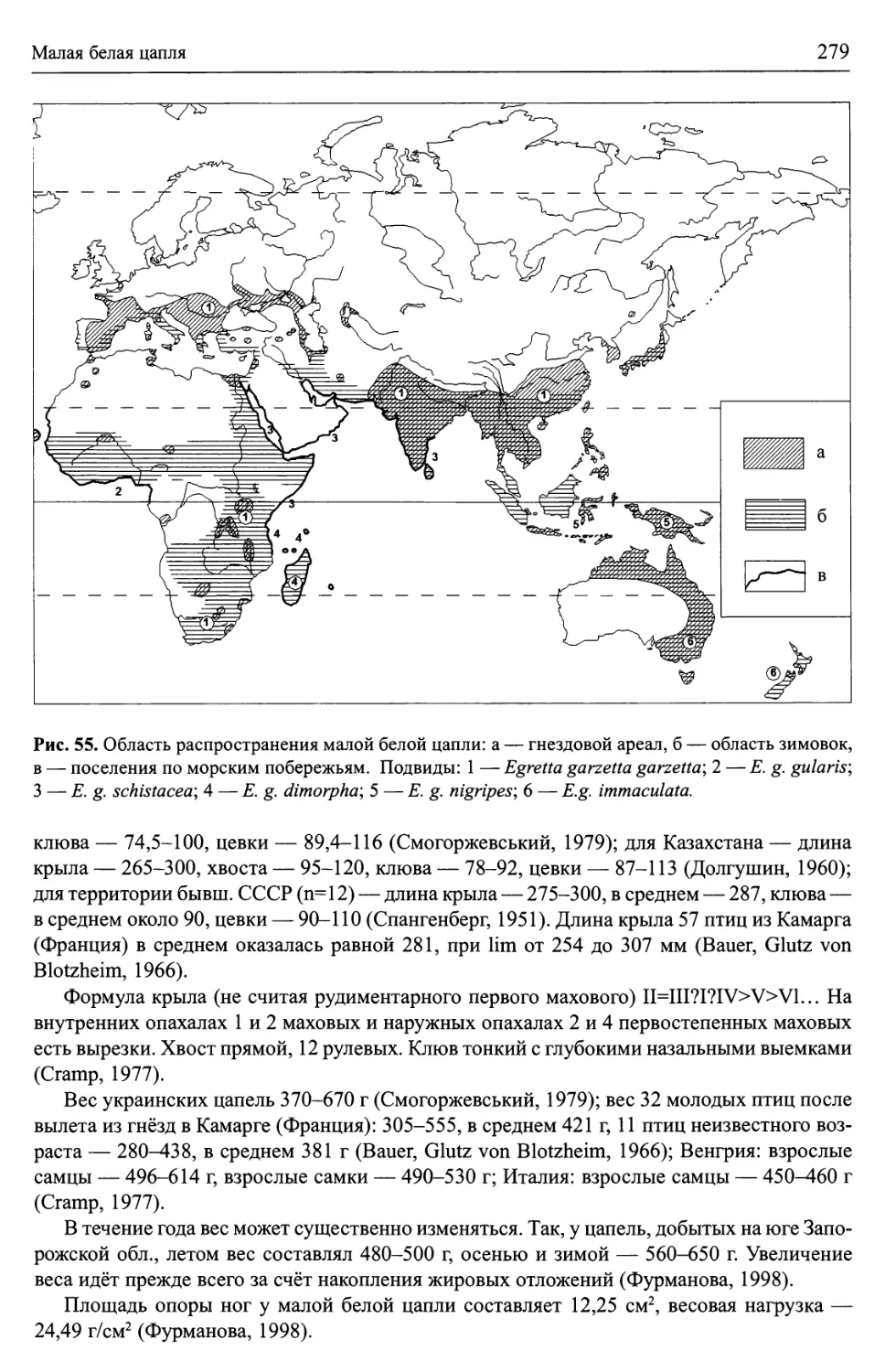
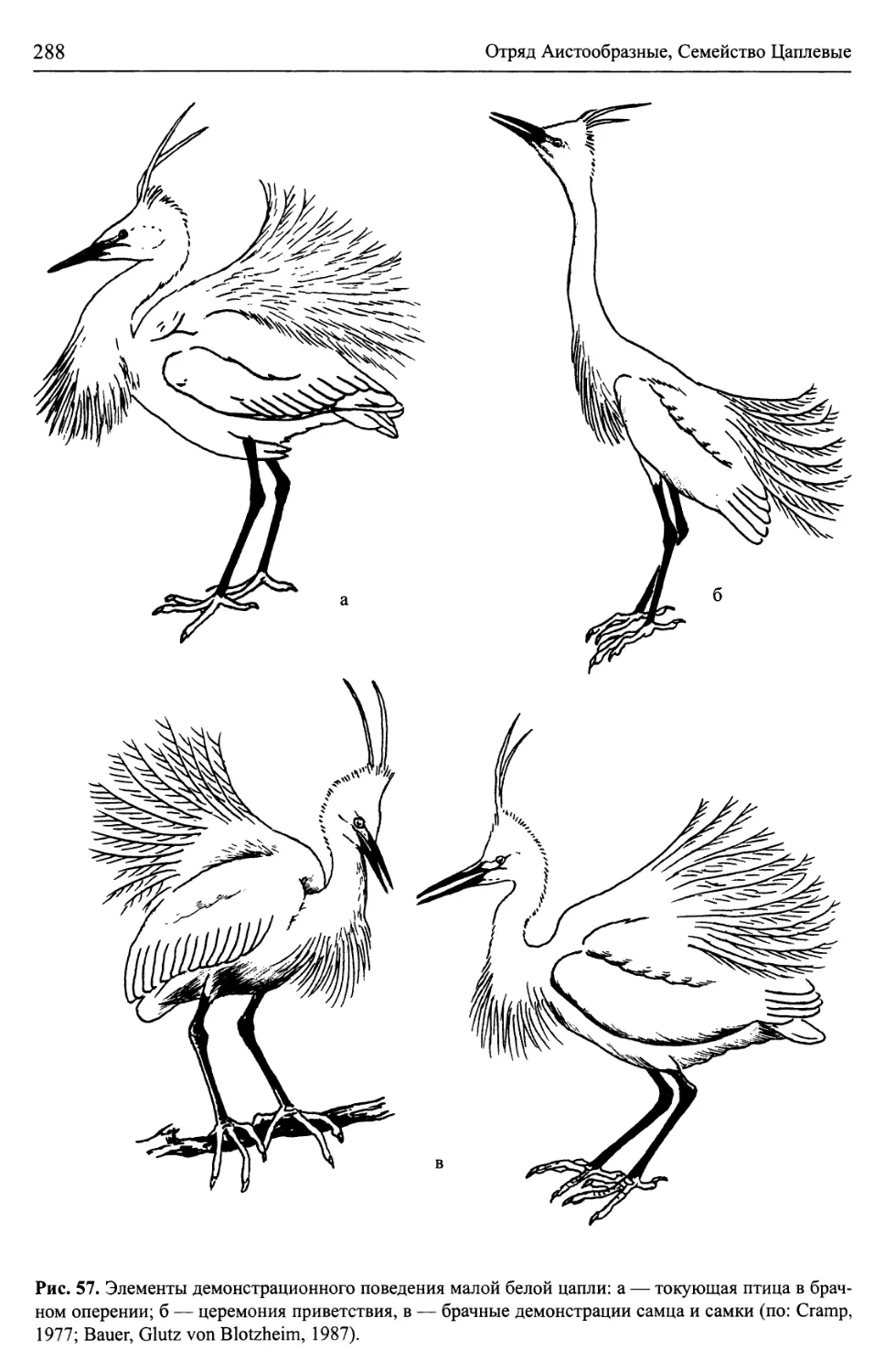
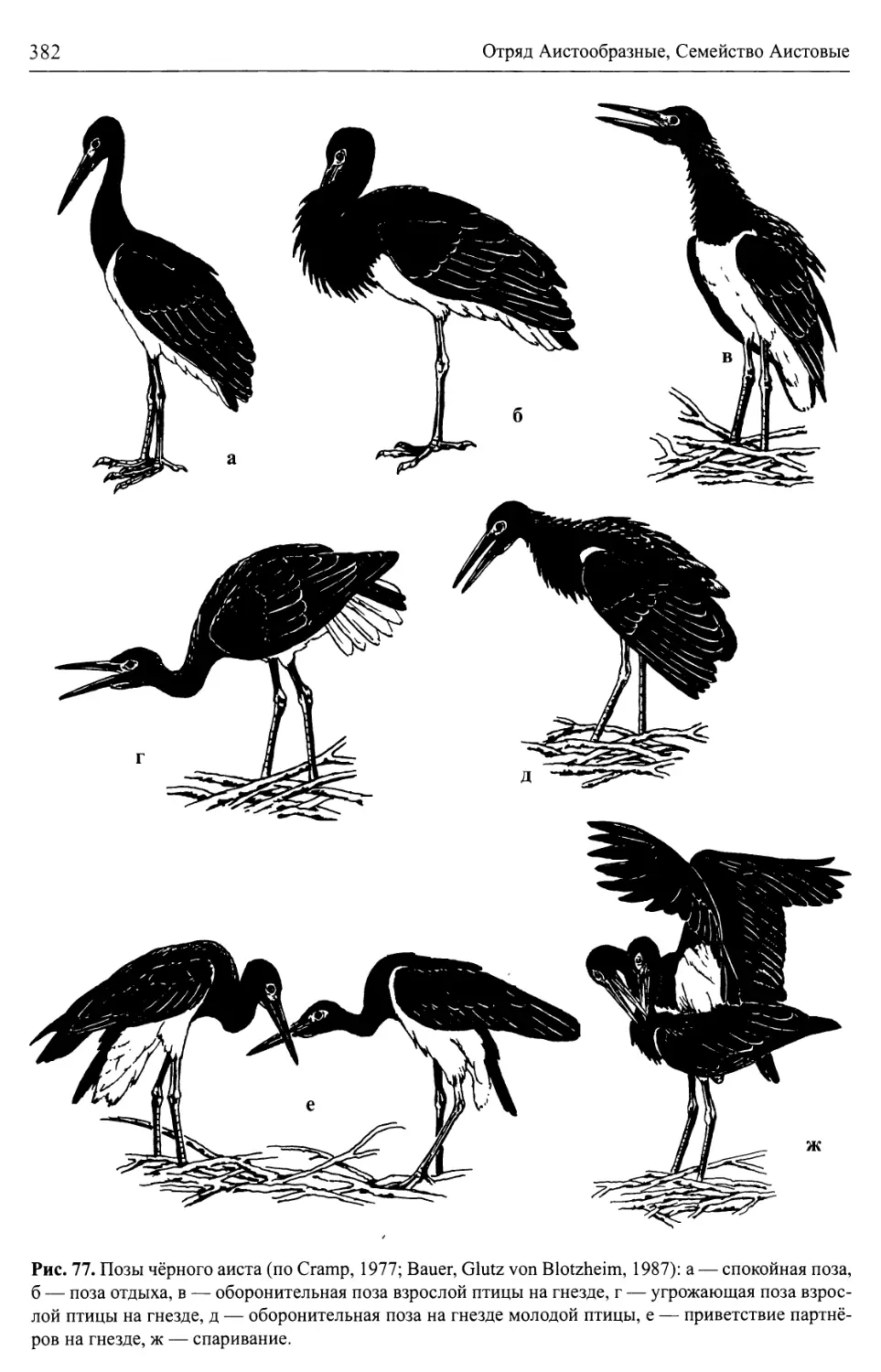
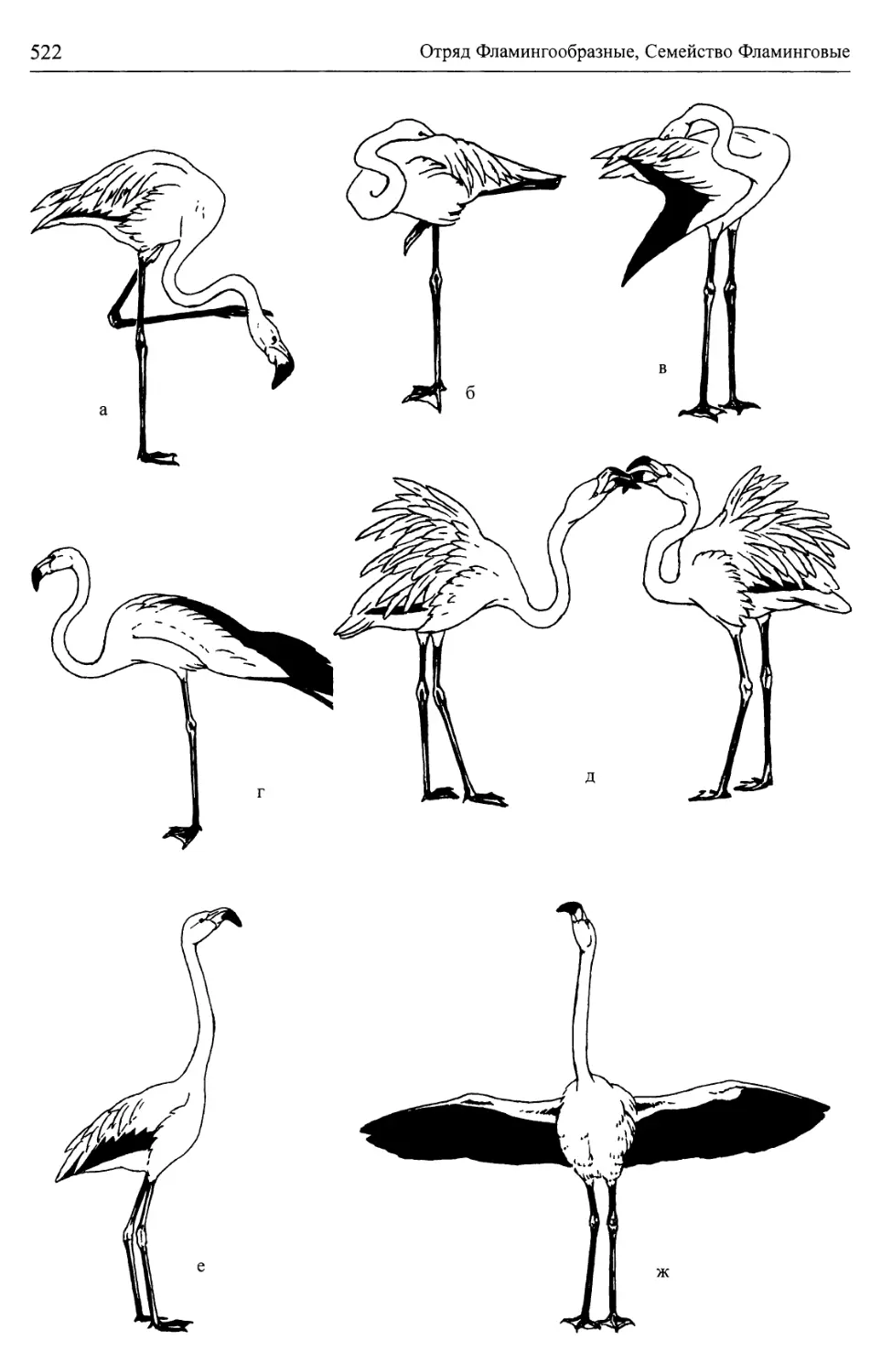
/














































































































































































































































































































































































































































































































































































































































Author: Андронов В.А. Ардамацкая Т.Б. Артюхин Ю.Б.
Tags: aves птицы в целом орнитология птицы география биология
ISBN: 978-5-87317-754-7
Year: 2011
Similar




Text
ПТИЦЫ РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
Пеликанообразные Аистообразные
Фламингообразные
<D
J
X
СП
tu
a; о
to s
co
x
X
e;
<v
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Программа фундаментальных исследований Президиума РАН
«Биологическое разнообразие»
Комиссия по сохранению биологического разнообразия
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Зоологический институт РАН
Мензбировское орнитологическое общество
Зоологический музей МГУ
Союз охраны птиц России
ПТИЦЫ РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
РЕДАКЦИОННАЯ КОЛЛЕГИЯ
В.Д. ИЛЬИЧЕВ
В.А. ЗУБАКИН
(ответственные редакторы)
В.М. ГАЛУШИН
М.В. КАЛЯКИН
Е.А. КОБЛИК
А.Ф. КОВШАРЬ
В.А. ПАЕВСКИЙ
С.Г. ПРИКЛОНСКИЙ
П.С. томкович
Товарищество научных изданий КМК
Москва ❖ 2011
ПТИЦЫ РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
ПЕЛИКАНООБРАЗНЫЕ
АИСТООБРАЗНЫЕ
ФЛАМИНГООБРАЗНЫЕ
Товарищество научных изданий КМК
Москва ❖ 2011
УДК 598.2
ББК 28.693.35(2)
П87
Авторы:
В.А. АНДРОНОВ, Т.Б. АРДАМАЦКАЯ, Ю.Б. АРТЮХИН, В.П. БЕЛИК,
М.М. БЕСКАРАВАЙНЫЙ, О.А. БУРКОВСКИЙ, Ю.Д. ГАЛЧЁНКОВ, ТВ. ГАМОВА,
Ю.Н. ГЛУЩЕНКО, В.Н. ГРИЩЕНКО, ГС. ДЖАМИРЗОЕВ, М.Г ДМИТРЕНОК,
В.А. ЗУБАКИН, Е.А. КОБЛИК, Н.Б. КОНЮХОВ, Ю.В. КОТЮКОВ, Ю.В. КРАСНОВ,
|Н.М. ЛИТВИНЕНКО!. А.Е. ЛУГОВОЙ, А.И. МАЦЫНА, Е.Л. МАЦЫНА,
А.Л. МИЩЕНКО, А.А. МОСАЛОВ.Гю.Н. НАЗАРОВ, В.А. НЕЧАЕВ,
С.Г ПРИКЛОНСКИЙ, И.Т. РУСЕВ, С.Г СУРМАЧ, Ю.В. ШИБАЕВ
Птицы России и сопредельных регионов: Пеликанообразные, Аистообразные,
Фламингообразные. / Андронов В.А., Ардамацкая Т.Б., Артюхин Ю.Б. и др. Отв.
ред.: С.Г. Приклонский, В.А. Зубакин, Е.А. Коблик. М.: Товарищество научных
изданий КМК. 2011. 602 с., 16 цв. вкл.
Книга представляет собой седьмой том многотомной сводки «Птицы России и сопре-
дельных регионов» (первые четыре тома носили название «Птицы СССР»), посвященный
описанию распространения и образа жизни всех видов отрядов Пеликанообразных, Аисто-
образных и Фламингообразных, отмеченных на территории бывшего СССР. В видовых очер-
ках приводятся современные данные о гнездовых ареалах, местах зимовок, путях пролёта,
местообитаниях, численности, экологии гнездования, поведении, питании и других аспек-
тах биологии. Для каждого вида описаны все наряды, географическая изменчивость окрас-
ки и размеров, приведены сведения о хозяйственном значении и проблемах охраны. Моно-
графия снабжена цветными таблицами, картосхемами и штриховыми рисунками.
Для орнитологов, зоологов широкого профиля, работников природоохранных служб и
охотничьего хозяйства, любителей природы.
Ответственные редакторы тома:
С.Г. ПРИКЛОНСКИЙ
В.А. ЗУБАКИН
Е.А. КОБЛИК
Рецензенты:
В.М. Константинов, К.Е. Литвин
На первой странице обложки — краснолицый баклан (фото: Ю.Б. Артюхин),
на последней — большая выпь (фото: И.П. Назаров)
ISBN 978-5-87317-754-7
© Коллектив авторов, 2011
© Российская академия наук, 2011
© Т-во научных изданий КМК, издание, 2011
ПРЕДИСЛОВИЕ
Данная книга представляет собой очередной, седьмой по порядку выхода в свет том
сводки «Птицы России и сопредельных регионов», посвященный описанию 44 видов птиц
из отрядов Пеликанообразных, или Веслоногих (15 видов), Аистообразных (28 видов) и
Фламингообразных (1 вид). Географический район, охваченный сводкой, остался пре-
жним — это территория бывшего СССР.
Сводка, первоначально носящая название «Птицы СССР», задумывалась в конце
1970-х гг. как новое фундаментальное руководство по птицам СССР, поскольку шеститом-
ник «Птицы Советского Союза», изданный в 1951-1954 гг. под редакцией Г.П. Дементьева
и Н.А. Гладкова, с одной стороны, стал библиографической редкостью, а с другой — нуж-
дался в значительных дополнениях (в ряде случаев, и исправлениях), поскольку за десяти-
летия, прошедшие с выхода в свет этой капитальной сводки, были собраны новые сведения
о распространении, численности, деталях биологии и поведения практически всех видов
птиц, населявших страну Во многом изменились представления об объёмах тех или иных
родов, видов и других систематических категорий птиц. Заметно изменилось и отношение
человека к птицам.
Первоначально сводка задумывалась в 10 томах, на издание которых отводилось около
10 лет и которые планировалось издавать последовательно, т.е. от первого к десятому тому:
1. История изучения птиц в СССР. Отряды Гагарообразных, Поганкообразных и Труб-
коносых.
2. Отряды Веслоногих (Пеликанообразных) и Голенастых (Аистообразных) (включая
Фламингообразных).
3. Отряды Пластинчатоклювых (Гусеобразных) и Дневных хищных птиц (Соколооб-
разных).
4. Отряды Курообразных и Журавлеобразных.
5. Отряд Ржанкообразных, подотряды чайковых и чистиковых.
6. Отряд Ржанкообразных, подотряд куликов.
7. Отряды Голубеобразных, Кукушкообразных, Совообразных, Козодоеобразных, Стри-
жеобраных, Ракшеобразных и Дятлообразных.
8-10. Отряд Воробьинообразных.
Однако в процессе работы над сводкой выяснилось, что за столь короткий срок издать
многотомник нереально. Ещё менее реально оказалось «втиснуть» все населяющие страну
виды птиц в объем десяти томов и обеспечить выход томов в последовательности, отража-
ющей систематический порядок расположения отрядов в классе птиц. Тома было решено
публиковать по мере их готовности; пришлось изменить и объем сводки.
Первые четыре тома вышли под названием «Птицы СССР», однако затем произошел
распад СССР, и первая часть следующего тома, посвященного отрядам Рябкообразных -
Дятлообразных и подготовленного ещё в советское время, была опубликована в 1993 г. уже
под названием «Птицы России и сопредельных регионов». Вторая часть тома должна была
выйти в свет вскоре после первой, но надежду на издание книги в 1990-х гг. перечеркнули
общий кризис в стране и безденежье российской науки. Возможность публикации появи-
лась лишь через десяток лет, и очередной этап работы над многотомником начался с 2003 г.
Вторая часть тома Рябкообразных - Дятлообразных вышла в свет в 2005 г. Естественно, её
пришлось основательно дополнить вновь появившимися материалами, а также внести кор-
рективы систематического и номенклатурного характера.
6
Предисловие
К настоящему времени издано 6 томов сводки:
«Птицы СССР. История изучения. Гагары, поганки, трубконосые» (1982)
«Птицы СССР. Курообразные, журавлеобразные»(1987)
«Птицы СССР. Чайковые» (1988)
«Птицы СССР. Чистиковые» (1990)
«Птицы России и сопредельных регионов. Рябкообразные, Голубеобразные, Кукушко-
образные, Совообразные (первая часть)» (1993)
«Птицы России и сопредельных регионов. Совообразные (заключительная часть), Ко-
зодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные» (2005).
История подготовки нынешнего тома сводки, посвященного веслоногим, голенастым и
фламинго, достаточно драматична. Эта книга должна была выйти в конце 1980-х - начале
1990-х гг., очерки по многим видам были уже написаны, однако работа затянулась. Безвре-
менная кончина организатора тома ГА. Кривоносова в 2001 г. прервала процесс подготов-
ки издания. Рукописи книги в архивах ГА. Кривоносова отыскать не удалось, и том при-
шлось собирать практически заново. Часть очерков удалось восстановить, дополнив их
современными данными. Однако к середине 2000-х гг., когда работа над данным томом
возобновилась, ряд авторов уже ушли из жизни, другие не захотели восстанавливать утра-
ченный текст двадцатилетней давности, поэтому авторский коллектив пришлось собирать
практически заново.
С изданием настоящей книги за российскими орнитологами остаётся «долг» в виде то-
мов по соколообразным, гусеобразным, куликам и воробьинообразным. Работа предстоит
большая, но процесс подготовки томов начался, особенно активно в отношении двух пер-
вых отрядов и некоторых семейств отряда воробьинообразных.
Многолетний интервал между изданиями первых и заключительных томов «Птиц Рос-
сии (СССР)», разумеется, должен учитываться пользователями и читателями. Тем не ме-
нее, мы полагаем, что описание состояния орнитофауны на территории бывшего СССР в
конце XX - начале XXI в. — дело стоящее. Прежде всего, потому, что такая сводка под-
ведёт итог орнитологическим исследованиям за все XX столетие и даст «точку отсчета»
для будущих поколений ученых. Наличие фактической информации, включая обширную
библиографию (которой не было в шеститомнике под редакцией Г.П. Дементьева и Н.А.
Гладкова), позволит нашим последователям уверенно ориентироваться в практически ве-
ковых (имея в виду и данные, опубликованные в упомянутом шеститомнике) изменениях
фауны птиц Северной Евразии, их биологии, обилия, биоценотических связей и степени
изученности.
Схема изложения видовых очерков в основном осталась прежней. Изменения, в некото-
рых случаях внесенные в нее, специально оговорены. Порядок расположения таксонов при-
нят по Л.С. Степаняну (2003), как это декларировалось для всей сводки, но с уточнениями
(del Hoyo et al., 1992; Dickinson, 2003; Коблик и др., 2006), которые оказалось необходимым
внести в связи с развитием орнитосистематики и в соответствии с современной номенкла-
турой.
Названия частей тела и оперения птиц, а также способы измерения приняты по «Опре-
делителю птиц СССР» (Гладков и др., 1964). Нумерация первостепенных маховых — от
дистального конца крыла к проксимальному; второстепенных маховых — от первостепен-
ных к проксимальной части крыла; рулевых — от центральной пары к краям хвоста.
Первостепенные маховые обозначаются римскими цифрами, второстепенные и руле-
вые — арабскими. Промеры сделаны авторами в полевых условиях при отлове птиц для
кольцевания, добыче, сборе материалов от охотников, а также по музейным экземплярам.
Широко использованы в этих целях и литературные данные. Длина клюва измерялась от
Предисловие
7
границы оперения лба, длина крыла — при прижатом к линейке и выпрямленном крыле,
длина хвоста — по самым длинным рулевым. В том случае, если измерения проводились
другими способами, это оговаривается в тексте.
Под географическим понятием «Восточная Европа и Северная Азия», или «Северная
Евразия», употребляемым в тексте, имеется в виду территория бывшего СССР. Границу
СССР на помещенных в книге картах следует отождествлять с границей региона, охвачен-
ного данной сводкой.
Цветные иллюстрации выполнены Е.А. Кобликом, штриховые рисунки — А.А. Моса-
ловым (частично по оригиналам, представленным авторами). Если рисунки были взяты из
других изданий, это специально оговорено в каждом конкретном случае. Картосхемы аре-
алов птиц по эскизам авторов и литературным данным выполнены Н.Б. Конюховым. Ха-
рактеристики отрядов и семейств, а также разделы видовых очерков, касающиеся полевых
признаков, описаний внешнего вида и окраски, систематики, географического распростра-
нения отредактированы и дополнены Е.А. Кобликом. Разделы видовых очерков, касаю-
щихся охраны птиц, в ряде случаев дополнены В.А. Зубакиным.
Редколлегия рекомендует при цитировании сводки «Птицы России...» ссылаться на ав-
торов конкретных видовых очерков, например: Нечаев В.А. 2011. Фрегат-ариель // С.Г.
Приклонский и др. (отв. ред.). Птицы России и сопредельных регионов. Пеликанообраз-
ные, Аистообразные, Фламингообразные. М.: Т-во научн. изданий КМК. С. 147-151. Ав-
торство очерков указано в «Содержании» данного тома (стр. 601—602).
Географические названия, насколько это возможно, приведены в современном русском
написании. При их цитировании — в написании, использовавшемся в соответствующие
годы. В тексте, помимо обычных физико-географических, использованы следующие со-
кращения: БПИ ДВО — Биолого-почвенный институт Дальневосточного отделения РАН,
Владивосток; ЗИН — Зоологический институт РАН, Санкт-Петербург; ЗМ МГУ — Зооло-
гический музей Московского государственного университета; ЗМ ДВГУ — Зоологический
музей Дальневосточного государственного университета, Владивосток; ЗМ КНУ — Зооло-
гический музей Киевского национального университета; обл. — область (административ-
ная); окр. — окрестности; ос. — особь (особи), р-н — район (административный); ЦКАН —
Центр кольцевания РАН, Москва.
Авторы и редакторы тома выражают искреннюю благодарность В.М. Лоскоту и П.С. Том-
ковичу за предоставленную возможность ознакомиться с орнитологическими коллекция-
ми ЗИН РАН и Зоомузея МГУ, К.Е. Литвину — за возможность получения информации по
миграциям птиц из Центра кольцевания ИПЭЭ РАН, научному сотруднику Окского запо-
ведника Л.С. Денис — за помощь в составлении списка литературы.
Редколлегия
Отряд ПЕЛИКАНООБРАЗНЫЕ, или ВЕСЛОНОГИЕ —
PELECANIFORMES
Внешний облик и экология очень разнообразны, но все представители отряда тесно
связаны с морскими побережьями и пресными водоёмами, питаются главным образом
рыбой. Размеры средние и крупные (масса 360 г - 15 кг). Все четыре пальца ноги соедине-
ны плавательной перепонкой, доходящей обычно до когтевых фаланг, что и отражено в
старом латинском и часто употребляемом втором русском названии отряда — веслоногие
(Steganopodes). Самый длинный — наружный четвёртый палец, самый короткий — пер-
вый, он направлен не назад, а внутрь (частичная помпродактилия), поэтому птицы «косола-
пят» при ходьбе. Подавляющее большинство веслоногих хорошо летают и плавают, по зем-
ле же передвигаются мало, медленно и неуклюже. Характерны удлинённое обтекаемое тело,
короткие ноги, длинная подвижная шея, мощные длинные крылья. Глаза относительно
маленькие.
Клюв длинный, разнообразной формы — от конического до плоского с крючком на
конце. Рамфотека сложная, состоит из нескольких роговых пластин, у старых птиц они
обычно сливаются в монолитный роговой покров. Ноздри маленькие, у некоторых групп
наружные отверстия ноздрей зарастают. Обычно язык и подъязычный аппарат редуциру-
ются, поскольку развит эластичный горловой мешок, переходящий в очень растяжимый
пищевод. Эти особенности, а также способность эффективно «распяливать» ветви ниж-
ней челюсти (стрептогнатия) позволяют веслоногим глотать очень крупные пищевые
объекты. Желудок тоже сильно растяжим, в железистом желудке обособлен пилоричес-
кий отдел, стенки мускульного желудка слабые, слепые кишки сильно редуцированы.
Зоба нет. Хорошо развиты надорбитальные солевыводящие железы. Череп переходного
типа между десмогнатическим и схизогнатическим, сошник редуцирован (сохранился у
фаэтонов и фрегатов). Шейных позвонков 12-20. Грудина мощная, широкая, с большим
килем. Таз обычно широкий, у хорошо ныряющих форм — суженный. Скелет крыла уд-
линён, особенно плечо и предплечье, степень пневматизации скелета варьирует (Карта-
шев, 1974; Коблик, 2001).
Оперение густое, обычно плотное, водоотталкивающее. Копчиковая железа хорошо
развита, оперена, имеет несколько выводных протоков. Аптерии узкие, пух равномерно
покрывает тело, контурные перья без побочного ствола. Первостепенных маховых перьев
11 (у олуш 10), рулевых — 6-12 пар. В окраске чаще всего сочетаются чёрные и белые
тона, половой диморфизм развит обычно слабо, возрастной, сезонный, индивидуальный
морфизм — сильно. Линька сильно растянута, маховые перья линяют полицентрично, без
утраты способности к полёту (есть исключения).
Пеликанообразные склонны к социальному образу жизни, образуют гнездовые коло-
нии, иногда поливидовые. В норме моногамны, образуют постоянные пары. Спариванию
обычно предшествуют длительные брачные демонстрации. Яйца относительно мелкие, с
довольно рыхлой скорлупой; как правило, одноцветные — белые или зеленовато-голубые,
нередко покрытые «известковым» налетом. Инкубация начинается с первого яйца и про-
должается 4-7 недель, насиживают оба партнёра. Птенцы вылупляются слепыми и голы-
ми, для них характерен птенцовый тип развития. Глаза у них открываются на 3-5-й день, к
6-10-му дню птенцы покрываются густым коротким пухом. Выкармливают оба родителя,
отрыгивая полупереваренную пищу, часто для получения корма птенцы проникают клю-
вом прямо в пищевод взрослой птицы. Подросшим птенцам родители приносят добычу в
клюве. В клюве и пищеводе приносят также воду птенцам — не только для питья, но и для
охлаждения поверхности тела. И взрослые птицы, и птенцы отводят избыточное тепло из
Отряд Пеликанообразные
9
организма обычно путём испарения влаги через горловой мешок и за счёт его охлаждения.
Во время дыхания горловой мешок часто вибрирует. В гнезде или поблизости от него птен-
цы остаются до 6-15 недель. Покинувшие гнездовья молодые птицы кочуют вместе со
взрослыми. Полный взрослый наряд веслоногие надевают, достигнув половой зрелости,
на 3-6-м году жизни. Большинство видов совершают сезонные миграции и кочёвки, не-
многие тропические — оседлы.
Распространены веслоногие всесветно, большинство видов приурочено к океаничес-
ким побережьям материков и архипелагов. Особенно многочисленны колонии веслоногих
и других морских птиц в зонах апвеллинга, в тех местах, где к берегу подходят высокопро-
дуктивные и богатые кислородом и кормом холодные течения. В хозяйственном отноше-
нии виды, селящиеся на морских островах, ценятся (наряду с трубконосыми), прежде все-
го, как производители первоклассного органического удобрения (гуано). В отличие от бу-
ревестникообразных многие представители отряда (пеликаны, бакланы, змеешейки) селятся
и на внутренних пресных водоёмах, богатых рыбой. Естественных врагов у взрослых птиц
немного, больше опасностей грозит кладкам и птенцам, отчего пеликанообразные стре-
мятся гнездиться в труднодоступных для наземных хищников местах — на островах, ска-
лах, деревьях. В исчезновении гнездовых колоний заметную роль играет фактор беспокой-
ства со стороны человека. В последнее время всё большее значение в сокращении числен-
ности морских пеликанообразных приобретает загрязнение акваторий отходами производ-
ства, нефтепродуктами и подрыв кормовой базы птиц в результате хищнического лова рыбы
в Мировом океане. 28 видов пеликанообразных мировой фауны стали глобально редкими,
занесены в Красную книгу МСОП и нуждаются в охране, а один (стеллеров баклан) был
полностью истреблён в историческое время.
Пеликанообразные — древняя группа птиц, возникшая ещё в конце мезозоя; наиболее
разнообразны и многочисленны они были, очевидно, в начале кайнозоя. Насчитывают до 8
ископаемых семейств. Некоторые вымершие формы были сходны с альбатросами (до 5 м в
размахе крыльев) и даже с пингвинами (ростом до 1 м и более). Края челюстных костей,
или только края рамфотеки «ложнозубых» веслоногих (подотряд Odontopteryges) были за-
зубрены, что расценивается как вторичное «приобретение» для лучшего удержания рыбы
(слегка зазубрены и края рамфотеки современных олуш и фаэтонов). Ни палеонтологичес-
кие находки, ни особенности современного распространения, не позволяют определить
единый центр возникновения отряда (Коблик, 2001).
Традиционно пеликанообразных сближали с буревестникообразными или аистообраз-
ными, а неоднородность отряда подчёркивалась наличием промежуточных таксонов —
выделяли 3 подотряда, 2 надсемейства. По некоторым молекулярно-генетическим данным
(Sibley, Ahlquist, 1990; Sibley, Monroe, 1990), все семейства веслоногих входят в огромный
отряд «аистообразных» (иногда с понижением таксономического ранга до подсемейств или
триб). В последнее время всё большее распространение получают взгляды на веслоногих,
как на сборную группу, не составляющую единого отряда. Например, фаэтоны (семейство
Phaethontidae, составляющее подотряд Phaethontes) резко отличаются от остальных пели-
канообразных обликом, рядом черт морфологии и онтогенеза. Внешне они напоминают
крупных крачек, имеют недлинный конический клюв со слитной рамфотекой и крупными
сквозными щелевидными ноздрями, хорошо развитый язык, отсутствие голого горлового
мешка и голых участков вокруг клюва и глаз. Они гнездятся закрыто, откладывают един-
ственное яйцо с гладкой красновато-коричневой скорлупой, испещрённой густыми тёмны-
ми пестринами, птенец вылупляется зрячий, покрытый густым светло-серым пухом, (тип
развития — полу выводковый). Из-за этих особенностей высказываются мнения об отряд-
ном ранге группы и большей близости фаэтонов к подотряду чайковых отряда ржанкооб-
10
Отряд Пеликанообразные
разных. По результатам недавних молекулярных исследований (Ericson, 2008; Hackett et al.,
2008) фаэтоны входят в состав парвкласса Metaves, противопоставленного парвклассу
Coronaves, который включает прочих веслоногих (а также всех аистообразных и ржанкооб-
разных). На основании сходства морфологии челюстного аппарата и участков ДНК пели-
канов нередко сближают с африканским китоглавом (Balaeniceps rex, Balae-nicipitidae) и
помещают в кладу аистообразных.
Шесть современных семейств пеликанообразных (в принятом здесь традиционном по-
нимании) включают 6-9 родов и 52-66 современных видов. На территории Северной Ев-
разии зарегистрированы 1 вымерший и 14 ныне живущих видов, которые принадлежат к
шести родам пяти семейств двух подотрядов.
Подотряд ПЕЛИКАНОВЫЕ — Pelecani
Семейство Пеликановые — Pelecanidae Rafinesque, 1815
Размеры крупнейшие в отряде (масса от 7 до 15 кг, размах крыльев до 3,6 м), облик
очень характерен. Птицы заметно крупнее гуся. Ноги короткие, с широкой перепонкой,
толстые, расположены ближе к середине туловища. Крылья очень длинные и широкие, не
заострённые, хвост короткий, закруглённый. Клюв сильно удлинён (до 47 см), надклювье с
щелевидными ноздрями, уплощено, заканчивается крючком. Симфиз гибких ветвей под-
клювья короткий, расположен на самой вершине, ветви соединены только эластичной ко-
жей объёмистого горлового мешка. Стрептогнатия развита лучше, чем у других птиц: при
открытом клюве подклювье с растянутым горловым мешком образует широкий и объёми-
стый сачок, используемый для лова рыбы. Шея длинная, подвижная, состоит из 17 позвон-
ков. Гетероцельные грудные позвонки сливаются в грудную кость, вилочка срастается с
вершиной киля грудины. Киль длинный и идёт почти вдоль всей грудины. Грудина широ-
кая, почти квадратная. Имеется os dorsale. Межглазничная перегородка полностью окосте-
невает и имеет одну фонтанель. Весь скелет лёгкий, пневматизированный, хорошо развита
сеть воздушных мешков, которые образуют воздухоносный слой, особенно хорошо выра-
женный на брюшной стороне тела, а также пенистая воздушно-пузырьковая подкожная
ткань. Голосовая мускулатура нижней гортани не развита, сохраняется только левая сонная
артерия, слепые кишки короткие (около 5 см). Зоб и обводящая мыщца отсутствуют. Коп-
чиковая железа крупная, с 6-8 выводными протоками (Дементьев, 1940; Судиловская, 1951;
Карташев, 1974).
Вершина крыла образована II и III маховыми, рулевых 10-12 пар (Судиловская, 1951).
В отличие от других веслоногих, оперение у пеликанов рыхлое, неплотно прилегает к телу.
Окраска взрослых птиц обычно светлая — белая, сероватая, часто с розовым отливом. Пос-
ледний, как и у фламинго, обусловлен питанием некоторыми видами ракообразных, в теле
которых присутствует соответствующий пигмент. Маховые перья обычно тёмные, контра-
стируют с остальным оперением, тёмными могут быть и хвост, участки мантии. Клюв,
голые участки «лица» окрашены обычно ярко, особенно в брачный период. Рассученные
перья на затылке нередко образуют хохол. Самки немного меньше и тусклее самцов.
На суше пеликаны ходят неуклюже, но хорошо плавают, на воде сидят высоко, почти не
погружаясь. Могут нырять (по крайней мере, бурые и розовые пеликаны). Из-за малого
удельного веса пеликаны испытывают трудности с нырянием, но зато хорошо летают, мо-
гут долго парить, с воды поднимаются после разбега. В полёте, из-за длинного тяжёлого
клюва держат шею сложенной буквой S, как цапли и марабу. Ноги у летящего пеликана не
Семейство Пеликановые
11
выдаются за обрез хвоста. Полет достаточно быстрый (50-60 км/час), с чередованием глу-
боких и редких взмахов крыльями и планирования. Для набора высоты в потоках восходя-
щего воздуха часто используется парение по спирали. Совершают перелеты, главным об-
разом днем, на высоте 50-3000 м. При дальних кормовых и миграционных перемещениях
образуют линейные построения в форме клина, углов, скосов, цепочек, шеренг, волнистых
рядов, дуг и кругов. Рыхлые и скученные стаи образуются только при перелетах на незна-
чительное расстояние (Молодовский, 1997, 2001).
Пеликаны предпочитают пресноводные водоёмы, даже во внегнездовой период дер-
жатся группами. Питаются рыбой, плавая по поверхности водоёма, периодически опуская
голову в воду и черпая добычу клювом. Особенно результативны коллективные охоты, ког-
да птицы, выстроившись полукругом, хлопая крыльями и шумя, загоняют рыбу на мелко-
водье. Иногда в совместных охотах принимают участие бакланы. Поймав рыбу, пеликан
выливает воду из горлового мешка (до 5 литров!) и проглатывает добычу; поверье о том,
что пеликаны складывают рыбу в горловом мешке, лишено основания. В день птица съеда-
ет 0,2-1 кг рыбы. Сравнительно некрупный (длина 105-152 см, масса 3,5-4,5 кг, 203-228
см в размахе крыльев) бурый пеликан (Р occidentalism — единственная морская птица се-
мейства, которая ловит рыбу, пикируя в воду с высоты до 20 м и погружаясь на 2-2,5 м. От
чувствительного удара о воду пеликана предохраняет воздушная подушка на груди. Счита-
ется, что размеры, тип окраски, способ охоты, биотопическая приуроченность бурого пе-
ликана были характерны для предков всей группы.
Гнездятся пеликаны колониями, насчитывающими сотни пар, нередко совместно с дру-
гими околсводными птицами. Крупные виды сооружают гнёзда на земле или заломах тро-
стника в виде высоких куч из веток и растительного мусора с неглубоким лотком. Мелкие
виды — бурый, пестроклювый (Р philippinensis), розовоспинный (Р rufescens) пеликаны
охотно гнездятся на деревьях, если они растут у облюбованных водоёмов. Известны коло-
нии пеликанов даже в черте городов. В кладке обычно 2-4 белых, желтоватых или голубо-
ватых яйца. Они продолговатые, довольно мелкие (до 200 г, 110x70 см), с рыхлым, неров-
ным наружным известковым слоем скорлупы. Насиживание начинается с момента отклад-
ки первого яйца, поэтому вылупление идёт асинхронно. Наседное пятно не образуется.
Инкубация длится 4-6 недель, самка насиживает более интенсивно. Птенцы покрываются
белым или тёмным пухом на 6-12-й день, к этому времени могут покидать гнездо, остава-
ясь поблизости, с этого же времени способны плавать, собираются в стайки и уплывают на
середину водоёма. К 20-му дню жизни начинает разворачиваться оперение на брюхе и кры-
льях. Учатся летать в возрасте 2-2,5 месяца. Смертность птенцов очень высока: от хищни-
ков, недоедания, климатических колебаний гибнет свыше половины вылупившихся птен-
цов. Молодые птицы окрашены в грязно-бурый или серый цвет, клюв и голые участки кожи
тусклые. С каждой линькой оперение светлеет, окончательный наряд надевают на 3-4-й год
жизни, в эти же сроки впервые приступают к размножению.
В ископаемом состоянии известны с нижнего эоцена (60 млн. лет назад, Франция), бо-
лее поздние находки — в различных частях современного ареала. Эти ископаемые остатки
относят к 5 ныне живущим и 10 вымершим видам, часть из них выделяют в род
Protopelicanus. В семействе единственный современный род Pelecanus, насчитывающий 7
видов. Глобально уязвимыми признаны 2 вида и 2 подвида пеликанов, они включены в
Красный список МСОП.
Современные пеликаны населяют все континенты: в Новом Свете и Африке по 2 вида, в
Австралии — 1, в Евразии — 3. Распространены очень спорадично, главным образом в
г. бтропиках и тропиках, но американский белый пеликан (Р. erythrorhynchos) гнездится на
внутренних озёрах Канады почти у Полярного круга, зимует же в прибрежных районах юга
12
Отряд Пеликанообразные, Семейство Пеликановые
США и Мексики. Перелётны и другие виды умеренных широт, в том числе оба вида, гнез-
дящихся в Северной Евразии.
Род Pelecanus Linnaeus, 1758
Розовый пеликан
Pelecanus onocrotalus Linnaeus, 1758
Pelecanus onocrotalus Linnaeus, 1758, Syst. Nat., ed. 10, с.132, Африка.
European [great, white] pelican (англ.); Rosapelikan, Gemainer Pelikan (нем.);
pelican blanc [commun] (фр.).
Син. (лат.): Pelecanus onocrotalus orientalis Linnaeus, 1766; Pelecanus roseus J.F. Gmelin,
1789; Onocrotalusphoenix Lesson, 1828; Pelecanus major C.L. Brehm, 1831; Pelecanus minor
Ruppell, 1837; Pelecanus sharpei Bosage, 1870; Pelecanus longirostris Hume, 1877.
Син. (рус.): пеликан, баба, баба-птица (народное, применяется как в отношении розового,
так и кудрявого пеликанов).
Статус. Гнездящийся, перелетный вид.
Общая характеристика и полевые признаки. Очень крупная птица своеобразной
внешности, размером с лебедя. Ярким отличительным признаком служит длинный клюв с
большим кожистым горловым мешком жёлтого цвета. Шея довольно длинная, S-образно
изогнута. В полете и во время отдыха на воде шея и голова лежит на спине. Оперение белое
с розоватым оттенком. На груди охристо-жёлтое пятно. В полете хорошо заметны чёрные
концы крыльев и чёрный задний край крыла. У сидящей птицы интеркарпальные суставы
сложенных крыльев сильно выдаются вперёд. На голове пучок из узких заострённых перь-
ев. Лоб неоперен, оперение макушки заходит на него в виде мыска. Ноги короткие, жёлтые
или розоватые. Радужина у взрослых птиц красно-бурая. По земле ходит вразвалку. Взлета-
ет после довольно большого разбега, отталкиваясь от воды или земли лапами. Обычно
молчалив, но на колонии можно слышать низкие глухие ворчащие, рычащие и похожие на
хрюканье звуки.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Наличие различных форм объясняется сильной индиви-
дуальной изменчивостью (Судиловская, 1951). Существование подвида Р.о.roseus в насто-
ящее время не признается. Само это название (roseus) является синонимом Р. philippensis,
но иногда всё ещё используется для обозначения африканской и восточноазиатской попу-
ляций P.onocrotalus (Cramp, 1977).
Описание. Окраска. Самцы и самки окрашены одинаково, но самцы имеют более
крупные размеры. Оперение взрослой птицы белое с розоватым оттенком, более интенсивно
выраженным на нижней стороне тела. Старые птицы имеют более интенсивную розовую
окраску. На груди охристо-жёлтое пятно. Первостепенные маховые и их кроющие чёрно-
бурые. Стержни первостепенных маховых белые. Наружные опахала второстепенных махо-
вых серебристо-серые. Внутренние второстепенные маховые светлее наружных. Голые учас-
тки вокруг глаз розовые. Горловой мешок жёлтый. Конек и основание надклювья и подклю-
вья серовато-голубые, края розовые или красные. Коготок на кончике клюва ярко-красный
Ноги жёлтые, или розоватые, на сгибах оранжевые. Радужина буро-красная.
Розовый пеликан
13
В брачный период все неоперённые участки головы окрашены более ярко — у самцов
розово-жёлтые, у самок интенсивно оранжевые (Cramp, 1977). На передней части лба у обо-
их полов образуется вздутие; горловой мешок жёлто-охристый. Ноги желтовато-красные.
Птенцы вылупляются голыми. В первые часы после вылупления, кожа птенцов розовая,
затем постепенно темнеет, становится чёрной, с синим отливом.
Пуховой наряд. Тёмно-коричневый пух развивается в возрасте 3-14 дней, после 28 дней
становится бледнее. Радужина свинцово-серая, клюв, ноги, горловой мешок синевато-чёр-
ные.
Гнездовой наряд. Голова, шея и остальные участки верхней части тела серовато-бурые,
светлее на крестце. Лоб и темя бледнее. Плечевые, большие и средние кроющие крыла
серо-бурые с более светлыми вершинами, малые кроющие светло-бурые с охристым оттен-
ком. Маховые чёрно-бурые со светлыми окаймлениями на первостепенных маховых и с
серебристым налётом на второстепенных. Рулевые светло-серые. Брюшная сторона гряз-
но-белого цвета. Радужина у неполовозрелых птиц бурая (Судиловская 1951). Горловой
мешок неяркий серовато-жёлтый, клюв рогового цвета с бурым коньком, ноги буровато-
розовые или телесные, голые участки вокруг глаз розоватые.
Строение и размеры. Первостепенных маховых 11. Первое маховое значительно
короче больших верхних кроющих первостепенных маховых. Формула крыла: П=Ш>
IV>I=V>VI (Судиловская, 1951). Размах крыльев 270-360 см. Длина тела 140-175 см (Cramp,
1977). Длина крыла пеликанов (без разделения на половые группы) по Казахстану — 640-
770 мм, клюва — 300-450, цевки — 130-150 мм (Долгушин, 1960). Названный автор числа
измеренных птиц не указывает. Размеры молодой самки, отловленной в Кировской обл.
были такими: длина крыла — 625, клюва — 320, а цевки — 110 мм (Сотников, 1999). Вес
взрослых птиц, отловленных в Юж. Африке (п=3) — 6 120-8 040, в среднем — 7 120 г
(Underhill et al., 1999). Вес взрослых птиц из Вост. Европы и Сев. Азии — до 10 кг (Бутур-
лин, Дементьев, 1935). Размерные показатели по половым группам приведены в табл. 1.
Линька. Плохо изучена; её продолжительность, по-видимому, варьирует. Имеет следу-
ющие особенности.
Послебрачная — полная; маховые линяют сериями, по три активных центра на крыле.
Смена первостепенных маховых и рулевых идёт от внутренних перьев к наружным. Линь-
ку контурного оперения и крыла регистрировали в период размножения — в мае-августе и
позднее — в июле-ноябре (Cramp, 1977). У пеликана, добытого 22 июня в дельте Волги
отмечена линька первостепенных маховых, а птица, добытая в начале января в Казалинске,
имела свежее оперение, линька была завершена (Судиловская, 1951).
Предбрачная линька взрослых', длинные перья на затылке развиваются перед сезоном
размножения.
Таблица 1. Размеры самцов и самок взрослых розовых пеликанов (мм)
Показатель п Самцы lim М п Самки lim М Источник
Восточная Европа и Северная Азия
Длина крыла 5 700-720 716 2 640-690 66,5 Судиловская, 1951
Западная Европа
Длина крыла 24 665-772 684 17 586-650 620 Cramp, 1977
Длина хвоста 5 155-188 176 7 138-178 162
Длина клюва 9 347^71 409 14 289^00 328
Длина цевки •з 145-149 146,7 6 125-145 132
14
Отряд Пеликанообразные, Семейство Пеликановые
Рис. 1. Места гнездовий и районы зимовки розового пеликана: а — места известных гнездовий, б —
районы зимовок, в — основные пути осенних миграций.
Линька молодых не изучена. Взрослый наряд появляется на 3-м году жизни. Постепен-
ная смена наряда может указывать на то, что линька молодых идёт медленно в течение
первых лет жизни (Cramp, 1977).
Распространение. Гнездовой ареал разобщён. Распространение спорадическое.
Существенное сокращение ареала наблюдается в Европе, где гнездовые колонии остались
в Греции (оз. Микра Преспа), Македонии и Румынии (дельта Дуная). В тропической и Юж.
Африке гнездится в Мавритании, в бассейнах Сенегала, Нигера, Нила и Конго, в Кении (в
районе оз. Виктория), в Ботсване (оз. Нгами и Суан Пан), на побережье Намибии, в Мо-
замбике и в ЮАР на островах в районе Кейптауна. В Азии колонии известны в Турции,
Иране, на побережье Персидского зал., на севере Индии и на юге Индокитая (Crivelli,
1997) (рис. 1).
На Чёрном море до середины XX века розовый пеликан гнездился в дельте Днестра,
затем долгое время встречался здесь только на пролёте. В последние годы постоянно дер-
жится в весенне-летний период на Днестровском лимане (Русев, 2004). В 1990-х гг. отмече-
но появление и устойчивый рост гнездовой численности вида в Черноморском заповедни-
ке (Яремченко, 2000, цит. по: Жмуд, 2000). С 1999 г. розовый пеликан начал гнездиться в
Крыму на Лебяжьих о-вах (Костин, Тарина, 2002). В 1960-1961 гг. гнездился в дельте Тере-
ка (Сапетин, 1968). В настоящее время в России гнездится в долине Маныча. На оз. Ма-
ныч-Гудило и Маныч гнездится регулярно, периодически и на Чограйском вдхр. (Кривен-
ко, Любаев, 1975). В 1969-1973 гг. отмечено гнездование на Пролетарском вдхр. наЧикал-
динских о-вах (Языкова, Казаков, 1975). Колонии розовых пеликанов наблюдали в Волж-
Розовый пеликан
15
ско-Уральском междуречье (Кривоносов, Бондарев, 1978; Русанов и др., 2000; Гисцов, 2003,
2005, 2006). В 1989-1992 гг. птицы гнездились в устье Эмбы совместно с кудрявыми пели-
канами, однако, в 1999-2000 гг. колония здесь не была обнаружена (Гисцов, 2001).
Вместе с кудрявыми пеликанами гнездятся на Наурзумских озёрах (Стоцкая, Кривенко,
1988; Блинова, Блинов, 1997). На оз. Сарыкопа после его наполнения в 2002 г. две колонии
розовых пеликанов располагались на тростниковых заломах вдоль восточного берега в 2003
и 2004 гг. (Брагин, Брагина, 2002). В 1987 г. в небольшом количестве гнездились на оз.
Сарыкамыш (Стоцкая, Кривенко, 1988). Известно гнездование на Чушкакульских озёрах в
низовьях р. Бугуни. Обычным и многочисленным вид был в районе Кзыл-Орды, на оз.
Майкуль, Берказан, озёрах Бийлюкульской группы, в низовьях р. Чу, до 1960-х гг. был мно-
гочислен в дельте р. Или и на оз. Балхаш (Долгушин, 1960). В Вост. Казахстане традицион-
ным местом гнездования розового пеликана были Алакольские озёра, однако в 1986 г. пе-
ликаны здесь не гнездились, но были обнаружены на гнездовании в 1987 г. на оз. Сасык-
коль (Стоцкая, Кривенко, 1988). В июле 1971 г. стаями по 12-30 птиц розовые пеликаны
наблюдались вместе с кудрявыми на оз. Кызыл-Коль. По устному сообщению В.С. Лобаче-
ва, в это же время существовала колония при впадении р. Тургай в оз. Челкар-Тенгиз (Ауэзов,
1977). В Узбекистане вид продолжает гнездиться в нижнем течении Аму-Дарьи, на озёрах
Тогызтар и Судочье, в Аральском море на архипелаге Ак-Петки. Гнездится на оз. Айдар и
Айдарских разливах северо-западнее Нуратинского хр. и на оз. Замонбобо на юго-западе
Бухарской обл. (Мекленбурцев и др., 1987). Отмечалось возобновление гнездования в дельте
Чёрного Итрыша (Стоцкая, Кривенко, 1988) (рис. 2).
Рис. 2. Ареал розового пеликана в Восточной Европе и Северной Азии: а — места гнездовий, б —
места зимовок в юго-западном Прикаспии, в — залёты.
16
Отряд Пеликанообразные, Семейство Пеликановые
Зимовки. В связи с недостатком данных кольцевания, сведения о зимовках и особен-
ностях миграций весьма ограничены. Часть птиц дунайской популяции остаются зимовать
на Балканах и в Турции, на юго-западном побережье Чёрного моря, о чем свидетельствуют
четыре возврата колец, полученные в год кольцевания, осенью и зимой (Нанкинов, Сапе-
тин, 1978). Часть пеликанов летят южнее через Турцию, Сирию, Ливан, Израиль. Переме-
щения птиц в южном направлении наблюдались в заливе Акаба. Места зимовочных скоп-
лений известны в Египте, на Суэцком канале, на Красном море, Израиле. На побережье
Красного моря в зимнее время более обычны с африканской стороны, нежели на стороне
Аравийского полуострова. Установить присутствие зимующих птиц на африканском кон-
тиненте сложно по той причине, что стаи прибывших на зимовку птиц сливаются с пред-
ставителями местной популяции (Cramp, 1977). Розовые пеликаны с оз. Микра Преспа (Гре-
ция) продвигаются на запад до Амвракийского залива, часть их зимует в Греции, оставша-
яся часть летит в Африку (Zelenka, 1963). Птицы, гнездящиеся в долине Маныча, мигриру-
ют на зимовку преимущественно в юго-восточном направлении. В период миграции розо-
вый пеликан встречается в Дагестане по низменным приморским районам и вдоль побере-
жья Каспия. Зимующих птиц отмечали на крупных водоёмах и на морском побережье в
Азербайджане, учитывали на зимовках в Кызыл-Агачском заповеднике (Олигер, 1967).
Существование здесь зимовок и, одновременно, пролётного пути было подтверждено дан-
ными кольцевания, проводившегося в дельте Терека в 1960-1961 гг. (Сапетин, 1968). Веро-
ятно, этим путем мигрировали и птицы, гнездившиеся в дельте Волги. Места зимовок птиц
из казахстанских колоний точно не установлены (Cramp, 1977). Однако, заметный пролёт
розовых пеликанов проходит в дельте и пойме Аму-Дарьи (Мекленбурцев и др., 1987).
Отмечено, что на весеннем пролёте птицы придерживаются строго северного направле-
ния, пересекая пустынные территории (Ажимуратов, Пиржанова, 1986). Наличие активно-
го пролёта осенью наблюдали на Айдарских разливах (Гончаров, 1981). Миграционные
перемещения наблюдались вдоль р. Тигр в Ираке с марта до начала апреля и с сентября до
начала ноября. (Cramp, 1977). Значительное увеличение численности розовых пеликанов в
зимний период в Индии позволяет предполагать, что мигрирующие птицы из казахстан-
ских колоний, вероятно, достигают данного региона (Cramp, 1977).
Миграции. Пеликаны европейских, черноморских и казахстанских популяций совер-
шают регулярные миграционные перемещения. Птицы, гнездящиеся в тропических облас-
тях, вне периода размножения могут предпринимать дальние и ближние кочёвки. По на-
блюдениям на Айдарских разливах, пролёт, как правило, проходит в ясные солнечные дни,
утром, через 3-4 часа после восхода солнца. Высоту птицы набирают по спирали, исполь-
зуя термальные потоки (Гончаров, 1981; Cramp, 1977).
Весной розовый пеликан появляется дней на 10 позднее кудрявого (Долгушин, 1960).
Весенний пролёт в Греции проходит в апреле, мае и продолжается до начала июня (Zelenka,
1963). В дельту Дуная пеликаны возвращаются в конце марта - апреле (Cramp, 1977; Пан-
ченко, Балацкий, 1991). В дельте Днестра встречались уже в начале марта (Стойловский,
2000). На территории Черноморского биосферного заповедника самая ранняя встреча за-
регистрирована 13 апреля (Руденко, Яремченко, 2005). В Крыму на Лебяжьих о-вах наибо-
лее ранняя дата первой регистрации вида — 3 марта (Костин, Тарина, 2002). На юге Арала
и в дельте Амударьи весенний пролёт проходит в последней декаде марта - первой полови-
не апреля (Ажимуратов, Пиржанова, 1986; Мекленбурцев и др., 1987). Стаи, пролётающие
в северо-восточном направлении у западного хребта Нуратау, зарегистрированы в первой
половине апреля (Салихбаев и др., 1970, цит. по: Мекленбурцев и др., 1987). В Алакольской
котловине пики пролёта розовых пеликанов приходились на 2 и 20 апреля (Сема, Ауэзов,
1984). В дельте Тентека (южная часть оз. Сасыкколь) передовые особи зарегистрированы
Розовый пеликан
17
20 марта (Березовиков, 20046). На Шардаринском вдхр., в районе Келеса и Голодной степи
розовые пеликаны начинают появляться с начала февраля (Долгушин, 1960).
Осенний отлёт почти повсеместно на описываемой территории начинается в сентябре и
продолжается до ноября, пик пролёта приходится на октябрь (Zelenka, 1963; Cramp, 1977;
Гончаров, 1981; Ажимуратов, Пиржанова, 1986); в теплые зимы птицы могут задерживать-
ся в местах летнего пребывания до декабря (Стойловский, 2000). В Казахстане пеликаны
задерживаются до первых заморозков и почти полностью исчезают здесь в начале — сере-
дине ноября (Долгушин, 1960).
В период кочёвок пеликаны могут встречатся довольно далеко от мест гнездования.
Известно несколько случаев залётов в Смоленскую (Судиловская, 1951) и Ленинградскую
обл. (Мальчевский, Пукинский, 1983). В мае 1935 г. молодая самка розового пеликана была
отловлена в Кировской обл. (Сотников, 1999). В Нижегородской обл. в 1982 г. раненая пти-
ца была подобрана рыбаками; в 1984 г. пара держалась летом не севере области; в 1985 г.
также наблюдали единичную особь на юге области. Известны две встречи одиночных птиц
в низовьях р. Белой в мае 1996 г. (Карякин, 1998а) и в апреле 1998 г. В Кемеровской области
осенью 1975 г. розовый пеликан был снят с проводов высоковольтной линии вблизи Мари-
инска, а в конце ноября 1988 г. молодой самец был отловлен в среднем течении р. Томь
(Гагина, Скалой, 2000). В мае 2000 г. две птицы встречены в Омской обл. на оз. Тенис. В
Челябинской обл. на оз. Тишки в 1996 г. 20 июня была встречена одиночная особь, а в июле
2001 г. 24 птицы наблюдались в колонии кудрявых пеликанов и больших бакланов на оз.
Курлады (Гордиенко, 2002, 2006). Дальние залёты свойственны, как правило, молодым не-
половозрелым особям.
Местообитания. Населяя районы степей, пустынь, саванн, розовый пеликан тесно свя-
зан с водными ассоциациями. В Африке обитает преимущественно в тропических райо-
нах, где сухой сезон приходится на зимнее время. Основным условием является наличие
поблизости водоёмов, богатых рыбой. Для гнездовий предпочитает не очень глубокие, теп-
лые, пресные водоёмы, болота, чаще в дельтах крупных рек. Реже населяет солоноватые
или солёные водоёмы — лагуны, эстуарии или побережья внутренних морей. Гнёзда поме-
щает в недоступных и малодоступных для наземных хищников местах: среди плотных за-
рослей околоводной растительности (тростника, камыша), на сплавинах, на каменистых,
ракушечных или песчаных островках и косах, часто, почти лишённых растительности
(Cramp, 1977; Багрикова, Костин, 2005).
Численность. С конца XIX в. наблюдается тенденция снижения численности. Мировая
численность этого вида оценивается приблизительно в 90 тыс. пар, до 80% из них гнездят-
ся в Африке (Crawford et al, 1995). Одна из крупнейших африканских колоний находится в
Мавритании, в национальном парке Даулинг, где в 1999 г. размножалось 11 480 пар (Diagana
et al., 2001). На оз. Св. Лючии на севере Квазулу-Натал гнездится 3 тыс. пар (Crawford et al.,
1995). По результатам учётов околоводных птиц в Юж. Азии 1997-2001 гг. численность
розовых пеликанов здесь составила от 15 до 30 тыс. особей (Li, Mundkur, 2004). На терри-
тории Турции размножается от 250 до 400 пар (Crivelli, 1997). В Палеарктике численность
данного вида составяет 6 660-11 550 пар (Линьков, 20006). По данным Всесоюзных учё-
тов колониальных околоводных птиц в 1973-1977 гг. в СССР численность розового пели-
кана составляла 2-2,2 тыс. гнездящихся пар, по учётам в 1986-1987 гг. — 2,3 тыс. (Флинт и
др., 1991). Колония на оз. Микра Преспа в Греции с 1989 г. продолжает оставаться относи-
тельно небольшой — от 40 до 100 пар; в 1983 г. эта колония насчитывала 116 гнездящихся
пар (Hadzilacos, 1986; Crivelli, 1997). Крупнейшая европейская колония располагается в
дельте Дуная и насчитывает около 3 500 гнездящихся пар. При этом, неразмножающаяся
часть дунайской популяции в конце 1990-х гг. только на украинской территории, включая
18
Отряд Пеликанообразные, Семейство Пеликановые
придунайские озёра и водохранилище Сасык, насчитывала 5 000 особей. Столь же значи-
тельные скопления неразмножающиеся пеликаны образовывали в румынской части дель-
ты, а также в молдавской части поймы Прута. С учётом всех перечисленных территорий,
общее число неразмножающихся розовых пеликанов дунайской популяции в конце 1990-х
гг. составляло около 10 тыс. особей, что более, чем в два раза превышает число гнездящих-
ся в дельте пар. По-видимому в результате роста численности дунайской популяции и про-
движения молодых особей на восток по черноморскому побережью в 1995 г. образовалась
колония в Черноморском заповеднике. В 1999 г. в ней насчитывалось уже 100 гнездящихся
пар (Жмуд, 2000). В том же году в Крыму на Лебяжьих о-вах была зарегистрирована по-
пытка гнездования 51 пары розовых пеликанов. Птицы пытались гнездиться и в последую-
щие годы, но их гнёзда разрушали рыбаки. Так, в 2002 г. на одном из островов было об-
наружено 19 гнёзд с кладками и 23 пустых. Рядом с колонией держалось 326 птиц, а по
краю острова — 218 (Костин, Тарина, 2002). На оз. Маныч-Гудило в 1972 г. гнездилось 60
пар; в настоящее время здесь насчитывается около 300 пар и наблюдается тенденция к
увеличению численности. Изредка гнездится на Чограйском вдхр. численностью от 2 до 8
пар (Кривенко, Любаев, 1975; Казаков, Кривенко, 2000). На Чокалдинских о-вах Пролетар-
ского вдхр. насчитывали 98 гнёзд (Языкова, Казаков, 1975). В дельте Волги в 1963 г. гнез-
дилось 7 пар, а в 1980 — 2 пары. С этого времени гнездование птиц этого вида здесь не
наблюдалось (Русанов и др., 2000).
Современная численность розового пеликана на территории Казахстана между Тургай-
ской низменностью и Джунгарским хр., Юж. Сибирью и Копетдагом по оценке В.В. Ви-
ноградова (Vinogradov, 1998) составляет 5 500 пар. В разливах нижнего течения рр. Иргиз
и Тургай в 1971-1972 гг. встречались стаи розовых пеликанов численностью до 300 осо-
бей. В смешанных стаях с кудрявыми пеликанами розовые пеликаны составляли 74-82%
(Варшавский и др., 1977). По данным авиаучётов, проведённых здесь в июле 1985 г., учтено
530 пеликанов, а в августе этого же года — 1 630 пеликанов, 80% из них составляли розо-
вые пеликаны (Ауэзов, Виноградов, 1986). На оз. Сарыкопа Кустанайской обл. в 2003 г.
обнаружено 2 колонии по 100-130 гнёзд в тростнковых заломах (Брагин, 2004). В Наурзум-
ском заповеднике розовый пеликан на гнездовье численно преобладает над кудрявым. В
1987 г. на озёрах было учтено 100 пар. Впервые птицы здесь загнездились в 1984 г. (Стоц-
кая, Кривенко, 1988; Блинова, Блинов, 1997). Встречаются здесь и летующие особи. В 2004 г.
на летовке было отмечено около 100 особей, по большей части, неполовозрелых птиц (Бра-
гин, 2005). На оз. Балхаш в дельте р. Или ещё в первой половине прошлого века пеликаны
были многочисленным видом. Совместные колонии кудрявых и розовых пеликанов насчи-
тывали до 2 500 гнёзд (Грачев, 1977а,б). В результате массового забоя птенцов, который
продолжался до 1956 г. (в 1930-е гг. охотники сдавали до 2 700 тушек), большие колонии
исчезли (Абрамова и др., 1975). Весной 1963 г. в районе оз. Балхаш насчитывалось около
1 800 пар пеликанов, 80%, которых составляли розовые пеликаны (Владимиров, Злобин,
1969). В мае 1968 г. в двух самых крупных колониях в дельте р. Или насчитывалось около
1 100-1 300 гнёзд, в которых численность кудрявого пеликана составляла не более 10-20%
(Грачев, 1977 а). В 1972 г. птицы переместились и разбились на несколько небольших коло-
ний, всего здесь было учтено в тот год около 820 пар (Страутман, Степанов, 1977). В 1960-е
гг. исчезли гнездовья в низовьях р. Чу (Ёлкин и др., 1975). На озёрах Алакольской группы
розовый пеликан был многочислен до 1940-х гг., затем численность сократилась из-за на-
чавшейся охоты на птиц. В 1947 г. колония насчитывала свыше 500 гнёзд, в 1948 г. — 150,
гнёзда были брошены (Слудский, 1953, цит. по: Ауэзов, Грачев, 1977). Из-за преследования
колония постоянно перемещалась. В 1962 г. в ней насчитывалось около 100 пар, к 1964—
1972 гг. — около 500 (Ауэзов, Грачев, 1977). В восточной части оз. Ссыкколь на оз. Тысячи
в 1999-2001 гг. гнездилось около 500-600 пар (Березовский, Левинский, 2004).
Розовый пеликан
19
По прогнозам В.В. Виноградова (1998) в связи с наступлением в начале XXI в. новой
климатической фазы — более сухой и тёплой — многие водоёмы Центр. Казахстана могут
либо пересохнуть, либо обмелеть и повысить свою соленость, в связи с чем к 2010 г. чис-
ленность розового пеликана здесь составит 1 500-2 000 пар.
Размножение. Начинают размножаться в возрасте трёх лет. Гнездятся большими плот-
ными колониями, которые могут состоять как из десятков, сотен, так и из десятков тысяч
пар. Отдельными парами поселяются в исключительных случаях. Внутри колоний, особен-
но крупных, выделяются отдельные субколонии, в которых сроки размножения синхрони-
зированы. Существование субколоний, вероятнее всего, обусловлено различными датами
подлёта стай (прилёт продолжается в большинстве мест гнездования с марта - апреля до
начала июня), поэтому сроки гнездования в больших колониях растянуты: на оз. Маныч-
Гудило 4-5-недельных птенцов отмечали в июне и августе (Zhatkanbayev, 1994). На оз.
Балхаш в начале июня встречали и свежие кладки, и начинающих летать молодых (Влади-
миров, Злобин, 1969).
В местах обитания обоих видов, как правило, образуют совместные колонии с кудря-
вым пеликаном в любых соотношениях.
На места гнездования прилетают стаями. Моногамы. Пары сохраняются, вероятнее всего,
в течение одного сезона и могут образовываться как в удалении, так и непосредственно на
месте расположения колонии. В период формирования пар холостые самцы собираются
вместе и устраивают брачные демонстрации — стоя вместе или прохаживаясь друг мимо
друга, то делают выпады в центр группы, хватая друг друга клювами, то поднимают клювы
вверх с мычанием. Действия беспорядочные, без определённой последовательности и ри-
сунка. Самки пристраиваются у края такой группы. Время от времени от группы самцов
отделяется один или несколько самцов и, пытаясь привлечь внимание самки, преследуют
её. Если самец не один, то вскоре в группе проявляется доминирующая особь, которая наи-
более активно отгоняет остальных. Оставшись с самкой вдвоём, самец плавает за ней и
может предпринимать попытки к спариванию — по всей видимости, неуспешные, поскольку
самка уходит под воду под весом самца. Птицы вновь образовавшейся пары могут совмес-
тно плавать поблизости друг от друга; самец старается держаться сбоку или позади самки.
Самцы из уже образовавшихся пар могут периодически приближаться к группе холостых
самцов и участвовать некоторое время в общих стычках, затем вновь удаляются и присое-
диняются к своим самкам. Охраняя гнездовой участок, пары принимают позу угрозы, стоя
рядом с поднятыми вверх клювами (рис. За). При этом они издают гортанные крики, напо-
минающие мычание. На суше самцы следуют за самками в демонстративной позе ухажива-
ния. Шея при этом вытянута, клюв полуопущен, а крылья полураскрыты (рис. 36, в). При
этом самец может покачивать телом из стороны в сторону.
Пара временами совершает небольшие совместные полёты, затем птицы садятся, трут-
ся клювами друг о друга. После периода ухаживания самка в сопровождении самца начи-
нает обходить территорию колонии, подыскивая место для гнёзда среди других пар, порой
сталкиваясь с проявлением агрессии со стороны птиц, уже занявших место. Выбрав место
для гнёзда, самка садится на землю. Здесь же происходит копуляция, во время которой
самец, стоя рядом, наклоняется вперёд, захватывает шею самки клювом и взбирается на
неё сбоку. Спаривание может повторяться несколько раз в день. Вскоре после копуляции
самка начинает строить гнездо, сгребая субстрат клювом. Самец принимает участие в стро-
ительстве, поднося гнездовой материал в горловом мешке и иногда воруя его в соседних
гнёздах. Принесённый материал самец кладёт перед самкой, которая, стоя на гнезде, укла-
дывает его вокруг и под себя, сооружая рыхлую, грубую на вид постройку. На строитель-
ство гнёзда уходит 2-3 дня (Судиловская, 1951; Долгушин, 1960; Cramp, 1977).
20
Отряд Пеликанообразные, Семейство Пеликановые
Рис. 3. Позы розовых пеликанов: а — угроза; б, в — ухаживание (б — самка, в — самец), г — поза
при смене партнёров на гнезде (по: Cramp, 1977).
К строительству гнёзд на оз. Микра Преспа птицы приступали в апреле (Hatzilacos,
1986). На Лебяжьих о-вах на Крымском п-ове 28 мая были обнаружены гнёзда с кладками
(Костин, Тарина, 2002). В дельте р. Волги постройка гнёзд начиналась 20 апреля, на Сыр-
Дарье — в конце апреля или начале мая (Судиловская, 1951). Основным условием для вы-
бора места колонии является недоступность её для наземных хищников. Поэтому пелика-
ны селятся либо на песчаных, ракушечных и каменистых островках на озёрах и у морских
побережий среди невысокой и редкой растительности или в дельтах рек, где часто поселя-
ются в прибрежных зарослях надводной растительности ближе к открытой воде. В зарос-
Розовый пеликан
21
лях тростника или камыша приминают растения, делая заломы, и на них строят гнёзда.
Гнёзда могут располагаться также на сплавинах на крупных болотах и озёрах.
Гнездовая постройка состоит из стеблей тростника, веточек и другой растительности,
иногда выстилается перьями (АН, 1960, цит. по: Cramp, 1977). Высота гнёзда 30-60 см,
диаметр у основания 70-100, диаметр у вершины 30-50 см (Terrasse et al., 1969а, цит. по:
Cramp, 1977). В кладке от 1 до 3, чаще 2 яйца. Форма яиц овальная, скорлупа белого
цвета с толстым известковым налётом (Судиловская, 1951). Размеры яиц: (п=150) 80-
104x52-64, в среднем 94x59 (Schonwetter, 1967); в дельте Волги 80-112 х 50-75 мм; на
Сыр-Дарье 90-106,3 х 58,9-60,5 мм. Вес яиц 155-195 г. (Судиловская, 1951). В дельте
Волги процесс откладки яиц начинался 20 апреля; на Сыр-Дарье — в конце апреля или в
начале мая (Спангенберг, 1936, цит. по: Судиловская, 1951). Откладка яиц идёт с интер-
валом в одни или двое суток. Насиживание начинается с откладки первого яйца, из-за
чего вылупление происходит неодновременно и птенцы в гнёздах иногда сильно отлича-
ются по размеру. В сезоне один цикл размножения. Замещение погибшей кладки проис-
ходит лишь в том случае, если от начала насиживания до момента разорения прошло не
более 10 дней (Судиловская, 1951). Процесс инкубации продолжается, по наблюдениям в
Казахстане, 33-39 дней (Долгушин, 1960), на оз. Микра Преспа — 33 дня (Hatzilacos,
1986), в Кении — 35-36 дней (Brown et. al., 1973, цит. по: Cramp, 1977). Насиживает
кладку преимущественно самка, самец сменяет её лишь на непродолжительное время
(Судиловская, 1951).
Вылупление происходит почти одновременно в гнёздах всей колонии (или в субколо-
ниях). Птенцы вылупляются голыми, без эмбрионального пуха, глаза и наружные слухо-
вые отверстия закрыты, кожа сначала бледно-розовая, но через несколько часов после
вылупления начинает темнеть (Мекленбурцев и др., 1987). Птенцов выкармливают оба
партнёра. В первое время после рождения птенцы не проявляют кормовой активности, и
родители стимулируют их к принятию первой порции корма, помещая свой клюв между
ногами птенца. Ярко-красный коготок на кончике клюва родителя возможно стимулирует
у птенцов «клевательный» рефлекс (Cramp, 1977). В последующие кормления взрослые
птицы побуждают птенцов к принятию пищи, слегка поклёвывая их в голову или тулови-
ще. Со временем птенцы начинают активно требовать корм, кричат при виде приближа-
ющегося родителя, толкают друг друга, стараясь придвинуться к краю гнёзда (Судиловс-
кая, 1951).
Птенцов в первые дни родители выкармливают полупереваренной пищей, отрыгивая
её птенцам прямо в рот. Позднее птенцы сами залезают за кормом родителям в горловой
мешок, порой очень глубоко, достигая пищевода (Cramp, 1977). В возрасте 5-8 дней птен-
цы начинают получать в корм мелкую рыбу, размеры приносимой добычи увеличивают-
ся по мере роста птенца. Одна порция корма в первые дни жизни птенцов составляет
около 30^40 г за одно кормление. В возрасте 10 дней птенец съедает 200-250 г (Судилов-
ская, 1951). По наблюдениям в Индии, птенцы заглатывали рыбу длиной до 25 см и весом
500-600 г (Aly, Ripley, 1968, цит. по: Cramp, 1977). Обычно взрослые птицы, прибыв с
новой порцией корма, отводят птенцов на удобное для кормления место, затем наклоня-
ются так, чтобы птенцу было удобно достать рыбу (Cramp, 1977). Кормовые биотопы
часто бывают удалены от места расположения колонии. Так, пеликаны, гнездящиеся на
оз. Св. Лючии, вынуждены совершать перёлеты до 100 км, чтобы преодолеть расстояние
между гнездовым и кормовым биотопами (Crawford et al., 1995). Пеликаны, гнездящиеся
на оз. Микра Преспа, кормятся на озёрах Зап. Македонии (Чимадитис, Вегорритис, Пет-
ров, Кастория и Зазари) (Hatzilacos, 1986), отстоящих более, чем на 50 км от мест гнездо-
вания.
22
Отряд Пеликанообразные, Семейство Пеликановые
В возрасте 20-30 дней птенцы покидают гнездо и объединяются в «ясли», где держатся
вместе плотной стайкой. В этот период родители, прилетая с кормом, опознают и кормят
только своих птенцов. Преследующих их чужих птенцов они довольно жестко отгоняют,
иногда хватают их за шею или туловище и оттаскивают в сторону или даже резко встряхи-
вают. С приобретением опыта птенцы, спустя некоторое время, преследуют уже только
своих родителей (Cramp, 1977). Плавать птенцы могут с раннего возраста и в случае опас-
ности сходят с гнёзда и стараются уплыть (Долгушин, 1960). В возрасте 55 дней они спо-
собны сами ловить рыбу. Лётными молодые становятся в возрасте 65-70 дней, почти в это
же время они становятся самостоятельными (Cramp, 1977). По данным В.Г. Кривенко, на
оз. Маныч-Гудило на одну гнездящуюся пару в среднем приходится от 0,7 до 1 птенца,
поднявшегося на крыло. В Черноморском биосферном заповеднике — 1 птенец на размно-
жающуюся пару (Руденко, Яремченко, 2005). Успешность размножения в популяции, гнез-
дящейся на оз. Микра Преспа в Греции, оценивается в 1,83 молодых на гнездо (Hatzilacos,
1986).
По данным кольцевания в Юж. Африке, максимальный возраст розового пеликана со-
ставлял 30 лет 1 месяц (Underhill et al., 1999), в Вост. Африке — 23 года 5 месяцев (Urban,
Ash, 2001).
Суточная активность, поведение. На протяжении всех фаз годового цикла держатся
группами разной величины. Стаи состоят как из взрослых, так и из молодых птиц.
Обычно миролюбивы, но случаются стычки между соседями по колонии и даже между
партнёрами; наблюдались агрессивные выпады и в адрес гнездящихся совместно с ними
бакланов. Самцы более агрессивны, чем самки. Агрессивное поведение проявляется наи-
более интенсивно в период формирования пар. У всех пеликанов, принимающих участие в
размножении, клюв обычно покрыт царапинами и шрамами. При типичной демонстрации
позы угрозы птица поднимает и вытягивает клюв вверх и к сопернику, иногда с нанесением
резких ударов, сопровождающихся повторяющимися звуками, похожими на хрюканье или
мычание. Соперник либо делает ответный выпад, либо удаляется с опущенным клювом;
атакующая птица редко преследует, его. Столкновения чаще заканчиваются вскоре после
начала, однако отмечается и иной финал: победитель захватывает верхнюю или нижнюю
челюсть противника, нагибает его голову вниз и держит так 1-3 сек. Насиживающая птица
обороняет только территорию вокруг гнезда, на расстоянии вытянутой шеи. На пути к гнез-
ду птицы приземляются у края колонии и дальше идут пешком. Мимо чужих гнёзд прохо-
дят, подняв клюв и вытянув шею вверх, крылья держат полуразвёрнутыми. По пути иногда
случаются стычки с соседями — взаимные выпады клювом в сторону друг друга. У гнезда
птицы приветствуют друг друга, издавая низкие, похожие на хрюканье звуки. Сидящая на
гнезде птица медленно поднимается и сходит с гнезда с опущенной головой и с движением,
напоминающим поклон (рис. Зг). Иногда в гнезде отмечаются агрессивные столкновения
между птенцами: более старшие птенцы жестоко бьют клювом младших без всякой види-
мой причины (Cramp, 1977; Zhatkanbayev, 2001).
Пеликаны мигрируют или перемещаются между кормовыми районами стаями по 5-500
птиц, состоящими из более мелких подгрупп — по 20-100 особей. С ослаблением действия
термальных потоков, мигрирующие стаи пеликанов спускаются на ночёвку, примерно за
1-2 ч до наступления темноты, и продолжают полет обычно через 1-3 ч после восхода
солнца (Cramp, 1977). На Айдарских разливах совместные стаи розовых и кудрявых пели-
канов стартовали через 3-4 ч после восхода (Гончаров, 1981).
Перёет от гнездовых к кормовым районам совершают небольшими группами по 4-50
особей; по прибытии в кормовые биотопы стаи разбиваются на более мелкие группы. Кор-
мовые территории не охраняются, в местах кормёжек птицы ведут себя мирно.
Розовый пеликан
23
Для ночёвок и дневного отдыха из года в год используются одни и те же места. Как
правило, это песчаные пляжи (преимущественно на островах), заросли тростника или ка-
менистые острова. Изредка птицы могут отдыхать на деревьях, в основном в тех случаях,
когда их застигает плохая погода, или во время перелётов. Могут покидать ночёвки и кор-
миться в ночное время, особенно в ясные лунные ночи. Свободные от насиживния партне-
ры могут ночевать как рядом с гнездом, так и около кормовых биотопов (Cramp, 1977).
Питание. Основу рациона составляет исключительно рыба, которую птицы вылавлива-
ют из воды, пользуясь клювом как черпаком; горловой мешок при этом сильно растягивает-
ся. Коготок на кончике надклювья помогает удерживать рыбу. Охотятся коллективно, раз-
деляясь на группы. При этом птицы совершают в высшей степени согласованные действия:
выстроившись цепочкой, начинают двигаться к берегу, хлопая крыльями, загоняя рыбу на
мелководья, затем все одновременно опускают головы под воду, либо ныряют (вопреки
существующему мнению, по данным киносъёмки, розовые пеликаны могут нырять).
Часто к «загонной цепочке» пеликанов присоединяются бакланы, которые плывут вме-
сте с ними, то и дело ныряя. Группу могут сопровождать и чайки, которые с криком носятся
вокруг, выхватывая из воды рыбу. Поднимаемый при этом шум способствует эффективно-
сти загона рыбы (Судиловская, 1951).
В дельте Волги в состав корма входит рыба средних размеров — сазан, лещ, вобла и
густера, реже — окунь. На Балхаше розовый пеликан поедает преимущественно сазана,
губача и балхашского окуня. Основу питания розовых пеликанов африканской популя-
ции составляют цихлиды рода Tilapia (Cramp, 1977). Взрослая особь потребляет в день
около 0,9-1,2 кг рыбы (Brown, Urban, 1969, Din, Eltrigham, 1974a, цит. no: Cramp, 1977).
Птицы, содержащиеся в неволе, могут съедать до 1,6 кг (Andone et al., 1969, цит. по:
Cramp, 1977).
Враги, неблагоприятные факторы. Пеликаны обычно селятся в местах, труднодос-
тупных для наземных хищников — на островах или в широких полосах надводной расти-
тельности ближе к открытой воде. Однако попадающие на острова, чаще всего в зимний
период, лисицы и енотовидные собаки могут наносить ощутимый урон колониям. Кладки
также может разорять чайка-хохотунья; кроме того, известны случаи клептопаразитизма на
птенцах пеликана (Костин, Тарина, 2004; Руденко, Яремченко, 2005). Наиболее существен-
ные факторы, влияющие на численность розового пеликана — деградация местообитаний
и усиление фактора беспокойства. Потревоженные людьми пеликаны могут покинуть гнёз-
да с кладками либо сменить местоположение колонии; даже если птицы возвращаются в
колонии, то за время их отсутствия яйца перегреваются на солнце и эмбрионы гибнут (Хох-
лов, Ильюх, 2002). Во многих местах пеликан до сих пор подвергается прямому преследо-
ванию со стороны человека — местные рыбаки и промысловики, усматривая в пеликане
нежелательного конкурента, разоряют гнёзда и ведут незаконный отстрел птиц (Жмуд, 2000;
Костин, Тарина, 2002; Хохлов, Ильюх, 2002).
Деградация местообитаний связана в первую очередь с изменением гидрологического
режима степных водоёмов в результате создания плотин и водохранилищ. Колонии пелика-
нов гибнут также из-за паводковых и нагонных вод. На успешность размножения на оз.
Маныч-Гудило отрицательное влияние оказывают ливневые дожди, затопляющие гнёзда.
В годы с высоким уровнем воды на этом водоёме происходит интенсивный размыв остро-
вов — мест гнездования. Кроме того, неконтролируемые сбросы воды из многочисленных
оросительных систем резко усиливают размывание островов волнобоем во время штормов
Кривенко, 2000). Серьёзным негативным фактором становится попадание в воду хлорор-
гзнических соединений, из-за которых изменяется толщина скорлупы и снижается успеш-
ность размножения птиц (Казаков, Кривенко, 2000).
24
Отряд Пеликанообразные, Семейство Пеликановые
Резкое уменьшение рыбных запасов в дельте Дуная, вызванное возросшей рыбопро-
мысловой нагрузкой на угодья, увеличением масштабов нелегальной добычи рыбы и рас-
ширением «чёрного рынка» в рыботорговле, особенно в украинском секторе дельты, мо-
жет привести к падению численности вида в этом регионе (Жмуд, 2000).
Хозяйственное значение, охрана. Численность розового пеликана настолько мала, что
реальной угрозы рыбному промыслу и рыборазводным хозяйствам он принести не может. В
1960-е гг. на Аральском море численность веслоногих была сокращена на треть, но ощутимого
эффекта для рыбного хозяйства в результате принятых мер получено не было (Марков, 1965).
До 1960-х гг. нелетающих птенцов охотники на лодках сгоняли в кучу вблизи колонии и
забивали палками. Из тушек добывали жир, которым смазывали оси телег, сапоги, оружие
и пр. Взрослых пеликанов отстреливали на шкурку или просто ради надклювья, из которо-
го изготавливали оригинальные ножны (Абрамова и др., 1975; Грачев, 1977а,б).
В настоящее время розовый пеликан повсеместно находится под охраной. Он занесён в
Красную книгу Российской Федерации, Красные книги Украины, Армении, Казахстана,
Узбекистана, Туркменистана и ряда других стран СНГ. Местам гнездовий этого вида, как
правило, присваивается статус охраняемых территорий. Вид включён в Приложение 1 Бон-
нской Конвенции, Приложение 2 Бернской Конвенции, Приложение Соглашения об охране
мигрирующих видов, заключённого между Россией и Индией. В России охраняется на оз.
Маныч-Гудило в филиале заповедника «Чёрные Земли», в Дагестанском и Астраханском
заповедниках (как потенциально гнездящийся и мигрирующий вид), в республиканском
заказнике Маныч-Гудило и заказнике Чограйский в Калмыкии. На путях миграции охраня-
ется в ряде заказников Ростовской области, в республиканских заказниках Приазовский
(Краснодарский край) и Самурский (Дагестан). На Украине охраняется в Дунайском и Чер-
номорском заповедниках, в заповеднике «Лебяжьи острова». В Казахстане — в Наурзум-
ском и Кургальджинском заповедниках.
Для сохранения вида необходимо осуществление комплекса мероприятий, направлен-
ных на улучшение условий его гнездования, а также проведение эколого-просветительской
работы среди населения и рыбаков-профессионалов. На численности вида положительно
могут сказаться биотехнические мероприятия, особенно специальная подготовка мест для
гнездования (Руденко, Яремченко, 2005).
Кудрявый пеликан
Pelekanus crispus Bruch, 1832
Pelecanus crispus Bruch, 1832, Isis, стб. 1109, Далмация.
Dalmatian pelican (англ.), Krauskopfpelikan (нем.), pelican frise (фр.)
Син. (лат.): Pelecanus patagiatus C.L. Brehm, 1855.
Син. (рус.): баба, баба-птица (используется в отношении как кудрявого, так и розового пе-
ликанов), серый пеликан.
Статус. Гнездящийся, перелётный вид, изредка зимующий в небольшом количестве.
Общая характеристика и полевые признаки. Очень крупная птица, размером с лебе-
дя, легко узнаваемая по массивному клюву с большим горловым мешком. Крылья длинные
и широкие, с тёмными концами. Шея в полёте изогнута, голова лежит на спине, как и при
отдыхе на воде. При быстром плавании пеликан вытягивает шею почти вертикально вверх.
Стараясь предохранить крылья от намокания, приподнимает их над водой. По земле пере-
Кудрявый пеликан
25
двигается неторопливо, вразвалку. Взлетает как с суши, так и с воды, несколько раз оттал-
киваясь от поверхности обеими ногами (Молодовский, 1997, 2001). Может присаживаться
на деревья, обхватывая ветку тремя пальцами спереди и одним сзади (Судиловская, 1951).
На голове и задней стороне шеи развита грива из удлинённых закрученных перьев; у моло-
дых птиц она развита слабо. В отличие от розового пеликана лоб полностью оперен, с
небольшой округлой выемкой посередине, куда вдаётся надклювье. Молчалив; голос напо-
минает низкий рёв или ворчание. Услышать его можно, как правило, только в гнездовой
колонии. Может издавать звуки, похожие на хрюканье, лай, шипение, фыркание, треск. В
делом, наблюдается большое сходство с голосовыми особенностями розовых пеликанов
(Cramp, 1977). В полёте отличим от розового пеликана по отсутствию резкого цветового
контраста между нижними кроющими крыла и маховыми и по тёмным ногам; взрослые
птицы в брачный период — по оранжево-красному, а не жёлтому, горловому мешку, отсут-
ствию розовых оттенков в оперении; вблизи — по светлой радужине, меньшей площади
неоперённых участков на голове, форме хохла. Молодые особи выглядят заметно светлее
молодых розовых пеликанов, с серыми а не розоватыми ногами и голыми участками кожи
на голове.
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Взрослые птицы. Самцы и самки выглядят одинаково. Опере-
ние взрослых птиц преимущественно белое. Спина, плечевые перья и верхние кроющие
крыла светло-серые с тёмными стволами и белыми вершинами. На голове и верхней стороне
шеи — удлиненные «курчавые» перья («грива»). Первостепенные маховые и их кроющие
буровато-чёрные, с тёмными стержнями. Второстепенные маховые — серые с белыми кай-
мами по внутреннему краю опахал. Клюв серый с красноватыми или жёлтыми краями,
ноготок оранжевый, горловой мешок светло-жёлтый. Неоперённые участки кожи вокруг
глаз жёлтые. Радужина бледно-жёлтая. Сезонные отличия в оперении взрослых птиц не-
значительны. В брачном наряде на зобе заметно жёлтое или рыжеватое пятно, кожа вокруг
глаз становится пурпурной, а горловой мешок — оранжевым, красновато-жёлтым или кро-
ваво-красным (Судиловская, 1951; Cramp, 1977).
Пуховой наряд. При вылуплении птенцы голые, с красновато-розовой кожей. Белый пу-
ховой наряд начинает развиваться на пятый день. Клюв буровато-чёрный, подклювье не-
сколько светлее.
Гнездовой наряд грязно-белый с заметным буровато-охристым или серым налетом. Кро-
ющие крыла охристые, плечевые буровато-чёрные с чёрными вершинами. Первостепен-
ные маховые бурые. Рулевые светло-серые с беловатыми каемками. Брюшная сторона тела
грязно-белая, бока светло-бурые. Удлинённые перья на голове и шее почти не развиты.
Радужина жемчужно-серая, клюв жёлто-серый, горловой мешок жёлтый, ноги свинцово-
серые (Судиловская, 1951; Долгушин, 1960; Cramp, 1977).
В промежуточных нарядах спинная сторона с дымчатыми или песочно-бурыми отме-
тинами. На голове и шее заметны серые, а на кроющих крыла и плечевых перьях — буро-
вато-охристые тона. «Грива» развита слабо. Окраску взрослых птицы приобретают, оче-
видно, к третьему году жизни, но точная последовательность смены нарядов по-прежнему
не изучена (Судиловская, 1951; Cramp, 1977).
Строение и размеры. Первостепенных маховых 11. Формула крыла
IV>III>II>V>I>VI. Хвост прямой, из 22 мягких рулевых (Судиловская, 1951). Самцы в сред-
нем крупнее самок (табл. 2). Размах крыльев 270-345 см, длина тела — 160-180 см (Суди-
ловская, 1951).
Короди-Гаал (Korodi-Gal, 1964, цит. по: Cramp, 1977) приводит ср. размеры неполовоз-
релых птиц из низовий Дуная: 3 сГсГ и 4 у которых длина крыла была соответственно
26
Отряд Пеликанообразные, Семейство Пеликановые
Таблица 2. Размеры взрослых кудрявых пеликанов (мм)
Показатель п Самцы lim Самки Источник
М n lim М
Длина крыла 7 Восточная Европа и Северная Азия 725-770 750 2 685-700 692 Судиловская, 1951
Длина крыла Дина клюва Длина цевки Длина крыла 19 720-800 400-450 115-125 690-800 Казахстан 680-720 Долгушин, 1960 320-380 Дельта Дуная, Румыния. 744 16 670-780 725 Dombrowski, 1912 и Korodi-GJ,
Длина хвоста 4 230-250 240 1 230 1961, (1964). Цит. по: Cramp,
Длина клюва 19 390-450 420 16 350-430 392 1977. Материалы объединены по
Длина цевки 4 130-135 131 1 130 первоисточникам.
равна 686 и 670, а клюва — 397 и 370 мм. Вес взрослых птиц колеблется от 9 до 12 кг и
может достигать 13 кг (Тугаринов, 1947; Cramp, 1977).
Линька. Постювенилъная. Не изучена. Постепенная смена нарядов может указывать
на то, что у молодых птиц линька проходит растянуто, в течение всех первых лет жизни
(Cramp, 1977).
Предбрачная. Длинные перья на затылке развиваются перед сезоном размножения; дру-
гие детали линьки не известны.
Послебрачная. У взрослых птиц полная послебрачная линька происходит после окон-
чания периода размножения. Сроки её начала сильно растянуты в различных частях гнез-
дового ареала (Судиловская, 1951). Линька оперения тела и крыла регистрировалась и во
время сезона размножения, в мае-августе. Смена первостепенных маховых идёт от внут-
ренних перьев к наружным, при этом на крыле может быть до трёх активных центров. К
декабрю смена оперения заканчивается (Cramp, 1977).
Распространение. Гнездовой ареал. Спорадически гнездится в умеренном и,
реже, в субтропическом поясах. Участки гнездового ареала разбросаны от Восточной Ев-
ропы до Монголии. В прошлом был широко распространён в Европе; в настоящее время
европейские колонии расположены в Албании (лагуна Караваста), в Болгарии (оз. Сребар-
на), в Черногории (оз. Скадар), в Греции (оз. Преспа и залив Амвракикос), в Румынии (дельта
Дуная) и в Турции (западная Анатолия, вдоль Черноморского побережья) (Peja et al., 1996;
Crivelli, 1997), на островах оз. Актас-Гулю на турецко-грузинской границе (Yarar, 1995). В
Монголии гнездится в западной её части на оз. Айраг Нуур (Collar et al., 2001) (рис. 4).
На территории Восточной Европы и Северной Азии гнездится на Украине, в Дагестане,
Азербайджане, Казахстане, Узбекистане и на территории России (рис. 5). Колонии найде-
ны на придунайских озёрах Кугурлуй и Картал. С 2000 г. отмечено гнездование на террито-
рии Дунайского биосферного заповедника (Жмуд, 2000). В дельте Днестра гнездился до
середины XX в.; в настоящее время наблюдается здесь в летнее время, но попыток гнездо-
вания не зарегистрировано (Русев, 2004). В Вост. Приазовье известны гнездовья на Кир-
пильских (Kazakov et al., 1994) и Кизилташских лиманах (Иваненко и др., 2000а), найдены
на оз. Ханском (Иваненко и др., 1995,20006; Динкевич и др., 1999; Белик, Динкевич, 2004).
Гнездится на островах озёр Маныч-Гудило (Миноранский, 1962; Кривенко, Любаев, 1975;
Хохлов, Ильюх, 2002), Сага-Бирючья, Дадынское, на Чограйском вдхр. (Хохлов, Ильюх,
2002), на островах Пролетарского вдхр. (Кривенко, Любаев, 1975; Языкова, Казаков, 1975),
на оз. Деед-Хулсун (Стоцкая, Кривенко, 1988; Сохина и др., 2000). В Дагестане на гнездо-
Кудрявый пеликан
27
Рис. 4. Область распространения кудрявого пеликана: а — места гнездования, б — районы зимовки.
вье распространён в Кизлярском и Бабаюртовском районах, в низовьях р. Терек (Прилуц-
кая, Пишванов, 1989; Вилков, Пишванов, 2000). В Азербайджане гнездился (вероятно, с
перерывами в отдельные годы) на оз. Акгёль и Сарысу (Судиловская, 1951; Султанов, 2002).
Гнездится в дельте Волги, у южных границ Дамчикского участка Астраханского заповед-
ника (Луговой, 1961; Бондарев, Кривоносов, 1977; Кривоносов, Живогляд, 1989; Русанов и
др., 2000). В С ев.-Во ст. Прикаспии в Казахстане колонии найдены в пойме Урала — на оз.
Челкар, Кушумских озёрах, на водохранилище бассейна р. М. Узень (Стоцкая, Кривенко,
1988) и в дельте р. Урал (Русанов, 1995), в районе Забурунского залива и Карабулака (Гис-
цов, 2001). При обследовании Убаган-Ишимского междуречья и Северного Тургая коло-
нии найдены на оз. Шошкалы, оз. Каракамыс и оз. Кулыколь (Брагин, 2001). Нерегулярно
гнездится на оз. Тенгиз и Кургальджин (Азаров, 1975; Кривицкий и др., 1985). В Наурзум-
ском заповеднике гнездится совместно с розовыми пеликанами (Блинова, Блинов, 1997). В
Кустанайской области на оз. Сарыкопа эти два вида также образуют совместные колонии
(Брагин, 2004). Гнездится на озёрах Балхаш-Алакольской котловины: на оз. Алаколь, Са-
рыкопа (на оз. Тысячи) (Ауэзов, Грачев, 1977; Березовиков, Левинский, 2004) и на оз.
Балхаш в дельте Или и у протоки Ир (Судиловская, 1951; Долгушин, 1960; Абрамова и
др., 1975).
В Узбекистане колонии были обнаружены в дельте Аму-Дарьи: в окрестностях Ушарым
на юго-западном побережье Муйнакского зал., на озёрах Карахожабах, Шегекуль, в урочи-
ще Узынкаир и в устье Кокдарьи на юго-восточном побережье Аральского моря (Кенжегу-
лов, 1974). Гнездится также в дельте Сыр-Дарьи (Марков, 1965; Мекленбурцев и др., 1987).
С 1984 г. начал гнездиться на оз. Айдаркуль (Шерназаров, 1988). В Туркмении колонии
располагались на Сарыкамышском озере (Стоцкая, Кривенко, 1988). На север кудрявый
пеликан распространён до Тоболо-Ишимского междуречья, где колонии располагаются на
озёрах Чёрное, Тундровое, Б. Белое (Молчанова, 1972; Тарасов, Примак, 2005), и оз. Сал-
28
Отряд Пеликанообразные, Семейство Пеликановые
Рис. 5. Распространение кудрявого пеликана в Восточной Европе и Северной Азии: а — места гнез-
дования, б — места зимовки.
таим-Тенис в Омской обл. (Якименко, 1998, 2006). Отмечен факт гнездования в Кемеровс-
кой обл. (Белянкин, 1997), на востоке Оренбургской обл.
Зимовки. Кудрявый пеликан зимует в прибрежных районах Средиземноморья, юж-
ной части Каспийского моря, в Персидском заливе, в Пакистане и в северной Индии (Crivelli,
1997; Van der Have et al., 2002). В целом, районы зимнего пребывания вида разобщены с
гнездовыми областями в меньшей степени, чем у розовых пеликанов. Птицы из албанской
колонии либо остаются в гнездовых районах, либо мигрируют и распределяются вдоль
Адриатического побережья и по внутренним озёрам, встречаются на зимовках также в ла-
гуне Нарта. Птицы, гнездящиеся в Греции, держатся в дельтах Эвроса и Аксиоса, в заливе
Амвракикос и Месолонги, встречаются на озёрах Лангада и Кастория. Из болгарской коло-
нии птицы в зимнее время перемещаются южнее по черноморскому побережью и по внут-
ренним водоёмам. Места зимовок пеликанов из дельты Дуная, включая украинскую часть
дельты, не выяснены. Вероятнее всего, они частично остаются в южной части черномор-
ского побережья, а частично присоединяются к птицам, мигрирующим в Турцию, Ливан,
Иран, Ирак. Пеликаны из колоний Турции распределяются, главным образом, по прибреж-
ным водоёмам в западной и южной части страны (Cramp, 1977; Crivelli, 1997). Первые
птицы, прибывшие на зимовку в Синд, регистрируются с конца августа (Collar et al., 2001).
На зимовках встречаются в январе-феврале в прибрежных водно-болотных угодьях Пер-
сидского залива (Van der Have et al., 2002). На оз. Денгизкуль их регулярно отмечали как на
осеннем пролёте, так и на зимовке (129 птиц в 1987-1988 гг.) (Лановенко и др., 2001).
Птицы, гнездящиеся в Дагестане, в тёплые зимы остаются в устье р. Терек и в Аграханском
зал., где их численность достигает 800 особей. При резком похолодании мигрируют в юго-
восточном направлении, иногда надолго задерживаясь на оз. Аджи. В суровые зимы часть
Кудрявый пеликан
29
особей погибает (Прилуцкая, Пишванов, 1989; Вилков, Пишванов, 2000). По данным коль-
цевания, кудрявые пеликаны из колоний в дельте р. Волги также проводят зиму в западном
и южном Прикаспии (Кищинский, 1978ж). В этих же районах, вероятно, зимуют пеликаны,
гнездящиеся на Маныче и в его окрестностях. Птицы из Казахстана и колоний юга Запад-
ной Сибири могут зимовать как в южной части Каспийского побережья на территории Турк-
мении и Ирана, так и мигрировать южнее, в Пакистан и сев. Индию (Cramp, 1977; Crivelli,
1997; Collar et al., 2001). Так, погибшая птица, окольцованная в Казахстане, была найдена
на северо-западе Индии в Гуджарате (Аджимуратов, Пиржанова, 1986).
Миграции. Пути миграций совпадают с таковым розовых пеликанов. Как и другие не-
многочисленные виды, летит широким фронтом (Аджимуратов, Пиржанова, 1986).
В послегнездовой период, когда птенцы становятся на крыло, кудрявые пеликаны кочу-
ют стаями (Кенжегулов, 1974). Осенний отлёт наблюдается в Дагестане в середине-конце
ноября (Прилуцкая, Пишванов, 1989). На осеннем пролёте у Чикишляра и Гасан-Кули (Юго-
Восточный Каспий) интенсивный пролёта наблюдали в октябре и ноябре (Караваев, Бело-
усов, 1983). В дельте Аму-Дарьи осенний пролёт проходит в основном в сентябре, продол-
жается до второй половины октября и к концу месяца заканчивается (Кенжегулов, 1974).
Отлёт кудрявого пеликана в Кургальджинском заповеднике проходит в основном в первой
половине октября и заканчивается к концу этого месяца. Последняя встреча — 27 октября
(Кривицкий и др., 1985). Около Самарканда кудрявого пеликана добывали в сентябре и
ноябре (Мекленбурцев и др., 1987). На Айдарских разливах пролёт начинается в конце
августа и продолжается до конца октября. В дельте Аму-Дарьи наиболее интенсивный про-
лёт наблюдали в октябре. Последняя дата встречи — 23 октября (Гончаров, 1981). В Ала-
кольской котловине кудрявого пеликана встречали на пролёте в значительно меньшем чис-
ле, чем розового; наибольшее число встреч зарегистрировано в сентябре (Сема, Ауэзов,
1984). На оз.Чёрное на юге Зап. Сибири отлёт заканчивается уже к концу августа (Блинова,
Блинов, 1997).
Весной в южные районы прилетают в середине марта, примерно на 10 дней раньше
розового пеликана. В дельтах Дуная и Днестра отмечался с начала апреля (Панченко, Ба-
лацкий, 1991). В Дагестане на местах гнездовий появляется в конце февраля (Прилуцкая,
Пишванов, 1989). В это же время появляются в Закавказье вдоль Черноморского побере-
жья. В дельте Волги первые птицы регистрируются в середине марта (Судиловская, 1951).
В низовьях р. Аму-Дарьи пеликаны появляются во второй половине марта, а заканчивается
прилёт в первой декаде апреля. Около Самарканда кудрявого пеликана добывали в марте
(Кенжегулов, 1974; Ажимуратов, Пиржанова, 1986; Мекленбурцев и др., 1987). В низовьях
Сыр-Дарьи пролёт отмечен с середины марта (Долгушин, 1960). На озёра Тенгиз и Кур-
гальджин начинает появляться с первых чисел апреля. Средняя дата прилёта по годам при-
ходится на 15 апреля (Кривицкий и др., 1985). Через Кураминскую долину миграция прохо-
дит с февраля до апреля. В Синде последние задержавшиеся на зимовках птицы регистри-
руются до апреля (Collar et al., 2001).
Случаев дальных залётов немного. Отмечались залёты кудрявого пеликана в Татарстан
(в XIX в. наблюдались даже случаи гнездования в устье р. Камы) (Сотников, 1999), в низо-
вья р. Белой (Маматов, 1998), на юго-восток Свердловской обл. (Карякин и др., 1999), в
Прибайкалье (Гагина, 1962; Жаров, Митейко, 1967; Мельников, 2000). Отмечались залёты
под Павлодаром, Омском, Барабинском (Долгушин, 1960).
Местообитания. Населяет материковые водоёмы: реки, пресные озёра, болота, соло-
новатые прибрежные лагуны внутренних морей, дельты крупных рек. Встречается на вы-
сотах до 850 м над уровнем моря (Cramp, 1977; Crivelli, Michev, 1997; Рогачева, Сыроеч-
ковский, 2003). Кудрявый пеликан по большей части гнездится на глубоких, окружённых
30
Отряд Пеликанообразные, Семейство Пеликановые
труднопроходимыми тростниковыми крепями озёрах, часто, на плавучих островках из от-
мершего тростника (Долгушин, 1960; Марков, 1965; Русанов, 1995), но может устраивать
гнёзда и на низких песчано-илистых или каменистых островах (Судиловскач, 1951; Мино-
ранский, 1962; Долгушин, 1960; Жмуд, 2000).
Численность. Уязвимый вид на всём протяжении своего ареала. Мировая численность
гнездящихся особей оценивается в 3 215-4 280 пар, более 80-84% их гнездится на терри-
тории бывшего СССР. В Греции обитает 6-8% мировой популяции. Общее количество птиц,
гнездящихся в Европе, в начале 1990-х гг. оценивалось в 886-1 204 пар (около 30% миро-
вой численности): в Албании 40-70 пар; в Болгарии — 70-80 пар; в Югославии — 10-20
пар; в Греции — 190-260 пар; в Румынии — 70-150 пар; на Украине — 6-14 пар; в России —
400-500 пар; в Турции — 100-150 пар (Crivelli, 1997).
По данным Всесоюзных учётов колониальных околоводных птиц на территории быв-
шего СССР в 1973-1977 гг. численность кудрявых пеликанов составляла 1,5-2 тыс. гнездя-
щихся пар, а в 1986-1987 гг. — 2,2 тыс. (Флинт и др., 1991). По данным В.В. Виноградова
(1998), современная численность только центрально-казахстанской популяции кудрявого
пеликана составляет 3 750 пар.
С 2000 г. отмечается постоянный рост численности вида в Причерноморском регионе.
В 2002 г. на территории всей Дунайской дельты в трёх поселениях учтено 419 пар кудрявых
пеликанов (Жмуд, 2004), с 2001 г. вид единично отмечается в дельте Днестра (Русев, 2004).
По материалам учётов колониальных птиц в 1986-1989 гг. на территории Краснодарского
края отмечено 3 колонии кудрявого пеликана общей численностью около 74 гнездящихся пар
(Емтыль и др., 1990). На настоящий момент здесь также насчитывается около 70-80 гнездя-
щихся пар в составе 3-5 колоний. Наиболее значимое поселение — 55-60 пар — зарегистри-
ровано на оз. Ханском (Динкевич, 2006). В Ставропольском крае в разные годы гнездится
до 40 пар, летует — от 80 до 200 особей (Хохлов, Ильюх, 2002). В Дагестане в двух колони-
ях в Аграханском заливе гнездилось около 55 пар (Прилуцкая, Пишванов, 1989), по более
поздним данным, в результате подъема уровня Каспия и регенерации Кизлярского и Агра-
ханского залива наблюдается некоторый рост численности вида. Сейчас здесь насчитыва-
ют 450-500 особей (Вилков, Пишванов, 2000). В 1974-1991 гг. в дельте Волги гнездилось
от 25 до 242 пар кудрявых пеликанов (Русанов, 1997). Численность колонии в Забурунье в
дельте Волги колеблется от 70 до 100 пар (Гисцов, 2002). В 1988 г. в дельте Урала была
обнаружена колония из 20 гнезд. В 1986 и 1987 гг. здесь наблюдали молодых птиц и группу
из 220 особей (Русанов 1995). На водоёмах Уральской обл. (оз. Челкар, Кушумские озёра и
водохранилище, бассейн р. М. Узень) гнездилось 160-200 пар (Стоцкая, Кривенко, 1988).
На юге Зап. Сибири на оз. Чёрном гнездится 180 пар, а на оз. Б. Белое — 90 пар (Тарасов,
Примак, 2005). В Кургальджинском заповеднике с 1975 по 1981 г. численность гнездящих-
ся пар возросла с 10 до 76 (Кривицкий и др., 1985), в 1987 г. здесь уже гнездилось 315 пар
(Стоцкая, Кривенко, 1988). В 2002 г. точное число гнёзд установить не удалось, но 25 июня
на оз. Есей насчитали 650 особей, а на оз. Шишалкар 23 июня — 146 особей (Кошкин,
2004). В период учётов, проведённых 1-10 июня 2000 г. на озёрах Убаган-Ишимского меж-
дуречья и Сев. Тургая, в колонии на оз. Каракамыс было найдено 20 пар кудрявых пелика-
нов, на оз. Шошкалы — 30^40 пар, оз. Кулыколь — 20 пар (Брагин, 2001). В Наурзумском
заповеднике на оз. Сары-Моин гнездилось 9 пар (Стоцкая, Кривенко, 1988). В 2004 г. на
Наурзумских озёрах на летовке было отмечено около 400 особей — по большей части,
неполовозрелых птиц (Брагин, 2005). На озёрах Сарыкопинской системы гнездилось 120—
160 пар (Брагин, 2004). На оз. Балхаш в 1968-1972 гг. численность кудрявого пеликана
составляла 150-Л00 гнездящихся пар (Грачев, 1977а,б). По учётам 1987 г. на островах в 3
колониях здесь гнездилось около 80 пар. В дельте р. Или в 1986 г. в 5 колониях поселилось
тэявый пеликан 31
5 ?*>-650 пар, а в 1987 г. обнаружено 4 колонии общей численностью 760-870 пар (Стоцкая,
’<гивенко, 1988). На озёрах Алакольской котловины, кроме совместного гнездования с розо-
ым пеликаном, селился и самостоятельно. В 1966 г. здесь существовало два поселения — на
: з. Пеликанья Курья в дельте р. Тентек (4 гнездящихся пары) и на оз. Закрытое (2 колонии
<з 16 пар) (Ауэзов, Грачев, 1977). В 1987 г. на этих озёрах гнездилось 250 пар (Стоцкая,
Гривенко, 1988), а в 2002 г. найдено всего 74 гнезда с кладками (Березовиков, 2002). На оз.
Алаколь в 1986 г. гнездилось 8 пар, на оз. Зайсан в дельте Чёрного Иртыша было обнару-
жено несколько больших колоний суммарной численностью около 250 пар (Стоцкая, Кри-
зенко, 1988). В Туркмении на Сарыкамышском озере гнездилось 287 пар. При авиаучё-
тдх на Аму-Дарье в пределах Чарджоузской обл. учтено 116 летующих особей (Стоцкая,
Кривенко, 1988). 13 пар гнездилось на оз. Айдаркуль (Шерназаров, 1988). В мае 2003 г.
тыла обнаружена колония на Сорбулаке порядка 50 пар (Ковшарь, 2004). В общей слож-
ности в районе Сорбулакской системы озёр насчитывали 309 птиц (Ашби, Анненкова,
1005).
В.В. Виноградов (Vinogradov, 1998) прогнозирует снижение общей численности этого
ззиа в Казахстане к 2010 г. до 300^400 пар в результате наступления новой, более теплой и
згсушливой климатической фазы. В этот период многие водоёмы на указанной террито-
рии. по его мнению, обмелеют, превратятся в гиперсолёные или полностью высохнут, что
приведёт к катастрофическому сокращению кормовой базы и деградации местообитаний,
пригодных для гнездования как кудрявого, так и розового пеликанов.
Размножение. Половой зрелости достигает в возрасте 3 лет. Гнездится колониями от 3-
. 0 до нескольких сотен пар, как изолированно, так и совместно с другими видами птиц,
наиболее часто с розовым пеликаном, большим бакланом и чайками-хохотуньями (Суди-
ловская, 1951; Петров, Миноранский, 1962; Молчанова, 1972; Кенжегулов, 1974; Шерназа-
гов. 1988; Русанов, 1995; Жмуд, 2000; Березовиков, 2002; Тарасов, Примак, 2005). Иногда
•удрявые пеликаны образуют смешанные колонии с голенастыми: серой и белой цаплями,
нваквами и колпицами. Известен случай мирного соседства с лебедем-шипуном в дельте
Волги — гнёзда располагались на расстоянии 0,5 м друг от друга (Кривоносов, Бондарев,
_978; Панченко, Балацкий, 1991). В смешанных колониях гнёзда кудрявых пеликанов рас-
положены обычно несколько обособленно от других видов птиц. Изредка селятся отдель-
ными парами. Внутри крупных колоний выделяются отдельные поселения, которые могут
. остоять из 5-20 пар. Гнёзда в группах иногда располагаются очень плотно, почти соприка-
:зются, особенно при гнездовании на островках. Такие группы гнёзд иногда представляют
:обой единые комплексные сооружения (Миноранский, 1962; Cramp, 1977; Кривицкий и
пр.. 1985; Тарасов, Примак, 2005).
Моногам. Пары сохраняются, как минимум, в течение одного сезона. В момент форми-
рования пары самец производит перед самкой серию брачных демонстраций: встает сбоку
: т неё, приопускает карпальные суставы при сложенных крыльях, поднимая первостепен-
ные вверх, широко раскрывает хвост и отвешивает многократные поклоны, после чего энер-
~лчно трепещет крыльями и перьями хвоста; одновременно громко захлопывает клюв и
держит его плотно закрытым, горловой мешок при этом оказывается сильно раздут, и са-
чец начинает порциями выпускать из него воздух, производя ритмичные шипящие звуки.
Гзмка в это время стоит тихо рядом. Позднее такие демонстрации могут повторяться и на
ездовом участке, который выбирает самка. Небольшая территория вокруг пары в период
• хдживания охраняется, чаще самцом. Позднее охраняемая территория ограничивается
чзстком непосредственно вокруг гнезда (Cramp, 1977). Копуляция может происходить до
' раз в день, иногда, с интервалом 10-15 мин, в том числе ночью. В период спаривания
1 дмка может подолгу сидеть на месте гнезда. Самец стоит рядом, периодически инициируя
32
Отряд Пеликанообразные, Семейство Пеликановые
копулятивный процесс, демонстрируя при этом специфическое поведение: ходит вокруг
самки, раскрыв крылья, приближаясь и отходя, периодически бросается в воду, отплывает
на небольшое расстояние и снова возвращается. Может при этом пытаться схватать самку
за перья на затылке. Самка первоначально отстраняется от него; самец предпринимает по-
вторные попытки, теребя самку клювом за перья затылка, шеи и спины, затем внезапно
крепко хватает её клювом и взбирается на нее. Копуляция происходит быстро, пока самец
удерживается наверху, энергично хлопая крыльями. Самка в этот момент вытягивает клюв
вверх и вперёд и наклоняет его набок, слегка приподнимая и опуская при этом крылья,
либо просто держит их приоткрытыми. Самец соскальзывает с самки в сторону, продолжая
хлопать крыльями.
Самка остаётся на гнездовом участке до откладки яиц, занимаясь строительством гнез-
да. Самец приносит ей гнездовой материал (25^40 раз за день), поднося его в клюве или
набивая им горловой мешок. Между самцами часто происходят драки из-за строительного
материала. Гнездо строится 3^4, иногда 5 дней (Судиловская, 1951; Cramp, 1977).
При гнездовании в тростниковых зарослях, пеликаны подминают под себя тростник и
на нём строят гнездо (Долгушин, 1960; Марков, 1965; Русанов, 1995). В таких местах раз-
мещение колоний кудрявых пеликанов тяготеет к границе распространения надводной ра-
стительности (Судиловская, 1951; Долгушин, 1960; Луговой, 1961; Жмуд, 2004). Подоб-
ный выбор места гарантирует относительную безопасность от наземных хищников. Окру-
жающие такую колонию участки воды обычно не зарастают плавающими водными расте-
ниями (чилимом, нимфейником и пр.), что играет существенную роль для обеспечения
безопасности подрастающего потомства, т.к. единственным способом укрыться от опасно-
сти для нелетных птенцов служит возможность быстро сойти с гнезда и уплыть, что труд-
ноосуществимо в условиях зарастающего водоёма. Заросли надводной растительности
имеют значение для пеликанов, в основном, как база и материал для постройки гнезд. На
оз. Кургальджин, где птицы заселялют берега проток в тростниковых массивах, гнёзда рас-
полагаются на вросших в дно тростниковых сплавинах (Кривицкий и др., 1985). Пеликаны
могут гнездиться и на открытых островах, почти или полностью лишенных растительнос-
ти — как, например, на оз. Маныч-Гудило (Миноранский, 1962) и Тенгиз (Кривицкий и др.,
1985). В Астраханском заповеднике, к примеру, в 1957 г. кудрявые пеликаны гнездились на
остове старого затонувшего рыбацкого судна (Луговой, 1961). На юго-восточном побере-
жье Аральского моря гнёзда располагались на берегах заливов и больших тихих озёр (Кен-
жегулов, 1974).
Гнёзда цилиндрической формы, чаще располагаются плотными группами. На оз.Чер-
ное смешанная колония, состоявшая из 29 гнёзд кудрявого пеликана и 35 гнёзд большого
баклана, находилась на сплавине площадью 42 м2 (Молчанова, 1972). Гнездовая плотность
на оз. Микра-Преспа варьировала от 0,12 до 1,2 гн./м2, в смешанных колониях (с розовым
пеликаном) — от 0,34 до 1,13 гн./м2 (Catsadorakis, Crivelli, 2001).
Одиночные гнёзда встречаются редко, преимущественно в тех случаях, когда распо-
лагаются среди густых зарослей тростника (Долгушин, 1960; Панченко, Балацкий, 1991).
Обычно для строительства используется тростник, рогоз, трава, сучья, камни, иногда палки
длиной до 1 м и диаметром 5-7 см; встречаются перья линяющих птиц и даже кости
павших животных. В основании гнёзд лежат толстые стебли и ветви, которые к вершине
гнёзда сменяются все более тонкими, лотки выстилаются тонкими стеблями и сухими
водорослями (Судиловская, 1951; Миноранский, 1962; Кривицкий и др., 1985); на оз.
Чёрное в качестве выстилки служил и мох (Молчанова, 1972). Средние размеры гнёзд (в
см) на оз. Маныч-Гудило (п=9): наружный диаметр — 68-88, диаметр лотка — 25^41,
высота — 18^42, глубина лотка — 6-11 (Миноранский, 1962). В Кургальджинском запо-
Кудрявый пеликан
33
веднике (п=15): наружный диаметр 62-95 (в среднем 72,4), диаметр лотка — 37-50 (в
среднем 42,6), высота 10-66 (в среднем 35,8), глубина лотка— 8-11 (в среднем 10) (Кри-
вицкий и др., 1985). На оз. Чёрное на юге Зап. Сибири (п=34): наружный диаметр — 57-
77 (в среднем 67), диаметр лотка — 31^43 (в среднем 36), глубина лотка 5-14 (в среднем
9); над уровнем сплавины гнёзда поднимались на 50-90 см (Молчанова, 1972). На юго-
восточном побережье Аральского моря (п=10): наружный диаметр 55-94 (в среднем 63,2),
внутренний диаметр 37-53 (в среднем 42,9), высота гнезда 16-27 (в среднем 21,5), глуби-
на лотка 8-15 (в среднем 12,1), высота гнезда от уровня воды 46-87 (Кенжегулов, 1974).
По мере подрастания птенцов гнездо заметно оседает и к завершению гнездового перио-
да обычно не превышает в высоту 50 см, иногда опускаясь до уровня воды (Судиловская,
1951).
Откладка яиц в колонии, особенно крупной, чаще происходит не одновременно, поэто-
му общие сроки гнездования порой оказываются сильно растянутыми. Свежие и слабо на-
сиженные яйца в отдельных гнёздах можно найти до середины июля, в то время как в дру-
гих гнёздах колонии могут быть уже вылупившиеся и даже оперившиеся птенцы (Судилов-
ская, 1951; Долгушин, 1960). 10 июля на оз. Черное наблюдали птенцов размером со взрос-
лую птицу, при этом в отдельных гнёздах колонии были обнаружены сильно насиженные
кладки (Тарасов, Примак, 2005).
На Маныче к откладке яиц приступают в конце апреля - начале мая, но в одной из
колоний свежие яйца находили 4 июня (Миноранский, 1962). В Кургальджинском заповед-
нике в дельте р.Тентек с 5 по 13 апреля уже были полные кладки (Березовиков, 2002). Пер-
вые кладки в дельте Волги и на Сыр-Дарье появляются в 3-й декаде апреля (Долгушин,
1960). В дельте Аму-Дарьи дата откладки первого яйца приходилась на 10 апреля (Кенже-
гулов, 1966).
Яйца белые, неравномерно покрытые известковым слоем, придающим скорлупе шеро-
ховатую поверхность. В процессе насиживания скорлупа, загрязняясь, жёлтеет или буреет
(Рябицев, 2001). Размеры яиц в мм: в дельте Дуная (п=3) - 91-92 х 60,4-62,4 (в среднем
91,1x61,5) (Панченко, Балацкий, 1991); в дельте Волги (величина выборки не указана) —
86-102 х 53-65 (Судиловская, 1951); на оз. Чёрном (п=21) — 85-99 х 45-67 (в среднем
93x56) (Молчанова, 1972); на оз. Айдаркуль (п=22) средние значения — 91,2x58,3 (Шерна-
заров, 1988); на юго-вост, побережье Аральского моря (п=9) — 87-98 х 59-61 (в среднем
92,9x59,6) (Кенжегулов, 1974); по данным из разных колоний — 78-106 х 53-64 (п=150), в
среднем 95x60 (Cramp, 1977). Размеры яиц в одной кладке сильно варьируют; так, в одном
из гнёзд размеры яиц были — 98x63 и 89x56, во втором — 95x50 и 85x45 (Молчанова,
1972). Вес яиц в дельте Волги — 143-195 г (Судиловская, 1951); на юго-восточном побере-
жье Аральского моря — от 147 до 184,7 г (п=9) (Кенжегулов, 1974).
Величина кладки неодинакова в разных местах (возможно, в зависимости от даты нача-
ла откладки) и колеблется от 1 до 6 яиц. В дельте р.Тентек в одиннадцати гнёздах было по
1 яйцу, в одном — 4, ещё в одном — 5 (Березовиков, 2002). На оз. Шегекуль 13 апреля из 21
гнёзда в четырех было по 1 яйцу, в шестнадцати — по 2 и в одном — 3. В дельте Амударьи
в урочище Узынкаир 7 мая в двух гнёздах было по 6 и в пяти — по 3 оперившихся птенца,
ещё в семи гнёздах было 10 голых зрячих птенцов (Кенжегулов, 1974). В дельте Или, на
озёрах Алакульской группы и на Балхаше кладка содержала чаще всего 2 яйца, в дельте
Волги — 2-3, на Сыр-Дарье обычно 3^4, реже 5 и, как исключение, 6 яиц (Долгушин,
1960). В Кургальджинском заповеднике в кладках было 1 (6 гнёзд) и 2 (5 гнёзд) яйца, в
среднем 1,5 яйца (Кривицкий и др., 1985).
Насиживание начинается сразу после откладки первого яйца, в результате чего вылуп-
ление происходит не .одновременно и птенцы в гнезде разновозрастные. Инкубация про-
34
Отряд Пеликанообразные, Семейство Пеликановые
должается от 30-32 (Cramp, 1977) до 39^40 дней (Судиловская, 1951); в дельте Аму-Да-
рьи — 33-34 дня (Кенжегулов, 1974). Насиживает преимущественно самка, самец сменяет
её только утром и вечером на время кормёжки. На гнезде птицы сидят очень плотно, потре-
воженные уходят с гнезда вплавь и так же возвращаются к нему. Если гнездо расположено
в зарослях тростника, найти его в период насиживания крайне трудно. С появлением птен-
цов сделать это значительно проще, ориентируясь на их голос (Судиловская, 1951).
Вылупившиеся птенцы совершенно голые, глаза и слуховые отверстия закрыты. Пер-
вые пушинки, равномерно покрывающие тело птенца, появляются на 5-й день. В 10-днев-
ном возрасте птенцы покрыты густым грязновато-белым пухом и уже способны плавать. В
возрасте 20 дней появляются пеньки маховых перьев, кроющих крыла и перья на нижней
стороне тела. На спине остатки пуха сохраняются дольше всего — до 2 месяцев.
По наблюдениям за развитием птенцов в дельте Волги, вес однодневного птенца— 110 г, на
2-й день — 202 г, на 6-й день — 480 г, на 12-й — 1 950 г, на 20-й — 3 500 г, в месячном
возрасте — 6 930 г и в возрасте 2 месяцев — 9 200 г. Таким образом, птенцы набирают вес
взрослых птиц раньше, чем заканчивается полное формирование перьевого покрова. В
возрасте двух с половиной недель птенцы собираются вместе, образуя «ясли» — плотные
птенцовые стаи, которые держатся обычно в центре колонии. В возрасте 4-5 недель стай-
ками перемещаются на воду поблизости от колонии (Cramp, 1977).
Сначала родители выкармливают потомство полупереваренной рыбой, для чего глубо-
ко захватывают в рот голову птенцов и отрыгивают содержимое желудка. Воду птенцам
также приносят в клюве. По мере роста птенцов масса и размеры приносимой добычи
увеличиваются; позднее молодые могут проглотить целых рыб весом 800-1 200 г, прине-
сённых родителями. Родители ловят рыбу в это время рядом с колонией и не совершают
дальних кормовых перелетов. Подлетая с кормом, всегда садятся на воду и подходят к гнез-
ду пешком, даже если оно расположено на значительном расстоянии от воды. При этом в
зарослях постепенно пробивается широкая тропа. Молодые встречают родителей глухим
ревом, толкают друг друга и соперничают из-за пищи.
Летными молодые пеликаны становятся примерно в 2,5-месячном возрасте, после чего
они сбиваются в стаи, покидают гнездовые колонии и до отлёта кочуют в поисках пищи по
более крупным, глубоким озёрам и морским побережьям.
Гибель потомства наблюдается чаще на стадии яиц. Успех вылупления варьирует от 35
до 70%. В колониях, расположенных на охраняемых территориях, пара кудрявых пелика-
нов довольно легко выращивает по два птенца. Успешность размножения может достигать
1,35 (Crivelli, 1996). Репродуктивный успех кудрявых пеликанов, гнездящихся в морских
лагунах, ниже, чем на пресноводных водоёмах, и составляют от 0 до 0,85 птенца на 1 гнез-
до (Peja et al., 1996). Однако на оз. Сребрана в Болгарии успешность размножения до 1993
г. не превышала 0,84 (Crivelli, 1996). Средний размер выводка (в возрасте 4 недель) на оз.
Маныч-Гудило не превышал 0,8 (Crivelli, Vizi, 1981). Довольно низкую успешность гнез-
дования наблюдали в Узбекистане на оз.Айдаркуль, где из 22 отложенных яиц вывелось
всего 3 птенца (13,6%); причиной гибели кладок послужило беспокойство птиц в период
насиживания (Шерназаров, 1988). Подсчитано, что при успешности размножения, равной
по меньшей мере 0,8, популяция кудрявого пеликана может находиться в стабильном со-
стоянии, а при успешности размножения свыше 1 возможен рост численности (Crivelli,
1997).
Суточная активность, поведение. Подобно розовым пеликанам, кудрявые пеликаны
держатся группами разной величины на протяжении всех фаз годового цикла. Семейные
группы, как правило, сохраняются и после подъёма молодых на крыло. Кормятся птицы
как в одиночку, так и группами. На зимовках держатся преимущественно относительно
??.дрявый пеликан
35
Рис. 6. Позы кудрявого пеликана: а — угроза, б — демонстрация поднятого клюва (по: Cramp, 1977).
крупными стаями. Для отдыха используют открытые острова и косы, сплавины, отдельные
крупные коряги, обеспечивающие хороший обзор и защиту от наземных хищников. В по-
лете нередко объединяются в стаи с другими видами птиц (иногда с большими бакланами).
Негнездящиеся взрослые птицы часто проводят лето в колонии. Неполовозрелые особи
держатся, в основном, в местах кормёжек, периодически посещают колонию и могут нахо-
диться там до нескольких дней, но взрослые гнездящиеся птицы рано или поздно их прого-
няют (Cramp, 1977).
Демонстрация угрозы по отношению к представителям своего и других видов птиц пред-
ставляет собой покачивание открытым клювом (рис. 6а). Поза с поднятым клювом (рис.
66) может быть адресована партнеру, птенцам и соседям и либо тоже относится к агрессив-
ным демонстрациям, либо служит для опознавания птицы (Cramp, 1977).
Ночуют стаями, иногда на деревьях вместе с бакланами. В пределах колонии обе взрос-
лые птицы отдыхают у своего гнезда и днем и ночью, пока не перестанут охранять моло-
дых. Когда молодые достигают возраста 4-5 недель, предпочитают отдыхать на берегу око-
ло колонии или на воде, перемещаясь полностью на воду к тому времени, когда птенцы
достигают возраста 7-8 недель. Молодые отдыхают в колонии, первоначально собираясь в
кучу, а затем распределяются поодиночке. Подросшие молодые наведываются в колонию
для дневного отдыха и на ночёвку, большую часть остального времени проводя на воде
-Cramp, 1977).
Питание. Ихтиофаг. Основу рациона составляет рыба средних размеров и молодь. Мо-
жет охотиться как в одиночку, медленно плавая и схватывая клювом рыбу, всплывающую к
ловерхности, так и небольшими группами. В отличие от розовых пеликанов, могут ловить
рыбу и на глубокой воде (Судиловская, 1951; Collar et al., 2001). Добывает пищу часто со-
вместно с большими бакланами и розовыми пеликанами, изредка — с большими поганка-
ми (Кенжегулов, 1974). После подъёма молодых на крыло для групповых охот часто объе-
диняются птицы из одного или нескольких выводков. Направляясь на охоту, птицы уже в
лолете выстраиваются в линию или углом. Опустившись на воду, они медленно продвига-
ются плотной шеренгой в сторону мелководья, хлопают крыльями по воде и попутно схва-
тывают замеченную рыбу, часто погружая в воду голову и шею. Нередко к ним присоединя-
ются розовые пеликаны, большие бакланы, чайки. Стая поднимает большой шум, заставля-
ющий рыбу двигаться в нужном направлении. Часть рыбы птицы поедают на месте, при
необходимости значительное количество добычи могут уносить в горловом мешке. На оз.
36
Отряд Пеликанообразные, Семейство Пеликановые
Чёрное пеликаны заплывали разреженной стаей в центр озёра, затем выстраивались в ше-
ренгу и, не спеша, подгоняли рыбу к берегу, где и вылавливали её (Тарасов, Примак, 2005).
Время охоты приурочено, как правило, к утренним часам или к концу дня. Кудрявые
пеликаны обычно не совершают длительных перелётов между местами кормёжки и отды-
ха. В дельте Дуная это расстояние составляет, как правило, 1-2 км. Кормовые перёлеты
гнездящихся птиц могут быть значительно протяжённее — до 10 км (Collar, 2001; Жмуд,
2004). В дельте Амударьи расстояние, которое пролётали гнездящиеся птицы от колонии
до кормовых участков, составляло 20-30 км (Марков, 1965).
При отсутствии корма птицы легко могут голодать в течение 3—4 дней. Но более про-
должительные голодовки — до 2 недель — сильно истощают птиц. По наблюдениям А.Т.
Ромашевой, за восемь месяцев пребывания в дельте Волги семья пеликанов из двух взрос-
лых и двух молодых птиц съедает около 1 080 кг рыбы (Судиловская, 1951). Среднесу-
точный рацион одной птицы при этом составляет около 1 100 г. Размер добычи колеблет-
ся от 30 до 1 200 г по весу и от 5 до 42 см в длину (Crivelli, Vizi, 1981), хотя регистриро-
вались случаи добычи щуки длиной 50 см (del Hoyo et al., 1992, цит. по: Collar et al.,
2001). Средний вес содержимого 12 обследованных желудков кудрявых пеликанов соста-
вил 1 289 г. В состав пищи в дельте Дуная входило 10 видов рыб; наиболее часто встреча-
емые — сазан (Cyprinus carpio) — 65,2%, окунь (Perea fluviatilis) — 6,3% и линь (Tinea
tinea) — 6,3% (Korodi, 1961). При анализе 14 желудков пеликанов, добытых в дельте
Аму-Дарьи, обнаружено, что 31,4% от общего числа встреч в них также составляет сазан,
6,2% — бычки (Gobius sp.). Кроме того, около гнёзд пеликана находили щук (Esox lucius),
усачей (Barbus barbus), чехонь (Pelecus cultratus), судаков (Sander lucioperca) длиной до
34 см, а сазанов — до 37 и 39 см (Кенжегулов, 1974). Основу питания кудрявого пеликана
в Средиземноморье составляет, главным образом, угорь (Anguilla anguilla), кефаль (Mugil
sp.), бычки (Gobius sp.) и атерина (Atherina sp.). Поскольку эти виды рыб имеют коммер-
ческое значение, кудрявого пеликана здесь воспринимают как конкурента человеку (Peja
et al., 1996).
Враги, неблагоприятные факторы. Те же, что и у розового пеликана: нарушение гид-
рологического режима и деградация мест гнездования и зимовок; загрязнение среды тяже-
лыми металлами и пестицидами; беспокойство на местах гнездования и разрушение коло-
ний рыбаками; отстрел; гибель на линиях электропередач.
Существование кудрявого пеликана в морских лагунах находится под угрозой в связи с
усилением антропогенного воздействия и развитием прибрежного туризма (Peja et al., 1996).
Хозяйственное значение, охрана. Так же, как и розовый пеликан, в прошлом был объек-
том промысла, но первоначально в меньшей степени — в силу своей малочисленности, а
также благодаря тому, что места расположения колоний кудрявого пеликана чаще приуро-
чены к более труднодопроходимым и глубоким участкам водоёмов. После резкого сокра-
щения численности розового пеликана чаще стал объектом промысла (Марков, 1965). Кро-
ме того, колонии разоряли рыбаки, считавшие его своим конкурентом.
В настоящее время вид находится под охраной, он занесён в Красные книги Российс-
кой Федерации, Украины, Армении, Казахстана, Узбекистана, Туркменистана и ряда дру-
гих стран СНГ. Внесен в Приложение 1 Конвенции СИТЕС, Приложение 2 Боннской Кон-
венции, Приложение 2 Бернской Конвенции. Места гнездования охраняются в Астраханс-
ком, Наурзумском, Кургальджинском и Арнасайском заповедниках; в заказниках Маныч-
Гудило, Тургайском, Чограйском и Светлинском. На юге Западной Сибири озёра Б. Белое и
Тундровое, где гнездятся пеликаны, находятся на территории заказников. Озеро Чёрное в
Тюменской части своей акватории объявлено памятником природы, однако реальной госу-
дарственной охраны оно в целом не имеет и используется для активного промысла рыбы и
Семейство Олушевые
37
в качестве охотничьего угодья. Все попытки организации здесь особо охраняемой природ-
ной территории, что совершенно необходимо для сохранения кудрявого пеликана, сталки-
ваются с серьёзным противодействием (Тарасов, Примак, 2005),
Для поддержания численности вида необходимо сохранить наиболее ценные водоёмы,
расположенные в пределах засушливых территорий, Так, колонию пеликанов на Кургальд-
жинском озере можно сохранить, повысив уровень воды за счет создания новых каналов
(Vinogradov, 1998). На оз. Маныч-Гудило следует восстановить небольшие пологие остро-
ва. В местах появления новых колоний пеликанов должны быть созданы особо охраняемые
природные территории. Необходимо также отказаться от отлова молодых птиц для зоопар-
ков и различных выставок.
Семейство Олушевые — Sulidae Reichenbach, 1849
Крупные морские птицы (длина 64-100 см, масса 1-3,5 кг, размах крыльев до 180 см) с
удлинённым коническим клювом, без крючка на конце. По краям как надклювья, так и
подклювья имеются мелкие зазубрины, позволяющие удерживать добычу. Анатомически
хорошо приспособлены к основному способу охоты — пикированию в воду с высоты 10-
30 м с погружением до 5 м. Наружные отверстия ноздрей полностью зарастают, что вызва-
но требованиями защиты от воды во время ныряния. Для дыхания с закрытым клювом
олуши, очевидно, приоткрывают дополнительный парный подвижный щиток рамфотеки,
расположенный у основания надклювья. Шея длинная, но относительно толстая, шейных
позвонков 16-18. Тело обтекаемое, грудина и таз удлинённые. Скелет сильно пневматичен,
хорошо развиты подкожные воздухоносные полости. Оперение плотное, густое, покрыто
водоотталкивающей смазкой (копчиковая железа хорошо развита, имеет 5 выводных про-
токов). Крылья длинные, довольно узкие, заострённые, первостепенных маховых 10; у не-
которых видов есть редуцированное первое первостепенное маховое. Хвост также длин-
ный, клиновидный, из 12-18 рулевых перьев. Машущий полёт очень быстрый (60-90 км/
ч), маневренный; хорошо планируют, могут долго находиться в воздухе. При пикировании
возле самой поверхности воды шея делает упреждающее движение вперёд и голова с ост-
рым клювом почти без сопротивления входит в воду, крылья же сильно отводятся назад.
Это помогает избежать чувствительного удара о водную поверхность и позволяет не слиш-
ком терять скорость при переходе из одной среды в другую. В воде олуша подгребает полу-
сложенными крыльями, устремляясь дальше вниз, поскольку инерции броска недостаточ-
но для глубокого погружения и вода стремится вытолкнуть тело с очень малым удельным
весом. По некоторым сведениям птицы попадались в рыболовецкие сети, поставленные на
глубине до 70 м. Олуши приспособлены к пикирующему полёту лучше бурого пеликана,
бакланов, фаэтонов, чайковых. Обычно охотятся стаями, высматривая косяки рыб при по-
мощи острого зрения. Глаза расположены на голове так, что область бинокулярного зрения
смещена книзу от оси клюва. Это явно охотничье приспособление, на суше же из-за этого
олушам приходится часто задирать голову, чтобы посмотреть вперёд. Плавательная пере-
понка у олуш очень велика, они хорошо перемещаются по поверхности воды, мощно заг-
ребая лапами. Питаются рыбой, головоногими моллюсками, значительную долю в рацио-
не тропических видов занимают летучие рыбы (Карташев, 1974; Коблик, 2001)
Наиболее часто встречающийся тип окраски — белый с тёмными концами (а иногда и
задним краем) крыльев, тёмным бывает также хвост. Верх тела и крыльев ряда видов —
бх-роватый или серый, часто с чешуйчатым рисунком. У тропических олуш нередко быва-
ют светлая, тёмная и.промежуточная фазы окраски. Клюв, ноги, неоперённые участки «лица»
38
Отряд Пеликанообразные. Семейство Олушевые
нередко бывают ярко окрашены и выполняют функцию видоспецифичных маркеров. Мо-
лодые птицы имеют тёмное оперение, испещрённое светлыми пятнами и пестринами; с
каждой линькой наряд приближается к наряду взрослых птиц, неоперённые части тела на-
чинают приобретать яркую окраску. Половой диморфизм в окраске и размерах практичес-
ки не выражен.
Гнездятся колониями в сотни и тысячи пар на морских побережьях, преимущественно
на островах. Спариванию предшествуют брачные игры с ритуальными позами, телодвиже-
ниями, криками дуэтом. Иногда исполняется нечто вроде парного танца с синхронными и
асинхронными па. Голоса у олуш грубые, громкие, каркающие (несмотря на то, что не
развита голосовая мускулатура нижней гортани). Большинство видов строит гнёзда — не-
брежные кучки из веток, водорослей — на ровной земле или уступах скал, некоторые тро-
пические виды предпочитают гнездиться на деревьях, что, очевидно, вызвано чрезмерным
нагревом грунта и приземного слоя воздуха. В кладке 1, реже 2-3 голубоватых или зелено-
ватых яйца. Насиживающая птица греет яйцо, обхватив его плавательными перепонками,
наседных пятен не образуется. Смена партнёров происходит иногда только через сутки (до
3 суток). Инкубация продолжается 6-7 недель. Птенцы вылупляются голые, с тёмной ко-
жей, на 7-10-й день жизни покрываются белым или светло-бурым густым пухом. Младший
птенец, оттесняемый во время кормёжки старшим, обычно погибает в первые дни жизни.
Подросшие птенцы в пуховом наряде имеют чёрные «лицо», клюв, радужину. Контурное
оперение начинает развиваться через 6-7 недель. Выкармливание длится 12-20 недель, до
появления способности к полёту. Некоторые виды олуш гнездятся не ежегодно, а через
каждые 1-2 года. Чрезвычайная доверчивость ряда видов по отношению к человеку во
время гнездования обусловила мнение о глупости птиц и нашла отражение в названиях —
устаревшие русские — «глупыш», «олух» (отсюда—«олуша»), английское «ЬооЬу» — «глу-
пец». Во внегнездовое время олуши совершают протяжённые кочёвки, районы кочёвок
молодых порой далеко отстоят от мест гнездования или кочёвок взрослых (Коблик, 2001).
Олуши распространены всесветно, за исключением Антарктики, Субантарктики, Се-
верной Пацифики. В Арктику заходит единственный североатлантический вид. По одному
виду обитает у южных побережий Африки и Австралии, 2 приурочены к тихоокеанскому
побережью Америки, 3 широко распространены в тропических водах всех океанов и их
популяции насчитывают миллионы пар. Наиболее угрожаемое положение — у олуши Эб-
ботта (Sula (Papasuld) abbotti Ridgway, 1893) — эндемика о-ва Рождества близ побережья
Явы. Популяция вида насчитывает не более тысячи пар, местам гнездования угрожают раз-
работки фосфатов, вид включён в Красный список МСОП. Остальные виды олуш не отно-
сятся к глобально угрожаемым, хотя численность многих снижается в результате загрязне-
ния акваторий, сокращения кормовой базы, промысла яиц и птенцов на колониях.
Древнейшие олуши (Sula ronzoni) известны из олигоценовых отложений Франции (око-
ло 30 млн. лет назад). Остатки из Северной Америки, Австралии и других районов отно-
сятся к 15 ископаемым и 6 современным видам. 9-10 современных видов семейства до
недавнего времени объединяли в единственный род Sula, сейчас обычно выделяют также
род Morus (3 вида), иногда — Papasula (1 вид) (Howard, Moore, 1980; del Hoyo et al., 1992;
Dickinson, 2003). В России 1 гнездящийся и 2 редких залётных вида олуш.
_ еверная олуша
39
Род Morus Vieillot, 1816
Северная олуша
Morus bassanus (Linnaeus, 1758)
Pelecanus bassanus Linnaeus, 1758, Syst. Nat., ed. 10, c. 133, «Шотландия, Америка».
Gannet (англ.); Basstolpel (нем.); Fou de Bassan (фр.)
Син. (лат.): Pelecanus maculatus J.F. Gmelin, 1789; Sula bassana (Linnaeus, 1758); Sula major
Lacepede, 1802; Sula alba Meyer, 1810; Sula melanura Gould, 1837, Sula americana Bonaparte,
1838; Sula lefevri Baldamus, 1851.
Син. (рус.): жаннита, олух, глупыш (устар.)
Статус. Гнездящийся, мигрирующий вид.
Общая характеристика и полевые признаки. Птица величиной с гуся; за счёт длин-
ных узких крыльев (размах 160-180 см) кажется ещё крупнее. Клюв мощный, конический,
стального цвета. В полевых условиях легко определяется по планирующей манере полёта,
чередующейся с сериями взмахов, и контрастной окраске взрослых особей. Белая окраска
кроющего оперения с ярко-охристой головой и шеей, чёрными вершинами крыльев позво-
ляют легко обнаруживать и уверенно определять взрослую северную олушу даже на боль-
ших расстояниях. От крупных чаек олуша в любом наряде хорошо отличается также силу-
этом — узкие заострённые крылья, клиновидный хвост, обтекаемое торпедообразное тело.
Молодые особи бурые с белыми пестринами, неполовозрелые имеют участки белого опе-
рения, степень развития которого зависит от возраста.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Иногда к М. bassanus в качестве подвидов относят также
остальные виды рода Morus — капскую (М. capensis (М.Н.К. Lichtenstein, 1823)) и авст-
ралийскую (М. serrator (G.R. Gray, 1843)) олуш. Они распространены, соответственно, у
южных и юго-западных побережий Африки, побережий южной Австралии и Новой Зелан-
дии, сходны с северной олушей обликом, размерами и биологией, а отличаются главным
образом деталями окраски. Ранее все три вида включали в род Sula sensu lato.
Описание. Окраска. Взрослые птицы имеют в целом белое оперение, за исключени-
ем первостепенных маховых и крылышка, окрашенных в тёмно-бурый цвет. Голова и шея
имеют сильный охристо-жёлтый налёт (наиболее яркий на задней стороне шеи), хорошо
заметный даже с большого расстояния. Неоперённая лицевая часть и узкие полоски на
горле — чёрного цвета, как и вся кожа. Кольцо вокруг глаз светло-голубое. Радужина свет-
лая, с кремовым оттенком. Клюв серо-стального цвета с чёрными узкими полосками вдоль
щитков рамфотеки. Ноги чёрноватые, с контрастными полосками на передней стороне цевки
и наружной стороне пальцев. Тип окраски этих полосок является одним из немногих при-
знаков полового диморфизма. По Дж. Б. Нельсону (Nelson, 1964) при значительных вариа-
циях оттенков в целом у самцов они всегда более жёлтые, у самок — сине-зелёные.
Пуховой наряд. Птенец появляется на свет почти голый; кожа чёрного цвета. К концу
первой декады он полностью покрыт густым белым пухом. Клюв, ноги и голая кожа «лица»
остаются чёрными. Радужина тёмная.
Окраска птиц первого года жизни и неполовозрелых особей сильно варьирует (Nelson,
1964). В гнездовом наряде птицы имеют в целом тёмно-бурую окраску. Сверху она более
насыщенная и заметно светлеет к низу. Варьирующие по размерам светлые пестрины ши-
роко распределены по всему телу, клюв тёмно-серый. К концу первого года жизни опере-
40
Отряд Пеликанообразные. Семейство Олушевые
ние приобретает более бурый оттенок и светлеет на брюхе. У некоторых особей светлеют
также верхняя часть головы, грудь и даже спина. Оперение второго года жизни ещё более
изменчиво. Многие особи в этом возрасте могут иметь белую голову, кроющие плеча, зад-
ней части спины и нижнюю часть тела. Белые перья могут встречаться и среди второсте-
пенных маховых. Постепенно светлеют клюв и радужина. На третий год жизни северные
олуши, как правило, напоминают обликом взрослых особей, отличаясь от них наличием
отдельных чёрных второстепенных маховых перьев и небольших участков тёмного опере-
ния на спине. Голова практически без охристого оттенка. Четырёхлетние птицы имеют
почти такое же оперение, как и взрослые особи, при наличии отдельных тёмных перьев
среди второстепенных и третьестепенных маховых и центральных рулевых. Охристый от-
тенок на голове проявляется сильнее.
Строение и размеры. Первостепенных маховых десять. Хвост хорошо выражен-
ной клиновидной формы с 12 рулевыми перьями. Масса взрослых птиц около 3,5 кг. Крю-
чок на конце клюва слабо выраженный; по краям надклювья и подклювья имеются внут-
ренние ряды зазубрин. Наружные отверстия ноздрей отсутствуют.
Три измеренные на острове Харлов (архипелаг Семь островов, Восточный Мурман)
взрослые птицы имели следующие размеры (мм): длина крыла — 490-497; длина хвоста —
246-355; длина головы с клювом— 183,5-188,9; длина клюва — 96,1-97,8; длина цевки —
61,6-63,8; длина среднего пальца с когтем — 105,8-107,8. Масса — 3 000-3 200 г.
По личному сообщению Р.Т. Баррета (R.T. Barrett), размеры северных олуш, отловлен-
ных на Лофотенских о-вах (Норвегия) были следующими (мм): длина крыла (п=91) —
491,1±8,8; длина головы с клювом (п=66) — 198,9±5,3; длина клюва (п=91) — 97,9±3,0.
Масса (п=91) — 3204,7±219,9 г.
Линька. В ходе годового цикла имеются две линьки. Частичная — в марте-апреле (смена
оперения головы и шеи) и полная — по окончании сезона размножения.
Распространение. Гнездовой ареал. Северная олуша широко распространена
на гнездовании в Северной Атлантике: на Британских о-вах, во Франции, в Исландии, Фарер-
ских о-вах, Ньюфаундленде, Новой Шотландии, островах залива Св. Лаврентия. Центром
распространения является Британия (рис. 7). Рост численности олуш обусловил не только
появление новых гнездовий внутри ареала (в Ирландии, Великобритании, Канаде), но и
привел к его значительному расширению. В 1946 г. было отмечено начало размножения
олуш на о. Рунде вблизи южного побережья Норвегии (Montevecchi et al., 1987). Вторая
колония северных олуш возникла вблизи границ СССР на побережье Баренцева моря в
Сюльт-фьорде (Syltefjord), где первые гнёзда были найдены в 1961 г. (Brun, 1972).
В первой половине XX века северная олуша рассматривалась в СССР как редкая, слу-
чайно залетавшая к берегам Мурмана птица (Судиловская, 1951). С начала 1970-х гг. дос-
товерные встречи птиц этого вида время от времени регистрировали на Мурмане (Коханов,
Скокова, 1967), а иногда и на разных участках акватории Белого моря (Шкляревич, Коха-
нов, 1980). Учитывая, что даже для гнездящихся особей этого вида характерны кормовые
полеты на удаление до 300 км, после возникновения колонии в Сюльт-фьорде появление
олуш вблизи берегов Мурмана было вполне предсказуемо. Начиная с 1977 г., северных
олуш стали относительно регулярно наблюдать в прибрежных водах Мурмана. В пределах
архипелага Семь островов до 1981 г. встречали исключительно одиночных особей, по-
зднее — небольшие группы из 2-3 птиц. В начале 1990-х гг. у Мурманского побережья
олуши держались в течение всего летнего периода, создавая порой крупные кормовые скоп-
ления (Краснов, Барретт, 1997). В 1993 г. было зарегистрировано появление пары северных
олуш в колонии кайр на о. Харлов, выбор ими территории и оборона её от других видов
птиц. Во второй половине лета 1995 г., после годичного перерыва, на данном участке по-
_ еверная олуша
41
Рис. 7. Область распространения северной олуши: а — места гнездовий, б — район встреч во вне-
гнездовое время.
явились уже 3 пары птиц, одна из которых построила гнездо и имитировала насиживание.
В 1996 г. здесь же наблюдали начало и ход размножения одной пары северных олуш, вклю-
чая откладку и насиживание яйца. Таким образом, начиная с 1996 г., гнездовой ареал олу-
ши включает северо-западные районы России, в частности, побережье Мурмана (Krasnov,
Barrett, 1997).
Миграции и зимние кочёвки. Районы зимних кочёвок северных олуш младших возра-
стных групп находятся гораздо южнее, чем у половозрелых и хотя бы уже однажды
размножавшихся птиц. На основании анализа возвратов от 40 тысяч окольцованных в 1904 г.
олуш из британских колоний впервые было установлено, что молодые птицы мигрируют в
атлантические воды Марокко и Сенегала, иногда достигая берегов Гвинеи-Бисау (Nelson,
1978). Некоторые особи проникают в воды Средиземного моря. Во второе и третье лето
своей жизни они нередко встречаются у берегов Испании. Незначительная часть птиц пер-
вого года жизни может оставаться в европейских водах и с началом летнего сезона перио-
дически посещать гнездовые колонии. Но большинство олуш вновь появляется у родных
берегов лишь на третий-четвертый год. Птицы более старших возрастных групп так дале-
ко на юг не откочёвывают.
Анализ миграционных связей северных олуш из колоний Британии и Норвегии проде-
монстрировал почти полное их сходство (Barrett, 1988). По данным кольцевания птиц в
норвежских колониях, в период с сентября по март основная масса половозрелых особей
встречается у побережий Норвежского и Северного морей (Barrett, Krasnov, 2000). Наблю-
дения с борта морского судна, выполненные А.В. Ежовым в 2005 г. (устн. сообщ.), показа-
ли, что в юго-западных районах Баренцева и юго-восточных районах Норвежского моря в
октябре-ноябре северные олуши полностью отсутствуют. Но у побережья южной Норве-
гии и в юго-восточных районах Северного моря они встречаются в значительных количе-
ствах, соответственно 0,4 и 1,0 особь на 1 км2.
В колониях северной олуши на побережье Норвегии, включая его баренцевоморскую
часть, были получены возвраты от птиц, окольцованных в Британии, на Нормандских о-вах
42
Отряд Пеликанообразные. Семейство Олушевые
и в Исландии. Это свидетельствует о продолжающемся процессе иммиграции птиц из цен-
тра ареала на его периферию (Barrett, Folkestad, 1996). В единственной российской коло-
нии также неоднократно встречали птиц с иностранными кольцами. При наблюдениях в
длиннофокусную оптику все они были определены как норвежские. В одном случае номер
кольца был прочитан полностью; оказалось, что северная олуша, размножавшаяся на о-ве
Харлов, была окольцована в колонии на баренцевоморском побережье Норвегии вблизи
мыса Нордкап (Krasnov, Barrett, 1997).
Местообитания. Годовой цикл большинства взрослых северных олуш проходит в пре-
делах континентального шельфа Северной Атлантики (Nelson, 1978). В Баренцевом море
на основе наблюдений с судов и самолетов установлено, что зона трофической активности
этих птиц ограничена акваторией прибрежных районов Финнмарка и Мурмана (не далее
100 км от берега). В гнездовой период размножающиеся особи держатся на скалистых
островах, для строительства гнёзд предпочитая скальные уступы. При дефиците пригод-
ных мест в старых и плотно заселённых участках, гнездятся на склонах или даже на плос-
ких, удаленных от моря вершинах островов. Известны случаи существования материковых
колоний северной олуши (Nelson, 1978). Обычно олуши устраивают гнёзда на участках,
давно освоенных другими видами птиц. На норвежском побережье они гнездятся среди
хохлатых и больших бакланов, в колониях моевок и кайр. Все случаи выбора гнездовых
территорий северной олуши на Мурмане имели место исключительно на участках, засе-
ленных кайрами. Чаще всего при этом новые хозяева птиц-резидентов активно изгоняли.
Численность. С начала XX столетия численность северных олуш увеличивается на 2-
3% в год (Nelson, 1978); общее их количество в мире превышает 260 тысяч пар (Lloyd et al.,
1991). Основная часть мировой популяции северной олуши размножается в Британии и
Ирландии (72%). Общая численность норвежской популяции (5 гнездовых поселений) со-
ставляет около 3 700 пар. В начале периода колонизации (1945-1965 гг.) рост численности
в Норвегии составлял 20-25% в год, снизившись до 7,5% в период 1975-1985 гг.; в первой
половине 1990-х гг. численность северной олуши здесь стабилизировалась. В России чис-
ленность гнездящейся группировки затри года, начиная с 1996, возросла до 35 пар. Обсле-
дования побережий Мурмана показали, что за последние десять лет новых колоний север-
ной олуши здесь не появилось, а численность гнездящихся и территориальных птиц на о.
Харлов до 2004 г. колебалась на уровне 50-60 пар (Мищенко и др., 2004). В 2005 г. она
возросла до 145-160 пар (Мельников, 2006).
Размножение. Моногамы. Сформировавшиеся пары существуют много лет (Nelson,
1978). К первому размножению северные олуши чаще всего приступают в возрасте 5 лет.
Территорию для гнёзда самцы подыскивают и завоевывают в предшествующие годы, в
возрасте от 3 до 5 лет. Самки, в конечном итоге, выбирают самца с территорией, после чего
происходит формирование пары и дальнейшая защита осуществляется совместно.
В период поиска участков, пригодных для гнездования, северные олуши регулярно по-
сещают колонии птиц. На Мурмане, где других колоний олуш не существует, они отвоевы-
вали участки в колониях кайр. Иногда олуши захватывали кайр за шею и сбрасывали их с
гнездового карниза. Оставшихся без надзора птенцов кайр олуши «усыновляли». Клювом
они продвигали кайрят под себя, размещая их либо на перепонках лап, либо под приспу-
щенными крыльями. Птенцы кайр постоянно предпринимали попытки побега, особенно
активные при призывном крике родителей. Если кайра при этом приносила своему птенцу
корм, то олуши пытались его отобрать. Но сами олуши кайрят никогда не кормили, хотя
такие случаи и наблюдали в других частях ареала (Fairhurst, 1976).
На выбранном участке олуша строит гнездо, однако к кладке приступает лишь в следу-
ющем году. В Британии большинство гнёзд многолетние и хорошо сохраняются в течение
Северная олуша
43
зимы. В России они, как правило, расположены не слишком высоко над уровнем моря и
большей частью смываются зимними штормами. В колонии о. Харлов олуши появляются в
апреле-мае. Большинство гнёзд в новом сезоне они полностью восстанавливают в течение
3-4 дней. Материалом для гнезда служат водоросли, подбираемые с воды на прилегающей
акватории. Попутно олуши приносят в гнездо разнообразный мусор, плавающий в море:
обрывки тралов, канатов, упаковочных лент и прочий бытовой хлам. Яркая расцветка этих
предметов часто придает гнезду своеобразный облик.
Копуляция происходит в ходе строительства гнезда и достигает пика за две недели до
начала кладки (Nelson, 1964). Кладка состоит из одного слегка голубоватого яйца массой
81-130 г, в среднем 104,5 г (Nelson, 1964). Размеры яиц составляют (п=100) 78,1x49,1 мм в
Британии, и (п=44) 77,6x47,0 мм в Канаде (Nelson, 2002). В России, по измерениям М.В
Мельникова (2006), — 77,6x47,0 мм (n= 121). Поверхность яйца в ходе насиживания быст-
ро пачкается, становится желтоватой, а иногда и бурой. Массовая кладка в Британии прохо-
дит в течении 3-4 дней в последних числах апреля (Nelson, 1964). В центральной и север-
ной Норвегии первые яйца появляются в середине - конце апреля, массовая кладка прохо-
дит в начале мая (Montevecchi et al., 1987). На о. Харлов — в третьей декаде мая. Как и у
многих других видов птиц, более старшие особи приступают к кладке яиц в более ранние
сроки.
Насиживание у северных олуш весьма своеобразное. Обогрев яйца происходит нижней
поверхностью перепонок лап. Фактически, олуша начинает насиживать, предварительно
установив лапы на скорлупу яйца. В конце насиживания, продолжающегося 43-^44 дня,
яйцо передвигается на верхнюю поверхность перепонок лап (Nelson, 1964). В насижива-
нии принимают участие оба пола. Время дежурства одного из родителей в период насижи-
вания или выкармливания птенцов достигает 24 ч. Птенцы появляются голыми и слепыми.
В 1998 г. в колонии на о. Харлов появление первых птенцов было отмечено 6 июля.
Взрослые олуши кормят птенцов, отрыгивая комочки пищи, при этом голова птенца
может проникать в пищевод родителя. В течение суток птенца кормят несколько раз. К
концу третьей декады появляются маховые и рулевые перья. В двухмесячном возрасте пте-
нец полностью оперяется. Выкармливание продолжается в течение 90-94 дней. Птенцы в
гнезде малоподвижны. Выпав за пределы гнезда, даже в присутствии родителей, как пра-
вило, остаются без помощи и погибают.
Начало схода птенцов на воду в центральной и северной Норвегии наблюдали в сентяб-
ре-октябре (Montevecchi et al., 1987). На Мурмане, вероятнее всего, он также происходит в
октябре. Период голодания в гнезде у слётков северной олуши отсутствует. Раскормленный
птенец, массой до 4 500 г, полностью оперённый, самостоятельно покидает карниз скалы,
ещё не владея техникой полета и, вероятно, ещё не имея такой возможности из-за излишне-
го веса. Спуск на воду представляет собой скорее управляемое падение, чем полёт. В море
птенец остаётся один без сопровождения родителей и некоторое время (до двух недель)
голодает до приобретения способности к полёту (Nelson, 1964).
Среднегодовая смертность северной олуши составляет 6,1%, причём смертность сам-
цов несколько выше: 7,6% против 4,7% у самок. Наибольшую смертность отмечают у не-
половозрелых птиц, из которых большинство погибает на первом году жизни. Средняя про-
должительность жизни — 16,2 года. Наибольший возраст — 21 год — зарегистрирован у
птицы из колонии Басс Рок (Nelson, 1966, цит. по: Cramp, 1977).
Суточная активность, поведение. В обычном полёте олуша чередует взмахи крыльев
:: скольжением, совершая от 170 до 190 взмахов в минуту (Nelson, 1978). Обнаружив до-
тычу, птицы ныряют с любой высоты, не превышающей 30 м. Погружение происходит
вследствие действия силы тяжести и при помощи взмахов крыльев. Олуша в броске за
44
Отряд Пеликанообразные. Семейство Олушевые
Рис. 8. Поведение северной олуши: а — церемония приветствия членов пары; б — совместная пост-
ройка гнезда самцом и самкой; в — рекламное поведение при ухаживании; г — спаривание; д —
рекламное поведение самца на гнезде и вытянутая поза; е — ритуализированный поклон на границе
гнездового участка; ж — угрожающие позы насиживающих самок (по Cramp, 1977; Nelson et al.,
2005).
_ еверная олуша
45
добычей способна изменять угол и направление падения, иногда ныряя почти отвесно. Счи-
тается, что олуши способны погружаться на глубину до 15 м (Nelson, 1978).
Неполовозрелые птицы много времени проводят в «клубах», существующих вблизи каж-
дой крупной колонии. В них оттачиваются социальные взаимодействия готовящихся к по-
иску и занятию гнездовой территории особей.
Территориальные особи в колониях ведут себя очень шумно, постоянно подтверждая
твое право на занятую территорию. При этом важным элементом является демонстрация
приподнятых крыльев, скрипучего голоса в сочетании с серией поклонов и энергичной
тряской головы из стороны в сторону (рис. 8). Появление самки вызывает взаимную де-
монстрацию. Самец проявляет по отношению к самке свою агрессивность, которую та га-
сит, отворачивая свой клюв в сторону от самца. Нередко такая демонстрация заканчивается
щипками самцом головы самки. Аналогично это происходит и при копуляции. При даль-
нейших посещениях территории формирование пар закрепляется строительством гнезда.
При появлении партнёра происходит взаимная демонстрация приветствия. Птицы вы-
тягивают вверх шеи, головы и клюв (рис. 8).
Питание. Обычный корм северной олуши — стайные пелагические виды рыб. У нор-
вежского побережья Баренцева моря в первой половине 1990-х гг. птицы чаще всего добы-
вали сельдь и мойву. В Норвежском море их корм состоял из скумбрии Scomber scombrus,
сайды, сельди, мойвы, песчанки и, отчасти, сёмги Salmo salar (Brun, 1972; Barrett, 1981,
Montevecchi, Barrett, 1987). Показано, что размещение гнездовий олуши в ряде районов
ареала коррелирует с распределением сельди Clupea harengus (Reinsch, 1969). Предполага-
ется, что данный вид рыб — предпочитаемый корм северных олуш на севере Норвегии
। Montevecchi, Barrett, 1987).
В поисках корма северные олуши способны удаляться от родных колоний на расстоя-
ние до 300 и более километров (Nelson, 1964). Начало проникновения северной олуши в
российскую часть Баренцева моря отмечено в период крупномасштабных изменений кор-
мовой базы птиц-ихтиофагов. Активная колонизация данным видом побережья Мурмана
проходила в годы появлений у его берегов крупных скоплений мойвы Mallotus villosus
и песчанки Ammodytes tobianus. Начало гнездования и рост колонии северной олуши на
о. Харлов (архипелаг Семь островов, Восточный Мурман) по времени совпадает с наличи-
ем у берегов архипелага скоплений молоди сельди. Анализ корма птенцов в 1996-1999 гг.
показал, что он состоял преимущественно из сельди (около 90% встреч) при незначитель-
ной примеси песчанки и мойвы. В результате наблюдений за трофическим поведением
взрослых олуш было установлено, что в эти же годы в водах Восточного Мурмана они
также добывали мелкую сайду Pollachius virens и даже крупную креветку Pandalus sp. Бо-
лее поздние наблюдения за северными олушами на о. Харлове продемонстрировали появ-
ление в их кормах незначительной примеси трески Gadus morhua (Мельников, 2006).
На большой части ареала северная олуша, обнаружив рыбу, ныряет за ней с большой
высоты, сложив крылья и вытянув шею. Кормодобывающее поведение птиц у берегов ар-
хипелага Семь островов заметно отличалось от поведения олуш в других районах ареала.
На Мурмане олуши меньше демонстрировали «ударное» ныряние — вероятно, из-за отсут-
ствия в годы наблюдений у берегов архипелага относительно крупной рыбы. Обычно, най-
дя косяк мелкой пелагической рыбы (или крупной креветки), олуша садилась на воду, пери-
одически резко опуская в не голову на всю длину шеи, выхватывая сразу по несколько рыб
/□и креветок.
Величина кормовой территории зависела от состояния трофических условий в прилега-
ющих к архипелагу Семь островов районах. В 1993 г., при постоянном наличии косяков
рыбы, олуши не удалялись от островов более чем на 1,5-2 км. С ухудшением кормовых
46
Отряд Пеликанообразные. Семейство Олушевые
условий в 1995 г. дальность кормовых полётов увеличилась настолько, что визуально опре-
делить её не удалось.
Враги, неблагоприятные факторы. Врагов у северной олуши немного. В колониях
олуш на норвежском побережье Баренцева и Норвежского морей известны случаи актив-
ного хищничества орлана-белохвоста. По мнению Р.Т. Барретта (личн. сообщ.) снижение
численности птиц в этих колониях обусловлено деятельностью данного хищника.
В гнёздах и в оперении северных олуш находили блоху Ceratophyllus borealis (Hicks,
1959).
На существование колоний северной олуши заметное негативное воздействие способ-
но оказывать антропогенное беспокойство. Предполагается, что именно по этой причине в
1978 г. исчезла одна из колоний на Лофотенских о-вах в Норвежском море (Barrett, 1979).
На о. Харлов, в единственной гнездовой колонии на территории России, также отмечены
случаи беспокойства насиживающих птиц и браконьерская добыча олуш местным населе-
нием. В последние годы беспокойство размножающихся птиц происходит в период сбора
научного материала по гнездовой биологии сотрудниками заповедника. Только этим мож-
но объяснить зарегистрированные случаи массового хищничества морских чаек (Larus
marinus) в колониях северной олуши (Мельников, 2006). Специальные исследования, про-
ведённые нами в первые годы существования колоний на о. Харлов, продемонстрировали
полное отсутствие влияния крупных чаек на успех размножения северных олуш при отсут-
ствии антропогенного беспокойства последних. В то же время, возможность снижения чис-
ленности российской гнездовой популяции северной олуши по данным причинам весьма
мала, так как численность постоянно увеличивается вследствие иммиграции птиц из коло-
ний норвежского побережья.
Общий долговременный успех заселения северной олушей Мурмана будет, вероятнее
всего, возможен при стабилизации существующих здесь кормовых условий.
Хозяйственное значение, охрана. Хозяйственного значения на территории России се-
верная олуша не имеет. В то же время, многолетнее неудовлетворительное состояние охра-
ны колоний морских птиц в Баренцевоморском отделе Кандалакшского заповедника про-
воцирует рост нелегальной добычи птиц, в том числе и северной олуши на о. Харлов. Вид
включён в Красную книгу Мурманской области как редкий или узколокальный, но в насто-
ящее время конкретных мер для его охраны не предпринимается.
Род Sula Brisson, 1760
Красноногая олуша
Sula sula (Linnaeus, 1766)
Pelecanus sula Linnaeus, 1766, Syst. Nat., ed. 12, vol. 1, c. 218, о-в Барбадос, Малые
Антильские о-ва (Grant, Mackworth-Praed, 1933, Bull. Brit. Omith. Club, 53, c. 118-119).
Red-footed booby (англ.); Rotfuatolpel (нем.); Fou a pieds rouges (фр.).
Статус. Редкий залётный вид.
Общая характеристика и полевые признаки. Крупная птица (длина тела в среднем
70 см, размах крыльев — 150 см) с сигарообразной обтекаемой формой тела, длинными
крыльями, длинным клиновидным хвостом и светлым коническим клювом. Окраска опе-
рения варьирует от полностью серовато-бурой у особей тёмной морфы до белой с чёрными
концами крыльев и чёрной полосой по заднему краю крыла у белой морфы. Специфичны
ноногая олуша 47
вида красные ноги и участок оголённой розовато-красной кожи вокруг основания клю-
Оперение молодых птиц бурое, с более светлым брюхом и тёмной размытой полосой на
“г? ди; клюв и кожа на лице у них тёмного цвета, ноги желтовато-серые. Как и другие виды
.-.глией, летает быстро и прямолинейно, чередуя скользящее планирование с редкими силь-
-ыми взмахами крыльев, может подолгу парить; охотясь за рыбой, стремительно пикирует
. полураскрытыми крыльями с высоты в воду.
Подвидовая систематика. Выделяют 3 подвида (Степанян, 2003; Dickinson, 2003).
изменчивость проявляется в общих размерах. Подвидовая принадлежность молодой осо-
: и. залетевшей на территорию России, установлена предположительно, на основании гео-
рафических соображений.
1. Sula sula rubripes
Sula rubripes Gould, 1838, Synops. Birds Austr., pt. 4, c. 7, о-в Рейн, сев. Квинсленд, Авст-
72_~ИЯ.
Самая крупная раса, распространенная на тропических островах Индийского, западной
центральной части Тихого океанов (1) (рис. 9).
Два других подвида распространены следующим образом: S. s. websteri Rothschild, 1898 —
:т побережья Зап. Мексики до о-в Галапагос (2); S. s. sula (Linnaeus, 1766) — острова Ка-
7ноского моря и юго-западной части Атлантического океана (3) (рис. 9).
Описание. Окраска. Взрослые птицы. Сезонных и половых различий в окраске
. церения нет. Окраска полиморфна, выделяют до 7 цветовых вариаций. Наиболее распро-
страненной является белая морфа: все оперение белое, за исключением первостепенных и
второстепенных маховых и больших верхних кроющих крыла, окрашенных в серебристо-
- ёрный цвет, и темного пятна на нижней стороне крыла у карпального сгиба. Самая темная
чорфа полностью бурая. Между этими крайними вариантами различают несколько проме-
жуточных типов окраски в зависимости от степени развития и топографии белого и бурого
цвета в оперении. Клюв светло-голубой; у его основания и вокруг глаза есть участок ого-
лённой кожи, цвет которой варьирует от розоватого до красного. Ноги ярко-красные.
Птенцы вылупляются голыми, кожа от серой до розоватой, клюв черновато-бурый, ноги
зерые. Пуховой наряд белого цвета. Гнездовой наряд в основном тёмно-бурый, более свет-
лый на груди и брюхе. Первый промежуточный наряд (белая морфа): голова и шея серова-
т ?-бурые; спина, крылья и нижняя сторона туловища белые с серо-бурыми пятнами, коли-
чество которых с возрастом сокращается. Второй промежуточный наряд (белая морфа):
цтова, шея и низ туловища белые; спина и верх крыльев белые, испещренные черноваты-
чи или бурыми перьями; поясница бурая; первостепенные и второстепенные маховые се-
7^е: рулевые серовато-бурые.
Строение и размеры. Первостепенных маховых 10 (не считая рудиментарного),
ц<рмула крыла I>II>III>IV; рулевых 14, форма хвоста клиновидная. Средние размеры — в
нм (±SD) самцов и самок S. s. rubripes (атолл Джонстон, Гавайские о-ва): длина крыла
3^8.0±14,3 (п = 283); длина хвоста 118,0±7,9 (п = 234); длина клюва 84,0±4,0 (п = 285);
”ина цевки 42,1±2,8 (п=281). Масса 1 110,0±131 г (п = 803). Самки несколько крупнее и
тяжелее самцов. Вес птиц белой морфы существенно больше, чем олушей с более тёмной
: краской оперения (Palmer, 1962; Carboneras, 1992; Schreiber et al., 1996).
Линька. Пуховой покров начинает активно развиваться на 10 сутки, остатки пуха со-
храняются до 15-недельного возраста, когда молодые в полностью сформировавшемся гнез-
ц: зом наряде поднимаются на крыло. Ювенильная линька первостепенных маховых начи-
-ается через 3,5-7 месяцев после вылета из гнезда. Окончательный наряд приобретается в
?-летнем возрасте. В связи с большой продолжительностью периода размножения взрос-
7 птицы не успевают сменить оперение в негнездовое время и активно линяют в ходе
48
Отряд Пеликанообразные. Семейство Олушевые
Рис. 9. Область распространения красноногой олуши: а — область кочёвок, б — места гнездования,
в — залёт на территорию России. Подвиды: 1 — S. s. rubripes, 2 — S. s. websteri, 3 — S. s. sula.
инкубации и выкармливания птенцов. Первостепенные маховые сменяются партиями од-
новременно в двух или трёх центрах линьки (Schreiber et al., 1996).
Распространение. Гнездовой ареал пантропический. Населяет острова Карибского моря,
Атлантического, Индийского и Тихого океанов. Ближайшие к границам российского Даль-
него Востока места размножения расположены на о-вах Рюкю; залётных птиц отмечали
несколько севернее — на о. Хонсю (Brazil, 1991; Ornithological Society of Japan, 2000).
В пределах России достоверно установлен единственный случай залёта: молодая особь
добыта в зал. Чихачева (Де-Кастри) в северной части Татарского пролива (Finsch, 1872).
Кроме того, птицу, похожую на красноногую олушу, наблюдали в июне 1960 г. в бух. Киев-
ка (Судзухэ) на юге Приморского края (Литвиненко, Шибаев, 1971) (рис. 9).
Миграции. Данных мало. По-видимому, совершают непродолжительные кочёвки. Наи-
более подвижны молодые птицы, которые могут удаляться от родных колоний более чем на
1 тыс. км.
Зимовки расположены в пределах гнездового ареала.
Местообитания. Гнездится на коралловых атоллах и вулканических островах, покры-
тых густой древесной и кустарниковой растительностью. На море встречается как в при-
брежных, так и в открытых водах в сотнях километров от суши.
Численность. Самый многочисленный вид олушей. В середине 1990-х гг. репродук-
тивная часть мировой популяции оценивалась в 300 тыс. пар. Большинство птиц этого вида
(250 тыс. пар) гнездятся на о-вах Галапагос. Общая численность красноногой олуши, по-
видимому, превышает 1 млн. особей (Carboneras, 1992; Schreiber et al., 1996).
Размножение. Моногам. Начинает размножаться обычно в 3^4-летнем, изредка в 2-лет-
нем возрасте. Колониальный вид. Почти половина взрослых птиц из года в год занимает для
гнездования одни и те же места. Гнёзда, как правило, устраивают на ветвях деревьев и кус-
тарников, иногда на земле среди травы. В кладке 1 яйцо. Скорлупа голубоватая, покрытая
меловым налетом. Средние размеры яиц (±SD) с атолла Джонстон, Гавайские о-ва (п = 179):
62,5±13,0 х 40,6±1,5 мм; масса 52,9±5,4 г. Инкубационный период длится в среднем 46 суток.
При благоприятных условиях птенцы проводят в гнёздах примерно 100 дней. После вылета
молодые посещают гнездо ещё в течение 1^1 месяцев, где продолжают подкармливаться ро-
дителями. Успех размножения старых опытных птиц почти вдвое выше (до 80%), чем у моло-
красноногая олуша, бурая олуша 49
zbix, гнездящихся в первый раз. В популяциях, населяющих Тихий океан, в годы активного
проявления Эль-Ниньо результативность гнездования резко снижается, так как из-за бескор-
^щы происходит массовая гибель потомства (Schreiber et al., 1996).
Суточная активность, поведение. На колониях активна в светлое время суток; кор-
мится как днём, так и ночью. Красноногая олуша — общественная птица, обычно держит-
ся группами и стаями, как на суше, так и в море. Часто подлетает к судам и садится на них.
Питание. Основу рациона составляют летучие рыбы Exocetidae и кальмары Отпта-
i:rephidae. Добычу хватает, пикируя вертикально с высоты в воду; летучих рыб может ло-
вить и в воздухе. Кормится обычно стаями до 100 и более особей, часто вместе с другими
видами морских птиц. Дальность кормовых перелетов составляет до 150 км.
Враги, неблагоприятные факторы. Колонии, расположенные по соседству с жильем
человека, активно эксплуатируются людьми, в результате чего численность птиц в них се-
рьезно сократилась. На многих островах вред наносят интродуцированные овцы, козы и
свиньи, которые уничтожают древесно-кустарниковую растительность в местах гнездова-
ния, а свиньи также — и самих птиц.
Хозяйственное значение и охрана. Будучи залётным видом, на территории России
хозяйственного значения не имеет. Специальных мер охраны не предусмотрено.
Бурая олуша
Sula leucogaster (Boddaert, 1783)
Pelecanus leucogaster Boddaert, 1783, Table Planches Enlum., c. 57, основано на табл. 973,
Daubenton, Le Fou de Cayenne. Terra typica не указана, предположительно Кайенна.
Brown booby (англ.); Brauntolpel, Weissbauchtolpel (нем.); Fou brun (фр.).
Син. (рус.): желтоногая олуша.
Статус. Редкий залётный вид.
Общая характеристика и полевые признаки. Крупная птица (длина тела в среднем
69 см, размах крыльев — 141 см) с характерным для олушей плотным сигарообразным те-
лосложением. Взрослые особи легко идентифицируются по шоколадно-бурой окраске голо-
вы, шеи и верхней стороны тела, резко контрастирующей с белым низом туловища и крыль-
ев. Светлая окраска клюва чётко выделяется на фоне тёмной головы. Специфична также жёл-
то-зелёная окраска ног. Молодые птицы похожи на взрослых, но бурые участки оперения у
них более тусклые, нижняя сторона туловища и крыльев не белая, а серо-бурая, светлеющая
по мере взросления. В море обычно летает низко над водой, чередуя сильные уверенные
взмахи крыльев с парящим скольжением; может подолгу парить в потоках воздуха.
Подвидовая систематика. Выделяют 4 подвида (Dickinson, 2003). Изменчивость про-
является в варьировании интенсивности окраски головы и шеи и в общих размерах. Заре-
гистрированные в России особи относятся к подвиду:
1. Sula leucogaster plotus
Pelecanus Plotus J. R. Forster, 1844, Descriptiones animalium quae in itinere ad Maris australis
Terras per annos 1772, 1773,1774 suscepto collegit, ed. A.A.H. Lichtenstein, p.278, близ Новой
Каледонии.
Наиболее крупная раса, у которой оперение на голове и шее по цвету не отличается от
соседних участков; распространена в тропической и субтропической зонах Индийского,
западной, центральной и южной частях Тихого океанов (1) (рис. 10).
50
Отряд Пеликанообразные. Семейство Олушевые
Рис. 10. Область распространения бурой олуши: а — область кочевок, б — места гнездования, в —
залеты на территорию России. Подвиды: 1 — S. I. plotus, 2 — S. I. brevsteri, 3 — S. I. etesiaca, 4 — S.
I. leucogaster, 5 — S. I. nesiotes.
Распространение остальных подвидов: 5. /. brewsteri Goss, 1888 — Калифорния и за-
падное побережье Мексики (2); S. L etesiaca Thayer et Bangs, 1905 — тихоокеанское побе-
режье от Гондураса до Колумбии (3); S. I. leucogaster (Boddaert, 1783) — Атлантический
океан от Мексиканского залива до побережья Африки (4). Прежде выделяли (Howard, Moore,
1980) и пятый подвид: 5. /. nesiotes Heller et Snodgrass, 1901, гнездящийся на о. Клиппертон
(5), (рис. 10), впоследствии сведенный в синонимы к S. I. brewsteri.
Замечания по систематике. Подвид S. I. brewsteri, хорошо отличающийся от прочих
рас светло-серой а не тёмно-бурой окраской головы, ранее считали самостоятельным ви-
дом.
Описание. О к р а с к a (S. I. plotus). Взрослые птицы. В окраске оперения сезонных и
половых различий нет. Голова, шея, верх груди, хвост, вся верхняя сторона туловища и
крыльев шоколадно-бурые. Низ груди, брюхо, бока и подхвостье белые. Подмышечные,
нижние большие и средние кроющие крыльев белые. Окраска неоперённых частей тела
варьирует в зависимости от пола птиц, времени года и географического положения. Клюв и
ноги чаще всего окрашены в зеленоватые или жёлтые тона. Участок оголённой кожи на
лице у самок желтоватый или зеленоватый; у самцов более яркий, обычно бирюзовый или
пурпурный; перед глазом тёмное пятно.
Птенцы вылупляются голыми, цвет кожи от розоватой до серой, клюв чёрный с жёл-
тым яйцевым зубом, ноги тёмно-серые. Пуховой наряд образован густым длинным белым
пухом, покрывающим все тело (менее развит на боках лицевой части и отсутствует на под-
бородке и горле). Гнездовой наряд бурого цвета, на нижней части груди, брюхе, подхвостье
и нижней стороне крыльев окраска более светлая, серо-бурая. Клюв серо-голубой, ноги
желтоватые или светло-оранжевые. Первый промежуточный наряд похож на предыдущий,
но низ груди и брюхо по мере линьки ещё больше светлеют, подмышечные беловатые, ниж-
ние большие и средние кроющие крыльев светлые серовато-бурые. Второй промежуточ-
ный наряд похож на обношенный окончательный (Palmer, 1962; Cramp, 1977; Schreiber,
Norton, 2002).
Строение и размеры. Первостепенных маховых 10 (не считая рудиментарного),
формула крыла обычно I>II>III>IV..., иногда II>I>III>IV...; рулевых 14, форма хвоста кли-
z-рая олуша 51
- : видная. Средние размеры (мм) самцов (п = 52) и самок (п = 54) S. I. plotus (атолл Джон-
ттон. Гавайские о-ва): длина крыла самцов 402,0±14,7 (±SD), самок — 418,04±12,8; длина
^доста самцов 123,0±14,6, самок — 125,0±8,9; длина клюва самцов 99,6±3,0, самок —
. >4.1 ±2,9; длина цевки самцов 53,2±3,5, самок — 56,5±2,9. Масса (г) самцов 1 187±126,
; амок — 1 525±153. Самки значительно крупнее и тяжелее самцов (Schreiber, Norton, 2002).
? 2змеры (мм) самки, добытой в российских водах: длина крыла — 420, длина хвоста —
230. длина клюва — 110, длина цевки — 50 (Нечаев, 2000).
Линька. Интенсивное развитие пухового наряда протекает со 2-й по 4-ю неделю жиз-
ни. остатки пуха сохраняются до конца 16-й недели. Гнездовой наряд полностью формиру-
ется на 13-14-й неделе. У молодых птиц линька первой партии первостепенных маховых
начинается в 10-месячном возрасте, второй — в 15-месячном. Смена контурного оперения
• них происходит одновременно с линькой маховых, протекает медленно и непрерывно в
течение 1,5 лет. Окончательный наряд птицы приобретают в 3-летнем возрасте. У взрос-
лых первостепенные маховые и рулевые начинают обновляться после вылупления птен-
дов. Маховые сменяются в дистальном направлении одновременно в двух или более цент-
тах линьки (Schreiber, Norton, 2002).
Распространение. Гнездовой ареал. Населяет острова тропического и субтро-
пического поясов Тихого, Индийского и Атлантического океанов и их окраинных морей
тис. 10). Ближайшие к границам российского Дальнего Востока места размножения нахо-
дятся в Японии у южного побережья о-вов Хонсю и Кюсю (Brazil, 1991; Ornithological
Society of Japan, 2000).
На территории России добыта залётная взрослая самка 2 июля 1976 г. в зал. Львиная
Пасть на охотоморском побережье о. Итуруп (Нечаев, 2000). Ещё одну взрослую самку на-
блюдали 25 июля 1997 г. восточное Малой Курильской гряды на границе исключительной
жономической зоны РФ в точке с координатами 43°05’ с.ш., 146° 15’ в.д. (Артюхин, 1997).
Миграции. Взрослые птицы круглый год встречаются в районах гнездования, молодые
весь период полового созревания проводят в море, кочуя в тропических водах.
Зимовки расположены в пределах гнездового ареала.
Местообитания. Гнездится обычно на лишённых растительности скалистых островах
и коралловых атоллах. На море держится, главным образом, в прибрежных водах.
Численность. Современная численность в мире оценивается в 221,2-275,4 тыс. пар, в
том числе 51-70 тыс. пар S. I. plotus. Большинство бурых олушей гнездятся на островах у
“обережий Австралии и Мексики. Численность этого вида сократилась в историческое время
в десятки раз из-за интенсивного промысла птиц и их яиц человеком (Schreiber, Norton,
Z002).
Размножение. Моногам. Большинство птиц впервые приступают к размножению в 3^4
года, реже с 2-летнего возраста. Гнездятся обычно неплотными группами из десятков или
готен пар, очень редко крупными колониями, иногда отдельными парами. Гнёзда располо-
жены открыто на земле либо на невысоких скалах. В кладке 1-3, обычно 2 яйца, со скорлу-
пой светло-голубого или зеленоватого цвета, покрытой меловым слоем. Средние размеры
•иц (±SD) с атолла Куре, Гавайские о-ва (п = 26): 62,1±3,5 х 43,3±3,3 мм. Масса яиц номи-
нативного подвида с о. Вознесения (п = 82) 39-65, в среднем 52 г. Кладку насиживают
:амец и самка примерно поровну в течение 4СМ-4 дней, в среднем 42 дня. Из двух птенцов,
как правило, выживает только один, так как старший изгоняет младшего из гнезда. Моло-
дые становятся летными в возрасте 95-120 дней, после чего ещё длительное время их про-
должают подкармливать родители. Из природных факторов успех размножения в наиболь-
шей степени зависит от проявления эффекта Эль-Ниньо, вызывающего массовую гибель
птенцов (Cramp, 1977; Schreiber, Norton, 2002).
52
Отряд Пеликанообразные, Семейство Баклановые
Суточная активность и поведение. По наблюдениям за гнездящимися птицами, бурая
олуша ведёт дневной образ жизни. На море держится в одиночку или небольшими стаями.
Питание. Рацион включает рыбу, кальмаров и креветок, величиной от 3 до 40 см. Важ-
ную роль в питании играют летучие рыбы Exocetidae. Обычно охотится, пикируя в воду с
небольшой высоты, реже хватает добычу с поверхности воды в полёте, иногда отнимает её
у других птиц. Кормится в одиночку либо небольшими группами, в основном в прибреж-
ных водах (Белопольский, 1989, 1994; Schreiber, Norton, 2002).
Враги, неблагоприятные факторы. Помимо промысла, негативные последствия дея-
тельности человека проявляются в виде разорения колоний бурых олушей интродуциро-
ванными домашними животными (крысами, кошками, свиньями и др.).
Хозяйственное значение, охрана. Будучи залётным видом, на территории России хо-
зяйственного значения не имеет. Специальных мер охраны не предусмотрено.
Семейство Баклановые — Phalacrocoracidae Reichenbach, 1836
Объединяет веслоногих птиц размером от утки до гуся (длина 45-100 см, масса 360-
4 000 г). Основная трофическая специализация бакланов — подводная погоня за рыбой —
наложила отпечаток на всё строение птиц. Проблема увеличения удельного веса, что необ-
ходимо для длительного нахождения под водой, решена за счёт непневматичности скелета,
отсутствия подкожных воздухоносных полостей и устранения воздушной прослойки из
оперения. Перья бакланов промокают, и птицы вынуждены долго их сушить, расправив
крылья на солнце и ветру. Контурное оперение короткое, плотное, крылья относительно
короткие, с тупой вершиной (первостпенных маховых — 11), иногда под водой находятся в
полусложенном состоянии и выполняют функцию рулей глубины. Хвост длинный, слабос-
тупенчатый, из 12-16 жёстких рулевых перьев, играет под водой ту же роль. Тело обтекае-
мое, удлинённое, таз суженый. Шея длинная, подвижная и гибкая, относительно тонкая.
Шейных позвонков 18. Клюв относительно тонкий и длинный, с большим острым крюч-
ком на конце надклювья, края надклювья и подклювья гладкие, режущие, но на нёбе разви-
ты направленные назад шипы, препятствующие выскальзыванию добычи. Размерами гор-
лового мешка и степенью стрептогнатии бакланы уступают пеликанам, но могут очень
широко раскрывать клюв из-за высокой степени подвижности верхней челюсти относи-
тельно черепа. Добавочные порции челюстных мышц, прикреплённые к своеобразной па-
лочковидной косточке, отходящей от верхнезатылочной кости, способствуют силовому
сжатию челюстей. Щелевидные наружные отверстия ноздрей имеются лишь у молодых
птиц, у взрослых они зарастают. Под водой глаза прикрыты прозрачным третьим веком,
выполняющим функцию «подводных очков», его кривизна «настроена» на преломление
лучей света в водной среде и обеспечивает чёткое видение.
Короткие сильные ноги отнесены далеко назад, в результате чего гребки оказываются
очень эффективными, но на суше эта черта строения всех хороших пловцов и ныряльщи-
ков приводит к вертикальному положению тела и неустойчивости при ходьбе. Коленная
чашечка имеет отверстие, через которое проходит сухожилие, малая берцовая кость до-
ходит до интертарзального сустава, что обеспечивает возможность вращательных дви-
жений лапы. Бакланы могут грести лапами по очереди и одновременно. Если баклан плы-
вёт на поверхности, для него характерна очень низкая посадка тела. Ныряет без всплес-
ка, иногда погружается, просто прижав оперение. Обычная продолжительность нахож-
дения под водой 1-2 мин, глубина преследования рыбы до 10 м, однако, в виде исключе-
Семейство Баклановые
53
ния, бывают гораздо более длительные и глубокие погружения. Практикуются группо-
вые загонные охоты. В питании преобладают рыбы длиной 15-25 см, встречаются и мол-
люски, ракообразные, иглокожие, земноводные. Бакланы считаются очень прожорливы-
ми, однако установлено, что средняя дневная норма крупных видов составляет всего 300—
500 г рыбы.
В связи с меньшей необходимостью смазывать оперение копчиковая железа у бакланов
небольшая, всего с двумя выводными протоками. Слепые кишки рудиментарны, функцио-
нируют обе сонные артерии.
Летают бакланы неплохо для столь специализированных к нырянию птиц, могут даже
планировать, но полёт маломаневренный, с воды взлетают после разбега. Иногда бакланы
охотятся с пикирующего полёта, как олуши. Во время миграций и дальних перемещений к
местам кормежки бакланам свойственно образование стай в виде верениц — усложнённых
линейных построений, состоящих из сочетания различных простых форм: клиньев, углов,
цепочек, косых шеренг, дуг, вытянутых в длину от нескольких сотен метров до нескольких
километров. В кормовых недальних перелетах характерны скученные плотные построения
(Молодовский, 2001). В семействе есть нелетающий вид — эндемик Галапагосских остро-
вов, где отсутствуют наземные хищники. В результате отказа от полёта крылья галапагос-
ского баклана (Nannopterum harrisi) укоротились до 25 см, но сам он стал самым крупным
в семействе и наиболее приспособленным к подводной охоте. Как и у других видов, отка-
завшихся от полёта, в его облике присутствуют педоморфные (ювенильные) черты: окрас-
ка оперения однотонно-бурая с чуть более светлым низом, нет сезонной смены наряда,
отсутствуют украшающие перья.
Окраска взрослых птиц у большинства представителей семейства тёмная, обычно с
металлическим блеском и чешуйчатым рисунком по спине и на крыльях. Ряд видов имеют
белый низ тела, иногда тёмная и двуцветная фазы существуют в пределах одного вида.
Нередко на голове развиты хохлы из тонких перьев, в брачный период развиваются волосо-
видные белые перья на голове или по всему телу. Бывает ярко окрашена радужина, голая
кожа «лица» и горлового мешка, орбитального кольца. Голоса бакланов грубые и низкие,
галапагосский баклан во время тока издаёт рычание или глухой барабанный стук, исполь-
зуя надувающийся эластичный пищевод в качестве резонатора. Нижняя гортань несёт сла-
боразвитую мускулатуру.
Смена оперения бывает дважды в году. В начале года, в сжатые сроки, происходит не-
полная предбрачная линька (в основном вырастают украшающие перья). Полная после-
брачная линька со сменой рулевых и маховых, ведущая к частичной потере лётных качеств,
начинается летом и тянется до поздней осени.
Бакланы населяют как морские побережья, так и внутренние водоёмы, гнездятся коло-
ниями от нескольких десятков до тысяч пар. Гнёзда строят оба пола, носит гнездовой мате-
риал преимущественно самец. Гнездо — груда веток, травы, водорослей, скреплённых по-
мётом, — помещается на уступах скал, в нишах, на заломах тростника, в кронах деревьев и
прямо на земле. Виды, гнездящиеся в местах, лишённых растительности (например, в Су-
бантарктике), строят гнёзда из камешков. Иногда колонии удалены от воды на 3-5 км. В
кладке обычно 3^ белых, зеленоватых, голубоватых яйца. Насиживание, в котором прини-
мают участие оба родителя, длится 28-50 дней. Птенцы обычно тёмные, с более светлой
головой, на 3—4-й день прозревают, на 10-14-й день покрываются тёмным или серым пу-
хом, а полностью раскрывшимся контурным пером — в возрасте около 50 дней. Способны
к полёту в возрасте от 7 недель, ещё несколько недель лётных молодых докармливают взрос-
лые. Половозрелы на 2^-й год, в природе доживают до 18-20 лет (Карташев, 1974; Коб-
лик, 2001).
54
Отряд Пеликанообразные, Семейство Баклановые
Распространены бакланы практически по всём морским побережьям мира, кроме вы-
сокой Арктики и ледового щита Антарктиды, обитают также на многих внутренних водо-
ёмах. Широко кочуют вне сезона гнездования, с внутренних водоёмов обычно перекочё-
вывают на морские акватории. Предполагается, что внутренние водоёмы бакланы освои-
ли вторично. Центр происхождения группы неизвестен, возможно, это бассейн Индийс-
кого океана. Ископаемые остатки обнаружены в разных районах, начиная с верхнего па-
леоцена или нижнего эоцена (Северная Америка, 60-70 млн. лет назад). Описано около
30 вымерших видов из 3 родов, в ископаемом состоянии найдено не менее 10 современ-
ных видов. Один вид истреблён в историческое время, в угрожаемом состоянии находит-
ся ещё 11 видов, в основном узкоареальные островные эндемики. Наибольшее современ-
ное видовое разнообразие наблюдается в умеренных и холодных широтах южного полу-
шария.
Все 33^0 современных видов семейства часто объединяют в единственный род Phala-
crocorax в широком понимании (del Hoyo et al., 1992; Dickinson, 2003), или выделяют из
него в самостоятельный род Nannopterum только галапагосского баклана (напр. Howard,
Moore, 1980). Согласно недавней ревизии (Siegel-Causey, 1988), семейство делят на две
эколого-морфологические группы, которым предлагается присвоить ранг подсемейств Phala-
crocoracinae (пресноводные и частично морские формы) и Leucocarboninae (морские бак-
ланы), а число родов увеличивают до 8-9. При этом в роде Phalacrocorax в узком понима-
нии остаётся всего 3—4 крупных, преимущественно пресноводных вида, остальные же рас-
пределяются между родами Hypoleucos, Leucocarbo, Euleucocarbo, Stictocarbo, Notocarbo,
Compsohalieus, Halietor (= Micro carbo).
В данной сводке, согласно Л.С. Степаняну (2003), к роду Phalacrocorax sensu lato отно-
сятся все 8 видов бакланов, зарегистрированных в России и сопредельных странах. Из них
6 видов гнездятся, 1 залетает, 1 вымер.
Род Phalacrocorax Brisson, 1760
Большой баклан
Phalacrocorax carbo (Linnaeus, 1758)1
Pelecanus carbo Linnaeus, 1758, Syst. Nat., ed. 10, c. 133, «Европа» = северная
Скандинавия (Vaurie, 1965, Birds pal. Fauna, c.48).
Great [black, common, European] cormorant (англ.), Kormoran (нем.),
grand cormoran, cormoran ordinaire (фр.)
Син (лат.): для подвида Ph. с. carbo: Carbo vulgaris Lacepede, 1790; Carbo cormoranus
Meyer et Wolf, 1810; Carbo glacialis C.L. Brehm, 1824; Phalacrocorax carbo major Nilsson,
1835; Graculus americanus Reichenbach, 1850; Phalacrocorax carbo macrorhynchus Bonaparte,
1857; Phalacrocorax carbo litoralis Schuster, 1905; для подвида Ph.c. sinensis: Pelecanus
sinensis Shaw et Nodder, 1802; Carbo subcormoranus C.L. Brehm, 1824; Phalacrocorax carbo
medius Nilsson, 1835; Phalacrocorax humilirostris C.L. Brehm, 1835.
Син (рус.): корморан, морской ворон (устар.)
1 Разделы, касающиеся японского подвида большого баклана Ph. с. hanedae, написаны В.А. Нечае-
вым (прим. ред.).
большой баклан
55
Статус. Гнездящийся оседлый и мигрирующий вид. Атлантический подвид Ph.c.carbo
ъ значительной мере оседлый, но в условиях Белого моря совершает регулярные перелеты.
Материковый подвид Ph.c.sinensis на значительной части гнездового ареала — мигрирую-
щий либо кочующий, на юге — частично оседлый.
Общая характеристика и полевые признаки. Один из самых крупных видов бакла-
нов, величиной с гуся, с темной окраской оперения. У взрослых оно почти сплошь чёрное,
блестящее. В бинокль видно жёлтое пятно у основания длинного крючковидного клюва,
эелые щеки и подбородок, а в брачном оперении также белые пятна по бокам тела над
эедром и голенью. Последние признаки, как и отсутствие заметного хохолка на голове,
позволяют отличать большого баклана от хохлатого (Ph. aristotelis). У молодых птиц опере-
ние тусклое, бурое с беловатым низом. От близкого вида — японского, или уссурийского
баклана (Ph. capillatus) отличается большей площадью голого участка кожи у основания
лодклювья (заходит назад дальше разреза рта) и меньшим развитием белого поля на боках
головы.
Ведёт стайный образ жизни. Летают бакланы скученными и упорядоченными линейны-
ми построениями (Молодовский, 1990, 2001), при этом большие стаи (до 1200 особей)
летят клином, углом, скосами или зигзагами, более мелкие (до 200 особей) — шеренгой
либо дугой, а небольшие (до 30 особей) — цепочкой или «змейкой». При ежедневных кор-
мовых полётах бакланы могут лететь невысоко над водой (0,5-5 м), при дальних перелётах
в том числе кормовых) набирают высоту до 100 и более м. При линейных построениях
характерно волнообразное движение птиц друг относительно друга в вертикальной плос-
кости. Полёт несколько тяжёлый, но быстрый (60-70 км/ч), с чередованием не очень час-
тых взмахов крыльев и скольжения (планирования). Ноги и шея в полете вытянуты гори-
зонтально. Для взлёта с воды бакланам нужен предварительный разбег. На земле бакланы
неуклюжи, но на деревьях держатся вполне уверенно. Нелетные птенцы прекрасно лазают
по веткам, цепляясь за них лапами и ещё неоперёнными крылями. Бакланы превосходно
плавают, глубоко погружая тело в воду; при этом они держат шею вертикально, а голову и
клюв приподнимают немного вверх. Часто ныряют за добычей. После охоты подолгу су-
шат оперение, сидя неподвижно на каком-нибудь островке, ветке или скале среди воды в
характерной позе: туловище вытянуто вертикально вверх, крылья наполовину развёрнуты.
Бакланы обычно молчаливы, но в колониях достаточно шумны, порой даже крикливы.
Около гнёзд при кормежке птенцов они глухо урчат на низких тонах, что отдаленно напо-
минает карканье. При подходе к гнездовой бакланьей колонии эти звуки, издаваемые де-
сятками особей, слышны издалека. Иногда птицы в полёте издают громкое и резкое кар-
кание.
Подвидовая систематика. Большой баклан образует 6-8 подвидов (Howard & Moore,
.980; del Hoyo et al., 1992; Степанян, 2003; Dickinson, 2003), отличающихся по окраске,
тональности металлического блеска оперения и размерами. В Вост. Европе и Сев. Азии
считают 3 подвида, все они гнездятся в России.
1. Phalacrocorax carbo carbo
Pelecanus carbo Linnaeus, 1758, Syst. Nat., ed. 10, c. 133, «Европа» = северная Скандина-
вия
Самый крупный из российских подвидов. Оперение взрослых птиц имеет фиолетово-
с?:ний металлический оттенок. Побережья северо-востока Сев. Америки, Гренландии и
северо-запада Европейского материка, включая Кольский п-ов.
2. Phalacrocorax carbo sinensis
Pelecanus sinensis Blumenbach, 1798 (1796-1810), Abbildungen naturhist. Gegenstande, N
25. таблица и текст, Китай.
56
Отряд Пеликанообразные, Семейство Баклановые
Рис. 11. Область распространения большого баклана: а — область гнездования, б — область зимо-
вок. Подвиды: 1 —Phalacrocorax с. carbo, 2 — Ph. с. sinensis, 3 —Ph. с. hanedae, 4—Ph. c. maroccanus,
5 — Ph. c. lucidus, 6 — Ph. c. novaehollandiae.
Чёрные участки оперения взрослых птиц имеют металлический фиолетово-зелёный от-
тенок. Населяет восточную Европу, Азию вплоть до восточных берегов материка.
3. Phalacrocorax carbo hanedae
Phalacrocoraxcarbo hanedaeN. Kuroda, Sr., 1925, Tori, vol. IV, N 20, pp. 336-350, «Haneda,
between Tokyo and Yokohama», Япония.
По окраске оперения не отличается от Ph.c.sinensis, но характеризуется меньшими раз-
мерами. Япония, Курильские о-ва (Коблик и др., 2006)
Прочие подвиды не связаны с описываемой территорией. Ph. с. maroccanus Е. Hartert,
1906 (4), и Ph. с. lucidus (М.Н.К. Lichtenstein, 1823) (5) присущи африканскому континенту,
a Ph. с. novaehollandiae Stephens, 1826 (6) — Австралии и Новой Зеландии (рис. 11).
Замечания по систематике. Подвид Ph. с. hanedae выделен из Ph. с. sinensis сравни-
тельно недавно (N. Kuroda, 1925, цит. по: Коблик и др., 2006) и ещё в 1940-1950-х гг. при-
знавался далеко не всеми (Судиловская, 1951). Начиная с 1960-х гг., «материковый» афри-
канский подвид Ph. с. lugubris Ruppell, 1845 систематики, как правило, переводят в сино-
нимы Ph. с. lucidus, а южноавстралийский Ph. с. carboides Gould 1838 и новозеландский
Ph. с. steadi Е. Hartert, 1920 — в синонимы Ph. с. novaehollandiae (Howard, Moore, 1980; del
Hoyo et al., 1992). Высказываются и мнения о видовой обособленности рас Ph.c.lucidus и
Ph. с. novaehollandiae (del Hoyo et al., 1992).
Описание. Окраска. Самец и самка в брачном наряде. Голова, шея до основания,
нижняя часть спины, поясница и верхние кроющие хвоста, а также весь низ — чёрные с
блестящим сине-фиолетовым (атлантический подвид Ph. с. carbo) либо зеленовато-фиоле-
товым (материковый подвид Ph. с. sinensis и японский подвид Ph. с. hanedae) отливом.
Перья лопаток, верхней части спины, кроющие крыльев бронзово-фиолетовые (Ph. с. carbo)
или бронзово-оливковые (Ph. с. sinensis, Ph. с. hanedae) с чёрными каймами по краю, обра-
зующими чешуйчатый рисунок. Горло и щеки белые. С января и в течение всего периода
гнездования (по май-июнь включительно) на затылке имеется пучок удлинённых перьев, а
по бокам брюха в бедренно-голенной области — чисто-белые крупные пятна. Первосте-
пенные маховые и рулевые перья буровато-чёрные со слабым металлическим блеском на
к.тылой баклан 57
г-левых. Уздечка, пространство вокруг глаза, основание нижней челюсти и бока горла —
злые, жёлтого цвета. Граница между участком голой кожи на боках головы и оперенной
-лстью идёт от угла рта вниз или вниз и назад. Оперение шеи не доходит до угла рта и в
i -Lie узкого клина протягивается вперёд посредине голого подбородка. Радужина глаз изум-
~ ино-зелёная, ноги чёрные.
У’ материкового большого баклана Ph. с. sinensis на голове, по бокам шеи и на горле в
:сачном оперении (с января по май-июнь включительно) вырастают длинные узкие белые
е ерья, чего не наблюдается у номинативного подвида. У некоторых самок японского под-
:-<а Ph.c.hanedae (вероятно, в возрасте старше трх лет) в брачном наряде на фоне лимон-
х i-жёлтой голой кожи по бокам головы вблизи задней стороны глаза выделяется треуголь-
— :е розовато-красное пятно; у самцов подобная окраска не отмечается (Kuroda, 1925). Яр-
ое красное пятно на боках головы баклана заметно, в частности, на цветной фотографии,
: о бликованной в монографии «Wild Birds of Japan» (1985).
Взрослая птица в зимнем (послебрачном) наряде имеет чёрное с металлическим блес-
<м оперение с тёмными окаймлениями отдельных перьев спины. Плечевые, лопаточные,
верхние кроющие крыла и второстепенные маховые зеленовато-бурые с широкими тёмно-
1 :<ними окаймлениями, в результате чего образуется «чешуйчатый» рисунок. Белых пятен в
'едренно-голенной области нет.
Птенцы вылупляются голыми, их кожа чёрная с розоватым оттенком на голове и в мес-
lx будущих птерилий. Пуховой наряд, появляющийся только через 2 недели после выкле-
буровато-чёрный, но с беловатым оттенком в средней части груди и брюха, а также по
переднему краю крыла. Голая кожа горлового мешка светлая, клюв светло-серый.
Гнездовой наряд. Голова, шея и верхняя часть зоба буровато-серые с мелкими белыми
запинками, которые особо выделяются на нижней стороне шеи. Грязно-белое полукольцо
: темными мелкими пестринами охватывает нижнюю челюсть до глаз. Брюшная сторона
'етая, иногда с крупными тёмно-бурыми продольными пятнами. Бока тела, спина и над-
-достье не блестящие, тёмно-бурые с более светлыми окаймлениями перьев; отдельные
т ерья верхних кроющих крыла, плечевых и лопаточных партий, наоборот, окаймлены бо-
ее тёмным. Радужина бурая, клюв тёмно-бурый, несколько светлее на подклювье. Голая
<жа горлового мешка жёлтая.
Первый брачный наряд. Голова и шея ещё сохраняют буровато-серую с белыми крапин-
12АШ окраску оперения. Грязно-белое полукольцо у основания подклювья теряет темные
естрины. Брюшная сторона туловища белая с крупными продольными бурыми пятнами,
:собенно заметными на груди и по бокам брюха. Подхвостье буровато-чёрное с редкими
'етыми пестринами. Спинная сторона, за исключением головы и шеи, уже чёрная, как и у
в трослых птиц, но металлический блеск ещё слабый.
Первый зимний наряд. Верх головы и шеи тёмно-бурые с сероватым налетом и редкими
ел о вато-бурыми пестринами. На нижней стороне шеи и её боках пестрин больше. Брюхо
"ёчно-бурое с редкими продольными пестринами. Спинная сторона, как и раньше — чёр-
-is со слабым металлическим блеском.
Второй брачный наряд. Похож на таковой у взрослых птиц. Оперение уже с выражен-
-Д13! металлическим блеском, но ещё не появился характерный для вполне взрослых птиц
-чок удлиненных перьев на затылке. В то же время на бедрах появляются пока ещё незна-
чительные белые пятна.
Строение и размеры. Первостепенных маховых 11. Формула крыла П>Ш>1>1 V...
*_т>: II=III>I=IV>V... Вершины передних первостепенных маховых на внутренних опахалах
глкены примерно до половины длины. Хвост округлый, состоит из 14 жёстких рулевых
- ер ь^в умеренной длины. Оперение птиц густое, плотно прилегающее к телу.
58
Отряд Пеликанообразные, Семейство Баклановые
Размеры (мм) и вес взрослых птиц атлантического подвида Ph. с. carbo по измерениям,
сделанным в Белом море согласно разным отечественным источникам, таковы: длина кры-
ла 335-370 (у самок до 365); длина клюва (от оперения лба) 65-85 (в среднем 70); длина
цевки 66-75. Размах крыльев 1 320. Вес может превышать 3 000 г. Размеры птиц из Запад-
ной Атлантики и северной Скандинавии согласно С. Крэмпу (Cramp, 1977) следующие: у
самцов (п = 6) длина крыла в среднем 357 (350-363); длина клюва — 69,6 (67-73); длина
цевки — 74,0 (68-78). У самок (п = 12), соответственно: 339 (318-351); 63,7 (59-68); 70,6
(67-74).
Размеры взрослых птиц (мм) материкового подвида Ph. с. sinensis согласно отечествен-
ным источникам: длина крыла 330-356; длина клюва (от оперения лба) 56-75; длина цевки
60-75; размах крыльев 1270-1560. У самок размеры несколько меньше, чем у самцов. По
С. Крэмпу (Cramp, 1977), у материкового большого баклана размеры таковы: у самцов
(п = 38) длина крыла в среднем 347 (338-364); длина хвоста — 155 (145-165); длина цевки —
69,4 (66-73). У самок (п = 18), соответственно: 325 (311-337); 144 (133-154); 66,1 (64-70).
Длина клюва самцов (п = 34) в среднем 62,6 (58-67); самок (п = 15) — 55,7 (50-58). Сред-
ний вес самцов (п = 55) 2 529 г, самок (п = 25) 2 025 г (однако в целом вес очень изменчив,
он колеблется в пределах 1 800-3 000 г). По месяцам средний вес самцов и самок (вес
последних приведен в скобках), обитающих в дельте Волги, составил: в марте 2 369 г (1 999 г),
в апреле 2 650 г (2158 г), в мае 2 421 г (2 112 г), в сентябре 2 570 г (1 882 г), в октябре 2 670
г (Судиловская, 1951).
Японский подвид Ph. с. hanedae характеризуется самыми мелкими размерами (мм). Сам-
цы (п = 6): длина крыла 326-340 (в среднем 330), длина клюва 57,5-69,0 (64,6). Самки (п =
8): длина крыла 314-326 (320), длина клюва 56,0-61,0 (58,0) (Kuroda. 1925).
Линька. Различают наряды: пуховой - гнездовый - первый брачный - первый зимний -
второй брачный - второй зимний - и т.д. Окончательный, дефинитивный наряд (третий
брачный и третий зимний) появляется на четвёртом году жизни. Линька проходит у атлан-
тического и материкового бакланов идентично.
У молодых птиц известны две линьки в году: частичная в августе-сентябре и полная с
апреля по август. Гнездовый наряд появляется у молодых птиц в августе. В таком наряде
они зимуют, а в апреле начинается линька рулевых, кроющих крыла, а затем маховых перь-
ев. Эта линька заканчивается к июлю.
У взрослых птиц — две линьки в году. Неполная, предбрачная начинается в декабре и
завершается в январе. Птицы одевают брачный наряд, в котором они находятся до мая, а то
и до июня включительно. Вторая, послебрачная линька полная. Она растянута и начинает-
ся в мае-июне. Вначале исчезают белые украшающие перья брачного наряда (на бедрах, на
голове и шее), затем наступает очередь мелкого оперения. Эта смена заканчивается к сен-
тябрю-октябрю. В августе и сентябре происходит смена рулевых (от центральных к край-
ним), а в конце августа - сентябре и по октябрь сменяются маховые перья (от внутренних к
наружным).
Распространение. Гнездовой ареал. Сев.-вост. часть Сев. Америки (побережья
Ньюфаунленда, залива Св. Лаврентия, о-ва Антикости, Микелон, Принца Эдуарда, пролуо-
стров Новая Шотландия), Западное побережье Гренландии, Сев.-Зап., Вост, и Юж. Афри-
ка, Юж. и Юго-Вост. Азия, Австралия, Тасмания, Новая Зеландия, о-ва Чатем. Евразия от
атлантического до тихоокеанского побережья (рис. 11).
В Вост. Европе и Сев. Азии в пределах бывш. СССР почти весь видовой ареал занимает
материковый большой баклан Ph. с. sinensis^ на побережье Кольского п-ова и островах Кан-
далакшского и Онежского заливов Белого моря гнездится атлантический подвид Ph. с. carbo,
>:_тыпой баклан
59
20 60 100 140 180
3Ис. 12. Область распространения большого баклана в Сев/ Евразии: а — область гнездования, б —
-Хасть зимовок.
-с Курильских о-вах и, возможно, на Сахалине — японский большой баклан Ph. с. hanedae
-гг. 12).
В Калининградской области вид обычен, даже многочислен (Гришанов, 1994, Гриша-
<5. Беляков, 2000); гнездится в Литве (Zydelis et al., 2002), а в последние годы появился
’ ^сже в Латвии и Эстонии (Приедниекс, Страздс и др., 1989; Hagemeijer, Blair, 1997; Я. Вик-
_~е. личн. сообщ.), хотя ещё в начале 1980-х гг. большой баклан в Латвии считался залёт-
-дсч видом (Тауринып, 19836). Начиная с 1980-х гг. отмечались все более учащающиеся
чггречи стаек бакланов на реках Тиса, Латорица, Уж и прочих водоёмах Закарпатской низ-
менности (Луговой, 1994); бакланы прилетали сюда в основном из Венгрии, где в конце
-X века гнездилось не менее 1 800 пар (Magyar et al., 1998). С 2006 г. стали известны
-большие (в 2-3 гнезда) поселения бакланов и в самой Закарпатской области (личное
. >?бщение охотоведа Л. Потиша). Далее на восток вид регулярно и давно гнездится в Дне-
-рювских лиманах (выше по Днестру бакланы не гнездятся — Когут и др., 1996), а начиная
. е?нца 1990-х гг. — спорадично в Ивано-Франковской обл. (Бучко,1998). Обычен баклан в
лгеллиманной части дельты Днепра, где вверх по течению реки доходит до южных днеп-
хзских каскадов (Мельничук, 2001). Выше Каховского вдхр. самая большая колония в 57
была отмечена в 1986 г. (Poluda et al., 1997). Обнаружена небольшая колония на р.
2гсне выше Чернигова, в 1986 г. 2 пары гнездились на Днепре близ Киева; возможно вид
останавливается и на других реках, таких как Сула на Полтавщине, р. Самара на Днепро-
стровщине, где бакланы гнездились в начале XX века (Poluda et al., 1997). Начиная с 1970-
— гнездится на Лебяжьих о-вах Крыма (Костин, 1983). Заселяет заросшие тростниками
’ южинские о-ва близ Скадовска и комплекс Устричных о-вов Джарылгачского залива Чёр-
-х: моря (Ардамацкая, 2001); стал обычен и на островах Молочного лимана на Азовском
ксе (Кошелев и др., 1990). С недавних пор вновь гнездится в Ставропольском крае (Хох-
.6. Бичеров, 19866). Обычен в плавнях Кубани, а также от устья Терека и Кизляра по
60
Отряд Пеликанообразные, Семейство Баклановые
Каспийскому побережью до дельты Волги, где многочислен (Луговой 1963). На север дохо-
дит до юга Саратовского Заволжья (Завьялов и др., 1997). В центральном Кавказе и по
прилежащему побережью Чёрного моря отсутствует. В Закавказье местами гнездится в
Грузии от Батуми до Поти; в Армении — в бассейне оз. Севан (Судиловская, 1951). В Азер-
байджане гнездится по бассейну Куры, начиная от Мингечаурского вдхр. (Васильев, 1969)
до Ленкоранской низемености (Туаев, 1962), а также по берегу и островам Каспия.
В центральной Калмыкии стал гнездиться с 1980-х гг. (Кукиш, 1984). В Волжско-Ураль-
ских степях к северу доходит до Камыш-Самарских озёр и далее на север до 49° с.ш. (Завь-
ялов и др., 1997). На северо-восточном Каспии гнездится в устьях рек Урала и Эмбы, к югу
до Устюрта, и, конкретно, по южному берегу п-ова Мангышлак (Молодовский, 1975). Вверх
по Уралу не доходит до Оренбурга. На Арале был известен в районе оз. Шомке-Куль, обра-
зованого одним из рукавов Аму-Дарьи у мыса Ак-Кала (Рашкевич, 1969). На Илеке встре-
чается только в низовьях. В Зап. Сибири обитает на оз. Чаны, далее на восток отмечен на
Алтае на Иртыше, на Кара-Иртыше и Зайсане. В восточной части Казахстана материковый
большой баклан вполне обычен в пойме р. Или выше Капчагайского вдхр. и по степным
озёрам (Tamm, 2006), а также в Алакульской котловине, где во второй половине 1960-х гг.
проводилось массовое кольцевание птенцов (Гаврилов, 1972). Гнездится на островах Бал-
хаша и в дельтах впадающих в него рек (Долгушин, 1960), а также по южному побережью
этого озёра от протоки Жидель до урочища Коржун; в Вост. Прибалхашье — в низовьях
р. Каратал (Владимиров, Злобин, 1969). В Тянь-Шане — на оз. Иссык-куль и на горном
озере Тюп. К востоку от Алтая отмечен на озёрах Джигетай-куль в Танну (Тува) (Суди-
ловская, 1951).
В Туркмении большой баклан распространён на юго-восточном побережье Каспия к
северу до Кара-Богаз-Гола, на Мургабе, Теджене и Аму-Дарье. Ранее гнездился в некото-
рых районах Таджикистана (Судиловская, 1951), но более поздние исследования птиц это-
го региона гнездования баклана не подтверждают. Большой баклан в Таджикистане отне-
сен к пролётным и зимующим видам (Абдусалямов, 1971).
В Саянах и в Минусинском крае отсутствует, в Туве отмечен к северу до 51 -й параллели.
До середины XX в. гнездился на оз. Байкал (Судиловская, 1951), затем оттуда исчез, хотя
продолжал гнездиться на соседнем монгольском озере Хубсугул, что всего в 200 км юго-
западнее (Скрябин, 1982). Об отсутствии большого балана на гнездовье в дельте Селенги,
куда отдельные особи лишь залетали, указывал также И.В. Фефелов (Фефелов и др., 2001).
Однако в 2006 г. большой баклан вновь загнездился на Байкале: два гнезда с птенцами
найдены на о. Шаргадагон в проливе Малое Море (Пыжьянов, 2007). В Забайкалье боль-
шой баклан гнездится на Торейских озёрах (Зубакин, 1981). Далее на восток вид распрос-
транён до Приморья. Гнездится на Аргуни, в верхнем и нижнем течении Амура, от устья и
до верховий Уссури.
Гнездовой ареал японского подвида Ph. с. hanedae — Японские острова (Хонсю, Сико-
ку, Кюсю, где птицы оседлы). Нерегулярно залетает на о-в Хоккайдо (возможно, гнездит-
ся). В России, по всей вероятности, гнездится; в гнездовой период бакланов отмечали на
Курильских о-вах: Шикотан (Kobayashi, 1933), Итуруп (Bergman, 1935) и Симушир (экзем-
пляр самца, добытого 8 августа 1955 г., сбор С.К. Клумова, колл. ЗМ МГУ). Указания А.Г.
Велижанина (1977а) о добыче самца 18 мая 1974 г. на о-ве Итуруп (тушка хранится в кол-
лекции ЗМ МГУ) ошибочны; при переопределении птица оказалась Ph. capillatus (=
filamentosus). Весной, 22 апреля 1992 г., стаю из 6 особей наблюдали в устье р. Сусуя, на
юге о-ва Сахалин (Нечаев, 2005).
Необходимо отметить, что в связи с выделением подвида Ph. с. hanedae, сведения о
подвидовой принадлежности больших бакланов, встречающихся по дальневосточному
Бшъгюй баклан 61
г гсережью, на Сахалине и Курильских о-вах подлежат серьезной ревизии. Если А.И. Ги-
еико (1955) вслед за А.М. Судиловской (1951) полагал, что там может обитать только ма-
-ет«псовый большой баклан, то В.А. Нечаев находил там и японский подвид (Нечаев, 19916;
-шаев, Фудзимаки, 1995).
Зимовки. Большие бакланы подвида Ph. с. carbo, гнездящиеся на Мурмане, а также
• *1зндалакшском и Онежском заливах Белого моря, зимуют, очевидно, на Балтийском море
Скокова, 1978а; Bianki et al., 1997).
При сравнении размещения мест зимовок трёх групп материковых больших бакланов
-: sinensis, гнездящихся на севере Чёрного моря, в Азово-Сивашском регионе и дельте
Белги, можно утверждать (если не брать во внимание особей, которые зимуют поблизости
- мест гнездования), что для черноморских птиц наиболее характерны места зимовок в
Балканских странах (Албания, Греция, Черногория, Болгария, Румыния) и в их окрестнос-
та. т.е. основные зимовки расположены к западу и юго-западу от места гнездования. Часть
—ш. зимующих в Дунайском бассейне, кочуют в Закарпатской обл. Украины по р. Тисе
1к. в декабре-январе 2002-2004 гг. встречены одиночки и стаи в Береговском р-не чис-
тенлюстью более 100 особей). Места зимовок птиц из дельты р. Волги более компактны —
. ни расположены на Каспийском море и, частично, в Персидском заливе (Азербайджан,
л.ркмения, Иран, Арабские эмираты), то есть находятся к югу от мест гнездования. Наи-
хлыпий «разброс» мест зимовок характерен для бакланов Азово-Сивашской группиров-
ки Зимовки у них расположены в равной степени как южнее (Израиль), так и восточнее
Краснодарский край России, Абхазия, Азербайджан) и западнее (Румыния, Болгария)
«ест гнездования. Это может быть связано с тем, что бакланы освоили данный регион
е главно, и азовская группировка бакланов формировалась из разных географических ПО-
H. ПЯЦИЙ.
Бакланы, гнездящиеся в различных частях Казахстана — на Капчагайском вдхр., в дель-
~z Или, в низовьях Таласа, на Аральском море и в низовьях Иргиза, зимуют в южных час-
-tx Таджикистана, в Туркмении и в Иране, на юж. Каспии (36-38° с.ш. и 49-69° в.д.).
Декабрьская добыча в низовьях Вахша окольцованной молодой птицы показала, что
некоторые бакланы с Аральского моря зимуют на юге Памиро-Алая. Бакланы из Балхаш-
-лакольской котловины проводят зиму значительно южнее — в Индии и, вероятно, в Юж.
?*>пае, откуда пока возвратов колец нет (Иванов, 1969; Гаврилов, Гисцов, 1978). В конце
XX столетия условия зимовки бакланов в южных районах Средней Азии значительно улуч-
шались в связи с тем, что здесь был создан ряд крупных водохранилищ на Сыр-Дарье,
Дуэгабе, Тенджене (Чардинское, Фархадское и Кайрак-Кумское, Султанбентское, Первое
- Второе Тендженские и ряд других).
Если говорить о северной, прибалтийской популяци Ph. с. sinensis, то по данным коль-
_ езания в Польше, Германии, а также в бывш. Чехословакии эти бакланы пересекают евро-
пйский континент и зимуют в Средиземноморье — в Италии, на Корсике, Сицилии, в
_~гснах бывш. Югославии, Греции (Скокова, Кищинский, 1978). К таким птицам, по-види-
< чу. относятся и бакланы, гнездящиеся на территории Калининградской обл. РФ, в Литве
лптих странах Прибалтики. Одиночки зимуют на Балтике, в открытом море (Тауринып,
Большие бакланы японского подвида Ph. с. hanedae зимой отмечаются на Японских о-
Цусима, Идзу, Огасавага, Дайто, Рюкю и др. (Check-list..., 2000).
Миграции. Для разных подвидов и географических популяций миграции проходят по-
? _:н?чу: по классификации Фесенко (2001), большой баклан — «ближний» мигрант.
^.антический подвид. В Англии часть птиц оседла, часть совершает перёлеты до юга
Хпен ейского п-ова и берегов Сев.-Зап. Африки. Большие бакланы, гнездящиеся на Белом
62
Отряд Пеликанообразные, Семейство Баклановые
Рис. 13. Миграции большого баклана,
гнездящегося на Семи Островах: а — ме-
сто мечения, б — сентябрьские и октябрь-
ские встречи молодых, в — ноябрьские и
декабрьские встречи молодых, г — встре-
ча взрослой птицы в октябре, д — встре-
ча на весеннем пролёте (по Скоковой,
1978а, с уточнениями).
море, летят осенью (начиная с конца августа) через Финляндию на Балтийское море и зи-
муют вдоль побережий Дании и юго-вост. Швеции (Bianki et al., 1997). Осенние миграции
здесь начинаются в конце августа и продолжается до октября-ноября. Молодые птицы с
Мурмана летят как непосредственно на юго-запад через систему озёр Финляндии, так и
огибая Кольский п-ов, через горло Белого моря (рис. 13). Над сушей бакланы летят, оста-
навливаясь на внутренних водоёмах, и встречаются здесь до ноября, единично до декабря.
Один молодой баклан 12 ноября найден уже на восточном побережье Швеции, в 1 600 км
от гнездовья; взрослая птица на четвёртый год после кольцевания добыта в октябре в Фин-
ляндии, на побережье Финского зал.
Н.Н. Скокова (1978а) считает, что широкий разброс находок окольцованных птиц пока-
зывает, что они летят над Карелией и Финляндией «веером», расширяющимся от Онежско-
го залива по направлению к Финскому зал. Балтийского моря. Путь от места кольцевания
до юж. Финляндии (800-1 200 км) популяция больших бакланов преодолевает за 2-3 меся-
ца. Добыча одной молодой птицы в апреле следующего года в Ленинградской обл. показы-
вает, что часть ещё негнездящихся птиц может летовать вне своей родины.
Материковый подвид. В Эстонии (п-ов Пурикаринеэм) пролёт в 1990-1992 гг. отмеча-
ли с 17 марта по 3 апреля весной и 17 июля, 14 августа — поздним летом (Домашевский,
2001). В регионе Рижского зал. весной бакланов наблюдают регулярно, хотя и в ограничен-
ном количестве, стайками до 5 особей. Незначительный весенний пролёт отмечали также в
заливе Хара-Лахт (Эстония) в 1990-1992 гг. (Домашевский, 2003). В южной части Латвии
регистрируются летние (июль-август) кочёвки одиночных особей и небольших групп на
рыбоводных прудах, оз. Лиелауцес и в других местах (Тауринып, 19836).
В Ивано-Франковская обл. Украины бакланы появляются в середине февраля (Бучко,
1998), в низовьях Днестра — в конце февраля, хотя массово — в середине марта (Ког-
zyukov, 1997). В дельту Днепра вид прилетает не раньше 1 марта (Ардамацкая, 1968). В
западной части дельты Волги бакланы появляются около 25 февраля (в среднем за 16 лет
наблюдений), сроки колеблются в пределах от 15 февраля (1952 г.) до 7 марта (1954 г.). В
восточную часть дельты прилетают на неделю позже, в среднем 4 марта — амплитуда коле-
баний дат от 25 февраля (1949 г.) до 14 марта (1954 г.) (Луговой, 1963). Массовый осенний
отлёт бакланов из дельты Волги приходится на конец первой - вторую декаду ноября. Уста-
новлено, что местная, «дельтовая» популяция бакланов к осени пополняется птицами с
востока Каспия (Мангистаусские о-ва) и Арала, которые устремляются в дельту на жиров-
>_ ’ьшой баклан 63
«Гладков, 1957, цит. по: Скокова, 1960а). Кроме того, сюда прилетают птицы и с запада,
-: стороны Калмыцких степей. Так, в Лиманском р-не Астраханской обл 4 октября 1959 г.
\гла отмечена стая, насчитывавшая 2 тыс. особей (Луговой, 1963).
Для Казахстана И.А. Долгушин (1960) приводит такие сроки весенного прилёта боль-
~-fx бакланов: в дельте р. Или они появляются в начале - второй половине марта, на Сыр-
дарье близ Чиназа — в середине марта, а ниже Джулека — в апреле. На Аральском море (о.
Дсрса-Кельмес) — в конце марта; на Зайсане около 10-15 апреля, тогда же становятся обыч-
-здми и под Семипалатинском. В Кустанайской обл. весенний пролёт приходится на тре-
~-ю декаду апреля (Гынгазов, Миловидов, 1977). Осенний пролёт бакланов в северной по-
• :зиние Казахстана идёт в конце сентября - октябре, в южной — в октябре-ноябре.
На восточном побережье Аральского моря в 1978 и 1979 гг. весенний пролёт больших
Ггчланов был несущественным, вид составлял менее 0,2% от учтённых там водных и око-
: водных птиц. В 1978 г. с марта по май было зафиксировано 295 пролетевших бакланов, в
:"9г. — 159; максимальное число птиц отмечено в апреле (Березовский, 1983).
Особняком стоят данные по перемещениям бакланов на Памиро-Алае. Здесь сезонные
грёлеты выражены очень слабо. Единично бакланов отмечали на весеннем пролёте меж-
10 марта и 20 апреля на юге региона (долина Вахша, на реках Кафирниган, Ханака,
Д.тиамбе). На осеннем пролёте — начиная с сентября и до 22 ноября; порой стайками до
:; особей. Летние кормовые кочёвки, в основном молодых «бродячих» птиц, регистриро-
вались преимущественно в горной части этого региона — на Памире (Аму-Дарья, озёра
' Дракуль, Яшилькуль и др.) (Иванов, 1969).
По данным возвратов колец, пути миграций бакланов из колоний Азово-Черноморского
тегиона выглядят следующим образом (Мациевская, Кошелев и др., 1998). Птицы, поме-
- еиные птенцами в дельте Днестра, летом оказываются в дельте Дуная — в районе Вилкова
з Румынии (в одном случае даже на 6-й день после кольцевания). Однако часть молодых
—щ летит на восток (встреча 26 августа в Джарылгачском заливе Чёрного моря), а часть
стаётся до глубокой осени в дельте Днестра (находки окольцованных птиц в октябре и
ервой половине ноября). 25 октября 1979 г. в Турции, в окрестностях оз. Измир был обна-
~жен «днестровский» баклан, погибший на пятый год после кольцевания, а 23 ноября 1978
- з Болгарии близ г. Бургас — сеголеток. Балканские страны служат основным местом
:имовок птиц, гнездящихся не только в дельте Днестра, но и в Тендровском зал. Об этом
.зидетельствуют находки птиц из обеих популяций в Греции (21 февраля и 20 марта), Чер-
- . гории (Скадарское озеро, 23 февраля), Албании (2 декабря), Болгарии (3 января). 28 мар-
1985 г. одна птица добыта уже на весеннем пролёте в Одесской обл.
Бакланы с Лебяжьих о-вов (сев.-зап. Крым) также частично зимуют на Балканах (по
. дной декабрькой, январской и февральской встрече в Румынии, Греции и Македонии), а
сдельные птицы остаются там и летовать (окольцованный 20 мая 1980 г. баклан был от-
.-делян 28 мая 1981 г. в Греции, в окр. Комотини). Однако большинство «лебяжьеостровс-
бакланов дают зимние возвраты из Краснодарского края (6 находок в Анапе, п-ов
"дмань, Темрюк, лиман Гнилой и др.) а также из Абхазии (Сухуми) и приморских мест
'г-дни (4 возврата). Во время летних послегнездовых кочёвок бакланы с Лебяжьих о-вов
- 2сто регистрируются близ Керчи и в других местах Крыма, в Запорожской и Херсонской
<л Встреча черноморского баклана 1 июля 1987 г. на седьмой год после кольцевания в
Венгрии (г. Чонград) позволяет предполагать, что часть птиц осваивает новые места гнез-
z. зания, далёкие от места своего рождения.
Птиц, окольцованных птенцами в сев. Крыму (на Сиваше), в период зимовок отстрели-
=а_ти в пределах Чёрного моря — юго-восточнее (4 марта — Анапа), южнее (1 февраля —
Турция, дельта р. Гексу) и юго-западнее (28 февраля — Болгария, Старозагорский округ)
64
Отряд Пеликанообразные, Семейство Баклановые
мест гнездования. Позднелетние (август) и осенние (сентябрь-октябрь) встречи отмечены
как в районе гнездования (Геническ), так и в разных местах Азовского моря (Бердянская
коса, Ачуевская коса, Молочный лиман).
В свою очередь бакланы, окольцованные птенцами на Молочном лимане, в летний пе-
риод были встречены в Азовском море (Бердянская коса), в Чёрном море (Херсонская об-
ласть, дельта Дуная) и в Каховском вдхр. Птицы этой популяции бакланов зимуют как дале-
ко к югу от мест гнездования (возвраты из Израиля 1 декабря 1994 г., 28 января 1996 г. и 1
февраля 1995 г.), так и в Краснодарском крае, в том числе на р. Кубань (18 декабря 1999 г.,
30 декабря 1989 г., 10 марта 1998 г.). Часть перезимовавших молодых птиц остаётся лето-
вать в Краснодарском крае (находка 21 мая 1991 г. близ г. Темрюк).
Среди птиц, окольцованных на Обиточной косе Азовского моря (северо-восточнее Мо-
лочного лимана), картина возвратов практически схожа. Обращают на себя внимание сле-
дующие две находки: одна птица, окольцованная птенцом 2 июня 1993 г., была обнаружена
через 4,5 года на зимовке в Швеции (Блекнинг); другая была застрелена 30 ноября 1986 г.
(через 5 месяцев после кольцевания) близ г. Пойлу, в горном районе верховий реки Куры,
относящейся уже к бассейну Каспийского моря.
79 возвратов колец от бакланов, помеченных в дельте Кубани, показали, что птицы этой
популяции в большинстве случаев не совершают далёких миграций (Скокова, 1978а). По-
чти все они были добыты в заливах и лиманах Азовского и Чёрного морей, причём 80% во
все сезоны — поблизости от места кольцевания в плавнях Кубани и прилежащих районах,
не далее 100-120 км. Максимальное удаление от места кольцевания — около 1000 км к
западу (пролив Босфор).
Молодые бакланы, появившиеся на свет в дельте Волги (п= 171), после вылета из гнёзд
держатся в течение 4-5 месяцев преимущественно вблизи мест гнездования (Скокова, 1960а).
С мест ночёвки бакланы совершают кормовые полёты в радиусе 10-30 км в зависимости от
мест скопления рыбы. 16 возвратов колец показывает, что бакланы западной части дельты
(Дамчикский участок Астраханского заповедника) осенью летают на кормёжку в район
Волжско-Каспийского канала, где в сентябре-октябре проходит предзимовочный миграци-
онный путь чехони, а в октябре-ноябре — воблы и леща. Это подтверждается и визуальны-
ми наблюденими. Во время утреннего учёта на Дамчикском участке заповденика 12 сен-
тября в сторону Волжско-Каспийского канала пролетело 7 тыс. бакланов — фактически
все, ночевавшие в данном районе.
В период позднелетних кочёвок (первая декада августа) баклан вполне обычен на Вол-
ге. На маршрутном учёте по руслу реки от с. Солодники (близ Волгограда) до Солёного
Займища (104 км) он составил почти 18% от всех зарегистрированных птиц, причём всте-
чался большими стаями, в среднем насчитывающими более 65 особей (Golawski, Kasprzy-
kowski, 2004).
Хотя подавляющее большинство бакланов остаётся в дельте Волги до середины ноября,
часть птиц уже летом откочёвывает вдоль западного побережья Каспия к югу. Так, окольцо-
ванный в мае баклан 18 августа оказался уже в Азербайджане. А в начале ноября зафикси-
ровано несколько возвратов колец из Махачкалы (Дагестан) и Порт-Ильича (Азербайджан).
Основной путь пролёта бакланов проходит вдоль западного берега Каспийского моря. На
восточном берегу Каспия пролёт бакланов весьма незначителен и, можно сказать, случаен.
В 1977 г. близ пос. Чикишляр пролётные особи отмечались от 15 августа до 9 ноября (Кара-
ваев, Белоусов, 1983).
Небольшая часть молодых птиц летом и ранней осенью, ещё до начала дальних мигра-
ций к югу, разлетаются из дельты Волги на восток, север и запад. Так, 4 «дамчикские»
птицы были пойманы осенью на Обжоровском участке Астраханского заповедника — в
г»: ~ьшой баклан 65
.<> км восточнее места кольцевания, но ещё в пределах дельты Волги. Три возврата полу-
- ены ещё восточнее — в Денгизском р-не Атырауской (бывш. Гурьевской) обл. Казахстана.
'-*2 север молодые птицы поднимаются вверх по течению проток на десятки и даже сотни
одометров. 3 августа в Волго-Ахтубинской пойме был добыт молодой баклан, окольцо-
*я-:ный птенцом 6 июня на Дамчикском участке заповедника, что лежит на 290 км южнее.
Второй, уже определённо лётный баклан, окольцованный там же 3 сентября, через 4 дня
: чгзался в 130 км севернее, в Красноярском р-не Астраханской области. О западном на-
ставлении разлета молодых птиц в послегнездовое время свидетельствуют одиночные
-сходки «дельтовых» бакланов в Ставропольском крае (23 августа), в западных подстеп-
-ях ильменях и т.д
В середине ноября численность бакланов в дельте Волги резко снижается, а на юго-
:гладе Каспия уже в конце октября — возрастает; здесь она достигает максимума в конце
-: чбря и в декабре. Часть бакланов, в связи с уходом воблы на большие глубины, в янва-
те-феврале откочёвывает с юга Каспия до Персидского залива, однако значительное ко-
лче ство птиц остаётся на зимовку в водах юго-западного, юж. и юго-вост. Каспия. В
~ёг_лые зимы часть бакланов может оставаться в пределах дельты Волги и Сев. Каспия.
Г: наблюдениям А.В. Молодовского (1981), в ноябре-декабре 1956-1957 гг. число бак-
енов в Кызыл-Агачском заповеднике исчислялось сотнями и тысячами птиц, в Красно-
ярском заповеднике и прилежащих акваториях — десятками, а на Юж. Мангышлаке —
синицами.
Весеннее движение бакланов на Каспии начинается в феврале. В районе Кызыл-Агач-
. < го заповедника это совпадает с подходом косячных частиковых рыб. Февральские и
«артовские возвраты птиц получены с о. Сара, Апшеронского п-ова. По восточному бере-
' Каспия, как и осенью, заметного пролёта нет.
Хотя большинство молодых бакланов на второй год жизни возвращаются в дельту
Ъ г лги (15 возвратов), некоторых птиц летом отмечали далеко от мест появления на
лег — в Саудовской Аравии и Иране. Так, баклан, окольцованный осенью (18 ноября
-54 г.) на востоке дельты Волги, спустя два года (19 июля 1956 г.) обнаружен на п-ове
ангышлак.
Данные Н.Н. Скоковой (1960а) по миграциям больших бакланов дельты Волги были
: ян ее подтверждены и дополнены новыми сведениями (Скокова, Кищинский, 1978). В
- ятности установлено, что дальность миграций этой популяции, как правило, не превы-
_тег 800-900 км, хотя есть случаи зимовок и в 2 000 км от места гнездования. При этом
:яы могут удаляться на значительные расстояния от морских берегов. Так, по р. Куре
'яяаны поднимаются вверх по течению на 150 км, достигая Муганской и Мильской сте-
; г Получены возвраты «волжских» бакланов из окрестостей Багдада, в 600 км от моря, с
г в Лурестане, из области Иранского нагорья в 500 км от моря и т.д. Выяснено, что
- ясльшая часть птиц из дельты Волги летит зимовать и в юго-восточном направлении, о
- м говорят встречи птиц у форта Шевченко и в Красноводском зал., что в 350 и 800 км от
• ггт гнездования.
Территориальные связи больших бакланов, населяющих Балхаш-Алакольскую котло-
• -г. и Аральское море, как в период пролёта, так и на зимовках, позволяет отнести их к
7 яличным популяциям (рис. 14). Наиболее тесная связь существует между каспийской и
л ягьской популяциями. Выведшихся на Аральском море бакланов неоднократно встреча-
42 восточном побережье Каспия, а одного отметили ещё западнее — в Краснодарском
<7 ±е. часть птиц проводит зиму совместно на южном Каспии. Балхаш-Алакульская популя-
__? эолее обособлена, птицы в западном направлении перемещаются исключительно ред-
Зи.мовки этих бакланов находятся вдвое дальше, чем у аральских. Пути пролёта бал-
66
Отряд Пеликанообразные, Семейство Баклановые
Рис. 14. Области миграций бакла-
нов, гнездящихся в дельте Волги
и в Казахстане (по книге «Мигра-
ции птиц Восточной Европы и
Северной Азии», 1978): а — бак-
ланы дельты Волги, б — бакланы
Аральского моря, в — бакланы
озёр Балхаш и Алакуль.
хаш-алакульских бакланов проходят через Тянь-Шань, Джунгарский Алатау, пустыни Центр.
Азии, Тибет и Гималаи. Это подтверждается отсутствием возвратов колец из Средней Азии,
Ирана и Афганистана, где бакланы данной популяции практически не пролетают. Тянь-
Шань и Джунгарский Алатау птицы пересекают по межгорным проходам и речным доли-
нам. Каким путём бакланы пересекают Тибет и Гималаи — пока не известно (Гаврилов,
Гисцов, 1978).
В период современного увеличения численности баклана и освоения им новых терри-
торий под гнездовья, расширяется и география кормовых кочевок, которые порой трудно
отличить от залётов. Так, участившиеся встречи одиночных птиц и стаек больших бакла-
нов в Оренбургской обл. по рекам Урал, Киндель, Боровка, на Ченкалинском вдхр. и т.д.,
которые ГМ. Самигуллин (1988) относит к разряду залётов, следует скорее всего считать
регулярными кормовыми кочёвками.
Залёты одиночных материковых бакланов или их групп известны из очень многих пун-
ктов. Они отмечены в Кировской (по р. Вятке) и Нижегородской областях (Попов, 1977),
неоднократно в Московской обл. (Калякин, Волцит, 2006), на Рыбинском вдхр. осенью 1946
г. (Зиновьев, 1981а), в среднегорье Зап. Кавказа 7 февраля 1992 г. (Мнацеканов, 1998). В
южных окраинах лесной зоны Сибири птиц отмечали в Новосибирске, Томске, Тюмени,
Зырянске на р.Чулыме (Гынгазов, Миловидов, 1977), в Приангарье (Липин, Сонин, 1986).
Местами эти залёты приняли регулярный характер. Так, в юго-западной части Чернигов-
ской обл. бакланов стайками до 12 особей отмечали в весеннее время в 1986, 1991, 1993,
1994 гг. (Бабко, 1995).
Залёты японского подвида зарегистрированы на п-ов Корея и о-в Тайвань (Check-list...,
2000), о-в Сахалин и Курильские о-ва (Нечаев, 2005).
Местообитания. Местом гнездования и отдыха атлантического подвида Ph. с. carbo
служат скалистые берега и острова, местами кормёжки — морские литоральные отмели.
Местообитания материкового подвида Ph. с. sinensis весьма разнообразны и зависят от
местных условий каждого конкретного региона. На затапливаемых участках речных дельт
материковый баклан гнездится преимущественно на деревьях и кустарниках (ветла, ольха,
Большой баклан
67
г>таи, турангии, заросли тамариска, тальник). На озёрах и в опреснённых заливах морей
предпочитает поселяться в тростниковых зарослях, хотя известны случаи наземного гнез-
дования на островах озёр Калмыкии, Забайкалья и Дальнего Востока (Зубакин, 1981; Ку-
киш, 1984; Глущенко и др., 2003). Наземное гнездование особенно характерно для морских
островов (Чёрное, Азовское и, в меньшей степени, Каспийское моря). Данный подвид мо-
жет гнездиться и на скалах либо на скалоподобных береговых обрывах. Это характерно для
птиц, обитающих на островах озёр Балхаш и Алакуль, а также на восточном побережье
Каспия (Долгушин, 1960; Молодовский, 1975; Андреев и др., 1977). На искусственных во-
дохранилищах — например, Бурштынском (Зап. Украина) — может поселяться на остро-
вках с небольшими группами и даже одиночными деревьями (Бучко, 1998).
Кормовыми биотопами материкового подвида служат морские мелководья, озёра, пру-
ды и крупные реки, богатые рыбой.
Численность. Численность вида со второй половины XIX в. подвергалась большим
колебаниям. Ещё в конце XIX в. атлантический большой баклан Ph. с. carbo на Мурмане
был обычным и даже многочисленным видом. Но в первой половине XX в. произошел
резкий спад численности. Это позволило А.М. Судиловской (1951), ссылаясь на других
исследователей, писать о том, что большой баклан на Мурмане редок и кое-где исчез со-
всем, а в районе архипелага Семь Островов не только больше не гнездится, но даже не
залетает. Однако в 1960 г. на о. Кувшин этого архипелага вновь загнездились 22 пары, а в
1972 г. — 10 пар (Корнеева, Рахилин, 1975). По данным Ю.В. Краснова с соавторами (1995),
первые три колонии численностью в 100 пар на Семи Островах появились в 1976 г., когда
вблизи островов отмечали большие скопления тресковых рыб. Но уже в 1978 г. косяки рыб
стали редкими, и количество бакланов на архипелаге уменьшилось. Следующий подъём
численности начался в 1990 г., что было связано с подходом нерестовых косяков мойвы, а
позднее различных тресковых рыб. В 1992 г. численность гнездящихся больших бакланов
на архипелаге Семь Островов достигла примерно 140 пар.
В Кандалакшком заливе Белого моря с 1950 по 1980 г. численность атлантических боль-
бакланов возросла в 20 раз, и этот рост продолжается. Первые семь пар загнездились
на о. Средняя Луда в 1967 г. (Bianki et al., 1997; Бианки, Бойко, 2001).
Что касается материкового большого баклана Ph. с. sinensis, то колабания численности
:<ого подвида были ещё более значительными. Некогда широко распространённый вид в
а . пинах крупных рек, большой баклан к первой половине XX в., в результате снижения
пбилия рыбы и преследования со стороны человека, оставался обычным в основном толь-
в дельтовых участках, причём преимущественно в заповедных зонах. Так, на Волге ещё
= середине XIX в. большой баклан гнездился вверх по течению вплоть до Самарской Луки
Богданов, 1871), а к началу XX в. он был обычным уже только в дельте этой реки, где
п'разовывал рассеянные, малочисленные колонии (Луговой, 1963). С организацией Астра-
дн с ко го заповедника (1919 г.) численность баклана в дельте Волги стабилизировалась, но
—иды были привязаны только к заповедным участкам. К началу XX столетия резко сокра-
~ _‘2сь численность бакланов также на Кубани, Дону (Белик, Данкевич, 2004) и в прочих
г^тдонах былого распространения вида.
2;ггуация резко изменилась в последней трети XX века, когда началось увеличение чис-
г-:-юсти вида и экспансия его в новые места обитания. Однако не везде экспансия была
__едной. Так, на Львовщине (заповедник «Росточье») встречаемость кочующих летне-
-1 -1них птиц сначала росла, а затем упала: 1986 г. — бакланов не отмечали; 1987 г. — 11
•- :<ей: 1992 г.—48 особей; 1994 г. — 15 особей; 1995 г. — 3 особи (Гузш, 1997). В Ивано-
-тднковской обл. (Бурштынское вдхр.), где ранее птицы вообще не гнездились, в 1989 г.
• >з?1лась колония из 11 гнёзд, которая к 1991 г. выросла до 44 гнёзд. Но уже в 1993 г. здесь
68
Отряд Пеликанообразные, Семейство Баклановые
оставалось не более 20 гнёзд, а в 1995 г. только одно, после чего бакланы на водохранили-
ще гнездиться перестали (Бучко, 1998).
В пригодных для бакланов угодьях экспансия оказалось более успешной. На Балтике и,
конкретно, в Калининградской обл., на территории которой бакланы не гнездились по край-
ней мере полтора столетия (последние сведения датированы 1787 г.), птицы появились вновь
только в 1980-е гг. (Гришанов, 1994). К концу XX столетия там уже гнездилось более 1 000
пар (Гришанов, Беляков, 2000). Рост численности бакланов на этом не остановился, и в
2005 г. в Калининградской обл. гнездилось порядка 8 тыс. пар (ГВ. Гришанов, in lit.). В
соседней Литве небольшие поселения бакланов в колониях серых цапель начали появлять-
ся с 1990-1991 гг., затем рост поголовья усилился, и в 2002 г. здесь насчитывали уже до 2
тыс. гнёзд (Zydelis et al., 2002).
В дельте Днестра, где в 1970-х гг. насчитывалось 100 гнездовых пар, к 1998 г. было уже
3 800 пар. Позже (2002-2004 гг.), в связи с упадком прудового хозяйства, численость снизи-
лась и держалась на уровне около 2 000 пар (Русев, 2004).
Крупнейшие поселения большого баклана находятся в дельте Дуная (Киш и др., 2003).
В 2002 г. здесь в 28 колониях насчитывалось 22 652 пары (23 колонии с 17 397 парами на
румынской стороне и 5 колоний с 5 255 парами на украинской стороне дельты).
История роста численности большого баклана в Черноморском заповеднике (Херсон-
ская область) подробно описана А.Г. Руденко и О.А. Яремченко (2004). Первое поселе-
ние бакланов здесь было отмечено в 1993 г. (11 гнезд), а максимальная численность в
период 1993-2002 гг. составляла уже 1 300-1 860 пар. В это время основные колонии на
территории заповедника размещались на Конских о-вах (Ягорлыцкий залив), о. Орлов и
Новых о-вах (Тендровский залив). Затем началось расселение вида на прочие заповед-
ные острова.
В 1984 г. на юге Украины суммарно насчитывалось 5 100 пар, а в 1985 г. — 3 500 пар
больших бакланов (Ардамацкая и др., 1988). О численности большого баклана на Азово-
Черноморском побережье Украины в 1998 г. даёт представление таблица 3.
На Лебяжьих о-вах северо-западного Крыма в начале 1980-х гг. в новообразовашихся
колониях гнездились сотни пар (Костин, 1983), большим успехом завершилось заселение
островов на Азовском море (Кошелев и др., 1990). На Каховском вдхр. (о-ва Большие и
Малые Кучугуры), где к 1960 г. бакланы были истреблены, поголовье начало восстанавли-
ваться в 1971 г. (появились 2 пары); уже в 1997 г. там гнездилось 1010 пар (Кошелев и др.,
1998). Общая численность больших бакланов на территории Украины, включая Крым, в
середине 1990-х гг. оценена в 24 000 пар (Poluda et al., 1997), что в пять и более раз превы-
шает их численность в середине 1980-х гг.
Таблица 3. Распределение и численность (количество пар) большого баклана
на Азово-Черноморском побережье Украины в 1998 г. (по: Сиохин и др., 2000)
123456789 10 Z
1590 3800 713 1200 1756 393 1534 4185 620 1233 17024
Примечание: цифрами в верхней строке обозначены водно-болотные угодья: 1 — дельта Дуная; 2 —
дельта Днестра; 3 — Ягорлыцкий зал.; 4 — Тендровский зал.; 5 — Джарылгачский зал.; 6 — Карки-
нитский залив; 7 — Центр. Сиваш; 8 — Вост. Сиваш; 9 — Молочный лиман; 10 — устье р. Берда и
Бердянская коса, залив и коса Белосарайская (в 1998 г. бакланы не гнездились на оз. Сасык, системе
озёр Шаганы-Алибей-Бурнас; Куяльницком лимане; Тилигульском лимане; заливе и косе Кривая).
большой баклан
69
Основными причинами роста численности большого баклана на Азовском море А.И. Ко-
шелев с соавторами (1990) считает обогащение кормовой базы за счёт сооружения многочис-
ленных рыборазводных прудов, доминирования в водоёмах мелкой сорной рыбы вслед-
ствие их эвтрофикации, опреснения Сиваша из-за сбросов воды с рисовых чеков; кроме
того, свою роль сыграло усиление природоохранной работы и, как следствие, ослабление
антропогенного пресса. Роль рыбхозов в увеличении численности баклана подчеркивал и
М.Х. Емтыль (1986). По его данным, в Краснодарском крае в многочисленных колониях в
978-1984 гг. гнездилось 7-10 тыс. бакланов (3,5-5,0 тыс. пар); введение в строй 10 тыс. га
трудовых хозяйств способстовало расширению кормовой базы этих птиц.
В дельте Дона, где на о-вах М. и Б. Дворян в 1990-х гг. гнездилось 900-1 000 пар (Мино-
ранский, Добринов, Маркитан и др., 1998), к 2003 г. учтено 1 800 пар (Белик, Данкевич,
2004). На косе в Кизилташском лимане Анапского р-на Краснодарского края в 2003 г. гнез-
дилось 1 820 пар. Такую же численность (около 2 тыс. пар) для черноморских лиманов
Таманского п-ва приводят и Ю.В. Лохман с соавторами (2005). По Краснодарскому краю в
телом в 16 колониях в 1990-2002 гг. насчитывалось 9 490 пар (Белик, Данкевич, 2004), что
в 3 раза больше, чем в 1950-е гг.
В западной части оз. Маныч-Гудило в 1990-х гг. была известна одна колония больших
такланов (25 гнездовых пар) на о-ве Прибрежном (Миноранский, Подгорная, 1998); на
калмыцком оз. Деед-Хулсун гнездилось 40 пар (Кукиш, 1984).
Следует подчеркнуть, что современный рост численности большого баклана во многих
местах происходит несмотря на активное «регулирование» поголовья этих птиц. Только в
Астраханском заповеднике с 1936 по 1960 г. в плановом порядке было уничтожено около
2СЮ тыс. больших бакланов. В те годы в дельте Волги их колониальные поселения были
: концентрированы практически только на заповедных участках, и общее количество бак-
ланов оценивалось в 10 тыс. гнездящихся пар (Луговой, 1963). Хотя в начале 1950-х гг. в
заповеднике наметилась тенденция к сокращению численности бакланов (Скокова, 1955),
: ни вскоре начали широко расселяться как на соседние участки собственно дельты, так и в
азанделыу Волги, на острова Сев. Каспия и на созданные землечерпалками острова вдоль
Вэджско-Каспийского канала. В итоге к 1973-1974 гг. численность больших бакланов в 17
заявленных колониях достигла почти 18 тыс. пар (Бондарев, 1975а). К началу XXI в. число
сдоний выросло до 27, и в них было учтено в 1999 г. 34 846 гнездовых пар, в 2000 г. —
?5 653, а в 2001 г. — 36 390 пар (Гаврилов и др., 2003). Следовательно, по сранению с
: срединой XX века численность бакланов увеличилась в три с половиной раза, и пока не
-лмечается тенденции к прекращению этого роста. Средняя плотность населения бакланов
-з водоёмах различного типа в пределах ключевой орнитологической территории «Дельта
Вс лги» в 2004 г. составляла: в феврале — 0,26; весной — 2,72; летом — 80,85; осенью —
--.49 особей на 100 га акватории (Русанов, 2005).
На северо-востоке Каспия, в труднодоступных тростниковых зарослях между Урало-
« Аспийским и Яицким каналами известна смешанная колония, в которой гнездится около
5 тыс. особей большого баклана (Гисцов, 2001), а на п-ове Мангышлак — моновидовая
<лония в 200 пар (Молодовский, 1975). В Дагестане вид в основном встречается на пролё-
при этом в Кизлярском и Аграханском заливах Каспия образует скопления порядка 15 000
с сбей (Джамирзоев, 1999а).
В Кызыл-Агачском заповеднике в первой половине XX в. колониальных поселений
з< слоногих и голенастых вообще не было. Тогда эти птицы гнездились в незаповедном
зз; с о ко ствольном лесу, удалённом на 3-5 км от моря. С падением уровня Каспия колонии
. - зли обычными на территории заповедника, но большой баклан в них составлял всего
5-2.0% от числа прочих гнездящихся видов. Исключением было поселение в Калинов-
70
Отряд Пеликанообразные, Семейство Баклановые
ском лимане, где в смешанной колонии с кваквой большой баклан составлял 18,5% чис-
ленности колонии. Общая численность гнездящихся больших бакланов в Кызыл-Агачс-
ком заповеднике на 1977 г. составляла 1 тыс. особей (Коновалова, 1979), что по крайней
мере в 10 раз меньше численности баклана, указанной для заповедника в 1958 г. В.С.
Грековым (1965).
В Аральском море и его бассейне в середине XX столетия были известны колонии по
Сыр-Дарье и на о. Барса-Кельмес (Долгушин, 1960); Аму-Дарьинская колония в 1960-е гг.
ещё насчитывала около 300 пар (Рашкевич, 1969). На оз. Сарыкамыш в 1975-1976 гг. учте-
но 1,5 тыс. гнездящихся особей (Великанов, Хохлов, 1979); однако позднее, в 1987 г., Д.Ю.
Кашкаров с соавторами (1989) на этом же озере, а также на оз. Айбигур у подножья Устюр-
та, выявили уже не более 330 особей. О существенном сокращении численности бакланов
в этом регионе по сравнению с началом XX в. писал Р.Н. Мекленбурцев (Мекленбурцев и
др., 1987). В связи с экологической катастрофой, постигшей Арал и впадающие в него
реки, сократились рыбные запасы в регионе, что обусловило уменьшение численности бак-
ланов. Возможно, в будущем численность бакланов в бассейне Аральского моря нормали-
зуется, что некоторые орнитологи связывают с осуществлением планов строительства в
Каракумах «Туркменского моря» (Сопыев, 2001).
В Казахстане большой баклан наиболее многочислен в Балхаш-Алакольской котловине
(Гаврилов, Гисцов, 1978). Так, на Алаколе в 1974 г. было учтено 200-250 пар; в дельте
Тентека на оз. Софроново в 1968-1969 гг. — около 1 000 пар. На р. Или, где бакланы заг-
нездились с образованием Капчагайского вдхр., в 1973 г. в трёх колониях учтено 700 пар.
Численность баклана на оз. Балхаш в 1963 г. Ю.В. Владимировым и Б.Д. Злобиным (1969)
оценена в 1 000-1 050 пар. Какова численность бакланов на юго-востоке Туркмении, где
согласно А.О. Ташлиеву (1973) бакланы населяют тугаи по Тенджену и Кара-Кумскому
каналу, не известно. В Узбекистане на Айдарских разливах после подъёма птиц на крыло
насчитывали до 10 тыс. особей (Мекленбурцев и др. 1987), т.е. численность здесь могла
составлять около 4 тыс. гнездящихся пар. В связи с прогрессирующим, начиная с 1996 г.,
усыханием Наурзумских озёр, возможности обитания водных и околоводных птиц там со-
кращаются. В июне 2000 г. веслоногие (пеликаны и бакланы) составляли здесь всего 3,4%
от 13 718 учтённых птиц (Брагин, 2001).
На Торейских озёрах в Забайкалье в 1976 г. учтено 1 160 гнёзд вида (Зубакин, 1981). В
районе оз. Ханка на Дальнем Востоке, где ещё 30 лет назад большой баклан встречался
нерегулярно, к 2000 г. гнездилось уже 250 пар (Глущенко, Мрикот, 2001). В целом в При-
ханкайской низменности Дальнего Востока численность больших бакланов стремительно
растет, и в начале XXI в. этот вид стал одним из наиболее многочисленных среди водных и
околоводных птиц (Глущенко, Коробов, Кальницкая, 2003). Если ранее здесь он гнездился
нерегулярно, то в 2002 г. в колониях насчитывалось порядка 1 050 гнездящихся пар, а в
2003 г. численность ещё более возросла. На крайнем юго-западе Приморья гнездится на
некоторых островах залива Петра Великого, где размножение впервые зарегистрировано в
1979 г. (не более 10 пар), а в 1989 г. обнаружено 202 гнезда (Литвиненко, Шибаев, 1996).
Подытоживая, можно утверждать, что суммарно в Черноморском и Каспийском бас-
сейнах в пределах описываемой территории в начале XXI в. гнездилось не менее 80
тыс. пар Ph. с. sinensis. Восточнее численность бакланов бесспорно меньше, и ситуация
там не столь ясна, особенно в связи с экологической катастрофой на Аральском море и
реках Аму-Дарье и Сыр-Дарье. Тем не менее, от Туркмении и до Дальнего Востока можно
ожидать гнездования примерно 9-10 тыс. пар данного подвида баклана. Таким образом,
общая численность материкового подвида большого баклана составляет ныне не менее
90 тыс. пар.
эольшой баклан
71
Говоря о численности бакланов в другие сезоны года, можно указать, что на Ниж-
ней Волге южнее Волгограда в августе встречается в среднем по 208,5 птиц на каждые
10 км, а размер наибольшей стаи оценен в 1 400 особей (Golawski, Kasprzykowski, 2004).
3 лиманах Таманского п-ва зимует порядка 10 тыс. больших бакланов (Лохман и др.,
2005).
В зарубежной Европе численность большого баклана также повсеместно возросла.
Этот процесс в странах Зап. Европы (например, в Австрии) был поначалу даже поддер-
жан проектами, предусматривающими эксперименты по реакклиматизации бакланов, со-
здание для них заповедных зон (Straka, 1988). Первоначальный круглогодичный запрет
на отстрел бакланов на Боденском оз. был отменён только в 1988 г., и то лишь частично,
поскольку запрет сохраняется в период с 1 января по 31 августа. Всё это способствовало
г-осту численности вида. Если в середине 1970-х гг. в Австрии зимовало порядка 100—
21'0 бакланов, то к концу 1980-х гг. их там было уже около 2 000, а на Боденском озере
число зимующих птиц за 20 лет (1970-1990 гг.) поднялось с нескольких сотен до 8 000
Siraka, 1991; Winkler, 1999).
В федеральной земле Гессен ФРГ большой баклан продолжает оставаться охраняе-
мым видом, несмотря на то, что количество гнездящихся пар с единиц в начале 1980-х гг.
• зеличилось до 300-350 пар к середине 1990-х гг. (Kreuziger et al., 1993-2000). Частич-
ный отстрел бакланов, разрешённый в связи с осуществлением проекта зарыбления р.
?1ссса (Земля Гессен) западно-европейским ельцом (Leuciscus souffid), вызвал бурю воз-
шения у природоохранителей; отдельные предложения по регулированию численнос-
ти бакланов на Дунае (Sturzlbaum, 1990) тоже подверглись жёсткой критике (Spitzenberger,
. -90) и т.д.
К началу 2000-х гг., согласно оценке Международной ассоциации охраны птиц BirdLife
li:emational (BirdLife International, 2004), численность больших бакланов в Европе достиг-
2 310-370 тыс. гнездящихся пар. В том числе в Дании обитало 36-^1 тыс. пар, в Норве-
жи — 20-25 тыс. пар, в Нидерландах — 18,4-19,5 тыс. пар, в Великобритании — 9,1 тыс.
"гг. в Швеции — 25-26 тыс. пар, в Германии — 16,8 тыс. пар, в Польше — 12,5 тыс. пар,
= Румынии — 18-20 тыс. пар, на Украине — 65-75 тыс. пар. В Европейской России, по
ей оценке, гнездилось 35-60 тыс. пар.
Размножение. Бакланы — колониально-гнездящиеся птицы, однако эта колониальность
- есколько своеобразна. По мнению Н.Н. Скоковой (1960а), бакланов нельзя в полном смысле
:ва отнести к общественным птицам. Колониальное гнездование не обеспечивает им той
_~ивной защиты гнездовья, как это имеет место у грачей, чаек и других подобных видов
-си. Колониальный тип гнездования дает баклану преимущества не в отношении лучшей
жжггы кладок и птенцов от хищников, а в отношении лучшего обеспечения пищей; тем
. гчым достигается уменьшение гибели птенцов.
Бакланы чаще гнездятся в смешанных колониях — с пеликанами, малыми бакланами,
: ieчастыми, ибисовыми и Чайковыми птицами (включая крачек). Но встречаются и «чис-
-=> г . одновидовые колонии. В случае гнездования на деревьях, бакланы «уживаются» с
~!ми видами птиц, занимая для своих гнёзд верхние ярусы и оставляя цаплям и ибисо-
более низкие. Большой баклан в таких колониях держится несколько обособленно,
ч тазуя моновидовые «островки»; под ними другие птицы обычно не гнездятся (Греков,
— ? •. В местах нового заселения бакланы могут гнездиться и отдельными парами, пре-
<мужественно в колониях других птиц (например, голенастых), что было отмечено в 1982 г.
• Гжзропольском крае А.Н. Хохловым и А.П. Бичеровым (19866). Ныне то же можно на-
ежзть в Закарпатье. Тип гнездования зависит от геоморфологических, гидрологических
гдботанических условий конкретной местности.
72
Отряд Пеликанообразные, Семейство Баклановые
Гнездование на скальных карнизах характерно в первую очередь для бакланов морских
побережий северных широт — Атлантики, Белого моря — т.е. присуще в основном подви-
ду Ph. с. carbo. Реже такое гнездование встречается и у Ph. с. sinensis в глубине материка.
Оно отмечено, например, для бакланов, гнездящися на островных скалах оз. Балхаш и
прибрежных скалах п-ова Мангышлак на Каспии (Долгушин, 1960, Молодовский, 1975).
И.А. Долгушин (1960) упоминает также о гнездовании бакланов на глинистых обрывах
некоторых рек Казахстана, что экологически близко к обитанию на скалах.
Наземные колонии на плоских островах характерны для материкового подвида Ph. с.
sinensis на Чёрном и Азовском морях и на ряде внутренних водоёмов. В случае наземного
гнездования бакланы занимают все типы островов, хотя аккумулятивным предпочитают
более крупные материковые. Причём птицы гнездятся на одном месте только 2-3 года кря-
ду, после чего начинают осваивать новые участки острова (Костин, 1983; Кошелев и др.,
1990). Гнёзда располагаются на возвышениях, ровных площадках высотой не менее 0,3—
0,5 м от уровня воды. При этом гнёзда хотя и располагаются близко друг от друга, однако не
«срастаются» в общий конгломерат. «Срастание» случается в старых, многолетних колони-
ях. На о. Комсомольский (Аральское море) в недалёком прошлом гнёзда, разрастаясь, со
временем образовывали общий «бугор» высотой в человеческий рост и включали в себя по
10-30 гнёзд. Вся эта масса была сцементирована экскрементами птиц (Гладков, цит. по:
Долгушин, 1960). «Срастание» гнёзд отмечал также А.В. Молодовский (1975) на Мангыш-
лаке. Видимо, такие гнездовья характерны в первую очередь для засушливых регионов, где
подобная конструкция не размывается дождями.
Весьма распространено гнездование бакланов на заломах тростника. Оно присуще бак-
ланам, которые селятся по берегам лиманов, озёр. В Казахстане такой тип гнездования
главенствует — в тростниковых крепях размешено до 95% колоний (Долгушин, 1960). В
схожих условиях гнездятся бакланы и на другой относительно невысокой растительнос-
ти — на кустах тамариска (Греков, 1965), на саксауле (Долгушин, 1960) и т.д.
Бакланам дельтовых популяций присуще гнездование на деревьях, преимущественно в
ивовых подтапливаемых лесах. Такое предпочтение легко объяснимо: пик половодья в
южных дельтах нашей страны приходится на период гнездования птиц (в дельте Волги —
на первую декаду июня). В подобных условиях наземное гнездование попросту невозмож-
но — все острова в этот период затоплены. В редких случаях отмечается гнездование на
хвойных деревьях — елях и лиственницах (Измайлов, Котова, 1973).
На деревьях располагаются и гнёзда японского подвида большого баклана Ph. с. hanedae',
он также поселяется колониями, нередко вместе с различными видами цапель — кваква-
ми, средними и малыми белыми цаплями, серыми цаплями (Austin, Kuroda, 1953).
При наземном типе гнездования бакланы активно вытесняют других птиц на перифе-
рию колонии, в менее удобные (например, подверженные затоплению) участки. Такая гнез-
довая агрессия по отношению к серебристым чайкам отмечена, например, в колониях Азо-
во-Черноморского региона (Кошелев и др., 1990). В организации новых колоний в трост-
никовых зарослях и ивовых лесах бакланы выступают в роли пионеров и основателей (Ко-
шелев и др., 2001).
Вопреки мнению И.А. Долгушина (1960), что бакланы упорно придерживаются посто-
янных мест гнездования даже в случае преследования их со стороны человека, птицы не-
редко вынуждены менять насиженные участки при смене гидрологического либо кормово-
го режима местности. Так, в дельте Волги в годы регрессии Каспийского моря, начавшейся
в 1930 г., колонии бакланов постепенно смещались из средней и нижней зоны в заливно-
култучную (Луговой, 1961), а позднее бакланы стали заселять ещё более дальние острова в
аванедельте, меняя при этом древесный способ гнездования на тростниково-наземный. С
Большой баклан
73
началом трансгрессии моря в 1980 г. процесс пошёл в обратном направлении: бакланы из
авандельты вновь стали селиться в култучнуой зоне и т.д. (Гаврилов и др., 2003). В отноше-
нии атлантического подвида Ю.В. Краснов с соавторами (1995) также подчёркивают, что у
бакланов мурманского региона нет выраженного гнездового консерватизма и птицы спо-
собны часто менять расположение колонии.
Считалось, что бакланы приступают к гнездованию в возрасте трёх лет, когда они оде-
вают брачное оперение (Судиловская, 1951). Однако освоение бакланами Лебяжьих остро-
вов Чёрного моря в 1970-х гг. было начато молодыми (белобрюхими) птицами. Среди ус-
пешно загнездившихся птиц в 1976-1977 гг. не было ни одной во взрослом (чернобрюхом)
наряде. Трёхлетние птицы стали гнездиться там только начиная с 1978 г. (Костин, 1983).
Вполне взрослые птицы в колонии отличались не только окраской оперения, но и феноло-
гией размножения: они приступали к откладке яиц значительно раньше молодых, белобрю-
хих птиц. В колониях на островах Молочного лимана доля годовалых размножающихся
бакланов составляла до 20-25%, вполне взрослых было только 10-15%, остальные птицы
были двух-трёхлетние (Кошелев и др., 1990). Скорее всего, «преждевременное» половое
созревание бакланов присуще периодам экспансии вида, что наблюдается в наши дни. При
?том пионерами в расширении гнездового ареала оказываются как раз молодые птицы.
Бакланы моногамы, могут образовывать постоянные пары (Долгушин, 1960). В начале
сезона гнездования происходят брачные демонстрации. Им предшествуют драки самцов,
которые хватают друг друга за клюв, пытаясь столкнуть или стянуть противника с места,
йамец, находящийся на месте гнезда, принимает своеобразные позы для привлечения сам-
(Мекленбурцев и др., 1987). Он демонстрирует специфическую позу (wing-waving), в
кгторой быстро поднимает и опускает концы крыльев, так что белые пятна на бёдрах с
частотой примерно дважды в секунду то заслоняются крыльями, то «вспыхивают» вновь
Cramp, 1977). При этом звуки не издаются, закрытый клюв направлен вверх и вперёд,
перья на темени и затылке слегка приподняты, шея изогнута S-образно, задняя часть туло-
вища приподнята, грудь опущена, хвот поднят и развёрнут, голова покачивается вверх-вниз,
трло и клоака пульсируют в ритме движения крыльев (рис. 15а). Данная демонстрация не
: гмечается после откладки яиц. Во время и после формирования пары самец, приветствуя
приближающуюся самку, поднимает закрытый клюв, голову и шею вертикально вверх, за-
тем. широко раскрыв клюв и издав громкий крик, отклоняет голову назад (рис. 156), после
г его касается закрытым клювом основания хвоста, быстро вращая головой из стороны в
.торону и издавая рокочуще-булькающие звуки (рис. 15в); в конце демострации птица бы-
стро перенаправляет голову и шею вперёд. Приветственная церемония у самки сходна с
гковой у самца и различается некоторыми деталями (Cramp, 1977). Ряд других демонстра-
ций и характерных поз большого баклана показаны на рис. 15 (г-к).
Свои гнёзда на островах Чёрного и Азовского морей бакланы располагают отдельными
группами — субколониями, удалёнными друг от друга на 150-600 и более м (Кошелев и
пр.. 1990). Внутри субколонии установлена высокая синхронность размножения (7-25 дней),
е то время как разница сроков размножения между субколониями доходит до 50 дней. Рас-
гттяние между гнёздами в субколонии на островах Молочного лимана составляет от 0,1—
.5 до 3,0-5,0 м.
Гнёзда на прибрежных островах Чёрного моря имеют вид «башенок», их основания
гл гжены из толстых сухих стеблей полыни, корневищ тростника и морской капусты. Лоток
з-; отдан сухими листьями взморника (зостеры), крупными перьями самих бакланов и дру-
~ -з. птиц, кусками хлорвиниловой плёнки, иногда зелёными листьями тростника. Размеры
5 гнёзд, приведённые Ю.В. Костиным (1983) для Лебяжьих о-вов: диаметр гнезда (в см) —
• (в среднем 48,9), диаметр лотка — 21-30 (26,3), высота гнезда — 8^2 (20,1), глуби-
74
Отряд Пеликанообразные, Семейство Баклановые
Рис. 15. Демонстрационное поведение большого баклана (по: Cramp, 1977; Bauer, Glutz von Blotzheim,
1987; Nelson et al., 2005): a — рекламное поведение самца, б, в — элементы поведения при привет-
ствии между членами пары, г — обвивание шеей партнёра на гнезде, д — спаривание, е — террито-
риальный конфликт в колонии, ж — ритуализованный ремонт гнезда, з — вытянутая поза на гнезде,
и — поза перед взлётом, к — поза после посадки.
эолыпой баклан
75
:-и лотка — 5-11 (7,9). На островах Черноморского заповедника высота гнёзд 14-42 см
отдельные до 80 см), в среднем (п=32) — 25,2±4,0 см. Диаметр гнёзда колеблется от 38 до
?8 см, в среднем (п=30) 50,3±3,5 см. Среднее растояние между центрами гнёзд (п=25)
”1.8 ±1,6 см (Руденко, Яремченко, 2004). Гнёзда в тростниковых зарослях устраиваются
исключительно из фрагментов этого растения и своими основаниями опираются на дно,
заломы тростника либо на кучи плавника. Постройка гнезда прочна, она сцементирована
помётом бакланов. Размер гнёзд, устроенных на деревьях, небольшой, в диаметре около
-0-50 см, лоток мелкий.
На оз. Ханка гнёзда имеют размеры (n=31): диаметр гнезда 380-780 мм (в среднем 600
мм), диаметр лотка 210-330 мм (в среднем 281 мм), толщина гнезда 210-640 мм (в среднем
321 мм), глубина лотка 30-140 мм (в среднем 77 мм). Расстояние между краями ближай-
ших гнёзд в отдельных случаях может составлять 15-20 см (Глущенко и др., 2006).
Начало откладки яиц зависит от возраста размножающейся пары, состояния прошло-
годнего гнёзда, климатических и кормовых условий конкретного года. Ряд исследовате-
лей указывает на значительный разрыв между временем прилёта птиц в колонию и нача-
лом кладки. Так, по данным В.В. Бианки с соавторами (Bianki et al., 1997), в Кандалакш-
ский залив Белого моря атлантические большие бакланы Ph. с. carbo прилетают весной
обычно в последних числах апреля (в разные годы от 18 апреля до 19 мая), а откладка яиц
начинается в первых числах июня. В дельте Днепра, куда бакланы подвида Ph. с. sinensis
прилетают около 1 марта, птицы приступают к кладке только в конце этого месяца или
даже в начале апреля (Ардамацкая, 1968). В Черноморском заповеднике бакланы присту-
пают к яйцекладке в конце марта - начале апреля; средняя дата за 13 лет наблюдений —
28 марта (Руденко, Яремченко, 2004), а в тугаях Туркмении начало откладки яиц отмече-
но даже в третьей декаде апреля (Ташлиев, 1973). В дельту Волги бакланы прилетают в
конце февраля - начале марта (Луговой, 1963), но начало яйцекладки датируется началом
длреля (Судиловская, 1951). На юго-западе Каспия, в Кызыл-Агаче, первые птицы появ-
ляются в феврале, первые кладки бывают в начале марта, а массовая кладка начинается
:л\стя 15-20 дней после первой (Греков, 1965). Вместе с тем известны случаи и очень
тайней яйцекладки, следующей сразу по прилету птиц в колонию. Так, на Лебяжьих о-
чах Чёрного моря уже 26 февраля 1978 г. было обнаружено 30 гнёзд с кладками из 1-5
гиц (Костин, Тарина, 2004). В целом, сроки откладки яиц материкового большого бакла-
на очень растянуты. Чёткую фенологию трудно проследить вследствие регулирования
численности бакланов, что заставляет птиц менять места гнездования и приступать к
пзвторным кладкам.
Откладка яиц японского подвида большого баклана Ph. с. hanedae начинается в декабре
на юге о-ва Хонсю) и марте (на севере острова); гнездовой период продолжается до июня
Austin, Kuroda, 1953).
Яйца бакланов бело-голубые либо бело-зелёные, покрытые сверху известковым нале-
77 м. матовые. Полная кладка материкового подвида Ph. с. sinensis в Узбекистане содер-
впгг 2-6 яиц (Мекленбурцев и др., 1987); в Казахстане — 3-5 яиц (Долгушин, 1960); на
: стровах Черномоского заповедника — 3-7 яиц, обычно 3-5 (Руденко, Яремченко, 2004),
= Прибалтике — 1-4 яйца (Zydelis et al., 2002), на Ханке в полной кладке от 2 до 5 яиц, в
шепнем (п=59) 3,1 яйца (Глущенко и др., 2006). Размеры яиц в Черноморском заповедни-
ке п=110): 63,3±0,28 х 39,65 ±0,62 мм. На Лебяжьих о-вах (Крым) размеры яиц (п=105):
5-8-70,5 х 35,6^12,4 мм, в среднем 62,2x39,8 мм. В дельте Волги (п=23): 51—61х 34-38
* v. на восточном берегу Аральского моря: 58-66 х 37—42,6 мм, в среднем 62,3x38,6 мм;
= пеньте Аму-Дарьи: 63,3x39,6 мм; в нижнем течении Сыр-Дарьи (п=4): 66,9x39,9 мм; в
шъе р. Урал: 61-70 х 36^15 мм, в среднем 65,02x40,7 мм; на оз. Севан (п=16): 63,5 х
76
Отряд Пеликанообразные, Семейство Баклановые
38,7 мм. Вес яиц в Крыму (п=75) составил 39,7-65,0 г, в среднем 58,5 г; в дельте Волги —
37-50 г; в Армении (п=16) — 46,2 г; в Узбекистане — 56,5 г (Долгушин, 1960; Костин,
1983; Мекленбурцев и др., 1987; Руденко, Яремченко, 2004). В Приханкайской низменно-
сти размеры яиц (п=65): 56,2-66,8 х 34,8-39,7 мм, в среднем 60,8x37,3 мм (Глущенко и
др., 2006).
В полной кладке атлантического подвидаP/z. с. carbo бывает 2-6 яиц (в среднем 3,5±0,1).
Их размеры (п = 54) в среднем 64,2±0,2 х 40,0±0,2 мм (59,0-70,0 х 37,2^4,0) (Bianki et al.,
1997). У японского подвида большого баклана Ph. с. hanedae в кладке 3-6 яиц, средние
размеры яиц 62x39 мм (Austin, Kuroda, 1953).
Как указывает А.И. Кошелев с соавторами (1990), при упрощенной пространственной
структуре колонии (отсутствие ярусности) нередко наблюдаются случаи гнездового пара-
зитизма. Смешанные кладки включают до 14 яиц, в том числе могут попадаться и яйца
других видов птиц (в 5 случаях отмечено по 1-2 яйцу чайки-хохотуньи).
Насиживание длится 28-30 дней (Судиловская, 1951), хотя для материкового подвида
С. Крэмп приводит и другие сведения, ссылаясь на Портилье: 23-24 дня (Portielje, 1927,
цит. по: Cramp, 1977). Инкубационный период японского подвида РА. с. hanedae продол-
жается в течение 47-50 суток (Austin, Kuroda, 1953). Насиживают оба родителя начиная с 1-го
яйца, поэтому птенцы разновозрастные. Они рождаются голыми, беспомощными. До окон-
чательного вылета молодых птиц проходит около двух месяцев, но уже трёхнедельные птен-
цы могут вылезать из гнезда, а месячные — плавать. Большинство взрослых птиц остаётся
в районе колонии и после выведения птенцов. Так, в дельте Днестра общая продолжитель-
ность пребывания бакланов на месте гнедования составляет 160-190 дней (Korzyukov, 1997),
т.е. 5-6 месяцев. Но часть птиц начинает кочевать сразу после вылета из гнезда, о чем
свидетельствуют находки окольцованных 3-5-месячных молодых в 100-290 км от родных
колоний (Скокова, 1960а).
В прибалтийских колониях вылупляется 2,66-2,72 птенца на гнездо (Zydelis et al., 2002),
в то время как у птиц из дельты Волги эта цифра равна примерно 3-3,5 птенца. Гибель
кладок и птенцов в Узбекистане в среднем состаляет 37%, однако известны и более круп-
ные потери. Н.А. Гладков отметил, что из 51 птенца в колонии на о. Комсомольском до
вылета из гнёзд выжило только 15 птиц (Мекленбурцев и др., 1987). Яйца и птенцы в коло-
ниях на ракушечных островах очень чувствительны к перегреву: голые птенцы гибнут от
солнечных полуденных лучей в течение 15 мин (Костин, 1983).
Смертность больших бакланов в первый год жизни, по данным казахстанских орнито-
логов, составляет 69,4%. Установлен факт продолжительности жизни одной птицы в 14 лет
и 2 месяца (помечена 10 июля 1959 г. в дельте р. Или, добыта 25 августа 1973 г. на Балхаше)
(Гаврилов, Гисцов, 1978).
По материалам кольцевания для бакланов из дельты Волги установлена средняя про-
должительность жизни, равная 1,9 года, а для переживших первый год — 3 года. Гибель
молодых бакланов на 1-м году жизни составляет 44,5%, на втором — 25,3%, на третьем —
12,3%; у птиц старше трёх лет — 7% (Скокова, Бондарев, 1978).
Суточная активность, поведение. Вид ведёт дневной образ жизни. Поскольку места
кормежки бакланов часто находятся в нескольких километрах от мест гнездования или от-
дыха, птицам приходится делать дальние перелеты. В разные периоды года эти перёлеты
совершаются по-разному. В период гнездования кормовые перёлеты носят челночный ха-
рактер: за светлое время суток родители не менее двух раз (а в некоторых случаях и 3-5
раз) вылетают в строну моря или другого рыбного водоёма и возвращаются обратно с кор-
мом для птенцов. Во внегнездовое время, особенно осенью, бакланы совершают лишь один,
утренний кормовой вылет с мест ночёвок. Если бакланов не беспокоят, места ночёвок по-
г-: :ьшой баклан 77
:т гянны на протяжении всего осеннего сезона. Подобным образом ведут себя бакланы и на
- естах зимовок.
Как правило, из колоний архипелага Семь Островов бакланы атлантического подвида
3 с с. carbo вылетают на кормёжку в одиночку или небольшими группами в 2-3 особи,
/дксимальная дальность полётов от 3 до 12 км. Достигнув района кормёжки, птицы по-
прежнему в одиночку ныряют за рыбой, не практикуя коллективной охоты, столь характер-
-:_ й для материкового подвида. Ситуация меняется лишь в случае появления скоплений
трагических рыб — песчанки или мойвы. Если такое скопление большое, бакланы могут
::ч5ираться и ловить рыбу группами до 10 особей. Концентрация птиц вне колонии для
: тдыха и обсушки проходит на небольших скалистых островах, но и здесь собирается обычно
5—6. максимум до 10 особей.
Бакланы материкового подвида P/z. с. sinensis в дельте Волги осенью значительную часть
времени проводят в местах постоянных ночёвок, которые расположены на ивах прибреж-
ных галерейных лесов, растущих вдоль ериков дельты. Утренний вылет на кормёжку пред-
ставляет собою впечатляющее зрелище — армады крупных птиц летят волна за волной
иглообразными шеренгами и цепочками в сторону Каспия, заполняя в течение 20-30 ми-
чп.т всё небо.
Своеобразно защитное поведение материковых бакланов в гнездовой колонии. Птицы
не в состоянии защитить свои кладки от нападений серых ворон. Поначалу насиживающая
птица пытается отогнать ворону от гнёзда, совершая выпады клювом в её сторону. В конце
концов птица приподнимается на гнезде, приоткрыв таким образом кладку, чем незамедли-
тельно пользуется другая ворона, караулящая сзади. Сидящий поблизости партнёр насижи-
вающей птицы не реагирует на действия ворон.
Питание. Все бакланы облигатные ихтиофаги, хотя в местах, бедных рыбой, частично
переходят и на другие виды кормов. Особенности пищеварения и других сторон трофики
большого баклана подробно описаны в работе Н.Н. Скоковой (1962). Работа была выпол-
нена в дельте Волги, объектом исследований был материковый подвид большого баклана
?h. с. sinensis, однако полученные выводы можно распространить на вид в целом.
По данным этого исследователя у ихтиофагов, с их железистым типом желудка, перева-
ривание пищи происходит медленнее, чем у птиц с хорошо выраженным мускульным же-
лудком. Опыты показали, что даже через 8 ч после приёма пищи в желудке рыбоядной
птицы можно обнаружить в среднем 1/4 кормов (по весу). У бакланов, отловленных осе-
нью на местах ночёвки в 20-21 ч, удавалось полностью восстановить (по глоточным зубам,
челюстям) видовой состав и количество рыб, съеденных утром, т.е. минимум 9-10 ч назад.
При отсутствии в регионе камешков (как, например, в дельте Волги), роль гастролитов
выполняют обломки старых раковин моллюсков (например, Dreissena polymorpha), скор-
лупа водяного ореха (Trapa natans) и песчаные конкреции, достигающие в поперечнике до
2-3 см. Однако подобные гастролиты обнаруживаются только у птенцов или у взрослых в
период выкармливания. Очевидно, гастролиты необходимы птенцам для компенсации ещё
недостаточно энергичной деятельности стенок желудка и пищеварительных ферментов.
Взрослые птицы отрыгивают гастролиты вместе с пищей и, частично, с погадками.
Поскольку пища у бакланов переваривается наполовину в среднем за 3-5 ч, птицы мо-
п>т задерживаться на местах кормёжки по 3-4 ч для поисков и добычи необходимой птен-
да.м порции корма, которая приносится в гнездо лишь частично переваренной. Баклан от-
рыгивает пищу птенцам из пищевода и желудка, так что на долю родителя приходится та
чисть корма, которая не успела перевариться до прилёта баклана к гнезду.
Суточная норма пищи у бакланов составляет в среднем 20,6% от веса тела птиц. Для
большого баклана в дельте Волги такая норма составляет около 465 г в сутки. Для этого
78
Отряд Пеликанообразные, Семейство Баклановые
баклан должен в день поймать, например, 13 вобл длиной в 12-13 см или 18 чехоней дли-
ной в 15 см.
Атлантический большой баклан Ph. с. carbo добывает свой корм преимущественно в
морских водах. Птицы этого подвида очень хорошо ныряют, на глубину 3-9 м, макси-
мально до 9,5 м (Cramp, 1977). Согласно Ю.В. Краснову и др. (1995), на Мурмане птицы
кормятся в местах с глубинами от 8 до 37 м. В связи с этим в рационе часто попадаются
донные и придонные рыбы литорали. В Атлантике рыбы поверхностных слоёв составля-
ют в питании не более 40%. Там в корме больших бакланов встречаются сельдевые, ло-
сосевые, угревые, тресковые, губановые и камбаловые рыбы. Перечень видов рыб, обна-
руженных в желудках Ph. с. carbo, которых добывали у берегов Шотландии, Исландии и
Скандинавии, таков (Cramp, 1977): атлантическая сельдь (Clupea harengus), шпрот (Sprat-
tus sprattus), сёмга (Salmo salar), кумжа (S. trutta), мальма (Salvelinus alpinus), речной
угорь (Anguilla anguilla), атлантическая треска (Gadus morhua), сайда (Pollachius virens),
мерланг (Merlangius merlangius), речная камбала (Platichthys flesus), анадромная камбала
лиманда (Limanda limanda). Кроме того, в питании отмечали таких пресноводных рыб,
как обыкновенная щука (Esox lucius) и обыкновенный окунь (Perea fluviatilis). Размеры
большинства поедаемых рыб 7-36 см, речных угрей — до 65 см. В качестве случайного
корма можно назвать также травяную лягушку (Rana temporaria), птенцов камышницы
(Gallinula chloropus) и обыкновенной пеганки (Tadorna tadorna), водяную полевку
(Arvicola terrestris). В питании беломорских бакланов по данным В.В. Бианки с соавтора-
ми (Bianki et al., 1997) 50-70% составляет атлантическая треска, далее идёт атлантичес-
кая сельдь и навага (Eleginus navaga). В питании птенцов больших бакланов на архипе-
лаге Семь Островов в первую очередь встречались треска (Gadus), песчанки (Ammodytes),
бычки (Gobiidae), мойва (Mallotus villosus), зубатка (Anarhichas) и, реже, маслюк (Pholis
gunnellus), пинагор (Cyclopterus lumpus), камбалы (Pleuronectidae), сайда (Pollchius virens),
пикша (Melanogrammus aeglefinus), девятииглая колюшка (Pungitiuspungitius). Впрочем,
это было характерно для 1985-1992 гг., когда отмечали депрессию рыбных ресурсов (Крас-
нов и др., 1995). Было отмечено, что при массовом питанаии мойвой бакланы добывали
исключительно отнерестившихся самцов. При благоприятных кормовых условиях в пи-
тании бакланов преобладают более крупные рыбы. Бакланы способны добыть и прогло-
тить треску и зубатку длиной около 40 см. В зависимости от подхода скоплений пелаги-
ческих рыб к местам обитания птиц спектр рациона большого баклана может изменяться
(Краснов и др., 1995).
Добычей материковым бакланам Ph. с. sinensis служит почти исключительно рыба,
обитающая в поверхностных слоях воды, поскольку эти бакланы, в отличие от номина-
тивного подвида, ныряют не глубже 3—4 м (Долгушин, 1960). Стайный во все периода
года образ жизни облегчает им добычу пищи с меньшей затратой энергии на поиски и,
главное, на ловлю рыбы. Это особенно проявляется на мелководье, когда одна часть птиц
кормится, а другая выполняет роль «невода» (загонщиков), после чего роли меняются
(Скокова, 1960а). Порой такая коллективная ловля рыбы выполняется в сообществе с пе-
ликанами.
Бакланы, населяющие дельты рек, кормятся в зависимости от концентрации рыбы то в
авандельте и море, то собственно в речных протоках, вблизи от гнездовых колоний. Рацион
состоит из рыб любых видов, которые доступны по размерам (обычно до 25 см длины) и
глубине обитания. Тем не менее, существует некоторая избирательность. Н.Н. Скокова (1955)
разбила корма бакланов дельты Волги на 4 группы по размерам.
В первую группу входят рыбы длиною от 2 до 10 см — бычки (Gobius kessleri, G. fluvia-
tilis, G. melanostomus), щиповка (Cobitis taenia), уклея (Alburnus alburnus), молодь карпо-
>:лыиой баклан 79
звлх (Cyprinus carpio, Scardinius erytrophtalmus, Leuciscus idus и т.д.). Значение этой группы
= литании бакланов невелико, и массово такие рыбы встречались лишь у единичных птиц
zo 34 особей молоди сазана в одном желудке).
Во вторую группу включены рыбы размером от 10 до 16 см. Это вобла (Rutilus rutilus
.jispicus), густера (Blicca bjoerkna), мелкий лещ (Abramis brama), сеголетки щуки и судака
Lucioperca lucioperca). Эта пища в кормах баклана преобладала (в разные годы от 54 до
-3%), причём здесь первое место занимала вобла (61-77%).
Третья группа — рыбы размером от 17 до 26 см — состояла из леща, сельдей (Caspiolosa
Lessleri volgensis, С. caspia), чехони (Pelecus cultratus), линя (Tinea tinea) и окуня, четвер-
тая — размерами от 27 до 40 см была представлена судаком и щукой. Рыбами последних
лвух групп бакланы питаются редко, в основном в периоды, когда в водоёмах дельты Вол-
ги нет заметных скоплений другой добычи. Тогда птицам «выгоднее» добывать более круп-
ные экземпляры рыб.
Из видов, не перечисленных в данных четырех группах, Н.Н. Скокова (1955) называет
ещё следующие виды, входящие в рацион бакланов дельты Волги: килька (Clupeonella
delicatula), карась (Carassius carassius), синец (Abramis ballerus), пескарь (Gobio gobio),
жерех (Aspius aspius), com (Silurus glanis), ёрш (Acerina cernua), берш (Lucioperca volgensis).
Из нерыбных кормов названы озёрная лягушка (Rana ridibunda), речной рак (Potamobius
yachipus) и ряд насекомых, а также моллюсков, встречаемость которых составляла от 0,1
го 1,8%.
Предпочитаемыми рыбами в кормах бакланов дельты Волги являются вобла и чехонь.
На питание окунем, судаком, лещом, густерой и краснопёркой бакланы переходят вынуж-
дено. Вобла и чехонь преобладают в пище не только из-за высокой концентрации, но и
вследствие особенностей формы тела, отсутствия жёстких лучей в плавниках и т.д. Изби-
рательность проявляется в том, что баклан предпочитает эти виды другим, столь же много-
численным. Так, при одновременных нерестовых миграциях воблы и леща в разные годы
вобла составляла в пище баклана 89-94%, в то время как лещом бакланы обычно пренебре-
гали.
В других регионах видовой состав рыб в корме бакланов различен. Так, у литовских
чтиц это преимущественно пресноводные рыбы, которые располагаются по убыванию зна-
чимости (биомассе) таким образом: плотва — 45,01%, судак — 29,85%, лещ — 12,95%,
окунь — 5,01%, рыбец (Vimba vimba) — 3,98%, чехонь — 3,71%, сазан — 3,05%, атланти-
ческая сельдь — 1,85%, карась — 1,24%, налим (Lota lota) — 0,81%, ёрш — 0,76%, густе-
ра — 0,75%, большеротая корюшка (Osmerus eperlanus) — 0,03% (Zydelis et al., 2002).
В 10 желудках больших бакланов, добытых в марте-июне в Херсонской и Одесской
областях, Л.О. Смогоржевський (1959) обнаружил бычков (ближе не опредёленных) — 40%,
морского языка (Solea nasuta) — 30%; из других видов встречены черноморская сельдь
Alosa kessleri), бычок-мартовик (Mesogobius batrachocephalus), лещ, плотва, чехонь, зо-
лотистый карась, речной рак, мидии (Midia sp.), прудовик (Limnea palustris). Согласно
Ю.В. Костину (1983), в апрельских отрыжках птенцов, гнездящихся на Лебяжьих остро-
вах. отмечены бычки и креветки, в июне — бычки, карпы, караси. А зимующие на юге
Крыма бакланы кормились преимущественно кефалью (Mugil sp.) длиной до 40 см и мас-
::й ло 950 г.
Для бакланов, гнездившихся в дельте Днепра и проводивших значительную часть свое-
времени в Тендровскоми и Ягорлыцком заливах Чёрного моря, Т.Б. Ардамацкая (1968)
созывает в числе кормов судака, тарань (Rutilus rutilus heckeli), густеру, сазана, чехонь,
тычков. А.Г. Руденко и О.Я. Яремченко (2004) уточняют видовой состав употребляемых
ггсанами бычков в Чёрном море. Это Neogobius melanostomus, N. fluviatilis, Zostersessor
80
Отряд Пеликанообразные, Семейство Баклановые
ophionalus, Mesogobius batrachocephalus, Gobius niger. Из морских рыб реже встречается
кефаль (вид не уточнён), европейский анчоус (Engraulis encrasicholus) и камбала глосса
(Platichthys flesus luscus). Пресноводные рыбы составляют менее 50% корма черноморских
бакланов.
В последние десятилетия бакланы Краснодарского края в значительной степени пере-
ключились с питания таранью, хамсой (Engraulis encrasicholusponticus), тюлькой (Clupeo-
nella delicatuld), сельдю-пузанком в Азовском море на питание прудовыми видами рыб —
карпом, амурским и пестрым толстолобиком (Hypophthalmichthys molitrix, Aristichthys по-
bilis\ белым и чёрным амуром {Ctenopharyngodon idella, Mylopharyngodon piceus). Культи-
вируемая в прудах рыба составляет почти 98% питания бакланов, остальное приходится на
малоценную «сорную» рыбу — окуня, краснопёрку и др., а также на водных беспозвоноч-
ных — раков, креветок (Емтыль, 1986).
В Туркмении в рационе больших бакланов обнаружены аральская плотва, елец (Leuciscus
latus), аральский усач {Barbus brachycephalus), маринка (Schizothorax sp.) храмуля
{Varicorhinus sp.), белый амур, сом, а также стрекозы (Odonata), прямокрылые (Orthoptera),
водолюбы (Hydrophilidae), совки (Noctuidae) и другие насекомые (Ташлиев, 1973).
И.А. Долгушин (1960) указывает, что бакланы в Казахстане поедают даже маринок
(Schizothorax sp.) с их ядовитой икрой, а кроме рыб иногда ловят также моллюсков, насеко-
мых и лягушек.
Бакланов отличает от таких ихтиофагов, как цаплевые, дружный переход на массовые
виды корма в периоды нерестовых и предзимовочных миграций. Так, в дельте Волги в
апреле-мае, когда идёт нерестовый ход воблы, в желудках бакланов преобладает именно
этот вид рыбы. Чехонь чаще всего служит кормом в октябре (встречаемость выше 50%).
Лещ, который в пище баклана не столь обилен, попадается главным образом в период
осеннего хода, в ноябре (Скокова, 1955). При этом уменьшаются затраты энергии на по-
иски пищи, что приводит к заметному увеличению веса птиц в эти периоды года (Скоко-
ва, 1962).
Враги, неблагоприятные факторы. В гнёздах больших бакланов (без уточнения под-
вида) выявлены следующие насекомые-сожители: Ceratophyllus vagabunda Bohem, С. bo-
realis Roths., Scydmaneus barnevillei Reiter, Cryptophagus pilosus Gyll., Apion cruentatum
Walt., Choleva watsoni Spence, Omosita colon L., Sunius lyonesius Joy, Oncocephalusplumicornis
Germ., Leptocera (Thoracochaeta') penteseta Richards, L. zosterae HaL, L.(Limosind) silvatica
Meigen, L. (Limosina) minutissima Zett., L.(Limosina) heteroneura Haliday, Pteromalusfucicola
Walk. (Hicks, 1959). Часть из них — кровососущие, большинство же потребляют несведен-
ные остатки пищи и опад.
Среди врагов бакланов, гнездящихся на деревьях, можно назвать серых ворон и некото-
рых других птиц, которые уничтожают кладки и маленьких птенцов. Как выяснилось, к
таковым можно отнести и квакв (Patrikeev, 2004). Для потомства птиц, гнездящияхся на
скальных карнизах, существенную опасность могут представлять крупные чайки. Причём,
большие бакланы, которые гнездятся не в скалистых нишах, как хохлатые бакланы, а от-
крыто, более уязвимы для воздействия этих неспециализированных хищников (Краснов и
др., 1995). Для наземногнездящихся популяций опасны крупные чайки (Кошелев и др., 2001),
а также млекопитающие (лисицы, енотовидные собаки и т.д.), сумевшие проникнуть на
острова, где расположены колонии бакланов. Так, в 1996 г. из-за поздней весны на о. Боль-
шой (Приазовье) проникла пара енотовидных собак, которая вывела там потомство. Еното-
видные собаки уничтожили большую часть кладок и птенцов; на крыло поднялось всего
800 молодых бакланов из 9 600 ожидаемых. Эти же авторы указывают на опасные послед-
ствия для наземногнездящихся бакланов нагонных вод. Примером может служить гибель
эельшой баклан
81
колонии больших бакланов на о. Долгом, случившаяся в результате продолжительных штор-
мов в 1995 и 1996 гг.
В литературе приводятся случаи массовой гибели бакланов от малоизученных причин.
Так. в 1994-1995 гг. в приазовских колониях была отмечена эпизоотия, в результате кото-
гой погибли сотни взрослых птиц и птенцов. Там же в 1997 г. от заболевания пищевари-
тельного тракта погибло более 350 птенцов в гнёздах (Кошелев и др., 1993) В черномор-
ских колониях на Каржинских островах в 1997 г. найдено 278 погибших птенцов, в 2000 г. —
310 птенцов, в основном старшего возраста (Ардамацкая, 2001). На территории Украины у
больших бакланов были выявлены такие гельминты, как Ligula intestinalis, Eustrongilides
^ergorum, E. exsicius, которые в личиночной стадии паразитируют у рыб, а также Apophallus
^ilhlingi, Metagonimus yokogawai, Cryptocotyle concavum, C. lingua, опасные и для млеко-
питающих (Смогоржевський, 1959). Согласно А.О. Ташлиеву (1973), у больших бакланов,
добытых в Туркмении, были найдены такие гельминты, как Diplostomum spathaceum,
Рагу pho stomum radiatum, Ligula intestinalis, Paradilepis scolecina, Capillaria carbonis,
Skrjabinocara squamata, Desmidocercella skrjabini, Contracaecum spiculigerum, C. micro-
yapillatum.
Из перьевых клещей у бакланов паразитирует Freyana largifolia largifolia.
На протяжении столетий главным врагом бакланов был и остаётся человек, который не
только истребляет этих птиц как конкурентов-рыболовов, но и уничтожает места гнездо-
вий либо кормовую базу птиц. Так, на юго-западе Каспия в Кызыл-Агачском заповеднике
лосле «залпового» сброса ядовитых коллекторных вод резко сократились площади тама-
рисковых зарослей, где гнездились бакланы (Литвинова, 1989а), а антропогенная экологи-
ческая катастрофа в бассейне Арала, приведшая к катастрофическому сокращению вод-
ных площадей, стала причиной уменьшения рыбных запасов в регионе, от чего пострадали
и рыбоядные птицы.
В прошлом, во время сбора яиц на птичьих базарах (у кайр и т.д.) мурманского архипе-
лага Семь Островов, для колониальных поселений больших бакланов очень ощутимым
был фактор беспокойства (Краснов и др., 1995).
Прямое преследование птиц также становится причиной существенного изменения чис-
ленности бакланов. По данным А.И. Кошелева с соавт. (1998), на Каховском водохранили-
ще к 1960-м гг. бакланы были практически полностью уничтожены в рамках специаль-
ной кампании. Они загнезилилсь здесь вновь, на о-вах Б. и М. Кучугуры, лишь в 1971 г.
Бреди причин гибели взрослых больших бакланов превалируют отстрел в целях ограниче-
ния численности и любительская охота (Скокова, Бондарев, 1978). Человек добывает бак-
щнов в том числе и ради мяса (в копчёном виде оно вполне съедобно), а также ради шку-
рок.
Хозяйственное значение, охрана. Выраженная ихтиофагия бакланов, встречающихся к
: му же большими скоплениями, вызывает среди рыбоводов и орнитологов постоянные спо-
тд как о степени ущерба, наносимого бакланами, так и о необходимости сокращения их
численности. Наиболее радикальные меры по сокращению численности бакланов в России
тедлагал А.И. Пахульский (1951, 1957). Л.О. Смогоржевський (1959) акцентировал внима-
-ие не негативной роли бакланов в распространении гельминтозных заболеваний у рыб.
В оценке хозяйственного значения больших бакланов, безусловно, необходим диффе-
тендированный подход. Что касается, например, значения атлантических больших бакла-
-: з как конкурентов человека в рыболовстве, то оно ничтожно, а при невысокой численно-
. бакланов практически равно нулю.
Бакланы материкового подвида, кормящиеся на рыборазводных водоёмах, несомнен-
- вносят ущерб рыбному хозяйству. Согласно исследованиям, проведённым на Ахтыр-
82
Отряд Пеликанообразные, Семейство Баклановые
ском рыбокомбинате, бакланы за сезон уничтожают 25% выращиваемой рыбы. При ин-
тенсивном отпугивании птиц удаётся снизить выедание годовиков на 10%, но полностью
избавиться от бакланов не удаётся (Емтыль, 1986). Однако в условиях естественных во-
доёмов, где человек занимается лишь рыболовным промыслом, не вкладывая средств и
труда в выращивание рыбы, бакланы съедают крайне незначительную долю рыбы (деся-
тые доли процента) от добытой человеком, не говоря уже о доли от общих запасов рыбы
(Скокова, 1955). Более того, при авиационных учётах на Северном Каспии мы неоднок-
ратно отмечали, как бакланы (и прочие рябоядные птицы) «объедают» «хвост» рыбьего
косяка и никогда не охотятся на тех рыб, что плывут в «голове» скопления. Таким образом,
птицы-ихтиофаги изымают из косяка рыб-«аутсайдеров», среди которых значительный
процент представлен неполноценными, больными особями, и к местам нереста в дельту
Волги приходит отборная рыба, от которой можно ждать хорошего потомства. Кроме того,
своей жизнедеятельностью (удобрение водоёмов экскрементами) птицы положительно вли-
яют на функционирование водных биоценозов. Биомасса зоопланктона в водах ёриков дель-
ты Волги, где скапливаются бакланы, в 5,5 раза выше, чем в водах ёриков, где бакланов нет
(Чуйков, Головкин, 1979). Уместно отметить и «подкормку» многих позвоночных и беспоз-
воночных животных оброненной бакланами рыбой и трупами птенцов в колониях (Коше-
лев и др., 2001).
Заслуживает внимание негативная роль бакланов в изменении зоот в местах нового рас-
селения. Так, при освоении островов Черноморского заповедника бакланы вытеснили на
менее пригодные для выведения птенцов участки кулика-сороку, а из чаек — хохотунью,
черноголового хохотуна, чеграву. В итоге количество гнездящихся пар двух последних
видов снизилось, соответственно, с 180 до 11, и со 180 до 30 пар (Руденко, Яремченко,
2004). Экспансия большого баклана на оз. Ханка (Большая Лефинская колония) привела к
вытеснению на периферию колонии цапель, в том числе редких в регионе — египетской, а
также южной, средней и малой белых цапель (Глущенко и др., 2003).
Роль птиц в распространении арбовирусов пока изучена недостаточно, однако среди
реальных носителей «клещевых» вирусов одними из первых названы колониальные бакла-
ны (Львов, Лебедев, 1972). Колония бакланов на Вост. Каспии была поражена клещами
Ornithodoros capensis (Андреев и др., 1977). Таким образом, бакланы могут становиться
переносчиками некоторых вирусных заболеваний человека и животных.
Если возникает острая необходимость сокращения численности бакланов, она должна
проводиться не путем поголовного истребления птиц и гнёзд, а путём уменьшения количе-
ства яиц в кладках до 1-2 — как это, например, практикуется в Черноморском заповеднике
(Руденко, Яремченко, 2004).
Уже упоминалось, что человек утилизирует мясо, шкуры и яйца бакланов для своих
нужд, в частности, в дельте Волги и в Дагестане (Джамирзоев, 1999а). Однако такое ис-
пользование бакланов не идёт ни в какое сравнение с подобным использованием водопла-
вающих птиц.
Атлантический большой баклан Ph. с. carbo занесён в Красные книги Мурманской об-
ласти и Республики Карелия (в последнем случае — без разделения на подвиды), материко-
вый подвид Ph. с. sinensis — в Красную книгу Армении, а в России — в Красные книги
Республики Алтай, Республики Карелия, Читинской и Иркутской областях, а также в спис-
ки охраняемых видов Республики Бурятия, Республики Хакасия, Тюменской области. Япон-
ский большой баклан Ph. с. hanedae занесён в Красную книгу птиц Японии (A Red Data
Book of Japanese Birds, 1975).
Японский, или уссурийский баклан
83
Японский, или уссурийский баклан
Phalacrocorax capillatus (Temminck et Schlegel, 1849)2
Carbo capillatus Temminck et Schlegel, 1849, Siebold, Fauna Japonica, Aves, табл. 83,
Япония.
Japanese Cormorant, Temminck’s Cormorant (англ.), Japan-Kormoran (нем.),
cormoran de Temminck (фр.)
Син. (лат.): Carbo filamentosus Temminck et Schlegel, 1850; Siebold, Fauna Japonica, Aves,
стр. 129, Carbo capillimentosus Bonaparte, 1855.
Статус. Гнездящийся и мигрирующий вид.
Общая характеристика и полевые признаки. Один из самых крупных бакланов. По
величине немного превосходит материкового большого баклана (птиц дальневосточных
популяций), встречающегося иногда вместе с ним на морских побережьях. В полевых ус-
ловиях виды не всегда различимы. У сидящего уссурийского баклана верх тела имеет зеле-
новатый металлический оттенок и чешуйчатый рисунок. У большого баклана — более чёт-
ная коричневато-бронзовая «чешуя». Затылок у уссурийского баклана гладкий; у взрослых
больших бакланов летом и осенью на затылке — невысокий и неровный, но хорошо замет-
ный чёрный хохол-гребень («гребешок»).
На близком расстоянии видна разница в конфигурации голых участков кожи на го-
лове — у уссурийского баклана граница оперения доходит до угла рта, а на нижней челю-
сти даже выдаётся от него вперёд, тогда как у большого баклана площадь голой кожи боль-
ше и оперение не доходит до угла рта, а ниже граница оперения углом смещается назад.
Клюв у уссурийского баклана выглядит более массивным. Помимо этих признаков, виды
различаются по характеру размещения — уссурийский баклан, в отличие от большого, очень
редко посещает пресные водоёмы и не удаляется от побережья моря; он никогда не садится
на деревья.
Голос взрослых уссурийских бакланов, как и у большинства бакланов, грубый, гортан-
ный: «кроо-ко ... кгоооо ...» (крик испуганного, слетающего с гнёзда баклана). В море —
молчаливая птица. Голос только что вылупившегося птенца — тихий, хрипловатый и не-
много скрипучий писк. Через несколько дней он становится громче, но по-прежнему оста-
ётся хриплым: «кхии... кхии...».
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Близок к большому баклану. Оба вида составляют номи-
нативный подрод рода Phalacrocorax в широком понимании (del Hoyo et al., 1992) и оста-
ется в составе указанного рода в узком понимании (Siegel-Causey, 1988). В таксономичес-
ких сводках есть разночтения по поводу года опубликования первоописания вида — 1850 г.
напр., Степанян, 2003) или 1848 г. (напр., Dickinson, 2003; Коблик и др., 2006). Недавние
исследования окончательно устанавливают год первоописания — 1849 (Morioka et al., 2005),
который принят здесь. Под названием filamentosus вид описан теми же авторами только в
850 г., поэтому это название должно фигуровать в качестве младшего синонима.
Описание. Окраска. Самец и самка в брачном наряде. Общая окраска оперения —
чёрная с зелёным металлическим блеском. На голове, шее, пояснице, подхвостье и нижней
части тела этот блеск более тёмный и имеет холодный синевато-зелёный оттенок. На верх-
: Очерк написан Н.М. Литвиненко в 1990-1991 гг. Дополнения и общее редактирование Ю.В. Шиба-
ева {прим. ред.).
84
Отряд Пеликанообразные, Семейство Баклановые
ней части тела он светлее, с чуть заметным бронзовым отливом. Бока головы и горла бе-
лые, в мелких чёрных (иногда с зелёным блеском) пестринах. Голову и верхнюю часть шеи
украшают длинные белые волосовидные перья. Перья лопаток, плечевые и верхние крою-
щие крыла с бронзово-зелёным оттенком; при определённом освещении на концах перьев
заметен фиолетовый оттенок. Перья окаймлены чёрными полосами, что создает на верх-
ней части тела заметный чешуйчатый рисунок. По бокам тела над голенью по большому
белому пятну. Маховые черновато-бурые; участки с металлическим блеском расположены
главным образом на наружной стороне опахал так, что на сложенном крыле образуется
равномерный зеленоватый оттенок. Испод крыла бурый. Рулевые сверху чёрные с зелено-
ватым отливом, снизу буровато-чёрные. Клюв буроватый, светлеющий в основании и ниж-
ней части подклювья; с боков он покрыт крупными роговыми чешуями, расположенными
черепицеобразно и создающими заметный поперечный рисунок. Ноги чёрные. Кожа лица
оранжево-жёлтая. Радужина изумрудно-зелёная.
Только что вылупившийся птенец покрыт полупрозрачной розовой кожей с лёгкой жел-
тизной у основания хвоста и на крыльях, спина с глянцем. Когти серые с белыми концами,
конец клюва тёмный. Сквозь кожу просвечивают внутренности. Темнеть и терять «про-
зрачность» кожа начинает через несколько часов. Однодневный птенец покрыт грифель-
ной с красноватым оттенком кожей. Голова, лапы и крылья бледно-розовые, местами с жел-
тизной. Сквозь кожу синеют глазные яблоки. От ноздрей через глаз идёт чуть заметная
темная полоса. Клюв бледно-розовый с прозрачной желтизной, конец его беловатый, перед
концом на надклювье и подклювье по чёрному пятну. Яйцевой зуб белый. Окраска зева —
сочетание тонов прозрачно-розовых и прозрачно-жёлтых. Когти серые с белыми концами.
Через несколько дней кожа птенца ещё более темнеет, становясь чёрной на спине и тёмно-
серой на брюхе. Грифельный глянец ещё хорошо заметен, он исчезает с появлением пуха.
По средней линии груди и брюха намечается узкая желтоватая полоса. Лапы становятся
серовато-жёлтыми сверху и бледно-жёлтыми снизу. На когтях появляются черноватые пред-
вершинные пятна.
Пуховой наряд. Птенец покрыт буровато-чёрным пухом; голыми остаются горловой
мешок и желтоватая полоса, идущая вдоль середины нижней части тела. У младших пухо-
вичков, у которых пух недавно появился, клюв в основном имеет жёлтый оттенок. Пере-
дняя половина надклювья — серая; чёрные предвершинные пятна, характерные для голых
птенцов, исчезают. Горловой мешок розовато-жёлтый. Ноги сверху бледно-бурые, пере-
понки серовато-жёлтые, снизу цевка и перепонки — бледно-жёлтые. Когти серые с чёрны-
ми концами. У старших пуховиков с полностью отросшим пухом клюв серый, за исключе-
нием жёлтого конца, основания и середины подклювья, а также полос, окаймляющих его
края. Горловой мешок желтоватый в темных пятнах. Ноги чёрные, перепонки и подошва —
жёлтые.
Гнездовой наряд. Общая окраска верха бурая. На лопатках, плечевых и кроющих крыла
перья окаймлены полосами, состоящими из светло-охристых или сероватых наружных и
тёмно-бурых внутренних каёмок. Низ бурый с грязно-белой грудью и средней частью жи-
вота. Маховые и рулевые тёмно-бурые. На перьях верхней части тела, а также на маховых
и рулевых уже заметен зеленоватый металлический блеск того же оттенка, что и у взрос-
лых птиц. Надклювье чёрное, к концу и краям — светло-розовое. Подклювье бледно-жёл-
тое с тёмными продольными пятнами в средней части верхнего края. Цевка спереди и с
боков чёрная, сзади (там, где нет щитков) грязновато-жёлтая. Пальцы сверху чёрные, пере-
понки тёмно-бурые, желтеющие возле пальцев. Снизу лапа серовато-жёлтая. Когти тёмно-
розовые, светлеющие к концу и острому нижнему краю. Голые участки кожи вокруг клюва
и нижней части «лица» лимонно-жёлтого оттенка, вокруг верхней — жёлто-бурого. От ноз-
Японский, или уссурийский баклан
85
Таблица 4. Размеры (мм) взрослых птиц (п, если известно, в скобках)
Место добычи Пол Размах крыльев Длина крыла Длина хвоста Длина цевки Длина клюва Источник
- - - 310-340 - 63-67 58-70 Тугаринов, 1947
— — — 320-332 (3) - - 61—67 О) Судиловская, 1951
Приморье сГ 1355 330 153 58 - Taczanowski, 1891-1893
_ к _ о 1366 326 164 60 — II
_ и _ ? 1400 330 155 64 — II
_ и _ ? - 315 148 65 60 Зоомузей ДВГУ
_ и _ О — 302 139 65 63 _ и
и сГ - 332 155 68 70 н
_ и _ - — 335 - 65 65 Зоомузей БПИ ДВО РАН
и — — 330 — 60 67 II
Япония - - 312-350 140-150 60-68 63-73 Kobayashi, 1962
Китай $ - 330 157 65 71 La Touche, 1931-1934
_ и _ сГ? - 375 175 58 - II
три через глаз до границы оперения проходит узкая тёмно-бурая полоса. Такая же полоса
ллёт параллельно ей вдоль края надклювья. Горловой мешок по ярко-жёлтому фону испещ-
рен чёрными продольными пятнами. Радужина серовато-бурая.
Строение и размеры. Размеры приведены в таблице 4.
Масса тела самцов (п=7) в среднем 3150 г, самок (п=8) — 2525 г (Kato et al., 2001).
Распространение. Гнездовой ареал вида лежит между 34 и 52° с.ш. Вид
-вселяет берега Японского, Жёлтого и Восточно-Китайского морей, южную часть Куриль-
ной гряды и Тихоокеанское побережье Японских о-вов.
В России гнездится на Курильских о-вах от Малой Курильской гряды и о. Кунашир до о.
Гимушир (Велижанин, 19776). Недавнее новое обследование Курильских о-вов показало,
-то в настоящее время вид не идёт севернее о. Уруп (Артюхин и др., 2001а). На Сахалине
-ездование известно только для крайнего юга острова. Это п-ов Крильон в прошлом (Ги-
те нко. 1955), Тонино-Анивский п-ов (Нечаев, 19916; Матюшков, 1996) и о. Монерон (Ги-
тенко. 1955; Беньковский, 1968; Нечаев, 1975а; Шибаев, Литвиненко, 1996).
На побережье материка в России гнездится практически от границы с КНДР до северной
-тети Татарского пролива (Litvinenko, Shibaev, 1991; Блохин, Тиунов, 2003). В Приморье наи-
s. лее крупная колония (около 1 500 гнездящихся птиц) находится в зал. Петра Великого на о.
tур>тельма. Кроме неё, в зал. Петра Великого есть 14-15 колоний поменьше — на о-вах
У-драмзина, Стенина, Большой Пелис, Гильдебрандта, а также на других островах и некото-
участках побережья (Шибаев, 1987). Далее к северо-востоку, начиная от мыса Пово-
тного, до м. Аукан небольшие поселения уссурийских бакланов встречаются вдоль всего
- .бережья, хотя распределение их неравномерно, что связано, главным образом, с характе-
7<м структуры береговых скал (авиаучёты 1982 и 1986 гг., данные автора очерка). Три ко-
: -лай известны из Лазовского заповедника и его окрестностей (Литвиненко, Шибаев, 1971),
_есть — найдены в районе зал. Ольги (Лабзюк, 1975), десять — обнаружены на отрезке
.бережья между м. Счастливый и устьем р. Пея (Елсуков, 1984).
86
Отряд Пеликанообразные, Семейство Баклановые
Рис. 16. Область гнездования японского, или уссурий-
ского баклана (без китайской части ареала): а — рай-
оны гнездовий на континенте, б — отдельные извест-
ные колонии на островах, в — район гнездовий на Ку-
рильских о-вах, г — область зимовки.
В Японии гнездовья этих бакланов размещаются в основном в северной части архипе-
лага. Наиболее известные колонии на Хоккайдо находятся на о-вах Юрури, Моюрури и
Дайкокудзима, п-ове Сиретоко, на мысах Отииси и Эсан, а также на о-вах Теури, Ягисири.
Возможно, гнездится на о. Ребун. На тихоокеанском побережье Хонсю известны две коло-
нии — на о-вах Санкандзима и Кинказан, на западном — на о-вах Ава, Садо, Нанацусима,
а также западнее зал. Вакаса вблизи г. Касуми. Самая южная колония этого вида в Японии
находится у северной оконечности о. Кюсю на о. Окиносима (Sato et al., 1968; Survey of
Birds in Japan, 1973, 1980; Hasegawa, 1984; Фудзимаки, 1986).
На Корейском п-ове известны 6 мест гнездования этого вида, все они находятся в север-
ных и центральных районах полуострова: о. Сонбонг Альсом (Арсом, Нандо), расположен-
ный на крайнем северо-востоке КНДР у пос. Унги, два острова в Восточно-Корейском за-
ливе (один из них о. Тхончхон Альсом у г. Тхончхон), о. Уллындо, о. Сончхонрап-то у севе-
ро-западного побережья п-ова в провинции Пхенан-Пукто и один из островов в зал. Че-
мульпо (Austin, 1948; Вон Хон Гу, 1963; Endangered bird species in the Korean Peninsula,
1987) (рис. 16).
В Китае гнездится на берегах Жёлтого моря: на Ляодунском п-ове и на морских берегах
провинции Хэбэй и Шаньдунского п-ова вблизи городов Яньтай, Вэйхай и Циндао (Meize,
1934; Cheng, 1976).
Зимовки. Область зимовок не поднимается выше 47° с.ш. Самая северная точка
зимней встречи — устье р. Самарга (Шульпин, 1936), где в январе 1926 г. была добыта
взрослая птица.
Зимой на дальневосточных побережьях России уссурийский баклан довольно редкая
или немногочисленная птица. В Приморье, кроме упомянутой выше встречи в устье Са-
марги, зарегистрирован ещё лишь несколько раз. 21 декабря 1867 г. в зал. Ольги двух бакла-
нов этого вида встретил Н.М. Пржевальский (Дневник по птицам Уссурийского путеше-
ствия 1867-1869 гг., цит. по: Шульпин, 1936). Шесть птиц отмечены вблизи пос. Терней
8 февраля 1958 г. (Рахилин, 1972). Ю.В. Шибаев, проводивший учёты морских птиц в При-
морье в зимние сезоны 1977/78 и 1978/79 гг., встретил уссурийских бакланов только в зал.
Петра Великого, причём, в очень небольшом числе. Вид зимует, кроме того, в незамерзаю-
щих водах Южно-Курильского пролива, где численность его также невелика (Нечаев, 1969).
Основные районы зимовки лежат южнее.
В Японии зимует на побережье центральных и южных районов страны (Check-list of
Japanese birds, 1974), причём в некоторых местах о. Хонсю концентрируется в больших
Японский, или уссурийский баклан
87
количествах. Так, на скалистом островке Кабесима в префектуре Ямагути (крайний юго-
запад Хонсю) с ноября по март скапливаются тысячи бакланов, покрывая за этот период
скалы толстым слоем гуано. Кроме того, большие скопления уссурийских бакланов встре-
чаются зимой на островах Оки, Ава (западная часть Хонсю) и на других островах в префек-
турах Айти, Хиого и Ибараки (Austin, Kuroda, 1953). Почти ежегодно зимует на о. Кинка-
ззн (Sato et al., 1968). Отмечен также в северной части архипелага Рюкю (Vaurie, 1965;
Brazil, 1991) и на о-вах Идзу. О. Миякодзима, например, служит местом ночлега для уссу-
г и неких бакланов; стаи по несколько десятков птиц собираются сюда вечером вплоть до
конца апреля (Moyer, 1957).
На Корейском п-ове зимует на восточном, западном и южном побережьях, встречен
также на о-вах Уллындо и Чечжудо (Вон Хон Гу, 1963; Vaurie, 1965; Check-List of Japanese
T.rds, 1974). Во время учётов морских птиц, проведённых на крайнем юго-западе Корей-
ского п-ова с января по март 1974-1975 гг., было зарегистрировано 167 уссурийских бакла-
нов. причём, на отрезке побережья от г. Мокпхо до о. Вандо этот вид составлял 8,64% от
=<ех встреченных птиц (Min Yoon & Won, 1976).
В КНР зимует от провинции Шаньдун до провинции Фуцзянь (Cheng, 1976).
Судя по материалам кольцевания (данные автора очерка; Рахилин, 1972), различные
т эпуляции этого вида мигрируют и зимуют в разных районах. Молодые птицы из коло-
ний Приморья продвигаются к югу вдоль материкового побережья и проводят зиму в
Корейском проливе, концентрируясь у южного побережья Корейского п-ова и на крайнем
< ro-западе о. Хонсю, где наблюдаются особенно крупные скопления этих бакланов (см.
зыше).
По-видимому, в Корейском проливе зимуют преимущественно птицы из материковых
". опуляций (Приморье, Корейский п-ов), так как бакланы, гнездящиеся в Японии, до сих
~?р не были здесь встречены (Japanese Bird Banding in Recent Years, 1961-1983, 1985).
Корейский пролив привлекает птиц тем, что в его водах в течение всего зимнего периода в
5: льшом количестве концентрируются стада сайры (Cololabis saira), иваси (Sardinops mela-
’ :<rictus) и анчоуса (Engraulis japonicus) (Шунтов, 1972).
Области миграций и зимовок бакланов о. Хоккайдо лежат немного севернее, причём
-:и также довольно четко распределены между птицами из различных поселений. Бакла-
-л с о. Теури, расположенного на северо-западе Хоккайдо, распространяются вдоль запад-
-zzx побережий Японских о-вов, не спускаясь южнее о. Садо. Птицы с о-вов Юрури и Мо-
<тури (восточный Хоккайдо) мигрируют главным образом вдоль восточных берегов
’ “.:нии, а их соседи с двух близлежащих островков больше тяготеют к западным берегам
\ ~ : нии (Japanese Bird Banding in Recent Years, 1961-1983,1985). Такое разграничение райо-
-: з зимовок и миграций отдельных популяций бакланов связано, по-видимому, с их пище-
ы й специализацией. Но при этом лимитирующим фактором в местах зимовок иногда мо-
я tr быть не столько пища, сколько наличие мест дневного отдыха (для сушки оперения) и
-: -ёвок.
Миграции. Судя по данным кольцевания, большинство молодых птиц, покинувших гнез-
: г ъ? колонию, остаются до конца октября (единицы — до середины ноября) в районе гнез-
:. мн>1я. Районы кочёвок птиц с о. Фуругельма и м. Островного, несмотря на относитель-
< близость этих мест гнездования, перекрываются лишь частично. Дальность разлета
а --.«тих бакланов невелика. Некоторые особи начинают продвигаться к югу раньше —
лтица с о. Фуругельма уже в октябре была встречена в Юж. Корее (окрестности г.
. 1ЧЧХ0К). В октябре-ноябре начинается заметное движение птиц к местам зимовок.
Зглётные птицы отмечены на о-вах Тайвань, Окинава, Куродзима (один из самых южных
•еттчевов архипелага Рюкю), Бонин и Торисима (Vaurie, 1965; Cheng, 1976; Brazil, 1991).
88
Отряд Пеликанообразные, Семейство Баклановые
Местообитания. Вид связан с морскими побережьями — скалистыми берегами мате-
рика и островов. Гнездится на уступах скал, предпочитая широкие карнизы, а также на
вершинах небольших скалистых островков. Может гнездиться на низких островках, если
они не затапливаются водой во время штормов. Кормится в море вблизи побережья. Изред-
ка залетает в устья рек и на прибрежные озёра.
Факт гнездования уссурийских бакланов совместно с большими в хвойных лесах
окрестностей Нагои и Токио (Jahn, 1942) последующими исследователями не был под-
твержден (Austin, Kuroda, 1953; Check-list of Japanese birds, 1974). Биологическая спе-
цифика уссурийского баклана свидетельствует о том, что гнездование его на деревьях
невозможно. В частности, он никогда не садится на деревья, стоящие на берегу (что
сразу выдает большого баклана). В то же время, совместное гнездование обоих видов
на материковом побережье — доказанный факт (Шибаев, 1987; Бабенко, 2000; Блохин,
Тиунов, 2003). Таким образом, уссурийский баклан, по сравнению с большим, более
стенотопен.
Численность. Несмотря на то, что имеющиеся оценки численности получены для раз-
ных регионов в разные годы, они позволяют с определённой долей условности судить о
состоянии вида в конце XX столетия.
На материковом побережье Японского моря в России по приблизительным подсчетам
гнездится не более 8 тыс. уссурийских бакланов. Около 3 тыс. (15 колоний) обитает в зал.
Петра Великого, на о. Фуругельма. В 1979 г. здесь гнездилось около 1 тыс. птиц, в 1989 г. —
1 450 птиц. В 1982 г. на о. Б. Пелис обитало 110 бакланов, о. Гильдебрандта — более 200,
о. Стенина — 110, на кекурах3 у п-ова Гамова — 434 (Шибаев, 1987).
Далее к северу от мыса Поворотного до северной части Татарского пролива было
обнаружено 38-40 небольших колоний, насчитывающих от 5 до 100 пар (преимуще-
ственно от 10 до 20 пар) (Литвиненко, Шибаев, 1971; Лабзюк, 1975; Елсуков, 1984;
авиаучёты Ю.В. Шибаева 1982 и 1986 гг.; Бабенко, 2000; Блохин, Тиунов, 2003).
На Курильских о-вах по А.Г Велижанину (19776) гнездится 7-8 тыс. уссурийских бак-
ланов. Однако Ю.Б. Артюхин и др. (2001а), проанализировав его материалы (учёт 1963 г.),
нашли эту оценку «несколько завышенной». Учёт численности вида, проведённый на рубе-
же столетий (Артюхин и др., 2001а), дал вдвое меньшую цифру — 3 600 особей.
Численность на Юж. Сахалине (о. Монерон) составила на этот период около 500 особей
(Нечаев, 1986; Шибаев, Литвиненко, 1996; Матюшков, 1999).
Общую численность репродуктивной части популяции в России таким образом можно
оценить в 12 000 особей (или несколько более).
В Японии число гнездящихся уссурийских бакланов оценивается примерно в 1 100 осо-
бей (Hasegawa, 1984). Однако, по мнению Ватануки в соавторами (Watanuki et al., 1988),
только на Хоккайдо в 1980-е гг. гнездилось 1 900 пар (3 800 особей). Общая же числен-
ность по стране была, по-видимому, не более 4 000 особей.
Численность вида на Корейском п-ове и в Китае неизвестна, но по имеющимся литера-
турным данным, колонии уссурийского баклана встречаются в этих странах нечасто (Gore,
Won, 1971; Tomek, 1999; Meize, 1934; Cheng, 1976). Видимо, птиц здесь немного, посколь-
ку это край гнездового ареала, включающий весьма населенные людьми территории.
Таким образом, общая численность репродуктивной группировки вида на конец XX
столетия вряд ли превышала 17 000 особей. В гнездовом ареале выделяются два участка с
повышенной плотностью вида — материковый (Приморье, юг Хабаровского края) и ост-
ровной: Юж. Курилы (от о. Итуруп) и о. Хоккайдо.
3 Кекур — местное название небольшого безлестного скалистого островка, морской скалы.
Японский, или уссурийский баклан
89
Самая крупная колония на материковом участке расположена на о. Фуругельма (725
пар, 1985 г.) (Шибаев, 1987). Самые крупные колонии на островном участке — на о. Хок-
кайдо (п-ов Сиретоко, более 500 пар, конец 1980-х гг.; о. Теури, 738 пар, 1987 г.) (Watanuki
et al., 1988).
В последние 60 лет численность уссурийского баклана в зал. Петра Великого подверга-
лась резким колебаниям. Это хорошо прослеживается на примере колонии о. Фуругельма.
В 1929-1930 гг. гнездовья этого баклана были полностью уничтожены завезенными на ос-
тров песцами. Через несколько лет, когда звери вымерли, бакланы вернулись на остров и
численность их начала увеличиваться. Однако в 1967 г. она по неизвестной причине резко
снизилась во всех колониях зал. Петра Великого. На о-вах Большой Пелис, Стенина, Ка-
рамзина и др. гнездилось в том году лишь несколько пар птиц (Лабзюк и др., 1971). На
следующий год численность их немного возросла, но лишь через несколько лет достигла
прежнего уровня. На о. Фуругельма в 1969 г. было зарегистрировано только 43 гнездящих-
ся пары уссурийских бакланов (Лабзюк и др., 1970). После 1970 г. число гнездящихся птиц
снова начало расти и к 1985 г. колония увеличилась примерно в 17 раз (по сравнению с
1969 г.), после чего численность стабилизировалась (Шибаев, 1987).
Росту численности способствовали, по крайней мере, два обстоятельства. В 1970-
1980-е гг. в Юж. Приморье несколько снизился антропогенный пресс по отношению к
морским птицам в связи с организацией в 1978 г. Дальневосточного морского заповедни-
ка. куда вошли основные колонии уссурийского баклана. Кроме того, в эти же годы зна-
чительно улучшилась кормовая обстановка для морских птиц. Изменения гидрологиче-
ских условий на севере Японского моря вызвали, в частности, мощную вспышку числен-
ности иваси (Шунтов, 1986), и некоторые морские птицы, в том числе и уссурийский
баклан, в значительной степени сменили спектр кормов, переключившись на питание этим
массовым видом.
Размножение. Колониальная птица, отдельными парами гнездится очень редко. Охот-
но селится рядом с другими видами колониальных птиц (Fujimaki, 1961; Вон Хон Гу, 1963
и др.), образуя смешанные поселения. В Юж. Приморье гнездится в непосредственной
близости (расстояние между гнёздами 2-3 м) с тонкоклювой кайрой (Uria aalge), черно-
хвостой чайкой (Larus crassriostris), серой цаплей, большим бакланом и др.
К размножению приступает, по-видимому, в возрасте около двух лет. Молодые негнез-
дящиеся птицы посещают гнездовые колонии, ночуют здесь, иногда пытаются строить гнёзда
?: совершать брачные демонстрации. Обычно они составляют не более 20% населения ко-
лонии. Молодые птицы появляются в колонии перед заходом солнца и улетают утром. Та-
ким образом, основной период, который они проводят здесь, приходится на темное время
суток.
Как и у других видов бакланов, у уссурийского наблюдается тенденция к смене места
~ездования (Литвиненко, Шибаев, 1971; Лабзюк, 1975; Yamamoto, 1967; Sato, Misuno,
Tikemaru, 1968). Часть большой колонии (обычно обособленная группа) или отдельная
- ебольшая колония в начале сезона размножения занимает новую территорию (зачастую
: на располагается поблизости от прежней, иногда в нескольких десятках метров). Через
сд-два бакланы могут снова вернуться на прежнее место. В некоторых случаях такое Пе-
ле с едение объясняется прямым вмешательством человека: в колонии проводился сбор яиц,
• дльцевание и т.п. Но часто птицы меняют место без видимой причины.
Самец, привлекающий самку, выполняет следующий ритуал: он сидит в гнезде, грудь
: лущена, поясница приподнята, шея и хвост подняты вверх. Находясь в такой позе, птица
взмахивает крыльями и забрасывает голову назад: 5-6 взмахов крыльями и затем 3-Л дви-
жения шеей. Одиноко сидящий в гнезде самец может неспешно помахивать крыльями в
90
Отряд Пеликанообразные, Семейство Баклановые
течение 1-2 ч, но при приближении самки движения его становятся быстрее, и он начинает
энергично забрасывать назад шею.
Похожую демонстрацию совершают оба партнёра во время ритуала смены на гнезде.
Она отличается рядом деталей. В частности птицы, сгибая шею назад, издают глухой кар-
кающий звук.
Ещё одна демонстрация, которую выполняют оба партнёра, совершается обычно перед
началом копуляции. Партнеры вытягивают навстречу друг другу шеи и скользящими дви-
жениями вперёд и вверх легко соприкасаются боками головы с одной стороны, потом —
с другой.
Ритуал смены на гнезде обычно происходит так: прилетевшая птица на 1-2 сек вытя-
гивает вверх шею, голова её при этом пригнута вниз, а подъязычная кость опущена. За-
тем птицы обмениваются криками и выполняют одну из описанных выше совместных
демонстраций, иногда — обе. Прибывшая птица усаживается рядом с гнездом или непо-
далеку от него и в течение 1-30 мин чистит и сушит оперение; только после этого направ-
ляется к гнезду. Насиживающая птица иногда сразу встает и отходит в сторону, а партнёр
занимает её место. Но в большинстве случаев прилетевшей птице приходится прибегать
к ритуальным «уловкам». Наиболее распространенная в таких случаях демонстрация —
подношение строительного материала. Порой достаточно бывает партнеру поднять с земли
первую попавшуюся веточку и предложить её сидящей на гнезде птице, как она уступает
ему место. Однако, как правило, эту демонстрацию приходится повторять несколько раз;
воздействие её усиливается, если партнёр не сразу предлагает веточку, а улетает с ней в
море и, сделав большой круг в пределах видимости супруга, возвращается и тут же бе-
жит с веточкой к гнезду. Но и этот ритуал приходится повторять иногда по 5-6 раз, преж-
де чем сидящая на гнезде птица уступает место. В любом случае родители во время сме-
ны на гнезде стараются ни на минуту не оставлять кладку или птенцов без надзора —
сменившаяся птица не улетает до тех пор, пока новая не усядется на яйца или не подой-
дёт к птенцам.
Территориальные конфликты в колониях уссурийских бакланов, как правило, разреша-
ются спокойно: хозяин гнёзда, резко вытянув шею с взъерошенными на ней перьями в
направлении к чужой птице и широко раскрыв клюв, издает хрипловатый крик «ко-ко-оо...»
или «ко-о-о...». Так же реагируют бакланы и на других «пришельцев», например, на близко
подлетающих к их гнёздам чернохвостых чаек. Драки у бакланов случаются крайне редко
и в основном на первых этапах размножения, до начала инкубационного периода.
На островах Юж. Приморья уссурийские бакланы появляются в гнездовых колониях к
началу апреля, но в этот период их бывает ещё очень мало и они не ночуют на территории
колонии. В десятых числах апреля бакланы приступают к постройке гнёзд: 13 апреля 1971
г. на о. Карамзина птицы уже начали строить и ремонтировать гнёзда, а к 25 апреля все
гнёзда были в разной степени готовности, но лишь некоторые из них имели законченную
форму и лоток. Большинство же представляло собой рыхлую постройку, напоминающую
скорее охапку прутьев. 3 мая 1968 г. почти у всех пар гнёзда были достроены.
Гнёзда, как было сказано выше, уссурийский баклан устраивает на уступах береговых
скал и на выступающих из моря камнях. В отличие от берингова и краснолицего бакланов,
он избегает отвесных участков и узких карнизов, предпочитая скалы с широкими уступа-
ми, иногда заросшими травой, или плоские верхушки небольших островков. Такой харак-
тер размещения гнёзд прослеживается у вида в разных частях его ареала: на Японских о-
вах (Yamamoto, 1967; Fujimaki et al., 1976), на Юж. Курилах (данные Ю.В. Шибаева) и в
Приморье. Он сохраняется и в местах совместного гнездования уссурийского баклана с
другими бакланами: верхнюю часть островов и широкие уступы всегда занимает уссурий-
Японский, или уссурийский баклан
91
с кий баклан, узкие карнизы и ниши на отвесных скалах — берингов и краснолицый. В то
же время в Приморье, где уссурийский баклан устраивает общие поселения с серой цап-
лей, он иногда вынужден уступать последней «лучшие места».
Гнёзда уссурийского баклана иногда располагаются в доступных для наземных хищни-
ков и человека местах — в отличие, например, от берингова баклана, у которого такие
гнёзда представляют собой исключение.
Гнездо не всегда имеет правильную округлую форму. Если карниз узок, оно может быть
овальным. Как и у большинства других бакланов, это массивная, не очень аккуратная пост-
ройка. Материал гнезд, как правило, «подручный». На одних островах Юж. Приморья (Фу-
ругельма, Большой Пелис) это чаще всего сухая прошлогодняя трава, растущая поблизости
от гнездовой колонии — стебли лебеды (Chenopodium sp.) диаметром до 1 см, реже — пру-
тики кустарниковой полыни (Artemisia sp.) и барбариса (Berberis amurensis); лоток высти-
лается измочаленным колосняком (Elymus mollis). На других островах (о. Карамзина) гнёз-
да были выстланы преимущественно зостерой (Zostera marina). В колониях вблизи бухты
Киевка (Судзухе) строительным материалом служили веточки плавника и также сухая зос-
тера (Литвиненко, Шибаев, 1971). В Японии на о. Кинказан (вост. Хонсю) гнёзда этого
баклана были устроены из веток сосны (Pinus densiflora), лоток же был выстлан большим
количеством игл этого дерева, а в некоторых гнёздах, кроме хвои, лоток выстилали стебли
чискантуса (Miscanthus sinensis) и, опять-таки, морская зостера (Sato et al., 1968). Таким
эбразом, среди небольшого набора видов строительного материала, характерного для гнёзд
?того баклана в различных частях ареала, зостера (как материал выстилки) — наиболее
распространённый.
Средние размеры гнёзд в Юж. Приморье (п=15): диаметр гнезда — 57,1 см, высота
гнезда — 20,3 см, диаметр лотка — 24,5 см, глубина лотка — 7,1 см. Максимальная высота
гнезда — 25 см. В Японии на о. Хонсю некоторые гнёзда достигают 1 м в высоту; обычно
?то старые, неоднократно ремонтированные постройки (Yamamoto, 1967). Большинство
гнёзд за зиму разрушаются, и птицам приходится возводить всю постройку заново.
Гнездо строят оба партнёра. На островах залива Петра Великого неоднократно прихо-
дилось видеть, как бакланы летали за строительным материалом на ближайший от колонии
склон и, усевшись там среди зарослей прошлогодней травы, ломали или подбирали с земли
д.хие стебли. Очень распространенным способом добычи строительного материала явля-
ется кража его из чужих гнёзд. Достройка гнезда продолжается и в период насиживания —
•темя от времени один из партнеров приносит прутики, а насиживающая птица, не вста-
вая. вставляет их в края гнезда.
Первые кладки у бакланов Юж. Приморья появляются в конце апреля, массовая отклад-
<д яиц приходится обычно на начало мая. В иные годы сроки гнездования запаздывают: 3
чая 1968 г. на о. Карамзина ни в одном из 50 осмотренных гнёзд яиц ещё не было, а в 1966
~ откладка яиц на этом острове началась только в середине мая (Нечаев, Юдаков, 1968).
2 днако яйца в гнёздах этого баклана можно увидеть здесь не только в мае, но и значитель-
г позже — вплоть до середины июля. Примерно в такое же время (начало мая) откладыва-
?: т яйца уссурийские бакланы в других частях ареала: на о. Монерон (Гизенко, 1955), на о-
згх Юрури и Моюрури у северо-восточного побережья Хоккайдо (Fujimaki et al., 1976) и на
а: стоке Хонсю в префектуре Ивате (Yamamoto, 1967). Однако, в соседней, более южной
ттефектуре Мияги (о. Кинказан), птицы приступают к размножению значительно раньше:
.. апреля 1965 г. в гнёздах уже были яйца (Sato et aL, 1968). По-видимому, такие ранние
:гх:ки гнездования определяются не столько южным положением острова, сколько близос-
теплого течения Куро-Сиво, омывающего его берега.
На Корейском п-ове этот вид образует гнездовые пары в апреле (Вон Хон Гу, 1963).
92
Отряд Пеликанообразные, Семейство Баклановые
У птиц из Юж. Приморья (зал. Петра Великого) величина кладки от 2 до 4 яиц, в сред-
нем — 3,35 (п=45). Такие же по размеру кладки в 2-А яйца у бакланов на Хоккайдо (Fujimaki,
1961) и на Корейском п-ове (Вон Хон Гу, 1963). Лишь по А.И. Гизенко (1955) на о. Монерон
количество яиц в кладках доходило до 6.
Яйца бледно-голубые, матовые, покрыты толстым известковым слоем. Форма — удли-
нённо-яйцевидная. Размер яиц из Юж. Приморья (п=34), мм: 59,9-66,2 х 36,5^40,2; сред-
нее 63,2x38,6. С Курильских о-вов (о-ва Демина, п=3): среднее — 60,4x41 (А.И. Кошелев,
коллекция Биологического института СО РАН)
Инкубационный период продолжается около 30 дней. Массовое вылупление в колониях
Юж. Приморья происходит обычно в конце мая - начале июня.
Голые птенцы беспомощны и первые дни лежат в гнезде, обогреваемые родителями.
Приблизительно на 4-5-й день у них открываются глаза, ещё через 2-3 дня появляется пух.
Родители по очереди кормят птенцов и в течение 35^40 дней постоянно их охраняют. В
жаркие дни они поят птенцов и «купают» их в гнезде, обливая струей воды. После затяж-
ных дождей сушат гнездо: энергичным движением клюва протыкают его в нескольких ме-
стах, временами вытаскивая пучок влажной подстилки и разбрасывая её в гнезде.
Примерно на третий день головы птенцов уже начинают показываться из-под греющей
птицы. Пуховики, особенно старшие, очень энергичны. Выпрашивая корм, они вскидыва-
ют голову и раскачивают ею из стороны в сторону, сопровождая это движение своеобраз-
ной прерывистой трелькой. При этом они выпячивают подъязычную кость, и жёлтая кожа
горлового мешка становится особенно хорошо заметной. Такие раскачивающиеся жёлтые
головки — характерная картина для колоний бакланов в начальный период выкармливания
птенцов. Движения голодного птенца настолько энергичны, что взрослой птице не сразу уда-
ётся накормить его. Выбрав удобный момент, баклан осторожно хватает клювом птенца за
голову и тот погружает её в горловой мешок родителя. Каждое кормление состоит из 5-6
таких «погружений». Интервалы между отдельными кормежками трехдневного птенца в ут-
ренние часы составляли от 15 до 30 мин. По мере роста птенцов интервалы увеличиваются.
Каждое кормление выводка из трёх 10-12-дневных птенцов продолжалось от 15 до 20 мин.
Период пребывания в гнезде птенцов (п = 11) составлял в 1983 г. (о. Фуругельма) от 50
до 57 дней (в среднем, 53,2 дня). Массовый вылет в Приморье обычно наблюдается в сере-
дине июля. Покинувшие гнёзда выводки спускаются к воде и собираются на пляжах и отло-
гих каменных мысах, образуя своеобразные «детские сады». В каждой отдельной группе
насчитывается примерно от 20 до 100 птенцов. Вся колония постепенно перемещается
сюда — родители кормят здесь своих птенцов, отгоняя чужих. Молодые птицы то и дело
спускаются на воду, учатся плавать, нырять, взлетать с воды. Они подхватывают любые
плавающие на воде предметы — веточки, водоросли и подолгу манипулируют ими. 8 авгу-
ста 1983 г. удалось наблюдать, как молодежь из такого «детского сада» в течение несколь-
ких минут успешно охотилась на стайку анчоусов, приблизившуюся к берегу. «Детские
сады» существуют на гнездовом острове около 1 месяца, постепенно перемещаясь с берега
на окружающие остров скалы. Меняется характер кормления — птенцы всё чаще получа-
ют корм в воде, встречая родителей в стороне от перенаселённых пляжей. В начале августа
в Приморье уже встречаются сеголетки, самостоятельно добывающие пищу. Они охотятся
поодиночке или небольшими группами.
Через две недели после массового вылета молодые птицы уже хорошо летают, хотя по-
прежнему тяжело поднимаются с воды и более часто, чем взрослые, взмахивают крыльями
в полете. Они собираются в группы по 10-20 птиц и рассаживаются на скалах и на воде
вблизи колонии, по-прежнему пытаются неловко ловить плавающий мусор — щепки, об-
рывки водорослей. Через месяц после вылета молодые птицы ещё держатся рядом с род-
Японский, или уссурийский баклан
93
ной колонией. Многие к этому времени уже приобретают навыки в ловле рыбы и кормятся
самостоятельно.
Суточная активность, поведение. Ведёт дневной образ жизни. Кормовая активность
в период выкармливания птенцов начинается довольно поздно. Утренний отлёт первых
птиц из колонии на о. Фуругельма в середине июля 1984 г. зарегистрирован около 8 ч, а
примерно через час начали прилетать первые птицы с кормом. После 9 ч 30 мин число
• летевших из колонии птиц резко возрастало. Пик активности (максимальное количество
сгкланов, курсирующих между колонией и районами кормодобывания) наблюдали с 10 до
3 ч. после чего активность птиц падала.
Бакланы, направляющиеся в колонию с мест кормежки, летят обычно низко над водой
с :одиночке или группами до 15 особей, выстраиваясь цепочкой, реже — клином. Полет их
'таелый, у многих приоткрыты клювы. Птицы, отправляющиеся на кормёжку, летят быст-
7 ее и выше, стаи иногда формируются вблизи гнездовой колонии. Приходилось наблюдать,
iiix одиночные птицы пытаются догнать стаю или другого отдельно летящего баклана и
"растроиться к ним.
После 18 ч и до наступления темноты в колонии на о. Фуругельма наблюдается хорошо
‘Сраженное увеличение количества направляющихся в колонию птиц — это взрослые бак-
'дны. прилетающие на ночёвку. Большая часть из них сразу же летит в колонию, другие
т досаживаются на выступающих из воды скалах, а с наступлением сумерек дружно снима-
йся и тоже летят ночевать на остров. В этот период в колонии наблюдается заметное ожив-
ание, усиливается шум, учащаются территориальные конфликты. Хотя многие ночующие
~пхы, по нашим наблюдениям, имеют в колонии постоянные места, им часто приходится
. твоёвывать» их снова и снова, а затем некоторое время охранять. Хозяева гнёзд также
встречают прилетевших птиц угрожающими демонстрациями и криками. Заняв, наконец,
:д?и места, бакланы принимаются энергично чиститься, сушить оперение и, закончив эти
процедуры, тут же засыпают.
Разница в размерах отражается на поведении полов при нырянии в процессе кормодо-
: звания (Watanuki et al., 1996; Kato et al., 2001). Различия касаются глубины ныряния, про-
должительности, количества погружений в течение дня. Самцы ныряют глубже, но реже.
Различия создают возможность маневра, как во времени, так и по охвату глубин.
Питание. Питается рыбой. Охотится в стаях или поодиночке в прибрежных водах и
отъях рек. Приходилось неоднократно видеть, как уссурийские бакланы наблюдали со скал
чернохвостыми чайками, занятыми поиском корма, и как только чайки обнаруживали
с сяк рыбы и начинали охоту, немедленно присоединялись к ним.
В корме птенцов из залива Петра Великого чаще всего встречали рыб из следующих
:емейств (встречаемость, соответственно, в 1976-1979 гг.; 1980 г.; 1983 г): Clupeidae (Clupea
Sardinops melanostictus, Clupanodon punctatus) — 2,5, 15,7, 29,5%; Engraulidae
Zrgraulis japonicus) — 2,5, 0, 40,8%; Gadidae (в основном Eleginus gracilis) — 2,5, 17,1,
~ Stichaeidae (Stichaeus grigorjewi, Opistocentrus zonope, O. dybowskii и др.) — 56,4, 2,8,
1 :. Hexagrammidae (Hexagrammus octogrammus, Pleurogrammus azonus) — 15,3, 5,7, 0%;
::~dae — 7,6, 15,7, 4,2%.
В указанный период (1976-1983 гг.) в составе корма уже намечалась тенденция к увели-
- - них? доли сельдевых и снижению значения других групп (в частности, стихеев). В после-
г чдие годы (1984 и 1985 гг.) в корме явно преобладала сардина иваси. Такая смена спек-
тлитания, безусловно, явилась результатом изменений состава ихтиофауны в водах Япон-
. с 70 моря, связанных с общим потеплением вод в северной Пацифике в период с середи-
-Дх *.9"0-х до конца 1980-х гг. Они вызвало рост численности некоторых видов рыб, в час-
--: сти. иваси (Шунтов, 1986).
94
Отряд Пеликанообразные, Семейство Баклановые
Состав корма, приносимого птенцам, изменяется в течение суток: в июле 1984 и 1985 гг.
на о. Фуругельма утренний рацион состоял из разнообразного набора преимущественно
донных рыб, вечером — главным образом из сардины иваси.
Враги, неблагоприятные факторы. Серьезных врагов у этого крупного баклана нет.
В Приморье в его колониях постоянно «дежурят» большеклювые вороны (Corvus тасго-
rhynchos), чёрные вороны (Corvus согопе) и чернохвостые чайки (Larus crassirostris). Чай-
ки то и дело низко пролетают над гнёздами бакланов, вороны наблюдают за колонией изда-
ли. При возникновении паники среди бакланов, вызванной, например, появлением челове-
ка, эти птицы немедленно прилетают в колонию, чтобы расклевать яйцо или утащить ма-
ленького птенца из покинутых хозяевами гнёзд. В июле, когда яиц нет, а птенцы слишком
большие, чайки и вороны по-прежнему посещают колонии, довольствуясь оброненным
птенцами кормом.
Отмечены редкие случаи нападения орлана-белохвоста (Haliaeetus albicilld) на моло-
дых бакланов и даже на гнёзда с птенцами. Ещё реже эти птицы становятся жертвами на-
земных хищников: в 1972 г. в одной из колоний, расположенных вблизи залива Ольги, не-
сколько гнёзд бакланов были разорены гималайским медведем (Лабзюк, 1975).
Гельминтологическое обследование уссурийских бакланов (п=35), проведенное в Юж.
Приморье с 1966 по 1969 г. (Сметанина, Алексеев, 1970), дало следующие результаты: тре-
матоды обнаружены у 35 птиц, цестоды — у 7, нематоды — у 35, скребни — у 31. Видовой
состав гельминтов, по данным З.Б. Сметаниной, следующий:
Trematoda — Petasiger phalacrocoracis, Galactosomum phalacrocoracis, Renicola umi-
garasu, Renicola vladica, Liliatrema skrjabini, Liliatrema sobolevi, Hysteromorphe triloba.
Cestoidea — Ligula colymbi, Schistocephalus solidus.
Acanthocephala — Polymorphus sp., Corynosomaphalacrocoracis, Corynosoma strumosum,
Hemiechinosoma ardese, Hemiechinosoma mergi, Prosthorhynchus sp.
Nematoda — Contracaecum spiculigerum, Desmidocercellia incognita.
Выяснен состав клещей, обитающих в гнёздах уссурийских бакланов из залива Петра
Великого. В шести гнёздах были обнаружены только гамазовые клещи. Общее число со-
бранных экземпляров — 1 430, из них — Macrochelespinicilliger составлял — 29%, Andro-
laelapspavlovskyi — 0,2%, Haemolaelaps casalis — 25,9%, Hypoaspis sp. — 45% (Беликова,
Литвиненко, 1973).
Из иксодовых клещей на уссурийских бакланах Приморья найден один вид — Ixodes
signatus.
Серьезное воздействие на популяции уссурийского баклана оказывает антропогенный
фактор. В последние 50-60 лет XX столетия численность вида в большинстве его колоний
в Приморье существенно изменилась, и причиной сокращения во многих случаях было
отрицательное антропогенное влияние. Многие колонии находятся вблизи населённых пун-
ктов, выросших за последние десятилетия. Посещение гнездовий людьми, сбор яиц и от-
стрел бакланов в колониях в 1950-1960-х гг., наряду с усиливающимся загрязнением вод,
не только сократило количество гнездящихся птиц, но, зачастую, приводило к полному унич-
тожению отдельных поселений. Так, исчезли колонии бакланов на островах залива Петра
Великого — Рейнеке, Рикорда и Попова (Лабзюк, и др., 1971), ещё раньше прекратила своё
существование колония на о. Аскольд, упоминаемая В. Тачановским (Taczanowski, 1891—
1893).
Пагубно сказалась на уссурийском баклане практика акклиматизации млекопитающих
на морских островах Дальнего Востока, интенсивно проводившаяся в 1930-е гг. Свидете-
лем катастрофически быстрой гибели колоний морских птиц (в том числе и уссурийского
Японский, или уссурийский баклан, Ушастый баклан
95
баклана) на о. Фуругельма оказался известный писатель М.М. Пришвин4. Осенью 1929 г.
на острове был организован питомник голубых песцов с полувольным содержанием и заве-
зена первая партия зверей (50 шт.), а через два года многотысячный птичий базар острова
перестал существовать. «Целыми остались только гнёзда бакланов, устроенные на отвес-
ных и совершенно неприступных скалах, — пишет М.М. Пришвин — этих последних птиц
работники питомника достреливали с лодок на корм песцам».
Хозяйственное значение, охрана. Хозяйственного значения не имеет. В Японии, в пре-
фектуре Гифу, до сих пор используется для ловли рыбы (традиция, заимствованная японца-
ми из Китая), но исключительно в рекламных целях и для развлечения туристов. Для этого
на р. Нагара устраиваются ежегодные праздники рыбной ловли с помощью обученных ус-
сурийских бакланов (Brazil, 1991).
Так как уссурийский баклан имеет невысокую численность и сравнительно узкий аре-
ал. в основном лежащий в районах с большой плотностью населения, он должен быть взят
под охрану. Вид занесён в Красную книгу Сахалинской области. Самая крупная в России
колония этих бакланов, расположенная на о. Фуругельма в заливе Петра Великого, и не-
сколько небольших колоний на соседних островах вошли в состав Дальневосточного госу-
дарственного морского заповедника, организованного в 1978 г. Кроме того, вид охраняется
з Лазовском и Курильском заповедниках. Эти меры частично решают проблему охраны
вида в нашей стране.
Ушастый баклан
Phalacrocorax auritus (Lesson, 1831)
Carbo auritus Lesson, 1831, Trailed Om. livr.8 с.605, «Новая Зеландия» (ошибочно) —
Северная Америка.
Double-crested cormorant, White-crested Cormorant (для западных подвидов) (англ.);
Ohrenscharbe (нем.); cormoran a aigrettes (фр ).
Син. (лат.): Phalacrocorax dilophus (Swainson et Richardson, 1831); Dilophalieus auritus (Lesson,
1831); Hypoleucus auritus (Lesson, 1831).
Син. (рус.): двухохлый баклан
Статус. Редкий залётный вид.
Общая характеристика и полевые признаки. Крупный баклан. Длина тела в сред-
нем 84 см, размах крыльев — 134 см. Оперение чёрно-бурое с зеленоватым или бронзо-
вым блеском. Характерна оранжево-жёлтая окраска кожи на лицевой части и горле. На
лицевой части есть пучки украшающих перьев — «уши», давшие название этому виду
имеются только в начальный период размножения). У молодых птиц оперение в основ-
ном бурое, более светлое на груди и шее. Кожа на «лице» — жёлтая. В районах вероятных
залетов на север Дальнего Востока ушастых бокланов можно спутать с молодыми берин-
товыми, от которых они отличаются более крупными размерами и специфической окрас-
ной участков голой кожи на «лице». Кроме того, у ушастого баклана в полёте шея слегка
прогнута вниз, а не выпрямлена в одну линию с туловищем. Во время отдыха бакланы
-того вида часто сидят в характерной позе, держа крылья для сушки оперения в расправ-
"енном состоянии.
<и1лвин М.М. 1956. Голубые песцы // Собр. соч. Т. 2. С. 649-679.
96
Отряд Пеликанообразные, Семейство Баклановые
Подвидовая систематика. Выделяют до 5 подвидов (del Hoyo et al., 1992; Dickinson.
2003), изменчивость которых проявляется в общих размерах и в разнице в окраске украша-
ющих перьев (хохлов) на голове. Подвидовая принадлежность особи, залетевшей на терри-
торию России, установлена предположительно, на основании географических соображе-
ний:
1. Phalacoracorax auritus cincinatus
Carbo cincinatus J.F. Brandt, 1837, Bull. Sci. Acad. Sci. St. Petersb., 3, No. 4, col. 55, Nov
16, о. Кодьяк, Аляска. Распространён от Аляски до северо-западной Канады. Типовой эк-
земпляр этого подвида находится в ЗИН РАН.
Другие подвиды распространены: Ph. a. albociliatus Ridgway, 1884 (2) — от северо-
западной Канады до западной Мексики; Ph. a. auritus (Lesson, 1831) (3) — центральная и
восточная Канада, восток США; Ph. a.floridanus (Audubon, 1835) (4) — юг США, Гонду-
рас; Ph. a. heuretus Watson, Olson et Miller, 1991 (5) — Багамские о-ва и, возможно, Куба.
Замечания по систематике. Ряд систематиков относит ушастого баклана к подроду
(при дроблении рода Phalacrocorax — к самостоятельному роду) Hypoleucos Reichenbach,
1852 (Siegel-Causey, 1988; del Hoyo et al., 1992).
Описание. Окраска. Половой диморфизм в окраске не выражен. Взрослые птицы в
брачном наряде. Голова, шея, задняя часть спины, поясница, надхвостье и вся нижняя сто-
рона туловища чёрные, с тусклым зеленоватым блеском. Перья передней части спины,
плечевые и верхние кроющие крыльев буровато-серые с бронзовым отливом, с широкими
чёрными краевыми каймами. Крылышко, маховые и рулевые буровато-чёрные. По бокам
темени пучки украшающих перьев, цвет которых варьирует от белого в западной части
ареала (включая Ph. a. cincinatus) до чёрного — в восточной. Участок голой кожи от осно-
вания клюва на подбородке ярко-оранжевого цвета. Надклювье темное, подклювье желто-
ватое. Ноги чёрные. Радужина яркая сине-зелёная.
В зимнем наряде хохлы отсутствуют, цвет оголённой кожи на голове — жёлтый.
Птенцы вылупляются голыми; кожа их при рождении буроватая, но вскоре чернеет.
Пуховой наряд образован коротким черным пухом.
Гнездовой наряд', голова буровато-серая, подбородок и горло — светлые. Верхняя сто-
рона туловища и крыльев — серовато-бурые с тёмным чешуйчатым рисунком. Низ тулови-
ща в буровато-белых размытых пятнах. Маховые и рулевые тёмно-бурые. Кожа на «лице»
желтоватая. Клюв бурый с более светлым подклювьем. Ноги чёрные.
Молодые птицы второго календарного года жизни похожи на птиц в гнездовом наряде,
но оперение у них в основном тёмно-бурое; к лету третьего года становится похожим на
изношенный дефинитивный наряд. Хохлы отсутствуют, либо развиты слабо.
Строение и размеры. Первостепенных маховых 11 (включая рудиментарное).
Рулевых — 12. Размеры (мм) самцов (п=9) и самок (n=4) Ph. a. cincinatus с Аляски. Длина
крыла самцов — 312-349, в среднем — 331, самок — 330-340, в среднем — 334; длина
клюва самцов — 52-63, в среднем — 59, самок — 54-61, в среднем — 58, длина цевки
самцов — 64—72, в среднем — 68, самок — 62-71, в среднем — 66 (Palmer, 1962; Johansgard,
1993; Hatch, Weseloh, 1999).
Линька. У птенцов тело покрывается пухом в течение второй недели жизни. Гнез-
довой наряд формируется в конце лета в возрасте около двух месяцев и сохраняется в
основном до следующей весны (частично до 13-месячного возраста). Возраст приоб-
ретения окончательного наряда варьирует. По-видимому, чаще всего это происходит на
третьем году жизни. У взрослых птиц частичная предбрачная линька головы, шеи и
передней части спины протекает в январе-марте, полная «послебрачная» — в основ-
ном, в конце лета. Но украшающие перья на голове выпадают раньше, в июне. Перво-
Ушастый баклан
97
Рис. 17. Область распространения ушастого баклана: а — гнездовой ареал, б — отдельные поселе-
ния вне сплошного ареала, в — районы зимовок, г — залёт на территорию России. Подвиды: 1 —
Ph. a. cincinatus\ 2 — Ph. a. albociliatus', 3 —Ph. a. auritus\ 4 — Ph. a. floridanus; 5 — Ph. a. heyretus.
степенные маховые сменяются «ступенчато» в дистальном направлении (Hatch, Weseloh,
1999).
Распространение. Гнездовой ареал. Эндемик Сев. Америки. Гнездится на
тихоокеанском побережье от о. Нунивак на севере до о-вов Ревилья-Хихедо — на юге. На
атлантическом побережье: от о. Ньюфаундленд до Белиза, а также во внутренних конти-
нентальных районах Канады и США. В России зарегистрирован единственный случай за-
лёта 25 августа 1991 г. Одиночную взрослую особь наблюдали возле пос. Эгвекинот в зали-
ве Креста на Чукотке (Уорд, Шмутц, 1993).
Зимовки расположены в пределах границ гнездового ареала на всём тихоокеанском
побережье Сев. Америки, в южной части континента, на островах и побережьях Мексикан-
ского зал. и западной части Карибского моря. Большинство птиц, гнездящихся на внутрен-
них водоёмах, проводят зиму в приморских районах (рис. 17).
Миграции. Фактически миграции совершают только птицы, населяющие центральные
районы Канады и США и север атлантического побережья Америки, которые на зиму отле-
тают на юг континента. На тихоокеанском побережье бакланы в основном ведут оседлый
образ жизни, но птицы с Аляски могут кочевать до Британской Колумбии. Южные популя-
ции оседлы. Пути перелётов пролегают вдоль морских побережий и по долинам рек.
Местообитания. Гнездятся как на морских побережьях, так и на пресноводных водо-
ёмах — реках, озёрах, прудах, болотах. На море держатся, как правило, невдалеке от суши.
Численность. Репродуктивная часть мировой популяции оценивается в 370 тыс. пар,
большинство которых (220 тыс.) гнездится во внутренней части континента. Численность
Ph. a. cincinatus — 20 тыс. пар (Hatch, Weseloh, 1999).
Размножение. Моногам. Большинство птиц приступают к размножению в 3-летнем
возрасте. Гнездится колониями. В зависимости от типа занимаемых местообитаний, гнёзда
сооружают или на земле, или на широких уступах скал, а также — на деревьях или кустар-
98
Отряд Пеликанообразные, Семейство Баклановые
никах. В кладке — 1-7, чаще — 3^4 яйца. Скорлупа яиц светло-голубая, часто с белым
меловым налётом. Размеры яиц (п=23) номинативного подвида: 56,9-69,0 х 35,7-41,1, в
среднем — 61,2x38,6 мм. Масса в среднем 44,9 г (п=448). На Аляске откладка яиц начина-
ется в конце мая - начале июня. Продолжительность инкубации — 25-29 дней. Птенцы
поднимаются на крыло на 6-8-й неделе, начинают самостоятельную жизнь в возрасте 10
недель.
Продуктивность составляет 1,2-2,4 слётка на гнездо (Gabrielson, Lincoln, 1959; Hatch,
Weseloh, 1999).
Суточная активность и поведение. Ведёт дневной образ жизни.
Питание. Основу рациона составляет рыба длиной от 3 до 40 см, Чаще всего это мало-
подвижные донные виды. Изредка в пробах пищи встречают ракообразных, насекомых,
амфибий, части растений. Кормится в одиночку или стаями, как правило, на прибрежных
мелководьях. Часто в массе посещает пруды рыборазводных хозяйств.
Враги, неблагоприятные факторы. Из биотических факторов наибольший урон гнёз-
дам и потомству наносят чайковые и врановые птицы, хищничество которых особенно
ощутимо при посещении колоний людьми. В местах массовых концентраций подвергается
преднамеренному истреблению в интересах сохранения рыбных запасов.
Хозяйственное значение и охрана. В России, будучи редким залётным видом, хозяй-
ственного значения не имеет. Специальные меры охраны не предусмотрены.
Стеллеров баклан
Phalacrocorax perspicillatus Pallas, 1811
Phalacrocoraxperspicillatus Pallas, 1811, Zoographia Rosso-Asiatica, 2 c. 305, о. Беринга.
Pallas’ [spectacled] cormorant (англ.); Brillemeerscharbe (нем.);
cormoran de Ра11а8(фр.).
Син. (рус.): очковый баклан, палласов баклан.
Статус. Вымерший вид.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Как и большинство «морских» бакланов северного полу-
шария, при дроблении рода Phalacrocorax стеллеров баклан, очевидно, должен включаться
в род Stictocarbo Bonaparte, 1855 (Siegel-Causey, 1988).
Описание (на основе описаний Ф.Ф. Брандта5, опубликованного Л. Штейнегером
(Stejneger, 1889), и П. Джонсгарда (Johnsgard, 1993)). Окраска. Взрослые птицы в брач-
ном наряде. Общая окраска оперения черная с выраженным металлическим блеском раз-
ных оттенков. Задняя часть головы и верх шеи с фиолетовым отливом, переходящим ниже
по шее в темный голубовато-зелёный. Основание шеи, межлопаточная область и грудь с
бронзово-зелёным оттенком; остальные части туловища с зеленовато-синим, за исключе-
нием крупных треугольных белых пятен на боках поясницы и бедрах, образованных длин-
ными редкими перьями. Плечевые и верхние кроющие крыльев с пурпуровато-фиолето-
вым блеском, с узкими чёрными каёмками. Первостепенные маховые и нижние кроющие
крыльев чёрные с буроватым отливом. Второстепенные маховые чёрные, с более или ме-
нее выраженным пурпуровато-фиолетовым блеском. Рулевые чёрные. На задней части лба
5 Перевод с латинского выполнен С.Л. Николаевым.
Стеллеров баклан
99
и темени — хохол, направленный назад, длиной 7,5 см и шириной до 5 см из расширенных
перьев Чёрного цвета, с пурпуровато-фиолетовым или, иногда, зеленоватым отблеском по
краям. На границе затылка и шеи веерообразный хохол, более узкий в основании, длиной 5
см и шириной 6,5 см, из довольно широких перьев чёрного цвета с зеленовато-пурпурным
оттенком по краям.
На голове и на боках верхней части шеи отдельные белые нитевидные перья, короткие
(2,5 см или немного больше) на лбу и более длинные (5-7,5 см) на шее, а также белые
короткие кистевидные перышки. Передняя часть лба, окологлазничное пространство, уз-
дечка, подбородок и верх горла полностью голые, киноварно-красного и лазурного цвета;
вокруг глаз широкое эллипсоидное кожистое кольцо — «очки», давшее одно из названий
этому виду.
Клюв чёрный, на конце беловато-розового цвета, у основания и по краям челюстей —
рогового цвета. Ноги угольно-чёрные с когтями тёмно-рогового цвета.
Согласно Г. Стеллеру, у самок, в отличие от самцов, отсутствуют хохлы и «очки» вокруг
глаз.
Прочие наряды не описаны.
Строение и размеры. Первостепенных маховых 10 (не считая рудиментарного),
формула крыла: II=III=IV>V>I>VL.. (Stegmann, 1936). Рулевых 12; хвост имеет форму паль-
мовой ветви, у основания суженный. Размеры (мм) четырёх птиц, половая принадлежность
которых не определена, из музеев Санкт-Петербурга, Лондона, Лейдена и Хельсинки (Stej-
neger, 1889; Palmgren, 1935): длина крыла 351, 356, 359 и 364, в среднем 357,5; длина хво-
ста 169 (рулевые сильно обношены), 189, 201 и 229, в среднем (по трём последним) —
206,3; длина клюва 74,5 и 95, в среднем 84,8; длина цевки 68 и 76, в среднем 72. Масса
птиц, по описанию Г. Стеллера, составляла 12-14 фунтов, или 5,44-6,35 кг.
Распространение. Эндемичный вид Командорских о-вов (рис. 18). Открыт Г. Стеллером
во время зимовки 1741/42 гг. на о. Беринга. Л. Штейнегер (Stejneger, 1883), прибывший на
Командоры в 1882 г, стеллеровых бакланов уже не нашел, но от местных жителей узнал, что
около 30 лет назад эти птицы ещё водились на о. Арий Камень. По опросным данным, бакла-
ны населяли и соседний о. Медный, но доказательств тому нет. Предположение о распрост-
ранении стеллерова баклана на камчатском побережье, основанное на информации из тру-
дов П.С. Палласа и О.Е. Коцебу (Siegel-Causey et al., 1991), представляется маловероят-
ным, так как описание этого вида отсутствует в основных академических монографиях
того времени, посвящённых Камчатке (Крашенинников, 1994; Стеллер, 1999). Более того, Г.
Стелл ер (Steller, 1781) определённо указывал, что никогда не видел эту птицу на Камчатке.
По Л. Штейнегеру (Stejneger, 1889), стеллеров баклан в прошлом мог населять Алеутс-
кие о-ва, но исчез оттуда из-за вулканических извержений. Сообщение о находке одной
кости (пряжки) этого вида на о. Амчитка в раскопках алеутской стоянки возрастом чуть
более 2 тыс. лет (Siegel-Causey et al., 1991) не подтвердилось: при повторном определении
её отнесли к ушастому баклану Ph. auritus cincinatus (Olson, 2005).
В коллекциях мира сохранились 6 экземпляров стеллерова баклана: по два в Санкт-
Петербурге и Лондоне, по одному в Лейдене и Хельсинки. Один из них поступил в ЗИН
РАН в 1833 г. от главного правителя Русской Америки Ф.П. Врангеля, в ведении которого
находились Командорские о-ва. Другие птицы происходят из коллекции его преемника
И.В. Купреянова. В 1839 г. он передал один экземпляр в Британский музей естественной
истории, остальные птицы в 1841 г. поступили в Зоологический музей АН, откуда они ра-
зошлись по другим европейским музеям (Нейфельдт, 1978). В коллекциях ЗИН РАН и На-
ционального музея США в Вашингтоне имеются, кроме того, ископаемые кости стеллеро-
ва баклана, собранные Л. Штейнегером и Н.А. Гребницким на м. Песчаный у северо-запад-
100
Отряд Пеликанообразные, Семейство Баклановые
Рис. 18. Гнездовой ареал (а) стеллерова бак-
лана.
ной оконечности о. Беринга (Stejneger, 1889; Плес-
ке, 1896). Кроме того, кости этого вида содержала
коллекция Н.П. Сокольникова, которая поступила
сначала в зоологический музей лорда Ротшильда
в г. Тринг (Hartert, 1920), а затем в Американский
музей естественной истории (Johansen, 1961).
Описание фрагментов скелета стеллерова бакла-
на выполнено Ф.А. Лукасом (Lucas, 1889, 1895).
Численность. Сведения о численности стел-
лерова баклана в период его открытия противо-
речивы. П.С. Паллас, использовавший материа-
лы Г. Стеллера при описании вида, отмечает его
многочисленность на о. Беринга (цит. по: Stejneger,
1885). Однако сам Г. Стеллер (Steller, 1781) пи-
шет, что этот баклан встречается на острове толь-
ко в одном месте — на скалах около пещеры, на-
званной именем Стеллера (расположение этого
места не определено — Ю. А.). В первой полови-
не XIX в., по опросным данным, этих птиц было
«много» на о-вах Беринга и Арий Камень; мест-
ные жители добывали их в большом количестве
для пропитания (Stejneger, 1883). Всё же вероятно, стеллеров баклан уступал в численнос-
ти другим видам бакланов, населявшим острова. Косвенное подтверждение тому содер-
жится в записках штурмана И.Ф. Васильева, посетившего Командоры в 1812 г. на бриге
«Финляндия». В них отмечено, что, запасаясь провизией, экипаж добыл на Арьем Камне
«множество урилов» (бакланов), среди которых, судя по крупным размерам, оказался толь-
ко один стеллеров (цит. по: Пасенюк, 1981).
Особенности биологии. Остались не изученными. Стеллеров баклан вёл оседлый образ
жизни. Вероятно, гнездился в основном на прибрежных островках и скалах, недоступных
для песцов, которых, как свидетельствуют участники экспедиции В. Беринга (Ваксель, 1940),
в то время было «неимоверное количество». Описание гнёзд и кладок отсутствует.
Причины вымирания. Главной причиной исчезновения стеллерова баклана считается
истребление его человеком. С момента своего открытия этот вид стал важным источником
питания для людей, чему способствовали простота охоты на эту доверчивую малоподвиж-
ную птицу, её крупные размеры и хорошие вкусовые качества мяса. По словам Г. Стеллера
(цит. по: Stejneger, 1889), одной птицы «с избытком хватало на троих измученных голодом
людей». Вслед за участниками экспедиции В. Беринга, бакланов добывали команды зверо-
промышленников, которые вели постоянный промысел на Командорах либо эпизодически
заходили сюда на пути в Русскую Америку, чтобы пополнить запасы продовольствия. По-
видимому, определяющим моментом в судьбе стеллерова баклана стало появление на ост-
ровах в 1820-х гг. постоянных алеутских поселений. Морские птицы — важный компонент
традиционного питания алеутов, поэтому, как известно (Stejneger, 1889), население добы-
вало бакланов в большом количестве, особенно в зимний период, когда другие источники
пропитания были ограничены.
С другой стороны, по мнению Э. Хартерта (Hartert, 1920), причиной вымирания этого
вида могла стать эпизоотия, подобная тем, что периодически распространяются в популя-
циях берингова и краснолицего бакланов.
Берингов баклан
101
Берингов баклан
Phalacrocorax pelagicus Pallas, 1811
Phalacrocoraxpelagicus Pallas, 1811, Zoographia Rosso-Asiatica, 2 c. 303, «море восточней
Камчатки и острова у Америки».
Pelagic cormorant, pelagic shag (англ.); Meerscharbe, Beringmeerscharbe (нем.);
cormoran pelagique (фр ).
Син. (лат.): Phalacrocorax [Stictocarbo] kenyoni Siegel-Causey, 1991
Син. (рус.): берингийский баклан, морской баклан.
Статус. Гнездящийся мигрирующий, в южных частях ареала — оседлый, пролётный и
зимующий вид.
Общая характеристика и полевые признаки. Имеет характерную для бакланов
валькообразную форму тела с длинной шеей и умеренно длинным хвостом. Самый мел-
кий из дальневосточных бакланов: длина тела составляет в среднем 73 см, размах кры-
льев — 98 см. Самцы несколько крупнее самок. Оперение однообразно черное с метал-
лическим зелёным блеском (на шее — с фиолетовым). У взрослых птиц в весенне-
летнее время на боках поясницы характерные белые пятна, в начальный период раз-
множения на шее и спине заметны отдельные узкие белые перья, на темени и затылке
два коротких черных хохла. Кожа вокруг глаз, вдоль углов разреза рта и на подбородке не
оперена, в брачном наряде — тёмно-красная, в зимнем — бурая. Оперение молодых птиц
в основном бурое.
Очень похож на краснолицего баклана, от которого отличается меньшими размерами и
стройным телосложением (особенно заметна более тонкая шея, относительно небольшая
голова, узкий тонкий клюв). В любом возрасте у берингова баклана клюв темнее; площадь
оголённой кожи на «лице»6 меньше, лоб всегда оперен; граница оперения на боках нижней
челюсти выдаётся углом вперёд, доходя, как минимум, до уровня глаза (у краснолицего
баклана край оперения ровный и лишь иногда достигает заднего края глаза). Кроме того, у
взрослых беринговых бакланов окраска лица не такая яркая и заметная, особенно на рас-
стоянии; основание клюва красное, а не синее. При хорошем освещении видно, что спин-
ная сторона у берингова баклана окрашена однообразно, в то время как у краснолицего
блеск оперения на спине и крыльях имеет разные оттенки.
Из-за относительно коротких и тупых крыльев беринговы бакланы летают только ак-
тивным полётом, на парение переходят лишь в момент приземления. В полете голова и шея
вытянуты в одну линию с туловищем. Сидящие птицы держатся почти вертикально. Пре-
восходно плавают и ныряют, под водой передвигаются с помощью ног. С воды взлетают,
как правило, с разбега. В море — молчаливая птица; на колониях издаёт разнообразные, в
основном глухие гортанные звуки.
Подвидовая систематика. Изменчивость проявляется в варьировании общих разме-
ров. Выделяют два подвида (Степанян, 2003; Dickinson, 2003). В России — один подвид.
1. Phalacrocorax pelagicus pelagicus.
Phalacrocoraxpelagicus Pallas, 1811, Zoographia Rosso-Asiatica, 2 c. 303, «море восточ-
ней Камчатки и острова у Америки».
Более крупная раса, занимающая всю азиатскую и американскую часть ареала вида к
югу до Британской Колумбии.
6 Ниже термин «лицо» приводится без кавычек.
102
Отряд Пеликанообразные, Семейство Баклановые
Другой подвид Ph. р. resplendens Audubon, 1838 (2) распространён на тихоокеанском
побережье Северной Америки от Британской Колумбии до юго-восточной границы аре-
ала — п-ова Калифорния (Palmer, 1962).
Замечания по систематике. В кладистической системе семейства баклановых, осно-
ванной на анализе остеологических признаков (Siegel-Causey, 1988), берингов баклан вклю-
чён в род Stictocarbo Bonaparte, 1855. Л.А. Портенко (1939) более крупных птиц с матери-
кового побережья Азии и Аляски выделил в подвид Ph. р. aeolus, но это не было признано
убедительным вследствие малочисленности выборки (Тугаринов, 1947). Однако новые дан-
ные также свидетельствуют, что птицы в центральной части Алеутской гряды мельче, чем
на о-вах Прибылова и побережье Аляски (Rohwer et al., 2000), что противоречит традици-
онной точке зрения.
Результаты повторного исследования типовых экземпляров баклана Кениона (Phalacro-
corax [Stictocarbo} kenyoni), описанного по трем скелетам и ископаемым костным остаткам
с о. Амчитка (Siegel-Causey, 1991), не подтвердили его валидность (Rohwer et al., 2000).
Данный таксон предложено считать синонимом берингова баклана, что находит отражение
в современных сводках (American Ornithologists’ Union, 1998; Dickinson, 2003). Очевидно,
это относится и к экземплярам с островов Уналашка, Атту и Беринга из фондов ЗИН РАН
(№ 8901, 8945 и 8955), которых Д. Сигел-Кози переопределил как бакланов Кениона, а
также к двум неполным скелетам из этой же коллекции и к костным остаткам из археологи-
ческих памятников Сахалина, описанным А.В. Пантелеевым (1997а, 2001,2002) в качестве
Ph. kenyoni.
Описание. Окраска. Половой диморфизм в окраске не выражен. Взрослые птицы в
брачном наряде. Оперение в основном черное с сильным металлическим блеском. Голова,
основание шеи, плечи, спина, поясница, надхвостье, зоб и вся нижняя сторона туловища —
с зелено-малахитовым блеском, большая часть шеи — с ультрамарино-фиолетовым, верх-
ние кроющие крыльев — с зеленовато-бронзовым, маховые — со слабым зелёным, руле-
вые — почти без блеска. На боках поясницы и наружной стороне бёдер пара белых пятен,
образованных узкими рыхлыми перьями. На шее, зобе, задней части спины, пояснице ред-
кие белые узкие перья длиной до 35 мм. На голове два чёрных хохла, один расположен на
темени, другой — на границе затылка и шеи. Уздечка, углы разреза рта и основание под-
клювья неоперённые, с кожей киноварно-красного цвета. Клюв черновато-бурый, в осно-
вании красноватый. Ноги чёрные. Радужина зелёная.
Взрослые птицы в зимнем наряде. Оперение отличается от предыдущего наряда отсут-
ствием белого цвета. Хохлов нет либо они слабо развиты. Голые части лица буроватого
цвета; основание клюва жёлто-бурое.
Пуховой наряд образован густым серо-бурым пухом длиной до 15 мм, более редким и
коротким (до 5 мм) на голове и шее; грудь и брюхо — в грязно-белых размытых пятнах.
Голые части лица светло-розовые. Клюв и ноги буровато-чёрные.
Гнездовой наряд. Оперение в основном бурое. Голова и шея серо-бурые, со светлыми
окончаниями перьев. Шея испещрена светлыми пятнышками, образованными нитевидными
перьями с рассученными вершинами. Перья мантии с тёмными предвершинными и светлы-
ми вершинными каймами, образующими чешуйчатый рисунок. Спина, поясница, надхвос-
тье, плечевые и верхние кроющие крыльев с лёгким зеленоватым блеском; у некоторых птиц
нижняя часть шеи с едва заметным фиолетовым блеском. Голая кожа на лице серая, на горле
розовая. Клюв черноватый; на конце надклювья, по краю верхней челюсти и у основания
подклювья бывает окрашен в роговый цвет. Ноги черноватые. Радужина светло-бурая.
Молодые птицы. Окраска оперения в промежуточных нарядах сильно варьирует вслед-
ствие значительной продолжительности и индивидуальных различий постювенильной линь-
Берингов баклан
103
ки. В первую зиму молодые остаются похожими на птиц в гнездовом наряде, только голова
и верхняя часть шеи у них темнеют. Весной второго календарного года наряд, изнашива-
ясь, светлеет, летом в результате линьки покровного оперения становится пятнистым чёр-
но-бурым с более выраженным металлическим блеском; голая кожа на лице темнеет. Летом
третьего календарного года молодые похожи на взрослых, но оперение с примесью бурого.
Блеск его не такой яркий; на боках поясницы вместо пятен только отдельные белые перья;
хохлы выражены слабо; голая кожа на лицевой части головы и основание клюва тёмные, с
едва заметным красноватым оттенком.
Строение и размеры. Первостепенных маховых 10 (не считая рудиментарного),
формула крыла II>III>IV>V=I>VI..., II>III>IV>I>V... или III>II=IV>I>V>VI... Рулевых 12,
форма хвоста округлая. Размеры (мм). Длина крыла самцов (п = 37) 256-296, в среднем
278; самок (п = 24) 244-285, в среднем 263. Длина хвоста самцов (п = 27) 135-161, в сред-
нем 146; самок (п = 21) 127-155, в среднем 141. Длина клюва самцов (п = 35) 46-55, в
среднем 51; самок (п = 24) 42-55, в среднем 49. Длина цевки самцов (п = 37) 52-61, в
среднем 57, самок (п = 24) 49-57, в среднем 54 (колл. ЗИН, ЗМ МГУ, БПИ, ДВГУ, ориг.).
Масса (г) взрослых птиц: самцов (п = 30) 1 040-2 295, в среднем 1 896; самок (п = 10)
1 300-2 200, в среднем 1 663 (Портенко, 1972; Кондратьев и др., 1992; Н. Герасимов, 1999;
С.В. Мараков, неопубл, данные; колл. ЗИН и ЗМ МГУ). Самцы заметно крупнее и тяжелее
самок.
Линька. Гнездовой наряд полностью формируется на седьмой неделе жизни (Van Tets,
1959). Линька молодых птиц до приобретения ими дефинитивного наряда в зиму третьего
календарного года протекает без чёткого разделения на периоды, с широкой индивидуаль-
ной изменчивостью сроков и полноты. Она начинается вскоре после вылета из гнезда со
смены мелких перьев на голове и шее, крупное покровное оперение активно линяет с июня-
июля второго календарного года, остатки ювенильных покровных перьев сохраняются до
осени третьего календарного года. Линька первостепенных маховых у птиц на восточном и
западном тихоокеанском побережье несколько различается, что предположительно связа-
но с оседлостью американских птиц и сезонными перелётами азиатских (Filardi, Rohwer,
2001). У молодых бакланов она обычно начинается в феврале-апреле второго календарно-
го года (на восточном побережье в среднем на месяц раньше, чем на западном). Линька
происходит двумя волнами, вторая из которых инициируется в летнее время после неболь-
шого перерыва (при запоздалом начале линьки — без перерыва). Обе дистально направ-
ленные волны начинаются с замены самого внутреннего пера, поэтому за период смены
всех ювенильных маховых, который заканчивается обычно в конце второй зимы, прокси-
мальные перья успевают обновиться дважды. Количество линяющих перьев в одной волне
у азиатских птиц больше (2-5, в среднем 3), чем у американских (1-3, в среднем 1,4). Часть
молодых бакланов с азиатского побережья не успевает обновить все маховые перья до вес-
ны (по-видимому, это птицы, совершающие миграции). Судя по экземплярам из коллекций
ЗМ МГУ и ДВГУ, иногда 1-2 крайних дистальных пера гнездового наряда остаются до
осени третьего календарного года. Рулевые сменяются, начиная с центральной пары; сро-
ки их линьки крайне вариабельны.
У взрослых птиц частичная предбрачная линька контурного оперения начинается зи-
мой. Белые украшающие перья появляются в феврале-марте. На шее они выпадают в
начале периода размножения (обычно в июне), пятна на боках поясницы исчезают в июле-
августе. Полная послебрачная линька начинается во время гнездования со смены мелко-
го покровного оперения передней части тела. Рулевые линяют обычно с августа с цент-
ральной пары. Замена первостепенных маховых начинается после окончания сезона раз-
множения — в августе-сентябре (неразмножавшиеся и неудачно гнездившиеся особи,
104
Отряд Пеликанообразные, Семейство Баклановые
по-видимому, линяют раньше). Она происходит одновременно двумя-тремя дистально
направленными волнами, начинаясь с первого внутреннего пера. Преобладающее боль-
шинство американских птиц успевают обновить все маховые к следующему гнездовому
циклу, а у азиатских — остаётся от 2 до 5 старых внешних перьев, замена которых про-
должается после перерыва на время размножения (Filardi, Rohwer, 2001; данные состави-
теля очерка).
Можно предположить, что у азиатских молодых и взрослых бакланов, которые не со-
вершают миграций и проводят зиму в районах гнездования, линька протекает примерно
так же, как у оседлых птиц с американского побережья.
Распространение. Гнездовой ареал. Берингов баклан — эндемичный вид Сев.
Пацифики, населяющий острова и морские побережья Азии и Сев. Америки (рис. 19). Ази-
атская часть гнездового ареала почти полностью находится в пределах Российской Феде-
рации. В Восточно-Сибирском и Чукотском морях распространён на о-вах Врангеля и Ге-
ральд (Стишов и др., 1991) и спорадично на континентальном побережье от западного бе-
рега Чаунской губы (окрестности м. Наглейнын) до о. Колючин (Кондратьев, 1986; Соловь-
ева, Дондуа, 2006). На Чукотском п-ове повсеместно гнездится от м. Сердце-Камень до м.
Беринга, включая о. Ратманова (Konyukhov et al., 1998). В глубине Анадырского залива
колонии известны в районе бух. Руд ера, в зал. Креста, у м. Поворотный и на о. Алюмка в
Анадырском лимане (Белопольский, 1934; Смирнов, Велижанин, 1986; Велижанин, 1987;
Е.Е. Сыроечковский-мл., личн. сообщ.). Гнездится повсеместно на побережье Корякского
нагорья от м. Гинтера (зал. Анадырский) до м. Олюторский и далее на запад до Камчатско-
го перешейка (Кищинский, 1980; Вяткин, 1986, 2000; Смирнов, Велижанин, 1986; Доро-
гой, 1992). Распространён по всей восточной стороне п-ова Камчатка до о. Гаврюшкин
Камень на юге (кроме участков с низинными берегами), в том числе на о-вах Верхоту-
рова и Карагинский. На крайнем юго-западном побережье Камчатки в 1983 г. гнездился
в районе м. Сивучий, но в дальнейшем здесь не найден (Вяткин, 1986,2000; Лобков, Алек-
сеев, 1987; Артюхин и др., 20016). Населяет все основные острова Командорского архипе-
лага — Беринга (в основном южную часть), Медный, Топорков и Арий Камень (Артюхин,
1999). На севере Охотского моря распространён по северо-западному берегу Камчатки (к
северу от м. Хайрюзова), на большинстве о-вов и побережьях зал. Шелихова, Пенжинской,
Гижигинской и Тауйской губы (Яхонтов, 1974, 19756; Вяткин, 1986; Голубова, Плещенко,
1997; Kondratyev et al., 2000). На западном континентальном побережье Охотского моря
гнездится спорадично, распространение здесь изучено плохо; населяет о-ва Нансикан, Ионы
и Шантарские (Яхонтов, 1975а; Харитонов, 1980; Росляков, Росляков, 1996). На о. Саха-
лин в последние годы существует только одна колония — на м. Анива (Нечаев, 1986,
19916); в прошлом гнездование было установлено на м. Терпения, на восточном побере-
жье Тонино-Анивского п-ова и на п-ове Шмидта (Гизенко, 1955). Более полувека назад вид
перестал гнездиться на близлежащем о. Тюлений (Трухин, Кузин, 1996). В Татарском про-
ливе и Японском море колонии известны на о. Монерон (Шибаев, Литвиненко, 1996), на
континентальном побережье от зал. Чихачева до зал. Ольги (Шульпин, 1936; Лабзюк, 1975;
Велижанин, 1978; Елсуков, 1984; Бабенко, 2000; Блохин, Тиунов, 2003), на островах зал.
Петра Великого: Русский, Верховского, Карамзина, Стенина, Большой Пелис, Матвеева,
Гильдебрандта и Фуругельма (Шибаев, 1987; Litvinenko, Shibaev, 2001; Катин и др.,
2004). На Курилах в прошлом населял все острова, за исключением самых низких (Ве-
лижанин, 1971). В последние годы гнездится в основном на северных островах — от
Атласова и Шумшу до Шиашкотана, а также в центральной части — от о. Кетой до север-
ной оконечности Урупа, и нерегулярно на островах Малой Курильской гряды — Лисьих и
Демина (Артюхин и др., 2001а; Артюхин, 2003; Ушакова, 2003).
Берингов баклан
105
Рис. 19. Область распространения берингова баклана: а — гнездовой ареал, б — районы зимовок,
в — залёты. Подвиды: 1 — Ph. р. pelagicus, 2 — Ph. р. resplendens.
За пределами России в Азии гнездится только на о. Хоккайдо (Ornithological Society of
Japan, 2000; Osa, Watanuki, 2002). В американской части ареала распространён на островах
Берингова моря и континентальном побережье от м. Лисберн на севере Аляски до о. Санта-
Барбара, Калифорния (Hobson, 1997).
Зимовки. В Азии охватывают часть гнездового ареала и лежащие южнее побережья
Японии, Кореи и Вост. Китая. В пределах российской территории беринговы бакланы ре-
гулярно зимуют в незамерзающих водах Командорских о-вов (Stejneger, 1885; Иогансен,
1934) и тихоокеанского побережья Камчатки к югу от Камчатского зал. (Лобков, 1986а),
вдоль всей гряды Курильских о-вов (Bergman, 1935; Гизенко, 1955; Подковыркин, 1955;
Нечаев, 1969), у юго-западной оконечности Сахалина (Нечаев, 19916), в южной части Та-
тарского пролива, вдоль Приморья и в зал. Петра Великого (Лабзюк и др., 1971; Елсуков,
1982; Шунтов, 1998). Кроме того, в незначительном количестве регулярно проводят зиму
на незамерзающей Сирениковской полынье у южных берегов Чукотки (Богословская, Вот-
рогов, 1981; Конюхов, 1990). В Японии зимой берингов баклан распространён на юг до о.
Танегасима (Ornithological Society of Japan, 2000), а вдоль континентального побережья
Кореи и Китая — до Гонконга (Siegel-Causey, Litvinenko, 1993). В американской части зи-
мует в основном в пределах гнездового ареала, кочуя на юг вдоль океанского побережья до
середины п-ова Калифорния (Hobson, 1997).
Залеты за границы области распространения известны на о. Беннетта, Новосибирские
о-ва (Успенский, 1964, цит. по: Шунтов, 1998), на о-ва Мидуэй и Лайсан Гавайского архи-
пелага (Hobson, 1997) (рис. 24).
Миграции. В районах зимовки на юге Японии весенние перемещения к северу начинаются
в марте (Austin, Kuroda, 1953; Brazil, 1991), в Кунаширском проливе — с середины апреля и
продолжаются в течение всего мая (Нечаев, 1969). На юго-восточном побережье Камчатки
первые пролётные птицы появляются 5-13 апреля (по наблюдениям 1974-1976 гг. в зал.
Кроноцкий); миграция продолжается здесь до первой декады июня включительно, но наи-
106
Отряд Пеликанообразные, Семейство Баклановые
более интенсивно она проходит в последних числах апреля и в мае (Лобков, 1980; Ю. Гера-
симов, 1999, 2001). Северной части п-ова Камчатка бакланы достигают в самом конце ап-
реля: весной 1971 г. на о. Карагинский их впервые увидели 29 апреля (Н. Герасимов, 1999).
Массовый прилёт бакланов на этот остров и соседний с ним о. Верхотурова в 1973 г. на-
блюдали 9 мая (Трухин, Косыгин, 1987). На южном побережье Корякского нагорья актив-
ную миграцию отмечали 13-22 мая 1960 г. (Кищинский, 1980), а на восточном побережье у
бух. Наталии — 20 мая 1973 г. (Трухин, Косыгин, 1987). К чукотским берегам Берингова
моря бакланы начинают прибывать в конце апреля - начале мая: в 1931 г. в зал. Креста
первых птиц зарегистрировали 1 мая, в 1932 г. в бух. Провидения — в первых числах мая,
в 1938 г. на м. Чаплина — 22 апреля (Портенко, 1972). На островах и побережьях Берингова
пролива их отмечали с третьей декады апреля (Портенко, 1972; Kessel, 1989). В районах
гнездования в Чукотском море бакланы появляются в первой или второй декадах мая: на о.
Колючин в 1953-1963 гг. — в период с 3 по 17 мая (Кречмар и др., 1978), на о. Врангеля в
1979-1987 гг. — с 8 по 20 мая (Стишов и др., 1991).
В Охотском море миграция проходит примерно в те же сроки, что и в прикамчатских
водах Тихого океана и Берингова моря. На юго-западе Камчатки активный пролёт наблю-
дается в течение всего мая (Герасимов, Калягина, 1997; Герасимов, 2003). Дальше на север
в районе устья р. Морошечная в 1977 г. прилёт зафиксирован 3 мая (Герасимов и др., 1992);
в 1990 г. миграция началась 5 мая, наиболее оживлённо она протекала в период с 11 по 21
мая, в небольшом количестве птицы продолжали лететь по первую декаду июня (Гераси-
мов, 1992). Бакланы, следующие континентальным побережьем, достигают Северного Охо-
томорья в самом конце апреля. Так, у берегов о. Талан, Тауйская губа, в 1987 г. первые
птицы появились 29 апреля (Кондратьев и др., 1992). В годы с тяжелой ледовой обстанов-
кой их прилет задерживается до образования в районах гнездования больших участков от-
крытой воды; например, в 1998 г. в такую весну первая встреча на о. Талан произошла
лишь 12 мая (Утехина, 1999).
Сроки осенней миграции на севере ареала зависят от ледовой обстановки и погодных
условий, поэтому сильно варьируют в разные годы. В целом, Чукотское море большинство
бакланов покидают в конце сентября - середине октября (Портенко, 1972; Кречмар и др.,
1978; Стишов и др., 1991). В Карагинском зал. бакланы весьма обычны до середины нояб-
ря; последних птиц здесь отмечали 3 декабря в 1970 г. и 6 декабря в 2003 г. (Н. Герасимов,
1999; данные автора очерка). На юго-восточном побережье Камчатки пролёт хорошо выра-
жен с середины августа до конца ноября, иногда и в первой половине декабря, но массовая
миграция обычно проходит в конце сентября - первой половине октября (Лобков, 1980). На
южной оконечности Камчатки на м. Лопатка в 1987 г. пролёт бакланов стал хорошо заме-
тен с 9 сентября, массовая миграция началась 19 сентября и ещё не прекратилась в день
окончания учётных работ 11 октября (Лобков, 20036). Север Охотского моря бакланы по-
кидают к концу сентября (Кондратьев и др., 1992). На северной оконечности Сахалина
(м. Елизаветы) крупные скопления пролётных бакланов держатся с сентября до ноября
(Гизенко, 1955). На материковом побережье Японского моря (Сихотэ-Алинский заповед-
ник) миграция начинается с первых чисел сентября и растягивается до первой декады но-
ября (Ел суков, 1982).
Наиболее многочисленный миграционный поток беринговых бакланов охватывает Ку-
рильские о-ва, Вост. Камчатку и континентальное побережье Берингова моря. Птицы, гнез-
дящиеся в северо-восточной части Охотского моря, мигрируют главным образом камчатс-
ким берегом. По осенним наблюдениям на м. Лопатка, где эти два миграционных потока
сходятся, большинство бакланов пересекают Первый Курильский пролив в направлении
охотоморской стороны о. Шу мшу, но в штиль и при попутных ветрах увеличивается коли-
Берингов баклан
107
чество птиц, продолжающих миграцию вдоль тихоокеанской стороны этого острова (Лоб-
ков, 20036). На западе Охотского моря основная миграционная трасса проходит по при-
брежной зоне с восточной стороны Сахалина (Тиунов, 2005). Водное пространство между
Сахалином и северным охотоморским побережьем часть птиц преодолевает, по-видимому,
напрямую (Шунтов, 1998).
Конкретные данные, позволяющие количественно оценить масштабы миграции, полу-
чены на Камчатке в ходе проведения специальных учётных работ. Интенсивность пролёта
на полуострове многократно выше на восточном побережье, чем на западном. Так, в устье
р. Ходутка на юго-востоке Камчатки с 30 апреля по 20 мая 1995 г. в прибрежной полосе
моря шириной 10 км пролетели 71,9 тыс. бакланов (Ю. Герасимов, 1999), в то время как на
западном побережье за весну учитывали лишь от 1,4 до 4,5 тыс. особей (Герасимов, 1992,
2003; Герасимов, Калягина, 1997). На осенней миграции на м. Лопатка за осень 1987 г.
пролётело не менее 100 тыс. беринговых и краснолицых бакланов, большинство которых
(92,2%) следовали тихоокеанским берегом (Лобков, 20036). Общая численность беринго-
вых бакланов, мигрировавших осенью 1975 г. вдоль юго-восточного побережья полуостро-
ва в районе пос. Жупаново, оценена в 29 тыс. особей (Лобков, 1980).
Пути миграций беринговых бакланов пролегают в основном вдоль морских побережий.
Фронт пролёта на участках прямого открытого берега довольно широкий: птицы летят на
дистанции от 300-400 м до 10 км и более от суши. Мигранты могут сокращать расстояние,
двигаясь от мыса к мысу напрямую. При этом они никогда не пересекают мысы над сушей,
за исключением южной оконечности Камчатки, где нередко пролётают над основанием м.
Лопатка. Во время весенней миграции бакланы избегают покрытых льдами акваторий, при-
держиваются прибрежных разводий. Одиночные, вероятно, сбившиеся с пути птицы, регу-
лярно встречаются в открытых водах на большом удалении от берега, особенно часто в
период осенней миграции. Иногда залетают вглубь суши на пресные водоёмы: такие слу-
чаи известны для оз. Ханка и Приханкайской равнины. Мигрирующие бакланы летят оди-
ночками, парами и стаями из десятков особей. В дни массового пролёта размеры стай увели-
чиваются до нескольких сотен особей; временами бывает, что птицы летят непрерывной ве-
реницей, в которой сложно выделить отдельные группы. Форма стаи — чаще всего цепочка,
реже клин; при перестроениях стая нередко рассыпается, превращаясь в беспорядочную
группу. Как правило, птицы летят низко над водой на высоте от 3 до 25 м, но при благопри-
ятных погодных условиях могут подниматься на 100-150 м. Мигрируют в светлое время
суток, особенно активно в первые часы после рассвета, но в период валового пролёта — в
течение всего дня (Поливанова, 1971; Велижанин, 1981; Глущенко и др., 1997; Шунтов,
1998; Ю. Герасимов, 1999; Лобков, 1980, 20036).
Местообитания. Круглый год обитает в прибрежной полосе моря. Гнездится на участ-
ках абразионного берега со скалистыми обрывами, островками и кекурами. Непременным
условием является наличие вблизи колоний невысоких надводных камней или рифов, на
которых бакланы собираются для сушки оперения и отдыха. Кормится на прибрежных мел-
ководьях в бухтах и заливах, на лиманах и лагунах, иногда залетает в низовья рек. Мигри-
рует над морем в основном в пределах нескольких километров от береговой линии. Зимой
держится на незамерзающих прибрежных акваториях, в том числе на полыньях.
Численность. Характерны значительные колебания численности, как в локальных по-
селениях, так и на больших участках ареала. На о-вах Врангеля и Геральд в 1987 г. на
гнездовании учтено 1,2 тыс. беринговых бакланов (Стишов и др., 1991). На арктическом
побережье от западной границы ареала до Колючинской губы обитает примерно 1 тыс.
особей. Здесь преобладают мелкие поселения. Единственная крупная колония — о. Колю-
чин; в 1938 г. в ней было 2-3 тыс. особей, в 1974 г. — 350, в 1985 г. — 800 особей (Портен-
108
Отряд Пеликанообразные, Семейство Баклановые
ко, 1972; Кондратьев, 1986; Кондратьев и др., 1987). На Чукотском п-ове от м. Сердце-
Камень до м. Беринга гнездится 16,1 тыс. бакланов, самые крупные колонии расположены
на м. Столетия и о. Нунэанган (3 и 2,8 тыс. особей соответственно; Konyukhov et al., 1998).
Крупные поселения общей численностью 3,8 тыс. особей обнаружены в 2004 г. на мысах
Чирикова и Бакланий в районе бух. Рудера; сотни птиц гнездились в зал. Креста и у м.
Поворотный (Е.Е. Сыроечковский-мл., личн. сообщ.). Далее на юг до административной
границы Корякского округа (м. Рубикон) сведения весьма неполные. Судя по имеющейся
информации (Кищинский, 1980; Смирнов, Велижанин, 1986; Дорогой, 1992; Е.Е. Сыроеч-
ковский-мл., личн. сообщ.), бакланы гнездятся здесь в значительном количестве на круп-
ных птичьих базарах, расположенных в районе м. Наварин и к северу и западу от него.
Предположительно, суммарная численность вида на всём этом побережье сопоставима с
таковой на Чукотском п-ове. На беринговоморском и тихоокеанском побережье Камчатки в
1994-95 гг. учтено 23,7 тыс. бакланов, в том числе 10 тыс. на о-вах Верхотурова и Карагин-
ский (Вяткин, 2000). В 1970-х - начале 1980-х гг. здесь их было вдвое больше — 46,3 тыс.
(Вяткин, 1986). Серьезно изменилась численность и на Командорских о-вах: с 18-20 тыс.
особей в 1960-1970-х гг. до 5,4 тыс. в 1986 г. и 6,2 тыс. в 1993-1994 гг. (Артюхин, 1991а,
1999). Примерно такой же она остаётся в последние годы (Зеленская, 2001а). Сокращение
численности в Камчатской области произошло вследствие распространения среди птиц
сильных эпизоотий (Мараков, 1972; Вяткин, 2000).
На северо-западном охотоморском побережье Камчатки (м. Хайрюзова - устье р. Пенжи-
на) в 1970-х гг. учтено 5,6 тыс. бакланов (Вяткин, 1986); принимая во внимание, что несколь-
ко колоний остались необследованными, всего здесь гнездилось не менее 6 тыс. особей.
Экспертная оценка численности на северном и западном континентальном побережье
Охотского моря — более 20 тыс. особей (Kondratyev et al., 2000). Эта территория обследо-
вана не полностью. Для западной стороны Пенжинской губы и Хабаровского края конкрет-
ных данных о численности бакланов нет. На побережье и близлежащих островах Магадан-
ской обл. (от западной границы до м. Поворотный на входе в Пенжинскую губу), по резуль-
татам последних учётов (Kondratyev, Mendenhall, 1995; Голубова, Плещенко, 1997; U. S.
Fish..., 2005; Зеленская, 2006; Утехина, 2006), гнездится примерно 6 тыс. особей. Больше
всего птиц в Тауйской губе, самые крупные поселения находятся на о-вах Умара (752 особи
в 2005 г.), Шеликан (724 — в 2000 г.) и Спафарьева (400 — в 1991 г.). В Гижигинской губе
наиболее крупная колония расположена на м. Тайночин (334 особи). Берингов баклан мно-
гочислен на о. Нансикан (Бабенко, 2000), но единственная имеющаяся оценка — 5 тыс. пар
(Яхонтов, 1975а) — в настоящее время представляется завышенной, учитывая тенденцию
сокращения численности данного вида, отмеченную в северной части Охотского моря в
1990-х гг. (Kondratyev et al., 2000). На о. Ионы гнездится около 60 особей (Харитонов,
1980). На Шантарских о-вах берингов баклан распространён в небольшом количестве (из-
вестны колонии из 5-35 пар); численность непостоянна, в отдельные годы его здесь вооб-
ще не бывает (Росляков, 1992; Росляков, Росляков, 1996).
На о. Сахалин в колонии на м. Анива гнездятся 50-60 бакланов (Нечаев, 1986), на о.
Монерон — 24 особи (Шибаев, Литвиненко, 1996).
В Японском море население берингова баклана оценивается в 2 тыс. особей. Около 350
птиц гнездятся на островах зал. Петра Великого, остальные — на континентальном побе-
режье Приморья и Хабаровского края (Шибаев, 1987; Litvinenko, Shibaev, 1991; Kondratyev
et al., 2000). Недавняя информация об увеличении численности на локальных участках зал.
Петра Великого (Litvinenko, Shibaev, 2001; Катин и др., 2004) является скорее следствием
перераспределения гнездящихся птиц между колониями, чем отражением общего тренда
популяции в регионе.
Берингов баклан
109
На Курильских о-вах в 1963 г. А.Г. Велижанин (1971) учел в колониях 38-42 тыс. берин-
говых и краснолицых бакланов и на этом основании оценил общую численность двух ви-
дов до 80 тыс. особей, в том числе 40-50 тыс. беринговых бакланов. Результаты недавних
учётов (Артюхин и др., 2001а; Артюхин, 2003) показывают результат на порядок ниже:
количество гнездящихся птиц сократилось до 4 тыс. особей. Преобладающее большинство
бакланов в настоящее время обитает на о-вах Шумшу и Парамушир (2,1 и 1,1 тыс. особей
соответственно). По-видимому, ухудшение состояния началось на рубеже 1970-х и 1980-х
гг. примерно в одно время с аналогичными изменениями на соседнем Камчатско-Коман-
дорском участке ареала. На Хоккайдо также исчезли колонии берингова баклана, извест-
ные в 1950-1960-х гг.; сейчас здесь осталось чуть более десятка размножающихся пар, в
основном на о. Теури в Японском море (Фудзимаки, 1986; Brazil, 1991; Osa, Watanuki, 2002).
Эти изменения в состоянии берингова баклана, как и некоторых других видов морских
птиц, возможно, обусловлены динамикой климато-океанологических и гидробиологичес-
ких условий, носящих эпохальный характер, а именно наступлением в дальневосточных
морях в 1970-1990-х гг. тёплого периода (Шунтов, 1998).
На основе приведенных выше данных можно утверждать, что на всём Дальнем Вос-
токе России (и в азиатской части ареала в целом) в настоящее время гнездится примерно
100 тыс. беринговых бакланов. Этот показатель ниже прежних оценок — более 300 тыс.
(Siegel-Causey, Litvinenko, 1993) и 200 тыс. (Шунтов, 1998), что в значительной мере объяс-
няется как существенным падением численности птиц, так и проведением новых деталь-
ных учётов в ряде районов гнездования. В американском секторе Пацифики репродук-
тивная часть популяции берингова баклана оценивается в 69 тыс. особей (Wetlands
International, 2006).
В пределах гнездового ареала в летнее время всегда держатся неразмножающиеся (в
основном неполовозрелые) птицы. Их особенно много в южных районах; вероятно, часть
молодых птиц из северных популяций остаётся здесь до наступления половой зрелости.
Так, на Чукотке доля неразмножающихся птиц составляет 20% (Konyukhov et al., 1998), а
на северном побережье Приморья — более 40% (Елсуков, 1984). Крупные летние скопле-
ния негнездящихся бакланов из сотен и даже тысяч особей наблюдали на юго-востоке Кам-
чатки, на восточном и южном побережьях Сахалина и на Юж. Курильских о-вах (Лобков,
1986а; Нечаев, 1991; Матюшков, 1999; Артюхин и др., 2001а; Блохин, Тиунов, 2005).
Размножение. Сведения о сроках наступления половой зрелости у беринговых бакла-
нов противоречивы. По мнению Л. Штейнегера (Stejneger, 1885), птицы с Командорских о-
вов начинают размножаться в возрасте двух лет; а по наблюдениям за помечеными особя-
ми в Британской Колумбии — с трёх (Van Tets, 1959).
Строительство гнезда инициируется самцом, но в этом процессе принимают участие
оба члена пары. До начала откладки яиц в колонии наблюдаются конфликты (рис. 20а) и
«саморекламирование» самцов — «Wing-Waving»: птица в характерной позе (шея закинута
на спину, голова и клюв направлены вертикально вверх, хвост опущен) ритмично двигает
головой из стороны в сторону и похлопывает крыльями, демонстрируя белые пятна на бо-
ках поясницы (рис. 206). При приближении самки партнеры начинают двигать вверх-вниз
головой с раскрытым клювом, издавая специфичные ритмичные горловые звуки («Gape
Recognition Display»). Копуляция обычно происходит на гнезде (Van Tets, 1959, 1965).
Берингов баклан — факультативно-колониальный вид. Преобладающее большинство
птиц гнездится колониями, средняя величина которых в разных частях ареала составляет
первые десятки пар, максимальная — 1,5 тыс. пар (м. Голенищева на о. Карагинский в годы
высокой численности; Герасимов, 1986). Вместе с тем не представляют редкости отдельно
гнездящиеся пары. Как правило, беринговы бакланы гнездятся совместно с другими вида-
110
Отряд Пеликанообразные, Семейство Баклановые
Рис. 20. Демонстрационное поведение берингова баклана (по Nelson et al, 2005): а — поза угрозы
при территориальном конфликте в колонии; б — рекламное поведение самца.
ми морских птиц (чаще всего с краснолицыми бакланами, кайрами, моевками), но нередко
образуют и моновидовые поселения. Местоположение колоний нередко сильно меняется
из года в год. Имеются два основных объяснения этому явлению. По мнению С.В. Марако-
ва (1972), переселяясь на новое место, птицы избавляются от кровососущих насекомых-
паразитов, в массе населяющих гнездовые постройки. По другой точке зрения, гнёзда за
время нахождения в них птенцов настолько сильно деформируются и пропитываются по-
мётом, что весной превращаются в промерзшие скользкие «горки», непригодные для по-
вторного гнездования (Н. Герасимов, 1999). Кроме того, на севере ареала причиной исчез-
новения колоний может стать позднее освобождение прилегающей морской акватории ото
льда (Томкович, Сорокин, 1983).
Для гнездования беринговы бакланы занимают места, недоступные наземным хищным
млекопитающим, в основном на скалистых обрывах и крутых склонах морского побере-
жья, реже на островках и кекурах вблизи берега (к примеру, на Командорских и Курильских
о-вах в колониях кекурного типа размещается 11-12% птиц). Гнёзда располагаются на
широких горизонтальных карнизах, уступах и площадках, как частично задернованных,
так и лишённых растительности. В первом случае основанием для гнездовой постройки
могут служить разросшиеся кочки колосняка. Пространственная структура поселений в
значительной мере определяется особенностями микрорельефа занимаемой поверхности
и стремлением этого вида к колониальному типу гнездования (Стишов, 1991). Крупные
колонии состоят из нескольких субколоний—так называемых «бакланьих городков». Гнёзда
Берингов баклан
111
нередко располагаются настолько близко друг к другу, что соприкасаются краями. По дан-
ным с Командорских о-вов, среднее расстояние между центрами ближайших соседних гнёзд
(п=95) в колониях на клифах составляет 3,2 м. На береговых скалах в большинстве случаев
бакланы гнездятся на высоте от 4 до нескольких десятков метров, иногда — более 100 м
над ур. моря. При отсутствии четвероногих хищников птицы могут занимать 2-3-метро-
вые обрывы, находящиеся выше зоны заплеска; изредка устраивают гнёзда на земле у ос-
нования обрыва (Артюхин, 1989). В Британской Колумбии в условиях перенаселенности
естественных гнездовых местообитаний наблюдали формирование колоний на техниче-
ских конструкциях — маяке, опорах моста, сваях (Hobson, Wilson. 1985).
Гнёзда массивные, обычно конусовидной формы. Старые гнёзда имеют мощное сце-
ментированное помётом основание, на котором возвышается свежая постройка. В каче-
стве строительного материала используют листья, стебли и корни произрастающих побли-
зости растений, мох, водоросли, почву, камешки. В выстилке лотка нередки перья, встреча-
ются куски капроновых веревок (от рыбацких сетей). Строительный материал птицы обычно
собирают на близлежащих участках вытаявших склонов и на берегу моря, реже вылавлива-
ют его в воде. Иногда разбирают прошлогодние гнёзда или похищают материал из постро-
ек соседних пар своего вида, других бакланов и моевок. Размеры (см) гнёзд с Командорс-
ких о-вов (п = 43): диаметр — 29-50, в среднем 38,6; высота — 7-33, в среднем 22,8;
диаметр лотка — 16,5-26, в среднем 20,8; глубина лотка — 3-10, в среднем 5,2 (Артюхин,
19916). На Чукотке находили гнёзда высотой до 70-73 см (Портенко, 1972).
Сроки размножения сильно растянуты. Например, на Командорских о-вах, где часть
беринговых бакланов, видимо, ведёт оседлый образ жизни, птицы начинают занимать гнёзда
в марте-апреле; откладку яиц наблюдали с начала мая по 27 июля, но в большинстве гнёзд
она проходит в первой половине июня. Столь длительный период послужил основанием
для ошибочного представления о бицикличности размножения берингова баклана на ост-
ровах (Stejneger, 1885). В южных частях ареала первые яйца в гнёздах появляются в конце
апреля, разгар кладки приходится на первую декаду мая (Гизенко, 1955; Велижанин, 1971;
Siegel-Causey, Litvinenko, 1993). В более суровых условиях Сев. Охотоморья бакланы начи-
нают нестись в конце мая (Кондратьев и др., 1992). На о. Карагинский первые яйца в гнёз-
дах наблюдали в начале июня, в массе — во второй декаде этого месяца (Н. Герасимов,
1999). На севере ареала в Беринговом и Чукотском морях кладки появляются в первой
половине июня (Портенко, 1972; Кречмар и др., 1978; Стишов и др., 1991), но в заливах,
поздно освобождающихся от ледового покрова, сроки откладки яиц сдвигаются на первую
декаду июля (Томкович, Сорокин, 1983). По наблюдениям на о. Врангеля (Придатко, 1986),
в среднем спаривание начинается через 19 дней после прилёта, а от спаривания до появле-
ния первого яйца проходит 6 дней. Ранние кладки нередко погибают. Пары, потерявшие
кладку в первую половину инкубационного периода, способны откладывать яйца повторно
(Харкевич, Вяткин, 1977).
В гнёздах находили до 7 яиц. Средний размер кладки на Командорских о-вах в 1988 г.
(п = 45) — 3,16 яйца (Артюхин, 19916), на о. Карагинский в 1968-1969 гг. (п=77) — 3,21 (Н.
Герасимов, 1999), на о. Верхотурова в 1975 г. (п=123) — 3,49 (П.С. Вяткин, личн. сообщ.),
на Чукотке (п=19) — 3,21 (Томкович, Сорокин, 1983), на о. Врангеля в 1981 г. (п=15) — 2,9 и
в 1984 г. (п=17) — 2,5 (Стишов и др., 1991), на о. Талан в 1993 г. (п=21) — 4,33 и в 1995 г.
(п=18) — 2,17 (Kondratyev et al., 2000). Размеры повторных кладок меньше, чем первых
(Hobson, 1997).
Яйца удлинённо-эллиптической формы. Скорлупа голубоватого либо бледно-зелёного
цвета, покрыта меловым слоем, скрывающим эту окраску. Яйца, отложенные первыми в
кладке, крупнее последующих, и они же самые «грязные», часто со следами крови на
112
Отряд Пеликанообразные, Семейство Баклановые
поверхности. Размеры яиц (мм) с Командорских о-вов: 52,4-60,4 х 32,5-38,8, в среднем
56,0 х35,6 (п=66; Артюхин, 19916); из зал. Кроноцкий: 55,2-62,0 х 34,3-39,1, в среднем
58,7х 37,2 (п=12; Лобков, 1986а); с о. Карагинский: 55,6-66,0 х 34,1-38,6, в среднем 58,6x36,5
(n=28; Н. Герасимов, 1999), с Чукотки: 53,0-66,0 х 32,0-39,2, в среднем 59,3x37,0 (п=43;
колл. ЗМ МГУ); с Аляски: 48,7-61,6 х 33,5-38,6, в среднем 56,3x35,9 (n=97; Hobson, 1997).
Толщина скорлупы — 0,317-0,596, в среднем 0,427 мм (Hobson, 1997). Масса свежих
яиц — 35,1^1,8, в среднем 38,7 г (п=8; колл. ЗМ МГУ).
Яйца откладывают обычно с интервалом в 2 дня (Hobson, 1997; П.С. Вяткин, личн. со-
общ.). Насиживание начинается с первого яйца. Продолжительность инкубационного пе-
риода на о. Врангеля 24-36 суток (Стишов и др., 1991), в зал. Аляска и Британской Колум-
бии — 28-32, в среднем 31 день (Hobson, 1997). В насиживании кладки, а также в обогре-
вании и кормлении птенцов принимают участие оба партнёра. Доли участия самца и самки
в этом процессе существенно варьируют у разных пар, но в среднем распределяются при-
мерно поровну.
Птенцы вылупляются асинхронно, голыми и слепыми. Кожа у них тёмно-серая, клюв
бурый с белым яйцевым зубом, ротовая полость светло-розовая, ноги бурые с коготками
рогового цвета. Масса новорожденных птенцов по одним данным (Анисимов и др., 1994)
составляет 37-39 г, по другим (Герасимов, 1979) — от 24 до 41,5, в среднем (п=7) 35,5 г. В
первые дни жизни птенцы малоактивны, издают слабый писк, плотно прижимаются друг к
другу. На 4-5-е сутки у них открываются глаза, и начинает пробиваться пух. По наблюдени-
ям на Командорских о-вах, развитие птенцов в гнёздах длится 53-60 дней (п=10; Фирсова
и др., 1986). После вылета они держатся на берегу поблизости от колонии и могут регуляр-
но возвращаться в гнездо, в том числе на ночёвку. Период вылета из гнёзд весьма продол-
жителен. Так, в северных частях ареала первые летающие молодые появляются с середины
августа, а последние покидают гнёзда в конце сентября.
Количество слетков в выводках варьирует: на о. Талан от 2,06 (п=38) в 1989 и 1995 гг. до
3,41 (п=29) в 1993 г. (Kondratyev et al., 2000); на Командорах от 2,39 (п=282) в 2007 г. до
2,63 (п=169) в 1988 г. (Артюхин, 19916, ориг.). На о. Беринга в 1992 г. успех размножения
составил 36,8 %, или 1,4 птенца на гнездо (п=5; Вяткин, Зеленская, 1993). На о. Умара в
1995 г. продуктивность была 1,20 слетка на гнездо (п=7), в 2005 г. — 2,67 (п=12) (Зелен-
ская, 2006); на о. Топорков (Командорские о-ва) в 2007 г. — 1,85 (п=431; ориг.); на сопре-
дельной американской территории (о. Булдырь) этот показатель значительно колеблется по
годам — от 0,6 до 1,8 птенца на пару размножающихся птиц (Williams et al., 2002).
Максимальная продолжительность жизни берингова баклана в природе, по данным коль-
цевания, — 17 лет 10 месяцев (Clapp et al., 1982).
Суточная активность, поведение. Берингов баклан ведёт дневной образ жизни. Одна-
ко в короткие зимние дни птицы не успевают насытиться засветло и продолжают кормить-
ся в темноте. На берег, в этом случае, они возвращаются нередко за полночь (Мараков,
1972). Взрослые птицы тратят времени на отдых в 3,3 раза больше, чем на поиски пищи, а
молодые — в 3,7 раза. Предполагают, что бакланы не могут находиться подолгу в холодной
воде из-за опасности переохлаждения вследствие быстрого намокания оперения (Hobson,
Sealy, 1985). В дневное время для отдыха и сушки оперения птицы собираются группами
на прибрежных невысоких камнях, окружённых водой. Ночуют на островках либо на ска-
листых обрывах на постоянных местах, хорошо распознаваемых по обильным пятнам по-
мёта. В колониях берингов баклан ведёт себя более осторожно, чем краснолицый, из-за
чего при беспокойстве людьми сильнее страдает от пернатых хищников.
Питание. Кормятся беринговы бакланы в основном на прибрежных мелководьях. В
период гнездования птицы обычно держатся в пределах нескольких километров от своих
Берингов баклан
113
колоний. Дальность кормовых перелётов может увеличиваться, если льды подступают близко
к берегу, либо когда птицы летают за кормом на удалённые от суши банки. Охотятся в
одиночку или мелкими дисперсными группами, периодически ныряя с поверхности воды и
затем методично обшаривая морское дно. По наблюдениям в Британской Колумбии, при
глубине 2-5 м средняя продолжительность пребывания под водой взрослых птиц составля-
ла 34,9 сек, а паузы между нырками были в 2,3 раза короче; это соотношение почти не
менялось до 20-метровой глубины (Hobson, Sealy, 1985).
Трофически беринговы бакланы связаны не с конкретными объектами питания, а с
кормовыми стациями. Рацион в значительной мере определяется глубиной, на которую
ныряют птицы, и характером донной поверхности. По этой причине спектры питания
даже в близлежащих колониях могут существенно различаться. Анализ погадок взрос-
лых птиц, собранных в колониях на Командорских о-вах и в Тауйской губе Охотского
моря, показал, что они кормятся преимущественно рыбой (всего зарегистрирован 41 вид),
чаще всего бентосными (63,4%) и придонными (22%) видами из семейств Gadidae, Sti-
chaeidae, Cottidae, Scorpaenidae. Кроме того, в их рацион входят донные беспозвоноч-
ные — в основном ракообразные (креветки, раки-отшельники) и полихеты, реже мол-
люски и иногда морские ежи. У птенцов, судя по их отрыжкам, состав кормов значитель-
но беднее видами рыб и почти не содержит беспозвоночных, за исключением десятино-
гих раков (Зеленская, 20016).
На о. Медный в 1950-х гг. в 24 из 25 осмотренных желудков была найдена рыба (рогат-
ковые — сем. Cottidae и мойва), в 5 — остатки крабов, креветки длиной до 45 мм и боко-
плавы (С.В. Мараков, неопубликованные данные). На Сахалине в желудках 8 птиц обнару-
жили остатки рыб (корюшки, наваги, мойвы, трёхиглой колюшки, песчанки, морского оку-
ня) и креветок (Нечаев, 19916). В колониях на юго-востоке Камчатки (о. Старичков) нахо-
дили в основном остатки рыб (бычков, песчанки, мойвы) и в меньшем количестве ракооб-
разных (Вяткин, 1983). На Чукотке желудки добытых в летнее время птиц чаще всего со-
держали рыб (в их числе — сайка и керчаки), ракообразных (бокоплавы, креветки), голово-
ногих моллюсков (Портенко, 1972). В желудке одной молодой птицы у пос. Уэлен было
найдено 10 бычков общим весом 520 г (Томкович, Сорокин, 1983). В колониях у м. Сердце-
Камень бакланы предпочитали питаться креветками длиной до 4 см (Головкин, Флинт, 1975).
На соседнем американском побережье Чукотского моря (м. Томпсон) в питании бакланов
наибольшее значение имеет рыба (сайка, стихеи) и десятиногие раки (Swartz, 1966 цит. по:
Hobson, 1997).
Враги, неблагоприятные факторы. Результативность гнездования в значительной мере
определяется хищничеством крупных чаек. Причиняемый ими ущерб особенно велик при
распугивании бакланов в колониях людьми. В тех местах, где проводят массовый сбор ча-
ячьих яиц, уровень хищничества чаек значительно возрастает, и они разоряют почти все
находящиеся по соседству гнёзда, в том числе и бакланьи. Часть потомства бакланов про-
падает из-за хищничества воронов (Corvus corax) и чёрных ворон. Подросшие птенцы не-
редко гибнут, падая с узких гнездовых карнизов. Слётки, намокшие в воде, даже выбрав-
шиеся на сушу, становятся добычей лисиц и росомах, когда пытаются сушить оперение на
береговых камнях (Харкевич, Вяткин, 1977; Зеленская, 1995, 1999, 2006; Н. Герасимов,
1999). На Сев. Охотоморье взрослые и молодые беринговы бакланы обычны в питании
белоплечих орланов, гнездящихся в колониях морских птиц (Utekhina et al., 2000; Зеленс-
кая, 2006). В Юж. Приморье останки этого вида находили в поедях сапсана и филина (Ко-
ломийцев, Поддубная, 1985; Назаров, Трухин, 1985). На Командорах бакланы найдены в
рационе песцов и белых сов. Известен случай нападения на птенцов баклана в гнезде инт-
родуцированной американской норки.
114
Отряд Пеликанообразные, Семейство Баклановые
Берингов баклан, вероятно, уступает в конкуренции за места гнездования более крупно-
му краснолицему. Последний в годы экспансии на юге Камчатки занял большинство коло-
ний, которые прежде принадлежали берингову баклану (Вяткин, 1986),
Отмечаемая в некоторые годы повышенная смертность птенцов в гнёздах и взрослых
птиц, возможно, обусловлена недостатком кормов вследствие изменения гидрологических
условий (Лобков, 19866; Артюхин, 1992). В колониях отдельные птицы и гнёзда гибнут под
камнепадами. Низко расположенные гнёзда смываются волнами во время штормов. На
Аляске известны случаи разрушения гнёзд сильными дождями (Hatch, Hatch, 1990). Моло-
дые неопытные птицы довольно часто получают смертельные травмы и переломы крыль-
ев, кормясь в полосе прибоя (Кондратьев и др., 1992).
Важным фактором, резко ограничивающим численность бакланов, являются периоди-
чески вспыхивающие эпизоотии, приводящие к массовой гибели птиц. На Командорских
о-вах с конца XIX в. зафиксированы, по крайней мере, 4 эпидемии среди бакланов: в 1876—
1877, 1923-1924, 1968-1969 и 1975-1976 гг. (Stejneger, 1885; Мараков, 1972, 1976, 1977).
Основным переносчиком инфекции служит иксодовый клещ Ixodes signatus — облигатный
паразит различных видов бакланов, распространённый в умеренных широтах Сев. Паци-
фики (Львов, Ильичев, 1979). Кроме того, гнёзда бакланов в большом количестве заселяют
блохи Myoctenopsylla sp. Все семь беринговых бакланов из зал. Петра Великого и с о. Та-
лан, вскрытые для паразитологического обследования, оказались заражены нематодами
Contracaecum spiculigerum и Desmidocercella skrjabini, трематодами Liliatrema sp. и скреб-
нями Andracantha sp. (Алексеев, Сметанина, 1968; Хоберг, 1992).
Негативное влияние оказывают факторы антропогенного происхождения: в прошлом —
прямое истребление, приведшее к исчезновению ряда колоний (Гизенко, 1955; Мараков,
1963; Нечаев, 1975), сейчас — беспокойство людьми в сезон размножения, которое спо-
собствует разорению гнёзд пернатыми хищниками. Отдельные птицы гибнут в рыболов-
ных снастях во время проведения промысла ярусами, дрифтерными и донными сетями
(Артюхин и др., 2000, 2006; Artyukhin, Burkanov, 2000; Мягких, 2002), а также из-за загряз-
нения прибрежных вод нефтепродуктами. В желудках нередко находят различные пласти-
ковые частицы, которые птицы заглатывают, принимая за пищевые объекты (Robards et al.,
1995); однако последствия этого вида загрязнения на состояние беринговых бакланов не
изучено.
Хозяйственное значение и охрана. В недалёком прошлом был традиционным объек-
том охоты коренного населения Дальнего Востока. Птиц употребляли в пищу, шкурки ис-
пользовали для изготовления верхней одежды (Портенко, 1972; Артюхин, 1991а; Стеллер,
1999). Судя по обилию костных остатков в археологических памятниках (Бурчак-Абрамо-
вич и др., 1987; Динесман и др., 1996; Савинецкий, Пташинский, 1999; Пантелеев, 2001),
беринговых бакланов часто добывали и древние люди. В настоящее время берингов баклан
как объект добычи потерял свое значение. В некоторых поселках Чукотки на бакланов про-
должают охотиться, но размеры добычи невелики: например, за 2002 и 2003 гг. жителями
посёлков Певек и Рыткучи всего добыто около 50 птиц (Е.Е. Сыроечковский-мл., личн.
сообщ.). Прежде берингов баклан признавался вредным видом, якобы наносящим урон
запасам ценных лососёвых рыб (Успенский, 1959), что, разумеется, абсурдно. Вид имеет
некоторое эпидемиологическое значение, так как участвует в циркуляции возбудителей
вирусных и бактериальных инфекций (Львов, Ильичев, 1979).
Берингов баклан занесён в Приложения двухсторонних соглашений, заклюённых Рос-
сией с США и Японией об охране мигрирующих птиц. Многие гнездовья, пути пролёта и
районы зимовок находятся в пределах особо охраняемых природных территорий. Наиболь-
шее значение в сохранении этого вида имеют заповедники «Остров Врангеля», «Командор-
Берингов баклан, Краснолицый баклан
115
ский» и «Дальневосточный морской», а также заказники: Южно-Камчатский федеральный
и местные — «Остров Верхотурова» и «Остров Карагинский».
Краснолицый баклан
Phalacrocorax urile (J.F. Gmelin, 1789)
Pelecanus urile J.F. Gmelin, 1789, Syst. Nat., 1, c. 575, Камчатка.
Red-faced cormorant, red-faced shag (англ.); Rotgesichtsscharbe,
Meerscharbe (нем.);согшогап a face rouge (фр ).
Син. (лат): Phalacrocorax bicristatus Pallas, 1811. •
Статус. Гнездящийся мигрирующий, на части ареала — оседлый, пролётный, зимую-
щий вид.
Общая характеристика и полевые признаки. Очень похож на берингова баклана, но
заметно крупнее (длина тела в среднем 79 см, размах крыльев — 117 см), более массивного
телосложения. Плечевые и внутренние маховые перья с пурпурно-фиолетовым блеском,
выделяющимся на зеленом фоне спины (у берингова баклана вся спинная сторона с одно-
образно зелёным отливом). Участок оголённой кожи на лицевой части головы развит силь-
нее, граница оперения на боках нижней челюсти ровная и, как правило, не доходит до
заднего края глаза (у берингова баклана—с далеко выступающим вперёд мыском). У взрос-
лых птиц лоб не оперён, лицо имеет характерную яркую оранжевую или красную окраску,
видимую издалека. Клюв заметно крупнее и светлее, чем у берингова баклана; в брачном
наряде его основание ярко-синее, а не красное. Поведением от берингова баклана не отли-
чается.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. В кладистической системе семейства баклановых, пост-
роенной на основе анализа остеологических признаков (Siegel-Causey, 1988), краснолицый
баклан, как и берингов, включён в род Stictocarbo Bonaparte, 1855, что, однако, как и в
отношении второго, современной систематикой не признается (Степанян, 2003; Dickinson,
2003; Коблик и др., 2006). Коллекционные сборы этого вида весьма малочисленны, поэто-
му его географическая изменчивость и популяционная структура не изучены. Отмечено,
что птицы с Аляски крупнее, чем из азиатской части ареала (Causey, 2002), что, однако,
может быть следствием ограниченности выборки.
Описание. Окраска. Половой диморфизм в окраске не выражен. Взрослые птицы в
брачном наряде. Оперение в основном черное с сильным металлическим блеском. Голова
и туловище — с зелено-малахитовым блеском, шея — с зеленовато-фиолетовым, внутрен-
ние маховые, плечевые и верхние кроющие крыльев — с пурпурно-фиолетовым, наружные
маховые и рулевые — почти без блеска. На боках поясницы и наружной стороне бёдер пара
белых пятен, образованных узкими рыхлыми перьями. На шее и иногда на спине редкие
белые узкие перья длиной до 45 мм. На темени и на границе затылка и шеи два черных
хохла. Вокруг основания клюва и глаза большой участок неоперенной кожи оранжевого
или красного цвета. Надклювье буровато-чёрное со светлой жёлто-роговой полоской по
краю челюсти. Подклювье жёлто-рогового цвета, тёмное на конце. Основание клюва опоя-
сано яркой лазурно-синей полосой. Ноги чёрные. Радужина бурая.
Взрослые птицы в зимнем наряде. Оперение отличается от предыдущего наряда отсут-
ствием белого цвета. На темени либо есть короткий хохол, либо его нет. Лоб частично зара-
116
Отряд Пеликанообразные, Семейство Баклановые
стает редкими мелкими чёрными перьями. Окраска основания клюва и оголённой кожи на
лице более тусклая.
Пуховой наряд серо-бурый, с грязно-белыми пятнами на груди, брюхе и боках тулови-
ща. Длина пушинок на туловище до 20 мм, на голове и шее до 10 мм. Лоб полностью
покрыт коротким редким пухом. Вдоль углов разреза рта и на подбородке кожа голая, серо-
го или светло-розового цвета. Клюв первоначально полностью буровато-чёрный, но посте-
пенно основание подклювья светлеет, приобретая серовато-голубую окраску. Ноги бурова-
то-чёрные; когти рогового цвета, на конце буро-чёрные.
Гнездовой наряд. Оперение в основном тёмно-бурое. Голова и верх шеи серо-бурые, с
многочисленными светлыми нитевидными перьями. Едва заметный металлический блеск:
на нижней части шеи — зелёно-фиолетовый, на спине, пояснице и надхвостье — зелёный,
на плечах и крыльях — пурпурно-фиолетовый. Лобная часть головы покрыта короткими
редкими серо-бурыми перьями. Голая кожа на лице серая, на горле — розовая. Клюв буро-
вато-чёрный, испещрённый желтоватыми пятнами; основание подклювья серо-голубое. Ноги
буровато-чёрные. Радужина светло-бурая.
Молодые птицы. Окраска оперения в промежуточных нарядах довольно вариабельна
вследствие значительных индивидуальных различий в продолжительности, объеме и пол-
ноте линьки. В первую зиму оперение остаётся от гнездового наряда, голова и верхняя
часть шеи темнеют. К лету второго календарного года наряд, изнашиваясь, становится в
целом светлее; клюв (особенно подклювье) тоже светлеет, пятнистый рисунок на нем раз-
мывается; голое пространство на лице несколько увеличивается (но лоб по-прежнему опе-
рён) и приобретает оранжевый оттенок. Летом в период интенсивной линьки покровного
оперения общая окраска становится пятнистой. Во вторую зиму наряд буровато-чёрный, с
более выраженным металлическим блеском, распределение которого аналогично таковому
у взрослых птиц, но менее интенсивным. Летом третьего календарного года молодые пти-
цы похожи на взрослых, но блеск оперения менее ярок; на боках поясницы вместо пятен
только отдельные белые перья; хохлы слабо развиты; оголённая кожа на лбу и около глаза
сравнительно небольшого размера, окрашена в бледно-оранжевый цвет; основание клюва
светло-синее.
Строение и размеры. Первостепенных маховых 10 (не считая рудиментарного),
формула крыла II>III>IV>I>V..., III>II>IV>I>V... или III>II>I>IV>V... Рулевых 12, форма
хвоста округлая. Размеры и масса тела приведены в таблице 5. Самцы заметно крупнее и
тяжелее самок.
Линька. Гнездовой наряд полностью формируется на 7-й неделе жизни. Линька моло-
дых и взрослых птиц изучена слабо. Судя по малочисленным экземплярам, имеющимся в
отечественных коллекциях, она протекает примерно так же, как у берингова баклана.
Распространение. Гнездовой ареал. Эндемик Сев. Пацифики, гнездящийся на
островах и побережьях Северо-Востока Азии и Сев. Америки (рис. 21). Азиатская часть
гнездового ареала почти полностью находится на территории Российской Федерации.
Населяет все основные острова Командорского архипелага — Беринга (в основном юж-
ную часть), Медный, Топорков и Арий Камень (Артюхин, 1999). На восточном побере-
жье Камчатки северная граница распространения проходит через о. Столбовой, где крас-
нолицего баклана впервые обнаружили в 1995 г. (Вяткин, 1999). Дальше к югу случай
гнездования известен в районе м. Шуберта, но основные места гнездования расположе-
ны южнее — от устья р. Жупанова до м. Лопатка (Вяткин, 2000; Лобков, 2003а). На за-
падном побережье Камчатки граница распространения в 1980-х гг. проходила в 3,5 км
севернее м. Сивучий (Вяткин, 1986; Лобков, Алексеев, 1987), но в последние годы крас-
нолицего баклана на охотоморской стороне полуострова нет (Артюхин и др., 20016). Гнез-
Краснолицый баклан
117
Таблица 5. Размеры (мм) и масса (г) взрослых особей краснолицего баклана
Показатель М Самцы lim п М Самки
lim п
Курильские и Командорские о-ва (колл. ЗИН, ЗМ МГУ, БПИ, ДВГУ, ориг.)
Длина Крыла 283 272-290 10 271 268-274 4
Хвоста 151 140-160 10 149 148-150 2
Цевки 59 54-61 9 58 56-61 4
Клюва 55 52-57 10 51 47-53 4
Командорские о-ва (Мараков, 1976)
Длина Крыла 291 275-310 13 268 265-270 4
Хвоста 161 150-170 12 152 140-163 4
Цевки 60 55-65 13 57 * 54-60 4
Клюва 53 51-56 13 53 52-56 4
Масса 2300 2000-2700 13 1900 1800-2000 4
Алеутские о-ва, зал. Аляска (Causey, 2002)
Длина Крыла 284 270-285 20 266 240-280 16
Хвоста 157 140-170 20 145 135-165 16
Цевки 58 54-61 20 58 50-60 16
Клюва 56 52-60 20 54 52-57 16
Масса 2428 2200-2500 20 1847 1650-2100 16
Аляска, о-ва Волрес и Кадьяк (Palmer, 1962)
Длина Крыла 277 269-288 6 272 255-296 9
Клюва 54 50-58 6 55 50-59 9
дится практически на всех островах Курильской гряды (Артюхин и др., 2001а; Артюхин,
2003; Ушакова, 2003).
За пределами России в Азии распространение ограничено о. Хоккайдо, где вид гнездит-
ся на островках в районе п-ова Немуро (Ornithological Society of Japan, 2000; Osa, Watanuki,
2002). Американская часть гнездового ареала охватывает о-ва Алеутские, Прибылова и по-
бережье Аляски от м. Ньюэнхем до о. Кадьяк (Causey, 2002; U. S. Fish..., 2005).
В России залёт за границы области распространения известен на о. Сахалин вблизи
г. Корсаков (Momiyama, 1928, цит. по: Нечаев, 1991). Кроме того, годовалая птица добыта
В.И. Лабзюком 2 июня 1976 г. на м. Маневского в зал. Ольга Приморского края (колл. БПИ
ДВО РАН). Самая южная находка на Японских о-вах — префектура Вакаяма, юго-западное
побережье о. Хонсю (Brazil, 1991). Есть информация о зимних находках вида на северо-
востоке Китая (Cheng, 1976 цит. по: Siegel-Causey, Litvinenko, 1993). На американском по-
бережье зарегистрирован залёт на о-ва Королевы Шарлотты в Британской Колумбии
(American Ornithologists’ Union, 1998).
Находки костных остатков краснолицего баклана в археологических памятниках на се-
вере Берингова моря (Causey, 2002) и на юго-западе Сахалина (Пантелеев, 19976) могут
свидетельствовать о том, что в позднеголоценовое время этот вид имел более широкий
ареал.
Зимовки. В Азии часть птиц остаётся зимовать в пределах гнездового ареала там, где
море не замерзает. Краснолицые бакланы зимой держатся в акватории Командорских о-
вов, у южной оконечности Камчатки и вдоль всей гряды Курильских о-вов (Мараков, 1972;
Нечаев, 2005; Вяткин, Артюхин, 2006). В Японии в это время южная граница распростра-
118
Отряд Пеликанообразные, Семейство Баклановые
Рис. 21. Область распространения краснолицего баклана: а — гнездовой ареал, б — районы зимо-
вок, в — залёты.
нения опускается до северной части о. Хонсю (Brazil, 1991; Ornithological Society of Japan,
2000). На Алеутах и Аляске зимует, не выходя за границы области гнездования (Causey,
2002) (рис. 21).
Миграции. На юге ареала краснолицые бакланы либо оседлы, либо совершают недалё-
кие кочёвки. Ярко выраженные перёлеты предпринимают птицы только из северных райо-
нов гнездования, главным образом с побережий Камчатки, Командор и Северных Курил.
При этом не все бакланы покидают острова; например, на о. Медный в 1950-х гг. на зиму
оставалась примерно треть птиц (Мараков, 1963).
В южной части Курильских о-вов (о. Уруп) весенний массовый пролёт краснолицых
бакланов в 1974 г. наблюдали 11-14 мая (Велижанин, 1975). Однако эта весна была затяж-
ной и холодной, поэтому, вероятно, миграция проходила позднее, чем обычно. В это же
время (11 мая 1974 г.) множество мигрирующих стай беринговых и краснолицых бакланов
отмечено на севере Курильской гряды между о-вами Шумшу и Парамушир (Остапенко и
др., 1975). На Камчатке и Командорских о-вах краснолицые бакланы появляются на местах
гнездования значительно раньше этих сроков. Так, в районе Кроноцкого зал. их регистри-
ровали с 31 марта (Лобков, 1986а), на Командорах в марте у них уже происходит строитель-
ство гнёзд (Мараков, 1976). Очевидно, эти наблюдения относятся к зимовавшим поблизос-
ти птицам, но не перелётным, которые прибывают сюда позднее. В конце апреля - мае на
юго-восточном побережье Камчатки наблюдали активный пролёт бакланов, считая, что все
Краснолицый баклан
119
они — беринговые (Ю. Герасимов, 1999,2001). Безусловно, здесь мигрируют и красноли-
цые бакланы, которых во время учётов на пролёте сложно выделить из массы беринговых
(Ю.Н. Герасимов, личн. сообщ.).
Сведения об осенней миграции ограничены наблюдениями на крайнем юге Камчатки
на м. Лопатка. В 1987 г. в конце августа - начале сентября здесь наблюдали только местные
кочёвки краснолицых бакланов, но с 9 сентября начался пролёт, который с 19 сентября
превратился в массовую миграцию краснолицых и беринговых бакланов (Лобков, 20036).
Оценку численности мигрирующих птиц проводил А.Г. Велижанин (1975), наблюдав-
ший весенний пролёт на о. Уруп. По его мнению, в 1974 г. за четыре дня массовой мигра-
ции вдоль берегов этого острова на север пролетело 9 тыс. особей именно краснолицего
баклана.
Мигрируют вдоль морских побережий; вдоль Курильских о-вов продвигаются как по
охотоморской, так и по тихоокеанской стороне. Как правило, птицы пролётают на удалении
не более 10 км от суши. Летят в светлое время суток одиночками, парами и стаями, состо-
ящими обычно из нескольких десятков птиц, но в дни массовой миграции наблюдали стаи
из сотен (до 800) особей. Стаи имеют форму клина, либо цепочки. Пролёт проходит на
высоте от 15 до 100 м над водой (Велижанин, 1975; Лобков, 20036).
Местообитания. Обитатель прибрежных вод. Гнездится на скалистых обрывистых уча-
стках морского побережья. Кормится на мелководьях вблизи берега, в бухтах и заливах, на
лиманах и лагунах, иногда залетает в низовья рек. Зимой держится на незамерзающих при-
брежных акваториях. Как правило, краснолицые бакланы не встречаются дальше 10-20 км
от суши, за исключением птиц с удалённых островов, которые во время миграций вынуж-
дены пересекать открытые морские пространства.
Численность. Область распространения и численность краснолицего баклана подвер-
жены серьезным колебаниям, имеющим, по-видимому, характер долговременной циклич-
ности, обусловленной изменениями климато-океанологических условий. Кроме того, осо-
бенности динамики населения этого вида трудно поддаются изучению из-за частой смены
мест расположения гнездовых колоний.
На Командорских о-вах в конце XIX в. этот вид был довольно редким, встречался толь-
ко на о-вах Медный и Арий Камень (Stejneger, 1885). В первой половине XX в. его гнездо-
вья имелись лишь на первом из этих островов; в начале 1950-х гг. здесь гнездилось не
более 1,5-2 тыс. пар (Иогансен, 1934; Мараков, 1963). В дальнейшем вид широко рассе-
лился по всём островам архипелага, и его численность к 1970 г. достигла 12 тыс. пар, но в
1975-1976 гг. резко сократилась из-за распространившейся в популяции эпизоотии (Мара-
ков, 1972, 1977). В период с 1986 по 1994 г. число размножающихся на Командорах птиц
колебалось от 850 до 1 100 пар (Артюхин, 1991а, 1999). Примерно столько же бакланов (не
менее 1 тыс. пар) гнездилось здесь в 1999 и 2000 гг. (Зеленская, 2001а).
На п-ове Камчатка этот вид впервые нашли на гнездовании на о. Уташуд в 1972 г. (Вят-
кин, 1981). Общая численность в 1970-х гг. составляла 1,5-2 тыс. пар, в 1983 г. — около 5,6
тыс. пар (Вяткин, 1986). Во время проведения следующих учётов в 1995 г. отмечено сокра-
щение численности до 3 тыс. пар (Вяткин, 2000). Современная численность на полуостро-
ве оценивается в 2 тыс. пар (Kondratyev et al., 2000; Вяткин, Артюхин, 2006). Возможно, в
последние годы численность вида на юго-востоке Камчатки находится на подъёме. Об этом
свидетельствуют наблюдения на о. Старичков, расположенном у входа в бух. Авачинскую,
где количество гнездящихся краснолицых бакланов увеличилось с 60 пар в 2002 г. до 400 —
в 2006 г. (Лобков, 2009).
На Курильских о-вах, по результатам наблюдений, выполненных до середины XX в.,
краснолицый баклан никем из исследователей не приводится как массовый вид (Stejneger,
120
Отряд Пеликанообразные, Семейство Баклановые
1898; Yamashina, 1931;Bergman, 1935;Гизенко, 1955;идр.).В 1950-егг. С.К.Клумов(1960)
нашел его здесь весьма обычным. В 1963 г. краснолицый баклан был распространён на
островах уже повсеместно; А.Г. Велижанин (1971, 1976) оценил его численность в 20-
30 тыс. особей. Однако по результатам следующего учёта в 2000 г., численность вида на
Курилах составила всего лишь 4,5 тыс. пар, из них 3 тыс. пар гнездились на о-вах Шумшу
и Парамушир (Артюхин и др., 2001а). Вероятно, численность упала к началу 1980-х гг., так
как некоторые наблюдатели (Ильяшенко и др., 1988; Анисимова, 1996; Артюхин и др., 2001а)
отмечали малочисленность вида, начиная с этого десятилетия. В это же время красноли-
цый баклан почти исчез на южной границе ареала в Японии; в последние годы на о. Хок-
кайдо гнездится примерно 25 пар (Brazil, 1991; Osa, Watanuki, 2002).
Таким образом, современная численность краснолицего баклана в азиатской части аре-
ала составляет 7,5 тыс. размножающихся пар. Это на порядок ниже прежних оценок (Siegel-
Causey, Litvinenko, 1993; Шунтов, 1998), которые выводились на основе данных, получен-
ных в период наивысшей численности вида в дальневосточных морях.
В американском секторе ареала в 1970-х гг. насчитывалось 130 тыс. особей красноли-
цых бакланов, большинство из них (88 тыс.) обитало на Ближних о-вах в западной части
Алеутской гряды (Sowls et al., 1978). В последние годы здесь отмечено значительное паде-
ние численности бакланов, но причины этого явления не установлены (Byrd, Williams,
2004; Causey et al., 2006). Современная оценка численности вида на Аляске снижена до
20 тыс. особей (Denlinger, 2006).
Размножение. Точных данных о сроках наступления половой зрелости у краснолицых
бакланов нет. Вероятно, они начинают размножаться в трёхлетием возрасте, подобно близ-
кородственному берингову баклану (Van Tets, 1959).
Факультативно-колониальный вид. Предпочитает гнездиться колониями, величина ко-
торых варьирует от нескольких пар до первых сотен пар; одиночные гнёзда встречаются
относительно редко. Селится как самостоятельными колониями, так и вместе с другими
видами морских птиц (чаще всего с беринговыми и уссурийскими бакланами, чайками,
кайрами). Расположение колоний сильно меняется из года в год, благодаря чему, по мне-
нию С.В. Маракова (1976), птицы ослабляют контакты с многочисленными гнездовыми
паразитами (клещами и блохами). Занимает недоступные для наземных хищников места на
скалистых обрывах и крутых склонах морского берега, на прибрежных островках и кеку-
рах. Значительно чаще, чем берингов баклан, образует колонии кекурного типа (на Куриль-
ских о-вах в них размещается 19% всех птиц, а на о. Медный — до 57%). На береговых
обрывах гнёзда обычно размещаются в средней либо верхней части клифа на высоте до
70-80 м, но иногда значительно выше (до 200 м); на островках и кекурах — начиная с 3^-
м над уровнем моря. Подмечено (Мараков, 1972; Nysewander, 1986; данные автора очерка),
что при совместном гнездовании краснолицый баклан предпочитает занимать более высо-
кие участки обрыва, чем берингов.
Брачное и территориальное поведение изучено недостаточно. Судя по имеющимся дан-
ным, оно сходно с таковым у берингова баклана (Van Tets, 1965; Kennedy et al., 1996).
Гнёзда устраивают на широких горизонтальных карнизах и уступах, на ровных площад-
ках и плоских вершинах кекуров. Гнездо представляет собой массивную постройку, возве-
денную из стеблей и листьев произрастающих поблизости растений (чаще злаков), в выс-
тилке лотка бывают отдельные перья. На Курилах в качестве материала птицы часто ис-
пользуют водоросли (Велижанин, 1976), чего ни разу не отмечалось на Командорах. Гнез-
до имеет конусовидную форму с широким основанием, которое сцементировано помётом.
Размеры гнёзд (см) с Командорских о-вов (п = 14): диаметр — 30-50, в среднем 43,7; высо-
та — 6-30, в среднем 15,8; диаметр лотка — 17-24, в среднем 21,1; глубина лотка — 3-7,5,
Краснолицый баклан
121
в среднем 4,9 (Артюхин, 19916). Максимальной высоты (до 70 см; Мараков, 1976) гнездо
достигает, если возводится на старом основании. Свежие постройки на ровных вершинах
кекуров бывают очень низкими, тарелкообразной формы. В строительстве гнёзда прини-
мают участие оба партнёра.
По срокам размножения краснолицые бакланы обычно опережают беринговых, за ис-
ключением отдельных случаев, отмеченных на юге Курильских о-вов. Строительство гнёзд
начинается в конце марта, первые яйца появляются во второй половине апреля, но массо-
вая откладка яиц происходит значительно позже — в конце мая - первой половине июня.
На Командорских о-вах незавершенные кладки находили до конца июня, а свежие яйца —
и в июле. Отмечено, что на о. Медный разница в сроках начала яйцекладки между северной
и южной оконечностями острова достигает 20-30 дней (Иогансен, 1934; Мараков, 1976;
Велижанин, 1976,1977; Фирсова и др., 1986; Артюхин, 19916). Судя по длительному пери-
оду яйцекладки, птицы способны откладывать яйца повторно взамен утерянных, как это
установлено для берингова баклана (Харкевич, Вяткин, 1977; Hobson, 1997), но прямых
доказательств тому нет.
В гнёздах бывает до 8 яиц (Фирсова и др., 1986). Размер кладки на Командорских о-вах
в 1989 г. (п = 42) — от 1 до 5, в среднем 2,83 яйца (Артюхин, 19916). На Аляске в зависимо-
сти от места и года наблюдений средние размеры кладок варьировали в пределах 2,1-3,1
яиц (Johnsgard, 1993). Яйца удлинённо-эллиптической формы. Скорлупа светло-голубая или
бледно-зелёная, покрытая сверху меловым слоем, который скрывает окраску. Размеры яиц
с Командорских о-вов: 56,8-62,8 х 35,5-40,8, в среднем (п = 37) 59,4x38,3 мм (Фирсова и
др., 1986; Артюхин, 19916; колл. ЗМ МГУ); масса — 45-49, в среднем (п = 13) 47,3 г (Ма-
раков, 1976). Л. Штейнегер (Stejneger, 1885) приводит более крупные размеры двух яиц с о.
Медный: 63,5x39 и 66x37 мм.
Кладку насиживают оба партнёра, начиная с появления первого яйца. По наблюдениям
на о-вах Прибылова длительность инкубации составляет в среднем 31 сутки (Hunt et al.,
1981), в зал. Аляска — 33 (Nysewander, 1986).
Вылупление птенцов происходит асинхронно. Они рождаются голыми, с буроватой ко-
жей, слепыми, малоподвижными. Выкармливанием птенцов занимаются оба родителя.
Известен случай, когда пара краснолицых бакланов «усыновила» двух птенцов берингова
баклана в возрасте 2 недель и успешно их выкормила (Зеленская, 1993). Птенцы прибавля-
ют в весе в среднем от 48,5 до 72,0 г в день (Hunt et al., 1981). Средняя продолжительность
птенцового периода составляет по одним данным 50 дней (Nysewander, 1986), по другим —
59 (Hunt et. al, 1981).
На Командорских о-вах начало появления птенцов в гнёздах регистрировали со второй
половины июня (например, 17 и 26 июня в 1986 и 1989 гг. соответственно), массовое вы-
лупление обычно приходится на начало июля. Большинство птенцов поднимается на кры-
ло в конце августа - начале сентября. На крайнем юго-западном участке ареала птенцы
покидают гнёзда с июля.
Количество слетков в выводках на Командорах в 1988 г. было в среднем 2,15 (п=13), в
1990 г. — 2,68 (п=19), в 2007 г. — 2,30 (п=106); на японских колониях в 1972 г. — 2,8 (п=37).
Успех размножения значительно варьирует, в отдельные годы наблюдается массовая смер-
тность потомства. Продуктивность краснолицего баклана на о. Топорков (Командорские о-
ва) в 2007 г. составила 1,22 птенца на пару размножавшихся птиц (п=196 гнёзд), в пунктах
мониторинга на Аляске этот показатель колеблется от 0 до 2,5 (Нечаев, 1969; Fujimaki et al.,
1976; Артюхин, 19916, 1992, ориг.; Dragoo et al., 2004).
Суточная активность, поведение. Краснолицый баклан активен в светлое время суток.
Однако в короткие зимние дни птицы нередко кормятся до 22-24 ч и возвращаются на берег
122
Отряд Пеликанообразные, Семейство Бакланов ые
в темноте (Мараков, 1976). Ночуют на недоступных для песцов скалистых обрывах и остро-
вках, днём для отдыха собираются группами на каменистых рифах. Малоосторожен, наси-
живающая птица нередко подпускает человека вплотную. При появлении людей в колониях,
краснолицые бакланы, в сравнении с беринговыми, позднее покидают гнёзда и быстрее воз-
вращаются обратно, поэтому меньше страдают от пернатых хищников, но в прошлом по
этой причине они попадали под выстрел в первую очередь (Мараков, 1963).
Питание. Кормовые биотопы краснолицых бакланов расположены в прибрежной зоне
моря, на лиманах, в лагунах и устьях рек. В период гнездования птицы кормятся поблизос-
ти от своих колоний. Так, в акватории Командорских о-вов в летнее время бакланы, как
правило, ищут корм в пределах 10 км от берега, но зимой нередко улетают мористее в
среднюю часть пролива между о-вами Беринга и Медный. Кормятся в одиночку или не-
большими дисперсными группами. Добычу ищут в донном и придонном слое воды, пери-
одически ныряя.
Основу питания составляет рыба (не только донных и придонных, но и пелагических
видов) и в меньшей степени ракообразные. На Командорских о-вах в 1950-х гг. в 16 из 20
осмотренных желудков была обнаружена рыба (мойва, песчанка, рогатковые), в 4 — мел-
кие крабы, креветки и бокоплавы (Мараков, 1976). В колониях на о. Старичков, юго-вос-
точное побережье Камчатки, в 1979 г. у гнёзд находили обычно остатки песчанки, бычков,
мойвы, колюшки (Вяткин, 1983). На сопредельной американской территории в западной
части Алеутской гряды (о. Агатту) в питании взрослых краснолицых бакланов доминиро-
вала песчанка, в малых количествах встречались люмпены, креветки и кальмары (Springer
et al., 1996). На о-вах Прибылова в составе рыбных кормов преобладали рогатковые, мин-
тай и песчанка; в рационе взрослых птиц наряду с рыбой важную роль играли ракообраз-
ные, в то время как птенцы выкармливались только рыбой (Hunt et al., 1981; Scheider, Hunt,
1984).
Враги, неблагоприятные факторы. Основными естественными врагами краснолице-
го баклана являются пернатые хищники — серокрылая и тихоокеанская чайки, ворон и
чёрная ворона, которые часто похищают яйца и маленьких птенцов. На Аляске, кроме того,
на бакланов охотятся сапсаны и белоголовые орланы (Murie, 1959). Из наземных хищников
доступные гнёзда разоряют песцы, лисицы и бурые медведи. Отмечаемая в некоторые годы
высокая смертность птенцов, по-видимому, происходит от истощения при недостатке кор-
мов. Отдельные гнёзда разрушаются ливнями и волнами во время сильных штормов. Пе-
риодически в популяциях краснолицего баклана вспыхивают эпизоотии, в результате кото-
рых погибает множество птиц, и численность вида существенно сокращается (Мараков,
1972, 1976). Переносчиками заболевания служат иксодовые клещи Ixodes signatus и блохи
Myoctenopsylla sp.
Из факторов антропогенного происхождения в недалёком прошлом наибольшее воз-
действие на состояние вида оказывала добыча птиц местным населением для пропитания.
На Командорах в период островного звероводства (1930-1950-е гг.) неумеренный промы-
сел стал причиной исчезновения многих колоний бакланов (Мараков, 1963). Отдельные
птицы гибнут в сетях во время дрифтерного промысла лососей (Артюхин и др., 1999). Вид
весьма чувствителен к локальным загрязнениям прибрежных вод нефтепродуктами.
Хозяйственное значение, охрана. На Командорах до середины 1960-х гг. красноли-
цый баклан являлся традиционным объектом промысла коренного алеутского населения. В
1950-х гг. здесь ежегодно добывали не менее 500 особей (Мараков, 1976). В настоящее
время он исключён из перечня видов, относящихся к объектам охоты. Краснолицый баклан
имеет некоторое эпидемиологическое значение, так как птицы этого вида участвуют в цир-
куляции арбовирусов (Львов, Ильичёв, 1979).
Краснолицый баклан, Хохлатый баклан
123
Занесён в Красную книгу Севера Дальнего Востока России (Вяткин, Артюхин, 1998),
Красную книгу Камчатки (Вяткин, Артюхин, 2006), приложения двухсторонних соглаше-
ний, заключённых Россией с США и Японией об охране мигрирующих птиц. На гнездова-
нии, пролёте и зимовке охраняется в Командорском, Кроноцком и Курильском заповедни-
ках, Южно-Камчатском федеральном заказнике, Южно-Камчатском природном парке, за-
казниках и природных памятниках областного подчинения.
Хохлатый баклан
Phalacrocorax aristotelis (Linnaeus, 1761)
Pelecanus aristotelis Linnaeus, 1761, Fauna Suecica, Ed. altera, N 146,c. 51, Швеция.
Common shag (англ.), Krahenscharbe, Mittelmeer-Kormoran (нем.),
cormoran huppe (фр.).
Син. (лат.): для подвида aristotelis: Pelecanus gracilus Linnaeus, 1766; Pelecanus cristatus J.F.
Gmelin, 1789; Pelecanus lophurus Graves, 1813; Carbo brachyurus C.L. Brehm, 1824; Graculus
linnaeii G.R. Gray, 1845; Phalacrocorax graculus (Мензбир, 1895); для подвида desmarestii'.
Carbo desmarestii Payraudeau, 1826; Phalacrocorax gracilus croaticus Brusina, 1891;
Phalacrocorax graculus riggenbachi Hartert,* 1923.
Син. (рус.): длинноносый, [средний] баклан.
Статус. На севере ареала гнездящийся мигрирующий вид, на юге в основном оседлый.
Общая характеристика и полевые признаки. Птица среднего размера, заметно меньше
большого баклана; тело более стройное, клюв тоньше и длиннее. Оперение взрослых пол-
ностью черное, с примесью металлически-зеленого оттенка. В брачном наряде (зимой и до
конца весны) на голове имеется хохол из тёмных перьев. Клюв и лапы тёмные, голый уча-
сток кожи вокруг клюва и основание подклювья жёлтые. Молодые сверху бурые, нижняя
часть туловища, шеи и подбородок более светлые.
Подвидовая систематика. Вид включает три подвида, которые различаются по длине
клюва, интенсивности окраски неоперенных частей головы и плавательных перепонок,
длине хохла (Судиловская, 1951; Cramp, 1977; Степанян, 2003). В пределах Вост. Европы и
Сев. Азии встречаются 2 подвида.
1. Phalacrocorax a. aristotelis.
Pelecanus aristotelis Linnaeus, 1761, Fauna Suecica, Ed. altera, N 146,c. 51, Швеция.
Плавательные перепонки чёрные, в окраске неоперённых частей головы более развит
чёрный цвет, хохол — до 50 мм. Северо-западные берега Европы до Испании и Португа-
лии; в России — северные берега Кольского п-ова.
2. Phalacrocorax a. desmarestii.
Carbo desmarestii Payraudeau, 1826, Annales Sci. Nat., 8, c. 464, острова западной части
Средиземного моря.
Несколько меньше предыдущего подвида, клюв длиннее. Плавательные перепонки жёл-
тые у молодых птиц, бурые у взрослых, неоперённые части головы более жёлтые, хохол —
менее 50 мм (Cramp, 1977; Степанян, 2003). Острова и северные берега Средиземного моря,
на Украине — черноморские берега Крыма, в России — у Таманского п-ова (Белик, 2003а).
Третий подвид — Ph. a. riggenbachi Е. Hartert, 1923 (3) — распространён в северо-запад-
ной Африке вдоль побережья Марокко (Howard, Moore, 1980). Размеры и окраска неоперен-
ных частей как у Ph. a. desmarestii, длина клюва — как у Ph. a. aristotelis (Cramp, 1977).
124
Отряд Пеликанообразные, Семейство Баклановые
Замечания по систематике. При дроблении рода Phalacrocorax хохлатого баклана от-
носят к роду Stictocarbo Bonaparte, 1855 (Siegel-Causey, 1988; del Hoyo et al., 1992), кото-
рый может рассматриваться в качестве подрода при широкой трактовке Phalacrocorax.
Описание. Окраска. Половой диморфизм в окраске не выражен. Взрослые птицы в
брачном наряде. Все оперение чёрное. Перья с заметным металлически-зелёным отливом
на туловище и фиолетово-зелёным — на голове и шее. Перья передней части спины, лопа-
ток, плечевые и верхние кроющие крыла с бархатно-чёрными окаймлениями. Маховые
чёрные, внешние опахала слегка зеленоватые, внутренние коричневатые. Рулевые чёрные
с фиолетово-зеленоватым отливом. Ноги и плавательные перепонки буровато-чёрные. На
голове хохол из тёмных загнутых вперёд перьев, более развитый у самцов. Клюв чёрный с
бледной вершиной; основная его часть и окружающие голые участки кожи жёлтые. Раду-
жина зелёная.
Взрослые птицы в зимнем наряде. То же, что и в брачном, но хохол отсутствует.
Пуховой наряд. Птенцы вылупляются голыми, с тёмно-буроватым цветом кожи, бледно-
жёлтыми подклювьем и перепонками лап. Подросшие птенцы покрыты чёрно-бурым пу-
хом. Голая кожа вокруг глаз тёмная, у углов рта и на подбородке грязно-жёлтая. Надклювье
черное, подклювье бледно-жёлтое в основной части, постепенно темнеющее к вершине.
Радужина светло-бурая. Ноги и плавательные перепонки тёмно-бурые.
Гнездовой наряд. Верх тела и бока тёмно-коричневые, с едва заметным металлически-
зелёным блеском; перья поясницы, лопаточные и плечевые с широкими неблестящими тем-
ными краями. Верхние кроющие крыла бурые с более светлыми окаймлениями. Нижняя
часть шеи и её бока бурые; подбородок, грудь, брюхо и бока тела грязно-белые, с редкими
светло-бурыми пятнами на груди и боках брюха, светлее у птиц южных популяций. Руле-
вые матово-чёрные, со светлыми краями, маховые как у взрослых, но более тусклые. Ноги
тёмно-розовые. Надклювье бурое, более тёмное по коньку, подклювье бледно-жёлтое, тем-
нее к вершинной части. Радужина светло-бурая.
Между первой зимой и второй осенью приобретает «переходный наряд» между гнездо-
вым и первым брачным (Cramp, 1977). В этом наряде верхние кроющие почти как у взрос-
лых, но в основаниях перьев головы имеется некоторая крапчатость, окаймления кроющих
спины, лопаток и крыльев шире и менее чёрные. Нижняя часть туловища темнее, особенно
на боках; передняя часть шеи тёмно-коричневая с более бледным подбородком.
Первый брачный наряд. Надевается между 2-й и 3-й осенью жизни. Верхняя сторона
головы, шея и спина с более сильным металлически-зелёным блеском. Мелкие кроющие
крыла и маховые тёмно-бурые с металлическим блеском на кроющих крыла и наружных
опахалах маховых. Нижняя сторона тела равномерно бурая без блеска, светлеющая к гряз-
но-белому подбородку. Ноги бурые.
Первый зимний наряд. Как у взрослых. Только низ тела тёмно-бурый с очень слабым
металлическим блеском, подбородок бурый с мелкими белыми пестринами (Судиловская,
1951; Cramp, 1977).
Строение и размеры. Первостепенных маховых 11. Хвост закруглённый,
рулевых 12. Клюв с небольшим крючком на конце. Средний вес 1 865 г (Судиловская, 1951).
Вес самца и самки, добытых на Мурмане, составил 1 700 и 2 050 г (Ю.В. Краснов, личн.
сообщ.). Размеры приведены в таблице 6.
Линька. Последовательность смены нарядов: пуховой - гнездовой - первый брачный -
первый зимний - второй брачный - второй зимний. У взрослых птиц 2 линьки в году. Час-
тичная предбрачная протекает с ноября по январь: происходит смена кроющих крыла и
мелкого пера (Судиловская, 1951). На третьем году жизни на голове вырастает хохол (Смо-
горжевський, 1979). Полная послебрачная линька (июнь-октябрь) начинается потерей пе-
Хгхлатый баклан
125
Таблица 6. Размеры хохлатого баклана. Ph. a. aristotelis — по Cramp, 1977.
Ph. a. desmarestii — по коллекциям зоомузеев ННПМ Украины и Киевского нац. ун-та
(ориг.) и по Cramp, 1977
Пол Д;1 ина крыла Длина хвоста Длина цевки Длина клюва
п lim М n lim М n lim М n lim М
12 Ph. a. aristotelis (Англия и Голландия; размеры клюва — только для ЮЗ Англии) 261-278 271 6 119-133 129 12 62-68 64,7 10 53-58 55,7
-- 18 251-269 258 10 114-125 119 18 58-65 62,0 5 55-58 56,2
Ph. a. aristotelis (Норвегия) 21 51-58 55,0
-- 18 51-59 55,3
Ph. a. aristotelis (Северная Шотландия) 14 56-61 58,9
-- 11 57-63 59,3
5 Ph. a. desmarestii (черноморская популяция, ориг.) 256-268 262 5 60-63,4 61,8 5 57,3-65,5 62,8
-- 8 236-251 245 8 57,4-61,6 59,5 8 56,8-65,4 60,6
12 Ph. a. desmarestii (западное Средиземноморье — по Cramp, 1977) 243-271 258 6 58-65 60,9
2 0 11 246-265 249 5 61-65 63,2
рьев хохла, частичной сменой мелкого оперения и в сентябре-октябре — линькой маховых
и рулевых (Судиловская, 1951). Пост-ювейильная линька полная, продолжается в течение
года. Голова, шея, туловище и мелкие кроющие крыла линяют в течение осени первого года
жизни, затем, после зимней задержки — с весны и до поздней осени; первостепенные ма-
ховые и крупные кроющие крыла — после мая второго календарного года и до декабря. В
продолжение второго календарного года линяют перья хвоста. Линька в первый брачный
наряд полная. Крылья линяют с лета 3-го календарного года, туловище — с июля до зимы.
Некоторые коричневые перья иногда сохраняются до 5-го года (Cramp, 1977).
Распространение. Гнездовой ареал — морские побережья Европы, западной
части Малой Азии, сев. Африки (рис. 22). В Европе гнездится по северным берегам Коль-
ского и Скандинавского п-овов, по берегам Великобритании, Ирландии, запада Исландии,
Франции, Испании и Португалии. Южная часть ареала охватывает берега и острова Среди-
земного, Адриатического, Эгейского и Чёрного морей. В Африке известен на северо-запад-
ном (Марокко), местами на северном побережье (Cramp, 1977).
В России распространён на Кольском п-ове от Варангер-фьорда до мыса Святой Нос;
самый восточный пункт гнездования — губа Дворовая. В Зап. Мурмане гнездится на юж-
ном берегу Варангер-фьорда (бухты Базарная и Печенгская) и на Айновых о-вах (Шкляре-
вич, Татарникова, 1986). На Вост. Мурмане (Семь Островов) первые гнёзда найдены в 1932
г. на о. Харлов и в 1937 г. — на о. Кувшин (Спангенберг, 1941; Модестов, 1967). В после-
дние годы наиболее крупные поселения существуют на архипелаге Семь Островов (о. Веш-
няк) и Гавриловских о-вах (Татарникова, 20016; Ю.В. Краснов, личн. сообщ.).
Самая крупная черноморская группировка находится на п-ове Тарханкут в зап. Крыму.
В юж. Крыму спорадично гнездится на побережье от м. Херсонес у Севастополя до м.
126
Отряд Пеликанообразные, Семейство Баклановые
Рис. 22. Места гнездования (а) и районы зимовок (б) хохлатого баклана. Подвиды: 1 —Ph. a. aristotelis,
2 — Ph. a. desmarestii, 3 — Ph. a. riggenbachi.
Киик-Атлама западнее Феодосии и в Опукском заповеднике на Керченском п-ове (Костин,
1983; Цвелих, 2002; Бескаравайный, 2004). В 2003 г. колония обнаружена в Керченском
проливе, у м. Панагия Таманского п-ова (Белик, 2003а). Ранее гнездился в северо-западной
части Чёрного моря: в конце XIX в. — на о. Березань и в 1929 г. — на о. Змеиный (Назарен-
ко и др., 1975).
Миграции. Регулярные сезонные перемещения характерны для птиц северной популя-
ции. После подъема на крыло молодых, бакланы кочуют у берегов Мурмана, смещаясь на
запад. Большая часть птиц зимует у сев. Норвегии, некоторые остаются на Мурмане (Моде-
стов, 1967; Кищинский, 1978д; Татарникова и др., 1983). Места зимовок удалены от мест
гнездования на 200-1 000 км, иногда до 1 500 км (Кищинский, 1978д).
Черноморская популяция в основном оседла: большинство птиц в негнездовое время
держится не далее 20 км от районов гнездования и сосредоточена в пределах Крымского
полуострова (Костин, 1983; Бескаравайный, 2004), дальние залёты нерегулярны. Известны
встречи на Сиваше (Никольский, 1891; Лысенко, Сиохин, 1991); как многочисленный вид
отмечался осенью у Геническа (Воронцов, 1937). На летних и зимних кочёвках встречается
у северных берегов Керченского п-ова (Кошелев и др., 2002; А.Б. Гринченко, личн. со-
общ.). Указанные пункты находятся на расстоянии от 45-50 до 150-170 км от районов гнез-
Хохлатый баклан
127
дования. Известна встреча у г. Гагра на Кавказе (Строков, 1974) — примерно в 300 км от
границы гнездового ареала.
Местообитания. Хохлатый баклан — стенотопный вид, связанный на гнездовании ис-
ключительно со скалистыми морскими берегами, наличие которых является одним из ос-
новных лимитирующих факторов его распространения.
В Крыму гнездовые поселения приурочены к выпуклым (в плане) участкам берега и
вершинам мысов, состоящих из прочных горных пород: известняков, конгломератов, вул-
канитов. Скальные поверхности, на которых располагаются гнёзда, полностью лишены
растительности. Используется два типа скально-береговых форм рельефа. Во-первых: эро-
дированные обрывы (клифы), нависающие над водой, реже — над берегом. Гнёзда здесь
располагаются в глубоких нишах, реже открыто, на скальных полках. Во-вторых: скальные
островки высотой от 3-5 до 30 м от морской поверхности, на удалении до 4 км от берега:
для них более характерно открытое гнездование. На Мурмане хохлатые бакланы гнездятся
в сходных стациях; здесь регистрировали также гнездование в нагромождениях крупных
камней и осыпях, образованных крупными скальными обломками (Модестов, 1967; Крас-
нов и др., 1995). В Крыму гнёзда распологались на высоте от 2 до 120 м, обычно от 5 до 40 м;
на о. Харлов — до 20 м.
В Баренцевом море зона жизнедеятельности ограничена трёхкилометровой прибреж-
ной полосой (Ю.В. Краснов, личн. сообщ.). В Крыму кормятся на расстоянии до 2 км,
обычно до 600 м от берега (Гринченко, 1994). В сев. Атлантике дальность кормовых ко-
чёвок в гнездовой период составляет 7,0=Ы ,9 (до 17) км от берега (Wanless, Burger, Harris,
1991).
Для дневного отдыха и ночёвки птицы используют выступы береговых обрывов, камни
в море, а при отсутствии фактора беспокойства — пологие участки берега. В Крыму часто
отдыхают на береговых сооружениях.
Численность. Современная численность вида в Европе (1980-е гг. - начало 2000-х гг.)
составляет по одним оценкам 75-81, по другим — 85-88 тыс. пар. Существенное её возра-
стание зарегистрировано в 1970-1990-х гг., однако в 1990-2000-х гг., отмечено некоторое
снижение численности (Hagemaeijer, Blair, 1997; BirdLife International, 2004).
На Вост. Мурмане наиболее крупные колонии располагаются на Семиостровском и Гав-
риловском участках Кандалакшского заповедника. С начала 1970-х гг. численность на Семи
Островах росла и достигла максимума в 1977 г. (80 пар), затем, после спада и стабилизации
(20-30 пар), к 1998 г. снова возросла до 54 пар (Шкляревич, Татарникова, 1986; Ю.В. Крас-
нов, личн. сообщ.). Рост численности зарегистрирован и на Гавриловских о-вах: в 1978 г.
здесь учтено 20, а в 1995 г. — 102 пары (Татарникова, 20016; Рев и др., 2003). В конце 1990-
х гг. количество гнёзд на Вост. Мурмане стало постепенно сокращаться (Ю.В. Краснов,
личн. сообщ.). На Зап. Мурмане колонии появились только в конце 1970-х гг.: в 1978-1982
гг. 30-40 пар гнездились на южном побережье Варангер-фиорда, с 1979 г — на Айновах о-
вах, где к середине 1980-х гг. численность достигла 40 пар; в 1997 г. на о. М. Айнов гнезди-
лось не менее 300, в 2002 г. на о. Б. Айнов — 30-35 пар (Шкляревич, Татарникова, 1986;
Краснов и др., 1995; Татарникова, 20016; Иваненко, 2005). В настоящее время общая
численность вида на Мурмане оценивается в 400-500 пар и продолжает нарастать, при
этом наблюдается перераспределение мест гнездования (Татарникова, 20016; Мищенко
и др., 2004).
Наиболее крупная черноморская гнездовая группировка известна на обрывах Тархан-
кута (запад Крыма). В 1957 г. здесь учтено 1 750 взрослых птиц, в 1962 г. — около 1 200
пар, а в 1970-е гг. — не более 500 пар (Смогоржевський, 1979; Костин, 1983). На том же
уровне или ниже численность оценивается здесь и в последние годы — 200-500 пар (Чер-
128
Отряд Пеликанообразные, Семейство Баклановые
ничко и др., 1993; Directory..., 2003). В разрозненных поселениях Юж. Крыма (м. Херсо-
нес — м. Киик-Атлама), включающих от 2-3 до нескольких десятков гнёзд (Цвелих, 2002;
Костин, 1983; Бескаравайный, 2004), в сумме гнездится не менее 300 пар. Самое крупное
поселение находится в Карадагском заповеднике, где за последние 25 лет численность воз-
росла от 2-4 (1980, 1981 гг.) до 128 (2005 г.) пар. На юге Керченского п-ова (Опукский
заповедник) учтено 55 пар (Бескаравайный, 2004). Общая численность в Крыму в 1980-х
гг. оценена примерно в 1 875 птиц и 600 гнёзд, современная — приблизительно 900 пар;
годовалые птицы в период размножения составляют до 30-50% (Костин, 1983; Ардамац-
кая и др., 1988; Гринченко, 1991; данные автора очерка). В недавно обнаруженной колонии
у Таманского п-ова гнездится 20-25 пар бакланов (Белик, 2003а).
Согласно оценке Международной ассоциации охраны птиц BirdLife International, к
2000 г. наибольшее количество хохлатых бакланов гнездилось в Великобритании (28,9 тыс.
пар) и Норвегии (15,5-16,5 тыс. пар); во Франции гнездились 6,3-6,4 тыс. пар, в Исландии —
6,2-7,0 тыс.пар, в Испании — 4,39 тыс. пар, в Хорватии — 2,5-5,0 тыс. пар. В других
странах численность этих птиц была менее значительной (BirdLife International, 2004).
Зимой вид обычен у черноморских берегов Крыма. Среднезимняя плотность колеблет-
ся от 1,9 (1981/82 гг.) до 11,1 (2003/04 гг.) экз./км, среднемноголетняя составляет 5,2±0,6
экз./км (п=115), а максимальная при разовом учёте — 28,6 экз./км (10.12.2003 г.). В локаль-
ных скоплениях насчитывали до 230 птиц.
Размножение. В размножении участвует около 50% взрослых птиц (Гринченко, 1991).
Размножаться начинают в возрасте 2 (самцы) и 3 (самки) лет. Хохлатый баклан — моногам,
но при дефиците гнездовых стаций среди 2-3% птиц отмечена бигамия (Cramp, 1977).
В районах гнездования на Мурмане первые птицы появляются в середине марта (в
1939 г. — 15 марта), массовый прилёт — в конце этого месяца (Белопольский, 1957). В
конце апреля образуют пары. С первой по третью декаду мая продолжается постройка гнёзд
(в 1939 г. — с 5 мая) (Модестов, 1967). В Крыму гнездовой цикл начинается значительно
раньше. Птицы держатся на гнездовых участках с января, к спариванию и постройке гнёзд
приступают в конце февраля. Гнездо без кладки отмечено 3 марта 2004 г. Гнездостроение
продолжается в апреле, иногда в мае (самая поздняя дата — 21 мая 1978 г.: Ю.В. Костин,
архив). В Опукском заповеднике 30 апреля 1997 г. на стадии постройки было 25-30% гнёзд.
Гнёзда располагаются поодиночке или группами разной величины и плотности, что обус-
ловлено наличием и распределением гнездопригодных элементов нанорельефа. Расстоя-
ние между гнёздами составляет иногда менее полуметра.
На постройку гнезда уходит 2-3 дня. Гнездовой материал бакланы собирают на суше и
в море, по наблюдениям в Крыму — на уступах береговых обрывов, в 50-100 м от гнезда.
Иногда крадут ветки из соседних гнёзд. Время полета за материалом составляет 1-4 (в
среднем 1,7±0,3) мин, интервал между стартами — 1,5-4,5 (в среднем 2,6±0,4) мин (п=9).
Подлетая к гнезду, птица часто делает 1-2 круга, затем садится и передаёт строительный
объект партнёру. «Достройка» гнезда продолжается и при насиживании, но в это время
полёты за материалом осуществляются реже — 1 раз в 8-45 мин. Насиживающая птица, не
вставая с кладки, регулярно подправляет гнездовую постройку, подбирает и укладывает
находящийся в пределах досягаемости материал.
В составе материала гнезда присутствуют объекты как наземного, так и морского про-
исхождения. На Мурмане это ветки ивы, можжевельника, слоевища ламинарии и фукуса,
стебли вороники и сухая трава, крупные перья чаек, мусор (Модестов, 1967; Шкляревич,
Татарникова, 1986). В Крыму в большинстве гнёзд, расположенных на материковых обры-
вах, преобладает наземный материал. Внешние стенки делаются из плавника, веток, жёст-
ких стеблей травянистых многолетников. В лотке присутствуют травы (часто злак Anisantha
Хохлатый баклан
129
lectorum), морская трава Zostera marina, реже мхи (в т.ч. Homalotecium sp.). Как в стенках,
так и в выстилке имеются фрагменты водорослей (обычно Cystoseira sp.), иногда крупные
перья птиц. На удалённых от берега скалах-островках все гнёзда состоят из водоросли
Cystoseira sp. (около 90% по составу) и морской травы Zostera marina (около 10%).
На Мурмане размеры гнёзд следующие (см): диаметр 40-55, высота до 15, диаметр
лотка 18-25, глубина лотка 5-7 (Модестов, 1967; Шкляревич, Татарникова, 1986). Близкие
значения получены в Крыму (n=l 1): диаметр 33-49 (41,3±1,7), высота 10-18 (12,2±1,2),
диаметр лотка 19-30 (23,3±0,9), глубина лотка— 5-9 (6,7±0,4). В Крыму многие открыто
расположенные гнезда разрушаются уже к концу лета.
Основные элементы демонстрационного поведения в колонии показаны на рис. 23. Спа-
риваются птицы рядом с гнездом или на нем. Копуляции предшествует чистка перьев на
голове и шее. Затем самец некоторое время удерживает самку клювом за шею, сидя у неё на
спине; самка запрокидывает голову и открывает клюв. Весь процесс занимает 6-7 мин,
копуляция — примерно 20 сек. Интервал между актами копуляции — около 4 мин. В Кры-
му наблюдали случаи, когда самка занимала положение на спине самца, что известно и для
других частей ареала (Ortega-Ruano, Graves, 1991).
Начало откладки яиц на Мурмане колеблется между 6 мая (в 1935 г.) и 2 июня (в 1948 г.),
в среднем 14 мая (Белопольский, 1957). Регистрировались и более ранние сроки — вторая
половина апреля (Шкляревич, Татарникова, 1986). Разница в сроках начала яйцекладки в
разных даже близко расположенных колониях составляет до 1-1,5 мес. В Крыму откладка
яиц начинается в 1-й декаде марта, а по данным А.Б. Гринченко (1994) — в конце февраля, и
продолжается 2-2,5 мес. Период, когда насиживает большинство птиц, колеблется в разные
годы между 2-й половиной марта и 1-й половиной мая. В Карадагском заповеднике насижи-
вание примерно в 1/4 гнёзд происходило 21 мая 1995 г. Яйца откладываются с интервалом
1-5 дней, в случае гибели кладки, повторная появляется через 16-20 дней. В полных клад-
ках на Мурмане 1-4 яйца (Бианки и др., 1993), средняя величина варьирует по годам от 2,1
до 2,7 и зависит от состояния кормовой базы (Краснов и др., 1995). Величина кладок крым-
ских птиц — 1-5 яиц (Костин, 1983; данные автора очерка), в среднем 3,0±0,2 (п=17).
Яйца удлинённой формы, скорлупа бледно-голубая, покрыта известковым слоем, кото-
рый со временем становится грязно-буроватым. Размеры на севере ареала (n= 121) следую-
щие (мм): длина 56,7-69,9, диаметр 34,1-41,1, средние 62,1 х 38,1 (Бианки и др., 1993).
Вес 36,2-50,2 г (Модестов, 1967), средний 42,4±1,0 г (п=17), по другим данным (п=35) —
50,2±0,5 г (Шкляревич, Татарникова, 1986). В Крыму (п=59) размеры яиц 56,5-74,6 х 31,0—
41,3 (Смогоржевський, 1979, данные автора очерка), в среднем 62,1±0,4 х 36,9±0,3. Вес —
35,5-53,0 г, скорлупы — 4,8-7,5 г (Смогоржевський, 1979).
Насиживание начинается после откладки первого яйца и продолжается от 30 до 32 (или
33) дней на Мурмане (Модестов, 1967; Шкляревич, Татарникова, 1986) и 28 дней в Крыму.
В нем участвуют оба партнёра. Смена на гнезде происходит приблизительно через 2-2,5 ч.
С интервалом примерно в полтора часа насиживающая птица переворачивает яйца. К момен-
ту вылупления вес яиц уменьшается в среднем на 8,8 г, или на 18,5% (Белопольский, 1957).
Птицы агрессивно реагируют на приблизившихся к гнезду особей своего вида, при этом
издают низкий хрипящий звук; насиживающая птица иногда выскакивает из гнезда, чтобы
отогнать чужака. Кульминация конфликта — сцепление птиц на короткое время клювами.
Отмечалась угрожающие выпады на пролетающих и находящихся рядом чаек-хохотуний.
Вылупление птенцов на Мурмане происходит в середине июня, средняя дата — 15 июня
(Белопольский, 1957). В Крыму оно начинается в начале апреля и обычно заканчивается в
конце мая. По наблюдениям М.А. Воинственского (дневник), 10 июня 1957 г. в гнёздах на
Тарханкуте были птенцы «от совсём маленьких, до порядочных...». Птенцы вылупляются
130
Отряд Пеликанообразные, Семейство Баклановые
Рис. 23. Основные элементы поведения хохлатого баклана в колонии (по Cramp, 1977; Bauer, Glutz
von Blotzheim, 1987; Nelson et al., 2005): a — демонстрация зева на гнезде, б — поза перед взлётом,
в — поза во время взлёта, г — поза умиротворения после посадки в колонию; д, е — элементы
рекламного поведения самца, ж — ритуализованный ремонт гнезда самцом и самкой, з — привет-
ствие: вытягивание шеи самкой и щелчки клювом самца; и, к — поклоны при ухаживании и реклам-
ном поведении.
Хохлатый баклан
131
голыми и слепыми, с белым яйцевым бугорком. Глаза открывают примерно через 2-3 дня
Карадагский заповедник). На севере ареала на 9-й день глаза намечаются в виде узких
щелей, а открываются на 12-й (Судиловская, 1951). Вес вылупившегося птенца 30,5-31,5 г
Модестов, 1967). В первые три дня средний прирост в день по сравнению с весом взрос-
лого составляет всего 0,05% (Белопольский, 1957). Терморегуляция устанавливается по-
здно — между 13 и 25-м днем после вылупления (Рольник, 1948). С 8-дневного возраста,
при весе 75 г, начинается быстрый рост, достигающий максимума интенсивности с 21 по
30-й день (3,1% в день от веса взрослого); на 42-й день вес достигает 1760 г. Затем проис-
ходит замедление роста, а иногда даже снижение веса. Пух появляется на 8-9-й день, на
10-й — покрывает все тело, на 20-й появляются трубочки маховых и рулевых, на 50-52-й
день птенец одет в гнездовый наряд. В 60-дневном возрасте, при весе 1 700-1 800 г, птенцы
слетают (Модестов, 1967). В Крыму развитие перьевого покрова идёт примерно так же
I Бескаравайный, 2004). 12-дневные птенцы проявляют агрессию по отношению к человеку
и пытаются покинуть гнездо.
На Мурмане величина выводка к моменту подъема на крыло составляет 1,9 (п=54), в
разные годы 1,8-2,5 птенцов (Шкляревич, Татаринкова, 1986). Величина выводка в Крыму
(птенцы размером 1/3 взрослой птицы и более) — 1-3, в среднем 2,0±0,1 (п=29).
Выкармливают птенцов оба родителя, за кормом летают дважды в день (Смогоржевсь-
кий, 1979). Корм (полупереваренную рыбу) птенец получает, засовывая голову в ротовую
полость взрослого. За один акт кормления это происходит до 3 раз с интервалом в несколь-
ко минут. Кормлению предшествует выпрашивание, которое продолжается до 3 мин и вы-
ражается в горизонтальном покачивании клювом и прикосновениях к клюву родителя.
На Мурмане вылет первых птенцов происходит во второй половине августа - начале
сентября (Шкляревич, Татаринкова, 1986), самая ранняя дата — 4 августа 1935 г., средняя —
15 августа (Белопольский, 1957). В Крыму в некоторые годы вылет начинается уже в 3
декаде мая; наиболее поздние сроки — июль и август (Гринченко, 1994). После подъема
птенцов на крыло взрослые продолжают докармливать их до конца лета.
Суточная активность, поведение. Деятельны днем. В Крыму птицы кормятся уже около
500 утра (июнь). Подлет на ночевку начинается до наступления сумерек и продолжается
примерно 2 ч — с 19-1930 до 2100 (июль). Ночуют на выступах береговых обрывов, камнях
в море, а в малодоступных местах — на пологих участках берега, группами до 110 особей.
По сравнению с большим бакланом менее склонны объединяться в стаи; значительно чаще
птицы держатся поодиночке. Взрослые и неполовозрелые птицы много времени проводят
в «клубах» вблизи крупных гнездовых поселений (Ю.В. Краснов, личн. сообщ.; данные
автора очерка).
Исключительно морские птицы. Летают только над акваторией, не поднимаясь выше
верхней кромки берегового обрыва и облетая вдающиеся в море участки суши (Смогор-
жевський, 1979; данные автора очерка). Ныряют на глубину до 4 м, по другим данным —
до 33-35 м; задерживаются под водой до 3^1 мин (Судиловская, 1951; Wanless, Harris, Morris,
1991).
По наблюдениям на Мурмане, при появлении человека птицы из открытых гнёзд надол-
го их покидают и лишь отдельные пары обороняют гнездо. В некоторых колониях Норве-
гии бакланы терпимее к появлению человека, многие продолжают находиться в гнезде,
что, возможно, объясняется регулярным присутствием людей (Ю.В. Краснов, личн. сообщ.).
На юге Крыма подпускают на расстояние 10-20, иногда 6-7 м, спугнутые садятся на воду
рядом с гнёздами, возвращаясь вскоре после ухода человека.
Питание. Вид относится к специализированным ихтиофагам, кормовые объекты помимо
рыбы играют в питании второстепенную или случайную роль. Обеспеченность кормовыми
132
Отряд Пеликанообразные, Семейство Баклановые
ресурсами в весенне-летний период является основным фактором, определяющим успеш-
ность размножения и распределение колоний у побережья Мурмана (Краснов и др., 1995).
Основные корма на севере Европы — песчанковые — Ammodytes tobianus, A. marinus
(29-51 % по частоте встречаемости и до 54,1 % по объему), тресковые—Merlangus merlangus,
Gadus morhua, Pollacris virens, сельдевые — Clupea harengus, Sprattus sprattus, корюшко-
вые — Mallotus villosus (Cramp, 1977; Barrett et al., 1990; Wanless et al., 1993). В питании
птенцов и молодых большое значение имеют песчанки (Harris, Wanless, 1991, 1993).
У Мурманских берегов основу рациона составляют тресковые, сельдь — Clupea harengus,
песчанка — Ammodytes tobianus (до 83,3%), мойва — Mallotus villosus, бычки (Модестов,
1967; Краснов и др., 1995). Единично регистрировали крабов — Hyas araneus (Модестов,
1967). Ориентация хохлатого баклана на поимку пелагической добычи позволяет ему сни-
жать уровень пищевой конкуренции с большим бакланом (Краснов и др., 1995).
В Крыму важную роль в питании играют бычки (встречаемость 71,2%), песчанка —
Gymnammodytes cicerellus (15,1 %), смарида — Spicara smaris (13,6%), губановые (12%). Реже
(встречаемость 1,5-6%) поедаются хамса — Engraulis encrasicolus, тюлька — Clupeonella
cultriventris, ставрида — Trachurus mediterraneus, ласкирь — Diplodus annularis, губаны —
Ctenolabrus rupestris и Symphodus ocellatus, мерланг — Merlangius merlangus, уточка —
Lepadogaster decandollei, собачка — Blennius sp., трубкорот — Syngnathus sp. У Тарханкута
осенью в питании возрастает роль ставриды, а в мае отмечены бычки. В незначительном
количестве в пробах встречали ракообразных: креветок — Leander sp., крабов — Brachiura
sp., равноногих — Idotea sp. (Смогоржевський, 1959, 1979; Ю.В. Костин, архив). На южном
берегу Крыма поедают мелкую кефаль—Mugil sp., песчанку, атеринку —Atherina sp., ласки-
ря, зеленушку — Symphodus sp., барабулю — Mullus barbatus, ставриду, морских собачек —
Parablennius sanguinolentus (Костин, 1983; данные автора очерка). Таким образом, в Крыму
пищевой рацион включает не менее 16 видов рыб, большинство из которых (не менее 13) —
придонные и прибрежные (бычки, песчанка, смарида, губановые, барабуля).
Вес дневного рациона — 200-246 г (Cramp, 1977; Смогоржевський, 1979). В Крыму
суточное изъятие рыбы всей популяцией хохлатого баклана составляет приблизительно 720
кг (Бескаравайный, 2004).
Враги, неблагоприятные факторы. Из позвоночных в числе врагов известны 4 вида
птиц. Хохотунья (Larus cachinnans): отмечено похищение ею яиц, птенцов и поедание рыбы,
отрыгиваемой потревоженными птенцами. Филин (Bubo bubo\. головы бакланов найдены
в местах кормежки филина на Тарханкуте (Смогоржевський, 1979). Серая ворона: хищни-
чество в период откладки яиц отмечено для Мурманского побережья (Шкляревич, Тата-
рникова, 1986) и Крыма. Ворон: уничтожение им кладок регистрировали на Мурмане (Шкля-
ревич, Татарникова, 1986), такое же возможно и у крымских берегов. От чаек и воронов
птенцов защищает скрытное гнездование: при опасности они перемещаются в глубину ук-
рытий и у них полностью отсутствует реакция на испуг, выраженная в отрыгивании корма
(Краснов и др., 1995). Уничтожение яиц и птенцов неадаптированными хищниками (чайки,
врановые), как правило, спровоцировано фактором беспокойства и практически не оказы-
вает влияния на эффективность размножения (Ю.В. Краснов, личн. сообщ.; данные автора
очерка).
Видовой состав экто- и эндопаразитов изучен слабо. Сильное заражение гельминтами
(полость тела и желудок) обнаружено у птиц, добытых в Крыму (Ю.В. Костин, архив). На
Мурмане выявлена высокая заражённость круглым червем Ascaris spiculigera, в Шотлан-
дии— скребнем Andracantha tunitae. Известно 3 вида эктопаразитов птенцов: пухоед Мепо-
роп breviapalpe, клещ Ixodes putus (Мурман) и пухоед Pectinopygus brevicornis (Испания)
(Модестов, 1967; Munro et al., 1995; Martin-Mateo, 1990).
Хохлатый баклан, Малый баклан
133
В гнёздах обнаружены блохи: Ceratophyllus vagabunda insularis, С. columbae, С. garei, а
также — Cercyon littoralis и Gnathoncus nanus (Hicks, 1959).
Угнетения птенцов или их гибели из-за паразитарного заражения не установлено.
На фоне регулярного распугивания птиц, существенной причиной смертности может
стать температурный фактор, что обусловлено поздними сроками установления терморе-
гуляции у птенцов: в первые две недели жизни птенцы погибают в течение 8-10 мин при
10 °C (Модестов, 1967). Высокая смертность птиц и гибель гнёзд имеют место при продол-
жительных зимних и весенних ветрах (Harris, Wanless, 1996; Aebischer, 1993).
К основным антропогенным причинам снижения численности относится загрязнение
моря нефтепродуктами (Гринченко, 1994; Munro et al., 1995) и фактор беспокойства. Ис-
чезновение колоний на Мурмане во время Великой Отечественной войны произошло вслед-
ствие распугивания птиц при сборе яиц на птичьих базарах (Шкляревич, Татаринкова, 1986;
Краснов и др., 1995). В последние годы антропогенное беспокойство, наблюдаемое в этом
регионе, лимитирует уровень успешности размножения, но не приводит к распаду и остав-
лению колоний птицами (Ю.В. Краснов, личн. сообщ.). На Тарханкуте (Крым) с 1962 по
1978 г. по этой причине отмечено более чем двукратное падение численности: в первую
очередь пострадали колонии, расположенные вблизи населенных пунктов (Костин, 1983).
В Юж. Крыму большинство колоний находится в активно эксплуатируемой курортной зоне.
Иногда бакланы погибают, попадаясь в рыболовные сети.
Хозяйственное значение, охрана. Хозяйственного значения в России и на Украине
хохлатый баклан не имеет. Учитывая преобладание в рационе непромысловых видов рыб
(Крым) и малочисленность вида, его следует считать безвредным для рыболовства.
Хохлатый баклан занесён в Красные книги Российской Федерации и Украины. В России
все крупные колонии охраняются в Кандалакшском заповеднике. В Крыму гнездится в Кара-
дагском и Опукском заповедниках, где охраняется около 20% крымской популяции. Районы
гнездования на Тарханкуте входят в состав территории, зарезервированной для включения в
природно-заповедный фонд Крыма7. Однако современный уровень охраны не обеспечивает
надлежащей защиты вида как на Мурмане (Ю.В. Краснов, личн. сообщ.), так и в Крыму.
Малый баклан
Phalacrocorax pygmaeus (Pallas, 1773)
Pelecanuspygmaeus Pallas, 1773, Reise versch. Prov. Russ. Reichs, 2. c. 712, табл. G,
Капийское море.
Little [pygmy] cormorant (англ.); Zwergscharbe (нем.); cormoran pygmee (фр ).
Син. (лат.): Graculus pygmaeus algeriensis Reichenbach, 1850; Carbo niepcii Malherbe, 1855.
Статус. Гнездящийся мигрирующий, частично оседлый вид.
Общая характеристика и полевые признаки. Размером с утку-широконоску. При
общей схожести конституции и повадок,* малый баклан существенно мельче и проворнее
прочих видов бакланов. Превосходно плавает и ныряет. В отличие от большого баклана,
плавающая птица держит клюв параллельно поверхности воды, а не приподнимает его.
7 Пост. Презид. ВС Крыма № 538-1 от 12.04.94: «О развитии перспект. сети терр. и объектов прир.-
запов. фонда Крыма».
134
Отряд Пеликанообразные, Семейство Баклановые
Полет легче, чем у других бакланов. Форма самой птицы в полёте, как и у всех бакланов,
крестообразная, но шея выглядит заметно короче. Часто держится одиночно или парами,
сбивается в стаи лишь осенью; максимальная величина стай до 250 особей, обычно мень-
ше. Типы построения летящих стай такие же, как и у большого баклана, причём строй
«цепочкой» преобладает (Молодовский, 2001).
Отдыхая, малые бакланы сидят в вертикальной позе на ветке или тростнике, изогнув
шею S-образно. Летом у обоих полов тёмная красно-коричневая голова, блестящее зелено-
вато-чёрное оперение, с заметными белыми каплевидными пестринами на теле. Вне брач-
ного периода эти пестрины исчезают, зато обращает на себя внимание беловатый зоб и
красновато-бурая грудь. Молодые птицы отличаются белым подбородком, коричневыми
зобом и грудью, буровато-белым низом и желтоватым клювом.
В период гнездования малые бакланы крикливы — голос напоминает «каркание» боль-
шого баклана, но глуше. В остальные периоды птицы молчаливы.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. При дроблении рода Phalacrocorax малого баклана отно-
сят к роду Halietor Heine, 1860 (Siegel-Causey, 1988; del Hoyo et al., 1992), который может
рассматриваться в качестве подрода при широкой трактовке Phalacrocorax. Иногда к этому
же виду в широком понимании относят яванского баклана {Phalacrocorax niger Vieillot,
1817), широко распространенного в тропической Азии (del Hoyo et al., 1992).
Описание. Окраска. Самцы и самки окрашены одинаково. Взрослые птицы в брач-
ном наряде. Большая часть головы и шеи тёмно-шоколадно-бурые (хотя попадаются особи
с черной окраской этих частей тела), на границе клюва оперение темнее, почти чёрное;
вокруг глаза частые мелкие белые пёрышки, а в основании лба короткие, жёсткие, щетини-
стые. Уздечка голая. Все остальное оперение чёрное с тусклым зеленоватым отливом, кро-
ме плечевых и верхних кроющих крыла, которые серо-аспидные с чёрными краями. В гнез-
довой период на спине и брюхе располагаются мелкие каплевидные белые перья («украша-
ющие» перышки в виде мелких белых кисточек на волосовидном стерженьке). Клюв тём-
но-роговой, ноги чёрные, радужина бурая.
Взрослые птицы в зимнем наряде. Белые каплевидные («украшающие») перья на теле
исчезают, голова и верхняя половина шеи тёмно-коричневые, кольцо вокруг клюва почти
чёрное. Плечевые, лопаточные и верхние кроющие крыла чёрные с сероватым оттенком и
с чёрными каймами на отдельных перьях, в результате чего образуется чешуйчатый рису-
нок.
Пуховой наряд. Птенец покрыт однотонным тёмно-бурым пухом.
Гнездовой наряд. Верх головы и шеи бурый, подбородок беловатый, нижняя сторона
шеи светло-бурая. Спина и бока тела тёмно-бурые, перья верхней части спины со светло-
бурыми окаймлениями. Плечевые и верхние кроющие крыла с сероватым налётом и с тём-
ными каймами. Брюшная сторона буровато-белая, в особенности посередине брюха, с ры-
жевато-бурыми размытыми пятнами на груди и зобе. На задней части брюха пятна чернова-
то-бурые, переходящие в тёмно-бурое подхвостье. Уже в этом наряде в январе появляются
на боках зоба несколько (3-5) белых каплевидных пёрышек.
Первый брачный наряд. Верх головы и шеи буровато-коричневый, нижняя сторона шеи
светлее, подбородок светло-бурый. Спина и надхвостье чёрные с зеленоватым блеском.
Кроющие крыла, плечевые и второстепенные маховые с сероватым налетом, с тёмными
предвершинными и бурыми вершинными каймами. Низ тела тёмный, коричнево-бурый с
крупными нерезкими светлыми пятнами, которые образуются каймами перьев. Бока тела и
подхвостье черновато-бурые. Весенних каплевидных белых пятен на боках зоба, шеи и на
бедре больше, чем в гнездовом наряде.
/2-7ЫЙ баклан 135
Второй брачный наряд. Верх головы, шеи и вся верхняя сторона тела окрашены как у
е зрослой птицы, нижняя сторона светлее. Подбородок светло-бурый, низ тела однообразно
' .ровато-чёрный без блеска. Весенние белые каплевидные пятна (появляются начиная с
января) хотя ещё и редки, но разбросаны уже по всему телу.
Второй зимний наряд. Почти не отличается от окраски оперения взрослой птицы. Ниж-
цяя сторона с металлическим блеском, но более слабым, чем у взрослой. Кроме того, на
нижней части брюха ещё имеются отдельные светлые перья. Этот наряд носится, по-види-
чому, до января (Судиловская, 1951).
Строение и размеры. Первостепенных маховых 11, с вырезкой на внутреннем
эпахале. Формула крыла II>III>IV>I>V... Хвост округлый, из 12 жёстких рулевых. Длина
крыла 185-216 мм; длина клюва (от оперения лба) 28-32 мм; длина цевки 36-38 мм. Вес
зимних экземпляров от 490 до 683 г (Иванов, 1969). А.М. Судиловская (1951) приводит
цифру 800 г, без указания сезона взвешивания. По С. Крэмпу, размеры малых бакланов
следующие (мм): длина крыла самцов (n=l 1) 195-217, в среднем 206, самок (п=7) 193-208,
в среднем 201; длина хвоста самцов (п=6) 137-145, в среднем 142, самок (п=5) 139-147, в
среднем 141; длина клюва самцов (п=6) 29-33, в среднем 30,5, самок (п=5) 27-31, в сред-
нем 29,2; длина цевки самцов (п=6) 37^40, в среднем 38,0, самок (п=5) 36-39, в среднем
37,8. Вес трёх самцов 650, 710, 870 г, трёх самок 565, 640, 640 (Cramp, 1977).
Линька. Последовательность нарядов соответствует таковой у большого баклана. У
взрослых птиц две линьки в году: частичная предбрачная в декабре-январе, во время кото-
рой птица одевает брачный наряд и в нем находится до июня, и полная послебрачная с
июня по октябрь-ноябрь. При этом происходит смена рулевых, маховых (заканчивается в
августе-сентябре) и мелкого оперения (заканчивается в октябре-ноябре).
Распространение. Гнезовой ареал. Северо-восточное побережье Алжира, побе-
режья и речные бассейны балканских стран, Румынии. Гнездится в дельтах Дуная, Днест-
ра, Днепра, в последние годы и на черноморских островах (Гринченко, 20046). Бытующее
представление о гнездовании малого баклана по всему Крыму — ошибочно (Костин, 1983).
Но вид гнездится на Сиваше (Гринченко, 2004а), а с 1980 г. в небольшом количестве стал
также гнездиться на Лебяжьих о-вах (Костин, Тарина 2004). Далее на восток малый баклан
заселяет побережья Азовского моря, дельты Волги (Гаврилов и др., 2003) и Терека (Бонда-
рев, 1975а), юго-западное побережье Каспийского моря (дельта Куры и Кызыл-Агач), а
также озёра Кура-Аракской и Самар-Дивичской низменностей Азербайджана (Туаев, 1962;
Туаев, Васильев, 1969, Patrikeev, 2004). Ранее обитал на Аральском море в устье Аму-Да-
рьи (Рашкевич, 1969); гнездится в долине Сыр-Дарьи вверх до Чиили и связанных с ней
озёрных систем, предположительно до долины Теджена и туркменского Мургаба. Малая
Азия, озёрные и речные системы Закавказья к югу до Ирака и сев.-зап. Ирана (Степанян,
2003) (рис. 24).
Зимовки. Область зимовок ряда популяций малого баклана лежит в Средиземном
море у Сев.-Зап. Африки, в Адриатическом и Эгейском морях, у побережья Греции, в Пале-
стине, Иране и т.д. (Судиловская, 1951). В целом малые бакланы, гнездящиеся в России и
на прилежащих территориях, в годы с нормальными погодными условиями могут обитать
в пределах этих стран в течение всего годового цикла, и лишь частично вылетать в более
дальние регионы (Нанкинов, Сапетин, 1978).
Многотысячные стаи остаются зимовать в низовьях Дуная и сотенные стаи — в Крыму,
на взморье (Гринченко, 20046). Зимние возвраты получены с оз. Карасу в Ширванской сте-
пи (12 декабря), с Малого Кызыл-Агачского зал. (13 января) и из юго-вост. Ирана, близ г.
Заболь (29 декабря). Три зимних возврата от кызыл-агачских взрослых птиц получены из
Юж. Азербайджана и Саудовской Аравии (Нанкинов, Сапетин, 1978).
136
Отряд Пеликанообразные, Семейство Баклановые
Рис. 24. Область распространения малого баклана: а — места гнездовий, б — районы зимовок.
По данным И.А. Долгушина (1960), в низовьях Сыр-Дарьи малый баклан не зимует, но
уже в районе Джулека попадается на глаза наблюдателям в заметном числе, особенно в
теплые зимы. Южнее зимует регулярно, и близ Ташкента в некоторые зимы бывает даже
многочислен. Однако Р.Н. Мекленбурцев с соавторами (1987), говоря о птицах Узбекиста-
на, сообщает, что малый баклан там зимует лишь изредка. Тем не менее, он же приводит
(ссылаясь на З.Л. Сатаева) сроки зимовки малого баклана в Узбекистане: с ноября по фев-
раль. Существенная концентрация малого баклана в период зимовок отмечается в системе
Айдар-Арнасайских водоёмов Узбекистана (Лановенко и др., 2001). В Туркмении малый
баклан зимует в заметном числе в районе Келифских озёр (Ташлиев, 1973), а также на
озёрах Делили по Атреку (Судиловская, 1951). Многочислен он также на зимовках в пой-
мах и на озёрах рек Вахш и Пяндж (Иванов, 1969).
Миграции. Откочёвка малых бакланов, гнездящихся на юге Украины, происходит уже
в августе. На пролёте птицы держатся стаями, набирая высоту в 250 м (Гринченко, 20046).
Три баклана, выведшихся в дельте Дуная, добыты: 19 ноября в Черногории и 22 декабря
(две птицы) — в Македонии, в 300-800 км от места кольцевания (Нанкинов, Сапетин, 1978).
В регионе Крыма в сентябре появляются птицы со стороны Кубани, которые затем концен-
трируются в массивах плавней до начала замерзания речных проток и каналов (обычно в
ноябре), а потом перекочевывают на взморье. К концу февраля крымские зимовки малого
баклана начинают распадаться. Часть птиц улетает на северо-запад в устье Днепра, и к
концу марта в Крыму остаются уже только местные особи (Гринченко, 20046).
Птенцы из колоний в дельте Терека после вылета из гнёзд довольно долго держатся
неподалеку от места появления на свет и только в октябре отлетают к относительно недале-
ким зимовкам. По данным кольцевания они летят сперва на юг вдоль западного берега
Каспия, и только поравнявшись с долиной бассейна Куры, устремляются на юго-запад, к
внутренним водоёмам Азербайджана, таким, как оз. Акгёль и озёра в Мильской степи. От-
дельные птицы могут до середины декабря задерживаться в Предкавказье (Сапетин, 1968).
Малый баклан
137
Малых бакланов, окольцованных в Кызыл-Агачском заповеднике (юго-запад Каспия), в
июле-ноябре находили, как правило, не далее 50 км от мест появления на свет. Но есть и
более дальние мигранты: одна молодая птица всего лишь через пол месяца после кольцева-
ния 8 июня оказалась в Иране (Мазандеран), в 450 км от места появления на свет (Нанки-
нов, Сапетин, 1978). Во время послегнездовых кочёвок, когда тысячи молодых особей
концентрируются в болотах Кура-Аракской низменности, небольшая часть птиц попадает
и в горную местность — например, на оз. Ноур хребта Куткашен (Patrikeev, 2004).
В Казахстане валовый весенний пролёт малых бакланов проходит в середине и конце
марта, в то время как осенний идёт малозаметно (Долгушин, 1960).
Залётных малых бакланов наблюдали во многих местах. 5 февраля 1995 г. вид встречен
на Бурштынском вдхр. Ивано-Франковской обл. Украины (Бучко,1998). Позднелетние (ав-
густ) залеты в Ставропольский край, порой стаями в 50 особей, отмечали в середине 1980-
х гг. на озёрах Дадынском, Бирючья Сага, Соленое (Хохлов, 1990). Известны залеты в стра-
ны Европы — например, в апреле 1926 г. в Вост. Словакию (Грабар, 1997), в ноябре 1864 г.
на р. Ольшу в Польше (Tomialojc, 1990) и т.п.
Местообитания. Гнездятся малые бакланы в болотистой местности, в дельтах рек, пре-
дустьевых лиманах, на пресноводных озёрах, в морских заливах. В горных странах, таких
как Армения и Азербайджан, заселяют низменные долины. Местами кормёжек служат пре-
имущественно большие озёра с глубинами до 3 м, с прозрачной водой и с древесно-кустар-
никовой растительностью по берегам, морские лиманы, рыбоводные пруды. Временно ме-
стами кормежек могут служить и оставшиеся после паводков мелководные, пересыхаю-
щие пойменные озерца — до того, как там истощатся рыбные запасы (Иванов, 1969).
Численность. Численность малого баклана, как и большого, претерпела в последних
десятилетиях XX в. существенные изменения (Белик, Данкевич, 2004; Русев, 2001, 2004 и
др.). В украинской части дельты Дуная в 1980-х гг. гнездилось не более 150-200 пар малых
бакланов, но в 1990-х гг. начался интенсивный рост поголовья, и в 2002 г. здесь в трёх
колониях насчитывалось уже 1 030 пар. Во всей дельте Дуная (включая румынскую сторо-
ну) в том же году было учтено почти 20 тысяч взрослых птиц (9 341 пара) в 14 колониях
(Киш и др., 2003).
В 2001 г. в дельте Днепра была обнаружена пока единственная для региона колония на
Кардашинских болотах (по дороге Голая Пристань - Цюрупинск), где вместе с цаплями и
ибисовыми гнездилось 350 пар малого баклана (Петрович, Щеголева, 2003; Гринченко,
20046). В это же время на Днестре гнездилось 700 пар, в Крыму — 35^10 пар (Гринченко,
20046). Касательно Днестра эти данные подтверждает также И.Т. Русев (2004), который
приводит такую динамику днестровской популяции: до конца 1980-х гг. малых бакланов
здесь было не более 20-30 пар, в 2000 г. их стало уже 600, в 2003 г. — 960 и в 2004 г.
численность остановилась на отметке 900 пар. Т.Б. Ардамацкая с соавторами (1988) уточ-
няет, что в 1984 г. в целом по всей дельте Днестра гнездилось свыше 100 пар малых
бакланов.
Общая численность гнездящихся малых бакланов на юге Украины, с учётом погреш-
ности подсчета, составляет сегодня примерно 11 000 пар (Гринченко, 20046). Эта цифра
намного выше указанной для этого вида в коллективной монографии «Численность и
размещение гнездящихся птиц в водно-болотных угодьях Азово-Черноморского побере-
жья Украины» (2000). Однако в названной книге даны сведения, собранные до макси-
мального всплеска численности бакланов; в ней отсутствуют также данные по дельте
Днепра. Тем не менее, эти сведения ценны, поскольку позволяют прослеживать динами-
ку изменения численности по отдельным водно-болотным угодьям региона. По данным
указанной монографии, в 1998 г. малые бакланы ещё не гнездились в таких местах, как
138
Отряд Пеликанообразные, Семейство Баклановые
Таблица 7. Распределение и численность малого баклана на Азово-Черноморском
побережье Украины в 1998 г. (количество пар)
Дельта Дуная Дельта Днестра Тилигульский Восточный В сумме
____________________________________лиман___________Сиваш___________________
730 315 1 1 1047
оз. Сасык, на системе озёр Шаганы-Алибей-Бурнас, на Куяльницком лимане, в заливах
Ягорлыцком, Тендровском, Джарылгачском, Каркинитском, на Зап. и Центр. Сиваше, Мо-
лочном лимане, заливе и косе Обиточной, Бердянской косе, Бесарайской косе, Кривой
косе. Тогда малые бакланы были отмечены на гнездовье только в четырёх пунктах реги-
она, что отражено в таблице 7.
Отметим, что ещё в начале 1990-х гг. вид в Приазовье считался залётным (Лысенко,
1992).
С 1998 г. малые бакланы стали появляться в низовьях Дона, и к 2004 г. их здесь гнезди-
лось 15-20 пар (на оз. Монастырском). В Краснодарском крае, где ещё в конце XX в. малые
бакланы не гнездились, теперь насчитывается 1 000 пар (Белик, Данкевич, 2004).
В дельте Волги, где раньше малые бакланы встречались случайно и единично, начиная
с 1957 г. они уже появлялись постоянно стайками до 10 птиц, причём в пределах Астрахан-
ского заповедника лишь на самом маловодном, заболоченном участке — Трёхизбинском
(Луговой, 1963). Затем их численность в дельте стала стремительно расти. В колонии «Кре-
стовая Черепашка» в Камызякском р-не Астраханской обл., где в 1995 г. насчитывалось 200
пар малых бакланов, в 2001 г. их стало уже 1 500 пар (Гаврилов и др., 2003). Всего в 4
колониях култучной зоны дельты Волги этими авторами выявлено в 1999 г. 670 гнездовых
пар; в 2001 г. — 1 650.
Колониальные поселения малого баклана известны также в дельте Терека и прилегаю-
щих водоёмах, где в 1973-1974 гг. в трёх колониях (Каракольская, Ачикольская, Дагестан-
ская) гнездилось 500, 150 и 40 пар малых бакланов. В Дагестане, в Кизлярском и Аграхан-
ском заливах Каспия, малый баклан (до 2 000 особей) скапливается преимущественно в
пролётный период (Джамирзоев, 1999а), а также частично зимует (Вилков, 1999), но на
ключевой орнитологической территории (КОТР) «Водохранилище Мехтеб» гнездится в
количестве 15-20 пар (Джамирзоев, 19996).
В Кызыл-Агачском заповеднике (Азербайджан) численность малого баклана весьма измен-
чива в зависимости от колебаний уровня моря и других факторов (Коновалова, 1979). Если в
1957 г. здесь отмечали 10 тыс. гнездящихся особей, которые составляли в колониях 15% всех
гнездящихся там птиц, то впоследствии (1972 г.) эти цифры уменьшались до 1,4 тыс. (6%),
а к 1974-1977 гг. численность стабилизировалась на уровне около 3 тыс. гнездящихся птиц.
Д.Г. Туаев и В.И. Васильев (1969) для заповедника называли цифру 5 тыс. особей. Подчер-
кнем, что в обоих случаях речь идёт не о гнездящихся парах, а об абсолютном числе учтён-
ных в гнездовое время птиц. Цифры, приводимые в двух упомянутых работах, значительно
ниже данных, представленных В.С. Грековым (1965). Последний писал об огромных 4-5-
миллионных смешанных колониях, где малый баклан составлял в 1958 г. 6% населения (т.е.
примерно 250-320 тыс. особей). Причиной таких расхождений может быть как деградация
тамарисковых зарослей — мест колониальных поселений — в результате залповых сбро-
сов токсичных вод на этом участке (Литвинова, 1989а), так и неточность в подсчётах. Ма-
лые бакланы гнездятся также в тростниковых зарослях озёр Б. и М. Акгёль (Виноградов,
Чернявская, 1969). Уточнённые сведения о численности вида в Азербайджане находим у
М. Патрикеева (Patrikeev, 2004). Согласно его данным, в конце 1980-х - начале 1990-х гг.
здесь гнездилось порядка 4 800-5 800 пар малых бакланов.
Малый баклан
139
Таким образом, в настоящее время на пространстве от западного побережья Чёрного
моря до Каспия включительно гнездится не менее 25-26 тыс. пар малых бакланов, а с учё-
том пограничной с Украиной румынской части дельты Дуная — 36 тыс. пар.
Восточнее Каспия данные по численности малого баклана скудны и противоречивы.
Прежние сведения по Аральскому морю теперь вряд ли соответствуют новым реалиям,
поскольку в результате антропогенной катастрофы это озеро-море вместе с впадающими в
него реками значительно обсохло. В 1960-х гг. в колонии на оз. Шомкекуль (дельта Аму-
Дарьи) гнездилось 200 пар малого баклана (Рашкевич, 1969). Уменьшение численности
этих птиц прослеживалось на другом водоёме данного региона — на оз. Карахожабах, где
в 1961 г. насчитывали 71, в 1962 г. — 70, а в 1964 г. — уже только 14 гнёзд малого баклана
। Мекленбурцев и др., 1987). Данные авторы подчеркивали, что в те годы численность ма-
лого баклана в Узбекистане была незначительной. В Казахстане вид ранее достоверно гнез-
дился только по восточному побережью Арала (Долгушин, 1960); в настоящее время эти
гнездовья вряд ли сохранились.
По внегнездовой численности есть следующие данные. Во второй декаде сентября 1984—
1985 гг. в пойме р. Прут в районе Кагульского рыбокомбината Молдавии было зарегистри-
ровано 2 500 малых бакланов (Куниченко, 1986). Зимой 1999/2000 гг. в украинской части
дельты Дуная держалось 1250 малых бакланов, а на участке Дуная в пределах бывшей
Югославии — до 5 тысяч. В Крыму, в зависимости от погоды, зимует 400-700 малых бак-
ланов (Гринченко, 20046).
Вдоль черноморского побережья Кавказа в 1950-е гг. зимовки малого баклана были
малочисленными, здесь наблюдали лишь пару десятков особей (Строков, 1974). В настоя-
щее время численность малых бакланов на зимовках в Черноморском бассейне увеличива-
ется. Так, в Крыму, близ Феодосии и Коктебеля, где ранее малые бакланы вообще не зимо-
вали, начиная с конца 1990-х гг. в декабре и январе стали появляться стайки этого вида в
20-70 особей. Здесь их регистрируют главным образом на заросших гидрофильной расти-
тельностью пресных водоёмах; встречаются они и на солёном озере Ачи близ Владисла-
вовки (Бескаравайный, 2001).
На туркменских зимовках (Келифские озёра) малый баклан в 1961 г. количественно
доминировал (40,8% от всех учтённых «водолюбивых» птиц) и встречался с частотой 64,1
особей за 1 ч наблюдений (Ташлиев, 1973).
На юге Памиро-Алая, на озёрах и в пойме рек Вахш и Пяндж вид был многочислен во
время зимовок, но его численность испытывала сильные колебания по годам. В годы с
высокими летними паводками бакланы были более многочисленны и прилетали к местам
зимовок раньше. По подсчётам Р.Л. Потапова (цит. по: Иванов, 1969), общая численность
малых бакланов на зимовке в низовьях р. Вахш в декабре 1958 г. была равной 6 000 особей;
основную массу зимующих здесь птиц составляли первогодки.
Размножение. Моногамы. Поселяются колониально, выбирая для гнездования тростнико-
вые крепи, кустарниковые (например, тамарисковые или тугайные) заросли, реже высокостволь-
ные леса, порой отстоящие от водоёма на 3-5 км (Спангенберг, Фейгин, 1936). В тамариско-
вых зарослях малые бакланы поселяются обычно совместно с другими колониальными
видами птиц (большими бакланами, каравайками, кваквами и т.д.). Гнездятся также назем-
но на морских побережьях и островах (Греков, 1965; Гринченко, 20046; Русев, 2004 и др.).
На юго-западном Каспии (Кызыл-Агачский заповедник) малые бакланы прилетают в
колонии в феврале-марте, на озёра в глубине Кура-Аракской низменности — только в кон-
це марта (Patrikeev, 2004). В условиях юга Украины малые бакланы прилетают в колонию в
конце марта, часто парами. Они не образуют моновидовых поселений, а гнездятся вместе с
другими птицами — голенастыми и ибисовыми, тяготея к группировкам мелких цапель.
140
Отряд Пеликанообразные, Семейство Баклановые
Предпочитают селиться на краю колонии. В многоярусных колониях малые бакланы зани-
мают верхний и средний уровни. Согласно данным 3.0. Петровича и И.В. Щеголева (2003),
в дельте Днепра гнездятся на кустах вербы на высоте 1,5-5 м, где верхний ярус делят вме-
сте с серыми цаплями. По А.Б. Гринченко (20046), расстояние между гнёздами в колонии
от 1-2 до 20-30 м, высота над водой — 0,8 - 5-7 м. Этот же автор указывает, что гнездо
представляет собой довольно рыхлую постройку. Для Казахстана, наоборот, подчеркивает-
ся прочность полушаровидного гнезда малого баклана (Долгушин, 1960). Здесь птицы строят
гнездо из тростника или мелких веток, лоток иногда выстлан ветошью. Если размеры рых-
лых гнёзд на Украине имеют показатели (п=2): 390-340 х 210-200, при глубине лотка 50
мм (Гринченко, 20046), то в Казахстане диаметр полушаровидных гнёзд редко достигает
25 см, при этом лоток сильно углублён (Долгушин, 1960). Размеры гнёзд на тамариске:
диаметр 220-280 мм; глубина лотка 70-129 мм (Patrikeev, 2004).
На Украине бакланы приступают к гнездо строению в начале апреля, а во второй декаде
этого месяца — к массовой откладке яиц. В первой декаде мая кладки в основном заверше-
ны, в гнёздах бывает по 5-6 яиц. В Азербайджане кладка состоит из 2-7, чаще 4-6 яиц (в
среднем 4,5 яйца); откладка яиц на оз. Акгёль приходится на середину - конец апреля, а в
бассейне р. Варвары — даже на начало мая. (Греков, 1965; Patrikeev, 2004). В случае утери
кладки, если это произошло в начале гнездового сезона, птицы приступают к повторному
гнездованию. Размеры яиц на Украине (n=l 1): 44,3^4-8,6 х 29,4-31,3 мм, в среднем 46,23х
30,56 мм (Гринченко, 20046). Для других регионов приводятся такие размеры: (п=40) 42,4-
53,5 х 28,7-31,3 мм (нижнее течение Сыр-Дарьи; Спангенберг, Фейгин, 1936); (п=16) 45,0-
47,3 х 28,0-34,0 мм (дельта Сыр-Дарьи; Зарудный, 1916, цит. по: Судиловская, 1951); 47,Ох
29,9 мм (Азербайджан; Patrikeev, 2004). Окраска яиц в украинских колониях бледно-голу-
бая, матовая, у казахстанских малых бакланов отмечена зеленоватая окраска яиц (Долгу-
шин, 1960). Известковый поверхностный слой яиц тоньше, чем у большого баклана.
Насиживают кладку оба родителя. Инкубация длится 23-28 дней (Patrikeev, 2004). На
Украине выклев птенцов происходит во второй половине мая, в дельте Сыр-Дарьи — в
середине июня. В целом, начало размножения малого баклана происходит заметно позже,
чем у большого баклана, но птенцы затем растут быстрее. По данным М.В. Патрикеева
(Patrikeev, 2004), птенцы остаются в гнезде 40-43 дня. Выкармливают птенцов оба родите-
ля — вначале полупереваренной пищей, которую отрыгивают в глотку птенца, позднее
мелкой рыбой, а в конце гнездового периода — более крупной, достигающей размеров 10-
12 см (Судиловская, 1951).
На Украине первых слётков наблюдали в 1-2 декадах июня, но птенцы из поздних и
повторных кладок становятся лётными лишь в июле. Подъем молодых на крыло в Азербай-
джане происходит с конца июня (в засушливые годы) до начала августа (Греков, 1965).
Поначалу молодые птицы держатся группами близ мест гнездования; в августе выводки
распадаются, и птицы кочуют в районе гнездования (Гринченко, 20046; Patrikeev, 2004).
Гибель птенцов в гнёздах бывает довольно высокой. Так, в колонии на оз. Шомке-Куль
(юг Арала), расположенной в заломах тростника, из 4,2 вылупившихся птенцов доживали до
вылета в среднем 3,1 (Рашкевич, 1969). В колониях Азербайджана потери яиц в период наси-
живания относительно невелики (6%), но гибель птенцов в гнёздах достигает более 60%.
Одна из главных причин высокой смертности птенцов — хищничество квакв, которые при
совместном поселении способны разорить более половины гнёзд малых бакланов. В коло-
ниях, где кваквы отсутствуют, потери бакланов значительно меньше (Patrikeev, 2004).
Максимальная известная продолжительность жизни в природе, по данным кольцева-
ния — 6 лет 7 месяцев. Средняя продолжительность жизни — 1,2 года, для птиц, пережив-
ших первый год, — 2,7 года (Сапетин, 1968).
Малый баклан
141
Суточная активность, поведение. Малые бакланы — птицы довольно скрытные. В
смешанных колониях они очень осторожны, при появлении людей улетают раньше цапель
и прочих обитателей этого сообщества; возвращаются же назад позже других. Летом и
осенью часто кормятся в скоплениях лысух. Полёты на кормежку от мест гнездования и
обратно совершают в одиночку, реже парами; летят на высоте 5^40 м. В летнее время ма-
лые бакланы активны всё светлое время суток, кроме жаркого периода с 12 до 15 ч. На
местах летних кормежек держатся рассредоточено, в 20-100 м друг от друга; зимой могут
группироваться по нескольку десятков особей (Гринченко, 20046). На большую осторож-
ность малого баклана по сравнению с большим указывает факт различной реакции на от-
пугивание этих двух видов. Малые бакланы полностью покидают пруды, на которых их
преследуют, в то время как от больших бакланов избавиться труднее (Куниченко, 1986).
У зимующих малых бакланов существует четкий суточный ритм жизни. Так, в долине р.
Вахш основная масса птиц ночует в густых зарослях тростника на больших озёрах, причём
места ночёвки постоянны. Перед восходом солнца бакланы вылетают на места кормёжек
стаями по 15-20 особей, летят на большой высоте (200-300 м) и очень дружно, так что через
15-20 мин перелёт, в основном, завершается. Обратный перелет не столь дружен, птицы
возвращаются вразброд, небольшими группами. На ночевку птицы устраиваются после захо-
да солнца, но время зависит и от погоды: в пасмурные дни это происходит уже начиная с 16 ч.
Тишина на зимних ночевках воцаряется лишь с наступлением полной темноты. При замерза-
нии водоёмов бакланы бывают вынуждены кормиться на полыньях глубоких озёр и усажи-
ваться для отдыха прямо на льду (Иванов, 1969). По иному описывает суточную актив-
ность зимующих в Туркмении малых бакланов А.О. Ташлиев (1973). Согласно этому авто-
ру, птицы в районе Келифских озёр (оз. Часкак) вечером снимаются с мест днёвок и улета-
ют на мелководья, богатые рыбой, откуда в 8 ч утра возвращаются на места днёвки. Утрен-
ний, как и вечерний, перёлеты продолжаются 30^40 мин. В стаях насчитывают от 100 до
300 птиц. Где конкретно располагаются места днёвок — не уточняется. Не исключено, что
автор ошибочно принял время дневной кормежки за период отдыха птиц (днёвки).
Питание. Малые бакланы преимущественно рыбояды, но в их рационе встречаются и
другие группы кормов. Кормятся на пресноводных озёрах, лиманах с чистой водой, охотно
посещают протоки в массивах плавней, а также каналы и рисовые чеки. В отличие от боль-
ших бакланов не используют загонного способа добычи корма. В питании малого баклана
в Черноморском бассейне преобладает сорная рыба (Гринченко, 20046). В Крыму основу
питания составляют караси (Carassius sp.) длиною 4-7 см (70% встреч). Кроме них в от-
рыжках птенцов отмечены (перечисляются по степени убывания встречаемости): трёхиг-
лые колюшки (Gasterosteus aculeatus), остатки водных насекомых, карпы (Cyprinus carpius),
краснопёрки (Scardinius erythrophthalmus).
По данным М. Патрикеева (Patrikeev, 2004), в Азербайджане малые бакланы кормятся
на затапливаемых паводками территориях, болотах и рисовых полях. В Кызыл-Агаче осно-
ву питания малых бакланов составляют карп (49%) и плотва (12,1%). Кроме того, отмече-
ны молодь щуки, взрослые гамбузии {Gambusia affinis), а также личинки насекомых.
На зимовках охотятся за рыбой обычно два раза в сутки — утром и вечером; в проме-
жутке отдыхают на прибрежных кустах и заломах тростника. Суточная норма съедаемой
на зимовках рыбы составляет 250 г (Иванов, 1969). В Туркмении в рационе малых бакла-
нов обнаружены аральская плотва (Rutilus rutilus), елец (Leuciscus latus), амурский чебачок
(Pseudorasbora parva), аральский усач {Barbus brachycephalus), маринка (Schizothorax sp.),
белый амур (Ctenopharyngodon idella), храмуля (Varicorhinus sp.), сом, а также стрекозы
। Odonata), прямокрылые (Orthoptera), жуки-водолюбы (Hydrophilidae), а из бабочек — со-
вки (Noctuidae). В пище найдены также другие насекомые (Ташлиев, 1973).
142
Отряд Пеликанообразные, Семейство Змеешейковые
Враги, неблагоприятные факторы. У малых бакланов Туркмении, согласно А.О. Таш-
лиеву (1973), выявлены следующие паразиты: из гельминтов — Clinostomum complanatum,
Paryphostomum testitrifolium, Ligula intestinalis, Schistocephalus solidus, Capillaria carbonis,
Desmidocercella skrjabini, Contracaecum microcephalum, C. micropapillatum; из перьевых
клещей — Scutomegninia phalacrocoracis, Alloptes suberassipes.
На юге Арала выпавшие из гнёзд птенцы малых бакланов становятся жертвой полу-
взрослых птенцов серебристых чаек, плавающих по плесам колонии (Рашкевич, 1969).
Малых бакланов довольно интенсивно отстреливают охотники-любители. По данным
Я.В. Сапетина (1968), в 1960-х гг. возврат колец от птиц старших возрастов, помеченных
птенцами в дельте Терека, достигал 40%. Среди добытых птиц 72% составляли особи пер-
вого года жизни; 12% — в возрасте от 1 до 2 лет; 8% — от 2 до 3 лет; 4% — более старые
птицы.
Хозяйственное значение, охрана. Как ихтиофаг, имеющий невысокую численность,
малый баклан может наносить лишь несущественный урон прудовому хозяйству.
Занесён в Красные книги Российской Федерации, Украины, Армении, Узбекистана.
Семейство Змеешейковые — Anhingidae Reichenbach, 1849
Тропическое семейство, морфологически и экологически близкое к бакланам, отличающе-
еся рядом анатомических особенностей. Сесамовидная косточка, которая у бакланов примыка-
ет к затылочному отделу черепа, у змеешеек отсутствует. Мускульное сухожилие не пронизыва-
ет коленную чашечку, как это характерно для бакланов, а проходит по желобку на поверхности.
В желудке обособлен пилорический отдел; пищеварительные железы лежат в углублении сте-
нок большого железистого желудка. У копчиковой железы два выводных протока. Мускулатура
нижней гортани развита слабо. Функционирует только левая сонная артерия.
Приспособления к нырянию, долгому нахождению под водой, подводной охоте за ры-
бой сходны с таковыми бакланов, но выражены ещё сильнее. Основные отличия от бакла-
нов — в строении клюва и шеи. Клюв длинный, тонкий, прямой, с конически заострённой
вершиной и слегка зазубренными краями, наружные отверстия ноздрей — в виде узких
щелей. Рыбу змеешейки могут не только хватать, но и протыкать клювом насквозь, как
копьём или гарпуном. Шея очень длинная и подвижная (о чём и говорит название птиц),
обычно изогнута буквой S. Шейных позвонков 20, их тела удлинённые, между 8-м и 9-м
позвонками развит специфический сустав, обеспечивающий возможность резкого выпрям-
ления шеи. При этом голова мгновенно выбрасывается вперёд, клюв схватывает или прон-
зает добычу. Глаза находятся на средней линии клюва, что обеспечивает точность прицела.
Подобные приспособления есть и у цапель. Змеешейки, как и цапли, могут подстерегать
добычу с берега, засекая её сквозь поверхность воды. С воздуха, подобно олушам, никогда
не охотятся. Посадка на воде очень низкая, порой над водой видна лишь голова и шея.
Пойманную добычу змеешейки глотают только над поверхностью воды. Помимо рыбы
охотятся на земноводных, водных беспозвоночных.
Хвост более длинный, чем у бакланов, закруглённый, опахала всех 6 пар рулевых перь-
ев «гофрированные», что помогает увеличить их жёсткость и делает хвост более эффектив-
ным в качестве руля в воде. Крылья довольно большие, притуплённые; взлетают змеешей-
ки тяжело, но летают хорошо, могут даже парить. Намокшее оперение подолгу сушат, рас-
кинув крылья и распустив хвост, сидя на берегу или корягах. В отличие от других веслоно-
гих, у змеешеек при полной линьке все маховые выпадают одновременно, и птицы на неко-
торое время теряют способность к полёту. Окраска оперения тёмная: чёрно-каштановая, на
Чернобрюхая змеешейка
143
крыльях развит красивый чёрно-серебристый сетчатый рисунок, плечевые перья удлинены
в виде чёрно-белых косиц. Эти косицы птицы топорщат во время тока. Клюв желтоватый,
голая кожа вокруг глаз зеленовато-жёлтая, радужина жёлтая или коричневая, лапы телесно-
го цвета или тёмные. В окраске развит половой диморфизм: самки американской змеешей-
ки, или анхинги, имеют охристо-коричневые шею и грудь, контрастирующие с тёмными
спиной и брюхом. У форм, обитающих в Старом Свете, низ головы, шеи, а порой и брюхо
самок тоже светлее, чем у самцов. Молодые птицы окрашены сходно с самками, не имеют
украшающих косиц.
Змеешейковые — единственное семейство веслоногих, не представленное морскими
формами; птицы населяют пресные и солоноватые внутренние водоёмы со стоячей или сла-
бопроточной водой и богатой водной растительностью. Лишь изредка встречаются на при-
морских лиманах, в мангровых зарослях. Больших колоний не образуют, гнездятся отдель-
ными парами или небольшими группами до десятка пар, часто в колониях других околовод-
ных птиц. Гнёзда—массивные сооружения из веток — строят на заломах тростника, кустах,
невысоко в кронах деревьев. В кладке 3-6 белых, с зеленоватым отливом яиц. Птенцы покры-
ваются не тёмным, как у бакланов, а беловатым или кремовым пухом. Инкубация длится
около месяца, выкармливание в гнезде — 6-8 недель, ещё 1-3 недели взрослые докармлива-
ют лётный выводок. Вне сезона размножения кочуют небольшими стайками по водоёмам
недалеко от мест гнездования. Змеешейки становятся половозрелыми на 3-м году жизни.
Ископаемые остатки минимум 6 видов змеешеек найдены на разных континентах, в том
числе и в Европе; наиболее древняя находка с Суматры датируется эоценом (около 30 млн.
лет назад). Современный ареал группы — тропики и субтропики Америки от юга США до
севера Аргентины, тропики восточного полушария от Африки до юга Австралии. В ряде
районов змеешейки считаются конкурентами рыбаков, из-за внешности (птица-змея) мес-
тами распространено суеверно-неприязненное отношение к этим пернатым. На большей
части ареала змеешейки — обычные птицы. Иногда группу считают подсемейством семей-
ства баклановых, но целый ряд морфологических отличий от последних и древность груп-
пы свидетельствуют о более высоком ранге змеешейковых. В семействе 1 современный
род с 2^1 видами (Карташёв, 1974; Коблик, 2001).
Род Anhinga Brisson, 1760
Чернобрюхая змеешейка
Anhinga melanogaster Pennant, 1769
Anhinga melanogaster Pennant, 1769, Indn.Zool. с.13, табл. 12, Цейлон и Ява
Darter, Oriental/Indian Darter, Snakebird (англ.), Schlangenhalsvogel (нем.),
Anhinga roux (фр.)
Син (рус.): индийская змеешейка, змеешейка.
Статус. Редкий залётный вид.
Общая характеристика и полевые признаки. Крупная птица, величиной с большого
баклана. Длина тела 85-97 см, размах крыльев 116-128 см, вес 1 058-1 815 г (del Hoyo et
al., 1992). Клюв длинный, заострённый, его длина 71-87 мм. Хвост заметно длиннее, чем у
бакланов. У взрослых самцов разных подвидов окраска головы и шеи варьирует — от чёр-
но-шоколадной до коричневато-рыжей с беловатыми продольными полосами по бокам шеи;
у самок оперение головы и шеи светлее. Остальное оперение чёрное с серебристо-серыми
144
Отряд Пеликанообразные, Семейство Змеешейковые
полосками на мантии. Плечевые перья удлинены в виде косиц. У молодых в оперении пре-
обладают более светлые, палевые тона; чёрный цвет заменён бурым.
Плавающая птица держит клюв поднятым наклонно вверх, туловище нередко полнос-
тью погружено в воду. Намокшее оперение змеешейки сушат, развернув крылья и хвост.
При взлете широкие крылья и длинный, веерообразно раскрытый хвост образуют как бы
общий полукруг. В отличие от бакланов, змеешейка способна парить.
Подвидовая систематика. Выделяют 4 подвида, различающиеся деталями окраски (del
Hoyo et al., 1992): A. m. melanogaster Pennant, 1769 (1), распространённый от западной
Индии до о. Сулавеси, А. т. rufa (Daudin, 1802) (2), населяющий Африку южнее Сахары и
Ближний Восток, А. т. vulsini Bangs, 1918 (3), обитающий на Мадагаскаре, и А. т. novae-
hollandiae (Gould, 1847) (4), распространённый в Австралии и Новой Гвинее. Нередко боль-
шинству этих подвидов придают видовой статус, выделяя три вида: A. melanogaster, A. rufa
(вместе с А. т. vulsini) и A. novaehollandiae.
Подвидовую принадлежность особи, залетевшей на территорию Узбекистана, устано-
вить не удалось, ближе всего к точке находки подходят границы ареала азиатского подвида
А. т. melanogaster.
Распространение. Африка южнее Сахары, Мадагаскар, Индия, Юго-Вост. Азия, вклю-
чая Филиппины и Индонезию, Новая Гвинея, Австралия. Изолированная гнездовая попу-
ляция существует в низовьях Тигра и Евфрата (Cramp, 1977; King, Dickinson, 1995) (рис.
25). В местах обитания змеешейка ведёт оседлый образ жизни.
Единственный залёт вида на территорию бывш. СССР зарегистрирован 6-7 апреля 2006 г.
Одиночную особь примерно годовалого возраста в течение двух дней наблюдали в юго-
западной части системы Айдарских озёр в точке с координатами 40°55.632' с.ш. и 65°57.672'
в.д. (Навоийская обл., Республика Узбекистан) (Митропольский и др., 2006).
Местообитания. Пресные водоёмы в тропической и субтропической зоне с древесной
растительностью по берегам: озёра, пруды, водохранилища, лиманы, медленно текущие
реки, эстуарии. Нуждается в местах для отдыха и просушки оперения — торчащих из воды
корягах, стволах деревьев, пнях и т.п. Несмотря на свойственную виду осторожность, в
местах судоходоходства может обитать вблизи поселений человека (Cramp, 1977).
Численность. Вид достаточно обычен, местами многочислен в Африке; обычен в Ав-
стралии. Численность на Ближнем Востоке заметно сократилась в течение XX столетия; в
настоящее время очаг размножения остался в юж. Ираке, где популяция пострадала во вре-
мя войны в Персидском заливе в 1991 г. В Юго-Вост. Азии в целом немногочислен, в ряде
мест здесь отмечено снижение численности (del Hoyo et al., 1992).
Размножение. Половой зрелости птицы достигают, по-видимому, не ранее двухлетнего
возраста. Гнездятся одиночными парами или рыхлыми колониями из 6-14 пар, часто вме-
сте с бакланами и голенастыми птицами. Гнёзда устраивают на невысоких деревьях, кус-
тах, изредка на тростнике, на высоте до 2 м над водой. Гнездо — платформа из веток и,
иногда, тростника, диаметром около 45 см. В кладке 2-6 (обычно 3-5) светло-зелёных или
голубоватых яиц. Насиживание длится 26-30 дней, в нем принимают участие оба партнёра.
Птенцы покрыты беловатым пухом; на крыло молодые поднимаются в возрасте около 7
недель. Наибольшая известная продолжительность жизни в природе — более 9 лет, в нево-
ле — более 16 лет (Cramp, 1977; del Hoyo et al., 1992).
Питание. Основу пищевого рациона составляет рыба, видовой состав которой разли-
чен в разных частях ареала. Кроме того, змеешейки поедают амфибий, водяных змей и
различных водных беспозвоночных — ракообразных, насекомых и моллюсков. Рыбу
пронзают или схватывают клювом под водой; активное преследование добычи не харак-
Чернобрюхая змеешейка
145
Рис. 25. Область распространения чернобрюхой змеешейки: а — область обитания, б — залёт на
территорию Северной Евразии. Номера подвидов — см. раздел подвидовой систематики.
терно. Могут также охотиться, подстерегая добычу с берега (del Hoyo et al., 1992; Коб-
лик, 2001).
Враги, неблагоприятные факторы. Наибольшую угрозу для вида представляют, ви-
димо, деградация местообитаний и загрязнение водоёмов.
Хозяйственное значение, охрана. В Северной Евразии, будучи залётным видом, хо-
зяйственного значения не имеет. Специальные меры охраны не предусмотрены.
Подотряд ФРЕГАТОВЫЕ — Fregatae
Семейство Фрегатовые — Fregatidae Degland et Gerbe, 1867
Крупные (длина тела 71-114 см, размах крыльев у Fregata minor — до 230 см, у F
magnificens — до 244 см) но при этом очень лёгкие (630-1 640 г) океанические птицы
(Harrison, 1983; Orta, 1992); по некоторым данным вес может достигать 2 кг. Скелет высо-
копневматизирован, но подкожных воздухоносных полостей нет. Шея короткая, из 14-15
позвонков, грудные позвонки не срастаются в спинную кость, они гетероцельные (разно-
вогнутые), как у фаэтонов и пеликанов, а не опистоцельные (задневогнутые), как у олуш,
бакланов, змеешеек. Грудина широкая и короткая, киль высокий, его основание занимает
лишь около половины длины грудины. Вилочка срастается с коракоидами, а её вершина
прирастает к вершине киля. Таз широкий и короткий. Сошник есть. Сонные артерии пар-
ные. В отличие от других веслоногих, в скелете крыла фрегата больше всего удлинена кисть.
Как и у крупных буревестникообразных, развиты сесамовидные косточки, заклинивающие
локтевой сустав, что позволяет без особой затраты мышечной энергии сохранять опреде-
лённое положение крыла при парении. Крылья очень большие относительно размеров тела,
заострённые, довольно широкие. Первостепенных маховых — 11. Хвост длинный, глубоко
вырезанный; рулевых — 6 пар. Клюв выглядит несоразмерно большим по сравнению с
головой. Он удлинёный, цилиндрической формы, с заметно сдавленным хребтом, заканчи-
вается большим крючком. Имеются щелевидные наружные ноздри. Ноги короткие, цевка
расширенная, с неполностью слившимися, как у пингвинов, метатарзалиями, оперена прак-
тически до пальцев. Пальцы длинные, но перепонка слаборазвита, выемчатая, не доходит
до концевых фаланг. Когти загнутые; средний из них зазубрен по внутреннему краю. Фре-
гаты практически не могут ходить по плоской поверхности, плохо плавают, не способны
нырять, но могут обхватывать пальцами ветки деревьев.
Оперение относительно рыхлое. Окраска в основном чёрная с металлическим блеском
(особенно на украшающих плечевых перьях), сверху на крыльях иногда развит более свет-
лый чешуйчатый рисунок. У самок грудь и брюхо белые, контрастируют с остальным опе-
рением; белые участки меньшего размера имеются и у самцов некоторых видов. У самок
Вознесенского фрегата (Fregata aquila) есть две цветовые морфы. Порой различается цвет
клюва и голого окологлазничного кольца, у самцов они свинцово-серые или тёмные, у са-
мок — розоватые или светлые. Радужина тёмная. Горловой мешок у самок слаборазвитый,
тёмный, у самцов же обширный, ярко-красный, в период тока раздувается огромным пузы-
рём-резонатором. Половой диморфизм проявляется и в размерах: самки крупнее самцов.
Молодые птицы имеют белые или охристые голову, шею, грудь, контрастирующие с ос-
тальным тёмным оперением.
Фрегаты — одни из лучших летунов среди птиц. Хотя они регулярно залетают в субтро-
пические и даже умеренные воды, но всё же в основном придерживаются тропических зон
Мирового океана, особенно тех районов, где с востока на запад дуют постоянные океани-
ческие ветры — пассаты. Силу постоянного ветра фрегаты используют для скольжения над
водой так же, как альбатросы используют постоянные западные ветры в Субантарктике.
Для фрегатов парение и скольжение, экономящие мышечную энергию, ещё более важны,
поскольку они неохотно плавают и не умеют нырять. У них слабо развита копчиковая желе-
за, поэтому перья быстро намокают. Мнение, что фрегаты не удаляются далеко от побере-
жий и архипелагов в открытое море, справедливо лишь для некоторых видов. Отдыхают
фрегаты на скалах или ветвях деревьев, поскольку с ровной поверхности взлетают с трудом
из-за слишком больших крыльев. Вместе с тем конструкция крыльев и хвоста обеспечива-
Фрегат-ариель
147
ет высокую маневренность в воздухе, что и позволяет фрегатам эффективно кормиться —
подбирать корм с поверхности воды, ловить летучих рыб, мелких кальмаров, также выле-
тающих из воды. Планируя над песчаными пляжами, хватают вылупившихся морских че-
репашат, выхватывают птенцов из гнёзд. Хвост во время маневров то разворачивают, то
складывают, изгибают и вращают в разных плоскостях. Чаще любых других птиц, кроме
некоторых поморников, фрегаты занимаются клептопаразитизмом — отбирают добычу у
других видов морских птиц, преследуя их, ударяя клювом и крыльями и заставляя отрыг-
нуть или бросить добычу, а затем подхватывая её на лету или с поверхности воды. Охотятся
фрегаты в одиночку или небольшими группами. Иногда преследуют рыболовецкие суда,
образуют скопления у рыбоперерабатывающих предприятий, питаясь отходами рыбного
промысла, оказавшимися в воде.
Гнездятся рассеянными колониями, гнёзда из сухих веток строят на деревьях, крупных
кустах, реже — на вершинах скал. Размножению предшествует ток, когда самцы, часами
сидя на деревьях и скалах, обозначают индивидуальные участки и привлекают самок, тряся
головой и распростёртыми крыльями, раздувая горловые мешки и издавая рокочущие и буль-
кающие крики. В кладке 1 белое яйцо, в виде исключения 2, но второй птенец обычно не
выживает. Инкубация длится 40-55 дней. Примерно к 12-14-му дню голые при вылуплении
птенцы покрываются светло-серым пухом. Полностью оперяются и становятся способными
к полёту они только в возрасте 4-5 месяцев. Птенцовая смертность очень высока, часть
птенцов съедается взрослыми птицами и птенцами старших возрастов. Взрослые, кочую-
щие вместе с молодыми в районе колоний, докармливают молодых ещё 2-18 месяцев. Се-
зонности в гнездовании нет, пары обычно гнездятся через год. Половая зрелость наступает
лишь на 5-10-й год, продолжительность жизни неизвестна, очевидно, высока.
Ископаемые остатки фрегатов известны с раннего эоцена (находки на западе США име-
ют возраст 50 млн. лет). В настоящее время в семействе сохранился всего один род Fregata
с 5 видами (Harrison, 1983; Dickinson, 2003). Два вида широко распространены, в основном
в Тихом и Индийском океанах, самый крупный — великолепный фрегат (Е magnificens
Mathews, 1914) — замещает их в американском секторе Тихого и Атлантического океанов.
Узкоареальные, занесённые в Красный список МСОП Вознесенский (F. aquila (Linnaeus,
1758)) и рождественский (F andrewsi Mathews, 1914) фрегаты гнездятся, соответственно,
на о-ве Вознесения на юге Атлантики и о-ве Рождества в Индийском океане, кочуют непо-
далёку от мест размножения. В России зарегистрирован один залётный вид; встречи боль-
шого фрегата (F. minor (J.F. Gmelin)) на Дальнем Востоке и великолепного фрегата на
Белом и Баренцевом морях вызывают сомнения в достоверности определения вида и нуж-
даются в дальнейших подтверждениях (Коблик и др., 2006).
Род Fregata Lacepede, 1799
Фрегат-ариель
Fregata ariel (G.R. Gray, 1845)
Atagen ariel G.R. Gray, 1845, The Genera of Birds., 3, табл. 185, северная Австралия;
о-в Рейн
Lesser frigatebird (англ.), Ariel-frigattvogel (нем.), fregate ariel (фр.)
Син. (лат.): к подвиду ariel: Fregata ariel tunnyi Mathews, 1914.
Син. (рус.): малый фрегат (Бутурлин, Дементьев, 1935).
148
Отряд Пеликанообразные, Семейство Фрегатовые
Назван в честь летающего персонажа восточной мифологии — Ариэля. В русской ор-
нитологической литературе ранее (смотри, например, Судиловская, 1951) ошибочно при-
водился под названием Fregata minor (Иванов, 1976).
Статус. Нерегулярно залётный вид.
Общая характеристика и полевые признаки. Фрегат-ариель по величине и тёмной
окраске оперения похож на баклана, но отличается длинными крыльями и длинным виль-
чатым хвостом. По сравнению с другими видами фрегатов, он меньших размеров (размах
крыльев до 193 см), и у него иное расположение белых полей на нижней стороне тела. В
отличие от большого фрегата, возможно залетавшего в воды дальневосточных морей Рос-
сии, у взрослых самцов фрегата-ариеля на нижней стороне тела (боках груди) выделяются
поперечные белые пятна, переходящие на подкрылья, а у самок — тёмные подбородок и
горло. У молодых фрегатов-ариелей, в отличие от больших фрегатов, также белые пятна на
подкрыльях и более тёмная голова. Птицы в промежуточных возрастных нарядах по окрас-
ке оперения похожи на больших фрегатов, но отличаются меньшими размерами и иной
конфигурацией белого рисунка на нижней стороне тела. Во всех возрастных нарядах отли-
чительный признак фрегата-ариель — наличие чётко выраженного белого пятна (в виде
скобки) на подкрыльях у основания нижней стороны крыльев. Молодые фрегаты-ариели и
более крупные виды этого рода в полёте трудно различимы.
Голос. В гнездовой сезон голос фрегатов напоминает квохтание и бормотание.
Подвидовая систематика. Изменчивость проявляется в оттенках окраски оперения и
общих размерах. Выделяют 3 подвида (Alexander, 1963; Степанян, 2003; Dickinson, 2003),
из которых в водах России отмечен только номинативный:
1. Fregata ariel ariel
Atagen ariel G.R. Gray, 1845, The Genera of Birds., 3, табл. 185, северная Австралия; о-в
Рейн (Mathews, 1914, Austr. Av. Rec., 2, c. 121). Распространён на островах и в акваториях
центральных и восточных районов Индийского и западных и центральных частей Тихого
океанов (Sibley, Clapp, 1967).
Подвид Е a. iredalei Mathews, 1914 (2) населяет острова и акватории западной части
Индийского океана, а подвид Е a. trinitatis Miranda-Ribeiro, 1919 (3) — острова и аквато-
рии тропической Атлантики (Harrison, 1983).
Замечания по систематике. Следует отметить, что фрегат, добытый в России (на р.
Амур) в 1926 г., был определён М.А. Мензбиром по фотографии как фрегат-ариель (Карда-
ков, 1928). Однако в дальнейшем в литературе за этой птицей укоренилось название —
большой фрегат F. minor (Johansen, 1927; Бутурлин, Дементьев, 1935; Шульпин, 1936;
Судиловская, 1951 и др.). В результате переопределения чучела этого фрегата, хранящего-
ся в Краеведческом музее г. Хабаровска, было установлено, что данная птица представляет
собой всё же фрегат-ариель (Нечаев, 1972). Фрегата-ариеля считают наиболее обособлен-
ным видом фрегатов и иногда выделяют в подрод (или даже самостоятельный род) Atagen
G.R. Gray, 1841 (Orta, 1992).
Описание. Окраска (по: Alexander, 1963; Harrison, 1983; коллекции Биолого-почвен-
ного института ДВО РАН и ДВГУ). Половой диморфизм взрослых птиц проявляется в
окраске оперения и размерах: самки немного крупнее самцов.
Самец в брачном наряде. Общая окраска оперения чёрная с синим, фиолетовым или
зелёным блеском на спине. На нижней стороне тела резко выражены два поперечных удли-
нённо-дельтовидной формы белых пятна, которые располагаются на боках груди (по одно-
му на каждой стороне) и протягиваются клином (“spur”, по: Harrison, 1983) на подкрылья
поперёк пазушных перьев (axillaries). Горло и зоб не оперены; кожистый горловой «ме-
шок» ярко-красный. Ноги красно-бурые или чёрные. Клюв серый или чёрный. Радужина
коричневая. Орбитальное кольцо чёрное.
Фрегат-ариель
149
Самка в брачном наряде. Общая окраска оперения чёрно-бурая с фиолетовым отливом
на спине. Голова, подбородок и горло чёрные; на горле участок кожи красного цвета. Заты-
лок и задняя сторона шеи светло-коричневые. Грудь, верх живота и участок на пазушных
перьях подкрыльев белые. На верхней стороне крыльев — продольная белая полоса. Ноги
розовые или красные. Клюв голубоватый или розовато-серый. Радужина коричневая. Ор-
битальное кольцо розовое, красное или синее.
Пуховой наряд. Светло-серый.
Первый ювенильный наряд. Самцы и самки не различаются. Окраска оперения в ос-
новном черно-бурая. Голова и верх шеи тёмно-рыжие (или тускло-жёлтые). Подборо-
док, горло и передняя сторона шеи белые с продольными рыжими полосками. Грудь бе-
лая с поперечной сплошной или разорванной посредине чёрно-бурой полосой («бантом»).
Верхняя часть брюшка, бока тела и участок на пазушных перьях белые. На верхней сто-
роне крыльев — продольная светлая полоса. Ноги розовато-белые. Клюв голубоватый.
Второй ювенильный наряд. Самцы и самки не различаются. Окраска оперения в основ-
ном черно-бурая. Голова серовато-белая, на темени и боках головы слабый рыжеватый
оттенок. Подбородок и горло белые с продольными рыжеватыми полосками. Тёмная попе-
речная полоса на груди («бант») отсутствует. На белом брюшке чёрные полоски.
Первый промежуточный наряд (immature). Окраска самцов и самок в основном чёр-
но-бурая. Черная поперечная полоса на белой груди отсутствует; у самца на груди узкие
чёрные полоски. Горло самцов чёрное, самок — белое.
Второй промежуточный наряд (immature). Похож на предыдущий, но у самца голова
черная. На белой груди протягивается поперечная чёрная полоса («бант»), от которой до
нижней границы белого поля проходит продольная чёрная полоса, разделяющая белую грудь
на две части в виде двух треугольников. У самки голова бурая с белыми штрихами, на
подбородке и горле чёрные полоски, поперечная полоса («бант») на груди отсутствует.
Третий промежуточный наряд (immature). Похож на предыдущий. У самца на белой
груди более резко выражены чёрные поперечная и продольная полосы и заметны пятна на
верхней части груди.
Предбрачный наряд (subadult). Похож на взрослый наряд. У самцов белые пятна на
боках груди шире, чем у птиц в брачном наряде, а перемычка между ними на середине
груди испещрена чёрными пятнами. Окраска самки неотличима от птиц в брачном на-
ряде.
Строение и размеры. Самый мелкий и относительно короткохвостый вид
фрегатов. Размеры (мм). Общая длина 710-810. Длина крыла самцов 503-549, самок 510-
560, длина клюва самцов 95-106, самок 98-110 (Orta, 1992; Степанян, 2003). Длина хвоста
самцов 270-337, самок 285-363 (Бутурлин, Дементьев, 1935). Длина крыла самцов и самок
527-555, хвоста 265-330 (Alexander, 1963). Размах крыльев 175-193 см (Harrison, 1983).
Масса самцов 625-875 г, самок — 760-955 г (Orta, 1992).
Линька. Последовательность смены нарядов: пуховой - первый ювенильный - второй
ювенильный - первый промежуточный - второй промежуточный - третий промежуточ-
ный - предбрачный - брачный. Окончательный наряд птицы надевают, вероятно, в возрас-
те 5 лет. Перья сменяются постепенно. Линька фрегатов изучена слабо.
Распространение. Гнездовой ареал. Острова в экваториальных водах океанов и
морей. В Индийском океане — в центральных и восточных районах, в частности на Ко-
косовых о-вах (Килинг) и островах у северо-западных берегов Австралии, в западных
частях — на о-вах Альдабра, Тромлен, Агалега, Чагос и Мальдивских (Harrison. 1983). В
Коралловом море на островах вблизи о-ва Новая Каледония. В Тихом океане — в западных
и центральных районах на восток до атолла Джонстон, включая о-ва Феникс, Лайн, Фид-
150
Отряд Пеликанообразные, Семейство Фрегатовые
Рис. 26. Область распространения фрегата-ариеля (по Harrison, 1983): а — район постоянных коче-
вок, б — места гнездования.
жи, Туамоту, а также Маркизские о-ва (Sibley, Clapp, 1967). В Атлантическом океане — о-
ва Тринидади и Мартин-Вас (Harrison, 1983) (рис. 26).
В XX веке было известно не менее 20 достоверных случаев залётов фрегатов-ариелей в
прибрежные воды России (рис. 27). Птиц регистрировали в основном в июне-августе; пре-
обладали молодые особи в ювенильном наряде и только дважды встречены птицы в брач-
ном наряде. Так, в долине р. Амур: вблизи г. Хабаровск молодую птицу добыли 1 июня
1926 г. (Johansen, 1927; Кардаков, 1928), у г. Комсомольск найдены полуживыми молодая
птица в августе 1962 г. и взрослый самец в августе 1963 г. (Яхонтов, 1966 в; данные В.А.
Поповича), у с. Тыр в устье Амура молодую птицу добыли 30 июня 1972 г. (Росляков,
1981). Кроме того, молодую птицу (вероятнее всего, фрегата-ариеля) наблюдали на р. Амур
у с. Малышево 2 июня 1987 г. (Вальчук, 1997). Молодая птица, окольцованная птенцом
11 ноября 1964 г. на о-ве Эндербери (о-ва Феникс) на юго-западе Тихого океана, была до-
быта через 8 месяцев, 28 июля 1965 г., вблизи г. Советская Гавань на побережье Татарского
пролива (Винокуров, 1967).
В Японском море у берегов Приморского края птиц встречали в заливе Петра Велико-
го: в Амурском заливе у п-ова Де-Фриз молодые птицы добыты 14 июня 1959 г. и 2 августа
1965 г.; молодая птица сфотографирована 29-30 июля 1969 г. (Омелько, 1962; Омелько
М.А., Омелько М.М., 1974). В устье р. Амба молодой фрегат был добыт В.К. Абрамовым
21 июля 1969 г. (Судиловская, 1974). В дельте р. Раздольная молодую птицу наблюдали 11
июня 1975 г. (Назаров, Куринный, 1981). Кроме того, двух птиц отмечали в полёте над г.
Владивосток 9 августа 1979 г. и одну около с. Рязановка Хасанского р-на — 5 июля 1978 г.
(Назаров, Куринный, 1981). В Уссурийском зал. в устье р. Артёмовка птица встречена 30
июля 1964 г. (Омелько М.А., Омелько М.М., 1974). На зал. Восток вблизи г. Находка моло-
дого фрегата наблюдали в первой декаде сентября 1997 г. (данные В.А. Нечаева), а птицу
(возраст не определён) на оз. Ханка — в июне 1998 г. (Глущенко, Нечаев, 2000). В водах о-
ва Сахалин фрегаты были встречены 12 июня 1982 г. на оз. Невское (Воронов, 1987) и 16
июля 2000 г. в бухте Лососей в зал. Анива (наблюдения американского орнитолога С. Ат-
кинсона; Нечаев, 2005). Не исключено, что некоторые визуальные наблюдения фрегатов
относятся к большому фрегату — Fregata minor.
С>регат-ариель
151
Рис. 27. Места встреч фрегата-ариеля в Рос-
сии: а — встречи залетных особей.
Миграции. После периода размножения
ррегаты разлетаются в основном в западном
направлении вдоль пассатных течений в пре-
делах тропических и субтропических зон (Шун-
тов, 1998). Однако отмечаются и дальние пере-
лёты, во время которых птицы залетают в уме-
ренные широты. Фрегатов, окольцованных в
гнездовой сезон на островах в центральных и
западных районах Тихого океана, в мае отмеча-
ли в водах Японских о-вов и в Жёлтом море у
берегов Китая и п-ова Корея (Sibley, Clapp, 1967).
Благодаря течению Куросиво (Цусимское), они
проникают в Японское море, достигая Татарс-
кого пролива, Амурского лимана и долины р.
Амур (Шунтов, 1998); залетают в Охотское море
(Шунтов, 1993).
Местообитания. Фрегаты — океанические
птицы. Поселяются колониями на небольших
скалистых островах, нередко вместе с большими фрегатами. В негнездовой период посто-
янно держатся в открытых водах океанов и морей.
Численность. Обычный вид у берегов Австралии, в других местах встречается реже
(Alexander, 1963). Гнездится многочисленными колониями. В период кочёвок обычен в
тропических и субтропических водах, редок в умеренных широтах.
Размножение. Гнездовой период — в течение всего года (Alexander, 1963). Моногамы.
Токуют поодиночке или группами. В период тока самцы раздувают горловой мешок в виде
большого ярко-красного шара, хлопают крыльями, топорщат перья на голове, издают квох-
тание и бормотание. Поселяются колониями. Гнездо строят на ветвях низких и густых кус-
тарников. Используют сухие прутья, которые обламывают клювом, вылавливают из воды
или воруют у соседей. В кладке 1, редко 2 яйца белой окраски. Насиживают оба гнездовых
партнёра. Инкубационный период около 2 месяцев. Птенцы находятся в гнёздах 4-6 меся-
цев; взрослые выкармливают их полупереваренной пищей, которую отрыгивают в рот птен-
ца. Затем взрослые птицы подкармливают молодых в течение 5-6 месяцев.
Суточная активность, поведение. Фрегаты — птицы дневного образа жизни. Ночью
отдыхают и спят на ветвях деревьев и уступах отвесных скал. Чаще всего фрегатов можно
наблюдать парящими над водами океанов и морей. Заметив добычу, с большой скоростью
настигают её. Плавают плохо и на воду не садятся. Не ныряют. Нередко отнимают добычу
у других птиц.
Питание. Пелагический вид. Объекты питания — рыбы, моллюски, главным образом
кальмары, ракообразные, медузы. Добычу выхватывают с поверхности воды. Нападают на
бакланов, пеликанов, олуш, чаек, крачек и других морских птиц, заставляя их отрыгнуть
добычу. В полёте ловят летучих рыб, качурок и других птиц. В колониях разоряют гнёзда
других фрегатов. В желудке фрегата, добытого у с. Тыр (Нижний Амур) обнаружены остат-
ки трёх косаток-скрипунов — Pelteobagrus fulvidraco (Росляков, 1981).
Враги, неблагоприятные факторы. Для взрослых птиц враги не известны. Отмечает-
ся высокая смертность птенцов от хищных птиц и сородичей, гнездящихся по соседству.
Нередко птенцы выпадают из гнёзд и погибают. Некоторые колонии разоряют люди.
Хозяйственное значение, охрана. Фрегат-ариель не относится к редким или охраняе-
мым видам птиц. Хозяйственного значения не имеет.
Отряд ЛИСТООБРАЗНЫЕ — CICONIIFORMES
Разнообразные по величине (массой от 0,1 до 9 кг) длинноклювые, длинноногие и длин-
ношейные околоводные птицы. Высота стоящей взрослой птицы некоторых видов дости-
гает 1,5 м. Эта группа (чаще всего с включением фламинго) раньше называлась «голенас-
тые».
Скелет хорошо пневматизирован. Грудина и таз широкие, обычно образуется спинная
кость. Череп десмогнатического типа; базиптеригоидные сочленения редуцированы. Шей-
ных позвонков 16-20. Ноги, четырехпалые, пальцы длинные (задний укорочен у аистов),
иногда с зачаточными перепонками, приспособлены к передвижению по зыбкому грунту,
лазанию по ветвям и стеблям. Ноздри щелевидные, сквозные (у форм с массивным или
уплощённым клювом — не сквозные). Глотка пищевод и желудок сильно растяжимы, что
помогает глотать крупную добычу. Слепые кишки рудиментарны (кроме ибисов), есть желч-
ный пузырь, у аистов сохраняется рудимент пениса (Карташев, 1974).
Оперение относительно рыхлое. Уздечка, кожа вокруг глаз, цевка и, как правило, ниж-
няя часть голени не оперены; у некоторых аистов и ибисов оголены также значительные
участки головы и шеи. В основании контурных перьев развита пуховая часть, обычно есть
добавочное пуховое перо. У цапель и китоглавов развиты пудретки. Первостепенных махо-
вых 10-12. Смена их при линьке идёт постепенно, в связи с чем большинство видов в этот
период не теряют способности к полету. Крылья большие, широкие, с тупой вершиной.
Листообразные хорошо, но медленно летают, часть видов может парить. Половой димор-
физм выражен слабо (исключение — некоторые волчки), у многих цапель и ибисов хорошо
развиты сезонные изменения наряда (включая неоперённые части тела). Молодые и перво-
годки до наступления половой зрелости обычно также заметно отличаются по окраске от
взрослых птиц.
Питаются разнообразным животным кормом, в основном добываемым в воде и у воды,
поедание растений носит случайный характер. Основной способ охоты — прицельный удар
острым клювом, но существует и другие стратегии охоты. В отличие от веслоногих, аисто-
образные не являются морскими птицами, для гнездования предпочитают берега внутрен-
них водоёмов, на морских побережьях (за исключением мангровых зарослей и защищён-
ных от волнения лагун) встречаются в основном во время сезонных миграций и послегнез-
довых кочёвок.
В норме моногамы, у многих видов пары постоянные. Большинство видов гнездится
колониями, нередко смешанными, где присутствуют представители не только аистообраз-
ных, но и других отрядов. Гнёзда на заломах прибрежной и болотной травяной раститель-
ности, на кустах, деревьях; иногда — на постройках человека или специально приспособ-
ленных для них сооружениях (белый аист). Редко — на земле или уступах скал. В кладке 2-
10 однотонных (белых, голубых, желтоватых) яиц. Продолжительность насиживания от 2,5
до 5 недель, насиживает в основном самка. Тип развития птенцов — птенцовый с перехо-
дом к полувыводковому. У многих видов потревоженные птенцы уже в 2,5-3-недельном
возрасте перебираются на соседние ветви, откуда могут возвращаться в гнездо. У одиноч-
но гнездящихся видов птенцы находятся в гнезде в течение всего гнездового периода. По-
томство выкармливают оба родителя, у некоторых видов они носят птенцам и воду. Корм
отрыгивают в гнездо, порой птенцы залезают клювами в пищевод родителей, как это дела-
ют и птенцы веслоногих. Постэмбриональное развитие обычно быстрое, подъём на крыло
происходит в возрасте 4-10 недель, дольше только у крупных аистов. Взрослый наряд оде-
вают по наступлению половозрелости, обычно в возрасте 2-3 лет. Живут долго, крупные
виды доживают в природе до 10-16 лет, в неволе до 30 и более лет.
Зцряд Листообразные, Семейство Цаплевые
153
Родственные связи и состав отряда аистообразных весьма дискуссионны. Традицион-
но в отряде выделяют 5 современных семейств, группируемых, как минимум, в 3 подо-
тряда — монотипичные Ardeae (цапли), Scopi (молотоглавы) и включающий три семей-
ства Ciconiae (китоглавы - аисты - ибисы), причём семейства последнего подотряда счи-
таются обособленными и на уровне надсемейств. Раньше к голенастым в ранге подотря-
да относили и семейство фламинго (Карташов, 1974; Howard, Moore, 1980), либо сбли-
жали фламинго с ибисами, а через эту группу всех голенастых считали родственниками
гусеобразных. В настоящее время считаются доказанными высокая степень обособлен-
ности и отрядный ранг фламинго. Через американских грифов, или катартид (семейство
Cathartidae) аистообразных сближали с отрядом соколообразных, однако морфологичес-
кими и молекулярными исследованиями доказано, что катартиды, действительно проис-
ходящие от древних аистов, к настоящим хищным птицам отношения не имеют и сходны
с ними лишь конвергентно. Вероятно, катартиды дожны включаться в Ciconiae. Исходя
из общности ряда анатомических и генетических признаков, Ardeae сближают с баклана-
ми и змеешейками, a Ciconiae с пеликанами (вплоть до включения в семейство пеликано-
вых китоглава Balaeniceps rex). В системе, основанной на сходстве структур ДНК, в от-
ряд аистообразных включают рябков и ржанкообразных (подотряд Charadrii), а также
всех дневных хищных птиц, трубконосых, веслоногих, пингвинов, гагар, поганок, фла-
минго (на правах разных семейств подотряда Ciconii sensu lato). В таком расширенном
объёме отряд объединяет около 30 семейств и более 1000 видов птиц (Sibley, Ahlquist,
1990; Sibley, Monroe, 1990). В наиболее «революционной» трактовке (электронная вер-
сия www.zoonomen.net 2009.07.18) цапель, ибисов, молотоглава и китоглава включают в
состав Pelecaniformes, а в отряд Ciconiiformes, помимо собственно аистов, помещают всех
соколообразных, ржанкообразных и рябков. Здесь принят более традиционный объём
отряда — 113-116 видов, относимых к 36 родам пяти семейств (del Hoyo et al., 1992;
Dickinson, 2003).
Центр происхождения аистообразных неясен, наиболее ранние представители датиру-
ются палеоценом (ранний кайнозой, около 60 млн. лет назад), хотя по данным молекуляр-
ного анализа группа обособилась ещё в конце мезозоя. Описано до 70 вымерших видов из
более чем 30 родов. Некоторые группы, которым иногда придаётся ранг семейств, пред-
ставлены только ископаемыми формами. Современные представители отряда наиболее
разнообразны и многочисленны в тропиках. Несколько подвидов и 2-3 островных вида
голенастых исчезли уже в историческое время по вине человека. Из-за загрязнения водо-
ёмов ядохимикатами и отходами производств, беспокойства на гнёздах и исчезновения
подходящих для гнездования прибрежных биотопов, ещё около 20 видов стали глобально
редкими и нуждаются в охране. В России и сопредельных государствах зарегистрированы
представители 28 видов из 18-19 родов, объединяемых в три семейства двух подотрядов
(Степанян, 2003; Коблик и др., 2006). Из них несомненно гнездится 21 вид.
Подотряд ЦАПЛЕВЫЕ — Ardeae
Семейство Цаплевые — Ardeidae Leach, 1820
Птицы характерного облика цапли или выпи. Размеры различны: индейский (Ixobrychus
exilis) и пестроспинный (I. involucris) волчки имеют длину 28-33 см и массу до 100 г при
размахе крыльев около 52-55 см, цапля голиаф (Ardea goliath) достигает 150 см, весит до 4
кг и имеет размах крыльев до 230 см.
154
Отряд Листообразные, Семейство Цаплевые
Клюв прямой, конический, обычно сжатый с боков, реже (челноклювы) утолщенный и
лодкообразно расширенный. Края надклювья и подклювья режущие, иногда слегка зазуб-
ренные. В отличие от других аистообразных язык длинный. Шея тонкая и длинная, из 19-
20 удлинённых позвонков. Обычно она S-образно изогнута, этот изгиб у выпей и некото-
рых ночных цапель может быть замаскирован длинными перьями, отчего они выглядят
короткошеими и сгорбленными. Сочленения некоторых позвонков, как и у змеешейки, рез-
ко ограничивают боковые движения, но обеспечивают резкий выпад вперёд и точный «гар-
пунный» удар клювом по добыче или врагу. Распрямиться до конца шея не может. Глаза
цапель расположены на линии, соединяющей основание черепа и кончик клюва, что тоже
увеличивает прицельность удара клювом. Область острого бинокулярного зрения находит-
ся перед клювом и ниже его (Коблик, 2001).
Нижняя часть голени не оперена (исключение — некоторые волчки). Пальцы более длин-
ные, чем у других аистообразных, задний палец прикреплён на одном уровне с остальными
и не укорочен, как у аистов и ибисов. Когти длинные, коготь среднего пальца зазубрен в
виде гребёнки, используется для ухода за оперением.
Редкий пух расположен только по аптериям, оперённая или голая копчиковая железа
развита слабо. На груди, надхвостье, иногда на брюхе развиты пудретки — участки непре-
рывно растущего ломкого пуха, продуцирующего роговую пудру, используемую для ухода
за оперением вместо жировой смазки. Именно для её нанесения и соскабливания и необхо-
дим «туалетный» коготь. Пудра белая (у видов с белым оперением) или сизоватая, придаёт
оперению нежные оттенки. В году одна полная линька, обычно после сезона размножения.
У большинства цапель в брачном оперении на голове, шее и спине развиваются украшаю-
щие перья в виде ниспадающих лент, ажурных эгреток и т.п. Крылья со слитной вершиной
(отчего цапли почти не могут парить). Хвост короткий, рулевых обычно 6 пар, первосте-
пенных маховых 11. Полёт с глубокими взмахами, вытянутые назад ноги у всех видов вы-
даются за прямой обрез хвоста. В полете характерно S-образный изгиб шеи, при котором
голова птицы как бы ложится на плечи. В периоды миграций крупные виды объединяются
в стаи, образующие разные типы «геометрически правильного» полётного строя (линия,
угол).
Из черт внутреннего строения характерны крупный сошник, чётко обособленный пило-
рический отдел желудка, одна слепая кишка, парные сонные артерии. Голосовая мускула-
тура обычно развита.
Питаются преимущественно рыбой, но в рационе присутствуют также земноводные,
пресмыкающиеся, мелкие млекопитающие (до суслика), беспозвоночные (среди после-
дних — главным образом, моллюски и насекомые). Основной способ охоты — затаивание
и выслеживание добычи при помощи зрения, а затем — удар сквозь воду. Иногда добыча не
схватывается клювом, а протыкается; впрочем, это случается реже, чем у змеешеек. При
ударе цапля делает поправку на искажение направления световых волн под водой и берёт
ниже видимой добычи. Иногда птицы бродят по мелководью, водной растительности или
по земле, покрытой травой, выпугивая и выискивая добычу. Для ряда видов известны груп-
повые охоты.
Большинство видов ведут дневной образ жизни, но некоторые — также сумеречный
или ночной. Дневные цапли гнездятся обычно колониями (до сотен пар), ночные — как
колониями, так и поодиночке. Часто в одной колонии гнездится несколько видов, суще-
ствует межвидовая иерархия по высоте расположения гнёзд. Гнёзда из веток, стеблей и
другого растительного материала строят на деревьях, кустах, заломах тростника, редко на
плавучей растительности или земле. Откладке яиц предшествуют брачные церемонии, де-
монстрации украшающих перьев, приобретающих яркую окраску клюва, ног, голой уздеч-
льшая выпь 155
:<и. У ночных видов в это время отмечена интенсивная вокализация. Голоса многих цапель
арой очень громкие, квакающие, каркающие, гогочущие, но некоторые виды могут изда-
вать лишь тихий треск или шипение. В кладке от 2 до 5 (у мелких видов до 9) голубоватых
?ши зеленоватых яиц, насиживают оба партнёра, но самка — больше по времени. Насижи-
вание начинается с первого-второго яйца и продолжается от 16-17 до 28-30 дней. Птенцы
вылупляются голыми или слабо опушенными, с прорезавшимися, но не полностью откры-
тыми глазами. В начале постэмбрионального периода самка согревает и защищает птен-
цов, а кормит их самец; в дальнейшем выкармливают оба партнёра. Выкармливание длится
—8 недель до полного оперения, но уже в возрасте около 15 дней при опасности птенцы
}аодят из гнёзда и затаиваются поблизости. Полностью оперяются и становятся способны-
ми к полёту в возрасте 4-5 (мелкие) или 7-8 (крупные виды) недель. У общественных
видов молодые после вылета из гнёзда кочуют вместе с взрослыми, у одиночных видов
держатся самостоятельно. Половая зрелость наступает в возрасте 2-3 лет, у мелких видов
। волчки) — вероятно, на следующий год после рождения.
Цапли известны уже из раннего эоцена (55 млн. лет назад). Описано около 35 вымер-
ших видов, в ископаемом состоянии найдено и более 20 современных видов. В семействе
насчитывают около 60 видов, относимых к 17-25 родам (Карташёв, 1974; Howard, Moore,
1980; Dickinson, 2003). Выделяют разное число подсемейств и триб, по наиболее рас-
пространённой классификации подсемейство Ardeinae объединяет большинство дневных
и некоторых сумеречных цапель (роды Syrigma, Pilherodius, Ardea, Egretta, Casmerodius,
Bubulcus, Ardeola, Butorides, Agamia, 34 вида), подсемейство Nycticoracinae включает квакв
(Nycticorax, Gorsachius, Cochlearius, 8 видов), в Tigrisomatinae помещают тропических
тигровых цапель или тигровых выпей (Tigrisoma, Zonerodius, Tigriornis, 5 видов), нако-
нец, к Botaurinae относят зебровых и настоящих выпей (Zebrilus, Ixobrychus, Botaurus, 13
видов) (Martinez-Vilalta, Motis, 1992). Ранее челноклювов (род Cochlearius), на основа-
нии некоторых морфологических особенностей, выделяли в самостоятельное семейство
Cochleariidae (смотри, например, Карташёв, 1974). Представители семейства распрост-
ранены по всему свету от умеренных и даже субарктических широт до тропиков, где со-
средоточено большинство видов; отсутствуют лишь в безводных пустынях и на некоторых
океанических архипелагах. На постсоветском пространстве зарегистрировано 19 видов (15
из них, безусловно, гнездятся), принадлежащих 9-10 родам (Степанян, 2003; Коблик и др.,
2006).
Род Botaurus Stephens, 1819
Большая выпь
Botaurus stellaris (Linnaeus, 1758)
Ardea stellaris Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Швеция.
Bittern, [eurasian, great] bittern (англ.), Rohrdommel (нем.), butor fttolift (фр.)
Син.(лат.): для подвида stellaris’. Botaurus lacustris C.L. Brehm, 1831; Botaurus arundinaceus
C.L. Brehm, 1831; Botaurus stellaris orientalis Buturlin, 1908.
Син.(рус.): выпь, бугай, водяной бык (народи.)
Статус: гнездящийся перелётный, в юго-западной части ареала — зимующий, оседлый
или совершающий недалёкие кочёвки вид.
156
Отряд Листообразные, Семейство Цаплевые
Общая характеристика и полевые признаки. Птица плотного сложения, несколько
мельче серой цапли. Голова довольно массивная, ноги относительно короткие, пальцы длин-
ные. Крылья широкие, округлые, хвост короткий. Покровительственная окраска оперения
желтовато-коричневая, с многочисленными чёрными и коричневыми пестринами, велико-
лепно маскирует птицу на фоне сухой растительности (тростника, рогоза и т.д.).
В полете большая выпь выглядит как типичная цапля, она несколько мельче, чем серая
цапля, силуэт её имеет более округлые очертания, голова меньше выступает за туловище,
ноги — за хвост. В полёте выпь несколько напоминает сову, так как полёт довольно плав-
ный и тихий.
Проще всего узнать о наличии выпи в исследуемом районе по характерному брачному
крику самца, который хорошо слышен на расстоянии 1-2 км, а в идеальных условиях —
5 и даже 6 км. Однако, в некоторых случаях (например, в ветреную погоду) его можно не
услышать на расстоянии всего 200-400 м.
Голос. «Песня» самца состоит из нескольких частей: щелканье клювом (некоторыми
авторами описывается как «клик», слышно только на очень близком расстоянии и не всегда
издается), затем серия звуков, похожих на вдох (слышны обычно в непосредственной бли-
зости от птицы), затем — серия из 3-9 слогов «уп-бум». В целом это слышится как: «клик-
клик-клик — уп-уп-уп-бум, уп-бум, уп-бум, уп-бум». Элемент «уп», ассоциируется с набо-
ром воздуха и является «функциональным» (Puglisi et al., 2001).
Перед тем, как издать крик, самец осматривается или прислушивается. Затем он опус-
кает шею вниз, держа её s-образно согнутой, перья по бокам шеи встопорщены, на тулови-
ще приподняты (птица как бы «надувается»), клюв чуть опущен, иногда почти касается
земли или воды. Потом самец начинает издавать первую часть крика «уп», при этом его шея
движется немного вперёд в горизонтальном направлении, он как бы делает хватательные
движения полураскрытым клювом. Затем клюв закрывается и следует «буханье». (Alessan-
dria et al., 2003, по видеоматериалам Ф. Сабина и Ф. Дешандоля (Ph. Sabine, F. Deschandol);
данные автора очерка).
В гнездовой период самцы кричат практически круглые сутки, но пик активности при-
ходится на 2 ч до восхода и 2 ч после захода солнца. В это время самец может издавать
крики примерно каждые 1-2 мин. В дневное и ночное время при высокой плотности насе-
ления самец издаёт крик каждые 20^40 мин (вероятно, самцы провоцируют друг друга),
при низкой — значительно реже. Во время активной вокализации самцы могут как нахо-
диться всё время примерно в одном и том же месте, так и довольно активно перемещаться.
Так, за 40 мин наблюдения один самец прошёл по своему участку около 150 м, при этом
регулярно издавая крики. Другой за 1 ч наблюдений дважды перелетал из одного островка
растительности в другой, также почти не переставая активно кричать (Белоруссия, данные
автора очерка).
Голос самца очень низкий, его частота лежит в пределах до 700 герц. Песня каждой
особи имеет индивидуальные признаки, благодаря чему птиц можно отличать друг от дру-
га в поле даже без применения звукозаписывающей техники (Gilbertet, McGregor, 1994;
данные автора очерка). Часто 1-2 элемента «бум» звучат очень глухо и тихо; песня некото-
рых самцов состоит только из таких звуков, и в этом случае их можно услышать только на
небольшом расстоянии (обычно не более 500 м в хорошую погоду). Исследования показа-
ли, что количество таких элементов, как и вообще количество «бухов» на одну песню, не
может служить отличительным признаком состояния и возраста птицы (Puglisi et al., 2001).
У самцов в весеннее время сильно развивается мускулатура шеи, необходимая для того,
чтобы издавать крик (Puglisi et al., 2001); у отловленных в Белоруссии самцов диаметр шеи
составлял 3-5,7 см, в среднем 4,7 см (п=7).
Большая выпь
157
В полете (вечерние полёты кругами и во время миграции) выпь издает хриплые гнуса-
вые крики «кау», или «кво», повторяющиеся с интервалом 6-50 сек. По мнению некоторых
специалистов, этот крик играет фундаментальную роль в формировании миграционных
групп (Puglisi, Baldaccini, 2000). Очень сходные звуки, но с большей частотой, издаёт самка
при беспокойстве у гнезда.
При беспокойстве пуховые птенцы издают тихие быстро повторяющиеся крики «ки-ки-
ки-ки-ки». И.А. Долгушин (1960) отмечал, что начавшие летать молодые как-то особенно
покрякивают, издавая нечто вроде «корк... корк».
Подвидовая систематика. Выделяют два подвида. На территории Северной Евразии
обитает один подвид:
1. Botaurus stellaris stellaris
Ardea stellaris Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Швеция.
Весь ареал вида в пределах бывш. СССР.
Подвид В. s. capensis Shlegel, 1863 (2) обитает в Юж. Африке (рис. 28), он отличается
меньшими размерами и менее интенсивной охристой окраской.
Замечания по систематике. Выпи с Дальнего Востока более густо исчерчены и могли
бы быть выделены в самостоятельный подвид. Однако различия между ними и другими
представителями вида могут быть отнесены и к обычным цветовым вариациям (Hancock,
Kushlan, 1984).
Описание. Окраска. Самцы и самки по окраске оперения не различаются. Окраска
взрослых птиц. Основные тона в окраске оперения — соломенно-жёлтый, охристый, чёр-
ный (буро-чёрный), коричневый разных оттенков. Голова песочно-жёлтая до светло-жёл-
то-коричневой (новое перо). Горло, вся нижняя сторона шеи и живот — палевые. На голове
черная или тёмно-бурая «шапочка», продолжающаяся полосой того же цвета по верхней
стороне примерно до первой трети или середины шеи. Перья на затылке образуют длин-
ную «гриву», которую птица встопорщивает в возбужденном состоянии. Длинные перья на
этой «гриве» имеют светлые соломенно-жёлтые кончики, которые со временем обламыва-
ются (изнашиваются). Верхняя часть шеи покрыта пуховым пером, которое прикрыто длин-
ными перьями «гривы» и перьями с боков шеи. От угла рта и нижнего века по границе
палевого и жёлтого идёт чёрный или тёмно-коричневый «ус». Его ширина и длина варьиру-
ют в широких пределах независимо от половой принадлежности птицы. У всех обследо-
ванных в Белоруссии оперённых молодых (п=4) ус был слабо выражен (очень короткий);
но небольшое количество обследованных птиц не позволяет утверждать, что этот признак
характерен для всех представителей данной возрастной группы.
Боковая сторона шеи и верхняя часть у её основания песочно-жёлтая или коричневатая,
перья с тонкой тёмной окантовкой. На нижней части горла и шеи по палевому фону идут длин-
ные коричневые полосы или пестрины, низ тела и испод крыла так же окрашен — светло-
жёлтый или палевый с мелкими чёрными или коричневыми пестринами. Спина покрыта чёр-
ными перьями с широкой жёлтой окантовкой. Перья поясницы мягкие, пуховые, охристые с
чёрным рисунком. Перья хвоста песочно-жёлтые, покрыты чёрными пятнами и разводами
различного рисунка. Надхвостье окрашено сходно, подхвостье очень светлое, почти белое.
Верхние кроющие крыла по окраске сходны с общим тоном шеи — жёлто-коричнева-
тые с чёрным рисунком, сильно обесцвечиваются при выгорании и износе. Маховые обыч-
но несколько темнее, чем кроющие, однако интенсивность окраски и количество чёрного
на них очень широко варьирует. Характерно преобладание тёмных тонов в окраске этих
перьев, достигающее 80-95% их поверхности.
Пропорции чёрного и светлого в оперении сильно варьируют у разных экземпляров,
форма и цвет пестрин так же весьма различны. Кроме того, интенсивность окраски опере-
158
Отряд Листообразные, Семейство Цаплевые
ния меняется по сезонам вследствие сильного выгорания, чёрный цвет становится корич-
неватым или бурым, жёлто-коричневый — песочным, светло-жёлтым.
Окраска голой уздечки, как и кожа вокруг глаз, у самок в сезон размножения буровато-
оливкового цвета, выступающие участки желтоватые. Подобная окраска отмечалась у в
всех отловленных на гнёздах самок (п = 31). У самцов в течение сезона размножения (март-
июнь) голая кожа на голове становится яркой светло-голубой, с розоватым оттенком из-за
просвечивающих под кожей кровеносных сосудов (п = 34). У одной ещё не приступавшей
к гнездованию самки, отловленной в Белоруссии 16 мая 2005 г., так же был отмечен голубо-
ватый оттенок уздечки, однако он был не столь ярким, как у самцов, выступающие части
были желтоватыми. Все птицы, пойманные вне сезона размножения (п=5, из них 1 $ и 4 сГсГ),
имели цвет уздечки буро-оливковый, независимо от пола (Dmitrenok et al., 2007; Hancock,
Kushlan, 1984).
Окраска радужины может быть различной: большей частью жёлто-оливковая вокруг
зрачка с оранжевым ободком по краю, с некоторым затемнением под зрачком; оранжево-
коричневая с небольшим жёлтым кругом вокруг зрачка; однотонно тёмно-бордовая, тем-
ная оранжево-красная или тёмно-каштановая. Окраска радужной оболочки глаза варьирует
индивидуально, но, как выявлено с помощью повторных отловов ранее окольцованных птиц,
с возрастом становится темнее. Последовательность смены цветов в ходе изменения окрас-
ки радужной оболочки пока точно не установлена (Dmitrenok et al., 2007).
Птенцовый наряд. Птенцы вылупляются в пуху. Эмбриональный пух коричневый, на
нижней стороне тела светлее. Встречаются вариации окраски от оранжево-коричневой до
тёмно (шоколадно)-коричневой. Ноги у однодневных птенцов телесно-розовые, затем се-
ровато (голубовато-) жёлтые, с возрастом становятся более жёлтыми. Голая уздечка олив-
ково-бурая посередине и желтоватая по разрезу клюва и в его верхней части. Клюв при
вылуплении телесно-розовый с чёрным кончиком, яйцевой зуб белый. Яйцевой зуб и чёр-
ная окраска кончика клюва сохраняется как минимум до 30-дневного возраста.
Окраска радужной оболочки глаза у птенцов оливково-жёлтая, тусклая, однотонная и
одинаковая у всех птенцов (более чем 300 обследованных на территории Белоруссии и
Франции птенцов) (Dmitrenok et al., 2007).
К 25-30 дням птенцы полностью покрыты перьями, гнездовой наряд мало отличается
от наряда взрослых птиц.
Последующие наряды неотличимы от наряда взрослых птиц.
Строение и размеры. Основные промеры взрослых больших выпей из Европы
приведены в таблице 8.
Длина тела самцов 740-790 мм, самок 680-776 мм. Размах крыльев самцов 1100-1365
мм, самок 1130-1340 мм (п=3 для сГс? и п=8 для $$, ЗМ МГУ). По данным А.И. Янушеви-
ча и др. (1960), для Киргизии длина тела 730-775, крыло 323-359, хвост 110-125, цевка
95-105, клюв 63-80 мм, вес в среднем до 1 000 г (п=6). Для Узбекистана: длина крыла
260-362 мм, клюва — 60-75 мм, цевка 90-102 мм, вес 820-1 000 г8 (Салихбаев, Богданов,
1961).
Первостепенных маховых 11 (первое рудиментарно), самое длинное из них третье; вто-
рое и четвёртое первостепенные почти равны ему (отличаются на 1-6 мм или равны тре-
тьему). На маховых перьях вырезки нет. Хвост короткий, рулевых перьев 10.
Клюв длинный, его длина от границы оперения равна или чуть больше длины головы от
основания клюва до затылка. На передней части надклювья и подклювья (немного не дохо-
дя до ноздри) расположены мелкие зубчики.
Количество промеров авторы не указывают (прим. ред.).
Большая выпь
Таблица 8. Размеры (мм) и масса (г) большой выпи (среднее ± стандартное отклонение, минимальные и максимальные значения,
размер выборки)
Регион, источник Масса тела Самцы Цевка Масса тела Самки Цевка
Длина крыла Клюв от пера Длина крыла Клюв от пера
Белоруссия, 1510 ± 194,72 351 ±8,98 72,4 ± 2,52 103,5 ± 8,00 844 ±157,25 317 ±5,98 68,5 ± 1,76 90,5 ± 2,77
Dmitrenok et all., 1195-1800 335-366 67,2-75,8 92,7-121,0 734-966 308-326 64,7-71,0 85,5-94,6
(2007) n=10 n=10 n=10 n=10 n=14 n=13 n=14 n=14
Зап. Европа, 1574 ±241,72 348 ± 9,94 72,4 ±3,13 103,5 ± 6,63 878 ± 102,03 316 ±8,55 67,3 ± 2,67 91.7 ±3,32
Dmitrenok et all., 990-2060 318-368 65,5-78,0 92,7-121,0 700-1145 290-330 61,0-71,1 85,5-99,7
(2007) n=43 n=39 n=43 n=36 n=30 n=29 n=30 n=28
Bauer & Glutz von 966-1940 377 80 125 867-1150
Blotzheim (1966) Зап. Европа Cramp (639 ±194,6 346 ± 6,67 69.1 ± 3,24 102 ±3,53 (556 ± 86,0 311 ±7,34 64,3 ±2,10 91,0 ±2,63
(1977) 462-867 335-357 61-74 97-109 430-750 296-327 60-68 87-95
n=12)* n=21 n=20 n=ll n=16)* n=16 n=16 n=9
Baker (1993) 333,4 70,8 98 306 66,4 91,6
310-347 66-75 88-105 278-336 60-71 83-98
n=20 n=20 n=18 n=15 n=15 n=14
* — масса тела истощенных птиц.
160
Отряд Листообразные, Семейство Цаплевые
Масса тела сильно колеблется в зависимости от сезона. В гнездовое время масса самок
(п=14) в Белоруссии составляет в среднем 844, самцов (п=7) —1468 г. К осени птицы «на-
бирают» жир, и масса их тела может возрасти почти вдвое (Dmitrenok et. al., 2005). Исто-
щенные птицы очень сильно теряют в весе. Так, самцы, умершие, как полагают авторы, от
голода, весили 600, 602, 702 г (Newton et al., 1994).
Линька. У взрослых птиц в сезоне одна послегнездовая линька, которая начинается
вскоре после окончания сезона размножения и заканчивается к середине осени. По некото-
рым данным (Witherby et al., 1939, цит. по: Cramp, 1977), у некоторых птиц она может
длиться до января, и даже до марта (Baker, 1993), но у большинства заканчивается к нояб-
рю. Взрослый самец выпи, пойманный на юге Белоруссии 5 октября 2002 г., имел полнос-
тью новое контурное оперение, линька была полностью завершена. У одной самки, успеш-
но выведшей птенцов, смена контурного и хвостового оперенья происходила в июле на её
гнездовом участке. Считается, что линька маховых перьев в семействе цаплевых, в том
числе у выпи, нерегулярная (Cramp, 1977; del Hoyo et al., 1992; Baker, 1993). Она начинает-
ся от нескольких центров, при этом в каждом есть восходящее или нисходящее направле-
ния. От одного года к следующему сохраняется различное количество перьев.
Некоторые данные по линьке маховых у большой выпи собраны на территории Бело-
руссии, Франции и Италии (Dmitrenok et al., 2007). Обследованные взрослые выпи имели
маховые перья разных генераций. Не выявлено явной очередности в смене маховых: линь-
ка может включать как удалённые друг от друга, так и серии последовательных перьев. У
48 отловленных взрослых птиц перья разных генераций выглядели как случайно распреде-
ленные.
В пяти случаях была возможность сравнить одно и то же крыло до и после послегнездо-
вой линьки (п=50 первостепенных маховых). В итоге получены следующие данные: 32%
первостепенных маховых перьев менялись через два последующих года, 62% линяло в те-
чение года, 6% не сменилось. Для второстепенных маховых эти показатели были, соответ-
ственно, 26, 60 и 14%. Следовательно, маховые перья заменялись в среднем каждые два
года, однако некоторые из них менялись чаще, другие сохранялись и после двух последова-
тельных линек. Некоторая часть взрослых птиц полностью меняет летом все маховые пе-
рья.
В 9 случаях была возможность сравнить 2 крыла одной птицы. Сравнение показало, что
линька шла не вполне симметрично, поскольку одно-три соответствующих пера оказыва-
лись разной генерации. При этом, кроющие первостепенных маховых были примерно того
же возраста, как и соответствующие маховые в 68% случаев (п=28 у 18 птиц), тогда как
большие кроющие второстепенных — никогда (Dmitrenok et al., 2007).
Частичная постювенильная линька ограничивается оперением туловища и хвоста. Она
начинается от головы и шеи, вскоре после подъёма птиц на крыло. Частично меняются
кроющие крыла и, возможно, некоторые маховые. Не исключено, что голова линяет снова
в середине зимы или ранней весной (Baker, 1993). Отдельные неполовозрелые особи могут
менять только часть маховых перьев (Cramp, 1977).
Распространение. Гнездовой ареал. Выпь можно назвать эндемиком Старого
Света. Ещё в середине прошлого века её гнездовой ареал занимал практически всю Запад-
ную Европу, за исключением большей части Британских о-вов, Исландии и северных рай-
онов Скандинавии, простирался по всей территории бывш. СССР южнее 58-62° с.ш. — до
Магаданской обл. и Сахалина, в Якутии местами по Колыме доходил до 68° с.ш. (Воробьев.
1963). При этом на территории Вост. Европы от южной Карелии до Крыма, в Закавказье и
республиках Средней Азии выпь отмечали на гнездовании во всех подходящих биотопах,
за исключением засушливых зон и горных районов (Воронцов, 1937; Иванов, 1940; Пуза-
Большая выпь
161
Рис. 28. Область распространения большой выпи: а — область гнездования, б — районы зимовки,
в — залёты, г — наиболее вероятные пути осенних миграций. Подвиды: 1 — Botaurus s. stellaris, 2 —
В. s. capensis. По Hancock, Kushlan, 1984 и Cramp, 1977, с уточнениями.
нов и др., 1955; Янушевич и др., 1959; Долгушин, 1960, 1969; Федюшин, Долбик, 1967;
Птушенко, Иноземцев, 1968; Костин, 1983; Мальчевский, Пукинский, 1983; Мекленбур-
цев и др., 1987; Астафьев, 1998; Сотников, 1999; Рябицев, 2001; Иванчев и др., 2003;
Patrikeev, 2004 и др.); её считали обычной также на Дальнем Востоке (Шульпин, 1936;
Яхонтов, 1942; Воробьев, 1954; Панов, 1973 и др.). На Сахалине выпь относили к редким
залётным (пролетающим) видам (Нечаев, 1991), гнездование её не было подтверждено.
Гнездование Botaurus s. stellaris регистрировали на средиземноморском побережье Ма-
рокко и Алжира, а В. s. capensis — на крайнем юге Африканского континента (Спангенберг,
1951).
К началу XXI в. положение существенно изменилось. Согласно С. Крэмпу (Cramp, 1977),
выпь в Европе уже в 1980-х гг. гнездилась спорадично (рис. 28), а обширные пространства,
занятые её ареалом на Азиатском континенте, объяснялись не повсеместным там обитани-
ем, а скорее относительно слабой изученностью этой территории. Тем не менее, выпь мож-
но считать здесь скорее обычным немногочисленным, чем редким на гнездовании видом,
хотя кое-где её относят к последним (например, в Белоруссии, по М.Е. Никифорову и др.,
1989). Два очага гнездования существуют на юго-востоке Африки (del Hoyo et al., 1992).
На просторах Вост. Европы и Сев. Азии северная граница распространения выпи про-
ходит следующим образом. Огибая Ладожское оз., она захватывает юг Онежского оз. и
вдоль 61° с.ш. устремляется, смещаясь немного к северу, к месту слияния Сухоны с Вычег-
дой. В этом районе регистрируют самые северные места встреч выпи на востоке Европы.
162
Отряд Листообразные, Семейство Цаплевые
Рис. 29. Ареал выпи в Вост. Европе и Сев. Азии: а — область гнездования; б — районы зимовки.
Затем граница несколько отклоняется к югу, пересекает Уральские горы и лишь около мес-
та впадения р. Вах в Обь вновь отклоняется к северу. Енисей пересекает ниже впадения в
него р. Вельмы. Идёт на восток примерно вдоль 63-й параллели, пересекает Лену на уров-
не Вилюя. Затем поднимается вновь севернее, к верховьям Яны и Индигирки. Пересекая
Колыму, резко сворачивает на юг и подходит к океанскому побережью вдоль 152° в.д. По
всей территории южнее очерченной линии большая выпь гнездится во всех подходящих
биотопах (Воробьев, 1963; Мальчевский, Пукинский, 1983; Сотников, 1999). В последние
годы гнездовой ареал вида был уточнён (рис. 29).
Зимовки. В середине XX века к местам зимовки причисляли как юг Европы, Закав-
казья, Средней Азии, так и обширные районы в Индии, Китае, Вост, и Центр. Африке (Спан-
генберг, 1951). В настоящее время ситуация несколько изменилась. В частности, область
зимовок в Африке значительно сократилась: если в середине XX столетия выпь регистри-
ровали почти на 90% этого континента (Спангенберг, 1951), то уже к концу 1980-х гг. счи-
талось, что Botaurus s. stellaris зимует там лишь в окрестностях оз. Танганьика (Cramp,
1977). Впрочем, оказалось, что это не совсём так (Hancock, Kushlan, 1984) — материалы
мечения, коллекционные сборы и отдельные наблюдения показывают, что в незначитель-
ном количестве выпь встречается зимой также в Египте, некоторых оазисах Сахары, в Ни-
герии, Судане, Эфиопии, Эритрее, северо-востоке Конго, Анголе, Намибии, на востоке ЮАР
в примыкающих районах Мозамбика, Зимбабве, Ботсваны (del Hoyo et al., 1992).
Необходимо отметить, что выпи, в отличие от многих других цапель, часто остаются зи-
мовать в местах гнездования, особенно на юге ареала. Зимовки их отмечены также на бере-
гах Балтийского моря и, в том числе, в Прибалтике. В Белоруссии зимуют лишь единичные
экземпляры на незамерзающих водоёмах (Дмитренок и др., 2005). В Великобритании, Фран-
ции, Италии большая выпь — оседлый и зимующий вид. Однако в неблагоприятные годы
при замерзании водоёмов многие птицы сильно истощаются, а часть из них гибнет.
эольшая выпь
163
Миграции. Редко регистрируются из-за того, что выпь не склонна к образованию боль-
ших стай. Весной на местах размножения птицы появляются с исчезновением снежного
покрова (в Казахстане, по данным И.А. Долгушина (1960), это происходит ещё при его
наличии). Для суждения о появлении выпи используют как визуальные встречи птиц, так и
их брачные крики — «буханье», по которым можно определить наличие прилетевших осо-
бей, не видя их, но находясь в соответствующих угодьях. Считают (Плесский, 1960), что
первое время после прилёта на места гнездования птицы молчат, что подтверждают на-
блюдения в Рязанской обл., где визуальные встречи и «буханье» отличались по срокам ре-
гистрации почти на месяц (Иванчев и др., 2003). Поэтому использование разных толкова-
ний термина «первая встреча» может привести к существенным погрешностям. Это следу-
ет иметь в виду при анализе приводимых разными авторами дат — в случае, если авторы не
указывают применённого ими оценочного критерия.
На юге Белоруссии первые крики выпей регистрируют в середине — конце марта (Дмит-
ренок и др., 2005). В Ленинградской области в наиболее ранние весны первые особи появ-
лялись 12-17 апреля, а разгар брачных криков приходится на май, хотя крики самцов не-
редко слышатся и в конце апреля (Мальчевский, Пукинский, 1983). По обширным терри-
ториям постсоветского пространства Вост. Европы прилет выпи происходит плавно, вслед
за наступлением весны. Ю.В. Костин (1983) наблюдал первые встречи этой птицы в Кры-
му с 14 марта по 27 апреля. В центральных регионах Европейской России (Рязанская обл.)
визуальные встречи регистрировали 1-9 апреля, а брачные крики — 6 мая (Иванчев и др.,
2003). В Нижегородской обл. и Татарстане выпь появляется в середине апреля (Пузанов и
др, 1955; Приклонский, 1977). Конкретные данные приводятся для Кировской обл.: это 14
апреля 1995 г. и 5 мая 1988 г., а в среднем за годы наблюдений (п=7) — 25 апреля (Сотни-
ков, 1999).
В ближнем Зауралье, Зап. Сибири и Средней Азии движение выпи весной к местам
гнездования происходит таким же образом. В Вост. Сибири и на Дальнем Востоке эти даты
смещаются на несколько более поздний срок (Воробьев, 1963; Иванов, 1969; Мекленбур-
цев и др., 1987; Салихбаев, Богданов, 1961; Янушевич и др., 1959 и т.д.). Весной наиболее
раннее появление на Ханке отмечено 28 марта 1978 г. и 30 марта 1973 и 1974 гг. (Глущенко
и др., 20066), а в долине р. Раздольная у Уссурийска наиболее раннее появление зарегист-
рировано 27 марта 2003 г. и 30 марта 2005 г. (Глущенко и др., 2006а).
Осенние перемещения выпи происходят столь же незаметно, но иногда удаётся наблю-
дать, как птицы собираются в группы, готовясь к миграции. Так, 17 сентября 1957 г. в месте
впадения р. Белой в Каму (Татарстан, дер. Каракулы) в 21.35 в наступающих сумерках заме-
тили выпь, взлетевшую с небольшого болотца и кругами устремившуюся вверх. Через мину-
ту оттуда же появилась вторая птица, которая также начала подниматься по восходящей спи-
рали. Чуть позднее из соседнего болотца появились ещё 3 выпи. Подъём происходил с
редкими гортанными вскриками птиц. Совершив несколько кругов над болотом на высоте
около 150-200 м, птицы выстроились в «линию» (по терминологии Молодовского, 2001) и
устремились на юго-восток. Удалось заметить, что примерно в 500 м к ним присоединилось
ещё 4-6 птиц. Точно такую же картину сбора птиц к осеннему перелёту наблюдали в Окском
заповеднике в середине сентября 1963 г., с той только разницей, что здесь выпи направи-
лись на запад (С.Г. Приклонский, устн. сообщ.). По наблюдениям В.П. Иванчева с соавто-
рами (2003), отлёт выпи в Рязанском крае происходит во второй половине сентября — на-
чале октября. Летящие в густых сумерках на юг или на юго-запад птицы отмечены 2 сен-
тября, 3, 19 и 21 октября 2001 г. в разных частях поймы в охранной зоне ОГЗ.
В Белоруссии осенний пролёт выпи растянут с августа по ноябрь. Уже в конце августа
на р. Припять удаётся увидеть одиночных птиц, пролетающих в западном или юго-запад-
164
Отряд Листообразные, Семейство Цаплевые
ном направлениях. Регистрируют в это время по одной птице за 4-5 дней. Наибольшее
количество мигрирующих птиц в Белоруссии отмечают в первой половине октября (Дмит-
ренок и др., 2007).
В Сокольском р-не Ивановской обл. удалось подметить массовое движение на юг и юго-
восток выпей как по одиночке, так и небольшими (до 4-5 особей) стайками на вечерних
зорях в начале октября 1953 г. (Н. Соболев, устн. сообщ.).
Приведенные наблюдения показывают, что осенняя миграция большой выпи в центре
Европейской части бывш. СССР начинается с подвижек отдельных особей в последней
декаде августа, продолжается в сентябре (с большей численностью участвующих в процес-
се птиц) и завершается массовым отлётом в первой половине октября. Однако часть птиц
может задерживаться до наступления морозов. Вероятно, таков же характер осеннего дви-
жения выпи на всём пространстве её ареала.
Местообитания Выпь населяет различные типы водно-болотных угодий, в которых
имеется достаточное количество высокой надводной растительности. Типичным для вида
гнездовым биотопом служат обширные заросли тростника, растущие по берегам стоячих
или медленно текущих водоёмов. Вид столь же успешно населяет и сильно мозаичные
места обитания, например, озёра или пруды с отдельными незначительными по площади (с
полгектара и меньше) зарослями растений, поймы рек с небольшими островками тростни-
ка по краю стариц и пойменных озёр. Гнездится также на низинных болотах. Рыбхозы и
зарастающие тростником и рогозом вторично заболачиваемые торфоразработки во многих
регионах (Белоруссия, Кировская, Нижегородская области и др.) также являются важными
гнездовыми и кормовыми биотопами вида, наряду с рисовыми чеками в Крыму (Костин,
1983) и водоёмами-отстойниками в Рязанской обл. (В.П. Иванчев, устн. сообщ.). В Якутии
выпь населяет многочисленные озёра, особенно, в долинах рек (Воробьев, 1963).
Во время пролёта и на зимовке выпь значительно менее требовательна к выбору мест
обитания, её можно встретить на морском побережье, по окраинам заболоченных участков
с кустарником и по берегам арыков с небольшими зарослями тростника (Дементьев, 1952),
по незамерзающим арыкам и разливам, на прудах, озёрах и речках (Долгушин, 1960), в
заболоченных местах с кустарниковой растительностью и по берегам оросительных кана-
лов с негустой высокоствольной растительностью (Мекленбурцев и др., 1987).
Места гнездования, в основном, располагаются не выше 200 м над ур. моря. Выпь пере-
носит солоноватую воду, может обитать в эстуариях или болотистых дельтах, но отсутству-
ет на морском побережье (Cramp, 1977). Избегает мест обитания с сильными перепадами
уровня воды и мест, где кислотность превышает pH 4,5 (Voisin, 1991).
Численность. Из-за одиночного образа жизни численность выпи трудно поддается оцен-
ке: в Восточной Европе её чаще относят к разряду обычных, хотя и немногочисленных
птиц, подчеркивая спорадичность её гнездования, тесно связанного с наличием необходи-
мых биотопов. Конкретные сведения по некоторым странам, опубликованные в Атласе птиц
Европы (Hagemeijer, Blair, 1997), таковы. На Украине гнездится 4 200 (4 000—4 300) пар, в
Белоруссии — 1 000 (950-1 200), в Эстонии и Латвии — по 250 пар. Здесь же сообщается,
что в европейской части России численность достигает 10-30 тыс. пар. По современной
оценке численность большой выпи в Европейской части России составляет 13-25 тыс. пар
(Мищенко, 2004). В некоторых других европейских странах количество выпей (гнездящих-
ся пар = «кричащих» самцов) следующее: в Польше — 4 100-4 800, Румынии — 1 500-2 000,
Германии — 360-620, Венгрии — 450, Франции — 210-317, Швеции — 360—400, а в сум-
ме в 25 странах Евросоюза — 7 900-10 000 (Hagemeijer, Blair, 1997). Если сопоставлять эти
данные с опубликованными прежде (Cramp, 1977), создаётся впечатление о стабильности в
целом численности выпи в Европе за последние полстолетия.
Большая выпь
165
Плотность размещения гнёзд на территории ареала весьма неоднородна: от 1 гнезда на
100 км2 до 3,9-6,5 пар на 1 км2 в характерных местах обитания вида (Спангенберг, 1951;
Дементьев, 1952; Зиновьев, 1982; Астафьев, 1998; Сотников, 1999; Рябицев, 2001; Иванчев
и др., 2003; Мищенко, 2004; Дмитренок, 2005 и др.). Замечено, что после суровых зим
плотность гнездования выпи уменьшалась в таких странах, как Британия, Швеция, Нидер-
ланды (Cramp, 1977).
Размножение. Вскоре после прилёта самцы начинают вокализировать. В регионах с
холодными зимами первые выпи начинают кричать, когда на водоёмах ещё достаточно много
льда. Например, оз. Нарочь (Белоруссия) в 2006 г. полностью освободилось ото льда толь-
ко в мае, а первые самцы выпи начали кричать здесь уже 12 апреля, когда появились пер-
вые небольшие полыньи в местах впадения ручьёв (О.А. Островский, устн. сообщ.). В.Н.
Сотников (1999) отмечал, что при возврате холодов самцы выпи надолго замолкают. Одна-
ко в Белоруссии в разгар гнездового сезона выпи продолжают активно кричать даже при
резких похолоданиях с небольшими ночными заморозками и снегопадами, которые иногда
здесь случаются в апреле-мае (данные автора очерка).
На юге Белоруссии быстрое увеличение числа самцов происходит в течение 14-20 дней
с момента начала вокализации первых из них. Самцы, появляющиеся после основной мас-
сы территориальных птиц (после первой декады апреля), как правило, занимают худшие
участки (меньшие по территории «острова» растительности или только что заполненные
водой пруды рыбхозов). В Кировской обл. интенсивность токования нарастает к середине
мая, когда самцы «гудят» почти круглые сутки с небольшим перерывом в полдень (Сотни-
ков, 1999). В начале июня в Белоруссии вокализационная активность самцов заметно сни-
жается, а у большинства птиц прекращается, что может свидетельствовать о снижении их
половой активности. К концу июня вокализация прекращается как в Белоруссии, так и в
Кировской обл. (Сотников, 1999; Дмитренок и др., 2005).
К размножению самки способны приступать в годовалом возрасте. Из 12 самок, отлов-
ленных на гнёздах в Белоруссии, 2 оказались птицами первого года жизни. Отловленные в
1999-2005 гг. территориальные самцы (п=9) были не менее чем двухлетнего возраста; дан-
ных о размножении годовалых самцов нет.
Размер участка растительности, где обитает или строит гнездо выпь, не имеет перво-
степенного значения при условии, если он расположен на значительном по площади озере,
болоте, заливаемой пойме и т.д. Гнёзда выпи могут располагаться как в центре большого
массива, более чем в 100 м от открытой воды, так и на маленьких островках растительно-
сти вблизи воды (в среднем на рыборазодных прудах рыбхозов в Белоруссии в 19 м от
воды, п=38). В пойме р. Припять гнёзда выпи ежегодно находили в полосе тростника ши-
риной 5-20 м. В Англии так же отмечено гнездование в очень маленьких островках расти-
тельности, оставшихся после выкашивания массива (Tyler 1994).
Среди растений, в зарослях которых гнездится выпь, наиболее предпочитаемый — тро-
стник, здесь были расположены подавляющее большинство (71%) из найденных гнёзд
(п=313). На втором месте стоит рогоз (20,3%), узколистный и широколистный. Кроме того,
вид может гнездиться в зарослях камыша озёрного, в посадках риса (Крым). В.Н. Сотников
(1999) находил гнёзда выпи на осоково-вейниковых закочкаренных участках, в заросшем
рогозом и хвощом водосборном непроточном канале. В Рязанской обл. (В.П. Иванчев, устн.
сообщ.) гнездо было найдено в куртине ивняка на заболоченном лугу. В Белоруссии также
было найдено гнездо, расположенное в основании ивового куста, однако такие случаи не
типичны. Исследования в Великобритании показали, что сукцессионный процесс, приво-
дящий к развитию кустарниковой и осоковой растительности, оказывает негативное воз-
действие на численность выпи (Tyler, 1997). Отметим, что растительность, в которой гнез-
166
Отряд Листообразные, Семейство Цаплевые
дится выпь, может быть как очень высокая (около 4 м в пойме р. Припять, белорусское
Полесье), так и низкая, сильно заломанная (данные автора очерка).
Одним из важнейших параметров среды для гнездования вида служит уровень воды.
Средняя глубина воды у основания гнёзда на рыбхозе в Белоруссии составила 34 см (от О
до 64 см; n=38). В.Н. Сотников (1999) отмечал находки гнёзд на сплавине в зарослях трос-
тника при глубине воды около 1 м. В Рязанской обл. (В.П. Иванчев устн. сообщ.) глубина
воды около гнёзд была 26-83 см. В пойме р. Припять при высоком паводке выпи строили
гнёзда при глубине воды 86-95 см. Следует заметить, что тростник в этих местах был очень
толстым и достаточно густым. В Англии выпь в основном гнездится при глубине воды
менее 15 см (Tyler, 1994). В Италии уровень воды у около гнёзд составляет около 30 см
(Adamo et al., 2004). Таким образом, глубина воды в районе постройки гнезда может быть
от 0 см (влажный ил, сплавина) до 1 м; предпочтение же отдается глубине около 35 см.
Наличие воды в зарослях растительности в момент прилёта может играть решающую роль
в выборе конкретного места, однако известны случаи, когда самцы занимали участок рас-
тительности на пруду со спущенной водой и оставались там в течение всего сезона (уже
после наполнения пруда водой). При сильном падении уровня воды самки часто бросают
гнёзда, оказавшиеся на сухом месте.
В брачный период самцы очень агрессивны по отношению друг к другу. При проигры-
вании фонограммы голоса выпи самцы начинают более активно кричать и медленно под-
ходить к «нарушителю» границ участка. При встрече двух самцов может возникнуть драка,
при которой соперники встопорщивают перья на шее и голове, крылья держат развернуты-
ми и в этой позиции наносят удары друг другу. Обычно драка заканчивается бегством одно-
го из них, но в некоторых случаях птицы сильно калечат друг друга, а иногда даже убивают
(Voisin, 1991). Описываются воздушные бои, когда оба самца, кружась в воздухи, пытают-
ся нанести удары друг другу (Cramp, 1977).
С помощью радиопрослеживания выявлено, что в Великобритании средний размер тер-
ритории самца в период размножения составляет 14,6 и 33,1 га зимой (Gilbert et al., 2005).
В Италии территория самца в гнездовой период составляет от 5,4 до 120 га (в среднем 34,4
га), причём 75% всего времени самцы проводят в участке от 2 до 16 га. Вокализация отме-
чалась не только в пределах участка, в котором самец находился 75% времени, но и за его
пределами. Из четырёх отслеженных самцов в сезон размножения 3 имели одинаковый
размер участка в течение обоих лет изучения, у одного самца на второй год участок увели-
чился. Участок единственной отслеженной самки составил 8,5 га, но 75% регистраций были
в пределах 0,5 га. Причём центр активности самки располагался между двумя центрами
активности самцов. Территории выпей типично «многоядерные», как в течение сезона раз-
множения, так и после него (Puglisi et al., 2003).
Брачные отношения выпей можно отнести к территориальной полигинии. На террито-
рии одного самца может гнездиться от одной до пяти самок (Hancock, Kushlan, 1984). В
Белоруссии на прудах рыбхоза «Белое» (1999-2001 гг.) на 7-8 самцов приходилось 7-10
гнёзд, в половине случаев на территории каждого самца было по 1 гнезду, в пяти случаях на
территории одного самца было по 2^1 гнезда, в двух — гнёзд не было найдено. В пойме р.
Припять на 4 самцов приходилось 4 гнезда, причём 3 из них — на территории одного сам-
ца. На оз. Черствятское в Витебской обл. 16 июня 2006 г. было найдено 9 гнёзд на площадке
320x100 м; расстояние между близлежащими гнёздами составляло 13-64, в среднем 31,8
м. Они располагались вдоль береговой полосы. По данным, собранным на рыбхозе «Бе-
лое», в случае, когда несколько самок гнездятся на участке одного самца, их гнёзда распо-
лагаются в 17-130 м друг от друга. Можно предположить, что самцы занимают любые
свободные участки с подходящим типом растительности и пытаются привлечь туда самок.
Большая выпь
167
тогда как самки выбирают наилучший из занятых самцами участков, и в особо привлека-
тельных местах селятся близко друг к другу. Вероятно, выбор места гнездования в первую
очередь зависит от наличия самца и от выбора места самцом, а не от типа растительности.
Самки не имеют индивидуального предпочтения к определённому виду растительности.
Так, по данным радиопрослеживания, одна самка через год после успешного гнездования в
рогозе широколистном, загнездилась в смешанных зарослях тростника и рогоза узколист-
ного, но — вблизи от прошлогоднего гнезда. Другая отслеженная с помощью радиопере-
датчика самка после прилёта некоторое время держалась на прошлогодней гнездовой тер-
ритории (гнездование в прошлом году было успешным), однако в этом году здесь не было
самца, и самка загнездилась в 500 м от прошлогоднего гнезда, на ближайшей территории
самца (возможно, другого).
Роль самцов в выращивании потомства до сих пор не вполне ясна. Вероятнее всего, они
не участвуют в выращивании птенцов и их основная роль — удержание границ участка и
привлечение самок. Все птицы, отловленные на гнёздах, были самками (n=21, Dmitrenok et
al., in litt.). Существует мнение, что моногамные самцы иногда помогают выкармливать
птенцов и кормят сидящих на гнезде самок, однако точных сведений на этот счет пока нет.
Н.А. Зарудный указывал на добычу самцов с наседными пятнами в Псковской обл. Самец с
голой сморщенной кожей на животе, очень напоминающей наседное пятно, был пойман в
мае в Белоруссии. На участке, на котором он в то время находился и постоянно кричал,
было найдено недавно построенное гнездо без кладки. Вместе с ним была отловлена сам-
ка, ещё не приступавшая к размножению.
Самки приступают к гнездованию примерно через 3 недели после начала вокализации
самцов (Дмитренок и др., 2005). Сроки гнездования сильно зависят от географического
положения региона. Предполагается, что во внутриконтинентальных регионах Евразии,
сроки откладки яиц у выпи в большей степени зависят от климатических факторов, чем в
странах Западной Европы с более мягким климатом.
Для России и территорий сопредельных государств данных по фенологии гнездования
выпи очень мало. Исходя из описаний степени насиженности яиц и гнездового наряда птен-
цов, мы попытались восстановить приблизительные даты начала кладок в гнёздах для раз-
ных регионов по опубликованным данным. На северо-востоке Украины (Астафьев, 1998)
отмечено, что начало массового гнездования приходится на первую декаду мая, две наибо-
лее ранние кладки здесь были начаты ориентировочно 28 и 30 апреля. В Псковской обл. 3
кладки были начаты ориентировочно 7-14 мая, одна — во второй декаде июня (Спанген-
берг, 1951). В Ленинградской обл. три кладки были начаты 12,15 мая и в последней декаде
июня (Мальчевский, Пукинский, 1983). В Липецкой обл. три кладки были начаты пример-
но в период с 19 по 22 мая. В Рязанской обл. (Иванчев и др., 2003) 11 кладок было начато в
период со 2 мая по 1 июня. В Кировской обл. начало кладок приходится на 20 мая — 9 июня
(п=5), в Татарстане — на первую декаду мая (Сотников, 1999). В Казахстане сроки начала
кладок в 6 обследованных кладках (Долгушин, 1960) пришлись на период с 19 апреля по 22
мая. В Туркмении (Дементьев, 1952) начало кладки в единственном обследованном гнезде
пришлось примерно на 12 апреля. Под Оренбургом две кладки были начаты 15 и 19 мая, на
юге Иркутской обл. (п=4) — 21, 23 мая, 16 и 25 июня (Спангенберг, 1951). Две находки
гнёзд в Якутии, к сожалению, были мало информативны с точки зрения сроков гнездова-
ния. В одном из гнёзд начало кладки, судя по всему, было примерно во второй половине
мая, в другом — во второй половине июня. В Приморском крае гнёзда с кладками регист-
рировались в мае (Глущенко и др., 2006а,б).
Гнёзда выпи могут располагаться как на какой-либо твёрдой опоре, так и непосред-
ственно на воде среди растительности. Большинство гнёзд на прудах рыбхоза «Белое» в
168
Отряд Листообразные, Семейство Цаплевые
Белоруссии (34 гнезда из 40 обследованных в 1999-2002 гг.) были сооружены без какой-
либо опоры. Глубина подводной части таких гнёзд была значительной, около 20-30 см.
Держались они за счёт того, что сухой строительный материал некоторое время сохранял
плавучесть, а также за счёт частичной опоры на вертикальные стебли растений, среди
которых гнёзда были построены. Среди 6 других гнёзд одно располагалось на заломе
рогоза, остальные опирались на грунт. Из 5 гнёзд, найденных на болоте Званец (Белоруссия,
2000-2001 гг.) одно было построено на кочке (А. Козулин, устн. сообщ.), одно — в находя-
щейся ниже уровня воды развилке куста ивы; в пойме р. Припять (2002-2006 гг.) 4 из 8
гнёзд были сооружены без опоры, все остальные опирались на дно. Из 12 гнёзд, обследо-
ванных в Рязанской обл., 10 опирались на сплавину, 1 на ивовые ветки, а одно было сделано
на сплавине рогоза узколистного и было приподнято над водой на 19 см (Иванчев и др.,
2003).
В качестве строительного материала используют сухие прошлогодние растения, собран-
ные в окрестностях гнезда (Иванчев и др., 2003). Чаще всего это сухие стебли тростника
или рогоза. В пойме р. Припять на заливном осоково-злаковом лугу было найдено гнездо,
построенное из стеблей злаков и осок.
Гнездо выпи имеет округлую или слегка эллиптическую форму. На стадии насиживания
гнёзда, построенные из тростника, имеют диаметр 28—43, в среднем 36 см (п=19), гнёзда из
рогоза узколистного — 32—46, в среднем 39 см (п=15), из рогоза широколистного — 40-50
см (п=2). Гнездо, построенное из камыша озёрного, имело диаметр 51 см. Последние три
гнёзда были массивнее остальных.
Лоток слабо выражен, часто встречаются совершенно плоские гнёзда. Из 34 гнёзд с
кладками, обследованных на юге Белоруссии, 18 имели углубление лотка (4 из 19 гнёзд из
тростника и 14 из 15 гнёзд из рогоза узколистного; глубина лотка составляла 0,5—4 см). В
Рязанской обл. из 12 гнёзд лоток был выражен только у двух, его глубина была до 2,5 см
(Иванчев и др., 2003).
Высота гнезда над водой колеблется от 3 до 30 см. Если его основание ни на что не
опирается, то вся постройка постепенно погружается в воду, и жилое гнездо постоянно
надстраивается сверху. В случае, когда гнездо расположено на твёрдой опоре, со временем
становится заметно его увеличение в высоту. Построенные на воде гнёзда, по той или иной
причине не ремонтируемые самкой, примерно за 2 недели полностью погружаются в воду.
Гнёзда, обнаруженные на Приханкайской низменности (Глущенко и др., 20066), имели
размеры (п=6): диаметр гнезда 35-80 см (в среднем 53,8 см), диаметр лотка 15-27 см (в
среднем 21,5 см), толщина гнезда 6-13 см (в среднем 8 см).
По литературным данным, найденные летом гнёзда бывают несоразмерно велики, дос-
тигая диаметра 90 см (Спангенберг,1951). Среди исследованных нами гнёзд максимальный
размер составил 74x46 см при высоте 5 см; гнездо имело форму цифры «8». Оно было
построено из рогоза узколистного, возраст птенцов в это время составлял несколько более
3 недель. На стадии насиживания размер этого гнезда был 41x43 см при высоте 8 см. В
большинстве же случаев диаметр жилых гнёзд мало изменяется в процессе выкармливания
птенцов; со временем он может и уменьшаться. В одном случае за неделю диаметр гнезда
сократился с 30x45 до 29x35, высота гнезда при этом увеличилась с 13 см до 18 см (данные
автора очерка).
В критических ситуациях при подъёме уровня воды самка может достаточно быстро
надстроить гнездо и, таким образом, спасти потомство. Так, 10 мая 2002 г. на одном из
прудов рыбхоза «Белое» было найдено гнездо, возвышавшееся над водой на 8 см. В нём
было два свежих яйца. При последующем визите (1 июня 2002 г.) уровень воды в пруду
оказался на 10-15 см выше, гнездо возвышалось над водой на 4 см и представляло собой
Большая выпь
169
очень рыхлую свежую платформу; в нем было 2 яйца и 1 птенец. Очевидно, гнездо быстро
надстраивалось в условиях повышающегося уровня воды.
Если гнездо построено на солнечном припёке, самка может защитить птенцов от пере-
грева, соорудив для них новое гнездо в более благоприятном месте (Cramp, 1977). На рыб-
хозе «Белое» нами был зафиксирован случай, когда самка, видимо, пытаясь спасти птен-
цов, построила 5 новых гнёзд (рыхлых и меньшего размера, чем обычное гнездо) в районе
старого гнезда, где хищник истребил часть выводка.
В Белоруссии в кладке 3-7 яиц, в среднем 5 (п=45). В условиях Приморского края в
полной кладке содержится от 3 до 6 яиц, в среднем (по 15 кладкам) 5,1 яйца на кладку
(сообщение Ю.Н. Глущенко). Средная величина кладки для Англии, Франции, Италии,
Польши и Белоруссии составляет 4,54±0,79 яйца (п=184). Выявлено, что размер кладки
несколько увеличивается от Атлантики к центру Европы (Bretagnolle et al., in litt.). В Казах-
стане величина кладки составляет 3-5, редко 6 яиц (Долгушин, 1960), в Туркменистане —
3-5 (Дементьев, 1952), в Кировской обл. в полной кладке — 4-5 яиц, в среднем 4,6 (п=5)
(Сотников, 1999). Откладывание яиц происходит с интервалом 2-3 дня (Cramp, 1977) или
1-3 дня (Demongin et al., 2007).
Скорлупа яиц матовая, окраска глинисто-оливково-серая, в разных кладках оттенок мо-
жет быть более коричневым, зеленоватым или серым. Изредка встречаются яйца, имею-
щие грязно-голубую окраску (одно такое яйцо найдено в кладке на р. Припять в 2006 г.).
Размеры яиц (мм) в Белоруссии составляют 46,2-59,3 х 34,3-41,1, в среднем 52,8±2,1 х
38,6±1,1 (п=189); в Рязанской области 47,1-54,7 х 36,1-39,9, в среднем 51,79±0,260 х
38,18±0,139 (п= 37) (Иванчев и др., 2003). По материалам, собранным в Приморском крае,
размеры яиц составляют 49,8-57,1 х 31,5-41,7, в среднем (п=58): 52,1x38,2 мм (данные
Ю.Н. Глущенко, И.Н. Кальницкой и Д.В. Коробова). Для Англии, Франции, Италии, Польши
и Белоруссии размер яиц составил 46,0-59,7 х 34,0—43,5, в среднем 52,6±2,2 х 38,7±1,3 мм
(153 кладки); существенных различий по размерам яиц в вышеназванных странах не выяв-
лено. Вес свежих яиц — 38,2±3,52 г (26—47 г) (n=163) (Demongin et al., 2006).
Насиживание может начинаться с первого, второго или третьего яйца (Cramp, 1977;
Спангенберг, 1951; Makatsh, 1974). Судя по различиям в размерах между птенцами (обсле-
довано 18 гнёзд), самки в основном начинают насиживать с первого яйца (61%) или со
второго (39%) Лишь в одном случае первые три птенца были практически неразличимы по
размерам; можно предположить, что здесь самка начала насиживание с третьего яйца.
Средняя продолжительность инкубации равна 25,3±1,24 дня (n=16) (Demongin et al.,
2006). Но возможны отклонения — от 22 до 30 дней в зависимости от режима насиживания
(Voisin, 1991; данные автора очерка). В результате насиживания ежедневная потеря веса
яйца (в г) составляет в среднем 0,195±0,32 (0,125-0,250; n=20, Demongin et al., 2007). При
искусственной инкубации одного яйца, изъятого из гнезда 12 июня, птенец проклюнулся 4
июля, а вывелся через 25 ч 5 июля. Срок инкубации, таким образом, составил минимум 23
дня, и яйцо за это время уменьшилось в весе на 2,55 г (6,3%). Обсохший птенец весил 29,2
г, что составляло 67,8% первоначального веса яйца (Сотников, 1999). Вылупление асинх-
ронное, растягивается на 4-13 дней в зависимости от количества яиц и, видимо, режима
насиживания (Ceutz, 1961, цит. по: Cramp, 1977; данные автора очерка).
Успех вылупления у выпи довольно высокий. На рыбхозе «Белое» (Белоруссия) из 33
обследованных гнёзд в двух было найдено по одному «болтуну», в одном — погибший
зародыш. Основной причиной гибели яиц оказалось хищничество. Из 9 гнёзд, в которых
отмечена полная или частичная гибель кладки, 24 яйца были уничтожено хищниками.
Одним из неясных моментов биологии большой выпи до недавних пор оставался воп-
рос о возможности повторной кладки. В 2000-2002 гг. с помощью радиопрослеживания в
170
Отряд Листообразные, Семейство Цаплевые
Белоруссии и Франции отслежено 9 самок, гнёзда которых были разорены на стадии наси-
живания или маленьких птенцов. Из них в трёх случаях птицы отложили повторную клад-
ку. Самки откладывали первое яйцо повторной кладки через 9-21 день после разорения
первого гнезда (Dmitrenok et al., 2005). Более того, английские орнитологи зарегистрирова-
ли два успешных выводка за один гнездовой сезон у одной самки; при этом птица присту-
пила ко второй кладке, когда возраст старшего птенца первого выводка был 55-60 дней.
Вероятно, это стало возможным благодаря очень раннему гнездованию: первая кладка была
начата 29 марта (Mallord et al., 2000).
Для птенцов выпи характерен полувыводковый тип развития, они вылупляются покры-
тыми пухом и с открытыми глазами. Уже в первые сутки после вылупления птенцы начина-
ют выпрашивать корм. Голодный птенец издаёт пищевой сигнал, представляющий собой
тихую булькающую трель (Пчелинцев, 1990). Одновременно птенцы хватают самку за клюв,
стимулируя её к кормлению. Самка отрыгивает пищу птенцам из клюва в клюв или на гнез-
до. Если пища крупная и птенцы не могут её съесть, взрослая птица снова заглатывает её.
Птенцы в возрасте 1-2 дней способны подбирать и заглатывать мелкий корм, лежащий на
гнезде (данные автора очерка).
Рост птенцов был подробно исследован на 200 птенцах из 99 разных гнёзд. В результате
удалось выстроить кривую их роста и построить уравнение, позволяющее определить воз-
раст птенцов по их промерам (длина цевки, клюва и масса тела) до достижения ими 25-
дневного возраста (Demongin et al., 2006). Данные по росту птенцов приведены в таблице 9.
До 10-14-дневного возраста птенцы, по-видимому, практически все время проводят в
гнезде. В возрасте 2-3 недель они уже часто покидают гнездо на 30 и более минут (И.
Бышнев, личн. сообщ.; данные автора очерка); вероятно, в этом возрасте птенцы уже час-
тично собирают корм самостоятельно. При опасности, начиная с возраста 10-14 дней, птен-
цы убегают из гнёзда и затаиваются в 1-5 м от него. Младшие при этом обычно сидят в
гнезде, и только при подходе человека вплотную 4-6-дневные птенцы могут сместиться на
его край и попытаться спрыгнуть в воду. Будучи «загнанными в угол», птенцы активно
защищаются: широко открывают клюв, клюются, отрыгивают полупереваренную пищу.
Как и взрослые птицы, они ловко лазают по стеблям растений или плывут, если раститель-
ность недостаточно густая (Иванчев и др., 2003; данные автора очерка).
В возрасте 26 дней некоторые птенцы по весу тела и длине цевки могут уже несколько
превышать размеры взрослых самок, к 25-30 дням они уже почти полностью оперены,
рост маховых перьев продолжается. Таким образом, в возрасте 1 месяца внешне они уже
почти не отличаются от взрослых птиц (данные автора очерка) (табл. 9).
По данным, полученным за 4 года исследований на юге Белоруссии, выживаемость птен-
цов (по Mayifeld, 1975, в интерпретации В.А. Паевского, когда за единицу берётся не гнез-
до, а индивидуум), оказалась равной 0,31. Основной причиной гибели птенцов в эти годы
было хищничество (не менее 52% от всех умерших птенцов убито хищниками в 2000 г. и
75% — в 2001 г.). Наибольшая смертность птенцов наблюдается в возрасте до 2 недель. На
этот возраст приходится 83% (п=44) всех случаев гибели, в том числе 70% — от хищников.
Отмечены случаи, когда птенцы погибали от переохлаждения в гнезде в результате беспо-
койства самки наблюдателем в дождливую погоду. В Англии одной из важных причин низ-
кого успеха размножения является гибель птенцов от истощения, причём вероятность ги-
бели значительно выше у младших птенцов в выводке (Gilbert et al., 2002).
На основании материалов радиопрослеживания, птенцы в 57-72-дневном возрасте (в
среднем 62,4 дня; п = 8) уже улетают на расстояние 1 200-3 500 м от гнезда, причём
птенцы из одного выводка разлетаются по разным прудам. Самки не кочуют вместе с
ними. Вылет основной массы птенцов на юге Белоруссии происходит с 8 по 30 июля
Большая выпь
171
Таблица 9. Изменение некоторых морфологических признаков птенцов большой выпи
в зависимости от возраста по данным, полученным на юге Белоруссии в 1999-2002 гг.
В верхней строке каждой графы приведено среднее значение и стандартное отклонение,
во второй строке — максимальные и минимальные значения (lim)
Возраст (сутки) Длина клюва от оперения лба (мм) Длина цевки (мм) Вес (г) Второе первосте- пенное маховое Развитие оперения
1 12,8±1,4 11,4-15,4 (п=8) 16,7±1,1 15-18,5 (п=8) 31,9±1,8 29-35 (п=8) Покрыты пухом
4-5 16,6±1,8 13,8-21,9 (п=21) 21,1±2,2 17,6-26,3 (п=20) 61,4±18,3 37-96,5 (п=21) Есть маленькие пеньки на лопатках
7-8 22,8±2,3 31,0±3,8 124,6±35,3 Пенек 0,5- Начинают появляться пеньки на
19,4-28,1 (п=22) 25,5-41,7 (п=22) 90-216 (п=19) 1,5 мм бедрах, в верхней части спины, на голове, на крыльях. Начинают расти маховые перья.
10-11 29,7±3,2 43,3±6,3 221,9±60,0 Пенек 4-16 Пеньки по всей поверхности тела,
21,9-36 (п=27) 27,2-58,5 (п=26) 98-370 (п=20) мм у части птенцов на лопатках и предплечьях из пеньков начинают выходить перья (до 6 мм составляет опахало).
14 36,9±1,8 56,6±3,9 370±34,3 Пенек 7-23 У части птенцов начинают
35,1-40,1 51-63,9 315-435 мм, у раскрываться пеньки на затылке.
(п=И) (п=11) (п=10) некото- рых уже раскрыты На лопатках перо выходит из пеньков на 15-20 мм, второстепенные маховые выходят из пеньков на 2-5 мм.
16-17 40,6±2,5 64,5±4,2 438,9±57,3 Пенек 10- Спина полностью покрыта
36,7-44,8 (п=17) 55,7-70,7 (п=17) 342-545 (п=16) 23 мм. опахалами перьев, но они продолжают расти; на голове пеньки раскрыты только на затылке.
21-24 50,7±3,6 82,0±4,5 674,4±107,9 24-31 мм, Предплечье и голова покрыта
46-56,2 75,9-90,7 540-840 из них перьями (перья продолжают
(п=10) (п=10) (п=8) опахало 4- 12 мм расти)
25-31 54,9±5,1 48-61,8 (п=5) 88,3±9,2 73,7-95,7 (п=5) 813,7±153,3 700-1030 (п=4) Птенцы полностью покрыты перьями, рост большинства кроющих завершен, рост маховых
продолжается
(Дмитренок и др., 2005). В Кировской обл. выводки распадаются с конца июля до конца
августа (Сотников, 1999).
Суточная активность, поведение. На содержащихся в неволе птицах установлено,
что в норме выпи активны в дневное время, а в ночное время спят примерно с 20 ч до
4.00 (Voisin, 1991). В природе в светлое время суток птицы кормятся, в вечерние и утрен-
ние сумерки в гнездовой период самцы активно кричат. С помощью радиопрослежива-
ния выявлено, что в ночное время самцы выпи большую часть времени остаются на од-
ном и том же месте, но так же зафиксированы их перемещения на 30-210 м (Gilbert,
172
Отряд Листообразные, Семейство Цаплевые
Tyler, Smith, 2005). Миграции также происходят в сумерках, ночью, изредка днём в пас-
мурную погоду.
Большая выпь мало летает и большую часть времени передвигается пешком по зарос-
лям растительности. Обычно птица ходит медленно, высоко поднимая ноги. На глубоких
местах перемещается, обхватывая длинными пальцами стебли растений. Может взбирать-
ся вверх по тростнику, ходить по заломам. При необходимости способна двигаться быстро
через открытые места и плавать по открытой воде. Устраивает присады в траве, на рогозе и
тростнике на высоте до 1 м над водой, которые представляют собой заломанную расти-
тельность, образующую небольшую круглую или вытянутую площадку. Здесь выпи отды-
хают (рис. 30а); под такими присадами часто можно найти выпавшие перья, частички пе-
рьевых чехликов, иногда погадки. В травянистой растительности можно заметить «тропы»
из примятой в одном направлении осоки или злаков на некоторой высоте над водой (Hancock,
Kushlan, 1984; данные автора очерка). При приближении человека птицы затаиваются,
принимая характерную «позу выпи»: тело, шея и клюв вытягиваются почти вертикально
вверх, глаза смотрят вперёд, при этом птицы могут слегка покачиваться, как бы имитируя
движения растительности под ветром (рис. 306). Нам часто удавалось наблюдать иную
позу затаивания: птица плотно сжималась в комок, втянув шею и выставив клюв вперёд и
вверх, при этом обычно нижняя часть тела выпи и лапы были погружены в воду. Вероят-
но, надеясь на свою покровительственную окраску, птицы часто подпускают человека
вплотную. «Загнанные в угол», выпи (особенно, самцы) активно защищаются, принимая
угрожающую позу (рис. ЗОв): раскрыв крылья и встопорщив перья на шее, птица широко
раскрывает клюв и делает резкие выпады, нанося удары агрессору (Voisin, 1991; данные
автора очерка).
В дневное время выпи совершают кормовые перелеты. Большинство таких перелетов
короткие, при этом птицы обычно летят низко над растительностью или водой и быстро
садятся. Особенно часто такие полёты можно наблюдать в период выкармливания птенцов.
Внутривидовые демонстрации мало изучены. При некоторых демонстрациях большое
значение имеют длинные светлые перья, растущие на плечевой птерилии, которые в спо-
койном состоянии обычно скрыты под крыльями, а в состоянии возбуждения поднимаются
и становятся очень заметными (Cramp, 1977; Voisin, 1991).
Брачное поведение большой выпи изучено недостаточно. Описано лишь несколько слу-
чаев ухаживания. В одном из них самец регулярно «бухал» в нескольких метрах от кормя-
щейся самки. Держа шею вытянутой, а клюв направленным вниз, он приблизился к самке и
прошелся около неё налево, затем направо. В это время самка держала тело параллельно
земле, шея и клюв были направлены вверх. Самец зашёл к ней сзади, взобрался на нее,
держа крылья слегка опущенными, после чего последовала копуляция. Затем самка приве-
ла в порядок оперение и взлетела; самец последовал в ту же сторону пешком. В другом
случае самец «бухал», когда две самки находились примерно в 10 и 30 м от него. Он подо-
шел к ближайшей самке, втянул шею в плечи, затем вытянул шею и, опустив клюв, двигал-
ся сначала с левой стороны самки, затем с правой, однако самка убежала. Самец перелетел
ко второй самке и некоторое время стоял рядом, опустив клюв вниз. Через некоторое время
обе птицы полетели туда, где приземлилась первая самка (Alessandria et al., 2003).
Одной из характерных особенностей поведения большой выпи служат полеты кругами
над гнездовым биотопом в сумерках или в темноте. Полет неравномерный, на высоте около
60 м, длится до 10 мин. Часто несколько выпей летают вместе, могут парить как чайки, а
затем резко «падают» в заросли растительности (Hancock, Kushlan, 1984). В полёте птицы
регулярно издают характерный крик «кау». Роль этих полётов пока точно не выявлена, но,
видимо, они довольно полифункциональны и связаны как с брачным, так и с территориаль-
Большая выпь
173
Рис. 30. Некоторые позы большой выпи (по: Bauer, Glutz von Blotzheim, 1966): a — поза покоя, б —
поза при затаивании, в — угрожающая поза.
ным поведением. В.Н. Сотников (1999) отмечает, что подобные полеты можно наблюдать
до конца мая, иногда в июне. Обычно по 2-3 птицы летают вечером, ночью, утром, иногда
и днем. В Италии отмечали антагонистическое поведение в полёте: одна выпь летела по
направлению к другой с шеей, вытянутой вертикально; перья на шее и затылке подняты,
клюв расположен горизонтально (Puglisi, Baldaccini, 2000).
Полеты кругами наблюдаются также в конце лета и осенью. В одном случае в середине
октября на рыбхозе «Локтыши» в Белорусском Полесье отмечено, как в глубоких сумерках
начали кричать, летая кругами, сразу несколько птиц. В стороне от них появилась ещё одна
выпь, которая спешно летела с криками «кау» в сторону отмеченной прежде группы. Нео-
днократно одиночных выпей и небольшие группы до 12 птиц наблюдали в августе-октябре
в разных районах Белоруссии. Птицы в сумерках начинали летать кругами над тростника-
ми, а затем улетали в одном направлении; при этом они постоянно издавали крик «кау».
Несомненно, такие полёты играют важную роль при формировании миграционных групп
(Puglisi, Baldaccini, 2000; Дмитренок и др., 2005; см. также раздел «Миграции»).
Отмечено, что в весеннее время крик «кау» стимулирует активизацию «буханья» сам-
цов (Gilbert, 1994). В Белоруссии в гнездовой период дважды удалось наблюдать, как до
этого молчавший самец начинал активно «бухать», услышав крик «кау» от пролетавшей
над его участком большой выпи (данные автора очерка).
Питание. Большая выпь охотится в мелководной зоне водоёмов и изредка на суше,
собирая в основном водных и околоводных животных. Её рацион достаточно разнообра-
зен. В настоящее время в питании выпи найдено 85 видов животных из 31 семейства; из
174
Отряд Листообразные, Семейство Цаплевые
них 40 видов — насекомые, 20 — рыбы, 6 — амфибии. Благодаря характерной для многих
цаплевых способности отрыгивать содержимое желудка при беспокойстве, материал по
питанию этого вида в большинстве случаев собирают прижизненным методом. Кроме того,
выпи отрыгивают непереваренные остатки пищи (в основном кусочки хитина от покровов
насекомых), образующие плотные погадки. Такие погадки можно найти на гнёздах или под
присадами.
Преобладание той или иной группы животных в питании выпи напрямую зависит от
видового состава потенциальных жертв в осваеваемом ею биотопе, и, возможно, от сезона.
В большинстве случаев основным кормом большой выпи является рыба (Спангенберг, 1951;
Cramp, 1977; Voisin 1991). Однако видовой состав потребляемых рыб широко варьирует в
зависимости от местных условий. В.Г. Пчелинцев (1990) отмечал, что в пищевом рационе
более 80% составляет рыба, в одном из гнёзд более половины всей принесённой рыбы
приходилось на вьюна (Misgurnus fossilis). В Белорусском Полесье в питании выпей, гнез-
дящихся на прудах рыбхозов, где основными выращиваемыми видами рыб являются карп и
карась (Cyprinus carpio, Carassius auratus), 61% сырой биомассы корма составляли эти два
вида рыб, а в пойме р. Припять и на низинном болоте в том же регионе среди рыб домини-
ровали окунь и щука (соответственно, 7,2 и 8,5% биомассы, данные автора очерка). Кроме
того, в питании отмечены щиповка Cobitis taenia, вьюн, судак Lucioperca lucioperca, ерш
Gymnocephalus сегпиа, ротан Perccottus glehni, краснопёрка Scardinis erythrophthalmus, a
так же представители родов Rutilus, Leuciseus, Cottus, Tinea, Gasterostidae, Salmo, Thymallus
(Cramp, 1977; Пчелинцев, 1990; Иванчев и др., 2003; данные автора очерка) и другие виды
рыб.
Не менее важными, а зачастую и основными кормовыми объектами выпи служат члени-
стоногие. Так, В.Н. Сотников (1999) отмечает, что питание выпи составляют в основном
насекомые (жуки, клопы, стрекозы) и их личинки, а также другие водные беспозвоночные
(пиявки, ракообразные). На юге Франции в Камарге интродуцированный из Америки крас-
ный болотный рак Procambarus clarkii составлял значительно больше половины биомассы
в пищевых пробах (п=57) выпи (Poulin et al., 2004).
При исследовании питания выпи в Белоруссии встречаемость беспозвоночных соста-
вила 87%, (86 проб), причём она оказалась одинаковой на прудах рыбхоза, где рыба значи-
тельно преобладала по биомассе в питании выпи, и в естественных биотопах. По биомассе
в пробах питания, собранных в пойме р. Припять и на низинном болоте Званец, наиболь-
шее значение имели личинки жука-плавунца Dytiscus circumcinctus (21% биомассы в изу-
ченных пробах, п=21). Всего беспозвоночные в естественных биотопах составили 42%
общей биомассы проб. На рыбхозе из беспозвоночных в рационе доминируют личинки
другого водного жука Cybister lateralimarginalis (4,7% биомассы, п=65).
Сухопутные насекомые в питании выпи не играют существенной роли и в основном
представлены видами, которые часто встречаются на надводной растительности: взрослые
стрекозы, жуки рода Donacia. Реже встречаются жужелицы и слоники. Несомненно, неко-
торая часть этих жуков попадает в желудок выпи из пищеварительных трактов переварен-
ных ими лягушек. На одном из исследовательских стационаров в Белоруссии часто в пита-
нии находили остатки пауков рода Dolomedes, единично отмечены медведка и даже коло-
радский жук (данные автора очерка). Кроме того, в литературе в качестве кормовых объек-
тов указывают кобылок, дождевых червей, пиявок (Воробьев, 1963; Пчелинцев, 1990; Gilbert
et al., 2003).
Роль амфибий в питании большой выпи, как правило, не столь велика, как рыб и члени-
стоногих, однако в некоторых случаях эта группа доминирует. Так, Н.А. Зарудный (1888,
цит. по: Спангенберг, 1951) считал, что головастики являются основной пищей птенцов.
Большая выпь
175
И.А. Долгушин (1960) указывал, что основную роль в питании выпи играют лягушки, их
головастики и рыбы. По данным А.А. Винокурова (1965), 95% биомассы в пище выпи в
Приазовье в летне-осенний период составляли земноводные, рыбы же были представлены
только в 2%.
В Белоруссии встречаемость амфибий составила 10% на рыбхозе и 23,5% в естествен-
ных биотопах (соответственно 4,6 и 21,9% биомассы). В основном это были зелёные ля-
гушки на рыбхозе и головастики чесночницы в пойме р. Припять.
Из млекопитающих в рационе выпи отмечены водяная полевка Arvicola terrestris, по-
левки родаЛ/zcrote и кутора Neomys fodiens (Долгушин, 1960; Cramp, 1977). В Белоруссии
на рыбхозе встречаемость мыши-малютки Micromys minutus в пробах питания составила
9,2%, но только на одном из 8 обследованных прудов, где этот вид встречался наиболее
часто.
Значительно реже из позвоночных животных в рационе выпи встречаются рептилии и
птицы. Среди пресмыкающихся в питании отмечены уж Natrix natrix, гадюка Vipera berus,
ящерицы рода Lacerta', из птиц упоминаются крапивник, усатая синица, а также «гнездя-
щиеся в тростнике птицы» без указания их видовой принадлежности (Дементьев, 1952;
Долгушин, 1960; Cramp, 1977; Hancock, Kushlan, 1984; Мекленбурцев и др., 1987; данные
автора очерка).
Потребление моллюсков, вероятно, носит случайный характер9. Среди 86 проб питания
из Белоруссии только в одной были найдены два моллюска из рода Succinea.
В пробах питания встречается так же кусочки побегов и семена растений. Вероятнее
всего, они попадают в желудок случайно, при захватывании корма. Не исключено, однако,
что эти кусочки могут играть определённую роль при образовании погадок, на что указы-
вает Н.Н. Скокова (1962).
Таким образом, выпь поедает наиболее доступные и многочисленные виды животных,
и значимость того или иного вида в её рационе зависит от доступности и обилия его в
кормовых биотопах.
Для птиц, содержащихся в неволе, суточный расход пищи оценён как одна пятая от веса
тела (Lundevall, 1953, цит. по: Cramp, 1977). Восстановленная биомасса проб питания,
собранных от птенцов выпи 2^1-дневного возраста на рыбхозе в Белоруссии, составляла 5-
27 г (в среднем 14,2 г, п=5), и 12-110 г (в среднем 50,5 г, п=11) — у птенцов 12-15-дневного
возраста. Самый крупный карп, отрыгнутый птенцом (возраст 10-13 дней), имел длину
140 мм (данные автора очерка). И.А. Долгушин (1960) отмечал, что выпь поедает главным
образом мелкую рыбу, однако в желудках находили щук до 35 см длиной (такая щука весит
не менее трети массы взрослой самки выпи). Количество жертв в одной пробе питания
изменяется от 1 до 43 экз. (в среднем 7,2 экз., п=86, Белоруссия, данные автора очерка).
Охотится выпь в светлое время суток (Puglisi et al., 2003; Gilbert et al., 2005; данные
автора очерка). Некоторые авторы считают, что изредка она может охотиться и кормить
птенцов и ночью (Voisin, 1991). Днём в хорошую погоду часто можно наблюдать кормовые
перелёты выпи. При этом птицы, как правило, летят низко над тростником или водой и
быстро «падают» в заросли растительности. Полёт прямолинейный и довольно тяжелый.
Самки садятся и взлетают несколько в стороне от гнезда. Обычно они совершают короткие
перелёты по 50-200 м, но иногда улетают и на большее расстояние (1 000-1 500 м). Инте-
ресно, что такие перелёты выпь совершает даже в местах гнездования, очень богатых ры-
бой, таких, как выростные пруды рыбхозов. В ветреную или дождливую погоду перелётов
9 Здесь следует заметить, что многие моллюски быстро перевариваются в пищеварительном тракте и
поэтому трудно учитываемы в составе пищи (прим. ред.).
176
Отряд Листообразные, Семейство Цаплевые
не наблюдали. По-видимому, в таком случае птицы предпочитают охотиться, передвигаясь
пешком по зарослям растительности. По данным, полученным с помощью видеозаписи,
самка может охотиться, и непосредственно находясь на гнезде (Adamo et al., 2004).
По нашим наблюдениям, самки приносят птенцам корм примерно каждые 30^40 мин.
Кормятся выпи в одиночку, но в особо кормных местах иногда можно спугнуть 2-3 птиц с
отрезка берега в 50 м.
Выпи могут собирать корм в зарослях различных растений — тростника, рогоза, осоки,
ежеголовника и т.д. Часто для охоты выбирают край сплавины, берега каналов, но так же
часто они охотятся и в зарослях надводной растительности в нескольких десятках метров
от открытой воды. Изредка выпь может кормиться и на берегу, далеко от воды (Пчелинцев,
1990; данные автора очерка).
Основной метод кормежки — медленное перемещение и выслеживание добычи. На мел-
ководье птица неспешно шагает почти по брюхо в воде, высоко поднимая ноги и часто оста-
навливаясь с вытянутой вперёд шеей, высматривая добычу, затем быстро выхватывает из
воды жертву. Время от времени она наполовину погружает клюв в воду, вытянув шею вперёд,
в такой позе застывает на несколько мгновений, затем следует короткий бросок. На глубоких
местах выпи охотятся, передвигаясь по стеблям растений, приминая зелёные листья. Добычу
заглатывает целиком (Cramp, 1977; Hancock, Kushlan, 1984; Voisin, 1991; видеоматериалы
Ф. Сабина и Ф. Дешандоля [Ph. Sabine, F. Deschandol]; данные автора очерка).
Охота выпи весьма успешна: за одно получасовое наблюдение птица 12 раз схватывала
что-то в воде и сразу заглатывала добычу (Пчелинцев, 1990), за 50 мин наблюдений выпь
поймала 42 маленьких рыбки (Voisin, 1991).
Враги, неблагоприятные факторы. Наибольший вред популяции выпи наносит унич-
тожение мест её обитания. Осушительная мелиорация была главной причиной сокращения
численности выпи в Европе. Весенние палы так же приводят к сокращению численности
выпи на гнездовании. Выгорание 50-60% сухой высокостебельной надводной растительнос-
ти на прудах «экспериментального» рыбхоза привело к снижению численности вокализиру-
ющих самцов в среднем на 65%, плотность гнёзд уменьшилась на 86% (Дмитренок, 2005). К
гибели гнёзд приводят также сильные перепады уровня воды в течение сезона размножения.
В Белоруссии за один сезон в окрестностях одного населенного пункта около 15 особей
большой выпи погибло в капканах, поставленных на ондатру (О. Парейко, личн. сообщ.).
Выпь нередко попадает под выстрел во время охоты на гусей, нелетающих птенцов часто
давят бродячие собаки (Сотников, 1999). Интенсивность добычи большой выпи во время
осенней охоты в Европейской части бывш. СССР, по данным из Тверской области, оцени-
вается как 3 особи на 100 человеко-дней охоты (Зиновьев, 1982). В Белоруссии на рыбхозе
«Белое» домашние собаки были основными разорителями гнёзд выпи. Гнёзда разоряют
серая ворона, ворон, возможно, и другие врановые, енотовидная собака, норка, вероятно, и
другие наземные хищники.
В питании больших подорликов в Белоруссии встречаемость большой выпи составила
около 1% (4% от всех птиц — В. Домбровский, устн. сообщ.). Возможно, молодых выпей
могут добывать и другие крупные хищники и совы — например, филин.
При обследовании 18 умерших в Англии выпей выявлено, что в крови у большинства
умерших по разным причинам птиц содержалась ртуть и пестициды, иногда с высокой
концентрацией (Newton et al., 1994). По мнению цитированных авторов, одна из этих 18
птиц умерла именно от отравления пестицидами.
Среди паразитов у выпи найдены гельминты: нематоды Thominx contorta, Contracaecum
microcephalum, Porrocaecum crassum, Subulura suctoria, Desportesius brevicaudatus, Desmi-
docercella numidica, Paronchocerca tonkinensis\ цестоды Ligula intestinalis, Schistocephalus
Большая выпь, Волчок
177
solidus', трематоды Echinostoma paraulum, Е. sudanense, Е. tschertkovi, Е. petrovi, Echino-
chasmus amphibolus, E. bursicola, Pegosomum spiniferum, Clinostomum complanatum, Prostho-
gonimus cuneatus, Codonocephalus urnigerus, Apharyngostrigea cornu, Posthodiplostomum
impraeputiatum, P cuticola (Зиновьев, 1982).
На территории Белоруссии в крови большой выпи впервые зарегистрированы парази-
ты рода Leucocytozoon (Sporozoa, Haemosporida), а также гельминты Microfilaria sp. Общая
экстенсивность заражения Leucocytozoon leboeufi (Mathis et Leger, 1911) составляет 29,4%.
Экстенсивность паразитемии Microfilaria sp. — 11,8%. Этих паразитов регистрировали в
смешанной инвазии с гемоспоридиями рода Leucocytozoon. Все отмеченные виды инвазии
характерны лишь для взрослых птиц (Бабушникова, 2006).
В гнёздах большой выпи, преимущественно в странах центральной части Западной
Европы, найдены Lynchia albipennis (=ardeae) (Say), Ornithoponus ardeae Macq, Ornithomyia
avicularia L. и Lynchia ardea (Macq) (Hicks, 1959).
Хозяйственное значение, охрана. Вид не имеет существенного хозяйственного значе-
ния. Плотность гнездования большой выпи значительно меньше, чем у других рыбоядных
птиц, поэтому ущерб для рыбхозов от выпи минимален. Кроме того, выпь во множестве
поедает личинок жука-плавунца, который, как полагают, является врагом молоди рыб.
Во многих странах Западной Европы вид в настоящее время может гнездиться только
благодаря специальным мерам по восстановлению мест его обитания. (Tyler, 1994).
Вид занесён в Красные книги Белоруссии, Литвы, Латвии; в России — в Красные кни-
ги или списки охраняемых видов многих субъектов федерации: республик Алтай, Башкор-
тостан, Карелия, Коми, Татарстан, Удмуртия и Хакасия; Красноярского, Пермского, При-
морского и Хабаровского краёв; областей Белгородской, Кировской, Ленинградской, Нов-
городской, Орловской, Сахалинской и Ульяновской; Еврейской автономной области.
Род Ixobrychus Billberg, 1828
Волчок — Ixobrychus minutus (Linnaeus, 1766)
Ardea minuta Linnaeus, 1766, Syst. Nat., ed. 12, c. 240, Швейцария.
Little bittern (англ.), Zwergdommel (нем.), biongios nain, butor [heron] biongios (фр.).
Син.(лат.): для подвида minutus'. Botaurus pusillus C.L. Brehm, 1831; Botaurus melanotos
C.L. Brehm, 1842.
Син. (рус.): малая выпь, бугайчик (народи).
Статус. Гнездящийся, мигрирующий вид; в южных регионах возможны нерегулярные
зимовки.
Общая характеристика, полевые признаки. По складу тела птица похожа на боль-
шую выпь, но гораздо мельче её, размером с галку. Вес около 130-160 г, длина 25-38 см.
Взрослый самец характеризуется контрастной, жёлто-охристой с чёрным окраской крыль-
ев; у самки чёрный цвет заменён тёмно-бурым, палевый — светлым жёлто-коричневым.
Скрытная птица, держится главным образом в заболоченном кустарнике, зарослях трост-
ника и другой околоводной растительности. Хорошо лазает по стеблям тростника и рогоза,
а также по веткам кустов. По суше ходит крупными шагами, при необходимости способна
к быстрому бегу. Взлетает неохотно, летит низко над растительностью быстрым для цапель
полетом с мелкими и поверхностными взмахами крыльев; ноги свисают, как у коростеля.
178
Отряд Листообразные, Семейство Цаплевые
При снижении некоторое время летит парящим полётом. От китайского волчка отличается
более крупными размерами, более коротким клювом, чёрной, а не коричневой спиной, се-
ровато-коричневым, а не жёлто-коричневым крыловым пятном. От охристого волчка отли-
чается тёмной спиной и головой.
Чаще всего птицу можно увидеть летящей над болотной растительностью или выпуг-
нуть из прибрежных зарослей. У гнёзда волчки сидят на ветви дерева или в густых зарос-
лях. Заметив приближение человека, птица вытягивает вверх шею и голову и подолгу оста-
ётся неподвижной.
Голос. Брачный крик самца — размеренное повторение низкочастотных, коротких сло-
гов «кохр.., кохр...» или «кхоу..., кхоу...» (иногда передают как «хок.., хок..» или хриплое
«вро... вро...»). Издали этот крик напоминает отдалённый собачий лай или карканье, он
довольно тихий, в обычных условиях хорошо и отчетливо слышен на расстоянии не более
200-300 м. Среди других криков — быстрые квакающие звуки «кве-кве-кве...», издавае-
мый в полёте громкий крик «квор» и крик тревоги — резкое отрывистое «гэк» или «кэ»,
«уик» (Snow, Perrins, 1998; Рябицев, 2001; Kushlan, Hancock, 2005).
Подвидовая систематика. Выделяют пять подвидов (Howard, Moore, 1980; del Hoyo et
al., 1992; Степанян, 2003), отличающихся друг от друга размерами и окраской головы, шеи
и кроющих крыла. Из них в Восто. Европе и Сев. Азии обитает только номинативный под-
вид.
1. Ixobrychus minutus minutus
Ardea minuta Linnaeus, 1766, Syst. Nat., ed. 12, c. 240, Швейцария.
Распространён от Зап. и Юж. Европы до Зап. Сибири, а также в Средней Азии, на Ближ-
нем Востоке и от Ирана до сев.-вост. Индии.
Другие подвиды в своем распространении не связаны с Северной Евразией: I. m.payesii
(Hartlaub, 1858) (2) обитает в Африке от Сенегала до Капской провинции; I. т. podiceps
(Bonaparte, 1855) (3) — на Мадагаскаре; I. т. dubius Mathews, 1912 (4) — в восточной и
юго-зап. Австралии, в Новой Гвинее (иногда восточноавстралийских птиц выделяют в са-
мостоятельный подвид I. т. alisteriy Подвиды незначительно различаются размерами и
окраской оперения. Крупный и тёмный I. т. novaezelandiae (Potts, 1871) (5), которого часто
считают самостоятельным видом, обитал в Новой Зеландии и вероятно вымер к 1900 г.
Замечания по систематике. Близок к китайскому и индейскому, или карликовому (I.
exilis (J.F. Gmelin, 1789)) волчкам, имеющим сходный тонкий клюв, однообразную дор-
зальную окраску и умеренный, выраженный в окраске оперения, половой диморфизм. Воз-
можно, составляет с ними группировку типа superspecies. Эти три вида, в отличие от всех
прочих голенастых, имеют оперённую до сочленения с цевкой голень.
Описание. Окраска (Спангенберг, 1951, с дополнениями). Сезонного диморфизма в
окраске нет. У взрослого самца верхняя часть головы, спина, плечевые, надхвостье — чёр-
ные с зеленоватым металлическим отливом; бока головы и верхняя часть шеи — серова-
тые, бока шеи и её нижняя сторона постепенно приобретают кремовый оттенок, на зобе —
прядь удлиненных перьев той же окраски. От подбородка до зоба тянется беловатая полоса
с охристой или коричневатой линией посередине, образованной продольными пестринами
(таких линий может быть несколько). Верхние кроющие крыла — желтоватые с кремовым
оттенком; большие кроющие могут быть светлее, до беловатых, нижние кроющие крыла
(подкрылья) белые, маховые и рулевые — чёрные. Брюшная сторона светло-охристая с
беловатыми основаниями перьев и бурым продольным рисунком на груди; подхвостье бе-
лое. Клюв жёлтый с тёмным коньком, голая уздечка жёлтая, от клюва через глаз4 идёт тём-
но-бурая полоса; ноги жёлто-оливковые или зеленоватые. В разгар брачного периода уз-
дечка и часть клюва приобретают более яркую (до оранжевой или красной) окраску, кото-
Волчок
179
рая исчезает, когда птица находится в спокойном состоянии. Радужина жёлтая, с оранже-
вым ободком по внешнему краю. Взрослая самка отличаются от самца тёмно-бурой (а не
черной) окраской оперения спины и плечевых; эти перья у них имеют желтовато-охристые
каемки. Бока головы и шеи рыжевато-бурые; на передней стороне тела от подбородка до
брюха на желтоватом фоне выделяется тёмный продольный рисунок из нескольких тёмных
продольных полос. Радужина глаз, клюв и уздечка окрашены, как у самца, окраска уздечки
в брачный период так же может стать более яркой, но не в такой степени, как у самцов
(Langley, 1983; данные М.Г. Дмитренок).
Пуховой птенец до полуторанедельного возраста покрыт густым светло-коричневым
пухом. На плечах пух рыжеватый, на брюшной стороне белый, на остальных частях тела
желтовато-рыжеватый. Вершины пушинок на спинной стороне шелковисто-блестящие. Глаза
и слуховые отверстия открыты. Клюв телесного цвета или буровато-розовый, ротовая по-
лость розовая, яйцевой зуб белый. Лапы и ноги оливково-серые с розовыми пальцами Ра-
дужина глаз тёмно-оливковая или тёмно-коричневая.
Птица в гнездовом наряде по окраске сходна с взрослой самкой, но удлиненных перьев
на зобе нет, и всюду, включая большие кроющие крыла, по светлому фону разбросаны тём-
ные пестрины. Темя буроватое с рыжеватыми отметинами; маховые и рулевые бурые. Кры-
ловое пятно в коричневых пятнах.
После частичной линьки в первом зимнем наряде молодые волчки отличаются от взрос-
лых большим развитием продольных отметин на брюшной стороне. У самцов серый цвет
на шее менее чист; маховые и рулевые сохраняются от гнездового наряда и имеют бурова-
тый цвет. Иногда сменяются только внутренние второстепенные маховые.
Строение и размеры. По строению — типичная цапля, со сжатым с боков телом,
узкой длинной головой, прямым острым клювом, коротким, слегка закругленным хвостом
и широкими крыльями, что хорошо видно в полёте. Первостепенных маховых 11, рулевых
10. Формула крыла: II>I?III>IV>V>VI>VII... I первостепенное короче II первостепенного
на 1,1-3,5 (2,2±1,2, п=3) мм, ПКП на 2,0-2,9 (2,5±0,5) мм, IV<III на 3,1-6,4 (4,8±1,7) мм,
V<IV на 5,1-6,0 (5,5±0,5) мм, VI<V на 5,6-6,7 (6,НО,6) мм, VIKVI на 5,2-7,3 (6,5±1,1) мм,
VIIKVII на 7,0-8,3 (7,8±0,7) мм. Крыло у волчка имеет более заострённую и ступенчатую
форму, чем у китайского волчка.
Длина крыла птиц с территории Вост. Европы и Сев. Азии: у самцов (п=19) в среднем
154,6 мм (150-160 мм), у самок (п=25), соответственно, 147,1 мм (140-150 мм) (Спанген-
берг, 1951). С. Крэмп (Cramp, 1977) приводит такие сведения по длине отдельных частей
тела взрослых птиц, коллектированных в Зап. Европе и хранящихся в музеях Лейпцига и
Антверпена (в мм): длина крыла у самцов (п=15) в среднем 153 (149-157), у самок (п=12) в
среднем 148 (142-153); длина хвоста у самцов (п=14) в среднем 49,6 (47-53), у самок (п=10)
в среднем 49 (47-53); длина клюва у самцов (п=14) в среднем 48,6 (46-53), у самок (n=l 1)
в среднем 46,5 (44-49); длина цевки у самцов (п=12) в среднем 46,2 (43-52), у самок (п=8)
в среднем 43 (44-47).
Вес птиц с территории бывш. СССР от 136 до 150 г; из Западной Европы: самцы —
145-150 г, самки — 140-150 г.
Линька. Данных мало. Вероятно, взрослые птицы имеют полную послегнездовую линь-
ку (Cramp, 1977). У обитающего в Африке подвида I. т. payesii линька полная, замена
первостепенных маховых происходит последовательно от XI к V, остальных — иррегуляр-
но; III первостепенное маховое линяет последним (Cramp, 1977, Baker, 1993) Вероятно,
линька номинативного подвида проходит сходным образом, однако данных недостаточно.
Сроки линьки, по-видимому, сильно растянуты. Некоторые птицы начинают линьку крою-
щих и некоторых маховых перьев в конце июля. Предполагается, что они прерывают линь-
180
Отряд Листообразные, Семейство Цаплевые
ку во время миграции и заканчивают её в Африке, на местах зимовки. Однако большинство
птиц из западной Европы не начинают линьку до прибытия на места зимовки (Pezzo, Gosler,
2005).
Некоторыми авторами предполагается наличие частичной предгнездовой линьки, кото-
рая может затрагивает контурное оперение туловища и часть внутренних второстепенных
маховых перьев, так как весной около 76% отловленных в Италии птиц имели маховые
перья разных генераций (Pezzo, Gosler, 2005). Однако, возможно, это есть следствие силь-
но растянутых сроков постгнездовой линьки — если второстепенные линяют в после-
днюю очередь и не успеют износиться.
Постювенильная линька частичная, начинается в ноябре-январе и включает контурное
оперение и часть внутренних второстепенных маховых. Весной у молодых птиц обычно
остаются старые перья в области лопаток и кроющих крыла (Pezzo, Gosler, 2005). Брачный
наряд молодые птицы одевают на зимовках в конце первого (возможно — второго) года
жизни.
Распространение. Гнездовой ареал. Евразия от Атлантического побережья и
Юж. Европы к востоку до Оби, зап. Синьцзяна, Непала, Синда. Северная граница ареала в
Вост. Европе проходит в пределах Ленинградской, Тверской, Нижегородской обл., по югу
Кировской обл. и Удмуртии; в Сев. Азии — примерно по 56-й параллели. На восток ареал
простирается примерно до 90° в.д., откуда восточная граница спускается на юг до Сев.
Индии. Затем ареал захватывает Юж. Иран, Ирак, Малую Азии; незаселенные видом оста-
ются обширные пустынные регионы Средней Азии и Казахстана. Волчок широко распрос-
транён в Африке к югу от Сахары, очагами встречается на юге и востоке Австралии, юге
Новой Гвинеи; в Новой Зеландии на гнездовании исчез (рис. 31, 32) (Cramp, 1977; del Hoyo
et al., 1992; Сотников, 1999; Степанян, 2003).
По-видимому, изолированное место гнездования нескольких пар волчков существует
на оз. Донты в южной части Республики Коми, севернее основного ареала вида (Красная
книга Республики Коми, 1998).
Совершенно неожиданной оказалась находка волчка на юге Приморского края. Гнездо-
вание отмечено на одном из озёр побережья Амурского залива в 2007-2009 гг. По данным
фото и видеосъёмки, выполненной С.Г. Сурмачем, в 2007 г. гнездилась пара, в которой
самец был волчком, а самка с некоторыми сомнениями (в пользу гибридного происхожде-
ния) была определена как китайский волчок (Гамова и др., 2007). В 2008-2009 гг. в том же
районе гнездилось по одной паре, в которых оба партнёра по окраске тела и форме клюва в
большей степени походили на волчка, но имели признаки, свойственные китайскому волч-
ку, что позволило идентифицировать их как гибридных особей (данные Т.В. Гамова и др.).
В 2007-2008 гг. гнездование пар было успешным, в 2009 г. птенцы погибли.
Зимовки. Зимовки номинативного подвида располагаются в Африке южнее Сахары
и в Индии. Остальные подвиды волчка оседлы. Европейские волчки (7. т. minutus) зимуют
в Алжире, Чаде, Судане, Эфиопии, Сенегале, Нигерии, Заире, Анголе, Замбии, Зимбабве,
Юго-Вост, и Юж. Африке, а также на о-ве Мадейра и Канарских о-вах (Нанкинов, 1999).
Часть молодых птиц остаётся на местах зимовки в Африке во второе лето жизни (Pezzo,
Gosler, 2005). Известны единичные случаи зимовки волчков в Зап. Европе (Нанкинов, 1999).
Нерегулярные зимние встречи одиночных птиц отмечались в Киргизии в западной части оз.
Иссык-Куль (Птицы Средней Азии, 2007), и, возможно, в Туркменистане (Спангенберг, 1951).
Миграции. Под Адлером в Краснодарском крае первые особи зарегистрированы 18
апреля 1982 г.; весенний пролёт здесь хорошо выражен до первой декады мая. Массовое
появление волчков в Вост. Приазовье и на Зап. Маныче отмечается во второй половине
апреля; в Ставропольском крае первых птиц отмечали в начале второй декады апреля; мае-
Волчок
181
Рис. 31. Область распространения волчка: а — область гнездования мигрирующих популяций и об-
ласти обитания оседлых; б — районы зимовки, установленные коллекционными сборами и, отчасти,
мечением. Подвиды: 1 — Ixobrichus minutus minutus, 2 — I. m. payesii, 3 — I. m. podiceps, 4 — I. m.
dubius, 5 — I. m. novaezelandiae.
Рис. 32. Ареал волчка в Вост. Европе и Сев. Азии: а — область гнездования; б — залёты, в — изоли-
рованные гнездования.
182
Отряд Листообразные, Семейство Цаплевые
совый пролёт наблюдается здесь в середине мая (Казаков и др., 2004). В Закавказье под
Ленкоранью волчки не представляют редкости уже в самом начале апреля (Спангенберг,
1951). В Крыму первых птиц отмечали с конца марта, но обычно пролёт идёт здесь в апре-
ле и первой половине мая, с пиком в последней декаде апреля (Костин, 1983); первые вол-
чки на юге Украины зарегистрированы в первых числах апреля, в северных областях Укра-
ины — во второй половине апреля (Смогоржевський, 1979). В Белоруссии первых птиц
регистрировали 20 апреля (Федюшин, Долбик, 1967). В Московской обл. волчок появляет-
ся в период с 11 апреля по 5 мая (Птушенко, Иноземцев, 1968), в Калужской обл. самая
ранняя встреча отмечена 5 мая 1977 г. (Марголин, 2000), в Нижегородской обл. и Татарста-
не первые встречи регистрируются в первую декаду мая (Попов, 1977), на юге Кировской
обл. прилет отмечается в конце апреля - начале мая (Сотников, 1999). В Туркмении волчки
появляются с середины марта, пролёт их отмечен здесь во второй половине марта; под Са-
маркандом птиц регистрируют в середине апреля, под Ташкентом — в конце второй или
начале третьей декады апреля. В Таджикистане массовый пролёт проходит в апреле, в Кир-
гизии — в начале мая; на юге Казахстана первых волчков отмечают в первой половине
апреля, пролёт длится до начала июня (Птицы Средней Азии, 2007). В Алтайский край
волчок прилетает в начале мая (Кучин, 1976).
Осенний отлёт начинается в августе и заканчивается в конце сентября - начале октября
(Спангенберг, 1951; Птушенко, Иноземцев, 1968; Костин, 1983; Марголин, 2000; Казаков и
др., 2004 и др.). Во время осеннего пролёта, как и весной, часть птиц пересекает Главный
Кавказский хр. (Казаков и др., 2004). В Средней Азии в большом количестве волчок летит
в первой половине сентября (Птицы Средней Азии, 2007), в южном Казахстане пролёт
наблюдали с конца сентября до 20-х чисел октября, но отдельные птицы могут задержи-
ваться до начала ноября (Долгушин, 1960). Азиатские птицы летят на юг, возможно на юго-
восток, а европейские — на юг и юго-запад (Нанкинов 1999).
Во время миграции волчки могут лететь поодиночке или небольшими группами от 2-3
до 10-15 особей. Дневные часы птицы проводят в зарослях тростника и кустарниках по
берегам водоёмов, в особо благоприятных местах останавливается до нескольких десятков
птиц (Cramp, 1977; Лопатин и др., 1992; Нанкинов, 1999; Казаков и др., 2004; Птицы Сред-
ней Азии, 2007).
Местообитания. Гнездится в зарослях тростника, камыша, рогоза, ольхи, ивы по бере-
гам различных водоёмов. В средней полосе Европейской России — это берега озёр и ста-
риц в поймах рек, а также заросли надводной растительности на различных искусственно
созданных водоёмах (карьеры старых торфоразработок, пруды рыбхозов, водохранилища,
фильтрационные пруды-отстойники). Так, в Московской обл. основная часть популяции
гнездится на искусственных водоёмах, после создания которых численность вида заметно
возросла (Суханова, 2008). Гнездится на заросших тростником степных озёрах, в плавнях
речных дельт, тугайных зарослях в долинах южных рек. Водоёмов, расположенных в глу-
бине лесных массивов, избегает (Спангенберг, 1951). В Белоруссии при выборе мест гнез-
дования предпочитает участки с глубиной воды около 40 см (от 0 до 120 см). Стабильность
уровня воды в течение всего года также играет большую роль, так как влияет на кормовую
базу, защитные свойства биотопа и т.д. (Дмитренок и др., 2009).
В горной местности Туркмении находили гнёзда на берегах горных ручьёв (Шестопё-
ров, 1936), в Зеравшанской долине волчок гнездится на рисовых полях (Птицы Средней
Азии, 2007). В целом, местообитания сильно варьируют на протяжении обширного ареала
вида. Не избегает волчок антропогенного ландшафта и может гнездиться в местах, регу-
лярно посещаемых людьми.
Для поисков пищи использует затопленные заросли околоводной растительности, мел-
ководья и различные отмели по берегам водоёмов, в том числе открытые грязевые пляжи
Волчок
183
среднерусских рек. Так, в июле 1956-1960 гг. этих птиц неоднократно удавалось отлавли-
вать для мечения по ночам автоматическими лучками на грязевых отмелях заливов в сред-
нем течении р. Оки (Приклонский и др., 1962).
На зимовках волчки придерживаются более открытых водоёмов, отмечаются в антро-
погенном ландшафте, на сухих злаковых полях и плантациях сахарного тростника (del Hoyo
et al., 1992).
Численность. Волчок многочислен в характерных для него биотопах на юге Украины,
в Приазовье, Средней Азии, но севернее только обычен, становясь редким к северной гра-
нице ареала. В то же время, из-за скрытного образа жизни впечатление о его немногочис-
ленности нередко ошибочно (Спангенберг, 1951; Долгушин, 1960; Смогоржевський, 1979;
Костин, 1983; Казаков и др., 2004; Птицы Средней Азии, 2007 и др.). По данным на конец
прошлого - начало нынешнего века в Европейской России гнездилось 15-50 тыс. пар этих
птиц (Мищенко и др., 2004). В Белорусском Полесье численность составляет не менее 400-
700 пар (Дмитренок, 2009), в северной части Белоруссии вид значительно более редок. В
Эстонии нерегулярно гнездятся отдельные пары (Leibak et al., 1994), численность в Латвии
оценивается в 5-20 пар, в Литве — 1-10 пар, Молдове — 1,8-2,3 тыс. пар, Украине —
13,2-22,3 тыс. пар, Армении — 1,0-2,5 тыс. пар, Азербайджане — 0,25-2,5 тыс. пар; из
других стран Европы волчок наиболее многочислен в Румынии (8,5-10 тыс. пар) и Венг-
рии (4-6 тыс. пар); в Турции обитает 6-9 тыс. пар. Общая численность волчка в Европе
оценивается в 60-120 тыс. пар (BirdLife International, 2004).
В Средней Азии волчок считается нередким видом, однако данные по его общей чис-
ленности здесь отсутствуют (Птицы Средней Азии, 2007); в Зап. Сибири вид редок (Крас-
ная книга Новосибирской обл., 2000; Красная книга Алтайского края, 1998).
В местах, где вид многочислен — например, в ряде мест Краснодарского края — его
численность может достигать 50 особей на 1 км2 (Казаков и др, 2004). В Белоруссии отме-
чены отдельные места обитания с очень высокой для региона плотностью — 15,7 пар на
1 км2 (вторично заболоченные торфоразработки, Брестская обл). В Липецкой обл. России
на 280 га рыборазводных прудов было выявлено 23-29 пар малой выпи (Кузнецова, 2006).
Однако столь высокая плотность встречается достаточно редко; например, в пойме р. При-
пять (Белорусское Полесье) она составляет только 1-2 пары на 1 км2 (Дмитренок, 2009). В
средней полосе Европейской России вид никогда не достигает высокой численности даже
в благоприятных для него местообитаниях. Так, на не расчищаемых от надводной расти-
тельности рыборазводных прудах Московской обл. плотность населения в 2007 г. составля-
ла 0,02 пары на 1 га акватории (Суханова, 2008).
Размножение. Половой зрелости волчок достигает в возрасте одного года, хотя часть
птиц, как и большинство других видов цапель, видимо приступают к размножению на вто-
рой-третий год жизни. Самки могут приступать к размножению в возрасте менее одного
года (Kushlan, Hancock, 2005). Часть молодых птиц остаётся на местах зимовки во второе
лето жизни (Pezzo, Gosler, 2005).
Моногам; пары образуются, как минимум, на один сезон. По данным большинства ав-
торов, для волчка характерно одиночное гнездование, и групповых поселений вид не обра-
зует. Некоторые авторы указывают, что хотя волчок и не колониальный вид, он иногда мо-
жет гнездиться небольшими группами по 2-3 пары в 50 м друг от друга (Cramp, Simmons,
1977). Однако, имеются и другие данные. Так, на Украине на Днестре отмечены 7 гнёзд
малой выпи, расположенные поблизости друг друга; в Каневе несколько лет подряд нахо-
дили по 2-3 гнезда, расположенные рядом (Смогоржевский, 1979). В богатых кормовыми
ресурсами прудах рыбхозов на Сев. Кавказе волчки могут образовывать подобия рыхлых
колоний, где расстояния между гнёздами составляют 2,3-36 м, в среднем 17-20 м (Казаков
184
Отряд Листообразные, Семейство Цаплевые
и др., 2004). Плотные гнездовые поселения отмечены также на юго-востоке Казахстана:
там найдены два гнезда, расстояние между которыми составляло менее 1 м; гнёзда были
расположены по обеим сторонам густого залома тростника (Лопатин и др., 1992). В Бело-
руссии в 2005-2007 гг. на территории заболоченного карьера в Бобруйском р-не в центре
массива рогоза располагалась группа из 6 гнёзд, кратчайшие расстояния между которыми
составляли 3,7-15 м. В Минске на Чижовском вдхр. все три найденных здесь в 2007 г.
гнёзда находились на расстоянии 2-20 м друг от друга (Островский и др., 2004). На вторич-
но заболоченных торфоразработках в Брестской обл. 10 гнёзд располагались на расстоянии
10-25 м одно от другого. Таким образом, у малой выпи имеется склонность к формирова-
нию агрегированных гнездовых поселений.
Подобные групповые поселения становятся возможными благодаря специфике терри-
ториального поведения самцов. Самцы отстаивают гнездовую территорию, с которой изго-
няют всех других самцов. Однако птицы, создавшие пару и закончившие кладку, становят-
ся менее агрессивными, защищают только гнездо и его ближайшие окрестности, что по-
зволяет поселиться по соседству с ними другой паре (Langley, 1983; Voisin, 1991). Эти дан-
ные косвенно подтверждаются наблюдениями в Белоруссии: здесь близко расположенные
гнёзда различались по срокам откладки яиц на 7-12 дней (Островский и др., 2004).
Интересно отметить, что в четырёх находившихся под наблюдением местах гнездова-
ния антропогенного происхождения в Белоруссии, в том числе и в наиболее плотных гнез-
довых поселениях, гнёзда волчков находились в непосредственной близости от колоний
озёрных чаек. Гнёзда волчков могут также тесно соседствовать с гнёздами хохлатой черне-
ти, чомги (Островский и др., 2004), дроздовидной камышевки, лысухи, красноносого ныр-
ка (Лопатин и др., 1992).
После прилёта самцы занимают гнездовые территории, при этом между ними нередко
происходят драки (Cramp, 1977; Птицы Средней Азии, 2007). Наряду с использованием
брачного крика, территории защищают с помощью наземных и воздушных атак и угрожа-
ющих демонстраций. В последнем случае птица, находясь сбоку от оппонента, разворачи-
вает крылья и затем одно крыло опускает, а другое поднимает. В период образования пар
самцы демонстрируют круговые полёты. Копуляция происходит на гнезде, ей предшеству-
ет церемония приветствия, наблюдаемая также во время смены партнёров на гнезде: парт-
нёры открывают и закрывают клювы, касаются клювами друг друга; перья на спине и груди
при этом приподняты, хохол на голове поднимается и опускается (Cramp, 1977; Kushlan,
Hancock, 2005). Некоторые позы волчка приведены на рисунке 33.
В строительстве гнезда, которое занимает 2-6 дней, участвуют оба члена пары (Дол-
гушин, 1960; Лопатин и др., 1992; Птицы Средней Азии, 2007). Излюбленные места гнез-
дования — затопленные кустарники, тростниковые и рогозовые крепи; часто гнездо распо-
лагается в развилках кустов, отмечены гнёзда на ольхе, шиповнике среди таких зарослей.
В Белорусси волчок явно предпочитает устраивать гнёзда в густом кустарнике — так,
чтобы гнездо невозможно было бы обнаружить сверху; в рогозе и тростнике он выбирает
наиболее загущённые участки, преимущественно с сухими прошлогодними стеблями. Все
обследованные в пойме р. Припять гнёзда располагались в смешанных зарослях ивняка с
тростником. Такое сочетание растительности обеспечивает максимальную маскировку для
гнёзд (Дмитренок и др., 2009). Однако иногда встречаются и совершенно открытые гнёзда.
В Предкавказье волчки строят гнёзда в густых рогозовых и тростниковых зарослях, на
заломах их стеблей или кочках; при этом птицы нередко используют естественные укры-
тия. Предпочтение укрытий нередко приводит к тому, что волчок поселяется под гнёздами
других цапель и даже в искусственных гнездовьях для уток (Казаков и др., 2004). На Укра-
ине (Смогоржевский, 1979) и в Белоруссии отмечены случаи гнездования в старых сорочь-
их гнёздах.
Волчок
185
Рис. 33. Некоторые позы волчка (Bauer, Glutz von Blotzheim, 1966): а-г — вертикальная поза при
нарастании возбуждения, д-ж — позы приветствия и угрозы птицы на гнезде при снижении степени
возбуждения, з — поза угрозы, направленная на приближающуюся птицу.
186
Отряд Листообразные, Семейство Цаплевые
Места расположения гнёзда очень разнообразны. Гнездо, как правило, расположено низко
над водой; иногда оно касается воды нижнем краем, может быть построено на сплавине, но
чаще приподнято на 14-40 см. В Казахстане отмечены случаи гнездования на деревьях на
высоте до 3 м (Лопатин и др., 1992), в Белоруссии — 1,5-2 м (в сорочьих гнёздах). Перво-
начально гнездо имеет форму перевёрнутого конуса, позднее растаптывается и может стать
плоским. Постройка на вид лёгкая (особенно если гнездо сооружено на ветвях куста), в
некоторых случаях яйца просматриваются сквозь стенки и дно гнезда. В качестве гнездо-
вого материала используются растения, которые преобладают в данном месте: стебли тро-
стника, рогоза (при гнездовании в тростниковых и рогозовых зарослях) или веточки (при
гнездовании на кустах). Лоток может выстилаться листьями и метёлками тростника, листь-
ями рогоза, сухими стеблями травы (Спангенберг, 1951; Долгушин, 1960; Казаков и др.,
2004 и др.); однако в Белоруссии при обследовании 40 гнёзд в большинстве из них выстил-
ки не было или она была минимальной. Размеры гнёзд из Предкавказья (п=42): диаметр
гнёзда 14-29 х 17,8-32 см, в среднем 22,1x23,6 см; диаметр лотка 9-17 х 10,4-17 см, в
среднем 12,2x13,2 см; высота гнезда 10-35 см, в среднем 15,8 см; глубина лотка 1,5-9 см,
в среднем 4,3 см (Казаков и др., 2004). В Белоруссии по промерам 28 гнёзд получены сход-
ные данные. По Средней Азии имеются следующие сведения (n= 10): диаметр гнезда 15-20
см, диаметр лотка 9,0-16,0 см, высота гнезда 6,3-21,0 см, глубина лотка 1,2—4,0 см. Гнездо
строится за 2-4- дня (Птицы Средней Азии, 2007). Первые яйца могут откладываться в ещё
недостроенное гнездо, оно постоянно достраивается в процессе откладки яиц, насижива-
ния и выкармливания птенцов.
Первые свежие кладки в Предкавказье отмечаются в середине мая, последние — 2-18
июня; сезон гнездования, таким образом, сильно растянут (Казаков и др., 2004). В более
северных регионах волчки приступают к откладке яиц позже. Так, в Калужской обл. кладки
отмечаются с начала июня, но свежие кладки могут быть встречены здесь и в первой дека-
де июля (Марголин, 2000). В Белоруссии гнездовой сезон также очень растянут, самая ран-
няя из обнаруженных кладок (Брестская обл.) была отложена ориентировочно 10-15 мая
(исходя из размеров найденных в июне птенцов). Но основная масса кладок появляется в
июне-июле. В Зап. Палеарктике обычно бывает один выводок за сезон; иногда, после гибе-
ли первой кладки, возможно повторное гнездование (Cramp, 1977; del Hoyo et al., 1992).
Высказывалось предположение о наличии у некоторых пар двух кладок в году (Марголин,
2000). Теоретически это возможно, так как птенцы малой выпи очень быстро растут, а
гнездовой сезон сильно растянут. В Белоруссии в гнезде, где 20 июня 2007 г. был найден
выводок уже почти полностью оперившихся птенцов, через 20 дней обнаружена новая,
слегка насиженная кладка. Не исключено, что это была вторая кладка одной и той же
пары. Для подвида I. т. payesii доказано, что в случае разорения кладки птицы отклады-
вают новую; отмечен случай повторного гнездования и даже третьего гнездования в году
(Langley, 1983).
В полной кладке от 3 до 9 яиц; в Предкавказье (п=56) в 4 гнёздах было по 3 яйца, в
одном — 4, в 23 — по 5, в 22 — по 6, в 5 — по 7 и в одном — 9 яиц; в среднем — 5,6 яиц на
гнездо. На Украине полная кладка обычно содержит 6 яиц, но встречаются кладки с 5-9
яйцами; в Средней Азии величина кладки составляет 4-9 яиц, в Рязанской обл. в 1960-1990-
х гг. (п=5) в кладках было от 3 до 7 яиц. В Белоруссии в насиженных кладках было 4-7 яиц,
в среднем 5,6 (п=9). Размеры яиц (мм) из дельты Кубани (п=14): 34,0-35,8 х 25,0-27,2, в
среднем 35,0x26,1; из Челбасских плавней (п=59): 31,1-39,0 х 23,6-27,9, в среднем 35,2x28,5;
из Рязанской обл. в среднем по двум кладкам: 37,6±0,52 х 26,57±0,29; из Средней Азии (п=20):
32,3-35,7 х 25,0-21,6; из Белоруссии (п=62): 31,6-38,5 х 24,6-28,0, в среднем 34,9x26,2. Вес
свежих яиц (п=10) — 13,2-13,9 г, в среднем 13,5 г. Яйца белые, изредка голубоватые или
Волчок
187
зеленоватые, блестящие или матовые, овальной или округлой формы (Долгушин, 1960; Snow,
Perrins, 1998). Яйца откладываются с разным интервалом: ежедневно (в 80% случаев), через
одни сутки, иногда даже через 2-3 суток (Лопатин и др., 1992). Насиживание начинается с
первого (по некоторым сведениям — с первого-второго) яйца и продолжается, по данным
разных авторов, 16-19 дней или 18-21 день. В насиживании принимают участие оба партнё-
ра, но больше времени на гнезде проводит самка (Смогоржевський, 1979; Нумеров и др,
1995; Казаков и др., 2004; Птицы Средней Азии, 2007). Предполагается, что у волчка суще-
ствует внутривидовой гнездовой паразитизм, так как дважды наблюдались случаи, когда в
пронумерованных кладках одно из яиц исчезало и появлялось новое. Описан случай под-
кладывания в гнездо волчка яйца камышницы (Лопатин и др., 1992).
В жаркую погоду родители могут вставать над гнездом, раскрывая одно из крыльев и
создавая тень для кладки или птенцов (Лопатин и др., 1992; данные М.Г. Дмитренок). Сме-
на партнеров на гнезде за 8 часов наблюдений происходила 6 раз во время откладки яиц и
12 раз на гнезде с маленькими птенцами (данные М.Г. Дмитренок). Насиживающие птицы
проявляют защитную реакцию по отношению к нежелательным визитерам — раскрывают
клюв, встопорщивают перья на голове (Voisin, ориг.). Застигнутые врасплох на гнезде вол-
чки могут атаковать даже человека, защищая своё потомство. Однако если птицы заранее
слышат приближение человека, они загодя покидают гнёзда.
Вылупление протекает в течение 3-5 дней. Средний вес однодневного птенца (п=10)
10,6=1=0,1 г. До 7-8-дневного возраста один из родителей остаётся с птенцами, другой при-
носит корм; позднее за кормом начинают летать оба родителя (Казаков и др., 2004).
Глаза у птенцов открываются через 3-Л часа после вылупления, по другим сведениям
птенцы вылупляются зрячими (Сагитов и др., 1973). Первые два дня жизни птенцы мало-
подвижны, но уже на четвёртый день они хорошо стоят на ногах и способны при опасности
принимать «позу выпи» — вытягивать, как взрослые, шею и клюв вверх (Сагитов и др.,
1973; данные М.Г. Дмитренок).
Общее число прилетов с кормом варьирует в разные дни от 12 до 17 раз в день (Казаков
и др., 2004). По данным Е.П. Спангенберга (1951), родители собирают корм для птенцов в
основном вблизи гнезда, но когда потомство подрастёт и требует больше пищи, они совер-
шают вылеты за кормом на более отдалённое расстояние, не превышающее, однако, 1 км
(по наблюдениям в Предкавказье волчок может улетать от гнезда на кормежку и на 1,5 км —
Казаков и др., 2004). Птицы летят низко над растительностью или водой.
В течение двух перых дней родители кладут корм для птенцов на дно гнезда. На третий
день птенцы уже выпрашивают корм у родителей, захватывая их клюв. Несмотря на разни-
цу в возрасте птенцов, соперничество у них за пищу выражено слабо. Птенцы растут и
развиваются быстро. Пеньки первостепенных маховых появляются на четвёртый день, ру-
левых — на пятый. Наиболее интенсивный рост крыла, клюва и цевки происходит в пер-
вые 2-8 дней жизни, веса — в первые 2-9 дней. Из гнезда птенцы начинают выходить в
возрасте 8-10 дней, а полностью покидают гнездо на 14-18 день. В возрасте 20 дней птен-
цы уже покрыты перьями; на крыло они поднимаются через месяц после вылупления (Ло-
патин и др., 1992; Казаков и др., 2004; Kushlan, Hancock, 2005). У птенцов хорошо изучен-
ного подвида I. т. payesii наиболее интенсивный рост крыла, клюва и цевки происходит в
первые 2-15 дней жизни, веса — в первые 2-20 дней (Langley, 1983).
Данные об успехе размножения скудны. Для Средней Азии имеются следующие сведе-
ния: в четырёх гнёздах из 16 яиц вылупилось 7 птенцов (43,8%), из которых до подъёма на
крыло дожили 5 молодых (71,4%). В.В. Лопатиным с соавторами (1992) получены следую-
щие показатели успеха размножения: отход яиц — 0,3-0,4 яиц на гнездо, птенцов — 0,1-
0,3 птенца на гнездо. Количество погибших гнёзд — 16,2-28,1%. Причины гибели: эмбри-
188
Отряд Листообразные, Семейство Цаплевые
ональная смертность, перепады уровня воды, разорение болотными лунями, змеями, лас-
кой (Лопатин и др., 1992; Птицы Средней Азии, 2007).
В Белоруссии успех гнездования в значительной степени различается в разных услови-
ях обитания, причём наиболее высокий процент выживания отмечен на антропогенных
водоёмах. Так, в пойме Припяти успех гнездования крайне низок — из 7 найденных в 2002-
2007 гг. гнёзд птенцы дожили до 10-дневного возраста только в одном, остальные были
разорены хищниками (куньими). На двух водоёмах антропогенного происхождения успех
гнездования был, наоборот, очень высок: из 9 гнёзд на Чижовском вдхр. (Минск) сохрани-
лись все, из 14 гнёзд на выработанных и залитых водой торфоразработках в Брестской обл.
сохранились 12. Как в Белоруссии, так и в Казахстане гибель потомства в основном проис-
ходит на стадии насиживания.
Максимальная продолжительное жизни волчков, по данным кольцевания — 21 год (Нан-
кинов, 1999).
Суточная активность, поведение. Большинство орнитологов считают волчка птицей
с сумеречной или сумеречной и ночной активностью. Дневная активность отмечается в
брачный сезон и в период выкармливания птенцов (Спангенберг, 1951; Казаков и др., 2004;
Птицы Средней Азии, 2007 и др.). Однако существует точка зрения, что волчок ведёт не
ночной, а скрытный дневной образ жизни (Марголин, 2000). Для обитающего в Африке
подвида I. т. payesii с помощью размещённого на гнезде датчика определено, что основное
количество посещений гнезда происходит в светлое время суток и в сумерках. Пик актив-
ности приходится на полдень, небольшая двигательная активность отмечена также в тече-
ние трёх предрассветных часов, ночью зарегистрированы единичные передвижения от гнезда
или к нему (Langley, 1983). Для номинативного подвида установлено, что в период выкар-
мливания птенцов наибольшую активность по сбору корма родители проявляют в раннеут-
ренние (4-6), полуденные (10-12) и вечерние (18-20) часы (Казаков и др., 2004).
Вне сезона размножения для волчка характерен преимущественно одиночный образ
жизни; в период миграции птицы могут объединяться в группы от 2-3 до 10-15 особей
(Cramp, 1977; Казаков и др., 2004; Птицы Средней Азии, 2007).
Питание. Питается животной пищей. В корме преобладают мелкие рыбешки, лягушки,
головастики, а также водные и наземные насекомые, пауки. В Челбасских плавнях основу
рациона волчка составляла мелкая рыба (38,2% по весу и 40% по встречаемости) — ко-
люшка, бычки, речной окунь, язь, карась и др. На втором месте стояли земноводные (34,1 %
и 28,4% соответственно) — главным образом, головастики озёрных лягушек, реже взрос-
лые лягушки и чесночницы. Третье место занимали насекомые (20,2% по весу и 43,3% по
встречаемости), 94,7% массы которых приходилось на водных насекомых (личинки 6 ви-
дов стрекоз, личинки жуков-плавунцов и жуков-водолюбов, различные водяные клопы). В
дельте Кубани основу питания составляли водные насекомые (62,2% по весу и 90,0% по
встречаемости) (Казаков и др., 2004).
В Белоруссии на 1 пищевую пробу в среднем приходится 3,7 экз. жертв (1-13 экз.), из
них 1,9 экз. беспозвоночных и 1,8 экз. рыб. Размер рыб в пробах питания был 2,6-8,1 см, в
среднем 4,6 (п=15). Среди беспозвоночных доминировали клопы рода Notonecta и Naucoris.
Общее количество этих насекомых составило 23,6% от всех жертв, тогда как остальных
беспозвоночные (включая пауков и мух) — 25%. Кроме того, в питании волчков отмечены
Cybister lateralimarginalis, Rhantus sp., Hydaticus, личинки стрекоз Aeschna, Corduliidae,
пауки. Из позвоночных зарегистрированы амфибии, рыбы Carassius auratus, Leucaspius
delineatus, Misgurnus fossilis, Rutilus rutilus, Perccottus glenii. На Украине в питании волчка
отмечены 8 видов земноводных, 15 видов рыб, водные и наземные жесткокрылые, водные
клопы, стрекозы, подёнки, прямокрылые, пауки, муравьи, моллюски, щитни и пиявки (Смо-
горжевський, 1979).
Волчок, Китайский волчок
189
В Казахстане и Средней Азии основу кормового рациона волчка составляют разнооб-
разные водные насекомые и их личинки, а также мелкая рыба и земноводные (Долгушин,
1960; Птицы Средней Азии, 2007). Подобный спектр питания характерен для вида в По-
волжье (Попов, 1977) и в средней полосе Европейской России (Птушенко, Иноземцев, 1968;
Марголин, 2000 и др.). Под Ленкоранью малые выпи, гнездящиеся на лесных водоёмах, в
большом количестве поедали древесных лягушек (квакш), реже рыбу и массу разнообраз-
ных насекомых (Спангенберг, 1951). Многие авторы, изучавшие питание вида, отмечают
нередкое разорение волчками гнёзд мелких птиц, из которых они забирают как птенцов,
так и яйца; изредка отмечалось и разорение утиных кладок. Иногда волчки почти полнос-
тью переключаются на питание сухопутными насекомыми. Так, по наблюдениям Е.П. Спан-
генберга (1951), на осеннем пролёте в Крыму желудки волчков были буквально набиты
остатками кобылок и, видимо, богомолов.
Добывая мелкую рыбу и водных насекомых, волчки подкарауливают добычу, подолгу
сидя на ветке, нависающей над водой, или на пучке тростника (Спангенберг, 1951). При
активном способе охоты птица быстро перемещается в сгорбленной позе с опущенной го-
ловой. Заметив добычу, поднимает голову и делает резкий выпад. Кормится в одиночку на
своей территории (Kushlan, Hancock, 2005).
Враги, неблагоприятные факторы. Специфических естественных врагов, по-види-
мому, не имеет. На южных реках и, особенно, в районах вновь созданных водохранилищ,
где перепады уровня воды весьма значительны, часто страдает от затопления гнёзд. В этом
случае отмечены повторные кладки. На прудах рыбхозов и некоторых других водоёмах
гнёзда нередко гибнут при выжигании сухого прошлогоднего тростника. Отмечено сниже-
ние численности из-за загрязнения водоёмов. Так, на оз. Муромском (Московская обл.) из-
за залповых сбросов загрязнённой воды Шатурской ГРЭС численность волчка снизилась с
2 особей на 1 км береговой линии в 1980 г. до 0,25 особей на 1 км береговой линии в 2007 г.
(Мищенко, 2008; Суханова, 2008). В пойме р. Припять отмечена высокая смертность кла-
док волчков в результате хищничества куньих (предположительно, куницы).
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Волчок занесён
в Красные книги, Латвии, Литвы, Республики Беларусь и в региональные Красные книги
ряда субъектов Российской Федерации: республик Коми, Марий-Эл, Мордовия, Татарстан,
Удмуртия; Алтайского и Пермского краёв; Владимирской, Калужской, Курганской, Липец-
кой, Московской, Нижегородской, Новосибирской, Омской, Орловской, Пензенской, Ря-
занской, Тверской, Ульяновской, Ярославской областей.
Китайский волчок
Ixobrychus sinensis (J.F. Gmelin, 1789)
Ardea sinensis J.F. Gmelin, 1789, Syst. Nat., 1, c. 642, Китай.
Yellow bittern [Chinese little, long-nosed] bittern (англ.), Chinesedommel (нем.),
biongios de Chine, biongios chinois (фр ).
Син. (лат.): Ardea lepida Horsfield, 1821; Ardetta luteola Stejneger, 1887; Ardetta bryani Seale,
1901; Ixobrychus sinensis astrologus Wetmore, 1918; Ixobrychus sinensis moorei Wetmore, 1918;
Ixobrychus sinensis p elewens is Momiyama, 1932; Ixobrychus sinensis yapensis Momiyama, 1932.
Син. (рус.): китайская выпь, длинноносый волчок, [охристая выпь — ошибочно].
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Одна из самых мелких цапель, сходная
по сложению и типу движений с амурским волчком, а в ещё большей степени — с волчком.
190
Отряд Листообразные, Семейство Цаплевые
Из всех видов малых выпей имеет наиболее тонкий и длинный клюв. У самца тёмные пе-
рья темени удлинены и образуют небольшой хохол. В полете хорошо отличается от самцов
волчка коричневой (а не чёрной) спиной и тёмно-жёлтым крыловым пятном. В окраске
преобладают охристые тона, поэтому птица кажется очень светлой по сравнению с амур-
ским и охристым волчками; у летящей птицы тёмные маховые перья резко контрастируют
с остальным светлым оперением. От амурского и охристого волчов отличается меньшими
размерами, более светлой окраской верха тела, оперённой до сочленения с голенью цевкой.
У самок и молодых птиц, в отличие от амурского волчка, вместо белых и охристых крапин
на спинной стороне и кроющих крыла имеются достаточно широкие охристые каёмки по
краю опахал, создающие общий «полосатый» фон верхней части тела. На нижней части
тела пятнистость менее выражена.
Подобно другим видам волчков, ведёт одиночный образ жизни. Держится главным об-
разом в тростниковых зарослях, кустарниках или в другой густой растительности, особен-
но вокруг полей, озёр, медленно текущих рек, мангровых болот. Этих скрытных птиц чаще
можно увидеть в коротком и низком полёте. Хорошо лазает по стеблям тростника и рогоза,
по веткам кустов, садится на деревья. Взлетает неохотно, летит низко над растительнос-
тью, прямым быстрым полетом.
Голос. Брачный крик самца — довольно громкое и частое повторение слогов «ох-ох-
ох...». Его чаще всего можно услышать в сумерки в начальный периода гнездования (Austin,
Kuroda, 1953). Три типа позывок — контактная, тревожная и крик бедствия — характерны
как для взрослых, так и для молодых птиц. Контактная позывка («кавканье») — короткий
(от 0,01 до 0,08 с у взрослых и от 0,1 до 0,3 с у молодых птиц) широкополосный звук с
шумоподобным спектром заполнения; при повышении уровня тревожности частотный ди-
апазон и длительность позывок увеличиваются. Тревожная позывка («гудок»), преобразо-
ванная из учащенной контактной позывки — короткий широкополосный тональный сиг-
нал с гармоническим спектром. Крик бедствия — продолжительный (0,3-0,6 с) широкопо-
лосный (в среднем 10,8-23,3 кГц) шумовой сигнал, используемый при крайней степени
тревоги. Специализированная тревожная позывка самки («кукареканье») — низкий по зву-
чанию, широкополосный, с наличием гармонических составляющих сигнал, используе-
мый совместно с тревожным «кавканьем» возле гнездовых птенцов в присутствии челове-
ка. Ювенильные позывки: «мяуканье» — первичная позывка для поддержания контакта
между молодыми одного выводка и взрослыми в момент кормления (плотная гомотопиче-
ская серия из широкополосных нот), которая позднее преобразуется в сугубо пищевой сиг-
нал; «шипение», или «гогот», имеющий совершенно иную структуру (тональные звуки с
шумоподобным или гармоническим спектром); контактно-тревожная «перекличка» между
молодыми птицами, покинувшими гнездо — широкополосный составной трелевый сиг-
нал, в котором происходит постепенное уменьшение интенсивности звучания посылок.
Помимо голосовых сигналов китайский волчок использует механические звуки — глу-
бокие удары крыльями в рекламирующих круговых полётах.
Подвидовая систематика. Монотипический вид. Существуют незначительные вариа-
ции в размерах и окраске между птицами разных частей ареала (Спангенберг, 1951). Пред-
полагалось выделение до пяти подвидов на основе различий в окраске, но из-за большой
индивидуальной изменчивости это предложение было отвергнуто (Дементьев и др., 1951;
Kushlan, Hancock, 2005). Образует одну надвидовую группу с волчком (Z. minutus) и карли-
ковым, или индейским волчком (Z. exilis) (del Hoyo et al., 1992).
Описание. Окраска. Самец в брачном наряде. Лоб и темя чёрные с сизым оттенком;
на лбу каймы перьев бурого цвета; задняя сторона шеи рыжеватая; плечи и верхняя часть
спины глинисто-буроватые с охристыми каймами перьев; нижняя часть спины серая; верх-
ние кроющие хвоста сизо-серые; первостепенные маховые, их кроющие и рулевые чёрные
Китайский волчок
191
с серым налётом; внутренние второстепенные маховые глинисто-охристые; верхние крою-
щие крыла светлые, глинисто-песочного цвета; подбородок и верхняя часть горла светлые
с охристой полосой; нижняя часть горла и грудь охристые; по бокам верхней части груди
расположены крупные пятна из чёрных с узкими охристыми краями перьев; брюхо, испод
крыла, подмышечные и нижние кроющие хвоста белые; клюв бледно-жёлтый с бурым конь-
ком надклювья; в основании надклювья и подклювья в разгар брачного сезона проступают
розоватые пятна. Голые части лица зеленоватые; радужина жёлтая; ноги зеленовато-жёл-
тые, задняя сторона цевки и нижняя сторона пальцев красновато-буроватые.
Самка в брачном наряде. У самки на лбу перья рыже-бурого цвета с тонкой централь-
ной чёрно-бурой полоской, на темени перья чёрно-буроватые; бока головы глинисто-охри-
стые; перья на спине рыжие с тонкой охристой каймой, мелкие кроющие и внутренние
второстепенные маховые каштановые с широкими охристым каймами; большие кроющие
крыла светло-охристые с бурым налётом по центральной части; первостепенные и второ-
степенные маховые чёрно-бурые с бурой вершинной частью, рулевые чёрно-бурые, подбо-
родок и горло светлые с продольной буровато-охристой полосой, по бокам верхней части
груди по светло-бурому пятну; на груди немногочисленные продольные буро-охристые пе-
стрины.
Гнездовой наряд. Лоб и темя каштановые с чёрными центрами перьев; перья на спин-
ной стороне и кроющих крыла рыже-бурые с широкой светлой окантовкой; брюшная сто-
рона каштаново-охристая с немногочисленными узкими продольными тёмно-бурыми пе-
стринами; на шее многочисленные рыжие пестрины; остальное оперение, как у взрослой
самки.
У птенца, готового покинуть гнездо (в возрасте около 13 суток), имеются перья на кры-
льях и спине, на остальных частях тела только пеньки: у первостепенных маховых (39 мм)
пеньки тёмно-серые, кисточки чёрные; у второстепенных маховых (34 мм) пеньки тёмно-
серые, кисточки тёмно-коричневые со светло-рыжими каемками; кисточки горловых перь-
ев коричнево-рыжие с тёмно-коричневым цветом по центру; кисточки спинных перьев тём-
но-коричневые с рыжими краями; бедренных — белые с охристыми краями. На всех пте-
рилиях сохраняются остатки пуха. Клюв телесный с серым коньком, на кончике клюва ещё
просматриваются следы яйцевого зуба в виде белого пятна. Лапы от светло-жёлтых до жёл-
тых с зеленоватым оттенком, когти серовато-коричневые. Радужина тёмно-зелёная. Пуд-
ретки жёлтые.
Пуховой птенец. Покрыт светло-рыжим длинным, с выраженным блеском пухом, на
брюшных и межчелюстных птерилиях пух светлее — светло-золотистый с белым в основа-
нии. Длина пуха на лобной, теменной и затылочной птерилиях 7-8 мм, на глазных птерили-
ях 2 мм, на надглазничных, межчелюстных, кистевых и локтевых птерилиях 4-6 мм, на
спинной и бедренных — 9-10 мм. Ноги, когти, клюв и ротовая полость розовато-телесные,
кожа вокруг глаз и на уздечке голубоватая, радужина тёмно-серая. Яйцевой зуб белый с
коричневым цветом в основании. Слуховые отверстия круглые, 2 мм; ноздри щелевидные
(Гамова и др., в печати).
Строение и размеры. Небольшая стройная цапля, 30^40 см в длину. Голова
относительно небольшая, узкая. Клюв длинный и относительно тонкий, значительно длин-
нее цевки. Голень оперена до сочленения с цевкой. Цевка относительно длинная, покрыта
щитками. Первая фаланга среднего пальца меньше половины длины второй фаланги (в
отличие от амурского волчка). Хвост короткий, из 8 рулевых перьев. Крылья короче 135
мм, широкие, закруглённые. Первостепенных маховых 10, второстепенных маховых 13.
Формула крыла: II>III>I?IV>V>VI>VII... I первостепенное короче II первостепенного
на 2,5^,8 (3,9±0,7, n=l 1) мм, ПКП на 0,3-2,0 (1,2±0,6) мм, IV<III на 0,5-5,0 (2,6±1,4) мм,
192
Отряд Листообразные, Семейство Цаплевые
V<IV на 1,2-5,8 (3,5±1,6) мм, VI<V на 1,2-5,8 (3,5± 1,6) мм, VIKVI на 2,7-6,5 (5,0±1,0) мм.
VIIKVII на 3,2-6,5 (5,4±0,9) мм, IX<VII на 4,5-8,3 (6,4±1,2) мм, Х<1Х на 4,9-7,3 (5,7±1,0)
мм.
Размеры (мм). Птицы из Китая, Японии, Приморского края (колл. ЗИН, ДВГУ, БПИ
ДВО РАН, Пекинского Зоологического института, Института Ямасины). Взрослые самцы
без учёта сезона: длина крыла (максимально выпрямленного на плоскости) (п=6) — 120,0-
141,3 (в среднем 131,7±6,9); длина хвоста (п=4) — 43,7—49,0 (46,4±3,7); длина клюва от
оперения (п=6) — 48,0-53,5 (50,3±2,0), длина клюва от переднего края ноздри (п=4) —
38,7-40,4 (39,6±1,2); высота клюва на уровне переднего края ноздри (п=4) — 8,5-8,8
(8,7±0,2); ширина клюва на уровне переднего края ноздри (п=4) — 6,4-7,0 (6,7±0,4); длина
цевки (п=6) — 45,0—46,8 (45,9±1,3).
Взрослые самки без учёта сезона: длина крыла (максимально выпрямленного на плос-
кости) (п=16) — 118,6-138,7 (в среднем 127,7±5,5); длина хвоста (п=7) — 45,5-53,0
(48,3±2,7); длина клюва от оперения (п=17) — 47,8-53,6 (51,0±1,6); длина клюва от пере-
днего края ноздри (п=5) — 39,2-51,6 (43,3±4,8); высота клюва на уровне переднего края
ноздри (п=5) — 7,9-9,7 (8,6±0,7); ширина клюва на уровне переднего края ноздри (п=4) —
6,0-7,5 (6,8±0,6); длина цевки (п=7) — 42,0^6,0 (44,9±1,3).
Размеры (мм) птиц (п=3) из Сев. Кореи: длина крыла -126-138 (в среднем 132±6,0),
длина клюва — 45-53 (48,5±4,1), длина цевки — 43—45 (44,3±1,2), длина хвоста — 48-60
(52,7±6,4) (Тошек, 1999).
Масса тела (г): самцов (п=18) — 63-115, в среднем 94; самок (п=10) — 75-120, в сред-
нем 95 (Shaw, 1936).
Линька. Информация отсутствует.
Распространение. Гнездовой ареал. Юж. и Юго-Вост. Азия к северу до Японии
(включая о. Хоккайдо), Корейского п-ова и Сев.-Вост. Китая, к югу — до островов Зондс-
кого архипелага. В Китае область распространения в целом совпадает с распространением
амурского волчка, она охватывает Вост, и Юж. Китай, Тайвань, на север доходит до г. Хар-
бин; западная граница проходит по провинциям Шеньси, Ганьсу и Сычуань (Cheng Tso-
hsin, 1987). На юге охватывает Индокитай, Большие и Малые Зондские о-ва, Филиппины,
Новую Британию, Каролинские и Марианские о-ва, на западе включает Индию, Шри-Лан-
ку, Мальдивские и Сейшельские о-ва. Залёты отмечены в юго-зап. Австралию и на о. Рож-
дества в Индийском океане (del Hoyo et al., 1992; Kushlan, Hancock, 2005) (рис. 34).
Неоднократно отмечался на юге Дальнего Востока России. Впервые самец был добыт
27 мая 1879 г. на о. Аскольд (Taczanowski, 1891-1893). На о. Большой Пелис (залив Петра
Великого) самец добыт 29 мая 1967 г., погибшая птица найдена 10 июня 1968 г. (Лабзюк и
др., 1971), взрослого самца наблюдали 1 июля 1989 г. Самца встретили 26 июля и 2 августа
1972 г. у пос. Хасан (Назаров, Лабзюк, 1975), взрослая самка добыта 20 сентября 1979 г. под
г. Уссурийск (Глущенко и др., 1986). Одна птица была добыта на северо-восточном побере-
жье Приморского края у пос. Терней 24 июня 1979 г. На побережье Уссурийского зал. в
дельте р. Раздольная у устья р. Грязной взрослый самец кормился 10 июня 1989 г. (Назаров,
2004), в том же районе встречен самец на гнездовом участке 26 и 30 июля 2000 г. (Нечаев,
2003). Самка добыта 29 мая, а самец — 8 июня 1980 г. на побережье зал. Ольга (колл. БПИ
ДВО РАН). На Приханкайской низменности отмечены две встречи этого вида: 6 июня 2003 г.
в окрестностях с. Новосельское отмечена самка и 23 июня 2009 г. — самец на озёрах вос-
точного побережья Ханки. Характерные токовые крики китайского волчка отмечены 29
мая 2004 г. в долине р. Уссури в окрестностях посёлка Кировский и 7 июля 2008 г. в окрес-
тностях озёра Утиное, в низовьях р. Раздольной (Волковская-Курдюкова, 2009; А.Б. Кур-
Китайский волчок
193
Рис. 34. Область распространения китайского волчка: а — гнездовой ареал, б — места зимовок, в —
недостаточно выясненная граница гнездового ареала.
дюков, личн. сооб.). В 2008 г. установлено гнездование на юге Приморского края, в приго-
роде г. Владивостока (Нечаев, Гамова, 2009; Гамова и др., в печати).
В гнездовой период птиц наблюдали на о. Сахалин вблизи пос. Стародубское, где были
добыты 4 птицы: один самец 24 июня, один самец и две самки 25 июня 1914 г. (Thayer, Bangs,
1916; Munsterhjelm, 1922), одна птица (без указания даты) — на оз. Невское (Takahashi, 1937а,
с, d). На о. Кунашир одну птицу наблюдали в августе 1929 г. (Yamashina, 1931) (рис. 35).
Зимовки. Зимует китайский волчок на юге Китая, в провинции Гуандун, на о-вах
Хайнань, Тайвань, на южных Японских о-вах, в Индии, Индокитае, Малайзии, Индонезии,
на Филиппинах, о-вах Новая Гвинея, Сулавеси и Микронезии (del Hoyo et al., 1992; Robson,
2000).
Миграции. Южные популяции, в том числе в Юж. Японии, Юж. Китае, Сев. Бирме и
Сев. Индокитае оседлые. Северные популяции совершают миграции. Птицы летят ночью.
Весной миграции в северном направлении проходят с середины апреля до конца мая. На о.
Хоккайдо китайский волчок весной прилетает в мае и держится здесь до октября, на севере
о. Хонсю в префектуре Акита птиц отмечали в конце декабря и середине февраля (Austin,
194
Отряд Листообразные, Семейство Цаплевые
Kuroda, 1953). В Сев.-Вост. Китай прилетает в
середине апреля - мае (La Touche, 1931-1934),
а улетает в первой половине сентября (Jones,
1911), по другим данным — в октябре (Kushlan,
Hancock, 2005). Через п-ов Малакка осенний
пролёт проходит в октябре-декабре, обратная
миграция совершается в марте-апреле (del
Hoyo et al., 1992).
Местообитания. Населяет тростниковые
плавни в устьях рек и по берегам озёр, заросли
бамбука, приречные кустарники, болота и ри-
совые поля. В Китае отмечен в мангровых за-
рослях. В Индии обитает на высоте до 900 м,
на Шри-Ланке—до 1 200 м, на Суматре — 1 500
м (del Hoyo et al., 1992). В России все встречи
вида отмечены в водно-болотных угодьях.
Численность. О размере популяций извес-
тно мало. В различных частях ареала указыва-
Рис. 35. Места встреч китайского волчка в
России: а — пункты регистрации птиц.
ется от сотен до тысяч особей. В Японии весьма обычен на всех крупных островах (Austin,
Kuroda, 1953; Check-List..., 1974). В Юж. и Сев. Корее редок (Moo-Boo Yoon, 1995; Тошек,
1999). Считается наиболее обычным видом среди остальных видов волчков в Китае. Обы-
чен в Пакистане, Таиланде, Филиппинах, на о. Борнео (зимой) (Cheng Tso-hsin, 1995; del
Hoyo et al., 1992).
Размножение. Китайские волчки — моногамы, но не исключается полиандрия, в пользу
которой свидетельствует слабое участие самок в насиживании кладок и обогреве птенцов.
Кроме того, самцы чаще самок остаются в гнёздах на ночь.
В Юго-Вост. Азии сезон размножения длится с марта по ноябрь (Robson, 2000). Начало
гнездовой активности совпадает с наступлением весны или с периодом дождей в субтропи-
ках, который приходится на июнь-сентябрь в Индии, июль-октябрь в Малайзии, май-ав-
густ в Японии и Китае и сентябрь-апрель на Соломоновых о-вах (del Hoyo et al., 1992;
Kushlan, Hancock, 2005).
Китайские волчки предпочитают гнездиться в густой околоводной растительности. Гнёз-
да устраивают как в травянистой растительности — на заломах тростника среди воды, в
рогозе, бамбуке, рисе, диком сахарном тростнике, камыше — так и на кустарниках и не-
больших деревьях в 1,5^4,5 м от земли. В отличие от других видов выпей, высокая плот-
ность травостоя при выборе места для устройства гнезда играет меньшую роль, но при
этом уровень воды является важным фактором.
Гнездятся птицы одиночными парами, при высокой численности — небольшими дис-
персными группами или колониально. На одном дереве может располагаться до 6 гнёзд. В
Японии в состав колонии входило 5 пар, среднее расстояние между гнёздами составляло
6,7 м, а между близкорасположенными гнёздами — до 2,5 м. При колониальном гнездова-
нии характерно синхронное строительство гнёзд (Uchida, Matsuda, 1990). Плотность гнез-
дования на орошаемых полях в Китае может достигать 31,2 пар/га, в среднем она составля-
ет 11,4 пары/га (Kushlan, Hancock, 2005).
Самец рекламирует территорию с помощью брачного крика, демонстративных поз и
полетов. Брачные демонстрации самцов происходят на вершинах кустарников и сопровож-
даются мягкими, монотонными криками «ох-ох-ох...». При этом самец принимает сутулую
позу, перья на груди распушены, основание клюва краснеет. Во время рекламирующих кру-
Китайский волчок
195
говых полетов самец взлетает с земли и медленно летит по кругу, делая неглубокие взмахи
крыльями, после чего приземляется на место рекламирования. В состоянии возбуждения
он поднимает хохол, вытягивает в сторону одно из крыльев, разводит хвост из стороны в
сторону (del Hoyo et al., 1992; Kushlan, Hancock, 2005).
Гнездо — небольшая, аккуратная конструкция в виде платформы, состоящая из стеблей
и листьев тростника и других трав и расположенная на высоте 0,1-3 м над водой. Часто
окружающая растительность смыкается над гнездом, формируя его крышу. Гнездо строит
самка, но на протяжении всего гнездового периода самец его обновляет и поправляет, при-
нося всё новые травинки. Размеры гнёзд: высота 12 см, диаметр 17-25 см, глубина 2,5-4 см
(Kushlan, Hancock, 2005).
В Приморье о возможном гнездовании китайского волчка свидетельствовали наблю-
дения В.А. Нечаева на побережье Уссурийского зал. в 2000 г. (Нечаев, 2003). Так, на
одном из озёр вблизи побережья залива 26 и 30 июля 2000 г. наблюдали, без сомнения,
одного и того же самца, который, судя по поведению, держался на гнездовом участке. Он
совершал регулярные (не реже 2-3 раз в течение часа) перелеты из топких зарослей рого-
за, занимавших центральную часть озера, где предположительно находилось гнездо, в
прибрежные заросли тростника и на мелководье вдоль берега; там он разыскивал пищу
(очевидно, для птенцов), а затем возвращался в обратном направлении. Обнаружить гнездо
не удалось. По-видимому, гнездовая пара была добыта В.И. Лабзюком на побережье за-
лива Ольга в 1980 г.: у самки, добытой 29 мая, размер самого крупного фолликула состав-
лял 3 мм, а у самца, добытого 8 июня, длина семенников достигала 9 мм (колл. БПИ ДВО
РАН). Репродуктивное состояние другого самца, добытого на юге Приморья 29 мая, так-
же свидетельствовало о подготовке к размножению: его семенники были увеличены и
достигали размеров 8x4 мм (колл. Зоомузея ДВГУ). В 2008 г. гнездование китайского
волчка на юге российского Дальнего Востока было доказано. На одном из озёр побере-
жья Амурского зал. в пригороде г. Владивостока обнаружено 2 гнезда китайского волчка
(Нечаев, Гамова, 2009; Гамова и др., в печати). Гнёзда располагались в густых зарослях
подтопленного прошлогоднего тростника, в нескольких метрах от кромки свободной воды
и в 15 м одно от другого. Они крепились с помощью нескольких стеблей тростника, про-
низывающих стенки и дно гнёзд в 3-6 местах. К постройке гнёзд птицы приступили 5 и
27 июня; продолжительность строительства составила около 3 дней. Размеры гнёзд (см):
диаметр 19,8 и 22,0, высота 8,0 и 10,0, диаметр лотка 19,8 и 22, глубина лотка 1,0 и 2,0.
Высота расположения гнёзд от воды 33 и 88 см, высота травостоя в районе гнезда 1,5-2,0
м, плотность травостоя 90-100%.
В Вост. Китае кладки находили в июне и в июле, в Юж. Китае — с начала мая до авгу-
ста, в Японии — с начала июня до середины августа. Кладка содержит 3-7, в среднем 4-6
яиц. В Приморье кладки содержали 5 и 7 яиц, к откладке птицы приступали на следующий
день после завершения строительства (Гамова и др., в печати). Яйца откладываются ежед-
невно или с однодневными интервалами спустя 1-3 дня после окончания строительства
гнезда. В условиях колониального гнездования откладка может быть растянута (в Японии,
например, с июня по август). Размеры яиц китайского волчка в среднем чуть меньше, чем
у амурского волчка. По данным разных исследователей они таковы: 27,5-33,7 х 22,2-25,0,
в среднем 30,9x23,7 мм (n=40) (Baker, 1929); 28,5-35,0 х 22,5-25,0, в среднем 32,0x24,5 мм
(n=30) (La Touche, 1931-1934); 29-37x22-27, в среднем 32x24 мм (n=180) (Shaw, 1936);
28,0-35,0 х 22,2-26,5, в среднем — 31,8x24,0 мм (n=60) (Schonwetter, 1960). Размеры яиц
из Приморья (п=12): длина 28-36 (32,2±2,0), диаметр 22-24,9 (23,9± 1,1) мм. Масса яиц на
. S-й день насиживания (п=7): 7,9-10,1 (9,3±0,7) г. Окраска однотонная. Цвет скорлупы ва-
рьирует от бледно-зеленоватого, голубовато-зеленоватого, бледно-голубого до белого. Яйца
196
Отряд Листообразные, Семейство Цаплевые
с голубоватым оттенком составили 66,7%. Форма яиц — овальная, яйцевидная, удлинённо-
овальная; яйца овальной формы составили 50% (Гамова и др., в печати).
Насиживание начинается сразу после откладки первого яйца и длится 17-22 (в среднем
18-19) дней. Насиживают и кормят птенцов оба партнёра. Ритуал смены партнёра на гнез-
де занимает 1 минуту. При смене самца самкой самец встречает её раскрыванием клюва и
киванием головы без поднятого хохла, самка проделывает тоже самое. После ухода самца
самка поднимает хохол во время кормления птенцов. При смене самки самцом самка не-
сколько раз наклоняет голову в сторону самца и издаёт писклявые звуки, такие же, как при
кормлении птенцов («верещание»). В момент встречи у обеих птиц хохлы прижаты, но
после ухода самки самец поднимает хохол, принося гнездовой материал и во время кормле-
ния птенцов.
Вылупление птенцов одного выводка занимает до 4 дней. В гнёздах из Приморья вылу-
пилось по 5 птенцов, в одном из гнёзд 2 яйца оказались неоплодотворёнными. До 5-суточ-
ного возраста птенцы беспомощны и малоподвижны. Родители кормят их, отрыгивая пищу
на дно гнёзда. В возрасте 5 суток птенцы во время кормления издают пищевую позывку
«шипение», к этому возрасту осуществляется «прямое» кормление из клюва родителя в
клюв птенцам, при этом птенцы обхватывают клюв родителя, как пинцетом. Птенцы уже
ходят в «туалет», отползая к краю гнезда. В возрасте 6-7 суток птенцы способны ненадолго
покидать гнездо, садясь ниже него на тростнике, когда уходят в «туалет», или при беспо-
койстве со стороны человека. В этом возрасте начинается активный рост перьев на всех
птерилиях; меняется окраска частей тела: лапы желтеют, когти становятся серовато-корич-
невыми, глаза — зелёными. 10-суточные птенцы уже покидают гнездо постоянно и поддер-
живают связь между собой и родителями при помощи контактной трелевой позывки. В
этом возрасте появляются пеньки рулевых и теменных перьев. 11-суточные птенцы могут
находиться в 1-2 м от гнезда, сидя на стеблях тростника, а 12-суточные — в 3 м от гнезда
возле кромки воды. В возрасте 15 дней молодые с проворством могут подниматься вверх
по растениям. В Приморье гнездовой период развития птенцов занимал 15-16 суток; все
птенцы были успешно выкормлены (Гамова и др., в печати). По литературным данным,
средний успех размножения составляет 86% (Uchida, Matsuda, 1990; Kushlan, Hancock, 2005).
Суточная активность, поведение. Принято считать, что китайский волчок ведёт суме-
речный и ночной образ жизни (del Hoyo et al., 1992). Однако, в гнездовой период он наибо-
лее активен в дневное время суток. Суточный ритм нахождения в гнезде родителей в пери-
од выкармливания птенцов у самца и самки отличается: продолжительность непрерывного
нахождения в гнезде у самца значительно выше, чем у самки — 1,7±2,9 и 0,3±0,6 ч соответ-
ственно. Однако средние интервалы между нахождением в гнезде примерно одинаковы:
0,5±0,8 ч у самки и 0,6±0,6 ч у самца. Максимальное время непрерывного нахождения в
гнезде с птенцами у самца — 10,4 ч, у самки — 2,4 ч. Активность выкармливания, выра-
женная в среднесуточном количестве кормлений/час, у обоих родителей примерно одина-
кова: 1,3-1,4 кормлений/час, но количество скормленных пищевых объектов за час и за
одно кормление у самца больше (2,3 и 1,9 у самца; 1,3 и 1,7 у самки). Кроме того, самец
кормит за одно кормление и за один час большее количество птенцов (самец в среднем —
1,9 и 2,2 птенцов; самка — 1,7 и 1,1). Выявлена отрицательная корреляция между време-
нем отсутствия в гнезде и количеством скормленных пищевых объектов после прилёта с
кормом — Sr=0,46, п=23, р=0,03 (Гамова и др., в печати).
Питание. Кормится китайский волчок на сырых лугах, болотах, по берегам зарастаю-
щих водоёмов, на рисовых полях. Нередко улетает за кормом от гнезда на расстояние более
500 м. Охотится в одиночку, наиболее активно на рассвете и в сумерках. Высматривает
добычу, медленно передвигаясь по воде, или же неподвижно сидит на возвышающейся над
Китайский волчок, Амурский волчок
197
водой растительностью. Во время активной охоты может бегать и прыгать (Kushlan, Hancock,
2005). В Индии в питании отмечены мелкие лягушки, водные насекомые, рыбы, моллюски,
стрекозы (Ali, Ripley, 1968), в Китае — мелкие рыбы и креветки (Shaw, 1936). В Японии
68% пищевых объектов, получаемых птенцами, составили гольяны длиной до 80 мм и весом
до 3 г, 14% — лягушки длиной до 30 мм и весом до 1 г, 12% — мелкая рыба Pseudorasbora
sp., остальные 6% — раки и пауки (Ueda, 1992). В желудке добытого в Приморье самца
была обнаружена квакша {Нуla japonica). Родители в Приморье кормили птенцов рыбой:
гольянами, ротанами, молодью карасей.
Враги, неблагоприятные факторы. Как и для всех других мелких цапель, неблаго-
приятным является изменения среды обитания, в том числе любая мелиорация водно-бо-
лотных угодий. Негативное влияние на состояние вида оказывает использование пестици-
дов, отравляющих пищу и самих волчков.
В оперении китайских волчков обнаружены блоха Blankaartia acuscutellaris и перьевой
клещ из рода Ardeacarus (McClure, Ratanaworabhan, 1973).
Хозяйственное значение, охрана. Хозяйственного значения не имеет, однако может
участвовать в контролировании численности насекомых-вредителей на рисовых полях.
Специальные меры охраны не предусмотрены; вид занесён в Аннотированный пере-
чень таксонов и популяций животных, нуждающихся в особом внимании к их состоянию в
природной среде (Приложение 3 к Красной книге РФ).
Необходимо уточнение распространения вида на территории России, дальнейшее вы-
явление мест гнездования и оценка численности.
Амурский волчок
Ixobrychus eurythmus (Swinhoe, 1873)
Ardetta eurhythma Swinhoe, 1873, Ibis (3) 3, c. 74, Амой и Шанхай, Китай.
Schrenk’s [little] bittern (англ.), Mandschuren-Dommel (нем.), biongios de Schrenk (фр ).
Син. (лат.): Ardetta riedeli Meyer et Wiglesworth, 1894.
Син. (рус.): амурская выпь.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Одна из самых мелких цапель; наибо-
лее обычный волчок на российском Дальнем Востоке. В окраске сочетаются каштановые и
охристые тона. Обычно птицу можно видеть внезапно взлетающей из под ног или летящей
на высоте несколько метров. Полёт относительно медленный, с резкими взмахами крыль-
ев. В полёте ноги висят вниз. Вспугнутый амурский волчок не складывает шею, летит низ-
ко над землёй и бросается в траву в нескольких десятках, реже — сотнях метров от места
взлёта.
От сходных по размеру и совместно с ним встречающихся китайского и охристого вол-
чков отличается окраской верхней части тела. В отличие от охристого волчка, у которого
спина, маховые и рулевые полностью каштановые, у амурского волчка каштановая спин-
ная сторона резко контрастирует с чёрно-сизыми маховыми перьями. От китайского волч-
ка отличается более крупными размерами, более мощным в основании клювом, тёмным
надклювьем. В полёте хорошо отличается тёмной окраской спинной стороны (у самки она
с чёткими светлыми или белыми крапинами), меньшим контрастом чёрного и тёмно-жёл-
того на крыльях и серыми подкрыльями. У кормящихся птиц заметно, что нижняя часть
голени на 1,5 см выше сочленения с цевкой не оперена (у китайского волчка голень опере-
198
Отряд Листообразные, Семейство Цаплевые
на полностью). Первая фаланга самого длинного среднего пальца равна по длине второй
фаланге.
Голос. По сравнению с другими видами волчков, амурский волчок молчалив. Самая
обычная позывка — контактно-тревожный отрывистый, глухой крик «квек» или «кав» —
полифункциональный тональный, узкополосный сигнал длительностью 0,02-0,03 с с бо-
лее продолжительными межслоговыми интервалами (0,5-0,6 с). Брачный крик самца —
это серия сдвоенных низкочастотных выкриков «уп-уп, уп-уп, уп-уп», которые произносят-
ся от 6 до 20 раз. Песня слышна за 1-1,5 км (Пукинский, 2003). Пищевая позывка («шипе-
ние») у 1-3-суточных птенцов имеет гармонический спектр и более широкий частотный
диапазон (до 6 кГц), чем у птенцов более старшего возраста (с шумоподобным спектром и
узким частотным диапазоном до 4 кГц). В возрасте 9 суток птенцы издают тревожную
позывку («гудок») с гармоническим спектром и диапазоном частот до 7 кГц.
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Самец в брачном наряде. Верх головы черновато-бурый с
каштановым налетом; задняя сторона шеи, бока головы, спина, плечи, внутренние второ-
степенные маховые и мелкие кроющие крыла каштановые; область крестца буровато-серая
с коричневым налётом. Большие и средние верхние кроющие крыла серовато-охристые;
маховые и рулевые от чёрно-серых до буровато-серых с сизым налётом; наружное опахало
первого махового рыжевато-охристое с более светлым наружным краем; внутренние пер-
востепенные и первые второстепенные маховые постепенно светлеют к вершине. Горло
светло-охристое, грудь и брюхо охристые; от подбородка вдоль горла тянется узкая бурая
полоса, соединяющая с чёрно-бурыми пестринами верхней части груди; под кроющими
уха назад идёт белая полоса. Испод крыла и подмышечные белые с охристым налётом;
нижние кроющие хвоста белые с лёгким охристым налётом. Надклювье и режущий край
подклювья тёмные, подклювье желтоватое. Голая кожа у основания клюва зеленоватая.
Радужина жёлтая с более или менее выраженным затемнённым сектором у заднего края,
который в сочетании со слегка эллипсовидным зрачком создаёт эффект «лягушачьего гла-
за». Ноги буровато-зелёные, задняя сторона цевки и подошва пальцев желтоватые.
Самка в брачном наряде. На голове, как и у самцов, тёмная шапочка; задняя сторона
шеи, плечевые каштаново-бурые; спинная сторона от чёрной с буроватым оттенком до чёр-
но-бурой (встречаются птицы с рыжей спиной). Спина, включая верхние кроющие крыла,
с многочисленными белыми и иногда охристыми пятнами. Брюшная сторона охристая с
крупными продольными бурыми пестринами, самая нижняя часть брюха и подхвостье свет-
ло-охристые с отдельными узкими светло-коричневыми наствольными пестринами. Боль-
шие кроющие крыла имеют светлую охристую кайму по вершине пера или же по наружно-
му опахалу, что создаёт чуть заметный осветлённый контраст со спиной, верхними крою-
щими и маховыми. Рулевые и маховые — как у самца. Клюв в основном жёлтый с тёмным
коньком. Ноги оливковые.
Зимний наряд самки и самца. Зимний наряд самок и самцов отличается насыщенно-
стью цвета нижней части тела. В зимнем наряде у самца на спине рыже-бурые перья со
слабой светлой соломенной окантовкой. У самок белая пятнистость на спине более разви-
та, и рулевые перья с тонкой белой каймой по краю.
Птенцовый наряд. У однодневного птенца клюв рогового цвета, конек и конец подклю-
вья светло-коричневые, конец надклювья более тёмный. Яйцевой зуб белый, круглый, диа-
метром 1 мм; участок в основании яйцевого зуба бурый или чёрный. Ноздри щелевидные,
слуховые отверстия овальные, диаметром мм. Ротовая полость и язык розовые или
бледно-розовые. Корни языка белые, а кончик светло-коричневый. Кожа тела, головы и
конечностей телесного цвета. Когти на пальцах ног молочно-белые, с бурыми кончиками.
Амурский волчок
199
Кромки век чёрного цвета. Длина пуха на теменной птерилии составляет 12,5 мм, затылоч-
ной — 12; боковых шейных — 5, плечевых и межлопаточной — 19,5, спинной и крестцо-
вой — 18, бедренных — 18,5, грудной и брюшных — 8, на месте первостепенных махо-
вых — 4, второстепенных — 11 мм. На большинстве птерилий пух чёрного цвета, с корич-
невым, светло-коричневым или белым основанием. На межчелюстной и каудальных частях
боковых шейных птерилий пух целиком белый. На грудной птерилии он белый с чёрными
кончиками (Ильяшенко, 1987; Гамова и др., 2005). Участок кожи вокруг глаз голый и имеет
голубовато-зеленоватую окраску. Радужина тёмно-бурая с фиолетовым оттенком (Пукинс-
кий, 2003).
Гнездовой наряд. Пуховой покров сменяется перьевым к концу первого месяца жизни
птенца (Пукинский, 2003). У самца задняя сторона шеи каштановая; спина и плечи с бело-
ватыми пестринами; верхние кроющие крыла как у взрослых птиц — охристо-сероватые
без пестрин; подбородок, полоса под кроющим уха и нижние кроющие хвоста белые; ма-
ховые и рулевые черновато-серые; клюв зеленовато-жёлтый с бурым коньком надклю-
вья; ноги желтовато-зелёные. У самки на спинной стороне, включая верхние кроющие
крыла, многочисленные желтоватые или светлоохристые и более редкие белые пестрины
и крапины.
Возрастная изменчивость в окраске невелика. В первую осень жизни молодые самки не
отличаются от взрослых птиц. У самцов молодые птицы в первом зимнем пере имеют свет-
лые или белые пятна на спине и иногда менее выраженные и частые, чем у самок, тёмные
настовольные пестрины по бокам тела и на груди. У части размножающихся самцов (по-
видимому, годовалых птиц) летом сохраняется слабая пятнистость на спине или на боль-
ших кроющих крыла.
Строение и размеры. Стройная цапля, сходная по размеру с волчком (малой
выпью), но более короткокрылая. Первостепенных маховых 11, рулевых 12. Хвост корот-
кий, закруглённый; клюв относительно массивный в основании. Формула крыла: П?Ш>
I>IV>V>VI>VII... I первостепенное короче II на 2,2-2,5 мм, Ш<П на 1,5 мм, IV<III на 4
мм, V<IV на 6 мм, VI<V на 4,5 мм, VII<VI на 6 мм, VIII<VII на 8 мм, IX<VIII на 8,5 мм,
Х<1Х на 8,3 мм.
Размеры (мм) птиц из Приморья (коллекционные материалы Зоомузея ДВГУ и БПИ
ДВО РАН). Самцы (n=l 1): длина крыла — 141-149, в среднем 146; длина клюва от опере-
ния — 42,1—46,7, в среднем 44,9; длина клюва от переднего края ноздри — 35,8-37,9, в
среднем 36,9; длина цевки — 43,5-50,2, в среднем 47,0; длина хвоста — 44—48, в среднем
46,5. Самки (n=l 1): длина крыла — 136-146, в среднем 139,8; длина клюва от оперения —
41,8—48,5, в среднем 44,94; длина клюва от переднего края ноздри — 35,7-38,5, в среднем
37,1; длина цевки — 42,СМ7,6, в среднем 47; длина хвоста — 41,4—49,0, в среднем 45,5.
Размеры (мм) самцов из Сев. Кореи (п=3): длина крыла — 140-153, в среднем 145;
длина клюва — 44-48, в среднем 45,3; длина цевки — 45—49, в среднем 46,7; длина хвоста —
49-59, в среднем 53 (Тошек, 1999). Размеры птиц из Вост. Китая (п=12): длина крыла —
142-153, в среднем 149; длина хвоста — 39—47; длина клюва (n=l 1) — 45-50, в среднем
47,2; длина цевки — 45-52 (La Touche, 1931-1934).
Масса самцов (п=12): 120-158 г, в среднем — 139 г (Shaw, 1936). Масса трёх птиц из Примо-
рья: 106, 115,9 и 125 г (колл, материал). Масса самки с о. Сахалин— 137,7 г (Нечаев, 1991).
Линька. Сведения по линьке амурского волчка отрывочны. Самки начинают линять в
июле, к началу августа маховые полностью сменяются, рулевые и мелкое оперение в эти
сроки продолжают интенсивно линять; в июле самцы ещё сохраняют брачное оперение
(Спангенберг, 1951). У самца, добытого 15 августа, заканчивался рост контурного пера.
Птицы, добытые на пролёте в Приморье в сентябре, были уже в свежем пере.
200
Отряд Листообразные, Семейство Цаплевые
Смена наряда молодых птиц: пуховой - гнездовой - первый годовой (переходный) с
ювенильными маховыми и рулевыми - второй годовой окончательный (Спангенберг, 1951).
Распространение. Гнездовой ареал. Ото. Сахалин, Японских о-вов (о. Хоккайдо
и сев. часть о. Хонсю) и Кореи до юго-вост. Забайкалья и Вост, и Юго-Вост. Китая, включая
Внутреннюю Монголию и Сычуань (Cheng Tso-hsin, 1987; Степанян, 2003; Yoon, 1995)
(рис. 36).
На Сахалине гнездование в северной части острова установлено на побережье Саха-
линского зал. и зал. Тык, в южной части острова — также и на оз. Невское (Нечаев, 19916).
В центральной части острова вид добывали в долине р. Тымь у с. Воскресеновка (Пекло,
1997).
В Приморье в свойственных виду типах местообитаний отмечен на всей территории, за
исключением горных и таёжных районов. Гнездится в бассейне оз. Ханка, на водно-болот-
ных угодьях южной части Приморья, в бассейне р. Уссури, на морском побережье и в доли-
нах рек восточного склона хр. Сихотэ-Алинь, на о. Большой Пелис (зал. Петра Великого)
(Спангенберг, 1965; Лабзюк и др., 1971; Лабзюк, 1990; Литвиненко, 2005а).
На Нижнем Амуре гнездится на побережье оз. Эворон (Нечаев, 1963); самые северные
находки — на оз. Мухтель, в Ульбанском зал., на р. Тугур (Воронов, Пронкевич, 1991;
Поярков, Будрис, 1991; Бабенко, 2000). В нижнем течении р. Амур отмечен до с. Софийск
и оз. Удыль (Кистяковский, Смогоржевский, 1973; Бабенко, 2000). В бассейне р. Бурея встре-
чается к северу до дер. Желунда и с. Островское (Кистяковский, Смогоржевский, 1964). По
р. Зея гнездо найдено в 80 км от устья (Дымин и др., 1970).
Рис. 36. Область распространения амурского волчка: а — гнездовой ареал, б — районы зимовок, в —
недостаточно выясненная граница ареала.
Амурский волчок
201
В юж. Забайкалье гнездится в долине р. Аргунь и в бассейне нижнего течения р. Онон
(Гагина, 1961).
Зимовки. Зимует на Суматре, Яве, Калимантане, Сулавеси и Филиппинах; отмечен
также в Бирме и на о-вах Палау. По-видимому, в небольшом количестве остаётся на зимов-
ку в Юж. Китае (о. Хайнань), в Таиланде и п-ове Малакка (Cheng Tso-hsin, 1987; del Hoyo et
al., 1992; Robson, 2000).
Миграции. Миграции амурского волчка проходят ночью; летящих птиц легко узнать
по характерному для этого вида крику.
По наблюдениям К.А. Воробьева (1954), появление птиц на юге Приморья происходит
между 23 и 29 мая, но немногочисленные находки гнёзд в конце мая (Лабзюк, 1990; Глу-
щенко и др., 20066) свидетельствуют о том, что часть птиц появляется в южной части При-
морья в конце апреля - начале мая. Весной в Приморье первые птицы отмечены на оз.
Ханка 20-25 мая (Шульпин, 1936; Глущенко и др., 20066), на р. Б. Уссурка — 22 и 29 мая
(Спангенберг, 1965). На о. Монерон пролётная птица встречена 6 июня (Нечаев, 19756). В
Нижнем Приамурье появление птиц отмечено во второй половине мая (Кистяковский,
Смогоржевский, 1973; Колбин и др., 1994).
Осенний отлёт в Нижнем Приамурье приходится на первую половину сентября, отдель-
ные птицы отмечены до конца сентября (Бабенко, 2000). Начало осенних перемещений на
юге Приморья трудно проследить; по-видимому, они начинаются с «местных» кочёвок мо-
лодых птиц в начале-середине августа. В этот период в ночное время можно услышать
крики летящих амурских волчков даже находясь в населённых пунктах (Ю.Н. Глущенко,
личн. сообщ.).
Пик пролёта, по-видимому, приходится на вторую декаду сентября; именно на этот пе-
риод времени приходится наибольшее количество разбившихся в разные годы птиц, проле-
тавших ночью через г. Владивосток (колл, материал). В Приморье наиболее поздние встре-
чи отмечены 26.09 и 2.10 (Воробьев, 1954, колл, материал). На побережье оз. Ханка после-
дняя осенняя встреча зарегистрирована 30 сентября (Ю.Н. Глущенко, личн. сообщ.). В осен-
ний период амурского волчка на Сахалине добывали 13 сентября и 21 ноября (Нечаев, 19916).
Через Гонконг пролёт проходит в сентябре-октябре, а на п-ове Малакка пролётные пти-
цы отмечены в октябре-ноябре (del Hoyo et al., 1992).
Известны два дальних залёта в Европу: в Германию и Италию (del Hoyo et al., 1992).
Местообитания. В Нижнем Приамурье встречается на сырых лугах, покрытых вейни-
ком, манником, сабельником, камышом, осоками с редкими кустами ивы, густыми зарос-
лями спиреи. Обитает также на заболоченных участках с невысокой травяной растительно-
стью, на заболоченных марях (с багульником, пушицей, мхом, редкими ивами, ольхой, ли-
ственницей) с обязательным наличием мелководных богатых рыбой водоёмов (Бабенко,
2000). На о. Сахалин амурского волчка наблюдали на топких берегах озёр, заросших вах-
той (Menyanthes), сабельником и осоками, а также в северной части острова на берегу эвт-
рофного озера, расположенного среди заболоченного лиственничного леса с кедровым стла-
ником (Нечаев, 19916). В Приморье встречается по берегам озёр, поросших тростником,
рогозом, диким рисом; по вейничковым или травяным болотам. Обитает также по сырым
луговинам морского побережья. В долинах рек придерживается сырых разнотравных или
осоковых лугов с отдельными кустами ивы и спиреи (Воробьев, 1954).
В Китае и Японии отмечен по травяным болотам, зарослям тростника, открытым учас-
ткам долин рек и морского побережья, ирригационным каналам полей, рисовым полям, по
берегам искусственных прудов. Помимо этого, в Японии гнездование отмечено на сухих
пастбищах и участках, покрытых кустарником (A field guide to the birds of Japan, 1982;
Cheng Tso-hsin, 1987; del Hoyo et al., 1992).
202
Отряд Листообразные, Семейство Цаплевые
Численность. Конкретных данных по численности немного. На Филиппинских о-вах
этот волчок — редкий зимующий вид. Немногочислен он и в Японии (del Hoyo et al., 1992:
A field guide to the birds of Japan, 1982). В Китае же амурский волчок считается обычным
видом (Cheng Tso-hsin, 1987).
На о. Сахалин — редкий гнездящийся вид (Нечаев, 19916), по р. Зея малочислен (Ды-
мин и др., 1970), в Юж. Забайкалье редок (Гагина, 1961). На Нижнем Амуре и в Приморье
его относят к обычным видам (Бабенко, 2000; Литвиненко, 2005).
В Приморье численность амурского волчка значительно колеблется по годам, отчасти в
зависимости от количества осадков в весенне-летний период. В сезоны, когда на лугах и
болотах стоит высокий уровень воды, численность вида низкая. В окрестностях ст. Хасан в
1976 г. плотность его населения достигала 7-8 пар на 1 км2 и была в 4-6 раз выше, чем в
1970-1972 гг., отличавшихся дождливой погодой (данные Ю.Н. Назарова). Колебания чис-
ленности наблюдаются и на побережье оз. Ханка, где в 2003-2004 гг. имел место всплеск
численности амурского волчка: с одной точки в вечернее время учитывали 4-5 кричавших
самцов, а за часовой маршрут регистрировали до 10 птиц (Глущенко и др., 20066). На При-
ханкайской низменности во второй половине лета и до отлёта птицы прилетают кормиться
на рисовые чеки, где за час в сумерки отмечали до 25 особей (Глущенко, 1979). В пойме р.
Бикин Ю.Б. Пукинский (2003) в первой декаде июня по брачным крикам учитывал 2-3
особи в радиусе около 1 км. Экстраполируя учётные данные на площадь поймы, он опреде-
лил ориентировочное число амурского волчка для низовий р. Бикин в 75-100 пар.
Плотность населения в Среднем Приамурье колеблется от 0,4-0,8 пар на 1 км2 на травя-
ных болотах до 2,7-5,9 пар на 1 км2 на влажных лугах (Смиренский, 1986). По данным В.Г
Бабенко (2000), в Нижнем Приамурье она составляла от 1,9 пары на 1 км2 на лугах в устье
р. Черной до 6 пар на лугах и марях в окрестностях оз. Эворон, с максимальными показате-
лями 13,3 пары на 1 км2 на о. Шарголь.
Размножение. Обычно брачный крик самца можно слышать ночью или на заре, иногда
днем в пасмурную погоду. При этом самец сидит на земле или на сухих ивовых кустах, при
новой серии криков может менять место. По наблюдениям Ю.Б. Пукинского (2003), брач-
ные крики присущи как самцу, так и самке. В Приморье, в окрестностях ст. Хасан брачный
крик самцов приходилось слышать только до середины июня, а на побережье Амурского
залива — до середины июля (до начала откладки яиц); один самец токовал до 25 июля (до
середины периода насиживания). На оз. Чукчагирское в Нижнем Приамурье брачные кри-
ки и токовые полёты пары амурских волчков были отмечены вечером 8 июня 1980 г. (Ба-
бенко, 2000; данные Ю.Н. Назарова).
Судя по находкам гнёзд амурского волчка, гнездовой период этого вида в Приморье
весьма растянут и зависит от состояния травостоя, пригодного для устройства гнёзд. Гнёз-
да с только что отложенными яйцами и кладки в различной степени насиженности находи-
ли здесь с 25 мая по 10 августа (Шульпин, 1936; Спангенберг, 1951; Нечаев, 1971; Глущен-
ко и др., 20066; данные Ю.Н. Назарова и др.). В Нижнем Приамурье гнёзда находили 10 и
И июня 1961 г. (Кистяковский, Смогоржевский, 1973), с насиженной кладкой — 2 июля
1978 (Бабенко, 2000), со слабо насиженной кладкой — 3 июля 1994 г. (Рябцев, 1997). На Зее
слабо насиженная кладка найдена 17 июня (Дымин и др., 1970). На Сахалине гнездо со
слабо насиженной кладкой найдено 3 июля, а 15 июля добыта самка, которая уже отложила
4 яйца (Нечаев, 1991).
Гнёзда устраивает среди густой травы или в зарослях кустарника (лещина, поросль дуба)
на сухих и влажных участках открытых пространств, по гривам с низкорослым густым
кустарником среди пониженных луговых участков. В отличие от других волчков, чаще ус-
траивает гнёзда на сухих лугах. При высокой численности птиц гнёзда соседних пар могут
Амурский волчок
203
располагаться в 150-200 м друг от друга (Литвиненко, 2005; Гамова и др., 2005; данные
Ю.Н. Назарова и др.).
Гнездо располагается на земле, иногда в неглубокой ямке, а в густой траве может под-
держиваться стеблями на высоте 10 см от земли. Одно гнездо в Приморье располагалось в
густых зарослях злака цицании среди озёра, где оно помещалось на плавающих корневи-
щах (Нечаев, 1971). Постройка хорошо скрыта высокой густой травой или кустом. От гнез-
да птицы протаптывают тропинку, которая ветвится, ведя в заросли и на небольшую утоп-
танную площадку в 0,5-1 м от гнезда. По форме гнездо напоминает перевёрнутый конус, в
котором стебельки расходятся от центра радиально (Спангенберг, 1951), или же оно чаше-
видное, со слабо выраженным лотком. Материалом служат сухие и зелёные стебли и лис-
тья злаков и осок иногда лоток выстилается метёлками тростника. Гнездо может представ-
лять собой ямку диаметром 13-14 см, выстланную незначительным числом прутиков, су-
хими листьями, осок, злаков или просто утоптанную площадку с несколькими сорванными
и примятыми травинками свежих злаков (Пукинский, 2003; Гамова и др., 2005).
Одно из гнёзд (п=7), обнаруженных в Приморье, располагалось на ровном участке зем-
ли диаметром 20x30 см и состояло из нескольких сорванных и примятых травинок колос-
няка и тимофеевки. Трава вокруг гнезда смыкалась почти полностью, образуя подобие
шалаша (Гамова и др., 2005). Остальные гнёзда были найдены во влажных условиях, среди
тростниково-осокового болота, на кочках или на земле между кочками в окружении 1-2-
метрового прошлогоднего и свежего тростника и осоки (доля которой может составлять до
90%). Размеры гнёзд (п=7): диаметр 11,0-31,5 (в среднем 25,5±7,0) см, высота 1,0-16,0
(9,3±5,0), диаметр лотка 11,0-31,5 (21,8±8,8), глубина лотка 0-2,0 (0,7±0,8) см. Высота рас-
положения гнёзд от земли или воды 0-5,0 (0,7±1,9) см, высота травостоя в районе гнёзд
0,4-2,0 (1,1±0,6) м, плотность травостоя 40-100 (65,7±23) % (данные Т.В. Гамовой и др.).
Размеры гнёзд, обнаруженных в Верхнем Приамурье (п=4): диаметр 17-22 см, высота
5,5-9 см, диаметр лотка 10-13 см, глубина 1,5-2,2 см; гнездо, расположенное в ямке, имело
глубину лотка 4 см (Дымин и др., 1970). В условиях Приханкайской низменности диаметр
гнезда составил (п=7) 12-30 см, в среднем 18,4 см; диаметр лотка 5,5-13 см, в среднем 9,2
см; глубина лотка 0,4-3 см, в среднем 1,4 см (Глущенко и др., 20066).
Строительство гнёзда продолжается 2-4 дня, в нём участвуют оба пола. Кроме этого, на
протяжении всего гнездового периода взрослые регулярно обновляют и ремонтируют гнездо,
принося новые веточки. Наблюдения за двумя парами волчков показали, что после силь-
ных дождей, когда уровень воды поднялся на 10 см, они построили новые гнёзда в 30 и 100
см от прежних: одно было почти точной копией старого, другое представляло собой при-
гнутую листву осоковой кочки (данные Т.В. Гамовой и др.).
Полная кладка содержит 3-6 яиц. В Юж. Приморье в найденных кладках (п=6) находи-
лось 4-6 (4,7±0,8) яиц. Размеры яиц (п=33): длина 31,5-37,2 (34,1±1,5) мм, диаметр 25,0-
29,1 (26,6±1,0) мм. Масса яиц на 13-й день насиживания (п=6) 10,5-12,1 (11,5±0,6) г, на
18-й день 9,2-12,6 (11,5±1,4) г. Яйца амурских волчков достоверно больше по длине (Р<0,05),
диаметру (Р<0,01) и массе (Р<0,05) яиц китайского и малого волчков (данные Т.В. Гамовой
и др.).
В кладке с р. Зея яйца имели следующие размеры (п=5): 36,0x27,0, 35,0x27,5, 34,2x28,0,
35,0x29,0, 36,0x29,0 мм (Дымин и др., 1970); из окрестностей Хасана (п=4): 33,0x30,0,
33,5x28,5,34,0x28,2,34,0x28,0 мм (Нечаев, 1971). Размеры яиц с побережья оз. Ханка (п=6):
31,5-35,7 х 25,4-28,0, в среднем — 33,9±1,4 х 27,0±1,0 мм (Гамова и др., 2005); по другим
данным (п=25): 32,0-37,9 х 25,2-27,4 мм, в среднем 33,7x26,5 мм (Глущенко и др., 20066).
Масса слабо насиженных яиц с о. Сахалин (п=4): 11,8; 12,0; 12,2; 12,3 г (Нечаев, 1991), из
Сев. Приморья (п=5) 13,5-13,7 г (Пукинский, 2003).
204
Отряд Листообразные, Семейство Цаплевые
Окраска яиц белая, реже голубоватая, матовая. Форма яиц овальная, редко (по одному
яйцу в кладке) округлая. Поверхность мелкозернистая, шероховатая. При насиживании скор-
лупа приобретает лёгкий буроватый налёт и слабый блеск (данные Т.В. Гамовой и др. по
Юж. Приморью).
По мнению Ю.Б. Пукинского (2003), птица приступает к насиживанию после откладки
3-4 яиц, по нашим данным — 1-3 яиц. Насиживание длится 16-18 дней (del Hoyo et al.,
1992; Пукинский, 2003), в Юж. Приморье (п=6) — 18-19 (18,3±0,5)дней (данные Т.В. Га-
мовой и др.). В нем принимают участие оба партнёра; самка насиживает преимущественно
в светлое время суток, самец меняет самку в сумерках и ночью (Пукинский, 2003). По
данным авторов очерка, больший вклад самцов наблюдается на ранних стадиях насижива-
ния; в дни перед вылуплением насиживание очень плотное (яйца почти не остаются без
обогрева) и осуществляется преимущественно самками. Так, на 16-19-й дни насиживания
в трёх гнёздах самки непрерывно оставались на гнёздах в течение 1-2,9 суток, изредка
покидая гнездо на несколько минут. В одном из гнёзд на 15-е сутки насиживания самец
ловил рыбу, не покидая гнезда (1 рыба в час; весь процесс занимал 2 мин), а самка проде-
лывала это на 17-е сутки насиживания (3 рыбы за 5 ч).
Ритуал смены партнёра на гнезде похож на подобный ритуал других волчков (китайско-
го и малой выпи), однако амурские волчки меньше контактируют друг с другом. Ожидая
прихода самки, самец часто принимает тревожную позу: беззвучно раскрывает клюв, при-
поднимает хохол. Во время смены самки самцом, она наклоняет голову вниз и в сторону и
редко раскрывает клюв. Самец при этом щёлкает клювом, слегка наклоняя голову и прика-
саясь клювом к спинному оперению самки, а самка — к оперению нижней стороны тела
самца. Церемония смены самца самкой часто происходила без участия самца — он убегал
из гнезда, попискивая или издавая позывку «кав» в ответ на протяжный крик самки «кии-и-
и...», которая демонстрировала элементы ритуала смены на гнезде и без участия самца.
Часто самка, побеспокоенная человеком, после повторного подхода к гнезду выжидала до
10 мин, замирая в характерной для выпи тревожной позе: вытягивала шею вверх, припод-
нявшись на ногах (Гамова и др., 2005; данные Т.В. Гамовой и др.).
Вылупление птенцов в восьми обследованных гнёздах происходило в течение 2-3 дней.
В Приморье самая ранняя находка гнезда с 2-3-дневными птенцами зарегистрирована 25
мая 1984 г. (Лабзюк, 1990). В окрестностях ст. Хасан появление птенцов отмечено 30 июня,
2 и 9 июля, 13 августа; на побережье оз. Ханка — 8 июля, на побережье Амурского зал. —
с 5 июля по 14 августа (Нечаев, 1971, Гамова и др., 2005; данные Ю.Н. Назарова). По
мнению В.Г. Бабенко (2000), появление птенцов в Нижнем Приамурье происходит в пер-
вой декаде июля, так как с этого времени взрослые птицы, начиная кормить птенцов, ста-
новятся хорошо заметными. У однодневного птенца длина клюва от края ноздри — 5,4 мм,
длина крыла от интертарзального сочленения до конца кисти — 12,6 мм, цевки — 12,7 мм
(Ильяшенко, 1987).
Наблюдения за семью парами амурских волчков в Приморье показали, что во время
насиживания птицы плотно сидят на гнезде, а потревоженные человеком не покидают гнёзда
до последнего момента, затаиваются, нахохлив оперение и приняв характерную для выпей
вертикальную позу. После вылупления птенцов взрослые становятся крайне осторожны-
ми. При обогревании птенцов птицы взлетали, увидев или услышав человека, уже в 10 м от
гнезда. По-видимому, самки в период выкармливания птенцов ведут себя осторожнее, чем
самцы. В окрестностях ст. Хасан в светлое время суток удавалось видеть только самцов,
приносящих корм молодым.
На обогрев птенцов в первые два дня жизни родители тратят много времени, сменяя
друг друга и принося пищу не чаще 3 раз в час. В возрасте двух суток птенцы самостоя-
Амурский волчок
205
тельно уходят в «туалет», отползая на край гнезда. В этом возрасте у них начинают расти
первостепенные маховые, на 3-4 сутки — второстепенные маховые, кроющие маховых и
теменные перья, на 6 сутки — рулевые перья. Голодные птенцы активно шевелятся под
обогревающими их родителями и выпрашивают корм, издавая характерную шипящую по-
зывку и принимая позу выпрашивания. На четвертые сутки птенцы способны ходить, они
1-2 раза в час ходят в «туалет», а также прячутся от солнца в траве в 15 см от гнезда. При
появлении человека птенцы затаиваются в позе, аналогичной «вертикальной позе» взрос-
лых птиц. 6-суточных птенцов взрослые всё чаще кормят вне гнезда. 7-суточные птенцы
уже ненадолго покидают гнездо, выбегая из него навстречу приближающимся родителям с
кормом, но сразу же после кормления возвращаются обратно. На 8-12-е сутки они в основ-
ном находятся вне гнезда, но ночуют в гнезде. Окончательно покидают гнездо птенцы в
13-14-суточном возрасте. Общая продолжительность гнездового периода развития птен-
цов (п=3) составляет 11-16 (13,3±2,5) суток (данные Т.В. Гамовой и др.). Летать молодые
начинают в месячном возрасте (Пукинский, 2003).
Активность выкармливания птенцов, выраженная в среднесуточном количестве корм-
лений в час, и количество скормленных пищевых объектов за час у обоих родителей одина-
ковы (0,3±0,2 и 0,5±0,6 соответственно), однако самец за одно кормление и за час кормит
больше птенцов (1,9 и 0,7 птенцов), чем самка (1,6 и 0,4 птенцов) (данные Гамовой и др.).
В Приморье в шести наблюдаемых гнёздах количество птенцов достигало 4-5 (4,3±0,5),
одно гнездо было разорено на поздней стадии насиживания, в двух птенцы погибли в пер-
вые дни жизни; успех гнездования составил 43,8%. Гнёзда часто разоряются хищниками
(енотовидной собакой и воронами) (Пукинский, 2003; Гамова и др., 2005; данные Т.В. Га-
мовой и др.).
Суточная активность, поведение. Амурский волчок деятелен круглые сутки, но, ис-
ключая период выкармливания птенцов, наиболее активен в сумерки и ночью. Отмечено,
что в период воспитания потомства вид ведёт дневной образ жизни, регулярно летая на
водоёмы за кормом (Спангенберг, 1951; Нечаев, 1963; Бабенко, 2000; данные Т.В. Гамовой
и др.). Ночные видеосъёмки гнёзд амурского волчка в Приморье показали, что родители в
ночное время птенцов не кормили; пара начинала кормить их в 630 и заканчивала в 2130.
Лишь в одном из гнёзд самец, обогревавший ночью 9-суточных птенцов, вёл себя на гнезде
очень беспокойно: птенцы, требуя корм, клевали его снизу. В результате самец был вынуж-
ден покинуть гнездо и покормить птенцов в 1 ч ночи (Гамова и др., 2005; данные Т.В.
Гамовой и др.). В другом гнезде птицы также были активны днем, а ночью в течение 2,5-3 ч
отдыхали у гнёзда; тем не менее, из 78 прилётов к гнезду самки и 42 самца 83% пришлось
на сумеречные и ночные часы (Пукинский, 2003). В течение суток наиболее активно роди-
тели кормят птенцов в утренние и вечерние часы. За один раз они приносят и отрыгивают
до нескольких пищевых объектов, которыми кормят одного или нескольких птенцов. Су-
точный ритм нахождения в гнезде родителей в период выкармливания птенцов у самца и
самки отличается: продолжительность непрерывного присутствия в гнезде у самки в сред-
нем выше (2,3±3,9 ч), чем у самца (1,6±2,8 ч), однако средние интервалы между нахожде-
нием в гнезде одинаковы (1,2±1,6 ч). Максимальное время непрерывного присутствия в
гнезде с птенцами у самца — 12 ч, у самки — 13,3 ч (данные Гамовой и др.).
Питание. Амурский волчок кормится по берегам пойменных озёр, заболоченных ручь-
ёв, на сырых лугах, болотах, рисовых полях.
В пищеводах и желудках Е.П. Спангенберг (1951) находил насекомых, мелких вьюнов
(до 76 мм), бычков (до 57 мм). В окрестностях ст. Хасан В.А. Нечаев (1971) обнаружил в
двух желудках этих птиц мелких ротанов и гольянов, остатки стрекозы, жука и личинок
водных насекомых. В двух отрыжках птенцов из этого же района оказалось 13 ротанов
206
Отряд Листообразные, Семейство Цаплевые
(2,5-5 см), 2 вьюна (7 см), 2 мелких жука. В желудке взрослой самки 7 июня на о. Б. Пелис
обнаружено 19 гусениц совок (данные автора очерка). В Приханкайской низменности из 58
определённых визуально кормовых объектов, принесённых птенцам одного гнезда, 86%
составили чернопятнистые лягушки, 14% пришлось на крупных жуков и мелкую рыбу (Га-
мова и др., в печати). В пойме р. Бикин основной корм птенцов (43 порции) составили
налимы, бычки и другая рыба длиной до 11 см, некрупные миноги, а также водные и около-
водные насекомые, преимущественно личинки стрекоз (Пукинский, 2003). На Сахалине в
питании одной птицы отмечены озёрные гольяны, девятииглые колюшки, личинка жука-
плавунца, пиявки (Нечаев, 19916). В Нижнем Приамурье среди объектов питания встрече-
ны чебаки, ротаны и гольяны. Наблюдали охоту птицы на саранчовых (Нечаев, 1963; Ба-
бенко, 2000). В Китае амурский волчок поедает креветок, равноногих ракообразных, насе-
комых, личинок стрекоз и других насекомых, однако основу питания составляют вьюны,
молодь карасей и других рыб (Shaw, 1936).
Враги, неблагоприятные факторы. В Приморье к числу основных врагов амурского
волчка относятся енотовидная собака, лисица и колонок, разоряющие гнёзда. Часть гнёзд
гибнет при сенокошении (данные авторов очерка; Ю.Н. Глущенко, личн. сообщ.). Амур-
ский волчок найден в питании большого подорлика в Хинганском заповеднике (Рябцев,
1997) и восточного болотного луня (Ю.Н. Глущенко, личн. сообщ.).
Вид занимает наиболее уязвимые, подверженные трансформации ландшафты. Осуше-
ние и распашка земель неминуемо ведут к деградации свойственных виду местообитаний.
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Занесён в Крас-
ные книги Приморского края (Литвиненко, 2005), Читинской области, Сахалинской облас-
ти и Хабаровского края, а также в Аннотированный перечень таксонов и популяций живот-
ных, нуждающихся в особом внимании к их состоянию в природной среде (Приложение 3
к Красной книге РФ).
Охристый волчок
Ixobrychus cinnamomeus (J.F. Gmelin, 1789)
Ardea cinnamomea J.F. Gmelin, 1789, Syst. Nat., 1, 2, c. 643, Китай.
Cinnamon [chestnut] bittern (англ.); Zimtdommel (нем.); biongios cannelle (фр ).
Син. (лат.): Ardea nebulosa Yjrsfield, 1821; Botaurus assimilis C.L. Brehm, 1842; Nannocnus
ijimae Ogawa, 1905.
Син. (рус.): рыжий волчок, охристая выпь, коричный волчок.
Статус. Нерегулярно залётный вид.
Общая характеристика и полевые признаки. Мелкая цапелька с обликом и повадка-
ми типичными для представителя рода Ixobrychus. По размерам примерно соответствует
чирку-трескунку или большому веретеннику (длина тела 39-41 см, размах крыльев 60-65
см). Общий тон окраски оперения рыжевато-коричневый или ржаво-шоколадный, низ тела
более светлый. От всех других волчков отличается яркой рыжевато-коричневой окраской
маховых и рулевых перьев, а также отсутствием контраста между верхними кроющими
крыла и остальными участками оперения верха тела. Птицы нередко поселяются на рисо-
вых полях и зачастую активны в присутствии людей, обрабатывающих посевы.
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Взрослый самец. Темя коричневое; бока головы и шеи, спина,
крылья, хвост яркие, рыжевато-коричневые. Нижняя сторона в основном бледная желтова-
Охристый волчок
207
то-коричневая. Горло и узкая полоска, идущая вдоль щек и по бокам шеи, беловатые. По-
средине шеи коричневая полоса. На боках груди тёмно-коричневые пятна. Клюв от зеле-
новато-жёлтого до жёлто-оранжевого с бурым коньком, уздечка жёлтая, ноги желтовато-
зелёные. В разгар брачного периода голая кожа уздечки и основания клюва становится
ярко-красной. Радужина жёлтая с затемнённым задним сектором (как и у амурского волч-
ка, это создает эффект «лягушачьего глаза»).
Взрослая самка окрашена в целом темнее и более тускло, чем самец: сверху коричнева-
то-бурая с черноватым теменем, верхние кроющие крыла, плечевые и третьестепенные
маховые с охристой каймой. Нижняя сторона бледная коричневато-охристая, бока тела с
бурыми полосками.
Пуховой наряд розовато-коричневый, более тёмный сверху, особенно на голове и спине;
низ, особенно живот, светлее.
Гнездовой наряд похож на наряд взрослой самки, отличаясь от него наличием охристой
каймы на перьях мантии и густых бурых пестрин на нижней стороне тела.
Стороение и размеры. Размеры (мм): длина крыла 137-158, цевки 45-50, клюва
43-55 (Бутурлин, 1935). Размеры самки: длина крыла 146, цевки 47, клюва от переднего
края ноздри 40, от границы оперения на лбу — 49.
Линька. Не изучена. Предположительно происходит, как и у других представителей
рода Ixobrychus. Взрослые птицы имеют две линьки в году: полную послебрачную и непол-
ную предбрачную. Первогодки претерпевают постювенильную линьку осенью и первую пред-
брачную в конце зимы. Вероятно, постювенильная линька неполная, о чём свидетельствует в
частности состояние оперения экземпляра от 20 октября 1976 (Лабзюк, Глущенко, 1984).
Распространение. Гнездовой ареал. Юж., Юго-Вост, и Вост. Азия. К западу до
Иранского нагорья, к северу до южного склона Гималаев, провинции Сычуань в центр.
Китае и провинции Ляонин в сев.-вост. Китае; к югу и востоку до океанического побере-
жья. О-ва Мальдивские, Шри-Ланка, Андаманские, Никобарские, Хайнань, Тайвань, Рюкю,
Филиппины, Большие и Малые Зондские к востоку до Сулавеси (рис. 37).
Несколько раз отмечен в России. В Приморском крае погибшая самка найдена 20 октяб-
ря 1976 г. в районе бухты Ольги (Лабзюк, Глущенко, 1984). 16 июня 1996 г. в окрестностях
пос. Хасан дважды встречен взрослый самец (Литвиненко, Шибаев, 1999). Что же касается
упоминавшейся в литературе встречи охристого волчка на юге Сахалина (Бутурлин, 1935),
то переопределение показало, что речь в данном случае идёт о китайском волчке (В.А.
Нечаев, личн. сообщ.).
Зимовки. Северные популяции зимуют в южной части гнездового ареала.
Миграции. Популяции, населяющие Индокитай, Филиппины и Индонезию оседлы,
остальные предпринимают сезонные миграции. На п-ове Малакка пролёт наблюдается в
сентябре-ноябре и в феврале-мае. Сезонные перемещения птиц п-ова Индостан носят в
основном локальный характер, и их масштабы зависят от состояния водоёмов.
Местообитания. Заполненные водой рисовые чеки, сырые луга и другие сельскохозяй-
ственные угодья. Селится также на пресноводных травянистых и тростниковых болотах,
на заросших канавах, в кустарниковых зарослях, тростниковых бордюрах озёр и прудов, в
придорожных зарослях тростника. В Индии отмечается иногда на побережье, в мангровых
зарослях. В соответствующих биотопах встречается как на равнинах, так и в горах на высо-
те до 2000 м.
Численность. На большей части ареала обычный вид. Хорошо приспособился к искус-
ственным (созданным в процессе хозяйственной деятельности человека) местообитаниям,
благодаря чему многие популяции вполне благополучны. В январе 1990 г. в округе Темпе
(южная часть о-ва Сулавеси) подсчитано 1 070 особей (Martinez-Vilalta, Motis, 1992).
208
Отряд Листообразные, Семейство Цаплевые
Рис. 37. Область распространения охристого волчка: а — гнездовой ареал; б — места зимовок; в —
залёты на территорию России.
Размножение. Гнездовой сезон в Индии начинается обычно в период муссонных дож-
дей и длится с июня по сентябрь. На Суматре гнёзда с кладками находили с июня по март
на равнине и в октябре-декабре в горах, на п-ове Малакка — в сентябре, декабре, феврале-
июле, на о-ве Ява — с октября по июнь. Птицы поселяются обычно небольшими группами,
иногда в колониях других цапель. Гнёзда устраивают на заломах тростника, в густых зарос-
лях камыша, на молодых деревцах или кустах, обычно в 1 м над водой (влажным грунтом)
или прямо на земле. Гнездо — плоская платформа, сложенная из маленьких кусочков ве-
ток, тростника и осоки, лоток выстилают листьями и травой. В кладке 2-6 (обычно 3-5)
яиц (Martinez-Vilalta, Mods, 1992).
Суточная активность, поведение. Характерна как дневная (особенно в период выкар-
мливания птенцов), так и сумеречная активность.
Питание. В состав кормов входят мелкая рыба длиной до 13 см, головастики и взрос-
лые лягушки, моллюски, ракообразные, насекомые и их личинки, а также другие живот-
ные. Добычу птицы подкарауливают на мелководье или в зарослях надводной растительно-
сти. Охотятся поодиночке, охотничью территорию охраняют.
Враги, неблагоприятные факторы. В гнездовой период враги — вороны, сороки, мел-
кие хищные млекопитающие. Не испытывает преследования со стороны человека и охотно
селится на созданных им влажных местообитаниях.
В оперении обнаружены блохи: Blankaartia acuscutellaris и Leptotrombidion sp., перье-
вой клещ Bicentralges orientalis и пухоеды: Ardeicola ixobrychae и Ciconiphilus decimfasciatus
(McClure, Ratanaworabhan, 1973).
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Особые меры
охраны не предусмотрены.
Японская кваква
209
Род Gorsachius Bonaparte, 1855
Японская кваква
Gorsachius goisagi (Temminck, 1836)
Nycticorax goisagi Temminck, 1836, Temminck et Laugier, Planaches Coloriees, livr. 98,
табл. 582, Япония.
Japanese [brown] night-heron, Japanese bittern (англ.), Rotscheitelreiher (нем.),
bihoreau goisagi (фр.)
Син. (рус.): японская выпь
Статус. Регулярно залётный вид.
Общая характеристика птицы и полевые признаки. Отдалённо напоминает обык-
новенную квакву, но мельче, более изящного сложения. Шея относительно короткая, ноги
умеренно длинные, пальцы, в отличие от других цапель, относительно короткие. Клюв ко-
роткий, толстый, конек надклювья слегка изогнут книзу. Общая окраска тёмная. В брачном
наряде перья затылка удлинены, образуют густой хохол; на нижней части горла длинные
заострённые перья. Повадками сходна с кваквами. Полёт плавный, не быстрый, с довольно
частыми неглубокими взмахами крыльев. Шею в полёте не складывает. Очень скрытна.
Взрослые птицы от близкой по размеру, но чуть более крупной обыкновенной кваквы
резко отличаются общим рыжим тоном окраски тела и коротким клювом. Молодые птицы
японской кваквы, в отличие от молодых квакв, у которых основной тон задаёт продольный
рисунок из тёмных и светлых пятен, более светлые со струйчатым поперечным рисунком
груди и верха тела.
Голос — негромкое «крэк» или «кхыу»; обычно молчалива.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Род лесных квакв Gorsachius, объединяющий 4 (а в узкой
трактовке — 2) вида, нередко включают на правах подрода в род Nycticorax. От настоящих
выпей эти цапли далеки, поэтому название «японская кваква», видимо, следует предпо-
честь названию «японская выпь», широко употреблявшемуся в отечественной литературе.
В таксономической литературе существуют разночтения по поводу года опубликования
первоописания вида — 1835 (традиционная трактовка) или 1836 (специальное исследова-
ние Dickinson, 2001). Последняя трактовка принята в «Списке птиц Российской Федера-
ции» (Коблик и др., 2006) и в последнем издании “The Howard and Moore Complete Checklist
of the Birds of the World” (Dickinson, 2003).
Описание. Окраска. Половой диморфизм отсутствует. Самец и самка в дефинитив-
ном наряде. Лоб и темя тёмно-коричневые, бока головы и шеи коричневато-рыжие, от под-
бородка вдоль горла тянется тёмная полоса; спинная сторона, включая кроющие крыла,
также тёмно-коричневая с тонким струйчатым рисунком. Первостепенные маховые аспид-
но-чёрные с широкими каштановыми вершинами и белым кончиком на первом маховом;
второстепенные маховые чёрно-бурые с широкими буровато-каштановыми вершинами,
испещрёнными мелкими бурыми крапинками и струйчатым рисунком; два пера крылышка
аспидно-чёрные с широкими белыми вершинами. Большие кроющие первостепенных ма-
ховых светло-каштановые; верхние кроющие хвоста и рулевые тёмно-аспидные. Брюшная
сторона бледно-охристая с бурыми продольными пятнами; нижние кроющие хвоста белые
с буроватыми пестринами. Испод крыла и подмышечные рыжевато-охристые с коричнева-
то-бурыми поперечными полосами. Клюв тёмно-роговой, уздечка зелёно-жёлтая, веки чёр-
ные, роговица жёлтая; ноги чёрные или зеленовато-бурые, когти бледно-роговые.
210
Отряд Листообразные, Семейство Цаплевые
Гнездовой наряд. Верх головы чёрно-бурый со светлыми пестринами; спинная сторона
темная, ржаво-бурая с чёрными пестринами; верхние кроющие крыла светлее; брюшная
сторона буроватая с пестринами; по бокам горла округлые тёмные пестрины. Клюв серый
с желтоватым оттенком; ноги яркие желтовато-зелёные, когти бледно-желтоватые.
У птиц первого года в отличие от птиц старших поколений ржаво-коричневый цвет ок-
раски оперения, особенно спинной стороны, кроющих маховых, шеи и головы, менее на-
сыщенный, на шее и голове сохраняется хотя и слабый, но заметный струйчатый попе-
речный рисунок.
Строение и размеры. Формула крыла I<II<III>IV>V...........Размеры (мм) самцов
(п=3): длина крыла — 267; 270; 250, длина клюва — 37,6; 37,7; 36,7, длина цевки — 70,6;
66,0; 64,3, длина хвоста 111,4; 113; 104. Длина крыла трёх погибших особей — 260; 274;
270 мм (колл. ДВГУ, БПИ ДВО РАН).
Линька. Не изучена.
Распространение. Гнездовой ареал. Японская кваква встречается на всей
территории Японии от п-ова Мацумоэ (Хоккайдо) до о-вов Цусимы, Идзу, Кадзан (Волка-
но) и Сикисима (Check-List ..., 1974), но гнездование доказано только на островах Идзу,
примыкающих частях о. Хонсю в префектурах Канагава, Сидзуока и южной Яманаси, а
также на о. Сикоку в префектуре Токусина (Brazil, 1991). В Корее добывали у г. Пусан 2.05
и 11.06 1966 г. (Gore, Won, 1971) и в окрестностях Сеула (без даты, сборы Мольтрехта, ЗИН
АН СССР). Регистрировали на о. Корор (Каролинские о-ва) (рис. 38).
В России отмечалась на о. Монерон, где была добыта 18 июня 1935 г. (Takahashi, 1937а),
там же остатки птицы найдены 7 июня 1954 г. и 22 июля 1973 г. (Нечаев, 1975); одна птица
добыта на юге о. Сахалин 15 марта 1937 г. (Takahashi, 19376).
Японскую квакву неоднократно отмечали также на юге Приморья в прибрежных райо-
нах материка (рис. 39): птица, добытая у Владивостока 12 мая 1921 г., хранится в Примор-
ском краевом музее (Омелько, Омелько, 1981); мёртвый взрослый самец найден в апреле
1961 г. в пригороде Владивостока (Лабзюк, 1965); В.И. Лабзюк (1981; 1990) наблюдал три
особи близ устья р. Аввакумовка 11 и 14 мая 1976 г., одну — в этом же районе на неболь-
шом лесном озере, одну — ус. Михайловка (Ольгинский р-н) 14 сентября 1977 г., группу из
4 птиц в зал. Ольги 23 мая 1980 г. и одну птицу 7 июня 1984 г. Неполовозрелый самец
держался в окрестностях с. Рязановка 25 и 26 мая и одна птица — 1 июня 1983 г. Там же
остатки одной птицы были найдены 26 мая 1981 г. (Назаров, 1986; 2004). В Лазовском
заповеднике в мае 1990 г. у оз. Чехуненко найдена погибшая птица и одна сильно истощен-
ная (Лаптев, Медведев, 1995). Японскую квакву дважды отмечали в мае 1999 г. на ключе
бух. Петрова (Хохряков, Шохрин, 2002); в низовьях р. Кедровой взрослая самка добыта 21
мая 1976 г. (Глущенко, Шибнев, 1977; Пекло, 1997), там же одиночных птиц наблюдали с
10 по 16 мая 1983 г., 3 мая 1988 г., 1-2 мая 1998 г (Шибнев, 1992; Курдюков, 2004). Одиноч-
ная птица встречена 25 апреля 2001 г. в верховьях р. Шкотовка (Курдюков, устн. сообщ.) и
26 июня 2003 г. в окрестностях пос. Андреевка (Глущенко, устн. сообщ.).
Японская кваква неоднократно регистрировалась на островах зал. Петра Великого, где
чаще находили остатки погибших птиц. На о. Б. Пелис эту птицу наблюдали 2 июня 1967 г.
(Лабзюк и др., 1971), одна птица держалась 9-10 июня 1979 г и 23 и 26 июня 1988 г.,
остатки трёх птиц найдены в мае 1979 г. и в 1980 г (Назаров, 2004). Свежие остатки япон-
ской выпи найдены 11 июня 1979 г. на о. Гильдебранта Ю.В. Шибаевым (устн. сообщ.) и 25
июля 1979 на о. Матвеева (Назаров, 2004). Погибшая птица найдена 7 мая 1972 г на о.
Фуругельма (Литвиненко, 2005).
Одна птица встречена 2 мая 1975 г. на восточном побережье оз. Ханка в окрестностях
с. Гайворон (Глущенко и др., 1992) (рис. 39).
Японская кваква
211
Рис. 38. Область распространения японской кваквы: а — область гнездования, б — районы зимовок.
Имея в виду регулярность встреч японской кваквы в южном Приморье, высказано пред-
положение о возможности гнездования этого вида на юге Дальнего Востока (Назаров, 2004;
Литвиненко, 2005). Появление птиц в Приморье весной, возможно, объясняется тем, что
часть птиц, зимующих в вост. Китае, пролетает вдоль морского побережья континента.
Зимовки. Зимует от о-вов Рюкю и Волкано до Филиппинских и сев. Сулавеси. В
небольшом количестве зимует на о. Кюсю (A Hand-List...., 1958). Отмечена на зимовке на
Тайване и в юж. Китае в провинции Гуандун в устье р. Чжуцзян (Cheng Tso-hsin, 1987).
Миграции. Весной появляется на о. Хонсю в начале апреля, но отдельных птиц добы-
вали здесь и в марте; отлёт в сентябре-октябре, последних регистрировали в ноябре (Austin,
Kuroda, 1953; del Hoyo et al., 1992). В Китае пролётные особи отмечены у г. Шанхай и в
провинции Фуцзянь (Cheng Tso-hsin, 1987). У Шанхая регистрировалась с конца марта до
конца мая (La Touche, 1912), в Фучжоу — в апреле (Caldwell, Caldwell, 1931). На Филиппи-
нах одна птица поймана в августе (McClure, 1974).
Местообитания. Населяет густые тенистые леса предгорных районов, не поднимаясь
выше 800 м (Jahn, 1942). В Приморье японских выпей отмечали на берегах лесных водо-
ёмов и на лесистых склонах островов.
Численность. Точных сведений нет. На местах гнездования немногочисленна или ред-
ка. С середины 1950 гг. отмечено сокращение численности (del Hoyo et al., 1992). Очень
редка на зимовках в Китае (Cheng Tso-hsin, 1987).
Размножение. Сведений мало. Гнездовой биотоп — преимущественно вечнозелёные
широколиственные густые леса в долинах небольших речек и ключей. Гнездовой сезон
212
Отряд Листообразные, Семейство Цаплевые
Рис. 39. Места встреч японской кваквы в Рос
сии: а — пункты регистрации вида.
растянут. Кладки находили с середины мая
до конца июля (Austin, Kuroda, 1953). Пары
поселяются на отдельных гнездовых участ-
ках, а в наиболее благоприятных условиях
могут гнездиться группами, но не колония-
ми. Брачный крик представляет собой серию
низких однотонно повторяющихся гудящих
звуков.
Гнездо японская кваква устраивает на вет-
вях деревьев, обычно вечнозелёных, в 7-20 м
от земли и в 1^4 м от ствола. Постройка гру-
бая, из прутьев. Похожа на гнездо восточной
горлицы, но крупнее. Полная кладка содер-
жит 3—4 белых яйца. Размеры яиц (п=7): 45,0-
51,0 х 36,7 х 38,3 мм, в среднем 47,8x37,8
мм (Schonwetter, 1960) и 47,5x37,0 мм (Austin,
Kuroda, 1953). Продолжительность насижи-
вания около 17-20 дней, птенцы приобрета-
ют способность к полёту в возрасте 35-37 дней. Самостоятельные молодые птицы встреча-
ются с начала июля. Вероятно, может быть повторная кладка (del Hoyo et al., 1992).
У самца, добытого 26 мая на юге Приморья, семенники были чуть увеличены, их раз-
мер составлял 8x3 мм (колл. ДВГУ).
Суточная активность и поведение. Ведёт ночной образ жизни. В течение дня спокой-
но сидит в кроне крупного дерева, но иногда в туманную дождливую погоду активна днем
(Austin, Kuroda, 1953).
Питание. Поедает крабов и других ракообразных, насекомых и мелких рыб, которых
вылавливает в маленьких лесных ручьях или на болотистых просеках. В желудках двух
птиц из Приморья обнаружены лягушки (Лабзюк, 1965) и дождевые черви (Глущенко, устн.
сообщ.). В дождливую погоду наблюдали птицу на аллее парка, кормящуюся дождевыми
червями и жуками-плавунцами (Назаров, 1986, 2004).
Враги и неблагоприятные факторы. Основной неблагоприятный фактор для японской
кваквы — освоение и трансформация мест её обитания человеком (del Hoyo et al., 1992).
Хозяйственное значение и охрана. Хозяйственного значения не имеет. Японская квак-
ва внесена в Красный список МСОП и Красную книгу Азии; в России занесена в Красные
книги Сахалинской области и Приморского края (Литвиненко, 20056).
Род Nycticorax Forster, 1817
Кваква
Nycticorax nycticorax (Linnaeus, 1758)
Ardea nycticorax Linnaeus, 1758, Syst. Nat., ed. 10, C. 142, Южная Европа.
Black-crowned night-heron (англ), Nachtreier (нем.), bihoreau gris (фр.).
Син. (лат.): для подвида nycticorax'. Ardea grisea Linnaeus, 1766; Ardea kwakwa S.G. Gmelin,
1771; Ardeaferruginea S.G. Gmelin, 1771; Ardea maculata J.F. Gmelin, 1789; Nycticorax infaustus
Forster, 1817; Nycticorax europaeus Stephens, 1819; Nycticorax orientalis C.L. Brehm, 1831;
Кваква
213
Nycticorax badus C.L. Brehm, 1831; Nycticorax meridionalis C.L. Brehm, 1831; Nycticorax
ardeola Temminck, 1840; Nycticorax aegiptius Gurney, 1872.
Син. (рус.): квак (народное, звукоподражательное), ночная цапля, обыкновенная кваква
I книжное), черноголовая кваква (переводное с англ.)
Статус. Гнездящийся, перелётный, в южных частях ареала — изредка зимующий вид.
Общая характеристика и полевые признаки. Средних размеров коренастая цапля
(несколько крупнее вороны) с относительно короткими ногами и клювом, закруглёнными
крыльями и хорошо различимыми чёрным, серым и белыми цветами окраски оперения.
Молодые бурые со светлыми продольными пестринами; их окраска во многом сходна с
окраской молодых больших выпей, однако последние окрашены преимущественно в золо-
тисто-коричневый цвет.
Кваква — типичная ночная птица. Повышенная активность начинается с наступлением
сумерек и заканчивается незадолго до восхода солнца. В дневные часы кваквы стайками
спокойно сидят на деревьях, в кустах или на тростнике, прячась от лучей солнца. Иногда их
удаётся видеть летящими днем, причём во время полёта они сильно втягивают и без того
короткую шею и обычно выстраиваются в поперечную линию. Будучи потревоженными в
тростниках, взлетают в воздух довольно быстро, но, поднявшись, летят медленно, легко,
плавно взмахивая крыльями.
Кваквы не очень осторожны, близко подпускают к себе человека, неожиданно взлетая
почти вертикально. Охотятся на мелководьях по окраинам тростниковых зарослей или сидя
на свисающих ветках над протоками. При добывании пищи птицы нередко медленно бро-
дят по мелководью или (чаще) неподвижно стоят, всматриваясь в воду (Спангенберг, 1951).
Голос. Кваква более криклива, чем другие цапли. Её обычные крики звучат как «квау,
квау» или «квак, квак, квак», повторяясь несколько раз со значительными интервалами.
Наиболее часто голос кваквы удаётся слышать с наступлением сумерек, когда птицы нахо-
дятся высоко в воздухе, или же при взлёте. В гнездовых колониях взрослые и молодые
кваквы чрезвычайно крикливы.
Подвидовая систематика. Четыре слабо различающихся морфологией и биологией
подвида.
1. Nycticorax nycticorax nycticorax
Ardea nycticorax Linnaeus, 1758, Syst. Nat., ed. 10, C. 142, Южная Европа.
Европейская, азиатская и африканская часть видового ареала.
Кроме номинативного, выделяют ещё три подвида: N. л. hoactli Gmelin, 1789 (2), кото-
рый обитает в американской части ареала от юго-востока Канады и северо-запада США
через Центр. Америку до вост. Аргентины, а также на Гавайских о-вах, N. п. obscurus,
Bonaparte, 1855 (3), распространённый в Юж. Америке на юге Перу, в Чили, в зап. Арген-
тине на юг до Огненной Земли, и N. п. falklandicus Е. Hartert, 1914 (4) с Фолклендских о-
вов (del Hoyo et al., 1992; Dickinson, 2003) (рис. 40). Две южные формы отличаются от
прочих сравнительно тёмной окраской и ведут практически оседлый образ жизни, совер-
шая лишь незначительные сезонные кочёвки даже на субантарктичекиом юге материка и
на озёрах Анд, располагающихся на высотах до 4 816 м н.у.м.
Описание. Окраска. Половой диморфизм в окраске отсутствует. Самец и самка в
брачном наряде. Окраска верха головы и спины чёрная; крылья, поясница и хвост пепель-
но-серые, низ тела и шея белые с сероватым оттенком, лоб и щёки чисто-белые. На затылке
развиты 2-А узких длинных белых украшающих пера-косицы. Клюв чёрный, ноги розовые
или жёлтые (в разгар брачного периода до красных), уздечка и кожа вокруг глаз тёмные с
синеватым оттенком, радужина красная.
214
Отряд Листообразные, Семейство Цаплевые
Зимний наряд взрослых птиц почти неотличим от брачного, обычно отсутствуют укра-
шающие косицы, чёрные партии оперения сохраняют матовый сероватый оттенок и не имеют
зелёного блеска.
Пуховой птенец покрыт светло-бурым пухом (особенно длинным на темени и спине),
сквозь который просвечивает серовато- или зеленовато-жёлтая кожа. На брюшной стороне
пух светлее — до белесого. Радужина тускло-жёлтая, клюв и ноги сероватые или серовато-
жёлтые.
Молодые птицы в гнездовом наряде сверху бурые со светлыми продольными пестрина-
ми на спине и беловатыми каплевидными пятнами на кроющих крыла. Голова, шея, низ
тела светло-бурые с охристым оттенком и многочисленными тёмными продольными пест-
ринами; радужина от жёлтой до жёлто-оранжевой, жёлто-бурой, клюв буровато-жёлтый с
тёмным коньком, ноги буровато- или зеленовато-жёлтые.
Первогодки (первый летний наряд) сохраняют от гнездового наряда бурые со светлыми
каплевидными пестринами крылья, и охристые оттенки низа тела. Шапочка и спина буро-
ватые, однотонные; изредка вырастают украшающие косицы. Продольные пестрины на
шее и груди по сравнению с гнездовым нарядом более размыты, лоб светлее. Клюв бурый
с желтоватым основанием подклювья, ноги тусклые, желтоватые, радужина от буровато-
оранжевой до красной.
У птиц во втором летнем наряде (второгодков) тип окраски в разной степени прибли-
жается к окраске взрослых, шапочка и спина, как правило, однотонные, буровато-серые,
сохраняется охристый налёт на серых и белых партиях оперения, на шее и кроющих крыла
ещё остаётся некоторое количество буроватых пестрин. Украшающие косицы, как прави-
ло, отсутствуют.
Строение и размеры. Длина тела 58-65 см, самцы в среднем чуть крупнее самок.
Размах крыльев 105-112 см. В среднем самцы несколько крупнее самок (табл. 10).
Линька. Схема линьки — как у других цапель. Пуховой наряд, образованный густым,
длинным и мягким пухом, сменяется на перьевой постепенно, начиная с возраста в 13-15
дней. Формирование гнездового наряда заканчивается к 40-дневному возрасту. Молодые
птицы в гнездовом наряде сменяют мелкое оперение в первую осень жизни между авгус-
Рис. 40. Область распространения кваквы: а — гнездовой ареал; б — места зимовок. Подвиды: 1—
N. п. nycticorax; 2 — N. п. hoactli; 3 — N. п. obscurus\ 4 — N. п. falklandicus.
Кваква
215
Таблица 10. Средние размеры и вес (масса) взрослых квакв
Пол Число особей (п) Длина (мм) Масса, г
клюва цевки крыла череп с клювом
Самки 35 69 75 287 133 580
Самцы 30 69 76 290 134 630
том и декабрем. Ювенильная линька происходит на местах зимовок. Весной молодые пти-
цы ещё отличаются от взрослых темной, буро-коричневой окраской с белыми пятнами на
маховых и покровных перьях крыла. В июле-августе начинается очередная линька «полу-
взрослых» птиц и, вероятно, частичная послебрачная — взрослых.
Линька взрослых птиц в пределах репродуктивной части ареала изучена недостаточ-
но. Применяемая методика осмотра отловленных взрослых птиц ночью на местах кор-
мёжек (Русев, 19836) и линяющих птиц в густых зарослях тростника не дает возможно-
сти полностью проследить это явление. Отмечается смена мелкого пера у отдельных осо-
бей, могут меняться и маховые. Такие птицы не способны летать, они держатся в густых
тростниковых зарослях, выходя на кормёжку к открытым плёсам. Например, у отловлен-
ных 27 июля 1980 г. в плавнях Днестра «полувзрослых» квакв имелось всего по 3 наруж-
ных первостепенных маховых пера; опахала остальных только начинали показываться из
пеньков.
У подавляющего большинства особей крупное перо полностью меняется, вероятно, на
местах зимовок, и взрослый наряд птицы приобретают там. Одновременно появляются и
«брачные» перья. Наблюдения за группировкой зимующих квакв в дельте Днестра показа-
ли, что уже в январе у многих особей имеется брачный наряд, хотя у некоторых он форми-
руется непосредственно перед гнездованием. После гнездования, обычно перед отлётом,
взрослые птицы теряют «брачные» перья и частично меняют мелкое перо.
Полный наряд, не отличимый по окраске от наряда взрослых птиц, кваквы одевают в
годовалом возрасте. Это второй зимний (а затем — второй брачный) наряд. Смена брачно-
го оперения начинается с выпадения украшающих перьев головы и мелкого оперения.
Распространение. Гнездовой ареал. Сев. и Юж. Америки, Азия от Малой
Азии, Сирии, Палестины, Месопотамии, Ирана и Средней Азии до Юж. Маньчжурии,
Китая (от Чжили до Юньнаня, Гуаньдуна, Фуцзяня), Юж. Приморья, Кореи и Японии (на
север до Хондо), на юг до Филиппинских, Зондских о-вов и Сулавеси. В Африке гнездит-
ся в Марокко, Тунисе, Алжире, Сенегале (Cramp, 1977; Fishpool, Evans, 2001), а также
очагами — по всей тропической части материка и на Мадагаскаре. В Зап. и Центр. Евро-
пе гнездится в юж. Португалии, Испании, Албании, Австрии, Бельгии, Боснии и Герцего-
вине, Болгарии, Хорватии, Чехии, Франции, Германии, Греции, Венгрии, Италии, Маке-
донии, Мальте, Польше, Сербии, Словакии, Словении, Румынии (Birdlife International,
2004) (рис. 40).
Область гнездования кваквы в Вост. Европе и Сев. Азии простирается от западных
границ бывшего СССР до оз. Балхаш, к северу до 52-53° с.ш. (рис. 41). В самой северной
части ареала кваква очень редка. На Украине встречается почти повсеместно (Смогор-
жевський, 1979), но на севере гнездится спорадически и изредка, в южной же части —
многочисленна. В Азово-Черноморском регионе ею заселены практически все водно-бо-
лотные угодья международного и национального значения (Микитюк, 1999; Сиохин и
др., 2000).
216
Отряд Листообразные, Семейство Цаплевые
В прошлом кваква была обычна в дельтах Терека, Кубани, в низовьях Бейсуга и Челбас
(Спангенберг, 1951), на Зап. Маныче. В 1970-х и 1980-х гг., в связи с интенсивным развити-
ем рисоводства и прудово-лиманного рыбоводства, география её поселений расширилась,
в ряде мест возникли новые колонии. В настоящее время кваква гнездится в долине Ниж-
него Дона, на Зап. и Вост. Маныче, в Вост. Приазовье, на водоёмах Ставрополья, Сев. Осе-
тии, в дельте Терека (Птицы Северного Кавказа, 2004).
Регулярно гнездится в устьях Волги и Урала. К северу достигает районов Орла, Вороне-
жа, на Волге — местностей несколько севернее Камышина. Северная граница в Азиатской
части доходит до низовьев Тургая, района Атбасара, р. Или. К югу отсюда широко распро-
странена по Средней Азии. В Туркмении гнездится в поймах Аму-Дарьи, Мургаба и Тед-
жена. Довольно широко гнездится в низменном Таджикистане, в частности у ст. Мельнико-
ве, в урочище Джили-куль на Вахше, в долине Аму-Дарьи и Пянджа от Пархара до Терме-
за, по-видимому, у Куляба в тугаях по Ях-су. В Таджикистане на гнездовье в горы, вероят-
но, не заходит, но в послегнездовое время кочующие птицы появляются в горах довольно
высоко. Птиц наблюдали также в Фергане, преимущественно около Андижана, близ стан-
ций Чиназ и Сыр-Дарьинской, а к северу она обыкновенно гнездится вниз по течению реки,
во всяком случае, до широты ст. Джусалы. Редко гнездятся на оз. Камышлы-баш. На восток
от Сыр-Дарьи в годы, богатые водой, населяет озёра Тели-куль и низовья Сары-су. Гнездит-
ся на Лепсе, найдена в низовьях Чу, по тугаям в дельте р. Или (Долгушин, 1939; цит. по
Спангенберг, 1951; Долгушин, 1960; Птицы Средней Азии, 2007). К северу отсюда ещё
гнездится в тростниковых зарослях дельты Тургая (Сушкин, 1908) и найдена в Адбасарс-
ком р-не Акмолинской обл. Как исключительный случай гнездование кваквы было отмече-
но Н.А. Зарудным на р. Мечетке около Буранной, но в целом названный исследователь
считал её редкозалётной птицей для всего бывшего Оренбургского края, что вероятней
(Зарудный, 1888). На Дальнем Востоке известно гнездование только на оз. Ханка (Глущен-
ко и др., 20066) (рис. 41).
Рис. 41. Гнездовой ареал кваквы в Сев. Евразии: а — область гнездования, б — места зимовок.
Кваква
217
Случаи залётов известны в Прибалтику, в район Пскова, в Московскую и Тверскую обл.
и в Башкирию. На севере Зап. Европы кваква залетает в Англию, Голландию, Данию, Фин-
ляндию, Швецию и даже в Гренландию. Залетает также на Азорские о-ва и Мадейру (Спан-
генберг, 1951; Птушенко, Иноземцев, 1968; Cramp, 1977).
Зимовки. В Африке и юге Азии кваква оседлая, кочующая и лишь частично мигриру-
ющая птица. Протяженность же пролётного маршрута квакв, гнездящихся в Вост. Европе и
Сев. Азии к местам зимовок, колеблется от 4 до 5 тыс. км. Основные места зимовок евро-
пейских квакв расположены в Африке южнее Сахары. Среднеазиатские кваквы, очевидно,
зимуют на полуострове Индостан.
В то же время, нерегулярные зимовки вида отмечаются и в более северных местах. В
дельте Днестра в отдельные теплые зимы кваквы встречаются небольшими стаями в пой-
менных лесах, вылетая на кормёжку на рыбоводные пруды (Русев, 1983а; Русев, Русева,
1983). В отдельные годы, когда погода резко меняется и пруды покрываются льдом, кваквы
переходят на питание мышевидными грызунами. Кваква частично зимует также в Закавка-
зье в Шемахинском р-не Азербайджана и под Ленкоранью. Зимовки здесь носят нерегуляр-
ный характер, хотя иногда птицы встречаются здесь зимой в большом количестве. Моло-
дых квакв зимой неоднократно отмечали в дельте Волги (Хлебников, 1928; цит. по: Спан-
берберг, 1951), но эта зимовка, как правило, кончалась гибелью птиц от голода.
Интересным является возврат от молодой птицы из Житомирской обл., полученный 10
ноября 1978 г., который может свидетельствовать о том, что некоторые птицы остаются на
зимовку в этих районах Украины, хотя достоверных наблюдений подобного нет. Однако
существуют возвраты от днестровских птиц, зимующих в Скадовском р-не Херсонской
обл. (добыта 28 января 1982 г.), и вблизи Красной косы и в Белгород-Днестровском р-не
Одесской обл. (29 декабря 1978 г.).
Основные зимовки днестровской популяции кваквы находятся в Зап. Африке, в основ-
ном в верхней части дельты р. Нигер, откуда получено 2 возврата колец. Одна из зимую-
щих в Африке птиц добыта на 13-м году жизни в Мали (окольцована 15 июня 1979 г.,
добыта 15 января 1992 г.). Остальные особи квакв добыты на р. Шари вблизи оз. Чад
(февраль), одна особь встречена в декабре во Франции на р. Дордоно у Бискайского зал.
Ещё одна особь, окольцованная в верховьях Тилигульского лимана, добыта в декабре в
заливе Бенин в Нигерии.
Зимой 1979/80 г. кваква впервые обнаружена на зимовке на Новотроицком вдхр. в Став-
рополье. В последующие годы там зимовало до 20 особей (Бичеров, Хохлов, 1981). В 1981 г.
6 декабря при положительной температуре 15-20 квакв наблюдали на Новомарьевском
лимане в 20 км западнее г. Ставрополя. 7 декабря здесь же на открытых местах держалось 5
взрослых и 3 молодые птицы. Видимо, кваквы, в 1980-х - 1990-х гг. остававшиеся на зиму
вблизи мест размножения, в критический период зимовки концентрировались на Новотро-
ицком вдхр., где имелась незамерзающая акватория у сбросов Ставропольской ГРЭС.
В первую зиму три молодые азербайджанские кваквы были обнаружены неподалеку от
места рождения, тоже в Азербайджане (Агджабединский р-н, Сальяны и Астара); одна пти-
ца — 17 декабря в Ливане, и ещё одна — в начале апреля в Ираке близ Басры. 8 мая кваква
из Кызыл-Агача была поймана в Туркмении, под Небит-Дагом.
Миграции. Места зимовки покидает в феврале или марте. В Азово-Черноморском ре-
гионе кваквы появляются в первой половине марта. Самые ранние встречи, в первых чис-
лах марта, отмечены в дельте Днестра. Интенсивный пролёт кваквы на Дунае наблюдал
З.А. Петрович в середине марта; массовый пролёт в Крыму проходил между 20 марта и 13
апреля; в дельте Днепра — в конце марта - начале апреля (Смогоржевський, 1979). Прилёт
к местам гнездовий очень растянут. Как правило, сначала появляются передовые группи-
218
Отряд Листообразные, Семейство Цаплевые
ровки по 100-150 особей. В последствии, в течение 2-3 недель прилетает основная масса
птиц. Обычно кваквы летят с заката солнца и до утренней зари, на рассвете останавлива-
ются в удобных для отдыха местах (Русев, Русева, 1983). Птицы летят небольшими стайка-
ми — до 50 особей. Построение стай имеет либо форму слабовыраженного клина, либо в
виде косой линии. Во время дневного отдыха стаи не распадаются. В период весеннего
пролёта квакв можно встретить почти на всех внутренних водоёмах региона. Но морское
побережье они пролетают без остановки и их здесь видят лишь изредка.
Прилет кваквы в дельту Дона, на Зап. Маныч и в Вост. Приазовье начинается обычно в
первой декаде апреля и продолжается до конца второй декады этого месяца. В ранние вес-
ны прилетает и раньше. В 1979 г., например, первые 27 особей отмечены в дельте Дона 22
марта (Казаков и др., 1986). По данным Е.П. Спангенберга (1951), на Черноморском побе-
режье Кавказа кваквы появляются 27 марта, а пролёт их затягивается обычно до конца
первой трети мая. На приморских низменностях в окрестностях Адлера в 1980 г. их отме-
чали с 11 апреля. Последние мигранты в районе р. Мзымты в 1985 г. встречены в конце мая,
а в 1987 г. — 17 июня. Обычно отмечаются одиночные птицы и пары, и только однажды
наблюдали стаю из 12 птиц. Во время пролёта птицы придерживаются участков древесной
растительности вблизи водоёмов или лесных массивов по берегам рек, граничащих с от-
крытыми пространствами. В Сев. Осетии на оз. Бекан птицы появляются в конце третьей
декады марта- первой половине апреля; в 1984-1986 гг. они отмечены 14-16 апреля (Пти-
цы Северного Кавказа, 2004).
Весной наиболее раннее появление на оз. Ханка отмечено в первой декаде апреля (Глу-
щенко и др., 20066), а в долине р. Раздольная в окрестностях г. Уссурийска наиболее ранняя
регистрация датирована 31 марта 2006 г. (Глущенко и др., 2006а).
Весенние возвраты взрослых птиц, родившихся в Азербайджане, поступили 29 марта
из Поти (Грузинская ССР), 28 марта — из низовий Терека, 10 апреля — из северного Азер-
байджана (г. Сиазань) и из-под Сочи, где труп погибшей «до 22 июня10» цапли был обнару-
жен в лесу.
Осенние перемещения кваквы можно наблюдать уже в июле. Сведения о сроках перехо-
да послегнездовых кочёвок в собственно миграцию отсутствуют. Однако перемещения не-
больших стай квакв отмечены уже в середине августа. Основная масса днестровских квакв
отлетает в сентябре: массовый отлёт — конец сентября - середина октября. На Дунае отлёт
происходит также в сентябре-октябре; аналогичны сроки и для днепровских квакв (Смо-
горжевський, 1979). Интенсивность осенней миграции несколько ниже весенней. Она бо-
лее растянута по срокам и стайки более рассредоточены по территории. К концу ноября
кваквы практически полностью покидают гнездовые территории.
Послегнездовые перемещения наиболее полно изучены на Днестре. Массовый подъём
на крыло молодых квакв происходит обычно в середине июля. В первую неделю молодые
находятся вблизи колоний. В последствии они совместно с взрослыми вылетают на кор-
мёжку и живут в одних стаях с ними (Русев, 1986). Спустя три недели после вылета связь
молодых с гнездовой колонией становится менее прочной. Птицы из разных колоний обра-
зуют смешанные стаи. Часть птиц держится до осени на территории рождения, но боль-
шинство улетает далеко от неё. Для молодых квакв характерен послегнездовой разлёт в
восточном, южном, северном и северо-западном направлениях. Возвраты от квакв, поме-
ченных в дельте Днестра, регистрировали в Черниговской, Ровенской, Житомирской, Львов-
ской, Черновицкой, Винницкой обл. Украины (рис. 42). Это наводит на мысль о том, что
птицы используют некие «экологические коридоры», в данном случае — бассейн р. Днестр,
10 Год не назван (прим. ред.)
Кваква
219
достигая Львовской обл. Возможно, они потом оттуда летят в Зап. Европу и вместе с птица-
ми европейских стран мигрируют через Средиземное море на юг в Африку либо остаются
зимовать в европейских странах.
Первые молодые птицы отмечены в Ровенской обл. уже 17 июля, на расстоянии более
чем в 500 км от мест рождения. Встречены они также в Житомирской, Черниговской и
других областях. Доля таких «бродяг» (по данным кольцевания) составляет 43%. Разлет в
северном направлении характерен и для квакв, обитающих на Днепре (Сапетин, 1978в).
Вероятно, в этот период и в таких местах формируются группировки из особей различных
популяций для отлёта к местам зимовок. Объединение небольших стай в послегнездовой
период в более крупные, сопровождающееся дисперсией вида по территории, вероятно
имеет существенное значение для благополучия вида (Юзефик, 1970).
Такую же картину можно наблюдать и у «нижнеднепровских» квакв (кольцевание в
Херсонской обл.). Но это явление распространяется здесь и на более взрослые группы птиц.
Лишь две из них (молодые) в начале сентября встречены невдалеке от гнездовий. Одна,
также первогодок, добыта у Днепропетровска (270 км к северо-востоку). 2-летняя же квак-
ва из этого района в октябре была убита в бассейне р. Припять у с. Хойники на юге Бело-
руссии (600 км от места рождения), а 3-летняя в ноябре — в Запорожской обл. (250 км).
В отличие от «западно-черноморских» квакв, область послегнездовых кочёвок птиц
того же вида из Вост. Приазовья сравнительно невелика. Встречи окольцованных здесь
птиц регистрировались в 20-100 км от мест рождения (Сапетин, 1977в). Так, в августе-
сентябре 23 молодые кваквы отмечены в Краснодарском крае и одна — в Крыму. Взрос-
лые птицы держались в основном здесь же, и лишь одна встречена 27 сентября в Грузии
у г. Поти. Не далее 50 км от места кольцевания найдена основная масса молодых квакв и
в октябре, хотя одна особь уже 2 сентября встречена в Сирии у г. Изра (Птицы Северного
Кавказа, 2004).
Рис. 42. Послегнездовой разлёт квакв из дельты Днестра (по данным мечения).
220
Отряд Листообразные, Семейство Цаплевые
Сам процесс осенней миграции, если его начало отсчитывать с момента вылета моло-
дых из гнезд, носит затяжной характер. Уже в конце июля - начале августа собравшиеся
стайки кваквы предпринимают широкие кочёвки, о чем было сказано выше. В конце авгу-
ста начинается слабо выраженное движение птиц к югу, которое постепенно усиливается,
приобретая характер настоящего отлёта. В конце сентября во многих северных районах
своего распространения численность кваквы резко сокращается, но на южных гнездовьях,
в местах, обильных пищей, они задерживаются чрезвычайно долго. Под Харьковом осен-
ний отлёт начинается в некоторые годы с последних чисел августа, но чаще с первых чисел
сентября. В конце сентября пролёт достигает наибольшей силы — птицы летят стаями в
сотни особей. Затем численность пролётных квакв убывает, и движение к югу совершенно
заканчивается в самом конце сентября или в первых числах октября. Одновременно начи-
нается движение птиц на юг в низовьях Днепра, достигая здесь наибольшей силы в середи-
не сентября и заканчиваясь в конце первой трети следующего месяца. Заметный осенний
пролёт в районе Ростова начинается в последней трети сентября. В низовьях Волги отлёт
происходит в сентябре, запоздавшая молодёжь задерживается до половины ноября; иногда
квакв наблюдают в приморской части Волги и в зимнее время. В центральной части Сев.
Кавказа пролёт происходит в сентябре-октябре, но на Каспийском побережье в Кизляр-
ском районе кваквы задерживаются на весь ноябрь и первую декаду декабря. На Черномор-
ском побережье Кавказа в районе Новороссийска и Сочи выраженный пролёт наблюдается
по ночам во второй половине сентября (Спангенберг, 1951).
Данные о сроках отлета для азиатской части бывшей территории Советского Союза
менее подробные. Н.А. Зарудный (1915) отмечал крайне раннее появление квакв в пустыне
Кызыл-Кум. 2 и 3 августа птицы летели прямо на юг. На нижней Сыр-Дарье Н.А. Северцов
слышал квакв 4 октября, но по наблюдениям Е.П. Спангенберга (1951) в середине октября
на Сыр-Дарье кваквы уже отсутствовали. В юж. Прибалхашье и сев. Киргизии пролёт про-
ходит во второй половине августа, заканчиваясь к середине сентября. В юго-зап. Бетпакда-
ле пролёт наблюдался в 1974 г. 22-24 сентября, на перевале Чокпак — в разные годы 31
августа - 22 сентября (Птицы Средней Азии, 2007). В Туркмении осенние кочёвки квакв
начинаются с конца июля - начала августа, а в сентябре и октябре идёт интенсивный осен-
ний пролёт (Дементьев, 1952). В низовьях Атрека осенний пролёт квакв происходит с тре-
тьей декады сентября до конца ноября (Караваев, 1991).
В осенний миграционный период одиночные птицы изредка встречаются на предгор-
ных и горных водоёмах. Так, 24 и 27 августа 1976 г. квакву регистрировали в окрестностях
пос. Гузерипль Кавказского заповедника, 16 сентября 1977 г. — в долине р. Теберда.
Пролёт квакв из Краснодарского края на места зимовок идёт через Зап. Кавказ и Малую
Азию. Возможно, часть птиц отлетает и на юг Европы. Молодые птицы на местах зимовки
встречены в центральной Африке (18 февраля), в Колхиде (19 марта); одна взрослая птица
добыта в феврале в Эчмиадзинском р-не Армении (Сапетин, 1978в). В 1987 г. одна кваква
добыта 7 февраля на Черноморском побережье в низовьях р. Псоу. Последние находки мо-
гут свидетельствовать как о местах зимовок, так и о начале движения квакв к районам
гнездования.
Из дельты Волги птицы летят через Дагестан и Ставропольский край в Экваториальную
Африку (оз. Чад, Камерун). Не исключён частичный отлёт и в Европу (взрослая птица 20
августа встречена в Днепропетровской обл.). Молодые кваквы из дельты Волги в августе и
сентябре держатся в пределах гнездовых территорий, откочевывая, однако, из приморской
полосы к западу и северо-западу (Скокова, 1959, цит. по: Сапетин, 1978в). Лишь 30 сентяб-
ря добыта одна птица, отлетевшая на юг, в северную часть дельты Терека. В октябре три
птицы были добыты близ мест кольцевания, а две — на пролёте: 29 октября на берегу
Кваква
221
Каспийского моря в районе Кизляра и 20 октября — в Ставропольском крае (480 км к юго-
западу). Ещё одна молодая кваква 14 октября уже достигла места зимовки на оз. Чад в
Африке; вторая найдена 15 января в Камеруне.
Все летне-осенние (август - ноябрь) возвраты колец от молодых птиц, помеченных в
дельте Терека (Дагестан), согласно данным Я.В. Сапетина (1978в), получены из самой дельты
или с близлежащего побережья, не далее 100-150 км от мест кольцевания. Но одна годова-
лая кваква добыта 27 августа в Карасукском р-не Новосибирской обл. в 2 500 км от места
рождения, другая — в декабре в Джелилабадском р-не Азербайджана. Из 11 возвратов 6
поступило из дельты Терека, не далее 50 км от места рождения, в период с 14 июня по 17
октября от птиц в возрасте 1, 2, 2, 5, 6 и 7 лет. Ещё три дагестанские кваквы были добыты
через 2, 2 и 6 лет во внутренних частях дельты Волги — соответственно, 12 августа, 27
августа и 23 октября. Поскольку дельта Волги не входит в область послегнездовых кочёвок
дагестанских молодых квакв, можно предположить, что часть этих птиц переселяется туда
для выбора мест будущих гнездовий.
По Азербайджану наиболее массовые материалы получены от квакв, окольцованных в
Кызыл-Агачском заповеднике. Молодые птицы в августе-сентябре (некоторые до ноября)
держатся в том же районе или откочёвывают в массив плавней р. Куры у устья Аракса.
Однако некоторые из них отлетают на зимовку очень рано. Так, уже 2 сентября одна птица
была поймана в Индийском океане в 800 км западнее южной оконечности Индостанского
п-ова. Не исключено, что она, находясь на пути к местам зимовки, была снесена туда воз-
душным потоком. В октябре получен 1 возврат из района кольцевания и два — из Ирака,
пров. Дивания (около 800 км на ЮЮЗ). На путях осеннего пролёта взрослых (2-8-летних)
квакв добывали: 19 августа в Италии близ Милана, 1 сентября в сев. Иране и в середине
ноября — в Ливане. В начале зимы (до 4 января) азербайджанскую квакву убили в Судане,
а 5 апреля — под Багдадом.
Не установлено существенных различий в путях миграций между популяциями Вост.
Приазовья и дельты Волги. Птицы, гнездящиеся здесь, совершенно не встречаются зимой
севернее Кавказского хр., но в небольшом количестве бывают в Закавказье. Основные же
зимовки их, несомненно, находятся в тропической Африке. Дагестанские кваквы также
летят через Кавказ и вост. Средиземноморье (Греция), иногда задерживаясь до декабря в
Азербайджане. Взрослые птицы, родившиеся в Азербайджане, наоборот, частично зимуют
близ родных колоний, а частично также отлетают в Африку через Зап. Азию (Иран, Ирак,
Ливан). Осенние встречи в Италии и Индийском океане позволяют предполагать ещё более
широкий «веер разлёта».
О путях миграций квакв из сев. Причерноморья мы можем судить по осенним возвра-
там из Греции, Мальты, Испании, Италии. Вероятно, попав на Балканский п-ов, они влива-
ются в общий миграционный поток дунайских квакв, летящих через Италию, Балканы и
Грецию в северную и экваториальную Африку. Возможно, птицы пересекают Сахару. Одна
из птиц, окольцованных в Венгрии, добыта в оазисе этой пустыни в весенний период (Snow,
Perrins, 1998). Вместе с тем, возвраты, полученные от квакв на Атлантическом побережье
Франции и в Испании, наталкивают на мысль о возможном пролёте части их популяции
вдоль Атлантического побережья Африки. Тем более, что некоторые европейские кваквы
используют, кроме долины Нила, ещё и западноатлантический путь пролёта (Snow, Perrins,
1998).
Кваквы, по-видимому, довольно консервативны в территориальном отношении и, как
правило, возвращаются для гнездования в места, где они родились. Переселение птиц, ког-
да оно бывает, идёт обычно в северном и северо-восточном направлениях. Наиболее ярко
это видно на птицах Кызыл-Агачской и в меньшей степени дагестанской популяций. Об
222 Отряд Листообразные, Семейство Цаплевые
этом свидетельствуют данные о встречах в дельте Волги квакв, окольцованных птенцами в
Кызыл-Агачском заповеднике. Прямых доказательств гнездования птиц, родившихся в
Кызыл-Агачском заповеднике и встреченных на гнездовьях в дельте Волги, нет. Однако
значительный процент возвратов колец из возможных мест следующего гнездования не
может не обратить на себя внимания. На меньшем материале, но похожую картину видим
для квакв дагестанского происхождения.
О том же свидетельствуют и некоторые возвраты колец от взрослых птиц из устья Днеп-
ра и дельты Днестра, а также от квакв, помеченных птенцами за пределами Вост. Европы и
Сев. Азии. 11 мая 1960 г. в Ейском районе Краснодарского края убита кваква, окольцован-
ная в 1959 г. птенцом на оз. Маньяс (Турция). 28 октября и 7 сентября в Астраханской обл.,
а 18 октября под Херсоном добыты птицы из Венгрии (оз. Кисбалатон) в возрасте соответ-
ственно 4, 2 и 8 лет. В Краснодарском крае у станицы Динская 20 октября на шестом году
жизни встречена кваква из Румынии (дельта Дуная). Там же, в Курганинском р-не у стани-
цы Павловская на 4-м году жизни обнаружена кваква из Украины (дельта Днестра). Не
исключено, что это птицы, не вернувшиеся в места своего рождения и гнездившиеся на
новых местах.
Численность. Численность гнездящихся квакв подвержена значительным несинхрон-
ным колебаниям, как и у многих других видов, трофически и топически связанных с водно-
болотными угодьями.
На Украине, в её западной части кваква гнездится спорадически (Страутман, 1963). В
бассейне верхнего Днестра численность незначительна и колеблется в пределах 20-100
пар (Бокотей и др., 2003). В Черновицкой обл. обнаружено более 400 пар и отмечена тен-
денция к росту численности (Скильский и др., 2000).
Наиболее западные поселения вида отмечены в плавнях оз. Кагул, где количество край-
не невелико — не более 30 пар. Более крупные гнездовые скопления кваква образует в
Кугурлуйских плавнях. Здесь, в смешанных колониях голенастых и веслоногих птиц её
число достигает 210 пар. Небольшие скопления птиц в гнездовой период отмечены и на
других озёрах Придунайской низменности, в частности, на оз. Китай и Катлабух. Периоди-
чески гнездится кваква на о. М. Татару и Б. Даллер. Однако численность её здесь невелика
и не превышает 80-100 пар. В Килийской дельте Дуная наиболее крупные поселения этого
вида сосредоточены в Дунайском биосферном заповеднике. Число гнездящихся пар кваквы
здесь колеблется в разные годы от 300 до 1 650 пар (Панченко, Чёрный, 1987; Жмуд, 1998).
Далее к востоку до дельты Днестра, кваква редка. В низовьях р. Сараты она гнездится
в небольших количествах в Саратском лесничестве, и численность её здесь колеблется от
60 до 112 (Гулай, 1975), а в устье р. Кагильник — 220 пар (Русев и др., 2005).
В дельте Днестра сосредоточено одно из наиболее крупных гнездовых поселений квак-
вы не только Азово-Черноморского региона, но и всей территории Европейского Юго-За-
пада (Русев, 2003). За период наблюдений здесь отмечены две депрессии численности — в
1974 и 1987 гг. (табл. 11). Первый спад численности произошел после относительно дли-
тельного, в течение одного сезона резкого естественного падения уровня воды, а второй —
в результате полного зарегулирования естественного стока реки плотиной Днестровской
ГЭС. В обоих случаях была подорвана кормовая база кваквы. Полностью исчезли имаго и
личинки водных насекомых — наиболее массовый и доступный корм мелких цапель (Ру-
сев, 2005).
Небольшие колонии периодически формируются вблизи Одессы в зарослях скумпии, а
также в верховьях Тилигульского лимана, где численность кваквы в смешанных поселени-
ях колеблется в пределах 200^117 пар (Русев, 2000а; Греков и др., 2005). Обычна кваква в
дельте Днепра. Здесь её численность колебалась в 1980-х гг. от 198 до 214 пар (Ардамац-
Кваква
223
кая, 2005), а в настоящее время не превышает 80 пар (Петрович, Щеголев, 2003). Несколь-
ко десятков пар гнездятся в Черноморском биосферном заповеднике на соснах в урочище
«Волыжин лес» (Яремченко, Руденко, 2004).
На Лебяжьих о-вах (Крым) численность нестабильна и составляет десятки пар (Костин,
Тарина, 2004). Многочисленные гнездовые поселения сосредоточены в тростниковых за-
рослях Восточного Сиваша — до 1201 пары (Гринченко, 2004а). В Сев.-Зап. Приазовье
кваквы гнездятся в плавнях р. Молочная в количестве до 400 пар в отдельные годы (Коше-
лев и др., 2004).
В пойме Дона на окраине Ростова в смешанной колонии на тополях в 1981 г. отмечено
около 300 гнёзд, в 1982 г. — 25 пар, в 1983 г. колония перестала существовать. На оз. Мона-
стырском в пойме Дона близ г. Аксай в 1982 г. в смешанной колонии на кустарниковых
ивах гнездилось около 10 пар, в 1986 г. — в тростниках 200-300 пар. В 1981 г. кваквы
гнездились в большом смешанном поселении в пойменном ветляннике на правом берегу
Дона у хут. Арпачин Багаевского р-на. В 1982 г. цапли бросили эту колонию, но с 1983 г.
квакв начали отмечать в смешанной колонии уже на землях Новочеркасского рыбхоза близ
того же хутора.
Поселения квакв отмечены также в смешанных колониях цапель в пойменном лесу близ
ст. Багаевской, где в 1985 г. учтено около 70-80 пар; в 10 км выше г. Константиновска, где
в 1986 г. на сухих деревьях в затоне реки гнездилось 150-300 пар; у станицы Задоно-Ка-
гальницкой в 1979 г. — 10 пар, а в 1980 г. — несколько пар; близ станицы Николаевской в
1980 г. — 600-700 пар, в 1986 г. — колония сохранялась; у станицы Мариинской в 1979 г. —
50 пар; у станицы Камышевской в 1979 г. — 5 гнёзд, в 1980 г. — больше; близ станицы
Каргальской в 1978 г. — более 50, а в 1979 г. — 60-80 пар. Предположительно кваквы
гнездились в пойме Дона в районе станицы Мелиховской, в устье р. Сухой в Цимлянском р-
не, в ветляннике на берегу залива близ хут. Балобанов на Цимлянском песчаном массиве в
Волгоградской обл. и в лесу на острове среди Цимлянского вдхр. в устье р. Чир. В 1993 г.
смешанная колония, в которой было не менее 50 пар квакв, обнаружена в заболоченных
тальниках на Цимлянском песчаном массиве.
Таблица 11. Численностиь кваквы в дельте Днестра в 1972-2005 гг.
Год Число пар Год Число пар Год Число пар
1972 1000 1986 1780 2000 2100
1973 1060 1987 650 2001 1850
1974 660 1988 1100 2002 ?
1975 1030 1989 1200 2003 1700
1976 2260 1990 1300 2004 1900
1977 2700 1991 1000 2005 1500
1978 2040 1992 1620
1979 2200 1993 1400
1980 2400 1994 1500
1981 2500 1995 9
1982 2500 1996 1550
1983 1300 1997 1500
1984 1350 1998 1570
1985 1000 1999 1840
Примечание'. ? — птицы гнездились но учёт не проводился.
224
Отряд Листообразные, Семейство Цаплевые
В пойме Северского Донца 10-20 пар квакв гнездились в 1977 г. вместе с серой цаплей
в старом дубовом лесу в урочище Огиб. Но в 1985 г. эта колония оказалась брошена, а
цапли были обнаружены в новой смешанной колонии в 8 км ниже по течению реки в забо-
лоченном ольшанике на окраине станицы Нижне-Кундрюченской, где учтено до 50 пар
квакв (в 2002 г. эта колония сохранялась, но её численность не определена).
На Зап. Маныче кваква встречается на водохранилищах с развитой жесткой надводной
растительностью. На Усть-Манычском вдхр. в 1948 г. гнездилась в значительном количе-
стве, но в последующие два года численность её сильно сократилась, а в 1951 г. — вновь
несколько увеличилась (Олейников, 1953). В 1966-1970 гг. здесь отмечено всего несколько
десятков особей (Языкова, 19706).
На Веселовском вдхр. в 1948-1950 гг. учтено около 120 пар (Олейников, 1953), но в
1966-1970 гг этот вид здесь не был обнаружен вовсе. В 1977 г. колонии кваквы отмечены в
балках Б. и М. Садковка и на приплотинном участке водохранилища, где они регулярно
гнездились в небольшом числе в смешанных поселениях с другими цаплями в тростниках.
Учёты проводили здесь с 1977 по 1987 г. Число птиц в колониях по годам изменялось от 40
(1981 г.) до 160-180 (1978 г.) пар.
На Пролетарском вдхр. гнездование квакв регистрировали в смешанных колониях в
тростниках на оз. Казинка, на водохранилище у места впадения р. Егорлык и близ охот-
станции «Удачная»: здесь в конце мая 1986 г. учтено 15, а в 1987 г. — 10 пар. Случай гнез-
дования 15 пар квакв зарегистрирован на Состинских озёрах близ г. Черноземельска (Па-
дутов и др., 1975).
Численность квакв на лиманах Вост. Приазовья в гнездовой период 1953-1954 гг., по
данным А.А. Винокурова (1959), составляла 1-1,5 тыс. особей. На р. Челбас поселение
квакв впервые было обнаружено в 1964 г. (Ломадзе, 1973). Эта колония сохранялась до
1980-х гг. На р. Бейсуг 50 пар этих птиц учтено в 1986 г. в смешанной колонии цапель на
территории одного из рыбхозов.
В дельте Кубани в 1964-1972 гг. квакв отмечали на гнездовье на лиманах Ахтарско-
Гривенской системы: на Ахтарском лимане — 27 пар, на Кочковатом — 40 пар (Ломадзе,
1973). В 1980 г. на лиманах этой системы зафиксировано 6 колоний квакв: на лимане Ряс-
ном — 15 гнёзд, на Б. Карпиевском — 30, на Солодко-Рясном — 100, Кочковатом — 39,
близ станицы Гривенской — 5 гнёзд. В 1987 г. они гнездились здесь в тростниках на Ясен-
ской (20 пар) и Ачуевской (30 пар) косах, Солодко-Рясном лимане (20 пар), на Солёных
озёрах близ станицы Некрасовской (50 пар) и Ахтарских озёрах (50 пар).
В Сладковско-Черноерковской группе лиманов колонии обнаружены южнее кубанского
рукава Протоки у места её впадения в Азовское море (10 гнёзд), на лимане Горьком (10),
лимане Лозоватом (10) и лимане Войсковом (10), кроме того — близ пос. Забойский (150
гнёзд). В 1987 г. их поселения отмечены также на лимане Мечетном (10 гнёзд).
С развитием рисосеяния на Кубани кваквы, вместе с другими цаплями, стали гнездить-
ся в лесных урочищах в «старой дельте». Так, в Красноармейском р-не в урочище Круглик
в 1970 г. учтено 304 гнёзда (колония возникла за 3^1 года до этого). В 1983 г. здесь обнару-
жено около I 500 гнёзд кваквы, в 1984 г. учтено 2 100 гнёзд. В искусственных, посаженных
в начале 1950-х гг. насаждениях Крымского лесхоза близ станицы Анастасиевской (Сла-
вянский р-н), поселение цапель возникло в 1971 г.; в тот год там было обнаружено несколь-
ко сотен гнёзд кваквы. В 1982 г. здесь гнездилось около 800 пар квакв; в 1983 г. — 2 400; в
1984 г. — 880; в 1987 г. — 1 230 пар. Таким образом, общая численность гнездящихся в
дельте Кубани квакв в 1980-1990-х гг. составляла 3-5 тыс. пар.
В 1977 г. кваквы обнаружены на гнездовье в долинах степных рек Краснодарского края:
в Крыловском лесничестве — 45-60 пар и в Новопокровском лесничестве — 40 пар. В
Кваква
225
пойменном лесу на р. Чамлык (приток р. Лабы) восточнее г. Лабинска в 1980 г. учтено 64
гнезда.
В 1970-1980-х гг. эти цапли обнаружены на водоёмах Ставрополья. Гнездование квак-
вы отмечено здесь в 10 смешанных колониях: в пойменном лесу между реками Кубань и
Барсучки в 10 км северо-западнее г. Невинномысска в 1980 г. — 23 гнезда и в 1983 г. — 29
гнёзд; на Новотроицком вдхр. в Изобильненском р-не в 1978 г. в смешанной колонии на
деревьях зарегистрировано 380 пар, а в 1984 г. — 200 пар квакв; в лесополосе между стани-
цей Новотроицкой и с. Птичье в северо-западной части Изобильненского р-на в 1981 г.
учтено 33, а в 1982 г. — 42 гнёзда квакв, В 1983 г. эта колония исчезла в связи со строитель-
ством здесь автотрассы. В 1983 г. поселение квакв обнаружено на рыборазводных прудах
близ с. Птичье (Птицы Северного Кавказа. 2004).
В 1981 г. два поселения этих цапель встречены в Кировском р-не на р. Кура: в районе с.
Пегушин — 40 гнёзд и западнее с. Новая Деревня — 107 гнёзд. В пойменном лесу у места
впадения р. Кумы в Отказненское вдхр. гнездование квакв зарегистрировано в Торосов-
ском лесничестве, где в 1981 г, учтено 188 гнёзд, и в лесу у с. Иргаклы, где учтено около 130
гнезд. В пойменном лесу у с. Владимировка Левокумского р-на число квакв в смешанном
цапельнике в 1985 г. было 160 пар. Общая численность квакв, гнездившихся на водоёмах
Ставрополья, достигала 900-1 000 пар (Птицы Северного Кавказа. 2004).
В Северной Осетии колония квакв отмечается с 1977 г. на искусственном оз. Бекан в Ар-
донском р-не: максимальное число гнёзд (75-80) зарегистрировано в 1978, 1982 и 1983 гг.,
минимальное (13 пар) — в 1986 г.; в 1979-1980 гг. в связи со спуском воды из озёра кваквы
здесь не гнездились. Кроме того, эти птицы гнездятся в Моздокском р-не в пойменном лесу
на р. Терек, предположительно — в окрестностях станицы Павлодольской.
В дельте Терека в 1973-1974 гг. кваквы гнездились только в смешанных колониях в
тростниках охотхозяйства «Кизлярское» — 50 пар, на Каракольских озёрах — 100 пар и на
деревьях в урочище «Латышский остров» — 170 пар (Бондарев, 1975). В 1985 г. 12 пар
квакв гнездились на оз. Прудском (Ачикольские озёра). В 1986-1987 гг. гнездования этого
вида в дельте Терека не отмечали.
Таким образом, общая численность квакв на водоёмах Северного Кавказа (Нижний Дон,
Зап. и Вост. Маныч, Вост. Приазовье, водоёмы Ставрополя, дельта Терека) согласно учёт-
ным данным, достигает 5 900-9 100 пар (Птицы Северного Кавказа, 2004).
В последние 10 лет, на фоне трансгрессии Каспийского моря и увеличения глубин в
дельте Волги, численность кваквы волжской дельты испытывала резкие колебания. До на-
чала интенсивного увеличения обводненности водоёмов в дельте Волги в середине 1970-х
гг. и до начала 1980-х гг. здесь насчитывалось до 4 300 пар квакв, но в 1987 г. их было
только 2 500 пар. По мере дальнейшего возрастания глубин, число гнёзд кваквы сократи-
лось до 1 000 — в 1991 г. (Бондарев, 19756; Кривенко, 2004). Однако в последние годы
численность кваквы в дельте Волги можно назвать относительно стабильной. Она изменя-
ется в пределах от 2 100 (2004 г.) до 3 010 пар (2003 г.) (Гаврилов, 2005).
Современная численность кваквы в Европейской России предполагается равной 10-15
тыс. пар (Мищенко и др., 2004).
На оз. Ханка первое гнездо было обнаружено в 1973 г. (Глущенко, 1981). С 1994 г. гнездо-
вание стало носить регулярный характер, численность поступательно нарастала: в 1994 г.
гнездилось 15 пар, в 2000 г. — 240 пар, в 2001 г. — около 300 пар, в 2002 г. — около 310 пар.
С 2003 по 2005 г. численность снизилась; в 2007 и 2008 гг. она составляла, соответственно,
около 360 и 380 пар (данные Ю.Н. Глущенко, И.Н. Кальницкой и Д.В. Коробова).
В Казахстане кваква обычна как в дельте Волги, так и в долине Сыр-Дарьи (на отрезке
от оз. Камышлыбаш до Чиили), малочисленна по р. Урал, на Теликульских озёрах, Балха-
226
Отряд Листообразные, Семейство Цаплевые
ше. Конкретные данные о численности здесь отсутствуют (Долгушин, 1960). В Средней
Азии, несмотря на падение численности, по-прежнему гнездится 12-15 тысяч пар (Птицы
Средней Азии, 2007).
В зарубежной Европе наибольшая численность кваквы зарегистрирована в Италии
(14 000 пар), Румынии (8 000), Франции (5 500), Испании (4 430), Венгрии (3 600), Сербии
(2 500), Болгарии (2 500), Греции (1 200), Хорватии (600), Чехии (500), Польше (513),
Словакии (400), Македонии (200 пар). В незначительном количестве гнездится в Южной
Португалии (120 пар), Албании (10), Австрии (70), Бельгии (14), Боснии и Герцеговине (5),
Германии (35), Мальте (2), Словении (5 пар). В большом числе гнездится в Турции (8000
пар) (Birdlife International, 2004). В целом европейская популяция кваквы в конце XX в
насчитывала 47,3 (44,9-51,8) тыс. пар (Fasola, Hudec, 1997).
Дж. Хегемейер и М. Блеер (Hagemeijer, Blair, 1997) приводят несколько иные сведения.
По их данным в Италии гнездится 20 тыс. пар кваквы, в Румынии, Венгрии и Франции —
немногим более чем по 4 тыс. пар в каждой из названных стран, в Испании и Болгарии —
по 2 тыс., Греции — 900, в Хорватии — до 1 000 пар, в Европейской России, в Молдавии и
на Украине — в общей сложности до 20 тыс. пар. По их оценке в Европе в целом гнездится
около 50 тыс. пар кваквы, что не сильно отличается от цифр, приведённых выше.
Местообитания. Необходимым условием возникновения колониальных поселений квакв
является наличие удобных мест для гнездования при весьма обильной кормовой базе. В
гнездовой период кваквы заселяют преимущественно пойменные биотопы, предпочитая
заросли кустарниковой ивы, а также тростниковые ассоциации. Отмечается заметное стрем-
ление птиц к труднодоступным для человека участкам околоводных ландшафтов, что обус-
ловлено все возрастающим антропогенным прессом: нередки пожары, велика рекреацион-
ная нагрузка. Так, в центральной части плавней, в районе оз. Квашино регулярно до 1981 г.
сосредотачивалось на гнездовании около 50% днестровской популяции квакв (более 1 000
пар). Гнёзда размещались на вкраплённых в тростники кустах ивы. В летний период этим
биотопам присущ весьма своеобразный микроклиматический режим — повышенная влаж-
ность воздуха, высокая температура с незначительной амплитудой суточных колебаний —
обеспечивающий нормальное размножение даже таких теплолюбивых птиц, как жёлтая
цапля и каравайка.
В пойме Нижнего Днестра периодически появляются колонии и на заломах тростника.
На заломах тростника гнездится кваква и в Кугурлуйских плавнях, в плавнях нижнего тече-
ния р. Тилигул, Вост. Сиваша.
В низовьях Днепра, в пойме малых рек Одесской обл. Сарата и Кагильник гнездовья
квакв локализованы на деревьях, в основном, акациях. Колонии у г. Одессы сформированы
на кустах скумпии (Cotinus).
Кормовые стации используются кваквой ещё более широко. Птицы отдают предпочте-
ние тростниковым окраинам мелководных плесов и пойменных озёр, где обычно много
корма. Нередки они на кормежке и по берегам лесных проток, по берегам рек. В половодья
на Днестре их можно наблюдать на мелководных заливаемых лугах. Здесь они встречаются
даже после спада воды. В отдельные годы в зимний период птицы часто посещают сельхо-
зугодья, особенно те их участки, где в массе обитают мышевидные грызуны, которые вхо-
дят в их рацион. И, наконец, кваква часто использует искусственные рыборазводные пру-
ды, особенно в случаях подрыва кормовой базы в естественных экосистемах.
Размножение. Половой зрелости кваквы достигают в годовалом возрасте, однако раз-
множаются в первый год жизни не все птицы (Птицы Средней Азии, 2007). Моногамы,
пары формируются каждый сезон (Cramp, 1977). Прилетевшие птицы не приступают сразу
к гнездованию, а образовывают небольшие скопления, придерживаясь богатых кормом
Кваква
227
биотопов невдалеке от будущих колоний. Участок гнездования выбирают самцы, предпо-
читая места прошлогодних поселений. Формирование пар длится в среднем 16 дней и
сопровождается разнообразными церемониями (рис. 43) (Cramp, 1977).
Первые группировки птиц приступают к гнездостроению уже в начале апреля, тогда
как последние — в конце этого месяца или даже в первой декаде мая. Весь репродуктив-
ный цикл Азово-Черноморской популяции квакв очень растянут. Так, в 1980 г. в дельте
Днестра он был необычайно продолжительным — размножение квакв закончилось в нача-
ле сентября. Растянутость сезона гнездования характерна для вида и в Средней Азии (Пти-
цы Средней Азии, 2007).
Колониальный вид, нередко выступающий в качестве организатора смешанной коло-
нии мелких аистообразных. Так, в условиях Днестра каравайка, малая белая и жёлтая цап-
ли не формируют моновидных колоний, тем более они никогда не являются «зачинателя-
ми» поливидовых сообществ. Они занимают для гнездования лишь те участки, которые
выбирает кваква. Последняя же способна как формировать самостоятельные, так и органи-
зовывать поливидовые колонии.
Численность в смешанных колониях может сильно варьировать — от 30-50 до 600-700
гнёзд; в моновидовых колониях число пар колеблется от 15 до 200. И.А. Долгушин (1960)
упоминал о колониях более 1 000 гнёзд. Величина колоний, прежде всего, зависит от пло-
щади гнездопригодного пространства. Как и в других колониальных поселениях птиц, в
колониях, где организатором является кваква, сильно развиты центростремительные тен-
денции. Птицы стараются загнездиться в центральных, наиболее защищенных от хищни-
ков участках, которые одновременно являются и местами с благоприятными микроклима-
тическими условиями. Однако такому стремлению противодействует наличие вокруг гнёз-
да каждой пары охраняемой территории. Поэтому позднее прилетающим другим кваквам
и остальным видам смешанной колонии приходится занимать ещё оставшиеся свободны-
ми гнездопригодные участки. Жёлтая цапля и каравайка в смешанной колонии с кваквой
занимают преимущественно нижний ярус. Малая белая цапля — средний, нижний и час-
тично верхний11.
Каждая пара квакв в колонии имеет свою определённую гнездовую территорию. Пара
охраняет пространство вокруг гнезда на расстоянии вытянутой шеи сидящей на гнезде птицы
(радиус не менее 50 см). Птицы обычно занимают старые гнёзда, достраивая их (Русев,
Арнаут, 1983). В период выбора места для гнезда проявляется агрессивность друг к другу
различных видов и особей одного вида, особенно в колониях с плотным поселением (Ру-
сев, 1983в).
К гнездостроению кваквы приступают вскоре после прилёта. Птицы, гнездящиеся на
деревьях, устраивают гнёзда на различной высоте, что в первую очередь связано с высотой
гнездовых деревьев и составом колониального поселения. В местах, где кваквы гнездятся
совместно с серыми цаплями на высоких ольхах, их гнёзда располагаются на высоте в пол-
дерева, тогда как вершины занимают более крупные виды. В смешанных колониях на боль-
ших деревьях под Ленкоранью основная масса гнёзд квакв сосредотачивалась несколько
выше среднего яруса деревьев, на одной высоте с гнёздами малых бакланов. Ниже гнезди-
лась жёлтая цапля, тогда как вершины были заняты крупными гнёздами серых цапель и
больших бакланов. В дельте Волги гнёзда квакв также помещаются относительно низко,
обычно на той же высоте, где гнездится каравайка. В одной из колоний на Сыр-Дарье, где
кваквы гнездились в роще карагача, их гнёзда занимали верхний ярус деревьев, чуть ниже
основной массы поселения грачей. В другой колонии, расположенной в тугае, постройки
11 Ярусность гнездования видов в разных частях Вост. Европы и Сев. Азии может быть различной —
смотри очерки по другим видам цапель (прим. ред.}
228
Отряд Листообразные, Семейство Цаплевые
Рис. 43. Элементы демонстрационного поведения кваквы (а-е) (по: Cramp, 1977, с изменениями).
Кваква
229
квакв густо покрывали вершины низкорослых деревьев джиды и тополя. Часть гнёзд была
выстроена на высоте не более двух метров. При гнездовании в камышовых и тростниковых
зарослях легкие постройки квакв, как правило, находятся выше гнёзд колпиц, больших
бакланов, иногда караваек. Гнёзда, устроенные среди зарослей тростника, могут распола-
гаться на различной высоте от воды, что в значительной мере зависит от состояния жест-
кой надводной растительности. На Зап. Маныче и в Вост. Приазовье (п=25) их находили на
высоте 40-50 см, в низовьях Дона (п=15) — в 1,2-2,5 м над водой. На деревьях гнёзда
укрепляются в развилках горизонтальных и вертикальных веток, под Ленкоранью часто
среди перепутанных колючих лиан; в тростниках и камышах — на прошлогоднем заломе
этих растений.
Гнездо у кваквы, как и у многих других цапель, имеет вид перевёрнутого конуса. Мате-
риалом служат сухие ветки кустарников и деревьев, обыкновенный тростник. Гнёзда, пост-
роенные из тростника, обычно несколько больше, и гнездовой материал уложен более плот-
но, тогда как сделанные из древесных веточек — меньше по размерам и рыхлее. Порой они
не имеют лотка. Вся постройка (особенно в тех случаях, когда занимаются и достраивают-
ся старые гнёзда) крупнее и грубее, чем у других мелких цапель, гнездящихся на деревьях.
В крупных смешанных колониях кваквы часто занимают старые гнёзда других мелких ца-
пель, перестраивая их. Вследствие этого гнёзда кваквы весьма изменчивы в размерах и
форме.
Размеры гнёзд кваквы (в см) из дельты Дона (п=97): диаметр гнезда — 34,4-63, в сред-
нем — 48; диаметр лотка — 15-39, в среднем — 26; высота гнезда — 13^3, в среднем —
30; глубина лотка — 1-17, в среднем — 8. Размеры гнёзд из леса Круглик в Краснодарском
крае (п=49) и в Анастасиевском лесхозе (п=29): диаметр гнезда — 25-52, в среднем — 39;
диаметр лотка — 12-28, в среднем — 20; высота гнезда — 12-35, в среднем -26; глубина
лотка — 3-10, в среднем — 6,5 см. На оз. Ханка гнёзда имеют размеры (п=16): диаметр
гнезда — 28-60 см, в среднем 43,1 см; диаметр лотка — 13-30 см, в среднем 23,3 см;
глубина лотка — 0,5-1,4 см, в среднем 1 см; толщина гнезда 17-30 см, в среднем 24,2 см
(Глущенко и др., 20066). В постройке гнезда принимают участие оба партнёра. Самец при-
носит материал, самка укладывает его и охраняет от других птиц. По наблюдениям в Аст-
раханском заповеднике иногда приносит материал и самка (Ромашова, 1938). Материал
птицы собирают обычно в непосредственной близости от колонии, причём часто вытаски-
вают (крадут) из гнёзд, расположенных по соседству.
Первые кладки у кваквы появляются в первой декаде апреля, последние — в середине
июня, а при повторном гнездовании — ив конце июля (наблюдения в дельте Днестра).
Массовые кладки появляются в конце апреля - начале мая, хотя эти сроки могут варьиро-
вать в зависимости от гидрологического режима и микроклиматических условий. Отклад-
ка яиц наиболее интенсивно происходит во второй половине апреля - начале мая (Ломадзе,
1982). Так, в колонии на оз. Бекан 22 апреля 1982 г. в 6 из 23 гнёзд было по 1 яйцу; в 7 — по
2; в 2 — по 3; в 6 — по 4; два гнезда ещё строились. 6 июня 1982 г. отмечены птенцы
разного возраста, свежие и насиженные кладки. В гнёздах, расположенных по периферии
колонии, происходила откладка яиц.
В полной кладке кваквы 2-5 яиц. В кладке днестровских квакв 3-5 яиц, в среднем —
3,5, в дельте Дуная — 3,3. Из 78 кладок в дельте Дона 55,2% содержали по 4 яйца; 30,8 % —
по 3; 7,6% — по 2; 6,4% — по 5 яиц. В условиях Ханки полная кладка содержит от 3 до 6
яиц, в среднем (п=20) 3,7 яйца на гнездо (Глущенко и др., 2006). Окраска яиц кваквы зеле-
новато-голубая. Форма — правильная яйцевидная. Размеры яиц из дельты Дона (п=21):
46,5-52,5 х 31,0-36,0, в среднем — 49,6x35,1 мм. Немногим отличаются размерами яйца
из колонии в низовьях Дона (п=29): 45,5-52,3 х 31,6-37,3, в среднем — 48,9x35,5 мм. Раз-
230
Отряд Листообразные, Семейство Цаплевые
меры яиц из дельты Кубани близки к первым: лес Круглик (п=85) — 43,5-56,5 х 33,2-37,4,
в среднем — 49,7x35,1 мм; лесхоз Анастасиевский (n= 12) — 45,4-51,0 х 32,9-36,2, в сред-
нем — 49,7x34,9 мм. Яйца из колонии на оз. Бекан несколько крупнее: 45,1-53,4 х 37,0-
38,8, в среднем -51,8x35,4 мм. Длина яиц днестровских квакв (п=120) изменяется в преде-
лах 45,5-57,0, в среднем 50,6 мм, ширина — 30,2-38,5, в среднем — 33,9 мм. В плавнях
Днепра (п=90) длина 43,9-52,2, ширина 32,6-36,5, в среднем 47,3-34,5 мм, на Дунае, соот-
ветственно, 49,3-53,5 и 32,5-35,5 мм (Смогоржевський, 1979). Размеры яиц из Средней
Азии (п=140): 42,5-54,0 х 32,0-38,1, в среднем 48,06x35,23 мм (Птицы Средней Азии, 2007).
На оз. Ханка яйца квакв (п=66) имеют размеры 42,5-56,2 х 31,8-39,4 мм, в среднем 49,8x34,2
мм (Глущенко и др., 20066).
Интервал в откладке яиц составляет около суток; насиживание начинается с первого
яйца. В нем участвуют оба родителя, но самец обогревает крадку меньше времени, чем
самка. Насиживающая птица каждые 40-90 мин встаёт, переворачивает яйца, поправляет
прутики на гнезде. Птицы меняются на гнезде от 2 до 4 раз в сутки, чаще 3 раза в сутки.
Продолжительность насиживания — 23 дня.
Перед вылуплением птенцов кваквы очень плотно сидят на гнезде и клювом часто «про-
щупывают» кладку. Вылупление птенцов начинается не раньше чем через 24 ч после про-
клёвывания яиц. После вылупления каждого птенца взрослые птицы выбрасывают скор-
лупу из гнёзда. Интервал в вылуплении птенцов составляет от 1 до 1,5 суток и аналогичен
интервалу откладки яиц. Разрыв в возрасте между первыми и последними птенцами одно-
го гнёзда может достигать 7-9 дней.
Птенцы вылупляются слепыми и беспомощными. Вскоре они обсыхают и через день
открывают глаза. В первые дни после вылупления птенцы кваквы пассивны, дремлют в
глубине лотка, оживляясь только во время кормёжки. Весят они в первый день 25 г, что
составляет 4,6% от веса взрослой птицы. Веса взрослых молодые достигают спустя 2 меся-
ца. Прирост веса у кваквы замедляется с середины третьей недели, что, возможно, связано
с труднностями выкармливания птенцов в условиях, когда один из родителей постоянно
находится на гнезде. Повторное увеличение этого показателя соответствует периоду, когда
кормить выводок начинают оба родителя.
В возрасте 5-6 дней птенцы ведут себя более активно, в случае опасности занимают
оборонительную позу. В двухнедельном возрасте, при беспокойстве их наблюдателем, пе-
реходят на стебли тростника или ветви дерева близ гнёзда; при этом они могут погибнуть,
падая в воду или на землю. В выкармливании птенцов принимают участие оба партнёра.
Птенцы первое время вскармливаются отрыгнутой полу переварившейся пищей. Молодые
в возрасте 5-7 дней получают пищу 10 раз в сутки, в возрасте 21-23 дней — 3^ раза.
Наиболее дружный вылет взрослых за кормом происходит в 4-6 ч утра. В середине дня
активность полётов за кормом падает и вновь возрастает к 17 ч. Наблюдения показывают,
что принесенный родителями корм удаётся получить только старшим птенцам, которые
отталкивают младших и иногда за одну кормёжку успевают получить пищу 2 раза, тогда,
как младшим её не достаётся вовсе. Ослабленные и отставшие в развитии малыши часто
гибнут; смертность младших птенцов кваквы в 2^1 раза выше, чем у старших. Отмечали
случаи, когда старшие птенцы поедали младших.
Постэмбриональное развитие птенцов кваквы длится 30-35 дней. Наиболее интенсив-
ный прирост веса, а также длины клюва и цевки наблюдается в первые 3-5 дней жизни
(табл. 12). Перо на теле птенцов появляется в недельном возрасте. Оно, в первую очередь,
приходит на смену пенькам на спинной и плечевой птерилиях — в 7-9-дневном возрасте
(табл. 13). На 24-27-й день полностью раскрываются все пеньки.
Характерная черта птенцов кваквы — появление и развитие на 15-17-й день пудреток.
За несколько дней до этого кожа на месте их появления слегка припухает.
Кваква
231
Обогревание и защита птенцов от перегрева взрослыми птицами отмечается до двух-
недельного возраста. Во время дождя и сильных ветров родители обогревают и более стар-
ших птенцов. Птенцы поднимаются на крыло в возрасте 5 недель. С этого времени они
начинают вместе с взрослыми кочевать поисках корма.
Постоянно существуя в условиях нестабильного гидрологического режима, кваквы,
как и другие виды мелких голенастых, выработали специальные адаптации, компенсирую-
щие гибель приплода. Птицы, как правило, не гнездятся в последующем году в том месте,
где в предшествующем гнездование было неудачным. В случае потери кладки или выводка,
птицы приступают к повторному, а иногда и к третьему гнездованию. Очень важно в этом
случае, на какой стадии выводкового процесса произошла потеря гнезда. Наблюдения за
днестровской популяцией квакв показали, что на поздних стадиях насиживания и при гибе-
ли гнёзд с птенцами птицы реже гнездятся повторно. При потере кладки в начале размно-
жения, повторная откладка яиц происходит достаточно быстро (не более, чем через 10 дней).
В случае гибели яиц на более поздних стадиях насиживания, птицы долго держатся у пре-
Таблица 12. Возрастные изменения веса тела, длины клюва, крыла и цевки птенцов
кваквы (размеры — в мм; вес — в г; усреднённые данные; п=18)
Возраст в днях Клюв Крыло Цевка Вес
0 13,6 14,7 15,8 23,5
1 14,6 15,5 16,3 31,6
2 16,0 16,8 18,2 48,4
3 17,5 18,6 19,6 57,8
4 19,9 21,9 22,9 83,2
5 22,3 24,7 24,9 104,3
6 24,2 26,3 27,2 114,3
7 26,5 32,3 31,5 145,4
8 29,1 35,5 34,3 159,1
10 33,9 47,9 40,3 217,9
11 35,9 52,1 42,5 248,3
12 37,6 58,4 45,6 289,3
13 39,8 72,4 48,1 293,1
14 41,2 83,5 51,0 316,1
15 42,5 94,1 54,3 360.4
16 43,9 105,2 57,5 380,8
17 45,7 121,7 60,2 399,3
19 47,8 135,3 63,0 412,8
21 49,5 147,0 66,5 478,8
23 51,6 159,7 67,9 480,1
24 52,8 165,1 68,9 481,2
25 53,6 171,2 70,8 490,6
26 54,7 . 176,6 71,9 512,0
28 55,9 187,0 74,7 527,5
232
Отряд Листообразные, Семейство Цаплевые
Таблица 13. Очередность появления и развертывания пеньков на теле птенцов кваквы
(дни от вылупления).
Птерилии Начало появления пеньков Сроки развертывания пеньков
Плечевая Спинная Маховые перья Шейная Крыловая Голень Рулевые перья 4-5 7-8 4-5 8-9 5-6 9-11 6-8 10-11 6-8 11-12 6-8 11-12 8-9 14-16
жних мест гнездования и приступают к повторной откладке яиц спустя 2 недели и более.
При этом уменьшается количество вновь загнездившихся пар.
Так, в 1980 г. в одном из поселений днестровских квакв вследствие стремительного
подъема воды во время половодья погибло около 600 гнёзд, построенных на низком кус-
тарнике и заломах тростника. Затем около 500 пар приступили к повторному гнездованию,
сформировав на небольшом удалении (до 600 м) новую колонию. Очередной паводок в
период последней стадии насиживания уничтожил около 60% гнёзд повторно загнездив-
шихся квакв. Около 250 пар приступили к третьей попытке выведения потомства. Птицы
заняли преимущественно верхний и, частично, средний ярусы кустарниковой ивы, и пер-
вые кладки здесь появились в конце июля. Стремительно поднявшийся в середине августа
уровень воды вновь уничтожил подавляющую часть поселения, а из не затронутых павод-
ком 28 гнёзд самого верхнего яруса успешно вывелись птенцы.
Менее эффективна достройка гнёзд по мере подъёма воды. При медленном подъёме
уровня воды, птицы нижнего яруса (поздно загнездившиеся группировки квакв, жёлтая
цапля, каравайка) стараются надстраивать гнёзда. Каравайка и жёлтая цапля достраивают
гнёзда быстрее по сравнению с кваквой. Для них это более важно, так как основная часть
их популяций гнездится именно в нижнем ярусе. Гнёзда же кваквы нижнего яруса являют-
ся как бы «резервными», и их потеря в меньшей степени чувствительна для вида.
Смертность птенцов в конкретных гнёздах определяется асинхронностю их вылупле-
ния и разновозрастностью. Как уже упоминалось, птенцы, вылупившиеся последними, редко
выживают, особенно в малокормные годы. Обычно они становятся жертвами старших со-
братьев.
Незначительна гибель кладок и птенцов кваквы от пернатых (болотный лунь, серая во-
рона) и наземных (енотовидная собака, серая крыса) хищников. Она становится более зна-
чимой во время посещения колоний человеком. В обычных условиях число разорённых
хищниками гнёзд для днестровской популяции не превышает 5%.
В случае посещения человеком колонии, смертность птенцов до подъёма их на крыло
может достигать 80%. Это происходит в тех случаях, когда птенцы, способные передви-
гаться, пытаясь спастись, уходят из гнёзд, а обратно возвратиться не могут. В результате
они падают в воду или погибают от переохлаждения, голода и хищников. В поселениях, не
подверженных влиянию антропогенных факторов, успешность размножения составляет 2,3
птенца на пару (дельта Днестра). В дельте Дуная этот показатель колеблется от 2,7 на запо-
ведной территории до 2,4 на оз. Кугурлуй.
На примере днестровской популяции хорошо видны экологические особенности вида,
которые дают квакве определённые преимущества по сравнению с другими видами аистооб-
разных (Русев, Арнаут, 1983). Кваква прилетает в дельту Днестра первой из мелких цапель и
Кваква
233
отлетает позже них. В отдельные, благоприятные в кормовом и климатическом отношениях
годы, часть её популяции остаётся зимовать и весь годовой цикл проводит в дельте. Более
раннему прилёту кваквы в дельту способствует её эврифагия (Русев, 1981, 1985), тогда как
малая белая и, тем более, жёлтая цапли и каравайка относятся к более узкоспециализирован-
ным в питании видам. Доступный им массовый корм появляется позднее. Кваква, в отличие
от других мелких аистообразных, также более эвритермна, особенно по сравнению с жёл-
той цаплей и каравайкой, которые весьма теплолюбивы (Назаренко, 1953). Она обладает
высокой экологической пластичностью в добывании корма, что позволяет ей реагировать
на количественные его изменения и переключаться в конкретной ситуации на более доступ-
ные и массовые корма или перемещаться в места их обилия. Кваква способна своевременно
и быстро реагировать на неблагоприятное воздействие гидрологического режима путём
повторного и даже третьего гнездования, что не столь выражено у малой белой, жёлтой
цапель и каравайки. Все эти экологические и поведенческие особенности позволяют квак-
ве компенсировать даже существенную гибель кладок и птенцов и способствуют быстро-
му, адекватно емкости среды, восстановлению своей численности.
Суточная активность, поведение. Кваквы особенно активны ночью и в сумерках, но
в гнездовой период птицы кормятся и в дневное время суток. Несмотря на общественный
характер поведения, больших стай, как правило, не образуют. Свободные от насиживания
птицы отдыхают вблизи гнезда. Первые вылеты за кормом отмечены перед заходом солн-
ца, однако наиболее массовый вылет из колонии можно наблюдать спустя полчаса после
его захода. Вылетая в сумерки и ночью на кормёжку, кваквы издают характерные квакаю-
щие крики. Птицы, уже занявшие место охоты, отвечают им. Такая перекличка, по-видимо-
му, помогает птицам находить ещё не занятые для охоты места или, наоборот, определять
наиболее привлекательные кормовые угодья.
В случае нападения хищника (болотного луня) сигналы бедствия издает конкретная особь
(или пара), чьё гнездо подверглось нападению. Реакция колонии выражается лишь в об-
щем беспокойстве соседних птиц (ориентировочное движение головой, прекращение чис-
тки оперения, кормления птенцов и др.). Общего взлёта, как правило, не происходит. Особь,
чьё гнездо подверглось нападению, либо издает тревожные крики, оставаясь на гнезде и
при этом пытаясь атаковать врага (на последних стадиях насиживания или при защите ма-
леньких птенцов), либо покидает гнездо (на первых стадиях насиживания). Каких-либо
совместных действий по защите кладок и птенцов от пернатых хищников члены колонии
не предпринимают.
При появлении человека в колонии птицы, первыми заметившие опасность, издают сиг-
налы беспокойства, сопровождающиеся взлётом особей с ближайших гнёзд. Характерны
при этом своеобразные пассивные защитные действия птиц: дефекация, отрыгивание пищи.
Сигналы беспокойства имеют только локальное значение. Никаких агрессивных действий
взлетевшие птицы не проявляют. Как только наблюдатель исчезает из поля зрения, птицы
успокаиваются и занимают свои гнёзда.
Питание. Морфофизиологические возможности кваквы, связанные с добыванием кор-
ма — высокая подвижность шейного отдела, ведущая роль зрения в поисках пищи в любое
время суток и не слишком специализированный набор локомоторных способностей — оп-
ределяют набор возможных кормовых объектов: они должны быть хорошо видимы и нахо-
диться недалеко от охотящихся птиц (Кокшайский, 1962). Из описанных для цапель 28
типов кормодобывания основным для квакв является подкарауливание. Во время охоты
кваква находится на низко расположенных над водой ветвях деревьев, торчащих из воды
предметах, а также стоит на берегу у кромки воды, на мелководье, среди зарослей водных
растений.
234
Отряд Листообразные, Семейство Цаплевые
Во время половодья и паводков, когда пойменные луга покрыты водой, подкараулива-
ние у квакв может сопровождаться медленным перемещением по облюбованному участку.
При этом птица осторожно двигается по мелководью, что противоречит мнению Н.Н. Ско-
ковой, считающей, что это не доступно квакве из-за её коротких ног. В то же время, морфо-
функциональные особенности квакв, по Н.Н. Скоковой, ставят её, в отношении добывания
корма, почти вне конкуренции с другими видами цапель (Скокова, 1954).
Кормовыми стациями кваквы служат протоки, заросшие мелководные лиманы, ерики и
гирла, рисовые чеки, сбросные каналы оросительных систем, рыборазводные пруды. При
наличии вблизи от колоний кормопригодных стаций, кваквы предпочитают кормиться имен-
но там. В случае отсутствия таковых летят на более удалённые места кормёжек. Макси-
мальная удалённость кормовых угодий от мест гнездования, по наблюдениям за птицами,
окольцованными цветными кольцами в условиях дельты Днестра, составляет 12 км. Кор-
мовой спектр составляют рыбы, земноводные, насекомые, низшие ракообразные, пиявки и
др. Встречаемость их в пище неодинакова (как в различных местах обитания, так и в раз-
ные годы на одних и тех же водоёмах) и находится в прямой зависимости от обилия и
доступности кормовых объектов. В целом, состав пищи у квакв широко варьирует. Он раз-
личается не только у птиц из различных микропопуляций, колоний, но даже у соседних
гнездовых пар.
В питании молодых и взрослых основное значение имеют рыба и лягушки. В дельте
Волги птенцы вскармливаются рыбой, лягушками, водяными крысами, личинками водо-
любов и других крупных насекомых. В районе Харькова кваквы ночью охотно посещают
илистые берега водоёмов в местах водопоев для скота. Здесь они отыскивают различных
червей и личинок лошадиных оводов. Зимой на юге Каспия кваквы питаются рыбой, пре-
имущественно семейства карповых, водными насекомыми (Macrodytes, Colymbetes и др.) и
какими-то растениями (Исаков, Воробьев, 1940, цит. по: Спангенберг, 1951). В Бессарабии
они поедают рыбу, водяных насекомых и семена различных водных растений (Остерман,
1912).
В пище квакв из дельты Днестра обнаружены кормовые объекты шести групп. Посто-
янными и наиболее Часто употребляемыми служат лишь четыре: амфибии, водные и на-
земные беспозвоночные и рыбы (Русев, 1981). В летний период (июнь, июль) доминируют
водные беспозвоночные, составляющие 56,2%, весной и осенью — рыба (70%). В дельте
Днестра в рационе кваквы отмечено 45 видов животных. Ряд видов используется как в
имагиальной так и в личиночной формах. Разнообразнее всего представлены наземные (24
вида) и водные беспозвоночные (8 видов), а также рыбы (8 видов). Самые постоянные из
кормовых объектов — карась, вьюн, умбра (сем. Umbridae), лягушка озёрная и её головас-
тики, большой и малый водолюбы, скоморох (р. Cybister) и их личинки. Такой широкий
диапазон в выборе кормов ставит квакву вне жёсткой зависимости от дефицита того или
иного потенциального кормового объекта. В период массового появления личинок водных
насекомых кваква может переключаться на питание в основном этими животными. В слу-
чае выхода на поверхность почвы дождевых червей после паводков или дождей, кваква
добывает их. В послепаводковый период, когда на лугах остаётся много мелких водоёмов,
кваква предпочитает кормиться именно там.
Сезонные изменения в питании кваквы, как и других голенастых, связаны с появлением
и исчезновением временных мелководий. Рацион квакв, остающихся на зимовку в дельте
Днестра, состоит, главным образом, из имагинальных форм водных насекомых и некото-
рых видов рыб — карася, вьюна, умбры. Некоторые особи охотятся на мышевидных гры-
зунов, вылетая на кормёжку в сельхозугодья. Весной основу питания составляют лягушки,
рыбы, а также животные, спасающиеся от поднимающейся воды (медведки, грызуны). Лет-
Кваква
235
нее питание состоит в основном из личинок водных насекомых, головастиков и мальков
рыб. В конце лета вплоть до отлёта кваква опять переходит на питание лягушками и рыбой.
Из 8 видов рыб, объектов питания днестровских квакв, один относится к промысловым
видам (сазан), пять — к малоценным и сорным (умбра, вьюн, краснопёрка, карась, колюш-
ка) и два — к хищным промысловым (судак и щука). Ведущее место в питании квакв зани-
мает вьюн (Misgurnus fossilis), его кваква ловит круглый год. Зимой он иногда становится
одним из основных кормовых объектов. В труднодоступных участках плавней, в слабо про-
точной воде, кваква добывает также карасей и умбру.
У квакв, добытых в мае-сентябре на выростных прудах рыбхоза в дельте Дона (п=30),
карп обнаружен в 60% проб, толстолобик — в 13,3%, земноводные — в 26,7%, насеко-
мые — в 40%, остатки птиц — в 6,6%. У квакв, добытых в мае-сентябре на рыборазводных
прудах на р. Сал (n=l 1), карп обнаружен в 81,8%, окунь — в 9,1%, неопределённые остатки
рыб — в 9,1%, земноводные — в 9,1%, насекомые — в 18,2% проб.
На Манычском вдхр. кваква также питается преимущественно рыбой. Среди съеден-
ных рыб обнаружены краснопёрка (22,5% по весу и 21,5% по встречаемости), язь — соот-
ветственно, 19,3 и 19%,плотва — би5%,карась — 2,3 и5%, сазан — 0,3 и 1,3%,щука — 1,1
и 1,3%). Заметную роль в питании играют лягушки (16,3 и 24% соответственно), а также
насекомые — 9,3 и 79,6%. Основная масса съеденных насекомых представлена личиноч-
ными стадиями водных жуков сем. Dytiscidae и Hydrophilidae — 7,8% по весу и 65,8% по
встречаемости (Олейников, 1953).
Основу рациона кваквы на лиманах Вост. Приазовья, как и в дельте Дона, составляли
водные насекомые (37% по весу и 76,4% по встречаемости), представленные главным об-
разом личинками жуков-плавунцов и водолюбов. Рыбы составляли одну четвёртую часть
рациона — соответственно, 26,4 и 27,3%, причём более половины рыбного корма (17,3%)
было представлено сорными видами и хищниками. Молодь промысловых рыб составляла
9,3% и встречалась редко (5%). Заметное место в пище кваквы занимали здесь земновод-
ные (16,7 и 22,7%) и пиявки (14,4 и 22,7%). Значение млекопитающих (5,1 и 4,5%), пауков
(0,1 и 4,5%), бокоплавов (0,1 и 9,1%) и наземных насекомых (0,2 и 32,2%) было небольшим
(Ломадзе, 1973, 1984, цит. по: Птицы Северного Кавказа, 2004).
Основу питания кваквы в местах рисосеяния в старой дельте Кубани (лес Круглик) со-
ставляли в разные годы ракообразные (22,4, 38 и 50,6% по весу и 63,9, 71,6% и 59% по
встречаемости, соответственно в 1970, 1971 и 1984 гг.), представленные в основном щит-
нями. Значительное место в её рационе здесь занимали также земноводные (22,1, 19,8 и
26,8% по весу и 67,4, 24,5 и 56% по встречаемости), представленные озёрными лягушками
и краснобрюхими жерлянками, а также рыба (22,1, 20,5 и 11,7% по весу и 47, 29,8 и 31,6%
по встречаемости). В составе рыбного корма встречались разводимые в прудах сеголетки
сазана и толстолобика, а также окунь, малая южная колюшка, щиповка, краснопёрка, се-
ребряный карась, уклея, обитающие в каналах оросительных систем. Из других кормовых
объектов в пище кваквы в местах рисосеяния важное место по встречаемости занимали
пиявки (20,5, 9,8, 31,6% в названные выше годы) и наземные насекомые (93,1, 67,7, 61,5%).
В число последних входили сверчки, медведки, земляные щитники, клопы-черепашки, зем-
ляные клопы, жужелицы, щелкуны, листоеды, чернотелки, пластинчатоусые, коровки, со-
вки и др. Водные же насекомые встречались реже (10,8, 9,8, 35,9%). Из них в пище кваквы
отмечали плавунчиков, вертячек, пискунов, личинок жуков-плавунцов и водолюбов. Мле-
копитающие были представлены обыкновенной полёвкой и полевой мышью, рептилии —
прыткой ящерицей и водяным ужом (Ломадзе, 1984, цит. по: Птицы Сев. Кавказа, 2004).
Кваквы Анастасиевского лесничества кормились не только на рисовых системах, но и в
поймах рек Протока и Кубань. Поэтому основу рациона у них составили земноводные (50,4%
236
Отряд Листообразные, Семейство Цаплевые
по весу и 60% по встречаемости) и рыба (29,4 и 44%). Значение других кормовых объектов
было значительно меньшим. Кроме видов рыб, встреченных в пище квакв в колонии в лесу
Круглик, здесь обнаружены также линь, тарань и лещ.
Кваквы в районе Ахтарского рыбозавода кормятся на выростных прудах у источников
подачи воды. Суточный рацион птицы составляет здесь 90 г. В пище доминирует карп (67%),
белый толстолобик (22%) и пёстрый толстолобик (11%) встречаются реже.
В Средней Азии основу пищевого рациона квакв, как и в Европе, составляет рыба, ам-
фибии и их личинки, водные и наземные беспозвоночные (Птицы Средней Азии, 2007).
Враги, неблагоприятные факторы. В качестве ведущего негативного фактора, ока-
зывающего воздействие на потомство квакв, выступает гидрологический режим местооби-
таний. Так, в отдельные годы, вследствие затопления в дельте Днестра до его зарегулирова-
ния, гибло до 70% гнёзд квакв, гнездившихся на кустах ивы и заломах тростника. Суще-
ственное влияние на численность гнездящихся птиц может оказывать также низкая темпе-
ратура предыдущей зимы.
Среди биотических факторов следует упомянуть хищничество, конкуренцию и канни-
бализм. Из типичных хищников и животных, проявляющих хищнические наклонности и
способных оказать существенное влияние на численность квакв, наибольшее отрицатель-
ное воздействие оказывают болотный лунь (наиболее значимый хищник), серая ворона,
енотовидная собака, серая крыса и шакал. Наибольший ущерб от наземных хищников око-
ловодные птицы испытывают в маловодные годы, когда многие гнездовья птиц вполне
доступны для этой группы животных. Как уже упоминалось, в случае недостатка корма
старшие птенцы могут поедать младших сородичей, выступая в качестве типичных кан-
нибалов.
В условиях оз. Ханка временное, но существенное снижение численности квакв было
вызвано конкуренцией за гнездовья с большим бакланом (данные Ю.Н. Глущенко).
Кроме естественных факторов, огромное влияние на квакв оказывают антропогенные
преобразования среды и прямое преследование птиц человеком (Krivenko, Crivelli, Vino-
gradov, 1994, цит. по: Lysenko, 1994). На успешность размножения квакв существенное
воздействие оказывают возникающие по вине отдыхающих пожары в плавнях и поймен-
ном лесу. Из-за них в дельте Днестра пропали многие колонии, формировавшиеся квак-
вой на протяжении десятка лет. Серьезно влияет на гибель потомства, как показано выше,
фактор беспокойства, провоцирующий, помимо прочего, массовое разорение гнёзд серой
вороной. В прошлом значительную опасность представлял довольно широко практиковав-
шийся неконтролируемый сбор яиц, что иногда приводило к исчезновению даже крупных
колоний в дельтах Днестра и Дона.
В гнёздах кваквы найдены блохи Ornithomya avicularia, Dermestes elongatus и D. nidum
(Hicks, 1959), в оперении — пухоеды: Ardeicola goisagi и Ciconiphilus decimfasciatus
(McClure, Ratanaworabhan, 1973).
Хозяйственное значение, охрана. В связи с относительной малочисленностью и пита-
нием преимущественно так называемыми «сорными» видами рыб, хозяйственного значе-
ния не имеет. Занесена в Красную книгу Республики Беларусь; в России — в Красные
книги Северной Осетии, Белгородской и Пензенской областей.
Зелёная кваква
237
Род Butorides Blyth, 1852
Зелёная кваква
Butorides striatus (Linnaeus, 1758)
Ardea striata Linnaeus, 1766., Syst. Nat., ed. X, 1:144, Суринам
Striated, [little green, green-backed] heron (англ.), Mangrovereiher (нем.),
heron vert (фр.)
Син. (лат): для подвида amurensis’. Butorides schrenckii Богданов, 1884.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Общая окраска тела темная, зеленова-
то-сизая, на брюшной стороне светлее. Удлинённые перья темени и затылка образуют зао-
стрённый хохол. На верхней части спины и плечах имеются узкие заострённые украшаю-
щие перья. Полёт довольно быстрый, с частыми глубокими взмахами крыльев, шею в поле-
те обычно складывает S-образно. Чаще зелёную квакву удаётся видеть пролётающей над
рекой или сидящей в тени деревьев у воды на берегу, коряге и т.п. В случае опасности
скрывается в кронах деревьев, открытых пространств избегает, но во время пролёта её
можно видеть на скалах и пляжах морского побережья.
По величине и сложению наиболее сходна с жёлтой и белокрылой цаплями. От других
мелких цапель фауны России хорошо отличается тёмной окраской и голосом. Голос — рез-
кий, звонкий или приглушенный крик, который можно передать как «тиуу».
Подвидовая систематика. Выделяют от 20 до 36 подвидов, различающихся размера-
ми и окраской (del Hoyo et al., 1992; Dickinson, 2003). На территории Северной Евразии
обитает один подвид:
1. Butorides striatus amurensis
Ardea (Butorides) virescens amurensis Schrenck, 1860, Reise...Amur-Lande, 1, Zweite
Lieferung, c. 441, область Амура.
Ареал вида в России, северо-восточном Китае, Кореи, Японии.
Прочие подвиды не связаны с территорией Восточной Европы и Северной Азии. Поми-
мо В. s. amurensis различают следующие основные подвиды (Dickinson, 2003):
В, s. striatus (Linnaeus, 1758) (2). От восточной Панамы до северной Аргентины и Боли-
вии.
В. s. atricapilla (Afzelius, 1804) (3). Африка к югу от Сахары, о-ва в Гвинейском заливе.
В. s. brevipes (Hemprich et Ehrenberg, 1833) (4). Сомали, побережье Красного моря.
В. s. rutenbergi (Hartlaub, 1880) (5). Мадагаскар, о. Реюньон
В. s. rhizophorae Salomonsen, 1934 (6). Коморские о-ва.
В. s. crawfordi Nicoll, 1906 (7). О-ва Альдабра и Амирантские.
В. s. degens Е. Hartert, 1920 (8). Сейшельские о-ва.
В. s. albolimbata Reichenow, 1900 (9). Архипелаг Чагос, Мальдивские о-ва.
В. s. chloriceps (Bonaparte, 1855) (10). Пакистан, Индия, Шри-Ланка, Лаккадивские о-ва.
В. s. spodiogaster Sharpe, 1894 (11). Андаманские и Никобарские о-ва, Зап. Суматра.
В. s. actophila Oberholser, 1912 (12). Юж. Китай, Сев. Вьетнам и Сев. Бирма.
В. s. javanica (Horsfield, 1821) (13). Бирма, Таиланд, Юж. Индокитай, о-ва Хайнань и
Тайвань, п-ов Малакка, М. и Б. Зондские о-ва, Сулавеси, Филлипины.
В. s. moluccarum Е. Hartert, 1920 (14). Молуккские о-ва, о-ва Кай.
В. s. papuensis Мауг, 1940 (15). Северо-западная область Новой Гвинеи.
238
Отряд Листообразные, Семейство Цаплевые
В. s. idenburgi Rand, 1941 (16). Сев., вост, и юго-вост, часть Новой Гвинеи.
В. s, macrorhyncha (Gould, 1848) (17). Вост, и Сев.-Вост. Австралия.
В. s. stagnatilis (Gould, 1848) (18). Сев.-Вост. Австралия.
В. s, solomonensis Мауг, 1940 (19). Соломоновы о-ва.
В. s. patruelis (Peale, 1848) (20). О. Таити.
Замечания по систематике. Род Butorides весьма далёк от настоящих квакв и сближа-
ется с цаплями рода Ardeola. Возможно, более правильным русским названием для вида
было бы «зелёная цапля», а не «зелёная кваква». Считающиеся подвидами зелёной кваквы
в широкой трактовке (В. striatus sensu lato) расы virescens, anthonyi, frazari, maculate, обита-
ющие от востока Канады и США до Панамы и Карибских о-вов, а также раса sundevalli с
Галапагосских о-вов, иногда выделяют (напр. Dickinson, 2003) в самостоятельные виды,
соответственно, зеленоспинная цапля (В. virescens (Linnaeus, 1758)) и галапагосская
лавовая цапля (В. sundevalli (Reichenow, 1877)).
Описание. Окраска. Половой диморфизм и сезонный морфизм отсутствуют. Самец
и самка в дефинитивном наряде. Лоб, темя и перья хохла чёрные с зеленоватым металли-
ческим блеском. Спинная сторона, включая кроющие крыла, буровато-серая с зелёным
металлическим блеском; у старых птиц украшающие перья на спине голубовато-серые.
Первостепенные маховые чёрные, второстепенные и их кроющие серовато-бурые с зелё-
ным оттенком и белыми, отчасти охристыми, каемками по краю опахал. Бока головы и шеи
тёмно-серые, от клюва под глазом идёт чёрная полоска; подбородок белый, верхняя часть
горла белая с редкими буроватыми пестринами. Низ тела, включая испод крыла, дымчато-
серый, светлеющий на брюхе и нижних кроющих хвоста. Надклювье и режущий край под-
клювья чёрные, нижняя часть подклювья желтоватая; голые участки «лица» зелёные; раду-
жина жёлтая. Ноги буровато-зеленоватые; задняя сторона цевки и подошва пальцев оран-
жево-желтоватые.
Птенцовый наряд. В возрасте 1-2 недель от роду пух на спинной стороне серый, иногда
с белыми кончиками или буроватым налётом; на брюшной стороне пух белый; длина пуха
на голове 11,0-14,4 мм, на остальных птерилиях — 7,2-11,4 мм. Кожа голых участков голо-
вы и шеи зеленовато-жёлтая или желтовато-оранжевая, клюв жёлтый или телесно-серый,
обычно с буроватой вершиной; яйцевой зуб белый. Стенки ротовой полости грязно-розо-
вые, полоски по бокам неба и кончик языка тёмно-серые. Веки жёлтые; радужина мутно-
жёлтая или зеленоватая. Лапы телесные с оранжево-жёлтым оттенком или жёлтые; когти
телесные или бледно-жёлтые.
Во втором пуховом наряде кожа всех участков тела с зеленоватым оттенком; клюв и
лапы с серым оттенком; радужина светлая, зеленовато-жёлтая.
Гнездовой наряд. Хохол на голове немного короче, чем у взрослой птицы; спинная сто-
рона буроватая с хорошо выраженным зелёным металлическим блеском и узкими рыжева-
то-охристыми каймами перьев; кончики маховых белые. Брюшная сторона беловатая с
рыжевато-охристым налётом и многочисленными крупными продольными тёмными пест-
ринами.
Возрастная изменчивость окраски проявляется в исчезновении пестрин на брюшной
стороне, по-видимому, к началу третьего года жизни. У годовалых птиц от подбородка до
средней линии и по бокам горла чёрные пестрины образуют три полосы; на груди размы-
тые продольные светлобурые пестрины; концы второстепенных маховых белые; остальное
оперение как у взрослых. У молодых птиц второго года жизни сохраняются пестрины на
подбородке и горле, а концы второстепенных маховых — белые.
Строение и размеры. Формула крыла, не учитывая первое рудиментарное
первостепенное маховое — I<II?III>IV>V.....Размеры (мм) птиц (п=15): длина крыла —
Зелёная кваква
239
186-205, в среднем 195,5; длина клюва 53,0-67,4, в среднем — 63,8; длина цевки 48,0-
55,8, в среднем 52,7; длина хвоста — 67,7-79,6, в среднем — 73,8. Промеры выполнены по
коллекционным материалам зоомузея ДВГУ и БПИ ДВО РАН.
Масса 266,0-370,9 г (Спангенберг, 1951).
Линька. Сведения по линьке практически отсутствуют. Известно, что она начинается в
конце июня (Шульпин, 1936) и проходит в течение июля и августа (Спангенберг, 1951).
Распространение. Гнездовой ареал. От Нижнего Амура на севере до северо-зап.
и вост. Австралии на юге, острова юж. Полинезии на восток до о. Таити, в юж. Азии на
запад включая Индию, тропические и субтропические острова Индийского океана, юго-
зап. побережье Аравийского п-ова, Африка от Сахары к югу до Заира и провинции Наталь
(ЮАР) включительно, Центр, и Юж. Америка до Перу, сев. Аргентины и Уругвая (рис. 44).
В России распространена в Приморье, на севере от устья р. Зея вниз по р. Амур до устья
р. Горин (Шульпин, 1936; Бабенко, 2000), по восточным склонам Сихотэ-Алиня на север
проникает до р. Ботчи (Иванов, 1976) (рис. 45). Отмечен залёт зелёной кваквы на о. Саха-
лин, но дата и точное место не приводятся (Check-List..., 1974).
Данных об исторических изменениях ареала нет. Локальные изменения связаны со сме-
ной пойменной растительности. В районах лесоразработок расселяется в верховья рек.
Зимовки. Большинство подвидов проводят зиму в местах гнездования. Гнездящийся
в России В. s. amurensis, как и подвид В. s. actophila, зимуют в Юж. Китае, на о. Тайвань, о-
вах Рюкю (самка от 17 декабря 1926 г., ЗИН), Ито, Идзу, в Индокитае, на Филиппинах,
Зондских о-вах и о. Сулавеси. Относительно распределения мест зимовок между этими
двумя подвидами в литературе существует противоречивая информация. Для В. s. amurensis
места зимовок указаны от Юж. Китая до о. Суматра и Филиппин, а для В. s. actophila —
Никобарские о-ва, о. Суматра и о. Борнео (del Hoyo et al., 1992). По другим источникам
места зимовок В. s. amurensis находятся на Филиппинских о-вах, о. Сулавеси и Больших
Зондских о-вах, а В. s. actophila — в Юго-Вост. Азии и на Б. Зондских о-вах (Dickinson,
2003).
Миграции. Весенняя и осенняя миграции известны только для двух подвидов — В. s.
amurensis и В. s. actophila. Для африканских популяций отмечены местные кочёвки, связан-
ные с сезоном дождей.
В Приморье пролёт зелёной кваквы протекает незаметно, ввиду ночной миграции этого
вида. Весной первые птицы появляются в конце апреля, массовый прилет приходится на
начало мая (Пукинский, 2003; Назаров, 2004). В этот период в ночное время суток в соот-
ветствующих угодьях можно часто слышать голос пролетающих зелёных квакв. На юге
Приморья прилёт затягивается до 10-11 июня. Осенью с мест гнездования улетает в сен-
тябре, но часть птиц летит позже. В устье р. Гур самец в гнездовом наряде добыт 2 октября
(колл. ЗИН РАН).
В Шанхае сроки миграций сходны — весной с 21 апреля, осенью до 13 октября (Wilkinson,
1935).
Местообитания. В бассейне р. Бикин придерживается приречной уремы (Пукинский,
2003), на юге Приморья встречается по долинам рек, слабопроточных озёр с обязательным
присутствием пойменных и прибрежных лесов, в период миграций встречается на морс-
ком побережье. Открытых мест избегает. Чаще её можно встретить в пойменных ивовых
лесах. Гнездится по долинам рек, сильно пострадавшим от антропогенного освоения, по
берегам искусственных водохранилищ, особенно с подтопленными участками леса (Горча-
ков, 1990; ориг.).
В других частях ареала также придерживается лесистых берегов рек, озёр, прудов, эсту-
ариев рек, а также встречается в мангровых зарослях. Отдельных птиц иногда можно уви-
240
Отряд Листообразные, Семейство Цаплевые
Рис. 44. Область распространения зелёной кваквы: а — гнездовой ареал; б — область зимовок. Ос-
новные подвиды (по Dickinson, 2003). 1 —В. s. amurensis; 2 — В. s. striatus', 3 —В. s. atricapilla', 4 —
В. s. brevipes', 5 — В. s. rutenbergi', 6 — B. s. rhizophorae', 7 — B. s. crawfordi’, 8 — B. s. degens', 9 —
B. s. albolimbata; 10 — B. s. chloriceps; 11 —B. s. spodiogaster', 12 — B. s. actophila; 13 —B. s.javanica;
14 — B. s. moluccarum’, 15 — B. s. papuensis', 16 — B. s. idenburgi', 17 — B. s. macrorhyncha; 18 — B. s.
stagnatilis', 19 — B. s. solomonensis’, 20 — B. s. partuelis.
деть на более открытых пространствах — на грязевых отмелях, в зоне приливов и отливов
на коралловых рифах, тростниковых зарослях, на рисовых полях, болотах, пастбищах (del
Hoyo et al., 1992). В гористых районах отмечена до высоты 1 400 м (Robson, 2000).
Численность. По численности B.s. amurensis конкретных данных немного. В южном
Приморье зелёная кваква обычна в низовьях рек, где на 10 км уремы может приходиться до
35 пар (ориг.). Сходная плотность гнездования наблюдается и на р. Б. Уссурка (Иман), где
Рис. 45. Гнездовой ареал зелёной кваквы в
России (а).
на 1 км приходится от 1 до 3 пар (Спанген-
берг, 1951). Обычна в нижнем течении р. Би-
кин, выше пос. Красный Яр. В зоне первич-
ных кедрово-широколиственных лесов редка.
Общая численность для всего бассейна р. Би-
кин оценивается в 350—400 гнездящихся пар
(Пукинский, 2003). На Амуре в районе Мало-
го Хингана обычна (Яхонтов, 1973), но в Боль-
шехехцирском заповеднике немногочисленна
(Казаринов, 1973), а у Комсомольска-на-Аму-
ре редка, хотя в прошлом была обычна. Осен-
няя численность в Хабаровском крае предпо-
лагается равной 2-3 тыс. особей (Красная
книга Хабаровского края, 2000). Обычна в
Японии и Китае (Check-List ..., 1974; Cheng
Tso-hsin, 1987).
В Таиланде, Малайзии, на юге Лаоса и
Вьетнама, в Сингапуре (подвид/?, s.javanica)
зелёная кваква немногочисленна, но местами
Зелёная кваква
241
обычна, особенно на побережье, в период же зимовки северных популяций этот вид стано-
вится весьма заметным (Robson, 2000).
Вырубка водоохранных лесов по Иману в 1930-1950 гг. и по р. Бикин в 1970-х гг. спо-
собствовала зарастанию поймы черёмухой и ивой, что в свою очередь привело к росту
численности этого вида (Спангенберг, 1965; Пукинский, 2003).
Размножение. Половая зрелость В. s. amurensis наступает в возрасте 2 лет. Гнездится
по берегам стариц и проток, захламлённых корягами, в зарослях невысоких, густых дере-
вьев со сплетающимися кронами. В высокоствольной чозениевой уреме встречается зна-
чительно реже. Местами селится в 100-200 м от водоёма, в небольших заболоченных ро-
щицах, состоящих из японской ольхи и берёзы. Обычно гнездится отдельными парами, в
благоприятных условиях (устье р. Раздольной, юг Приморья) гнёзда соседних пар распола-
гаются в 6-15 м, так что образуются как бы скопления из 3-7 пар. По Бикину на островах
площадью 0,2-0,5 га отмечены колониальные поселения от 3^4- до 12 пар (Пукинский, 2003).
В Японии известны колонии до 10 пар (Jahn, 1942). Для других подвидов отмечены коло-
нии до 300-500 пар (del Hoyo et al., 1992). В колониях в начале сезона размножения можно
видеть разнообразные формы демонстрационного поведния (рис. 46).
Сроки размножения зеленой кваквы в разных частях ареала сильно варьируют; в тропи-
ческих областях они приурочены периоду дождей (del Hoyo et al., 1992). Размножение B.s.
amurensis растянуто с мая по август. Первые птицы приступают к строительству гнёзд на
юге Приморья в начале мая. Наиболее ранняя полная кладка (6 яиц) найдена здесь 18 мая,
одна кладка (5 яиц) закончена 19 мая. Поздние кладки здесь же найдены 24 июня (три; одна
из них только что завершена) и 7 июля; они содержали по 3, реже 4 яйца (ориг.). Сходные
сроки отмечены и на северо-западе Приморья. На Бикине первые брачные крики слышны
в сумерках и по ночам в первой декаде мая. Начало кладки отмечено в 6 из 9 гнёзд 9-13
мая, а также начало кладок зарегистрировано 14, 17 и 21 июня и 1-3 июля (Пукинский,
2003). Предполагается, что поздние кладки могут быть повторными.
Гнёзда сооружают на невысоких густых раскидистых деревьях (ива, приземистая фор-
ма чозении, черёмуха, яблоня, ольха, ильм и т.д.), очень редко на сухих, подтопленных
водой деревьях. Часто птицы используют старые постройки в течение нескольких лет, дос-
траивая их. При повторной кладке нередко занимают старые гнёзда. Обычно гнездо устра-
ивают в развилках, на скрещивающихся или расположенных параллельно горизонтальных
ветвях в 2-3 м от ствола, гораздо реже в основании ветви у главного ствола. Высота распо-
ложения гнёзд над землей или водой — от 2 до 7,7, в среднем 3,75 м (по 23 измерениям);
изредка 10-12 м (Спангенберг, 1951). Часто гнёзда труднодоступны (для исследователя),
особенно когда расположены на тонких ветвях, склонившихся над водой. По форме хоро-
шо различаются гнездовые постройки двух типов. Один из них представляет собой неболь-
шую площадку из небрежно наложенных друг на друга веток. При этом лоток выражен
слабо. Содержимое такого гнезда можно разглядеть снизу. В другом случае гнёзда более
массивные, плотные и аккуратные, по форме напоминающие перевернутый конус, в кото-
ром часть веток располагается радиально. Рыхлые постройки принадлежат, по-видимому,
молодым птицам, гнездящимся впервые12. Более плотные конусообразные могут быть, как
и первой постройки, так и достроенными прошлогодними. Основным материалом для гнёзд
обычно служат сухие веточки дерева, на котором они построены. В гнёздах второго типа
лоток выстилается прямыми концевыми веточками ивы или чозении. Размеры гнёзд варьи-
руют. Постройки, обследованные в устье р. Раздольной (п=22), имели «внешний» диаметр
(мм) — 200-380, в среднем — 302; высоту 50-170, в среднем — 102; диаметр лотка 100—
200, в среднем — 165; глубину лотка — 20-55, в среднем — 40 мм (Назаров, 2004).
12 Это не подтверждается по наблюдениям за др. видами цапель (прим. ред.).
242
Отряд Листообразные, Семейство Цаплевые
Рис. 46. Элементы демонстрационного поведения зелёной кваквы (по Meyerriecks, 1960): а — агрес-
сивная демонстрация, б — демонстрация щёлканья клюва, в — вытянутая поза, г — хлопанье крыль-
ями в полёте.
Гнездо птица строит за 3-5 дней. По мере насиживания и выкармливания птенцов сам-
ка его достраивает, часть веток приносит самец. К откладке яиц птицы приступают сразу
после окончания строительства (Пукинский, 2003).
Первые три яйца откладываются ежедневно, последующие — с большим промежутком
(Спангенберг, 1951). В устье р. Раздольная во всех гнёздах самки откладывали по одному
яйцу через день. Полная кладка содержит 2^4- яйца (Африка), 3-5 (Юго-Вост. Азия) (Robson,
2000) или 3-6 (Приморье) (Назаров, 1979). Количество яиц в гнёздах (п=15): 3 (2 гнезда), 4
(6), 5 (6) и 6 (1 гнездо) (Пукинский, 2003). Нередко в период насиживания яйца выпадают
из гнёзда. При полной гибели кладки откладывают новые яйца, но их уже не бывает больше
четырёх. Размеры яиц из разных частей ареала (п=216): 32,3^4-5,0 х 23,8-33,0 мм (Schon-
wetter, 1960); из Приморья (п=21): 37,4^43,0 х 29,0-31,1 мм (Спангенберг, 1965) (п=82):
37,8^4-3,4 х 28,5-32,2 мм, в среднем 30,6x40,8 мм (Назаров, 1979) (п=22): в среднем —
Зелёная кваква
243
40,5x30,4 мм (Пукинский, 2003). Масса свежеотложенных яиц (п=8): 22,0-25,5, в среднем —
24,4 г (Пукинский, 2003). Форма эллипсовидная различной удлинённости, реже правиль-
ная яйцевидная. Окраска однотонная. Цвет скорлупы бледно-голубой, на просвет голубова-
тый или зеленовато-голубоватый. Поверхность мелкозернистая, шероховатая, матовая. Во
время насиживании скорлупа приобретает грязно-белёсые тона.
К насиживанию зелёная кваква приступает после откладки первого яйца (Спангенберг,
1951; ориг.). По мнению Ю.Б. Пукинского (2003) насиживание обычно начинается с 3^4-
яйца. В насиживании принимают участие оба партнёра. Через 19-20 дней после заверше-
ния кладки в гнезде появляется первый птенец; остальные — в течение последующих 1-2
суток. На юге Приморья наиболее раннее вылупление отмечено 4 июня, массовое — 7-9
июня. В одном гнезде птенцы появились 20 июля; сроки вылупления могут быть и более
поздними. Основная смертность птенцов в гнёздах приходится на первые дни их жизни.
Эмбриональная смертность (п=7) составила около 32%, смертность птенцов — 10%. При-
чины гибели отдельных яиц в кладках — продавливание скорлупы веточками лотка и выпа-
дение из гнёзд.
Пеньки маховых перьев начинают прорезываться на 3-5-й день жизни, на одиннадца-
тый день прорезаются кисточки маховых, на 13-й — кисточки рулевых перьев. Одноднев-
ные птенцы весят 11,6-15,9 г; в первые три-четыре дня вес увеличивается на 3-5 г ежед-
невно, через 10 дней их вес составляет уже 105 г. Выкармливают птенцов оба партнёра. По
данным Ю.Б. Пукинского (2003) самец, особенно в первую половину гнездового периода,
приносит корм значительно чаще, самка в это время практически всё время находится с
птенцами.
В первые дни жизни птенцы спокойно лежат в гнезде, на четвёртый день при попытке
взять их в руки они шипят и пытаются защищаться. Взрослые птицы затаиваются непода-
леку от гнёзда, но если птенцы подают голос, то родители с громким тревожным криком
перелетают с дерева на дерево, иногда усаживаются близ гнезда и издают устрашающие
каркающие звуки. 6-7-дневные птенцы при приближении человека приподнимаются в гнезде
и затаиваются, вытягивая шеи вверх. Через неделю после вылупления птенцы выходят из
гнезда и отдыхают на ветвях в 1 м от него, двухнедельные бродят по ветвям гнездового
дерева, могут перебираться на соседние, а в случае опасности ловко скрываются в кроне и
затаиваются, но на ночь возвращаются в гнездо. В возрасте 27-32 дней поднимаются на
крыло и способны летать на небольшие расстояния. В это время они держатся выводками в
районе гнезда, но заметить их бывает очень трудно. В конце июля - августе можно встре-
тить самостоятельно кормящихся молодых птиц (Назаров, 2004).
Суточная активность, поведение. Зелёная кваква деятельна круглые сутки. Днем можно
видеть птиц, строящих гнёзда, не только в глубине древесных зарослей, но и на опушках
или перелетающих через открытые участки к водоёмам. Особенно часто встречаются кор-
мящиеся птицы в полуденное время. В солнечную погоду их активность снижается. Ночью
часто можно услышать крик пролетающей над рекой птицы.
Питание. В Приморье питается мелкой рыбой (Шульпин, 1936; Спангенберг, 1965;
Панов, 1973; Пукинский, 2003). В одном желудке обнаружены стрекозы (Воробьев, 1954),
в устье р. Раздольной в одном желудке и трёх отрыжках обнаружены щиповки и вьюны
длинной 5-9 см.
В Индии пищей служит рыба, в том числе илистые прыгуны (р. Periophthalmus) и сарга-
ны (р. Belone), длиной 16,5-20 см, лягушки, креветки, крабы, водяные жуки (Ali, Ripley,
1968); в Африке — морские черви и моллюски (Mackworth-Praed, Grant, 1952). Обычно
кормятся на затененных участках лесных речек и стариц, реже на озёрах и морском побере-
жье. Подкарауливает добычу неподвижно сидя на берегу или на коряге, иногда медленно
244
Отряд Листообразные, Семейство Цаплевые
прогуливаясь у кромки воды. На речных перекатах подкрадывается к добыче (Панов, 1973).
Ю.Б. Пукинский (2003) наблюдал, как охотящиеся у кромки воды птицы для привлечения
молоди рыб бросали в воду палочки, кусочки коры и после этого хватали подманившихся
на эту уловку рыб.
Враги, неблагоприятные факторы. В Приморье врагами являются сорока, черная и
большеклювая вороны, разоряющие гнёзда. Вороны нападают на взрослых зелёных квакв
(о-ва Римского-Корсакова). В августе отмечена гибель молодых птиц в рыборазводных сад-
ках (ориг.). Локальные снижения численности наблюдаются при уничтожении прибрежной
растительности, а также при постоянном беспокойстве гнездящихся птиц со стороны чело-
века.
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Занесена в Крас-
ную книгу Хабаровского края (2000). В дальневосточных заповедниках и заказниках на
гнездовье немногочисленна.
Род Ardeola Boie, 1822
Жёлтая цапля
Ardeola ralloides (Scopoli, 1769).
Ardea ralloides Scopoli, 1769, Annus 1 Historico-Naturalis, c. 88, Крайна.
Squacco heron (англ.), Rallenreiher (нем.), crabier chevelu (фр.).
Син. (лат.): Ardea pumila Lepechin, 1770; Ardea castanea S.G. Gmelin, 1771; Ardea comata
Pallas, 1773; Ardea squaiotta J.F. Gmelin, 1789; Ardea senegalensis J.F. Gmelin, 1789; Ardea
audax Picot de la Peirouse, 1794; Ardea botaurulis Schrenk, 1798; Ardea deaurata Merrem,
1820; Buphus illyricus C.L. Brehm, 1831; Bubulcus luteus Doderlein, 1872.
Син. (рус.): косматая цапля.
Статус. Гнездящийся, перелётный вид.
Общая характеристика и полевые признаки. Небольшая (слегка крупнее вороны)
коренастая цапля с относительно короткими ногами и клювом, безошибочно отличается от
других цапель по размерам и общему сложению. Сидящую птицу легко отличить от других
и по окраске оперения. Напротив, во время полета жёлтая цапля кажется белой и на боль-
шом расстоянии может быть легко спутана с другими видами — например, с египетской
цаплей. На лету отличается от них особенно широкими и тупыми крыльями. Видеть её
удаётся в одиночку, спокойно сидящей на берегу водоёма, на поваленном с краю уреза
воды сухом камыше, на кочке или коряге, при этом птица втягивает шею, сжимается и ка-
жется значительно меньше, чем есть на самом деле. Реже жёлтые цапли кормятся неболь-
шими группами, но и в этих случаях птицы держатся поодаль друг от друга.
Иногда утрами и на заходе солнца приходится наблюдать цапель, летящих маленькими
стайками. Полет их медленный, но удивительно лёгкий с редкими взмахами широких кры-
льев. В одних случаях цапли летят низко беспорядочной стайкой, в других — выстроив-
шись углом или в косую линию. Такие перелеты стайками затягиваются до поздних суме-
рек; вообще же жёлтую цаплю надо считать дневной птицей. Несмотря на светлую окраску
оперения, делающую её заметной, жёлтая цапля отличается тем, что она не очень пуглива и
близко подпускает к себе человека.
Голос — что-то вроде грубого «карр», его удаётся слышать очень редко, чаще — при
испуге и взлёте.
Жёлтая цапля
245
Подвидовая систематика. Монотипический вид.
Описание. Окраска (Спангенберг, 1951). Половой диморфизм в окраске не выра-
жен. Самец и самка в брачном наряде. Бока головы и шея взрослой птицы — жёлтые или
кремово-жёлтые. На голове длинный хохол, перья зоба и плечевые удлинены и рассучены.
Нижняя часть шеи, зоб и грудь ярко-охристые, горло белое. На затылке и задней части шеи
по желтоватому фону — тонкие продольные, почти чёрные полоски. Наиболее длинные
лентовидные перья хохла белые с чёрными краями. Спина и украшающие перья спины —
винного цвета, несколько желтеющие на концах. Крылья, подхвостье, хвост и вся нижняя
сторона тела от зоба, грудь и брюхо — белые, местами со слабым желтоватым налетом.
Голая кожа вокруг глаз зеленоватая, клюв голубой с почти чёрной вершиной, ноги жёлто-
оранжевые, а в разгар токования ярко-красные, подошвы жёлтые, когти чёрные. Радужина
жёлтая.
Самец и самка в зимнем наряде. В зимнем наряде украшающие перья отсутствуют, го-
лова и шея с черновато-бурым продольным рисунком. Спина, плечевые и внутренние вто-
ростепенные маховые — землисто-бурые. Ноги зеленовато- или серовато-жёлтые. Клюв
рогового или желтовато-охристого цвета с тёмным концом, уздечка зеленовато-бурая.
Пуховой наряд. Смотри табл. 17.
Гнездовой наряд. Молодые птицы похожи на взрослых в зимнем оперении, отличаясь от
них серо-бурым продольным рисунком на кроющих крыла; маховые — с тёмными ствола-
ми, вершинами и наружными опахалами. Ноги, уздечка, клюв тусклые, серовато- или буро-
вато-жёлтые. Радужина бледно-жёлтая или белесая.
В первом брачном наряде стволы первостепенных маховых бурые (у взрослых белые).
Строение и размеры. На наружных опахалах II и IV махового имеются
неглубокие вырезки. Размеры (мм): длина крыла самцов (п=9) 208-234, в среднем 225;
длина хвоста (п=8) 73-84, в среднем 81,1; длина цевки (п=9) 51-62, в среднем 58; длина
клюва (п=8) 62-70, в средн. 64,8. У самок (п=9), соответственно, крыло 209-228 (216),
хвост 66-84 (73,4), цевка 54-59 (56,2), клюв 58-65 (61,8) мм (Cramp, 1977).
Взрослые самцы и самки по внешнему виду не различаются. Однако при детальном
изучении можно заметить, что самцы несколько крупнее самок, а украшающие перья хохла
и спины у последних менее развиты (табл. 14).
Линька. Плохо выяснена. Общая схема линьки — примерно как у других цапель. Мо-
лодые птицы сменяют мелкое оперение между июлем и декабрем. У взрослых две линьки
в году: предбрачная между январем и маем, послебрачная полная, между июлем и декаб-
рем. Возрастные изменения в окраске оперения незначительны.
Пуховой наряд сменяется на перьевой, начиная с 10-12-го дня. Формирование юве-
нильного наряда заканчивается к 25-30-дневному возрасту. Молодые птицы в гнездовом
наряде сменяют мелкое оперение в первую осень жизни между августом и декабрем. Юве-
нильная линька происходит на местах зимовок. У подавляющей части особей крупное перо
полностью меняется, вероятно, на местах зимовок и птицы приобретают там взрослый
наряд. После гнездования, обычно перед отлётом, взрослые птицы теряют «брачные» пе-
рья и частично меняют мелкое перо.
Распространение. Гнездовой ареал. Состоит из изолированных очагов. В Европе
гнездится в юж. Португалии и юж. Испании, в дельте Роны во Франции, в Италии, Венг-
рии, в Хорватии, Словакии, Болгарии, Румынии. В Африке распространена севернее Саха-
ры от Марокко до Туниса, южнее — локальными очагами по всему континенту; Мадагас-
кар, Азия от Малой Азии, Сирии, Ирака до Ирана и Средней Азии. Изредка встречается в
Чехословакии, Великобритании, Голландии, Дании, Финляндии, Польше, Австрии, Швей-
царии, на Азорских о-вах, на Мадейре, на Канарских о-вах (Cramp, 1977) (рис. 47).
246
Отряд Листообразные, Семейство Цаплевые
Таблица 14. Основные размеры и вес самцов и самок жёлтых цапель (дельта Днестра)
Промеры, Молодые (п=20) Взрослые
в мм’ Самцы (п= 12) Самки (п= 15)
вес в г lim М lim М lim М
Крыло 195-230 205 225-245 240 205-227 215
Клюв 53-65 58 64-70 67 55-67 64
Цевка 54-67 59 55-67 62 53-67 59
Вес 195-312 238 245-400 285 220-302 238
Рис. 47. Область распространения жёлтой цапли: а — гнездовой ареал; б — места зимовки.
Подробности распространения жёлтой цапли в странах Вост. Европы и Сев. Азии тако-
вы. Периодически гнездится в Молдавии (Birdlife International, 2004). На Украине гнездит-
ся в дельте Дуная и на придунайских водоёмах Украины — озёрах Картал, Китай, о-вах
Малый Даллер, в Стенцовско-Жебрияновских плавнях и в Дунайском биосферном запо-
веднике, в низовьях р. Кагильник (Жмуд, 2000; Русев, Корзюков, Яковлев, 2005). В дельте
Днестра гнездилась на протяжении всего XX столетия (Русев, 2003), в дельте Днепра изве-
стна на гнездовании давно, однако в последние годы гнездовые поселения исчезли (Арда-
мацкая, 2005). Изредка гнездится в Черноморском биосферном заповеднике (Яремченко,
Руденко, 2004). Периодически гнездится в верховьях Тилигульского лимана (Русев, 2000а).
Жёлтая цапля
247
Рис. 48. Распространение жёлтой цапли в Сев. Евразии: а — области гнездования; б — места зимовки.
С 1972 г. гнездится на Лебяжьих о-вах в Крыму, однако никогда не была здесь многочислен-
на. Позднее заселила и другие районы в зоне рисосеяния (Костин, Тарина, 2004). Гнездит-
ся в плавневых массивах Вост. Сиваша (Гринченко, 2004а). Периодически жёлтые цапли
гнездятся также в плавнях Молочного лимана в Запорожской обл. (Кошелев, 2004, Коше-
лев и др., 2004) (рис. 48).
Во второй половине XIX в. жёлтая цапля была редкой птицей Сев.-Вост. Приазовья
(Алфераки, 1910, цит. по: Птицы Северного Кавказа, 2004), в начале XX в. неоднократно
отмечалась в Приазовье, где, по-видимому, гнездилась; встречалась в плавнях Кубани и
низменной Осетии, отмечена в районе Кизляра (Спангенберг, 1951). В 1950-х гг. в плавнях
Кубани найдена на гнездовье (Винокуров, I960). В 1970-1980-е гг. в связи с развитием
рисосеяния и прудово-лиманного рыбоводства область её поселений на Сев. Кавказе зна-
чительно расширилась. В настоящее время гнездится в долине Нижнего Дона, на Западном
Маныче, в дельтах Кубани и Терека, на Вост. Маныче и водоёмах Ставрополья (Птицы
Северного Кавказа, 2004). В последние годы обычна в Армении и, возможно, гнездится в
Грузии (Birdlife International, 2004).
На северном берегу Каспия регулярно гнездится в дельте Волги и редко в устье Урала.
В 1870-х гг. гнездилась на Сарпинских озёрах (Мензбир, 1918). В Средней Азии гнездова-
ние жёлтой цапли приурочено к устьям рек, впадающих в Каспийское и Аральское море:
Аму-Дарьи, Сыр-Дарьи, Атрека, а также к дельтам Теджена и Мургаба. Из приустьевых
участков цапли распространяются вверх по долинам рек, не удаляясь, однако, далеко от их
нижнего течения (Птицы Средней Азии, 2007).
Зимовки. Часть жёлтых цапель из Азербайджана, так же, как часть кубанских и
дагестанских цапель, в тёплые зимы остаются в южных районах области гнездования, в
248
Отряд Листообразные, Семейство Цаплевые
том числе поблизости от своих гнездовий. Однако, общее количество птиц, проводящих
здесь зиму, невелико (Сапетин, 19786). Регулярная зимовка, по-видимому, существует на
юго-восточном (иранском) побережье Каспия. Основная же масса азербайджанских цапель
через Иран, Двуречье и Аравию улетает на зимовки в Экваториальную Африку. В Тропи-
ческой Африке, вероятно, зимуют и дунайские жёлтые цапли (Нанкинов, 1978).
Сведения о зимовках взрослых жёлтых цапель, родившихся в Азербайджане, получен-
ные в результате их мечения, таковы. 13 сентября годовалая птица обнаружена в Эфиопии.
Во второй половине января взрослая цапля убита под Нахичеванью; в феврале — в дельте
Тигра-Евфрата и, как ни странно, в Дании; в марте — невдалеке от г. Сари в Иране. Отме-
чены нерегулярные зимовки жёлтых цапель на западе Марокко, в Ираке, Иране, Персид-
ском заливе.
Миграции. В низовья Дона, на Зап. Маныч и в Вост. Приазовье жёлтые цапли прилета-
ют во второй половине апреля — начале мая. Первые птицы на лиманах Ахтарско-Гривен-
ской системы в 1964 г. отмечены 16 апреля, в 1969 г. — 25 апреля, в лесу Круглик в 1970 г. —
27 аперля, в 1971 г. — 8 мая, на Веселовском вдхр. в 1986 г. — 16 апреля. На приморских
низменностях в районе Адлера их регистрировали в конце апреля. Пролёт здесь завершает-
ся обычно к началу июня (8 июня 1982 г.). В первой декаде июня отдельные особи встреча-
ются в среднегорье: 10 июня 1935 г. и 7 июня 1981 г. одиночки встречены близ Гузерипля.
Возможно, эти особи пересекли Главный Кавказский хр. со стороны Черноморского побе-
режья (Птицы Северного Кавказа, 2004).
В Ставропольском крае первых птиц тоже встречали в конце апреля - начале мая. Стай-
ки по 5-7 птиц отмечали в окрестностях Ставрополя на озёрах Вшивом и Кравцовом, на
Новомарьевском лимане в 1978-1980 гг. с 20 апреля по 15 мая. В 1981 г. 12 птиц встречены
5-7 мая на оз. Б. Тамбукан в окрестностях Пятигорска; в 1982 г. — 26 сентября на рыбораз-
водных прудах у с. Птичье. Весенний пролёт жёлтых цапель в дельте Терека наблюдали с
конца марта до конца апреля. Наиболее ранние встречи цапель в местах гнездовий в дельте
Днестра относятся к середине апреля. У Лебяжьих о-вов наиболее ранних птиц наблюдали
20 апреля (Костин, 1983).
Во время весенних миграций основная масса приазовских и северо-кавказских жёлтых
цапель летит вдоль Черноморского и Каспийского побережий Кавказа, часть птиц пересе-
кает Кавказский хребет. Жёлтая цапля, окольцованная птенцом в Вост. Приазовье, в возра-
сте трёх лет встречена 3 мая у Апшеронска; взрослая птица, окольцованная птенцом в дельте
Терека, добыта 13 мая в Грузии (Сапетин, 19786).
В Средней Азии первые жёлтые цапли появляются в последней декаде марта, однако
регулярно начинают встречаться с конца апреля. Один из миграционных путей идет, види-
мо, из юго-восточного Каспия по долине Сумбара и далее через Копетдаг; возможно, имен-
но таким путем цапли попадают в низовья Аму-Дарьи и Сыр-Дарьи. Пролёт идёт также
через п-ов Мангышлак, куда птицы попадают, пересекая Каспийское море либо с дельты
Терека, либо прямо из Азербайджана; первые птицы отмечаются на Мангышлаке в пер-
вой-второй декаде мая (Птицы Средней Азии, 2007).
В период осенней миграции жёлтые цапли из Азово-Черноморского региона, вероятно,
летят через Средиземноморье, как вдоль побережья, так и пересекая его акваторию. Впол-
не вероятно, что птицы затем пересекают Сахару, направляясь в Экваториальную Африку
(Cramp, 1977). Послегнездовые кочёвки начинаются сразу после вылета из гнёзд. С 14-17
августа до 8-12 сентября птицы регулярно встречаются по всей пойме низовий Дона. В
1979 г. в районе станицы Большовской (Волгодонский р-н) последняя жёлтая цапля встре-
чена 29 сентября. В Вост. Приазовье осенний пролёт наблюдали с конца августа до середи-
ны сентября (Винокуров, 1960). По данным кольцевания, молодые птицы из дельты Кубани
Жёлтая цапля
249
в течение всей осени держатся в основном близ мест своего рождения, улетая на юг через
Кавказ, Турцию, Ливан. Дальнейший их путь на зимовки неизвестен (Сапетин, 19786). На
осеннем пролёте на приморских низменностях в районе Адлера наблюдалась всего 2 раза:
8 сентября 1984 г. и 4 октября 1989 г. Осенний отлёт жёлтых цапель в дельте Терека растя-
нут и продолжается с конца августа до середины октября, а массовый — проходит в сентяб-
ре. Отдельных особей встречали в дельте Терека и в декабре. Со 2 сентября по 8 октября
молодых цапель отмечали в разных районах Сев. Кавказа, Армении и Азербайджана. Изве-
стен случай добычи жёлтой цапли зимой: 27 февраля в дельте Кубани в 60 км от места
рождения (Сапетин, 19786).
Большая часть среднеазиатских жёлтых цапель осенью мигрируют вверх по долинам
Мургаба и Теджена или через низовья Атрека, где первые мигрирующие к югу птицы на-
блюдаются уже с конца июля; осенний пролёт наблюдается также на Мангышлаке (Птицы
Средней Азии, 2007).
Местообитания. Жёлтые цапли предпочитают теплый климат, преимущественно тро-
пический и субтропический. Хотя гнездование регистрировалось на горных озёрах на уровне
до 2000 м, эти птицы все же предпочитают водно-болотные угодья в низменностях (Cramp,
1977). В гнездовой период жёлтые цапли заселяют преимущественно пойменные экосис-
темы, отдавая предпочтение зарослям кустарниковой ивы. Используют также тростнико-
вые ассоциации и древесные насаждения в пойменных лесах. Изредка гнездится в сухо-
дольных лесопосадках на акации (Русев и др., 2005). В последние годы отмечается замет-
ное стремление птиц к труднодоступным для человека участкам околоводных ландшафтов,
что обусловлено возрастающим антропогенным прессом (Русев и др., 2005). Кормовой
биотоп — берега и мелководья разнообразных стоячих водоёмов, краевые участки зарос-
лей околоводных растений, плавающие водные растения, берега проток и ериков; отмеча-
ются кормежки и на сухих лугах (Спангенберг, 1951; данные составителя очерка).
Численность. Число мест гнездовий и численность гнездовых пар жёлтой цапли на
протяжении двух последних столетий подвергались значительным колебаниям с общей
тенденцией к снижению. Изменения численности с середины XIX века по вторую полови-
ну XX века в палеарктической зоне ареала жёлтой цапли можно разбить на 4 периода (Jozefic,
1962):
- прямое сокращение численности. Снижение числа мест гнездовий было наиболее за-
метным в 1850-1920 гг. В это время сильно была развита мода на перья этих птиц. Числен-
ность в результате промысла упала не менее, чем на 85%;
- 1921-1940 гг. Продолжающийся количественный регресс в этот период обуславли-
вался главным образом мелиоративными мероприятиями;
- 1940-1960 гг. Стабилизация численности вида. Она происходила благодаря созданию
сети заповедных территорий, охране вида и началу его синантропизации;
- 1961-1969 гг. Бурный рост численности, одновременное восстановление ранее исчез-
нувших мест гнездования и возникновение новых. Это объяснялось внедрением мер по
охране экосистем и самого вида, а также адаптацией вида к жизни в антропогенных ланд-
шафтах — как в местах гнездования, так и на зимовках.
Наиболее значительные вековые колебания численности и распространения жёлтой
цапли происходили лишь вдоль широтной оси ареала. Максимальная концентрация вида (в
меридиональном разрезе) отмечается между 44 и 48° с.ш.; при этом более чем в 50% мест
гнездится только 6% особей вида (Jozefik, 1970). По сравнению с 1850 г., количество жёл-
тых цапель здесь увеличилось на 480%. За последние 100 лет область наибольшей плотнос-
ти населения вида переместилась из Венгерской низменности к северному побережью Чёр-
ного моря, а во второй половине XX столетия — в Ленкоранскую низменность (Jozefik, 1971).
250
Отряд Листообразные, Семейство Цаплевые
Определённая различными методами численность жёлтой цапли в Палеарктике даёт
следующие цифры: 16 400 пар в 165 пунктах гнездования в 1850-1900 гг., 6 800 пар в 115
пунктах в 1900-1920 гг., 6 000 пар в 80 местах гнездования в 1920-1940 гг. и 8 200 пар в 71
пункте гнездования в 1940-1960 гг. (Cramp, 1977). Согласно Хагемайеру и Блейру (Hage-
meijer, Blair, 1997), число гнездовых пар жёлтых цапель в последнем десятилетии XX в.
было таким: в Румынии — 1 900, Испании — 500, Италии — 400, Албании — 300, Греции —
280, Хорватии — 200, Франции — 150, прочих европейских странах — 50-70 пар. По рас-
четам этих авторов, на Украине гнездилось 270 пар, в России — 5 745 (5,5-6 тыс.) и в
Турции — 5 477 (3-10 тыс.).
В последнее десятилетие в Зап. Европе наибольшая численность жёлтой цапли зареги-
стрирована в Румынии (6 500 пар), Испании (1 000 пар), Сербии и Черногории (900 пар),
Греции (700 пар), Италии (650 пар), Болгарии (650 пар), Хорватии (500 пар), Венгрии (410
пар), Франции (130 пар), Македонии (150 пар). В незначительном количестве вид гнездит-
ся в юж. Португалии (10 пар), Албании (48 пар), Боснии и Герцеговине (12 пар), Словакии
(5 пар). Пока ещё в большом количестве гнездится в Турции (6 000 пар), (Birdlife International,
2004). По мнению Дилейни и Скотта (Delany, Scott, 2002), численность жёлтой цапли нача-
ла заметно возрастать во Франции, Испании, Италии только к 1990-м гг.
Численность жёлтой цапли на территории Вост. Европы и Сев. Азии испытывает значи-
тельные колебания по годам и отдельным районам. На Украине наиболее западные поселе-
ния вида отмечены в плавнях оз. Кагул, где численность цапли крайне низка и гнездится
она, количеством не более 10 пар, лишь в отдельные годы. Более крупные гнездовые скоп-
ления жёлтая цапля образует на оз. Картал. Здесь, в смешанных колониях голенастых и
веслоногих птиц, её численность иногда достигает 30 пар. Регулярно гнездится в небольшом
количестве (до 15 пар) на о. Б. Даллер в Измаильском р-не Одесской обл. (Потапов, 2001).
В Килийской дельте р. Дунай наиболее крупные поселения этого вида сосредоточены в
Дунайском биосферном заповеднике на о-вах Лебединка и Гнеушев. Число гнездовых пар
было самым высоким в 1990 г. — 125 пар (Жмуд, 1999). Далее к востоку до дельты р.
Таблица 15. Численность жёлтой цапли в дельте Днестра
(по: Русев и др., 2005)
Год Число пар Год Число пар Год Число пар
1972 ? 1986 100 2000 280
1973 312 1987 14 2001 350
1974 90 1988 90 2002 ?
1975 153 1989 140 2003 310
1976 370 1990 190 2004 210
1977 370 1991 190 2005 150
1978 400 1992 210
1979 505 1993 190
1980 500 1994 200
1981 510 1995 ?
1982 123 1996 250
1983 ? 1997 300 /
1984 ? 1998 215
1985 ? 1999 220
Примечание'. ? — птицы гнездились но учёт не проводился.
Жёлтая цапля
251
Днестр жёлтая цапля редка. В низовьях р. Кагильник её численность не превышает 10 пар
(Русев и др., 2005).
В дельте Днестра находятся одно из наиболее крупных гнездовых поселений жёлтой
цапли в Азово-Черноморского регионе Украины (Русев, 2003). В начале XX столетия чис-
ленность жёлтой цапли здесь была крайне низкой из-за её преследования с целью добычи
перьев. В середине XX века эти цапли стали уже довольно обычными (Назаренко, 1953). За
период наблюдений в последние 30 лет XX столетия (табл. 15) отмечены 2 спада численно-
сти — в 1974 и 1987 гг. Первый произошел после резкого, в течение одного сезона, есте-
ственного падения уровня воды, второй — в результате полного зарегулирования стока
плотиной Днестровской ГЭС. В обоих случаях была подорвана кормовая база вида (Русев
и др., 2005).
Небольшие колонии периодически формируются в тростниковых зарослях и на деревь-
ях в верховьях Тилигульского лимана, где количество жёлтых цапель в смешанных поселе-
ниях измеряется 5-10 парами (Русев, 2000а). Обычной была жёлтая цапля в дельте Днеп-
ра, где её численность колебалась в 1980-х гг. от 16 до 196 пар (Ардамацкая, 2005). В насто-
ящее время здесь насчитывают лишь 6 пар (Петрович, Щеголев, 2003).
В дельте Дона жёлтые цапли селятся на островах в устье протоки Б. Кутерьма, устраи-
вая гнёзда на деревьях в смешанной колонии цапель, больших бакланов и грачей. В 1976 г.
18 пар гнездилось в смешанной колонии квакв и серых цапель на оз. Лебяжьем близ г.
Азова, но в последующие годы это поселение исчезло. Его гибель нельзя не связать с по-
вреждением местообитаний птиц при проведении строительных работ. В 1981 г. около 100
пар жёлтых цапель гнездились в смешанной колонии цапель в пойменных лесонасаждени-
ях на юго-восточной окраине Ростова, а в 1983 г. около 60 пар поселились в тростниковых
зарослях одного из озёр в 5 км южнее, однако позже эта колония исчезла в связи со строи-
тельством здесь рыбхоза. В 1982 г. 10 гнёзд жёлтых цапель обнаружены в пойме Дона близ
г. Аксай в смешанной колонии на ивах в пределах оз. Монастырского. В 1986 г. здесь в
тростниках гнездилось 40-70 пар. В 1981 г. 5 пар этих цапель найдено в смешанной коло-
нии на прудах Новочеркасского рыбхоза, но позже этих птиц здесь не встречали. Случаи
гнездования жёлтых цапель в разные годы фиксировались в смешанных колониях в пой-
менных лесах в окрестностях станиц Багаевской, Николаевской (в 1979-80 гг. учтено около
100 пар), Мариинской (в 1979 г. учтено 10 пар, в 1980 г. — более 10 пар), Каргальской (в
1978 г. гнездилось 10 пар, в 1979 г. — до 20 пар) и, возможно, на одном из озёр близ стани-
цы Мелиховской (Птицы Северного Кавказа, 2004).
На Усть-Манычском вдхр. в прежние годы жёлтые цапли гнездились не ежегодно, одна-
ко их численность была довольно высокой — около 150 гнёзд, но в мае 1970 г. здесь встре-
чено только 2 птицы. На Веселовском вдхр. эта цапля тоже была немногочисленной: в 1977 г.
гнездилось 150 пар, в 1978 г. — 50-60, в 1979 г. — 16, в 1981 г. — 2, в 1982 г. — 5-6 пар, в
1987 г. — 10 пар.
До 1976 г. жёлтая цапля регулярно гнездилась на оз. Казинка, где в 1976 г. размножа-
лось 10-12 пар. В 1987 г. одна пара встречена в смешанной колонии в западной части Про-
летарского вдхр. В Калмыкии жёлтые цапли найдены на южных водоёмах: на Состинских
оз., в пойме Вост. Маныча, на оз. Лысый Лиман и в устье р. Калаус. Число гнездящихся
птиц здесь не превышало 40-60 пар. На Состинских озёрах, Лысом лимане и в пойме р.
Калаус жёлтые цапли гнездились в смешанных колониях с другими голенастыми, а в пойме
Вост. Маныча держались обособленной колонией (Птицы Северного Кавказа, 2004).
В Вост. Приазовье в 1953-1954 гг. жёлтые цапли гнездились в небольшом количестве.
В 1964-1969 гг. на Кочковатом и Ахтарском лиманах в дельте Кубани в тростниках обитало
от 32 до 72 пар, а на Черкасском лимане в Челбасских плавнях — от 32 до 110 пар. В 1979
252
Отряд Листообразные, Семейство Цаплевые
г. на лимане Горьком в Сладковско-Черноерковской системе водоёмов дельты Кубани гнез-
дилось около 250 пар, а в 1980 г. на пяти лиманах Ахтарско-Гривенской системы (Ахтарс-
ком, Бойкиевском, Солодко-Рясном, Кочковатом и Круглом) учтено 80 пар. В 1987 г. в сме-
шанных колониях на лиманах Ахтарско-Гривенской системы зарегистрировано 560 пар: на
Орлином — 50, Лимане Чайкины Плёсы — 30, Рясном — 300, Бакланьем — 30, Солодко-
Рясном — 150, а на водоёмах Сладковско-Черноерковской группы — 90 пар: на Прорвенс-
ком лимане — 60, Мечетном — 30. Всего здесь в 1980-1983 гг. обитало 470-500, а в 1987
г. — около 650 пар.
В условиях рисосеяния жёлтая цапля найдена также в лесных урочищах старой дельты
Кубани, где она гнездилась в смешанных колониях на деревьях. В лесу Круглик в 1970—
1971 гг. учтено 12 и 20, в 1983 г. — около 100, в 1984 г. — 20 гнезд; в лесу близ станицы
Анастасиевской в 1983 г. гнездилось 10 пар. Кроме того, в 1977 г. 2 гнезда этих птиц найде-
ны в колонии рыжих цапель на заросшем озере в пойме Кубани близ г. Кропоткина (Птицы
Северного Кавказа, 2004).
На Ставрополье поселения жёлтых цапель отмечены в 5 известных цапельниках: в Не-
винномысском пойменном лесу в междуречье Кубань - Барсучки в 1983 г. — 22 гнезда; в
Новотроицком пойменном лесу в смешанной колонии в 1978 г. — 32 гнезда, в 1985 г. — 113
пар; в Новодеревенском в 1984 г. — 14 гнёзд; в Кировском и Торосовском лесничествах в
1978 г. — 30, в 1981 г. — 17 гнёзд. В 1979 г. поселение жёлтых цапель из 11 гнёзд отмечено
в тростниковых зарослях на рыборазводных прудах близ с. Птичье, в 1981 г. здесь учтено
26, а в 1984 г. — 20 гнёзд (Бичерев, Хохлов, 1981).
В дельте Терека в 1973-1974 гг. этот вид зарегистрирован в 3 колониях, где гнездилось
около 1 300 пар, но в 1985 г. здесь не обнаружено ни одного поселения этих птиц. Согласно
учётным данным 1987 г., около 100 пар гнездилось в смешанных колониях в низовьях Те-
река, Средней и Прорвы, в Аграханском зал., на Ачикольских озёрах и оз. Караколь (Бонда-
рев, 19756; Птицы Севверного Кавказа, 2004).
Численность жёлтой цапли в дельте Волги в последние десятилетия испытывала резкие
колебания. До интенсивного увеличения обводненности водоёмов в дельте Волги насчиты-
вали до 5 200 пар жёлтых цапель (1987 г.). По мере дальнейшего возрастания здесь глубин,
её численность сократилась до 100 пар в 1994 г. (Кривенко, 1991). Однако в последние
годы количество жёлтой цапли относительно стабильно и колеблется в пределах от 400
(2004 г.) до 700 пар (2003 г.) (Гаврилов, 2005).
Современная численность вида в России оценивается в 1 500-2 000 пар (Мищенко и
др., 2004), из которых на Сев. Кавказе обитает 1 000-1 200 пар (Птицы Северного Кавказа,
2004). В Средней Азии численность жёлтой цапли невелика, причём за последнее столетие
она значительно снизилась (Птицы Средней Азии, 2007).
Размножение. Моногам; пары формируются на один сезон (Cramp, 1977). Гнездится
колониями, причём моновидовых колоний, по-видимому, не образует, поселяясь в смешан-
ных поселениях других цапель и ибисов. В Средней Азии только в немногих смешанных
поселениях доминирует по численности, в большинстве же случаев гнездится в таких по-
селениях небольшими группами (Птицы Средней Азии, 2004).
После прилёта птицы не сразу приступают к гнездованию, а формируют небольшие
подвижные скопления, которые держатся в богатых кормом биотопах невдалеке от буду-
щих колоний. Необходимыми условиями возникновения колониальных поселений жёлтых
цапель являются наличие удобных мест для гнездования и присутствие вида-организатора
смешанной колонии; таким видом-организатором, по крайней мере в условиях Сев. При-
черноморья, служит кваква. Жёлтые цапли устраивают гнёзда как на деревьях, так и в тро-
стниковых зарослях. Формирование пар сопровождается демонстрационным поведением
(рис. 49).
Жёлтая цапля
253
Материалом для постройки гнёзда на
деревьях обычно служат сухие веточки ивы
и тополя, стебли и листья тростника (если
деревья растут среди тростника). На лима-
нах эти птицы устраивают свои гнёзда на
заломах тростника, используя в качестве
строительного материала стебли, листья и
метелки последнего.
Гнёзда на деревьях, как правило, устра-
иваются в развилках ветвей не ближе 1 м
друг от друга, иногда на одной ветке птицы
строят несколько гнёзд. В строительстве
принимают участие оба партнёра, однако
веточки в основном поставляет самец, а сам-
ка их укладывает (Cramp, 1977). Материал
для гнезда птицы собирают недалеко от ко-
лонии. Лоток, если гнездо расположено на
дереве или в кустарнике, выстлан незначи-
тельным количеством тонких веточек ивы.
Постройка гнезда длится 6-8 дней (Sterbetz,
1960).
В дельте Волги жёлтая цапля устраива- „ ~ ...
Рис. 49. Самец желтой цапли в брачном опере-
ет гнёзда обычно в нижнем ярусе деревьев, нии в токовой позе (по Baue|. Glutz von Blotzheim>
ниже всех других цапель, но иногда на од- 1966).
ном уровне с кваквой, малой белой цаплей
и каравайкой (Доброхотов, 1961). В дельте Днестра жёлтая цапля занимает в основном
средний ярус кустарниковой колонии (1,2-1,5 м от уровня воды), хотя довольно часто мож-
но наблюдать гнёзда и в верхнем ярусе на высоте 2,5-3 м. В низовьях Кагальника (Татарбу-
нарский р-н Одесской обл.) цапли гнездятся на акации, на высоте 3,5-5 м. В дельте Дона
гнёзда располагаются на старых ивах во втором-третьем ярусах, чаще всего на одном уровне
с гнёздами малых белых цапель на высоте 3,5-6,5 м. Гнёзда жёлтых цапель в лесу Круглик
находили в нижних ярусах крон на высоте 4,5-6,3 м, изредка на одном уровне с гнёздами
квакв и малых белых цапель. Гнёзда в тростниках располагаются обычно ниже гнёзд дру-
гих видов цапель; гнездо имеет вид пологой чаши, расположенной чаще всего на высоте
20-60 см от поверхности воды.
Гнездо жёлтой цапли отличается от гнёзд всех других колониальных цапель небольши-
ми размерами. Диаметр гнёзд из дельты Волги 26-36 см, в среднем 30 см; высота гнёзда
14-31, в среднем 19 см; диаметр лотка 14-23, в среднем 19 см; глубина лотка 3-6, в сред-
нем 4,5 см (Доброхотов, 1961). В дельте Днестра размеры гнёзд (п=21) составляли: диа-
метр 22-32, в среднем 27 см; высота гнезда 12-29, в среднем 17 см (ориг.). На Лебяжьих о-
вах (п=29) диаметр гнезда составлял 20-23, в среднем 25,9 см; высота — 8-30, в среднем
13,5 см (Костин, Тарина, 2004). В дельте Дона (п=33) диаметр гнезда был равным 11-40, в
среднем 28 см; диаметр лотка — 10-19, в среднем 14 см; высота гнезда — 15-28, в сред-
нем 22 см; глубина лотка — 1-12, в среднем 7 см. Размеры гнёзд в древесной колонии
около Новотроицкого вдхр. (п=8): диаметр гнезда 27-37, в среднем 31,5 см; диаметр лотка
17-29, в среднем 21,8 см; высота гнезда 25-35, в среднем 28,3 см; глубина лотка 4-8, в
среднем 5,6 см. Размеры гнёзд жёлтых цапель из Вост. Приазовья (п=36): диаметр гнезда
18,7-25,3; диаметр лотка 8-10 см. Размеры гнёзд с оз. Лебяжьего (n=l 1): диаметр гнезда
254
Отряд Листообразные, Семейство Цаплевые
20,3-32, в среднем 24,7 см; диаметр лотка 9-22, в среднем 12,2 см; высота гнезда 10,3—
21,5, в среднем 16,6 см; глубина лотка 2,2-7, в среднем 5,5 см (Птицы Северного Кавказа,
2004).
Начало кладки яиц у жёлтых цапель в дельте Дона, на Веселовском вдхр. и в Вост.
Приазовье наблюдали во второй-третьей декадах мая, в некоторые годы — в начале мая.
Гнездовой период растянут, так как сроки прилёта разных групп птиц в одних и те же коло-
ниях не совпадают: в колониях одновременно встречаются строящиеся гнёзда, гнёзда со
свежими и насиженными яйцами, птенцами разного возраста. Так, 5 июня 1976 г. в коло-
нии на оз. Лебяжьем птенцы в возрасте 1-5 дней отмечены в 5 из 18 гнёзд; в одном — 2
только что вылупившихся птенца и 3 яйца; в 10 гнёздах было по 3-5 яиц разной степени
насиженности, в том числе в одном — наклюнутые яйца; в 2 гнёздах было 1 и 2 свежих
яйца; одно гнездо строилось. На о. Б. Дворян в дельте Дона 30 мая 1979 г. в гнёздах зареги-
стрированы первые птенцы и кладки разной степени насиженности. То же наблюдали 9-10
июня в колониях на Веселовском вдхр. В гнёздах на Ахтарском лимане в 1964 г. 20 июня
были кладки из 4-5 яиц, в некоторых гнёздах — птенцы в возрасте 3-7 дней. В 1965 г. в
этой же колонии жёлтые цапли загнездились позже — 25-30 июня; 30 июня в 6 из 12 гнёзд
было по 2, в двух — по 1 яйцу, а 4 гнёзда ещё строились. В 1966 г. начало откладки яиц в
этой колонии зарегистрировано 12 мая. В лесу Круглик в 1970-1971 гг. кладка началась 15-
30 мая (Птицы Северного Кавказа, 2004).
Число яиц в насиживаемых кладках на Лебяжьих о-вах — 5-6, в дельте Днестра — 4-5,
в Зап. Предкавказье — 3-7. Из 64 просмотренных кладок в 6,2 % было по 3 яйца, в 25% по
4, в 46,9% — по 5, в 18,8% — по 6 и в 3,1 % — по 7 яиц. Полные кладки этих птиц в Центр.
Предкавказье состояли из 4-5 яиц, в среднем 4,8 яйца на гнездо.
Яйца жёлтой цапли довольно мелкие, они обычно удлинённой формы, их окраска зеле-
новато-голубая, со временем значительно выцветающая. В дельте Волги (п=25) они имели
Таблица 16. Возрастные изменения размеров (мм) и веса (г) птенцов жёлтой цапли
(усредненные данные; п= 20) (по Доброхотову, 1961)
Возраст в днях Вес Длина тела Длина клюва Длина цевки Длина крыла
1 16,5 36,2 12,0 12,3 12,2
2 18,2 38,4 12,4 15,9 14,1
3 28,4 41,5 14,6 20,8 19,8
4 40,2 53,1 18,9 23,2 22,6
5 60,5 57,3 22,1 25,3 23,0
6 80,1 60,9 22,7 29,1 28,4
7 85,4 63,4 24,1 31,0 36,7
8 103,8 68,2 25,2 33,1 45,4
9 109,6 72,5 29,1 37,3 55,1
10 110,3 76,5 31,2 40,2 60,0
11 121,5 78,4 31,3 43,4 67,1
12 126,7 80,9 32,5 44,1 75,2
13 130,4 84,1 32,9 44,8 84,1
14 145,2 88,8 36,1 49,3 92,5
15 155,1 92,4 39,1 53,0 110,2
16 164,2 95,1 43,5 55,1 135,2
17 171,6 96,1 44,1 56,0 140,5
18 178,3 96,8 45,2 56,6 152,1
19 180,4 98,1 45,9 56,9 158,6
20 185,7 98,5 46,5 57,0 165,5
21 (слетки) 225 120,5 57,1 59,0 205,5
Жёлтая цапля
255
длину 43,3-37,0 (в среднем 38,9) мм и ширину 31,5-27,2 (в среднем 28,4) мм. На Северном
Кавказе в дельте Дона (п=46) размеры яиц составляли 35,2-42,0 х 26,5-30,0, в среднем
38,7x28,3 мм; на лиманах Вост. Приазовья (п=49) — 35-41x24-31, в среднем 38x29,7 мм;
в лесу Круглик (п=24) — 32,0-40,3 х 28,3-29,5, в среднем 38,2x28,9 мм; на Ставрополье
(п=19) — 36,0-42,1 х 26,0-29,1, в среднем 38,7x28,0 мм. На Лебяжьих о-вах в Крыму
(п=152) — 32,8x41,3, в среднем 36,2x28,2 мм; в дельте Днестра (Одесская обл.) (п=30) —
35,7x41,1, в среднем 37,2x28,1 мм (ориг.). В Туркмении (п=8) — 37-42,3 х 26-28,5, в сред-
нем 39,5x27,6 мм (Эминов и др., 1984). Вес яиц в среднем 10-13 г.
Продолжительность насиживания в среднем составляет 24 дня. Насиживают оба парт-
нёра. Насиживающая птица сидит на гнезде довольно плотно и слетает лишь при значи-
тельном шуме и беспокойстве. Массовое вылупление птенцов в дельте Днестра происхо-
дит во второй половине июня. Вылупившиеся птенцы совершенно беспомощны. Скорлупа
яиц сбрасывается родителями прямо под гнездо. Изменение размеров и оперения птенцов
по мере роста и развития последних представлены в таблицах 16 и 17.
До 17-20-дневного возраста, т.е. до тех пор, пока птенцы находятся в гнезде, там же
происходит и их кормление. В выкармливании птенцов принимают участие как самка, так
и самец. Подлетевшая взрослая птица садится на дерево рядом и некоторое время осматри-
вается, после чего медленно спускается в гнездо, перебираясь с ветки на ветку. Птенцы
Таблица 17. Возрастные изменения оперения, окраски
и поведения птенцов жёлтой цапли
Возраст, Описание птенцов:
в днях___________________________________________________________________________
1-3 Верхняя часть спины покрыта редким буроватым пухом, на голове буро-
коричневый хохолок. Окраска голых частей тела зелёная. Поясница и надхвостье
покрыты белым пухом. Клюв серо-розовый; плюсна и голень зеленовато-жёлтая.
Птенцы беспомощно лежат в гнезде; постоянно пищат.
4-6 На шее и темени коричневый пух. На спинной и плечевой птерилиях пеньки 2
мм; появляются пеньки маховых. Брюхо тёмно-зелёное, покрыто очень редким
белым пухом. Птенцы молча лежат в гнезде, на ноги ещё не встают.
7-9 Пробиваются пеньки на голени и шее (до 2 мм). Начинается развертывание пера
на шейной, спинной и плечевой птерилиях. Пробиваются пеньки рулевых;
пудреток ещё не видно. Птенцы становятся на ноги, но из гнезда не выходят.
10-12 Пеньки голени пробились на 4 мм, перо развернуто на шейной птерилии до 2 мм,
перо кроющих крыла — до 3 мм, перо на спинной и плечевой птерилии — до 7
мм. Маховые развернуты до 2 мм. На спине, плечах и шее коричневое перо, на
пояснице — белое, пуховое. Яйцевой бугорок есть. Пудреток еще не видно.
Потревоженные птенцы пытаются убежать или защищаются.
13-15 Продолжается развертывание пеньков на головной, шейной, брюшной
птерилиях. Маховые перья до 1 см. На спине сплошной покров из перьев. Кожа
— тёмно-зелёная, лапы — жёлто-зелёные. Радужна глаз - буро-зелёная. Места,
где появляются пудретки — более светлые и слегка припухшие. Птенцы все
время стоят в гнезде или на ближайших ветках.
16-18 Весь птенец покрыт ровным перьевым покровом. Маховые белые.
Первостепенные маховые с серыми окончаниями. Рулевые перья развёрнуты,
брюхо, грудь и поясница покрыты буроватым пером. Пудретки уже появляются.
Птенцы сидят на ветках у гнезда.
19-25 Все перья хорошо развёрнуты, кроме нижних кроющих крыла. Яйцевой бугорок
отделяется. Пудретки функционируют. Птенцы могут перепархивать с ветки на
ветку. Птенец отличается от взрослой птицы более тёмной буро-коричневой
окраской. Потревоженные птенцы могут улететь из колонии.
256
Отряд Листообразные, Семейство Цаплевые
поднимаются на ноги, пищат и теребят клюв родителя. После этого взрослая птица запус-
кает свой клюв в клюв птенца (иногда наоборот) и отрыгивает пищу. Освободившись от
принесенного корма, взрослая птица отлетает на ближайшие открытые ветви и чистится ми-
нут 20, иногда улетает сразу. После ухода родителя с гнезда, птенцы быстро успокаиваются.
Количество приносимой пищи изменяется в зависимости от возраста птенцов. Птенец
в возрасте 1-3 дней получает 1,5-2,5 г пищи за одно кормление, в возрасте 6-9 дней — 3-
5 г, а в возрасте 11-15 дней — 6-8 г пищи. Отрыжки полулетных птенцов весят 20-25 г.
Видовой состав пищи не зависит от возраста птенцов, а изменяется только в зависимости
от наличия многочисленного и доступного корма. Суточная активность взрослых птиц в
период выкармливания птенцов имеет три пика: утром в 6-7, днем в 14-15 и вечером в 19-
20 ч. При этом каждый из родителей совершает около трёх кормлений в день. За один
прилет взрослая птица кормит, как правило, не более трёх птенцов.
Птенцы прекрасно приспособлены к жизни в зарослях: они очень цепки, подвижны.
Однако если птенец по какой либо причине падает в воду или на землю, ему не всегда
удаётся взобраться в своё гнездо. Птенцы, сидящие в гнезде, делают попытки защиты (от
наблюдателя) только в возрасте 9-10 дней; более молодые защищаться ещё не могут, стар-
шие — стремятся убежать. Защищаясь, птенец с резким гортанным криком выбрасывает
вперёд голову, пытаясь ударить врага клювом.
Вылет птенцов происходит в возрасте 25-35 дней, начинается во второй декаде июля и
тянется до середины августа.
Успешность размножения жёлтых цапель в условиях Лебяжьих о-вов достаточно высо-
кая — 90,6-95,6% при показателях эмбриональной смертности 1,75-9,38% и ювенильной —
2,67-9,09%. Большая часть погибших яиц (40-70%) приходится на неоплодотворённые.
Часть яиц расклевывают чайки-хохотуньи, а птенцы гибнут, сходя с гнёзд вследствие бес-
покойства птиц в колонии людьми (Костин, Тарина, 2004). Гибель птенцов в первую неде-
лю жизни в дельте Днестра много выше и составляет 25-30%. Они погибают от нападения
ворон и болотного луня. Гибнут преимущественно младшие птенцы, поскольку они полу-
чают меньше пищи и погибают от истощения или бывают задавлены старшими. По этой
причине гибнет от 40 до 50% птенцов, родившихся последними в выводке. Ещё одной
важной причиной гибели птенцов является выпадение их из гнезда. При этом, падая, они
повреждают себе неокрепшие кости, попадают в развилки ветвей, в воду. Однако с птенца-
ми жёлтой цапли это случается реже, чем с другими видами цапель, так как гнёзда их рас-
положены преимущественно в среднем, нижнем ярусах и реже всего — в верхнем. К тому
же птенцы жёлтой цапли довольно цепкие, что позволяет им эффективно взбираться в гнездо.
Из гнезда с 5 яйцами в дельте Днестра в среднем вылетает 3 птенца (ориг.).
По результатам меченья выяснено, что смертность молодых до 1 года составляет около
68%, более взрослых — 21-29%. При этом относительная смертность на 2-6-м годах жиз-
ни составляет около 24%, а впоследствии снижается. Максимальная продолжительность
жизни 13-14 лет. Средняя продолжительность жизни 1,5 года, а для птиц, переживших
первый год — 4 года (Сапетин, 19786).
Суточная активность, поведение. Жёлтые цапли активны преимущественно в свет-
лое время суток, но кормятся также и в сумерках. Общественная птица во все сезоны, хотя
вне периода размножения и в местах кормежки нередко встречается поодиночке. Свобод-
ные от насиживания птицы ночуют вблизи своих гнёзд. Первые вылеты за кормом отмече-
ны с восходом солнца, однако наиболее активно вылет из колонии можно наблюдать спустя
час после его восхода, когда за кормом улетает более 50% птиц данной колонии.
Питание. Важная кормовая стация вида, по крайней мере в дельте Днестра — плаваю-
щие водные растения (кувшинки, кубышки, водяной орех др.), заломы тростника и ежего-
Жёлтая цапля
257
ловника. На плавающих растениях жёлтые цапли не встречают конкурентов среди других
видов, так как подобный субстрат не выдерживает веса более крупных цапель. Во время
кормежки жёлтые цапли держатся как одиночно, так и небольшими группами до 20-30
особей. В таких скоплениях птицы придерживаются определённой дистанции — от двух и
более метров друг от друга. В период половодья предпочитают пойменные луга; сюда их
привлекает множество мальков рыб и головастиков, личинок водных насекомых, дожде-
вых червей. Активно используются массивы рисовых чеков и других мелководий глубиной
10-15 см, где скапливается много молоди рыб, водных насекомых, головастиков, лягушек.
Кормовыми стациями служат также берега проток и ериков, небольшие островки, появив-
шиеся после спада воды, берега пересыхающих озёр и стариц, пруды рыбхозов, сырые
луга. Подстерегающая добычу птица обычно стоит неподвижно, реже медленно бродит,
выслеживая её. Схватывает цапля добычу быстрым колющим движением.
Жёлтые цапли предпочитают кормиться недалеко от колонии — на расстоянии в 1,5-2
км. Заметно реже они встречаются в отдалении от гнездовий. Однако в период маловодных
лет и при искусственном регулировании стока жёлтые цапли из-за отсутствия кормовых
биотопов могут улетать за 8 и более километров от колонии.
В питании жёлтых цапель можно выделить четыре группы кормов: рыбы; земновод-
ные, водные насекомые и наземные насекомые. В дельте Волги рыбы составляют 29,76%
от общего количества добытых пищевых объектов, земноводные (в основном лягушки) —
20,16%, водные насекомые — 48,69%, наземные насекомые — 1,39%. В рацион жёлтых
цапель, по данным Доброхотова (1961), входит 34 вида животных. В дельте Днестра вод-
ные насекомые в питании жёлтых цапель в гнездовой период доминируют и составляют
70% общего числа жертв (в основном это личинки водолюбов и плавунцов). На втором
месте — головастики и мелкие лягушки — 21%; незначительную долю занимают мальки
рыб (6%), дождевые черви (2%) и наземные насекомые (1%). Во внегнездовой период ос-
нову питания составляют головастики и мальки рыб. Имаго же и личинки водных насеко-
мых занимают подчиненное положение.
В дельте Дона значение каждой из групп кормов в рационе жёлтых цапель в разные
годы неодинаково, что определяется доступностью и обилием кормов (табл. 18).
Из анализа 56 пищевых проб в дельте Дона и 57 на лиманах Вост. Приазовья следует,
что птицы, гнездящиеся на естественных водоёмах, отдают предпочтение земноводным,
водным насекомым и их личинкам. Из водных насекомых на обоих водоёмах они поедали
личинок и имаго жуков-плавунцов и водолюбов, водных клопов-плавтов и гладышей, ли-
чинок стрекоз. Видовой состав рыб в рационе жёлтых цапель из дельты Дона и лиманов
Вост. Приазовья несколько отличался. В дельте Дона цапли поедали как сорных, так и про-
мысловых рыб. Доля промысловых в их рационе в 1981 г. составила 60%, в 1982 г. — 38,8%
Таблица 18. Состав пищи жёлтой цапли в дельте Дона (Птицы Северного Кавказа, 2004)
Кормовые 1981 г. (п=23) 1982 г. (п=33) 1981- (п=56)
объекты По весу Встреча- По весу Встреча- 1982 гг. По весу Встреча-
(%) емость (%) (%) емость (%) (%) емость (%)
Земноводные 48,9 86,8 54,6 81,8 51,8 83,9
Рыбы Насекомые и 20,1 65,1 10,6 54,5 15,4 58,9
их личинки 31,0 100,0 34,8 93,9 32,9 96,4
258
Отряд Листообразные, Семейство Цаплевые
от всех съеденных рыб. На лиманах в пищевых пробах этих цапель обнаружены красно-
пёрка, линь, малая южная колюшка, щиповка, бычки.
Основой корма жёлтой цапли в местах рисосеяния составляют наземные насекомые и
низшие ракообразные (в основном, щитень). Из насекомых в пробах пищи были встречены
стрекозы, сверчки, саранчовые, долгоносики, жужелицы, листоеды, коровки, щелкуны,
муравьи. Однако среди них преобладали медведки (30,5% по весу), число которых в одной
пробе было от 1 до 17 (в среднем, 4). Из водных насекомых встречали личинок жуков-
плавунцов и водолюбов. Земноводные были представлены краснобрюхими жерлянками и
озёрными лягушками. Из рыб в составе корма встречена только щиповка. Рисовые зерна
(1% встречаемости) были, видимо, проглочены случайно. В желудках жёлтых цапель из
Аграханского залива в июне 1966 и 1967 гг. встречены медведки, уховертки, жуки, кобыл-
ки, божья коровка (Птицы Северного Кавказа, 2004).
В юж. Туркмении по данным анализа содержимого 16 желудков, рыбы (9 видов) состав-
ляли 25,82% от числа съеденных животных, земноводные (головастики и озёрные лягуш-
ки) — 12,64%, насекомые — 61,54%. Среди рыб преобладала гамбузия, среди насекомых —
водяные жуки и их личинки, а также личинки стрекоз (Ташлиев, 1973).
Враги, неблагоприятные факторы. По данным кольцевания, важнейший фактор смер-
тности лётных птиц — охота: из 90 возвратов колец от жёлтых цапель, помеченных в бывш.
СССР, 67 (три четверти) убито охотниками, 8 — «найдено», 4 — поймано, 1 погибла, раз-
бившись о провода. Обстоятельства гибели остальных 20 птиц не указаны. Основная масса
возвратов колец получена в августе-сентябре, т.е. в период наиболее интенсивной охоты
(Сапетин, 19786).
Пернатые хищники также способны значительно увеличить гибель жёлтых цапель. На-
блюдения показали, что в поселениях, не подверженных влиянию антропогенных факто-
ров, успешность размножения составляет 3,5 птенца на пару. Из типичных хищников и
животных, проявляющих хищнические наклонности и способных оказать существенное
влияние на численность вида, следует отметить болотного луня, серую ворону, енотовид-
ную собаку, серую крысу (дельта Днестра, ориг.).
Наибольшее количество гнёзд подвергается разорению пернатыми хищниками (вклю-
чая серую ворону) после возникшей тревоги, особенно во время посещения колоний чело-
веком, когда часть птиц долго не возвращаются на свои гнёзда. Тогда смертность птенцов
до подъёма на крыло может достигать 80%, в том числе и по причине того, что птенцы, уже
способные передвигаться, пытаясь спастись, падают в воду и не могут возвратится в гнездо.
На успешность размножения существенное влияние оказывают возникающие по вине
отдыхающих пожары в плавнях и пойменных лесах, а также аномальные половодья. На-
пример, в 1998 г. в дельте Днестра из-за повторяющихся высоких и нерегулярных подъе-
мов воды несколько сотен цапель разных видов были вынуждены гнездиться повторно и
даже в третий раз. В результате отрицательного влияния половодья больше всего гнёзд
пострадало у жёлтой цапли (82,1%) и каравайки (75,0%) (Русев и др., 2005). Эти два вида
гнездятся преимущественно в нижнем ярусе колоний, расположенных на кустарниковых
ивах, и поэтому первыми подвергаются негативному влиянию подтопления. Повторные же
гнездования значительно менее результативны.
В прошлом существовал довольно широко практиковавшийся неконтролируемый сбор
яиц цапель, что иногда приводило к исчезновению даже крупных колоний в дельтах Днес-
тра и Дона.
В случае недостатка естественного корма, взрослые птенцы жёлтых цапель могут по-
едать сородичей, выступая в качестве типичных каннибалов.
Хозяйственное значение, охрана. Прямого хозяйственного значения не имеет. Явля-
ясь сочленом водно-болотных экосистем, играет в них весьма заметную роль, представляя
Белокрылая цапля
259
собой важнейшее звено в трофических цепях. Поскольку жёлтая цапля очень чутко реаги-
рует даже на незначительные перемены в осваиваемых ею угодьях, она может быть ис-
пользована как индикатор таких изменений.
Устраивая гнёзда в плотных, многовидовых и многочисленных колониях голенастых
птиц, жёлтая цапля играет ведущую роль в циркуляции ряда арбовирусных инфекций, пе-
редаваемых от особи к особи через комаров и трансовариально (Львов, Ильичёв, 1979;
Русев, 2004; Греков и др., 2005). Немаловажна её роль как промежуточного звена в разви-
тии гельминтозов. У жёлтой цапли найдено 20 видов паразитических червей (Корнюшин и
др., 2004).
Образуя единое поливидовое сообщество, все виды мелких цапель, включая жёлтую,
подлежат безусловной охране. Режим охраны должен предусматривать минимальное ант-
ропогенное вмешательство. Места гнездовий жёлтых цапель целесообразно охранять в
рамках охраны ландшафтов и типичных местообитаний.
Жёлтая цапля занесена в Красную книгу Украины, Казахстана; в России — в Красные
книги Астраханской области и Республики Ингушетия. Охраняется в заповедниках Азер-
байджана, России и Украины.
Белокрылая цапля
Ardeola bacchus (Bonaparte, 1855)
Buphus bacchus Bonaparte, 1857, Consp. Generum Avium, 2, c. 127, п-ов Малакка.
Chinese pond-heron (англ.), Bacchusreiher (нем.), crabier chinois (фр.).
Син.: Ardeolaprasinosceles Swinhoe, 1860.
Статус. Залётный вид. Возможно нерегулярное гнездование.
Общая характеристика и полевые признаки. Мелкая цапля, сходная по сложению с
жёлтой цаплей, от которой отличается более контрастной окраской из сочетания тёмных и
белых тонов. В брачном наряде несколько длинных узких перьев затылка образуют хохол;
на плечах и верхней части спины хорошо заметны украшающие волосовидные перья, дос-
тигающие вершины хвоста. На нижней части горла и по бокам верхней части груди —
длинные рассученные украшающие перья. Кормящаяся птица движениями напоминает еги-
петскую цаплю. В случае опасности скрывается в кронах деревьев или в высокой траве, где
и затаивается, низко приседая и втягивая голову в плечи. Полёт довольно быстрый, с резки-
ми взмахами крыльев.
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Половой диморфизм отсутствует. Самец и самка в брачном
наряде. Голова с затылочным хохлом, шея и украшающие перья горла рыже-каштановые;
верхняя часть спины и плечевые, включая украшающие перья чёрно-аспидные с фиолето-
во-зелёным металлическим блеском; удлинённые перья по бокам верхней части груди чёр-
ные. Большие верхние кроющие крыла дымчатые; подбородок, полоса вдоль горла и ос-
тальное оперение белое с жёлтым оттенком. Клюв жёлтый с чёрной вершиной и голубова-
тым основанием надклювья; голая кожа лица и основание подклювья зеленовато-жёлтые;
радужина жёлтая. Ноги бледно-оранжевые с розоватой голенью и буровато-зеленоватыми
когтями.
260
Отряд Листообразные, Семейство Цаплевые
Самец и самка в зимнем наряде. Зимой украшающие перья на затылке, плечах, спине,
горле и боках груди отсутствуют. Голова, шея и горло буровато-охристые с широкими чёр-
но-бурыми краями опахал; плечи и верхняя часть спины коричнево-бурые, на плечах с уз-
кими беловатыми наствольными полосками; на боках верхней части груди по рыжевато-
бурому пятну с узкими охристыми наствольными полосками; внутренние второстепенные
маховые бурые. Ноги оливково- или зеленовато-жёлтые, клюв желтоватый с более тёмным
коньком, радужина тускло-жёлтая.
Гнездовой наряд. Как у взрослых зимой, но стержни первостепенных маховых перьев
тёмные, наружные опахала и концы маховых с дымчатым налётом и пестринами. Надклю-
вье, режущий край и вершина подклювья тёмнорогового цвета, остальная часть подклювья
желтоватая; ноги зеленовато-жёлтые.
Возрастная изменчивость окраски проявляется в исчезновении пестрин на голове и шее
к первой весне жизни; клюв становится желтоватым с бурой вершиной. На вторую весну
жизни от взрослой в брачном наряде отличается буроватым налётом на маховых перьях
крыла.
Строение и размеры. Формула крыла I<II>III>IV>V............Размеры (мм) самцов
и самок: длина крыла — 185-238, длина клюва — 54-69, длина цевки — 57-64; масса
(п=9): 220-380 г (Shaw, 1936); по А.И. Иванову (1961), масса (п=9): 170-300 г.
Размеры (мм) взрослых птиц (п=3) из Приморья: длина крыла — 204,214,228, клюва —
59,7, 62,8, 63,2, цевки — 58,6, 64,0, 63,2, хвоста — 84,6, 78,6, 90,2. Масса самца из Примо-
рья — 229,5 г (по материалам коллекции зоомузея ДВГУ).
Рис. 50. Область распространения белокрылой цапли: а — область гнездования, б — районы зимо-
вок, в — недостоточно выясненная граница распространения, г — дальные залёты.
Белокрылая цапля
261
Линька. Две линьки в году; вероятно, пол-
ная линька в зимний наряд и весенняя линька в
брачный наряд проходит на местах зимовок и
частично в период пролёта. В это время сменя-
ется контурное перо.
Птицы, добытые в декабре, январе и марте
на Малайском п-ове (Robinson, Kloss, 1911), а
также 3 и 6 марта в Лаосе (Robinson, 1931),
были в зимнем наряде. Самец от 31 марта из
Юньнани (колл. ЗИН РАН) интенсивно линял,
на голове, шее и спине преобладали перья зим-
него наряда. У самца от 20 мая из Приморья
интенсивно росли перья головы, шеи, спины;
от зимнего наряда сохранились единичные пе-
рья, а у самца от 12 мая заканчивался рост не-
которых эгреток на груди, а на верхней части
спины сохранилась небольшая группа перьев
прежнего наряда (Назаров, 2004).
Рис. 51. Распространение белокрылой цапли в
России: а — область регулярных встреч птиц;
б — залёты.
Распространение. Гнездовой ареал. Гнездится в Индокитае, Ассаме, Бирме и
Андаманских о-вах (del Hoyo et al., 1992), в Китае — от верхнего течения р. Сунгари, цен-
тральной части Внутренней Монголии до южных границ, на запад — включая провинции
Шаньси, Сычуань, Юньнань (Cheng Tso-hsin, 1987). Отмечена в Юж. и Сев. Корее без ука-
зания на факты гнездования (Wong, 1996; Томек, 1999) (рис. 50).
В России регулярно встречается в южной части Приморья и по восточному побережью
края (рис. 51). Одна птица добыта 27 мая у с. Сидеми (Taczanowski, 1891-1893), две — 1
июня 1911 г. у п. Славянка (Бутурлин, 1911), одна 20 мая 1926 г. в устье р. Илистой (Иоган-
зен, 1928), молодая самка добыта 30 мая 1967 г. в зал. Петра Великого на о. Б. Пелис (Лаб-
зюк и др., 1971). Три одиночные птицы отмечены в мае 1972 г. и 1973 г. у пос. Хасан, по
одной — 18 июня 1957 г., 18 мая 1958 г. и 5 июня 1972 г. у пос. Ольги (Назаров, Лабзюк,
1975), одна птица — 8 июня 1976 г. на р. Змеиной и одна — 24 июня 1976 г. на рисовых
полях восточного побережья оз. Ханка (Глущенко, Шибнев, 1977). Группа из 3 птиц встре-
чена в бухте Нарва 17 апреля и одиночка — 4 августа 1976 г. (Глущенко, 1979). Одна бело-
крылая цапля встречена 25 мая в долине р. Джигитовки, одна — 29 и 31 мая у пос. Пластун,
одна — 29 мая и 6 июня 1976 г. у пос. Терней (Елсуков, 1977). Остатки взрослой птицы
найдены в 1971 г. в бассейне р. Бикин (Поливанова, Глущенко, 1977). В окрестностях бух-
ты Ольга одиночек отмечали в приустьевых участках лесных речек и на морском побере-
жье 15-24 мая 1976 г., 13-14 июня 1977 г., 12-17 июня 1978 г., 11 и 13 июня 1980 г. и трёх
птиц — 23 мая 1980 г. (Лабзюк, 1981). У пос. Преображение пара встречена 15 июня 1981 г.,
в одной из бухт Лазовского заповедника одиночку наблюдали 26 июня 1981 г. (Коломийцев,
1986). В районе с. Рязановка одна птица в промежуточном наряде кормилась на солонова-
том озере 20 июня и одна в брачном — на лугу 25-28 июня 1983 г. (Назаров, 1986); здесь же
7 июня 1984 г. держалась на лугу одна птица и пара пролётела вечером на юго-восток, а 8
июня 1984 г. пролетела группа из 9 птиц. В зал. Петра Великого на о. Б. Пелис одиночных
особей регистрировали 19-20 мая, 31 мая и 10 июня 1979 г., одна птица пролетела над о.
Де-Ливрона 15 июня 1980 г., на о. Рикорда две птицы кормились на болоте 7 мая 1990 г., а
на ночь улетели в лес. Одна птица отмечена у ст. Рязановка — 26 мая 1981 г. (Назаров,
2004). Одиночные птицы в промежуточном наряде отмечены 20 июля 1999 г. в устье р.
Кедровая и 4 июля 2004 г. у п. Пограничный (Курдюков, 2004; О.А. Бурковский, ориг.).
262
Отряд Листообразные, Семейство Цаплевые
За пределами Приморья белокрылая цапля отмечена в долине р. Амур: в Амурском ли-
мане (Спангенберг, 1951), по одной птице добыто у Хабаровска в 1984 г., 18 мая 1975 г. и у
с. Малышево 13 октября 1983 г. (Смиренский, 1986; Бабенко, 2000). Самец-второгодок до-
быт на оз. Сопочное (север о. Итуруп, Курильские о-ва) 9 июля 2004 г (коллекция ЗМ МГУ)
(рис. 51). Указывается залёт в район Кяхты (Спангенберг, 1951), что вызывает некоторые
сомнения (Иванов, 1976).
Наконец, две молодые особи встречены в октябре 2004 г. на южных склонах хр. Боркол-
дой в Центр. Тянь-Шане (Киргизия), одна из них добыта (Остащенко, 2005; Рустамов, Ков-
шарь, 2007).
Зимовки. Зимует в Юж. Китае, Индокитае, на Малаккском п-ове, на о-вах Борнео,
Суматре, в Японии, о-вах Рюкю (del Hoyo et al., 1992).
Залёт зарегистрирован в долину р. Бура на границе Монголии и СССР (Фомин, Болд,
1991).
Миграции. Северные популяции совершают миграции. В Китае на местах гнездования
белокрылые цапли появляются в марте-апреле, и улетают в сентябре-октябре. В Индии
совершает локальные кочёвки (del Hoyo et al., 1992). Судя по встречам птиц на морском
побережье и островах зал. Петра Великого в Приморье, весенняя миграция белокрылой
цапли проходит с середины апреля до начала июня, наибольшее количество птиц встрече-
но во второй половине мая.
Местообитания. В Юго-Вост. Азии придерживается преимущественно культурного
ландшафта у стоячих водоёмов, держится на болотах, по берегам рек, на влажных участках
листопадных лесов. Не избегает мангровых зарослей. В горы поднимается до высоты 1400
м над уровнем моря (del Hoyo et al., 1992; Robson, 2000). В Приморье её отмечали в доли-
нах рек и на лугах морского побережья.
Численность и её изменения. Точных сведений по численности нет. В середине XIX
столетия и в начале 1900-х гг. в местах гнездования была многочисленна или обычна (Irby,
1861; Lloyd, 1873; Kershaw, 1904; La Touche, Rickett, 1905; La Touche 1907, 1924; Delacour,
Jabouille, 1925; Delacour, 1928, 1929; Deignan, 1945). В настоящее время в местах гнездова-
ния обычна, что позволяет считать, что её численность остается, по-видимому, на прежнем
довольно высоком уровне. Одна из наиболее обычных цапель в Вост, и Юж. Китае (Cheng
Tso-hsin, 1987). В Гонконге отмечено 100-150 гнездящихся пар, обычна во Вьетнаме и Та-
иланде. На о. Борнео в последние годы зарегистрировано сокращение численности этих
птиц (del Hoyo et al., 1992).
Размножение. Гнездится на деревьях, часто в парках, на высоких бамбуковых изгоро-
дях в деревнях, в бамбуковых зарослях. Селится в смешанных колониях совместно с други-
ми видами цапель. Сроки размножения растянуты с мая по август. Образование пар и нача-
ло строительства гнёзд в Китае приходится на первую декаду мая (Li Yang-hsin, Liu Xi-ye,
1963). В строительстве гнезда, которое продолжается около недели, участвуют оба партнё-
ра. Гнездо располагается на вершине дерева или вблизи от нее в 16-19 м от земли, на бам-
буке в 3 м от земли и ниже. Гнездо представляет собой подобие блюдца. Материалом для
него служат сухие ветки, тонкий бамбук, иногда лоток выстилается небольшим количе-
ством свежих листьев и травы. Размеры (мм) гнёзд (п=4): диаметр гнезда 270-400, высота
65-160, диаметр лотка 140-280, глубина лотка 25-55.
Самка откладывает по одному яйцу через день. Кладка содержит 3-6 яиц. Размеры яиц
(п=50): 34,0-39,2 х 26,7-30,1, в среднем 37,7x28,4 мм (Baker, 1929); (п=17): 37,5-42,0 х
29,0-30,0, в среднем 39,5x29,5 мм (La Tauche, 1931-1934); (n=14): 38,0-41,8 х 29,6-31,5, в
среднем 39,5x30,7 мм (Li Yang-hsin, Liu Xi-ye, 1963); (n=70): 36,5—42,0 x 27,0-30,2, в сред-
нем 38,4x29,0 мм (Schonwetter, 1960). Окраска яиц однотонная, цвет скорлупы зеленовато-
Индийская прудовая цапля
263
голубой. К насиживанию белокрылая цапля приступает после откладки первого яйца. Птенцы
появляются через 18-22 дня с начала насиживания. Выкармливают птенцов оба партнёра.
В юго-вост. Китае наиболее поздних молодых птиц встречали в гнёздах до 4 сентября
(Vaughan, Jones, 1913). Молодые приобретают способность к полёту в возрасте 30-33 дней.
В Приморье у двух самцов, добытых 12 и 20 мая, семенники были сильно увеличены и
достигали размеров 10x6 и 18x9 мм соответственно (ориг.). Регулярность встреч белокры-
лый цапли в южной части Приморья на протяжении всего периода размножения, встречи
пар на пролёте, а также состояние взрослых птиц (готовы к размножению), позволяет пред-
полагать возможность гнездования этого вида в Приморье.
Суточная активность. Деятельна в светлое время суток. На кормежку птицы улетают
за 3-5 км от места ночёвки. В период выкармливания птенцов наиболее активна в утренние
часы и после полудня.
Питание. Корм собирает на берегах водоёмов и на низкотравных лугах. В питании птен-
цов преобладают лягушата, головастики, мелкая рыба, реже встречаются водные насеко-
мые и прямокрылые (Li Yang-hsin, Liu Xi-ye, 1963). В Приморье на о. Б. Пелис в желудке
одной птицы обнаружены 4 медведки (Gryllotalpa africana), у другой — остатки рыбки (не
определена) и жука-листоеда.
Хозяйственное значение и охрана. В Китае птицы и яйца традиционно используются
в пищу (Vaughan, Jones, 1913; del Hoyo et al., 1992). Специально не охраняется.
Индийская прудовая цапля
Ardeola grayii (Sykes, 1832)
Ardea Grayii Sykes, 1832, Proc. Zool. Soc. London, Pt.2, no 23, c. 158, Декан, Индия
Indian Pond Heron (англ.), Paddyreiher (нем.), Heron crabier de Gray (фр.)
Син. (рус.): восточная жёлтая цапля
Статус. Редкий, нерегулярно залётный вид.
Общая характеристика и полевые признаки. Небольшая цапля (длина тела 18-20
см, размах крыльев 75-90 см), почти идентична по размерам жёлтой цапле, на которую
внешне похожа. Во всех нарядах отличается от жёлтой цапли более тёмной, тёмно-корич-
невой спиной и плечевыми перьями, которые в полёте резко контрастируют с белыми кры-
льями. У сидящей птицы голова светлее спины, особенно это заметно у птиц во взрослом
оперении. В брачном наряде на голове длинные белые перья-косицы, которые ниспадают
на спину и резко выделяются на фоне тёмной спины. Клюв, в отличие от жёлтой цапли, не
голубой, а жёлтый с тёмной вершиной. Окраска самцов и самок сходная (Cramp, 1977).
Держится одиночно или неплотными группами. В целом, индийская прудовая цапля
менее скрытная, чем жёлтая цапля, и для неё характерен более широкий спектр местооби-
таний. К человеку относится толерантно (Cramp, 1977; del Hoyo et al., 1992).
Голос — резкий, высокий по тону крик. В колониях, кроме того, постоянно слышны
крики «ува-куу», перемежающиеся коротким кваканьем (Hancock, Kushlan, 1984).
Подвидовая систематика. Монотипический вид.
Распространение. Юг Ирана, Пакистан, Индия, Непал, Шри-Ланка, Бангладеш, Мьян-
ма, Мальдивские, Никобарские и Андаманские о-ва. В местах обитания ведёт оседлый об-
раз жизни (Cramp, 1977; del Hoyo et al., 1992) (рис. 52).
264
Отряд Листообразные, Семейство Цаплевые
Рис. 52. Область распространения индийской прудовой цапли: а — область обитания; б — залёт на
территорию Сев. Евразии.
Единственный залёт вида на территорию бывш. СССР зарегистрирован 16 июля 2009 г.
Взрослая птица в брачном, судя по имеющимся фотографиям Андрея Коваленко, наряде
встречена на озере-накопителе сточных вод Сорбулак в 80 км северо-западнее Алма-Аты,
Республика Казахстан (Коваленко, Дякин, 2009; www//birds.kz/Ardeola grayii/index.html).
Местообитания. Разнообразные пресные и солёные водоёмы в тропической и субтро-
пической зонах: реки, озёра, водохранилища, мангровые заросли; предпочитает, однако,
небольшие стоячие водоёмы. Населяет главным образом низменности, однако может под-
ниматься в горы до абсолютной высоты 2 150 м. Нередко поселяется в населённых пунктах
и даже в городах (Cramp, 1977; del Hoyo et al., 1992).
Численность. Вид обычен на всём протяжении ареала. В Индии эта цапля многочис-
ленна и относится к самым обыкновенным околоводным птицам (del Hoyo et al., 1992).
Размножение. Сезон размножения на севере Индии продолжается с мая по сентябрь,
на юге Индии и в Шри-Ланке — с декабря по май.
Колониальный вид, поселяется небольшими моновидовыми колониями или совместно
с другими цаплями и бакланами; в последнем случае обычно занимает краевую зону сме-
шанной колонии. Иногда гнездится отдельными парами. Гнёзда устраивает на деревьях
или кустах на высоте от 2 до 16 м. В местах, где птиц не беспокоят, цапли гнездятся из года
в год в одних и тех же колониях.
В кладке 3-6 яиц, их окраска при откладке сине-зелёная, но быстро выцветает. Размеры
яиц в среднем 38x28,5 мм. Насиживают оба партнёра, длительность инкубации 24 дня.
Длительность периода выкармливания птенцов не известна. Птенцов родители первое вре-
мя кормят, отрыгивая пищу (Hancock, Kushlan, 1984; del Hoyo et al., 1992).
Суточная активность, поведение. По-видимому, птицы активны как в светлое, так и в
темное время суток — по крайней мере, кормиться этот вид может как днём, так и ночью
(del Hoyo et al., 1992). Полёт и поведение сходно с таковым у жёлтой цапли, однако соци-
альность у индийской прудовой цапли выражена в большей степени, и для этого вида ха-
рактерны коллективные ночёвки (Cramp, 1977).
Египетская цапля
265
Питание. В спектре питания отмечены мелкая рыба, лягушки, крабы и другие ракооб-
разные, водные и наземные насекомые. Кормятся цапли на илистых берегах прудов и ка-
нав, на рисовых полях и грязевых литоралях, в том числе, в мангровых зарослях. Цапли
подстерегают добычу, стоя неподвижно на грязи, мелководье или сидя на присаде над во-
дой, либо перемещаются медленными шагами (Hancock, Kushlan, 1984; del Hoyo et al., 1992).
Враги, неблагоприятные факторы. Нет данных.
Хозяйственное значение, охрана. Как залётный вид, в Сев. Евразии хозяйственного
значения не имеет. Специальные меры охраны не предусмотрены.
Bubulcus Bonaparte, 1855
Египетская цапля
Bubulcus ibis (Linnaeus, 1758)
Ardea ibis Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Египет.
Cattle egret (англ.); Kuhreiher (нем.); heron garde-boeuf (фр.).
Син. (лат.): для подвида ibis'. Ardea lucida Rafinesque, 1810; Ardea veranii Roux, 1825; Ardea
bubulcus Audouin, 1826; Ardea russata Wagler, 1827; Bubulcus ruficrista Bonaparte, 1885;
Ardeola ibis seychellarum Salomonsen, 1934; для подвида coromandus'. Cancroma coromanda
Boddaert, 1783; Ardea coromandelensis Stephens, 1819; Ardea affinis Horsfield, 1820; Ardea
flavirostris Vieillot, 1823; Ardea coromandelica M.H.K. Lichtenstein, 1823; Ardea caboga
Franklin, 1832.
Син. (рус.): рыжеголовая цапля (устар.), коровья цапля (перевод с англ.)
Статус. Гнездящийся, мигрирующий, изредка зимующий вид.
Общая характеристика и полевые признаки. Некрупная цапля, несколько крупнее
вороны и жёлтой цапли, но меньше кваквы. В полете примерно одинакового размера с
малой белой цаплей. Клюв и шея относительно короткие. Голова большая. Общая окраска
белая, с желтовато-рыжими, охристыми перьями на голове, спине и груди. Ноги бледно-
оранжевые. Клюв яркий, оранжево-жёлтый, с малозаметным красноватым оттенком снизу.
Молодые и не размножающиеся птицы целиком белые, с тёмно-бурыми ногами. Клюв у
молодых птиц серовато-чёрный, с жёлтым кончиком, у не размножающихся особей — це-
ликом жёлтый.
Малопуглива (доверчива). Часто держится на влажных лугах по окраинам деревень, в
местах выпаса коров и буйволов. Полёт лёгкий, сравнительно медленный, как правило,
невысоко над землей.
Голос. Египетская цапля сравнительно молчалива. Голос — глухие каркающие звуки
«корр», «коок», «коак»; реже — сравнительно звонкие «керр», «кхее». Молодые птицы бо-
лее крикливы и издают разнообразные звуки.
От малой белой цапли отличается короткой шеей, массивной головой и светлым клю-
вом. От жёлтой — обилием белого цвета в оперении, отсутствием тёмных перьев на голове
и светлым, ярким клювом.
Подвидовая систематика. Чаще всего выделяют два подвида, различающихся деталя-
ми окраски оперения и общими размерами (Vaurie, 1963; Степанян, 2003).
266
Отряд Листообразные, Семейство Цаплевые
1. Bubulcus ibis ibis
Ardea ibis Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Египет.
Западная часть ареала — Европа, Африка, Передняя Азия. В России и прилегающих
странах — низовья Волги и Урала, Предкавказье, Закавказье. В оперении верхней части
головы, шеи, спины и груди присутствует охристый оттенок; наиболее насыщенная окрас-
ка на темени.
2. В. L coromandus
Cancroma coromanda Boddaert, 1783, Table Planches Enlum., с.54, Коромандельский бе-
рег, Индия.
Восточная часть ареала — Индия, Индокитай и обширная зона от южной части Япон-
ского архипелага до Зондских о-вов и центрального побережья Австралии. В России —
Юж. Приморье. Голова, шея и верхняя часть груди оранжево-золотистые; спина винно-
золотистая. Размерами незначительно крупнее предыдущего (табл. 19).
Замечания по систематике. В последние годы восточному подвиду порой придается
ранг самостоятельного вида В. coromandus — восточной коровьей цапли (напр. Brazil,
2009). Иногда в качестве самостоятельного подвида B.L seychellarum (3) выделяют птиц с
Сейшельских о-вов, отличающихся от первых двух меньшими размерами и более золотис-
той окраской украшающих перьев (Cramp, 1977; Dickinson, 2003). Признания в системати-
ке, в том числе и отечественной, третий подвид не получил (Степанян, 2003).
Описание. Окраска. Самец и самка в брачном наряде белые, с желтовато-оранжевы-
ми, охристыми верхней частью головы, спиной и зобом. Эти участки тела покрыты удли-
нёнными, рассученными перьями, более длинными у самцов. Наиболее интенсивно окра-
шена верхняя часть головы. Клюв оранжево-жёлтый, с красноватым оттенком на нижней
части и у угла рта. Уздечка зеленовато-жёлтая. Радужина жёлтая с оранжевой каймой по
внешнему краю. Общая окраска ног желтовато-оранжевая. Цевка оранжевая, пальцы не-
сколько темнее. Неоперённые части голени желтоватые. В разгар брачного периода цвет
клюва меняется до ярко-красного с жёлтым кончиком, ноги становятся тёмно-красными,
уздечка меняет цвет до фиолетового, а радужина — до алого.
Окраска восточного подвида в брачном наряде отличается распространением оранже-
во-золотистого цвета на всю голову, шею и зоб (без границ). Украшающие перья на спине
яркие, винно-золотистые, ноги более тёмные.
Таблица 19. Размеры египетских цапель (мм)
Параметр п Самцы Самки Источник
lim М n lim М
Длина крыла Bubulcus ibis ibis (для всего ареала подвида) 230-257 Степанян, 2003
Длина цевки Длина крыла 3 70-85 Низовья Аракса 235-255 247 2 242-249 245 Adamian & Klem, 1999
Длина цевки 3 81-83 82 2 74-81 77
Длина клюва 3 51-55 54 2 56-57 56,5
Длина крыла 20 Западная Европа 241-266 253 20 240-258 248 Vaure, 1963
Длина цевки 20 70-85 77 20 70-81 76
Длина клюва 12 Западная Европа 52-60 52-58 Witherby et al., 1939
Длина крыла Bubulcus ibis coromandus (для всего ареала подвида) 240-265 Степанян, 2003
Длина цевки 80-91
Египетская цапля
267
Самец и самка в зимнем наряде. Оперение снежно-белое, на голове сохраняется неболь-
шое бледно-рыжее пятно. Ноги становятся темными, буроватыми с жёлтыми участками на
голени и верхней части цевки, жёлтыми подошвами (Спангенберг, 1951), клюв неяркий
желтоватый, радужина жёлтая.
Гнездовой наряд. Молодая птица вся чисто белая, с проявляющимся позже на голове
оранжевым оттенком. Клюв серовато-чёрный, с жёлтой вершиной. Ноги темные, почти
чёрные, за исключением более светлых, с зеленоватым оттенком сочленений голени с цев-
кой. Кожа вокруг глаз свинцово-серая, радужина грязно-белая.
Кожа вылупившихся птенцов оливково-зелёная, позже становится серой. Птенцы в гнёз-
дах покрыты белым пухом (Спангенберг, 1951), клюв телесного цвета, ноги сероватые.
Строение и размеры. В сводке Стэнли Крэмпа (Cramp, 1977) приведены
сведения о размерах птиц номинативного подвида для Зап. Европы без разделения на поло-
вые группы, суммарно для подросших молодых и взрослых птиц. Они таковы: длина крыла
(п=23), в среднем 265 (244-281) мм; хвоста (n=l 1) в среднем 91,4 (82-101) мм; длина
клюва (п=21) в среднем 83,8 (79-89) мм; цевки (п=19) в среднем 89,8 (82-94) мм. Размеры
для отдельных половых групп и для обоих подвидов, встречающихся на пространстве Во-
сточной Европы и Сев. Азии, представлены в табл. 19.
Вес самцов и самок в Зап. Палеарктике колеблется в пределах 300^400 г. На юге Афри-
ки — от 325 до 387 г. У молодых птиц (40 дней) вес достигает 350 г (Cramp, 1977).
Линька. Полная послебрачная линька проходит с июля по ноябрь, реже продолжается
до декабря. Смена мелкого оперения взрослых особей начинается у гнёзд в 10-х числах
июля, в это же время птицы начинают терять украшающие перья. Затем, в течение осени,
происходит смена крупного пера. Частичная предбрачная смена пера происходит с февра-
ля до мая. К середине февраля у птиц появляются украшающие перья на голове. У молодых
частичная линька происходит в первую календарную осень жизни — мелкое оперение сме-
няется между сентябрем и декабрем. Первые украшающие перья появляются весной вто-
рого календарного года (Спангенберг, 1951; Cramp, 1977).
Распространение. Гнездовой ареал. Западный подвид’, южная Европа (Португа-
лия, Испания, Франция, о-ва Сев. Италии) и Сев. Африка (Марокко, Тунис, Египет). Вся
тропическая и субтропическая Африка к югу и востоку от Сахары, Мадагаскар, Юж. и Зап.
Аравия, Малая Азия (Палестина, Израиль, Турция, Иран) (рис. 53).
Из Суринама, где впервые вид был отмечен между 1877 и 1882 г., широко расселилась
на север по территории Центр. Америки, США и южных провинций Канады. На юг египет-
ская цапля проникла здесь до юго-вост. Бразилии и северной половины Аргентины (Weber,
1972; Hilty, Brown, 1986).
В Азербайджане спорадично гнездится в низовьях Куры и Аракса, в Ленкоранской низ-
менности (Patrikeev, 2004). Встречается и на водоёмах вдоль побережья Каспия, но гнездо-
вание здесь не доказано.
В Армении гнездится в долине Аракса (Ляйстер, Соснин, 1942) и по его притокам иног-
да встречается до бассейна Севана (Даль, 1954). В последних сводках египетская цапля
указывается только для низовьев Аракса (Adamian, Klem, 1999).
В Туркмении первый залёт отмечен под Ашхабадом осенью 1927 г., а птица с наседным
пятном и в брачном наряде была добыта в июне 1950 г. на оз. М. Делили (Дементьев и др.,
1955; Караваев, 1991). Впервые гнездование вида установлено в 1964 г., в долине Теджена
(Эминов, 2002). В настоящее время египетская цапля нерегулярно гнездится в низовьях
Атрека, долине Теджена и в окрестностях Ашхабада (Эминов, 2002).
В Казахстане до второй половины 1970-х гг. были известны лишь единичные залеты
(Долгушин, 1960), а в настоящее время египетская цапля несомненно гнездится в низовьях
Урала. Впервые гнездование отмечено здесь в 1990 г. (Березовиков, Гисцов, 1999).
268
Отряд Листообразные, Семейство Цаплевые
Рис. 53. Область распространения египетской цапли в Восточном полушарии: а — области гнездова-
ния; б — места зимовок. Подвиды: 1 — В. i. ibis, 2 — В. i. coromandus, 3 — В. i. seichellarum.
В России поселения египетской цапли известны для нижнего течения Терека и дельты
Волги. На Латышском о-ве Нового Терека египетская цапля была найдена в смешанной
колонии голенастых. В 1970 г. здесь, вместе с кваквой, малой белой и жёлтой цаплей гнез-
дилось 56 пар египетской цапли, однако в 1973 г. обнаружено только 3 гнезда (Бондарев,
1975, 1977; Пишванов, 1975), а в 2003 г. вид здесь не встречен (ориг.). В Дагестане в насто-
ящее время египетская цапля, предположительно, гнездится на оз. Юж. Аграхан (окрест-
ности с. Нов. Терек) и на Ачикольских озёрах (недалеко от станицы Александрийской).
Возможно гнездование также на Каракольских озёрах и в устье р. Средней, в южной части
Кизлярского зал. В гнездовое время одиночных птиц встречали также в низовьях р. Кумы.
В дельте Волги гнездится на Трёхизбенском участке Астраханского заповедника. Первые
данные о гнездовании вида в дельте поступали в 1950 и 1951 гг. С 1976 г. встречи этих
цапель в Астраханском заповеднике стали регулярными (Русанов, 1999; Русанов и др., 2000).
В 2004 г. египетская цапля в гнездовой обстановке отмечена в низовьях Кубани, на По-
нурском лимане (Р.А. Мнацеканов, личн. сообщ.).
Залёты египетской цапли известны для Сев. Осетии (Спангенберг, 1951) и Чечни. В
северо-западной части Крыма одиночная птица, сидевшая в поле, встречена 14 мая 1949 г.
(Костин, Тарина, 2005). Известен случай залёта в Таджикистан. 12-13 июля 1960 г. Дваж-
ды встречена цапля на берегу реки Ак-Су в районе Кизыл-Рабата на Памире (Абдусалямов,
1971).
В Зап. Палеарктике залёты египетской цапли отмечены по всей Европе, включая Анг-
лию, скандинавские страны, Прибалтику, Белоруссию и Украину (Snow, Perrins, 1998). Пти-
Египетская цапля
269
цы, гнездящиеся в Зап. Палеарктике, зимуют в Передней Азии, Африке и южной части
Пиренейского п-ова.
Восточный подвид гнездится в Индии (к западу включительно до Сеистана), Индоки-
тае (до Сев.-Вост. Китая), в Японии, на Филиппинских и Зондских о-вах. В последнее вре-
мя он заселил северные и центральные районы побережья Австралии (Mckillgan, 1990b;
Snow, Perrins, 1998). В пределах России встречается только в Юж. Приморье (рис. 53).
Ранее были известны залеты египетской цапли, в частности, на р. Сунгаче и в устье р.
Тачингоуза (Спангенберг, 1951). В Лазовском заповеднике одиночную египетскую цаплю
встретили в мае-июне 1960 г. в долине р. Судзухе около с. Киевка. Здесь она в течение
недели собирала корм, держась около табуна лошадей. Другая встреча произошла уже в
1962 г. в окрестностях п. Валентин. В 1960-1970-х гг. её четырежды регистрировали в
Сихоте-Алинском заповеднике (Суворов, 2004, стр. 176, 185). Впервые для территории
России гнездование восточного подвида египетской цапли было зарегистрировано в 1999
г. в Б. Лефинской колонии на оз. Ханка (Мрикот, Глущенко, 2000).
Зимовки. В северной части ареала, в том числе на юге России, по всему Прикаспию
и в Закавказье — типично перелётная птица. Основные зимовки египетских цапель Запад-
ной Палеарктики расположены на юге Пиренейского п-ова (Испания, Португалия), в Сев.
Африке (Алжир, Марокко, Египет, Эфиопия) и на юге Передней Азии (Иран, Ирак). В от-
дельные годы птицы могут зимовать на юго-востоке Азербайджана. В частности, 4 цапли
отмечены в январе-феврале 1996 г. в Кызыл-Агачском заповеднике (Patrikeev, 2004). В ни-
зовьях Куры, на оз. Сарысу 16 января 2004 г. держалось 6 птиц (Султанов, Джаббарова,
2006). Указание на встречи зимующей птицы под Ленкоранью есть также у Г.И. Радде (1885).
Северные популяции восточного подвида зимуют, видимо, в восточном Китае и Юго-
Вост. Азии (Cramp, 1977).
Миграции. Даты весеннего прилёта и отлёта птиц точно не установлены. По опросным
данным на юге Азербайджана появляются в конце марта и, реже, в самом начале апреля, но
достоверно первые птицы отмечены с 11 апреля и позднее. Осенний отлет, предположи-
тельно, ранний, и в октябре в Азербайджане египетская цапля не встречается (Спангенберг,
1951). В Кызыл-Агачском заповеднике появляется в марте-апреле, а отлетает в основной
массе в сентябре. Отдельные птицы задерживаются до ноября-декабря и, вероятно, зимуют
(Греков, 1965). На оз. Акгёль отмечается весной с апреля (Туаев, 1975). В Армении наиболее
ранние встречи египетской цапли зарегистрированы 11 апреля 1994 г. у р. Ахуриан, а самая
поздняя — 10 октября 1994 г. около Арташана (Adamian, Klem, 1999). В Юго-Вост. Прикас-
пии первых птиц отмечали весной в конце апреля. В августе-сентябре цапли отлетают, но в
1981 г. они задержались здесь до конца октября, что было связано с высокой численностью
саранчовых и непересыханием разливов в летние месяцы (Караваев, 1991). По данным Е.
Вилкова (2000) в окрестности г. Махачкалы египетскую цаплю отмечали на весеннем про-
лёте с третьей декады марта до первой декадя мая включительно. Но указания автора на
встречи вида в марте, возможно, ошибочны. Численность мигрирующих птиц по тем же
данным колеблется от 12 до 120 особей. В устье р. Самур за время наблюдений в 1970-
1980-х гг. она отмечена только один раз: 21 мая 1989 г. Одиночная птица кормилась на
залитом водой поле с редкими посадками сосны недалеко от берега моря (Бутьев и др.,
1990). В Астраханском заповеднике встречается с апреля по сентябрь (Русанов и др., 1999).
Можно предположить, что в Закавказье первые птицы появляются в конце марта - нача-
ле апреля, а в Юго-Вост. Прикаспии, Дагестане и дельте Волги — в середине апреля. Из
Сев. Прикаспия осенью птицы отлетают до конца сентября. В Юго-Вост. Прикаспии мест-
ные и пролётные птицы могут задерживаться до второй половины октября, а в Закавказье
одиночные египетские цапли в тёплые годы зимуют.
270
Отряд Листообразные, Семейство Цаплевые
Местообитания. В континентальной части Азербайджана гнездовой биотоп — неболь-
шие группы высокоствольной ветлы, чаще перелески и рощи широколиственных пород
(дуб, грецкий орех, тутовник, карагач), обвитые колючими и другими лианами и разбро-
санные среди рисовых полей, болот или водохранилищ. Нередко птицы гнездятся среди
крупных участков широколиственного леса, расположенных поблизости от кормовых ста-
ций. Последними служат, главным образом, рисовые поля и, в меньшей степени, луга, бо-
лота, подсыхающие водохранилища, берега озёр и речек, протекающих в открытой мест-
ности. В период размножения, да и в другое время, птица не избегает близкого соседства с
человеком, гнездясь и кормясь у самых селений. В поисках пищи охотно держится среди
стад буйволов, часто отдыхая и кормясь на спинах этих животных (Спангенберг, 1951). В
Кызыл-Агачском заповеднике гнездятся в смешанных колониях голенастых и веслоногих в
зарослях тамариска вдоль побережья и на мелководьях опресненных участков залива, к
которым прилегают обширные полупустынные пространства и заболоченные земли (Гре-
ков, 1965).
В Армении населяют мелководные и слабопроточные участки речных долин, окрестно-
сти рыборазводных прудов и прочие водно-болотные угодья долины Аракса, с редкой дре-
весной растительностью и обширными лугами (Adamian, Klem, 1999).
В Дагестане — небольшие широколиственные леса по берегам и на островах нижнего
течения Терека, водоёмы с широкими плавнями и древесно-кустарниковыми зарослями.
Обязательное условие — наличие недалеко от мест гнездования обширных приплавневых
лугов и солончаковых полупустынных участков (пастбищ) или рисовых полей.
В дельте Волги цапли придерживаются ивовых лесов по берегам проток в нижней (при-
морской) части дельты, где гнездятся в смешанных колониях веслоногих и голенастых (Г.М.
Русанов, личн. сообщ.). В Юго-Вост. Прикаспии предпочитают озёра с хорошо развитой
надводной растительностью и затопленными зарослями, с прилегающими влажными со-
лончаковыми и сухими лугами, разливами Атрека (Караваев, 1991).
В Юж. Приморье — берега озёр с лесной растительностью и обширными сырыми лугами
и пастбищами (Глущенко и др., 2003; Суворов, 2004). В Японии — небольшие водоёмы с
лесистыми островами или берегами, окружённые рисовыми полями (Fujioka, Yamagishi, 1981).
Численность. На большей части ареала, от экватора до субтропиков, египетская цапля
обычный, местами многочисленный вид. Однако на северных границах ареала, в умерен-
ной зоне, она распространена спорадично, больших колоний не образует и является ред-
ким видом. Численность вида в Европе Хагемайером и Блейером (Hagemeijer, Blair, 1997)
оценивалась в 69,5-87,3 (в среднем 77,8) тыс. пар. Из них в Испании гнездилось 60 000,
Португалии — 17 500, Франции — около 350, Италии — 50 пар. Количество египетских
цапель, гнездившихся в те же годы в России было 30-50 (в среднем 39), в Турции — 1-50
(в среднем 7) пар. Данные численности, приводимые Европейским Советом по учёту птиц
таковы: в Европе в начале XXI в. было 88-95 тыс. пар египетских цапель. Из них до 69 тыс.
пар гнездилось в Испании, 15-20 тыс. в Португалии, 4-5 тыс. пар во Франции и от 2 до 500
пар в Италии. В Турции — до 50 пар, в Греции и Румынии — единичные пары (Bird Life
International, 2000).
На юге России численность может быть оценена в 50-100 пар (Бондарев, 1977; Русанов
и др., 1991, 2000; Джамирзоев и др., 2000). Из них в Астраханском заповеднике (Трехиз-
бенский участок) гнездится до 16 пар; обычно от 2 до 6 пар (Русанов и др., 2000; Г.М.
Русанов, личн. сообщ.). В Дагестане в 1970 г. только на одной колонии в низовьях Терека
гнездилось 56 пар, в 1973 г. там гнездилось всего 3 пары (Бондарев, 1977). В 2001 г. в
окрестностях с. Новая Коса, на юго-западной окраине Аграханского зал., учтено 17 птиц, а
гнездовая численность египетской цапли на Юж. Аграхане оценена в 8-10 пар. Столько же
Египетская цапля
271
может гнездиться на Ачикольских озёрах (Джамирзоев, Исмаилов, 2004). Небольшие ко-
лонии вида могут также размещаться в сев. части дельты Терека — на Каракольских озёрах
и южной половине Кизлярского зал. Численность вида в Дагестане может быть оценена в
20-25 пар, хотя реально она подвержена сильным колебаниям, и, вероятно, птицы гнездят-
ся не ежегодно (Джамирзоев и др., 2000).
В Азербайджане современная численность составляет 3-6 тыс. пар, сосредоточенных
главным образом в Кызыл-Агачском заповеднике и на озёрах Махмуд-Чал а и Аггёль. По-
скольку в 1956-1958 гг. только в трёх колониях Кызыл-Агачского заповедника насчитыва-
ли около 4-5 млн. веслоногих и голенастых птиц, из которых 19% составляли египетские
цапли (Греков, 1965), численность этого вида в середине 1950-х гг. могла быть оценена в
4CW5 тыс. пар (Patrikeev, 2004). Следовательно, за 50 лет произошло её десятикратное
снижение.
В Армении гнездовая численность не известна. Судя по данным встреч вида в 1994 г., в
долине Аракса может гнездиться от 10-15 до 50 пар. В частности, 4 июля 1994 г. около
Шахомяна было отмечено 204 птицы (Adamian, Klein, 1999).
В Туркмении гнездится, вероятно, не более 100 пар. В Юго-Вост. Прикаспии египетс-
кая цапля более обычна в многоводные для Атрека годы. Так, на оз. М. Делили в 1980-1982
гг. гнездилось, соответственно, 15, 50 и 28 пар. В другие годы цапля была редка или же
птиц на гнездовании не отмечали вовсе (Караваев, 1991). На Ашхабадском озере в 1982 и
2001 гг. гнездились единичные пары (Эминов, 2002).
Численность восточного подвида в пределах России (на Приханкайской низменности)
варьирует. В 1999 г. Б. Лефинской колонии обнаружено 3 гнезда (Мрикот, Глущенко, 2000).
В 2000 г. здесь размножалось 2 пары (Глущенко, Мрикот, 2001), в 2001 г. — 8-10 пар, а в
2002 г. их число сократилось до 6 пар (данные Ю.Н. Глущенко). В 2003-2005 гг. как в выше
указанной, так и в других колониях птицы достоверно не наблюдались, хотя многократно
регистрировались в гнездовой период у южной оконечности оз. Ханка. В 2007 г. египет-
ские цапли гнездились уже в другой колонии в количестве 15-17 пар (Коробов, Глущенко,
2008).
Размножение. Египетская цапля приступает к размножению в возрасте 2 лет (Литвино-
ва, 2001). Моногам, но для вида характерны внебрачные и промискуитетные связи самок и
самцов (Fujioka, Yamagishi, 1981; McKillgan, 1990b). У 19 индивидуально помеченных пар
в колонии на юго-востоке Квинсленда (Австралия) приблизительно 30% успешных копуля-
ций были «внебрачными» (McKillgan, 1990b). Моногамность поддерживается благодаря
тому, что до начала кладки яиц самец более 78,8% времени находится около гнезда и защи-
щает самку, которая склонна к спариванию с любым самцом, появляющимся около гнезда.
Лишь после откладки яиц, когда самки слабо реагируют на самцов, их напарники больше
времени проводят вне колонии. В это время увеличивается частота спариваний с «чужими»
самцами. Более агрессивные и доминирующие самцы находятся у гнёзд дольше, и интен-
сивнее спариваются со своими партнёрами и соседними самками (Fujioka, Yamagishi, 1981).
Формирование пары сопровождается разнообразными демонстрациями (рис. 54).
Египетская цапля — колониальная птица, преимущественно гнездящаяся на деревьях,
реже — в тростниковых крепях и на кустах тамариска. Предпочитает лиственные породы,
но известно гнездование и на хвойных деревьях, в частности, на японской чёрной сосне
(Pinus thunbergii) в центр. Японии (Fujioka, Yamagishi, 1981). Обычно гнездится в смешан-
ных колониях голенастых и веслоногих (с малой белой и жёлтой цаплями, кваквой, серой
цаплей, каравайкой, колпицей, а также с большим и малым бакланами).
В дельте Терека гнездилась совместно с малой белой и жёлтой цаплями, кваквой. В
Астраханском заповеднике кроме цапель, в колонии гнездятся и бакланы. На юге Азербай-
272
Отряд Листообразные, Семейство Цаплевые
Рис. 54. Демонстрационное поведение египетской цапли (по Cramp, 1977): а, б — элементы ритуаль-
ного поведния при ухаживании, в — рекламное поведение самца, г — взаимная чистка оперения
(аллопрининг).
джана, в смешанной колонии вместе с цаплями гнездились также колпицы и каравайки. На
Ширванской равнине и Ленкоранской низменности, известны моновидовые колонии еги-
петской цапли.
В смешанных колониях на деревьях египетская цапля устраивается в среднем и нижнем
ярусах, где гнёзда располагаются бок о бок с гнёздами малой белой и жёлтой цапель, тогда
как основная масса других видов в колонии гнездится выше (Спангенберг, 1951). В коло-
нии голенастых и веслоногих в Кызыл-Агачском заповеднике гнездится на кустах тамарис-
ка в среднем, реже в верхнем ярусе (Греков, 1965). В смешанной колонии в тростниках на
оз. Махмуд-Чала (Юж. Азербайджан) египетские цапли строили гнёзда на разной высоте, не
образовывая моновидовых скоплений и не занимая какую-то определённую часть колонии.
Они, вместе с жёлтыми цаплями, селились как по периферии колонии, так и в её центре.
На деревьях гнёзда располагаются на высоте до 8-10 м от земли, количеством от 3-5 до
50 на одном дереве (Сатунин, 1907). На заломах тростника гнёзда могут находиться на
высоте до 1,5 м. Расстояние между ними составляет от 20-50 см до 1 м (Патрикеев, 19916).
Египетская цапля
273
Особенно густо заселёнными оказываются деревья, стволы и ветви которых обвиты
колючими лианами, что позволяет птицам укреплять гнёзда не только на ветвях, но и среди
лиан. Трава под такими деревьями, их стволы, ветви и листья густо покрыты помётом.
Такие деревья редко выдерживают два-три гнездовых сезона и, как правило, гибнут. Гнёз-
да птицы строят в разветвлении трёх и более веток. Вновь построенное гнездо имеет ха-
рактерную для цапель форму перевёрнутого конуса с радиально расходящимися из верши-
ны веточками. Оно настолько рыхло, что яйца видны с боков и снизу. Материалом служат
тонкие сухие веточки от 0,1 до 0,5 см толщиной. Обычно материал для гнезда птицы соби-
рают на земле в той же роще или вытаскивают из пустующих (а иногда и из жилых) гнёзд.
В результате этого среди птиц колонии происходят частые ссоры (Сатунин, 1907; Мензбир,
1918; Спангенберг, 1951).
Многие пары занимают прошлогодние постройки своего вида или гнёзда жёлтых и ма-
лых белых цапель. В таких случаях гнёзда «приводятся в порядок» и несколько достраива-
ются. Гнездо сооружается обеими птицами гнездовой пары, причём самец, как правило,
приносит материал, а самка укладывает его и защищает гнездо от соседей (Спангенберг,
1951).
Размеры гнёзд варьируют чрезвычайно сильно. Свежевыстроенное гнездо бывает срав-
нительно мало в диаметре, имеет большую высоту и глубокий лоток. С подрастанием птен-
цов гнездо сильно «растаптывается» ими, его стенки становятся пологими, поперечный
диаметр увеличивается, а лоток «мелеет». Особенно крупных размеров бывают прошло-
годние, достроенные в текущем году, гнёзда. Подстилки в гнёздах (лотках) нет.
Размеры гнёзд, промеренных под Ленкоранью, таковы: диаметр вновь выстроенного
гнезда с незаконченной кладкой или с кладкой со свежими яйцами около 30 см, диаметр
лотка около 17 см, глубина лотка до 12 см (Спангенберг, 1951). В районе Агдаша «боль-
шой» диаметр гнёзд составлял 40-50 см, высота 25-30 см. Лоток — глубиной 5 см и диа-
метром 20 см (Сатунин, 1907).
Полная кладка содержит от 4 до 9 яиц, причём число яиц в кладках сильно колеблется в
различные годы. Более 200 гнёзд, осмотренных под Ленкоранью в 1929 г., содержали пол-
ные кладки из 4-5 яиц, а в 1933 г. в большинстве гнёзд было от 5 до 7 яиц, изредка до 8-9
яиц (Спангенберг, 1951). В кладках птиц, гнездившихся в Кызыл-Агачском заповеднике,
было 4-5, реже 2 или 9 яиц (Греков, 1965). От 7 до 9 яиц содержали гнёзда, осмотренные у
Агдаша в 1894 г. А.Б. Шелковниковым (Сатунин, 1907). В южных частях ареала, в частно-
сти, в Африке, число яиц в кладках никогда не бывает столь большим — обычно 2-3 яйца.
Во Флориде размер кладок колебался от 2 до 5 яиц, в среднем составляя 3±0,13 яйца (41
кладка) (Maxwell, Kale, 1977).
Массовая откладка яиц под Ленкоранью происходит в первой половине мая. В 1929 г. 1
мая постройка многих гнёзд заканчивалась, и только одно гнездо из осмотренных содержа-
ло первое яйцо. Там же 10 мая в гнёздах обнаружено 1-2 яйца, а 15 мая — законченные
кладки (Спангенберг, 1951). У Агдаша откладка яиц начинается во второй половине апре-
ля, но в конце первой декады мая яйца оказались частью свежими, частью насиженными.
Свежие и частично насиженные яйца обнаружены в гнёздах в том же районе Смирновым
(1915) 11 июня. Из Армении известны кладки в 3 и 6 яиц, взятые 3 мая (Ляйстер, Соснин,
1942).
Сроки начала кладки у различных самок сильно варьируют. Насиженные яйца в гнёздах
удаётся встречать даже в первой половине июля (Сатунин, 1907). Промежуток времени
между откладкой отдельных яиц кладки в Вост. Европе и Сев. Азии не выяснен, во Флори-
де среднее время между откладкой первого и последующих яиц составляло около 2,2 дня
(Maxwell, Kale, 1977).
in
Отряд Листообразные, Семейство Цаплевые
Яйца египетской цапли формой и окраской хорошо отличаются от яиц других цапель.
Их форма правильно яйцевидная, с хорошо выраженными острым и тупым концами. По-
верхность яиц шероховатая. Окраска скорлупы бледно-голубая.
Размер яиц (мм): из-под Ленкорани (п=21) — 43,9-50,0 х 32,8-34,8, в среднем 47,4х
33,8 (Спангенберг, 1951); из Азербайджана (п=7) — 43,8^47,4 х 31,1-34,7, в среднем 45,1х
33,0 (Adamian, Klein, 1999); на оз. Ханка (п=37) — 41,4-48,7 х 31,4-36,0, в среднем 45,81х
33,91 (данные Ю.Н. Глущенко, И.Н. Кальницкой и Д.В. Коробова).
Масса яиц (г): из Азербайджана (п=7) — 22,2-27,5; в среднем 25,1 (Adamian, Klein,
1999); из центр. Флориды — 19-26,5; в среднем 24,0 (Weber, 1972).
Насиживают оба родителя, продолжительность насиживания 21-24 дня (Уайзерби, 1939,
цит. по Спангенберг, 1951). Во Флориде инкубационный период длится 22-23 дня, в сред-
нем 22,7±0,15 дней (10 кладок). В 1973 г. в 78% случаев инкубация начиналась с первого
яйца, в 15% — после второго и 7% — после откладки третьего яйца (Maxwell, Kale, 1977).
Как правило, в сезон бывает один выводок, в южных районах могут быть 2 и даже 3
выводка в год (Литвинова, 2001). Во Франции установлены случаи повторного гнездова-
ния за одно лето, с вылетом птенцов в сентябре-октябре (Hafner, 1970). Доставка пищи,
пока птенцы малы, лежит на самце, самка же остаётся на гнезде. Позднее, когда птенцы
подрастут, в добывании пищи принимают участие оба родителя.
По наблюдениям А.Б. Шелковникова (Сатунин, 1907; Мензбир, 1918), у Агдаша первые
птенцы вылуплялись в середине мая, а в начале июля некоторые молодые уже летали.
В кладках египетских цапель на оз. Махмуд-Чала 7-8 июня 1990 г. были слабо и средне
насиженные яйца. 23 июня птенцы вывелись лишь в отдельных гнёздах. Однако, по дан-
ным опроса, в 1989 г. в конце июня в гнёздах находились уже хорошо оперённые птенцы
(Патрикеев, 1991).
Подросшие, но ещё нелётные птенцы покидают гнёзда и, цепляясь ногами, клювами и
шеями, лазают по ветвям деревьев и лианам. Особенно охотно они занимают сухие ветви
по окраине колонии, покрывая их сплошной массой, откуда криками встречают взрослых
птиц, прилетевших с пищей. Летные птенцы совместно с родителями образуют стаи, раз-
мер которых возрастает с каждым днем. Эти стаи первое время кочуют по рисовым полям
и болотам близ колоний, возвращаясь ночевать на гнездовые деревья. Затем птицы покида-
ют колонию и, продолжая кочевать, медленно подвигаются к югу (Сатунин, 1907). В разгар
размножения южнее Ленкорани удаётся встречать холостых египетских цапель. Числен-
ность их весьма ограничена — в стайке бывает 4-6 особей (Спангенберг, 1951).
Успешность гнездования в колониях в Камарге (Франция) очень высокая — более трёх
вылетевших молодых на одно гнездо (Hafner, 1970). На юге США она составляет 71% (Weber,
1972). Во Флориде в 1973 г. успешность вылупления составляла 78% от числа отлиженных
яиц, до 10-дневного возраста доживали 89% вылупившихся птенцов (Maxwell, Kale, 1977).
В Юж. Америке, по данным наблюдений на Рио-Гранде в 1999-2001 гг., успех гнездования
колебался от 40 до 45% (Petry, Fonseca, 2005). В колониях на юго-западе Капской провин-
ции покинувшие гнёзда птенцы составляли в среднем 34% от числа отложенных яиц. Са-
мые большие потери происходят во время откладки яиц, гибель во время инкубационного
периода вдвое меньше. Хищничество не играет большой роли в гибели яиц и птенцов в тех
гнёздах, за которыми уже постоянно присматривают родители. Основная причина смерт-
ности птенцов — голод (Siegfried, 1972).
Суточная активность, поведение Египетская цапля, как правило, очень молчалива.
Лишь на гнезде взрослые птицы издают резкие крики. Молодые птицы, наоборот, крикли-
вы. Издаваемые ими звуки очень разнообразны и трудно поддаются описанию (Спанген-
берг, 1951).
Египетская цапля
275
Для вида характерна защита гнезда от пернатых хищников и агрессивность по отноше-
нию к другим членам колонии, что обусловливает высокую продуктивность вида (Литви-
нова, 2001).
Максимальное расстояние от колонии, на которое птицы удаляются для добывания пищи,
для континентальных колоний в штате Алабама равно 28,2 км, а для прибрежных колоний
на атлантическом побережье в Новой Каролине — 20,1 км. Средняя дальность разлета птиц
на кормежку от колонии во время гнездования составила в последнем случае 5,68 км (Custer,
Osborn, 1978).
Египетскм цаплям свойственно групповое кормление. Примерно в 30% случаев птицы
прилетают в места кормёжки в сопровождении одной или нескольких особей своего вида.
Предполагается, что колонии служат своеобразными «информационными центрами» и пти-
цы имеют возможность быстрее найти оптимальные места для кормодобывания. Косвенно
это подтверждается наблюдениями, когда «неудачные» особи при следующем вылете из
колонии летели очень медленно, как бы «поджидая» других птиц, чтобы присоединиться к
ним (Custer, Osborn, 1978).
Вне гнездовий египетские цапли большими стаями, до 200-300 особей, держатся на
чалтыках, или же группами в 15-20 особей сопровождают пасущихся на лугах и болотах
буйволов, отыскивая на их спинах приставших в грязи червяков и личинок (Сатунин, 1907;
Спангенберг, 1951). В Дагестане, в низовьях Терека, египетские цапли небольшими груп-
пами также сопровождают пасущихся на лугах и окраинах плавней коров и буйволов. Ве-
роятно, сопровождая крупный рогатый скот, цапли получают больше возможностей для
добычи пищи на лугах и в приплавневых зарослях.
Питание. Обычный способ добывания пищи — пробежки или проходы, чередующиеся
с ударами клювом для ловли добычи. Кормовая активность уменьшается к середине дня
(Siegfried, 1971). Эти наблюдения выполнены в Капской провинции (Африка), но сходное
поведение характерно для вида на всём пространстве его ареала.
Основная пища — беспозвоночные (кузнечики, саранчовые, стрекозы, слепни, водя-
ные жуки и их личинки, пауки, сколопендры, медведки, моллюски) и, в меньшей степени,
лягушки и мелкая рыба. Ещё реже в рацион входят пресмыкающиеся, яйца, мелкие грызу-
ны и птенцы.
У вскрытых в районе Агдаша египетских цапель в желудках обнаружены кузнечики,
пауки, живущие в воде личинки насекомых, жуки и мелкие лягушата. Однако в наиболь-
шем количестве цапли поедали кузнечиков, которых в желудках бывало до 100 и более, и
которые, видимо, предпочитались птицами (Сатунин, 1907). В желудке добытой 8 июня
1963 г. египетской цапли содержалось большое количество кузнечиков, 1 целая разноцвет-
ная ящурка, фрагмент озёрной лягушки, а также личинки жука-плавунца (Туаев, 1975).
По данным В.И. Васильева (1975), на территории Азербайджана соотношение различ-
ных кормов в питании египетской цапли следующее: земноводные (озёрная лягушка) —
67,2%; рыбы (уклея) — 8,4%; насекомые (медведки, слепни, личинки плавунца) — 22,0%;
паукообразные — 2,4%.
В Кызыл-Агачском заповедникеке основу питания гнездовых птенцов египетской цап-
ли составляют личинки стрекоз и другие беспозвоночные (63%) и лягушки (32%). По дру-
гим данным, птенцы питаются головастиками и молодыми лягушками (Мустафаев и др.,
1968). Взрослые птицы здесь же питаются главным образом саранчой, кузнечиками, мед-
ведками и другими сухопутными насекомыми, редко — личинками стрекоз и лягушками
(Греков, 1965).
В местах «вторичного» расселения пристрастие к названным выше группам живот-
ных сохраняется. На севере центр. Флориды в летнем рационе встречаемость прямокры-
276 Отряд Листообразные, Семейство Цаплевые
лых составила 96,8%, а по объёму — 80,5% от всей съеденной пищи (Fogarty, Hetrick,
1973).
Враги, неблагоприятные факторы. Смертность молодых птиц египетской цапли на
юге Африки, по данным кольцевания, составляет в первый год 37%, в последующие годы в
среднем 25%. Максимальная продолжительность жизни 13,5 лет, средняя — 3-3,5 года
(Siegfried, 1970).
В дельте Волги отрицательное влияние на гнездовую группировку, вероятно, оказывает
неустойчивый гидрологический режим, связанный с нерегулярными паводками и происхо-
дящим в настоящее время повышением уровня Каспийского моря. На оз. Ханка египетская
цапля страдает от конкурентных взаимоотношений в колонии с большим бакланом (дан-
ные Ю.Н. Глущенко).
Антропогенные факторы, сдерживающие распространение или лимитирующие числен-
ность вида в современных условиях, достоверно не установлены. Возможно, отрицательно
сказывается вырубка пойменных зарослей и рощ в низовьях больших рек бассейна Кас-
пийского моря и по берегам пресных водоёмов.
Распространение этой южной цапли на юге России и в западных районах Казахстана,
вероятно, ограничивается климатическими условиями и отсутствием достаточного коли-
чества легкодоступной пищи (прямокрылых, личинок водных насекомых и лягушек) в пе-
риод кормления птенцов в мае-июне.
В оперении египетской цапли найдены пухоеды: Ardeicola expallidus, Ciconiphilus decim-
fasciatus и Comatomenopon ibis (McClure, Ratanaworabhan, 1973).
Хозяйственное значение, охрана. Специально охотниками не добывается и случаев
отстрела на территории юга России не известно. Как редкий, «краеареальный» вид, египет-
ская цапля нуждается в адресной охране в местах гнездования. Вне колоний для этого вида
специальных мер охраны не требуется. Египетская цапля занесена в Красные книги Турк-
менистана и Российской Федерации, а в России также в Красные книги Республики
Дагестан, Астраханской области, Сахалинской области и Приморского края. Включена в
Приложение 2 Бернской конвенции и Приложение соглашения, заключённого Россией с
Республикой Корея об охране мигрирующих птиц.
В России охраняется в Астраханском заповеднике. Формально охраняется в заповедни-
ке «Дагестанский», но здесь отмечены только единичные встречи этого вида. В Аграхан-
ском заказнике вид встречается в летнее время, однако гнездование пока не доказано. В
Азербайджане колонии египетских цапель охраняются в Кызыл-Агачском и Акгёльском
заповедниках.
Род Egretta Т. Forster, 1817
Малая белая цапля
Egretta garzetta (Linnaeus, 1766)
Ardea garzetta Linnaeus, 1766, Syst. Nat., ed. 12, c. 237, «Восток»; северо-восток Италии
(Grant, Mackworth-Praed, 1933, Bull. Brit. Omith. Club, 53).
Little egret (англ.), Seidenreiher (нем.),
aigrette [blanche, petite aigrette, heron] garzette (фр.)
Син. (лат.): для подвидаgarzetta'. Ardea nivea S.G. Gmelin, 1770; Ardea orientalis Gray, 1831;
Ardea xanthodactyla Rafinesque, 1810; Garzetta garzetta Sharpe, Herodias garzetta L., Herodias
Малая белая цапля
277
lindermayeri, C.L. Brehm, 1855; Ardea (Herodias) procerula Cabanis, 1868.
Син. (рус.): чепура-нужда.
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Некрупная цапля с длинной тонкой шеей,
длинными клювом и ногами, заметно меньше большой белой. Длина тела 55-65 см, размах
крыльев 90-105 см, вес до 500-600 г. Оперение чисто белое, клюв и ноги чёрные, пальцы
жёлтые. В брачном наряде на затылке длинный «чуб» из 2 ланцетовидных перьев. В полёте
«складывает» S-образно шею и вытягивает ноги. Встречается, обычно, на берегах водо-
ёмов, лугах, пастбищах, болотах.
От большой белой цапли отличается значительно меньшими размерами, черной окрас-
кой клюва во все сезоны, жёлтым цветом пальцев у взрослых птиц. Совершенно различ-
ный у этих цапель голос: у большой белой — хриплый треск, у малой — «карканье» или
«кваканье». Полет более легкий, чем у большой белой цапли, с большей частотой взмахов.
Египетские цапли в зимнем наряде имеют близкий размер и сходную окраску с малыми
белыми, но отличаются светлым клювом и чёрным цветом верха пальцев. У молодых еги-
петских цапель клюв хотя и тёмный, но не чёрный, к тому же он имеет другую форму
(более короткий и толстый). У средней белой цапли также более короткий и толстый клюв,
жёлтый в зимнем наряде, и чёрные пальцы.
Голос. За пределами гнездовой колонии кричит редко. У вспугнутой птицы можно ус-
лышать характерный каркающий крик, похожий на «арк» или «орк». В местах гнездования
вокальный репертуар довольно богат. Д. Блейкер (Blaker, 1969) описывает 8 типов криков
малой белой цапли, издаваемых в различных ситуациях: «ааах» — при взлёте; «да-вах» —
в разных случаях, но прежде всего при подлёте к гнезду и во время приветственной цере-
монии; «доу» — демонстративный крик самца без пары; щебечущий крик издаётся во вре-
мя токования и церемонии приветствия; «ггроу» и некоторые другие звуки можно услы-
шать во время токования. Демонстративный крик самца напоминает бульканье при полос-
кании горла. Крик выпрашивающих корм птенцов похож на «хе-хе-хе-хе» (Bauer, Glutz von
Blotzheim, 1966).
Подвидовая систематика. Разные авторы выделяют 2 (Howard, Moore, 1994), 3 (Степанян,
2003), 4 (Спангенберг, 1951; Cramp, 1977; Snow, Perrins, 1998) и до 6 (Martinez-Vilalta, Motis,
1992) подвидов, в зависимости от принимаемого объёма вида Egretta garzetta. Подвиды раз-
личаются размерами и окраской. В Вост. Европе и Сев. Азии распространён один подвид:
1. Egretta garzetta garzetta.
Ardea garzetta Linnaeus, 1766, Syst. Nat., ed. 12, c. 237, «Восток»; северо-восток Италии
(Grant, Mackworth-Praed, 1933, Bull. Brit. Omith. Club, 53).
Неоперённые участки кожи на голове голубоватого или зеленовато-желтоватого цвета,
пальцы желтоватые. Подвид широко распространён в Палеарктике от Португалии, Ирлан-
дии и Сев.-Зап. Африки до Юж. Приморья, Кореи и Японии. На юг доходит до материково-
го побережья Юж. и Юго-Вост, (исключая п-ов Малакка) Азии, Шри-Ланки, сев. Филип-
пин. Местами гнездится также в Африке к югу от Сахары.
Распространение других подвидов не связано с территорией Вост. Европы и Сев. Азии:
К g. gularis (Bose, 1792) (2) обитает на побережье Зап. Африки от Мавритании до Габо-
на; Е. g. schistacea (Ehrenberg, 1828) (3) — на побережье Вост. Африки до Красного моря,
Аравийского п-ова, Персидского залива, а также на Шри-Ланке, западе и юге Индии; Е. g.
dimorpha Е. Hartert, 1914 (4) — на Мадагаскаре, Коморских и Сейшельских о-вах, встре-
чается также на побережье Вост. Африки от юга Кении до Северного Мозамбика; Е. g.
nigripes (Temminck, 1840) (5) — на островах Юго-Вост. Азии от Зондских о-вов и юж.
278
Отряд Листообразные, Семейство Цаплевые
Филиппин до Новой Гвинеи; Е. g. immaculata (Gould, 1846) (6) — в Сев. и Вост. Австра-
лии, спорадично — в Новой Зеландии (Martinez-Vilalta, Motis, 1992; Howard, Moore, 1994)
(рис. 55).
Замечания по систематике. Формы gularis/schistacea и dimorpha, обитающие на мор-
ских побережьях восточного полушария, частью систематиков (напр. Dickinson, 2003)
рассматриваются как два самостоятельных вида — западная рифовая цапля (Egretta
gularis (Bose, 1792)) и мадагаскарская рифовая цапля (Egretta dimorpha Е. Hartert, 1914).
Они отличаются от «типичной» малой белой цапли более светлыми клювом и ногами и
частой встречаемостью в популяциях тёмной цветовой морфы с голубовато-серым опе-
рением.
Описание. Окраска. Самец и самка в брачном наряде. Оперение снежно-белое. На
затылке «коса» из 2 (иногда 3) удлинённых до 15-20 см ланцетовидных перьев. На зобу
перья также удлинённые и узкие, образуют «гриву», на спине развиты эгретки — длинные
рассученные перья длиной до 24 см, на конце слегка загнутые вверх. Эгретки на спине
образуют шлейф, заходящий по длине за конец хвоста. Клюв чёрный с серовато-зелёным
основанием подклювья. Голая кожа на лице серая с зеленоватым оттенком; в начале сезона
гнездования она становится жёлтой, а в разгар брачных демонстраций — оранжевой или
розовой (у дальневосточных птиц — до фиолетово-лиловой). Радужина глаза жёлтая. Ноги
чёрные или зеленовато-чёрные, пальцы ярко-жёлтые, в разгар брачных демонстраций —
до оранжевых, красноватых). Самец несколько крупнее самки, украшающие перья у после-
дней развиты меньше.
Самец и самка в зимнем наряде. Отличаются отсутствием длинных украшающих перь-
ев. Основание подклювья сероватое, а не чёрное, голая кожа на лице голубовато-серая (у
дальневосточных птиц — зеленоватая). Пальцы менее ярко окрашенные, чем в брачный
сезон.
Пуховой птенец. Пух чисто белый, волосовидный, на темени он немного удлинен. У
недавно вылупившегося птенца кожа желтовато-зеленоватая, лапы жёлтые, клюв желтова-
тый с розовым или серым оттенком. На шестой день кожа становится светло-зеленоватой,
на 15-20-й — на ней появляется голубоватый оттенок. Перья начинают развиваться с 10
дня. Лапы становятся светло-зелёными, на клюве появляются чёрные пятна, позже он тем-
неет. Глаза обычно ярко-жёлтые. Остатки пуха, особенно на голове, сохраняются до вылета
(Cramp, 1977; Смогоржевський, 1979).
Гнездовой наряд. По окраске сходен с зимним нарядом взрослой птицы. Клюв и ноги
более тускло окрашенные, чем у взрослых птиц — зеленоватые или сероватые, а не чёр-
ные. Пальцы также могут быть сероватыми, а не жёлтыми, слабо контрастирующими с
цветом цевки. Основание нижней челюсти жёлтое.
Первый зимний наряд. Оперение как в гнездовом наряде, но перья на зобу немного уд-
линены и заострены, хотя и меньшей длины, чем у взрослых птиц (Bauer, Glutz von Blotzheim,
1966).
У некоторых подвидов малой белой цапли в тропиках встречаются темные морфы. Пти-
цы имеют черноватую или серую окраску оперения; горло, обычно, белое. Существует так-
же большое количество переходных состояний между белой и тёмной морфами. Изредка
залеты таких птиц регистрируют в Европе.
Строение и размеры. Размеры малой белой цапли приводятся в таблице 20.
Самец несколько крупнее самки. Достоверные различия по выборке из ЗМ КНУ получены
для длины крыла и клюва (при р<0,002) и цевки (р<0,02).
В литературе приводятся следующие размеры малых белых цапель в мм (без разделе-
ния на половые группы): для Украины — длина крыла — 251-296, хвоста — 83-138,
Малая белая цапля
279
Рис. 55. Область распространения малой белой цапли: а — гнездовой ареал, б — область зимовок,
в — поселения по морским побережьям. Подвиды: 1 — Egretta garzetta garzetta', 2 — Е. g. gularis;
3 — E. g. schistacea; 4 — E. g. dimorpha; 5 — E. g. nigripes; 6 — E.g. immaculata.
клюва — 74,5-100, цевки — 89,4-116 (Смогоржевський, 1979); для Казахстана — длина
крыла — 265-300, хвоста — 95-120, клюва — 78-92, цевки — 87-113 (Долгушин, 1960);
для территории бывш. СССР (п=12) — длина крыла — 275-300, в среднем — 287, клюва —
в среднем около 90, цевки — 90-110 (Спангенберг, 1951). Длина крыла 57 птиц из Камарга
(Франция) в среднем оказалась равной 281, при lim от 254 до 307 мм (Bauer, Glutz von
Blotzheim, 1966).
Формула крыла (не считая рудиментарного первого махового) II=III?I?IV>V>V1... На
внутренних опахалах 1 и 2 маховых и наружных опахалах 2 и 4 первостепенных маховых
есть вырезки. Хвост прямой, 12 рулевых. Клюв тонкий с глубокими назальными выемками
(Cramp, 1977).
Вес украинских цапель 370-670 г (Смогоржевський, 1979); вес 32 молодых птиц после
вылета из гнёзд в Камарге (Франция): 305-555, в среднем 421 г, 11 птиц неизвестного воз-
раста — 280^138, в среднем 381 г (Bauer, Glutz von Blotzheim, 1966); Венгрия: взрослые
самцы — 496-614 г, взрослые самки — 490-530 г; Италия: взрослые самцы — 450^460 г
(Cramp, 1977).
В течение года вес может существенно изменяться. Так, у цапель, добытых на юге Запо-
рожской обл., летом вес составлял 480-500 г, осенью и зимой — 560-650 г. Увеличение
веса идёт прежде всего за счёт накопления жировых отложений (Фурманова, 1998).
Площадь опоры ног у малой белой цапли составляет 12,25 см2, весовая нагрузка —
24,49 г/см2 (Фурманова, 1998).
280
Отряд Листообразные, Семейство Цаплевые
Таблица 20. Размеры (длина крыла, длина хвоста, длина клюва и длина цевки) самцов
и самок малых белых цапель в разных регионах (в мм)
Часть тела п Самцы Самки Источник
lim М п lim М
У к р айна
Крыло 23 253-304 282,3±2,4 12 249-284 269,1±2,9 Колл. ЗМ КНУ,
Хвост 23 87-114 94,2±1,2 11 83-105 92,0±2,0 ориг.
Клюв 22 78,1-93,0 87,15±0,92 12 77,1-87,5 82,69±0,95
Цевка 23 93-116 105,3±1,3 12 94-110 99,9±1,5
Евр опа
Крыло 24 260-295 240-282 Bauer & Glutz von
Хвост 90-110 90-110 Blotzheim, 1966
Клюв 83-92 78-90
Цевка 100-110 97-105
Западная Палеарктика
Крыло 17 245-303 280 17 251-297 272 Cramp, 1977
Хвост 16 84-113 97,9 14 81-101 93,6
Клюв 17 97-103 84,4 17 68-89 79,5
Цевка 17 78-112 101 17 88-110 97,3
Линька. У молодых птиц частичная постювенильная линька после отлета с мест гнез-
дования в августе - ноябре. В это время сменяется мелкое оперение. Следующая линька
полная, она начинается несколько раньше, чем послегнездовая взрослых птиц — весной
второго года жизни. У взрослых птиц частичная предгнездовая линька в январе-апреле,
реже до мая. Начинается она ещё на зимовках, заканчивается уже после прилёта на места
гнездования. Украшающие перья, особенно на затылке, отрастают у разных птиц в разное
время — с февраля по май. Полная послегнездовая линька начинается в июне ещё во время
гнездования, заканчивается уже на зимовках в ноябре, реже в декабре. Сначала выпадают
украшающие перья на затылке, затем на спине (Bauer, Glutz von Blotzheim, 1966; Stresemann,
Stresemann, 1966; Cramp, 1977).
Распространение. Гнездовой ареал. Малая белая цапля широко распространена
в Восточном полушарии (рис. 55). В Евразии она гнездится от Пиренейского п-ова и Ир-
ландии до Вост. Китая, Юж. Приморья, Тайваня, Кореи и Японии. На север распростране-
на до Великобритании, Голландии, Германии, Польши. В Вост. Европе и Сев. Азии север-
ная граница ареала проходит через Черновицкую обл., север Молдавии, Одесскую, Пол-
тавскую, Харьковскую, Луганскую обл. Украины; Ростовскую, Волгоградскую обл. Рос-
сии; устье Урала в Казахстане, низовья Аму-Дарьи и южное побережье Аральского моря в
Узбекистане. Дальше северная граница ареала проходит по восточным окраинам Алайской
системы, южным подножьям Гималаев, через Китай, Корею и о. Хонсю в Японии (Martinez-
Vilalta, Motis, 1992; Степанян, 2003). С 2001 г. периодическое гнездование регистрируется
на оз. Ханка (Глущенко и др., 20066; Коробов, Глущенко, 2008), возможно гнездование
также на крайнем юге Приморья в низовьях р. Туманган на стыке границ России, КНДР и
Китая (Литвиненко, Шибаев, 1999); в различных частях Приморья, за пределами оз. Ханка,
малая белая цапля — нередкий летующий вид (Глущенко, Шибнев, 1977; Бурковский, 1998;
Литвиненко, Шибаев, 1999; Шохрин, 2001). В Азии малая белая цапля распространена на
юг до долины Евфрата, Персидского залива, побережья Юж. и Юго-Вост. Азии. Гнездится
на Б. и М. Зондских о-вах, Филиппинах, Молуккских о-вах, Новой Гвинее, на севере и
востоке Австралии, в Новой Зеландии. В Африке встречается на о-вах Зелёного мыса, в
Марокко, на севере Алжира, в Тунисе, на северо-западном побережье Ливии, в дельте Нила,
Малая белая цапля
281
в Вост, и Юж. Африке (Bauer, Glutz von Blotzheim, 1966; Cramp, 1977; Martinez-Vilalta,
Motis, 1992; Fasola, Hafner, 1997; Степанян, 2003).
Более детальное распространение малой белой цапли в Восточной Европе и Северной
Азии таково (рис. 56). В Молдавии она спорадически распространена на всей территории
(Аверин и др., 1971; Зубков и др., 2001, 2004). На западе Украины первая гнездовая коло-
ния была обнаружена в 1974 г. на острове р. Прут в Черновицкой обл. (Смогоржевський,
1979). В 1977 г. ниже по течению найдена другая колония на острове у с. Зелёный Гай. С
1991 г. 2 пары гнездились в колонии кваквы и серой цапли в прижелезнодорожной лесопо-
лосе поблизости от рыборазводных прудов в окрестностях с. Кливодин на севере области.
Колонии на р. Прут впоследствии исчезли (Годованець и др., 1996). Гнездование отмечено
также на Дунае, Днестре, в Черноморском заповеднике (Смогоржевський, 1959).
В Приднепровье малая белая цапля спорадично и нерегулярно гнездилась на север до
Полтавской (Гавриленко, 1929) и Киевской (Кирпиченко, 1932) областей. После создания
каскада водохранилищ в 1950-1970-х гг. птицы расселились вверх по Днепру, а затем и по
долинам левобережных притоков — Сулы, Пела, Ворсклы. В настоящее время малые бе-
лые цапли гнездятся на север до Сульского залива Кременчугского вдхр., низовий Пела,
Ворсклы, Орели (Гаврись, 1994). В 1983 г. одно гнездо было обнаружено в колонии серой
цапли на левом берегу Каневского вдхр. южнее Киева (Фесенко, 1984), однако вскоре ма-
лая белая цапля перестала там гнездиться. По долине р. Орели птицы поднялись до южных
районов Харьковской обл. (Лисецкий и др., 1980; Ткаченко, 1987; Есилевская и др., 1988).
Гнездование отмечали также в Луганской обл. (Ветров и др., 1991; В.В. Ветров, личн. со-
общ.).
С 1961 г. малая белая цапля начала регулярно гнездиться на Лебяжьих о-вах у северо-
западного побережья Крыма, до этого в Крыму она была известна лишь как пролётная и
летующая птица (Костин, 1983). В начале 1980-х гг. малая белая цапля уже гнездилась на
Рис. 56. Ареал малой белой цапли в Вост. Европе и Сев. Азии: а — область гнездования, б — районы
зимовки.
282
Отряд Листообразные, Семейство Цаплевые
Сиваше, что связано с его опреснением вследствие интенсивного орошения полей. Извес-
тны колонии и на севере Крыма (Костин, Тарина, 2004).
В Сев. Приазовье малая белая цапля впервые загнездилась в 1986 г. в верховьях Молоч-
ного лимана и на о-вах у Обиточной косы (Лысенко, Сиохин, 1991). В последующие годы
она появилась на гнездовании и в других местах на юге Запорожской обл. (Кошелев и др.,
2004). В 1980-е гг. дважды отмечали гнездование её в Донецкой обл. (Завгородний и др.,
1989).
На Северном Кавказе гнездится на нижнем Дону до Цимлянского вдхр., в низовьях Се-
верского Донца, на водоёмах Зап. и Вост. Маныча, в дельте Кубани, в низовьях рек Бейсуг
и Челбас, в дельте Терека. В 1970-1980-е гг. отмечено дальнейшее расселение в связи с
развитием рисосеяния и прудового рыбоводства (Казаков и др., 2004). В Азербайджане
малая белая цапля многочисленна в Кызыл-Агачском заповеднике, дельте Куры, на оз. Ах-
чала в Пушкинском р-не (Литвинова, 1988, 19896). Гнездится в Армении (Спангенберг,
1951).
В Поволжье малая белая цапля гнездится на север до Волгоградской обл. Гнездование
известно на Сарпинских озёрах (Чернобай и др., 2000), Кисловской оросительной системе
(Чернобай, 2000), Цимлянском вдхр. (Казаков и др., 2004). В Саратовской обл. это уже
лишь редкий залётный вид (Завьялов и др., 1997, 2005).
В Казахстане малая белая цапля сейчас гнездится только в низовьях Урала (Ковшарь,
1999). В Узбекистане — редкий гнездящийся вид низовьев Аму-Дарьи и южного побере-
жья Аральского моря (Сагитов, 1987; Лукашевич, 1990). Изолированная популяция суще-
ствовала на нижнем и среднем течении Аму-Дарьи вверх до Чарджоу, в низовьях Сыр-
Дарьи и на побережье Аральского моря. Малая белая цапля гнездилась также в Мервском
оазисе и в низовьях Теджена на востоке Туркмении (Спангенберг, 1951; Долгушин, 1960).
В настоящее время в Вост. Туркмении не гнездится; изредка встречается на весеннем про-
лёте (Пославский, Ширеков, 1990).
Зимовки. Основные места зимовки евразийских популяций малой белой цапли рас-
положены в Средиземноморье, тропической Африке, на Ближнем Востоке, в Закавказье, на
юге Туркмении, в районе Персидского залива, на юге Китая, в Юго-Вост. Азии (Спанген-
берг, 1951; Bauer, Glutz von Blotzheim, 1966; Cramp, 1977; Martinez-Vilalta, Motis, 1992).
Большая часть E. g. garzetta зимует в Африке (рис. 55). Тропические популяции оседлы.
Балканские цапли обнаружены на зимовке в Тунисе, Нигерии, Сьерра-Леоне (Нанкинов,
Кищинский, 1978); нижнеднепровские — в долине Нигера; приазовские — в Ливане, Ниге-
рии, Гане; волжские — в Иране; дагестанские — в Кувейте. Часть азербайджанских цапель
остаётся на зимовку на юге Азербайджана, другие улетают к иранскому побережью Кас-
пия, низовьям Тигра и Евфрата, побережью Персидского зал. (Сапетин, 1978д). Зимой 2003/
04 гг. на территории Азербайджана было учтено 697 зимующих малых белых цапель (Сул-
танов, Джаббарова, 2006). Птицы, окольцованные на Лебяжьих о-вах, найдены в Зап. и
Центр. Африке (Костин, Тарина, 2005).
С 1950-х гг. все больше малых белых цапель остаётся зимовать в Юж. Европе. В насто-
ящее время в Европейской части Средиземноморья зимует около 10% европейской популя-
ции. Это можно объяснить общим потеплением климата (Fasola, Hafner, 1997).
Случаи зимовки известны также в Молдавии (Аверин и др., 1971), на юге Украины — в
Крыму (Бескаравайный, Костин, 1999), Одесской обл. (Архипов, 1996; Фесенко, Архипов,
1996; Русев и др., 2002), Сев. Приазовье (Кошелев, 2000). В районе заповедника «Аскания-
Нова» в Херсонской обл. 21-24.01.2001 г. учтены 183 птицы (Гавриленко и др., 2002). От-
дельные случаи зимовки отмечали и севернее — в Ивано-Франковской обл. (Бучко, 1998),
а также на очистных сооружениях Тернополя (Страшнюк, 2004). Регулярно зимуют малые
Малая белая цапля
283
белые цапли на Сев. Кавказе — на Веселовском вдхр., в дельте Кубани, на водоёмах Став-
рополья (Казаков и др., 2004), в Сев. Осетии (Комаров, 1991).
Миграции. По данным кольцевания, среднедунайские птицы осенью мигрируют через
Балканы, Италию и Сицилию в западные части тропической Африки. Цапли из Сев. При-
черноморья на зимовку летят также на юго-запад — через Балканский п-ов, южную
половину Италии и Тунис в бассейн Нигера. Из вост. Приазовья миграция идёт через Чер-
ное море, Кавказ и Восточное Средиземноморье к побережью Гвинейского зал. Прикас-
пийская популяция мигрирует вдоль западного побережья Каспийского моря в Иран и к
северо-западной оконечности Персидского зал. (Кищинский, Сапетин, 1978).
Фенология. Весной малая белая цапля прилетает позднее большой. Первые птицы по-
являются в различных регионах европейской части бывшего СССР во второй половине мар-
та - апреле. Средние сроки приходятся на первую декаду апреля (табл. 21). В Молдавии
прилетает во второй половине марта, массовый прилет на Кучурганском лимане наблюда-
ли в конце марта - начале апреля. Появление первых птиц отмечено здесь 17 марта 1960 г.
и 28 марта 1968 г. (Аверин и др., 1971). На юге Крыма пролётных птиц чаще всего наблю-
дают с середины апреля. Пролёт обычно заканчивается в третьей декаде мая. Отдельные
птицы могут задерживаться до июня, самая поздняя встреча — 25 июня 1984 г. Максималь-
ная продолжительность миграционного периода — 52 дня. В пролетающих стаях насчиты-
вается до 28 особей (Бескаравайный, 2005). В Вост. Приазовье прилёт малых белых цапель
наблюдается в первую и вторую декаду апреля. На приморских низменностях возле Адлера
пролёт отмечали с первой декады апреля. Здесь видели в основном одиночных птиц, из-
редка стайки по 8-10 особей. Завершается пролёт к концу мая. На Ставрополье малые бе-
лые цапли появляются в середине апреля. В дельте Терека прилет отмечается с первой
декады марта до начала апреля. Летят птицы, как правило, небольшими стаями (Казаков и
др., 2004). В Туркменистане весенняя миграция начиналась в середине марта и продолжа-
лась весь апрель (Спангенберг, 1951). В дельте Аму-Дарьи в Узбекистане малая белая цап-
ля прилетает в начале апреля (Лукашевич, 1990).
Осенняя миграция начинается в августе. В западных и центральных областях Украины
последних птиц наблюдают с конца августа до октября, в южных — с конца сентября до
ноября (табл. 22). В Молдавии отлёт начинается со второй декады августа, массовая мигра-
ция идёт в сентябре, частично — в октябре. Отдельные особи могут задерживаться до кон-
ца октября и даже ноября (Аверин и др., 1971). На Сев. Кавказе первые особи покидают
область летне-осенних кочёвок в конце августа - первой половине сентября. Массовый
Таблица 21. Сроки прилёта (первых встреч) малой белой цапли
в Вост. Европе и Сев. Азии
Регион п М SD lim Источник
Южная Украина 11 3 апреля и,з 16 марта - 20 апреля Лысенко, 1992; Корзюков, 1996; Бескаравайный, 1999; 2005; Костин, Тарина, 2005
Центральная Украина 8 4 апреля 12,2 17 марта - 22 апреля Шакула, Ткаченко, 1986; Грищенко, Гаврилюк, 2000; Кныш, 2001
Ростовская обл. 9 9 апреля 12,4 20 марта - 24 апреля Сапетин, 1978д; Ломадзе и др., 1981; Белик, 1999; Казаков и др., 2004
Дельта Волги 10 1 апреля 6,6 22 марта - 13 апреля Луговой, 1963
284
Отряд Листообразные, Семейство Цаплевые
Таблица 22. Даты последних встреч малой белой цапли в Вост. Европе и Сев. Азии
Регион п М SD lim Источник
Западная Украина 13 16 сентября 17,2 26 августа - 21 октября Орштолопчш спостереження, 1994; Матер1али, 1995а, б, 1996; Годованець и др., 1996; Капелюх, Гузш, 2000; Новак, 2002
Центральная Украина 4 14 сентября 19,2 25 августа - 10 октября Роговий, 1996; Грищенко, Гаврилюк, 2000; Грищенко (ориг.)
Южная Украина 6 18 октября 17,9 27 сентября - 18 ноября Грищенко, 1995; Бескаравайный, 1999, 2005; Костин, Тарина, 2005.
пролёт начинается в середине сентября и обычно заканчивается в первой половине октяб-
ря. Наиболее поздняя встреча — 3 ноября 1964 г. в Ахтарско-Гривенских плавнях (Казаков
и др., 2004). В дельте Волги малые белые цапли отлетают в сентябре - первых числах ок-
тября, отдельные особи могут задерживаться до ноября (Луговой, 1963). На северо-запад-
ном побережье Каспийского моря пролёт идёт с конца августа до середины ноября. Наибо-
лее интенсивен он в конце августа - сентябре. Миграция проходит в основном утром и
вечером на высотах от 50 до 80 м, иногда до 300 м. Птицы летят стайками по 10-20 особей,
одиночные цапли встречаются редко. Чаще, чем у крупных видов цапель, пролёт идёт над
морем (Михеев и др., 1975). На юго-восточном побережье Каспия миграция идёт весь но-
ябрь и первую половину декабря (Спангенберг, 1951).
В Зап. и Центр. Европе отлёт начинается в конце августа - сентябре, на юге Европы
может продолжаться до ноября и даже декабря (Bauer, Glutz von Blotzheim, 1966).
Послегнездовые кочёвки. Для малой белой цапли характерны широкие послегнездовые
кочёвки. В это время птицы появляются за пределами гнездового ареала. Кочёвки начина-
ются вскоре после вылета птенцов и к концу августа - сентябрю постепенно переходят в
осеннюю миграцию. В благоприятных местах могут образовываться скопления из десят-
ков и сотен особей. По наблюдениям на Лебяжьих о-вах, после вылета из гнёзд молодые
птицы держатся по соседству с колонией в составе поливидовых скоплений — «клубов»,
откуда летают на кормёжку и куда возвращаются на ночевку. Затем происходит постепен-
ный разлет, переходящий в кочёвки и миграции. В августе цапли кочуют по Крыму в преде-
лах 100 км от колоний (Костин, Тарина, 2005). По данным мечения на Балканах, в этот
период молодые цапли разлетаются в северном, восточном и юго-восточном направлени-
ях, обычно не далее 100 км, иногда до 180-360 км (Нанкинов, Кищинский, 1978). Птицы,
окольцованные на нижнем Днепре, во время послегнездовых кочёвок также в основном
держались за 20-150 км от гнездовий. В Вост. Приазовье цапли разлетаются в разных на-
правлениях. В июле и августе большинство молодых особей найдены не далее 160 км от
колоний, среднее удаление в это время до 100 км. Отдельные птицы улетели на 420 км.
Осенняя миграция к местам зимовок начинается в первой половине сентября (Сапетин,
1978д). В низовьях Волги молодые птицы в послегнездовой период широко кочуют по дельте,
преимущественно к северу, вверх по течению, удаляясь по Волго-Ахтубинской пойме на
200-250 км от моря (Скокова, 1959а). В дельте Терека, по всей вероятности, большая часть
молодых цапель после вылета из гнёзд откочёвывает в дельту Волги, проводит там август,
а в сентябре возвращается обратно. В Азербайджане молодые птицы с конца июля по сен-
тябрь держатся обычно не далее 100 км от гнездовых колоний, в низменностях к югу от
низовий Куры и Аракса, однако в августе отдельные особи улетают уже за 200-230 км к
западу и северо-западу. В сентябре отмечены два дальних залета в северном и северо-за-
падном направлениях — в Гурьевскую обл. (ок. 900 км) и в устье Дона (1 170 км) (Сапетин,
Малая белая цапля
285
1978д). Испанские малые белые цапли во время кочёвок могут удаляться до 400 км к северу
(Bauer, Glutz von Blotzheim, 1966).
Залёты. Для малой белой цапли они достаточно обычны, хотя птицы отмечаются и не
так далеко, как в случае с большой белой цаплей.
В Европе залёты отмечались на север до Ирландии, Великобритании (Cramp, 1977;
Combridge, Parr, 1992), Голландии, Сев. Германии, Швеции (Bauer, Glutz von Blotzheim,
1966), Польше (Tomialojc, Stawarczyk, 2003). Известны залёты малой белой цапли даже в
Америку — этих птиц встречали на о-вах Ньюфаундленд, Тринидад и Мартиника (Cramp,
1977), а также в США (Kain, 1994, 1995; Rohrbacher, 2000). В странах Вост. Европы залёты
отмечали в Белоруссии (Никифоров и др., 1997; Никифоров, 2001; Абрамчук и др., 2003;
Абрамчук, Абрамчук, 2004), в России на север до Калужской (Галченков и др., 1998), Ли-
пецкой, Тамбовской (Климов и др., 2004), Пензенской (Спангенберг, 1951), Саратовской
(Завьялов и др., 2005) обл. В Казахстане залёты малой белой цапли регистрировали в раз-
личных регионах, удаленных от мест гнездования — в низовьях Илека (Долгушин, 1960), в
Наурзумском заповеднике (Гордиенко, 1987), в районе Кургальджинского заповедника (Кош-
кин, 2002), восточнее Алма-Аты (Ковшарь, 2002), на Шошкакольских озёрах (Хроков, Бек-
баев, 2002). Залётных птиц наблюдали в Оренбургской обл. и в районе Кузнецка (Спанген-
берг, 1951). В Хабаровском крае впервые найдена (занесённая ветром) птица 18.05.1986 г.
на Амуре (Вальчук, 1997). Залётных птиц отмечали также на Сахалине (Нечаев, 1991а, б).
Местообитания. Обитает в разнообразных влажных биотопах — в поймах, плавнях и
дельтах рек, по берегам озёр и прудов, морских лагун, эстуариев, в тростниковых зарослях
по берегам водоёмов и т.п. Предпочитает водоёмы со стоячей водой или с медленным тече-
нием. Нередко можно увидеть колонии этого вида в подтопляемых пойменных лесах. Охот-
но селится возле рисовых полей. На севере Италии в зоне рисосеяния колонии из сотен гнёзд
расположены через каждые 4-10 км (Fasola, Hafner, 1997). Обычно гнездится на равнинах, но
может подниматься и в горы. В Армении обитает на высоте до 2 000 м н.у.м. (Cramp, 1977).
В Молдавии населяет мелководные водоёмы, заросшие надводной растительностью, с
затопляемыми древесными и кустарниковыми зарослями. Раньше основным местом оби-
тания были пойменные леса (Аверин и др., 1971).
В Центр. Предкавказье 44,1% колоний находились в подтопленных пойменных лесах,
остальные — в тростниково-рогозовых зарослях: 46,9% — на обводнённых солончаках,
9,0% — на пересыхавших до обводнения реках и балках (Бичерев, 1988). В дельте Волги
малая белая цапля гнездится в древесных зарослях култучной зоны в смешанных колониях
с другими видами цапель (Луговой, 1963).
Кормится на мелководьях водоёмов различного типа, по берегам, на лугах и пастбищах,
болотах. Можно встретить кормящихся птиц в сухих местах довольно далеко от воды. Не
избегает близости человека. В Ленкорани гнездится возле самых населённых пунктов, охотно
кормится среди стад пасущихся буйволов. Нередко можно увидеть птиц, сидящих на их
спинах (Спангенберг, 1951).
Распределение колоний малой белой цапли и их численность зависят как от наличия
кормовых биотопов, так и пригодных для гнездования мест. В странах с сильно изменен-
ным ландшафтом последний фактор может быть определяющим. Так, на севере Италии на
рисовых полях кормовая база вполне достаточна, но количество пригодных для гнездова-
ния мест ограничено из-за интенсивного сельскохозяйственного производства (Fasola, Alieri,
1992).
Численность. Общая численность в Европе оценивается в 68-94 тыс. пар. Из них: 8-12
тыс. пар в Азербайджане, 7-10 тыс. пар в России, 4,1-4,6 тыс. пар на Украине, 400-500 пар
в Молдавии, 50-250 пар в Армении. Численность зимующих в Европе птиц составляет
286
Отряд Листообразные, Семейство Цаплевые
более 28 тыс. особей. В большинстве стран численность стабильна или медленно растет
(Birds in Europe, 2004).
На Украине основные места гнездования — низовья Дуная, Днестра, Днепра, север и
северо-восток Крыма, лиманы и косы Сев. Приазовья, водохранилища южной части Днеп-
ровского каскада. В большинстве мест отмечаются колебания или рост численности. В
Дунайском заповеднике в 1984-1997 гг. общее количество гнездящихся пар колебалось от
14 до 429 (Жмуд, 1999). В дельте Днестра численность малой белой цапли в 1952-2004 гг.
изменялась в пределах 50-513 пар, но обычно составляла от 200 до 400 пар (Русев, 2004).
На Лебяжьих о-вах численность непрерывно повышалась с 4 пар в 1961 г. до 682 в 1975 г.
В 1976-1993 гг. она колебалась в пределах 211-905 пар. Затем последовало снижение чис-
ленности, в 1994-2003 гг. она составляла всего 23-96 пар (Костин, Тарина, 2004). На Вост.
Сиваше в начале 1980-х гг. в общей сложности гнездилось 270 пар. Численность достигла
максимума в 1989 г. — 770 пар, затем последовал спад до 190 пар в начале 1990-х гг. После
2000 г. численность стабилизировалась на уровне 350-450 пар (Гринченко, 2004).
Общая численность на Сев. Кавказе оценивается в 3 300^4 100 пар (Казаков и др., 2004).
В Вост. Приазовье в 1990-х гг. обнаружено 19 колоний, в которых гнездились 3 142 пары
малой белой цапли (Емтыль и др., 2003). В низовьях Дона общая численность в настоящее
время составляет примерно 500-700 пар (Белик, Динкевич, 2004), по другим данным —
1 100-1 400 пар (Казаков и др., 2004). 300^400 пар гнездится на Зап. и Вост. Маныче (Каза-
ков и др., 2004). Общая численность малой белой цапли в Ставропольском крае в 1990-х гг.
оценивалась в 100-150 пар (Хохлов, 1993). В дельте Терека в 1985 г. было учтено 430 пар,
в 1986 г. — 380 (Казаков и др., 2004). В дельте Волги численность снизилась с 3,6 тыс. пар
в 1980 г. до 1,4 тыс. пар в 1994 г. (Кривенко и др., 1998). В 1999 г. в дельте Волги и западном
ильменно-бугровом районе учтено 3 170 гнездящихся пар малой белой цапли, в 2000 г. —
1 407, в 2001 г. — 1 650 (Гаврилов и др., 2003). Плотность населения малой белой цапли на
Дамчикском участке Астраханского заповедника весной 2003 г. составляла 0,02 ос./100 га,
летом — 0,19 ос./100 га, осенью — 0,09 ос./100 га (Русанов, 2004). Численность в Кизляр-
ском зал. в 1996 г. оценивалась в 250-300 пар (Вилков, Близнюк, Джамирзоев, 2000). В
Кызыл-Агачском заповеднике в 1976-1986 гг. она колебалась в пределах 1,65-5,55 тыс. пар
(Литвинова, 1988).
На оз. Ханка гнездование носит нерегулярный характер. Судя по поведению, от 1 до 5
пар гнездилось колониях дельты р. Илистая в период с 2001 по 2003 г. (Глущенко и др.,
2003), а в 2007 г. в трёх колониях цапель, расположенных в дельте р. Илистая, суммарно
гнездилось 8 или 9 пар (было осмотрено 5 гнёзд с яйцами или птенцами) (Коробов, Глу-
щенко, 2008).
Размножение. Возраст первого размножения точно не установлен, вероятно, он состав-
ляет 1 год (Cramp, 1977). Однако, как показывают данные кольцевания, значительная часть
годовалых птиц не возвращаются в места гнездования, проводя лето значительно южнее
(Сапетин, 1978д). Моногамы. Пары образуются, по всей видимости, только на время гнез-
дового сезона, связь партнеров ограничена гнездом и его окрестностями.
Гнездится колониями, обычно совместно с другими голенастыми птицами — большой
белой, рыжей, серой цаплями, кваквой, колпицей. Может гнездиться в общих колониях
также с грачами и некоторыми хищными птицами. По данным анкетного учёта, в 1986 г. на
территории Украины всего лишь 9,7% колоний малой белой цапли были моновидовыми, в
них находилось 3,6% учтённых гнёзд (Серебряков, Грищенко, 1992). В благоприятных ме-
стах колонии могут достигать огромных размеров — до сотен и тысяч гнёзд.
В начале гнездового сезона самцы без пары часто посещают свободные гнездовые уча-
стки и токуют на них. Позже пары энергично охраняют территорию в 3-4 м вокруг гнезда,
Малая белая цапля
287
но при плотном гнездовании её радиус может быть и меньше 1 м. Самец, привлекающий
самку, часто издает характерный крик, взлетает, хлопая крыльями, совершает круговые
полеты с вытянутой шеей на 30-300 м, возвращаясь на прежнюю присаду или садясь около
нее. Токующий самец привлекает как самок, так и других самцов. Все вместе они с крика-
ми летают вокруг колонии. Несколько самок могут садиться поблизости от токующих сам-
цов, приглядываться, вытягивая шеи, подлетать к ним. Пары образуются через несколько
дней. Токовое поведение очень разнообразно (рис. 57 а, в): птицы соприкасаются крылья-
ми, вытягивают шеи, расправляют эгретки, взлетают, хлопая крыльями, трещат клювами.
Вскоре после образования пары начинается постройка гнезда и копуляции. Каждый раз,
когда один из партнеров возвращается к другому на гнездо, происходит церемония привет-
ствия: подлетающая птица издаёт особый крик, другая ей отвечает и поднимается в гнезде,
вертикально вытянув шею (рис. 576). Затем обе цапли издают щебечущий крик. Через не-
сколько секунд после посадки прилетевшей птицы оба партнёра принимают сходные токо-
вые позы, поднимая украшающие перья; затем нередко можно услышать трещание клювом
(Blaker, 1969; Cramp, 1977).
В отличие от большой белой цапли, малая чаще гнездится на деревьях и кустарниках,
но устраивает гнёзда и на заломах тростника. Из деревьев предпочитает те, которые растут
в поймах рек, по берегам водоёмов, среди болот. Это ива, ольха, дуб, вяз, сосна. В кустар-
никах чаще всего гнездится на ивах и тамариске. В агроландшафте может поселяться в
лесополосах или рощах среди орошаемых земель, возле рисовых полей или рыборазвод-
ных прудов. По наблюдениям в Сев. Приазовье, из 120 гнёзд в тростниковых колониях
66,6% находились в средних по густоте зарослях тростника, 30% — в густом заломанном
тростнике, 3,4% — в очень густом мозаичном тростнике (Кошелев и др., 2004). Может
гнездиться и на земле. В Испании и Португалии известны случаи гнездования на скалах
(Bauer, Glutz von Blotzheim, 1966).
Гнёзда на деревьях располагаются, как правило, в средней части кроны среди густых
ветвей, часто на горизонтальных ветках далеко от ствола. На крупных деревьях может на-
ходиться до нескольких десятков гнёзд. Под Ленкоранью большое количество гнёзд соору-
жается среди густой сети перепутанных колючих лиан. Гнёзда как бы висят между ветвей
стоящих рядом деревьев (Спангенберг, 1951). В Молдавии в поливидовых колониях на де-
ревьях малая белая цапля устраивает гнёзда во всех ярусах, но большая часть гнёзд нахо-
дится в среднем ярусе (Журминский, 1984; Журминский, Куниченко, 1986). На о. Городки
на Днепродзержинском вдхр. смешанная колония цапель располагалась в загущенной со-
сновой посадке на чахлых деревьях со слабо развитой кроной. Центральную часть колонии
занимали гнёзда серой и большой белой цапель, малая белая цапля и кваква располагались
кольцом по периферии. 70% гнёзд цапель находятся в развилках ветвей деревьев, 25% —
около стволов, остальные — на боковых ветках (Харченко и др., 1986).
Высота расположения гнёзд на деревьях зависит от высоты последних и архитектоники
кроны. Средняя высота расположения гнёзд в колонии в низовьях Днестра составляет
7,8±0,3 м (Журминский, Куниченко, 1986). На о. Городки большинство гнёзд построены на
высоте 7-11 м (Харченко и др., 1986). В Черноморском заповеднике гнёзда на дубах были
на высоте 3,5-8 м (Смогоржевський, 1979), в колонии у с. Кливодин в Черновицкой обл. —
на вязах и ясенях на высоте 14-22 м, в среднем — 17,3 м (Годованец и др., 1996). На Сев.
Кавказе гнёзда на деревьях птицы строят на высоте от 2 до 20, чаще всего, в — 4-8 м от
субстрата. Гнёзда в колониях могут располагаться как редко, так и скученно на расстоянии
0,5^4 м одно от другого (Казаков и др., 2004). В Астраханском заповеднике гнёзда были на
вершинах низкорослых верб, кустарниковых ив или во второй трети высоты деревьев на
высоте 7-10 м, иногда и ниже (Спангенберг, 1951).
288
Отряд Аистообразные, Семейство Цаплевые
Рис. 57. Элементы демонстрационного поведения малой белой цапли: а — токующая птица в брач-
ном оперении; б — церемония приветствия, в — брачные демонстрации самца и самки (по: Cramp,
1977; Bauer, Glutz von Blotzheim, 1987).
Малая белая цапля
289
Гнёзда в тростнике располагаются на небольшой высоте — от 0,5 до 1,5 м (Спанген-
берг, 1951). На Сев. Кавказе гнёзда в зарослях находили на высоте от 0,4 до 1,2 м (Казаков
и др., 2004). В колонии в плавнях Молочного лимана (Запорожская обл.) из 300 гнёзд малой
белой цапли 20% были построены на высоте 9-50 см над водой, 75% — в 51-100 см, 5% —
в 101-150 см; средняя высота — 60 см. По мере насиживания и роста птенцов гнёзда по-
степенно оседают, опорные стебли пригибаются, часть их ложится на воду, поэтому гнёзда
постепенно опускаются почти до самой воды. Среднее расстояние между гнёздами своего
вида в разных колониях Запорожской обл. составляет 2,5-3,5 м, до гнёзд других цапель —
2,1-3,3 м (Кошелев и др., 2004). В дельте Аму-Дарьи малая белая цапля строит гнёзда на
небольших куртинах тростника на высоте 0,4-1,4 м вблизи небольшого окна открытой воды
или у тропы, что, по-видимому, обеспечивает условия для взлёта и посадки птиц в высоком
тростнике (Лукашевич, 1990). В смешанных колониях в тростниковых зарослях малая бе-
лая цапля обычно занимает нижний ярус.
Интересно, что в разные годы, а иногда и за один сезон, малые белые цапли могут ме-
нять не только места гнездования, но и тип колоний — в тростниках или на деревьях. Так,
в Запорожской обл. в 1990-1994 гг. кваквы и малые белые цапли из тростниковых плавней
у с. Светлодолинское переселились в искусственные лесопосадки у с. Орлово, а затем в
свежепосаженный лес у с. Тихоновка. Подобные перемещения наблюдали и на Обиточной
косе в 2001-2003 гг. (Кошелев и др., 2004).
Гнёзда на деревьях цапли строят обычно из тонких длинных прутиков, реже сухих стеб-
лей тростника и корешков. Гнёзда в зарослях — из стеблей тростника и камыша. Такие
гнёзда с подрастанием птенцов «растаптываются», стенки их становятся более пологими,
лоток мелким, диаметр гнезда увеличивается (Казаков и др., 2004). В дельте Аму-Дарьи
птицы строили гнёзда из тонких стеблей тростника, не более 1 см в диаметре (Лукашевич,
1990).
Прилетевшие на место гнездования птицы прежде всего занимают старые гнёзда, при
их недостатке — строят новые. Малая белая цапля может занимать и старые гнёзда других
птиц, причём не только других цапель, но и грачей. Вновь построенное гнездо представля-
ет собой типичный для цапель перевёрнутый конус с довольно рыхлыми стенками. Строят
его оба партнёра. Самец, как правило, приносит гнездовой материал, который собирает в
непосредственной близости от колонии, а укладывает его самка. Обе птицы охраняют гнез-
до от других цапель колонии, которые могут растаскивать принесённый материал (Спан-
генберг, 1951; Bauer, Glutz von Blotzheim, 1966; Смогоржевський, 1979; Казаков и др., 2004).
Нередко первые яйца птицы откладывают в не готовые ещё гнёзда, которые затем достраи-
ваются и подновляются (Костин, 1983). Размеры гнёзд приводятся в табл. 23.
К «ремонту» и постройке гнёзд птицы приступают вскоре после прилёта. Сроки гнездо-
вания растянуты. В наибольшей степени это характерно для колоний, в которых гнёзда
часто разоряют хищники. Так, на о. Городки свежие повторные кладки встречались до пер-
вой декады июля (Харченко и др., 1986). В низовьях Днестра в Одесской обл. откладка яиц
начиналась с 12 по 29 апреля, в среднем за 6 лет — 22 апреля (Смогоржевський, 1979). В
Молдавии на нижнем Днестре малые белые цапли начинают гнездиться в первой - второй
декадах апреля, первые птенцы появляются в начале мая (основная масса — в конце меся-
ца), большая часть птенцов покидает гнёзда в конце июня, последних птенцов можно встре-
тить на колонии в третьей декаде июля (Журминский, Куниченко, 1986). На Лебяжьих о-
вах откладка яиц начинается со второй декады апреля, но обычно происходит в третьей
декаде апреля - первой декаде мая. Разница между первыми и поздними кладками за 12 лет
наблюдений составляет от 5 до 44 дней, в среднем 25±11 дней. Пик вылупления приходит-
ся на последние числа мая - первую декаду июня. Наиболее ранняя дата появления птен-
290
Отряд Листообразные, Семейство Цаплевые
Таблица 23. Размеры гнёзд (см) малой белой цапли в разных частях ареала Вост. Европы и Сев. Азии
Регион п Диаметр гнезда Высота гнезда Диаметр лотка Глубина лотка Источник
Низовья Днестра, Молдавия 34±6 22±8 17±4 6±1 Журминский, Куниченко, 1986
Лебяжьи о-ва, Крым* О. Городки, Полтавская обл. 120 22-65 (36,5) 30-42 х 27- 39 15-67 (35,3) 10-25 16^15 (26,8) 13-25х12-17 4-15 (7,7) Костин, 1983 Харченко и др., 1986
Северо-Западное 9 36-52 х 33- 11-25 18,5-30 х 17-25 4-10 Кошелев и
Приазовье 46 (42,3±2,1 х37,8±1,2) (15,8±1,3) (23,8±1,3 х 21,7±1,1) (5,9±0,8) др., 2005
Дельта Дона 123 15-56 (37) 6-39 (24) 11-30 (21) 1-15(9) Казаков и др., 2004
Ур. Круглик, дельта Кубани 58 22-37(31,6) - 14-25 (23,6) 6-10(7,6) Казаков и др., 2004
Оз. Казинка, Ростовская обл.* 45 28-46 (38,3) 15-50 (23,3) 15-37 (25,6) 3-8 (4,8) Казаков и др., 2004
* — гнёзда на заломах тростника.
цов - 6 мая 1986 г. Разница в сроках колеблется от 5 до 35 дней, в среднем составляет 24±9
дней. Вылет молодых происходит в третьей декаде июля - августе. Продолжительность
гнездового цикла от 66 до 92 дней, в среднем 80±8 дней (Костин, Тарина, 2004). В Сев.-Зап.
Приазовье в 1988-2002 гг. постройка гнёзд отмечалась с 23 апреля по 15 мая, откладка яиц —
с 29 апреля по 26 мая, вылупление птенцов — с 24 мая по 14 июня, вылет птенцов — с 23
июня по 30 июля. Гнездовая жизнь пары составляла 60-65 дней (Кошелев и др., 2005). На
о. Городки основной период кладки яиц приходится на конец апреля - начало июня, массо-
вая откладка наблюдается в начале мая (Харченко и др., 1986). На Сев. Кавказе к откладке
яиц малые белые цапли приступают в конце апреля - начале мая, массовая яйцекладка
наблюдается во второй половине мая. Свежие кладки находили до начала июля. Вылупле-
ние птенцов начинается во второй-третьей декадах мая и продолжается до конца июля.
Вылет молодых начинается в третьей декаде июня и продолжается до середины августа
(Казаков и др., 2004). В дельте Аму-Дарьи откладка яиц начинается в конце апреля - начале
мая (Лукашевич, 1990).
В год одна кладка, после разорения откладывается повторная. В полной кладке от 2 до 7
яиц, чаще всего 3-5. Известны кладки даже из 8 яиц (Bauer, Glutz von Blotzheim, 1966). На
Обиточное косе (Запорожская обл.) в 1992-1997 гг. из 192 полных кладок 4,2% имели 2
яйца, 34,9% — 3, 39,6% — 4, 18,2% — 5, 3,1% — 6 яиц. В различных колониях Сев. При-
азовья (выборка по 35-100 гнёзд) в 1988-2002 гг. средняя величина кладки колебалась от
3,6 до 4,8 яиц, в среднем составляя 4,3±0,2 яйца (п=80) (Кошелев и др., 2005). В колонии на
Каржинских о-вах в Джарылгачском зал. (Херсонская обл.) в 1997 г. средний размер клад-
ки составлял 4,3 яйца (Ардамацкая, 1999). На Сев. Кавказе из 125 насиживаемых кладок
10,4% содержали 3 яйца, 28% — 4, 45,6% — 5, 12,8% — 6, 3,2% — 7 яиц. Средний размер
кладки в низовьях Дона, на Зап. Маныче и в Вост. Приазовье был равен 4,7 яйца, в колонии
на Новотроицком вдхр. — 5,1 яиц (Казаков и др., 2004). В дельте Аму-Дарьи из 9 осмотрен-
ных в 1989 г. кладок по 44,4% имели 3 и 4 яйца, 11,1% — 5, средний размер кладки состав-
лял 3,7 яйца (Лукашевич, 1990).
Малая белая цапля
291
В Испании из 199 кладок 3% имели по 3 яйца, 24% — 4, 61% — 5, 12% — 6, 0,5% —
7 яиц; средний размер кладки был равен 4,8 яйца (Cramp, 1977).
Яйцо малой белой цапли зеленовато-голубого цвета, эллипсовидное, реже яйцевидное.
По мере насиживания окраска становится более тусклой (Спангенберг, 1951; Cramp, 1977;
Смогоржевский, 1979). Изредка встречаются яйца и чисто белого цвета (Bauer, Glutz von
Blotzheim, 1966). Размеры яиц указаны в табл. 24. Вес свежеснесённых яиц из колонии
Лебяжьих о-вов (п=4) — 28,0-28,7 г (Костин, 1983), средний вес (п=13) слабонасиженных
яиц из дельты Аму-Дарьи — 25,2±1,3 г (Лукашевич, 1990). Вес скорлупы яиц из Македо-
нии 1,44-2,36, в среднем 1,90 г (Bauer, Glutz von Blotzheim, 1966).
Яйца откладываются с интервалом в 1, реже 2 дня. По наблюдениям на Лебяжьих о-вах
(Крым), на откладку 4 яиц уходит от 4,5 до 6,5 сут. (Костин, Тарина, 2004). Насиживание
начинается с первого яйца. Насиживают оба партнёра 21-25 дней, по другим данным — до
28 дней (Смогоржевський, 1979; Костин, Тарина, 2004). Время насиживания больше при
ветреной и пасмурной погоде. В начале периода насиживания птицы довольно часто поки-
дают гнёзда. Смена партнёров чаще всего происходит в утренние часы (с 5 до 11) и в вечер-
ние (с 16 до 19) (Харченко и др., 1986). Последовательность вылупления птенцов, за редкими
исключениями, повторяет порядок откладки яиц. Вылупляется обычно 1 птенец в сутки, в
кладках из 5-6 яиц вылупление может длиться на сутки дольше (Костин, Тарина, 2004).
Птенцы полуптенцового типа (Cramp, 1977). Кормят их родители в гнезде, отрыгивая
пищу в рот птенцу или в лоток гнезда. Вылупившихся птенцов родители кормят полупере-
варенной пищей; в возрасте 1,5-2,5 недель птенцы получают частично переваренный корм,
большая часть которого состоит из личинок насекомых. С 3—4-недельного возраста родите-
ли кормят птенцов уже всеми объектами своего («взрослого») рациона (Костин, Тарина,
2004). Пока птенцы маленькие, один из родителей постоянно остаётся в гнезде. Только
когда птенцы оперяются, за кормом начинают летать оба партнёра. В это время птенцы уже
вылезают на ветки возле гнезда, где ожидают родителей (Смогоржевський, 1979).
Прилетая с кормом к гнезду, птицы занимают одно и то же место на соседней ветке или
на самом гнезде. Птенцы сразу же реагируют на прилет родителей — издают звуковые
сигналы, взмахивают крыльями, передвигаются в гнезде. Количество кормлений обычно
не превышает 2-3 в сутки (Харченко и др., 1986). Птенцы остаются в гнезде 30-36 дней
(Bauer, Glutz von Blotzheim, 1966; Ткаченко, 1996; Казаков и др., 2004).
Наибольшая скорость роста птенцов в первые 3-6 дней жизни, затем она снижается, а
позднее снова увеличивается и снова снижается перед вылетом. Вес птенцов увеличивает-
ся с 20-25 г при рождении до 420-470 г перед вылетом. В возрасте 13-20 дней начинается
Таблица 24. Размеры яиц (мм) малой белой цапли в Вост. Европе и Сев. Азии
Регион и м lim Источник
Македония 160 45,8 х 33,8 41,1-51,8 x 30,6-36,0 Bauer, Glutz von Blotzheim, 1966
Лебяжьи о-ва, Крым 513 45,4 х 33,3 41,0-55,7 х 30,5-35,5 Костин, 1983
Черноморский заповедник 108 45,9 x 33,1 42,8-51,1 х 31,2^13,4 Смогоржевський, 1979
Ур. Круглик, дельта Кубани 29 44,7 x 33,1 42,5^6,5 х 32,2-35,4 Казаков и др., 2004
Оз. Казинка, Ростовская обл. 144 46,6 х 33,4 41,3-52,5 x 30,8-38,4 Казаков и др., 2004
Ставрополье 27 46,7 х 33,9 43,5—49,5 х 33,0-34,5 Казаков и др., 2004
Ленкорань Дельта Аму-Дарьи, Узбекистан 20 33 46,5 х 32,6 44,6 х 32,5 41,6-50,1 х 30,0-34,8 Спангенберг, 1951 Лукашевич, 1990
292
Отряд Листообразные, Семейство Цаплевые
интенсивный рост пера, вследствие чего замедляется прирост веса тела. Средняя суточная
потребность птенца в энергии составляет 527,8 кДж (Ткаченко, 1994).
Оба родителя продолжают заботиться о птенцах после вылета их из гнезд. Молодые
птицы продолжают держаться вместе, даже совместно защищаются от опасности.
На Лебяжьих о-вах успешность размножения на контрольных участках колоний с опти-
мальными условиями варьирует от 79,9 до 94,4%, эмбриональная смертность составляет
1,9-11,8%, ювенильная — 2,2-18,5% (Костин, Тарина, 2004). В Сев. Приазовье успеш-
ность размножения в 1998-2002 гг. составляла 79,7%, гибель кладок — 5,0%, выводков —
15,3%, средний размер выводка был равен 3,4±0,1 птенца (Кошелев и др., 2005). В Кызыл-
Агачском заповеднике успешность размножения малой белой цапли колеблется в пределах
60,2-74,7%. Определяется она там в основном погодными условиями года, влияющими на
кормовую базу (Литвинова, 1988).
Анализ многолетних данных по успешности размножения в Камарге на юге Франции
показал зависимость успешности размножения от численности цапель — она уменьша-
лась с увеличением количества гнездящихся птиц; величина кладки увеличивалась в более
влажные годы, успешность гнездования уменьшалась с возрастанием скорости ветра
(Bennetts et al., 2000).
По данным мечения в Азово-Черноморском и Каспийском бассейнах, средняя продол-
жительность жизни малых белых цапель равна 1,2 года, а птиц, переживших первый год, —
3,9 года. Смертность в первый год жизни составляет 77%, во второй — 40%, в последую-
щие — в среднем около 25%. Показатели различаются для разных популяций. Максималь-
ный возраст — 14 лет 10 месяцев (Сапетин, 1978д).
Суточная активность, поведение. Дневная птица (Bauer, Glutz von Blotzheim, 1966).
Во время миграции летит в основном днем. В полёте малые белые цапли образуют рыхлые
скученные стаи численностью в 10-50 особей или же линейные построения — цепочки,
клин, углы и т.п. — до 10 особей (Молодовский, 2001).
Ведёт как одиночный, так и стайный образ жизни. Во все времена года кормится обыч-
но поодиночке, скопления на кормёжке образуются лишь временно. Часто кормится вместе
с другими цаплями.
Отдыхают цапли на деревьях и кустарниках. Отдыхающих молодых птиц нередко мож-
но увидеть на плавающих островках водных растений. На ночёвку птицы слетаются в су-
мерках, при этом образуются постоянные скопления, часто с другими цаплями.
Питание. Питается малая белая цапля различными мелкими водными и околоводными
животными: рыбой, земноводными, пресмыкающимися, насекомыми и их личинками, ра-
кообразными, пауками, червями, моллюсками, пиявками. Может поедать грызунов, мелких
воробьиных птиц. Обычный размер добычи — от 1 до 15 см.
Состав пищи значительно варьирует в зависимости от места, сезона и условий года.
Рацион может отличаться даже в разные дни одного месяца (Казаков и др., 2004). На о.
Городки на Днепродзержинском вдхр. в 1981-1983 гг. рыба составляла 96,4% рациона ма-
лой белой цапли, насекомые — 2,7%, земноводные — 0,6%, пиявки — 0,3%, моллюски —
0,08%, пресмыкающиеся — 0,07% (Ткаченко, 1987). В Сев. Приазовье рыба составляла
всего 25%, земноводные — 40%, беспозвоночные — 35% (Кошелев, 2004). В Кызыл-Агач-
ском заповеднике малые белые цапли также добывают большое количество земноводных,
в основном неполовозрелых особей озёрной лягушки. В июне лягушата составляют 57,1%
по весу среди земноводных. На долю рыбы приходится 47,5% по весу в мае, 30,6% — в
июне, 41,6% — в июле. Птицы поедали чаще всего атерину и гамбузию (Литвинова, 1988).
На Лебяжьих о-вах в рационе преобладали насекомые — 62,6%, рыбы составляли лишь
16,2%, лягушки — 15,6%. 5,5% приходилось на долю полихет, моллюсков, ракообразных,
Малая белая цапля
293
пауков и пресмыкающихся. Из насекомых чаще всего в пробах питания встречали личинок
стрекоз (21,6%), медведок (20,8%), личинок водяных жуков (19,2%) и двукрылых (16,4%).
Состав пищи изменяется по годам. В 1983 г. земноводных было 70%, а рыб — 28%, в
1984 г. эти показатели почти сравнялись — 48 и 50%, в 1987 г. разрыв был очень большим —
62 и 20% соответственно (Костин, Тарина, 2004).
В дельте Дуная в мае в желудках добытых цапель находили в основном рыбу, прежде
всего колюшек — до 199 экз. у одной птицы. В плавнях Днепра в Запорожской обл. в пер-
вое время после прилёта малые белые цапли кормились в основном насекомыми, ракооб-
разными, рыбой и земноводными. Среди насекомых преобладали плавунцы и их личинки,
а также клопы. Ракообразные были представлены почти исключительно щитнем (отр.
Notostraca). Из рыб преобладали щуки; обнаружены также карась, язь и другие карповые.
Из земноводных выявлены озёрные и прудовые лягушки. В гнездовой период (июнь-июль)
в низовьях Днепра и в верховьях Днепровского лимана малые белые цапли выкармливали
птенцов и питались сами в основном рыбой, земноводными и насекомыми. Из рыб в боль-
шом количестве поедали бычков, колюшек, молодых сазанов. В пищеводах и желудках птиц,
добытых в местах нереста и роста мальков сазана, были обнаружены почти исключительно
мальки этих рыб. У одной птицы их количество колебалось от 28 до 82 экз. Из земновод-
ных цапли в большом числе поедали головастиков, чесночниц и в меньшей мере — озёр-
ных лягушек. Из насекомых преобладали личинки стрекоз и плавунцов (Смогоржевський,
1959). Анализ содержимого желудков и пищеводов 39 малых белых цапель, добытых в мае -
августе в 6 областях Украины, показывает, что из 1805 обнаруженных кормовых объектов
45,8% составляли насекомые, 40,6% — рыбы, 8,3% — ракообразные, 5,2% — земновод-
ные. Из рыб преобладали сазан и колюшка, в большом количестве встречались также щука,
плотва, бычки, атеринка. Из насекомых чаще всего встречались личинки стрекоз и плавун-
цов, клопы (Смогоржевський, 1979).
В Вост. Приазовье на лиманах основу рациона малой белой цапли составляли водные
насекомые, преимущественно личинки жуков плавунцов и водолюбов. Рыбы занимали вто-
рое место. Удельный вес промысловых рыб относительно невелик — 7,8%. Это мальки
тарани, сазана, судака, бычка-кругляка. Земноводные (взрослые особи и головастики озёр-
ной лягушки и жерлянки) составляли 22,9% от общего веса пищи. В колонии в ур. Круглик
в дельте Кубани цапли питались в основном низшими раками (щитни, жаброноги, эсте-
рии), которых добывали на рисовых полях, земноводными (взрослые особи и головастики
озёрной лягушки и жерлянки), рыбами, насекомыми. Доля последних зависела от доступ-
ности их в разные годы. Из рыб птицы поедали мелких толстолобиков, сазанов, тарань,
окуней, карасей и др. Наземные насекомые были представлены в основном медведками, из
жуков встречены жужелицы, листоеды, долгоносики, божьи коровки. Из водных насеко-
мых обнаружены личинки и имаго водных жуков, полужесткокрылые, личинки стрекоз.
Среди кормовых объектов отмечены также дождевые черви, пиявки, уж, скворец (птенец),
обыкновенная полевка, домовая мышь. На водоёмах Ставрополья основу питания малых
белых цапель в гнездовой период составляет рыба: окунь — 23,9%, сазан — 19,5%, уклея —
19,2%, пескарь — 6,5%. Роль земноводных невелика — 6,3%. В дельте Дона, на Усть-Ма-
нычском вдхр. и оз. Казинка в Ростовской обл. в питании также преобладает рыба. В раци-
оне птиц из дельты Дона встречены окунь, судак, сазан, уклея, верховка, тарань, густера.
Как по весу (38,5%), так и по встречаемости (62,1 %) преобладает молодь сазана, реже встре-
чаются уклея (8,6 и 57,5%) и верховка (4,2 и 25,7%). Второе место в рационе занимали
водные насекомые — личинки стрекоз, водных жуков и полужесткокрылые. Из земновод-
ных цапли поедали взрослых озёрных лягушек и их головастиков. Из ракообразных встре-
чены гаммарусы и речной рак (Казаков и др., 2004).
294
Отряд Листообразные, Семейство Цаплевые
В питании отмечены также кузнечики, саранча, сверчки. Во второй половине лета у
добытых птиц часто попадаются наземные насекомые (Спангенберг, 1951).
Обычно малая белая цапля кормится на мелководье и берегах водоёмов, на рисовых
полях, затопленных лугах и т.п. Тактика охоты разнообразна — от активного преследова-
ния добычи до подкарауливания. Чаще всего птица бродит по мелководью, схватывая быс-
трыми движениями клюва мелких животных в воде или на её поверхности. Малая белая
цапля более подвижна, чем большая белая цапля. Нередко можно увидеть, как птица быст-
ро семенит по воде, активно преследуя добычу. Может выгонять мелкую придонную жив-
ность, взмучивая воду и ил быстрыми движениями одной ноги. Отмечались и необычные
способы охоты: кормежка с плавающих растений (Parasharya, Bhat, 1987) и даже ловля
рыбы с зависанием и посадкой на воду (Olioso, 2002).
В районе Лебяжьих о-вов кормовыми биотопами служат мелкие пресноводные и соло-
новодные разливы, рисовые чеки, протоки, каналы; часто птицы собирают корм по кромке
берега у зарослей тростников (Костин, Тарина, 2004). В Узбекистане после вылета птенцов
стаи малых белых цапель сопровождают пасущийся на лугах скот (Лукашевич, 1990).
На о. Городки на Днепродзержинском вдхр. отмечена групповая охота, чаще всего груп-
пами из 3 птиц. Цапли охотились близко одна к другой на одном участке, так что живот-
ное, вспугнутое одной птицей и ускользнувшее от неё, становилось добычей другой осо-
би. Какой-либо согласованности в действиях птиц при этом не наблюдалось (Харченко и
др., 1986).
Во время гнездового сезона малые белые цапли могут улетать за кормом на расстояние
до 20 км от колонии, а иногда и дальше (Bauer, Glutz von Blotzheim, 1966).
Враги, неблагоприятные факторы. Малая белая цапля может становиться жертвой
крупных хищных птиц — ястребов, орлов, орланов, из млекопитающих — лисиц, волков,
бродячих собак. Гнёзда разоряют болотные луни, врановые, крупные чайки, четвероногие
хищники — лисица, енотовидная собака, шакал, а также крысы. Гнёзда на мелководье мо-
гут страдать от кабанов. Так, в одной из колоний возле Лебяжьих о-вов в результате прохо-
да семьи кабанов было разрушено 11 гнёзд малой белой цапли (Костин, Тарина, 2004).
Существенный ущерб колониям малой белой цапли могут причинять врановые, прежде
всего серые вороны и грачи. На о. Городки хищническая деятельность врановых начина-
лась с появлением первых кладок у цапель и заканчивалась после вылета птенцов из гнёзд.
Вороны и грачи группами летали над колонией. Обнаружив незащищенное гнездо с яйца-
ми или птенцами, они с криками атаковали его. При этом кладка уничтожалась, само гнез-
до также в большинстве случаев разрушалось. Чаще всего врановые нападали на гнёзда с
2-3 птенцами в возрасте 1-2 недель. Птенцов серые вороны и грачи хватали клювом за
область желудка, и если они были маленькими — поднимались с ними и улетали. Более
крупные птенцы либо падали на землю, либо оставались в гнезде с пробитым желудком;
большинство из них впоследствии погибали. Появление ворон или грачей в колонии вызы-
вает бурную реакцию у цапель: поднимается шум, птицы слетают с гнёзд, садятся на край
гнёзда, издавая крики тревоги. Однако при этом они защищают только свои гнёзда, но не
соседние, подвергшиеся атаке (Харченко и др., 1986). На Сиваше значительный ущерб ко-
лониям цапель наносят сороки; некоторые из них даже гнездится в тростниках недалеко от
колоний. Гибель потомства увеличивается, если цапель кто-то вспугивает с гнёзд (Гринчен-
ко, 2004).
Из абиотических факторов наибольшее отрицательное влияние имеют экстремальные
погодные условия. Во время сильных ветров часть гнёзд может разрушаться или затапли-
ваться. Из-за ливней, особенно с градом, нередко погибают кладки и птенцы. Отрицатель-
но сказываются и резкие колебания уровня воды на реках с зарегулированным стоком, а
Малая белая цапля, Желтоклювая цапля
295
также летние паводки. Так, в дельте Днестра необычный паводок летом 1998 г. уничтожил
46,8% гнёзд малых белых цапель (Русев, 2004).
Во многих местах фактором, лимитирующим численность малых белых цапель, служит
разрушение среды обитания человеком — вырубка пойменных и прибрежных лесов, осу-
шение болот, затопление плавней вследствие гидростроительства, выкашивание и выжига-
ние тростника, нарушение нормального гидрорежима рек и т.п. Отрицательно сказывается
и фактор беспокойства.
На юге Украины у малой белой цапли зарегистрировано 33 вида гельминтов: Grypo-
rhynchuspusillus, Dendrouterina herodiae, Clinostomum complanatum, Pegosomum spiniferum,
Echinochasmus amphibolus, E. beleocephalus, E. euryporus, E. ruficapensis, Saakotrema
metatestis, Ignavia ardeae, Cryptocotyle concavum, C. lingua, Ciureana cryptocotyloides, Pygi-
diopsis genata, Prostogonimus ovatus, Opisthorchis simulans, Hepatiarius longissimus, Renicola
sudaricovi, Strigea falconis, Apharyngostrigea cornu, Codonocephalus urnigerus, Ophiosoma
patagiatum, Posthodiplostomum cuticola, Tylodelphys clavata, Bilharziella p о Ionic a, Polymorphus
minutus, P striatus, Souths ell ina hispida, Contracaecum micro cephalum, Tetrameres fissispina,
Tetrameres sp., Desmidocercella numidica, Desportesius invaginatus. Ядро гельминтофауны
включает 4 вида трематод: Cryptocotyle concavum, С. lingua, Pygidiopsis genata, Apharyn-
gostrigea cornu. Роль малой белой цапли в поддержании численности этих трематод доста-
точно велика, но всё же уступает серой цапле. Экстенсивность заражения нематодами дос-
тигает 40%, найдены 2 вида, из которых Desportesius invaginatus относится к числу доми-
нирующих и входит в состав ядра гельминтофауны. У других хозяев этот вид не найден
(Корнюшин и др., 2004).
В оперении малой белой цапли найдены пухоеды: Ardeicola expallidus и Ciconiphilus
decimlineatus (McClure, Rotanaworabhan, 1973).
Хозяйственное значение, охрана. Может приносить некоторый ущерб рыбному хо-
зяйству, особенно на рыборазводных прудах, поедая мальков; служит также распространи-
телем ряда гельминтов, вызывающих опасные заболевания у рыб. Однако этот ущерб ком-
пенсируется пользой, которую малая белая цапля приносит, уничтожая вредителей рыбно-
го хозяйства. На Украине они составляют 59,1% общего количества обнаруженных живот-
ных (Смогоржевський, 1979). Кроме того, поедает вредителей сельского хозяйства — мы-
шевидных грызунов, саранчовых, медведок и т.п. Как и другие белые цапли, вид заслужи-
вает охраны и как эстетически ценный природный объект.
Малая белая цапля занесена в Красные книги Казахстана и Узбекистана, а в России —
в Красные книги Республики Ингушетия и Сахалинской области.
Желтоклювая цапля
Egretta eulophotes (Swinhoe, 1860)
Herodias eulophotes Swinhoe, 1860, Ibis, c. 64, Амой, Китай.
Chinese [Swinhoes] egret (агл.), Schneereier (нем.), aigrette de Chine (фр.)
Син. (рус.): жёлтоклювая чепура-нужда.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Некрупная (несколько крупнее малой
белой) изящная цапля белой окраски. Взрослая птица в брачном оперении имеет густой
296
Отряд Листообразные, Семейство Цаплевые
хохол, хорошо отличающий её от средней белой цапли (не имеющей хохла) и от малой
белой, у которой хохол иного типа. В конце лета и осенью, когда перья хохлов утрачены,
различить эти виды сложнее. В целом желтоклювая цапля выглядит более толстоклювой и
коротконогой по сравнению с малой белой. У взрослой желтоклювой цапли в брачный
период — жёлтый клюв, в отличие от целиком (или главным образом) чёрного клюва ма-
лой и средней белых цапель. У средней белой цапли, кроме того, ноги полностью чёрные,
и она несколько крупнее.
Желтоклювая цапля, как правило, не встречается вне прибрежной полосы моря; она
никогда не кормится в траве и, тем более, следуя за скотом, как египетская цапля. Избегает
пресных озёр, заросших околоводной растительностью.
Очень похожи и трудно различаются молодые птицы (первогодки) желтоклювой и ма-
лой белых цапель. Однако у этих видов разное отношение к морю. Если желтоклювые, в
том числе, и молодые, могут в тихую погоду спокойно стоять и кормиться в зоне заплеска,
то для малой белой цапли это не характерно.
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Самец и самка в брачном наряде. Оперение белое, на затылке
хохол из удлинённых перьев, развиты удлинённые перья на зобу и эгретки на лопатках.
Клюв жёлтый, цевка чёрная, пальцы зеленовато-жёлтые (как и у малой белой цапли), обна-
жённая кожа «лица» тёмно-голубая.
Самец и самка в зимнем наряде. Длинные украшающие перья отсутствуют. Клюв чёр-
ный или тёмный, причём подклювье чаще всего жёлтое. Цевка от желтовато-зелёного до
голубовато-зелёного цвета, пальцы серовато-жёлтые, обнаженная кожа на лицевой части
зеленоватая (King, Dickinson, 1980).
Строение и размеры. Приводим размеры (мм) 1 экз. взрослой желтоклювой
цапли в брачном наряде, который был в распоряжении авторов. Добыт В.И. Лабзюком в
июне 1970 г. в бухте Ольга (Приморье) хранится в Зоологическом музее ДГУ, Владивосток.
Пол не указан. Длина крыла — 235; хвоста — 88; клюва — 73; цевки — 89. Длина перьев
хохла 64 мм. Хохол неполный, концы первостепенных маховых слегка обтрёпаны, сохра-
нились эгретки и украшающие перья на груди.
Размеры (мм) по справочным изданиям: крыло — 235-282, цевка — 71-100, средний
палец с когтем — 51-73.
Распространение. Гнездовой ареал. Восточноазиатский вид. Основная часть
популяции гнездится на нескольких островах Жёлтого моря у берегов Кореи и юго-вост.
Китая. В качестве пролётной птицы отмечена в Юж. Корее, Японии, в Вост. Китае на о-вах
Хайнань, Тайвань, Суматра, Борнео и Сулавеси.
В России единственная гнездовая колония располагается на о. Фуругельма в Японском
море. Впервые на территории России залётная птица была найдена на м. Олимпиады 28
июня 1915 г. (Спангенберг, 1951). Во второй половине XX в. залётных особей неоднократ-
но отмечали в разных местах Приморского края, в том числе на берегах и о-вах зал. Петра
Великого (Литвиненко, 2005) (рис. 58).
Зимовки. Прибрежные болота в Юго-Вост. Азии к югу до Филиппин, Борнео и
Сингапура.
Численность. Репродуктивную часть мировой популяции» в 1980-х гг. оценивали в 960
пар (Lansdown, 1990; Litvinenko, Shibaev, 2000), однако позднее — уже в 2,6-3,4 тыс. осо-
бей (Bird Life Intern., 2001). В колонии на о. Фуругельма гнездится 35^4-0 пар этих цапель
(Литвиненко, 2005). В 2006 г. на острове гнездилось 20 пар, а к концу гнездового сезона
учтено около 100 птиц (в основном, на ночевках).
Местообитания. Весь жизненный цикл вида связан с морскими побережьями. Гнездо-
вой биотоп — небольшие морские острова вблизи материкового побережья. Кормовые ста-
Желтоклювая цапля
297
ции — различного рода мелководья на материковом побережье (солоноватые лагуны, мел-
ководные заливы, эстуарии с отмелями и пр.). На о. Фуругельма в первые годы после вселе-
ния вид гнездился исключительно на кустах, расположенных на склонах, обращенных к
открытому морю.
Размножение. На о. Фуругельма большинство гнёзд построено на кустах бузины (Sambu-
cus latipinna) и барбариса (Berberis amurensis). Предпочтение отдаётся густым, плотным
группам кустов, в которых гнездо было замаскировано и хорошо укрыто сверху листвой.
Однако некоторые гнёзда на кустах расположены почти открыто. В последние годы были
обнаружены гнёзда в пустотах между глыбами камней, хорошо защищавших взрослых и
молодых птиц от непогоды. Величина кладки (п=6) — 3 яйца. Величина выводка (п=22):
1 (4,5%), 2 (31,8%) и 3 (63,6%) птенца. Сроки размножения растянуты. 10 июля 1998 г. в 14
гнёздах были разновозрастные оперяющиеся птенцы, некоторые из которых уже пробова-
ли летать, а в 6 гнёздах — яйца или 2-5 суточные птенцы (Litvinenko, Shibaev, 2000). Все 35
птенцов из 14 выводков, обнаруженных 10 июля, благополучно покинули остров (Литви-
ненко, Шибаев, 1999).
Питание. Мелкие водные беспозвоночные, обитающие как в солоноватой, так и в пре-
сной воде, реже — рыба, ракообразные. Места кормежки цапель из колонии на о. Фуру-
гельма находились в 15 км от гнездовий.
Рис. 58. Область распространения желтоклювой цапли: а — область гнездования и отдельные посе-
ления; б — места зимовок.
298
Отряд Листообразные, Семейство Цаплевые
Враги, неблагоприятные факторы. Единственным существенным фактором, ограни-
чивавшим численность птиц, признается охота на них (в прошлом) из-за эгреток. Взаимо-
отношения между желтоклювой цаплей и другими животными, как и воздействие погод-
ных условий, изучены слабо.
Хозяйственное значение, охрана. В связи с малочисленностью, хозяйственного значе-
ния не имеет. В настоящее время включена в Красный список МСОП как вид, находящийся
под глобальной угрозой исчезновения, и взята под охрану во всех странах, где гнездится.
Занесена в Красную книгу РФ и Красную книгу Приморского края. Остров Фуругельма,
где находится единственная гнездовая колония желтоклювой цапли в России, входит в со-
став Дальневосточного морского заповедника.
Для исключения случайной гибели цапель в местах кормёжки в сезон охоты, необходи-
мо запретить охоту на этих территориях или включить их в состав названного выше запо-
ведника.
Средняя белая цапля
Egretta intermedia (Wagler, 1829)
Ardea intermedia Wagler, 1829, Isis, стб. 659, Ява.
Intermediate [yellow-billed, median] egret (англ.); Mittelreiher, Edelreiher (нем.),
aigrette intermediate [moyene] (фр ).
Син. (лат.): для подвида intermedia’. Ardea melanopus Wagler, 1829; Ardea nigrirostris Gray,
1831; Mesophoyx intermedia Sharpe, 1898; Egretta intermedia palleuca Deignan, 1947.
Статус. Нерегулярно гнездящийся перелётный вид.
Общая характеристика и полевые признаки. Стройная цапля средних размеров (длина
тела около 70 см) и типичного «цаплевого» сложения. Оперение рыхлое, чисто белой ок-
раски. В брачном наряде на спине выделяются длинные рассученные перья (эгретки), выс-
тупающие за вершину хвоста на 10-15 см. На нижней стороне шеи и зобе расположены
рассученные перья длиной до 20 см, образующие «гриву». Удлинённые перья на голове
отсутствуют. Характерны сезонные изменения окраски клюва.
По размерам крупнее малой и желтоклювой, но мельче южной и большой белых ца-
пель. По полёту и характеру передвижения по земле также занимает промежуточное поло-
жение между относительно мелкими (малой, желтоклювой) и крупными (большой и юж-
ной) белыми цаплями. Кроме того, от желтоклювой и малой белых цапель отличается од-
нотонными с цевкой пальцами (чёрными, а не жёлтыми), а в брачном наряде — отсутстви-
ем удлинённых перьев на затылке и относительно более длинными эгретками, далеко вы-
ходящими за вершину хвоста. От южной и большой белых цапель, помимо более мелких
размеров, отличается целиком чёрными ногами, высоким у основания и укороченным клю-
вом, а в брачном наряде — наличием на зобе длинных рассученных перьев. На близком
расстоянии заметно, что у средней белой цапли угол рта заканчивается прямо под глазом, а
не заходит намного дальше, как у большой белой (Cramp, 1977; Beaman, Madge, 1998).
Полёт спокойный и прямолинейный, с глубокими взмахами широких крыльев. Взлетает
легко и быстро. Во время полёта ноги вытянуты за вершину хвоста, а шея согнута в верти-
кальной плоскости и втянута в плечи. Населяет водно-болотные угодья. Поселяется совме-
стно с другими видами цапель. В период кочёвок и миграций предпочитает держаться груп-
пами, зачастую объединяясь с другими видами белых цапель, образуя на кормёжке разре-
Средняя белая цапля
299
женные скопления. Неполовозрелые птицы в летний период ведут кочевой образ жизни,
встречаясь далеко за пределами гнездового ареала.
Голос. Молчаливая птица, изредка издает при тревоге громкий и резкий звук: «кроа-кр,
кроа-кр ...».
Подвидовая систематика. Политипичный вид, географическая изменчивость которо-
го проявляется в общих размерах и варьировании окраски неоперённых частей тела (клюв,
ноги, уздечка). Три подвида, из которых в пределах России известен лишь номинативный.
1. Egretta intermedia intermedia,
Ardea intermedia Wagler, 1829, Isis, стб. 659, Ява.
Общие размеры несколько крупнее, а окраска неоперённой части голени чёрная, а не
желтовато-оранжевая, как у двух других подвидов. Во внегнездовой сезон клюв жёлтый с
чёрной вершиной. Населяет Юж., Юго-Вост, и, отчасти, Вост. Азию.
Подвид Е. L plumifera (Gould, 1848) (2) распространён в Австралии, на о. Новая Гвинея
и прилежащих островах, имеет более мелкие размеры, а по окраске неоперённых частей
тела приближается к африканской расе. Подвид Е. L brachyrhyncha (А.Е. Brehm, 1854) (3)
распространён в Центр, и Юж. Африке, по размерам приближается к номинативному под-
виду, но имеет желтовато-оранжевые клюв и неоперённую часть цевки.
Замечания по систематике. Среднюю белую цаплю иногда относят к монотипичному
роду Mesophoyx Sharpe, 1894 (чаще трактуется как подрод рода Egretta) или помещают в
род Casmerodius Gloger, 1842, вместе с большой белой цаплей. Исследования гибридиза-
ции ДНК показали более близкое родство средней и большой белой цапель с родом Ardeay
чем с Egretta (Sheldon, 1987). Таким образом, систематическое положение средней белой
цапли требует дальнейшего изучения и уточнения.
Описание (по: A field guide to the waterbirds of Asia, 1993; King at al., 1995; Mackinnon,
Phillipps, 2000; коллекции Биолого-почвенного института ДВО РАН и Дальневосточного
государственного университета, г. Владивосток).
Окраска. Половой диморфизм в окраске отсутствует, однако, у самцов украшающие
перья немного длиннее, чем у самок.
Взрослые самец и самка. Оперение белое. Ноги целиком чёрные; голая кожа «лица» в
гнездовой период желтовато-зелёная, в остальное время — жёлтая. Радужина бледно-жёл-
тая. В брачном наряде клюв чёрный с жёлтым основанием, в остальное время жёлтый с
чёрным концом. В брачный период имеются украшающие перья на зобе и спине (эгретки).
Первый пуховой наряд. Пух белый. Клюв телесно-розовый с чёрным кончиком.
Второй пуховой наряд. Пух белый. Клюв желтоватый с чёрным кончиком.
Гнездовой наряд. Оперение белое. Клюв жёлтый с чёрным кончиком. Голая кожа в углах
клюва, на уздечке и вокруг глаза желтоватая. Ноги черноватые.
Первый годовой наряд. Оперение белое. Украшающие перья на спине и нижней части
шеи отсутствуют. Клюв жёлтый с темной вершиной. Голая кожа на уздечке и вокруг глаза
желтоватая. Ноги чёрные.
Строение и размеры. Стройная птица. Шея длинная и тонкая, но выглядит
заметно толще и короче, чем у других белых цапель, ноги длинные. Клюв относительно
несколько короче и выше, чем у других белых цапель, встречающихся в пределах России.
Размеры (мм). Длина крыла самцов 290-325, цевки 110-130, клюва 70-96 (Степанян,
2003). Птица, добытая в Приморье (пол не установлен) имела длину крыла 307, длину цев-
ки 101 и длину клюва 96 (Бутурлин, Дементьев, 1935). Размеры самцов и самок из Китая:
длина крыла 280-330, длина цевки 98-100, длина клюва 67,5-100 (Иванов, 1961). Особи из
Сев. Кореи: самка — крыло 313, цевка 114, хвост 122, клюв 74; птица неопределённого
пола — крыло 308, цевка 111, хвост 118, клюв 71 (Tomek, 1999). Птицы, добытые на Саха-
300
Отряд Листообразные, Семейство Цаплевые
лине: самцы (п=2) — крыло 303 и 313, цевка 117-118, клюв 76 и 76,5 (Takahashi, 1937);
самка — крыло 290, цевка 105, хвост 123, клюв 71 (Нечаев, 19916).
Птицы, добытые в Приморском крае: самцы (п=2) — длина крыла 300 и 300, длина
цевки 105 и 115, длина клюва 75 и 75; самки (п=3) — длина крыла 295, 300 и 300, длина
цевки 100, 103 и 105, длина клюва 70, 74 и 75; птицы, пол которых не установлен (п=3) —
длина крыла 290,295 и 300, длина цевки 103,108 и 110, длина клюва 70,71 и 72 (колл. БПИ
ДВО РАН и ДВГУ, г. Владивосток).
Размеры самцов и самок (мм) подвида Е. i. intermedia (Cramp, 1977): длина крыла в
среднем 299 мм (275-327, п=13), длина хвоста— 118 (103-135, п=7), длина клюва — 72,8
(66-76, п=14), длина цевки — 106 (93-111, п=7).
Масса птиц из Китая: самцы (п=2) — 470 г и 642 г, самка — 600 г, птица, пол которой не
установлен — 700 г (Иванов, 1961). Самка, добытая на Юж. Сахалине, весила 458 г (Неча-
ев, 19916).
Линька. Полная годовая линька взрослых птиц протекает с июля по ноябрь. Частичная
предбрачная линька происходит на зимовках. Молодые птицы в гнездовом наряде начина-
ют смену мелких перьев в августе, продолжают осенью и заканчивают во время зимовки;
на втором году жизни они проходят полную годовую линьку.
Две птицы, добытые в районе зал. Ольги (Приморский край) 20 мая 1980 г., были в
брачном наряде, однако, клюв самца был жёлтым с тёмной вершиной, а у самки — чёрным
с жёлтым основанием. Из трёх птиц, добытых на оз. Ханка, одна особь от 30 июня 1978 г.
не имела следов линьки; у экз. от 15 июля 1977 г. начиналась линька мелкого оперения;
взрослая самка, добытая 25 июля 1980 г. находилась в разгаре полной линьки (примерно
треть маховых и рулевых перьев оказались утерянными либо растущими, происходила ин-
тенсивная линька мелкого оперения).
Распространение. Гнездовой ареал. Центр, и Юж. Африка, Шри Ланка, Бирма,
Индонезия, Китай, Япония, Корейский п-ов, Молуккские о-ва, Сев. и Вост. Австралия (Спан-
генберг, 1951; Степанян, 2003; Vaurie, 1965; Dickinson, 2003; и др.). Средние белые цапли
номинативного подвида гнездятся в центральных и южных районах Китая, на о-вах Тай-
вань и Хайнань (Mackinnon, Phillipps, 2000), в Гонконге (Carey et al., 2001), в центральных
и южных районах п-ова Корея (Won Pyong-oh, 1996; Tomek, 1999), в Японии — на о-вах
Хонсю, Сикоку, Кюсю и Садо (Check-list of Japanese birds, 2000), а также в Индии и на Шри
Ланка (рис. 59).
На юге Дальнего Востока России гнездование зарегистрировано на оз. Ханка (Полива-
нова, Глущенко, 1977; Глущенко, Мрикот, 2000) и на о. Шикотан, Курильские о-ва (Динец,
1996). Попытку гнездования наблюдали в районе залива Ольги, Приморский край (Лаб-
зюк, 1981). Кроме того, гнездование предполагается на крайнем юго-западе Приморья (Лит-
виненко, Шибаев, 1999) (рис. 60).
Зимовки. Птицы номинативного подвида зимуют в Юго-Вост. Азии: в южных райо-
нах Китая, на о-вах Тайвань и Хайнань (Cheng Tso-Hsin, 1987; Mackinnon, Phillipps, 2000),
Филиппинах, Калимантане, в Индонезии (Vaurie, 1965), Вьетнаме (Во Кви, 1983), Таилан-
де (Lekagul, Round, 1991), в южных районах Японии (о. Кюсю и острова, расположенные
южнее) (Check-ist of Japanese birds, 2000), Гонконге (Carey et al., 2001).
Первые залёты на территорию Приморского края были зарегистрированы в начале
XX века (Бутурлин, Дементьев, 1935; Белопольский, 1955). Начиная с 1960-х гг. они уча-
стились и стали регулярными (Литвиненко, Шибаев, 1965; Лабзюк и др., 1971; Елсуков,
1974; Глущенко, 1981; Лабзюк, 1981, 1990). Отмечены залёты в Нижнее Приамурье (Ба-
бенко, 2000), на Сахалин (Нечаев, 19916), Монерон (Нечаев, 1975), Юж. Курильские о-
ва: Кунашир (Нечаев, 1969) и Шикотан (Дыхан, 1990) и Камчатку (Артюхин и др., 2000).
Средняя белая цапля
301
Рис. 59. Область распространения средней белой цапли: а — гнездовой ареал, б — установленные
места обособленного гнездования, в — районы зимовок северных популяций. Подвиды: 1 — Egretta
i. intermedia, 2 — Е. i. plumifera, 3 — E. i. brachyrhyncha.
В северной части Японии отмечены залёты
на о. Хоккайдо (Check-list of Japanese birds,
2000).
Миграции. В Юж. Приморье весенние
миграции протекают в конце апреля и в мае.
Наиболее раннее появление зарегистрирова-
но 14 апреля 1993 г. на крайнем юге Примо-
рья в устье р. Туманная (данные Ю.Н. Глущен-
ко), 26 апреля 2004 г. в окрестностях Уссурий-
ска (Глущенко и др., 2006а), 27 апреля 1994 г.
на оз. Ханка (Глущенко и др., 20066) и 27 ап-
реля 1979 г. в зал. Ольги (Лабзюк, 1981). Пос-
легнездовые кочёвки на оз. Ханка происходят
в августе, а самая поздняя достоверная встре-
ча осенью отмечена 17 сентября 1973 г. В ок-
рестностях Владивостока (устье р. Шмитов-
ка) одна особь наблюдалась 16 сентября 2007
г. (данные Ю.Н. Глущенко). На о. Шикотан
двух птиц наблюдали ещё 16 октября 1986 г.
(Дыхан, 1990).
Рис. 60. Ареал средней белой цапли в России,
а — установленное место гнездования, б —
район встреч птиц во время сезонных мигра-
ций и летних кочевок, в — предполагаемое
место гнездования, г — залёты.
302
Отряд Листообразные, Семейство Цаплевые
Местообитания. На Ханке две колонии, в которых гнездились средние белые цапли,
размещались в приустьевой части р. Илистая в полосе затопленных ивняков, окружённых
озёрно-болотным массивом. Местами кормёжки здесь служат сырые луга, травяные боло-
та, мелководья озёр и рисовые поля. На о. Шикотан птицы гнездились в заболоченной пой-
ме ручья, в тростниковых зарослях с куртинами курильского бамбука и группами деревьев
(Динец, 1996). В период сезонных миграций и летних кочёвок средние белые цапли встре-
чаются по берегам озёр, рек и водохранилищ, на рисовых полях, сырых лугах и травяных
болотах, как во внутренних районах, так и на побережье.
В Китае, на Корейском п-ове и в Японии средние белые цапли населяют травяные боло-
та, сырые луга, марши, грязевые отмели и рисовые поля (A field guide to the birds of Asia,
1993). В Японии они устраивают гнёзда на соснах и в бамбуковых зарослях (Jahn, 1942), на
Корейском п-ове — на деревьях, окружающих рисовые поля (Gore, Won Pyong-oh, 1971).
Численность. На оз. Ханка гнездится не регулярно и с переменной численностью. Впер-
вые два гнёзда были обнаружены в приустьевой части р. Илистая в 1971 г. (Поливанова,
Глущенко, 1977). В 1973-1980 гг. вид почти ежегодно регистрировали в летнее время в
южных и восточных районах Приханкайской низменности, а в 1976-1977 гг. отмечали мо-
лодых особей, что позволяет предполагать их нерегулярное гнездование в данный период
(Глущенко, 1981). В 1999-2002 гг. средние белые цапли вновь обнаружены гнездящимися в
той же колонии, однако их численность, определённая в 1999 и 2000 гг. соответственно в
20-30 и 30-40 гнездящихся пар (Глущенко, Мрикот, 2000), оказалась завышенной. Реально
здесь могло гнездиться от 7 до 10 пар (Глущенко и др., 2003). В 2000 г. гнездилось 8 пар, а три
осмотренных гнезда содержали 1,3 и 4 яйца. В 2002 г. вид вовсе не был здесь обнаружен, а в
июне 2003 г. от 1 до 3 птиц наблюдали в одной из субколоний дельты р. Илистая. Одиночек
также видели кормящимися на лугах в окрестностях с. Сиваковка (Глущенко и др., 2003).
На побережье зал. Ольги (Приморский край) в устье р. Аввакумовка отмечена неудач-
ная попытка гнездования: птицы начали строить гнездо, но позднее оно оказалось брошен-
ным (Лабзюк, 1981). Кроме размножающейся популяции в южных и восточных районах
Приморского края в тёплое время года (преимущественно с мая по июль) регулярно встре-
чали отдельных холостых птиц и их группы, насчитывавшие до 10 и более птиц (Литвинен-
ко, Шибаев, 1965, 1999; Лабзюк и др., 1971; Елсуков, 1974; Волошина и др., 1999; Лабзюк,
1981, 1990; Глущенко, Назаров, ориг.). На о. Шикотан в 1988 г. найдено гнездо с двумя
птенцами (Динец, 1996).
В Японии средняя белая цапля немногочисленна летом и малочисленна зимой на край-
нем юге (A field guide to the birds of Japan, 1982). В Китае это обычный вид (Mackinnon,
Phillipps, 2000); в Гонконге — обычен как летом, так и зимой (Carey et al., 2001); в Сев.
Корее — редок на гнездовании (Тошек, 1999), а в Юж. Корее — немногочислен в гнездо-
вой сезон (Won Pyong-oh, 1996).
Размножение. Колониальная птица. Моногам. Формирование пар сопровождается раз-
нообразными демонстрациями (рис. 61). Гнездовая биология в условиях Приморского края
изучена крайне слабо. Гнездовой сезон длится здесь с мая по июль. На оз. Ханка гнездится
в поливидовой колонии цапель и больших бакланов. Гнёзда располагаются на кустах ивня-
ка и построены почти исключительно из сухих прутьев ив. Размеры (см) гнёзд (п=3): диа-
метр гнезда 27-38 (в среднем 30,7); диаметр лотка 15-23 (в среднем 18); толщина гнезда 9-
21 (в среднем 16). В кладке 3-5 однотонных, слегка зеленовато-голубоватых яиц. Размеры
(мм) яиц с побережья Ханки (п=8): 45,9-50,9 х 33,9-37,7 (в среднем 48,3x35,1) (Глущенко
и др., 20066); из Китая: 43-53 х 33-39 (в среднем 48x36) (Zhao Zheng-jie, 2001); из Индии
(п=60) 42,6-52,8 х 33,1-38,6 (в среднем 47,6x35,8) (Baker, 1929). Кладку насиживают оба
партнёра в течение 21-27 суток.
Средняя белая цапля
303
В Китае птенцы вылупляются в июне,
в июле встречаются слётки (Латуш, 1934,
цит. по: Спангенберг, 1951).
Гнездо, найденное на о. Шикотан, по-
мещалось в развилке ствола ивы на вы-
соте около 5 м над землёй и было пост-
роено из ивовых веточек, стеблей трав и
листьев курильского бамбука. Толщина
гнезда составила около 30 см, а диаметр —
не менее 60 см; 12 и 13 июля 1988 г. в
нём находилось 2 птенца (Динец, 1996).
Для птенцов средней белой цапли ха-
рактерен сиблицид, как и для птенцов
большой белой цапли (McKilligan,
1990b).
Суточная активность, поведение.
Ведёт дневной образ жизни. Кормятся
цапли обычно поодиночке, но иногда
могут собираться в стаи до десятков и
даже сотен птиц (Martinez-Vilalta, Mods,
1992). Поведение вида на территории
России не изучено.
Питание. Основными объектами пи-
тания служат водные и наземные беспоз-
воночные (моллюски, пауки, насекомые Рис- 61. Токовая поза самца средней белой цапли (по
и их личинки) и позвоночные (рыбы, ам- фотографиям Ong Long-Shin).
фибии) животные. Желудок птицы, добы-
той на о. Сахалин 26 мая 1974 г., содержал остатки личинок водных насекомых (Нечаев,
19916). В желудке птицы, добытой 30 июня 1978 г. на оз. Ханка, оказались головешка-
ротан (Perccottus glenii) и три личинки стрекозы, а в желудке особи, добытой там же 25
июля 1980 г., обнаружены три личинки плавунцов, паук и остатки хитина насекомых (Глу-
щенко, ориг.).
Враги, неблагоприятные факторы. В колонии на побережье оз. Ханка цапли испыты-
вают мощный пресс со стороны основного конкурента — большого баклана. Другой суще-
ственный негативный фактор здесь — высокий уровень беспокойства со стороны людей и
скота. Он особенно велик в маловодные годы, когда болотистая низменность становится
легко доступной (Гусаков, Виноградов, 1998). Катастрофические последствия для гнездя-
щихся на оз. Ханка птиц может иметь уничтожение зарослей ивняка пожарами в месте
размещения колонии во время засушливой погоды (Глущенко, 2005).
В Японии средняя белая цапля была многочисленной в прошлом, однако загрязнение
среды обитания и беспокойство птиц в колониях привело к существенному сокращению
численности, начиная с 1960-х гг. (Martinez-Vilalta, Mods, 1992).
Хозяйственное значение и охрана. Как очень редкий вид хозяйственного значения не
имеет. Занесена в Красную книгу Российской Федерации (2001) и Красную книгу Примор-
ского края (2005). Место размещения колонии на оз. Ханка входит в состав охранной зоны
Ханкайского государственного заповедника. Рекомендуется увеличение площади данного
заповедника за счёт включения территории указанной колонии в его состав.
304
Отряд Листообразные, Семейство Цаплевые
Род Casmerodius Gloger, 1842
Большая белая цапля
Casmerodius albus (Linnaeus, 1758)
Ardea alba Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Швеция.
Great white heron, great egret (англ.), Silberreiher (нем.), grande aigrette (фр ).
Син. (лат.): для подвида albus\ Ardea egrettoides S.G. Gmelin., 1774; Ardea garzette major
Boddaert, 1783; Ardea egretta Bechstein, 1793, nec Linnaeus; Herodius Candida C.L. Brehm,
1824; Lepterodas flavirostris Hemprich et Ehrenberg, 1828; Egretta alba (Linnaeus, 1758); Egretta
orientalis Bonaparte, 1832; Erodius victoriae MacGillivray, 1832.
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Крупная птица размером с серую цап-
лю с длинной тонкой шеей, длинными клювом и ногами. Длина тела 85-102 см, размах
крыльев — 140-170 см, вес до 1,5-2 кг. Оперение чисто белое, клюв в гнездовой период
чёрный с жёлтым основанием, в зимнем наряде — жёлтый. Голень желтоватая, цевка и
пальцы чёрно-бурые. В полете, как и большинство других цапель, складывает шею. Молча-
лива. От вспугнутой цапли можно услышать характерный раскатистый треск «крр-рр-ра».
Встречается, обычно, на берегах водоёмов, лугах, пастбищах, болотах.
От серой цапли отличается белой окраской оперения, более «изящными» контурами, от
малой белой — величиной (почти вдвое крупнее). На большом расстоянии по размеру лег-
ко спутать с другими белыми цаплями. От малой белой можно отличить по более тяжёлому
полету с меньшей частотой взмахов крыльев, более стройному телу и длинной шее, чёр-
ным пальцам; в негнездовой период также по жёлтой окраске клюва. От средней — по
более длинным клюву и шее. От обоих этих видов, а также желтоклювой цапли — по отсут-
ствию в брачный период гривы из узких перьев на зобе (перья здесь лишь немного удлине-
ны). У большой белой цапли, в отличие от средней, угол рта заканчивается не под глазом, а
заходит намного дальше. В полёте ноги выступают назад дальше, чем у более мелких бе-
лых цапель. Хорошо отличается по голосу: у малой и средней белых цапель это не треск, их
позывы больше похожи на «кваканье» кваквы. От изредка встречающихся альбиносов се-
рой цапли большая белая отличается более длинной и тонкой шеей, менее массивной голо-
вой с более длинным и тонким клювом, более тёмными ногами (Cramp, 1977; Beaman,
Madge, 1998).
Голос. Летящие или кормящиеся птицы кричат редко. Крик похож на раскатистое «крр-
рр-ра» или «крраак». На колонии можно услышать громкие каркающие звуки: «рра», «аррп».
«Приветствие» на гнезде звучит как «рррооо-рррооо» (Bauer, Glutz von Blotzheim, 1966).
Крик птенцов в гнезде, выпрашивающих корм, напоминает «чечётку» птенцов серой цап-
ли: «ке-ке-ке» или «че-че-че».
Подвидовая систематика. Выделяют 3-4 подвида (Martinez-Vilalta, Motis, 1992; Howard,
Moore, 1998; Dickinson, 2003; Степанян, 2003), различающихся размерами, особенностями
оперения, окраской клюва и ног. В Вост. Европе и Сев. Азии обитает один:
1. Casmerodius albus albus
Ardea alba Linnaeus, 1758, Syst. Nat., ed. 10, c. 144, Швеция.
Наиболее крупная форма. Длина крыла самцов 410-485, цевки 170-215, клюва 117-140
мм (Степанян, 2003). Встречается от Юж. Европы и Голландии до Приморья и Сев. Япо-
нии, на юг доходит до Турции, Ирана, Афганистана, Монголии, Китая и Японии.
Большая белая цапля
305
Для прочих регионов описаны также G a. melanorhynchos (Wagler, 1827) (2) — встре-
чается в Африке к югу от Сахары и на Мадагаскаре, С. a. egretta (J.F. Gmelin, 1789) (3) —
подвид распространён в Америке от юга Канады до Патагонии (Спангенберг, 1951; Bauer,
Glutz von Blotzheim, 1966; Howard, Moore, 1980). Формы, обитающие в тропической и Вост.
Азии, в Австралии и Океании и традиционно включаемые в состав этого вида, отнесены в
настоящем издании к виду Casmerodius modestus (см. след, очерк).
Замечания по систематике. Традиционно большую белую цаплю относили к роду
Egretta (напр., Спангенберг, 1951; Cramp, 1977; Howard, Moore, 1980; del Hoyo et al., 1992;
Степанян, 2003). Внутри этого рода большая белая цапля обособлялась на уровне подрода
Casmerodius Gloger, 1842 (Степанян, 2003). Однако было показано, что по ряду признаков,
в частности, строению скелета, структуре ДНК, поведению, большая белая цапля стоит
ближе к роду Ardea, чем Egretta (Bauer, Glutz von Blotzheim, 1966; Sheldon, 1987; McCracken,
Sheldon, 1998), поэтому многие систематики повышают ранг подрода Casmerodius са-
мостоятельного рода, (напр. Grimett et al., 1998; Robson, 2000; Brazil, 2009) или даже вклю-
чают этот вид в род Ardea (напр. Dickinson, 2003). Учитывая ряд существенных отличий
большой белой цапли от цапель рода Ardea (характерные для цапель рода Egretta белое
оперение, сезонная смена окраски неоперённых частей тела, наличие рассученных эгре-
ток, а также характер вокализации, непохожий на вокализацию цапель обоих родов), в на-
стоящем издании принято «компромиссное» решение, и этот вид выделен в род Casmerodius
(со сменой женского рода видового названия alba на мужской albus). В широком смысле
вид С. albus включает также азиатско-австралийскую форму modesta (modestus), однако
симпатрия этой формы и номинативной albus на юге Приморья при разнице в сроках и
особенностях гнездования, а также отсутствии видимой гибридизации не позволяет счи-
тать данные формы конспецифичными. Ряд исследователей выделяют азиатско-австралий-
скую форму в самостоятельный вид Casmerodius modestus (Egretta modesta) (Глущенко и
др., 2003; Коблик и др., 2006). Очевидно, следует также принимать во внимание различия в
окраске ног западных и дальневосточных популяций С. albus и различия в размерах и ок-
раске ног тропических и японско-дальневосточных популяций С. modestus. Исследования
по установлению таксономического статуса отдельных форм, а также родовой принадлеж-
ности всего комплекса «больших белых цапель» должны быть продолжены.
Описание. Окраска. Самец и самка в брачном наряде. Оперение чисто белое. Перья
на затылке и зобе немного удлинены. На спине до 30-50 рассученных и сильно удлинённых
плечевых перьев (эгреток) длиной до 50 см, они заходят за хвост и несколько свисают с
боков тела. Клюв чёрный с жёлтым, реже — красновато-жёлтым основанием. Голая уздеч-
ка и кольцо вокруг глаза зелёные или жёлто-зелёные. Голень жёлтая или розовато-жёлтая,
по бокам этот цвет заходит и на бока цевки в виде узких полос. В разгар брачного периода
жёлтые участки ног становятся красноватыми (у дальневосточных птиц до ярко-красных).
Цевка и пальцы обычно черноватые, но их окраска может варьировать от зеленовато-серой
до чёрной. Окраска ног в целом может изменяться от преимущественно чёрной (западные
особи) до преимущественно жёлтой (дальневосточные особи). Радужина жёлтая. У самки
удлинённые перья несколько короче, чем у самца.
Самец и самка в зимнем наряде. Отличаются отсутствием эгреток и удлиненных перьев
на затылке и зобе. Некоторые плечевые перья имеют эгреткообразные рассученные вер-
хушки. Клюв жёлтый.
Пуховой наряд. Пух чисто белый, на затылке несколько удлинённый, образует подобие
хохолка. Кожа вокруг глаза, подбородок и горло голые, серовато-телесного цвета, клюв и
ноги телесного или рогового цвета, иногда с желтоватым оттенком.
Гнездовой наряд. По окраске сходен со взрослым зимним нарядом.
306
Отряд Листообразные, Семейство Цаплевые
Строение и размеры. Приведены в таблице 25.
Размеры птиц (мм) без разделения на половые группы таковы. По Украине: длина кры-
ла от 403 до 440, хвоста — 158-184, клюва — 121-131, цевки — 180-209 (Смогоржевсь-
кий, 1979); по Казахстану: длина крыла — 410-470, хвоста— 175-200, клюва— 115-143,
цевки — 165-212 (Долгушин, 1960). В целом по прежней территории СССР (п=14): длина
крыла — от 412 до 465, в среднем — 437,8 (Спангенберг, 1951).
По Украине (на основании коллекционных материалов Зоологического музея Киевско-
го национального ун-та) (п=9): длина крыла — 410-458, в среднем — 436,4±5,2; хвоста —
149-181, в среднем — 166,3±4,3; клюва — 121-131,4, в среднем — 125,6±1,1; цевки —
185-213, в среднем — 197±3,7 мм (ориг.).
Молодые особи имеют несколько более короткие крылья и хвост, дорастание маховых и
рулевых перьев идёт в течение первых 5-10 месяцев жизни (Фурманова, 1998).
Формула крыла (не считая рудиментарного первого махового) II>III>IV>I>V>VI. На
внешних опахалах II—IV и на внутренних опахалах I—III первостепенных маховых есть вы-
резки. Хвост слегка закруглён. Рулевых — 12. Клюв крепкий, заострённый на конце. Режу-
щий край его слегка зазубрен. Ноги относительно длиннее, чем у видов Egretta и Ardea
(Cramp, 1977). Вес 3 особей подвида С. a. albus — 960-1 680 г (Bauer, Glutz von Blotzheim,
1966). В течение года вес может существенно изменяться. Так, у цапель, добытых на юге
Запорожской обл., летом вес составлял 1 600-1 700 г, осенью и зимой — 2 000-2 100 г. Увели-
чение веса идёт прежде всего за счёт накопления жировых отложений (Фурманова, 1998).
Площадь опоры ног у большой белой цапли составляет 20,86 см2, весовая нагрузка —
43,14 г/см2 (Фурманова, 1998).
Линька. У молодых птиц частичная постювенильная линька происходит в августе-но-
ябре первого года жизни. В это время сменяется мелкое оперение. Маховые перья линяют
весной следующего года. У взрослых птиц частичная предгнездовая линька в декабре-мае
и полная послегнездовая — в августе-ноябре, первостепенные маховые сменяются нерегу-
лярно. Украшающие перья начинают выпадать уже в начале гнездового сезона (Bauer, Glutz
von Blotzheim, 1966; Stresemann, Stresemann, 1966; Cramp, 1977). В Крыму эгретки сбрасы-
вают в мае. Линька мелкого оперения начинается в конце июня (Костин, 1983).
Распространение. Гнездовой ареал. Большая белая цапля — вид-космополит,
широко распространённый в тропическом и умеренном поясах. Она встречается на всех
континентах, кроме Антарктиды и Австралии; в тропической Азии, очевидно, только зиму-
ет, будучи замещена здесь на гнездовье близким видом — южной белой цаплей (рис. 62).
Таблица 25. Размеры (длина крыла, хвоста, клюва и цевки) самцов и самок больших
белых цапель из разных регионов (в мм)
Часть тела Самцы Самки Источник
п lim М n lim М
Европа
Крыло 12 410-465 400-450 Witherby, 1943
Хвост 140-185 140-185
Клюв 110-135 110-132
Цевка 170-215 170-215
Украина, Юг Запорожской области. Молодые (imm.) птицы
Крыло 2 440,0 5 418,0 Фурманова, 1998
Хвост 2 164,0 5 157,8
Клюв 2 123,5 5 113,6
Большая белая цапля
307
Рис. 62. Область обитания большой белой цапли: а — районы гнездования, б — места зимовок.
Подвиды: 1 — С. a. albus, 2 — С. a. melanorhynchus, 3 — С. a. egretta.
В Евразии распространена от юга Испании до Приморья и Японии13. В Зап. и Центр.
Европе спорадически встречается на гнездовании на север до Голландии и Польши. В Вост.
Европе северная граница её ареала простирается до южной Белоруссии, Брянской, Там-
бовской, Самарской обл.; далее на восток она проходит примерно по 56° с.ш., затем откло-
няется южнее к 54° с.ш., проходит вдоль восточных границ Казахстана, через Сев. Монго-
лию, Приамурье. На юг в Азии доходит до Турции и Юж. Ирана. В Африке распространена
от границы Сахары до ЮАР и Мадагаскара. В Америке обитает от юга Аргентины до юга
Канады, а также на Б. и М. Антильских о-вах (Cramp, 1977; Martinez-Vilalta, Motis, 1992;
Степанян, 2003).
Ареал большой белой цапли на протяжении последних 150 лет претерпел значительные
изменения. В прошлом эта птица была распространена достаточно широко. В середине
XIX в. её ареал доходил на север до нынешней Львовской (Dzieduszycki, 1896; Страутман,
1963) и Тернопольской (Бокотей, Соколов, 2000) обл., районов Умани (Goebel, 1879), Харь-
ковской губернии (Сомов, 1897), Сарпы, низовий Урала и Эмбы (Мензбир, 1895).
Из-за моды на украшения из перьев во второй половине XIX в. началось истребление
многих видов белых цапель, достигшее пика на рубеже веков. В результате на значитель-
ной части своего ареала большая белая цапля была полностью истреблена. В Сев. Евразии
немногочисленные поселения сохранились лишь в самых труднодоступных местах, в ос-
новном в плавнях низовий крупных рек. Так, в Молдавии считанные особи уцелели только
в низовьях Днестра и Прута (Аверин и др., 1971). Почти полностью были истреблены бе-
лые цапли на юге Украины, в дельте Волги и в Казахстане.
После запрета добычи белых цапель в СССР в 1920-е гг. численность и ареал начали
постепенно восстанавливаться, однако на протяжении нескольких десятилетий это проис-
ходило весьма медленно. Ещё Е.П. Спангенберг (1951) отмечал, что во многих частях её
прежнего ареала в СССР большая белая цапля остаётся редкой птицей. В середине XX в. в
Молдавии она гнездилась только в низовьях Прута и Днестра, на территории Украины от-
носительно крупные колонии существовали в низовьях Дуная, Днестра, Днепра, на Молоч-
13 Выделение вида С. modestus усложняет описание ареала, поскольку нет чёткого представления о
существовании границ, разделяющих области обитания его и С. albus в Приморье (Глущенко и др.,
2003) (прим. ред.}.
308
Отряд Листообразные, Семейство Цаплевые
ном лимане в Запорожской обл. Севернее отмечали лишь редкие случаи гнездования, в
основном отдельных пар (Смогоржевский, 1959). Дальше на восток граница ареала прохо-
дила через низовья Дона, Сарпинские озёра в Поволжье, низовья Урала (Спангенберг, 1951).
В Казахстане — от Камыш-Самарских озёр через Тургай и Кургальджинскую впадину к
Зайсанской котловине (Долгушин, 1960; Ковшарь, Березовиков, 2001). В Сибири большая
белая цапля гнездилась в Юж. Забайкалье, в Приморье. Но была обычной только в южной
его части (Спангенберг, 1951).
По Л.С. Степаняну (1975), северная граница ареала большой белой цапли проходила в
Центр. Европе примерно по 48-й параллели, в европейской части СССР она гнездилась до
49-й параллели, в долинах Волги и Урала, Волго-Уральском междуречье, бассейнах Иргиза
и Тургая — до 50-й, в Сев. Казахстане — до 51 -й широты, восточнее — до Зайсана, долины
Чёрного Иртыша, сев.-зап. Монголии, долины Аргуни, среднего течения Амура. В Примо-
рье предположительно гнездилась до 46-й параллели.
Во второй половине XX столетия наметились как рост численности, так и расселение
большой белой цапли. В начале 1970-х гг. отмечен рост численности и расселение на но-
вые территории в Венгрии (Schmidt, 1977). Число гнёзд здесь возросло с 5-6 до 700 (Bauer,
Berthold, 1997). В 1978 г. зарегистрирован первый случай гнездования этой птицы в Нидер-
ландах (Munteanu, Ranner, 1997). В 1993 г. пара гнездилась в Баварии (Bezzel, 1994).
В Латвии пара предположительно гнездилась в 1977 г. на оз. Энгуре (Тауринын, 1983),
однако в 1980-84 гг. в гнездовой период здесь наблюдали лишь одиночных птиц (Приедни-
екс и др., 1989). Попытки гнездования возобновились через 20 лет: в 1997 и 1999 гг. Отме-
чали гнездование смешанной пары с серой цаплей (Baumanis, Kalnips 1997; Lipsbergs, Roze
2001), в 2000 г. загнездилась уже нормальная пара (Lipsbergs et al., 2001). В 1997 г. гибрид-
ная пара вывела 4 птенцов (Baumanis, 1997). В это же время происходит рост численности
и расселение вида в странах СНГ. Причём идёт как расселение внутри ареала, так и про-
движение его границы на север.
В Крыму до 1970-х гг. большая белая цапля была редкой пролётной, спорадически лету-
ющей и зимующей птицей. Ситуация изменилась после строительства Северо-Крымского
канала в 1960-х гг. Уже в 1967-1968 гг. на опресняемых мелководьях возле Лебяжьих о-вов
у северо-западного побережья Крыма летовки белых цапель стали регулярными, число птиц
на послегнездовых кочёвках выросло от единиц до десятков. В 1970 г. на Лебяжьих о-вах
загнездилась первая пара. Численность гнездящихся птиц всё время увеличивалась, уже в
1976 г. тут насчитывали 424 пары. Цапли расселялись и по другим районам Крыма (Кос-
тин, 1983). Для орошения полей началось отведение дренажных вод в Сиваш, что вызвало
коренное преобразование биотопов. На месте солоноводных и гиперсолёных водоёмов с
крайне бедной растительностью образовались пресноводные болота с плавневым комп-
лексом растений. Первая колония большой белой цапли на Вост. Сиваше была обнаружена
в 1978 г., в 1983 г. было уже 7 колоний, в которых гнездилось 153 пары (Гринченко, 2004а).
В Молдавии с 1981 г. вид регулярно стал гнездиться на прудах Кагульского рыбокомби-
ната (Куниченко, Гусан, 1992). Сейчас цапля гнездится также на прудах в южной и цент-
ральной частях страны (Куниченко, Гусан, 2001) и на среднем Днестре (Зубков и др., 2001).
В западных областях Украины большая белая цапля гнездилась в XIX в. (Dzieduszycki,
1896; Страутман, 1963; Грабар, 1997; Бокотей, Соколов, 2000). В последний раз она была
обнаружена на гнездовании в 1925 г. в Тернопольской обл. (Храневич, 1926). В дальней-
шем более полувека встречались лишь залётные и летующие особи (Страутман, 1963; Скиль-
ский и др., 2001а). В 1985 г. 4-5 пар загнездились в колонии серой цапли и кваквы возле
одного из рыборазводных прудов в Черновицкой обл. Во второй половине 1980-х - начале
1990-х гг. были обнаружены ещё две колонии в Черновицкой обл. (Скильский и др., 2001а,
Большая белая цапля
309
20016). В 1994 г. найдена колония на пруду в Львовской обл. (Чорненька, 1998), в 1990 г. 3
пары загнездились в колонии серой цапли на оз. Любязь в Волынской обл., в 1994 г. здесь
было уже 11 пар, в 1997 г. — 23-25 пар (Горбань, Фладе, 2000).
В Приднепровье первый этап расселения большой белой цапли на север начался после
создания каскада водохранилищ в 1950-70-х гг. Область её обитания продвинулась до вер-
ховий Киевского вдхр., где в тростнике гнездилось более 100 пар. В 1980-х гг. началось
расселение вверх по долинам левобережных притоков Днепра — Сулы, Пела, Ворсклы,
Десны (Гаврись, 1994). Несколько раньше, после создания Днепродзержинского вдхр. в
начале 1960-х гг., птицы поднялись по долине р. Орели до южных районов Харьковской
обл. (Лисецкий и др., 1980; Ткаченко, 1987; Есилевская и др., 1988). В пойме Десны в
середине 1980-х гг. появились небольшие поселения в колониях серых цапель на севере
Сумской и Черниговской обл. (Афанасьев, 1998). В Сумской обл. большую белую цаплю
добывали в XIX в. у г. Сумы, но с тех пор её здесь не встречали вплоть до конца 1970-х гг.
С 1980-х гг. начали появляться очаги гнездования в поймах Сулы, Ворсклы, Сейма и других
рек (Кныш, Сыпко, 1994). С 1984 г. стали отмечаться ежегодные залёты в междуречье Дес-
ны и Неруссы на юге Брянской обл., а в 1994 г. найдены два гнезда в колонии серой цапли
в низовьях Неруссы (Лозов, 1998). В Среднем Приднепровье в последние годы продолжа-
ется расселение, отмечено появление новых колоний (Гаврилюк и др., 2005).
В Белоруссии единичные случаи гнездования регистрировали в 1970-1980-х гг. в Брес-
тской и Гомельской обл. (Долбик, 1985; Чырвоная кшга, 1993); впрочем, достоверность
этих данных подвергалась сомнению (Самусенко, Журавлев, 2000). В 1994 г. была обнару-
жена колония из 8 гнёзд в Гомельской обл. на р. Припять у границы с Украиной (Никифо-
ров, 2001), в 1997 г. — две колонии в Брестской и Гомельской обл. (Самусенко, 1999). В
последующие годы в бассейне Припяти выявлено ещё несколько колоний и предполагае-
мых мест гнездования (Абрамчук, 2003; Абрамчук, Абрамчук, 2005). Таким образом, гра-
ница ареала продвинулась значительно дальше на север.
На востоке Украины в конце XIX - первой половине XX в. большая белая цапля была
распространена весьма спорадично. Одиночные гнёзда или небольшие поселения её отме-
чали лишь в бассейнах рек Орель и Самара, а также на Днепре. Регулярное гнездование
наблюдается с 1970-х гг., происходит быстрое расселение вида (Гудина, 2003а). В Донец-
ком Приазовье большая белая цапля начала гнездиться после длительного перерыва в нача-
ле 1980-х гг. (Завгородний и др., 1989). На Обиточной косе первое гнездование отмечено в
1981 г., когда в колонии серой цапли загнездились 2 пары. В 1983 г. здесь учтено уже 6 пар
(Лысенко, Сиохин, 1991). В 1980-е гг. цапли расселяются в бассейне Северского Донца в
Луганской обл. (Ветров и др., 1991). Вскоре после этого они появляются на гнездовании в
черноземных областях России.
В Курской обл., после того как в 1926 г. под Курском было разорено одно из последних
гнезд, большая белая цапля встречалась лишь как залетный вид в поймах рек Сейм и Псёл.
Только в 1990-х гг. она вновь появилась на гнездовании на рыборазводных прудах (Миро-
нов, 1996, 1998). В Воронежской обл. в 1975-78 гг. две пары регулярно регистрировали в
гнездовой период в устье р. Воронеж, в 1980-е гг. неоднократно отмечали залёты в разных
частях области (Воробьев, Лихацкий, 1987; Соколов, 1999). В 1997 г. зарегистрировано
гнездование двух пар в окрестностях г. Боброва (Соколов, Простаков, 1997), в последую-
щие годы выявлено ещё несколько предполагаемых мест гнездования в поймах рек и на
искусственных водоёмах (Соколов, 2005). В 1997 г. 1-2 пары гнездились в пойме Хопра у
оз. Ильмень на северо-востоке Воронежской обл. (Венгеров и др., 2000). В 2002 г. около 10
пар отмечены на гнездовании на пруду совхоза «Победа» в Мучкапском р-не Тамбовской
:бл. (Гудина, 20036). Возможно гнездование также в Липецкой обл. (Климов и др., 2004).
310
Отряд Листообразные, Семейство Цаплевые
В Поволжье ещё в середине XX в. большая белая цапля не гнездилась севернее Сарпин-
ских озёр (Спангенберг, 1951). Однако уже в 1970-е и 1980-е гг. вид расселяется значитель-
но дальше на север. В 1978 г. отмечено гнездование пары на Селитьбенском озере, а в
1982 г. — на Сурском вдхр. в Пензенской обл. (Денисов, Фролов, 1991; Фролов и др., 2001).
Позднее, поселения появляются в верхней зоне Волгоградского вдхр. и на малых реках
Саратовской обл. (Шляхтин и др., 1994; Беляченко и др., 2000; Морозов, 2000; Завьялов и
др., 2005). Отмечено гнездование пары в Сусканском заливе Куйбышевского вдхр. в Са-
марской обл. (Лебедева, Пантелеев, 2000).
На Урале и в Зауралье с конца 1970-х гг. отмечаются залётные птицы, а через некоторое
время белые цапли появляются на гнездовании. В Оренбургской обл. с 1979 г. с апреля по
сентябрь регулярно встречали залётных и летующих птиц. Возможно, белые цапли уже
тогда гнездились на некоторых озёрах на востоке области (Самигуллин, 1988). В 1990-х гг.
гнездование было зарегистрировано уже в разных частях области (Самигуллин, 1990; Ков-
шарь, Березовиков, 2001). В Башкортостане гнездо в колонии серых цапель было найдено
в 1996 г. в устье р. Белой (Карякин, 1998а). Появляются гнездовья в Челябинской обл. — в
Октябрьском (Захаров и др., 2000) и Чесменском (Морозов, Корнев, 2002а) р-нах. Совре-
менная численность вида в области оценивается в 20 пар (Гордиенко, 2006). В Тургайской
депрессии на севере Казахстана по В.Ф. Рябову (1982) встречали лишь «бродячих» особей,
но уже в 1981 г. белая цапля была обнаружена на гнездовании в Наурзумском заповеднике.
(Гордиенко, 1987; Блинова, Блинов, 1997). В Курганской обл. большая белая цапля появи-
лась на гнездовании в конце 1990-х гг. В настоящее время она обычна на границе с Тюмен-
ской обл. и на юге лесостепи, на остальной территории встречается спорадично, в северо-
западных районах отсутствует (Тарасов, Поляков, 2006).
В Западной Сибири также отмечено продвижение гнездового ареала на север. Большая
белая цапля появилась на гнездовании на юге Тюменской обл. В 2001 и 2002 гг. гнездящих-
ся и предположительно гнездящихся птиц отмечали на озёрах в Бердюжском и Казанском
р-нах (Морозов, Корнев, 20026; Гашев и др, 2003а, б). Возможно гнездование на оз. Алабо-
та в Омской обл. (Морозов, Корнев, 20026). В Новосибирской обл. известно гнездование в
районе системы озёр Чаны: с 1970-х гг. от одной до 12 особей почти ежегодно наблюдали
на оз. М. Чаны и в его окрестностях, в 1998 г. две пары успешно гнездились на озере у с.
Шелчиха в Здвинском р-не. В 1990-е гг. птиц неоднократно наблюдали на юге Новосибир-
ского вдхр. (Юрлов, 2000). Возможно гнездование также на западе Алтайского края. Белые
цапли встречаются по тростниковым займищам р. Кулунда, но характер их пребывания
здесь пока не ясен (Петров, Торопов, 2000).
Дальше на восток в Сибири известны лишь залёты этих птиц. В верховьях Оби большая
белая цапля — редкий залётный вид (Ирисова и др., 1999). Залетнх белых цапель отмечали
в Томской обл. (Шастовский, Селевин, 1928; Гагина, Скалой, 2000), возле Анжеро-Суджен-
ска (Белянин, 2000) и Таштагола (Гагина, Скалой, 2000) в Кемеровской обл. В 1957 г. белая
цапля добыта в верховьях Подкаменной Тунгуски (Сыроечковский, 1959; Сыроечковский,
Рогачева, 1995). Залётных птиц наблюдали на оз. Беле в Хакасском заповеднике (Девяткин
и др., 2000).
На востоке Казахстана большая белая цапля — редкий, но регулярно гнездящийся вид
Зайсанской котловины и поймы Чёрного Иртыша (Березовиков, 2004а), гнездится также в
Алакольской котловине.
На Байкале и в Забайкалье большая белая цапля в настоящее время также лишь залёт-
ный вид. Залетных птиц отмечали в дельте Селенги и на оз. Таглей в Бурятии (Попов, Мат-
веев, 2005). По данным О.А. Горошко (личн. сообщ.), в благоприятные годы возможно
гнездится на р. Аргунь. В Вост. Монголии гнездится на оз. Буйр-нуур, вероятно гнездится
также на сопредельных территориях Китая.
Большая белая цапля
311
Вероятно гнездится в Верхнем Приамурье (Дугинцов, Панькин, 1993). В настоящее время
большая белая цапля встречается в Амурской обл. по Амуру и Зее, возможно гнездование
3-5 пар (В.А. Дугинцов, личн. сообщ.). По данным В.Г. Бабенко (2000), в Нижнем Приаму-
рье большая белая цапля совсем недавно была лишь редким залётным видом. В настоящее
время в Еврейской АО она спорадически гнездится по Амуру. В 1997 г. гнездо было найде-
но ниже устья р. Биры, в 1999 г. в колонии было уже 3 гнезда. В 1999 г. пара обнаружена в
колонии серой цапли ниже с. Амурзет (Антонов, 1999). В 2004 г. 3 гнезда найдены в коло-
нии серых цапель и больших бакланов в Ленинском р-не в истоках р. Венцелевская. Всего
в Еврейской АО возможно гнездование нескольких десятков особей, численность увеличи-
вается, вид расселяется к северу (А.А. Аверин, личн. сообщ.). В 1997 г. отмечали гнездова-
ние в Хабаровском крае в Комсомольском заповеднике (Росляков, 20006).
На о. Сахалин, по данным П.И. Супруненко (1890, цит. по: Нечаев, 19916), большая
белая цапля когда-то гнездилась на болотах и в долинах некоторых рек на юге острова. В
настоящее время встречаются лишь залётные птицы (Нечаев, 19916).
В Закавказье большая белая цапля обычна лишь в Азербайджане. Гнездится в Грузии
(Birds in Europe, 2004). В Армении встречается только во время миграций и на зимовке.
Таким образом, современная северная граница гнездового ареала большой белой цапли
в Вост. Европе и Сев. Азии проходит через Юж. Белоруссию близ сев. границы с Украиной.
Севернее есть небольшой изолированный очаг гнездования в Латвии. В России граница
ареала идёт через юг Брянской обл. (низовья р. Неруссы), север Воронежской обл., юго-
восток Тамбовской обл., северо-восток Пензенской обл., Сусканский залив Куйбышевско-
го вдхр. в Самарской обл. В Предуралье — примерно по 56° с.ш.: устье р. Белой в Башкор-
тостане, южнее Челябинска. На Урале и в Зап. Сибири — по северу Курганской обл., Бер-
дюжскому и Казанскому р-нам Тюменской обл., затем отклоняется к 54° с.ш. — оз. Алабо-
та в Омской обл. и район оз. Чаны в Новосибирской обл. Дальше граница ареала проходит
на юго-восток через оз. Зайсан и Чёрный Иртыш, затем по границе с Монголией, по Аргу-
ни и Амуру (рис. 63). В Нижнем Приамурье ареал доходит на север до 51° с.ш. С ареалом
южной белой цапли в Приморье она граничит примерно по 44° с.ш., хотя не исключено
здесь её аллопатрическое обитание с последней (Глущенко и др., 2003). Внутри обширной
очерченной выше территории большая белая цапля распространена очень спорадично; ос-
таются довольно большие участки, где вид на гнездовании не обнаружен.
Заметим, что в ходе расселения вид не только восстановил прежний ареал, но и значи-
тельно его расширил. Благоприятными для белых цапель оказались многие антропогенные
изменения среды — создание водохранилищ и запруд на реках, рыборазводных прудов,
прокладка каналов и обводнение аридных земель, расширение рисосеяния. Так, вверх по
течению рек расселение началось после создания каскадов водохранилищ — как на Днеп-
ре (Клестов, 1991; Гаврись, 1994), так и на Волге (Рахимов, Уленгов, 2003). В Крыму боль-
шие белые цапли быстро распространились вдоль трассы Северо-Крымского канала уже
во время проведения там работ (Костин, 1983). В Ставропольском крае создание обводни-
тельно-оросительной системы значительно улучшило условия обитания многих водно-бо-
лотных птиц, в частности белых цапель (Бичерев, 1988). В Узбекистане благодаря образо-
ванию нескольких искусственных озёр большая белая цапля появилась на гнездовании в
центральных и южных районах, где раньше встречалась только во время пролёта и зимовок
(Шахназаров, 1995).
Рост численности и расселение большой белой цапли наблюдается не только в Евразии,
но и в Сев. Америке (Martinez-Vilalta, Motis, 1992; Crozier, Gawlik, 2003).
Зимовки. Основные места зимовки евразийских популяций большой белой цапли
находятся в Сев. Африке, на юге Европы, на Ближнем Востоке, в Юж. Азии, Китае (Спан-
312
Отряд Листообразные, Семейство Цаплевые
Рис. 63. Область распространения большой белой цапли в Вост. Европе и Сев. Азии: а — гнездовой
ареал, б — зимовки, в — изолированные гнездования.
генберг, 1951; Bauer, Glutz von Blotzheim, 1966; Cramp, 1977). Птицы из США зимуют в
Калифорнии, Мексике, Центр. Америке, на островах Карибского моря (Martinez-Vilalta,
Motis, 1992). Тропические популяции оседлы.
Птицы, окольцованные на Лебяжьих о-вах, в зимние месяцы обнаружены в Болгарии,
Турции, Греции, Италии, Югославии, Грузии, а также в Краснодарском крае, Одесской обл.
и в Крыму (Костин, Тарина, 2005). Цапли, окольцованные в Вост. Приазовье, найдены на
зимовке в Греции, Сев. Ираке, Грузии, Израиле, Египте, Центральноафриканской Респуб-
лике. Птицы из Ростовской обл. — в Дагестане, Краснодарском крае, Иране. Цапли из Да-
гестана и Азербайджана зимовали недалеко от мест гнездования. Две дагестанские особи
обнаружены в 150 и 240 км южнее от места кольцевания, из 6 возвратов цапель из Кызыл-
Агачского заповедника, 5 приходится на долину р. Куры в Азербайджане, одна птица най-
дена в низовьях Атрека в Туркмении (Сапетин, 1978а). Зимующих птиц отмечали также в
ряде стран Центр, и Зап. Европы — в Румынии, Хорватии, Австрии, Нидерландах (Munteanu,
Ranner, 1997), Словении (Vogrin, 1999), Польше (Piotrowska, 2003), Германии (Flore, Komer,
Marzimeier, 1999).
В южных частях Вост. Европы и Сев. Азии большая белая цапля — обычный, хотя и
немногочисленный, зимующий вид.
На юге Украины она достаточно обычна в мягкие теплые зимы. Так, в январе 2001 г. 28
особей было учтено в Сев. Приазовье (Кошелев и др., 2002), 318 — на Сиваше (Андрю-
щенко и др., 2003), 6 — в районе заповедника Аскания-Нова (Гавриленко, Думенко, Лопу-
шанский, 2002), 4 — в районе Черноморского заповедника (Руденко, Яремченко, Моска-
ленко , 2002), 194 — в Сев.-Зап. Причерноморье (Русев и др., 2002). В Крыму на зимовку
остаются одиночные птицы или небольшие группы (Костин, 1983). На юж. берегу Крыма
большая белая цапля зимует на заросших участках ручьёв и речек; обычно наблюдают оди-
Большая белая цапля
313
ночных птиц или группы до 5 особей (Бескаравайный, Костин, 1999). При значительных
похолоданиях цапли отлетают южнее. В Молдавии белые цапли зимуют на незамерзающих
участках Днестра, Прута, Турунчука и других рек, на Кучурганском лимане, прудах рыбхо-
зов (Куниченко, Гусан, 1992; Куниченко, Тищенков, 1999; Зубков и др., 2004; Тищенков,
2004).
На территории Украины зимовки отмечаются и значительно севернее: в Летичевском р-
не Хмельницкой обл. в 2000-2003 гг. от 5 до 25 особей зимовали в долине р. Юж. Буг и её
притоков (Новак, 2003), одиночных особей и небольшие группы птиц наблюдали зимой
возле г. Тернополя (Страшнюк, 2004), в Черкасской обл. (Гаврилюк, Грищенко, 2001), в
окрестностях Киева (Давиденко, Сыпко, 2002; Костюшин, Полуда, 2005), в Луганской обл.
(Панченко, Сулик, 1978).
Одну птицу наблюдали в феврале 2002 г. на р. Муховец в черте г. Бреста (Абрамчук,
Абрамчук, 2005).
На Северном Кавказе большая белая цапля регулярно зимует на приморских низменно-
стях у Адлера, реже по берегам рек. Изредка в зимнее время птиц отмечают на Зап. Маны-
че. Одиночки и небольшие группы цапель, особенно в тёплые зимы, регулярно встречают-
ся в бассейнах Кумы, Терека, Сулака, Самура, у лесных родников в Кизилюртовском р-не
Дагестана, на водоёмах Чечни и Ингушетии. В Ставропольском крае, начиная с 1977 г.
почти ежегодно в разных точках отмечали одиночных птиц и группы до 30^0 ос. Обычна
белая цапля зимой в Сев. Осетии. Зимовка расположена на Брутских озёрах, где постоян-
но держится 80-140 ос. Одиночные встречаются и на других водоёмах, поднимаясь в пред-
горья. Изредка зимой белых цапель видели в Тебер дине ком и Кавказском заповедниках
(Казаков и др., 2004). В январе 2004 г. на берегу Каспия в Дагестане и в приморских водо-
ёмах учтено 155 больших белых цапель (Джамирзоев, Исмаилов, 2004). Отмечали и случаи
зимовки в дельте р. Урал (Березовиков, Гисцов, 2001).
Большая белая цапля обычна на зимовке в Азербайджане, встречается на многих водо-
ёмах страны (Спангенберг, 1951; Смогоржевський, 1979; Султанов, Джаббарова, 2006). В
Армении зимующих птиц отмечали в долине р. Араке и в бассейне оз. Севан. Всего в Ара-
ратской равнине, по данным М.С. Адамяна, зимует 250-1 000 этих птиц.
В Туркменистане белые цапли зимуют в небольшом количестве на р. Атрек и в предгор-
ной полосе возле Ашхабада (Спангенберг, 1951). В 1978-1986 гг. численность зимующих
на юго-востоке Туркменистана птиц колебалась от 71 до 499 особей (Молева, Рустамов,
1986). Зимующих белых цапель отмечали на юге Казахстана (Долгушин, 1960; Белялов,
Карпов, 2004), в Узбекистане (Сагитов, 1987; Лукашевич, 1990), Кыргызстане (Шукуров,
1981; Кулагин, 2005), Таджикистане (Иванов, 1940), в бассейне р. Тарим в Китае (Судилов-
ская, 1968).
В зимний период белых цапель отмечали и значительно севернее границы гнездового
ареала. Так, одна из них зимовала на незамерзающем участке Шершневского вдхр. у г.
Челябинска с 19 декабря 1985 г. до 31 марта 1986 г. (Гордиенко, 1987). В декабре 1996 г.
одна птица появилась на незамерзающих Выбутских порогах на р. Великой выше Пскова и
оставалась там до третьей декады февраля. В марте 1997 г. одна особь две недели держа-
лась в черте Пскова на незамерзающих участках рек Мирожки и Псковы. Здесь, ослабев-
шая за зиму птица, была разорвана бродячими собаками (Фетисов, 1998).
Миграции. По данным кольцевания, направления миграций неодинаковы у разных
популяций. Большие белые цапли из Приазовья мигрируют как на юго-запад через Балка-
ны и Сицилию, так и на юго-восток через Зап. Закавказье и Переднюю Азию. Цапли с
Маныча частично улетают через Предкавказье в область Каспийского моря и в Иран (Сапе-
тин, 1978а).
314
Отряд Листообразные, Семейство Цаплевые
Фенология. Большая белая цапля принадлежит к рано прилетающим видам. Первые
птицы могут появляться, ещё когда водоёмы покрыты льдом, но в целом прилет довольно
растянут. В Вост. Европе его регистрируют с конца февраля до первой половины апреля. В
южных районах первые особи могут регистрироваться и раньше. Так, на Лебяжьих о-вах в
2003 г. прилёт отмечен ещё 7 февраля (Костин, Тарина, 2004). В 1958 г. на Маныче первые
цапли были встречены 18 февраля (Олейников, Гончаров, 1967). Средние сроки прилёта в
разных регионах приходятся на март - первую декаду апреля. Отклонение от средних дат
составляет обычно 11-14 дней (табл. 26). По мере продвижения на север и в глубь конти-
нента сроки прилёта смещаются на более поздние. Так, в Саратовской обл. цапли прилета-
ют лишь в первой половине апреля (Завьялов и др., 2005). В южной части Белорусии пер-
вых птиц отмечают в конце марта - начале апреля, наиболее ранняя регистрация — 6 марта
2002 г. (Абрамчук, Абрамчук, 2005). В дельте Волги первые птицы появляются в конце
февраля, в некоторые годы даже раньше. В первой декаде марта идёт интенсивный пролёт
стаями до 200 особей (Луговой, 1963). По наблюдениям Е.П. Спангенберга (1951), под
Ленкоранью численность цапель начинает возрастать с конца февраля, интенсивный про-
лёт идёт в марте и заканчивается к концу этого месяца.
Отлёт зимующих птиц с приморских низменностей у Адлера наблюдается в конце апре-
ля (Казаков и др., 2004).
В азиатской части ареала сроки прилёта сильно различаются в зависимости от региона.
В Узбекистане весеннее движение цапель к местам гнездования начинается в конце февра-
ля - начале марта (Шахназаров, 1995). Наиболее ранняя дата прилёта в дельте Аму-Да-
рьи — 22 февраля 1989 г. (Лукашевич, 1990). В Казахстане в Илийской долине белая цапля
появляется в некоторые годы уже в конце февраля, но чаще в первой половине марта; в
дельте Или и низовьях Сыр-Дарьи — во второй половине марта, в низовьях Иргиза — в
конце марта - апреле (Долгушин, 1960). По восточному побережью Каспия пролёт наблю-
дается в марте (Спангенберг, 1951). На Нижнем Амуре весной большие белые цапли встре-
чаются со второй декады апреля (Бабенко, 2000). В Сихотэ-Алиньском заповеднике, пер-
вых птиц наблюдают в третьей декаде марта - апреле. Наиболее ранние встречи на Ханке
датированы 9 марта 2004 г., 14 марта 1992 г., 15 марта 2001 г., 16 марта 1994 г., 18 марта
1998 г. и 23 марта 1973 г.; основное население местных колоний пребывает в последних
числах марта и в первой декаде апреля, а транзитный пролёт продолжается до середины
апреля (сообщение Ю.Н. Глущенко).
Отлетает большая белая цапля поздно. В разных областях Украины, Молдавии, Казах-
стана последних птиц наблюдают в сентябре-ноябре. Средние сроки последних встреч
приходятся на конец сентября - начало ноября. Отклонение от средних дат осенью значи-
тельно больше, чем весной — 16-20 дней (табл. 27). В Белоруссии последних птиц наблю-
дают в конце сентября-октябре. Наиболее поздняя регистрация — 21 октябя 2003 г. (Аб-
рамчук, Абрамчук, 2005). На Сев. Кавказе отлёт на зимовку начинается в сентябре, закан-
чивается в октябре-ноябре (Казаков и др., 2004). В Саратовской обл. откочёвка большей
части популяции происходит в первой декаде августа, отдельные птицы встречаются до
первой декады октября (Завьялов и др., 2005).
В дельте Волги осенний отлёт начинается с 25 сентября и тянется весь октябрь, запоз-
давших особей наблюдали до 8 ноября. На разливах Терека, в Аграханском заливе и по
озёрам в районе Кизляра стаи пролётных птиц отмечали на протяжении всего ноября и до
6 декабря, уже при сильных ночных заморозках. В дельте Урала пролёт можно наблюдать
уже в конце сентября, а отдельные особи задерживаются до конца октября. В юго-восточ-
ной части Каспия пролёт идёт в течение всего октября и ноября. В низовьях Сыр-Дарьи
отлёт начинается в конце сентября, к двадцатым числам октября цапли становятся уже редки-
Большая белая цапля
315
Таблица 26. Сроки прилёта большой белой цапли в Вост. Европе и Сев. Азии
Регион п М SD Lim Источник
Южная Украина 9 5 марта 14,3 7 февраля - 25 марта Лысенко, 1992; Корзюков, 1996; Бескаравайный, 1999; Костин, Тарина, 2004
Западная Украина 16 23 марта 13,5 24 февраля - 12 апреля Орнгголопчш спостереження..., 1994; Матер1али..., 1996; Скильский и др., 20016; Капелюх, Гузш, 2000; Новак, 2002
Центральная Украина 32 22 марта 12,8 27 февраля - 17 апреля Шакула, Ткаченко, 1986; Роговой, 1992; Очеретный, 1998; Грищенко, Гаврилюк, 2000; Слюсар, 2000; Гаврилюк, 2002; (ориг.)
Северная Украина 16 2 апреля 14,6 26 февраля - 23 апреля Марисова и др., 1991; Головушкин, 1992; Книш, 1992; Афанасьев, 1998; Полюшкевич, 1998; Домашевский, 2002
Молдавия 8 20 марта 3,4 17 марта - 25 марта Аверин и др., 1971, Тищенков, 2004
Северный Кавказ 9 13 марта 11,8 18 февраля - 2 апреля Олейников, Гончаров, 1967; Казаков и др., 2004
Кургальджино, Казахстан 8 8 апреля 9,4 Сема, 1989
Окр. Алма-Аты, Казахстан 4 10 марта 6,5 Сема, 1989
Сихотэ-Алинский заповедник, Приморский край 9 13 апреля 11,9 21 марта- 25 апреля наблюдения С.В. Елсукова
ми, одиночные птицы задерживаются до первых чисел ноября, а в среднем течении реки —
до ноября (Спангенберг, 1951).
По данным А.В. Михеева и его соавторов (1975), пролёт по западному побережью Кас-
пия начинается в конце августа. До конца октября и начала ноября он выражен слабо. В
конце ноября и начале декабря пролетает наибольшее количество птиц, пролёт продолжа-
ется до середины или конца декабря. Летят птицы на высоте 100-150 м, реже 200-300 м,
над морем ниже — иногда до 10-20 м. Пролётные стаи насчитывают десятки и даже сотни
особей. Миграция идёт как ночью, так и в светлое время суток.
В Сихотэ-Алиньском заповеднике последних птиц наблюдали 7 ноября 1995 г. и 9 нояб-
ря 1998 г. (данные Летописи природы заповедника). На Приханкайской низменности пос-
ледние птицы отмечены 28 октября 1986 г., 29 октября 1989 г., 30 октября 1982 г., 5 ноября
1995 г., 16 ноября 1993 г. и 19 ноября 2004 г. (Глущенко и др., 20066). В долине р. Раздоль-
ная наиболее поздние встречи зарегистрированы в окрестностях Уссурийска 24 ноября 2002
г. и 18 декабря 2003 г., а одна особь наблюдалась в середине января 1993 г. (Глущенко и др.,
2006а). Единичные случаи зимнего пребывания данного вида в Приморье известны также
для окрестностей Владивостока (Воробьёв, 1954) и р. Нарва (Сидими) в Хасанском р-не
(Панов, 1973).
Послегнездовые кочёвки. Для цапель характерен так называемый промежуточный пере-
лет в разнообразных направлениях, который постепенно переходит в осеннюю миграцию.
Кочующие белые цапли появляются значительно севернее границы гнездового ареала. В
богатых кормом местах в августе-сентябре могут образовываться большие скопления из
десятков и даже сотен птиц.
316
Отряд Листообразные, Семейство Цаплевые
Таблица 27. Сроки последних встреч большой белой цапли в Вост. Европе и Сев. Азии
Регион п М SD lim Источник
Западная 32 17 20,2 12 сентября - Орштолопчш спостереження, 1993;
Украина октября 28 ноября 1994; Матер1али, 1994; 1995а; 19956; 1996; Новак, 2002; Скшьський и др., 2001
Центральная 34 18 20,5 15 сентября - Роговий, 1994; Очеретный, 1998;
Украина октября 27 ноября Грищенко, Гаврилюк, 2000; Бредб1ер, 2002; Гаврилюк, 2002; Шевцов и др., 2004; (ориг.)
Северная 16 27 16,6 7 сентября - Марисова и др., 1991; Книш, 1994;
Украина сентября 5 ноября Домашевский, 1996; Афанасьев, 1998; Полюшкевич, 1998
Молдавия 11 2 ноября 16,4 30 сентября - Аверин и др., 1971; Куниченко, Гусан,
29 ноября 1992; Тищенков, 2004; А.А. Тищенков (личн. сообщ.)
Низовья 3 13 Сема, 1989
Тургая, Казахстан октября
Кургальджино, Казахстан 3 6 октября Сема, 1989
Окрестности 4 28 Сема, 1989
Алма-Аты, Казахстан октября
Дальность разлёта по материалам меченья обычно не превышает 200-250 км, иногда
достигает 400-600 км (Сапетин, 1978а). Птицы, окольцованные на Лебежьих о-вах, кочуют
в августе-сентябре по северу Крыма и в Херсонской обл. Но уже через месяц первые особи
достигают Греции (Костин, Тарина, 2005).
В Каневском заповеднике (Черкасская обл.), на островах Днепра, расположенных ниже
плотины Каневской ГЭС, первые кочующие белые цапли появляются в конце июня - пер-
вой половине июля. Численность их постепенно возрастает, достигая максимума во второй
половине августа - сентябре, затем снижается вплоть до середины октября. Сроки появле-
ния и численность кочующих птиц зависят от уровня воды в Днепре. Когда мелководья во
второй половине лета затоплены, птицы появляются позже и больших скоплений не обра-
зуют. Максимальное количество белых цапель наблюдали в 2001 г. — около 200 особей
(Грищенко, 2001). На юге Брестской обл. в августе — ноябре 2002 г. на 6 водоёмах в общей
сложности учтено 1011 особей. Наиболее высокая численность вида отмечена на рыбхозе
«Новоселки», где 6 октября встречено не менее 519 птиц (Абрамчук и др., 2003).
В Крымском Присивашье в 1978-80 гг. во время послегнездовых кочёвок с середины
августа до конца октября держалось не менее 5-6 тыс. больших белых цапель. Отдельные
скопления кормящихся птиц включали до 700 особей (Костин, 1983).
На юге Приморья одиночных белых цапель и их стаи до 15 особей наблюдали на рисо-
вых полях с июля по август (Глущенко, 1979).
Залеты. Для большой белой цапли характерны дальние залёты. Чаще всего залётных
птиц наблюдают во время миграций и послегнездовых кочевок, но могут встречаться и
летующие особи в гнездовой период. Известны залёты для Эстонии (Leibak, Lillerht, Vero-
mann, 1994). В России залётных белых цапель отмечали во многих областях, расположен-
ных гораздо севернее границы гнездового ареала — от Псковской (Фетисов, 1998; Фети-
Большая белая цапля
317
сов, Иванов, 2003) и Ленинградской обл. (Мальчевский, Пукинский, 1983; Ковалев, 2001)
до Камчатки (Lobkov, 1997). На север птицы могут проникать до Северного полярного
круга. Наиболее северные находки залётных белых цапель — оз. Лача в Каргопольском р-
не Архангельской обл. (Андреев, 2004), р. Печора в Республике Коми (Нейфельд, Теплов,
2000), окрестности г. Салехарда (Азаров, 1996), Беринговский р-н Чукотки.
На западе Украины залеты белых цапель заметно участились с конца 1970-х гг. (Гор-
бань, Фладе, 2000). В Латвии в 1990-х гг. ежегодно отмечали лишь единичные случаи залё-
тов, в 2000 г. учтено 13 птиц, а в последующие три года — уже десятки особей. В 2004 г.
здесь зарегистрировали 342 птицы, большинство из них — во второй половине лета (Celmips,
2004). Повышение частоты встреч залётных птиц отмечается и во многих регионах России
(Lobkov, 1997; Приклонский и др., 1998; Рахимов, Уленгов, 2003).
В Зап. Европе время от времени зимой регистрируют залёты больших белых цапель с
красными ногами и чёрной окраской клюва. Считается, что это могут быть Е. modesta.
Поскольку визуальные наблюдения не дают возможности однозначно судить об их видо-
вой принадлежности, такие встречи относят к группе «modesta-type». Отмечались такие
птицы и в Вост. Европе. Так, О.А. Форманюк (личн. сообщ.) 6 ноября 2005 г. в низовьях
Тилигульского лимана в Одесской обл. в группе больших белых цапель наблюдал одну особь
с красными ногами и чёрным клювом.
Местообитания. Населяет разнообразные типы влажных биотопов — плавни рек, при-
брежные заросли водоёмов, заросшие пруды, поймы рек, тростниковые заросли на остро-
вах, косах и болотах, мангровые заросли и т.п. В аграрном ландшафте может селиться на
рисовых чеках, каналах, искусственных водоёмах. Известны случаи гнездования даже на
деревьях, растущих среди сухих пастбищ (Martinez-Vilalta, Motis, 1992). Оптимальный гнез-
довой биотоп — обширные заросли, чередующиеся с открытыми мелководными водоёма-
ми или плёсами.
В Белоруссии все обнаруженные колонии находились в рогозово-тростниковых и трос-
тниковых зарослях на реках, водохранилищах, рыбхозах и болотах (Абрамчук, Абрамчук,
2005). В Центр. Предкавказье все колонии большой белой цапли находились в тростнико-
во-рогозовых зарослях: 63,8% — на обводненных солончаках, по 18,1% — на пересыхав-
ших до обводнения реках и балках и на водохранилищах (Бичерев, 1988).
Большая белая цапля предпочитает равнинный ландшафт, в горы поднимается редко,
хотя может гнездиться и на значительных высотах. В Грузии найдена на гнездовании на
высоте 1800 м н.у.м. (Чхиквишвили, 1930), а в Перу отмечались попытки гнездования на
высоте 4080 м н.у.м. (Martinez-Vilalta, Motis, 1992).
Неоднократно повторявшиеся утверждения, что большая белая цапля избегает селиться
вблизи человека и предпочитает самые глухие и труднодоступные уголки зарослей (Спан-
генберг, 1951; Долгушин, 1960 и др.), в последнее время подвергаются сомнению. Колонии
возле населённых пунктов сейчас уже не редкость. Например, в окрестностях г. Кременчу-
га (Полтавская обл.) была обнаружена колония из 25-30 пар на тростниковом болоте возле
железнодорожной станции между железной дорогой и автострадой. Другая колония нахо-
дилась за 300 м от отстойника нефтеперерабатывающего завода (Гаврись, 1994).
Кормится большая белая цапля на мелководьях по берегам водоёмов, в поймах рек, на
лугах и пастбищах, болотах. В большом количестве птицы собираются вместе с другими
рыбоядными птицами на спущенных для облова рыборазводных прудах, на участках рек
ниже плотин ГЭС, работающих в пиковом режиме. Из-за постоянных перепадов уровня
воды здесь создаются хорошие условия для рыбоядных птиц — после очередного спада
воды остаётся большое количество мелководных временных водоёмов, что весьма удобно
для кормёжки. В глубоких местах можно увидеть подстерегающих добычу птиц на корягах
318
Отряд Листообразные, Семейство Цаплевые
и затопленных кустах. Охотно кормится на рисовых чеках (Спангенберг, 1951; Глущенко,
1979; Костин, 1983; Сагитов, 1987; Казаков и др., 2004).
Зимой встречается на незамерзающих участках водоёмов, техногенных прудах, кана-
лах, арыках, горных речках, возле родников. В Узбекистане зимующих белых цапель отме-
чали на распаханных полях хлопчатника, когда проводилась промывка почвы (Шахназа-
ров, 1995).
Численность. Общая численность в Европе оценивается в 11-24 тыс. пар. Из них 3-10
тыс. гнездится в европейской части России, 4,5-7,3 тыс. на Украине, 250-800 пар в Азер-
байджане. Численность зимующих птиц составляет более 8600 особей, из них 1-2 тыс. —
в Азербайджане. Для всех европейских стран, где вид гнездится, кроме Албании и Турции,
отмечены положительные тренды численности. В 1970-1990 гг. происходило увеличение
численности, в 1990-2000 гг. — значительное её увеличение (BirdLife International, 2004).
На Украине основные колонии расположены в низовьях Дуная, Днестра, Днепра, на
севере и северо-востоке Крыма, на лиманах и косах Сев. Приазовья, водохранилищах Днеп-
ровского каскада. В большинстве мест отмечаются как незначительные флуктуации, так и
рост численности. В Дунайском биосферном заповеднике в 1984-1997 гг. общее количе-
ство гнездящихся пар колебалось от 0 до 170 (Жмуд, 1999). В дельте Днестра численность
большой белой цапли в 1978-2004 гг. изменялась в пределах 120^30 пар (Русев, 2004).
Пик численности большой белой цапли на Лебяжьих о-вах был отмечен в 1981 г. — 830
пар; в дальнейшем численность колебалась в пределах 200-778 пар (Костин, Тарина, 2004).
Общая численность в колониях на Вост. Сиваше за последние 30 лет изменялась в преде-
лах 130-260 пар (Гринченко, 2004а).
В Молдавии по приблизительной оценке гнездится в разные годы от 30 до 150 пар. Из
них от 17 до 120 пар в 1-5 колониях на землях Катульского рыбокомбината и от 10 до 50
пар на прудах в центральной и южной частях страны (Куниченко, Гусан, 2001). В Белорус-
сии известно не менее 8 колоний, в которых гнездится, как минимум, 110-140 пар; с учётом
предполагаемых мест гнездования общая численность может быть определена в 200-300
пар (Абрамчук, Абрамчук, 2005).
Общая численность в Юж. России оценивается в 3-9 тыс. пар (Белик, 2005). В Красно-
дарском крае в 1980-х гг. было известно 10 колоний общим числом 757 пар, а в 1990-х гг. —
уже, соответственно, 14 и 1347 (Емтыль и др., 1989,2003). В дельте Кубани к концу 1980-х гг.
численность достигла 500-600 пар (Казаков и др., 2004). В низовьях Дона, по учётам 1990—
91 гг., общая численность составляла 230-250 пар (Казаков, Ломадзе, 1991,1992). На Весе-
ловском вдхр. в 1990-1991 гг. было учтено 400-500 пар (Казаков, 1998, 2000а). Общая
численность больших белых цапель на Зап. и Вост. Маныче в настоящее время достигает
500-600 пар (Казаков и др., 2004). На водоёмах Ставрополья в 1990-е гг. гнездилось при-
мерно 100 пар (Хохлов, 1993).
В традиционных местах гнездования на Каспии численность в последние десятилетия
снижается из-за трансгрессии моря и повышения глубин в низовьях рек. В дельте Волги
вследствие этого общая численность большой белой цапли сократилась с 6 тыс. пар в 1980 г.
до 1 тыс. в 1994 г. (Кривенко и др., 1998), в дельте Терека — с 1290 пар в 1973 г. (Бондарев,
1975) до 175 в 1986 г. (Казаков и др., 2004). В 1999 г. в дельте Волги и западном ильменно-
бугровом районе учтено 3090 гнездящихся пар большой белой цапли, в 2000 г. — 995, в
2001 г. — 1850 (Гаврилов и др., 2003). Численность в Кизлярском зал. в 1996 г. оценивалась
в 300^100 пар (Вилков и др., 2000). 6 ноября 2003 г. в угодьях предустьевого взморья Волги
(Астраханский заповедник) учтено 1 790 особей (Русанов, 2004).
Довольно крупные поселения есть в Приуралье. Так, на оз. Шалкар-Ега-Кара на восто-
ке Оренбургской обл. гнездится около 100 пар (Коршиков, Корнев, 2000).
Большая белая цапля
319
В Узбекистане в 1987-1989 гг. учтено 1 892 особей на гнездовании, 4 275 особей — во
время миграций и 1 773 особей — на зимовке. Основными местами гнездования служат
озёра Айдаркуль, Судочье, Думалак, Караджар, залив Жылтырбас. Наибольшие миграци-
онные скопления отмечены на озёрах Айдаркуль и Закир куль. Наибольшее количество ца-
пель зимует на озёрах Денгизкуль, Айдаркуль, Султандаг и в Кашкадарьинском прудовом
хозяйстве (Шахназаров, 1995).
В западноевропейских странах наибольшее количество большой белой цапли зарегист-
рировано в Венгрии (500 пар), Австрии (250 пар) и Румынии (200 пар). Общее число гнез-
дящихся птиц в других странах, где она отмечена (Греция, Хорватия, Болгария, Словакия и
др.), не превышает 50 пар.
В Приморье на оз Ханка в начале 1960-х гг. гнездилось 200-300 пар14 (Поливанова,
1971). Позднее численность отмечалась в пределах 170-520 гнездящихся пар (Глущенко и
др., 1992).
Размножение. Моногам. Пары сезонные, в Европе образуются весной во время возвра-
щения с зимовок. Возраст первого размножения — вероятно, 2 года (Cramp, 1977).
В брачном и антагонистическом поведении широко используются удлинённые перья
(эгретки). Самцы обычно появляются в колонии первыми, занимают гнездовые участки и
активно их защищают. Они строят из гнездового материала небольшие платформы, кото-
рые служат для тока. Токующая птица распрямляет эгретки широким веером, трясёт ими,
делая волнообразные движения шеей и двигая хвостом. Шея и клюв поднимаются вверх,
затем опускаются вниз (рис. 64). Приветственная церемония партнеров на гнезде также
происходит с поднятием эгреток (Cramp, 1977; Martinez-Vilalta, Motis, 1992).
Гнездится большая белая цапля обычно колониями, часто совместно с другими голена-
стыми птицами — малой белой, серой и рыжей цаплями, каравайкой, колпицей и др. Чис-
ленность в крупных колониях может достигать сотен пар. Там, где плотность населения
невысока, чаще гнездится отдельными парами или небольшими группами. По данным ан-
кетного учёта, в 1986 г. на территории Украины всего лишь 9,3% колоний большой белой
цапли были моновидовыми, в них находилось 15,2% учтённых гнёзд (Серебряков, Грищен-
ко, 1992). В Предкавказье большая белая цапля также чаще гнездится в смешанных коло-
ниях с другими видами, образуя в них моновидовые «микропоселения» из 7-20 гнёзд (Ка-
заков и др., 2004). Расселяющиеся на север птицы на новых территориях чаще всего гнез-
дятся в колониях серой цапли; известны случаи гибридизации этих видов. Из 8 обнаружен-
ных на юге Белоруссии колоний, только одна была моновидовой, остальные — смешанные
с серой цаплей (Абрамчук, Абрамчук, 2005).
В дельтах рек, имеющих обширные пространства тростниковых крепей, колонии рас-
полагаются обычно в глубине зарослей тростника, камыша, рогоза. По наблюдениям в Се-
верном Приазовье, большая белая цапля — один из наиболее пластичных видов среди ца-
пель в выборе гнездовых стаций, хотя предпочтение отдает средним по густоте зарослям
тростника. В них располагалось 80% из 275 обнаруженных гнёзд, 14,5% — в густом зало-
манном тростнике, 4,4% — в очень густом мозаичном тростнике и 1,1% — в очень густом
сплошном (Кошелев, Кошелев, Пересядько, 2004). На оз. Ханка из 39 осмотренных гнёзд
87,2% располагались в густых зарослях тростника, находящегося на плаву, 12,8% — на
кустах тальника (Поливанова, 1971).
Гнёзда птицы устраивают на заломах прошлогодних стеблей, растущих среди зарослей
кустах или низких деревьях. Они могут находиться как возле самой воды, так и на высоте
до 4-5 м. В Прикарпатье (2 колонии в Черновицкой и Ивано-Франковской обл.) гнёзда
14 По крайней мере часть из них принадлежала виду С. modestus (прим. ред.).
320
Отряд Листообразные, Семейство Цаплевые
Рис. 64. Элементы демонстрационного поведения большой белой цапли (по фотографиям J. Zonneveld.
J. Kille, М. Drummond, S. Ingraham, H. Rufe, C.Lockwood): a-e — последовательность токовых поз
самца, ж — конфликт двух особей, з — спаривание.
Таблица I. Пеликаны
1-6 — Розовый пеликан (1-2 — в брачном наряде; 3 — птица третьего года жизни в предбрачном
наряде: 4 — в послебрачном наряде; 5 — молодая птица; 6 — птенец); 7-11 — Кудрявый пеликан
(7-8 — в брачном наряде; 9-10 — в послебрачном наряде; И — птенец).
2
Таблица П. Олуши
1-6 — Северная олуша (1-2 — взрослая; 3 — птица третьего года жизни; 4 — птица второго года
жизни; 5 — молодая; 6 — птенец); 7-10 — Красноногая олуша (7 — 8 — взрослая белой морфы;
9 — взрослая бурой морфы; 10 — молодая).
Таблица III. Олуши, Фрегаты и Змеешейки
1-2 — Бурая олуша (1 — самец; 2 — молодая птица); 3-5 — Фрегат-ариель (3 — самец; 4 — самка;
5 — молодая птица); 6-8 — Чернобрюхая змеешейка (6 — самец; 7 — самка; 8 — молодая птица).
Таблица IV. Бакланы
1-7 — Большой баклан (1 — в брачном наряде, подвид carboy 2 — в брачном наряде,
подвид sinensis^ 3 — в брачном наряде в полете; 4 — в послебрачном наряде; 5 — молодой:
6 — птицы в промежуточных нарядах; 7 — яйцо и птенцы разных возрастов);
8-9 — Японский баклан (8 — в брачном наряде; 9 — в послебрачном наряде).
5
Таблица V. Бакланы
1-3 — Ушастый баклан (1-2 — в брачном наряд, подвид cincinatus', 3 — молодой); 4-7 — Берингов
баклан (4-5 — в брачном наряде; 6 — в послебрачном наряде; 7 — молодой); 8-12 — Красноли-
цый баклан (8-9 — в брачном наряде; 10 — в послебрачном наряде; 11 — молодой; 12 — птенец);
13 — Стеллеров баклан в брачном наряде.
Таблица VI. Бакланы
1-5 — Хохлатый баклан (1-2 — в брачном наряде; 3 — в послебрачном наряде; 4 — годовалая
птица; 5 — молодой); 6-9 — Малый баклан (6 — в брачном наряде; 7-8 — в послебрачном на-
ряде; 9 — молодой; 10 — птенец); 11-17 — Головы бакланов (11 — малый в послебрачном на-
ряде; 12 — большой в послебрачном наряде; 13 — японский в послебрачном наряде;
14 — хохлатый, подвид aristotelis в брачном наряде; 15 — хохлатый, подвид desmarestii в брачном
наряде; 16 — берингов в брачном наряде; 17 — краснолицый в брачном наряде).
3
Таблица VII. Цапли
1-7 — Волчок (1-2 — самец; 3^4 — самка; 5-6 — молодая птица; 7 — птенец); 8-10 — Китайский
волчок (8-9 — взрослый; 10 — молодой); 11-15 — Амурский волчок (11-12 — самец;
13-14 — самка/молодая птица; 15 — птенец).
Таблица VIII. Цапли
1-3 — Охристый волчок (1-2 — самец; 3 — самка/молодая птица); 4-8 — Зелёная кваква
(4-6 — взрослая, варианты окраски; 7 — молодая; 8 — птенец); 9-11 — Японская кваква
(9-10 — взрослая; 11 — молодая).
Таблица IX. Цапли
1^4 — Большая выпь (1-2 — взрослая/молодая; 3 — поза затаившейся птицы; 4 — птенец);
5-10 — Кваква (5-6 — взрослая; 7 — птица второго года жизни; 8-9 — молодая; 10 — птенец).
1-7 — Желтая цапля (1-2 — в брачном наряде; 3 — самец в разгар тока; 4-5 — в небрачном на-
ряде; 6 — молодая; 7 — птенец); 8-11 — Белокрылая цапля (8-9 — в брачном наряде;
10-11 — в небрачном наряде); 12-13 — Индийская прудовая цапля в брачном наряде.
Таблица XL Цапли
1-8 — Египетская цапля (1 — в брачном наряде, подвид ibis; 2 — самец в разгар тока, подвид ibis;
3 — в брачном наряде, подвид coromandus; 4-5 — в послебрачном наряде; 6 — голова в начале
брачного сезона; 7 — молодая; 8 — птенец); 9-17 — Малая белая цапля (9 — пара в разгар тока;
10-11 — в небрачном наряде; 12 — голова птицы из западной Евразии в послебрачном наряде;
13 — она же в брачном наряде; 14 — голова птицы с Дальнего Востока в послебрачном наряде;
15 — она же в брачном наряде; 16 — голова молодой птицы; 17 — птенец);
18-19 — Желтоклювая цапля (18 — в брачном наряд; 19 — голова птицы в небрачном наряде).
1-3 — Средняя белая цапля (1 — в брачном наряде; 2 — в послебрачном наряде; 3 — голова в по-
слебрачном наряде); 4-10 — Большая белая цапля (4 — в брачном наряде; 5 — самец дальневос-
точной популяции в разгар тока; 6 — голова в брачном наряде; 7-8 — в послебрачном наряде;
9 — молодая; 10 — птенец); 11-12 — Южная белая цапля (11 — голова птицы в брачном наряде;
12 — в послебрачном наряде).
Таблица XIII. Цапли
1-6 — Серая цапля (1-3 — взрослая; 4 — голова в разгар тока; 5 — молодая; 6 — птенец);
7-12 — Рыжая цапля (7-9 — взрослая; 10-11 — молодая; 12 — птенец).
Таблица XIV. Аисты
1-5 — Черный аист (1-3 — взрослый; 4 — молодой; 5 — птенец); 6-11 — Белый аист (6-8 — взрос-
лый; 9 — ток у гнезда; 10 — молодой; 11 — птенец); 12-13 — Дальневосточный аист, взрослый.
Таблица XV. Ибисы
1-3 — Красноногий ибис (1 — в брачном наряде; 2-3 — в послебрачном наряде); 4-6 — Священ-
ный ибис (4-5 — взрослый; 6 — молодой); 7-8 — Черноголовый ибис, взрослый; 9-13 — Кара-
вайка (9 — в брачном наряде; 10 — летящая, в сравнении с большим кроншнепом;
11 — в послебрачном наряде; 12 — молодая; 13 — птенец).
Таблица XVI. Колпицы и Фламинго
1-7 — Колпица (1 — в брачном наряде; 2-3 — в небрачном наряде; 4 — летящая, в сравнении с
большой белой цаплей; 5-6 — молодая; 7 — птенец); 8-12 — Малая колпица (8 — в брачном на-
ряде; 9-10 — в послебрачном наряде; 11 — молодая; 12 — птенец); 13-19 — Розовый фламинго
(13-15 взрослый; 16 — птица третьего года жизни; 17 — птица второго года жизни; 18 — моло-
дой; 19 — птенец).
Большая белая цапля
321
располагались в тростнике на высоте от 0 (2 гнезда находились на поверхности воды, воз-
можно построены они были на заломах тростника, но упали под своей тяжестью) до 65 см,
средняя высота — 35,5±7,5 см (п=10, по: Скильський и др., 20016; неопубл, данные В.В.
Бучко). В колонии в плавнях Молочного лимана (Запорожская обл.) из 300 гнёзд большой
белой цапли 2% располагались на высоте 9-50 см над поверхностью воды, 3% — в 51-100
см, 30% — в 101-150 см, 60% — в 151-250 см, 5% — более чем в 250 см. Ср. высота— 170
см (Кошелев и др., 2004). В Молдавии (п=400) гнёзда располагались на высоте от 95 см до
2,3 м, в среднем 1,6 м (Куниченко, Гусан, 2001). На Усть-Манычском вдхр. гнёзда были на
высоте от 1 до 2,3 м, в среднем 1,5 м от поверхности воды, в дельте Кубани — на высоте от
0,6 до 2 м (Казаков и др., 2004). В колонии в устье р. Лань на юге Белоруссии в 1998 г.
гнёзда (п=28), построенные на затопленных кустах ивы, находились на высоте от 0,3 до 2,5
м, в среднем 1,4 м. Глубина воды под гнёздами варьировала от 0 до 30 см (Самусенко,
Журавлев, 2000). Размещение колоний цапель в тростнике определяется уровнем воды и
состоянием зарослей. В маловодные засушливые сезоны происходит смещение колоний на
сохранившиеся обводнённые участки и их укрупнение (Кошелев и др., 2005). В разрежен-
ном тростнике гнёзда устраиваются на меньшей высоте, чем в плотных зарослях (Куничен-
ко, Гусан, 2001).
Место расположения колоний в тростниковых зарослях каждый год меняется, что свя-
зано с недолговечностью этих стаций: стебли тростника легко ломаются, влияют на состо-
яние зарослей и птицы. Участок зарослей с колонией вскоре становится непригодным для
гнездования. Уже следующей весной цапли вынуждены устраивать новую колонию за 50-
200 м от прошлогодней (Кошелев, Кошелев, 1999). Возможны поселения и на деревьях в
сухих местах. Так, на севере Крыма в 2003 г. обнаружена поливидовая колония голенастых
на деревьях 12-рядной лесополосы в пределах колонии грача из примерно тысячи пар. Здесь
гнездилось около 100 пар большой белой цапли (Костин, Тарина, 2004).
В центре колонии гнёзда располагаются более скученно, на расстоянии 1-2 м одно от
другого, а на периферии удалены на 15-30 м (Казаков и др., 2004). В Молдавии расстояние
между гнёздами составляет от 3,8 до 12,2 м, в среднем 6,1 м (п=400) (Куниченко, Гусан,
2001). Гнёзда в тростнике могут строиться так близко друг к другу, что к концу гнездового
сезона образуется общая сплошная платформа (Bauer, Glutz von Blotzheim, 1966).
Новое, только что отстроенное гнездо представляет собой перевернутый конус. Пост-
ройка довольно рыхлая, с просвечивающими стенками. Гнёзда в зарослях строятся, как
правило, из сухих стеблей тростника, на деревьях — из веток. Лоток выстилается листьями
тростника, изредка перьями (Спангенберг, 1951; Долгушин, 1960; Смогоржевский, 1979;
Абрамчук, Абрамчук, 2005).
Размеры гнёзд сильно варьируют. Гнёзда из тростника больше по диаметру, чем постро-
енные из веток, что связано с большей длиной его стеблей (Кошелев и др., 2005). Размеры
гнёзд (см). Лебяжьи о-ва (п=22): диаметр гнезда — 45-74 (в среднем, 56,4), диаметр лот-
ка — 21-35 (26,6), высота гнезда — 13-70 (25,8), глубина лотка — 4,5-13,5 (8,2) (Костин,
1983); Сев. Приазовье (п=22): диаметр гнезда min — 60-120 (83,3±3,9), диаметр гнезда
max — 75-200 (100,7±5,9), высота гнезда — 14-30 (23,4±1,1), диаметр лотка min — 25-50
(34,9±1,4), диаметр лотка max — 30-50 (37,5±1,4), глубина лотка — 4-16 (9,4±0,7) (Коше-
лев и др., 2005); Прикарпатье (п=10): диаметр гнезда — 49-80 (63,9±3,4), диаметр лотка —
18^4-7 (31,2±3,1), высота гнезда — 15^4-0 (28,8±2,4), глубина лотка — 3-16 (8,1±1,4) (по:
Скильський и др., 20016; неопубл, данные В.В. Бучко); дельта Кубани (п=27): диаметр
гнезда — 60-75, высота гнезда — 30-50, глубина лотка — 5-6 (Казаков и др., 2004); сред-
ние размеры гнёзд в юж. Белоруссии (п=14): диаметр гнезда — 59,5, диаметр лотка — 26,2,
зысота гнезда — 51,2, глубина лотка — 2,8 (Абрамчук, Абрамчук, 2005).
322
Отряд Листообразные, Семейство Цаплевые
В сооружении гнёзда принимают участие оба партнёра. Приносит гнездовой материал
чаще самец, а укладывает и утаптывает его самка. Достройка гнезда продолжается во вре-
мя насиживания и выкармливания птенцов (Спангенберг, 1951; Bauer, Glutz von Blotzheim,
1966). По мере роста птенцов, диаметр гнезда постепенно увеличивается, т.к. птицы зала-
мывают в гнездо близко растущий зелёный тростник и приносят обломки сухих стеблей.
Из этого же материала вокруг гнезда на высоте 20-50 см от него цапли образуют настил
шириной 20-30 см, на который садятся прилетевшие с кормом родители. С ростом птенцов
гнездо уплотняется и несколько оседает (Казаков и др., 2004).
Сроки начала гнездо строения регистрируются преимущественно с марта по апрель.
Однако, по данным Ю.В. Костина (1983), в 1970-х гг. на Лебяжьих о-вах птиц на гнёздах
отмечали уже с первыми признаками потепления — конце января - феврале. К ремонту
гнёзд цапли приступали в конце марта, первые кладки в колонии появлялись во второй
декаде апреля, а пик яйцекладки приходился на вторую половину этого месяца. В начале
мая появлялись первые птенцы, которые поднимались на крыло к концу июня. В 1977 и
1978 гг. цапли загнездились необычайно рано, отложив яйца в начале марта. В последнее
время сроки гнездования на Лебяжьих о-вах стали ещё более ранними. Период между при-
летом и появлением первых кладок в настоящее время колеблется в широких пределах —
от 12 до 60 дней. Начало откладки яиц приходится, обычно, на вторую декаду марта, массо-
вая яйцекладка — на первую декаду апреля. Сроки гнездования сильно растянуты. Наибо-
лее поздняя встреча слабо насиженных кладок — 20 июня 1986 г. Первые птенцы в гнёздах
появляются в разные годы с последних чисел марта до второй декады мая, обычно во вто-
рой декаде апреля. Разгар вылупления чаще всего приходится на первую декаду мая, а
поздние птенцы появляются в десятых числах июня. Покидают гнёзда птенцы с начала
второй декады мая до последних чисел августа - начала сентября. Разница крайних дат
фенофаз за 11 лет в среднем составляла (в днях): яйцекладки — 69±29 (11-104), появления
птенцов — 70±27 (16-104), подъёма на крыло — 69±28 (16-108). Средняя продолжитель-
ность гнездового цикла — 140±35 (73-176) дней (Костин, Тарина, 2004). В северо-запад-
ном Приазовье в 1988-2002 гг. постройка гнёзд отмечалась с 5 по 20 апреля, откладка яиц —
с 10 апреля по 15 июня, вылупление птенцов — с 5 мая по 20 июня, вылет — с 15 по 25
июля. Общая продолжительность гнездовой жизни одной пары — 100-111 дней (Кошелев
и др., 2005). На Сев. Кавказе откладка яиц обычно начинается во второй декаде апреля,
хотя полные кладки можно встретить и раньше. Первые птенцы появляются, как правило, в
конце апреля - начале мая, в ранние весны — раньше. Вылупление птенцов продолжается
около месяца. Вылет молодых происходит с конца июня до середины июля (Казаков и др.,
2004). На юге Белоруссии белые цапли приступают к гнездованию в конце апреля - мае.
Птенцы появляются в конце мая - начале июня (Абрамчук, Абрамчук, 2005). В Астрахан-
ском заповеднике откладку яиц отмечали между 15 и 25 апреля, вылупление первых птен-
цов — 1-10 июня, массовое появление птенцов — 15-25 июня, вылет первых птенцов —
5-15 июля, массовый вылет — 20-30 июля (Спангенберг, 1951). По данным А.Е. Лугового
(1963), откладка яиц в Астраханском заповеднике начинается в конце марта, гнёзда с клад-
ками могут встречаться до июня. В дельте Аму-Дарьи в Узбекистане большие белые цапли
приступают к размножению во второй половине марта, большинство кладок появляется в
третьей декаде марта - первой декаде апреля. Свежие кладки (вероятно, повторные) встре-
чаются до конца мая (Лукашевич, 1990).
Яйцо зеленовато-голубого цвета. Форма яиц разнообразна. Они могут быть яйцевид-
ными, слегка удлинёнными или вздутыми. Во время насиживания скорлупа становится
блестящей (Спангенберг, 1951; Bauer, Glutz von Blotzheim, 1966; Cramp, 1977; Смогор-
жевський, 1979). Размеры яиц приводятся в табл. 28. Вес свежеснесённых яиц (г): Лебя-
z.: пылая белая цапля 323
Таблица 28. Размеры яиц (мм) большой белой цапли в Вост. Европе и Сев. Азии
Регион п М lim Источник
е-: =жьи о-ва, Крым 120 62,4 х 42,9 56,9-69,9 х 34,2^6,7 Костин, 1983
вья Днепра Ге верное Приазовье 74 335 61,5 х 43,5 61,2x42,8 58,0-64,0 х 42,0-44,8 Смогоржевський, 1979 по: Кошелев и др., 2005
Прикарпатье 25 62,7 х 42,3 59,1-68,7 х 40,3—44,4 Скильський и др., 20016; неопубл, данные В.В. Бучко
Юж. Белоруссия 52 61,2x42,1 56,2-65,0 х 40,0—43,9 Абрамчук, Абрамчук, 2005
Усть-Манычское вдхр., Ростовская обл. 46 61,3 х 43,3 58,1-65,0 х 41,3—45,2 Языкова, 1970
Оз. Казинка, Ростовская обл. Дельта Волги 73 60,2 х 43,0 61,3 х 42,2 54,0-66,8 х 37,6—45,0 59,0-64,0x41,0-43,0 Казаков и др., 2004 Спангенберг, 1951
Оз. Ханка, Приморский край 25 62,6 х 42,7 58,9-70,1 х 39,0^17,8 неопубл, данные Ю.Н. Глущенко
жьи о-ва (п=100) — 47,7-71,5 (в среднем 58,7) (Костин, 1983); Сумское Полесье (п=5) —
50,3-52,7 (51,4) (Афанасьев, 1998); Усть-Манычскоевдхр. (п=19)— 49,0-58,8 (54,1) (Язы-
кова, 1970).
В году бывает одна кладка, возможны повторные после гибели основной (Cramp, 1977).
В полной кладке от 2 до 7 яиц, обычно 3-5. На Обиточное косе (Запорожская обл.) в
1992-1997 гг. из 117 полных кладок по 2 яйца было в 5,1% гнёзд, по 3, как и по 4, в 41,9%,
по 5 — в 9,4%, по 6 и 7 — в 1,8% гнёзд. В различных колониях Сев. Приазовья (выборки по
35-100 гнёзд) в 1988-2002 гг. средняя величина кладки колебалась от 2,9 до 4,1 яиц, в
среднем составляя 3,8±0,1 яйца (п=64) (Кошелев и др., 2005). На Сев. Кавказе из 107 пол-
ных кладок 2,8% содержали по 2 яйца, 14% — по 3, 66,4% — по 4, 16,8% — по 5, средняя
величина кладки составляла 3,3 яйца (Казаков и др., 2004). Величина ранних кладок в сред-
нем больше, чем поздних. Так, на Лебяжьих о-вах в 1984 г. в ранних кладках было 3-6 яиц
(в среднем 4,5; п=100), а в поздних — 2^4- (в среднем, 2,9; п=50) (Костин, Тарина, 2004). То
же отмечено и в Сев. Приазовье (Кошелев и др., 2005). По наблюдениям в центр. Калифор-
нии (США) на протяжении 13 лет, число яиц в кладке большой белой цапли колебалось от
1 до 6, средняя величина кладки составляла 2,87±0,3 яйца, изменяясь в разные годы от 2,7
до 3,07 (Pratt, Winkler, 1985).
Насиживание начинается после откладки первого яйца, но плотно птицы на кладку са-
дятся примерно после откладки третьего. Яйца откладываются с интервалом в 1-2 дня.
Насиживают оба партнёра 25-28 дней. Птенцы вылупляются асинхронно, также с интерва-
лом в 1-2 дня. Последовательность их появления соответствует порядку откладки яиц, но
птенцы из первых двух яиц могут появляться одновременно. От стадии наклюнутого яйца
до вылупления птенца проходит около 1,5 сут. (Warga, 1938; Спангенберг, 1951; Bauer, Glutz
von Blotzheim, 1966; Cramp, 1977; Смогоржевський, 1979; Костин, Тарина, 2004). Как по-
казали исследования в Техасе (США), средняя продолжительность инкубации яиц 27,3±0,3
дня (п=18), при этом отложенное первым яйцо насиживалось достоверно дольше, чем вто-
рое и третье (Custer et al., 1992).
Птенцы полуптенцового типа (Cramp, 1977). Кормят их родители в гнезде, отрыгивая
пищу. Птенцы стимулируют отрыжку, теребя родителей за клюв. В больших колониях с
плотно размещёнными гнёздами подросшие птенцы могут свободно переходить в другие
324
Отряд Листообразные, Семейство Цаплевые
гнёзда, ко времени вылета теряя связь с собственными. В такой колонии может образовы-
ваться общая платформа для всех выводков (Bauer, Glutz von Blotzheim, 1966).
По наблюдениям на Усть-Манычском вдхр., первые 10-12 дней после вылупления
один из родителей постоянно остаётся с птенцами, защищая их от хищников, переохлаж-
дения и перегрева. Вторая птица (преимущественно самец) летает за кормом. На 15-17-й
день птенцы уже могут оставлять гнездо и выходить на образовавшийся вокруг него на-
стил из зелёного тростника. К этому времени потребность их в пище возрастает, летать
за кормом начинают обе взрослые птицы. Число прилётов взрослых птиц с кормом варь-
ирует в зависимости от количества и возраста птенцов от 6 до 11 в день (Олейников,
Гончаров, 1967).
Птенцы остаются в гнезде 40-45 дней (Warga, 1938; Смогоржевський, 1979), по другим
данным — 52-56 (Олейников, Гончаров, 1967). После вылета птенцы держатся с родителя-
ми до периода осенней миграции (Cramp, 1977). Первые дни молодые птицы на ночь воз-
вращаются к родным гнёздам, примерно через неделю они группируются в небольшие стай-
ки и начинают кочевать по водоёмам (Олейников, Гончаров, 1967).
На Лебяжьих о-вах успешность размножения в разные годы варьирует от 79,0 до 93,1%,
эмбриональная смертность составляет 1,8-9,8%, ювенильная — 2,8-14,1% (Костин, Тари-
на, 2004). В Сев. Приазовье успешность размножения в 1998-2002 гг. составляла 80,2%,
гибель кладок — 6,5%, выводков — 13,3%, средний размер выводка— 3,2±0,1 птенца (Ко-
шелев и др., 2005). В центр. Калифорнии (США) на протяжении 13 лет средний размер
выводка был 0,9±0,04 птенца на гнездо, составляя в разные годы от 0,03 до 2,04; успеш-
ность размножения была равной 33%. Наиболее продуктивными оказались кладки из 4
яиц (Pratt, Winkler, 1985).
На уровень смертности птенцов существенное влияние оказывает конкуренция между
ними из-за разновозрастности. При недостатке пищи, принесенной родителями, она дос-
таётся преимущественно старшим птенцам. Младшие постепенно слабеют и погибают или
их заклевывают другие птенцы. Они выживают лишь при особо благоприятных условиях
или при гибели старших, представляя собой своеобразную «страховку» на случай смерти
последних. На Лебяжьих о-вах из-за асинхронности вылупления ежегодно гибнет до 20-
30% птенцов (Костин, Тарина, 2004). Д. Мок (Моск, 1985) предполагает, что высокий уро-
вень сиблицида у большой белой цапли связан с кормлением птенцов мелкой пищей. До-
минирующие птенцы при этом стремятся «монополизировать» весь принесённый корм. У
тех же цапель, которые приносят птенцам более крупный корм, этого не наблюдается.
По данным кольцевания на юге европейской части бывшего СССР, средняя продолжи-
тельность жизни больших белых цапель составляет 1,7 года, а птиц, переживших первый
год, — 4,5 года. Смертность молодых птиц, рассчитанная по данным мечения — 68%, птиц
старше года — 25%. Максимальный возраст — 14 лет 10 месяцев (Сапетин, 1978а). У ца-
пель с Лебяжьих о-вов в первый год жизни погибает 55,6% особей, средняя продолжитель-
ность жизни составляет 1,7 года, а у птиц, переживших первый год жизни — 3,1 года (Ко-
стин, Тарина, 2005). Данные кольцевания в США дали близкие результаты: в первый год
жизни погибает 76% птиц, ежегодная смертность цапель старше года — 26%, средняя
продолжительность жизни — 1,4 года, начиная со 2-го года — 3,3 года, максимальный воз-
раст — 22 года 10 месяцев (Cramp, 1977).
Суточная активность, поведение. Большая белая — чисто дневная цапля (Bauer, Glutz
von Blotzheim, 1966; Cramp, 1977; Martinez-Vilalta, Motis, 1992). Во время миграции она
также летит в основном в светлое время суток. В полёте образует рыхлые скученные стаи
численностью в 10-150 особей или же линейные построения — цепочка, клин, углы и
т.п. —до 50 ос. (Молодовский, 2001).
Большая белая цапля
325
В колониях взрослые птицы начинают полеты за кормом ещё на рассвете и продолжают
их с разной активностью почти всю светлую часть суток. Наибольшая активность проявля-
ется с 3 до 8-9 ч, затем она ослабевает и в полуденные часы (особенно в первую половину
периода выкармливания птенцов) почти прекращается. Второй пик активности, меньший,
чем утром, отмечается с 15-16 до 19-20 ч (Олейников, Гончаров, 1967).
Ведёт как одиночный, так и стайный образ жизни. После вылета птенцов семейные
группы широко кочуют. В местах с высокой численностью они могут объединяться в боль-
шие стаи. Образовавшиеся стаи обычно не распадаются даже на зимовках. Птицы днём
разлетаются на кормёжку, но вновь собираются на коллективную ночёвку.
При совместных кормёжках с другими цаплями большая белая доминирует над более
мелкими видами. С серой она примерно равна «по силе». Поэтому в конфликтах может
побеждать как один, так и другой вид (Exner, Schubert, 2005).
Многие орнитологи отмечают большую осторожность большой белой цапли по сравне-
нию с серой. Л.М. Шульпин (1931, цит. по: Ганя, Зубков, 1989) связывал это с тем, что
животные белого цвета лучше заметны хищникам и вынуждены раньше реагировать на
опасность. Действительно, по проведенным на Украине замерам, средняя дистанция вспу-
гивания большой белой цапли составляет 173,1 ±5,6 м (n=l 18), а серой —124,2±2,9 м (n=231)
(Грищенко, ориг.). Дистанции вспугивания обоих видов достоверно возрастают после на-
чала осенней охоты (Грищенко, 2003).
Питание. Основу питания большой белой цали составляют рыба и водные насекомые.
Помимо этого она поедает также большое количество других мелких животных — земно-
водных, мелких пресмыкающихся, грызунов, мелких птиц и их птенцов, ракообразных,
моллюсков, других водных беспозвоночных и их личинок.
Состав пищи сильно варьирует в зависимости от места и сезона. Так, на о. Городки
(Днепродзержинское вдхр., Полтавская обл.) в 1981-1983 гг. рыба составляла 96,6% пита-
ния большой белой цапли (из них 66,7% — промысловые виды), насекомые — 2%, земно-
водные — 0,7%, пресмыкающиеся — 0,3%, пиявки — 0,2%, моллюски — 0,2%, млекопи-
тающие — 0,04% (Ткаченко, 1987). В Сев. Приазовье рыба составляла 47,1 % (из них промыс-
ловая всего 10,3%), пресмыкающиеся — 5,4%, земноводные — 1,5%, беспозвоночные —
46,0% (Кошелев, 2004). На Лебяжьих о-вах в гнездовой период рыба составляет 78,9%,
насекомые — 9,0%, озёрные лягушки — 7,2%, ракообразные — 1,3%. Из насекомых цапли
чаще поедают медведок, личинок большого водолюба и стрекоз. Основу осенне-зимнего
рациона составляет рыба — караси (84%), бычки (14%), карпы (8%). До 46% возрастает
доля лягушек. Насекомые встречены в 20% проб (Костин, Тарина, 2004). На Катульском
рыбкомбинате в Молдавии до 30% корма большой белой цапли составляют беспозвоноч-
ные, 10-12% — земноводные и мышевидные грызуны, не менее 30% — сорная рыба из
сбросных и мелиоративных каналов (Куниченко, Гусан, 1992).
На Усть-Манычском вдхр., по анализу 340 отрыжек птенцов и содержимого 25 желуд-
ков взрослых птиц, рыба составляла по весу 60,9% пищи, насекомые — 29,0%, земновод-
ные — 8,5% (в основном головастики озерной лягушки). Среди рыб преобладали щука
(19,4% общего веса пищи), сазан, плотва, окунь, уклея, лещ, карась и др. В основном это
была молодь длиной от 3 до 16 см, а в отдельных случаях (ранней весной) — до 20-24 см.
68,3% рыбного корма приходилось на долю хищных и сорных видов. Из насекомых пре-
обладали жуки-плавунцы — взрослые особи и личинки, водолюбы, крупные водяные
клопы, личинки стрекоз. Из наземных позвоночных в небольшом количестве встреча-
лись обыкновенные полёвки и прыткие ящерицы. Соотношение кормов изменялось на
протяжении сезона. Весной резко преобладала рыба, причём в первую половину весны
цапли добывали относительно крупных рыб, а в мае — преимущественно мальков и се-
326
Отряд Листообразные, Семейство Цаплевые
голеток. В первую половину периода выкармливания птенцов (16 мая - 4 июня) вес рыбы
в пище птенцов составлял 89,3%, а насекомых — 2,2%, а во вторую половину (5-27 июня)
доля рыбы уменьшилась до 48,4%, а насекомых возросла до 50,5%. После вылета птен-
цов процент рыбы в их корме мог снова возрасти. Соотношение основных компонентов
пищи изменялось и по годам, в зависимости от доступности кормовых объектов (Олей-
ников, Гончаров, 1967).
На лиманах Вост. Приазовья в летне-осенние месяцы 1953-1954 гг. белые цапли пита-
лись в основном рыбой (55,4% по весу), хищными водными насекомыми (22,2%), озёрны-
ми лягушками и их головастиками (22,1%). Наибольшая встречаемость рыб наблюдалась в
конце июня - начале июля, когда мелели и пересыхали лиманы. Из рыб в пище преоблада-
ли молодь сазана, щука, окунь, краснопёрка. На Ахтарском рыбкомбинате большая белая
цапля охотилась в основном на выростных и частично на нагульных прудах. Вес суточного
рациона достигал 250 г. Из прудовых рыб на долю карпа приходилось 75%, белого толсто-
лобика — 22%, пёстрого толстолобика — 3% (Казаков и др., 2004).
На оз. Ханка в 16 пробах бычки составляли 41,1%, вьюны — 30,4%, девятииглая ко-
люшка — 9,7%, жуки-плавунцы — 8,8%, стрекозы — 4,0%, дальневосточная полёвка —
4,0% (Поливанова, 1971).
В желудках птиц, добытых зимой на родниках, находили лягушек и мышевидных гры-
зунов (Казаков и др., 2004). В негнездовое время в сухих местах большая белая цапля мо-
жет питаться преимущественно мышевидными грызунами и наземными насекомыми (Спан-
генберг, 1951; Cramp, 1977). Неоднократно в питании отмечали саранчу (Спангенберг, 1951;
Олейников, Гончаров, 1967).
Кормится цапля преимущественно на открытых мелководных водоёмах и их берегах. На
мелководье охотится, бродя по воде и высматривая добычу, или подкарауливает её, стоя на
месте. Если вода слишком глубокая, птица стоит на берегу, наклонив голову к воде. Добычу
хватает быстрым ударом клюва. Может охотиться также в сухих местах, активно разыскивая
или подкарауливая добычу. Охотится обычно поодиночке, агрессивно защищая кормовую
территорию от конкурентов. В местах, обильных пищей, может собираться в большие стаи.
Наблюдения на оз. Нойзидлерзее в Австрии показали, что распределение белых цапель
на водоёме зависит от обилия рыбы. Птицы обычно держатся там, где плотность её наи-
большая (Nemeth, Schuster, 2005). При этом цапли могут наблюдать за охотой других рыбо-
ядных птиц, например крачек, и перемещаться туда, где она была успешной (Peter, 1994). В
США большие белые цапли часто кормятся вместе со снежными цаплями. Последние бо-
лее подвижны, они вспугивают добычу, которая становится доступнее для более медли-
тельных больших белых цапель (Brzorad et al., 2004).
Враги, неблагоприятные факторы. Начиная со второй половине XIX в. и в первую
половину XX в. большая белая цапля подвергалась хищническому истреблению из-за моды
на украшения из перьев. Особым спросом пользовались эгретки. У одной большой белой
цапли в брачном наряде было 30-50 этих перьев. 1 кг эгреток получали примерно от 150
особей этих птиц. О масштабах истребления говорят следующие цифры. В 1902 г. только
одна лондонская фирма, специализировавшаяся на торговле перьями, продала 48 240 ун-
ций эгреток. Поскольку на одну унцию этого товара требовалось, как минимум, 4 птицы,
для такого количества эгреток было убито по меньшей мере 190 тыс. больших белых ца-
пель. В 1899-1912 гг. из Аргентины, Бразилии и Венесуэлы было экспортировано 15 т.
перьев, прежде всего больших белых и снежных (Egretta thuld) цапель. Белых цапель добы-
вали в разгар их гнездового сезона, обычно прямо на колониях. При этом в массе гибли
яйца и птенцы, родители которых были уничтожены (Martinez-Vilalta, Motis, 1992). В Вене-
суэле в 1898 г. было отстреляно 1,5 млн. больших белых цапель (Sedlag, 1983). Ажиотаж
Большая белая цапля
327
подогревался очень высокими ценами на эгретки. В 1903 г. «охотники за перьями» в США
получали по 30 долларов за унцию перьев, унция золота была в то время вдвое дешевле.
Через некоторое время цена поднялась до $80 (Martinez-Vilalta, Motis, 1992). В 1912 г. в
России эгретки скупались по цене 600 руб. золотом за фунт (Шнитников, 1957). Торговля
перьями птиц пошла на убыль лишь после Первой мировой войны и прекратилась после
Второй (Дорст, 1968). Огромный ущерб причиняла и заготовка яиц, которые вывозились с
крупных колоний птиц «лодками», в смысле — объёма (Поливанова, 1971; Белик, 2000;
Русев, 2004).
В настоящее время основной фактор, лимитировавший численность — интенсивное
истребление человеком — перестал действовать. Целенаправленно преследовать белых
цапель сейчас могут разве что в некоторых рыборазводных хозяйствах. Население же отно-
сится к этой красивой, изящной птице вполне дружелюбно.
Успешность размножения больших белых цапель зависит от погоды, кормности угодий,
деятельности хищников, беспокойства. При пересыхании водоёмов гнёзда становятся дос-
тупными для четвероногих хищников, которые их разоряют. Взрослые птицы успешно за-
щищают кладки и маленьких птенцов от хищных птиц и врановых, но если люди часто
беспокоят цапель в колонии, они надолго покидают гнёзда, чем активно пользуются серые
вороны (Кошелев и др., 2005).
Взрослых больших белых цапель могут добывать крупные хищники — орланы, орлы,
ястреб-тетеревятник, из млекопитающих — лисицы, волки, бродячие собаки. Анализ на-
ходок окольцованных птиц показал, что основными причинами гибели вне мест гнездова-
ния являются неблагоприятные условия зимовки (57,8%) и преследование человеком (39,4%)
(Костин, Тарина, 2005). Гнёзда разоряют болотные луни, врановые, крупные чайки, четве-
роногие хищники — лисица, енотовидная собака, шакал, а также крысы. Гнёзда на мелко-
водье могут страдать в результате проходов кабанов.
Как раноприлетный вид, большая белая цапля страдает от продолжительных возвратов
холодов весной. При этом часть птиц может погибнуть, другие ослабевают, что в итоге
приводит к снижению результативности размножения.
Главный отрицательный фактор для гнездования белых цапель в настоящее время —
разрушение среды обитания человеком вследствие осушения заболоченных территорий,
затопления плавней и прибрежных зарослей водохранилищами застройки побережий и т.п.
Негативно влияет также заготовка тростника. Существенный отрицательный фактор —
выжигание зарослей. Так, в 2004 г. в дельте р. Или в Казахстане во время пожара, охватив-
шего более 1000 га тростников, погибли колонии большой белой цапли и других птиц.
Кроме того, цапли не гнездятся в молодых зарослях тростника, которые возникают на сле-
дующий год после выкашивания или пожаров (Кошелев, Кошелев, 1999). Отрицательное
влияние оказывает беспокойство птиц в колониях людьми, особенно в начале гнездования.
Достаточно 1-2-кратного посещения колонии во время постройки гнёзд или в начале от-
кладки яиц, чтобы цапли вообще покинули это место (Кошелев, Кошелев, 1999). В период
насиживания и выкармливания птенцов яйца и маленькие птенцы в таких ситуациях могут
погибнуть от хищников и переохлаждения. В активно посещавшихся людьми колониях на
Усть-Манычском вдхр. в 1958 г. отход яиц и птенцов составил 55%; в гнёздах перед выле-
том насчитывалось всего 1,6 птенца (Олейников, Гончаров, 1967).
Численность колоний зависит от уровня воды в водоёмах. Причём отрицательное воз-
действие оказывает как слишком низкий, так и слишком высокий уровень. Во многих мес-
тах отмечалось снижение численности вплоть до полного исчезновения колоний при сни-
жении уровня воды — в Венгрии (Furi, Vidra, 2002), на Украине (Кошелев и др., 2004;
Кошелев, 2004), в Казахстане (Дебело, 2002). Причём влияние гидрологических условий
328
Отряд Листообразные, Семейство Цаплевые
чаще и резче проявляется на малых водоёмах (Кошелев и др., 2004). Исследования на оз.
Нойзидлерзее в Австрии дали несколько иной результат — размер колоний зависел не от
глубины воды, а от ширины полосы тростниковых зарослей (Griill, Ranner, 1998).
В дельте Волги в последнее время происходит непрерывное увеличение глубин из-за
трансгрессии Каспийского моря, что привело к сокращению численности белых цапель
(Кривенко и др., 1998). С другой стороны, в низовьях Урала сформировалась обширная
дельта с водно-болотным орнитокомплексом, близким к волжскому (Березовиков, Гисцов,
1999).
Существенный ущерб колониям в плавнях рек могут приносить поздневесенние и лет-
ние паводки. Так, два наводнения в июне и июле 1998 г. уничтожили 69,6% гнёзд в колонии
на Горелых озёрах в дельте Днестра (Русев, 2004). В том же году отмечалась высокая смер-
тность птенцов во время паводка и на Волге (Завьялов и др., 2005). Кладки и птенцы могут
погибать во время сильных ливней, особенно, сопровождающихся градом.
К неблагоприятным факторам относится накопление пестицидов и тяжелых металлов в
органах и тканях вследствие загрязнения среды обитания. Так, в Кызыл-Агачском заповед-
нике у 7 из 10 обследованных больших белых цапель содержание хлорорганических пести-
цидов в печени колебалось от 0,06 до 0,3 мг/кг, в жировой ткани — от 0,06 до 0,35 мг/кг
(Литвинова, 1988). На Лебяжьих о-вах в 1989-1992 гг. в тканях птенцов обнаружено высо-
кое содержание ДДТ и ДДЕ. В 1991 г. у цапель наблюдалось снижение успешности размно-
жения из-за значительного количества неоплодотворённых и аномально мелких (безжёлт-
ковых) яиц, 3% гнёзд содержали по одному аномальному яйцу (Костин, Тарина, 2004). На
юге Флориды в перьях большой белой цапли обнаружено 7,1 частей на миллион ртути,
концентрация её увеличивалась с возрастом птиц. Такое содержание ртути уже может быть
достаточным для снижения продуктивности (Beyer et al., 1997). По наблюдениям в Кали-
форнии, после запрета использования ДДТ у белых цапель достоверно увеличились раз-
мер кладки и успешность размножения (Pratt, Winkler, 1985).
Белые цапли могут становиться жертвами клептопаразитизма, подвергаясь нападениям
различных птиц от крупных чаек до орланов. Интересный случай клептопаразитизма заре-
гистрирован в Венгрии: на летевшую цаплю с пойманной на поле мышью напал балобан и
отобрал добычу (Kovacs, 2004).
На юге Украины у большой белой цапли зарегистрировано 35 видов гельминтов: Schisto-
cephalus pungtii, Gryporhynchus pusillus, Clinostomum complanatum, Pegosomum saginatum,
P spiniferum, Echinochasmus beleocephalus, E. coaxatus, E. euryporus, E. militaris, E. mordax,
E. bursicola, Saakotrema metatestis, Philophtalmus palpebrarum, Cryptocotyle concavum, C.
lingua, Pygidiopsis genata, Galastosomum sp., Prostogonimus anatinus, Strigea falconis, Apha-
ryngostrigea cornu, Hysteromorpha triloba, Histeromorphoides sp., Posthodiplostomum cuticola,
Bilharziella polonica, Polymorphus minutus^ P striatus, Southwellina hispida, Contracaecum
microcephalum, Porrocaecum reticulatum, Desmidocercella numidica, Tetrameres fissispina,
Desportesius brevicaudatus, Desportesius sp., Cosmocephalus oblevatus, Avioserpens galliardi.
Цестоды встречаются редко, трематодофауна достаточно богата. Доминируют Cryptocotyle
convacum и Pygidiopsis genata, для которых большая белая цапля является основным хозя-
ином в регионе (Корнюшин и др., 2004).
Хозяйственное значение, охрана. Большая белая цапля заслуживает всяческой охра-
ны хотя бы как эстетически ценный вид, украшающий любой ландшафт. Она может прино-
сить некоторый ущерб рыбному хозяйству, особенно на рыборазводных прудах. Однако,
как показали специальные исследования, он с лихвой компенсируется истреблением вре-
дителей рыбного хозяйства — хищных водных насекомых (имаго и личинок), лягушек и
головастиков, поедающих икру и мальков. Поедают эти птицы также большое количество
Большая белая цапля, Южная белая цапля
329
хищных и сорных рыб. На Усть-Манычском вдхр. они составляли около 12% веса пищи
(Олейников, Гончаров, 1967). Кроме того, белые цапли уничтожают мышевидных грызу-
нов, саранчовых, медведок, считающихся вредителями сельского хозяйства.
Белые цапли могут быть распространителями ряда опасных инфекций, экологически
связанных с птицами — орнитоза, гриппа, лептоспироза и др. (Греков и др., 2004).
Большая белая цапля занесена в Красные книги Армении, Белоруссии, Грузии, Молда-
вии, а в России — в Красные книги республик Ингушетии, Мордовии, Татарстана, Тывы,
Хакасии, Алтайского и Хабаровского краев, Брянской, Курганской, Липецкой, Новосибир-
ской, Омской, Пензенской, Саратовской, Сахалинской, Смоленской и Тюменской областей,
Еврейской АО.
Южная белая цапля
Casmerodius modestus (J.E. Gray, 1831)
Ardea modesta J.E. Gray, 1831, Zool. Miscellany, c. 19, Индия
Southern Egret, Southern White Egret, Eastern/Oriental White Egret (англ.);
Siidlicherreicher (нем.); sud aigrette (фр.).
Син. (лат.): Ardea alba modesta J.E. Gray, 1831; Egretta alba modesta (Gray, 1831); Ardea torra
Franklin, 1831; Ardea putea Franklin, 18.31; Ardea timoriensis Lesson, 1831; Ardea longicollis
Meyen, 1834; Herodias syrmatophorus Gould, 1846; Ardea syrmatophora Taczanowski, 1876;
Egretta alba neglecta Mathews, 1912; Herodias alba maoriana Iredale et Mathews, 1913.
Син. (рус.): восточная белая цапля.
Статус. Нерегулярно гнездящийся, перелётный вид.
Общая характеристика и полевые признаки. Довольно крупная цапля: заметно мельче
большой белой цапли, но крупнее средней. Длина тела около 90 см, размах крыльев немно-
гим более 1 м. Оперение рыхлое, чисто белой окраски. В брачном наряде на спине выделя-
ются длинные и рассученные перья (эгретки), заметно выступающие за обрез хвоста. На
нижней стороне шеи и зобе расположены гораздо менее длинные и рассученные перья,
образующие «гриву». Удлинённые перья на голове отсутствуют. Характерны сезонные из-
менения окраски клюва и голых участков кожи на «уздечке» и вокруг глаз. От большой
белой цапли отличается меньшими размерами, несколько более частыми взмахами крыль-
ев в полёте, а также голубой или голубовато-зелёной (а не зеленовато-жёлтой) окраской
голой кожи на «уздечке» и вокруг глаз в брачном наряде и практически целиком чёрными
ногами как в гнездовой, так и в негнездовой период. От средней белой цапли отличается
более крупными размерами, несколько менее частыми взмахами крыльев (в полете), менее
длинными и слабо рассученными украшающими перьями на нижней стороне шеи и зобе,
относительно более длинным и тонким клювом, а также голубой или голубовато-зелёной (а
не жёлтой) окраской кожи на «уздечке».
Полёт спокойный и прямолинейный, с глубокими взмахами крыльев, иногда скользя-
щий. При тревоге круто взлетает вверх, быстро взмахивая крыльями и набирая высоту. Во
время полёта ноги вытянуты за вершину хвоста, а шея согнута в вертикальной плоскости и
втянута в плечи. По твёрдому субстрату передвигается большими шагами, по дну мелко-
водных водоёмов — медленно, опустив голову вниз. Во время отдыха нередко стоит на
одной ноге. Поселяется в колониях других видов цапель (влючая больших белых). В пери-
330
Отряд Листообразные, Семейство Цаплевые
од кочёвок и миграций предпочитает держаться группами, зачастую объединяясь с други-
ми видами белых цапель, образуя на кормёжке разреженные скопления.
Голос. Молчалива. При тревоге издаёт хриплый каркающий звук «кра-а, кра-а ...»).
Подвидовая систематика. Монотипический вид. Иногда в качестве отдельного подви-
да рассматривают новозеландскую форму maorianus.
Замечания по систематике. Южная белая цапля была описана в 1831 г. из Индии как
Ardea modesta J.E. Gray. В 1913 г. она была вновь описана, но уже в качестве подвида боль-
шой белой цапли Herodias alba maoriana Iredale et Mathews. В российской орнитологичес-
кой литературе её традиционно считали подвидом большой белой цапли, хотя и указыва-
лось, что таксономические отношения форм alba и modesta выяснены недостаточно (Сте-
панян, 2003).
Следует отметить, что до последнего времени южная и большая белые цапли рассмат-
ривались как аллопатрические формы, линия разделения гнездовых ареалов которых про-
ходит по 44-й параллели (Воробьёв, 1954; Степанян, 2003). Реально же в Приморье (и на
Приханкайской низменности, в частности) почти до самого конца XX столетия было изве-
стно гнездование лишь Egretta alba alba (Casmerodius albus albus) (Шульпин, 1936; Поли-
ванова, 1971; Глущенко и др., 1992). Гнездование Е. modesta (С. modestus) здесь впервые
было установлено лишь в 1999 г. (Глущенко и др., 2003).
Морфологические отличия между albus и modestus проявляются, прежде всего, в общих
размерах, которые не перекрываются. Значительная разница в размерах и выявленные отли-
чия в брачном поведении недавно позволили вновь возвратить форме modesta видовой статус
(Inskipp et al., 1996). Установление факта совместного гнездования обеих рассматриваемых
форм в одной и той же колонии на оз. Ханка в 1999-2003 гг. показало их локальную симпат-
рию. Обнаруженные при этом отличия в фенологии, особенностях гнездостроения, разме-
рах гнёзд и яиц (Глущенко, Коробов, 2005) явно свидетельствуют о видовой самостоятель-
ности южной белой цапли. Согласно результатам молекулярного анализа, американская
большая белая цапля (С. albus garzetta) и С. modestus из австралийских популяций различа-
ются по генотипу на видовом уровне (Christidis, Boles, 2008). О выделении «комплекса
больших белых цапель» в самостоятельный род Casmerodius см. предыдущий очерк.
Русское название «южная белая цапля», впервые предложенное ещё С.А. Бутурлиным
(Бутурлин, Дементьев, 1935), удачно характеризует особенности её географического рас-
пространения по сравнению с космополитом — большой белой цаплей, подвидом которой
она считалась. Однако, учитывая, что в Америке и Африке большая белая цапля обитает на
тех же широтах и даже южнее (до Патагонии), чем «южная белая цапля» в Азии и Океании,
название «восточная белая цапля» (Коблик и др., 2006) выглядит не менее правомерным, а
может быть и более точным.
Описание (по: Sonobe, Usui, 1993; Mackinnon, Phillipps, 2000; Коллекции Биолого-по-
чвенного института ДВО РАН и Дальневосточного государственного университета, г. Вла-
дивосток). Окраска. Половой диморфизм в окраске отсутствует, однако у самцов укра-
шающие перья немного длиннее, чем у самок.
Взрослые самец и самка. Оперение белое. Клюв и ноги чёрные, но в брачный период на
нижней части голени может просматриваться красно-розовый оттенок (King, Dickinson,
1995; данные В.А. Нечаева, — ориг.); во внегнездовой период клюв жёлтый. Окраска голой
кожи на «уздечке» и вокруг глаз в брачный период голубая или голубовато-зелёная, а в
остальное время — зелёная или жёлтая. Радужина жёлтая. В гнездовой период на спине
развиты эгретки, как и у большой белой цапли.
Первый пуховой наряд. Пух белый. Клюв телесно-розовый с чёрным кончиком.
Второй пуховой наряд. Пух белый. Клюв желтоватый с чёрным кончиком.
Южная белая цапля
331
Гнездовой наряд. Оперение белое. Клюв жёлтый с чёрным кончиком надклювья. Голая
кожа в углах клюва, на «уздечке» и вокруг глаза жёлто-зеленоватые. Ноги черноватые.
Первый годовой наряд. Оперение белое. Украшающие перья отсутствуют. Клюв жёл-
тый с тёмным кончиком надклювья. Голая кожа на «уздечке» и вокруг глаза желтоватая с
зелёным оттенком. Ноги чёрные.
Строение и размеры. Стройная птица, обликом очень напоминающая большую
белую цаплю. Шея длинная и тонкая, клюв и ноги длинные. Размеры (мм). Птицы (самцы
и самки), добытые на юге Приморья (п=8) имели длину крыла 343-385 (в среднем 357,3),
длину цевки 138-158 (в среднем 149,8), длину клюва 92-109 (в среднем 96 мм) (Дубинин,
Горчаковская 1951; Спангенберг, 1951). Размеры двух цапель, добытых в окрестностях
Лазовского заповедника: птица, пол которой не установлен, имела длину крыла 358, цев-
ки — 153, клюва — 113 (Воробьёв, 1954); самка имела длину крыла 375, хвоста — 145
клюва — 97 (Белопольский, 1955). Экземпляры из Юж. Приморья, хранящиеся в коллек-
ции Зоомузея ДВГУ: самец — крыло 370, цевка 165, клюв 115; самка — соответственно
350, 145 и 105; птица, пол которой не установлен — соответственно 355, 155 и 105. Самцы
(n=l 1) из Индии: крыло 345-378 (в среднем 361), цевка 141-172 (в среднем 157), клюв
101-115 (в среднем 108); самцы (п=10) из Китая: крыло 340-380 (в среднем 366,4), цевка
135-167 (в среднем 153,8), клюв 98-125 (в среднем 112) (Vaurie, 1965). Две особи из Сев.
Кореи (пол не установлен): крыло 350 и 360, цевка 152 и 155, хвост 100 и 148, клюв 112 и
128 (Тошек, 1999).
Масса птиц (п=8), добытых в Приморском крае, составляла 792-1 108 г, в среднем 908,3
г (Спангенберг, 1951); самка имела массу 815 г (Белопольский, 1955).
Линька. Взрослые особи проходят полную годовую линьку в период с июля по ноябрь.
На местах зимовок у взрослых птиц происходит частичная предбрачная линька: вырастают
украшающие перья. Цапли в гнездовом наряде начинают смену мелких перьев уже в авгу-
сте, продолжают осенью и заканчивают во время зимовки; на втором году жизни они, по-
добно взрослым птицам, проходят полную годовую линьку. Самка, добытая на крайнем
юге Приморского края 6 сентября 1946 г. начала линьку мелкого пера на шее (Дубинин,
Горчаковская, 1951).
Распространение. Гнездовой ареал. Юж., Юго-Вост, и Вост. Азия, Индонезия,
Новая Гвинея, Австралия и Новая Зеландия (Спангенберг, 1951; Glenister, 1959; Vaurie, 1965;
King, Dickinson, 1995; Степанян, 2003; Dickinson, 2003; и др.) (рис. 65).
В Китае южная белая цапля распространена локально: гнездится в провинциях Гирин,
Ляонин, Хэбэй, Фуцзянь и юго-западных районах провинции Юньнань (Cheng Tso-Hsin,
1987; Mackinnon, Phillipps, 2000); нерегулярное гнездование отмечено в Гонконге (Carey et
al., 2001). Известны колонии на юге Таиланда (Lekagul, Round, 1991). На п-ове Корея эта
цапля гнездится в центральных и южных районах (Won Pyong-oh, 1996; Tomek, 1999), а в
Японии — на о-вах Хонсю, Сикоку и Кюсю (Check-list of Japanese birds, 2000).
В России достоверное гнездование известно только для южного побережья оз. Ханка в
приустьевой части р. Илистая (Глущенко и др., 2003; Глущенко, Коробов, 2005) (рис. 66).
Предполагается гнездование двух пар именно этого вида белых цапель на о. Фуругельма в
зал. Петра Великого (Литвиненко, Шибаев, 1999). Следует отметить, что серия птиц данно-
го вида, собранная в Юж. Приморье в летний период 1945-46 гг., позволила отнести их к
категории гнездящихся (Дубинин, Горчаковская, 1951), однако, как справедливо заметил
Е.Н. Панов (1973), такое утверждение тогда было голословным.
Зимовки. Область зимовок птиц северных популяций — Юго-Вост, и Вост. Азия:
южные районы Китая, о-ва Тайвань и Хайнань (Cheng Tso-Hsin, 1987; Mackinnon, Phillipps,
2000), Гонконг (Carey et al., 2001), центральные и южные районы п-ова Корея (Won Pyong-
332
Отряд Листообразные, Семейство Цаплевые
Рис. 65. Область распространения южной белой цапли: а — гнездовой ареал, б — установленные
гнездовые поселения, в — район зимовок.
oh, 1996; Tomek, 1999), южная часть о-ва Хонсю и острова, расположенные южнее (Check-
list of Japanese birds, 2000), страны Юго-Вост. Азии, в частности, Таиланд (Lekagul, Round,
1991), Вьетнам (Во Кви, 1983), Малайзия (Glenister, 1959) и другие. Южные популяции по-
видимому, оседлы (рис. 65).
К.А. Воробьёв (1954) указывает на находку одного экземпляра формы modesta в Юж-
ном Приморье (п-ов Муравьёва-Амурского, район Ляньчихэ) 25 января 1948 г. В коллекции
Зоологического музея БПИ ДВО РАН хранится крыло белой цапли с указанием упомянуто-
го места и датированное январём 1948 г., однако, его длина составляет 400 мм, что соответ-
ствует предельно мелкому экземпляру С. albus albus.
Залеты на юге Приморского края были известны с последней четверти XIX века (Tacza-
nowski, 1893; Шульпин, 1936). Кроме того, на Дальнем Востоке России в качестве залётной
птицы вид известен для Сахалина, Юж. Курильских о-вов: Кунашир, Итуруп, Анучина (Не-
чаев, 2005) и Камчатки (Артюхин и др., 2000). Залёты птиц, похожих на С. modestus, заре-
гистрированы в Европу (del Hoyo et al., 1992), однако там их относят к форме «modesta
type» (см. раздел по миграциям очерка о большой белой цапле).
Миграции. В Южном Приморье наиболее раннее появление весной зарегистрировано
9 апреля 2005 г. на побережье Уссурийского зал. (данные В.А. Нечаева); 20 марта 2007 г. и
31 марта 2006 г. в окрестностях г. Уссурийска; 7 апреля 2006 г. на оз. Ханка (данные Ю.Н.
Глущенко). В окрестностях г. Уссурийск транзитный весенний пролёт проходит в апреле и
первой половине мая (Глущенко, Коробов, 2005). Птицы обычно летят совместно с други-
ми видами цапель. Послегнездовые кочёвки начинаются в августе, а осенний пролёт завер-
шается в октябре. Сроки осеннего отлёта гнездящейся популяции не известны.
Южная белая цапля
333
Холостые птицы регулярно отмечаются в
различных районах Юж. Приморья, гл. обр. с
мая по август. Здесь они образуют разрознен-
ные смешанные группировки с другими ви-
дами цапель. Такие скопления, в которых до-
минировали южные белые цапли, встречены
на юге Хасанского р-на (крайний юго-запад
Приморья) со второй половины августа до
первой декады октября 1946 г. (Дубинин, Гор-
чаковская, 1951; Воробьёв, 1954), а также в
июле-августе 1996 г. (Литвиненко, Шибаев,
1999).
Местообитания. На оз. Ханка колонии, в
которой гнездились южные белые цапли, раз-
мещались в полосе затопленных ивняков, ок-
ружённых озёрно-болотным массивом. Мес-
тами кормёжки здесь служат сырые луга, тра-
вяные болота, мелководья озёр и рисовые поля.
Рис. 66. Ареал южной белой цапли в России:
а — установленные места гнездования, б —
район встреч, в — залёты.
Кочующие в Юж. Приморье птицы придерживаются сырых луговин, травяных болот, мел-
ководных побережий озёр и рек, а также морских побережий (заболоченные участки на
приморской низменности, лагуны, литораль).
За пределами России птицы гнездятся на открытых пространствах, занятых озёрами,
реками, травяными болотами и затопленными лесами (Sonobe, Usui, 1993). В Таиланде и
Малайзии, кроме того, птицы населяют мангровые леса (Lekagul, Round, 1991; Glenister,
1959), а в Японии (о. Хонсю) одна из крупных колоний была расположена в лесу из крипто-
мерий, причём гнёзда помещались на верхних ветвях этих хвойных растений (Jahn, 1942).
Численность. В Приморском крае это очень редкий, нерегулярно гнездящийся вид. По
приблизительной оценке численность южной белой цапли в колонии, расположенной на
южном побережье оз. Ханка в 1999-2000 гг., находилась в пределах от 15 до 20 пар, а в
2001 г. она составляла 30-40 гнездящихся пар (данные Ю.Н. Глущенко). Краткосрочное
посещение колонии в 2002 г. выявило лишь 8 пар южных белых цапель, причём снижение
их численности, по-видимому, было связано с экспансией большого баклана, вытеснивше-
го цапель. В 2003 г. основная часть цапель оставила данную колонию, сформировав в её
окрестностях серию дочерних поселений, в одном из которых отмечено несколько особей
южной белой цапли, которые, судя по поведению, здесь гнездились. В 2004 г. наблюдения
не проводились, а в 2005 г. рассматриваемый вид, несмотря на специальные поиски, обна-
ружен не был (Глущенко, Коробов, 2005). В 2008 г. птиц наблюдали в двух колониях, где,
вероятно, гнездилось от 9 до 13 пар и было осмотрено 6 жилых гнёзд (данные Ю.Н. Глу-
щенко, И.Н. Кальницкой и Д.В. Коробова).
Холостые птицы, кочующие в Юж. Приморье, по суммарной численности заметно пре-
восходят гнездящуюся на Ханке популяцию и в своей основе, безусловно, состоят из птиц,
которые вывелись за пределами России.
В Китае южная белая цапля местами обычна в колониях и немногочисленна во внегнез-
довой период (Cheng Tso-Hsin, 1987; Mackinnon, Phillipps, 2000). В Японии вид немного-
числен в гнездовой сезон и малочислен зимой (A field guide to the birds of Japan, 1982). Ha
Корейском п-ове эта цапля обычна летом и немногочисленна зимой (Won Pyong-oh, 1996),
а в Гонконге — редка в гнездовой сезон (в 1997 г. было учтено 30 пар), но обычна и даже
многочисленна в другие времена года (Carey et al., 2001).
334
Отряд Листообразные, Семейство Цаплевые
Размножение. Колониальная птица. Моногам. Гнездовая биология в условиях Примор-
ского края изучена крайне слабо. На местах размножения птицы, вероятно, появляются в
первой-второй декадах апреля. Гнездовой сезон длится со второй половины апреля по июль
и начинается на 10-15 дней позднее, чем у большой белой цапли (Глущенко, Коробов, 2005).
В многовидовом поселении цапель и больших, бакланов на побережье оз. Ханка южные
белые цапли тяготеют к формированию самостоятельной группировки, в состав которой
также входят средняя и малая белые цапли. Гнёзда размещаются на ветвях затопленных ив,
и сооружаются почти исключительно из ивовых прутьев с редким включением стеблей
тростника. Гнёзда меньшего размера и более компактны, чем у больших белых цапель.
Размеры (см) гнёзд (п=12): диаметр гнезда 29-64 (в среднем 47,9); диаметр лотка 20-34 (в
среднем 26,1), высота гнезда 14-33 (в среднем 23,2); глубина лотка 4,2-9,3 (в среднем 6,8).
Откладка яиц, судя по всему, происходит в конце апреля и в первой половине мая. В
кладке от 3 до 6 яиц. Окраска яиц однотонная, слегка голубовато-зеленоватая. Размеры
(мм) яиц (п=39): 53,3-58,3 х 35,8-41,1 (в среднем 55,5x38,4). Насиживают оба партнёра в
течение 25-26 суток. В 2001 г. вылупление птенцов отмечено 21 мая (в двух из 8 гнёздах
были пуховые птенцы, в остальных — яйца). 29 мая 2008 г. в двух гнёздах было 3 и 5
ненасиженных яиц, в трёх гнёздах содержалось по 2, 3 и 3 пуховых птенца, а в одном — 3
оперённых птенца (данные Ю.Н. Глущенко, И.Н. Кальницкой и Д.В. Коробова). Птенцы
покидают гнёзда в июле, а в августе уже начинаются кочёвки.
Суточная активность, поведение. Активны преимущественно в дневное время, одна-
ко транзитные миграции отмечались и в сумерках. Поведение в условиях Приморского
края не изучено.
Питание. В целом для вида характерно питание как водными, так и наземными позво-
ночными и беспозвоночными животными (моллюсками, пауками, насекомыми и их личин-
ками, рыбами, амфибиями и т.д.). В желудке самки, добытой в Лазовском заповеднике 9
мая 1944 г., были обнаружены остатки лягушки и рыб: колюшки и бычка.
Враги, неблагоприятные факторы. В условиях оз. Ханка известен случай острой кон-
курентной борьбы с большим бакланом, вытеснившим птиц из гнездовой колонии (Глу-
щенко, Коробов, 2005).
Хозяйственное значение и охрана. Ввиду крайней редкости в пределах России хозяй-
ственного значения южная белая цапля не имеет. Занесена в Красную книгу Приморского
края (2005) и подлежит включению в последующее издание Красной книги Российской
Федерации. Одна из смешанных колоний цапель с участием южной белой цапли находится
в Ханкайском государственном природном биосферном заповеднике, другая — в его ох-
ранной зоне. Для улучшения охраны вида необходимо увеличение площади данного запо-
ведника за счёт включения в его состав участка охранной зоны с колонией цапель.
Род Ardea Linnaeus, 1758
Серая цапля
Ardea cinerea Linnaeus, 1758
Ardea cinerea Linnaeus, 1758, Syst. Nat., ed. 10, c. 134, Швеция
Gray [common] heron (англ.), Fischreiher, Graureiher (нем.), heron cendre (фр.)
Син.: для подвида cinerea'. Ardea major Linnaeus, 1766; Ardea vulgaris Bechstein, 1803 (1802);
Ardea cineracea C.L. Brehm, 1831; Ardea rectirostris Gould, 1843; Ardea leucoptera Gould,
1848; Ardea brag Saint-Hilaire, 1844.
Серая цапля
335
Статус: гнездящийся, перелётный, в южных регионах зимующий вид.
Общая характеристика и полевые признаки. Крупная птица с длинной шеей, пря-
мым длинным клювом и высокими ногами. В полёте шея изогнута S-образно и втянута в
плечи. От всех других цапель региона отличается светло-серой окраской верха, в полете —
контрастом светлых (кроющие) и тёмных (маховые) тонов на крыльях. По этим признакам
похожа на серого журавля, но несколько меньше его; кроме того, у стоящей цапли отсут-
ствуют свисающие над хвостом перья, характерные для журавля, а в полёте она складыва-
ет, а не вытягивает шею. Полёт серой цапли спокойный, с медленными взмахами широких
крыльев. Летят птицы обычно небольшими стаями клином или шеренгой, где расстояние
между отдельными особями составляет 10-15, иногда до 25-30 м (Кокшайский, 1959).
Ночью цапли летят рыхлой стаей, перекликаясь между собой (Молодовский, 1990).
Голос серой цапли похож на голос рыжей цапли, но более громкий, резкий и скрипучий
(Спангенберг, 1951; Смогоржевський, 1979); его можно передать звуками «кря-а-а-к». В
колонии взрослые издают хриплое «хрёк». Птенцы малоактивны и молчаливы только в
первые дни, когда они лежат в гнезде, тесно прижавшись друг к другу, но к 5-6-му дню
становятся крикливыми: бесконечно «чакают» или урчат, издавая звуки, похожие на про-
тяжное «ка-а-ка-ка-ка», умолкая лишь при приближении опасности.
Подвидовая систематика. Выделяют от 3 до 5 подвидов, различающихся окраской и
размерами (Спангенберг, 1951; Степанян, 2003; Dickinson, 2003). На территории бывшего
СССР обитает, по-видимому, один подвид.
1. Ardea cinerea cinerea
Ardea cinerea Linnaeus, 1758, Syst. Nat., ed. 10, c. 134, Швеция
Этот же подвид населяет Зап. Европу, Африку и внетропическую Азию, за исключени-
ем её востока. Вне пределов указанной территории выделяют подвиды: А. с. monicae Jouanin
et Roux, 1963 (2), оседло обитающий на о-вах Банк д’Аркуин у побережья Мавритании;
А. с. jouyi (rectirostris) Clark, 1907 (3) из континентальной тропической Азии, Японии, Ко-
реи, Вост. Китая и Тайваня; А. с. firasa Е. Hartert, 1917 (4), населяющий Мадагаскар, Ко-
морские о-ва и о-ва Альдебра; А. с. altirostris Mees, 1971 (5), обитающий на Яве и Суматре
(выделяется не всеми).
Замечания по систематике. Некоторые систематики относят серых цапель Забайкалья
и Приморья к подвиду jouyi. Однако большинство склонны всех серых цапель, обитающих
на этом пространстве, относить к номинативному подвиду или, в крайнем случае, считать
их таксономически промежуточными между cinerea и jouyi.
Описание. Окраска. Самец и самка в брачном наряде. Голова белая, за глазами
серовато-чёрные полосы, которые на затылке соединяются и переходят в чёрный хохол из
нескольких удлинённых перьев; шея серовато-белая, на ней чёрные пятна, образующие 2-
3 продольные полосы. Спина и покровные перья второстепенных маховых сизовато-серые,
лопаточные перья удлинены в виде узких косиц, бахромой свисающих на крылья и спину.
Цвет косиц обычно светлее, чем спины и кроющих крыльев (от светло-серого до белесого).
Удлинённые и заострённые перья зоба свисают вниз. По бокам тела проходит по широкой
чёрной полосе, при сложенных крыльях район кистевого сгиба прикрыт рассученными
перьями боков груди, образующими выделяющееся чёрно-белое пятно. Хвост серый, ма-
ховые чёрные с сизым налётом порошкового пуха, грудь, брюхо и подхвостье серовато-
белые. На груди, брюхе и в паху имеется пух, окончания которого при трении образуют
порошок, наподобие талька. Клюв желтовато-бурый с более тёмным коньком, голое пятно
на уздечке желтоватое. Радужина жёлтая с чуть зеленоватым оттенком, кольцо вокруг глаза
зеленоватое. Ноги оливково-бурые. В разгар брачных демонстраций неоперённые участки
становятся ярче: клюв и ноги — до красно-оранжевых, уздечка и кольцо вокруг глаза — до
голубых, радужина — до оранжево-жёлтой.
336
Отряд Листообразные, Семейство Цаплевые
Взрослая птица в зимнем (послебрачном) наряде. Украшающие, удлинённые перья зоба
и на плечах снашиваются.
Пуховой наряд. Верх пепельно-серый, низ беловатый, горло и брюшко голые, кожа се-
рая с зеленоватым оттенком, радужина белесая или светло-голубовато-серая.
Гнездовой наряд. Общая окраска верха серая. Голова светло-серая с едва развитым се-
ровато-черным хохлом; шея светло-серая с тёмным рисунком спереди; низ беловатый с
широкими бурыми полосами по бокам, чёрные полосы по бокам тела не развиты. Маховые
тёмно-серые, рулевые серые; украшающие перья зоба и спины отсутствуют. Клюв двуцвет-
ный: надклювье тёмно-серое, подклювье — жёлтое. Радужина едва голубоватая. Ноги серо-
бурые.
Строение и размеры. Несмотря на многочисленность серой цапли, данных о её
размерах в печати на удивление мало (табл. 29).
Кроме того, И.А. Долгушин (1960), приводит следующие сведения о параметрах этих
птиц обоего пола (в мм), не указывая числа измеренных экземпляров: длина крыла — 445-
478; длина хвоста — 144-188; длина цевки — 144-170; длина клюва — 109-135.
Масса серых цапель (г), добытых в период зимовки на юго-востоке Каспия: самцы (п=22):
1350 — 1770, самки (n=21): 1 100-1 470 г (Воробьев, Исаков, 1940). Максимальный вес
серой цапли, по Е.П. Спангенбергу (1951), достигает 2 кг. В Нидерландах средний вес (г)
самцов (п=17) составляет 1 503 (1 071-2 073), самок (п=13) — 1 361 (1 020-1 783) (Cramp,
1977). Вес вылупившегося птенца равен 45 г, а 3,5-недельных птенцов — 1 200-1 600 г
(Owen, 1960, цит. по: Cramp, 1977).
Линька. Данных мало. По-видимому, полная линька происходит один раз в году (Спан-
генберг, 1951). На юге Украины линька взрослых птиц начинается в июне, когда у основ-
ной части популяции заканчивается период размножения. Выпавшие первостепенные ма-
ховые можно найти на земле под деревьями в местах отдыха и сбора птиц на ночлег и в
колониях, в течение всего лета и начала сентября. Линька контурного пера протекает мед-
ленно и, вероятно, заканчивается уже в районах зимовок.
Распространение. Гнездовой ареал. Серая цапля распространена в лесном и
пустынно-степном поясе Палеарктики, в Африке (кроме Сахары) и Юго-Вост. Азии (Спан-
герберг, 1951; Степанян, 2003). В Зап. Европе на гнездовании отсутствует севернее 65° с.ш.
(Смогоржевський, 1979), хотя по северному побережью Норвегии поднимается до 68° с.ш.
(Cramp, 1977). В Великобритании на гнездовании обычна по всей территории этой страны
(Stafford, 1963). В континентальной Европе распространение спорадично (Cramp, 1977),
осваивая все пригодные для нее территории. Населяет Малую Азию, Месопотамию, Иран,
Пакистан, Индию, Сев. Китай, ряд островов Индонезийского архипелага (рис. 67).
Таблица 29. Размерные показатели серой цапли для разных территорий (мм)
п Самцы Самки М lim n М lim Источник
Крыло 20 Территория бывшего СССР 472 430-490 14 458 440-475 Спангенберг, 1951
Нидерланды
Крыло 20 467 440 - 485 12 443 428 - 463 Cramp, 1977
Хвост 20 174 161-187 12 166 157-174
Цевка 23 151 136-172 16 141 132-153
Клюв 26 120 110-131 19 112 101-123
Серая цапля
337
Рис. 67. Область распространения серой цапли: а — гнездовой ареал, б — районы зимовок. Подви-
ды: 1 —Ardea cinerea cinerea; 2 —А. с. monicae; 3 —A. c.jouyi; 4 — A. c.firasa; 5 —А. с. altirostris.
В Вост. Европе и Сев. Азии северная граница распространения серой цапли проходит
по линии: Санкт-Петербург - Вологда - Пермь - низовья Амура - Сахалин. Изолирован-
ный участок ареала находится в районе Якутска (рис. 68). На Украине, в Белоруссии, в
южных районах и средней полосе Европейской территории России вид встречается в под-
ходящих местах повсюду. Как и другие виды цапель, серая цапля тесно связана с устьями
таких рек, как Волга, Терек, Урал, Дон, Днепр, Днестр, Дунай и др. Охотно заселяет поймы
Северного Донца, Кубани, лиманы и водохранилища.
Зимовки. Европейские и западносибирские популяции зимуют преимущественно в
Африке. Среднеазиатские цапли отлетают зимовать в Индию и Китай. Подвиды, распрост-
ранённые на островах, оседлы. Цапли востока России встречаются на зимовках в Индии,
Японии, Юж. Китае (Спангенберг, 1951; Смогоржевський, 1979).
Область зимовок серой цапли Вост. Европы и Сев. Азии очень широка. Птицы регуляр-
но остаются на зиму в приморских низменностях и низовьях рек, впадающих в Черное
море, в Сев. Причерноморье и Крыму, встречаются на зимовке в дельте Кубани. Серая
цапля — обычный зимующий вид на Ставрополье (Бичеров, 1988). Птицы, гнездящиеся в
дельте Днепра, в основном зимуют в Италии, Греции, Западной Африке (Ардамацкая, 1975);
гнездящиеся на Лебяжьих о-вах в Крыму — зимуют в Средиземноморье, в Африке вплоть
до Нигерии (Костин, 1983). Молодые птицы из дельты Терека встречены в декабре близ
Махачкалы (Скокова, 1978г).
Зимовки цапель, гнездящихся на Рыбинском вдхр., расположены в пределах Восточной
Европы от устья Кубани, по черноморскому побережью Кавказа до долины р. Риони, а
также в Сев. Германии, Юж. Польше, Венгрии, Болгарии, Югославии, Австрии, Юж. Фран-
338
Отряд Аистообразные, Семейство Цаплевые
Рис. 68. Распространение серой цапли в Сец. Евразии: а — область гнездования вида; б — зимовки.
ции, Италии, в Сев. Африке от Алжира до Египта, в Зап. Африке по долине р. Нигер и
берегам Гвинейского залива.
Наиболее ограниченные районы зимовки и вполне определённые пути миграций име-
ют норвежские цапли. Большое количество птиц из этого региона проводит зиму на юго-
западе самой Норвегии, основная масса отлетает к Британским о-вам, где концентрируется
в пределах Шотландии, немногие в зимние месяцы посещают Данию и Исландию.
В Приморском крае одиночные особи наблюдались на незамерзающих участках р. Ра-
ковка в городской черте Уссурийска 6 января 2005 г. и 27 января 2008 г. (данные Ю.Н. Глу-
щенко, И.Н. Кальницкой и Д.В. Коробова). Одна зимующая птица периодически наблюда-
лась в декабре 1997 и январе 1998 г. в долине р. Михайловка (данные Е.А. Волковской-
Курдюковой). В окрестностях Владивостока одна серая цапля отмечена 7 февраля 1989 г.
(Назаров, 2004); в Хасанском р-не одна особь наблюдалась 14 и 30 января 1962 г. в устье р.
Кедровая и две птицы зарегистрированы 6 февраля того же года у оз. Хасан (Панов, 1973).
Миграции. После окончания гнездового периода начинаются кочёвки. В первые дни
после вылета молодые держатся вместе со взрослыми вблизи колоний. В этот период в
центральных районах материка они кормятся на озёрах, старицах, мелководьях рек и водо-
хранилищ. Приморские популяции тяготеют к лиманам, устьям крупных рек, мелковод-
ным частям заливов. В это время птицы часто ночуют в колонии, куда залетают и днём.
Однако вскоре цапли порывают связь с местами гнездования и уже с конца июня — начала
июля держатся в наиболее кормных местах. Если цапель не беспокоить, то они и на ночь
остаются здесь же, устраиваясь на отдых на тростнике или деревьях. Постепенно кормо-
вые кочёвки превращаются в разлёт в различных направлениях — зачастую, прямо проти-
воположных путям осенних миграций. Окольцованных на Лебяжьих о-вах птиц добывали
в Луганской, Ровенской, Курской обл. (Костин, 1978). Молодые цапли из дельты Днепра в
июле встречаются также в северных областях Украины, хотя часть птиц вплоть до октября
Серая цапля
339
обитает как в Херсонской, так и в Николаевской обл. Птицы из Эстонии направляются в
сторону Ленинградской обл. — на восток (Кищинский, 1978в). Цапли, окольцованные в
Дарвинском заповеднике, разлетаются по всему Рыбинскому вдхр.; значительная часть их
направляется на север — по долинам рек Мологи, Суды, Шексны, к озёрам Белому и Ку-
бенскому в Вологодской обл. (Скокова, 1978в). Для молодых цапель, помеченных в Рязан-
ской обл., более характерно западное и южное направления миграции, хотя значительная
часть молодежи движется на север (в Ярославскую, Владимирскую области) и северо-вос-
ток — вниз по Оке и Волге, вплоть до устья Суры (Приклонский, Сапетин, 1978). По дан-
ным кольцевания настоящая осенняя миграция у днепровской популяции начинается в сен-
тябре. Птицы летят на юг и юго-запад, в основном придерживаясь прибрежных районов.
Немногие уже во второй половине августа долетают до Молдавии, большинство в сентябре
регистрируются в Болгарии, где некоторые задерживаются на зимовку. Массовый отлёт с
юга Украины отмечается с 10 по 30 сентября. Серая цапля в это время летит в основном
ночью (Спангенберг, 1951; Смогоржевський, 1979; Creutz, 1981), широким фронтом, чаще
всего — на высоте 150-400 м. По данным, полученным В.Н. Грищенко и В.В. Серебряко-
вым (1993), в 1975-1989 гг. для осенней миграции серой цапли на Украине были характер-
ны два основных направления. На западе страны миграция шла на юго-запад двумя широ-
кими ветвями: через Волынскую и Львовскую обл. и от Житомирской обл. на севере до
Ивано-Франковской и Винницкой на юге. В восточной части Украины миграция шла на юг:
от Харьковской обл. на севере до Запорожской и Донецкой на юге. Центральные и юго-
западные области Украины занимает обширная «область запаздывания», где цапли задер-
живаются дольше всего. Осенняя миграция заканчивается на Украине в среднем в октябре,
но разброс дат по отдельным годам очень большой: от первой декады сентября (Харьковс-
кая, Черниговская, Сумская обл.) до третьей декады ноября (Николаевская, Одесская обл.).
Скорость осенней миграции — 30-40 км/сут., что согласуется с литературными данными,
полученными при помощи кольцевания: 30-35 км/сут. (Creutz, 1981).
«Промежуточная миграция» (с июля по сентябрь), когда птицы разлетаются во все сто-
роны от мест рождения по кормным угодьям, существует повсеместно. Например, цапли,
окольцованные в Приморско-Ахтарском р-не Краснодарского края, в августе того же года
встречены в Днепропетровской обл. (400 км к северо-западу) и в Калмыкии (530 км к вос-
току). Однако, нельзя игнорировать и то обстоятельство, что уже в августе идёт направлен-
ная осенняя миграция. Так, Н.Н. Скокова (1978в) сообщает, что 11 августа взрослая птица
обнаружена в Нигерии, более чем в 5 000 км от места рождения. По данным этого же авто-
ра, часть молодых птиц в это время ещё держится в приазовских плавнях, хотя всё же чаще
отлетает к местам зимовок — вдоль Черноморского побережья Кавказа. В октябре они
зарегистрированы в 400 км к востоку от места рождения, а также на побережье Египта.
Перемещение прибалтийских цапель (Латвия, Эстония, Калининградская обл. РФ) к райо-
нам зимовки начинается тоже в сентябре, а в ноябре почти все особи достигают мест зи-
мовки — территории бывшей Югославии, Венгрии, о. Сардиния, Алжира и Туниса (Лиепа,
1966; Кищинский, 19786, в, г).
Наглядно иллюстрирует современные представления о распределении во время осен-
них миграций и на зимовке отдельных популяций серой цапли, гнездящихся в Восточной
Европе и Сев. Азии, картосхема, предложенная А.А. Кищинским (1978а) (рис. 69). Несмот-
ря на то, что она основана на материалах мечения этого вида в начальный период второй
половины XX века, использованные данные позволяют составить представление и о со-
временном положении дел. Наблюдения, выполненные в последние годы, убеждают нас,
что существенных изменений миграций и размещения зимовок у серой цапли за прошед-
шее время не произошло.
340
Отряд Листообразные, Семейство Цаплевые
Массовый отлёт цапель с Рыбинского вдхр. начинается 15 сентября и заканчивается 5
октября. Последних одиночных птиц видели вплоть до 16 ноября. В сентябре близ мест
кольцевания остаётся не более 29% всей популяции этого вида. Птицы, откочевавшие вна-
чале к северу, поворачивают в эти сроки к югу и западу. Около 8% из них в сентябре и
октябре находится на территории северной и центральной Украины, ещё 4% достигает её
южных причерноморских районов. Наконец, 10% цапель появляется на основных местах
зимовки этой популяции — во Франции, Италии и Сев. Африке.
Молодые птицы, вылетевшие из колоний в Литве и Калининградской обл., как и латвий-
ские цапли, частично летят на север и северо-восток, залетают в Латвию, а некоторые по
Зап. Двине — в Витебскую обл. Однако другие птицы из этих мест уже в августе достигают
юго-вост. Польши, Чехословакии и даже сев. Италии. Зимой основная масса птиц отмеча-
ется в Югославии и Италии (Исаков, 1957). Залетают птицы в Грецию, проникают в Афри-
ку до р. Нигер, зимуют и в Юж. Франции. Наиболее северные места зимовки расположены
в Баварии.
По данным кольцевания, основных мест зимовки в Африки днепровская популяция се-
рых цапель достигает иногда уже в конце октября. В ноябре и декабре птиц встречают в
Италии, Греции, Экваториальной Африке.
Размещение на зимовках цапель, наделяющих Рыбинское вдхр., имеет некоторые прин-
ципиальные отличия от птиц других популяций. Процент особей, достигающих африкан-
Рис. 69. Области миграций и зимовок различных географических популяций серых цапель (по Ки-
щинскому, 1978а). Границы областей миграции и зимовок популяций: а — атлантической, южнока-
захстанской и дальневосточной; б — балтийско-средиземноморской; в — верхневолжской; г — чер-
номорской; д — нижневолжской; е — область зимовки балтийско-средиземноморских, верхневолж-
ских и черноморских цапель в тропической Зап. Африке (по данным индивидуального мечения).
Серая цапля
341
ских зимовок, у них выше, а путь, который проделывают эти птицы во время своих перелё-
тов, намного превосходит среднюю длину миграционного пути цапель, населяющих райо-
ны, расположенные южнее. Например, птицы, окольцованные в дельте Волги, не покидают
её и во время кочевок: лишь некоторые обнаружены за её пределами в устье р. Кубани.
Птицы, гнездящиеся по берегам Кызыл-Агачского зал. Каспийского моря, остаются, по-
видимому, зимовать на Каспии (в декабре цапля добыта на Апшеронском п-ове, другая 20
февраля на восточном побережье Каспийского моря).
Отмечены случаи появления на территории бывшего СССР птиц, гнездящихся и поме-
ченных за рубежом. Чаще всего залетают польские цапли. Их добывали в Литве, Белорус-
сии, Латвии, Эстонии, Калининградской обл., в западных областях Украины. Преимуще-
ственно это молодые птицы, попавшие сюда в период осеннего разлёта и «ненаправлен-
ных» миграций. Зимовки у польских цапель расположены на берегах Средиземного моря,
некоторые долетают до Туниса, а часть зимует на юге Германии и Чехословакии.
Весенняя миграция серых цапель начинается с конца февраля, продолжается в марте,
но особой интенсивности достигает в апреле. Цапли в эти месяцы появляются на западе и
в средней полосе европейской территории России близ мест гнездовий. Однако часть птиц
прилетает и в начале мая, когда у птиц, появившихся раньше, откладка яиц и, зачастую,
насиживание уже в самом разгаре. Молодые птицы после первой зимовки возвращаются в
места гнездования далеко не все (вероятно, менее половины), они часто задерживаются по
пути на родину или в районах зимовки.
В Приморском крае весной отдельные птицы появляются на оз. Ханка уже в первой
половине марта: 2 марта 1995 г., 8 марта 2004 г., 9 марта 1992,1998 и 2004 гг., 10 марта 1996
г., 11 марта 1992 г., 13 марта 1993 г. и 14 марта 2001 г., однако, основная часть популяции
прибывает в места размножения обычно в третьей декаде этого месяца (данные Ю.Н. Глу-
щенко). В долине р. Раздольная весенний пролёт хорошо выражен. Наиболее раннее появ-
ление цапель в окрестностях с. Раздольное В.А. Нечаев (2006) отметил 9 марта 1990 г. и 12
марта 1994 г., а в окрестностях Уссурийска первые птицы наблюдались 12 марта 2004 г., 14
марта 2005 и 2007 гг., 15 марта 2006 и 2008 гг. и 16 марта 2003 г. Массовый пролёт здесь
проходит в последней декаде марта или в первых числах апреля; слабая миграция продол-
жается во второй половине апреля и в первой половине мая (Глущенко и др., 2006а).
Пролёт весной на Украине идёт по трем основным направлениям. Раньше всего цапли
появляются в Крыму. Отсюда они летят на север широким фронтом через Херсонскую,
Днепропетровскую, Полтавскую, Сумскую обл. Второй поток проходит на север-северо-
восток от Одесской обл. на юге до Киевской и Черниговской обл. на севере. Третий — на
северо-восток через Волынскую, Ровенскую, Львовскую обл. Может быть, существует ещё
один миграционный путь — вдоль побережья Азовского моря по южным районам Запо-
рожской, Донецкой и Луганской обл. Средняя скорость весенней миграции составляет 60-
80 км/сут. Она проходит в более сжатые сроки, чем осенняя.
Местообитания. Для серой цапли при выборе мест гнездования характерна чрезвы-
чайно высокая экологическая пластичность. Эти птицы гнездятся одинаково охотно как на
деревьях, так и в тростниковых зарослях; наиболее многочисленны они в плавнях больших
рек, на крупных водохранилищах. Часто занимают острова, расположенные в мелковод-
ных заливах, с зарослями тростника, полыни, лебеды татарской и других травянистых рас-
тений (Ардамацкая, 1999, 2000, 2001; Костин, 2004). Цапли предпочитают лесные насаж-
дения поблизости от водоёма, устраивая гнёзда в старых ольховых рощах, в сосновых вы-
соких насаждениях, на дубах.
Большинство птиц поселяются в отдалении от человеческих поселков, однако, посколь-
ку при выборе места для гнездования серые цапли предпочитают поселяться вблизи кор-
342
Отряд Листообразные, Семейство Цаплевые
мового биотопа, колонии иногда размещаются на рыборазводных прудах (как в тростни-
ках, так и в рощах), вблизи хуторов или на окраине городов. На Северном Кавказе основ-
ными местами концентрации серой цапли на гнездовье в настоящее время и являются рай-
оны прудового рыболоводства и рисосеяния (Казаков и др., 2004). В дельте Днестра цапля
занимает в основном высокоствольные деревья (Русев, 2004).
Численность. Конкретные данные о численности серой цапли на территории бывшего
СССР в 1930-1940-х гг. отсутствуют. Численность была различна в разных частях ареала,
наиболее многочисленны цапли были по рекам низинного Кавказа и в плавнях рек, впада-
ющих в Чёрное и Каспийское моря (Спангенберг, 1951). Единственные учётные данные
приводятся для Астраханского заповедника: 18,5 тыс. особей — в 1934 г. и 23,5 тыс.
особей — в 1935 г. (Ромашова, 1938).
По ряду стран Зап. Европы имеются сведения о количестве гнездящихся пар в 1960-
1970-х и даже 1950-х гг., что дает возможность судить о динамике численности вида. В
1954 г. был проведен наиболее полный учёт цапель на Британских о-вах, где найдена 521
колония с 6 193 гнёздами. По данным выборочных учётов (с последующей экстраполяци-
ей) в Англии, Уэльсе, Шотландии и Ирландии в I960 г. предполагалось наличие 211 коло-
ний с 3 522 гнёздами, в 1961 г. численность оценивалась в 3 702 гнезда, в 1962 г. — в 2946,
в 1963 г. — в 1 863 гнезда. Таким образом, по сравнению с 1954 г. численность снизилась
на 55,4%. Основными причинами этого считаются суровые зимы с возвратами холодов
весной и отравление птиц ядохимикатами (Stafford, 1969). В 1970-х гг. численность раз-
множающихся серых цапель в Великобритании составляла 4 500 пар. В 2000 г. в Великоб-
ритании гнездилось 14 800 пар (BirdLife International, 2004).
В Германии регулярные учёты проводились в Вестфалии (Stichmann, 1968), в прави-
тельственным округе Люнебург (Kirsch, 1963), в Зап. Германии (Stichmann, 1962b) и в коло-
нии у Потсдама. В Вестфалии в 1957 г. гнездилось 295, в 1962 г. — 422, в 1967 г. — 395 и в
1968 г. — 349 пар. Динамику численности здесь связывают с погодными условиями. В
разных местах округа Люнебург в 1885-1965 гг. ежегодно насчитывали 17 колоний, в кото-
рых гнездилось до 550 пар, но к 1966-1970 гг. общая численность снизилась до 81 гнездя-
щейся пары; большая часть колоний исчезла. Причиной послужила регуляция численности
цапель из-за ущерба, наносимого рыбному хозяйству. В Зап. Германии в 1960 г. сохрани-
лось 3 400 гнёзд; в колонии у Потсдама в 1957 г. насчитывалось 60 гнёзд, в 1970 г. — 121
гнездо. Во второй половине 1990-х гг. общая численность гнездящихся пар на территории
Германии составляла 24 000-27 500 пар (BirdLife International, 2004).
С начала 1950-х гг. в СССР в наиболее труднодоступных районах с многочисленными
колониями цапель стали применять авиаучёты. В 1953-1954 гг. в Вост. Приазовье при авиа-
учётах выявлено около 3-3,5 тыс. особей серых цапель (Винокуров, 1959а). На рубеже
нового тысячелетия общая численность здесь составила около 2-3 тыс. пар (Белик, 2004).
Общая численность серой цапли в низовьях Дона, по данным авиаучётов 1990-1991 гг. и
учётов на взморье, оценивается в 600-900 пар (Казаков, Ломадзе, 1991, 1992).
В центре Европейской России в 1950-х и 1970-х гг. учёт проводили с помощью анкет-
ных обследований. В 1958 г. в 15 субъектах Федерации этим способом было обнаружено 55
колоний серой цапли с 700 гнёздами. При повторном учёте тем же методом в том же реги-
оне в 1978 г. обнаружено 153 колонии с 1 430 гнёздами. При этом меньшее число птиц
было учтено только в Нижегородской, Рязанской и Ульяновской обл. (Маркин, Приклонс-
кий, 1995). Увеличение числа учтённых птиц авторы объясняют как лучшим качеством
последнего учёта, перед которым была проведена специальная подготовку среди коррес-
пондентов, так и фактическим ростом количества цапель в районе исследования. Исключе-
ние составила самая крупная из известных колоний в этом регионе — Тереховская (Рязан-
Серая цапля
343
ская обл.). Здесь из-за гибели в 1970-х гг. большинства деревьев, на которых птицы гнезди-
лись ещё в 1956 г. (Сапетин, Галушин, 1958), число гнёзд реально сократилось. Если в 1956
г. было обнаружено 266 гнёзд, то в 1975 г. — 115, а в 1978, по анкетным сведениям, — около
100 (по данным Летописи природы ОГЗ — 210 гнёзд; двукратная разница объясняется при-
менением различной методики учёта гнёзд: при анкетном учёте — «на глаз», а при кольце-
вании — подсчёт на каждом дереве). В 1980-х гг. количество учитываемых гнёзд здесь
колебалось от 182 до 241, в 1990-х гг. — от 164 до 170, а в 2001 г. оно составило 272
(Летописи природы ОГЗ, данные С.Г. Приклонского).
В Астраханском заповеднике, участки которого служат эталоном местообитаний весло-
ногих и голенастых птиц, гнездящихся колониями в древесных зарослях, в конце 1970-х гг.
наметилась тенденция к сокращению численности серой цапли (Кривоносов, Гаврилов, 1981).
В 1974 г. было учтено 963 пары, в 1975 г. — 938, в 1977 г. — 890, в 1978 г. — 479 и в 1979 г. —
444 пары. В связи с резким ухудшением условий гнездования и кормодобывания в авандельте
(сократилась гнездопригодная площадь на островах авандельты из-за вымокания и гибели
лесов) у аистообразных (в том числе, у серой цапли) происходит уменьшение плотности
населения, как в «речной» части дельты Волги, так и в авандельте (Русанов, 2006).
В последней четверти XX столетия на Приханкайской низменности в разные годы гнез-
дилось 1 200-1 700 пар (Глущенко и др., 2003). На территории бывшего СССР, включая
азиатскую часть, по данным Всесоюзного учёта околоводных птиц, проведённого в 1986—
1987 гг., выявлено 70 000 гнездящихся пар серой цапли (Кривенко, 1991). По сравнению с
данными учётов 1970-х гг. (Шкуратова, 1981), отмечено заметное возрастание численнос-
ти у всех видов в среднем на 20-30%. На Украине основные колонии аистообразных сосре-
доточены в южных регионах, где весной 1984-1985 гг. членами Азово-Черноморской ор-
нитологической группы было учтено 2 400-3 100 пар серой цапли (Ардамацкая и др., 1988).
Общая численность вида на территории Украины по учётам 1986 г. составила 16 000-20 000
пар (Серебряков, Грищенко, 1992; Микитюк, 1999 и др.). В 1998 г. при проведении едино-
временного учёта околоводных птиц на юге Украины учтено 1 628 пар, т.е. численность по
сравнению с 1984-1985 гг. несколько уменьшилась (Численность и размещение..., 2000).
Атлас птиц Европы (Hagemejier, Blair, 1997) приводит для конца 1980-х гг. следующие
данные: общее число серых цапель в Европе оценено в 122 тыс. пар (117,5-128,5 тыс. пар),
при этом численность вида в Европейской России по данным Атласа составляет 20-25 тыс.
пар, на Украине — 18 000, в Белоруссии — 5 000 пар.
По данным Международной ассоциации по охране птиц (BirdLife International, 2004), в
1990-2000 гг. в Европейской России гнездилось 35 000-60 000 пар серых цапель (отмечена
тенденция роста численности); вся европейская гнездящаяся популяция по тем же данным
насчитывала в эти годы более 210 тыс. пар, причём заметное увеличение численности про-
изошло между 1970 и 1990 гг. На территории Украины гнездится 23 800-32 900 пар; в
течение 1990-2000 гг. здесь отмечено сокращение численности. В Белоруссии гнездящая-
ся популяция составляет 4,5-5,5 тыс. пар. Значительно увеличилась численность цапель в
Латвии, где в 1970-1979 гг. гнездилось 600-650 пар (Липсберг, 1981), а в 1990-2000 гг. —
1,1-1,5 тыс. пар. В Эстонии в 1970-х гг. насчитывалось в общей сложности не более 230
пар (Вероманн, Роотсмяэ, 1981), а в конце 1990-х гг. — 1,2-1,5 тыс. пар. В Литве гнездовая
популяция насчитывает от 3 до 6 тыс. пар, в Армении — 50-250 пар, в Азербайджане —
250-1 000 пар; по Грузии конкретные цифры отсутствуют.
Резюмируя, можно утверждать, что, несмотря на снижение численности серых цапель в
некоторых регионах, её общая численность имела тенденцию к увеличению. Многочис-
ленность или, по крайней мере, обычность вида на территории Вост. Европы и Сев. Азии
убеждает нас, что вид здесь находится в оптимальных условиях существования.
344
Отряд Листообразные, Семейство Цаплевые
Размножение. Половая зрелость наступает обычно на второй, иногда — на третий год
жизни, хотя по данным Хавершмидта (Haverschmidt, 1961) в Англии размножение самок в
годовалом возрасте — явление обычное. Серая цапля гнездится поливидовыми и монови-
довыми колониями, реже встречаются одиночные гнёзда. Размер колоний варьирует от 5
до 200 гнезд, зредка встречаются и более крупные колонии. Например, в пойме р. Сев.
Донец обнаружили колонию из 326 гнёзд (Сулик, 1981), в дельте Днепра — из 500 гнёзд на
древесных насаждениях (Ардамацкая, ориг.); колония из 500 гнёзд располагалась на скалах
о. Фуругельма (Стоцкая, 1984). Прилёт на место гнездования происходит в разные сроки в
зависимости от характера весны и географического положения. На юге Украины (Арда-
мацкая, 1999) и в Крыму (Костин , 1983) первые особи появляются в конце февраля - нача-
ле марта, в Казахстане в низовьях Сыр-Дарьи — в первой половине марта (Долгушин, I960).
В северных регионах и в Сибири — в конце апреля - начале мая (Ларионов, Винокуров и
др., 1976), в Московской обл. — в конце марта - начале апреля (Птушенко, Иноземцев,
1968), на Рыбинском вдхр. — 5-10 апреля (Исаков, 1957).
Серые цапли гнездятся на деревьях, на заломах тростника или на островах, прямо на
земле. В глубине материка цапли часто выбирают для гнездования высокоствольные ели и
сосны до 30-35 м высоты, в прошлом веокрестностях Москвы — даже до 40 м (Мензбир,
1918); в пойменных лесах и затопленных ольшаниках гнездятся на высоте 12-25 м. На
северной границе ареала в Ленинградской обл. серая цапля строит гнёзда только на дере-
вьях, в частности, — на усыхающих дубах на берегу рек (Мальчевский, Пукинский, 1983).
В Московской и Калужской обл. гнёзда серых цапель тоже располагаются только на дере-
вьях — соснах, елях, ольхе, дубах, реже на берёзах и осинах (Зубакин и др., 1986; Коваль-
ковский и др., 1986; Марголин, 2000). По данным Штихмана (Stichmann, 1962а), в Вестфа-
лии цапли гнездятся на дубах, буках, лиственницах, елях, ольхе и ясене. В Якутии в таёж-
ной части центральных районов колонии серых цапель также располаются на деревьях
(Ларионов и др., 1976). В Латвии гнёзда серых цапель обычно находили на деревьях, и
только в 1970 г. птицы впервые загнездились на заломах тростника на оз. Энгуре (Lipsbergs,
1971).
В Тереховской колонии (Рязанская обл.) гнёзда размещались преимущественно на ду-
бах и редко — на вязах, периферийные же были построены на ольхах. За годы наблюдения
за колонией (1956-2001) многие дубы в центральной части колонии погибли. Цапли про-
должали строить гнёзда на высохших деревьях, хотя их число на таких деревьях, по срав-
нению с живыми, резко сократилось. В то же время, более интенсивно начали заселяться
ольхи по краям колонии. На дубах, многие из которых имели весьма обширную крону,
число гнёзд на одном дереве достигало 17-20 (до 25), тогда как на ольхах в самом благо-
приятном случае оно не превышало четырёх. В другой колонии на Рязанщине, в Шацком
р-не, гнёзда цапель располагались исключительно на соснах. Первоначально (в 1940-1960-х
гг.) колония находилась на территории поселка Ямбирнского лесничества, на 120-170-лет-
них строевых соснах. Затем, по мере их гибели, она переместилась в сторону от поселка,
но тоже в 70-100-летний сосняк (Нумеров и др, 1995). Число гнёзд на соснах обычно не
превышало 2-3 на одном дереве, что, вероятно, связано со строением крон сосен.
На Каржинских о-вах (Костин, 1983; Ардамацкая, 2000) серые цапли гнездятся в высо-
корослом тростнике, и их новые гнёзда имеют типичную для цапель форму «перевернутого
конуса». Однако часть птиц здесь же поселяется на земле среди лебеды и полыни. Тогда
гнёзда низкие и плоские, как и в зарослях тамариска в Кызыл-Агаче (Греков, 1965).
В Казахстане серые цапли гнездятся преимущественно в тростниках, значительно реже
устраивают гнёзда на деревьях — джигдах, турангах, ветлах. На деревьях цапли гнездятся
на высоте 6 м, а в тростниках гнёзда устраивают на заломах тростника, в 20-80 см над
Серая цапля
345
водой (Долгушин, I960). И.А. Долгушин также сообщает о необычных местах устройства
гнёзд — на вершине стога, стоявшего в тростниках, и в старом гнезде белого аиста.
На о. Хухан (юг Читинской обл.) серые цапли устраивают гнёзда на каменистой повер-
хности, покрытой редкой галофитной растительностью (Зубакин, 1981). На о. Фуругельма
(Стоцкая, 1983) цапли гнездятся на уступах скал.
В поливидовых колониях именно серые цапли раньше других птиц приступают к гнез-
дованию, на 1-2 недели опережая большую белую цаплю. Они являются основателями
таких колоний, и хотя в норме, при наличии гнездопригодной площади, цапли не поселя-
ются на одних и тех же участках из года в год (Костин, Тарина, 2004), зачастую встречают-
ся многолетние поселения на одной и той же территории. В высокоствольной дубраве в
«Лесу на Ворскле» постоянно занимаются те гнёзда, в которых в предыдущем году гнездо-
вание прошло успешно (Корякин, 1981). В Вост. Приазовье цапли так же из года в год
занимают одни и те же гнёзда, восстанавливая их после зимы (Казаков и др., 2004).
Появившись в колонии, цапли приступают к «ремонту» или постройке гнезда. В этом
деле принимают участие оба партнёра, образовавшие пару. В качестве строительного ма-
териала используется тростник, ветки и сучки деревьев и кустарников. В это время в коло-
нии можно наблюдать разнообразное демонстрационное поведение цапель (рис. 70).
Размеры гнёзд зависят от их «возраста» и месторасположения. В поливидовых колони-
ях серая цапля занимает верхний ярус дерева, а ниже неё устраиваются кваква, малая белая
цапля, иногда — каравайка. Гнездовые постройки на деревьях птицы используют в тече-
ние многих лет.
Размеры гнёзд из дельты Дона: диаметр гнезда — 59-112, в среднем 87 см, диаметр
лотка — 23-51, в среднем 41 см, высота гнезда — 37-70, в среднем 49 см, глубина лотка —
2-15, в среднем 8 см (Казаков и др., 2004). С Лебяжьих о-вов: диаметр гнезда — 43-90, в
среднем 67,3 см, диаметр лотка — 30-61, в среднем 46 см, высота гнезда — 39-89, в сред-
нем 59,9 см, глубина лотка — 5,5-23, в среднем 11,9 см. (Костин, 1983). На скалистом о.
Хухан средний диаметр гнёзд — 74 см, лотка — 40 см (Зубакин, 1981). На оз. Ханка гнёзда
имеют размеры (п=14): диаметр гнезда 37-82 см, в среднем 69,6 см; диаметр лотка 24-46
см, в среднем 32,8 см; глубина лотка 9-15 см, в среднем 12,6 см; толщина гнезда 20-50 см,
в среднем 32,3 см (данные Ю.Н. Глущенко).
Откладка яиц начинается на юге Украины и в Крыму в первой или второй декадах марта
(Костин, Тарина, 2004), на оз. Маныч-Гудило — в конце марта - начале апреля, а в ранние
весны — в двадцатых числах марта (Казаков и др., 2004), на Сыр-Дарье — в первой декаде
апреля (Долгушин, I960), в Верхневолжье — во второй декаде апреля (Зиновьев, 19816), в
лесостепном Зауралье — с 23 апреля (Тарасов, Поляков, 2006), на Братском вдхр. — 10-15
мая (Мельникова и др., 1981), в Приморье на о. Фуругельма — в первой декаде апреля
(Стоцкая, 1984). По наблюдениям Е.Э. Стоцкой (1984), самка откладывает первое яйцо на
4-5-й день после первого спаривания.
Гнездовой период очень растянут. Птицы, прилетающие позже (возможно, младшие по
возрасту), приступают к откладке яиц и в мае и в июне. Самая поздняя встреча ненасижен-
ных яиц на Лебяжьих о-вах — 28 июля (Костин, 1983).
Яйца светло-голубые, часто с белыми «мазками»; по Е.П. Спангенбергу (1951) они зеле-
новато-голубого цвета. Скорлупа свежих яиц без блеска, насиженных — со слабым блес-
ком. Форма яиц изменчива, они или правильной яйцевидной формы, или эллипсовидной.
Размеры яиц характеризуются существенной изменчивостью. Е.П. Спангенберг (1951) при-
водит такие данные о размерах яиц серой цапли (п=43): 55,3-60,4 х 41,8-46,3 мм. На Лебя-
жьих о-вах (п=400) средние размеры были 60,3x42,6 мм (Костин, Тарина, 2004); на Усть-
Манычском вдхр. (п=13) — 49-81 х 38-48, в среднем 63,5x41,6 мм; в Пензенской обл.
346
Отряд Листообразные, Семейство Цаплевые
Рис. 70. Позы серой цапли (по Bauer, Glutz von Blotzheim, 1987): a — поза отдыха, б — почёсывание,
в — принятие солнечных ванн, г-д — элементы брачного поведения (вытянутая поза), е — элементы
брачного поведения (щелканье клювом), ж — спаривание, з — приветствие подлетающего к гнезду
партнёра, и — оборонительная поза, к — поза угрозы.
Серая цапля
347
(п=413) — 50,4-68,0 х 39,0-46, в среднем 60,17x42,56 мм (Муравьев, Иванов, 2006); в
Тереховской колонии (Рязанская обл.) (п=20) средние размеры были 60,7±0,44 х 41,95±0,21
(Нумеров и др, 1995); в Курганской обл. (п=96) — 52,1-67,4 х 39,6-44,7, в среднем 59,1х
42,6 мм (Тарасов, Поляков, 2006); на оз. Ханка (п=80) — 52,1-65,1x37,9-45,2, в среднем
59,8x42,1 мм (данные Ю.Н. Глущенко). Вес свежих яиц на Лебяжьих о-вах (п=40) от 49,8
до 66,5 г, в среднем 57,6 г; на Усть-Манычском вдхр. (п=13) несколько меньше — от 42 до
56, в среднем 45 г.
В кладке от 3 до 9, чаще всего 5-6 яиц (Спангенберг, 1951 ;Скокова, 1954; Костин, 1983;
Казаков и др., 2004), изредка — 2. На оз. Ханка полная кладка содержит от 3 до 6 яиц, в
среднем (п=45) 3,9 яйца на гнездо (данные Ю.Н. Глущенко). Величина кладки варьирует по
годам. В 1990-х гг. она составляла в средн. 4,8 яйца на Каржинских о-вах (Ардамацкая,
1999). На Лебяжьих о-вах в 1984 г. в ранних кладках — 4,48 яйца, в поздних 2,98; в Пензен-
ской обл. в 2002-2005 гг. средняя величина ранних кладок была 4,25, в поздних — 4,12
яйца. Яйца откладываются с интервалом в 1,5-3 суток (Спангерберг, 1951; Долгушин, I960;
Костин, Тарина, 2004; Казаков и др., 2004). Общая продолжительность откладывания яиц в
гнезде составляет 6-12 суток (Бичеров, 1988). Наиболее высокая ритмичность отмечена у
цапель, рано приступивших к размножению.
Насиживание начинается с момента откладки первого яйца. В инкубации принимают
участие оба родителя, днём обычно насиживает самец, ночью самка (Смогоржевський, 1979),
на о. Фуругельма смена партнёров происходила чаще — дважды в сутки (Стоцкая, 1984).
Инкубационный период длится 25-29 суток (Тугаринов, 1947; Спангерберг, 1951; Ско-
кова, 1954; Мельникова, 1981; Казаков и др., 2004; Костин, Тарина, 2004). Как и у других
цапель, у серой в кладках бывают неоплодотворённые яйца; в ранних кладках их меньше, в
ддздних — больше. Обычно число неоплодотворённых яиц невелико, по данным С.Г.
Лриклонского (1975) в Рязанской обл. оно составляет 4,1%, в окрестностях Рыбинского
вдхр. — 2,0-3,7% (Скокова, 1954), на оз. Ханка — 2-4% (Поливанова, 1971), однако на о.
Фуругельма оно оказалось равным 19,3% (Стоцкая, 1984). Большое воздействие на количе-
ство неоплодотворённых яиц оказывают погодные условия. После суровой зимы 1984/85
гг. на Лебяжьих о-вах отметили 36,1% кладок с неоплодотворёнными яйцами, причём в не-
которых гнёздах было до двух безжелтковых яиц, а 8 гнёзд содержали только 4-6 аномально
мелких яиц, тогда как яиц нормального размера в них не было (Костин, Тарина, 2004).
В период насиживания цапли регулярно переворачивают яйца и много времени прово-
дят, ухаживая за оперением: перебирают рассученные перья на груди, распределяя поро-
шок пудреток по всему оперению (Стоцкая, 1984).
Первые птенцы появляются в Крыму, в плавнях Днепра и на островах Джарылгачского
залива в конце марта - первой декаде апреля, массовое вылупление отмечается в конце
апреля - начале мая. В Вост. Приазовье птенцы появляются во второй декаде апреля, в
Верхневолжье — с середины мая, в лесостепном Зауралье — в 20-х числах мая. Одноднев-
ные птенцы встречаются в колониях до конца июня и позже. На Лебяжьих о-вах в 1986 г.
последние птенцы вылупились 21 июля. Только что вылупившиеся птенцы беспомощны,
но зрячие, с щелевидными глазами; их средний вес (п=17) — 41г, длина клюва — 18 мм,
длина крыла — 22 мм, длина цевки — 19 мм (Бичеров, 1988). Первые перья появляются на
"-9-й день. На ноги птенцы начинают подниматься после двух недель жизни. Постэмбри-
гнальное развитие в гнёздах продолжается 45-50 (Олейников, 1953; Ардамацкая, ориг.), по
другим данным — 50-55 дней (Ломадзе, 1973); молодые становятся лётными на 53-56-й
день. Наиболее ранняя дата подъёма первых птенцов на крыло на Лебяжьих о-вах — 13 мая
'^*89 г.; птенцы из поздних кладок вылетают в третьей декаде августа (Костин, Тарина,
2004).
348
Отряд Листообразные, Семейство Цаплевые
Выкармливание птенцов начинается с однодневного возраста, добычу родители в этот
период ловят как днём, так и в сумерках и ночью. По данным Е.Э. Стоцкой (1984), до двух
недель птенцы получают полупереваренную пищу, а более старшие — почти всегда све-
жую. Маленьким птенцам родители отрыгивают пищу прямо в клюв. Подросшие птенцы
не всегда могут справиться с принесённой крупной рыбой, и родители нередко подъедают
пищу за птенцами. После вылупления птенцов значительную часть времени родители тра-
тят на их обогрев. Маленьких птенцов взрослые птицы обогревают постоянно; нерегуляр-
ный обогрев продолжается до 18-22-дневного возраста птенцов (максимально до 25 дней).
После прекращения обогрева родители перестают постоянно находиться около птенцов и
прилетают только, чтобы их накормить. Большая часть времени серых цапель, не занятых
уходом за потомством, приходится на отдых, сон и чистку оперения. Когда появляется сол-
нце (на о. Фурунгельма лето в основном туманное и дождливое), цапли принимают солнеч-
ные ванны, изредка подновляют гнездо. Скорлупа от яиц обычно выбрасывается, неопло-
дотворённые яйца поедаются птенцами или их соседями либо лежат в гнёздах, из которых
птенцы уже ушли (то же отмечалось в Тереховской колонии цапель в Рязанской обл.). Пос-
ле смены на гнезде или кормления птенцов цапли не сразу улетают за кормом на материк, а
некоторое время сидят на каменистых вершинах хребтов, ограничивающих колонию. За
пределами колонии на о. Фуругельма находилось два небольших «клуба», где в течение дня
постоянно отдыхало до 60 взрослых птиц, свободных от дежурства и покормивших птен-
цов. В основном они спали или стояли, отдыхая (Стоцкая, 1984).
Из-за растянутости гнездового периода вылет молодых на юге начинается со второй
декады мая - начала июня, достигает пика в первых числах июля и продолжается до конца
августа, когда вылетают птенцы из поздних кладок. В северных регионах этот процесс
тянется до начала сентября. Сразу после вылета молодых семьи держатся вместе, прилетая
ночевать в колонию, затем собираются большими стаями, кочуя по кормным местам.
Эффективность размножения серых цапель в разных колониях и в разных регионах раз-
лична и находится под влиянием различных факторов, как природных, так и антропоген-
ных. В Юж. Приморье на о. Фуругельма среднее количество слётков на гнездо составило
1,25, преобладали выводки из двух птенцов, а 36,4% пар не вырастили ни одного птенца
(Стоцкая, 1984). На Братском вдхр. отход яиц составлял 25-30%. Основной причиной гибе-
ли яиц здесь оказалось беспокойство колонии людьми и/или хищниками и непосредствен-
ное разорение гнёзд чёрной вороной (Мельникова, 1981). На о. Хухан в 1976 г. в разных
колониях было разорено от 10,8 до 17,8% гнёзд, по-видимому, серебристыми чайками (Зу-
бакин, 1981). На Ставрополье в 1980-х гг. самая высокая эффективность размножения от-
мечена у птиц, гнездившихся на прудах в тростниково-рогозовых крепях — 3 птенца на
одно гнездо, самая низкая — в пойменных поливидовых колониях, расположенных на де-
ревьях: от 1,8 до 2 птенцов (Бичерев, 1988). В Астраханском заповеднике у цапель, гнез-
дящихся в колониях на деревьях, отмечается значительная гибель оперяющихся молодых
птиц как от падения с веток дерева, куда они вылезают из гнезда, так и в результате застре-
вания их крылев, шей и лап в ветвях и развилках стволов (Кашин, 1976). Однако основным
фактором постэмбриональной смертности служит хищничество серой вороны, болотного
луня, камышевого кота, в меньшей степени — шакала и волка (Греков, 1965). В Терехов-
ской колонии цапель (Рязанская обл.) в 1957-1987 гг. обследовано 499 гнёзд. Из них с 1
птенцом было 6, с двумя — 34, с 3 — 98, с 4 — 197, с 5 — 143 и с 6 — 21 гнездо. Среднее
число птенцов в гнезде на момент кольцевания, т.е. в середине июня, оказалось равным
4,00±0,05 (Нумеров и др., 1995).
В оптимальных условиях на Лебяжьих о-вах в середине 1980-х гг. успешность размно-
жения рано загнездившихся пар составляла 77,5-95,6%. В 1986 г. этот показатель у рано
Серая цапля
349
загнездившихся птиц (п=100) был равен 86,5% (при эмбриональной смертности — 4,42%,
ювенильной — 9,08%), у поздних, (п=87) — 64,48% (эмбриональная смертность при этом
была 16,99%, ювенильная — 18,53%). После суровой зимы 1984/85 гг. успех размножения
на Лебяжьих о-вах был очень мал в поздних кладках и составил всего 34,98% (эмбриональ-
ная смертность — 23,68%, ювенильная — 39,34%); из 101 гнёзда 24 были брошены в нача-
ле насиживания (Костин, Тарина, 2004), на крыло поднялось в среднем 1,14 птенца на гнез-
до. На р. Ворскле на крыло поднималось, по наблюдениям А.С. Корякина (1981), 2,8-3,6
птенца из гнезда.
В Германии в колонии у Потсдама в старом сосняке в 1969-1973 гг. отмечали в среднем
по 2,0-2,5 вылетевших птенцов (Rainer Merhout, 1974), в Вестфалии успешно заканчивали
развитие менее 3 птенцов в пересчёте на одно гнездо (Stichmann, 1968). В Великобритании
в 1970-х гг. серые цапли выкармливали, как правило, одного птенца на гнездо, реже боль-
ше. Наибольшая гибель (60%) происходила в период насиживания и в первые три месяца
жизни (Prestt, Bell, 1973).
По данным В. Штихмана (Stichmann, 1968) до половозрелости доживает 15-18% при-
плода. По данным кольцевания отдельные особи живут до 18-24 лет; наибольшая зарегис-
трированная продолжительность жизни — 25 лет 4 мес. (Rydzewski, 1973).
Суточная активность, поведение. Серую цаплю нельзя назвать ни дневным, ни суме-
речным, ни ночным видом. Взрослые кормятся сами и добывают пищу для потомства и в
ранние утренние часы, улетая за кормом с 2 ч 30 мин до 5 ч 35 мин, и в поздневечерние —
с 21 до 23 ч. Днём они собирают добычу в степи или на мелководье; при этом цапли не
улетают далеко от гнездовой колонии. В сев.-зап. Йоркшире (Великобритания) зимующие
серые цапли собираются в дневное время в полях. Прилетают поодиночке, спустя час пос-
ле рассвета, большинство в 9-10 ч. Отлёт за кормом начинается в 15-16 ч — за час до
наступления темноты. Активность у цапель распределяется следующим образом: 77,5%
времени они бодрствуют (стоя) и охотятся (непосредственно на процесс охоты приходится
всего лишь 0,7% времени), 5,9% — спят (отдыхают), 16,6% — чистят оперение (Birkhead,
1973).
Питание. Плотоядный вид; пищевой рацион очень разнообразен, меняясь в разные се-
зоны и годы в зависимости от наличия массовых кормов. В 1959 г. Л.А. Смогоржевский
классифицировал серую цаплю как «в основном рыбоядную». В Астраханском заповедни-
ке и в районе Рыбинского вдхр. цапли также питались преимущественно рыбой (Скокова,
1954). Такую же картину наблюдали в таежной части центральных районов Якутии (Лари-
онов, Винокуров и др., 1976), где среди рыб по встречаемости в корме преобладали елец (р.
Leuciscus) и плотва (41,3%), а также окунь (14,8%). Серые цапли, гнездящиеся на о. Фуру-
гельма в заливе Петра Великого, в основном приносят птенцам морскую рыбу (80%), а
20% рациона птенцов составляют креветки. Однажды цаплей был добыт узорчатый полоз
(Elaphe dione) (Стоцкая, 1984). Основным кормом в период размножения на оз. Ханка, судя
по отрыжкам потревоженных птенцов, служат рыбы, различные беспозвоночные живот-
ные и мелкие млекопитающие (преимущественно дальневосточная полёвка), соответственно
составляющие в разные годы от 70,5 до 95%, от 4,7 до 29,1% и от 0,4 до 1,2% от общего
объёма потреблённой пищи (Поливанова, 1971).
В Казахстане набор кормов более разнообразен: это мелкая рыба, лягушки и их голова-
стики, ящерицы, небольшие змеи, водяные крысы. При затоплении весной колоний песча-
нок, цапли в массе их ловили; весной, после прилёта, птицы также охотно поедают сусли-
ков, полёвок и пеструшек. Во время массового размножения саранчи серые цапли почти
исключительно питаются ею (Долгушин, 1960). В Вост. Приазовье основным кормом явля-
ется рыба, но значительную часть составляют земноводные и пресмыкающиеся. В районах
350
Отряд Листообразные, Семейство Цаплевые
рисосеяния возрастает доля ракообразных, наземных насекомых, дождевых червей, пия-
вок, водных насекомых, земноводных, млекопитающих. Из-за почти полного отсутствия
рыбы в некоторые годы на оз. Маныч-Гудило, цапли были вынуждены кормиться преиму-
щественно в степи. Здесь основу их питания составляли наземные животные (58,3%). Наи-
большее значение при этом имели медведки (21 % по встречаемости), малые суслики (15,4%),
пластинчатоусые жуки (7,7%), жужелицы — 4,3%; на долю рыб приходилось всего 17,4%
(Языкова, 1970).
Изучение кормового рациона в двух колониях цапель, расположенных в разных биото-
пах на Ставрополье, показало что в «сухопутной» колонии в корме преобладали наземные
животные, доля же рыб была невелика. В «водно-болотной» основу питания составляли
именно рыбы. Значительно реже встречались земноводные и млекопитающие, роль же бес-
позвоночных оказалась ничтожной (Бичерев, Сигида, 1984). Анализ желудков и погадок
цапель в Черниговской, Черкасской, Киевской, Одесской и Херсонской обл. показал чрез-
вычайно широкий спектр питания: мышевидные млекопитающие (от водяной полёвки до
обыкновенной бурозубки), пресмыкающиеся (гадюка, прыткая ящерица), земноводные (пре-
обладает чесночница обыкновенная, особенно в плавнях рек, а также озёрная, остромор-
дая и прудовая лягушки) и 16 видов пресноводных рыб (доминирует карась). Кроме того,
обнаружено 10 видов морских рыб (главенствуют бычки), встречаются моллюски, ракооб-
разные (креветки, бокоплавы и др.), наземные и водные насекомые (Смогоржевський, 1979).
В Крыму, по данным А.И. Гизенко (1957) и Ю.В. Костина (1983), в годы с высокой числен-
ностью грызунов цапли питались и выкармливали птенцов в основном ими. При дефиците
грызунов птенцы на Лебяжьих о-вах получали в качестве корма преимущественно бычков
и креветок, пресноводных рыб и крупных наземных жуков. В дальнейшем, при увеличении
площади пресноводных биотопов и создании прудового рыбного хозяйства, ведущее место
в питании серой цапли заняли пресноводные позвоночные (лягушки, рыбы) и крупные на-
секомые (Костин, Тарина, 2004). В осенне-зимний период серые цапли в Крыму и Причер-
номорье питаются в основном рыбой. В Припятском охотхозяйстве Брестской обл. в лет-
ний период в пище серой цапли рыба отсутствует, здесь преобладают мышевидные грызу-
ны; доминирует водяная полевка, лягушки и водные насекомые: плавунцы, плавунчики,
водолюбы (Самусенко, 1981). На п-ове Сара (Кызыл-Агачский заповедник) зимой серые
цапли едят ежевику (Греков, 1965).
В Германии в районах, где широко развито прудовое хозяйство, серая цапля добывает
69,5% «ценных» видов рыб, 24% «малоценных» и 6,5% других групп животных (по весу)
(Creutz, 1964). В Румынии исследователи считают, что рыбоядность серой цапли преувели-
чена, и её корм состоит в основном из мышей, лягушек, моллюсков, ужей (Catuneanu, 1963).
Корм серые цапли добывают по-разному. На мелководье они медленно бродят или по-
долгу неподвижно стоят, поджидая добычу; иногда сидят на какой-нибудь торчащей из воды
коряге. Рыбу хватают клювом, молниеносно выбрасывают шею и голову вперёд и вниз. В
степи, подкарауливая жертву, цапли также очень часто стоят неподвижно и взлетают лишь
при тревоге. Иногда птицы добывают рыбу, плавая с поднятыми вверх крыльями (что ха-
рактерно только для этого вида); схватив её, летят к берегу, где и съедают.
Враги, неблагоприятные факторы. Из абиотических факторов наиболее значимый —
экстремальные погодные условия. Продолжительное весеннее похолодание с обильными
снегопадами, штормовые ветры и буреломы ведут к массовому разрушению старых мас-
сивных гнёзд на деревьях, срезают или размывают часть берега островов, где гнездятся
цапли, сокращая гнездопригодную площадь. Поздняя, холодная весна задерживает сроки
прилёта птиц в колонию; по этой причине начало гнездования сдвигается на 1-2 недели,
что отрицательно сказывается на успешности размножения. Анализ динамики численное-
Серая цапля
351
Рис. 71. Изменения численности серой цапли в 1928-1973 гг. в Англии и Уэльсе в зависимости от
суровости зим. Стрелками обозначены суровые зимы (по Cramp, 1977).
ти гнездящихся серых цапель в Англии и Уэльсе в 1928-1973 гг. показал, что сокращение
количества гнёзд происходит после неблагоприятных (суровых) зим, а подъёмы численно-
сти — после мягких. Особенно показательными в этом отношении были холодные зимы
1929, 1939-1941, 1947 и 1962-1963 гг., после которых количество приступивших к гнездо-
ванию птиц сокращалось, соответственно, на 12, 38, 45 и 53%15. После таких депрессий
численность восстанавливалась в течение 2-3 лет (рис. 71).
Из биотических факторов на первое место выступает воздействие хищников. В плавнях
южных рек в поливидовых колониях аистообразных часто поселяется серая ворона, кото-
рая питается и выкармливает своих птенцов яйцами и маленькими птенцами цапель. Бо-
лотный лунь и сорока постоянно гнездятся в тростниковых зарослях вблизи колоний, в их
рационе существенную роль также играют яйца и подрастающее потомство цапель. Из
млекопитающих основными врагами являются енотовидная собака, лисица, серая крыса,
шакал. Наземные хищники, в отличие от пернатых, главным образом подбирают выпав-
ших из гнёзда птенцов или ловят ещё плохо летающих, но уже пытающихся передвигаться
по колонии молодых. Считается, что наземных хищников привлекают к колониям обильно
падающие из гнёзд остатки принесённой пищи, которую не успели съесть молодые цапли.
Каннибализм не носит массового характера, но при недостатке пищи старшие птенцы мо-
гут поедать младших. В некоторых местах (о. Фуругельма) обычными явлениями стано-
вятся «братоубийство» и «нахлебничество» (Стоцкая, 1983). Младшие птенцы часто поги-
бают от истощения и побоев. Конкуренция за пищу проявляется у птенцов рано и отмечает-
ся до 35^46-дневного возраста. В результате драк при кормлении отмечали гибель молодых
15 Расчёт автора очерка по данным Cramp, 1977 (прим. ред.).
352
Отряд Листообразные, Семейство Цаплевые
птиц (Скокова, 1954). «Нахлебничество», когда на прилетевшую с кормом птицу набрасы-
ваются помимо своих ещё до 10 чужих птенцов, тоже может приводить к истощению и
гибели потомства, поскольку собственные птенцы (особенно, если они моложе) при этом
остаются голодными. Хотя у серой цапли отмечается высокий уровень агрессивности, взрос-
лые особи пугливы и при тревоге оставляют гнездо. Для них менее, чем для других птиц
этой группы, характерны как индивидуальная, так и коллективная защита гнездовий и по-
томства.
Что касается антропогенных факторов, то в конце XIX - начале XX вв. на серых цапель
оказывалось необычайно сильное промысловое воздействие. Отстреливали взрослых птиц,
разоряли гнёзда, собирали яйца. Объяснялось это негативным отношением к цапле со сто-
роны человека, считавшего её одним из главных своих конкурентов в рыболовстве. С дру-
гой стороны, в Германии (Вестфалия), а также во Франции серую цаплю в течение многих
столетий использовали как наиболее удобный объект для охоты с ловчими птицами (для
тренировки, «натаскивания» последних). Поэтому её поселения находилась под особой
охраной. Их специально оберегали в соответствии с распоряжениями феодалов, иногда, —
согласно королевским указам. С падением моды на такую охоту цапель огульно причисли-
ли к вредным рыбоядным птицам, в результате большинство колоний в Европе было унич-
тожено, а численность птиц резко сократилась (Stichmann, 1968). В России, как и на Украине
и в Белоруссии, на серую цапля специально не охотятся, но она может попадать под выстрел
попутно, вместе с «прочей» водно-болотной дичью. Первый всероссийский учёт добычи
пернатой дичи, проведённый в 1960 г. (Иванов и др., 1965), показал, что в числе этой, так
называемой «прочей» дичи, серых цапель было добыто около 40-50 тыс. особей. В последу-
ющие годы результативность охоты падала, причём на «прочую» дичь за 20 лет — в 4 раза
(Priklonski, Sapetina, 1990). Поэтому современные размеры добычи серых цапель охотника-
ми никак не превышают 15-30 тыс. особей для всей территории Вост. Европы и Сев. Азии.
Возрастание антропогенного пресса в последние десятилетия сказывается на цаплях
по-разному на разных этапах годового цикла. Наиболее уязвимы цапли к фактору беспо-
койства в гнездовой период на начальных стадиях гнездостроения, откладки яиц, начала
насиживания (Ардамацкая, 1981; Микитюк, 1999 и др.). Потревоженные в это время птицы
бросают гнёзда, кладки и даже колонии.
В XX в. возросла смертность цапель от отравления загрязнёнными объектами питания.
Увеличилось и само загрязнение водоёмов различными ядохимикатами. В тканях птенцов
серой цапли на Лебяжьих о-вах (Крым) была обнаружена высокая концентрация хлорорга-
нических пестицидов (Жерко, 1998). В 1999 г. здесь наблюдали снижение успешности гнез-
дования из-за значительного количества неоплодотворённых яиц, а также случаи откладки
самками аномально мелких яиц (Костин, Тарина, 2004).
В Великобритании в органах серой цапли в середине XX в. найдено больше остатков
хлорорганических пестицидов, чем в тканях других видов птиц. Серая цапля выступала в
этом отношении неким аккумулятором вредных веществ, поскольку она употребляла в пищу
обширный спектр кормов, обитающих в самых различных стациях осваиваемого ею реги-
она. Практически все взрослые особи, яйца и птенцы серой цапли содержали ОДЦТ (в
виде метаболита рр-ДЦЕ и дильдрина), в тканях большинства особей находили полихлор-
бифенилы — основной компонент сточных вод предприятий. Предполагается, что в связи
с этим примерно за 30 лет индекс толщины скорлупы яиц серой цапли уменьшился на 20%,
средний размер кладки — до 3,9, а количество выкормленных птенцов на одну пару — до
1,9. Кроме того, более трети гнездящихся пар, подвергшихся химическому загрязнению,
умышленно расклёвывали свои яйца, и почти каждое отложенное ими яйцо имело ненор-
мально тонкую скорлупу (Prestt, 1970).
Серая цапля
353
В период с 1952 по 2001 г. на юге Украины была изучена циркуляция гельминтов в
колониях голенастых птиц (Корнюшин и др., 2004); выяснилось, что все исследованные
серые цапли заражены паразитическими червями. Цестодофауна у них оказалась богаче,
чем у других голенастых, а трематодами были заражены все обследованные особи. У зна-
чительно меньшего числа птиц (35%) были найдены нематоды. Общая экстенсивность за-
ражения цестодами молодых птиц, покинувших колонию, составляет 60%, взрослых —
70%. Трематодофауна молодых цапель разнообразнее, чем у взрослых. Нематодами зара-
жено 40% молодых и 30% взрослых. Однако, несмотря на высокое заражение гельминта-
ми, не было установлено достоверных случаев гибели птиц по этой причине.
Фауна наружных паразитов и обитателей гнёзд серой цапли достаточно обширна. В
оперении птиц и в гнёздах обнаружено более 60 видов членистоногих. Многими авторами
среди прочих найдены кровососущие Ornithomyia avicularia и Ornitheza metallica (Hicks,
1959). В Средней Азии на серых цаплях выявлен пухоед Cicnoiphilus decimfasciatus.
Хозяйственное значение, охрана. Из-за своей многочисленности, широкого распрос-
транения и рыбоядности серая цапля имеет большое и неоднозначное хозяйственное зна-
чение. В середине - второй половине XX века мнения о «вреде» и «пользе» этого вида
широко обсуждались в литературе. С одной стороны, в степных районах в гнездовой пери-
од основной пищей серой цапле служат суслики, полёвки и другие мышевидные грызуны,
наносящие ущерб сельскому хозяйству (Спангенберг, 1951; Костин, 1983; Костин, Тарина,
2004 и др.). На естественных водоёмах цапли в значительном числе поедают сорную и
малоценную рыбу, а также большое количество водных насекомых и их личинок (водолю-
бов, плавунцов, клопов), в свою очередь поедающих мальков рыб; в плавнях рек цапли
выкармливают птенцов, помимо рыб и водных насекомых, также медведками и лягушка-
ми. Животные, которые в середине XX столетия считались несомненными вредителями,
составляют здесь 22,6% её рациона.
С другой стороны серая цапля наносит ущерб рыбному хозяйству, особенно в после-
гнездовой период, когда начинаются кормовые миграции, и птицы в значительном количе-
стве собираются на специальных рыборазводных прудах, поедая молодь разводимых здесь
рыб. По этой причине Л.А. Смогоржевський (1979) считал серую цаплю вредным видом
для рыборазводных водоёмов Украины. В южных районах в основном страдают карповые
и кефальные хозяйства, где цапля охотится с наступлением темноты, что затрудняет борьбу
с нею. В дельте Дона птицы кормятся по мелководьям основного русла, но часто посещают и
прудовые хозяйства (Казаков и др., 1986). В дельте Кубани в добыче серой цапли из прудовых
рыб 84% приходится на долю карпа, 12% — на белого и 4% — на пёстрого толстолобика.
В средней полосе России значение вида также далеко не однозначно. В одной из коло-
ний под Казанью питание серой цапли состояло почти исключительно из рыб, добываемых
на Волге и её притоках, но не на рыборазводных прудах (судак, щука, окунь, язь и др.). В
Тереховской колонии цапель во время работ по кольцеванию неоднократно находили среди
принесённой птенцам пищи довольно больших (до 25-30 см в длину) щучек, лещей и круп-
ную плотву.
Полагают, что цапли могут наносить ущерб рыборазводным хозяйствам по всему Евро-
пейскому центру РФ, поедая молодь, выпущенную для разведения. Однако нельзя не заме-
тить, что это актуально при плохой охране прудов и отсутствии там людей. При появлении
на прудах рыбхозов людей, даже без оружия, цапли немедленно улетают. Охота цапель на
мальков менее результативна, если последние жизнеспособны и активны. Как и всякие
хищники, серые цапли прежде всего вылавливают малоподвижных и нежизнеспособных
особей. Поэтому их деятельность и значение в рыборазводных хозяйствах следует оцени-
вать и в этом контексте.
354
Отряд Листообразные, Семейство Цаплевые
После многолетних специальных исследований хозяйственного значения рыбоядных птиц
(Скокова, 1965; Винокуров, 1959; Языкова, 1970; Чельцов-Бебутов, 1982; Ткаченко, 1987 и
мн. др.), орнитологи пришли к выводу, что мнение о вредоносности всех видов цапель на
рыборазводных прудах, высказанное рядом авторов в начале-середине XX в. (Браунер, 1923;
Пахульский, 1951, 1957; Смогоржевський, 1959), было ошибочным. Цапли съедают менее
0,0045% от общего запаса в водоёмах сазана и менее 0,0045% тарани, а молодь промысловых
видов рыб занимает в их рационе не более 35% (Скокова, 1965; Чельцов-Бебутов, 1982). Под-
считано, что огромное количество хищных насекомых, лягушек и сорной рыбы, съеденных
цаплями, истребили бы в 1 500 раз больше мальков, чем съедают птицы (Скокова, 1965).
Поэтому рыбоядных цапель нельзя считать вредными не только на естественных водоёмах,
но и в нерестово-вырастных хозяйствах (Чельцов-Бебутов, 1982; Кошелев, 2000 и др.).
По исследованиям на рыбоводных прудах в районе Оберлаузица в Германии, серая цап-
ля приносит значительный ущерб рыбному хозяйству, но в то же время она является «тупи-
ком» в циркуляции лигулеза. Поедая рыб, цапля поглощает в большом количестве яйца и
личинок лигул, и этим уравновешивается приносимы ею ущерб от изъятия ценных пород.
Для регуляции численности цапли здесь применяют различные способы отпугивания, в
зависимости от типа водоёма (Creutz, 1964).
В некоторых округах Германии традиционно существовала охота на серую цаплю, по-
этому, наряду с регуляцией её численности на рыбоводных прудах, одновременно создава-
лись охранные зоны вокруг сохранившихся поселений, в которых ограничивалось прове-
дение сельскохозяйственных и лесохозяйственных работ в гнездовой период. Регулирова-
лась и сама охота на цапель; она разрешалась только с 1 ноября по 31 декабря (Kirsch,
1965). В последние годы сроки изменялись, но незначительно.
Крупные многолетние поселения серых цапель отрицательно влияют на состояние дре-
востоя. Под тяжестью массивных гнёзд часто обламываются ветви у деревьев, особенно
если на одном дереве размещается до 20-30 гнездовий, как, например, в Припятском охот-
хозяйстве. Предположение, что помёт цапель и остатки пищи, падающие на землю из гнезд,
улучшают питательную ценность субстрата под деревьями, достаточно спорно.
Серая цапля играет определённую роль в распространении гельминтов рыб. Из 26 вы-
явленных видов, 11 встречаются в личиночной стадии и у рыб. Один из них (Posthodiplo-
stomus cuticola) вызывает чернопятнистую болезнь у рыб и является опасным паразитом,
особенно на юге страны (Смогоржевський, 1979).
Несмотря на необдозначность в оценке хозяйственного значения серой цапли, этот вид,
безусловно, подлежит охране как характерный представитель околоводных экосистем, игра-
ющий важную роль в циркуляции биогенных веществ между водной средой и сушей. В мес-
тах, где их присутствие нежелательно, цапель необходимо отпугивать, но не отстреливать.
Интересен опыт привлечения серой цапли на искусственные гнездовья. Так, старший
научный сотрудник Дарвинского заповедника В.В. Немцов рассказывал С.Г. Приклонско-
му о попытке привлечь цапель в искусственные гнёзда на Рыбинском вдхр. (за год до экспе-
римента осенними штормами было уничтожено большинство деревьев, на которых ранее
гнездились цапли). Для этого в сохранившихся массивах крупных деревьев в развилках
сучьев и на отдельных ветвях помещали груды хвороста, обильно политые сверху извес-
тью, имитировавшей помёт цапель. Особого успеха опыт не имел. Первой весной несколь-
ко птиц пытались гнездиться, но безрезультатно. На второй год цапли вообще искусствен-
ную «колонию» не посещали. Отрицательный результат был получен и при попытке при-
влечения серых цапель на гнездовья в Окском заповеднике, где в пойменной Окской дубра-
ве (никогда до этого не посещавшейся цаплями в целях гнездования) на вершинах дубов в
1957 г. В.М. Галушин по указанию В.П. Теплова разместил около 30 искусственных гнёзд,
Серая цапля, Рыжая цапля
355
сплетённых из ивовых прутьев, наподобие плоских корзин (подсадные птицы и окрашива-
ние гнёзд известью в этом эксперименте не применялись). Негативный опыт учтён в Ниж-
ней Саксонии, где в 1972 г. на деревьях было построено 7 искусственных гнёзд. Для «под-
страховки» здесь содержали 3 приманочных птицы. Через год в этой искусственной коло-
нии уже поселились 7 пар цапель, которые благополучно выкормили 22 птенца (Finkenstaedt,
Heckenroth, 1974).
В Российской Федерации серая цапля занесена в ряд региональных Красных книг: Рес-
публики Саха (Якутия), Республики Алтай, Кировской, Кемеровской, Нижегородской, Том-
ской областей.
Рыжая цапля
Ardea purpurea Linnaeus 1766
Ardea purpurea Linnaeus, 1766, Syst. Nat., ed. 12, c. 236, «на Востоке»; Франция
(Stresemann, 1920, Avifauna Macedonica, c. 226)
Purple heron (англ.), Purpurreiher (нем.), heron pourpre (фр.).
Син.: для подвида purpurea: Ardea rufa Scopoli, 1769; Ardea variegata Scopoli; 1769; Ardea
caspia S.G. Gmelin, 1774; Ardea purpurata J.F. Gmelin, 1789; Ardea badia J.F. Gmelin, 1789;
Ardea monticola Picot de la Peirouse, 1799; Ardea purpuracens C.L. Brehm, 1831; Ardea
pharaonica Bonaparte, 1855; для подвида manilensis: Phouxpurpurea ussuriana Shulpin, 1928.
Статус: гнездящийся, перелётный, изредка зимующий вид.
Общая характеристика и полевые признаки. Внешне походит на серую цаплю, с
которой может быть спутана на большом расстоянии или при недостаточной освещённос-
ти; однако размеры рыжей цапли несколько меньше, а шея выглядит заметно тоньше. Про-
порционально голове клюв тоньше и длиннее, представляет собой копьевидное продолже-
ние линии лба и всей узкой головы. В полете шея, как и у других цапель, согнута подобно
латинской букве «S», благодаря чему голова птицы прижата к спине. Полет напоминает
таковой серой цапли, но взмахи крыльев более резкие, «судорожные», из-за меньшей мас-
сы тела. Летящую рыжую цаплю можно отличить от серой по изящным контурам тела и
несколько меньшей длине вытянутых назад ног, а также по заметно более тёмной окраске
верха, без резкого контраста на крыльях между кроющими и маховыми. Существенным
отличительным признаком рыжей цапли служит то обстоятельство, что средний палец (с
когтем) всегда длиннее цевки; это, однако, можно увидеть, только держа птицу в руках. На
земле отличается «крадущейся» походкой, чему способствуют более короткая цевка и от-
носительно более длинные пальцы. Корм значительно чаще, чем серая цапля, добывает,
преследуя добычу пешком. Останавливаясь, подолгу стоит в одном положении, позволяю-
щем нанести быстрый нацеленный удар.
По мнению Е.П. Спангенберга (1951), менее осторожна по отношению к человеку, чем
серая цапля; это связывается им с сумеречным (и даже ночным) образом жизни. В отличие
от серой цапли, редко садится на деревья, предпочитая скрываться в тростниковых зарос-
лях.
Голос (крик в полете) похож на голос серой цапли, но менее громок (Спангенберг, 1951).
Птенцы «урчат». Их голос в гнезде можно передать словами: «пью-пью, кхью-кхью», иног-
да: «грау-рау-рау» и «ко-ко-ко» (ориг.). Иным исследователям крик взрослой птицы напо-
356
Отряд Листообразные, Семейство Цаплевые
минает карканье, и они передают его как громкое «крак-краак» или «клек-клек-клак» — у
гнезда (Bauer, Glutz von Blotzheim, 1966).
Подвидовая систематика. Л.С. Степанян (2003), вслед за Е.П. Спангенбергом (1951)
выделяет 3 подвида, из которых на территории Вост. Европы и Сев. Азии обитает два.
1. Ardea purpurea purpurea
Ardea purpurea Linnaeus, 1766, Syst. Nat., ed. 12, c. 236, «на Востоке»; Франция
(Stresemann, 1920, Avifauna Macedonica, c. 226)
Западная часть ареала в Вост. Европе и Сев. Азии.
2. Ardea purpurea manilensis
Ardea purpurea manilensis Meyen, 1834, Nova Acta Acad. Caes. Leop.Carol., 16.suppl.,
с.102, Филиппины.
Восточный подвид; в пределах бывш. СССР населяет Приморье и бассейн нижнего
Амура. От номинативного подвида он отличается несколько более крупными размерами,
не развитым пятнистым рисунком в нижней части шеи, более тёмными, серовато-буланы-
ми перьями груди, более насыщенной чёрной окраской живота, примесью свинцово-серых
перьев в украшающих косицах на спине (Степанян, 2003).
Не связан с описываемой территорией подвид А. р. madagascariensis Oort, 1910 (3),
оседло обитающий на Мадагаскаре. Ряд орнитологов (Cramp, 1977; Howard, Moore, 1980)
выделяют также ещё один подвид Л. р. bournei de Naurois, 1966 (4), оседло живущий на о-
вах Зелёного Мыса (рис. 72).
Замечания по систематике. В составе рода Ardea рыжая цапля занимает обособлен-
ное положение и большинством систематиков выделяется в подрод Pyrrherodia Finch et
Hartlaub, 1870. Форму bournei, заметно отличающуюся от остальных рас светло-рыжей
окраской верха и слабым развитием чёрного рисунка, иногда считают самостоятельным
видом — кабовердесской цаплей (A. bournei de Naurois, 1966).
Описание. Окраска. Взрослая птица в брачном наряде. На затылке развита узкая
косица из 3-5 чёрных удлинённых перьев (более длинная у самцов). Шапочка и задняя
сторона шеи чёрные, бока шеи и головы охристо-рыжие (до рыже-коричневых) с продоль-
ными чёрными полосами от угла рта назад к затылку и вдоль боковой стороны шеи. Подбо-
родок, горло, передняя сторона шеи белые, по передней стороне шеи тоже проходят узкие
темные продольные полосы или ряды продольных пестрин. На зобе развиты удлинённые и
заострённые перья, светло-охристые с примесью тёмных пестрин. Верхняя сторона тела,
от зашейка до хвоста, включая верхние кроющие крыльев, аспидно-серая (иногда с бурова-
тым налётом в верхней части крыльев), маховые перья тёмные, подмышечные — аспид-
ные. На лопаточных партиях оперения развиты узкие удлинённые украшающие перья, бах-
ромой свешивающиеся на крылья и заднюю часть спины; их цвет варьирует от светло-
серого до охристого. Бока груди и брюха а также оперённые части голени и нижние крою-
щие крыльев интенсивного каштанового цвета, при сложенном крыле кистевой сгиб при-
крыт рыхлым оперением, образующим сбоку каштановое пятно. Передний край крыла
рыжеватый. Середина груди и брюха, подхвостье — чёрные. Клюв жёлтый с тёмным конь-
ком, неоперённые участки кожи на голове зеленовато-жёлтые, радужина жёлтая. Ноги жёл-
тые или оливково-жёлтые, передняя сторона цевки и пальцы тёмно-коричневые.
Зимний наряд взрослой птицы отличается от брачного меньшей выраженностью укра-
шающих перьев (косицы, бахромы на спине, «гривы» на зобе), более тусклым цветом нео-
перённых частей тела.
Птенец. Первый и второй пуховые наряды на спинной стороне буровато-серые, на брюш-
ной — беловатые, на голове развит беловатый хохол из удлинённых пушинок. Неоперён-
Рыжая цапля
357
ные участки кожи тёмные с интенсивной пятнистостью на голове и шее. Клюв с тёмным
надклювьем и желтоватым подклювьем, ноги сероватые, радужина светлая.
Гнездовой наряд. Косица на голове и украшающие перья на спине отсутствуют, на зобе —
развиты слабо. Верх глинисто-бурый с более темными размытыми центрами перьев, ша-
почка тёмно-бурая, бока головы, шея и низ тела рыжевато-охристые с рядами бурых пест-
рин, подхвостье беловатое. Каштановые и аспидно-серые тона развиты слабо. Клюв у мо-
лодых желтовато-бурый, короче (тупее), чем у взрослых. Ноги оливково- или зеленовато-
жёлтые, радужина бледно-жёлтая. Двухлетние птицы по сравнению с более старыми окра-
шены светлее, и имеют рыжеватые каймы на верхних кроющих крыла.
Строение и размеры. Размеры (мм) рыжих цапель номинативного подвида
(самцов и самок) для территории бывш. СССР (п=25): длина крыла — 340-390, в среднем
367,2; длина цевки — 115-130; длина среднего пальца с когтем — 125-135 (Спангенберг,
1951); в Казахстане и Узбекистане: длина крыла — 340-390; длина хвоста — 125-140;
длина клюва— 112-146; длина цевки — 108-145 (Долгушин, 1960; Салихбаев, Богданов,
1961). Размеры для разных подвидов и половых групп приведены в табл. 30.
Данные по весу тела таковы. Для А. р. purpurea с территории бывш. СССР вес двух
взрослых птиц (пол не указан) — 1 220 и 1 225 г (Спангенберг, 1951). По данным Н.А.
Зарудного (Зарудный, 1888, цит. по: Спангенберг, 1951) — до 1 650 г. Вес самцов из Голлан-
дии: 617-1 218 г; самок: 525-1 035 г (Cramp, 1977). Вес птенца при вылуплении — 35 г, в
возрасте 50 дней — 1 100-1 200 г (Tomlinson, 1978, цит. по: Cramp, 1977). Для двух самцов
А. р. manilensis с территории бывш. СССР — 1 310 и 1 358 г (Спангенберг, 1951).
Рис. 72. Область распространения рыжей цапли: а — гнездовой ареал; б — область зимовки. Подви-
ды: 1 —А. р. purpurea; 2 —А. р. manilensis; 3 —А. р. madagascariensis; 4 — А. р. bournei.
358
Отряд Листообразные, Семейство Цаплевые
Таблица 30. Размеры рыжих цапель (мм) из разных участков ареала
Название части тела п Самцы Самки Источник М lim n М lim
Длина крыла 10 Ardea purpurea purpurea, для территории бывш.СССР 370-395 9 340-370 Спангенберг, 1951
Длина крыла 13 Ardea purpurea purpurea. Палеарктика 371 357-383 9 355 337-372 Cramp, 1977
Длина хвоста 13 125 118-136 8 119 112-127
Длина клюва 13 126 120-131 8 116 109-123
Длина цевки 13 122 113-131 8 118 112-123
Длина сред- 13 132 121-139 7 126 118-131
него пальца Длина крыла Длина крыла 2 Ardea purpurea purpurea, Палеарктика 330 290 Witherby et al. (1939), цит. no Cramp, 1977) Ardea purpurea manilensis, для территории бывш.СССР 370,370 Спангенберг, 1951
Длина крыла Длина клюва Длина цевки Ardea purpurea manilensis. Дальний Восток 370-401 360-388 Шульпин (1927), цит. 125—137,3 117,6-119,5 по Спангенберг, 1951 128-142 125,3-129
Линька. Данных мало. У взрослеющих (не достигших годовалого возраста) птиц час-
тичная смена контурного оперения и некоторых кроющих крыла происходит в ноябре-
апреле, во время зимовки. У птиц, возраст которых приближается к двум годам, она начи-
нается уже в мае. Линька взрослых птиц, завершивших сезон размножения, начинается в
июне и длится до ноября, часто до февраля. Замена первостепенных маховых происходит
не регулярно, по-видимому, на протяжении всего летнего периода, до начала миграций.
При описании линьки, которое приведено выше, как Е.П. Спангенберг (1951), так и Стэнли
Крэмп (Cramp, 1977) ссылаются только на мнение Уайзерби с соавторами (Witherby et al,
1939). Более поздние сведения по линьке вида нам не известны, что свидетельствует о том,
что этот процесс у рыжих цапель изучен совершенно недостаточно.
Распространение. Гнездовой ареал. На постсоветском пространстве — южный
вид, в своём распространении связанный с устьями таких европейских рек, как Дунай,
Днепр, Кубань, Волга, Терек, Урал. Гнездится в тростниковых зарослях как по западному,
так и восточному побережьям Каспийского моря. В Средней Азии — по долинам Аму-
Дарьи и Сыр-Дарьи. На Дальнем Востоке населяет самый юг Приморского и Хабаровского
краёв. Встречается в устье р. Горин, в пойме Нижнего Амура, на озёрах Эворон, Болонь,
Чукагирское и Удыль; на Листвяном и Б. Уссурийском о-вах под Хабаровском (Росляков,
2000а). В Приморье — обычный вид оз. Ханка и его окрестностей.
За пределами территории бывш. СССР перелётные популяции номинативного подвида
обитают также в Испании, Франции, Бельгии, Австрии, Румынии, Болгарии, Сев. Греции,
Турции и Сев. Иране. Оседлые популяции известны из районов Центр. Африки, располо-
женных южнее экватора, а так же на самом юге ЮАР; на Мадагаскаре и о-вах Зелёного
Мыса обитают особые подвиды (см. выше).
Зимовки. Места зимовок птиц номинативного подвида располагаются в Экватори-
альной Африке севернее экватора, а также в юго-восточной части этого континента. Изред-
ка в зимнее время рыжих цапель отмечали на юго-вост. Каспии. Места зимовок подвида Л.
Рыжая цапля
359
р. manilensis, гнездящегося в России, не были установлены до конца 1950-х гг. Однако,
благодаря кольцеванию птенцов на оз. Ханка, выяснено, что область их зимовок располага-
ется на всей территории Индокитая. Помеченые на оз. Ханка птицы найдены в Юж. Китае,
во Вьетнаме, Таиланде, островах Индонезии, на северном побережье п-ова Малакка (рис.
72, 73).
В Азии оседлые популяции А. р. manilensis обитают в Пакистане, Индии, Юж. Китае и
в Индонезии.
Миграции. Расселение молодых птиц с мест гнездовий выглядит следующим образом.
После того, как птенцы оставляют гнездо, уже с конца июня и в июле, они разлетаются в
его окрестностях на удаление до 20-50 км. В июле и в августе происходит дальнейшее
расселение птиц, когда их встречают на расстояниях от 150 до 300 км от места гнездова-
ния. В этот период многие из птиц перемещаются в направлениях, противоположных об-
щему руслу осеннего перелёта: в северном и северо-восточном направлениях (Schenk, 1934;
Сапетин, 1978ж). Только с начала сентября начинается «планомерный» отлёт рыжих ца-
пель на юг и юго-запад (Кищинский, 1978е). У перелётных популяций, населяющих евро-
пейские, северо-африканские и среднеазиатские страны, даты последних встреч в районах
гнездования регистрируют в конце августа - начале сентября (Страутман, 1954; Cramp,
1977). А уже 15 сентября пролётных рыжих цапель наблюдали в Вост. Иране (Зарудный,
1903).
Я. Шенк (Schenk, 1934), анализируя разлёт молодых рыжих цапель, окольцованных в
Югославии, называет время с июля по сентябрь промежуточной миграцией. В этот период
птиц, родившихся в плавнях Дуная, отмечали севернее мест рождения (37%), юго-западнее
(30%), южнее и юго-восточнее (27%). И лишь 6% птиц летят на запад (п = 70), тогда как в
дальнейшем последнее и южное направления будут основными. В сущности, таков же ха-
рактер разлёта молодых цапель, гнездящихся в дельте Кубани и на сев. Каспии (Сапетин,
Рис. 73. Ареал рыжей цапли в Вост. Европе и Сев. Азии: а — гнездовой ареал; б — залёты.
360
Отряд Листообразные, Семейство Цаплевые
1978ж). В ноябре-декабре возвраты как дунайских, так и азово-каспийских птиц лежат
уже к западу и юго-западу от мест мечения. Их регистрируют в Италии, Франции, на Сици-
лии, Мальте, в Юж. Тунисе. Это позволяет утверждать, что основная область их осенне-
зимней концентрации находится в странах Зап. Средиземноморья. Приазовские цапли ос-
ваивают Восточное Средиземноморье. Возвраты колец от них получены из Греции, с Кип-
ра, из Турции, Египта. При дальнейшем движении на юг они достигают стран Западной и
Центр. Африки, причём дунайские птицы занимают более западные области зимовок. Окон-
чательно «освобождают» районы гнездовий европейские популяции в конце октября - но-
ябре (Schenk, 1934; Сапетин, 1978ж).
Для полноты картины перемещения западных рыжих цапель и их размещения во вне-
гнездовой период можно привести несколько данных, не входящих в рамки предложенной
выше концепции. Так, цапля, помеченная в дельте Волги птенцом, в возрасте 2 лет в ноябре
была обнаружена на юго-западе Камеруна (5 300 км от места кольцевания). Птица, родив-
шаяся в дельте Терека, на третьем году жизни, 7 мая, была убита в Телавском р-не Грузии.
Наконец, третья птица, родившаяся в низовье Маныча, на первом году жизни в октябре
добыта в Калмыкии, в 600 км юго-восточнее места рождения (Сапетин, 1978ж).
Миграции птиц, гнездящихся в Средней Азии, с помощью кольцевания не изучали.
По аналогии с близкими видами цапель можно предположить, что у них тоже существует
«промежуточный перелёт», когда эти птицы разлетаются во все стороны от мест рожде-
ния по пригодным для добычи пищи угодьям. Затем, при наступлении осеннего похоло-
дания, цапли устремляются к югу и юго-западу. По-видимому, уже зимующих птиц на-
блюдали на юго-восточном берегу Каспийского моря в районе Гассан-Кулийского запо-
ведника, а также в долине Атрека и его притоков, в Сев. Иране. Документированных
сведений о более дальних перемещениях среднеазиатских рыжих цапель во время мигра-
ции не имеется.
От 2 780 птенцов, окольцованных на оз. Ханка в 1960-1973 гг., к 1975 г. получено 42
возврата колец (1,5%), т.е. возврат колец был почти вчетверо ниже, чем для подвида А. р.
purpurea. В.М. Поливанов (1978) объясняет это отсутствием традиции охоты на цапель в
этом регионе и весьма неполным возвращением колец. Для птенцов восточного подвида,
как и для западного, зарегистрирован ранне-осенний разлёт молодых (в июле-сентябре) по
всём направлениям, с последующей миграцией на юго-юго-запад вдоль восточного побе-
режья Азии вплоть до п-ова Малакка (рис. 74).
Залеты в Европейской части бывш. СССР отмечаются до 60-й параллели. Так, напри-
мер, в октябре 1957 г. одна особь, пойманная в Баварии и содержавшаяся там 2 года в
неволе, добыта около пос. Вожега Вологодской обл. (Шалаков, 1958). Отдельные встречи
рыжих цапель отмечали на всём протяжении XX в. в Литве, Латвии, Белоруссии, а в Рос-
сийской Федерации — в Псковской, Новгородской, Ивановской, Нижегородской, Тамбов-
ской, Пензенской, Саратовской обл., в Мордовии и Татарстане (Вадкоусю, 1964; Федю-
шин, Долбик, 1967; Попов, 1977; Пузанов и др., 2005 и др.).
Местообитания. Рыжая цапля предпочитает обширные открытые пространства, в зна-
чительной степени занятые водоёмами, с обширными зарослями тростника и камыша и,
много реже, низкорослыми кустарниками ивы (Спангенберг, 1951). Такие угодья распола-
гаются в Вост. Европе и Сев. Азии главным образом в устьевых участках южных рек, по
побережьям южных морей и озёрных систем. На облесненных территориях, в горной мес-
тности и в пустынях рыжая цапля редка или не встречается вовсе. Правда, существуют
наблюдения, что в пустынные районы Казахстана рыжие цапли залетают иногда довольно
далеко в поисках корма (Костин, 1936, цит. по: Салихбаев, Богданов, 1961).
Рыжая цапля
361
Рис. 74. Миграции рыжих цапель, установленные путем кольцевания (по: Поливанов, 1978; Сапетин,
1978ж, с дополнениями по материалам Центра кольцевания РАН до 2004 г.): а — районы «промежу-
точной» миграции; б — границы территорий разлёта птиц разных популяций. Популяции: I — «ду-
найская», II - «азово-каспийская», III — «приморско-тихоокеанская».
Гнездовья располагаются в отдалении от человеческих поселений; напротив, во время
кормежки сами птицы часто встречаются рядом с рыбаками и в сельскохозяйственных уго-
дьях (Спангенберг, 1951).
Наиболее типичный биотоп для устройства гнёзд — тростниковые крепи в дельтах и
долинах южных рек. Здесь эти птицы находят как удаленность от человеческих поселений,
так и недоступность самих гнездовий для человека и четвероногих хищников.
Кормовые стации разнообразны. Например, на Сыр-Дарье многие птицы добывают пищу
вблизи от гнёзд, стоя на краю маленького плёса. Другие же залетают далеко от гнездовых
колоний, углубляясь в несвойственные им стации. Добывают пищу также повсюду по по-
логим речным берегам, иногда на свежевспаханных полях, в степных участках.
Численность. Конкретные данные о численности рыжей цапли по всему гнездовому
ареалу на территории бывшего СССР в 1960-1980-х гг. отсутствуют. Авторы региональ-
ных орнитологических сводок чаще всего характеризуют её в качестве «обычного», «не
редкого» или, наоборот, «редкого», а иногда и «случайно залётного» вида (Воробьев, 1954;
Страутман, 1954; Долгушин, 1960; Панов, 1973; Костин, 1983; Сагитов, 1987; Patrikeev, 2004
и др.). Е.П. Спангенберг (1951), основываясь на данных, первой половины XX века, относит
рыжую цаплю к разряду «довольно многочисленных видов» тогдашней территории СССР.
На Сев. Каспии в 1960-1970-х гг. подсчёт гнёзд проводился силами Каспийской орнито-
логической станции. По результатам этих учётов, в 1973-1974 гг. из 46,6 тыс. гнёзд колони-
альных веслоногих и аистообразных, рыжей цапле принадлежало 3 010 гнёзд. При этом в
дельте Волги обнаружено 2450, в дельте Терека — 510 и на Прикумских озёрах — 50 гнёзд
(Бондарев, 19756).
По сведениям, собранным М. Патрикеевым (Patrikeev, 2004), в 1950-1960-х гг. в окрес-
тностях Кызыл-Агачского заповедника гнездились 600-700 пар этих птиц, в Калиновском
лимане в 1983-1984 гг. — 52-440 пар, в прочих участках Азербайджана (Махмуд-хала,
дельта р. Куры, оз. Шоргель, долины рек Варвара и Самур и др.) в 1980-1998 гг. — около
60-80 пар.
362
Отряд Листообразные, Семейство Цаплевые
Крупный очаг гнездования голенастых существует в Вост. Приазовье, в частности, в
устье Кубани. В результате летних учётов в этом районе в 1953-1954 гг. количество рыжих
цапель, кормившихся на лиманах, оценено в 6,5-7 тыс. особей, что составляло тогда около
четверти всего обилия птиц названной группы (Винокуров, 1960).
Целесообразно привести данные о конкретном количестве гнездящихся пар рыжей цап-
ли с некоторыми сведениями о динамике численности вида в ряде стран Зап. Европы по
материалам, собранным до середины 1970-х гг. Большей численностью вида отличались
страны Средиземноморья, где рыжая цапля находит более подходящие условия для обита-
ния в плавнях устьев рек, впадающих в Средиземное море. В Испании общее число гнездя-
щихся пар приближается к 1 000 (Fernandez Cruz, 1975). Во Франции для 1964-1968 гг. его
оценивали в 2 100-2 500, а в 1974 г. в 1 350-1 500 гнездящихся пар. В Италии в 1975 г. — в 1
500-2 400. В Греции — около 300. В странах, расположенных севернее, численность вида
значительно ниже. В Зап. Германии рыжая цапля гнездилась в долине Рейна. Здесь она
впервые обнаружена в 1967 г.; в 1968 г. учтено не менее 9, в 1970 — до 18-20 пар. В сере-
дине 1970-х гг. возможно гнездилось 20-30 пар. В Швейцарии в 1941 г. учтено 3 пары, к
1955 г. количество их возросло до 54. Затем последовал спад, и в 1961 г. отмечено 40-45, а
в 1972 г. — лишь 8-10 пар. В Австрии в 1950-1955 гг. обнаружили 240-350 пар. Для терри-
тории прежней Чехословакии известно гнездование с 1947 г. 4-20 пар. В Венгрии на одной
из колоний (Кисбалатон) в период с 1968 по 1972 гг. численность сократилась со 107 до 37
пар. На о-вах Зелёного Мыса гнездилось не более 200 пар (Cramp, 1977). Таким образом,
в середине 1970-х гг. в странах европейского Средиземноморья популяцию рыжей цапли
оценивали в 5 300, а в более северных странах Зап. Европы — примерно в 400 гнездя-
щихся пар.
Рассматривая эти данные в целом, видим, что численность рыжей цапли в 1930-1970-е гг.
в Западной Европе колебалась в значительных пределах. В южных регионах, как правило, в
этот период отмечали почти двукратное сокращении её количества, в северных — рост изве-
стных поселений и появление новых мест гнездования. По всей вероятности, сходным обра-
зом проходили изменения численности рыжих цапель в европейской части бывшего СССР.
В Атласе гнездящихся птиц Европы для 1990-х гг. приводятся следующие данные: Фран-
ция — 2 750, Испания — 1 100, Румыния — 1 000, Венгрия — 900, Италия — 600, Нидер-
ланды и Португалия — по 250, Греция — 150, прочие страны Европы, за исключением
России, — 70 пар. При этом западно-европейская популяция рыжей цапли оценивается в
7 766-8 556 (в среднем 8 090) пар. Считается, что на Украине в те же годы гнездилось 800,
Молдавии — 150 пар, а в России — 40-90 тыс. пар (в среднем 60 тыс. пар) (Bankovics,
1997). Никаких конкретных данных, на которых основывается эта явно завышенная чис-
ленность цапель в России, не приводится.
По данным, опубликованным Международной ассоциацией охраны птиц BirdLife
International (Bird Life International, 2004) и, по-видимому, наиболее полным на сегодняш-
ний день, на рубеже XX-XXI веков общая численность европейской популяции рыжей цапли
составляла 29-42 тыс. гнездящихся пар, из которых в России гнездилось 10-15 тыс. пар, на
Украине — 6,7-11,9 тыс. пар, в Молдове — 250-350 пар, в Армении — 80-330, в Азербай-
джане — 250-500 пар. Общая тенденция европейской популяции — умеренное снижение
численности; та же тенденция прослеживается и в России (Мищенко и др., 2004).
Конкретных сведений о численности рыжих цапель в Средней Азии и Казахстане нет.
Судя по данным, приведенным И.А. Долгушиным (1960), в Казахстане даже в типичных
местообитаниях этот вид скорее немногочислен. Этот же автор полагал, что на восточном
побережье Аральского моря цапля обычна лишь на отрезке долины Сыр-Дарьи между
Джусалами и Чиили, т.е. примерно на 300-километровом участке. В Узбекистане довольно
Рыжая цапля
363
обычна в дельте р. Аму-Дарьи и юж. Приаралье (Сагитов, 1987). В остальных местах гнез-
дится редко, спорадично. В нижнем течении Зеравшана и в Ферганской долине на гнездо-
вании не обнаружена (Салихбаев, Богданов, 1961). В Туркменистане на гнездовании редка,
отмечена главным образом на Аму-Дарье, на Ташаузских озёрах (т.е. опять-таки в долине
Аму-Дарьи) и в низовьях Мургаба (Дементьев, 1952). В связи с резкими негативными из-
менениями экологических условий в Средней Азии, численность рыжей цапли в этом реги-
оне, по-видимому, сильно сократилась (Птицы Средней Азии, 2007).
В Приморье рыжая цапля гнездится по берегам озёр Ханка и Дорицени, а также у зал.
Посьет (Воробьев, 1954; Панов, 1973). В 1960-1970-х гг. здесь она была весьма обычной, а
гнездовья её вполне доступными (Поливанов, 1965; Поливанов, Поливанова, 1971). Н.Н.
Поливанова (1971) считает, что в районе оз. Ханка гнездится 1-1,5 тыс. пар. Высокая чис-
ленность и доступность колоний рыжей цапли на оз. Ханка в те годы подтверждается тем,
что в 1950-х гг. там было окольцовано вчетверо больше птенцов, чем в Приазовье, дельте
Терека и в Астраханском заповеднике вместе взятых, при примерно одинаковой активнос-
ти метчиков.
Ю.Н. Глущенко, Д.В. Коробов и И.Н. Кальницкая (2003) оценивают современную чис-
ленность вида в Приханкайской низменности как крайне непостоянную. Они связывают
увеличение количества гнездящихся птиц с периодами высокого уровня воды в оз. Ханка и
максимального увлажнения плавней южного и восточного побережий. В такие годы рыжая
цапля, по их мнению, на гнездовании преобладает среди других видов семейства Ardeidae.
В 2002 г. здесь было обнаружено 20 гнездящихся пар; названные авторы полагают, что это
лишь десятая часть их истинного числа. Кроме того, до 50 пар гнездится в Хабаровском
крае (Росляков, 2000). Приведенные данные можно расценивать как признак сокращения
численности рыжих цапель в Приморье к началу XXI в.
К. А. Воробьев (1954) обращает внимание на тот факт, что эта, по его мнению, «хорошо
заметная птица» не была обнаружена здесь Н.М. Пржевальским в 1860-х гг., современни-
ками Н.М. Пржевальского и последующими исследователями вплоть до конца XIX столе-
тия. Первые (хранящиеся в коллекциях) экземпляры рыжих цапель на Дальнем Востоке
добыты 9 и 12 сентября 1912 г. Это заставляет его предположить, что этот вид проник в
Приморье лишь в начале XX столетия. Е.Н. Панов (1973) о времени проникновения рыжей
цапли в Юж. Приморье сообщает уклончиво, ссылаясь лишь на наблюдения Л.М. Шульпи-
на в 1930-х гг., зафиксированные в дневниках последнего. По-видимому, можно допустить,
что проникновение рыжей цапли на русский Дальний Восток действительно произошло в
конце XIX - начале XX столетий.
Размножение. Половая зрелость наступает на 3-4 год жизни. В.М. Поливанов (1978) на
основании данных по мечению, высказывает предположение о том, что молодые птицы до
наступления половой зрелости держатся в местах зимовок. По крайней мере, так выглядит
это для восточного подвида.
Гнездятся, как правило, моновидовыми колониями (Винокуров, 19596). Правда, И.А.
Долгушин (1960) встречал в Казахстане как отдельно располагавшиеся гнёзда рыжих ца-
пель, так и отмечал их присоединение к колониям серых, белых цапель и колпиц. Поблизо-
сти от колоний рыжих цапель встречаются гнёзда других видов цапель и птиц иных систе-
матических групп. Отношение цапель к птицам, живущим в районе колоний, преимуще-
ственно безразличное. Они лишь изредка нападают на болотных луней и всегда отгоняют
появляющихся вблизи серых ворон; на остальных птиц, по-видимому, просто не обращают
внимания (Винокуров, 19596). Размеры колоний от нескольких (2-5) до 200-250 гнёзд (Ви-
нокуров, 19596; Glutz von Blotzheim, 1962). Однако в большинстве случаев колонии состо-
ят из 10-20 (35% колоний) или 50-60 (29%) гнёзд (Винокуров, 19596).
364
Отряд Листообразные, Семейство Цаплевые
Расстояние между гнёздами в дельте Кубани составляло 1,5-2 м. Располагались они
небольшими группами по 3-4 гнезда, удаленными одна от другой на 10-20 м, а иногда и на
большее расстояние. Поэтому колония состоит как бы из нескольких «элементарных коло-
ний» (Винокуров, 1959). Другая колония может находиться в 500 м от первой или на боль-
шем расстоянии (Долгушин, 1960; Cramp, 1977; Костин, 1983 и др.).
Гнёзда размещаются в зарослях тростника высотой 2,5-3 м с диаметром стеблей от 5 до
10 мм, при плотности тростника в таких местах до 340 стеблей на 1 м2 (в среднем 250).
Глубина воды у гнёзд от 0,3-0,6 до 0,8-1,3, местами до 1,5 м.
Основанием гнёзда служат не только так называемые «заломы» или кучи тростника, но
и сгущения стеблей этого вида. Удалось подсчитать, что в последнем случае птицы строят
гнездо на 27-29 кучно стоящих растениях (Кенжегулов, 1965, цит. по: Сагитов, 1987). В
качестве строительного материала используются стебли того же тростника. В Средней Азии
цапли часто строят гнёзда в переплетениях ветвей мелких ив и колючих кустарников. Не
было найдено гнёзд на высоких деревьях, уступах скал и на земле (на болотных кочках),
хотя П. Патев (1950) такую возможность для Болгарии не исключает. Есть указание на
гнездование цапель небольшими колониями на невысоких деревьях, иногда, по нескольку
гнёзд на одном дереве (Мензбир, 1918). По Е.П. Спангенбергу (1951), такие случаи не харак-
терны для птиц, обитающих в нашей стране, и должны рассматриваться как исключение.
В постройке гнезда принимают участие оба партнёра. По наблюдениям в устье Терека
их участие в самом строительстве равное. Спаривание осуществляется на гнезде или на
соседних ветвях растений и происходит до 3-7 раз в дневное время до появления первых
яиц и по 4-6 раз во время их откладки.
Архитектура гнёзда подобна строениям других цапель. Его форма во время откладки
яиц похожа на перевёрнутый конус с «просвечивающими» стенками (Спангенберг, 1951).
Размеры: 35-90 см в диаметре и от 16 до 55 см в высоту (Спангенберг, 1951; Долгушин,
1960; Винокуров, 19596; Костин, 1983). Характерен глубокий лоток гнезда, где помещают-
ся яйца. Конкретные сведения по 18 промеренным гнёздам на оз. Блан (в Узбекистане)
сообщает К. Кенжегулов (Кенжегулов, 1960, цит. по: Сагитов, 1987). Их наружный диаметр
был от 45 до 52 см, диаметр лотка — 27-31 см, глубина лотка — 8 см, высота расположе-
ния основания гнезда над уровнем воды — 1,5-1,6 м. По А. А. Винокурову (19596), в дельте
Кубани наружный диаметр гнёзд во время насиживания составляет 38-65 см, глубина лот-
ка — 3-7 см, высота над водой — 75-137 см (и = 14).
Размер кладки от 3 до 8 яиц, чаще всего 4-5 яиц (Спангенберг, 1951; Долгушин, 1960;
Cramp, 1977; Костин, 1983; Климов и др., 1990). На Ханке полная кладка содержит от 3 до
7 яиц, в среднем (по 289 кладкам) 3,9 яйца на кладку (Поливанова, 1971). По результатам
обследования 151 кладки в Швейцарии, по 2 яйца было в 6% гнёзд, по 3 — в 21 %, по 4 —
в 31%, по 5 — в 36%, по 6 — в 7% (Bauer, Glutz von Blotzheim, 1966).
Окраска яиц зеленовато-голубая при снесении, позднее сильно выцветающая (Спанген-
берг, 1951). Размеры яиц (мм) в коллекциях СССР до 1950 г. (п = 17): 53-57,5 х 37,9-39,2, в
среднем 54,8x38,5. В сев. Крыму (п=56) в 1976-1978 гг.: 51,1-58,5 х 37,5-43,4, в среднем
54,7x40,5 (Костин, 1983). На Днепре (п=56): 54,8-58 х 40,6-44,5 мм (Ардамацкая, личн.
сообщ.). На оз. Ханка (п=24): 53,8-60,5 х 39,0-43,4 мм, в среднем 56,9x41,6 мм (данные
Ю.Н. Глущенко). В зарубежной Европе (Schonwetter, 1967) (п = 300): 57x41. Масса яйца (г)
в зарубежной Европе (п=42) 37-60, в среднем 50 (Cramp, 1977); по Ю.В. Костину (1983),
масса 5 яиц — от 40,8 до 45,6, в среднем — 45,3 г; по Т.Б. Ардамацкой (личн. сообщ.) в
устье Днепра (п=56) 43,4-59,2, в среднем 50,7.
Яйца откладывают через один или два дня (Спангенберг, 1951), иногда интервалы меж-
ду откладкой очередного яйца составляют 2-3 дня, но никогда не превышают 5-7 дней
Рыжая цапля
365
(Cramp, 1977). Насиживание начинается с момента откладки первого яйца. Насиживают
оба родителя (Долгушин, 1960); время насиживания делится поровну между партнёрами.
Только в последние дни перед вылуплением птенцов на гнезде больше времени (примерно
65%) проводит самка (дельта Терека, ориг.). Насиживание длится 25-27 дней.
Как и у всех цапель, одно-два яйца из кладки (особенно, в крупных кладках) как прави-
ло оказываются неоплодотворёнными или погибают во время насиживания. Исключение
из этого правила отметил А.А. Винокуров (19596), который в 1953 г. в Чумаковском лимане
(Вост. Приазовье) в 10 гнёздах с общим числом 35 яиц не обнаружено ни одного неопло-
дотворённого яйца. Число птенцов в выводках — от 1 до 5, чаще всего 3-4. Время пребы-
вания птенцов в гнёздах — немногим более месяца. Однако по свидетельству многих орни-
тологов (Сапетин, 1978ж; Климов и др., 1990), уже в возрасте 25 дней птенцы часто пере-
бираются на соседние кусты и в другие гнёзда. Мы склонны объяснять это явление прежде
всего беспокойством подрастающего поколения со стороны наблюдателя. По данным С.
Крэмпа (Cramp, 1977), время нахождения птенцов в гнезде, соответствующее их полному
оперению, составляет 45-50, но иногда растягивается до 55-60 дней.
Пока птенцы малы, пищу приносит только самец, позднее к этому процессу присоеди-
няется самка. Впечатление от вида крупной (монолитной) колонии рыжих цапель в период,
когда основная масса птенцов достигла примерно 3-недельного возраста, незабываемо. По
словам Т.Б. Ардамацкой (личн. сообщ.) одна из таких колоний, разместившаяся в 1970 г. на
заломах тростника на Днепре под Херсоном, представляла собой «сплошной ковёр из под-
росших уже птенцов рыжих цапель, лишь по периферии украшенный вкраплениями дру-
гих видов: большой и малой белых цапель, кваквы и каравайки».
По данным большинства исследователей, изучавших этот вид, вылетевшие из гнёзд мо-
лодые по крайней мере в течение месяца держатся семьями, а затем объединяются в более
крупные стаи, состоящие из нескольких выводков. Такими группами они отлетают на зи-
мовки.
По наблюдениям в устьях Кубани и Терека, из 161 яйца, найденного в 41 полной кладке,
вылупилось 122 птенца и были окольцованы в возрасте 2-3 недель 114 сохранившихся.
Следовательно, эмбриональная смертность составила 24,2%, а гибель птенцов в гнёздах в
возрасте до 3 недель — 6,6% (устн. сообщ. В.Н. Карповича, ориг.). О размерах общей убы-
ли потомства до момента вылета птенцов из гнёзд данных не имеется. Но она составляет
никак не менее трети от числа отложенных яиц. В числе причин гибели гнёзд — разруше-
ние их в непогоду (ветер), затопление водой в периоды штормов. Однако от этих факторов
больше страдают гнёзда, расположенные по краям зарослей тростника. В глубине зарос-
лей основания гнёзд прочнее. А уже трёхнедельные птенцы спасаются от наводнений, пе-
ребираясь на более высокие расположенные рядом ветви и кустарники.
70% молодых птиц западного подвида, по данным мечения, погибает в течение первого
года жизни (Сапетин, 1978ж). Максимальная продолжительность жизни в естественных
условиях, как и у прочих крупных цапель, превышает 20 лет. Из 118 проанализированных
(окольцованных) цапель, одна прожила не менее 16, другая — 13 лет (Сапетин, 1978ж).
Причём первая в этом возрасте ещё гнездилась (район Белье, Австрия) (Schenk, 1934). Дж.
Деджон и М. Чайковский (Dejonghe, Czajkowski, 1983) сообщают о трёх птицах, окольцо-
ванных в Германии и Франции, одна из которых прожила 25 лет, 5 месяцев и 17 дней,
вторая — 23 года и 2 месяца, а третья — 20 лет и 2 месяца.
Суточная активность, поведение. Рыжая цапля — сумеречный вид, она кормится и
собирает пищу для потомства в вечерние, предрассветные и раннеутренние часы, редко
днем. Е.П. Спангенберг (1951) предполагал даже ночной образ жизни этого вида, однако С.
Крэмп (Cramp, 1977) определённо пишет о сумеречном образе жизни рыжей цапли, сооб-
366
Отряд Листообразные, Семейство Цаплевые
щая как о ночёвочных, так и днёвочные скоплениях отдыхающих птиц. В период размно-
жения взрослые птицы держатся днём рядом с гнездом. Вспугнутые улетают недалеко и по
завершении тревоги возвращаются к выводку.
Перелётные стаи главным образом моновидовые. Но иногда рыжие цапли во время
миграций объединяются с гнездившимися по соседству кваквами, реже — с серыми цапля-
ми и большими выпями. Птицы летят строем типа «угла» или «прямой линии».
Питание. Плотоядный вид. О находке растений в желудках птиц (кусочки тростника,
рогоза, рдеста, хары) есть сведения лишь у К. Кенжегулова (цит. по: Сагитов, 1987); по
всей вероятности, растительные объекты были случайно проглочены цаплями вместе с ос-
новной добычей. В порциях пищи, принесённой на гнездо птенцам, растения не отмеча-
лись вовсе. Все исследователи подтверждают, что основой корма рыжей цапли служит рыба,
в меньшей степени — лягушки (Патев, 1950; Спангенберг, 1951; Воробьёв, 1954 и др.).
Анализ 208 желудков рыжих цапель в Вост. Приазовье дал следующие результаты. Рыба
составила 52% веса их содержимого, насекомые и их личинки — 23,1%, земноводные —
18%, млекопитающие — 3,7%. Прочие объекты, включая птиц, пресмыкающихся и расте-
ний — 2,3% (Винокуров, 1965). При этом масса суточного рациона рыжей цапли в летне-
осенний период достигала 160-220 г (Винокуров, Дубровский, 1957). В Азербайджане рыба
(преимущественно промысловых пород) составляла 27,4%, земноводные — 46,7%, члени-
стоногие — 23-30% (Туаев, Васильев, 1971). В желудках птиц, добытых в Крыму весной,
находили жуков, медведок, озёрных лягушек, крымских ящериц (Щербак, 1966). Здесь же
в степной местности — общественных полёвок, ящериц. В отрыжках птенцов (Крым) при-
сутствовали караси, краснопёрки, медведки, прыткие ящерицы (Костин, 1983). С. Крэмп
(Cramp, 1977) называет в числе объектов питания также ужей, ракообразных, изредка —
птиц, моллюсков и пауков. В результате вскрытия 113 желудков рыжих цапель, отстрелян-
ных в Венгрии, установлено, что первостепенное значение в рационе там составляла рыба
(найдена в 52% желудков) и насекомые: наземные жуки (46%), водные жуки и их личин-
ки (38,9%), личинки стрекоз (30,9%), водяные клопы (28,3%), а также мелкие млекопита-
ющие (обыкновенные полёвки и, редко, землеройки) (24,7%) и земноводные (22,1%)
(Vasvari, 1938). В числе поедаемых цаплей рыб называют наиболее распространенных в
той или иной местности мелких (длиной от 5 до максимум 22 см) карповых (плотва, язь,
краснопёрка, лещ, сазан, карась), щуку, окуня, бычков. Встречаемость отдельных видов
рыб (число проб пищи, в которых найден данный вид в процентах от суммы) в рационе
цапель Приазовья составила: сазан — 36,3%, щука — 13,1%, окунь — 7,1-11,1% (Ви-
нокуров, 1965).
Изменения, происходящие в видовом составе ихтиофауны водоёмов, не могут не сказы-
ваться на рационе цапель. Так, в Таганрогском зал. Азовского моря в пище всех птиц-ихти-
офагов (включая и рыжую цаплю) в последние годы заметное место занимает серебряный
карась (Carassius auratus gibelio), резко увеличивший свою численность. Среди объектов
питания стал встречаться также недавно акклиматизированный на Азове пелингас (Mugil
soiuy) (Лебедева и др., 2006).
Враги, неблагоприятные факторы. Из абиотических факторов наибольшее воздей-
ствия оказывают штормовые ветра и затопление, приводящие к гибели гнёзд с кладками и
птенцами младших возрастов. В качестве основных врагов цапель называют млекопитаю-
щих среднего размера — таких, как шакал, лисица — способных разорить главным обра-
зом одиночные гнёзда. На Дальнем Востоке ущерб цаплям наносит акклиматизированный
там енот (JProcyon lotor).
В гнёздах рыжих цапель в 1930-1940-х гг. обнаружены мухи — кровососки (сем. Hippo-
boscidae) Ornithoponus (Lynchia) ardeae и клоп, ближе не определённый (Hicks, 1959).
Рыжая цапля
367
Хозяйственное значение, охрана. В силу рыбоядности рыжей цапли, оценка хозяй-
ственного значения этого вида, как и других цапель, в свое время была предметом острых
дискуссий. В середине XX в. многие исследователи обращали внимание на негативное зна-
чение рыжей цапли для рыбного хозяйства, особенно в условиях нерестово-выростных
прудов, куда слетаются цапли, привлеченные обилием и доступностью корма (Олейников,
1953; Пахульский, 1957; Олейников, Данилов, 1958; Винокуров, 19596; Назаренко, Попо-
ва, 1959; Смогоржевський, 1959). Правда, многие исследователи обращали и обращают
внимание на то, что цапли выбирают главным образом неполноценных и больных рыб,
осуществляя, таким образом, санитарные функции. Предположения о «вреде» рыбоядных
птиц часто тенденциозны или основаны на «сомнительных данных или неправильных рас-
четах» (Гладков, 1964).
При всей противоречивости мнений о действительном влиянии цапель на продуктив-
ность рыбоводных хозяйств, надо полагать, что такое влияние заметно в периоды массово-
го выпуска мальков в водоёмы. Наблюдения специалистов показали, что поедание рыбы
особенно существенно на водоёмах, обильно заросших тростником. В качестве способа
борьбы с этим явлением А.А. Винокуров (1965) предлагал ликвидацию на водоёмах рыб-
хозов зарослей надводной растительности, которая привлекает сюда голенастых птиц. По
его данным, когда осенью 1953 г. в районе колонии цапель в Чумаковском лимане были
прокошены тропинки и «ерики», оставшийся по краям прокосов тростник за зиму упал,
намок и утонул. В результате массив зарослей оказался как бы разорванным, и цапли были
вынуждены покинуть это место.
Рыжая цапля имеет определённое значение как переносчик и хозяин паразитических
червей рыб (Смогоржевская, 1956).
Во время охоты на водоплавающих рыжая цапля нередко попутно отстреливается охот-
никами-любителями. Именно благодаря охотникам, добывшим окольцованных цапель, уда-
лось получить сведения о послегнездовых перемещениях, миграциях и зимовках этого вида.
В 1950-1970-х гг. по инициативе общественной комиссии содействия учётам животных
(Львов, Ильичев, 1979) проведены учёты добычи дичи во всех республиках тогдашнего
СССР (Приклонский, 1968; Долбик, 1984; Полякова, 1984, 1984а; Кашенцева, 1984; и др.).
Суммируя эти данные, можно предполагать, что в 1960-1970-х гг. на территории бывш.
СССР добывали в год около 900 тыс. особей так называемой «прочей вводно-болотной
дичи», куда относили в том числе и цапель. Конечно, в это количество входили не только
рыжие цапли, но также каравайка, колпица, другие цапли и многие пастушковые. Однако
можно полагать, что несколько десятков тысяч рыжих цапель в те годы служили непремен-
ной добычей российских и среднеазиатских охотников. Поскольку к настоящему времени
результативность охоты значительно снизилась, объём добычи цапель, безусловно, замет-
но сократился. Но, несмотря на это, рыжую цаплю, очевидно, по-прежнему добывают в
южных районах исследуемой нами области (Priklonski, Sapetina, 1990).
В настоящее время, в связи с невысокой численностью, рыжая цапля заметного хозяй-
ственного значения не имеет и подлежит охране как характерный представитель околовод-
ных экосистем. В местах, где присутствие цапель нежелательно (например, на прудах рыб-
хозов), необходимо проводить мероприятия по отпугиванию, а не отстрелу этих птиц.
Рыжая цапля занесена в Красную книгу Республики Беларусь, а в Российской Федера-
ции — в Красные книги Хабаровского края, Курской, Липецкой, Сахалинской, Тамбовс-
кой, Читинской областей.
Подотряд АИСТОВЫЕ — Ciconiae
Семейство Аистовые — Ciconiidae Sundevall, 1836
Наиболее крупные представители отряда Листообразных, весят от 1 кг (африканский
аист-разиня Anastomus lamelligerus) до 9 кг (африканский марабу Leptoptilos crumeniferus).
Шея несколько короче, чем у цапель, не столь подвижна, в полёте обычно вытянута. Шей-
ных позвонков 17. Клюв массивнее, чем у цапель, сжатый с боков, прямой, конический,
реже с загнутым вниз кончиком. У ряда видов при почти прямом коньке надклювья про-
филь подклювья заметно изогнут (скошен) кверху. Ноздри сквозные. Затылочных фонтане-
лей нет, задний край грудины с одной парой вырезок, конец вилочки иногда срастается с
вершиной киля. Язык и голосовая мускулатура сильно редуцированы, иногда отсутствует и
голосовая мембрана. Многие виды способны лишь шипеть и щёлкать клювом, только неко-
торые издают сложные громкие звуки, при этом резонатором служат петли длинной трахеи
и разделённый на камеры воздушный мешок, расположенный под кожей шеи. Глаза менее
крупные, чем у цапель, с тёмной или светлой радужиной. В желудке обособлен пилоричес-
кий отдел, парные слепые кишки сильно редуцированы. Сонные артерии парные, есть ру-
димент пениса. Ноги очень длинные с относительно короткими пальцами с притуплённы-
ми когтями. Гребёнка на среднем пальце отсутствует (как и пудретки), задний палец распо-
ложен выше остальных и совсем короткий. Между основаниями передних пальцев есть
небольшие перепонки. На первом пальце кисти имеется маленький коготок (Карташев, 1974).
Оперение плотное, прилегающее, копчиковая железа оперена. Контурные перья обыч-
но без побочных стволов, пух распространён по всему телу, украшающих перьев нет. Хвост
короткий, обычно из 6 пар рулевых, крылья большие, широкие, первостепенных маховых
11-12, они имеют вырезки и суженные вершины, при полёте расставлены в виде «паль-
цев». Прекрасно летают, парят (в отличие от цапель), в полете держат шею вытянутой (только
марабу складывают её, как цапли). Окраска оперения обычно контрастная — сочетание
белого и чёрного с металлическим отливом. Иногда в брачный период развиваются розова-
тые оттенки. Ноги, клюв, оголённые участки кожи могут быть ярко окрашены: от красного
до жёлтого, буроватого, голубовато-серого цветов. У многих видов оголены лишь уздечка,
кольцо вокруг глаза, у других — «лицо» или вся голова, иногда и шея. Самцы несколько
крупнее самок, чуть ярче окрашены, более длинноклювые.
У большинства видов пары сохраняются постоянно. Гнездятся чаще одиночными пара-
ми, реже небольшими рассеяными группами, лишь немногие — многочисленными плот-
ными колониями. В смешанных поселениях гнездятся выше других видов голенастых и
веслоногих. При одиночном гнездовании гнёзда часто располагаются в десятках километ-
ров одно от другого. Гнездовые постройки — массивные платформы, диаметром от 1 до
1,5 м, располагаются в центральных развилках, реже на боковых ветвях крупных деревьев,
на сооружениях человека (крыши домов, водонапорные башни, столбы электропередач и
пр.), скалах, реже на земле или зарослях околоводной растительности. Строительный мате-
риал — ветви деревьев, сухая трава. Многие гнёзда используются десятки лет, не одним
поколением, и порой достигают внушительных размеров. В кладке 3-8 белых яиц, инкуба-
ция 4-6 недель, редко больше. Насиживают и кормят птенцов оба партнёра, у ряда видов
более интенсивно насиживает самец. Вылупившиеся птенцы зрячие, покрыты коротким,
довольно редким сероватым пухом. У многих видов первый пуховой наряд сменяется вто-
рым — более длинным, густым и светлым. В гнезде птенцы остаются до приобретения
способности к полёту — около 70 дней (у крупных аистов — до 100 дней и дольше). После
Чёрный аист
369
вылета из гнёзда выводок держится вместе, часто семьи объединяются в крупные монови-
довые стаи. Половозрелы с 3-4 лет.
В отличие от многих цапель, аисты активны обычно днём. По пищевым предпочтениям
и способам кормодобывания можно выделить несколько экологических групп. Большин-
ство некрупных аистов (Ciconia) — собиратели мелкого разнообразного корма (рыба, ам-
фибии, рептилии, грызуны, насекомые), как на мелководьях, так и на лугах, в саваннах, в
других травянистых стациях. Крупные аисты (Ephippiorhynchus, Jabiru, Leptoptilos) повы-
сили мощь гарпунного удара за счёт увеличения массивности клюва, а чтобы конёк над-
клювья оставался прямым (что важно для прицеливания по добыче), усилился скос под-
клювья кверху. Эти преобразования позволяют справляться с объектами крупного размера,
вплоть до молодых крокодилов, водоплавающих птиц размером с утку, водяных млекопи-
тающих крупнее крысы. Некоторые из крупных аистов время от времени питаются пада-
лью, а два вида марабу стали специализированными падальщиками и мусорщиками. Аис-
ты-клювачи (Mycteria) имеют относительно самые длинные клюв и ноги и обычно нашари-
вают добычу в мутной воде или на илистом дне, погрузив в воду весь клюв, а иногда и
голову. Такой способ кормодобывания, при котором снижена роль зрения и возрастает роль
осязания, больше характерен для ибисов. Наконец, аисты-разини (Anastomus) имеют край-
не специализированный челюстной аппарат, позволяющий им раскрывать раковины дву-
створчатых моллюсков, которыми они питаются (Коблик, 2001).
Аистовые известны начиная с верхнего эоцена. Самые древние ископаемые находки
сделаны во Франции, наибольшего разнообразия семейство достигало в олигоцене. В на-
стоящее время описано более 30 ископаемых видов и 9 родов. Три современных рода по-
явились уже в эоцене, 7 современных видов известно в ископаемом состоянии только из
плейстоцена. Некоторые вымершие аисты были гораздо крупнее современных.
В семействе обычно насчитывают 19 современных видов, группируемых в 6 родов (del
Hoyo et al., 1992; Dickinson, 2003), иногда количество родов доводят до 11. Порой выделя-
ют 3 трибы или подсемейства, но, скорее всего, это не систематические единицы, а эколо-
гические группировки. Аисты распространены преимущественно в тропиках, где они оседлы
или номадны; виды умеренных широт перелётны. Пять азиатских видов находятся под гло-
бальной угрозой исчезновения и занесены в Красный список МСОП. В России и сопре-
дельных регионах гнездится 3 вида аистов рода Ciconia.
Род Ciconia Brisson, 1760
Чёрный аист
Ciconia nigra (Linnaeus, 1758)
Ardea nigra Linnaeus, 1758. Syst. Nat., ed. 10,c. 142, Швеция
Black Stork (англ.); Schwarzstorch (нем.); Cigogne noigre (фр ).
Син.: Ardea atra J.F. Gmelin, 1789; Ardea crysopelargus A.A.H. Lichtenstein, 1793; Ciconia
fusca C.L. Brehm, 1831.
Статус. Гнездящийся, мигрирующий вид.
Общая характеристика и полевые признаки. Крупная птица; встречается в гнездо-
вой период в лесных стациях, как правило, в самых глухих, редко посещаемых человеком
участках леса, в прибрежной полосе лесных стариц и других водоёмов, на открытых вод-
370
Отряд Листообразные, Семейство Аистовые
ных участках лесных болот; в горных местностях отмечается до 3000 м над у.м., в период
миграции — над открытыми пространствами.
От белого аиста отличается большей стройностью телосложения, несколько меньшими
размерами, преобладанием чёрных тонов в оперении, заметно более скошенным к верши-
не профилем клюва. Встречи в одних и тех же скоплениях вместе с белым аистом в период
зимовки регистрируют редко (Спангенберг, 1951). В полёте шея вытянута и чуть прогнута
дугой вниз; птица легко лавирует в переплетении ветвей. В конце гнездового сезона харак-
терно парение парами, а перед отлетом — семьями (по 4-6 особей), когда птицы, почти не
взмахивая крыльями, набирают высоту, используя восходящие токи воздуха (ориг.).
Голос. Предположение о «немоте» чёрного аиста из-за отсутствия у него сиринкса (Voigt,
1950) безосновательно, однако чёрный аист заметно молчаливее белого. Изредка от него
можно услышать протяжные вскрики, сходные с криком сарыча (Buteo buteo\ чаще всего —
при испуге (неожиданное появление опасности при кормежке) или во время тревоги у гнёз-
да, главным образом во время нахождения в нем птенцов (но не кладки). Птенцы в гнезде
издают глухое «утробное» бормотание: «ххьии-ххью». Иным наблюдателям это напомина-
ет пищание (Спангенберг, 1951) или повторяющиеся звуки, похожие на «хьюйии-йи» (Bauer,
Glutz von Blotzheim, 1966). Обороняясь, птенцы щёлкают клювами, пытаются ударить ими
«нападающего» (Приклонский, Галушин, 1959).
Подвидовая систематика. Монотипический вид.
Описание. Окраска. Птицы во взрослом наряде. Полового диморфизма в окраске
нет. Оперение головы, шеи, зоба, спины, надхвостья, а также маховые перья, все верхние
кроющие крыла и рулевые — чёрные с металлическим отливом (цвета воронова крыла),
зелено-оливковым в сочетании с фиолетово-малиновым. При поворачивании тушки птицы
из коллекции в определённых направлениях к источнику освещения, создается впечатле-
ние, что верх оперения покрыт тёмно-синим или тёмно-фиолетовым налётом. При рас-
сматривании отдельных перьев это впечатление создается только когда перо находится под
определённым углом к свету. Подобная окраска сохраняется на коллекционных материалах
не менее 100 лет (колл. П.П. Сушкина из собрания ЗИН РАН). Основания контурных перь-
ев чаще бурые, иногда тёмно-серые. Нижняя часть тела — все брюхо, подмышечные перья
и подхвостье — белые. Клюв и ноги, лишенные перьев до верхней трети голени — мясо-
красного цвета. Голая кожа около клюва и глаз также красная, в редких случаях — бурова-
то-зелёная. По Е.П. Спангенбергу (1951), уздечка и кожа вокруг глаз — ярко-красные. Ра-
дужина бурая или красно-коричневая.
Первый и второй пуховые наряды белые. По наблюдениям в Окском заповеднике, пух
только что вылупившегося из яйца аистенка со слегка желтоватым налетом, исчезающим к
концу первой недели жизни. Ноги у пуховых птенцов сероватые, радужина и кожа уздечки
темные, клюв жёлтый.
Гнездовой наряд. К моменту вылета из гнёзда (в 65-70-дневном возрасте) все в будущем
чёрные отделы оперения тёмно-бурого цвета без металлического блеска. Концы контур-
ных перьев окаймлены отчётливой более светлой, иногда — коричневой узкой полоской.
На концах перьев — белые пушинки. Начиная с 3-недельного возраста, оперение брюха —
грязновато белое, отчасти из-за того, что гнездо к концу периода пребывания в нем птенцов,
становится неопрятным. Клюв и ноги птенцов, вылетающих из гнёзда, тёмные, зеленовато-
бурые; неоперённые участки кожи вокруг глаз и уздечка — более светлые, желтовато-зелё-
ные или желтовато-бурые. По М.И. Лебедевой (1959), клюв у основания — оранжеватый.
Различия в окраске оперения птиц в возрасте от двух месяцев до одного года и более
старших групп — не установлены. Считают, что линька годовалых особей начинается со
второго календарного года (Bauer, Glutz von Blotzheim, 1966).
Чёрный аист
371
Строение и размеры. Длина крыла самцов и самок (п=8) 539 мм (520-580 мм)
(Спангенберг, 1951), хвоста— 190-240; клюва— 160-190; цевки — 180-200 мм (Witherby
et al., 1939, по: Cramp, 1977). Размеры аистов из Белоруссии несколько выше. По А.В. Фе-
дюшину и М.С. Долбику (1967), крыло самцов достигает длины 580 мм, хвост — 255 мм,
цевка — 225 мм, клюв — 193 мм; общая длина тела у самцов равна 790-1 090, в среднем
973 мм. Средний вес самцов (п=8) составляет 2 987 г (от 2 800 до 3 200 г), самок (число
исследованных птиц не указано) — в среднем 2 900 г (Федюшин, Долбик, 1967).
Линька. Происходит один раз в году. Начинается весной в феврале и кончается в мае-
июне. Птица, добытая 8 июля, почти закончила линьку. Старые перья сохранились только
на лбу, темени и в незначительном количестве среди маховых и предплечевых. Белые перья
брюха значительно обносились и приняли грязный оттенок (Сомов, 1897). Вслед за Е.П.
Спангенбергом (1951) и С. Крэмпом (Cramp, 1977) приходится констатировать, что линька
Чёрного аиста изучена слабо.
Распространение. Гнездовой ареал. В конце XIX - начале XX вв. считалось, что
в Евразии вид гнездится от южной части Пиренейского п-ова, юга Скандинавии (южная
Швеция), Прибалтики и Балкан по всей лесной зоне до юж. Якутии, юга Магаданской обл.,
Хабаровского и Приморского краев, включая Сахалин и Курилы. Последующими исследо-
ваниями границы были уточнены. Так, К.А. Воробьев (1963) считал аиста в Якутии в ос-
новном залётной птицей, «так как местному населению он не известен и нам во время
поездки по Вилюю не попадался» Однако на основании приводимых тем же автором дан-
ных о встречах одиночных аистов (в конце мая 1927 г.) близ Вилюйска и ссылки на наблю-
дения А. А. Меженного, видевшего летом 1964 г. двух (возможно, пару) чёрных аистов близ
устья р. Токко, а также принимая во внимание скрытность этого вида даже в тех районах,
где он относительно многочислен, северную границу распространения вида можно прове-
сти по югу Якутии. На Курильских о-вах и Сахалине чёрный аист — редкая залётная птица,
и сведения о его гнездовании здесь, приведённые Е.П. Спангенбергом (1951), ошибочны
(Гизенко, 1955). Южная граница гнездового ареала проходит на востоке вдоль государ-
ственной границы России, захватывая лесные и горные участки от Алтая (Кучин, 1988) до
Сев. Кореи и Сев. Китая (Маньчжурии). По А.И. Иванову (1969), аист регулярно гнездится
в пределах Памиро-Алая, и, следовательно, в этом районе он распространён и южнее пре-
делов бывшего СССР. Далее на запад южная граница обитания проходит по центральным
районам Афганистана, сев. Ирану и центр. Турции, затем огибает с юга практически все
страны Балканского п-ова. В этих пределах в Сев. Палеарктике в настоящее время извест-
но, по крайней мере, два региона, где чёрный аист в настоящее время безусловно не гнез-
дится. Это Зап. Европа — от Австрии и Германии до юга Пиренеев, включая Британские о-
ва — и степные, лесостепные и пустынные области России и сопредельных стран на вос-
ток до оз. Маркаколь. Точная граница северных пределов зоны, где аист не гнездится, не
установлена. По всей вероятности, она совпадает с границей лесной зоны, лишь в некото-
рых местах чуть смещаясь к северу. Южная же граница области негнездования проходит
чуть севернее горных массивов Тянь-Шаня и Памира, по югу пустынь Средней Азии и
южному побережью Каспийского моря. В Турции, Болгарии, Греции, Румынии, Польше и
Германии места гнездования чёрного аиста четко приурочены к сохранившимся лесным
массивам, в связи с чем его распространение здесь носит «пятнистый» характер (рис. 75).
Небольшой участок оседлого обитания чёрного аиста существует на западе Испании по
границе с Португалией (Cramp, 1977).
Необходимо отметить, что из-за скрытного образа жизни аиста и его незначительной
численности, сведения о нем на пространстве ареала весьма скудны. По дошедшим до нас
свидетельствам специалистов, работавших в конце XIX и начале XX в., на которых в ос-
372
Отряд Листообразные, Семейство Аистовые
Рис. 75. Область распространения чёрного аиста в конце XX в.: а — область гнездования, б — рай-
оны зимовок.
новном ссылается Е.П. Спангенберг (1951), распространение вида было повсеместно спо-
радичным. Это заставляло орнитологов чаще всего высказывать лишь предположения о
гнездовании аиста при редких обнаружениях самих гнёзд. Так, М.А. Мензбир (1895) счи-
тал аиста возможно гнездящимся от Петербургской губернии до Тверской (Вышний Воло-
чек) и далее на восток — в Вологодской губернии и, вероятно, до Урала, где определённо
находили его «отдельные» гнёзда (Карамзин, 1901). Южнее, в Центр. России, по М.А. Менз-
биру, чёрный аист безусловно гнездился, но был малочислен и спорадично распространен.
Этот же автор вскользь упоминает о гнездовании чёрного аиста в горных районах Крыма.
Документальные свидетельства об этом мы находим у И.И. Пузанова (1931), который сооб-
щает о находке гнезда чёрного аиста в 1917 г. на сухом дубе в пределах нынешнего Крым-
ского заповедника в урочище Япалах. Весьма существенно замечание М.А. Мензбира о не
редком гнездовании чёрного аиста в Новгородской губернии. В отношении южных преде-
лов гнездового распространения вида в центре Европейской России М.А. Мензбир пола-
гал, что аист определённо селится вплоть до запада Харьковской губернии и на восток до
Юж. Урала. Сейчас эти пределы можно считать южной границей ареала вида в Вост. Евро-
пе (исключая Закавказье). М.А. Мензбир (1895) выделял также особенно густо населенную
черным аистом территорию в пределах Ленкорани и Закавказья, где, по его выражению,
эта птица не редка (стр. 778). О распространении аиста на просторах Сибири для конца
XIX в. конкретные сведения отсутствуют.
Современные знания о распространении чёрного аиста на территории России и сопре-
дельных государств более определённые (рис. 76). Достоверно известно, что вид гнездится
во всех подходящих местообитаниях Калининградской обл. РФ, в странах Балтии. Восточ-
Чёрный аист
373
Рис. 76. Ареал чёрного аиста в Вост. Европе и Сев. Азии: а — территория гнездования; б — места
встреч молодых птиц и возможного гнездования.
нее — в Ленинградской обл. — его распространение носит уже более спорадичный харак-
тер (Мальчевский, Пукинский, 1983). Что же касается Белоруссии и центра Европейской
части России, то далеко не во всех областях этого региона гнездование чёрного аиста заре-
гистрировано документально. Исключением является территория Урала, где в каждом из 4
субъектов федерации — Башкирии, Пермской, Свердловской и Челябинской обл. — к кон-
цу XX столетия было известно по 4-6 гнёзд (Давыгора, 1984; Карякин, 2000). Исследовате-
ли, работавшие в центре Европейской России, чаще всего сообщают об одной-двух наход-
ках (добыче) птиц в исследуемом районе или ссылаются на немногочисленные шкурки,
хранящиеся в местных краеведческих музеях или при зоологических кафедрах вузов. А
затем на этом основании и с учётом подходящих местообитаний предполагают гнездова-
ние вида по аналогии с «соседними территориями» (Пузанов и др., 2005 — для Нижегород-
ской обл.; Б.В. Зимин — для Карелии; Шапошников и др., 1959 — для Самарской обл.;
Кузнецов, Макковеева, 1959 — для Ярославской обл.; Авданин, 1984 — для Тверской обл.
в районе Центрально-Лесного заповедника и др.). В Московской обл. вид на гнездовье не
отмечался с 1920-х гг. и занесён в областную Красную книгу как вид, вероятно исчезнув-
ший на гнездовании (Калякин, 2008).
Показательна динамика сведений о чёрном аисте в Рязанской области, с конца XIX в.
находившейся под пристальным вниманием орнитологов. Несмотря на предположение М. А.
Мензбира (1895) о гнездовании этого вида в Рязанской губернии, орнитологи, работавшие
там в позапрошлом столетии и в начале XX в. (Павлов, 1879; Хомяков, 1900; Туров, 1918,
1925; Бекштрем, 1927), ничего об этом виде не пишут, что явно свидетельствует об отсут-
ствии встреч ими чёрного аиста на Рязанщине. Позднее Е.С. Птушенко (рукопись), сослав-
шись на свидетельство С.С. Турова (1925) о находке гнезда аиста в 1914 г. близ с. Кельцы
Рязанского уезда, сообщает, что в районе Окского заповедника чёрный аист «не представ-
ляет редкости» по крайней мере с 1935 г. Основываясь, по-видимому, на своих записях в
дневниках, он далее пишет, что в летнее время здесь регистрируют 18—40 встреч этого вида
374
Отряд Листообразные, Семейство Аистовые
в год. О находках гнёзд аиста в этом районе Е.С. Птушенко известно с 1938 г. (Птушенко,
рукопись, 1958; Птушенко, Иноземцев, 1968), а С.Г. Приклонскому (1958) — с 1953 г. Е.С.
Птушенко сообщает также о единственной пока находке гнеда аиста на правобережье р.
Оки напротив заповедника у с. Нармушадь в 1938 г. Позднее гнёзда чёрных аистов находи-
ли и по р. Пре выше заповедника, в окрестностях д. Деулино. Здесь их количество в 1983-
1987 гг. оценивали в 2-5 (Пегова и др., 1990, И.П. Назаров, устн. сообщ. и др.). И если
прежде Окский заповедник считался единственным местом гнездования чёрного аиста в
Рязанском крае, то нынче приходится признавать, что этот вид был распространён и обитал
здесь, по крайней мере в середине XX в., намного шире. На основании сказанного можно
сделать обоснованное предположение, что аист, несомненно, гнездился в Рязанской обл. в
XIX столетии, но не был обнаружен из-за особенностей своего скрытного поведения в гнез-
довой сезон. Гнездование чёрного аиста, таким образом, можно выявить только при специ-
альных исследованиях — в первую очередь там, где численность этих птиц хотя бы немно-
го выше обычной, или там, где ведутся более тщательные наблюдения.
Очаг гнездования есть на юго-востоке Африки, от Малави, Зимбабве и Намибии до ЮАР.
Численность оседлой африканской популяции оценивается в более чем 200 пар, в Драконо-
вых горах в пределах Трансвааля и Лесото вид местами нередок, имеет стабильную числен-
ность, гнездится с сентября (del Hoyo et al., 1992). Вероятно, эта популяция ведёт начало от
мигрантов из Евразии, по каким-либо причинам начавших размножаться на зимовках.
Зимовки. Зимуют европейские чёрные аисты главным образом в Вост, и Юж. Афри-
ке; небольшие скопления на зимовках отмечены также в отдельных точках Центр, и Зап.
Африки. Азиатские аисты зимуют на юге Пакистана, севере Индии, в центр, и юж. Китае,
а также на самом юге Японии.
Миграции. Осенняя миграция проходит незаметно. Она начинается уже в августе,
почти сразу после того, как птенцы покинут гнездо. Так, молодой чёрный аист, который
ещё 4 августа находился в своем гнезде в Окском заповеднике, 15 числа того же месяца
добыт в Пензенской обл., а другой птенец из того же гнезда — 25 августа в Балашовском
р-не Саратовской обл. Все авторы, описывающие перелет аистов, отмечают, что птицы
летят небольшими, возможно семейными группами. Судя по возвратам колец от окольцо-
ванных птенцов, осенний перелёт европейских чёрных аистов происходит в южном на-
правлении. По данным мечения, птенцов 2-3-месячного возраста из Прибалтийских стран
добывали в августе и в сентябре в Белоруссии, Польше, на Зап. Украине, в Молдавии,
Болгарии, Румынии, Греции, Турции. А птиц той же возрастной группы, окольцованных
в Окском заповеднике — в Пензенской, Саратовской, Тамбовской и Курской обл., а также
в Грузии. Дальнейший путь «прибалтийских» аистов тянется по восточному побережью
Средиземного моря и вдоль долины р. Нил. Аисты из центра Европейской России огиба-
ют Чёрное море с востока, а затем также оказываются на нильском пролётном пути (Ле-
бедева, 1979в). Самый дальний возврат окольцованной птицы зарегистрирован из Юж.
Африки (птенец был помечен в Окском заповеднике). Во время миграций чёрные аисты
редко присоединяются к стаям более многочисленных белых аистов. Материалов по ме-
чению сибирских и дальневосточных чёрных аистов не имеется. Судя по наблюдениям
орнитологов, аисты в горных районах Средней Азии движутся вместе с другими мигран-
тами, придерживаясь горных разрезов и долин. Не исключено, что часть птиц совершает
перелеты в более крупных стаях на большой высоте (Дольник, 1981). Однако в связи с
отсутствием конкретных данных, это можно считать пока только предположением, более
или менее логичным по аналогии с другими видами птиц. Наконец, аисты, гнездящиеся в
Вост. Сибири и на Дальнем Востоке, следуют к местам зимовок через горные районы
Маньчжурии и Сев. Корею.
Чёрный аист
375
В 2002-2004 гг. чешским орнитологам с помощью радиослежения за птицами, снаб-
женными специальными передатчиками, удалось установить путь осенней миграции чер-
ных аистов, гнездящихся в районе Новосибирска. Оказалось, что миграционный маршрут
проходит через нижнее течение Сыр-Дарьи и прилегающие Кызыл-Кумы, пересекает Центр.
Узбекистан и далее направляется к Центр, и Зап. Индии (Ковшарь, 2007).
О весеннем перелёте к местам гнездовий сведения и вовсе отсутствуют. Известны лишь
даты появления на местах гнездовий, в своем большинстве указанные ещё Е.П. Спанген-
бергом (1951). Он пишет, что в азиатской части страны пролёт сильно растянут и местами
происходит очень поздно. Под Ашхабадом в 1923 г. птиц впервые видели 21 апреля, а в
1924 г. — 7 апреля. В более северных местах их впервые отмечали как в начале апреля, так
и 10 июня. У Зайсана аистов регистрировали 26 марта. На западе Алтая птицы появляются
в конце апреля, но в центре Алтая и в Монголии «значительно позднее». В горах на высоте
2000 м н.у.м. у оз. Кара-Гель стайка впервые была отмечена 5 июня (Сушкин, 1938, цит. по
Спангенбергу, 1951) (возможно, это были неполовозрелые птицы). Для Сибири приводятся
даты первых встреч весной от середины апреля до первых чисел мая, для Приморья — во
второй половине марта.
Если судить по срокам вылупления птенцов, принимая продолжительность насижива-
ния равной примерно месяцу, то в Ленкорань аисты прилетают в конце марта - первых
числах апреля, а по всему Закавказью распределяются не позднее 10 апреля.
Даты первых встреч чёрных аистов в Европейской части исследуемого региона таковы.
В Белоруссии аисты появляются уже в первых числах апреля, хотя конкретные данные сви-
детельствуют о середине этого месяца. Однако А.В. Федюшин и М.С. Долбик (1967), осно-
вываясь на материалах из Пинского и Лепельского р-нов, приходят к выводу, что птицы
появляются здесь между 27 марта и 15 апреля, в среднем 4-7 апреля. В Окском заповедни-
ке дат появления аистов в непосредственной близости от гнёзд за 75-летний период суще-
ствования заповедника не зафиксировано. Имеется лишь единственное свидетельство, что
следы чёрного аиста обнаружены на мокром снегу частично оттаявшего мохового болота
30 марта 1986 г. Все остальные наблюдения относятся уже к птицам, заканчивающим про-
цесс насиживания (конец мая) или к гнёздам, в которых уже появились птенцы (первая
половина июня).
Численность. Выше мы отмечали, что ещё М.А. Мензбир в позапрошлом веке предпо-
ложил относительно высокую численность аиста на гнездовании в Новгородской губер-
нии. В настоящее время, благодаря усилиям орнитологов северо-западной части террито-
рии бывш. СССР, установлено, что в Калининградской обл., Литве, Латвии и Эстонии чёр-
ный аист вполне обычен. По периферии этого района с юга и востока (Новгородская, Псков-
ская, Ленинградская обл. и Белоруссия) лежит полоса более редкого, спорадичного, но так-
же обычного гнездования этой птицы. Дальше к востоку и югу (разумеется, только в преде-
лах лесной зоны) он все более редок. В Трубчевском и Суземском р-нах Брянской обл. в
начале 1980-х гг., по данным И.П. Шпиленка, на небольшом участке (около 1 200 га) орга-
низованного по его инициативе местного заказника, впоследствии вошедшего в состав
заповедника «Брянский лес», ежегодно обитало до 6 пар чёрного аиста.
Другим очагом более высокой численности чёрного аиста в Европе является, вероятно,
Закавказье. К сожалению, орнитологические исследования в этом районе проводятся в пос-
ледние годы нерегулярно. Поэтому свои заключения о высокой численности гнездования
аиста в Закавказье от Ленкорани до, по крайней мере, восточной части Краснодарского
края мы можем строить только на основании данных исследователей позапрошлого и пер-
вой половины прошлого века и лишь отдельных сообщений современников (Пекло, Оча-
повский, 1984; Тильба, 1984). Ещё М.А. Мензбир (1895) считал, что чёрный аист «особен-
316
Отряд Листообразные, Семейство Аистовые
но обилен в лесах под Ленкоранью». В том же районе Г.И. Радде (1884) удалось увидеть
три гнёзда этой птицы на одном дереве (случай единственный и уникальный для этого
вида; правда, вышеназванный автор не сообщает, все ли они были жилыми). По Е.П. Спан-
генбергу (1951), между устьем р. Куры и Иранской границей «в соответствующих биотопах
в 1934 г. гнездо от гнезда редко располагалось на расстоянии более 1 км» (стр. 380). В 2000
г. в урочище Красный лес Краснодарского края на осеннем пролёте было зарегистрировано
350 чёрных аистов (Букреев, 2001).
В бассейне р. Бикин в 1970-х годах общая численность определялась в 80-100 пар (Пу-
кинский, 2003), а по другим оценкам в 1990-х гг. здесь размножалось не более 10 пар (Ми-
хайлов и др., 1998). Численность пролётных птиц на Ханкайско-Раздольненской равнине с
начала текущего столетия значительно сократилась (сообщение Ю.Н. Глущенко).
На остальной территории гнездового ареала, составляющей не менее 90% ареала, чёр-
ный аист редок. Хотя полностью исключить возможности находки других очагов его более
высокой численности при проведении более тщательных исследований нельзя.
Как уже упоминалось, для чёрного аиста характерна скрытность в сезон размножения,
приводящая к тому, что его видят очень редко. В 1959 и 1970 гг. Центральная орнитологи-
ческая станция проводила анкетные обследования распространения чёрного аиста в лесах
15 субъектов федерации европейской части России. В 1959 г. получено 802 ответов из лес-
ничеств, причём только 6 корреспондентов сообщили о наличии чёрных аистов (в Горьков-
ской, Ульяновской обл., Марийской и Татарской автономных республиках). В 1970 г. о наход-
ке гнёзда аиста из 650 ответов пришло сообщение только для 1 пункта во Владимирской
обл. (Приклонский, Теплов, 1962; Попов, 1977).
Фактические данные для европейской части ареала чёрного аиста по материалам более
позних лет таковы. В Калининградской обл. на начало 1980-х гг. гнездилось не менее 50
пар (Белопольский, 1984; Гришанов, 1988). В Литве в 1958 г. учтено 1 610 птиц, а в 1968 г. —
1 185 (Флинт, 1984). В этом случае, вероятно, имеются в виду как взрослые птицы, так и
птенцы; таким образом общее число пар могло быть, соответственно, 300 и 200. В Латвии
численность чёрного аиста в 1930-х гг. оценивали примерно в 300-400 пар (Transche, 1965),
в 1970 г. — в 150-200 пар (Балтвилкс, 1972), хотя фактическая численность, как предпола-
гает Ю.К. Липсберг (1984), могла быть и выше; в 1977-1980 гг. получены сведения о 107
жилых гнёздах аиста (Липсберг, 1984). Для конца 1990-х гг. Петер Сакл и Марис Страздс
(Sackl, Strazds, 1997) оценивают численность чёрного аиста в Латвии в 1 075 гнездящихся
пар. В Эстонии в 1962 г. насчитывали 150 пар, в 1970 г. — не более 80, а в начале 1980-х гг. —
до 170 пар (Манк, 1963; Юсси, Рандла, 1970; Рандла, Ыун, 1984). В Ленинградской обл.
гнездится не менее 10 пар (Мальчевский, Пукинский, 1983).
В Белоруссии чёрный аист на гнездовании в общем обычен и встречается во всех обла-
стях (Шнитников, 1913; Мензбир, 1928). Зарегистрирован здесь он и во всех заповедниках.
Так, в Беловежской пуще (Попенко, Дацкевич, 1984) с 1949 по 1981 г. найдено 47 гнёзд
Чёрного аиста, хотя из них использовалась птицами только половина (из текста не ясно —
в общей сложности или ежегодно). В 1949 г. здесь одновременно гнездилось 20 пар, к 1969 г.
число жилых гнёзд сократилось до 5. В 1979 г. в Беловежье зарегистрировано 20 пар чёр-
ного аиста (Приклонский, Постельных, 1984). Есть чёрный аист и в других заповедниках
Белоруссии: в Березинском — до 16 пар (Волков, 1977) и в Припятском — 4-5 пар (Ю.А.
Вязович, устн. сообщ.).
Вообще же чёрный аист на особо охраняемых природных территориях более обычен,
чем за их пределами. Анализ статистической отчётности заповедников и специальных оп-
росов, выполненных в 1979 и 1982 гг., показал следующее. В 1979 г. из 140 особо охраняе-
мых природных территорий страны (ООПТ), составлявших 11,2 млн. га (0,51% площади
Чёрный аист
377
тогдашнего СССР), чёрный аист отмечен в 28 из них. Это 86% ООПТ, расположенных в
пределах ареала чёрного аиста. В 1982 г. в СССР был уже 151 заповедник. Чёрный аист
зарегистрирован в 24 из них, причём в 6 из тех, где не был отмечен во время первого учёта.
Число учтённых пар составило не менее 96 (Приклонский, Постельных, 1984).
На основании всех приведенных данных представляется возможным дать ориентиро-
вочную оценку суммарной численности чёрного аиста для описываемой территории в пер-
вое пятилетие XXI в. На северо-западе Европейской части бывшего СССР предполагается
гнездование 700 пар, на Сев. Кавказе и в Закавказье — 300 пар, в Приморье — 250 пар, в
горных районах Средней Азии — 400 пар, на остальной территории ареала — 2 000 пар.
Это дает минимальный среднегодовой показатель в пределах 3 650 пар.
Экстраполируя сведения по заповедникам на всю площадь ареала аиста в пределах опи-
сываемой территории, при разных расчетах получаем от 3 870 до 23 140 (в среднем 6 000)
пар гнездящихся ежегодно птиц. К сходным выводам приходит В.Е. Флинт (1984), оцени-
вающий численность этого вида для территории бывш. СССР в 3-5 тыс. гнездовых пар.
Данные о численности чёрного аиста в некоторых странах Европы для первых трёх
четвертей XX в. таковы. В Польше зарегистрирован рост от 40 пар в 1914 до 500-530 в
1966 г. (Keller, Profus, 1992). В Германии (включая Западную и Восточную её части) в
среднем в первой половине XX в. обитало 20-22 пары. На территории прежней Чехосло-
вакии аист гнездится преимущественно в Словакии, где его численность в период с 1940
по 1960 гг. возросла до 100 пар. В Австрии первое гнездо было обнаружено в 1939 г.; в
1951 г. найдено уже 5 гнёзд, и количество аистов продолжало увеличиваться. В Венгрии
число пар в 1941 г. составляло 50-60, в Греции — менее 20. В Испании зарегистрировано
50-60 пар, всего же в пределах Пиренейского п-ова гнездится в обычные годы до 150
пар. В Югославии отмечают уменьшение числа гнездовий, особенно на севере (Cramp,
1977).
На конец XX столетия (Hagemeijer, Blair, 1997) число гнездящихся пар в ряде европей-
ских стран таково: в Белоруссии — 1 100, Латвии — 1 050, Польше — 1 000, Словакии и
Литве — по 370, Украине — 240, Хорватии и Чехии по 220, Испании — 200 и Венгрии —
175; в прочих странах, за исключением России — 675 пар. Названные авторы предполага-
ют, что в России (на её европейской территории) может обитать от 1 до 10 тыс. пар (в
среднем 3 162 пары) чёрных аистов, а в Турции — в среднем до 1 тыс. пар. Плотность
населения в этом случае в Вост. Европе составляет от 1,3 до 1,8 гнездящейся пары на 100
км2 лесных местообитаний. Особо отмечено, что в некоторых лесах Польши плотность
населения достигает 8,4 пары на 100 км2 (Keller, Profus, 1992).
Названные выше цифры, так же как и мнение ряда исследователей (Белопольский, 1984;
Кривоносов, 1984; Флинт, 1984 и др.), как будто бы подтверждают представление о сокра-
щении численности чёрного аиста, происходившем в конце XX столетия. В то же время,
нельзя не обратить внимания на мнение Б.В. Щербакова (1984), Т. Рандлы и А. Ыуна (1984)
и ряда других орнитологов, отметивших рост численности вида. Увеличивается количе-
ство чёрных аистов и во многих странах Зап. Европы (Hagemeijer, Blair, 1997). Нельзя,
однако, забывать о том, что «рост» количества обнаруженных гнёзд иногда напрямую свя-
зан с усилением интенсивности исследований. Можно предположить, что наблюдающиеся
изменения численности не выходят за пределы естественных флуктуаций, свойственных
данному виду. Таким образом, приведенные цифры и рассуждения настраивают нас в отно-
шении будущего чёрного аиста весьма оптимистично.
Местообитания. В лесной зоне чёрный аист выбирает для гнездования наиболее глу-
хие и удалённые от жилья человека участки леса, «уремы», черноолыпаники, долины не-
больших лесных рек (Мальчевский, Пукинский, 1983), окраины вырубок и гарей в спелых
378
Отряд Листообразные, Семейство Аистовые
и перестойных сосняках (Приклонский, 1958). В районе гнездового участка аиста должны
находиться заболоченные лесные водоёмы — чаще всего небольшие озёра и старицы, уча-
стки главным образом низовых болот с открытым зеркалом воды, где птицы добывают
корм. В горных районах в Средней Азии, Закавказье, Приморье отмечены случаи гнездова-
ния на уступах скал (Панов, 1973; Красная книга Узбекской ССР, 1983). Устройство гнёзд
на скалах характерно для этих птиц также в пределах Средиземноморья, Карпат, горных
районов Центр. Европы. Так, в Испании на скалах располагалось 61% найденных гнёзд
аиста, а в Австрии — 28% (Sackl, Strazds, 1997).
В некоторых районах, например, в Закавказье чёрные аисты обитают рядом с поселка-
ми, иногда селятся прямо в аулах (Спангенберг, 1951). Не всегда сторонится жилья и при-
сутствия человека чёрный аист и в лесной зоне средней полосы России — в местах более
массового гнездования. Отдельные гнёзда вида в 1970-х гг. на территории будущего запо-
ведника «Брянский лес» найдены И.П. Шпилёнком всего в 1,5 км от обитаемых лесных
кордонов (Ф.Р. Штильмарк, письм. сообщ.). Однако такие случаи не правило, а лишь ред-
кое исключение. Нам удалось найти сведения о них лишь в 3 публикациях из примерно 80
проанализированных. При этом заметим, что близкие к жилью гнёзда отыскать много лег-
че, чем удалённые от него.
Осенью, перед началом миграции, аистов можно наблюдать в пойменных лугах. Во вре-
мя миграций, как было сказано выше, они встречаются над обширными безлесными про-
странствами, в степях и пустынях.
Размножение. Считается, что половая зрелость у чёрного аиста наступает в трехлетием
возрасте (Спангенберг, 1951; Cramp, 1977). Однако, в свете последних данных, большая
часть птиц скорее всего достигает зрелости к 4-6 годам жизни. Нельзя исключать, что об-
разование пар происходит раньше, ещё на зимовках или в скоплениях неполовозрелых осо-
бей (о которых, в сущности, ничего не известно). Возможно, после этого пары занимают
гнездовые участки и делают попытки гнездования, строя или надстраивая прошлогодние
незанятые гнёзда, но размножаться начинают только после достижения половой зрелости
обоими партнёрами. В пользу этого свидетельствует находка трёх пустовавших гнёзд аис-
тов в Окском заповеднике в 1955, 1973 и 1978 гг. Все они имели вполне пригодное для
гнездования состояние. Аисты в мае и июне держались около этих гнёзд; отмечена попыт-
ка «насиживания» одним из них еловой шишки. Однако никаких признаков откладки яиц
не наблюдали. Не встречали аистов около таких гнёзд и со второй половины июня. Следует
заметить, что пары чёрных аистов образуются на продолжительный срок, чему способ-
ствует относительно невысокая численность птиц. В пользу этого говорит следующее на-
блюдение. На одном из гнёзд в Окском заповеднике, где успешно прошло гнездование в
1967 г., на следующий год наблюдали только одну птицу. Вероятно эта же птица держалась
в районе гнездового участка и на третий год, и только на четвёртый не была здесь встрече-
на. К весне пятого года гнездо разрушилось. По-видимому, партнёр после гибели супруга
не нашёл ему замены, но в течение двух лет «охранял» гнездовой участок.
Гнездо аисты строят сами; оно, как правило, располагается в средней части дерева. Ча-
сто основанием гнезду служит большая развилка в кроне (Приклонский, 1958; Мальчевс-
кий, Пукинский 1983; Пекло, 1984) или мощный, отходящий в сторону сук. Основанием
гнезду может послужить «ведьмина метла» — заболевание, чаще всего свойственное хвой-
ным деревьям. Однако иногда гнёзда бывают расположены на относительно тонких и, на
человеческий взгляд, непрочных ветках. Особенно это характерно для гнёзд, помещаемых
в верхней части кроны или на боковых сучьях.
Вблизи гнезда обычно находятся обширные прогалины — зарастающие вырубки или
поляны, где в период насиживания можно видеть одного из партнеров. Птица стоит на
Чёрный аист
379
сухом суку открыто стоящего дерева и взлетает, как только наблюдатель или другой трево-
жащий объект оказывается поблизости.
Для гнездования используют наиболее распространенные породы крупных деревьев с
массивными ветвями, произрастающие в данной местности. В Белоруссии это огромные
ольхи, на Черниговщине — тополя, на Алтае — столетние дубы. Под Ленкоранью для стро-
ительства гнёзд аисты избирают дубы, ольхи, старые тутовники, у которых сломана верши-
на; это место как раз и служит основанием гнёзда (Спангенберг, 1951). В Окском заповед-
нике гнезда (п = 56) были построены на соснах и лишь одно, расположенное в пойменном
ольшанике — на старом дубе.
Как уже упоминалось, в горных районах и в Приморье чёрные аисты могут устраивать
гнёзда на скалах. Так, Е.Н. Панов (1973) 15 июля 1961 г. нашёл гнездо аиста в верховьях р.
Нарва (ныне р. Сидими). Оно было расположено в средней части отвесной каменной сте-
ны, в небольшой пещере-нише. Гнездо было недоступно как снизу, так и сверху. Гнездова-
ние на скалах, сообщает далее Е.Н. Панов, вообще характерно для корейской популяции
чёрных аистов. То же свойственно виду в Вост. Казахстане (Щербаков, 1984).
Диаметр гнездовой постройки достигает одного метра, но иногда бывает и больше —
до 1,5-1,8 м (Березовиков, Воробьёв, 1984; Николаев, Керданов, 1984; Полозов, 1984; По-
пенко, Дашкевич, 1984). Конкретные данные, приводимые М.Е. Никифоровым с соавтора-
ми (1989), свидетельствуют о том, что диаметр гнёзд (п = 5) составляет в среднем 110 см
(от 80 до 132 см), высота гнёзда — в среднем 37 см (30—40 см). Так как гнездо обычно
используется в течение многих лет, высота постройки может достигать одного метра. В
Белоруссии известен случай, когда гнездо использовалось аистами непрерывно в течение
14 лет (Федюшин, Долбик, 1967). Нижние слои веток, перемешанных с выстилкой и помё-
том птенцов, с годами перепревают, превращаются в труху и опадают на землю. Ежегодно
аисты достраивают гнездо, принося на него новые ветки, иногда до 3-4 см толщиной в
месте облома. В строительстве всегда используются сухие сучья, которые собираются на
земле или обламываются с близ стоящих деревьев. Лоток выстилается корневищами, стеб-
лями плауна (в сосняках в средней полосе Европейской России) и лиан (в Приморье), зелё-
ным мхом, ободранным с оснований деревьев (Мальчевский, Пукинский, 1983), сфагну-
мом, собранным на болотах, травой (Березовиков, Воробьев, 1983). Все это скрепляется
влажным песком и землей. Чёрный аист никогда не выстилает лоток пухом и перьями, хотя
к концу насиживания они в небольшом количестве накапливаются здесь — вероятно, в
результате чистки аистами своего оперения (Попов и др., 1977). В этой «вторичной» выс-
тилке преобладают контурные перья аиста, которые впоследствии смешиваются с рыбьей
чешуей, испражнениями птенцов, остатками их пищи.
К концу гнездового сезона птенцы основательно утрамбовывают центр гнёзда и оно
становится почти плоским, а в некоторых случаях — покатым к краям или в одну сторону.
Однако, поскольку птенцы либо лежат в гнезде, либо стоят на голенях, падения птенцов из
гнёзд не регистрировали. Встают «на ноги» птенцы в трёхнедельном-месячном возрасте.
Кладка состоит из 2-6, редко из 1 или 7 яиц. Наиболее часто отмечают 4-5 яиц в кладке.
Скорлупа яиц грязновато-белая, ненасиженные яйца зеленоватые на просвет. Из 19 кладок,
осмотренных в Окском заповеднике, с 2 яйцами было 2 гнезда, с 3 — 6 гнёзд, с 4 - 4, с 5
яйцами — 5 гнёзд. В 2 гнёздах было по 6 яиц. Средние (средневзвешенные) размеры клад-
ки — 3,95 яйца (Приклонский, 1984). Средние размеры 9 яиц из двух кладок — 66,61±
0,44x50,19±0,26 мм, индексы формы яиц: I = 75,36±0,45; К = 1,33±0,008. Коэффициент
вариации длины яиц — 2,01%, диаметра — 1,55% (Нумеров и др., 1995). По В. Макачу
(Makatsch, 1974), минимальные размеры яйца — 60,3x45,2, максимальные — 74,3x47,5
мм, средний вес — 81,3 г. По М.Е. Никифорову с соавторами (1989), средняя масса йена-
380
Отряд Листообразные, Семейство Аистовые
сиженных яиц (п = 33) равна 81,3 г, средний вес скорлупы яиц — 8,1 (6,8-9,66) г; размеры
яиц — 75,9x42,6 мм. Интересные данные приводят А.С. Мальчевский и Ю.Б. Пукинский
(1983), которые в Ленинградской области обнаружили самые мелкие, насколько известно,
яйца аиста. Размеры 4 яиц были: 49,5-51,0 х 33,9-34,9 мм, а вес, при средней степени
насиженности, 84-89 г. Вес «непродуктивных» яиц (неоплодотворённых или с погибшими
эмбрионами) (п = 7) из Окского заповедника — 72-80 г (Приклонский, 1958).
Как правило, одно из яиц в кладке (по Е.П. Спангенбергу, 1951 — одно-два), бывает
неоплодотворённым. Поэтому птенцов в гнезде в среднем на одного меньше, чем яиц. В
1957 г. в одном из гнёзд чёрного аиста в Окском заповеднике было обнаружено 2 неразвив-
шихся яйца. При вскрытии одно из них оказалось неоплодотворённым, а во втором было
обнаружено 2 погибших аистёнка. Зародыши погибли на стадии образования пуха на ос-
новных птерилиях.
Насиживание у чёрного аиста продолжается 28-30 дней. По другим данным, длитель-
ность насиживания составляет 32-38 дней, а по некоторым сведениям (Уайзерби, 1939,
цит. по Спангенберг, 1951) — до 46 дней. Мы склонны сомневаться в интерпретации на-
блюдений последнего автора, поскольку считаем процесс инкубации строго «запрограм-
мированным» в онтогенезе вида. У дальневосточного аиста в питомнике Московского зоо-
парка (п=2) инкубация продолжалась 32 дня (Т.В. Лавровская, устн. сообщ.). Следователь-
но, мы можем предположить, что и у чёрного аиста она в природных условиях длится не
более 30-32 дней.
Участие партнеров в насиживании примерно равное, хотя один из них (по-видимому,
самка), насиживает кладку плотнее и проводит на гнезде ночное время. Вторая птица в это
время держится поблизости, стоя на ветви какого-нибудь большого дерева в пределах ви-
димости гнёзда, как бы «контролируя» происходящее. Вылупление птенцов происходит в
течение 1-2 суток (данные автора очерка), из чего можно предположить, что насиживание,
в сущности, начинается после завершения кладки. По Е.П. Спангенбергу (1951), насижи-
вание начинается с первого яйца, но взрослые аисты сидят на нем сначала короткое время,
хотя и не отлучаются от него надолго. В это время одна из птиц постоянно стоит на гнезде,
оберегая кладку от хищников.
Процесс вылупления птенца в одном из гнёзд в Окском заповеднике наблюдал В.М.
Галушин. По его данным, птенец сначала проделал щель в скорлупе яйца, а затем повора-
чивался в нем в одном направлении, стараясь занять положение клювом вниз. При этом
взрослая птица поворачивала яйцо в противоположном направлении. В результате образо-
вывалась щель, опоясывающая яйцо ближе к тупому концу. В конце этого процесса мень-
шая часть скорлупы откинулась, как «крышечка».
Масса вылупившегося птенца обычно не превышает 50 г. Птенец только что вылупив-
шегося чёрного аиста в гнезде близ г. Казани (сейчас это территория Волжско-Камского
заповедника) 17 июня 1947 г. весил 48,6 г (Попов и др., 1977).
Из 39 гнёзд с птенцами в Окском заповеднике в четырёх было по 1 птенцу, в шести — по
2, в восьми — по 3, в тринадцати — по 4, в восьми — по 5 птенцов. Средний размер
выводка— 3,4 птенца (Приклонский, 1984).
В первые дни рост птенцов мало заметен. Однако уже через неделю-другую птенцы
начинают внушительно прибавлять в весе. К моменту вылета из гнёзда их масса достигает
2,5-3 кг.
В первые один-два дня после вылупления птенцов взрослые их практически не кормят.
Одна из взрослых птиц в это время стоит на гнезде, как бы заслоняя птенцов от солнечных
лучей. Попытки кормления начинаются на третий-четвёртый день жизни птенцов и стиму-
лируются активностью последних. Чаще всего аисты приносят пищу в зобу (возможно, и в
Чёрный аист
381
пищеводе), отрыгивают её в центр гнезда, после чего начинают раздавать корм птенцам, не
разрывая добычу на куски. В первые дни аистята получают полупереваренные части добы-
чи. В двухнедельном-месячном возрасте птенцы начинают требовать пищу более активно.
При появлении взрослой птицы они открывают клюв, пищат, пытаются ухватить на лету
отрыгиваемую пищу. В это время корм доставляется на гнездо сразу после охоты, и поэто-
му часть добычи (иногда более половины улова) бывает ещё живой. В 30-50-дневном воз-
расте аистят родители, отрыгнув пищу на гнездо, сразу же улетают за новой порцией кор-
ма; птенцы сами подбирают корм (в первую очередь, ещё двигающихся животных) и загла-
тывают их.
Места охоты взрослых аистов располагаются как поблизости от гнезда, так и на некото-
ром отдалении от него. М.И. Лебедева (1959), по своим наблюдениям в Беловежье, опреде-
ляет радиус охотничьего участка аиста в 6 км. По её мнению, это подтверждается тем об-
стоятельством, что она не находила гнезда аистов ближе, чем в 6 км друг от друга. Впро-
чем, А.В. Федюшин и М.С. Долбик (1967), ссылаясь на наблюдения В.Ф. Гаврина на При-
пяти, приводят данные о находках гнёзд аистов на расстоянии от 1 до 1,5 км друг од друга.
По данным литовских орнитологов места охоты чёрных аистов могут отстоять на удалении
до 9 км от их гнёзд (Ivanauskas, 1959).
Вылет в нормальных условиях, то есть слёт с гнезда не слишком потревоженных на-
блюдателем аистят, происходит на 60-70-й день с момента вылупления. Потревоженные
исследователем аистята могут слететь с гнезда уже на 50-55-е сутки. В этом случае полёт
молодых неровный, птенцы чаще всего не могут уверенно приземлиться на ноги и падают
на землю грудью. При этом они могут получить повреждения, вызывающие впоследствии
гибель, особенно если травмированы ноги. Но обычно повреждается только кожный по-
кров груди. Наблюдателю нередко приходилось при подходе к гнёздам вспугивать 65-70-
дневных птенцов. В этом возрасте они чаще всего отлетают и садятся на землю в 100 или
более метрах от гнезда; иногда им удаётся сесть на соседние деревья. Однако, первые полё-
ты (по крайней мере, спугнутых птенцов) идут со снижением. Спугнутые с гнезда птенцы
некоторое время держатся по соседству. Кольцевание показало, что в этих случаях вероят-
ность гибели их невелика.
Как было сказано выше, в 75-80-дневном возрасте молодые птицы приступают к миг-
рации. В средней полосе Европейской России это происходит в 10-20-х числах августа.
Материалов о семейной жизни чёрного аиста после окончания гнездового сезона нет.
Нельзя исключить (по аналогии с другими видами того же отряда), что миграции начина-
ются с перелетов семейных групп, объединяющихся затем в стаи. В редких случаях образо-
вываются совместные стаи с белым аистом.
Суточная активность, поведение. Чёрный аист — в гораздо меньшей степени обще-
ственная птица, чем другие виды аистов, он никогда не гнездится колониально. В сезон
размножения и на зимовках держится одиночно или отдельными парами, стаи (до 100 осо-
бей) образует только в период миграций (Cramp, 1977). Брачное поведение изучено плохо.
По данным некоторых наблюдателей, видевших его, оно напоминает поведение белого аиста.
Отличие состоит в том, что чёрный аист более скрытен, его брачный «танец» происходит в
гнезде, причём в то время, когда к гнезду трудно подобраться. По словам И.П. Шпиленка
(устн. сообщ.), взрослые птицы, находясь на гнезде и стоя один против другого, запроки-
дывают головы к спине и постоянно трещат (щелкают) клювами. Такие действия происхо-
дят неоднократно, с 10-15-минутными перерывами, в моменты между активным строи-
тельством гнезда (Брянская обл.). Спаривание происходит на гнезде, что наблюдал Е.С.
Птушенко в Ленкорани (Спангенберг, 1951). Некоторые позы чёрного аиста показаны на
рис. 77.
382
Отряд Листообразные, Семейство Аистовые
Рис. 77. Позы чёрного аиста (по Cramp, 1977; Bauer, Glutz von Blotzheim, 1987): a — спокойная поза,
б — поза отдыха, в — оборонительная поза взрослой птицы на гнезде, г — угрожающая поза взрос-
лой птицы на гнезде, д — оборонительная поза на гнезде молодой птицы, е — приветствие партнё-
ров на гнезде, ж — спаривание.
Чёрный аист
383
Питание. Со времени Е.П. Спангенберга (1951), считавшего, что данных по питанию
чёрного аиста мало, накоплены внушительные материалы по этому вопросу. Прежде всего,
установлено и доказано, что чёрный аист — плотоядный вид. Луковицы водных растений,
равно как корневища луговых или семена в его рационе отсутствуют либо присутствуют
как примесь к приносимым на гнездо животным кормам.
Характер кормовых стаций определяет состав кормов чёрного аиста на территории все-
го исследуемого пространства. Выше было сказано, что аист кормится по берегам водо-
ёмов, в том числе на мелководных берегах стариц, на неглубоких болотцах, лужах, высыха-
ющих (и, следовательно, мелеющих на протяжении лета) участках травяных (низовых) бо-
лот. Здесь он подбирает наиболее массовые виды живности, обитающей в воде. В корме
птенцов чёрного аиста в условиях Окского заповедника в 1954-1956 гг. (п = 474) преобла-
дали вьюн (Misgurnus fossilis) — 67,5% и щука (Esox lucius) — 30%. Оставшиеся 2,5%
пришлись на три вида лягушек и единственного окуня (Perea fluviatilis). Заметим, что в
пище не найдено водных насекомых и их личинок. Вьюны, принесённые родителями птен-
цам, были от 12 до 25 см длиной и массой до 60 г. Щурята — от 9 до 17 см и массой 6^40 г.
Общий вес пищи, принесённой одним аистом на гнездо за один раз, составлял 220-600 г.
Однажды, 13 июля 1954 г., четырём полуторамесячным птенцам пищу на гнездо принесли
одновременно оба родителя. Вес 58 обнаруженных там вьюнов, составил 1 120 г (Приклон-
ский, 1958). В Беловежье 20-дневным птенцам аисты за три раза принесли 387 пищевых
объектов, общим весом 1 127 г (Крапивный, 1957). Е.П. Спангенберг (1951) упоминает в
числе объектов питания аиста водных насекомых, рыбу, лягушек, змей. Как первые, так и
последние кормовые объекты в пище птенцов чёрного аиста в Окском заповеднике не встре-
чены, что свидетельствует как об обилии в местах охоты аистов в заповеднике основных
для этих птиц кормов, так и об определённом постоянстве в выборе корма, по крайней мере
в этом районе. По материалам М.И. Лебедевой (1959), спектр питания птенцов аиста в
Беловежской пуще иной. Амфибии (лягушки, включая жерлянок, и тритоны) в 1948 г. со-
ставили там 65,2%, а в 1949 г. — 78,1%; рыбы, соответственно, — 26,3 и 15,6%; рептилии
(живородящая ящерица) — 3,2 и 0,9%; насекомые (стрекозы, жуки-водолюбы и их личин-
ки) — 3,5 и 4,6%; растения — 0,0 и 0,9%.
В Ленинградской обл. среди остатков пищи у гнезда были обнаружены 7 водяных поле-
вок, 4 лягушки (вид не указан), 3 серых полёвки рода Microtus, молодая ондатра и неболь-
шая щука (Мальчевский, Пукинский, 1983). Следует заметить, что в последние полвека,
учитывая редкость аиста, изучали питание только его птенцов путём анализа проб пищи,
принесённой на гнездо, в то время, как в довоенные годы применялось исследование со-
держимого желудков отстрелянных птиц. Логично, что в желудках находили более разно-
образный состав кормов.
Из приведенных данных понятно, что и взрослые птицы питаются животной пищей.
Поэтому нельзя исключить, что редкость гнездования и спорадичность распространения
этой птицы определяется не только малым числом подходящих для самого гнезда биото-
пов, но и ограниченностью мест с необходимым виду обилием пищи.
В период пролёта аисты днюют в основном на сходных с гнездовыми биотопами мес-
тах, что позволяет им использовать в основном такую же пищу, как в сезон размножения.
Однако в более южных местностях и на зимовках эти птицы чаще используют насекомых,
особенно, если последние находятся в массе в районах стоянок, а также пресмыкающихся.
Судя по С. Крэмпу (Cramp, 1977), на зимовках рацион этих птиц мало отличается от раци-
она в местах гнездования. Здесь в пище аиста преобладает мелкая, но обильная в местах
охоты рыба, а при необходимости остановок в более засушливых местах — пресмыкающи-
еся, насекомые, грызуны.
384
Отряд Листообразные, Семейство Аистовые
Враги, неблагоприятные факторы. Пернатых врагов чёрный аист практически не име-
ет, хотя во время миграций и на зимовках не исключено нападение на аистов крупных хищ-
ных птиц. В принципе возможно похищение яиц из гнёзд аистов серой вороной, но практи-
ческое отсутствие её в местах гнездования аиста и, к тому же, упоминавшееся постоянное
нахождение взрослых при кладке как будто бы исключает такую вероятность. Птенцов,
покинувших гнездо (особенно, если это произошло до обычного срока их подъёма на кры-
ло) из-за неосторожности человека, в первые часы их нахождения на земле могут поймать
четвероногие хищники: лисица, волк, барсук, енотовидная собака, возможно куница. Од-
нако уже в 35^0-дневном возрасте аистята отчаянно сопротивляются при попытках чело-
века взять их в руки для исследования или кольцевания. Удар клювом таких птенцов доста-
точно болезнен, поэтому добыча аистёнка хищником сопряжена с немалой опасностью для
последнего.
Благополучие вида зависит в первую очередь от сохранности и состояния гнездовых
биотопов, а они пока ещё используются человеком в относительно малой степени, по край-
ней мере на основной территории ареала. В иных же местах — таких, как Калининградская
обл. РФ и страны Прибалтики — для охраны местообитаний вида приняты строгие меры.
О болезнях чёрного аиста данных не имеется. Сведения о паразитах и гнездовых сожи-
телях чёрного аиста также отсутствуют (Hicks, 1969; McClure, Ratanaworabhan, 1973).
Практическое значение, охрана. Практического значения в обычном смысле этого
понятия, т.е. в хозяйстве человека, чёрный аист не имеет. С другой стороны, этот вид обла-
дает исключительной эстетической привлекательностью. Встреча одинокого чёрного аис-
та на берегу лесного водоёма или на поляне близ гнёзда не может оставить равнодушным
любителя природы, она запоминается надолго и многие годы затем хранится в его памяти.
Чёрный аист занесён в Красную книгу Российской Федерации (2001), в большинство
Красных книг субъектов Российской Федерации и государств, прежде входивших в состав
СССР. Основной формой охраны вида следует считать сбережение мест его гнездования.
Следует заметить, что много чёрных аистов, особенно в первую осень жизни, отстрелива-
ют охотники. Этот негативный фактор можно снизить путём повышения культуры охоты, а
возможно также и запрещая охоту в местах постоянных остановок этих птиц на осеннем
перелёте. Последнее потребует от орнитологов усиления целенаправленных исследований
осенней миграции аистов.
Белый аист
Ciconia ciconia (Linnaeus, 1758)
Ardea ciconia Linnaeus, 1758. Syst. Nat., ed. 10, c. 142, Швеция.
White stork (англ.); WeiBstorch (нем.), cigogne blanche (фр ).
Син. (лат.): для подвида ciconia'. Ciconia alba Bechstein, 1793; Ciconia albescens C.L. Brehm,
1831; Ciconia nivea C.L. Brehm, 1831; Ciconia Candida C.L. Brehm, 1831; Ciconia major C.L.
Brehm, 1855; для подвида asiatica'. Ciconia asiatica Severtzov, 1873; Ciconia orientalis
Severtzov, 1875.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Крупная птица с длинными ногами,
шеей и клювом. Длина тела 100-115 см, размах крыльев 155-165 см, вес взрослой птицы
Белый аист
385
от 2,5 до 4,5 кг. Самцы несколько крупнее самок, но внешне они практически неотличимы.
Оперение белое, маховые чёрные. Клюв и ноги красные. При наблюдении за летящей пти-
цей, обращают на себя внимание вытянутые шея и ноги, контрастное чёрно-белое опере-
ние. Ходит по земле, немного покачивая головой в такт движению. На гнёздах или приса-
дах может подолгу стоять на одной ноге, втянув шею в оперение туловища. Часто использу-
ет парящий полёт, практически без взмахов крыльев способен подниматься в восходящих
потоках воздуха. При резком снижении и посадке — немного прижимает крылья к тулови-
щу и выставляет ноги вперёд. Стаи образуются во время перелёта, их формируют также
негнездящиеся птицы в ходе кочёвок в поздневесенний и летний периоды. Строго порядка
в летящих стаях нет. При снижении из восходящего потока птицы одна за другой скользят
вниз. От чёрного аиста отличается белым оперением, от журавлей и цапель — красным
цветом клюва и ног. В отличие от цапель, в полете вытягивает, а не складывает шею.
Голос. Основа звукового общения белых аистов — трещание клювом. Изредка можно
услышать шипение. Звуковой репертуар птенцов более разнообразен. Крик выпрашиваю-
щего корм аистёнка напоминает протяжное мяуканье. Первая часть этого крика имеет бо-
лее высокий тон, вторая — низкий. У птенцов на гнезде можно услышать также громкий
писк и шипение; уже в первые недели жизни птенцы пытаются трещать клювом.
Подвидовая систематика. Выделяют 2 подвида, отличающихся размерами и формой
клюва:
1. Ciconia ciconia ciconia
Ardea ciconia Linnaeus, 1758, Syst. Nat., ed. 10, c. 142, Швеция.
Более мелкая форма. Длина крыла самцов 545-600 мм, длина цевки 188-226 мм, длина
клюва 150-200 мм. Клюв менее резко скошен к вершине (Степанян, 2003). Распространён
в Европе, Сев. Африке, Зап. Азии.
2. Ciconia ciconia asiatica
Ciconia alba asiatica Severtzov, 1873, Изв. Имп. о-ва любителей естествознания, антро-
пологии и этнографии, 8, вып. 2, с. 145, Туркестан.
Более крупная форма. Длина крыла самцов 580-630 мм, длина цевки 200-240 мм, дли-
на клюва 184-235 мм. Клюв, в особенности подклювье, более резко скошен к вершине
(Степанян, 2003). Обитает на территории Узбекистана, Казахстана, Таджикистана и Кир-
гизии.
Замечания по систематике. Ранее дальневосточного аиста (Ciconia boyciana) считали
подвидом белого аиста, однако в настоящее время большинством систематиков он рассмат-
ривается как самостоятельный вид. Специальные исследования показали значительные
морфологические и поведенческие различия, достаточные для разделения видов (Hancock
et al., 1992). Специального изучения требует подвидовая принадлежность популяции бело-
го аиста из Закавказья.
Описание. Окраска. Взрослые самец и самка. Сезонных различий в окраске нет.
Большая часть оперения белая, первостепенные маховые, наружные второстепенные, пле-
чевые и часть кроющих перьев предплечья чёрные с металлическим блеском. Наружные
опахала второстепенных маховых имеют сероватые поля вдоль ствола (признак варьирует,
виден обычно только с близкого расстояния). Перья на шее и груди несколько удлинены;
возбуждённые птицы (например, во время токования) часто их распушивают. Клюв и ноги
ярко-красные. Голая кожа вокруг глаза и передняя часть кожи подбородка чёрные. Радужи-
на глаза бурая.
Первый пуховой наряд. Птенец после вылупления покрыт редким и коротким серовато-
белым пухом. Ноги розоватые, через несколько дней становятся серовато-чёрными. Клюв
и кожа вокруг глаз чёрные, на подбородке кожа красноватая, радужина тёмная.
386
Отряд Листообразные, Семейство Аистовые
Второй пуховой наряд. Пух чисто белый, более густой и длинный. Сменяет первый
примерно через неделю.
Гнездовой наряд. Молодая птица по окраске похожа на взрослую, но чёрный цвет в опере-
нии заменен буроватым, без блеска. Клюв и ноги тёмно-бурые; ко времени вылета птенцов из
гнёзд они обычно становятся красновато-бурыми, но нередко можно увидеть и летающих
слетков с чёрным клювом или бурым с черноватой вершиной. Радужина глаза серая.
Строение и размеры. Как правило, публикуют промеры различных частей тела
аистов, не разделяя выборку на половые группы. Длина крыла номинативного подвида бе-
лого аиста при таком подходе для территории бывш. СССР составляет, по 6 особям, 585—
605 мм (Спангенберг, 1951), для Украины (Смогоржевський, 1979) — 534-574 мм. После-
дний автор сообщает также, что длина хвоста колеблется в пределах 206-232 мм, клюва -
156-195, а цевки — 193-227 мм. Ревизия коллекций Зоомузея Киевского национального
университета и Национального научно-природоведческого музея Украины дал следующие
результаты: длина крыла (п=14) — 513-587 мм, при среднем значении 559,9±5,8 мм; хвос-
та (n=l 1) — 201-232, в среднем 222,5±4,2; клюва (п=12) — 150-192, в среднем 166,4±3,5;
цевки (п= 14) — 187-217, в среднем 201,4±2,5 мм (ориг.). Для азиатского белого аиста
длина крыла по 9 измеренным особям оказалась 550-640, в среднем 589 мм
Размеры белого аиста по половым группам и подвидам для различных территорий при-
водятся в табл. 31.
Формула крыла (не считая рудиментарного первого махового) IV?III?V>I>VI... Наруж-
ные опахала II и IV первостепенных маховых имеют вырезки. Хвост слегка закруглён, ру-
левых перьев 12. Клюв длинный, прямой, суживающийся к вершине. Ноздри длинные,
щелевидные. Вес 41 самца из Вост. Пруссии 2 900-4 400 г (в среднем 3 571), 27 самок — 2 700-
3 900 г (3 325). Вес на протяжении лета немного увеличивается. Средний вес 14 самцов в
июне 3 341 г, 14 самок — 3 150 г; в июле-августе 12 самцов весили в среднем 3 970 г, 12
самок — 3 521 г (Steinbacher, 1936).
Самец, таким образом, несколько крупнее самки, имеет более длинный и массивный
клюв. Кроме того, клюв у самца имеет несколько иную форму: подклювье перед вершиной
слегка изогнуто вверх, у самки же клюв прямой (Bauer, Glutz von Blotzheim, 1966; Creutz,
Таблица 31. Размеры (мм) различных половых групп и подвидов белого аиста
Параметр Самцы Самки Источник
п lim M n lim M
Ciconia ciconia ciconia. Европа
Длина крыла 530-630 530-590 Witherby et al., 1939
Длина хвоста 215-240 215-240
Длина клюва 150-190 140-170
Длина цевки 195-240 195-240
Длина крыла 18 556-598 576 15 543-582 558 Hancock et al., 1992
Длина хвоста 18 221-268 247 15 218-256 237
Длина клюва 18 157-198 179 15 155-180 164
Длина цевки 18 191-230 214 15 184-211 197
Ciconia ciconia asiatica. Средняя Азия
Длина крыла 18 581-615 596 9 548-596 577 Hancock et al., 1992
Длина клюва 18 188-223 204 9 178-196 187
Длина цевки 18 213-247 234 9 211-234 220
Белый аист
387
1988). По длине клюва можно определить пол 67% птиц с вероятностью ошибки не более
5% (Post et al., 1991). Возможно также индивидуальное узнавание птиц по рисунку чёрного
пятна на подбородке (Fangrath, Helb, 2005).
Линька. Изучена недостаточно. У молодых птиц полная постювенильная линька начи-
нается, в зависимости от обстоятельств, с декабря по май первого года жизни. У взрослых
птиц полная линька занимает большую часть года. Первостепенные маховые сменяются в
нерегулярной последовательности на протяжении гнездового периода, некоторые зимой
(Stresemann, Stresemann, 1966).
Более детально линька маховых перьев прослежена на 5 аистах, содержавшихся в пи-
томнике в Швейцарии (Bloesch et al., 1977). Рост пера происходит с линейной скоростью.
Первостепенные маховые растут по 8-9 мм в день, второстепенные — 6,5-6,9 мм. На сме-
ну махового пера уходит от 50-55 до 65-75 дней. У находившихся под наблюдением птиц в
год сменялось по 6 первостепенных маховых и 13 второстепенных на обоих крыльях. Про-
должительность ношения различных перьев разная; для первостепенных маховых она ко-
лебалась от 1,2 до 2,5 лет. Смена перьев идёт ступенчато. У первостепенных маховых она
начинается с XI, у второстепенных — с нескольких точек. Циклы линьки начинаются со
второго года жизни, их окончательный ход устанавливается лишь к 4-5 году. Во время пер-
вой-третьей линьки смена перьев начиналась в марте-апреле, в дальнейшем — в середине
мая и продолжалась до начала ноября. Большая часть перьев сменялась в летние месяцы
между началом насиживания и отлётом.
Совмещение линьки и гнездования может быть связано с тем, что у белого аиста в это
время нагрузка на крылья гораздо меньше, чем во время продолжительной миграции или
при кочевой жизни на зимовках (Creutz, 1988).
Распространение. Гнездовой ареал. Европа, Сев.-Зап. Африка, Зап. и Средняя
Азия (рис. 78).
Европейский подвид распространён на большей части Европы от Пиренейского п-ова
до Поволжья и Закавказья. На север его ареал доходит до Дании, Юж. Швеции, Эстонии,
северо-запада России. Во Франции аисты обитают лишь в нескольких провинциях, поэто-
му места гнездования в Испании, Португалии, Зап. Франции и Сев.-Зап. Африке оказыва-
ются оторванными от основного европейского ареала. Однако, в связи с продолжающимся
расселением, вполне вероятно смыкание этих двух частей ареала. В Сев.-Зап. Африке бе-
лый аист гнездится в Марокко, на севере Алжира и Туниса. В Зап. Азии — в Турции, Си-
рии, Ливане, Израиле, Ираке, Иране, в Закавказье — на юге Грузии, в Армении, Азербайд-
жане, а также в Республике Дагестан РФ. Известны случаи размножения также на местах
зимовки в Юж. Африке (Broekhuysen, 1965, 1971; Broekhuysen, Uys, 1966; Hancock et al.,
1992). В 2004 г. отмечена попытка гнездования на северо-востоке Англии в графстве Йор-
кшир. Это первый случай размножения белого аиста в Великобритании после 1416 г., когда
птицы гнездились на кафедральном соборе в Эдинбурге.
В России белый аист издавна населяет территорию Калиниградской обл. В других обла-
стях он появился сравнительно недавно, расширяя гнездовой ареал в восточном и северо-
восточном направлениях. Первые случаи гнездования в современных границах Ленинг-
радской и Московской обл. отмечались ещё в конце XIX в. (Мальчевский, Пукинский, 1983;
Зубакин и др., 1992). К началу XX в. белый аист начал гнездиться в Псковской, Тверской и
Калужской обл. (Зарудный, 1910; Филатов, 1915; Бианки, 1922). К этому времени он уже
был достаточно обычным в западных районах Смоленской (Граве, 1912, 1926) и южной
части Брянской обл. (Федосов, 1959). Расселение на новые территории имело волнообраз-
ный характер. Особенно интенсивным освоение новых местностей отмечено в 1970-1990-х
гг. В настоящее время на территории России северную и восточную границу регулярного
388
Отряд Листообразные, Семейство Аистовые
Рис. 78. Область распространения белого аиста: а — область гнездования, б — районы зимовок, в —
основные направления осеннего пролёта, г — направления экспансии.
гнездования восточноевропейской популяции можно условно провести по линии Санкт-
Петербург - Волхов - Тихвин - Ярославль - Липецк - Воронеж - граница Ростовской обл.
и Украины (рис. 79).
Периодические выселения отдельных пар отмечены далеко за пределами указанной гра-
ницы: в Юж. Карелии, Костромской, Нижегородской, Кировской, Пермской, Ульяновской,
Пензенской, Саратовской, Волгоградской и Ростовской обл., Краснодарском крае (Лапшин,
1997, 2000; Бакка и др., 2000; Бородин, 2000; Дылюк, Галченков, 2000; Карякин, 2000;
Комлев, 2000; Мнацеканов, 2000; Пискунов, Беляченко, 2000; Сотников, 2000; Фролов и
др., 2000; Чернобай, 2000а и др.). Западноазиатская популяция номинативного подвида
распространена в Терско-Сулакской низменности Дагестана (Бабаюртовский, Хасавюртов-
ский, Кизлярский, Тарумовский р-ны), периодически гнёзда появлялись за пределами Да-
гестана — в Ставропольском крае, Карачаево-Черкесии, Пролетарском р-не Ростовской
обл. (Хохлов, 1988а; Бичерев, Скиба, 1990). Регистрировали белых аистов и в предгорьях
Сев. Осетии (Комаров, 1986). Ростовская обл., очевидно, является территорией, где с раз-
ных направлений практически сходятся восточноевропейская и западноазиатская популя-
ции. Первая проникает сюда с севера по Дону и с запада — с Украины, вторая — с юго-
востока по Кумо-Манычкой впадине. Подтверждением последнего, наиболее плохо выяс-
ненного направления перемещения птиц, может служить встреча 13 мая 1996 г. в районе оз.
Дадинское, на крайнем северо-востоке Ставропольского края, стаи из 18 птиц, мигриро-
вавших на большой высоте в северо-западном направлении (Дылюк, Галченков, 2000).
На Украине современная граница ареала проходит через Сев. и Сев.-Вост. Крым, юж-
ные части Запорожской и Донецкой обл., Луганскую обл. (Грищенко, 2005). В 2006 г. отме-
Белый аист
389
чен первый случай гнездования
белого аиста на юго-востоке Кры-
ма возле Феодосии (М.М. Беска-
равайный, личн. сообщ.).
Туркестанский белый аист рас-
пространён в Средней Азии — на
юго-востоке Узбекистана, в Тад-
жикистане, Киргизии, на юге Ка-
захстана. Ранее ареал доходил до
Чарджоу в Туркмении, низовьев
Аму-Дарьи; отмечались случаи
гнездования и на западе Китая —
в Кашгарии (Спангенберг, 1951;
Долгушин, 1960; Сагитов, 1987;
Шерназаров и др., 1992). Изредка
попытки гнездования — очевид-
но, уже европейского подвида —
отмечаются на юго-востоке Турк-
менистана (Белоусов, 1990).
Небольшой очаг гнездования
белого аиста (около 10 пар) воз-
ник на крайнем юге Африки. Пти-
цы здесь приступают к гнездова-
нию в сентябре-ноябре — в пору
прилёта за зимовки аистов север-
ных популяций (del Hoyo et al.,
Рис. 79. Ареал белого аиста в Вост. Европе и Сев. Азии: а —
регулярное гнездование, б — недостаточно выясненная гра-
ница гнездового ареала, в — нерегулярное гнездование.
Подвиды: 1 — С. с. ciconia, 2 — С. с. asiatica.
1992). Как и в случае с чёрным аистом, эта микропопуляция ведёт начало от мигрантов, по
каким-то причинам начавших размножаться на зимовках.
Зимовки. Основные места зимовки западной популяции европейского подвида —
саванны к югу от Сахары от Сенегала на западе до Камеруна на востоке. Важнейшие места
концентрации зимующих птиц — долины рек Сенегал, Нигер и район оз. Чад. Здесь же
зимуют и аисты, гнездящиеся на северо-западе Африки. Восточная популяция зимует в
Вост, и Юж. Африке от Судана, Эфиопии и Сомали до ЮАР. Большая часть птиц проводит
зимние месяцы в Танзании, Замбии, Зимбабве, ЮАР. Аисты из Зап. Азии зимуют частично
в Африке, частично — на юге Азии. Азиатский подвид зимует в основном в Индии на юг до
Шри-Ланки. На восток эти птицы могут встречаться до Таиланда (Schulz, 1988, 1998; Ash,
1989; Hancock et al., 1992). В Индии основными местами зимовки аистов служат штаты
Бихар на северо-востоке и Гуджарат на западе (Majumdar, 1989). Интересно, что в Индии
находили и птиц, окольцованных в Европе (Лебедева, 1979а). Видимо, это аисты, сбивши-
еся с пути в районе залива Искандера — не свернувшие на юг, а продолжившие миграцию
на юго-восток.
Часть птиц зимует в южной части гнездового ареала. В Испании в зимние сезоны 1991
и 1992 гг. в дельте р. Гвадалквивир и на побережье Андалузии было учтено около 3 000
особей (Tortosa et al., 1995). В Португалии зимой 1994/95 гг. зимовало 1 187 аистов (Rosa et
al., 1999). Тысячи аистов остаются на зиму в Израиле (Schulz, 1998). В Армении в долине
Аракса ежегодно зимуют сотни птиц (Адамян, 1990). В Болгарии аисты оставались на зи-
мовку ещё в конце XIX в., сейчас их количество значительно возросло. Отмечаются стаи
до 10 особей (Nankinov, 1994). Известны случаи зимовки и в более северных широтах — на
390
Отряд Листообразные, Семейство Аистовые
Украине (Грищенко, 1992), в Чехии (Tichy, 1996), Германии, Дании (Schulz, 1998). На тер-
ритории России зимовки белых аистов отмечались в Дагестане (Т.К. Умаханова, В.Ф. Ма-
матаева, личн. сообщ.). В Средней Азии аисты зимуют в небольшом количестве в Ферган-
ской долине (Третьяков, 1974, 1990). Здесь в районе Пунгана - Ургенча в зимние месяцы
1989 г. регистрировали до 250 птиц. Полагают, что частичная оседлость белых аистов в
Ферганской долине способствовала общему увеличению их количества в регионе. Зимов-
ки, носящие нерегулярный характер, наблюдали в долине Сыр-Дарьи и на р. Пяндж в Юж.
Таджикистане (Митропольский, 2007).
Окольцованные в бывш. СССР белые аисты обнаружены на зимовке в основном в ЮАР,
отдельные птицы — в Эфиопии, Судане, Уганде, Кении, Намибии, Зап. Африке (Лебедева,
1979а; Смогоржевський, 1979).
Как установил X. Шульц (Schulz, 1988), распределение аистов на местах зимовки в
Африке определяется прежде всего запасами корма. В первую очередь птицы выбирают
влажные биотопы, но могут держаться и в богатых кормом засушливых местах. Большие
стаи встречаются даже в пустынях и горах. В Лесото в 1987 г. была обнаружена стая из 200
аистов на высоте ок. 2 000 м. Птицы кормились на водоёмах, изобиловавших земноводны-
ми. В местах, богатых кормом, аисты могут скапливаться в огромных количествах. В янва-
ре 1987 г. в Танзании на участке в 25 км2 было учтеноюколо 100 тыс. особей. Птицы корми-
лись на люцерновых полях, где массово размножились гусеницы одной из местных бабо-
чек. В Юж. Африке в этот сезон белые аисты почти не встречались.
Благодаря результатам кольцевания и спутниковой телеметрии установлено, что места
зимовки западной и восточной популяций не изолированы друг от друга. В Центр. Африке
существует зона смешанных зимовок, где встречаются птицы из обеих популяций. Здесь
особи из одной популяции могут увлекаться стаями аистов из другой популяции и возвра-
щаться весной другим путем и в другие места гнездования (Berthold et al., 1997; Brouwer et
al., 2003).
Миграции. Белый аист — дальний мигрант. Птицы из северо-восточной части ареала
совершают перелеты более 10 тыс. км. Существуют две основные географические популя-
ции европейского подвида, различающиеся путями пролёта и местами зимовки. Линия раз-
дела между ними проходит через Голландию, Гарц, Баварию, Альпы (Schiiz, 1953, 1962;
Creutz, 1988; Schulz, 1988, 1998). Птицы, гнездящиеся к западу от нее, мигрируют осенью
на юго-запад через Францию, Испанию, Гибралтар. Далее пролёт идёт через Марокко,
Мавританию, западную часть Сахары. Зимуют эти птицы в Зап. Африке. Аисты, гнездящи-
еся к востоку от этой разделительной линии, летят осенью в юго-восточном, а из России,
Украины, Белоруссии и Прибалтики — в южном направлениях. Через территорию Украи-
ны осенью проходит три основных пролётных пути, которые сливаются в мощный мигра-
ционный поток, проходящий по западному побережью Чёрного моря (Грищенко, Серебря-
ков, 1992; Grischtschenko et al., 1995). Далее аисты летят через Балканы и Турцию, через
Босфор, Малую Азию. От Искандера они выходят к средиземноморскому побережью, где
снова поворачивают на юг и узким потоком мигрируют через Ливан, Израиль, Синайский
п-ов к долине Нила. Вдоль этой реки и Рифтовой долины идёт дальнейшая миграция к
основным местам зимовки в Вост, и Юж. Африке. В Вост. Судане аисты делают длитель-
ную остановку на 4-6 недель и интенсивно кормятся, чтобы восстановить жировые резер-
вы для продолжения миграции (Schulz, 1988, 1998).
Аист, как сухопутный паритель, избегает длительного перелета над морем, поэтому об-
разуются миграционные потоки вдоль побережий. Аисты из западных, северных и цент-
ральных областей Украины мигрируют вдоль западного побережья Чёрного моря и через
Босфор, а птицы из Вост. Украины летят на юго-восток к восточному побережью Чёрного
Белый аист
391
моря. Сюда же летят и аисты из восточной части ареала в России. Часть аистов, хотя и
незначительная, все же летит напрямую через море. Существует «промежуточный» про-
лётный путь через Италию и Сицилию в Тунис. В 1990-1992 гг. на мысе Бон в Тунисе
учтено 1 378 мигрировавших аистов и 67 — возле Мессины в Сицилии (Kisling, Horst,
1999). Предполагается, что этот путь используют птицы как из западной, так и из восточ-
ной популяций (Schulz, 1998). Особь, окольцованная в Латвии, обнаружена в сентябре у
Неаполя (Лебедева, 1979а). А один аист со спутниковым передатчиком перелетел через
Средиземное море напрямую от г. Сан-Тропе во Франции до Туниса; путь через море со-
ставил как минимум 752 км (Chernetsov et al., 2005). Возможно, часть аистов перелетает
через Чёрное море, пересекая Крым.
Миграции аистов из Закавказья, Ирака и Ирана изучены недостаточно. Предполагается,
что они улетают на юго-восток в Юж. Азию (Schiiz, 1963; Schulz, 1998). Птица, окольцо-
ванная в Армении, была найдена в Нахичеванской АССР в 160 км к юго-востоку (Лебедева,
1979а). Линия раздела между популяциями, мигрирующими в Африку и в Азию пока наи-
звестна. Видимо, она проходит где-то на востоке Турции. По крайней мере, в этом реги-
оне осенью наблюдают стаи птиц, мигрирующих как на юго-восток, так и на запад (Schiiz,
1963).
Туркестанские аисты осенью улетают на юг через Афганистан в Индию, пересекая Гин-
дукуш через перевал Саланг (Schiiz, 1963; Schulz, 1998). Окольцованных в Узбекистане
аистов добывали весной в Афганистане и Пакистане (Лебедева, 19796).
Анализ спутникового прослеживания 140 немецких аистов показал, что у этих птиц
могут в довольно широких пределах изменяться пути и сроки миграции, места зимовки и
остановок, но, при возможности, они остаются постоянными. Изменения вызываются при-
родными факторами, прежде всего условиями кормежки (Berthold et al., 2004). Сроки отле-
та с мест зимовки зависят от метеорологической ситуации. При неблагоприятных условиях
птицы могут задерживаться. Так, в крайне неблагоприятном 1997 г. аисты стартовали с
мест зимовки на месяц позже обычных сроков (Косарев, 2006). К этому добавилась задер-
жка из-за длительных холодов на Ближнем Востоке. Снабженные передатчиками аисты
совершали длительные остановки в Сирии и Турции. Отмечался обратный пролёт (Kaatz,
1999). В итоге в 1997 г. только 20% птиц из восточной популяции прилетели в нормальные
сроки, большинство же — с задержкой в 4-6 недель (Schulz, 1998).
С мест зимовки массовое движение в обратном направлении происходит в конце января
или в феврале. В Израиле начало весенней миграции взрослых птиц становится заметным
в середине февраля, пик пролёта приходится на вторую половину марта, заканчиваются
особенно заметные перемещения в конце апреля; молодые птицы мигрируют через Изра-
иль в апреле-мае (van den Bossche et al., 2002). В местах гнездования на севере Африки
аисты появляются ещё в декабре-феврале. Пик пролёта над Гибралтаром отмечают в фев-
рале-марте, над Босфором — с конца марта до конца апреля (Schulz, 1998).
В Молдавии прилетевшие аисты отмечаются с первой декады марта (Аверин и др., 1971).
На территории Украины прилет регистрируется с первых чисел марта до второй половины
апреля, средние даты прилёта приходятся на третью декаду марта - начало апреля. Раньше
всего птицы появляются во Львовской и Черновицкой обл., облетая Карпаты; затем мигра-
ция идёт двумя потоками: одни птицы летят на северо-восток, другие — на восток по юж-
ным областям Украины. Позже всего аисты появляются в восточных областях и в Крыму
(Грищенко, Серебряков, 1992; Grischtschenko et al., 1995). На севере Сумской обл. прилёт
регистрировали с 18 марта по 26 апреля, средняя дата за 16 лет — 30 марта (Афанасьев,
1998). На юго-западе Белоруссии прилёт аистов отмечается в третьей декаде марта - пер-
вой половине апреля (Шокайо, Шокало, 1992).
392
Отряд Листообразные, Семейство Аистовые
Аисты, гнездящиеся в европейской части России, достигают родины в начале марта -
первой половине мая. На территории Калиниградской обл. в первой половине XX в. пер-
вые птицы появлялись на гнёздах в период с 19 марта по 12 апреля (данные за 23 года,
Tischler, 1941). В 1970-е гг. прилет аистов происходил с начала марта (Беляков, Яковчик,
1980). В 1990 г. первые птицы на гнёздах в Калининградской обл. отмечены 18 марта (Гри-
шанов, Савчук, 1992). В Себежском р-не Псковской обл. прилет наблюдали в конце марта -
первой декаде апреля (Фетисов и др., 1986). За период с 1989 по 1999 гг. наиболее ранняя
регистрация в Калужской обл. зафиксирована 20 марта (1990 г.), наиболее поздняя — 8 апре-
ля (1991 и 1997 гг.), в среднем 30 марта. В отдельные годы самые ранние птицы появляются
весной при высоте снегового покрова на полях 30^40 см. Пик прилёта первых аистов на
гнёзда в Калужской обл. приходится на вторую пятидневку апреля (1990-1999 гг.) (Галчен-
ков, 20006). В Воронежской обл. первых аистов наблюдали в те же сроки: с 19 марта по 8
апреля, в среднем 30 марта (1995-1998 гг.) (Нумеров, Макагонова, 2000). К северо-восточ-
ной границе ареала аисты прилетают на 2^4 недели позже. В Ярославскую обл. птицы
прилетали 22-26 апреля (1994 г.), 16 апреля (1996 г.), 2 мая (1995 г.) (Голубев, 2000). В
восточных районах Лениградской обл. самый ранний прилет отмечен 20 апреля 1999 г.
(Тихвинский р-н), обычные же сроки — с 1 по 8 мая (1983-1999 гг.) (Храбрый, 2000). В
южных районах Карелии первые птицы появляются в конце апреля - середине мая; в очень
раннюю весну 1990 г. одиночную птицу видели в начале второй декады апреля (Лапшин,
2000). В Кировской обл. наиболее ранняя регистрация белого аиста — 17 апреля 1992 г.
(Сотников, 2000). На черноморском побережье Сев. Кавказа весенний пролёт отмечается с
первой декады марта до второй половины апреля, в Ростовской обл. и Краснодарском крае
первых птиц отмечали в апреле (Казаков и др., 2004). В Дагестане первые особи появляют-
ся в начале и середине марта (Маматаева, Умаханова, 2000).
Появление белых аистов весной в Средней Азии приходится на конец февраля - начало
марта и на большей части территории отмечается практически одновременно (Дементьев,
1952; Митропольский, 2007). На перевале Чокпак их регистрировали 11-14 марта 1974 г.
(Гаврилов, Гисцов, 1985), интенсивный пролёт отмечен 24 марта (Сема, 1989).
В Калужской обл. в 69% случаев прилёт белого аиста проходил по схеме 1+1: сначала
прилетала одна птица из пары, через некоторое время после нее — вторая. Первая особь
появляется с 20 марта по 18 мая, в среднем (п=176) — 10 апреля, вторая — с 25 марта по 26
мая, в среднем (п=150) — 14 апреля. Запаздывание второй птицы происходит в интервале
от нескольких часов до 31 дня, в среднем, на 4 дня. В указанной схеме прилёта существуют
редко встречающиеся варианты: при первом каждая из особей пары подлетает с одной или
двумя другими птицами, которые на гнезде не остаются, а летят дальше; при втором — к
одиночному аисту прилетает пара и изгоняет его. В 31% случаев на гнездо прилетали сразу
две птицы.
Гнездящиеся птицы восточноевропейской популяции улетают в августе. Молодые, как
правило, отлетают раньше взрослых птиц. В Калужской обл. молодые покидали гнёзда на-
чиная с 8 августа, чаще во второй декаде этого месяца. Взрослые птицы оставляют родину
позже, отлёт последних особей заканчивается в среднем 30 августа (1985-1999 гг.) (Гал-
ченков, 20006). В Тверской обл. аисты отлетают 28 августа - 5 сентября (Николаев, 2000).
В Ярославской обл. птицы отлетали 23 августа (1996 г.) и 29 августа (1995 г.) (Голубев,
2000). Отдельные особи и пары задерживаются до сентября - октября. В юго-западных
регионах России перед отлётом образуют скопления в десятки и до 100 и более особей, как
например, в Смоленской обл. (Бичерев, Барнев, 1998). На Сев. Кавказе осенняя миграция
отмечается с первой половины августа до конца сентября (Казаков и др., 2004). Не выясне-
ны пути миграции и районы зимовки дагестанских аистов: известно, что гнездовую об-
Белый аист
393
ласть последние из них покидают с 25 октября по 10 ноября, иногда задерживаясь до сере-
дины-конца этого месяца (25 ноября 2003 г. и 15 ноября 2004) г. Вероятнее всего аисты,
гнездящиеся в Терско-Сунженской низменности, следуют вдоль западного берега Каспий-
ского моря, где птицы этого вида были отмечены 23 октября 1998 г. в районе г. Каспийск
(Е.В. Вилков, личн. сообщ.).
В Молдавии отлёт начинается в конце августа и продолжается до середины сентября.
Отдельные птицы могут задерживаться до первой половины октября. Наиболее поздняя
встреча — 9 ноября 1964 г. (Аверин, Ганя, Успенский, 1971). На Украине первые пролёт-
ные стаи наблюдаются с первой декады августа до сентября и начала октября. Средние
даты начала отлёта приходятся на третью декаду августа - первую декаду сентября. Раньше
всего пролёт начинается во Львовской, Житомирской и Полтавской обл. Последних птиц
наблюдали со второй половины августа до октября. Средние даты последнего наблюдения
в большинстве областей Украины приходятся на первую и вторую декады сентября. Доль-
ше всего аисты задерживаются в Запорожской обл. и в Крыму (Грищенко, Серебряков, 1992;
Grischtschenko et al., 1995). Отдельные запоздавшие особи могут отмечаться и в ноябре.
Иногда очень поздно можно встретить целые стаи. Так, 4 декабря 1985 г. над Ивано-Фран-
ковском наблюдали стаю из нескольких десятков аистов (Штыркало, 1990). 5 ноября 1997 г.
над Брестом видели стаю из 40 особей (Шокайо, Шокало, 1992). Пролёт по восточному побе-
режью Чёрного моря отмечали с 29 августа по 4 октября (Абуладзе, Элигулашвили, 1986).
Среднеазиатские аисты отлетают с конца августа до средины октября (Долгушин, 1960;
Третьяков, 1990).
Перелет трёх молодых аистов, помеченных на гнёздах в Зеленоградском и Гурьевском
р-нах Калининградской обл. спутниковыми передатчиками, был прослежен в 2000 г. Одна
птица отправилась на зимовку 10 августа, две другие — 14-го. Пролётный путь прошёл
через северо-восточную Польшу, крайний юго-запад Белоруссии, западную часть Украи-
ны, восток Румынии и Болгарии, далее через пролив Босфор, Турцию, Палестину, Синайс-
кий п-ов. Пролива Босфор аисты достигли, соответственно, 23, 25 и 26 августа, т.е. через
13, 11 и 12 дней после начала миграции. На южной оконечности Синайского п-ова аисты
были 29, 31 августа и 1 сентября соответственно (через 19, 17 и 18 дней после начала
миграции, или через 6 дней после пересечения Босфора каждой из птиц); здесь аисты сде-
лали остановку. Далее аисты проследовали вдоль долины Нила в континентальном Египте.
Быстрое движение к югу птицы прекратили 6, 7 и 10 сентября, к этому моменту две из них
находились в центр. Судане, одна — на востоке Чада поблизости от суданской границы
(Chernetsov et al., 2004).
В ходе осенних миграций средняя протяженность суточных перемещений аистов вос-
точной популяции по данным телеметрии составляет: в Европе — 218 км (для взрослых
птиц от 52 до 504, для молодых — от 51 до 475 км), на Ближнем Востоке — 275 км (для
взрослых птиц от 52 до 490, для молодых — от 55 до 408 км), в Сев. Африке — 288 км
(для взрослых птиц от 70 до 503, для молодых — от 108 до 403 км) (van den Bossche et al.,
1999).
Комплексное исследование миграций белого аиста показало, что у этого вида, по край-
ней мере его восточной популяции, совершенно особый тип миграции, пока не известный
для других птиц. Для него характерен очень быстрый перелет от мест гнездования к зоне
отдыха в Вост. Африке. Расстояние в 4 600 км, как взрослые, так и молодые птицы преодо-
левают в среднем за 18-19 дней. При нормальных условиях аисты летят ежедневно, прово-
дя в пути по 8-10 ч. Длительные, особенно многодневные остановки, встречаются лишь
как исключение и связаны прежде всего с неблагоприятными погодными условиями. У
аистов, в отличие от других перелетных птиц, жировые запасы во время миграции незначи-
394
Отряд Листообразные, Семейство Аистовые
тельны. Заметной гиперфагии во время перелета не наблюдается. Аисты практически не
восстанавливают вес до самой Африки (Berthold et al., 2001).
Большинство неполовозрелых аистов проводят летние месяцы вдали от мест гнездова-
ния. После первой зимовки птицы мигрируют в направлении области гнездования, но дос-
тигают её очень редко. Только треть однолетних аистов была обнаружена ближе 1000 км от
места кольцевания. С возрастом доля «невозвращенцев» быстро уменьшается. Значитель-
ная часть 1-2-годовалых аистов проводят лето южнее Сахары, но 3-летние птицы в гнездо-
вой период там не встречаются вообще. Кольцевание показало, что в большинстве случаев
аисты впервые появляются в местах гнездования в 3-летнем возрасте (Libbert, 1954; Kania,
1985; Bairlein, 1992).
Залётные аисты могут встречаться значительно севернее и восточнее границы гнездо-
вого ареала. В России они отмечались на берегу Белого моря в Мурманской обл. (Коханов,
1987), у с. Холмогоры в Архангельской обл. (Плешак, 1987), в Башкирии (Карякин, 1998а),
Татарстане (Аськеев, Аськеев, 1999), Пермской обл. (Демидова, 1997; Карякин, 19986),
Свердловской обл. (Зеленцов, 1995), в степях Юж. Урала (Давыгора, 2006). По недостаточ-
но надёжным данным, двух птиц наблюдали в августе в Курганской обл. (Тарасов и др.,
2003). Залёты белых аистов регистрируют также в Финляндии, Швеции, Норвегии, Вели-
кобритании, Ирландии, Исландии (Hancock et al., 1992; Бирина, 2003). Во время миграции
могут отмечаться настоящие инвазии, когда большие стаи оказываются вдали от основных
пролётных путей. Так, 15 сентября 1984 г. возле г. Абу-Даби на востоке Аравийского п-ова
появилась стая из 3 000 аистов (Reza Khan, 1989). 27-29 августа 2000 г. скопление из 300-
400 особей держалось в долине р. Теберда на Сев. Кавказе (Поливанов и др., 2001). Иногда
пролётные стаи аистов заносит ветром далеко в море. Такие птицы отмечаются даже на
Сейшельских о-вах, лежащих за 1 000 км от африканского побережья (Stork, 1999).
Местообитания. Белый аист — типичный обитатель открытых ландшафтов; сплош-
ных лесов и заросших болот избегает. Предпочитает территории с влажными биотопами —
лугами, болотами, пастбищами, поливными землями, рисовыми полями и т.п. Встречается
также в степях и саваннах с одиночными крупными деревьями или сооружениями челове-
ка. Оптимальным биотопом в наших условиях являются обширные поймы рек с нормаль-
ным гидрорежимом и экстенсивным сельскохозяйственным использованием. В таких мес-
тах плотность населения может достигать десятков пар на 100 км2. Населяет, как правило,
равнинные территории, но может гнездиться и невысоко в горах с подходящими условия-
ми. В Центр. Европе белые аисты редко гнездятся выше 500 м н.у.м (Schulz, 1998). В Кар-
патах они поднимаются до 700-900 м (Смогоржевський, 1979; Rejman, 1989; Stollmann,
1989), в Армении и Грузии — до 2 000 м н.у.м. (Адамян, 1990; Gavashelishvili, 1999), в
Турции до 2 300 м (Creutz, 1988), а в Марокко даже до 2 500 м н.у.м (Sauter, Schiiz, 1954). В
Болгарии 78,8% аистиных пар гнездится на высотах от 50 до 499 м н.у.м. и только 0,2% —
от 1 000 до 1 300 м (Petrov et al., 1999). В Польше отмечено расселение аистов на более
значительные высоты в ходе роста численности (Tryjanowski et al., 2005). Кормиться белый
аист предпочитает на открытых участках с невысокой травянистой растительностью, на
мелководье стоячих и медленнопроточных водоёмов. Реже встречается на берегах круп-
ных рек, горных ручьёв. Пахотные земли и интенсивно культивируемые луга и поля много-
летних трав также используются аистами для кормёжки, однако благоприятный период для
сбора корма в таких местах очень короткий — сразу после вспашки или сбора урожая.
Гнёзда аиста встречаются на периферии колоний цапель и других голенастых птиц.
Но чаще всего он гнездится в населённых пунктах. Может селиться даже среди плотной
застройки в больших городах, откуда ему приходится летать за кормом за 2-3 км. Поки-
нутые людьми села белый аист обычно со временем оставляет. Так, перестали эти птицы
Белый аист
395
гнездиться в большинстве выселенных сёл Чернобыльской зоны (Samusenko, 2000; Га-
шек, 2002).
Во время миграции белый аист также предпочитает открытый ландшафт; большие вод-
ные пространства и лесные массивы старается облетать, поскольку, как мы считаем, полет
над ними у специализированного парителя требует большего расхода энергии.
Численность. Общую численность белого аиста по результатам V Международного
учёта в 1994-1995 гг. можно оценить минимум в 170-180 тыс. пар, из них на восточную
популяцию приходится 140-150 тыс. пар (Грищенко, 2000а). По сравнению с предыдущей
переписью в 1984 г., общая численность выросла на 23%. Причём численность западной
популяции увеличилась значительно больше — на 75%, восточной — на 15% (Schulz, 1999).
Наибольшее количество белого аиста отмечено в Польше. В 1995 г. там учтено около 40900
пар, на 34% больше, чем в 1984 г. Средняя плотность гнездования в Польше составляет
13,1 пар/100 км2 (Guziak, Jakubiec, 1999). В Испании, где гнездится основная часть запад-
ной популяции, в 1996 г. численность оценивалась в 18 000 пар. В этой стране отмечен
наибольший её рост: между двумя международными переписями она увеличилась здесь
более, чем вдвое (Marti, 1999).
По предварительным итогам VI Международного учёта, проходившего в 2004-2005 гг.,
общая численность белого аиста оценивается в 230 тыс. пар. Наибольшая численность в
Польше — 52,5 тыс. пар, далее идут Испания — 33,2 тыс. пар, Украина — ок. 30 тыс. пар,
Белоруссия — 20,3 тыс. пар, Литва— 13 тыс. пар, Латвия — 10,7 тыс. пар, Россия — 10,2
тыс. пар. Наибольший прирост численности отмечен во Франции — 209%, Швеции —
164%, Португалии— 133%, Италии— 117%, Испании— 100%. Снизилась (вдвое) числен-
ность только в Дании. Там осталось всего 3 гнезда. По азиатскому подвиду данные приво-
дятся только для Узбекистана, где учтено 745 пар; численность снизилась на 49%.
По материалам, собранным в России в 1994-1997 гг., а также экспертным оценкам для
территорий, где учёт не был проведён или оказался неполным, общая численность гнездя-
щейся группировки составила минимум 7 100-8 400 пар (Cherevichko et al., 1999). Наибо-
лее плотно заселены аистами Калининградская и Псковская обл. — соответственно, 2 371
и 1 910 пар. В Брянской обл. учтено около 600 пар, однако, скорее всего здесь гнездилось от
800 до 1 000 пар; не менее 600 пар размножались в Смоленской обл. (в 12 западных из 25
административных р-нов области отмечено 449 пар). В Курской обл. учтено 325 пар, в
Новгородской — 316, в Тверской 200-230, в Калужской — около 200, в Ленинградской —
не менее 100 пар. От нескольких десятков до 100 пар обитали в Орловской и Белгородской
обл., в Московской обл. учтено 23 пары, в Воронежской — 10, Ярославской — 15-20, Ли-
пецкой — 5, Рязанской — 216, Кировской — 1, Мордовии — 1 пара (Галчёнков, 2000а;
Голубев, 2000; Дылюк, 2000).
В ходе VI Международного учёта, по предварительным данным, учтено: Курская обл. —
929 пар (+186% по сравнению с V Международным учётом, данные В.И. Миронова), Брян-
ская обл. — 844 (+31%, С.М. Косенко), Калужская обл. — 285 (+58%, Ю.Д. Галчёнков),
Ленинградская обл. — 160 (+344%, В.Г. Пчелинцев), Орловская обл. — 129 (С.В. Недосе-
кин), Московская обл. — 80 (+248%, М.В. Калякин).
Современная численность в Армении оценивается в 1-1,5 тыс. пар, в Азербайджане —
1-5 тыс. пар, в Молдавии — 400-600 пар (Birds in Europe, 2004).
На протяжении XX столетия численность белого аиста подвергалась значительным из-
менениям (см. Грищенко, 2000а). В первой половине века (а местами ещё раньше) нача-
лось быстрое её сокращение во многих странах Европы. К концу 1940-х гг. в Средней Ев-
16 В 2002 г. в Рязанской обл. по данным сотрудников Окского заповедника гнездилось не менее 4 пар
белых аистов (прим. ред.).
396
Отряд Листообразные, Семейство Аистовые
ропе она уменьшилась почти вдвое. Проводившиеся в 1934, 1958, 1974, 1984 гг. междуна-
родные учёты белого аиста показывали неуклонное снижение числа занятых гнезд. Так,
если в 1907 г. в Германии насчитывалось 7-8 тыс. гнездящихся пар (Wassmann, 1984), то к
1984 г. их количество уменьшилось до 649 в ФРГ (Heckenroth, 1986) и 2 724 в ГДР (Creutz,
1985). В Нидерландах в XIX в. белый аист был одной из обычных птиц, в стране были
тысячи гнёзд. Но уже в 1910 г. оставалось всего 500 гнездящихся пар, численность продол-
жала стремительно уменьшаться: 209 пар в 1929 г., 85 — в 1950 г., 5 — в 1985 г. (Jonkers,
1989). После 1991 г. не осталось вообще ни одной «дикой» пары, гнездились только птицы,
выпущенные из специальных питомников (Vos, 1995). Перестали аисты гнездиться в Бель-
гии, Швейцарии, Швеции, подошли к грани исчезновения во Франции, Дании и некоторых
других странах. Наиболее уязвимой оказалась западная популяция белого аиста. По дан-
ным IV Международного учёта в 1984 г., только за 10 лет численность её сократилась на
20%, восточной популяции — на 12% (Rheinwald, 1989).
Радикальное изменение ситуации началось в 1980-х гг., раньше всего в Испании. При-
мерно с 1987 г. начинается рост количества аистов. За 11 лет оно увеличилось более чем в
2,5 раза и вскоре превысило уровень полувековой давности (Gomez Manzaneque, 1992;
Martinez Rodriguez, 1995). Более чем в 2 раза выросла численность и в Португалии (Rosa et
al., 1999). Все это объяснялось прежде всего климатическими причинами. Во второй поло-
вине 1980-х гг. наконец прекратился длительный период засухи в зоне Сахеля, значительно
ухудшивший условия зимовки западной популяции белого аиста. Способствовало росту
численности и значительное улучшение кормовой базы на местах гнездования. В Испании,
например, увеличилась площадь орошаемых земель; кроме того в каналах прижился юж-
ноамериканский рак Procambarus clarkii, которого охотно поедают аисты (Schulz, 1994;
1999). В Испании и Португалии гораздо больше птиц стало оставаться на зимовку, что
также снизило смертность (Gomez Manzaneque, 1992; Rosa et al., 1999). Скачок численнос-
ти белого аиста на Пиренейском п-ове способствовал быстрому росту всей западной попу-
ляции. Вскоре подъем численности и расселение этих птиц начались во Франции, причём
связь с процессами, происходившими в Испании, была доказана: в 1990 и 1991 гг. нашли
аистов, гнездившихся на Атлантическом побережье Франции, а окольцованных в Испании.
Предполагается, что часть аистов, загнездившихся в департаментах по побережью Бискай-
ского залива, расселилась из Испании. На северо-востоке и в центре Франции появились
аисты из Эльзаса, Швейцарии и Нидерландов. В департаменте Приморская Шаранта в
1995 г. гнездился аист, окольцованный в 1986 г. птенцом в Польше. Отмечено также быст-
рое расселение аистов в Голландии, Швейцарии, Италии, Германии и других странах. Во
Франции с 1984 по 1995 гг. численность увеличилась на 830% (Duquet, 1999).
У восточной популяции не было таких резких скачков численности, как у западной, но
отмечен её положительный тренд. Подчеркнем, что при общем снижении численности
продолжалось расселение аистов на восток в России и на Украине и её рост у границы
ареала. Увеличение численности восточной популяции началось примерно в те же сроки,
что и западной, хотя темпы роста были гораздо ниже. Практически синхронно изменилась
ситуация и у азиатского подвида. С 1984 до 1994 г. численность белого аиста в Средней
Азии выросла более чем в 7 раз (Shemazarov, 1999), а к 2005 г. количество этих птиц здесь
оценивают в 700-1 000 гнездящихся пар (Митропольский, 2007).
По данным мониторинга на постоянных пробных участках на Украине, в 1990 гг. про-
шла волна роста численности. Она наметилась уже в первой половине 1990-х гг., несколько
раньше на северо-востоке Украины, позже всего — в западных областях. В 1992-1994 гг. в
селах по р. Сейм в Сумской обл. отмечалось увеличение численности на 25-30% ежегодно
(Грищенко, 1995а, 20006). С 1994 г. средний по Украине прирост все время увеличивался
Белый аист
397
(спад отмечен лишь в 1997 г., крайне неблагоприятном для белого аиста по всей Европе),
достигнув максимума в 1996 и 1998 гг. — соответственно 13,7 ± 2,9 и 16,3 ± 3,6 %. Затем
темпы роста начали уменьшаться, а в 2001-2003 гг. популяция стабилизировалась. (Гри-
щенко, 2004).
В этот же период активизировалось расселение на восток в восточных областях Украи-
ны и в России. В Харьковской обл. к 1994 г. отмечено смещение границы ареала на восток
по сравнению с распространением в 1974-1987 гг., в 1998 г. обнаружены гнёзда на правом
берегу р. Оскол (Атемасова, Атемасов, 2003). В Луганской обл., где белый аист встречался
на восток до р. Айдар, в 1998 г. найдены 2 гнезда в пойме р. Деркул на границе с Россией
(Ветров, 1998). В Ростовской обл. в 1996 г. аисты снова загнездились после 5-летнего пере-
рыва — было обнаружено гнездо в долине Маныча (Казаков и др., 1997). В Краснодарском
крае аисты начали гнездиться в середине 1990-х гг. (Мнацеканов, 2000). В 1993 г. гнездова-
ние впервые отмечено в Кировской обл. (Сотников, 1997, 1998), в 1994 г. — в Тамбовской
обл. (Евдокишин, 1999), в 1995 г. — в Мордовии (Лапшин, Лысенков, 1997,2000), в 1996 г. —
в Вологодской обл. (Дылюк, 2000). В 1996 г. отметили резкое увеличение количества птиц
(на 20,1%) в Калужской обл. (Галченков, 2000а).
Размножение. Белые аисты начинают размножаться обычно в возрасте от 3 до 7 лет,
иногда в 2 года. Соотношение возрастов первого размножения различается в разных реги-
онах. Так, на северо-западе Германии 1,3% аистов начинали гнездиться в возрасте 2 лет,
12,0% — 3 лет, 26,6% — 4,19,0% — 5,17,7% — 6,23,4% — 7 лет; на юго-западе Германии:
5,9% — в возрасте 2 лет, 45,4% — 3, 31,9% — 4, 13,4% — 5, 3,4% — 6 лет; в Эльзасе во
Франции: 3,2% — в возрасте 2 лет, 50,5% — 3, 36,6% — 4, 9,7% — 5 лет. Успешность
размножения повышается с возрастом, по крайней мере в промежутке между 6 и 10 года-
ми. Большинство размножающихся птиц имеют возраст 7 лет (Meybohm, Dahms, 1975). На
юго-западе Германии средний возраст первого размножения в 1961-1976 гг. составлял 3,78±
0,94 года (Bairlein, Zink, 1979).
В Баварии 5% птиц местной популяции имели возраст 2-3 года, 51% — 4-7 лет, 27% —
8-11, 17%— 12 лети более; в Шлезвиг-Гольштейне: 7% — 2-3 года, 43% — 4-7 лет, 31% —
8-11, 19% — 12 и более (Schulz, 1998). В Сев.-Зап. Европе птицы до 5 лет составляют 21%,
в возрасте 6-8 лет — 33%, 9-11 — 27%, 12-26 — 19% (Meybohm, Dahms, 1975).
Белый аист — моногам, пары образуются только на период гнездования. Центром соци-
альной активности для размножающихся птиц служит гнездо, где можно видеть разнооб-
разные демонстрации аистов (рис. 80а-н). За пределами гнездовой территории партнеры
могут вообще игнорировать друг друга (Kahl, 1972). Как правило, самец весной возвраща-
ется первым и занимает гнездо, защищая его от других аистов. Пара образуется на гнезде.
Приближающихся других аистов хозяин гнёзда встречает трещанием клюва, запрокидыва-
ет назад и опускает голову, распуская при этом крылья, поднимая хвост и топорща перья на
шее (up-down display, рис. 80в-з).
Это универсальная демонстрация, она используется постоянно обоими партнёрами и
имеет различное развитие в зависимости от ситуации. Если прилетевший на гнездо аист
оказывается самкой, через некоторое время демонстрации переходят в приветственную це-
ремонию. Самец ложится в гнездо, топорщит перья на шее и энергично качает головой из
стороны в сторону. При приближении самцов токовое поведение переходит в угрожаю-
щее: хозяин гнезда опускает крылья и распускает веером хвост, громко треща клювом.
Если чужой самец пытается сесть на гнездо, птица принимает позу угрозы: аист стоит
неподвижно на полусогнутых ногах с опущенными и разведёнными в стороны крыльями,
задранным хвостом и встопорщенными перьями на шее; голова и шея вытянуты вперёд
(рис. 80л).
398
Отряд Аистообразные, Семейство Аистовые
Рис. 80.
Белый аист
399
Рис. 80. Элементы демонстрационного и иного поведения белого аиста (по Cramp, 1977; Bauer,
Glutz von Blotzheim, 1987; Schulz, 1993): a — спокойная поза; б — тревога, низкий уровень опаснос-
ти; в-з — последовательность поз во время токования (up-down display); и-к — совместное токова-
ние на гнезде пары белых аистов; л — поза угрозы на гнезде; м — спаривание; н — защита птенцов
от солнечных лучей.
400
Отряд Листообразные, Семейство Аистовые
Из этой позиции птица может садиться на гнездо с широко расправленными крыльями,
как бы закрывая его. Если чужак все же приземляется на гнездо, следует атака. В драке
аисты используют удары клювом и крыльями. При этом хозяин старается столкнуть при-
шельца с края гнезда. Драки могут продолжаться часами, нередко заканчиваясь серьезны-
ми травмами и смертью (см. выше). Преследуя чужака, хозяин может демонстрировать
угрозу и в полёте: он летит немного выше и сзади, треща клювом, направленным на сопер-
ника. После образования пары аисты приветствуют подлетающего к гнезду партнёра опи-
санными выше токовыми демонстрациями. На гнезде обе птицы нередко токуют «дуэтом»
(рис. 80 и, к). Партнеры охотно перебирают друг другу перья, особенно на голове и шее
(Kahl, 1972; Cramp, 1977; Hancock et al., 1992).
Гнёзда белый аист чаще всего устраивает на деревьях и различных сооружениях чело-
века— жилых и хозяйственных постройках, электрических и телефонных столбах, водона-
порных башнях (чаще всего на так называемых башнях Рожновского с плоской вершиной)
и т.п. На Украине, по данным VI Международного учёта аистов в 2004-2005 гг., 40,8%
гнёзд находились на столбах электролиний, 34,0% — на деревьях, 13,4% — на водонапор-
ных башнях, 9,4% — на постройках. Расположение гнёзд различается в разных природных
зонах. Наибольшая доля гнёзд на деревьях отмечается в лесной зоне, на столбах — в степ-
ной. Есть различия, связанные с культурными традициями. Так, больше всего гнёзд на по-
стройках находится в западных областях Украины. Общая тенденция — повсеместно по-
вышается доля гнёзд на столбах: в 1931 г. их не было вовсе, в 1987-1988 гг. в целом по
Украине на столбах располагалось 25,6% гнёзд (Грищенко и др., 1992), в 1994-1995 гг. —
33,2% (Grishchenko, 1999). Сейчас доля гнёзд на столбах в некоторых северных областях
уже почти такая же, как на юге. Количество гнёзд на постройках, напротив, все уменьшает-
ся: в 1931 г. их было на тогдашней территории УССР 68%, в 1987-1988 гг. — 20% (Грищен-
ко и др., 1992), в 1994-1995 гг. — 12,9% (Grishchenko, 1999). В настоящее время увеличи-
вается также доля гнёзд на водонапорных башнях, особенно в восточных областях. Места-
ми их количество превышает треть от общего числа.
В Белоруссии в 1994-1996 гг. 50,8% гнёзд располагались на деревьях, 17,5% — на водо-
напорных башнях, 17% — на постройках, 14,4% — на столбах (Samusenko, 1999). Здесь
отмечена та же тенденция изменения расположения гнёзд. В 1967 г. в Белоруссии 59,5%
аистов гнездились на деревьях и 40,5% — на крышах строений. Гнёзда на столбах и водо-
качках не были отмечены. В 1985 г. количество гнёзд на крышах сократилось до 28,1%,
7,5% гнёзд находились на столбах и 7,3% — на водонапорных башнях (Самусенко, 1992). В
Латвии доля гнёзд на деревьях уменьшилась с 91% в 1934 г. до 30% в 1995 г., на столбах —
увеличилась с 1% в 1974 г., до 43% в 1995 г. (Janaus, Stopniece, 1999). Аналогичные тенден-
ции отмечены в Армении (Адамян, 1990), Болгарии (Petrov et al., 1999), Венгрии (Fenyxsi,
1994, Lovaszi, 2004), Польше (Bogucki, Oi'go, 1999) и других странах. В Литве в 1994-1995
гг. 52% гнёзд были построены на деревьях, 20% — на столбах, 16% — на водонапорных
башнях, 12% — на постройках (Malinauskas, Zurba, 1999). В Польше в 1995 г. 36,7% гнёзд
размещались на столбах, 32,0% — на постройках, 26,0% — на деревьях; расположение
гнёзд было различным в разных регионах (Guziak, Jakubiec, 1999). В Узбекистане в 1994 г.
75% гнёзд находились на столбах, 22,1% — на деревьях, 2,9% — на водонапорных башнях
(Shemazarov, 1999). Гнездятся аисты и на скалах. Так, в Португалии в 1994 г. на них находи-
лось 2,3% гнёзд (Rosa et al., 1999). Помимо этого, белые аисты могут устраивать гнёзда на
развалинах, памятниках, различных вышках, скирдах соломы, кучах навоза и хвороста,
стрелах автокранов и т.п. (Creutz, 1988; Schulz, 1998; Грищенко, 2005). Известно немало
случаев гнездования и на земле (Dahms, 1973, 1999; Antipow, Domaszewicz, 1997).
Высота расположения гнёзд зависит от высоты выбранной для гнездования опоры и
может варьировать от 0 (гнёзда на земле) до десятков метров (гнёзда на трубах и других
Белый аист
401
сооружениях человека). В Испании гнездо на одной из военных вышек связи находилось
на стометровой высоте (Schulz, 1998). Большинство гнёзд строится на высоте от 5 до 20 м.
В Черниговской обл. 38,8% гнёзд были расположены на высоте 8-10 м, 24,3% — 3-7 м,
22,7% — 11-16, 7,9% — 17-20, 6,2% — выше 20 м (Марисова, Макаренко и др., 1992).
Как показывают наблюдения в зоне роста численности, аисты используют все суще-
ствующие возможности для постройки гнёзд в равной мере, и их размещение отражает не
столько предпочтения птиц, сколько соотношение этих возможностей (Грищенко, 1998а;
2005). Нет также особых предпочтений к разным вариантам искусственных гнездовий (Гри-
щенко, 1999). Пластичность птиц в выборе мест гнездования подтверждается и тем, что
аисты охотно переходят на искусственные гнездовья при массовой их постройке. Так, в
Словакии удалось сократить число гнёзд на столбах электролиний путём постройки, при
поддержке государства, искусственных гнездовий. В 1995 г. на столбах ЛЭП располагалось
37% гнёзд, а в 1984 г. — всего 2,5% (Fulin, 1999).
В Калиниградской обл. в качестве гнездового субстрата преобладают водонапорные
башни: на них размещены 32% гнезд, далее следуют черепичные и шиферные крыши —
26%, столбы и опоры — 21%, деревья — 16%, трубы — 5% (Гришанов, Савчук, 1992). В
Ленинградской обл. 47% гнёзд размещены на водонапорных башнях, на деревьях — 43%
(Пчелинцев, Ильинский, 2000). В Тверской обл. на водонапорных башнях селятся 70% пар,
на деревьях — 29% (Николаев, 2000). В Калужской обл. на водонапорных башнях распо-
лагается 73% гнёзд, на деревьях — 17%, на различных столбах и опорах ЛЭП — 5%,
церквях — 4%, на крышах и трубах зданий и сооружений — менее 1% (Галченков, 20006).
В Воронежской обл. в 1992-1999 гг. на водонапорных башнях находилось 50% гнёзд, на
различных строениях 31%, на ЛЭП — около 13% и на деревьях — около 6% (Нумеров,
Макагонова, 2000). В Липецкой обл. все известные гнёзда размещались на сооружениях
человека (7 — на водонапорных башнях и 1 — на церкви) (Сарычев, 2000). В Курской обл.
в 2004 г. на водонапорных башнях находилось 48% гнёзд, 40% располагалось на деревьях,
9% — на опорах ЛЭП и 3% — на других сооружениях (В.И. Миронов, личн. сообщ.).
Большинство аистов селится не далее 50-100 км от места рождения. На северо-западе
Германии 3,7% аистов гнездились на расстоянии до 1 км от места рождения, 1,1% — 2-10
км, 23,8% — 11-25 км, 17,9% — 26-50 км, 23,6% — 51-100 км, 10,7% — 101-200 км,
3,2% — 201-700 км (Meybohm, Dahms, 1975). На юго-западе Германии 70% птиц селились
в радиусе до 50 км от места рождения, среднее расстояние для самцов составляло 33 км,
для самок — 61 км (Zink, 1967). Эти цифры могут различаться в разных регионах. Так, в
Баварии только 30% аистов гнездились не далее 50 км от родительского гнезда, в Дании —
85,6%, на востоке Польши — 81%, в различных регионах Вост. Германии — 43,7-65,6%
(Schulz, 1998). Известен случай переселения на 910 км (Meybohm, Fiedler, 1983). Наблюде-
ния за 93 окольцованными аистами в Вост. Германии показали, что 20 из них занимали
одно и то же гнездо 2 года подряд, 13 — 3 года, 7 — 4 года, 3 — 5 лет, 2 — 6 лет, 3 — 8 лет,
по 1 паре — 11, 14 и 19 лет. В Эльзасе один аист занимал то же самое гнездо 16 лет
подряд (Creutz, 1988). При создании водохранилищ на Днепре, аисты некоторое время
возвращались на затопленные территории, пока уровень воды был небольшим. Птицы
гнездились на оставшихся среди воды постройках, холмиках, пнях, выброшенных дере-
вьях. Оставили они старые места лишь после полного их затопления (Гавриленко, 1968;
Клестов, 1983).
Белые аисты гнездятся обычно отдельными парами, но могут образовывать и довольно
крупные колонии. Иногда можно увидеть больше десятка гнёзд на одном дереве. В Арме-
нии есть несколько колоний на железнодорожных станциях численностью до нескольких
десятков пар, где аисты гнездятся на опорах контактной сети (Адамян, 1990). Крупнейшая
402
Отряд Листообразные, Семейство Аистовые
колония аистов на одном здании известна в г. Альфаро в Испании: в 1994 г. на крыше собо-
ра Св. Мигеля гнездилось 60 пар (Schulz, 1996).
Гнездо белого аиста — крупная постройка из древесных веток толщиной до нескольких
сантиметров; внутренняя часть гнезда выложена более тонкими ветками. В стенках гнёзда
попадается также земля, дерн, стебли растений, сено, солома, навоз. Лоток выстилается
толстым слоем мягкого материала — сухой травой, листьями, мхом, сеном, шерстью и т.п.
В гнезде можно найти и различный мусор — куски бумаги и полихлорвиниловой пленки,
тряпки, обрывки верёвок. Вновь построенное гнездо меньше размерами, особенно в высо-
ту. Но т.к. гнёзда занимаются много лет, птицы постепенно их надстраивают. Старые гнёзда
могут достигать 2 м в диаметре, 3-4 м в высоту и веса 1-2 т (Bauer, Glutz von Blotzheim,
1966; Creutz, 1988). В Черниговской обл. 36,5% гнёзд использовались птицами от 6 до 10
лет, 22,9% — 3-5, 16,2% — 11-20, 13,3% — более 20, 11,0% — всего 1-2 года (Марисова,
Макаренко и др., 1992). Возраст наиболее старых гнёзд в Калужской и Тверской обл. —
30-35 лет. В Польше, Венгрии, Германии известны гнёзда «возрастом» более 100 лет (Creutz,
1988; Jakubiec, Szymocski, 2000).
Как показали наблюдения в населенных пунктах, в первую очередь занимаются старые
многолетние гнёзда, тогда как построенные 1-2 года назад — как правило, к концу периода
гнездостроения. Новые гнёзда птицы строят обычно лишь после того, как уже заняты все
существующие (Грищенко, 1998а, 2005). Новые гнёзда чаще всего строят молодые аисты,
поэтому успешность размножения в них в среднем ниже (Вероман, 1981; Грищенко 20006;
Lovaszi, 2005).
Строят гнездо оба партнёра, но самцы принимают в этом больше участия. Нередко один
из аистов сторожит постройку, пока другой собирает материал. Новое гнездо обычно стро-
ится в течение 8 дней (Bauer, Glutz von Blotzheim, 1966; Schulz, 1998). По наблюдениям в
Армении, постройка гнезда занимает от 4 до 15-20 дней (Адамян, 1992). Быстрее гнёзда
строят птицы, начавшие гнездиться в более поздние сроки, либо взамен разрушенных. Стро-
ительный материал аисты собирают на расстоянии до 500 м от гнезда, реже дальше. Приле-
тают с ним каждые 10^40 мин, проявляя наибольшую активность в утренние часы (Ада-
мян, 1992). Достройка гнёзда продолжается практически весь гнездовой сезон.
В крупных гнёздах белого аиста довольно часто селятся другие птицы. Обычно это до-
мовые и полевые воробьи, скворцы. Известны случаи гнездования и таких крупных видов
как галка, домовый сыч, пустельга, сизоворонка и даже серая неясыть (Creutz, 1988; Schulz,
1998; Brix, 2005).
Размеры гнёзд (см). В Киевской и Винницкой обл. (п = 16): диаметр гнезда— 106-230
(в среднем 186), диаметр лотка — 76-172 (115), высота гнезда — 53-108; диаметр трёх
вновь построенных гнёзд составлял 129-134 см, диаметр лотка — 66-72 см (Смогоржевсь-
кий, 1979). В Белоруссии (п = 7): диаметр гнезда — 70-280 (130), диаметр лотка — 35^40
(37), высота гнезда — 40-115 (77), глубина лотка — 8-12 (10) (Никифоров и др., 1989). В
Армении: (п = 3): диаметр гнезда — 150-206, диаметр лотка — 75-102, высота гнезда —
30-58, глубина лотка — 10-15 (Адамян, 1992). В России (п = 12, Калужская, Тверская и
Ярославская обл.): диаметр гнезда — 110-255, диаметр лотка — 50-200, высота гнезда —
10-150, глубина лотка— 12-15.
Средние размеры гнёзд туркестанского белого аиста в Узбекистане (см): диаметр гнез-
да — 110, диаметр лотка — 94, высота гнезда — 54, глубина лотка — 11 (Сагитов, 1987).
На гнёздах аисты появляются вскоре после прилёта. Как правило, первым занимает гнез-
до самец, реже самка, иногда появляются обе птицы сразу. Пара образуется на гнезде, при-
чём возможна неоднократная смена партнёров. Так, на одном из гнёзд в Польше в 1994 г.
после прилёта сменились, по крайней мере, три самца и одна самка. Первая пара продер-
Белый аист
403
жалась 5-8 дней, вторая — всего один, и лишь третья комбинация партнеров оказалась
устойчивой (Wuczycski, 2005) в течение всего сезона. Известны и случаи бигамии. В одном
из гнёзд в Германии в 1957 г. самец и две самки успешно вывели 5 птенцов (Schmidt, 1989).
Хозяева активно защищают гнездо от других аистов. При этом на гнёздах нередко воз-
никают жестокие драки, которые иногда заканчиваются гибелью или серьезными ранения-
ми птиц; могут также выбрасываться отложенные яйца и даже маленькие птенцы. В Вост.
Пруссии в 1931 г. на 8 835 гнёздах отмечено 2 660 драк, 77 из которых окончились смер-
тью птиц. Наиболее жестокие драки возникают между взрослыми птицами одного пола
(Schiiz, 1944).
Сроки размножения белого аиста довольно растянуты. Одновременно можно увидеть и
насиживающих птиц, и только занимающих гнёзда. Связано это с растянутостью сроков
прилёта и с тем, что не всём птицам удаётся сразу занять гнёзда. Кроме того, часть отло-
женных кладок гибнет. Потерявшие их аисты откладывают яйца повторно. Повторные кладки
могут появляться и после гибели птенцов (Anderegg, 2000). Занятие гнёзд растягивается со
второй половины марта до мая. Так, на юге Польши первая птица появлялась на гнезде в
5,8% случаев до 20 марта, в 31% — 21-31 марта, в 43,9% — 1-10 апреля, в 13,2% — 11-20
апреля, в 4,1% — 21-30 апреля, в 2% — в мае. Второй партнёр появлялся на гнезде в сред-
нем через 4,5 дня после первого (от 0 до 25 дней) (Profus, 1991).
Откладка яиц начинается в апреле, а в южной части Европы — даже в марте. В Армении
первые яйца птицы откладывают уже в конце марта. В долине Аракса самый ранний вылет
птенцов отмечен в первой половине июля, массовый вылет происходит в августе. Птенцы
из поздних кладок покидают гнёзда в начале сентября (Адамян, 1992). В Молдавии первые
яйца появляются в гнёздах уже в начале апреля, а полные кладки обычно отмечаются с
конца второй декады. Птенцы вылупляются во второй половине мая, вылет их происходит
во второй половине июля (Аверин и др., 1975). Под Ленкоранью в Азербайджане откладка
яиц отмечалась в первую декаду апреля, птенцы вылуплялись в начале мая (Спангенберг,
1951). На Украине яйца откладывают во второй половине апреля - первой половине мая,
(хотя запоздалые пары приступают к размножению лишь к концу мая). Птенцы появляются
в конце мая - первой половине июня, покидают гнёзда во второй половине июля — начале
августа (Страутман, 1963; Смогоржевський, 1979; Стадницкий, 1992; Грищенко, 2005). На
севере Сумской обл. средняя дата начала массовой откладки яиц — 24 апреля (Афанасьев,
1998). В Белоруссии откладка яиц начинается в третьей декаде апреля - начале мая (Ники-
форов и др., 1989). В Эстонии аисты откладывают яйца в конце апреля - первой половине
мая. Птенцы начинают вылупляться в конце мая, вылетают из гнёзд во второй половине
июля - в августе (Вероман, 1976).
В Калининградской обл. яйцекладка происходит с 11 по 20 апреля (Беляков, Яковчик,
1980). В Воронежской обл. откладка яиц завершается к 24 апреля - 19 мая, в среднем 8 мая
(1995-1998 гг.) (Нумеров, Макагонова, 2000). Вылупление птенцов происходит в Калинин-
градской обл. с 10 мая (Tischler, 1941) с пиком на третью декаду мая, в Воронежской обл. —
с 22 мая по 24 июня, в среднем — 10 июня. Птенцы покидают гнёзда с середины июля по
конец августа. В Калужской обл. наиболее ранняя дата вылета молодых из гнезда — 15
июля, наиболее поздняя — третья декада августа. В Воронежской обл. последние молодые
слетали с гнёзд 20 июля - 8 августа, в среднем 29 июля (1995-1998 гг.). В Псковской обл.
птенцы вылетают во второй половине июля - начале августа (Фетисов и др., 1986).
В Узбекистане туркестанские белые аисты начинают откладку яиц в третьей декаде
марта - первой половине апреля (Сагитов, 1987; Третьяков, 1990). На юге Казахстана пер-
вые яйца появляются уже в середине марта, к началу апреля попадаются уже насиженные
кладки (Долгушин, 1960).
404
Отряд Листообразные, Семейство Аистовые
В полной кладке обычно 3-5 яиц, реже 1-2 или 6-8. В Молдавии из 453 обследованных
кладок 62,5% содержали по 4 яйца, 23,4% — по 5, 11,3% — по 3, 1,8% — по 2, 0,9% — по
6, 0,2% — 7 яиц (Аверин и др., 1971); в Белоруссии (Беловежская пуща) из 236 кладок
47,5% содержали 4 яйца, 27,1% — 3, 13,1% — 2, 11,0% — 5, 0,8% — 1, 0,4% — 6 яиц
(Голодушко, 1958); в Эстонии из 36 гнёзд с полными кладками в 38,9% было 3 яйца, в
33,3% — 4, в 13,9% — 5, в 5,6% — 2, в 2,8% — по 6, 7 и 8 яиц (Вероман, 1976); в Армении
из 22 гнёзд 50% содержали 4 яйца, 40,9% — 5, 4,5% — по 6 и 7 яиц (Адамян, 1992); в
Польше из 90 кладок в 54,5% было по 4 яйца, в 23,3% — по 5, в 11,1% — по 3, в 7,8% — по
2, в 3,3% — по 6 яиц (Profus, 1986). Размер 8 полных кладок в Воронежской обл. составил
от 2 до 5 яиц, в среднем 3,38±0,37 яиц. Средняя величина кладки в одном из сел Волынской
обл. в 1978-1989 гг. (по 3-8 гнёзд в год) изменялась от 4 до 5,5 (Шкаран, 1990). Средняя
многолетняя величина кладки в европейских странах варьирует в пределах 3,5-5,02 (Creutz,
1988; Profus, 1991).
В 75% обследованных кладок туркестанского белого аиста в Узбекистане (п = 24) было
по 4 яйца, в 25% — по 5 (Сагитов, 1987).
Яйца чисто белого цвета, после начала насиживания они часто приобретают желтова-
тый оттенок и покрываются коричневатыми пятнышками. Свежее яйцо на просвет зелено-
ватое. Форма яиц довольно разнообразна, обычно близка к эллиптической. Зёрна скорлупы
имеют характерную штриховку (Schonwetter, 1967; Creutz, 1988).
Размеры яиц приводятся в табл. 32. Размеры яиц из гнезда в Кировской обл. на северо-
восточном краю ареала (д. Вершинята Уржумского района, 1996 г.): 67,0 х 52,7 и 71,0 х
52,6 мм, вес: 95,2 и 102,0 г (яйца слабонасиженные). Средний вес 84 свежих яиц в Польше
115 г (90-139,9 г) (Profus, 1991). Вес 7 яиц из Черниговской обл.: 86,7-110,6 г, в среднем —
102,7 г (Смогоржевський, 1979), 6 яиц из Сумской обл.: 96,5-104,8 г, в среднем — 101,1г
(Афанасьев, 1998). Вес скорлупы (п = 150) 8,6-13,6 г, в среднем — 11,1 г. Толщина скорлу-
пы — 0,53 мм (Creutz, 1988). В ходе насиживания яйцо теряет около 13% веса (Creutz,
1988). Яйца белого аиста из Сев. Африки и Юж. Европы немного мельче, чем в Сев. и Вост.
Европе (Profus, 1986).
Таблица 32. Размеры яиц (мм) белого аиста в разных частях ареала
Регион п М Lim Источник
Ciconia ciconia ciconia
Территория бывш. СССР 16 73,8 x 53,8 71,0-78,2 x 47,9-57,0 Спангенберг, 1951
Армения 102 72,5 x 53,7 63-79 x 50-58 по: Адамян, 1992
Белоруссия 65 72,68 x 52,25 67,2-79,1 x 47,0-55,3 Никифоров и др., 1989
Молдавия 15 73,5 x 52,9 Аверин и др., 1971
Россия (Псковская обл.) 4 73,5 x 53,2 72,0-74,9 x 52,3-54,6 Фетисов и др., 1986
Венгрия 42 72,44 x 51,82 64,2-78,8 x 46,9-55,3 Profus, 1991
Польша (Силезия) 125 73,17 x 52,02 66,6-88,6 x 46,3-57,1 Profus, 1991
Чехословакия 53 72,8 x 52,9 65,6-78,7 x 42,7-55,0 Ciconia ciconia asiatica Hudec, Иегпу, 1972
Узбекистан (Зеравшан) 29 74,7 x 54,1 Сагитов, Гульмурадов, 1972
Узбекистан 19 71,3 x 51,8 70,5-72,0 x 51,0-52,5 Мамашукуров и др., 1998
Узбекистан (Ферганская долина) 6 72,4-78,5 x 50,7-52,9 Третьяков, 1990
Белый аист
405
У туркестанского белого аиста (п = 19) средний индекс удлинённости яйца 72,5±0,2%,
средний вес — 102,2 г (Мамашукуров и др., 1998). Вес 6 яиц из Ферганской долины 95,0-
112,2 г (Третьяков, 1990).
Яйца аист откладывает с интервалом от 1 до 4 дней, обычно 2 дня. Чаще всего это
происходит ночью. Насиживание начинается после откладки 2-го, реже — 3-го яйца. Инку-
бация продолжается от 29 до 34 дней, в среднем — 32. Насиживают обе птицы, но самка
проводит на гнезде больше времени, особенно ночью. Птенцы вылупляются асинхронно с
интервалом от 6 ч до 2,5 дней. Скорлупу яиц родители обычно перекладывают на край
гнёзда, где она через некоторое время накрывается приносимым гнездовым материалом
(Creutz, 1988; Schulz, 1998). Успешность вылупления зависит от величины кладки и после-
довательности откладки яиц. В больших кладках реже вылупляются все птенцы из-за боль-
шего количества неоплодотворённых яиц (Profus, 1991; Адамян, 1992). В Швейцарии наи-
меньшая вылупляемость была у 1-го, 5-го и 6-го яиц, наибольшая — у 3-го и 4-го (Bloesch,
1982).
К моменту вылупления у птенцов сохраняются довольно большие жёлточные мешки, за
счет которых они могут существовать первые сутки (Смогоржевський, 1979). Птенцы рож-
даются слепыми, но глаза у них открываются уже через несколько часов. Вес они набирают
очень быстро. Вылупившийся птенец весит 68-88 г, пятидневный — 160 г, в возрасте 9-13
дней — 500 г, 14-18 дней — 1 000 г, 18-24 дней — 1 500 г (Creutz, 1988; Schulz, 1998). Вес
птенца перед вылетом достигает 3,5-3,9 кг (Смогоржевський, 1979). Увеличение массы
тела лучше всего описывается логистическим уравнением, длины тела — линейным. Наи-
больший рост происходит первые 10 дней. Масса тела примерно на 45-й день достигает
асимптотического значения в 3,4 кг. Длина клюва линейно увеличивается на протяжении
51 дня (Tsachalidis et al., 2005).
В первые дни после вылупления птенцов аисты кормят их в основном мелкой и мягкой
пищей — червями, головастиками, личинками насекомых. С 3-й недели размер добычи
увеличивается, а после 5-й появляются грызуны, пресмыкающиеся и другие сравнительно
крупные животные (Крапивный, 1957; Schulz, 1998).
До 3 недель один из родителей постоянно остаётся с птенцами, защищая и обогревая
их. Птенцы с первого дня едят самостоятельно. Взрослые аисты приносят пищу в клюве
или глотке и отрыгивают её в середину гнезда. Они обеспечивают птенцов также водой,
выливая её из клюва. Частота прилётов зависит от возраста птенцов и расстояния полётов
за кормом. Обычно корм приносится несколько раз в день каждые 1-2 ч. Подросшие птен-
цы принимают характерные позы и пищат, выпрашивая корм, могут хватать родителей за
клюв. В возрасте 6-7 недель они уже начинают махать крыльями. Птенцы покидают гнез-
до в возрасте от 54 до 68 дней. Первое время родители подкармливают их. Выводок дер-
жится поблизости от гнезда на лугах или пастбищах, возвращаясь к нему на ночевку.
Полностью самостоятельными молодые аисты становятся в возрасте около 3 мес. (Creutz,
1988; Schulz, 1998).
Гнёзда покидают обычно от 2 до 4 птенцов, реже — 1 или 5, совсем редко — 6. Извест-
но несколько случаев выкармливания 7 птенцов — в Венгрии (Radvanyi, 1943; Satori 1943),
Дании (Skov, 1999), на Украине (Смогоржевський, 1979; Грищенко, 1996а). Абсолютный
рекорд — 8 слётков в гнезде белого аиста у одной из пар в Венгрии в 1999 г. (Kovacs,
Kovacs, 1999). На Украине, по данным мониторинговых наблюдений на постоянных проб-
ных участках в 22 областях в 1992-2006 гг. (п=5 781), 41,3% выводков перед вылетом име-
ли 3 птенцов, 29,3% — 2, 20,5% — 4, 4,8% — 1, 4,3% — 5, 0,3% — 6, 0,02% — 7 птенцов
(единственный случай за 15 лет наблюдений). В благоприятные годы увеличивается коли-
чество больших выводков, доля гнёзд с 3 и 4 птенцами почти выравнивается, а в отдельных
406
Отряд Листообразные, Семейство Аистовые
регионах последние могут даже преобладать. В неуспешные же годы максимум смещается
на выводки из 2 птенцов. На западе Польши в 1971-1992 гг. (п = 9 200) 38,2% выводков
имели по 3 птенца, 37,5% — 2, 14,6% — 4, 8,0% — 1, 1,5% — 5, 0,03% — 6 птенцов
(Bogucki, 1994).
Репродуктивные показатели сильно варьируют в зависимости от года и региона. Так, на
Украине, по данным 1992-2006 гг., среднее количество вылетевших птенцов на размно-
жавшуюся пару составляло 2,51±0,03, на успешную пару — 2,86±0,03; средняя доля неус-
пешных пар была равной 12,9±0,6%. Наименее успешным был 1997 г. —1,87±0,12,2,49±0,10
и 24,9±3,0% соответственно; наибольшее среднее количество птенцов на размножавшую-
ся пару составило 2,94±0,10 (1995 г.), на успешную пару — 3,34±0,13 (1996 г.); наимень-
шая доля неуспешных пар отмечена в 1992 г. — 5,6±2,0%. Обнаружены существенные ре-
гиональные отличия. Успешность размножения оказалась выше в северо-восточных обла-
стях, Среднем Приднепровье и на юге Украины, чем в западных и центральных областях.
Наиболее высокие показатели отмечены в Полтавской и Харьковской обл. (Грищенко, 2005).
В Центр. Европе размножавшаяся пара выводит в среднем около 2 птенцов. Этот показа-
тель колеблется от 0,8 до 3,4 (Bauer, Glutz von Blotzheim, 1966). Средние многолетние пока-
затели для разных частей ареала приводятся в табл. 33.
В России в Калужской обл. на 1 успешно размножавшуюся пару поднимается на крыло
в разные годы от 2,70 (2005 г.) до 3,73 (1999 г.) молодых (1988-2006 гг.); преобладают
выводки из 3 или 4 птенцов — по 34%, выводки из 2 птенцов составляют 15%, из 5 — 13%,
1 птенца выращивают 4% пар. В благоприятные годы могут преобладать выводки из боль-
шего количества птенцов: в 1999 г. 5 птенцов были у 32% пар, 4 — у 29%. В неблагоприят-
ном 1997 г. выводков из 5 птенцов было только 2%. Шесть птенцов пара аистов вывела в
Таблица 33. Средние репродуктивные показатели белого аиста в разных частях ареала
Регион Период JZa JZm %НРо Источник
Латвия 1989-1999 1,98 2,29 13 Janaus, 2000
Эстония 1939-1984 1,96 2,66 26,6 Veromann, 1989
Белоруссия 1975-1996 2,42 2,66 8,0 по: Samusenko, 1999
Украина 1992-2006 2,51 2,86 12,9 В.Н. Грищенко, ориг.
Украина, Львовская обл.. 1992-2005 2,24 2,53 14,7 Грищенко, 2005
Украина, Полтавская обл. 1992-2005 3,17 3,54 10,3 Грищенко, 2005
Россия, Псковская обл. 1982-1984 2,00 2,27 11,9 по: Фетисов и др., 1986
Россия, Калужская обл. 1988-2006 2,67 3,27 18,9 Ю.Д. Галчёнков, ориг.
Дания 1952-1996 1,86 2,72 32 Skov, 1999
Испания, 1983-2002 1,84 2,42 26,2 по: Martin, 2002
г. Алькала-де-Энарес
Центральная Европа 1973-1978 1,87 2,62 28,5 Profus, 1991
Германия, Оберлаузиц 1966-1983 1,93 2,69 28,4 Creutz, 1988
Германия, Саксония 1986-1995 1,91 2,56 26,6 по: BaBler, Schimkat, 1999
Германия, Шлезвиг- 1930-2005 1,68 2,45 31,5 U. Peterson, личн. сообщ.
Г ольштейн
Польша, Верхняя Силезия 1973-1978 1,91 2,50 23,4 Profus, 1986
Польша, Лешновское 1974-1990 1,96 2,61 22,9 Kuzniak, 1994
воеводство
Венгрия 1958-2002 2,46 2,73 14,6 Lovaszi, 2004
Словакия 1951-1984 2,09 2,56 18,3 Stollmann, 1989
Чехия 1984-1995 2,20 2,74 19,9 no: Rejman, 1996, 1999
Примечание'. JZa — среднее число птенцов на размножавшуюся пару, JZm — среднее число птенцов
на успешную пару, %Нро — средняя доля неуспешных пар в %.
Белый аист
407
одном случае из более чем 1 100 находившихся под наблюдением выводков (Боровский р-
н, 2001 г.) (Галченков, 20006). Средний размер выводка в Тверской обл. составил 3,2 слетка
на успешно гнездившуюся пару (п=35) (Николаев, 2000), в Воронежской — 2,29 (п=31)
(Нумеров, Макагонова, 2000), в Липецкой обл. — 3,50 (п=8) (Сарычев, 2000). Наименьший
репродуктивный успех отмечен у пар, гнездящихся на краю ареала (в Ярославской обл.) —
1,79 молодых на 1 успешно гнездившуюся пару (Голубев, 2000).
На репродуктивные показатели влияет много различных факторов, но решающим явля-
ются условия зимовки. Установлено, что в основном они определяют как успешность раз-
множения, так и выживаемость взрослых птиц (Dallinga, Schoenmakers, 1987,1989; Kanya-
mibwa et al., 1990; Schaub et al., 2005). Так называемые катастрофические годы, когда на
значительной части гнездового ареала белого аиста резко снижаются и численность, и реп-
родуктивные показатели, связаны именно с неблагоприятными условиями зимовки.
Такими в последнее время были 1997 и 2005 гг. Исследования на Украине показали, что
колебания численности и репродуктивных показателей у белого аиста имеют тесную связь,
т.е. определяются одними и теми же факторами (Грищенко, 2004).
В местах гнездования наиболее существенным фактором, влияющим на репродуктив-
ный успех, служит качество кормовых биотопов. Так, в Голландии обнаружена связь между
успешностью размножения и уровнем грунтовых вод на гнездовом участке, от чего зависит
обилие корма (van der Have et al, 1999). Репродуктивный успех зависит также от сроков
гнездования. Птицы, приступившие к размножению раньше, выводят в среднем больше
птенцов (Profus, 1986, 1991; Адамян, 1990; Goutner, Tsachalidis, 1995; Tryjanowski et al.,
2004). Объясняют это тем, что прилетевшие раньше аисты занимают лучшие гнездовые
участки; кроме того, у них остаётся больше времени для восстановления сил после мигра-
ции. В ряде стран выявлена отрицательная корреляция между успешностью размножения
белого аиста и плотностью его населения из-за возрастания внутривидовой конкуренции
(Profus, 1986, 1991; Barbraud et al., 1999). Больше всего птенцов выращивают аисты в воз-
расте 10-16 лет (Meybohm, Dahms, 1975).
Смертность на юго-западе Германии в 1950-1959 гг. для первогодков составляла 60,4%,
для аистов, старше 1 года, — 25,8%. В 1960-1976 гг., когда произошло резкое сокращение
численности западной популяции, смертность была равной, соответственно, 74,2 и 26,5%
(Bairlein, Zink, 1979). В Баварии смертность однолетних птиц составляет 60,0%, двухлет-
них — 26,5%, трёхлетних 19,9%, затем она стабилизируется на уровне около 18% (Schulz,
1998).
В Швейцарии содержавшаяся в питомнике свободно летающая птица прожила 33 года
и 8 месяцев (Creutz, 1988). К. Брувер (Brouwer, 1990) приводит для белого аиста макси-
мальный возраст 35 лет. В природе обнаружена окольцованная птица, прожившая 29 лет и
3 месяца (Rydzewski, 1973). Максимальный возраст размножения полудикой немигриро-
вавшей птицы — 30 лет (Creutz, 1988), в дикой природе — 25 лет (Meybohm, Fiedler, 1983).
Средний возраст птиц, переживших первый год жизни: в Оберлаузице в Германии — 8,22
года (Creutz, 1988), в Нижней Саксонии — 8,9 года (Meybohm, Dahms, 1975), в Баварии —
8,66 лет (Schulz, 1998).
Суточная активность, поведение. Белый аист — дневная птица, однако известны слу-
чаи кормления птенцов и в светлые ночи (Schuz, Schuz, 1932). Ночью птицы могут прояв-
лять активность на гнезде: отмечались копуляции, уход за оперением, смена насиживаю-
щих партнёров и т.п. Во время миграции аист летит днём, но на северо-западе Африки при
высокой дневной температуре пролётные стаи отмечались и ночью (Bauer, Glutz von
Blotzheim, 1966). Большие стаи чаще всего скученные, неупорядоченные; птицы летят на
разной высоте (Молодовский, 2001).
408
Отряд Листообразные, Семейство Аистовые
По земле белый аист передвигается шагами, реже бегает. Активный полёт довольно
тяжелый, с медленными взмахами крыльев. При благоприятных условиях предпочитает
парение, особенно при полёте на большие расстояния. В восходящих потоках нередко об-
разуются скопления набирающих высоту птиц. Белый аист может плавать, хотя делает это
неохотно. При благоприятном ветре способен взлетать с поверхности воды (Bauer, Glutz
von Blotzheim, 1966; Creutz, 1988).
Во внегнездовой период белый аист ведёт стайный образ жизни. Во время гнездования
также могут образовываться колонии и скопления в местах кормежки. Неразмножающиеся
птицы летом держатся стаями, численность которых достигает десятков и даже сотен осо-
бей. Они держатся в богатых кормом местах, ведя бродячий образ жизни. Численность
таких стай постепенно увеличивается от мая к июню, в июле их размеры заметно возраста-
ют; ещё более многочисленными они становятся в августе, в связи с образованием предот-
летных скоплений. По наблюдениям в Калужской обл. в 1990-е гг., средняя численность
птиц в летних стаях составляла: в мае — 3,4 особи, в июне — 4,0, в июле — 7,8, в августе —
10,5 (п=50). Выводки после вылета объединяются в стаи, которые постепенно укрупняют-
ся по ходу миграции. Так, если на Украине обычный размер пролётных стай осенью —
десятки, реже сотни особей, то уже на черноморском побережье Болгарии средний размер
стаи составляет 577,5 особи (Michev, Profirov, 1989). На Ближнем Востоке и в Сев.-Вост.
Африке нередко отмечают огромные скопления, превышающие 100 тыс. особей (Schulz,
1988, 1998). Установлено, что эффективность миграции (скорость передвижения, компен-
сация сноса ветром и т.п.) выше в больших стаях (численностью в несколько тысяч осо-
бей), чем в мелких группах или у отдельных птиц (Liechti et al., 1996).
Отдыхают аисты преимущественно ночью. В гнездовой период количество времени,
которое остаётся для отдыха и чистки оперения, зависит от обилия пищи и количества
птенцов. При её изобилии, аисты могут и днём часами отдыхать или чистить оперение.
Поза отдыхающей птицы весьма характерна: аист стоит чаще всего на одной ноге, втянув
голову в плечи и пряча клюв в распушенное оперение шеи. Как правило, отдыхают аисты
на высоких присадах с хорошим обзором — на сухих деревьях, столбах, крышах.
Белые аисты используют довольно необычный способ терморегуляции — испражняют-
ся на ноги. В жаркий день можно увидеть многих птиц с белыми «чулками» на лапах. По-
видимому, жидкая мочевая кислота испаряется, охлаждая поверхность цевки. Кожа её обиль-
но пронизана кровеносными сосудами, проходя через которые кровь охлаждается (Prinzin-
ger, Hund, 1982; Schulz, 1987). Как показали эксперименты на американском лесном аисте
(Mycteria americand), при интенсивном испражнении на ноги температура тела снижается
(Kahl, 1972). X. Шульц (Schulz, 1987), наблюдая за аистами в Африке, установил, что час-
тота испражнений зависит от температуры воздуха. Температурный порог, после которого
птицы с забрызганными помётом ногами начинают встречаться регулярно, равен пример-
но 28 °C. При 40° частота испражнений доходит уже до 1,5 раз в минуту. Белый помёт,
кроме того, экранирует ноги от палящих лучей солнца. При облачной погоде частота ис-
пражнений снижается. Наблюдения на Украине показали, что в местах гнездования аисты
также начинают использовать такой способ терморегуляции при температуре около 30 °C
(Grischtschenko, 1992).
При совместной кормежке белого и чёрного аистов и цапель доминирует белый аист
(Kozulin, 1996).
Питание. Питание белого аиста очень разнообразно. Он поедает различных мелких
животных от дождевых червей до грызунов и мелких птиц: пиявок, моллюсков, пауков,
ракообразных, насекомых и их личинок, рыб, земноводных, пресмыкающихся и т.п. Может
разорять гнёзда наземногнездящихся птиц или поймать зайчонка. В питании отмечены даже
Белый аист
409
мелкие хищники, такие как ласка (Mustela nivalis) (Lohmer et al, 1980; Штыркало, 1990).
Размер добычи ограничивается лишь возможностью её проглотить. Рацион зависит от ус-
ловий местности и численности объектов охоты. В сухих местах он почти полностью мо-
жет состоять из насекомых, на лугах и болотах их доля значительно меньше. Так, по дан-
ным Э.Г. Самусенко (1994), в Белоруссии удельный вес различных групп животных в пита-
нии белого аиста колеблется в значительных пределах. В поймах Сожа и Березины беспоз-
воночные составляли по частоте встреч 51,6-56,8%, во внепойменных биотопах — до 99%.
Добычу аисты глотают целиком. Мелких животных проглатывают сразу, крупных насе-
комых и грызунов сначала убивают ударами клюва. Иногда можно видеть, как белый аист
некоторое время как бы «жуёт» клювом пойманную полёвку или крота. Крупную сухую
добычу при наличии поблизости воды птица некоторое время прополаскивает, пока её можно
будет легко проглотить. Таким же образом аисты отмывают запачканных в иле лягушек или
рыбу (Creutz, 1988).
Непереваренные остатки пищи отрыгиваются в виде погадок. Погадки формируются
на протяжении 36-48 ч. Состоят они из хитиновых остатков насекомых, шерсти и костей
млекопитающих, чешуй рыб и пресмыкающихся, щетинок червей и т.п. Размер погадок
20-100 х 20-60 мм, вес 16-65 г. У птенцов они несколько мельче — 20^15 х 20-25 мм
(Creutz, 1988; Muzinic, Rasajski, 1992; Schulz, 1998).
Кормятся аисты в разнообразных открытых биотопах — на лугах, пастбищах, болотах,
берегах водоёмов, полях, огородах и т.п. Излюбленные места кормёжки — участки с нару-
шенной растительностью или почвенным слоем, где лишённые убежищ мелкие животные
становятся лёгкой добычей. Эффективность охоты в таких ситуациях может быть весьма
значительной. Так, в Польше кормившийся за убиравшим пшеницу комбайном аист за 84
мин поймал 33 грызуна (Pinowski et al., 1991). По наблюдениям в пойме Эльбы в Германии,
наибольшая эффективность охоты (в среднем 5 г добычи в минуту) была во время сеноко-
шения или сразу после него (Dziewiaty, 1992). Поэтому скопления кормящихся аистов мож-
но увидеть на свежих покосах, на обрабатываемых полях и даже среди горящей травы. В
Африке эти птицы собираются там, где местные жители выжигают саванну в сухой сезон.
Достаточно им увидеть дым, как аисты слетаются отовсюду, концентрируясь за стеной огня.
Они вышагивают по ещё дымящимся стеблям и вылавливают насекомых. Иногда на таких
пожарищах собираются сотни особей (Creutz, 1988). Охотно аисты сопровождают стада
домашнего скота или диких животных на пастбищах. Копытные вспугивают мелких жи-
вотных, облегчая их добычу. На лугах аисты чаще всего кормятся на участках с низкой
травой или на мелководье водоёмов. Глубже 20-30 см забредают редко. Дождевых червей
аисты собирают чаще всего после дождей, когда те вылезают на поверхность, или на све-
жевспаханных полях. Охотно кормятся на поливных полях, изобилующих дождевыми чер-
вями. Хотя в высокой растительности численность насекомых выше, эффективность охоты
белого аиста при этом снижается. Так, в Австрии она составляла 61% в растительности
высотой 25 см и 52% при высоте растений 25-30 см (Schulz, 1998).
Основной способ охоты белого аиста — активный поиск добычи. Птица размеренно
вышагивает по траве или мелководью, то замедляя, то ускоряя темп; может делать резкие
броски или замирать на месте. Реже аисты подкарауливают жертву, прежде всего грызунов
и крупных насекомых. Корм птицы собирают на земле, на мелководье, реже — на растени-
ях. Могут они ловить клювом и пролетающих животных — стрекоз, жуков и других насе-
комых. Иногда даже сбивают их крыльями. Содержащиеся в неволе аисты быстро обуча-
ются хватать на лету клювом брошенную им пищу. Описаны даже случаи успешной охоты
аистов на пролетающих мимо воробьёв и других мелких птиц (Niethammer, 1967; Creutz,
1988; Berthold, 2004). Дождевых червей и других почвенных беспозвоночных птица нашу-
410
Отряд Листообразные, Семейство Аистовые
пывает клювом, погружая его в землю на несколько сантиметров (Schulz, 1998). Наблюда-
ли также, что аисты в полете схватывали рыбу с поверхности воды (Neuschulz, 1981; Schulz,
1998).
По исследованиям П. Закла (Sackl, 1985, цит. по: Schulz, 1998) в Австрии, средняя ско-
рость передвижения аиста во время кормёжки — 1,7 км/ч. При этом он делает от 1 до 90
шагов в минуту, в среднем 39,3. Время подкарауливания добычи варьирует от 10,5 до 720
сек, в среднем составляя 151,8 сек. Изредка птицы могут замирать на месте до 12 и даже 20
мин. Кормящийся аист делает в среднем 5,3 клевков в мин, из них успешных — 4,0. При
кормежке головастиками и молодыми лягушками на мелководье в пойме р. Сава в Хорва-
тии частота клевков составляла 5,9 в мин, из них 2,9 — успешных (Schulz, 1998).
Добычу птица обнаруживает чаще всего визуально. Иногда в мутной воде на мелково-
дье белые аисты используют также тактолокацию, аналогично аистам родаА/ус/епя (Ьц11г1,
1957; Резанов, 2001). По наблюдениям А.Г. Резанова (2001) на юге Украины, зондирование
мутной воды и илистого дна осуществлялось безостановочно слегка приоткрытым клю-
вом. Аисты ходили по мелководью, делая по 43-89 шагов в мин, постоянно прощупывая
участки дна перед собой. 98,9% клевков были разовыми тактильными зондированиями.
Успешность кормёжки составила 2,3%.
Аисты могут поедать и мертвых животных, например, снулую рыбу или погибших во
время сенокошения птенцов, и даже питаться отбросами. В Испании в 1990-х гг. они осво-
или свалки и теперь кормятся там вместе с чайками и врановыми. Часть птиц даже зимует
на свалках (Martin, 2002; Tortosa et al., 2002).
Описаны случаи клептопаразитизма. Так, однажды наблюдали аиста, который пресле-
довал в воздухе серую ворону, пытаясь отобрать пойманную мышь. Предполагается, что
такое поведение связано с недостатком пищи (Creutz, 1988). Могут аисты отбирать добычу
и у озёрных чаек (Ranner, Szinovatz, 1987).
Кормятся аисты как поодиночке, так и стаями. В местах, богатых кормом, могут образо-
вываться огромные скопления, которые на зимовках иногда достигают десятков тысяч осо-
бей. Причём в скоплениях эффективность кормления аистов повышается, поскольку они
лучше защищены от хищников и меньше времени тратят на осматривание (Carrascal et al.,
1990).
В гнездовой период аисты добывают корм, как правило, поблизости от гнёзда, но могут
летать за пищей и за несколько километров. От расстояния к основным кормовым угодьям
в значительной степени зависит успешность размножения. Исследования на Эльбе в Гер-
мании показали, что среднее расстояние от гнезда до мест сбора корма обратно пропорци-
онально количеству выращенных птенцов (Dziewiaty, 1999). Обнаружена достоверная кор-
реляция между числом вылетевших птенцов и долей влажных лугов, болот и водоёмов на
гнездовой территории (Nowakowski, 2003). По данным наблюдений за одним из гнёзд в
Силезии в Польше, птицы летали за едой чаще всего к нескольким предпочитаемым мес-
там, расположенным на расстоянии от 500 до 3 375 м, в среднем 1 900 м (Jakubiec, Szymocski,
2000). Наблюдения за другой парой в Померании на севере Польши показали, что аисты
кормились на территории около 250 га. Больше чем в половине случаев они искали добычу
на нескольких предпочитаемых участках, которые составляли лишь 12% общей площади.
65% времени они кормились на лугах и пастбищах, 24% — на полях и 11% — на пруду.
Максимальное расстояние полётов за добычей — 3 600 м, среднее — 826 м. В 53% случаев
аисты кормились не далее 800 м от гнезда. Дальше всего они летали, когда уже подросли
птенцы. Интересно, что самец и самка различались в своих предпочтениях, кормясь пре-
имущественно в разных местах (Oigo, Bogucki, 1999). На Эльбе в 80% случаев аисты соби-
рали корм не далее 1 км от гнезда (Dziewiaty, 1992).
Белый аист
411
Максимальное расстояние полётов за кормом, определённое для окольцованных птиц в
Зап. Европе, составляет 10 км (Lakeberg, 1995).
Анализ 242 образцов пищи, собранных в негнездовой период на Украине, показал, что
весной наибольшее значение имеют земноводные и щитни, в августе — прямокрылые и раз-
личные жуки. Птенцов аисты выкармливают в основном земноводными и насекомыми на
различных стадиях развития. Из насекомых наибольшее значение имеют прямокрылые и жуки,
всего в питании обнаружены представители 19 семейств 3 отрядов (Смогоржевський, 1979).
В погадках, собранных в верховьях Киевского вдхр. в Черниговской обл., 96,1% фраг-
ментов от общего числа принадлежали остаткам членистоногих. Причём питание аистов
было очень разнообразным: в одной погадке находили до 130 видов животных, в том числе
таких мелких, как муравьи. Среди насекомых преобладали жесткокрылые (35,3%), пере-
пончатокрылые (21,0%) и ручейники (19,6%). Позвоночные в питании играли лишь
незначительную роль (Марисова, Самофалов, Сердюк, 1992).
По данным анализа 337 погадок, собранных в южной и центральной частях Белоруссии
в 1986-1992 гг., основу питания белого аиста составляли беспозвоночные — 99% общего
числа определённых экземпляров кормовых объектов. Преобладали водные жуки и клопы,
массовые виды жужелиц, населяющие преимущественно увлажнённые местообитания,
моллюски. В населенных пунктах повышается доля мелких млекопитающих и насекомых,
характерных для сухих биотопов (Самусенко, 1994). М.И. Лебедева (1960) в погадках, со-
бранных в Беловежской пуще, обнаружила среди 187 кормовых объектов 80 экз. моллюс-
ков, 75 — насекомых, 24 лягушки, 8 живородящих ящериц. Из насекомых найдены 42 стре-
козы, 20 личинок жуков-плавунцов и водолюбов, 9 медведок, 2 кузнечика, 1 гусеница. По
данным А.П. Крапивного (1957), в питании птенцов белого аиста в Беловежской пуще 72,5%
по весу составляли позвоночные, из них 60,6% — лягушки. Доля дождевых червей была
равной всего 1%.
В Калужской обл. энтомологический анализ погадок показал наличие представителей
17 видов, относящихся к 7 семействам отряда жесткокрылых (Coleoptera). Наиболее обыч-
ными оказались представители семейства жужелиц (Carabidae) — 41%. Далее идут плас-
тинчатоусые жуки (Scarabaeidae) — 22%, водолюбы (Hydrophilidae) — 15%, листоеды (Chry-
somelidae) и стафилины (Staphylinidae) — по 7%, плавунцы (Dytiscidae) и ложнослоники
(Anthribidae) — по 4%. Представленные виды жуков в основном являлись обитателями
умеренно влажных и сухих лугов, а также антропогенных ландшафтов, и были характерны
для поверхности почвы — 44%, населяли небольшие пруды и лужи или являлись навозни-
ками — по 19%; за ними следовали жесткокрылые, населяющие поля и обитающие на рас-
тительности, а также населяющие смешанные леса и живущие на коре и листьях — по 7%.
В Тверской обл. в пище отмечены представители 7 семейств жуков, большинство из кото-
рых составляют пластинчатоусые и жужелицы (61,3%) (Николаев, 2000).
В Мазурии в Польше из 669 собранных погадок 97,3% содержали остатки насекомых
(преобладали представители семейств Carabidae, Silphidae, Dytiscidae, Scarabeidae), 72,2% —
мелких млекопитающих (в основном кроты, мыши и полевки), 1,6% — моллюсков, 1,0% —
мелких птиц, 0,7% — земноводных. Доля насекомых в питании была наибольшей на полях
во время роста зерновых и люцерны и на скошенных лугах и полях после уборки урожая,
довольно высокой — на вспаханных полях (Pinowski et al., 1991). В Австрии во время гнез-
дового периода по количеству в пище преобладали прямокрылые (67,7%) и жуки (24,1%),
по весу — позвоночные (55,5%), прежде всего мелкие грызуны (33,2%). Среди насекомых
аисты предпочитали саранчовых, жужелиц, листоедов и пластинчатоусых жуков. В апре-
ле-июне рацион был более разнообразным, с преобладанием мелких грызунов, в июле-
августе преобладали прямокрылые (Sackl, 1987). В питании летних стай неразмножаю-
412
Отряд Листообразные, Семейство Аистовые
щихся птиц на лугах в Польше по количеству преобладали насекомые (83%), прежде всего
жуки, по биомассе — мелкие млекопитающие, в основном полевки (58%), насекомые (22%)
и дождевые черви (11,5%) (Antczak et al., 2002). Исследования в Греции показали широкую
вариабельность рациона в различных местообитаниях, но в погадках везде преобладали
остатки насекомых, прежде всего прямокрылых и жуков (Tsachalidis, Goutner, 2002).
Рацион аистов может изменяться по годам, в зависимости от погодных условий. Так, на
севере Германии в 1990 г., когда отмечался всплеск численности мышевидных грызунов,
последние составляли на двух участках, где проводились исследования, 59,1 и 68% веса
пищи, а в 1991 г. — всего лишь 3,6 и 3,8%. В очень влажный 1991 г. в питании преобладали
дождевые черви — 50 и 61,6% по весу (Thomsen, Struwe, 1994). На юге Германии в разные
годы весовая доля дождевых червей в питании белого аиста колебалась от 28,9 до 84%,
членистоногих — от 8,9 до 28,5%, пиявок — 0 до 51,9%, грызунов — от 1,5 до 55,2%,
лягушек — от 1,2 до 5,4% (Lakeberg, 1995).
Одна из основных групп насекомых, которыми питается белый аист — прямокрылые,
прежде всего саранча. Наибольшее значение в рационе она имеет на местах зимовки в
Африке, в связи с чем в языках некоторых африканских народов белый аист называется
«саранчовой птицей». Аисты могут поедать огромное количество саранчи, объедаясь иног-
да так, что не могут взлететь. Во время налёта саранчи на Хортобадь в Венгрии в 1907 г. в
пищеварительном тракте одного из добытых аистов обнаружили около 1 000 экз. саранчи.
Желудок и пищевод птицы были набиты до самой глотки. В одной из аистиных погадок
нашли 1 600 мандибул саранчи (Schenk, 1907). По оценке последнего автора, стая из 100
аистов способна за день уничтожить 100 тыс. экз. этих опасных вредителей. В местах гнез-
дования белый аист также уничтожает большое количество вредителей сельского хозяй-
ства, прежде всего медведок (Gryllotalpa gryllotalpa), долгоносиков, проволочников. По
данным А.П. Крапивного (1957), в Беловежской пуще в питании птенцов медведки состав-
ляли 8% по количеству и почти 14% по весу. В Мазурском Поозерье в Польше 31% погадок
содержал остатки проволочников, 14% — долгоносиков, 16% — медведок (Pinowska et al.,
1991). В Зап. Франции в пище, которую аисты приносили птенцам, преобладали жуки-
водолюбы и медведки (Barbraud, Barbraud, 1998).
При содержании в неволе суточная потребность в пище взрослого аиста колеблется от
300 г в теплое время года до 500 г зимой. В год птице необходимо 110-130 кг (Bloesch,
1982). Суточная потребность в энергии пары аистов, выкармливающих птенцов, оценива-
ется в 4 660 кДж. Такое её количество дает потребление 1,4 кг дождевых червей, 1 044 г
лягушек или 742 г мелких грызунов (Profus, 1986). По другим данным, пара с 1-2 птенцами
потребляет около 5200 кДж (Bohning-Gaese, 1992). На р. Сава в Хорватии пара аистов ежед-
невно приносила птенцам в возрасте 3-6 недель в среднем 1,4 кг пищи на птенца (Schulz,
1998), на севере Германии (возраст птенцов 3-8 недель) — 1,2 кг (Struwe, Thomsen, 1991).
Для белого аиста наиболее выгодной в энергетическом плане пищей являются позво-
ночные животные. Во влажных местообитаниях это обычно земноводные. Однако из-за
мелиоративных и гидротехнических работ численность их во многих странах значительно
снизилась. Так, питание находившейся под наблюдением пары аистов в швейцарской Юре
на 2/3 состояло из дождевых червей, позвоночные составляли лишь 0,4% (Wermeille, Biber,
2003). В таких условиях все большее значение для аистов приобретают грызуны. Наблюде-
ния в долине р. Обры на западе Польши показали, что успешность размножения и даже
количество заселённых гнёзд были выше в годы с высокой численностью обыкновенной
полевки (Microtus arvalis) (Tryjanowski, Kuzniak, 2002).
Враги, неблагоприятные факторы. Естественных врагов у белого аиста мало. Гнёзда
могут разорять крупные хищные птицы, врановые, куницы. Взрослые птицы становятся
Белый аист
413
жертвами нападений орлов, орланов, крупных четвероногих хищников — лисиц, бродячих
собак, волков и т.п. Однако гибель большинства взрослых белых аистов прямо или косвен-
но связана с человеком.
В наибольшей степени за смертность ответственны линии электропередачи. В 1986-
1989 гг. на Украине из 489 случаев гибели взрослых аистов с известной причиной 64,0%
пришлось на ЛЭП. Среди жертв электролиний 80,8% погибли на опорах от поражения то-
ком и 19,2% разбились о провода. Наибольшую опасность электролинии представляют для
молодых плохо летающих птиц: 72,8% случаев гибели приходится на аистов, недавно по-
кинувших гнездо. На втором месте оказалось прямое уничтожение людьми — 12,7%. 8,8%
аистов погибли в результате драк на гнёздах и при формировании стай перед отлётом, 7,6% —
из-за неблагоприятных погодных условий, 2,9% — из-за отравления пестицидами, 1,6% —
из-за столкновений с транспортом, 1,2% — из-за болезней, 0,8% — от хищников, 0,4% —
из-за падения в большие трубы. Таким образом, всего по не связанным с деятельностью
человека причинам погибли лишь 18,4% аистов. Основная причина гибели птенцов (742
случая с известной причиной) — выбрасывание птенцов родителями из гнёзд. На нее прихо-
дится 41,9%. 20,2% птенцов погибли из-за неблагоприятных погодных условий, 12,9% — из-
за падения гнёзд, 7% — при драках между взрослыми аистами на гнёздах, 6,2% — уничто-
жены человеком, 4,5% — из-за сгорания гнёзд, 2,7% — в результате гибели родителей,
2,0% погибли от хищников, 1,5% — отравились, 1,1% — погибли из-за принесённых в
гнездо материалов (Грищенко, Габер, 1990).
В Калужской обл. картина несколько иная. По данным, собранным в 1960-99 гг., основ-
ной причиной гибели взрослых птиц служит браконьерский отстрел. На его долю прихо-
дится 74% случаев с установленной причиной гибели (п = 19). В 21% случаев птицы гибли
на ЛЭП, 1 раз взрослая птица погибла во время драки за гнездо с другими аистами. Основ-
ной причиной гибели слетков является контакт с электрическими коммуникациями: от уда-
ра током на открытых трансформаторах и опорах ЛЭП, а также при столкновении с прово-
дами. Часть случаев пропажи молодых птиц вскоре после вылета из гнёзд, вероятно, следу-
ет отнести к браконьерскому отстрелу. Связаны такие различия с тем, что на территориях,
недавно заселённых аистами, отношение людей к ним гораздо менее благожелательное.
Известны даже случаи уничтожения появившихся гнёзд. Так, первое гнездо в Мордовии
было разрушено местными жителями из-за опасений, что аисты повредят посевы огурцов
(Лапшин, Лысенков, 1997). В Нижегородской обл. основная причина гибели гнёзд — пре-
следование человеком (Бакка, Бакка, Киселева, 2000). Уничтожение взрослых птиц и разо-
рение гнёзд отмечалось на юго-востоке Туркменистана, где аисты пытались гнездиться в
1980-х гг. (Белоусов, 1990). Однако и в тех регионах, где белый аист обитает уже давно,
отношение к нему местного населения изменилось в худшую сторону. Об этом свидетель-
ствует хотя бы высокий процент уничтожения птиц людьми среди причин гибели и разру-
шение гнёзд на опорах линий электропередач.
Среди причин гибели птенцов, как уже упоминалось выше, на первом месте стоит роди-
тельский инфантицид. Существенная часть птенцов выбрасывается из гнёзд или даже съе-
дается взрослыми аистами. Так, в Беловежской пуще выбрасывали птенцов почти 30% пар,
причём иногда уничтожались даже все птенцы выводка (Федюшин, Долбик, 1967). В Испа-
нии инфантицид отмечен в 18,9% находившихся под наблюдением гнезд. Во всех случаях
выбрасывался самый слабый птенец. Средний возраст выброшенных аистят—7,3 дня (Тог-
tosa, Redondo, 1992). Обычно такое поведение связывают с нехваткой корма. По Д. Лэку
(1957), инстинкт абортирования части отложенных яиц или вылупившихся птенцов — при-
способление, позволяющее привести размер семьи в соответствие с количеством наличной
пищи. Предполагается, что распространённость инфантицида у белого аиста связана с от-
414
Отряд Листообразные, Семейство Аистовые
сутствием сиблицида и конкуренции за пищу в выводках. Родители приносят большое ко-
личество мелкого корма, и более крупные птенцы не могут монополизировать его. Поскольку
наиболее слабые птенцы не погибают сами, их «приходится» уничтожать родителям (Тог-
tosa, Redondo, 1992; Zielicski, 2002).
Аналогичная ситуация отмечается не только на территории бывш. СССР, но и в других
странах. Больше всего взрослых аистов погибает на ЛЭП, наибольшую опасность линии
электропередач представляют для молодых, ещё плохо летающих птиц. Это отмечено в
Болгарии (Нанкинов, 1992), Германии (Riegel, Winkel, 1971; Fiedler, Wissner, 1980), Испа-
нии (Garrido, Fernandez-Cruz, 2003), Польше (Jakubiec, 1991), Словакии (Fulin, 1984), Швей-
царии (Moritzi, Spaar, Biber, 2001). В округе Росток в Восточной Германии из 116 погиб-
ших птенцов белого аиста 55,2% были выброшены родителями, 20,7% погибли из-за паде-
ния гнёзд, 9,5% — от переохлаждения (Zollick, 1986). На путях пролёта и в местах зимовки
основными причинами гибели аистов служат отстрел и другие формы преследования людь-
ми, гибель на ЛЭП, отравление пестицидами (Schulz, 1988). Если многотысячный поток
мигрирующих аистов пересекает район с густой сетью электролиний, одновременно гиб-
нут десятки особей (Нанкинов, 1992).
Во многих странах Африки белый аист — традиционно охотничий вид. По данным
возвратов колец, в Сев. и Зап. Африке около 80% случаев гибели приходится на отстрел. По
подсчётам X. Шульца (1988), в 1980-х гг. на восточном пролётном пути ежегодно отстрели-
валось 5-10 тыс. аистов, из них 4-6 тыс. — в Ливане.
Массовая гибель аистов может вызываться катастрофическими погодными явлениями —
бурями, крупным градом и т.п. 5 августа 1932 г. возле одного из сел на севере Болгарии во
время небывалого градобоя (с неба падали куски льда до полукилограмма весом!) погибло
около 200 аистов и ещё около сотни остались с поломанными ногами и крыльями (Schumann,
1932). В 1998 г. в двух сёлах Львовской обл. почти все птенцы в 19 находившихся под
наблюдением гнёздах погибли во время сильных дождей (Горбулшська и др., 2004). Боль-
шой урон может причинять возвращение холодов после прилёта аистов. Так, в 1962 г. во
Львовской обл. сотни особей погибли из-за морозов и снегопадов в третьей декаде марта
(Черкащенко, 1963).
Иногда птенцы погибают, пытаясь проглотить принесенную родителями слишком круп-
ную добычу. Например, отмечен случай гибели аистёнка, подавившегося ужом (Kuppler,
2001). Опасность птенцам представляют и некоторые принесённые родителями в гнездо
материалы — куски шпагата, пакли, в которых аистята могут запутываться; обрывки плен-
ки или клеёнки в лотке, на которых собирается вода.
Белый аист может становиться жертвой клептопаразитизма. Например, на Днепре в
Черкасской обл. наблюдали случай нападения на пролетавшего над рекой аиста орлана-
белохвоста (Haliaeetus albicilla). Аист отрыгнул добычу, две рыбёшки из которой орлан
подобрал с поверхности воды (Лопарев, 1997).
К неблагоприятным факторам принадлежат происходящие в последние десятилетия
изменения среды обитания. Постройки с соломенными и тростниковыми крышами, на ко-
торых охотно гнездились аисты, практически исчезли из деревень. Уменьшается и количе-
ство пригодных для гнездования старых деревьев в населённых пунктах. Чрезмерная мели-
орация, затопление пойм рек водохранилищами, нарушение нормального гидрорежима
водоёмов приводит к обеднению кормовой базы. Особенно актуально это для многих стран
Зап. Европы, где приходится специально разводить земноводных на корм аистам. В после-
днее время добавилась ещё одна проблема — сокращение площадей традиционно исполь-
зующихся лугов и пастбищ во многих регионах Вост. Европы и Сев. Азии из-за экономи-
ческого спада. Всё увеличивающаяся химизация сельского хозяйства вызывает накопление
Белый аист
415
пестицидов в цепях питания, что вызывает отравления и заболевания птиц. В наибольшей
степени это проявляется на местах зимовки, где проводится активная борьба с саранчой и
другими вредителями сельского хозяйства, которые служат основным кормом аистов.
В Средней Азии наиболее важным фактором, повлиявшим на изменения ареала и чис-
ленности, стало освоение новых земель под посевы сельскохозяйственных культур с пре-
обладанием монокультуры хлопчатника, вырубка деревьев в долинах рек, осушение болот,
уменьшение площади рисовых полей. Благодаря укрупнению полей было вырублено мно-
го лесополос. Современная архитектура и градостроительные тенденции не способствуют
гнездованию белого аиста в населенных пунктах (Сагитов, 1990; Шерназаров и др., 1992).
В России существенным фактором, ограничивающим численность гнездящихся пар,
служит разрушение гнёзд на церквях в связи с их реставрацией, на телеграфных столбах и
опорах ЛЭП при обслуживании электрических коммуникаций, а также демонтаж водона-
порных башен для установки на новое место или сдачи на металлолом. Последний фактор
представляется особенно угрожающим, так как на водонапорных башнях гнездится более
половины российской группировки белого аиста.
К неблагоприятным факторам следует отнести и ухудшение положительного отноше-
ния к белому аисту местного населения, утрата давних народных традиций. Так, проведен-
ный в Киевской обл. опрос показал, что значительная часть сельских жителей не только не
знает, как привлечь белого аиста на гнездование, но и не хочет иметь гнездо на усадьбе
(Грищенко и др., 1992). Это при том, что наличие гнезда раньше считалось большим бла-
гом; привлечение белого аиста на гнездование было одним из элементов древней аграр-
ной магии (Грищенко, 19986, 2005). В Узбекистане белый аист считался священной пти-
цей, но теперь население в некоторых местах занимается разорением гнёзд и сбором яиц
(Сагитов, 1990).
На юге Украины у белого аиста зарегистрированы 4 вида гельминтов: Dyctimetra disco-
idea, Chaunocephalus ferox, Tylodelphys excavata, Histriorchis tricolor (Корнюшин и др., 2004).
В гнёздах белого аиста обнаружено около 70 представителей разных видов насекомых,
главным образом жуков (Coleoptera) (Hicks, 1959).
Хозяйственное значение, охрана. Белый аист уничтожает большое количество вреди-
телей сельского хозяйства, прежде всего насекомых и грызунов. Он широко известен как
один из наиболее активных истребителей саранчи. Аист может приносить некоторый ущерб
рыбному и охотничьему хозяйству, поедая рыбу, птенцов, зайчат и т.п., однако подобное
носит лишь случайный характер, и такие кормовые объекты не занимают сколько-нибудь
заметного места в рационе белого аиста. Более или менее существенный ущерб рыбному
хозяйству возникает лишь там, где образуются большие скопления аистов и практически
отсутствует другая пища (например, на рыборазводных хозяйствах в Израиле). В странах
же Вост. Европы и Сев. Азии это случается редко.
Белый аист — давний спутник человека, он имеет огромное эстетическое значение,
считается одной из наиболее любимых и почитаемых у многих народов птиц. Культ его
сформировался в глубокой древности, вполне вероятно вскоре после возникновения про-
изводящего хозяйства (Грищенко, 19986, 2005). Аист — прекрасный объект для экологи-
ческого образования и воспитания, принимает помощь человека, позитивно воздействует
на эмоции живущих рядом людей. Для охраны аиста необходима активная пропагандист-
ская и разъяснительная работа, возрождение давних народных традиций помощи этой пти-
це. При этом благодаря большой популярности белого аиста удаётся привлечь к природо-
охранной деятельности значительное количество людей. Большую эффективность имеют
широкомасштабные научно-пропагандистские акции, например, проводившиеся на Укра-
ине операции «Лелека» («Аист») и «Год белого аиста» (Грищенко, 1991,19966; Грищенко и
416
Отряд Листообразные, Семейство Аистовые
др., 1992). Особенно важны как пропагандистская работа, так и практическая помощь в
зоне расселения — для закрепления птиц на новых местах гнездования.
Белый аист занесён в Красные книги Казахстана, Узбекистана, а в Российской Федера-
ции — в Красные книги Карелии, Мордовии, Чечни, Краснодарского и Ставропольского
краев, Белгородской, Волгоградской, Калужской, Кировской, Липецкой, Московской, Ни-
жегородской, Пензенской, Ростовской, Рязанской, Тамбовской, Тверской и некоторых дру-
гих областей.
Дальневосточный аист
Ciconia boyciana (Swinhoe, 1873)
Ciconia boyciana Swinoe, 1873, Proc. Zool. Soc. London, c. 513, Иокогама, Япония
Oriental [eastern Japanese, blackbilled, Chinese] White Stork (англ.);
Swarzschnabelstorch (нем.); cigogne orientale (фр.)
Син. (рус): дальневосточный [черноклювый] белый аист, белый китайский аист.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Белоснежная птица с массивным чер-
ным клювом, яркими красно-оранжевыми ногами и такого же цвета небольшими оголен-
ными участками кожи вокруг глаз и под клювом. Чёрные крылья стоящей птицы прикрыва-
ют хвостовое оперение и создают обманчивое впечатление, что хвост у аиста чёрного цве-
та. Летящий аист имеет силуэт лука с настроенной к полёту стрелой: ноги и шея вытянуты
горизонтально, линия крыла очерчена характерным дугообразным изгибом, первостепен-
ные маховые раскрыты пальцеобразно.
Самый крупный аист из обитающих на территории России. Размерами превышает боль-
шую белую цаплю (шею в полёте, в отличие от цапель, аист вытягивает), но уступает япон-
скому (уссурийскому) журавлю, с которым в полевых условиях имеет относительное внеш-
нее сходство (оба вида практически полностью белые с чёрными маховыми перьями). От
белого аиста отличается более крупным телосложением, длинным и массивным, заметно
скошенным вверх клювом и деталями окраски — клюв взрослых птиц чёрный, радужина
светлая, уздечка, орбитальное кольцо и горловой мешок красные, широкими светлые поля
на чёрных второстепенных маховых шире и хорошо заметны. В противоположность бело-
му аисту для дальневосточного аиста характерно также чрезвычайно осторожное, пугли-
вое поведение. Совместно эти виды не встречаются.
Полет дальневосточного аиста машущий, не скоростной, с промежуточным коротким
планированием; на большой высоте парящий. Посадка на гнездо, помимо стандартных ва-
риантов, может осуществляться практически по вертикали с непродолжительным зависа-
нием в воздухе. Большую часть времени птица пребывает на земле, деревья использует в
основном для отдыха и гнездования.
Типичный представитель открытых пространств, в первую очередь'болот и увлажнён-
ных лугов, расположенных среди релочных лесов и характерных для природных экосистем
юга Дальнего Востока.
Голос. У взрослых птиц голос отсутствует, его заменяет дробь, издаваемая быстрым
щёлканьем клюва; во время исполнения дроби птица, как правило, описывает клювом в
воздухе дугу, поднимая его от земли и запрокидывая на спину. Вне сезона размножения
дальневосточные аисты обычно молчаливы.
Дальневосточный аист
417
Подвидовая систематика. Монотипический вид.
Замечания по систематике. До конца 1970-х гг. считался подвидом европейского аис-
та — Ciconia ciconia boyciana, хотя эти виды между собой существенно отличаются по
морфологии, экологии и поведению, а также географически разделены расстоянием более
чем в 5000 км (Archibald, Schmitt, 1991).
Описание. Окраска. Взрослые птицы. Половой диморфизм отсутствует. Самец и
самка во взрослом наряде имеют белоснежное оперение. Маховые, перья крылышка и боль-
шие верхние кроющие крыла чёрные. Наружная сторона опахала, стержни второстепен-
ных и части первостепенных маховых, как и наружное опахало больших верхних кроющих
крыла с бело-серебристым отливом, хорошо заметным в сложенном крыле. Рулевые перья
белые. В гнездовой сезон оперение на шее, груди и животе имеет грязно-охристый оттенок
из-за окислов железа, содержащихся в дальневосточных осоковых болотах. Неоперённая
кожа вокруг глаз к основанию клюва и под горлом по краю нижней челюсти красно-оран-
жевая. Кожа на теле под оперением тонкая с жёлто-оранжевым оттенком. Ноги красно-
кирпичного цвета, не выступающие закругленные когти на пальцах темнее. Клюв массив-
ный, тёмно-коричневого, почти чёрного цвета с невыраженным красноватым отливом по
верху надклювья. Язык немного мясистый, короткий, бледно-розовый. Радужина светло-
голубая, с возрастом почти белёсая.
Первый пуховой наряд образован серовато-молочным пухом длиной 10-15 мм на спине,
5-10 мм на груди, животе, по бокам тела и 2-3 мм на голове. Рудиментарный короткий пух
имеется на внутренней стороне бедер, вокруг глаз и по нижнему краю подклювья, на кры-
льях. Аптерии располагаются на спине в межлопаточной области с переходом в узкую ап-
терию на дорзальной стороне шеи, также по бокам тела и на брюхе; обширная брюшная
аптерия переходит в грудную и узкую по всей длине на передней стороне шеи. Кожа на
аптериях светлая с желтоватым оттенком. Небольшие аптерии имеются на внешней и внут-
ренней стороне крыла. Клюв серовато-розоватый, к концу с зелено-коричневым оттенком;
вершина клюва с преобладанием жёлтого цвета. Яйцевой зуб величиной не более 3 мм,
молочного цвета; сохраняется до трёхнедельного возраста. Кожный участок под клювом и
кожа вокруг глаз яркого оранжевого цвета. Глазные щели и слуховые проходы открыты.
Радужина глаз коричневая, зрачок тёмно-серый, кожа вокруг век коричневая. Ротовая по-
лость и язык бледно-розовые. Ноги и когти на пальцах розовые с коричневатым оттенком.
Второй пуховой наряд начинает формироваться в возрасте 6-8 дней. Всё тело равно-
мерно покрывается белым пухом длиной 10-15 мм другой генерации и ювенальным пе-
ром, на вершинах которого сохраняются эмбриональные пушины. Смена происходит на
дорзальных птерилиях: верхнешейной, межлопаточной, крестцовой, околокопчиковой, око-
лоанальной, на голенях и боковой стороне тела. Появляются пеньки первостепенных и вто-
ростепенных маховых, верхние и нижние кроющие крыла. Растёт перо на лопатках. Пер-
вый пуховой наряд сохраняется на лицевой части головы, спереди на шее, брюхе и внут-
ренней стороне голени. В 15-дневном возрасте тело птенца полностью покрыто густым
белым пухом и пуховидными перьями, за исключением небольшого размера оголённых
аптерий на спине, груди, шее и брюхе. Кончики растущих маховых и перьев на межлопа-
точной птерилии чуть освобождаются от чехликов, растут кроющие крыла. Меняется цвет
клюва на серо-зелёный, в вершине клюв жёлто-зелёный. Цвет оголённых кожных участков
на голове становится оранжевым. Светлеет кожа век, а ноги, наоборот, темнеют; на них
чётче проявляется рисунок пластин рогового покрытия.
В мезоптилъном наряде птенцы пребывают с 10-дневного до 1,5-месячного возраста.
Все тело птенца покрыто оперением с мезоптилем, включая растущие маховые и рулевые
перья. В месячном возрасте интенсивный рост юношеского пера наблюдается на крыле
418
Отряд Листообразные, Семейство Аистовые
(маховые и кроющие крыла), спине, между лопаток, крестце и шее. Клюв ещё больше тем-
неет: основание и вершина клюва серо-зелёного цвета, гребень — с красно-зелёным отли-
вом. Кожа вокруг глаз и под клювом вновь приобретает яркий красно-оранжевый цвет.
Размер кожного оголённого участка вокруг глаз сокращается и имеет обратно-яйцевидный
контур, суживающийся к основанию клюва. Кожный участок под горлом покрывается мел-
ким белым пером и неоперённым остаётся только по краю нижней челюсти и вдоль крыль-
ев подклювья. Светлеет радужина глаз и кожа вокруг век. Мезоптильный наряд у птенцов
сохраняется непродолжительное время и изнашивается ещё до слёта с гнезда.
Гнездовой (ювенильный) наряд начинает развиваться в возрасте 15-20 дней и заканчи-
вает формирование около 2-месячного возраста. Молодая птица в этом возрасте внешне
уже выглядит взрослой. Всё тело равномерно покрывает белоснежное перо и белый пух,
более длинный на спине и животе. Кожа прозрачная, розовато-оранжевая. Рост маховых
практически завершён, чёрный цвет матовый. Клюв зелёно-коричневый с красным оттен-
ком; он много короче, чем у взрослой птицы (рост клюва медленно продолжается ещё не
менее года). Участки кожи вокруг глаз, в углах ротовой полости и под клювом насыщенно-
го красно-оранжевого цвета, их размер становится меньше; они представляют собой неши-
рокое неправильное кольцо и узкие полоски по нижнему краю подклювья. Радужина глаз
меняется до жёлто-серого цвета, зрачок тёмный. Ротовая полость и язык остаются бледно-
розовыми. Ноги розово-кирпичного цвета. Признак, отличающий молодых птиц от взрос-
лых, по мнению С.В. Винтера (1978) — тёмно-серая либо чёрная окраска верхних крою-
щих кисти. Однако подобное обнаруживается у небольшого числа птенцов, и нельзя ис-
ключить, что это вызвано нарушением обмена веществ в организме.
Строение и размеры. Самцы, в основном, крупнее самок, но ни морфометрия,
ни различия в поведении не являются гарантами правильного визуального определения
пола (Kahl, 1971; Archibald, 1985). Контурное перо у взрослых птиц по большей части сред-
него и крупного размера, с нешироким, чуть заостряющимся в вершине, мягким рыхлым
опахалом и не жёстким стержнем. Пух, покрывающий все тело, в большинстве своем круп-
ный, с длинными бородками первого и второго порядка. Спереди на шее и груди удлинен-
ные и слегка рассученные перья образуют воротник, который у возбужденных или, наобо-
рот, отдыхающих птиц приподнят. Перо на темени и затылке длиннее, чем на других учас-
тках головы, при выражении агрессии оно встает дыбом в виде невысокой короткой гривы.
Крылья длинные, неширокие и даже узкие; их ширина практически не меняется по всей
длине. Вершины первостепенных маховых перьев слегка заострённые, с невыраженным
срезом по наружному краю; внутреннее опахало шире наружного. Первостепенных махо-
вых 11, рулевых 12. Рулевые перья средней длины, не жёсткие, закруглённые в верхней
части. Хвост с почти прямым срезом, у сидящей птицы полностью прикрыт сложенными
крыльями, в полёте раскрыт веером. По данным А.И. Иванова и Б.К. Штегмана (1978),
длина крыла дальневосточного аиста составляет 620-670 мм. Крыло имеет пропорции,
характерные для птиц, преимущественно перемещающихся полётом (Познании, 1979); в
сложенном состоянии вершины первостепенных маховых совмещаются с внутренними
второстепенными маховыми. Длина скелетных отделов крыла (мм): плеча (п=14) 230-300,
в среднем 252,7; предплечья (п=14) 220-310, в среднем 279,6; кисти (п=12) 100-170, в
среднем 130,4.
Голова некрупная округлой формы, затылок немного скошен вниз. Лобная часть прак-
тически без границы переходит в основание клюва. Клюв конической формы, чуть сплюс-
нутый с боков с выраженным гребнем на надклювье, подклювье имеет 3-6 см скоса вверх.
Ноздри в виде узких щелей в первой четверти клюва. Края клюва очень острые, режущие;
на конце надклювья имеется роговой зубчик. Длина клюва у годовалых птиц (мм) (п=16)
Дальневосточный аист
419
200-272, в среднем 221,4; ширина у основания (мм) (п=16) 30,8-37,5, в среднем 34,5. Счи-
тается, что форма и длина клюва различаются у разных полов (Murata et al., 1988).
Ноги длинные, тонкие, с недлинными смотрящими вперёд тремя пальцами; четвёртый
палец короткий и расположен сзади на одной линии с центральным пальцем. Неоперенная
часть голени, цевка и пальцы покрыты роговыми пластинками спереди и мелкими чешуй-
ками сзади и с боков. Первые три пальца расположены друг к другу под острым углом и
имеют короткую оторочку. Размеры бедра (мм) (п=14) 110-155, в среднем 128,6; голени
(мм) (п=13) 220-335, в среднем 295,4; цевки (мм) (п=12) 240-280, в среднем 261,3.
Форма туловища яйцевидная; в спокойном состоянии шея опущена вниз в виде буквы S,
клюв упирается в шею или лежит сбоку на шее. Общая длина от кончика клюва до копчика
(мм) (п=14) 915-1 050, в среднем 957,2. Вес (г) (п=20) 3 600-5 900, в среднем 4 940.
Линька. Практически не изучена. По наблюдениям за сменой оперения у птиц в нево-
ле, в ежегодном цикле линьки можно выделить 3 периода: осенний, зимний и летний (Ан-
дронова, 2000). Начало осенней линьки приходится на конец октября и продолжается до
середины декабря у взрослых птиц. У молодых аистов первая ювенальная линька начина-
ется раньше, в сентябре - начале октября. Смена контурного, пухового и полупухового
пера на шее, груди, животе происходит пассивно, при перепадах температуры воздуха она
прерывается на несколько дней, в последнюю очередь меняется мелкий пух. Как у моло-
дых, так и у взрослых птиц выпавшее перо не имеет обношенного вида. Зимняя линька у
неполовозрелых птиц начинается в первой декаде января и продолжается до конца апреля.
Замена пера начинается на спине и брюхе, меняется полупуховое и пуховое перо, обломан-
ное и изношенное контурное. В феврале и до конца марта происходит смена контурного
пера на крыле и замена 1-2 третьестепенных маховых. В конце мая наступает летняя линь-
ка, которая начинается со смены контурного оперения на крыле. В это же время выпадают
первостепенные и второстепенные маховые, но не более 2, реже 4 перьев с крыла в месяц.
Из первостепенных первым выпадает одно из центральных маховых. Далее замена проис-
ходит центробежно. Обновление маховых на каждом крыле происходит поочерёдно. По-
врежденные маховые перья заменяются в первую очередь. Относительно интенсивная лет-
няя линька контурного пера проходит с июля по середину сентября. Смена маховых и руле-
вых перьев без нарастания интенсивности продолжается до августа. Ближе к осени вместе
с контурными обношенными перьями начинают выпадать полупуховые с груди, шеи, голо-
вы, нижних частей тела. Выпавшее перо в большинстве имеет обношенный вид. К середи-
не сентября, перед отлётом на зимовку, линька прекращается.
Линька по срокам соотносится с важными моментами жизненного цикла: летняя совпа-
дает со временем появления и выкармливания птенцов и наличием наилучшей кормовой
обеспеченности охотничьих биотопов. Осенняя и зимняя линьки позволяют птицам иметь
оптимальные летные и теплоизоляционные кондиции оперения. Вообще, линька не при-
водит к обильному выпадению пера, что, в целом, характерно для всех аистовых (Туга-
ринов, 1947) и, видимо, больше связана с поддержанием оперения в удовлетворительном
состоянии.
Распространение. Гнездовой ареал. Почти весь современный гнездовой ареал
дальневосточного аиста находится на территории российского Дальнего Востока и при-
урочен главным образом к бассейну реки Амур и его притоков Уссури и Зея (рис. 81). На
гнездовании зарегистрирован в Амурской обл. и Еврейской автономной области, в Хаба-
ровском и Приморском краях (Смиренский и др., 1987; Андронов, 1988, 1989, 2001). Заре-
гистрировано гнездование одной пары аистов в Читинской обл., на Шилке (Golovushkin,
Goroshko, 1995).
В Амурской обл. находится самый важный участок гнездования дальневосточного аис-
та в России; высокая плотность гнездовий отмечается здесь на Зейско-Буреинской равни-
420
Отряд Листообразные, Семейство Аистовые
Рис. 81. Ареал дальневосточного аиста: а — область гнездования; б — зимовки.
не, на Архаринской низменности и на севере Амурской обл., в поймах среднего и верхнего
течения рек Ульма, Ташина, Томь (Андронов, 1983, 1987; Дарман и др., 20006).
В Еврейской АО аист гнездится по заболоченным равнинам рек Урми, Тунгуска, Бира,
Биджан, Добрая и их притокам (Росляков, 1981а; Горобейко, Крюков, 2000).
На территории Хабаровского края северные пределы гнездования дальневосточного
аиста ограничены р. Амгунь. Но в период осенних пролётов птицы регистрировались и
севернее — в долине Нижнего Амура, что указывает на вероятность гнездования вида в
районе озёр Дальжа, Орель и Чля (Росляков, 1987). Встречается аист практически на всех
равнинных территориях в бассейнах озёр Чукчагирское, Эворон, Удыль, Болонь, в между-
речье Кур-Урми, в долинах рек Тунгуска, Амур, Уссури, а также на сопредельной террито-
рии Еврейской автономной области (Росляков и др., 2000).
В Приморье гнездится на Приханкайской низменности и в долине рек Уссури, Бикин,
Иман (Большая Уссурка) (Глущенко и др., 1995; Сурмач, Шибаев, 2000; Глущенко, Мрикот,
2000). В вост. Приморье единичное гнездование отмечено в Ольгинском р-не (в 5 км выше
устья р. Аввакумовка) 20 июня 1970 г. (Назаров, Лабзюк, 1975). Залёт одиночной особи
отмечен на юге Сахалина (Нечаев, 19916).
Вне России гнездится в сев.-вост. Китае, в основном в провинции Хэйлунзцян
(Heilongiiang) в пойме реки Сунгари (Sanjiang), а также в небольшом количестве в провин-
циях Гирин (Jilin) и Внутренняя Монголия (Jiangying, 1995; Threatened birds of Asia, 2001).
Зимовки. Основные зимовки вида находятся в Центр. Китае, в бассейне р. Янцзы
(Jianzhang et al., 1995), на оз. Поянху (Poyang Hu), Ченху (Chen Hu), Шенгжинху (Shengjin
Hu) и Дунтинху (Dongting Hu) (Changhu et al., 1995; Renzhu, Kemin, 1995; Hu, 1995; Qishan,
1995a). Некоторых особей также видели и южнее, вплоть до Гонконга (Chan, 1995). В от-
Дальневосточный аист
421
дельные годы дальневосточных аистов отмечали на территории Японии и Корейского п-
ова (Simba Chan, устное сообщение).
Миграции. Движение аистов с мест зимовок начинается с середины февраля. Весен-
ний и осенний пролётные пути, по данным спутникового мечения, практически одинако-
вы, но весной птицы летят ускоренными темпами (Higuchi et al., 2000; Tamura et al., 2000;
Van den Bossche et al., 2001). На места размножения аисты прилетают одиночками, парами
или небольшими группами, в конце марта - начале апреля. За последние 20 лет первая
встреча аистов на Архаринской низменности приходится на 19 марта, среднемноголетняя
дата прилёта в этот район — 30 марта. На территории Хабаровского края и Еврейской АО
аисты появляются 20-22 марта (Росляков и др., 2000). Самый ранний прилёт на Прихан-
кайской низменности отмечен 10 марта 2004 г. (Глущенко и др., 2006), а в районе г. Хаба-
ровска зарегистрирован 12 марта 2004 г.
Скопления аистов в несколько десятков особей (неполовозрелые и холостые особи) вес-
ной начинают регистрировать с 15 апреля по 31 мая, число птиц в таких скоплениях посте-
пенно нарастает, а затем снижается. Первые скопления аистов в поелегнездовой период
появляются в начале августа, их обнаруживают по берегам крупных водоёмов, на заливных
лугах. Осенние скопления формируются, в первую очередь, из молодых птиц и насчитыва-
ют от 6 до 36 особей. Постепенно небольшие группы сливаются и образуют довольно круп-
ные стаи в 50 и более птиц.
Покидают места гнездования аисты в августе-сентябре небольшими стаями или оди-
ночно и достигают зимовочных участков в ноябре-декабре. Последние регистрации оди-
ночных птиц в ареале гнездования на Приханкайской низменности приходятся на 14-28
октября (Глущенко и др., 2006), а в долине р. Амур 8-18 октября, до установления стабиль-
но низких температур воздуха; массовый отлёт происходит 17-30 сентября. Осенью птицы
медленно движутся на юг, часто останавливаясь в определённых, наиболее кормных мес-
тах; в среднем в пути они находятся 117±36 дней. По данным мечения спутниковыми пере-
датчиками (12 особей), протяжённость их осеннего миграционного пути от места гнездо-
вания (южная часть Амурской обл. и Еврейской АО и Хабаровского края) до мест зимовок
в Китае (р. Янцзы) составляет в среднем 3 256±802 км. Скорость передвижения нарастает
от 1-10 км/день до более 50 км/день. На пролёте аисты из различных участков российско-
го ареала собираются вместе в районе городов Цицикар и Харбин (долина р. Сунгари),
позднее их путь лежит вдоль берега Жёлтого моря, где они концентрируются над заливом
Бохай и, пролетая через устье р. Жёлтая, приземляются на зимовку в долине р. Янцзы.
Численность. Общая численность дальневосточного аиста в мире, по экспертным оцен-
кам, составляет 2 500-3 000 особей (Qishan, Coulter, 1995, данные составителя очерка). На
территории сев.-вост. Китая возможно нахождение до нескольких десятков гнёзд аистов. В
1994 г. в Амурской обл. обнаружено 99 жилых гнёзд (Andronov, 1994). В 1998 г. здесь же
был проведён анкетный опрос населения, на основании которого осуществлены полевые
исследования и авиаучёт дальневосточного аиста на большей части его гнездового ареала
(Andronov, Darman, 1998; Андронов и др., 2001). В 1999 г. в Амурской обл. исследованиями
была охвачена только Архаринская низменность, где численность аиста в 1999 г. увеличи-
лась по сравнению с прошлым сезоном на 28,5%. Расчётная численность аиста в Амурской
обл. в 1998 г. составила 530 особей, в 1999 г. — не менее 650 птиц (Дарман и др., 2000а;
Masatomi et al., 2002).
Для выяснения состояния популяции дальневосточного аиста на территории Еврейской
АО в 1999 г. проведен анкетный опрос охотников и работников природоохранных служб.
Затем осуществлено авиаобследование и проведено 4 экспедиции для проверки анкетных
данных. Расчётная численность аистов в области составила 405 особей, из них: гнездящих-
422
Отряд Листообразные, Семейство Аистовые
ся — 120, не размножающихся — 101, птенцов — 184 (средний размер выводка — 2,96
птенца) (Горобейко, Крюков, 2000).
По данным комплексных учётов аистов (авиаучёт, анкетный опрос, авиационное и на-
земное обследования гнездовий) в 1998-2000 гг., проведённых практически по всему гнез-
довому ареалу, численность аистов на территории Зейско-Буреинской равнины, на Средне-
Амурской низменности и в бассейне р. Уссури можно оценить в 380^430 размножающих
пар. При этом в бассейне р. Уссури численность популяции оказалась примерно на том же
уровне, что и в середине 1970-х гг. Однако на Зейско-Буреинской равнине и Средне-Амур-
ской низменности численность дальневосточного аиста значительно сократилась.
В Приморском крае в конце прошлого столетия (1999-2000 гг.) гнездилось от 67 до 71
пары, из них 17-18 пар на Приханкайской низменности, 5-8 пар на правобережье р. Сунга-
ча и 45 пар на правобережье Уссури, включая низовья р. Б. Уссурка (Дарман и др., 2000). В
бассейне р. Бикин в 1970-х гг. размножалось 6-8 пар (Пукинский, 2003), в то время как в
1996 г. достоверно было известно одно жилое гнездо, а в 1997 г. отмечены только бродячие
птицы (Михайлов и др., 1998). На оз. Ханка в 80-е годы прошлого столетия гнездилось в
среднем 7 пар; в 1990-е гг. около 14 пар (Глущенко, Мрикот, 2000); в 2007 г. было обнаруже-
но 20 жилых гнёзд, а всего в указанном году здесь предположительно гнездилось 27-29 пар
(Коробов, Глущенко, 2008).
Изменение количества жилых гнёзд имеет естественную выраженную 4-летниюю цик-
личную динамику спадов и подъёмов. Есть опасение, что динамика численности жилых
гнёзд всё больше определяется долговременным негативным влиянием антропогенных
факторов и имеет тенденцию к убыванию с ярко выраженной циклической составляющей
(Андронов, 1986; Андронов, Кастрикин, 2000).
Местообитания. Основное требование к местообитанию в гнездовом ареале — нали-
чие разреженных участков леса с крупными деревьями и обводнённые пространства или
водоёмы в качестве кормовых угодий. Аисты селятся вблизи водно-болотных угодий в ре-
лочных или в прирусловых лесах, избегая близкого соседства с человеком. Наиболее плот-
но гнездовья расположены на Архаринской низменности и в междуречье Ульмы и Ташины
(Амурская обл.), в пойме р. Тунгуски и в бассейне оз. Болонь (Хабаровский край), на При-
ханкайской низменности (Приморский край), где аисты строят гнёзда в островках релоч-
ных лесов, разбросанных среди обширных осоковых и осоко-вейниковых болот, с включе-
нием озёр и речек.
Состав древесных пород в релочных лесах невелик; в основном, в них произрастают
лиственница, плосколистная белая и чёрная березы, осина, монгольский дуб и, реже, амур-
ская липа. Гнёзда аисты чаще делают на лиственнице, дубе монгольском, плосколистной
березе и осине. Анализ распределения гнёзд дальневосточных аистов по авиаучётным ра-
ботам в 1998-1999 г.г. на территории Амурской обл., Еврейской АО и Хабаровского края
показал, что для устройства гнезда аисты в большинстве случаев используют деревья
(91,1 %), и абсолютным доминантом по размещению гнёзд является лиственница — 29,55%
от численности всех обнаруженных гнёзд на деревьях (Парилов и др., 2000). Кормовая
насыщенность осоковых болот зависит от гидрологического режима, который определяют
атмосферные осадки и паводки. Во время разливов рыба из рек, небольших и крупных озёр
проникает в болота, облегчая аистам поиск и добывание корма. По северу ареала, а также в
Хабаровском и Приморском краях, аисты занимают под гнездовья преимущественно ши-
рокие поймы рек с многочисленными заливами, озёрами, болотами, старицами, где пред-
почитают поселяться вдоль опушек леса, лесных релок с разреженным древостоем, неред-
ко занимают одиночно стоящие деревья (Воронов, 1985; Бабенко, Поярков, 2000; Сурмач,
Шибаев, 2000).
Дальневосточный аист
423
С конца 1980-х гг. при дефиците гнездопригодных деревьев на безлесных пространствах
аист стал строить гнёзда на опорах линий электропередач и триангуляционных вышках.
Для кормления и отдыха во время миграций и зимовки птицы выбирают только увлаж-
ненные места (болота, берега озёр, рисовые поля и т.д.).
Размножение. О сроках наступления половой зрелости можно судить только по пти-
цам, выращенным в неволе. Половозрелыми аисты становятся в возрасте не менее трёх
лет, но обычно в вольерных условиях самки начинают размножаться в возрасте 3-5, а сам-
цы — в возрасте 5-7 лет; продолжительность жизни в неволе составляет до 48 лет (King,
1990; Topola, 2004). Считается, что дальневосточный аист — моногамный вид, хотя в паре
птицы живут только в сезон размножения; на зимовках аисты не проявляют ни партнерс-
ких, ни семейных отношений и держатся одиночно или небольшими группами (Qishan,
1991). В места гнездования прилетают одиночно. Сразу после прилёта зрелые особи зани-
мают гнездовые деревья, в то время как молодые аисты ещё только ищут подходящие дере-
вья для устройства гнезда. Формирование новых пар происходит сразу после прилёта и
продолжается, в среднем, до середины мая. Одиночный аист (по поведению — предполо-
жительно, самец) выбирает подходящее для устройства гнезда дерево и начинает строи-
тельство. Известно, что в неволе занятие гнездового дерева, строительство гнезда и его
охрану также осуществляет самец (Archibald, 1991; Rozdina et al., 1991; H.R. Park, устн.
сообщ.). В весенний период одиночная птица на дереве, по нашим визуальным наблюдени-
ям, часто сама по себе привлекает внимание других аистов. Кроме того, инициирующая
создание пары птица привлекает к себе внимание, исполняя клювом отрывистую дробь, с
демонстрацией наклонов головы или без них. В неволе инициатором создания пары высту-
пает самка, она садится рядом с самцом на гнездо или ближнюю ветку, поднимает голову и
клюв немного вверх и производит неглубокие наклоны головы, издавая при этом клювом
отрывистую дробь; звук усиливается с помощью подгорлового резонирующего мешка.
Самец повторяет позу самки, но в наклонах описывает клювом большую дугу — от спины
до земли. Соединившиеся в пару аисты исполняют дуэтом дробный треск клювами, стоя
бок о бок; глубокие наклоны головы и закидывание клюва на спину птицы производят по-
очередно. В нижнем положении головы аисты, не прекращая треска, водят клювом из сто-
роны в сторону. В природе инициировать создание пары могут и самцы. Имеются свиде-
тельства (п=6), когда холостые самцы пытались отбить самку в устоявшихся парах, в том
числе даже когда в гнезде уже были яйца и птенцы.
Дальневосточные аисты не гнездятся колониями, что подтверждается наблюдениями за
размножением вида в разных частях гнездового ареала (Austin, 1948; Шибаев и др., 1976;
Винтер, 1976; Fujimaki et al., 1990; Qishan, 1995в). Однако нам приходилось встречать гнез-
дование аистов в виде компактных группировок или «полуколоний» (до 10 гнёзд), когда
расстояние между соседними гнёздами не превышало 100-200 м. В своем отчёте Y. Yama-
shina (1977с) тоже описывает рыхлую колонию, которая включала несколько гнёзд, отстоя-
щих друг от друга не более чем на 100 м. Довольно часто в природе попадаются небольшие
смешанные колонии аистов и цапель; как правило, в такой колонии одно гнездо аиста окру-
жено многочисленными гнёздами серой цапли.
Обычно гнездовое дерево располагается по кромке релочного леса, реже в глубине. Для
устройства гнёзда птицы выбирают крупные живые или мертвые деревья разных пород:
лиственницу, монгольский дуб, осину, плосколистную белую и чёрную берёзы, иву и неко-
торые другие. Гнездо располагается в верхней части кроны на высоте (п=143) 3,4-15 м, в
среднем 8,4 м, в развилке самых крупных живых или мертвых веток или на сломанной
вершине. На Приханкайской низменности при жёстком дефиците крупных деревьев гнёзда
дальневосточного аиста располагаются на высоте от 1,8 до 11 м, в среднем (п=12) 4,2 м
424
Отряд Листообразные, Семейство Аистовые
(Глущенко и др., 2006). Минимальное расстояние между гнёздами (в пределах 200-300 м)
отмечено на территории Архаринской низменности, максимальная удалённость гнёзд друг
от друга до 15 км наблюдается в северной части ареала и в Приморье. Аисты используют
свои гнёзда на протяжении длительного времени, иногда более 10 лет (Austin, 1948). Для
вида характерен гнездовой консерватизм. Причины, из-за которых птицы могут покинуть
старое гнездо — это постоянное беспокойство пары в период размножения, сбор или унич-
тожение потомства человеком, падение или разрушение гнезда и другие подобные причи-
ны. При наличии подходящих деревьев новое гнездо аисты чаще всего устраивают недале-
ко от прежнего.
Строят гнездо или подстраивают старое оба партнёра. Строительным материалом слу-
жат сухие длинные тонкие и толстые ветки, которые птицы сначала собирают на гнездовом
дереве, а потом приносят с некоторого расстояния. Лоток выстилают прошлогодней осо-
кой, мхом и растительной ветошью, приносимой с близлежащих болот. Форма гнезда зави-
сит от места его постройки, но в общих чертах оно напоминает чашу самой разнообразной
конфигурации. Размеры гнезда (в см) (п=29): диаметр 100-250; высота 136,8-200; диаметр
лотка 55-60; глубина лотка 5-14. На протяжении всего гнездового периода, да и после
слета птенцов, аисты достраивают и поправляют гнездо. Больше времени на это занятие
затрачивает самец. Строительство гнезда продолжается не более недели, новое гнездо пара
способна построить за 2-3 дня. Ежегодное подновление и достройка гнезда приводят к тому,
что высота старых гнёзд превышает один метр, а масса всей гнездовой конструкции увеличи-
вается до сотен килограммов. Это становится основной причиной падения старых гнёзд при
сильном и даже средней силы ветре. Помимо больших размеров и веса гнезда, на сохран-
ность и продолжительную эксплуатацию гнездовых деревьев влияют пожары, глубина по-
чвенного слоя до слоя мерзлоты под гнездовым деревом и попадание на дерево экскрементов
аистов, которые «сжигают» и иссушают древесину. Весна на юге Дальнего Востока сопро-
вождается сильными ветрами, и когда корневая система гнездового дерева неустойчива, ве-
роятность падения такого дерева очень высока. Влияние пожаров на гнездовые деревья сво-
дится в основном к повреждению древесины у основания ствола, что ускоряет гибель дерева.
Брачное поведение аистов можно наблюдать только на гнезде. Партнеры чистят друг
другу оперение на голове и шее, самка кладет голову самцу на крыло или на спину; пары
исполняют (синхронно с наклонами головы и закидыванием клюва на спину) барабанную
дробь, которую слышно на расстоянии сотен метров. В брачный период самец демонстри-
рует особое поведение и издает клювом «дробь ухаживания», что не отмечено у самцов
европейского белого аиста (Archibald, 1991). Спаривание происходит на гнезде в дневное
время суток. Инициирует его самка (Dianjin, 1991); она становится перед самцом в спокой-
ной позе, хвост при этом приподнят вверх. Самец поднимается ей на спину и, балансируя
крыльями, приседает на цевки; хвост при этом опущен, и самец полностью закрывает сво-
им телом спину самки. Копуляция продолжается 10-30 сек (Fulin, 1991). По завершению
спаривания самка первой исполняет клювом дробь, оперение на шее и груди у нее припод-
нимается. Спаривания продолжаются во время и после откладки яиц, но могут происхо-
дить и в конце гнездового сезона. Аисты возобновляют размножение даже после падения
гнезда с хорошо оперёнными птенцами, что говорит о большом репродуктивном потенци-
але этого вида (Андронов, Андронова, 1992). По нашим наблюдениям (1982-2005 гг.) за
размножением самки дальневосточного аиста в Хинганском заповеднике, индивидуально
опознаваемой по травмированной ноге, хорошие репродуктивные способности у неё со-
храняются уже на протяжении, как минимум, 23 лет.
В норме пара аистов в сезон имеет только одну кладку, однако при потере потомства она
способна повторить размножение и отложить вторую кладку. Насиживание начинается в
Дальневосточный аист
425
зависимости от погодных условий, как правило, в период с 4 по 12 апреля, через 8-12 дней
после прилета. Так, 9 апреля 1989 г. в одном из гнёзд аистов на Архаринской низменности
уже имелось два яйца. В северной части ареала откладка яиц приходится на 17-18 апреля,
а вылупление птенцов — на конец мая - первую половину июня (Росляков, 1981а). Отклад-
ка яиц происходит с интервалом в 2-5 дней. В кладке (п=178) 1-6 яиц, средний размер
кладки колеблется по годам от 2,79 до 3,75 яиц. Яйца однородного молочного цвета с мато-
вой поверхностью и без рисунка, овальной формы. Размеры яиц (в том числе с использова-
нием данных С.В. Винтера, 1978) (п=30): 71,1-80,4 х 51,5-58,3 мм; в среднем 75,6x54,7
мм; вес (г) (п=4): 101,5-105,0, в среднем 102,9. На Приханкайской низменности размеры
19 яиц находились в пределах 74,4-80,0 х 53,5-58,3 мм, составляя в среднем 77,3x56,3 мм
(Глущенко, Шибнев, Волковская-Курдюкова, 2006). Максимальный размер кладки (6 яиц)
фиксировали на протяжении четырёх лет только в одном гнезде (видимо, одна семейная
пара); после падения этого гнезда с птенцами в тот же год пара отложила вторую кладку из
5 яиц. Процент оплодотворенных яиц в кладке очень высокий. Так, по данным о размноже-
нии аистов в Цицикаре (КНР), из 22 яиц в природных гнёздах 18 (81,8%) было оплодотво-
ренными (Dianjin, 1991); в неволе оплодотворёнными оказались 45 из 48 яиц (93,8%), из
всех этих яиц вылупились птенцы (Е. King, устн. сообщ.). По сведениям С.В. Винтера (1978),
в 12 гнёздах на Архаринской низменности из 45 яиц вылупилось 33 (73,3%) птенца.
Насиживать кладку аисты начинают после снесения третьего яйца, до этого времени
птицы на гнезде проводят мало времени. Насиживают оба партнёра, соотношение време-
ни, проведенного на гнезде самцом и самкой, может отличаться у разных пар. В целом,
самка проводит на гнезде больше времени; она же насиживает и в ночное время. Инкуба-
ционный период продолжается 32-34 дня. Вылупление отмечается с 4 мая по 18 июня,
массовое появление птенцов приходится на 10-14 мая. В выводке 1-5 птенцов, средний
размер выводка (п=191) по годам варьирует от 1,7 до 3,41 птенцов. Для Приханкайской
низменности число птенцов в среднем (по 26 гнёздам) составляет 2,8 особей на гнездо (Глу-
щенко и др., 2006). Гибель птенцов вызывается недостатком корма, погодными условиями и
беспокойством пары; она составляет в разные годы 7,0-27,3%. Успешность размножения у
зрелых пар выше, чем у молодых. Вылет молодняка происходит по среднемноголетним
данным с 11 июня по 14 июля, массовый слёт птенцов с гнёзд приходится на 10-15 июля.
Выкармливают птенцов оба родителя. Так, из зарегистрированных 113 посещений гнёзда
на р. Бикин, на самца пришлось 46, на самку — 62 прилёта (Fujimaki et al., 1990). Первые
10 дней с момента появления птенцов самка продолжает плотно сидеть на гнезде, обогре-
вая аистят. Ночью оба родителя остаются на гнезде или рядом на соседних ветках. Моло-
дые пары и пары с малым количеством птенцов проводят на гнезде больше времени. С
ростом птенцов время пребывания родителей на гнезде сокращается. До 20-дневного воз-
раста птенцы остаются под постоянным присмотром родителей, в 30-дневном возрасте
отлучки взрослых птиц составляют от 3 до 12 мин, в 40-50-дневном возрасте — 133 мин, в
60-дневном — 304 мин в течение светлого времени суток. Пары, имеющие большие вывод-
ки, вынуждены много времени расходовать на сбор корма, поэтому длительность пребыва-
ния на гнезде у них составляет не более 10 мин, т.е. они остаются на время, необходимое
для кормления и осмотра гнездовой постройки. Как правило, на гнезде находится только
один из родителей; одновременно самец и самка находятся там во время смены или в днев-
ное время на отдыхе. Смена происходит, в среднем, через 3 ч. Сменяемый аист задержива-
ется на гнезде на 1-7 мин, в редких случаях слетает сразу при появлении партнёра; отдыха-
ющий аист может оставаться на гнезде свыше 20 мин. Находясь на гнезде, взрослые птицы
чистят себя и птенцов, кормят птенцов, защищают их от непогоды и перегрева, поправляют
и охраняют гнездо. После месячного возраста птенцы всё чаще остаются в дневное время
426
Отряд Листообразные, Семейство Аистовые
одни, особенно в ясные, тёплые дни, однако поначалу один из аистов добывает корм в
непосредственной близости от гнезда.
Кормят птенцов родители 4-6 раз в сутки; по мере роста птенцов количество кормежек
увеличивается до 12-15 порций, но при нехватке пищи в природе кормежка может сокра-
титься до 1 раза в сутки. Перед слетом птенцов родители сокращают количество кормле-
ний почти в два раза. Самых маленьких птенцов родители кормят отрыжкой прямо в клюв,
старшим птенцам отрыгивают корм на гнездо, и птенцы его тут же разбирают. Подросшие
аистята прилет родителя с кормом встречают, сидя на цевках, с визгом, треском клювами,
хлопаньем крыльями. Голодные птенцы выхватывают корм из клюва взрослой птицы и
друг у друга. Весь процесс кормления после появления на гнезде взрослой птицы заверша-
ется в считанные минуты, остатки корма выбрасываются из гнезда или взрослый аист съе-
дает их сам. У сытого птенца пищевод полностью заполнен кормом, что не позволяет ему
согнуть шею. В жаркие дни родители приносят птенцам воду в клюве и защищают их от
палящих лучей солнца, стоя над птенцами с раскрытыми крыльями. Подрастающим птен-
цам родители всё чаще приносят корневища, пучки осоки, ветки или другой строительный
материал, который используется для игр и починки гнезда.
Хорошая возможность проводить наблюдения за потомством появляется с 15-дневного
возраста, когда аистята начинают активно перемещаться по гнезду и видны с большого
расстояния. Растут и развиваются птенцы очень быстро. До месячного возраста общий
рост массы тела постоянно увеличивается, следующие 10 дней наблюдаются колебания
прироста, но относительный прирост остаётся ещё высоким и составляет, в среднем, 20%.
В 40-50-дневном возрасте прирост массы тела не превышает 2-3%. Перед слётом с гнезда
масса тела птенца после стабилизации падает на 3,5-4,3%, что, видимо, необходимо для
снижения весовой нагрузки на крыло у летной молодой птицы. Формирование скелета за-
вершается в возрасте 60-70 дней.
У вылупившегося птенца задние конечности значительно длиннее передних — соотно-
шение составляет 1,4. В процессе постэмбрионального развития крылья «догоняют» по
длине задние конечности и в возрасте 30-40 дней опережают их в росте. Конечный индекс
соотношения длины задних и передних конечностей составляет 1,1; окончательная длина
конечности больше первоначальной для ног в 3,6 раза, для крыла — в 4,5 раза.
В период активного роста птенец съедает в сутки 800-1 500 г корма. Основу питания
птенцов составляет вьюн, речная и озёрная рыба. В Корее пара аистов выкармливала птен-
цов змеями (Austin, 1948). В целом, спектр питания птенцов чрезвычайно разнообразен и
включает разных представителей беспозвоночных и позвоночных животных, а также рас-
тительные объекты (Винтер, 1978).
По мере роста у аистят изменяется вокализация. Вылупившиеся птенцы до месячного
возраста издают хрюкающие, лающие и визжащие звуки, потом появляются звуки, напоми-
нающие поросячье повизгивание. Первую дробь клювом аистят можно слышать приблизи-
тельно в месячном возрасте, а голос молодые птицы теряют в первую осень жизни.
Гнездо птенцы покидают в возрасте 55-70 дней. Взрослые аисты участия в обучении
птенцов полету не принимают. Слет аистят с гнезда происходит самостоятельно, молодые
оставляют его одновременно или поодиночке. Беспокойство может спровоцировать ран-
ний слёт птенцов, вследствие чего они становятся лёгкой добычей для наземных хищни-
ков. На протяжении следующих после подъёма на крыло 14-30 дней лётный молодняк дер-
жатся вблизи гнездового дерева, иногда ночуя на нём. До откочёвки птенцов и распада
семейной группы родители подкармливают выводок и подправляют гнездо. По нашим на-
блюдениям, в одном случае самка продолжала держаться около птенца, который недоста-
точно хорошо летал, до сентября.
Дальневосточный аист
427
Суточная активность, поведение. Дальневосточные аисты ведут дневной образ жиз-
ни. В гнездовой период первый слёт с гнезда происходит в 400 утра, последний прилет про-
исходит с темнотой, после 2300. В поведении аистов прослеживаются два пика активности:
в утренние часы с 500 до 1200 и вечером с 1600 до 2100. Большую часть светового дня аисты
заняты сбором корма, особенно в период выкармливания птенцов. На протяжении всего
периода выкармливания во всех сферах деятельности чувствуется приоритет самки, будь
то нахождение на гнезде, прилёты с кормом или со строительным материалом, защита птен-
цов от дождя или лучей солнца.
В гнездовой сезон чаще всего можно встретить либо одиночную птицу в кормовом био-
топе, либо малочисленные скопления молодых птиц. Во время поиска корма аист подолгу
топчется на одном месте, переходы делает небольшие, пешим перемещениям предпочита-
ет полет. На чистку оперения взрослая птица затрачивает в среднем 15-70 мин в день, 6-30
мин на уход за оперением птенцов, 5-10 мин на ремонт гнезда. Находясь на гнезде, аисты
много времени затрачивают на осмотр окрестностей или просто спокойно стоят (Андро-
нов, Артемова, 1988).
На зимовке птицы с рассветом улетают на кормовые участки и остаются там до 1700.
Сбором корма зимой аисты заняты с 700 и до 1000 и после обеда с 1400 до 1700 (Qishan, 1991;
Zerong, 1991), во время дневного отдыха они не покидают кормовых зон. Ночуют в стаях
на мелководье, но очень часто меняют и кормовую зону и, соответственно, стаю. Во время
дневного отдыха занимаются чисткой оперения.
Питание. При наличии подходящих кормовых биотопов места кормежки располагают-
ся нередко в непосредственной близости от гнезда. У каждой пары и даже особи существу-
ют собственные предпочитаемые кормовые участки. При затяжных дождях, когда уровень
воды в марях повышается, птицы значительную часть времени проводят рядом с гнездом,
не улетая на дальние участки. При недостатке корма в сухие годы улетают на кормежку за
два и более км от гнезда. Предпочитают мелководье по берегам рек и озёр, заливные луга,
мочажины и канавы с водой. Уровень воды на кормовом участке обычно не превышает 50
см, оптимальная глубина воды 15-20 см. Спектр кормов чрезвычайно разнообразен и во
многом зависит от обилия того или иного компонента в биотопе. Аисты всеядны; тем не
менее, основу питания как в гнездовом ареале, так и на зимовке составляет, в первую оче-
редь, рыба, потом земноводные, моллюски, мышевидные грызуны, ракообразные (Пань-
кин, Нейфельдт, 1976; Винтер, 1978; Dianjin, 1991). Пищевод взрослой птицы, убитой бра-
коньерами 29.04.1975 г., содержал 39 ротанов (от 30 до 115 мм длиной) и 13 сибирских
лягушек (35-50 мм), а желудок — остатки 7 ротанов и 7 лягушек, хитин водных жуков
(Винтер, 1978).
В поиске и добывании корма птицы используют зондирование и хватание, мелкие объек-
ты склевывают с поверхности. При зондировании аисты погружают клюв в воду или грунт
и частыми пинцетообразными движениями клюва нащупывают съедобные объекты. Пери-
одически птица быстро вынимают из воды клюв и тут же опускает его рядом, постоянно
прощупывая кормовой участок и медленно продвигаясь по нему. Аисты никогда не добы-
вают корм в высокой воде, им не свойственно погружать голову в воду. Насекомых и назем-
ных позвоночных ловят бросками головы вперёд или склевывают с субстрата. Пойманный
объект помогает удерживать зубец на надклювье; посредством быстрых боковых движений
клюва птица продвигает добычу от кончика клюва к его середине, а затем заглатывает её
целиком, не расчленяя. Иногда аист довольно долго перебирает добычу клювом, умерщв-
ляет и разминает её; рыбу, прежде чем проглотить, разворачивает головой к глотке. Однако
голодная птица добычу проглатывает сразу, не перебирая её клювом и не разбирая положе-
ние головной и хвостовой части. Взрослый аист и лётный птенец способны заглатывать
428
Отряд Листообразные, Семейство Аистовые
очень крупную рыбу, до 25 см длиной. Корм накапливается в пищеводе до полного его
заполнения. Отводом головы вбок и вниз аист проталкивает и утрамбовывает корм; шея, по
мере наполнения пищевода, раздувается и принимает «палкообразный» вид. После пер-
вичной обработки ферментами и частичного переваривания, корм из пищевода попадает в
желудок. Неперевариваемые остатки пищи вместе с растительной массой удаляются из
организма в виде погадок. Взрослый аист в сутки добывает не менее 3 кг корма, но пита-
тельную его часть, видимо, составляет не более 1,5 кг. Сытая птица на 2-3 ч погружается в
состояние покоя (отдыхает, чистится), таким образом, экономно расходуя энергию.
Враги, неблагоприятные факторы. В природе у здоровой взрослой птицы врагов нет.
Кроме естественных причин, на аистов и среду их обитания влияют антропогенные факто-
ры (разрушение местообитаний, отстрел, отлов и т.д.), особенно в местах зимовки.
Повышенная смертность наблюдается в период миграций и зимовок. Отмечено доста-
точно много случаев гибели аистов при столкновении с высоковольтными линиями элект-
ропередач. Существенными причинами смертности являются также браконьерство (с при-
менением оружия и отравленных приманок) и использование химикатов в сельском хозяй-
стве. Накапливаясь в кормовых объектах, ядохимикаты представляют реальную угрозу для
жизни птицы и будущего потомства. О влиянии браконьерства в местах зимовок дальнево-
сточного аиста можно судить по находкам мёртвых аистов (Jong-jun, Zhou, 1989), обнару-
женных на одной территории в течение нескольких лет: 1980 г. — 40 особей, 1981 г. — 73,
1982 г. — 85, 1983 г. — 30, 1984 г. — 23, 1985 г. — 10, 1986 г. — 8, 1987 г. — 38 особей.
Известны также случаи гибели птиц от бескормицы.
В настоящее время местообитания аистов сильно трансформируются вследствие хозяй-
ственной деятельности (мелиорация, вырубка леса, выпас скота и т.д.), что на сегодняшний
день представляет самую серьезную угрозу для вида. С начала 1980-х гг. в Амурской обл. (а
затем и на других участках ареала в России) на безлесных территориях аисты стали стро-
ить гнёзда на опорах телеграфных линий, линий электропередач и триангуляционных пун-
ктах, а кормиться на открытых озёрах вблизи населенных пунктов. Там, где их не беспоко-
ят люди, аисты благополучно выводят птенцов, и подобные гнездовые платформы на ис-
кусственных сооружениях могут существовать довольно долго. Однако при гнездовании
на ЛЭП случаются замыкания электрической сети — вероятно, в результате контакта про-
водов либо с гнездовым материалом, либо с самой птицей, либо со струёй экскрементов.
Устраняя неисправность, электрики стремятся ликвидировать и её причину, для чего сбра-
сывают гнёзда аистов с опор ЛЭП. Данная проблема существует практически во всех реги-
онах юга Дальнего Востока, где обитает дальневосточный аист. В ближайшее время мес-
там обитания аистов серьёзную угрозу будет представлять также гидроэлектростанция на
реке Бурея (Буреинская ГЭС).
Другим лимитирующим фактором, наносящим большой ущерб гнездовой популяции,
служат лесостепные пожары. У местного населения существует полуторавековая традиция
весеннего выжигания сухой растительности. Этот бесконтрольный, законодательно запре-
щенный прием улучшения сенокосов применяется по сей день в местах обитания аистов.
Площади пожарищ достигают значительных размеров, особенно в весенний период с ха-
рактерными для этих мест сильными ветрами. Аисты часто предпочитают старые, сухие
деревья, а весенние пожары не только повреждают прикомлевую часть дерева, но иногда и
сжигают гнездовое дерево целиком. Повреждённые пожаром деревья с гнёздами чаще все-
го падают именно в сезон размножения аистов, когда в гнёздах уже имеются яйца или
птенцы.
Воздействие других лимитирующих факторов — таких, как сильные ветра, заморы рыб
на озёрах, разорение гнёзд белогрудым медведем и т.п. — в целом незначительно.
Дальневосточный аист
429
Хозяйственное значение, охрана. Как вид, находящийся под угрозой исчезновения,
дальневосточный аист занесён в Красный список МСОП (IUCN Red list) и Красную книгу
Российской Федерации, включён в Приложение 1 к Конвенции СИТЕС, в Приложения к
двусторонним соглашениям, заключённым Россией с Японией, Республикой Корея и КНДР
об охране перелётных птиц и среды их обитания. Занесён в региональные Красные книги
субъектов федерации юга Дальнего Востока. Охраняется в природных заповедниках Бас-
так, Болоньский, Норский, Ханкайский и Хинганский, а также в некоторых заказниках
Приморского и Хабаровского краев, Амурской области и Еврейской АО. Для сохранения
вида в странах Азии (Япония, Южная Корея, Китай) и Европы (Германия, Россия) разраба-
тываются и реализуются программы по вольерному содержанию и разведению птиц в це-
лях их последующей реинтродукции. На Приханкайской низменности проводится уста-
новка искусственных опор для размещения гнёзд. В заповеднике «Ханкайский» и его ох-
ранной зоне было установлено 12 опор, 10 из которых к настоящему моменту сохранили
функциональность, причём 8 из них в 2007 г. были заняты аистами (Коробов, Глущенко,
2008). Ещё задолго до установки первой опоры, когда местная гнездовая группировка на-
ходилась в критическом состоянии, предполагалось, что при наличии достаточного коли-
чества опор численность вида на Приханкайской низменности может быть доведена до 30-
40 гнездящихся пар (Глущенко, 1985).
Семейство Ибисовые - Threskiornithidae Poche, 1904
Листообразные средних размеров: от 47 см и 500 г у пятнистогрудого ибиса (Bostrychia
(Lampribis) гага) из Экваториальной Африки до 104 см и 2 кг у южноазиатского гигантско-
го ибиса (Pseudibis (Thaumatibis) gigantea). Тело довольно плотное, ноги от длинных до
довольно коротких, пальцы средней длины, задний короче остальных, но прикрепляется на
одном уровне с ними (в отличие от аистов). Основания передних пальцев соединены не-
большой перепонкой, когти заострённые, коготь среднего пальца гладкий. Клюв длинный,
обычно изогнут книзу дугой, у колпиц прямой, уплощённый, с лопатообразным расшире-
нием на конце. От несквозных щелевидных или овальных наружных ноздрей к концу клю-
ва идут чёткие борозды, функция которых не до конца ясна (предполагается, что она связа-
на с осязанием). На нижней челюсти хорошо развит задний угловой отросток (processus
angularis posterior), сближающий эту группу с фламинго (но, очевидно, это результат кон-
вергенции, вызванной сходством некоторых приёмов кормодобывания). Язык редуциро-
ван, имеется небольшой горловой мешок. Голосовая мускулатура нижней гортани развита
слабо или отсутствует, у части видов нет и голосовой мембраны, они немые. У немногих
видов с громким голосом (например, у индийского чёрного ибиса Pseudibispapillosa) длин-
ная трахея образует петли, лежащие в углублении грудины и играющие роль резонаторов.
Подкожного воздушного мешка на шее, характерного для аистов, нет. Глаза небольшие,
возможности бинокулярного зрения снижены. Шея обычно не такая длинная, как у цапель
и аистов, иногда выглядит совсем короткой, шейных позвонков 17-18. По заднему краю
грудины есть 2 пары вырезок, конец вилочки обычно не доходит до вершины киля и соеди-
нён с ним эластичной связкой. Первый палец кисти не несёт коготка. Сонные артерии пар-
ные. Слепые кишки парные, недлинные, но функционирующие, рудимента пениса нет. Кон-
турное перо со слаборазвитым побочным стержнем, пух растёт по птерилиям и аптериям,
копчиковая железа оперена, пудреток нет. У большинства видов не оперён участок между
глазами и клювом, у других голая большая часть головы, у немногих — вся голова и боль-
шая часть шеи. Первостепенных маховых 11, рулевых 12 (Карташов, 1974). Хвост от ко-
роткого до длинного, клиновидного, крылья имеют форму, промежуточную между цапля-
ми и аистами. Ибисы, особенно крупные виды, способны парить, шею в полёте не склады-
вают. Окраска очень разнообразна, часто с металлическим блеском, нередко развиваются
удлинённые украшающие перья на затылке, шее, нижней части спины. Яркую окраску обыч-
но имеют и неоперённые части тела. Для ряда видов характерна сезонная смена нарядов.
Полная линька происходит в конце лета - осенью (Коблик, 2001).
По сравнению с цаплями и аистами в питании ибисов более заметную роль играют
водные беспозвоночные. В рационе присутствуют черви, ракообразные (креветки, бокоп-
лавы и др.), различные насекомые и их личинки, амфибии на разных стадиях развития,
рыбы и др. Максимальный размер пищевых объектов обычно не превышает в длину 4-6
см. Можно выделить два основных способа кормодобывания. Первый — это зондирование
клювом грязевых отмелей или «кошение» клювом в мутной воде, обычно группами. Иног-
да под водой или в слое ила оказывается вся голова и шея. При этом способе охоты сильно
возрастает роль осязания, роль зрения падает. Изогнутый книзу клюв лучше, чем прямой,
обеспечивает схватывание придонных организмов наощупь. Участки головы и шеи ибисов
оголились, поскольку постоянно мокрое и грязное оперение на них причиняло бы неудоб-
ства. Некоторые ибисы, ведущие более сухопутный образ жизни во влажных лесах, хвата-
ют добычу с поверхности земли и с веток.
Ещё более специализированы в питании колпицы, имеющие цедильный аппарат из мел-
ких зубчиков по краям уплощённого клюва, особенно на концевом расширении. Они ловят
мелких ракообразных и другой зоопланктон в верхних слоях воды.
Священный ибис
431
Гнездятся как колониями разной величины, так и отдельными парами, чаще всего вме-
сте с другими голенастыми и веслоногими, редко — в колониях грачей. Гнездо — плоская
платформа из небрежно набросанных веток, или стеблей тростника, располагается на кус-
тах, ветвях деревьев, заломах околоводной растительности, изредка на уступах скал. Мо-
ногамны. В кладке 2-3, редко 4 белых, или голубоватых яйца, у немногих видов окраска
скорлупы слабопятнистая. Насиживание продолжается 28-35 дней. Птенцы вылупляются
слепыми, но прозревают в первый же день жизни. Пуховой наряд от беловатого до бурого.
Ещё не научившись летать, птенцы покидают гнездо и держатся поблизости в ветвях или
тростнике. Молодые полностью оперяются и приобретают способность к полёту на 6-7
неделе жизни. Половой зрелости достигают в возрасте 2-3 лет, хотя известно, что у некото-
рых видов самки становятся способными к гнездованию уже после первой зимовки. Вне
периода размножения держатся стайками. Активны как днём, так и в сумерках, ночью.
Некоторые виды ибисов, видимо с давних пор, терпимо относятся к присутствию челове-
ка, охотно добывают корм на полях, особенно на затопленных рисовых чеках, концентри-
руются вокруг деревенских свалок, выполняя роль мусорщиков. В ряде стран ибисы счита-
ются охотничьей дичью, истребляются из-за вкусного мяса. К глобально редким и исчеза-
ющим относятся 6 современных видов ибисов. Вероятно, уже в историческое время на о-
ве Реюньон (Маскаренские о-ва) вымер эндемичный Borbonibis (Threskiornis) latipes, изве-
стный только по остаткам костей.
В ископаемом состоянии ибисы известны с раннего эоцена (около 60 млн. лет назад;
находки в Англии), возможно, наиболее близкая к ним группа — верхнемеловые Plegador-
nithidae. Известно более 10 ископаемых видов (3 рода), с плиоцена в ископаемом состоя-
нии найдены несколько современных родов и, как минимум, 8 современных видов. Группа
распространена в тропиках и субтропиках всего земного шара, лишь немногие виды захо-
дят в южную часть умеренных широт. По разным современным источникам (Howard, Moore,
1980; Matheu, del Hoyo, 1992; Dickinson, 2003) семейство насчитывает 14—20 родов, включа-
ющих 32-33 современных вида. Традиционно семейство разделяют на два подсемейства —
собственно ибисовых (Threskiomithinae Poche, 1904), включающее роды Threskiornis, Pseud-
ibis, Geronticus, Nipponia, Bostrychia, Theristicus, Cercibis, Mesembrinibis, Phimosus, Eudo-
cimus, Plegadis, Lophotibis c 26-27 видами и колпицевых (Plataleinae Bonaparte, 1838), вклю-
чающее роды Platalea и Ajaia с 6 видами. В пределах Восточной Европы и Северной Азии
отмечено 6 видов, относящихся к 4 родам и обоим подсемействам. Из них три вида гнез-
дятся (один из них — только в Дальневосточном морском заповеднике), гнездование чет-
вёртого отмечалось в прошлом, и в настоящее время он исчез из региона (Ильяшенко, 2001;
Коблик и др., 2006), а два вида относятся к случайно залётным.
Род Threskiornis G.R. Gray, 1842
Священный ибис
Threskiornis aethiopicus (Latham, 1790)
Tantalus aethiopicus Latham, 1790, Index Omith., 2. с.706, «Эфиопия»
Sacred ibis (англ.), Heiliger Ibis (нем.), Ibis sacre (фр.)
Син. (лат.): для подвида aethiopicus'. Numenius ibis Cuvier, 1804; Ibis religiosa Cuvier, 1817;
Ibis egretta Temminck, 1840; Thereschiornis minor C.L. Brehm, 1855; Thereschiornis alba C.L.
Brehm, 1855; Ibis sacer Bohm, 1886.
432
Отряд Листообразные, Семейство Ибисовые
Статус. Редкий залётный вид.
Общая характеристика и полевые признаки. Крупная голенастая птица характерно-
го облика ибиса, величиной почти с гуся, с размахом крыльев до 124 см. Длинным, загну-
тым вниз клювом священный ибис несколько напоминает каравайку, но крупнее её, отно-
сительно более коротконогий и отличается общей белой окраской оперения. Лишь ноги,
голая голова и большая часть шеи окрашены в чёрный цвет. У взрослых птиц длинные
рассученные плечевые перья, свешивающиеся поверх хвоста (эгретки), тоже имеют чёр-
ный цвет. В полёте выделяются также чёрные кончики маховых перьев (как у молодых
колпиц) и полоска красноватой голой кожи ближе переднему краю испода крыла.
Кормятся ибисы обычно группами или стаями на болотах или мелководьях. Ночуют
обычно на деревьях. При агрессивном поведении широко раскрывают крылья и топорщат
чёрные эгретки, поднимая их высоко над спиной. Молодые птицы отличаются отсутствием
эгреток и оперенной головой и шеей.
Название «священный» почти на всех языках этот ибис получил, поскольку почитался
древними египтянами в качестве символа бога мудрости Тота (последний обычно изобра-
жался в виде человека с головой ибиса) и во множестве фигурировал на фресках и барель-
ефах.
Подвидовая систематика. Обычно выделяют 3 подвида: номинативный Th. ае. aethio-
picus (1), занимающий почти весь видовой ареал; Th. ае. bernieri (Bonaparte, 1855) (2), засе-
ляющий Мадагаскар, и Th. ае. abotti (Ridgway, 1893) (3), обитающий на о-вах Альдабра
(Спангенберг, 1951; del Hoyo et al., 1992; Степанян, 2003; Dickinson, 2003) (рис. 82).
1. Threskiornis aethiopicus aethiopicus
Tantalus aethiopicus Latham, 1790, Index Omith., 2. с.706, «Эфиопия»
Отличается более толстым клювом и широкой чёрной каймой по заднему краю крыла, а
также тёмно-бурой окраской радужины глаз (del Hoyo et al., 1992).
Предполагают, что на территорию Вост. Европы и Сев. Азии залетали птицы номина-
тивного подвида, распространенного к северу до Ирака (Степанян, 2003). Но описания
ибисов, добытых в Азербайджане в 1874—1875 гг. (Сатунин, 1907; Бутурлин, Дементьев,
1935), позволяют допускать возможность залётов сюда и мадагаскарской формы Th. ае.
bernieri — без чёрных пятен на концах маховых и со светлыми глазами.
Замечания по систематике. Маргинальные островные расы очень сходны между со-
бой, отличаясь, в основном, окраской радужины: белой — у Th. ае. bernieri и синеватой —
у Th. ае. abotti. Поэтому, возможно, их целесообразнее объединять в один подвид. Иногда
островные расы считают самостоятельным видом Th. bernieri (Bonaparte, 1855) (напр.,
Cramp, 1977). Ибисы Th. aethiopicus, южноазиатский Th. melanocephalus и австралийский
Th. molucca (Cuvier, 1829), формируют один надвидовой комплекс с викарирующими вида-
ми; иногда последние два вида рассматривают как подвиды Th. aethiopicus (см.: Cramp,
1977; del Hoyo, 1992).
Описание. Окраска. Половых и сезонных различий в окраске нет. Взрослые птицы имеют
белое оперение с большими чёрными пятнами на вершинах всех маховых перьев. Снизу
часто развит желтоватый оттенок. Плечевые перья очень длинные, рассученные, при осно-
вании — пепельно-серые, к концам — почти чёрные, с фиолетово-пурпурным отливом.
Голова и шея — голые, морщинистые, матово-чёрного цвета. Клюв блестяще-чёрный, ноги
чёрные в красноватых колечках (Сатунин, 1907; Мензбир, 1918). Голая кожа аптерий (ее
можно увидеть, если раздвинуть перья на соседних птерилиях) по бокам груди и под кры-
льями— красная.
Птенцы покрыты белым пухом, и лишь голова — в чёрном пуху. Клюв короткий, пря-
мой, «мясного» цвета.
Священный ибис
433
Рис. 82. Область обитания священного ибиса: а — граница гнездового ареала, б — гнездование на
берегу Бискайского залива, в — места встреч залетных птиц в Вост. Европе. Подвиды: 1 — Th. ае.
aethiopicus, 2 — Th. ае. bernieri, 3 — Th. ае. abotti.
Гнездовой наряд. Молодые птицы окрашены сходно со взрослыми, но цвет оперения
более тусклый, грязно-белый. Плечевые перья удлинённые, но не рассученые, и чёрные
эгретки поэтому отсутствуют. Голова и шея покрыты мелким пером; внизу перья белые, а
на верхней части и боках головы и зашейке — черновато-бурые с мелкими беловатыми
пестринами.
Строение и размеры. Крылья сравнительно короткие, тупые; первостепенных махо-
вых — 11; вершину крыла составляют III или III и IV первостепенные маховые. Первое
маховое редуцировано, оно короткое и узкое, скрыто под кроющими крыла. Хвост корот-
кий, прямой, из 6 пар рулевых. Голень и цевка сетчатые, клюв очень толстый, сильно сжат
с боков, с узкими длинными бороздами по бокам надклювья, в основании которых лежат
щелевидные ноздри.
Длина тела (см) — 65-89, размах крыльев — 112-124, длина крыла — 36-40, хвоста —
14-16, цевки — 9-11, клюва (от лба) — 13,5-17. Вес — около 1 500 г (Бутурлин, Дементь-
ев, 1935; Тугаринов, 1947; Cramp, 1977; del Hoyo, 1992). Самки заметно меньше самцов;
длина крыла самок и самцов равны, соответственно, 336-362 и 362-385 мм (Snow, Perrins,
1998). Залётная птица, пойманная в Юж. Литве, имела следующие размеры: размах крыль-
ев — 1019, крыло — 340, клюв — 140, цевка— 102, хвост— 125 мм. Вес её равнялся 1 400 г
(Nedzinskas, 1987).
Линька. Полная линька проходит вскоре после завершения гнездования, но маховые
последовательно сменяются в течение всего негнездового периода, иногда с перерывом
на время миграций. Перед началом сезона размножения происходит частичная линька
434
Отряд Листообразные, Семейство Ибисовые
верхних плечевых перьев и некоторых партий оперения на нижней стороне тела (Cramp,
1977).
Распространение. Гнездовой ареал. Заселяет почти всю Эфиопскую область к
югу от Сахары, а также побережья Мадагаскара и о-ва Альдабра (рис. 82). Изолированный
участок гнездового ареала сохранился, по-видимому, в Ираке — в низовьях Тигра и Евф-
рата. До XIX в. гнездился в Египте, вплоть до дельты Нила. Кроме того, в 1980-е гг. за счет
птиц, улетевших из зоопарка, во Франции на северном побережье Бискайского залива сфор-
мировалась искусственная популяция из свободно живущих особей. Гнездование ибисов
отмечается там в цапельниках с начала 1990-х гг. (Snow, Perrins, 1998). Возможно, именно
к этой популяции принадлежал залётный священный ибис, появившийся в сентябре 1986 г.
в Юж. Литве на озёрах Жувинтас и Симнас и державшийся там около двух недель до 29
сентября (Nedzinskas, 1987).
В основном же ибисы залетали в Вост. Европу, по-видимому, из Ирака. В XIX в. они
были хорошо известны жителям Азербайджана, которые называли их «птицами пророка
Али», причём в начале века ибисы появлялись там регулярно и были нередки. В начале
августа 1874 и 1875 гг. в заболоченных районах Ленкоранской низменности были добыты 2
одиночные птицы (Сатунин, 1907). Последний раз стайка из 7 птиц встречена там 4 апреля
1944 г. в 40 км к югу от г. Баку (Бурчак-Абрамович, 1946). Весной 1924 г. группу из 2^1
птиц наблюдали в дельте Волги (Тугаринов, 1947). П.С. Паллас (1811, цит. по: Спанген-
берг, 1951) наблюдал будто бы этих птиц на Сарпинских озёрах и сообщал, что ибиса
Numenius ibis изредка отмечали также на Каспийском и Черноморском побережье до устья
Кубани и Волги. Но, судя по комментариям М. Богданова (1879, с. 169), это сообщение
относится, скорее всего, не к ибису, а к каравайке.
Исчезновение священного ибиса из современной фауны России и сопредельных регио-
нов связано, несомненно, с сокращением численности птиц, гнездившихся на северной
границе ареала — в Египте и Ираке близ границ Азербайджана.
Зимовки. Священный ибис — оседлый или кочующий вид, покидающий гнездовые
районы в сухие периоды и возвращающийся обратно к сезону дождей. Размах кочёвок до-
стигает сотен километров, а в Юж. Африке, по данным мечения, ибисы могут мигрировать
на расстояние до 1 000-1 500 км. Часть птиц, гнездящихся к северу от экватора, откочёвы-
вает обычно в северном направлении; более южные популяции мигрируют на юг. В Ираке
ибисы встречались весь год, а в соседнем Иране, Кувейте и Йемене — лишь изредка на
кочёвках или зимой (del Hoyo, 1992).
Местообитания. Кормится преимущественно на мелководьях лагун, озёр, болот, на ри-
совых полях, по берегам других водоёмов. Нередко посещает свалки, скотобойни, фермер-
ские подворья, иногда встречается далеко от воды — на выгоревших травянистых равни-
нах. Гнездится в лесах или кустарниках, а также на земле на скалистых островах или среди
болот; иногда селится в городах (Гладков, 1970; del Hoyo, 1992).
Численность. В Африке — широко распространенный, обычный и многочисленный
вид. Общая численность там сейчас стабильна и насчитывает не менее 200 тыс. особей
(Rose, Scott, 1994). В 1970-е гг. священный ибис был ещё довольно обычен в Ираке; в 1990-е
гг. местная популяция оценивалась в 200 птиц (Rose, Scott, 1994), но сейчас статус пребы-
вания ибиса там неясен (Snow, Perrins, 1998). В Египте в прошлом тоже был очень обычен,
о чем свидетельствует захоронение 1,5 млн. мумий священного ибиса в катакомбах Сахары
(del Hoyo, 1992). Ибис встречался в Египте до начала XIX в. и к 1850 г. практически исчез.
Популяция во Франции в 1994 г. насчитывала 280 пар (Snow, Perrins, 1998).
Размножение. Священный ибис имеет один выводок в году. Его размножение приуро-
чено к периоду дождей, но в заболоченных районах он может гнездиться и в сухие сезоны.
Священный ибис, Черноголовый ибис
435
В Ираке откладка яиц происходит в апреле-мае, в Эфиопии — в марте-апреле. Селится
обычно плотными колониями от 50 до 2 000 пар, часто вместе с цаплями, бакланами и
другими колониальными видами. Гнёзда делает на высоких деревьях. Высота гнезда до 20
см, диаметр максимально до 42 см (Cramp, 1977); средний диаметр гнезда составляет 34
см. Гнездо сооружается из сучьев и веток и выстилается травой, листьями, иногда перьями.
Как и у других родственных видов, сучки и травинки приносит самец, а строительством
занимается самка. На островах гнёзда могут располагаться на низких кустах или на земле
среди скал, иногда — в тростниках среди болот. В кладке обычно 2^4, иногда до 5 яиц
овальной или слегка округлой формы с шероховатой скорлупой. Их окраска тусклая, бело-
ватая, с бледно-голубым или бледно-зелёным оттенком, иногда с мелкими красно-бурыми
пестринами. Размеры яиц: 57-65 х 39,5^15 мм (Спангенберг, 1951); 58-68 х 39^16, в сред-
нем 63x43 мм (Cramp, 1977). Насиживают обе птицы, инкубация длится 28-29 дней. Моло-
дые в возрасте 2-3 недель покидают гнёзда и собираются в группы; в 35^10 дней они опе-
ряются, а к 44^18-му дню оставляют колонии. Успех размножения невысок; в Эфиопии
успешное гнездование наблюдается, в среднем, у 35% пар. Из выводка часто поднимается
на крыло, в среднем, менее 1 слётка. Продолжительность жизни, по данным кольцевания,
более 21 года (Cramp, 1977; del Hoyo, 1992; Snow, Perrins, 1998).
Питание. Кормовые объекты ибиса — преимущественно насекомые (саранча, сверчки,
водяные жуки), а также пауки, ракообразные, черви, моллюски, рыба, лягушки, ящерицы и
мелкие млекопитающие, иногда — птенцы и яйца птиц. Местами ибисы специализируются
на некрофагии, поедая на свалках падаль, отбросы и т.п. (Гладков, 1970; Cramp, 1977; del
Hoyo, 1992; Snow, Perrins, 1998). Кормятся днем, группами по 2-20, иногда до 300 особей.
Медленно шагая, ибисы зондируют клювом ил или собирают живую добычу с поверхности
почвы. На ночёвку собираются обычно на деревьях у водоёмов. Кормовые перемещения из
колоний достигают 10 км и более (Cramp, 1977).
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Специальные
меры охраны не предусмотрены.
Черноголовый ибис
Threskiornis melanocephalus (Latham, 1790)
Tantalus melanocephalus Latham, 1790, Index Omith., 2, c. 709, Индия.
Oriental [Oriental Asian, White, Black-headed, Indian Black-headed] ibis (англ.);
Schwarzhalsibis (нем.); ibis a tete noire (фр.).
Син. (лат.): Ibis macei Wagler, 1790; Ibis leucon Temminck, 1829; Ibis bengala Cuvier, 1829;
Ibis propiqua Swinoe, 1870.
Син. (рус.): Индийский [белый, азиатский] ибис.
Статус. Залётный вид.
Общая характеристика и полевые признаки. Ибис средних размеров, меньше свя-
щенного ибиса, но крупнее каравайки. Длина 65-76 см. У птиц относительно длинные шея
и ноги, длинный серповидно изогнутый книзу клюв, короткий слабо округленный хвост.
Оперение белого цвета. У взрослых птиц вся голова и верхняя часть (более половины) шеи
голые (без перьев), кожа чёрного цвета. У молодых птиц голова и шея покрыты перьями.
Третьестепенные маховые и плечевые перья у птиц в брачном наряде удлинённые и рассу-
ченные.
436
Отряд Листообразные, Семейство Ибисовые
По внешнему виду и окраске похож на священного ибиса, но отличается от него тем,
что «украшающие» перья серого цвета, а не пурпурно-синевато-чёрные; кроме того, у чер-
ноголового ибиса имеются узкие ланцетовидные перья на зобе, а граница голой кожи на
шее проходит выше. В полете взрослые птицы отличаются от взрослых священных ибисов
также отсутствием чёрных пятен на вершинах маховых. От красноногого ибиса, помимо
окраски, отличается чуть большей величиной, более длинными чёрными ногами и отсут-
ствием красного цвета на «лице».
Полёт спокойный, прямолинейный, с глубокими взмахами широких крыльев; чередует-
ся с планированием. Во время полёта шея вытянута вперёд, ноги выступают за вершину
хвоста. У пролетающих птиц на нижней стороне развернутых крыльев заметна узкая про-
дольная полоса красного цвета. Ибисы населяют водно-болотные угодья в основном в тро-
пических и субтропических широтах. Поселяются колониями. Птицы, не участвующие в
размножении, в летний период ведут кочевой образ жизни и встречаются нередко за преде-
лами гнездового ареала. Во время миграций летят поодиночке или небольшими стаями.
Голос. Молчалив. В брачный сезон птицы издают хриплые и ворчащие звуки, отдалённо
напоминающие хрюканье.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Некоторые орнитологи считают черноголового ибиса под-
видом священного ибиса.
Описание (по: Sonobe, Usui, 1993; Mackinnon, Phillipps, 2000 и др.). Окраска. Поло-
вой диморфизм в окраске оперения отсутствует. Брачный наряд. Оперение белое с желто-
вато-серым налётом на спине, верхней стороне крыльев и груди. Голова и большая часть
шеи не оперены; голая кожа чёрного цвета. Третьестепенные маховые и плечевые перья в
вершинной части серого цвета, они удлинены и рассучены. На зобе пучок узких ланцето-
видных перьев. На нижней стороне крыльев протягивается узкая полоса голой кожи крас-
ного цвета. Ноги и клюв чёрные. Радужина красновато-бурая.
Пуховой наряд. Пух на голове коричневый, на туловище белый.
Гнездовой наряд. Оперение серое. Перья на голове и шее коричневые, на передней сто-
роне шеи серовато-белые. Первостепенные маховые имеют снаружи чёрные вершины. Го-
лые участки на нижней стороне крыльев черноватые.
Первый годовой наряд. Оперение белое. Голова и задняя часть шеи покрыты буроваты-
ми, серыми и черными, а горло, подбородок и передняя часть шеи — белыми перьями.
Вершины первостепенных маховых бурые. Продольная (голая) полоса на нижней стороне
крыльев розового цвета. Ноги и клюв чёрные. Нет «украшающих» и ланцетовидных (на
зобе) перьев.
Самец, найденный 9 октября 1972 г. в Приморском крае, был белой окраски. У него
верх и бока головы, а также верхняя сторона шеи светло-бурые, на лбу и темени разброса-
ны редкие чёрные перышки. Подбородок и горло белые. Стержни первостепенных махо-
вых, включая первое, почти целиком чёрные, стержни остальных — светло-бурые. Концы
первостепенных и третьестепенных маховых, большие кроющие кисти и крылышко свет-
ло-бурые (Докучаев, Лаптев, 1974). У птицы, добытой в мае 1972 г. в Якутии, в оперении
преобладала белая окраска, голова была серая, дистальная половина второстепенных махо-
вых — светло-серая; голый участок кожи на голове был незначителен и не захватывал уш-
ные отверстия (Дегтярев и др., 1990).
Строение и размеры (мм). Первостепенных маховых 11, рулевых 12. II или II и
III маховые самые длинные. Размер птиц обоего пола (мм): длина крыла 340-370, хвоста
130-145, цевки 95-115, клюва 135-170 (Бутурлин, 1935). Размеры самца: длина крыла 370,
хвоста 135, цевки 110, клюва 145 мм (Докучаев, Лаптев, 1974); молодой птицы (пол не
Черноголовый ибис
437
установлен): длина крыла 341, хвоста 138, цевки 95, клюва 148 мм (Дегтярев, Ларионов,
Винокуров, 1990).
Линька. Две линьки в году: неполная предбрачная и полная послебрачная. Окончатель-
ный наряд птицы надевают, вероятно, на втором году жизни.
Распространение. Гнездовой ареал: Пакистан, Непал, Индия, к западу до
бассейна р. Инд, Шри Ланка, юг Вьетнама, Индонезия (о. Ява и, возможно, о. Суматра),
Сев.-Вост. Китай (Степанян, 2003; Dickinson, 2003; del Hoyo et al., 1992; Sonobe, Usui, 1993
и др.) (рис. 83). В XIX-XX вв. был широко распространён в Юж., Юго-Вост, и Вост. Азии.
В настоящее время возможно гнездится в Сев.-Вост. Китае, в провинции Хэйлунцзян; в
других районах Китая на гнездовании отсутствует (Mackinnon, Phillipps, 2000).
Зимовки в Юж. и Юго-Вост. Азии (на Филиппинских о-вах, в Бирме, Таиланде,
Индонезии, на юго-восточном и южном побережье Китая, о. Тайвань и в других регионах).
Кроме того, зимой птиц регистрировали во внутренних районах Китая (Сычуань, Юньнань
и Тибет), а также на Японских о-вах — в центральных и южных частях о-ва Хонсю и остро-
вах, расположенных южнее (del Hoyo et al., 1992; Mackinnon, Phillipps, 2000; Dickinson,
2003 и др.).
В России известны следующие случаи зал тов. В Приморском крае зарегистрированы
3 встречи: молодой самец добыт 9 октября 1972 г. в окрестностях с. Щербаковка Ольгинс-
кого р-на (Докучаев, Лаптев, 1974); птиц наблюдали 15 июня 1984 г. вблизи пос. Рязановка
Хасанского р-на (Nazarov et al., 2001) и 27 марта 2003 г. на оз. Тростниковое у побережья
оз. Ханка (Глущенко, Волковская-Курдюкова, 2005). В Якутии молодого ибиса добыли из
стаи в 7 птиц между 20 и 25 мая 1972 г. на озере в окрестностях пос. Тюнгюлю в 60 км к
северо-востоку от г. Якутска (Дегтярев и др., 1990). Ибисов встречали в Бурятии в окрест-
ностях г. Улан-Удэ 11 мая 1990 г. (Болд и др., 1991), в Иркутской обл. на западном берегу оз.
Байкал в окрестностях пос. Зама 22 июня 2003 г. (Попов, 2004) и в Амурской обл. (Дугин-
Рис. 83. Область распространения черноголового ибиса: а — гнездовой ареал, б — места изолиро-
ванного гнездования, в — места зимовок, г — места встреч залётных особей в России.
438
Отряд Листообразные, Семейство Ибисовые
цов, Панькин, 1993) (рис. 83). В периоды сезонных миграций и летних кочёвок залетных
птиц регистрировали в Китае, на Корейском п-ове, в Японии (на о. Хоккайдо), в странах
Юго-Вост. Азии (Mackinnon, Phillipps, 2000; Won, 1996а; Check-list of Japanese birds, 2000;
и др.). Вид включён в список птиц Узбекистана (Митропольский, Митропольский, 2009),
однако конкретные данные о встречах черноголового ибиса в этой работе не приведены.
Миграции. В Индии, Пакистане и других регионах Юж. Азии ибисы оседлы; кочёвки
отмечаются лишь в засушливые сезоны года. Из Сев.-Вост. Китая птицы мигрируют на
зимовки в южные районы страны, на о. Тайвань и страны Юго-Вост. Азии.
Местообитания. Мелководные пресные и солоноватые водоёмы (реки, озёра, водохра-
нилища, лагуны, заливы и т.д.) и их берега, затопленные леса и древесно-кустарниковые
заросли, травянистые болота, влажные луга, мангровые заросли, рисовые поля и другие
сельскохозяйственные угодья, литораль мелководных заливов и т.д. Нередко птицы посе-
ляются вблизи жилищ человека.
Численность. Немногочисленный вид. В большинстве регионов, где гнездится, отно-
сится к редким птицам. Редок в Пакистане; в 1965 г. наиболее крупная стая состояла из 100
особей. По данным на январь 1991 г., в Индии учтено 2,5 тысячи, в Шри-Ланке — 685, в
Бангладеш — 252, в Бирме — 730 птиц. В прошлые годы был многочисленным во Вьетна-
ме, в 4 колониях насчитывали от 500 до 2 000 особей; сейчас здесь редок. Стал также ред-
ким в Индонезии на о. Ява. На Суматре в 1990-е гг. было учтено около 2 тысяч птиц, но
гнездование не доказано (del Hoyo et al., 1992).
Размножение. Гнездовой сезон в северной Индии длится с июня по октябрь, в юж.
Индии и Шри-Ланке — с ноября по март. Сроки гнездования зависят от состояния уровня
воды после окончания сезона дождей. Моногам. Птицы поселяются колониями, обычно
в поселениях других аистообразных и бакланов. Гнездо - чашеобразная платформа из
сухих веток и стеблей трав, располагается на ветвях деревьев над водой или вблизи от
воды. В кладке 2-3, редко 4 яйца белой окраски с зеленоватым или голубоватым оттен-
ком. Средние размеры яиц (п=112): 53,5-71,5 х 38-45,6 мм (Hartert, 1920а). Инкубацион-
ный период длится 23-25 суток. Насиживают оба партнёра. Молодые птицы находятся в
гнёздах до 40 суток.
Суточная активность, поведение. Характерна дневная и сумеречная активность. По-
ведение не изучено.
Питание. Основные объекты питания — водные, околоводные и наземные беспозво-
ночные (моллюски, ракообразные, черви, насекомые, их личинки и др.) и позвоночные (ля-
гушки, рыбы и др.) животные. Иногда птицы поедают растительную пищу. Добычу отыс-
кивают на мелководьях, зондируя клювом мягкий ил или влажную почву; часто погружают
в воду голову и шею.
Враги, неблагоприятные факторы. В гнездовой сезон враги — вороны, хищные пти-
цы и человек. В настоящее время основная причина сокращения численности птиц—транс-
формация водно-болотных угодий под влиянием хозяйственной деятельности человека, охота
и использование пестицидов.
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Занесён в спи-
сок охраняемых видов животных Приморского края России.
Красноногий ибис
439
Род Nipponia Reichenbach, 1853
Красноногий ибис
Nipponia nippon (Temminck, 1835)
Ibis nippon Temminck, 1835, Temminck et Laugier, Planches Coloriees, livr. 93, табл. 551,
Япония.
Japanese [crested] ibis (англ.); Nipponibis (нем.); ibis nippon (фр.).
Син. (лат.): Nipponia temminckii Reichenbach, 1852 (1853); Ibis sinensis David, 1872.
Син. (рус.): Японский [китайский, белый] ибис (книжн.), хохлатый ибис.
Статус: вид, исчезнувший в России на гнездовье.
Общая характеристика и полевые признаки. Птица размером с крупную домашнюю
курицу. Вне периода размножения окраска целиком белая. Ноги, «лицо» (оголённая кожа
на лбу, в основании клюва и вокруг глаз) и кончик клюва красные, остальная часть клюва
чёрная. На голове густой, довольно длинный хохол из перьев разной длины, ниспадающий
на шею в спокойном состоянии. У летящей птицы за пределы хвоста выдаются только кон-
чики пальцев вытянутых лап (по цветным фотографиям из Yamashina, Nakanishi, 1983).
Л.М. Шульпин (1936) описывал красноногого ибиса следующим образом: «Ибис низок
на ногах, плотен, коренаст, со сравнительно короткой шеей при длинном клюве. Он не
избегает садиться на деревья и даже на тальник, и, сидя на сучке, обычно прижимает шею,
так что из сгорбленной фигуры тела птицы странно торчит длинный тонкий клюв. На поле-
те птица медленно и плавно машет крыльями, вытягивает шею и ноги. Клюв держит почти
горизонтально».
Голос похож на карканье вороны, но гораздо громче, ниже по тону и грубее: «ка-га, ка-
га, кага ....»; иногда звучит как «каг, каг, каг» или «кагаак, кагаак» (Н.М. Пржевальский,
дневник; цит. по: Шульпин, 1936).
Птица очень осторожна.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. В таксономической литературе существуют разночтения
по поводу года опубликования первоописания вида— 1835 (традиционная трактовка) или
1836 (специальное исследование Dekker et al., 2001). Последняя трактовка принята в «Списке
птиц Российской Федерации» (Коблик и др., 2006). В то же время Э. Дикинсон, один из
соавторов упомянутого исследования, в дальнейших работах (Dickinson, 2003 и выпуски
дополнений и исправлений к этой работе) продолжает считать годом описания вида 1835.
Таким образом, вопрос остаётся открытым. Здесь принята традиционная трактовка.
Описание. Окраска. Взрослая птица. Маховые имеют жёлто-оранжевые (или охри-
сто-оранжевые) стержни. Интенсивность их окраски возрастает к очину. Опахала — свет-
ло-оранжевые вдоль стержня, светлеют до белого к краям. Стоящая птица выглядит белой.
Летящая — сочетание белого и тёплого оранжевого (не очень интенсивного) цвета (по цвет-
ным фотографиям из Yamashina, Nakanishi, 1983). В конце зимы одновременно с началом
брачных демонстраций окраска головы, хохла, шейных и плечевых перьев меняется от свет-
ло-розовой до тёмно-серой. Источник этого тёмного пигмента (меланина) — кожа у осно-
вания перьев, окружающих голое «лицо» ибиса. После купания птица в течение 20-30 мин
потирает головой о «плечи» («daubing behaviour»). По мере повторения операции перья на
шее, плечах, а также хохол темнеют до тёмно-серого цвета. После летней линьки птицы
снова становятся белыми.
440
Отряд Листообразные, Семейство Ибисовые
Радужина взрослой птицы желтоватая или желтовато-оранжевая. Лоб, бока головы, кожа
горла (на подклювье), глотка — красные. Ноги красные, клюв чёрный с красным кончиком.
Ушное отверстие расположено на оголенном участке кожи (не прикрыто перьями) (по цвет-
ным фотографиям из Yamashina and Nakanishi, 1983).
Однодневный птенец. Тело покрыто светло-серым пухом. «Лицо» и конец клюва —
розовые, основная часть клюва чёрная, ноги оранжево-красные (по Арчибальд, Лантис,
1981 и цветным фотографиям из Yamashina, Nakanishi, 1983).
Гнездовой наряд. Оперение в целом белое, шея и уже довольно длинный хохол — свет-
ло-серые. Два первых первостепенных маховых имеют частично чёрные (тёмно-серые) опа-
хала. «Лицо» и голени жёлтые, цевка и пальцы тёмные. Клюв чёрный, светлеющий к кон-
цу. Радужина коричневая (по цветным фотографиям из Yamashina, Nakanishi, 1983).
Строение и размеры. Птица плотного сложения, по сравнению с большинством
ибисов большеголовая, с короткой толстой шеей и корткими ногами, недлинными крылья-
ми с закруглёнными вершинами. Общая длина 550-785 мм, крыло 375^110 мм, цевка 75-
83 мм, клюв от лба 150-183 мм (Тугаринов, 1947).
Распространение. Гнездовой ареал. В прошлом на материке гнездился от
долины Амура на севере до Центр, и Вост. Китая на юге, а также на п-ове Корея и на
Тайване. На Японских о-вах обитал от юга Хоккайдо до Кюсю (BirdLife International.
Threatend Birds..., 2001) (рис. 84 A).
Имеющаяся информация о прошлом гнездовом распространении в северной части аре-
ла (Россия, крайний северо-восток Китая) очень скудна. На севере ареала в известных ме-
стах гнездования красноногий ибис был редок уже в 1850-1860-е гг. Об этом пишет Л.М.
Шульпин (1936), цитируя Р. Маака (1861) и Н.М. Пржевальского (1870). Речь при этом идёт
о долине р. Уссури, р. Сунгача и оз. Ханка. Н.М. Пржевальский так оценивает численность
ибиса в Ханкайском бассейне: «Много-много, если здесь остаётся для вывода десятка два
экземпляров». В. Тачановский (1875, 1893) пишет о редкости вида в устье Уссури, ссыла-
ясь на Б. Дыбовского и В. Годлевского (они работали там в 1874 г.).
Для всех этих районов гнездование ибиса было строго доказано. Встречу ибисов в 60
км к востоку от Бурей (3 экз., 17 апреля 1858 г.) (Radde, 1863) Л.М. Шульпин также оцени-
вает как свидетельство гнездования. При этом он пишет: «Тунгусы знают её и имеют для
неё специальное название (ургун), так что птица, вероятно, нормально гнездится здесь».
Все перечисленные районы находятся на периферии гнездового ареала, и низкая числен-
ность вида там ожидаема и объяснима.
Что касается левобережья Уссури, то прямая информация о гнездовании вида здесь прак-
тически отсутствует, не считая упоминания Н.М. Пржевальского о гнездовании ибиса на р.
Мурени (р. Мулинхэ, левый приток р. Уссури) (Дневник; цит. по: Шульпин, 1936). Тем не
менее, Л.М. Шульпин считал, и, безусловно, обоснованно, что основные гнездовья нахо-
дятся на китайской стороне.
Информация из предполагаемого района зимовок уссурийских ибисов дополняет наши
представления о состоянии популяции. Численность ибиса в конце XIX - начале XX столе-
тий была, по-видимому, невысокой, но до катастрофы дело ещё не дошло. Уссурийско-
корейская популяция на тот период насчитывала по крайней мере несколько тысяч особей.
Это максимальное количество птиц, которое наблюдали на южнокорейских зимовках в
1911 г. (Shimokoriyama, 1913; цит. по: Austin, 1948). Затем началось падение численности,
которое завершилось исчезновением уссурийско-корейской популяции.
В 1929 г. в провинции Чхунчхон-намдо (юго-запад Корейского п-ова) ещё учли более
100 птиц; весной 1934 г. возле Инчхона наблюдали 20 птиц (Yamashina, Nakanishi, 1983;
цит. по: BirdLife International.Threatend Birds..., 2001). В период 1966-1978 гг. с Корейского
Красноногий ибис
441
Рис. 84. Область обитания красноногого ибиса. А. Распространение: а — известные места гнездовий
в прошлом (пункт западнее оз. Ханка в районе р. Мурень [Мулинхэ] — предположителен), б —
район современного гнездования вида (Китай, 1980-е гг.), в — район гнездования в XIX - начале XX
вв.; г — районы известных зимовок. Б. Вероятные пути миграций уссурийско-корейской популяции:
д — точно установленные пункты встреч, е — предполагаемые пункты встреч птиц в миграционный
период (по: Шульпин, 1936; BirdLife International. Threatend Birds..., 2001; с ориг. добавлениями).
п-ова известны несколько встреч (от десяти до одной птицы), последний раз ибисов (снача-
ла двух, потом одну птицу) видели здесь в 1978 г. (Арчибальд, Лантис, 1981).
Л.М. Шульпин (1936) в годы своей работы в Приморье (1926-1928 гг.) отметил сниже-
ние численности красноногого ибиса: «В настоящее время ибис решительно исчезает, он
уже не гнездится, как раньше на средней Лефу (р. Илистая), в Сунгачинских болотах стал
ещё более редкой птицей, чем был раньше, на пролёте реже встречается на юге страны, а в
связи с общим высыханием края сокращается и его стация, независимо от его боязни чело-
века. Только за нашей границей, по болотам левых притоков Уссури и на Сунгаче, вероят-
но, плотность его населения более или менее сохраняется близ прежнего уровня. С расту-
щим заселением края, осушением болот и запашкой их — ибис совершенно исчезнет».
Как показал дальнейший ход событий, Л.М. Шугльпин был абсолютно прав. Начавшее-
ся в первой половине XX столетия заселение и освоение равнины Саньцзян (междуречье
Уссури, Сунгари, Амура) полностью трансформировали этот регион. За период 1949—
2000 гг. площадь водно-болотных угодий уменьшилась здесь более чем в 5 раз (с 5 340 000
до 889 786 га) (Liu, Ма, 2002). По-видимому, этот процесс поставил точку в судьбе уссу-
рийско-корейской популяции красноногого ибиса.
Имеются косвенные свидетельства о том, что многие из птиц, встреченных в России в
1960-1980-х гг., находились в стадии размножения (Ильяшенко, 2001). По данным того же
автора, с 1990 г. в пределах России вид более не отмечался.
442
Отряд Листообразные, Семейство Ибисовые
В Японии красноногий ибис был широко распространён и обычен до конца XIX столе-
тия (Austin, Kuroda, 1953). Однако с конца XIX столетия традиционная охрана перестала
действовать, и в результате чрезмерной охоты численность вида быстро уменьшилась. К
концу 1908 г. красноногий ибис почти исчез (Yasuda, 1983; цит. по: BirdLife International.
Threatend Birds..., 2001). К 1950-м гг. уцелели две небольшие группы птиц (на п-ове Ното и
о. Садо). Их пытались сохранить, однако, несмотря на все прилагаемые усилия, это не уда-
лось. Последний самец умер в неволе в 1995 г.
В Центр. Китае ибис местами был обычен до начала XX столетия в провинциях Чжец-
зян, Шэньси и Ганьсу. В Нинбо (Чжецзян) наблюдались «большие партии» птиц в конце
августа и очень большие стаи в середине ноября (Swinhoe, 1873, цит. по: BirdLife International.
Threatend Birds..., 2001); в 1904 г. в долине Вэйхэ (провинция Шэньси) красноногий ибис
был одной из обычнейших птиц (Blackwelder, 1907; цит. по: BirdLife International. Threatend
Birds..., 2001). В провинции Шэньси в начале 1940-х гг. наблюдали сотни красноногих иби-
сов в местах отдыха на деревьях. Позднее там же было найдено более 30 гнёзд (Li Bin, Tian
Qing, 1998; цит. no: BirdLife International. Threatend Birds..., 2001). Вид был обычен в Гань-
су ещё в 1955 г., однако затем численность неожиданно резко упала, и к 1957 г. ибис почти
исчез (Wang Xiangting, 1991; цит. по: BirdLife International. Threatend Birds..., 2001).
Небольшая сохранившаяся популяция была обнаружена в 1981 г. в провинции Шэньси
на южных склонах хребта Циньлин (Liu Yinzeng, 1981; цит. по: BirdLife International.
Threatend Birds..., 2001). В результате предпринятых усилий численность этой группиров-
ки удалось увеличить. В 2000 г. из 15 гнёзд вылетело 33 птенца. Общая численность дикой
популяции составила к концу сезона более 120 птиц (Ding Changqing, устное сообщение,
2001; цит. по: BirdLife International. Threatend Birds..., 2001). Кроме того, в Центре по разве-
дению и охране (Янсянь) содержалось в 1998 г. 52 птицы (Cao Yonghan et aL, 1999; цит. по:
BirdLife International. Threatend Birds..., 2001).
Зимовки. Птицы из северных частей ареала были мигрирующими, гнездившиеся на
юге — оседлыми или кочующими (BirdLife Intemational.Threatend Birds..., 2001). Л.П. Шуль-
пин (1936) и Е.М. Спангенберг (1951) полагали, что гнездящиеся у нас птицы летят зимо-
вать в Китай. Иное мнение говорит в пользу корейских зимовок (Арчибальд, Лантис, 1981;
BirdLife Intemational.Threatend Birds..., 2001). В пользу второго варианта свидетельствует и
размещение встреч мигрирующих птиц (рис. 84 Б). При этом крайний юго-запад Примо-
рья, побережье зал. Петра Великого, судя по количеству встреч (Шульпин, 1936), был мес-
том заметного пролёта красноногого ибиса (правда следует иметь в виду, что здесь, види-
мо, чаще бывали орнитологи).
Имеющаяся информация о численности красноногого ибиса в двух основных местах
зимовок на Корейском п-ове свидетельствует об их значительности в конце XIX и первые
десятилетия XX века. Ибис на восточном побережье полуострова, по свидетельству Л. Та-
чановского, был обычен, встречаясь стаями до 50 особей (Taczanowski, 1888, цит. по: Austin,
1948). На юго-западе Корейского п-ова в провинции Чхунчхон-Намдо видели около 100
птиц в 1929 г. (Yamashina, Nakanishi, 1983). Наконец, там же, в районе границы двух про-
винций Чхунчхон-Намдо и Чолла-Пукто, в конце декабря 1911 г. наблюдали скопление от-
дыхающих на соснах ибисов численностью в несколько тысяч особей (Shimokoriyama, 1913,
цит. по: Austin, 1948).
Миграции. О возможных маршрутах миграций можно судить на основании некоторой
имеющейся конкретной информации. Известна серия встреч в период весенней миграции
во второй половине XIX - начале XX столетия вдоль долины Уссури, в бассейне оз. Ханка
и вдоль побережья зал. Петра Великого (Шульпин, 1936). Весной, осенью и зимой птиц
отмечали в Сев. Корее (на Корейском п-ове) в провинции Хамген-Пукто, Хамген-Намдо,
Красноногий ибис
443
Канвондо (р-н Вонсана) (Taczanowski, 1888, цит. по: Tomek, 1999; Янковский, 1898; Austin,
1948). Цепочка точек регистрации прочерчивает пролётный путь красноногого ибиса от
корейских зимовок до р. Уссури. Это один из вероятных миграционных маршрутов (рис.
84 Б).
Другой маршрут, видимо, проходил западнее. Основные зимовки ибиса на Корейском
п-ове располагались в юго-западной его части на границе провинций Чхунчхон-Намдо и
Чолла-Пукто (BirdLife International. Threatend Birds..., 2001).
В пользу реальности существовавших миграционных маршрутов говорит и тот факт,
что ряд видов, в частности японский, чёрный и даурский журавли, гнездящиеся в бассейне
Уссури, по наблюдениям в 1870-х гг. летели на корейские зимовки и в Японию теми же
путями. По поводу же прилёта ибиса на о. Кюсю сообщается, что он держится во время
перелета вместе с журавлями (Yasuda, 1983, цит. по: Bird Life International, 2001).
Численность. На рубеже XIX и XX вв. красноногий ибис был многочисленным в севе-
ро-восточном Китае, обычным в Японии и на Дальнем Востоке России (Ильяшенко, 2001).
На протяжении XX в. численность значительно сократилась в результате охоты, деграда-
ции мест гнездования и загрязнения пестицидами (смотри раздел «Распространение»).
Конкретные цифры сохранившихся в настоящее время на гнездовании пар японский иби-
сов назвать трудно. Благодаря учётам в Китае в 1981—1988 гг. в лесах у подножия Циньлиня
зарегистрировано до 40 гнёзд красноногого ибиса (Luthin, 1987); в некоторых из них раз-
множение прошло успешно. В Японии же отмечали только неразмножавшихся птиц; рез-
кое сокращение численности красноногих ибисов там произошло в конце второй полови-
ны XX в. Так, на о. Садо в 1930-х гг. обитало около 100 птиц, а в 1950-х гг. зарегистрирова-
но всего 4 особи. Во второй половине 1960-х гг. эти птицы, несомненно, размножались в
горных лесах о. Садо, так как молодых видели осенью во время кормёжки на водоёмах. В
результате предпринятых мер по охране вида к 1974 г. их количество достигло 15 особей.
Однако в 1975 г. численность вновь снизилось до 8 птиц (Yamashina, 19776).
На основании перечисленных сведений можно предполагать, что общая численность
красноногого ибиса в естественных местообитаниях в настоящее время не превышает 200-
500 особей. Тренд численности, несмотря на принятые меры (см. ниже), отрицательный, и
вид безусловно находится в угрожаемом положении.
Местообитания. Гнёзда ибис устраивает на высоких деревьях, главным образом в уча-
стках перестойных лесов. Кормовые стации представляют собою заболоченные широкие
долины рек в невысоких горах, имеющих низменные участки. Охотно осваивает рисовые
чеки, кормится на берегах озёр, стариц. На севере области распространения зимует на не-
замерзающих участках рек, заброшенных, затопленных и не замерзающих в зимние меся-
цы рисовых полях. Районы обитания, особенно в зимнее время, тесно связаны с террито-
риями, в разной степени освоенными человеком.
Размножение. Красноногий ибис гнездится на высоких соснах и дубах в средней части
кроны. Гнёзда располагаются как в непосредственной близости от ствола, так и на боковых
ветвях на удалении до 3-5 м. При выборе места для гнезда предпочитает «укрытые» про-
странства с несколькими путями подлёта к ним (Yamashina, 1977а). Яиц в полной кладке 2-
5. Нормальная толщина скорлупы яиц в естественных условиях — 0,189-0,315 мм (Yama-
shina, 19776). В 1981 г. в провинции Циньлинь проведено наблюдение за двумя гнёздами.
Одно, в котором было 4 яйца, погибло. Во втором вывелось и вылетело 3 птенца.
Питание. Наиболее тщательно изучалось на о. Садо в Японии (Yamashina, 19776). До-
бычу ибис собирает в ручьях, прудах и других водоёмах. В рационе отмечены: из пресно-
водных рыб — вьюн Misgurnus anguillicaudatus, карась Carassius carassius, речной сомик
Parasilurus asotus; из амфибий — лягушка Rhacophorus viridis, тритон Cynops pyrrhogaster,
444
Отряд Листообразные, Семейство Ибисовые
углозуб Hynobius nigrescens\ из ракообразных — речной краб Potamon dehaani, рак Camba-
roides japonicus', из насекомых — водолюб Hydrophilus acuminatus, японский плавунец
Cybister japonicus, личинки жуков, а также представители прямокрылых Acridioidea sp.; из
пресноводных моллюсков — улитки Viviparus japonica и Cristaria plicata.
В условиях клеточного содержания нормальный вес дневного рациона составляет SOO-
SOO г.
Враги, неблагоприятные факторы. Считается, что основным фактором, вызвавшим
резкое сокращение численности красноногого ибиса в 1950-х гг. в Японии (да и по всему
ареалу) было отравление его пестицидами, которые он получал в основном через объекты
питания на рисовых полях. В числе причин снижения численности вида называется также
охота и вырубка высокоствольных лесов в пределах гнездового ареала вида; эти два факто-
ра главенствовали в России и в Китае. О том, что ибис был желанной добычей для русских
поселенцев и китайцев (с Мурени), писал Н.М. Пржевальский (Дневник; цит по: Шульпин,
1936). Охота частично сыграла роль также в снижении численности вида в Корее в начале
XX века (Austin, 1948). Что же касается деградации мест гнездования вида, то особенно
болезненно отразилась на его судьбе вырубка в 1958 г. массива высокоствольных лесов в
восточной части Зап. Китая на хр. Циньлинь, где красноногий ибис до этого успешно
гнездился.
Яйца в кладках нередко повреждают сойки Garrulus glandarius, из-за чего эмбрионы
погибают на ранних стадиях развития (Yamashina, 19776).
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет. Занесён в
списки особо охраняемых животных и Красные книги всех стран в пределах своего ареала,
в том числе в Красную книгу Российской Федерации. Правительством Японии на о. Садо в
1970 г. создан специальный рефугиум площадью 1 000 га в особо привлекательных кормо-
вых угодьях этого вида. Возможно, именно благодаря принятым мерам в 1972-1974 гг.
местная популяция ибиса продуцировала не менее 2 птенца ежегодно, и общее число птиц
к 1974 г. составило 15 особей. Здесь же с 1965 г. проводились опыты клеточного разведе-
ния красноногого ибиса: отдельных птиц, опасаясь за их судьбу в зимние месяцы, отлавли-
вали для содержания в неволе. Неудачу подобных экспериментов (погибло 5 птиц) Я. Яма-
шина (Yamashina, 19776) связывает с недоброкачественным кормом; этот же автор высоко
оценивает достижения Базельского зоопарка (Швейцария), где, по его мнению, разработан
близкий к естественному состав кормов для ибисов.
Род Plegadis Каир, 1829
Каравайка
Plegadis falcinellus (Linnaeus, 1766)
Tantalus falcinellus Linnaeus, 1766, Syst. Nat., ed.12, с.241, «Австрия, Италия».
Glossy ibis (англ.), Braunsichler, Brauner Sichler (нем.), ibis falcinelle (фр ).
Син. (лат.): Scolopax rufa Scopoli, 1769; Numenius igneus S.G. Gmelin, 1770; Numenius viridis
S.G. Gmelin, 1770; Tantalus castaneus P.L.S. Miiller, 1776; Ibis sacra Temminck, 1815; Ibis
cuprea C.L. Brehm, 1831; Ibis ordi Bonaparte, 1838; Ibisperegrina Bonaparte, 1855; Falcinellus
bengalensis Bonaparte, 1855; Falcinellus erithrorhynchus Cabanis, 1856.
Каравайка
445
Статус. Гнездящийся перелётный, в южных частях ареала нерегулярно зимующий вид.
Общая характеристика и полевые признаки. Крупная птица, величиной больше во-
роны, весьма характерного облика, позволяющего легко отличать каравайку от сходных
видов. Это типичная голенастая птица с длинными ногами и длинной тонкой шеей. В отли-
чие от большинства наших голенастых, имеет общую тёмно-каштановую или рыжевато-
бурую окраску оперения с металлическим отливом разных оттенков и тонкий длинный клюв,
дугообразно загнутый на конце книзу. В полёте шею и ноги вытягивает в одну линию, час-
тые взмахи крыльев периодически чередует с непродолжительным планированием. Дер-
жится преимущественно стаями, иногда очень большими, близ открытых пустынно-степ-
ных водоёмов. Гнездится в тростниковых плавнях, местами в пойменных древесно-кустар-
никовых зарослях. Кормится небольшими группами или в одиночку в основном на зарос-
ших травой мелководьях, иногда на лугах, изредка также на полях или в степи. При перелё-
тах птицы выстраиваются косой, волнистой колеблющейся линией. Летают обычно невы-
соко над землей.
Сравнительно молчалива; голос — негромкое, грубое «гррр-гррр» — слышен обычно в
колониях при спаривании или беспокойстве птиц (Аскаров, 1938). У гнёзд при ссорах пе-
риодически раздается глухое гортанное гоготание или кряканье, при смене партнёров на
кладке слышны своеобразные воркующие звуки (Смогоржевський, 1979; Черничко и др.,
1991), а при внезапном взлёте птицы иногда издают негромкое хриплое «кррраа, крра».
Каравайка — народное название, происходит от казахского «карабай» (чёрный богач);
на юге России местное население называет птицу также чёрным кроншнепом.
Подвидовая систематика. В настоящее время рассматривается как монотипический
вид (Степанян, 2003).
Замечания по систематике. Австралийские популяции каравайки, обитающие также
на Филиппинах, Целебесе и Яве, иногда выделяют в качестве отдельного подвида Р f реге-
grinus Bonaparte, 1855 (Спангенберг, 1951; Cramp, 1977; Howard, Moore, 1980; Rose, Scott,
1994). Ранее в данный вид в качестве подвида включали американскую белолицую кара-
вайку Р chihi Vieillot, 1817 (син.: Р guarauna (Linnaeus, 1766); Р mexicana (Ord, 1817))
(Спангенберг, 1951; Wolters, 1982), незначительно отличающуюся окраской и образом жиз-
ни. Сейчас оба вида трактуют как группировку типа superspecies. Порой в эту же группи-
ровку включают и третий вид рода — тонкоклювую каравайку, или каравайку пуны (Р
ridgwayi J.A. Allen, 1876) — эндемика южноамериканских Анд.
Описание. Окраска. Самец в брачном наряде имеет общее каштаново-бурое или
красновато-коричневое оперение. Маховые и рулевые, а также большие кроющие крыла
чёрно-бурые с ярким зеленовато-синим или сине-зелёным металлическим отливом. На лбу,
темени, спине и плечевых перьях — контрастный фиолетовый или пурпурный отлив. Из-за
интенсивно ирридизирующей поверхности оперения его окраска у караваек воспринима-
ется довольно субъективно, поэтому в разных описаниях этих птиц имеется много проти-
воречий. Длинный загнутый на конце клюв и ноги оливково-бурые. Неоперенная уздечка,
кольцо вокруг глаз и подбородок зеленовато-бурого цвета, вдоль границ треугольного уча-
стка голой кожи, идущих от глаза к клюву, выделяются тонкие белые полоски. Радужина
бурая.
Ряд авторов (Аскаров, 1938; Долгушин, 1960) отмечают, что на голове у самцов имеется
слабо выраженный хохолок, который при агрессивном возбуждении может подниматься в
виде султана.
Самка в брачном наряде слегка отличается от самца чуть более тусклым оперением,
более мелкими размерами и более коротким, круто загнутым вниз клювом (Спангенберг,
Фейгин, 1936; Аскаров, 1938).
446
Отряд Листообразные, Семейство Ибисовые
В зимнем наряде у взрослых птиц голова и шея черновато-бурые, без следов металли-
ческого блеска. Отдельные перья имеют здесь узкие беловатые каёмки, в результате чего
вся шея и голова испещрены мелкой продольной штриховкой. Низ тела тусклый чёрно-
бурый или землисто-бурый. Нет светлых полосок, идущих вдоль границ неоперённого
«лица». Остальное оперение — как летом.
Птенцы покрыты матово-черным пухом. Со временем на горле, нередко также на теме-
ни у них появляются 1-5 белых пятна или поперечные полоски. Иногда эти пятна отсут-
ствуют вовсе. Ноги у пуховичков жёлто-розовые, чернеющие на 3-й день. Когти чернеют
через 3-4 ч после вылупления. Клюв при вылуплении тоже жёлто-розовый, но в течение
первых суток становится чёрным с широкими желтоватыми или оранжево-розовыми пере-
вязями в основании, на конце и посередине. В 5-7-дневном возрасте исчезает светлое коль-
цо в основании клюва, на 27-30-й день — следующее, а иногда и последнее (Аскаров,
1938; Туаев, Исрафилов, 1978).
Гнездовой наряд похож на зимний наряд взрослых птиц, но окраска более тусклая, свет-
ло-бурая, особенно на голове и шее, со значительно более слабым металлическим блеском.
Кроме того, нередко встречаются птицы, у которых на шее, горле и иногда на темени оста-
ются от птенцового наряда белые пятна. Клюв розовато-бурый с чёрной поперечной поло-
сой посередине и чёрным концом, значительно более короткий, чем у взрослых птиц.
Строение и размеры. Некрупный изящный ибис. Крылья сравнительно короткие,
закругленные, так что внутренние второстепенные почти достигают вершины сложенного
крыла. Первостепенных маховых 11. Формула крыла: II>III>I>IV>V... Вырезки располо-
жены на внутренних опахалах I и II и на внешних опахалах III и IV первостепенных. Хвост
короткий, прямой, из 12 рулевых перьев. Пудретки отсутствуют.
Щелевидные ноздри расположены в основании клюва в узких длинных бороздках. Го-
лень оперена до середины. Цевка спереди покрыта поперечными щитками, сзади сетчатая.
Основания 3-го и 4-го пальцев ног соединены зачаточной перепонкой. Когти довольно длин-
ные, тупые, на среднем пальце — с 2-3 зазубринами на внутренней стороне.
Имеющиеся в литературе фрагментарные морфометрические показатели каравайки
(табл. 34) подтверждают различия самца и самки в размерах. Средняя длина самцов превы-
шает длину самок на 82, а размах крыльев — на 97 мм (Аскаров, 1938; Спангенберг, 1951).
Длина крыла самца больше, чем у самки, в среднем, на 15 мм (Симеонов, Мичев, Нанки-
нов, 1992). Весьма заметны различия также в длине клюва и цевки (Аскаров, 1938; Долгу-
шин, 1960; Симеонов и др., 1992).
Размеры птиц из Вост. Европы и Сев. Азии (мм) без деления по полу таковы: длина —
559; размах крыльев — 919-1029; длина крыла — 273-292, хвоста — 121, клюва — 132,
цевки — 102 (Мензбир, 1918). Длина крыла — 248-305, клюва — 100-145, цевки — 95-
120 (Бутурлин, 1935). Длина крыла — 230-305, клюва — 100-142, цевки — 85-112 (Туга-
ринов, 1947; 1951). По Украине получены следующие данные: длина крыла — 250-302,
клюва — 100-142, цевки — 82-112 мм (Вошственський, Юстяювський, 1962); длина
крыла — 353-310, хвоста — 94-101, клюва — 102-148, цевки — 71-109 мм (Смогор-
жевський, 1979).
Вес птиц, по данным всех цитированных авторов, изменяется в пределах 530-770 г. Он
заметно снижается в период насиживания и выкармливания птенцов (в среднем, на 110 г у
самцов и на 150 г у самок). После вылета птенцов, когда взрослые прекращают кормление
молодняка и начинают активно питаться сами, вес быстро восстанавливается (Аскаров,
1938).
Линька. Линяет дважды в году: в конце лета и осенью проходит полная послебрачная
линька и надевается зимний наряд, а уже с ноября по май идёт частичная предбрачная
Каравайка
447
Таблица 34. Морфометрические показатели (размеры — мм, вес — г) караваек
из разных регионов
Параметр п Самцы п Самки Источник
lim М lim М
Восточная Европа и Северная Азия
Длина тела 14 608-660 635 12 516-588 553 Аскаров,
Размах крыльев 14 1000-1068 1014 12 870-968 917 1938;
Длина крыла* 6 292-305 297,3 5 265-285 Спанген-
Длина цевки (плюсны) 14 90-123 112 12 81-96 90 берг, 1951
Вес** 7 556-768 673,3 7 530-679,5 599,8
А р м е н и я
Длина тела 2 531-588 559,5 2 540-590 565 Adamian,
Длина крыла 3 289-295 3 261-266 Kleim, 1999
Длина хвоста 3 106-126 3 108-127
Длина клюва 2 130-132 131 3 105-117
Длина цевки (плюсны) 3 96-107 3 82-95
К а з а х с т а н
Длина крыла 290-306 265-280 Долгушин,
Длина клюва 126-150 100-125 1960
Длина цевки (плюсны) 90-123 80-96
Б о . л г а р и я
Длина крыла 8 281-305 290 2 271-280 275,5 Симеонов и
Длина хвоста 8 108-140 2 111-116 113,5 др., 1992
Длина клюва 8 112-140 2 111-122 116,5
Длина цевки (плюсны) 8 102-117 2 89-95 92
3 а п а д н а я П а л е а Р к тика
Длина крыла 280-306 267-281 Snow,
Вес 557-768 530-680 Perrins,
1998
Западная Европа, (гл. образом - - Нидерланды)
Длина крыла 8 281-305 297 7 267-281 273 Cramp,
Длина хвоста 6 96-111 106 7 90-99 94 1977
Длина клюва И 126-141 132 8 106-114 110
Длина цевки (плюсны) 18 101-113 107 13 82-90 86
Примечания. Размеры птиц Е.П. Спангенбергом (1951) взяты в основном из работы Г. Аскарова
(1938), но в текст первого «прокрались» опечатки, которые по возможности здесь исправлены. Сред-
ние показатели для ?? из Болгарии рассчитаны нами. Сведения о весе птиц в сводке С. Крэмпа
(Cramp, 1977) даны по Е.П. Спангенбергу (1951) и поэтому не приводятся. * — только по Е.П. Спан-
генбергу (1951). ** — средние показатели рассчитаны по материалам Г. Аскарова (1938) нами. —
Ред.
линька, в ходе которой сменяется мелкое оперение и формируется брачный наряд. Взрос-
лый наряд надевается на втором году жизни в ходе первой послебрачной линьки. Детали
линьки неизвестны.
Распространение. Гнездовой ареал. По своему современному распростране-
нию — пантропический вид, гнездящийся в Америке, Африке, по югу Евразии и в Австра-
лии (рис. 85). В настоящее время это единственный вид семейства ибисовых, распростра-
нённый как в восточном, так и в западном полушарии. Однако в Америку, на её атлантичес-
кое побережье, каравайка проникла лишь в конце XIX в., примерно в 1880 г. Расселяясь, к
1986 г. она достигла Канады, в 1988 г. обнаружена на гнездовье в Мексике. Сейчас каравай-
ка гнездится в Америке только на восточном побережье от Канады до Флориды, на Б. Ан-
448
Отряд Листообразные, Семейство Ибисовые
Рис. 85. Область распространения каравайки: а — примерная граница области обитания в Вост. Ев-
ропе и Сев. Азии в XIX в., б — современные области гнездования, в — область зимовок.
тильских о-вах и в Венесуэле на самом севере Юж. Америки (del Hoyo et al., 1992). В Афри-
ке распространена по востоку континента от Капской провинции до Мадагаскара и Кении,
а также на северо-западе — на побережьях Марокко, Алжира и Туниса.
В Юж. Европе обитает места-
Рис. 86. Ареал каравайки в Вост. Европе и Сев. Азии: а —
области современного гнездования, б — области гнездова-
ния, исчезнувшие во второй половине XX в.
ми на Пиренейском п-ове, во Фран-
ции, Италии, Венгрии, Югославии,
Албании, Греции; регулярно гнез-
дится в Причерноморских странах.
Далее на восток обычна в Предкав-
казье и Прикаспии, местами встре-
чается в Казахстане и Средней
Азии. По югу Азиатского матери-
ка локально гнездится в Турции,
Сирии и Израиле; обычна в Ираке,
к востоку распространена до Ин-
дии (до южных подножий Гимала-
ев) и океанических побережий
Юго-Вост. Азии, а также до Боль-
ших Зондских о-вов, Юж. Филип-
пин и Новой Гвинеи.
На территории Вост. Европы и
Сев. Азии северная граница ареа-
ла проходит через низовья Дуная,
Днестра, Днепра, Дона, Волги,
Урала, а также Иргиза и Тургая в
Центр. Казахстане. По Дону под-
нимается вверх, возможно, до Цим-
лянского вдхр., по Волге — по-
чти до г. Волгограда (Спангенберг,
Каравайка
449
1951; Чернобай, 2004). В Волго-Уральском междуречье встречается в районе Урды, в низо-
вьях Узеней и Кушума, но гнездование здесь не подтверждено (Шевченко и др., 1993; Дебе-
ло, 1996) (рис. 86).
В указанных выше пределах каравайка эпизодически гнездится на Тилигульском и Мо-
лочном лиманах в Сев. Причерноморье и Приазовье, обычна на северном побережье Кры-
ма в Каркинитском зал. и на Сивашах (Сиохин, 2000), многочисленна в Восточном Приазо-
вье, гнездится по долине Маныча, в низовьях Кумы и Терека (Казаков и др., 2004), много-
численна в дельте Волги и в Азербайджане (Султанов, 2000; Гаврилов, 2005), в небольшом
числе сохранилась в Армении (Adamian, Klem, 1999).
В Средней Азии в середине XX в. гнездилась на равнинных участках долин рек Аму-
Дарья и Сыр-Дарья, кроме того — в низовьях рек Талас, Чу и Или в южном Прибалхашье
(Дементьев, 1952; Долгушин, 1960; Сагитов, 1987). В прошлом колонии были известны
также в северных предгорьях Копетдага — в низовьях р. Теджен и по р. Мургаб (Спанген-
берг, 1951). В настоящее время в Казахстане каравайка практически полностью исчезла
(Ковшарь, 1999). Здесь уже в 1980-е гг. в разных районах — от дельты Урала до Балхаша —
эпизодически встречались лишь кочующие одиночки и стайки (Ковшарь, 1991). Резко со-
кратилась её численность в дельте Аму-Дарьи (Крейцберг-Мухина, Лановенко, 2003), где
ещё в 1988-89 гг. находились огромные колонии (Лукашевич, 1990). Но в это же время
гнездовья начали формироваться на искусственных сбросных водоёмах среди пустыни
Каракум, например, на оз. Каракыр близ г. Газли (Сагитов и др., 1989). В Туркмении эпизо-
дически гнездится на оз. Делили в низовьях р. Атрек (Венгеров, 1975; Караваев, 1991). В
горные районы Киргизии и Таджикистана каравайка изредка залетает лишь во время кочё-
вок и миграций (Северцов, 1873; Абдусалямов, 1971).
Прежде границы ареала каравайки проходили значительно севернее и восточнее. Так, в
начале XIX в. она была обычна в низовьях Северского Донца и по всему Нижнему Дону
(Кондратьев, 1885, 2004). В Заволжье и Приуралье, в пределах бывшего Оренбургского
края, в середине XIX в. была нередка к северу до 50° с.ш., изредка залетая до 52° с.ш.
(Эверсманн, 1866). Гнездилась каравайка также в низовьях р. Илек в среднем течении р.
Урал (Зарудный, 1888). На востоке Казахстана обитала в Семиречье, встречаясь вплоть до
Алакольской котловины (Шнитников, 1949).
Позже, в результате неумеренного промысла птиц и сбора яиц в колониях, а также, ве-
роятно, из-за интоксикации пестицидами в годы борьбы с азиатской саранчой (Белик, 1997),
ареал каравайки значительно сократился. Глубокую депрессию численности вида, в ре-
зультате которой птицы исчезли на степных озёрах по югу Украины, на Дону, по Уралу, на
Алаколе, наблюдали в первой половине XX в., когда проводилось массовое опыление плав-
ней южных рек остротоксичными препаратами (Скалов, 1928; Соколов, 1928; и др.). Во
второй половине, особенно — в конце XX в. численность и ареал каравайки в Вост. Европе
в значительной мере восстановились. Это было вызвано отчасти мерами по её охране, от-
части — благодаря широкой ирригации и обводнению степных районов, а в последние
годы — из-за упадка сельскохозяйственного производства в этих районах. В то же время, в
Зап. Европе популяции каравайки сейчас по-прежнему находятся в состоянии депрессии,
её ареал там очень фрагментирован, а в некоторых странах (Румыния, Болгария, Сербия)
~о сих пор продолжается падение численности и сокращение распространения вида (Thucker,
Heath, 1994; Snow, Perrins, 1998; BirdLife International, 2004). Исчезают каравайки в после-
днее время также на востоке ареала — в Казахстане и странах Средней Азии.
Следует отметить непостоянство мест гнездования каравайки. Эпизодически эти птицы
могут группами и целыми колониями спонтанно переселяться с места на место, меняя гнез-
довые районы, как под воздействием антропогенных факторов, так и без видимых причин
450
Отряд Листообразные, Семейство Ибисовые
(Кеве, 1978; Сапетин, 1978е). Это приводит к сильному перемешиванию популяций; массо-
вое перераспределение птиц между соседними районами гнездовий может влиять и на ди-
намику видового ареала.
Зимовки. Судя по данным кольцевания, у караваек в пределах Сев. Евразии суще-
ствует несколько популяций, различающихся районами своих зимовок и пролётными путя-
ми. Птицы из дунайской популяции в своем большинстве улетают на зимовку через Гре-
цию в Вост. Африку — в бассейн Верхнего Нила; частично они летят через Италию и Фран-
цию на юго-запад и запад — вероятно, в Сев. и Зап. Африку (Кеве, 1978). Возможно, к этой
же популяции относятся и каравайки, гнездящиеся в Сев. Причерноморье и Вост. Приазо-
вье, а осенью улетающие, в основном, на запад к Дунаю и дальше через Балканы и Италию
в Зап. Африку — в бассейн р. Нигер (Сапетин, 1978е; Щеголев, 1992; Schogolev, 1996b;
Корзюков, 2001; Русев, 2004; Костин, Тарина, 2005). На запад в основном перемещаются
большие стаи птиц, кочующих в июле в Сев. Крыму (Гринченко, 2004).
Птицы из Азербайджана мигрируют преимущественно на юго-запад через Иран и Ирак
в Вост. Африку — в бассейн Верхнего Нила, но частично остаются на зимовку в Ираке —
в низовьях Тигра и Евфрата. Волжские же каравайки зимуют, в большинстве, в Индии,
однако пути их миграций прослежены слабо. Скорее всего, они летят туда вдоль восточно-
го побережья Каспия (Смирнов, 1916; Дементьев, 1952) или через Аральское море, Сыр-
Дарью и Аму-Дарью (Сапетин, 1978е). Возможно, эти птицы совершают кольцевой пере-
лёт, поскольку интенсивность их миграций в разных районах резко различается по сезо-
нам. Так, на юго-восточном побережье Каспия в 1930-х гг. каравайки появлялись в боль-
шом числе лишь осенью, а в 1980-х гг. — в основном, весной (Исаков, Воробьев, 1940;
Караваев, 1991). Каравайки из Казахстана и Средней Азии зимуют, вероятно, тоже в Индии
(Спангенберг, 1951; Долгушин, 1960).
Пролёт караваек вдоль западного побережья Каспия сравнительно слабый (Воробье-
ва, 1979; Михеев, 1991) и характерен, скорее всего, лишь для дагестанских птиц. Сама же
популяция из дельты Терека имеет, по-видимому, смешанное происхождение, поскольку
основная часть птиц улетает оттуда на зимовку в Ирак и, отчасти, в Вост. Африку, некото-
рые каравайки вместе с волжскими птицами летят в Индию, а отдельные вместе с кубан-
скими мигрируют на запад, долетая до Франции. Об этом свидетельствуют и направле-
ния послегнездовых кочёвок молодняка из дельты Терека, часть которого улетает осенью
на север, в дельту Волги, а часть — на юг, в Азербайджан. Подтверждают это и эпизоди-
ческие массовые переселения караваек из Азербайджана в более северные районы (Са-
петин, 1978е).
Каравайки изредка остаются на зиму в субтропических районах Юж. Европы, а также в
Азербайджане (Радде, 1884; Греков, 1965; Сапетин, 1978е; Патрикеев, 1991а) и Туркмении
(Житников, 1900). В последние десятилетия, в связи с потеплением климата, приемлемые
условия для их зимовок складываются местами и в умеренном поясе. Так, в 1967/68 г. в
дельте Кубани зазимовали 6 птиц (Олейников и др., 1975), а с 1973 г., одиночки и стайки до
20 особей регулярно отмечались там зимой на убранных рисовых полях. На незамерзаю-
щих же водоёмах у насосных станций птицы держались даже при температуре -14 °C.
Кроме того, в январе 1988 г. одиночных караваек наблюдали в Каневском р-не на севере
Краснодарского края. В середине декабря каравайки встречены однажды на родниках в
Кизилюртовском р-не Дагестана (Заболотный, Хохлов, 1989а; Казаков и др., 2004). В де-
кабре 1973 и 1980 гг. одиночек отмечали также в степной части Крыма (Костин, 1983).
Миграции. Каравайка, как и колпица, относится к типичным «погодным» мигрантам, и
поэтому время её возвращения с зимовок во многом зависит от синоптических условий в
местах гнездования; в Сев. Евразии в разные годы птицы появляются с начала марта до
Каравайка
451
конца мая. Кроме того, прилет отдельных групп птиц в конкретные колонии тоже идёт нео-
дновременно.
В юго-западных районах Украины первые каравайки появляются в конце марта — нача-
ле апреля. На Днестре их прилёт отмечали с 19 марта по 6 апреля, в среднем за 6 лет (1972-
1977) — 27 марта (Смогоржевський, 1979), массовый прилёт в 1953-1973 гг. приходился в
среднем на 8 апреля, а наиболее ранний — 31 марта 1970 (Назаренко и др., 1975). На Тили-
гульский лиман каравайки прилетали в период с 25 марта (1981 г.) по 18 апреля (1980 г.), в
среднем за 1978-1983 гг. — 8 апреля (Черничко и др., 1991). На Лебяжьих о-вах в северо-
западном Крыму прилет первых птиц регистрировали с 4 по 24 апреля, в среднем за 16 лет
(1987-2002 гг.) — 15 апреля, однако в 1980-е гг. они появлялись здесь позже, в начале мая
(Костин, Тарина, 2002, 2004). Примерно в эти же сроки (9 апреля - 10 мая), отставая в
среднем на 10 дней, пролёт каравайки начинается в Николаевской обл. (Рединов, 1999). На
южном же берегу Крыма первые пролётные птицы появляются значительно раньше — в
середине марта (14 марта 1998 г., 13 марта 2002 г.), иногда даже в начале этого месяца (3
марта 1952 г.), но обычно с середины апреля. Пик пролёта регистрируется здесь в 3-й дека-
де апреля - 1-й декаде мая, последних пролётных караваек отмечают в конце мая (24 мая
1983 г., 23 мая 2002 г.). Причём в 1-й декаде мая на водоёмах Юж. Крыма нередко встречают-
ся скопления до нескольких десятков особей (Костин, 1983; Бескаравайный, 2001; 2005).
В Вост. Приазовье первые каравайки прилетают обычно в конце марта — с 14 марта по
4 апреля, в среднем за 6 лет — 25 марта (Сарандинаки, 1909; Казаков и др., 2004). Массо-
вый прилёт идёт во 2-й половине апреля, но стаи могут появляться в гнездовых районах
вплоть до середины мая. Например, около 300 пролётных птиц видели в дельте Кубани 17
мая 1965 г. (Казаков и др., 2004). В дельте Дона в конце XIX в. однажды наблюдали «гро-
мадное количество стай», летевших на восток в конце апреля (ст. ст.) (С.[Алфераки], 1890;
Алфераки, 1910). В низовьях Дона, где весной идёт слабо выраженный пролёт на восток, в
1978-2001 гг. первых караваек отмечали с 31 марта по 29 апреля, в среднем за 8 лет — 15
апреля. Всего здесь учтено 15 пролётных стай численностью от 4 до 120, в среднем — по
23 особей (В.П. Белик, ориг.). На Маныче же каравайки появляются с 24 марта по 6 апреля,
но в основной массе они прилетают туда к середине мая (Олейников и др., 1975; Казаков и
др., 2004).
Выраженный пролёт караваек на северо-запад идёт весной вдоль Черноморского побе-
режья Кавказа. В Аджарии его наблюдали с 17 марта по 10 апреля (Вильконский, 1897). В
районе Адлера птиц регистрировали с 11 апреля до 26 мая, но более обычны они здесь с
конца апреля до середины мая (Тильба, 1999). На Ставрополье каравайки летят с 3-й дека-
ды апреля, но в основном во 2-й половине мая, причём часть птиц мигрирует на восток, а
часть — на северо-запад (Казаков и др., 2004).
В Азербайджан каравайки прилетают в середине марта - начале апреля и регистриру-
ются на миграциях до 21 мая (Греков, 1965; Туаев, Исрафилов, 1978; Патрикеев, 19916). В
Дагестане они мигрируют вдоль Каспия на северо-запад со 2-й пятидневки апреля до сере-
дины мая, причём пик пролёта (более 50 % особей) падает на 3-ю пятидневку апреля (Буть-
ев и др., 1989; Михеев, 1991; Казаков и др., 2004). В дельте Волги сроки прилёта первых
птиц, по данным А.Е. Лугового (1963), колеблются в интервале около полутора месяцев —
с 17 марта до 29 апреля, в среднем за 23 года — 8 апреля. При этом в 1920-1930-е гг. прилёт
проходил в среднем на 11 дней позже, чем в 1940-1950-е гг. (соответственно 15 апреля и 4
апреля). Время появления караваек в восточной части дельты, по сравнению с западной, в
первый период запаздывало на 1-3 дня, тогда как во второй период, наоборот, на востоке
птиц отмечали на 2-3 дня раньше; возможно, это свидетельствует о смене весенних мигра-
ционных маршрутов караваек.
452
Отряд Аистообразные, Семейство Ибисовые
В Туркмении пролёт идёт с конца 1-й декады марта до 3-й декады мая (Дементьев,
1952), но в юго-вост. Прикаспии птицы отмечаются лишь с 22 марта, обычно же с 1-й декады
апреля, и летят до середины мая, задерживаясь в небольшом числе, судя по учётным дан-
ным, до июня (Караваев, 1991). В Юж. Казахстане каравайки появлялись иногда уже в на-
чале марта, но чаще в конце этого месяца. На Сыр-Дарье в районе Кзыл-Орды первых птиц
отмечали 15-25 марта, массовый пролёт наблюдали во 2-й декаде апреля; миграция закан-
чивалась к концу этого месяца (Долгушин, 1960).
Летом, вскоре после вылета из гнёзд, каравайки сбиваются в стаи и с середины июля
начинают кочёвки в поисках кормных мест, в которых задерживаются затем до отлёта на
зимовку. Гнездовые районы у северной границы ареала (Средний Дунай, дельта Днестра,
Нижний Дон) большинство птиц покидает весьма рано, в течение июля-августа. При этом
их перемещения к югу иногда приобретают характер массового пролёта (Молчанов, 1912;
Зарудный, 1916). Южнее — в низовьях Дуная, в дельте Кубани, в Крыму и Ленкорани —
они могут оставаться до октября. Но некоторые птицы из Азербайджана и дельты Кубани
уже в конце августа - середине сентября оказываются на местах зимовок в Двуречье (Ирак)
или в Экваториальной Африке (Сапетин, 1978е).
На Тилигульском лимане кочёвки отмечаются с середины июля, во второй половине
августа заметна миграция птиц на запад, а в сентябре они исчезают (Черничко и др., 1991).
На Лебяжьих о-вах в Крыму каравайки в августе вполне обычны, в сентябре встречаются
реже, но ещё регулярно, а пик их пролёта наблюдается в сентябре - начале октября (Кос-
тин, Тарина, 2005). В дельте Кубани выраженный пролёт на юго-запад регистрируется в
августе, основная масса птиц отлетает в середине сентября (Винокуров, 1963; Олейников и
др., 1975). В дельте Волги массовый отлёт продолжается с конца августа до середины сен-
тября; в 1926-1929 гг. его регистрировали 6,7,15 и 17 сентября (Аскаров, 1938; В.Б. и Н.М.
Дубинины, 1940). В Дагестане в дельте Терека массовый отлёт птиц на юго-восток идёт с
конца августа до конца сентября (Казаков и др., 2004), а в устье р. Самур пик пролёта (око-
ло 75% особей) наблюдается в последней пятидневке августа. Однако в отдельные годы
основная масса птиц мигрирует в конце сентября - начале октября, что связано, вероятно,
с особенностями гнездовых сезонов (Бутьев и др., 1989; Михеев, 1991). В Ленкорани, на
юго-западном побережье Каспия, массовые кочёвки птиц начинаются в середине августа и
продолжаются до конца августа - сентября, а с середины сентября наблюдается их выра-
женный пролёт на юг (Спангенберг, 1951; Patrikeev, 2004). На юго-восточном берегу Кас-
пия, в низовьях р. Атрек, пролёт караваек идёт в основном в августе и в первой половине
сентября (Исаков, Воробьев, 1940; Караваев, 1991). В низовьях Сыр-Дарьи в начале сен-
тября каравайки были ещё многочисленны, а в начале октября их уже не встречали (Спан-
генберг, Фейгин, 1936).
В октябре в гнездовых районах кочуют обычно лишь случайные одиночки и небольшие
стайки; последние каравайки исчезают в ноябре. На Днестре их отмечали 21 ноября 1974 г.
и 20 ноября 1976 г. (Смогоржевський, 1979), в Крыму последних птиц регистрировали до
30 ноября, в среднем за 16 лет (1987-2002 гг.) — до 1 ноября (Костин, Тарина, 2002). Вплоть
до конца ноября одиночных караваек регулярно встречали в дельте Волги (Луговой, 1963).
В Аграханском заливе в Дагестане очень тёплой осенью 1974 г. каравайки держались даже
до 6 декабря (Михеев, Орлов, Резанов, 1975). В Азербайджане последнюю стайку видели 6
ноября 1956 г. (Греков, 1965).
Следует отметить отсутствие выраженных осенних миграций на Черноморском побе-
режье Кавказа и Крыма (Тильба, 1999; Бескаравайный, 2005). Это позволяет предполагать,
что весной каравайки попадают сюда случайно, отклоняясь к гористым берегам, по всей
видимости, при перелётах через Чёрное море (Корзюков, 2001).
Каравайка
453
Во время послегнездовых кочёвок молодняк обычно держится отдельно от взрослых
птиц, которые дружно отлетают на зимовки на месяц раньше молодых (Щеголев, 1992). В
период кочёвок молодые каравайки разлетаются весьма широко, улетая иногда за 780-1100-
1900 км на северо-запад - север - северо-восток от гнездовых колоний, в направлениях,
противоположных основным миграционным путям (Сапетин, 1978е). Однажды молодая
каравайка из дельты Днестра была добыта 11 сентября 1982 г. на оз. Кулундинском в Ал-
тайском крае, преодолев за 2 месяца 3,5 тыс. км (Щеголев, 1992; Schogolev, 1996b). Такие
кочёвки можно рассматривать как «промежуточный перелёт», который затем переходит в
миграцию к местам зимовки. Однако, скорее всего, в основной массе это «изгои» или пти-
цы, потерявшие ориентацию; в своем большинстве они, по-видимому, обречены в незнако-
мых местах на гибель. Так, 8 из 12 птиц, залетевших в октябре 1981 г. в Пензенскую обл.,
были вскоре добыты на прудах местного рыбхоза (Фролов и др., 2001). Хотя нельзя исклю-
чить, что некоторые выжившие особи могут «запоминать» районы кочёвок и в последую-
щие годы образовывать там новые колонии.
Дальние залёты изредка отмечали и у взрослых птиц (Юрлов и др., 1977; Сотников,
1999; Марголин, 2000). Встречи залётных караваек регистрируются практически по всей
Европе на север до Исландии и Норвегии (Snow, Perrins, 1998), до Мурмана (Бианки и др.,
1993), Московской, Кировской и Пермской обл. (Ушков, 1927; Спангенберг, 1951; Сотни-
ков, 1999; Зубакин, 2007). В Сибири их отмечали к северу до Томской и Кемеровской обл.
(Рябицев и др., 2001). Большинство залётов приходится обычно на август (Hudec, Сету,
1972), когда начинаются кочёвки молодняка.
Мечение показывает, что каравайки из дельты Волги в основном мигрируют на зимовки
в юго-восточном направлении, птицы из Азербайджана — в юго-юго-западном. В своем
движении к местам зимовок они достигают верховий Нила и Ливии. Каравайки, помечен-
ные в устье Кубани и на Украине, стремятся в Юж. Европу, пересекают Средиземное море
и оказываются в Центр. Африке южнее Сахары (рис. 87).
Миграции караваек идут в основном вдоль экологических русел и направляющих ли-
ний, но эти птицы способны пересекать моря (Сапетин, 1978е; Корзюков, 2001) и иногда
появляются на высокогорных озёрах Кавказа (Казаков и др., 2004). Миграции наблюдают-
ся обычно в ранние утренние часы; более слабый пролёт идёт во второй половине дня
(Михеев, 1991). Во время осеннего пролёта птицы могут собираться в стаи, достигающие
500-1000 и более особей; весной стаи, как правило, меньше: до 200-300 птиц (Спанген-
берг, 1951; Смогоржевський, 1979; Казаков и др., 2004).
Местообитания. Гнездовые и кормовые местообитания караваек существенно разли-
чаются, поэтому птицы регулярно совершают кормовые миграции, иногда на расстояние
до 20 км и больше (Спангенберг, Фейгин, 1936).
Основными местами кормежки служат опресненные мелководья до 20-25 см глубиной
(Скокова, 19606), покрытые мозаичной надводной растительностью. Прежде всего, это
весенне-летние разливы рек (полой), травянистые злаково-осоковые болота и сырые, забо-
лоченные луга, а также усыхающие временные водоёмы в луговых понижениях. В дельте
Волги с весны кормятся на полоях и ильменях среди лугов, а с середины лета, когда паво-
док спадает и пойма обсыхает, птицы перемещаются на взморье, собираясь стаями на или-
стых заливах (култуках) (Аскаров, 1938). В течение всего гнездового периода каравайки
очень охотно посещают рисовые поля с мелкой прогретой водой, где кормятся, зачастую,
полностью скрываясь в зарослях риса. Птицы кормятся также по топким, илистым берегам
лиманов, рыборазводных прудов, водохранилищ, промышленных фильтрационных полей,
однако обширных водоёмов, полностью лишённых надводной растительности, они, как
правило, избегают. При этом охотно используют залитые водой тростниковые гари в плав-
454
Отряд Листообразные, Семейство Ибисовые
Рис. 87. Распределение отдельных популяций караваек Вост. Европы и Сев. Азии по областям пролё-
та: а — места кольцевания; б — места встреч птиц, окольцованных в Приазовье; в — встречи птиц,
окольцованных в Астрахани; г — встречи птиц, окольцованных в Дагестане; д — встречи птиц, околь-
цованных в Азербайджане (по: Сапетин, 1978е, с дополнениями).
нях (Лукашевич, 1995). Иногда птицы собирают появляющихся в массе насекомых в траве
на суходольных лугах или в степи, местами кормятся на убранных хлебных полях (Хохлов,
1987; 1993).
Гнездятся каравайки преимущественно в тростниковых зарослях среди плавневых мас-
сивов, изредка в прибрежных бордюрных тростниках или на тростниковых островах. Тро-
стниковые колонии распространены по всему ареалу каравайки, а в некоторых районах
(например, в дельтах Кубани и Терека) они преобладают. Для гнездования птицы предпо-
читают старые высокие заросли тростника с диаметром стеблей у воды 1,5-2,2 см. В тон-
ких, разреженных зарослях селятся только вынужденно. Избегают и сильно загущенных
зарослей тростника, где трудно ломать стебли для строительства гнёзд (Черничко и др.,
1991).
Местами, например, в дельтах Волги, Дуная, на Сыр-Дарье, колонии нередко распола-
гаются в пойменных лесах из ветлы (Salix alba), туранги (Populus diversifolia), лоха, или
джиды (Elaeagnus angustifolia), растущих по берегам проток и озёр. В дельтах Дуная, Дне-
стра, Днепра, на Дону, Волге и в Азербайджане каравайки селятся также в зарослях мощ-
ных кустарников (различные виды ивы, тамариск и др.) среди болот. Особого предпочте-
ния древесным или кустарниковым ивам, по наблюдениям в дельте Волги, каравайки не
отдавали (Аскаров, 1938). Здесь в 1973-1974 гг. в 10 «лесных» колониях было сосредоточе-
но более 96% гнёзд местной популяции караваек (Бондарев, 19756, 1979).
В Азербайджане колонии голенастых на побережье моря в Кызыл-Агачском заповедни-
ке сформировались лишь в 1950-е гг., когда там развились обширные массивы заболочен-
ных тамарисковых зарослей, труднодоступные для наземных хищников. Гнездовья разме-
щались на самых глубоких участках болот и не ближе 2 км от берега (Греков, 1965; Муста-
Каравайка
455
фаев, Кязимов, 1965). В дельте Днестра около 1,5 тыс. пар караваек в 1970-1973 гг. гнезди-
лись, в основном, на кустах ивы; в 1974-1977 гг. из-за низкого уровня воды в плавнях и
обсыхания этих ивняков каравайки переселились в тростники. Доля птиц, гнездившихся в
тростниках, менялась с 25% в 1972 г. до 48% в 1973 г., 97% в 1974 г. и 100% в 1975 г. Всего
в дельте Днестра было известно 5 колоний в кустарниках, 3 — в тростниках и 1 смешанное
поселение на древовидных ивах (Щеголев, 1977; Schogolev, 1996а).
Как исключение, на Лебяжьих о-вах в Крыму однажды отмечено поселение караваек в
колонии цапель на земле среди низкорослой полыни в 5-12 м от берега (Смогоржевський,
1979; Костин, Тарина, 2004).
Численность. Будучи колониальным, особо охраняемым видом, каравайка в большин-
стве регионов находится под регулярным контролем, и в европейских странах её числен-
ность оценена сейчас достаточно точно.
Общая численность мировой популяции, без австралийской формы Р. f. peregrinus, со-
ставляет более 60-100 тыс. особей (Rose, Scott, 1994). В Европе, по оценкам, сделанным в
начале 1990-х гг. и в начале XXI в., гнездилось, соответственно, 21-23 и 16-22 тыс. пар, в
том числе на Украине — 5,5-7,0 и 1,7-3,6, в Европейской части России — 10,3 и 7-8 тыс.
пар и в Молдавии — 1-5 и 2-5 пар. Кроме того, в начале XXI в. 20-80 пар было учтено в
Армении и 4-6 тыс. пар — в Азербайджане. Остальные птицы гнездились, в основном, в
Румынии (4,0 и 2,5-2,8 тыс. пар), Болгарии (200-300 и 50-130 пар), Греции (50-70 и 150—
200), Албании (100-300 и 0), Испании (1-4 и 393-423 пары), а также в Турции (0,5-1,4 и
0,5-1,0 тыс. пар) (BirdLife International..., 2000; BirdLife International, 2004).
В бывшем СССР в 1973-1977 гг. насчитывалось 15 тыс., а в 1986-1987 гг. — 28 тыс.
пар. Заметный рост популяций объяснялся развитием в Палеарктике благоприятной для
лимнофилов прохладно-влажной климатической фазы, а также улучшением охраны птиц
и, отчасти, большей полнотой учёта (Флинт и др., 1991). В 1980-е гг. в России было учтено
около 6,6 тыс. пар, на Украине — 2,7-2,9 тыс., в Азербайджане — 18,7 тыс. пар, в Казахста-
не — 100, в Узбекистане — 20 пар (Стоцкая, Кривенко, 1988). В 1990-е гг., по данным
Красных книг, в России (2000) насчитывалось 10-11 тыс. пар, на Украине (1994) — 2,7-3,6
тыс. пар, в Узбекистане (2003) — 700-800 пар. В Казахстане, кроме восточной части дель-
ты Волги, жилые колонии в тот период не были известны (Красная книга Казахстана, 1996;
Ковшарь, 1999).
В Молдавии небольшие колонии эпизодически отмечали в 1960-е гг. в плавнях низовий р.
Прут ниже г. Кагул (Аверин и др., 1971). На Украине каравайка гнездится в дельтах Дуная,
Днестра и Днепра, в верховьях Тилигульского лимана в Одесской обл., на Лебяжьих о-вах и
недалеко от них в Кумовских плавнях на северо-западном побережье Крыма, а также на
Сивашах в сев.-вост. Крыму, где известны 2 большие, относительно стабильные колонии и
7-8 непостоянных поселений (табл. 35). Эпизодически она появляется на островах Джа-
рылгачского зал., возможно также в верховьях Молочного лимана (Сиохин, 2000).
В украинской части дельты Дуная каравайка в массе гнездились в середине XX в. Так, в
1970-е гг. до 3,5 тыс. пар обитало там в тростниковых зарослях и около 100 пар — на дере-
вьях (Смогоржевський, 1979); однако в 1980-е гг. на оз. Кугурлай и в Стенсовских плавнях
осталось лишь 190-300 пар (Панченко, Балацкий, 1991), по другим данным — всего 1-10
пар (Platteeuw et al., 2004). В 1990-е гг. каравайки исчезли там вовсе и гнездились только в
Румынии, где в 2001-2002 гг. в 10-12 колониях насчитывалось 2 055-3 340 пар. Однако в
2002 г. на украинском оз. Кугурлай вновь найдены 2 колонии из 160 и 320 пар (Platteeuw et
al., 2004).
В дельте Днестра в начале 1970-х гг. численность составляла 1,4-1,6 тыс. пар, но в
1976-1978 гг. она катастрофически снизилась и в 1980-е гг. не превышала 200-900, в
456
Отряд Листообразные, Семейство Ибисовые
<р 75 ад о . д-s ° а о о к & * Б £ g s 3 в s 2 Д со к ’8 8 8 Д о ОО ? 2 £ X й Д (N S. * <D Й Г 2 к к S СО Си
cd [_/ so
си 77 о
о
CQ Н-( О гч
uQ О
К (N §
S • ~
СО сч Д
CU о сО
► X о > ГЧ i
х 2 к S s а <с
•2 си 2- rcd § g S Й Ж * СО • - CQ —< со О си О S 04 к S м н 2 О Е О СО а £ к <D П ч . - о о S о * о со a sc S ® 3 X а ° а $ £ Os Св Os Е — S £ се Н
1990-е — 340-925, а в начале XXI в. — 260^180
пар (Щеголев, 1977; Schogolev, 1996а; Русев,
2004). На Тилигульском лимане в начале 1980-х
гг. гнездилось до 200 пар, но затем численность
резко снизилась и в 1998 г. насчитывала всего 15
пар (Черничко и др., 1991; Сиохин, 2000).
В дельте Днепра в 1952 г. впервые отмечена 1
пара, в 1959 г. найдено уже 107 гнёзд, а в 1970-е
гг. там только в двух известных колониях гнез-
дилось от 300 до 600 пар (Смогоржевский, 1953;
Гизенко, 1963; Смогоржевський, 1979; Ардамац-
кая, 2006). Однако в 1980-е гг. из-за отстрела
больших бакланов, а также из-за засух и пожа-
ров в тростниках, связанных с длительным от-
сутствием паводков, колонии эти распались, и
сейчас в дельте Днепра гнездится лишь около 20
пар караваек (Ардамацкая, 2006).
На Лебяжьих о-вах в Крыму каравайки впер-
вые загнездились в 1967 г. (7 пар), в середине
1970-х гг. их численность поднялась до 300 пар,
но в дальнейшем колебалась на уровне 10-100
пар. В некоторые годы они исчезали вовсе (Кос-
тин, 1983; Костин, Тарина, 2004). На Сивашах
птицы загнездились в конце 1970-х гг. (Костин,
1983), переселившись туда, очевидно, с Дуная
(Гринченко, 2004). В 1980-е гг., на начальных
стадиях сукцессии водной растительности, выз-
ванной антропогенными факторами, популяция
караваек здесь резко увеличилась (до 2,2-2,7 тыс.
пар), но в 1990-е гг., в связи со старением биото-
пов, последовал быстрый спад численности (до
140-630 гнездящихся пар). Однако летом на Си-
вашах в последние годы регулярно держится до
1 500-2 500 холостых кочующих особей (Грин-
ченко, 2004). Всего на Украине в 1998 г. было
учтено 1 648 пар этих птиц (Сиохин, 2000).
В России основные гнездовья сосредоточены
в Вост. Приазовье, на Нижнем Дону, в дельтах
Волги и Терека. В Приазовье в 1953-1954 гг. в
двух колониях было учтено 150-200 особей (Ви-
нокуров, 1959), в 1961-1972 гг. было известно три
колонии из 20-70 пар (Олейников и др., 1975; Бе-
лик, Динкевич, 2004). В 1980-е гг. в 4 колониях
было учтено около 1 300 пар, к 1992 г. выявлено 6
колоний, общей численностью 5 500 пар, в том
числе самое крупное гнездовье из 4,0-4,5 тыс. пар
на Понурском лимане в Калининских плавнях, а
к 2002 г. зарегистрировано уже 8 колоний с 5 610
Каравайка
457
парами (Емтыль и др., 1989, 2003; Емтыль, Лохман, 1994). Кроме того, в этот перечень не
вошел ещё ряд колоний в дельте Кубани (100-600 пар) и в плавнях р. Ея (250-400 пар) (Бе-
лик, Динкевич, 2004). Всего же в Краснодарском крае — от Кубани до Ей — во второй
половине XX в. гнездовья каравайки отмечались на 20 водоёмах, которые заселялись пти-
цами попеременно, вследствие выгорания или естественного старения тростников. Общая
численность караваек составляет там сейчас, очевидно, не менее 6-7 тыс. пар.
В дельте Волги, где находится вторая по численности группировка в России, популяция
каравайки после глубокой депрессии в начале XX в. восстановилась к середине 1930-х гг.,
составив около 3,0 тыс. пар (табл. 36). В 1945 г. там насчитывалось уже 5,0 тыс. пар, а затем
численность колебалась из-за перераспределения птиц от 2,3 в 1968 г. до 6,1 тыс. пар в
1985 г. (Кривоносов, Гаврилов, 1981; Кривенко, 1983). В 1990-е гг. в связи с подъёмом
уровня Каспия и ухудшением кормовых и гнездовых условий, численность птиц в дельте
резко снизилась (до 316 пар в 1995 г.), и в настоящее время она колеблется на уровне 660-
1 825 пар (Гаврилов, 2002, 2005; Гаврилов и др., 2003).
Выше дельты каравайка гнездится сейчас на водоёмах Волго-Ахтубинского междуре-
чья почти до г. Волгограда (6-20 пар в Волгоградской обл.), а также до Сарпинских озёр,
где в последние десятилетия отмечается не более 5-10 пар (Чернобай, 2004).
На Нижнем Дону сотенные колонии были известны в разное время на озёрах и прудах в
окрестностях Азова, Ростова, Аксая, Новочеркасска и близ устья р. Сал. Птицы переселя-
лись с места на место в связи с гидромелиоративным строительством или выжиганием
тростников. Единичные пары эпизодически отмечали в колониях голенастых в верховьях
Цимлянского вдхр. (Чернобай, 2004). Всего в низовьях Дона в 1970-е гг. гнездилось 450-
600 пар, в 1980-е их численность сократилась вдвое, составив 200-250 пар, а в 1991 г. при
авиаучётах было выявлено 110-120 пар (Казаков, 20006). Сейчас колония из 300-500 пар
известна только на оз. Монастырское близ г. Аксай, где каравайки постоянно гнездятся с
1986 г. (Белик, Динкевич, 2004).
На Зап. Маныче каравайки обитали на Усть-Манычском вдхр., где в 1948 г. выявлены 3
колонии общим числом около 1 500 пар. С 1967 г. несколько колоний известно в нижней
части Веселовского вдхр., но численность птиц в них, как и в других местах, значительно
колеблется по годам. В течение 1977-1988 гг. там учитывали от 6 до 72 пар, иногда кара-
вайки отсутствовали вовсе (1980, 1985 гг.). На Пролетарском вдхр. они гнездятся лишь в
опресненной западной части близ устья р. Егорлык, где сохранились заросли тростника.
В прошлом каравайки обитали также в устье балки Тройной (Миноранский, 1963), но
после осолонения Маныча и гибели тростников исчезли там, так же как и на переоборудо-
ванном под рыборазводные пруды оз. Казинке, где гнездились до 1976-1977 гг. (Казаков и
др., 2004).
В верховьях Зап. Маныча, близ устья р. Калаус, колония из 8-23 пар была известна в
1978-1981 гг. на оз. Лысый Лиман. На Вост. Маныче 8-18 пар гнездились в 1973-1977 гг.
на Состинских озёрах близ г. Черноземельска. Всего в долине Маныча в 1990-1991 гг. было
учтено 250-300 пар караваек (Казаков и др., 2004). По имеющимся сведениям, до 300-400
пар гнездится на Бурукшунских лиманах в Ипатовском р-не на севере Ставропольского
края (Рудницкий, 2003).
В дельте Терека каравайка была многочисленна до середины XX в., но позже её числен-
ность значительно сократилась в связи с хозяйственным освоением водоёмов. В 1971-1974
гг. в 3 колониях гнездилось всего 400-450 пар (Бондарев, 1975; Пишванов, 1975), в 1985-
1986 гг. в 2-4 колониях учтено 430-480 пар (Казаков и др., 2004). В 1990-е гг. там было
известно 4 места гнездования 125-130 пар. Всего же в Дагестане, по экспертной оценке,
сейчас гнездится 250-300 пар (Джамирзоев и др., 2000).
458
Отряд Листообразные, Семейство Ибисовые
Таблица 36. Динамика численности каравайки в дельте Волги
Годы Астраханский заповедник Дельта пары Источники цитирования
взрослые осоои пары
1934 3870 * Сыроечковский, 2005
1935 4960 3,0 тыс. Кривенко, 1975; Сыроечковский, 2005
1939 1490 Сыроечковский, 2005
1945 5,0 тыс. Кривенко, 1975
1948 11153 Сыроечковский, 2005
1950 2355 4710 ** Скокова, 19606; Сыроечковский, 2005
1951 3542 Скокова, 19606
1952 1048 524 Скокова, 19606; Луговой, 1963
1953 1358 1358 679 -2000 *** Скокова, 19606; Луговой, 1963; Сыроечковский, 2005
1954 642 321 Скокова, 19606; Луговой, 1963
1955 672 336 3,5 тыс. Скокова, 19606; Луговой, 1963; Кривенко, 1975
1956 278 139 Скокова, 19606; Луговой, 1963
1957 381 Скокова, 19606
1961 550 Луговой, 1963
1968 2,3 тыс. Кривенко, 1975
1973- 4528 Бондарев, 1975, 1979, 2005
74
1974 678 Кривоносов, Гаврилов, 1981
1975 453 Кривоносов, Гаврилов, 1981
1976 418 Кривоносов, Гаврилов, 1981
1977 477 Кривоносов, Гаврилов, 1981
1978 314 Кривоносов, Гаврилов, 1981
1979 933 Кривоносов, Гаврилов, 1981
1980 4459 Гаврилов, 1984, 2005
1983 1443 Кривоносов и др., 1991
1984 995 Кривоносов и др., 1991
1985 941 6063 Кривоносов и др., 1991; Гаврилов, 2002, 2005
1986 865 Кривоносов и др., 1991
1987 778 Кривоносов и др., 1991
1991 1024 Гаврилов, 2002, 2005
1995 316 Гаврилов, 2002, 2005
1999 1372 Гаврилов и др., 2003
2000 660 Гаврилов, 2002, 2005
2001 1825 Гаврилов и др., 2003; Гаврилов, 2005
2002 1050 Гаврилов, 2002
2003 1400 Гаврилов, 2005
Примечания. * В начале 1930-х гг. почти все каравайки гнездились в заповеднике (Ромашова, 1938;
Сыроечковский, 2005); **Снижение численности птиц в заповеднике в 1950-е гг. происходило за счёт
выселения караваек на взморье (Скокова, 19606); *** В начале 1950-х гг. в дельте было известно 11
колоний каравайки (Сыроечковский, 2005).
Таким образом, общая численность каравайки в России на начало XXI в. составляла
около 8-10 тыс. пар.
В Казахстане из-за антропогенных изменений в гнездовых и кормовых биотопах, про-
исходивших в середине XX в., каравайка к 1980-м гг. полностью покинула крупные гнездо-
вья в пойме Сыр-Дарьи (Спангенберг, Фейгин, 1936), после чего исчезли и подпитывавши-
Каравайка
459
еся, вероятно, за их счет колонии в степной зоне (Ковшарь, 1991; 1999). В последние деся-
тилетия до 100-300-800 пар эпизодически гнездятся лишь в дельте р. Урал (Гисцов, Ауэзов,
1991; Русанов, 1992; Березовиков, Гисцов, 2001; Гаврилов, 2005) и на восточной окраине
Волжской дельты (Жатканбаев, 1996; Ковшарь, 1999). В 1990-е гг. каравайки практически
исчезли также в Узбекистане, где сейчас 700-800 пар гнездится лишь в дельте Аму-Дарьи
(Крейцберг-Мухина, 2003).
В Азербайджане в начале XX в. каравайки в большом числе гнездились на озёрах и
разливах, а также в галерейных лесах вдоль Куры, Аракса и в Ленкоранской низм. (Сату-
нин, 1907; Мустафаев, 1975). К середине XX в. птицы стали там редки, и лишь появление
обширных тамарисковых зарослей на обсыхающих берегах Каспия в Кызыл-Агачском за-
поведнике позволило сформироваться в начале 1950-х гг. очень крупному гнездовью голе-
настых и веслоногих птиц общей численностью до 337,5 тыс. пар. В 1957 г. в нем было
более 77 тыс. пар (23 %) караваек (Дюнин, 1960а; Греков, 1965). Однако в середине 1960-х
гг., возможно из-за интоксикации птиц пестицидами на орошаемых полях, их количество
резко снизилось (до 150 особей). В 1972 г. там насчитывалось 900 караваек; в дальнейшем,
в 1973-1995 гг., их численность колебалась на уровне 2-6 тыс. гнездящихся особей (Васи-
льев и др., 1972; Морозкин, 1975; Коновалова, 1977, 1979; Туаев, Исрафилов, 1978; Литви-
нова, 1981; Султанов, 2000).
Кроме Кызыл-Агача, в 1990-е гг. крупные колонии каравайки находились также в сме-
шанных поселениях голенастых и веслоногих птиц на оз. Акгёль (2 900 пар) и на оз. Мах-
мудчала-Ахчала (250 пар, или 10% общей численности колонии). В 1986 г. на упомянутом
оз. Махмудчала-Ахчала численность каравайки была определена в 10 тыс. пар, или 60%
общей численности поселения, в 1990 г. — 40% общей численности (Литвинова, 1989;
Патрикеев, 1991 в). Ещё 4-5 колоний каравайки известны на Дивичинском лимане, в дельте
р. Куры, на оз. Сарысу, Варваринском вдхр. (Султанов, 2000).
Для Средней Азии (как географического региона) известны данные только по южной
части Караджарской равнины, где Р.В. Лукашевич в 1988-1989 гг. оценил численность ка-
раваек в 700-800 особей (Птицы Средней Азии, 2007).
Дать оценку общей численности каравайки в Сев. Евразии без более точной информа-
ции из Азербайджана, где сосредоточена, вероятно, основная часть популяции, в настоя-
щее время достаточно сложно. Но, учитывая экспертные оценки по Азербайджану в 4-6
тыс. пар и Украине в 1,7-3,6 тыс. пар (BirdLife International, 2004), она может составлять,
очевидно, не менее 15-20 тыс. пар.
Размножение. Срок наступления половой зрелости не установлен. Известно, что неко-
торые годовалые птицы остаются на лето в местах зимовки или на путях пролёта, очевидно
не приступая к размножению (Сапетин, 1978е). Связь с гнездовой территорией относи-
тельно слабая, и птицы могут спонтанно переселяться с места на место целыми колония-
ми. Особенно лабильны молодые генерации птиц. Впрочем, при сохранении благоприят-
ных кормовых и защитных условий каравайки гнездятся в постоянных местах обычно в
течение довольно длительного времени; к переселению же птиц вынуждают пожары в тро-
стниках, беспокойство в колониях, пересыхание водоёмов и другие факторы. Смешанные
поселения в тростниках обычно ежегодно меняют места расположения в пределах гнездо-
вого водоёма; птицы переселяются на соседние участки зарослей из-за сплошного выла-
мывания сухого тростника в колониях при строительстве гнёзд (Щеголев, 1977; Черничко
и др., 1991).
Каравайка — моногам; пары ежегодно формируются в местах размножения после возвра-
щения птиц с зимовок. В парах самцы доминируют над самками (Щеголев, 1975). Каравайки
образуют поселения, насчитывающие от нескольких пар до нескольких сотен и тысяч пар. Наи-
460
Отряд Листообразные, Семейство Ибисовые
более крупные, многотысячные колонии известны на Кубани (Емтыль, Лохман, 1994; Мна-
цеканов и др., 2004), а также в Азербайджане (Дюнин, 1960а; Виноградов, 1967; Литвино-
ва, 1989; Султанов, 2000 и др.). Величина колоний определяется в первую очередь кормо-
выми ресурсами, зависящими от гидрологического режима водоёмов. Численность птиц
заметно увеличивается при периодических мощных паводках, заливающих мелководья (Чер-
ничко и др., 1991), но при устойчивом повышении уровня воды, как это наблюдалось в
1990-е гг. в дельте Волги, каравайки могут лишаться мелководных кормовых угодий и мно-
гих гнездовых стаций из-за вымокания древесно-кустарниковой растительности.
Каравайки гнездятся только в смешанных колониях голенастых и веслоногих птиц, из-
редка также вместе с грачами в лесонасаждениях вдоль водоёмов. Возвращаясь на места
гнездования с зимовок одной из последних среди голенастых птиц, каравайка поселяется,
как правило, в уже сформировавшихся колониях различных видов цапель, колпиц, бакла-
нов, обычно размещая свои гнёзда небольшими группами или цепочками по окраинам за-
нятых цаплями и бакланами тростниковых или древесно-кустарниковых куртин. Имея тем-
ную окраску, каравайка занимает, в основном, самые затенённые нижние ярусы древесно-
кустарниковых зарослей, прячась от солнца под кронами, а в тростниках располагается
обычно у самой воды, иногда — под гнёздами цапель, колпиц и др.
Как облигатно колониальный вид, каравайка не имеет гнездовых территорий. Она охра-
няет лишь сами гнёзда, которые часто располагаются плотными агрегациями (до несколь-
ких десятков построек) с минимальным расстоянием между центрами гнёзд около 53 см. В
этом случае, при среднем диаметре гнёзд в 37 см, промежутки между их краями составля-
ют 16 см, что является нижним пределом «нейтральной» зоны. Сокращение последней ве-
дёт к разрушению построек соседними парами (Щеголев, 1975; Черничко и др., 1991).
Обычное же расстояние между гнёздами в тростниках (п=12) составляет 0,8-1,0 м (Лука-
шевич, 1991, 1995).
Каравайки активно защищают свое гнездо. Нарушителя обычно предупреждают хрип-
лым каркающим криком и демонстративными позами. Тело при этом принимает горизон-
тальное положение, крылья раскрываются, птица приседает и совершает стремительные
выпады головой в сторону врага. Если демонстраций оказывается недостаточно, происхо-
дит короткая драка. Особенно достаётся чужим птенцам, бродящим по колонии. Своих
птенцов каравайки начинают различать с 10-12-дневного возраста (Щеголев, 1975; Чер-
ничко и др., 1991).
Гнездо строят оба партнёра. По наблюдениям в Азербайджане, самец приносит строи-
тельный материал, а самка в основном укладывает его. Птицы собирают материал для гнёзда
обычно в колонии или в её окрестностях, но иногда носят его в клюве за несколько кило-
метров от гнездовий. На строительство гнезда требуется 3-5, в среднем (п=7) 4 дня, но его
ремонт и достройка продолжаются весь период насиживания и выкармливания птенцов
(Мустафаев, Кязимов, 1965,1966). По наблюдениям в дельте Волги, в период насиживания
самец и самка время от времени проводят срочный ремонт гнезда, в течение 15-20 мин
обновляя выстилку лотка и укрепляя его борта (Аскаров, 1938). О непрекращающемся об-
новлении гнёзд свидетельствует и тот факт, что брошенные гнездовые постройки караваек
растаскиваются членами колонии в течение 2-3 дней (Черничко и др., 1991).
В тростниковых плавнях каравайки гнездятся обычно на невысоких тростниковых коч-
ках или заломах тростника у самой воды, иногда в 0,5-1,5 м над водой. В Азербайджане на
оз. Акгёль расстояние от воды до основания гнезда составляло, в среднем (п=214), всего 16
см (Туаев, Исрафилов, 1978). Изредка птицы устраивают гнёзда на старых хатках ондатры
или занимают гнёзда цапель, перестраивая их (Спангенберг, 1951). При повышении уровня
воды каравайки надстраивают гнездо, которое может достигать 40 см в высоту (Черничко и
Каравайка
461
др., 1991). Гнёзда в тростниках обычно ежегодно делаются заново, но местами птицы гнез-
дятся в них несколько лет подряд, достраивая сверху (Олейников и др., 1975; Туаев, Исра-
филов, 1978). Материалом для гнезда служат, в основном, сухие стебли тростника диамет-
ром до 1,0-1,7 см, иногда с примесью тины и грязи (Костин, 1983; Черничко и др., 1991).
Лоток выстилается сухими и зелёными листьями тростника или рогоза, на дно нередко
укладываются сухие метёлки тростника, размочаленные листья и другая мягкая раститель-
ная ветошь. В отличие от цапель и колпиц, гнёзда каравайки более плотные, аккуратные,
округлые, с глубоким, хорошо выраженным лотком. После появления птенцов лоток посте-
пенно растаптывается, гнездо проседает, намокает и часто приобретает неряшливый вид
полусгнившей кочки. Из-за специфического корма и своеобразного помёта, от гнезда в это
время исходит характерный для колоний караваек запах (Аскаров, 1938).
Размеры гнёзд, устраиваемых в тростниковых зарослях на Тилигульском лимане в Одес-
ской обл., в среднем (п=25) составляли: диаметр гнезда — 37; высота гнезда — 20,3; диа-
метр лотка — 21,7 см (Черничко и др., 1991). На Лебяжьих о-вах в Крыму диаметр гнезда
был равным 27-43 см; высота гнезда — 11-46; диаметр лотка — 13-23; глубина лотка —
3-9,5 см, в среднем (п=34), соответственно, 33,5; 29,0; 19,2; 7,1 см (Костин, 1983). В дельте
Кубани гнёзда были построены на заломах камыша и рогоза в 40-50 см над водой. Их
средние размеры составляли: диаметр гнезда — 37; высота гнезда — 31; диаметр лотка —
14; глубина лотка — 4,1 см (Заболотный, Хохлов, 1989а). В Вост. Приазовье в Челбасских
плавнях гнёзда (п=31) располагались в 28-61 см над водой (в среднем, 41 см), но в процес-
се насиживания осели до 15-60 см (в среднем, 27 см). Их размеры (п=51) составляли: диа-
метр гнезда — 24-46; диаметр лотка — 14-21; глубина лотка -3-10, в среднем, соответ-
ственно, 33; 17 и 6 см. Высота свежих гнёзд равнялась 15-25 см, а прошлогодних, достро-
енных — 35-60 см. В долине Зап. Маныча на оз. Казинка гнёзда (п=49) строились в 20-140
см над водой (в среднем, 63 см). Их размеры равнялись: диаметр гнезда — 25-44; высота
гнезда — 15-24; диаметр лотка — 22-40; глубина лотка -3-8, в среднем, соответственно,
33,7; 17,5; 26,6; 6,5 см (Олейников и др., 1975; Казаков и др., 2004). В дельте Аму-Дарьи
птицы гнездятся, в среднем, на 0,58±0,27 м над водой. Диаметр их гнёзд составляет 30-40
см, диаметр лотка— 12-15 см (Лукашевич, 1991, 1995).
Древесно-кустарниковые колонии располагаются обычно на болотах с глубиной воды
от 0,5 до 1 м и более (Аскаров, 1938; Мустафаев, Кязимов, 1965). При обсыхании колоний
птицы бросают их из-за появления у гнёзд наземных хищников (Виноградов, Чернявская,
1965). На древесно-кустарниковой растительности каравайки гнездятся на высоте до 6,2 м.
Гнёзда размещаются преимущественно в наиболее густых зарослях, которые лучше защи-
щают кладки и птенцов от хищных птиц (Аскаров, 1938; Сыроечковский, 2005). Здесь гнёзда
более рассредоточены, чем в тростниках, распределяясь обычно среди построек мелких
видов цапель, но, как и в тростниковых плавнях, находятся, в основном, в самом нижнем
ярусе, в тени листвы. Лишь в джидовых тугаях на островах Сыр-Дарьи каравайки гнезди-
лись в средних и верхних ярусах крон невысоких, но густых и колючих деревьев (Спанген-
берг, 1951).
Гнёзда на деревьях и кустах каравайки занимают обычно несколько лет подряд, достра-
ивая их весной. Сооружаются они из сухих прутьев ивы, тамариска, лоха длиной 10-82 см
и толщиной от 0,3 до 2 см (Аскаров, 1938). Нередко птицы используют также сухие стебли
тростника. Лоток выстилают листьями ивы, тростника, рогоза, клубнекамыша и др. Как и в
тростниковых зарослях, гнёзда каравайки на деревьях отличаются аккуратным, плотным
строением, округлой формой и глубоким лотком. В дельте Волги их размеры составляли:
диаметр гнезда — 30x30 см; глубина лотка — 5-8 см (Аскаров, 1938). В Азербайджане
гнёзда, размещавшиеся в зарослях тамариска на высоте от нескольких сантиметров до 1,0-
462
Отряд Листообразные, Семейство Ибисовые
1,9 м над водой, имели примерно такие же размеры: диаметр гнезда — 28-35; глубина
лотка — 4-6 см (Греков, 1965).
Период размножения оседлых популяций каравайки в тропических районах приурочен
обычно к сезону дождей (del Hoyo et al., 1992). В Сев. Евразии каравайки приступают к
гнездованию весной после возвращения с зимовок. В дельте Волги, на северной границе
ареала, они начинают гнездостроение вскоре после прилёта, откладывая первые яйца с 28
апреля, но в массе — с 10 мая. При гибели первых кладок птицы гнездятся повторно до 1-
12 июля, а птенцы из поздних гнёзд вылетают иногда в конце августа (Воробьев, 1936;
Аскаров, 1938; Луговой, 1963). В более южных районах гнездование начинается обычно в
середине - конце апреля, через 2-5 недель после появления первых птиц. До этого каравай-
ки стаями кочуют по кормовым стациям в районе колоний, возвращаясь к ним на ночевку
(Назаренко и др., 1975; Черничко и др., 1991).
В дельте Дуная начало строительства гнёзд отмечено 20 апреля 1977 г., в конце апреля
начинается кладка яиц. На Днестре начало кладки яиц наблюдали с 16 апреля (1972 г.) до 3
мая (1974 г.), в среднем за 1970-1977 гг. — 23 апреля (Смогоржевський, 1979). В дельте
Днепра 25 мая 1959 г. найдены первые кладки, 10 июня обнаружено уже 107 гнёзд с яйцам,
а 10 июля в них находились птенцы перед вылетом (Гизенко, 1963).
На Тилигульском лимане строительство первых гнёзд отмечено 20 апреля 1980 г., 27
апреля 1982 г. и 22 апреля 1983 г.; их основная масса сооружалась в период 26 апреля - 1 мая
1980 г., 21 апреля - 3 мая 1981 г., 28 апреля - 3 мая 1982 г., 23-28 апреля 1983 г. Яйцекладка
тоже проходила очень дружно. Первые яйца появлялись 30 апреля - 4 мая 1980 г., 25 апреля -
7 мая 1981 г. и 1-5 мая 1982 г. Первые птенцы отмечены 22 мая 1982 г. и, по расчетным
данным, 16 мая 1981 г.; массовое вылупление проходило 20-29 мая 1981 г. и 23-29 мая
1982 г. Слётки встречались с конца 2-й декады июня, в конце 3-й декады наблюдался массо-
вый вылет молодых (Черничко и др., 1991).
На Лебяжьих о-вах в Крыму фенология гнездования варьирует значительно сильнее.
Там в 1990-х - начале 2000-х гг. самые ранние кладки отмечали 4 мая 1997 г. и 8 мая 1994
г., а последние — 18 июня 1997 г. Первые кладки появлялись через 12^11 день после при-
лёта птиц, в среднем за 12 лет — через 32±9 дней. Продолжительность периода яйцекладки
в колонии колебалась от 4 (1998 и 2002 гг.) до 45 (1997 г.) дней, в среднем за 12 лет— 14±12
дней. Вылупление птенцов проходило обычно в 2-3-й декадах июня, иногда раньше: 16
мая 1992 г., 26 мая 1997 г., 2 июня 1994 г. Разница в сроках вылупления птенцов из ранних
и поздних кладок составляла 3^13 дня, в среднем за 12 лет — 16±12 дней. Вылет птенцов
проходил в июле, с 2 июля (1996 г.) - 5 июля (2000 г.) до 29 июля (1998 г.) - 7 августа (1997
г.). Весь гнездовой цикл от начала яйцекладки до вылета птенцов длился от 55 дней (2002
г.) до 95 дней (1997 г.), в среднем за 12 лет — 71± 13 дней (Костин, Тарина, 2004). Однако в
1970-х гг. каравайки начинали яйцекладку в Крыму в конце апреля - начале мая и лишь в
1971 и 1973 гг., когда птицы поздно вернулись с зимовок, откладка яиц началась в середине
и конце мая. В 1975 г. у большинства пар 25 мая оказались уже летающие птенцы, хотя в
другой части той же колонии десятки гнёзд были ещё с кладками (Костин, 1983). Таким
образом, в Крыму каравайки могут приступать к гнездованию в отдельные годы уже в кон-
це марта.
В Вост. Приазовье в колонии на р. Челбас массовое вылупление в 1972 г. наблюдалось с
31 мая до 15 июня, а в отдельных гнёздах продолжалось до 24 июля. В колонии же в дельте
Кубани 28 мая 1961 г. были ещё только свежие кладки, а в колонии на р. Бейсуг 24 июня
1961 г. почти все птенцы уже поднялись на крыло (Олейников и др., 1975). На Зап. Маныче
каравайки приступают к откладке яиц во 2-3-й декадах мая, в дельте Терека начало гнездо-
вания приходится на 1-ю декаду мая (Казаков и др., 2004).
Каравайка
463
В Азербайджане в Кызыл-Агачском заповеднике в 1964 г. продолжительность периода
размножения от начала строительства гнёзд до вылета птенцов составляла 59 дней у от-
дельных пар и 83 дня (с 16 мая по 7 августа) в целом по колонии (Мустафаев, Кязимов,
1966). На оз. Акгёль 15 мая 1964 г. уже шла массовая яйцекладка (Виноградов, 1967). По
наблюдениям в 1973-1976 гг., откладка яиц на этом озере продолжалась с конца апреля до
конца июня, а наиболее ранняя находка яйца в яйцеводе добытой птицы отмечена 26 апре-
ля 1975 г. (Туаев, Исрафилов, 1978). В дельте Аму-Дарьи первые кладки появлялись в кон-
це апреля, массовая яйцекладка проходила в 1-й декаде мая (Лукашевич, 1991,1995). Нако-
нец, в низовьях Сыр-Дарьи первые яйца тоже отмечались в конце апреля, в 1-й декаде мая
шла массовая яйцекладка, 28 мая 1925 г. в большинстве гнёзд были слабо насиженные
яйца, а 20 июля 1926 г. все птенцы караваек уже покинули гнёзда (Спангенберг, Фейгин,
1936; Спангенберг, 1951).
Копуляция у караваек происходит после завершения строительства гнезда на соседних
с гнездом ветвях, на кучах слежавшегося камыша или на самих гнёздах (Аскаров, 1938;
Спангенберг, 1951). Насиживание начинается с откладки первого яйца, но в период яйцек-
ладки оно обычно «неплотное», так как большую часть дня птицы часто лишь стоят над
кладкой. От плотности насиживания в период откладки яиц зависит длительность инкуба-
ционного периода различных яиц в одном и том же гнезде. Если насиживание вначале было
неплотным, а родители приступили к нему уже после откладки всех яиц, птенцы появляют-
ся в течение суток. Если же птицы с первого яйца плотно сидели на кладке, процесс вылуп-
ления растягивается на 5-6 дней.
Таким образом, время начала откладки яиц и гнездового периода в целом у каравайки
достаточно продолжительно, хотя завершается первая его «волна» примерно в одни и те же
сроки на разных широтах Сев. Евразии. В норме репродуктивный период продолжается
55-59 дней, но птицы разных возрастных генераций, прилетающих с зимовки и приступа-
ющих к гнездованию в разное время, могут «удлинять» его (в том числе и из-за повторных
кладок) до июля, а у поздних пар — до конца лета (Аскаров, 1938; Мустафаев, Кязимов,
1965 и др.). Инкубация, по наблюдениям в Азербайджане, продолжается 19-23, в среднем
(п=18 яиц) — 21,2 дня (Мустафаев, Кязимов, 1966). Продолжительность инкубации в 21
день экспериментально определена также в дельте Кубани (Олейников и др., 1975). Инку-
бационный период в Крыму составляет 23-25 дней (Костин, Тарина, 2004). Откладка яиц
происходит, как правило, ежедневно. При средней величине кладки в 3,2 яйца, яйцекладка
продолжалась, в среднем (п=5), 3,5 дня, а вылупление — 1-5 дней, в среднем 2,3 дня. Вы-
лет птенцов происходил на 30-32-й, в среднем на 31-й день их развития (Мустафаев, Кязи-
мов, 1966).
Насиживают оба партнёра: самка — ночью, самец — днем, сменяясь обычно утром (в
7-8 ч) и вечером (в 16-19 ч) (Спангенберг, Фейгин, 1936; Аскаров, 1938; Мустафаев, Кязи-
мов, 1965). Если в период насиживания погибает один из партнёров, птицы, как правило,
гнездо бросают (Аскаров, 1938). Смена караваек на гнезде сопровождается звуками, напо-
минающими воркование. В начале периода насиживания партнеры неохотно уступают
место на яйцах, и второй птице приходится «сгонять» другую с кладки (Черничко и др.,
1991). Величина кладки в норме составляет 3-6 яиц. Размер её зависит, по-видимому, от
обеспеченности птиц кормами, особенно в зимний период (Гизенко, 1963; Мустафаев, Кя-
зимов, 1966). Нередко встречающиеся кладки из 1-2 яиц появляются, очевидно, при по-
вторном гнездовании или вследствие хищения яиц из гнёзд (Аскаров, 1938). Изредка реги-
стрируемые гнёзда с 7-9 яйцами представляют собой, в большинстве случаев, сдвоенные
кладки. На Днестре, например, в 1955 г. было обнаружено 6 сдвоенных кладок из 6 (2
гнезда), 7 (1 гнездо) и 8 яиц (3 гнезда), борьба за которые велась между двумя самками,
464
Отряд Листообразные, Семейство Ибисовые
тогда как самцы участия в этих конфликтах не принимали, одинаково относясь к обеим
самкам. Сдвоенные кладки возникли здесь после того, как готовые к яйцекладке самки
потеряли в результате шторма свои гнездовые постройки и вынуждены были захватывать
соседние уцелевшие гнёзда более мелких, «слабых» самок, только приступивших к отклад-
ке яиц (Jozefik, 1960). Обычная кладка в дельте Днестра состоит из 3-4, изредка — 5 яиц, в
среднем 3,6 яйца (Смогоржевський, 1979). В дельте Днепра в 1959 г. кладки были значи-
тельно крупнее, в среднем (п=107) по 4,4 яйца; гнёзда с 6 яйцами составляли 18,7% (Гизен-
ко, 1963). На Тилигульском лимане в кладках было от 2 до 5 яиц, в среднем (п=62) 3,7 яйца
(Черничко и др., 1991). На Лебяжьих о-вах кладки состояли из 3-5 яиц (Костин, Тарина,
2004), а по более ранним данным — из 2-7, чаще 4-5 яиц (Костин, 1983).
В Вост. Приазовье в гнёздах регистрировалось от 2 до 8 яиц, в среднем (п=27) 3,8 яйца,
но самая крупная кладка была, по-видимому, сдвоенная (Олейников и др., 1975). В дельте
Кубани кладки состояли из 2-4, в среднем (п=26) 3,6 яиц; кроме того, здесь же обнаружено
6 расклёванных яиц (Заболотный, Хохлов, 1989). По более поздним данным этих же авто-
ров, в кладках было 2-5, в среднем (п=60) 3,5 яиц (Хохлов и др., 1997). На Зап. Маныче в
гнёздах отмечалось по 3-5 яиц (по 3 яйца — в 4, по 4 — в 14, по 5 — в 2 кладках), в среднем
(п=20) 3,9 яиц (Казаков и др., 2004). На Вост. Маныче — тоже по 3-5 яиц (по 3 яйца — в 5,
по 4 — в 10 по 5 — 2 кладках), в среднем (п=17) 3,8 яиц (Ильюх, Хохлов, 2006). В дельте
Волги в гнёздах было обычно 3—4, очень редко 5 яиц (Воробьёв, 1936), а по данным Г. Аска-
рова (1938) — 3-4, изредка 2, иногда 1 яйцо, что было вызвано разорением гнёзд серыми
воронами. В Азербайджане на оз. Акгёль в 1964 г. абсолютно преобладали кладки из 3-4,
изредка 5 яиц, в среднем 3,6 яйца на гнездо (Виноградов, 1967); в 1973-1976 гг. средний
размер кладок здесь колебался от 2,5 (1973 г., п=37) до 3,8 (1976 г., п=24) яиц (Туаев, Исра-
филов, 1978). В Кызыл-Агачском заповеднике в 1964-1965 гг. кладки состояли из 1-4, в
среднем 3,2 (п=48) - 3,1 (п=100) яиц (Мустафаев, Кязимов, 1966); в 1975-1976 гг. там же
было, в среднем, по 3,5-3,7 яиц (Коновалова, 1977), в 1958 г. отмечались кладки, в основ-
ном, из 4-6, иногда до 9 яиц (Греков, 1965).
В дельте Аму-Дарьи в 1988 г. кладки состояли из 2-6, в среднем (п=12) 3,67 яиц (из 2
яиц — 3 гнезда, из 3 — 2, из 4 — 4, из 5 — 2 и из 6 — 1 гнездо), но в 1989 г. средний размер
кладок значимо снизился — до 3,12 яиц (Лукашевич, 1991, 1995). Наконец, в низовьях
Сыр-Дарьи в кладках было 3^4, редко 5 яиц (Спангенберг, Фейгин, 1936).
Яйца каравайки яркого сине-зелёного, зеленовато-синего или голубого цвета, более тём-
ного и более стойкого к выцветанию, чем у цапель. Яйца имеют обычно правильную яйце-
видную форму; изредка встречаются закругленные, коротко-эллиптические и удлинённо-
эллиптические с заострёнными полюсами (Спангенберг, Фейгин, 1936; Мустафаев, Кязи-
мов, 1966).
Величина яиц, как и кладок, зависит, вероятно, от обеспеченности птиц кормами (Ги-
зенко, 1963), и между этими параметрами прослеживаются положительные корреляции
(табл. 37).
По наблюдениям в дельте Волги, более крупные яйца появляются в поздних кладках,
когда в воде развивается много насекомых (Аскаров, 1938). В Вост. Приазовье самыми
крупными в кладках были первые яйца (53,2x37,9 мм; CV=4,8 % и 7,7 %; п=10), у которых
сильно варьирует диаметр, определяя больший объём и вес яиц. Вторые и третьи яйца
заметно меньше, и у них сильнее колеблется длина (2-е яйцо: 51,2x36,8 мм; CV=6,3 % и
3,9 %; п=10; 3-е яйцо: 51,2x36,9 мм; CV=4,4 % и 3,9 %; п=10). Последние, четвертые яйца
несколько крупнее и равномернее предыдущих (52,3x36,8 мм; CV=3,7 % и 2,5 %; п=10)
(Олейников и др., 1975). Эти особенности тоже связаны, вероятно, с трофикой птиц. Повы-
шение упитанности самок перед яйцекладкой позволяет им продуцировать более крупные
Каравайка
465
Таблица 37. Размеры кладок и яиц каравайки в Вост. Европе и Сев. Азии
Регион Величина кладки Масса яиц, г Сред, размеры яиц (мм) Пределы размеров яиц (мм) Источник
Тилигульск. лиман 3,7 37,1-26,5 51,8x36,3 (п=35) 48,3-55,0x33,0-39,1 Черничко и др., 1991
Крым 4-5 26,7-43,1; в ср. 35,9 52,2x35,8 (п=122) 45,2-58,3x30,5-39,0 Костин, 1983
Вост. Приазовье 3,8 52,5x36,7 (п=168) 48,0-61,0x31,0-46,0 Олейников и др., 1975
Дельта Кубани 3,6 30,6-42,2; в ср. 36,3 (п=93) 51,6x36,4 (п=93) 46,0-55,7x33,3-38,5 Заболотный, Хохлов, 1989
Дельта Кубани Зап. Маныч 3,5 51,3x36,3 (п=82) 52,0x36,9 (п=10) 44,0-55,7x33,3-37,9 46,4-58,0x35,2-38,5 Хохлов и др., 1997 Олейников и др., 1975
Зап. Маныч 3,9 35,9-42,4 (п=16) 52,3x36,7 (п=58) 48,6-56,6x33,0-38,9 Казаков и др., 2004
Вост. Маныч 3,8 52,5x36,2 (п=29) 44,0-58,6x32,3-38,4 Ильюх, Хохлов, 2006
Дельта Волги 3-4 51,2x36,5 (п=27) 48,2-54,2x34,6-39,0 Воробьев, 1936
Дельта Волги 2-4 31,4-38,9; в ср. 35,4 (п=20) 53,3x36,7 (п=20) 50,0-57,1x34,6-40,5 Аскаров, 1938
Азербайджан 3,1-3,2 52,4x35,8 (п=51) 47,5-56,5x34,0-38,5 Мустафаев, Кязимов, 1966
Азербайджан 2,5-3,8 50,1x35,0 (п=51) 46,9-58,7x32,7-39,0 Туаев, Исрафилов, 1978
Аму-Дарья 3,1-3,7 36,8 (п=75) 52,4x36,2 (п=151) Лукашевич, 1995
Сыр-Дарья 3-5 52,0x35,9 (п=32) 49,0-54,5x33,1-37,5 Спангенберг, 1951
первые яйца, используя основной запас питательных ресурсов. Но это приводит затем к
уменьшению размеров 2-го и 3-го яиц. Увеличение 4-го яйца может быть связано как с
сокращением энергозатрат самки после начала насиживания, так и с увеличением интерва-
ла между откладкой яиц.
Масса свежих яиц указана в таблице 37. За период насиживания яйца теряют 1,2-2,0 г,
или 3,2-5,6% первоначальной массы. Вес только что вылупившегося птенца составляет
20,2-25,8 г (Аскаров, 1938; Туаев, Исрафилов, 1978).
Птенцы вылупляются слепыми, покрытыми тёмным пухом. В первые дни они беспомощ-
ны и неподвижно лежат в гнезде. В этот период самка постоянно находится в гнезде, согревая
птенцов и защищая их от перегрева на солнце. Глаза у птенцов открываются через 2-3 - 5-6 ч
после вылупления (Аскаров, 1938; Черничко и др., 1991). Кормление начинается на 2-3-й
день (Туаев, Исрафилов, 1978; Черничко и др., 1991). Корм птенцы получают из глотки роди-
телей, засовывая туда свой клюв и голову до самых глаз. К 4-му дню исчезает яйцевой зуб. На
4-5-й день птенцы пытаются подняться на ноги, а на 7-й день уже довольно уверенно стоят в
гнезде. В период с 6-7-го до 11-12-го дней взрослые птицы начинают принудительно стиму-
лировать птенцов к тренировкам крыльев. В это время птенцы уже могут склевывать пищу,
отрыгнутую родителями в гнездо. С 9-10-го дня у них проявляются признаки полового ди-
466
Отряд Листообразные, Семейство Ибисовые
морфизма, прежде всего в длине клюва и цевки (Аскаров, 1938). До 9 дней птенцы находят-
ся в гнезде и при опасности затаиваются, позже они по тревоге покидают гнездо, а с 11-13-
го дня постоянно бродят по колонии, независимо от беспокойства, при опасности сбивают-
ся в табунки (Черничко и др., 1991). Однако в лесных колониях на Волге птенцы сидят в
гнезде до приобретения способности перелетать с одного дерева на другое (около 24 дней);
лишь в присутствии родителей двухнедельные птенцы могут покидать гнездо и ходить по-
близости по ветвям. К 27-му дню молодые летают уже относительно свободно, а на ЗО-й
день улетают за родителями на места кормежки (Аскаров, 1938).
Пеньки маховых перьев появляются у птенцов на 4-5-й день, на 8-9-й день начинают
разворачиваться их опахала. В возрасте 15 дней тело покрывается пером, пух остаётся только
на голове и шее. В 20 дней птенцы полностью оперяются, а в 26 дней почти достигают
размеров взрослых птиц, хорошо отличаясь коротким клювом. В 27-30 дней на клюве ис-
чезают жёлтые перевязи, и он весь становится чёрным.
В развитии птенцов выделяются 3 периода: до 3-4 дней у них идёт наиболее интенсив-
ный прирост массы тела, длины крыла, клюва и цевки; до 12-15 дней наиболее интенсивно
растёт оперяющееся крыло; после 15 дней интенсивно развивается контурное оперение.
Перед вылетом птенцы достигают 440-635 г, или 80-90% массы взрослых птиц (Аскаров,
1938; Черничко и др., 1991; Казаков и др., 2004).
С маленькими птенцами постоянно находится самка, которую самец сменяет лишь на
2-4 ч в полдень (Спангенберг, Фейгин, 1936). Кормят птенцов оба родителя, но самец толь-
ко приносит корм в гнездо и передаёт его самке, постоянно находящейся с птенцами млад-
ших возрастов, а та распределяет корм птенцам. Поэтому если в паре в это время погибает
самец, то самка может выкормить выводок одна, однако если исчезает самка, то птенцы,
как считает Г. Аскаров (1938), обречены на гибель. Для подросших 8-10-дневных птенцов
корм начинает собирать и самка, вылетающая на добычу корма утром, днем и вечером.
Продолжительность добывания корма составляет от 30 мин до 1 ч 30 мин (в среднем, 1 ч),
кормёжка птенцов длится 10-15 мин при одном приеме пищи и 25 мин. при двух приёмах,
повторяющихся через 15-20 мин. До 5-дневного возраста родители кормят птенцов 8-12
раз в сутки, 5-10-дневных — 6-10 раз, 10-15-дневных — 5-8 раз, а более старших — 5-7
раз в день (Аскаров, 1938). По другим наблюдениям, число кормёжек увеличивается с 5-8
раз в 15-дневном возрасте до 14 перед вылетом птенцов из гнёзд (Туаев, Исрафилов, 1978).
После вылета молодые ещё около недели выпрашивают (и получают) корм у взрослых птиц.
Успех размножения каравайки изучен слабо. На Дунае из яиц вылуплялось 77% птен-
цов (Смогоржевський, 1979). В дельте Днестра в благоприятные годы выживало, в сред-
нем, 2,5 птенца на гнездо (при кладке в 3,6 яиц), а при летних паводках или штормах —
всего 0,9 птенца (Щеголев, 1977). На Лебяжьих о-вах в Крыму успешность размножения
достигала 79,2-94,2%. Эмбриональная смертность составляла 3,4-8,3%, а ювенильная —
2,5-13,6%. Количество неоплодотворённых яиц колебалось здесь от 0,3 до 2,1% в разные
годы (Костин, Тарина, 2004).
В дельте Волги погибли 3 из 21 гнезда, находившегося под контролем (2 — с яйцами и
1 — с птенцами), а из 78 яиц в контрольных гнёздах выросло 50 птенцов (64,1%). Эмбрио-
нальная и постэмбриональная смертность составила 35,9%, в том числе 4 яйца оказались
неоплодотворенными (Аскаров, 1938).
На оз. Акгёль вылетало, в среднем, по 3 птенца, при средней кладке в 3,6 яйца (Виногра-
дов, 1967). В Кызыл-Агачском заповеднике в 1964 г. из 154 яиц в 48 гнёздах вылупилось 134,
а вылетело 124 птенца, т.е. отход яиц составил 13%, а птенцов — 7,5%. Таким образом, об-
щая эффективность размножения равнялась 79,8% (Мустафаев, Кязимов, 1966). В 1975 г. там
же эмбриональная смертность составила 13,8%, а постэмбриональная — 3,4%, в целом же —
Каравайка
467
16,8%- Но в 1976 г., при невысоком уровне воды, у караваек, приступивших к гнездованию
до середины мая, погибло 44,7% яиц, а у птиц, загнездившихся позже, — 16,2%. Постэмб-
риональная смертность составила 5,7%, а общая — 52,2% (Коновалова, 1977).
Следует отметить, что в дельте Днестра при наступлении паводка в период насижива-
ния почти все гнёзда каравайки погибают, но гибель гнёзд с птенцами, достигшими 10-
дневного возраста и способными к лазанию по кустарникам, не превышает 30% (Назарен-
ко, Николаевский и др., 1975).
Таким образом, наиболее успешно гнездование протекает на Лебяжьих о-вах (80-95%),
где хищничество ворон и других животных сведено к минимуму. На степных озёрах Азер-
байджана эффективность гнездования несколько ниже (80-85%), но при неблагоприятных
гидрологических условиях она может снижаться до 50%. В дельтовых же экосистемах, для
которых характерна нестабильность гидрологического режима и где обитает много ворон и
других хищников, успешность размножения самая низкая (65-70%), снижаясь при паводках
до 25%. Постоянство дельтовых колоний может объясняться, поэтому, лишь высокими кор-
мовыми качествами пойменных угодий и многочисленными повторными кладками.
Максимальная продолжительность жизни, по данным мечения, составляет 20-21 год
(Сапетин, 1978е; del Hoyo et al., 1992). В результате кольцевания было установлено, что для
азербайджанских птиц она равнялась 10 лет и 4 месяца, дагестанских — 8 лет и 4 месяца, а
для краснодарских — лишь 4 года и 2 месяца. Это было обусловлено мощным промысло-
вым прессом в Приазовье в период работ по кольцеванию (Сапетин, 1978е). Соответствен-
но различались также средняя продолжительность жизни и ежегодная смертность каравай-
ки (табл. 38).
До занесения вида в Красную книгу России (1983) основная масса караваек (около 40%
взрослых птиц и 65% молодых) гибла в августе, главным образом (более 95%) от выстре-
лов охотников. В настоящее время промысловый пресс на неё снизился, и она стала посте-
пенно восстанавливать свой ареал и численность, особенно в Вост. Приазовье.
Суточная активность, поведение. Суточная активность караваек в период выкармли-
вания птенцов начинается ещё до рассвета и продолжается до густых сумерек. Утренний
вылет происходит недружно и растягивается с 3-4 до 10 ч утра. Второй пик активности
начинается около полудня, третий — с 16-18 ч и продолжается до 22 ч. На кормёжку птицы
летят поодиночке и небольшими группами. Размер таких групп при кормовых перелётах в
период гнездования составляет, в среднем, 2,1-2,4 особи (Лукашевич, 1995). После вылета
птенцов из гнезда характер суточной активности меняется: птицы в массе покидают коло-
нию в 4-5 ч утра и возвращаются назад только в 19-21 ч (Аскаров, 1938; Скокова, 19606;
Казаков и др., 2004).
Таблица 38. Смертность и средняя продолжительность жизни караваек из разных
регионов Вост. Европы по данным кольцевания (по: Сапетин, 1978е)
Регион Средняя продолжительность жизни, годы Ежегодная смертность, %
средняя для всех птиц для птиц, переживших 1 -й год в 1-и год жизни птиц старше 1 года
Восточное Приазовье (Краснодарский край) 0,4 2,4 92 42
Дельта Терека (Дагестан) Южный 1,3 4,5 75 23
1,2 5,2
Азербайджан 68 21
468
Отряд Листообразные, Семейство Ибисовые
В период выкармливания птенцов на ночь у гнезда остаётся одна взрослая птица, а дру-
гая улетает на кормежку, возвращаясь к утру (Спангенберг, Фейгин, 1936).
Питание. Кормятся каравайки, в основном, на мелководьях или илистых литоралях,
как днем, так и ночью. В корме преобладают водные животные, преимущественно беспоз-
воночные: личинки стрекоз и жуков-плавунцов, моллюски, нередко встречаются мальки
рыб, а летом — появляющиеся в массе головастики и мелкие лягушата. Обычны также
жуки, сверчки и другие прибрежные насекомые (Аскаров, 1938). Корм в желудке перевари-
вается очень быстро. Этому способствует сильное развитие мышц мускулистого желудка, а
также гастролиты — обломки раковин дрейссены (Скокова, 19606).
В дельте Дуная в 33 желудках выявлено 435 экз. добычи, среди которых 65% составляли
насекомые, 20% — растения, 6% — моллюски, 4% — амфибии и 2% — рыбы (Platteeuw et
al., 2004). В дельте Днестра и Днепра в корме (19 желудков) преобладали личинки мух-
львинок и других двукрылых (325 экз.), личинки стрекоз (276 экз.), а также личинки водо-
любов и плавунцов (149 экз.) и сами эти жуки (89 экз.) (Смогоржевський, 1959, 1979).
Анализ обширных материалов по питанию каравайки, собранных в Вост. Приазовье и
на Зап. Маныче (Казаков и др., 2004), свидетельствует, что здесь в корме тоже абсолютно
преобладают водные насекомые (жуки, клопы, стрекозы, двукрылые); временами добыва-
ется много пиявок. Рыбы и земноводные поедаются регулярно, но в сравнительно неболь-
ших количествах. Во время жестокой засухи 1972 г., когда пересыхали Челбасские плавни,
каравайки вынуждены были носить птенцам также растительную пищу
В дельте Волги, по данным Скоковой (19606, 1965), в 1949-1954 гг. среди добычи кара-
вайки от 50,7 до 94,1% составляли водные беспозвоночные, 0,8-18,9% — наземные бес-
позвоночные, 4,9-21,3% — амфибии и 0,2-9,1 % — рыбы. В 1953 г. водные членистоногие,
преимущественно личинки водолюбов, составляли 73,8% добытых экземпляров животных,
моллюски — 8,5%, наземные членистоногие — 8,5%, амфибии — 8,3%, рыба (в основном
мальки сазана) — лишь 0,9%. Спектр питания каравайки состоял из более чем 60 видов
животных.
Аналогичные данные приводятся и для других регионов. Рацион может меняться в те-
чение сезона в зависимости от массовости и доступности кормов. Так, летом основным
кормом часто становятся головастики и лягушата. При массовом появлении саранчи кара-
вайки могут переходить на питание ею (Спангенберг, 1951; Танюшкин, 1958).
Враги, неблагоприятные факторы. В лесных колониях основной враг — серая воро-
на, активно хищничающая на кладках и птенцах каравайки. В дельте Волги на долю воро-
ны приходится не менее 20 из 36% общего отхода яиц и птенцов (Аскаров, 1938). Особенно
сильные опустошения вороны производят в колониях при их посещении людьми, распуги-
вающими птиц. Определённое негативное воздействие оказывает горностай, разоряющий
гнёзда. Изредка птенцов в гнёздах добывает болотный лунь. В смешанных поселениях го-
ленастых яйца и небольших птенцов каравайки может поедать кваква (Аскаров, 1938; Му-
стафаев, Кязимов, 1965). В плавневых колониях существенный вред гнёздам каравайки,
расположенным на небольшой высоте, иногда наносят дикие кабаны.
В северных районах гнездового ареала на численность каравайки существенное воз-
действие оказывает суровость зимних сезонов. Например, в 1976, 1978 и 1985 гг. после
холодных зим популяция резко сокращалась в дельте Днестра. Из других абиотических
факторов основными являются сильные колебания уровня воды во время летних паводков
и штормов. Флуктуации летней погоды, особенно сильные засухи, сказываются на обеспе-
ченности взрослых птиц и птенцов кормами, т.е. на эффективности размножения каравай-
ки. Синоптические условия зимних и весенних периодов определяют выживаемость и реп-
родуктивное состояние взрослых птиц, возвращающихся на места гнездования. Нередко
Каравайка
469
каравайки вынуждены бросать колонии из-за пожаров в тростниковых плавнях и на остро-
вах дельты Волги (Кривоносов и др., 1991).
В конце XIX - начале XX вв. на численности всех колониальных голенастых, в том
числе каравайки, мощное влияние оказывал, вероятно, массовый сбор яиц и промысловый
отстрел птиц на шкурки (Кривоносов, 1981; Белик, 1998; Русанов, 2004; Сыроечковский,
2005 и др.). Каравайка интенсивно добывалась до 1980-х гг. в качестве охотничьей дичи;
она и сейчас, несмотря на занесение её в Красную книгу России (1983, 2000) и других
соседних государств, по-прежнему попадает под выстрел.
Катастрофические последствия для каравайки должны были иметь истребительные
мероприятия против азиатской саранчи, охватившие в 1920-е гг. плавни всех южных рек.
Авиационное распыление над тростниковыми массивами чрезвычайно токсичных париж-
ской зелени и арсенита натрия послужило, очевидно, основной причиной глубокой депрес-
сии популяций каравайки и других околоводных птиц в начале XX в. (Белик, 1997, 20006).
Очень много саранчи, которой кормились стаи караваек, появилось в дельте Волги также в
мае 1950 г. Для борьбы с нею было проведено опыление плавней с самолёта и вручную
(Ганюшкин, 1958). В последующие годы численность караваек здесь вновь значительно
снизилась, что тоже было связано, очевидно, именно с этим мероприятием.
Снижение численности каравайки в середине XX в. могло быть обусловлено также заг-
рязнением пестицидами (ДДТ и др.) многих водоёмов, а через цепи питания — и всех
лимнофильных птиц. Наиболее сильно это загрязнение должно было отразиться на состо-
янии популяций каравайки в Азербайджане, где в тот период широко внедрялось хлопко-
водство, требовавшее массового применения различных химических препаратов.
Каравайка имеет достаточно специфичную паразитофауну, характерную для ибисовых
и отличающую их от других голенастых птиц. В дельте Волги для неё характерны из гель-
минтов Plagiorchis uhlwormii, Ophthalmophagus skrjabini, Hymenolepis microcephala. У всех
без исключения караваек отмечаются пухоеды ардеиколы, интенсивность заражения кото-
рыми доходит до 10 тыс. особей на одну птицу. Общим для каравайки, колпицы и цапель
является пухоед С. nyctardeae (Семенова, Иванова, 1991).
В колониях Кызыл-Агачского заповедника 100% караваек заражены трематодой Patagifer
bilobus (Саилов, 1965). На Днепре у двух из трёх птенцов обнаружены 2 цестоды и трема-
тода Plagiorchis laricola (в кишечнике) и нематода (в пищеводе). В кишечнике взрослой
птицы было 5 трематод Patagifer bilobus и 2 цестоды, а под кожей — около 15 цист тремато-
ды Tetracotyle ardeae (Смогоржевский, 1953).
В гнёздах караваек в большом количестве размножаются ложно скорпионы (Chelifer),
жуки-карапузики, сверчки, разнообразные гусеницы и сосальщики (Аскаров, 1938).
Хозяйственное значение, охрана. Из-за своей оригинальной внешности, напоминаю-
щей священного ибиса, каравайка в южных районах имеет определённое эстетическое зна-
чение. В большом количестве поедает водных насекомых и их личинок, считающихся вред-
ными. В дельте Волги за одну кормежку (2-2,5 ч) каравайка добывает, в среднем, 40-50,
иногда до 250 экз. животных, а за 4 месяца гнездового периода — более 15 тыс. обитателей
водоёмов и их побережий, в том числе более 10 тыс. водных насекомых и 2,5 тыс. назем-
ных беспозвоночных. Доля рыбы в рационе весьма ограничена, так что вся популяция ка-
раваек в дельте Волги, например, добывает за лето не более 0,5% мальков сазана, выращи-
ваемого в местных рыбхозах, или 0,0009% его молоди, отрождающейся во всей дельте Волги.
При этом одна каравайка за лето поедает около 8,5 тыс. хищных водных насекомых, кото-
рые за 2 месяца летнего половодья способны выловить более 5 млн. мальков рыбы. Кроме
того, каравайки в большом количестве добывают земноводных, тоже питающихся малька-
ми рыб (Скокова, 19606, 1965).
Mb
Отряд Листообразные, Семейство Ибисовые
В прошлом каравайку относили к охотничьим видам птиц (Мензбир, 1900, Шульпин,
1952; Назаренко и др., 1975; Смогоржевський, 1979 и др.). Уничтожение этих птиц и массо-
вый сбор яиц в колониях в XIX - начале XX в. привели к резкому снижению численности и
сокращению ареала каравайки по всей степной зоне. В Прикаспии, по-видимому, только
организация в 1919 г. Астраханского заповедника спасла её от окончательного истребле-
ния.' К концу 1920-х гг. основная часть гнездовий в дельте Волги сосредоточилась в запо-
веднике, под охраной которого каравайки стали интенсивно размножаться. Важное значе-
ние для охраны каравайки имела также организация в 1929 г. Кызыл-Агачского заповедни-
ка, на территории которого в середине XX в. сформировались крупнейшие гнездовья этого
вида.
В настоящее время каравайка занесена в Красные книги Российской Федерации, Арме-
нии, Украины, Казахстана, Узбекистана и ряда других государств Средней Азии.
Род Platalea Linnaeus, 1758
Колпица
Platalea leucorodia Linnaeus, 1758
Platalea leucorodia Linnaeus, 1758, Syst. Nat., ed.10, с.139, Европа, южная Швеция.
Spoonbill (англ.), Loffler (нем.), spatule blanche (фр.)
Син. (лат.): для подвида leucorodia'. Platalea leucopodius S.G. Gmelin, 1770; Platalea nivea
Cuvier, 1817; Platalea major Temminck et Schlegel, 1849.
Син. (рус.): обыкновенная колпица; колпик — народное (на юге России), от праславянско-
го къ1рь — лебедь, очевидно ошибочно перенесенное позже на другую крупную белую
птицу.
Статус. Гнездящийся перелётный, в южных частях ареала нерегулярно зимующий вид.
Общая характеристика и полевые признаки. Крупная птица величиной с гуся, с те-
лом длиной 75-90 см и размахом крыльев 115-145 см. Имеет очень характерный облик,
позволяющий легко отличать колпицу от сходных видов. Это типичная голенастая птица с
оперением общей белой окраски, с длинными ногами, длинной тонкой шеей и длинным,
уплощённым клювом, расширяющимся на конце в виде ложки. В полёте, в отличие от ца-
пель, шею вытягивает вперёд. Летает обычно гребным полётом с размеренными, сильны-
ми взмахами крыльев, периодически планирует, может парить, особенно в восходящих
потоках теплого воздуха.
Держится преимущественно группами или небольшими стаями на открытых пустын-
но-степных водоёмах. Гнездится колониями в тростниках, на деревьях или открытых ост-
ровах. Кормится обычно на илистых солоноватых мелководьях. Охотясь на слабопроточ-
ных водоёмах, колпицы часто выстраиваются шеренгой и медленно идут вперёд, держа
клюв опущенным в воду и двигая им при каждом шаге то вправо, то влево, как бы «косят»
тину (одно из народных названий колпицы — «косарь»). На заиленных стоячих озерках
могут кормиться в одиночку. При кормёжке, в отличие от цапель, ведут себя значительно
подвижнее. При перелётах птицы часто выстраиваются развернутым фронтом, нередко
поднимаются на значительную высоту.
Голос. Молчаливы; лишь в колониях можно слышать различные негромкие ворчащие
секвенции: «ху-гур», «ку-гу-гуг», «хо-хо-хок» и другие гортанные звуки. Птенцы, выпра-
Колпица
471
шивая у родителей корм, нередко издают своеобразные дребезжащие трели «хю-ррюк, хюр-
рю-рюк».
Сезонные различия в окраске оперения и габитусе незначительны; самец отличается от
самки более длинными ногами и клювом, что заметно лишь при сравнении. Молодые пти-
цы хорошо отличаются чёрными вершинами маховых перьев.
Подвидовая систематика. Обычно выделяют 2 подвида: (1) номинативный Р. I. leuco-
rodia Linnaeus, 1758, и (2) Р. I. archeri Neumann, 1928, гнездящийся на южном побережье и
островах Красного моря (Спангенберг, 1951; Степанян, 2003). Сейчас уверенно выделяют
также подвид (3) PL balsaci Naurois et Roux, 1974 с островов у побережья Мавритании (del
Hoyo et al., 1992; Dickinson, 2003). На территории России и в сопредельных регионах оби-
тает только номинативный подвид:
1. Platalea leucorodia leucorodia
Platalea leucorodia Linnaeus, 1758, Syst. Nat., ed.10, с.139, Европа, южная Швеция.
От Р /. archeri отличается несколько более крупными размерами: длина крыла у Р. I.
leucorodia — 360-410 мм против 325-360 мм у Р. I. archeri, длина клюва — 173-228 против
145-170 мм, цевки — 120-168 против 102-118 мм (Cramp, 1977; Степанян, 2003). К номи-
нативной форме большинство систематиков относят и популяции населяющие простран-
ство от Центр. Азии до Кореи, включая Египет и Индию, иногда выделявшиеся в подвид Р
I. major Temminck et Schlegel, 1849 (4).
Замечания по систематике. Крупная популяция (до 1 000-2 000 пар), найденная на
труднодоступных островах у атлантического побережья Мавритании, сравнительно недав-
но выделена в отдельный подвид Р I. balsaci de Naurois et Roux, 1974, отличающийся ок-
раской (Brouwer, 1964; Cramp, 1977; del Hoyo et al., 1992; Snow, Perrins, 1998). Эта форма
по размерам занимает промежуточное положение между Р. I. leucorodia и Р I. archeri, но
клюв у нее полностью чёрный, а жёлтое пятно на груди практически отсутствует (Cramp,
1977).
Следует также отметить, что у молодых колпиц из восточных частей ареала (Средняя
Азия, Монголия, Джунгария) описан особый морфологический признак, не встречающий-
ся у птиц из Европейской части России, Украины и Кавказа (Тугаринов, 1947). Их голое
пятно кожи на горле не вдаётся, как у взрослых птиц, клином вниз вдоль шеи, а имеет W-
образную форму, такую же, как и у взрослых особей аллопатричной малой колпицы (Platalea
minor), обитающей в Корее. Возможно, эта характерная особенность, помимо жёлтой, а не
черной вершины подклювья и чуть больших размеров птиц (длина цевки — 130-165 мм,
длина клюва — до 235 мм против 120-145 и 183-223 мм у западных популяций), отличает
азиатскую расу колпицы Р I. major Temminck et Schlegel, 1849, которую выделяли некото-
рые орнитологи (Бутурлин, 1901; Бутурлин, Дементьев, 1935; Rose, Scott, 1994 и др.).
Колпицу Р. leucorodia, вместе с восточноазиатской малой колпицей (Р. minor Temminck
et Schlegel, 1849), африканской белой колпицей (Р. alba Scopoli, 1786) и австралийской
королевской колпицей (Р. regia Gould, 1838), обычно объединяют в составе одного надви-
дового комплекса (del Hoyo et al., 1992) или отдельного подрода Platalea (Wolters, 1982).
Последняя форма (regia) иногда рассматривается также в качестве подвида Р leucorodia
(del Hoyo et al., 1992).
Описание. Окраска. Самец в брачном наряде имеет общую белую окраску оперения.
Спереди на нижней части шеи располагается большое желтовато-охристое пятно. Жёлто-
ватый оттенок имеют также удлинённые перья хохла. Голая кожа вокруг глаз и в основании
клюва жёлтая, на уздечке — с чёрным пятном. Большое эластичное кожистое пятно на
горле красновато-жёлтого цвета. Участок голой жёлтой кожи находится также на зобе. Клюв
в основании окрашен в чёрный цвет, а у вершины постепенно желтеет. Величина и конфи-
472
Отряд Листообразные, Семейство Ибисовые
гурация дистального жёлтого пятна имеет индивидуальную изменчивость, позволяющую
распознавать большинство особей (Петрович, 2000). Ноги чёрные. Радужина красная. Самка
слегка отличается от самца чуть меньшей величиной и менее развитым хохлом на голове.
Зимний наряд, одеваемый колпицами после осенней линьки, сходен у обоих полов и
характеризуется отсутствием хохла на голове и охристого пятна на зобе.
Птенцы вылупляются покрытыми коротким густым эмбриональным пухом белого или
розоватого цвета, сквозь который просвечивает ярко-жёлтая кожа (Тугаринов, 1947). На
лицевых частях и брюхе пуховой покров разрежен. Второй ювенильный пуховой наряд,
появляющийся примерно на 10 день, белый с охристым оттенком. Клюв и ноги у пуховиков
розовато-жёлтые (Спангенберг, Фейгин, 1936).
Гнездовой наряд сходен с зимним нарядом взрослых птиц, но первостепенные маховые
с дымчатым оттенком и темными стержнями, а на вершинах маховых расположено черное
пятно, величина которого сокращается по направлению от первого пера к последующим.
Эти пятна на перьях сохраняются и в первом брачном наряде. Клюв молодых птиц гладкий,
голубовато-серого или розовато-серого цвета, ноги тёмные, почти чёрные, радужина буро-
вато-жёлтая.
Строение и размеры. Крылья сравнительно короткие, так что внутренние
второстепенные маховые почти достигают вершины сложенного крыла. Первостепенных
маховых— 11. Формула крыла: III>II>IV>V>VI... или III>IV>II>V>VI..., I маховое укоро-
чено и скрыто под верхними кроющими крыла. На внутренних опахалах III—V и на наруж-
ных опахалах II—IV первостепенных маховых имеются вырезки. Хвост короткий, прямой,
из 12 рулевых перьев. На затылке длинные узкие перья образуют свисающий вниз хохол,
который может разворачиваться над головой веером во время брачных демонстраций. У
самца длина этих перьев — 120-150 мм, у самки — 105-135 мм. Пудретки отсутствуют.
«Чешуя» цевки и голени сетчатая. Голень не оперена примерно на половину длины.
Между передними пальцами ног — зачаточные перепонки. Коготь среднего пальца без
зазубрин. Длинный плоский, резко расширяющийся на конце клюв сверху слегка морщи-
нистый за счёт поперечных валиков. Ноздри узкие, вытянутые вдоль клюва, расположены в
канавках в его основании.
Развитие типичного для колпиц расширенного на конце клюва продолжается у птенцов
более 2 месяцев. У пуховиков он короткий, толстый, слегка приплюснутый и прямой, на
конце закруглённый и чуть загнут вниз. В это время он больше напоминает клюв уток.
Лишь в месячном возрасте он начинает удлиняться, а к 1,5 месяцам, к вылету птенцов из
гнезд, расширяется на конце. Но только через год он приобретает размеры и форму, харак-
терные для взрослых птиц (Тугаринов, 1947). Расширенная часть клюва у колпиц богата
чувствительными нервными окончаниями, помогающими птицам при кормёжке обнару-
живать добычу в мутной воде.
Из-за недостатка коллекционного музейного материала морфометрия колпицы изучена
слабо. Размеры птиц (мм) по разным сводкам без разделения их по полу в целом для Вост.
Европы и Сев. Азии таковы. Длина крыла — 368-381, хвоста -127, клюва — 165-183,
цевки — 127-140 (Мензбир, 1918); длина крыла — 350^105, клюва — 180-228, цевки —
120-165 (Бутурлин, Дементьев, 1935); крыла — 355-400, клюва— 180-222 (Тугаринов,
1947; 1951). Для Казахстана И.А. Долгушин (1960) приводит следующие данные: длина
крыла — 360-397, клюва — 183-223, цевки — 120-145. Для Украины (Смогоржевський,
1979): длина крыла — 342^105, хвоста — 112-129, клюва — 173-226, цевки — 120-190
мм. Последние цифры вызывают сомнения, поскольку они, по-видимому, ошибочны (ско-
рее всего, типографская опечатка: спутаны 6 и 9), но эти данные «перекочевали» в разные
отечественные сводки (Воинственский, 1962; Смогоржевський, 1979) ещё из работы А.Я.
Колпица
473
Таблица 39. Морфометрические показатели колпиц (мм) из разных регионов.
Параметр п Самцы lim М п Самки Источник
lim М
Восточная Европа и Северная Азия
Общая длина 3 777-830 802,3 Спангенберг, 1951
Размах крыльев 3 1258-1450 1369,3
Армения
Общая длина 3 750-810 1 764 Adamian, Kleim,
Длина крыла 3 370-388 2 361-369 1999
Длина хвоста 3 130-141 2 120-121
Длина клюва 3 207-212 2 178-187
Длина цевки 3 155-158 2 137-140
Болгар и я
Длина крыла 3 378—400 2 365-370 Симеонов и др.,
Длина хвоста 3 132-134 2 138-150 1992
Длина клюва 3 197-215 2 176-186
Длина цевки 3 159-168 2 143-150
Зап адная Пал еарктика
Длина крыла 386—412 360-377 Snow, Perrins, 1998
Нидерланды
Длина крыла 13 386—412 394 10 360-370 370 Cramp, 1977
Длина хвоста 25 108-126 117 24 108-118 ИЗ
Длина клюва 15 195-231 213 14 168-191 172
Длина цевки 24 140-163 149 19 123-141 131
Тугаринова (1947). Причём последний автор, в следующей по хронологии своей статье,
указывает максимальный размер плюсны колпицы уже 160 мм. Л.С. Степанян (2003), по-
дытоживший все данные с 1940-х гг. до начала XXI в., пишет, что длина крыла колпиц
составляет 360^410, клюва — 180-223 и цевки — 120-160 мм.
Масса птиц, по С.А. Бутурлину (1935) — 1 700 г; по С. Крэмпу (Cramp, 1977) — самца
1960, самки 1 130 г (в обоих случаях взвешено по 1 птице). Средний вес птиц из дельты
Волги составляет 1 950 г, колеблясь в пределах ±4,1% у самцов и ±7,2% у самок (Скокова,
19596, 1962). В таблице 39 приведены размеры птиц различных половых групп по данным
различных авторов. Судя по этим фрагментарным сведениям, самцы имеют заметно более
длинное крыло, клюв и цевку.
По Дель Хойо с соавторами (del. Hoyo et al., 1992) длина колпиц составляет 700-900
мм, размах крыльев — 1 150-1 350 мм, вес — 1 130-1 960 г.
Линька. Колпица имеет одну полную послебрачную и частичную предбрачную линьку.
Последняя протекает в январе-марте и захватывает мелкое оперение головы и шеи, в ре-
зультате чего у птиц развивается хохол и охристое пятно на шее. Полная послебрачная
линька идёт очень медленно и растягивается с августа до марта. Во время осенней мигра-
ции линька, вероятно, приостанавливается и заканчивается на зимовках (Cramp, 1977).
Последовательность смены нарядов следующая: первый (эмбриональный) пуховой наряд -
второй (ювенильный) пуховой наряд - гнездовой наряд - первый брачный наряд - первый
послебрачный наряд - второй брачный наряд и т.д. Взрослый наряд птицы приобретают,
по-видимому, на втором году жизни, после первой послебрачной линьки и смены маховых
с чёрными вершинами (Спангенберг, 1951). Детали линьки изучены слабо.
Распространение. Гнездовой ареал. Ареалогически колпица — в основном
пустынно-степной палеарктический вид, краями ареала заходящий на север тропическо-
474
Отряд Листообразные, Семейство Ибисовые
Рис. 88. Область распространения колпицы: а — примерная северная граница ареала в XVIII-XIX вв.,
б — современный гнездовой ареал и известные места гнездовий, в — основные места зимовок. Под-
виды: 1 — Р. I. leucorodia, 2 — Pl. archeri, 3 — Р I. balsaci, 4 — P. I. major.
го пояса и в умеренные широты Зап. Европы. Гнездится от Сев. Африки (берега Маври-
тании, Марокко, Алжира) и Атлантического побережья Европы, где спорадичные коло-
нии встречаются в разных странах к северу эпизодически до Англии, Германии и Дании
(сотни пар — в Испании и Нидерландах, десятки пар — в Португалии и Франции), к
востоку до Индостана и Кореи (Спангенберг, 1951; Cramp, 1977; del Hoyo et al., 1992;
Snow, Perrins, 1998) (рис. 88).
В Вост. Европе и Сев. Азии северная граница ареала проходит в настоящее время через
дельту Дуная, низовья Днестра, Дона, Волги и Урала. В Сев.-Зап. Причерноморье периоди-
чески гнездится также в верховьях Тилигульского лимана, до середины XIX в. в неболь-
шом числе обитала в дельте Днепра (Браунер, 1894; Мензбир, 1918). Единичный случай
гнездования отмечен в 1980 г. в Латвии (Виксне, 1983), куда проникают, вероятно, птицы
западноевропейской популяции. По Волге вверх поднимается до г. Волгограда (Чернобай,
2004). В 1984 г. гнездование колпицы впервые отмечено в Новоузенском р-не на юго-вос-
токе Саратовской обл. (Хрустов и др., 1995).
По северу Казахстана эпизодически гнездится на степных озёрах в окрестностях Урды,
а также в низовьях Узеней, Балыкты и Кушума в Волго-Уральском междуречье, на оз. Чел-
кар и в низовьях р. Уил в Зауралье (Волчанецкий, 1937; Шевченко и др., 1993; Дебело,
1996,2002), на озёрах к югу от р. Илек, в Наурзуме, в низовьях Иргиза и Тургая, по р. Нура
у Кургальджина, а также на водоёмах Алакольской и Зайсанской котловин (Зарудный, 1888;
Сушкин, 1908; Шнитников, 1949; Долгушин, 1960). Кроме того, в 1999-2001 гг. колпиц
неоднократно отмечали на востоке Оренбургской обл. на озёрах Светлинского р-на, а в
Колпица
475
2002 г. там были встречены 2 выводка (Коршиков, Корнев, 1999; Коршиков, 2001; Корти-
ков, Шубин, 2002).
По югу Сибири колпица, возможно, ещё сохранилась на степных озёрах Тувы и в Дау-
рии (Stegmann, 1929; Янушевич, 1952; Баранов, 1991; Забелин, 2002; Горошко, Кирилюк,
2003), заходя сюда краем ареала из Монголии, где она гнездится от оз. Убсу-нур до оз.
Орог-нур (Фомин, Болд, 1991; Мельников, 2000а). Восточнее эпизодически встречается по
долине Амура: в низовьях р. Зея, у М. Хингана, в районе устья р. Сунгари и ниже устья р.
Уссури вплоть до оз. Болонь (Stegmann, 1930; Шульпин, 1936; Росляков, 19816; Панькин,
1984; Нечаев, 1989; Бабенко, 2000). В XIX в. была многочисленна на оз. Ханка в Приморье
(Пржевальский, 1870), до 10 пар гнездилось там ещё в 1970-е гг. (Глущенко, 1981; Нечаев,
1989).
К югу от очерченной границы колпица гнездится в сев. Крыму — на Лебяжьих о-вах, в
Каркинитском заливе и на Сиваше (Сиохин, 2000). Обычна в Вост. Приазовье, по долине
Маныча, в дельтах Волги и Терека (Бондарев, 1979; Казаков и др., 2004; Гаврилов, 2005;
Сыроечковский, 2005), а также на равнинных водоёмах Азербайджана (Виноградов, 1967;
Коновалова, 1979; Султанов, 2000; Patrikeev, 2004 и др.). В первой половине XX в. размно-
жалась на оз. Севан и в долине р. Араке в Армении (Ляйстер, Соснин, 1942; Даль, 1954), но
позже гнездование там не отмечали, хотя в 1980-1990-е гг. летом регулярно регистрирова-
ли одиночек и группы птиц (Adamian, Klem, 1999).
В Средней Азии гнездится местами на озёрах в дельте Аму-Дарьи, по Сыр-Дарье, в
долинах Чу, Или, Каратала и других рек в Юж. Прибалхашье (Долгушин, 1960; Сагитов,
1987). В Узбекистане сейчас заселила также водохранилища, фильтрационные и сбросные
озёра вдоль Каршинского и Аму-Бухарского каналов и в других местах (Шерназаров, 2003).
В Туркмении прежде обитала по долинам Мургаба, Теджена, Атрека (Дементьев, 1952), в
последнее же время эпизодически гнездится, по-видимому, лишь на оз. Делили в низовьях
р. Атрек (Венгеров, 1975; Караваев, 1991). В 1984 г. первые гнездовья появились на искус-
ственном сбросном оз. Сарыкамыш на севере Каракумов (Чернов, 1990; Эминов, 1999). В
Таджикистане в начале XX в., по наблюдениям Н.А.Зарудного, возможно размножалась в
тугаях по р. Кафирниган, Вахш и Пяндж, затем птицы исчезли оттуда, но с 1987-1990 гг.
они вновь регулярно гнездятся в Тигровой балке в низовьях Вахша (Иванов, 1969; Абдуса-
лямов, 1971). В Киргизии колпица изредка встречается на пролёте в верхней части Чуйской
долины и в котловине оз. Иссык-Куль (Шнитников, 1949; Янушевич и др., 1959; Умрихина,
1970; Федянина, 1990) (рис. 89).
В прошлом граница ареала проходила значительно севернее. Так, до XVII в. колпица
регулярно гнездилась в Великобритании (Мензбир, 1918; Cramp, 1977; Snow, Perrins, 1998),
в конце XIX в. отмечалось её гнездование в Украинском Закарпатье (Страутман, 1963; Гра-
бар, 1997), тогда же нередко она встречалась на Пинских болотах в верховьях р. Припять в
Белоруссии (Шнитников, 1913; Мензбир, 1918). О гнездовании колпицы в Челябинском и
Шадринском уездах Зауралья сообщал Л.П.Сабанеев (1874). В первой половине XX в. кол-
пиц отмечали на гнездовье на оз. Кушмурун, а также на оз. Тениз в низовьях р. Убаган на
самом севере Кустанайской обл. (Деревягин, 1930, 1934). Вероятно на эти же находки ссы-
лался Е.П. Спангенберг (1951), приводя колпицу для района Кургана на юге Зап. Сибири. В
начале XX в. она возможно гнездилась в устье р. Абакан в Минусинских степях Хакасии
(Сушкин, 1914), а П.С. Паллас (Pallas, 1811, цит. по: Мензбир, 1918) включал в ареал кол-
пицы также Барабинские озёра и р. Селенгу в Забайкалье, где позже эту птицу на гнездовье
никто не регистрировал. В XIX в. Пржевальский (1870) отмечал большие колонии близ оз.
Ханка, но к концу XX в. колпица там, по-видимому, почти полностью исчезла (Шульпин,
1936; Воробьёв, 1954; Панов, 1965; Глущенко, 1979, 1981; Нечаев, 1989; и др.).
476
Отряд Листообразные, Семейство Ибисовые
Рис. 89. Ареал колпицы в Вост. Европе и Сев. Азии: а — современная область гнездования, б —
области гнездования, исчезнувшие во второй половине XX века, в — места зимовок.
Периодическая перекочёвка колоний с места на место или полное исчезновение колпиц
из отдельных районов вызываются обычно изменениями в их гнездовых или кормовых
местообитаниях; эти изменения связаны с пересыханием степных озёр или затоплением
островов вследствие синоптических флуктуаций, с выгоранием тростниковых крепей (Суш-
кин, 1908) или их естественным старением (Казаков, Ломадзе, 1990; Гринченко, 2004), а
также с антропогенной трансформацией ландшафтов. Но при восстановлении благоприят-
ных условий колпицы могут вновь возвращаться на прежние места гнездования (Шевченко
и др., 1993; Дебело, 2002; и др.).
Сокращение ареала и численности, наблюдавшееся до 1980-х гг. во многих странах
Европы, включая Украину (Tucker, Heath, 1994; Bird Life International..., 2000), сейчас сме-
нилось там, судя по последним данным, ростом численности и расселением птиц (Сиохин,
2000; BirdLife International, 2004). Колпица вновь загнездилась, например, в Великобрита-
нии (BirdLife International, 2004), появилась в нескольких колониях в Чехии и Словакии
(Hagemeijer, Blair, 1997; Snow, Perrins, 1998), где до этого лишь в 1949 г. было отмечено 1
гнездо (Hudec, Сету, 1972). Возможно, с расселением колпиц на север, связано недавнее
появление их гнездовий в Заволжье и Зауралье (Хрустов и др., 1995; Коршиков, Шубин,
2002). Расселение было обусловлено, вероятно, как охраной птиц, так и потеплением и
увлажнением климата, улучшившим кормовую базу и позволившим продлить гнездовой
период и повысить репродуктивный успех птиц за счет повторных попыток размножения.
Зимовки. Западноевропейские популяции зимуют почти исключительно на побере-
жье Мавритании, мигрируя туда, в основном, вдоль берегов Атлантики, редко удаляясь от
них более чем на 100 км в глубь суши (Brouwer, 1964). Центральноевропейские популяции
из бассейна Дуная зимуют, судя по материалам кольцевания (Харашти, 1978; Muller, 1984),
в основном, на северном побережье Африки, а также на юге Италии и Греции, изредка
залетая в Экваториальную Африку (Нигерия, Судан). По всей видимости, к этой же популя-
ции относятся и птицы из юго-запада Украины. Судя по отсутствию выраженных миграций
Колпица
477
на северном берегу Азовского моря и в низовьях Дона, а также на Черноморском побере-
жье Кавказа (Тильба, 1999; ориг.), колпицы с Сиваша, вероятно, тоже улетают через дельту
Дуная на запад, а затем на юг. Об этом же свидетельствует западное направление сентябрь-
ского пролёта птиц в районе г. Джанкой в Крыму (Гринченко, 1991).
Птицы из Юж. России (Приазовье, Маныч) зимуют, по данным мечения, в основном на
юге Пакистана и Ирана, улетая с мест гнездования сначала на юго-восток, а затем вдоль
берегов Каспия — на юг. Прикаспийские птицы (из дельты Волги, а также — из Дагестана
и Азербайджана) мигрируют вдоль западного берега Каспийского моря, а затем через Иран
в Индию (Сапетин, 1978г). Птицы из Сев. Казахстана летят, вероятно, вдоль восточного
берега Каспия (Исаков, Воробьёв, 1940; Караваев, 1991), а также по долинам крупных рек
(Дементьев, 1952; Пославский, Ширеков, 1990), тоже направляясь, по-видимому, в Индию.
Наконец, птицы из Вост. Сибири и Монголии зимуют в восточном Китае (Спангенберг,
1951). До 1870 г. они регулярно зимовали в Японии, но сейчас встречаются там редко,
вероятно в связи с сокращением численности восточных популяций (Панов, 1973; del Hoyo
et al., 1992).
Колпицы изредка встречаются зимой также и на территории Сев. Евразии. Так, их отме-
чали в Ленкорани (Loudon, 1909), хотя позже зимовки этих птиц в Вост. Закавказье отрица-
лись (Спангенберг, 1951; Смогоржевський, 1979). Однако на оз. Акгёль 8 из 11 зимовавших
птиц благополучно выжили даже в суровую зиму 1963/64 г. (Виноградов, 1967). Кроме
того, однажды, по данным мечения, взрослая колпица была добыта 1 декабря в Азербайд-
жане близ места кольцевания (Сапетин, 1978г). В декабре и январе (1988-1992 гг.) оди-
ночки и группы птиц до 3-6 особей несколько раз регистрировались также в Армении в
долине р. Араке (Adamian, Klem, 1999). А зимой 2005/06 г., по сведениям В.Н. Мосейки-
на, группа из 10 колпиц благополучно перезимовала на Новотроицком вдхр. близ г. Став-
рополя.
В середине декабря 1967 г. одиночных колпиц наблюдали в верховьях Сыр-Дарьи на
севере Таджикистана — на островах Кайраккумского вдхр. (Абдусалямов, 1971). Одиноч-
ная птица была отмечена 28 декабря в низовьях р. Вахш (Потапов, 1958). Имеются сведе-
ния о нерегулярных зимовках колпиц в Узбекистане, где их отмечали в декабре и феврале
(Сагитов, 1987), а в долине р. Сурхан-Дарья в первой декаде февраля наблюдали будто бы
их «отлёт с зимовок» (Остапенко, 1974, цит. по: Сагитов, 1987). В последние десятилетия,
с появлением лиманов в низовьях р. Атрек, колпицы изредка стали оставаться на зиму так-
же в Туркмении. Например, у пос. Гасан-Кули 9 птиц были учтены в январе 1987 г., 55
особей — в декабре 1988 г. и 24 колпицы — в январе 1989 г. (Караваев, 1991).
Миграции. Сроки появления первых птиц весной определяются, в основном, погодны-
ми условиями в местах гнездования. В отдельных регионах Сев. Евразии прилёт регистри-
руется с середины февраля до начала мая. В целом период миграции весьма продолжите-
лен, поскольку отдельные группы птиц — вероятно, разного возраста (Brouwer, 1964) —
прилетают в колонии асинхронно, и их возвращение с зимовок растягивается иногда на 2-
3 месяца. Так, в долине Зап. Маныча на оз. Казинка колония колпиц формировалась в 1977
г. с конца марта до июня, а в 1979 г. на Веселовском вдхр. — с середины марта до июня
(Казаков и др., 2004). На Сивашах подселение колпиц в колонии продолжается с конца
марта до начала мая (Гринченко, 2004а).
В дельту Дуная колпицы прилетают в феврале-марте. В 1953 г. первая птица отмечена
там 17 февраля, но основная масса появилась к 27 марта (Смогоржевський, 1979), в 1976—
1978 гг. прилёт отмечали, соответственно, 18, 6 и 9 марта (Петрович, 2000), а в 1981—
1983 гг. — 26, 28 и 30 марта (Панченко, Балацкий, 1991). В Румынии прилёт колпиц реги-
стрировали с 20 марта, но обычно он происходит 10-15 апреля (Vespremeanu, 1968).
478
Отряд Листообразные, Семейство Ибисовые
На Лебяжьих о-вах в сев.-зап. Крыму прилёт первых птиц отмечался с начала февраля
до начала мая, причём в последние годы колпицы появлялись там всё раньше (Костин,
Тарина, 2004). По данным этих же авторов (Костин, Тарина, 2002,2005), средняя дата при-
лёта в 1989-2003 гг. (п=14) приходилась на 21 марта (3 февраля 1991 — 3 мая 1992). Основ-
ной же пролёт на северо-западе Крыма идёт в конце марта - начале апреля, а последние
пролётные стаи встречаются до мая (11 мая 1992 г., 26 мая 1993 г.).
В Азербайджане под Ленкоранью первых птиц отмечали уже 19 и 27 февраля (Radde,
1884, цит. по: Мензбир, 1895), а выраженный пролёт идёт там с начала апреля (Исаков,
Воробьев, 1940). В Кызыл-Агачском заповеднике появление первых птиц регистрировали
31 марта 1958 г. и 14 апреля 1959 г., а на оз. Акгёль — 27 марта 1962 г. и 19 марта 1963 г.
(Виноградов, Чернявская, 1965; Виноградов, 1967). В устье р. Самур на юге Дагестана
весенний пролёт, направленный на северо-запад вдоль берега моря, в 1980 г. начался в
конце марта и продолжался до 5-й пятидневки апреля (до окончания наблюдений); наибо-
лее интенсивно он шел во 2-й пятидневке апреля. Однако в целом численность мигрантов
была невелика — 372 особи за 3 года наблюдений (Михеев, 1991). По данным В.Т. Бутьева
с соавторами (1989), пролёт там начинается во второй декаде марта, в конце марта - начале
апреля наблюдается первый пик миграции, затем в конце апреля — второй; активный же
пролёт продолжается до 9 мая.
В Предкавказье (Кубань, Маныч, дельта Терека) колпицы появляются обычно вскоре
после того, как водоёмы освобождаются ото льда: в 3-й, реже — во 2-й декадах марта,
иногда — в конце февраля, а в годы с холодной весной — в апреле (Олейников и др., 1975;
Хохлов, 1990в). По данным Б.А. Казакова с соавторами (2004), в течение 1960-1980-х гг. в
Восточном Приазовье и на Зап. Маныче прилёт колпиц дважды был отмечен во 2-й декаде
марта (1979,1984 гг.), в 3-й декаде наблюдался 12 раз, а в 1-й декаде апреля — 2 раза (1980,
1982 гг.). Лишь однажды птицы появились в начале 3-й декады апреля (1987 г. — Веселов-
ское вдхр.). Массовый же прилёт колпиц в Предкавказье регистрируется обычно в первой
декаде апреля.
В дельту Волги колпицы прилетают обычно в начале апреля, но в годы с мягкой зимой
они могут появляться и раньше. В 1938-1959 гг. даты их прилёта колебались между 15
марта (1955 г.) и 15 апреля (1959 г.), в среднем (п=14) — 4 апреля. Массовый же прилёт
наблюдался примерно через 2 недели после появления первых птиц, т.е. в середине апреля
(Луговой, 1963).
В Туркмении в прошлом колпицы мигрировали с середины марта до мая (Дементьев,
1952), причём в устьях р. Атрек они держались в большом количестве вплоть до середины
мая (Radde, Walter, 1889). Но к концу XX в. их прилёт стал отмечаться там значительно
раньше — с конца февраля - начала марта. Заметное увеличение числа птиц наблюдали во
2-й половине марта, а пик пролёта (до 100-1 100 особей, или, в среднем, 361 птица за 1
учётный день) приходился на апрель. Небольшие группы задерживаются на Атреке до се-
редины мая, а иногда их встречали там в июне и даже в июле (Караваев, 1991).
В Узбекистане мигрирующих колпиц изредка регистрируют с марта (Сагитов, 1987;
Лукашевич, 1995). На юге Казахстана первые птицы появляются иногда уже в начале мар-
та, но чаще — в конце этого месяца; прилет наблюдается в период перехода средних темпе-
ратур воздуха от 5 до 10 °C (Сема, 1989). В низовья Сыр-Дарьи колпицы прилетают между
15 и 25 марта, массовый прилёт наблюдается во 2-й декаде апреля, заканчиваясь к концу
апреля. В низовьях Тургая передовых птиц встречали 27-28 апреля, на оз. Сорбулак близ
Алма-Аты — 4 апреля (Сема, 1989). В Алакольской котловине и на оз. Зайсан колпица
появляется в конце марта - начале апреля. На соседнем оз. Маркаколь в горах Юж. Алтая
залетных птиц регистрировали 3 апреля 1983 г. (Березовиков, 1989). В Центр. Казахстане
Колпица 479
на Кургальджинских озёрах наиболее ранняя встреча зафиксирована 21 апреля 1971 г. (Кри-
вицкий и др., 1985). В устье р. Урал колпицы появлялись в конце марта, а на Мангышлаке
последних мигрантов видели вплоть до середины мая (Смирнов, 1916; Спангенберг, 1951;
Долгушин, 1960). В Приморье, на оз. Ханка, прилет колпиц Н.М. Пржевальский в 1868-
1869 гг. отмечал 3-14 апреля. В долине р. Раздольная у г. Уссурийска наиболее ранние
встречи в разные годы отмечались с 23 марта по 1 апреля, а наиболее поздние — 18 и 29
апреля (Глущенко, Липатова, Мартыненко, 2006).
В целом по фенологическим данным хорошо прослеживается поступательное направ-
ление весеннего пролёта на восток вдоль северо-западного побережья Чёрного моря и на
север — вдоль берегов Каспия. На Маныч и Кубань колпицы прилетают, вероятно, с восто-
ка, но фактические наблюдения над континентальным перелётом птиц через Предкавказье
весьма ограничены (Бичерев, Хохлов, 1990). Собранный материал чётко демонстрирует
также сдвиг сроков весеннего прилёта в последние десятилетия на более ранний период,
что связано, вероятно, с потеплением климата.
Последние весеннепролётные колпицы составляют, по-видимому, волну дисперсантов,
состоящую из молодых птиц, которые в ходе миграций часто залетают за пределы гнездо-
вого ареала. В Чехословакии, например, наибольшее количество залётных колпиц (около
50%) отмечалось именно в мае-июне (Hudec, Сету, 1972). В ходе этой дисперсии птицы,
очевидно, импринтингуют районы, подходящие для гнездования, чтобы в последующем
возвратиться туда на размножение. Так, в дельте Днестра, где колпицы размножались до
1952 г., их майские залёты начали регулярно регистрировать в 1970-е гг. (5 особей — в 1972
и 1973 гг., 10 — в 1974 г., 2 — в 1975 г.), в 1976 г., после 20-летнего перерыва, было найдено
их первое гнездо с кладкой, а с 1983 г. птицы стали гнездиться там постоянно (Смогор-
жевський, 1979; Русев, 2004).
Летом, после вылета птенцов, основная масса колпиц долгое время — с середины июля
до августа-сентября — кочует в районах своих гнездовий. В сентябре часть молодняка
разлетается, улетая иногда за 300-700 км на северо-запад - север - северо-восток от коло-
ний. В это же время нередко отмечаются дальние залеты и у взрослых колпиц (Сапетин,
1978г). В Западной Европе залётные птицы регистрировались к северу до Исландии и Нор-
вегии (Snow, Perrins, 1998), в Европейской России — до Архангельской обл. (Спангенберг,
1951) и Татарстана (Рахимов, 1998), а в Сибири — до широты Екатеринбурга (Сабанеев,
1874; Тарасов, 2004), Омской и Томской обл. (Иоганзен, 1914; Гынгазов, Миловидов, 1977;
Якименко, 1995; Рябицев и др., 2001 и др.), до среднего течения Енисея (Архипов, 2002;
Бурский и др., 2003), Среднего Байкала (Мельников, 2000а) и о. Итуруп Курильской гряды
(Bergman, 1935, цит. по: Нечаев, 1969).
Отлёт колпиц на зимовку из Зап. Причерноморья происходит в сентябре, последние
птицы отмечались там 10 сентября 1952 г. и 24 сентября 1953 г. (Смогоржевський, 1979)
или в начале октября (Петрович, 2000). В Румынии они исчезают 15-25 октября (Vespremeanu,
1968). В Крыму запоздавших птиц наблюдали 18 ноября 1965 г. и 17 ноября 1978 г. (Костин,
1983; Костин, Тарина, 2005). Обычно же их отлёт заканчивается там к концу сентября - нача-
лу октября, а последних колпиц регистрировали с 11 сентября (1995 г.) по 4 ноября (1994 г.),
в среднем за 1994-2002 гг. (п=9) — 4 октября (Костин, Тарина, 2002, 2005).
В дельте Кубани в сентябре колпицы ещё обычны, встречаясь стаями до 100-150 осо-
бей. К концу этого месяца их численность заметно сокращается, а в начале октября они
улетают (Винокуров, 1960). На Зап. Маныче их регулярно наблюдают в сентябре, после-
дние встречи отмечали 20 октября 1989 г. и 12 октября 1987 г. (Казаков и др., 2004).
Из дельты Волги, по данным А.Е. Лугового (1963), отлёт идёт в сентябре, а к началу
октября, в основном, заканчивается. Но отдельные группы и особи встречаются вплоть до
480
Отряд Листообразные, Семейство Ибисовые
ноября: 21 ноября 1934 г. (17 птиц), 18 ноября 1957 г. (10), 8 ноября 1958 г. (4). В дельте
Терека птицы начинают выраженные миграции с конца августа, но иногда задерживаются
до 18-22 ноября (Казаков и др., 2004). В устье р. Самур пролёт птиц на юго-восток регис-
трируется с середины августа, пик миграций наблюдается в конце этого месяца (65% мигран-
тов), а затем в начале октября (32%), последние же колпицы отмечены 21 ноября 1981 г.
(Бутьев и др., 1989; Михеев, 1991). В Вост. Закавказье у Хармандали появление первых
пролётных стай прослежено в конце августа (Спангенберг, 1951), последних птиц наблю-
дали 8 ноября 1957 г., 9 октября 1958 г. и 23 октября 1959 г. — в Кызыл-Агаче и 22 ноября —
на оз. Акгёль (Виноградов, Чернявская, 1965; Греков, 1965; Виноградов, 1967). В Индии
большие скопления колпиц появляются в октябре (Cramp, 1977).
В Казахстане отлёт начинается в августе и приобретает массовый характер в сентябре.
В нижнем течении Урала миграцию наблюдали с 9 по 28 сентября (Сема, 1989). Устье р.
Урал птицы покидают в сентябре-октябре (Карелин, 1875; Бостанжогло, 1911; Березови-
ков, Гисцов, 2001), но отдельные особи задерживаются до 23 ноября (Спангенберг, 1951).
На Сыр-Дарье слабый пролёт идёт между 2 сентября и 17 октября (Сема, 1989), а на юге
Казахстана последние птицы исчезают в конце октября - начале ноября (Долгушин, 1960).
В низовьях р.Сарысу последних птиц встречали 17 сентября, на р. Тургай — между 17
сентября и 12 октября (Сема, 1989), в дельте р. Или — 10 октября (Мищенко, 1991). В
Центр. Казахстане на Кургальджинских озёрах наиболее поздние встречи зафиксированы 9
сентября 1977 г., 23 сентября 1975 г., 26 сентября 1969 г. (Кривицкий и др., 1985). В Ала-
кольской котловине последние встречи приходятся на 5 октября, на оз. Сорбулак близ Алма-
Аты— 8-14 октября (Сема, 1989; Хроков, 1991).
В Узбекистане колпицы задерживаются обычно до начала октября, редко — до середи-
ны этого месяца (Сагитов, 1987). В Туркмении в низовьях р. Атрек первые стаи появляют-
ся в конце августа, пик миграции приходится на сентябрь (до 150-180 особей за 1 учётный
день); во 2-й половине октября колпицы встречаются реже, а последние отмечаются до
конца 2-й декады ноября (Караваев, 1991). В некоторые годы птицы бывают там многочис-
ленны во 2-й половине октября (Исаков, Воробьев, 1940).
В общем, в последние десятилетия, в связи с более ранним прилетом и гнездованием,
прослеживается и несколько более ранний отлёт основной массы птиц на зимовки. Но за-
поздавшие особи (возможно, из повторных выводков) задерживаются на местах гнездовий
дольше прежнего. Чаще они стали оставаться и на зиму. Взрослые колпицы осенью улета-
ют обычно раньше молодых (Скокова, 19596). Их миграции наблюдаются преимуществен-
но во второй половине дня. Стаи летят обычно на высоте 40-200 м. Размеры их сравни-
тельно небольшие — 5-1CM0, редко до 60 особей (Бутьев и др., 1989; Михеев, 1991). Над
морскими акваториями птиц наблюдают очень редко (Кошелев и др., 1991).
Местообитания. Гнездовые и кормовые стации у колпиц, как правило, разобщены и
существенно различаются. В результате птицы вынуждены совершать регулярные кормо-
вые миграции, иногда на расстояние до 10-15-25 км (Спангенберг, 1951; Лукашевич, 1995;
Platteeuw et al., 2004).
Кормятся птицы обычно на илистых мелководьях глубиной до 35 см, доступных им по
длине ног, но чаще — на отмелях до 20-25 см, соответствующих длине клюва (Скокова,
19596). Предпочитают различные открытые, лишённые надводной растительности водо-
ёмы: лагуны, заливы и берега морей, озёр, прудов, равнинных рек. Очень охотно использу-
ют временные пересыхающие водоёмы, в которых концентрируются мальки рыб, головас-
тики, различные водные беспозвоночные. Нередко кормятся на рисовых полях до их зара-
стания. Во время весенне-летних паводков птицы часто посещают залитые луга с мелкой,
прогретой водой. В дельте Волги прослеживается сезонная смена кормовых стаций: вес-
Колпица
481
ной — в заливах на взморье, а летом, когда глубины в море из-за паводка повышаются,
колпицы перемещаются на луга. В связи с этим сенокосы и палы тростника на пойменных
островах могут значительно расширять и улучшать кормовые местообитания колпицы (Лу-
говой, 1963). В августе, с обсыханием полоев, колпицы вновь откочевывают на взморье
(Скокова, 19596). На зимовках они придерживаются аналогичных местообитаний (del Hoyo
et al., 1992).
Гнездится колпица в трёх различных типах биотопов. Основным из них являются, веро-
ятно, тростниковые массивы на водоёмах, обычно — плавневого типа, реже — небольшие
тростниковые острова, иногда прибрежные бордюрные заросли. Поселения колпицы в та-
ких условиях встречаются по всему её ареалу, а в безлесных дельтовых районах — в част-
ности, в Вост. Приазовье — они преобладают (Белик, Динкевич, 2004). Основные гнездо-
вья колпиц в Казахстане сейчас тоже приурочены к тростниковым зарослям на трудноступ-
ных водоёмах в дельтах рек (Урал, Или, Тентек, Чёрный Иртыш) или на замкнутых систе-
мах степных озёр (Кургальджино, Наурзум).
Второй тип биотопов — древесные формации. Это преимущественно ветловые (с Salix
alba\ местами туранговые (с Populus diversifolid) или джидовые (с Elaeagnus angustifolia)
леса по берегам речных проток и озер, в которых колпицы гнездятся в дельте Волги, на
Среднем Дунае, иногда — по Сыр-Дарье и др. В начале XIX в. гнездование на деревьях
отмечалось также для бассейна Дона (Кондратьев, 1885). В дельте же Волги (Бондарев,
1979), в 1973-1974 гг. насчитывалось 12 лесных колоний, в которых гнездилось 1 157 пар
колпицы (96% местной популяции). Промежуточное положение между лесными и трост-
никовыми колониями занимают кустарниковые поселения, размещающиеся обычно среди
болот на густых высоких кустах различных видов ивы (Salix cinerea, S. triandra) или тама-
риска (Tamarix sp.). Такие колонии изредка встречаются в разных районах (Азербайджан,
дельта Волги, Приморье и др.). Иногда колпицы гнездятся на одиночных кустах или дере-
вьях близ водоёмов — например, на саксауле в Сарыкамышской впадине (Чернов, 1990).
Третий тип гнездовых биотопов — небольшие открытые сухие острова, где колпицы
селятся среди зарослей лебеды, мари, крестовников и другого разнотравья. Подобные ко-
лонии наиболее характерны для островов оз. Маныч-Гудило, на которых колпицы начали
гнездиться в 1960-е гг. вскоре после создания Пролетарского вдхр. на р. Зап. Маныч. На-
блюдаются они также на Лебяжьих о-вах в Крыму. В большинстве случаев на такие острова
колпицы переселялись вынужденно из-за исчезновения тростниковых зарослей вследствие
засоления водоёмов или размыва и затопления прибрежных кос. Поэтому тростниковые
островки используются колпицами в этих случаях, как правило, до последней возможнос-
ти. На островах они придерживаются обычно экотонных сообществ между сухими злако-
выми плакорами и периодически заливаемыми понижениями, где обильно разрастается
грубое высокотравье, необходимое птицам для строительства гнёзд (Линьков, 1988).
Гнездятся колпицы обычно в смешанных поселениях голенастых и веслоногих птиц,
изредка также вместе с грачами в пойменных лесонасаждениях. Иногда они формируют
самостоятельные колонии. В смешанных поселениях колпицы образуют, как правило, не-
сколько обособленные довольно плотные агрегации гнезд, нередко соприкасающихся друг
с другом.
Численность. Общая численность мировой популяции составляет до 40-60 тыс. осо-
бей (Rose, Scott, 1994). В Европе, по экспертным оценкам и учётным данным, в 1980-е и
1990-е гг. гнездились, соответственно, 6-10 и 8,9-15,0 тыс. пар, в том числе в европейской
части России — 2 300-2 800 и 2 500-3 000, на Украине — 300-800 и 200-250 и в Молда-
вии — 10-20 и 0-20 пар. Кроме того, в начале XXI в. 800-2 500 пар обитало в Азербайджа-
не. Остальные птицы гнездились, в основном, в Румынии (750 и 1 100-1 500 пар), Венгрии
482
Отряд Листообразные, Семейство Ибисовые
(600-750 и 850-1 200), Нидерландах (400-530 и 1 000-1 270), Испании (675 и 1 000-2 500),
а также в Турции (500-3 000 и 600-1 200 пар) (BirdLife International..., 2000, 2004). В Ин-
дии, по учётам 1987-1989 гг., обитало 1 765-5 017 особей (Пракаш, 1990).
В бывш. СССР в 1973-1977 гг. насчитывалось 3,7, а в 1986-1987 гг. — 4,5 тыс. пар.
Отмеченное увеличение численности связывали с развитием в Палеарктике благоприят-
ной для лимнофилов прохладно-влажной климатической фазы, а также с улучшением ох-
раны птиц, отчасти — с большей полнотой учёта (Флинт и др., 1991). В России в 1980-е гг.
было учтено около 3 тыс. пар, на Украине — 500-750, в Казахстане — 550-580, в Азербай-
джане — 411, в Молдавии — 12 пар (Стоцкая, Кривенко, 1988). В 1990-е гг., по данным
республиканских Красных книг, в России (2000) гнездилось 1,6-2,4 тыс. пар, на Украине
(1994) — около 250 пар, в Казахстане (1996) — 400-650 пар, в Узбекистане (2003) — около
400 пар.
Всего на Украине в 1984-1985 гг. было учтено 230 и 750 пар, в основном в дельте Дуная
(Ардамацкая и др., 1988), а в 1998 г. — 213 пар, преимущественно в Северном Крыму
(Сиохин, 2000). Сейчас колпица гнездится на Украине в 5 районах (табл. 40): в дельтах
Дуная и Днестра, нерегулярно на Тилигульском лимане в Одесской обл., на Лебяжьих о-вах
в Крыму и на Сивашах (Смогоржевський, 1979; Костин, 1983; Сиохин, 2000; Костин, Тари-
на, 2002, 2004; Русев, 2004; и др.). В украинской части дельты Дуная она сохранилась сей-
час лишь в одной колонии на оз. Кугурлуй близ г. Измаил, где в 2002 г., по сообщению И.И.
Черничко, насчитывалось около 130 пар, хотя в 1978-1979 гг. на Дунае в 3-6 колониях
гнездилось 180-250 пар, в 1985-1986 гг. — 100-160 пар (Жмуд, 2000; Петрович, 1981,
2000; Потапов, 2001), а по данным других авторов — даже до 630-800 пар (Панченко,
Чёрный, 1986; Панченко, Балацкий, 1991; Кошелев и др., 1991). После 1990 г. большинство
этих поселений распалось, и в настоящее время в дельте Дуная основная часть гнездовий
сосредоточена в Румынии, где в 2001-2002 гг. было известно 7-9 колоний общим числом
218-339 пар (Platteeuw et al., 2004)17.
В дельте Днестра в 4 колониях сейчас гнездится 10-70 пар. Там они размножались до
1952 г., затем надолго исчезли, вновь появившись в 1976 г., и с 1983 г. гнездятся постоянно,
испытывая лишь депрессии численности после морозных зим. В верховьях Тилигульского
лимана в 1998 г. в 2 колониях голенастых обнаружены 24 пары этих птиц. На Лебяжьих о-
вах первые единичные пары загнездились в 1976 г., затем — в 1987-1988 гг., а с 1992 г. их
численность начала увеличиваться и к началу XXI в. достигла 102-116 пар. На Сивашах в
Крыму колпицы регулярно гнездятся с 1983 г. Там в Джанкойском заливе численность с 4-
5 пар в 1989 г. постепенно увеличилась до 100 пар в 1998 г. Кроме того, в 1990-е гг. неболь-
шие поселения эпизодически отмечались ещё в 6-7 колониях голенастых на Сивашах (Си-
охин, 2000).
В России основные гнездовья сосредоточены сейчас в долине Маныча — на многочис-
ленных островах оз. Маныч-Гудило, образовавшихся в 1950-е гг. в связи с созданием Про-
летарского вдхр. (табл. 41). Здесь в 1968-1969 гг. было учтено 208-349 пар, в 1970-е гг. —
240-360 пар, в 1980-е гг. — 400-925 пар, а в 1990-1991 гг. — 400-660 пар (Языкова, Каза-
ков, 1975; Кривенко, 1981; Казаков и др., 1988; Линьков, 1988; Казаков, Ломадзе, 1991,
1992; Казаков и др., 2004). В последние годы полные учёты птиц на Маныче не проводи-
лись, но судя по эпизодическому мониторингу отдельных колоний, общая численность кол-
пиц, колеблясь из-за их переселения с острова на остров, остаётся, по-видимому, относи-
тельно стабильной (Белик, 2004).
17 В это число ошибочно включена, по-видимому, и колония на украинском оз. Кугурлуй, которая
была обследована, судя по приведенным в монографии данным и сведениям И.И. Черничко (см.
выше), в 2002 г.
Колпица
Таблица 40. Динамика численности колпицы на водоёмах Украины в 1983-2004 гг., в парах (по: Панченко, Чёрный, 1986; Жмуд, 1999;
Сиохин, 2000; Потапов, 2001; Костин, Тарина, 2002, 2004; Гринченко, 2004; Русев, 2004; Platteeuw et al., 2004)
Годы: 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03 04
Дунай, дельта 230 630 160 2 20 10 359 0 0 0 0 0 5 0 0 0 0 0 130
Днестр, дельта Тилигул, лиман 4 0 15 0 18 0 2 6 8 8 12 10 9 10 14 25 10 30 24 65 55 40 70 12 3
Лебяжьи о-ва Зап. Сиваш Вост. Сиваш 0 1 0 0 0 0 4 1 1 0 9 0 0 2 2 18 3 16 5 16 24 0 19 17 52 72 41 5 118 81 82 102 109 20 116 40
Примечание. На Вост. Сиваше в указанные годы обследовались не все пригодные для гнездования районы
Таблица 41. Минимальная численность гнездящихся колпиц в долине Маныча в 1968-1991 гг., в парах (по: Падутов и др., 1975;
Языкова, Казаков, 1975; Кривенко, 1981; Казаков и др., 1988; Линьков, 1988; Казаков, Ломадзе, 1991, 1992). В 1971 и 1988-1989 гг.
учёты не проводились
Водоем / год 68 69 70 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 90 91
Веселовское вд. вдхр. 63 185 135 55 53 84 35 80 28 44 61 80 9
оз. Казинка 120 93 38 9 44 36 42 0 0 0 0 4 40
Запад Маныча 198 281 316 59 280
Центр Маныча 349 208 103 300 250 324 210 220 200 291 240 288 247 235 316 420 376 368 13 140 110
Восток Маныча 38 66 91 41 69 100 123 191 167 220 165 273 276 194 200 270
Лысый Лиман 97 49 117
Чограйское вд. вдхр. 5
оз. Соста-Дада 92 92 103 112 135
484
Отряд Листообразные, Семейство Ибисовые
Большие колонии известны местами также на других водохранилищах Манычской до-
лины, на озёрах и лиманах в прилежащих к Манычу степях. Так, на Усть-Манычском вдхр.
в 1940-1960-е гг. гнездилось 25-50 пар (Олейников и др., 1975), в низовьях Веселовского
вдхр. в 1960-е гг. обитало 250-300, а в 1977-1990 гг. — от 28 до 185 пар (Языкова, 19706;
Казаков, Ломадзе, 1991; Казаков и др., 2004). В 2001-2003 гг. 20-30 пар размножалось
также на Курниковом лимане в Ремонтненском р-не Ростовской обл. (Белик, 2004).
В Калмыкии, кроме оз. Маныч-Гудило, колпица гнездится на Чограйском вдхр. (р. Вос-
точный Маныч), на Сарпинских и Состинских озёрах и др. Общую численность колпицы
на этих водоёмах А.Б. Линьков (2000) оценивал в 400-550 пар, из которых 150-200 пар
размножается на оз. Деед-Хулсун (Свиридова, Зубакин, 2000; Музаев и др., 2004). В 6 посе-
лениях на внутренних водоёмах Ставропольского края (озёра Дадынские, Солёное, Хунд-
Хур, Бурукшунские лиманы, устье р. Калаус и др.) гнездится 120-170 пар (Хохлов, Биче-
рев, 1986а; Хохлов, 1990в; 1993; Хохлов и др., 2005).
В Восточном Приазовье на многочисленных лиманах в низовьях Кубани, Бейсуга, Чел-
баса, Ей и других рек в 1953-1954 гг. в 4 колониях было учтено 250-300 особей (Виноку-
ров, 1959а), в 1986-1989 гг. выявлено 7 колоний общей численностью 235 пар, а в 1990—
1992 гг. — 10 колоний с 323 парами (Емтыль и др., 1989,2003). Однако размещение гнездо-
вий здесь непостоянное, и контролировать их в труднодоступных плавнях весьма трудно.
Поэтому общая численность колпиц в Приазовье, несомненно, занижена. Так, в 1966-1970
гг. их популяция на Кубани резко снизилась (Очаповский, 1971), однако в эти же годы толь-
ко на Ахтарском лимане было обнаружено 200 пар (Борисов, 1977). Ещё одна неизвестная
колония из 150-200 пар выявлена при авиаучётах в 1999 г. на лимане близ с. Могукоровка
в Калининском р-не Краснодарского края (Белик, Динкевич, 2004).
На Нижнем Дону, у северной границы ареала, колпица с 1986 г. регулярно гнездится на
оз. Монастырское близ г. Аксай (до 3-5 пар), в 1980-е гг. несколько лет отмечалась в коло-
ниях голенастых птиц на прудах Новочерскасского рыбхоза (до 4-16 пар), в 1984 г. найдена
на озере в низовьях р. Сал (1 пара), а в 1991 г. — в верховьях этой реки (до 10 гнёзд) (Белик,
20036; Белик, Динкевич, 2004; Казаков и др., 2004). В июне 1975 г. 9 колпиц наблюдали у
цапельника на Цимлянских песках на северном берегу Цимлянского вдхр. (Чернобай, 2004).
В бассейне Волги самые северные колонии располагаются на оз. Сарпа (20-35 пар), на
Большом лимане близ г. Волжский (5-10 пар) и в верхней части Волго-Ахтубинской поймы
(до 10-20 пар) (Коленов, 1981; Чернобай, Букреев, Сохина, 2000; Чернобай, 2004). Всего в
Волго-Ахтубинском междуречье в 1970-е гг. гнездилось 200 пар, а в 1990-е гг. — 100-150
пар (Кривенко, 1983; Линьков, 2000), но в настоящее время колония в южной части Волго-
Ахтубинской поймы исчезла в связи с вырубкой леса (Русанов, 2004).
В дельте Волги после глубокой депрессии в начале XX в. численность колпиц к 1930-м
гг. увеличилась до 500 пар и к середине 1940-х гг. достигла 2,5 тыс. пар. В 1955 г. там
гнездилось 1,5 тыс. пар, в 1968 г. — 0,9 тыс. пар, в 1973-1974 гг. — 1,2 тыс. пар в 14
колониях, а в 1980 г. — 500 пар (Кривенко, 1975, 1983). Затем в течение 1980-х гг. числен-
ность птиц снизилась до 200-Л00 пар, гнездившихся в 8 колониях, а в 1995 г. в дельте Волги
было учтено лишь 55 пар (табл. 42). В последующие годы наблюдался рост численности до
200—400 пар, в том числе 50-100 пар гнездилось в 3-5 лесных колониях в островной дельте
и 150-200 пар — в одной колонии среди западных «подстепных» ильменей (Гаврилов, 2002).
Сейчас в дельте Волги известны всего 4 колонии, в которых в 2001 г. насчитывалось 285, а
в 2003 г. — 140 пар (Гаврилов и др., 2003; Гаврилов, 2005).
В Дагестане в первой половине XX в. колпица гнездилась в низовьях Терека, Сулака,
Аксая и Акташа, на Темиргоевских озёрах и в низовьях Кумы. В 1939 г. лишь в дельте
Терека было известно не менее 5 многосотенных колоний, но к началу 1950-х гг. там оста-
Колпица
485
Таблица 42. Динамика численности колпицы в дельте Волги
Годы Астраханский заповедник Дельта пары Источник
взрослые особи пары
1934 240* Сыроечковский, 2005
1935 610 0,5 тыс. Кривенко, 1975; Сыроечковский, 2005
1939 1542 Сыроечковский, 2005
1945 2,5 тыс. Кривенко, 1975
1948 2957 Сыроечковский, 2005
1950 1395 2790 Сыроечковский, 2005; Скокова, 19596
1951 4862 Скокова, 19596
1952 1670 Скокова, 19596
1953 1140 1140 -1700** Сыроечковский, 2005; Скокова, 19596
1954 850 Скокова, 19596
1955 646 1,5 тыс. Скокова, 19596; Кривенко, 1975
1956 400 Скокова, 19596
1958 808*** Скокова, 19596
1968 0,9 тыс. Кривенко, 1975
1973-74 1206 Бондарев, 2005
1974 164 Кривоносов, Гаврилов, 1981
1975-79 25-46 Кривоносов, Гаврилов, 1981
1980 416 Гаврилов, 1984, 2005
1985 357 Гаврилов, 2002, 2005
1987 310 Русанов, 2004
1991 183**** Гаврилов, 2002, 2005
1995 55 Гаврилов, 2002, 2005
1997-03 135-410 Гаврилов, 2002, 2005; Гаврилов и др.
2003; Русанов, 2004
Примечания-. * В начале 1930-х гг. почти все колпицы гнездились в заповеднике (Сыроечковский,
2005); ** В начале 1950-х гг. в дельте было известно 12 колоний колпицы (Сыроечковский, 2005);
*** Снижение численности в заповеднике происходило в 1950-е гг. за счёт выселения колпиц на
взморье, а их общая численность оставалась относительно стабильной (Скокова, 19596);**** В 1980-е гг.
численность колоний в дельте снизилась с 15 до 8 (Гаврилов, 2005).
лись лишь 3 поселения, в которых было примерно 200, 50 и 25 гнёзд (Сыроечковский,
2005). Сейчас её гнездовья сохранились только в дельте Терека, где в 1973-1974 гг. в 3
колониях гнездилось 370 пар, в 1985-1986 гг., после осушения дельты, в 2 колониях было
учтено лишь 40 пар (Бондарев, 1977, 2005; Прилуцкая, Пишванов, 1989; Казаков и др.,
2004), а в 1990-е гг. были известны 3 места гнездования с общим числом в 45 пар (Джамир-
зоев и др., 2000).
В Юж. Сибири и на Дальнем Востоке А.Б. Линьков (2000) предполагает гнездование
20-50 и 2СМ0 пар, соответственно. Всего, таким образом, современная численность кол-
пицы в России может составлять 1 700-2 900 пар, а принимая во внимание возможный
недоучёт — 2-3 тыс. пар.
В Азербайджане в колониях, сформировавшихся в 1950-е гг. в Кызыл-Агачском запо-
веднике, колпица была относительно немногочисленна (45-300 особей). По разным оцен-
кам, она составляла от 0,02 до 2% общего числа голенастых и веслоногих, насчитывавших
в 1957 г. до 337,5 тыс. пар или 2 млн. особей в конце лета (Дюнин, 1960а; Виноградов,
Чернявская, 1965; Греков, 1965). Однако, эта оценка была, возможно, сильно завышена,
поскольку в 1959 г. общая численность данных колоний была оценена всего в 15 020 пар
(Чернявская, 1961). В 1960-е гг. колония голенастых и веслоногих в Кызыл-Агаче умень-
шилась в целом до 300-Л00 тыс, а в 1970-е — до 20-80 тыс. особей. При этом колпица
486
Отряд Листообразные, Семейство Ибисовые
вовсе выпала из гнездовой фауны и только с 1975 г. там вновь появились 1-2 пары (Муста-
фаев, Кязымов, 1965; Васильев и др., 1972; Морозкин, 1975а; Коновалова, 1979; Литвино-
ва, 1981). Но в 1985-1986 гг. там насчитывали 35^46 пар, а в 1995 г. —до 800 гнездящихся
птиц (Султанов, 2000; Patrikeev, 2004).
В 1960-е гг. более 2 000 пар гнездились также на оз. Акгёль в Кура-Араксинской низ-
менности (Виноградов, 1967; Васильев, 1968; Виноградов, Чернявская, 1969). В последу-
ющие годы их численность там резко сократилась: с 5 320 особей в 1964 г. до 2 200 в 1968 г.
и всего 100 в 1976 г. (Туаев, Исрафилов, 1977). Однако затем популяция на оз. Акгёль вос-
становилась до 200-700 пар в 1988-1990 гг. и до 1 750 пар — в 1990-е гг. (Султанов, 2000;
Patrikeev, 2004).
Всего в Азербайджане в 1980-е гг. было учтено лишь 411 пар, в том числе 360 — на оз.
Махмудчала-Ахчала в Муганской степи (Стоцкая, Кривенко, 1988; Литвинова, 1989). К концу
1980-х - началу 1990-х гг. численность колпицы в Азербайджане увеличилась до 500-1 200
пар (Patrikeev, 2004), а в конце 1990-х гг. она была оценена в 800-2 500 пар (Султанов,
2000; BirdLife International, 2004).
В Казахстане в середине XIX в. колпица в большом количестве гнездилась на озёрах
среди «киргизских» степей, на нижнем Урале, по берегу Каспия от р. Урал до Мертвого
Култука: на Гогольских о-вах, по Черепнинскому култуку, близ устья р. Эмбы (Эверсманн,
1866; Карелин, 1883, цит. по: Сыроечковский, 2005). Большинство этих колоний исчезло,
очевидно, уже к концу XIX в., но в устье Урала колпицы оставались обычными ещё в нача-
ле XX в. (Бостанжогло, 1911) и были уничтожены, по-видимому, лишь в начале 1930-х гг.
(Сыроечковский, 2005). Однако в 1990-е гг. в дельте Урала вновь обитало около 200 птиц
(Березовиков, Гисцов, 2001).
В Западно-Казахстанской обл. в 1980-е гг. в 20-25 поселениях гнездилось 300^450 пар
(Стоцкая, Кривенко, 1988; Дебело, 1996). Ещё 7-15 пар было учтено в Наурзуме, до 23 — в
Кургальджино, около 45 — в низовьях р. Тургай, 80-100 — в дельте р. Или, 90-100 пар —
на оз. Зайсан (Стоцкая, Кривенко, 1988; Ковшарь, 1991). Кроме того, в 1971 г. колония (12
гнезд) отмечена в дельте р. Эмба (Неручев, 1991), в 1974-1981 гг. — на оз. Караколь у
Капчагайского водохранилища (60-70 пар) (Хроков, Гаврилов, 1991), в 1980-83 гг. — на
оз. Сорколь в 90 км восточнее Джаныбека (6-20 пар) (Линдеман, 1991), в 1984-85 гг. — в
долине р. Чу на Ташуткольском вдхр. (2^4 пары) (Березовиков, 1991). В 1985-1986 гг. 49 и
32 пары гнездились также в дельте р. Чёрный Иртыш на Зайсане (Березовиков, Самусев,
2003) и до 11-12 пар — в дельте р. Тентек в Алакольской впадине, однако последняя коло-
ния к 2004 г. распалась из-за беспокойства браконьерами (Березовиков, 20046). Большие
колонии колпиц, существовавшие в первой половине XX в. в низовьях Сыр-Дарьи (Заруд-
ный, 1915; Спангенберг, Фейгин, 1936), исчезли уже к 1980-м гг. (Пославский, 1991).
В 1990-е гг., по данным А.Ж. Жатканбаева (1996) и А.Ф. Ковшаря (1999), в Казахстане
обитало 400-650 пар, в том числе около 150 — в низовьях Тургая, 100 — в дельте р. Или,
60-70 пар — у Капчагайского вдхр., 100-150 — на Чушкакольских озёрах в среднем тече-
нии Сыр-Дарьи, 10 пар — в Наурзумском и Кургальджинском заповедниках. В Наурзум-
ском заповеднике колпицы гнездились в 1983-1992 гг., а в последующие годы (период низ-
кого уровня озёр) наблюдались единично (Брагин, Брагина, 2002). В Кургальджино сейчас
постоянно держится около 40-50 птиц (Кошкин, Кошкина, 2003).
В Узбекистане в дельте Аму-Дарьи колония из 321 гнезда была найдена в 1959 г. близ
мыса Аккала и 84 гнезда — на оз. Б. Дауткуль (Рашкевич, 1961,1969). До 1975 г. колония из
235 гнёзд располагалась на Дауткульском вдхр., но позже она исчезла из-за выжигания
тростников, и лишь в 1984 г. эти птицы опять здесь появились. В 1988-1989 гг. там же в
дельте Аму-Дарьи в 4 колониях учтено уже не менее 500 пар (Лукашевич, 1990, 1995), но к
Колпица
487
2000 г., в связи с обсыханием дельты, численность птиц вновь сократилась до 200 пар. В
1990-е гг. ещё около 100 пар гнездилось вдоль Аму-Бухарского и Каршинского каналов,
около 50 пар — на Айдар-Арнасайских озёрах по Сыр-Дарье, где колпицы впервые появи-
лись в 1987 г., и около 50 пар — в Ферганской долине на Кайраккумском вдхр. (Шерназа-
ров, 2003).
В целом в Вост. Европе и Сев. Азии популяции колпицы насчитывают сейчас, ориенти-
ровочно, от 3,5 до 6,5 тыс. пар, по-видимому, несколько увеличившись, по сравнению с
1980-ми гг. (Флинт и др., 1991). Численность колпиц испытала резкую депрессию в начале
XX в., но к его середине восстановилась. Однако с середины 1960-х гг. вновь было отмече-
но резкое сокращение популяций. Сейчас численность вида в Европе постепенно восста-
навливается, хотя в азиатской части ареала тенденция динамики численности остаётся в
общем отрицательной.
Отмеченные флуктуации численности колпицы отчасти объясняются, вероятно, пере-
распределением птиц по разным регионам, в частности, из дельты Волги — на озёра Вол-
го-Уральского междуречья или на Маныч. В основном же они обусловлены прямым (пес-
тицидное воздействие, отстрел птиц) или косвенным (ирригация, осушение пойм, выжига-
ние тростников) влиянием человека, а также воздействием естественных факторов (изме-
нение климата, обводненность, сукцессии древесной растительности и др.).
Размножение. Колпицы начинают размножаться, по-видимому, с 3^4-летнего возраста
(Гладков, 1970; Cramp, 1977; del Hoyo et al., 1992). Часть неполовозрелых птиц прилетает
весной на родину и кочует в гнездовых районах, а некоторые проводят своё первое лето в
районах зимовок (Сапетин, 1978г; Харашти, 1978). Судя по материалам кольцевания (Са-
петин, 1978г; Казаков и др., 2004), колпицы отличаются выраженным территориализмом,
обычно возвращаясь в места расположения своих старых колоний и оставляя их лишь при
очевидном ухудшении гнездовых или кормовых условий. Местные переселения птиц выз-
ваны их беспокойством или разорением гнездовий.
Колпица гнездится моногамными, по-видимому, сезонными парами (Cramp, 1977). Это
типичный колониальный вид, селящийся сообществами от нескольких до нескольких со-
тен пар. Так, на открытых сухих островах оз. Маныч-Гудило находили колонии до 160 пар
(Линьков, 1988), в лесах в дельте Волги колонии насчитывали до 350 пар (Бондарев, 1979),
а в плавневых колониях в дельтах Терека и Аму-Дарьи и на озёрах Азербайджана были
известны колонии до 300-360 пар (Рашкевич, 1969; Бондарев, 1975а; Литвинова, 1989); на
оз. Акгёль — даже до 1 047 гнёзд на 1 га (Виноградов, 1967). Максимальная величина
колоний определяется в первую очередь, по-видимому, размерами гнездопригодных мес-
тообитаний, зачастую также наличием гнездостроительного материала и пространствен-
но-социальной организацией поселений (Олейников и др., 1975; Линьков, 1988).
В плавневых массивах местами встречаются моновидовые колонии колпиц. В дельте
Волги в 1973-1974 гг., например, по данным Д.В. Бондарева (1979) все колонии колпиц
были именно такого типа. Но гораздо чаще птицы присоединяются к смешанным поселе-
ниям, в которых можно увидеть разные виды цапель, бакланов, пеликанов, а также кара-
ваек. Иногда колпицы гнездятся одиночными парами, но только в составе поливидовых
колоний. В смешанных поселениях колпицы обычно подселяются к бакланам, боль-
шим белым и серым цаплям, начинающим гнездиться раньше их. На сухих островах с
колпицами нередко соседствуют также различные чайки, прежде всего — многочислен-
ные хохотуньи, часто расклёвывающие яйца колпиц, особенно при наличии фактора бес-
покойства.
Как облигатно колониальный вид, колпица не имеет и не защищает гнездовых террито-
рий. Охраняются лишь сами гнёзда, которые могут соприкасаться своими краями, нередко
488
Отряд Листообразные, Семейство Ибисовые
Рис. 90. Элементы брачного и гнездового поведения колпицы (по: Bauer, Glutz von Blotzheim, 1966;
Cramp, 1977): a — взаимная чистка оперения, б — смена партнёров на гнезде, в — кормление моло-
дой птицы.
образуя в колониях очень плотные агрегации. Борта гнёзд уже в период насиживания яиц
покрываются белыми фекалиями.
Место будущего гнезда, по наблюдениям 3.0. Петровича (2000) в дельте Дуная, занима-
ют и защищают самцы. Демонстрациями клюва, охристой груди и хохла они привлекают
на гнездо самку. Тут же происходит сближение партнёров и формируется пара (рис. 90 а,б).
Гнездо птицы строят вместе: самец собирает поблизости и носит строительный материал,
а самка укладывает его (Петрович, 2000). Строительство гнезда в тростниках продолжает-
ся 4-6 дней (Туаев, Исрафилов, 1977; Смогоржевський, 1979). Затем оно подновляется и
достраивается в течение всего гнездового периода (Скокова, 19596).
В плавневых колониях колпицы гнездятся обычно на низких тростниковых кочках или
заломах тростника, иногда — на сплавинах или мелководье. Гнёзда размещаются, как пра-
вило, плотными группами (часто соприкасаясь друг с другом) в небольших прогалах, по
окраинам тростниковых куртин или на открытой воде. Между такими гнездовьями могут
формироваться тропинки в виде длинных, узких, утоптанных и покрытых помётом прохо-
дов в зарослях (Бостанжогло, 1911; Спангенберг, Фейгин, 1936). Одиночные постройки
Колпица
489
колпиц встречаются иногда в гуще высоких тростниковых зарослей среди гнёзд цапель.
Типичные гнёзда колпиц довольно крупные, массивные. Они занимают, обычно, вто-
рой после цапель ярус и основанием, как правило, касаются воды, но иногда размещаются
на заломах мощного тростника на высоте до 1,5-3,5 м (Смогоржевський, 1979; Сагитов,
1987; Лукашевич, 1995). При этом, в отличие от гнёзд цапель, они открыты с боков в ре-
зультате обламывания птицами всех окружающих стеблей тростника (Лукашевич, 1995).
Гнёзда птицы строят из сухих стеблей тростника, иногда с примесью сухих веток деревьев
и кустарников, и выстилают сухими листьями тростника и другой растительной ветошью.
Размеры (см) таких гнёзд (п=20) в дельте Дуная: диаметр гнезда — 50-90; высота гнезда —
21-30; диаметр лотка — 25-30; глубина лотка — 5 (Петрович, 2000), а по данным Смогор-
жевского (1979): диаметр гнезда — 63-110; высота гнезда — 50-80; диаметр лотка — 26-
35; глубина лотка — 7-8. В плавнях Кубани диаметр гнезда — 40-55; высота гнезда — 10-
20; глубина лотка — 3-10, в среднем 7,4 см (Винокуров, 1960). В Азербайджане на оз.
Акгёль гнёзда (п=16) имели следующие размеры (см): диаметр гнезда — 40-106; высота
гнезда — 11-30; диаметр лотка — 20-32; глубина лотка — 5-10 (Туаев, Исрафилов, 1977).
В Узбекистане, в дельте Аму-Дарьи, средние размеры гнёзд (п=6) составляли: диаметр
гнезда — 42,5; диаметр лотка — 28,3; глубина лотка — 7,3 см (Кенжегулов, 1972).
На Усть-Манычском и Веселовском водохранилищах в мощных зарослях тростника
гнёзда заметно крупнее (диаметр гнезда — 80-140 и высота гнезда — 60-100 см), чем в
изреженных зарослях на солоноватом оз. Казинка в долине оз. Маныч-Гудило: диаметр
гнезда — 38-78; высота гнезда — 23-100; диаметр лотка — 20-33; глубина лотка — 2-8, в
среднем (п=39), соответственно — 58; 62; 27; 7,5 см (Казаков и др., 2004). Занимая старые
гнёзда в тростниках в течение ряда лет, колпицы достраивают их в виде многослойных куч
(Спангенберг, Фейгин, 1936).
На открытых островах оз. Маныч-Гудило гнёзда обычно группируются среди высоких
зарослей сухой лебеды четко выраженными субколониями. В них гнёзда располагаются на
расстоянии от нескольких десятков см до нескольких метров друг от друга (по измерениям
составителя очерка в двух субколониях по 7 гнёзд — в 0,5-4-8 м, в среднем — 1,8 и 3,1 м
между центрами гнезд) или плотными «монолитами», слившимися в виде изломанных ли-
ний или каре, в состав которых входит от 2 до 9-13, чаще всего 4-8 гнёзд (Линьков, 1988).
На открытых сухих островах колпицы строят гнёзда в форме рыхлых, широких кочек, тумб
или башенок из грубых сухих стеблей лебеды с примесью древесных прутьев и прочей
ветоши, а выстилают тонкими сухими стеблями лебеды и других травянистых растений,
укладываемыми обычно, в отличие от цапель, концентрическими слоями. Кроме того, в
лотках нередко присутствуют зелёные побеги и листья лебеды, злаков и других растений,
иногда встречаются комки сухих водорослей, сухие листья и стебли кукурузы, птичьи пе-
рья и другая ветошь. Размеры таких гнёзд (см): диаметр гнезда — 37-117; высота гнезда —
17-73; диаметр лотка — 25-36; глубина лотка — 2-9, в среднем (п=20), соответственно, 73,
44, 29 и 5 (Казаков и др., 2004). По данным составителя очерка, полученным в 2004 г.,
гнёзда в одной из колоний имели следующие параметры: диаметр гнезда — 60-99; высота
гнезда — 25-65; диаметр лотка — 24^43; глубина лотка — 4,5-10, в среднем (п=27) —
78,5±6,7; 47,5±11,8; 33,0±3,8; 6,9±1,1 см.
В дельте Волги гнёзда обычно располагаются открыто на обособленных деревьях —
либо в развилках ветвей на усыхающих вершинах в 5-6 м над землей или водой, либо на
концах длинных боковых ветвей в 2,5-3 м от воды. Это облегчает слабо маневренным пти-
цам посадку и слёт с гнезда в древесных зарослях. На концах боковых ветвей размещают-
ся, в основном, небольшие однолетние постройки, тогда как на вершинах деревьев и кус-
тарника они существуют в течение ряда лет, ежегодно подновляясь и становясь очень мае-
490
Отряд Листообразные, Семейство Ибисовые
сивными. Их размеры: диаметр гнезда — 63-79; диаметр лотка — 30-35; глубина лотка —
5-7 см. Гнёзда строятся из веток ивы, лоток выстилается мелкими веточками и сухими
листьями ивы и тростника, располагающимися концентрически (Скокова, 19596).
В тамарисковых зарослях Кызыл-Агача колпицы гнездятся небольшими группами в
центре смешанных колоний или в одиночку по их окраинам. Громадные гнёзда колпиц,
сделанные из веток тамариска, располагаются примерно в половину высоты кустов. Диа-
метр лотка в таких гнёздах равен 26-30, а его глубина — 7-8 см. Выстилается лоток листь-
ями тростника и стеблями клубнекамыша морского (Греков, 1965). На отмирающих трост-
никовых островках в заливах оз. Маныч-Гудило гнёзда часто строятся из гнилых корневищ
тростника, собираемых поблизости на отмелях.
Оседлые птицы на юге ареала размножаются обычно в сезон дождей. Так, в северных
районах Индии гнездовой период приурочен к июлю-октябрю, а в южных районах — к
ноябрю-январю (Пракаш, 1990).
В Сев. Евразии колпицы приступают к гнездованию сразу же после возвращения с зи-
мовок. Однако из-за сильной растянутости сроков миграций, гнездовой период тоже очень
растянут. Первыми начинают размножаться старые птицы, а затем, по мере прилёта, более
молодые (Brouwer, 1964; Cramp, 1977). Однако на Маныче однажды наблюдалось поселе-
ние 4-летних колпиц, приступивших к размножению почти на 1 месяц раньше 6-7-летних
птиц в соседней микроколонии (Линьков, 1992).
На Лебяжьих о-вах в Крыму первые кладки появляются в период с середины марта (16
марта 2002 г.) до конца апреля (26 апреля 1997г.; 29 апреля 1996 г.), в среднем за 9 лет —
через 23 дня, а иногда уже через 9 дней после прилёта первых птиц. Разница же в сроках
появления первых и последних кладок составляет, в среднем за 9 лет, 53 дня, а максималь-
ная — 85 дней. Вылупление первых птенцов отмечается с 11 апреля (2002 г.) по 24 мая
(1996 г.). Весь период вылупления продолжается, в среднем, 48 дней, максимально же —
84 дня. До 1999 г. первые птенцы вылетали из гнёзд в 3-й декаде июня (20 июня 1997 г.; 26
июня 1996 г.), но в последние годы — значительно раньше: с середины мая - начала июня
(14 мая 2002 г.; 23 мая 2001 г.; 31 мая 1999 г.; 6 июня 2003 г.). Последние птенцы покида-
ют гнёзда в июле - августе (24 августа 1999 г.). Общая продолжительность гнездового
периода в колонии составляет 64-142, в среднем (п=9 лет) — 108±26 дней (Костин, Тари-
на, 2004).
В дельте Дуная начало гнездования в 1977-1978 гг. отмечено 17 и 19 марта. Оно прохо-
дило тремя волнами: очень ранняя кладка (18% пар), ранняя (15%) и основная (67%). Из 6
поздних, повторных кладок птенцы вылупились лишь в середине июля, так что 17 июля
1977 г. и 24 июля 1978 г. в гнёздах наблюдались ещё 2-7-дневные птенцы (Петрович, 2000).
На оз. Маныч-Гудило первые кладки отмечены 5 апреля 1969 г. и 5-10 апреля 1977 г., а
на Веселовском вдхр. — в конце марта 1977 г. и в начале апреля 1979 г. Однако вследствие
очень растянутых сроков прилета, гнездостроение и свежие кладки регистрируются вплоть
до середины июня. Например, на Веселовском вдхр. 19 июня 1979 г. в колонии с уже лёт-
ными птенцами обнаружены 4 свежие кладки. В колонии на оз. Маныч-Гудило 27 мая 1986
г., кроме гнёзд с птенцами, было 12 ещё пустых гнёзд, а 25 мая 1987 г. — 13 пустых гнёзд и
13 гнёзд с неполными кладками из 1-2 яиц. Свежие и слабонасиженные кладки встреча-
лись там иногда вплоть до начала-середины июля (например, 11 июля 1969 г.), что объяс-
няется также нередкими случаями повторного гнездования колпиц после гибели первых
кладок (Казаков и др., 2004).
По данным А.Б. Линькова (1992), общая продолжительность периода яйцекладки на
Маныче составляет около 70 дней. В нормальные годы колпицы имеют там 2 пика массо-
вой кладки (в 3-й декаде апреля и в 1-2-й декадах мая), разделенные интервалом в 10-35
Колпица
491
дней, в среднем (п=6) — 21 день. Соотношение числа птиц, приступающих к размножению
в эти сроки, может существенно меняться по годам, но в целом оно относительно выровне-
но. Птицы, размножающиеся в разные сроки, формируют на островах, как правило, обо-
собленные микроколонии с синхронизированным в пределах 5-8 дней началом откладки
яиц. Хотя иногда в эти субколонии птицы могут подселяться и позже — возможно, при
повторном гнездовании.
Первые птенцы в колониях на Маныче появляются иногда в 3-й и даже во 2-й декаде
апреля (Линьков, 1992). Появление лётных молодых в 1977-1986 гг. отмечали там 9-25
июня, в среднем за 4 года — 16 июня. Массовый же вылет птенцов наблюдался в июле.
Последние птицы покидают колонии в сентябре (Казаков и др., 2004).
В дельте Волги в первой половине XX в. массовая откладка яиц начиналась 15-25 мая,
но в некоторые годы — с начала мая (Воробьев, 1936; Дубинин, Дубинина, 1940). Массо-
вый вылет птенцов наблюдали в конце июля - начале августа; последние птенцы (вероят-
но, из повторных кладок) вылетели в 1952 г. в первой половине сентября, тогда как в 1953
г. во многих гнёздах в августе ещё были яйца (Скокова, 19596). На Кургальджинских озё-
рах в Центр. Казахстане самка с полностью сформировавшимся яйцом добыта 7 мая 1969 г.
(Кривицкий и др., 1985).
В Азербайджане на оз. Акгёль в 1962 г. начало строительства гнёзд отмечено 18 апреля,
а к 20 июня часть выводков уже разлетелась, но в 2 гнёздах ещё оставались пуховики. В
1963 г. вылупление птенцов началось лишь в первой декаде июня; в 1964 г. колпицы начали
гнездостроение уже 2 апреля, но массовое появление пуховиков отмечено только 3 июня
(Виноградов, 1967).
В дельте Аму-Дарьи первые слётки зарегистрированы 5 июня 1989 г., однако большин-
ство птенцов поднимается на крыло к началу июля; некоторые из них могут задерживаться
в гнёздах до конца сентября - начала октября. Например, 12 сентября 1959 г. там были ещё
пуховики в возрасте 10-12 дней (Рашкевич, 1961; Лукашевич, 1995). На оз. Айдаркуль
колпицы приступают к яйцекладке в начале апреля, но большинство птиц — в конце ап-
реля - мае, а некоторые — в июне (вплоть до 22 июня). Вылупление птенцов отмечали с
начала мая до 10-17 июля, первых слётков видели с 15 июня (Шерназаров, 1991).
В целом прослеживается запаздывание начала размножения колпиц в более северных
районах в соответствии с более поздним возвращением птиц с зимовок (Спангенберг, 1951).
В последние десятилетия наблюдается также заметный сдвиг всех репродуктивных фено-
фаз на более ранний период (возможно, в связи с потеплением климата). Хотя общее тече-
ние процессов размножения в значительной мере определяется синоптическими условия-
ми отдельных лет. Так, например, в Венгрии в 1961 г. часть птиц 29 марта уже насиживала
кладки, а в 1962 г. в той же колонии в конце марта 38 колпиц только появились с зимовок, и
36 из них погибли затем из-за возврата холодов (Sovago, 1964, цит. по: Харашти, 1978).
В случае гибели первых кладок колпицы приступают, как правило, к повторному гнез-
дованию, в результате чего часть птиц заканчивает размножение на 1-2 месяца позже обыч-
ного — к концу лета или началу осени.
Величина кладки у колпиц довольно строго детерминирована и составляет обычно 3-4
яйца. Размер её колеблется по годам и регионам, возможно в соответствии с обеспеченно-
стью птиц кормами. По наблюдениям Е.П. Спангенберга (1951) на Сыр-Дарье, в одни годы
в гнёздах абсолютно преобладают кладки из 3 яиц, в другие — из 4. Кладки из 5-6 яиц
(имеются сведения о кладках даже из 7 яиц; Гладков, 1970; Cramp, 1977) в некоторых слу-
чаях могут быть сдвоенными; мелкие кладки из 1-2 яиц появляются, видимо, при повтор-
ном гнездовании, а также, что вероятнее, в результате хищничества ворон, сорок и чаек или
могут быть просто незаконченными.
492
Отряд Листообразные, Семейство Ибисовые
В дельте Дуная в 1976-1979 гг. кладки из 4 яиц составляли 43,1%, из 3 — 41,4%, из 2 —
14,4%, а из 5 — 1,1%, в среднем (п=181 гнездо) кладка составляла 3,3 яйца (Петрович,
2000). В 1958-1963 гг. в Румынии преобладали кладки из 3 яиц (53,8% от 415 гнезд), а с 2
и 4 яйцами было по 23,1% гнёзд (Vespremeanu, 1968).
В дельте Волги характерны кладки из 3 яиц (Скокова, 19596), в Азербайджане — из 2-
3 яиц (Греков, 1965; Виноградов, 1967), в дельте Аму-Дарьи — из 3 (33,3% кладок) и 4
(53,3%) яиц, в среднем (n= 12-15) — 3,3 яйца (Лукашевич, 1991; 1995), на оз. Айдаркуль —
из 3 (38,7%) и 4 (58%) яиц, в среднем (п=31) — 3,6 яйца (Шерназаров, 1991), на Капчагай-
ском водохранилище в юго-восточном Казахстане — тоже из 3 (33,3%) и 4 (52,4%) яиц
(Хроков, Гаврилов, 1991).
На Маныче, по данным Б. А. Казакова с соавторами (2004), преобладают кладки из 4 яиц
(58,5% гнёзд), реже встречаются 3 яйца (34,4%). Кладки из 2 яиц составляют всего 3,4%, а
из 5 яиц — 2,4%. Лишь 1 кладка содержала 6 яиц и 3 кладки — по 1 яйцу. Средний размер
кладки (п=294) составил 3,6 яиц. По данным составителя очерка, полученным в 2004 г.,
кладки из 4 яиц были в 86,2% гнезд. Кроме того, учтено по 1 гнезду с 1, 2, 3 и 6 яйцами,
причём в последней кладке 2 яйца заметно отличались по размерам и форме. Средний раз-
мер кладки (п=29) составил 3,9±0,39 яиц.
По данным А.Б. Линькова (1992), средний размер кладки на Маныче (п=771) равен
3,54±0,03 яйца, варьируя от 1-2 до 5-6, но в основном составляя 3—4 яйца. При этом в
первый, апрельский пик гнездования средняя величина кладки (3,59-3,69 яйца) была дос-
товерно выше (р < 0,001), чем в майский пик (3,27-3,31 яйца).
Яйца колпицы имеют необычную для большинства голенастых пятнистую окраску, боль-
ше напоминая яйца хищных птиц. Их матовая грязновато-белая или известково-белая скор-
лупа покрыта чёткими (лишь иногда расплывчатыми) редкими поверхностными коричне-
ватыми пятнами и мазками разной величины и густоты, концентрирующимися обычно вок-
руг тупого полюса яйца. Эти пятна могут быть как плотными, тёмными, так и бледными,
светлыми, изредка — с охристым или сиреневым оттенком.
Форма яиц обычно овальная, иногда — короткоовальная. Изредка встречаются удли-
нённо-овальные или удлинённо-элипсоидные яйца. У некоторых яиц клоакальный полюс
может быть заострён, придавая им слегка грушевидную форму (Makatsch, 1974).
Размеры яиц колпицы из Сев. Евразии варьируют в пределах 51,7-80,7 х 36,0-53,0 мм
(табл. 43). При этом прослеживается тенденция к увеличению средних размеров яиц в бо-
лее восточных популяциях (Смогоржевський, 1979). Величина яиц может зависеть также
от размера кладки: в больших кладках они меньше, и наоборот. Так, в дельте Дуная в 2-
яйцевых кладках размеры яиц составили 72,3-77,1 х 41,9^7,9 мм (75,4x45,4; п=36), в 3-
яйцевых — 54,0-69,6 х 37,0^16,3 мм (62,8x43,2; п=63), а в 4-яйцевых — 51,7-57,1 х 36,0-
38,7 мм (54,0x37,2; n=6) (Vespremeanu, 1968). Отчасти этим фактором, возможно, обуслов-
лена и географическая вариация средних размеров яиц.
Однако по нашим расчетам, проведенным на основе данных М.П. Ильюха и А.Н. Хох-
лова (2006), различия в размерах яиц из разных кладок в Манычской популяции колпицы
оказались не выражены: в 2-яйцевых кладках, в среднем (п=16 яиц) — 68,0±2,1х 45,8±1,5
мм, в 3-яйцевых (п=64) — 69,8±2,2 х 46,1±1,3 мм; в 4-яйцевых (п=154) — 68,1±2,6 х 46,3±1,3
мм, а в 5-яйцевых (п=35) — 68,1±1,9 х 47,3±1,2 мм.
Масса свежих яиц из колоний на оз. Маныч-Гудило варьирует в пределах 92-96 г, а
слабонасиженных — 74,9-84,5 г (Казаков и др., 2004). На Аму-Дарье средний вес 18 нена-
сиженных яиц составил 73,0±6,3 г (Лукашевич, 1995), а 12 других — 78,7 г (Кенжегулов,
1966, цит. по: Смогоржевський, 1979). Вес яиц (п=3) перед вылуплением снижается до 69,3 г
(Шерназаров, 1991). В Азербайджане вес яиц составлял 67,75-91,2, в среднем (п=100) —
Колпица
493
Таблица 43. Размеры яиц колпицы из разных регионов (в мм)
Регион п Lim М Источник
Венгрия 12 66,4-75,3 х 43,7—47,7 70,5 х 45,2 Makatsch, 1974
Югославия 250 58,5-76,4 х 40,8^19,2 67,6 х 45,9 Makatsch, 1974
Румыния 50 52,7-74,6 x 36,8^17,1 65,8 х 45,1 Lintia, 1955
Румыния 105 51,7-77,1 х 36ДМ7,9 61,2x42,4 Vespremeanu, 1968
Дунай, дельта 62,3-70,8 х 43,6—47,6 Смогоржевський, 1979
Дунай, дельта 101 61,0-75,4 x41,5-50,0 69,3 х 46,2 Петрович, 2000
Кубань, дельта 20 65,5-73,0 х 43,7—49,5 68,2 х 47,5 Хохлов и др., 1997
Крым, Лебяжьи о-ва 25 59,2-79,3 х 42,5^19,9 Костин, Тарина, 2004
Долина Маныча, оз. Казинка 83 62,8-80,7 х 42,7—48,6 70,3 х 46,6 Казаков и др., 2004
Озеро Маныч-Гудило 103 54,6-79,5 х 43,6-53,0 68,5 х 46,6 Казаков и др., 2004
Озеро Маныч-Гудило 95 60,9-75,2 x42,3-50,1 67,9 х 46,7 Ориг. данные
Озеро Маныч 224 60,3-78,4 x41,2-50,4 68,2 х 46,5 Ильюх, Хохлов, 2006
Восточный Маныч 50 60,8-75,3 х 41,8-50,2 70,1 х 45,8 Ильюх, Хохлов, 2006
Волга, дельта 62,6-72,8 х 42,6—47,5 Скокова, 19596
Азербайджан, оз. Акгёль 100 59,9-77,9 х 42,2^19,0 70,4 х 46,6 Туаев, Исрафилов, 1977
Азербайджан, Ленкорань 3 63,7-66,6 х 43,8—45,0 65,6 х 44,3 Makatsch, 1974
Аму-Дарья, дельта 15 71,0x45,7 Кенжегулов, 1966
Аму-Дарья, дельта 42 68,6 х 45,4 Лукашевич, 1995
Сыр-Дарья, оз. Айдаркуль 45 66,0-76,2 х 43,0-^19,0 69,5 х 46,1 Шерназаров, 1991
Сыр-Дарья 16 65,2-72,5 х 43,0^18,9 Спангенберг, Фейгин, 1936
Сыр-Дарья (?) 17 65,1-73,0 x 43,2—49,0 69,5 х 45,0 Спангенберг, 1951
79,91 г (Туаев, Исрафилов, 1977). Масса скорлупы — 4,36-8,79, в среднем (п=176) — 6,95 г
(Makatsch, 1974).
Откладка яиц идёт с интервалами в 2-3 дня (Cramp, 1977). Насиживание начинается с 1-
го или 2-го яйца (Туаев, Исрафилов, 1977; Петрович, 2000), но в период откладки яиц оно
«неплотное», и самки часть времени только стоят на гнезде (Спангенберг, 1951). По наблю-
дениям на Маныче (Линьков, 1992), в период массовой яйцекладки в апреле и мае колпицы
начинают плотное насиживание фактически лишь после завершения кладки, и все птенцы
у них вылупляются обычно в течение 3^1 ч. Яйцекладка между пиками идет, вероятно,
прерывисто, насиживание начинается до завершения кладки, и птенцы у таких пар (п=111)
вылупляются в среднем в течение 5,67 дней. В некоторых гнёздах часть птенцов появляет-
ся синхронно, а последние запаздывают. В целом же синхронное вылупление наблюдается
в 79,7%, асинхронное — в 20,3% гнёзд.
Продолжительность инкубации составляет 24-25 дней (Тугаринов, 1947; Туаев, Исра-
филов, 1977; Петрович, 2000), по другим данным — 20-25 дней (Спангенберг, 1951; Cramp,
1977; Смогоржевський, 1979; Костин, Тарина, 2004). Насиживает, в основном, самка, са-
мец сменяет её лишь днём на непродолжительное время, нужное ей для кормежки (впро-
чем, сменённая самка большую часть времени находится на гнезде, охраняя его от конку-
рентов) (Петрович, 2000). Одна из птиц постоянно дежурит на гнезде и при птенцах, зак-
рывая их от солнечных лучей, к которым пуховики очень чувствительны.
Птенцы вылупляются беспомощными, слепыми, но глаза открываются в первые же сут-
ки. Вес только что вылупившихся пуховичков составляет 48,5-60 г (Тугаринов, 1947; Туа-
ев, Исрафилов, 1977). Максимальный прирост массы тела (+153,4%) идёт в первую неделю
жизни (Денисова, 1958), затем замедляется. Перед вылетом из гнезда масса птенцов на
494
Отряд Листообразные, Семейство Ибисовые
550-700 г меньше, чем у взрослых птиц. До отлёта на зимовку молодняк так и не успевает
набрать вес, характерный для взрослых (Скокова, 19596).
Пеньки маховых перьев появляются к 10-12-му дню (Петрович, 2000), по другим дан-
ным — на 4-5-й день, а рулевые пробиваются на 9-10-й день (Скокова, 19596). К 20 дням
птенцы оперяются, поднимаются с пяток на цевки и могут перемещаться по колонии. По
сравнению с каравайками и цаплями, птенцы колпицы сравнительно малоподвижны, так
как благодаря белому оперению не особенно нуждаются в поисках тени (Кокшайский, 1976).
В возрасте 36 дней птенцы уже тренируют крылья, а в 45^17 — летают с родителями на
места кормежки (Петрович, 2000). По данным других авторов, вылет птенцов происходит в
возрасте 45-50 (Скокова, 19596; Cramp, 1977), иногда — 50-55-60 дней (Греков, 1965;
Vespremeanu, 1968; Смогоржевський, 1979). После подъема молодых на крыло родители
могут подкармливать птенцов до 70-дневного возраста, хотя молодые сами постепенно
начинают добывать себе корм (Cramp, 1977). К этому времени у птенцов удлиняется клюв и
формируется расширение на его конце, характерное для взрослых птиц, однако окончатель-
ные размеры и форму клюв приобретает лишь к концу первого года жизни (Тугаринов, 1947).
Птенцов кормят оба родителя, но чаще — самка. Сначала птицы кормят их 2 раза в день
(утром и вечером), а подросших птенцов — 3 раза в день. Самец приносит корм в основном
по утрам, самка — днем и вечером (Скокова, 19596; Петрович, 2000). Птенцы получают
отрыгиваемую полупереваренную пищу небольшими порциями по несколько раз за кор-
межку, для чего засовывают голову в раскрытый клюв родителей, доставая корм из глуби-
ны глотки (рис. 90 в).
По наблюдениям в 1977-1978 гг. в украинской части дельты Дуная, к месячному возра-
сту доживает около 75% птенцов от числа отложенных яиц, а средний размер выводка со-
ставляет 2,5 молодых (Петрович, 2000). В соседней Румынии в 1958-1963 гг. отход яиц
(п=1234) составлял всего 3,7%, в основном из-за гибели эмбрионов (2,7%). Гибель птенцов
достигала 6,4% в первые 2 дня жизни, 24,8% — к 10 дням, 28,9% — к 40 дням и 29,7% —
к 50 дням развития. Причём в гнёздах с 2 птенцами их смертность была ничтожной; поги-
бали преимущественно самые младшие в выводках, состоявших из 4 птенцов. Колебания
уровня смертности птенцов по годам, связанные в основном с погодными факторами, со-
ставляли от 17,1 до 100%, в среднем - 29,7% (Vespremeanu, 1968).
В колониях на оз. Маныч-Гудило, по данным А.Б. Линькова (1989, 1992), средний раз-
мер кладок (п=771) составляет 3,54 яйца, а 3-недельные выводки (п=178) состоят из 2,96
птенцов, т.е. эмбриональная и постэмбриональная смертность там равна 16,4%. При этом в
асинхронных выводках младшие птенцы, появляющиеся на 5 и более дней позже, не в со-
стоянии конкурировать за корм со старшими и в большинстве случаев (п=29) затаптывают-
ся в первые 2-5 дней. Поэтому у таких пар до 3-недельного возраста доживает, как прави-
ло, лишь 1-2 птенца. В колониях на оз. Акгёль в Азербайджане, при средней кладке в 3
яйца, средняя величина выводка оценена в 2 слётка (Виноградов, 1967; Виноградов, Чер-
нявская, 1969), т.е. отход яиц и птенцов составил около 33%.
В Узбекистане, на оз. Айдаркуль, отход яиц в 20 гнёздах составил 25% (17 яиц из 68, в
том числе 11 неоплодотворённых и 1 расклёванное чёрной вороной); постэмбриональная
смертность в 6 гнёздах равнялась всего 9,2% (встали на крыло 12 из 13 птенцов) (Шерназа-
ров, 1991). В дельте Аму-Дарьи, при средней кладке 3,3 яйца (Лукашевич, 1991, 1995),
вылуплялись, в среднем, 3,2 птенца, а вылетали 2,6 слётка на 1 гнездо (Рашкевич, 1969),
т.е. отход яиц составлял 3,0%, а птенцов — 18,8%. При этом общие потери равнялись 21,2%,
т.е. примерно в тех же пределах, что и в Европе. О сравнительно незначительной гибели
яиц колпицы в естественных условиях свидетельствуют и данные В.В. Хрокова и Э.И. Гав-
рилова (1991), обнаруживших всего 3 расклёванных воронами яйца возле 11 гнёзд с клад-
Колпица
495
ками (отход — 7,7%). Но при беспокойстве колпиц человеком оставшиеся без защиты кладки
часто уничтожаются врановыми, чайками или другими пернатыми хищниками. Так, в При-
азовских плавнях 4 обнаруженных в 1953 г. гнезда были уничтожены на глазах исследова-
телей одной серой вороной, а в 1954 г. кладку в единственном оставшемся там гнезде рас-
клевал степной лунь, после чего колпицы бросили колонию (Винокуров, 1960).
Большое негативное воздействие серых ворон на успешность гнездования голенастых
птиц отмечали в лесных колониях дельты Волги (Воробьев, 1936; Луговой, 1963; Кривен-
ко, 1983), хотя при этом нельзя не обратить внимания на отсутствие колпицы в приводимых
списках жертв вороны. Аналогичная ситуация наблюдается и на островах оз. Маныч-Гуди-
ло, где многочисленные хохотуньи при беспокойстве колоний человеком активно расклё-
вывают яйца в гнёздах цапель (серой и малой белой) и большого баклана, но значительно
реже нападают на гнёзда колпицы.
Болотные луни, поселяющиеся в тростниках рядом с колониями голенастых, служат
для них своеобразной защитой от врановых, значительно снижая отход яиц и птенцов (Би-
черев, Хохлов, 1984).
До занесения колпицы в Красную книгу СССР (1978 г.) смертность молодых птиц в 1-й
год жизни составляла 82%, от 1 до 3 лет — 29%, а половозрелых — 21,5%. При этом не
менее 76% из этих птиц добывалось охотниками (Сапетин, 1978г). В среднедунайской по-
пуляции смертность колпиц в 1-й год жизни составляла 72% (Харашти, 1978), на оз. Ма-
ныч-Гудило она не превышала 60% (Линьков, 1989, 2000). В настоящее время охотничий
пресс на колпиц снижен, но на рыбхозах Предкавказья местами продолжается незаконный
отстрел этих птиц вместе с цаплями (Хохлов, 1990в; Хохлов и др., 2005). Продолжительность
жизни колпицы, по данным кольцевания, составляет более 28 лет (Cramp, 1977; del Hoyo et
al., 1992); средний возраст птиц, размножающихся в колониях на оз. Маныч-Гудило (п=99),
равен 6,25 лет, а в некоторых небольших поселениях — 4,2 года (Линьков, 1992, 2000).
Суточная активность, поведение. Колпицы деятельны в течение круглых суток. Ле-
том они кормятся преимущественно в сумерках и ночью, но в гнездовой период активны и
днем. На кормёжку вылетают главным образом в 4-6 и 17-20 ч после смены партнеров на
гнёздах, а летом, при выкармливании подросших птенцов, также и в 11-13 ч. Птицы при-
держиваются обычно постоянных трасс, летя на кормёжку в одиночку и группами по 5-7,
редко до 20 особей. В колонию они возвращаются в те же часы, но более крупными стая-
ми — до 30-35 особей (Скокова, 19596). Днем значительную часть времени птицы стаями
отдыхают на отмелях; ночуют они обычно на заломах тростника близ кормовых угодий
(Спангенберг, Фейгин, 1936; Скокова, 19596).
Питание. Пищу колпицы собирают почти исключительно в воде; во время сбора корма
они перемещаются по мелководью, опустив клюв в воду и двигая им из стороны в сторону.
Корм состоит из водных беспозвоночных и мелких позвоночных животных. Нередко в пи-
щеварительном тракте встречаются водные растения, иногда колпицы собирают также на-
земных беспозвоночных, случайно попадающих в воду (например, саранчу), или медве-
док, живущих на илистых литоралях (Спангенберг, 1951; Скокова, 19596; Языкова, 19706).
В дельте Волги основным кормом служат мальки рыб (сазан, вобла, краснопёрка и др.)
длиной от 1 до 14 см, обычно — 2-8 см, а также водные насекомые (малый водолюб, ли-
чинки жука-скомороха, плавт, личинки стрекоз). Колпицы постоянно добывают амфибий
(головастиков и лягушат) и различных моллюсков (Скокова, 19596). На Усть-Манычском
вдхр. в добыче по весу абсолютно доминирует молодь рыб (98%), часто встречаются вод-
ные насекомые (жуки и клопы), но в весовом отношении они занимают лишь доли процен-
та (Олейников, 1953). На солёном, бедном рыбой оз. Маныч-Гудило, наоборот, и по встре-
чаемости, и по массе преобладали водные насекомые (соответственно — 90 и 46,6%), в
496
Отряд Листообразные, Семейство Ибисовые
основном личинки крупных плавунцов. Рыба занимала второе место: 57,7% по встречае-
мости и 43,8% по весу, а затем шли земноводные: 16,4 и 9,6% (Языкова, 1970). Аналогич-
ные или близкие пищевые спектры приводятся также для Сыр-Дарьи, Аму-Дарьи, дельты
Кубани, Терека, Дуная (Спангенберг, 1951; Смогоржевський, 1959,1979; Кенжегулов, 1972;
Казаков и др., 2004; Platteeuw et al., 2004). Спектры питания могут резко меняться в зависи-
мости от сезона и, следовательно, от обилия тех или иных кормов в опромышляемых пти-
цами водоёмах (Скокова, 19596).
Вес потребляемой пищи одной особью по расчётным данным должен составлять 331г
рыбы в сутки (Platteeuw et al., 2004). За одну кормежку колпицы вылавливают обычно 30-
50 мелких животных (максимум — 260 экз.) общим весом, в среднем, 100-150 г. Как ис-
ключение, восстановленный первоначальный вес пищи может достигать 300-500 г, что
следует считать, очевидно, полной суточной нормой. Такое же количество корма (до 500 г)
составляет среднесуточную норму для птиц, содержащихся в неволе. Это соответствует
15,5-25,6% средней массы тела колпицы (Скокова, 19596).
Корм в желудке колпицы за 2 ч переваривается на 50%. Перевариванию способствуют
своеобразные «гастролиты»: костянки рдестов, резухи, ежеголовника, семена ириса, арбу-
зов, плоды дурнишника, скорлупа чилима, кусочки стеблей тростника, раковины моллюс-
ков и т.п. (Скокова, 1962).
Враги, неблагоприятные факторы. Среди врагов прежде всего следует указать серую
и чёрную ворону и чайку-хохотунью — неспециализированных хищников, уничтожающих
кладки и птенцов в колониях голенастых при высоком уровне фактора беспокойства (см.
выше). Лимитирующее воздействие на размножение и численность колониальных гнездо-
вий оказывает, несомненно, и орлан-белохвост, численность которого заметно увеличи-
лась в последние десятилетия в лесистых дельтах южных рек. Из наземных хищников гнёз-
да колпиц в тростниках могут разорять кабаны (Воробьев, 1936).
Среди лимитирующих абиотических факторов важное значение имеют синоптические
условия зимних и весенних периодов, определяющие выживаемость и репродуктивное со-
стояние взрослых птиц, возвращающихся на места гнездования, и резкие флуктуации лет-
ней погоды, сказывающиеся на гибели кладок и птенцов, а также на обеспеченности мо-
лодняка кормом. Неожиданные весенне-летние паводки, например, иногда приводят к пол-
ной гибели колоний (Воробьёв, 1936; Vespremeanu, 1968); крайне опасны также штормо-
вые ветры и ливни с градом (Кривенко, 1983; Линьков, 2000).
Из антропогенных факторов большое значение имеют обширные пожары в тростнико-
вых плавнях, лишающие колпиц гнездовых местообитаний (Сушкин, 1908; Заболотный,
Хохлов, 19896). Кроме того, в XIX - начале XX вв. на численности птиц сказывались, веро-
ятно, промысловый сбор яиц в колониях и отстрел птиц на шкурки (Кривоносов, 1981;
Белик, 1998; Русанов, 2004; Сыроечковский, 2005; и др.). Катастрофические последствия
должны были иметь истребительные мероприятия против азиатской саранчи, охватившие
в 1920-е гг. плавни всех южных рек. Авиационное распыление над тростниковыми масси-
вами чрезвычайно токсичных парижской зелени и арсенита натрия, очевидно, и послужило
основной причиной глубокой депрессии популяций колпицы (и других околоводных птиц)
в начале XX в. (Белик, 1997, 20006). Снижение численности в середине XX в. могло было
быть связано с загрязнением водоёмов пестицидами (ДДТ и др.), которое сказалось на
состоянии популяций очень многих видов птиц-ихтиофагов во всём мире.
Во второй половине XX в. чрезвычайное влияние на размещение гнездовий колпицы и
других голенастых оказали гидромелиоративные работы в аридной зоне, которые привели
к перераспределению птиц из обсыхавших пойменных угодий на сбросные и фильтраци-
онные озёра и водохранилища в орошаемых районах.
Колпица, Малая колпица 497
В гнёздах колпицы обнаружены паразиты: Ornithoponus massonati и Lynhia palustris
(Hicks, 1959).
Хозяйственное значение, охрана. Колпица имеет, несомненно, большое эстетическое
значение — как из-за своей оригинальной внешности, так и из-за особенностей охотничь-
его поведения. Она издавна привлекала к себе внимание людей, давших ей целый ряд ха-
рактерных народных названий («косарь», «лопатень» и др.).
Будучи рыбоядным видом, колпица в значительном количестве добывает в водоёмах
молодь рыб, но в силу своего ограниченного распространения и невысокой численности
не оказывает существенного влияния на их ресурсы. По расчётам для дельты Волги, общее
изъятие всеми колпицами молоди рыб составляет за лето от 0,015 до 2,7% их поголовья. В
то же время, уничтожение колпицами хищных водных насекомых, питающихся мальками
рыб, позволяет сохранить в 210 раз больше рыбьей молоди, чем колпицы добывают сами
(Скокова, 19596, 1965).
В прошлом колпицу относили к охотничьим птицам (Мензбир, 1900; Шульпин, 1952;
Смогоржевський, 1979; и др.). Например, в августе-сентябре 1953 г. на нескольких озёрах
в центральной части дельты Волги были убиты 42 колпицы, что составило 8% добытой
охотниками дичи. Колпица занимала среди нетрадиционно охотничьих птиц («прочей дичи»)
пятое место, пропустив вперёд лишь квакву, серощекую поганку, лысуху и серую цаплю
(Сыроечковский, 2005).
В 1978 г. колпица была занесена в Красную книгу СССР, а затем и республиканские
Красные книги, и с тех пор охота на неё в России и сопредельных регионах полностью
запрещена. В настоящее время вид занесён в Красные книги Российской Федерации, Ар-
мении, Украины, Казахстана и ряда государств Средней Азии.
Малая колпица
Platalea minor Temminck et Schlegel, 1849
Platalea minor Temminck et Schlegel, 1849, Fauna Japonica, Aves, c. 120, табл. 76, Япония
Black-faced spoonbill (англ.), Schwarzstirnloffler (нем.), petite spatula (фр.).
Син.: Platalea japonica Reichenow, 1877.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Малая колпица — самая мелкая из кол-
пиц восточного полушария, её длина 73 см (Sonobe, Usui, 1993). По внешнему облику и
поведению это типичная колпица; во время кормёжки она, как и предыдущий вид, как бы
«косит» опущенным в воду клювом, помахивая им из стороны в сторону. В целом похожа
на колпицу (Platalea leucorodia), с которой во время миграций и кочёвок последней встре-
чается совместно в прибрежных районах Юж. Приморья. Взрослые птицы в брачном наря-
де, особенно стоящие рядом, различаются достаточно хорошо. У малой колпицы — чёрное
«лицо» (пространство за клювом, захватывающее лоб и глаза), и глаз расположен на чёр-
ном фоне. У колпицы глаз на белом фоне, хорошо выделяется (узкая «уздечка» — не в
счет). Бросается в глаза разная окраска клюва. Различие в окраске (площадь чёрного в пе-
редней части головы) заметно и у птиц, летящих в смешанной группе, если они не очень
далеко от наблюдателя.
498
Отряд Листообразные, Семейство Ибисовые
Стоящие рядом птицы отличаются высотой и общими размерами. Колпица — более
высокая и длинноногая, рядом с ней малая колпица выглядит относительно коротконогой и
даже коренастой.
Почти все перечисленные признаки (исключая окраску клюва) «работают» и в случае
птиц в промежуточных нарядах, если особи двух видов находятся рядом. В противном слу-
чае вероятность ошибки возрастает.
Кроме того, эти виды различаются по своим биотопическим предпочтениям. Весь жиз-
ненный цикл малой колпицы связан с морскими побережьями. Гнездится вид исключи-
тельно на скалистых прибрежных островках, в многовидовых колониях морских и назем-
ных колониальных птиц; его кормовые биотопы — прибрежные мелководья (морские за-
ливы, приустьевые участки, лагуны).
Известны достоверные встречи малой колпицы внутри материка в Китае (Bird Life
International. Threatend Birds..., 2001), но это скорее исключение, нежели правило.
Подвидовая систематика. Монотипический вид.
Описание. Описание сделано по цветным фотографиям и птицам, наблюдавшимся в
природе (не исключены некоторые неточности). Кроме того, в нашем распоряжении имел-
ся один экземпляр вида из Юж. Приморья от 5 августа 1995 г. (залив Голубиный, или лагу-
на Вудунупты), хранящийся в Зоологическом музее Дальневосточного государственного
университета, г. Владивосток.
Окраска. Взрослые самец и самка в брачном наряде. Половой диморфизм в окраске
не выражен. Окраска белая. Основание шеи желтовато-охристое. Окрашенная часть замы-
кается сзади, образуя ошейник; края и задняя часть его бледнее. Длинный свисающий хо-
хол окрашен так же, но менее интенсивно. Голая кожа на передней части головы вокруг
клюва («лицо») чёрного цвета. На чёрном фоне «лица» выделяются жёлтые веки: верхнее —
узкое, нижнее — шире (спереди от глаза цветовое поле века часто продолжается в виде
жёлтого пятна). Поперечный рисунок надклювья напоминает по расцветке шкуру зебры в
миниатюре, он складывается из чёрных и белых узких полос примерно одинаковой шири-
ны и издали кажется рельефным. Края надклювья и всё подклювье — чёрные. Радужина
красная, неяркая. Ноги чёрные.
Оперение взрослых птиц зимой (после осенней линьки) полностью белое, хохол отсут-
ствует. «Лицо», клюв, ноги — полностью чёрные.
Пуховой наряд. Информация отсутствует.
Гнездовой наряд. Оперение белое, хохла нет. Концы первостепенных маховых и стерж-
ни маховых — чёрные. «Лицо» чёрное. Клюв в основании чёрный или почти чёрный, по-
степенно светлеющий к концу. Ноги чёрные.
Приводим описание самки в возрасте 1 года и 2-3 месяцев (судя по оперению). Птица
белая, с загрязненным оперением. Уже имеется хохол (длина перьев от 25 до 30 мм). Пер-
востепенные маховые (дистальные) имеют значительные участки чёрного цвета на наруж-
ных опахалах (до 185 мм в длину). Чёрное поле есть на концах второстепенных маховых и
на части стержней третьестепенных маховых. При раскрытом крыле образуется заметный
участок чёрного на конце крыла, как у птицы в гнездовом наряде. Концы перьев крылышка
и стержни также чёрные. Нижние кроющие крыла — с чёрными кончиками и чёрными
стержнями. «Лицо» чёрное. Лапы чёрные. Клюв постепенно светлеет к концу.
Строение и размеры. Ниже приведены результаты измерений 13 особей,
пойманных для мечения в феврале 1998 г. в Гонконге (Melville et al., 2000). Пол птиц неиз-
вестен; возраст, по мнению авторов, 2-5 лет. Данные промеров (мм) таковы (среднее опре-
делено авторами очерка): длина крыла — 346-371 (среднее 361,9); длина клюва (до грани-
цы оперения) — 163-207 (среднее 183,3); длина цевки — 129-152 (среднее 139,3); длина
хвоста— 103-116 (среднее 108,8). Вес (г)—1 470-1 900 (среднее 1 660).
Малая колпица
499
Экземпляр из Юж. Приморья: длина крыла — 328 мм; длина клюва — 178 мм (от гра-
ницы оперения лба) и 166 мм (от границы с голой кожей); длина цевки — 122 мм; длина
хвоста — 121 мм (от основания рулевых перьев).
Количество рулевых перьев — 12.
Линька. На протяжении гнездового сезона у взрослых птиц идёт постепенное изнаши-
вание пера. Издали это заметно, в частности, по снашиванию оперения хохла. К моменту,
когда семья покидает гнездовую колонию, от хохлов у взрослых птиц-родителей остаются
отдельные перья или их небольшие пучки. Тускнеет и почти исчезает жёлто-охристый от-
тенок на шее, загрязняется оперение в целом. Клюв полностью чернеет. Послебрачная и
предбрачная линька проходят в основном на местах зимовки.
У птицы-«прошлогодка» (см. выше), по-видимому, всё крупное оперение осталось от
гнездового наряда; концы первостепенных, а также третьестепенные маховые были истрё-
паны. Сменилась проксимальная часть первостепенных маховых (несколько свежих перь-
ев). Чёрный цвет везде выгорел до светло-бурого.
Распространение. Ареал вида в целом — восточная и юго-восточная периферия Ази-
атского континента. Птицы встречаются на пространстве от о. Хоккайдо и Юж. Приморья
до п-ова Малакка и о. Калимантан (рис. 91).
Гнездовой ареал. Основные гнездовья (более 90% популяции) расположены на
восточном побережье Жёлтого моря на территории двух корейских государств. Известно
также о гнездовании малой колпицы на одном острове у побережья Ляодунского п-ова в
Китае (BirdLife International. Threatend Birds..., 2001). С недавнего времени (2003 г.) вид
гнездится в России на о. Фуругельма (Юж. Приморье) (Литвиненко, Шибаев, 2005). Это
северный предел гнездования вида, единственное место гнездования малой колпицы вне
основного гнездового района, удалённое от него более чем на 600 км, и единственное изве-
стное поселение малой колпицы на побережье Японского моря.
Этапы вселения малой колпицы в Юж. Приморье выглядят следующим образом:
1995 г. — первые встречи вида на побережьях зал. Петра Великого (Нечаев, Шибаев,
1996);
1996-2002 гг. — период относительно регулярных встреч (Литвиненко, Шибаев, 1999;
Нечаев, 2003; Литвиненко, Шибаев, 2005; С.Г. Сурмач, личн. сообщ.);
2003 г. — первая (неудачная) попытка гнездования на о. Фуругельма;
2004-2009 гг. — ежегодное успешное гнездование там же.
От первого появления птиц до начала гнездования прошло 8 лет. До начала успешного
гнездования — 9 лет. В течение 15 лет вселения вида (1995-2009 гг.) наблюдается позитив-
ная тенденция в освоении им нового гнездового района: появление, регулярные встречи,
первая попытка гнездования, стабильное размножение.
Зимовки. Основные зимовки располагаются в прибрежной зоне на Тайване, в Гон-
конге, в Юж. Китае, Вьетнаме, Японии, Юж. Корее (BirdLife International. Threatend Birds...,
2001; Dahmer, Felley, 1995; Yu, 2005; Yu, Swennen, 2005) (рис. 91).
Миграции. Два птенца, окольцованные на о-ве Токдо (Сев. Корея) в июле 1995 г.,
улетели на зимовку в разные регионы. Одного видели в Идзуми (о. Кюсю, Япония) в
ноябре 1995 г., другого - в дельте Красной реки (Вьетнам) в декабре того же года (BirdLife
International. Threatend Birds..., 2001). В 1998 и 1999 гг. на Тайване и в Гонконге помёти-
ли спутниковыми передатчиками 18 зимовавших взрослых и неполовозрелых малых кол-
пиц (Ueta et al., 2002). В результате прослеживания выяснилось, что взрослые птицы
двигались к северу вдоль побережья Китая, далее пересекли Жёлтое море и прибыли на
острова, находящиеся на границе Сев. и Юж. Кореи, где и провели гнездовой сезон. Не-
половозрелые птицы держались летом либо на местах зимовки (1 птица), либо в преде-
500
Отряд Листообразные, Семейство Ибисовые
Рис. 91. Область распространения малой колпицы: а — известные места гнездовий, б — районы
зимовки, в — отдельные встречи зимой (по: BirdLife International. Threatend Birds..., 2001 с ориг.
уточнениями).
лах пролётного пути. Одна из этих птиц долетела до гнездовий, где и провела лето (вне
гнездовых колоний).
В результате цветного мечения молодых птиц на о. Фуругельма (2006-2009 гг.) было
выяснено, что птицы южно-приморской «группировки» во время миграций и зимовки по-
сещают значительную (вероятно, большую) часть пространства, которое осваивается ви-
дом в этот период (рис. 92). Наши птицы были встречены в зимнее и в осеннее время в
Японии (на юго-западе о. Хонсю, на юге архипелага Рюкю), на материковом побережье юго-
восточного Китая (у г. Ханчжоу и Хайфын), в Гонконге, во Вьетнаме (устье Красной реки).
О маршрутах движения наших мигрантов пока можно судить только в общих чертах. Их
два — материковый (вдоль восточного и юго-восточного побережья Китая) и островной
(от юго-западного Хонсю вдоль архипелага Рюкю). Одна из птиц, помеченная цветными
метками зимой на о. Окинава (К. Озаки, личн. сообщ.), была встречена нами летом на о.
Фуругельма.
Численность. Начиная с 1989 г. и по настоящее время проводятся ежегодные согласо-
ванные международные учёты абсолютной численности малой колпицы на зимовках
(Dahmer, Felley, 1995; Yu, 2005; Yu, Swennen, 2005).Тенденция изменения численности, ко-
Малая колпица
501
Рис. 92. Разлёт (места осенних и зимних встреч) малых колпиц, окольцованных на о. Фуругельма
(Юж. Приморье). Цифры — количество встреч в каждой точке.
торая выстраивается по результатам учётов, строго положительная: от 288 экз. зимой 1989/
90 гг. (Kennerley, 1990, цит. по: Dahmer, Felley, 1995) до 2041 экз. зимой 2008/09 гг. (Yu,
2010). Однако существует точка зрения (Dahmer, Felley, 1995), что, поскольку условия зи-
мовки ухудшаются повсеместно, данная позитивная картина отражает не реальный рост
численности вида, а есть следствие перераспределения птиц между разными зимовочными
районами и результат более полного обследования зимовок. Так, за 4 года число обследо-
ванных точек (мест зимовки) выросло с 27 до 61 (Yu, 2005). Обнаружены новые места
зимовки, в частности, в Юж. Китае. Число участников учётов с 1993 г. выросло с 20 до 100
(Dahmer et al., 2007). В настоящее время, однако, авторы считают, что позитивная динами-
ка учётов отражает и реальный рост численности вида.
Численность гнездовой группировки из Юж. Приморья находится примерно на одном
уровне. В 2004-2009 гг. здесь ежегодно успешно гнездились от 1 до 3 пар малых колпиц.
Общая численность птиц в гнездовой сезон, с учётом вставших на крыло молодых птиц, в
последние годы достигает 15-16 особей, что составляет менее 1% от численности вида.
В 2008 г. период с 20 по 22 августа вблизи корейской границы учтено 16 особей, боль-
шая часть которых держались на островках приустьевой части р. Туманная (данные Ю.Н.
Глущенко и Д.В. Коробова).
Местообитания. В гнездовое время — небольших скалистые островки, расположен-
ные не очень далеко от материкового побережья; птицы поселяются здесь в смешанных
колониях (BirdLife International. Threatend Birds..., 2001). Кормовые стации — мелководья,
в частности, в приливно-отливной зоне на материке. Основной район гнездования — мел-
ководное Жёлтое море — изначально предоставлял виду идеальные условия для обитания:
502
Отряд Листообразные, Семейство Ибисовые
большое количество островов в прибрежной зоне и различные варианты мелководий. По-
добных условий нет нигде более в пределах ареала вида. В Юж. Приморье кормовые ста-
ции — солоноватые мелководные части лагун, приустьевые участки рек.
Большинство мест зимовки — солоноватые водно-болотные угодья вблизи устьев рек, с
грязевыми отмелями, которые открываются при отливе. В более южных районах — манг-
ровые болота. Кроме того, это песчаные отмели и острова в низовьях рек (например, Крас-
ная река во Вьетнаме) или рисовые поля в холодные зимы (на о. Чеджудо, Юж. Корея)
(BirdLife International. Threatend Birds..., 2001).
Размножение. Малая колпица гнездится в многовидовых колониях, состоящих как из
морских птиц (трубконосые, бакланы, чайки, чистики), так и из колониальных птиц, тяго-
теющих к материку (голенастые и др.) (Chong et al., 1996; BirdLife International. Threatend
Birds..., 2001). На о. Фуругельма (Юж. Приморье) малые колпицы поселились в колонии,
состоящей из бакланов (три вида), чаек (чернохвостая чайка и др.), голенастых (серая
цапля, желтоклювая цапля и др.) и чистиковых (два вида). В непосредственной близости от
гнёзд малых колпиц находились гнёзда чернохвостой и тихоокеанской чаек.
Колониальный вид, однако нередко (и даже обычно) отмечается одиночное гнездование
в колониях других птиц, что, возможно, есть следствие антропогенного воздействия. Из 15
проанализированных поселений птицы гнездились одиночно в 53,3% случаев, поселения
из 2 гнёзд найдены в 20% случаев, а числом более 2 гнёзд — в 26,6% (BirdLife International.
Threatend Birds..., 2001).
Численность в основном районе гнездования (пространство, прилегающее к демилита-
ризованной зоне на границе Сев. и Юж. Кореи) оценивалось в 2009 г. в 443-515 пар. Три
самые крупные колонии насчитывали от 100 до 150-200 пар, однако преимущественно это
были поселения от нескольких до 20-25 гнёзд (Lee, 2010).
Прилет птиц на гнездовья растянут. В Жёлтом море он приходится в основном на ап-
рель (Ueta et al., 2002). Ниже приводится время прилёта в район гнездования (на острова в
демилитаризованной зоне между двумя корейскими государствами) шести взрослых птиц,
помеченных спутниковыми передатчиками: 31 марта — 1 птица; 9-10 апреля — 3 птицы;
22 апреля и 4 мая — 2 птицы. Птицы были помечены в Гонконге и на Тайване.
В Юж. Приморье первые малые колпицы прилетают, по-видимому, где-то в начале ап-
реля. Наиболее ранняя регистрация на гнездовом участке — 11 апреля 200? г. (4 взрослые
птицы в брачном пере).
По данным Chong et al. (2000), половая зрелость у малых колпиц наступает в условиях
неволи в возрасте 2 года 8 месяцев у самок и годом-двумя позднее у самцов. В условиях о.
Фуругельма в размножении реально участвуют примерно 36% птиц в возрасте около 3 лет
(2 года 10-11 мес.; по данным 14 гнездившихся в 2004-2009 гг. пар). У этих птиц сохраня-
ются ювенильные признаки в окраске оперения, «лица», клюва, радужины, а также в разме-
рах хохла (он примерно вдвое короче). Их возраст, возраст начала размножения в наших
условиях, определен по меченой и повторно наблюдавшейся птице. Остальные размножа-
ющиеся птицы находятся в окончательном брачном наряде. Возраст этих особей, видимо,
не менее 4 лет.
В колониях, расположенных в основной части гнездового ареала, птицы с ювенильны-
ми признаками также присутствуют. Однако все участвующие в размножении особи, на-
сколько можно судить по опубликованным фотографиям — это взрослые птицы в оконча-
тельном брачном наряде (Chong et al., 1996; Won Pyong-Oh, 1996; Lee, Kim, 2007). Возмож-
но, ситуация на о. Фуругельма специфична — в Приморье мы имеем дело с начальным
периодом колонизации, и более молодые половозрелые птицы получают возможность для
более ранней самореализации.
Малая колпица
503
Судя по косвенным признакам, малой колпице в определённой степени присущ гнездо-
вой консерватизм. На о. Фуругельма после первой попытки гнездования (2003 г.) птицы
прилетали в последующие три года на одну и ту же небольшую площадку. Скорее всего, это
были одни и те же особи (по крайней мере, старшие из них). Впоследствии поселение
сместилось примерно на 20 м в сторону.
Сведения о демонстрационном поведении отрывочны. На зимовках отмечена демонст-
рация «треск клювом» (bill-clappering) (Severinghaus et al., 1995). Ухаживание на террито-
рии колонии состоит во взаимной чистке оперения (allopreening). Наблюдали, как птицы
перебирали клювом перья друг у друга на голове и шее. Это продолжалось 4-5 мин, после
чего самец встопорщил хохол и, удерживая самку за среднюю часть клюва, инициировал
копуляцию, длившуюся 7-8 сек. Ухаживание и спаривания продолжались до дня перед от-
кладкой последнего яйца. Спаривание происходило в гнезде, птицы при этом сидели (Chong
et al., 1996). На о. Фуругельма приходилось изредка наблюдать элементы ухаживания
(allopreening) и позже, уже при наличии оперяющихся птенцов. Некоторые примеры пове-
дения птиц в колонии приведены на рисунке 93.
О. Токдо, где гнездились 5 пар малых колпиц, при высоте 85 м имеет протяженность
береговой линии 1,2 км. Гнёзда колпиц располагались на крутом скалистом обрыве на вы-
соте 42,4 м над ур. моря. В двух случаях колпицы использовали старые гнёзда серой цапли,
в двух — старые гнёзда птиц своего вида, одно гнездо построили заново. Гнёзда находи-
лись на расстоянии 1,2-1,5 м одно от другого. Размеры вновь построенного гнезда были
50x20 см. Все пары достраивали гнёзда в процессе инкубации и в период выкармливания
птенцов. Самец приносил строительный материал и передавал самке, которая его уклады-
вала (Chong et al., 1996).
На о. Фуругельма птицы также гнездились на скалистой стороне, обращенной в сторо-
ну открытого моря. Гнёзда в 2003-2006 гг. располагались на одном и том же небольшом,
относительно ровном горизонтальном участке скалистого гребня размерами 6-7 на 2-3 м.
Высота гнездовой площадки над уровнем моря составляла около 10 м. Расстояние между
двумя гнёздами (2005 г.) равнялось 5 м. Только один раз гнездо (повторное, 2003 г.) было
выстроено непосредственно у края обрыва на уступе, в выемке; в остальных пяти случаях
гнёзда размещались на ровном участке и несколько в стороне от обрыва. Материалом слу-
жили сухие стебли полыни, выстилкой — более мягкие стебли злаков. Материал для гнез-
да, видимо, собирался поблизости. Размеры одного из гнёзд с яйцами — 52x20 см. Глубина
лотка — 5-7 см. Пространство вокруг гнёзд летом зарастало травой (до 30 см), по которой
бродили и прятались здесь от чаек-соседей подросшие птенцы колпиц.
Откладка яиц на о. Токдо (1995 г.) происходила в 4 гнёздах в период с 25 мая по 8 июня.
В одном гнезде было 2 яйца, в остальных — по 3. Две повторные кладки (после гибели
первых) состояли из 3 и 2 яиц. Птицы откладывали яйца каждый день. Яйца имели оваль-
ную форму и были белого цвета с маленькими коричневыми точками (Chong et al., 1996).
Начало откладки яиц на о. Фуругельма точно не установлено. 6 мая 2006 г. в гнезде
была полная кладка из 3 яиц, которую птица насиживала. Яйцо с о. Фуругельма (коллекти-
ровано 7 июня 2003 г.) имеет белый фон с разбросанными в беспорядке светло-коричневы-
ми и зеленовато-коричневыми пятнами, мелкими и более крупными. Один конец яйца зао-
стрён. Три другие яйца выглядели грязновато-белыми с бледно-бурыми, не очень много-
численными пятнами. Размеры яиц (п=4): 63,9-66,0 х 42,6-45,3, в среднем — 64,9x44,0 мм
(ориг.).
Продолжительность насиживания, по наблюдениям в пяти гнёздах на о. Токдо — 26
дней. Ночью насиживает самка, днем — в основном самец (70% времени). Две пары птиц,
у которых кладки были уничтожены чайками, загнездились повторно через две недели.
504
Отряд Листообразные, Семейство Ибисовые
Малая колпица
505
Рис. 93. Элементы поведения малой колпицы; рисунки выполнены по фотографиям, сделанным ав-
торами очерка (кроме рисунков «г» и «д»): а — тактильный контакт двух молодых птиц, б, в —
элементы ухаживания неполовозрелых особей, г — ухаживание взрослых половозрелых особей на
гнезде (по фотографиям Chong et al., 1996, 2000), д — спаривание (по фотографиям Chong et al.,
1996, 2000), е-з — выпрашивание корма (begging) птенцами разных возрастов; и — кормление птен-
ца; к, л — комфортные движения; м — предполётная тренировка крыльев (молодая птица); н — сон;
о, п — конфликт между соседними парами (о — агрессивная демонстрация, п — защитная реакция
пары, на которую направлена агрессия).
506
Отряд Листообразные, Семейство Ибисовые
Взрослые птицы постоянно находились при птенцах до 11-го дня после вылупления. Птен-
цы полностью оперяются и начинают летать в возрасте около 40 дней; в это время отмеча-
ются их совместные полёты со взрослыми (Chong et al., 1996). В 1995 г. в пяти гнёздах на о.
Токдо до состояния полной оперённости дожили 6 птенцов, что составило 42,8% от числа
отложенных яиц и 66,6% от вылупившихся птенцов. Причины гибели яиц — хищничество
чаек; из 5 яиц птенцы не вылупились. Причина гибели птенцов — падение из гнёзд (в
возрасте 18, 20 и 26 дней).
В 2003 г. на о. Фуругельма погибли обе кладки, первая и повторная. Наиболее вероятная
причина — отсутствие партнёра (вторая птица не была отмечена ни разу). Насиживающая
птица (самка) в конце концов бросила кладку, сначала первую, потом — повторную. В 2004
и 2005 гг. в трёх гнёздах вылупились суммарно за 2 года 7 птенцов (2 и 5 соответственно).
До вылета дожили 4 птенца (57% от числа вылупившихся), три птенца погибли: два пухо-
вика выпали из гнёзд (лежали рядом), один, уже оперяющийся, птенец был найден мёрт-
вым в гнезде. Причина гибели не установлена.
Самостоятельными молодые птицы становятся в середине августа (Severinghaus et al.,
1995; J.R. Chong, личн. сообщ.). Однако в Юж. Корее видели молодых птиц, выпрашивав-
ших корм во второй половине сентября, т.е. незадолго до начала осенней миграции (Yu,
Swennen, 2005). Те же авторы наблюдали как выпрашивание, так и кормление взрослыми
птицами молодой (одной и той же особи несколько раз) уже на месте зимовки в Гонконге (в
ноябре и декабре 2004 г.). Правда, это был единственный случай, отмеченный с начала
наблюдений в 1995 г. (Yu, Swennen, 2005)
В 2006 г. одна из размножающихся на о. Фуругельма пар была смешанной, состоящей
из взрослой особи малой колпицы и колпицы (Pl. leucorodia), ещё не имеющей окончатель-
ного брачного наряда. В течение второй половины июля неоднократно наблюдалось корм-
ление оперённого птенца обеими птицами, вплоть до его благополучного отлёта на мате-
рик (птенец был помечен цветными кольцами). Внешне птенец выглядел как типичная мо-
лодая малая колпица. Впоследствии он был зафиксирован на зимовке в Японии. В тот се-
зон в мае и июле о. Фуругельма посещали ещё не менее четырёх особей колпицы (Р1.
leucorodia) разного возраста. В 2007-2008 гг. на острове регистрировались только малые
колпицы, а в 2009 г. появилась сексуально активная, явно половозрелая самка PL leucorodia,
имеющая, однако ювенильные черты оперения. До размножения дело не дошло, хотя по-
пытки спаривания были. Птица посещала гнездовье около 40 дней.
Питание. Для малой колпицы обычна кормёжка в ночное время, хотя это не всегда
обязательно и зависит от конкретной ситуации. Так, в гнездовой период самцы кормятся в
основном ночью (самка насиживает вечером и ночью, самец — в дневное время). На зи-
мовках в Японии птицы кормятся наиболее активно во время прилива (Chong et al., 1996;
BirdLife International. Threatend Birds..., 2001); кормёжка на зимовках происходит в основ-
ном в стаях. Размер добычи обычно менее 5 см в длину (С. Swennen, in litt, 1999; Yu Yat-
tung, in litt, 1999; цит. no: BirdLife International. Threatend Birds..., 2001). Объекты пита-
ния — рыба, ракообразные, моллюски.
Непосредственных сведений о кормовых объектах малой колпицы в Юж. Приморье
нет. На о. Токдо (Сев. Корея) главным объектом питания птиц в период размножения была
рыба, в основном золотистый бычок, Acanthogobius flavimanus (Chong et al., 1996) — вид,
зарегистрированный и в солоноватоводных водоёмах Юж. Приморья (Спур, Николаева,
2004).
Враги, неблагоприятные факторы. Малая колпица — вид, обитающий в одном из
наиболее населенных районов мира; прибрежные мелководья здесь — зона активной чело-
веческой деятельности. Поэтому основные негативные факторы, воздействующие на ма-
Малая колпица
507
лую колпицу, имеют антропогенный характер. В их числе трансформация местообитаний
во всех основных районах зимовки (Япония, южная часть материкового Китая, Гонконг,
Тайвань, Вьетнам), где преобразованные мелководья используются под застройку или ак-
вакультуру; все формы загрязнения водной среды; охота на зимующих мигрантов; повсе-
местно высокий уровень беспокойства. На гнездовьях в ряде мест (Ляонин, Китай) мест-
ным населением практикуется сбор яиц колониальных видов птиц, в том числе и малых
колпиц (BirdLife International. Threatend Birds..., 2001).
К естественным врагам малой колпицы относятся крупные чайки. Так, в колонии на о.
Токдо (Сев. Корея) две кладки колпиц были уничтожены серебристыми чайками (Larus
argentatus) (Chong et al., 1996).
Хозяйственное значение, охрана. Анализ географической и экологической ситуации в
прибрежной зоне Приморья показал, что в южной его части имеются места, теоретически
пригодные для обитания вида (подходящие острова и кормовые биотопы). Однако тща-
тельное двойное обследование прибрежной зоны залива Петра Великого (с материка, ле-
том 2009 г.) подтвердило присутствие малой колпицы на побережье только в непосред-
ственной близости от известной точки гнездования на о. Фуругельма. Вся прибрежная по-
лоса очень активно осваивается человеком и трансформируется в зону отдыха. Недостаток
кормовых угодий — основное препятствие для формирования в перспективе новых гнез-
довых поселений.
Вид занесён в Красный список МСОП. Поселение малой колпицы на о. Фуругельма
находится в относительной безопасности, поскольку остров входит в состав Дальневосточ-
ного морского заповедника. Потенциальная опасность здесь может исходить от туристов и
фотографов (фактор беспокойства в сочетании с хищничеством крупных чаек). Однако глав-
ная проблема состоит в том, что кормовые биотопы малой колпицы не охраняются, здесь
ведется интенсивная весенняя и осенняя охота на водоплавающих, и нередки случаи брако-
ньерского отстрела птиц вне охотничьего сезона. Первая зарегистрированная в Южном
Приморье птица была найдена 5 августа 1995 г. убитой именно здесь.
Условие сохранения вида в Приморье — охрана водно-болотного угодья «Туманган».
Здесь необходимо создать особо охраняемую природную территорию федерального значе-
ния и придать этой территории статус Рамсарского угодья (Литвиненко, Шибаев, 1996,
Шибаев, Литвиненко, 2007; Litvinenko, Shibaev, 2001). Существующий природный парк
«Хасанский» подобной охраны не обеспечивает, являясь, по сути, далеко не лучшим вари-
антом охотничьего угодья.
Сохранение водно-болотного угодья «Туманган» важно не только для малой колпицы,
но и для целого комплекса мигрирующих и гнездящихся видов, в том числе редких и нахо-
дящихся под угрозой исчезновения (два вида журавлей, желтоклювая цапля и др).
Отряд ФЛАМИНГООБРАЗНЫЕ — PHOENICOPTERIFORMES
Взгляды на положение фламинго в системе класса птиц противоречивы (Карташев, 1974).
Это связано с тем, что по одним признакам фламинго близки к гусеобразным (Anseriformes),
тогда как по другим — к аистообразным (Ciconiiformes). С первыми их связывают общие
признаки строения клюва взрослых птиц, языка, редукция заднего пальца, наличие плава-
тельных перепонок, одновременное выпадение маховых перьев, что на две недели лишает
их способности к полету. Со вторыми — размеры и форма ног (особенно длина цевки),
форма клюва птенцов при вылуплении, затылочные фонтанели, форма и строение грудины
и таза, особенности мускулатуры, безусловно связанные с общей конституцией птицы. Ре-
зультаты серологических и иммунологических исследований позволяли сближать фламин-
го с аистообразными, а исследования вокализации — с гусеобразными. Считалось таким
образом, что группа занимает промежуточное положение между аистообразными и гусе-
образными. Некоторыми авторами (Карташёв, 1974; Иванов, 1976; Cramp, 1977 и мн. др.)
она помещалась в ранге подотряда в отряд аистообразных. Ранее, так же в ранге подотря-
да — в отряд гусеобразных (Мензбир, 1895; Холодковский, Силантьев, 1901; Матвеев, 1935).
Некоторые ископаемые находки в сочетании с чертами кормового поведения, локомо-
ции, гнездовой биологии современных фламинго дают основание говорить о родстве этой
группы с ржанкообразными, а именно с семейством шилоклювковых (Recirvirostridae). Дан-
ные исследований генома однозначно исключают гусеобразных из числа близких родствен-
ников фламинго (и других неогнат), в остальном они также весьма противоречивы. В сис-
теме Ч. Сибли и Б. Монро (Sibley, Monroe, 1990), фламинго — лишь одно из 18 семейств
подотряда Ciconii огромного отряда Ciconiiformes sensu lato. По результатам других моле-
кулярно-генетических исследований, аистообразные и фламинго разделены на уровне пар-
вклассов Coronaves и Metaves; наиболее близкой (сестринской) группой фламинго счита-
ются поганки, а другими соседями по Metaves — фаэтоны, козодои, стрижи, колибри, голу-
би, рябки, некоторые журавлеобразные и гоацин (Ericson, 2008; Hackett et al., 2008 и др.).
К настоящему времени наиболее обоснованным выглядит выделение фламинго в само-
стоятельный отряд, представители которого имеют лишь конвергентное сходство с аисто-
образными и гусеобразными (Степанян, 1990, 2003; del Hoyo et al., 1992; Коблик и др.,
2006). Это реликтовая группа птиц, расцвет которой пришёлся на ранний кайнозой. Поми-
мо единственного современного семейства в отряд, очевидно, следует включить ископае-
мые семейства Agnopteridae, Palaeolodidae, Scaniomithidae. Они были представлены срав-
нительно примитивными формами (известно более 10 видов) с более короткими ногами,
длинными пальцами, прямым или слабоизогнутым клювом. С учётом ископаемых семейств
фламингообразные относятся к самым древним отрядам птиц, обособившимся ещё в верх-
нем мелу. Единственное современное семейство в составе нескольких родов и более чем 11
видов (включая рецентные) известно начиная со среднего эоцена — более 50 млн. лет. Уже
30 млн лет назад появились формы, относящиеся к современным родам. Центр происхож-
дения отряда неизвестен, наибольшее современное разнообразие наблюдается в Юж. Аме-
рике (Коблик, 2001).
Семейство Фламинговые — Phoenicopteridae Bonaparte, 1831
Птицы крупных размеров, весом от 2 до 4,5 кг, длиной тела 90-155 см и размахом кры-
льев 95-165 см. Ноги и шея очень длинные относительно размеров тела (Долгушин, 19606).
Скелет лёгкий, пневматичный, рёбер пять пар, строение грудины и таза, мускулатура крыла
и ноги сходны с таковыми аистов. Череп десмогнатический, есть затылочные фонтанели.
Семейство Фламинговые
509
Голова сравнительно небольшая, клюв же массивный, примерно посередине он перегнут
вниз почти под прямым углом. Большая часть надклювья образована верхнечелюстной ко-
стью (у гусеобразных — предчелюстной). На заднем краю нижней челюсти хорошо развит
processus angularis posterior, длинный и крючкообразный (как у ибисов). Ноздри сквозные,
голоринальные, хорошо развиты надорбитальные солевыводящие железы — фламинго без
ущерба пьют солёную и щелочную воду. Тонкая шея с 19 позвонками (последний входит в
состав спинной кости) в спокойном состоянии изогнута в форме буквы S. Цевка значитель-
но длиннее голени и слегка прогнута вперёд для лучшей остойчивости короткого корпуса с
высоко поднятым центром тяжести. Пальцы короткие, с тупыми когтями, три направлены
вперёд и соединены хорошо развитой перепонкой. Задний палец мал, прикреплён выше
ступни, у видов рода Phoenicoparrus редуцирован. Сонные артерии парные, соединяющи-
еся в проксимальной части шеи. Нижняя гортань трахеобронхиального типа. Под кожей
шеи развит разделённый на четыре камеры доли воздушный мешок, служащий резонато-
ром. Мускульный желудок объёмист и имеет мощную мускулатуру, функционируют круп-
ные парные слепые кишки. Имеется обводящая мышца. Есть рудимент пениса (Дементьев,
1940; Карташёв, 1974; Коблик, 2001).
Челюстной аппарат высоко специализирован в процеживании воды в поисках мелкого
корма. По краям плоского надклювья и объёмного вогнутого подклювья, а также по краям
большого, мясистого языка расположены роговые пластины, которые образуют фильтро-
вальный аппарат (функционально схожий с таковым колпиц и уток).
Крыло относительно короткое, заострённое. При сложенном крыле внутренние второ-
степенные маховые длиннее первостепенных. Первостепенных маховых 12, при полной
послебрачной линьке первостепенные маховые выпадают одновременно и птица на 2-3
недели теряет способность к полёту (как и многие гусеобразные). Хвост короткий, закруг-
лённый и состоит из 12-16 перьев, средняя пара рулевых самая длинная. Оперение доволь-
но мягкое и рыхлое, у стоящей птицы крылья и короткий хвост полускрыты плечевыми
перьями и перьями надхвостья, ниспадающими в виде эффектной «бахромы». Аптерии и
птерилии покрыты пухом. На перьях хорошо развиты побочные стволы. Копчиковая желе-
за довольно крупная, оперённая. Секрет копчиковой железы не предохраняет оперение
фламинго от намокания
Голень на большей своей части не оперена. Кольцо вокруг глаз, уздечка и подбородок
также лишены оперения. Цевка и основная часть пальцев покрыты поперечными пластин-
ками. Окраска взрослых преимущественно белая, с более или менее развитым розовым
оттенком, или красная, маховые чёрные, цвет клюва, ног, неоперённых участков кожи, ра-
дужины разнообразен и видоспецифичен. Розово-красные оттенки оперения обусловлены
пигментом астаксантином из группы липохромов-каротиноидов, близким к красным пиг-
ментам ракообразных. Этот пигмент поступает в организм птиц с кормом, в составе кото-
рого различные рачки занимают ведущее место. Пигмент очень нестойкий и быстро разру-
шается на свету (Карташев, 1974; Коблик, 2001). Полового и сезонного диморфизма в ок-
раске нет.
Фламинго населяют высокосолёные и щелочные озёра, нередко с высоким содержани-
ем едкого натра, гипса и сероводорода, а также прогреваемые мелководные морские лагу-
ны, солёные прибрежные болота и дельты с высокой биологической продуктивностью. При
кормлении на мелководье птица опускает голову так, что надклювье находится под повер-
хностью субстрата. Поводя головой в разные стороны, а языком действуя как поршнем,
фламинго процеживает воду и ил. На больших глубинах в субстрат погружается вся голова,
а иногда и шея до плеч. В состав пищи входит различный фито- и зоопланктон (сине-зелё-
ные водоросли, мелкие ракообразные, черви, моллюски). Очевидно, скопления пищи нахо-
510
Отряд Фламингообразные, Семейство Фламинговые
дят при помощи зрения, поэтому активны днём. По земле и мелководью передвигаются
обычно медленно, но при необходимости довольно быстро бегают. Часто стоят на одной
ноге. Довольно хорошо плавают, несмотря на намокаемое оперение. Полёт ровный с раз-
меренными взмахами; парить фламинго не могут, взлетают после разбега. Во время полета
шею и ноги птицы вытягивают в одну линию (Долгушин, 1960а). Голос фламинго похож на
гоготание гусей.
Строго колониальные птицы, гнездование отдельными парами неизвестно. Иногда сме-
шанные колонии образуют 2-3 вида фламинго, у которых во избежание пищевой конкурен-
ции выработались цедильные аппараты, «нацеленные» на разный размерный класс добы-
чи. Фламинго моногамны, пары образуются на один сезон. Гнёзда представляют собой стол-
бообразные или конусовидные кучи из ила и ракушечника, высотой до 50-60 см, распола-
гаются на мелководьях и островах в непосредственной близости друг от друга. Они еже-
годно строятся заново, хотя расположение колонии не меняется. В кладке 1-2, редко 3 круп-
ных белых яйца. В насиживании и выкармливании птенцов участвуют оба партнёра, инку-
бация длится 27-33 дня, в жарких местностях она превращается скорее в создание тени над
кладкой и защиту от окружающих экстремальных температур. Птенцы выводкового типа,
вылупляются зрячими, с прямым коротким клювом, короткими толстыми ногами, покры-
тые густым белым пухом. Родители выкармливают птенцов полупереваренной пищей, сме-
шанной с выделениями стенок пищевода и желудка, причём продолжают подкармливать
молодых вплоть до подъёма на крыло. К началу второго месяца жизни птенцы надевают
второй пуховый наряд — серого цвета, ещё через две недели у них начинает развиваться
оперение. Птенцы в ювенильном наряде нередко сбиваются в группы, образуя в гнездовой
колонии своеобразные «детские сады». Способными к полёту становятся на 65-75-й день
жизни, к этому времени у них окончательно формируется и фильтровальный аппарат клю-
ва. Молодые птицы долгое время остаются серыми, с тёмными ногами и клювом. Взрос-
лый наряд надевают лишь на 3-м году жизни, вероятно, одновременно с наступлением
половой зрелости, хотя, по некоторым данным, крупные виды впервые приступают к раз-
множению лишь на 5-6-й год, а мелкие — на 3^4-й год жизни. Медленный темп воспро-
изводства и позднее созревание компенсируются долгожительством: по данным кольце-
вания, многие птицы живут свыше 20-30 лет, отдельные особи, вероятно, более 50 лет
(Коблик, 2001).
Семейство включает три рода {Phoenicopterus, Phoeniconaias, Phoenicoparrus) с 5-6
видами, населяющими главным образом тропические и субтропические зоны Евразии,
Африки, Центр, и Юж. Америки (в Австралии вымерли в историческое время). Короткок-
лювые фламинго {Phoenicoparrus, 2 вида) — узкоареальные эндемики пояса альтиплано
или пуны (3500—4500 м) в Андах. Птицы, гнездящиеся у северных границ ареала, соверша-
ют регулярные сезонные миграции. В тропических зонах живут оседло или совершают
небольшие перемещения, в горах — вертикальные кочёвки. Видов, находящихся под гло-
бальной угрозой исчезновения, нет. На территории Сев. Евразии семейство представлено
единственным видом.
Розовый фламинго
511
Род Phoenicopterus Linnaeus, 1758
Розовый фламинго
Phoenicopterus roseus Pallas, 1811
Phoenicopterus roseus Pallas, 1811, Zoographia Rosso-Asiatica, 2, c. 207, устье Волги.
Greater flamingo (англ.), Flamingo (нем.), Flamant rose (фр ).
Син. (лат.): Phoenicopterus major Dumont, 1820; Phoenicopterus europaeus Swainson, 1837;
Phoenicopterus plinii Reichenbach, 1852 (1853); Phoenicopterus platyrhynchus C.L. Brehm,
1855; Phoenicopterus andersoni Brooks, 1875.
Син. (рус.): обыкновенный фламинго, краснокрыл, красный гусь.
Статус. Гнездящийся перелётный вид, в южных частях региона — зимующий.
Общая характеристика и полевые признаки. Крупные птицы с длинной, S-образно
изогнутой шеей, очень высокие на ногах. По размерам тела сравнимы с гусями. Голень
заметно длиннее цевки. Стоящая на земле с вытянутой шеей птица достигает в высоту
1,5 м (Долгушин, 1960а). Голова маленькая, с массивным, коленообразно изогнутым вниз
клювом. Глаза небольшие, радужина жёлтая или свинцово-серая (у молодых). Взрослые
птицы окрашены в белый цвет с розовым оттенком. Верхняя и нижняя поверхности крыла
окрашены наиболее интенсивно — от ярко-розового до красного, что хорошо заметно в
полете. Концы крыльев и их задний край чёрные. Клюв розовый с чёрным кончиком. Ноги
ярко-розовые.
Полёт ровный и быстрый (до 60 км/ч) с размеренными взмахами крыльев, без планиро-
вания и парения. В полёте шея и ноги вытянуты в одну линию. Ноги далеко выступают за
обрез короткого хвоста. Летают как небольшими группами в несколько особей, так и боль-
шими стаями, чаще всего до 250 особей, как правило, обособленно от других видов птиц.
Полёт совершают как днём, так и ночью (особенно во время миграции). Небольшие и сред-
ние по размеру группы фламинго (15-120 особей) образуют в полёте различные линейные
построения — углы, клинья, цепочки и линии. При незначительных местных перелётах
стая фламинго часто меняет форму, может образовывать скученные или рыхлые неупоря-
доченные построения (Молодовский, 1997, 2001). Кормятся фламинго, медленно передви-
гаясь по мелководью и погружая клюв в воду. Хорошо плавают. Молодые при особой опас-
ности могут и нырять, но очень быстро выныривают (Долгушин, 1960а).
Держатся стаями или группами на мелководьях солёных озёр, лиманов и на морских
отмелях. Гнездятся плотными колониями.
Голос — различные варианты гоготания, при большом скоплении переходящего в не-
громкий гул.
Сходных видов в фауне России и сопредельных регионов нет. В природе легко отличим
от других птиц благодаря своеобразным пропорциям тела и окраске.
Подвидовая систематика. Монотипический вид
Замечания по систематике. В состав рода Phoenicopterus (Linnaeus, 175 8) входит над-
видовая группировка типа superspecies с двумя allospecies: розовым фламинго (Phoenicopterus
roseus Pallas, 1811) и красным фламинго (Phoenicopterus ruber Linnaeus, 1758). В этой трак-
товке, принятой здесь, обе формы считаются монотипическими. Розовый фламинго при-
надлежит к фауне Старого Света, красный — к фауне Нового Света. Ареалы их разобщены
и отстоят друг от друга на очень большом расстоянии (Степанян, 2003). Вместе с тем розо-
вый фламинго чаще рассматривается систематиками как один из двух подвидов Phoenico-
pterus ruber — Ph. ruber roseus (Спангенберг, 1951; Карташёв, 1974; Cramp, 1986; del Hoyo
512
Отряд Фламингообразные, Семейство Фламинговые
et al., 1992; Johnson, 1997; Svensson et al., 2002; Dickinson, 2003). В этом случае фламинго
Нового Света, населяющих Карибский бассейн и Галапагосские о-ва, относят к номина-
тивному подвиду — Ph.ruber ruber. Реже в состав Ph. ruber sensu lato включают и третий
вид рода Phoenicopterus — чилийского фламинго (Ph. chilensis Molina, 1782).
Описание. Окраска. Половой диморфизм в окраске оперения отсутствует. Оперение
птиц во взрослом наряде белое с нежным розовым оттенком, который с возрастом стано-
вится интенсивнее (Спангенберг, 1951; Долгушин, 1960а). Подмышечные и часть верхних
и нижних кроющих крыла ярко-розовые (до красного); первостепенные и ближайшие вто-
ростепенные маховые чёрные. Ноги интенсивно розовые. Голые места на голове и большая
часть клюва розоватые. Конец клюва чёрный. Радужина жёлтая. Самцы крупнее самок.
Сезонной смены нарядов нет, но у птиц в свежем пере розовые тона выражены более ин-
тенсивно (Cramp, 1977).
Первый пуховой наряд. Птенцы похожи на гусят. Они рождаются зрячими, с прямым
коротким клювом и короткими толстыми ногами (Johnson, 1987). Первые 2-3 недели птен-
цы покрыты густым белым пухом, который затем сменяется на второй пуховой наряд серо-
го цвета. На пятой неделе начинается рост пера первого гнездового наряда (Спангенберг,
1951). Клюв у птенцов в двухнедельном возрасте начинает постепенно искривляться. У
двухмесячных птенцов клюв похож по своему строению на клюв взрослых птиц (Кустано-
вич, 1986). Вес птенцов в возрасте около месяца 680-1 450 г (среднее по 11 экземплярам —
960 г; середина июля, оз. Тенгиз) (Долгушин, 1960а).
Гнездовой наряд. Молодые птицы снизу белые, с рыжеватым налетом, сверху серовато-
бурые, с жёлтыми пятнами. Голова и шея серо-коричневые, с белым крапом; мантия и ло-
патки коричневатые; испод крыла розоватый; концы крыльев чёрные; кроющие крыла бу-
ровато-серые с белым рисунком. Клюв свинцово-серый, ноги почти чёрные, радужина чер-
новатая. Шея и ноги у молодых несколько короче, чем у взрослых.
Взрослый наряд молодые надевают в возрасте от 3,5 до 5,5 лет (Johnson, 1987).
Строение и размеры. Основные промеры взрослых особей приведены в таблице
44.
Размеры взрослого самца, добытого в Татарстане 30 сентября 1958 г. (мм): длина
крыла — 423, клюва — 126, цевки — 367. Неполовозрелая особь, залетевшая в мае на
Таблица 44. Размеры взрослых фламинго, в мм
Пол Длина крыла Длина клюва Длина цевки Источник
п lim М п lim М п lim М
самцы 8 406-464 428 7 117-125 121 8 291-373 323 Cramp, 1977
самки 6 360-396 380 7 112-121 116 7 232-280 259 Cramp, 1977
самцы 12 360-445 12 120-135 12 240-365 Witherby et.al. цит.: no Cramp, 1977
самки — 355^125 Witherby et.al. цит.: no Cramp, 1977
самцы 1 420 132 265 Adamian, Klem, 1999
самки 1 390 118 250 Adamian, Klem, 1999
самцы 21 435^190 459 Спангенберг,
и самки 1951
самцы 350-470 111-135 260-370 Долгушин,
и самки 1960a
Розовый фламинго
513
Средний Амур, имела длину крыла — 424, хвоста — 148, цевки — 252, клюва — 146 мм
(Дугинцев, Панькин, 1994).
Вес взрослых самцов в мае в зал. Кара-Богаз составлял 3 900-4 400 г; самок — 3 050-
3 400. Все птицы, добытые в конце мая в заливе, имели большие жировые запасы. Жир
слоем 10 мм покрывал всё туловище, включая спину и грудные мышцы (Исаков, 1948а).
Вес самки в июле на оз. Кургальджино составил 2 100 г; самцов в июле на оз. Тенгиз —
2 700-4 100 г, в среднем 3 500 г (п=16), вес двух самок там же — по 3 100 г. Вес отдельных
взрослых особей достигает 4 500 г (Долгушин, 1960а). В Камарге фламинго имели следую-
щий вес: самцы — 3 000-4 100 г, в среднем 3 579 г (п=13), самки — 2 100-3 300 г, в среднем
2 525 г (n=12) (Gallet, 1950, цит. по: Cramp, 1977).
Неполовозрелые птицы имеют в среднем меньший вес: в мае 1938 г. в Кара-Богаз-Голе —
2 600 г; в Ленкорани в феврале 1931 г. хорошо упитанные птицы — от 1 960 до 2 400 г
(Исаков, 1948а).
Линька. Изучена недостаточно и, вероятно, характеризуется высокой степенью измен-
чивости.
Известно, что у взрослых фламинго в году две линьки: полная послебрачная начинается
в конце периода размножения в июле и продолжается, по одним данным, до конца августа
(Кустанович, 1986), по другим — до ноября-декабря (Спангенберг, 1951). Вторая линька —
предбрачная, охватывает лишь мелкое оперение. Она начинается в январе и идёт до мая
(Спангенберг, 1951).
Время смены маховых перьев зависит от возраста и гнездового статуса птиц (Shannon,
2000а). Гнездящиеся птицы имеют последовательную смену полетного оперения. Перво-
степенные маховые выпадают поочередно и симметрично, парами, по одному на каждом
крыле, что позволяет птицам сохранить способность к полёту. При смене дистальных ма-
ховых фламинго взлетают с трудом. По-видимому, одновременное выпадение маховых пе-
рьев, приводящее к временной потере способности к полёту, свойственно молодым и не
участвующим в размножении фламинго (Андрусенко, 1981; Кустанович, 1986).
Оперение на одних птерилиях меняется 3 раза в году, на других — 1 раз в 2 года.
Каждая птерилия имеет определённые периоды смены, которые не всегда соответствуют
циклам других. В результате круглый год в стае есть линяющие птицы (Shannon, 2000а).
Отгнездившиеся фламинго или имевшие неудачную попытку гнездования держатся во
время линьки группами поблизости от основной колонии (Cramp, 1977). Значительные
скопления линяющих фламинго известны на оз. Тенгиз (Долгушин, 1960). Неразмножа-
ющиеся птицы, как правило, линяют далеко от колоний, в местах линочных скоплений.
Наиболее известным ранее местом массовой линьки фламинго был залив Мертвый Кул-
тук на восточном побережье Каспийского моря, где с половины июля фламинго собира-
лись в несметном количестве (Мензбир, 1918). В настоящее время скопления линяющих
фламинго регулярно отмечаются у побережий Сев. Прикаспия от пос. Прорва до залива
Комсомолец (Русанов, 2004). Численность линяющих птиц в разные годы сильно колеб-
лется. В 1984 г. она составила 65,5 тыс. особей, а в 1988 — только 1,1 тыс. По мнению
Г.М. Русанова (20046), это связано с тем, что в годы с благоприятным ходом размноже-
ния большая часть птиц держится в местах гнездования за пределами побережий Кас-
пийского моря.
Первую линную птицу на оз. Тенгиз отлавливали 22 июля. В июле наблюдали массовую
линьку фламинго. Последние линные птицы, не успевшие полностью потерять старые ма-
ховые, отмечались 16 августа (Андрусенко, Жулий, 1977).
Распространение. Гнездовой ареал разобщён (рис. 94). Распространение
прерывистое, спорадическое в тропических и субтропических регионах Европы, Африки и
514
Отряд Фламингообразные, Семейство Фламинговые
Рис. 94. Область распространения фламинго: а — места гнездования; б — места кочёвок в зимний
сезон. Цифрами обозначены отдельные популяции этих птиц (см. раздел «Численность»).
Азии. Гнездится отдельными колониями, далеко отстоящими одна от другой. В Африке
поселения известны на островах Зелёного Мыса, по побережьям и на внутренних водо-
ёмах в Сев. Мавритании, на юге Марокко, в юж. Тунисе, Египте, Алжире, Кении, Танза-
нии, Намибии, Ботсване, ЮАР. На востоке и юге Африки розовый фламинго нередко
образует смешанные колонии с малым фламинго (Phoeniconaias minor Geoffroy, 1798),
питающимся в основном сине-зелёными водорослями и не составляющим ему трофи-
ческой конкуренции.
В Европе наиболее значительные гнездовья находятся в Средиземноморье, во Фран-
ции в дельте р. Роны (природный парк Камарг) и в дельте р. Вар; в Испании в дельте р.
Эбро, в Андалусии в Кадисском зал. (Уэльв и дельта Гвадалквивира) и в районе Малаги; на
португальском побережье в устье р. Тежу. Известны также гнездовья на юге Сардинии
(Johnson, 1997; Gurssu, 2000). С 1984 года зарегистрирована колония в лагуне Орбетелло,
Италия (Dall’Antonia et al., 1996). Около 25 лет назад в прибрежных водах Нидерландов
(Lensink, 1996), Германии и Дании начали гнездиться улетевшие из зоопарков розовый,
чилийский, и красный фламинго. Эти свободноживущие группы фламинго хорошо адапти-
ровались к местным условиям. Гнездятся в удалённых от моря колониях, зимуют в дельтах
рек Рейн, Мез, Шельда (Тгеер, 2000).
Розовый фламинго
515
В зарубежной Азии колонии известны в западных и центральных районах Турции, в
Иране на оз. Урмия, на побережье северной части Персидского зал., на озёрах Юж. Афга-
нистана, в сев.-зап. Индии и в Шри-Ланке.
На территории бывшего СССР основным местом постоянного гнездования фламинго
служат горько-солёные бессточные озёра Центр. Казахстана (рис. 95). Оз. Тенгиз (50°30'
с.ш.) — самая северная точка распространения этого вида и единственный водоём, на кото-
ром гнездование птиц центрально-казахстанской части популяции наблюдается довольно
регулярно (Спангенберг, 1951; Долгушин, 1960; Елкин, 1969; Волков, 1972,19756,1977а,б;
Андрусенко, Жулий, 1977; Андрусенко, 1979; Стоцкая, Кривенко, 1988; Кошкин, 2004). В
периоды трансгрессии колонии перемещаются с оз. Тенгиз на оз. Челкар-Тениз (Сушкин,
1908; Спангенберг, 1951; Чельцов-Бебутов, 1958; Волков, 19756; Варшавский и др., 1977;
Кривицкий и др., 1985), однако, в отдельные годы это озеро полностью пересыхает (Елкин,
1969; Волков, 19756, 1977а). Фламинго с нелётными молодыми наблюдали на оз. Ащитаз-
тысор в июне 1954 г. М.Д. Бирюков (Долгушин, 1960а) и А.А. Слудский (1960). В 1957 г.
это озеро было обнаружено пересохшим (Чельцов-Бебутов, 1958), и в последующие годы
как место расположения колоний фламинго не упоминается. По данным Е.Н. Волкова
(1977а,б), в 1969-1975 гг. оно оставалось сухим. Оз. Жаман-Акколь, на котором была обна-
ружена колония в 1956 г. (Чельцов-Бебутов, 1958), также время от времени подвержено
полному пересыханию (Долушин, 1960а; Волков, 1972; Даниленко и др., 1977). Периоди-
чески появлялись данные о гнездовании фламинго на северо-восточном побережье Каспия
от устья Эмбы до п-ова Бузачи (Исаков, 19486; Спангенберг, 1951; Долгушин, 1960а; Неру-
чев, Худяков, 1967; Пославский и др., 1977). Предположительно фламинго гнездились здесь
в 1985 г. (Стоцкая, Кривоносов, 1988). В 1939 г. фламинго гнездились в Туркмении в зал.
Рис. 95. Область распространения фламинго в Вост. Европе и Сев. Азии: а — места гнездования; б —
граница области регулярных встреч в сезон гнездования; в — места зимовок.
516
Отряд Фламингообразные, Семейство Фламинговые
Кара-Богаз-Гол, но с 1939 г. гнездование этих птиц в заливе прекратилось, видимо, вслед-
ствие резкого повышения солености воды залива (Исаков, 19486). Значительные колонии
располагались в 1954-1956 гг. и в более поздние годы в районе зал. Сев. Челекен в Красно-
водском заповеднике (Залетаев, 1960). В 1970-1971 гг. Б.В. Сабиневский наблюдал попытку
гнездования фламинго на оз. Сарыкамыш, на островах в районе впадения Дарьялка. Коло-
ния погибла в результате браконьерства и повышения уровня воды в озере. Хотя сейчас на
оз. Сарыкамыш нет подходящих условий для гнездования фламинго, птицы могут появить-
ся здесь при понижении уровня озера и появлении мелководий с топкими островами (Чер-
нов, 1990а, 19906, цит. по: Букреев, 1997). В настоящее время фламинго не гнездится на
территории Туркменистана (Букреев, 1997).
В Азербайджане в 1952 г. фламинго гнездились на оз. Ax-Чала в Муганской степи, но в
дальнейшем озеро пересохло (Patrikeev, 2004). В 1982-1984 гг. фламинго гнездились в
Кызыл-Агачском заповеднике на юго-западном побережье Каспия (Литвинова и др., 1984;
Стоцкая, Кривенко, 1988), однако поселение исчезло из-за хищничества шакалов (Canis
aureus). В 1985 г. колония здесь не была найдена, но в июле наблюдали взрослых птиц с
молодыми (Стоцкая, Кривенко, 1988).
Негнездящиеся птицы летуют преимущественно у северо-восточного побережья Кас-
пия (Исаков, 19486; Молодовский, 1963; Гисцов, 2003а). Обычны встречи фламинго в лет-
нее время и на восточном побережье в районе зал. Кара-Богаз-Гол, в меньшем количестве -
в Красноводском зал. (Пославский и др., 1977). На территории Центр. Казахстана в гнездо-
вой период размещается в подавляющем большинстве половозрелая часть популяции.
Молодые фламинго, рассредоточиваясь по пути пролёта и на зимовках, в область гнездова-
ния проникают в небольшом количестве. Неполовозрелые птицы отмечались также в низо-
вьях Тургая, на оз. Жаман-Акколь, оз. Кызыл-Коль и в южной части оз. Сарыкопа (Волков,
1975а; Ауэзов, 1977; Ауэзов, Виноградов, 1986).
В летнее время единичные особи отмечались в Предкавказье (долине оз. Маныч-Гуди-
ло) и Дагестане (в низовьях рек Терек, Сулак и Шура-Озень, на оз. Аджи и в Рутульском р-
не в высокогорьях Вост. Кавказа) (Красовский, 1932; Пишванов, Прилуцкая, 1988; Хохлов,
19886). В конце лета встречи фламинго здесь и на западном побережье Каспийского моря
более часты, что, вероятно, связано с разлётом птиц из районов гнездования (Казаков и др.,
2004). Небольшие стайки фламинго встречались в июле и августе в Армении (Adamian,
Klem, 1999).
В Узбекистане встречается не ежегодно, в различных местах, небольшими группами.
Чаще всего наблюдается на Аральском море. Имеются сведения о встречах фламинго в
Амударьинском заповеднике, в Ферганской долине, долинах Зеравшана и Сыр-Дарьи (Мек-
ленбурцев и др., 1987). Весной 1987 г. при авиаучёте по Аму-Дарье наблюдали 58 птиц в
районе Чарджоу (Стоцкая, Кривоносов, 1988).
Довольно часто фламинго залетают далеко от мест гнездования. Веер разлёта фламинго
довольно широк. В 1935 г. было несколько залетов в Полтавскую обл. (Гавриленко, 1958).
Несколько раз залетали фламинго в Ленинградскую обл. (в 1933, 1946, 1947 и 1972 гг.), в
Эстонию и Латвию (Мальчевский, Пукинский, 1983). Два года подряд в августе и сентябре
(в 1965 и 1966 гг.) наблюдались фламинго на оз. Ильмень (Леус, 1972). В ноябре 1959 и
октябре 1969 гг. фламинго встречали и добывали в Кировской обл. (Плесский, 1960; Сотни-
ков, 1999). Имеются сведения о залетах этих птиц в Нижегородскую обл. — в сентябре
1921 г. и августе 1951 г. В 1921 г. фламинго был добыт в Татарии (Пузанов и др. 2005). В
августе 1959 г. в Татарии было встречено 6 фламинго (Попов, 1977). В октябре 1990 г.
молодой фламинго был пойман в Пермской обл. (Лапушкин и др., 1995). Поздней осенью
1959 г. фламинго был обнаружен в Тарском р-не, а в 1987 г. так же поздней осенью отмечен
Розовый фламинго
517
залёт в Тюменскую обл. (Якименко, 1998). В ноябре 1998 г. отловлена птица этого вида в
Томской обл. (Блинова и др., 2001). Во второй половине прошлого века эти птицы неоднок-
ратно залетали в Кемеровскую обл.: в 1963 г. была добыта одна птица из небольшой груп-
пы фламинго; в 1965 г. пара держалась всё лето на границе с Новосибирской обл.; в 1966 г.
там же наблюдали стайку фламинго; осенью 1968 г. отмечено около 10 особей; в январе
1972 г. найдена 1 погибшая птица (Гагина, Скалой, 2000). В 1988 г. двухлетний фламинго в
течение лета держался в низовьях р. Зеи на Среднем Амуре (Дугинцев, Панькин, 1994). В
Приморском крае одиночные особи наблюдались в устье р. Киевка в конце апреля и в нача-
ле мая 1977 г. (Назаров, Куринный, 1981), а затем в первой декаде апреля 2004 г. (Шохрин,
2007); стая, численностью около 20 особей, встречена в районе устья р. Туманная в конце
октября 1984 г. (Литвиненко, Шибаев, 1999). Отмечались залёты фламинго на берега Бай-
кала, на р. Лену (Усть-Кутский район) и в Иркутскую обл. (Гагина, 1962).
Подавляющее большинство случаев дальних залетов фламинго приходится на время
осенней миграции. Среди залётных птиц большую часть составляют молодые особи (Иса-
ков, Формозов, 1946). Во время залётов птицы встречаются в несвойственных им биото-
пах: на песчаных отмелях рек, влажных травяных лугах, в поймах среди кустарников и
близ жилья человека. Основная масса птиц, попавших в непривычные условия, как прави-
ло, погибает (Долгушин, 1960а).
Зимовки. Область зимовок европейской популяции фламинго лежит в Средиземно-
морье, Сев. и Сев.-Запа. Африке (Волков, 1977в). В Египте на охраняемой территории За-
раник численность зимующих фламинго составляет около 10 тыс. (Salama, Grieve, 1996).
Зимовки центрально-казахстанских фламинго располагаются на Азербайджанском (Кы-
зыл-Агачский заповедник), Туркменистанском (Красноводский заповедник) и Иранском
побережьях Каспийского моря (Спангенберг, 1951; Долгушин, 1960а), но большая часть
птиц отлетает южнее - в Ирак, Афганистан, Индию, Пакистан и Египет (Долгушин, 1960;
Волков, 19776,в; 1979).
Численность птиц на зимовках сильно колеблется по годам в зависимости от погодных
условий. Так, численность фламинго, учитываемых на зимовках в Кызыл-Агачском запо-
веднике, составила: в 1963 г. — 1 700 особей (Олигер, 1967), 1984/85 г. — 13 500 особей,
1989/90 г. — 3 300,1990/91 г. — 12 970,1994/95 г. — 8 900,1995/96 г. — 5 200 птиц (Patrikeev,
2004). В январе 2004 г. в Кызыл-Агачском зал. зимовало около 6 000 птиц, на оз. Аджино-
ур — 3 500, на Акгёле — 1 200 (Султанов, Джаббарова, 2006). В последние годы зимовки
вида отмечены на восточном побережье Среднего Каспия, на оз. Караколь у города Актау
(Гисцов, 2003а).
В 1980-1987 гг. фламинго отмечались на зимовке на Осетинской равнине (Комарова,
Комаров, 1988). Одиночные птицы изредка задерживаются на зимовке на юге Дагестана.
2 января 2006 г. птица, летящая на юг, отмечена на побережье Каспия в устье реки Самур.
5 января 2006 одиночный фламинго держался на мелководье оз. Аджи.
Места зимовок одних и тех же особей фламинго постоянны, и птицы могут менять
зимовочные районы только в связи с резкими изменениями условий среды обитания или
под воздействием фактора беспокойства. Птиц, окольцованных на каспийских зимовках,
находили в дальнейшем либо в местах мечения, либо в непосредственной близости от
них. Не смотря на такую привязанность к местам зимовок, в суровые зимы, которые пе-
риодически случаются на Каспии, происходит откочёвка фламинго в южном направле-
нии (Волков, 1979).
Миграции. Фламинго, принадлежащие к популяциям, гнездящимся в умеренном по-
ясе, совершают регулярные сезонные миграции. К ним относятся и птицы центрально-
казахстанских колоний.
518
Отряд Фламингообразные, Семейство Фламинговые
Распадение зимовок на юго-западе Каспия происходит обычно в марте, но иногда про-
движение птиц к северу начинается уже в феврале. Весенний отлёт фламинго из Кызыл-
Агачского заповедника заканчивается в конце апреля (Греков, 1962; Морозкин, 19756). В
конце марта и начале апреля отмечались крупные скопления фламинго на оз. Аджи (Каза-
ков и др., 2004). В устье р.Самур на юге Дагестана весенний пролёт проходит с конца марта
до начала мая, причём основная масса птиц пролётает в марте и начале апреля (Бутьев и
др., 1989; Михеев, 1997). В марте же начинают появляться первые пролётные фламинго на
Юж. Мангышлаке. Максимальное число мигрирующих птиц этого вида в день наблюдали
здесь со второй-третьей декады апреля до конца мая (Молодовский, 1963).
Таким образом, весенний прилёт фламинго, гнездящихся на озёрах Центрального Ка-
захстана, довольно сильно растянут и может продолжаться около двух месяцев. Первых
птиц на оз.Тенгиз регистрировали с конца марта - начала апреля. Прилёт птиц продолжает-
ся до конца мая. После тяжелых зимовок появление первых особей регистрируется в более
поздние сроки (Андрусенко, 1979).
Большая часть зимующих в Азербайджане фламинго пересекают Каспийское море в
южной и средней его части. Лишь небольшая часть птиц мигрируют вдоль западного побе-
режья Каспия до Юж. Дагестана и в отдельные годы до Аграханского зал., откуда они на-
правляются к восточному побережью Каспийского моря. Весной некоторые птицы, оче-
видно, уклоняются от традиционного пути пролёта и проникают по водоёмам Маныча до
восточных границ Ростовской обл. (Казаков и др., 2004). В устье Урала пролётающих фла-
минго наблюдали в течение всего мая (Долгушин, 1960а). В это же время наблюдается
активный пролёт птиц в районе северного побережья Аральского моря (Спангенберг, 1951;
Долгушин, 1960а; Варшавский и др., 1977) вдоль северного чинка Устюрта (Сушкин, 1908;
Бостанжогло, 1911; Варшавский и др., 1977). В 1950-1960 гг. в районе г. Аральска регуляр-
но регистрировали пролётные стаи фламинго от 160 до 770 особей. Преимущественное
направление пролёта птиц здесь во время весенней миграции — северо-восток. Весной
1972 г мигрирующие стаи фламинго (до 60 птиц) наблюдали на плато между Б. и М. Барсу-
ками (Варшавский и др., 1977). Слабый пролёт отмечен на восточном берегу Аральского
моря в районе Кызылкумов (Березовский, 1983) и на Сыр-Дарье (Волков, 19776).
Осенние миграции начинаются ненаправленными летними кочёвками молодых и поте-
рявших кладки птиц во второй-третьей декадах июля (Андрусенко, 1986). Часть птиц ле-
тит от оз. Тенгиз в западном направлении — через Челкар-Тенгиз и Жаман-Акколь, Сред-
нее течение Эмбы, Сев.-Вост. Каспий, Мангышлак. Отсюда часть птиц летит вдоль восточ-
ного побережья к югу (Андрусенко, 1986), остальные пересекают Каспий в средней части
и попадают на западное побережье моря в районе оз. Аджи (на юге Дагестана). После
короткого отдыха, птицы, по-видимому, транзитно летят до Кызыл-Агачского зал., поскольку
пик осеннего пролёта и прилёта там также приходится на конец октября (Морозкин, 19756;
Бутьев и др., 1989). Осенний пролёт на юге Дагестана (в устье р. Самур) проходит в октяб-
ре-ноябре, но большая часть фламинго пролётает во второй половине октября. Птицы здесь
не останавливаются, летят почти исключительно над морем, стаями от 10-15 до 500 осо-
бей (Бутьев и др., 1989).
Иногда одиночные особи и небольшие стаи летят через Сев. Прикаспий; основной про-
лёт завершается здесь в октябре (Русанов, 2002). Эти птицы останавливаются осенью на
водоёмах Калмыкии, Ставрополья и Сев. Дагестана (Хохлов, 1988; Пишванов, Прилуцкая,
1988). В редких случаях здесь отмечаются массовые скопления мигрирующих птиц. Так,
стая из молодых и взрослых птиц, насчитывающая более 1 тыс. особей, держалась на мел-
ководьях Чограйского вдхр. (Аргзирский р-н Ставропольского края) с конца августа до
середины октября 1977 г. (Хохлов, 1988).
Розовый фламинго
519
Данные, полученные в результате кольцевания и визуальных наблюдений, дают основа-
ния предполагать наличие прямого пролётного пути от низовий Тургая через область Араль-
ского моря к юго-восточной оконечности Каспия и в Иран (Волков, 1979; Караваев, Бело-
усов, 1983; Андрусенко, 1986).
Часть птиц (вероятно, небольшая) летит от оз. Тенгиз на юг и юго-восток в Афганис-
тан, Пакистан и Индию, о чем свидетельствуют многочисленные встречи на данном на-
правлении небольших групп и одиночных особей фламинго (Абдусалямов, 1971; Гисцов и
др., 1977; Андрусенко, 1986).
Местообитания. Жизнь всех 6 видов фламинго связана с водоёмами с высокой концен-
трацией солей (от 40 до 280 промилле). Разнообразие водных живых организмов в них
крайне ограничено. Фламинго используют эти водоёмы как для кормежки, так и для гнез-
дования (Goesner, 1987). Тем не менее, птицы из тех европейских колоний, которые образо-
ваны улетевшими из зоопарков особями, несмотря на обилие солоноводных местообита-
ний, предпочитают пресноводные водоёмы (Тгеер, 2000). Местами фламинго гнездятся
высоко в горах в холодных климатических зонах (Cramp, 1977).
Птицы поселяются, как правило, на топких илистых или песчаных труднодоступных
островах и отмелях, отделенных от берега. Наличие влажного ила или заиленного песка
для постройки гнёзда также определяет гнездование фламинго в пониженных частях рель-
ефа, в которых грунт весной бывает влажным, и где временами стоит вода. По этой же
причине колонии чаще всего вытянуты узкой лентой вдоль края воды (Исаков, 1948а).
Значительные годовые и многолетние колебания уровня воды, свойственные пустын-
ным водоёмам, влияют на наличие или отсутствие гнездопригодных участков, а также ска-
зываются на концентрации солей, что, в свою очередь, влияет на состав и количество гид-
робионтов (Исаков, 19486; Чельцов-Бебутов, 1958; Долгушин, 1960а; Волков, 1977а). Все
это служит причиной периодического исчезновения колоний фламинго на одних озёрах и
появления их на других, имеющих более подходящий гидрологический режим (Чельцов-
Бебутов, 1958; Чекменев, 1964а,б; Волков, 1977а,б).
В условиях Челкар-Тениза фламинго гнездятся на песчаных островах (Волков, 19776;
Андрусенко, Жулий, 1977) в годы, когда глубина воды в озере не превышает 3 м (Андрусен-
ко, Жулий, 1977).
Численность. В настоящее время можно выделить пять популяций обыкновенного фла-
минго (Johnson, 1997; 1998):
1. Популяция, гнездящаяся в Юго-Зап. Азии от Центр. Турции до Казахстана и севе-
ро-западной Индии. В зимнее время широко разлетается от восточной части Средизем-
ного моря, Сев.-Вост. Африки и востока Аравийского п-ова до Юж. Индии и Шри-Ланки.
Численность её оценивается в 500 тыс. особей; серьёзных изменений численности не
наблюдается.
2. Популяция национального парка Банк д’Арквин в Мавритании, в зимнее время ши-
роко распределяющаяся по побережью Мавритании, Сенегала и Гамбии, изредка до Либе-
рии. Численность этой популяции — около 40 тыс. особей. Тенденция изменения числен-
ности неизвестна.
3. Восточно-африканская популяция, гнездящаяся, главным образом, в Эфиопии и
до Юж. Танзании; величина этой популяции — около 35 тыс. особей, и численность её
падает.
4. Южно-африканская популяция, состоящая из колоний, гнездящихся в националь-
ном парке Этоша в Намибии и в Ботсване. Вне брачного периода разлетается от Анголы и
Замбии на юг до ЮАР; численность популяции составляет около 50 тыс. особей (55 тыс.
по зимним учётам) и имеет тенденцию к понижению.
520
Отряд Фламингообразные, Семейство Фламинговые
5. Популяция, обитающая в западной части Средиземноморья, оценивается в 80 тыс.
особей. Численность её возрастает.
В Вост. Европе и Сев. Азии фламинго — редкая птица. Численность взрослых особей
на территории Центр. Казахстана в 1970-1975 гг. составляла 9-13,4 тыс. (Волков, 1977в). В
1953 г. на оз. Тенгиз насчитывалось 15 тыс. гнёзд (Чекменев, 1962); в течение 1958-1975 гг.
количество гнёзд фламинго на этом озере снизилось с 24 тыс. до 1,43 тыс. (в 1972 гнездова-
ния не наблюдалось), а общее количество взрослых птиц сократилось с 50 до 9 тыс. особей
(Волков, 19776). В дальнейшем отмечено постепенное восстановление численности фла-
минго. В сентябре 1982 г. на побережье Каспия от устья Урала до Эмбы встречали до 20
тыс. особей вида, в 1983 г. только в устье Урала отмечено около 50 тыс. этих птиц (Гисцов,
2001). По результатам учёта в 1986 г. на территории Казахстана обитало 22 179 фламинго
(Стоцкая, Кривоносов, 1988). В 1980-х гг. в период летней линьки общая численность фла-
минго у северо-восточных побережий Каспия достигала 50 тыс. Общая численность про-
лётных скоплений фламинго на Сев. Каспии в период максимальной концентрации птиц в
сентябре 1983-1986 и 1988-1989 гг. составила соответственно 73,0; 70,4; 27,5; 51,2; 35,6 и
17,9 тыс. птиц (Русанов, 2004г). В районе зал. Комсомолец с апреля по октябрь держится
до 20 тыс. птиц, а на пролёте встречается до 40-50 тыс. особей. На авиамаршруте 13-14
апреля 2002 года в Мангышлакском зал., зал. Комсомолец и соре Мертвый Култук учтено
20,6 тыс. особей (Гисцов, 2004). Там же 17-18 апреля 2004 г. отмечено 17,3 тыс. фламинго
(Гисцов, 2005). В январе 2004 в Азербайджане зимовало около 11 тыс. фламинго (Султа-
нов, Джаббарова, 2006).
Численность колоний фламинго, обитающих во Франции в Камарге, оценивалась в 1997 г.
в 14 тыс. птиц, испанская колония составляла около 2 тыс., итальянская — около 200 осо-
бей (Johnson, 1997). В Мавритании в 1998 г. с использованием ГИС учтено 8 545 фламинго
(Diagana et al., 2001). В Португалии по данным зимних учётов в устье р. Тежу плотность
фламинго составляла 0,83 особи на га (Rosa et al., 2003). На о. Сардиния в 1999 г. на соле-
вых озёрах успешно размножались 1 200-1 400 пар фламинго, вырастившие около 860
птенцов (Gurssu, 2000).
Уместно привести некоторые сведения о численности других близких видов фламинго.
В Кении на оз. Накуру, наряду с 30 тыс. обыкновенных фламинго обитает и более миллиона
малых (Nasirwa, 2000). Там, после увеличения их численности в 1990-1993 гг. с десятков
тыс. до 1,5 млн. (Bennun, 1993), в связи с усыханием этого озера отмечается общая тенден-
ция уменьшения поголовья (Nasirwa, 2000). В результате учётов, проведенных в январе
1997 г. на 100 высокогорных солёных озёрах в Юж. Америке, было обнаружено 47 619
фламинго Джеймса и 33 927 андских фламинго; в январе 1998 г. на 133 озёрах соответ-
ственно — 63 143 и 25 643 особей, а в период зимовки в августе там же — 31 657 и 13 944
особей фламинго этих видов соответственно (Valqui et al., 2000). Численность красного
фламинго (Ph. ruber) составляла в 1952-1954 гг. 5 600-6 057 особей, в 1972 г. — 12 ПО, в
1986 г. — 26 000 особей (Arengo, 1995).
На зимовках в Египте насчитывается около 10 тыс. обыкновенных фламинго (Salama,
1996).
Размножение. Возраст первого гнездования — 2-3 года (Cramp, 1977). По данным коль-
цевания в Камарге, размножаться фламинго начинают в возрасте 3 лет. Большинство птиц
возвращается на гнездование в места рождения, однако, они могут размножаться и в дру-
гих колониях (Johnson, 1987). В норме фламинго гнездятся колониями с высокой плотнос-
тью. Размеры колоний могут колебаться от нескольких десятков до нескольких сотен и
тысяч особей. Так, например, в колониях фламинго в зал. Кара-Богаз-Гол насчитывалось
184, 300 и 419 гнёзд. На Мёртвом Култуке — от 50 до 500 гнёзд (Исаков, 1948а). На остро-
Розовый фламинго
521
вах оз. Тенгиз в разные годы в колониях насчитывалось от 2 до 24 тыс. гнёзд (Долгушин,
1960а; Сурков, 1969; Андрусенко, Жулий, 1977; Волков, 19776; Кривицкий и др., 1985).
Колонии состоят из многочисленных дискретных групп, каждая из которых отличается
высоко синхронизированными сроками откладки яиц (Cramp, 1977). На оз. Тенгиз наблю-
далось одновременное начало откладки яиц более чем в 15 тыс. гнёзд (Чекменев, 1964).
Птицы обороняют только место расположения гнезда; враждебность между членами
своей группировки ниже, чем между птицами из разных групп (Cramp, 1977). Моногамы,
образуют постоянные пары, которые сохраняются, возможно, на протяжении всей жизни.
Наблюдения в Одюбоновском зоопарке в Новом Орлеане выявили, что у красных фламин-
го некоторые пары сохраняются до 13 лет. Из 108 гнездовых пар 49 существовали в тече-
ние одного сезона, 31 — в течение 2-3 последовательных сезонов, остальные пары остава-
лись постоянными 4-12 лет. После начала гнездостроения 7,8% пар склонны к промискуи-
тету (Shannon, 20006).
Фламинго свойственны сложные формы социального поведения. За 8-10 недель до гнез-
дования групповые демонстрации, которые обычно происходят нерегулярно и слабо выра-
жены в течение всего года, становятся более частыми. В некоторых группах брачные де-
монстрации наблюдаются в течение всего дня, обычно поблизости от островов, где птицы
в конце концов загнездятся. Принимать участие в демонстрациях может довольно много
особей (часто 15-30 или 40 птиц). Фламинго собираются вместе, формируется группа из
самцов и самок, хотя ритуальные движения не имеют направленности к конкретной птице
или полу. Движения в процессе групповой демонстрации совершаются в заданной после-
довательности синхронно среди особей группы. Сюда входят: покачивание шеей из сторо-
ны в сторону, вытягивание шеи, определённые двусложные крики, синхронные перемеще-
ния. Выделяются несколько фиксированных поз: вытягивание шеи вверх с последующим
раскрыванием крыльев; демонстративная чистка оперения, когда крыло вытянуто в сторо-
ну, а клюв касается внутренних кроющих крыла с одной стороны; поклон, при котором
голова и шея вытянуты вперёд, крылья раскрыты и приподняты позади тела; вытягивание
крыла и ноги и пр. (рис. 96). Повторения демонстраций интенсивнее в группах по 20-30
птиц. Крайняя степень агрессии выражается изгибом шеи в виде крючка вниз, одновремен-
но с её наклоном, клюв при этом направлен к низу шеи. Хотя период интенсивных демонст-
раций в колонии может длиться от недели до месяца, отдельные особи принимают участие
в этом ритуале несколько дней, но не больше 2-3 недель до начала копуляций и проявления
интереса к гнездованию (Studer-Thiersch, 2000). Сходные формы демонстрационного по-
ведения наблюдались и у андских фламинго (Lindgren, Pickering, 1998).
Формирование пар происходит незаметно; возможно, это случается во время кормле-
ния, когда самцы и самки, стоя рядом, время от времени поднимают головы вверх и издают
повторяющиеся мягкие звуки. Отмечен специальный ритуал — ложное кормление, когда
птицы быстро прохаживаются бок о бок с шеями, опущенными вниз; время от времени
одна птица или обе чистят клювом свои верхние кроющие крыла. Когда пара образована,
птицы помогают друг другу во время стычек, имеют короткие взаимные индивидуальные
расстояния и синхронно выполняют обычные действия; кроме того, они кричат дуэтом,
вытянув шеи, иногда после драки (как знак триумфа), после копуляции и в некоторых дру-
гих случаях.
Копуляции обычно начинаются через несколько дней после того, как пара перестаёт
принимать участие в групповых демонстрациях. Приглашение к спариванию может исхо-
дить от обоих партнёров, но чаще самка прогуливается вокруг, а самец следует за ней с
вытянутой вперёд шеей, иногда дотрагиваясь до её спины. Когда самка останавливается,
часто с немного опущенными крыльями, самец взлетает ей на спину, быстро взмахивая
522
Отряд Фламингообразные, Семейство Фламинговые
Розовый фламинго
523
Рис. 96. Элементы брачного, агрессивного, группового и комфортного поведения фламинго (по: Bauer,
Glutz von Blotzheim, 1966; Cramp, 1977): a — почёсывание, 6 — сон, в — ритуализованная чистка
оперения, г — потягивание, д — агрессивное поведение при территориальном конфликте, е — пока-
чивание головой (“head-flag”), ж — «салют крыльями» в вертикальной позе, з — «салют крыльями»
с поклоном, и — сгибание шеи крючком (“hooking-posture”), к-н — последовательность поз при
спаривании.
524
Отряд Фламингообразные, Семейство Фламинговые
крыльями, затем опускает шею и вытягивает голову наклонно вниз. После эякуляции, са-
мец встает на спине самки и спрыгивает впереди нее. Последующие действия могут быть
разными, они могут включать мягкие взаимные позывы, чистку оперения друг у друга,
когда птицы, стоят рядом (бок о бок), кормёжку и проявление угрозы по отношению к
другим птицам. Случающиеся копуляции с посторонними птицами не влияют на прочность
пары (Studer-Thiersch, 1974, цит. по: Cramp, 1977; Андрусенко, 1998).
Поведение, предшествующее гнездованию — такое, как интенсивные групповые де-
монстрации и копуляция — могут происходить не только в районах гнездования. Данное
поведение приводит фламинго в гнездовую готовность за несколько недель до гнездования
и часто в местах, далеких от возможного места гнездования. Даже демонстрация подгото-
вительных действий, предшествующих строительству гнезда, может происходить до при-
лёта на гнездовые участки (Brown, 1958; Brown et al., 1973, цит. no: Cramp, 1981). Подобная
готовность к гнездованию позволяет группе птиц начать кладку в течение нескольких дней
после их появления в местах, благоприятных для размножения, или при улучшении усло-
вий гнездования (Studer-Thiersch, 2000).
По прибытии в район гнездования птицы продолжают держаться отдельными группа-
ми, которые сохраняются и во время гнездования (Studer-Thiersch, 1974, цит. по: Cramp,
1977). Постоянные пары обычно не участвуют в брачном поведении, а партнеры почти все
время находятся в непосредственной близости друг от друга. В групповом брачном поведе-
нии чаще участвуют птицы, меняющие партнеров регулярно, или которым свойствен про-
мискуитет (Shannon, 20006).
Прилет фламинго на места гнездовий сильно растянут. Первыми прилетают наиболее
старые и, возможно, наиболее сильные птицы. Передовые особи на оз.Тенгиз регистриру-
ются с конца марта - начала апреля. Подлет птиц продолжается до конца мая. Таким обра-
зом, сроки начала гнездования в колонии растянуты почти на два месяца (Андрусенко, 1979).
Отмечена зависимость успешности гнездования от сроков прилёта: среди фламинго, при-
летающих позднее, процент загнездившихся пар значительно ниже. По-видимому, это свя-
зано с тем, что последними подлетают ослабленные, неполовозрелые и впервые гнездящи-
еся особи (Андрусенко, 1979). Сроки размножения у фламинго также сильно растянуты.
Так, в одной из двух колоний оз. Тенгиз в 1970 г. 31 июля пуховые птенцы уже покинули
гнёзда, а во второй колонии 500 птиц ещё продолжали насиживание (Волков, 1972). Наибо-
лее ранние сроки откладки яиц наблюдали 1-2 мая 1970, наиболее поздние — 2-7 июля
того же года. По данным Н.Н. Андрусенко (1980), в некоторые годы откладка яиц начинает-
ся с третьей декады апреля (Кривицкий и др., 1985).
Старые гнёзда не используются (Спангенберг, 1951), гнездо всегда строится заново,
обоими партнёрами; строительство часто начинается за много дней до начала откладки яиц
(Cramp, 1977). Так, на оз. Тенгиз 8 мая наблюдалось строительство гнёзд, а к гнездованию
птицы приступили лишь в начале июня (Андрусенко, 1979). Однако иногда яйца отклады-
ваются в рудиментальное гнездо, которое затем быстро достраивается (Cramp, 1977). Гнёз-
да фламинго имеют форму усечённого конуса с чашеобразным углублением наверху. Выс-
тилки в лотке нет. Высота гнезда зависит от влажности места, в котором находится коло-
ния. Наиболее высоки те гнёзда, основания которых находятся в воде (Исаков, 1948а). Гнёзда,
построенные из ила, могут достигать в высоту 61 см, из песка — имеют небольшую высо-
ту, могут быть всего 10-15 см (Cramp, 1977). Состоит гнездо из того грунта, на котором
расположено (Долгушин, 1960а). На Багамских о-вах фламинго устраивали свои колонии в
мангровых зарослях; гнёзда их состояли, в равной мере, как из ила, так и из растительных
остатков. В Камарге подводная часть гнёзд состоит из чёрного ила, надводная — из ила с
вязкой глиной (Swift, 1960). На Кара-Богаз-Голе гнёзда состояли только из солёного ила с
Розовый фламинго
525
примесью мелких створок Cardium. Большинство гнёзд имело в основании большие шап-
ки осевшего гипса. Птицы брали вокруг влажный ил и укладывали его в гипсовое основа-
ние, утрамбовывая материал клювом и ногами (Исаков, 1948а). Гнёзда на Тенгизе состояли
из песка и водорослей (Долгушин, 1960а). Иногда, при строительстве гнёзд из песка, фла-
минго скрепляют их в верхней трети своими перьями, преимущественно плечевыми (Вол-
ков, 1972).
Плотность расположения гнёзд в колонии очень велика. В колонии в Кара-Богаз-Голе
на одно гнездо приходилось в среднем 1,14 кв.м (Исаков, 1948а). Ещё более скученное
гнездование наблюдалось на оз. Тенгиз, средняя плотность расположения гнёзд в разных
колониях колебалась здесь от 2 до 3,7 гнёзд на м2 (Долгушин, 1960а; Кривицкий и др.,
1985). В Камарге гнёзда были расположены группами по 4-5; на 1 м2 приходилось в сред-
нем 1,3 гнезда (Swift, 1960). В центре колонии гнёзда расположены плотнее, чем на пери-
ферии (Кривицкий и др.. 1985).
По данным, приведенным Ю.А. Исаковым (19486), 8 гнёзд на о. Тараба (зал. Кара-Бо-
газ-Гол) имели размеры: высота — 12-32 см, в среднем 21,9 см; диаметр основания — 41-
48 см, в среднем 43.6 см: диаметр лотка — 21-23, в среднем 21,9 см; размеры 6 гнёзд на
косе Сары-Тамчи (зал. Кара-Богаз-Гол) были следующими: высота — 10-12 см, в среднем
11,2 см; диаметр основания — 40-44 см, в среднем 41,8 см; диаметр лотка — 21-23, в
среднем 22,0 см.
Размеры 11 гнёзд на оз. Тенгиз, построенных из песка, но хорошо сохранившихся после
гнездования: высота — 15-20 см, в среднем 17,6 см, диаметр основания — 35-55 см, в
среднем 45,8 см, наружный диаметр у вершины — 32-40 см, в среднем 35,2 см, диаметр
лотка— 19-28 см, в среднем 22,5 см (Волков, 1972). Гнёзда, построенные на солончаковых
отмелях, массивнее (п=25): высота — 17-35 см, в среднем 25,5 см, диаметр основания —
40-59, в среднем 47 см, наружный диаметр у вершины — 32-50 см, в среднем 38,1 см,
диаметр лотка — 18-27 см, в среднем 23,5 см. В период инкубации птицы поправляют
гнёзда, при необходимости наращивают их (Кривицкий и др., 1985).
В кладке фламинго 1-2, изредка 3 яйца. Причём, в кладках казахстанской популяции
нередко встречается 2 яйца и иногда бывает до 3 (Исаков, 1948а), а у европейских, афри-
канских и американских птиц — чаще всего 1 и, редко, 2 яйца. Так, в Камарге только в 40
гнёздах из 2 тыс. обнаружено по 2 яйца (Gallet, 1950, цит. по: Cramp, 1977), в Кении — в 5
из 900 гнёзд (Brown, 1958, цит. по: Cramp, 1977), в Юж. Африке — в 1 из 400 гнёзд (Uys et
al., 1963 цит. по: Cramp, 1977).
Основной цвет скорлупы яиц оливково-зелёный, с обильным мягким меловым налётом
(Долгушин, 1960а). В процессе инкубации налёт стирается, яйца пачкаются, изменяя, та-
ким образом, свой первоначальный цвет (Cramp, 1977).
Размеры яиц 77-103 х 48-60 мм, в среднем 90x55 мм (п=133). Средний вес яйца 140 г
(Schonwetter, 1967, цит. по: Cramp, 1977). В Камарге описано два типа яиц: 89x58 мм (ко-
роткие, правильной формы) и 94x54 мм (удлинённые, узкие). Яйца первого типа более
обычные, второго — встречаются реже (Gallet, 1950, цит. по: Cramp, 1977). Размеры яиц
фламинго на оз. Тенгиз: 80,1-101,0 х 52,6-58,0 мм, в среднем 88,24x55,62 мм (п=29) (Ан-
друсенко, Жулий, 1977).
В насиживании участвуют оба партнёра. Инкубация длится 28-31 (27-36) день. Скор-
лупа выбрасывается из гнезда или остаётся в нем (Cramp, 1977).
О птенцах заботятся оба партнёра. Сначала птенцы находятся в гнезде, затем родители
водят их, кормят и защищают небольшую территорию вокруг них до тех пор, пока птенцы
не присоединятся к группе остальных птенцов, так называемым «яслям»; это происходит в
возрасте 8-10 дней. Птенцы в это время совершенно незащищены. Первое время родители
526
Отряд Фламингообразные, Семейство Фламинговые
часто возвращаются, чтобы накормить их. В период инкубации и нахождения птенцов в
«яслях» родители летают на кормёжку в разные, иногда весьма удаленные, места. На оз.
Фуэнте-Де-Пьедра (Юж. Испания) пока птенцы были маленькие и требовали немного кор-
ма, некоторые родители прилетали кормить их вечером, а затем снова улетали в места кор-
мёжки. При этом каждую ночь они пролетали около 300 км (Rendon-Martos et al., 2000).
Птенцы в первые недели после вылупления получают пищевой сок из клюва родителей.
В начале выкармливания сок имеет жидкую консистенцию и яркий кроваво-красный цвет;
интенсивность окраски по мере роста птенцов ослабевает. Пищевой сок имеет высокое
содержание жира (17,5%) и глюкозы (190 мг/100 мл), а также протеина, кальция и фосфо-
ра. Алая окраска вызвана присутствием пигментов p-каротина, ксантофила и кантаксанти-
на (основной пигмент оперения). На характер окраски влияет также наличие в соке фор-
менных элементов крови — эритроцитов, лейкоцитов и др. Пищевой сок продуцируется
многочисленными ацинозными железами с мерокринным типом секреции, расположенны-
ми по всей длине пищевода от глотки до железистого желудка. Железы богато снабжаются
кровью из густой сети тесно оплетающих их капилляров. Пищевой сок, продуцируемый эти-
ми железами, изливается в просвет пищевода. Сокообразование у взрослых птиц идёт непре-
рывно, так что каждая пара даёт ежедневно 200 мл высококалорийного корма, и вес птенцов
за первые 2 месяца жизни увеличивается со 100 г до 2-4 кг (Lang, 1963; Wackemagel, 1964).
Родители отличают своего птенца и кормят именно его. Только что вылупившиеся птен-
цы сначала выпрашивают корм и у своих соседей, но вскоре после оставления гнёзда они,
видимо, начинают опознавать своих родителей. Старшие птенцы, разыскивая родителей,
иногда толкают спящих птиц, провоцируя их подавать голос. Родители предлагают пищу
крошечным птенцам, дотрагиваясь до них несколько раз клювом. Позднее птенец выпра-
шивает корм криком и иногда пощипывает перья на шее родителя. Подросшие птенцы вып-
рашивают корм, приближаясь к родителю и издавая при этом мягкий звук. Ноги птенца при
этом широко расставлены, шея опущена к земле; птенец стремится занять место под гру-
дью у взрослой птицы, затем вытягивает шею и клюв вверх и кричит уже более громко и
часто. Стоит взрослой птице побежать, птенец устремляется за ней, затем припадает к зем-
ле впереди неё. Стычки между молодыми в «яслях» редки.
Птенцы держатся в «яслях» долго, иногда до 100 дней, затем «ясли» постепенно распа-
даются (Cramp, 1977). Лётными молодые становятся в возрасте 90-120 дней, когда длина
крыла достигает 95-100% длины крыла взрослых птиц (Studer-Thiersch, 2000). По данным,
приводимым С. Крэмпом (Cramp, 1977), период выкармливания птенцов длится 70-75 (65-
90) дней.
Успешность размножения фламинго в Камарге в разные годы колебалась от 0 до 95%,
составляя в среднем 47%.
Смертность молодых фламинго в первые годы — 40% (Johnson, 1987). По данным коль-
цевания в Камарге, максимальный возраст фламинго составляет 30,5 лет (Johnson, 1987).
Имеются также сведения о четырёх птицах, одна из которых прожила 21 год, две — 22
года, и одна — 27 лет (Dejonge, Czajkowski, 1983).
Суточная активность, поведение. Фламинго — очень осторожные птицы. Кормятся
днём и ночью, на рассвете и в сумерках. Постоянно держатся большими скоплениями, кон-
центрируясь тысячами в местах, богатых пищей, и около источников пресной воды. Совер-
шают кормовые перемещения на большие расстояния, часто в вечернее время; перемеще-
ния из гнездовой колонии в Фуэнте-де-Пьедра к местам кормежки происходили ночью
(Rendon-Martos et al., 2000). Собираются для сна в мелководных участках, часто плотными
группами, как в дневное, так и в ночное время. Расстояние между спящими птицами на-
столько мало, что если одна из птиц теряет равновесие, то задевает несколько других.
Розовый фламинго
527
Действия стаи более или менее синхронны, особенно в пределах группы соседей по
гнездованию в течение сезона размножения. Не известно, поддерживается ли такая же спло-
ченность группы в стае после гнездового сезона. В кормовых угодьях птицы рассеиваются
довольно равномерно на большой территории. У пресных источников фламинго скаплива-
ются в огромных количествах и держатся плотной группой.
Отдельные стычки внутри группы возникают нередко, но настоящие нападения и драки
довольно редки (Studer-Thiersch, 1972, цит. по: Cramp, 1977). Низкий уровень угрозы пти-
цы демонстрируют, приподнимая плечевые перья (Aly 1945; Brown 1958, цит. по Cramp,
1977) и издавая одновременно низкий, короткий, повторяющийся звук; при нарастании ин-
тенсивности угрозы начинается покачивание шеей, вытягивание головы и шеи горизон-
тально с их движениями из стороны в сторону, подчас с клевками противника. Во время
драки две птицы бессистемно клюют и треплют друг друга клювами, но без нанесения
серьезных повреждений (Brown, 1958, цит. по: Cramp, 1977). Подобные драки нечасты,
случаются они только во время перераспределения гнездовых участков, когда самка, пользу-
ясь поддержкой самца, становится очень агрессивной (Studer-Thiersch, 1967, цит. по: Cramp,
1977). Умеренная агрессия наблюдалась в начале периода интенсивных групповых демон-
страций, во время строительства гнёзд и вылупления молодых. Агрессия в этом случае
связана с сохранением индивидуальной дистанции. Партнёры поддерживают друг друга
во время драк. Реакция бегства слабо выражена, хотя иногда после стычки птицы уходят
или убегают, взмахивая крыльями; фламинго могут и преследовать друг друга. В кормо-
вых биотопах угроза с приподниманием плечевых перьев часто удерживает других птиц
от приближения. Перемещаясь среди других плотно стоящих птиц, фламинго часто на-
клоняют шею и голову, как бы принимая позу покорности (Studer-Thiersch, 1967, цит. по:
Cramp, 1977).
Питание. Для вида характерен фильтрационный тип питания, сходный с таковым у
пластинчатоклювых. Циркуляцию воды обуславливают синхронные движения мясистого
языка и челюстей. Фламинго прогоняют воду возвратно-поступательно, вытесняя её нару-
жу на всём протяжении разреза рта, и фильтруя её через пластинки на краях челюстей и
языка. Для задержания пищевых частиц используют только пластинки челюстей. Изменяя
взаимное расположение пластинок, птицы могут настраивать фильтр на задержание объек-
тов определённого размера. Фламинго способен эффективно отфильтровывать объекты раз-
мером 0,8-5 мм (при оптимуме 3 мм), производительность фильтрации до 30 таких объек-
тов в секунду (Berkhoudt, Zweer, 1994).
Кормятся птицы как в светлое время (в основном на рассвете и в вечерних сумерках),
так и ночью (Дементьев и др., 1955; Cramp, 1977), почти исключительно на мелководьях
солёных озер, обычно с вязким дном. На пресных водоёмах встречаются редко (Долгушин,
1960а). Птицы медленно переступают ногами и зачёрпывают взмученный при этом со дна
корм (Исаков, 1948а). В момент фильтрации клюв, часто вместе с головой, погружен в
воду так, что верхняя челюсть оказывается внизу. В одних случаях (например, в зал. Кара-
Богаз-Гол) птицы погружают в воду только клюв, что, по предположению Ю.А. Исакова
(1948а), может быть реакцией на высокую солёность воды, действующей на слизистую
оболочку глаза. В других (на Ленкоранских зимовках, в районе станции Челкар в Казахста-
не и в ряде других пунктов) птицы погружают в воду не только голову, но и часть шеи
(Исаков, 1948а; Спангенберг, 1951).
В состав корма входят личинки Chironomidae и Ephidridae; гребляки (Corixidae) — Sigaria,
Micronecta, Notonecta', водяные жуки (Coleoptera); муравьи (Formicidae); моллюски —
Paludestrina, Neritina, Cerithium, Cardium, Venus, Mytilus, Tapes и Tympanotomus', мелкие
ракообразные — Sphaeroma, Gammarus, Asellus и Paradiaptomus', водоросли Spirulina,
528
Отряд Фламингообразные, Семейство Фламинговые
Arthrospira и Oscillatoria\ семена осок, кисточки руппии Ruppia и тростника Juncus (Ridley
et al., 1955; Allen 1956; Palmer, 1962, цит. no: Cramp, 1981). В водах умеренной солёности
большое значение в питании фламинго имеет рачок Artemia salina и его икра (Максимович,
1896, цит. по: Исаков, 1948а) Во всех желудках фламинго (п=7), добытых Ю.А. Исаковым
(1948) в 1938 г. на восточном берегу Каспийского моря, содержались одноклеточные водо-
росли Apanothence salina и семена рупии Ruppia maritima. У добытых 12 июля на оз. Тен-
гиз 3 взрослых фламинго в желудках содержались исключительно семена рупии и рдеста.
В желудках 2 птиц, добытых здесь же 15 июля, основную массу пищевого комка составля-
ли личинки Tendipedidae и небольшое количество личинок Ephydridae, семян рупии и рде-
ста. В желудке фламинго, добытого В.Ф. Гавриным на солёном озере близ Кургальджина,
масса пищевого комка составила 84 г, 95% содержимого было представлено личинками
Ephydra sp. (всего 1 890 особей), в небольшом количестве присутствовали рачки-бокопла-
вы (Долгушин, 1960а).
Враги, неблагоприятные факторы. Одним из основных факторов, снижающих чис-
ленность фламинго, населяющих Центр. Казахстан, является ухудшение условий обитания
этих птиц как в границах гнездового ареала, так и на местах их летних скоплений и зимо-
вок. Серьёзную угрозу для численности фламинго создаёт проведение работ по освоению
и разведке углеводородного сырья, ведущихся на всём побережье Каспия от Мангышлака и
Бузачей до дельт Урала и Волги (Русанов, Кривоносов, 1991; Русанов, 2002).
Отрицательно сказываются на численности фламинго экстремально суровые зимы, пе-
риодически, раз в 5-8 лет, наступающие на Юж. Каспии (Греков, 1962; Кривоносов и др.,
1977). По-видимому, часть птиц в таких случаях откочёвывает в более южные области — в
Иран, Турцию, Египет (Волков, 19776, 1979; Кривоносов и др., 1977), однако, некоторое
количество птиц остается, и значительная часть оставшихся особей погибает (Дюнин, 19606;
Греков, 1962; Кривоносов и др., 1977). Даже в европейских популяциях, где условия зимо-
вок гораздо мягче, чем на Каспии, на уровень выживаемости фламинго сильно влияет су-
ровость зимы. В годы с обычной зимой выживаемость европейской популяции составляла
93%, тогда как после суровой зимы 1984/85 г. — 76% (Cezilly et al., 1996). В Кызыл-Агач-
ском заповеднике в наиболее суровые зимы погибает большая часть оставшихся фламинго.
Так, в 1950 г., когда зимние температуры опускались до -20 °C, а глубина снежного покро-
ва достигала 60-80 см, численность зимовавших там фламинго упала с 15 тыс. до 200-300
особей. В 1957 г. от зимовавших здесь 5 тыс. особей осталось 30СМ100 крайне истощённых
птиц (Греков, 1962). Тем не менее, степень консерватизма фламинго в отношении избран-
ных мест зимовки и гнездования чрезвычайно велика (Исаков, 19486; Спангенберг, 1951;
Греков, 1962).
Приуроченность колоний фламинго к солёным пустынным озёрам и морским мелково-
дьям является основной причиной нерегулярности их гнездования. Для этих водоёмов ха-
рактерно постоянное изменение уровня и, как следствие, солёности воды. Как чрезмерное
повышение, так и падение концентрации солей негативно влияет на кормовые достоинства
водоёма для фламинго. Кроме того, сильное падение уровня воды способствует соедине-
нию островов, на которых гнездятся эти птицы, с берегом, что способствует доступности
колоний для хищников и человека. Сильное же повышение уровня вод приводит к затопле-
нию мест, пригодных для гнездования (Исаков, 19486), а также к разрушению гнёзд и гибе-
ли кладок (Кустанович, 1986).
В местах гнездования у фламинго естественных врагов немного. И в Камарге, и на оз.
Тенгиз большой вред колониям наносит хохотунья (Larus cachinnans), уничтожающая как
яйца, так и птенцов фламинго (Кустанович, 1986; Cramp, 1977). В местах зимовок на фла-
минго охотятся шакалы, гиены, орлан-белохвост (Кустанович, 1986).
Розовый фламинго
529
Мигрирующие в ночное время птицы погибают при столкновении с высоковольтными
ЛЭП (Кустанович, 1986) и проводами телефонных линий (Сушкин, 1908; Волков, 19776).
Гибнут фламинго и под выстрелами браконьеров.
Вид очень чувствителен к фактору беспокойства в сезон гнездования (Азаров, 1975;
Волков, 19776). После вторжения людей колонии распадаются или перемещаются на дру-
гое место, но, как правило, в данном сезоне потревоженные колонии уже не восстанавлива-
ются.
Для фламинго, гнездящихся в Кении, где наблюдается заметное падение численности,
главные факторы, провоцирующие этот процесс — неудачное размножение, потери птен-
цов в результате хищничества марабу, усыхание озёр, антропогенный пресс, загрязнение
среды обитания сточными водами и тяжёлыми металлами, а также болезни (туберкулез и
сепсис) (Nasirwa, 2000).
Хозяйственное значение, охрана. В недавнем прошлом проводился массовый сбор
яиц и варварское уничтожение фламинго в местах скоплений на линьку (Долгушин, 1960а).
Сейчас этот вид повсеместно находится под охраной. Места скоплений и гнездования вида,
как правило, входят в состав охраняемых территорий. Так, неоднократно упоминавшееся
оз. Тенгиз расположено на территории Кургальджинского заповедника. В период пролёта и
зимовок в России вид охраняется в заповеднике «Дагестанский», Аграханском и Самур-
ском заказниках, в Азербайджане — в Акгёльском, Ширванском и Кызыл-Агачском запо-
ведниках, в Туркменистане — в Хазарском (Красноводском) заповеднике (Северный и Гас-
санкулийский участки). Сарыкамышском и Огурчинском заказниках. Однако это не исклю-
чает случаев браконьерства и беспокойства птиц в колониях. Необходим строгий контроль
над соблюдением охранного режима в местах размножения, линьки, зимовки и остановок
на пролёте.
Возможно, имеет смысл осуществить некоторые гидротехнические мероприятия для
оптимизации гидрологического режима водоёмов, где гнездятся фламинго (Андрусенко,
1986) и где они гнездились в недавнем прошлом.
Во Франции и Италии фламинго в 100% случаев гнездятся на территории особо охраня-
емых природных территорий (Gustin et al., 2003). В Камарге и в лагуне Фуэнте-де-Пьедра
применялись технические средства для противодействия размыванию островов, пригод-
ных для успешного гнездования фламинго. Предполагается создание новых искусствен-
ных островов, а также применение специальных платформ для размещения гнёзд (Martos,
Johnson, 1996).
Фламинго занесён в Красные книги России, Азербайджана, Казахстана и других стран,
на территории которых он встречается; включён в Приложение 2 СИТЕС, Приложение 2
Бернской Конвенции, Приложение 2 Боннской Конвенции, Приложение Соглашения об
охране мигрирующих птиц, заключённого между Россией и Индией.
Вид содержится во многих зоопарках мира и при создании соответствующих условий
успешно там размножается.
Литература
Абдусалямов И.А. 1971. Птицы // Фауна Таджикской ССР. Душанбе. Т. 19. Ч. 1. 403 с.
Абрамова Р.Ю., Васильев А.Л., Воронин А.А., Минаев Л.С., Тимошин Е.В. 1975. Пеликаны в запад-
ной части дельты Или // Колон, гнездовья околоводн. птиц и их охрана. М. С. 187-189.
Абрамчук А.В. 2003. Новые находки гнездовых колоний большой белой цапли в Брестском полесье
// Subbuteo. Вып. 6. С. 3-5.
Абрамчук А.В., Абрамчук С.В. 2004. Наблюдения за редкими видами птиц на Днепре на границе
Беларуси и Украины И Беркут. Т. 13. Вип. 1. С. 132-134.
Абрамчук А.В., Абрамчук С.В. 2005. Большая белая цапля в Беларуси: распространение и экология //
Беркут. Т. 14. Вип. 1. С. 50-55.
Абрамчук А.В., Абрамчук С.В., Сербун А.А., Богданович И.А. 2003. Осенние скопления водно-бо-
лотных птиц на водоёмах Брестского полесья в 2002 г. И Беркут. Т. 12. Вып. 1-2. С. 128-142.
Абуладзе А.В., Элигулашвили В.Э. 1986. Белый аист (Ciconia ciconia L.) в Грузии // Сообщ. АН
ГрузССР. Т. 124. № 2. С. 417—420.
Аверин Ю.В., Ганя И.М., Успенский ГА. 1971. Птицы Молдавии. Кишинев. Т. 2. 236 с.
Авданин В.Д. 1984. Краткое сообщение о Чёрном аисте в Центрально-Лесном зап-ке // Исследования
в обл. запов. дела. М.: ВНИИприрода. С. 110.
Адамян М.С. 1990. Численность и некоторые особенности популяции белого аиста в Армении //
Аисты: распростр., экол., охрана. Минск. С. 46-51.
Адамян М.С. 1992. Биология белого аиста в Армении // Совр. орнитол. 1991. М. С. 22-30.
Ажимуратов X., Пиржанова Р. 1986. Влияние синоптических процессов на динамику осенней мигра-
ции некот. массовых видов птиц в дельте Аму-Дарьи И Мигр. птиц в Азии. Новосибирск. С. 7-11.
Азаров С.И. 1975. Гнездовые колонии околоводных птиц на озёрах Кургальджинского заповедника //
Колон, гнездовья околоводн. птиц и их охрана. Тез. совещ. М. С. 119-121.
Азаров И.В. 1996. Редкие животные Тюменской области и их охрана. Тюмень: Вектор Бук. 238 с.
Алексеев В.М., Сметанина З.Б. 1968. Нематоды рыбоядных птиц островов Римского-Корсакова //
Гельминты животных Тихого океана. М.: Наука. С. 97-104.
Алфераки С.Н. 1910. Птицы Восточного Приазовья // Орнитол. вести. № 1. С. 11-35.
Андреев В.А. 2004. Встречи в Архангельской обл. новых для её территории птиц // Русск. орнитол.
ж., Экспр.-вып. № 249. С. 20-21.
Андреев В.П., Аристова В.А., Щербина А.А. 1977. Зараженность гнездовых колоний морских птиц
иксодовыми клещами на Восточном Каспии // Тез. докл. 7-й Всес. орнитол. конф. Ч. 2. Киев:
Наукова думка. С. 63.
Андронов В.А. 1983. Состояние популяций журавлей и аистов в Хинганском заповеднике в 1982 г. //
Охр. жив. прир. М. С. 8-10.
Андронов В.А. 1986. Зависимость гнездования дальневосточного белого аиста от условий сезона //
Изуч. птиц СССР, их охрана и рац. использ. Л. Ч. 1. С. 31-32.
Андронов В.А. 1987. Редкие птицы юга Амурской области // Пробл. охраны редк. животн. М. С. 117-
121.
Андронов В.А. 1988. Численность и распространение дальневосточного аиста - Ciconia boyciana
Swinh - в Амурской области // Редк. птицы Дальн. Востока и их охрана. Владивосток. С. 60-61.
Андронов В.А. 1989. Стратегия сохранения редких видов птиц в Среднем Приамурье // Геол, и экол.
басе. Амура. Благовещенск. Ч. 3(1). С. 5-6.
Андронов В.А. 2001. Дальневосточный аист Ciconia boyciana Swinhoe, 1873 II Красная книга РФ.
Животные. М.: ACT, Астрель. С. 388-389.
Андронова Р.С. 2000. Заметки о линьке у дальневосточного аиста Ciconia boyciana Swinh. II Дальне-
вост. аист в России. Владивосток: Дальнаука. С. 86-89.
Андронов В.А., Андронова Р.С. 1992. Получение повторных кладок у дальневосточного белого аиста
// Аисты. Минск. С. 176-180.
Андронов В.А., Артемова Е.В. 1988. Некоторые стороны гнездовой жизни дальневосточного белого
аиста — Ciconia boyciana Swinhoe//Редк. птицы Дальн. Востока и их охрана. Владивосток. С. 33-
34.
Литература
531
Андронов В.А., Дарман Ю.А., Парилов М.П. и др. 2001. Состояние популяций журавлей и аистов в
Амурской области в 1998 г. // Орнитология. Вып. 29. С. 314-316.
Андронов В.А., Кастрикин В.А. 2000. Динамика численности жилых гнёзд дальневосточного аиста в
Хинганском заповеднике // Дальневост, аист в России. Владивосток: Дальнаука. С. 25-30.
Андрусенко Н.Н. 1979. Сроки гнездования фламинго на оз. Тенгиз и их зависимость от времени
весеннего прилёта // Тез. конф.: «Экол. гнездования птиц и методы её изуч.». Самарканд. С. 16-
17.
Андрусенко Н.Н. 1981. О последоват. линьке первостеп. маховых у обыкн. фламинго // Тез. докл. 8-й
Всесоюзн. орнитол. конф. Кишинев. С. 10.
Андрусенко Н.Н. 1986. Осенние миграции фламинго в СССР // Мигр. птиц в Азии. Новосибирск.
С.150-158.
Андрусенко Н.Н. 1998. Некоторые аспекты социального поведения фламинго // Орнитология. Вып. 28.
С. 236.
Андрусенко Н.Н., Жулий В.А. 1977. О гнездовании и линьке фламинго на оз. Тенгиз в 1976 г. // Тез.
докл. 7-й Всес. орнитол. конф. Киев. Ч. 2. С. 189-190.
Андрюшенко Ю.А., Попенко В.М., Черничко И.И., Арсиевич Н.Г., Олейник Д.С. 2003. Результаты
среднезимних учётов птиц на Сиваше в 2001 г. // Бранта. Вып. 6. С. 173-178.
Анисимов В.Д., Барсова Л.И.. Голубева Т.Б. 1994. Гнездование птиц на островах залива Лаврентия
Берингова моря ' Орнитология. Вып. 26. С. 174-175.
Анисимова О.А. 1996. О гнездовании морских колониальных птиц на Южных Курильских остро-
вах // Птицы пресных вод и морсчких побережий юга ДВ России и их охрана. Владивосток:
Дальнаука. С. 228-231.
Антонов А.И. 1999. Материалы по редким птицам Еврейской автономной области // Русск. орнитол.
ж., Экспр.-вып. № 86. С. 17-20.
Ардамацкая Т.Б. 1967. Материалы по экологии голенастых птиц района Черноморского заповедни-
ка // Мат-лы конф., посвящ. 40-летию Черном, зап-ка. Киев. С. 9-12.
Ардамацкая Т.Б. 1968. Материалы по биологии большого баклана // Мат-лы научн. сессии, посвящ.
50-летию Астраханск. гос. зап-ка. Астрахань. С. 176-177.
Ардамацкая Т.Б. 1975. Сезонная миграция птиц Черноморского заповедника // Ориент. и мигр. птиц.
М.: Наука. С. 59-64.
Ардамацкая Т.Б. 1981. Методика обследования колониальных гнездовий околоводных птиц Сев.
Причерноморья // Научн. основы обслед. колон, гнездовий околоводных птиц. М.: МОИП. С. 26-
36.
Ардамацкая Т.Б. 1999. Структура поселений птиц водно-болотного орнитокомплекса на Каржинских
островах // Фауна, экол. и охр. птиц Азово-Черном, per. Симферополь. С. 9-11.
Ардамацкая Т.Б. 2000. Орнитокомплексы о. Джарылгач и их современное состояние. // Вести, зоол.,
спецвып. «Биоразнообр. Джарылгача». С. 76-83.
Ардамацкая Т.Б. 2001. Современное состояние орнитокомплексов островов Джарылгачского залива
Чёрного моря // Актуальн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 45-47.
Ардамацкая Т.Б. 2005. Динамика численности колониальных аистообразных и пеликанообразных
птиц в дельте Днестра // Геоэкол. и биоэкол. пробл. Сев. Причерноморья. Тирасполь. С. 6-8.
Ардамацкая Т.Б. 2006. Изменения численности и статуса каравайки в районе Черноморского био-
сферного заповедника // Стрепет. Т. 4. Вып. 2. С. 71-77.
Ардамацкая Т.Б., Гринченко А.Б., Щеголев И.В. 1988. Численность колониальных голенастых и вес-
лоногих на юге Украины по данным учётов 1985 г. // Орнитология. Вып. 23. С. 198-199.
Артюхин Ю.Б. 1989. Морские колониальные птицы о. Топорков (Командорские о-ва) // Пром, фауна
Сев. Пацифики. Киров: ВНИИОЗ. С. 25-31.
Артюхин Ю.Б. 1991а. Гнездовая авифауна Командорских о-вов и влияние человека на её состояние //
Прир. рес. Командорских о-вов. М.: изд-во МГУ. С. 99-137.
Артюхин Ю.Б. 19916. Гнездовая авифауна Командорских о-вов (современное состояние и динамика,
охрана и перспективы использ.): Дис. ... канд. биол.н. М.: МГУ. 163 с.
Артюхин Ю.Б. 1992. О гнездовании моевки и красноногой говорушки на Командорских о-вах в
1990 г. // Изуч. морск. колониальных птиц в СССР. Магадан: ИБПС ДВО РАН. С. 13-15.
532
Литература
Артюхин Ю.Б. 1997. Залёт бурой олуши Sula leucogaster к о-вам Малой Курильской гряды // Русск.
орнитол. ж., Экспр.-вып. № 27. С. 13-14.
Артюхин Ю.Б. 1999. Кадастр колоний морских птиц Командорских о-вов // Биол. и охрана птиц
Камчатки. М.: Диалог МГУ. Вып. 1. С. 25-35; 139-144.
Артюхин Ю.Б. 2003. Дополнения к «Кадастру колоний морских птиц Командорских о-вов» // Биол. и
охр. птиц Камчатки. М.: ЦОДП. Вып. 5. С. 10-12.
Артюхин Ю.Б., Бурканов В.Н. 1999. Морские птицы и млекопитающие юга Дальнего Востока Рос-
сии. М.: ACT. 215 с.
Артюхин Ю.Б., Бурканов В.Н., Вяткин П.С. 1999. Случайная гибель морских птиц в дрифтерных
сетях на промысле лосося японскими судами в исключительной экономической зоне России в
1993-98 гг. // Биол. и охр. птиц Камчатки. М.: Диалог МГУ. Вып. 1. С. 93-108.
Артюхин Ю.Б., Бурканов В.Н., Заочный А.Н., Никулин В.С. 2000. Смертность морских птиц в дрифтер-
ных сетях на японском промысле лососей в российских водах Берингова моря в 1993-99 гг. // Биол.
и охр. птиц Камчатки. М. Вып. 2. С. 110-126.
Артюхин Ю.Б., Винников А.В., Терентьев Д.А. 2006. Морские птицы и донное ярусное рыболовство
в Камчатском регионе. М.: отд. WWF в России. 56 с.
Артюхин Ю.Б., Трухин А.М., Корнев С.И., Пуртов С.Ю. 2001а. Кадастр колоний морских птиц Ку-
рильских о-вов // Биол. и охр. птиц Камчатки. М.: ЦОДП. Вып. 3. С. 3-59.
Артюхин Ю.Б., Трухин А.М., Корнев С.И., Пуртов С.Ю. 20016. Численность морских колониальных
птиц на юге Камчатки в 2000 г. // Там же. С. 62-63.
Архипов В.Ю. 2002. Залеты некоторых видов птиц в район экологической станции «Мирное» (Сред-
не-таежный Енисей) // Мат-лы к распр. птиц на Урале. Екатеринбург. С. 10-11.
Аскаров Г. 1938. Материалы по биологии каравайки // Тр. Астраханск. зап-ка. Вып. 2. С. 59-77.
Астафьев В.Т. 1998. Материалы по биологии выпи и болотного луня на северо-востоке Украины //
Авифауна Украины. Вып. 1. С. 16-23.
Аськеев И.В., Аськеев О.В. 1999. Орнитофауна республики Татарстан (конспект совр. состояния).
Казань. 124 с.
Атемасова Т.А., Атемасов А.А. 2003. Белый аист в бассейне р. Северский Донец // Птицы басе. Се-
верского Донца. Харьков. Вып. 8. С. 57-68.
Ауэзов Э.М. 1977. Материалы по исчезающим и редким птицам озёр Тенгиз Тургайской депрессии //
Редк. и исчез, звери и птицы Казахстана. Алма-Ата. С. 131-134.
Ауэзов Э.М., Виноградов В.Г. 1986. Летний авиаучёт водоплавающих птиц на водоёмах Тургайской
депрессии // Тез. докл. 9-й Всесоюзн. орнитол. конф. Л. Ч. 1. С. 46-47.
Ауэзов Э.М., Грачев В.А. 1977. Исчезающие и редкие птицы Алакольской котловины // Редк. и исчез,
звери и птицы Казахстана. Алма-Ата. С. 135-138.
Афанасьев В.Т. 1998. Птицы Сумщины. Киев. 93 с.
Ашби В., Анненкова С. 2005. Орнитологические наблюдения с группами «Birdfinder» // Казахстанск.
орнитол. бюлл. 2004 г. Алматы. С. 133-136.
Бабенко В.Г. 2000. Птицы Нижнего Приамурья. М.: Прометей. 725 с.
Бабенко В.Г, Поярков Н.Д. 2000. Дальневосточный аист Ciconia boyciana Swinoe в нижнем Приаму-
рье // Дальневост, аист в России. Владивосток. С. 55-60.
Бабко В.М. 1995. Наблюдения большого баклана и белой совы на юго-западе Черниговской облас-
ти // Беркут. Т. 4. Вып. 1-2. С. 162.
Багрикова Н.А., Костин С.Ю. 2005. Биоценотические связи растит, и колон, гнездящихся веслоногих
и голенастых птиц на Лебяжьих о-вах // Бранта. Сб. тр. Азово-Черноморск. орнитол. станции.
Вып. 8. С. 27-43.
Бака С.В., Бака А.И., Киселева Н.Ю. 2000. Белый аист в Нижегородской области // Белый аист в
России: дальше на восток. Калуга. С. 110-111.
Балтвилкс Я. 1972. Результаты учёта гнездящихся черных аистов в Латвии // Тез. докл. 8-й Прибалт,
орнитол. конф, по изуч. мигр. птиц. Таллин. С. 7-8.
Баранов А.А. 1984. Кр. сообщ. о Чёрном аисте на юге Тувинской АССР // Иссл. в обл. запов. дела. М.:
ВНИИ прир. С. 118-119.
Баранов А.А. 1991. Редкие и малоизученные птицы Тувы. Красноярск: Красноярск, ун-т. 320 с.
Литература
533
Бекштрем Э.А. 1927. О фауне зверей и птиц Рязанской Мещеры // Мат-лы к изуч. флоры и фауны
Центр.-пром. обл. М.: изд. Гос. музея ЦПО. С. 32-33.
Белик В.П. 1997. Некоторые последствия использования пестицидов для степных птиц Восточной
Европы // Беркут. Т. 6. Вип. 1-2. С. 70-82.
Белик В.П. 1998. Исторические изменения и современная динамика авифауны Степного Придонья //
Соврем, орнитол. М.: Наука. С. 39-66.
Белик В.П. 1999. Авифауна Нижнекундрюченского песчаного массива и его окрестностей // Инвент.,
мониторинг и охр. Ключевых орнитол. терр. России. М. С. 15-37.
Белик В.П. 2000а. Птицы Северного Придонья. Ростов-на-Дону. 260 с.
Белик В.П. 20006. Птицы Степного Придонья: формирование фауны, её антропогенная трансформа-
ция и вопросы охраны. Ростов-на-Дону. 376 с.
Белик В.П. 2003а. Гнездовая колония хохлатого баклана на юге России // Стрепет. Вып. 1. С. 67-71.
Белик В.П. 20036. Имя из «Красной книги». Наземн. позвоночн. животн., нуждающиеся в особой
охране. Ростов-на-Дону: изд-во «Донской издат. дом». 432 с.
Белик В.П. 2004. Птицы долины оз. Маныч-Гудило: Non-Passeriformes // Тр. зап-ка «Ростовский».
Ростов-на-Дону. Вып. 3. С. 111-177.
Белик В.П. 2005. Кадастр гнездовой орнитофауны Южной России // Стрепет. Т. 3. Вып. 1-2. С. 5-37.
Белик В.П., Динкевич М.А. 2004. Колониальные веслоногие и голенастые птицы Восточного При-
азовья // Бранта. Со. тр. Азово-Черноморск. орнитол. станции. Вып. 7. С. 131-157.
Беликова Н.П., Литвиненко Н.М. 1973. Эколого-фаунистические связи гамазовых клещей с колони-
альными птицами островов зал. Петра Великого // Мед. паразитол. № 2. С. 204-210.
Белопольский Л.О. 1934. К орнитофауне Анадырского края // Тр. Арктич. ин-та. Т. 11. С. 23-44.
Белопольский Л.О. 1955. Птицы Судзухинского заповедника, ч. 2 // Тр. Зоол. ин-та СССР. Л. Т. 17.
С. 224-265.
Белопольский Л.О. 1957. Экология морских колониальных птиц Баренцева моря. М.-Л.: изд-во АН
СССР. 454 с.
Белопольский Л.О. 1984. Краткие сообщения о Чёрном аисте в Калининградской области // Иссл. в
обл. запов. дела. М.: ВНИИ прир.
Белопольский Л.О. 1989. Новые данные о распределении и питании олуш на кочёвках в тропиках
восточной части Тихого океана Вести, зоол. № 3. С. 59-61.
Белопольский Л.О. 1994. Обзор некоторых морфоэкологических особенностей олуш (Sulidae) Ми-
рового океана // Совр. орнитол. 1992. М.: Наука. С. 75-85.
Белоусов Е.М. 1990. Материалы о новых гнездящихся видах птиц низовьев р. Атрек (ЮВ Прикас-
пий) // Орнитология. Вып. 24. С. 103-107.
Беляков В.В., Яковчик Н.Л. 1980. Результаты учёта численности белого аиста на территории Кали-
нинградской области в 1974-75 гг. Фауна Нечерноземья, её охр., воспр., и использование. Ка-
линин. С. 1-18.
Белялов О.В., Карпов Ф.Ф. 2004. Орнитологические наблюдения на Сорбулаке в 2003 г. // Казах-
станок. орнитол. бюл. 2003. С. 81-84.
Белянин А.Ф. 1984. Кр. сообщ. о Чёрном аисте в Кузнецком Алатау, Кузнецкой котловине и в Горной
Шории // Иссл. в обл. запов. дела. М.: ВНИИ прир. С. 117-118.
Белянин А.Ф. 1997. О случае гнездования кудрявого пеликана в Кемеровской обл. // Пробл. сохранен,
биоразнообр. Юж. Сибири. С. 25-26.
Белянин А.Ф. 2000. К орнитофауне равнинной части Кемеровской обл. и сопредельных территорий
Новосибирской обл. // Мат-лы к распр. птиц на Урале, Екатеринбург. С. 18-21.
Беляченко А.В., Пискунов В.В., Антончиков А.Н., Варламов А.Г 2000. Чербаевская пойма // Ключе-
вые орнитол. терр. России. Т. 1. Ключевые орнитол. терр. междунар. знач. в Европ. России. М.
С. 469.
Беньковский Л.М. 1968. Заметки о птицах островов Тюлений и Монерон // Орнитология. № 9. С. 222-
227.
Березовиков Н.Н. 1989. Птицы Маркакольской котловины (Юж. Алтай). Алма-Ата: Наука. 200 с.
Березовиков Н.Н. 1991. Колпица в долине р. Чу // Редк. птицы и звери Казахстана. Алма-Ата: Гылым.
С. 45.
534
Литература
Березовиков Н.Н. 2002. О неудачном гнездовании кудр. пеликана в дельте р. Тентек // Казахстанск.
орнитол. бюл. 2002 г. Алматы. С. 51-52.
Березовиков Н.Н. 2004а. Гнездящиеся птицы ключевых орнитологических терр. Зайсанской котло-
вины // Selevinia. С. 112-133.
БерезовиковН.Н. 20046. Птицы Алакольского заповедника//Тр. Алакольск. зап-ка. Алматы: Мектеп.
Вып. 1. С. 199-257.
Березовиков Н.Н. 2004в. Наблюдения за пролётом птиц в дельте реки Тентека в марте 2003 г. // Казах-
станск. орнитол. бюл. 2003 г. Алматы. С. 67-68.
Березовиков Н.Н., Воробьев И.С. 1984. Краткие сообщения о Чёрном аисте в южн. и зап. Алтае //
Иссл. в области заповеди, дела. М.: ВНИИ природы. С. 116-117.
Березовиков Н.Н., Гисцов А.П. 1999. Орнитологические комплексы реки Урал и их изменение в свя-
зи с очередной трансгрессией Каспийского моря // Selevinia. 1996-1997. С. 79-87.
Березовиков Н.Н., Гисцов А.П. 2001. Птицы дельты реки Урал // Русск. орнитол. ж., Экспр.-вып.
№ 153. С. 635-649.
Березовиков Н.Н., Левинский Ю.П. 2004. Орнитологич. наблюдения в Алакольской котловине в 2004
г. // Казахстанск. орнитол. бюл. 2003 г. Алматы. С. 72-79.
Березовиков Н.Н., Самусев И.Ф. 2003. Птицы Зайсанской котловины. 1. Gaviformes, - Phoenicopteri-
formes // Русск. орнитол. ж., Экспр.-вып. № 210. С. 71-86.
Березовский В.Г. 1983. Весенний пролёт водоплавающих и околоводных птиц на восточном побере-
жье Аральского моря в 1978-1979 гг. // Мигр. птиц в Азии. Алма-Ата: Наука. С. 26-34.
Бескаравайный М.М. 1999. Некоторые особенности миграции птиц в Восточном Крыму // Фауна,
экол. и охрана птиц Азово-Черноморск. региона. Симферополь. С. 12-17.
Бескаравайный М.М. 2001. Современное состояние и некоторые тенденции динамики численности
редких видов птиц юго-восточного Крыма // Беркут. Т. 10. Вип. 2. С. 125-139.
Бескаравайный М.М. 2004. Хохлатый баклан на юге Украины // Бранта. № 7. С. 172-192.
Бескаравайный М.М. 2005. Видовой состав, сезонная динамика и биотопическое распределение аис-
тообразных птиц в Южном Крыму // Бранта. Сб. тр. Азово-Черноморск. орнитол. станции. Вып. 8.
С.114-127.
Бескаравайный М.М., Костин С.Ю. 1999. Структура и распределение зимней гидрофильной орнито-
фауны южного берега Крыма // Пробл. изуч. фауны юга Украины. Мелитополь-Одесса. С. 19-33.
Бианки В.В., Бойко Н.С. 2001. Осенний пролёт прибрежных птиц в Кандалакшских шхерах // Акту-
альн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 92-93.
Бианки В.В., Коханов В.Д., Корякин А.С. и др. 1993. Птицы Кольско-Беломорского региона // Русск.
орнитол. ж. Т. 2. Вып. 2. С. 491-586.
Бианки В.Л. 1922. Распространение птиц в СЗ части Европейской России // Ежегодн. Зоомуз. АН.
Т. 23. №2. С. 97-128.
Бирина У.А. 2003. Поздняя встреча белого аиста в Финляндии // Русск. орнитол. ж., Экспр.-вып.
№222. С. 531-532.
Бичеров А.П. 1988. Биология аистообразных птиц Центрального Предкавказья и сопредельных тер-
риторий. Автореф. дис. ... канд. биол. н. М. 18 с.
Бичеров А.П., Барнев В.В. 1998. О гнездовании белого аиста на Смоленщине // Редк. виды птиц
Нечерноземн. центра России. М.: МГПУ им. В.И. Ленина. С. 162-163.
Бичеров А.П., Бичерова М.А. 2001. Белый аист Смоленщины: численность, пространственная струк-
тура // Мат-лы 11-й (Всес.) орнитол. конф. Казань. С. 94-95.
Бичеров А.П, Сигида С.И. 1984. К оценке роли серой цапли в биоценозах Ставропольской возвы-
шенности // Фауна и экол. животн. Калмыкии. Элиста. С. 9-14.
Бичеров А.П., Скиба С.А. 1990. Заметки по редким и малоизученным птицам Ставрополья // Мало-
изуч. птицы Сев. Кавказа. Ставрополь. С. 160-163.
Бичеров А.П., Хохлов А.Н. 1979. К биологии серой цапли на Ставрополье // Нов. пробл. зоол. науки
и их отраж. в вузовск. Преподавании. Тез. докл. конф, зоологов педвузов. Ставрополь.Ч. 2. С. 212-
213.
Бичеров А.П., Хохлов А.Н. 1981. Кваква - новый зимующий вид на территории Европейской части
СССР // Природа. № 8. С. 118-119.
Литература
535
Бичеров А.П., Хохлов А.Н. 1984. О взаимоотношениях врановых и голенастых птиц в Предкавка-
зье // Экол., биоценотич. и хоз. знач. врановых птиц. М.: Наука. С. 173-174.
Бичеров А.П., Хохлов А.Н. 1990. Послегнездовая жизнь аистообразных Предкавказья // Мигр. и зи-
мовки птиц Кавказа. Ставрополь. С. 120-134.
Блинова Т.К., Блинов В.Н. 1997. Птицы Южного Зауралья: Лесостепь и степь. Т. 1. Фаунистич. обзор
и охрана птиц. Новосибирск: Наука. 296 с.
Блинова Т.К., Мухачева М.М., Романова Н.А. и др. 2001. Птицы Томского Причулымья // Мат-лы к
распростр. птиц на Урале. Екатеринбург. С. 40^13.
Блохин А.Ю., Тиунов И.М. 2003. Колониально гнездящиеся птицы залива Лихачева (Татарский про-
лив, Японское море) // Биол. моря. Т. 29. № 6. С. 405-408.
Блохин А.Ю., Тиунов И.М. 2005. Мыс Делиль-де-ла-Кройера (о-в. Сахалин) - новая точка гнездова-
ния морских птиц // Биол. моря. Т. 31. № 1. С. 61-62.
Богданов М.Н. 1871. Птицы и звери черноземной полосы Поволжья и долины Средней и Нижней
Волги. Казань. 226 с.
Богословская В.С., Вотрогов Л.М. 1981. Массовые зимовки птиц и китов в полыньях Берингова
моря // Природа. № 1. С. 42-43.
Бокотей А.А., Дзюбенко Н.В., Соколов Н.Ю., Кучинская И.В. 2003. Значение рыборазводных прудов
для видового разнообразия водно-болотных орнитокомплексов бассейна верхнего Днестра // Мат-
лы докл. 8-й научн. конф, орнитол. запада Украины, Кам.-Подольск. С. 101-103.
Болд А, Дорджиев Ц.З., Юмов Б.О., Цэвэмядаг Н. 1991. Фауна птиц бассейна озёра Байкал // Экол. и
фауна птиц Вост. Сибири. Улан-Удэ. С. 3-24.
Бондарев Д.В. 1975а. Колониальные гнездовья веслоногих и голенастых птиц на северном Каспии.
Астрахань. 30 с.
Бондарев Д.В. 19756. Околоводные колониально-гнездящиеся птицы и изменение условий среды //
Колон, гнездовья околоводн. птиц и их охрана. М. С. 11-13.
Бондарев Д.В. 1977. Колониальные гнездовья голенастых и веслоногих птиц на западном побережье
Каспия // Ресурсы пернатой дичи побер. Каспия и прилег, районов. Астрахань. С. 83-86.
Бондарев Д.В. 1979. Колониальные гнездовья околоводных птиц в дельте Волги // Тр. Кызыл-Агачск.
зап-ка. Баку. Вып. 1. С. 202-219.
Бондарев Д.В. 2005. Колониальные гнездовья голенастых и веслоногих птиц на СЗ Каспии в 1970-е
гг. (период максимального снижения уровня Каспийского моря) // Долговр. мониторинг и сохр.
колон, водн. птиц Сев. Каспия в связи с колеб. ур-ня Касп. моря. М.-Астрахань. С. 186-214.
Бондарев Д.В., Кривоносов Г.А. 1977. Кудрявый пеликан в дельте Волги // Тез. докл. 7-й Всес. орни-
тол. конф. Киев. Ч. 2. С. 193-194.
Бондарев Д.В., Скокова Н.Н. 1978. Миграции серых цапель дельты Волги Мигр. птиц Вост. Европы
и Сев. Азии. Гагарообр. Аистообр. М.: Наука. С. 192-194.
Борисов А.М. 1977. Водные угодья и водоплавающие птицы Приморско-Ахтарских лиманов // Мат-
лы 4-го Всесоюзн. совещ. по водопл. птицам. М. С. 47^19.
Бородин О.В. 2000. Белый аист (Ciconia ciconia) в Ульяновской области // Белый аист в России:
дальше на восток. Калуга. С. 179-180.
Бостанжогло В.Н. 1911. Орнитологическая фауна Арало-Каспийских степей // Мат-лы к познан, фа-
уны и флоры Росс. Имп. Отд. зоол. Вып. П.С. 1^4-10.
Брагин Е.А. 2001. Динамика озёр и водоплавающие птицы Убаган-Ишимского междуречья и Се-
верного Тургая // Актуальн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 113—
115.
Брагин Е.А. 2004. Орнитол. исследования в Кустанайской обл. в 2003 г. // Казахстанск. орнитол. бюл.
2003 г. Алматы.
Брагин Е.А. 2005. Наурзумские озёра весной 2004 г. // Казахстанск. орнитол. бюл. 2004 г. Алматы.
С. 24-25.
Брагин Е.А., Брагина Т.М. 2002. Фауна Наурзумского заповедника (аннот. списки позвоночн.). Коста-
най. 56 с.
Браунер А.А. 1894. Заметки о птицах Херсонской губернии // Зап. Новоросс. о-ва естествоиспытат.
Т. 19. Вып. 1. С. 39-93.
536
Литература
Браунер А.А. 1899. О вредных и полезных животных Херсонской губ.// Зап. о-ва с/х Юж. России. №
4 и 5-6. С. 99-122.
Браунер А.А. 1923. Сельскохозяйственная зоология. Одесса: Укргиз. 436 с.
Бредб1ер П. 2002. Дншропетровськ - Micro i птахи // Свята справа. Вип. 1. С. 41-46.
Букреев С.А. 1997. Орнитофауна и заповедное дело Туркменистана. М.: ЦОДП. 160 с.
Букреев С.А. 2001. Мониторинг КОТР - 2000 // Инф. бюлл. «Ключевые орнитологические террито-
рии России». М.: СОПР. № 13. С. 4-21.
Бурковский О.А1998. Некоторые интересные встречи птиц в Приморье // Русск. орнитол. ж., Экспр.-
вып. №43. С. 13-15.
Бурский О.В., Пагенкопф К., Форстмайер В. 2003. Птицы Среднего Енисея // Мат-лы к распростр.
птиц на Урале. Екатеринбург. С. 48-71.
Бурчак-Абрамович Н.И. 1946. О залетах священных ибисов на побережье Каспия // Природа. № 4.
С. 62.
Бурчак-Абрамович Н.И., Лобков Е.Г., Пономаренко А.К. 1987. К изучению исторического прошлого
авифауны Камчатки // Бюл. МОИП. Отд. биол. Т. 92. Вып. 3. С. 42-53.
Бутурлин С.А. 1901. Синоптические таблицы охотничьих птиц Российской империи. СПб. 126 с.
Бутурлин С.А. 1911. Белокрылая цапля в Уссурийском крае // Орнитол. вести. № 3-4. С. 289-290.
Бутурлин С.А., Дементьев Г.П. 1935. Птицы СССР. Полный определитель птиц СССР. М.-Л.: КОИЗ.
Т. 2. 280 с. (также - Бутурлин, 1935).
Бутьев В.Т., Михеев А.В., Костин А.Б. и др. 1989. Заметки о редких видах птиц Кавказского побере-
жья Каспия // Орнитол. ресурсы Сев. Кавказа. Ставрополь. С. 137-152.
Бучко В.В. 1998. Птахи Галицького регюнального ландшафтного парку та його околиць // Заповщна
справа в Украпп. Т. 4. Вып. 2. С. 32-41.
Ваксель С. 1940. Вторая Камчатская экспедиция Витуса Беринга. Л.-М.: изд. Главсевморпути. 175 с.
Вальчук О.П. 1997. Случаи заносов птиц ветром в Приамурье // Русск. орнитол. ж., Экспр.-вып.
№ 30. С. 6-8.
Варшавский С.Н., Варшавский Б.С., Гарбузов В.Е. 1977. Некоторые редкие и исчезающих птицы
Северного Приаралья // Редкие и исчезающие звери и птицы Казахстана. Алма-Ата. С. Мб-
153.
Васильев В.И. 1968. Рыбоядные птицы Азербайджана. Автореф. дис. ... канд. биол. н. Баку. 20 с.
Васильев В.И. 1969. О факторах, определяющих размещение и численность рыбоядных птиц в Азер-
байджане // Орнитол. в СССР. Ашхабад. Кн. 2. С. 102-105.
Васильев В.И. 1975. Трофические связи рыбоядных птиц и их хозяйственное значение в условиях
Азербайджана // Мат-лы по фауне и экол. наземн. позв. Азерб. Баку. С. 189-216.
Васильев В.И., Мирзоева Н.М., Соколова Э.И. 1972. Колония голенастых и веслоногих птиц, как
возможный природный очаг арбовирусных инфекций // Трансконтинент, связи перел. птиц и их
роль в распростр. арбовирусов. Новосибирск: Наука. С. 314-316.
Велижанин А.Г. 1971. Морские колониальные птицы Курильских о-вов. Дис... канд. биол.наук. Но-
восибирск: Зап.-Сиб. проекта, изыскат. эксп. Главохоты РСФСР. 249 с.
Велижанин А.Г. 1976. Краснолицый баклан на Курильских о-вах // Тр. Окск. заповеди. Рязань. Вып. 13.
С. 12-13.
Велижанин А.Г. 1977а. Новые сведения о морских птицах Дальнего Востока // Зоол. ж. Т. 56. Вып. 7.
С. 1077-1084.
Велижанин А.Г. 19776. О некоторых редких и малоизвестных птицах Курильских островов // Орни-
тология. Вып. 13. С. 25-32.
Велижанин А.Г. 1978. Размещение и состояние численности колоний морских птиц на Дальнем Восто-
ке // Актуальн. вопр. охр. прир. на Дальн. Востоке. Владивосток: ДВО АН СССР. С. 39-42.
Велижанин А.Г. 1981. Наблюдения беринговых бакланов Phalacracorax pelagicus Pall, на Прихан-
кайской равнине // Редк. птицы Дальн. Востока. Владивосток: ДВНЦ АН СССР. С. 123-124.
Велижанин А.Г. 1987. Заметки о морских птицах о. Алюмка (Чукотский АО) // Распростр. и биол.
морск. птиц Дальн. Востока. Владивосток: ДВО АН СССР. С. 39-42.
Великанов В.П., Хохлов А.Н. 1979. Об орнитофауне и особенностях биологии водоплавающих и
околоводных птиц озёра Сары-Камыш // Тр. Кызыл-Агачского зап-ка. Баку. Вып. 1. С. 236-240.
Литература
537
Венгеров М.П. 1975. О колониально гнездящихся птицах заповедного оз. Делили (ЮЗ Туркмения) //
Колон, гнездовья околоводн. птиц и их охрана. М.: Наука. С. 29.
Венгеров П.Д., Воробьев И.И., Нумеров А.Д., Соколов А.Ю. 2000. Пойма Хопра у оз. Ильмень //
Ключевые орнитологические территории России. М. Т. 1. С. 298-299.
Вероман X. 1976. Некоторые данные по экологии белого аиста в Эстонии // Мат-лы 9-й Прибалт,
орнитол. конф. Вильнюс. С. 48-50.
Вероман X. 1981. Результаты гнездования впервые гнездящихся и старых пар белого аиста. // Экол. и
охр. птиц. Кишинев. С. 39-40.
Вероманн X., Роотемяэ Л. 1981. Краткий обзор о распространении колоний серых цапель и грачей в
Эстонии // Сообщ. Прибалт, комис. по изуч. мигр. птиц. Тарту. С. 169-173.
Ветров В.В. 1998. К распространению белого аиста в Луганской области // Беркут. Т. 7. Вип. 1-2.
С. 69.
Ветров В.В., Самчук Н.Д., Литвиненко С.П. 1991. О находках редких птиц на территории Луганской
обл. // Мат-лы 10-й Всес. орнитол. конф., Минск. Ч. 2. Кн. 1. С. 107-109.
Виксне Я. 1983. Птицы Латвии. Рига: Зинатне. 224 с.
Вилков Е.В. 1999. Новые ключевые орнитологические территории Дагестана: Туралинская и Сулак-
ская лагуны // Инвент.. мониторинг и охр. Ключ, орнитол. терр. России. М. С. 54-60.
Вилков Е.В., Близнюк А.И.. Джамирзоев Г.С. 2000. Кизлярский залив // Ключевые орнитологические
территории России. М. Т. 1. С. 384-385.
Вилков Е.В., Пишванов Ю.В. 2000. Редкие и малочисленные птицы Дагестана // Редк., исчез, и мало-
изуч. птицы России. Со. научн. статей. Махачкала. С. 13-29.
Вильконский Ф.В. 1897. Орнитологическая фауна Аджарии, Гурии и СВ части Лазистана // Мат-лы к
познан, фауны и флоры Росс. имп. Отд. зоол. Вып. 3. С. 1-121.
Виноградов В.В. 1967. Биологические ресурсы водно-болотных охотугодий Мильской степи, их про-
изводительность и преспективы хозяйственного использования // Тр. зап-ков Азерб. М.: Лесн.
пром-сть. Вып. 2. С. 1-144.
Виноградов В.В., Чернявская С.П. 1965. Материалы по орнитофауне Кызыл-Агачского заповедни-
ка // Тр. зап-ков Азерб. М.: Лесн. пром-сть. Вып. 1. С. 22-52.
Виноградов В.В., Чернявская С.П. 1969. Материалы к изучению гнездовых колоний голенастых и
веслоногих птиц на озере Акгель в Азербайджане // Орнитол. в СССР. Ашхабад. Кн. 2. С. 108—
112.
Винокуров А.А. 1959а. Распределение и численность голенастых птиц в Приазовских лиманах //
Зоол. ж. Т. 38. Вып. 6. С. 939-941
Винокуров А.А. 19596. Рыжая цапля в юго-восточном Приазовье // Орнитология. Вып. 2. С. 256-
261.
Винокуров А.А. 1960. Материалы по биологии некоторых голенастых птиц Приазовских лиманов //
Охрана природы и озеленение. Вып. 4. С. 18-22.
Винокуров А.А. 1963. Аэровизуальный абсолютный учёт голенастых птиц в Плавнях дельты Куба-
ни // Организ. и методы учёта птиц и вредн. грызунов. М. С. 53-57.
Винокуров А.А. 1965. Голенастые птицы в плавнях Кубани и их значение в рыбном хозяйстве Севе-
ро-Западного Предкавказья // Рыбоядн. птицы и их знач. в рыбн. хоз-ве. М.: Наука. С. 151-155.
Винокуров А.А. 1967. О залете в Приморье фрегата Ариеля // Орнитология. Вып. 8. С. 339.
Винокуров А.А., Дубровский Э.Б. 1957. О значении некоторых рыбоядных птиц в рыбном хозяйстве
юго-восточного Приазовья // Вопр. ихтиологии. Вып. 8. С. 91-96.
Винтер С.В. 1976. О биологии черноклювого белого аиста (Ciconia boyciana Swinhoe) в Среднем
Приамурье // Пробл. зоол. Л. С. 21-23.
Винтер С.В. 1978. Гнездование черноклювого белого аиста, Ciconia boyciana Swinhoe, в Среднем
Приамурье // Тр. Зоол. ин-та АН СССР. Л. Т. 76. С. 9-23.
Владимиров Ю.В., Злобин Б.Д. 1969. Размещение и численность колониальных птиц на озере Бал-
хаш // Орнитол. в СССР. Ашхабад. Кн. 2. С. 122-125.
Вошственский М.А., Кютяювський О.Б. 1962. Визначник птах!в УРСР. Вид. 2-е. Кшв: Рад. школа.
371 с.
Во Кви. 1983. Каталог птиц Вьетнама // Фауна и экол. животн. Вьетнама. М.: Наука. С. 12-43.
538
Литература
Волков Е.И. 1972. Гнездование фламинго в Центр. Казахстане // Ресурсы водопл. птиц СССР, их
воспроизв. и использ. М. Кн. 2. С. 17-19.
Волков Е.Н. 1975а. О сезонном размещении, численности и мерах охраны фламинго центрально-
казахстанской популяции // Мат-лы Всесоюзн. конф, по мигр. птиц. М. Ч. 2. С. 257-259.
Волков Е.Н. 19756. Динамика численности и размещения птиц на оз. Тенгиз // Колон, гнездовья
околоводн. птиц и их охрана. М. С. 125-129.
Волков Е.Н. 1977а. Динамика обводненности озёр и размещение центрально-казахстанской популя-
ции фламинго // Фауна и биол. гусеобр. птиц. С. 82-85.
Волков Е.Н. 19776. О размещении и численности центрально-казахстанской популяции фламинго //
Редк. и исчез, звери и птицы Казахст. Алма-Ата. С. 153-157.
Волков Е.Н. 1977в. О зимовочном ареале фламинго // Тез. докл. 7-й Всесоюзн. орнитол. конф. Киев:
Наукова думка. Ч. 2. С. 201.
Волков Н.И. 1977г. Чёрный аист в Березинском заповеднике // Мат-лы 7-й Всесоюзн. орнитол. конф.
Киев: Наукова думка. Ч. 2. С. 202.
Волков Е.Н. 1979. Миграции фламинго - Phoenicopterus roseus Pall // Миграции птиц Вост. Европы и
Сев. Азии. Аистообр. - Пластинчатою!. М.: Наука. С. 31-37.
Волковская-Курдюкова Е.А. 2009. Редкие и малоизученные птицы Приморского края: новые матери-
алы за 1997-2009 годы // Русск. орнитол. ж., Экспр.-вып. 18 (494). С. 1103-1114.
Волошина И.В., Елсуков С.В., Вдовин А.Н. 1999. Кадастр позвоночных животных Сихотэ-Алинско-
го заповедника и Сев. Приморья. Владивосток: Дальнаука. 91 с.
Волчанецкий И.Б. 1937. К орнитофауне Волжско-Уральской степи // Пращ наук.-доел, зоол.-бюл. ш-
ту Харк. ун-ту. Т. 4. С. 21-81.
Вон Хон Гу. 1963. Птицы Кореи. Пхеньян. Т. 1. 129 с. (на кор. яз.).
Воробьев Г.П., Лихацкий Ю.П. 1987. Новые данные по редким птицам Воронежской области // Ор-
нитология. Вып. 22. С. 176-177.
Воробьев К.А. 1936. Материалы к орнитологической фауне дельты Волги и прилежащих степей //
Научн. тр. зап-ков, сер. 1: Астраханск. зап-к. Вып. 1. С. 3-60.
Воробьев К.А. 1954. Птицы Уссурийского края. М.: изд. АН СССР. 360 с.
Воробьев К.А. 1963. Птицы Якутии. М.: изд. АН СССР. 336 с.
Воробьева Т.Д. 1979. Материалы по миграциям птиц в Кызыл-Агачском заповеднике // Тр. Кызыл-
Агачск. зап-ка. Баку. Вып. 1. С. 41-61.
Воронов Б.А. 1985. О встречах и гнездовании некоторых редких птиц в Приамурье // Редк. и исчез,
птицы Дальн. Востока. Владивосток. С. 24-26.
Воронов Б.А., Пронкевич В.В. 1991. О некоторых орнитологических находках в Хабаровском крае //
Бюл. МОИП. Отд. биол. Т. 96. № 5. С. 23-28.
Воронов ГВ. 1987. О встречах редких и экзотических птиц на Сахалине и Курильских островах //
Итоги иссл. по вопр. рац. использ. и охраны водных, земельных и биологич. рес. Сахалина и
Курильск, о-вов. Южно-Сахалинск. С. 69-71.
Воронцов Е.М. 1937. До шзнання орштофауни Присивашшя i Сиваппв // Пращ н.-д. Зоол.-бюл. ш-ту
Харк. ун-ту. Т. 4. С. 83-124.
Вяткин П.С. 1981. Новые сведения о распространении морских птиц на побережьях Камчатки //
Бюл. МОИП. Отд. биол. Т. 86. Вып. 4. С. 14-19.
Вяткин П.С. 1983. Материалы по орнитофауне о. Старичкова // Биол. и промысел охотн. животн.
Пермь: Пермск. с/х ин-т. С. 8-14.
Вяткин П.С. 1986. Кадастр гнездовий колониальных птиц Камчатской области // Морск. птицы Дальн.
Востока. Владивосток: ДВНЦ АН СССР. С. 20-36.
Вяткин П.С. 1999. Морские колониальные птицы острова Столбовой // Морск. птицы Берингии.
Вып. 4. Магадан: ИБПС ДВО РАН. С. 28-30.
Вяткин П.С. 2000. Кадастр гнездовий колониальных морских птиц Корякского нагорья и восточ-
ного побережья Камчатки // Биол. и охрана птиц Камчатки. Вып. 2. М.: Тип. Россельхозакад.
С. 7-15.
Вяткин П.С., Артюхин Ю.Б. 1998. Краснолицый баклан Phalacrocorax urile (Gmelin, 1789) // Кр. кн.
севера Дальн. Востока России. Животные. М.: ТОО «Пента». С. 101-103.
Литература
539
Вяткин П.С., Артюхин Ю.Б. 2006. Краснолицый баклан // Кр. книга Камчатки. Т. 1. Животные. Пет-
ропавловск-Камчатский: Камч. печ. двор. С. 102-103.
Гавриленко Н.И. 1929. Птицы Полтавщины. Полтава. 133 с.
Гавриленко Н.И. 1958. Залеты некоторых птиц на Полтавщину // Уч. зап. Полтавск. гос. пед. ин-та.
Вып. 197. Орнитология.
Гавриленко Н.И. 1968. Экологические заметки о птицах Украины // Орнитология. Вып. 9. С. 343.
Гавриленко Н.С., Думенко В.П., Лопушанский Е.А. 2002. Зимовки птиц в районе заповедника «Аска-
ния-Нова» в 2000-2001 гг. // Мониторинг зимующ. птиц в Азово-Черном, регионе Украины. Одес-
са-Киев. С. 9-15.
Гаврилов Н.Н. 1984. Динамика колониальных гнездовий околоводных птиц в дельте Волги // 8-я
Всес. зоогеогр. конф., М.. С. 31-33.
Гаврилов Н.Н. 2002. Динамика численности колониальных околоводных птиц дельты Волги // Мат-
лы конф. «Птицы Юж. России». Ростов-на-Дону. С. 61-66.
Гаврилов Н.Н. 2005. Сохранение колониальных гнездовий веслоногих и голенастых птиц в дельте
Волги и на Сев. Каспии в 1975-2003 гг. (период подъема ур-ня Каспия) //Долговрем. мониторинг
и сохр. колон, водных птиц Сев. Каспия в связи с колебанием уровня Касп. моря. М.-Астрахань.
С. 214-257.
Гаврилов Н.Н., Русанов Г.М.. Бондарев Д.В. 2003. Состояние колониальных гнездовий птиц в дельте
Волги, западном ильмено-бугровом районе и на о. Малый Жемчужный (1999-2001 гг.) // Ключе-
вые орнитол. терр. России. Информ, бюл. № 2. М. С. 30-42.
Гаврилов Э.И. 1972. Предварительные итоги массового кольцевания птиц на юго-востоке Казах-
стана // Трансконт. связи перед, птиц и их роль в распростр. арбовирусов. Новосибирск: Наука.
С.108-109.
Гаврилов Э.И.. Гисцов А.П. 1978. Миграции больших бакланов в Казахстане // Мигр. птиц Вост.
Европы и Сев. Азии. Гагарообразные-аистообразные. М.: Наука. С. 88-101.
Гаврилов Э.И., Гисцов А.П. 1985. Сезонные перелеты птиц в предгорьях Западного Тянь-Шаня. Алма-
Ата. 223 с.
Гаврилюк М.Н. 2002. Строки сезонних м!гращй птах!в у Черкаському Подншров! в 1991-2002 рр. //
Ав!фауна Украши. Вып. 2. С. 86-96.
Гаврилюк М.Н., Грищенко В.Н. 2001. Современная зимняя орнитофауна Восточной Черкасщины //
Беркут. Т. 10. Вип. 2. С. 184-195.
Гаврилюк М.Н., Грищенко В.Н.. Яблоновська-Грищенко С.Д. 2005. Нов! дан! про рщюсних та мало-
вивчених птах!в Центрально! Украши // Беркут. Т. 14. Вип. 1. С. 28-37.
Гаврись ГГ 1994. Расселение большой и малой белых цапель на Левобережье Украины // Вести,
зоол. № 1. С. 80-83.
Гагина Т.Н. 1961. Птицы Восточной Сибири (Список и распространение) // Тр. Баргузинск. зап-ка.
М. Вып. 3. С. 99-123.
Гагина Т.Н. 1962. Залётные птицы Восточной Сибири // Орнитология. Вып. 4. С. 367-372.
Гагина Т.Н., Скалой Н.В. 2000. Залеты редких видов птиц на территорию Кемеровской области //
Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 57-59.
Галченков Ю.Д. 2000а. История формирования населения белого аиста на территории Калужской
области // Белый аист в России: дальше на восток. Калуга. С. 132-151.
Галченков Ю.Д. 20006. Мониторинг популяции белого аиста в Калужской области // Там же. С. 132—
151.
Гамова ТВ., Рыжов А.В., Сурмач С.Г. 2005. Материалы к гнездовой биологии амурского волчка
Ixobrychus eurhythmus в Приморье // Русск. орнитол. ж., Экспр.-вып. 14 (303). С. 990-995.
Гамова Т.В., Сурмач С.Г, Бурковский О.А. Первое свидетельство гнездования китайского волчка
Ixobrychus sinensis (J. F. Gmelin, 1789) на юге Дальнего Востока России // Birdland.ru, в печати.
Танюшкин М.А. 1958. Из фенологических дневников // Тр. Астраханск. зап-ка. Т. 4. С. 353-366.
Ганя И.М., Зубков Н.И. 1989. Редкие и исчезающие виды птиц Молдавии. Кишинев: Штиница. 152
с.
Гашев С.Н., Некрасов А.Л., Низовцев Д.С. и др. 2003. Интересные авифаунистические находки в
Тюменской области // Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 74-76.
540
Литература
Гашев С.Н., Шаповалов С.И., Попов Н.Я. и др. 2003. Результаты обследования КОТР на лесостепном
юге Тюменской области в 2002 году // Ключевые орнитол. терр. России. Вып. 18. С. 42-44.
Гашек С.П. 2002. Нотатки про деяких рщюсних птах!в з територш Чорнобильськоп зони вщчуження //
Беркут. Т. 11. Вип 2. С. 141-147.
Герасимов Н.Н. 1986. Морские колониальные птицы острова Карагинского // Морск. птицы Дальн.
Востока. Владивосток: ДВНЦ АН СССР. С. 57-65.
Герасимов Н.Н. 1999. Берингов баклан Phalacrocorax pelagicus на Карагинском острове // Биол. и
охр. птиц Камчатки. М.: Диалог-МГУ. Вып 1. С. 83-86.
Герасимов Н.Н., Соколов А.М., Томкович П.С. 1992. Птицы орнитологического заказника «Река
Морошечная», Зап. Камчатка // Русск. орнитол. ж. Т. 1. Вып. 2. С. 157-208.
Герасимов Ю.Н. 1992. Миграция бакланов и чистиковых вдоль западного побережья Камчатки //
Изуч. морск. колон, птиц в СССР. Магадан: ИБПС ДВО РАН. С. 25-27.
Герасимов Ю.Н. 1999. Наблюдения за весенней миграцией птиц в устье р. Ходутки (Юго-Вост. Кам-
чатка) // Биол. и охр. птиц Камчатки. М.: Диалог-МГУ. Вып. 1. С. 69-71.
Герасимов Ю.Н. 2001. Наблюдения за миграцией водных и околоводных птиц вдоль юго-вост, побе-
режья Камчатки // Биол. и охр. птиц Камчатки. М.: ЦОДП. Вып. 3. С. 86-95.
Герасимов Ю.Н. 2003. Мат-лы по весенней миграции некот. видов птиц вдоль юго-зап. побережья
Камчатки // Биол. и охр. птиц Камчатки. М.: ЦОДП. Вып. 5. С. 55-56.
Герасимов Ю.Н., Калягина Е.Е. 1997. Мат-лы по весенней миграции гагар, поганок и бакланов в
прибрежной полосе Юго-Зап. Камчатки, // Рус. орнитол. журн., Экспр.-вып. № 28. С. 21-22.
Гизенко А.И. 1955. Птицы Сахалинской области. М.: изд-во АН СССР. 328 с.
Гизенко А.И. 1957. Материалы по экологии и численности птиц Лебяжьих островов // Тр. Крымск,
зап-ка. Вып. 4. С. 167-184.
Гизенко А.И. 1963. О гнездовании каравайки на Днепре // Орнитология. Вып. 6. С. 469.
Гисцов А.П. 2001. Численность птиц водно-болотного комплекса на северо-восточном побережье
Каспия // Актуальн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 174-176, 177—
178.
Гисцов А.П. 2003. Особенности летней орнитофауны на побережье Каспия // Совр. пробл. орнитол.
Сибири и Центр. Азии. Улан-Удэ. Ч. 1. С. 81-85.
Гисцов А.П. 2003а. Фламинго в Казахстанской части Каспия // Казахстанск. орнитол. бюл. Алматы.
С.133.
Гисцов А.П. 2004. Мониторинг состояния водоплавающих и околоводных птиц Сев. Каспия на рубе-
же столетий // Тр. Ин-та зоол. Алматы. Т. 48. Орнитология. С. 260-278.
Гисцов А.П. 2005. Мониторинг состояния птиц северной части Каспия в 2004 г. // Казахстанск. орни-
тол. бюл. Алматы. С. 7-8.
Гисцов А.П. 2006. Водно-болотные птицы СВ побережья Каспия // Тез. 12-й Межд. орнитол. конф.
Сев. Евразии. Ставрополь. С. 141-142.
Гисцов А.П., Ауэзов Э.М. 1991. Численность и размещение фоновых и редких околоводных птиц СВ
побережья Каспийского моря // Мат-лы 10-й Всес. орнитол. конф., Минск: Навука i тэхшка. Ч. 2.
Кн. 1. С. 147-148.
Гисцов А.П., Гаврилов Э.И., Бородихин И.Ф, 1977. Дополнение к орнитофауне Зап. Тянь-Шаня //
Мигр. птиц в Азии. Новосибирск. С. 215-218.
Гладков Н. 1964. Стрелять или не стрелять? Как охотнику относиться к рыбоядным птицам // Охота
и охотн. х-во. № 10. С. 23-25.
Гладков Н.А. 1970. Отряд Голенастые // Жизнь животных. Т. 5. М.: Просвещ.. С. 87-106.
Гладков Н.А., Дементьев Г.П., Птушенко Е.С., Судиловская А.М. 1964. Определитель птиц СССР.
М.: Высш, школа. 370 с.
Глущенко Ю.Н. 1979. О птицах рисовых полей Приханкайской низменности // Биология птиц юга
Дальн. Востока СССР. Владивосток: БПИ ДВНЦ АН СССР. С. 55-66.
Глущенко Ю.Н. 1981. К фауне гнездящихся птиц Приханкайской низменности // Редк. птицы Дальн.
Вост. Владивосток: ДВНЦ АН СССР. С. 25-33.
Глущенко Ю.Н. 1985. Проблемы охраны и привлечения дальневосточного аиста на Приханкайской
низменности // Редкие птицы Дальнего Востока. Владивосток: ДВНЦ АН СССР. С. 138-139.
Литература
541
Глущенко Ю.Н. 2005. Средняя белая цапля // Кр. книга Приморск, края. Животн. Владивосток. С. 201—
202.
Глущенко Ю.Н., Волковская-Курдюкова Е.А. 2005. Птицы // Позвон. животн. зап-ка «Ханкайский»
(аннот. списки видов). Спасск-Дальний. С. 30-85.
Глущенко Ю.Н., Коробов Д.В., Кальницкая И.Н. 2003. Численность и размещение колоний около-
водных и водоплавающих птиц на Приханкайской низменности в 2002 г. // Животн. и растит, мир
Дальн. Востока. Уссурийск. Вып. 7. С. 54-65.
Глущенко Ю.Н., Коробов Д.В. 2005. Южная белая цапля Egretta modesta (J.E. Gray, 1831) в Примор-
ском крае: таксономический статус в свете новых данных // Русск. орнитол. ж., Экспр.-вып. № 304.
С.1028-1032.
Глущенко Ю.Н., Липатова Н.Н., Мартыненко А.Б. 2006. Птицы города Уссурийска: фауна и динамика
населения. Владивосток. 264 с.
Глущенко Ю.Н., Мрикот К.Н. 2000а. Динамика популяции дальневосточного аиста в российском
секторе Приханкайской низменности // Дальневост, аист в России. Владивосток: Дальнаука. С. 77-
85.
Глущенко Ю.Н., Мрикот К.Н. 20006. Новые данные о гнездовании средней белой цапли Egretta
intermedia на озере Ханка Русск. орнитол. ж., Экспр.-вып. № 114. С. 20-22.
Глущенко Ю.Н., Мрикот К.Н. 2001. Динамика колониальных гнездовий голенастых и веслоногих
птиц российского сектора Приханкайской низменности в последней трети XX столетия // Акту-
альн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 179-180.
Глущенко Ю.Н., Нечаев В.А. 2000. Новые встречи фрегата-ариеля в Приморском крае // Животн. и
растит, мир Дальнего Востока. Уссурийск: Изд-во Уссурийск, гос. пед. ин-та. Вып. 4. С. 109-110.
Глущенко Ю.Н., Поливанова Н.Н., Шибнев Ю.Б. 1992. Цапли Приханкайской низменности // Жи-
вотн. и растит, мир Дальн. Востока. Уссурийск: УГПИ. С. 27-36.
Глущенко Ю.Н., Шибаев Ю.В. Лебяжинская И.П. 1995. Современное состояние популяций некото-
рых редких видов птиц Приханкайской низменности // Тр. междунар. науч.-практ. конф. «Пробл.
сохр. ВБУ междунар. знач.: озеро Ханка». Спасск-Дальний. С. 45-50.
Глущенко Ю.Н., Шибнев Ю.Б. 1977. Новые данные о редких птицах Приморья // Тез. докл. 7-й Все-
союзн. орнитол. конф. Киев. Ч. 1. С. 49-50.
Глущенко Ю.Н., Шибнев Ю.Б.. Медведев В.Н. 1986. Заметки о новых для Приморья и редких птицах //
Распростр. и биол. птиц .Атгая и Дальн. Востока. Тр. Зоол. ин-та АН СССР. Л.: Наука. Т. 150.
С. 83-84.
Глущенко Ю.Н., Шибнев Ю.Б.. Волковская-Курдюкова Е.А. 2006. Птицы // Позвоночные животные
заповедника «Ханкайский» и Приханкайской низменности. Владивосток. С. 77-233.
Годованець Б.Й., Скыьський I.B.. Бучко В.В. и др. 1996. Мала бита чапля в швшчно-захщнш частиш
Прут-Дшстровського межир!ччя на прилеглих територ!ях Украпнських Карпат // Беркут. Т. 5.
Вип. 1. С. 26-30.
Головкин А.Н., Флинт В.Е. 1975. Колониальные гнездовья морских птиц в районе мыса Сердце-
Камень (Чукотка) // Колон, гнездовья околоводн. птиц и их охрана. М.: Наука. С. 171-173.
Головушкин М.И. 1992. Фенология весеннего прилёта птиц в окрестностях Киева // Сез. мигр. птиц
на терр. Украины. Киев: Наукова думка. С. 242-249.
Голодушко Б.З. 1958. К биологии белого аиста // Тр. заповедно-охотн. хоз-ва «Беловежская пуща».
Вып. 1. С. 110-119.
Голубев С.В. 2000. Белый аист в Ярославской области: история и совр. состояние // Белый аист в
России: дальше на восток. Калуга. С. 100-105.
Голубова Е.Ю., Плещенко С.В. 1997. Колонии морских птиц северной части Охотского моря // Видо-
вое разнообразие и сост. популяций околоводн. птиц Северо-Востока Азии. Магадан: СВНЦ ДВО
РАН. С. 141-162.
Гончаров ГФ. 1981. Осенний пролёт веслоногих на Айдарских разливах // Миграции птиц в Азии.
Ашхабад. С. 276-278.
Горбань И., Фладе М. 2000. Значение Верхней Припяти (Украина) ля охраны птиц // Экол. и охрана
птиц низинных болот Полесья. Минск. С. 32-39.
542
Литература
Горбулшська С., Сребродолська С., Бадецька К. 2004. Мошторинг за популящею быого лелеки (Ciconia
ciconia L.) в селах Монастырець та Поляна Городоцького району Лв1вськоп облает! // Вюник
Львюк. ун-ту. Сер. бюл. Вип. 35. С. 154-158.
Гордиенко Н.С. 1987. Новые и редкие виды птиц Наурзумского заповедника // Орнитология. Вып. 22.
С. 177-179.
Гордиенко Н.С. 2002. Новые материалы по фауне и распространению птиц в Челябинской обл. //
Матер, к распростр. птиц на Урале, в Приуралье и Зап. Сибири. Екатеринбург. С. 104-108.
Гордиенко Н.С. 2006. Состояние редких видов водно-болотных птиц на Южном Урале // Орнитол.
иссл. в Сев. Евразии. Ставрополь. С. 148-149.
Горобейко В.В., Крюков В.Х. 2000. Состояние популяции дальневосточного аиста в Еврейской авто-
номной области в 1999 г. // Дальневост, аист в России. Владивосток, Дальнаука. С. 31-33.
Горошко О.А., Кирилюк В.Е. 2003. Птицы зап-ка «Даурский» // Наземн. позв. Даурии. Чита: Поиск.
С. 20-32.
Горчаков Г.А. 1990. К фауне гнездящихся птиц антропогенных ландшафтов юга Приморского края //
Экол. и распростр. птиц Дальн. Востока. Владивосток: ДВО АН СССР. С. 69-73.
Грабар А. 1997. Птицы Подкарпатской Руси (Avifauna Carpathorossica) в обработке А.Е. Лугового с
переводом с чешского и русинского языков // Беркут. Т. 6. Вип. 1-2. С. 91-102.
Граве Г.Л. 1912. Несколько слов к вопросу о стремлении птиц расширить свою гнездовую область //
Орнитол. вести. № 1. С. 58-59.
Граве Г.Л. 1926. Очерк авифауны Смоленской губернии // Тр. о-ва изуч. Смоленск, края. Т. 3. Смо-
ленск. С. 3-64.
Грачев В.А. 1977а. Редкие и исчезающие птицы дельты р. Или // Редк. и исчез, звери и птицы Казах-
стана. Алма-Ата. С. 175-177.
Грачев В.А. 19776. Современное распространение и численность пеликанов на оз. Балхаш // Там же.
С.178-179.
Греков В.С. 1962.0 зимовке фламинго в Кызыл-Агачском заповеднике // Орнитология. Вып. 5. С. 356-
261.
Греков В.С. 1965. Колонии голенастых и веслоногих птиц в Кызыл-Агачском заповеднике // Орнито-
логия. Вып. 7. С. 258-265.
Греков В.С., Русев И.Т., Нехороших З.Н. и др. 2004. Обзор эпизоотологического значения веслоно-
гих и голенастых птиц на примере Азово-Черноморского региона // Бранта. Сб. тр. Азово-Чер-
ном. орнитол. станции. Вып. 7. С. 215-240.
Гринченко А.Б. 1991. Новые данные о редких и исчезающих птицах Крыма // Редк. птицы Причерно-
морья. Киев-Одесса: Лыбидь. С. 78-90.
Гринченко А.Б. 1994. Баклан довгоносий // Червона книга Украши. Тваринний св!т. Ки!в: «Украшьс-
ка енциклопед!я» iM. М.П. Бажана. 306 с.
Гринченко А.Б. 2004а. История и динамика колониальных поселений аистообразных птиц в восточ-
ных районах Крыма в связи с антропогенной сукцессией Вост. Сиваша и Присивашья // Бранта.
Сб. тр. Азово-Черноморск. орнитол. ст. Вып. 7. С. 61-81.
Гринченко А.Б. 20046. Размещение, численность и экология малого баклана (Phalacracoraxpygmaeus)
на юге Украины // Там же. Вып. 7. С. 167-171.
Гришанов Г.В. 1988. Чёрный аист в Калининградской области // Рес. редк. жив. РСФСР, их охрана и
воспроизв. М.: ЦНИЛ Главохоты РСФСР. С. 61.
Гришанов Г.В. 1994. Гнездящиеся птицы Калининградской области: территориальное размещение и
динамика численности в XIX-XX вв. 1. Non-Passeriformes // Рус. орнитол. журн. Т. 3. Вып. 1.
С. 83-116.
Гришанов Г.В., Беляков В.В. 2000. Наземные позвоночные Калининградской области. Справ, посо-
бие. Калининград: Калинингр. ун-т. 69 с.
Гришанов Г.В., Савчук М.М. 1992. Изменения гнездования биологии белого аиста в Калининградс-
кой области за последние десятилетия // Аисты: распростр., экол., охр. Минск. С. 79-81.
Грищенко В.М. 1991. Проведение «девизных годов» по охране редких птиц на Украине // Мат-лы
Всесоюзн. орнитол. конф. Ч. 2. Кн. 1. С. 170-171.
Грищенко В.М. 1992.0 зимовках белого аиста на Украине // Аисты: распростр., экол., охрана. Минск.
С. 82-85.
Литература
543
Грищенко В.М. 1995а. До питания про динам!ку чисельности быого лелеки на швшчному сход! Ук-
рапни (м. Дрогобич. 1-3.02. 1995 р.). Льв!в-Чершвщ. С. 37.
Грищенко В.М. 19956. Фенолопчш спостереження за осшньою м!гращею птах!в у Миколапвськш
облает! у 1983 рощ // Пробл. вивч. та охор. птах!в. Льв!в-Черн!вщ. С. 36-37.
Грищенко В.М. 2003. Вплив полювання на дистанщю сплохування птах!в // Прюр. орштол. доел.
Льв!в-Кам’янець-Под!льський. С. 118-120.
Грищенко В.М. 2005. Чар!вний св!т быого лелеки. Чершвщ. 160 с.
Грищенко В.М., Гаврилюк М.Н. 2000. Фенология миграций птиц в районе Каневского заповеднике
во второй половине XX в. // Заповщна справа в Укради. Т. 6. Вип. 1-2. С. 67-76.
Грищенко В.Н. 1996а. Семь вылетевших птенцов в гнезде белого аиста // Беркут. Т. 5. Вип. 1. С. 20.
Грищенко В.Н. 19966. Быий лелека. Чершвщ. 127 с.
Грищенко В.Н. 1998а. До екологш быого лелеки в зон! росту чисельност! // Мат-ли 3-й конф, моло-
дих орштолопв Украши. Чершвщ. С. 31-34.
Грищенко В.Н. 19986. Белый аист в мифологии европейских народов и современные представления
о происхождении индоевропейцев // Беркут. Т. 7. Вип. 1-2. С. 120-131.
Грищенко В.Н. 1999. Постройка и заселение искусственных гнездовий для белого аиста в Украине //
Беркут. Т. 8. Вип. 2. С. 203-212.
Грищенко В.Н. 2000а. Современное состояние мировой популяции белого аиста// Б. аист: дальше на
восток. Калуга. С. 13-20.
Грищенко В.Н. 20006. Белый аист в Сумском Посеймье // Там же. С. 202-205.
Грищенко В.Н. 2004. Динамика численности белого аиста в Украине в 1994-2004 гг. // Беркут. Т. 13.
Вип. 1. С. 38-61.
Грищенко В.Н., Борейко В.Е., Дремлюга Г.Н. и др. 1992. Опыт проведения операции «Лелека» в
Киевской области Аисты: распр., экол., охр. Минск. С. 85-95.
Грищенко В.Н., Габер Н.А. 1990. Соотношение причин гибели взрослых белых аистов и их птенцов
на Украине // Аисты: распростр., экол., охр. Минск. С. 90-93.
Грищенко В.Н., Серебряков В.В. 1992. Миграция белого аиста на Украине по данным фенологичес-
ких наблюдений // Сез. мигр. птиц на терр. Украины. Киев. С. 258-273.
Грищенко В.Н., Серебряков В.В. 1993. Миграция серой цапли на Украине по данным фенологичес-
ких наблюдений // Бюл. МОИП. Отд. биол. Т. 98. Вып. 5. С. 33-36.
Грищенко В.Н., Серебряков В.В., Борейко В.Е., Грищенко И.А. 1992. Современное состояние попу-
ляции белого аиста (Ciconia ciconia) на Украине // Русск. орнитол. ж. Т. 1. Вып. 2. С. 147-156.
Губский В.С. 1960. Гельминтофауна кваквы Нижнего Днестра// Научн. ежег. Биол. ф-та ОГУ. Одесса.
С. 94-96.
Гудина А.Н. 2003а. Птицы Русской равнины на границе степной и лесостепной зон. Т. 1. Птицы
Восточной Украины. Вып. 1. История изучения, обзор раритетов. Воронеж: ВГУ. 216 с.
Гудина А.Н. 20036. Редкие и малоизученные птицы среднего течения реки Ворона // Экол. и эволю-
ция животн. Рязань. С. 41-45.
Гузш A.I. 1997. Фауна i населения хребетних Захщного репону Украши. Кшв. Т. 1. 147 с.
Гулай В.И. 1975. О колонии квакв и малых белых цапель в низовьях рп. Сарата // Колон, гнезд,
околоводн. птиц и их охрана. М.: Наука. С. 32-33.
Гусаков Е.С., Виноградов В.Г. 1998. Озеро Ханка // Водно-бол. угодья России. М. Т. 1. С. 237-245.
Гынгазов А.М., Миловидов С.П. 1977. Орнитофауна Запапдно-Сибирской равнины. Томск. 350 с.
Давиденко И.В., Сыпко А.В. 2002. Зимовка птиц в районе очистных сооружений г. Киева зимой 2000/
01 гг. // Ав!фауна Украш!. Вип. 2. С. 70-73.
Давыгора А.В. 1984. Кр. сообщ. о Чёрном аисте в Бузулукском бору // Иссл. в обл. запов. дела. М.:
ВНИИприроды. С. 111.
Давыгора А.В. 2006. Залеты птиц и их значение в познании истории и долговременной динамике
познания авифаун // Тез. 12-й Межд. орнитол. конф. Сев. Евразии. Ставрополь: СГУ. С. 170-171.
Даль С.К. 1954. Животный мир Армянской ССР. Ереван. Т. 1. 415 с.
Даниленко У.Ф., Даниленко А.К., Чибисова В.В. 1977. О распределении фламинго в послегнездовое
время на юге Тургайской ложбины // Орнитология. Вып. 13. С. 202-204.
544
Литература
Дарман Ю.А., Андронов В.А., Парилов М.П., Хигучи X., Нагендран М., Кириченко Ю.И. 2000а.
Авиаучёт редких видов птиц на Средне-Амурской низменности в 1999 г. // Дальневост, аист в
России. Владивосток: Дальнаука. С. 13-19.
Дарман Ю.А., Андронов В.А., Парилов М.П., Хигучи X., Нагендран М., Кириченко Ю.И. 20006.
Состояние популяции дальневосточного аиста в Амурской области // Там же. С. 20-25.
Дарман Ю.А., Шибаев Ю.В., Сурмач С.Г. 2000. Результаты учёта дальневосточного аиста в России в
1998-2000 гг. // Там же. С. 107-109.
Дебело П.В. 1996. О состоянии редких и исчезающих животных Западно- Казахстанской области //
Краеведч. пробл. экологии Зап. Казахст. Уральск. С. 98-100.
Дебело П.В. 2002. Материалы по орнитофауне оз. Шалкар (неворобьиные) // Мат-лы к распростр.
птиц на Урале. Екатеринбург. С. 108-126.
Девяткин Г.В., Кустов Ю.И., Липаткина О.О. и др. 2000. Хакасский заповедник // Заповедники Рос-
сии. Заповедники Сибири. Т. 2. М.: Логата. С. 129-143.
Дегтярев В.Г., Ларионов Т.П., Винокуров В.Н. 1990. Второй факт залета индийского ибиса в СССР //
Орнитология. Вып. 24. С. 148.
Дементьев Т.П. 1940. Руководство по зоологии. Позвоночные. Птицы. Т. 6. 856 с.
Дементьев Т.П. 1952. Птицы Туркменистана. Ашхабад: изд-во АН Туркм. ССР. 517 с.
Дементьев Т.П., Гладков Н.А. (ред.). 1951-54. Птицы Советского Союза. Тт. 1-6. М.: Сов. наука.
Дементьев Г.П., Караев М.К., Карташов Н.Н. 1955. Птицы Юго-Зап. Туркмении // Уч. зап. МГУ.
Вып. 171, биология. С. 53-172.
Демидова М.И. 1997. О залете белого аиста в Пермскую область // Мат-лы к распр. птиц на Урале.
Екатеринбург. С. 56.
Денисов В.П., Фролов В.В. 1991. Новые данные о распространении птиц в Пензенской обл. // Орни-
тология. Вып. 25. С. 155-156.
Денисова М.Н. 1958. Особенности роста птенцовых, полувыводковых и выводковых птиц // Уч. зап.
МГУ. Вып. 197. С. 165-182.
Деревягин П.Я. 1930. Орнитофауна оз. Куш-Мирун и его окрестностей // Тр. Кустанайск. краеведч. о-
ва. Вып. 1. С. 21-39.
Деревягин П.Я. 1934. Материалы к проблеме использования системы водоёмов Арало-Тургайско-
Убаганской депрессии // Арало-Тургайский пролив (автор: Шиллингер Ф.Ф.). М. С. 112-136.
Джамирзоев Г.С. 1999а. Территориальная охрана мигрирующих птиц в Дагестане // Инвентар., мо-
ниторинг и охр. Ключевых орнитол. терр. России. М. С. 83-89.
Джамирзоев Г.С. 19996. Некоторые ключевые орнитологические территории Дагестана // Там же.
С. 50-53.
Джамирзоев Г.С., Исмаилов Х.Н. 2004. Результаты зимних учётов птиц в республике Дагестан //
Ключевые орнитологические территории России. Инф. бюл. Вып. 19. С. 34-36.
Джамирзоев Г.С., Хохлов А.Н., Ильюх М.П. 2000. Редкие и исчезающие птицы Дагестана и их охра-
на. Ставрополь. 145 с.
Динесман Л.Г, Киселева Н.К., Савинецкий А.Б., Хасанов Б.Ф. 1996. Вековая динамика прибрежных
экосистем северо-востока Чукотки. М.: Аргус. 189 с.
Динец В.Л. 1996.0 гнездовании средней белой цапли на Курильских островах // Орнитология. Вып. 27.
С. 273.
Динкевич М. А. 2006. Современное состояние и размещение редких гнездящихся лимнофильных птиц
в Краснодарском крае // Тез. 12-й Межд. орнитол. конф. Сев. Евразии. Ставрополь. С. 176-178.
Динкевич М.А., Иваненко А.М., Лохман Ю.В. и др. 1999. О реализации проекта «Пеликан-99» и его
результатах // Мат-лы Всерос. науч.-практич. конф. «Человек и ноосфера». Геленджик-Красно-
дар. С. 98-100.
Дмитренок М.Г. 2005. Мониторинг большой выпи // Мониторинг животн. мира Беларуси (Осн. прин-
ципы и результаты).
Дмитренок М.Г. 2009. Оценка численности и популяционная динамика большой выпи (Botaurus
stellaris) и малой выпи (Ixobrychus minutus) в пойме р. Припять (Беларусь). Збереження та
вщтворення бюр!зноманптя природно-заповщних територш. Матер1али м!жнародно1 науково-прак-
тично! конференцп, присвячено! 10-р1ччя Р1вненьского природного заповедника. Р1вне. С. 405-409.
Литература
545
Дмитренок М.Г., Журавлев Д.В., Демонжан Л. 2005. Фенология большой выпи в Белоруссии // Орни-
тология. № 32. С. 79-86.
Дмитренок М.Г., Лундышев Д.С., Гаращук Ю.В. 2009. Влияние биотических и абиотических факто-
ров среды на выбор гнездовых биотопов малой выпью (Ixobrychus minutus) II Проблемы сохране-
ния биологического разнообразия и использования биологических ресурсов. Мат-лы межд. науч.-
практич. конф, и 10-й зоол. конф. Минск. Ч. 2. С. 425-427.
Доброхотов Б.П. 1961. К экологии жёлтой цапли в дельте Волги // Тр. Астрах, зап-ка. Вып. 5. С. 250-
261.
Докучаев Н.Е., Лаптев А.А. 1974. Два новых вида птиц в фауне Приморья // Зоол. ж. Т. 53. Вып. 3.
С. 484-485.
Долбик М.С. 1978. Использование пернатой дичи в Белоруссии // Тр. Окского госуд. зап-ка. Вып. 14.
Научн. осн. охраны и рац. использ. птиц. Рязань: Рязанск. отд. изд-ва «Московский рабочий».
С. 303-311.
Долбик М.С. 1985. Рэв1з1я саставу i размеркавания арштафауны Беларус! // Весщ АН БССР. Сер.
б!ял. навук. № 2. С. 85-89.
Долгушин И.А. 1960. Птицы Казахстана. Алма-Ата: изд-во АН Каз. ССР. Т. 1. 469 с.
Долгушин И.А. 1960а. О фламинго на озере Тенгиз (Центральный Казахстан) // Тр. Ин-та зоол. АН
КазССР. Т. 13. С. 125-132.
Дольник В.Р. 1981. Динамическая модель прогноза миграции птиц; Прогноз осеннего дневного про-
лёта птиц на основе динамической модели // Методы обнаружения и учёта миграции птиц. Тр.
Зоол. ин-та АН СССР. Л. Т. 140. С. 123-133, 133-143.
Домашевский С.В. 2001. Материалы по орнитофауне полуострова Пурикаринэм (Эстония) // Беркут.
Т. 10. Вип. 1. С. 3 М2.
Домашевский С.В. 2002. Наблюдения за миграциями хищных и околоводных птиц в нижнем течении
р. Десна // Ав1ф. Украши. Вип. 2. С. 52-59.
Домашевский С.В. 2003. Видовой состав и динамика мигрирующих птиц в заливе Хаара-Лахт на
севере Эстонии // Беркут. Т. 12. Вип. 1-2. С. 143-152.
Дорогой И.В. 1992. Орнитологические наблюдения на птичьих базарах Вост. Чукотки // Изуч. морск.
колониальн. птиц в СССР. Магадан: ИБПС ДВО РАН. С. 7-8.
Дорст Ж. 1968. До того, как умрет природа. М. 415 с.
Дубинин В.Б., Горчаковская Н.Н. 1951. Редкие птицы Советского Приморья // Природа. № 2. С. 75-
76.
Дубинин В.Б., Дубинина М.Н. 1940. Паразитофауна колониальных птиц Астраханского заповедни-
ка // Тр. Астраханск. зап-ка. Вып. 3. С. 190-298.
Дугинцев В.А., Панькин Н.С. 1993. Список птиц Верхнего и Среднего Приамурья в административ-
ных границах Амурской области // Пробл. экол. Верхи. Приамурья. Благовещенск. С. 25-145.
Дугинцев В.А., Панькин В.С. 1994. Залёт фламинго на Зейско-Буреинскую равнину // Соврем, орни-
тол. 1992. М. С. 228.
Дылюк С.А. 2000. Состояние популяции белого аиста в Европейской части России по данным все-
российского учёта 1994-1997 гг. // Белый аист в России: дальше на восток. Калуга. С. 42-52.
Дылюк С.А., Галченков Ю.Д. 2000. История расселения белого аиста в России // Там же. С. 21-41.
Дымин В.А., Ефремов В.Ф., Панькин Н.С. 1970. О гнездовании некоторых птиц верхнего Приамурья
// Вопр. геогр. Дальн. Востока. Благовещенск. Т. 11. С. 26-29.
Дымин В.А., Панькин Н.С. 1975. О гнездовании и пролёте аистов (Ciconidae) и журавлей (Gruidae) в
Верхнем Приамурье // Орнитол. исслед. на Дальн. Востоке. Владивосток. С. 253-267.
Дыхан М.Б. 1990. Новые сведения о птицах острова Шикотан // Экол. и распр. птиц юга Дальн.
Востока. Владивосток: ДВО АН СССР. С. 83-86.
Дюнин А.П. 1960а. Изменение состава орнитофауны Малого Кызыл-Агачского залива в связи с его
опреснением // Охр. прир. и озеленен. М. Вып. 4: Птицы водоёмов. С. 53-54.
Дюнин А.П. 19606. Поведение птиц на зимовке в период похолодания // Охр. природы и озеленен. М.
Вып. 2. С. 39-41.
Евдошкин С.А. 1999. Новые виды птиц Тамбовской области // Экол. животн. Экол. человека. Мичу-
ринск. С. 15-16.
546
Литература
Елкин К.Ф. 1969.0 фламинго в Центральном Казахстане // Орнитол. в СССР. Ашхабад. Кн. 2. С. 221—
223.
Елкин К.Ф., Азаров В.И., Сотников В.Л. 1975. Хозяйственная деятельность и колониальные гнездо-
вья околоводных птиц в низовьях реки Чу (Южн. Казахстан) // Колон, гнездовья околоводных
птиц и их охрана. М. С. 134-135.
Елсуков С.Е. 1974. К авифауне ВС Приморья // Мат-лы 6-й Всесоюзн. орнитол. конф. М.: МГУ. Ч. 1.
С. 199-200.
Елсуков С.В. 1977. О редких птицах северо-востока Приморья // Тез. докл. 7-й Всесоюзн. орнитол.
конф. Киев.Ч. 1. С. 58-59.
Елсуков С.В. 1982. Птицы // Растит, и животн. мир Сихотэ-Алинского заповеди. М.: Наука. С. 171-
173.
Елсуков С.В. 1984. К орнитофауне морского побережья Сев. Приморья // Фаунистика и биол. птиц
юга Дальнего Востока. Владивосток: ДВНЦ АН СССР. С. 34-43.
Емтыль М.Х. 1986. Значение большого баклана в прудовом рыбоводстве // Изуч. птиц СССР, их охр.
и рац. использ. Тез. докл. 1-го съезда Всесоюзн. орнитол. о-ва. Л. Ч. 1. С. 218-219.
Емтыль М.Х., Лохман Ю.В. 1994. К весенней орнитофауне Калининских плавней // Тез. межресп.
науч.-практ. конф.: «Актуальн. вопр. экол. и охр. прир.степных экосистем и сопред. терр.». Крас-
нодар. Ч. 1. С. 170-173.
Емтыль М.Х., Лохман Ю.В., Иваненко А.М., Емтыль А.М., Короткий Т.В. 2003. Гидрофильные коло-
ниальные птицы в Западном Предкавказье // Актуальн. вопр. экол. и охр. прир. экосист. южн.
регионов России и сопред. терр. Краснодар. С. 181-187.
Емтыль М.Х., Тильба П.А., Плотников Г.К., Мнацеканов Р.А. 1989. Численность и распределение
колоний околоводных птиц в Краснодарском крае // Актуальн. вопр. экол. и охр. прир. Азовск.
моря и Вост. Приазовья. Краснодар. Ч. 1. С. 165-168.
Есилевская М.А., Кривицкий И.А., Лисецкий А.С., Кныш Н.П. 1988. О новых орнитологических
находках в СВ оконечности Украины. Сообщ. 1 // Вести. Харьковск. ун-та. № 313. С. 84-85.
Жаров О., Митейко В. 1967. Залёт кудрявого пеликана на Байкал // Орнитология. Вып. 8. С. 350.
Жатканбаев А.Ж. 1996. Колпица. Каравайка // Кр кн. Казахст. Т. 1.4. 1, Алматы. С. 100-102.
Житников М. 1900. Орнитологические наблюдения на реке Атрек (зима 1898 и весна 1899) // Псовая
и руж. охота. Кн. 10. С. 1-16; кн. 11. С. 17-32; кн. 12. С. 33-57.
Жерко Н.В. 1998. Геохимический фоновый мониторинг заповедника «Лебяжьи острова» // Сост. прир.
компл. Крымск, зап-ка и др. запов. терр. Украины. Алушта. С. 26-28.
Жмуд М.Е. 2000. Дельта Дуная // Числ. и размещ. гнездящ. околоводн. птиц в ВБУ Азово-Черно-
морск. побер. Украины. Киев. С. 27-41.
Жмуд М.Е. 2000а. Изменение статуса пеликанов в Северном Причерноморье на рубеже тысячелетий //
Бранта. Сб. тр. Азово-Черноморск. орнитол. станции. Мелитополь. Вып. 3. С. 112-117.
Жмуд М.Е. 2004. Кудрявый пеликан (Pelecanus crispus Bruch.) на юге Украины // Бранта. Сб. Тр.
Азово-Черноморск. орнитол. станции. Вып. 7. С. 158-166.
Жмуд М.С. 1999. Птахи // Бюразн. Дунайськ. зап-ка. Кшв: Наук, думка. С. 146-149.
Журминский С.Д. 1984. Ярусное распределение гнёзд цапель в смешанной колонии в зависимости от
вида // Тез. конф, молодых ученых АН МССР, посвящ. 60-летию Молдавской ССР и созд. Комм,
партии Молдавии. Кишинев: Штиница. С. 104.
Журминеский С.Д., Куниченко А.А. 1986. Гнездование и структура колоний цапель в низовьях Дне-
стра // Млекопит. и птицы атнропог. ландшафта Молдавии и их практич. знач. Кишинев: Штини-
ца. С. 60-69.
Забелин В.И. 2002. Колпица // Кр.кн. респ. Тыва, Животн. Новосибирск. С. 62.
Заболотный Н.Л., Хохлов А.Н. 1989а. Каравайка в низовьях Кубани // Науч. тр. ЦНИЛ Главохоты
РСФСР. М. С. 54-56.
Заболотный Н.Л., Хохлов А.Н. 19896. Колпица в низовьях Кубани // Науч. тр. ЦНИЛ Главохоты РСФСР.
М. С. 54.
Завгородний В.И., Молодан Г.Н., Чернышов А.И. 1989. Голенастые птицы сев. побер. Азовского
моря // Всес. совещ. по пробл. кадастра и учёта животн. мира. Уфа. Ч. 3. С. 86-87.
Завьялов Е.В., Шляхтин Г.В., Капранова Т.Р и др. 1997. Водоплавающие и околоводные птицы Сара-
товской области (Gaviiformes, Podicipediformes, Pelecaniformes, Ciconiiformes, Phoenicopteriformes,
Anseriformes) // Беркут. T. 6. Вип. 1-2. C. 3-18.
Литература
547
Завьялов Е.В., Шляхтин Г.В., Табачишин В.Г. и др. 2005. Птицы севера Нижнего Поволжья. Кн 1.
История изуч., общ. характ. и состав орнитофауны. Саратов: изд-во СГУ. 296 с.
Залетаев В.С. 1960. Новые сведения о распространении фламинго в Зап. Туркмении // Охр. прир. и
озеленен. М. Вып. 4. С. 99-101.
Зарудный Н.А. 1888. Орнитологическая фауна Оренбургского края // Зап. Имп. Акад. наук. СПб.
Т. 57. № 1. С. 1-338.
Зарудный Н.А. 1903. Птицы Восточной Персии. Орнитологические результаты экскурсии по Восточ-
ной Персии в 1898 г. // Зап. РГО по общ. геогр. СПб. Т. 36. № 2. 468 с.
Зарудный Н.А. 1910. Птицы Псковской губ. // Зап. Имп. Акад, наук по физ-мат. отд. СПб. Сер. 8.
Т. 25. №2. С. 1-181.
Зарудный Н.А. 1915. Птицы пустыни Кизыл-Кум // Мат-лы к познан, фауны и флоры Росс. Имп. Отд.
зоол. Вып. 14. С. 1-149.
Зарудный Н.А. 1916. Птицы Аральского моря // Изв. Туркест. отд. РГО. Т. 12. Вып. 1. С. 1-229.
Зеленская Л.А. 1993. Усыновление птенцов берингова баклана парой краснолицых бакланов // Морск.
птицы Берингии. Вып. 1. Магадан: ИБПС ДВО РАН; Анкоридж: Упр. дикой природы США. С. 51-
52.
Зеленская Л.А. 1995. Состояние морских колониальных птиц на о. Умара в 1995 г. // Морск. птицы
Берингии. Вып. 3. Магадан: ИБПС ДВО РАН. С. 46-47.
Зеленская Л.А. 1999. Состояние колонии о. Топорков (Командорские о-ва) в 1998 г. // Морск. птицы
Берингии. Вып. 4. Магадан: ИБПС ДВО РАН. С. 39-42.
Зеленская Л.А. 2001а. Распределение и численность морских колониальных птиц на Командорских
о-вах в 1999-2000 гг. Биол. и охрана птиц Камчатки. М.: ЦОДП. Вып. 3. С. 64-71.
Зеленская Л.А. 20016. Экология питания Берингова баклана, Phalacrocorax pelagicus в гнездовой
период // Зоол. ж. Т. 80. № 9. С. 1120-1130.
Зеленская Л.А. 2006. Динамика изменений в колонии морских птиц о. Умара, Северное Охотморье //
Биол. и охрана птиц Камчатки. М.: ЦОДП. Вып. 7. С. 100-106.
Зеленцов Л.С. 1995. Некоторые орнитологические находки в окрестностях Красноуфимска (Сверд-
ловская обл.) Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 28.
Зиновьев В.И. 1981а. Дополнение к списку видов птиц Калининской области // Геогр. и экол. наземн.
позвон. Нечерноземья. Владимир. С. 3-4.
Зиновьев В.И. 19816. Серая цапля в Верхневолжье // Размещ. и сост. гнездовий околоводных птиц на
терр. СССР. М.: Наука. С. 52-53.
Зиновьев В.И. 1982. Птицы лесной зоны Европейской части СССР. Аистообразные, дрофообразные,
голубеобразные, кукушкообразные // Животн. мир центра лесн. зоны Евр. части СССР. С. 62-91.
Зубакин В.А. 1981. Колониальные птицы Торейских озёр // Размещ. и сост. гнездовий околоводн.
птиц на терр. СССР. М.: Наука. С. 132-134.
Зубакин В.А. 2007. Первый залёт каравайки в Московскую область // Московка. Новости программы
«Птицы Москвы и Подмосковья». № 6, сентябрь 2007 г. С. 2-3.
Зубакин В.А., Иванов А.В., Смирнова Е.В. 1992. Белый аист в Московской области // Аисты: рас-
простр., экол., охр. Минск. С. 97-101.
Зубакин В.А., Мищенко А.Л., Абоносимова Е.В., Волошина О.Н. и др. 1986. Современное состояние неко-
торых редких видов птиц Московской области. Неворобьиные // Орнитология. Вып. 21. С. 77-93.
Зубков Н., Журминский С., Манторов О. 2004. Managementul integral al resulselor naturale din bazinul
transformatlier al fluviului Nistru Мат-лы conf, int. Кишинев. C. 125-128.
Зубков H., Журминский С., Штирбу В. и др. 2001. Современное состояние водно-болотных птиц
Среднего Днестра // Diversitatea. valorificarea rational §i protectia lumii animale. C. 81-82.
Иваненко A.M., Лохман Ю.В., Тильба П.А., Емтыль М.Х. 1995. О новом месте гнездования кудряво-
го пеликана (Pelecanus crispus Bruch) в Восточном Приазовье // Актуальн. вопр. экологии и охр.
природы водных экосистем и сопред. терр. Мат-лы науч. конф. Краснодар. Ч. 1. С. 137-140.
Иваненко А.М., Емтыль М.Х., Динкевич М.А. и др. 2000а. Кизилташские лиманы - новое место
гнездования кудрявого пеликана (Pelecanus crispus Bruch) на Северном Кавказе // Мат-лы 13-й
межд. научно-практич. конф. «Актуальн. вопр. экологии и охр. природн. экосист. юж. регионов
России и сопред. терр.». Краснодар. С. 105-106.
548
Литература
Иваненко А.М., Емтыль М.Х., Динкевич М.А. и др. 20006. Современное состояние колонии кудряво-
го пеликана на озере Ханском // Там же. С. 106-107.
Иваненко Н.Ю. 2005. Современное состояние орнитофауны о. Большой Айнов (Варангер-фьорд) в
2002 г. // Мат-лы 23-й конф, молодых ученых, посвящ. 70-летию МБС-ММБИ (май 2005 г.). Апа-
титы: Кольский НЦ РАН. С. 31-36.
Иванов А.И. 1940. Птицы Таджикистана. М.-Л.: изд-во АН СССР. 300 с.
Иванов А.И. 1961. Материалы по птицам автономной области Сишуаньбанна и смежных частей про-
винции Юньнань. Сообщ. 2-е // Acta zoologica sinica. Vol. 13. No. 1-4. C. 70-96.
Иванов А.И. 1969. Птицы Памиро-Алая. Л.: Наука. 448 с.
Иванов А.И. 1976. Каталог птиц СССР. Л.: Наука. 275 с.
Иванов А.И., Козлова Е.В., Портенко Л.А., Тугаринов А.Я. 1951. Птицы СССР. Ч. 1. М.-Л.: изд-во
АН СССР. 281 с.
Иванов А.И., Штегман Б.К. 1978. Краткий определитель птиц СССР. М.
Иванов Ф.В., Приклонский С.Г., Теплое В.П. 1965. Опыт количественного учёта пернатой дичи, до-
бываемой на территории РСФСР // Тр. Окск. гос. зап-ка. Воронеж: Центр.-Черноземн. кн. изд-во.
Вып. 6. С. 5-49.
Иванчев В.П., Котюков Ю.В., Николаев Н.Н., Лавровский В.В. 2003. Птицы долины Оки в пределах
Рязанской области // Тр. Окск. биосферн. гос. зап-ка. Вып. 22. С. 47-147.
Измайлов И.В., Боровицкая Г.К. 1973. Птицы Юго-Западного Забайкалья. Владимир. 303 с.
Ильинский И.В., Фетисов С.А. 1986. Характеристика демонстративного «пения» большой выпи //
Мат-лы 9-й Всесоюзн. орнитол. конф. Л. Ч. 1. С. 264-266.
Ильюх М.П., Хохлов А.Н. 2006. Кладки и размеры яиц птиц Центрального Предкавказья. Ставро-
поль. 220 с.
Ильяшенко В.Ю. 1987. Пуховой птенец амурского волчка// Орнитология. Вып. 22. С. 211-212.
Ильяшенко В.Ю. 2001. Красноногий ибис // Красная книга РФ (животн.). М.: Астрель. С. 385-387.
Ильяшенко В.Ю., Калякин М.В., Соколов Е.П., Соколов А.М. 1988. Некоторые результаты орнитоло-
гических исследований на Кунашире и Шикотане // Вопр. экол., фаунист, и системат. птиц Пале-
арктики. Л.: Зоол. ин-т АН СССР. С. 70-88.
Иоганзен Г.Х. 1914. Материалы по орнитологии Томской губ. // Орнитол. вести. № 3.
Иоганзен Г.Х. 1928. Замечания по списку птиц из Южноуссурийского края // Uragus. No. 3. С. 19-28.
Иогансен Г.Х. 1934. Птицы Командорских островов // Тр. Томск, ун-та. Т. 86. С. 222-266.
Ирисова Н.Л., Гармс О.Я., Вотинов А.Г. и др. 1999. Птицы Вехнего Приобья (Алтайский край) //
Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 98-108.
Исаков Ю.А. 1948а. Заметки по биологии фламинго // Чайка-хохотунья и фламинго на Касп. море. М.
С. 45-54.
Исаков Ю.А. 19486. К вопросу о распростр. фламинго в СССР // Там же. С. 55-61.
Исаков Ю.А. 1957. Сезонные миграции серых цапель (Ardea cinerea) по данным кольцевания в СССР //
Тр. Бюро кольц. М.: изд-во Мин-ва сельск. хоз-ва СССР. Вып. 9. С. 46-85.
Исаков Ю.А., Воробьев К.А. 1940. Обзор зимовок и пролёта птиц на Юж. Каспии // Тр. Всес. орни-
тол. зап-ка «Гассан-Кули». Вып. 1. С. 5-159.
Исаков Ю.А., Формозов А.Н. 1946. Непериодические миграции (залеты) фламинго в СССР // Зоол.
ж. Т. 25. Вып. 5. С. 473-480.
Казаков Б.А. 2000а. Веселовское водохранилище // Ключевые орнитол. терр. России. М. Т. 1. С. 314—
315.
Казаков Б.А. 20006. Каравайка Plegadis falcinellus (Linneaus, 1766) // Красная книга РФ. С. 384-385.
Казаков Б.А., Кривенко В.Г. 2000. Озера Маныч-Гудило и Восточный Маныч // Водно-бол. угодья
России. Т. 3. С. 188-193.
Казаков Б.А., Ломадзе Н.Х. 1991. Результаты авиаобследования поселений колониальных околовод-
ных птиц в долинах Дона и Зап. Маныча // Соврем, свед. по сост., распростр. и экол. птиц Сев.
Кавказа. Ставрополь. С. 84-87.
Казаков Б.А., Ломадзе Н.Х. 1992. Результаты авиаобследования поселений колониальных околовод-
ных птиц в долинах Дона и Зап. Маныча // Кавказск. орнитол. вести. Ставрополь. Вып. 4. Ч. 1.
С. 104-109.
Литература
549
Казаков Б.А., Ломадзе Н.Х., Белик В.П. 1986. Колонии голенастых в бассейне Нижнего Дона // Тез.
докл. Всесоюзн. совещ. по пробл. кадастра и учёта животн. мира. М. Ч. 2. С. 306-307.
Казаков Б.А., Ломадзе Н.Х., Белик В.П., Бичеров А.П. и др. 1988. Колпица на Сев. Кавказе // Рес.
редк. животн. РСФСР, их охрана и воспроизв. М. С. 56-59.
Казаков Б.А., Ломадзе Н.Х., Белик В.П. и др. 2004. Птицы Северного Кавказа. Т. 1. Ростов-на-Дону.
398 с.
Казаков Б., Ломадзе Н., Миноранский В., Белик В. 1997. Белый аист в Ростовской области // Стрепет.
Вып. 1. С. 9.
Казаринов А.П. 1973. Фауна позвоночных Болынехехцирского заповедника // Вопр. геогр. Дальн.
Востока. Хабаровск. № 11. С. 3-29.
Калякин М.В., Волцит О.В. 2006. Атлас. Птицы Москвы и Подмосковья. София-М.: Pensoft. 372 с.
Капелюх Я.1., Гузий A.I. 2000. Орштофауна заповедника «Медобори» // Заповщна справа в Украпп.
Т. 6. Вип. 1-2. С. 59-67.
Караваев А.А. 1991. Численность и размещение водно-болотных птиц в Юго-Вост. Прикаспии //
Прир. среда и животн. мир ЮВ Прикаспия. Тр. Красноводск. зап-ка. М. Вып. 2. С. 37-143.
Караваев А.А., Белоусов Е.М. 1983. Миграции птиц на ЮВ Каспии осенью 1977 г. // Мигр. птиц в
Азии. Алма-Ата: Наука. С. 52-65.
Карамзин А.Н. 1901. Птицы Бугурусланского и сопредельных с ним частей Бугульминского, Бузулук-
ского уездов Самарской губернии и Белебейского уезда Уфимской губернии // Мат-лы к познан,
фауны и флоры Росс. Имп. Отд. зоол. СПб. Вып. 5. С. 274-284.
Кардаков А.И. 1928. Фрегат Fregata ariel Gould // Охотник. № 7. С. 24
Карелин Г.С. 1875. Разбор статьи А. Рябинина «Естественные произведения земель Уральского каза-
чьего войска» /; Тр. СПб о-ва естествоиспытат. Т. 6. С. 186-298.
Карташов Н.Н. 1974. Систематика птиц. М.: Высшая школа. 342 с.
Карякин И.В. 1998а. Конспект фауны птиц республики Башкортостан. Пермь. 253 с.
Карякин И.В. 19986. Конспект фауны Пермской области. Пермь. 261 с.
Карякин И.В. 2000. Чёрный аист в Уральском регионе и на прилежащих территориях // Редкие, исчеза-
ющие и малоизученные птицы России. Сб. науч, статей. М.: Союз охр. птиц России. С. 144-148.
Катин И.О., Семенова О.А., Тюрин А.Н. и др. 2004. Биота островов: распределение, состав и структу-
ра // Дальневост, морской биосферный заповеди. Биота. Т. 2. Владивосток: Дальнаука. С. 673-786.
Кашенцева ТА. 1984. Итоги учёта добычи охотничьих птиц в Казахстане в 1974 г. // Тр. Окского зап-
ка. Рязань: Рязанск. отд. изд-ва «Московский рабочий». Вып. 15. С. 128-136.
Кашин Т.Н. 1976. О гибели молодых цапель на деревьях // Орнитология. Вып. 12. С. 231-234.
Кашкаров Д.Ю., Филатов А.К., Зинченко Е.И. 1989. Весенние скопления водоплавающих птиц в ЮЗ
Приаралье // Тез. докл. Всесоюзн. совещ. по пробл. кадастра животн. мира. Уфа. Ч. 2. С. 350-
352.
Кеве А. 1978. Результаты кольцевания каравайки в Венгрии // Мигр. птиц Вост. Европы и Сев. Азии:
Гагарообр. - Аистообр. М. С. 243-244.
Кенжегулов К. 1966. Экология и значение рыбоядных птиц в низовьях р. Аму-Дарьи и юга Аральско-
го моря. Автореф. дис. ... канд. биол. н. Ташкент. 20 с.
Кенжегулов К.К. 1972. К экологии голенастых и чайковых птиц дельты Аму-Дарьи // Экол. млекопит.
и птиц Каракалпакии. Ташкент. С. 153-193.
Кенжегулов К. 1974. О распространении и биологии кудрявого пеликана в дельте Аму-Дарьи // Ор-
нитология. Вып. 11. С. 378-380.
Кирпиченко М. 1932. Матер1али до орштофауни Быоцерювщини // Журн. бю-зоол. циклу ВУАН.
№4. С. 91-104.
Кистяковский А.Б., Смогоржевский Л.А. 1964. О границе китайского орнитофаунистического комп-
лекса по реке Бурее // Науч. докл. высш. шк. Биол. н. № 3. С. 26-29.
Кистяковский А.Б., Смогоржевский Л.А. 1973. Материалы по фауне птиц Нижнего Амура // Вопр.
геогр. Дальн. Востока. Зоогеогр. Хабаровск. Сб. И. С. 182-224.
Киш Я.Б., Жмуд М.Е., Платтэу М. 2003. Колониальные веслоногие (Pelecaniformes) и аистообразные
(Ciconiiformes) птицы дельты Дуная - полный учёт 2002 г. // Мат-лы 2-го съезда и научн. конф.
АЧОС. Николаев. С. 30-34.
550
Литература
Кищинский А.А. 1978а. Заключение (к обзору мигр. серой цапли) // Миграции птиц восточн. Европы
и сев. Азии. Гагарообразные - аистообразные. М.: Наука. С. 206.
Кищинский А.А. 19786. Миграции серых цапель, гнездящихся в Калининградской области (СССР) и
в СВ Польше // Там же. С. 166—171.
Кищинский А.А. 1978в. Результаты кольцевания в Эстонской ССР // Там же. С. 175-176.
Кищинский А.А. 1978г. Результаты кольцевания серых цапель в Латвийской ССР // Там же. С. 171—
175.
Кищинский А.А. 1978д. Длинноносый баклан // Там же. С. 103-104.
Кищинский А.А. 1978е. Заключение (к обзору миграции рыжей цапли) // Там же. С. 231-232.
Кищинский А.А. 1978ж. Кудрявый пеликан // Там же. С. 67-68.
Кищинский А.А. 1980. Птицы Корякского нагорья. М.: Наука. 335 с.
Клестов Н.Л. 1983. Орнитофауна Сев. Днепра и её изменение под влиянием гидростроительства.
Дис. ... канд. биол. н. Киев: КГУ. 251 с.
Клестов Н.Л. 1991. Формирование околоводных комплексов под влиянием гидростроительства (на
примере р. Днепр). Киев. 70 с. (Перепр. / АН УССР; 91.3).
Климов С.М., Сарычев В.С., Мельников М.В., Землянухин А.И. 2004. Фауна птиц бассейна Верхнего
Дона. Неворобьиные. Липецк. 224 с.
Климов С.М., Сарычев В.С., Недосекин В.Ю. 1990. Современное состояние редких птиц Липецкой
области // Редкие виды птиц центра Нечерноземья. М.: изд-во ЦНИЛ Главохоты РСФСР. С. 45-
47.
Клумов С.К. 1960. Промысловые животные Курильских островов // Охрана полезн. зверей. М. Вып. 3.
С. 9-29.
Книш М.П. 1992. Фенолопя весняно! м!грацп птах!в в околицях м. Суми за даними спостержень
1967-1992 рр. // Пробл. oxop. i рац. використ. прир. рес. Сумщини. Суми. С. 95-112.
Кныш Н.П., Сыпко А.В. 1994. Большая белая цапля в Сумской области // Мат-ли 1-й конф, молодих
орштолопв Украши. Чершвщ. С. 15-17.
Коблик Е.А. 2001. Разнообразие птиц. М.: изд-во МГУ. Ч. 1. 384 с.
Коблик Е.А., Редькин Я.А., Архипов В.Ю. 2006. Список птиц Российской Федерации. М.: Т-во на-
учн. изд. КМК. 256 с.
Ковалев В.А. 2001. Орнитологические находки в Лодейнопольском р-не Ленинградской обл. в 1998—
2000 гг. // Русск. орнитол. ж., Экспр.-вып. № 137. С. 248-251.
Коваленко А.В., Дякин Г.Ю. 2009. Индийская прудовая цапля (Ardeola grayii) - новый вид в фауне
Казахстана // Селевиния. Казахстанск. зоол. ежегодн. 2009. С. 237.
Ковальковский С.Ю., Мищенко А.Л., Суханова О.В. 1986. Колонии серой цапли в Московской обла-
сти // Бюл. МОИП. Отд. биол. Т. 91. Вып. 4. С. 31-33.
Ковшарь А.Ф. (ред.). 1991. Редкие птицы и звери Казахстана. Мат-лы ко второму изд. Кр. кн. Каз.
ССР. Алма-Ата: Гылым. 334 с.
Ковшарь А.Ф. 1999. Редкие, ичезающие и уязвимые птицы Казахстана (Состояние и перспективы
территориальной охраны) // Терр. аспекты охраны птиц в Ср. Азии и Казахстане. М. С. 77-84.
Ковшарь А.Ф. 2002. Малая белая цапля // Казахск. орнитол. бюл. Алматы. С. 53.
Ковшарь А.Ф. 2007. Чёрный аист // Птицы Средней Азии. Алматы. Т. 1. С. 124-130.
Ковшарь А.Ф., Березовиков Н.Н. 2001. Тенденции изменения границ ареалов птиц в Казахстане во
второй половине XX столетия // Достиж. и пробл. орнитол. Сев. Евразии на рубеже веков. Ка-
зань: Магариф. С. 250-270.
Когут I.B., Бокотей А.А., Соколов Н.Ю. 1996. Особливосп бютошчного розподыу птах!в Верхнього
Дшстра // Мат-ли 2-й конф, молодих орштол. Украши. Чершвщ. С. 86-90.
Козарь Ф.В. 1985. Трофические связи озерной лягушки в Приднестровье и их сезонная динамика //
Изв. АН МССР. Сер. биол. и хим. н. № 2. С. 64-66.
Кокшайский Н.В. 1959. О морфо-функциональных различиях летательного аппарата ибисов и ца-
пель // Тр. 1-й конф, молод, науч. сотр. московск. морфологич. лабор. М. С. 124-126.
Кокшайский Н.В. 1962. Морфо-функциональные исследования полета птиц (на примере голенастых:
ибисы, цапли). Автореф. дис. ... канд биол. н. М. 23 с.
Кокшайский Н.В. 1976. О гибели молодых цапель на деревьях // Орнитология. Вып. 12. С. 231-234.
Литература
551
Колбин В.А., Бабенко В.Г., Бачурин Г.Н. 1994. Птицы Комсомольского заповедника // Позвон. жи-
вота. Комсомольск, зап-ка. Флора и фауна заповедников. М. Вып. 57. С. 13^4-1.
Коленов В.В. 1981. Материалы по размещению околоводных птиц в Волгоградской области И Раз-
мещ. и сост. гнездовий околоводн. птиц на терр. СССР. М.: Наука. С. 98-99.
Коломийцев Н.П. 1986. Некот. интересн. залеты птиц в район Лазовского заповедника И Распред, и
биол. птиц Алтая и Дальн. Востока. Тр. Зоол. ин-та АН СССР. Л.: Наука. Т. 150. С. 84-85.
Коломийцев Н.П., Поддубная Н.Я. 1985. Материалы к биологии филина -Bubo bubo (L.) в Лазовском
заповеднике (Юж. Приморье) // Редкие и исчезающие птицы Дальнего Востока. Владивосток:
ДВНЦ АН СССР. С. 81-84.
Комарова Н.А., Комаров Ю.Н. 1988. О зимовке водоплавающих и околоводных птиц на Осетинской
равнине И Ресурсы животн. мира Сев. Кавказа. Ставрополь. С. 90-93.
Комаров Ю.Е. 1986. О встречах редких видов птиц на Северо-Осетинской равнине // Редк. и исчез,
виды раст. и животн., флорист, и фаунистич. компл. Сев. Кавказа, нужд, в охране. Ставрополь.
С.101-103.
Комаров Ю.Е. 1991. Список птиц Сев. Осетии И Кавказск. орнитол. веста. Вып. 2. С. 25-32.
Комлев Е.П. 2000. Гнездование белого аиста в Костромской области И Белый аист в России: дальше
на восток. Калуга. С. 99.
Кондратьев А.Я. 1986. Колонии морских птиц на арктическом побережье крайнего Северо-Востака
СССР // Морск. птицы Дальнего Востока. Владивосток: ДВО АН СССР. С. 37^4-7.
Кондратьев А.Я.. Зубакин В.А.. Голубова Е.Ю. и др. 1992. Фауна наземных позвоночных животных
о. Талан // Приор, экосист. Сев. Охотморья. Остров Талан. Магадан: ИБПС ДВО РАН. С. 72-108.
Кондратьев А.Я., Коровин М.М., Бодяк Н.Д. 1987. Численность и структура колоний морских птиц
на о. Колючин (Чукотское море) // Распростр. и биол. морских птиц Дальнего Востока. Владиво-
сток: ДВО АН СССР. С. 22-35.
Кондратьев В. 1885. Систематическое описание животных в войске Донском, составленное в 1882
году // Газ. «Казачий вестник». Новочеркасск. № 48-59.
Кондратьев В. 2004. Систематическое описание животных в войске Донском, составленное в 1882
году // Стрепет: Фауна, экол. и охрана птиц Юж. Палеаркт. Т. 2. Вып. 1. С. 12-33.
Коновалова Н.А. 19^7. У спешность размножения караваек в условиях Кызыл-Агачского заповедника
И 7-я Всес. орнитол. конф. Киев. Ч. 1. С. 261.
Коновалова Н.А. 1979. Состояние гнездовых колоний голенастых и веслоногих птиц в Кызыл-Агач-
ском заповеднике Тр. Кызыл-Агачск. зап-ка. Баку. Вып. 1. С. 83-88.
Конюхов Н.Б. 1990. Зимовки птиц в Смрениковской полынье И Изуч. морских колониальн. птиц в
СССР. Магадан: ИБПС ДВО АН СССР. С. 36-38.
Корзюков А.И. 1996. Фенология весеннего прилёта птиц в СЗ Причерноморье (по мат-лам 1995—
1996 гг.) И Экосистемы дикой природы. Вып. 5. С. 24-27.
Корзюков А.И. 2001. Географические связи птиц придунайских водоёмов Украины И Бранта. Вып. 4.
С. 101-108.
Корнеева Т.М., Рахилин В.К. 1975. Веслоногие архипелага «Семь островов» И Колон, гнездовья око-
ловодн. птиц и их охрана. М. С. 192-194.
Корнюшин В.В., Смогоржевская Л.А., Исакова Н.И. 2004. Циркуляция гельминтов в колониях весло-
ногих и голенастых птиц юга Украины И Бранта. Сб. тр. Азово-Черноморск. орнитол. станции.
Вып. 7. С. 241-278.
Коробов Д.В., Глущенко Ю.Н. 2008. Новые сведения о некоторых редких видах аистообразных
(Ciconiiformes, Aves) заповедника «Ханкайский» и Приханкайской низменности И Чистый
Амур - долгая жизнь: материалы международной научной конференции. Хабаровск. С. 1 Об-
ll 1.
Коршиков Л.В. 2001. Новости орнитологического сезона 2001 г. в Оренбуржье И Мат-лы к распростр.
птиц на Урале. Екатеринбург. С. 121-122.
Коршиков Л.В., Корнев С.В. 1999. Новости орнитологического сезона 1999 г. в Оренбуржье И Мат-
лы к распростр. птиц на Урале, в Приуралье и Зап. Сибири. Екатеринбург. С. 140-142.
Коршиков Л.В., Корнев С.В. 2000. Дополнительные заметки и комментарии к «Орнитологической
фауне Оренбургской области» // Мат-лы к распр. птиц на Урале. Екатеринбург. С. 118-122.
552
Литература
Коршиков Л.В., Шубин А.О. 2002. Новости орнитологического сезона 2002 года в Оренбуржье И
Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 147-149.
Корякин А.С. 1981. Колония серых цапель на р. Ворскле И Размещ. и сост. гнездовий околоводн.
птиц на терр. СССР. (МОИП). М.: Наука. С. 60-61.
Косарев В.В. 2006. Влияние метеорологической ситуации в Африке на начало миграции белых аис-
тов, помеченных спутниковыми передатчиками И Тез. 12-й Межд. орнитол. конф. Сев. Евразии.
Ставрополь. С. 282-283.
Костин С.Ю., Тарина Н.А. 2002. Редкие птицы заповедника «Лебяжьи острова» и прилегающих тер-
риторий И Бранта. Вып. 5. С. 113-128.
Костин С.Ю., Тарина Н.А. 2004. Распределение и биология размножения веслоногих и голенастых
птиц на Лебяжьих о-вах и сопредельных территориях И Бранта. Сб. тр. Азово-Черноморск. орни-
тол. станции. Вып. 7. С. 82-110.
Костин С.Ю., Тарина Н.А. 2005. Послегнездовое распределение и миграции веслоногих и голенас-
тых птиц северо-западной части Крыма И Бранта. Сб. тр. Азово-Черноморск. орнитол. станции.
№ 8. С. 80-95.
Костин Ю.В. 1978. Результаты кольцевания серой цапли в Северном Крыму И Мигр. птиц Вост. Ев-
ропы и Сев. Азии: гагарообр. - аистообр. М.: Наука. С. 197-202.
Костин Ю.В. 1983. Птицы Крыма. М.: Наука. 293 с.
Костюшин В.А., Полуда А.М. 2005. Учёт водно-болотных птиц в окрестностях Киева зимой 2000/
2001 гг. И Вести, зоол. Т. 39. Вып 2. С. 85-87.
Коханов В.Д. 1987. Обзор изменений, отмеченных в орнитофауне Мурманской области за последнее
столетие И Пробл. изуч. и охр. природы Прибеломорья. Мурманск. С. 20-37.
Кошелев А.И. 2000. Новое место зимовки цапель и бакланов - Сев. Приазовье И Птицы Азово-Чер-
номорск. региона на рубеже тысячелетий. Одесса: Астро-принт. С. 10.
Кошелев А.И., Корзюков А.И., Лобков В.А., Пересадько Л.В. 1991. Состояние численности редких и
исчезающих видов птиц И Редк. птицы Причерноморья. Киев. С. 9-36.
Кошелев А.И., Кошелев В.А. 1999. Гнездование цапель в зарослях тростника на водоёмах Сев. При-
азовья И Бранта. Вып. 2. С. 39^49.
Кошелев А.И., Кошелев В.А., Пересядько Л.В. 2004. Динамика видового состава и численности гнез-
дящихся цапель (Ardeidae) в СЗ Приазовье (1988-2004) И Бранта. Сб. тр. Азово-Черноморск.
орнитол. станции. Вып. 7. С. 111-130.
Кошелев А.И., Кошелев В.А., Пересядько Л.В., Покуса Р.В. 2005. Репродуктивные показатели цапель
(Ardeidae) в СЗ Приазовье И Бранта. Сб. тр. Азово-Черноморск. орнитол. станции. Вып. 8. С. 96-
113.
Кошелев А.И., Кошелев В.А. 2004. Состояние изученности цапель на юге Украины И Бранта. Сб. тр.
Азово-Черноморск. орнитол. станции. Вып. 7. С. 8-22.
Кошелев А.И., Попенко В.М., Пересадько Л.В. и др. 1990. Особенности колониального типа гнездо-
вания большого баклана в свете его экспансии на островах Молочного лимана в Приазовье И
Совр. пробл. изуч. колониальное™ у птиц, Симферополь-Мелитополь: «Сонат». С. 71-77.
Кошелев А.И., Сиохин В.Д., Покуса Р.В., Белашков И.Д. 1998. Динамика локальных гнездовых посе-
лений большого баклана в Сев. Приазовье (Запорожская обл.) И Мат-ли 3-й конф, молодих
орштолопв Украши. Чершвщ. С. 91-95.
Кошелев В.А. 2000. Новое место зимовок цапель и бакланов (Сев. Приазовье) И Птицы Азово-Чер-
номорск. региона на рубеже тысячелетий. Одесса. С. 10.
Кошелев В.А., Кошелев А.И., Покуса Р.В. 2001. Консервативные связи в колониях цапель и бакланов
Сев. Приазовья И Акт. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 331-332.
Кошелев В.О. 2004. Чаши (Ardeidae) як структурний елемент бюгеоценоз!в Швшчного Приазов’я
(вщновлення, охорона, управлшня. Автореф. дис. ... канд. бюл. н. Дншропетровськ. 20 с.
Кошелев О., Черничко I., Черничко Р. и др. 2002. Нацюнальна доповщь про стан IBA територш
УкраТни. КиТв. С. 36-39.
Кошкин А.В. 2002. Дополнения к фауне птиц Кургальджинского заповедника И Selevinia. С. 127—
130.
Кошкин А.В. 2004. Орнитологические наблюдения в Кургальджинском заповеднике И Казахстанск.
орнитол. бюл. Алматы. С. 54-56.
Литература
553
Кошкин А.В., Кошкина О.И. 2003. Краткий обзор состояния редких видов птиц в Тенгизском регио-
не (Центр. Казахстан) И Selevinia. С. 209-210.
Крашуны А.П. 1957. Харчаванне пушанят белага бусла у Белавежскай пушчы И Весщ АН БССР. Сер.
б!ял. навук. № 1. С. 91-98.
Красная книга Алтайского края. 1998. Т. 2. Животные. Барнаул. 238 с.
Красная книга Казахстана. 1996. Т. 1. Животные. Ч.. 1. Позвон. животн. Алма-Ата. 326 с.
Красная книга Новосибирской области. Животные. 2000. Новосибирск. 316 с.
Красная книга Приморского края. Животные. 2005. Владивосток. 408 с.
Красная книга Российской Федерации. Животные. 2001. М.: изд. МПР РФ и РАН, Аст., Астрель. 862 с.
Красная книга Республики Коми. 1998. М.-Сыктывкар. 528 с.
Красная книга Республики Узбекистан.. Т. 2, Животные. 2003. Ташкент. 328 с.
Красная книга РСФСР. Животные. 1983. М.: Россельхозиздат. 454 с.
Красная книга Узбекской ССР. 1983. Т. 1. Позвоночные животные. Ташкент: изд-во ФАН. 128 с.
Красная книга Хабаровского края. 2000. Хабаровск: ИВЭП ДВО РАН. 464 с.
Краснов Ю.В., Матишов Г.Г.. Галактионов К.В., Савинова Т.Н. 1995. Морские колониальные птицы
Мурмана. СПб.: Наука. 224 с.
Красовский Д.Б. 1932. Материалы к познанию фауны наземных позвоночных Рутульского кантона
ДАССР И Изв. 2-го Сев.-Кавказск. пед. ин-та. Орджоникидзе. Т. 9. С. 186-219.
Крашенинников С.П. 1994. Описание земли Камчатки. Т. 1. СПб.: Наука; Петропавловск-Камчатс-
кий: Камшат. 438 с. (Репринт изд. 1755 г.).
Крейцберг-Мухина Е.А. 2003. Каравайка И Красная книга Респ. Узбекистан. Ташкент. Т. 2. С. 156-157.
Крейцберг-Мухина Е.А.. Лановенко Е.Н. 2003. Влияние сукцессий и реабилитации водно-болотных
угодий системы Судочье на состояние орнитокомплексов в дельте Амударьи И Selevinia. С. 169-
181.
Кречмар А.В.. Андреев А.В., Кондратьев А.Я. 1978. Экология и распространение птиц на Северо-
Востоке СССР. М.: Наука. 196 с.
Кривенко В.Г. 1975. Влияние экологических сукцессий на численность колониально гнездящихся
птиц в дельте Волги Колон, гнездовья околоводн. птиц и их охрана. М.: Наука. С. 18-20.
Кривенко В.Г. 1981. Закономерности динамики численности гнездящихся птиц на водоёмах р. Ма-
ныча // Научн. осн. обслед. гнездовий околоводн. птиц. М.. С. 68-75.
Кривенко В.Г. 1983. Колпица. Каравайка И Кр. кн. РСФСР, Животные. М. С. 165-169.
Кривенко В.Г. 2000. Чограйское водохранилище (река Вост. Маныч) И Водно-бол. угодья. М. Т. 3.
С. 193-198.
Кривенко В.Г. 2004. Водоплавающие птицы и их охрана. М.: Агропромиздат. 272 с.
Кривенко В.Г., Любаев В.Л. 1975. Колониально гнездящиеся птицы Восточного Маныча И Колон,
гнездовья околоводн. птиц и их охрана. С. 142-143.
Кривицкий И.А., Хроков В.В., Волков Е.Н., Жулий В.А. 1985. Птицы Кургальджинского заповедни-
ка. Алма-Ата: Наука. 194 с.
Кривоносов Г.А. 1984. Краткие сообщения о Чёрном аисте в Астраханском заповеднике И Иссл. в
области заповеди, дела. М.: ВНИИ природы МСХ СССР. С. 111.
Кривоносов Г.А., Бондарев Д.В. 1978. Судьба пеликанов в дельте Волги И Бюл. МОИП. Отд. биол.
Т. 83. Вып. 5. С. 42^8.
Кривоносов Г.А., Васильев В.И., Морозкин Н.И. 1977. Фламинго на Каспийском море И Тез. докл. 7-й
Всес. орнитол. конф. Киев. Ч. 2. С. 222-223.
Кривоносов Г.А., Гаврилов Н.Н. 1981. Современное состояние численности голенастых и веслоно-
гих птиц в Астраханском заповеднике И Размещ. и сост. гнездовий околоводных птиц на терр.
СССР. (МОИП). М.: Наука. С. 99-103.
Кривоносов Г.А., Живогляд А.Ф. 1989. Астраханский заповедник И Заповедники СССР. Зап-ки Ев-
роп. части РСФСР. Ч. 1. М.: Мысль. С. 264-292.
Кузнецов Н.В., Макковеева И.И. 1959. Животный мир Ярославской области. Ярославль: кн. изд-во.
228 с.
Кузнецова Е.Н. 2006. Относительная численность и распределение волчка на территории Добро-
вского зонального рыбопитомника (Липецкая область) И Орнитологические исследования в се-
верной Евразии. Тез. 12-й межд. орнитол. конф, северной Евразии. Ставрополь. С. 300-301.
554
Литература
Кукиш А.И. 1984. Влияние обводнения на распространение околоводных колониально-гнездящихся
птиц в Калмыкии И Пробл. регион, экологии животных в цикле зоол. дисциплин педвуза. Ви-
тебск. Ч. 1. С. 97-98.
Кулагин С.В. 2005. Зимний учёт птиц на оз. Иссык-Куль в январе 2004 г. И Казахстанск. орнитол.
бюл. 2004. С. 126-127.
Куниченко А.А. 1986. Изменение в поведении пеликанов и бакланов под воздействием антропоген-
ного фактора И Тез. докл. 1-го съезда Всесоюзн. орнитол. о-ва. Л. Ч. 1. С. 346-347.
Куниченко А.А., Гусан Г.З. 1992. Редкие виды птиц низовий Прута И Экол. и охрана птиц и млекопит.
в антроп. ландшафте. Кишинев: Штиница. С. 70-80.
Куниченко А.А., Гусан Г.З. 2001. Гнездовая экология большой белой цапли в Молдове И Геоэкол и
биоэкол.пробл. Сев. Причерноморья. Тирасполь: РИО ПГУ. С. 144-145.
Куниченко А.А., Тищенков А.А. 1999. Необычные и редкие случаи зимовок птиц на Кучурганском
лимане и др. водоёмах Южн. Приднестровья И Сохранен, биоразнообр. басе. Днестра. Кишинев.
С. 117-119.
Курдюков А.Б. 2004. К орнитофауне заповедника «Кедровая падь» и сопредельных территорий. Но-
вые сведения за 1998-2000, 2003 гг. И Мат-лы конф. «Научн. иссл. в зап-ках». Хабаровск. Ч. 1.
С. 150-154.
Курочкин Е.Н. 1991. Отряд Пеликанообразные И Фауна мира. Птицы. М. С. 42^49.
Кустанович С.Д. 1986. Жар-птица из Красной книги (обыкновенный фламинго). М. 80 с.
Кучин А.П. 1976. Птицы Алтая. Барнаул: Алтайское книжное изд-во. 232 с.
Кучин А.П. 1988. Чёрный аист на Алтае И Ресурсы редк. животн. РСФСР, их охрана и воспроизв. М.:
ЦНИЛ Главохоты РСФСР. С. 63-65.
Лабзюк В.И. 1965. Редкий случай залета японской выпи в южное Приморье И Зап. Приморск. фил.>
Геогр. о-ва СССР. Владивосток. Вып. 1 (24). С. 158-160.
Лабзюк В.И. 1975. Летняя авифауна морского побережья в районе зал. Ольги // Орнитол. иссл. на
Дальн. Вост. Владивосток: ДВНЦ АН СССР. С. 279-284.
Лабзюк В.И. 1981. Встречи редких птиц в заливе Ольги (Южное Приморье) И Редкие птицы Дальн.
Востока. Владивосток: ДВНЦ АН СССР. С. 122-123.
Лабзюк В.И. 1990. Цапли в Ольгинском районе Приморского края И Экол. и распростр. птиц юга
Дальн. Востока. Владивосток: ДВО АН СССР. С. 75-76.
Лабзюк В.И., Назаров Ю.Н., Нечаев В.А. 1971. Птицы островов северо-западной части залива Петра
Великого И Орнитол. иссл. на юге Дальн. Вост. Владивосток: ДВНЦ АН СССР. Т. 6. С. 52-78.
Лабзюк В.И., Назаров Ю.Н., Остапенко В.А. 1970. Птичий базар на о. Фуругельма (Юж. Приморье) И
Мат-лы 15-й науч. конф, проф.-преп. состава Биолого-почв. ф-та ДВГУ. Владивосток. С. 64-67.
Лановенко Е.Н., Филатов А.К., Загребин С.В. 2001. Значение водоёмов Узбекистана для сохранения
биоразнообразия зимующих птиц И Актуальн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии.
Казань. С. 358-359.
Лаптев А.А., Медведев В.Н. 1995. Птицы И Кадастр наземн. позвон. животн. Лазовского зап-ка. Вла-
дивосток: Дальнаука. С. 10-42.
Лапушкин В.А., Шепель А.И., Фишер С.В., Казаков В.П. 1995. Новые виды птиц Пермской облас-
ти И Мат-лы к распростр. птиц на Урале. Екатеринбург. С. 43^45.
Лапшин Н.В. 1997. Белый аист в Карелии И Русск. орнитол. ж., Экспр.-вып. № 6. С. 3-4.
Лапшин Н.В. 2000. Встречи и статус белого аиста в Карелии // Белый аист в России: дальше на
восток. Калуга. С. 61-62.
Лапшин Н.В., Лысенков Е.В. 1997. Белый аист {Ciconia ciconia) и луговой конек {Anthuspratensis) -
гнездящиеся виды Мордовии И Фауна, экол. и охр. птиц Ср. Поволжья. Саранск. С. 80-81.
Лапшин Н.В., Лысенков Е.В. 2000. Белый аист в Мордовии И Белый аист в России: дальше на восток.
Калуга. С. 176-177.
Ларионов Г.В., Винокуров В.Н., Гермогенов Н.И., Дегтярев А.Г. 1976. Гнездовые поселения серой
цапли в Центр. Якутии И Прир. рес. Якутии их использ. и охрана. Якутск. С. 44-46.
Лебедева Г.П., Пантелеев И.В. 2000. Сусканский заказник И Ключевые орнитол. терр. России. М.
Т. 1. С. 451.
Лебедева М.И. 1959. К биологии Чёрного аиста в Беловежской пуще // Орнитология. Вып. 2. С. 138—
142.
Литература
555
Лебедева М.И. 1960. Материалы по питанию белого аиста И Охр. прир. и озеленен., № 4. С. 111-116.
Лебедева М.И. 1979а. Миграции европейского белого аиста по данным кольцевания в СССР И Мигр.
птиц Вост. Европы и Сев. Азии. Аистообр. - пластинчатокл. М. С. 18-27.
Лебедева М.И. 19796. Миграции туркестанского белого аиста И Там же. С. 27-28.
Лебедева М.И. 1979в. Миграции Чёрного аиста - Ciconia nigra (L.) И Там же. С. 28-31.
Лебедева Н.В., Савицкий Р.М., Цветкова Ю.А. 2006. Динамика трофических ниш рыбоядных птиц
Азовского моря И Орнитол. иссл. в Сев. Евразии. Тез. 12-й межд. орнитол. конф. Сев. Евразии.
Ставрополь. С. 310-311.
Леус С.И. 1972. Залеты фламинго на озеро Ильмень И Орнитология. Вып. 10. С. 361.
Лиепа В.К. 1966. Сезонное размещение и миграции Латвийских серых цапель - Ardea cinerea, по
данным кольцевания И Орнитол. иссл. Рига: Зинатне. № 4. С. 32^4-2.
Линдеман Г.В. 1991. Колпица в западной части Волжско-Уральского междуречья И Редк. звери и
птицы Казахстана. Алма-Ата: Гылым. С. 42.
Линьков А.Б. 1988. Стациальное распределение и динамика численности колпиц, гнездящихся в
Центральном Предкавказье // Тр. ЦНИЛ Главохоты РСФСР. М. С. 43-55.
Линьков А.Б. 1989. Биология размножения, территориальные связи и охрана водных и околоводных
птиц (на примере Центрального Предкавказья). Автореф. дис. ... канд. биол. н. М. 20 с.
Линьков А.Б. 1992. Фенологическая структурированность и возможный механизм авторегуляции у
размножающихся колпиц 7 Тр. ЦНИЛ Главохоты РСФСР. М. С. 87-98.
Линьков А.Б. 2000. Розовый пеликан, Кудрявый пеликан, Колпица И Кр. кн. РФ (Животные). М.: ACT
Астрель. С. 371-375: 381-383.
Липин С.И., Дурнев Ю.А.. Сонин В.Д. и др. 1984. Кр. сообщ. о чёрном аисте в Восточной Сибири И
Иссл. в обл. запов. дела. М.: ВНИИ прир. С. 119-120.
Липин С.И., Сонин В.Д. 1989. Редкие виды в динамике фауны птиц И Тез. докл. 1-го съезда Всесо-
юзн. орнитол. о-ва. Л. Ч. 2. С. 26-27.
Липсберг Ю.К. 1981. Гнездовые колонии серой цапли - Ardea cinerea в Латвии И Сообщ. Прибалт,
комиссии по изуч. мигр. птиц. Тарту. Вып. 12. С. 133-147.
Липсберг Ю.К. 1984. Краткие сообщ. о Чёрном аисте в Латвии И Иссл. в обл. запов. дела. М.: ВНИ-
Иприроды. С. 108.
Лисецкий А.С., Кривицкий И.А., Есилевская М.А. и др. 1980. Материалы по голенастым птицам
Харьковщины / Вести. Харьковск. ун-та. № 195. С. 83-89.
Литвиненко Н.М. 2005а. Амурская выпь И Кр. кн. Приморск, края. Животные. Владивосток. С. 205-
207.
Литвиненко Н.М. 20056. Жёлтоклювая цапля, Японская выпь И Там же. С. 202-205.
Литвиненко Н.М., Шибаев Ю.В. 1965. О некоторых редких птицах Южного Приморья И Орнитоло-
гия. Вып. 7. С. 115-121.
Литвиненко Н.М., Шибаев Ю.В. 1971. К орнитофауне Судзухинского заповедника и долины р. Суд-
зухэ И Экол. и фауна птиц юга Дальн. Вост. Владивосток: ДВНЦ АН СССР: С. 127-186.
Литвиненко Н.М., Шибаев Ю.В. 1992. Миграции и зимовка молодых японских бакланов (по матери-
алам кольцевания) И Изуч. морск. колон, птиц в СССР. Информ, мат-лы. Магадан. С. 20-23.
Литвиненко Н.М., Шибаев Ю.В. 1996. Значение низовий реки Туманган для поддержания разнооб-
разия птиц (Материалы для создания национального парка и представления нового водно-болот-
ного угодья международного значения) И Птицы пресных вод и морских побережий юга Дальне-
го Востока России и их охрана. Владивосток: Дальнаука. С. 49-75.
Литвиненко Н.М., Шибаев Ю.В. 1999. Новые орнитологические находки и наблюдения на крайнем
юго-западее Приморья // Русск. орнитол. ж., Экспр. вып. № 71. С. 9-16.
Литвиненко Н.М., Шибаев Ю.В. 2005. Малая колпица // Красная книга Приморск, кр., Животные.
Владивосток. С. 209.
Литвинова Н.А. 1981. Колонии цапель и бакланов в Кызыл-Агачском заповеднике И Размещ. и сост.
гнездовий околоводн. птиц на терр. СССР. М.: Наука. С. 57-59.
Литвинова Н.А. 1988. Биология голенастых птиц Кызыл-Агачского заповедника и пути сохранения
колониальных гнездовий. Автореф. дис.....канд. биол. наук. М. 16 с.
Литвинова Н.А. 1989а. Проблемы сохранения гнездовых колоний цапель, бакланов и ибисов в Кы-
зыл-Агачском заповеднике И Тез. докл. 1-го съезда Всесоюзн. орнитол. о-ва. Л. Ч. 2. С. 31-32.
556
Литература
Литвинова Н.А. 19896. Размещение смешанных колоний голенастых и веслоногих птиц в юго-вос-
точном Азербайджане И Всес. совещ. по пробл. кадастра и учёта животн. мира. Уфа. Ч. 3. С. 146.
Литвинова Н.А. 2001. Египетская цапля И Красная книга Российской Федерации. М.: Астрель. С. 378—
379.
Литвинова Н.А., Ткаченко Е.Э., Литвинов В.П. 1984. Гнездование фламинго в Кызылагаджском за-
поведнике И Орнитология. Вып. 19. С. 182-183.
Лобков Е.Г. 1980. Мат-лы по миграции гагар, поганок, бакланов, чаек и чистиков вдоль восточного
побережья Камчатки И Экол. птиц морских побережий М.: ЦНИЛ Главохоты РСФСР. С. 74-86.
Лобков Е.Г. 1986а. Гнездящиеся птицы Камчатки. Владивосток: ДВНЦ АН СССР. 304 с.
Лобков Е.Г. 19866. О случаях массовой гибели птиц в летнее время на побережьях Камчатки И Морск.
птицы Дальнего Востока. Владивосток: ДВО АН СССР. С. 88-95.
Лобков Е.Г. 2003а. Гнездящиеся птицы м. Копыто (устье р. Жупанова) И Биол. и охр. птиц Камчатки.
М.: ЦОДП. Вып. 5. С. 117-118.
Лобков Е.Г. 20036. Осенняя миграция водных и околоводных птиц на м. Лопатка И Там же. С. 27-54.
Лобков Е. Г. 2009. Фауна, население птиц и их роль в экосистеме острова Старичков И Тр. Камчатск.
ф-ла Тихоокеанск. ин-та геогр. ДВО РАН. Вып. 8. Петропавловск-Камчатский: Камчатпресс.
С. 280-340.
Лобков Е.Г., Алексеев С.А. 1987. Размещение и численность морских колониальных птиц на юге п-
ова Камчатка // Распростр. и биол. морск. птиц Дальнего Востока. Владивосток: ДВО АН СССР.
С. 36-38.
Лозов Б.Ю. 1998. Новые виды редких птиц Центральной России и Брянской обл. И Редк. виды птиц
Нечерноземн. центра России. М. С. 94-97.
Ломадзе Н.Х. 1973. Рыбоядные птицы Приазовья и их хозяйственное значение. Автореф. дис. v.
канд. биол. н. Ростов-на-Дону. 32 с.
Ломадзе Н.Х. Казаков Б.А., Гончаров В.Т. 1981. О колонии больших бакланов и голенастых в дельте
Дона И Научн. осн. обслед. гнездовий околоводн. птиц. М. С. 84-86.
Лопарев С.А. 1997. Интересный случай клептопаразитизма И Беркут. Т.6. Вип. 1-2. С. 69.
Лопатин В.В., Карпов Ф.Ф., Клейменов С.В. 1992. Биология волчка (Ixobrychus minutus) на юго-
востоке Казахстана // Русск. орнитол. ж. Т. 1. Вып. 2. С. 227-234.
Лохман Н.В., Емтыль М.Х., Фадеев И.В. и др. 2005. Орнитофауна черноморских лиманов России и
прилегающих территорий И Инвент., мониторинг и охр. ключ, орнитол. терр. России. М. Вып. 5.
С. 72-96.
Луговой А.Е. 1961. Динамика гнездового распределения колониальных птиц низовьев дельты Волги И
Тр. Астрах, зап-ка. Астрахань. Вып. 5. С. 211-219.
Луговой А.Е. 1963. Птицы дельты Волги И Тр. Астрах, зап-ка. Астрахань. Вып. 8. С. 9-185.
Луговой А.Е. 1994. Змши населения птах!в твденно-захщно! частини Закарпаття за останш роки И
Наук. в1сн. Ужгородськ. ун-ту, сер. бюл. Вип. 1. С. 76-79.
Луговой А.Е., Потиш Л.А. 2004. Современное состояние и численность Чёрного аиста в Закарптской
обл. И Беркут. Т. 13. Вип. 1. С. 62-66.
Лукашевич Р.В. 1990. Некоторые редкие виды голенастых птиц дельты Аму-Дарьи и вопросы их
охраны И Редк. и малоизуч. птицы Ср. Азии. Бухара. С. 54-57.
Лукашевич Р.В. 1991. Колпица и каравайка в дельте Аму-Дарьи И Орнитология. Вып. 25. С. 162-163.
Лукашевич Р.В. 1995. Колпица и каравайка в дельте Аму-Дарьи И Орнитология. Вып. 26. С. 189-190.
Лысенко В.И. 1992. Особенности видимых миграций гидрофильных птиц в Северо-Западном При-
азовье И Сезонные мигр. птиц на терр. Украины. Киев: Наукова думка. С. 188-210.
Лысенко В.И., Сиохин В.Д. 1991. Современное состояние численности и распределения редких ви-
дов птиц Северного Приазовья И Редк. птицы Причерноморья. Киев-Одесса: Лыбидь. С. 69-78.
Львов Д.К., Ильичев В.Д. 1979. Миграция птиц и перенос возбудителей инфекции. М.: Наука. 271 с.
Львов Д.К., Лебедев А.Д. 1972. Закономерности географического распространения арбовирусов И
Трансконт. связи перел. птиц и их роль в распростр. арбовирусов. Новосибирск: Наука. С. 145—
151.
Лэк Д. 1957. Численность животных и её регуляция в природе. М.: Иностр, литер. 403 с.
Ляйстер А.Ф., Соснин Г.В. 1942. Материалы по орнитофауне Армянской ССР (Omis Armenica). Ере-
ван. 402 с.
Литература
557
Мальчевский А.С., Пукинский Ю.Б. 1983. Птицы Ленинградской области и сопредельных террито-
рий. Л.: изд-во ЛГУ. Т. 1. 480 с.
Маматаева В.Ф., Умаханова Т.К. 2000. Распространение и гнездование белого аиста в Бабаюртовс-
ком районе Республики Дагестан // Белый аист в России: дальше на восток. Калуга. С. 184-186.
Маматов А.Ф. 1989. Чёрный аист в Башкирии И Редк. и нуждающиеся в охране животн. М.: ЦНИЛ
Главохоты РСФСР. С. 56.
Мамашукуров А.У, Джаббаров А.Р., Фундукчиев С.Э. 1998. Оологическая характеристика яиц тур-
кестанского белого аиста (Ciconia с. asiatica) И Мат-лы 2-й Межд. конф, стран СНГ: «Актуальн.
прорбл. оологии». Липецк. С. 67.
Мараков С.В. 1963. Птичьи базары о. Медного и возможности их практического использования И Сб.
н.-т. информ. ВНИИЖП. Киров. Вып. 5. С. 51-65.
Мараков С.В. 1972. Природа и животный мир Командор. М.: Наука. 184 с.
Мараков С.В. 1976. Материалы по биологии и распространен, краснолицего баклана в СССР И Тр.
Окского гос. запов. Рязань. Вып. 13. С. 7-11.
Мараков С.В. 1977. Загадочный мир островов. М.: Мысль. 125 с.
Марголин В.А. 2000. Птицы Калужской области. Калуга. Ч. 1. Неворобьиные. 336 с.
Марисова И.В., Макаренко М.М., Бабко В.М. и др. 1992. К экологии аистов на Черниговщине И
Аисты: распр., экол., охрана. Минск. С. 222-224.
Марисова И.В., Самофалов М.Ф., Бабко В.М. и др. 1991. Изучение миграций птиц на Черниговщине.
Рукоп. деп. в УкрНИИНТИ 21.05.91. № 725-Ук-91. 39 с.
Марисова И.В., Самофалов М.В., Сердюк В.А. 1992. О питании белого аиста в Черниговской облас-
ти И Аисты: распр., экол., охрана. Минск. С. 111-114.
Маркин Ю.М., Приклонский С.Г. 1995. Изменения численности и размещения глухаря, серого жу-
равля и серой цапли за 20-летний период в центре Европейской части России И Тр. Окского зап-
ка. Рязань. Вып. 19. С. 160-181.
Марков В.И. 1965. Заметки о пеликанах и бакланах Аральского моря И Рыбоядн. птицы и их знач. в
рыбн. х-ве. С. 156-165.
Матвеев Б.С. 1935. Курс зоологии. М. 653 с.
Матер1али орштолопчних спостержень..., 1994 И Troglodytes. Луцьк. Вип. 4. С. 4-9.
Матер1али орштолопчних спостержень..., 1995а И Troglodytes. Луцьк. Вип. 5. С. 6-16.
Матер1али орштолопчних спостержень..., 19956 И Troglodytes. Луцьк. Вип. 5. С. 17-43.
Матер1али орштолопчних спостержень..., 1996 И Troglodytes. Луцьк. Вип. 6. С. 9^12.
Матюшков Г.В. 1999. О гнездовании морских колониальных птиц на побережье Тонино-Анивского
полуострова (о-в Сахалин) И Вести. Сахалинск, музея. Южно-Сахалинск. № 6. С. 285-292.
Мациевская Н.Б., Кошелев А.И., Сиохин В.Д. и др. 1998. Материалы по численности птиц в дельте
Дона И Кавказск. орнитол. вести. Ставрополь. Вып. 10. С. 109-114.
Мекленбурцев Р.Н., Сагитов А.К., Кашкаров Д.Ю. и др. 1987. Птицы Узбекистана. Ташкент. Т. 1. 291
с.
Мельников Ю.И. 2000. О встречах кудрявого пеликана Pelecanus crispus на территории Прибайкалья И
Русск. орнитол. журн., Экспр.-вып. № ПО. С. 21-22.
Мельников Ю.И. 2000а. Колпица Platalea leucorodia в Прибайкалье И Русск. орнитол. ж., Экспр.-
вып. № 104. С. 22-23.
Мельничук А.В. 2001. Орнитологическая ситуация на водохранилищах Днепра И Актуальн. пробл.
изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 417—419.
Мензбир М.А. 1895. Птицы России. М.: Типолит. т-ва Кушнерев и Ко. Т. 1, 836 с.; Т 2, 1120 с.
Мензбир М.А. 1900. Охотничьи и промысловые птицы Европейской России и Кавказа. М. Т. 1.478 с.
Мензбир М.А. 1918. Птицы России. (Европейская Россия, Сибирь, Туркестан, Закаспийская область
и Кавказ). Вып. 1. М.: Изд-во М.С. и С. Сабашниковых. 224 с.
Миграции птиц Вост. Европы и Сев. Азии. Гагарообр.-Аистообр. 1978. М.: Наука. 270 с.
Микитюк О.Ю. 1999. Территории Украины важные для сохранения видового разнообразия и количе-
ственного богатства птиц. Киев: СофтАРТ. 420 с.
Миноранский В.А. 1962. О гнездовании кудрявого пеликана на озере Маныч-Гудило И Зоол. ж. Т. 41.
Вып. 7. С. 1107-1108.
Миноранский В.А. 1963. Ещё об орнитофауне Маныч-Гудило И Орнитология. Вып. 6. С. 475^76.
558
Литература
Миноранский В.А., Добринов А.В., Маркитан Л.В., Подгорная Я.Ю. 1998. Материалы по численно-
сти птиц в дельте Дона И Кавказск. орнитол. вести. Ставрополь. Вып. 10. С. 86-96.
Миноранский В.А., Подгорная Я.Ю. 1998. Орнитологические наблюдения в западной части оз. Ма-
ныч-Гудило И Кавказск. орнитол. вести. Ставрополь. Вып. 10. С. 96-109.
Миронов В.И. 1996. Влияние антропогенных факторов на изменение состава Курской области за
последние 100 лет И Птицы бас. Сев. Донца. Харьков. Вып. 3. С. 26-28.
Миронов В.И. 1998. Редкие птицы Курской области И Редк. виды птиц Нечерноз. центра России. М.
С. 304-306.
Митропольский О.В. 2007. Белый аист И Птицы Средней Азии. Алматы. Т. 1. С. 114-123.
Митропольский О.В., Митропольский М.Г. 2009. Список птиц Узбекистана. Ташкент. 15 с.
Митропольский О.В., Митропольский М.Г., Коршунова Е.Н. 2006. Находка индийской змеешейки в
Узбекистане И Орнитология. Вып. 33. С. 185.
Михеев А.В. 1991. Пролёт по западному побережью Каспийского моря некоторых редких околовод-
ных птиц И Кавказск. орнитол. вести. М. Вып. 2. С. 33—40.
Михеев А.В. 1997. Видимый дневной пролёт водных и околоводных птиц по западному побережью
Каспийского моря. Ставрополь. 156 с.
Михеев А.В., Орлов В.И., Резанов А.Г. 1975. Пролёт аистообразных по Зап. побережью Каспия И
Мат-лы Всесоюзн. конф, по мигр. птиц. Ч. ЕС. 133-136.
Михайлов К.Е., Шибнев Ю.Б., Коблик Е.А. 1998. Гнездящиеся птицы бассейна Бикина (аннотиро-
ванный список видов) И Русск. орнитол. ж., Экспр.-вып. № 46. С. 3-19.
Мищенко А.Л. 2008. Современное состояние редких видов водоплавающих и аистообразных в Не-
черноземном центре И Редкие виды птиц Нечерноземного центра. Мат-лы 3-го совещ. «Редкие
птицы центра Европейской части России» (Москва, 1-3 декабря 2000 г.). М. С. 20-26.
Мищенко А.Л., Белик В.П., Равкин Е.С. и др. 2004. Оценка численности и её динамики для птиц
Европейской части России (Птицы Европы - II). М.: СОПР. 44 с.
Мищенко В.П. 1991. Колпица в Куртинском районе И Редк. птицы и звери Казахст. Алма-Ата: Гылым.
С. 45.
Мнацеканов Р.А. 1998. К авифауне среднегорий центральной части Зап. Кавказа И Кавказск. орнитол.
вести. Ставрополь. Вып. 10. С. 109-114.
Мнацеканов Р.А. 2000. К гнездованию белого аиста в Краснодарском крае И Кавказск. орнитол. вести.
Вып. 12. С. 146-147.
Мнацеканов Р.А. 2001. Чёрный аист в Краснодарском крае И Мат. межд. конф. «Актуальн. пробл.
изуч. и охр. птиц Вост. Европы и Сев. Азии». Казань. С. 434^36.
Мнацеканов Р.А., Динкевич М.А., Тильба П.А., Короткий Т.В. 2004. Новые сведения о регистрации
египетской цапли в Краснодарском крае И Стрепет. Т. 2. Вып. 1. С. 134-137.
Модестов В.М. 1967. Экология колониально гнездящихся птиц (по наблюдениям на Восточном Мур-
мане и в дельте Волги) И Тр. Кандалакшею зап-ка. Вып. 5. С. 49-150.
Молева Э.Б., Рустамов Э.А. 1986. О численности серой и большой белой цапель в Юго-Восточном
Туркменистане И Тез. докл. 1-го съезда Всесоюзн. орнитол. о-ва и 9-й Всесоюзн. орнитол. конф.
Л. Ч. 2. С. 78-79.
Молодовский А.В. 1963. Фламинго на Ю. Мангышлаке И Зоол. ж. Т. 42. Вып. 12. С. 1885-1886.
Молодовский А.В. 1975. О гнездовании большого баклана на Южном Мангышлаке И Колон, гнездо-
вья околоводн. птиц и их охр. М. С. 193.
Молодовский А.В. 1981. Зоогеографические аспекты стайных зимовок птиц на Каспии И Вести, зоол.
№ 5. С. 88-91.
Молодовский А.В. 1990. О стайном пролёте веслоногих, голенастых и фламинго И Вести, зоол. № 3.
С. 53-60.
Молодовский А.В. 1997. Полевой определитель стайных птиц. Ниж. Новгород. 310 с.
Молодовский А.В. 2001. Эколого-морфологические основы построения стайных птиц в полете.
Ниж. Новгород. 391 с.
Молчанов Л.А. 1912. Летняя орнитофауна дельты Аму-Дарьи // Орнитол. вести. № 4. С. 261-286.
Молчанова Т.В. 1972. К авифауне лесной зоны Зап. Сибири И Орнитология. Вып. 10. С. 370-372.
Морозкин Н.И. 1975а. Материалы по численности и экологии колониальных гнездовий голенастых и
веслоногих птиц в Кызыл-Агачском заповеднике И Мат-лы совещ.: «Колон, гнезд, околоводн.
птиц и их охрана». М. С. 147-148.
Литература
559
Морозкин Н.И. 1975b. Пролёт и зимовка водоплавающих птиц в Кызыл-Агачском заповеднике в
1971-1974 гг. И Мат-лы Всес. конф, по мигр. птиц. М. С. 221-224.
Морозов В.В. 2000. Долина р. Сафаровки // Ключевые орнитол. терр. России. М. Т. 1. С. 471—472.
Морозов В.В., Корнев С.В. 2002а. К орнитофауне Южного Зауралья И Мат-лы к распростр. птиц на
Урале. Екатеринбург. С. 161-164.
Морозов В.В., Корнев С.В. 20026. Дополнения к орнитофауне юга Западной Сибири И Там же. С. 164-
169.
Мрикот К.Н., Глущенко Ю.Н. 2000. Египетская цапля Bubulcus ibis гнездится на озере Ханка И Русск.
орнитол. ж., Экспр.-вып. 99. С. 10-11.
Музаев В.М., Бамбеева В.И., Меджидов Р.А. 2004. Материалы по гнездованию некоторых колони-
альных и околоводных птиц на озере Деед-Хулсун (Республика Калмыкия) в 1999-2001 гг. И Мат-
лы 2-й Межд. заочн. научн. конф. Элиста. С. 75-78.
Муравьев И.В., Иванов С.В. 2006. Оологоморфологические особенности кладок серой цапли (Ardea
cinerea) в Пензенской области // Тез. докл. 12-й межд. орнитол. конф, по Сев. Евразии. Ставро-
поль. С. 382-383.
Мустафаев Г.Т., Кязымов К.Д. 1965. Жизнь гнездовой колонии рыбоядных птиц в Кызыл-Агачском
заповеднике И Уч. зап. Азерб. ун-та, сер. биол. № 2. С. 31-35.
Мустафаев Г.Т., Кязымов К.Д. 1966. Результаты статистического изучения экологии рыбоядных птиц
на ЮЗ Каспии // Уч. зап. Азерб. ун-та, сер. биол. № 1. С. 27-32.
Мустафаев Г.Т., Кязымов К.Д., Аббасов Г.М. 1968. Питание гнездовых птенцов голенастых и весло-
ногих птиц в Кызыл-Агачском заповеднике И Изв. АН АзССР, сер. биол. № 5. С. 49-54.
Мягких К. А. 2002. Видовой состав уловов донных жаберных сетей в Авачинском заливе в зависимо-
сти от размеров их ячеи // Сохр. биоразнообр. Камчатки и прилег, морей: Мат-лы 3 науч, конф.,
Петропавловск-Камчатский: КамчатНИРО. С. 190-192.
Назаренко Л.Ф. 1953. Колониально гнездящиеся птицы Днестра и их хозяйственное значение И Мат-
лы по гидробиол. и рыболовству лиманов Сев.-Зап. Причерноморья. Киев: КГУ С. 151-163.
Назаренко Л.Ф., Назаренко М.Ф., Корзюков А.И., Чуйко В.П. 1975. Колониальные гнездовья около-
водных птиц Северо-Западного Причерноморья И Колон, гнездовья околоводн. птиц и их охрана.
М.: Наука. С. 149.
Назаренко Л.Ф., Николаевский О.В., Корзюков А.И., Щеголев И.В. 1975. Экологическая характери-
стика каравайки и динамика её колониальных гнездовий в дельте Днестра И Мат-лы совещ.: «Ко-
лон. гнездовья околоводн. птиц и их охрана». М. С. 45^16.
Назаренко Л.Ф., Попова М.Ф. 1957. О рыбоядности цапель в низовьях Днестра и их практическое
значение И Тез. докл. 3-й Прибалт, орнитол. конф. Вильнюс. С. 55-57.
Назаренко Л.Ф., Попова М.Ф. 1959. О рыбоядности цапель в низовьях Днестра и их практическом
значении И Тр. 3-й Прибалт, орнитол. конф. Вильнюс. С. 201-203.
Назаров Ю.Н. 1979. К биологии зеленой кваквы И Тез. докл. конф. «Миграции и экология птиц Сиби-
ри». Якутск. С. 198-202.
Назаров Ю.Н. 1986. Встречи редких птиц в Приморском крае И Тр. Зоол. ин-та АН СССР. Л.: изд-во
АН СССР. Т. 150. С. 81-83.
Назаров Ю.Н. 2000. Средняя белая цапля И Кр. книга РФ (животн.). М. С. 379-380.
Назаров Ю.Н. 2004. Птицы города Владивостока и его окрестностей. Владивосток: изд-во ДГУ 276 с.
Назаров Ю.Н., Куринный В.Н. 1981. Новые встречи редких птиц в Приморском крае И Филогения и
систематика птиц. Тр. Зоол. ин-та АН СССР. Л. Т. 102. С. 110-111.
Назаров Ю.Н., Лабзюк В.И. 1975. К авифауне южного Приморья И Орнитол. иссл. на Дальн. Восто-
ке. Владивосток. С. 268-276.
Назаров Ю.Н., Трухин А.М. 1985. К биологии сапсана и филина на островах зал. Петра Великого
(Южн. Приморье) И Редк. и исчезающ. птицы Дальнего Востока, Владивосток: ДВНЦ АН СССР.
С. 70-76.
Нанкинов Д.Н. 1978. Миграции жёлтых цапель на Балканах И Миграции птиц Вост. Европы и Сев.
Азии. Гагарообразные - аистообразные. М.: Наука. С. 125-126.
Нанкинов Д.Н. 1992. Картина гибели белых аистов от электропроводов в Болгарии И Аисты: распр.,
экол., охр. Минск. С. 114-120.
560
Литература
Нанкинов Д.Н. 1999. К вопросу о распространении и миграциях малой выпи И Беркут. Т. 8. Вип. 1.
С.15-20.
Нанкинов Д.Н., Кищинский А.А. 1978. Миграции малых белых цапель на Балканах И Миграции птиц
Вост. Европы и Сев. Азии. Гагарообр. - аистообр. М. С. 140-142.
Нанкинов Д.Н., Сапетин Я.В. 1978. Малый баклан - Phalacrocoraxpygmaeus (Pall.) И Там же. С. 104—
107.
Нанкинов Д.Н., Сапетин Я.В. 1978а. Розовый пеликан Pelecanus onocrotalus L. И Там же. С. 66-67.
Нейфельдт И.А. 1978. Вымершие птицы в коллекции ЗИН АН СССР И Сист. и биол. редких и мало-
изученных птиц. Л.: Зоол. ин-т АН СССР. С. 101-110.
Нейфельд Н.Д., Теплов В.В. 2000. Птицы ЮВ части республики Коми И Мат-лы к распростр. птиц на
Урале. Екатеринбург. С. 132-154.
Неручев В.В. 1991. Колпица на востоке дельты Волги И Редк. птицы и звери Казахст. Алма-Ата:
Гылым. С. 42.
Неручев В.В., Худяков И.В. 1967. Новые данные о фламинго на СВ побережье Каспия И Орнитоло-
гия. Вып. 8. С. 375-377.
Нечаев В.А. 1963. Новые данные о птицах Нижнего Амура И Орнитология. Вып. 6. С. 177-183.
Нечаев В.А. 1969. Птицы Южных Курильских о-вов. Л.: Наука. 246 с.
Нечаев В.А. 1971. К распространению и биологии некоторых птиц Южного Приморья И Орнитол.
иссл. на юге Дальн. Востока. Владивосток: Изд-во ДВНЦ АН СССР. Т. 6. С. 5-25.
Нечаев В.А. 1972. Какой вид фрегата добывали в СССР И Зоол. ж. Т. 51. Вып. 6. С. 936-938.
Нечаев В.А. 1975а. Колониально гнездящиеся птицы острова Монерон И Колон, гнездовья около-
водн. птиц и их охрана. М.: Наука. С. 181-183.
Нечаев В.А. 19756. Птицы острова Монерон И Орнитол. иссл. на Дальн. Востоке. Тр. Биол.-почв, ин-
та. Нов. сер. Т. 29 (132). Владивосток: ДВНЦ АН СССР. С. 5-25, 158-160.
Нечаев В.А. 1986. Новые сведения о птицах острова Сахалин И Морск. птицы Дальнего Востока.
Владивосток: ДВНЦ АН СССР. С. 71-81.
Нечаев В.А. 1989. Колпица И Редк. позвон. животн. Советск. Дальнего Востока и их охрана. Л.:
Наука. С. 51-52.
Нечаев В.А. 1991а. Встречи новых и редких птиц на о. Сахалин И Орнитология. Вып. 25. С. 167-168.
Нечаев В.А. 19916 (1991). Птицы острова Сахалин. Владивосток. 748 с.
Нечаев В.А. 2000. Встреча бурой олуши Sula leucogaster вблизи острова Итуруп (Курильские остро-
ва) И Русск. орнитол. журн., Экспр.-вып. № 122. С. 18.
Нечаев В.А. 2003. Новые сведения о птицах Южного Приморья И Русск. орнитол. журн., Экспр.-вып.
№210. С. 86-89.
Нечаев В.А. 2005. Обзор фауны птиц (Aves) Сахалинской области И Животн. и растит, мир о. Саха-
лин. Владивосток: Дальнаука. Ч. 2. С. 246-327.
Нечаев В.А., Гамова Т.В. 2009. Птицы Дальнего Востока России (аннотированный каталог). Влади-
восток: Дальнаука. 564 с.
Нечаев В.А., Фудзимаки Ю, 1995. Птицы Южных Курильских островов (Кунашир, Итуруп, Шико-
тан, Хабомаи). Владивосток-Токио. 123 с.
Нечаев В.А., Шибаев Ю.В. 1996. Малая колпица Platalea minor Temminck et Schlegel - новый вид
фауны России И Птицы пресн. вод и морск. побережий юга Дальн. Вост. России и их охрана.
С. 231-232.
Нечаев В.А., Юдаков В.Г. 1968. О гнездовании морских птиц на островах зал. Петра Великого (Юж.
Приморье) И Изв. Сиб. отд. АН СССР, сер. биол.-мед. наук. Т. 15. Вып. 3. С. 93-97.
Никифоров М.Е. 2001. Белорусская орнито-фаунистическая комиссия: обзор сообщений о наиболее
редких находках за 1990-1999 гг. И Subbuteo. Вып. 4. С. 25^0.
Никифоров М.Е., Козулин А.В., Гричик В.В., Тишечкин А.К. 1997. Птицы Беларуси на рубеже XXI
века. Минск. 188 с.
Никифоров М.Е., Яминский Б.В., Шкляров Л.П. 1989. Птицы Белоруссии. Справочник - определи-
тель гнёзд и яиц. Минск: Вышэйшая школа. 480 с.
Николаев В.И. 2000. Расселение белого аиста в Тверской области И Белый аист в России: дальше на
восток. Калуга. С. 95-98.
Литература
561
Николаев В.И., Керданов Д.А. 1984. Краткие сообщения о Чёрном аисте в Калининской области И
Иссл. в области заповеди, дела. М.: ВНИИ природы. С. 109.
Никольский А.М. 1891. Позвоночные животные Крыма // Прилож. к 68 т. Зап. Имп. акад, наук, СПб.
С. 1-484.
Новак В.О. 2002. Матер1али по фенологи мщращй птах!в на Подыл! 1. Non-Passeriformes // Ав1фауна
Украши. Вип. 2. С. 73-86.
Новак В.О. 2003. Зимова орштофауна схщних райошв Подшля // Беркут. Т. 12. Вип. 1-2. С. 14-20.
Нумеров А.Д. 2000. Численность и распространение белого аиста в Воронежской области И Белый
аист в России: дальше на восток. Калуга. С. 171-173.
Нумеров А.Д., Макагонова О.Б. 2000. Особенности гнездования белого аиста в Воронежской облас-
ти // Там же. С. 174-175.
Нумеров А.Д., Приклонский С.Г., Иванчев В.П. и др. 1995. Кладки и размеры яиц птиц юго-востока
Мещерской низменности // Тр. Окск. гос. биосферн. зап-ка. М.: ЦНИЛ Главохоты России. Вып. 18.
168 с.
Олейников Н.С. 1953. Рыбоядные птицы и их влияние на рыбное хозяйство манычских водохрани-
лищ И Уч. зап. Ростовск. ун-та. Т. 19. Вып. 3. С. 127-137.
Олейников Н.С., Гончаров А.И. 1967. Большая белая цапля на Усть-Манычском водохранилище И
Орнитология. Вып. 8. С. 280-284.
Олейников Н.С., Данилов Г.М. 1958. Материалы к биологии рыжей цапли // Уч. зап. Ростовск. ун-та.
Вып. 38. С. 57-71.
Олейников Н.С., Казаков Б.А., Ломадзе Н.Х., Языкова И.М. 1975. Семейство ибисовых в Предкавка-
зье // Изв. Сев.-Кавказск. науч. ц. высш шк. Сер. естеств. наук. № 3. С. 51-54.
Олигер Т.И. 1967. Ещё о численности птиц, зимующих в Кызыл-Агачском заповеднике // Орнитоло-
гия. Вып. 8. С. 378-379.
Омелько М.А. 1962. Новые данные о птицах Южного Приморья // Сообщ. Дальневост. фил. АН
СССР. Вып. 16. С. 119-123.
Омелько М.А., Омелько М.М. 1974. О новых редких птицах Южного Приморья И Фауна и экол.
наземн. позвон. юга Дальнего Востока СССР. Владивосток: изд-во ДВНЦ АН СССР. С. 200-203.
Омелько М.А., Омелько М.М. 1981. Наблюдения редких птиц в Южном Приморье, преимуществен-
но на п-ове Де-Фриза // Редк. птицы Дальн. Востока. Владивосток: изд-во ДВНЦ АН СССР. С. 117—
120.
Орштолопчш спостереження (1993) за 1991-1992 рр. // Troglodytes, 1993. Луцьк., Вип. 3. С. 1-69.
Орштолопчш спостереження (1994) на територп захщних областей Украши за 1993 рпс // Troglodytes,
1994. Луцьк. Вип. 4. С. 10-28.
Остапенко В.А., Гаврилов В.М., Ефремов В.Д. 1975. Характер и особенности весенних миграций
птиц Западной Камчатки И Мат-лы Всесоюзн. конф, по мигр. птиц. М.: изд-во МГУ. Ч. 2. С. 32-
35.
Остащенко А.Н. 2005. Первая в Средней Азии встреча китайской белокрылой цапли (Ardeola bacchus)
II Казахстанск. орнитол. бюл. 2004. Алматы: Tethys. С. 166-167.
Остерман А.И. 1912. Заметки о птицах Бессарабии И Тр. Бессарабск. о-ва естествоиспыт. и любит,
естествозн. Т. 2. Вып. 2. С. 165-191.
Остерман А.И. 1914. Заметки о птицах Бессарабии // Там же. Т. 5. С. 159-178.
Островский О.А, Дмитренок М.Г, Юрко В.В. 2004. Новые находки гнёзд малой выпи (Ixobrychus
minutus) в городе Минске И Subbuteo. Т. 7. С. 23-24.
Очаповский В.С. 1971. Редкие птицы Восточного Приазовья // Вести, зоол. № 5. С. 54-59.
Очеретный Д.Г 1998. Материалы по фенологии миграций птиц в Тульчинском районе Винницкой
области И Ав1фауна Украши. Вип. 1. С. 74-82.
Павлов П.П. 1879. Орнитологические наблюдения в Рязанской губернии // Тр. СПб о-ва естествоис-
пыт. Т. 10. С. 1-41.
Падутов Е.Е., Савицкий Б.П., Аршиев Н.А. 1975. Видовой состав, численность и питание колониаль-
но гнездящихся голенастых птиц средней части Восточного Маныча (КАССР) И Колон, гнездо-
вья околоводн. птиц и их охр. М. С. 151.
Панов Е.Н. 1965. Находки птиц на оз. Ханка // Орнитология. Вып. 7. С. 483-484.
562
Литература
Панов Е.Н. 1973. Птицы Южного Приморья (фауна, биология и поведение). Новосибирск: Сиб. отд.
изд-ва «Наука». 420 с.
Пантелеев А.В. 1997а. Кости птиц из поселений древнего человека на о. Сахалин // Вести. Саха-
линск. музея. Южно-Сахалинск. № 4. С. 281-285.
Пантелеев А.В. 19976. Остатки птиц из археологического памятника Ивановка (Сахалин) // Русск.
орнитол. ж., Экспр.-вып. № 9. С. 8-10.
Пантелеев А.В. 2001. Изучение четвертичной орнитофауны о-ва Сахалин И Актуальн. пробл. изуч. и
охр. птиц Вост. Европы и Сев. Азии. Казань. С. 486.
Пантелеев А.В. 2002. Особенности распространения некоторых видов птиц в Северной, Централь-
ной и Восточной Азии в четвертичном периоде // Русск. орнитол. журн., Экспр.-вып. № 186.
С. 471-484.
Панченко В.А., Балацкий К.Л. 1991. Редкие и исчезающие птицы дельт Дуная, Днестра и прилежа-
щих районов // Редк. птицы Причерноморья. Киев-Одесса: Лыбидь. С. 37-54.
Панченко В.А., Чёрный С.А. 1987. Современное состояние и численность колониальных веслоногих
и аистообразных птиц в Советской части дельты Дуная // Тез. докл. Всесоюзн. совещ. по пробл.
кадастра и учёта животн. мира. М. С. 371-373.
Панченко С.Г., Сулик В.Г. 1978. О зимовке перелетных птиц на юго-востоке Украины // Тез. сообщ. 2-
й Всесоюзн. конф, по мигр. птиц. Алма-Ата: Наука. Ч. ЕС. 49-51.
Панькин Н.С. 1984. Состояние и вопросы охраны редких птиц Верхнего и Среднего Приамурья //
Тез. докл. 3-й Всесоюзн. конф, зоологов пед. ин-тов. Ч. 1. Витебск. С. 131-132.
Панькин Н.С., Нейфельдт И.А. 1976. Дальневосточный белый аист в Амурской области // Тр. Окск.
зап-ка. Рязань. Вып. 13. С. 19-31.
Парилов М.П., Андронов В.А., Дарман Ю.А. 2000. Опыт создания базы данных по гнёздам дальне-
восточного аиста Ciconia boyciana Swinh. в Приамурье // Дальневост, аист в России. Владивос-
ток: Дальнаука. С. 89-95.
Паровщиков В., Ларионов В. 1958. Залеты голенастых птиц на Архангельский север И Охота и охотн.
хоз-во. № 5. С. 24.
Пасенюк Л.М. 1981. В одиночку на острове Беринга. М.: Мысль. 143 с.
Патев П. 1950. Птиците в България. София: изд. на Българската Академия Науките. 364 с.
Патрикеев М.В. 1991а. К весеннее-летней авифауне юго-восточной Ширвани и сопредельных терри-
торий // Мат-лы научно-пракг. конф.: «Фауна, насел, и экол. птиц Сев. Кавказа». Ставрополь.
С. 30-55.
Патрикеев М.В. 19916. О колонии голенастых и веслоногих птиц на озере Махмуд-Чала (Юж. Азер-
байджан) // Совр. сведения по составу, распростр. и экол. птиц Сев. Кавказа. Ставрополь. С. 88-
89.
Патрикеев М.В. 1991 в. Савка (Oxyura leucocephala) и другие редкие птицы на озере Аггель в январе
1991 г. // Мат-лы научно-пракг. конф. «Актуальн. вопр. экол. и охр. прир. Ставроп. кр. и сопред.
терр.». Ставрополь. С. 160-161.
Пахульский А.Н. 1951. Рыбоядные птицы южных морей СССР и их вред. М.: изд. МОИП. 220 с.
Пахульский А.Н. 1957. Рыбоядные птицы южных морей СССР и их вред. Киев. 156 с.
Пегова А.Н., Мокиевский В.О., Пономаренко С.В. и др. 1990. Новые данные о статусе некоторых
редких птиц Рязанской области // Редкие виды птиц центра Нечерноземья. М.: ЦНИЛ Главохоты
РСФСР. С. 95-98.
Пекло А.М. 1984. Краткое сообщение о Чёрном аисте в Юж. Приморье И Иссл. в обл. запов. дела. М.:
ВНИИприроды. С. 109-110.
Пекло А.М., Очаповский В.С. 1984. Краткое сообщение о Чёрном аисте в Зап. Предкавказье И Там
же. С. 110-111.
Петров В.С., Миноранский В.А. 1962. Летняя орнитофауна озёра Маныч-Гудило и прилежащих сте-
пей И Орнитология. Вып. 5. С. 266-275.
Петров В.Ю., Торопов К.В. 2000. Птицы Кулундинского озёра и его окрестностей // Мат-лы к рас-
простр. птиц на Урале. Екатеринбург. С. 157-163.
Петрович З.И., Щеголев И.В. 2003. Смешанная колония малого баклана на Кардашинских болотах И
Птицы Азово-Черноморск. региона. Николаев. С. 52.
Литература
563
Петрович 3.0. 1981. Колония околоводных птиц в советской части дельты р. Дунай И Размещ. и сост.
гнездовий околоводных на терр. СССР (МОИП). М.: Наука. С. 106-109.
Петрович 3.0. 2000. Косар в Украшський частиш дельти Дунаю И Бранта. Вып. 3. С. 117-121.
Пискунов В.В., Беляченко А.В. 2000. Статус белого аиста в Саратовской области И Белый аист в
России: дальше на восток. Калуга. С. 182.
Пишванов Ю.В. 1975. Некоторые данные к вопросу о колониальных гнездовьях околоводных птиц в
Дагестане // Колон, гнездовья околоводн. птиц и их охрана. М. С. 151-153.
Пишванов Ю.В., Прилуцкая Л.И. 1988. Фламинго в Дагестане // Ресурсы редких животных РСФСР,
их охрана и воспроизводсво. М. С. 67.
Плеске Ф.Д. 1896. Кости Phalacracoraxperspicillatus И Ежегодн. Зоол. музея Акад. наук. СПб. Т. 1. №
1-2.
Плесский П.В. 1960. Наземные позвоночные Кировской области // Природа Кировск, обл. Киров.
С. 192-216.
Плешак Т.В. 1987. О некоторых орнитологических находках в Архангельской области // Орнитоло-
гия. Вып. 22. С. 191.
Подковыркин Б.А. 1955. Список птиц северной части Курильской гряды // Зоол. ж. Т. 34. № 6. С. 1379—
1385.
Познании Л.П. 1979. Эколого-морфологический анализ онтогенеза птенцовых птиц (общий рост и
развитие пропорций тела в постэмбриогенезе). М. 296 с.
Поливанов В.М. 1965. О миграциях цапель, гнездящихся на оз. Ханка // Новости орнитол. Алма-Ата:
Наука. С. 76-87.
Поливанов В.М. 1978. Миграции рыжих цапель оз. Ханка (Приморский край, СССР) // Миграции
птиц Вост. Европы и Сев. Азии. Гагарообразные - аистообразные. М.: Наука. С. 229-231.
Поливанов В.М., Поливанова Н.Н. 1971. О миграциях цапель озёра Ханка // Экол. и фауна птиц юга
Дальнего Востока. Владивосток: Дальневост. кн. изд-во. С. 5-61.
Поливанов В.М., Ткаченко И.В., Поливанова Н.Н. 2001. Необычный массовый залёт белых аистов
Ciconia ciconia в долину реки Теберды // Русск. орнитол. ж., Экспр.-вып. № 130. С. 73-75.
Поливанова Н.Н. 1971. Птицы озёра Ханка. Владивосток: ДВНЦ АН СССР. 240 с.
Поливанова Н.Н., Глущенко Ю.Н. 1977. Новые данные о некот. редких и малочисл. птицах Приморья И
Тез. докл. 7-й Всесоюзн. орнитол. конф. Киев. Ч. ЕС. 95-96.
Полозов С.А. 1984. Краткие сообщения о Чёрном аисте в Западном Копетдаге // Иссл. в обл. запов.
дела. М.: ВНИИ природы. С. 112-114.
Полюшкевич I.M. 1998. Матер!али по фенологи м!гращй птах!в у КоросНшивському район! Жито-
мирськоТ облает! // Ав!фауна Укра!ни. Вип. 1. С. 62-74.
Полякова А.Д. 1984. Ориентировочные данные по добыче птиц в Средней Азии // Тр. Окск. гос. зап-
ка. Рязань: Рязанск. отд. изд-ва «Московский рабочий». Вып. 15. Научн. основы охраны и рац.
использ. птиц. С. 118-127.
Полякова А.Д. 1984а. Результаты учёта добычи охотничьих птиц в Азербайджане // Там же. С. 107-
117.
Попенко В.М., Дацевич В.А. 1984. Краткие сообщения о Чёрном аисте в Беловежской пуще // Иссл.
в области заповеди, дела. М.: ВНИИприроды. С. 108.
Попов В.А. (отв. ред.). 1977. Птицы Волжско-Камского края. Неворобьиные. М.: Наука. 296 с.
Попов В.В. 2004. Залёт индийского ибиса Threskiornis melanocephalus на Байкал // Русск. орнитол.
журн., Экспр.-вып. № 257. С. 322-323.
Попов В.В., Матвеев А.Н. 2005. Позвоночные животные Байкальского региона: видовой состав и
правовой статус. Иркутск: изд-во Иркутск, ун-та. 85 с.
Портенко Л.А. 1939. Фауна Анадырского края. Птицы. Ч. 1 и 2 // Тр. НИИ полярн. земледелия, жи-
вотноводства и промысл, хоз-ва. Сер. Промысл, хоз-во. Вып. 5, С. 5-211; вып. 6, С. 4-198.
Портенко Л.А. 1792. Птицы Чукотского полуострова и о. Врангеля. Ч. 1. Л.: Наука. 424 с.
Пославский А.Н. 1991. Колпица в низовьях Сыр-Дарьи // Редкие птицы и звери Казахст. Алма-Ата:
Гылым. С. 44.
Пославский А.Н., Сабиневский Б.В., Лури В.Н. 1977. Фламинго в Северо-Восточном Прикаспии //
Редк. и исчез, звери и птицы Казахст. Алма-Ата. С. 209-215.
564
Литература
Пославский А.Н., Ширеков Р.Ш. 1990. Редкие виды веслоногих, голенастых и фламинго в Вост. Тур-
кмении // Редк. и малоизуч. птицы Ср. Азии. Бухара. С. 35-38.
Потапов О.В. 1995. Птицы оз. Кугурлуй и прилегающих территорий // Экосистемы дикой природы.
Одесса. Вып. 2. С. 13-30.
Потапов О.В. 2001. Орнитофауна регионального ландшафтного парка «Измаильские острова» И Бран-
та. Вып. 4. С. 25-41.
Потапов РЛ. 1958. Новые данные о птицах, зимующих в Ю. Таджикистане И Докл. АН Тадж. ССР.
Т. 1. №3. С. 41-44.
Поярков Н.Д., Бурдис РР. 1991. Заметки о птицах оз. Мухтель // Орнитология. Вып. 25. С. 172-174.
Пракаш Гоул. 1990. Материалы по птицам из Красной книги РСФСР в Индии // Итоги изуч. редк.
животн.: мат-лы к Кр. книге. М. С. 34-40.
Пржевальский Н.М. 1870. Путешествие в Уссурийском крае в 1867—69 гг. СПб. 298 с.
Придатко В.И. 1986. Закономерности чередования сезонных явлений в колониях морских птиц на о. Вран-
геля И Экосист. экстрем, усл. среды в зап-ках РСФСР. М.: ЦНИЛ Главохоты РСФСР. С. 160-166.
Приедниекс Я., Страздс М., Страздс А., Петриньш А. 1989. Атлас гнездящихся птиц Латвии. Рига:
Зинатне. 352 с.
Приклонский С.Г 1958. Материалы по экологии Чёрного аиста в Окском заповеднике // Тр. Окск.
зап-ка. М. Вып. 2. С. 102-115.
Приклонский С.Г. 1968. Результаты первого учёта добычи пернатой дичи в Казахстане И Орнитоло-
гия. Вып. 2. С. 188-207.
Приклонский С.Г. 1975. Тереховская колония серых цапель близ Окского заповедника // Колон.^гнез-
довья околоводн. птиц и их охрана. М.: МОИП.
Приклонский С.Г. 1977. Отряд аистообразные Ciconiiformes И Птицы Волжско-Камск. края. Неворо-
бьиные. М.: Наука. С. 29-43.
Приклонский С.Г. 1984. Численность и успешность гнездования чёрного аиста в Окском заповедни-
ке // Исслед. в обл. запов. дела. М.: ВНИИприр. С. 105-107.
Приклонский С.Г, Бианки В.В., Карпович В.Н. и др. 1962. Отлов птиц автоматическими лучками И
Тр. Окск. зап-ка. Вологда: Волог. кн. изд.. Вып. 4. С. 402-424.
Приклонский С.Г, Галушин В.М. 1959. Новые данные по экологии Чёрного аиста // Тр. 3-й Прибалт,
орнитол. конф. Вильнюс. С. 231-236.
Приклонский С.Г, Котюков Ю.В., Сапетина И.М. и др. 1998. Редкие, малочисленные и малоизучен-
ные птицы Мещеры (голенастые, пластинчатоклювые и др. околоводные) И Редк. виды птиц Не-
черноз. центра России. М. С. 204-207.
Приклонский С.Г, Постельных А.В. 1984. Сведения о численности и распространении Чёрного аис-
та в СССР по данным различных обследований // Иссл. в обл. запов. дела. М.: изд. ВНИИприро-
ды. С. 100-105.
Приклонский С.Г, Сапетин Я.В. 1978. Результаты кольцевания (серой цапли) в Рязанской области И
Мигр. птиц Вост. Европы и Сев. Азии. Гагарообр. - Аистообр. М.: Наука. С. 188-191.
Приклонский С.Г., Теплов В.П. 1962. Опыт учёта численности глухаря, журавля и серой цапли в
лесах центральных областей Европейской части РСФСР И Тр. Окск. гос. зап-ка. Вологда: Воло-
годск. кн. изд-во. Вып. 4. Работы Центральной орнитол. станции. Вып. 2. С. 33-64.
Прилуцкая Л.И., Пишванов Ю.В. 1989. К кадастру редких и исчезающих птиц Дагестана И Всесо-
юзн. совещ. по пробл. кадастра и учёта животн. мира. Уфа. Ч. 3. С. 181-184.
Прилуцкая Л.И., Пишванов Ю.В. 1989а. Кудрявый пеликан в Дагестане И Тез. науч.-пракг. конф.
«Орнитол. рес. Сев. Кавказа». Ставрополь. С. 71-73.
Прокофьев С.М. 1984. Краткое сообщение о Чёрном аисте на юге Красноярского края И Иссл. в обл.
заповедного дела. М.: ВНИИприроды. С. 129-130.
Птицы Волжско-Камского края. 1977. - см. Попов В.А. 1977.
Птицы Северного Кавказа. 2004. Ростов-на-Дону: изд-во РГПУ Т. 1. Гагарообр. - гусеобр. 398 с.
Птицы Средней Азии. 2007. Алматы. Т. 1. 574 с.
Птицы Узбекистана, 1987. - см. Мекленбурцев PH. и др. 1987.
Птушенко Е.С. 1958. Список птиц Окского заповедника и Рязанской области // Тр. Окск. гос. зап-ка.
М. Вып. 2. Работы Окской орнитол. станции. Вып. 1. С. 192-206.
Литература
565
Птушенко Е.С., Иноземцев А.А. 1968. Биология и хозяйственное значение птиц Московской области
и сопредельных территорий. М.: изд-во МГУ. 461 с.
Пузанов И.И. 1931. Предварительные итоги изучения птиц Крымского заповедника // Сб. работ по
изуч. фауны Крымск, гос. зап-ка. С. 32-59.
Пузанов И.И., Козлов В.И., Кипарисов Г.П. 2005. Позвоночные животные Нижегородской области.
3-е изд. Ниж. Новгород. 543 с.
Пукинский Ю.Б. 2003. Гнездовая жизнь птиц бассейна р. Бикин. СПб.: Изд-во СПб. ун-та. 316 с.
Пчелинцев В.Г. 1990. К вопросу о питании большой выпи (Botaurus stellaris L.) И Тр. Зоол. ин-та АН
СССР. Т. 213. С. 91-93.
Пчелинцев В.Г., Ильинский И.В. 2000. Белый аист в Ленинградской области: результат 30-летнего
расселения И Белый аист в России: дальше на восток: Калуга. С. 63-65.
Пыжьянов С.В. 2007. Список птиц побережья Малого моря и прилегающих территорий // Тр. При-
байкальск. нац. парка. Иркутск. Вып. 2. С. 218-229.
Радде Г.И. 1884. Орнитологическая фауна Кавказа (Omis Caucasica). Систематическое и биолого-
географическое описание кавказских птиц. Тифлис. 451 с.
Рандла Т, Ыун А. 1984. Краткие сообщения о Чёрном аисте в Эстонии // Иссл. в обл. запов. дела. М.:
ВНИИприроды. С. 107-108.
Рахилин В.К. 1972. Характер миграций веслоногих на Дальнем Востоке // Итоги 6-го симп. по изуч.
вирусов, эколог, связанных с птицами. М. С. 69-70.
Рахимов И.И. 1998. О распространении некоторых видов птиц Татарстана // Мат-лы к распр. птиц на
Урале. Екатеринбург. С. 138-141.
Рахимов И.И., Уленгов Р.А. 2003. Изменения в авифауне Среднего Поволжья в связи с созданием
водохранилищ Волжско-Камского каскада // Русск. орнитол. ж., Экспр.-вып. № 245. С. 1352—
1357.
Рашкевич Н.А. 1961. Гнездовая жизнь птиц. Обзоры // Природа. № 5. С. 75-78.
Рашкевич Н.А. 1969. Колония рыбоядных птиц на южном берегу Аральского моря И Мат-лы 5-й
Всесоюзн. орнитол. конф. Ашхабад. Кн. 2. С. 530-533.
Рев Н., Татаринкова И.П., Панеева Т.Д. 2003. Хохлатый баклан Phalacrocorax aristotelis И Сост. по-
пул. морск. птиц, гнездящихся в регионе Баренцова моря. Норв. полярн. ин-т. С. 34-36.
Рединов К.А. 1999. Материалы по редким и малочисленным видам птиц Николаевской области И
Бранта. Мелитополь. Вып. 2. С. 152-158.
Резанов А.Г. 2001. Тактолокация и визуальное обнаружение добычи аистами Ciconia ciconia и С.
nigra при разыскивании корма на мелководье И Русск. орнитол. ж., Экспр.-вып. № 133. С. 152—
153.
Рогачева Э.В., Сыроечковский Е.Е. 2003. Атлас гнездящихся птиц Европы Европ. Совета по учётам
птиц [Хагемайер, Блейер, 1997]. Сокращ. версия текстовой части на русском языке. М.: ИПЭЭ
РАН. 338 с.
Роговий Ю.Ф. 1994. До фенологи м!гращй птах!в у долин! р. Кагамлик // Мат-ли 1-й конф, молодих
оршт. Украши. Чершвщ. С. 104-106.
Роговой Ю.Ф. 1992. Особенности пролёта птиц в долине реки Сухой Кагамлык // Сез. мигр. птиц на
терр. Украины. Киев. С. 127-133.
Рольник В.В. 1948. Развитие терморегуляции у некоторых птиц севера // Зоол. ж. Т. 27. Вып. 7. С. 535—
546.
Ромашова А.Т. 1938. Количественное изучение гнездовых колоний Астраханского заповедника И Тр.
Астраханск. зап-ка. Вып. 2. С. 78-111.
Росляков Г.Е. 1981а. Дальневосточный аист Ciconia boyciana в Нижнем Приамурье // Редк. птицы
Дальн. Востока. Владивосток: изд-во ДВНЦ АН СССР. С. 97-99.
Росляков Г.Е. 19816. Краткие сведения о некоторых редких и малоизученных птицах Нижнего При-
амурья И Там же. С. 112-115.
Росляков Г.Е. 1981 в. Редкие птицы Хабаровского края, нуждающиеся в особой охране // Там же.
С.141-144.
Росляков Г.Е. 1987. Дальневосточный аист // Они нуждаются в защите. Редк. животн. Хабаровск,
края. Хабаровск. С. 71-73.
566
Литература
Росляков ЕЕ. 1992. О состоянии морских птиц на Шантарских о-вах в 1991 г. И Изуч. морск. колон,
птиц в СССР. Магадан: ИБПС ДВО РАН. С. 11-12.
Росляков А.Г. 2000а. Рыжая цапля Ardea purpurea (Linnaeus, 1766) И Красная книга Хабаровского
края. 2-е изд. Хабаровск. С. 284-285.
Росляков А.Г. 20006. Состояние редких видов птиц, внесенных в Красную книгу Хабаровского края //
Инвент., монит. и охрана Ключевых орнитол. терр. Вып. 2. С. 120—127.
Росляков А.Г, Воронов Б.А., Сапаев В.М. 2000. Дальневосточный аист в Хабаровском крае // Даль-
невост. аист в России. Владивосток: Дальнаука. С. 34-43.
Росляков Г.Е., Росляков А.Г. 1996. К орнитофауне создаваемого национального парка «Шантарские
острова» // Птицы пресн. вод и морск. побер. югаДальн. Востока и их охрана. Владивосток: Даль-
наука. С. 86-92.
Руденко А.Г, Яремченко О.А. 2004. История гнездования и динамика колониальных поселений боль-
шого баклана {Phalacracorax carbo) в Черноморском биосферном заповеднике // Бранта. Сб. тр.
Азово-Черноморск. орнитол. станции. Вып. 7. С. 193-207.
Руденко А.Г, Яремченко О.А. 2005. Динамика численности и распределение скоплений розового
пеликана {Pelecanus onocrotalus) // Там же. Вып. 8. С. 16-26.
Руденко А.Г, Яремченко О.А., Москаленко Ю.А. 2002. Мониторинг зимующих птиц водно-болот-
ных угодий Черноморского зап-ка в 2000-2001 гг. И Мониторинг зимующих птиц в Азово-Чер-
ном. регионе Украины. Одесса-Киев. С. 40-54.
Рудницкий С.В. 2003. Из Ставропольского края // Ключевые орнитол. терр. России, инф. бюл. № 17.
С. 42.
Русанов ГМ. 1992. Редкие виды птиц дельты Урала // Редк. виды раст. и животных. Оренбург. С. 56-
58.
Русанов ГМ. 1995. Гнездование кудрявых пеликанов в дельте Урала и у СВ побережья Каспия И
Орнитология. Вып. 26. С. 194.
Русанов ГМ. 1997. Пеликаны на Северном Каспии И Бюл. МОИП. Отд. биол. Т. 102. № 5. С. 27-30.
Русанов ГМ. 2002. Фламинго на Сев. Каспии И Инф. бюлл. «Волга». Вып. 9.
Русанов ГМ. 2005. Мониторинг КОТР «Дельта Волги» (2004) // Инвент., монит. и охр. Ключевых
орнитол. терр. России. Вып. 5. С. 119-133.
Русанов ГМ. 2006. Изменение птичьего населения дельты Волги во второй половине XX столетия И
Орнитол. иссл. в Сев. Евразии. Ставрополь. С. 450-451.
Русанов Г.М., Кривоносов ГА. 1988. Фламинго {Ph. roseus Pall.) на севере и северо-востоке Каспии в
условиях современного повышения уровня моря // Бюл. МОИП. Отд. биол. Т. 93. Вып. 3. С. 13.
Русанов Г.М., Кривоносов ГА. 1991. Водоплавающие птицы и добыча нефти в СВ Прикаспии И Мат-
лы 10-й Всесоюзн. орнитол. конф. Минск. Ч. 1. С. 136-138.
Русанов Г.М., Реуцкий Н.Д., Кривоносов ГА. и др. 1999. Птицы // Позвон. животн. Астраханск. зап-
ка. Флора и фауна зап-ков. М. Вып. 75. С. 27-64.
Русанов Г.М., Реуцкий И.Д., Литвинова Н.А., Гаврилов Н.Н. 2000. Редкие птицы дельты Волги и
прилежащих территорий // Редкие, исчез, и малоизуч. птицы России. М. С. 64-68.
Русев И.Т. 1981. Хозяйственное значение кваквы в дельте Днестра // Тез. 7-й Всесоюзн. орнитол.
конф. Кишинев. С. 196-197.
Русев И.Т. 1983а. Зимовки кваквы в дельте Днестра И Бюл. МОИП. Отд. биол. Т. 85. Вып. 5. С. 58.
Русев И.Т. 19836. Способ отлова квакв с помощью света // Вести, зоол. Вып. 5. С. 8.
Русев И.Т. 1983в. Некоторые черты поведения кваквы в гнездовой период // Мат-лы 3-й Всесоюзн.
конф, по повед. животн. М. Т. 3. С. 187.
Русев И.Т. 1985. Значение птиц-ихтиофагов, посещающих рыборазводные пруды в низовьях Днестра И
Бюл. МОИП. Отд. биол. Т. 90. Вып. 6. С. 23-26.
Русев И.Т. 1986. Территориальные связи кваквы в послегнездовой период И Тез. докл. 1-го съезда
Всесоюзн. орнитол. о-ва и 9-й Всесоюзн. орнитол. конф. Л. Ч. 2. С. 205-206.
Русев И.Т. 2000а. Гнездование колониальных аистообразных птиц в верховьях Тилигульского лима-
на в 1999 г. И Птицы Азово-Черном, региона на рубеже тысячелетий. Одесса: Астропринт. С. 55.
Русев И.Т. 20006. Трансконтинентальные и экологические связи птиц в природных очагах арбовиру-
сов, как фактор эпидемического риска // Там же. С. 88.
Литература
567
Русев И.Т. 2000в. Численность и размещение околоводных птиц в водно-болотных угодиях Азово-
Черноморского побережья Украины. Киев. С. 66-98.
Русев И.Т. 2001. Динамика численности основных орнитокомплексов и доминирующих видов // Ак-
туальн. пробл. изуч. и охр. птиц Вост. Европы и Сев. Азии, Казань. С. 537-538.
Русев И.Т. 2003. Дельта Днестра. Одесса: Астропринт. 760 с.
Русев И.Т. 2004. Видовой состав и численность веслоногих и голенастых птиц в дельте Днестра //
Бранта. Сб. тр. Азово-Черноморск. орнитол. станции. Вып. 7. С. 23-52.
Русев И.Т., Арнаут И.Н. 1983. Некоторые аспекты колониальное™ аистообразных в дельте Днестра //
Мат-лы 3-й Всесоюзн. конф, по повед. животн. М. Т. 2. С. 129.
Русев И.Т., Корзюков А.И., Яковлев М.В. 2005. Низовья реки Кагильник - территория важная для
птиц // Мат-лы конф., посвящ. 140-летию Одесск. нац. ун-та. Одесса. С. 243-245.
Русев И.Т., Корзюков А.И., Форманюк О.А., Панченко П.С. 2002. Зимовки водоплавающих и водно-
болотных птиц в Северо-Западном Причерноморье в 2000-2001 гг. // Мониторинг зимующ. птиц
в Азово-Черном, регионе Украины. Одесса-Киев. С. 54-74.
Русев И.Т., Русева Т.Д. 1983. Некоторые особенности поведения ночных цапель в дельте Днестра //
Мат-лы 3-й Всесоюзн. конф, по повед. животн. М.: Наука. Т. 1. С. 238-239.
Рустамов А.К., Ковшарь А.Ф. 2007. От редакторов // Птицы Ср. Азии. Алматы. Т. 1. С. 11-13.
Рябцев В.В. 1997. Амурский волчек в Хинганском заповеднике // Русск. орнитол. ж., Экспр.-вып. №
8. С. 9-10.
Рябицев В.К. 2001. Птицы Урала, Приуралья и Зап. Сибири. Екатеринбург. 608 с.
Рябицев В.К, Бойко Г.В.. Москвитин С.С. и др, 2001. Фауна птиц регионов Зап. Сибири И Инвента-
рю., мониторинг и охр. Ключевых орнитол. терр. России. М. Вып. 3. С. 140-168.
Рябов В.Ф. 1982. Авифауна степей Сев. Казахстана. М.: Наука. 176 с.
Сабанеев Л. 1874. Позвоночные Среднего Урала: географическое распространение в Пермской и
Оренбургской губ. М. 204 с.
Савинецкий А.Б., Пташинский А.В. 1999. Промысловая добыча древних приморских жителей Кам-
чатки // М.: Диалог-МГУ Вып. 1. С. 109-111.
Сагитов А.К. 1987. Отряд Аистообразные Ciconiiformes // Птицы Узбекистана. Ташкент: изд-во “ФАН”
Узбекской ССР. Т. 1. С. 29-54.
Сагитов А.К. 1990. Динамика численности белого аиста (Ciconia asiatica Sev.) в Узбекистане // Аис-
ты: распранение, экология, охрана. Минск. С. 124-128.
Сагитов А.К., Гульмурадов С. 1972. Биология размножения белого аиста на Зарафшане // Тр. Самар-
канде^ ун-та. Нов. сер. Зоол. Вып. 211. С. 1-7.
Сагитов А.К., Салимов X. 1973. К биологии малой выпи Ixobrychus minutus minutus L. в пойме реки
Зарафшан // Вопросы экологии и морфологии животных. Тр. Самаркандск. гос. ун-та им. Алише-
ра Навои. Вып. 228 (зоол. позв.). Самарканд. С. 18-25.
Саилов Р.А., Пукинский Ю.Б., Пукинская М.В. 1989. Современное поведение веслоногих (Peleca-
niformes) и голенастых (Ciconiiformes) птиц на озёрах Каракыр Бухарской области // Фауна и
экол. птиц Узбекистана. Самарканд. С. 193-205.
Салихбаев Х.С., Богданов А.Н. 1961. Птицы // Фауна Узбекской ССР. Ташкент: Изд-во АН Узб. ССР.
Т. 2. 272 с.
С. (Алфераки). 1890. Летят ли птицы против ветра или за ветром // Природа и охота. Кн. 7. С. 1-13.
Самигуллин Г.М. 1988. Залеты большого баклана и большой белой цапли в Оренбургскую область И
Орнитология. Вып. 23. С. 220-221.
Самигуллин Г.М. 1990. Цаплевые Оренбургской области И Живота, мир Юж. Урала. Оренбург. С. 90-92.
Самусенко И.Э. 1992. Некоторые особенности распространения и экологии белого аиста как ланд-
шафтно-экологические индикаторы // Аисты: распранение, экология, охрана. Минск. С. 124-132.
Самусенко И.Э. 1994. Питание белого аиста (Ciconia ciconia') в Беларуси // Тез. докл. 7-й зоол. конф.
Минск. С. 312-314.
Самусенко И.Э. 1999. Первые подтверждения находки большой белой цапли (Egretta alba) на гнездо-
вании в Беларуси // Тез. докл. 8-й зоол. конф. Беларуси. Минск. С. 141-143.
Самусенко И.Э., Журавлев Д.В. 2000. К гнездованию большой белой цапли (Egretta alba) в Беларуси
// Subbuteo. Вып. 3. С. 14-16.
568
Литература
Сапетин Я.В. 1968. Материалы по кольцеванию розового пеликана и малого баклана в дельте Терека
// Миграции животн. Л.: АН СССР. Вып. 5. С. 113-115.
Сапетин Я.В. 1978а. Миграции больших белых цапель юга европейской части СССР И Миграции
птиц Вост. Европы и Сев. Азии. Гагарообразные - аистообразные. М.: Наука. С. 132-140.
Сапетин Я.В. 19786. Миграции жёлтых цапель Азово-Черноморского и Каспийского бассейнов
(СССР) // Там же. С. 127-132.
Сапетин Я.В. 1978в. Миграции квакв в СССР // Там же. С. 114-123.
Сапетин Я.В. 1978г. Миграции колпиц, гнездящихся в СССР // Там же. С. 236-242.
Сапетин Я.В. 1978д. Миграции малых белых цапель Азово-Черноморского и Каспийского бассейнов
(СССР) // Там же. С. 142-149.
Сапетин Я.В. 1978е. Результаты кольцевания каравайки в СССР // Там же. С. 245-255.
Сапетин Я.В. 1978ж. Результаты кольцевания рыжих цапель на юге европейской части СССР И Там
же. С. 220-229.
Сапетин Я.В., Галушин В.М. 1958. Крупная колония серой цапли И Тр. Окск. зап-ка. М. Вып. 2.
С. 168-169.
Сарандинаки Г. 1909. Некоторые данные для орнитологии Ростовского н/Д округа Донской области //
Сб. студ. научн. кружка при Новоросс. ун-те. № 4. С. 1-75.
Сарычев В.С. 2000. Распространение и численность белого аиста в Липецкой области И Белый аист
в России: дальше на восток. Калуга. С. 167-170.
Сатунин К.А. 1907. Материалы к познанию птиц Кавказского края И Зап. Кавказск. отд. РГО. Тиф-
лис. Кн. 26. Вып. 3. 144 с.
Свиридова Т.В., Зубакин В.А. (ред.). 2000. Ключевые орнитологические территории России. Т. 1.
Ключевые орнитологические территории международного значения в Европейской России. М.:
Союз охраны птиц России. 702 с.
Северцев Н.А. 1873. Путешествия по Туркестанскому краю и исследование горной страны Тянь-
Шань. СПб. 461 с.
Селлис У.Ю. 2001. Чёрный аист (Ciconia nigra) в Эстонии в конце XX в // Актуальн. пробл. изуч. и
охр. птиц Вост. Европы и Сев. Азии. Казань. С. 555.
Сема А.М. 1989. Фенология перелетов птиц в Казахстане. Алма-Ата: Наука. 152 с.
Сема А.М., Ауэзов Э.М. 1984. Материалы количественных учётов пролёта птиц в Алакольской кот-
ловине // Миграции птиц в Азии. Ташкент. С. 53-56.
Семенова Н.Н., Иванова В.М. 1991. Паразиты животных // Астрах, заповедник. М. С. 77-91.
Серебряков В.В., Грищенко В.Н. 1992. Распределение и численность колоний цапель на территории
Украины по данным анкетного учёта в 1986 г. И Беркут. Т. 1. С. 52-56.
Симеонов С.Д., Мичев Т.М., Нанкинов Д.Н. 1992. Фауна на България. Т. 20. Aves. Ч. 1. София: изд-во
Бългр. АН. 350 с.
Сиохин В.Д. (ред.). 2000. Численность и размещение гнездящихся околоводных птиц в водно-болот-
ных угодьях Азово-Черноморского побережья Украины. Мелитополь-Киев: Бранта. 475 с.
Скалов Ю. 1928. Перелетная саранча (Locusta migratoria L.) и меры борьбы с нею на Кубани за
период с 1874 по 1927 гг. // Изв. Сев.-Кавк. краев, станц. защ. раст. Вып. 4. С. 71-116.
Скильский И.В., Годованец Б.И., Бучко В.В. 2000. Гнездовая экология кваквы в регионе Украинских
Карпат // Беркут. Т. 9. Вып. 1-2. С. 12-22.
Скильский И.В., Годованец Б.И., Бучко В.В. 2001а., Большая белая цапля в регионе Украинских Кар-
пат: территориальное размещение и численность // Актуальн. пробл. изуч. и охраны птиц Вост.
Европы и Сев. Азии. Казань. С. 556-557.
Сюльський I.B., Годованець Б.Й., Бучко В.В. 20016. Чепура велика в репош Украшських Карпат //
Зелена Буковина. Вип. 1-2. С. 53-61.
Скокова Н.Н. 1954. Очерки экологии серой цапли в районе Рыбинского водохранилища // Уч. зап.
МГПИ им. В.П. Потемкина. Т. 28. Вып. 2. С. 89-153.
Скокова Н.Н. 1955. Питание большого баклана в дельте Волги // Вопр. ихтиол. Вып. 5. С. 170-185.
Скокова Н.Н. 1959а. Результаты кольцевания ибисовых (Ibididae) и цаплевых (Ardeidae) в СССР И
Мигр. животных. Вып. 1. С. 67-94.
Скокова Н.Н. 19596. Экология и хозяйственное значение колпицы в дельте Волги И Орнитология.
Вып. 2. С. 262-270.
Литература
569
Скокова Н.Н. 1960а. О сезонном размещении и миграции большого баклана на Каспийском море И
Мигр. животн. М.: АН СССР. Вып. 2. С. 76-99.
Скокова Н.Н. 19606. О хозяйственном значении каравайки в дельте Волги // Охр. и озеленение. М.:
ВООП. Вып. 4. Птицы водоёмов. С. 45-51.
Скокова Н.Н. 1962. О количественном изучении питания птиц-ихтиофагов И Орнитология. Вып. 4.
С. 288-296.
Скокова Н.Н. 1965.0 характере воздействия большого баклана и голенастых птиц на фауну водоёмов
дельты Волги и их хозяйственное значение // Рыбоядн. птицы и их знач. в рыбн. хозяйстве. М.:
Наука. С. 55-70.
Скокова Н.Н. 1978а. Атлантический большой баклан - Phalacracorax carbo carbo (L.) И Миграции,
птиц Вост. Европы и Сев. Азии. Гагарообр. - аистообразные. М.: Наука. С. 68-70.
Скокова Н.Н. 19786. Миграции большого баклана в дельте Кубани // Там же. С. 86-88.
Скокова Н.Н. 1978в. Миграции серых цапель, гнездящихся на Рыбинском водохранилище (СССР) И
Там же. С. 179-188.
Скокова Н.Н. 1978г. Результаты кольцевания серых цапель в бассейне Азовского моря // Там же.
С. 195-197.
Скокова Н.Н., Бондарев Д.В. 1978. Миграции больших бакланов дельты Волги // Там же. С. 80-86.
Скокова Н.Н., Кищинский А.А. 1978. Заключение (по большому баклану) // Там же. С. 101-102.
Скрябин Н.Г. 1982. Околоводные птицы экосистем крупнейших озёр Азии (на примере оз. Байкал и
оз. Хабсугул МНР) // Тез. докл. 18-го Междр. орнитол. конгр. М.: Наука. С. 233.
Слудский А.А. 1960. Фламинго в СССР и задачи его охраны // Охр. природы и озелененение. М.
Вып. 4. С. 95-98.
Слюсар М.В. 2000. Анотований список наземних чотириногих хребетних околиць бюстацюнару
«Лучки» Полтавського педагопчного ушверситету. Полтава. 47 с.
Сметанина З.Б., Алексеев В.М. 1970. Итоги гельминтологических исследований рыбоядных птиц
Приморского края // Мат-лы 15-й науч. конф, проф.-преп. состава Биол. почв, фак-та ДВГУ. Вла-
дивосток. С. 164-168.
Смиренский С.М. 1986. Эколого-географич. анализ авифауны средн. Приамурья. Дисс. на соиск. уч.
степ. канд. биол. н. М. 364 с.
Смиренский С.М., Андронов В.А., Былков А.Ф. 1987. Первые результаты цветного мечения дальне-
восточного аиста // Орнитология. Вып. 22. С. 219-220.
Смирнов Г.П., Велижанин А.Г. 1986. Колонии морских птиц Чукотского нац. округа И Бюлл. МОИП.
Отд. биол. Т. 91. Вып. 3. С. 29-35.
Смирнов Н.А. 1916. Наблюдения над весенним пролётом некоторых видов птиц в Мангышлаке //
Изв. Кавказск. муз. Т. 10. Вып. 3. С. 217-228.
Смогоржевський Л.О. 1951. Фауна Украши. Птахи. Т. 5. Вип. 1. 188 с.
Смогоржевский Л.А. 1953. К биологии и распространению каравайки (Plegadis falcinellus L.) на Ук-
раине // Уч. зап. Киевск. ун-та. Т. 12. Вып. 7. Тр. биол.-почв. ф-та. № 9. С. 173-177.
Смогоржевський Л.О. 1959. Рибощш птахи Украши. Киев. 122 с.
Смогоржевський Л.О. 1979. Гагари, норщ, трубконос!, веслоног!, голшаст!, фламшго // Фауна Укра!-
ни. Т. 5. Птахи. Вип. 1. Кшв: Наук, думка. С. 1-188.
Смогоржевская Л.А. 1956. Гельминтофауна рыбоядных птиц долины Днепра // Тр. 2-й паразитол.
конф. М. С. 124-126.
Соколов А.Ю. 1999. Встречи редких видов птиц в Хреновском бору и на сопред. территориях // Редк.
виды птиц и ценные орнитол. терр. Центр. Черноземья. Липецк. С. 60-63.
Соколов А.Ю. 2005. О тенденциях изменения численности некоторых видов птиц в фауне Бобровс-
кого Прибитюжья // Стрепет. Т. 3. Вып. 1-2. С. 51-56.
Соколов А.Ю., Простаков Н.И. 1997. Новые данные о встречах редких видов птиц в центр, части
Прибитюжья // Сост. и пробл. экосист. Средн. Подонья. Воронеж. Вып. 10. С. 45-47.
Соколов П.Т. 1928. К вопросу о химической борьбе с саранчей И Изв. Сев.-Кавказск. краев, станции
защ. раст. Вып. 4. С. 122-126.
Соловьева Д.В., Дондуа А.Г. 2006. Поселения морских птиц в Чаунской губе Восточно-Сибирского
моря и их обусловленность геолого-геоморфологическими факторами // Биол. и охр. птиц Кам-
чатки. М.: ЦОДП. Вып. 7. С. 125-126.
570
Литература
Сомов Н.Н. 1897. Орнитологическая фауна Харьковской губернии. Харьков: Типогр. А. Даре. 680 с.
Сопыев О.С. 2001. Новый проект «Использование возвратных вод» и перспективы улучшения усло-
вий пролётов и зимовок водно-болотных птиц в Туркменистане И Актуальн. пробл. изуч. и охр.
птиц Вост. Европы и Сев. Азии. Казань. С. 568-569.
Сотников В.Н. 1997. Редкие птицы Кировской области // Фауна, экол. и охр. птиц Ср. Поволжья.
Саранск. С. 42-46.
Сотников В.Н. 1998. Редкие птицы Кировской области И Редк. виды птиц Нечерноз. центра России.
М. С. 309-316.
Сотников В.Н. 1999. Птицы Кировской области и сопредельных территорий. Т. 1: Неворобьиные.
Ч. 1. Киров. 432 с.
Сотников В.Н. 2000. Белый аист в Кировской области // Белый аист в России: дальше на восток.
Калуга. С. 112-114.
Сохина Э.Н., Чернобай В.Ф., Линьков А.Б. 2000. Сарпинские озёра и озеро Деед-Хулсун // Водно-
бол. угодья России. М. Т. 3. С. 198-204.
Спангенберг Е.П. 1941. Состав авифауны острова Харлова и прилегающего побережья в гнездовой
период 1932 г. // Тр. зап-ка «Семь островов». Вып. 1. С. 74-84.
Спангенберг Е.П. 1951. Отряды фламинго и голенастых // Т.П. Дементьев, Н.А. Гладков (ред.). Пти-
цы Советского Союза. М.: Сов. наука. Т. 2. С. 341-475.
Спангенберг Е.П. 1965. Птицы бассейна реки Имана И Иссл. по фауне Советск. Союза (птицы). Сб.
тр. Зоол. музея МГУ. Т. 9. М.: Изд. МГУ. С. 98-202.
Спангенберг Е.П., Фейгин ГА. 1936. Птицы нижней Сыр-Дарьи и прилегающих районов И Сб. тр.
Зоол. муз. при МГУ Т. 3. С. 41-184.
Спур И.В., Николаева И.В. 2004. Морские солоновато-водные и пресноводные рыбы залива Голуби-
ного // Дальневост, морск. биосф. зап-к., Биота. Владивосток. Т. 2. С. 669-672.
Спурис З.Д. 1962. Кольцевание птиц в Латвийской ССР в 1950-1960 гг. И Тр. ин-та Биол. АН ЛатвССР.
Рига. Вып. 22. С. 5-21.
Стадницкий И.М. 1992. Роль школы в охране и изучении белых аистов // Аисты: распространение,
экология, охрана. Минск. С. 138-141.
Стеллер ГВ. 1999. Описание земли Камчатки. Петропавловск-Камчатский: Камч. печ. двор. 287 с.
(Пер. с нем. изд. 1774 г.).
Степанян Л.С. 1975. Состав и распределение птиц фауны СССР. Неворобьиные Non-Passeriformes.
М.: Наука. 370 с.
Степанян Л.С. 1990. Конспект орнитологической фауны СССР. М.: Наука. 728 с.
Степанян Л.С. 2003. Конспект орнитологической фауны России и сопредельных территорий (в гра-
ницах СССР как исторической области). М.: ИКЦ «Академкнига». 808 с.
Стишов М.С. 1991. Распределение и типы населения гнездящихся морских птиц на о. Врангеля и
Геральд // Попул. и сообщ. животн. о. Врангеля. М.: ЦНИЛ Главохоты РСФСР. С. 143-160.
Стишов М.С., Придатко В.И., Баранюк В.В. 1991. Птицы острова Врангеля. Новосибирск: Наука. 254 с.
Стойловский В.П. 2000. Птицы озёра Кагул // Бранта. Сб. тр. Азово-Черном, орнитол. станции. Вып. 3.
С. 78-86.
Стоцкая Е.Э. 1983. О колониальное™ голенастых на примере серой цапли И Колониальное™ у птиц:
структура, функция, эволюция. Куйбышев. С. 104-121.
Стоцкая Е.Э. 1984. Родительское поведение и некоторые особенности гнездования серой цапли на
острове Фуругельма // Орнитология. Вып. 19. С. 128-143.
Стоцкая Е.Э, Кривенко В.Г. 1988. Редкие виды по материалам Всесоюзного учёта колониальных
гнездовий околоводных и морских птиц // Ресурсы редких животных РСФСР, их охрана и вос-
производство. М.: ЦНИЛ Главохоты РСФСР. С. 39-46.
Страутман Е.Н., Степанов Ю.В. 1977. Численность лебедей и пеликанов на основных водоёмах Ка-
захстана и мероприятия по их охране // Редк. и исчез, звери и птицы Казахст. Алма-Ата: «Наука»
Каз. ССР. С. 226-229.
Страутман Ф.И. 1954. Птицы Советских Карпат. Киев: изд-во АН УССР. 331 с.
Страутман Ф.И. 1963. Птицы западных областей Украины. Львов: изд-во Львовск. ун-та. Т. 1. 200 с.
Страшнюк Д.В. 2004. Кшыбсна ощнка зимово! орнпофауни очисних споруд Тернополя // Облж птах!в:
шдходи, методики, результата. Житомир. С. 85-87.
Литература
571
Строков В.В. 1974. Зимовки водоплавающих птиц черноморских берегов Кавказа // Орнитология.
Вып. 11. С. 274-278.
Суворов Е. 2004. Заповедные хроники. Документальная повесть. Владивосток: изд-во ДВГУ. 378 с.
Судиловская А.М. 1951. Отряд Веслоногие - Pelecaniformes // Г.П. Дементьев, Н.А. Гладков (ред.).
Птицы Советского Союза. Т. 1. М.: Сов. наука. С. 13-69.
Судиловская А.М. 1968. Зимняя авифауна Таримского бассейна и окружающих его горных хребтов //
Орнитология. Вып. 9. С. 289-294.
Судиловская А.М. 1974. О залетах фрегатов в Советский Союз // Орнитология. Вып. 11. С. 307-310.
Султанов Э. (ред.). 2000. Потенциальные Рамсарские угодья Азербайджана. Баку: Wetland Intemat.,
АЕМЕ. 152 с.
Султанов Э.Г., Джаббарова А.Ф. 2006. Зимнее распределение аистообразных и фламингообразных в
Азербайджане // Кавказск. орнитол. вести. Ставрополь. Вып. 18. С. 212-222.
Сурков М. 1969. Розовый фламинго на озере Тенгиз // Охота и охотн. хоз-во. № 7. С. 15.
Сурмач С.Г., Шибаев Ю.В. 2000. Дальневосточный аист в российской части бассейна р. Уссури (1999—
2000) // Дальневост, аист в России. Владивосток: Дальнаука. С. 61-76.
Суханова О.В. 2008. Малая выпь, или волчок // Красная книга Московской области. 2-е изд. М.: Т-во
научн. изданий КМК. С. 45.
Сушкин П.П. 1908. Птицы Средней Киргизской степи (Тургайская область и вост, часть Уральской) //
Мат-лы к познан, фауны и флоры Росс. имп. Отд. зоол. Вып. 8. С. 1-803.
Сушкин П.П. 1914. Птицы Минусинского края, Западного Саяна и Урянхайской земли // Мат-лы к
познан, фауны и флоры Росс. имп. Отд. зоол. Вып. 13. С. 1-551.
Сыроечковский Е.Е. 1959. Новые материалы по орнитофауне Ср. Сибири (бассейн Подкаменной
Тунгуски) /7 Уч. зап. Красноярск, гос. пед. ин-та. Т. 15. С. 225-239.
Сыроечковский Е.Е. 2005. Географическе распределение колониальных водных птиц Северного Кас-
пия в период снижения его уровня (1950-е годы) // Долговрем. мониторинги сохран. колон, вод-
ных птиц Сев. Каспия в связи с колебанием уровня Каспийского моря. М.: Астрель. С. 17-186.
Сыроечковский Е.Е., Рогачева Э.В. 1995. Красная книга Красноярского края. Красноярск. 408 с.
Тарасов В.В. 2004. Залёт колпицы на Средний Урал // Мат-лы к распростр. птиц на Урале. Екатерин-
бург. С. 150.
Тарасов В.В., Поляков В.Е. 2006. К биологии большой белой и серой цапель в лесостепном Заура-
лье // Орнитол. иссл. в Сев. Евразии. Ставрополь. С. 509-510.
Тарасов В.В., Примак И.В. 2005. Распространение и численность кудрявого пеликана в лесостепной
части Тоболо-Ишимского междуречья // Инвент., мониторинг и охрана Ключевых орнитол. терр
России. М.: Союз охраны птиц России. Вып. 5. С. 167-174.
Тарасов В.В., Рябицев В.К., Примак И.В. и др. 2003.Орнитофауна северо-восточных районов Кур-
ганской области // Мат-лы к распр. птиц на Урале. Екатеринбург. С. 193-213.
Татарникова И.П. 2001а. Гнездование бакланов на Айновых островах // Биол. осн. устойч. развития
прибр. морск. экосистем. Апатиты: Кольск. НЦ РАН. С. 224-225.
Татарникова И.П. 20016. Хохлатый баклан Phalacrocorax aristotelis (Linnaeus, 1761) // Красная книга
РФ (Животные). М.: изд-во Астрель. С. 375-376.
Татарникова И.П., Карпович В.Н., Краснов Ю.В., Шкляревич Ф.Н. 1983. Результаты кольцевания и
территориальные связи птиц на Мурмане // Сообщ. Прибалт, ком. по изуч. мигр. птиц АН ЭССР.
Тарту. № 16. С. 38-54.
Тауринып Э. 1983а. Большая белая цапля // Птицы Латвии. Территориальное размещение и числен-
ность. Рига: Зинатне. С. 24.
Тауринып Э. 19836. Большой баклан Phalacrocorax carbo (L.) // Там же. С. 21-22.
Ташлиев А.О. 1973. Орнитологические комплексы юго-восточной Туркмении. Ашхабад: Ылым. 269 с.
Тильба П.А. 1984. Краткие сообщения о Чёрном аисте в Краснодарском крае // Исследования в обла-
сти заповедного дела. М.: ВНИИ охраны природы и заповедного дела.
Тильба П.А. 1999. Авифауна Имеретинской низменности. Сообщ. 1. Неворобьиные // Кавказск. ор-
нитол. вести. Ставрополь. Вып. 11. С. 166-204.
Тиунов И.М. 2005. Водно-болотные и морские птицы Северного Сахалина и материкового побере-
жья пролива Невельского. Автореф. дис. ... канд. биол. н. Владивосток: БПИ ДВО РАН. 22 с.
572
Литература
Тищенков А.А. 2004. Редкие виды птиц Приднестровья в заповеднике «Ягорлык» // Managementul
integral al resulselor naturale din bazinul transfrontalier al fluviululi Nistru. Кишинев. C. 319-323.
Ткаченко A.A. 1987. Колониальные голенастые птицы Лесостепи Левобережной Украины и их хо-
зяйственное значение. Автореф. дис. ... канд. биол. н. Харьков. 20 с.
Ткаченко О.О. 1994. Особливосп росту та розвитку пташенят малоп бигоп, cipon чапель та квака //
Беркут. Т. 3. Вип. 2. С. 96-99.
Томкович П.С., Сорокин А.Г. 1983. Фауна птиц Восточной Чукотки // Распростр. и системат. птиц.
М.: Изд-во МГУ. С. 77-159.
Третьяков Г.П. 1974. Зимовки белого аиста в Ферганской долине // Мат-лы 6-й Всесоюзн. орнитол.
конф. М.: Изд-во МГУ. Ч. 2. С. 145-146.
Третьяков Г.П. 1990. Белый аист в Ферганской долине // Редк. и малоизуч. птицы Ср. Азии. Бухара.
С. 74-77.
Трухин А.М., Косыгин Г.М. 1987. Распределение морских птиц во льдах западной части Берингова и
Чукотского морей // Распростр. и биол. морск. птиц Дальнего Востока. Владивосток: ДВО АН
СССР. С. 6-21.
Трухин А.М., Кузин А.Е. 1996. Многолетняя динамика видового состава и численности морских
птиц, гнездящихся на о. Тюлений (Охотское море) // Птицы пресн. вод и морск. побер. юга Дальн.
Востока России и их охрана. Владивосток: Дальнаука. С. 214-221.
Туаев Д.Г. 1962. Современное состояние водоёмов Прикаспийской низменности северной части Азер-
байджана и их роль в жизни водоплавающей и болотной птицы // Мат-лы 3-й Всесоюзн. орнитол.
конф. Львов: изд-во Львовск. ун-та. Кн. 2. С. 215-216.
Туаев Д.Г. 1975. Материалы по фауне и экологии водоплавающих и болотных птиц Кура-Араксинс-
кой низменности в современных условиях // Мат-лы по фауне и экол. наземн. позвон. Азербайд-
жана. Баку. С. 151-188.
Туаев Д.Г., Васильев В.И. 1969. Колониально гнездящиеся птицы водоёмов Азербайджана // Орнито-
логия в СССР. Ашхабад. Кн. 2. С. 649-652.
Туаев Д.Г., Васильев В.И. 1971. Питание, пищевые отношения и хозяйственное значение некоторых
птиц отряда голенастых в Азербайджане // Изв. АН АзССР, сер. биол. н. № 2. С. 86-93.
Туаев Д.Г., Исрафилов С.А. 1977. К биологии европейской колпицы в Азербайджане // Тез. докл. 7-й
Всесоюзн. орнитол. конф. Ч. 1. Киев: Наук, думка. С. 106.
Туаев Д.Г., Исрафилов С.А. 1978. Биология гнездовой жизни Platalea leucorodia leucorodia и Plegadis
falcinellusfalcinellus в Кура-Араксинской низменности // Изв. АН АзССР. Сер. биол. н. № 6. С. 70-
75.
Тугаринов А.Я. 1947. Веслоногие, аистообразные, фламинго // Фауна СССР. Еов. серия, № 33. Пти-
цы. Т. 1. Вып. 3. М.-Л.: изд-во АН СССР. С. 125-317.
Тугаринов А.Я. 1951. Отряд Ciconiiformes Аистообразные // Птицы СССР. Ч. 1. М.-Л. С. 66-88.
Туров С.С. 1918. К орнитофауне Рязанской губернии (1913-1915 гг.) // Ежегодн. Зоол. муз. АН. Пг.
Т. 22 (1918-1922). С. 64-78.
Туров С.С. 1925. Некоторые новые данные о фауне птиц Рязанской губернии // Тр. о-ва исслед. Ря-
занск. края. Рязань. С. 65-73.
Умрихина Г.С. 1970. Птицы Чуйской долины. Фрунзе: Илим, 133 с.
Уорд Д.Х., Шмутц Д.А. 1993. Двуххохлый баклан (Phalacrocorax auritus) в заливе Креста // Морск.
птицы Берингии. Вып. 1. Магадан ИБПС ДВО РАН; Анкоридж, Упр. дикой прир. США. С. 50.
Успенский С.М. 1959. Морские колониально гнездящиеся птицы северных и дальневосточных мо-
рей СССР, их размещение, численность и роль как потребителей планктона и бентоса // Бюл.
МОИП. Отд. биол. Т. 64. Вып. 2. С. 39-52.
Успенский С.М., Шапошников Л.К., Залетаев В.С. и др. 1959. Первые результаты исследования зи-
мовки водоплавающих птиц на Азовском море и северном побережье Чёрного моря // Мигр.
животн. М. Вып. 1. С. 48-58.
Утехина И.Г. 1999. Заметки о пролёте птиц на о. Талан весной 1998 г. // Морск. птицы Беринги.
Вып. 4. Магадан: ИБПС ДВО РАН. С. 16-19.
Утехина И.Г. 2006. К распределению колоний морских птиц на северном побережье Охотского моря //
Биология и охрана птиц Камчатки. М.: ЦОДП. Вып. 7. С. 95-99.
Литература
573
Ушакова М.В. 2003. К авифауне Малой Курильской гряды // Там же. Вып. 5. С. 105-109.
Ушков С.Л. 1927. Список птиц Пермского округа Уральской области // Бюл. МОИП. Нов. сер. Отд.
биол. Т. 36. Вып. 1/2. С. 68-116.
Федосов А.В. 1959. Распространение белого аиста в Брянской области // Орнитология. Вып. 2. С. 147—
149.
Федюшин А.В., Долбик М.С. 1967. Птицы Белоруссии. Минск. 520 с.
Федянина Т.Ф. 1990. Данные о численности некоторых редких водно-болотных птиц Чуйской доли-
ны // Редк. и малоизуч. птицы Ср. Азии. Ташкент: Фан. С. 82-83.
Фесенко Г.В. 1984. Гнездование малой белой цапли (Egretta garzetta L.) в Киевской области // Вести,
зоол. № 2. С. 88.
Фесенко Г.В. 2001. К вопросу о разделении птиц на ближних и дальних мигрантов // Актуальн. пробл.
изуч. и охр. птиц Вост. Европы и Сев. Азии. Казань. С. 602-603.
Фетисов С.А. 1998. Залёт большой белой цапли Egretta alba в Псковскую область // Русск. орнитол.
ж., Экспр.-вып. № 31. С. 8-10.
Фетисов С.А., Иванов И.Д. 2003. Залёт большой белой цапли Egretta alba в Себежское Поозерье //
Русск. орнитол. ж., Экспр.-вып. № 241. С. 1218-1219.
Фетисов С.А., Ильинский И.В., Пинигина Т.В. 1986. Материалы к биологии размножения белого
аиста (в Себежском р-не Псковской обл.) // Экол. и размнож. птиц. Л. С. 89-95.
Фетисов С.А., Федоров Ю.В.. Сагитов Р.А. 2000. Результаты пятого международного учёта белого
аиста в Новгородской области И Белый аист в России: дальше на восток. Калуга. С. 78-94.
Фетисов С.А., Черевичко В.И. 2000. Экспансия вида и географические различия в экологии белого
аиста в Псковской области Там же. С. 68-77.
Фефелов И.В., Тупицын В.А.. Подковыров В.А., Журавлев В.Е. 2001. Птицы дельты Селенги. Ир-
кутск. 320 с.
Филатов В.А. 1915. Птицы Калужской губернии // Мат-лы к познан, фауны и флоры Росс. Имп. Отд.
Зоол. М. Вып. 14. С. 194-379.
Фирсова Л.В., Михтарьянц Э.А., Андреев А.В. 1986. Фаунистические заметки о некоторых птицах
Командорских островов // Распростр. и биол. птиц Алтая и Дальнего Востока. Л.: Зоол. ин-т АН
СССР. С. 89-96.
Флинт В.Е. 1984. Чёрный аист // Красная книга СССР. Редкие и находящиеся под угрозой исчезнове-
ния виды животных и растений. 2-е изд. М.: Лесная промышленность. Т. 1. С. 109.
Флинт В.Е., Стоцкая Е.Э, Ардамацкая Т.Б., Виксне А.Я. и др. 1991. Современное состояние числен-
ности околоводных птиц в СССР // Мат-лы 10-й Всесоюзн. орнитол. конф. Минск.Ч. 2. Кн. 2.
С. 269-270.
Фомин В.Е., Болд А. 1991. Каталог птиц Монгольской Народной Республики. М.: Наука. 125 с.
Фролов В.В., Коркина С.А., Фролов А.В. и др. 2001. Анализ состояния фауны неворобьиных птиц
юго-лесостепн. зоны Правобер. Поволжья в XX в. // Беркут. Т. 10. Вип. 2. С. 156-183.
Фролов В.В., Муравьев И.В., Коркина С.А., Фролов А.В. 2000. Белый аист в Пензенской области //
Белый аист в России: дальше на восток. Калуга. С. 181.
Фудзимаки Ю. 1986. Колонии морских птиц на о. Хоккайдо // Морск. птицы Дальн. Востока. Влади-
восток: ДВНЦ АН СССР. С. 82-87.
Фурманова В.П. 1998. Морфо-экологическая характеристика цапель Сев. Приазовья // Мат-ли 3-й
конф, молодих орн1т. Украши. Чершвщ. С. 152-155.
Харашти Л. 1978. Миграции колпиц, гнездящихся в Венгрии // Мигр. птиц Вост. Европы и Сев.
Азии. Гагарообр. - Аистообр. М. С. 232-236.
Харитонов С.П. 1980. Материалы по птицам о. Ионы // Орнитология. Вып. 15. С. 10-15.
Харкевич С.С., Вяткин П.С. 1977. Остров Верхотурова в Беринговом море // Природа. № 4. С. 84-92.
Харченко Л.П., Зноменко С.К., Логвиненко Т.Л. 1986. Экология малой белой цапли в смешанной
колонии голенастых в гнездовой период // Экол. и размн. птиц. Л. С. 84-89.
Хоберг Э.П. 1992. Экология гельминтов морских птиц острова Талан (предварит, обзор) // Прибрежн.
экосист. Сев. Охотоморья. О. Талан. Магадан: ИБПС ДВО РАН. С. 43-59.
Холодковский Н.А., Силантьев А.А. 1901. Птицы Европы. 4.2. Специальная орнитология. СПб.:
изд-во А.А. Девриена. 636 с.
574
Литература
Хомяков М.М. 1900. Птицы Рязанской губернии // Мат-лы к познан, фауны и флоры Росс. Имп. М.
С.1-102.
Хонякина З.П. 1978. Материалы по питанию птиц Дагестана // Иссл. по зоол. и паразитол. Дагестана.
Махачкала. С. 166-179.
Хохлов А.Н. 1987. Каравайка в Ставропольском крае // Пробл. охр. редк. животн. Сб. научн. тр.
ЦНИЛ Главохоты РСФСР. М. С. 129-131.
Хохлов А.Н. 1988а. Аисты на Ставрополье в период сезонных перемещений и гнездования // Сезон,
перемещ. и структ. попул. наземн. позв. животн. М.: МГПИ им В.И. Ленина. С. 33-37.
Хохлов А.Н. 19886. Фламинго в Предкавказье // Ресурсы редк. животн. в РСФСР, их охрана и воспро-
изв. М. С. 65-67.
Хохлов А.Н. 1990в. Колпица в Ставропольском крае // Итоги изуч. редких животн. Мат-лы к Кр.
книге. Тр. ЦНИЛ Главохоты РСФСР. М. С. 75.
Хохлов А.Н. 1990г. Малый баклан // Там же. С. 74.
Хохлов А.Н. 1993. Животный мир Ставрополья. Ставрополь. 165 с.
Хохлов А.Н., Бичеров А.П. 1986а. Колпица в Ставрополье // Природа. № 11. С. 94-95.
Хохлов А.Н., Бичеров А.П. 19866. Новое место гнездования и зимовки большого баклана в СССР //
Орнитология. Вып. 21. С. 143.
Хохлов А.Н., Заболотный Н.Л., Ильюх М.П. 1997. Кладки и размеры яиц птиц низовий Кубани. Став-
рополь. 30 с.
Хохлов А.Н., Ильюх М.П. 2002. Розовый пеликан (Pelecanus onocrotalus Linnaeus, 1758) // Красная
книга Ставропольск. края. Т. 2. Животные. Ставрополь. С. 113.
Хохлов А.Н., Ильюх М.П., Казиев У.З. 2005. Редкие позвоночные животные Ставропольского края.
Ставрополь. 216 с.
Хохряков С.А., Шохрин В.П. 2002. Земноводные, пресмыкающиеся, птицы и млекопитающие Ла-
зовского заповедника (аннотированные списки видов). Лазо. 60 с.
Храбрый В.М. 2000. Белый аист в Ленинградской области // Белый аист в России: дальше на восток.
Калуга. С. 66-67.
Храневич В.П. 1926. Огляд фавни птах!в Подыля // Нарис фавни Под шля. Вшниця. С. 34-90.
Хроков В.В. 1991. Колпица на оз. Сорбулак // Редк. птицы и звери Казахст. Алма-Ата. С. 46.
Хроков В.В., Бекбаев Е.З. 2002. Летняя фауна птиц Шошкакольской системы озёр (Ю. Казахстан) //
Русск. орнитол. ж., Экспр.-вып. № 194. С. 747-752.
Хроков В.В., Гаврилов Э.И. 1991. Колпица на оз. Караколь // Редк. птицы и звери Казахст. Алма-Ата:
Гылым. С. 45-46.
Хрустов А.В., Подольский А.Л., Завьялов Е.В. и др. 1995. Редкие и исчезающие птицы Саратовской
области // Русск. орнитол. ж. Т. 4. Вып. 3/4. С. 125-142.
Цвелих О. 2002. Херсонесько-Фюлентсысе набережжя // Нац. доповщь про стан IBA територш Ук-
раши. 2002. Кшв. С. 34.
Чекменев Д.И. 1962. Размещение и численность колониально гнездящихся птиц на оз. Тенгиз (Центр.
Казахстан) // Мат-лы 3-й Всесоюзн. орнитол. конф. Ч. 2. Львов. С. 229-230.
Чекменев Д.И. 1964а. Очерки по биологии колониально гнездящихся видов птиц оз. Тенгиз // Тр. Ин-
та зоол. АН КазССР. С. 65-82.
Чекменев Д.И. 19646. Очерки по колониально гнездящимся видам птиц оз. Тенгиз (Центр. Казахст.) //
Мат-лы 3-й Всесоюзн. орнитол. конф Львов.. Ч. 2. С. 229-230.
Чельцов-Бебутов А.М. 1958. Новое гнездовье фламинго в Советском Союзе // Уч. зап. МГУ. Вып. 197.
Орнитология. С. 95-101.
Чельцов-Бебутов А.М. 1982. Экология птиц. М.: изд-во МГУ. 128 с.
Червона книга Украши: Тваринний св!т. 1994. Кшв: Укр. Енциклопед1я. 464 с.
Черкащенко M.I. 1963. Еколопчна характеристика гшздових водо плавких, лучних та болотних птах!в
долини верхнього Дшстра // Сичасна та минула фауна зах. обл-й Украпни. Кипв. С. 47-63.
Черничко И.И., Гринченко А.Б., Дядичева Е.А. 1993. Тарханкутское побережье // Инвент. и кадастр,
характ. водно-бол. угодий юга Украины. Бюл. № 1. Бранта. С. 66-68.
Черничко И.И., Черная А.Н., Чёрный С.А. 1991. Гнездование каравайки в верховьях Тилигульского
лимана // Редк. птицы Причерноморья. Киев-Одесса: Лыбидь. С. 113-128.
Литература
575
Чернобай В.Ф. 2000а. Белый аист в Волгоградской оласти // Белый аист: дальше на восток. Калуга.
С.183.
Чернобай В.Ф. 20006. Степновский угол // Ключевые орнитол. терр. России. М.: Союз охраны птиц
России. Т. 1. С. 490.
Чернобай В.Ф. 2004. Птицы Волгоградской области. Волгоград: Перемена. 287 с.
Чернобай В.Ф., Букреев С.А., Сохина Э.Н. 2000 Сарпинские озёра // Ключевые орнитол. терр. Рос-
сии. М.: Союз охраны птиц России. Т. 1. С. 485-486.
Чернов В.Ю. 1990. Редкие околоводные птицы озёра Сарыкамыш // Редк. и малоизуч. птицы Сред-
ней Азии. Ташкент: Фан. С. 46-49.
Чернявская С. 1961. Колпица // Охота и охотн. хоз-во. № 8.
Численность и размещение гнездящихся околоводных птиц в водно-болотных угодьях Азово-Черно-
морского побережья Украины, 2000 /Колл, авторов под общ. ред. В.Д. Сиохина. Мелитополь-
Киев: Бранта. 476 с.
Чорненька О.Б. 1998. Гшздування чапель у зм!шанш колони бшя с. Отиневич! на Льв1вщиш // Мат-
ли 2-й конф, молодих opHir. Украши. Чершвщ. С. 156-158.
Чуйков Ю.С., Головкин А.Н. 1979. Влияние поселений больших бакланов на гидрохимический ре-
жим и зоопланктон водоёмов дельты Волги // Тр. Кызыл-Агачск. зап-ка. Баку. Вып. 1. С. 220-231.
Чхикишвили И.Д. 1930. Материалы по орнитофауне Кахетии (Телавский уезд) // Закавказск. крае-
ведч. сб., сер. А. Естествознание. Тифлис. Т. 1. С. 105-128.
Чырвоная KHira Рэспублш Беларусь. 1993. Мшск: Бел. энц. 560 с.
Шакула В.Ф., Ткаченко А.А. 1986. Сроки прилёта и синхронизация процессов размножения в сме-
шанной колонии цапель // Изуч. птиц СССР, их охр. и рац. использ. Л. Ч. 2. С. 325-326.
Шалаков А. 1958. Судьба рыжей цапли // Охота и охотн. хоз-во. № 8. С. 61.
Шапошников Л.. Головин О., Сорокин М., Тараканов А. 1959. Животный мир Калининской области.
Калинин.
Шастовский П.А.. Селевин В.А. 1928. Залёт большой белой цапли в окрестности Томска // Uragus.
Т. 3. Вып. 3-4. С. 24
Шахназаров Э. 1995. Распространение и численность большой белой цапли в Узбекистане // Орнито-
логия. Вып. 26. С. 220-222.
Шевченко В.Л., Дебело П.В., Гаврилов Э.И. и др. 1993. Об орнитофауне Волжско-Уральского между-
речья И Фауна и биол. птиц Казахст. Алматы. С. 7-103.
Шевцов А.О., Санжаровський Ю.О.. Copiin Р.В., Сфремов В.Л. 2004. Нов! рщкгсш та малочисельш
птахи Юровоградсько! облает! // Беркут. Т. 13. Вип. 1. С. 13-17.
Шерназаров Э. 1988. Новое место гнездования кудрявых пеликанов в Узбекистане // Орнитология.
Вып. 23. С. 225.
Шерназаров Э. 1991. К биологии колпиц на оз. Айдаркуль // Орнитология. Вып. 25. С. 212-213.
Шерназаров Э.Ш. 2003. Колпица // Красная кн. Респ. Узбекистан. Тошкент. Т. 2. С. 154-155.
Шерназаров Э., Тураев М.М., Кучкаров А.Х. 1992. Некоторые моменты изменения ареала и числен-
ности белого аиста в Узбекистане // Аисты: распростр., экол., охрана. Минск. С. 174-175.
Шестоперов Е.Л. 1936. Фаунистические обследования окрестностей Тенджена // Бюл. Туркменск.
зоол. станции. № 1. С. 173-190.
Шибаев Ю.В. 1987. Кадастр колоний и мониторинг некоторых видов птиц зал. Петра Великого (Япон-
ское море) // Распростр. и биол. морск. птиц Дальн. Востока. Владивосток: ДВО АН СССР. С. 43-
59.
Шибаев Ю.В., Литвиненко Н.М. 1996. Современное состояние и проблемы охраны морских колони-
альных птиц о. Монерон (Юж. Сахалин) // Птицы пресн. вод и морск. побер. юга Дальн. Востока
России и их охрана. Владивосток: Дальнаука. С. 93-102.
Шибаев Ю.В., Литвиненко Н.М. 2007. Низовья реки Туманган - водно-болотное угодье международ-
ного значения. Ещё раз о проблеме сохранения // Проблемы устойчивого природопользования в
нижнем течении реки Туманная. Матер, межд. конф. Владивосток. С. 90-98.
Шибаев Ю.В., Семенченко Н.Г., Лимин В.А. 1976. Гнездовья белого аиста на правобережье Уссури ,/
Охр. прир. на Дальн. Востоке. Владивосток. С. 190-191.
Шибнев Ю.Б. 1992. Некоторые обобщения наблюдений и новые материалы по птицам заповедника
«Кедровая падь» // Совр. сост. флоры и фауны зап-ка «Кедровая падь». Владивосток: ДВО РАН.
С. 145-163.
576
Литература
Шкаран В.И. 1990. Динамика численности и успешность размножения белого аиста в селе Датынь
Волынской области в 1978-89 гг. // Аисты: распр., экол, охр. Минск. С. 169-170.
Шкляревич Ф.И., Татарникова И.П. 1986. Численность и некоторые стороны экологии хохлатого
баклана на Мурмане // Редкие, исчезающие и малоизуч. птицы СССР. М. С. 4-8.
Шкуратова Т.И. 1981. Современное состояние колониальных околоводных птиц в СССР и за грани-
цей // Научн. осн. обслед. колон, гнездовий околоводн. птиц. М.: Наука. С. 3-13.
Шляхтин Г.В., Беляченко А.В., Каширская Е.В., Завьялов Е.В. 1994. Генезис и пространственно-
временная структура экотонов верхней зоны Волгоградского водохранилища // Биология, эколо-
гия, биотехнология и почвоведение. М.: МГУ. С. 76-91.
Шнитников В.Н. 1913. Птицы Минской губернии // Мат-лы к познан, фауйы и флоры Росс. Имп. Отд.
зоол. Вып. 12. С. 1-475.
Шнитников В.Н. 1949. Птицы Семиречья. М.-Л. 665 с.
Шнитников В.Н. 1957. Звери и птицы нашей страны. М.: Молодая гвардия. 254 с.
Шокало С.И., Шокало Б.И. 1992. Сроки прилёта и отлета белого аиста в ЮЗ Беларуси // Аисты:
распр., экол., охр. Минск. С. 162-164.
Шохрин В.П. 2001. Редкие птицы Лазовского зап-ка и сопред. территорий: современное состояние //
Актуальные пробл. изуч. птиц Вост. Европы и Сев. Азии. Мат-лы 11-й Всесоюзн. орнитол. конф.
Казань. С. 651-653.
Шохрин В.П. 2007. Дополнения к орнитофауне Лазовского заповедника // 8-я Дальневост. конф, по
заповедному делу. Благовещенск. Т. 2. С. 85-89.
Штыркало Я.Е. 1990. Заметки по экологии белого аиста // Аисты: распр., экол., охр. Минск. С. 174—
175.
Шукуров Э.Д. 1981. Птицы Киргизии. Фрунзе: Мектеп. Ч. 1.
Шульпин Л.М. 1936. Промысловые, охотничьи и хищные птицы Приморья. Владивосток: ДВФ АН
СССР. 436 с.
Шульпин Л.М. 1952. Отряд Аистообразные Ciconiiformes // Атлас охот.-пром. птиц и зверей СССР.
М. Т. 1. Птицы. С. 35-46.
Шунтов В.П. 1972. Морские птицы и биологическая структура океана. Владивосток. 376 с.
Шунтов В.П. 1986. Состояние изученности многолетних циклических изменений численности рыб
дальневосточных морей // Биол. моря. № 3. С. 3-14.
Шунтов В.П. 1993. Трудная профессия альбатроса. М.: Наука. 224 с.
Шунтов В.П. 1998. Птицы дальневосточных морей России. Т. 1. Владивосток: ТИНРО. 423 с.
Щеголев И.В. 1975. Поведенческие особенности каравайки в дельте Днестра // Колон, гнездовья
околоводн. птиц и их охрана. М. С. 26.
Щеголев И.В. 1977. Динамика популяции каравайки и других аистообразных птиц в дельте Днестра //
Тез. докл. 7-й Всесоюзн. орнитол. конф. Киев. Ч. ЕС. 346.
Щеголев И.В. 1992. Результаты кольцевания каравайки в дельте Днестра // Сезон, мигр. птиц на терр.
Украины. Киев. С. 279-282.
Щербак Н.Н. 1966. Земноводные и пресмыкающиеся Крыма. Киев. 239 с.
Эверсман Э.А. 1866. Естественная история птиц Оренбургского края. Казань. Ч. 3. 622 с.
Эминов А. 1999. Колпица // Красная книга Туркменистана. Ашгабат. Т. ЕС. 212-213.
Эминов А. 2002. Новая гнездовая колония цапель в предгорьях Центрального Копетдага // Вопр.
орнитол. Туркменистана. М.: Союз охраны птиц России. С. 77-79.
Эминов А., Шамурадов А., Сапармурадов Д. 1984. Новое место гнездования цапель в предгорьях
Копетдага // Миграции птиц в Азии. Ташкент. С. 134-137.
Юзефик М. 1970. Изучение жёлтой цапли. Внутривидовая изоляция и экологическая зона ареала //
Acta omithol. Vol. 12. No. 11. С. 445-504.
Юрлов A.K. 2000. Цапля большая белая // Красная книга Новосибирской области Новосибирск. С. 53-
54.
Юрлов К.Т., Чернышов В.М., Кошелев А.И. и др, 1977. Новые и редкие птицы в районе озёра Чаны
(Зап. Сибирь) // Мигр. птиц в Азии. Новосибирск: Наука. С. 205-209.
Юсси Ф., Рандла Т. 1970. О некоторых редких видах птиц в Эстонии // Мат-лы 7-й Прибалтийск.
орнитол. конф. Рига. Вып. 3.
Литература
577
Языкова И.М. 1970а. Питание рыбоядных птиц на оз. Маныч-Гудило // Мат-лы 4-й научн. конф,
зоологов пед. ин-тов. Горький. С. 403-404.
Языкова И.М. 19706. Рыбоядные птицы манычских водохранилищ и их хоз. Значение. Автореф. дис.
... канд. биол. н. Ростов-на-Дону. 31 с.
Языкова И.М., Казаков Б.А. 1975. Пеликаны и голенастые Пролетарского водохранилища // Колон,
гнездовья околоводн. птиц и их охрана. М.. С. 164-165.
Якименко В.В. 1995. Регистрация залетов птиц на территории Омской области // Мат-лы к распростр.
птиц на Урале. Екатеринбург. С. 79-80.
Якименко В.В. 1998. Материалы к распространению птиц в Омской области // Мат-лы к распростр.
птиц на Урале. Екатеринбург. С. 192-196.
Якименко В.В. 2006. Омская область И Ключевые орнитол. терр. России. М. Т. 2. Ключевые орнитол.
терр. межд. значения в Зап. Сибири. С. 161-174.
Янушевич А.И. 1952. Фауна позвоночных Тувинской области. Новосибирск. 142 с.
Янушевич А.И., Тюрин П.С., Яковлев И.Д. и др. 1959. Птицы Киргизии. Фрунзе: АН Кирг.ССР. Т. 1.
229 с.
Яремченко О.А., Руденко А.Г. 1997. О возобновлении гнездования розового пеликана в Черноморс-
ком заповеднике // Заповтд. справа в Украпи. Т. 3. № 2. С. 56-57.
Яремченко О.А., Руденко А.Г. 2004. Характеристика современных гнездовых поселений голенастых
птиц в Черноморском биосферном заповеднике // Бранта. Сб. тр. Азово-Черноморск. орнитол.
станции. Вып. 7. С. 53-60.
Яремченко О.А., Руденко А.Г. 2005. Динамика численности и распределение скоплений розового
пеликана (Pelecanus onocrotalus) на Тендровском и Ягорлыцком лиманах Черноморского запо-
ведника // Там же. Вып. 8. С. 16-26.
Яхонтов В.Д. 1966. Ещё один фрегат // Природа. № 7. С. 67.
Яхонтов В.Д. 1973. Орнитологический комплекс Малого Хингана // Вопр. геогр. Дальн. Востока.
Хабаровск. № 11. С. 225-245.
Яхонтов В.Д. 1974. Птичьи базары Пенжинской губы и их состояние // Мат-лы 6 Всес. орнитол.
конф., ч.1, М.: Изд-во МГУ. С. 251-252.
Яхонтов В.Д. 1975а. Остров Нансикан // Колон, гнездовья околоводн. птиц и их охрана. М.: Наука.
С. 186-187.
Яхонтов В.Д. 19756. Птичьи базары Пенжинской губы и их состояние // Там же. С. 185-186.
Adamian M.S., Klem D.Jr. 1999. Handbook of the Birds of Armenia. California (USA). 649 p.
Adamo M.C., Puglisi L.. Baldaccini N.E. 2004. Factors affecting Bittern Botaurus stellaris distribution in a
Mediterranian wetland // Bird Conserv. Intemat. Vol. 14. P. 153-164.
Aebischer N.J. 1993. Immediate and delayed effects of a gale in late spring on the breeding of the Shag
Phalacracorax aristotelis 11 Ibis. Vol. 135. No. 3. P. 225-232.
A field guide to the birds of Japan. 1982. / Tokyo: Wild birds society Japan. 336 p.
A Hand-list of the Japanese birds. 1932. Tokyo. 211 p. (1942 - 238 p. 1958 - 254 p.).
Alessandria G., Carpegna F., Della Toffola M. 2003. Vocalisation and courtship displays of the Bittern
Botaurus stellaris 11 Bird study. Vol. 50. P. 182-184.
Alexander W.B. 1963. Birds of the Ocean. N.Y. 306 p.
Ali S., Ripley S.D. 1968. Handbook of the Birds of India and Pakistan. Vol. 1. Bombay - London - N.Y. 380
P-
American Ornithologists’ Union, 1998. Check-list of North American birds. 7th ed. Washington, D.C.: Amer.
Omithol. Union. 829 p.
Anderegg K. 2000. Weissstorch-Paar erbriitete zweites Gelege // Om. Beob. Bd. 97. H. 2. S. 153-155.
Andronov V 1994. The status and preservation of Oriental White Storks in Russia 1994. Toyoka. Jap.
Andronov V, Darman Y. 1998. The Oriental White Stork action // The Sixth northeast Asia and north
Pacific environmental broom workshop. Abstr. of artics. Peking. P. 4-6.
Antczak M., Konwerski S., Grobelny S., Tryjanowski P. 2002. The food composition of immature and non-
breeding White Storks in Poland // Waterbirds. Vol. 25. Is. 4. P. 424-428.
Antipow J., Domaszewicz A. 1997. L$gi dziko zyjacych bocianow bialych Ciconia ciconia w gniazdach
umiejscowionych na ziemi (Europa Srodkowa) // Chrohmy Przyr. Ojcz. R. 53. Z. 5. P. 111-118.
578
Литература
Archibald К. 1985. Captive management of the Eastern White Stork Ciconia boyciana 11 Avic. Mag. Vol. 91.
No. 1/2. P. 92-101.
Archibald К. 1991. Recommendations for captive management of the Oriental White Stork (Ciconia boyciana)
11 M.C. Coulter et al. (eds.). Biol, and Conserv. of the Oriental White Stork Ciconia boyciana. P. 193-
203.
Archibald K., Schmitt B. 1991. Comparison between the Oriental White Stork Ciconia c. boyciana and the
European White Stork Ciconia c. ciconia 11 Ibid. P. 3-17.
A Red Data Book of Japanese Birds. 1975. 285 p.
Arengo Felicity. 1995. American flamingos and ecotourism on the Yucatan Peninsula, Mexico // Integr.
People and Wildlio Sustainable Fut. P. 207-210.
Artyukhin Y.B., Burkanov VN. 2000. Incidental mortality of seabirds in the driftnet salmon fishery by
Japanese vessels in the Russian exclusive economic zone, 1993-1997 // Seabirds of the Russian Far
East. Ottawa: Can. Wildl. Serv. Soec. Publ. P. 105-115.
Ash J.S. 1989. The status of the White Stork (Ciconia ciconia) in Ethiopia, Somalia and Djibouti // WeiBstorch
- White Stork. Proc. I Int. Stork Conserv. Symp. Schriftenr. des DDA. Bd. 10. P. 341-353.
Austin O.L. 1948. The birds of Korea // Bull. Mus. Comp. Zool., Harvard Univ. Vol. 101. No. 1. 301 p.
Austin O.L., Jr., Kuroda N. 1953. The birds of Japan, their status and distribution // Bull. Mus. Comp. Zool.,
Harvard Univ. Vol. 109. No. 4. P. 279-637.
Bairlein F. 1992. Zugwege, Winterquartiere und Sommerverbreitung mittel europaischer WeiBstorche // Les
cigogn. d’Europe. Metz. S. 191-205.
Bairlein F., Zink G. 1979. Der Bestand des WeiBstorches Ciconia ciconia in Siidwestdeutschland: eine
Analyse der Bestandsentwicklung // J. Om. Bd. 120. H. 1. S. 1-11.
Baker K. 1993. Identification Guide to European Non-Passerines. BTO, Guide 24. Thetford.
Baker S.E.C. 1929. The fauna of British India, including Ceylon and Burma. Birds. London. Vol. 4. 499 p.
Bankovics A. 1997. Ardea purpurea', Purple Heron // Hagemaijer & Blair (eds.). The EBCC Atlas of European
breeding Birds: Their Distribution and Abundance. London: T. & A.D. Poiser. P. 52-53.
Barbraud C., Barbraud J.C. 1998. La regime alimentaire des poussins de Cigogne blanche Ciconia ciconia,
en Charente-Maritime: importance des insectes// Alauda. Vol. 65. P. 259-262.
Barbraud C., Barbraud J.-C., Barbraud M. 1999. Population dynamics of the White Stork Ciconia ciconia in
western France // Ibis. Vol. 141. Is. 3. P. 469-479.
Barret R.T., Rorv N., Loen J., Montevecchi W.A. 1990. Diets of shags Phalacrocorax aristotelis and
cormorants P carbo in Norway and possible implications for gadoid stock recruitment // Mar. ecol.
prog. ser. Vol. 66. No. 3. P. 205-218.
Bauer K.M., Glutz von Blotzheim U.N. 1966. Handbuch der Vogel Mitteleuropas. Band 1. Gaviiformes -
Phoenicopteriformes. Frankfurt am Main. 483 S.
Bauer K.M., Glutz von Blotzheim U. N. 1987. Handbuch der Vogel Mitteleuropas. Band 1. Gaviiformes -
Phoenicopteriformes. Wiesbaden: Aula-Verlag. 484 S.
Bauer N.G., Berthold P. 1997. Die Brutvogel Mitteleuropas: Bestand und Gefahrung. Wiesbaden: AULA-
Verlag. 715 S.
Baumanis J. 1997. Hibridisierung von Siberreiher und Grauereiher in Lettland // Omith. Mitt. Bd. 49. Nr. 11.
S. 296.
Baumanis J., Kalinips M. 1997. Pieradits liela balta gama un zivju ganja hibridizacijas gadijums Latvija //
Putni daba. Vol. 7. Is. 2. P. 1-3.
BaBler R., Schimkat J. 1999. Bestandsentwicklung und Schutz des WeiBstorches in Sachsen(Deutschland) //
WeiBstorch im Aufwind? - White Stork on the up? Hamburg, 1996. Bonn: NABU. S. 369-373.
Beaman M., Madge S. 1998. Handbook der Vbgelbestimmung: Europa und Westpalearktics. Stuttgart: Ulmer.
867 S.
Benmergui M. 1997. Premier cas de reproduction de la Grande Aigrette Egretta alba en Domes (Ain) //
Omithos. Vol. 4. Is. 4. P. 185-186.
Bennets R.E., Fasola M., Hafner H., Kayser Y. 2000. Influence of environmental and density-dependent
factors on reproduction of Little Egrets // Auk. Vol. 117. No. 3. P. 634-639.
Bennun L.A. 1993.Training water bird counters to monitor wetlands in Kenya: Progress and prospects //
Proc. Afr. Crane and Wetland Train. Worksh. Maun. P. 183-189.
Литература
579
Bergman S. 1935. Zur Kentniss Nordostasiatischer Vogel. Ein zur Systematik, Biologie und Verbreitung.
Kamchatkas und der Kurilen. Stokholm: Alb. Bonn. Forlag. 268 S.
Berkhoudt H., Zweer G. 1994. Feltremachanisms: design, perfomance and evolution in Flamigoes and
Anceriformes: [Pap.] 21st Int. Omithol.Congr., Vienna, 20-25 Aug. 1994/ Berkhoudt H., Zweer G. 11 J.
Om. Bd. 135. No. 3. S. 434.
Berthold P., Kaatz M., Quemer U. 2004. Long-term satellite tracking of White Stork (Ciconia ciconia)
migration: constancy versus variability // J. Om. Vol. 145. Is. 4. P. 356-359.
Berthold P., van den Bossche W, Fiedler W., et al. 2001. Der Zug des WeiBstorchs (Ciconia ciconia): eine
besondere Zugform auf Grund neuer Ergebnisse// J. Om. Bd. 142. Nr. 1. S. 73-92.
Berthold P, van den Bossche W., Leshem Y., et al. 1997. Satelliten-Telemetrie beim WeiBstorch Ciconia
ciconia: Wanderung eines Ost-Storchs westlich bis Nigeria // J. Om. Bd. 138. Nr. 3. S. 331-334.
Beyer W.N., Spalding M., Morrison D. 1997. Mercury concentrations in feathers of wading birds from
Florida // Ambio. Vol. 26. P. 97-100.
Bezzel E. 1994. Artenliste der Vogel Bayems // Garm. Vogelkundl. Berichte. Bd. 23. S. 1-65.
Bianki V, Boiko N., Kokhanov V 1997. The cormorant Phalacrocorax carbo in Kandalak-sha bay (White
Sea, Russia) // Ekologia Polska. Vol. 45. No. 1. P. 15.
Bibby C. 1981. Wintering bitterns in Britain // Brit. Birds. Vol. 74. No. 1. P. 1-10.
Billinger K. 2003. Erstbrutnachweis des Seidenreichers (Egretta garzetta) fur Oberdsterreich // Vogelkund/
Nachrichten aus Oberdsterreich, Naturschutz aktuell. Bd. 11. Nr. 1-2. S. 1-4.
BirdLife International. 2004. Birds in Europe: population estimates, trends and conservation status. Cambridge,
UK: BirdLife International (BirdLife Conservation Series No. 12). 374 p.
BirdLife International European Bird Census Council. 2000. European bird populations: estimates and trends.
Cambridge, UK: BirdLife International (BirdLife Conservation Series No. 10). 160 p.
BirdLife Intemational.Threatend Birds of Asia. 2001. Cambridge, UK. 3030 p.
Birkhead T.R. 1973. A wintre roost of Grey Herons // Brit. Birds. Vol. 66. No. 4. P. 147-156.
Blaker D. 1969. The Behaviour of Egretta garzetta and E. intermedia // Ostrich. Vol. 40. P. 150-155.
Bloesch M. 1982. Sechser Gelege beim WeiBstorch Ciconia ciconia // Om. Beob. Bd. 79. Nr. 1. S. 39—44.
Bloesch M., Dizerens M., Sutter E. 1977. Die Mauser der Schwungfedem beim WeiBstorch Ciconia ciconia //
Om. Beob. Bd. 74. Nr. 2. S. 161-188.
Blum V, Jacoby H. 1989. Kormorane am Bodensee zum AbschuB freigegeben // Vbgelschutz in Osterreich.
Nr. 3. S. 35-36.
Bogucki Z. 1994. Wielkosc l$gu bociana bialego (Ciconia ciconia) w Wielkopolsce w latach 1971-1992 H
Bocian bialy (Ciconia ciconia) w Wielkopolsce. Prace Zakl. Biol, i Ekol. Ptakow UAM. Poznan. P. 165—
176.
Bogucki Z., Ozgo M. 1999. Changes in nest selection of the White Stork Ciconia ciconia in Wielkopolska
(Western Poland) over the period 1974-1994 // WeiBstorch im Aufwind? - White Stork on the up? Proc.
Intern. Symp. on the White Stork, Hamburg, 1996. Bonn: NABU. P. 443-450.
Bohning-Gaese K. 1992. Zur Nahrungsokologie des WeiBstorchs (Ciconia ciconia) in Oberschwaben:
Beobachtungen an zwei Paaren // J. Om. Bd. 133. Nr. 1. S. 61-71.
Brasil M.A. 1991. The birds of Japan. London: A & C Black. Ltd. 466 p.
Brazil M.A. 2009. Birds of East Asia. London: A & C Black. Ltd. 528 p.
Brix M. 2005. Turmfalken als langjahrige Untermieter von WeiBstorchen // Om. Mitt. Bd. 57. Nr. 1. S. 27.
Broekhuysen G. 1965. Nesting of the White Stork (Ciconia ciconia (L.)) in South Africa // Vogelwarte.
Bd. 23. Nr. l.P. 5-11.
Broekhuysen G. 1971. White Storks breeding in the Bredasdorp District, most southern Part of the Wintering
Quarters // Vogelwarte. Bd. 26. Nr. 2. P. 164-169.
Broekhuysen G., Uys D. 1966. Uber das Bruten des WeiBstorchs in Siidafrika in der Brutzeit 1965/66 //
Vogelwarte. Bd. 23. Nr. 4. S. 235.
Brouwer J., Mullie W.C., Scholte P. 2003. White Storks Ciconia ciconia wintering in Chad, northern Cameroon
and Niger: a comment on Berthold et al. (2001) // Ibis. Vol. 145. Is. 3. P. 499-501.
Brouwer G.A. 1964. Some data on the Spoonbill, Platalea leucorodia L., in Europe, especially in the
Netherlands // Zool. meded. Vol. 39. P. 481-521.
Brouwer K. 1990. Longevity of storks // SIS-Group Newslet. Vol. 3. Is. 1-2. P. 5-6.
580
Литература
Brzorad J.N., Maccarone A.D., Konleyec K.J. 2004. Foraging energetics of Great Egrets and Snowy Egrets //
J. Field. Om. Vol. 75. Is. 3. P. 266-280.
Bufield I., Bommel F. 2004. Birds in Europae. Popul., trends and conserv. status. Cambridge. 374 p.
Byrd G.V, Williams J.C. 2004. Cormorant survey in the Near Island Group, Aleutian Islands, Alaska, in
July 2003 with notes on other species. U.S. Fish and Wildl. Serv. Rep. AMNWR 03/13. Homer, AK. 19
P-
Caldwell H.R., Caldwell G.C. 1931. South China Birds. Shanghai. 447 p.
Carboneras C. 1992. Family Sulidae (gannets and boobies) // Handbook of the Birds of the World. Barcelona:
Lynx Ed. Vol. 1. P. 312-325.
Carey G.J., Chalmers M.L., Diskin D.A. et al. 2001. The avifauna of Hong Kong. Hong Kong. 563 p.
Carrascal L.M., Alonso J.C., Alonso J.A. 1990. Aggregation size and foraging behaviour of White Storks
Ciconia ciconia during the breeding season // Ardea. Vol. 78. Is. 3. P. 399-404.
Catsodorakis G., Crivelli A.I. 2001. Nesting habitat characteristics and breeding performance of Dalmatian
Pelicans in Lake Mikri Prespa, NW Grees // Waterbirds. Vol. 24. P. 386-393.
Catuneau I. 1963. “Ihtiofagia” sticului cenu§iu (Ardea cinerea) H Ocrot. Naturii. Vol. 7. P. 145-147.
Causey D. 2002. Red-faced Cormorant (Phalacrocorax urile) // The Birds of North America. No 617.
Philadelphia, PA: The Birds of North America, Inc.P. 1-16.
Causey D., Byrd G.V, Williams J. 2006. Profound declines in Aleutian populations of cormorants indicate
possible ecosystem disequilibrium in the Near Island // Pacific Seabird Group: 33rd Annual Meeting.
Program and abstracts. Girdwood, AK. P. 47.
Celmins A. 2004. Liela balta garpa Egretta alba invazija Latvija 2004. gada vasara // Putni daba. Vol. 14.
Is. 4. P. 12-16.
Chan S. 1995. Report on Ciconia boyciana in Hong Kong // Cranes and Storks of the Amur river. The Proc,
of the Intern. Workshop. Moscow: Arts. Literature Publ. P. 107-109.
Changhu L., Fulan H., Jiachuan C. 1995. Distribution of the White and Black Storks in China // Ibid. P. 92-
100.
Check-List of Japanese Birds. 1974. 5-th and Rev. Ed. Tokyo, 364 p., 6-th and Rev. Ed., (2000) Tokyo, 345 p.
Cheng Tso Hsing. 1976. Distributional list of Chinese birds. 2-nd rev. ed. Peking. 1280 p.
Cheng Tso-hsin. 1987. A synopsis of avifauna of China. Beijing: Science Press. 1222 p.
Cherevichko V.I., Fedorov Y.V, Fetisov S.A. et al. 1999. The historical status, present-day distribution and
numbers of the White Stork (Ciconia ciconia) in North-western Russia // WeiЯstorch im Aufwind? -
White Storks on the up? Proc. Intern. Symp. on the White Stork, Hamburg 1996. Bonn: NABU. P. 305-
317.
Chernetsov N., Berthold P, Quemer U. 2004. Migratory orientation of first-year white storks (Ciconia
ciconia)'. inherited information and social interactions // J. Experim. Biol. Vol. 207. P. 937-943.
Chernetsov N., Kaatz M., Quemer U., Berthold P. 2005. Vieijahrige Satelliten-Telemetrie eines WeiBstorchs
Ciconia ciconia vom Selbstandigwerden an - Beschreibung einer Odyssee // Vogelwarte. Bd. 43. Nr. 1.
S. 39-42.
Chong J-R., Isumi T., Heizo S. 2000. Captive breeding of Black-faced Spoonbill // Conserv. and res. of
Black-faced Spoonbills and their habitat. Proc, of the Intern, workshop in Tokyo, 12-16 June, 1997.
Second edition. 2000. P. 47-53.
Chong J.R., Pak U.I., Rim C.Y., Kim T.S. 1996. Breeding biology of Black-faced Spoonbill Platalea minor
// Strix. Vol. 14. P. 1-10.
Christidis L., Boles W. E. 2008. Systematics and taxonomy of Australian birds. Collingwood, VIC, Australia:
CSIRO Publishing. 277 p.
Cezilly F., Viallefont A., Boy V, Johnson A.R. 1996. Annual variation in survival and breeding probability
in greater flamingos // Ecology. Vol. 77. No. 4. P. 1143-1150.
Clapp R.B., Klimkiewicz M.K., Kennard J.H. 1982. Longevity records of North American birds: Gaviidae
through Alcidae // J. Field Omithol. Vol. 53. No. 2. P. 81-208.
Collar N.J., Andreev A.V, Chan S. et al. 2001. Treatened birds of Asia: the BirdLife International Red Data
Book. Cambridge, UK: BirdLife Intemat. P. 54-67.
Combridge P, Parr C. 1992. Ibflux of Little Egrets in Britain and Ireland in 1989 // Brit. Birds. Vol. 85.
P. 16-21.
Литература
581
Cramp St. (ed.). 1977. [Cramp S., & Simmons K.E.L., eds. 1977,1978,1980,1982,1986 = Repr.] Handbook
of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic. Vol. 1.
Ostrich to Ducks. Oxford Univ. Press. 722 p.
Crawford R.J.M., Cooper J., Dyer B.M. 1995. Conservation of an increasing population of Great White
Pelicans Pelecanus onocrotalus in South Africa’s Western Cape H S. Afr. J. Marine Science. Vol. 15.
P. 32—42.
Creutz G. 1964. Emahrungsweise, Nahrungsauswachl und Abwehr des Graureihers (Ardea cinerea) 11 Zool.
Abhandl., Staatl Museum Tierkunde Dresden. Bd. 27. H. 1-10. S. 29-64.
Creutz G. 1981. Der Graureiher H Die Neue Brehm-Bucherei. Nr. 530. 195 S.
Creutz G. 1985. Die Entwicklung des Storchbestandes in der DDR 1958 bis 1984 П Vogelwarte. Bd. 106.
S. 211-214.
Creutz G. 1988. Der WeiBstorch H Neue Brehm-Bucherei. Nr. 375. Wittenberg Lutherstadt. 236 S.
Crivelli A.J. 1996. Action plan for the Dalmatian pelican (Pelecanus crispus) in Europe. 53 pp.
Crivelli A. 1997. White Pelican Pelecanus onocrotalus 11 Hagemeijer & Blair (eds.). The ABCC Atlas of
European breeding birds. London. P. 32.
Crivelli A., Michev T. 1997. Dalmatian Pelican Pelecanus crispus 11 Ibid. P. 33.
Crivelli A. J., Vizi 0.1981. The Dalmatian Pelican Pelecanus crispus Bruch, 1832, a recently world endangered
bird species H Biol, conserv. Vol. 20. P. 297-310.
Crozier G.E., Gawlik D.E. 2003. Wading birds nesting effort as an index to wetland ecosystem integrity H
Waterbirds. Vol. 26. Is. 3. P. 303-324.
Custer Th., Osborn R. 1978. Feeding habitat use colonially-breeding herons, egrets and ibises in North
Carolina // Auk. Vol. 95. P. 733-743.
Custer T.W., Pendleton G.W., Roach R.W. 1992. Determination of hatching date for eggs of Black-crowned
Night Herons, Egrets and Greats Egrets H J. Field Omith. Vol. 63. Is. 2. P. 145-154.
Dahmer T.D., Felley M.L. 1995. Report on Winter Range Survey of Black-faced Spoonbill, 1994-1995.
World Nature Association. P. 24.
Dahmer T.D., Yu Y-T., Felley M.L. 2007. Change in Black-faced Spoonbill Platalea minor global population
from 1988-2006 H Proceedings: Keeping Asia’s Spoonbills Airborne Intern. Symp. on Res, and Conserv.
of the Black-faced Spoonbill. Hong Kong. P. 29-38.
Dahms G. 1973. Bodenbruten beim WeiBstorch (Ciconia ciconia) inNiedersachsen H Vogelk. Ber. Niedersach.
Bd. 5. Nr. 1. S. 16-20.
Dahms G. 1999. Bodenbruten beim WeiBstorch (Ciconia ciconia) in freier Natur H WeiBstorch im Aufwind?
- White Stork on the up? Bonn, NABU. P. 459^161.
Dall’Antonia P, Baccetti N., Cianchi F. 1996. Origine, fenologia e movimenti flamingo dei Fenicotteri della
Laguna di Orbetello H Riv. ital. omitol. T. 66. No. 2. P. 97-117.
Dallinga, J. H., Schoenmakers S. 1987. Regional differences in the number of White Storks (Ciconia c.
ciconia) in relation to food resources H Colon. Waterbirds. Vol. 10. P. 167-177.
Dallinga J.H., Schoenmakers S. 1989. Population changes of the White Stork since the 1850s in relation to
food resources // White Stork. Proc. I Int. Stork Conserv. Symp. Schriftenreihe des DDA. Bd. 10. P. 231-
262.
Day J.C.U. 1981. Status of Bitterns in Europae since 1976 H Brit. Birds. Vol. 74. P. 10-16.
Dejonge J.F., Czajkowski M.A. 1983. Sur la longevite des Oileaux bagues en France Metropolitaine, dans
les Deportements d’outre-mer et dans les pays d’influence Francaise H Alauda. Vol. 51. No. 1. P. 27^17.
Deignan N.G. 1945. The birds of the northern Thailand H US Nat. Mus. Bull. Vol. 186. 616 p.
Dekker R.W.R.J., Dickinson E.J., Morioka H. 2001. Systematic notes on Asian birds. 18. Some nomenclatural
issues relating to Japanese taxa described in Planches Coloriees (1820-1839) and Fauna Japonica, Aves
(1844-1850) H Zool. Verh. Leiden. Vol. 335. P. 199-214.
Delany S., Scott D. 2002. Western Populations Estimates. 3-rd Ed. WI global series No. 12. 226 p.
Delacour J. 1928. On the Birds collected during the Third Expedition to Indo-China with notes by P. Jabonille,
M.B.O.U., and W.P. Lowe, M.B.O.U. // Ibis. Vol. 4. No. 1. P. 23-51.
Delacour J. 1929. On the Birds collected during the Fourth Expedition to French Indo-China П Ibis. Vol. 5.
No. 2. P. 193-220.
Delacour J., Jabouille P. 1925. On Birds of Quangtri, Central Annam: with Notes on others from other Parts
of Central French Indo-China П Ibis. Vol. 1. No. 1. P. 209-260.
582
Литература
del Hoyo J., Elliott A., Sargatal J. (eds.). 1992. Handbook of the Birds of the World. Barcelona: Lynx Ed.
Vol. 1. 696 p.
Demongin L., Dmitrenok M., Bretagonolle V. 2006. Determining Great Bittern Botaurus stellaris laying
date from egg and and chick biometrics // Bird study.
Denlinger L., comp. 2006. Alaska Seabird Information Series: Draft. Anchorage: US Fish and Wildl. Sefv.
85 p.
Diagana Cheikh Hamallah, Ould Messaoud Brahim, Diawara Yelli. 2001. The use of the Geograph. Inform.
Syst. as a tool for decision mak. the field of manag. of bird populat. in protect, areas. The Diawling Nat.
Park (Mauritania) as example И OMPO, Newsl. No. 23. P. 19-26.
Dianjin F. 1991. Observations on the breeding of the Oriental White Stork (Ciconia boyciana) near Qiqihar,
Heilongjiang province, China H M.C. Coulter et al. (eds.). Biol, and Conserv. of the Oriental White
Stork Ciconia boyciana. P. 21-30.
Dickinson E.J. 2001. Systematic notes on Asian birds. 9. The “Nouveau recueil de planches coloriees” of
Temminck& Laugier (1820-1839) and Fauna Japonica, Aves (1844-1850) 11 Zool. Verh. Leiden. Vol. 335.
P. 7-54.
Dickinson E.C. (ed.). 2003. The Howard and Moore Complete Checklist of the Birds of the World. 3rd ed.
London: Christopher Helm. 1039 p.
Directory of Azov-Black Sea Costal Wetlands. 2003. Rev. and adapt. Kyiv. 235 p.
Diagna CH. Hamallach et al. 2001.The use of the Geographical Inform. Systems, as a tool for dec. mak. in
the field of manag. of bird popul.areas П OMPO Newsl. No. 21. P. 19-26
Dmitrenok M., Demongin L., Zhurauliov D. 2005. Three cases of replacement clutches in the Great Bittern
Botaurus stellaris 11 Ardea. Vol. 93. No. 2. P. 271-274.
Dmitrenok M., Puglisi L., Demongin L. et al. 2007. Geographic variation, sex and age in Great Bittern
Botaurus stellaris using cjljuration and morphometries П Ibis.
Dragoo D.E., Byrd G.V., Irons D.B. 2004. Breeding status, population trends and diets of seabirds in Alaska,
2002., US Fish and Wildl. Serv. Rep. AMNWR 04/15. Homer, AK. 91 p.
Duquet M. 1999. Der WeiBstorch (Ciconia ciconia) in Frankreich 1995 - Ergebnisse des 5. Intemationalen
WeiBstorchzensus H WeiBstorch im Aufwind? Proc. Intern. Symp. on the White Stork, Bonn: NABU.
S. 97-102.
Duquet M., Seriot J. 1995. La Cigogne blance en France de 1975 a 1993 H Proc, of the Intern. Symp. on the
White Stork (Western Population). Basel. P. 53-56.
Dvorak M., Ranner A., Berg H.-M. 1993. Atlas der Brutvogel Osterreihes. Wien. 522 S.
Dyrcz A., Graibinski W., Stawarczyk T., Witkowski J. 1991. Ptaki Slaska. Wroclaw. 525 p.
Dzieduszycki W. 1896. Fuhrer durch das graflich Dzieduszyckische Museum in Lemberg. Lemberg. 234 S.
Dziewiaty K. 1992. Nahrungsokologische Untersuchungen am WeiBstorch Ciconia ciconia in der Dannen-
berger Elbmarsch (Niedersachsen) П Vogelwelt. Bd. 113. Nr. 3. S. 133-144.
Dziewiaty K. 1999. Die Elbtalaue als Lebensraum des WeiBstorchs - Bedeutung und Bewertung anhand
nahrungsokologischer und populationsdynamischer Untersuchungen П WeiBstorch Intern. Symp. Bonn,
NABU. S. 463—470.
Eisner J. 1997. Cormoran Phalacrocorax carbo sinensis H Vogelkundliche Nachrichten aus Oberdsterreich.
Naturschutz aktuell. Linz. Sonderband. S. 15-16.
Endangered Bird Species in the Korean Penisula. 1987. 75 p.
Ericson P.G.P. 2008. Current perspectives on the evolution of birds H Contributions to Zoology. Vol. 77.
No. 2. P. 109-116.
Exner S., Schubert P. 2005. Konflikte zwischen Silberreier Ardea alba und Graureier A. cinerea'. Die Dom-
inaz wechselt H Om. Mitt. Bd. 57. Nr. 6. S. 207-208.
Fangrath M., Helb H.-W. 2005. Die Kehlmusterung des WeiBstorchs - ein individuelles Oder geschlechtsdi-
morphes Merkmal? П Om. Mitt. Bd. 57. Nr. 5. S. 148-153.
Fasola M., Hafner H. 1997. Little Egret П The EBCC Atlas of European Birds. London. P. 46-47.
Fenyosi L. 1994. Data on the nesting of White Storks (Ciconia ciconia) in Somogy county H Aquila. Vol. 101.
P. 226-227.
Fernandez Cruz M. 1975. On the numbers of Ardea purpurea 11 Ardeola. Vol. 21. P. 65-126.
Fiedler G., Wissner A. 1980. Freileitungen als todliche Gefahr fur Storche Ciconia ciconia 11 Okol. Vogel.
Bd. 2. S. 59-109.
Литература
583
Filardi С.Е., Rohwer S. 2001. Life history inplications of complete and incomplete primary molts in Pelagic
Cormorants 11 Condor. Vol. 103:
Finsch O. 1872. Uber eine Vogelsammlung aus den Kustenlandem der chinesisch-japanichen Meere 11
Verhandlungen d. k.-k. Zool.-botan. Gesellschaft in Wien. S. 253-272.
Finkenstaedt Ch., Heckenrouth H. 1974. Eine Kiistliche Koloniegriindung beim Graureiher (Ardea cine-
red) 11 Vogelwelt. Bd. 95. No. 6. S. 227-231.
Fishpool L.D.,Evans M.I. 2001. Important bird Areas in Africa and associated islands Priority sites for
conservation. Newbury and Cambridge, UK: Pisces Publicat. and Birdlife Intern. (Birdlife Conserv.
series No. 11). 1144 p.
Flore B.-O., Komer F., Marzmeier U. 1999. Uberwintemde Silberreier (Egretta alba) in siidwestlichen
Niedersachsen 1998/99 H Vogelk. Ber. Niedersachs. Bd. 31. Nr. 1. S. 52-54.
Fogarty M., Hetrick W. 1973. Summer foods of Cattle Egrets in north central Florida H Auk. Vol. 90. No. 2.
P. 268-280.
Fujimaki Yu. 1961. Observations on sea birds of Moyururi Island eastern Hokkaido H Tori. Vol. 16. No. 78.
P.387-398.
Fujimaki Y, Fujita T., Yunoki O. et al. 1990. Breeding ecology of the Oriental White Stork Ciconia ciconia
boyciana in the Bikin-Alchan Mar in the Far East of the USSR П Strix. Vol. 9. P. 139-157.
Fujimaki Y, Hyakutake T., Matsuoka S. 1976. Birds of Yruri and Moyururi Islands, eastern Hokkaido in
summer. 1. Cormorants and anserine birds H Misc. Rep. Yamashina Inst. Omithol. Vol. 8. No. 1. P. 68-
88.
Fujioka J., Yamagishi S. 1981. Extramartial and pair copulations in the Cattle Egret П Auk. Vol. 98. P. 134—
144.
Fulin J. 1991. Preliminary observations on captive breeding of Oriental White Storks H M.C. Coulter et al.
(eds.). Biol, and Conserv. of the Oriental White Stork Ciconia boyciana. P. 213-217.
Fulin M. 1984. Straty bocianov bielych (Ciconia ciconia) na hnieziskach v rokoch 1977-1984 vo vycho-
doslovenskom kraji Zb. Vychodosl. omitol. klubu ZK ROH. Vol. 1. P. 29-35.
Fulin M. 1999. The White Stork in Slovakia in 1994 and 1995 H WeiBstorch. Intern. Symp. Hamburg.
P. 199-202.
Fiiri A., Vidra T. 2002. Effect of years rich in high level inland waters on the nesting bird life of the Tapio-
Hajta region H Aquila. Vol. 107-108. P. 117-118.
Gabrielson I.N., Lincoln F.C. 1959. The birds of Alaska. Harrisburd: Stackpole Co.; Washington D.C.:
WildL Manag. Inst. 922 p.
Garrido J.R., Fernandez-Cruz M. 2003. Effects of power lines on a White Stork Ciconia ciconia population
in Central Spain H Ardeola. Vol. 50. Is. 2. P. 191-200.
Gauckler A., Kraus M. 1965. Zur Brutbiologie der Grossen Rohrdommel (Botaurus stellaris) 11 Vogelwelt.
Bd. 86. S. 129-146.
Gavashelishvili A. 1999. The 1996 census of the White Stork (Ciconia ciconia) in Georgia H WeiBstorch
Intern. Symp. Bonn: NABU. P. 323-325.
Gilbert G., McGregor P. 1994. Vocal individuality as a census tool: practical considerations illustrated by a
study of two rare species П Field Omith. Vol. 65. No. 3. P. 335-348.
Gilbert G., Tyler G., Smith K.W. 2002. Local annual survival of booming male Great Bittern Botaurus
stellaris in Britain, in period 1990-1999 П Ibis. Vol. 144. P. 51-61.
Gilbert G., Tyler G., Smith K.W. 2005. Behaviour, home-range size and habitat use by male Great Bittern
Botaurus stellaris in Britain П Ibis. Vol. 147. P. 533-543.
Glenister A.Gr. 1959. The birds of the Malay Penisula, Singapore and Pennang. London: Oxf. Univ. Press.
282 p.
Goebel H. 1879. Die Vogel des Kreises Uman, Gouwemment Kiew, mit besonderer Riicksicht auf ihre
Zugverhaltnisse ihr Brutgeschaft 11 Beitr. zur Kenntn. des Russ. Reiches. Bd. 2. S. 1-124.
Goesner Kenneth L. 1987. Salt ponds and flamingos H Underwater Natur. Vol. 16. No. 4. P. 3-7.
Golawski A., Kasprzykowski. 2004. Comparison of bird fauna of the Volga and the Vistula rivers during
autumn migration И Беркут. T. 13. Вип. 1. P. 103-109.
Golovushkin M.I., Goroshko O.A. 1995. Cranes and Storks in South-Western Transbaikalia H Cranes and
Storks of the Amur River. The Proc, of the Intemat. Workshop. Moscow: Arts. Literat. Publish. P. 39-
40.
584
Литература
Gomez Manzaneque A. 1992. Situation actual de la poblation de Cigiiena Blanca en Espana 11 Les Cigognes
d’Europe. Metz. P. 184-188.
Goutner V., Tsachalidis E.P. 1995. Time of breeding and brood size of White Storks Ciconia ciconia in
North-eastern Greece H Vogelwarte. Vol. 38. Is. 2. P. 89-95.
Gore M.E.J., Won Pyong-Oh. 1971. The birds of Korea. Seoul-Tokyo. 450 p.
Grimett R., Inskipp C., Inskipp T. 1998. Birds of the Indian Subcontinent. London: Cristopher Helm A& C
Black Ltd. 888 p.
Grischtschenko V. 1992. Beinkoten beim WeiBstorch (Ciconia ciconia) dient der Abkiihlung H Om. Mitt.
Bd. 44. Nr. 9. S. 221-222.
Grischtschenko V, Serebryakov V, Galinska I. 1995. Phanologie des WeiBstorchzuges (Ciconia ciconia) in
der Ukraine H Vogelwarte. Bd. 38. Nr. 1. S. 24-34.
Grishchenko V. 1999. Die Situation des WeiBstorchs Ciconia ciconia in der Ukraine H WeiBstorch Intern.
Symp. Bonn, NABU. S. 289-303.
Griill A., Ranner A. 1998. Populations of the Great Egret and Purple Heron in relation to ecological factors
in the reed belt of the Neusieder See H Col. Waterbirds. Vol. 21. P. 328-334.
Gurssu M. 2000. Nuovo sito di nidificazione di Fenicottero, Phoenicopterus roseus, in Sardegna H Riv. ital.
omitol. Ser. 2. No. 1. T. 70. P. 84-86.
Gustin M., Brunner A., Rossi P., Celada C. 2003. Efficacia del sistema delle IBA e della rete delle ZPS per
la conservazione di alcune specie di non passeriformi nidifcanti in Italia H Riv. ital. omitol. Ser. 2. No 2.
T. 72. P. 151-159.
Guziak R., Jakubiec Z. 1999. Der WeiBstorch Ciconia ciconia in Polen im Jahr 1995 - Verbreitung, Bestand
und Schutzstatus H WeiBstorch Intern. Symp. Bonn, NABU. S. 171-187.
Hackett Sh.J., Kimball R.T., Reddy S., Bowie R.C.K., Braun E.L., Braun M.J., Chojnowski J.L., Cox W.A.,
Han K-L., Harshman J., Huddleston Ch.J., Marks B.D., Miglia K.J., Moore W.S., Sheldon F.H., Stead-
man D.W., Witt Ch.C., Yuri T. 2008. A Phylogenomic Study of Birds Reveals Their Evolutionary His-
tory 11 Science. Vol. 320. P. 1763-1768.
Hagemeijer W.J.M., Blair M.J. 1997. The EBCC Atlas of European breeding Birds: Their Distribution and
Abundance. London: T. & A.D. Poiser. 903 p.
Hafner H. 1970. A propos d’une population de herons garde-boeufs Ardeola ibis en Camargue H Alauda.
Vol. 38a. No. 4. P. 249-254.
Hancock J. A., Kushlan J. A., Kahl M.P. 1992. Storks, Ibises and Spoonbills of the World. Academic Press.
385 p.
Hancock J., Kushlan J. 1984. The herons handbook. N.Y.: Harper & Row.
Hand-List of Japanese Birds. 1958. Fifth and Revision Ed. Tokyo. 264 p.
Harris M.P., Wanless S. 1991. The importans of the lesser sandeel Ammodytes marinus in the diet of the
shag Phalacrocorax aristotelis 11 Omis-scand. Vol. 22, No.4. P. 375-382.
Harris M.P., Wanless S. 1993. The diet of shags Phalacrocorax aristotelis during the chick-rearing period
assessed by three methods H Bird-study. Vol. 40. No. 2. P. 135-139.
Harris M.P., Wanless S. 1996. Differential responses of guillemot Uria aalge and shag Phalacrocorax
aristotelis to a late winter wreck 11 Bird-study. Vol. 43. No. 2. P. 220-230.
Harrison P. 1983. Seabirds an identification guide. Boston: Haughton Mifflin Company. 448 p.
Hartert E. 1910. Die Vogel der Palaarktischen Fauna. Berlin: Fridlander. Bd. 1. 832 S.
Hartert E. 1920. The birds of Commander Islands H Nov. Zool. Vol. 27. No 1. P. 128-158.
Hartert E. 1920a. Ordnung Gressores H Die Vogel der Palaarktischen Fauna. Berlin. H. 10 (Bd. 2. H. 4).
S. 1213-1264.
Hasegawa H. 1984. Status and conservation of seabirds in Japan, with special attention to the Short-tailed
Albatross H Stat, and Conserv. of the World’s Seabirds. ICBP Techn. Publ. No. 2. P. 487-500.
Hatch J. J., Weseloh D.V. 1999. Double-creasted Cormorant (Phalacrocorax auritus) 11 The Birds of North
America. Philadelphia, PA: The Dirds of North America. No. 441. Inc. P. 1-36.
Hatch S.A., Hatch M.F. 1990. Components of breeding productivity in a marine bird community: key fac-
tors and concordance П Can. J. Zool. Vol. 68. P. 1680-1690.
Hatzilacos D. 1986. Pereliminary data on the breeding and feeding biology of the white pelican (Pelecanus
onocrotalus L.) at Lace Mikra Prespa H Biol. Gallo-hellen. 1986 (1987). No. 12. P. 497-506.
Литература
585
Haverschmidt F. 1961. The breeding of the common heron in immature plumage 11 Ardea. Vol. 49. No. 1-2.
P. 72-74.
Heckenroth H. 1986. Zur Situation des WeiBstorchs (Ciconia ciconia) in der Bundesrepublik Deutschland,
Stand 1984 H Artenschutzsymposium WeiBstorch. Beih. Veroff. Landschaftspflege Baden-Wurttem-
berg. Karlsruhe. Bd. 43. S. 111-120.
Hicks E.A. 1959. Check-list and bibliographyon the occurence of insects in birds nests. Ames. Iowa state
college press. 681 p.
Higuchi H., Nagendran M., Darman Y. et al. 2000. Migration and Habitat Use of Oriental White Storks from
Satellite Tracking Studies П Global Environment Res. Vol. 4. No. 2. P. 169-182.
Hilty S.L., Brown W.L. 1986. A Guide to the Birds of Colombia. Princeton: Princ.Univ. Press. 836 p.
Hobson K.A. 1997. Pelagic Cormorant (Phalacrocorax pelagicus) 11 The Birds of North America. Philadel-
phia: The Academy of Natural Sciences; Washington, DC: The American Ornithologists’ Union. No. 282.
P. 1-28.
Hobson K.F., Sealy S.G. 1985. Diving rhythms and diurnal roosting times of Pelagic Cormorants H Wilson
Bull. Vol. 97. No. 1. P. 116-119.
Hobson K.A., Wilson D. 1985. Colony establishment by Pelagic Cormorants on man-made structures in
southwest coastal British Columbia // Murrelet. Vol. 66. P. 84-86.
Howard R., Moore A. 1980. A complete checklist of the Birds of the World. Oxford: Oxford Univ. Press.
701 p. (Howard, Moore, 1998, - 2 ed., 630 p.).
Hudec K., Cerny W. (eds.). 1972. Fauna CSSR. Praha. T. 19. Ptaci - Aves. Pt. 1. 536 S.
Hu H. 1995. Status and Conservation of the Rare Waterfowl in Hubei, China H Cranes and Storks of the
Amur River. The Proc, of the Intemat. Worksh. Moscow: Arts. Literat. Publ. P. 102-104.
Hunt G.L., Jr., Eppley Z.A., Burgeson B., Squibb R.C. 1981. Reproductive ecology, foods and foraging
areas of seabirds nesting on the Pribilof Islands, 1975-1979. Environ. Assess. Alaskan Cont. Shelf,
Final Rep. Boulder, CO: NOAA. 257 p.
Inskipp T., Lindsey N., Duckworth W. 1996. An annotated checklist of the birds of the Oriental Region. 294
P-
Irby L.N. 1851. Notes on Birds observed in Oudh and Kumaon П The Ibis. Vol. 3. No 11. P. 217-251.
Ivanauskas T. 1959. Lietuvos paukscias. Vilnius. Vol. 2. 283 s.
Jahn H. 1942. Zur Oekologie und Biologie der Vogel Japans П J. Om. Bd. 90. H. 1/2. 302 S.
Jakubiec Z. 1991. Causes of breeding losses and adult mortality in White Stork Ciconia ciconia (L.) in
Poland H Population of White Stork Ciconia ciconia (L.) in Poland. Part II. Some aspects of the biology
and ecology of White Stork. Krakow. P. 107-124.
Jakubiec Z., Szymohski P. 2000. Bociany i bocky. Wroclaw: PTPP “pro Natura”. 144 S.
Janaus M. 2000. Balto star^u ligzdosanas sekmes Latvija no 1989. lTdz 1999. gadam П Putni daba. Vol. 10.
Is. 2. P. 14-21.
Janaus M., StTpniece A. 1999. The White Stork in Latvia: 1994-1995 11 WeiBstorch Bonn: NABU. P. 253-
264.
Japanese Bird Bunding in Recent Years (1961-1983). 1985. Bird Migration Res. Center, Yamashina Inst,
for Ornithology. 202 p.
Jiangying C. 1995. Distribution and Protection of Cranes and Storks in Inner Mongolia, China H Cranes and
Storks of the Amur River. The Proc, of the Intern. Worksh. Moscow: Arts. Literat. Publ. P. 48-50.
Jianzhang M., Jihong G., Liyang L. 1995. Status of Reserch on the Oriental White Stork in China 11 Ibid.
P. 88-92.
Johansen H. 1927. Fregata minor (Gm.) in Ostsibirien erlegt H Omithol. Monatsberichte. Jg. 35. Nr. 1.
S. 20.
Johansen H. 1961. Revised list of the birds of the Commander Islands 11 Auk. Vol. 78. No. 1. P. 44-56.
Johnsgard P.A. 1993. Cormorants, darters and pelicans of the world. Washington, D.C.; London: Smithson
Inst. Press. 445 p.
Johnson A. 1987. Etude de la population de flamants roses de la Camargue et de la Mediterranee occiden-
tale. H Rev. ecol. Vol. 42. No. 4, Suppl. P. 122.
Johnson A. 1997. Greater Flamingo // The EBCC Atlas of European Breeding Birds: Their Distribution and
Abundence. P. 62.
586
Литература
Johnson A.R. (ed.). 1998. Flamingo Specialist Group Newsletter. No. 8. Annual Reports 1995-1996 11
Wetl. Intemat. and lUCN-Spec. Surviv. Comm. P. Ml.
Jones S. - S.K.H. 1911. On some Birds observed in the Vicinity of Wei Hai Wei, North-East China 11 Ibis.
Vol. 5. No. 20. P. 657-695.
Jong-jun W., Zhou W. 1989. Hunting effects on the number variation of wintering White Storks in Cenhu
Lake 11 Chinese wildlife (Bimonthly). No. 4. July. P. 16-21.
Jonkers D.A. 1989. Status and conservation of the White Stork in the Netherlands: a review 11 Wei?storch
Intern. Conserv. Symp. Schriftenreihe des DDA. Bd. 10. P. 45-54.
Jozefik M. 1960. Legi podwojne ibisow u kasztanowatych , Plegadis falcinellus falcinellus (L.) w delcie
Dniestru 11 Acta omithol. Vol. 5. No. 14. P. 367-377.
Jozefik M. 1962. Studies on the squacco heron, Ardeola ralloides (Scop.), Part 11. Secular changes in the
numbers and distribution in the Palearctic range 11 Acta omith. Vol. 11. No. 7. P. 135-264.
Jozefik M. 1970. Studies on the squacco heron, Ardeola ralloides (Scop.), Part 111. Geographical variability
in the critical and mean micropopulation size H Acta omith. Vol. 12. No. 3. P. 57-102.
Jozefik M. 1971. Wplyw czynnikow antropogenych na structure rozmiezezenia gatunku c naprzykladzie
Ard. rail. П Wiad. ecol. Vol. 17. No. 2. P. 118-126.
Kaatz M. 1999. Warum sich 1997 WeiBstorchheimkehr so verzogerte? Die Satelliten-Telemetrie gibt Auskunft
П 6. und 7. Sachsen-Anhalt. S. 27-31.
Kahl M.P. 1971. Social behavior and taxonomic relationships of the storks H Liv. Bird. Vol. 10. P. 151-170.
Kahl M.P. 1972. Comparative ethology of the Ciconiidae. Part 4. The “typical” storks (genera Ciconia,
Sphenorhynchus, Dissoura and Euxenurd) 11 Z. Tierpsychol. Vol. 30. P. 225-252.
Kain T. 1994. 1994 report of the VSO Records Committee H Raven. Vol. 65. P. 84-87.
Kania W. 1985. Wyniki obrqczkowania ptakow w Polsce. W^drowki bocianow bialych Ciconia ciconia 11
Acta omithol. R. 21. Z. 1. P. 1-41.
Kanyamibwa S., Schierer A., Pradel R., Lebreton J.D. 1990. Changes in adult annual survival rates in a
western European population of the White Stork Ciconia ciconia 11 Ibis. Vol. 132. Is. 1. P. 27-35.
Kato A., Watanuki Y., Naito Y. 2001. Foraging and breeding performance of Japanese cormorants in rela-
tion to prey type H Ecol. Res. Vol. 16. P. 475M58.
Kazakov B.A., Khokhlov A.N., Pishvanov Y.V., Yemtyl M.K. 1994. Pelicans in wetlands of Predkavkaz’e
region (north of the Caucasus) H Pelicans in the former USSR. IWRB Publ. No. 27. P. 9-16.
Keller M., Profus P. 1992. Presentsituation, reproduction and food of the Black Stork in Poland H Les
Cigognes d'Europe. Mutz: Inst. Europeen d'Ecologie. P. 227-236.
Kennedy M., Hamish G.S., Russell D.G. 1996. Hop, step and gape: do the social displays of the Pelecani-
formes reflect phylogeny? H Anim. Behav. Vol. 51. P. 273-291.
Kershaw J.C. 1904. List of the Birds of the Quangutung Coast, China 11 The Ibis. Vol. 4. No. 14. P. 235-
248.
Kessel B. 1989. Birds of the Seward Penisula, Alaska. Fairbanks: Univ, of Alaska Press. 330 p.
King B., Dickinson E.C. 1995. Birds of South-east Asia. Hong Kong: Harp.Coll. Publ. 480 p. (London, 480
p)
King C. 1990. Reproductive management of the Oriental White Stork Ciconia boyciana 11 Int. Zoo Yrb.
Vol. 29. P. 85-90.
Kirsch Karl-Wilh. 1973. Der Gesembestand des Graureihers (Ardea cinerea} im Regierungsbezirk Luneb-
urg ab 1965 П Vogelskund. Ber. Niedersachsen. Bd. 5. Nr. 2. S. 33-37.
Kisling M., Horst B. 1999. Die “mittlere Zugroute” des WeiBstorchs uber Italien - Beobachtungen am Cap
Bon/Tunesien und bei Messina/Sizilien H Proc. Intern. Symp. on the White Stork, Hamburg, 1996.
Bonn: NABU. S. 529-534.
Kobayashi K. 1933. Summer birds in Shikotan Island, South Kurilen П Tori. Vol. 8. No. 36. P. 10-21.
Kobayashi K. 1962. Birds of Japan in Natural Colours. Osaka : Hoikusha. 204 p.
Kondratyev A.Y, Litvinenko N.M., Shibaev YV. et al. 2000. The breeding seabirds of the Russian Far East
H Seabirds of the Russian Far East., Ottawa: Can. Wildl. Serv. Spec. Publ. P. 37-81.
Kondratyev A.Y, Mendenhall V.M. 1995. A survey of seabird colonies in the northern Sea of Okhotsk H
Pacific Seabirds. Vol. 22. No. 1. P. 35.
Konyukhov N.B., Bogoslavskaya L.S., Zvonov B.M., van Pelt T.I. 1998. Seabirds of the Chukotka Penisu-
la, Russia H Arctic. Vol. 51. No 4. P. 315-329.
Литература
587
Korodi Gal I. 1961. Contributii la cunoa§terea biometrici hranei pelecanului cret (Pelecanus crispus
Bruch) din delta Dunarii П Bui. Inst, ceratari proiect. Pisocole. Vol. 20. No. 2. P. 90-95.
Korzyukov A.I. 1997. Seasonal distribution of the Cormorant Phalacrocorax carbo in the North-Western
part of the Black Sea region П Ekologia Polska. Vol. 45. No. 1. P. 169.
Kovacs G. 2004. Saker Falcon (Falco cherrug) tacing prey from Great Egret (Egretta alba) 11 Aquila. Vol. 111.
P. 208.
Kovacs G., Kovacs G. 1999. Nyolc fehergolya-fioka (Ciconia ciconia) egy feszekben П Tuzok. Vol. 4. Is. 4.
P. 125-126.
Kozulin A. 1996. Nahrungsverhalten des Schwarzstorches Ciconia nigra 11 Om. Mitt. Bd. 48. Nr. 7. S. 170—
172.
Kuppler O. 2001. WeiBstorch-Nestling erstickt an Ringelnatter П Om. Mitt. Bd. 53. Nr. 1. S. 32.
Kuroda N. 1925. On the Japanese Cormorant П Tori. Vol. 4. No. 20. P. 336-350.
Kushlan J. A., Hancock J. A. 2005. The Herons. Bird Families of the World. N.Y.: Oxford Univ. Press Inc.
433 p.
Kuzniak S. 1994. Bocian bialy (Ciconia ciconia) w wojwodztwie leszczyhskim w latach 1974-1990 П
Bocian bialy (Ciconia ciconia) w Wielkopolsce. Prace Zakl. Biol, i Ekol. Ptakow UAM. Poznan. P. 69-89.
Lakeberg H. 1995. Zur Nahrungsokologie des WeiBstorchs Ciconia ciconia in Oberschwaben (S-Deut-
schland): Raum-Zeit-Nutzungsmuster, Nestlingsentwicklung und Territorialverhalten П Okol. Vogel.
Bd. 17. Sonderheft. 87 S.
Lang E.M. 1963. Flamingoes raise their young on a liquid containing blood П Experientia. Vol. 19. No. 10.
P. 532-533.
Langley C.H., 1983. Biology of the Little Bittern in the Southwestern Cape H Ostrich. Vol. 54. P. 83-94.
Lansdown R. 1990. Little known oriental bird: Chinese egrett // Bull, of the Oriental Club. Vol. 11. P. 27-30.
La Touche J.D.D. 1907. Field-Notes on the Birds of Chinkiang, Lower Yangtse Basin., Part 3 П Ibis. Vol. 1.
No. 1. P. 1-30.
La Touche J.D.D. 1912. List of the species of birds collected and observed on the Island of Shaweishan П
Bull. B.O.S. Vol. 29. P. 124-160.
La Touche J.D.D. 1924. On the birds of South-East Yunnan, S.W. China. Part 4 П Ibis. Vol. 6. No. 2. P. 284-
307.
La Touche J.D.D. 1925-1930. A handbook of the Birds of Eastern China, 1. London.
La Touche J.D.D. 1931-1934. A handbook of the Birds of Eastern China, 2. London. 566 p.
La Touche J.D.D., Rickett C.B. 1905. Furter Notes on the Nesting of Birds in the Province of Funkien, S.E.
China H Ibis. Vol. 5. No. 17. P. 25-67.
Lee K. 2010. Breeding Situation and Survival Rate of Black-faced Spoonbills in Korea by the re-sighting of
color bands П Ecology, Migration and Conservation of the Black-faced Spoonbill. Glob. COE Intern.
Symp. 2010. P. 15-18.
Lee K., Kim I. 2007. Status and breeding biology of Black-faced Spoonbills П Proceedings: Keeping Asia’s
Spoonbills Airborne Intern. Symp. on Res, and Conserv. of the Black-faced Spoonbill. Hong Kong.
P. 56-62.
Leibak E., Lillelrht V, Veromann H. 1994. Birds of Estonia. Tallinn. 287 p.
Lekagul B., Round Ph.D. 1991. A guide to the birds of Thailand. Bangkok. 457 p.
Lensink Rob. 1996. Vreemde vogels in de Nederlandse avifauna: Verleden, heden en wat voor een toekomst
П Vogeljaar. Jg. 44. H. 4. S. 145-164.
Libbert W. 1954., Wo verbleiben die WeiBstorche aller Altersstufen in den Brutmonaten? П Vogelwarte.
Bd. 17. Nr. 2. S. 100-113.
Liechti F., Ehrich D., Bruderer B. 1996. Flight behaviour of White Storks Ciconia ciconia on their migra-
tion over southern Israel H Ardea. Vol. 84. Is. 1. P. 3-13.
Lindgren Cory J., Pickering S.P.C. 1998. Phoenicoparrus andinus. Ritualised displays and display frequen-
cies of Andean Flamingos Phoenicoparrus andinus 11 Wildfowl. No. 48. P. 194-201.
Lintia D. 1955. Pasarile din RPR. Vol. 3. P. 14-18.
Lipsbergs J. 1971. Ziviu gamie (Ardea cinerea) pirmas ligzdoDa - nas gadiums niedrayos Latvya // Zool
muz. raksti Latv. un-t. P. 87-89.
Lipsbergs J., Opermanis O., StTpniece A., StTpnieks A. 2001. Pirmais pieradTtais liela balta garna Egretta
alba ligzdosans gadljums Latvija П Punti daba. Vol. 11. Is. 1. P. 22-24.
588
Литература
Lipsbergs J., Roze V. 2001. Liela balta gama Egretta alba un zivju gama Ardea cinerea hibridpara otrais •
zinamais ligzdosans gadijumus Latvija H Punti daba. 1 pielikums. P. 51-53.
Litvinenko N., Shibaev Yu. 1991. Status and conservation of the seabirds nesting in southeast USSR П
ICBP Technical Publ. No. 11. Cambridge. P. 175-204.
Litvinenko N.M., Shibaev Yu.V. 2000. Importance of Furugelm island birds: the first record of a breedeng
colony of Chinese egret Egretta eulophotes 11 Oryx. Vol. 34. No. 4. P. 335-337.
Litvinenko N., Shibaev Yu. 2001. Birds of the wetland «Tumangan» (biodiversity and problems of protec-
tion) П The state of environment and biota of the southwestern part of Peter the Great Bay and the
Tumen River mouth. Vladivostok: Dalnauka. Vol. 2. P. 5-19.
Li Yung-hsin, Liu Xi-ye. 1963. On the breeding behaviour of the Pond Heron H Acta zool. Sinica. Vol. 15.
No. 2. P. 203-210.
Li Z.W.D., Mundkur T. 2000. Numbers and distribution of waterbirds and wetlands in the Asia-Pacific
region. Results of the Asian Waterbird Census: 1997-2001. Kuala Lumpur, Malaysia: Wetl. Intern.
Lloyd J.H. 1873. On the Birds of the Province of Kattiawar in Western India H Ibis. Vol. 3. No. 12. P. 397-
421.
Lobkov E. 1997. Die Vogelwelt Kamtschatkas H Acta omithoecologica. Bd. 3. Nr. 4. S. 1-451.
Lohmer R., Jaster P., Reck F.-G. 1980. Untersuchungen zur Emahrung und NahrungsraumgroBe des WeiBs-
torches (Ciconia ciconia) 11 Beitr. Naturk. Niedersachs. Bd. 33. S. 117-129.
Lohrl H. 1957. WeiBstorch “erschnabelt” Beute im Wasser H Vogelwarte. Bd. 19. Nr. 1. S. 52-53.
Loudon H. 1909. Meine dritte Reise nach Zentral-Asien und ihre jmithologische Ausbeute H J. Om. Jg. 57.
S. 505-573.
Lovaszi P. 2004. A feher golya (Ciconia ciconia) helyzete Magyarorszagon 1941-2002 kozott П Aquila.
Vol. 111. P. 11-18.
Lovaszi P. 2005. A feher golya (Ciconia ciconia) feszekrakohely-valasztasanak termeszetvedelmi vonat-
kozasai Magyarorszagon H Aquila. Vol. 112. P. 9-14.
Lucas F.A. 1889. Contributions to the natural history of the Commander Islands. B. Description of some
bones of Pallas’ cormorant (Phalacrocorax perspicillatus) 11 Proc. US Nat. Mus. Vol. 12. P. 88-94 + 3
pl-
Lucas F.A. 1995. Contributions to the natural history of the Commander Islands. XL, The cranium of Pal-
las’s cjrmorant H Proc. US Nat. Mus. Vol. 18. P. 717-719 + 2 pl.
Luthin C.S. 1987. Rare visit to Oriental Creasted ibis H Flying Free. Vol. 5. No. 2. P. 4-5.
Lysenko VI. 1994. Pelecanus crispus in the Ukraine H Pelecanus in the formed USSR. IWRB publ. No. 27.
P. 5.
Magyar G., Haderics T., Waliczky Z. et al. 1998. Magzarorszag madarainek nevjegyzeke. Budapest-Szeged.
202 S.
Majumdar N. 1989. Past and present wintering distribution of the White Stork (Ciconia ciconia) in India
and suggestions for its conservation 11 Wei?storch - White Stork. Proc. I Intern. Stork Conserv. Symp.
Schriftenreihe des DDA. Bd. 10. P. 373-378.
Makatsch W. 1974. Die Eier der Vogel Europas. Eine Darstlung der Brutbiologie aller in Europa briitenden
Vogelarten. Neuman Verlag. Bd. 1. 468 S.
Makinnon J., Phillipps K. 2000. A fild guide to the birds of China. Oxford. 586 p.
Mackworth-Praed C.W., Grant C.H.B. 1952. African Handbook of Birds. Ser. 1. Birds of eastern and north-
eastern Africa. Vol. 1. London - N.Y. - Toronto. 836 p.
Malinauskas V, Zurba M. 1999. White Stork - the national bird of Lithuania, results of the census 1994/
1995 П Proc. Intern. Symp. on the White Stork, Hamburg, 1996. Bonn: NABU. P. 265-275.
Mallord J.W., Tyler G.A., Gilbert G. & Smith K.W. 2000. The first case of successful double brooding in the
Great Bittern Botaurus stellaris H Ibis. Vol. 142. P. 672-675.
Marion L., Marion P. 1994. Premieres nidifications reussies de la Grande Aigrette Egretta alba en France,
au lac de Grand-Lieu П Alauda. Vol. 62. Is. 3. P. 149-152.
Martines-Vilalta A., Motis A. 1992. Family Ardeidae (Herons) H Handbook of the Birds of the World. Vol
1 (Ostrich to Ducks). Barcelona: Lynx Ed. P. 376-429.
Martinez Rodriguez E. 1995. Situation actual de la Cigiiena Blanca en Espana H Proc, of the Intern. Symp.
on the White Stork (Western Population). Basel 1994. P. 57-63.
Martin J.P. 2002. Las cigiienas de Alcala. 120 p.
Литература
589
Martin-Mateo M.P. 1990. Malofagos de aves marinas. Especies parasitas daves Pelecaniformes (Insecta,
Mallophaga) 11 Garcia-de-orta. zool. Vol. 17. No. 1-2. P. 37-51.
Marti R. 1999. Results of the 5th International White Stork Census (1994) in Spain 11 Proc. Intern. Symp. on
the White Stork, Hamburg, 1996. Bonn: NABU. P. 61-68.
Martos M.R., Johnson A.R. 1996. Management of nesting sites for Greater Flamingos 11 Colon. Waterbirds.
No. 19. P. 167-183.
Masatomi H., Smirenski S., Momose K. et al. 2002. Status of cranes and storks breeding in the middle Amur
River basin, Russia, in the summer of 1997 11 Biosph. Conserv. Vol. 4. No. 2. P. 87-102.
Matheu E., del Hoyo J. 1992. Family Threskiomithidae (Ibisses and Spoonbils). Handbook of the Birds of
the World. Vol. 1. Barcelona: Lynx Ed. P. 472-506.
Maxwell J., Kale H. 1977. Breeding biology of five species of herons in costal Florida H Auk. Vol. 94
(Oktober). P. 689-700.
McClure H.E. 1974. Migration and survival of the Birds of Asia. Bangkok. 476 p.
McClure H.E., Ratanaworabhan N. 1973. Some ectoparasites of the birds of Asia. San Francisco, Bangkok.
219 pp.
McCracken K.G., Sheldon F.H. 1998. Molecular and osteological heron phylogenies: sources of incongru-
ence//Auk. Vol. 115. Is. 1. P. 127-141.
McKilligan N.G. 1990a. The breeding biology of the Intermediate Egret. Part 1, The phisical and behav-
ioural development of chik, with special reference to sibling aggression and food intake 11 Corella.
Vol. 14. P. 162-169.
McKillgan N. 1990b. Promiscuity in Cattle Egret H Auk. Vol. 107 (April). P. 334-341.
Meise W. 1934. Die Vogelwelt der Mandshurei 11 Abhandl. und Berichte Mus. Tierkunde Dresden. Bd. 18.
Nr. 2. 186 S.
Melville D.S., Leader P.J., Carey G.J. 2000. Movements and biometriks of Black-faced Spoonbills Platalea
minor at Mai Po, Hong Kong in spring 1998 H Conserv. and res. of Black-faced Spoonbills and their
Habitat. Proc, of the Intern, workshop in Tokyo, 12-16 June, 1997. Second edition. 2000. P. 19-26.
Meybohm E., Dahms G. 1975. Uber Altersaufbau, Reifeadler und Ansiedlung beim WeiBstorch (C. ciconia)
im Nordsee-Kiistenbereich // Vogelwarte. Bd. 32. Nr. 1. S. 44-61.
Meybohm E., Fiedler G. 1983. Neue Faile von hohen Alter, Ortstreue, Um- und Femsiedlung und anderen
brutbiologischen Befunden beim WeiBstorch (C. ciconia) // Vogelwarte. Bd. 32. Nr. 1. S. 14-22.
Meyerriecks A. J. 1960. Comparative breeding bechavior of four species of North American herons H Nuttal
Ornithological Club Publication. No. 2. P. 1-158
Michev T., Profirov L. 1989. Der Herbstzug des WeiBstorchs (Ciconia ciconia L.) entlang der bulgarischen
Schwarzmeerkiiste H WeiBstorch - White Stork. Proc. I Int. Stork Conserv. Symposium. Schriftenreihe
des DDA. Bd. 10. S. 281-295.
Min Byung Yoon, Won Pyong Oh. 1976. A winter seabird survey on the shore south coast of Korean
Penisula (Mogro-Wandj-Yeosu area) П Misc. Rep. Yamashina Inst. Omithol. Vol. 18. No. 1 (45). P. 53-
67.
Mock D.W. 1985. Siblicidal brood reduction: the prey-size hypothesis H Amer. Nat. Vol. 125. P. 327-343.
Moo-Boo Yoon. 1995. Wild birds of Korea. Seoul. 548 p.
Morioka H., Dickinson E.C., Hiraoka T., Allen D., Yamasaki T. 2005. Types of Japanese birds. H Nai. Sci.
Mus. Monographs (Tokyo). No. 28. P. 1-154.
Moyer J.T. 1955. Further notes on Korea birds H Auk. Vol. 72. No. 4. P. 428^131.
Moyer J.T. 1957. The birds of Miyake-jima, Japan H Auk. Vol. 74. P. 215-228.
Muire O.J. 1959. Fauna of the Aleutian Islands and Alaska Penisula H North Am. Fauna. Vol. 61. P. 1-406.
Muller Ch.Y 1984. Bestandsentwicklung und Zugverhalten der Loffler (Platalea leucorodia L.) im osterre-
ichisch und ungarischen Raum H Egretta. Bd. 27. Nr. 2. S. 45-67.
Munisterhjelm L. 1922. Some ornithological notes from a journey to Saghalin in 1914 H Meddelanden fran
Goteborgs Musei Zoologiska Avdelning. Nr. 23. 112 p.
Munro J.F., Crompton D.W.T., Stoddart R.C. 1995. Andracantha tunitae (Acanthocephala) in the shag
(Phalacrocorax aristotelis) from Shetland, Scotland H J. Parasitol. Vol. 81. No. 3. P. 496-498.
Munteanu D., Ranner A. 1997. Great White Egret H The EBCC Atlas of European Breeding Birds: Their
Distrib. and Abund. London: T & A.D. Poiser. P. 48^19.
590
Литература
Murata К., Miyashita М., Nagase К., Komiya Т., Matsushima К. 1988. Sex determination in the Eastern ,
White Stork, Ciconia c. boyciana by bill measurements and disriminant analysis H J. Jpn. Assoc. Zoos
and Aquariums. Vol. 30. P. 43^17.
Muzinic J., Rasajski J. 1992. On food and feeding of the White Stork, Ciconia c. ciconia, in the Central
Balkan H Okol. Vogel. Vol. 14. Is. 2. P. 211-223.
Nankinov D.N. 1994. Wintering of the White Storks in Bulgaria П Research notes on avian biology: selected
contribut. from the 21st Intemat. Omithol. Congr. J. Om. Vol. 135. P. 208.
Nasirwa O. 2000. Conservation status of flamingos in Kenya H Waterbirds. Vol. 23. P. 47-51.
Nazarov Y.N., Shibaev Y.V., Litvinenko N.M. 2001. Birds of the Far State Marine Reserve (South Pri-
morye) 11 The Status of Env. and biota of the SW part of Peter the Great Bay and the Tumen Riv. Mouth.
Vladivostok. Vol. 3. P. 163-199.
Nechaev V.A. 1998. Nachweis des Schwarzstimlofflers Platalea minor bei Wladivostok H Omithol. Mitt.
Bd. 50. Nr. 5.
Nedzinskas V. 1987. Sventasis ibis H Miisu gamta. No. 1. P. 28.
Nelson J.B., Mackay A., Busby J. 2005. Pelicans, Cormorants, and their Relatives: Pelecanidae, Sulidae,
Phalacrocoracidae, Anhingidae, Fregatidae, Phaethontidae. Bird Families of the World. Oxford Univer-
sity Press, USA. 661 p.
Nemeth E., Schuster A. 2005. Spatial and temporal variation of habitat and prey utilization in the Great
Wtite Egret et Lake Neusiedl, Austria H Bird Study. Vol. 52. Is. 2. P. 129-136.
Neuschulz F. 1981. WeiBstorche {Ciconia ciconia) holen im Flug Nahrung aus dem Wasser 11 Omithol. Mitt.
Bd. 33. Nr. 1. S. 22-24.
Newton L, Wyllie I., Asher A. 1994. Populations in Great Bitterns // Brit. Birds. Vol. 87. No. 1. P. 22-25.
Niethammer J. 1967. Storche in Afghanistan H Vogelwarte. Bd. 24. Nr. 1. S. 42^14.
Nowakowski J. 2003 Habitat structure and breeding parameters of the White Stork Ciconia ciconia in the
Kolno Upland (NE Poland) H Acta Om. Vol. 38. Is. 1. P. 39^16.
Nysewander D.R. 1986. Cormorants {Phalacrocorax spp.) H The breeding biology and feeding ecology of
marine birds in the Gulf of Alaska. Environ. Assess. Alaskan Cont. Shelf, Final Rep. Anchorage: NOAA.
P. 207-236.
Olioso G. 2002. Unusual fishing behaviour of Little and Western Reef Egrets H Omithos. Vol. 9. P. 218—
219.
Ornithological Society of Japan. 2000. Check-list of Japanese birds. 6-th ed. Tokyo. 345 p.
Orta J. 1992. Family Fregatidae (Frigatebirds) H J. del Hoyo, A. Elliot, J. Sargatal (eds.). Handbook of the
Birds of the World. Vol. 1. Barcelona: Lynx Ed. P. 362-374.
Ortego-Ruano J., Graves J. A. 1991. Reverse mounting during the courtship of the European shag Phalacro-
corax aristotelis 11 Condor. Vol. 93. No. 4. P. 859-863.
Osa Y, Watanuki Y. 2002. Status of seabirds breeding in Hokkaido H J. Yamashina Inst. Omithol. Vol. 33.
No. 2. P. 107-141.
Ozgo M., Bogucki Z. 1999. Home range and intersexual differences in the foraging habitat use of a White
Stork {Ciconia ciconia) breeding pair Proc. Intern. Symp. on the White Stork, Hamburg, 1996. Bonn:
NABU. P. 48 W92.
Palmer R.S. (ed.). 1962. Handbook of North American birds. Vol. 1. Loons through flamingos. New Haven;
London: Yale Univ. Press. 567 p.
Palmgren P. 1935. Ein Exemplar von Phalacracoraxperspicillatus Pall, in den Sammlungen des zoologis-
chen Museums in Helsingfors H Omis Fenn. Vol. 12. P. 78-80.
Parasharya B.M., Bhat H.R. 1987. Unusal feeding strategies of the Little Egret and Pond Heron 11 Pavo.
Vol. 25. P. 13-15.
Patrikeev M. 2004. The birds of Azerbaijan. Sofia-Moscow. 380 p.
PejaN., Sarigul G., Siki M., Crivelli A.J. 1996. The Dalmatian pelican, Pelecanus crispus, nesting in Med-
iterranien Lagoons in Albania and Turkey H Colon. Waterbirds. Vol. 19. P. 184-189.
Peter J.M. 1994. A feeding interaction between Great Egret Ardea alba and Fairy Tern Sterna nereis 11
Austral. Bird Watcher. Vol. 15. P. 228.
Petrov T., lankov P., Georgiev D. 1999. Population status of the White Stork {Ciconia ciconia) in Bulgaria
in the years 1994/95 H Proc. Intern. Symp. on the White Stork, Hamburg, 1996. Bonn: NABU. P. 241-247.
Литература
591
Petry М.А., Fonseca V.S.D.S. 2005. Breeding success of the colonist species Bubulcus ibis (Linnaeus,
1758) and four native species // Acta Zool., Stockholm. Vol. 86. P. 217-221.
Pezzo, F., Gosler, A.G. 2005. Evidence of prenuptial moult in the Little Bittern Ixobrychus minutus II Ring-
ing & Migration. Vol. 22. P. 129-132.
Pinowska B., Buchholz L., Grobelny S. et al. 1991. Skipjacks Elateridae, weevils Curculionidae, orthopter-
ans Orthoptera earwigs Dermaptera in the food of White Stork Ciconia ciconia (L.) from the Mazurian
Lakeland. // Population of White Stork in Poland. Part 2. Some asp. of the biol. and ecol. of White Stork.
Krakow. P. 87-106.
Pinowski J., Pinowska B., de Graaf R. et al. 1991. Influence of feeding habitat on prey capture rate and diet
composition of White Stork Ciconia ciconia (L.) // Population of White Stork Ciconia ciconia (L.) in
Poland. Part II. Some asp. of the biol. and ecol. of White Stork. Krakow. P. 59-85.
Piotrowska M. 2003. Zimowanie ptakiiw na lubelskim odcinku Wisiy w latach 1985-2000 // Notatki Omi-
tol. Vol. 44. Is. 2. P. 141-149.
Platteeuw M., Kiss J.B., Zhmud M.Ye., Sadoul N. 2004. Colonial waterbirdsand their habitat use in the
Danube Delta, as an example of a large-scale natural wetlands // RIZA report, 002. 168 p.
Poluda A.M., Chemichko I.I., Serebryakov V.V. et al. 1997. The Cormorant Phalacrocorax carbo in Ukraine
// Ekologia Polska. Vol. 45. No. 1. P. 105-110.
Post H., Rueb F., Belterman F. 1991. Geschlechtsbestimmung beim WeiBstorch (Ciconia ciconia) И J. Om.
Bd. 132. Nr. 1. S. 99-101.
Pratt H.M., Winkler D.W 1985. Clutch size, timing of laying, and reproductive success in a colony of Great
Blue Herons and Great Egrets // Auk. Vol. 102. Is. 1. P. 49-63.
Prestt I. 1970. Organochlorine pollution of rivers and the Heron (Ardea cinerea) И IUCH Pubis New Ser.
No. 17. P. 95-102.
Prestt L, Bell T. 1973. Life in a heronry // Birds. Vol. 4. No. 12. P. 316-321.
Priklonski S.G., Sapetina I.M. 1990. Game statistics in the USSR // Manag. Waterf. Populat., Proc, of an
IWRB Symp., (Astrakhan, USSR, 2-5 Oct. 1989), IWRB Spec. publ. No. 12. P. 113-114.
Prinzinger R., Hund K. 1982. Beinkoten beim WeiBstorch (Ciconia ciconia) - eine thermoregulatorische
Verhaltensweise zur Warmeabgabe durch die Beine // Okol. Vogel. Bd. 4. Nr. 1. S. 82-83.
Profus P. 1986. Zur Brutbiologie und Bioenergetik des WeiBstorchs in Polen // Artenschutzsymposium
WeiBstorch. Beih. Veroff. Landschaftspflege Baden-Wwttemberg. Karlsruhe. Bd. 43. S. 205-220.
Profus P. 1991. The breeding biology of White Stork Ciconia ciconia (L.) in the selected area of Southern
Poland // Population of White Stork Ciconia ciconia (L.) in Poland. Part 2. Some asp. of the biol. and
ecol. of White Stork. Krakow. P. 11-57.
Puglisi L., Baldaccini E. 2000. Nocturnal flights during spring migration in the Bittern (Botaurus stellaris)
H Die Vogelwarte. Bd. 40. Nr. 3. P. 235-238.
Puglisi L., Bretagnolle V, in press. New data on the breeding biology of the secretive and declining Eur-
asian Bittern, and a synthesis of the current state of knoledge // Waterbirds.
Puglisi L., Cima O., Baldaccini E. 1997. A study of the sesonal booming activity of the Bittern Botaurus
stellaris’, what is the biological significance of the booms? // Ibis. Vol. 139. P. 638-645.
Puglisi L., Pagni M., Bulgarelli C., Baldaccini N.E. 2001. The possible function of calls organization in the
bittern (Botaurus stellaris) П Ital. J. Zool. Vol. 68. P. 315-321.
Qishan W. 1991. Wintering ecology of Oriental White Storks in the lower reaches of the Changjiang river,
central China // M.C. Coulter et al. (eds.). Biol, and Conserv. of the Oriental White Stork Ciconia
boyciana. P. 99-107.
Qishan W. 1995f. Oriental White Storks of Shengjin Lake and Strategies for their Protection // Cranes and
Storks of the Amur River. The Proc, of the Intern. Worksh. Moscow: Arts. Literat. Publ. P. 104-107.
Qishan W. 1995a. Current Studies on Oriental White Stork // J. of Anhui Univ. Nat. Sci. Edition # 1. P. 82-
99.
Qishan W., Coulter M.C. 1995. Statement of the Working group on Oriental White Storks // Cranes and
Storks of the Amur River. The Proc, of the Intern. Worksh. Moscow: Arts. Literat. Publ. P. 202-204.
Ison S.L. 2005. Correction of erroneous records of cormorants from archeological sites in Alaska // Condor.
Vol. 107. P. 930-933.
Radde G. 1884. Omis Caucasica. Kassel. 592 S.
592
Литература
Radde G., Walter A. 1889. Die Vogel Transcaspiens // Omis. Jg. 5. S. 1-128.
Radvanyi O. 1943. Ein siebener Gelege des Weissen Storches // Aquila. Bd. 50. S. 418.
Rainer Merhout. 1974. Bestandsuntersuchung in der Graureiherkolonie bei Potsdam-Geltow // Naturschutzarb.
Berlin und Brandenburg. Bd. 10. H. 3. S. 78-81.
Ranner A., Szinovatz H. 1987. Nahrungsschmarotzen beim WeiBstorch (Ciconia ciconia) И Egretta. Bd. 30.
S. 89-90.
Rejman B. 1989. Der WeiBstorch in der Tschechei 1934-1984 // WeiBstorch - White Stork. Proc. I Int. Stork
Conserv. Symposium. Schriftenreihe des DDA. Bd. 10. S. 69-72.
Rejman B. 1996. Die WeiBstorchbestande in der Tschechischen Republik nach dem 4. Intemationalen Zen-
sus von 1984 bis 1994 // Jubilawnsband WeiBstorchs - Jubilee Edition White Stork. Tagungsbandreihe
des Storchenhofes Loburg im MRLU-LSA. Bd. 3. S. 25-28.
Rejman B. 1999. Der Bestand des WeiBstorchs (Ciconia ciconia) in der Tschechischen Republik in den
Jahren 1994 und 1995 // Proc. Intern. Symp. on the White Stork, Hamburg, 1996. Bonn: NABU. S. 189-
197.
Rendon-Martos M., Vargas J.M., Rendon M.A. et al. 2000. Nocturnal movements of breeding Greater Fla-
mingos in southern Spain. // Waterbirds. Vol. 23. P. 9-19.
Renzhu P., Kemin F. 1995. Oriental White Stork in Sanjiang Plain // Cranes and Storks of the Amur river.
The Proc, of the Intemat. Worksh. Moscow: Arts. Literat. Publ. P. 100-102.
Reza Khan M.A. 1989. Migratory status of the White Stork (Ciconia ciconia L.) in the United Arab Emir-
ates // WeiBstorch - White Stork. Proc. I Intern. Stork Conserv. Symp. Schriftenreihe des DDA. Bd. 10.
P. 325-326.
Rheinwald G. 1989. Versuch einer Bilanz // WeiBstorch - White Stork. Proc. I Intern. Stork Conserv. Symp.
Schriftenreihe des DDA. Bd. 10. S. 221-227.
Riegel M., Winkel W. 1971. Uber Todesursachen beim WeiBstorch (C. ciconia) an Hand von Ringftmdang-
aben // Vogelwarte. Bd. 26. Nr. 1. S. 128-135.
Robards M.D., Piatt J.F., Wohl R.D. 1995. Increasing frequency of plastic particles ingested by seabirds in
the subarctic North Pacific // Mar. Poll. Bull. Vol. 30. No. 2. P. 151-157.
Robinson H.C. 1931. Some Birds from Siam and Laos (Middle Mekong). With Preface and Notes by C.
Boden Kloss // Ibis. Vol. 1. No. 2. P. 319-334.
Robinson H.C., Kloss C.B. 1911. The birds from the Northern Portion of the Malay Penisula, including the
Islands of Langkawi and Terutan: with Notes on other rare Malayan Species from the Southern Districts
// Ibis. Vol. 5. No. 17. P. 10-80.
Robson G. 2000. A field guide of the birds of South-East Asia / N. Collar (ed.). Singapore. 504 p.
Rohrbacher F. 2000. Delaware Records Committee Annual Repopt for 2000 // Delaware Ornithologist.
Vol. 32. P. 55-59.
Rohwer S., Filardi C.E., Bostwick K.S., Peterson A.T. 2000. A critical evaluation of Kenyon’s Shag (Pha-
lacrocorax [Stictocarbo] kenyoni) II Auk. Vol. 117. No. 2. P. 308-320.
Rosa G., Araujo A., Martins J. 1999. The present situation of the White Stork Ciconia ciconia in Portugal //
Proc. Intern. Symp. on the White Stork, Hamburg, 1996. Bonn: NABU. P. 49-59.
Rosa S., Palmeirim J.M., Moreira F. 2003. Factors affecting waterbird abundance and species richness in an
increasingly urbanized area of the tagus estuary in Portugal // Waterbirds. Vol. 26. No. 2. P. 226-232.
Rose P.M., Scott D.A. 1994. Waterfowl population estimates // IWRB Publ. No. 29. 102 p.
Rose P.M., Scott D.A. 1997. Waterfowl Population Estimates. 2nd ed. // Wetland Intemat. publicat. Vol. 44.
106 p.
Rozdina O.I., Sotnikova E.I., Skakunova E.N. 1991. Notes on Oriental White Storks, Ciconia boyciana, in
the Moscow Zoo, 1985-88 // M.C. Coulter et al. (eds.). Biol, and Conserv. of the Oriental White Stork
Ciconia boyciana. P. 203-207.
Rydzewski W. 1973. Longevity records. Ill // Ring. Vol. 76. P. 63-70.
Sackl P. 1987. Uber saisonale und regionale Unterschiede in der Emahrung und Nahrungswahl des WeiBs-
torches (Ciconia c. ciconia) im Verlauf der Brutperiode // Egretta. Bd. 30. Nr. 2. S. 49-80.
Sackl P., Strazds M. 1997. Ciconia nigra Black Stork // W.J.M. Hagemeijer, M.J. Blair (eds.). The EBCC
Atlas of European Breeding Birds: Their Distribution and Abundance. London: T & A.D. Poyser. P. 56-
57.
Литература
593
Salama W., Grieve A. 1996. The Zaranik experience // Sandgrouse. Vol. 18. No. 1. P. 14-17.
Samusenko I. 1999. Population development, current population status and biology of the White Stork in
Belarus // Proc. Intern. Symp. on the White Stork. Bonn: NABU. P. 277-278.
Samusenko I. 2000. Some aspects of White Stork Ciconia ciconia population dynamics in the region of
Chernobyl’s accident // Proc. Int. Confer, and 13th Meet. Eur. Bird Census Council, Pamu, Estonia /
Bird Census News. Vol. 13. Is. 1-2. P. 157-160.
Sato M., Mizuno N., Takemaru K., Matsuki K. 1968. Avifauna of Kinkazan Island, Miyagi Prefecture.
Especially winter birds // Tori. P. 356-378.
Satori J. 1943. Siebener Gelege des Weissen Storches // Aquila. Bd. 50. S. 419.
Sauter U., Schiiz E. 1954. Bestandsveranderungen beim WeiBstorch. Dritte Ubersicht, 1939-1953 // Vogel-
warte. Bd. 17. Nr. 1. S. 81-100.
Schaub M., Kania W., Koppen U. 2005. Variation of primary production during winter induces synchrony in
survival rates in migratory white storks Ciconia ciconia 11 J. Animal Ecol. Vol. 74. Is. 4. P. 656-666.
Schenk J. 1907. Die Heuschreckenplage auf dem Hortobagy im J. 1907 und die Vogelwelt // Aquila. Bd. 14.
S. 223-251.
Schenk J. 1934. Die Vogelberingungen des KonioL Ungarischen Omithologischen Institutes in den Jahren
1931-1932 // Aquila. Nr. 38-39. S. 216-224.
Schmidt E. 1977. Auffalende Zunahme des Silberreiers (Casmerodius albus) in Ungam im Jahre 1976 //
Egretta. Bd. 20. S. 68-70.
Schmidt K. 1989. Bigamie beim WeiBstorch (Ciconia ciconia) im Werratal // Om. Mitt. Bd. 41. Nr. 9.
S. 221-222.
Schneider D.C., Hunt G.L., Jr. 1984. A comparison of seabirds diets and foraging distribution around the
Pribilof Islands, Alasrf // Marine birds: their feeding ecology and commercial fisheries relationships.
Ottawa: Can. Wildl. Serv. Spec. Publ. P. 86-95.
Schogolev I.V. 1996a. Fluctuations and trends in breeding populations of colonial waterbirds in Dnestr
delta, Ukraine, Black Sea // Colonial Waterbirds. Vol. 19 (Spec. publ. No. 1). P. 91-97.
Schogolev I.V. 1996b. Migration and wintering grounds of Glossy Ibises (Plegadis falcinellus) ringed at the
colonies of Dnestr delta, Ukraine, Black Sea // Ibid. P. 152-158.
Schonwetter M. 1960. Handbuch der Oologie. Bd. 1 // Zool. Staatsinst. und Zool. Mus. Hamburg Berlin.
S. 65-128.
Schonwetter M. 1967. Handbuch der Oologie. Berlin. Bd. 1. 929 S.
Schreiber E.A., Norton R.L. 2002. Brown Booby (Sula leucogaster) И The Birds of North America. Phila-
delphia, PA. No. 649. P. 1-28.
Schreiber E.A., Schreiber R.W., Schenk G.A. 1996. Red-footed Booby (Sula sula) // The Birds of North
America. Philadelphia: The Academy of Natural Sciences; Washington, D.C.: The American Ornithol-
ogists’ Union. No. 241. P. 1-24.
Schulz H. 1987. Thermoregulatorisches Beinkoten des WeiBstorchs (Ciconia ciconia). Analyse des Verhalt-
ens und seiner Bedeutung fur Verluste bei beringten Storchen im afrikanischen Winterquartier // Vogel-
warte. Bd. 34. Nr. 2. S. 107-117.
Schulz H. 1988. WeiBstorchzug - Okologie, Gefahrdung und Schutz des WeiBstorchs in Afrika und Nahost.
Konigslutter-Lelm. 460 S.
Schulz H. 1993. Der WeiBstorch: Lebensweise und Schutz. Augsburg: Naturbuch-Verl. 64 S.
Schulz H. 1994. Zur Bestandssituation des WeiBstorchs - Neue Perspektiven fur den “Vogel des Jahres
1994”? // Berichte zum Vogelschutz. Bd. 32. S. 7-18.
Schulz H. 1996. Alfaro - die groBte urbane WeiBstorchansiedlung der Welt // Jubilaumsband WeiBstorch -
Jubilee Edition White Stork. Tagungsbandreihe des Storchenhofes Loburg in MRLU-LSA. Bd. 3. S. 51.
Schulz H. 1998. Ciconia ciconia White Stork // BWP Update. Vol. 2. Is. 2. P. 69-105.
Schulz H. 1999. Der Weltbestand des WeiBstorchs (Ciconia ciconia) - Ergebnisse des 5. Intemationalen
WeiBstorchzensus 1994/95 // Proc. Intern. Symp. on the White Stork, Bonn: NABU. S. 335-350.
Schumann A. 1932. Vemichtung einer grossen Anzahl Storche und Wildtauben durch ein ausserordentlich
Hagelwetter // Kocsag. Bd. 32. Nr. 3^4. S. 116-117.
Schuster A., Nemetz E., Grull A., Rossler M. 1998. Der Seidenreiher (Egretta garzetta) - ein neuer Brutvo-
gel fur Osterreich // Egretta. S. 61-66.
594
Литература
Schiiz Е. 1944. Nesterwerb und Nestbesitz beim WeiBen Storche // Z. Tierpsychol. Bd. 6. S. 1-5.
Schiiz E. 1953. Die Zugscheide des WeiBstorches nach den Beringungsergebnissen // Bonner zool. Beitr.
Bd. 4. S. 31-72.
Schiiz E. 1962. Uber die nordwestlichen Zugscheide des WeiBen Storchs // Vogelwarte. Bd. 21. Nr. 4. S. 269-
290.
Schiiz E. 1963. Uber die Zugscheiden des WeiBstorchs in Afrika, Ukraine und Asien// Vogelwarte. Bd. 22.
Nr. 2. S. 65-70.
Schiiz E., Schiiz T. 1932. Brutbeobachtungen am Storch (Ciconia ciconia) // Beitr. Fortpflanzungsbiol.
Vogel. Bd. 8. S. 18-21.
Sedlag U. 1983. Vom Aussterben der Tiere. Leipzig, Jena, Berlin: Urania-Verlag. 216 S.
Severinghaus L.L., Brouwer K., Chan S., Chong J.R., Coulter M.C., Poorter E.P.R., Wang Y. 1995. Action
plan for the Black-faced Spoonbill Platalea minor. Taipei: Wild Bird Society of Taiwan (Bird Conserva-
tion Research Series No. 10).
Shannon P.W. 2000a. Plumages and molt patterns in captive Caribbean flamingos // Waterbirds. Vol. 23.
P. 160-172.
Shannon P.W. 2000b. Social and reproductive relationships of captive Caribbean Flamingos // Waterbirds.
Vol. 23. P. 173-178.
Shaw Tsen-Hwang. 1936. The birds of Hoppei province // Biol. Sinica. Beijing. Vol. 15. No. 1. 529 p.
Sheldon F.H. 1987. Phylogeny of herons estimated from DNA-DNA hybridization data // Auk. Vol. 104.
Is. 1. P. 97-108.
Shemazarov E. 1999. Distribution and numbers of Ciconia c. asiatica in Central Asia // Proc. Intern. Symp.
on the White Stork, Hamburg, 1996. Bonn: NABU. P. 331-334.
Sibley F.C., Clapp R.B. 1967. Distribution and dispersal of Central Pacific Lesser Fregatebirds, Fergata
ariel // Ibis. Vol. 109. No. 3. P. 328-337.
Sibley Ch.G., Ahlquist J.E. 1990. Phylogeny and classification of birds a study in molecular evolution. New
Haven-London: Yale Univ. Press. 976 p.
Sibley Ch.G., Monroe B.L., Jr. 1990. Distribution and Taxonomy of Birds of the World. New Haven, Con-
necticut: Yale University Press, i-xxiv; 1111 p.
Siegel-Causey D. 1988. Phylogeny of Phalacrocracidae // Condor. Vol. 90. P. 885-905.
Siegel-Causey D. 1991. Systematics and biogeography of North Pacific shags, with a description of a new
species // Occas. Pap. Mus. Nat. Hist. Univ. Kans. No. 140. P. 1-17.
Siegel-Causey D., Lefevre C., Savinetskii A.B. 1991. Historical diversity of cormorants and shags from
Amchitka Island, Alaska // Condor. Vol. 93. P. 840-852.
Siegel-Causey D., Litvinenko N.M. 1993. Status, ecology and conservation of shags and cormorants of the
temperate North Pacific // The status, ecology and conservation of marine birds of the North Pacific.,
Ottawa: Can. Wildl. Serv. Spec. Publ. P. 122-130.
Siegfrid W.R. 1970. Mortality and dispersal of ringed cattle egrets // Ostrich. Vol. 41. No. 1-2. P. 122-135.
Siegfrid W.R. 1971. Feeding activity of cattle egret // Ardea. Vol. 59. No. 1-2. P. 38^46.
Siegfrid W.R. 1972. Breeding success and reproduction outputs of the Cattle Egret // Ostrich. Vol. 43.
No. 1. P. 43-55.
Skov H. 1999. The White Stork (Ciconia ciconia) in Denmark // Proc. Intern. Symp. on the White Stork,
Hamburg, 1996. Bonn: NABU. P. 111-131.
Snow D.W., Perrins C.M. (eds.). 1998. The birds of the Western Palearctic. Vol. 1. - Non-Passeriformes.
Oxford - N.Y: Oxford Univ. Press. 1008 p.
Sonobe K., Usui S. (eds.). 1993. A field Guide to the Waterbirds of Asia. Tokyo: Wild Bird Society of Japan.
224 p.
Sowls A.L., Hatch S.A., Lensink C.J. 1978. Catalog of Alascan Seabird Colonies., Anchorage: US Fish and
Wildl. Serv. 32 p + 153 maps and tables.
Spitzenberger F. 1990. Stellungnahme zu “Kormoranproblematik in den Fischenvieren” BN Komeufung,
DY Sturzlbaum. 3.1. 1990 // Vogelschutz in Osterreich. Nr. 6. S. 92-94.
Springer A.M., Piatt J.F., van Vliet G. 1996. Seabirds as proxies of marine habitats and food webs in the
western Aleutian Arc // Fish. Oceanogr. Vol. 5. No. 1. P. 45-55.
Stafford J. 1963. The census of heronries 1960-1961 // Bird St. Vol. 10. No. 2. P. 29-33.
Stafford J. 1969. The census of heronries 1962-1963 // Bird St. Vol. 16. No. 2. P. 83-88.
Литература
595
Starka U. 1988. Das Projekt zur Wiederansiedlung des Kormorans im Tullen Feld // Vogelschutz in Osterre-
ich. Nr. 2. S. 83-84.
Starka U. 1991. Verbreitung sommerliche und winterliche Bestandsentwicklung des Kormorans in Oster-
reich // Ibid. Nr. 6. S. 48-63.
Stegmann B. 1929. Die Vogel Sud-Ost Transbaikalens // Ежегодн. Зоол. муз. АН СССР, Т. 29. С. 83-242.
Stegmann В. 1930. Die Vogel des dauro-mandschurischen Ubergangsgebites // J. Om. Jg. 78. H. 4. S. 389-
471.
Stegmann B. 1936. Uber das Flugvermogen der ausgestorbenen Scharbe Phalacrocorax perspicillatus Pall. //
Omithol. Monatsber. Jg. 44. Nr. 5. S. 140-153.
Steinbacher J. 1936. Untersuchungen fiber die Nahrungsbiologie des WeiBen Storchs (Ciconia c. ciconia
(L.)) in OstpreuBen 1933 und 1934 // Schr. phys.-dkon. Ges. Konigsberg. Bd. 69. S. 23-36.
Stejniger L. 1883. Contributions to the history of the Commander Islands // Proc. U.S. Natl. Mus. Vol. 6.
P. 58-89.
Stejniger L. 1885. Results of ornithological explorations in the Commander Islands and in Kamtchatka //
Bull. U.S. Natl. Mus. No. 29. P. 1-382.
Stejniger L. 1889. Contributions to the history of the Commander Islands A. Contributions to the history of
Pallas’ cormorant // Proc. U.S. Natl. Mus. Vol. 12. P. 83-88.
Steijniger L. 1898. The birds of the Kuril Islands // Proc. U.S. Natl. Mus. Vol. 21. P. 269-296.
Steller G.V 1781. Topographishe und physikalische Beschreibung der Beringsinsel, welche im ostlichen
Weltmeer an der Kiiste von Kamtchatka liegt // Neue Nordische Beytrage [zur Physikalischen und Geog-
raphischen Erd- und Volkerbeschreibung, Naturgeschichte, und Oekonomie]. Bd. 2. St.-Petersb.; Leipzig.
S. 255-301.
Sterbetz I. 1960. Az ustokosgem a saseri rezervatumban // Aquila. P. 67-68.
Stichmann W. 1962a. Neues fiber den Graureiher in Westfalien // Natur und Heimat (BRD). Bd. 22. H. 1.
S. 22-26.
Stichmann W. 1962b. Der Graurehierbestand der Bundesrepublik Deutschland // Ibid. S. 26-29.
Stichmann W. 1968. Der Graureher, der groBte Koloniebrfiter Westfalens / Drei fur den Unterreicht bedeut-
same Aspecte // Naturkunde Westfalen. Bd. 4. H. 2-3. S. 57-62.
Stollmann A. 1989. Der WeiBstorch in der Slowakischen Sozialistischen Republik // WeiBstorch - White
Stork. Proc. I Int. Stork Conserv. Symp. Schriftenr. des DDA. Bd. 10. S. 73-76.
Stork H.-J. 1999. WeiBstorche auf den Seychellen // Proc. Int. Symp. on the White Stork. Bonn: NABU.
S. 557-558.
Stresemann E., Stresemann V. 1966. Die Mauser der Vogel // J. Om. Bd. 107. Sonderheft. S. 1^448.
Struwe B., Thomsen K.-M. 1991. Untersuchungen zur Nahrungsokologie des WeiBstorches (Ciconia cico-
nia, L. 1758) in Bergenhusen 1989 // Corax. Bd. 14. Nr. 3. S. 210-238.
Studer-Thiersch A. 2000. What 19 years of observation on captive Greater Flamingos suggests about adap-
tations to breeding under irregular conditions // Waterbirds. Vol. 23. P. 150-159.
Sturzlbaum H. 1990. Kormoranproblematik in den Fischrevieren // Vogelschutz in Osterreich. Nr. 6. S. 90-92.
Survey of Birds in Japan. 1973. Wild Bird Society of Japan. Env. Agency.
Survey of Birds in Japan. 1980. Wild Bird Society of Japan. Env. Agency. 556 p.
Svensson L., Grant P., Mullamey K., Zetterstrom D. 2002. Collins Bird guide. London. 392 p.
Swift J.J. 1960. Densite des nids et notion de territoire chez le flamant de Camargue // Alauda. Vol. 28.
No. 1. P. 1-14.
Taczanowski L. 1891-1893. Faune omithologique de la Siberia orientale. Pt. 1-2. St.-Petersb. 1278 p.
Takahashi T. 1937a. On the unrecordet bird from South Saghalien (1) // Toti. Vol. 9. No. 45. P. 464-472.
Takahashi T. 1937b. A list of the birds from Saghalien, depended on the discription to arrangement of birds
on the speciments in the Saghalien locality Museum. Repr. from the Transactions of the Saghalin Island
//Auk. Vol. l.No. l.P. 1-122
Tamm J. 2006. Zur Avifauna der Wiisten und Halbwtisten der Ostlichen Kasachstan // Omithol. Mitt. Nr. 1.
S. 6-17.
Tamura M., Higuchi H., Shimazaki H. et al. 2000. Satellite observation of movements and habitat condi-
tions of Red-crowned Cranes and Oriental White Storks in East Asia // Global Environm. Res. Vol. 4.
P. 207-217.
596
Литература
Thayar J.E., Bangs О. 1916. A collection of birds from Saghalin Island // Auk. Vol. 33. No. 1. P. 43^48.
The EBCC Atlas of European Birds. 1997. (See Hagemeijer & Blair, 1997).
Thiede W. 1996. Bemerkenswerte faunistische Feststellungen in Europa 1992/1993 // Omithol. Mitt. Bd. 48.
Nr. 3. S. 59-68.
Thomsen K.-M., Struwe B. 1994. Vergleichende nahrungsokologische Untersuchungen an WeiBstorch-Bmt-
paaren (Ciconia ciconia) in Stapelhohn und im Kreis Herzogtum Lauenburg // Corax. Bd. 15. Nr. 4.
S. 293-308.
Threatened birds of Asia. 2001 // Oriental Stork. P. 194-222.
Tichy H. 1996. Zimni vyskyt capu, Ciconia ciconia, Ciconia nigra, na Lounsku // Sb. Okres. mus. Moste. R.
prirodoved. Vol. 18. P. 76.
Tischler F. 1941. Die Vogel OstpreuBens und seiner Nachbargebiete. Konigsberg; Berlin. Bd.2. S. 721—
1304.
Tomek T. 1999. The birds of North Korea. Nonpasseriformes // Acta Zool. Cracov. Vol. 42. No. 1. 217 p.
Tomialojc L. 1990. Ptaki Polski, Rozmieszczenie i liczebnose. Warszawa: PWN. 462 s.
Tomialojc L., Stawarczyk T. 2003. Awifauna Polski. Rozmieszczenie, liczebnose i zmiany. Wroclaw: “pro
Natura”. T. 1. 439 s.
Topola R. 2004. Oriental White Stork (Ciconia boyciana). European Studbook #2., Lodz Zoo. 32 p.
Tortosa F.S., Manez M., Barcell M. 1995. Wintering White Storks (Ciconia ciconia) in South West Spain in
the years 1991 and 1992 // Vogelwarte. Vol. 38. Is. 1. P. 41—45.
Tortosa F.S., Redondo T. 1992. Motives for parental infanticide in White Storks Ciconia ciconia // Omis
Scand. Vol. 23. Is. 2. P. 185-189.
Transche N. von. 1965. Die Vogelwelt Lettlands. Hannower-Dohen. 516 S.
Treep J.M. 2000. Flamingos presumably escaped from captivity find suitable habitat in Western Europe //
Waterbirds. Vol. 23. P. 32-37.
Tryjanowski P., Kuzniak S. 2002. Population size and productivity of the White Stork Ciconia ciconia in
relation to Common Vole Microtus arvalis density // Ardea. Vol. 90. Is. 2. P. 213-217.
Tryjanowski P., Sparks Т.Н., Profus P. 2005. Uphill shifts in the distribution of the white stork Ciconia
ciconia in southern Poland: the importance of nest quality // Diversity and Distributions. Vol. 11. P. 219-
223.
Tryjanowski P., Sparks Т.Н., Ptaszyk J., Kosicki J. 2004. Do White Storks Ciconia ciconia always profit
from an early return to their breeding grounds? // Bird Study. Vol. 51. Is. 3. P. 222-227.
Tsachalidis E.P., Goutner V. 2002. Diet of the White Stork in Grece in relation to habitat // Waterbirds.
Vol. 25. Is. 4. P. 417—423.
Tsachalidis E.P., Liordos V, Goutner V. 2005. Growth of White Stork Ciconia ciconia nestlings // Ardea.
Vol. 93. Is. 1. P. 133-137.
Tucker G.M., Heath M.F. (eds.). 1994. Birds in Europe: their conservation status. Cambridge, UK. (BirdLife
Conserv. Ser., No. 3). 600 p.
Tyler G.A. 1994. Management of reedbeds for Bitterns and opportunities for Reedbed creation // RSPB
Conserv. Rev. Vol. 8. P. 57-62.
Tyler G.A., Smith K.W., Burges D.J. 1998. Reedbed management and breeding bitterns Botaurus stellaris in
the UK // Biol.Cons. Vol. 86: 257-266.
Uchida H., Matsuda T. 1990. Colonial Breeding of the Yellow Bittern Ixobrychus sinensis // Jap. J. Omithol.
Vol. 39. No. 2. P. 53-61.
Ueda K. 1992. Food of the Yellow Bittern Ixobrychus sinensis in boluses regurgitated by nestlings // J.
Yamashina Inst. Omithol. Vol. 24. P. 13-17.
Ueta M., Melwille D.S., Wang Y. et al. 2002. Discovery of the breeding sites and migration routes of Black-
faced Spoonbills Platalea minor II Ibis. Vol. 14. P. 340-342.
Underhill L.G., Tree A.G., Oschadleus H.D., Parker V. 1999. Review of Ring Recoveries of Waterbirds in
Southern Africa. Cape Town.
Urban E.K., Ash J.S. 2001. Longevity record of a Great White Pelican Pelecanus onocrotalus, from Lace
Shala, Ethiopia // Ostrich. Vol. 72. P. 123.
US Fish and Wildlife Service. 2005. Beringian seabird colony catalog: Computer database and colony status
record archives. Anchorage, AK.
Литература
597
Utekhina LG., Potapov E.R., McGrady M.J. 2000. Diet of the Steller’s sea eagle in the Northern Sea of
Okhotsk // First symposium on Steller’s and white-teiled sea eagles in East Asia. Tokyo: Wild Bird
Society of Japan. P. 71-82.
Valqui M., Caziani S.M., Rocha-O.O., Rodriguez-R.Ed. 2000. Abundance and distribution of the South
American altiplano flamingos // Waterbirds. Vol. 23. P. 110-113.
van den Bossche W., Berthold P., Darman Yu. et al. 2001. Satellite-tracking helps to discover stopover sites
of the threatened Oriental white stork (Ciconia boyciana) // www.microwavetelemetry.com, June 2001.
P. 3^4.
van den Bossche W., Berthold P., Kaatz C. et al. 1999. Preliminary results of satellite tracing White Stork
(Ciconia ciconia) migration // WeiBstorch im Aufwind? - White Storks on the up? - Proc. Intern. Symp.
on the White Stork, Hamburg 1996. Bonn: NABU. P. 517-524.
van den Bossche W., Berthold P, Kaatz M., at al. 2002. Eastern European White Stork Populations: Migra-
tion Studies and Elaboration of Conservation Measures / Final Report of the R+D-Project. Bonn. 198 p.
van der Have T.M., Enters A., Harte M. et al. 1999. The return of the White Stork to the Netherlands:
population size and breeding success in 1995 // WeiBstorch Intern. Symp. Bonn, NABU. P. 103-110.
van der Have T.M., Keijl G.O., Mansoori J., Mansoori V. 2002. Waterbirds in coastal wetlands along the
Persian Gulf cjast of Iran, January-February 2000 // Zool. Middle East. No. 26. P. 71-88.
van Tets G.F. 1959. A comparative study of the reproductive behaviour and natural history of three sympa-
tric species of cormorants (Phalacrocorax auritus, P penicillatus, P pelagicus) at Mandarte Island,
B.C.: M.A. thesis. Vancouver: Univ, of British Columbia. 86 p.
van Tets G.F. 1965. A comporative study of some social communication patterns in the Pelecaniformes //
Omithol. Monogr. Washington D.C.: Amer. Omithol. Union. No. 2. P. 1-88.
Vasvari M. 1938. Birds of Hungari // Aquila. Bd. 36-37. S. 231-293.
Vaughan R.E., Jones K.H. 1913. The Birds of Hong Kong. Macao and West River or Si Kiang in South-
Eastern China, with special reference to their Nidification and Seasonal Movements. Part 3 // Ibis.
Vol. 1. No. 3. P. 351-384.
Vaurie Ch. 1965. The birds of Palearctic fauna. A systematic reference Non-Passeriformes. London. 763 p.
Veromann H. 1989. Thirty-two year population trends of the White Stork in Estonian S.S.R. // WeiBstorch.
Proc. 1st Int. Stork Conserv. Symp. Schriftenreihe des DDA. Bd. 10. P. 153-158.
Vespermenau E.E. 1968. Distribution and biology of the Spoonbill in Roumania // Ardea. Vol. 56. No. 1-2.
P. 160-177.
Vinogradov V.V 1998. The outlook for the central Asian populations of pelicans // Ostrich. Vol. 69. No. 3-
4. P. 459.
Vogrin M. 1999. The Great White Egret Egretta alba during autumn and winter on Dravsko polje in north-
eastern Slovenia // Omis Hungarica. Vol. 8-9. No. 1-2. P. 33-37.
Voigt A. 1950. Excursionsbuch zum Studium der Vogelstimmen. Heidelberg. 218 S.
Voisin C. 1991. The herons of Europe. London. 364 p.
Vos K. 1995. Decline and reintroduction of the White Stork in central and northern parts of western Europe
// Proc, of the Intern. Symp. on the White Stork (Western Population). Basel. P. 49-51.
Wackemagel H. 1964. Was futtem die Flamingos ihren Jungen? // Z. Vitaminforsch. Bd. 34. H. 1. S. 141—
143.
Wanless S., Burger A.E., Harris M.P. 1991. Diving depths of Shags Phalacrocorax aristotelis breeding on
the Isle of May // Ibis. Vol. 133. No. 1. P. 37^42.
Wanless S., Harris M.P., Morris J.A. 1991. Foraging range and feeding locations of shags Phalacrocorax
aristotelis during chick rearing // Ibis. Vol. 133. No. 1. P. 30-36.
Wanless S., Harris M.P., Russel A.F. 1993. Factors influencing food-load brought in by shags Phalacroco-
rax aristotelis during chick rearing // Ibis. Vol. 135. No. 1. P. 19-24.
Warga K. 1938. Phanologische und Nidobiologische Daten aus den Kolonien von Egretta alba alba (L.) am
Kisbalaton // Proc. 8th Int. Om. Congr., Oxford. P. 655-663.
Wassmann R. 1984. Der WeiBstorchs - Vogel des Jahres 1984 // Voliere. Bd. 7. Nr. 1. S. 29-31.
Watanuki Y, Kato A., Naito Y. 1996. Diving preformance of male and female Japanese cormorants // Can.
J. Zool. Vol. 74. P. 1098-1109.
Watanuki Y, Kondj N., Nakagawa H. 1988. Status of seabirds breeding in Hokkaido // Jap. J. Omithol.
No. 37. P. 17-32.
598
Литература
Weber W. 1972. A new world for the cattle egret // Nature History. Vol. 81. No. 2. P. 56-62.
Wetlands International. 2002. Waterbird Population Estimates. 3-rd Ed. Wageningen, Netherlands: Wet-
lands Intemat. Glob. Ser. No. 12. 226 p.
Wetlands International. 2006. Waterbird Population Estimates. 4th Ed. Wageningen, Netherlands: Wetlands
International. 239 p.
Wermeille E., Biber O. 2003. Activite, utilisation de 1’habitat et regime alimentaire d’un couple de Cigognes
blanches Ciconia ciconia a Damphreux (Jura, Suisse) en premiere annee de reinstallation // Nos oiseaux.
Vol. 50. Is. 3. P. 147-158.
Wild Birds of Japan. 1955. Tokyo. 591 p.
Wilkinson E.S. 1935. The Shanghai bird year. A calendar of bird life in the country around Shanghai. 219 p.
Williams J.C., Sommer E., Brenneman K., Syria S., Moore H. 2002. Biological monitoring at Buldir Island,
Alaska in 2000 and 2002: Summary appendices. US Fish and Wildl. Serv. Rep. AMNWR 02/08., Adak,
AK. 146 p.
Winkler R. 1999. Avifauna der Schweiz // Omithol. Beobachter. Basel. Beiheft 10. 252 S.
Witherby H.F., Jourdian F.C.R., Ticehurst N.E., Tucker B.W. 1939. The handbook of British Birds. London.
Vol. 3. 765 p.
Wolters H.E. 1982. Die Vogelarten der Erde. Hamburg-Berlin. 745 S.
Wong Pyong-Oh. 1996. A Fild Guide to the Birds of Korea. Seoul-Korea: Korea Ass. of Wildl. conserv. 453
P-
Won Pyong-Oh. 1996a. Check-list of the birds of Korea // Bull. Kor. Omithol. Vol. 5. No. 1. P. 39-58.
Wuczynski A. 2005. The turnover of White Storks Ciconia ciconia on nests during spring migration // Acta
Om. Vol. 40. Is. 1. P. 83-85.
Yamashina Y. 1931. Die Vogel der Kurilen // J. Om. Bd. 79. H. 4. S. 491-541.
Yamashina Y. 1977a. Japanese creasted ibis // Red Data Book in Jap. Birds. Tokyo. P. 62-66.
Yamashina Y. 1977b. The Feeding of Japanese Creasted Ibises // Endand. Birds. Wisconsin-Madison (USA).
P. 161-164.
Yamashina Y. 1977. The present status of the Japanese Crested Ibis Nipponia nippon and the Japanese
White Stork Ciconia ciconia boyciana in continental Asia // Misc. Rep. Yamashina Inst. Omith. Vol. 9.
P. 131-142.
Yamamoto H. 1967. Phalacrocorax capillatus as a breeding bird on Iwate coast, Honshu // Misc. Rep.
Yamashina Inst. Omithol. Vol. 5. No. 1. P. 48-60.
Yoon M.-B. 1995. Wild birds of Korea. Seoul. 548 p.
Yu Yat-tung. 2005. The International Black-faced Spoonbill Census: 21-23 January 2005. 28 p.
Yu Yat-tung. 2010. World population and important sites of the endangered Black-faced Spoonbill // Ecol-
ogy, Migration and Conservation of the Black-faced Spoonbill. Glob. COE Intern. Symp. 2010. 143 p.
Yu Yat-tung, Swennen C. 2005. Food-beginning by a Juvenile Black-faced Spoonbill at the wintering site //
J. Yamashina Inst, for Omith. Vol. 37. P. 43^44.
Zerong S. 1991. Preliminary observations on the winter foraging ecology of Oriental White Storks (Ciconia
boyciana) in Jiangsu Province, China // M.C. Coulter et al. (eds.). Biol, and Conserv. of the Oriental
White Stork Ciconia boyciana. P. 107-114.
Zhao Zheng jie. 2001 A Handbook of the birds of China. Vol. 1. Non-Passeriformes. Jilin Science and
Technology Press. 797 p.
Zhatkanbayev A.Zh. 1994. Present status of pelicans in the Delta // Pelicans in the former USSR. IWRB
Publ. No. 27. P. 75-78.
Zhatkanbayev A.Zh. 2001. Colonial behaviour of the Dalmatian and Great White pelicans (Pelecanus cris-
pus, P onocrotalus) at the breeding season in Kazakstan // Adv. Ethol. No. 36. P. 292.
Zelenka G. 1963. Les mouvments saisonniers des pelicans en Grece // Alauda. Vol. 31. No. 3. P. 212-217.
Zielinski P. 2002. Brood reduction and parental infanticide - are the White Stork Ciconia ciconia and the
Black Stork C. nigra exceptional? // Acta Om. Vol. 37. Is. 2. P. 113-119.
Zimmermann R. 1931. Zur Fortpflanzungsbiologie der GroBen Rohrdommel, Botaurus stellaris L. // J. Om.
Bd. 79. S. 324-332.
Zink G. 1967. Populationsdynamik des Weissen Storches, Ciconia ciconia, in Mitteleuropa // Proc. 14th
Intern. Om. Congr. Oxford: Blackwell Scientific Publ. P. 191-215.
Литература
599
Zollick Н. 1986. Ergebnisse der vierten intemationalen Bestandserfassung vom WeiBstorch im Bezirk Ros-
tok 1984 // Falke. Bd. 33. Nr. 3. S. 82-83.
Zydelis S., Grazulevicius G., Zarankaite J., Mecionis R. 2002. Expansion of the Cormorant (Phalacrocorax
carbo sinensis) population in western Lithuania // Acta Zoologica Lituanica. Vol. 12. No. 3. P. 283-
287.
СПИСОК АВТОРОВ
Андронов Владимир Андреевич. Отдел по особо охраняемым природным территориям Депар-
тамента природных ресурсов по Дальневосточному федеральному округу. Россия, 689020, г. Хаба-
ровск, а/я 2098. E-mail: animal@pop.redcom.ru
Ардамацкая Татьяна Борисовна. Украинское общество охраны птиц. Украина, 75600, г. Голая
Пристань Херсонской области, ул. Кирова, д. 17, кв. 2.
Артюхин Юрий Борисович. Камчатский филиал Тихоокеанского института географии ДВО РАН.
Россия, 683028, г. Петропавловск-Камчатский, пр. Рыбаков, 19а. E-mail: artukhin@mail.kamchatka.ru
Белик Виктор Павлович. Ростовский педагогический государственный университет. Россия,
344091, г. Ростов-на-Дону, Коммунистический просп., д. 46, кв. 118. E-mail: vpbelik@mail.ru
Бескаравайный Михаил Михайлович. Карадагский заповедник. Украина, 98188, АО Крым, п/
о Курортное, Феодосия. E-mail: mbesk@pochta.ru
Бурковский Олег Александрович. Зоологический музей Дальневосточного федерального уни-
верситета. Россия, 690950, г. Владивосток, Океанский просп., д. 37. E-mail: oleg@museum.dvgu.ru
Галченков Юрий Дмитриевич. Центр «Кадастр». Россия, 284001, г. Калуга, ул. Мира, д. 4/1, кв.
9. E-mail: galchyonkov@mail.ru
Гамова Татьяна Владимировна. Биолого-почвенный институт ДВО РАН. Россия, 690022, г.
Владивосток, просп. 100-летия Владивостока, 159. E-mail: birds@.dvo.ru
Глущенко Юрий Николаевич. Уссурийский государственный педагогический институт. Рос-
сия, 692500, г. Уссурийск Приморского края, ул. Некрасова, д. 35. E-mail: root@uspi.ussuriisk.ru
Грищенко Виталий Николаевич. Каневский заповедник. Украина, 19000, г. Канев Черкасской
обл. E-mail: vgrishchenko@mail.ru
Джамирзоев Гаджибек Сефибекович. Заповедник «Дагестанский». Россия, 367010, Республика
Дагестан, г. Махачкала, ул. Гагарина, д. 120. E-mail: dzamir@mail.ru
Дмитренок Марина Григорьевна. Институт зоологии НАН Беларуси. Республика Беларусь,
220072, г. Минск, ул. Академическая, 27. E-mail: marinabittem@rambler.ru
Зубакин Виктор Анатольевич. Институт проблем экологии и эволюции РАН. Россия, 119071,
Москва, Ленинский просп., д. 33. E-mail: vzubakin@yandex.ru
Коблик Евгений Александрович. Зоологический музей МГУ. Россия, 125009, Москва, ул. Боль-
шая Никитская, д.6. E-mail: koblik@zmmu.msu.ru
Конюхов Николай Борисович. Институт проблем экологии и эволюции РАН, Центр кольцева-
ния. Россия, 119071, Москва, Ленинский просп., д. 33. E-mail: konyukhov@gmail.com
Котюков Юрий Валентинович. Окский заповедник. Россия, 391072, Рязанская обл., Спасский
р-н, п/о Лакаш. E-mail: br.bor@rambler.ru
Краснов Юрий Владимирович. Мурманский морской биологический институт КНЦ РАН. Рос-
сия, 183010, г. Мурманск, ул. Владимирская, 17. E-mail: kharlov51@mail.ru
Литвиненко Наталья Михайловна f
Луговой Алексей Евгеньевич. Закарпатский филиал Киевского славистического университета.
Украина, 88002, г. Ужгород, ул. Островная, д. 20, кв. 21. E-mail: lugovojalexej@mail.ru
Мацына Александр Иванович. Нижегородский университет. Россия, 603089, г. Нижний Новго-
род, Высоковский пр., д. 11/35. E-mail: mai-68@mail.ru
Мацына Екатерина Леонидовна. Нижегородский университет. Россия, 603089, г. Нижний Нов-
город, Высоковский пр., д. 11/35. E-mail: kaira-100@mail.ru
Мищенко Александр Леонидович. Институт проблем экологии и эволюции РАН. Россия, 119071,
Москва, Ленинский просп., д. 33. E-mail: almovs@mail.ru
Мосалов Алексей Александрович. Московский педагогический государственный университет.
Россия, 129278, Москва, ул. Кибальчича, д.6, корп. 5. Биолого-химический факультет МПГУ E-mail:
rallus@yandex.ru
Назаров Юрий Николаевич f
Нечаев Виталий Андреевич. Биолого-почвенный институт ДВО РАН. Россия, 690022, г. Вла-
дивосток, просп. 100-летия Владивостока, д. 159. E-mail: birds@ibss.dvo.ru
Приклонский Святослав Георгиевич. Окский заповедник. Россия, 391072, Рязанская обл, Спас-
ский р-н, п/о Лакаш. E-mail: svet-0632@mail.ru
Русев Иван Трифонович. Украинский научно-исследовательский противочумный институт им.
Мечникова. Украина, г. Одесса, ул. Пастера, д. 42, кв. 21а. E-mail: wildlife@paco.net
Сурмач Сергей Григорьевич. Биолого-почвенный институт ДВО РАН. Россия, 690022. г. Вла-
дивосток, просп.ЮО-летия Владивостока, 159. E-mail: birds@.dvo.ru
Шибаев Юрий Викторович. Биолого-почвенный институт ДВО РАН. Россия, 690022, г. Влади-
восток, просп. 100-летия Владивостока, 159. E-mail: birds@.dvo.ru
СОДЕРЖАНИЕ
Предисловие.........................................................................5
Отряд ПЕЛИКАНООБРАЗНЫЕ, или ВЕСЛОНОГИЕ — PELECANIFORMES.............................8
Подотряд PELECANI............................................................... 10
Семейство ПЕЛИКАНОВЫЕ — PELECANIDAE Rafinesque, 1815................. 10
Род Pelecanus Linnaeus, 1758.............................................. 12
Розовый пеликан Pelecanus onocrotalus Linnaeus, 1758, Е.Л. Мацына....... 12
Кудрявый пеликан Pelecanus crispus Bruch, 1832, А.И. Мацына, Е.Л. Мацына.24
Семейство ОЛУШЕВЫЕ — SULIDAE Reichenbach, 1849............................... 37
Род Morus Vieillot, 1816...................................................39
Северная олуша Morus bassanus (Linnaeus, 1758), Ю.В. Краснов.............39
Род Sula Brisson, 1760.....................................................46
Красноногая олуша Sula sula (Linnaeus, 1766), Ю.Б. Артюхин...............46
Бурая олуша Sula leucogaster (Boddaert, 1783), Ю.Б. Артюхин..............49
Семейство БАКЛАНОВЫЕ — PHALACROCORACIDAE Reichenbach, 1836................... 52
Род Phalacrocorax, Brisson, 1760 ......................................... 54
Большой баклан Phalacrocorax carbo (Linnaeus, 1758), А.Е. Луговой........54
Японский, или уссурийский баклан Phalacrocorax capillatus (Temminck
et Schlegel, 1849)j Н.М. Литвиненко\.................................... 83
Ушастый баклан Phalacracorax auritus (Lesson, 1831), Ю.Б. Артюхин........95
Стеллеров баклан Phalacracorax perspicillatus Pallas, 1811, Ю.Б. Артюхин.98
Берингов баклан Phalacracorax pelagicus Pallas, 1811, Ю.Б. Артюхин..... 101
Краснолицый баклан Phalacracorax urile (J.F. Gmelin, 1789), Ю.Б. Артюхин. 115
Хохлатый баклан Phalacracorax aristotelis (Linnaeus, 1761), М.М. Бескаравайный 123
Малый баклан Phalacracoraxpygmaeus (Pallas, 1773), А.Е. Луговой........ 133
Семейство ЗМЕЕШЕЙКОВЫЕ — ANHINGIDAE Reichenbach, 1849....................... 142
Род Anhinga, Brisson, 1760............................................... 143
Чернобрюхая змеешейка Anhinga melanogaster, Pennant, 1769,
В.А. Зубакин, С.Г. Приклонский......................................... 143
Подотряд FREGATAE.............................................................. 146
Семейство ФРЕГАТОВЫЕ — FREGATIDAE Degland et Gerbe, 1867.................... 146
Vor Fregata Lacepede, 1799............................................... 147
Фрегат-ариель Fregata ariel (G.R.Gray, 1845), В.А. Нечаев.............. 147
Отряд АИСТООБРАЗНЫЕ — CICONIIFORMES.............................................. 152
Подотряд ARDEAE................................................................ 153
Семейство ЦАПЛЕВЫЕ — ARDEIDAE Leach, 1820 .................................. 153
Род Botaurus, Stephens, 1819............................................. 155
Выпь Botaurus stellaris (Linnaeus, 1758), М.Г. Дмитренок............... 155
Род Ixobrychus Billberg, 1828............................................ 177
Волчок Ixobrychus minutus (Linnaeus, 1766), С.Г. Приклонский, М.Г. Дмитренок,
В.А. Зубакин, А.Л. Мищенко............................................. 177
Китайский волчок Ixobrychus sinensis (J.F. Gmelin, 1789)| А9.Я. Назаров^
Т.В. Гамова, С.Г. Сурмач, О.А. Бурковский........................... 189
Амурский волчок Ixobrychus eurhythmus (Swinhoe, 1873),[ю./7. Назаров^
Т.В. Гамова, С.Г. Сурмач, О.А. Бурковский.............................. 197
Охристый волчок Ixobrychus cinnamomeus (J.F. Gmelin, 1789), Ю.В. Котюков.206
Род Gorsachius Bonaparte, 1857........................................ 209
Японская кваква Gorsachius goisagi (Temminck, 1835)] Ю.Н. Назаров\
О.А. Бурковский.........................................................209
Род Nycticorax Т. Forster, 1817...........................................212
Кваква Nycticorax nycticorax (Linnaeus, 1758), И. Т. Русев..............212
Род Butorides Blyth, 1849 .............................................. 237
Зеленая кваква Butorides striatus (Linnaeus, \766\\Ю.Н.Назаров\
О.А. Бурковский.........................................................237
Род Ardeola Boie, 1822 ..................................................... 244
Желтая цапля Ardeola ralloides (Scopoli, 1769), И. T. Русев........... 244
Белокрылая цапля Ardeola bacchus (Bonaparte, 1 Ъ57)\ЮАГНазаров\
О.А. Бурковский...........................................................259
Индийская прудовая цапля Ardeola grayii (Sykes, 1832), В.А. Зубакин.....263
Род Bubulcus Bonaparte, 1857................................................265
Египетская цапля Bubulcus ibis (Linnaeus, 1758), Г.С. Джамирзоев........265
Род Egretta Т. Forster, 1817................................................276
Малая белая цапля Egretta garzetta (Linnaeus, 1766), В.Н. Грищенко......276
Желтоклювая цапля Egretta eulophotes (Swinhoe, 1860), Ю.В. Шибаев.......295
Средняя белая цапля Egretta intermedia (Wagler, 1829) В.Н. Грищенко,
В.А. Нечаев.............................................................298
Род Casmerodius Gloger, 1842................................................304
Большая белая цапля Casmerodius albus (Linnaeus, 1758), В.Н. Грищенко.....304
Южная белая цапля Casmerodius modestus (J.E.Gray, 1831), В.А. Нечаев,
Ю.Н. Глущенко...........................................................329
РодЛпЯя Linnaeus, 1758 ..................................................... 334
Серая цапля Ardea cinerea Linnaeus, 1758, Т.Б. Ардамацкая...............334
Рыжая цапля Ardea purpurea Linnaeus, 1766, С.Г. Приклонский.............355
Подотряд CICONIAE...............................................................368
Семейство АИСТОВЫЕ — CICONIIDAE Sundevall, 1836..............................368
Род Ciconia Brisson, 1760................................................ 369
Черный аист Ciconia nigra (Linnaeus, 1758), С.Г. Приклонский............369
Белый аист Ciconia ciconia (Linnaeus, 1758), В.Н.Грищенко, Ю.Д. Галчёнков.384
Дальневосточный аист Ciconia boyciana Swinhoe, 1873, В.А. Андронов......416
Семейство ИБИСОВЫЕ — THRESKIORNITHOIDEA Poche, 1904..........................430
Род Threskiornis G.R.Gray, 1842 ......................................... 431
Священный ибис Threskiornis aephiopicus (Latham, 1790), В.П. Белик......431
Черноголовый ибис Threskiornis melanocephalus (Latham, 1790), В.А. Нечаев.435
Род Nipponia Reichenbach, 1853............................................439
Красноногий ибис Nipponia nippon (Temminck, 1835)jH.M Литвиненко, I
Ю.В.Шибаев..............................................................439
Род Plegadis Каир, 1829...................................................444
Каравайка Plegadis falcinellus (Linnaeus, 1766), В.П. Белик.............444
Род Platalea Linnaeus, 1758...............................................470
Колпица Platalea leucorodia Linnaeus, 1758, В.П. Белик................. 470
Малая колпица Platalea minor Temminck et Schlegel, 1849,|Я.Л7. Литвиненко^
Ю.В. Шибаев.............................................................497
Отряд ФЛАМИНГООБРАЗНЫЕ — PHOENICOPTERIFORMES......................................508
Семейство ФЛАМИНГОВЫЕ — PHOENICOPTERIDAE Bonaparte, 1831 ................... 508
Род Phoenicopterus Linnaeus, 1758........................................ 511
Розовый фламинго Phoenicopterus roseus Pallas, 1811, Е.Л. Мацына,
Г.С. Джамирзоев........................................................ 511
Литература........................................................................530
Список авторов....................................................................600
Книги Товарищества научных изданий КМК
БИОЛОГИЯ
СЕРИЯ «ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ РОССИИ»
Деревья и кустарники зимой. 2-е изд. [Вып.9]. Е.Т. Валягина-Малютина. ISBN 978-5-
87317-388-4. 2007. 268 с. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб.
Булавоусые чешуекрылые Восточной Европы. [Вып.8]. А.Л. Львовский, Д.В. Моргун.
ISBN 978-5-87317-362-4. 2007. 443 с., 8 цв.вкл. Формат 170 х 240 мм. Те. перепл. — Цена
400 руб.
Флора Северо-Западного Кавказа. [Вып.7]. А.С. Зернов. ISBN 5-87317-338-9. 2006. 664
с. Формат 170 х 240 мм. Те. перепл. — Цена 500 руб.
Флора Нижнего Поволжья. Том 1 (споровые, голосеменные, однодольные). [Вып.6]. А.К.
Скворцов (ped.). ISBN 5-87317-285-4. 2006. 435 с. Формат 170 х 240 мм. Те. перепл. —
Цена 400 руб.
Планируется: Е.Т. Валягина-Малютина. Деревья и кустарники средней полосы европейской
части России (иллюстрированный определитель).
ПРОЧИЕ ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ
Атлас молоди пресноводных рыб России. А.П. Макеева, Д.С. Павлов, Д.А. Павлов. ISBN
978-5-87317-714-1. 2011. 383 с., ил. Формат 215 х 300 мм. В обл. — Цена 700 руб.
Краткий определитель мохообразных Подмосковья. Е.А. Игнатова, М.С. Игнатов, В.Э.
Федосов, Н.А. Константинова. ISBN 978-5-87317-746-2. 2011. 272 с., ил., 48 цв. вкл. Фор-
мат 165 х 240 мм. В обл. — Цена 350 руб.
Иллюстрированные определители свободноживущих беспозвоночных евразийских
морей и прилегающих глубоководных частей Арктики. Том 2. Немертины, головохо-
ботные, малощетинковые черви, пиявки, погонофоры, эхиуры, сипункулы, форони-
ды, плеченогие. Б.И. Сиренко (ped.). ISBN 978-5-87317-723-3. 2010. 186 с., ил. Формат
170 х 240 мм. Те. перепл. — Цена 300 руб.
Определитель зоопланктона и зообентоса пресных вод Европейской России. Том 1.
Зоопланктон. В.Р. Алексеев (ped.). ISBN 978-5-87317-684-7. 2010. 495 с., ил. Формат 170
х 240 мм. Те. перепл. — Цена 600 руб.
Флора и фауна Белого моря. Иллюстрированный атлас. А.Б. Цетлин, А.Э.Жадан, Н.Н.
Марфенин (ped.). ISBN 978-5-87317-672-4. 2010. 471 с., цв. фотоатлас. Формат 175 х
245 мм. Те. перепл. — Цена 1500 руб.
Растения Российского Западного Кавказа. Полевой атлас. А.С. Зернов. ISBN 978-5-
87317-587-1. 2010. 448 с., цв. фотоатлас. Формат 155 х 220 мм. Те. перепл. — Цена 600
руб.
Стрекозы Восточной Европы и Кавказа: Атлас-определитель. В.Э. Скворцов. ISBN 978-
5-87317-657-1. 2010. 623 с., ил. Формат 205 х 145 мм. Те. перепл. — Цена 400 руб.
Определитель наземных моллюсков лесостепи Правобережного Поволжья. Т.П Стой-
ко, О.В. Булавкина. ISBN 978-5-87317-639-7. 2010. 96 с., ил., 17 цв.табл. Формат 165 х
235 мм. В обл. — Цена 220 руб.
Иллюстрированные определители свободноживущих беспозвоночных евразийских
морей и прилегающих глубоководных частей Арктики. Том 1. Коловратки, морские
пауки, ракообразные: усоногие, тонкопанцирные, эуфаузииды, неполнохвостные,
мизиды, капреллиды. Б.И. Сиренко (ped.). ISBN 978-5-87317-626-7. 2009. 189 с., ил. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 300 руб.
Атлас дикорастущих растений Ленинградской области. ISBN 978-5-87317-577-2. 2010.
664 с., цв. фотоатлас. Бум. мелов. Формат 130 х 170 мм. Те. перепл. — Цена 600 руб.
Определитель сосудистых растений Оренбургской области. З.Н. Рябинина, М.С. Кня-
зев. ISBN 978-5-87317-572-7. 2009. 758 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена
600 руб.
Растения средней полосы Европейской России. Полевой атлас. И.А. Шанцер. 3-е изд.
ISBN 978-5-87317-591-8. 2009. 470 с., цв. фотоатлас. Бум. мелов. Формат 130х 170 мм.
Те. перепл. — Цена 500 руб.
Флора водоемов Волжского бассейна. Определитель сосудистых растений. Л.И. Ли-
сицына, В.Г. Папченков, В.И. Артеменко. ISBN 978-5-87317-554-3. 2009. 219 с., ил. Фор-
мат 143 х 210 мм. В обл. — Цена 150 руб.
Определитель грибов России. Порядок Афиллофоровые. Вып.З. И.В. Змитрович. ISBN
978-5-87317-561-1. 2008. 278 с. Формат 145 х 218 мм. Те. перепл. — Цена 300 руб.
Иллюстрированный атлас беспозвоночных Белого моря. Н.Н. Марфенин, С.А. Бело-
русцева (ред.). ISBN 5-87317-252-8. 2006. 312 с., цветной фотоатлас. Бум. мелов. Фор-
мат 150 х 220 мм. Те. перепл. — Цена 500 руб.
Определитель сосудистых растений Соловецкого архипелага. К.В. Киселёва, В.С.
Новиков, Н.Б. Октябрёва, А.Е. Черенков. ISBN 5-87317-230-7. 2004. 175 с., цв. фото.
Формат 170 х 240 мм. Те. перепл. — Цена 220 руб.
Иллюстрированное руководство для ботанических практик и экскурсий в Средней
России. В.Э. Скворцов. ISBN 5-87317-170-Х. 2004. 506 с. Формат 170 х 240 мм. Те. пе-
репл. — Цена 300 руб.
Иллюстрированный определитель растений Средней России. Том 3. И.А. Губанов,
К.В. Киселева, В.С. Новиков, В.Н. Тихомиров. ISBN 5-87317-163-7. 2004. 520 с. Формат
210 х 295 мм. Те. перепл. — Цена 350 руб. Том 2 (распродан; готовится 2-е издание) —
Том 1. ISBN 5-87317-091-6. 2002. 526 с. с портр. — Цена 350 руб.
Флора мхов средней части Европейской России. Том 2. М.С. Игнатов, Е.А. Игнатова.
2004. С.609-944. ISBN 5-87317-149-1. Бум. мелов. Формат 195 х 270 мм. Те. перепл. —
Цена 300 руб. Том 1. 2003. С. 1-608, илл. ISBN 5-87317-104-1. — Цена 500 руб.
Определитель грибов России. Дискомицеты. Вып.1. Копротрофные виды. В.П. Про-
хоров. ISBN 5-87317-136-Х. 2004. 255 с. Формат 145 х 218 мм. Те. перепл. — Цена 200
руб.
Планируется-. Ан.А. Шилейко, Т.С. Рымжанов. Фауна наземных моллюсков Казахстана и сопре-
дельных территорий.
СЕРИЯ «РАЗНООБРАЗИЕ ЖИВОТНЫХ»
Повелители бездны [Вып.5]. А.Н. Островский. ISBN 978-5-87317-569-7. 2009. 216 с., 16
с. цв. вкл. Формат 145 х 205 мм. — Цена 220 руб.
Мамонт [Вып.З]. А.Н. Тихонов. ISBN 5-87317-209-9. 2005. 90 с., цв. вкл. Формат 145х 205
мм. — Цена 100 руб.
Гидра: от Абраама Трамбле до наших дней [Вып.1]. С.Д. Степаньянц, В.Г. Кузнецова,
Б.А. Анохин. ISBN 5-87317-144-0. 2003. 101 с. + цв.вкл. Формат 145 х 205 мм. — Цена 100
руб.
Планируется’. А.Н. Островский. Подводный мир (в печати).
УЧЕБНИКИ ДЛЯ ВУЗОВ
Общая арахнология. Краткий курс. Часть 1. Введение. Малые отряды. КГ Михайлов.
ISBN 978-5-87317-749-3. 2011. 65 с. Формат 145 х 203 мм. В обл. — Цена 60 руб.
Протистология: Руководство. К. Хаусман, Н. Хюльсман, Р. Радек. Пер. с англ. ISBN 978-
5-87317-662-5. 2010. 495 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена 600 руб.
Жизнь на дне. Био-экология и био-география бентоса. И.А. Жирков. ISBN 978-5-87317-
613-7. 2010. 453 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена 600 руб.
Морская биогеоценология. И.В. Бурковский. ISBN 5-87317-341-9. 2006. 285 с. Формат
170 х 240 мм. Те. перепл. — Цена 220 руб.
Основы микологии (морфология и систематика грибов и грибоподобных организ-
мов). Л.В. Гарибова, С.Н. Лекомцева. ISBN 5-87317-265-Х. 2005. 220 с. Формат 170 х 240
мм. Те. перепл. — Цена 220 руб.
Малый практикум по зоологии беспозвоночных. Часть 1. 2-е изд. И.А. Тихомиров, А.А.
Добровольский, А.И. Гранович. ISBN 978-5-87317-442-3. 2008. 304 с., 14 ч/б вкл. Формат
170 х 240 мм. Те. перепл. — Цена 250 руб.
Основы биогеографии. В.Г. Мордкович. ISBN 5-87317-189-0. 2005. 236 с., 1 цв. вкл. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 220 руб.
Планируется-. И.А. Тихомиров (СПбГУ). Малый практикум по зоологии беспозвоночных (ч. 2).
СЕРИЯ «СОВРЕМЕННАЯ ОТЕЧЕСТВЕННАЯ БИОЛОГИЯ»
Экология и биогеография. Избранные работы. Ю.И. Чернов. ISBN 978-5-87317-528-4.
2008. 580 с. Формат 170 х 240 мм. Те. перепл. — Цена 500 руб.
Избранные труды. Н.В. Кокшайский. ISBN 978-5-87317-489-8. 2008. 410 с. с портр. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 400 руб.
Избранные труды. В.В. Кучерук. ISBN 5-87317-279-Х. 2006. 523 с. спортр. Формат 170х
240 мм. Те. перепл. — Цена 300 руб.
Избранные труды. Е.Н. Матюшкин. ISBN 5-87317-266-8. 2005. 658 с. с портр. Формат
170 х 240 мм. Те. перепл. — Цена 400 руб.
Избранные труды по эволюционной биологии. А.П. Расницын. ISBN 5-87317-243-9. 2005.
iv + 347 с. с портр., 16 фототаблиц. Формат 170 х 240 мм. Те. перепл. — Цена 250 руб.
Избранные труды. Организм, геном, язык. Б.М. Медников. ISBN 5-87317-197-1. 2005.
452 с. с портр. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб.
Планируется: С.М. Разумовский. Избранные труды.
НАУЧНО-ПОПУЛЯРНЫЕ ИЗДАНИЯ
Невероятная зоология. В.Н. Танасийчук. 2-е изд. ISBN 978-5-87317-735-6. 2011. 372 с., в
обл. Формат 140 х 213 мм. — Цена 220 руб.
Моя биосфера: научно-художественное изложение биосферных знаний. Г.И. Флёрова.
ISBN 978-5-87317-679-3. 2010. 247 с., ил. Формат 170 x 240 мм. Те. перепл. —Цена 300руб.
Киты и история китобойного промысла: взгляд из Японии. М. Комацу, С. Мисаки. ISBN
5-87317-262-5. 2005. 142 с., в те. перепл., цв. вкл. Формат 145 х 215 мм. — Цена 200 руб.
Планируется: В.Н. Танасийчук. Цокотуха ли муха.
СПРАВОЧНЫЕ ИЗДАНИЯ, ИСТОРИЯ БИОЛОГИИ
Столовые корнеплоды в России. В.И. Леунов. ISBN 978-5-87317-744-8. 2011. 272 с., ил.,
цв. книга. В те. перепл. Формат 210 х 290 мм. — Цена 900 руб.
Промысловые, перспективные для промысла и кормовые беспозвоночные Россий-
ских морей. Э.Н. Егорова, Б.И. Сиренко. ISBN 978-5-87317-724-0. 2010. 285 с., 8 вкл., 16
цв. вкл. В те. перепл. Формат 170 х 240 мм. — Цена 350 руб.
Методы исследования сообществ микроартропод. М.Б. Потапов, Н.А. Кузнецова. ISBN
978-5-87317-739-4. 2011. 84 с., ил., 4 цв. вкл., в обл. Формат 145 х 215 мм. — Цена 60 руб.
Методические подходы к экологической оценке лесного покрова в бассейне малой
реки. Л.Б. Заугольнова, Т.Ю. Браславская (отв. ред.). ISBN 978-5-87317-718-9. 2010. 383
с., ил., 14 цв. вкл., в те. перепл. Формат 170 х 240 мм. — Цена 500 руб.
Биологическая систематика: эволюция идей. И.Я. Павлинов, Г.Ю. Любарский. Сборник
трудов Зоологического музея МГУ. Том 51. ISBN 978-5-87317-685-4. 2011. 667 с., в те.
перепл. Формат 170 х 240 мм. — Цена 600 руб.
Как невозможно жили мы. В.В. Меншуткин (сост.). ISBN 978-5-87317-725-7. 2011. 219
с., 11 с. вкл., в обл. Формат 140 х 200 мм. — Цена 270 руб.
Каталог типовых образцов сосудистых растений Восточной Азии, хранящихся в Гер-
барии Ботанического института им. В.Л. Комарова (LE). часть 2 (Китай). А.Е. Грабовс-
кая-Бородина (отв. ред.). ISBN 978-5-87317-712-7. 2010. 517 с., в те. перепл. Формат
150 х 210 мм. — Цена 500 руб.
Культурные растения Главного ботанического сада им. Н.В. Цицина Российской ака-
демии наук: 60 лет интродукции. А.С. Демидов (отв. ред.). ISBN 978-5-87317-707-3. 2011.
511 с., 50 цв. вкл., в те. перепл., суперобл. Формат 170 х 240 мм. — Цена 700 руб.
Растительные ресурсы России. Дикорастущие цветковые растения, их компонент-
ный состав и биологическая активность. Том 3. А.Л. Буданцев (отв. ред.). ISBN 978-5-
87317-693-9. 2010. 601 с., в те. перепл. Формат 170 х 240 мм. — Цена 600 руб.
Большая энциклопедия цветочных многолетников. М.М. Диев. ISBN 978-5-87317-464-
5. 2011. 515 с., цв. фотоатлас. Формат 210 х 295 мм. — Цена 1500 руб.
«Философия зоологии» Жана-Батиста Ламарка: взгляд из XXI века. А.И. Шаталкин. ISBN
978-5-87317-642-7. 2009. 606 с., ил., в те. перепл. Формат 150 х 220 мм. — Цена 500 руб.
Справочник по семеноведению. В. Броувер, А. Штелин. Пер. с. нем. ISBN 978-5-87317-
633-5. 2010. 694 с. в те. перепл. Формат 210 х 290 мм. — Цена 1100 руб.
Эволюция и систематика: Ламарк и Дарвин в современных исследованиях. Сборник
трудов Зоологического музея МГУ. Том 50. А.В. Свиридов, А.И. Шаталкин (отв. ред.). ISBN
978-5-87317-617-5. 2009. 366 с., 40 с. вкл., в те. перепл. Формат 150 х 220 мм. — Цена
300 руб.
Растительные ресурсы России. Дикорастущие цветковые растения, их компонент-
ный состав и биологическая активность. Том 2. А.Л. Буданцев (отв. ред.). ISBN 978-5-
87317-600-7. 2009. 513 с., в те. перепл. Формат 170 х 240 мм. — Цена 600 руб.
История Зоологического музея МГУ: Идеи, люди, структуры. Г.Ю. Любарский. ISBN
978-5-87317-605-2. 2009. 744 с., ил., в те. перепл. Формат 21 Ох 290 мм. — Цена 900руб.
Вид и видообразование. Анализ новых взглядов и тенденций. (Труды Зоологического
института РАН, Приложение № 1). ISBN 978-5-87317-589-5. 2009. 298 с., в те. перепл.
Формат 210 х 267 мм. — Цена 500 руб.
Международный кодекс ботанической номенклатуры. ISBN 978-5-87317-588-8. 2009.
282 с., в те. перепл. Формат 145 х 220 мм. — Цена 300 руб.
Виды и формы хвойных, культивируемые в России. Часть 2. Picea, Thuja. Д.Л. Матю-
хин, О.С. Манина, Е.С. Сысоева. ISBN 978-5-87317-584-0. 2009. 288 с., бум. мелов., цв.
фото. Те. пер. Формат 170 х 240 мм. — Цена 700 руб.
Виды и формы хвойных, культивируемые в России. Часть 1. Juniperus, Cephalotaxus,
Taxus, Torreya. 2-е изд. Д.Л. Матюхин, О.С. Манина, Н.С. Королева. ISBN 978-5-87317-
590-1.2009. 259 с., бум. мелов., цв. фото. Те. пер. Формат 170 х 240 мм. — Цена 700 руб.
Учёные в вихре времени. Воспоминания об орнитологах, защитниках природы и
других натуралистах. Е. Новак. Пер с нем. ISBN 978-5-87317-575-8. 2009. 473 с., в те.
перепл. Формат 150 х 207 мм. — Цена 500 руб.
Каталог биоты Беломорской биологической станции МГУ. А.В. Чесунов, Н.М. Каляки-
на, Е.Н. Бубнова (ped.). ISBN 978-5-87317-485-0. 2008. 384 с., в те. перепл. Формат 170 х
240 мм. — Цена 500 руб.
Страна ББС. Е. Каликинская. ISBN 978-5-87317-479-9. 2008. 534 с., в те. перепл. Формат
170 х 245 мм. — Цена 500 руб.
Каталог чешуекрылых (Lepidoptera) России. С.Ю. Синёв (ped.). ISBN 978-5-87317-457-
7. 2008. 424 с., в те. перепл. Формат 220 х 290 мм. — Цена 620 руб.
Растительные ресурсы России. Дикорастущие цветковые растения, их компонент-
ный состав и биологическая активность. Том 1. А.Л. Буданцев (отв. ped.). ISBN 978-5-
87317-472-0. 2008. 421 с., в те. перепл. Формат 170 х 240 мм. — Цена 500 руб.
Введение в палеоэнтомологию. В.В. Жерихин, А.Г. Пономаренко, А.П. Расницын. ISBN
978-5-87317-454-6. 2008. 371 с., в те. перепл. Формат 170 х 240 мм. — Цена 300 руб.
Мировая коррелятивная база почвенных ресурсов: основа для международной клас-
сификации и корреляции почв. Пер. с англ. ISBN 978-5-87317-381-5. 2007. 278 с., в обл.
Формат 145 х 215 мм. — Цена 150 руб.
Мозаика судеб биофаковцев МГУ 1930-1960 годов поступления. Том II. 1951-1960-е
годы. Л.И. Лебедева (cocm.). ISBN 978-5-87317-372-3. 2007. 640 с., те. перепл. Формат
145 х 220 мм. — Цена 300 руб.
Мозаика судеб биофаковцев МГУ 1930-1960 годов поступления. Том I. 1930-1950-е
годы. Л.И. Лебедева (cocm.). ISBN 978-5-87317-372-3. 2007. 479 с., те. перепл. Формат
145 х 220 мм. — Цена 200 руб.
Ваш любящий Валя. Валентин Александрович Догель (1882-1955). Письма домой.
С.И. Фокин (ped.). ISBN 978-5-87317-369-3. 2007. 266 с., + 40 с., ил., в обл. Формат 145 х
210 мм. — Цена 150 руб.
Эрнст Майр и современный эволюционный синтез. Э.И. Колчинский. ISBN 5-87317-
245-5. 2006. 149 с., ч/б вкл., в обл. Формат 145 х 215 мм. — Цена 100 руб.
Александр Николаевич Формозов: Жизнь русского натуралиста. А.А. Формозов. ISBN
5-87317-331-7. 2006. 208 с., в обл. Формат 135 х 203 мм. — Цена 100 руб.
Пока горит свеча... Очерки по истории кафедры зоологии беспозвоночных МГУ. 2-е изд.
В.В. Малахов. ISBN 5-87317-317-6. 2006. 153 с., бум. мелов., в обл. Формат 145 х 210 мм.
— Цена 120 руб.
Конспект фауны земноводных и пресмыкающихся России. С.Л. Кузьмин, Д.В. Семенов.
ISBN 5-87317-303-6. 2006. 139 с., в те. перепл. Формат 145 х 210 мм. — Цена 150 руб.
Список птиц Российской Федерации. Е.А. Коблик, Я.А. Редькин, В.Ю. Архипов. ISBN 5-
87317-263-3. 2006. 281 с., бум. мелов., печать двухцветная, в обл. Формат 145 х 215 мм.
— Цена 220 руб.
Биологический факультет МГУ. А.И. Нетрусов и др. (ped.). ISBN 5-87317-257-9. 2005.
242 с., в те. перепл. Формат 170 х 240 мм. — Цена 220 руб.
Эволюционные факторы формирования разнообразия животного мира. Э.И. Воро-
бьева, Б.Р. Стриганова (ped.). ISBN 5-87317-199-8. 2005. 308 с., в те. перепл. Формат
170 х 240 мм. — Цена 220 руб.
Каталог типовых образцов сосудистых растений Восточной Азии, хранящихся в Гер-
барии Ботанического института им. В.Л. Комарова (LE). часть 1 (Япония и Корея).
В.И. Грубов (ред.). ISBN 5-87317-155-6. 2004. 188 с., ил. Формат 140 х 205 мм. В обл. —
Цена 80 руб.
Биология гидротермальных систем. А.В. Гебрук (ред.). ISBN 5-87317-099-1.2002. 543 с.
с цв. вкл., в те. перепл. Формат 210 х 260 мм. — Цена 400 руб.
Планируется: В.Б. Голуб, М.Н. Цурикое, А.А. Прокин. Коллекции насекомых: сбор, обработка и
хранение материала. — Ю.М. Марусик, Н.М. Коеблюк. Пауки Сибири и Дальнего Востока Рос-
сии (в печати). — В. Хенниг. Филогенетическая систематика (пер. с англ.).
НАУЧНЫЕ МОНОГРАФИИ
Новости систематики высших растений. Том 42. Н.Н. Цвелёв (отв. ред.). ISBN 978-5-
87317-759-2. 2011. 250 с. Формат 145 х 213 мм. Те. перепл. — Цена 300 руб.
Ботанические сады в современном мире: теоретические и прикладные исследова-
ния. А. С. Демидов (отв. ред.). ISBN 978-5-87317-756-1.2011. 765с., ил. Формат 210 х 295
мм. Те. перепл. — Цена 900 руб.
Восстановление и мониторинг природной флоры. Б.Р. Стриганова, А.А. Маслов (отв.
ред.). ISBN 978-5-87317-755-4. 2010. 116с., ил. Формат 145x210мм. Вобл. — Цена 100руб.
Онтогенетическая систематика и новая модель эволюции Bilateria. А.В. Мартынов.
ISBN 978-5-87317-750-9. 2011. 286 с., ил., 16 цв. вкл. Формат 170 х 240 мм. Те. перепл. —
Цена 350 руб.
Конспект фауны жуков южной тайги Западной Сибири (в бассейне Нижнего Ирты-
ша). С.П. Бухкало, Д.Е. Галич, Е.В. Сергеева, Н.В. Алемасова. ISBN 978-5-87317-745-5.
2011. 267 с. Формат 145 х 215 мм. В обл. — Цена 180 руб.
Орхидные северо-запада Европейской России (Ленинградская, Псковская, Новго-
родская области). П.Г. Ефимов. ISBN 978-5-87317-696-0. 2011. 211 с., цв. фотоатлас.
Формат 170 х 240 мм. Те. перепл. — Цена 600 руб.
Сравнительная анатомия, структурные преобразования и адаптивная эволюция ап-
парата двуногой локомоции птиц. А.В. Зиновьев. ISBN 978-5-87317-713-4. 2010. 285 с.,
ил., 2 цв. блока. Формат 210 х 290 мм. Те. перепл. — Цена 500 руб.
Особенности экспонирования коллекций декоративных растений. Вып. 2. А.С. Деми-
дов (отв. ред.). ISBN 978-5-87317-729-5. 2011. 128 с., ил. Формат 140 х 213 мм. В обл. —
Цена 100 руб.
Серебряноборское опытное лесничество: 65 лет мониторинга. Б.Р. Стриганова, А.А.
Сирин (отв. ред.). ISBN 978-5-87317-716-5. 2010. 260 с., ил., 8 цв. вкл., 6 карт. Формат
170 х 240 мм. Те. перепл. — Цена 400 руб.
Тараканы. Биология, экология, санитарно-эпидемиологическое значение, контроль
численности синантропных тараканов. В.П. Дремова, Н.А. Алешо. ISBN 978-5-87317-
708-0. 2011. 306 с., ил. Формат 145 х 210 мм. В обл. — Цена 300 руб.
Панцирные клещи Монголии. Б. Баяртогтох. ISBN 978-5-87317-703-5. 2010. 371 с., ил.
Формат 210 х 290 мм. Те. перепл. — Цена 500 руб.
Миграция воробьиных птиц: остановки и полёт. Н.С. Чернецов. ISBN 978-5-87317-698-
4. 2010. 173 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб.
Новости систематики низших растений. Том 44. А.Д. Потёмкин (отв. ped). ISBN 978-5-
87317-701-1. 2010. 378 с., ил. Формат 150 х 220 мм. Те. перепл. — Цена 300 руб.
Диатропика, эволюция и систематика. Ю.В. Чайковский. ISBN 978-5-87317-695-3. 2010.
407 с., ил. Формат 150 х 215 мм. Те. перепл. — Цена 400 руб.
Родентология. П.А. Пантелеев. ISBN 978-5-87317-688-5. 2010. 221 с., ил. Формат 145х
220 мм. Те. перепл. — Цена 360 руб.
Заказать эти и другие издания изд-ва КМК (биология, география, история, медицина) мож-
но по адресу:
123100 Москва, а/я 16 изд-во КМК, Михайлову Кириллу Глебовичу
Комп, почта: mikhailov2000@gmail.com
Интернет: http://avtor-kmk.ru (аннотации изданных книг)
Факс: (495) 629-4825 Тел. (495) 692-5894 раб.
Научное издание
Птицы России и сопредельных регионов: Пеликанообразные,
Аистообразные, Фламингообразные.
/ Андронов В.А., Ардамацкая Т.Б., Артюхин Ю.Б. и др.
Отв. ред.: С.Г. Приклонский, В.А. Зубакин, Е.А. Коблик.
М.: Товарищество научных изданий КМК. 2011. 602 с., 16 цв. вкл.
При участии ИП Михайлова К.Г.
Главный редактор издательства: К.Г. Михайлов
Вёрстка: Св.В. Найденко, М.В. Скороходова, К.Г. Михайлов
Для заявок:
123100, Москва, а/я 16, Издательство КМК
электронный адрес mikhailov2000@gmail.com
http://avtor-kmk.ru
Подписано в печать 12.07.2011. Заказ № 3521
Формат 70x100/16. Объём 38+1 п.л. Бумага офсетн. и мелов. Тираж 1200 экз.
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН,
121099, Москва, Шубинский пер., 6
4XVK>
---