi-
dium draba, в водоемах — Helodea canadense, на влажных берегах и про-
чих местах — североамериканские виды родов Aster и Solidago. Нельзя
еще говорить о внедрении многих новых пришельцев последнего времени,
изредка встречающихся то здесь, то там, поскольку не известно, удер-
жатся ли они на новых местах.
Даже у давно акклиматизировавшихся растений внедрение может быть только
кажущимся. Еще Валенберг указывал, что многие из таких растений удерживаются
только потому, что их семена непреднамеренно собираются и снова высеваются вместе
с другими. Без помощи человека они погибли бы от зимних холодов. Процессы засе-
ления и распространения пришельцев Крэ (Kreh, 1950, 1951) исследовал на примере
«флоры развалин» разрушенных городов.
Путем планомерного разведения или в ходе специальных хозяйст-
венных мероприятий человек изменил конституционные свойства многих
видов и создал новые расы или виды. Примером планомерного разведения
являются расы хлебных злаков с коротким периодом вегетации, которые
позволили продвинуть хлебопашество далеко к северу На сухих лугах
запада США растет опунция, стебли которой способны накапливать
воду в больших количествах. Пасущийся скот охотно поедает эти расте-
ния. Для лучшего использования их животными были выведены формы,
лишенные колючек.
Непреднамеренно, «с помощью косы», выведены сезонно-диморфные
расы (Wettstein, 1900) однолетних луговых растений, например из родов
Euphrasia и Alectorolophus, морфологически отличающиеся пары рас или
172
Единицы растительности в ландшафте
Рис. 42. Формы сезонного деморфизма у
очанки (Euphasia) (по Веттштейну).
I — рано цветущая разветвленная только в верх-
ней части форма, плодоносящая до скашивания;
II — разветвленная от основания форма; низко-
рослая вначале, она цветет только после покоса.
АВ — высота срезания при скашивании.
видов, из которых хотя бы одна систематическая единица в качестве
жизненной формы первого покоса заканчивает свое развитие до первого
скашивания, тогда как вторая начинает развиваться позднее (рис. 42).
В результате разведения некоторые культурные растения потеряли спо-
собность существовать самостоятельно, без помощи человека.
ЕСТЕСТВЕННОЕ ПРОСТРАНСТВЕННОЕ
РАЗМЕЩЕНИЕ РАСТИТЕЛЬНОГО ПОКРОВА
В ЛАНДШАФТЕ
КУЛЬТУРНЫЙ ЛАНДШАФТ И ПРИРОДА
Пространственное размещение растительности культурного ланд-
шафта обусловливается совместным влиянием деятельности человека
и сил природы. Эти два фактора взаимно связаны с самого момента возник-
новения. Многие антропогенные растительные формации, например луга,
пастбища и часть окультуренных лесов, состоят из спонтанных растений.
Подбор их определяется совместным влиянием человека и природных
факторов.
Человек, разрабатывая план хозяйственного использования данной
местности, прежде всего сталкивается с уже существующим естественным
пространственным подразделением ландшафта. Элементы ландшафта,
«флизе», не в одинаковой степени приспособлены для предполагаемого
использования. Поэтому, как правило, но, конечно, не в ущерб экономи-
ческим принципам полезные площади распределяются в соответствии
с определенным набором различных местообитаний естественных расти-
тельных сообществ. Разумеется, в этом нет никакой безусловной автома-
тичности, потому что от самого человека зависит, будет ли он и как именно
использовать предоставляемые ему природой возможности.
Наметилась связь между решением проблем истории ранних средне-
европейских поселений и выявлением форм использования растительности
населением или хозяйственной пригодности растительных сообществ для
Растительность в ландшафте
173
человека на различных этапах общественного развития. Уже самое общее
распределение открытых участков и леса и противоположность между
давно и недавно заселенными участками тесно связаны с природным деле-
нием пространства. Градман создал свою историю поселений на основа-
нии того, что области с доисторическим и раннеисторическим заселением
Рис. 43. Досредневековое заселение южной Германии и распространение «пустошно-
степной флоры» по Градману. Здесь объединены обе карты Градмана, ясно показы-
вающие совпадение участков.
1 — области старых поселений; 2 — старые поселения1 распределенные неравномерно;
з — старые поселения, периодически, 0flHaK0t пустовавшие; 4 — области с богатой пустош-
но-степной флорой; 5 — области с рассеяннной пустошно-степной флорой: 6 — области с
исключительно или почти исключительно лесной и болотной флорой.
в южной Германии совпадают с областями распространения определенных
растительных сообществ (рис. 43). С тех пор обнаружены многие подоб-
ные же зависимости и для других районов.
При сравнении карт областей доисторического заселения с естест-
венными ландшафтами всегда приходится удивляться, как ясно распо-
знаются по естественной растительности пространства, постоянно засе-
ленные еще с начала каменного века.
Можно привести большое количество таких примеров из южной и западной Гер-
мании, а также северной Швейцарии, где область естественного распространения дубо-
во-грабового леса совпадает с современными земледельческими ландшафтами, заселен-
ными еще в неолите, а впоследствии занятыми главным образом алеманнами 1 (Stamm,
1 Алеманны — группа племен, населявших западную Германию с III по VI в. н. э.
Ленточная керамика (Brandkeramik) — древнейшая земледельческая культура
племен, населявших юго-восток Средней Европы в эпоху неолита, за 4—3 тыс. лет
до н. э. Основное занятие — охота, скотоводство и мотыжное земледелие. Названа так
по характерному для нее ленточному рельефному орнаменту на глиняной посуде.
Мегалитическая культура — культура конца неолита (3 тыс. лет до н. э.), назван-
ная так по каменным сооружениям из крупных камней, мегалитам. Характерна для
племен скотоводов и земледельцев, населявших Западную Европу.— Прим. ред.
174
Единицы растительности в ландшафте
1938). В Нижней Саксонии поселения племен культуры ленточной керамики не выхо-
дят за пределы дубово-грабовых лесов, в то время как мегалитическая культура была
связана с областями дубово-березовых лесов (Тйхеп, 1931). Нитш попытался своей кар-
той распространения лесов в начале каменного века показать, что как мегалитические
сельские поселения, так и освоение земли племенами культуры ленточной керамики
были связаны с дубом. Желуди и лесная трава послужили основой для развития
скотоводства, позволившего в свою очередь заняться возделыванием под посевы тех
площадей, которые освободились от лесов в результате выпаса и затем были расширены
путем корчевания. Элленберг (Ellenberg, 1954) дополняет это положение, указывая
на различную сопротивляемость естественных лесных сообществ пастьбе скота.
Многие обстоятельства, подмеченные еще Градманом, находят сейчас
полное подтверждение, например резкое разграничение доисторического
ландшафта на заселенные участки и нетронутые лесные массивы, а также
приуроченность обжитых участков к площадям, занятым определенной
растительностью. С освоенными еще в начале каменного века областями
в южной Германии совпадает распространение местообитаний костровых
лугов. По новейшим данным, полученным при исследовании пустынь
и лугов, положение о постоянстве противоречий между лесными областями
и открытыми пространствами требует проверки.
Физиономически границы естественных участков местообитания могут
быть в культурном ландшафте более заметны, чем в первобытной расти-
тельности. Так, в естественном ландшафте болотистый лес, граничащий
с лесным сообществом на сухой почве, не так сильно отличается от послед-
него, как луга, занимающие в культурном ландшафте место болотистого
леса, отличаются от леса, сохранившегося на сухом местообитании.
Противоположность пашни и луга на границе местообитаний также
больше бросается в глаза, чем различие между двумя природными лес-
ными сообществами.
В результате неодинаковой пригодности местобитаний для деятель-
ности человека в культурном ландшафте некоторые единицы естественной
растительности часто в значительной степени, а иногда и совсем вытес-
няются хозяйственными формациями, тогда как другие сохраняются или
подвергаются лишь незначительному влиянию. Так, многие местообита-
ния вне территории Средней Европы, первоначально занятые типичными
дубово-грабовыми лесами, в настоящее время распаханы. Некоторые откры-
тые участки доходят сейчас почти до внешней границы местообитаний этих
дубово-грабовых лесов. Сомкнутые дубово-грабовые леса встречаются
поэтому весьма редко. Их находят лишь в виде небольших остатков по
краю открытых пространств илина площадях, не затронутых корчеванием.
С другой стороны, существуют лесные сообщества, местообитания кото-
рых почти или вовсе не подвергались воздействию человека, например
ненаселенный пояс влажных дубово-березовых лесов на северных склонах
горного хребта Высокий Венн и область естественных еловых лесов
в Северном Шварцвальде.
В Люнебургской Пустоши, в непосредственной близости от поселе-
ний, сохранился сырой дубово-грабовый лес, представлявший ценность
в качестве лесного пастбища. Сухой дубово-грабовый лес, напротив, почти
без остатка вытеснен со своих местообитаний пашнями. Заболоченные
ольховые леса остаются нетронутыми, если, конечно, в них не производит-
ся усиленный выпас скота. На месте сухого дубово-грабового леса в резуль-
тате выгона овец возникли луга, которые после прекращения выпаса
заросли сосняком.
Таблица 5
Сопротивляемость некоторых естественных лесных сообществ выпасу
и экстенсивному использованию древесины (по Элленбергу)
Сопротивляе- мость Среднеевропейская и за- падноевропейская области лиственных лесов Восточно- и североевро- пейская области хвойных лесов Альпы и среднегорья
1 Очень слабая Сухой дубово-бере- зовый лес на бедных питательными веще- ствами песках; тепло- любивый смешанный дубовый лес на плос- ких известняках или щебне или в сухих областях на лёссе Сухие дубовые ле- са в лесостепной об- ласти; сосновый лес на дюнах Лиственичный и кед- ровый лес субальпий- ских высот на легко- доступных участках
2 Слабая Сухой дубово-бере- зовый лес на лучших почвах (скальнодубо- во-березовый лес); бу- ковые и дубово-грабо- вые леса на плоских известняках Сосновые леса на сухом песке, камени- стых почвах или на окраинах болот Заросли горной сос- ны и еловые леса вблизи границы лесов; реликтовые сосновые леса центральных аль- пийских долин
3 Средняя Б у ково-ду бово-гра- бовые леса (типичные дубово-грабовые леса); сырой дубово-березо- вый лес; болотистый березовый лес Смешанные сосно- вые леса на разных местообитаниях; бо- лотистый сосновый лес Названные выше со- общества на более труд- нодоступных участках; заросли зеленой ольхи
4 Большая Буковый лес на глу- боких почвах; черно- ольховая топь Еловые и смешан- ные еловые леса на местообитаниях, бо- гатых питательными веществами Буково-пихтовый лес и еловый лес; серо-оль- ховые леса в долинах
5 Очень большая Сырой дубово-грабо- вый лес на глинистой почве с близкими грун- товыми водами; долин- ные леса Еловые болотистые леса и долинные леса Субальпийский ело- вый лес на труднодо- ступных склонах
176
Единицы растительности в ландшафте
Изменение хозяйственной оценки различных типов местообитаний
в процессе исторического развития можно проследить на примере сырых
дубово-грабовых лесов в Люксембурге. Поскольку здесь эти леса распо-
ложены на тяжелых почвах, они первоначально были сохранены, тогда
как местообитания типичных дубово-грабовых лесов были распаханы
еще в начале каменного века. С ростом населения в областях сырых дубо-
во-грабовых лесов возникли отдельные угодья с пастбищным хозяйством.
После введения усовершенствованных плугов оказалось возможным и на
этих почвах перейти к хлебопашеству. Позже, однако, пахотные земли
вновь сменились лугами и лесами, так как искусственные удобрения сде-
лали пригодными под посевы более легкообрабатываемые, но неплодород-
ные песчаные почвы, а недостаток в рабочей силе, возникший в результате
индустриализации страны, способствовал развитию пастбищного хозяй-
ства (Schmithusen, 1940).
Представление, будто при менее развитом хозяйстве формации полез-
ных растений теснее связаны с природными местообитаниями, чем при
современном уровне его развития, можно считать лишь условно правиль-
ным. Если технико-экономические возможности достаточно многосторонни,
а виды разводимых растений многочисленны, то более тонкие различия
участков местообитания могут оказаться заметными по распространению
«группировок полевых растений» даже там, где такие различия ранее не
обращали на себя внимания. Так, новое плодовое хозяйство может вызвать'
дробление в подразделении ландшафта, потому что виды или сорта
плодовых деревьев по своим требованиям занимают местообитания только
определенной растительности. Другой результат получается, если с по-
мощью технических средств изменять факторы местности, а вместе с тем
и самые границы естественных участков местообитания.
.ЗАМЕЩАЮЩИЕ СООБЩЕСТВА
Как естественные, так и антропогенные раститель-
ные сообщества дифференцируются по местообитаниям, и по
их размещению можно определить их отношение к потенциальной есте-
ственной растительности. Это выяснилось при сельскохозяйственном
и лесохозяйственном исследованиях растительности и подтвердилось при
фитоценологическом картировании. Растительные сообщества, возникшие
под влиянием человека, учитывая их принадлежность к определенным
естественным сообществам, называются «замещающими сооб-
ществами» (Тйхеп, 1932).
Замещающее сообщество — это понятие из области динамики сооб-
ществ. Оно обозначает антропогенную единицу растительности и приоб-
ретает собственный смысл только при расположении этой единицы в серии
сообществ, где она занимает место определенного растительного сообще-
ства потенциальной естественной растительности.
Одно и то же хозяйственное воздействие на различные природные
лесные сообщества приводит к возникновению на их месте разных заме-
щающих сообществ. Сенокошение, выпас или распашка приводят в каж-
дом случае к образованию на местообитаниях заболоченного ольшаника,
типичного дубово-грабового леса и сухого дубово-березового леса, фло-
ристически различных луговых или пастбищных сообществ и сообществ
полевых сорняков. Даже при близком родстве исходных естественных
сообществ, например при субассоциациях одного из названных лесных
сообществ, замещающие сообщества отличаются друг от друга.
Растительность в ландшафте
177
Экологически это легко объяснимо. Дело в том, что хозяйственное
воздействие, в результате которого возникают замещающие сообщества,
является дополнительным фактором в общем комплексе условий место-
обитания. Оно не может исключить действия всех прочих факторов. Даже
искусственно созданные путем посева сообщества, чуждые данному место-
обитанию, со временем самостоятельно превращаются в соответствующие
ему замещающие сообщества. Кнолль экспериментально доказал, и каж-
дый работник сельского хозяйства знает это по собственному опыту, что
выбранная при посеве смесь семян не определяет более позднего сочетания
видов в составе устойчивого луга. Для основных типов луговых место-
обитаний Швабии Кнолль составил четыре соответствующие им смеси
семян и высеял их для сравнения в двух областях, а именно в области
дубово-грабовых лесов, где луга из высокого райграса (Arrhenatheretum
elatioris) являются свойственными данному местообитанию свежими
и тучными лугами, и в горной области буковых лесов, где ту же роль
выполняет луг из желтоватого трищетинника (Trisetetum flavescentis).
Наибольший урожай был снят с тех опытных площадей, где возделыва-
лась смесь семян, соответствующая почвам данных лугов, но в обеих
областях на всех четырех опытных площадях позднее самостоятельно
возникли луга с таким составом, который соответствует замещающему
сообществу, обусловленному характером местообитания.
Менее удачно, чем понятие «замещающее сообщество», введенное при лесохозяй-
ственном картировании местообитании, понятие «лесокультурные сообщества», с по-
мощью которого древостои группируются по доминирующей древесной породе. Если
в сосновые «лесокультурные сообщества» объединить те замещающие сообщества, кото-
рые возникли в результате посадки сосен, то это не будет ни систематическим, ни дина-
мическим группированием сообществ, а только лишь объединением по характеру
использования. Слово «сообщество» здесь неуместно. Сосновые насаждения могут
относиться к разным единицам сообществ и динамически являться замещающими
сообществами весьма различных заключительных сообществ (Meisel-Jahn, 1955).
В лесохозяйствах, созданных путем подсадки видов деревьев, не
свойственных данному местообитанию, уже в первом поколении чуждых
древесных пород часть видов первоначального лесного сообщества выпа-
дает, тогда как другие виды добавляются. Сосновые лесонасаждения,
которые в течение двух-трех поколений растут на местообитании естест-
венного лиственного леса, уже весьма значительно отличаются по своему
составу и часто приобретают признаки, отражающие окисление почвы
и общее ухудшение местообитания. Исследования отдельных фаз дегене-
рации позволяют, однако, установить принадлежность заме-
щающего сообщества к потенциальному есте-
ственному лесному со об щ е с т в у даже в тех случаях,
когда уже нельзя обнаружить никаких характерных для него видов
(Тюксен). Последовательно сменяющиеся поколения хозяйственно ис-
пользуемых лесонасаждений являются стадиями регрессивной сукцессии.
Чем богаче местообитание основаниями, тем медленнее протекает процесс дегра-
дации под влиянием чуждого данному местообитанию древесного вида. Так, сосновые
насаждения на местообитании сухого черешчатодубово-березового леса в северо-запад-
ной Германии достигают уже во втором поколении характерной фазы, отличающейся
кислотолюбивыми видами, которая в сосновых насаждениях на местообитании скально-
дубово-березового леса с обилием фиалок возникает только в третьем или более позд-
нем поколении. Отнесение лесохозяйственно обусловленных замещающих сообществ
к потенциальным естественным лесным сообществам может быть подтверждено иссле-
дованиями истории лесного хозяйства и результатами пыльцевого анализа.
178
Единицы растительности в ландшафте
ГОМОЛОГИЧНЫЕ СООБЩЕСТВА
На местообитаниях с одинаковой потенциальной естественной расти-
тельностью в результате разнородных воздействий (пахота, выпас скота,
луговое или лесное хозяйство) и вследствие разной длительности и интен-
сивности этих воздействий вместо одного заключительного сообщества
могут возникать различные серии замещающих сообществ (регрессивная
сукцессия).
Если в конце концов поле забрасывается, если луг больше не ска-
шивается или если какое-либо другое замещающее сообщество перестает
подвергаться влиянию человека, то на этих площадях естественные сооб-
щества возникают вторично. Эта прогрессивная сукцессия ведет к обра-
зованию заключительного сообщества, обусловленного экологическим по-
тенциалом местообитания.
Все сообщества прогрессивных и регрессивных серий сукцессий
одного и того же естественного заключительного сообщества
являются гомологичными по динамике развития (vegetationsdynamisch
homolog).
ПОНЯТИЕ КРУГА СООБЩЕСТВ
Совокупность единиц растительных сообществ, которые встречаются
или могут встретиться на местообитаниях определенного типа, называют
«кругом сообществ» (Gesellschaftsring) данного типа местообитаний
(Schwickerath, 1954). Это понятие соответствует «местному ряду типов»
Куялы (Kujala, 1945). Круг сообществ охватывает заключительные сооб-
щества соответствующего местообитания со всеми замещающими сообще-
ствами, которые могут возникнуть на их месте, и всеми начальными и по-
Таблица 6
Пример круга сообществ, составленного по описанию виноградника,
заложенного в долине реки Майн на местообитании теплолюбивого смешанного
дубового леса и впоследствии частично снова заброшенного (Welte, 1934)
ТЕПЛОЛЮБИВЫЙ ДУБОВЫЙ ЛЕС
как исходное лесное сообщество, сохра- нившееся по исторической случайности Лесное сообщество, измененное уничтожением древесных пород (Корчевание) Сообщество сорняков виноградника как заключительное сообщество вторичной сукцессии t Кустарниковое терново-шиповниковое сообщество Сообщество сухого дерновинного луга Первичное сообщество сухого дерно- винного луга —> Остатки сообщества сорняков в забро- шенном винограднике
Растительность в ландшафте
179
следующими стадиями первичных и вторичных сукцессий, ведущими
к его образованию. Составные элементы круга сообществ являются одно-
временно гомологичными сообществами с точки зрения экологии место-
обитаний.
ПОНЯТ ИЕ «КЛИМАХ С-КОМ ИЛЕ К С»
Круг сообществ — это набор сообществ одного типа «флизе». Соот-
ветствующим понятием для набора сообществ всего ареала заключитель-
ного сообщества является «климакс-комплекс» (Braun-Blanquet, 1928).
Р и с. 44. Стадии деградации южпофранцузского каменнодубового
леса и их почвенные профили (по Браун-Блапке).
А. На компактном юрском известняке: 1 — Quercetum ilicis; 2 — Quercetum
cocciferae brachypodietosum; з — Brachypodietum ramosi; 4 —стадия после-
пастбищной Euphorbia charicia.
Б. На третичном мергеле: 1 — Quercetum ilicis', 2 — Quercetum coccijerae
rosmarinetosum; 3 — Rosmarineto — Litho spermetum; 4 — Aphyllanthion;
5 — голая хрящеватая почва; 6 — Deschampsietum mediae на осушенном
глинистом мелкоземе.
Климакс-комплекс охватывает все серии развития заключительных сооб-
ществ, гомологичных по динамике развития. Он может состоять из боль-
шого числа кругов сообществ. Как на начальных стадиях первичных
сукцессий, так и в деградированных замещающих сообществах внутри
ареала того же заключительного сообщества большее значение получают
различия местообитаний, которые обусловливают различие в сообще-
ствах.
На это указывают, например, серии регрессивных сукцессий леса из каменного
дуба на двух различных типах «флизе» (рис. 44). Не только потенциальное заключи-
180
Единицы растительности в ландшафте
тельное сообщество, но главным образом распределение серий по двум различным
кругам сообществ позволят охарактеризовать в этом случае неодинаковую произво-
дительность обоих местообитаний. В естественном ландшафте, по Браун-Бланке, на
обоих местообитаниях должны были бы находиться сходные растительные сообщества
(лес из каменного дуба). Их потенциальная экологическая дифференциация была бы
заметна только по замещающим сообществам, следовательно, в состоянии естественного
ландшафта определить и разграничить несовпадающие признаки обоих местообитаний
возможно лишь по различию почв. Таким образом, не только растительное сооб-
щество, но вся экосистема в целом являлась бы в этом случае критерием различного
качества естественных пространств.
ПРОСТРАНСТВО МЕСТООБИТАНИЙ КАК ПОТЕНЦИАЛЬНАЯ
БИОХОРА ИЛИ КАК ПРОСТРАНСТВЕННАЯ ЕДИНИЦА
ПОТЕНЦИАЛЬНОЙ ЕСТЕСТВЕННОЙ РАСТИТЕЛЬНОСТИ
Если известна местная динамика сообществ, а тем самым и круг сооб-
ществ, то по распространению входящих в него отдельных сообществ
можно выделить пространство местообитаний как потен-
циальную биохору определенного заключительного сообщества. На этом
методе основано составление специальных карт восста-
буковый лес
на известняках -------------1
'Ф'
5. Сообщество красавки на
участках сплошной вырубки
7 Точное пастбище
|—{разновидности сред-
ЗДцбобо-грабовый лес
с обилием первоцвета
него подорожника)
♦
♦
2. Терновый кустарник
на богатых
известью почвах
8. Тучный луг с обилием
первоцвета
♦
♦
I —< 9. Полевые
I культуры
Ф А _________________
6. Полусухие дернины
на известняках
1 Начальные стадии полусухих
________дернин на известняках____
А/С-Карбонат- А/С-Карбонатные почвы,
ные почвы перекрытые глиной
> --------------> 1
>---------------> 2
>--------------->3
-О 4
.........~»5
> ^6
Рис. 45. Круг сообществ на плоских известняковых склонах с глубоким
почвенным слоем па равнине у подножия Высокого Вейна (Штольберг)
(по Швикерату).
1 — естественное развитие; 2 — сплошная вырубка; 3 — нерегулярный покос или
выпас скота; 4 — регулярный выпас скота с внесением удобрений; 5 — регуляр-
ный покос с внесением удобрения; 6 — распашка.
Растительность в ландшафте
181
новленной (потенциальной) естественной расти-
тельности1. Они являются одновременно картами пространств
местообитаний и поэтому успешно применяются в сельском, лесном, вод-
ном хозяйствах и т. д.
Типы пространств местообитания одинакового качества (Fliesentypen)
являются элементарными растительными пространственными едини-
цами. Их растительный потенциал характеризуется кругом сообществ,
образованным гомологичными с точки зрения экологии местообитания
единицами растительности.
Воспроизведение круга сообществ в виде схемы сукцессии (рис. 45)
дает представление обо всех сообществах, входящих во «флизе» соответ-
ствующего типа по динамике их взаимоотношений, а также о процессах,
которые привели к их образованию.
В естественном ландшафте отдельные участки этого типа «флизе»
имеют ту же растительность, что и заключительное сообщество. В куль-
турном ландшафте они реагируют одинаково на сходные формы воздейст-
вия, оказываемого человеком. Поэтому знание их может иметь большое
значение для практического использования почв.
ИЗМЕНЕНИЕ ПОТЕНЦИАЛЬНОЙ ЕСТЕСТВЕННОЙ
РАСТИТЕЛЬНОСТИ ЧЕЛОВЕКОМ.
Еще недостаточно ясно, в какой степени мероприятия, проводимые
человеком, могут изменить потенциальную естественную растительность.
Необратимой деградацией почвы, происходящей под влиянием человека,
можно считать образование ортштейна под пустошами северо-западной
Германии на первоначальных местообитаниях дубово-березового леса. Но
поскольку на таких землях не применяется глубокая пахота и к тому же
в условиях равнинного рельефа эрозионные процессы весьма замедлены,
естественной регенерации почв не происходит и эти участки выпадают из
круга сообществ дубово-березового леса.
Подобные воздействия человека могут сказываться и косвенно, на-
пример в частичном изменении климата в окрестностях большого озера
после его осушения.
Некоторые непосредственные или косвенные изменения почв имеют
лишь преходящее влияние, как, например, обогащение почв лесных
окраин и как результат — изменение состава минеральных солей в почве
наносом пыли с пашен. Это явление может привести к изменению в серии
сукцессий, однако едва ли изменит потенциальную естественную расти-
тельность, поскольку решающее -влияние климата на почвообразование
снова надолго сгладит это отклонение. Напротив, вызванная и ускоренная
распашкой эрозия почвы способна настолько сильно преобразить мест-
ность, что в культурном ландшафте могут возникнуть новые границы поч-
венных разностей и с ними потенциальной естественной растительности,
которых в условиях естественного ландшафта не было бы. Наоборот,
в результате осушения болот, например, возможно исчезновение перво-
начальных различий.
1 В СССР карты потенциальной естественной растительности носят название
карт восстановленного растительного покрова, поскольку па них наносится суще-
ствовавшая на данных местообитаниях естественная растительность до видоизменения
ее хозяйственной деятельностью человека.— Прим, ред.
182
Единицы растительности в ландшафте
ИСПОЛЬЗОВАНИЕ ПОЧВ НА ОСНОВЕ
ГЕОГРАФИИ РАСТ ИТЕЛЬНОСТ И
Сельское, лесное и водное хозяйства, а также другие отрасли, свя-
занные с эксплуатацией природных ресурсов, строительством дорог
и населенных пунктов, все в большей степени опираются на результаты
изучения растительности с ценологической точки зрения, ибо методы
фитоценологии позволяют (и это самое главное) выделить отдельные
местообитания. Их качественные соотношения могут быть затем уточ-
нены с помощью других методов исследования (анализ почвы, микрокли-
матические измерения и т. д.), картирование же местообитаний помогает
перенести хозяйственный опыт в пределы тех областей, где он окажется
полезным.
Исходя из флористического состава сообществ, можно установить
степень различия определенных качеств местообитаний (например, уро-
вень грунтовых вод) (Tuxen, 1954); последняя в свою очередь обусловли-
вает значимость экологических единиц, которые в узком смысле являются
собственно низшими единицами в ассоциациях, вплоть до «ступеней»,
характеризуемых отдельными экологическими факторами.
В лесоводстве на основе фитоценологического картирования лесов и точного
исследования местообитаний можно установить, в каких сообществах и при какой доле
участия в составе древостоя данный вид деревьев будет расти быстрее и даст наиболь-
ший доход, поскольку интенсивность роста одного и того же вида в разных раститель-
ных сообществах может быть весьма различной.
Лесовод, стремящийся создать необходимые для хозяйства древесные
виды, должен учитывать «закон границы, переход через которую приводит
к результату, прямо противоположному желаемому» (МбПег, 1922).
Недостаточное внимание к природным условиям приводит в лесоводстве
к массовому развитию вредителей, резкому ухудшению почвы и умень-
шению производства древесины.
Цель современного научного лесоводства — создание лесных хозяйств, соответ-
ствующих природным условиям и содержащих максимально возможную долю полез-
ных древесных пород, без нарушения равновесия между растительностью и местооби-
танием. В практике лесоразведения удалось достичь успеха также путем подражания
той последовательности сообществ, которая наблюдается при естественном лесовозоб-
новлении на соответствующих местообитаниях, например при облесении пустошей,
и при попытках вторичного облесения в европейском Средиземноморье.
В странах высокоразвитого лесоводства, насколько это осуществимо
в пределах лесного сообщества, переданного в ведение хозяйства, часто
стремятся создать на отдельном местообитании насаждение, соответствую-
щее потенциальному естественному заключительному сообществу при
заранее определенном предпочтении полезным породам, если они успешно
растут в соответствующем лесном сообществе. Таким путем на длитель-
ное время достигают оптимальной хозяйственной производительности.
Виды, чуждые данному сообществу, вполне допустимы в виде ограничен-
ной примеси, если они происходят из экологически «аналогичных» расти-
тельных сообществ.
Куяла совершил путешествие по провинциям Квебек и Британская Колумбия,
исследуя биологию лесов, чтобы выработать научные основы для внедрения северо-
американских видов деревьев в Финляндию. Исходя из основного вопроса, при каких
Растительность в ландшафте
183
условиях можно перенести лесоводческий опыт и виды деревьев из одной части света
в другую, он прежде всего пытался установить, какие местности и местообитания
обеих стран являются аналогичными с точки зрения биологии леса, то есть экологи-
чески равноценными или хотя бы немного сходными.
ПОТЕНЦИАЛЬНЫЙ ЕСТЕСТВЕННЫЙ ЛАНДШАФТ
И ПЕРВОБЫТНЫЙ ЛАНДШАФТ
УЧЕНИЕ О КЛИМАКСЕ
Теория моноклимакса. Многократно оспариваемая гипотеза, что
региональный климат определяет климаксную растительность (теория
моноклимакса), которая выдвигалась еще Коулсом, Купером и Клементсом
и нашла поддержку у Браун-Бланке, утверждала в своей первоначаль-
ной форме следующее: в климатически однородной области естественное
развитие растительности вместе с зависящим от климата и обусловленным
растительностью развитием почвы приводит к сглаживанию первоначаль-
ных различий местообитаний и к образованию на всем пространстве
климатической области одинакового растительного сообщества как кли-
матически обусловленного заключительного звена (климакса) развития
растительности.
Такое представление появилось под влиянием учения о климатиче-
ских типах почв. Решительная формулировка теории моноклимакса
и то влияние, которым она сначала обладала, были основаны частично
на том, что авторы опирались прежде всего на наблюдения, произведен-
ные в районах, где сукцессия почв под естественной растительностью
приводит к напластованию ила, сглаживающего влияние на раститель-
ность первоначальных различий почвообразующих пород. Так, Купер
(Cooper, 1913) для облесенного лишь в послеледниковый период остро-
ва Айл-Ройал показал такие серии сукцессий на различных первич-
ных породах и на местообитаниях, относящихся к серии береговых отме-
лей, которые все привели к одинаковому заключительному звену, а именно
к растущему на 25—60-сантиметровом слое ила смешанному лесу из Abies
balsamea, Betula papyrifera и Picea canadensis.
Аналогично под осоковым дерном (Curvuletwri), который в горах
Центральных Альп считается заключительным сообществом на горных
породах всех видов (Braun-Blanquet, Jenny, 1926), обнаружена почва
с кислым гумусным горизонтом. Основываясь на этом факте, Браун-
Бланке пришел к выводу, что в каждой климатически однородной области
будет господствовать в качестве «климакса» только одна окончательная
стадия развития растительности. Это воззрение, однако, не нашло под-
тверждения при исследованиях, проведенных в других областях.
Проблема вековых (sakularen) сукцессий. Теория моноклимакса
исходит из представления, что в одинаковом климате при достаточно дли-
тельном развитии различия местообитаний, обусловленные горными поро-
дами и рельефом, оказываются сглаженными в ходе вековой сукцессии.
Между тем еще Никольс (Nichols, 1917) заметил, что развитие почвы под
влиянием климата и растительности вовсе не всюду приводит к полному
уравнению различий местообитаний, обусловленных составом горных
пород. Он противопоставлял региональному, обусловленному климатом,
184
Единицы растительности в ландшафте
заключительному сообществу («климатический климакс»—«climatic cli-
max») понятие почвенного климакса («эдафический климакс» — «edafic cli-
max»), который определял как «заключительное звено какой-либо особой
серии сукцессий».
Перрье де ля Бати (Perrier de la Bathie, 1921) обнаружил на Мада-
гаскаре в одном и том же климатическом поясе (муссоновых лесов) кли-
максные леса, разнящиеся именно по подстилающей горной породе. Тюк-
сен и Димонт (Tuxen, Diemont, 1937) установили то же самое для северо-
западной Германии, Вайбель (Waibel, 1948)— для Бразильского плато.
Это же было подтверждено и в почвоведении. Почвенные профили, кото-
рые могли бы служить доказательством сглаживания различий, вызван-
ных подстилающими горными породами, в ходе климатически обусловлен-
ной вековой сукцессии следует считать всего лишь исключениями из
общего правила.
Столь же мало, как и различия, определяемые подстилающими гор-
ными породами, сглаживаются климатом и различия, обусловленные
экспозицией. Напротив, в некоторых климатах они сохраняются именно
в результате климатических процессов (Budel, 1958).
Вопрос о заключительной стадии естественного развития раститель-
ности, то есть климаксе экосистемы, имеет смысл только
при рассмотрении его применительно к человеческому масштабу вре-
мени. В геологических масштабах, судя по данным геоморфологии и почво-
ведения, различия, обусловленные экспозицией и спецификой почвенных
условий, а также местными особенностями в стоянии грунтовых вод,
характере приливов, отливов и речных паводков, ни в коем случае не
могут быть настолько сглажены влиянием однородного климата, чтобы
в пределах определенной климатической области вековая сукцессия
привела бы на всех местообитаниях к одному и тому же заключительному
сообществу.
Свойственное местообитанию потенциальное естественное заключи-
тельное сообщество. Тюксен и Димонт (Tuxen, Diemont, 1937) выделяли
среди заключительных сообществ в пределах климатически однородной
области «группы», связывая их со спецификой подстилающих горных пород,
и «ряды» (Schwann), обязанные различиям в экспозиции. Теория моно-
климакса тем самым отпадает, однако новая система понятий пока еще
не создана.
Во избежание недоразумений слово «климакс» в новейшей литературе
многими фитоценологами не употребляется. Вместо этого говорят о «есте-
ственной растительности» или, точнее, о «потенциальном естественном
заключительном сообществе отдельного местообитания» (Tuxen, 1956).
Это сообщество, а тем самым и климакс всей экосистемы зависят от ком-
бинации всех факторов местности и могут быть определены лишь индук-
тивно для каждого типа местообитаний. Обобщение, распространенное на
другие типы местообитаний той же климатической области, будет непра-
вомочным. Это, однако, не означает, что многие типы местообитаний не
могут иметь одинаковых климаксов. В этом случае, помимо регионального
климата, предполагается одновременное и сходное воздействие и других
факторов, как подстилающей горной породы, экспозиций или грунтовых
вод.
Область климакса (Klimaxgebiet). Этим термином Браун-Бланке
(Braun-Blanquet, 1951) обозначает «область, занимаемую одним климакс-
Растительность в ландшафте
185
комплексом». «Пределы области климакса совпадают с теоретически воз-
можным ареалом климаксного сообщества».
Это понятие области климакса как ареала потенциального естествен-
ного заключительного сообщества можно сохранить и при нашей трак-
товке учения о климаксе. Однако «возможный ареал климаксного сооб-
щества» нельзя отождествлять с областью, определяемой исключительно
региональным климатом. Это может быть и дизъюнктивный (разорванный)
ареал, который тесно переплетается с ареалами других заключительных
сообществ.
Все высказывания о заключительном сообществе являются весьма
относительными. Строго говоря, ни развитие растительного сообщества,
ни развитие почвы никогда не приходят к завершению, ибо с течением
времени изменяется не только климат, изменяется также и комплекс
видов, составляющих эти сообщества. Некоторые систематические еди-
ницы широко распространяются, другие вымирают, и в результате воз-
никают новые формы.
Однако если даже флористический состав заключительного сооб-
щества может меняться, то пространственные единицы способны все же
оставаться одинаковыми в своих границах. На этом предположении,
которое учитывает возможные за более длительные отрезки времени изме-
нения форм поверхности, но которое можно считать лишь условно спра-
ведливым, основаны попытки — с привлечением методов палеонтологии,
пыльцевого анализа и др. — составления карт растительности прошлого
(например, карты Нитша, Фирбаса, Клютэ, Тролля и Френцеля).
ПОТЕНЦИАЛЬНЫЙ ЕСТЕСТВЕННЫЙ ЛАНДШАФТ,
ПЕРВОБЫТНЫЙ ЛАНДШАФТ И ИСТОРИЯ РАСТИТЕЛЬНОСТИ
Проблема климакса и потенциальной естественной растительности
на отдельных местообитаниях затрагивает круг вопросов, связанных
с исследованием ландшафта, приводящих к истории растительности
и исторической географии растительности. Ссылка на эти области науки
необходима здесь лишь для пояснения некоторых основных понятий
и круга задач, сведенных в табл. 7.
Естественный и культурный ландшафты можно различать не только
для настоящего времени, но для любых периодов прошлого. «Первобыт-
ный ландшафт» означает ландшафт, который предшествовал развитию
культурного ландшафта или существовал в самом начале его. Однако
это понятие часто применяется не только для «естественного ландшафта
в момент времени, непосредственно предшествующий его превращению
в культурный ландшафт», но и в более широком смысле для ранних (боль-
шей частью доисторических) стадий развития культурного ландшафта.
При исследовании растительности желательно понимание его в более
узком смысле. Реконструкция картины растительности первобытного ланд-
шафта (праландшафта) интересует прежде всего историю культуры, исто-
рию населенных пунктов и историческую географию. Методы исследова-
ния различны в зависимости от возраста того культурного ландшафта,
первобытный ландшафт которого должен быть воссоздан. При изучении
молодых культурных ландшафтов характеристику растительности пред-
шествовавших первобытных ландшафтов еще можно найти в путевых днев-
никах, описаниях путешествий или других источниках того времени.
Реконструкция более древних ландшафтов, например первобытного
ландшафта донеолитического заселения Европы, должна основываться на
186
Единицы растительности в ландшафте
Таблица 7
Понятия естественного, культурного и первобытного ландшафтов
Геологическая история Земли —> Современность
Бывший естественный ландшафт в разные периоды Современный естественный ландшафт
Первобытный ландшафт (естественный ландшафт перед его преобразова- нием в культурный ландшафт) Ранний культурный ландшафт (доисторический, исторический) Современный культурный ландшафт Реальный ландшафт
Ранний Современный Потенциальный естественный ланд- шафт (теоретический естественный ландшафт для определенного мо- мента времени, прошлого или настоящего) Восстановленный (теоретически возможный) естественный ландшафт области реального культурного ландшафта
Рис. 46. Распределение растительно-
сти Вестфалии в период от 500 г. до
и. э. до 1000 г. н. э. Попытка рекон-
струкции на основе пыльцевого анализа
(Budde, 1950).
1 — богатый видами буковый лес с участ-
ками остепненных пустошей; 2 — бедный
видами буковый лес иа западе с большой
примесью дуба; з — богатый видами буко-
вый лес, но беднее, чем 1\ 4 — дубово-гра-
бовый лес; 5 — дубово-березовый лес;
6 — верховое болото, окруженное низинным
болотом; 7 — заболоченный лес и низинные
болота в долине; 8 — пойменный лес.
методах палеоботанических исследований, пыльцевых диаграммах, дати-
ровании растительных остатков. Последние, однако, могут осветить лишь
отдельные моменты географии растительности. Поэтому для решения
Таблица 8
История среднеевропейской растительности и сопоставление ее с этапами
развития материальной культуры (по Фирбасу)
Господствующая раститель- ность Климатические пери- оды поздне- и после- ледниковой истории Средней Европы Абсолют- ная хро- нология Доисторический период
Культурные и полу куль- турные сообщества, ин- тенсивно используемые леса X Позднее «послетеп- лое» время (субат- лантический пе- _„.РИОД) + 1000
Буковые леса и смешан- ные леса с преобладани- ем бука (время бука) IX Древнее «послетеп- лое» время (суб- атлантический пе- риод) Начало н. э. Железный век
Превращение смешанных дубовых и горных пих- товых лесов в буковые леса (время буковых и смешанных дубовых ле- сов) Смешанные дубовые леса, в горах частично с пих- той (время смешанных дубовых лесов) й о н VIII Позднее теплое вре- мя- (суббореаль- ный период) —1000 —2 000 Бронзовый век
со К S о н й ф ч ф ч VII Среднее теплое вре- мя, поздняя часть (атлантический период) —3 000 —4 000 —5 000 —6 000 Неолитические культуры Поздние мезо- литические культуры Древние мезо- литические культуры
Богатые лещиной сосно- вые и смешанные дубо- вые леса или кустар- о К VI Среднее теплое вре- мя, древняя часть (атлантический период) V Раннее теплое вре- мя (бореальный период)
ники (время лещины) Березовые и сосновые леса (время березы и сосны) । IV «Предтеплое» время (предбореальный период) —7 000 —8 000 Культуры
Безлесные сообщества и светлые березовые и сос- III Поздний субаркти- ческий период —9 000 —10 000 Аренсбурга и Лингби
новые леса (новое дриа- довое и тундровое время) Сосновые и березовые леса ;ая эпоха II Средний субаркти- ческий период
Безлесное сообщество, к концу опять светлые березовые и сосновые леса (древнее дриадовое и тундровое время) Безлесные сообщества и О а й ф ч ф и tc со О К 16 Древний субаркти- ческий период 1а Древнейшее безлес- ное время —11000 —12 000 —13 000 —14 000 —15 000 Магдаленские культуры (Гамбургская ступень и > т. д.)
188
Единицы растительности в ландшафте
вопроса о пространственных различиях в первобытном ландшафте необ-
ходимо применять исследовательские методы самых различных отраслей.
Определенные контуры размещения, в частности положение границ, не
меняются на протяжении длительного времени и могут быть реконструи-
рованы, исходя из современного естественного размещения в пространстве
и данных из области истории климата, почв и растительности. Немаловаж-
ную роль при воссоздании первобытного ландшафта играет гипотеза
(рис. 46). История растительности послеледникового периода лучше
всего известна для тех областей, где широко распространены болота,
что облегчает применение пыльцевого анализа. Фирбас (Firbas, 1949)
попытался реконструировать историю растительности Средней Европы,
сопоставив ее с этапами развития среднеевропейского ландшафта.
Потенциальным естественным ландшафтом
(ранее — «теоретический естественный ландшафт») мы называем естест-
венный ландшафт, который занимал бы место современного культурного
ландшафта, если бы последний был полностью предоставлен природе
и человек не оказывал бы на него никакого воздействия. Вопрос о потен-
циальном естественном ландшафте в этом смысле может быть разрешен
лишь экспериментально на небольших примерах и имеет немаловажное
значение для исследования растительности и ее практического использо-
вания.
Часть третья
ПРОСТРАНСТВЕННОЕ ПОДРАЗДЕЛЕНИЕ
РАСТИТЕЛЬНОСТИ
Распространение единиц
растительности
ТОПОГРАФИЯ И КРУПНОМАСШТАБНОЕ
КАРТИРОВАНИЕ РАСТИТЕЛЬНОСТИ
Если не учитывать небольшого числа предшественников (Sendtner, 1854; Flahault,
1894; Drude, 1907), крупномасштабное картирование растительности начало развивать-
ся только после появления возможности идентифицировать единицы растительности
на фитоценологической основе. Вслед за изображением более мелких участков мест-
ности, например отдельных болот в масштабе от! 1 : 5000 до 1 : 25 000 (Gross, 1914;
Bertsch, 1918; Hueck, 1925, 1928; Gams, Ruoff, 1929) последовали попытки обработать
целые мензульные листы в виде карт растительности, например Hueck, 1931, 1932,
1934; Fischer, 1933; Oberdorfer, 1936, 1938; Faber, Heupel, 1938; Schlenker, 1940, и мно-
гочисленные неизданные листы Федерального управления картирования раститель-
ности.
В последнее время крупномасштабное картирование растительности было начато
и в некоторых других странах, и в отдельных случаях оно сделало большие успехи
(Ogilvie, 1937; Troll, 1939; Braun-Blanquet, Emberger, Molinier, 1947, INEAC, 1954
и след.). Библиография (Tuxen, Hentschel, 1955) обработанных в ФРГ до 1954 г. карт
растительности охватывает 216 напечатанных и еще большее число неопубликованных
карт. К сожалению, еще не существует обзора всех имеющихся в настоящее время в раз-
личных странах карт растительности.
Крупномасштабное картирование растительности осуществляется раз-
ными способами. На большинстве старых карт не делались различия
между реальной и потенциальной естественной растительностью. Часто
на одной и той же карте леса представлены с иных точек зрения, чем все
другие растительные сообщества, а некоторые единицы растительности,
например сообщества сорняков на используемых в сельском хозяйстве
площадях, не рассматривались вовсе. В последнее время стараются нано-
сить на карты все единицы растительности равномерно, насколько это
допускает масштаб (рис. 47).
Наряду с крупномасштабными картами все чаще встречаются и карты в масшта-
бах от 1 : 50 000 до 1 : 200 000, на которых частично еще допускается топографическое
разграничение (Troll, 1926; Тйхеп, 1931; Scamoni, 1935; Schwickerath, 1937, 1938;
Buker, 1939; Schmid, 1939). Однако эти карты и карты более мелкого масштаба, как пра-
вило, сильно генерализованы и ограничиваются общим воспроизведением потенциаль-
ной естественной растительности (Troll, 1926; Тйхеп, 1934; Oberdorfer, 1937; Hueck,
1937, 1938; Scharfetter, 1938; Louis, 1939; Bobek, 1951).
192
Пространственное подразделение растительности
Топографическое изображение реальных единиц растительности воз-
можно, как правило, лишь в масштабах, более крупных, чем 1 : 100 000,
как исключение — в масштабе 1 : 200 000.
Рис. 47. Карта растительности по профилю через Зенне (по неопубликованному
цветному оригиналу Ломейера, снятому в масштабе 1 : 25 000).
1 — ольховая топь; 2 — ольха — кочедыжник — сосновый лес; 3 — злаки — жимолость — со-
сновый лес; 4 — злаки, щитовник австрийский — сосновый лес; 5 — клюква — сосновый
лес; 6 — злаки — сосновый лес; 7 — ягодные кустарнички — сосновый лес; 8 — лишайни-
ки — вереск — сосновый лес; 9 — сухая песчаная пустошь; ю — жабник — вереск;
11 — сухая песчаная пустошь со злаками; 12 — типчаково-тимьяновый дерн; 13 — типчаково-
жабниковый дерн; 14 — заросли болотной калужницы — злаковые луга; 15 — лесные сит-
никовые болота; 16 — сообщество росички; 17 — сообщество паслена; 18 — сообщество
куриного проса; 19—влажное сообщество куриного проса.
Рис. 48. Растительность острова Бальтрум (Тйхеп, 1956).
1 — передовые дюны, поросшие ситниковым пыреем; 2 — основные дюны, поросшие пес-
колюбом, и дюны, покрытые шлемником; 3 — мелко-злаковые дюны, покрытые булавоно-
сцем и дерном из полевицы; 4 — кустарники из облепихи и дюнных ив; 5 — болотные сообще-
ства в междюнных понижениях; 6 —ситниковые прибрежные отложения (ватты); 7 —луга из
ситника жерарда; 8 — бескильницевые луга.
Специальные фитоценологические карты, воспроизводящие распро-
странение всех растительных сообществ реально существующей расти-
тельности, являются незаменимой основой как для научных исследований
в области географии растительности, так и для практического использова-
ния результатов изучения растительности при освоении почв. Поэтому
Распространение единиц растительности
193
в последние годы подобное картирование получило широкое распро-
странение (рис. 48).
По крупномасштабным картам растительных сообществ можно соста-
вить представление о размещении растительных сообществ и потенциаль-
ной естественной растительности, если растительные сообщества хорошо
описаны с точки зрения их формационного характера, а динамика их
настолько изучена, что могут быть установлены потенциальные заключи-
тельные сообщества.
В качестве примера современной карты тропической растительности на рис. 52
приводится несколько стилизованная и уменьшенная часть цветной карты окрестностей
Каниамы, снятой Муллендером в оригинале в масштабе 1 : 50 000.
АРЕАЛЫ, ГРАНИЦЫ АРЕАЛОВ,
ПРИЧИНЫ ИХ ВОЗНИКНОВЕНИЯ И ФУНКЦИОНАЛЬНЫЕ
СООТНОШЕНИЯ
Среди работ по географии растительности особенно многочисленны
работы, посвященные изучению отдельных единиц растительности и выяв-
лению причин их распространения. Подобные аналитические иссле-
дования являются обязательной предпосылкой для получения синте-
зированного воспроизведения растительности отдельных стран или
ландшафтов и для подразделения земной поверхности по характеру ее
растительного покрова. Неотъемлемой частью этих работ является также
изучение пространственного распределения единиц растительности с их
последующим картированием.
Исследование закономерностей распространения растительности начи-
нается с изучения отдельных единиц растительности (растительных
сообществ, формаций). Однако в центре внимания можно при этом поста-
вить и внешние факторы или другие моменты.
ГРАНИЦЫ РАСПРОСТРАНЕНИЯ ОТДЕЛЬНЫХ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ ИЛИ ФОРМАЦИЙ
Вопрос о распространении определенных единиц растительности
часто оказывается поводом для исследования причинных связей про-
странственного подразделения растительности. Натолкнуть на это могут
всевозможные специальные вопросы, например некоторые геологиче-
ские проблемы при изучении верховых болот (Keilhack, 1915; Krenkel,
1920; Post, 1926).
Монографическое описание растительного сообщества неизбежно
приводит к вопросу о его распространении (например, Braun-Blanquet,
1925, 1936), причем проблемы распространения часто выдвигаются на
первый план (Lindquist, 1931; Riibel, 1932; Meusel, 1941, 1942, 1943,
1951 —1952). Аналогичное положение создается при рассмотрении отдель-
ных растительных формаций (Waibel, 1921; Troll, 1936; Richards, 1952)
и господствующих на данном пространстве потенциальных естественных
растительных формаций более обширных районов земной поверхности
(Louis, 1939; Bobek, 1951). Тесно связаны с этим проблемы высотных
поясов (Volkens, 1897; Du Rietz, 1925; Troll, 1941, 1948; Tschermak,
1942; Markgraf, 1949), высотных пределов и общие вопросы границ расти-
тельности (Ratzel, 1889; Scharfetter, 1924; Markus, 1932).
Объяснение положения границы леса и отдельных древостоев отно-
сится к классическим проблемам географии растительности и вызвало
194
Пространственное подразделение растительности
появление обширной литературы (Kerner, 1964; Танфильев 1895, 1911;
Rikli, 1904; Brockmann-Jerosch, 1919; Кбрреп, 1919, Regel, 1920; Hjelm-
quist, 1940; Bliithgen, 1942, 1943; Hermes, 1955).
Рис. 49. Высота верхней границы леса (Hermes, 1955).
Аналогично положение и с вопросом о причинах возникновения и распростране-
ния травянистых пространств. «Степная проблема» (Miller, 1892; Краснов, 1894;
Танфильев, 1905; Woodard, 1924; Jones,1927; Schmieder, 1929; Stocker, 1929; Walter,
1939; Высоцкий, 1932; Wilhelmy, 1950), «проблема саванн» (Schimper, 1898; Perrier
de la Bathie, 1921; Troll, 1936; Roseveare, 1948; Duvigneaud, 1953; Walter, 1954; Dev-
red, 1956) и «проблема лугов» (Танфильев, 1925; Gradmann, 1932; Meusel, 1940; Zoller,
1954) — все они способствовали проведению многих исследований в области геобота-
ники и географии растительности.
ПРИЧИНЫ ПРОСТРАНСТВЕННОГО РАЗМЕЩЕНИЯ
РАСТИТЕЛЬНОСТИ
Пространственное распределение растительности обусловливается
в основном тремя группами сил, или тремя причинами, о которых частично
уже шла речь выше:
а) дифференцированием растительного мира, обусловленным биогене-
тическими причинами, связанными с историей Земли;
б) пространственным размещением местообитаний и различиями потен-
циальных пространств произрастания растительности, обусловленными
биотическими факторами;
в) воздействием человека, благодаря которому местная раститель-
ность часто приобретает особые черты, связанные с историей человеческой
культуры.
Причины распространения растительности рассматриваются в общем
виде в учебниках и справочниках по геоботанике и географии раститель-
ности, а со специальных точек зрения — в многочисленных монографиях
(Wallace, 1892;Ratzel, 1901; Kraus, 1911; Markus, 1936; Friederichs, 1937;
Распространение единиц растительности
195
Тйхеп, 1939; Thienemann, 1941; Bocher, 1949; Krause, 1950; Dansereau,
1952, 1956).
Большинство работ посвящено исследованию влияния климата на
растительность; здесь преимущественный упор сделан или на общий
характер климата (Grisebach, 1838; Philippson, 1903; Weberbauer, 1906;
Livingston, Schreve, 1921; Ruhle, 1928), или на его отдельные элементы:
температуру (Passarge, 1921), количество осадков и их распределение
(Hettner, 1893), силу и направление ветров (Bernbeck, 1911; Saxton, 1924;
Troll, 1952), испарение (Fuller, 1912), или же на комплексное влияние
нескольких факторов: например, океанический или континентальный
характер климата (Brockmann-Jerosch, 1913, 1919; Troll, 1925, 1951;
Teleki, 1930; Tschermak, 1944; Philippson, 1947; Bocher, 1954), ритм
климата (Scharfetter, 1922; Lauer, 1952; Bunning, 1955; Lautensach,
1955; Troll, 1955; Schwabe, 1956), непериодические крайности климата
(Ernst, 1934; Huber, 1947; Ludi, Zoller, 1948; Schmithusen, 1948; Geiger,
1951; Horn, 1952; Knapp, 1952; Kroner, 1952), или на климатически обус-
ловленное образование зон и их границ (Werth, 1927; Wissmann, 1948;
Bobek, 1952). Исследования различий экспозиций (Huttenlocher, 1923;
Тйхеп, 1954; Raabe, 1955) в конце концов также сводятся к изучению кли-
матических влияний.
Значению почвенных факторов для пространственных различий
растительности посвящены многочисленные исследования и монографии
(Schimper, 1891; Cowles, 1899; Frodin, 1919; Keller, 1925—1926; Baeyens,
1938; Vageler, 1938; Braun-Blanquet, 1947; Braun-Blanquet, Palmann,
Bach, 1954), освещающие вопрос с разных точек зрения. Однако общего
всеобъемлющего географического представления о взаимосвязях между
почвами и растительностью в современном почвоведении еще нет.
ПРОСТРАНСТВЕННОЕ РАЗМЕЩЕНИЕ РАСТИТЕЛЬНОСТИ
В ЗАВИСИМОСТИ ОТ ПРОЧИХ ЯВЛЕНИЙ
Очень часто ботанико-географические исследования определяются
вопросами, которые касаются не только самой растительности, но и других
объектов, зависящих от нее или поддающихся изучению только через
знание растительности. Это прежде всего вопросы хозяйственного про-
изводства и практического использования растений, которые «свой гео-
графический аспект приобретают только благодаря тому, что органиче-
ское сырье добывается не всюду и прежде всего — не везде в одинаковом
количестве и одним и тем же способом...это уже моменты, относящиеся
частично к географии растений и животных... Без знания экологии расте-
ний невозможно правильно объяснить картину распространения куль-
турных растений, или важного с точки зрения хозяйственных послед-
ствий положения естественной границы лесов, или размещения планта-
ций, хлебных культур, скотоводства. Климат во всех этих случаях являет-
ся начальным звеном в цепи причин и воздействий, а форма хозяйства —
звено заключительное; связывает их география растений и животных.
Без этого среднего звена цепь распадется (Gradmann, 1919).
Исследования такого рода касаются, например, центров происхож-
дения и распространения полезных растений (рис. 50) (Hock, 1890, 1899—
1900; Вавилов, 1928; Wissmann, 1957), культурной ценности определен-
ных единиц растительности, ботанико-географических зон (Schroter,
1901; Sapper, 1915, 1923; Troll, 1925; Obst, 1932; Waibel, 1937), естест-
венных основ производительности сельского хозяйства (Neumann, 1908;
196
Пространственное подразделение растительности
Troll, Gams, 1931; Domes, 1936; Gradmann, 1936; Valkenburg, 1938; Tuxen,
1940; Pallmann, Frei, 1949; Filzer, 1951; Ellenberg, 1954) и лесного хозяй-
ства (Jentsch, Biisgen, 1909; Rubner, 1934; Кostler, 1950). При исследо-
вании культурного ландшафта также возникают аналогичные вопросы
(Schmithiisen, 1934, 1942; Welte, 1934; Barenscher, 1939; Baasen, 1940).
С точки зрения исследований культурного ландшафта особенный интерес
Рис. 50. Основные центры происхождения культурных растений (по Вавилову).
представляют те направления и интенсивность воздействия человека на
растительность, выявление которых может оказаться решающим для
понимания особенностей какого-либо участка земной поверхности.
Многие вопросы, как, например, проблемы пустошей, пампасов и саванн, прохо-
дящие красной нитью через исследование ландшафтов, не могут быть решены с помощью
только чисто эколого-физиономического рассмотрения. В решении некоторых из них
нередко помогали исторические и географические старые описания той или иной мест-
ности. Но такие случаи — скорее исключение. Исторический метод редко удается при-
менить к большим площадям. «Социологическая геоботаника» в сочетании с почвове-
дением предоставляет возможность методами естествознания прийти к более надежным
результатам при решении подобных проблем.
Поэтому во многих специальных работах в области географии расти-
тельности заметно стремление определить потенциальную естественную
растительность по характеру формаций. С помощью одних только эколого-
физиономических методов достичь этой цели зачастую не удается. Тре-
буется одновременно флористико-фитоценологическое исследование дина-
мики растительности.
Влияние растительности на климат часто оспаривается, при этом
в более ранних работах влияние леса на макроклимат нередко переоце-
нивалось (ср. Geiger, 1932; Liidi, Zoller, 1948). В последнее время придают
большое значение воздействию растительности на процессы почвообра-
зования и эрозии (Erichson, 1923; Beadle, 1948; Haase, 1948; Mensching,
1951). Связям растительности с водным режимом ландшафта тоже посвя-
Распространение единиц растительности
197
щен ряд исследований (Reindl, 1903; Roll, 1942; Burger, 1948; Keller, 1948;
Berger-Landefeldt, 1949; Tuxen, 1954; Kausch, 1957).
Знание основ взаимоотношения растительности с местообитанием
нередко помогает использовать растительность как индикатор особен-
ностей местообитания (Valle, 1929; Tuxen, 1935, 1951; Ellenberg, 1948;
Krause, 1950) для изучения характера или интенсивности процессов
Р и с. 51. Фронт наступления весны 27 апреля 1934 г. в Люк-
сембурге как пример синоптической фенологической карты
(Schmithusen, 1940).
1 — цветет конский каштан; 2 — цветет сирень; з — цветет яблоня;
4 — цветет терн; 5 — цветет вишня; 6 — зацветание вишни.
эрозии (Schmid, 1925; Friedel, 1935; Hueck, 1951), а также для установле-
ния прежнего положения ледников и датировки отступания их (Friedel,
1934). Решению вопроса о том, обусловлена ли современная граница леса
естественными причинами или влиянием человека, тоже могут с успехом
помочь используемые в качестве свидетелей растительные сообщества
и почвенные биоценозы (Aichinger, 1930; Franz, 1950).
Часто проблемы пространственного распределения растительности
исследуются главным образом с целью получения критериев для разгра-
ничения климатических областей и климатических высотных ступеней
(Корреп, 1903; Troll, 1925, 1950, 1955). «Высотные ступени в горах клима-
тически никак нельзя охарактеризовать иначе, как сменой поясов расти-
198 Пространственное подразделение растительности
тельности» (Gradmann, 1919). Фенология растений преследует
частично сходные цели, прежде всего с точки зрения выявления юнких
микроклиматических различий (Hiltner, 1926; Schmithusen, 1940; Rosen-
kranz, 1948; Schnelle, 1949, 1950, 1955; Morgen, 1952; Lauscher, 1955).
Фенология наблюдает, регистрирует и описывает пространственные
различия в процессе сезонной смены явлений в жизни растений, живот-
ных и в сельском хозяйстве. С помощью изохронных карт, дающих начало
периодических явлений, например начало весны (Ihne, 1905; Schrepfer,
1923), или синоптических фенологических карт, отражающих различия
в развитии растений к определенному моменту времени (Schmithusen,
1940; Ellenberg, 1955), можно наглядно воспроизвести еще более тонкие
градации климата (рис. 51). Растения служат при этом в некоторой сте-
пени измерительными приборами, которые имеются всюду и чувствитель-
но реагируют на совокупное воздействие погоды.
Изучение климатов прошлого помогает также глубже понять особен-
ности распространения современной растительности (Dansereau, 1944;
Wilhelmy, 1952; Schmithusen, 1956).
Структура пространственного
распределения растительности
(Фитоценологически обоснованная хорология растительности)
ОБЩАЯ ПРОБЛЕМА. ПРИНЦИПЫ И ВОЗМОЖНОСТИ
ПРОСТРАНСТВЕННОГО ДЕЛЕНИЯ
С ТОЧКИ ЗРЕНИЯ ГЕОГРАФИИ РАСТИТЕЛЬНОСТИ
Существуют две принципиально различные возможности простран-
ственного расчленения растительности, а именно:
а) по критериям, не связанным непосредственно с растительностью,
например по климатическим зонам или отдельным факторам местообита-
ния (растительность побережий, растительность скал, заболоченных
почв и др,);
б) по самой растительности, например по растительным сообще-
ствам или по характеру формаций реально существующей потенциальной
естественной растительности.
Для охвата возможно большего пространства земной поверхности
в каждом отдельном случае приходится прибегать к обобщению. Генера-
лизация может быть достигнута путем: а) учета лишь некоторых домини-
рующих черт при отбрасывании всех других; б) создания комплексных
понятий для обозначения совокупности сообществ или формаций и в) ог-
раниченного изображения потенциальной естественной растительности
без учета некоторых реально существующих пространственных раз-
личий.
География склоняет исследователя от рассмотрения охватывающих
всю Землю климатических растительных зон и растительных формаций,
преобладающих в ландшафтах на обширных площадях, перейти к выделе-
нию пространственно более узких областей растительности. Всякая
попытка представить реально существующую растительность на мелкомас-
штабных обзорных картах требует сильной генерализации, основанной
только на самых общих понятиях; в результате нарушается принцип
единства изображения. Так, например, на составленной Мауллем карте
растительности Земли понятие «культурные земли» распространяется
на обширные территории без выделения сохранившейся в пределах этих
территорий, хотя бы]в виде остатков, естественной растительности.
Фитоценология, наоборот, исходит из растительных сообществ и их
локального распространения и с помощью крупномасштабного картирова-
ния растительности постепенно приходит тоже к выделению более обшир-
ных участков. Если рассматривается растительность всей Земли, то
и геоботаника (например, Брокман-Ерош) использует для разграничения
зон растительности формации, определяемые эколого-физиономическими
критериями, С другой стороны, география при крупномасштабных иссле-
200 Пространственное подразделение растительности
дованиях часто вынуждена обращаться к отдельным, определяемым флори-
стическими критериями, растительным сообществам. Поэтому должен быть
принят такой принцип структурного подразделения растительности
и найдена такая система понятий, которая была бы пригодной и для
решения географических задач. Следует стремиться к тому, чтобы обе
противоположные исходные точки зрения проявляли тенденцию к сбли-
жению и объединились бы в единой системе. Однако в настоящее время
делаются еще только первые попытки такого подхода.
ДЕЛЕНИЕ РЕАЛЬНО СУЩЕСТВУЮЩЕЙ РАСТИТЕЛЬНОСТИ
НО РАСТИТЕЛЬНЫМ СООБЩЕСТВАМ
Там, где комбинация растительных сообществ, характерная для
данной местности, уступает место иной комбинации, начинается с точки
зрения географии растительности уже другая пространственная единица.
Как растительные сообщества разграничиваются одно от другого на осно-
вании характерной для них совокупности видов, так можно различать
и растительные области по доминирующим, ведущим1
и постоянным сообществам. Характерный для данной области набор
сообществ (Gesellschaftsinventar) образует «ландшафтный расти-
тельный комплекс». Он состоит прежде всего из растительных
сообществ, но по эколого-физиономическому характеру последних может
восприниматься и как «комплекс растительных формаций».
Однако для разграничения областей растительности и прежде всего
в культурных ландшафтах целесообразно и даже необходимо исходить из
комплекса сообществ. По набору сообществ и его пространственному раз-
мещению мы различаем мельчайшие естественные пространственные еди-
ницы растительности —«р а с т и т е л ь н ы е районы» (Wuchsdistri-
kte), которые объединяются в более крупные единицы: округа (Bezir-
ke), провинции (Provinzen), регионы (Region), царства (Reich).
С увеличением ранга пространственных единиц увеличивается и набор
сообществ. Усложняется пространственная структура и, соответственно,
ее воспроизведение. Чем мельче масштаб рассмотрения, тем большего
обобщения требуют структурные элементы, определяющие разграничения.
При этом весьма важно выбрать именно тот признак, который наиболее
характерен для единицы данного порядка. Эти методы в подробностях
и частностях еще не разработаны. В зависимости от масштаба на первом
плане может оказаться тот или другой признак, причем для определенных
элементов структуры можно прибегнуть к критериям, которые использо-
вались для соседних величин высшего или низшего порядка. Пространст-
венные единицы более высокого ранга (округа, провинции, регионы,
царства) всегда состоят из нескольких смежных единиц низшего ранга,
родственных по составу сообществ. Они характеризуются присущими дан-
ным подразделениям единицами сообществ; округа — ассоциациями, про-
винции — союзами сообществ, регионы — порядками, а растительные
царства — классами.
1 Ведущие сообщества (Lcitgesellschaften), в понимании зарубежных геоботани-
ков (Шретцермайер),— группа лесных сообществ, флористически и экологически
сходных и имеющих одинаковый характер во всем, кроме основной древесной породы.
Ведущие сообщества в географически ограниченной области (например, в раститель-
ном округе) занимают часто такую большую площадь, что могут являться определяю-
щими для нее.— Прим. ред.
Структура пространственного распределения растительности
201
ДЕЛЕНИЕ РЕАЛЬНО СУЩЕСТВУЮЩИХ РАСТИТЕЛЬНЫХ
СООБЩЕСТВ НО ХАРАКТЕРУ ФОРМАЦИЙ
В естественном ландшафте расчленение по характеру формаций про-
извести, как правило, нетрудно. Кроме понятия «растительные формации»,
здесь применяют также понятие «комплексы растительных формаций»,
причем названия отдельных растительных формаций часто являются сим-
волом таких комплексов.
Р и с. 52. Растительные сообщества в поясе саванн, па северо-западной
границе Катанги. Вырезки из карты Капиамы (7°30' ю. ш., 24° в. д.).
Схематизировано. Подлинник содержит на том же участке 14 обозначений
(Mui lenders, 1955).
1 — вечнозеленый тропический дождевой лес (галерейный лес); 2 — дождезеленый
сухой лес (в области саванн — как остатки потенциальной естественной раститель-
ности); з — затопляемые саванны; / — влажные варианты Hyparrhenion conf inis-
саванн; .5 - Hyparrhenion con/inis-саванны, оптимальные варианты (с островками клн-
максного леса); 6 — слабо облесенные Andiopogonion schirt'nsis-саваппы (с переходом
в Hyparrhenion con/гпгя-саванны); 7 — IIyparrhenion con /inis-саванны, различные субас-
соцнацшт; 8 — возделываемые площади.
Для культурных ландшафтов тоже существуют понятия, которые,
подчеркивая одну физиономически выдающуюся черту, характеризуют
комплекс растительности определенного ландшафта. Такие типовые поня-
тия могут быть очень емкими по содержанию и поэтому пригодными для
широкого обозрения, когда важнее краткость выражения, чем подробность
описания.
Па примере Капиамы (рис. 52) типизацию выделенных сообществ по характеру
формаций дает комплекс галерейных лесов, состоящий из высокозлаковых сообществ
(саванна), сухих лесов, тропического дождевого леса и болот разного вида. Обобщение
понятий приводит к типу растительного комплекса —«влажная саванна» (Егер). Однако
202
Пространственное подразделение растительности
подобная генерализация возможна лишь при обзоре растительности целого континента
или даже всей земной поверхности. Для крупномасштабного деления растительности
оно значения почти не имеет, потому что при таком понятии вся область относится
к одному климатическому поясу растительности.
ДЕЛЕНИЕ ПО ПОТЕНЦИАЛЬНОЙ ЕСТЕСТВЕННОЙ
РА СТИТЕЛЬНОСТ И
Естественные жизненные пространства растительности могут быть
охарактеризованы и разграничены или фитоценологически по потенциаль-
ному естественному заключительному сообществу, выделенному на
основании реально существующей растительности, или же эколого-физио-
номически по характеру формаций этого заключительного сообщества.
Большинство мелкомасштабных карт, которые воспроизводят «обла-
сти естественной растительности», является — поскольку речь идет о куль-
турном ландшафте — картами потенциальной естественной растительности.
Это относится и к обзорным картам поясов растительных формаций или
климатических зон растительности Земли, если в последних не имеет
места чисто климатическое разграничение.
Представленные на примере Каниамы (рис. 52) единицы растительности могут
быть объединены или по экологической гомологии сообществ, или по их отношению
к потенциальной естественной растительности в зависимости от характера динамиче-
ских взаимосвязей растительности. Исследование сукцессии сухого леса, сохранивше-
гося в наше время только в виде остатков, показывает, что он представляет собой заклю-
чительное сообщество, замещающими сообществами которого являются травянистые
сообщества (злаковники). Те и другие вместе относятся, следовательно, к одной и той
же пространственной единице растительности.
ДЕЛЕНИЕ РАСТИТЕЛЬНОСТИ,
ОСНОВАННОЕ НА КОМПЛЕКСАХ СООБЩЕСТВ
ИЗОБРАЖЕНИЕ ПРОСТРАНСТВЕННОЙ СТРУКТУРЫ
С ТОЧКИ ЗРЕНИЯ ДИНАМИКИ СООБЩЕСТВ
Структуре «флизе» соответствует размещение основных единиц про-
странства, представленных динамикой сообществ,слагающих ландшафт.
Карта структуры «флизе» вместе с кругами сообществ, воспроизве-
денными в легенде в качестве отдельных типов «флизе», дает полную
и наглядную картину пространственной структуры с точки зрения динами-
ки сообществ (см. цветную карту в конце книги). На примере равнины
у подножия хребта Венн (Schwickerath, 1954) показано размещение в про-
странстве восьми типов «флизе», отмечены отдельные типы «флизе» по
характеру их местообитаний и показаны относящиеся к ним восемь кру-
гов сообществ от 1 до 6 с 9, 17, 9, 28, 8, 12, 4 и 7 сообществами соответ-
ственно. Участки 2а, 3, 4 и 5, строго говоря, уже являются структурами
«флизе», соответственно упорядоченными комбинациями схем сук-
цессии нескольких кругов сообществ с различными заключительными
сообществами. Поэтому общая структура динамики сообществ рассматри-
ваемой области содержит в целом одиннадцать заключительных сооб-
ществ, а именно четыре сообщества дубово-грабовых лесов, три варианта
гальмейных сообществ, два варианта ольховой топи и по одному сообще-
ству букового леса и леса из серебристой ивы.
Структура пространственного распределения растительностей
203
ОБЛАСТИ ЕСТЕСТВЕННОЙ РАСТИТЕЛЬНОСТИ
РАЗНОГО РАНГА, ОСНОВАННЫЕ НА НАБОРЕ СООБЩЕСТВ
Растительный район
Растительным районом мы называем самую низшую пространственную
единицу растительности, характеризующуюся собственной структурой
динамики сообществ. Предгорная равнина Высокого Венна и является
с точки зрения географии растительности именно таким растительным
Рис, 53, Карта потенциальной естественной растительности по
среднему течению реки Везер с границами растительных районов:
(I) — Дипхольцская болотистая низина и (IT) — Песчаная равнина
Сике (по Ломсйеру).
1 — верховые болота и березовые болота; 2 — ольховая топь с участками
березовых болот и ольхово-дубовым лесом; з — сырой дубово-березовый
лес с участками ольховой топи; 4 — сухой и сырой дубово-березовый лес;
5 — буково-дубовый лес и др.; 6 — сырой дубово-грабовый лес, чередую-
щийся с влажным дубово-березовым лесом; 7 — орехово-вязовый лес и др;
8 — влажные буковые смешанные леса.
районом. Изображение типичной структуры «флизе» вместе с относящи-
мися к ней кругами сообществ (см. цветную карту) дает нам в этом случае
временное пространственное строение растительности и может рассмат-
риваться как формула растительно-динамической структуры пространства
и набора сообществ данного растительного района.
204
Пространственное подразделение растительности
Весь комплекс набора сообществ данного растительного района
с характерной для него пространственной структурой мы называем «ком-
бинацией сообществ» (Gesellschaftspulk). Она является, таким образом,
комплексом кругов сообществ, пространственно ассоциированных в дан-
ном ландшафте. Нижняя карта схемы воспроизводит в северо-западном
углу пространственную структуру «комбинации сообществ» на равнине
у подножия хребта Венн, состоящую из восьми изображенных здесь кругов
сообществ, а в юго-восточном углу — структуру комбинации сообществ
склонов хребта Венн, состоящую из шести кругов сообществ ступенчатого
склона и четырех кругов сообществ облесенных долин (Schwickerath,
1954). Между ними проходит граница растительных районов. Сравнивая
«комбинации сообществ», можно установить и охарактеризовать степень
родства растительных районов.
Районы обычно настолько малы, что они не содержат свойственных
только им ассоциаций, а имеют в крайнем случае лишь эндемичные субас-
социации или местные варианты сообществ.
Строение «комбинации сообществ» можно пояснить на примере двух раститель-
ных районов по карте, составленной Ломейером (рис. 53 и табл. 9).
Пример I. «Комбинация сообществ» растительного района Дипхольцская боло-
тистая низина (Нижняя Саксония).
Характерная пространственная структура складывается из пяти типов «флизе»
с одним кругом сообществ в каждом; эти типы различаются на карте и названы по их
потенциальным естественным растительным сообществам. Три из пяти кругов сообществ
господствуют попеременно на больших площадях, а именно:
1. Круг сообществ дубово-березового леса со следующими характерными сообще-
ствами, ограниченными в пределах «комбинации сообществ» этим типом «флизе»:
а) естественные сообщества:
Querceto roboris-Betuletum,
Sparganium angustifolium-Sphagnum obesum (ass.),
Spergula vernalis-Corynephoretum canescentis,
Illecebretum verticillati,
Rubus gratus (ass.);
б) антропогенные сообщества:
сосновые насаждения (на месте Querceto roboris-Betuletum), Panicum ischaemum
(ass.).
2. Круг сообществ березовых болот и верховых болот с характерными сообще-
ствами:
Betuletum pubescentis typicum,
S phagnetum papillosi,
Sphagnetum medii subatlanticum,
Scheuchzerietum palustris,
а также дальнейшими 13 естественными сообществами и следующими замещающими
сообществами:
Calluneto-Genistetum,
Panicum crus galii-Spergula arvensis (ass.),
Teesdalio nudicaulis-Arnoseretum minimae.
3. Круг сообществ ольховой топи (на небольших площадях чередуется с набором
кругов сообществ, указанных в пункте 2) со следующими естественными сообществами,
ограниченными в пределах «комбинации сообществ» этим типом «флизе»:
Cariceto elongatae-Alnetum medioeuropaeum,
Еilipenduleto-Geranietum palustris, >
Cicuta virosa-Carex pseudocyperus (ass.),
Cariceto injlatae-vesicariae,
Структура пространственного распределения растительности
205
Таблица 9
Сопоставление растительных районов — (I) Дппхольцская болотистая низина
и (II) Песчаная равнина Сике — по классам сообществ,
участвующих в их «комбинациях сообществ»
Классы сообществ Число ассоциаций из соответствующих классов
только в I только во II в I и во II всего ассоциа- ций
Corynephoretea canescentis 1 — — 1
Vaccinietea uliginosi 1 — — 1
Oxy cocco-Sphagnetea 2 — 1 3
Epilobietea angustifolii 1 — 1 2
Quercetea robori-petraeae 1 1 — 2
Phragmitetea — 5 5 10
Potametea — 4 1 5
Nardo-Callunetea — 2 2 4
Alnetea glutinosae — 2 2 4
Querceto-F agetea — 1 1 2
Bidentetea tripartiti — 1 1 2
Artemisietea vulgaris — 1 2 3
Molinio-Arrhenatheretea — 1 5 6
Stellarietea mediae 1 2 6 9
Scheuchzerio-Caricetea fuscae 1 2 4 7
Litorelletea 1 1 4 G
I soeto-Nanojuncetea 1 1 1 «>
Plantaginetea maioris — — 5 5
Festuco-Brometea — — 2 2
Lemnetea — — 1 1
Asplenetea rupestris — — 1 1
10 24 45 79
Glycerietum maximae,
Scirpeto-Phragmitetum medioeuropaeuan (subass. Typha angustifolia),
Hottonietum palustris,
Lemna minor-Lemna trisulca сообщ.,
Polygonum hydropiper-Bidens tripartitus (ass.)
и замещающими сообществами:
Bromus racemosus-Senecio aquaticus (ass.),
J unce to-Mo lin ietum,
Oxaleto-Chenopodietum polyspermii subatlanticum.
Сюда же относятся следующие два круга сообществ на типах «флизе», которые
беспорядочно вкраплены в виде небольших площадей, а именно:
4. Круг сообществ буково-скальнодубового леса (на небольших площадях иногда
чередуется с характерным сообществом):
Fageto-Quercetum petraeae violetum.
206 Пространственное подразделение растительности
5. Круг сообществ дубово-грабового леса с Querceto-Carpinetum stachyetosum.
Все пять кругов сообществ в их характерном построении образуют «комбинацию-
сообществ» данного района и определяют ландшафтный характер его растительности.
Пример II. «Комбинации сообществ» растительного района Песчаная равни-
на Сике, граничащего с предыдущим.
Господствующим на большей части его площади является:
1. Круг сообществ Fageto-Quercetum (частью на небольших площадях чередуется
с Querceto roboris-Betuletum) с характерными сообществами:
Fageto-Quercetum petraeae violetum,
Sagittaria sagittifolia-Sparganium simplex (ass.), с сосновыми и пихтовыми насаж-
дениями на месте естественных лесных сообществ, а также с разделяющими сообщест-
вами (вместо дубово-березового леса и вместо дубово-грабового леса):
Juncetum acutiflorae,
Sarothamnion scopariae frag.
Эту структуру прорезывают узкие полосы долинных лугов:
2. Круг сообществ Cariceto elongatae-Alnetum.
3. Круг сообществ Querceto-Carpinetum stachyetsoum.
Круг сообществ II, 3 включает те же сообщества, что и I, 3, однако он богаче-
разделяющими сообществами, которые отсутствуют в комбинации сообществ I, а именно:
Ну dr о cha re to -S tra tiote turn,
Potameton lucentis,
Myriophylleto-Nupharetum,
Eleocharetum ovatae,
Runiecetum maritimi,
(Caricetum paniculatae),
Caricetum appropinquatae,
Caricetum gracilis,
Caricetum diandrae
В Дипхольцской болотистой низине встречаются 55 ассоциаций, среди них 10,
которых нет на равнине Сике. На равнине Сике имеются 69 ассоциаций, среди них 24,
отсутствующих в Дипхольцской болотистой низине; 45 ассоциаций одинаковы в обоих
участках.
Растительный округ
К пространственным единицам растительности этого ранга относятся
такие единицы, которые различаются по присущим им (эндемичным) ассо-
циациям. Например, округ Северного моря характеризуется набором
сообществ Querceto roboris-Betuletum, а рассмотренные выше районы
являются частью растительного округа, к которому среди других отно-
сится и Люнебургская Пустошь. В горных местностях растительные
округа иногда совпадают с опеределенными высотными поясами. В много-
численных районах, образующих округ, могут частично повторяться сход-
ные комбинации сообществ. Вместе с другими комбинациями они составля-
ют «с о ю з комбинаций» округа. Ведущие сообщества округа имеют
ранг ассоциаций и субассоциаций; следует отметить также наличие опре-
деленных контактирующих сообществ.
Растительная провинция
Характерными для набора сообществ являются союзы (или порядки),
как, например, Quercion robori-petraeae для западноевропейско-атланти-
ческой провинции («атлантические домены» Браун-Бланке), Fraxino-Carpi-
nion для среднеевропейской провинции, Quercion ilicis и Rosmarineta-
Структура пространственного распределения растительности
207
Рис, 54, Растительность Европы и Западной Сибири, расчлененная на комплексы
растительных сообществ, названные по господствующим или характерным сообще-
ствам, Выделенные области соответствуют по величине провинциям растительности.
В отдельных частностях этот набросок требует критического подхода (Krause, 1950)
Восточная Европа и Западная Сибирь: 1 — полынные (Artemisia) или саксауловые (Hatoxyton)
пустыни; 2 — полынные (Artemisia) степи; з — низкозлаковые и крупнозлаковые степи
(Festuca-Stipa), богаторазнотравные луговые степи; 4 — лесостепь (область, проникающая в з,
5 и б)\ 5 — континентальный смешанный дубовый лес (Querceto-Carpinetum typicum, на востоке
без Carpinus)’, 6 — березово-осиновые леса (Betuta-Poputus tremuta), в Западной Сибири
замещают 5; 7 — теплолюбивые сосновые и сосново-дубовые леса (Pinus и Pinus-Quci cus)
(олиготрофные параллели с 5); 8 — североевропейские смешанные хвойно-лиственные леса
(Querceto-Piceetum и др.); 9 — североевропейские хвойные леса. Возможно, частично в ком-
плексе с эвтрофными еловыми (Picea) и олиготрофными сосновыми (Pinus) лесами; 10 — тундра.
Средняя Европа: 11 — смешанный лиственный лес холмовой ступени (субассоциацин
Querceto-Carpinetum). Большая доля Fagus sitvatica: частичные комплексы: эвтрофный с Dentaiia-
Fagetum, олиготрофный c Querceto-Luzutetum nemorosae; 12 — балтийский смешанный буковый
лес (Querceto-Carpinetum etymetosum). Возможен частичный комплекс западный, приближаю-
щийся к 13, и восточный, близкий к 5. Встречается также содержащийся в 17 комплекс
чистого Fagetum silvaticae.
Западная Европа: 13 — атлантический дубово-ясеневый лес (Querceto-Fraxinr turn);
14 — атлантический дубово-березовый лес и сходные сообщества (Querceto-Betuletum, Blech по-
Quercetum и др.).
Юго-Западная Европа: 15 —листопадные субсредиземноморские леса (Querceto-Buxet нт,
Ometo-Ostryon и др.); 16 — вечнозеленые средиземноморские леса (Quericon ilicis).
Горные районы’, 17 — пояс Fagus-Abies\ пояс Picea', пояс Larix-Pinus cembra', пояс Vaccinium
uliginosum-Loiseleuria’, пояс Carex-El упа.
На для западносредиземноморской провинции. При установлении набора
сообществ, характеризующего провинцию, часто оказывается целесообраз-
ным выделить некоторые горные районы в подпровинцию с преобладанием
в ней бореальных единиц растительности (пихтовая ступень Германского
среднегорья) (рис, 54),
Растительная область, или регион
Характерными для региона являются порядки сообществ (а части-
чно и классы), как, например, для западноевропейского региона — Fage-
talia silvaticae и Arrhenatheretalia, а для средиземноморского региона —
Quercetalia ilicis, Т hero-Brachypodietalia и класс Cisto-Lavanduletea,
208
Пространственное подразделение растительности
Высокогорные округа, набор сообществ в которых не имеет ничего
общего с набором сообществ окружающего их растительного региона,
можно рассматривать как эксклавы другого растительного региона, так,
например, среднеевропейские высотные округа с сообществами Vaccinio —
Piceetea — как эксклавы североевропейского растительного региона. Харак-
тер набора сообществ и прежде всего принадлежность отдельных единиц
системы сообществ к определенным областям можно, таким образом, уста-
новить с большей точностью.
Растительное царство
Высшие хорологические единицы, основанные на систематике сооб-
ществ, растительные царства, пространственно могут совпа-
дать с флористическими царствами. Они характеризуются присущими им
классами сообществ.
Р и с. 35. Карта растительности бассейна Озегахара (Япония)
(Suzuki, 1954).
Верховые болота и топи: 1 — Sphagneta', 2 — Oxycocceto — Caricetum mid-
dendorfii; 3 — Motinietum japonicae; 4 — Phragmitetum о seanum и Comareto-
M enyanthetum.
Леса от горного до субальпийского пояса: 3 — Utmetum davidianac
(сырой вязовый смешанный лес); 6 — Fagetum crenatae (буковый смешанный
лес); 7 — Thujetum standishii (туевый лес).
Леса от субальпийского до альпийского пояса; 8 — Abictum marie sii
(пихтово-березовый лес); 9 — Nanoquercetum (Quercus crispula — карликовый
лес); 10 — Pinetum pumilae (криволесье из карликовых сосен).
Часть этих растительных сообществ близкородственна среднеевропей-
ским и западноевропейским единицам растительности.
Широко распространенными классами сообществ голарктического
растительного царства являются, например, Asplenietea rupestris, Аттор-
hiletea, Molinio-Arrhenat here tea, Scheuchzerio-Caricetea fuscae, Vaccinio-Pi-
ceetea, Querceto-Fagetea (рис. 55). Лишь некоторые классы растительных
Структура пространственного распределения растительности 209
сообществ распространены в нескольких растительных царствах одновре-
менно, как, например, Potametea (сообщества растений с плавающими ли-
стьями), Phragmitetea (тростниковые сообщества), Salicornietea (сообще-
ства солероса), Sphagnetea (сфагновые верховые болота).
Нетрудно видеть, что разделение флоры на царства и провинции должно совпа-
дать с фитоценологическим делением больших пространств. Разграничение флористи-
ческих царств обосновано флористико-статистически и построено с точки зрения флоро-
генеза. Границы проведены частью так, как нанесли их флористы еще в прошлом столе;
тии. Фитоценологический метод классификации растительности является в конечном
счете не чем иным как более подробным флористико-статистическим методом, так как
растительные сообщества определяются по их флористическому составу, который одно-
временно отражает генезис растительности и историю флоры.
Остается один вопрос, касающийся методов исследования и устано-
вления понятий,— согласовать деление на флористические царства и про-
винции с фитоценологическим подразделением больших пространств.
Ландшафтные растительные комплексы
География склонна слишком упрощенно представлять ландшафтные
комплексы растительности. Географы часто ограничиваются лишь характе-
ристикой доминирующей в естественном растительном покрове ландшафта
(то есть в его потенциальной естественной растительности) раститель-
ной формации и заменяющих ее, а в большинстве случаев и определяю-
щих физиономию ландшафта антропогенных формаций; кроме того, назы-
вается одна или несколько особенно характерных форм роста или видов
растений.
Однако для более детального и глубокого познания растительного
покрова этот метод является недостаточным, так как не учитывает все те
многочисленные и разнообразные признаки, по которым только и можно
составить полное описание ландшафтного растительного комплекса.
АНАЛИТИЧЕСКИЕ ПРИЗНАКИ РАСТИТЕЛЬНОСТИ
ЛАНДШАФТА ИЛИ РАСТИТЕЛЬНОЙ ОБЛАСТИ
ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ПРОСТ РАНСТ ВЕННОЕ
РАЗМЕЩЕНИЕ РАСТИТЕЛЬНОСТИ,
И ИХ СОВМЕСТНОЕ ВОЗДЕЙСТВИЕ
а) Естественные факторы местности, обусловливающие образование
единиц местообитания, и факторы, вызванные к жизни хозяйственной дея-
тельностью человека.
б) Пространственное деление растительности (карты потенциальной
естественной растительности).
в) Возможное изменение пространственной структуры во времени
в результате естественных процессов или воздействия человека.
СОСТАВ РАСТИТЕЛЬНЫХ ФОРМАЦИЙ
(ХАРАКТЕР ФОРМАЦИЙ ВСЕХ БИОЦЕНОЗОВ,
ОБУСЛОВЛИВАЮЩИХ ОБЩИЙ ОБЛИК РАСТИТЕЛЬНОСТИ)
а) Связь отдельных формаций с местообитаниями и их свойствами или
с определенными формами воздействия человека (антропогенные формации).
б) Характерные связи формаций с пространственными комплексами.
в) Изменение в распределении формаций (например, смена различных
формаций на одном и том же местообитании, дальнейшее распространение
или полное исчезновение определенных формаций и т. д.).
Ландшафтные растительные комплексы 211
НАБОР РАСТИТЕЛЬНЫХ СООБЩЕСТВ
а) Группировка сообществ по местообитаниям. Сообщества, гомоло-
гичные с точки зрения экологии местообитаний (круг сообществ).
б) Группировка растительных сообществ по их распространению
и распределению: доминирующие и локальные сообщества. Пространст-
венные комплексы сообществ по экологии местообитаний, самостоятельно
развивающиеся или антропогенные: ряды, рои, пояса или зоны. В связи
с этим интересны возникающие на границах типичные контакты сооб-
ществ или же особые контактирующие сообщества.
в) Порядок и характеристика набора сообществ по динамическому
характеру сообществ: первичные сообщества, последующие сообщества
(последовательные стадии серий сукцессии), замещающие сообщества,
замещающе-последующие сообщества, устойчивые и заключительные сооб-
щества и их принадлежность и распределение по пространственным еди-
ницам растительности данной области.
НАБОР СИСТЕМАТИЧЕСКИХ ЕДИНИЦ
а) Экологические группы видов.
б) Распространение характерных форм роста или важных в других
отношениях видов растений (полезных и культурных растений) и распре-
деление элементов флоры, входящих в набор таксономических единиц,
по растительным сообществам и единицам местообитания.
в) Изменения видового состава: старые и новые поселенцы, адвентив-
ная флора (занесенные и пришлые виды), реликты.
ПРИЗНАКИ, ОТЛИЧАЮЩИЕ
ДАННУЮ РАСТИТЕЛЬНУЮ ОБЛАСТЬ ОТ ДРУГИХ
ОБЛАСТЕЙ РАСТИТЕЛЬНОСТИ
ПОЛОЖЕНИЕ И ПРОСТРАНСТВЕННЫЕ СООТНОШЕНИЯ
а) Естественное и культурное положение в пространстве.
б) Характеристика области по типам ареалов растительных сообществ,
участвующих в построении растительности: спектр типов ареалов расти-
тельных сообществ.
в) Флористико-географическая характеристика по типам ареалов эле-
ментов флоры, входящих в набор таксономических единиц.
г) Классификация области по структурному размещению раститель-
ности в пространстве (растительный район, округ, провинция).
ПРИЗНАКИ, ОСНОВАННЫЕ НА СРАВНЕНИИ
С ДРУГИМИ РАСТИТЕЛЬНЫМИ ОБЛАСТЯМИ
а) Свойственные области виды, растительные сообщества или фор-
мации.
б) Симптоматичные, характеризующие ботанико-географические осо-
бенности области полезные растения, возделываемые растения, полевые
группировки растений и основанные на видовом составе растений формы
использования.
в) Количественные и качественные признаки общего набора единиц
растительности. Качественная оценка дается на основе эндемизма видов
212
Пространственное подразделение растительности
или сообществ, характерных и отличающих от соседних областей состав-
ных элементов (видов, сообществ, формаций), а также с помощью харак-
терных комплексов сообществ или формаций (естественных или антропо-
генных). Количественная оценка производится по величине площади,
занятой формациями, сообществами или кругами сообществ, и по доле,
занимаемой высшими единицами системы сообществ (например, классами
сообществ) в общем наборе сообществ ландшафтных комплексов раститель-
ности (см. табл. 9).
г) Общие, а также экологические или динамические признаки про-
странственной структуры растительности, как, например, сочетание есте-
ственных и антропогенных принципов подразделения, фенологический
характер (специфическая последовательность сообществ в смысле насту-
пления определенных фенологических фаз, их распределение во времени,
характерные для области градиенты запаздывания фаз в зависимости от
высотных ступеней), исторические особенности растительной области (ста-
рые и молодые сообщества или формации, возраст культурного ландшафта
и т. д.), а также некоторые общие критерии, как, например, наличие форпо-
стов или эксклавов для видов или сообществ других близких или отдален-
ных областей.
«ДИВИЗИОНЫ РАСТИТЕЛЬНОСТИ»
(VEGETATIONSDIVISIONEN)
Особый интерес для широкого сравнительного географического рас-
смотрения растительности представляют те «области растительности»,
которые охватывают пограничные местности с более или менее одинаковы-
ми комплексами растительных формаций.
Такими пространствами, определяемыми эколого-физиономическим
характером растительности, являются, например, область среднеевро-
пейских летпезеленых лиственных лесов или средиземноморская область,
потенциальная естественная растительность которой представлена жестко-
листными формациями. В географии при этом часто ограничиваются
абстрактной характеристикой по господствующему классу формаций
потенциальной естественной растительности. Нельзя, однако, упускать
из виду, что такие области в естественном ландшафте могут иметь даже на
небольшом пространстве весьма дифференцированный комплекс отличаю-
щихся друг от друга формаций вследствие локальных различий в рельефе,
типах почв, уровне грунтовых вод и возрасте местообитаний.
Так, например, в среднеевропейской области, если даже исключить
субальпийские высотные пояса, которые можно рассматривать как остров-
ные эксклавы другой растительной области, наряду с летнезелеными лист-
венными лесами в составе потенциальной естественной растительности
имеются хвойные леса, кустарниковое криволесье, солончаковые луга,
заросли тростника, осоковые сообщества, моховые болота, скальная,
осыпная и дюнная растительность и другие формации. В средиземноморской
жестколистной области естественный комплекс формаций не менее разно-
образен. В культурном ландшафте это разнообразие усиливается появле-
нием антропогенных формаций. Однако в ландшафтах, подверженных
интенсивному влиянию человека, например в средиземноморской жестко-
листной области, растительная формация, давшая название этой области,
может оказаться почти полностью вытесненной или быть ограниченной
столь малыми участками, что практически почти невозможно выделить
Ландшафтные растительные комплексы
213
эту область непосредственно по этим остаткам. Поэтому более целесообраз-
но такие «области» рассматривать не только с точки зрения гипотетическо-
го при данных обстоятельствах характера формаций потенциальной есте-
ственной растительности, но и с точки зрения всей фактически имеющейся
растительности и ее строения в фитоценологическом смысле. Естественные
растительные области в этом отношении оказываются, несмотря на силь-
ное или почти полное изменение их облика под воздействием человека,
конкретными пространственными единицами, потому что они, характе-
ризуемые господствующей естественной формацией, обусловленной преж-
де всего климатом, являются, собственно, естественнопространственными
растительными областями. Даже если естественная растительность здесь
уже отсутствует, в них остается характерный набор сообществ, состоящий
из замещающих и последующих сообществ, который за пределами области
заменяется другим. Соответственно с естественным распределением форма-
ций в пространстве находится и их флористический состав, обусловленный
типами ареалов слагающих его таксономических единиц.
Такие растительные области обладают, кроме того, как это было пока-
зано на примере чилийской жестколистной области (Шмитхюзен, Доклад
на географической сессии 1957 г.), собственным фитоценогенетическим
и флористико-генетическим характером. Этим и объясняются некоторые
особенности растительных сообществ, которые не могут быть поняты с точ-
ки зрения только современных экологических условий.
Характерные черты динамики растительности и проблемы развития
культурных ландшафтов также связаны со степенью сопротивляемости
растительных сообществ различным воздействиям человека, способностью
флоры создавать из местных видов псевдоестественные замещающие сооб-
щества, такие, как луга или пастбища, или способностью противостоять
внедрению пришлой флоры.
Таким образом растительная область характеризуется не только расти-
тельной формацией, дающей ей наименование, но всем растительным ком-
плексом, который, в частности, охватывает и группировки культурных
растений.
Такой широко распространенный ландшафтный комплекс раститель-
ности с определенным, регионально господствующим характером форма-
ций потенциальной естественной растительности мы и называем «дивизио-
ном растительности».
Понятие «дивизион растительности» охватывает весь набор сообществ,
который можно охарактеризовать всеми вышеупомянутыми признаками.
На основе признаков «дивизиона растительности» теперь можно уточнить
понятие «растительной области» как пространство, занятое «дивизионом
растительности». Иначе ее называют «областью дивизиона растительно-
сти». По структуре входящих в эту область сообществ она относится боль-
шей частью к рангу регионов или провинций растительности. Понятие
«дивизиона растительности» облегчает разделение областей на обширные
эколого-физиономические зоны или пояса.
В пространствах, занимаемых «дивизионами растительности», сталки-
ваются подразделения, исходящие из общего облика Земли и предпочитае-
мые географами, со структурным фитоценологическим подразделением,
исходящим из отдельных частностей. Здесь-то и скрыт ключ для составле-
ния карты растительности Земли, в которой учитывались бы одновремен-
но эколого-физиономическая, фитоценологическая и флористическая точ-
ки зрения (табл. 10).
214
Пространственное подразделение растительности
Таблица 10
Основные хорологические понятия в географии растительности
Структурная классификация Эколого-физиономическая классификация
растительности растительности
Растительный район-------------------1___> | Доминирующий тип формаций или
; У комплекс формаций потенциаль-
Растительный округ-------------------!___> I ной естественной растительности
Растительная про- винция ! V «Область дивизиона —> Ареал «областей Климатическая растительности» дивизионов зона раститель- ► (пространственная растительности» ности
Растительная об- ласть (регион) единица с индиви- с одинаковым дуальным общим эколого-физионо-
Растительное цар- ство набором и более мическим типом или менее однород- ным комплексом формаций)
ТИПЫ «ДИВИЗИОНОВ РАСТИТЕЛЬНОСТИ» ,
КЛИМАТИЧЕСКИЕ ЗОНЫ РАСТИТЕЛЬНОСТИ
И ПРОБЛЕМА СОСТАВЛЕНИЯ ОБЗОРНОЙ КАРТЫ
РАСТИТЕЛЬНОСТИ ЗЕМЛИ
Отдельные «дивизионы растительности» имеют свой собственный набор
сообществ. Однако и без учета их флористического состава их можно срав-
нить по характерным формациям или комплексам формаций их потенци-
альной естественной растительности и разделить на типы, например тип
«жестколистных дивизионов», «тундровых дивизионов» и т. д. Каждый из
этих типов объединяет «дивизионы растительности» с физиономически
аналогичными (конвергентными), но флористически различными комплек-
сами сообществ. Например, чилийский «жестколистный дивизион» по
структуре сообществ и составу таксономических единиц отличается от сре-
диземноморского или от калифорнийского «жестколистных дивизионов».
Распространение отдельных типов «дивизионов» вторгается в деление зем-
ной поверхности на климатические зоны (климатические пояса раститель-
ности). Пространственное расчленение растительного покрова Земли и его
картирование до настоящего времени производилось одновременно с трех
различных точек зрения:
1. С точки зрения географии флоры — на царства и провинции.
2. По эколого-физиономическому характеру потенциальной естествен-
ной растительности — на пояса растительных формаций и высотные сту-
пени (например, классификация Брокман-Ероша).
3. С точки зрения фитоценологии — на регионы сообществ (Braun-
Blanquet, 1951; Lebrun, 1947).
В результате одна и та же действительность воспроизводилась с трех
различных позиций и единой картины распределения растительности
на Земле не получалось. Используя пространства «дивизионов раститель-
ности», можно составить карту растительности Земли, в которой будут
Ландшафтные растительные комплексы
215
сочетаться все три принципа классификации — эколого-физиономический,
флористический и ценологический.
Структурное подразделение по комплексам сообществ является объек-
тивным отражением самого существа растительного покрова. Границы
районов, округов, провинций, регионов и царств должны быть взяты за
основу при составлении карты растительности Земли. Уже было доказа-
но, что флористико-географическое деление соответствует пространствен-
ному делению, вытекающему из структуры сообществ. Чисто генети-
ческое подразделение флоры представляло бы собой нечто совсем другое,
Рис. 56. Обзор растительных поясов Земли (Mattick, 1958).
1 — вечнозеленый дождевой лес; 2 — саванна и дождезеленый лес; 3 — колючая саванна,
колючие и суккулентные леса; 4 — мангры; 5 — высокогорные формации; 6 — полупустыни,
сухие и холодные пустыни; 7 — степь; 8 — жестколистные леса; 9 — лиственный и смешан-
ный лес умеренно холодного пояса; 10 — хвойный лес с бореальным березовым лесом; 11 —
тундра; 12 — культурные земли.
но оно, кроме того, не могло бы быть воспроизведено в пространственном
делении. Для этого необходимо найти другую форму воспроизведения,
которая отражала бы динамику пространственной дифференциации.
Если рассматривать всю схему снизу, то по крайней мере до ранга
провинции, и даже до региона, возможно однородное подразделение сточ-
ки зрения хорологии растительности (во всех масштабах), которое будет
соответствовать как фитоценологическим и флористическим, так и геогра-
фическим треб ованиям.
Более крупные пространственные единицы этой структуры, основан-
ные на флористическом характере сообществ (царства и частично — регио-
ны, а в виде исключения, возможно, и провинции) объединяют, напротив,
пространства Земли, которые по своему эколого-физиономическому харак-
теру (комплексы формаций) настолько различны, что с точки зрения гео-
графии оказывается необходимым воспроизвести эти различия на более
мелкомасштабной карте земной поверхности. Здесь получает значение
216
Пространственное подразделение растительности
понятие «типов дивизионов». Оно позволяет объединить эколого-физионо-
мически аналогичные «области дивизионов» (в ранге провинций или регио-
нов) путем одинаковой штриховки или окраски площадей за пределами
царств и кругов в пояса, которые соответствуют климатическим поясам
растительности.
Деление на пояса эколого-физиономических типов «дивизионов
растительности» перекрещивается, следовательно, с высшими простран-
ственными единицами структуры, а именно с царствами или регионами
(ср. табл. 10). Их границы должны, однако, укладываться в контуры,
предписанные структурным подразделением (в провинции и т. д.).
Это, конечно, не касается того, что обе пересекающиеся классифика-
ции — флористически и фитоценологически обоснованные структурные
пространства (царства, регионы, провинции и т. д.) и эколого-физионо-
мические «пояса типов дивизионов»— могут быть воспроизведены и раз-
дельно друг от друга. Однако, если воспроизведение соответствует дей-
ствительности, то при совмещении обеих карт положение границ на кар-
тах должно совпадать. В настоящее время это является лишь теоретическим
требованием, потому что данные, достаточные для точного разграничения
«областей дивизионов», имеются только для части земной поверхности.
В связи с этим мы пока ограничиваемся обзором «климатических поясов
растительности», базирующихся на общих представлениях о распростра-
нении характерных формаций (или комплексов формаций) (рис. 56).
Наименование этих поясов связано, как правило, с доминирующей
в потенциальном естественном ландшафте растительной формацией. Раз-
граничение основывается, кроме того, еще со времен Гризебаха на сравне-
нии ареалов характерных систематических единиц и на распространении
определенных сообществ.
Географический интерес к этим «поясам растительности» основывает-
ся не только на самой их растительности, но также и на их общей ланд-
шафтной экологии. Пояса комплексов формаций, или «типы дивизионов»,
являются важными отправными моментами при охватывающих всю
поверхность Земли хозяйственно-географических исследованиях и земле-
ведческих сопоставлениях.
Климатические зоны растительности
ПОЯСА РАСТИТЕЛЬНОСТИ ТРОПИКОВ
И СУБТРОПИКОВ
Рассматривая тропическую растительность, можно в общих чертах
заметить структуру ее распределения по климатическим зонам или поя-
сам. Но эта структура не так проста и очевидна, как иногда пытаются
представить. Тезис Шимпера о выделении трех типов климата — леса,
травянистых пространств и пустынь — не выдержал строгой критики.
Физиономическое сходство саванн, особенно подчеркиваемое Шимпером,
больше основано не на аналогии климатов, а на том обстоятельстве, что
мы, как правило, имеем дело с форма-
циями, обязанными своим возникнове-
нием пожарам. Эти формации, однако,
могут относиться к экологически раз-
личным типам растительности. Распро-
странение саванн не является поэтому
подходящей основой для разграничения
климатических поясов растительности.
Последнее можно все же осущест-
вить, используя имеющиеся еще остатки
климаксной растительности, если суметь
отнести каждое из различных раститель-
ных сообществ травянистых пространств
к его естественному заключительному
сообществу. Поэтому все встречающиеся
в области саванн единицы раститель-
ности, не затронутые пожарами, имеют
особое значение для исследований, даже
если они мало распространены и едва
выделяются в общей картине ландшафта.
Вопрос об исходной или потенци-
альной естественной растительности
некоторых участков в тропиках до сих пор еще не нашел определенного
решения. Как показал Вайбель на примере Бразильского плато, естествен-
ная растительность и в тропиках может быть при одинаковом климате
весьма различной в зависимости от характера горных пород, уровня грун-
товых вод, возраста поверхности, степени зрелости почв и т. д.
География, которая прежде всего должна дать общую картину, боль-
ше занималась разделением по климатическим зонам растительности, а не
Рис. 57. Распространение саван-
ного злака Andropogon schirensis
(общий судано-замбезийский ареал)
(по Лебрену).
218
Пространственное подразделение растительности
исследованием фактического распространения растительных форма-
ций.
Вместе с Шантцем, Вайбелем, Шевалье и др. Егер различал в тропи-
ках по продолжительности сухих времен года шесть климатических поя-
сов растительности. Менее удачно выбрано наименование трех из этих
поясов по типам саванн. Саванны встречаются во всех тропических кли-
матах, допускающих существование сомкнутого травяного покрова и име-
ющих продолжительность сухого периода, достаточную для возникновения
пожаров на обширных площадях (рис. 57). Даже в поясе дождевых лесов,
где самовозгорание происходит с трудом, на месте легкозагораю-
щихся вторичных кустарниковых сообществ («savoka») могут возникнуть
саванны.
В табл. 11 дается обзор распределения главнейших широко распро-
страненных групп формаций тропических низменностей по шести клима-
тическим поясам, установленным Егером.
Таблица 11
Климатические зоны растительности в тропиках и распространение формаций
Число
сухих
меся-
цев
Выделение климати-
ческих зон тропиче-
ских равнин по про-
должительности
сухого периода
Распределение важнейших широко распространенных
групп формаций по климатическим поясам
тропических низменностей
О
1
2
3
4
5
6
7
8
9
10
И
12
Зона тропических
дождевых’лесов
Зона влажных
саванн
Зона
сухих саванн
Зона колючих
саванн
Зона полупустынь
Зона пустынь
Тропические дожде-
” вые леса
, Сезонные дождевые
леса
> Муссоновые леса
«Campos cerrados» и
г листопадные сухие
леса
J Колючие и сук-
кулентные леса
J
Формации колючих и
ников, колючие саванны
Полупустыни
} Пустыни
Затопляемые и влажные
саванны (с вечнозеле-
ными галерейными
лесами)
Сухие саванны (без
вечнозеленых галерей-
ных лесов)
с листопадными сухими
или колючими лесами
суккулептных кустар-
ПОЯС ТРОПИЧЕСКИХ ДОЖДЕВЫХ ЛЕСОВ
В пояс тропических дождевых лесов мы объединяем территории,
в естественной потенциальной растительности которых господствуют
растительные сообщества, относящиеся по своему эколого-физиономическо-
му характеру к формациям вечнозеленых тропических дождевых лесов
(рис. 58).
Этот пояс растительности включает низменности и нижние высотные
пояса постоянно влажных тропиков, где в течение всего года имеются бла-
гоприятные условия для произрастания растений. Он простирается по обе
Р и С; 58. Распространение тропических дождевых лесов (по Ричардсу)^
стороны экватора во всех трех тропических частях, и только в Восточной
Африке его распространение нарушается большим разрывом, а в Южной
Америке — высокогориями Анд Эквадора и Колумбии.
Распространение участвующих в этом поясе групп формаций извест-
но лишь в общих чертах. В особенности недостаточно представление
о положении границ этого пояса, с одной стороны, с муссоновыми лесами
Южной Америки и Африки, и с другой — саваннами, расположенными
в лесных областях.
Настоящая, не измененная человеком первичная растительность зани-
мает в постоянно влажных тропиках меньшую часть всей площади. В бо-
лее плотно заселенных и интенсивно обрабатываемых частях пояса дожде-
вых лесов Азии и Африки преобладают возделываемые участки и вторич-
ные формации, так что границы областей потенциальной естественной
растительности дождевого леса определить бывает очень трудно. За послед-
ние 50—100 лет площадь девственных лесов сильно сократилась. Во мно-
гих областях, как, например, в южной Нигерии, от сплошных первобыт-
ных лесов сохранились лишь отдельные разъединенные остатки.
220
Пространственное подразделение растительности
Предположение, что кроме воздействия человека на сокращение площади под
девственными лесами влияют и климатические причины, до сих пор не находит под-
тверждения.
Только в некоторых местах, например на юге Конго и в Уганде (Robyns, 1838;
Eggeling, 1947) тропический дождевой лес, по-видимому, начинает занимать новые
площади.
Наиболее протяженная сплошная область дождевых лесов — это
южноамериканская «гилея». Она занимает почти весь бассейн Амазонки,
за исключением областей ее устья, и большую (южную и восточную) часть
нагорья Гвианы. В Венесуэле гилеями покрыты верховья Ориноко,
в Колумбии — территория восточных равнин вплоть до Гуавьяры на севе-
ре; на юге лес простирается до бассейнов рек Мадре-де-Диос и нижнего
течения реки Маморе и далее в глубь северо-восточной Боливии с послед-
ними форпостами у подножия Анд до Санта-Крус.
На тихоокеанских склонах Анд, почти от самого экватора, через
Эквадор и Колумбию до Панамы, тянется сплошная область дождевых лесов.
Она захватывает атлантическое побережье Центральной Америки, прохо-
дит через Коста-Рику, Никарагуа, Гондурас, Гватемалу до Мексики,
заканчиваясь у тропика Рака близ подножия восточных отрогов Сьерра-
Мадры. Большие Антильские острова с наветренной стороны гор тоже
покрыты небольшими участками дождевых лесов.
Этим выступам в поясе пассатов северного полушария соответствует
в южном полушарии длинная узкая полоса дождевых лесов на восточном
и южном побережье Бразилии к югу от 6° ю. ш., а также полоса горного
дождевого леса на восточных склонах Анд южнее Санта-Крус.
В Африке сомкнутая область дождевого леса простирается в бассейне
Конго, на юге Экваториальной Африки (среднее Конго, Габон, Камерун)
до южной Нигерии и после небольшого разрыва продолжается к западу
в прибрежных районах Верхней Гвинеи (Гана, Берег Слоновой Кости,
Либерия, Сьерра-Леоне).
В тропической Восточной Африке вечнозеленые дождевые леса огра-
ничены побережьем и горными наветренными склонами, которые открыты
юго-восточным пассатам или азиатским юго-западным муссонам. Более
обширная область, возникшая благодаря дождям при юго-восточных пас-
сатах, находится в восточной части Мадагаскара.
Третья главная область дождевых лесов простирается на пересекае-
мых экватором Зондских островах, через полуостров Малакку, Филип-
пинские острова, Новую Гвинею и ряд островов в западной части Тихого
океана, архипелаг Бисмарка, Соломоновы острова, Новые Гебридские
острова, острова Фиджи, Самоа и др. Небольшими выступами дождевые
леса вклиниваются в северное полушарие, простираясь через юго-восток
Индокитая и юго-западную или западную окраины обоих полуостровов
(Нижняя Бирма, Ассам, юго-западный Цейлон, Западные Гаты).
В Австралии тропический дождевой лес занимает лишь небольшой
участок на северо-восточном побережье Квинсленда, примерно от 16 до
19° ю. ш.
Описанные три главные области дождевых лесов расположены в посто-
янно влажном экваториальном климате внутритропической зоны конвер-
генции. В этих низких широтах, где продолжительность дня всегда почти
одинакова, нет резких сезонных колебаний температуры. Средние темпе-
ратуры самого холодного и самого жаркого месяца различаются только
на 1—4°. В более прохладных высотах леса тропических низменностей
Климатические зоны растительности
221
сменяются тропическими горными лесами (рис. 59). Они ограничены жар-
кой высотной ступенью («tierra caliente»), которая поднимается в среднем
до высоты 1000 м над уровнем моря.
Годовое количество осадков в большей части пояса дождевых лесов
составляет 1500—4000 мм, но в некоторых местностях оно вдвое больше.
Для существования вечнозеленых дождевых лесов, однако, более важно
не общее количество осадков, а распределение их в году. Экваториальный
дождевой климат имеет два пика осадков в течение 1 или 2 месяцев после
стояния солнца в зените. Эти зенитальные дожди вызывают резкий подъем
уровня воды в реках и затопление низменностей. Дождевой лес ограни-
чен в общем областями с максимум двумя сухими месяцами. Однако при
условии особенно большого количества осадков он может, как, например,
в Либерии, переносить и более продолжительный сухой период. Внутрен-
няя часть леса остается при этом сырой даже в сухое время года.
Средние месячные температуры находятся в пределах 25—28°. Макси-
мальная температура воздуха не поднимается выше 30°. Ночные миниму-
мы опускаются ниже 20э. Средние суточные температуры лежат большей
частью между 24 и 30°; средняя амплитуда температур составляет 6—8°.
Изменение погоды в течение суток — не говоря о различиях в коли-
честве осадков — происходит довольно однообразно на протяжении всего
года. В утренние, обычно туманные часы растения влажны от росы. В чис-
том воздухе царит приятная свежесть. Животное население в эти часы
особенно оживлено. Ближе к полудню жара усиливается и лишь вблизи
моря она смягчается дневным бризом. Безоблачное утром небо постепенно
становится молочно-голубым. Облачность все больше увеличивается;
наступает гнетущий зной. В послеполуденное время — в некоторых райо-
нах почти ежедневно, а в других менее регулярно — начинаются пролив-
ные дожди, часто сопровождаемые грозами. К вечеру небо обычно снова
бывает ясным. Ночью дожди выпадают лишь изредка.
Климат под сводом дождевого леса не без оснований сравнивается
с климатом в парниках; он еще более равномерен, чем климат открытых
районов. Воздух почти неподвижен. Суточные и сезонные колебания
температуры весьма незначительны. Вблизи почвы влажность воздуха
не меняется. Защита от испарения растениям здесь не требуется. На-
против, у них есть ряд приспособлений, которые облегчают транспи-
рацию, например тонкие листья с большой поверхностью и большим коли-
чеством устьиц или листья, в которых отдача влаги в уже насыщенный ею
воздух обеспечивается другим способом — капельными устьицами (на
концах листовых жилок), из которых выдавливаются капли воды.
Сухость воздуха в период минимума осадков сказывается только
на поверхности древесных крон, где листва большей частью состоит из бо-
лее мелких, более твердых и зачастую глянцевых листьев.
Внеэкваториальные тропические области дождевых лесов на постоян-
но влажных наветренных берегах, открытых пассатам или муссонам, име-
ют такое же количество осадков, столь же равномерное их распределение
и такую же температуру, как и другие области. Отличие заключается лишь
в более значительных сезонных колебаниях длины дня и температуры,
а также в несколько другом ритме изменения погоды: дожди слабее, дли-
тельнее и не приурочены к определенному времени суток.
Пояс дождевых лесов обладает большим избытком осадков, сильно
влажными почвами, частой сетью водоемов и полноводными реками. Сом-
кнутый древесный полог резко снижает силу осадков. Лес препятствует
денудации почвы, но не приостанавливает процессы вымывания и сноса
a)
м
35
30
25
20
15
10
5
О
м
35
30
25
20
15
10
5
О
и
35
30
25
20
15
10
5
О
О 10 20 30 40м
Р и с. 59. Профильная диаграмма вечнозеленого тропического
дождевого леса различных высотных ступеней на острове Тринидад.
Буквами обозначены различные виды деревьев (по Берду п Ричардсу).
а — горный лес па высотах 800 —1000 м; б — дождевой лес на 300—700 м‘,
в — дождевой лес на равнине до 300 Л1 над уровнем моря.
Климатические зоны растительности
223
почвы, о чем свидетельствует наличие наносного материала на затопляе-
мых низменностях. После сведения или уничтожения леса ураганом эро-
зия усиливается. На оголенных склонах часто возникают оползни или
промоины.
Тепло и влага обусловливают глубоко проникающее химическое выве-
тривание. Гидраты глинозема, имеющие благодаря наличию железа более
или менее красноватую окраску, при этом пассивно обогащаются в верх-
них горизонтах.
Почвы под тропическим дождевым лесом большей частью исключи-
тельно глубокие, вязкие, сильно выщелоченные (красные и бурые глины
Кубиены). Климат способствует жизнедеятельности микроорганизмов,
которые совместно с термитами, муравьями и другими животными быстро
уничтожают мертвые органические вещества. В лесах мало опавшей лист-
вы и нередко видны пятна оголенной земли. Если исключить почвы на
сильно щелочных вулканических породах, то обычно под дождевым лесом
гумус интенсивно разлагается и инфильтруется в почву в виде почти бес-
цветного органического вещества и возвращается частью в растворенном ви-
де в реки, вода которых благодаря этому темнеет («реки с черной водой»).
Под впечатлением роскошной растительности девственных лесов рань-
ше многие считали, что тропические лесные почвы неисчерпаемо плодо-
родны. Это заблуждение привело к гибели многих плантаций. В условиях
девственного леса плодородие почв сохраняется достаточно долго. Однако
оно быстро иссякает при уничтожении естественной растительности.
В результате смыва и выщелачивания, а также возделывания полезных
растений качество почвы ухудшается, если этому не препятствовать с помо-
щью интенсивного удобрения.
По составу флоры области дождевых лесов всех трех тропических
частей в корне различны. Однако растительные формации во многих отно-
шениях построены аналогично. Это относится как к их разделению по есте-
ственным типам местообитаний и стадиям сукцессионных серий, так и к
формам вторичной растительности, обусловленной влиянием человека.
Естественные комплексы растительности в поясе дождевых лесов
содержат целый ряд широко распространенных типов формаций, приспо-
собленных к определенным особенностям местности. От чаще всего описы-
ваемого «нормального» типа равнинных лесов на незатопляемых землях
с высоким уровнем грунтовых вод, например от леса «ete» на «terra firma»
в бассейне Амазонки, отличаются формации речных берегов, затопляемых
низин, болота и лесные топи главным образом меньшим обилием видов,
разнообразием форм роста, приспособленных к данным местообитаниям,
и меньшим числом ярусов древесного полога.
Леса, затопляемые при периодических паводках рек (varzea, vargem),
имеют довольно разнообразный состав с богатой крупнолистной напочвен-
ной растительностью, но с небольшим числом древесных великанов.
Леса на берегах рек, которые регулярно затопляются и при среднем
уровне воды как, например, амазонские «игапо», под которыми почвы
бывают сухими только несколько месяцев, бедны подлеском и лианами.
В болотистых лесах часто доминирует лишь небольшое число видов с осо-
быми формами роста (например, пальмовые или бамбуковые болота).
Для сообществ на берегах зарастающих водоемов во влажных тро-
пиках — в отличие от соответствующих формаций других климатов —
характерна большая доля древесных растений. То же относится к первич-
ным сообществам на твердых почвах и обусловлено непрерывной вегета-
ционной деятельностью растений.
224 Пространственное подразделение растительности
Соответственно сообщества плоских морских побережий во влажных
тропиках также являются древесными формациями, мангровой раститель-
ностью. Она развивается в основном в мелководных бухтах и лагунах
и может проникать в устья рек до верхней границы приливов. Обусловлен-
ное мангровыми зарослями образование суши часто облегчается и уско-
ряется с помощью развивающихся на этих же побережьях коралловых
рифов.
Сплошные пояса мангров распространены главным образом в поясе
дождевых лесов. Однако и за его пределами встречаются их изолирован-
ные участки, определяемые особыми местными условиями, например
на южной оконечности полуострова Калифорния, на побережье Флориды,
юге Ирана, в дельте Ганга и побережье Южного Китая вплоть до Гуанчжоу.
По флористическому составу различают западные и восточные ман-
гровые заросли. Первые, на тихоокеанском побережье Америки и по
обоим берегам Атлантического океана (восточное побережье Америки
и западное побережье Африки), имеют лишь четыре характерных вида,
тогда как вторые, на берегах Индийского и западной части Тихого океана,
более богаты видами (до 24 видов из десяти семейств). Только роды Rhiso-
phora и A vtcennia встречаются во всех областях. Мангровые деревья бла-
годаря содержащимся в них дубильным веществам находят разнообраз-
ное применение.
Наряду с древесными формациями в пределах пояса дождевых лесов
выделяются затопляемые саванны как естественные травянистые простран-
ства. Еще не достаточно ясно, в какой степени эти саванны, встречающиеся
и вне затопляемых областей среди дождевых лесов, являются естествен-
ными сообществами, возникшими вследствие особых эдафических условий.
Возможно, они имеют антропогенное происхождение, как это еще сто лет
назад предположил Юнгхун относительно алапговых саванн Индонезии.
Вторичные леса, широко распространившиеся к настоящему времени
на месте девственных лесов, сильно отличаются от последних, особенно
на первых стадиях обычно быстро протекающего повторного облесения.
Вторичная растительность состоит большей частью из других видов, неже-
ли девственные леса, и более однотонна. Вторичный лес имеет густой под-
лесок, но гораздо менее выраженную ярусность древесного полога. В какой
степени потенциальная естественная растительность совпадает с девствен-
ным лесом, в отдельном случае бывает трудно установить.
Тропические дождевые леса относятся к наименее населенным участ-
кам Земли и до настоящего времени еще остаются недоступными для чело-
века. Аборигены или занимаются сбором плодов, охотой и рыбной ловлей,
не возделывая почвы и не имея постоянных поселений, или примитивным
земледелием. Они обрабатывают почву мотыгой, предварительно очистив
ее от леса путем корчевания и выжигания. Когда плодородие почвы бывает
исчерпано, маленькие поля забрасываются и все племя переселяется на
новое место жительства. В результате на больших площадях возникли
вторичные леса, причем возобновление леса происходит обычно очень
быстро.
Разводимые плодовые культуры — это обычно клубневые растения,
в Старом Свете первоначально ямс и таро, в Америке — арроурут, батат
п маниока, из которых последние частично с помощью европейцев распро-
странились во всех трех тропических областях. Сюда же относятся много-
численные дикорастущие или даже саженые полезные деревья, дающие
фрукты или масличные плоды (кокосовая и масличная пальмы), соки
(пальмовое вино), саго, возбуждающие средства (орехи кола и бетель),
Климатические зоны растительности
225
строительный материал (дерево, бамбук, испанский тростник, пальмовый
лист), волокнистые вещества для изготовления одежды и других целей,
далее лианы, которые находят разнообразное применение, и, наконец,
широко распространенные в настоящее время бананы (плоды, мука, воло-
кно) и маис.
Природными путями сообщения служат реки. На их берегах, на кра-
ях прогалин и во вторичных сообществах лес обычно наиболее густ и труд-
нопроходим. Это создавало впечатление о непроходимости всего лесного
пояса, в то время как внутренняя часть девственных лесов на сухих поч-
вах является непроходимой скорее не из-за густого подлеска, а из-за
валежника и лиан, упавших на землю вместе со стволами деревьев.
Растения дождевого леса, приобретшие значение в мировой экономи-
ке, являются почти исключительно древесными растениями. Большей
частью это деревья, но наряду с этим распространены и другие формы,
например лианы (ваниль, черный перец, каучуконосные лианы), кустар-
ники (коричник), древовидные злаки (бамбук) и наиболее важная недре-
весная форма с травянистым стволом — бананы.
В зависимости от использования полезные растения тропического дождевого
леса разделяются на следующие группы:
Деревья, лианы и кустарники, дающие^каучук, гумми-соки или смолы: Hevea-
каучук, гуттаперча, гумми-балата, Ficus., различные каучуконосные лианы и др.
Деревья и кустарники, дающие продукты потребления:
какао, кофе, чай, кола.
Маслосодержащие плоды: масличная пальма, кокосовая пальма, орехи пара.
Пряности: гвоздики, мускат, перец, гвоздичный перец Pimenta^ корица, ваниль.
Ценная древесина: цветная и твердая древесина и древесина, содержащая дубиль-
ные вещества.
В каждой из трех тропических областей используются разные расте-
ния. Однако для большинства этих растений, культивируемых на планта-
циях, основной центр возделывания находится не на их родине. Исклю-
чением являются чай, масличная пальма, кокосовая пальма. Кокосовая
пальма с доисторических времен распространена на всех тропических
побережьях и главным образом на островах Полинезии. Настоящая роди-
на ее неизвестна. Из Африки происходят масличная пальма, кофе, орехи
кола; из Азии — перец, корица, гвоздика, мускат, гвоздичный перец, чай,
плодовые и текстильные бананы, джут и др.; из Америки — гевея, какао,
ваниль и др.
Древесина деревьев тропического леса различна по качеству. Неко-
торые породы весьма тверды, хрупки и тяжелы. Большинство, однако,
имеют легкую, губчатую и малоценную древесину. Их хозяйственное
использование является задачей будущего.
Некоторые ценные породы деревьев уже давно играют значительную роль в ми-
ровой торговле. К таким относятся:
1) породы, древесина которых благодаря отсутствию годичных колец имеет
одинаковую плотность, а также ценные породы для полировки и поделок; эбеновое
дерево, махагони, гуаята и др.;
2) породы, древесина которых окрашена в различные цвета (красное дерево,
синее дерево, желтое дерево) или содержит масла (розовое дерево) и дубильные веще-
ства (мангровые).
Пояс дождевых лесов отличается самым большим годовым приро-
стом древесины и огромными еще не тронутыми ее резервами, которые,
226
Пространственное подразделение растительности
однако, в результате бессмысленного уничтожения год от года уменьшают-
ся. Хотя половина площади земной поверхности, покрытой лесами, нахо-
дится в тропиках, их доля в мировом экспорте леса составляет всего 2%.
Это объясняется не столько их географическим положением, сколько
структурой лесов и общими условиями тропического пояса. Богатство
видами и мощность развития лесов затрудняют их рациональное исполь-
зование. На небольшом пространстве размещены тысячи деревьев различ-
ного качества, поэтому часто деревья одного вида приходится буквально
разыскивать среди других. Вырубку леса затрудняют лианы. Транспорт-
ные пути, за исключением водных артерий, отсутствуют, и к тому же
сплав некоторых наиболее ценных деревьев затруднен из-за их тяжести.
Лесное хозяйство в тропическом дождевом лесу до сих пор является чисто
хищническим хозяйством. Научное лесоводство там еще только начинается
или находится в стадии испытания.
ПЕРИОДИЧЕСКИ СУХИЕ ПОЯСА ТРОПИЧЕСКОЙ
РАСТИТЕЛЬНОСТИ С МУССОНОВЫМИ, СУХИМИ
И КОЛЮЧИМИ ЛЕСАМИ И САВАННАМИ
Разделение периодически сухих тропиков по характеру растительно-
сти еще не нашло удовлетворительного решения. Проведенное Егером
расчленение на три части по продолжительности сухого периода, исходя
из условий, существующих в Африке, а именно на климатические ланд-
шафтные пояса «влажных саванн», «сухих саванн» и «колючих саванн»,
для других частей света (например, муссонной Азии, Бразилии, Австра-
лии) пригодно лишь с известными ограничениями. Это связано частично
с тем, что наряду с продолжительностью сухого периода образование
растительных формаций обусловливается также количеством осадков
и некоторыми другими особенностями климата на каждом континенте.
Учитывая эти моменты, можно принять за основу деление по трем степеням
влажности:
а) Самый влажный пояс характеризуется тропическими муссоновыми
лесами, имеющими наибольшую протяженность в азиатских тропиках,
а также районами распространения «campos cerrados» и высокозлаковых
саванн с вечнозелеными галерейными лесами (влажные саванны).
б) В поясе средней влажности имеются листопадные сухие леса, бедные
суккулентами колючие леса и саванны со злаковым покровом от средней
высоты до низкотравья, без вечнозеленых галерейных лесов (сухие
саванны).
в) В третий пояс («колючекустарниковые степи» Вайбеля) входят
формации колючих и суккулентных кустарников и лесов (каатинга) и су-
хие травянистые пространства с большим количеством колючих кустарни-
ков (колючая саванна).
Два первых пояса охватывают главным образом саванны, как они
понимались в старых литературных источниках. В обоих поясах саванны,
возникшие после пожаров, и другие преимущественно или исключительно
антропогенные формации занимают более обширные пространства, чем
древесные формации, которые, видимо, следует частично рассматривать
как потенциальную естественную растительность.
Поскольку ни распространение саванн, ни соотношение их сообществ
с сукцессионными сериями комплексов сообществ муссоновых или сухих
лесов изучены еще недостаточно, нельзя четко разграничить оба эти
пояса.
Климатические зоны растительности
227
Вообще, не говоря об областях, изученных в последнее время новей-
шими методами (например, часть Конго), нельзя определенно решить, осо-
бенно в переходных полосах, находятся ли возделанные площади на месте
саванн или муссоновых лесов и какая часть саванн может рассматриваться
как эдафически обусловленная естественная формация.
К этому добавим, что часто, например, в обширных районах Индоки-
тая или Восточной Африки климаксные области обеих групп лесных фор-
маций не образуют резко отделенных друг от друга замкнутых поясов,
а вследствие климатических различий, обусловленных характером релье-
фа, попеременно чередуются на небольших пространствах или взаимно
проникают одна в другую.
Только в районах Центральной и Западной Африки севернее и южнее
области дождевых лесов оба эти пояса имеют ярко выраженное зональное
расположение. Но и здесь их разграничение затруднено, так как, напри-
мер, в Судане преобладают в основном культурные земли и саванны и часто
нелегко отличить влажные саванны от сухих. Дело в том, что и галерей-
ные леса все больше уничтожаются человеком, а травянистые простран-
ства изменены в такой степени, что их отнесение к определенной климакс-
ной растительности становится возможным разве только после основатель-
ного фитоценологического изучения.
Как в поясе тропических листопадных лесов, так и в поясе саванн
имеются, с одной стороны, еще обширные сильно облесенные области
(муссоновые леса Южной Азии, «campos cerrados» в Бразилии, миомбовые
леса в Южной Африке), а с другой — большие пространства, занятые
саваннами, тогда как леса, не говоря о галерейных лесах влажных саванн,
встречаются лишь в виде остатков. Последнее явление имеет место, напри-
мер, в Судане, на стыке обоих поясов,— эти саванны как травянистые
пространства физиономически более родственны друг другу, чем два лес-
ных района в одном и том же поясе. Физиономическое деление на травя-
нистые пространства и леса здесь не совпадает с климатическими поясами
растительности или с делением по потенциальной естественной раститель-
ности. Правда, современное распределение лесов и саванн на больших
пространствах все же зависит от условий местности. Например, лес может
возникнуть там, где из-за условий рельефа или характера подстилающих
пород почвы оказываются неблагоприятными для ведения сельского
хозяйства. Нередко, однако, гораздо более решающими оказываются
исторические и экономические факторы.
Особое значение имеет бытовавшее на некоторых территориях подсеч-
но-огневое земледелие. На землях, не знавших подобной обработки, как,
например, в древних культурных районах Южной Азии, леса сохранялись
гораздо дольше, несмотря на высокую плотность населения. В странах,
где пашенному земледелию предшествовало мотыжное, например в Китае,
леса сведены на значительной территории. В пределах Судана, севернее
пояса дождевых лесов, где первоначальное мотыжное земледелие кочевых
туземцев сочеталось с выжиганием лесов, ввиду небольшого количества
скота или даже вследствие полного его отсутствия из-за мухи цеце травя-
нистые пространства (злаковники) заняли место дождезеленых лесов.
Влажные саванны здесь имеют резко выраженную границу и примыкают
к дождевому лесу или даже вторгаются в него.
Плотность населения тоже приходится иметь в виду, например при
сравнении саванновых областей восточной Индии или Судана, «campos
cerrados» Бразилии.
228
Пространственное подразделение растительности
Пояс муссоновых лесов и влажных саванн
Муссоновые леса имеют наибольшую протяженность в области дей-
ствия азиатского муссона, от которого они и получили свое наименование
и где впервые были описаны. В северном полушарии муссоновые леса
занимают прежде всего обширные районы Индокитая. За исключением
пространств, занятых тропическим дождевым лесом, они покрывают бере-
говые горные склоны и наветренные относительно летнего муссона внут-
ренние районы, тогда как на подветренных сторонах и в долинах рек,
например в Таиланде, их часто сменяют сухие леса, саванны или даже фор-
мации колючих кустарников. На западном побережье Бирмы и северной
оконечности полуострова Малакка муссоновый лес с полувечнозелеными
переходными формациями примыкает к вечнозеленому дождевому лесу.
В Передней Индии, которая бедна лесами и покрыта большей частью
саваннами, леса встречаются преимущественно в горных районах Запад-
ных и Восточных Гатов и близ южных склонов Восточных Гималаев,
причем и здесь часто наблюдается постепенный переход к вечнозеленым
дождевым лесам. Эти более влажные полувечнозеленые муссоновые леса
при количестве осадков свыше 2000 мм зачастую богаты лианами, эпифи-
тами и бамбуком, тогда как напочвенный травянистый покров отсутствует.
Для гораздо шире распространенных типичных и частично также для
сухих муссоновых лесов характерно тиковое дерево (Tectona grandis,
Verbenaceae). В естественных лесах оно, однако, редко составляет более
1/ю всего древостоя. Эти леса остаются безлистными на протяжении 3—4
месяцев. С наступлением сухого периода они постепенно теряют листву
с разницей во времени, зависящей от погоды данного года, и одеваются
листвой снова в мае, незадолго до наступления сезона дождей. Некото-
рые виды деревьев, и среди них Tectona, цветут в середине дождливого
периода. Типы сухих муссоновых лесов (с количеством осадков 1000—
1500 мм), состоящие сплошь из разных видов деревьев, сильнее, чем все
другие леса, дифференцированы в зависимости от эдафических условий.
Южнее экватора муссоновые леса, частично в сочетании с саванна-
ми, встречаются на Яве, за исключением западной части острова, на Малых
Зондских островах и на юге островов Целебес и Новая Гвинея. На Яве
леса Tectona (называемые здесь лесами «djati») уже давно эксплуатируют-
ся как лесные хозяйства, причем они многократно превращались в чистые
тиковые насаждения. Они теряют листву обычно в конце июня и остаются
без листьев до октября.
В Австралии, где климатические предпосылки для возникновения
муссоновых лесов существуют на севере и северо-востоке, эти леса имеют
несколько отличный экологический габитус из-за господства характерных
вечнозеленых видов эвкалипта. Однако все же имеется определенное физи-
ономическое сходство этих «вечнозеленых муссоновых лесов» с листопад-
ными. Древесный полог в них двухъярусный и светлый. Деревья большей
частью прямоствольны, а на почве развит травяной покров, тогда как
кустарниковый подлесок отсутствует. Влажные саванны занимают более
обширные площади, главным образом в северной Австралии и в глубине
Квинсленда, западнее полосы прибрежных гор.
В американских тропиках листопадные муссоновые леса встречаются
у восточного подножия Анд, между южными выступами амазонских дож-
девых лесов и сухих лесов области Гран-Чако, в западных горных районах
Гвианы, в Венесуэле и Колумбии по краю льяносов и на северных отро-
гах Анд, а также на тихоокеанской стороне Центральной Америки и на
Климатические зоны растительности
229
Кубе. И здесь можно различить формации с разной степенью влажности;
так, на Тринидаде при количестве осадков от 800 до 1300 мм произрастают
леса только частично листопадного типа (рис. 60). Наиболее типичны
для северного пояса саванн описанные Гумбольдтом льяносы Ориноко.
Рис. 60. Профильная диаграмма последовательности климатически обу-
словленных формаций низменностей острова Тринидад и прилегающих частей
Венесуэлы (по Берду).
1 — вечнозеленый (сезонный) дождевой лес; 2 — влажный муссоновый лес; 3 — сухой
муссоновый лес и сухой лес; 4 — колючий и суккулентный лес; 5 — колючие и
суккулентные кустарники.
большей частью затопляемые саванны с высокозлаковым покровом
и пальмами. В основном на юге и в бассейне реки Мета это ти-
пичные влажные саванны с пышными вечнозелеными галерейными
лесами. Часть площадей, описанных Гумбольдтом как безлесная, покры-
лась с тех пор древесной порослью: это области с малой плот-
ностью населения и весьма экстенсивным пастбищным хозяйством.
Пояс муссоновых лесов и влажных саванн, прерываемый дождевыми
лесами, простирается до окраин Гвиан-
ского нагорья. Саванны расположены
здесь в венесуэльской части плоскогорья
на высоте 1000 м и доходят до района
верховий Риу-Бранку и западной части
Британской Гвианы (рис. 61). На широ-
те льяносов, несколько западнее, вплоть
до реки Магдалены, вновь появляются
саванны.
Северные выступы пояса влажных
саванн располагаются на тихоокеанской
стороне Центральной Америки, захва-
тывая небольшую часть известнякового
плато на Юкатане и подветренные
склоны Вест-Индских островов, где с
целью улучшения качества пастбищ
местами произведен подсев ввезенных
из Африки злаков.
Рис. 61. Растительные формации Британской
Гвианы (Fanshave, 1952).
1 — тропический дождевой лес; 2 — муссоновый
лес; з — тропический горный лес; 4 — сухой лес
и лавровый горный лес; 5 — болотистый и мангро-
вый лес; 6 — саванны; 7 — культурные земли.
230
Пространственное подразделение растительности
Южнее амазонской гилеи, в кампосах центральной Бразилии, про-
стирается самый крупный и в то же время наименее по сравнению с дож-
девыми лесами исследованный участок пояса муссоновых лесов и влаж-
ных саванн. Листопадные леса, прорезанные травянистыми пространства-
ми (злаковниками), муссоновые леса, сухие муссоновые леса и особенно
обширные «campos cerrados» перемежаются здесь травянистыми простран-
ствами, иногда с небольшим числом деревьев («сатро sujo» и «сатро Итро»)
и пересекающими их полосами вечнозеленых галерейных лесов. В сухое
время года часть травянистых пространств регулярно выгорает. Некото-
рые горные саванны по богатству видов и наличию эндемичной флоры
можно рассматривать как естественную климаксную раститель-
ность.
В Африке широкие пояса саванн тянутся севернее и южнее экватори-
ального дождевого леса и смыкаются на востоке, у западного края области
восточноафриканских озер. Поскольку вечнозеленый дождевой лес запол-
няет также долины и ущелья, а в виде галерейных лесов пересекает и высо-
котравные злаковники, то эти участки саванн мы относим к «влажной
саванне». Переход к граничащему с ними поясу сухих саванн и сухих
лесов часто нерезко выражен и положение границ во многих областях тре-
бует еще уточнения.
Характерным деревом влажных саванн Судана можно считать крупно-
листный бутироспермум (Butyrospermum parkii, Sapotaceae). Благодаря
распространению в этом поясе подсечно-огневого земледелия более круп-
ные лесные массивы встречаются настолько редко, что долгое время счи-
тали, будто саванны здесь примыкают непосредственно к дождевому лесу
как естественная формация. Остатки мезофитных листопадных или полу-
вечнозеленых лесов, однако, теперь и здесь известны на многих окраинных
участках «влажных саванн». Они встречаются на юге Судана, территории
Ганы, восточной Нигерии, Камеруна, а также на нижнем Конго, в различ-
ных районах Восточной Африки (Эфиопия, Танзания, Мозамбик) и во
внутренних районах Мадагаскара. Изучение данных аэрофотосъемок
(Meulenberg, 1949) показало, что в саваннах нижнего Конго, не считая
галерейных лесов, которые, кстати, сильно истреблены человеком, остатки
лесов встречаются значительно чаще, чем это предполагалось ранее. Эти
естественные сукцессии на оставшихся нетронутыми местообитаниях позво-
ляют считать муссоновые листопадные леса климаксной растительностью.
На площадях, которые охраняются от пожаров ради охоты или в целях
защиты поселений, образуются настоящие древесные заросли («nkunku»),
как и на месте заброшенных поселений («nvaka»).
Для более влажных участков восточноафриканских нагорий харак-
терны описанные Троллем (Troll, 1936) термитные саванны, которые
встречаются на плато Иринга на высоте 1500 м, а на юге (провинция
Наталь) несколько ниже. В области возвышенных саванн на равнинах На-
ционального парка Альберт все серии развития неиспользуемых травяни-
стых злаковых пространств приводят к образованию листопадного сухого
леса со склеро-ксероморфным и суккулентным составом (Лебрен). Внут-
ренняя часть Мадагаскара на всем протяжении пересекается поясом влаж-
ных саванн.
Средневлажный, в понимании Пенка, тропический климат этого
пояса растительности характеризуется примерно 3—5 сухими месяцами.
Остальные месяцы могут быть умеренно или даже исключительно дождли-
выми. Годовое количество осадков составляет 800—2000 мм, однако может
быть и значительно выше.
Климатические зоны, растительности
231
Периодичность в режиме влажности возникает в близких к экватору
широтах преимущественно вследствие смены осадков внутри тропической
конвергенции сухим периодом пассатов.Здесь господствует температур-
ный режим внутритропического типа с небольшими сезонными и суточны-
ми амплитудами. В более высоких широтах, и прежде всего в Южной
Азии, муссоны обусловливают смену влажного лета сухой зимой.
В зависимости от ветровой экспозиции возникают местные отклоне-
ния в режиме осадков, а с высотой количество осадков иногда резко возра-
стает. С широтой сезонные температурные различия углубляются, а на
внешних границах пояса температурный режим носит уже субтропи-
ческий характер. Для растительности при этом является определяющим
то обстоятельство, что период дождей совпадает с наиболее жарким вре-
менем. Реки пояса муссоновых лесов текут в основном круглый год; толь-
ко небольшие ручьи пересыхают в сухое время года. Зенитальные дожди
обусловливают прохождение по рекам паводков два раза в году.
Условия выветривания в поясе муссоновых лесов сходны во влажное
время с условиями тропического дождевого леса. В сухое время почвен-
ная вода с растворенными в ней солями поднимается к поверхности, бла-
годаря чему процесс выщелачивания замедляется. Вынос оснований
задерживается, и почвы, несмотря на пассивное обогащение альферитным
материалом, сохраняют рыхлую структуру. Эти тропические красно-
земы на породах, бедных железом, имеющие серый или желтоватый цвет,
менее пластичны, чем красные тропические глины дождевого леса и более
ценны для сельского хозяйства. Гумусом они беднее, чем красные глины.
Муссоновый лес, по Вагелеру, образует в год 50 т органического
вещества на гектар, а тропический дождевой лес в два и даже в четыре раза
больше. Листва муссонового леса, опадающая в сухое время года, быстро
разлагается без образования на поверхности гумусной подстилки. Однако
верхние горизонты почвы пропитаны б ^цветными органическими веще-
ствами и поэтому она не так бедна гумусом, как кажется на первый взгляд.
Саванна создает ненамного меньше органического вещества, чем мус-
соновый лес (30 т). Однако разложение растительных остатков и выщела-
чивание почвы происходят здесь гораздо быстрее. Так, на нижнем Конго
на одинаковых сланцевых породах под лесом обнаружены красные доста-
точно плодородные почвы, а под саванной — желтые и бедные (Бейенс).
Когда выщелачивание достигает определенной степени, начинается про-
цесс латеритизации, который завершается образованием плотной железной
корки, препятствующей существованию растительности.
Саванновые почвы обычно бедны калием и азотом. С возрастом они
претерпевают существенные качественные изменения. Более молодые поч-
вы, занятые большей частью злаковым: покровом со значительным числом
деревьев, являются более щелочными и более плодородными. Их верхний
горизонт, как и у лесных почв, пропитан бесцветными органическими
веществами. Более старые почвы тяжелее, плотнее, глинистее и с точки
зрения сельского хозяйства менее ценны. Они находятся преимуществен-
но под злаковым покровом, лишенным деревьев. Постройки термитов
местами хорошо вентилируют почву и улучшают ее водный режим. На
них часто встречаются лесистые островки («термитные саванны»).
Пожары в саваннах способствуют обеднению почв. Оголение от расти-
тельности перед периодом дождей облегчает смыв верхнего гумусного
слоя почвы. Минеральные вещества выщелачиваются и уносятся водой.
Стадиям обусловленной пожарами регрессивной сукцессии раститель-
ности соответствуют весьма определенные этапы состояния почвенных
232
Пространственное подразделение растительности
профилей, которые, как иногда и степень облесения, могут являться
мерилом плодородия различных саванн.
Чем сильнее деградирован почвенный профиль и чем глубже почва
смыта, тем больше при прочих равных условиях доля ксероморфных ком-
понентов в растительности. Чем реже покров из высоких злаков, тем
больше площадь обнаженных почв, которые после пожаров будут смыты,
как только начнется период дождей.
Пожары совместно с влиянием ряда крайне засушливых лет могут
навсегда воспрепятствовать поселению здесь деревьев. Чем мезофитнее
растительность и чем больше питательных веществ сохранила почва, тем
скорее может начаться возобновление деревьев, если, конечно, последую-
щие пожары опять не окажут своего губительного воздействия. После
6 или 10 лет отсутствия пожаров почва улучшается. Появляются древес-
ные проростки, и деревья могут быстро достигнуть высоты в несколько
метров. В других случаях, особенно на очень тяжелых почвах, должно
пройти 20—25 лет, прежде чем деревья достигнут такой высоты после
последнего пожара. Вновь появляются виды деревьев, отличные от тех,
которые росли в саваннах до пожара. В конце концов саванна сменяется
лесом.
Пояс тропических сухих лесов и сухих саванн
(«Суданская зона» Шевалье)
Области сухих лесов и сухих саванн находятся в основном в периоди-
чески сухом климате внешних тропиков, где сухой период продолжается
примерно полгода (от 5 до 7 месяцев). Лишь сравнительно недавно они
были признаны самостоятельными климатическими поясами растительно-
сти (Waibel, 1921), располагающимися между влажными саваннами и фор-
мациями колючих кустарников (рис. 62). Это область с годовым количест-
вом осадков от 500 до 1200 мм. Высокая температура и переменная влаж-
ность при энергичном передвижении почвенных растворов вверх по про-
филю во время сухих периодов обусловливают сильное химическое вывет-
ривание с тенденцией к латеритизации, при которой на поверхности горных
пород образуется сплошная железная корка, предотвращающая их даль-
нейшее разрушение. Возвышающиеся над почвой постройки термитов
тоже приобретают такую наружную защитную корку. Как по своему про-
странственному расположению, так и по характеру формаций эти пояса
растительности в разных частях света имеют разное строение. Частично
это обусловливается неодинаковой степенью воздействия человека, а так-
же тем, что различие почвообразующих горных пород оказывает на расти-
тельность здесь более сильное дифференцирующее влияние, чем в других
климатических поясах. С точки зрения типологии формаций, сухие леса,
рассматриваемые преимущественно как климаксная растительность, еще
недостаточно исследованы. Их связь с сухими саваннами, которые боль-
шей частью являются устойчивыми сообществами, возникшими в резуль-
тате пожаров, окончательно выяснена лишь для некоторых районов.
На неблагоприятных почвах встречаются местами формации колючих
кустарников и суккулентов (рис. 63); на заштрихованной части Трансва-
аля дополнительно нанесено два крестика по Обет — Кайзеру (Obst —
Kaiser, 1949).
Сухие леса на всем пространстве Африки к югу от экватора сохрани-
лись на больших площадях. К ним прежде всего относятся миомбовые леса^
Климатические зоны растительности
233
состоящие в основном из дождезеленых бобовых деревьев с перистой лист-
вой разной величины, а частью из мелколистных деревьев с широкими
зонтиковидными кронами. Они простираются через поднятие Лунда (вос-
точная Ангола, Замбия, Малави, южное Конго), северный Мозамбик
и другие районы Восточной Африки, а кроме того, по западной окраине
Мадагаскара.
В Судане леса сохранились лишь в незначительном объеме. Здесь
господствуют сухие саванны со средневысокими кустистыми злаками
Рис. 62. Судано-замбезийский
ареал Canthium vulgare (Rubiaceae)
(по Лебрену).
Рис. 63. Ареал «хлебного дерева
обезьян» (Adansonia digttata)
(Duvigneaud, 1953).
Точками отмечены точно описанные
местонахождения, крестиками — не-
уточненные местонахождения. В место-
нахождениях вне заштрихованного
ареала растение, как правило, занесено
человеком.
и местами более густой, а большей частью лишь скудной древесной расти-
тельностью, состоящей главным образом из дождезеленых неколючих
видов, а частью из колючих зонтичных деревьев, отдельных стеблесукку-
лентных деревьев или даже из пальм.
В Южной Америке соответствующие сухие леса и саванны не имеют
столь широкого распространения. Основная область их с перистолистным
сухим лесом типа «Piptadenia» (Paffen, 1957) и «Mimosos» (Лютцельбург)
тянется в виде сравнительно узкой и несплошной полосы в направлении
с севера на юг от штата Сеара до Минас-Жераис между «campos cer-
rados» и каатингой на северо-востоке Бразилии. Вторая, также узкая
и прерывистая полоса, включающая листопадные бобовые деревья
«chapadas», обрамляет северную часть области Гран-Чако и продолжается,
захватывая богатые злаками сообщества Acacia cebil, вплоть до аргентин-
ской провинции Тукуман. На севере Венесуэлы аналогичный пояс расти-
тельности окаймляет озеро Маракайбо.
В Азии к этому поясу растительности можно отнести районы восточ-
ного Декана, большинство которых в настоящее время окультурены,
а также сухие леса Верхней Бирмы и бассейна Менам; в Австралии —
редкие эвкалиптовые сухие леса и сухие саванны во внутренних частях
Северной территории и в западном и среднем Квинсленде.
234
Пространственное подразделение растительности
Пояс тропических и субтропических колючих и суккулентных
древесных формаций и колючих саванн (колючекустарниковых
степей)
Области с растительностью этого типа встречаются в различных кли-
матических условиях:
1. В сухих областях внутренних тропиков на востоке континентов
(северо-восток Бразилии, Восточная Африка).
2. В полусухих районах пояса пассатов с периодическими тропиче-
скими летними дождями и примерно 8—10 сухими месяцами (северная
и восточная Австралия, пояс Калахари, Гран-Чако, Мексика, тихоокеан-
ское побережье Центральной Америки, полуостров Юкатан, северное
побережье Венесуэлы, зона Сахель в Африке).
3. На обращенных к экватору субтропических окраинах с зимними
дождями, в переходных областях со скудными зимними дождями и сухим
периодом на протяжении 9 месяцев в году (полуостров Калифорния,
северное Чили, юго-западная Австралия).
4. В бассейнах тропических рек и межгорных долинах (например,
район Анд, Мексика и Индия).
Общим для этих областей является тропическая летняя жара, безмо-
розные зимы и прежде всего значительная продолжительность сухого
периода (от 8 до 10 месяцев) при колеблющемся количестве годовых осад-
ков от 250 до 750 мм. Большинство водотоков, за исключением рек, теку-
щих из других поясов, имеют периодический сток. Сопровождающие
речные долины древесные сообщества, если таковые встречаются, пред-
ставлены либо листопадными, либо ярко выраженными ксероморфными
лесами. В результате значительных колебаний температуры, обуслов-
ленных широтным положением, и прямого облучения поверхности, имею-
щего место из-за несомкнутости растительных формаций, происходит
интенсивное физическое выветривание с последующим сносом выветрив-
шихся слоев. Почвы в общем бедны гумусом, но в остальном довольно
различны, в зависимости от вида почвообразующих пород. Для некоторых
областей (например, Калахари) характерно образование поверхностных
известковых корок. Растительные формации в отдельных случаях довольно
разнообразны. Чем эпизодичнее осадки и чем больше сухость климата,
тем меньше злаковых сообществ и тем больше участие суккулентов.
Растительные формации можно объединить в три группы:
1) Колючие саванны (колючекустарниковые степи), ксероморф-
ные травянистые пространства, покрытые разреженными кустами жестко-
листных злаков высотой от 0,05—1 м и более или менее рассеян-
ными колючими деревьями со всеми переходами к сомкнутым сухим
лесам.
2) Колючие и суккулентные леса типа лесов каатинги и австралий-
ского скрэба или относительно редко встречающиеся чистые стеблесук-
кулентные сообщества (Мексика).
3) Бедные злаками формации колючих и суккулентных кустарни-
ков с преобладанием колючих кустарников или же стеблевых и листовых
суккулентов (Cactaceae, Euphorbiaceae, Bromeliaceae) или и тех и других
в самых разнообразных соотношениях.
Эти формации, в зависимости от эдафических или местных климати-
ческих условий, могут взаимно перемежаться.
В Африке злаковые сообщества занимают большую часть территории,
тогда как в Америке преобладают суккулентные деревья и кустарники,
Климатические зоны растительности
235
отличающиеся особенным богатством форм. В Америке отдельные расти-
тельные области расположены на значительном расстоянии друг от друга,
тогда как в Африке и в Австралии они образуют сплошные пояса.
Австралийский пояс имеет наибольшую протяженность и почти со
всех сторон охватывает внутренние полупустыни и пустыни. Здесь широко
распространены прежде всего кустарниковые формации с обилием акаций
(«мёлга-скрэб» и «бригелоу-скрэб»). В восточных и северных районах
страны простирается богатая злаками колючая саванна (овечьи паст-
бища).
В Африке колючекустарниковая степь пояса Сахель тянется от
широты Сенегала и озера Чад по южной окраине Сахары, пересекая
континент, в направлении Кордофана и северной Эфиопии. Восточно-
африканский участок, частично с древесными сообществами, богатыми
суккулентами, а частью тоже со злаковыми пространствами и колючими
кустарниками, простирается от внутренних районов полуострова Сомали
через большую часть восточной Кении до степи Масаи и далее на юг.
Южноафриканский пояс тянется от юго-западной Анголы через терри-
торию Амбо, Дамараленд и Калахари до южных окраин Южной Родезии,
земли Газа, захватывая юго-запад Мадагаскара. На юге, от Трансвааля
до нагорий Басутоленда, колючекустарниковая степь примыкает к летне-
влажным субтропическим высокозлаковым степям (Высокий Велд).
В Америке северная область пространственно наиболее расчленена
и по характеру формаций сильно дифференцирована. Основная ее часть
на плоскогорьях Мексики включает отдельные участки богатых злаками
колючих саванн, а также разнообразные суккулентные формации или
чистые заросли колючих кустарников без злакового покрова. Аналогичные
формации встречаются в Соноре и на полуострове Калифорния, на тихо-
океанском побережье южной Мексики, Юкатане, Антильских островах,
а также на северном побережье Венесуэлы и расположенных вблизи него
островах.
Каатинга засушливой северной Бразилии тоже делится на несколько
формаций. Каатинга буквально означает «белый лес» и названа так по
светлому тону деревьев, приобретаемому ими в сухое время, когда они
сбрасывают листву. Для этих лесов характерны «бутылочные» и «боченко-
видные деревья», а также древесные растения с водонакапливающими
подземными клубнями. Кроме того, здесь растут колючие деревья и кустар-
ники, листовые суккуленты и безлистные стеблевые суккуленты. Послед-
ние могут преобладать и образовывать отдельные формации. Наряду
с каатингой существуют кустарниковые формации («carascal»), которые
в самых засушливых местностях с осадками менее 350 мм переходят в сук-
кулентные полупустыни. Некоторые формации возникли под влиянием
человека, например «serido», представляющие собой обедненную фацию
повторного выгорания колючих кустарников каатанги.
Растительность области Гран-Чако в Парагвае и северной Аргентине
также не является однородной формацией. Она состоит частью из сухих
лесов с квебрахо (S chinop sis) и злаковым покровом на отдельных участках,
частью из колючих и суккулентных лесов с «бутылочными» деревьями,
канделябровидными стеблевыми суккулентами и колючими карликовыми
пальмами, частью из чистых зарослей колючих кустарников семейства
бобовых (Acacia, Prosopis и др.), а также из низкорослых формаций колю-
чих кустарников и суккулентов.
Пояс «монте» и «чаньяр», с востока и юга примыкающий к Чако,
может в большей своей части быть причислен к тому же комплексу форма-
236
Пространственное подразделение растительности
ций; сюда же можно отнести и растительность колючих зарослей чрезвы-
чайно сухих межгорных долин, глубоко врезанных в восточную окраину
Боливийских Анд,
С другой стороны Высоких Анд, покрытых сухой растительностью
пуны, на севере Чили в районе с редкими зимними дождями снова появ-
ляется узкий пояс формаций колючих кустарников и суккулентов, частич-
но с аналогичными видами, как, например, «альггарробо» (Prosopis juli-
flora) и «чаньяр» (Gourliera decorticans).
ПОЯС ТРОПИЧЕСКИХ И СУБТРОПИЧЕСКИХ
ПОЛУПУСТЫНЬ И ПУСТЫНЬ
Четко разграничить пустыни и полупустыни сухих субтропических и тропиче-
ских областей удается не везде. В зависимости от эдафических причин они могут тесно
переплетаться друг с другом на узком пространстве; зачастую они недостаточно иссле-
дованы, поэтому границы между ними воспринимаются разными авторами по-разному.
Замкнутые местами обширные области полупустынь окружают крайне аридные про-
странства и всюду образуют переход от пустынь к растительным формациям, требую-
щим большей влажности.
Кроме того, полупустыни в виде больших или меньших участков встречаются
и в поясе собственно пустынь. Резких границ провести здесь нельзя; сообщества
крайне ксероморфных растений постепенно становятся все более низкорослыми и раз-
реженными, так что даже непосредственному наблюдателю трудно бывает определить,
находится ли он в полупустыне или уже в пустыне.
Наибольшее протяжение и ярко - выраженное зональное распростра-
нение пустыни и полупустыни имеют в континентальном сухом поясе
северного полушария Старого Света, Эта зона простирается от восточных
Канарских островов через Сахару, юг Эритреи и Сомали, Аравийский
полуостров и южный Иран до северо-западной Индии,
В большей части Сахары годовое количество осадков, выпадающих
в виде редких, кратких, но сильных ливней (на юге летом, а на севере
обычно зимой), не превышает 50 мм. В обширных районах дождя не
бывает годами. На плоскогорьях Сахары полупустыни располагаются на
высотах до 3000 м. Здесь характерны значительные сезонные и суточные
колебания температуры; зимой нередки ночные заморозки. Среди расти-
тельных форм Сахары мало стеблевых суккулентов, В остальном расти-
тельность сильно варьирует в зависимости от качества субстрата. Наиболее
богата растениями каменистая пустыня (гамада); преобладают виды,
преимущественно сохраняющиеся в период покоя; карликовые древесные
виды, укореняющиеся в трещинах скал. Галечниковые пустыни (серир)
большей частью крайне бедны растениями, В песчаных пустынях, где
пески находятся в движении, нет вообще никакой растительности (эрг,
Ливийская пустыня), однако в условиях неподвижного субстрата, как
в некоторых районах внутренней аравийской пустыни, развивается
довольно обильная растительность. На засоленных глинистых низинах
встречаются разреженные галофиты. Сплошные соляные корки обычно
безжизненны, В местах близкого к поверхности залегания грунтовых
вод развиваются многолетние древесные растения с глубокими корнями,
карликовые кустарники или, особенно во многих «вади», деревья и ку-
старники (финиковые пальмы, олеандры). На ряде участков абсолют-
ных пустынь после необычно сильных дождей, которые выпадают,
может быть, раз в десятилетие, почва покрывается на короткое время
Климатические зоны, растительности
237
ковром зеленеющих эфемерных трав. Где осадки не столь редки, гео-
фиты сохраняются в почве многие годы, чтобы после первого же дождя
появиться на поверхности. Сахара является пограничной областью между
голарктикой и палеотрописом. В ее южной части встречаются африкан-
ские систематические единицы, а на севере — обедненная средиземно-
морская флора. Эндемы из систематических единиц более высокого ранга
здесь вряд ли возможны. С юга Сахару сплошной полосой окаймляют
полупустыни. Это преимущественно формации кустарничков, которые
произрастают здесь при 10—11 сухих месяцах и эфемерных летних осад-
ках 50—250 мм. На западе пояс полупустынь продвигается вдоль побе-
режья к северу и переходит в пояс, который в сходных климатических
условиях, однако с зимними осадками прослеживается вдоль северного
края пустыни. Растительность состоит из жестких дерновинных злаков,
которые в более влажных окраинных частях примыкают к субтропическим
зимнезеленым сухим степям. Альфа-альфа (Stipa tenactssima} возделы-
вается здесь как сырье для изготовления волокна. В Марокко и на Канар-
ских островах встречаются суккулентные кустарниковые формации,
а в полупустынях и пустынях полуострова Сомали характерны стебле-
вые суккуленты из эндемичных видов и родов. Дерновиннозлаковые полу-
пустыни и степи на северной окраине Сахары продолжаются в Южном
Атласе, сходные формации имеются и в юго-восточной Испании, и на Ближ-
нем Востоке (Палестина — южный Иран). Злаковые полупустыни —
основа кочевого скотоводства. С пустынными пространствами резко
контрастируют оазисы — участки среди пустыни или полупустыни, где
благодаря наличию открытой воды или близких грунтовых вод разви-
вается пышная растительность. В естественных оазисах североафрикан-
скоп засушливой области растут тростники, колючие кустарники и де-
ревья, частично суданского, частично ближневосточного происхождения.
Большинство оазисов превращено в культурные земли.
По характеру обводнения мы различаем: речные оазисы с частично регулярным
затоплением берегов (долина Нила, Месопотамия); оазисы у подножия гор на выходах
грунтовых вод (окраины Атласа, Тибет, Иран); оазисы у источников (или близ колодцев),
образующие либо сплошную полосу вдоль долины, либо беспорядочно рассеянные
по пустыне, а также оазисы на грунтовых водах, находящихся на глубине, доступной
для растений. В оазисах с помощью искусственной подачи воды можно увеличить оро-
шаемую площадь, а также регулировать степень и время увлажнения. Важнейшее
дерево североафриканских оазисов — это финиковая пальма. Она хорошо переносит
легкие заморозки, однако для созревания плодов требует пустынной жары. Ее можно
считать характерным деревом оазисов континентальных пустынь.
В пределах североамериканских субтропиков встречаются обшир-
ные пространства полупустынь в Мексике (Сонора, полуостров Калифор-
ния), в южной Аризоне, южной Калифорнии, а также в других районах
при наличии особых местных условий (подветренное положение, замкну-
тые внутриматериковые бассейны, наличие водопроницаемых пород,
антропогенное нарушение растительности и почв). Абсолютные пустыни
на больших пространствах здесь не встречаются. «Пустыни» южной Кали-
форнии (Мохаве, Долина Смерти) и южной Аризоны (Хила) по степени
сомкнутости растительного покрова в большей части следует уже отнести
к полупустыням. Американские ботаники понимают термин «пустыня»
гораздо шире, нежели европейские. Даже ксероморфные кустарниковые
формации и колючие суккулентные леса типа каатинги ими часто относятся
238
Пространственное подразделение растительности
к пустыням. Североамериканская сухая область только на некоторых
участках получает в год около 80 мм осадков. Обычно же здесь выпадает
более 100 и до 250 мм. Область, следовательно, далеко не столь же сухая,
как большая часть Сахары или прибрежные пустыни Южной Америки
и Южной Африки.
По данным климатической станции «Лаборатория в пустыне» (в Таксоне, южная
Аризона), среднее годовое количество осадков на высоте 740 м над уровнем моря состав-
ляет 293 мм. Нижняя высотная граница горных хвойных лесов расположена в южной
Аризоне уже примерно на высоте 1600 м. Особенность низинных сухих областей Север-
ной Америки с жарким летом заключается в исключительном обилии стеблевых сук-
кулентов.
Прибрежные пустыни южного полушария не так жарки, как конти-
нентальные пустыни северного полушария. Однако большая часть их
территории крайне суха. Если на земной поверхности и нет такой точки,
в которой наверняка никогда не бывает дождей, то все же в южноамери-
канской прибрежной пустыне на севере Чили имеются климатические стан-
ции, которые до сих пор не замерили даже малейшего количества осадков.
Под влиянием холодных морских течений, с одной стороны, и высокой
гряды Анд — с другой, климат пустыни на перуанском берегу доходит
почти до самого экватора (4° ю. ш.). Большая часть перуанско-чилийской
прибрежной пустыни, простирающейся до 24° ю. ш., является каменистой
пустыней. Однако встречаются и галечниковые пустыни и большие песча-
ные массивы. Сухой север Чили должен был бы иметь самые обширные
на Земле области, сплошь лишенные растительности, если не считать ледя-
ных пустынь. Только в виде узких и несплошных полос на береговых
склонах встречается кратковременная богатая травянистая растительность
и сохраняющиеся в период покоя заросли суккулентов, подушек и гео-
фитов (формация «лома»). Эти лишенные растений пространства ожив-
ляют лишь ленты оазисов, окаймляющих русла вдоль редких рек, прихо-
дящих сюда с гор. В северном Чили и южной Боливии, где пустыня заходит
высоко на плоскогорья, на окраинах обширных солончаков (салары)
также можно обнаружить кое-какую растительность, а на отдельных участ-
ках даже деревья (Prosopis tamarugd), которые, однако, большей частью
уже уничтожены. Во внутренних кордильерах Анд пустыня примыкает
на востоке к сухой пуне с формациями карликовых кустарников и дерно-
винных злаков, а на более высоких позициях — с характерными
растениями-подушками («llareta»). На юге чилийская пустыня примерно
у 27°ю. ш. переходит постепенно в очень однородную на обширном простран-
стве формацию карликовых кустарников, которая становится сомкнутее
и выше по направлению к югу. Она поднимается и на большие высоты
и доходит на юго-востоке до аргентинских склонов Анд.
Южноафриканская прибрежная пустыня (18—28° ю. ш.) не про-
стирается так далеко на север, как перуанская.
Речных оазисов здесь нет, вследствие отсутствия прилегающих высо-
когорий, откуда могли бы стекать живительные потоки. Количество
осадков в пустыне Намиб, доходящее иногда до 15—20 мм, почти столь
же мало, как и в Атакаме. Обе обладают большим числом эндемичных
групп, среди которых особенно характерным является древнее растение
Welivitschia. На окраинах Намиб окаймляют кустарничковые полупустыни
со скудными осадками на севере летом, а на юге зимой. В Карру эти
формации относятся к капскому флористическому царству, к полупусты-
ням, богатым суккулентами, с исключительным обилием видов, с эндемич-
Климатические зоны растительности
23$
ной и экологически весьма специально приспособленной флорой (например,
галечниковые и оконные1 растения). Во внутренних районах Южной
Африки с востока к ним примыкают высокозлаковые степи плоскогорий
с летними дождями. Сходная формация существует и на юго-востоке
Австралии и, возможно, в некоторых районах пампы, тогда как в север-
ном полушарии аналогичные формации, видимо, отсутствуют. На западе
Австралии прибрежная пустыня отсутствует. В этой части света лишь
в некоторых районах выпадает меньше 100 мм осадков и настоящих
пустынь мало. Каменистые пустыни имеют ограниченное распространение
во внутренних частях западной и южной Австралии, а кроме того, встре-
чаются эдафически обусловленные и локально ограниченные галечнико-
вые пустыни и песчаные пространства. Более обширные лишенные расти-
тельности солончаки, в которых теряются периодически текущие реки,,
берущие начало вне этих участков, распространены в основном вокруг
озера Эйр и окружены галофитными полупустынями. Большая часть
внутренних районов Австралии, ранее считавшихся пустыней, является,,
скорее, полупустынями с жесткими злаками (Spinijex — Halbwiiste).
На северо-западе и на юге они местами достигают побережья. Часто они
перемежаются кустарниками из акаций и казуариновых и через самые
различные формы переходят в замкнутый скрэб.
Л.ОЯС СУБТРОПИЧЕСКИХ ВЛАЖНЫХ ЛЕСОВ
И ЖЕСТКОЛИСТНЫХ ФОРМАЦИЙ
К сухому поясу континентальных внутренних районов и западных
побережий в субтропических широтах с их пустынями, полупустынями,
формациями колючих кустарников, суккулентов и сухих лесов примыкают
в западных областях континентов, там, где выпадают зимние дожди,
области с жестколистными лесами, а на высокогорных участках — с хвой-
ными лесами. Во влажных климатах восточных склонов с летними дождями
встречаются наряду с листопадными муссоновыми и сухими лесами прежде
всего вечнозеленые лавровые леса и жестколистные формации, а на боль-
ших высотах — вечнозеленые лиственные и хвойные смешанные леса.
Места, где дожди выпадают в течение всего года, особенно на восточном
побережье с пассатными дождями и на наветренных склонах гор, покрыты
вечнозелеными дождевыми и горными лесами. Горные дождевые и туман-
ные леса высоких позиций напоминают аналогичные леса внутренних
тропиков или принимают характер пышных лавровых или горных хвой-
ных лесов. Классификация этих типов растительности еще не во всех
случаях достаточно разработана.
В субтропиках растительность по климатическим причинам весьма
разнообразна. Это выражается не только в противоположности между
восточными и западными склонами, но сказывается и на небольших
участках в местных, порой крайне резких различиях растительности,
обусловленных особенностями экспозиции. Дело в том, что в этих широ-
тах разница в интенсивности солнечного облучения на солнечных и тене-
вых склонах особенно велика. Кроме того, исключительно резкая верти-
кальная дифференциация рельефа в особенности в Евразии ведет к тому,
что здесь нередко перемежаются полосы растительности умеренной зоны
1 Окопными растениями (Fensterpflanzen) называют некоторые виды Mesem-
bryanthemum (М, optirum, М, rhopalaphyllus), у которых все вегетативные органы
погружены в землю, па свет выставляется только верхушка листа, покрытая про-
зрачной водоносной тканью.— Прим, ред.
240
Пространственное подразделение растительности
(хвойные леса, летнезеленые лиственные леса, альпийские кустарнико-
вые формации и матты) с тропической растительностью.
На южной окраине Гималаев это явление особенно ярко выражено,
здесь растительность сильно дифференцирована одновременно как по
высоте, так и по направлению горных склонов. У подножия гор на востоке
Ассама располагаются вечнозеленые тропические дождевые леса, а на
западе (в Пенджабе) — субтропические формации колючих кустарников.
В Ассаме выше вечнозеленых субтропических горных дождевых лесов
на высоте 3000—4000 м проходит высотная ступень лаврового леса. Он
характеризуется здесь в основном видами Rhododendron и наличием
хвойных деревьев. Выше к нему примыкают влажные субальпийские
кустарники и матты. В Непале, от подножия и до отметки 1100 м, место
дождевых лесов занимает листопадный влажный муссоновый лес, тогда
как на больших высотах деление на высотные ступени примерно такое же,
как в Ассаме. В Западных Гималаях, напротив, горные леса состоят из
хвойных деревьев, а на высотах 2000—3500 м они представлены листвен-
ными и смешанно-хвойными породами. Выше леса и здесь сменяются
влажными альпийскими кустарниками и маттами. У подножия гор на
78° в. д. сухой муссоновый лес достигает высоты 1500 м. К северо-западу
на меньших высотах (до 600 м) его место занимают формации колючих
кустарников. Над Пенджабом между этими ступенями сухих кустарников
и хвойного леса вклинивается высотный пояс жестоколистного леса
(600—1100 м). Далее к северо-западу хвойный лес на высоте 600 м сле-
дует непосредственно за сухим кустарником; что касается обширной во-
сточноазиатской области распространения субтропических влажных лесов,
то в настоящее время известно очень мало определенного об облике и про-
странственном распределении первичных растительных формаций вслед-
ствие почти полного обезлесения этого района под воздействием человека.
С весьма неточными границами как между муссоновыми и дождевыми
лесами южного побережья, так и с частично вечнозелеными умеренными
лиственными лесами, примыкающими к ним севернее реки Янцзы, тя-
нется от гор Юньнани до восточного побережья южного Китая пояс шири-
ной почти 1000 км, естественная растительность которого, видимо, боль-
шей частью состояла из широколиственных лавровых лесов (типа магно-
лиевых) и частью, возможно, из жестколистных лесов с примесью листо-
падных компонентов. К этому же поясу относятся южные острова япон-
ской группы и южная оконечность Кореи.
В западной части евразийского континента субтропические влажные леса суще-
ствуют только на немногих участках, отличающихся особенностью своего располо-
жения. За исключением находящихся в полосе туманов атлантических островов с их
лавровыми лесами, это прежде всего северное подножие Понтийских гор, Колхида
и южное побережье Каспийского моря с пышными влажными лесами, отличающимися
богатством видов и состоящими из древесных пород, чувствительных к холоду, частью
листопадных, а частью вечнозеленых.
На юго-востоке Северной Америки субтропические влажные леса
встречаются во Флориде и на низменностях примыкающих к ней побе-
режий (побережье Мексиканского залива, Джорджия, Южная Каролина).
По мере продвижения в глубь страны они переходят в умеренные леса.
Только на юге Флориды не бывает заморозков, имеются вечнозеле-
ные широколиственные леса, богатые эпифитами, в которых наряду
с вечнозелеными дубами и магнолиями встречаются пальмы и другие
роды тропических деревьев, а также болотистые леса, травяные болота,
Климатические зоны растительности
241
а на побережье — мангровые заросли. На севере полуострова уже бы-
вают довольно сильные морозы, именно поэтому окончилась неудачей
попытка возделывания здесь цитрусовых. Эта область относится к поясу
«юго-восточных сосновых лесов», который распространяется к северу по
равнинам Пидмонта, а к западу через прибрежные низменности Мекси-
канского залива к долине Миссисипи. Леса состоят преимущественно
из длиннохвойных сосен субтропического типа. На влажных почвах
побережья и нижнего течения Миссисипи они чередуются с обширными
лиственными лесами с богатым видовым составом. Последние нередко
содержат в своем составе и вечнозеленые породы. Особенно ярко выра-
женный тропический характер имеют обширные затопляемые низмен-
ности («flats») и лесные болота («hammocks»). На месте речных стариц эти
низменности заросли лиственными деревьями, а на илистой почве реч-
ных лагун, лишенной торфа («glades»),— знаменитыми болотными кипа-
рисами {Taxodium distichum).
Их совершенно прямые, лишенные ветвей стволы, в противоположность сопро-
вождающим их лиственным деревьям, очень устойчивы против ураганов торнадо.
Деревья имеют сильно вздутое основание ствола, поднимающееся до высоты прибоя, и,
кроме того, удерживаются опорными корнями, изгибающимися над почвой на высоте
до 3-х м. Эти древостои используются в лесном хозяйстве, однако эксплуатировать
их в условиях болот чрезвычайно трудно,так что обширные массивы лесов еще сохра-
няются нетронутыми.
В южном полушарии вечнозеленые влажные леса произрастают
в субтропических восточных частях всех трех континентов. В восточной
Австралии они тянутся от 25° ю. ш. через весь Новый Южный Уэльс
сплошной полосой, имеющей, однако, на восточных склонах ширину
всего лишь в несколько десятков километров. На высоте более 800 м
равнинный лес переходит в горный дождевой лес, богатый папоротни-
ками. Этот последний во внутренних горных районах резко граничит
с сухими эвкалиптовыми лесами подветренных склонов. В наиболее бога-
той осадками юго-западной части Австралии тоже имеются океаниче-
ские влажные леса (карри), примыкающие к типу лавровых лесов.
На Северном острове Новой Зеландии, как и в восточной Австралии,
дождевые леса встречаются наряду с жестколистными формациями и сооб-
ществами хвойных деревьев (новозеландский агатис, Agathis australis).
Они могут быть отнесены к тому же поясу. Напротив, влажные леса
Тасмании и Южного острова Новой Зеландии относятся к поясу уме-
ренных дождевых лесов,
В Южной Африке восточные склоны гор между 24 и 32° ю. ш., где
наблюдается равномерное распределение годовых осадков, покрыты суб-
тропическим дождевым лесом. В сходных условиях, однако изолированно
от них, на южных склонах Капских гор растут пышные лавровые леса,
занимающие незначительный участок, примыкающий к области зимних
дождей.
Весьма обширны субтропические влажные леса Южной Америки.
На севере границу их с тропическими дождевыми и горными лесами
провести трудно. Переход наблюдается примерно на широте тропика
Козерога и происходит очень постепенно, что обусловлено широтным
простиранием побережья. В южной Бразилии, восточном Парагвае
и северной части Уругвая сильно разреженный в настоящее время лес
простирается далеко в глубь континента. Характер его в пределах Бра-
242
Пространственное подразделение растительности
зильского нагорья определяется частью лавролистными кустарниками
(Ilex paraguayensis), а частью хвойными породами (Araucaria angusti-
jolia). К субтропическим влажным лесам можно отнести и горные ту-
манные леса на восточных обрывистых склонах Боливийского плоско-
горья в пределах южного Юнгаса, а также изолированно встречаю-
щиеся на островах Хуан-Фернандес и в Малом Северном Чили лавровые
леса, обусловленные прибрежными туманами в горах Фрэй-Хорге и
Талинай.
К субтропическому поясу летневлажных лесов, видимо, следует отнести и часть
аргентинской и южноуругвайской пампы, первоначальный лесной или степной харак-
тер которой сейчас неясен. Лёссовые равнины, тянущиеся от побережья в глубь стра-
ны, превращены сейчас в сельскохозяйственные угодья. В области Ла-Платы выбор
возделываемых культур определяется субтропическим климатом и близостью моря.
Далее во внутренних районах страны посевы культурных злаков (маис, пшеница) смы-
каются с посевами люцерны и обширными пастбищами. Многолетние злаки на этих
участках, близких к побережью и влажных на протяжении всего года, остаются зеле-
ными и во время мягкой зимы. На западе, напротив, господствуют сухие пастбища
с формациями дерновинных злаков и полукустарников, чередующиеся с колючими
кустарниками (монте) и солеными озерами. Травянистые пространства более влажных
восточных районов необоснованно считались ранее равноценными прериям-степям
северного полушария. Злаковые сообщества пампы — явно вторичные формации, так
как состоят большей частью из чужеродных видов. То, что по крайней мере в централь-
ной пампе возможно произрастание деревьев, показывают рощи плодовых деревьев
и отдельные деревья близ поселений, а также участки искусственных насаждений.
Кроме того, существуют исторические свидетельства о наличии сообществ «monte»
уже после их лесохозяйственного использования переселенцами из Европы (Schmieder,
1929). С другой стороны, среди местных видов деревьев неизвестны такие, которые
могли бы входить в состав первоначально существовавшего здесь леса. Поэтому
у нас нет ясного представления о том, как выглядела первобытная растительность
пампы.
Все области распространения жестколистной растительности в запад-
ных районах континентов находятся на одинаковых широтах (около 35°).
Однако основная их часть в северном полушарии (от 37 до 38°) лежит
примерно на 3° ближе к полюсу, чем в южном полушарии. Средиземно-
морская область и ее ответвления в западных районах Атласских гор
и южной Франции имеют самую большую протяженность с севера на юг
(от 31 до 44°), а южноафриканская — наименьшую (от 30 до 35°). За исклю-
чением района Средиземного моря и Австралии, жестколистные области
в общем ограничены узкими участками на западных окраинах континен-
тов и имеют резкие границы с соседними областями растительности.
В Австралии имеются две жестколистные области; одна — на юго-
западе, по положению аналогичная южноафриканской, а другая —
в прибрежных районах юго-восточной Австралии, примерно от полу-
острова Эйре до южной оконечности Виктории. Обе выражены недоста-
точно четко, потому что по мере продвижения в глубь страны они пере-
ходят в сухие леса, в которых, как и в жестколистных лесах, преобладают
вечнозеленые виды Eucalyptus. Так же трудно выделить на юго-западе
Австралии жестколистные леса из лаврового леса «карри», уже упоминав-
шегося при описании влажных лесов.
Средиземноморская область, послужившая основной моделью при
обосновании типа областей жестколистных лесов, сама отнюдь не является
Климатические зоны растительности
243
характерным представителем этого типа. Это единственная из всех жест-
колистная область, проникающая далеко в глубь суши и вследствие
крайней изрезанности береговой линии и расчлененности рельефа являю-
щаяся и по горизонтальному протяжению и по вертикальному строению
исключительно сложной. На своих южных и юго-восточных границах
она перемежается с растительностью субтропических сухих областей,
на севере — с летнезелеными лиственными лесами Средней Европы,
Р и с, 64, Типичные ареалы средиземноморских видов растений (по Рикли и Мей-
зелю),
а — каменный дуб (Qucrcus ilex); б — пиния (Pinus pinta); черные площади внутри ареала
означают скопления, полузатепепные кружки вне ареала — посаженные и перезимовывающие
па открытом воздухе насаждения; в — эспарто (Lygevm spartum) — светлая штриховка, альфа-
альфа (Stipa tenacissima) — темная штриховка; г — иудино дерево (Cercis siliquastrum).
а на востоке — с сухими лесами и сухими степями Анатолийской возвы-
шенности, которые в свою очередь занимают промежуточное положение
между типами растительности субтропической и умеренной зон.
Калифорнийская и средиземноморская жестколистные области, рас-
положенные обе в голарктическом флористическом царстве, имеют
в составе таксономических групп большое число общих родов, как,
например, Quercus, Arbutus, Pinus, Juniperus. Флоры жестколистных
областей в южном полушарии весьма разнятся как между собой, так
и с флорой аналогичных областей северного полушария. Для них харак-
терно резко самостоятельное флористическое развитие с большим числом
эндемичных видов, родов и даже семейств. Наиболее четко выражена
самостоятельность южноафриканской области — основной части капского
флористического царства.
244
Пространственное подразделение растительности
Вечнозеленая жестколистная растительность субтропических обла-
стей с зимними дождями — один из лучших примеров климатически
обусловленной конвергенции. Здесь характерны сухое лето и мягкие
Р п с. 65. Карта и высотный профиль растительных поясов на острове
Гомера (Западные Канарские острова) (Ceballos, Ortuno, 1951).
1 — пояс «crassicauletum» с ксерофптной местами богатой суккулентами кустарни-
ковой формацией и участками сообщества Phoenicia canariensis', 2 — пояс
«sabinar» с кустарниковыми зарослями Juniperus phoenicea (маквисы); 3 — пояс
«laurisilva», туманный лавровый лес с Laurus canariensis, Ilex canariensis, Persea
indica и др.; 4—пояс — «fayal-brezal» с жестколистным лесом из Myrica faya и
Erica arborea.
или прохладные зимы, не " прерывающие периода вегетации. Однако
морозы ниже >—10°, случающиеся иногда в Средиземноморье, оказы-
ваются губительными для части местных жестколистных видов, а тем
более для цитрусовых. Летняя жара в большинстве жестколистных обла-
Климатические зоны растительности
245
стей довольно умеренна (средняя температура самого жаркого месяца
около 20°). Только во многих районах европейского Средиземноморья
лето бывает исключительно жарким. Количество осадков может значи-
тельно варьировать.
Там, где зимние дожди сочетаются с летними туманами, как это
наблюдается на открытых пассатам больших высотах западных Канар-
ских островов и на островах Хуан-Фернандес, жестколистная раститель-
ность сменяется или пышными лавровыми лесами (рис. 65), или экологи-
чески сходными хвойными лесами, например на калифорнийских при-
брежных Кордильерах и на Тенерифе.
Растительные сообщества, составляющие «дивизионы растительно-
сти» отдельных жестколистных областей, флористически совершенно раз-
личны, так как расположены в четырех различных флористических
царствах. Физиономически они, наоборот, на аналогичных местообита-
ниях совпадают. Не касаясь конституционно обусловленных различий
(хвойные в Калифорнии, эвкалипты в Австралии, пальмы в Чили), можно
сказать, что все жестколистные области обладают почти одинаковыми
сериями формаций: жестколистные леса, формации жестколистных кустар-
ников (маквисы, чапараль), гариги, на влажных местообитаниях —
частично листопадные леса и др. Антропогенные замещающие сообщества
при аналогичных условиях (распашка, косьба, выпас, пожары, ирри-
гация и т. д.) имеют такую же явно выраженную конвергенцию, а кроме
того, вследствие очень широкого антропогенного распространения многих
сорняков, часто и значительное флористическое сходство. Так, напри-
мер, ряд сообществ сорняков в чилийской жестколистной области почти
идентичен с сообществами сорняков европейского Средиземноморья. Эта
аналогия наблюдается и в других областях. К сожалению, планомерные
сравнительные исследования здесь не проводились. Конвергенция рас-
пространяется и на типы почв (средиземноморские желтоземы и красно-
земы), и на характер эрозии почвы, вызываемой хозяйственной деятель-
ностью человека. Однако степень разрушения почв в отдельных областях
по историческим причинам неодинакова. С этим связаны различия в рас-
пространении формаций. Так, в Средиземноморье районы с наиболее
деградированными почвами, покрытые гаригами или аналогичной им
растительностью, занимают уже на протяжении нескольких столетий
большую часть необрабатываемых земель. Наполовину естественные
участки каменнодубовых лесов, являющиеся на значительной части
западного Средиземноморья потенциальной естественной растительностью,
также представляют собой исключительную редкость. В других обла-
стях процесс обезлесения, связанный с разрушением или по крайней
мере с нарушением первоначальной почвы, закончился значительно
позже или происходит только сейчас, на наших глазах. Поэтому в таких
местностях еще можно найти на обширных площадях естественный жестко-
листный лес, хотя бы и в измененной форме. Связь широкого распростра-
нения почти обнаженных каменистых эродированных участков с уничто-
жением лесов, как это сначала в виде гипотезы принималось для Среди-
земноморья, совершенно безусловна там, где можно непосредственно
наблюдать последовательность всех стадий этого процесса, как, напри-
мер, в среднем Чили. Для более сухих районов жестколистных областей,
где характерны гариги или фриганы или — в юго-восточной Испании —
дерновиннозлаковые степи, состояние исследований еще не позволяет
сделать точного заключения, насколько далеко проникли первобытные
формации жестколистных лесов или кустарников и какой тип формации
246
Пространственное подразделение растительности
следует принять за потенциальную естественную растительность. В отно-
шении возделываемых растений — включая парковую и садовую флору,
придорожные деревья и т. д. — и рудеральной растительности в селениях
и на путях сообщения, а также на неиспользуемых участках жестколист-
ные области как по фактическому, так и по потенциальному видовому
составу в значительной степени совпадают. Различия в антропогенной
флоре больше зависят от трудности освоения новых местообитаний и от
давности, интенсивности и методов земледелия, существующих в данной
стране, чем от различий в природных условиях, которые, однако, тоже
имеют значение.
ПОЯСА РАСТИТЕЛЬНОСТИ УМЕРЕННЫХ ЗОН
ПОЯС ПУСТЫНЬ, ПОЛУПУСТЫНЬ
И СТЕПЕЙ УМЕРЕННЫХ ЗОН
Сухие пустыни в умеренных широтах ограничены засушливыми вну-
триконтинентальными районами. Обширные настоящие пустыни сущест-
вуют только в Азии (Западная Гоби, бассейн Тарима, Средняя Азия,
Иран). Однако полупустыни и сухие степи занимают в этих областях
также обширные пространства. На Иранском нагорье наблюдается пере-
ход к субтропическим сухим областям, которые вместе с зимнехолодными
сухими областями Внутренней Азии примыкают к обширному засушли-
вому поясу Старого Света. На западе Северной Америки внутренние высо-
когорные полупустыни и сухие степи у восточного подножия Скалистых
гор южнее переходят непосредственно в субтропический сухой пояс.
Важнейшие различия по сравнению с аналогичными формациями
субтропиков обусловлены зимними холодами. Так, например, во вне-
тропических полупустынях и пустынях отсутствуют стеблевые сук-
куленты. Не считая хвойных вечнозеленых кустарников (например, Juni-
perus), древесные растения здесь в основном представлены безлистными
зимой кустарничками или полукустарниками. Большую роль вместе
с тем играют дождезеленые многолетние дерновинные злаки. Солончако-
вые формации с галофитными и суккулентнолистными полукустарниками
и кустарниками сходны с соответствующими формациями субтропиков.
Особенно резкие различия наблюдаются в оазисах, поскольку здесь,
в зимнехолодных областях, все тропические растения отсутствуют (на-
пример, пальмы). В речных оазисах Тарима растут тополя и ивы, в Сред-
ней Азии, кроме того, лохи и высокие тростниковые заросли. Облик
культурного оазиса определяют листопадные деревья и прочие полезные
растения умеренных широт. Вечнозеленые плодовые деревья, такие, как
цитрусовые, выращиваются лишь там, где местный климат приближается
к субтропическому.
Примыкающие к полупустыням степные пояса обладают аналогичны-
ми для обоих континентов типами формаций, различающимися лишь по
степени влажности климата. Однако в то время как в Евразии полосы
однородной растительности в общем тянутся с запада на восток, в Север-
ной Америке они имеют меридиональное протяжение. К полупустыням
района Скалистых гор примыкают с последовательно уменьшающейся
сухостью по направлению к востоку:
а) сухие степи на бурых почвах с разреженным покровом из полу-
кустарников и жестких дерновинных злаков;
Климатические зоны растительности
247
б) более сомкнутые низкозлаковые сообщества на каштановых поч-
вах, по климатическим причинам лишенные деревьев (осадков менее
500 мм);
в) высокотравные прерии с обилием разнотравья и мягких злаков
на черноземе, почти полностью распаханные с середины XIX в.
г) лесостепи пояса гикори с чередующимися участками леса и пре-
рии на деградированных черноземах и черноземовидных почвах.
Для двух последних поясов остается открытым вопрос, в какой сте-
пени человек, выжигая степной травостой, способствовал распростране-
нию степных площадей за счет леса. В настоящее время первобытная
растительность, в основном в восточных поясах, почти полностью исчезла;
ее заменили сельскохозяйственные угодья, рудеральные сообщества и дру-
гая спонтанная вторичная растительность. В самых сухих поясах вне
границы богарного земледелия растительность сильно изменена выпа-
сом скота. Таким образом, приведенная выше последовательность поясов
растительности характеризует только потенциальную естественную расти-
тельность. Реально это деление проявляется в замещающих сообществах,
в возможности хозяйственного использования земли в соответствии
с таким поясным подразделением и различной степени уменьшения пло-
дородия почвы.
В степях Западной Сибири и Украины последовательность типов
формаций в направлении на север и северо-запад от среднеазиатских
пустынь Каракум и Кызылкум и окружающих их полупустынь такая же,
как и указанная для Северной Америки. Полупустыни представляют
собой в основном галофитные формации на засоленных почвах. К ним при-
мыкают открытые, зеленеющие короткой весной сообщества кустарнич-
ков и полукустарников, частью с низкорослыми дерновинными злаками
на серых или желтоватых почвах, обогащенных основаниями в верхних
горизонтах, или на бурых, но еще очень бедных гумусом почвах. Затем
следуют степи из перистых ковылей (Stipa — сухие степи) на каштановых
почвах и частью на малогумусных черноземах (южные черноземы).
Описание этой формации Кернером (Kerner, 1863), незначительно сокращенное,
может быть приведено здесь в качестве классического примера изображения им расти-
тельности: «Родиной перистого ковыля («сироткины волосы»— Waisenmadchenhaar)
являются степные просторы юга европейской России и Венгерской пушты. Его дер-
новины определяют физиономию ландшафта, а сам ковыль — характерное растение
разнотравной степи. Стебель и листья ковыля выглядят жесткими и хрупкими, причем
листья по форме напоминают щетину и из-за своей матовой серо-зеленой окраски
не могут придать той местности, на которой растение произрастает, свежий, живой вид.
Развитие новых листьев и стеблей начинается уже поздней весной, когда первые цветы
пушты — маленький нежный гусиный лук — уже давно отцвели, а весенние анемоны
несут на себе шарообразные покрытые волосками плоды. Как на континентальном
востоке все фазы развития растений следуют одна за другой, так и у этого степного
злака стадии распускания, цветения и увядания сменяют друг друга в течение корот-
кого времени. Через несколько дней после того, как из старой, словно бы высохшей
дернины пробьются молодые листья и стебли, начинает распускаться цветочная метел-
ка. Из самого верхнего листового влагалища вылезает целый пучок гибких волосовид-
ных остей, который сначала кажется серебристым блестящим пучком, но постепенно
удлиняется и превращается в белые, развевающиеся на ветру нити, каждую из которых
можно сравнить с тонким нежным пером. К этому времени вся растительная формация,
поскольку распустилось ее фоновое растение «сироткины волосы», достигает кульми-
национного пункта своего развития. Лютики, гвоздики, орхидеи, карликовые желтые
248
Пространственное подразделение растительности
касатики, прижимающиеся к почве пучки желтоцветных Astragalus и обладающая
мускусным запахом J иг inea, чьи репейниковые пурпурно-красные головки качаются над
белыми нитями Stipa, образуют красный ковер цветов, над которым возвышают свои
соцветия столь характерные для пушты перистолистные мотыльковые растения. Только
слишком уж быстро проходят эти картины. В июне все желтеет и высыхает под лучами
солнца, и если даже в мягкие дни осени еще распускаются кое-какие, цветы, они уже
не могут оживить ландшафта, и в то время, когда на северогерманском верещатнике
по белому фону разливается мерцающий красноватый тон, образованный миллионами
цветков, пушта остается пустынной, почти лишенной цветов землей с высохшими
кустиками травы и палевыми стеблями, качающимися на осеннем ветру».
Между тем степь уже отступила в связи с развитием сельского хозяйства. От пуш-
ты до Западной Сибири большие участки целинной степи теперь можно встретить только
в заповедниках.
К перистоковыльным степям примыкает следующая, более влаж-
ная ступень — «луговая степь», более богатая видами и совпадающая
с поясом оптимального развития чернозема. Пышное, метровой высоты
разнотравье, преобладающее здесь над злаками, охарактеризовал еще
Гумбольдт, когда он писал, что на низких повозках только стоя можно
ориентироваться в этих «травяных степях». С первых теплых дней после
таяния снегов до конца июля в богатой разнотравьем луговой степи
почти каждую неделю меняется аспект. Осенью она высыхает, и часть
корневой массы, образовавшейся в почве за лето, отмирает, способствуя
накоплению гумуса мощностью до 1,5 м. Благоприятствуют образованию
перегноя зимние холода и большая сухость летом, задерживающие про-
цессы бактериального распада, умеренное вследствие периодически направ-
ленного кверху движения почвенных вод выщелачивание оснований,
а также участие животного мира почвы в разложении остатков растений
и связывании органических веществ в почве. Благодаря большим резер-
вам гумуса эти почвы относятся к наиболее плодородным на Земле.
К безлесной степи примыкает следующий широкий степной пояс с лес-
ными островками на деградированных черноземах. Открытые участки заня-
ты степной растительностью, которая по флористическому составу близка
к лугово-степным сообществам, по эдафическим или местным климатическим
причинам проникающим далеко на запад в область среднеевропейских
летнезеленых лесов. Так как считается, что формирование чернозема
возможно только под безлесной степью, то пояс современной лесостепи
следует, видимо, рассматривать как бывший климатический степной пояс,
в котором чернозем образовался в период более континентального кли-
мата, когда граница леса проходила значительно севернее. Только после
окончания ледникового периода в этом поясе возникли условия, доста-
точные для произрастания леса. Но поскольку в России именно лесостеп-
ной пояс является областью наиболее раннего и интенсивного земледелия,
что можно доказать археологически, то предполагается, что это явилось
частичным препятствием к распространению леса на бывшие степные
почвы. Поэтому граница леса здесь обусловлена скорее не климатом, а хо-
зяйственной деятельностью человека (Wilhelmy, 1950). Однако на этот
счет существуют и другие мнения. Безлесие южных степей, напротив,
при естественных условиях и в настоящее время обусловлено климатиче-
скими причинами. Проростки деревьев и молодая поросль не в состоянии
выдержать естественной конкуренции со степными растениями. Однако
возможность роста здесь деревьев, если в раннем возрасте их охранять
от конкуренции степных растений, а в засушливые периоды поливать,
доказана существованием многочисленных лесных посадок.
Климатические зоны растительности
249
В южном полушарии нет формаций, соответствующих прериям-сте-
пям Евразии или Северной Америки, и нет настоящих черноземных почв.
В восточной Патагонии степной пояс в «дождевой тени» Анд содержит
сухие степи с гнездообразно растущими жесткими злаками, кустарнич-
ками и подушковидными растениями, которые могут переходить, с одной
стороны, в полустепные открытые формации, а с другой, на южной границе
с летнезелеными субантарктическими лиственными лесами,— в сомкну-
тую летнезеленую кустарниковую формацию. В Новой Зеландии на под-
ветренных склонах Южного острова наблюдаются сходные условия.
Наряду с сообществами разреженно растущих дерновинных злаков
существуют пустошные кустарничковые формации с подушковидными
жесткими злаками, кустарничками, кустарниками и разнотравьем.
ПОЯС ЛИСТВЕННЫХ ЛЕСОВ УМЕРЕННЫХ ЗОН
Область летнезеленых лиственных лесов умеренной зоны
Летнезеленый лиственный лес умеренной зоны, не считая Чили,
на обширных пространствах распространен только в северном полу-
шарии. Как потенциальная естественная растительность он господствует
в поясе, который широко раскинулся в Средней Европе, Восточной Азии
и восточной части Северной Америки, сильно сужаясь у северной окраины
прерии-степи и совершенно исчезая в горных областях Центральной
Азии и тихоокеанских районах Северной Америки.
Наибольшей протяженности с севера на юг пояс лиственных лесов
достигает на атлантическом побережье Европы, где он со своими самыми
северными ответвлениями доходит до северной границы леса, а на юге —
до северных районов Португалии. В Западной и Средней Европе основ-
ные массивы лиственных лесов расположены примерно между 45 и 55° с. ш.
а на восточных окраинах континентов, напротив, южнее 45° с. ш., что
обусловлено влиянием морских течений на климат. Почти на всех участ-
ках пояса к областям чистых лиственных лесов с севера примыкает пере-
ходный пояс из лиственных и хвойных лесов (рис. 66).
Этот последний в Восточной Азии (Камчатка, бассейн Амура) и на тер-
ритории Европы от средней Швеции до средней России особенно широк,
однако столь же отчетливо выражен он и в районе Великих озер Северной
Америки, и в северном лесном окружении канадских прерий. Далее на
юге хвойные леса встречаются в глубине областей лиственных лесов,
главным образом только в горном поясе, нижняя высотная граница
которого повышается по направлению к югу. Таким же образом в Европе
и Восточной Азии летнезеленый лиственный лес вторгается как высотная
ступень в жестколистную область лесов, примыкающую к нему на юге
(рис. 67).
Пояс лиственных лесов сильно изменен человеком, и в обширных
районах США, Средней Европы и Восточной Азии лес, исключая разве
только крутые горные склоны, почти полностью вытеснен сельскохо-
зяйственными угодьями, городскими и промышленными застройками.
Основные области летнезеленого лиственного леса расположены
в окраинных частях континентов с умеренно прохладным климатом,
с осадками, равномерно распределенными в течение года. Определяю-
щим является здесь, с одной стороны, ежегодный перерыв вегетацион-
ной деятельности зимними холодами (со снегом и морозами), а с дру-
250
Пространственное подразделение растительности
гой — периоды вегетации с обильными дождями или по крайней мере
достаточно влажные, умеренно или вполне теплые, длящиеся не менее
пяти месяцев. По типичному климату этого пояса растительности и сло-
жилось наше общее представление о четырех временах года.
Р и с. 66. Общее распространение Мajanthemum bifollium (по Липп-
маа).
Рис. 67. Схематические высотные профили распространения Fagus
silvatica, Abies alba и Acer pseudoplatanus в Средней и Южной Европе
(Meusel, 1942).
Южная граница летнезеленых лиственных лесов и их контакты с вну-
триматериковыми степями определяются главным образом сухостью
летнего периода, а граница с северным поясом хвойных лесов — преиму-
щественно поздними заморозками, губительными для цветения и молодых
побегов лиственных деревьев, и сокращением периода вегетации. В общем
же постепенное изменение климата пояса лиственных лесов определяется
удаленностью от океана.
Приморские части атлантической Европы (Ирландия, юго-запад
Великобритании, Бретань и северо-запад Пиренейского полуострова)
при прохладном и продолжительном лете отличаются исключительно
мягкой зимой (средняя температура наиболее холодных месяцев -j-2° С),
Климатические зоны растительности
251
так что полного зимнего периода покоя в этих широтах не бывает. Многие
факультативно зимнезеленые растения, как большая часть луговых злаков
и разнотравья, остаются зелеными в течение всего годы. Ареалы вечно-
зеленых растений южных широт («средиземно-атлантические» виды)
здесь продвигаются дальше всего на север (рис. 68). Полярная граница
Рис. 68. Типичные ареалы различных западпо- и среднеевропейских флористиче-
ских комплексов.
а — Arbutus uru’do, по Рикли и Сили; б — Genista angtica, по Хаиииигу; е — Dentaria but-
biiera, но Мейзелю; г — Titia cordata (пунктир), по Жаккарду и Фрею; Abies atba (штри-
ховка), по Маттфельду.
распространения некоторых из этих видов зависит от суммы летних тем-
ператур, необходимых для их плодоношения.
Еще более влажная ступень, переходящая в вечнозеленые дождевые
леса умеренной зоны южного полушария, примыкает здесь к типу кли-
мата южноамериканских летнезеленых лиственных лесов южной части
среднего Чили, отличающихся богатством вечнозеленых древесных расте-
ний. Субантарктические летнезеленые лиственные леса Огненной Земли
и верхнего высотного пояса южных Анд физиономически и по климати-
ческим условиям скорее всего можно сравнить с березовыми лесами Ислан-
дии и северной Норвегии.
В субокеаническом климате областей Средней Европы, примыкаю-
щих к атлантическому побережью, лето тоже умеренно жаркое, а зима
252
Пространственное подразделение растительности
сравнительно мягкая (средняя температура самого холодного месяца
от 4-2 до —3° С); продолжительность вегетационного периода 5—7 меся-
цев. Однако годовые колебания температуры здесь резче, чем на побе-
режье (средняя годовая амплитуда крайних температур составляет 15—
30°), а главное, что может оказаться определяющим для состава расти-
тельности, это случающиеся время от времени сильные зимние морозы или
летние засухи. Недостаток влаги приводит иногда к преждевременному
листопаду. Дифференцированность местообитаний по виду почв, разли-
чию экспозиций и высотному положению может здесь оказывать гораздо
более сильное влияние на распределение растительности, чем это наблю-
дается в других климатах.
В глубине континента влияние океана на климат ослабляется. Лето
становится теплее (средняя температура самого теплого месяца более
4-20°), зима холоднее (средняя температура самого холодного месяца
—6° С). Одновременно уменьшается количество осадков, особенно в юж-
ных районах, что может стать критическим моментом для роста деревьев.
Сходный климат, но с еще более резкими амплитудами и более жарким
летом (средняя температура самого теплого месяца до 4-26° С) имеют
области лиственного леса континентальной Восточной Азии и востока
Соединенных Штатов. Для внутренних равнин, расположенных запад-
нее Аппалачей, характерна сухая после относительно обильных летних
осадков осень, известная под названием «индейского лета» из-за красоч-
ной расцветки листвы на деревьях.
Сильная летняя жара, но более мягкая зима (средняя температура
самого холодного месяца выше -4-2° С) характерны для климата лесных
областей северной Японии и Кореи.
Почвы весьма разнообразны в зависимости от характера породы
и условий влажности. Преобладают всевозможные разновидности бурых
лесных почв (среднеевропейские буроземы), сменяющиеся местами более
или менее оподзоленными почвами, рендзинами или псевдоглеевыми
и глеевыми. На территории Средней Европы можно различить множество
почвенных разностей этого типа почв, которым, как это уже показало
картирование лесной растительности, соответствует столь же богатое
разнообразие природных единиц растительности.
Сильная подзолистость — невключение и устраняется часто только
при хозяйственном использовании этих почв. Большинство природных
лесных почв богато минеральными солями и гумусом и поэтому весьма
плодородно. Однако после корчевания и распашки достаточное содержа-
ние минеральных и органических веществ в почве сохраняется только
в случае внесения удобрений и правильной обработки почвы. За тысяче-
летия и века многие сельскохозяйственные почвы сильно изменились,
что привело к образованию их нового, антропогенного, профиля.
Большая часть пояса летнезеленых лиственных лесов — это области
с интенсивным земледелием и плотным населением, выросшим благодаря
наличию городов и промышленности. Лес на обширнейших простран-
ствах большей частью сведен, а остатки его с точки зрения хозяйственного
использования малоценны. При этом сохраняются различия в зависимости
от времени сведения леса и уровня культуры земледелия в соответствую-
щий период. В Средней Европе, например, где большинство лесов было
выкорчевано еще в доисторическое время (неолит) и средние века (VIII—
XIII вв.) это избирательное влияние сильнее, чем в Северной Америке,
где основное выкорчевывание леса начиная с XIX в. падает на эпоху искус-
ственных удобрений и механизированного сельского хозяйства.
Климатические зоны растительности
253
Аналогичные различия сохраняются и нри смене форм лесопользова-
ния. Так, нанример, современное распределение лиственных и хвойных
деревьев в Германии является но преимуществу отражением истории
хозяйства. Исследование растительности в таких областях богато пробле-
мами и довольно затруднительно; поэтому просто удивительно, что еще
Р и с. 69. Растительные сообщества лесных’площадей близ Беловежа как пример
европейской естественной растительности (Matuszkiewicz, 1954).
1 — сосновое верховое болото (Sphagnetum medii pinetosum)', 2 — ивово-крушиновая топь
(Salix aurita-Frangula alnus)\ 3 — ольховая топь (Alnetum glutinosae); 4 — дубово-грабовый
лес (Querceto-Carpinetum medioeuropaeum)', 5 — ольхово-ясеневый пойменный лес (Circaeo-Alne-
tum)', 6 — дубово-березовый лес (Querceto-Betuletum)', 7 — еловый и сосновый лес (Pineto-Vac-
cinietum myrtilliY, 8 — сосновый заболоченный лес (РгпМо-УассгпгеЩт uliginosi).
оказалось возможным, несмотря на сильное антропогенное воздействие,
достаточно полно охватить естественную пространственную дифферен-
циацию растительности.
Ниже приводится несколько обобщенное перечисление богатого
разнообразия единиц растительности на примере почти полностью окуль-
туренных ландшафтов Средней Европы.
Несмотря на бедность видов древесной флоры по сравнению с Восточ-
ной Азией и Северной Америкой, что вызвано последствиями ледникового
254
Пространственное подразделение растительности
периода, в лиственных лесах можно все же различить большое коли-
чество растительных сообществ, которые фитоценологически объеди-
няются в следующие союзы: черешчатодубово-березовые и буково-скально-
дубовые леса (преимущественно на западе Европы), буковые леса и
требовательные к условиям существования смешанные лиственные леса
(лещинные леса), дубово-грабовые леса, теплолюбивые смешанные дубовые
леса, пойменные леса, ольховые и березовые заболоченные леса. Сюда же
Р и с. 70. Примеры пространственного распределения атлантических и континен-
тальных элементов флоры в Баден-Вюртемберге (по Эйхлеру, Градману и Мейгену).
а — Digitalis purpurea (атл.) растет в сообществе сплошных вырубок (Atropiony б — Aster
amellus (конт.) на сухих лугах (Festuco-Brometea).
относятся северные и альпийские еловые и сосновые лесные сообщества
и хвойные заболоченные леса (рис. 69).
В более или менее закономерной связи с лесными формациями нахо-
дятся частью естественные, а частью антропогенные сообщества: лесо-
полосы, сообщества лесных опушек и лесосек, дюнные кустарники, фор-
мации лиственного и хвойного криволесья (на верхней границе леса),
горные луга, приатлантические луга, дернины на скалах, альпийские
тощие луга и пастбища, сообщества на каменных осыпях и в трещинах
скал, верховые и низинные болота, заросли камыша и крупноосоковые
сообщества, высокотравье, сообщества у ключей и по берегам прудов,
прибрежные сообщества, береговые и на намывных почвах, задернованные
дюны, тучные и тощие луга и пастбища, сухие луга, полевые и садовые
сообщества сорняков и рудеральные сообщества, многочисленные расти-
тельные сообщества текучих и стоячих вод. Большую часть сообществ
этих формаций можно с точки зрения типологии ареала отнести к вышеупо-
мянутым лесным сообществам. При таком условии Среднюю Европу
следует рассматривать как область с основным составом из среднеевропей-
ских сообществ со значительным проникновением атлантических и конти-
нентальных (рис. 70), северных и южных (субсредиземноморских) сооб-
ществ.
Климатические зоны растительности
255
Аналогично дифференцирован и состав сообществ североамерикан-
ской области лиственных лесов. Здесь можно различить по крайней
мере четыре основных компонента с различно расположенными центрами
распространения, которые, однако, смотря по локальным условиям место-
обитания, могут взаимно проникать друг в друга, а именно сообщества:
1) области смешанных лесов района Великих озер;
2) аппалачской области лиственных лесов;
3) пояса гикори западнее Аппалачей;
4) области смешанных и хвойных лесов южного Пидмонта и субтропического юга.
1) Леса района Великих озер в северной части представлены смешан-
ными лесами из различных лиственных деревьев с веймутовой сосной
и елью, в остальной же части — в основном буком, кленом и березой
с канадской тсугой, а также другими хвойными и лиственными породами
на более высоко расположенных участках.
2) Аппалачская область лиственных лесов на сухих склонах южной
экспозиции имела главным образом дубово-каштановые леса, которые
в настоящее время уже почти все уничтожены занесенной сюда грибковой
болезнью, а на теневой стороне — кленово-буковые леса и желтый тополь
на заливных поймах рек.
3) Богатые видами леса пояса гикори тянутся на запад до границы
с прериями. Они большей частью были уничтожены пожарами. Наряду
со встречавшимся в девственном лесу белокорым дубом (Quercus alba)
высотой до 45 м, который в своем кроваво-красном осеннем одеянии счи-
тался «царем лиственных деревьев» (Шенк), здесь росло около дюжины
других видов дуба, полдюжины видов гикори (Сагуа), а также вязы, ясени,
ореховые деревья, тюльпанное дерево и т. д. В лесах была развита бога-
тая напочвенная флора.
4) Леса Пидмонта и южной зоны хвойных лесов характеризуются
некоторыми видами сосны и дуба, а также большим числом других, частич-
но явно субтропических видов.
Вечнозеленые дождевые леса умеренной зоны
Этот тип формаций ограничен в основном резкоокеаническим дожде-
вым климатом зоны западных ветров в южном полушарии. Основные
области распространения — западная Патагония, Южная Новая Зелан-
дия, Тасмания и горные районы юго-восточной Австралии — родственны
между собой и по флористическому составу. В северном полушарии
в основном из-за сильных зимних холодов вечнозеленые лиственные леса
не столь обширны, а на Тихоокеанском побережье Северной Америки
встречаются лишь особенно пышные здесь хвойные леса (дождевые хвой-
ные леса). В Европе для умеренного вечнозеленого лиственного леса на
юго-западном побережье Ирландии характерна значительная примесь заве-
зенных сюда средиземноморских видов деревьев и кустарников. В Чили,
где пояс умеренных дождевых лесов имеет наибольшую сплошную протя-
женность с севера на юг, он смыкается на севере около 40° ю. ш. непо-
средственно с областью летнезеленых лиственных лесов и простирается
по наветренной стороне Анд через всю Патагонию до Огненной Земли.
Для него характерен климат зоны западных ветров с обильными (более
2000 мм) осадками, ближе к океану с умеренными, к югу со все более
низкими температурами, в глубинных же районах даже с частыми замо-
розками. В северной части, где лето достаточно теплое, растительность
256
Пространственное подразделение растительности
достигает большой пышности и представлена широколиственными дере-
вьями типа лавровых и магнолий, лианами, бамбуковым подлеском
и большим разнообразием эпифитных мхов и папоротников. К югу пыш-
ность леса, высота деревьев и богатство видов постепенно уменьшаются.
Листья становятся меньше и в составе древостоя все больше появляется
хвойных. Наконец на крайнем юге прохладное лето допускает только
умеренный рост растений. Число видов сокращается. Лес становится
однотонным, мелколистным и по высоте не достигает и трети той высоты,
которой обладает в Вальдивии дождевой лес на северной границе пояса.
ПОЯС БОРЕАЛЬНЫХ ХВОЙНЫХ ЛЕСОВ
Это единственный пояс растительности, кроме граничащих с ним
на севере тундр, который простирается по земной поверхности в виде
сплошной зоны, прерываемой лишь океанами. Он включает крупнейшие
лесные области Земли и достигает в среднем ширины более 1500 км. Он
охватывает в Европе 10—12, а в Азии и Америке 10—25 градусов по
широте. В Евразии его протяженность с севера на юг увеличивается по
направлению к востоку и достигает максимума между Енисеем и Леной,
в Америке она возрастает, наоборот, с востока на запад и максимальна
на Тихоокеанском побережье Америки.
(По Кёстлеру, хвойные леса, покрывающие несколько больше трети всей занятой
лесами площади на Земле, распределяются по частям света следующим образом:
Евразия —55%, Северная Америка — 40, Южная Америка — 4, Австралия —
Океания — Африка — 1%.)
В глубине континентов хвойный лес граничит с более или менее
широким переходным поясом к прериям и сухим степям, ближе к океану
(в Европе, Восточной Азии, районе Великих озер Северной Америки)—
с областями летнезеленых лиственных и смешанных лесов. Только на
Тихоокеанском побережье Америки южные ответвления пояса хвойных
лесов по не вполне выясненным причинам, вероятно связанным с историей
развития растительности, примыкают непосредственно к жестколистной
растительности области зимних дождей.
Большая часть зоны бореальных хвойных лесов простирается через
области континентальных климатов с холодными зимами, сильными моро-
зами (только в северном полушарии) и длительным снежным покровом.
Самые низкие, за исключением Антарктики, температуры (ниже —60° С)
отмечены в этом поясе растительности: в области нижнего течения Макен-
зи, а также в Восточной Сибири (до —78° С). Средняя температура янва-
ря в Канаде и в Евразии западнее Енисея составляет —10 до —30° С,
а в Восточной Сибири —30 до —50° С.
Лето здесь короткое, но теплое (средняя температура самого теплого
месяца 10—20° С), продолжительность вегетационного периода от 3 до
5 месяцев. В северной части Скалистых гор хвойный лес растет в условиях
с менее чем 90 безморозными днями. Амплитуда температуры между
самым теплым и самым холодным месяцами в большей части пояса хвой-
ных лесов равна примерно 30—50°, а в Восточной Сибири — более 50°;
в океанических окраинных областях эта величина снижается до 30
и даже 15° (на севере Скандинавии и Тихоокеанском побережье Британ-
ской Колумбии).
Годовое количество осадков составляет в среднем 250—750 мм. Оно
уменьшается на севере Восточной Сибири и увеличивается в океанических
Климатические зоны растительности
257
окраинных областях, прежде всего на Тихоокеанском побережье Север-
ной Америки, где оно превышает 3000 мм. Осадки выпадают во все времена
года, преимущественно, однако, летом. Поэтому при низких в течение
большей части года температурах климат даже в областях с малым коли-
чеством осадков остается все-таки влажным.
Тихоокеанская прибрежная область Северной Америки в северной
своей части (50—60° с. ш.), где хвойные леса наветренных склонов по
средней высоте деревьев и интенсивности роста относятся к наиболее
пышноразвитым лесам континента, обладает ярко выраженным океани-
ческим дождевым климатом с мягкой, но снежной зимой, прохладным
летом, продолжительным периодом вегетации и очень большим количеством
осадков, равномерно распределенных в течение всего года. Южнее этой
области при аналогичных температурных условиях количество летних
осадков постепенно уменьшается, и леса все с большей резкостью диф-
ференцируются в зависимости от экспозиции и высоты положения. В поло-
се южнее Орегона регулярные летние туманы на побережье допускают
проникновение пышных хвойных (из вечнозеленой секвойи) и смешанных
лесов далеко на юг (до 35° с. ш.) в область дождевых лесов, тогда как
в глубине континента, в горном окружении долины Сакраменто, сухой
сосновый лес встречается только на больших высотах.
Огромные (две трети пояса) площади хвойных лесов находятся в зоне
вечной мерзлоты; если в Европе это всего лишь узкая полоска на северной
окраине континента, то в Западной Сибири — добрая треть, в Канаде —
больше половины, а в Восточной Сибири — большая ее часть. Южная
граница вечной мерзлоты доходит в Монголии до 48° с. ш., южнее Гуд-
зонова залива — до 52° с. ш., в Норвегии же — только до 70° с. ш.
Оттаявший верхний слой почвы на больших участках в летнее время
остается заболоченным.
Ближе к северной границе лесного пояса определяющими подчас
становятся различия в типах почв. Песчаные почвы, которые легче нагре-
ваются и поэтому раньше и глубже оттаивают, здесь более благоприятны
для произрастания леса, чем, например, торф или тяжелые глинистые почвы.
Недостаток тепла на протяжении большей части года препятствует
деятельности микроорганизмов и в результате задерживается разложе-
ние опавшей хвои. Образующийся на поверхности грубый гумус резко
граничит с минеральной почвой, так как перемешивания горизонтов
земляными червями здесь не происходит. Почвы, которые летом лежат
выше уровня грунтовых вод, несмотря на относительно небольшое коли-
чество осадков, интенсивно выщелачиваются и оподзоливаются с обра-
зованием в нижней части профиля ортштейнового горизонта. Деревья
формируют в таких почвах, как и в оттаявших на небольшую глубину
мерзлых грунтах, плоскую очень широкую поверхностную корневую
систему. С этим связан здесь светлый характер лесов при господствующей
узкой форме кроны.
На склонах и участках, подвергшихся действию льда, часты камени-
стые маломощные почвы с неразвитым профилем. Сюда же относятся
почвы, затопляемые грунтовыми водами, аллювиальные и болотные
почвы и различные торфяники.
Физиономически преобладающими и господствующими на большей
части площади являются вечнозеленые хвойные леса с хорошо выраженным
пологом и умеренной высотой, уменьшающейся к северу.
Наличие хвои вместо листьев обеспечивает полное использование
периода вегетации и избавляет дерево от высыхания зимой. Форма хвои
258
Пространственное подразделение растительности
наряду с узкой кроной уменьшает, кроме того, опасность облома под
тяжестью снега, а почти колоннообразная форма кроны позволяет лучше
использовать свет при низком стоянии солнца.
Деревья растут медленно, с узкими годичными кольцами, однако на
северных границах распространения они достигают более старого воз-
раста, чем в южнее расположенных широтах. Европейская ель, которая,
например, в Средней Европе доживает до 200 лет, если ее не срубают
в 120 лет или раньше, на северной границе лесов достигает 300—400-лет-
него возраста и более.
Из тысячи распространенных по земной поверхности видов деревьев
только некоторые и большей частью хвойные приспособились к условиям
Таблица 12
Распространение древесных пород пояса бореальных хвойных лесов
Сканди- навия Европейская часть СССР и Западная Сибирь Восточная Сибирь Канада
Picea P, abies Р. obovata Р. canadensis Р. mariana
Larix L. sibirica L. gmelini L. laricina
Abies A. sibirica A. balsamea
Pinus P. si llvestris | Р. sibirica Р. pumila Р. banksiana
Betula B. papyri/era B. glandulosa B. glandulifera
d . puoesc B. verrucosa ens ' B. exilis
Populus P. tremuloides P. balsamifera
ли P. suaveolens
Alnus Л ->
Л
A. У1. ULI fruticosa A. sibirica A. hirsuta A. mitcheliana
Salix S. caprea S. vi S. tr\ minalis iandra S. sachalinensis S. bebbiana S. interior
Климатические зоны растительности
259
бореального климата с холодной зимой. Поэтому леса здесь очень бедны
видами и на огромных пространствах удивительно однородны. Только
в океанических и южных окраинных областях число видов хвойных
деревьев увеличивается и появляется обильная примесь лиственных
деревьев. В остальном же леса пояса бореальных хвойных лесов состоят
в основном из нескольких видов четырех родов хвойных — Picea, Larix,
Abies и Pinus,— и из занимающих подчиненное положение видов,
относящихся к родам лиственных деревьев — Betula, Populus, Alnus и
Salix.
В отдельных областях пояса роды древесных пород представлены раз-
личными, часто близко родственными викарирующими видами (см.
табл. 12).
Естественные пожары приводят обычно к временной смене видов.
На севере России, например, на месте сгоревших еловых лесов вырастает
сосна как растение, обладающее лучшей всхожестью на свету. Однако
под защитой соснового леса вырастает молодая еловая поросль, и в конце
концов ель снова сменяет сосну.
Типы лесов дифференцируются по местообитаниям. В Сибири ель
предпочитает свежие мощные минеральные почвы, тогда как сухие бедные
песчаные почвы и торфяники верховых болот остаются сосне, образующей
на них низкорослые, богатые лишайниками сосновые боры или лесные
болота. Сибирский еловый и пихтовый лес, поскольку молодая поросль
Picea obovata и Abies sibirica чувствительны к морозам, избегает склонов
и долин, подверженных ночным заморозкам. Здесь преобладают лист-
венные породы (береза, осина, ива), образующие участки заболоченной
тайги.
В обширных лесных болотах («muskegs») на внутреннем бессточном
плато Канадского щита господствует черная ель (Picea mariana) в виде
редкой поросли с малой интенсивностью прироста. Лиственница здесь —
также болотное дерево.
Наряду с лесами, лесными и верховыми болотами и островами тундры,
занимающими северную часть лесного пояса, для формационной структуры
пояса бореальных лесов характерны естественные луга. Весеннее поло-
водье, затопляющее на длительное время обширные низины, и препят-
ствующие нормальной аэрации почв, ледоход и длительный снежный
покров можно рассматривать как причины возникновения этих безлес-
ных злаковых и разнотравных сообществ (Танфильев, 1925).
В глубине континентов по южной окраине лесного пояса узкой пере-
ходной к степям полосой протягивается лесостепь, представляющая собой
полуоткрытую формацию с рассыпанными среди травянистых пространств
лесными островами.
Особое положение занимают лесные области североамериканских
Кордильер. Канадская территория относится почти полностью к лесному
поясу, тогда как по направлению к югу, исключая район Тихоокеанского
побережья, хвойные леса встречаются только в высотных ступенях гор-
ного обрамления засушливого Большого Бассейна.
Во внутриматериковых областях господствует в основном сосна,
являющаяся здесь вторичной породой, как, например, скрученная^сосна
(Pinus contorta), распространившаяся после пожаров. Верхний лесной
пояс в горах образован пихтами и елями (Picea engelmanni, Abies lasio-
carpa и др.)- Примерно на 45° с. ш. в Скалистых горах на высоте 1500 м
проходит нижняя засушливая граница леса. За поясом дугласовых жел-
тых сосен (до 1900 м) следует пояс скрученных сосен и наконец (до 2500 м)
260
Пространственное подразделение растительности
елово-пихтовый пояс с сосновым криволесьем (Pinus albicaulis) на верх-
ней границе леса.
Лесная область в районе Тихоокеанского побережья характеризуется
в северной части тсугой западной (Tsuga heterophylla) и ситхинской
елью (Picea sitchensis). В Британской Колумбии к ним добавляются Thuja
plicata, Pseudotsuga taxijolia, а южнее — достигающий больших горных
высот нутканский кипарисовик (Chamaecyparis nootkaensis). Из легко
раскалывающихся, противостоящих благодаря содержанию эфирных
масел гниению стволов Thuja plicata индейцы изготовляли резные тотем-
ные столбы и челны, теперь из них делают кровельный гонт, а из срублен-
ной при прочистке леса молодой поросли — телеграфные столбы. Такие
древостои со стволами высотой 60 м и диаметром до 6 м достигают местами
1000-летнего возраста. Кипарисовик Лавсона, встречающийся главным
образом на побережье штата Орегон, в нижней лесной ступени достигает
такой же высоты. Девственные участки этих лесов имеют двухъярусный
полог, с нижним древесным ярусом из тсуги западной, а иногда и из
ситхинской ели с обильным подлеском из вечнозеленых лиственных
деревьев и папоротников.
Сходный характер имеют примыкающие с юга леса из секвойи
вечнозеленой (Sequoia semper virens), однако деревья в них в два раза
выше.
Пояс бореальных хвойных лесов в большей его части по характеру
хозяйственного использования относительно однороден и поэтому в этом
отношении ему легче дать обобщающую характеристику, чем большинству
других поясов растительности. Это единственная на Земле зона, в которой
добыча древесины имеет доминирующее значение. Причина лежит в осо-
бенностях климата, которые, с одной стороны, обусловливают характер
леса, а с другой — резко ограничивают возможность использования его
в сельском хозяйстве, за исключением участков, представляющих интерес
для горной промышленности, прибрежных районов и южных окраин.
Весь пояс в общем населен очень редко. Поэтому леса здесь хорошо сохра-
няются, если они не уничтожаются в целях получения крепежного леса,
как это происходит, например, на Аляске. В обширных северных районах
пояса основными формами хозяйства редкого местного населения были
охота и рыболовство, дополняемые сбором ягод, грибов и добычей мел-
кого зверя. В Старом Свете, кроме того, на границе между лесом и тун-
дрой имело место кочевое пастбищное оленеводство, которое в более
позднее время было перенесено и на Аляску. Южная часть пояса хвойных
лесов относится с точки зрения его сельскохозяйственного использова-
ния к «субантарктической ячменной зоне». За исключением южной
Скандинавии и некоторых других точек, полярная граница земледелия
проходит примерно на 3—5, а в Восточной Сибири на 10 градусов по
широте ниже северной границы леса, если, конечно, не говорить о фор-
постах земледелия, которые на основе новейших достижений в этой обла-
сти продвигаются все дальше на север. В зоне хвойных лесов земледелию
препятствует короткий вегетационный период, бедность почв питатель-
ными веществами и необходимость в большом количестве удобрений.
Более благоприятны для возделывания щебенчатые почвы со слаборазви-
тым профилем, и прежде всего молодые почвы, на месте зарастающих озер.
В странах, где возделыванием сельскохозяйственных культур занимаются уже
на протяжении столетий, как, например в Финляндии, оно первоначально велось
на основе искусственного выжигания лесов под пашню, которое позднее было заменено
Климатические зоны растительности
261
постоянным землепользованием. Однако последствия прежних методов сказываются
на состоянии лесов и по сию пору. В южной Финляндии, где до конца прошлого столетия
широко практиковалось выжигание лесов, только десятая часть лесных массивов (тогда
как в северной Финляндии — более 2/3) имеет возраст свыше 100 лет. Несмотря на это,
средние запасы древесины на гектар на юге не меньше, чем на севере, потому что годич-
ный прирост по климатическим причинам здесь в среднем почти в два раза больше.
В конечном счете климат обусловил и прежнее обилие ценных пушных зверей
в хвойных лесах. Добыча пушнины, которая и до сих пор в некоторых районах, как,
например, в северных территориях Канады и в Восточной Сибири, играет значитель-
ную роль в экономике, является одной из основных причин освоения хвойных лесов.
Использование лесов на большей части их площади производилось до
настоящего времени только в европейских странах. В Канаде и Сибири
оно ранее было ограничено областями, расположенными более удобно
с точки зрения транспортировки древесины; в результате обширные мас-
сивы хвойных лесов почти совершенно нетронуты. Северные хвойные леса
являются основой всего лесного хозяйства на Земле. Более 9/10 пиленой
древесины, поступающей на мировой рынок, дают хвойные породы, такая
же доля хвойных идет на нужды бумажной промышленности.
Эксплуатация хвойных лесов облегчается их однородным составом
и бедностью видового состава. Это сильно упрощает заготовку, обработку
и сбыт леса, а также снижает затраты на лесохозяйственное восстановле-
ние лесов. Многие из наиболее распространенных видов деревьев имеют
прямые и ровные стволы с весьма стойкой и упругой ценной плотной дре-
весиной с узкими годичными кольцами. Менее ценные виды используются
в бумажной промышленности. Белая ель (Picea canadensis) — доминирую-
щий вид пояса бореальных лесов в Америке — дает сырья для бумажной
промышленности больше, чем какая-либо другая древесная порода. Кажу-
щиеся очень светлыми благодаря узкой форме крон более северные леса
имеют значительную густоту стволов и поэтому, несмотря на сравнительно
слабый годовой прирост древесины, дают относительно больше готовой про-
дукции. В районах с более благоприятными климатическими условиями,
как правило, происходит естественное возобновление леса или повторное
облесение обнаженных площадей при котором часто наблюдается смена
древесных пород.
Однако эксплуатационные преимущества хвойных лесов подчас сво-
дятся на нет трудностями транспортировки и малой плотностью населения
в районах лесоразработок. Это создает большие региональные различия
в степени освоения лесных богатств.
Если раньше заготовленная древесина вывозилась из леса в виде сырья (за исклю-
чением добычи древесного угля, сажи, дегтя и т. п.), то в последнее время все больше
развивается разделка и переработка ее на месте для экспорта или внутреннего потреб-
ления. В Канаде лес — основа экономики страны. Третья часть вырубки идет на лесо-
пильни, сосредоточенные главным образом на западе, остальное — в бумажную и цел-
люлозную промышленность. В Советском Союзе деревообрабатывающая промышлен-
ность — одна из ведущих отраслей хозяйства, продукты которой частично идут на
экспорт. Общий запас леса в Советском Союзе составляет примерно х/4 всех лесных бо-
гатств на Земле. Швеция, имеющая среди Скандинавских стран абсолютно наибольшую
площадь под лесом, опережает Финляндию по производству бумажной массы. Исклю-
чительно богата лесом Финляндия: 71% ее территории зан лесами, а па единицу
населения в Финляндии приходится в десять раз больше леса, чем в Средней и Западной
Европе.
262
Пространственное подразделение растительности
СУБПОЛЯРНЫЕ И ПОЛЯРНЫЕ ЗОНЫ
СУБАРКТИЧЕСКАЯ ЗОНА И АРКТИКА
Северная граница леса и деревьев и лесотундра
Границы леса и отдельных деревьев в природе не выражены непре-
рывными линиями. Между северной границей распространения сплош-
ных сомкнутых лесов и мысленно проведенной «границей деревьев», соеди-
няющей наиболее продвинутые на север форпосты одиночных деревьев
и групп, лежит пояс со средней шириной 100 км. На юге его еще господ-
ствует лес, чередующийся с открытыми тундровыми участками, а в север-
ной части — тундра либо с отдельными лесными островками, либо поло-
сками леса в долинах или отдельными группами деревьев. За исключе-
нием Исландии и Скандинавии, где на северном пределе лесов встречается
береза бородавчатая, эту границу повсюду образуют хвойные виды: в Скан-
динавии наряду с березой — лесная сосна, в Европейской части СССР,
на Кольском полуострове,— извилистая береза и сосна, от Белого моря
до Урала — сибирская ель, далее на восток, до Енисея, — сибирская
лиственница, а от Енисея до Берингова моря — даурская лиственница.
В Северной Америке это главным образом ели (Ptcea canadensis и Picea
mariana), а в восточных районах — лиственница и пихта. Сопровождаю-
щими видами являются различные представители рода березы, ольхи,
тополя и ивы, а также во многих местах вплоть до самой границы леса —
рябина.
Положение северной границы леса колеблется в пределах 20 граду-
сов по широте. Самая северная точка находится у устья Хатанги, а юж-
ная — на южном крае Алеутской дуги. Граница в общем определяется
степенью континентальности климата, однако возможны отклонения под
влиянием каких-либо других факторов. Самые северные леса Америки
и Евразии находятся на крупнейших массивах суши, а именно у устья
Хатанги на 72° с. ш. и в западном полушарии — у мыса Батерст на 69° с. ш.
Дальше всего на юг границы отступают в прибрежных областях кон-
тинентов. На восточных окраинах, омываемых холодными морскими тече-
ниями, пограничные линии достигают наиболее низких широт (восточное
побережье Лабрадора близ 53°30' с. ш., Камчатка — около 59° 30' с. ш.).
В глубине континентов самое южное положение границы леса отмечено
у Гудзонова залива (около 55°30' с. ш.). Гренландия находится вне гра-
ницы леса, тогда как граница деревьев проходит здесь на восточном побе-
режье у 66°30' с. ш. В южной Гренландии более теплые подветренные
участки заняты березовыми рощами высотой 3—6 м с примесью ивы,
ольхи и рябины. Относительно Исландии мнения разделяются: одни
помещают остров в пределы пограничного пояса, другие проводят гра-
ницу леса севернее острова. За исключением небольших площадей,
покрытых березовым кустарником, остров фактически безлесен.
Вопрос о том, какие факторы следует рассматривать в качестве основ-
ной причины того или иного положения северной границы леса, занимал
умы многих исследователей начиная с XVIII в. и вплоть до настоящего
времени. Еще Миддендорф обратил внимание на значение летнего тепла,
исходя из проникновения леса далеко на север в континентальных обла-
стях (полуостров Таймыр). Гризебах же подчеркивал, что северная гра-
ница леса не может определяться одним лишь климатическим фактором
и что решающее значение могут иметь как недостаток тепла, так и продол-
Климатические зоны растительности
263
жительность вегетационного периода. Давно уже было замечено и влияние
иссушающего действия ветров, особенно вблизи побережья, где в защи-
щенных от ветра низинах встречаются форпосты леса с деревьями, имею-
щими столообразно подрезанную форму.
Все попытки отразить многочисленные причины, обусловливающие
различное положение северного предела леса в единой климатической
формуле, остались безуспешными (см. литературу, Hennes, 1955).
Многие, и прежде всего финские, русские и шведские исследователи
показали, что на отдельных участках границы собственно экологические
причины в зависимости от видов, образующих эту границу, могут быть
очень различны, однако при этом основную роль, видимо, играет спо-
собность к восстановлению (задержка созревания семян, прорастания
или развития проростков).
У хвойных деревьев образование семян требует двух вегетационных
периодов и зависит от тепла двух последующих летних сезонов (оплодо-
творение и созревание семян), а тем самым от условий, которые создаются
за значительный промежуток времени. Поэтому древесные сообщества
близ границы леса состоят частью из деревьев одного вида лишь немно-
гих возрастных градаций, причем у сосны в Северной Европе могут ока-
заться промежутки в возрастных классах до 100 лет (Renvall, 1913).
На основании этих фактов в последнее время пришли к выводу, что
(не учитывая воздействия человека) на северной границе леса наблю-
дается значительная динамика наступания и отступания леса (Kujala,
1929; Hustich, 1937; Aario, 1940, 1941; Bluthgen, 1942).
В Лапландии, например, уже в течение столетий сосновый лес надви-
гается на тундру, заходя местами за границу леса, которая прежде была
образована березой. Семенные годы сосны наступают теперь не через
столетия, как раньше, а почти через десятилетние промежутки, что Блют-
ген объясняет удлинением вегетационного периода в сторону осени,
благодаря общему смягчению климата. Сосна, требующая более конти-
нентального климата, получает поэтому преимущества перед березой.
Для большей части открытых площадей пояса лесотундры характерен
тип крупнобугристой тундры. Покров из сухих мхов и лишайников на тор-
фяных буграх, поднимающихся до высоты 3 м («palsen»), сменяется боло-
тистыми осоковыми лугами и низкими кустарниками. Чисто лишайнико-
вая тундра в пограничной области часто перемежается лесом (зимние
оленьи пастбища).
Пояс тундры
Обширные безлесные тундровые пояса Евразии и Северной Америки
(«barren-grounds»), покрытые главным образом кустарничками и хамефит-
ными мхами и лишайниками, примыкают к субполярной континенталь-
ной климатической зоне, южная граница которой, по Кеппену, совпадает
с границей леса (рис. 71). Низкорослая растительность, бедная видами,
приспособлена к длительному периоду зимнего покоя с низкими и крайне
низкими температурами. Еще Кильман (Kihlmann, 1890) подчеркивал, что
характер тундры, кроме всего прочего, определяется в значительной сте-
пени тем, что она должна переносить большие и резкие скачки темпера-
туры и частые переходы температуры через нуль. Поэтому прирост здесь
замедлен, с длиной годичных побегов всего в несколько миллиметров.
Кильман насчитал на стволе можжевельника тощиной всего 83 мм
544 годичных кольца. «Карликовая форма роста, склеро- и микрофиллия,
преобладание мхов и лишайников, неполное покрытие почвы — таковы
264
Пространственное подразделение растительности
повсюду признаки тундры» (Шимпер — Фабер). Характерные формы
роста — это кустарнички с поверхностной корневой системой, с большей
частью вечнозелеными жесткими или чешуйчатыми маленькими листоч-
ками, которые полностью используют короткий период вегетации и хорошо
140,_________________150______________160____________170___________180 170 160 160 140
Рис. 71. Вечная мерзлота, тундра, холодная пустыня и полярные льды в северном
полушарии (по Троллю и др.).
1 — арктическая холодная пустыня; 2 — тундра; з — граница вечной мерзлоты; 4 — мате-
риковый лед; ,5 — полярный пак; 6 — замерзание в сентябре; 7 — замерзание в марте; 8 — пла-
вучие льды.
переносят периоды с неблагоприятным водным режимом. Широко распро-
страненные лишайники и мхи даже после полного высыхания способны
быстро вновь набирать воду.
Встречаются также полукустарники и гемикриптофитное разно-
травье, тогда как геофиты и терофиты отсутствуют.
Средняя температура самого холодного месяца, как правило, ниже — 8° С, в Си-
бири и Северной Америке даже ниже —20° С. Вегетационный период продолжается
Климатические зоны растительности
265-
не более 2—3 месяцев. Короткое лето начинается неожиданно теплыми днями, однако
ночью могут быть заморозки; из-за недостатка летнего тепла (средняя месячная темпе-
ратура самого теплого месяца 5—10° С) оттаивает только верхний слой почвы глуби-
ной до 1 м. Зимние снега и летние дожди приносят в среднем 200—300 мм осадков, кото-
Р и с. 72. Ареалы арктических родов вересковых (по Гуду).
1 — Bryanthus; 2 — Harrimanella', 3 — Cassiope\ 4 — Phyllodoce.
рых при недостатке тепла и промерзшем нижнем слое вполне достаточно, чтобы на
плоских позициях возникли стоячие озерца и болота. На возвышенных участках
песчаные и галечные почвы и скалистые склоны образуют сухие местообитания.
В зависимости от особенностей рельефа, типов почв и уровня грун-
товых вод, обусловленного глубиной оттаивания, на больших площадях
сменяются различные формации тундровой растительности (кустарнич-
ковые пустоши, лишайниково-моховые тундры, низинные болота) или же
они образуют на небольших пространствах пеструю мозаику раститель-
ности разного состава. В общем же преобладание той или иной формации
чередуется в зональной последовательности. На границе с лесотундрой,
в самой южной части, на сухих плоских возвышениях и на песчаных поч-
вах господствует лишайниковая тундра, прерываемая часто участками
кустарничковых пустошей с преобладанием Ericaceae (рис. 72). Последние
проникают сюда в результате стравливания. Во влажных низинах ветре-
266
Пространственное подразделение растительности
чаются еще лиственные кустарники вплоть до карликового криволесья
вдоль рек. Далее к северу кустарничковые формации все чаще переме-
жаются болотами с осокой и пушицей, а также другими формациями
низинных болот. Болотистая тундра Западно-Сибирской низменности весной
обычно затопляется талыми водами. Наконец, в северном поясе наряду
с моховыми и лишайниковыми тундрами, встречающимися и южнее, рас-
пространен тип полигональных тундр, в котором только края трещин
покрыты лишайниками, мхами и немногими цветковыми растениями, тогда
как большая часть поверхности почвы остается оголенной. Самые север-
ные окраины Азии носят уже арктический характер, здесь преобладают
каменистые тундры с подушковидными растениями, имеющими форму
полушарий.
Верховые болота располагаются в основном в южной части тундры,
вблизи границы леса, где летнее тепло еще допускает богатое развитие
растительности и быстрое образование торфа.
Лишайниковая тундра — основа пастбищного оленеводства с сезон-
ными перегонами между лесотундрой зимой и прибрежными районами
Ледовитого океана в середине лета.
Пояс субарктических маттов
В океанической части субарктического климатического пояса с более
мягкими зимами и меньшими годовыми колебаниями температуры, на
Алеутских островах и на подверженных влиянию Гольфстрима северо-
атлантических островах, где недостаток летнего тепла тормозит рост
деревьев, развиваются разнотравные матты («субполярные луга»). В Ислан-
дии, например, большая часть их используется под пастбища для овец.
Здесь преобладают гемикриптофиты, составляющие 53% флоры Ислан-
дии, 15,2% приходится на долю хамефитов, причем их число возрастает
с широтой и высотой над уровнем моря (34,2% видов встречаются на
высоте более 800 м). В спектре жизненных форм пояса субарктических
маттов в отличие от континентальной тундры немаловажную роль играют
терофиты и геофиты (в Исландии — 11,5 или 10,6%; к ним добавляются
водные и болотные растения — 9,2%).
Пространственная дифференциация растительности определяется про-
должительностью и мощностью снежного покрова, влажностью почвы
и глубиной грунтовых вод, а также экспозицией относительно ветра.
Одновременно формации разделяются и по высотным ступеням.
Р и с. 73. Общее распространение Cornus suedca (по Регелю).
Климатические зоны растительности
267
В Исландии можно различить следующие ступени:
1. Нижняя ступень, соответствующая лесотундре и простирающаяся до высоты
300 м, с пышными луговинами или маттами, болотами, а на защищенных от ветра
участках — с остатками субарктических березовых рощ высотой 2—3 м (Betula pubes-
cens, виды Salix и Sorbus aucuparia). Две трети видов этой ступени проникают и дальше
на юг, за пределы субарктических областей. Густое и богатое видами гемикриптофитное
разнотравье («jadar») приурочено к почвам, влажным зимой и сухим летом, и к участ-
кам, где по орографическим причинам снежный покров защищает растительность («ge-
iri») с осени до весны. На переувлажненных участках, где вода не покрывает почву
полностью, появляются геофиты и особенно многочисленными становятся хамефиты
(растительность «myri»).
2. Ступень фьельдов (300—700 м) преимущественно с кустарничковыми пусто-
шами, где больше половины видов имеют чисто субарктическое и арктическое распро-
странение.
3. Ступень исландской тундры с преобладанием лишайниковой и моховой тундры
и лишь с небольшим участием некоторых сосудистых растений, которые имеют почти
исключительно арктическое распространение. Последние вблизи снеговой линии вхо-
дят в состав каменистой тундры с подушковидной растительностью.
На самой нижней ступени, при особых условиях местообитания могут возникнуть
формации, свойственные верхним высотным ступеням. Так, например, на сухой, нахо-
дящейся вне влияния грунтовых вод почве при нормальном снежном покрове встре-
чается растительность «то»— кустарничковая пустошь с преобладанием травянистой
ивы (Salix herbacea) и водяники, с вкраплениями осок и злаков, а на каменистых поч-
вах наветренных участков, с которых снег сдувается ветром,— отдельные пятна расти-
тельности «melar»— кустарничковая пустошь или болотистые луга с немногими семен-
ными растениями, широко распространенными в высоких арктических широтах.
Высокая Арктика
На некоторых островах, расположенных в высокоарктических широ-
тах, температура летом тоже бывает достаточна для произрастания скуд-
ной растительности. Только в
глубине Гренландии господству-
ет абсолютная ледяная пустыня.
На южной оконечности
Г ренландии, в защищенных
фьордах встречается светлое
березовое криволесье высотой
до 3 м с несколькими видами
Betula и сопровождающими ее
древесными видами из родов
Salix, Alnus и Sorbus. До 68°
с. ш. проникает ивовый кустар-
ник высотой до полутора мет-
ров, а до 73° — высотой лишь
в полметра. В остальном расти-
тельность свободных ото льда
^Хамефиты ’
20\
80'с. ш.
60'
Рис. 74. Изменения в составе флоры с уве-
личением географической широты на запад-
ном побережье Гренландии (по Остенфельду
и Хансену).
65
70
75
берегов состоит из субполярных
лугов, ковровых и шпалерных кустарников и каменистой тундры с подушко-
видными растениями (рис. 74). Доля хамефитов возрастает с географической
широтой. На юге они составляют только 18,5%, на 82—83° с. ш. восточ-
ного побережья — 43, а на западной стороне — 32% флоры семенных
растений. Преобладание хамефитов на восточном побережье обусловлено
401
268
Пространственное подразделение растительности
влиянием холодного Восточно-Гренландского течения. На восточной бере-
говой полосе от 72 до 75° с. ш. еще встречаются 170 видов высших расте-
ний, из них на высоте более 600 м — 51, более 1000 м — 18 и на высоте
1275 м — еще 4 вида. Вся флора Гренландии насчитывает 400 сосудистых
растений, из них 80% североамериканского происхождения и только 8 эн-
демичных видов. Остальные виды европейского происхождения, главным
образом луговые растения, занесенные большей частью человеком. Обра-
зования верхового торфа в условиях высокой Арктики не наблюдается.
Самые северные торфяные болота на Новой Земле, вкрапленные среди
господствующих кустарничков и арктической каменистой тундры, следует
рассматривать как разрушающиеся реликты некогда широко распростра-
ненной субарктической тундры.
Шпицберген только в маленьких «оазисах», на отдельных благоприят-
ных участках имеет сомкнутый растительный покров. Его открытая
каменистая тундра с преимущественно подушковидной растительностью
весьма напоминает формацию высокогорий.
Как и в высокогорных районах, в Арктике число насекомоопыляемых растений
уменьшается по направлению к северу. На Шпицбергене наблюдается несоответствие
между насекомоопыляемыми растениями и фауной. Здесь нет ни пчел, ни шмелей
и едва ли встречаются бабочки. Две трети растений, опыляемых обычно насекомыми,
сохраняются только благодаря самоопылению. У некоторых видов образование цвет-
ков удачно заменяется вегетативным размножением. Как и в высокогорных районах,
здесь характерно наличие живородящих видов.
На Земле Франца-Иосифа, где ледники покрывают большую часть
площади, существуют еще 36 видов цветковых растений, подушковидное
разнотравье и кустарнички. Лишь вблизи птичьих базаров обычно весьма
скудная растительность становится несколько плотнее.
СУБАНТАРКТИКА И АНТАРКТИКА
Субполярная зона простирается в южном полушарии на значительно
более низких широтах, нежели в северном. При ярко выраженном океани-
ческом климате и малом размере островов в некоторых случаях трудно
бывает решить, определяет ли положение южной границы леса и отдель-
ных деревьев, недостаток тепла в равномерно прохладном, сыром и бес-
снежном климате или же влияние ветров. Эта граница проходит от южной
оконечности Южной Америки (56°) севернее Фолклендских островов и
далее в южной Атлантике примерно по 40-й параллели, а в Индийском
океане — между островами Сен-Поль и Амстердам даже по 38-й. Затем она
следует вдоль 53-й параллели южнее островов Окленд, на которых встре-
чаются низкорослые леса (миртовые и древовидный папоротник), а у
49-й — севернее островов Антиподов.
Растительность на островах, вне южной полярной границы древес-
ной растительности, состоит главным образом из формаций жестких поду-
шек, субполярных пустошей, пространств, покрытых подушковидными
злаками и разнотравьем. Некоторые исследователи называют весь ком-
плекс этих формаций «антарктической тундрой». Другие ограничивают
тундру формациями, образованными мхами и лишайниками, которые в юж-
ном полушарии имеют весьма незначительное распространение (например,
на высоком плато острова Южная Георгия).
Жесткоподушечники часто из изолированных, растущих на голой
скалистой почве, а частью из плотно сомкнутых подушек Azorella selago.
Климатические зоны растительности
269
достигающих высоты в полметра, на подветренной стороне которых
поселяются другие растения, господствуют в основном в растительности
Кергелена и на наветренных склонах островов Маку ори. Резко океани-
ческий прохладный климат с почти постоянными низкими температурами,
частыми переходами температуры через нуль, а также механическим и ис-
сушающим влиянием сильных ветров определяет условия существова-
ния этой формы растительности.
Варминг обратил внимание на сходство с климатом и раститель-
ностью тропических высокогорий. Климат островов Макуори вследствие
их небольшой величины почти изотермичен с разницей лишь в 5° между
максимальной и минимальной среднесуточной температурой в году
Р и с. 75. Диаграмма термоизоплет, харакеризующих ход сред-
ней суточной температуры воздуха для островов Макуори
(54°30' ю. ш., 158°57' в. д.) (по Троллю).
(рис. 75). На Кергелене в среднем в течение 2/3 всех дней термометр пока-
зывает выше нуля, поэтому почва здесь глубоко не промерзает. Шимпер
подчеркнул прежде всего воздействие ветра, которое проявляется в том,
что формация жестких подушек покрывает главным образом наветренные
склоны и вершины, с подветренной же стороны господствуют полукустар-
никовые пустоши с Асаепа adscendes (Rosaceae), а в понижениях и на влаж-
ных участках — разнотравье частью с крупнолистными, частью с густо
опушенными травами и папоротниками, и если почва не слишком забо-
лочена, также и злаками.
Своеобразным эндемичным растением с формой роста, напоминаю-
щей растения тропических плоскогорий, является кергеленская капуста
(по Троллю, «ствольно-метелковое растение», Stamm-Schopf-Pflanze).
Подушковые злаковые луга из «туссоковых злаков» разных видов гос-
подствуют на Южной Георгии и на островах Антиподов, широко распрост-
ранены на холмах островов Макуори, а также в восточной части Южного
острова Новой Зеландии и на Фолклендском архипелаге, где, однако, эта
формация сильно вытеснена пастбищным хозяйством. К туссоковым
формациям могут примыкать, с одной стороны, сухие дерновиннозлако-
вые степи, а с другой — болотистые разнотравные формации. Там, где
270
Пространственное подразделение растительности
их сообщества являются более или менее сомкнутыми, туссоковые злаки
образуют отложения торфа.
Полярный климат высоких широт охватывает в южном полушарии
около 20 градусов по широте. Антарктический континент, на котором,
за исключением отдельных районов Земли Грейама, средняя температура
самого теплого месяца остается ниже нуля, представляет собой ледяную
пустыню. Здесь известны только два вида цветковых растений, которые
встречаются вместе со скудной моховой тундрой на берегах западной
Антарктиды севернее 68° ю. ш. В более высоких широтах на скалах, лишен-
ных снега, растут только мхи, лишайники и водоросли с очень малым чис-
лом видов. Самые южные известные пока что местонахождения мхов и ли-
шайников расположены на 83° ю. ш.
* *
*
В заключение приводим схему, показывающую ассиметрию горизон-
тального и вертикального распределения растительности обоих полуша-
рий в условиях постоянно влажного климата (рис. 76).
Южн. Бразилия
,,, Юго-Восточная Тропические Юго-вост. Африка
шпиЧ~ Европа____________Азия Анды Юго-вост. Австралия
р * ^Rnnm' ' * ' ' Зап. Патагония Антарктика
Гш™°“ Нов Зеландия ' '
Мексика
Л7
60001-
5000 —
4000--
3000 —
2000-—4
0
С 60 70 60 50 40 30 20 10 0 10 20 30 40 50 60 70 ЫГ Ю
Рис. 76. Схематический вертикальный профиль Земли от Арктики до
Антарктики.
Приняты во внимание только постоянно влажные климаты, за исключением гра-
ницы снега п района пуны. Сходная растительность тропических высот и более
высоких широт характеризуется одинаковыми значками.
Литпература
А а г i о L., 1940. Waldgrenzen und subrezente Pollenspektren in Petsamo-Lappland,
«Ann. Akad. Scient. Fenn. A.», 54.
Abromeit J., 1892. Kartchen mit Verbreitungsgrenzen von Arten der preussischen
Flora, «Schr. Phys.-окоп. Ges. Konigsberg», 33.
Abromeit J., 1900. Uber die Erforschung der Flora von Ost- und Westpreussen
und die Anfertigung von Arealkarten von pflanzengeographisch wichtigen Arten
in unserem Gebiet, Konigsberg.
Aichinger E., 1930. Fichtenbestand, Latschenbestand und Biirstlingrasen im
Karawankengebiet und ihre almwirtschaftliche Bedeutung, «Carinthia», II, Klagen-
furt.
Aichinger E., 1933. Vegetationskunde der Karawanken, «Pflanzensoziologie»,
2, Jena.
A 1 e c h i n W. W., 1932. Die vegetationsanalytischen^Methoden der Moskauer Step-
penforscher, «Hdb. d. biol. Arbeitsmeth.», 11, 6, Berlin— Wien.
Ansiaux J. R., 1948. La notion de Mesoclimat et de Microclimat en Phytosociolo-
gie, Gembloux.
В a a s e n C., 1940. Wald und Bauerntum, Leipzig.
Baeyens J., 1938. Les sols de I’Afrique Centrale, specialement du Congo Beige,
Gembloux.
Barenscher F., 1939. Siedlungskundliches aus der siidlichen Liineburger Heide,
Oldenburg.
Beadle N. C. W., 1948. The vegetation and pastures of western New South Wales,
Sidney.
Beard J. S., 1944. Climax vegetation in tropical America, «Ecology», 25.
Beard J. S., 1946. The natural vegetation of Trinidad, «Oxford Forestry Memoires»,
20.
Beck v. Managetta G., 1902. Uber die Umgrenzung der Pflanzenformatio-
nen, «Osterr. Bot. Z.», LXII.
Berger-Landefeldt U., 1949. Zum Wasserverbrauch von Pflanzenbestiin-
den, «Forsch. u. Fortschr.», 25.
Bernatzki J., 1904. Anordnung der Formationen nach ihrer Beeinflussung sei-
tens der menschlischen Kultur und der Weidetiere, «Englers Bot. Jb.», 34.
Bernbeck O., 1911. Der Wind als pflanzengeographischer Faktor, Bonn.
Bertsch K. u. F., 1947. Geschichte unserer Kulturpflanzen, Stuttgart.
Black G. A., Dobzhansky T. and P a v a n C., 1950. Some attempts to
estimate species diversity and population density of trees in Amazonian forests,
«Bot. Gaz.», 111.
Bliithgen J., 1942. Die polare Baumgrenze in Lappland, «Veroff. Dt. wiss. Inst,
zu Kopenhagen», 10, 1, Berlin — Zehlendorf.
272
Литература
Bliithgen J., 1943. Zur Dynamik der polaren Baumgrenze in Lappland, «Forsch.
u. Fortschr.», 19.
В о b e к H., 1951. Die natiirlichen Walder und Geholzfluren Irans, «Bonner Geogr.
Abb.», 8.
Bobek H., 1952. Beitrage zur klima-okologischen Gliederung Irans, «Erdkun-
de», 6.
Bobek H., Schmithiisen J., 1949. Die Laudschaft im logischen System
der Geographic, «Erdkunde», 3.
Bock H., 1539. New Kreutter Buch, Strassburg.
Bodeux A., 1955. Alnetum glutinosae, «Mitt. Flor.— soz. Arbeitsgem.», N. F. 5.
В 6 c h e r T. W., 1949. Climate, soil and lakes in continental West Greenland in rela-
tion to plant life, Kopenhagen.
Bocher T. W., 1954. Oceanic and continental vegetatioual complexes in Southwest
Greenland, Kopenhagen.
Boer A. C., 1942. Plantensociologische beschrijving van de orde der Phragmitetalia,
Nederl. Kruidkunding Arcief.
Braun-Blanquet J., 1919. Essai sur les notions d’element et de territoire
phytogeografiques, «Archives Sci. Phys. Nat.», 124, Genf.
Braun-Blanquet J., 1921. Prinzipien einer Systematik der Pflanzengesell-
schaften auf floristischer Grundlage, «Jb. d. St. Gallischen Naturw. Ges.», 57.
Braun-Blanquet J., 1925. Zur Wertung der Gesellschaftstreue in der Pflan-
zensoziologie, «Vierteljahrsschr. Naturf. Ges. Zurich.», 70.
Braun-Blanquet J., 1928. Pflanzensoziologie, Grundziige der Vegetationskun-
de, «Biologische Studienbiicher», 7, Berlin; 2. Aufl., 1951, Wien.
Braun-Blanquet J., 1936. La chenaie d’yeuse mediterraneenne (Quercion
ilicis), «Comm. S.I.G.M.A.», 45.
Braun-Blanquet J., 1947. Le Tapis vegetal de la Region de Montpellier et
ses rapports avec les sols.
Braun-Blanquet J., 1951. Pflanzensoziologische Einheiten und ihre Klassi-
fizierung, «Vegetation», 3.
Braun-Blanquet J., 1952. Les groupements vegetaux de la France Mediter-
raneenne, Vaison-la-Romaine (Vaucluse).
Braun-Blanquet J., 1955. Die Vegetation des Piz Languard, ein Massstab
fur Klimaanderungen, Montpellier.
Braun-Blanquet J., 1956. Clef ecologique pour determiner les classes, ordres
et alliances phytosociologiques du midi mediterraneen, Montpellier.
Braun-Blanquet J.u. Mitarbeiter, 1933 и след., Prodromus der Pflan-
zengesellschafteii, 1—7.
Braun-Blanquet J., Emberger L., Molinier R., 1947. Instructi-
ons pour i’etablissement de la carte des groupements vegetaux, Montpellier.
Braun-Blanquet J., Jenny H., 1926. Vegetationsentwicklung und Boden-
bildung in der alpinen Stufe der Zentralalpen, «Denkschr. d. Schweiz. Nat. Ges.»,
63.
Braun-Blanquet J., Pallman n H., Bach R., 1954. Pflanzensozio-
logische und bodenkundliche Untersuchungen im schweizerischen Nationalpark und
seinen Nachbargebieten, II, Liestal.
Braun-Blanquet J., Tuxen R., 1949. Irische Pflanzengesellschaften,
«Veroff. Geobot. Inst. Riibel», 25, Zurich.
Brockmann-J.erosch H., 1913. Der Einfluss des Klimacharakters auf die Ver-
breitung der Pflanzen und Pflanzengesellschaften, «Englers Bot. Jb.» 49, Beibl.
Brockmann-Jerosch H., 1919. Baumgrenze und Klimacharakter, «Beitr.
Geobot. Landesaufn.», 6, Zurich.
Brunfelsius 0., 1530. Herbarum vivae icones, Strassburg.
Литература
273
В u d d е H., 1950. Versuch eines Rekonstruktion der Vegetation Westfalens in der alte-
ren Nachwarmezeit von 500 v. Chr. bis 1000 n. Ch., «Natur und Heimat», 3.
В iidel J., 1950. Das System der klimatischen Morphologie, «Dt. Geographentag
Munchen 1948».
В й к e r R., 1939. Die Pflanzengesellschaften des Messtischblattes Lengerich in West
falen, Munster.
В tinning E., 1949. Jahres- und tagesperiodische Vorgange in der Pflanze, «Stud.
Gen.», 2.
В tinning E., 1955. Jahreszeiten und Pflanzenleben, «Stud. Gen.», 8.
В ii n n i n g E., 1956. Der tropische Regenwald, «Verstandl. Wissensch.», 56, Berlin —
Gottingen — Heidelberg.
Burger H., 1948. Der Einfluss des Waldes auf den Wasserhaushalt, Zurich.
Busse W., 1908. Die periodischen Grasbrande im tropischen Afrika, «Mitt. a. d. Dt.
Schutzgeb.», 21.
Cain A. St., 1944. Foundations of plant geography, New York and London.
C a j a n d e r A. K., 1909. Uber Waldtypen, «Fennia», 28.
Campbell D. H., 1926. An outline of plant geography, New York.
Cannon W. A., 1921. Plant habits in the arid portions of South Australia,
Washington.
Ceballos L., Ortuno F., 1951. Estudio sobre la vegetacion у la flora forestal
de las Canarias occidentales, Madrid.
Cheval lier A., 1925. Essai d’une classification biogeografique des principaux
systemes de culture, «Rev. Int. de Renseign. agric.», 3.
Che val lier A., 1933. Le territoire geobotanique de 1’Afrique tropicale nord-
occidentale, «Bull. Soc. Bot.», 80.
Christ H., 1867. Uber die Verbreitung der Pflanzen der alpinen Region der euro-
paischen Alpenkette, Zurich.
Christ H., 1879. Das Pflanzenleben der Schweiz, Basel.
Christ H., 1910. Geographic der Fame, Jena.
Clements F. E., 1904. The development and structure of vegetation, Lin-
coln.
Clements F. E., 1905. Research methods in ecology, Lincoln.
Clements F. E., 1916. Plant succession, Washington.
Clements F. E., 1928. Plant succession and indicators, New York.
Clements F. E., Shelford V. E., 1939. Bioecology, New York.
Cooper W. S., 1913. The climax forest of Isle Royale. Lake Superieur and its develop-
ment, «Bot. Gas.», 55.
Cooper W. S., 1916. Plant succession in the Mount Robson region, British Colum-
bia, «Plant World», 19.
Cowles II. C., 1899. The ecological relations of the vegetation of the sand dunes of
the lake Michigan, «Bot. Gas.», 27.
Cowles H. C., 1911. The causes of vegetation cycles, «Bot. Gas.», 52.
Credner W., 1943. Die deutsche Agrarlandschaft im Kartenbild, Sitz. Ber. euro-
paischer Geographen, Leipzig.
Creutzburg N., 1928. Uber den Werdegang der Kulturlandschaften, «Sonderband
Z. Ges. f. Erdk. zu Berlin».
Diiniker A. U., 1928. Ein okologisches Prinzip zur Einteilung der Pflanzengesell-
schaften, Zurich.
Dancau B., Lutz J. C., 1956. Pflanzensoziologische Karte des Aischgrundes bei
Uhlfeld als Beispiel fur die Vegetationsverhaltnisse eines ungestorten frankischen
Flusstales, «Вауг. Landw. Jb.», 33.
274
Литература
Dansereau Р., 1943—1946. L’erabliere laurentienne I. u. II., «Contrib. Inst.
Bot. de 1’Univ. Montreal»., 45 et. 60.
Dansereau P., 1944. Les erablieres de la Gaspesie et le flcutuations du climat,
Montreal.
Dansereau P., 1944. Zonation et succession sur la restinga de Rio de Janeiro,
«I. Halosere. Rev. Canad. Biol.», 6.
Dansereau P., 1952. The varieties of evolutionary opportunity, «Rev. Canad.
Biol.», 11.
Dansereau P., 1956. Le regime climatique regional de la vegetation et le contro-
les edapliques, «Rev. Canad. Biol.», 15.
Dansereau P., 1957. Biogeography and ecological perspective, New York.
Darwin Ch., 1844—1845. A journal of researches in natural history kept during
a voyage round the world, London.
Darwin Ch., 1859. On the origin of species by means of natural selection, London.
De Candolle A., 1855. Geographic botanique raisonnee, Paris.
De Candolle A., 1874. Constitution dans le regne vegetal de groupes physiolo-
giques, «Archives Sci. Phys. Nat.», 50, Genf.
Delfs J., 1955. Die Niederschlagszuriickhaltung im Walde, «Mitt. Arbeitskr. Wald
mid Wasser», Nr. 2, Koblenz.
Devred R., 1956. Les savannes herbeuses de la region de Mvuazi (Bas-Congo)
«I.N.E.A.C.», 65, Bruxelles.
De Vries D. M., 1957. Overzicht van de ontwikkeling van het botanische grasland-
onderzoeg in Nederland, Wageningen.
Diels L., 1906. Die Pflanzenwelt von Westaustralien siidlich des Wendekreises,
«Vegetation d. Erde», 7, Leipzig.
Diels L., 1908. Pflanzengeographie. Sammlung Goschen, Leipzig, 5. Aufl. bearb.
von F. Mattick, Berlin, 1958. [Имеется русский перевод: Л. Дильс, 1916.
Ботаническая география. Пб.— Ред.]
Diemont W. Н., 1937. De beteekenis van de Plantensociologie voor den Boschbo-
uw, Nederl. Boschbouw Tijdschr.
Dodonaeus R., 1608. Herbarius oft Cruydt — Bock, Amsterdam (1 изд. в
1553 г.).
Doing Kraft H., Over de invloed van de mens op de plantengroi, «Levende
Natuur», 58, 6.
Domes N., 1936. Die klimatisch bedingte Abnahme des Ertrages von Wald und Wei-
de im Gebirge, Wien/Leipzig.
Dove H. W., 1844. Ober den Zusammenhang der Warmeveranderungen der Atmos-
phare mit der Entwicklung der Pflanzen, «Abh. Akad. Berlin Phys. К1.»
D r u d e O., 1876. Die Anwendung physiologischer Gesetze zur Erkliirung der Vegeta-
tionslinien, Gottingen.
D r u d e O., 1884. Die Florenreiche der Erde, «Р. M.», 30.
D r u d e O., 1887. Atlas der Pflanzenverbreitung, Gotha.
D r u d e 0., 1888. Pflanzengeographie. Neumayer, Anleitung zu wissensc haft-
lichen Beobachtungen auf Reisen, 2, Berlin.
Drude 0., 1890. Handbuch der Pflanzengeographie, Bibl. Geogr. Handbiicher,
Stuttgart.
Drude 0., 1902. Der hercynische Florenbezirk, «Vegetation der Erde», 6, Leipzig .
Drude 0., 1907. Die kartographische Darstellung mitteldeutscher Vegetationsforma-
tionen, «Вег. d. Freien Ver. syst. Bot. u. Pfl.-Geogr .»
Drude O., 1919. Die Elementarassoziation im Formationsbilde, «Вег. d. Freien Ver.
Pfl.-Geogr. u. syst. Bot.»
Du Rietz G. E., 1921. Zur methodologischen Grundlage der modernen Pflanzen-
soziologie, Upsala.
Литература
275
Du Rietz G. E., 1925. Die regionale Gliederung der skandinavischen Vegetation,
«Sv. Viixtsoc. Sallsk. Handl.», 8, Uppsala ooh Stockholm.
Du Rietz G. E., 1925. Studien fiber die Hohengrenzen der hochalpinen Gefass-
pflanzen im nordlichen Lappland, Feslschr. Schrdter, «Veroff. Geobot. Inst. Riibel»,
3, Zurich.
Du Rietz G. E., 1931. Life forms of terrestrial flowering plants, «Acta phytogeogr.
suecica», III.
Du Rietz G. E., Fries Th. С. E., T e n g wall T. A., 1918. Vorschlag zur
Nomenklatur dor soziologischen Pllanzengeographie, «Svensk. Bot. Tidskr.», 12.
Duvigneaud P., 1953. Les savannes du Bas-Congo, Liege.
Duvigneaud P., 1955. Etudes ecologiques de la vegetation en Afrique tropicale,
«Ann. Bot.». 31.
Ebermaier J.C., 1802. Von den Standorteu der Pflanzen im allgemeinen und
denen der Arzncigewachse besonders, Munster.
E b 1 i n B., 1901. Die Vegetationsgrenzen der Alpenrosen als unmittelbare Anhalte
zur Festsetzung friiherer bzw. moglicher Waldgrenzen in der Schweiz, «Schweiz.
Z. f. Forstwesen».
E h w a 1 d E., 1953. Der forstliche Wuchsbezirk als Mosaik von Standertseinheiten,
Dt. Akad. d. Landwirtschaftswiss. zu Berlin.
Eichler-Gradmann-Meigen, 1905—1927. Ergebnisse der pflanzengeo-
graphischen Durchforschung von Wurttemberg, Baden und Hohenzollern I bis
V, Stuttgart.
Ellenberg H., 1940. Mensch und Landschaft an Beispielen aus Nordwest-
Deutschland, «Natur und Volk», 70.
Ellenberg H., 1948. Unkrautgesellschaften als Mass fur den Sauregrad, die Ver-
dicbtung und audere Eigenschaften des Ackerbodens, «Вег. fiber Landtechnik»,
4, MiiDcheu.
Ellenberg H., 1954. Steppenhcide und Waldweide, «Erdkunde», 8.
Ellenberg H., 1955. Wuchsklimakarte Siidwestdeutschlands, Stuttgart.
Ellenberg H., 1956. Aufgaben und Methoden der Vegetationskunde. W a 1 t e r H.,
«Phytologie», IV, 1, Stuttgart.
Elster H. J., 1954. Einige Gedanken zur Systematik, Terminologie und Zielsetzung
der dynamischen Limnologie, «Arch. f. Hydrobiol.».
Emberger L., 1942. Un projet d’une classification des climats du point de vue
phytogeographique, Toulouse.
Emberger L., 1948. La carte des groupements vegetaux de la France, «Rev. gen.
Bot.», 55.
Emberger L., 1955. Une classification biogeograpbique des climats, «Recueil
, des Travaux des Laboratoires de Bot., Geol., et Zool. Univ. Montpellier», 7.
Engler A., 1879—1882. Versuch einer Entwicklungsgeschichte der Pflanzenwelt,
Leipzig.
Engler A., 1899. Die Entwicklung der Pflanzengeographie in den letzten bundert
Jahren und weitere Aufgaben derselben. Humboldt Centenar Schrift, Berlin.
Engler A., D r u d e O., 1896 ff. Die Vegetation der Erde, Leipzig.
Erichson R., 1923. Die Mangrovevegetation, Halle.
Ernst F., 1934. Die Bedeutung der Klimaextreme fur den Waldbau in Mitteleuropa,
«Forstw. C.», 56.
Eskuche U., 1955. Vergleichende Standortsuntersuchungen an Wiesen im Donau-
ried bei Herbertingen, «Jb. Ver. vaterl. Naturk. Wiirtt.», 109.
Etter H., 1954. Grundsatzliche Betrachtungen zur Beschreibung und Kennzeichnung
der Biochore, «Schweiz. Z. f. Forstwesen».
276
Литература
Faber А., 1937—1939. Pflanzensoziologisches Kartenblatt des mittleren Neckar-
und Ammertalgebiets, Stuttgart.
Fanshawe D. B., 1952. The vegetation of British Guiana, «Institute Paper
Nr. 29», Oxford.
F e 1 s E., 1935. Der Mensch als Gestalter der Erde, Leipzig.
F i 1 z e r P., 1936. Pflanzengemeinschaft und Umwelt, Stuttgart.
F i 1 z e r P., 1951. Die natiirlichen Grundlagen des Pflanzenertrags in Mitteleuropa,
Stuttgart.
Firbas F., 1949—1952. Waldgeschichte Mitteleuropas, Jena.
Fischer H., 1933. Pflanzengeographische Aufnahme des Blattes Grunwald, Mun-
chen.
Fitting H., 1922. Aufgaben und Ziele einer vergleichenden Physiologie auf geogra-
phischer Grundlage, Jena.
Fitting H., 1926. Okologische Morphologie der Pflanzen, Jena.
Fitting, Schumacher, Harder, Firbas, 1947. Lehrbuch der Botanik
fur Hochschulen, 23/24. Aufl., Jena.
F 1 a h a u 1 t M. Ch., 1894. Projet de carte botanique, forestiere et agricole de la Fran-
ce, «Bull. Soc. Bot. De france», 41.
Flahault M. Ch., Schrdter C.., 1910. Phytogeographische Nomenklatur,
Congr. Int. Bot. Bruxelles.
Flohn H., 1941. Die Tatigkeit des Menschen als Klimafaktor, «Z. f. Erdk.», 9.
Forbes E., 1846. On the connexion between the distribution of the existing fauna
and flora of the British Isles with the geological changes, «Memoirs Geol. Survey
of England», V.
Franz H., 1950. Bodenzoologie als Grundlage der Bodenpflege, Berlin.
Frei M., 1941. Der Anteil der einzelnen Tier- und Pflanzengruppen am Aufbau der
Buchenbiozonosen in Mitteleuropa, Zurich.
Frenzel B., Troll C., 1952. Die Vegetationszonen des nordlichen Eurasiens
wahrend der letzten Eiszeit, «Eiszeitalter und Gegenwart», 2.
Friedel H., 1934. Boden- und Vegetationsentwicklung am Pasterzenufer, «Carin- .
thia», II.
Friedel H., 1935. Beobachtungen an den Schutthalden der Karawanken, «Carin-
thia», II.
Friederichs K., 1937. Okologie als Wissenschaft von der Natur, «Bios», 7, Leip-
zig.
Friederichs K., 1943. Uber den Begriff «Umwelt» in der Biologie, «Acta bio-
theoretica», VII.
Fries Th. С. E., 1925. Uber primare und sekundiire Standortsbedingungen, «Svensk.
Bot. Tidskr.», 19.
Fro di n J., 1919. Uber nitrophile Pflanzenformationeu auf der Almen Jamtlands,
Bot. Notiser Lund.
Friih J., Schrdter C., 1904. Die Moore der Schweiz, Bern.
Fuchs L., 1542. Historia stirpium, Basel.
Fuller G. D., 1912. Evaporation and the stratification of vegetation, «Bot. Gaz.»,
54.
F u г о ii R., 1958. Causes de la repartition des etres vivants, Paris.
Furrer E., 1922. Begriff und System der Pflanzensukzession, Zurich.
Gams II., 1918. Prinzipienfragen der Vegetationsforschung, «Vierteljahrsschr. Naturf.
Ges. Zurich», 63.
Gams H., 1924. Die klimatische Begrenzung der Pflanzenareale, «G. Z.», 30.
Gams Ы., 1935. Zur Geschichte, klimatischen Begrenzung und Gliederung der immer-
griinen Mittelmeerstufe, «Veroff. Geobot. Inst. Riibel», 12.
Литература
277
Gams Н., 1940—1942. Pflanzengesellschaften der Alpen., 1—3. Jb. Ver. z. Schutze
d. Alpenpfl. u. -tiere.
Gams H., 1942. Der Zirmberg bei Innsbruck, «Naturschutz», 23.
Gams H., Nordhagen R., 1913. Postglaciale Klimaanderungen und Erdkru-
stenbewegungen in Mitteleuropa, Munchen.
Gams H., Ruoff S., 1928. Geschichte, Aufbau und Pflanzendecke des Zehlaub-
ruches, «Schr. Phys.— oekon. Ges. Konigsberg», 66.
Garner W. W., Allard H. A., 1920. Effect of relative length of day and night
and other factors of environment on growth and reproduction of plants, «Journ.
Agr. Res.», 18.
Gauss P., 1926. Vegetation und Anbau im Stromberg- und Zabergaugebiet, Heidel-
berg.
Gaussen H., 1933. Geographic des plantes, «Coll. Armand Colin», 152, Paris.
Gaussen H., 1938. La cartographic biogeographique. Congr. Int. de Geogr. Amster-
dam.
Gaussen H., 1952. Le dynamisme des biocenoses vegetates, «CoRoques Intern.
Centre Nat. de la Rech. Scient.», 33.
Geiger R., 1927. Das Klima der bodennahen Luftschicht, Braunschweig. [Имеется
русский перевод: P. Гейгер, 1960. Климат приземного слоя воздуха, ИЛ,
М. (перевод с 3-го немецкого издания).— РедД
Geiger R., 1930. Mikroklima und Pflaiizeukleid, «Hd. d. Klimatologie», I, Berlin.
Geiger R., 1932. Der Einfluss des Waldes auf das Klima, Berlin.
Geiger R., 1951. Die Waldgefahrdung dutch den Diirresommer 1947, «Forstw. Cen-
tralbl.», 70.
G e n s 1 e r G. A., 1946. Der Begriff der Vegetatioqszeit, Zurich.
Germain R., 1945. Note sur les premiers stades de la reforestation natureRe des
Savanes du Bas — Congo, «BuR. Agr. du Congo Beige», 36.
Germain R., 1952. Les Associationes vegetales de la Plaine de le Ruzizi (Congo
Beige), «Publ. de i’Inst. Nat. pour i’Etude agr. du Congo Beige», 52.
Germain R., Evrard C., 1956. Etudes ecologiques et phytosociologiques de
la foret a Brachystegia laurentii, «I.N.E.A.C. Serie scientifique», 67, Brussel.
G e s n e r C., 1555. Descriptio Montis Fractis sive Montis Pilati. Приложение к: De
Paris et Admirandis Herbis..., Zurich.
Gessner F., o. J. Pflanzengeographie, «Hdb. d. Biologic», 4, Potsdam.
Gessner F., 1955. Hydrobotanik, BerRn.
Ginzberger A., 1939. Pflanzengeographisches Hilfsbuch, Wien.
Глинка К. Д., 1913. Почвообразование, характер почвенных типов и география
почв. Пб.
G m е 1 i n J. G. 1741 —1752. Reisen dutch Sibirien, Gottingen.
Goppert H. R., 1868. Skizzen zur Kenntnis der Urwalder Schlesiens und Bohmens,
Bonn.
Good R., 1953. The geography of flowering plants, 2. AufL, London — New York —
Toronto.
Gradman n R., 1898. Das Pflanzenleben der Schwabischen Alb, Tubingen.
Gradmann R., 1901. Das mitteleuropaische Landschaftsbild nach seiner geschicht-
Rchen Entwicklung, «G. Z.», 7.
Gradmann R., 1909. Uber Begriffsbildung in der Lehre von den Pflanzenformatio-
nen, «Englers Bot. Jb.», 43.
Gradmann R., 1916. Wuste und Steppe, «G. Z.», 22.
Gradmann R., 1919. Pflanzen und Tiere im Lehrgebaude der Geographic, «Geogr.
Abende», 4, Berlin.
Gradmann R., 1932. Unsere Flusstaler im Urzustand, «Z. Ges. f. Erdk. zu Ber-
lin».
278
Литература
Gradmann R., 1934. Die Steppen des Morgenlandes, «Geogr. Abb.», III. 6, Stutt-
gart.
Gradmann R., 1936. Vorgescliichtliche Landwirtschaft und Besiedlung, «G. Z.»,
42.
Gradmann R., 1939 Mein Beitrag zur Urlandschaftsforschung, «Z. f. Erdk.», 7.
Grisebach A., 1838. Uber deu Einfluss des Klimas auf die Begrenzung der natiir-
lichen Floren, «Linnaea», 12.
Grisebach A., 1847. Gesammelte Abhandlungen und kleine Schriften, Leipzig.
Grisebach A., 1866. Die Vegetationsgebiete der Erde, «Р. M.», 11. [Имеется рус-
ский перевод: А. Гризебах, 1874—1877. Растительность земного шара.—
РедД.
Grisebach А. 1872. Die Vegetation der Erde nach ihrer klimatischen Anordnung,
Leipzig.
Gross H., 1914. Zwei bemerkenswerte Moore in Konigsberg Umgebung, Leipzig
u. Berlin.
Haase H., 1948. Die Folgen der Entwaldung des Harzes auf die Wasserfiihrung und
die Erosion, «Das Gas- und Wasserfach», 89.
Haberlandt G., 1893. Eine botanische Tropenreise, Leipzig.
Hahn E., 1910. Die Brandwirtschaft in der Bodenkultur, «Nachr. d. Klubs der Land-
wirte», Berlin.
Haller A., 1742. Emiumeratio niethodica stirpium Helvetiae indigenarum, Got-
tingen.
H a n n i g E., Winkler H. u. a., 1926—1938. Die Pflanzenareale, «Reihe», 1—4,
Jena.
Hansen H. M., 1930. Studies on the vegetation of Iceland, Copenhagen.
Hansen H. M., 1956. Life forms as age indicators, Ringkjobing.
Harder R., 1941. Einige neue Ergebnisse der Untersuchungen an Lang- und Kurztags-
pflanzen, «Forsch. u. Fortschr», 17.
Hardy M., 1925. The geography of plants, Oxford.
Hare F. K., 1950. Climate and zonal divisions of the boreal forest in Eastern Canada,
«Amer. Geogr. Soc. Spec. Publ.», 32, New York.
H a r n i s c h 0., ‘1929. Die Biologic der Moore, Stuttgart.
Harshberger J. W., 1911. Pliyt©geographic survey of North America, «Vegeta-
tion d. Erde», 13, Leipzig.
Hartmann F. K., 1941. Uber den waldbaulichen Wert des Grundwassers, «Mitt.
Forstwirtsch. u. Forstwiss.»
Hausrath H., 1911. Pflanzengeographische Wandlungen der deutschen Landschaft,
«Wissenschaft und Hypothcse», XIII, Leipzig — Berlin.
Hayek A., 1926. Allgemeine Pflanzengeographie, Berlin.
Heer 0. 1835. Die Vegetationsverhaltnisse des sudostlichen Teiles des Kantons Gla-
rus, «Mitt. a. d. Gebiet d. Theoret. Erdk.», Zurich.
H e g i G., 1935. Illustrierte Flora von Mitteleuropa, Munchen.
Heikinheimo 0., 1915. Der Einfluss der Brandwirtschaft auf die Walder Finn-
lands, «Acta Forest. Fennica», 4.
H e m s 1 e у W. B., 1888. Botany. Biologica centrali-americana.
Hermes K., 1955. Die Lage der oberen Waldgrenze und ihr Abstand zur Schnee-
grenze, «Kolner Geogr. Arb.», 5.
Herzog Th., 1923. Die Pflanzenwelt der bolivianischen Auden, «Vegetation der Erde»,
15, Leipzig.
Herzog Th., 1926. Geographic der Moose, Jena.
Herzog Th., o. J. (nach 1930); Pflanzengeographie, «Hdb. d. geogr. Wiss.», Ber-
lin. ’
Литература
279
Н е t t п е г А., 1893. Regenverteilung, Pflanzendecke und Besiedlung der tropischen
Anden, Leipzig.
Hettner A., 1927. Die Geographic, ihre Geschichte, ihr Wesen und ihre Methoden,
Breslau. [Имеется русский перевод: А. Геттнер, 1930. География, ее история,
сущность и методы, М.— Л.— Ред.].
Hettner А., 1935. Die Pflanzenwelt, «Vergleichende Landerkunde», 4, Leip-
zig.
Hiltner E., 1926. Die Phanologie und ihre Bedeutung, Munchen.
Hjelmquist H., 1940. Studien uber die Abhangigkeit der Baumgrenzen von
den Temper aturverhaltnissen, Lund.
Hock F., 1890. Nahrpflanzen Mitteleuropas, «Forsch. z. dt. Landes- u. Volkskunde»,
V, 1.
Hock F., 1899 u. 1900. Der gegenwartige Stand unserer Kenntnis von der urspriing-
lichen Verbreitung der angebauten Nutzpflanzen, «G. Z.», 5 и 6.
Hoff К. E. A. v., 1822—1841. Geschichte der natiirlichen Veranderungen der Erdo-
berflache, Gotha.
Hoffman H. К. H., 1857. Witterung und Wachstum oder Grundziige der Pflanzen
klimatologie, Leipzig.
Hoffman H. К. H., 1860. Vergleichende Studien zur Lehre von der Bodenstetig-
keit der Pflanzen, Giessen.
Hoffmann H. К. H., 1865. Untersuchungen zur Klima- und Bodenkunde mit
Rucksicht auf die Vegetation, «Bot. Ztg.», 23.
Hoffmann H. К. H., 1869. Pflanzenarealstudien in den Mittelrheingegenden, Gies-
sen.
Hooker J. D., 1847. On the vegetation of the Galapagos Archipelago, Transact.
Linn. Soc.
Hoops J., 1905. Waldbaume und Kulturpflanzen im germanischen Altertum, Strass-
burg.
H о r n s t e i n F. v., 1951. Wald und Mensch, Ravensburg.
H о r v a t J., 1929. Die Verbreitung und die Geschichte der mediterranen, illyrischen
und pontischen Florenelemente in Nordkroatien und Slovenien, «Acta Botanica»,
4, Zagreb.
Hosseus С. C., 1912. Durch Konigs Tschulalongkonrs Reich, Stuttgart.
Huber B., 1947. Die mutmasslichen Auswirkungen der heutigen Diirre auf den Wald,
«AHg. Forstzeitschr.», 2. Jg., Munchen.
Hueck K., 1925. Vegetationsstudien auf brandenburgischen Hochmooren, «Beitr.
Naturdenkmalpflege», 10, 5.
Hueck K., 1931. Vegetationskundliche Karte des Endmoranengebiets von Chlorin,
Berlin.
Hueck K., 1932. Erlauterung zur vegetationskundlichen Karte der Lebanehrung,
Berlin.
Hueck K., 1934. Erlauterung zur vegetationskundlichen Karte des Memeldeltas,
Neudamm.
Hueck K., 1937. Pflanzengeographie Deutschlands, Berlin.
Hueck K., 1938. Die natiirliche Vegetation 1 : 3 000 000.
Hueck K., 1951. Die biologische Methode zum Messen der erodiereden Tatigkeit
des Windes und des Wassers, «Вег. Dt. Bot. Ges.».
Hueck K., 1953. Urlandschaft, Raublandschaft und Kulturlandschaft in der Pro-
vinz Tucuman im nordwestlichen Argentinien, «Bonner Geogr. Abh.», 10.
Hueck K., 1957. Las regiones forestales de Sur America, «Boletin Inst. Forest.
Latino Americano», 2, Merida.
H u I t R., 1881. Forsok till analystik behandling af vaxtformationerna, «Meddel.
Soc. Faun. Flor. Fenn», VIII.
280
Литература
Н u 1 t ё n Е., 1950. Atlas of the distribution of vascular plants in NW Europe, Stock-
holm.
Humboldt A. v., 1805. Essai sur la geographic des plantes, Paris.
Humboldt A. v., 1806. Ideen zu einer Physiognomik der Gewachse, Tubin-
gen.
Humboldt A. v., 1808. Ansichten der Natur, Stuttgart. [Имеется русский пере-
вод: А. Гумбольдт, 1862. Картины природы, М.— РедД
Humboldt A. v., 1816. Sur les lois que Гоп observe dans la distribution des for-
mes vegetales, Paris.
Humboldt A. v., 1817. De distributione geographica plantarum secundum coeli
temperiem et altitudinem montium. Einleitung zu 6. Abt. in: Nova genera et spe-
cies plantarum partim adumbraverunt A. Bonpland et Alexander de Humboldt.
1815—1825.
Humboldt A. v., 1817. Des lignes isothermes et de la distribution de la chaleur
sur le globe, «Mem. de la Soc. d’Arcueil».
Humboldt A. v., 1845—1862. Kosmos, Entwurf einer physischen Weltbeschrei-
bung. [Имеется русский перевод: А. Гумбольдт, 1848—1863. Космос,
ч. 1—5, М. — РедД
Humboldt A. v., Bonpland А., 1814—1819. Voyages aux regions equinoxia-
les du nouveau continent, Paris.
Huttenlocher F., 1923. Sonnen- und Schattenlage, «Erdgesch. u. landeskun.
Abh. aus Schwaben u. Franken», 7, Ohringen.
Ihne E., 1884. Geschichte der pflanzenphanologischen Beobachtuugen in Europa,
Giessen.
Ihne E., 1905. Phanologische Karte des Friihlingseinzuges in Mitteleuropa, «Р. M.»,
51.
«I.N.E.A.C.», 1954 и след. Carte des sols et de la vegetation du Congo Beige et du Ruanda-
Urundi.
1. Kaniama (Haut-Lomami) 1955
2. Mvuazi (Bas-Congo) 1954
3. Vallee de la Ruzizi 1955
4. Nioka (Ituri) 1954
5. Mosso (Urundi) 1955
6. Yangambi Pl. 1 : Weko 1954
7. Bugesara-Mayaga (Ruanda) 1956
Brussel.
Iversen J., 1936. Biologische Pflanzentypen als Hilfsmittel in der Vegetationsfor-
schung, Kopenhagen.
Iversen J., 1949. The influence of prehistoric man on vegetation, «Danmarks
Geol. Unders.» IV, 3 Nr 6. Kopenhagen.
Jaeger F., 1945. Zur Gliederung und Benennung des tropischen Graslandgiirtels,
«Verb. Naturf. Ges. Basel», 56, 2.
Jentsch u. Biisgen M., 1909. Forstwirtschaftliche und forstbotanische Expe-
dition nach Kamerun und Togo. Tropenpflanzer.
Jessen O., 1933. Natur- Raub- und Kulturlandschaft in Angola, «Z. Ges. f. Erdk.
zu Berlin».
Jones Ph. M., 1927. The origin of prarie, «Science», 66.
Junghuhn F. W., 1852. Java, Leipzig.]
Kaiser E., 1951. Arbeitziele und Methoden der Biogeographie erlautert an Bei-
spielen der ober- und mitteldeutschen Steppenheide, «Р. M.», 95.
Литература
281
Karsten G., 1891. Uber die Mangrovevegetation im malayischen Archipel, «Bibl.
Bot.», 12.
Karsten G., Schenck H., 1903 и след. Vegetationbilder, Jena.
К e i 1 h а с к К., 1915. Uber tropische und subtropische Torfmoore auf der Insel Cey-
lon, «Jb. d. Pr. Geol. Landesanst», Berlin.
Келлер А., Сукачев В. H., 1932. Основные направления геоботанических
исследований, Изд-во Бот. ин-та АН СССР.
Келлер Б. А., 1923. Растительный мир русских степей, полупустынь и пустынь,
вып. 1, Воронеж.
Keller R., 1948. Zum Wasserverbrauch von Vegetation und Wirtschaft, «Erdkunde»,
2.
Kerner von Marilaun A., 1855. Uber den Einfluss der Temperatur des Quell-
wassers auf die im Rinnsale der Quellen vorkommenden Pflanzen, «Verh. Zool.—
Bot. Ges. Wien», V.
Kerner von Marilaun A., 1861. Zeitliche Umwandlung der Pflanzenfor-
mationen, «Verh. Zool.— Bot. Ges. Wien», XI.
Kerner von Marilaun A., 1863. Das Pflanzenleben der Donaulander, Inns-
bruck.
К e r n e r von Revue», I. Marilaun A. , 1864. Osterreichs waldlose Gebiete, «Osterr.
К erner von Marilaun A., 1869. Die Abhiingigkeit der Pflanzengestalt von
Klima und Boden, Innsbruck.
К i li 1 m a u n A. 0., 1890. Pflanzenbiologische Studien aus Russisch-Lappland,
«Acta Soc. Faun. Flor. Fenn.», VI.
Kirchhoff A., 1899. Pflanzen- und Tierverbreitung, «Allgemeine Erdkunde», 3.
К 1 a p p E., 1938. Wiesen und Weiden, Berlin.
К 1 a p p E., 1950. Pflanzensoziologische Auswirkungen verschiedener Weidetechnik,
«Mitt. Flor.— soz. Arbeitsgem.», N. F. 2.
Klapp E., S t a h 1 i n A., 1936. Standorte, Pflanzengesellschaften und Leistuug
des Grunlandes, Stuttgart.
К 1 a u s i n g 0., 1956. Untersuchungen uber den Mineralumsatz in Buchenwiildern,
«Forstw. Centralbl.», 75.
Клеонов Я. Д., 1941. Флористический анализ лиственных лесов Восточной
Европы, Харьков.
Knapp R., 1948. Einfuhrung in die Pflanzensoziologie, Stuttgart.
Knapp G. u. R., 1952. Beobachtungen uber Mikroklima und Spatfrosterscheinungen
im uordostlichen Vogelsberg, «Wetter u. Leben», 4.
К n о c h K., 1949. Die Gelandeklimatologie, «Вег. z. dt. Landesk.», 7.
Knoll J. G., 1925. Die Pflanzenbestandesverhaltnisse der Dauerwiesen in der Umge-
bung von Ulm, Hohenheim.
Koch W., 1926. Die Vegetationseinheiten der Linthebene, «Jb. d. St. Gallischeu
Naturw. Ges.», 61.
К о e g e 1 L., 1914. Das Urwaldphanomen Amazoniens, Munchen.
Koppen W., 1900. Versuch einer Klassifikation der Klimate vorzugsweise nach
ihren Beziehungen zur Pflanzenwelt, «G. Z.», 6.
Koppen W., 1919. Baumgrenze und Lufttemperatur, «Р. M.», 65.
Koppen W., 1926. Der jahrliche Temperaturgang und die Vegetationsperiode, «Met.
Zeitschr.»
К 6 s t 1 e r J., 1950. Waldbau, Berlin u. Hamburg.
Koorders S. H., 1895. Beobachtungen uber spontane Neubewaldung auf Java,
«Forstl. naturwiss. Z.»
Krassnow A. J., 1898. Die Grassteppen der nordlichen Halbkugel, «Englers Bot.
Jb.», XXVI.
282
Литература
Kraus G., 1900. Aus der Pflanzenwelt Unterfrankens I, «Verh. Phys.— Med. Ges
Wurzburg».
Kraus G., 1911. Boden und Klima auf kleinstem Raum, Jena.
Krause W., 1950. Uber Vegetationskarten als Hilfsmittel kausalanalytischer Un-
tersuchung der Pflanzendecke, «Planta», 38.
Krause W., 1952. Das Mosaik der Pflanzengesellschaften, «Planta», 41.
Krauss G., 1936. Aufgaben der Standortskunde, Jahresber. d. dt. Forstvereins.
Krauss G., H artel F., Muller K., Gartner G., Schanz H., 1939.
Stand ortsgemasse Durchfuhrung der Abkebr von der Fichtenwirtschaft im nordwest-
sachsischen Niederland, Tharandter Forstl. Jb.
Krebs N., 1923. Natur- und Kulturlandschaft, «Z. Ges. f. Erdk. zu. Berlin».
К r e h W., 1950. Verlust und Gewinn der Stuttgarter Flora im letzten Jahrhundert,
«Jb. Ver. vaterl. Naturk. Wiirtt.», 106;
Kreh W., 1951. Die Besiedlung der Trummerschutts durch Pflanzenwelt, «Naturw.
Rundschau».
Kreh W., 1956. Die Erhaltung des Lebens auf der Erde, «Naturw. Rund-
schau», 9.
Kreh W., 1957. Zur Begriffsbildung und Namengebung in der Adventivfloristik,
«Mitt. Flor.-soz. Arbeitsgem.», N. F. 6/7.
К r e n к e 1 E., 1920. Moorbildungen im tropischen Afrika, «Centralbl. f. Min. Geo].
Pal.»
Kubiena W. L., 1948. Entwicklungslehre des Bodens, Wien.
К u c h 1 e r A. W., 1951. The relation between classifying and mapping vegetation,
«Ecology», 32.
К u j a 1 a V., 1945. Waldvegetationsuntersuchungen in Kanada, «Ann. Acad, scient.
Fennicae», Helsinki.
Laatsch W., 1938. Dynamik der deutschen Acker- und Waldboden, Leipzig.
Lamprecht H., 1957. Sobre unos resultados de estudios estructurales en varios
tipos de bosques venezolanos, «Universitas Emeritensis», 4, Merida.
Lang R., 1920. Verwitterung und Bodenbildung, Stuttgart.
Lauer W., 1952. Humide und aride Jahreszeiten in Afrika und Siidamerika und ihre
Beziehungen zu den Vegetationsgiirteln, «Bonner Geogr. Abh.», 9.
Lauscher F. u. A.,Printz H., 1955—1959. Die Plianologie Norwegens, Oslo.
Lautensach H., 1938. Uber die Erfassung und zVbgrenzung von Landschafts-
raumen, Congr. Int. Geogr., Amsterdam.
Lautensach H., 1955. Der Rhytmus der Jahreszeiten auf der Iberischen Hal-
binsel, «Geogr. Rundschau», 7.
Лавренко E. M., С о ч а в а В. Б. (ред.), 1956. Растительный покров СССР,
Л. (с геоботанической картой СССР 1 : 4 000 000).
Lebrun J., 1947. La vegetation de la plaine alluviale au sud du Lac Edouard, Brus-
sel.
Ledebour K. F., 1829. Reise durch das Altaigebirge und die dsungarische Kir-
gisensteppe, Berlin.
L e i c k E., 1930. Die Pflanzendecke, Supan — Obst, «Grundziige der phys. Erdkun-
de», 7. AufL, II, 2, Berlin u. Leipzig.
Lindquist B., 1931. Den skandinaviska Bokskogens biologi, Uppsala.
L i n k о 1 a K., 1922. Zur Kenntnis der Verteilung, der landwirtschaftlichen Siedlungen
auf die Boden verschiedener Waldtypen in Finnland, «Acta Forest. Fennica», 22.
L i n k о 1 a K., 1941. Die Kartierung der Flora und Vegetation Finnlands, «Sitz.—
Ber. Finn. Ak. Wiss.», 1938, Helsinki.
Linne C. v., 1737. Flora lapponica, Amsterdam.
Linne C. v., 1745. Flora suecica, Stockholm.
Литература
283
L i п и ё C. v., 1751. Philisophia botanica, Stockholm. [Имеется русский перевод:
К. Линне ii, 1805. Философия ботаники..., Спб.— РедД
L i nn ё С. v., 1754. Stationes plantarum, «Amoenitates Acad.», IV.
L i ns tow 0. v., 1929. Bodenanzeigende Pflanzen, 2. Aufl., Berlin.
Livingston В. E., Shreve Forrest, 1921. The distribution of Vegetation
in the USA as relates to Climatic conditions, Washington.
Lohmeyer W., 1952. Naturlandschaftskarte des Gebietes beiderseits der Mittel-
weser zwischen Dummer, Steinhuder Meer und Bremen, 1 : 300 ООО, «Mitt. Flor.-
soz. Arbeitsgem.», N. F. 3.
Lorenz J. R., 1858. Allgemeine Resultate aus der pflanzengeographischen und gene-
tischen Untersucliung der Moore im praalpinen Hiigelland Salzburgs, «Flora», 16.
Louis H., 1939. Das natiirliche Pflanzenkleid Anatoliens, «Geogr. Abh.», Ill, 12.
Stuttgart.
L ii d i W., 1921. Die Pflanzengesellschaften des Lauterbrunnentals und ihre Sukzes-
siou, «Beitr. z. geobot. Landesaufn.», 9.
L ii d i W., Zoller H., 1948. Einige Beobachtuugen fiber die Diirreschaden des Som-
mers 1947 in der Nordschweiz und am schweizerischen Juraland, «Вег. Geobot.
Forsch.— Inst. Rubel», Zurich.
Luetzelburg P. v., 1925—1926. Estudio botanico do No^ste, Rio de Janeiro.
Lundegardh H., 1925. Klima und Boden in ihrer Wirkung auf das Pflanzenleben,
Jena und Neuauflagen 1930, 1949. 1954. [Имеется русский перевод: Г. Л ю н д е-
г о р т, 1937. Влияние климата и почвы на жизнь растений.— РедД
Lutz J., 1941. Die ehemaligen Eisenhammer und Hiittenwerke und die Waldentwi-
cklung im norddstlichen Bayern, «Mitt. Forstwirtsch. u. Forstwiss.», 12.
Magdefrau K., I960. Flechtenvegetation und Stadtklima, «Naturwiss. Rundschau».
Mager F., 1930. Entwicklungsgeschichte der Kulturlandschaft des Herzogtums
Schleswig in historischer Zeit, Bd. I., Breslau, 1930: Bd 2., Kiel, 1937.
M a r k g r a f F., 1949. Eine neue Hohenstufenkarte der Vegetation Albaniens, «Bor.
Geobot. Forsch.— Inst. Rubel».
Markus E., 1932. Chorogenese und Grenzverschiebung, Tartu.
Markus E., 1936. Geographisclie Kausalitiit, Tartu.
M a r 1 о t h R., 1908. Das Kapland, insonderheit das Reich der Kapflora, das Wald-
gebiet und die Karroo, Wiss. Erg. d. dt. Tiefsee-Exped. Valdivia.
M a r t i u s K. F. Ph., 1824. Die Pliysiognomie des Pflanzenreiches in Brasilien,
Munchen.
M a r t i u s K. F. Ph., S p i x J. B., 1823—1831. Reise nach Brasilien, Munchen.
M a r t о n n с E. de, 1925. ТгаПё de gёographie physique, 4. Aufl., 3 Bd — «Biogeo-
graphic» (avec la collaboration de A. Chevaliere et L. Сиёпо!), Paris. [Имеется рус-
ский перевод: Э. М а р т о н н, 1940. Основы физической географии, Т. III —
«Биогеография», М.— РедД
Matuszkiewicz A. u. W., 1954. Die Verbreitung der Waldassoziationen
des Nationalparks von Bialowieza, «Ekologia Polska», II.
Me. Dougall W. B., 1941. Plant ecology, Philadelphia. [Имеется русский пере-
вод первого издания: В.Д. Мак-Дугол, 1935. Экология растений.— РедД
М е i g е и F., 1896. Uber die Besiedlung der Reblausherde in der Provinz Sachsen,
«Englers Bot. Jb.», XX.
Meisel-Jahn S., 1955. Die Kiefern — Forstgesellschaften des nordwestdeu-
tschen Flachlandes, «Angew. Pflanzensoz.», 11, Stolzenau/Weser.
Melchior H., Werdermann E., 1954. A. Englers Syllabus der Pflanzeu-
familicn, 12. Aufl., 1, Berlin— Nikolassee.
Me ns chi ng H., 1951. Die kulturgeographische Bedeutung der Auelehmbildung,
Dt. Geographentag. Frankfurt a. M.
284
Литература
Merxmiiller Н., 1952. Untersuchungen zur Sippengliederung und Arealbildung
in den Alpen, «Jb. Ver. z. Schutze der Alpenpfl. u.-tiere.», 17.
Me us el H., 1937—1944. Verbreitungskarten mitteldeutscher Leitpflanzen I bis 6.
Reihe, «Hercynia», I bis III, Halle — Berlin.
M e u s e 1 H., 1940. Die Grasheiden Mitteleuropas. «Bot. Arch.», 41.
M e u s e 1 H., 1941. Liber die pflanzengeographische Stellung des nordwestdeutschen
Eichen — Birkenwaldes, «Вег. Dt. Bot. Ges.», 59.
Meusel H., 1942. Das Buchenwald als Vegetationstypus, «Bot. Arch.», 43.
M e u s e 1 H., 1943. Vergleichende Arealkunde, Berlin.
Meusel H., 1952. Liber Wuchsformen, Verbreitung und Phylogenie einiger mediter-
ran-mitteleuropaischer Angiospermen — Gattungen, «Flora», 139.
Meusel H., 1953—1954 u. 1955. Verbreitungskarten mitteldeutscher Leitpflanzen.
7. u. 8. Reihe, «Wiss. Z. d. Martin-Luther-Universitat Halle—Wittenberg», III u. V.
Meusel H., 1957. Fortschritte der pflanzengeographischen Forschung in China,
«Wiss. Z. Univ. Halle, Math — Nat.», 6.
Meynen E., Schmithiisen J., 1953 и след. Handbuch der naturraumlichen
Gliederung Deutschlands, Remagen.
Miller Ch., 1892. Why are the prairies treeless? Proceedings R. G. Society, London.
Mobius K., 1877. Die Auster und die Austernwirtschaft, Berlin.
Mobius M., 1937. Geschichte der Botanik von den ersten Anfiingen bis zur Gegen-
wart, Jena.
Moller A., 1922. Der Dauerwaldgedanke, Berlin.
Montfort C., 1918. Die Xeromorphie der Hochmoorpflanzen, «Z. f. Bot.», 10.
Morgen A., 1952. Die Bedeutung der Phanologie fiir die Landeskunde, «Вег. z. dt.
Landesk.»
Морозов Г. Ф., 1949. Учение о лесе, М.-Л.
Miickenhausen Е., 1936. Die Bodentypenwandlungen des norddeutschen Flach-
landes, «Jb. Pr. Geol. Landesanst,», 56.
Muller M., 1951. Der Verfall der Wiesenvegetation im Altmiihltal im Vergleich
mit der Diirreeinwirkung 1947 und 1949 in Wiesen anderer Flusstaler, Landw. Jb. f.
Bayern.
Myers J. G., 1933. Notes of the vegetation of the Venezuelan Llanos, «Journal of
Ecology», 23.
Myers J. G., 1936. Savannah and forest vegetation of the interior Guiana Plateau,
«Journal of Ecology», 24.
Neumann J., 1908. Die Grasfluren der Erde, «Z. f. Infektionskrankheiten der Haus-
tiere», I.
Nichols G. E., 1917. The interpretation and application of certain terms and con-
cepts in the ecological classification of plant communities, «Plant World», 20.
N i e t s c h H., 1939. Wald und Siedlung in vorgeschichtlichen Mitteleuropa, Leipzig.
Noirfalise A., 1952. La Frenaie a Carex, «Verh. Konikl. Belg. Inst. Naturwiss.»,
122, Brussel.
Oberdorfer E., 1936. Erlauterung zur vegetationskundlichen Karte des Ober-
rheingebiets bei Bruchsal, «Beitr. z. Naturdenkmalpflege», 16.
Oberdorfer E., 1938. Ein Beitrag zur Vegetationskunde des Nordschwarzwaldes,
«Beitr. z. naturkundll. Forsch. in Siidwestdeutschland», 3.
Oberdorfer E., 1948. Gliederung und Umgrenzung der Mittelmeervegetation auf
der Balkanhalbinsel, «Вег. Geobot. Forsch.-Inst. Riibel».
Oberdorfer E., 1949. Pflanzensoziologische Exkursionsflora fiir Siidwestdeutsch-
land, Stuttgart.
Oberdorfer E., 1957. Siiddeutsche Pflanzengesellschaften, Jena.
Литература
285
Obst Е., 1932. Die Gliederung der Tropenzone nach ihren Pflanzenerzeugnissen,
Festschr. f. Carl Uhlig. Ohringen.
Obst E., Kaiser K., 1949. Die grosse Randstufe auf der Ostseite Siidafrikas
und ihr Vorland, Hannover.
Ogilvie A. G., 1937. Natural and cultivated vegetation in the eastern Dora Baltea
Basin, «Scottish Geogr. Mag.», 53.
Oosting H. J., 1948. The study of plant-communities, San Francisco.
О s v a 1 d H., 1923. Die Vegetation des Hochmoores Kommosse, Svenska vaxtsocial
sallsk, Landl.
Paczoski J. K., 1896. Das soziale Leben der Pflanzen, «Wszeschawiat», 15.
P a f f en К. H., 1957. Caatinga, Campos und Urwald in Ostbrasilien, Dt. Geogra-
phentag Hamburg, Wiesbaden.
Pallas P. S., 1771—1776. Reisen durch verschiedene Provinzen des russischen Reiches,
Petersburg.
Pallmann H., 1947. Pedologie et Phytosociologie, C. R. Congr. Int. de Pedologie,
Alger.
Pallmann H., Frei E., 1949. Die natiirlichen Produktionsgrundlagen der schwei-
zerischen Milchwirtschaft, Die Scweiz. Milchwirtschaft.
Pallmann H., Haffter P., 1933. Pflanzensoziologische und bodenkundliche
Untersuchungen im Oberengadin, «Вег. Schweiz. Bot. Ges.», 42.
Palmgren A., 1921. Die Entfernung als pflanzengeographischer Faktor, «Acta
Soc. Faun. Flor. Fenn.», 49.
Palmgren A., 1927. Die Artenzahl als pflanzengeographischer Charakter sowie
der Zufall und die sakulare Landhebung als pflanzengeographische Faktoren, «Acta
Bot. Fenn.»
Passarge S., 1921. Kaltewiisten und Kaltesteppen, Berlin.
Penck A., 1910. Versuch einer Klimaklassifikation auf physiogeograpliischer Grund-
lage, «Sitz.-Вег. d. Pr. Akad. d. Wiss.», 12, Berlin.
Penck A., 1928. Neuere Geographic, Sonderband Z. Ges. f. Erdk. zu Berlin.
Perrier de la Bat hie H., 1921. La vegetation malgache, «Ann. Musee Colo-
nial Marseille».
Philippson A., 1895. Vegetationskarte des Peleponnes, «Р. N.», 41.
Philippson A., 1903. Uber Klima und Vegetation im westlichen Kleinasien,
Sitz.-Вег. d. niederrh. Ges. f. Natur- u. Heilkunde, Bonn.
Philippson A.,
63.
1919. Die Vegetation des westlichen Kleinasien,
«Р. M.»,
Philippson
Philippson
Philippson
A., 1922. Das Mittelmeergebiet, 4. Aufl., Leipzig.
A., 1929. Grundziige der Allgemeinen Geographic, Leipzig.
A., 1947. Griechenlands zwei Seiten, «Erdkunde», I.
P i t t i e r H., 1918. Forest formation of the Isthmus of Panama, «Journ. of Fore-
stry», 16.
P i t t i о n i B., 1949. Die klimaokologische Formel als Hilfsmittel der biogeographi-
schen Forschung, «Wetter u. Leben», 2.
Plochmann R., 1956. Bestockungsaufbau und Baumartenwandel nordischer
Urwalder, «Forstwiss. Forschungen», 6, Hamburg.
P о e p p i g E., 1835—1836. Reise in Chile-, Peru und auf dem Amazonenstrome 1827—
1832, Leipzig.
Post L. v., 1926. Einige Aufgaben der regionalen Moorforschung, Stockholm.
Potonie H., 1912. Die rezenten Kaustobiolithe, Abb. Preuss. Geol. Lande-
sanst.
Precht H., Christoph er sen J., Hensel N., 1955. Temperatur und
Leben, Berlin-Gottingen-Heidelberg.
286
Литература
Р г е i s i n g E., 1954. Ubersicht uber die wichtigen Acker- und Grunlandgesellschaf-
ten NW — Deutschlands unter Beriicksichtigung ihrer Abhangigkeit vom Wasser
und ihres Wirtschaftswertes, «Angew. Pflanzensoz.», 8.
Raabe E. W., 1955. Uber die Verarmung der Landschaft, «Schr. d. Naturw. Ver.
f. Schleswig-Holstein», XXVII.
R abeler W., 1937. Die planmassige Untersuchung der Soziologie, Okologie und
Geographie der heimischen Tiere, «Mitt. Flor.-soz. Arbeitsgem. Niedersachsen», 3.
R a b e 1 er W., 1951. Systematik der Vogelgemeinschaften im Hinblick auf Biozd-
notik und Pflanzensoziologie, «Ornithol. Abh.», 9.
R abeler W., 1955. Zur Okologie und Systematik von Heuschreckenbestanden
nordwestdeutscher Pflanzengesellschaften, «Mitt. Flor.-soz. Arbeitsgem», N. F. 5.
R a p a i e s R., 1931. Versuch einer Gesellschaftslehre der Pflanzen, «Forsch. z. Vol-
kerpsych. u. Soziologie», X.
R a t z e b u r g J. T. C., 1859. Die Standortsgewachse und Unkrauter Deutschlands,
Osterreicbs und der Schweiz, Berlin.
R a t z e 1 F. 1889. Hohengrenzen und Hohengurtel, Z. D. 0. Alpenver., 20.
Raunkiaer Ch., 1905. Types biologiques pour la geographie botanique, «Bull.
Acad. R. Sc. Kopenhagen».
Raunkiaer Ch., 1910. Statistik der Lebensformen als Grundlage fur die biologi-
sche Geographie, «Beih. Bot. Zentralbl.», 27.
Raunkiaer Ch., 1934. The life-forms of plants and statistical plant geography,
Oxford.
Ray J. (Rajus), 1693. Historia plantarum generalis, London.
R e g e 1 C., 1920. Zur Kenntnis des Baumwucbses an den polaren Waldgrenze, «Sitz-
ber. Naturf. Ges. Dorpat».
R e g e 1 C., 1950. Dynamik von Klima und Pflanzendecke in Nordeuropa, «Вег. Geo-
bot. Forsch.- Inst. Rubel f. 1949».
R e i n d 1 J., 1903. Die schwarzen Fliisse Siidamerikas, Miinclien.
Reiter H., 1885. Die Konsolidation der Physiognomik, Graz.
R e m a n e A., 1939. Die Gemeinschaft als Lebensform in der Natur. «Kieler Bl.», 2.
R en v a 1 1 A., 1913. Die periodischen Erscheinungen der Reproduktion der Kiefer
an den polaren Waldgrenzen, «Acta Forest. Fenn.»
Richards P. W., 1952. The tropical rain forest, Cambridge. [Имеется русский
перевод: П. Ричардс, 1961. Тропический дождевой лес, ПЛ, М.— РедД
Rikli М., 1904. Versuch einer pflanzengeographischen Gliederung der arktischen
Wald- und Baumgrenzen, Zurich.
Rikli M., 1913. Die Florenreiche, «Hdwb. d. Naturwiss.», IV, Jena.
Rikli M., 1948. Das Pflanzenkleid der Mittelmeerlander, Bern.
Robbins C. R., 1934. Northern Rhodesia, an experiment in the classification of
land with the use of aerial photographs, «Journal of Ecology», 22.
Robyns W., 1938. Considerations sur les aspects biologiques du probleme des feux
de brousse au Congo beige et au Ruanda — Urundi, «Bull. Inst. Roy. Colon. Belge>,
9, 2.
Rochow M. v., 1951. Die Pflanzengesellschaften des Kaiserstuhls, «Pflanzensozio-
logie», 8, Jena.
Rodenwaldt U., 1941. Forstliche Probleme in den Trockengebieten Afrikas unter
besonderer Berucksichtigung der Tsetsebekampfung, «Tropenhygien. Schriftenr. >,
2, Stuttgart.
Roll H., 1942. Zonation und Sukzession, «Biol. Gen.», 1.
Roll H., 1942. Pflanzensoziologische Gewiisserkunde, «Вег. Dt. Bot. Ges.»
Rosenkranz F., 1948. Die thermische Begunstigung gewisser Hohenstufen, «Wet-
ter u. Leben», 1.
Литература
287
Rosenthal G. E., 1784. Versuche, die zum Wachstum der Pflanzen benotigte
Warme zu bestimmen, Erfurt.
Roseveare G. M., 1948. The grasslands of Latin America, «Bull. 36», Imp. Bureau
of Pastures and Field Crops, Aberystwyth.
R ossm assler E. A., 1855. Die vier Jahreszeiten, Gotha.
Rossmassler E. A., 1863. Der Wald, Leipzig. [Имеется русский перевод:
E. Россмеслер, 1866. Лес, Спб.— РедД
R u b n е г Н., 1956. Die Walder in der Inn — Salzach — Platte, «Mitt. Geogr. Ges.
Miinchen», 41.
Rubner K., 1934. Die pflanzengeographisch-okologischen Grundlagen des Waid-
haus, 3. Aufl., Neudamm.
Rubner K., Reinhold F., 1953. Das natiirliche Waldbild Europas, Ham-
burg u. Berlin.
R ii b e 1 E., 1917. Anfiinge und Ziele der Geobotanik, «Vierteljahrsschr. Naturf. Ges.
Zurich», 62.
Riibel E., 1920. Uber die Entwicklung der Gesellschaftsmorphologie, «Mitt. Geo-
bot. Inst. Riibel».
R ii b e 1 E., 1922. Geobotanische Untersuchungsmethoden, Berlin.
R ii b e 1 E., 1930. Pflanzengesellschaften der Erde, Bern.
R ii h 1 e K., 1928. Die Vegetationsformationen Sudamerikas in ihrer klimatischen
Bediiurtheit, «Р. M.», 74.
Runge F., 1940. Die Waldgesellschaften des Inneren der Miinsterschen Bucht, Mini-
ster.
Russell E. J., 1921. Soil Conditions and Plant Growth, London.
R u t t n e r F., 1952. Grundriss der Limnologie, 2. Aufl., Berlin.
S a i n t - H i 1 a i r e A. de, 1831. Tableau de la vegetation primitive dans la province
de Minas Geraes, «Ann. Sciences Naturelles», 24, Paris.
S a i n t - H i 1 a i r e A. de, 1847—1848. Voyage aux sources du Rio de San Francisco
et dans la province de Goyaz, Paris.
Salisbury E. J., 1926. The geographical distribution of plants in relation to cli-
matic factors, «Geogr. Journal».
Sampson A. W., 1919. Plant succession in relation to range management.
Sapper K., 1915. Ober den Kulturwert der tropischen Vegetationsformationen,
«G. Z.», 21.
Sapper K., 1923. Die Tropen, Stuttgart.
Sauberer A., 1951. Die Verteilung rindenbewohnender Flechten in Wien, «Wetter
u. Leben», 3.
Sauer C., 1938. Destructive exploitation in modern colonial expansion, Congr. Int.
Geogr. Amsterdam.
S a u s s u r e H. B. de, 1779—1796. Voyage dans les Alpes, Genf.
Saxton W. T., 1924. Phases of vegetation under monsoon-conditions, «Journ. of
Ecology», 12.
Scamoni A., 1935. Vegetationsstudien im Forst Sarnow «Z. f. Forst-u. Jagdwesen»,
67.
Scamoni A., 1955. Einfiihrung in die praktische Vegetationskunde, Berlin.
Scharfetter R., 1922. Klimarhythmik, Vegetationsrhythmik und Formations-
rhythmik, «Osterr. Bot. Z.»
Scharfetter R., 1924. Die Grenzen der Pflanzenvereine, Sieger-Festscbr.,
Wien.
Scharfetter R., 1928. Die kartographische Darstellung der Pflanzengesellschaf-
ten, «Hdb. d. biol. Arbeitsmeth.»
Scharfetter R., 1938. Das Pflanzenleben der Ostalpen, Wien.
288
Литература
Schenck С. А., 1939. Fremdlandische Wald- und Parkbaume, Berlin.
Schi emenz F., 1935. Binnenfischerei und natiirliche Landschaft in Niedersachsen,
Berlin.
S c h i m p e r A. F. W., 1891. Die indomalayische Strandflora, «Bot. Mitt. a. d. Tro-
pen».
Schi m per A. F. W., 1898. Pflanzengeographie auf physiologischer Grundlage,
Jena; 3. Aufl. hrsg. v. F. C. Faber, Jena, 1935.
Schindewolf 0., 1955. Die Entfaltung des Lebens im Rahmen der geologischen
Zeit, «Stud. Gen.», 8.
S c h i n z H., 1893. Die Vegetation des deutschen Schutzgebiets in Siidwest — Afrika,
«Kolon. Jb.», VI.
Schlenker G., 1940. Erlauterungen zum pflanzensoziologischen Kartenblatt
Bietigheim, Tubingen.
Schluter H., 1957. Ein Beitrag zur Frage okologischer und soziologischer Arten-
gruppen, «Arch. f. Forstwesen», 6.
Schmid E., 1939. Die Vegetationskartierung der Schweiz 1 : 200000, Zurich.
Schmid E., 1948. Die Vegetationskarte der Erde im Schweizerischen Mittelschul-
atlas, «Geogr. Helv.», 3.
Schmid E., 1957. Ein Vergleich der Wuchsformen im illyrischen Buchen- und Laub-
mischwald, «Вег. Geobot. Forsch.— Inst. Rubel f. 1956».
Schmieder O., 1929. Das Pampaproblem, «Р. M.», 73.
Schmithusen J., 1934. Zur Vegetations- und Bodenkunde Siid — Frankreichs
und des ostlichen Spanien, «G. Z.», 40.
Schmithusen J., 1934. Der Niederwald des linksrheinischen Schiefergebirges,
«Beitr. z. Landesk. d. Rheinl.», II, 4, Bonn.
Schmithusen J., 1940. Das Luxemburger Land, «Forsch. z. dt. Landesk.», 34,
Leipzig.
Schmithusen L., 1942. Vegetationsforschung und okologische Standortslehre
in ihrer Bedeutung fiir die Geographic der Kulturlandschaft, «Z. Ges. f. Erdk. zu
Berlin».
Schmithusen J., 1948. Fliesengefiige der Landschaft und Okotop, «Вег. z. dt.
Landeskunde», 5.
Schmithusen J., 1948. Die Wirkungen des trockenen Sommers 1947 als For-
schungsaufgabe, «Вег. z. dt. Landesk.», 5.
Schmithusen J., 1950. Das Klimaxproblem vom Standpunkt der Landschafts-
forschung aus betrachtet, «Mitt. Flor.-soz. Arbeitsgem.», N. F. 2.
Schmithusen J., 1950. Die Diirreempfindlichkeit der mitteleuropaischen Wirt-
schaftslandschaft in Vergangenheit und Gegenwart, «Dt. Geographentag», 27.
Schmithusen J., 1951. Deutschlands Waldgebiete, Geogr. Taschenbuch.
Schmithusen J., 1952. Leo Waibel, «Die Erde», 4.
Schmithusen J., 1953. Die Grenzen der chilenischen Vegetationsgebiete, «Dt.
Geographentag Essen 1953», Wiesbaden.
Schmithusen J., 1954. Waldgesellschaften des nordlichen Mittelchile, «Vegeta-
tio», V—VI.
Schmithusen J., 1956. Die raumliche Ordnung der chilenischen Vegetation,
«Bonner Geogr. Abh.», 17.
Schmithusen J., 1957. Anfange und Zieleder Vegetationsgeographie, «Р. M.», 101.
Schmithusen J., 1958. Probleme der Vegetationsgeographie, «Dt. Geographen-
tag Wurzburg 1957», Wiesbaden.
Schmithusen J., 1959. Das System der geographischen Wissenschaft, Fest-
schrift Theodor Kraus, Bad Godesberg.
Schmithusen J., 1960. Die Nadelholzer in den Waldgesellschaften der siidli-
chen Anden, «Vegetatio», IX.
Литература
289
Schmucker W., 1942. Silvae orbis, Berlin.
Schnelle F., 1949. Phanologische Weltkarte: Beginn der Weizenernte, Verteilung
der Weizenanbauflachen und der Weizenausfuhr, «Met. Rundschau», 2.
Schnelle F., 1950. Kleinklimatische Gelandeaufnahme am Beispiel der Frost-
schaden im Obstbau, Dt. Wetterdienst.
Schnelle F., 1955. Pflanzen—Phanologie, Leipzig.
SchnizleinA., Frickhinger A., 1848. Die Vegetations — Verhaltnisse
der Jura- und Keuperformation in den Flussgebieten der Wornitz und Altmiihl,
Nordlingen.
Schomburgk R., 1847. Reisen in Britisch Guayana in den Jahren 1840—44,
Leipzig.
Schott C., 1951. Das Heideproblem in Schleswig— Holstein, Dt. Geographentag
Frankfurt.
Schouw J. F., 1822. Grundtrag til en almindelig Plantegeografie. Kopenhagen.
Dazu Atlas 1824.— Dt. Ausg., Berlin, 1823; Dazu Atlas 1823.
Schrepfer F., 1923. Das phanologische Jahr der deutschen Landschaften, «G. Z.»,
29.
Schrdter C., 1901. Die Palmen und ihre Bedeutung fiir die Tropenbewohner,
Zurich.
Schroter C., 1932. Kleiner Fuhrer durch die Pflanzenwelt der Alpen, Zurich.
Schultze W., 1937. Uber die pflanzengeographische Verhaltnisse der sogenannten
Lausitzer Heide, Berliner Geogr. Arb. Stuttgart.
Schulz A., 1887. Die Vegetationsverhaltnisse der Umgebung von Halle, Halle.
Schwabe G. H., 1956. Die okologischen Jahreszeiten im Klima von Mininco (Chile),
«Bonner Geogr. Abb.», 17.
Schweinfurth G., 1875. Im Herzen von Afrika, Leipzig.
Schweinfurth U., 1957. Die horizontale und vertikale Verbreitung der Vegeta-
tion im Himalaya, «Bonner Geogr. Abb.», 20.
Schwendener S., 1856. Die periodischen Erscheinungen der Natur, Zurich.
Schwickerath M., 1931. Die Gruppenabundanz, «Englers Bot. Jb.», 64.
Schwickerath M., 1937. Die nacheiszeitliche Waldgeschichte des Hohen Venns,
«Abb. Preuss. Geol. Landesanst.», N. F. 184.
Schwickerath M., 1938. Walder und Waldboden des Hohen Venns, Hannover.
Schwickerath M., 1940. Die Artmachtigkeit, «Fedde Rep. Beih.», 54.
Schwickerath M., 1942. Bedeutung und Gliederung des Differentialartenbegriffs,
Beih. Bot. Zentralbl.
Schwickerath M., 1944. Das Hohe Venn und seine Randgebiete, «Pflanzenso-
ziologie», 6, Jena.
Schwickerath M., 1954. Die Landschaft und ihre Wandlung auf geobotanischer
und geographischer Grundlage entwickelt und erliiutert im Bereich des Messtischblat-
tes Stolberg, Aachen.
Sendtner O., 1854. Die Vegetationsverhaltnisse Siidbayerns, Munchen.
Sendtner O., 1860. Die Vegetationsverhaltnisse des Bayerischen Waldes, Munchen.
Senft F., 1865. Die Vegetationsverhaltnisse der Umgebung von Eisenach, Eisenach.
Shantz H. L., Marbut C. F., 1923. Vegetation and soild of Africa, New York.
Shantz H. L., Zon R., 1923. The natural vegetation of the United States, Atlas
of American Agriculture, Washington.
Shreve F., 1914. A montane rain-forest (Jamaika), Washington.
S i s s i n g h G., 1939. Het Exotenvragstuk en de Plantensociologie, Nederl. Boschbo-
uw. Tijdschrift.
Skottsberg C., 1909. Pflanzenphysiognomische В eobach tungen aus dem Feuerlande,
Wiss. Erg. d. Schwed. Sudpolar — Exped. 1901—1909, Stockholm.
Sorge E., 1930. Die Trockengrenze Siidamerikas, «Z. Ges. f. Erdk. zu Berlin».
290
Литература
S о u 1 a v i е (J. L. Giraud), 1783. Histoire naturelle de la France meridionale, II, 1,
Paris.
Stamm E., 1938. Die_ Eichen — Hainbuchen — Walder der Nordschweiz,
Bern.
Stebler F. G., 1897. Die Streuwiesen der Schweiz, «Landw. Jb. d. Schweiz», 11.
Stebler F. G., Schrdter G., 1892. Versuch einer Ubersicht fiber die Wiesen-
typen der Schweiz, «Landw. Jb. d. Schweiz», 6.
Steffen H., 1939. Uber die floristischen Beziehungen der beiden Polargebieten zuein-
ander, Dresden.
Stocker 0., 1923. Die Transpiration und Wasserdkologie nordwestdeutscher
Heide- und Moorpflanzen am Standort, «Z. f. Bot».
Stocker 0., 1929. Ungarische Steppenprobleme, «Die Naturwissenschaften».
Stocker 0., 1950. Das Umweltproblem der Pflanze, «Stud. Gen.», 3.
Stocker 0., 1952. Grundriss der Botanik, Berlin-Gottingen-Heidelberg.
Stocker 0., 1954. Die Trockenresistenz der Pflanzen, VIII. Gongr. Int. de Botanique.
Stocker 0., 1957. Grundlagen, Methoden und Probleme der Okologie, «Вег. Dt.
Bot. Ges.», 70.
Str aka H., 1957. Pollenanalyse und Vegeta tionsgeschichte, «Die neue Brehm-Bii-
cherei», 202, Wittenberg.
Sukatschow W. N., 1934. Uber einige Grundbegriffe der Phytocoenologie, «Изд.
АН СССР», 0MEH, Ns 7 (резюме на русском языке).
Сукачев В. Н., 1950. Биогеоценоз, БСЭ, Т. 5, 2-е изд., М.
Sup ап А., 1915. Die Pflanzendecke und die geographische Verbreitung der Tiere.
B. «Grundziige der Phys. Erdkunde», Berlin.
Suzuki T., 1954. Forest and bog vegetation within Ozegahara Basin, Tokyo.
Tanfiljew G. J., 1895. Die Waldgrenze in Siidrussland, «Р. M.», 41.
Tanfiljew G. J., 1905. Die siidrussischen Steppen, Gongr. Int. Bot. Vienne.
Танфильев Г. H., 1911. Пределы лесов в полярной России, Одесса.
Tanfiljew G. J., 1925. Natiirliche Wiesen in Russland, Schrdter-Festschr.
T a n s 1 e у A. G., 1939. The British Islands and their vegetation, Cambridge.
Tansley A. G., Ghipp T. F., 1926. Aims and methods in the study of vegeta-
tion, London.
Teleki G. P., Nagy Z. de, 1930. Oceanic, continental, mediterranian and boreal
climatic influences and mountain climat in Europe as synthetised and represented
by characteristic plants, «Publ. Geogr. Inst. Budapest», I, Budapest.
Thienemann A., 1920. Untersuchungen uber die Beziehungen zwischen dem
Sauerstoffgehalt des Wassers und der Zusammensetzung der Fauna in norddeutschen
Seen, «Arch. f. Hydrobiol.», 12.
Thienemann A., 1931, Limnologie, «Handworterbuch d. Naturwiss», VI.
Thienemann A., 1941. Leben und Umwelt, «Bios», 12, Leipzig.
Thienemann A., 1955. Die Binnengewasser in Natur und Kultur, Berlin-Got-
tingen-Heidelberg.
Thurmann J., 1849. Essai de Phytostatique applique a la chaine du Jura et aux
con trees voisines, Bern.
Tischler W., 1951. Der biozdnotische Konnex, «Biol. Zentralbl.», 70.
Tischler W., 1955. 1st der Begriff «Kultursteppe» in Mitteleuropa berechtigt?
«Forsch. u. Fortschr.», 29.
Tournefort J. P. de, 1717. Voyage au Levant, Paris.
Troll G., 1925. Ozeanische Ziige im Pflanzenkleid Mitteleuropas, Drygalski-Festschr
Munchen-Berlin.
Troll G., 1936. Termiten-Savannen, Landerkundl. Forsch. Festschr. N. Krebs, Stutt-
gart.
Литература
291
Troll С., 1939. Das Pflanzenkleid des Nanga Parbat, «Wiss. Veroff. Dt. Mus. f.
Landesk.», N. F. 7.
Troll C., 1941. Studien zur vergleichenden Geographie der Hochgebirge, «Вег. Ges.
Frd. u. Ford. Univ. Bonn».
Troll C., 1948. Der asymmetrische Aufbau der Vegetationszonen und Vegetations-
stufen auf der Nord- und Siidhalbkugel, «Вег. Geobot. Forsch.— Inst. Rubel f.
1947.»
Troll C., 1950. Savannentypen und das Problem der Primarsavannen, Proceedings 7.
Int. Bot. Congr., Stockholm.
Troll C., 1951. Heckenlandschaften im maritimen Griinlandgiirtel und im Grimland
Mitteleuropas, «Erdkunde», 5.
Troll C., 1952. Die Lokalwinde der Tropengebirge und ihr Einfluss auf Niederschlag
und Vegetation, «Bonner Geogr. Abh.», 9.
Troll C., 1952. Das Pflanzenkleid der Tropen in seiner Abhangigkeit von Klima,
Boden und Mensch, Dt. Geographentag Frankfurt 1951, Remagen.
Troll C., 1955. Der jahreszeitliche Ablauf des Naturgeschehens in den verschiedenen
Klimagiirtel der Erde, «Stud. Gen.», 8.
Troll C., 1956. Das Wasser als pflanzengeograpliischer Faktor, «Hdb. d. Pflanzen-
physiologie», 3, Berlin.
Troll C., 1957. Forschungen in Zentralmexiko 1954, Dt. Geographentag Hamburg,
Wiesbaden.
Troll W., 1926. Die natiirlichen Walder in dem Gebiete des Isarvorlandgletschers,
«Landesk. Forsch. Geogr. Ges.», Munchen, 27.
Trommer C., 1853. Die Bonitierung des Bodens vermittels wildwachsender Pflan-
zen, Greifswald.
Tschermak L., 1944. Ozeanitiit und Waldklein im Gebirge, «Z. f. d. Ges. Forst-
wesen», 1.
Tschulok S., 1910. Das System der Biologie in Forschung und Lehre, Jena.
T ii x e n J., 1958. Stufen, Standorte und Entwicklung von Hackfrucht- und Garten-
unkrautgesellschaften und deren Bedeutung fiir Ur- und Siedlungsgeschichte, «Angew.
Pflanzensoz.», 16.
T ii x e n R., 1930. Uber einige nordwestdeutsche Waldassoziationen von regionaler
Verbreitung, «Jb. Geogr. Ges. Hannover».
T ii x e n R., 1931. Die Grundlagen der Urlandschaftsforschung, «Nachr. aus Nieder-
sachsens Urgesch.», 5.
T ii x e n R., 1932. Wald- und Bodenentwicklung in Nordwestdeutschland, Ber. 37.
Wandervers. d. Nordwestd. Forstvereins, Hannover.
T ii x e n R., 1933. Klimaxprobleme des nordwesteuropaischen Festlandes, «Nederl.
Kruidkundig Archief», 43.
T ii x e n R., 1934. Niedersachsen, Vegetation 1:800 000, Atlas Niedersachsen, 13,
Oldenburg.
T ii x e n R., 1935. Uber die Bedeutung der Pflanzensoziologie in Forschung, Wirt-
schaft und Lehre, «Der Biologe», IV.
T ii x e n R., 1937, Die Pflanzengesellschaften Nordwestdeutschlands, «Mitt. Flor.-
soz. Arbeitsgem. in Niedersachsen», 3.
T ii x e n R., 1939. Die Pflanzendecke Nordwestdeutschlands in ihren Beziehungen
zu Klima, Gestein, Boden und Mensch, «Dt. Geogr. Blatter», 42, Bremen.
T ii x e n R., 1951. Wasserversorgung und Pflanzensoziologie, «Gas- und Wasserfach»,
92.
T ii x e n R., 1952. Hecken und Gebiische, «Mitt. Geogr. Ges. Hamburg», 50.
Tiixen R., 1954. Die Wasserstufenkarte und ihre Bedeutung fiir die nachtragliche
Feststellung von Anderungen im Wasserhaushalt einer Landschaft, «Angew. Pflan-
zensoz.», 8.
292 Лите рат ура
Тйхеп R., 1954. Uber die raumliche, durch Relief und Gestein bedingte Ordnung
der nattirlichen Waldgesellschaften am nordlichen Rande des Harzes, «Vegetatio»,
V—VI.
Тйхеп R., 1954. Pflanzengesellschaften und Bodenprofil, 8. Congr. Int. Bot., Paris.
Тйхеп R., 1955. Das System der nordwestdeutschen Pflanzengesellschaften, «Mitt.
Flor.-soz. Arbeitsgem», N. F. 5.
Тйхеп R., 1956. Die heutige potentielle naturliche Vegetation als Gegenstand
der Vegetationskartierung, «Angew. Pflanzensoz.», 13.
T ti x e n R., 1956. Vegetationskarte Baltrum, Stolzenau/Weser.
Тйхеп R., Diemont H., 1937. Klimaxgruppe und Klimaxschwarm, «Jahresber.
d. Naturhist. Ges. Hannover», 88/89.
Тйхеп R., Hentschel G., 1955. Bibliographic der Vegetationskarten West-
deutschlands, «Mitt. Flor.-soz. Arbeitsgem.», N. F. 5.
Тйхеп R., Preising E., 1942. Grundbegriffe und Methoden zum Studium
der Wasser- und Sumpfpflanzen-Gesellschaften, «Deutsche Wasserwirtschaft»,
37.
Tuomikowski R., 1942. Untersuchungen йЬег die Untervegetation der Bruch-
moore in Ostfinnland, Helsinki.
Turesson G., 1922. The species and the variety as ecological units, Hereditas.
Turesson G., 1926. Die Bedeutung der Rassenokologie ftir die Systematik und Geo-
graphic der Gewiichse, «Rep. spec. nov. Beih.», 41.
Unger F., 1836. Ober den Einfluss des Bodens auf die Verteilung der Gewachse,
Wien.
U p h о f, 1920. Cold-resistence as an ecological factor in the geographical distribution
of cacti, «Journal of Ecology», 8.
V a gel er P., 1930. Grundriss der tropischen und subtropischen Bodenkunde, Ber-
lin — 2. Aufl. 1938.
Valle K. J., 1929. Konnen die stidfinnischen Seen vermittels der umgebenden Vege-
tation und Flora bonitiert werden? «Acta forest. Fennica», 34.
V a r e s c h i V., 1951. Zur Frage der Oberflachenentwicklung von Pflanzengesell-
schaften der Alpen und Subtropeu, «Planta», 40.
Вавилов H. И., 1926. Центры происхождения культурных растений, «Тр. по
прикл. бот. и сел.», Т. XVI, вып. 2.
V о 1 k е n s G., 1887. Die Flora der agyptisch-arabischen Wtiste, Berlin.
V о 1 к e n s G., 1897. Der Kilimandscharo, Berlin.
Volz W., 1921. Im Dammer des Rimba, Breslau.
Wagner H., 1923. Lehrbuch der Geographic. III. Biologische Geographic, 10.
Aufl., Hannover.
Wahlenberg G., 1804. Geographisk och economisk beskrifning om Kemi Lapp-
mark, Stockholm.
Wahlenberg G., 1812. Flora lapponica, Berlin.
Wahlenberg G., 1814. Flora Carpatorum principalium, Gottingen.
W a i b e 1 L., 1913. Lebensformen und Lebensweise der Tierwelt im tropischen Afri-
ka, «Mitt. d. Geogr. Ges. Hamburg», 27.
W a i b e 1 L., 1921. Urwald, Feld, Wtiste, Breslau.
W a i b e 1 L., 1922. Die periodisch-trockenen Vegetationsgebiete des tropischen Afri-
ka, Verb. 20. Dt. Geographentag Leipzig, Berlin.
W a i b e 1 L., 1933. Die Sierra Madre de Chiapas, «Mitt. d. Geogr. Ges. Hamburg», 43.
W a i b e 1 L., 1933. Probleme der Landwirtschaftsgeographie, Wirtschaftsgeogr.
Abh., Breslau.
Литература
293
W a i b е 1 L., 1935. Probleme der Landwirtschaftsgeographie, Verb. Dt. Geographen-
tag Bad Nauheim. Breslau.
W a i b e 1 L., 1937. Die Rohstoffgebiete des tropischen Afrika, Leipzig.
W a i b e 1 L., 1943. Place names as an aid in the reconstruction of the original vegeta-
tion of Cuba. «Geogr. Rev.», 33.
W a i b e 1 L., 1948. Vegetation and land use in the Planalto Central of Brazil, «Geogr
Rev.», 38.
Wallace A. R., 1892. Island life, London.
Walter H., 1927. Einfiihrung in die allgemeine Pflanzengeographie Deutschlands
Jena.
Walter H., 1931. Die Hydratur der Pflanzen, Jena.
Walter H., 1939. Grasland, Savanne und Busch der arideren Teile Afrikas in ihrer
okologischen Bedeutung, «Jb. wiss. Bot».
Walter H., 1942. Die Vegetation des europaischen Russlands, Berlin.
Walter H., 1949—1956. Einfiihrung in die Phytologie, Bd. I bis IV, 1, Stutt-
gart.
Walter H., 1954. Die Verbuschung, «Vegetatio», V—VI.
Wangerin W., 1915. Vorlaufige Beitrage zur kartographischen Darstellung der Ve-
getationsformationen im nordostdeutschen Flachland, Ber. Dt. Bot. Ges.
Wangerin W., 1932. Florenelemente und Arealtypen, «Beih. Bot. Zentralbl.», 49
Drude-Festschrift.
Wangerin W., 1934. Die Ordnungsbegriffe der chorologischen Pflanzengeographie,
«Die Naturwissenschaften».
Warburg O., 1893. Vegetationsschilderungen aus Ostasien, «Englers Bot. Jb.»,
17.
Warming E., 1895. Plantesamfund, Kopenhagen.
Warming E., Graebner P., 1933. Lehrbuch der okologischen Pflanzengeo-
graphie, 4. Aufl., Berlin.
Weaver J. E., Clements F. E., 1929. Plant Ecology, New York.— 2. Aufl.
1938.
Weber C. A., 1907. Aufbau und Vegetation der Moore Norddeutsch lands, «Englers
Bot. Jb.», 40.
Weberbauer A., 1906. Grundziige von Klima und Pflanzenverteilung in den pe-
ruanischen Anden, «Р. M.», 52.
Weberbauer A., 1911. Die Pflanzenwelt der peruanischen Anden, Leipzig.
Week J., 1947. Neue Unterlagen fiir eine Waldbrandbekampfung, «Mitt. Reichs-
inst. f. Forst-u. Holzwirtsch.», 1.
Week J., 1956. Entwicklungsstufen und Gefiigetypen von Baumbestanden, «Forstw.
Zentralbl.», 75.
Week J., 1957. Die Walder der Erde, «Verstandl. Wissensch.», 67, Berlin-Gottigen-
Heidelberg.
Weischet W., 1953. Zur systematischen Beobachtung von Baumkronendeformatio-
nen mit klimatologischer Zielsetzung, «Met. Rundschau», 6.
W e 1 t e A., 1934. Der Weinbau des mittleren Mainlandes, Forsch. z. Dt. Landesk.,
Stuttgart.
Werth E., 1927. Klima und Vegetationsgliederung in Deutschland, Berlin.
Wettstein R. v., 1900. Untersuchungen iiber die Saisondimorphismus, Wien.
Wettstein R. v., 1904. Die Biologie unsere Wiesenpflanzen, Wien.
Whittaker R.H., 1953. A consideration of climax thory. «Ecological Monographs»,
23.
Whittaker R. H., 1957. The kingdoms of the living world, «Ecology», 38.
W i 1 h e 1 m у H., 1950. Das Alter der Schwarzerde und der Steppen Mittel- und Osteu-
ropas, «Erdkunde», 4.
294
Литература
Wilhelmy Н., 1952. Die eiszeitliche und nacheiszeitliche Verschiebung der Klima-
und Vegetationszonen in Siidamerika, Dt. Geographentag Frankfurt, Remagen.
Willdenow C. L., 1792. Grundriss der Krauterkunde, Berlin.— 5. Aufl., Berlin,
1810.
W i r t g e n Ph., 1864. Die Schneifel, «Bot. Ztg.», 22.
W i r t g e n Ph., 1865. Ober die Vegetation der hohen und der vulkanischen Eifel,
«Verb. Naturhistor. Ver. Rh. u. Westf.»
Wissmann H.v., 1948. Pflanzenklimatische Grenzen der warmen Tropen, «Erdkun-
de», 2.
Wissmann H.v., 1957. Ursprungsherde und Ausbreitungswege von Pflanzen-
und Tierzucht und ihre Abhangigkeit von der Klimageschichte, «Erdkunde», 11.
Wittich W., 1952. Der heutige Stand unseres Wissens von Humus und neue Wege
zur Losiing des Rohhumusproblems im Walde, Frankfurt.
Woodard. J., 1924. Origin of prairies in Illinois, Botanical Gazette.
Вульф E. В., 1933. Введение в историческую географию растений, изд. 2-е, М.-Л.
Wyssotzky G. N., 1932. Bodenfeuchtigkeitsuntersuchungen in Waldbestanden
der ukrainischen Steppen- und Waldsteppenzone. «Tharandter Forstl. Jb.», LXXXIII,
9—10.
Zoller H., 1954. Die Arten der Bromus-Erectus-Wiesen des Schweizer Juras,
«Veroff. Geobot. Inst. Rubel in Zurich», 28, Bern.
Zuccarini J. G., 1833. Die Vegetationsgruppen in Bayern, Munchen.
Указатель русских терминов
Автотрофные организмы 11
Аллелопатия 101
Альпийские матты 145, 146
Ареал 27, 73, 193
биполярный 33
викарирующий 30
границы 74, 193
дизъюнктивный 28
замкнутый 28
изменение человеком 94
круги типов 32
минимальный ИЗ
«областей дивизионов растительно-
сти» 214
пояса 31
причины возникновения 193
разорванный 28, 90, 91
сокращение 94
сплошной 28
тип 32
увеличение 94, 95
Арктогейское царство 35
Аспект 111
смена 144
Ассоциация 114, 116, 200
общественно живущих видов 20
Биогеография 102
Биогеоценоз 105
Биология 14
Биосфера 9, И, 12
Биотип 46
Биотоп 46, 102, 103, 159
потенциальный 102
Биоформация 124
Биохора 102, 151
потенциальная 102, 152, 180
Биохорология 14
Биоценоз 99, 102, 108, 109, 159
тип формы 124
Биоценология 109
Биоценотическая связь 100
Биоценотическое равновесие 107, 108
.Болото 159
бамбуковое 223
березовое 204
верховое 148, 159
низинное 159
пальмовое 223
с жесткими подушками 148
сосновое 148
сфагновое 148, фото 31
тундровое 148
Ботанико-географический элемент 33
Борригудос, см. Деревья бутылочные
Бурые глины 223
Вариант ассоциации 115
Ветер, воздействие 79
Вивипария 57, 134
Вид 25, 26
верность 111, ИЗ
жизненность 111, 112
мощность 111, 120
общественность 111, 112
оценка покрытия 120
экологическая конституция 72, 73
Виды
абсолютное число ИЗ
благосклонные ИЗ
верные ИЗ
дифференциальные ИЗ, 114
зимнезеленые 133
летнезеленые 133
постоянные ИЗ
прочные ИЗ
разделяющие 114
сопутствующие ИЗ, 117
характерные ИЗ, 114
чуждые ИЗ
экологические группы 75
Водные устьица 50
Водный режим 169
Водный фактор 47
Водоросли 26
прикрепленные 69
сине-зеленые 12
эндолитные 76
Возделывание 165
Воздушное пространство 12
Выборочная рубка леса 168
Выветривание 88
аллитное 89
сиаллитное 89
Выжигание растительности 162
Выпас 166, 175
Высокотравье 67
Высотные ступени, пояса 18, 197
Галофиты 80
Гаммада 236
Указатель латинских названий растений
и растительных сообществ
Abies 26, 258, 259
alba 250, 251
balsamea 183, 258
lasiocarpa 259
sibirica 258, 259
Abietoideae 26
Abietum mariesii 208
Acacia 235
sebil 233
Acaena 43
adscendens 269
Acer 92
pseudoplatanus 250
Асегасеае 36
Aconitum napellus 68
Acorus calamus (ass.) 117
Adansonia digitata 233
Adesmia subterranea 66
Adiantetea 122, 123
Aesculus 92
Aextoxicum 29
Agathis australis 56, 241
Aizoaceae 43, 45, 65
Alectorolophus 171
Alisma plantago-aqatica 117
var. lanceolatum 112
Allium 67, 68
Alnetea glutinosae 120, 122, 205
Alnetum glutinosae 120
Alnion glutinosae 120
Alnus 258, 259, 267
fruticosa 258
hirsuta 258
incana 258
mitcheliana 258
sibirica 258
viridis 258
Amaryllidaceae 43, 68
Amaryllis 43, 68
Ammophiletea 122, 123, 208
Anacardiaceae 57
Ancistrocladaceae 39
Ancistrocladus 39
Andropogon schirensis 217
Andropogonion schirensis 201, 217
Angiospermae 26
Aphyllantion 117
Aquilegia vulgaris, ssp. atrata 119
Araceae 63, 65
Araliaceae 61
Araucaria angustifolia 242
araucana фото 13
Araucariaceae 41
Arbutus 243
unedo 251
Arctostaphylos 64
Aristotelia 43
Armerion maritimi 116
Arnoseris minima 69
Arrhenatheretalia 207
Arrhenatheretum elatioris 177
Artemisia 65, 207
Artemisietea vulgaris 122, 205
Artocarpus 92
Arum maculatum 68
Asclepiadaceae 39, 43, 45, 63, 65
Asphodelus 68
Asplenietea rupestris 122, 146, 208
Astelia 43
Aster 171
amellus 254
Astragalus 167, 248
Athyrium filix femina 36
Atropion 254
Bacteriophyta 26
Begonia 63
Bellis 67
Betula 36, 258, 259, 267
exilis 258
glandulifera 258
glandulosa 258
papyrifera 183
pubescens 258, 267
verrucosa 258
Betula-Populus tremula (ass.) 217
Betuleto-Adenostyletea 122
Betuletum pubescentis typicum 204
Bidentetea tripartiti (ass.) 122, 205
Bignoniaceae 61
Blechno-Quercetum 207
В lechnum spicant 118
Bolax 147
Bombacaceae 59
Bombax 92
Borassus 140
flabelliformis 60
Brachypodietum ramosi 179
Bromeliaceae 38, 62, 234, фото 2
Bromus racernosus-Senecio aquaticus
(ass.) 205
304
Указатель латинских названий растений и растительных сообществ
Brugiera conjugata фото 14
typ. 58
Bryanthus 265
Bupleurum spinosum 167
Butomus umbellatus 117
Butyrospermum parkii 230
Cactaceae 38, 45, 65, 234
Caesalpiniaceae 61
Cakiletea maritimae 122, 123
Calamagrostis villosa 118
Calandrinea 69
Callitriche 69
Calluna vulgaris 147
Calluneto-Genistetum 204
Caltha 43
Campanulaceae 36
«Campo limpo» 143, 230
«Campo sujo» 143, 230
«Campo cerrado» 165, 226
Canthium vulgare 233
Cardamine pratensis 36
Carex 66
gracilis 117
riparia 117
strigosa 121
Carex-Elyna (пояс) 207
Caricetea curvulae 122, 123
Cariceto elongatae — Alnetum 206
medioeuropaeum 120, 204
boreale 120
Cariceto inflatae-vesicariae 204
Cariceto laevigatae — Alnetum 120
Cariceto remotae — Fraxinetum atlanti-
cum 120, 121
oxidento-balticum 121
Caricetum appropinquatae 206
diandrae 206
gracilis 206
paniculatae 206
Carnegia gigantea 77
Carpinus 207
Carya 255
Caryophyllaceae 36, 45
Cassiope 265
Castanea dentata 170
Catharinea hausknechtii 119
Cedrus 26
Centaurea cyanus 69
Cercis siliquastrum 243
Chamaecyparis nootkaensis 260
Chamaerops humilis 62
Characeae 160
Charophyta 26
Chlorophyta 26
Chrysophyta 26
Cicuta virosa-Carex pseudocyperus (ass.)
204
Cinnamomum 39, 92
Circaea alpina 119
Circaeo-Alnetum 253
Cirsium 67
Cisto-Lavanduletea 207
Cladonia uncialis 120
Clematis 134
vitalba 61
Clivia 43
Comareto-Menyanthetum 208
Compositae 29, 41, 45, 59, 60
Coniferae 26, 45
Coniferopsida 26
Convallaria 68
Copernicia cerifera 140
Coprosma 43
Cordaitales 26
Cornus suecica 266
Corynephoretea canescentis 122, 205
Corypha umbraculifera 60
Crassulaceae 37, 45, 65
Crocus albiflorus 68
Cruciferae 36, 45
Cunnoniaceae 43
Cupresseceae 59
Cupuliferae 36
Curvuletum 183
Cuscuta 69
Cyanophyta 26
Cyatheaceae 59
Cycadaceae 50, 60
Cycas 60
Cyclamen 68
Cyperaceae 145
Dasylirion 60
Datura stramonium 171
Dentaria bulbifera 251
Dentaria-Fagetum (ass.) 207
Deschampsia flexuosa 118
Deschampsietum mediae 179
Dicranum scoparium 118
Digitalis 67
purpurea 254
Dioscorea 67
Distichia muscoides 148
«Djati» 228
Donatia 43
Draba verna 69
Dracaena 55, 59
Drapetes 43
Drosera 148
Dryopterideta cristatae — Alnetum 120
Ebenaceae 37
Echium 67
Eichhornia 69
Eliocharetum ovatae 206
Elymo-Seslerietea 122, 123
Empetrum 32, 33, 43
Encephalartos 60
Epacridaceae 43
Ephedra 62, 64
Epilobietea angustifolii 122, 205
Epilobium 112
Epipogium aphyllum 119
Equisetum limosum 117
Erica arborea 28
tetralyx 29
Ericaceae 265
Ericeta 99
Eschweilera sagotana 127
Espeletia 60
Указатель латинских названий растений и растительных сообществ
305
grandiflora фото 30 Homogyne alpina 118
Eucsdyptus 41, 242 regnans 56 Eucryphiaceae 43 Euglenophyta 26 Euphorbia charicias 179 Euphorbiaceae 37, 45, 59, 65, 234 Euphrasia 171, 172 Eupitys 25 Hottonietum palustris 205 Hydrochareto-Stratiotetum 206 Hydrostachyaceae 43 Hylocomium loreum 119 proliferum 118 Hymenophyllaceae 37 Hymenophyllum 50 Hyphaene 140, 164 tebaica 60
Fagaceae 43 Fagetalia silvaticae 207 Fageto-Quercetum 206 Fageto-Quercetum petreae violetum 205, 206 Fagetum crenatae 208 silvaticae 207 Fagus-Abies (пояс) 207 Fagus 92 silvatica 207, 250 Festuca 66 Festusa-Stipa (ass.) 207 Festuco-Brometea 122, 205, 254 Ficus 60, 61, 63, 92, 225 Filices 59 Filipenduleto-Geranietum palustris 204 Fitzroya 56 Fraxino-Carpinion 206 Fraxinus 92 Freesia 43 Fungi 26 Hypparrhenion confinis 201 Ilex 92 aquifolium 134 paraguayensis 242 Illecebretum verticillati 204 Impatiens 63 Iridaceae 43, 68 Iris pseudacorus 112, 117 Isoeto-Nanojuncetea 122, 149, 205 Isothecium myurum 119 Ixia 43 «Jadar» 267 Jubaea spectabilis 30, фото 10 Juglans 92 Juncaceae 148 Juncetalia maritimi 116 Juncetea maritimi 116, 122, 123 Junceto-Molinietum 205 Juncetum acutiflorae 206
Galium palustre 117 rotundifolium 119 triflorum 119 Geissolomaceae 43 Genista anglica 251 Gentiana purpurea 118 Geraniaceae 65 Gesneriaceae 37 Ginkgo 57, 90, 92 biloba 30 Gladiolus 63 Glaucophyta 26 Glyceria aquatica 117 Glycerietum maximae 205 Gourliera decorticans 236 Gramineae 29 Gunnera 43 Gymnospermae 10, 26, 59 gerardi 116 juncetosum maritimi 116 Juniperus 246 communis 44 Jurinea 248 Kingia 60 Kissenia 38 Laricoideae 26 Larix 26, 31, 57, 258 Gmelini 258 laricina 258 sibirica 258 Larix-Pinus cembra (пояс) 207 Lauraceae 57 Lavendula 65 Lemna minor-Lemna trisulca (сообщ.) 205
Haastia 62 pulvinaris 147 Haloxylon 207 Harrimanella 265 Hawortia 43, 65 Hedera 92 Helodea 69 canadense 96, 171 Hevea 225 Heteropogon contortus 142 Hieracium alpinum, ssp. halleri 120 murorum 118 vulgatum 118 Lemnetea 122, 205 Lepidium draba 171 Leucodendron 42 Leucoium 68 Lichenes 26 Liliaceae 29, 41, 43, 45, 59, 60, 65, 68 Linnaea borealis 91, 119 Liriodendron 90, 92 Listera cordata 119 Litorelletea 122, 205 «Llaretta» 147 Loasaceae 38 Lobelia 59, 60
306 Указатель латинских названий растений и растительных сообществ
Loiseleuria 64
Lonicera coerulea 119
nigra 119
Lophozia lycopodioides 118
Loranthaceae 63, 136
Lucuma paradisea фото 19
Luzula luzulina 118, 119
nivea 119.
silvatica, ssp. Sieberi 118
Luzuragoideae 43
Lycopodium alpinum 120
annotinum 119
Lygeum spartum 243
Lysimachia vulgaris 117
Lythrum salicaria 112, 117
Macrosamia 60
Magnolia 92
Majanthemum bifoliuin 118, 250
Malvaceae, 45
Marcgraviaceae 38
«Mato seco» 137
Mauritia vinifera 140
Melaleuca 41
Melampyrum silvaticum 118
«Melar» 147, 267
Mentha aquatica 117
Mercurialis annua 171
Mesembryanthemum 43, 65
Metrosideros 43
Minuartia peploides 30
Mnium spinosum 119
«Мо» 267
Molinietum japonicae 208
Molinio-Arrhenatheretea 122, 205, 208
Montio-Cardaminetea 122
Mora excelsa 127
gonggriipii 127
Moraceae 37
Musa 63, 95
paradisiaca ssp. seminifera 95
Myriophylleto-Nupharetum 206
«Myri» 267
Myrtaceae 41, 43, 57
Myxophyta 26
Myzodendraceae 43
Myzodendron 43
Nanoquercetum 208
Narcissus 68
Nardo-Callunetea 122, 205
Nepenthes 39, 40
Nipa 39, 92
«Nkunku» 230
Nolanaceae 38, 65
Nothofagus 43
antarctica фото 22
Oenanthe aquatica 117
Oenothera biennis 171
Oinaceae 43
Olea 42
cuspidata фото 5
europaea 56
Ononis 65
Orchidaceae 61, 63
Orchis 68
Orneto-Ostryon 207
Orobanchaceae 68
Orobanche rapum-genistae 81
Osmundaceae 42
Oxalis 67, 68
carnosa 65
corniculata 171
stricta 171
Oxaleto-Chenopodietum polyspermii su-
batlanticum 205
Oxycocceto-Caricetum middendorfii 208
Oxycocco-Sphagnetea 122, 205
Paliurus 67
Pandanaceae 59, 60
Pandanus 60
Panicum crus galli-Spergula arvensis
(ass.) 204
Panicum ischaemum (ass.) 204
Papaver 36
Papilionaceae 45
Palmae 45, 49
Paris 68
Pelargonium 65
Penaceaceae 43
Pernet tya 43
Peumus boldo фото 8
Phalaridetosum 117
Phalaris arundinacea 117
Phragmites communis 112, 117
Phragmitetea 117, 122, 205
Phragmitetum oseanum 208
Phragmition euro-sibiricum 117
Phylica 43
Phyllodoce 265
Picea 26, 207, 259
abies 258
canadensis 183, 258, 261, 262
engelmanni 259
excelsa 118
mariana 258, 259, 262
obovata 259
omorica 93
sitchensis 260
Piceetum montanum subass. galietosum
119
melicetosum 119
subalpinum 118, 119
transalpinum 119
Pinaceae 25
Pineta 99
Pinetum pumilae 208
Pineto-Vaccinietum myrtilli 253
uliginosi 253
Pinoideae 26
Pinus 25, 26, 259
albicaulis 260
banksiana 258
cembra 119
contorta 259
mugo 25
nigra 25
Указатель латинских названий растений и растительных сообществ
307
рейсе 93
pinea 243
pumila 258
sibirica 258
silvestris 25, 258
Pinus-Quercus 207
Piper 63
Plagiothecium undulatum 118
Plantaginetea maioris 122, 205
Pleurozium schreberi 118
Poa 66
Polygonatum 68
Polygonum amphibium terrestre 112, 117
convolvulus 69
Polygonum hydropiper-Bidens tripartitus
(ass.) 205
Polypodium 29, 63
Polyporus 81
Polytrichosum attenuatum 118
Polytrichum 148
Populus 92, 259
balsamifera 258
suaveolens 258
tremula 258
tremuloides 258
Potametea 122, 205, 209
Potameton lucentis 206
Potamogeton 69
Poterium spinosum 167
Pratia 43
Primula 67
Primulaceae 45
Pringlea antiscorbutica 62
Prosopis 235
juliflora 236, фото 18
tamarugo 238
Protea 42
Proteaceae 41, 42, 43
Pseudopanax laetivirens фото 7
Pseudotsuga 92
taxifolia 260
Pteridophyta 26
Ptilium crista-castrensis 118, 119
Puya 62
Pyrola uniflora 119
Pyrolaceae 68
Pyrrophyta 26
Quercetalia ilicis 207
Quercetea robori-petraeae 123, 205
Querceto-Betuletum 207, 253
Querceto-Buxetum 207
Querceto-Carpinetum 207
elymetosum 207
medioeuropaeum 253
stachyetosum 206
typicum 207
Querceto-Fagetea 122, 205
Querceto-Fraxinetum 207
Querceto-Luzuletum nemorosae 207
Querceto-Piceetum 207
Querceto roboris-Betuletum 204, 206
Quercetum cocciferae brachypodietosum
179
rosmarinetosum 179
Quercetum ilicis 179
Quercion ilicis 206, 207
Quercion robori-petraeae 206
Quercus 92, 293
alba 255
crispula 208
Quercus sp. фото 9
Racomitrium 148
Rafflesia arnoldi 68
Ranunculaceae 36
Ranunculus arvensis 69
ficaria 68
glacialis 32
lingua (subass.) 117
Raoulia 62
extmia 147
Restionaceae 42
Rhamnaceae 43
Rhisophora 58, 224
mangle 58
Rhododendron 240
ferrugineum 119
Rhodophyta 26
Rhodoreto-Vaccinietum 119
Rhytidiadelphus triquetrus 118
Rosaceae 36, 45, 269
Rosmarinetalia 206
Rosmarineto-Lithospermetum 179
Rubus gratus (ass.) 204
Rumecetum maritimi 206
Rumex hydrolapathum 117
Ruscus 49, 85
Rutaceae 41
Sagittaria sagittifolia (subass.) 117
Sparganium simplex (ass.) 206
Salicaceae 36
Salicetea herbaceae 122, 123
Salicornietea 209
Salix 92 258, 259, 267
aurita-Frangula alnus (ass.) 253
bebbiana 258
caprea 258
herbacea 66, 267
interior 258
retusa 80
sachalinensis 258
triandra 258
viminalis 258
Salvia 65
Salvinia 69
Sanguisorba officinalis 67
Sapindaceae 61
Sapium 37
Sapotaceae 230
Sarothamnion scopariae 206
Saxifraga cuneifolia 119
Saxifragaceae 36, 45
Scheuchzerietum palustris 204
Scheuchzerio-Caricetea fuscae 122, 205, 208
Schinopsis 235
Schoenoplectus lacustris 112, 117
triquetrus 117
308
Указатель латинских названий растений и растительных сообществ
Scilla 68
Scirpeto-Phragmitetum 112, 117, 205
Scirpus maritimus 117
Scrophylariaceae 68
Sedum 65
Sequoia 56, 90, 92
gigantea 57
sempervirens 56, 260
Sempervivum 65
Senecio 59, 60
smithii фото 25
vernalis 171
Sium latifolium 117
Smilax kraussiana 164
Solanum dulcamara 117
Solidago 171
Sorbus 267
aucuparia 118, 267
Sparganiaceae 36
Sparganium angustifolium-Sphagnum obe-
sum (ass.) 204
erectum ssp. neglectum 112
polyedrum (ass.) 117
Spergula vernalis-Gorynephoretum cane-
scentis 204
Sphagneta 208
Sphagnetea 209
Sphagnetum medii subatlanticum 204
pinetosum 253
papillosi 204
Sphagnum 80, 148
acutifolium 118
Spinifex 238
Stapelia 43
Stapelieae 39
Stellaria media 69
Stellarietea mediae 122, 205
sterculiaceae 59
Stipa 247
ichu 49
tenacissima 243
Stratiotes 69
Stylidaceae 43
Thero-Brachypodietalia 207
Thero-Salicornietea 122
Thlaspetea rotundifolii 122
Thuja plicata 260
Thujetum standishii 208
«Tierra caliente» 221
Tilia 92
cordata 251
Tillandsia 63
usneoides фото 11
Todea barbara 42
Trientalis europaea 27
Trisetum flavescentis 177
Tropaeolaceae 37
Tsuga heterophylla 260
Typha angustifolia 112, 117
latifolia 112, 117
Typhetosum angustifoliae 117
Ulmus 92
Ulmetum davidianae 208
Umbelliferae 36, 45, 67
Vaccinietea uliginosi 123, 205
Vaccinio-Piceetalia 118
Piceetea 123, 208
Piceion 118, 119
Vaccinium myrtillus 118
uliginosum-Loiseleuria 207
vitis-idaea 118
«Vargem» 223
«Varzea» 223
Verbenaceae 228
Veronica 43
tournefortii 171
Vicia hirsuta 69
Viola 33, 69
silvestris 67
Welwitschia 29, 62, 239
Taraxacum 67
Taxodium 92
distichum 241
Tectona grandis 228
Teesdalia nudicaulis-Arnoseretum
mae 204
Tepualia 43
Themeda triandra — Heteropogon
tus (ass.) 142
Theobroma cacao 56
Xanthorrhoea 41, 60
Yucca 59, 60
contor- Zingiberaceae 37
Zostera marina 36
Zosteretea marinae 122, 123
Оглавление
Предисловие к русскому изданию........................................... 5
Предисловие к немецкому изданию.......................................... 7
Введение
Растительность как предмет исследования географии........................ 9
Биосфера ............................................................ 9
Цели и задачи географии растительности.............................. 14
История географии растительности.................................... 17
Часть первая
СОСТАВНЫЕ ЭЛЕМЕНТЫ РАСТИТЕЛЬНОСТИ
И ПРИНЦИПЫ ИХ РАСПРОСТРАНЕНИЯ
Систематические единицы и их распространение............................ 25
Единицы систематики растений........................................ 25
Ареалы систематических единиц....................................... 26
Флористическое деление Земли........................................ 33
Формы роста растений.................................................... 44
Проблема типологии растительных форм................................ 44
Конвергенция и приспособляемость.................................... 45
Формы приспособления к водному фактору.............................. 47
Приспособленность наземных растений к свету, теплу и другим факторам
и система жизненных форм по Раункиеру .............................. 51
Классификация форм роста............................................ 52
Обзор форм роста.................................................... 55
Причины пространственного распределения систематических единиц и форм
роста .............................................................. 71
Биогенетические причины современного распространения растений .... 71
Экологические группы................................................ 75
Причины пространственного распределения систематических единиц и ра-
стительных форм..................................................... 81
Исторические факторы................................................ 89
Воздействие человека на распространение растений.................... 94
Часть вторая
ЕДИНИЦЫ РАСТИТЕЛЬНОСТИ В ЛАНДШАФТЕ
Единицы растительности.................................................. 99
Сообщества живых существ............................................ 99
Биоценоз и место существования....................................... 101
Голоцен и экосистема................................................. 104
310
Оглавление
Разграничение и классификация биоценозов............................. 108
Единицы растительности и растительные сообщества..................... 109
Типология и классификация единиц растительности (фитоценозов) по их
внешнему облику ..................................................... 123
Растительность в ландшафте............................................... 150
Естественный и культурный ландшафт................................... 150
Биохора и пространство местообитания................................. 151
Структура «флизе» и естественное распределение растительности но
условиям местообитания ............................................... 155
Независимая пространственная дифференциация растительности и взаимо-
связи биоценозов вне местообитаний.................................... 159
Деятельность человека как формирующий фактор..................... 161
Естественное пространственное размещение растительного покрова в ланд-
шафте ................................................................ 172
Потенциальный естественный ландшафт и первобытный ландшафт .... 183
Часть третья
ПРОСТРАНСТВЕННОЕ ПОДРАЗДЕЛЕНИЕ РАСТИТЕЛЬНОСТИ
Распространение единиц растительности..................................... 191
Топография и крупномасштабное картирование растительности............. 191
Ареалы, границы ареалов, причины их возникновения и функциональные
соотношения........................................................... 193
Структура пространственного распределения растительности (Фптоценологи-
чески обоснованная хорология растительности)............................. 199
Общая проблема. Принципы и возможности пространственного деления
с точки зрения географии растительности............................... 199
Деление растительности, основанное на комплексах сообществ.......... 202
Ландшафтные растительные комплексы........................................ 210
Аналитические признаки растительности ландшафта или растительной
области............................................................... 210
Признаки, отличающие данную растительную область от других обла-
стей растительности................................................... 211
«Дивизионы растительности» (Vegetationsdivisionen).................... 212
Типы «дивизионов растительности», климатические зоны растительности
и проблема составления обзорной карты растительности Земли............ 214
Климатические зоны растительности......................................... 217
Пояса растительности тропиков и субтропиков........................... 217
Пояса растительности умеренных зон.................................... 246
Субполярные и полярные зоны........................................... 262
Литература 271
Указатель русских терминов................................................ 295
Указатель латинских названий растений и растительных сообществ .... * 303
И. Шмитхюзен
ОБЩАЯ ГЕОГРАФИЯ
РАСТИТЕЛЬНОСТИ
Редактор Л. Б. Бабинцева
Художник Г. Л. Ларский
Художественный редактор Л. Ф. Шканов
Технические редакторы Ф. X. Джатиева
и Г. Е. Каледина
Сдано в производство 31/VIII 1965 г.
Подписано к печати 6/V 1966 г.
Бумага 70х 1081/16=103/8 бум. л.
27,30 печ. л. Уч.-изд. л. 28,06 в т. ч.
8 вклеек + 1 цв. вкладка Изд. № 11/2601
Цена 2 р. 12 к. Зак. 1
ИЗДАТЕЛЬСТВО «ПРОГРЕСС»
Комитета по печати
при Совете Министров СССР
Москва Г-21, Зубовский бульвар, 21
Московская типография №16
Главполиграфпрома Комитета по печати
при Совете Министров СССР
Москва, Трехпрудный пер., 9