/























































































Text
VOGELZUG
UND VOGELZUGFORSCHUNG
von
J. STEINBACHER
1951
И. ШТЕЙНБАХЕР
ПЕРЕЛЕТЫ ПТИЦ
И ИХ ИЗУЧЕНИЕ
Перевод с немецкого
К. Э. ФАБРИ
Под редакцией и с предисловием
проф. Г..П. ДЕМЕНТЬЕВА
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва — 1 9 56
Fl й
; ‘ ZOOME'l’GD.NAROD.RU
АННОТАЦИЯ
Автор на большом фактическом материале всесторонне, хотя и
в очень сжатой форме, разбирает перелеты — это интереснейшее яв-
ление в жизни птиц. Он касается вопроса о происхождении переле-
тов, об их движущих силах и направлениях, об ориентации во вре-
мя перелетов и об их физиологических основах. Книга предназначена
для биологов всех специальностей и для широких кругов читате-
лей-неспеци а листов.
Редакция биологической литературы
Заведующий редакцией — проф. А. А. НИЧИПОРОВИЧ
ПРЕДИСЛОВИЕ
К РУССКОМУ ИЗДАНИЮ
Вопрос о перелетах птиц представляет собой одну из
интереснейших общебиологических проблем. Ему посвя-
щена большая литература. Исследования в этой области
энергично продолжаются. Нельзя не отметить, в частности,
большое развитие, которое они получили в последние два
десятилетия в Советском Союзе, главным образом в ре-
зультате широкого и планомерного проведения кольцева-
ния птиц. Накопился огромный фактический материал.
Естественно, что дальнейшее изучение проблемы требует
обобщения этого материала. У нас немало обзорных работ
по перелетам птиц: книги академиков Миддендорфа и
Мензбира, монография Промптова, популярные книги
Житкова и Турова. Имеется по этому вопросу и иностран-
ная литература: книги Диксона, Дункера (переведена
на русский язык), Лендсборо-Томсона, Лукануса, Лин-
кольна, Бубье. Однако все эти работы теперь уже не охва-
тывают вопроса с достаточной полнотой. И в этом отноше-
нии небольшая книга Штейнбахера, несомненно, вполне
своевременна.
Штейнбахер всесторонне разбирает вопрос о перелетах
птиц. В первой главе дается определение понятия переле-
тов и краткий обзор истории вопроса. Вторая глава посвя-
щена современным методам изучения перелетов: полевым
исследованиям и наблюдениям, кольцеванию, эксперимен-
тальным работам по проблеме ориентации и выяснению
причин явления в целом. Третья — центральная = глава
6
Предисловие к русскому изданию
содержит описание самого явления перелета птиц. Разби-
раются вопрос о расположении мест зимовок по отноше-
нию к области гнездований, направления и «пути» проле-
та, особенности поведения пролетных птиц, сезонность
(время) пролетов, высота и скорость передвижения про-
летных птиц. Особые резделы посвящены вопросу о тех
физиологических требованиях, которые «предъявляют» пе-
релеты к организму птиц, вопросу о связи перелетов с по-
годой, ориентации перелетных птиц, причинам перелетов.
В качестве приложения дан подробный разбор перелетов
некоторых видов — скворца, сорокопута-жулана, белой
трясогузки, серебристой чайки, журавля и чибиса.
Автору удалось в сжатой и популярной форме осве-
тить проблему миграций птиц во всем ее многообразии.
А многообразие это очень велико: вопрос следует рас-
сматривать и с экологической, и с биогеографической,
и с физиологической, и с эволюционной точек зрения.
Для решения его необходимы и полевые зоологические
исследования, и эксперименты. Все это с разной степенью
полноты затронуто в книге Штейнбахера и притом на со-
временном научном уровне.
Кроме того, достоинство' книги состоит в насыщенности
ее фактическим материалом. Все положения подтвержда-
ются хорошо подобранными и свежими фактами, относя-
щимися, в основном, к европейским видам птиц. Книга,
таким образом, может быть использована и для самостоя-
тельного ознакомления с предметом, и для преподавания.
Из частных вопросов надо отметить следующее. Автор
совершенно правильно' вновь подвергает критике еще не-
давно широко распространенный в литературе взгляд о так
называемых «пролетных путях». Он приходит к правильно-
му выводу о наличии у перелетных видов птиц особых,'не
сводящихся только к зрительным способов ориентации
(хотя физиологическая сторона этого процесса остается,
Предисловие к русскому изданию 7
что отмечает и Штейнбахер, еще невыясненной). Автор
справедливо отвергает одностороннее объяснение явления
перелетов причинами гормонального порядка.
Существенный недостаток книги состоит в слабом ис-
пользовании русской и советской литературы. О Мензбире
автор даже не упоминает. Как известно, наше Централь-
ное бюро кольцевания издало несколько' выпусков своих
работ, широко освещающих результаты мечения птиц в
СССР. Они, как и другие исследования советских зоологов
по вопросу о перелетах птиц, остались Штейнбахеру, оче-
видно, неизвестными. В результате некоторые сообщаемые
автором сведения устарели или неточны. Слабо1 освещена
теория происхождения перелетов. На стр. 18 Штейнбахер,
в сущности сводит возникновение перелетов к событиям
ледникового времени и их последствиям. А между тем пе-
релеты, вероятно, существовали и в третичном времени;
существуют они и в местностях, не подвергавшихся оле-
денению, в частности в субтропиках и тропиках. В основе
происхождения перелетов птиц лежат, очевидно, реакции
их организма на значительные сезонные изменения усло-
вий существования, и привлекать сюда вопрос о леднико-
вом периоде, расселении или переселении, прародине — в
сущности нет оснований (за исключением отдельных част-
ных случаев).
Книга переведена полностью, без сокращений; опуще-
но лишь предисловие автора. Ввиду запутанности термино-
логии, в ряде случаев во избежание возможных недоразу-
мений принятый в переводе русский термин сопровож-
дается в скобках немецким термином.
Проф. Г. П. Дементьев.
Глава I
ВВЕДЕНИЕ
Что понимается под перелетом птиц?
Многие люди, утверждающие, что они интересуются
всеми вопросами жизни животных, не различают понятий
«полет птиц» и «перелет птиц». Само' по себе каждое из
них совершенно ясно. Однако из-за сходного звучания слов
или просто небрежного употребления эти термины неред-
ко смешивают, что, конечно, не может способствовать луч-
шему пониманию этого явления: не каждый сразу дога-
дается, о чем идет речь. Различие понятий «полет птиц»
и «перелет птиц» становится особенно ясным при сравне-
нии с аналогичными явлениями в других группах жи-
вотного мира, например плаванием рыб (в отличие от
основанного на иных принципах плавания медуз и раков)
и поразительными миграциями некоторых видов рыб,
одно явление только обусловливает другое. Поэтому, что-
бы предотвратить всякого рода недоразумения, оба тер-
мина следует употреблять только в их прямом значении.
Понятие «перелет птиц» почти такое же древнее, как и
знания о птицах вообще; оно упоминалось еще в античных
преданиях. Но впервые слово «Zug» (перелет) было
употреблено по отношению к птицам императором из ди-
настии Гогенштауфенов Фридрихом II около 1250 г. С тех
пор оно регулярно встречается в немецкой орнитологиче-
ской литературе. Один из виднейших орнитологов Брэм
указывал, что термин «Vogelzug» или «Zug der Vogel»
(«перелет птиц») почти всегда понимался в том же смысле,
что и понятие «миграции птиц» («Wanderungen der Vo-
gel»). Сравнительно недавно Вахе (1920) предложил
употреблять лишь последний термин. Термин же «перелет
птиц» он отклонил, поскольку он относится только к про-
лету в определенном месте и в определенное время. Одна-
ко такой подход к общепринятому термину слишком
сложен.
10
Глава I
Можно было бы также возразить, что всякого рода ми-
грации имеют место и в многочисленных группах живот-
, ных, что они обусловлены совершенно различными причи-
нами и часто совсем не похожи на миграции птиц. В этом
отношении следует особенно указать на миграции рыб,
прежде всего угря, сельди и лосося. Если не считать пере-
леты птиц, эти миграции относятся к наиболее поразитель-
ным примерам совершаемых животными перемещений, для
изучения которых также пытаются применить метод ин-
дивидуального мечения. Новейшими исследованиями
установлено удивительное сходство миграции североамери-
канского лосося с перелетами птиц (постоянство распро-
странения, ориентация и т. п.). Известны также массовые
миграции некоторых насекомых, например стрекоз и саран-
чи, которые передвигаются на сотни километров не только
в поисках пиши, но и побуждаемые неизвестными внут-
ренними стимулами. Миграции пеструшек на Крайнем Се-
вере, вероятно, обусловлены массовым размножением, но
и это явление не получило еще исчерпывающего объясне-
ния. Периодические переселения северных оленей, очевид-
но, связаны с изменениями климатических условий и с
питанием. Очень сходны с перелетами птиц миграции се-
верных летучих мышей, часть которых улетает вместе с
птицами в общей смешанной стае. Летучая мышь северный
кожанок (Eptesicus nilssoni} проводит лето и рождает
своих детенышей между 54 и 58° с. ш. К концу лета она
проникает еще дальше на север, а затем мигрирует вместе
с ласточками на юг на зимовки в Среднюю и Северную.
Германию. Некоторые летучие мыши Северной Америки
(Lasiurus cinereus) перелетают даже через открытое море
до Бермудских островов.
Таким образом, миграции некоторых животных не от-
личаются от миграций птиц, но тем не менее их очень
редко называют перелетом. Этот термин употребляется
только по отношению к птицам. Он обозначает не столько
способ осуществления миграции, который следует назы-
вать пролетом, сколько все явление миграции в целом,
представляющее собой такую же важную жизненную
функцию, как размножение или линька у птиц.
Не так давно термин «перелет птиц» было предложено
считать более узким по отношению к общему ( понятию
Введение
11
«миграции птиц» (ван Оордт, 1943). Согласно этому опре-
делению, под перелетом понимают те перемещения птиц,
которые они дважды ежегодно совершают между областью
гнездования (своей родиной) и областью зимовок в зави-
симости от периодической смены времен года. Такое огра-
ничение понятия, несомненно, имеет много преимуществ и
будет способствовать ясности будущих специальных работ.
Это следует особенно подчеркнуть, так как широкие рамки
данной книги не позволяют строго проводить такие тонкие
различия. Однако в наиболее ответственных местах изло-
жения на это будет обращено особое внимание и вместо
таких выражений, как «перелет» и «перелетные птицы»,
будут употреблены термины настоящий перелет» и «соб-
ственно перелетные птицы».
Как узнают перелеты птиц?
Наши знания о жизненных проявлениях у птиц основа-
ны на наблюдениях и их регистрации. В этом отношении
определение перелета птиц кажется простым и легким де-
лом. Так, если мы видим осенью стайки пролетающих над
полями мелких птиц или следим за воронами и галками,
кочующими по пашням и рощам, если иногда по серому
ноябрьскому небу с громким зовом »тянет выстроившаяся
клином стая журавлей или же доносятся крики пролетаю-
щих ночью диких гусей, то мы уже полагаем, что смогли
уловить часть того большого явления в природе, которое
именуется «перелетом птиц». Однако это ничтожно мало
по сравнению с той картиной, которая открывается нашим
взорам в нескольких пунктах земли. Так, например, на
Курской косе — узком перешейке между заливом и морем
в северной части б. Восточной Пруссии — можно наблю-
дать огромные массы птиц во время их путешествия на
юг. Непрерывным потоком, бесконечными вереницами или
бесчисленными стаями пролетают вьюрки, жаворонки,
скворцы, вороны, песочники, голуби и хищные птицы. Мно-
гие навсегда сохранят в памяти ночь у маяка на побережье
Северного моря во время перелета птиц: в белом свете
маяка, как искры, внезапно начинают сверкать птицы, их
становится все больше; громкие и мелодичные, как звуки
флейты, раздаются голоса ночных путешественников. А ко-
гда магическое сияние над темным морем привлечет свои-
12
Глава /
ми чарами тысячи пернатых, их крики сливаются в много-
голосый хор, способный привести в замешательство. Эта
картина производит сильное впечатление, и не надо быть
любителем природы, чтобы оказаться во власти такого
подлинного «чуда перелета птиц».
Помимо прямого опознавания перелета птиц, возможно
и косвенное — определение изменений в составе авифауны.
Такие возможности представляются и зимой, но особенно
во время перелета. Конечно, для этого необходимы некото-
рые знания о составе птичьего населения района в опре-
деленное время года. Таких крупных птиц, как аист или
журавль, заметит каждый; поэтому их возвращение на ро;
дину весной или отлет осенью установить нетрудно. Ла-
сточки и стрижи также слишком хорошо известны, чтобы
их появление или исчезновение могло остаться незамечен-
ным. Другие птицы, например кукушка или соловей, во
всяком случае весной, выдают свое присутствие голосами.
Но большинство птиц, перелет которых дважды в год ока-
зывает коренное влияние на общую картину жизни перна-
тых, известно далеко не всем; их не так просто узнать, за
ними не так легко вести наблюдения. С этой работой спра-
вится не каждый, но зато это наиболее надежный метод,
точно устанавливающий перелет птиц.
Таким путем можно получить представление о типич-
ных перелетных птицах и научиться отличать их от других
видов птиц, которые могут (но не обязательно должны)
совершать дальние путешествия. Их перелет без опреде-
ленной цели длится с осени до весны. Хотя птицы при этом
и придерживаются в основном южного направления, они
часто отклоняются то в одну, то в другую сторону. Это
скорее кочевки или скитания по тем местам, где в холод-
ное время года еще можно найти достаточно пищи. Поэто-
му такие виды птиц называют кочующими. К ним относит-
ся большая часть остающихся на зиму в Германии птиц
(синицы, дятлы, зяблики, зеленушки, черные дрозды, ще-
глы и чижи), а также некоторые из так называемых зим-
них гостей, т. е. птиц из северных и восточных областей
Европы, зимующих в Германии (например вьюрки, че-
четки и северные снегири).
Некоторые виды птиц в иные годы или вообще не ми-
грируют, или же перемещаются на совершенно незначи-
ЁведеНиё
13
тельные расстояния; зато в последующие годы они путеше-
ствуют в таком поразительном количестве и на таком про-
странстве, что в прежние времена с этим были связаны
многие суеверия. Время, так же как и конечные пункты
странствования птиц, совершенно различны, и их нельзя
предугадать. Часто птицы попадают в области, которые им
совершенно не подходят и где они большинству населения
неизвестны. Однако они без длительных остановок пересе-
кают и такие местности, которые им вполне подошли бы.
Эти миграции, объединяющие характерные черты как ко-
чевок, так и перелетов, называют залетами. Примерами
залетных птиц, появляющихся в Германии через неболь-
шие промежутки, от одного до нескольких лет, могут слу-
жить клесты, свиристели, сибирские ореховки, северные пе-
стрые дятлы и мохноногие канюки. Значительно менее ре-
гулярны вторжения саджи из Средней Азии или розового
скворца из Туркестана и Закаспийского края; такие залеты
обычно ограничиваются только югом СССР, но в отдель-
ных случаях птицы залетают значительно дальше на за-
пад. Так, например, в 1925 г. они в большой массе добра-
лись до Венгрии, а часть из них —даже до> Бельгии и
Шотландии. При этом иногда наблюдаются и попытки
гнездования. Впрочем, это относится и к залетам среди-
земноморских форм в Центральную Европу: ходулочника
в Голландию и фламинго в Швейцарию.
В противоположность перелетным, кочующим и залет-
ным птицам, оседлые птицы в течение всего года пребыва-
ют в своей родной местности: это — воробьи, сороки, куро-
патки, ястреб-тетеревятник, обыкновенная неясыть, кра-
пивник, хохлатый жаворонок и поползень. Они почти
никогда не удаляются на большие расстояния от своей
области гнездования; это правило иногда нарушают лишь
одиночки
Приведенная выше классификация птиц по их отноше-
нию к месту обитания произведена еще в начале XVIII в.
Фришем и с тех пор применяется во всей орнитологической
литературе. Ею еще широко пользуются и в настоящее
1 Попутно отметим, что при этом возможны и исключения, как
это иногда наблюдается у куропаток, например, в Восточной Евро-
пе—массовое появление куропаток у Курского залива (Позингис
и Шюц, 1938).
14
Глава I
время, хотя, строго говоря, она уже устарела. Опыт по-
следних десятилетий показывает, что и в данном случае
природа противопоставляет любой попытке схематизации
обилие переходов, которые нарушают всякий искусственно
установленный порядок. Так, в различных областях одни
и те же виды птиц могут по-разному относиться к своей
родной местности: в зависимости от географического' по-
ложения последней они могут быть либо оседлыми, либо
кочующими, либо перелетными птицами. Такие факты из-
вестны, в частности, о граче, серой вороне, скворце, зарян-
ке, зеленушке, черном дрозде, канюке и крякве. Но неред-
ко отношение вида к гнездовью зависит от возраста и по-
ла; это относится главным образом к зябликам, у которых
самки и молодые особи в подавляющем большинстве уле-
тают, в то время как старые самцы зимуют на родине. По-
добная зависимость наблюдается и у других видов, напри-
мер у многих хищных птиц. Помимо вышеупомянутых раз-
личий, встречаются и индивидуальные—у особей разных
выводков. Если к этому добавить, что в сравнительно ко-
роткий срок у одного и того же вида птиц может изме-
ниться отношение к миграции и перелетная птица может
стать кочующей, а кочующая — оседлой (исследования по-
следних десятилетий подтверждают этот вывод в отноше-
нии черного дрозда, скворца, канареечного вьюрка, луго-
вого конька и горной трясогузки), то невозможность про-
вести строгое разделение оседлых, кочующих и перелетных
птиц становится совершенно ясной. При употреблении этих
обозначений, которых иногда очень трудно избежать, сле-
дует иметь в виду указанные ограничения.
Наличие постепенных переходов между оседлыми, ко-
чующими, перелетными и залетными птицами доказывает
влияние факторов внешней среды, отчасти действующих
и поныне на размеры миграций.
В дальнейшем мы увидим, что эти факторы частично
определяют и направление миграций. Но целый ряд явле-
ний, относящихся прежде всего к самому перелету, можно
объяснить только исходя из предположения, что влияние
прошедших периодов истории Земли сохранилось в на-
следственности, и в настоящее время выступает в такой
форме, которую современными условиями уже объяснить
нельзя.
Введение
15
Где происходят перелеты птиц?
Широко распространено мнение, что подлинные пере-
леты птиц существуют лишь в умеренных и холодных зо-
нах и что они ограничиваются только северным полуша-
рием. И то и другое неверно. Правда, из-за наличия боль-
ших массивов суши и значительно более близкого располо-
жения их к полюсу перелеты в северном полушарии
выражены гораздно сильнее и более заметны, чем в юж-
ном полушарии. Но, например, и в Южной Америке есть
такие виды птиц, которые гнездятся в Патагонии, а зимуют
далеко на севере. Это относится и к некоторым пингвинам,
которые покидают свои гнездовья в Антарктике и вплавь
мигрируют к берегам Южной Америки. В Южной Африке
известно более двадцати видов типичных перелетных
птиц. В Австралии й Новой Зеландии многие птицы также
совершают регулярные миграции. Они простираются от
южного побережья Австралии через весь континент вплоть
до Индо-Малайских островов и, следовательно, вполне мо-
гут сравниваться с самыми дальними миграциями евро-
пейских перелетных птиц. Можно указать и на два вида
буревестников Puffinus gravis и Oceanites oceanicus, ко-
торые летят от своих гнездовий на Тристан-да-Кунья (для
одного- вида) и островах Кергелен, Южная Георгия и
Фальклендских (для другого вида) до широт Германии,
причем летом их можно не так уж редко встретить на Ат-
лантическом побережье Западной Европы и Северной
Америки. Типичные перелетные птицы есть и в тропиках.
В этом отношении лучше всего изучена Африка. Так, бело-
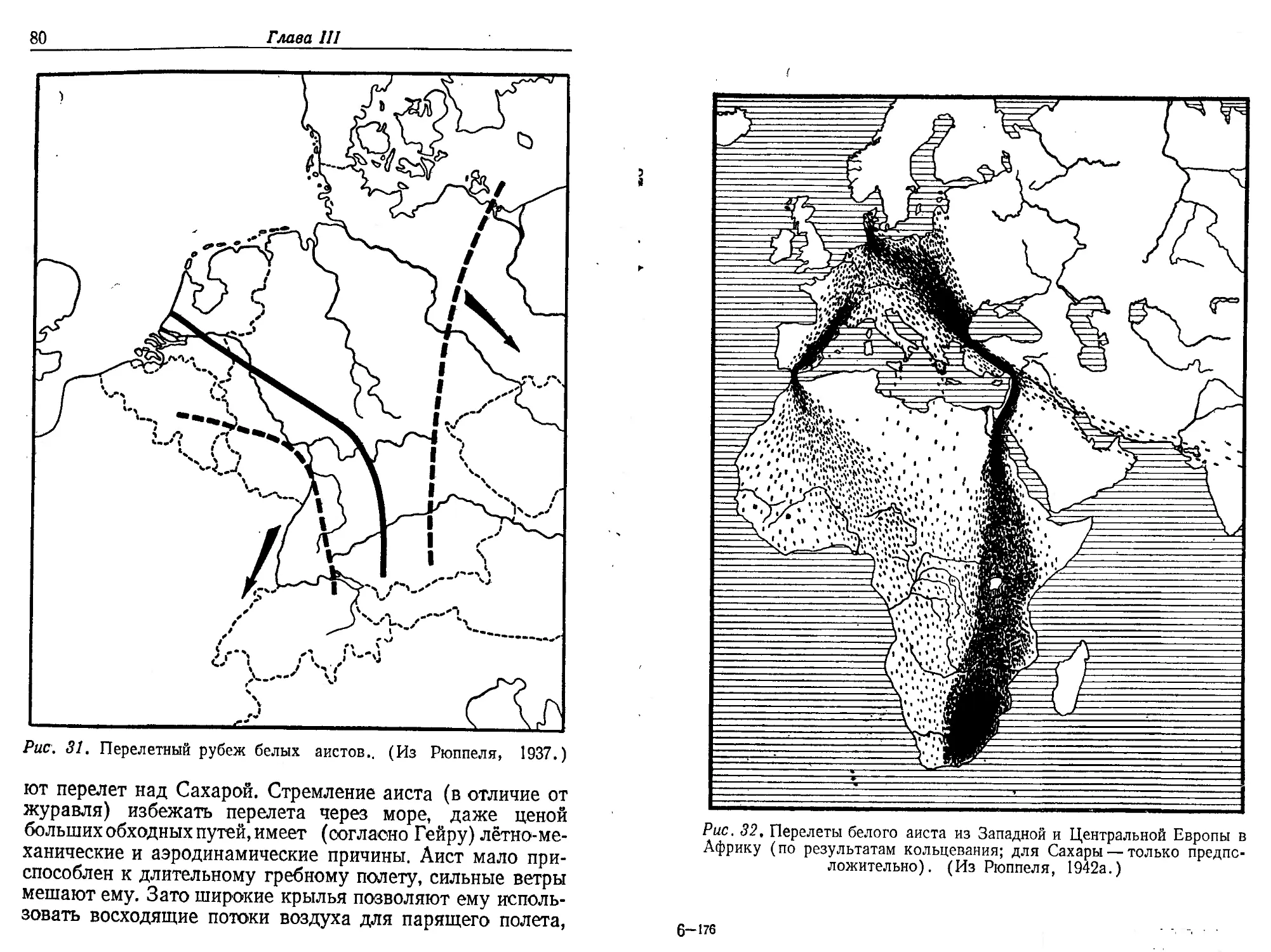
брюхий аист регулярно перелетает в начале южного- лета
из гнездовой области, находящейся в Судане, в Анголу,
Родезию и Мозамбик, вследствие чего он все время нахо-
дится в дождливой зоне, что важно при его специализиро-
ванном питании. Африканский ко-зодой, напротив, мигри-
рует из Родезии и Анголы через область Конго на свои
зимовки в Судан. Тем самым его пребывание в этих райо-
нах приурочивается ко времени года, наиболее благопри-
ятному для летающих термитов — его основного корма
(рис. 1). Ряд птиц перелетает к началу засушливого време-
ни года с острова Мадагаскар на Африканский континент,
где находит более благоприятные условия жизни, пока
16
Глава 1
Рис. 1. Перелеты вымпелового козодоя (Cosmetornis vexillarius) из
южной гнездовой области на знмовки на севере. (Из Штреземана,
1927—1934, по Чепину, 1932.)
наступающий период дождей не заставляет их вернуться
обратно.
Эти немногие примеры доказывают, правда, только са-
мый факт существования перелетов у тропических птиц1.
В целом авифауна тропиков состоит в подавляющем боль-
шинстве из оседлых и кочующих птиц. Миграции птиц в
тропиках обусловлены периодическими изменениями влаж-
ности воздуха и уровнем осадков, которые в определенных
1 Изучение перелетов птиц в Африке еще только начато и по-
ка ограничивается небольшими районами Восточной и Южной
Африки.
Введение
17
районах могут привести к существенным колебаниям в ко-
личестве пищи. В умеренной и холодной зонах периодиче-
ские изменения длины дня и температуры, вызывающие
колебания в количестве пищи, оказывают несравненно бо-
лее сильное влияние, которое усиливается внутренними
факторами, обусловленными наследственностью и при-
вычкой.
Поэтому мы встречаемся с перелетами птиц, прежде
всего, в северных частях Европы, Азии и Америки, где
преобладают перелетные или кочующие птицы и очень
мало оседлых. Но чем дальше мы продвигаемся на юг и
запад, а для некоторых видов и на восток, тем больше
изменяется это соотношение в пользу оседлых птиц.
Во всяком случае, для всех зон на Земле справедливо
следующее положение: чем менее выражена сезонная
периодичность климатических явлений, тем меньше число
перелетных птиц в. определенной области, и наоборот.
Как возник перелет птиц?
Едва ли существует другой вопрос, связанный с ми-
грациями птиц, на который даются столь различные отве-
ты. Причина этого заключается в отсутствии неопровержи-
мых фактов. Пытаясь увязать сведения из области палео-
географии, палеоклиматологии и палеозоологии с данными
о современных перелетах птиц, исследователь вынужден
полагаться только на свою проницательность и опыт. По-
этому, сколько бы ни обсуждался этот основной вопрос яв-
ления перелета птиц и сколько бы о нем ни писалось, все
заключения останутся более или менее вероятными теория-
ми, и вряд ли мы когда-либо при решении этого вопроса
сможем обойтись совсем без гипотез.
Некоторые из существующих многочисленных теорий
о возникновении перелетов считаются в общем приемлемы-
ми, Согласно Томсону (1926), их в настоящее время мож-
но свести к трем гипотезам:
L 1. Родиной перелетных птиц была их современная об-
ласть гнездования. Наступление ледника вынудило птиц
отступить на юг. Каждое лето они возвращались (насколь-
ко это было возможно) на север для выведения птенцов и
стали перелетными птицами.
18
Глава J
2. Родиной перелетных птиц была современная область
гнездования. Оттесненные ледником на юг, птицы засели-
ли небольшую территорию и превратились в оседлых
птиц. Но потребность возвращения на родину сохранилась
в наследственности. Поэтому по окончании ледникового
периода у них развились перелеты.
3. Родиной перелетных птиц была современная область
зимовки и другие южные местности. Массовое размноже-
ние в этих районах вынудило птиц расселяться на терри-
тории, освобождавшейся ото льда.
Согласно указанным гипотезам, причиной возникнове-
ния перелетов птиц является влияние ледникового перио-
да. При этом исключается возможность существования
перелетов до ледникового периода, как это допускал
Экардт. Перелеты в условиях равномерного теплого кли-
мата третичного периода, вероятно, напоминали переме-
щения наших современных кочующих птиц. В дальнейшем
ухудшение условий существования в начале ледникового
периода вынудило птиц все больше и больше расширять
свои миграции по мере того, как северные зимы станови-
лись все продолжительнее и суровее. Соответственно уко-
рачивались сроки летнего пребывания птиц на родине.
Когда северные материки были в большой степени охва-
чены оледенением, эти перелеты совсем прекратились.
После отступления льдов птицы вновь заселили освобо-
дившуюся область и так постепенно проникли в гнездовья
своей древней родины. Но ежегодно они были вынуждены
покидать ее на климатически неблагоприятное время года
и искать убежища в тех районах, в которые они отступа-
ли во время ледникового периода. Эти повторявшиеся в
течение тысячелетий отступления птиц осенью и ежегод-
ные новые или, точнее, повторные, заселения покинутых
областей весной как результат сезонных изменений клима-
тических условий вследствие наступления ледника и рас-
сматриваются в настоящее время как основная причина
возникновения перелетов птиц, если вообще можно гово-
рить об едином мнении в таком вопросе. В этом отноше-
нии мы придерживаемся взглядов Майра и Мейзе (1930),
которым мы обязаны новейшей разработкой этой пробле-
мы. Хотя их доказательства в общем ясны, все же многие
частности еще остаются сомнительными. Так, например,
Введение 19
почему поведение оседлых и кочующих птиц в Германии
во многом отлично от перелетных птиц? Вселились ли осед-
лые и кочующие птицы лишь после ледникового периода?
Почему так много видов птиц отлетают значительно даль-
ше, чем когда-либо могло простираться влияние леднико-
вого периода? И, наконец, все гипотезы о значении ледни-
кового периода в возникновении перелетов птиц не учиты-
вают возможного влияния последнего межледникового пе-
риода, когда, по крайней мере в Европе, на несколько ты-
сячелетий установились почти третичные климатические
условия. В результате субтропические растения и живот-
ные могли проникнуть глубоко в Центральную Европу. Не-
ужели это «время орешника» (Haselgipfel) от которого
нас отделяют приблизительно 8000 лет, не оказало на от-
ношение птиц к их месту обитания более стойкого влия-
ния, чем последнее оледенение, бывшее 23 000 лет назад?
Далее, если в течение тысячелетий сезонная периодичность
климатических явлений привела, в конце концов, к возник-
новению перелетов птиц, и эта приобретенная привычка
стала даже передаваться по наследству, так что в настоя-
щее время перелеты совершаются в значительной степени
независимо от влияния внешней среды, то чем можно объ-
яснить утрату этой привычки в течение столь непродолжи-
тельного времени, как несколько лет или даже десятиле-
тий, когда перелетные птицы вновь становятся оседлыми?
Мы уже приводили несколько соответствующих примеров,
когда говорили о разделении птиц на перелетных, кочую-
щих и оседлых, и еще вернемся к этому ниже.
Таковы некоторые из тех возражений и сомнений, кото-
рые можно выдвинуть против признанных гипотез о про-
исхождении перелетов птиц. Это не облегчает разрешения
вопроса, но и не должно отпугивать тех, кто хочет деталь-
нее заняться его разработкой. Мы сознательно указываем
здесь на эти трудности, чтобы поощрить критическое отно-
шение к теоретической части всей проблемы, так как хоро-
1 Haselgipfel — «время орешника» — начало послеледникового
потепления с преобладанием в растительности орешника. По систе-
ме Блитта — Сернандера это время называется «бореальным», у дру-
гих авторов — «атлантическим». Оба термина неудачны. — Прим,
ред.
2»
20
Глава 1
шо обоснованные гипотезы могут способствовать ее разви-
тию, а догадки, не подкрепленные научными данными,
только нанесут ей ущерб.
К истории изучения перелетов птиц
Началом каждой науки является первая попытка объ-
яснить наблюдаемые закономерности. Истоки изучения
естественнонаучных проблем связаны с трудами греческо-
го философа Аристотеля; его имя должно стоять и в на-
чале этого очерка. Ведь он первый в своей «Истории жи-
вотных» задумался над тем, где зимуют птицы, и пытался
объяснить ряд явлений перелета птиц. Неважно, что его
объяснения впоследствии оказались неверными; их надо
оценивать с учетом его времени. Сделанные им наблюде-
ния не утрачивали своего значения до средних веков.
Вполне правильно замечание Аристотеля о том, что после
осеннего равноденствия птицы улетают из холодных стран,
чтобы избежать зимы, направляясь в соседние, а иногда в
более отдаленные местности (например, журавли улетают
в Египет), а весной вновь возвращаются на север. Он упо-
минает даже о том, что отдельные виды появляются в раз-
личное время и что в период осеннего перелета они более
упитаны, чем во время весеннего. Его указания на то, что
некоторые птицы впадают в зимнюю спячку подобно зем-
новодным и пресмыкающимся, в такой форме, конечно, не-
верны. Но они делают особенно замечательным неодно-
кратно установленный в последнее время факт, что ласточ-
ки и стрижи иногда впадают в состояние, подобное зимней
спячке, которое при неблагоприятных условиях погоды мо-
жет продлиться много дней. Но едва ли можно допустить,
чтобы Аристотель знал об этом редком исключительном
явлении. Приблизительно 400 лет спустя римский писа-
тель Плиний в основном повторил указания Аристотеля в
своей «Истории природы» («Historia naturalis»).
С тех пор, вплоть до XIII в., эта область науки была
забыта. Только Фридрих II, император из династии Гоген-
штауфенов, который в изучении природы опередил свое
время на столетия, описал в своем капитальном труде
«De arte venandi cum avibus» перелеты птиц. Он не
только пытался объяснить отлет разных видов птиц в раз-
Введение
21
личное время и под влиянием различных внешних усло-
вий, но уже точно различал «перелеты» и «кочевки», упо-
минал о линьке и скоплениях птиц в определенных местах
перед началом перелета, отмечал влияние погоды и даже
сообщал о порядке в стае во время перелета. Эта работа
Фридриха II, написанная преимущественно на основе
личных наблюдений, значительно превосходила подобные
труды средних веков и должна считаться исходным пунк-
том научного изучения перелетов птиц.
После этого прошло еше два с половиной столетия,
прежде чем великий преобразователь естественных наук
Карл Линней продолжил изучение перелетов птиц. Он пер-
вый пытался создать сеть наблюдателей для регистрации
перелетов птиц, став, таким образом, основателем совре-
менных фенологических исследований, не утративших сво-
его значения и в наше время. Все позднейшие исследова-
тели придерживались главным образом взглядов Линнея,
так что до конца XIX в. авифенология была единственной
основой исследования перелетов птиц.
Минуя Бехштейна и видного орнитолога Наумана, ко-
торый четко различал оседлых, кочующих и перелетных
птиц, особенно отмечая влияние на перелет птиц погоды,
пищи и географических условий, а также дал вполне вер-
ные сведения о направлении миграций и о некоторых дру-
гих деталях перелета, мы переходим к труду Брэма «Пе-
релет птиц», в котором приводятся точные данные о днев-
ном и ночном перелете, определенных пролетных путях,
различном поведении старых и молодых особей одного и
того же вида и высказываются предположения о причинах
осеннего перелета. Ряд позднейших трудов, в том числе
Экстрёма, Фабера, Кесслера и Блогера, посвященных пе-
релетам птиц в Швеции, Исландии и на юге России, ука-
зывает на быстрый прогресс исследований в этой области
в первой половине XIX в. Если Экстрём впервые сопоста-
вил большое число дат прилета и отлета перелетных птиц,
то труд Миддендорфа «Изопиптезы России» («Isopiptesen
Rufilands») представляет собой серьезную попытку воссоз-
дать на основе авифенологических наблюдений картину пе-
релета и одновременно вскрыть его причины. Изопиптеза-
ми он назвал линии, соединяющие местности одновремен-
ного прилета весной определенных видов птиц. Мидден-
22
Глава I
дорф считал, что направление перелета определяется свой-
ственным перелетным птицам чувством северного магнит-
ного полюса Земли. Ниже мы подробнее остановимся на
содержании этой важной работы, не утратившей своего
значения и сегодня.
Несколько позднее финский исследователь Пальмен
(1876) опубликовал труд «О пролетных путях птиц» («Uber
die Zugstrafien der Vogel»). Он установил для 19 видов
птиц Крайнего Севера миграции по определенным узко
ограниченным пролетным путям, которые у водоплаваю-
щих и болотных птиц проходят по берегам материков.и
руслам рек и поэтому были обозначены как морские, при-
брежные и речные пути. Не будучи в состоянии объяснить
возвращение сухопутных птиц к старым гнездовьям, Паль-
мен прибегнул к допущению, что все птицы должны поль-
зоваться пролетными путями, соответствующими первона-
чально найденным. Именно из-за этого обобщения работа
Пальмена очень скоро подверглась резкой критике. Ее и,
прежде всего, пролетные пути как неправдоподобные до-
мыслы решительно отверг Гомейер. Он высказался за
веерообразный перелет, который впоследствии был назван
«перелетом широким фронтом». В течение последующих
десятилетий расхождение во мнениях о пролетных путях и
перелетах широким фронтом красной нитью проходит че-
рез все исследования о перелетах птиц. Только работы
новейшего времени положили конец этому спору, причем
совершенно иначе, чем это предполагали ученые того вре-
мени.
Как Пальмен, так и Гомейер уже указывали на то,
как важно было бы, помимо дат прилета и отлета птиц в
определенных районах, узнать также, откуда они приле-
тели и куда держат путь. Но они еще не понимали, что
это возможно только путем мечения отдельных особей пе-
релетных птиц. Этому открытию суждено было стать ре-
шающим для тех огромных успехов, которые сделала нау-
ка о перелетах птиц с начала XX в. и которые теснейшим
образом связаны с введением метода кольцевания. Ему
мы обязаны знанием множества фактов, ставших в настоя-
щее время достоянием зоологической науки: это — точные
знания о местах зимовок и пролетных путях, о «верности»
родине и порядке в стае во время перелета, о ночных и
Введение
23
дневных перелетах, о ведущих направлениях (Leitlinien)
и скорости перелета многих видов птиц. Частично резуль-
таты этих исследований мы найдем в таких трудах, как
«Атлас перелетов птиц» Шюца и Вейгольда (1931), или
«Росситтен», или «О перелетах птиц в Росситтене» Тине-
мана (1931). Мы еще неоднократно будем возвращаться
к этим работам.
Возраст новейшей страницы науки о перелетах птиц
исчисляется двумя десятилетиями. Ее характеризует при-
менение физиологических методов исследования, при по-
мощи которых изучается «пусковой механизм» перелета и
его внутренние причины. В этой области, так же как и в
вопросе об ориентации птиц, которому в течение послед-
них 25 лет также уделялось особое внимание, результаты
исследований и экспериментов еще недостаточны.
Глава II
МЕТОДЫ ИЗУЧЕНИЯ ПЕРЕЛЕТОВ ПТИЦ
Каждому научному исследованию свойственны свои пу-
ти и методы, более или менее многочисленные, которые с
различной скоростью приводят к цели. Это в равной мере
зависит как от накопленного в данной области науки опы-
та, так и от предмета исследования.
Точно так же и в науке о перелетах птиц соответствен-
но многообразию стоящих перед ней задач существую!
различные направления исследования. Если рассматривать
каждое в отдельности, то у них как будто мало общего
Но всех их можно, тем не менее, свести к двум основным
формам: наблюдению и эксперименту, которые связаны
между собой различными переходами.
Мы уже видели, что развитие науки о перелетах птиц
шло от случайных наблюдений явлений к систематической
регистрации наблюдаемых фактов и затем переросло в по-
пытки анализа фаунистического материала. Большим ша-
гом вперед явилось введение кольцевания как вспомога-
тельного метода изучения авифенологии. Наконец, стали
применяться физиологические методы исследования, кото-
рые должны были дать ответ на основные вопросы, до тех
пор рассматривавшиеся лишь умозрительно.
Такая историческая смена направлений исследования,
однако, отнюдь не означает, что одно из них должно было
заменяться другим по той причине, что оно устарело в
свете новых знаний. Большей частью оказывалось необхо-
димым применение всех этих методов, не утративших свое-
го значения и по сегодняшний день. Ниже мы познако-
мимся с объемом и значением этих методов, с теми вопро-
сами, для решения которых они используются, и с их
практическим применением.
Методы изучения перелетов птиц
25
Полевые наблюдения
В качестве первого метода исследования рассмотрим
полевые наблюдения за перелетом птиц.
Значение метода научного исследования полевые на-
блюдения приобретают лишь тогда, когда они проводятся
систематически и по хорошо продуманному плану. Для
этого необходимо сделать точные записи о видах, привести
хотя бы приблизительные цифры количества птиц, уста-
новить высоту и направление перелета и охарактеризовать
погоду. Кроме того, надо попытаться отметить первое по-
явление перелетных птиц, увеличение и уменьшение коли-
чества прилетных птиц и окончание перелета у отдельных
видов в определенной области. На основе этих данных со-
ставляются календари перелета для различных видов
птиц, по которым в течение нескольких последующих лет
можно вычислять средние показатели. Такие календари пе-
релета в ограниченных, экологически замкнутых областях
при нормальных сезонных условиях погоды точно отража-
ют ход перелета во времени. Вейгольд (1930) разработал
для острова Гельголанд пространный календарь переле-
тов в виде графических схем, охватывавший десятилетний
период и почти все встречавшиеся там виды птиц (рис. 2).
Для других наблюдательных пунктов также составлены
подобные календари, хотя бы для определенных видов
птиц и для менее продолжительного времени (например,
для Росситтена и Курской косы или Вассенаара в Гол-
ландии). В любой другой местности такие данные полу-
чить значительно труднее, чем на Гельголанде. При срав-
нении таблиц, кривых или графических схем можно сде-
лать ценные выводы об особенностях перелетов, обуслов-
ленных географическим положением, особенностями ланд-
шафта и погодой. Дальнейшее развитие этого направления
работ заключается в объединении наблюдений над пере-
летом, проводимых одновременно в более обширных райо-
нах специально подготовленными орнитологами, которые
образуют сеть наблюдателей, работающих под единым ру-
ководством. Как уже упоминалось, Линней еще в 1750 г.
организовал такую сеть наблюдателей. В дальнейшем его
примеру последовали многие ученые прошлого века. В
итоге этих наблюдений был накоплен огромный материал,
26
Глава II
Рис. 2. Количественная характеристика хода весеннего (Д) и
осеннего (5) пролетов полевого жаворонка (Alauda arvensis)
на Гельголанде. (Из Вейгольда, 1930.)
который, однако', в связи с неодинаковой надежностью
лиц, участвовавших в его сборе, имел ограниченное значе-
ние и до сих пор едва подвергся обработке; далеко не
полные данные изложены в «Сообщениях комитета наблю-
дательных станций Германии», «Орнитологическом Жур-
Методы изучения перелетов птиц
27
Рис. 3. Схема направлений осенних перелетов птиц в районе восточ-
ной части Северного моря. (Из ван Доббена и Маккинка, 1934.)
нале» («Journal fiir Ornithologie») за 1875—1893 гг. и в
журнале «Аквила» («Aquila») Венгерского орнитологиче-
ского центра. Бесспорно, наибольших успехов в этом отно-
шении достиг Венгерский орнитологический институт, ко-
торый в 1875—1914 гг. с помощью 120—150 подготовлен-
ных наблюдателей провел ценную работу и дал сведения по
многочисленным вопросам перелетов птиц в Венгрии. Так,
Шенк, ставший впоследствии руководителем этого инсти-
тута, составил карты возвращения различных перелетных
птиц. При помощи этих карт можно проследить последо-
вательность ежегодного расселения птиц в зависимости
от климатических условий и ландшафта. Сходные карты
28
Глава 11
перелетов для некоторых местностей Юго-западной Гер-
мании и для Швейцарии составлены Бретшером, уделяв-
шим особое внимание изучению влияния климатических ус-
ловий. Кук установил, что в Северной Америке изопиптезы
Рис. 4. Изопиптезы (жирные линии) пеночки-веснички (Phylloscopus
trochilus) и изотерма 9°С (пунктир) в Европе; весной совпа-
дение этих линий почти полное. (Из Саутерна, 1938.)
канадской казарки приблизительно соответствуют изотер-
ме 35°F. Этот факт он поставил в связь с улучшением ус-
ловий питания. Позже Саутерн (1938) сравнил изопипте-
зы деревенской ласточки и пеночки-веснички с данными о
распределении тепла весной (рис. 4), в то время как
Сливинский (1938) ограничился только изображением изо-
пиптез некоторых видов птиц. Эти авифенологические кар-
ты частично берут свое начало от работы Миддендорфа
«Изопиптезы России», о которой мы уже упоминали выше.
В настоящее время, несмотря на всякого рода затруд-
нения, также создаются сети наблюдателей, охватываю-
щие обширные пространства. Так, Дрост организовал
Методы, изучения перелетов птиц,29
такую сеть к осеннему перелету 1930 г. по всему району
Северного моря и получил ценные сведения о деталях пере-
лета на различных пунктах побережья. Осенью 1929 г.
сетью наблюдателей в менее обширном, но крайне важном
для перелета птиц районе Курского залива руководил
Шюц. При этом удалось установить поразительное, но для
данной местности весьма характерное распределение про-
летных путей. В этом же направлении работали в 1933 г.
и голландские наблюдатели, сумевшие получить ценные
сведения о перелетах в западной части Северного моря и
в его отдельных районах.
Конечно, не все местности в равной степени пригодны
для наблюдений над перелетом птиц. В некоторых из них
он еле заметен, другие же места являются прямо-таки
фокусами перелета птиц, и наблюдения в этих районах
обещают дать весьма ценные результаты. Эти местности
обычно отличаются особым географическим расположени-
ем (горные проходы, горы, места, богатые озерами, побе-
режья морей, русла рек). Поэтому в таких пунктах орни-
тологи многих стран организовали постоянные наблюда-
тельные станции, которые впоследствии стали центрами
научно-исследовательской работы по изучению перелетов
птиц. Это относится, в первую очередь, к Германии, где
обе орнитологические станции, Гельголанд и Росситтен,
расположенные в центре пролетных путей, сыграли исклю-
чительно важную роль в изучении перелетов птиц.
Первая в Германии орнитологическая станция была
основана в 1901 г. в маленьком рыбачьем поселке Россит-
тен на Курской косе, которая тянется слегка выпуклой ду-
гой от Клайпеды на юг по направлению к Земландскому
(Калининградскому) полуострову. Ее длина составляет
97 км, а ширина равна в среднем лишь немногим сотням
метров. Тинеман оценил значение для перелета птиц этой
узкой косы, расположенной между Балтийским морем и
Курским заливом, и указал на необычайно благоприятные
возможности этого места для наблюдений и исследований.
Тогда Германское орнитологическое общество начало кон-
тролировать деятельность этой орнитологической станции
(которая вначале была просто наблюдательным пунктом)
и назначило Тинемана ее руководителем. В 1929 г. он был
.менен Хейнротом и его постоянным заместителем ШюЦем.
30
Глава II
Последний оставался в Росситтене почти до конца второй
мировой войны. С 1923 г. орнитологическая станция была
прикреплена к Обществу покровительства наукам импе-
ратора Вильгельма (рис. 5) ’.
Чем объяснить значение Курской косы для перелетов
птиц? Ответим на этот вопрос при помощи прилагаемой
Рис. 5. Орнитологическая станция Росситтен.
Выставочный павильон и открытые сооружения.
схемы. Осенью перелетные птицы из обширных террито-
рий к востоку от Балтики (вплоть до Сибири) в боль-
шом количестве направляются к побережью, которое, на-
чиная от Эстонии, тянется преимущественно в южном
направлении. Все сухопутные птицы избегают полета над
морем, в связи с чем они сперва скапливаются на берегу,
а затем мигрируют на юг; в пути к ним присоединяются
птицы, летящие с севера и востока. Таким образом, этот
поток вскоре сильно разрастается, пока возле Клайпеды
1 Следует указать на коренное различие понятий «орнитологи-
ческая станция» (Vogelwarte) — научная станция по изучению пе-
релетов птиц — и «станция по охране птиц» (Vogelschutzwarte) —
учреждение по охране птиц, - - которые нередко путают.
Методы изучения перелетов птиц
31
Рис. 6. Осенний пролет птиц в районе Курского залива. (Из
Шюца, 1929.)
Большое скопление пролетных птиц приводит к образованию направ-
ляющих линий как на Курской косе, так и на восточном берегу этого
залива,
не достигает оконечности косы. Суша здесь несколько
отклоняется на юго-восток, а коса проходит в юго-запад-
ном направлении, которое больше соответствует направ-
лению перелета птиц. Поэтому птицы в основной массе
покидают в этом месте берег и продолжают путь над
узкой косой, причем и здесь к ним из внутренних райо-
32
Глава 11
нов Балтики примыкают стаи птиц, стремящихся к косе
через вначале еще узкий залив. Коса образует своего ро-
да мост, разделяющий водные пространства (отсюда и его
народное название «мост перелета птиц»), на котором тес-
нятся стаи птиц, пока они не увидят далеко на юге Зем-
ландский (Калининградский) полуостров, после чего пти-
цы вновь рассеиваются (рис. 6).
4
Рис. 7. Наблюдательный пункт у Ульменхорст, расположен-
ного южнее Росситтена в самом узком месте Курской косы.
Особые географические условия в сочетании с преиму-
ществами дюнного ландшафта с его большей частью ред-
кой растительностью (и соответственно благоприятными
возможностями для наблюдений) позволили выявить здесь
такие массовые перелеты птиц, какие не удавалось наблю-
дать ни в одном другом пункте Европы. Вот несколько
цифр для иллюстрации: в отдельные благоприятные для
перелета дни число пролетающих здесь птиц достигало
500 000; только' за три утренних часа, с 6 до 9 час., их бы-
ло около 200 000. В иные дни это были на 99% зяблики,
которых на этом основании считают особенно типичными
представителями массовых перелетов. В тесной связи с
Методы изучения перелетов птиц 33
этим обстоятельством стоит обилие полученных орнитоло-
гической станцией Росситтен данных по многим частным
вопросам изучения перелетов птиц.
Совершенно иную картину мы наблюдаем, рассматри-
вая положение и значение бывшей орнитологической стан-
ции Гельголанд (рис. 8). Она была основана в 1909 г. как
отделение Государственного биологического института. За
Рис. 8. Орнитологическая станция Гельголанд.
Служебное помещение и квартира руководителя станции.
исключением времени первой мировой войны ею образцо-
во руководил Вейгольд, пока в 1924 г. его не сменил
Дрост. Станция стала известной после работ Гетке, кото-
рый в течение десятилетий проводил на ней наблюдения
и впервые указал в своей книге «Орнитологическая стан-
ция Гельголанд» («Die Vogelwarte Helgoland») на значе-
ние острова для перелетов птиц. Расположенный более
чем в 40 км от ближайшего берега круто поднимающийся
утес из песчаника площадью всего 0,5 км2 является для
3-176
34
Глава II
перелетающих через море птиц единственной возможно-
стью для отдыха, которой особенно пользуются плохо ле-
тающие, а при туманной погоде — и другие птицы. Но
особое значение для бесчисленных ночных путешественни-
ков имеют далеко видимые огни маяка, они служат одно-
временно и указателем пути, и магнитом, который в пол-
ной темноте магически притягивает к себе птиц, так что
Рис. 9. Гельголанд, вид с северо-запада.
Справа утес, на котором гнездятся кайры.
часто огромные массы птиц вторгаются на остров, кото-
рый иногда кажется сплошь покрытым ими *. Едва ли в
каком-нибудь другом месте можно так хорошо проследить
именно ночной пролет птиц, который в общем плохо под-
дается наблюдениям. Конечно, и другие маяки и брандеры
предоставляют подобные возможности для наблюдений,
так как и они привлекают многочисленных птиц, которые
1 После выключения гельголандского маяка в начале второй
мировой войны Дрост все же отмечал попрежнему мощный ноч-
ной перелет над островом.
Методы изучения перелетов птиц 35
приближаются к ним с характерными криками, а иногда,
ослепленные, влетают в яркую полосу света и нередко по-
гибают, ударяясь о стекла или постройки, но гельголанд-
ский маяк был настолько же не сравнимым в своем влия-
нии, насколько невообразимо мощным был около него
ночной пролет птиц.
Рис. 10. Орнитологическая станция Гельголанд,
Участок для лова птиц.
Иногда при особых условиях погоды в восточных райо-
нах Северного моря к Гельголанду устремлялись такие
массы птиц, что1 по сравнению с ними обычные пролеты
над островом казались незначительными. Так, дни массо-
вых пролетов отмечались в октябре 1940 г. и октябре
1942 г. в связи с похолоданием, распространившимся с
севера, и плохими условиями видимости в Гельголандской
бухте.
Такого рода ограниченные во времени массовые и ноч-
ные пролеты, характерные для Гельголанда, определяли
его значение как орнитологической станции, тогда как в
Росситтене особую ценность представляла прежде всего
регулярность перелетов. На Гельголанде не так уж редко
О*
36
Глава II
появлялись залетные птицы из Сибири, Гренландии, Се-
верной Америки и Средиземноморской области, что дока-
зывает неоднородность условий перелетов в этом месте,
которые в большой степени обусловливались местоположе-
нием острова и метеорологическими условиями во всем
районе Северного моря (рис. 11).
Рис. 11. Орнитологическая станция Гельголанд.
Большая сеть на участке для лова птиц.
Третья германская орнитологическая станция Хидден-
зее (Hiddensee), расположенная перед островом Рюген,
была создана в 1936 г. Поставленные перед нею задачи
были вначале преимущественно фаунистического и фено-
логического характера, что соответствовало1 местоположе-
нию станции. Первый ее руководитель, Штади, подчерки-
вал, однако, физиологическое направление работ, и его пре-
емник Шильдмахер, видимо, собирается их продолжить '.
В этой связи необходимо еще вкратце остановиться на
возможностях определения перелетов птиц в ночное вре-
1 Работы по изучению физиологических сторон явления переле-
та птиц проводятся в настоящее время в Хиддензее проф. Шильд-
махером.— Прим. ред.
Методы изучения перелетов птиц
37
мя. О наблюдениях возле маяков и о гибели около них
птиц уже было сказано. Опасность для птиц еще больше
увеличивается, если маяки соединены с радиостанциями,
так как тонкие провода антенн, так же, как и телеграфные
провода, ночью невидимы. Вследствие этого после ночно-
го пролета около маяков часто находят много мертвых или
Рис. 12. Пролет птиц у Гельголандского маяка.
А — Гельголандский маяк ночью. Предохранительные лампы не
включены. В темные ночи перелетные птицы носятся вокруг ослеп-
ляющих лучей маяка. Многие из них ударяются при этом о башню
и погибают. Б — Гельголандский маяк ночью с включенными предо-
хранительными лампами. Ночью эти лампы горят непрерывно и ос-
вещают верхнюю часть маяка. Птицы видят ее и держатся на боль-
шом расстоянии от маяка.
раненых птиц. В Дании их уже в течение многих лет ре-
гистрируют и таким образом получают важные сведения
о перелетах птиц (см. сообщения Херринга «Fuglene ved
de danske Fyr»). Вейгольд предложил лампы, которые
слегка освещали также верхнюю часть маяка; тем самым
были предотвращены удары о них птиц, которые, будучи
ослеплены прожектором, не замечали верхней части баш-
ни, Эти лампы оправдали себя и на германском побе-
режье введены повсюду (рис. 12).
Во внутренних районах суши роль береговых маяков
Глава II______________________
выполняют различные яркие источники света: прожекторы,
дуговые лампы, сигнальные огни для самолетов, освещен-
ные площади и дома и т. д. Хотя они и не привлекают летя-
щих ночью птиц в такой степени, как равномерный огонь
сильных ламп маяков, все же часто заставляют их лететь
ниже и издавать характерные крики. Крики эти, вероят-
но, способствуют сплочению стаи, образуя своего рода
«звуковой контакт», но возможно также, что они только
выражают состояние возбуждения, которое охватывает
птиц при виде внезапно появившихся огней. В результа-
те оказывается возможным определять пролетающих птиц
по голосам. До сих пор речь шла только о наблюдениях
над перелетами птиц в ограниченных районах. Если же та-
кие наблюдения проводятся по всей области распростра-
нения какого-либо вида птиц (что отнюдь не обязательно
делать одновременно и для чего можно использовать раз-
личные литературные данные), то становится возможным
составить монографии перелетов, содержащих сведения
о пролетных путях, скорости перелета, зимовках и осо-
бенностях поведения во время перелета представителей
данного вида. Лишь в последние годы для этого стали
планомерно использовать фенологические наблюдения.
Первыми это сделали венгерские орнитологи, затем немец-
кий исследователь Гейр и североамериканские ученые, а
в последнее время Гроте и Штреземан.
Шагом вперед в описании перелетов в монографиях на
основе фенологических наблюдений является попытка ис-
пользовать при этом данные современной систематики.
Она делит многие виды птиц на легко различимые геогра-
фические расы, т. е. различные популяции. Если мы, на-
пример, обнаружим ясно отличимую от нашей белой трясо-
гузки британскую расу или британскую форму клуши,
или гренландскую каменку за пределами их области гнез-
дования, то мы можем сделать вывод относительно их
пролетных путей и зимовок. Так Штреземану (1927—1934)
удалось установить, что часто даже близкородственные
расы птиц по-разному относятся к перелету, мигрируют
различными путями и зимуют далеко друг от друга, хотя
никаких вынуждающих их к этому причин, казалось бы,
нет. Поэтому исследования по систематике, например
измерения величины и веса, определение состояния one-
Методы изучения перелетов птиц
39
рения, возраста и пола, также составляют часть работ по
изучению перелетов птиц, а следовательно, являются за-
дачей орнитологических станций.
Кольцевание
Кольцевание как метод изучения перелетов птиц за-
нимает промежуточное положение между наблюдением и
экспериментом. Вместе с тем кольцевание приходится счи-
тать одним из вспомогательных средств фенологии, так
как оно не отвечает условиям научного эксперимента, ко-
торый, помимо постановки определенной задачи, требует
изучения влияния внешних и внутренних условий жизни
живого существа и планомерных наблюдений за его от-
ветными реакциями. Если поэтому раньше говорили об
экспериментах с кольцеванием, то это было, по
меньшей мере, методически неверно, и лучше вообще не
употреблять этого термина.
Применение метода кольцевания, введенного 40 лет
назад, означало огромный прогресс. Можно без преувели-
чения сказать, что только благодаря ему были заложены
основы современных исследований перелетов птиц. Мече-
ние птиц, производимое с тем, чтобы впоследствии узнать
что-либо об их судьбе, казалось, напрашивалось само со-
бой, и все же многие ученые, занимавшиеся вопросами
перелетов птиц, были далеки от этой мысли. Примеры
мечения птиц были известны из прошлого. Так, еще в
средние века на пойманных во время охоты цапель на-
девали пластинки или кольца, после чего их выпускали
на волю. В 1740 г. Фриш привязывал улетающим ласточ-
кам на лапки красные нитки, которые сохранялись у них
и после возвращения. Можно сообщить о многих попыт-
ках мечения птиц, произведенных в прошлом столетии,
например, о случае с журавлем из Аскания-Нова, кото-
рый чуть было не стал роковым для Слатин-Паши
в 1892 г., когда он был взят в плен в Судане восставши-
ми махдистами. Но все эти попытки оставались опытами
любителей до тех пор, пока преподаватель гимназии
Мортенсен в Виборге не поднял их до уровня научного
метода. В 1890 г. он метил скворцов цинковыми пласти-
ночками, а в 1899 г. перешел к мечению птиц при по-
40
Глава II
мощи ножных колец из легкого металла. На этих коль-
цах был обозначен порядковый номер и адрес Мортен-
сена. Мортенсен надевал их скворцам, аистам, уткам и
морским птицам в надежде, что некоторые из них будут
вновь пойманы.
Три года спустя Тинеман определил кольцевание как
одну из задач незадолго перед тем созданной орнитоло-
гической станции Росситтен. В 1902 г. им был окольцован
Рис. 13. Ловля ворон на Курской косе. Привязанные ман-
ные птицы на «Черной горе».
первый пролетавший сарыч, затем его внимание привлек-
ли пойманные в большом количестве на Курской косе
серые вороны. Благодаря интенсивной агитационной дея-
тельности Тинемана, метод кольцевания преодолел все
препятствия и в относительно короткое время завоевал
себе равноправное место в ряду методов и способов изу-
чения перелетов птиц. Наука, находясь в плену косного
формализма, пыталась вначале осмеять этот метод. Лица,
занятые охраной животных, ополчились против него, и да-
же бессмертный певец степи Лёнс в своем злобно-сатири-
Методы изучения перелетов птиц
41
ческом «Разговоре ворон» раскритиковал «сумасшедшего
профессора». Но ученый-орнитолог из Росситтена не дал
себя смутить. Своей упорной устной и письменной разъ-
яснительной работой он привлек к этому делу общест-
венность, без помощи которой кольцевание не смогло бы
достичь тех успехов, которые ему очень скоро принесли
признание и со стороны широких научных кругов.
Успех нового метода исследования нашел свое выра-
жение в создании таких же центров по кольцеванию, ка-
ким все больше и больше становился Росситтен. Уже в
1903 г. Бартш в США приступил к работе по кольцева-
нию через Смитсоновский институтВ 1908 г. по иници-
ативе Шейка этому примеру последовал Венгерский
орнитологический центр в Будапеште. Затем в 1909 г.
была создана вторая германская орнитологическая стан-
ция на Гельголанде, которая, так же как и Росситтен,
очень скоро стала одним из центров кольцевания птиц.
В 1910 г. кольцеванием занялись университет в Абердине
(Шотландия), английский журнал «British Birds» под
руководством Уайзерби и естественнонаучное общество в
Риге под руководством Штолля. В работу по кольцеванию
быстро включились и другие станции и государства, напри-
мер в 1911 г. — Швейцария и Швеция, в 1912 г. — государ-
ственный музей в Лейдене (Голландия), в 1914 г. — Чехия
и Россия, в 1915 г. — Финляндия. После временного пре-
кращения всякой деятельности по кольцеванию во время
первой мировой войны, в 1930 г. существовало почти
30 центров по кольцеванию птиц приблизительно в 20
государствах Европы, Азии и Северной Америки. Число
окольцованных этими станциями и сотнями их сотруд-
ников птиц превысило в 1927 г. 1 млн., а несколько лет
спустя — даже 2 млн. К 1939 г. столько птиц было околь-
цовано уже в одной только Германии. В 1949 г. США и
Канада сообщили о кольцевании в общей сложности
5,5 млн. птиц; число занятых этой работой лиц составля-
ло около 1000 (Дэй, 1949).
В чем же заключается сущность кольцевания, чем об-
условливается его столь неожиданный успех и каковы пер-
1 Учреждение в США, соответствующее европейским академиям
наук.— Прим. ред.
Глава II
спективы этого метода? Для мечения употребляются на-
деваемые птице на ногу кольца из особо прочного спла-
ва алюминия. Размеры колец соответствуют величине
представителей данного вида, а их вес составляет 0,03—
0,04% веса птиц. Окольцованные птицы выделяются в
Рис. 14. Окольцованная кукушка.
дальнейшем из общей массы. На кольце, которое после его
прикрепления птица уже не может потерять, нанесены на-
звание станции или центра кольцевания, порядковое число
или буква (в зависимости от размера) и соответствующий
текущий номер. Каждая орнитологическая станция или
учреждение, выдающее кольца, имеет широкий круг до-
бровольных сотрудников и помощников, которым они по-
стоянно высылают кольца с точными указаниями относи-
тельно регистрации их деятельности по кольцеванию. Эти
сотрудники — каждая из обеих германских орнитологиче-
ских станций Росситтен и Гельголанд имела их около 1000
Методы изучения перелетов птиц
43
человек 1— выполняют основную работу по кольцеванию,
сами же орнитологические станции с их ограниченными
возможностями в отношении отлова птиц из-за недостат-
ка рабочих рук выполняют всегда только часть всей рабо-
ты. (До 1914 г. обе орнитологические станции ежегодно
Рис. 15. Орнитологическая станция Гельголанд. Рабочее помещение
на участке для лова птиц.
В транспортировочных ящиках (на столе слева) птиц приносят из сетей. При
массовом лове птиц часть из них помещают в одиночные садки (у стены
слева). Птиц взвешивают, измеряют и кольцуют. На столе справа —шта-
тив с кольцами всех видов, перед ним — измерительные приборы и блокноты.
Через проход (слева от окна) обработанных птиц выпускают на волю.
производили около 180 000—200 000 кольцеваний.) Коль-
цеванию подвергаются в первую очередь перелетные птицы
всех видов, причем из-за высокой смертности молодых осо-
бей (приблизительно 80% на первом году жизни) значи-
1 Новые германские орнитологические станции в Вильгельмс-
гафене и Радольфцелле, вероятно, еще не имеют стольких сотруд-
ников, хотя можно с удовлетворением отметить, что после войны
общий интерес к кольцеванию опять очень возрос.
44
Глава II
тельно интереснее, хотя и гораздо труднее, метить старых
птиц. Необходимо также кольцевать и некоторое число ко-
чующих и даже оседлых птиц, но с несколько иной целью,
чем перелетных. У последних кольцевание и последующие
находки, связанные большей частью со смертью птиц
(а также с повторной поимкой!), дают сведения о том,
куда и как быстро они улетели от места кольцевания (об
этом можно судить по времени, прошедшему между коль-
цеванием и находкой птицы), равно как и об избранном
птицей пути и местах ее зимовки. С большей достоверно-
стью об этом можно говорить на основании многочислен-
ных находок окольцованных птиц одного и того же вида,
имеющих одинаковую гнездовую территорию (родину), На
основании этих данных можно делать широкие выводы о
господствующем направлении перелетов для каждого ви-
да, о характере перелетов в зависимости от экологических
и географических факторов, о распределении популяции
одного и того же вида во время перелета и на зимовках, о
неодинаковом отношении к перелету особей различного
возраста, о «верности» родине и о постоянстве пролетных
путей и областей зимовок.
Кольцевание открывает, однако, и другие возможности
планомерных исследований, например в области экологии,
где благодаря мечению птиц, гнездящихся на ограничен-
ной территории, можно получить ценные сведения об отно-
шении разных видов к окружающим условиям, изучить
размножение и уточнить другие вопросы биологии птиц
(их этологии). Кольцевание дает точные данные о продол-
жительности жизни птиц, их половом созревании и разви-
тии оперения, об ориентации во время перелетов, а также
способствует разрешению многочисленных практических
вопросов, относящихся к охоте, рыболовству и охране
природы. Особенно большую пользу приносит кольцевание
охране птиц. Во многих случаях основы целесообразного
применения определенных мероприятий можно создать
только при помощи кольцевания. Поэтому нет ничего вред-
нее, чем бороться против кольцевания доводами об охра-
не птиц, как это иногда делают некоторые недостаточно
осведомленные люди. Немаловажным является, наконец, и
воспитательное значение серьезной работы по кольцева-
нию птиц, тесно связывающей орнитологические станции с
Методы изучения перелетов птиц
45
ее сотрудниками, которые, в свою очередь, оказывают про-
светительное влияние на другие круги населения.
За последние 25 лет кольцевание получило беспример-
ное в истории методов биологических исследований разви-
тие, причины которого наряду с отмеченными моментами
заключаются и в том, что каждый участник кольцевания
получает наглядное представление об успехах своей рабо-
ты, так как орнитологические станции постоянно сообща-
ют ему о всех случаях находок окольцованных им птиц;
кроме того, он получает большое удовлетворение, прини-
мая участие в работе над научной проблемой, в сферу ко-
торой вовлечен весь мир. Немалое значение имеет и то
ощущение радости, которое вызывают глубокие исследова-
ния природы и ее наиболее поэтических созданий — птиц.
Применение кольцевания ограничивается не столько
возможностями человека, сколько крайне неравномерным
распределением мест находок окольцованных птиц, как по
видам, так и по распространению. Дело в том, что про-
цент находок окольцованных мелких птиц остается более
или менее постоянно' значительно ниже 1, особенно в от-
даленных и малонаселенных местностях. Зато очень ча-
сто и в большом количестве находят окольцованных охот-
ничьих и особенно заметных или крупных птиц: вальд-
шнепов— 9%, черных аистов — 24%, ястребов в одном
случае — 45%, в другом — почти 70% 1. Большую роль
при этом играет, конечно, и то обстоятельство, что сооб-
щения о находках этих видов поступают из густонаселен-
ных мест. О нецелесообразности кольцевания молодых
особей из-за их высокой смертности уже говорилось вы-
ше. При этом напрасно затрачивается много усилий и тру-
да, так как заранее можно рассчитывать лишь на ничтож-
ный успех по сравнению с кольцеванием старых птиц.
Однако для решения определенных вопросов может ока-
заться совершенно необходимым кольцевание именно мо-
лодых гнездарей.
Особенно успешными оказались возможно полное коль-
цевание особей одного вида в ограниченной области или
же на зимовках, регулярное кольцевание в течение более
1 Согласно Дэю (1949), из 400 000 находок окольцованных аме-
риканских птиц 30% приходится на лысух, уток и гусей.
46
Глава 11
длительных отрезков времени пролетающих в определен-
ных местах птиц, поголовное кольцевание на границах
соприкосновения рас и видов и специальные планомерные
исследования по вопросам экологии, биологии размноже-
ния и этологии отдельных видов. Орнитологические стан-
ции уже давно направляют своих сотрудников на выпол-
нение именно такой планомерной работы, чтобы совер-
шенно исключить бесцельное массовое кольцевание птиц,
при котором едва ли возможно получение новых знаний.
Изучение ориентации птиц
Как уже говорилось, научный эксперимент является
наиболее современным методом изучения перелетов птиц.
Описывая кольцевание, мы уже указывали на сущность
научного эксперимента, не имеющего ничего общего со
всякого рода умозрительными заключениями и гипотеза-
ми. Он коренным образом отличается от наблюдений и
кольцевания, особенно в части изучения причин пере-
летов. Промежуточное положение между научным экспе-
риментом и кольцеванием занимает изучение ориентации
птиц, к рассмотрению которой мы и переходим.
Вопрос об ориентации птиц во время перелета относит-
ся к наиболее трудным в науке о перелетах птиц. Он был
поставлен уже сотни лет назад и обсуждался в связи с
другими проблемами, так как в ряде отношений он имеет
значение и для высших животных и даже для человека.
Достаточно часто путешественники и ученые сообщали об
удивительной способности местных жителей без карты и
компаса находить правильный путь в бесконечных сибир-
ских тундрах или в однообразных просторах пустынь. Мно-
го фактов правильной ориентации на больших расстояни-
ях известны также и в отношении собаки, кошки и лошади.
Однако нет ничего проще, чем довольствоваться
утверждением, что как у животных, так и у первобытных
людей многое из того, чему цивилизованный человек мо-
жет научиться только с трудом и чего он никогда полно-
стью не постигнет, является прирожденным. Наука этим
ограничиться не может, ее интересуют основы проблемы,
и она ищет возможности и методы их изучения. Одни
только наблюдения в данном случае не приводят к цели,
они должны, по меньшей мере, подкрепляться и допол-
Методы изучения перелетов птиц 47
мяться экспериментами. При решении вопроса о суще-
ствовании органа, ведающего способностью к ориентации,
необходимо прибегнуть к анатомическим исследованиям.
Специалисты по разведению почтовых голубей поста-
вили бесчисленное количество опытов по изучению возвра-
щения этих птиц домой. В результате этих опытов в науч-
ных кругах возникло мнение, будто ориентация птиц осно-
вана главным образом на зрительной памяти. Первые опы-
ты с дикими птицами, проведенные приблизительно1 так
же, как и с почтовыми голубями, а именно, путем пере-
возки птиц на большие расстояния от родных мест, как
будто подтвердили это мнение. Планомерное применение
метода кольцевания и в данном случае изменило существо-
вавшие взгляды и дало совершенно иные результаты, на
которых мы пока подробно останавливаться не будем.
Отметим лишь, что ориентация перелетных птиц при по-
мощи одного только зрения невозможна, что оседлые и
кочующие птицы, очевидно, не обладают столь выражен-
ной способностью к ориентации, как перелетные птицы, и
что сущность этой способности остается в настоящее вре-
мя, как и раньше, неразгаданной ’.
Проведенные до сих пор в этом направлении опыты
ограничивались перемещением гнездящихся птиц на боль-
шие или меньшие расстояния и наблюдениями за их воз-
вращением. Подобные опыты проводились и с птицами, ко-
торые только что закончили перелет или собираются при-
ступить к нему или же кочуют зимой. Опыты с перевозкой
молодых слетков также доказали их способность к ориен-
тации, равно как и эксперименты по переселению и аккли-
матизации, проведенные в различное время (европей-
ские виды птиц в Северной Америке, Южной Африке и
Австралии, а также экзотические птицы в Германии; опыты
с выпуском на волю аистов в Средней и Западной Герма-
нии, сизых чаек — в Силезии и в Росситтене и квакв —
в Бранденбурге). Все данные говорят о чрезвычайно боль-
шой изменчивости способности к ориентации и о быстрой
ее приспособляемости к условиям внешней среды.
Коснемся еще некоторых опытов и соображений, кото-
1 Работа Штейнбахера написана до опубликования весьма важ-
ных исследований Крамера (Труды X Международного орнитологи-
ческого конгресса, 1950) об ориентации птиц. — Прим. ред.
48
Глава II
рые являются предметами споров. Речь идет о ранних
предположениях Миддендорфа по поводу направляющей
роли земного магнетизма в перелетах птиц. Дрост (1934),
напротив, приписывает это влияние различному распреде-
лению тепла в пространстве и во времени. Другие мне-
ния, согласно которым направляющими факторами явля-
ются положение солнца, давление воздуха или электриче-
ские волны, мы упоминаем лишь ради полноты изложения.
До сих пор почти не проводилось анатомических ис-
следований в целях установления органа-рецептора, слу-
жащего для определения направления перелета или для
восприятия поступающих извне раздражений.
При изучении ориентации птиц необходимо прежде
всего отделить случаи, когда известное влияние на направ-
ление перелета может иметь традиция, где, следовательно,
возможно вождение молодых птиц старыми, от тех слу-
чаев, в отношении которых доказано, что старые и моло-
дые птицы улетают в разное время. Последние более уди-
вительны, но и значительно труднее объяснимы. Нахожде-
ние цели, т. е. мест зимовок, имеет для птиц подчиненное
значение. Это получается само собой, когда они достаточно
долго летели в определенном направлении, и стремление
к перелету угасло. Основным вопросом всегда остается на-
хождение пути, что предполагает знание определенного
направления, различного для каждого вида птиц (а не-
редко и для рас и популяций одного и того же вида).
Поэтому часто говорят о «чувстве направления», кото-
рое, как компас, помогает птицам ориентироваться, но ко-
торое вместе с тем принуждает их придерживаться во
время перелета определенного направления. Штреземан
определяет это чувство направления как «свойство пере-
летных птиц, о котором мы до сих пор знаем не многим
больше, чем то, что оно существует». Это свойство, неви-
димому, является наследственной «видовой памятью», так
как большинство птиц мигрирует без руководства со сто-
роны старых, опытных птиц. Предположение о существо-
вании особого чувства в обычном понимании, как, напри-
мер, зрение и слух, в отношении «чувства направления»
излишне. Птица (согласно Шюцу) как будто обладает
«чувством географического положения», но какие органы
ведают им, остается еще не ясным.
Методы изучения перелетов птиц
49
Изучение причин перелетов
В заключение мы переходим к объяснению основного
вопроса о перелетах птиц — причин перелетов. Почему
птицы совершают перелеты? Что побуждает их покинуть
родину или места зимовок? Почему они не остаются там,
где обилие пищи и тепло обеспечивают им наилучшие
условия жизни? Вызывается ли это ежегодно повторяю-
щееся событие факторами окружающей среды или же
внутренними стимулами? Эти вопросы ставит наука; ответ
на них может дать только эксперимент.
Ранние попытки объяснения причин перелетов в основ-
ном ограничивались изучением влияния смен времен года.
Особенно большое значение придавалось при этом возра-
стающему похолоданию осенью в сочетании с недостат-
ком пищи. Это объяснение исходит из уже обсужденного в
другом разделе предположения об историческом проис-
хождении перелетов птиц. Первоначально периодическая
нехватка пищи, наверное, каждый раз к началу зимы из-
гоняла птиц из их гнездовой области. На этой стадии раз-
вития «привычки» к перелетам остались до настоящего
времени многочисленные виды кочующих и перелетных
птиц, которых мы поэтому образно называем «бежен-
цами» от зимы. Они заранее, подобно барометру, чувствуют
перемену погоды. В соответствии с этим на их миграции в
большой степени влияют климатические условия. Появле-
ние перелетных и кочующих птиц можно ожидать при
определенном общем состоянии погоды (при котором име-
ют значение многие факторы: давление, температура воз-
духа, характер ветров, осадки). Конечно, при этом суще-
ствуют определенные отклонения, которые обычно тем
больше, чем раньше возвращается соответствующий вид.
В дальнейшем мы еще вернемся к этому вопросу.
Как особенно хороший пример такой зависимости при-
лета от погоды упомянем вальдшнепа. Охотнику известно
значение «вальдшнепиной погоды» весной, когда длин-
ноклювые птицы возвращаются на родину и когда на них
лучше всего охотиться перед началом сумерек. Венгерские
исследователи в результате многолетних предварительных
работ сумели настолько уточнить для местных условий
связь погоды с прилетом вальдшнепа, что могли на осно-
Л— 17В
50
Глава II
вании метеорологических сводок довольно точно предска-
зать начало прилета вальдшнепа.
Подчеркнем еще раз: существуют птицы, перелет кото-
рых в большой степени зависит от погоды («Wettervogel»),
т. е. их перелет регулируется внешними факторами. В от-
личие от них птицы, у которых перелет в большой степени
зависит от внутренних стимулов и в значительной мере
совершается независимо от внешних факторов, получили
название «Instinktvogel». В качестве примеров таких птиц
можно привести иволгу, соловья или черного стрижа, ко-
торые появляются у нас весной в определенное время,
иногда с небольшими отклонениями. Такое схематическое
разделение птиц по типу перелета, произведенное Вейголь-
дом (1924), возможно, разумеется, лишь при условии при-
знания многочисленных переходов. Об этом речь будет
впереди. Пока нам важно установить, что погода оказыва-
ет решающее влияние на перелет многих птиц и даже мо-
жет быть его причиной. В то же время на других птиц она
оказывает относительно слабое действие, изменяющееся во
времени и пространстве. Отношение к перелету последних
голландский орнитолог ван Оордт метко сравнивает с рас-
пусканием почек и опаданием листвы весной и осенью у
лиственных деревьев, которые также лишь незначительно
сдвигаются в ту или иную сторону внешними влияниями.
Согласно нашему определению, недостаток пищи также
может играть роль лишь для «Wettervogel», поскольку
«Instinktvogel» обычно никогда его не испытывают. Подав-
ляющее их число улетает еще в такое время, когда пища,
напротив, очень обильна. Из наблюдений над пойманными
перелетными птицами, которых содержали в клетках, из-
вестно, что они даже при наилучшем уходе и неизменной
температуре начинают проявлять беспокойство и целыми
днями и ночами машут крыльями и бьются в клетках,
иногда серьезно повреждая себя, в тот период, когда на
воле другие представители их вида отправляются в путь.
Этот факт ясно доказывает, во всяком случае для
«Instinktvogel», что причиной перелета является внутрен-
ний стимул. В определенные периоды времени он обретает
такую власть над птицей, что перекрывает и выключает
все другие инстинкты: спаривания, гнездостроения, выси-
живания и кормления птенцов, общественного сплочения и
Методы изучения перелетов птиц
51
даже инстинкт самосохранения. У очень многих видов птиц
этот «перелетный инстинкт» способен перестроить дневной
и ночной ритмы, т. е. сделать на время перелета дневных
птиц ночными. «Перелетный инстинкт» обусловлен наслед-
ственностью, но он может проявляться и с индивидуаль-
ными различиями, что подтверждает его зависимость от
других внутренних процессов. Об этом известно еще очень
немного, но все же нельзя сомневаться в тесной связи трех
властвующих над жизнью птиц периодических процессов:
размножения, линьки и перелета.
Эти знания были направляющими при постановке ис-
следований с целью изучения возникновения «перелетного
инстинкта». Задача заключалась в том, чтобы отграни-
чить значение различных факторов внешней среды от
внутренних причин, вызывающих перелеты, путем их
целенаправленного видоизменения. Далее предстояло
изучить изменения в состоянии накопленных в организме
перелетных птиц энергетических запасов и с этой целью
собрать данные о приеме пищи, весе и температуре тела.
Для сравнения анализ зависимости между обменом ве-
ществ и перелетом следует распространить на процессы
смены оперения и половой функции. При этом напра-
шивалось предположение о влиянии желез внутренней
секреции на явление перелета. Чтобы выявить влияние не-
которых гормонов на перелеты птиц были поставлены мно-
гочисленные опыты; кроме того, проводились морфологи-
ческие и гистологические исследования состояния желез
внутренней секреции в разное время года. Эти исследова-
ния уже дали ценные результаты, но исчерпывающих све-
дений пока еще нет. Это необходимо подчеркнуть в про-
тивовес фантастическим сообщениям о разгадке тайны
перелетов птиц, в которых говорится об обнаружении ка-
кого-то определенного «гормона перелета». Подобное
объяснение вряд ли когда-либо подтвердится, так как
воздействия всех гормонов слишком взаимосвязаны.
Вообще при таких опытах очень часто за первыми успе-
хами следует неудача, когда влияние тех же условий про-
веряется на более обширном материале. Поэтому и в дан-
ном случае обобщить полученные результаты невозмож-
но. Это относится не только к изучению причин переле-
тов, но и ко всем вопросам перелетов птиц.
!
Глава III
РЕЗУЛЬТАТЫ ИЗУЧЕНИЯ ПЕРЕЛЕТОВ ПТИЦ
Из предыдущих глав мы составили себе представление
о масштабе и разнообразии вопросов, связанных с переле-
тами птиц. Это делает понятным затруднения, связанные
с попыткой обобщения наших знаний в этой области. Ма-
териал, накопленный поколениями исследователей при
участии целой армии сотрудников, наблюдателей и работ-
ников по кольцеванию в течение многих лет исключительно
усердной работы, нельзя изложить даже приблизительно
полно в том ограниченном объеме, которым мы здесь рас-
полагаем. Поэтому для общего обзора, который мы наме-
рены дать, необходим жесткий отбор материала и ограни-
чение лишь самым существенным. Кое-что, вполне заслу-
живающее более подробного рассмотрения, будет лишь
упомянуто. Вместе с тем другие факты, если они хорошо
иллюстрируют различные вопросы, будут приводиться не-
однократно, особенно в качестве примеров. Тем самым об-
легчается обзор как всей проблемы в целом, так и частных
вопросов.
Все это нужно иметь в виду при анализе результатов
изучения перелетов птиц, полученных двумя основными
методами: наблюдением и экспериментом. Результаты по-
левых наблюдений и кольцевания группируются при этом
главным образом вокруг следующих вопросов: места
зимовок птиц, характер и сроки перелетов у различных
видов, скорость и высота полета и их физиологическое
объяснение, связь перелетов птиц с погодой. Во вто-
рую и третью очередь будут обсуждаться результаты изу-
чения ориентации птиц и причин перелетов. Деталей мы,
однако, при этом коснемся лишь в той мере, в какой это
будет возможно или целесообразно, так как пока еще нет
таких фактов, которые позволили бы сделать окончатель-
ные выводы.
Результаты изучения перелетов птиц
53
Цели перелетов (места зимовок)
Общепринято называть местности, в которых перелет-
ные птицы прекращают осенью свои миграции, местами зи-
мовок, или зимовками Гейр (1934) предложил заменить
термин «место зимовок» («Winterquartier»), пригодный
лишь для перелетных птиц умеренных широт, более
общим «цель отдыха» («Ruheziel») или «цель перелета»
(«Zugziel»). Тем самым удалось бы охарактеризовать
миграции птиц тропических областей и резче подчеркнуть
единую сущность перелетов птиц во всех климатических
зонах земного шара. Эта точка зрения логична и вполне
оправдана. Поэтому в каждом обобщающем труде о
перелетах птиц ее следовало бы придерживаться.
Однако пока новый термин войдет в употребление и за-
менит старый, несомненно, пройдет некоторое время. Во
избежание всяких недоразумений в этой книге попрежне-
му применяется термин «зимовка», тем более, что речь бу-
дет идти только о таких птицах, в отношении которых он
может употребляться недвусмысленно. Но в заголовках и
в некоторых особых случаях будет использована новая
терминология.
Термин «зимнее убежище» («Winterherberge») равно-
значен понятию «место зимовки». Это не относится к ча-
сто употребляемому термину «область зимовки» («Uber-
winterungsgebiet»), за исключением тех случаев, когда им
пользуются при общем объяснении явления перелета птиц.
Место зимовок — это географически четко ограниченная
область, в которой птицы остаются более или менее про-
должительное время и во многих случаях меняют оперение
или часть его, после чего вновь отправляются в обратный
путь к гнездовьям, на родину. Область зимовок, напротив,
редко бывает четко ограничена и часто весьма различна по
величине. Она в основном определяется факторами внеш-
ней среды: частично условиями питания, частично клима-
тическими условиями. Так, некоторые морские птицы (глу-
пыши) перемешаются с осени до весны с места на место,
следуя за периодическими миграциями животных, служа-
щих им пищей. Многочисленных перелетных птиц умерен-
ных, широт смена дождей и засухи в тропиках вынуждает
вести кочевую жизнь, Аналогично ведут себя и некоторые
Глава. Ill
Рис. 16. Находки окольцованных зябликов (Fringilla с. coelebs L.).
(Из Бохмана, 1934.)
Зяблики из Северной Европы, окольцованные во время пролета на Гельголанде,
зимуют в Северо-западной Германии, Бельгии, Голландии (места зимовок .обо-.
значены квадратами). Зяблики из Германии зимуют во Франции и Испании
(Места зимовок обозначены кружками и точками). Цифры — месяцы, в которые
птицы были окольцованы и вновь встречены. Черточки под цифрами — число
лет, прошедших после кольцевания птиц.
из зимующих в центральной Европе птиц, гнездовья кото-
рых расположены на севере, например мохноногий канюк,
свиристель, ореховка и клест, Все .она. пребывают зимой в
Результаты изучения перелетов птиц 55
различных по площади областях и не придерживаются
постоянных мест зимовок.
Где же расположены места зимовок европейских пере-
летных птиц? В какой стране зимует каждый вид? Неиску-
шенный в этом вопросе человек большей частью убежден,
что при этом речь может идти лишь о юге, в частности об
Италии или Африке. Но это правильно лишь в отношении
части перелетных птиц, особенно когда под «Африкой»
подразумевается тропическая зона континента. Многие
птицы, гнездящиеся на севере и востоке Европы, зимуют в
пределах Германии. Большая их часть, а также многие
виды, гнездящиеся в Германии, зимуют в Голландии, Бель-
гии, Северо-западной Франции и на Британских островах,
но главным образом в Юго-Западной Франции, на Пире-
нейском полуострове и в Северо-Западной Африке. Ита-
лия, Балканский полуостров и Малая Азия сильно уступа-
ют указанным странам по масштабу зимовок европейских
перелетных птиц. Это выражается как в количестве особей,
так и в видовом составе зимующих птиц. Много птиц
скапливается в долине Нила и экваториальной восточной
Африке, меньше — в центральных областях Африканского
континента и особенно на его западе, в то время как юг,
вплоть до Капской провинции, во время северной зимы за-
селяется птицами более густо. Данные, установленные для
Центральной Африки, требуют, однако, уточнения. Факты,
основанные на находках окольцованных птиц в этих без-
людных и неосвоенных местностях, настолько скудны и
случайны, что часто не могут служить основанием для
. определенных выводов.
Предпочтение западноевропейских стран в качестве
мест зимовок многочисленными европейскими перелетны-
ми птицами объясняется влиянием Гольфстрима на климат
этих районов. Он характеризуется небольшими сезонными
колебаниями температуры, относительно прохладным ле-
том и мягкой зимой. Такой климат называют морским или
океаническим в отличие от континентального климата
внутренних районов материков, где короткое жаркое лето
сменяется продолжительной холодной зимой. Перелетные
птицы, очевидно, стремятся попасть не столько в теплые
местности, сколько в такие, где наблюдаются небольшие
колебания температуры, оказывающие благоприятное вли-
56
Глава III
Рис. 17. Различия в перелетах разных популяций канюка (Buteo b.
buteoL.). (Из Бурра, 1936.)
Круги указывают «родину» каждой популяции. Районы пролетов (по данным
кольцевания) примыкают к ним с запада и юга. Данные за август 1935 г.
яние на жизнь растений и мелких животных. Это’ обеспе-
чивает птицам в любое время достаточно корма, во всяком
случае он обильнее, чем в еще более теплых местностях
с периодическими засухами.
Под влиянием морского климата, простирающимся да-
леко вглубь Центральной Германии, многочисленные виды
птиц этих районов зимуют в гнездовой области. На не-
сколько сотен километров севернее или восточнее те же
виды являются перелетными, а в этой местности они толь-
ко откочевывают при более продолжительных морозах на
юг и запад. Таким образом, у многих видов птиц наблю-
дается различное отношение к перелету в зависимости от
местонахождения их гнездовий, и решение вопроса о том,
имеем ли мы в данном случае дело с оседлыми, кочующи-
ми или перелетными птицами, целиком зависит от их
-происхождения, Но. даже когда оно.нам известно, возмож-
Результаты изучения перелетов птиц 57
ность ошибок не исключена. Мы, правда, можем в общей
форме утверждать, что та или иная популяция какого-ли-
бо вида зимует в гнездовой области или, напротив, отле-
гает, но при этом всегда нужно помнить, что это относит-
ся не ко всем особям данного вида и что многие из них со-
ставляют исключение. В связи с этим для некоторых мест-
ностей нельзя сказать с уверенностью, больше ли особей
данного вида зимует на месте или отлетает. Обычно более
выносливые старые птицы чаще пытаются зимовать и не
так далеко отлетают. На все это уже указывалось при
разборе понятий оседлых, кочующих и перелетных птиц,
так же как и на различное отношение к перелетам некото-
рых видов в зависимости от пола и на возрастающую
склонность к зимовкам у некоторых бывших ранее пере-
летными видов птиц. Причину этих явлений усматривают в
усилении связи птиц с населенными пунктами и благопри-
ятными возможностями добывания пищи, а также с по-
теплением зим, по крайней мере в Северо-Восточной Евро-
пе. Финские орнитологи, в частности Сиивонен и Калела
(1942, 1943), особенно отмечали наблюдающееся уже в те-
чение нескольких десятилетий изменение климата их ро-
дины в сторону морского и соответствующее увеличение
сухости климата в Центральной Европе. Это вызвало за-
метное смещение северных границ распространения мно-
гих видов птиц и изменение их отношения к перелету.
О возможностях такого изменения в результате расселе-
ния птиц в другие географические широты речь будет идти
еще неоднократно.
Теперь становится понятнее тот факт, что многие пере-
летные птицы встречаются в Германии зимой в большем
количестве, чем в период размножения, хотя они осенью
отлетают массами. Дело в том, что небольшое число
остающихся птиц увеличивается за счет птиц, прилетаю-
щих из северных гнездовий, которые часто в очень боль-
шом количестве зимуют в Центральной Европе. Это отно-
сится, в частности, к черному дрозду, скворцу, зарянке,
зяблику, коноплянке, зеленушке, вяхирю, вальдшнепу и
уткам. Правда, часть птиц всегда остается также и в
шведских, финских или балтийских гнездовых областях, но
.большинство отлетает. В Германии названные виды птиц
зимуют в значительно большем количестве, чем на севере.
58
Глава III
Рис. 18. Область распространения пролетающих над Росситтеном се-
рых ворон. (Из Грёббельса, 1932, по Тинеману.)
К северу от Росситтена расположена гнездовая область, к югу — область
зимовок.
По направлению с востока на запад это соотношение воз-
растает. Добавляются другие зимующие виды, например
полевые жаворонки, канареечные вьюрки, трясогузки, лу-
говые коньки, клинтухи и бекасы, которые в мягкие зимы
в небольшом числе довольно регулярно остаются в Рейн-
ской области, в то время как в Центральной Германии
только в исключительных случаях зимуют одиночные особи.
Наконец, на Британских островах, зимние температуры
которых приблизительно соответствуют североитальян-
ским, очень многие виды, перелетные в других местностях,
оказываются преимущественно оседлыми или кочующими.
Даже такие типичные перелетные птицы, как чибисы, се-
рые цапли, пеночки-кузнечики, чирки-свистунки или водя-
ные пастушки, остаются в большинстве йа зиму. Кроме то-
го, численность птичьего населения Англии и Ирландии
пополняется с севера и востока, так как большая часть
скворцов, вальдшнепов, дроздов, больших кроншнепов и
ворон из Скандинавии, Финляндии, Прибалтики и Герма-
нии перелетает в эти страны, а также в Голландию и
Результаты изучения перелетов птиц
59
Бельгию на зимовку. Птицы, которые частично зимуют в
Центральной Европе, летят также во Францию, Испанию и
Португалию. Перелеты этих птиц иногда захватывают и
Северную Африку. В эти места отправляются, кроме того,
трясогузки, горихвостки, луговые жаворонки, славки, пев-
чие дрозды, обыкновенные чайки, чибисы и серые цапли.
В Италии мы находим зимой зарянок, горихвосток-черну-
шек, пеночек-кузнечиков, трясогузок, горных трясогузок,
луговых и полевых коньков и луговых жаворонков, но не
так часто, как в западной части Средиземноморской обла-
сти. В Греции и Малой Азии перелетные птицы из Север-
ной и Центральной Европы бывают почти исключительно
во время пролета, но иногда вместе с другими видами из
Восточной Европы и Сибири в этих областях зимуют вер-
тишейка, черный коршун и малый подорлик. Долина Ни-
ла и граничащие с ней области Судана, Абиссинии и Во-
сточной Африки, как уже упоминалось, представляют со-
бой места массовых зимовок европейских перелетных птиц.
В огромных количествах в этих областях встречаются раз-
личные виды уток, прежде всего шилохвость и широконос-
ка; большими стаями теснятся на берегах Белого Нила и
его притоков журавли со всей Европы; бекасы, песочники,
ржанки, чибисы и большие улиты населяют совместно с
трясогузками, варакушками и другими мелкими птицами
районы половодья. В степях и саваннах зимуют западные
и восточные соловьи, иволги, удоды, мухоловки, славки,
пеночки, овсянки, желтые трясогузки, лесные коньки, лу-
говые чеканы, каменки и другие типичные перелетные пти-
цы. Некоторые из этих видов встречаются также в Восточ-
ной и Западной Экваториальной Африке, преимуществен-
но на западе, например мухоловка-пеструшка, соловей и
осоед. Через тропики в Южную Африку проникают также
серые мухоловки, ласточки, большинство сорокопутов, бе-
лые и черные аисты, чеглоки, кукушки, камышевые и луго-
вые луни, а также обыкновенные, пестроносые и поляр-
ные крачки.
Согласно Шюцу (1929), из 90 видов зимующих в Афри-
ке европейских перелетных птиц 6 встречаются только в
Западной Африке, 50 — только в Восточной и 34 — как на
западе, так и на востоке. Причины этого очевидны: долина
Нила с ее богатством кормов в течение тысячелетий при-
60
Глава Ill
Рис. 19. Пролетные пути азиатских жуланов. (Из Штрез?мана, 1927.)
Области гнездования (очерчены пунктиром) и зимовки различных рас жулана
(Lantus cristatus L.)t в том числе обыкновенного жулана (L. с. collurio L.);
--------------------------вероятные пролетные пути.
влекает множество птиц, а также служит путем, по кото-
рому они без больших опасностей следуют из Средиземно-
морья в тропики, например в Сенегамбию и отдаленные
местности побережья Гвинеи, которых с севера можно до-
стигнуть лишь после длительного и опасного пролета над
безводными песчаными и каменистыми пустынями.
Бросается в глаза, что1 целая группа азиатских перелет-
ных птиц также зимует в Восточной и Южной Африке.
Чтобы попасть туда, они должны совершить далекий
путь — через Персию и Аравию, в то время как располо-
женных прямо на юге тропических областей Северной Ин-
дии они могли бы достигнуть значительно1 быстрее. Дру-
гие виды также часто: летят на зимовки странными околь-
ными путями. Повидимому, птицы следуют по путям свое-
го исторического расселения, по которым они когда-то
продвигались из определенного центра. Штреземан
.(1927—1934) приводит.аналогичный пример: гнездящаяся
Результаты изучения перелетов птиц 61
в Северной Скандинавии пеночка-таловка пролетает через
всю Сибирь до Малайского архипелага, где расположены
места ее зимовок, вместо того чтобы сразу же отправить-
ся на юг, что значительно быстрее привело бы ее в тропи-
ки (в Африку). Эта птица только относительно недавно
проникла в Европу из Сибири и каждый раз совершает
свой перелет соответственно прежнему расселению. Грен-
ландская каменка, которая из Европы расселилась до
Лабрадора и в Америке нигде больше не обнаружена,
также придерживается при перелетах исторического пути
своего расселения. Это, очевидно, относится и к галстуш-
нику и к чирку-свистунку, который проник из Азии в
Аляску, но пролетный путь которого неизменно ведет об-
ратно в Азию. Аналогичным образом объясняются, неви-
димому, еще более странные перелеты полярных крачек.
Северной Америки: через Европу в Южную Африку и Юж-
ную Америку (рис. 20). Штреземан обращает внимание на
то, что продвижение какого-либо вида на север часто со-
пряжено с удлинением пролетного пути в южном направ-
лении, что приводит к перемещению мест зимовок на юг.
Во всяком случае, установлено, что птицы могут в тече-
ние длительного времени как менять места своих зимовок,
так и зимовать в определенных областях. Это доказывают
многочисленные случаи возврата колец. Следует указать
также на некоторые работы (Ренш, 1938; Кипп, 1936,
1942), посвященные вопросу связи между формой крыла
и перелетом. Перелетные птицы имеют более острые
крылья и поэтому кажутся более способными к дальним
перелетам, чем близкие им в систематическом отношении
оседлые птицы. Однако до сих пор нет точных выводов от-
носительно истории развития строения крыла и миграции.
Различия в отношении характера перелетов у различ-
ных рас и популяций одного и того же вида, о чем уже
неоднократно шла речь, не исключают того, что различные
расы очень часто зимуют в одной и той же области; это
можно отметить у многих певчих птиц в Восточной и Юж-
ной Африке. Однако отдельные расы нередко имеют и
строго разграниченные места зимовок, причем северные
формы перелетают дальше, чем южные. В этом отношении
характерен пример североамериканского' вьюрка Passerella
iliaca, шесть различных рас которого гнездятся на тихо-
Рис. 20. Пролет североамериканских полярных крачек на зимовки в
южной части Атлантического океана вдоль берегов Южной Европы и
Африки. (Из ван Оордта, 1943, по Линкольну, 1935.)
Рис. 21. Перелеты некоторых рас вьюрка Passerella iliaca у запад-
ного побережья Северной Америки (северные расы перелетают через
зимовки рас, обитающих южнее). (Из Линкольна, 1935.)
Гнездовые области обведены замкнутой линией, места зимовок обозначены точ-
ками. У расы 6 гнездовая область совпадает с областью зимовки.
Рис. 22. Ареалы соловья и восточного соловья. (Из Нитхаммера,
1937.)
Гнездовая область и места зимовок у первого обозначены точками, у второго —
заштрихованы. Пролетные пути обозначены стрелками.
Результаты изучения перелетов птиц
65
океанском побережье. Из них три северные отлетают юж-
нее, в то время как представители самой южной расы —
оседлые птицы (рис. 21). С соответствующими поправ-
ками это относится и к некоторым видам одного и того
же рода, например к славкам, пересмешкам, пеночкам, а
также к восточному и западному соловьям (рис. 22).
Разумеется, птицы предпочитаюттакиеобласти зимовок,
которые по характеру ландшафта в некотором отношении
соответствуют их гнездовой области. Во время перелета
птицы также по возможности придерживаются условий,
которые соответствуют особенностям отдельных форм.
Направления перелетов
При обсуждении вопроса о местах зимовок уже упоми-
налось вкратце, что распространенное представление о
южном направлении перелетов верно для европейских пе-
релетных птиц лишь в самом общем виде. Истинные на-
правления перелетов определяются, по крайней мере у
очень многих видов, расположением мест зимовок по от-
ношению к областям гнездования. У других видов направ-
ления перелетов зависят от экологических отношений или
определяются географическими условиями, в некоторых
же случаях они до сих пор не поддаются никакому объяс-
нению.
Очертания европейского континента заставляют при-
держиваться большинство перелетных птиц из Исландии и
Великобритании и многих птиц из Голландии, Бельгии и
Франции строго южного направления. Однако и мохно-
ногий канюк летит из Скандинавии на юг; ласточки в сво-
ем полете прямо на юг пересекают Средиземное море и
Сахару. Гнездящиеся в Италии представители различных
видов также мигрируют на юг. С последними, вероятно,
следуют и некоторые певчие птицы, зимующие в Тропиче-
ской Африке, например соловей. Но точных данных об
этом нет из-за недостаточных находок колец в Сахаре,
хотя Гейр и Моро, а в последнее время и другие англий-
ские исследователи во время войны наблюдали регуляр-
ные пересечения пустыни перелетными птицами (рис. 23,
24) «.
1 Советскими исследователями это установлено для средне-
азиатских пустынь (в частности, для Кара-Кумов). — Прим. ред.
К-176
66
Глава III
Рис. 23. Ареал мохноногого канюка как пример вида, перелет кото-
рого проходит только в южном направлении. (Из Шюца, 1930.)
Кружки — места находок окольцованных птиц.
В юго-западном направлении летят большинство сред-
неевропейских и многочисленные северо- и восточноевро-
пейские виды перелетных птиц, зимующих в Западной Ев-
ропе и Северо-западной Африке. В этой связи можно ука-
зать на приведенный при описании мест зимовок пере-
чень отдельных видов, который следует пополнить многи-
ми видами хищных птиц, уток, чаек и куликов. Целый
ряд финских, прибалтийских и северогерманских птиц ле-
тит почти строго в восточно-западном направлении на
зимовки в Западную Европу, о чем также уже говорилось.
На юго-восток направляются лишь немногие виды, из
числа которых мы укажем на иволгу, кукушку, жулана,
чернолобого сорокопута, чернозобую гагару, аиста из об-
ласти, расположенной восточнее Везера и Эмса, и на
большую часть больших ночных и обыкновенных горлиц
из восточных районов их гнездовой области. У последних,
трех видов разные популяции ведут себя- различно: часть
из них летит на юго-запад, часть — на юго-восток.
Результаты изучения перелетов птиц
67
Рис. 24. Места находок в Африке окольцованных европейских дере-
венских ласточек (Hirundo г. rustica) с указанием области гнездова-
ния. (Из Шюца, 1937.)
Цифры — месяцы кольцевания, черточки под цифрами — число лет, прошедших
после кольцевания птиц.
Как будто «неразумны» встречающиеся иногда переле-
ты в северном направлении, например у гагарок с британ-
ских островов, кайр с Гельголанда, западноевропейских
серебристых чаек и некоторых популяций обыкновенных
чаек. Последние летят осенью из Чехии и отчасти из Мо-
равии и Силезии, следуя по Эльбе и Одеру к побережью
моря, где они проводят зиму, или же устремляются даль-
ше на запад и юг. Серебристые чайки мигрируют как в се-
верном, так и в южном направлении.' Британские гагарки
и гельголандские кайры встречаются в особенно больших
5*
Рис. 25. Миграции в северном направлении молодых кайр с острова
Гельголанда с последующим распространением на юг и запад.
(Из Штехова, 1938.)
Результаты изучения перелетов птиц 69
количествах ранней осенью у южного побережья Норвегии
и только впоследствии они рассеиваются южнее по всему
району Северного моря. Во всех этих случаях причиной
подобной необычной картины перелета (это, впрочем,
относится не ко всему времени перелета, а лишь к его на-
чалу) являются, повидимому, экологические факторы (бо-
гатство кормов в прибрежных водах),. К этому надо еще
добавить, что большое количество находок окольцованных
птиц осенью и зимой нетрудно объяснить интенсивной
охотой.
Более или менее значительные отклонения от опреде-
ленного направления перелетов наблюдаются также у
многих других перелетных птиц, хотя они, конечно', не
столь заметны, как в приведенных выше двух примерах.
Иногда даже нельзя установить единое направление пе-
релета, когда какой-нибудь вид или популяция летит с
очень большим «рассеянием» («Streuung»). Большой
«угол рассеяния» («Streuungswinkel») встречается, как
правило, при отлете у очень многих видов перелетных птиц.
В таких случаях каждая птица летит не точно в том на-
правлении, в каком другие представители того же вида и
даже ближайшие соседи по гнездовью. Каждая особь мо-
жет в соответствии с внутренними и внешними факторами
выбрать (в известных пределах) «свое» направление. Та-
ким образом, угол рассеяния вида или популяции во мно-
гих случаях достигает 90° (деревенские ласточки, кукуш-
ки, горихвостки-чернушки и другие могут улетать как в
юго-западном и южном направлениях, так и на юго-во-
сток!), а иногда даже почти 180°. Как особое исключение
это удалось установить даже у птенцов одного выводка
(датские черные аисты).
Однако Гейр с полным основанием указывает на то,
что. почти все перелетные птицы придерживаются доволь-
но постоянного, в известной мере свойственного виду на-
правления перелета, которое, правда, неодинаково для
всей области распространения, но четко выявляется у
определенных популяций. Это направление перелета,
свойственное целому виду или соответственно популяции,
он называет «нормальным направлением» («Normalzug-
richtung») или «первичным направлением» («Primarrich-
tung») перелета.
Рис. 26. Рассеяние от запада — юго-запада до' юго-востока перелетных горихвосток-черну-
шек. (Из Дроста и Дессельбергера, 1932.)
Цифры — месяцы кольцевания, пунктир — находки окольцованных птиц спустя длительное время после
кольцевания.
Результаты изучения перелетов птиц
71
Рис. 27. Различия в перелетах разных популяций серой мухоловки.
(Из Крейца, 1941.)
Соответственно положению области гнездования (на западе или на востоке)
отлет совершается или в юго-западном, или в юго — юго-восточном направле-
нии. Цифры — месяцы кольцевания, пунктир — находки окольцованных птиц
спустя длительное время после кольцевания.
Оба приведенных выше взгляда не противоречат друг
другу, а лишь отображают то огромное многообразие яв-
лений, с которыми мы встречаемся при изучении перелета
птиц.
Необходимо еще вкратце упомянуть особый вид пере-
летов птиц, который Гейр (1930) обозначил как «промежу-
точный перелет» («Zwischenzug»), так как его направле-
ние совершенно отлично от обсуждавшихся выше направ-
лений перелетов, а иногда прямо противоположно на-
правлению отлета. Это направление непостоянно и может
изменяться. Некоторые виды птиц, а именно: все виды ца-
пель, каравайка, в меньшей мере — ласточки и, вероятно,
72
Глава 111
еще целый ряд других форм, рассеиваются вскоре после
периода размножения по самым различным направлениям,
часто на сотни километров. В результате эти птицы появ-
ляются иногда в таких местностях, где они едва ли ко-
му-нибудь известны и куда они никогда не залетали. Это
«бесцельное», ненаправленное перемещение не имеет ни-
чего общего с истинным перелетом, который начинается
много времени спустя в определенном направлении. «Про-
межуточный перелет» является, вероятно, способом рассе-
ления молодых птиц, стремящихся захватить новые жиз-
ненные пространства. Эти перемещения продолжаются до
достижения «промежуточной цели» («Zwischenziel»), кото-
рая, как уже упоминалось, нередко находится в противо-
положном направлении по отношению к цели осеннего
перелета. Иногда «промежуточная цель» располагается
вблизи последней. Во всяком случае «промежуточный пе-
релет» прекращается через несколько недель, когда начи-
нается истинный перелет на зимовки.
Пути перелетов
Ни одна птица не летит из гнездовой области на зи-
мовки совершенно прямым путем. Это следует учитывать
при изучении карт перелетов птиц, -на которых общепри-
нято места кольцевания и возврата колец какой-либо пти-
цы соединять прямой линией. Эта линия представляет
собой теоретический, или идеальный, путь перелета. В луч-
шем случае она только приблизительно соответствует дей-
ствительности. Строение поверхности земли, формы ланд-
шафта и растительность, условия питания и погода влия-
ют на перелет птиц и обусловливают различные отклонения
от идеального пролетного1 пути, который в основном ука-
зывает лишь на направление миграции. Направление пу-
ти может и изменяться, но чаще птицы строго придержи-
ваются раз избранного пролетного пути, несмотря на то,
что условия окружающей среды вынуждают их на от-
дельных участках к большим или меньшим отклонениям.
Рано или поздно птицы вновь возвращаются к первона-
чальному направлению.
Летят ли птицы на зимовки по определенным путям
или же они придерживаются лишь общего направления,
Результаты изучения перелетов птиц
73
влияют ли на направление перелета факторы внешней
среды или же внутренние импульсы, обусловленные тради-
цией и передаваемые по наследству? На все эти вопросы
нельзя дать общего ответа, ибо видовые различия в луч-
шем случае позволяют только объединять птиц в большие
или меньшие группы в соответствии с определенными фор-
мами перелетов и поведения при миграциях.
Основными формами перелетов являются перелеты уз-
ким и широким фронтом. Перелет узким фронтом прибли-
зительно соответствует понятию «пролетные пути» («Zug-
strasse»), которое употреблялось при ранних изучениях
перелетов птиц и которое до сих пор проникает недоста-
точно критически в литературу по этим вопросам.
Вторая форма перелетов характеризуется тем, что зна-
чительные массы птиц летят на большом пространстве в
одном и том же или приблизительно в одинаковом направ-
лении. При изложении истории изучения перелетов птиц
мы упоминали о многолетнем споре двух ученых о поня-
тиях «пролетные пути» и «перелет широким фронтом»,
мнения которых оказались в одно и то же время и вер-
ными и неверными, так как обе формы перелета встреча-
ются у различных видов птиц. Их связывает целый ряд
переходов, в связи с чем точно установить, в какой форме
совершает перелет данный вид птиц, часто затруднитель-
но. Нередко эти понятия употребляются в различном смыс-
ле, вследствие чего возникают дальнейшие недоразумения.
Окончательная ясность в отношении этих терминов была
внесена Гейром (1930). По его определению, птицы како-'
го-либо вида летят узким фронтом в том случае, когда
они населяют обширную область, а их пролетные пути на
большом протяжении так сближены, что по отношению к
размерам этой области они кажутся узкими и напоминают
«дороги». Перелеты широким фронтом имеют место тогда,
когда пути перелетов птиц, населяющих обширную об-
ласть, расположены далеко друг от друга, так что по от-
ношению к размерам этой области они кажутся широки-
ми и не похожими на «дороги». Однако на небольшом
протяжении широкий фронт может значительно сужи-
ваться.
Это .уточнение терминологии позволяет охарактеризо-
вать форму перелета каждого вида птиц, если имеется
74
Глава HI
Рис. 28. Значение Гельголанда как точки, направляющей перелеты
птиц в восточных районах Северного моря. Основные линии пролетов.
достаточное количество наблюдений и возврата колец.
Правда, при этом возможны субъективные суждения, на-
пример при наличии переходов между перелетами узким и
широким фронтом. Но практически только изредка нельзя
будет достигнуть ясности в решении этих вопросов. В та-
ких случаях мы должны будем ограничиться определени-
ем поведения птиц различных популяций. ....
Результаты изучения перелетов птиц 75
Если мы рассмотрим картины перелетов отдельных ви-
дов птиц, то придем к заключению, что в Европе и Азии
подавляющее большинство сухопутных птиц летит широ-
ким фронтом. Для некоторых видов это еще не установле-
но из-за отсутствия соответствующих данных. Однако при
равномерной расстановке наблюдателей выявилось бы и
приблизительно равномерное распределение птиц во вре-
мя перелета, причем, конечно, необходимо учесть влияние
особенностей ландшафта. При многочисленных возвратах
колец эти отношения выступают с полной ясностью. Пере-
леты узким фронтом встречаются относительно редко.
В качестве примеров можно назвать белого и черного аис-
тов, журавля, жулана, чернолобого сорокопута и крачек.
При определении понятия «перелет широким фронтом»
отмечалось, что вследствие сужения этого фронта на опре-
деленных участках перелет может приобретать характер
узкого фронта, обозначаемого в этом случае как путь
массового пролета птиц («Massenzugweg»). Это наблю-
дается тогда, когда рельеф поверхности земли создает пре-
пятствия на пути перелетов. Эти препятствия сходно влияют
на перелет многих видов птиц, поэтому путь массового
пролета представляет собой местное явление в противопо-
ложность узкому фронту, свойственному определенным
видам птиц и отличающемуся большой протяженностью.
Такими препятствиями бывают большей частью горы или
обширные водные пространства. В таких местах, на бере-
гах морей и по окраинам горных хребтов, наблюдается бо-
лее Тесное сближение пролетных путей при перелете ши-
роким фронтом, так как птицы не могут изменить направ-
ления и продолжать перелет иными, более благоприятны-
ми путями. Они стараются обойти препятствие и оставля-
ют его, как только оно оказывается вне направления их
перелета. Чем значительнее протяженность препятствия,
тем большее число птиц скопляется около него и затем
восстанавливает широкий фронт перелета. Берега морей
и окраины горных местностей определяют направление
перелета многочисленных видов и таким образом пред-
ставляют собой пути массового пролета. Подробнее об
этом говорилось в одном из предыдущих разделов в отно-
шении Курской косы и восточного берега Курского заливав
Там же указывалось и на значение для перелетов птиц
76
Глава III
острова Гельголанд. Подобно тому как берега служат на-
правляющими линиями (ведущими направлениями),
острова являются направляющими точками, которые вре-
менно сгущают широкий фронт перелетов на узком про-
странстве. О «воронкообразном» («Trichterzug») и «вееро-
образном» («Facherzug») перелетах говорят в тех случа-
ях, когда пролетные пути широкого фронта сближаются на
узком участке, а затем вновь расширяются.
Значение направляющих линий (ведущих направле-
ний) для отдельных видов и групп птиц весьма различно.
Большинство сухопутных птиц, избегающих перелета че-
рез обширные водные пространства, придерживается бере-
гов, расположение которых не всегда соответствует на-
правлению их перелета. Прибрежные, а отчасти и болот-
ные птицы из внутренних районов также часто летят
огромными стаями вдоль берегов, так как находят там
обильный корм. Но большинство болотных птиц при
своих миграциях тесно придерживается внутренних водо-
емов, причем направляющими линиями являются озера и
русла рек. Причины этого легко объясняются биологией
питания птиц.
Морские птицы, например чайки и крачки, как прави-
ло, пролетают частично вдоль берегов, частично — в глу-
бине суши; различные виды птиц ведут себя в этом отно-
шении по-разному. Долгое время считали, что утки летят
по ограниченным прибрежным путям, которые направляют
их перелет, например у Северных Фризских островов, так
как в этом месте во время перелета ловилось бесчислен-
ное множество уток в так называемые коечные ловушки.
Результаты кольцевания как будто подтверждали это
предположение. Но полевые наблюдения, произведенные
в глубине материка, доказали, что, по крайней мере, часть
северных уток не избегает даже дальних перелетов над
сушей. Эти птицы пересекают Центральную Европу или же
проникают глубоко на юг СССР, несомненно, используя
при этом озера и реки в качестве направляющих линий
(ведущих направлений). Кольцеванием уток в Астрахан-
ском заповеднике удалось установить, что эти птицы со-
вершают перелеты с юга на север на очень большие рас-
стояния над внутренними частями суши (Вучетич и Туга-
ринов, 1941) (рис. 29).
Результаты изучения перелетов птиц
77
Рис. 29. Карта пролета шилохвости над внутренними районами
материка из мест зимовок на юге в северную гнездовую область.
(Из Вучетича и Тугаринова, 1941.)
Подобный перелет широким фронтом, находящийся из-
за особенностей биологии питания птиц в тесной зависимо-
сти от направляющих линий (ведущих направлений),
Гейр и Шюц назвали перелетом «направленным широким
фронтом» («geleitete Breitfront»). Это обозначение осо-
бенно применимо к перелету обыкновенных чаек. До по-
следнего времени считалось, что они совершают миграции
по четко выраженным пролетным путям, или узким фрон-
том. Это мнение как будто подтверждалось и данными
кольцевания. Однако кольцевание многочисленных попу-
ляций показало, что это относится только к большинству
гнездящихся на побережье чаек, в то время как птицы,
гнездящиеся во внутренних районах суши, совершают пе-
релеты преимущественно' широким фронтом. Последние
предпочитают следовать вдоль рек и озер, но все же встре-
чаются и в отдалении от них. Так, например, популяция
западнобалтийских обыкновенных чаек отклоняется при
РйСг 30. Перелеты обыкновенных чаек направленным широким фронтом из Обер-
лаузица и Чехии. (Из Шюца и Венгольда, 1931.)
Цифры — месяцы кольцевания, черточки под ними — число лет, прошедших со времени коль-
Результаты. Изучения перелетов птиц
79
перелете Вислой и Одером на большие расстояния от при-
брежного пролетного пути и летит над участками, бедны-
ми водой, на зимовки к швейцарским озерам. В качестве
примера изменения направления перелета уже упомина-
лось о том, что летящие из Чехии чайки направляются по
Эльбе к побережью Северного моря, по которому они за-
тем следуют на запад и на юг. Из Моравии и Силезии, на-
против, птицы летят преимущественно в южном направ-
лении, к Дунаю и дальше к Адриатическому морю, следо-
вательно, отчасти над высокими горными хребтами. Эти
немногие примеры достаточно ясно характеризуют пере-
лет «направленным широким фронтом» и определяют
его отличия от перелета узким фронтом. Для первого ти-
пично разнообразие пролетных путей отдельных особей и
широкое рассеяние популяции, доходящее до 180°. Обо
всем этом уже говорилось.
Как уже упоминалось, журавли летят на зимовки огра-
ниченными пролетными путями, узким фронтом. Места их
зимовок расположены в Северо-Западной Африке и в Во-
сточном Судане, у Нила. Перелеты регулярно совершают-
ся над областями, в которых журавли не гнездятся и
поэтому во время пролетов особенно заметны: с одной
стороны — из Центральной Европы через Испанию и из
Восточной Европы через Южную Италию на юго-запад, с
другой стороны — из Восточной Европы через Эгейское
море и остров Крит на юг (см. приложение) .
Особенным своеобразием отличаются перелеты белого
аиста, летящего узким фронтом. Его популяции придер-
живаются при перелете двух-различных направлений; меж-
ду ними существует рубеж, который обычно представляют
проходящим через область, расположенную между Эльбой
и Зёйдер-зе. Гнездящиеся к востоку от этой местности
аисты улетают преимущественно на юго-восток через Ма,
лую Азию, Палестину и Египет. Гнездящиеся западнее и
южнее Зёйдер-зе улетают на юго-запад, через Францию;
Испанию и Марокко. Многочисленные находки колец да-
ют возможность точно проследить перелеты восточных по-
пуляций вплоть до южноафриканских мест зимовок; для
западных аистов точные данные на участке южнее Марок-
ко вплоть до озера Чад и Судана отсутствуют. Поэтому
пока можно лишь предположить, что эти птицы продолжа-
80
Глава III
Рис. 31. Перелетный рубеж белых аистов.. (Из Рюппеля, 1937.)
ют перелет над Сахарой. Стремление аиста (в отличие от
журавля) избежать перелета через море, даже ценой
больших обходных путей, имеет (согласно Гейру) лётно-ме-
ханические и аэродинамические причины. Аист мало при-
способлен к длительному гребному полету, сильные ветры
мешают ему. Зато широкие крылья позволяют ему исполь-
зовать восходящие потоки воздуха для парящего полета,
Рис. 32, Перелеты белого аиста из Западной и Центральной Европы в
Африку (по результатам кольцевания; для Сахары — только предпо-
ложительно). (Из Рюппеля, 1942а.)
6-176
82
Глава III
что дает большую экономию сил. Но при полете через
Средиземное море опасны сильные ветры, а восходящих
воздушных потоков нет (восходящие потоки воздуха над
морем, которые используют буревестники, слишком незна-
чительны). Поэтому аисты предпочитают сухопутную до-
рогу; большее расстояние компенсируется экономией сил.
Однако не все виды птиц при пролетах обходят боль-
шие водные пространства и горы. Многочисленные наблю-
дения показали, что даже над такими высокими горными
системами, как Альпы и Кавказ, проходят интенсивные
пролеты, причем не только над перевалами, но и над самы-
ми высокими хребтами и вершинами. Даже горные пусты-
ни Центральной Азии не являются непреодолимыми пре-
градами; об этом говорят находки сибирских птиц в Север-
ной Индии. Пустыни вообще значительно меньше препят-
ствуют пролетам, чем это было принято считать. Гейр.
Хартерт и английские орнитологи установили в Сахаре и
Ливийской пустыне интенсивный пролет многих видов мел-
ких птиц, а наблюдения и возврат колец в Судане позволи-
ли сделать заключение о регулярном пролете через пусты-
ню. Во время весеннего и осеннего перелетов птицы пере-
секают даже моря и океаны. Подробнее об этом будет
сказано ниже. По сравнению с- ними перелеты через Се-
верное или Средиземное моря едва ли заслуживают упо-
минания. Поэтому боязнь пролета даже над небольшими
водными пространствами представляется скорее как «пси-
хологический» момент, так как физически это невозможно
для птиц только при сильном ухудшении погоды.
Причины возникновения еще одной формы пролетных
путей заключаются в различных условиях погоды и пита-
ния весной и осенью в определенных областях. Пролет-
ный путь, который в одно время года вполне благоприятен,
может в другое время оказаться крайне неподходящим
(например, области весеннего половодья, которые в конце
лета высыхают и не доставляют больше пищи болотным
птицам). Некоторые виды птиц приспосабливаются к та-
ким меняющимся условиям и летят на родину и на зимов-
ки различными путями. Таким образом возник «петлеоб-
разный перелет» («Schleifenzug»), например, у трех севе-
ро-восточных сибирских уток, которые осенью направляют-
ся на юг вдоль тихоокеанского побережья, а весной
Рис. 33. Перелеты североамериканской золотистой ржанки (Charad-
rius dominicus). (Из Линкольна, 1935.)
Восточная раса летит на зимовки и обратно «петлеобразным перелетом». За-
падная раса летит из Аляски к Гавайским и Маркизским островам и обратно
по одному и тому же пути.
6*
84
Глава III
возвращаются к гнездовьям через материк. Подобные при-
меры дают нам пеночки, славки, сорокопуты и мухоловки,
которые .пролетают через Египет частично только осенью,
частично только весной. В этом отношении особенно ха-
рактерен перелет золотистых ржанок из Северной Амери-
ки, которые гнездятся на севере материка и летят (по
крайней мере старые птицы) на зимовку через Лабрадор
и открытое море в Южную Америку. На обратном пути
они следуют на север вдоль западной части Южной Аме-
рики, через Центральную Америку и по долине р. Мисси-
сипи, где весной условия жизни для них значительно более
благоприятны, чем во время осеннего перелета. Причину
ясно выраженного петлеобразного перелета других птиц,
например иволги и сизоворонки (они осенью летят через
восточную часть Средиземного моря к восточноафрикан-
ским местам зимовок, а весной — через центральные райо-
ны Средиземноморской области обратно на родину), Штре-
земан (1944, 1948) видит в том, что птицы придержива-
ются путей своего исторического расселения. Эти птицы,
повидимому, происходят из Восточной Евразии и распро-
странились в Западную Европу лишь в относительно не-
давнее геологическое время. Видовая «память» заставляет
их оставаться верными первоначальным местам зимовок
в Восточной Африке и добираться до них обходными путя-
ми; весной они следуют более прямым путем и преимуще-
ственно в северном направлении.
В Германии также можно иногда наблюдать, как в за-
висимости от времени года меняются пролетные пути
птиц.
Незадолго до второй мировой войны подобные из-
менения были установлены у журавля в Средней Герма-
нии. То же самое известно о перелетах сибирских черно-
зобых гагар (над Западной Балтикой) и различных песоч-
ников. Установить такие факты, конечно, трудно; для это-
го необходимы многолетние исследования в одной и той
же местности.
В тех случаях, когда птицы по внутренним или внеш-
ним причинам сбиваются с правильного направления про-
лета и появляются в местах, куда они обычно не залетают,
конечно, нельзя говорить о пролетных путях. В этом слу-
чае птицы летят по направлениям, которые не имеют ни-
Результаты изучения перелетов птиц
85
какого отношения к их нормальным пролетным путям, а
отчасти служат «продолжением» последних, и перелетают
через места зимовок. Таких птиц, случайно появившихся в
какой-либо местности, называют «залетными». Примером
могут служить сибирские виды дроздов: в прошлом сто-
летии, когда была распространена ловля силками, их от-
мечали в Германии (отдельные виды — до 18 раз). Это до-
казывает, что залетных птиц можно встретить повсюду.
Но обычно такие находки редки, за исключением тех мест,
где регулярно и тщательно учитываются изменения в ави-
фауне. Такими местами являются, например, остров Гель-
голанд, небольшая площадь которого и хорошая обозри-
мость особенно облегчают регистрацию авифауны, Россит-
тен, охраняемые колонии морских птиц и другие заповед-
ники, где проводятся постоянные наблюдения. На Гельго-
ланде находили залетных птиц из Средиземноморской
области, например пестрого каменного дрозда, обыкновен-
ную каменку, бледную пересмешку и журавля-красавку,
из Передней Азии — бледного сорокопута, черного жаво-
ронка, черноголовую овсянку и рыжехвостую славку, из
Сибири — одноцветного и краснозобого дроздов, несколь-
ко видов пеночек и овсянок, а также вилохвостых и розо-
вых чаек.
На маленьком скалистом острове не так редко
появляются залетные птицы даже из Северной Америки:
Тurdus migratorius и Тurdus minimus, а также американ-
ский перевозчик и золотистая ржанка. Европейские птицы
иногда залетают в Северную Америку. Для морских птиц
такие перелеты через океан не представляют труда. Но
этого нельзя сказать в отношении обыкновенных чаек, ко-
торые обычно не вылетают далеко в море. Однако околь-
цованных в Росситтене чаек находили в Мексике и на
островах Вест-Индии. Но совершенно удивительным был
залет английских чибисов, которые в декабре 1927 г. ста-
ей появились на Ньюфаундленде. Восточные бури оттес-
нили их от пролетного пути в Ирландию и пронесли за 24
часа через океан на расстояние 3600 км. Подобные зим-
ние бури являются также причиной появления различных
типичных морских птиц в глубине материков, где эти за-
летные птииы, обычно очень истощенные, как правило,
быстро погибают.
86
Глава III
Формы перелетов
Этот раздел посвящен обсуждению различий в по-
рядке перелетов и последовательности этого явления, об-
разованию перелетных стай, а также дневным и ночным
пролетам.
Нам уже знакомы многие различия в отношении к пе-
релету отдельных видов птиц, поэтому можно предпола-
гать, что мы встретимся с ними и в дальнейшем. Во вся-
ком случае, мы не должны удивляться, когда узнаем, что
птицы часто придерживаются особого порядка в сроках
перелета в соответствии с возрастными и половыми раз-
личиями. Так Вейгольд сообщает, что над Гельголандом у
зябликов, камышовых овсянок, полевых жаворонков, го-
рихвосток-чернушек и пустельги осенний перелет первыми
начинают молодые птицы, вслед за которыми летят ста-
рые самки и в последнюю очередь — старые самцы. Моро,
напротив, установил, что в Египте у кукушек, жуланов,
черноголовых славок и пестрых каменных дроздов старые
особи пролетают раньше молодых. Над Курской косой мо-
лодые серые вороны также пролетают раньше старых.
У скворцов и у вальдшнепов также установлен очень ран-
ний пролет молодых особей, в то время как старые следу-
ют за ними значительно позже. Молодые птицы соверша-
ют отчасти «промежуточный перелет», о котором говори-
лось выше, отчасти — направленный «раннелетний пере-
лет». У большого кроншнепа и некоторых других болотных
птиц миграцию к месту зимовок задолго до нормального
перелета начинают не молодые, а старые птицы, которые
не приступали к размножению или уже закончили его. До
недавнего времени считалось, что у черных дроздов и си-
ниц молодые птицы улетают, а старые остаются на зиму
на месте. Однако новые данные кольцевания выявили мно-
гочисленные исключения. У полевых луней старые самцы
как будто улетают (по крайней мере в земле Бранденбург,
но не в Западной Прибалтике), в то время как молодые
особи и самки большей частью остаются зимовать. В неко-
торых местностях на месте зимуют самцы зябликов, а уле-
тают только самки. У плавунчиков Крайнего Севера (Pha-
laropus), у которых в насиживании основную роль играют
самцы, самки улетают первыми, а самцы и молодые пти-
Результаты изучения перелетов птиц
87
цы значительно позднее. По данным голландских авторов,
это как будто имеет место и у полевых жаворонков (не-
смотря на то, что в насиживании участвуют обе птицы).
Очень часто (особенно у певчих птиц) самцы весной воз-
вращаются раньше самок. Это имеет биологическое зна-
чение, ибо самцы занимают гнездовые участки и защища-
ют их до прилета самок.
Многим, вероятно, известен особый строй пролетных
стай крупных птиц. Журавли и дикие гуси часто летят
клином. Этот же порядок пролета наблюдается и у уток.
Кулики-сороки образуют при перелете длинные, широко
растянувшиеся цепочки. Эта форма пролета дает извест-
ные аэродинамические преимущества; «ведущий» при
этом время от времени сменяется. Другим строем птиц
на пролете являются густые замкнутые стаи; так летят,
например, скворцы и кулики, которые своим четким манев-
рированием создают впечатление единой массы. Не так
крепко сплочены, особенно зимой, стаи мигрирующих вя-
хирей и клинтухов или мелких певчих птиц. «Рыхлыми»
стаями пролетают вороны, а также многочисленные виды
мелких птиц. Раздельно, на довольно больших расстояни-
ях, а также поодиночке летят хищные птицы над Курской
косой. Перепелятники соблюдают такое расстояние, чтобы
не терять друг друга из вида. Это относится, в частности,
к Южной Швеции и Босфору.
Указанные наблюдения поставили перед нами вопрос
об образовании стай при перелете; его нужно рассмотреть
подробнее.
Не только у большинства перелетных птиц, но и у
очень многих кочующих и оседлых после окончания перио-
да размножения пары разъединяются (мы не касаемся
птиц, гнездящихся колониями, например чаек, крачек, гра-
чей, береговых ласточек и др.). Вместе с птицами того же
или родственных видов они образуют небольшие или круп-
ные сообщества, которые, как правило, сохраняются до на-
чала следующего периода размножения. Это особенно за-
метно осенью и зимой, когда смешанные стайки жаворон-
ков, луговых коньков, зябликов, коноплянок, щеглов и
других птиц беспокойно носятся по полям или держатся
вблизи населенных пунктов. Соединения перелетных птиц
нередко, бросаются в глаза еще резче. Вспомним скопле-
88
Глава Ill
ния ласточек на телеграфных проводах незадолго до от-
лета, места сбора журавлей и аистов, откуда эти птицы
начинают миграции. Другие же виды, например лебеди и
дикие гуси, летят семьями. Об образовании стай у сквор-
цов и других мелких птиц (куликов) уже говорилось, так
же как и о «рыхлом» перелете ворон. Последние часто
летят вместе с галками, певчие дрозды — с дроздами-бело-
бровиками, зяблики — с зеленушками или чижами, вяхи-
ри — с клинтухами. Поодиночке пролетают очень немно-
гие виды птиц: кукушка, удод, зимородок, вертишейка,
большинство хищных птиц, так что стайный пролет сле-
дует считать правилом. В одной из своих новых работ
Гейр (1949) подчеркивает коренное различие между «со-
обществом по отдыху» и истинным «сообществом по про-
лету». Часто состав отдыхающих в пути стай птиц совер-
шенно' иной, чем непосредственно во время перелета. По
интенсивно используемым направляющим линиям вре-
менно может иметь место и «ложное» сообщество, кото-
рое в дальнейшем распадается. Большинство стайных пе-
релетных птиц летит только с представителями того же
вида, т. е. в видовом, или «чистом», сообществе. Другие
птицы предпочитают «смешанные» стаи. К ним относятся
клинтух, вяхирь, зяблик, вьюрок, галка, грач, перепел и
коростель.
Преимущества образования на пролете стай до сих пор
еще не ясны. Когда мигрируют совместно старые и моло-
дые птицы, то первые, вероятно, играют роль «учителей»
в перелете. Но и объединение старых особей, возможно,
выгодно для нормальных координированных действий.
Браун применил в данном случае термин «инстинктив-
ное корректирование поведения путем образования стаи».
Этот фактор играет значительную роль и в других
инстинктивных действиях птиц.
Почти все виды птиц, пролетающих стаями, во время
пролета постоянно издают характерные звуки, по которым
без труда можно установить их присутствие («перелетные
звуки»). Это особенно относится к ночному времени, ко-
гда, как отмечалось выше, пролетающие стаи реагируют
на световые «сигналы» усилением криков. Но и днем пти-
цы издают призывные крики, повидимому, способствую-
щие сплочению стаи (перелет с «голосовым контактом»).
Результаты изучения перелетов птиц
89
Птицы, летящие молча, часто образуют очень плотные
стаи или же пролетают только днем.
Поэтому необходимо различать ночные и дневные про-
леты; перелетные птицы могут, следовательно, и в этом от-
ношении вести себя различно, Днем летят главным обра-
зом крупные птицы, например журавли, аисты, хищные
птицы и вороны. И днем и ночью пролетают лишь очень
немногие виды птиц: чибисы, скворцы и полевые жаво-
ронки. Подавляющее большинство перелетных птиц мигри-
рует ночью, особенно мелкие насекомоядные певчие птицы,
но также и многие зерноядные певчие птицы, почти все
болотные, кулики и многие виды уток. Днем они кормятся
и пополняют запасы, необходимые им для перелета. Кроме
того, в это время они лучше защищены от хищных птиц,
пролетающих днем, так как в период пролета последние
не занимаются поисками пищи. Нахождение пути проле-
тающими ночью птицами не так трудно, как это может по-
казаться, так как даже в безлунные ночи земная поверх-
ность выделяется достаточно отчетливо благодаря отсвету
облаков, звезд или отражающих поверхностей воды.
В очень темные ночи, вероятно, не бывает никаких проле-
тов. Птицы, пролетающие ночью, в общем покрывают
большие расстояния, чем летящие днем, пролет которых
большей частью ограничивается только утренними часами.
В многочисленных случаях это подтвердили и данные
кольцевания.
Время перелетов
Обсуждение времени перелетов может на первый взгляд
показаться излишним, поскольку их сроки явно «непо-
движны». «Промежуточный» и «раннелетний» перелеты
происходят не в период собственно весенних и осенних
перелетов. К этому необходимо еще добавить иногда до-
вольно существенные миграции зимой, которые нельзя
считать только следствием изменившихся условий пита-
ния и погоды. Кроме того, прилет и отлет мигрирующих
птиц также продолжается в течение нескольких недель и
месяцев. У каждого вида — свое время прилета и отлета,
которое к тому же различно для каждой гнездовой об-
ласти, а следовательно, и для каждой популяции.
90
Глава 111
Прежде всего, можно сделать общее заключение, что
начало перелетов зависит от наступления неблагоприят-
ных условий: в тропиках — от периодов засухи, в умерен-
ных и холодных зонах — от проявлений «зимней» погоды.
Следствием различных требований, предъявляемых пере-
летными птицами к питанию и размножению, является
соответствующее различное отношение этих птиц к пере-
летам, так что из двух видов раньше улетает и позже воз-
вращается тот, который оказывается более восприимчи-
вым и требовательным. Время прибытия на родину
зависит у многих видов от наличия корма; у других же
видов—от возможности освоить гнездовую территорию.
Ясно, что перелетные птицы Крайнего Севера смогут вер-
нуться на родину только тогда, когда почва освободится
от снега и оттает. Некоторые из этих видов возвращаются
к гнездовьям в конце мая — начале июня и вынуждены
спешить с устройством гнезд и насиживанием, чтобы еще
до наступления холодной погоды подготовиться к отлету.
Они появляются в области гнездования иногда уже пара-
ми и отлетают даже позже, чем гнездящиеся южнее пред-
ставители того же вида. Это было установлено для чер-
ных стрижей (прибывают в Германию около 1 мая, в
Финляндию — в конце мая — начале июня; отлет из Гер-
мании — около 1 августа, из Финляндии — в конце авгу-
ста — начале сентября).
Как правило, позже всех прилетают на родину те
перелетные птицы, которые совершают наиболее дальние
миграции. Они раньше всех и отлетают. Все без исключе-
ния певчие птицы, возвращающиеся уже в марте — сере-
дине апреля, зимуют севернее Сахары, в то время как
прилетающие позже — почти все в Тропической или
Южной Африке. Последним свойственна большая
регулярность во времени отлета и прилета, хотя много-
летними наблюдениями установлены значительные откло-
нения.
Приведем некоторые примеры. Согласно многолетним
наблюдениям, проведенным Гагеном в Любеке и Тишле-
ром в центральной части б. Восточной Пруссии, колеба-
ния в сроках прилета птиц составляют для пеночки-пере-
смешки соответственно 11 и 14 дней, садовой славки— 13
и 17 дней, иволги — 14 и 10 дней, черного стрижа —17 и
Результаты изучения перелетов птиц
91
15 дней, кукушки — 19 и 22 дня, пеночки-веснички — 27
и 24 дня, мухоловки-пеструшки — 30 и 25 дней, певчего
дрозда — 37 и 30 дней и для аиста — 55 и 30 дней. В каж-
дом случае высчитывалось среднее число дней за период
свыше 20 или, соответственно, почти 40 лет.
Однако невозможно связать даты прилета некоторых
видов птиц с каким-либо определенным днем, как это
практиковалось раньше при использовании народных при-
мет для составления календарей. Это неверно ни для рано
ни даже для поздно возвращающихся птиц. Народные
приметы отражают только тот факт, что часто птицы ре-
гулярно и независимо от условий погоды придержи-
ваются обычных сроков миграций. Перелетные пти-
цы заселяют любой район весной (или покидают его
осенью) неравномерно. Произойдет ли это раньше или
позже — зависит от того, насколько благоприятны условия
на месте. Большое значение имеют высота местности над
уровнем моря и климатические факторы, так что в суро-
вых горных местностях в отличие от низменностей с мяг-
ким климатом иногда наблюдается опоздание на несколь-
ко недель. Используя многочисленные данные о датах
прилета птиц, собранные на большой территории, можно
составить фенологические карты для различных видов
птиц. Такие карты составлены Бретшером, Сливинским,
Саузерном и др. Если наблюдения проводились продол-
жительное время, то при помощи таких карт пролета мож-
но с уверенностью определить или предсказать для каж-
дой местности сроки прибытия соответствующего вида.
Этот метод основан на ранее упомянутой работе Мидден-
дорфа, который впервые соединил прямыми линиями ме-
ста одновременного прилета весной различных видов
птиц. Эти линии он назвал изопиптезами (по аналогии с
линиями, соединяющими места с одинаковой температу-
рой — изотермами, или с одинаковым давлением возду-
ха— изобарами). Позднее английские наблюдатели обо-
значили их как изохроны. Штреземан употребил термин
изофены. Эти линии показывают, как различные виды
продвигаются весной к своему гнездовью, где их задержи-
вают препятствия и в каких направлениях они расселя-
ются. Из этого вытекают противоположные выводы об их
образе жизни и распространении, Подобные выводы были
Рис. 31. Линии одновременного прилета весной (изопиптезы) черного стрижа
{Micropus apus) в Европу. (Из Сливинского, 1938.)
Результаты изучения перелетов птиц
93
сделаны многочисленными орнитологами в отношении ев-
ропейских и североамериканских видов птиц.
Можно легко установить, что не все птицы одного вида
одновременно прилетают на свою родину и отлетают, а,
следовательно, они в различное время покидают или со-
ответственно достигают мест зимовок. Обычно весной в
Германии появляются сначала лишь несколько предста-
вителей какого-либо вида, которых можно рассматривать
как «разведчиков». Лишь спустя некоторое время, в тече-
ние которого «разведчиков» часто совершенно невозмож-
но обнаружить, прибывает основная масса птиц, прилет
которых обычно не растягивается надолго. Затем в тече-
ние довольно продолжительного времени следуют запаз-
дывающие птицы. Количественное распределение по трем
названным группам меняется от вида к виду и из года в
год. Иногда большинство птиц прилетает или отлетает
сейчас же вслед за «разведчиком», в других же случаях
они следуют за ним значительно позже.
Мы уже упоминали о том, что живущие на севере
популяции мигрируют весной и осенью позже, чем популя-
ции того же вида, гнездящиеся южнее. Северные попу-
ляции пролетают через гнездовую область последних, в
связи с чем пролет часто значительно растягивается. Если,
например, ласточки в средиземноморской области уже при-
ступили к размножению, то в Германии они в это время
только начали появляться. Когда скандинавские пеночки
или желтые трясогузки пролетают над Германией, гер-
манские представители того же вида уже приступили к
размножению.
Таким образом, нельзя просто констатировать, что тот
или иной вид закончил перелет и приступил к размноже-
нию. Необходимо всегда учитывать различия между пти-
цами данного вида, гнездящимися в Германии и на севе-
ре. Что правильно для одних, неверно для других. Поэтому
определить, когда в Германии появляются первые пред-
ставители какого-либо вида и когда прилетит основная
масса перелетных птиц, нетрудно. Но очень сложно, а
иногда и невозможно установить конец прилета местных
популяций определенного вида и начало пролета северных
популяций. Поэтому некоторые наблюдатели легко впада-
ют в заблуждение, считая время пролета какого-либо
94
Глава III
вида значительно более продолжительным, чем оно бы-
вает в действительности. Это может быть правильным
только в тех случаях, когда они имеют в виду всю область
распространения данного вида, а не ареал одной или не-
многих рас или, соответственно, популяций, как это обыч-
но принято, и который можно определить при помощи
наблюдений в одном пункте (например на Гельголанде и
в Росситтене). Штреземан (1944) сравнил даты прилета
некоторых видов в средиземноморской области и в Цен-
тральной Европе и обнаружил совершенно иную последо-
вательность для этих районов, что едва ли вызвано разли-
чиями в скорости миграций, а скорее связано с особыми
требованиями, предъявляемыми различными видами к
условиям существования. Так, например, черные стрижи
и деревенские ласточки очень рано прилетают в Южную
Европу (начало — середина марта). При дальнейшем
продвижении на север их обгоняют другие виды, напри-
мер мухоловки-пеструшки и горихвостки-лысушки, кото-
рые появляются у Средиземного моря только в конце мар-
та, а в конце апреля прилетают в б. Восточную Пруссию,
в то время как стрижи и ласточки прибывают в эти места
лишь в конце апреля, а иногда даже в середине мая.
Последние предъявляют более высокие требования к ко-
личеству корма (насекомые) и поэтому медленно, по мере
потепления, продвигаются на север.
Скорость перелета
Едва ли по какому-нибудь вопросу, связанному с пере-
летами птиц, распространены столь ошибочные взгляды,
как по вопросу о скорости перелета. Мнения большинства
людей о скорости, с которой пролетают птицы, основаны
на случайных кратковременных наблюдениях, и поэтому
она обычно сильно преувеличена. Другие сравнивают ско-
рость пролета птиц со скоростью автомобиля, поезда или
самолета. Однако таких скоростей они не найдут даже у
наиболее быстрых из известных нам летунов. Так, напри-
мер, стрижи пролетают со скоростью 40 — 50 м/сек (не-
зависимо от ветра), что соответствует приблизительно
150 — 160 км/час. (Сравните: максимальная скорость экс-
пресса— 39 м/сек, или 140 км/час.) Это, конечно, не озна-
Результаты изучения перелетов птиц
95
чает, что птицы вообще не умеют летать быстрее. Гоняю-
щиеся друг за другом стрижи развивают скорость до
200 км/час, а сокол бросается на жертву со скоростью
70 м/сек, т. е. 250 км/час. Но эти предельные скорости в
течение очень короткого времени являются исключениями:
они в лучшем случае характеризуют способность к полету
некоторых видов, но их нельзя использовать для оценки
скорости полета при миграциях, когда необходимо' дли-
тельное напряжение.
При длительных миграциях имеют значение не только
способность к полету, но также и ветер. В зависимости от
его направления и силы скорость птиц может значительно
уменьшаться или увеличиваться. Особенно большие ско-
рости в полете можно объяснить только при учете под-
держки со стороны ветра. Так, в приведенном выше при-
мере скорость английских чибисов при полете через Ат-
лантический океан, равная приблизительно 70 км/час,
увеличилась до 150 км/час благодаря попутному ветру,
скорость которого достигала 90 км/час.
Учитывая задерживающее или ускоряющее влияние
ветра, можно точно измерить собственную скорость птиц
на коротких расстояниях и в соответствии с этим вычис-
лить истинную скорость пролета. Впервые такие расчеты
произвел Тинеман на Курской косе. Впоследствии они
были сделаны Мейнертцхагеном, Гаррисоном и др.
Таблица 8
СКОРОСТЬ ПОЛЕТА ПТИЦ ВО ВРЕМЯ ПЕРЕЛЕТА, км’час
Вид По Тинсману По Мейнер- тцхагену
Ястреб-перепелятник . . . 41,4
Серебристая чайка .... 49,7
Большая морская чайка 50
Ворона 50—52,2 51—59
Вьюрки 52,5 32—59
Сокол-сапсан 59,2
Галка 61,5
Скворец 74,1 63—81
Сокол 66—79
Кулики 66—85
Гуси .... 69—91
Утки 72—97
Ласточки 100—120
96
Глава II!
Цифры, приведенные в таблице, дают ясное предста-
вление о предельных скоростях перелета птиц. В общем
она, очевидно, равна 40 — 80 км/час, причем скорость
мелких певчих птиц приближается к низшим цифрам.
Птицы, совершающие перелет ночью, летят, повидимому,
быстрее, чем мигрирующие днем. Бросается в глаза ма-
лая скорость миграции хищных и других крупных птиц.
Одни и те же виды птиц летают в области гнездования
обычно значительно медленнее, чем на пролете, если во-
обще эти скорости можно сравнивать.
Как ни мала обычно скорость перелета птиц, вернее
какой малой она нам ни кажется, она вполне достаточна
для того, чтобы некоторые виды за несколько дней и но-
чей достигли мест зимовок. Больше того, с такой ско-
ростью при условии попутного ветра (как, например, при
перелете чибисами океана) многие перелетные птицы в
течение нескольких дней или ночей могли бы долететь в
тропики. Однако птицы не могут сохранить указанную
скорость пролета дольше, чем на несколько часов; они
почти никогда не летят несколько дней или ночей подряд;
как правило, их полет прерывается для короткого отдыха
или для более длительных остановок; последние придают
перелету в целом характер неторопливой «прогулки». Так
возникают длительные миграции.
При рассмотрении точно установленных кольцеванием
средних скоростей дневного или ночного перелета отдель-
ных видов всегда нужно иметь в виду, что они не характе-
ризуют способность к полету и развиваемую на пролете
скорость, а указывают лишь на длительность пролета и
расстояние между местами кольцевания и находок околь-
цованных птиц в пересчете на один день. Многочисленные
находки окольцованных птиц доказывают, что птицы бы-
стро пролетают большую часть пути, а остальное время
используют для отдыха в богатых кормом местах. Этот
вид пролета встречается наиболее часто. Значительно
реже наблюдается равномерное распределение нагрузки
и отдыха.
У птиц, летящих на большое расстояние, средний днев-
ной путь равен примерно 150—200 км, в то время как
летящие не так далеко не покрывают за это же время и
100 км. С этими данными согласуется продолжительность
Результаты изучения перелетов птиц
97
перелета в 2—3 или 3—4 мес. многих видов, которые зи-
муют в Тропической и Южной Африке. Так, например,
аист, обычно отлетающий из Германии в конце августа,
достигает зимовок в Южной Африке только в конце но-
ября или в декабре. Эти же сроки относятся и к жулану.
Ласточки мигрируют быстрее — с сентября до начала но-
ября. Сколь, однако, велики в данном случае индивиду-
альные различия, можно видеть на примере 3 окольцован-
ных горихвосток-лысушек, одна из которых покрывала
ежедневно 167 км, другая — 61 км и третья — лишь 44 км,
причем эти числа уменьшаются по мере увеличения отрезка
времени, для которого их вычисляют (6, 30 и 47 дней).
Основываясь на этих результатах, можно сделать вывод,
что дневная скорость более всего соответствует истинной
скорости перелета в том случае, когда она вычисляется на
основе общих показателей за короткий срок. Этот вывод
лучше всего доказывают следующие примеры скорости про-
летов отдельных птиц: аист покрыл за 2 дня 610 км, слав-
ка-черноголовка за 10 дней — 2200 км, лысуха за 7 дней—
1300 км, другая лысуха за 2 дня — 525 км, кряква за
5 дней— 1600 км. Этим данным можно противопоставить
дневную скорость певчего дрозда — 40 км (вычислена за
56 дней перелета), зяблика — 17,4 км (вычислена за 23дня
перелета) и ястреба-перепелятника — 12,5 км (вычислена
за 30 дней перелета). Эти данные сравнимы с приведен-
ными выше сведениями о горихвостках, на средние скоро-
сти которых при увеличении продолжительности пролета
сильно влияют длительные остановки для отдыха.
При оценке дневного пути и скорости пролета нельзя
упускать из виду и другой важный фактор: любые цифро-
вые данные можно вычислить только для идеального пути
перелета, т. е. для прямой линии, соединяющей места
кольцевания и находки окольцованной птицы. В действи-
тельности пролетный путь всегда больше, отклонения от
.прямой часто весьма значительны и выполненная работа
и скорость значительно выше вычисленных. Эти ошибки
практически невозможно исключить, и поэтому их необхо-
димо учитывать, особенно при очень длительных пере-
летах.
Кроме того, следует обращать внимание и на то, когда
были получены эти данные. Дело в том, что при весеннем
7-176
98
Глава Ilf
перелете показатели во многих случаях значительно выше,
чем при осеннем. В единичных случаях можно было' бы с
уверенностью доказать, что весенний перелет проходит
вдвое быстрее осеннего, например у аиста, американского
веретенника и жулана.
Штреземан (1944) точно установил, что весной про-
лет жулана длится приблизительно 60 дней, а осенью —
около 100 дней. В среднем эти птицы пролетают около
200 км в сутки. Однако они летят только ночью в течение
10 час. со скоростью 50 км/час. После такого пролета они
всегда отдыхают, так что расстояние 1000 км покрывается
ими за 5 суток: миграция — 2 ночи, сои — 3 ночи, кормеж-
ка — 5 дней.
Еще несколько слов о максимальных скоростях и дли-
тельности пролета, характеризующих возможности пере-
летных птиц: камнешарка, небольшая прибрежная птица,
окольцованная на Гельголанде, была найдена через
25 час. в Северной Франции, на 820 км южнее. Многочис-
ленные мелкие певчие птицы регулярно перелетают за
12 — 15 час. Мексиканский залив шириной 750 — 1000 км.
Согласно Моро (1938), некоторые небольшие сокола
/Falco concolor и F. amurensis), а также азиатские щурки
(Merops ' persicus и М. apiaster), зимующие на побе*
режье Южной Африки, также пролетают не меньше
3000 км над морем. Гавайские острова служат местом
зимовки для ряда северных куликов, которые, мигрируя
от Алеутских островов и Аляски, где расположены их гнез-
довья, вынуждены пролетать 3300 км над открытым мо-
рем. Золотистой ржанке, особенно сильному летуну, для
покрытия этого расстояния при скорости около 90 км/час
потребовалось бы приблизительно 35 час. Более высокие
скорости отмечены у другого вида ржанок, пролетающих
от Новой Шотландии до северной оконечности Южной Аме-
рики 3600 км над морем. Почти невероятным кажется
пролет одного из гнездяпщхся в Японии бекасов, который
зимует в Восточной Австралии и должен покрыть почти
5000 км, чтобы достигнуть зимовок. В пути он, вероятно,
совсем не отдыхает, так как в других местах его никогда
не отмечали.
К- пролету над водными пространствами можно при-
равнять пролет над большими пустынями. Такой пролет
Результаты изучения перелетов птиц
99
также, несомненно', проходит без перерыва, например про-
лет над Западной Сахарой мелких певчих птиц, трясогу-
зок и коньков, требующий 30—40 час. непрерывной рабо-
ты, если скорость их пролета считать равной приблизи-
тельно 50 км/час.
( Высота пролета
При обзоре направлений пролета уже указывалось, что
перелеты над высокими горными хребтами встречаются не
так редко, как можно было' бы ожидать на основании от-
клонений в перелете многочисленных видов на границах
больших возвышенностей. Кольцеванием и наблюдениями
удалось во многих случаях точно доказать наличие регу-
лярного, хотя и не очень интенсивного пролета над Аль-
пами, Кавказом и даже над мощными горными цепями
Гималаев. Следует особенно подчеркнуть, что птицы дале-
ко не всегда предпочитают долины и перевалы, а проле-
тают и над горными хребтами (например, в Тянь-Шане),
на высоте 6000 м. В сравнении с ними высоты, преодоле-
ваемые в Альпах, кажутся незначительными
Гейр впервые указал на необходимость различать от-
носительную и абсолютную высоту пролета. • Поэтому
очень важно, из какой точки наблюдатель ее регистрирует.
В дальнейшем речь будет идти большей частью об относи-
тельной высоте, т. е. о расстоянии от поверхности земли,
независимо от того, будет ли это берег моря, холмистая или
гористая местности. Во всяком случае, абсолютная высота
сама по себе как будто не оказывает на высоту пролета
птиц существенного влияния. Одни и те же виды птиц ле-
тят на равнине на такой же высоте, как и в горах, если
условия ветра и погоды существенно не различаются.
Долгое время считали, что перелеты птиц проходят
преимущественно на большой высоте и поэтому мало, до-
ступны человеческому глазу. Предполагали, что на боль^
шом расстоянии от поверхности земли полет облегчают
воздушные течения, а также лучшая ориентация. По дан-
ным Гэтке для Гельголанда, пролет обычно проходит на
1 Наибольшей высоты 9500 м, на которую когда-либо поднима-
лись птицы, достигла стая гусей над Эверестом. Высоту удалось
точно определить, стая гусей была сфотографирована с самолета
(Гаррисон, 1931).
100
Глава III
высоте приблизительно 2000 м, а у некоторых видов —
даже 3000 м и выше (у грачей на высоте приблизительно
4500 л!). Против этих взглядов в начале XX в. выступил
Луканус, Он поставил опыты о пределах видимости для
человека различных птиц, поднимая на аэростатах наби-
тые чучела с распростертыми крыльями. Оказалось, что на
высоте 800 м грачи имели вид точек, а на высоте 1000 м
они исчезали. Для перепелятников эти высоты составляли
соответственно 640 и 850 м, а такие крупные птицы, как
канюк и бородач, на высоте 1500—2000 м были еле
заметны.
Предположительная оценка высоты пролета обычно
ошибочна и дает преувеличенные результаты, так как для
сравнения нет никаких вспомогательных средств, анало-
гичных тем, которые мы имеем на земле. Основной источ-
ник ошибок заключается в различной прозрачности возду-
ха при безоблачном небе, небольшой и сплошной облач-
ности. В настоящее время точные данные о высоте
пролета птиц можно получить при помощи совершенных
приборов, созданных техникой для военных целей. В тех
случаях, когда необходимо доказать наличие пролетных
птиц на больших высотах, могут оказаться полезными
наблюдения летчиков и воздухоплавателей. Такие наблю-
дения собирали Вейгольд и Луканус. В последние годы к
ним прибавился еще ряд наблюдений, проведенных во
время полетов на планерах, и некоторые другие данные
(до сих пор не опубликованные, но уже общеизвестные
«тайны»). При помощи современных авиационных изме-
рительных приборов во время второй мировой войны ча-
сто удавалось установить перелеты птиц на больших вы-
сотах, например вяхиря на высоте 2500 м. Подводя итог,
можно, однако, утверждать, что пролеты птиц на высоте
более 1000 л сравнительно редки. В норме пролеты прохо-
дят на высоте нескольких сот метров, а часто, особенно у
мелких птиц, ниже 100 м. При благоприятной погоде и хо-
рошей видимости и не слишком сильном ветре птицы летят
значительно выше, чем при низкой облачности, дожде, ту-
мане или более сильном встречном ветре. Чем сильнее ве-
тер, тем ниже летят птицы, используя каждую возвышен-
ность, опушки лесов и долины рек, где сила ветра несколь-
ко слабее. Над морем птицы также летят большей частью
Результаты изучения перелетов птиц 101
над поверхностью воды. Они по возможности избегают ту-
манов и полетов в облаках. Над сплошным слоем облаков
редко можно встретить пролетных птиц, но иногда над
густой облачк>остью летят крупные птицы, например гуси
и журавли. Необходимо, однако, упомянуть о недавнем
сообщении Липпенса (1943), который у побережья Бель-
гии над двумя слоями облаков на высоте 150 и 500 м, где
было чистое небо, наблюдал оживленный перелет ржанок,
цапель, бакланов, гусей, скворцов, ворон, дроздов и вьюр-
ков. Но вообще птицы, безусловно, стремятся при проле-
те не терять из вида землю.
Предположения о том, что высота облегчает птицам
ориентацию, не подтвердились. Воздух редко бывает на-
столько прозрачен, чтобы птицы, находящиеся на высоте
более 1000 м, могли воспользоваться расширением поля
зрения; их-зрительные возможности далеко не безгранич-
ны. В этой области наши знания еще недостаточны, и
можно только предположить, что зрению птиц благодаря
особенностям строения их глаза и, в частности, сетчатки
туманы мешают меньше, чем зрению человека. В этой свя-
зи имеют значение сравнительные данные Триба (1939),
который обратил внимание на включенные в сетчатку
птиц желтые и красные маслянистые шарики, функция
которых до сих пор еще недостаточно выяснена. Триб
указывал на улучшение видимости на дальние
расстояния при закате солнца, а также на хорошую види-
мость красных сигнальных огней в туманную погоду. Это
явление объясняется тем, что длинноволновые желтые и
красные лучи лучше проникают через туманную атмосфе-
ру, чем коротковолновые зеленые, синие и фиолетовые.
При этом безразлично, рассматривается ли местность при
красноватом свете или же так, чтобы желтые и красные лу-
чи оказывались особенно действенными. Так, например,
поступает фотограф, который при съемке ландшафта с за-
туманенной далью экспонирует светочувствительную пла-
стинку через оранжевый светофильтр. Если исключить все
лучи, кроме наиболее длинных в инфракрасной части
спектра, то можно фотографировать даже на расстоянии
сотен километров. Подобное действие оказывают и крас-
но-желтые шарики в глазах птиц. Таким образом, перелет-
ные птицы могут даже при туманной погоде из Сицилии
102
Глава III
видеть берега Африки. Тем не менее создается впечатле-
ние, что птицы ориентируются главным образом по осо-
бым признакам ландшафта, а не по общим очертаниям по-
верхности земли. Если бы это было иначе, то некоторые
острова не играли бы роли направляющих линий и направ-
ляющих пунктов, исчезли бы массовые пролетные пути, а
многочисленные обходные пути сократились бы. Однако
все указанные условия существуют и оказывают влияние
на перелеты птиц, являясь доказательством того, что пе-
релеты проходят в среднем на небольшой высоте. Часто
птицам достаточно подняться лишь немного выше, чтобы
найти ориентиры, которые, казалось бы, делают излишним
следование по направляющим линиям. Дункер (1905) со-
ставил сводку дальности видимости (вне зависимости от
атмосферных условий) с различных высот. На основании
формулы г = У 2Rk = 113 V h получены следующие
цифры:
Высота, м Поле зрения, км
100 35,7
200 50,5
300 61,8
400 71,4
500 79,8
750 97,6
1000 113,0
2000 159,7
Согласно этим данным, птицы, находящиеся, например,
на высоте всего 100—200 м над Гельголандом, при бла-
гоприятной погоде увидели бы как на востоке, так и на
юге берега материка и не были бы вынуждены садиться
на остров для отдыха. То же самое относится к Курской
косе, к берегу Курского залива и ко многим другим мест-
ностям, значение которых в качестве направляющих ли-
ний при полете на больших высотах было бы уменьшено.
Перелет как физиологическая проблема
Как было установлено, проблема полета птиц на боль-
шой высоте требует более близкого рассмотрения. Прове-
денные ранее опыты относительно способности некоторых
Результаты изучения перелетов птиц 103
видов мелких птиц лететь на большой высоте кажутся бес-
полезными. Их результаты весьма незначительны, так как
изучение реакции организма на уменьшение давления
воздуха и понижение температуры возможно только в
том случае, если животное находится в состоянии покоя,
а это исключает сравнение с естественными условиями.
Напротив, при длительных пролетах птиц мы должны
определять физиологические основы усиленной мышечной
работы и ее источники. Это относится главным образом к
таким видам, которые совершают тысячекилометровые пе-
релеты над морем или в отношении которых удалось дока-
зать, что они летят непрерывно в течение 12—15 час. и
даже дольше. При 35-часовом пролете от Алеутских ДО' Га-
вайских островов американская золотистая ржанка долж-
на при двух ударах крыльев в секунду поднимать и опу-
скать их 252 000 раз, совершая при этом огромную мы-
шечную работу. Пролетный путь уже упоминавшейся
полярной крачки, очевидно, самый продолжительный. Она
следует из наиболее северной части своей гнездовой обла-
сти, расположенной всего в 7° от Северного полюса, через
Атлантический океан к берегам Европы и Африки и даль-
ше к месту зимовок в Антарктике. Эти 17 000 км птица
должна покрыть в течение приблизительно 10 недель;
следовательно, она должна ежедневно пролетать 225 км.
Другие примеры продолжительных перелетов от 10 000 до
15 000 км у куликов также уже упоминались.
Когда были получены точные сведения о длительных
перелетах, физиологи попытались согласовать их с дан-
ными об изменении обмена веществ. Однако до сих пор ис-
черпывающих результатов еще нет, и многие вопросы
остались открытыми. Некоторые ученые считали, что не-
прерывный полет на тысячи километров и продолжитель-
ностью более 15—20 час. физиологически невозможен.
Другие, напротив, пытались доказать, что необходимая для
пролета энергия лишь незначительно превышает энергию,
затрачиваемую птицей при ежедневных полетах в период
размножения. Никольсон приводит несколько примеров:
лазоревка, которая прилетает к гнезду только 35 раз в
час, отлетая в поисках пищи на 100 м, покрывает 100 км
в день. Ласточки или стрижи, весь день летающие со ско-
ростью более 60—80 км/час, несомненно покрывают около
104
Глава III
1000 км. По мнению этого автора, энергия, необходимая
для миграции, соответствует энергии, затрачиваемой в по-
вседневной жизни, но только направлена на определенную
цель.
К некоторым видам особенно сильных летунов эти рас-
суждения, возможно, вполне подходят, но общего значе-
ния они, очевидно, иметь не могут, ибо физиологическая
перестройка организма многих и даже большинства птиц
Рис. 35. Зависимость между весом тела, количеством пищи и «пе-
релетным беспокойством» у зарянки Erithacus rubecuia L. (Из Мер-
келя, 1940.)
------вес тела;--------количество пищи.
незадолго до перелета и во время миграции указывает на
то, что организм рассчитывает на совершенно исключитель-
ную нагрузку, в связи с чем изменяется его обмен веществ.
Наиболее наглядно это проявляется в сильном накоплении
жира, начинающемся вскоре после летней линьки и закан-
чивающемся за несколько дней или недель до начала
осеннего перелета. Подобное явление наблюдается и к на-
чалу весеннего перелета. В общем птицы становятся тем
более упитанными, чем длительнее предстоящая миграция
и чем напряженнее она будет проходить. (В отношении
крупных птиц данных еще нет.)
Это быстрое накопление жира до и в период перелета
происходит за счет усиленного приема пищи. Однако од-
Результаты изучения перелетов птиц
105
кого этого было бы недостаточно — питательные вещества
так же быстро расходовались бы. Физиологическая пере-
стройка организма в сторону замедления обмена энерге-
тических ресурсов и образования запасов, прежде всего —
накопления жира, создает необходимые источники энер-
гии. Если запасы жира после длительного полета израсхо-
дованы, птицы нё в состоянии лететь дальше. Они остают-
ся на богатых кормом местах до тех пор (иногда всего не-
сколько дней), пока энергетические запасы вновь не будут
пополнены.
Известно, что домашнего гуся особенно легко откор-
мить осенью. Он является потомком совершающего даль-
ние перелеты дикого серого гуся, у которого в периоды
перелетов наблюдается склонность к накоплению жира.
Став домашней птицей, гусь сохранил эту бесполезную для
него физиологическую особенность перелетной птицы.
Еще несколько слов о зависимости между перелетом и
сменой оперения у птиц. Оба процесса одинаково глубоко
влияют на жизнь птицы, оба они в известной мере зави-
сят друг от друга — один следует за другим,— о чем мы
подробнее скажем ниже. Линька требует большой затраты
энергии и поглощает все ресурсы организма. Это делает
понятным, почему большинство птиц, особенно мелкие ви-
ды, не приступает к миграции во время линьки. Более
крупные птицы в этом отношении менее чувствительны.
Певчие птицы меняют маховые и рулевые перья до и после
перелета. У них, помимо физиологических причин, решаю-
щее значение имеют летно-механические факторы (изъяны
в несущих поверхностях). Рано отлетающие виды линяют
обычно только на зимовках, поздно отлетающие — в гнез-
довой области. Часто мелкие перья сменяются на родине,
крупные — только на зимовках (например, у городской
ласточки, камышовок, пересмешек и др.). У крупных птиц
эта зависимость между сменой оперения и перелетом в
общем выражена менее четко, т. е. линька проходит мед-
леннее и с большими промежутками.
Перелет и погода
Мы уже несколько раз разбирали этот вопрос, устанав-
ливая различия между птицами, перелеты которых совер-
106
Глава III
шаются в большей зависимости от погоды («Wettervogeb)
или же от внутренних стимулов («Instinktvogel»), при об-
суждении периодов перелета, особенно времени прилета
различных видов весной, и, наконец, в связи со скоростью
перелетов и их высотой. Поэтому отпадает необходимость
останавливаться на всех подробностях влияния погоды на
перелет. Здесь будет приведен лишь обзор тех факторов
погоды, влияние которых на перелет можно считать до-
казанным.
Такие факторы, несомненно, существуют, но издавна их
либо недооценивали, либо преувеличивали главным обра-
зом из-за того, что авторы, исходя из отдельных наблюде-
ний, делали слишком общие выводы. Они учитывали лишь
немногие виды птиц или лишь определенные ландшафты и
исходили из очень коротких отрезков времени, ограничи-
ваясь только одним из многих метеорологических факто-
ров, которые в момент их наблюдения, возможно, были
определяющими для перелета. Неизбежным следствием
всех этих недостатков является отсутствие до сих пор
единого мнения О' зависимости между миграциями и
погодой. Можно с уверенностью сказать, что едва ли в ка-
кой-нибудь другой области науки о перелетах птиц суще-
ствуют столь разноречивые взгляды.
При характеристике птиц, перелет которых существенно
зависит от погоды («Wettervogeb), мы принимали, что со-
стояние погоды оказывает непосредственное влияние не
только на ход перелета, но и .на его «пусковой механизм»,
однако подчеркивали, что это имеет значение лишь для
некоторых видов птиц. К таким видам относятся, напри-
мер, скворец, полевой жаворонок, вальдшнеп, чибис, бе-
кас и несколько видов дроздов. Зимой или ранней весной
при приближении волны холода эти птицы в большом чис-
ле отлетают по направлению к более теплым массам воз-
духа. Явление «возвратного перелета» («Umkehrzug»)
можно наблюдать довольно регулярно весной в Германии.
Внезапное вторжение холодных масс воздуха во вре-
мя перелета может, однако, вызвать подобные отступле-
ния или отклонения и у птиц, перелет которых зависит
главным образом от внутренних стимулов. Согласно Греб-
бельсу, птицы в это время находятся в так называемом
«критическом» периоде, когда у них под влиянием внут-
Результаты изучения перелетов птиц
107
ренних причин появляется предрасположение к перелету,
но реализуется оно лишь под влиянием внешних факторов.
Другими словами, для того чтобы вызвать периодические
миграции, перелетный инстинкт должен подкрепляться
определенным, действующим извне раздражением. Таким
раздражением часто является понижение температуры.
Когда же «критический период» закончился, миграции
прекратились и перелетный инстинкт угас, перелетное на-
строение не может быть вызвано вновь, и поэтому птицы
не реагируют на изменение погоды. Это объясняет те не-
редкие случаи, когда поздней весной при вторжении масс
холодного воздуха в определенной местности погибает
почти все поголовье гнездящихся птиц этой группы, т. е.
типичных перелетных птиц, которые рано вернулись на ро-
дину с зимовок (например, пеночка-кузнечик, пеночка-
весничка, горихвостка-чернушка, горихвостка-лысушка,
черноголовая славка и черноголовый чекан). Поголовье
этих птиц затем пополняется за счет птиц соседних обла-
стей. Но поскольку такие, вызванные погодой, катастро-
фы происходят большей частью только на ограниченных
пространствах, общая потеря редко бывает очень тяжелой.
Иногда резкая перемена погоды наступает так внезап-
но, что перелетные птицы уже не могут избежать ее. Тогда
град, дождь и оледенение оказывают губительное действие
и приводят, особенно при пролетах птиц над морем, к
огромным потерям. После таких стихийных бедствий бе-
рег моря иногда усыпан тысячами трупов птиц.
В противоположность этой группе, у птиц, перелет ко-
торых определяется в основном условиями погоды, кри-
тический период охватывает не только период перелета, но
значительно более длительный отрезок времени: от начала
осеннего и до конца весеннего перелета, так что и в се-
редине зимы перелетный инстинкт не угасает и может
быть возбужден температурными воздействиями. Таким
образом возникают наблюдавшиеся Дростом на Гельго-
ланде зимние миграции, носящие характер типичного
перелета. Такие «зимние перелеты» можно установить
и во всех других местностях, где наблюдаются резкие,
внезапные смены температур.
Дрост (1934) объяснил направления перелетов осенью
у многих видов различиями в распределении тепла. Рано
108
Глава III
улетающие птицы, например чернолобый сорокопут, чече-
вица, малая мухоловка, пеночка-пересмешка, иволга, жу-
лан, кукушка и другие, покидающие гнездовья в августе
или немного позднее, почти без исключения летят на зи-
мовки в южном и юго-восточном направлении. Более позд-
но улетающие скворцы, вороны, вьюрки, жаворонки и
дрозды, напротив, всегда стремятся на юго-запад или за-
пад. Соответственно изотермы проходят в августе с запада
на восток или с юга-запада на северо-восток, а в сентяб-
ре-октябре — с северо-запада на юго-восток. Во время
осеннего перелета распределение тепла в Европе вплоть
до Средиземноморской области происходит, следователь-
но, с востока на запад. В соответствии с этим сдвигаются
на запад и направления пролетов, так как птицы стремят-
ся лететь в сторону большего тепла, т. е. в такие области,
из которых, как им по опыту известно, обычно распростра-
няются теплые воздушные течения.
В связи с этим укажем еще и на то, о чем при анализе
времени перелетов упоминалось лишь вкратце, а именно,
что у некоторых видов перелетных птиц линии, соединяю-
щие места одновременного прилета весной (изопиптезы),
частично параллельны изотермам.
Неоднократно предполагалось, что изменения атмо-
сферного давления оказывают прямое влияние на ход пере-
летов, но доказать это удавалось лишь в редких случаях.
То же относится и к влажности воздуха. Туманы, несо-
мненно, задерживают и меняют направление пролета, точ-
но так же, как низкая облачность смещает его по верти-
кали. Слабый дождь, напротив, не мешает перелету, а в
соединении с попутным ветром часто даже содействует
ему.
Несколько подробнее необходимо остановиться на
влиянии ветра на перелеты.
Господствовавшее ранее мнение, что пролеты птиц
возможны лишь при легком встречном ветре, в последнее
десятилетие было отвергнуто. Доказано, что пролет может
совершаться при любом направлении ветра. Наблюдения
о пролетах при встречном ветре опубликовали Гейр
(1931), Дрост (1931) и Кох (1934). Эти наблюдения под-
твердили, что птицы иногда даже меняют направление пе-
релета, чтобы иметь возможность лететь против ветра
Результаты изучения перелетов птиц 109
(возвратный перелет). Проводя наблюдения на побережье
Северного моря, Кох установил, что для зяблика, вьюрка
и полевого воробья характерен выраженный полет против
ветра (положительный анемотаксис). К таким птицам от-
носятся также полевой и лесной жаворонки, луговой конек,
желтая трясогузка, скворец, дрозд-деряба и белобровик.
У других видов особи ведут себя различно, так что обще-
го правила вывести нельзя. Названные виды могут совер-
шать перелет и при боковом ветре, а иногда и при попут-
ном ветре, когда этого требуют обстоятельства. Наблюда-
ются и индивидуальные отклонения, но все-таки они всегда
предпочитают встречный ветер. То, что сила ветра в за-
висимости от направления может как препятствовать, так
и способствовать перелету, уже отмечалось в другом месте.
Совершенно прекратить перелет, вероятно, не может даже
сильный шторм.
Здесь следует указать на явление так называемого
«перелетного» или «голодного сна» («Zug- oder Hunger -
schlaf»), которое вкратце упоминалось при обсуждении
исторических основ изучения перелетов птиц. Речь идет о
наблюдениях, которые раньше неоднократно ставились под
сомнение, но в последнее время подтвердились многочис-
ленными данными, о похожем на зимнюю спячку состоя-
нии, наблюдаемом у ласточек и стрижей (реже у пищух и
других птиц). При продолжительной неблагоприятной по-
годе указанные виды прерывают на несколько дней свой
осенний перелет, собираются в большом числе в сухих
теплых местах, подобно пчелиному рою, и впадают в со-
стояние летаргии, которое можно сравнить только с зим-
ней спячкой различных мелких млекопитающих, напри-
мер летучих мышей, орешниковых и садовых сонь и т. д.
В этот период температура тела и обмен веществ сильно
понижаются, активность сводится к минимуму, потребно-
сти в пище почти нет, и трудное время переносится легче.
В настоящее время еще нет объяснения физиологических
основ подобного приспособления у видов с очень интен-
сивным обменом веществ. В особенно большом масштабе
такой «перелетный сон» наблюдался осенью 1931 г. в
предгорьях Альп, где многие тысячи ласточек укрывались
от непогоды. Проведенные организациями по охране жи-
вотных спасательные мероприятия в форме подкармлива-
110
Глава III
ния и переброски спящих птиц на самолетах в Италию и
Грецию имели, вероятно, скорее теоретическое, чем прак-
тическое значение. Позже Менниг (1940) сообщил о «го-
лодном сне» черных стрижей в башне замка в Каннах и
указал также на то, что у Лазурного Берега зимуют гор-
ные ласточки, которые при хорошей погоде летят у бе-
рега и в горах на высоте до 2000 м, но при плохой погоде
исчезают и, вероятно, переживают холодные дни в пеще-
рах в состоянии сна. В известной «Книге Сан Мишеля»
Мунте это своеобразное явление нашло даже литератур-
ное отражение. В Швейцарии орнитологи недавно вторич-
но наблюдали «голодный сон» у черных и белобрюхих
стрижей: птицы собирались группами в 100—200 особей и
пытались переждать тяжелое время, что им, очевидно,
удалось только отчасти (Буркгардт, 1948).
Лишь редко отклонения в направлениях перелетов мож-
но объяснить воздействием погоды, как в данном случае.
Большей частью, например при ранее описанном массовом
пролете над Гельголандом, возникают затруднения из-за
незнания состояния погоды в областях отлета птиц. Осо-
бенности поведения при пролете только отражают послед-
ствия воздействия факторов, господствующих в месте отле-
та. Пока мы не будем точно знать метеорологических усло-
вий на обширных пространствах, всякое объяснение зави-
симости перелетов от погоды остается несовершенным.
В заключение еще несколько слов о влиянии различ-
ной интенсивности освещения на перелеты. Этот вопрос
также уже затрагивался, когда говорилось о влиянии сиг-
нальных огней и освещенных городов на пролетающих
ночью птиц. Как ни распространена привычка лететь
ночью и как ни многочисленны пролетающие ночью птицы;
всё же ни одна птица не совершает перелет в абсолютной
темноте. Дрост установил, что у многих видов пробуждение
отдыхающих перелетных птиц во время вечерних сумерек
зависит от захода солнца. Птицы, летящие ночью, отправ-
ляются в полет лишь при определенной интенсивности
освещения (измеряёмой фотометром): вид за видом, пти-
ца за птицей одного и того же вида с меньшими или боль-
шими промежутками. По другим наблюдениям, это отно-
сится также к птицам, летящим днем; они начинают ми-
грации еще до рассвета (например, журавли).
Результаты изучения перелетов птиц
111
Ориентация при перелете
При изложении методов изучения перелетов птиц по-
дробно разбирались общие вопросы этой проблемы, теоре-
тические возможности ее решения и опыты в этом направ-
лении. Мы будем неоднократно ссылаться на них, когда пе-
рейдем к более близкому рассмотрению путей, которые
привели к этим общим соображениям, и познакомимся не-
посредственно с проведенными в этом направлении экспе-
риментами.
Начнем с ранее широко распространенного мнения о
том, что молодым птицам перелетные пути показывают
старые птицы. Это относится к тем видам, которые мигри-
руют семьями или еще более крупными сообществами, глав-
ным образом ко многим крупным птицам, например ле-
бедям, аистам, журавлям и гусям. У последних молодые
птицы настолько нуждаются в «руководстве» старых птиц,
что не решаются отлетать без них. Ряд опытов, поставлен-
ных Тинеманом в 1926—1931 гг. и Шюцем в 1933 г., ука-
зывает на то, что молодые аисты сами могут найти пра-
вильное направление к местам зимовок. Выпущенные на
волю после отлета старых птиц молодые аисты летели в
основном правильно: из б. Восточной Пруссии на юго-во-
сток. Правда, отклонения от нормального пролетного пути
встречались при этом чаще, чем у других окольцованных
аистов. Таким образом, руководство над молодыми пти-
цами со стороны старых в данном случае, невидимому,
имеет значение лишь дополнительного фактора, уменьша-
ющего опасность сбиться с дороги и гарантирующего со-
блюдение строго ограниченных пролетных путей. Он опре-
деляет так называемую тонкую ориентацию, в то вр^мя
как знание направления перелета и общего «правильного»
поведения во время перелета (грубая ориентация), ве-
роятно, закреплено наследственностью.
Опыты с аистами были продолжены Шюцем на орни-
тологической станции Росситтен в 1934 г. с целью провер-
ки, постоянно ли «знание» птицами направления перелета
и в тех случаях, когда различные популяции ведут себя не-
одинаково, или оно изменяется в соответствии с условиями
внешней среды. Как указывалось выше, все без исключе-
ния восточногерманские аисты улетают в юго-восточном
112
Глава III
направлении, а западногерманские — в юго-западном.
Учитывая это, молодых аистов из б. Восточной Пруссии пе-
ревезли в Рейнскую область, где их вырастили и выпусти-
ли на волю, когда старые птицы уже отлетели. Большая
часть молодых аистов придерживалась юго-восточного на-
правления (меньшая часть отклонилась на юго-запад) и,
перелетев через Альпы, достигла долины р. По. Пови-
димому, наследственно закрепленное направление переле-
та сохраняется и в измененной обстановке, а влияние на-
следственности оказывается сильнее влияния ландшафта,
если о таковом вообще можно говорить.
В том же 1933/34 г. и позже орнитологическая стан-
ция Росситтен перевезла молодых аистов из б. Восточной
Пруссии, где они многочисленны, в Центральную и Запад-
ную Германию; там их содержали в условиях, по возмож-
ности близких к естественным. Это мероприятие проводи-
лось для решения следующих вопросов: возможности ак-
климатизации аистов, выяснения направления, в котором
произойдет отлет, и для проверки «приверженности» птиц
к гнездовой территории. При этом были получены резуль-
таты, противоречащие данным изложенного выше опыта с
аистами. Большинство выращенных в Центральной и За-
падной Германии аистов было обнаружено на пролете в
юго-западном направлении — во Франции. Можно пред-
положить, что, следуя примеру выросших на свободе
аистов, они при отлете направились по этому не свойствен-
ному для них пути «против воли». Факты возврата этих
аистов на родину (Южная Франция) и гнездования на
западе (только в одном случае) говорят в пользу пере-
селения этих птиц на запад.
Подобный же результат дала попытка выращивания в
Финляндии молодых крякв из Англии. Яйца этих птиц вы-
сиживались в Финляндии. Вылупившиеся особи акклима-
тизировались, полностью восприняли поведение финских
крякв, совершали длительные миграции и большей
частью возвращались следующей весной на свою но-
вую родину.
То же произошло и с сизыми чайками, когда их яйца
были привезены из Хиддензе в Росситтен и Силезию. Их
высиживали в колониях обыкновенные чайки, которые
также вскармливали птенцов. Впоследствии часть этих
Результаты изучения перелетов птиц
1'13
Рис. 36. Переселение английских крякв в Финляндию. (Из Великан-
гаса, 1933.)
На своей родине кряквы были оседлыми птицами. На новом месте они стали
перелетными птицами. На карте изображены дальние перелеты этих птиц уже
на первом году их жизни.
переселенных птиц вернулась на новую родину и даже
гнездилась там.
Таким образом, отношение птиц к перелету далеко не
неизменно, и они могут быстро акклиматизироваться в не-
известных для них местностях. К этому выводу мы пришли
и при ознакомлении с опытами по перевозке европейских
птиц за океан. Приспособление к новым условиям суще-
ствования может привести даже к неожиданному увеличе-
нию численности, расселению и вытеснению местных ви-
дов, как это отмечено у скворца в США и домового во-
робья во многих частях света.
Противоречащие результаты, полученные при описан-
114
Глава III
ных выше опытах с аистами, могут быть объяснены тем,
что ни юго-восточное, ни юго-западное направления, оче-
видно, наследственно у них не закреплены. Миграции мо-
лодых аистов в общем направлены к югу, и лишь в ре-
зультате влияния рельефа земной поверхности, экологиче-
ских условий и следования молодых птиц за старыми они
в дальнейшем летят на восток или на запад (Путциг, 1939).
При описании опытов с переселением аистов мы гово-
рили о возможном источнике ошибок, связанных с тем, что ♦
чуждые данной местности птицы как бы увлекаются про-
летающими стаями местных представителей данного
вида. Это затруднение, которое едва ли можно устра-
нить, не позволило получить точные результаты и в
опыте с переселением птиц, проведенном орнитологиче-
ской станцией Росситтен. В этом опыте 3000 скворцов
из Прибалтики были перевезены в Силезию и Саксонию.
Большая часть этих птиц, очевидно, под влиянием си-
лезских и саксонских скворцов отлетела на юго-запад,
в то время как прибалтийские скворцы обычно мигри-
руют преимущественно в западном направлении. Одна-
ко они перелетели за северные границы зимовок силез-
ских и саксонских скворцов и достигли своей прежней
области распространения (Кретциг и Шюц, 1936).
Объяснение ’ ориентации как результата традиции
(следование молодых особей за старыми птицами или
совместный перелет стаями) невозможно для птиц, у
которых различные по возрасту и полу особи мигрируют
отдельными группами, а также для многочисленных ви-
дов, совершающих перелеты поодиночке и ночью. Моло-
дая кукушка, которую воспитывали приемные родители,
принадлежащие к другому виду, совершает перелет одна
и тем не менее находит правильное направление к месту
зимовки. Жуланы достигают места зимовок также само-
стоятельно очень сложными и часто окольными путя-
ми. Не менее удивителен перелет молодых новозеланд-
ских кукушек Chalcites lucidus, которые значительно
позже старых птиц направляются к своим зимовкам на
Соломоновых островах и островах Бисмарка, пролетая
при этом над восточноавстралийским побережьем, т. е. ле-
тят сначала в северо-западном, а затем в северо-восточ-
ном направлении. Известно, что некоторые виды (на-
Результаты изучения перелетов птиц
115
пример, пеночки, гренландская каменка, галстушник)
осенью отлетают в направлении тех мест, откуда они
когда-то вселились в современную гнездовую область.
(Об этом уже упоминалось при анализе направлений пе-
релетов.) Как находит нужное направление подавляю-
щее большинство перелетных птиц — мы не знаем и по-
этому вынуждены пользоваться такими абстрактными
понятиями, как «чувство направления» и «восприятие
географического положения», о которых речь шла выше.
Для исследования этих загадочных для человека
способностей птиц пытались использовать данные опы-
тов с почтовыми голубями. Как известно, эти птицы на-
ходят обратный путь к своей голубятне даже из очень
отдаленных местностей. Способность этих птиц находить
обратный путь можно развить при помощи дрессировки
и тренировки, учебных полетов и тщательного отбора.
Возвращение голубей с небольших расстояний можно
очень просто объяснить зрительной ориентацией. Но не
так легко понять, как голуби находят обратный путь с
больших расстояний, превышающих иногда несколько
сот километров; это трудно объяснить даже огромной
зрительной памятью почтового голубя. Поэтому голубе-
воды приписывали птицам особое «чувство ориентации»,
позволяющее им найти голубятню. В качестве раздра-
жителей, воздействующих на это чувство ориентации,
они называли самые различные факторы — влияние
магнитного поля Земли, электрические волны, космиче-
ские лучи, метеорологические условия — или же пред-
полагали наличие врожденного чувства направления.
Однако все эти доводы были опровергнуты точной про-
веркой при помощи физических или биологических ме-
тодов исследования (насколько подобная проверка вооб-
ще была возможна).
Прошло довольно много времени, прежде чем О. и
К. Хейнротам (1941) удалось точно доказать, что спо-
собность голубей находить дорогу домой основана ис-
ключительно на зрении. С присущей им смелостью в
полете птицы долго кружатся над незнакомой местно-
стью и ищут до тех пор, пока вновь не попадут в такие
места, над которыми они когда-то пролетали. Хорошая
память облегчает им ориентацию. Это накопление и за-
9-176
116
Глава III
печатление «зрительных восприятий в памяти», сохра-
няющиеся в течение очень длительных промежутков
времени, определяют поразительные способности голу-
бей находить правильный обратный путь.
Таким образом, отпадает предположение о существо-
вании врожденного или развившегося в результате об-
учения особого чувства направления. Подводя итог сво-
им исследованиям, О. и К. Хейнроты отмечают, что осно-
ва поведения перелетных птиц совершенно отлична от
этого чувства направления у почтовых голубей. До них
это различие никогда так ясно не формулировалось.
Поэтому его следует особенно подчеркнуть теперь, когда
мы переходим к сравнению сведений, полученных при
изучении поведения голубей, с соответствующими данны-
ми о перелетных птицах
Уже давно известно, что поведение птиц в период
размножения подобно поведению почтовых голубей, т. е.
после насильственного удаления с гнезда они всегда
вновь возвращаются к нему даже с большого расстоя-
ния. Это свойство впервые использовал Лоос для провер-
ки способности птиц к ориентации. В дальнейшем его
примеру последовали многие другие орнитологи. Лоос
экспериментировал с ласточками и скворцами, Уотсон и
Лешли (1915) —с американскими крачками, которые
были переброшены на 800—1200 км от своих гнездовий
в Мексиканском заливе и, тем не менее, несколько дней
спустя вновь возвратились к ним. Подобные результаты
были получены Дирксеном (1932) для полярных и пе-
строносых крачек из района Халлиг Нордероог (из груп-
пы Северофризских островов). В это же время братья
Штиммельмайр установили, что варакушки и горихвостки-
лысушки, увезенные на несколько сот километров от гнез-
довых участков, возвращались через 2—3 недели. Вскоре
после этого Водзицкий и Войтусяк (1934) предприняли
подобные опыты с деревенскими и городскими ласточ-
ками. Одновременно Рюппель приступил к постановке
многочисленных опытов, продолжавшихся несколько
1 Согласно неопубликованным сообщениям, Крамер получил
недавно экспериментальные данные, противоречащие результатам
О. и К. Хейнротов и доказывающие возможность развития у поч-
товых голубей чувства направления в результате обучения
Рис. 37. Опыты Рюппеля по изучению ориентации у птиц.
Цифры — число выпущенных подопытных птиц по отношению к числу вновь найденных. (Из ван Оордта, 1934, по Рюппелю.)
118
Глава III
Рис. 38. Опыты Гриффина по перевозке северных качурок в откры-
тое море. (Из Ван Оордта, 1943, по Гриффину.)
лет. Существенно новым в этих опытах было большое
число подопытных птиц (преимущественно ласточки и
скворцы, но также и вертишейки, жуланы, тетеревятни-
ки, обыкновенные чайки, серые вороны и лысухи) и за-
воз их в самые различные места, в том числе и в на-
Результаты изучения перелетов птиц
119
правлениях, противоположных нормальному направле-
нию перелета данного вида (например, в опытах с жула-
нами, которых завозили на север, запад и юго-запад
вместо юго-востока). Кроме того, эти опыты производи-
лись с птицами, перелет которых в норме происходит
ночью; были учтены и другие специальные проблемы, на
которых мы остановимся особо. В результате в боль-
шинстве случаев было установлено, что птицы находят
обратный путь к гнезду; правда, это выражалось в раз-
личной степени, но вместе с тем не зависело от каких-
либо поддающихся учету внешних факторов, например
погоды, времени суток, длительности перевозок или
средств транспорта; необходимым условием являлось
лишь достаточно хорошее физическое состояние птицы.
Из других недавно проведенных опытов по перевозке
птиц назовем следующие: опыты Лэка и Локли (1938)
и Гриффина (1940) с морскими птицами, из которых
один буревестник (Puffinus puffinus) за 14 дней возвра-
тился из Венеции к гнезду, находящемуся в юго-западной
Англии. Эта птица, следуя вдоль берегов, покрыла, оче-
видно, 6000 км. Из числа Oceanodroma leucorrhoa, гнез-
дящихся в Новой Шотландии и выпущенных над откры-
тым морем в сотнях километров от суши, обратную до-
рогу нашли 75%. Водзицкий, Пухальский и Лихе (1939)
перевозили аистов на самолете из Львова в Палестину,
находящуюся на пути пролета этого вида. Даже с рас-
стояния, составляющего приблизительно 'А часть пути
их миграции на зимовку, куда они все равно должны
были бы лететь через 1—2 месяца, аисты вернулись за
12 дней обратно к гнездам. Шифферли (1942) отметил
возвращение в трехдневный срок белобрюхих стрижей,
перевезенных из Швейцарии в Лиссабон (1620 км). На-
конец, следовало бы упомянуть опыт Гриффина (1943)
по изучению ориентации у серебристых чаек и обыкно-
венных крачек, хотя его данные о возвращении много-
численных перевезенных птиц с большого расстояния
(1200 км) из неизвестных им местностей не представля-
ют чего-то принципиально нового.
Мейзе считал основой ориентации прилетающих в
гнездовую область перелетных птиц так называемые ки-
нестетические ощущения. Согласно его представлениям,
120
Глава 111
в памяти птиц должно фиксироваться направление поле-
та, а может быть, даже и все их движения во время
осеннего перелета. В таком случае весной птицам при-
шлось бы только повторить все движения в обратном
направлении, и, таким образом, летя как бы вдоль неви-
димой нити, они достигли бы родины. Исходя из этого
предположения, можно было бы легко объяснить нахож-
дение обратной дороги подопытными птицами даже в
тех случаях, когда они были завезены в местность, путь
которой противоположен направлению их перелета.
Чтобы исключить возможность всяких возражений,
Рюппель при перевозке непрерывно вращал подопытных
скворцов, а Клюйвер наркотизировал их, прежде чем
отправить в дорогу. Результат был в обоих случаях один
и тот же: птицы находили обратный путь так же хоро-
шо, как и контрольные. Гриффин (1940) также вращал
в дороге некоторых из перевозимых птиц, а других по-
мещал на короткое время в сильное магнитное поле. Эти
птицы, несмотря на такое воздействие, возвращались на-
зад так же уверенно и быстро, как и контрольные.
Мы слишком бы отвлеклись, если бы стали подробно
описывать все опыты по перемещению насиживающих
птиц. Поэтому мы ограничимся приведенными примера-
ми и вытекающими из них выводами. Следует отметить
общий для всех видов факт, что во многих случаях пти-
цы во время гнездования вновь находят обратную доро-
гу к гнездовому участку, даже когда их перевозят в не-
известные местности, лежащие вне области их переле-
тов. При этом исключается возможность зрительной
ориентации, так же как и кинестетического управления
полетом.
Так как эти опыты ограничивались периодом г:, заде-
вания, то невольно возникает предположение, что спо-
собность птиц к возвращению находится в тесной связи
с инстинктом размножения. Для проверки этого вопро-
са Дрост (1938) перевозил многочисленных ястребов-пе-
репелятников, пойманных во время осеннего пролета из
Скандинавии на острове Гельголанд, в Силезию и уста-
новил, что старые птицы направились по «правильному
пути» на свои зимовки, т. е. отклонились сильно на за-
пад, пока не достигли обычного района пролетов.
Рис. 39. Опыт по перевозке пролетающих над Гельголандом пере-
пелятников Accipiter nisus L. (Из Дроста, 1938.)
------ареал пролетающих над Гельголандом птиц; восточная граница
ареала окольцованных скандинавских птиц; молодые птицы, обнару-
женные в том же периоде перелета, + + + + взрослые птицы обнаруженные
в том__же периоде перелета; ••••молодые и взрослые птицы, обнаруженные
в более позднее время.
122
Глава III
Рис. 40. Перемещение серых ворон из Прибалтийской области гнездо-
вания в Южную Швецию (известный опыт Рюппеля).(Из Гриффина,
1949, по Рюппелю.)
Молодые перепелятники полетели в обычном направле-
нии и попали на новые места зимовок, к которым в по-
следующие поды приспособились и некоторые старые пе-
репелятники. Аналогичные результаты дал знаменитый
опыт Рюппеля (1942), в котором 900 серых ворон были
перевезены из Росситтена в Фленсбург. И в этом случае
произошло переселение птиц в новую гнездовую область,
так как после перевозки они придерживались обычного
направления перелета (рис. 40).
Чтобы исключить возражения о том, что способность
находить обратный путь свойственна только перелетным
птицам, Рюппель (1937, 1940) перевез тетеревятников
на большое расстояние (600 км), после чего птицы вер-
нулись если и не в самую гнездовую область, то при-
мерно в ту же местность. Когда же этих птиц удаляли
Результаты, изучения перелетов птиц 123
на небольшие расстояния (до 200 км), они, как прави-
ло, возвращались. Гете (1937) установил, что серебри-
стые чайки (которые, будучи кочующими птицами, не
улетают в каком-либо определенном направлении от
гнездовья) в подавляющем числе случаев находят обрат-
ный путь к месту гнездования. Хильпрехт (1935) пере-
возил зимой на значительные расстояния многочислен-
ных черных дроздов, зябликов, зеленушек, коноплянок и
больших синиц из района Магдебурга, где эти виды яв-
ляются большей частью оседлыми, и также очень часто
наблюдал их возвращение к прежнему месту обитания.
В опытах Крейца (1942) перевезенные зеленушки воз-
вращались зимой даже с расстояния более 790 км.
В 1939 г. Рюппель и Шифферли производили различные
испытания с обыкновенными чайками и лысухами, ко-
торых они перевозили с их берлинских или, соответ-
ственно, швейцарских мест зимовок, после чего птицы
вновь к ним возвращались (рис. 41).
Эти данные говорят о том, что способность птиц на-
ходить обратный путь не зависит от периода размноже-
ния и что зимой их приверженность к местообитанию
так же сильна, как и летом. Связана ли эта «верность»
с особенностями биологии питания, как склонны считать
Рюппель и Шифферли, или же в основе ее лежат более
общие причины — неизвестно. Примечательно, однако,
что перевозка птиц в период перелетов может привести
к смене одной гнездовой территории на другую, так же
как одного места зимовки на другое. Это относится
прежде всего к молодым птицам, которые в данном слу-
чае ведут себя несколько' иначе, чем старые. К такому
же выводу пришли Рюппель и Шейн (1941) в опытах с
молодыми выращенными в неволе скворцами, которые
в отличие от птиц, в течение года содержавшихся в
клетке, не возвращались на «родину» после перевозки
на большие расстояния. Таким образом, можно сделать
вывод, что у молодых птиц способность находить обрат-
ную дорогу еще не выражена в такой степени, как у
старых, и что для ее развития требуется известная
сноровка или опыт при перелете, которые невозможно
приобрести ни в течение первого года жизни, ни в
неволе.
124
Глава III
Рус. 41. Места зимовок средне- и североевропейских обыкно-
венных чаек (Lar us ridibundus) в Швейцарии. После перевозки птицы
возвращаются к прежним зимовкам. (Из Рюппеля, 1938.)
♦
В этой связи приведем некоторые данные относитель-
но общей верности птиц родной местности. Кольцевани-
ем удалось доказать, что подавляющее большинство пе-
релетных птиц возвращается к месту своего рождения.
Многочисленные виды птиц занимают даже то же самое
гнездо, что и в предыдущем году, или хотя бы то место,
где оно находилось. Так поступает, во всяком случае,
один из партнеров прежней насиживавшей пары, к ко-
торому затем часто вновь присоединяется и второй
Результаты изучения перелетов птиц 125
партнер. В общем можно сказать, что приблизительно в
80% случаев верность родине является правилом, осо-
бенно у старых птиц. Молодые птицы чаще расселяются
в более широких пределах гнездового ареала и нередко
впоследствии вновь сближаются. У форм, живущих ко-
лониями, привязанность к месту рождения резко выра-
жена уже на первом году жизни. Эти особенности в пове-
дении молодых и старых птиц вполне соответствуют тем
различиям, которые выявились в опытах по нахождению
птицами обратного пути.
Наблюдающаяся в норме привязанность к местности
не исключает, однако, возможности «добровольного» пе-
реселения в очень отдаленные районы. В таких случаях
мы говорим об «абмиграции» («Auswanderung»), кото-
рая чаще всего наблюдается у уток. Причина кроется в
свойственном этим птицам раннем образовании пар,
происходящем еще во время осеннего перелета или на
зимовке. Весной одна из птиц следует за своим партне-
ром (самец может последовать за самкой, и наоборот)
на его родину. При этом возможны переселения из
Англии в Германию, Финляндию или СССР и из Ислан-
дии в СССР.
Понятие постоянства местообитания включает и при-
верженность птиц к одному и тому же месту зимовки. Это
удалось установить для многих видов птиц при помощи
метода кольцевания. Подобная «верность» может выра-
зиться и в том, что одни и те же птицы в течение ряда
лет гнездятся не только в одной и той же местности, но
даже в одних и тех же определенных местах (водные
птицы — у прудов, чайки — в руслах рек, мелкие певчие
птицы — у мест кормления и в садах и даже у опреде-
ленных окон с кормушками). Сюда же относится и
отыскивание птицами одних и тех же мест отдыха, сбор-
ных пунктов и т. д.
Вернемся к обсуждению способности птиц находить
обратный путь. Опыты с перевозкой птиц дали ясные ре-
зультаты только у старых особей, которые в это время
высиживали, готовились к высиживанию или уже выве-
ли птенцов. Молодые птицы, очевидно, не смогли разо-
браться в изменившихся условиях, поэтому их способ-
ность к ориентации кажется менее развитой. Отсюда вы-
126
Глава III
текает возможность развития этой способности, хотя мы
и не знаем, чем она определяется.
Поэтому попрежнему остается загадочным, как птицы
впервые находят свои зимовки и обратный путь на ро-
дину. Не ясно также, чем руководствуются старые птицы
при своих повторных миграциях из гнездовой области и
обратно, особенно когда их перевозят в совершенно
чуждые им местности в направлении, противоположном
нормальному направлению их перелета. При этом не-
вольно возникает предположение о существовании «чув-
ства географического положения» («Gefiihl fflr die geogra-
phische Lage»), своего рода компаса, который может быть
установлен на определенные направления. Примечательно,
что пролетающие птицы сохраняют прежнее направление
перелета даже после их перемещения на большие рас-
стояния на несколько градусов широты и долготы.
Таким образом, предварительные результаты много-
численных экспериментов с перевозкой птиц сводятся к
тому, что при отсутствии опыта перелетная птица летит
сначала бесцельно, придерживаясь лишь определенного
направления, которое свойственно всему виду или всей
популяции и всегда передается по наследству. Крамер
(1949) поставил опыты с выращенными в неволе жула-
нами и черноголовыми славками, перелеты которых про-
исходят ночью, и установил, что эти птицы придержи-
ваются определенного направления перелета и при
отсутствии зрительной ориентации. Правда, направление,
избранное указанными особями, не вполне соответство-
вало' направлению перелетов их видов.
Как уже упоминалось выше и повторно отмечалось
при разборе вопроса об ориентации почтовых голубей,
для объяснения этой удивительной способности птиц вы-
двигали самые разнообразные теории, которые подкупа-
ли своей простотой, хотя и не отличались убедительно-
стью. На первый взгляд, многие факты как будто под-
тверждали их, но ни одна из этих теорий не выдержала
более тщательной проверки. Кое-что в них, возможно,
правильно, многое, повидимому, заслуживает дальней-
шего изучения, но сделанные выводы поспешны и оши-
бочны. Мы не беремся здесь оценивать эти теории, а
лишь кратко излагаем важнейшие из них.
Результаты изучения перелетов птиц
127
В связи с опытами по перевозке птиц братья Штим-
мельмайр установили, что на перелеты птиц, их время и
направление влияет изменение положения солнца при
перемещении с севера на юг. Это влияние осуществляет-
ся будто бы через электрические и магнетические явле-
ния в атмосфере. Именно поэтому летом птицы чувству-
ют себя хорошо только в гнездовой области, а зимой —
только на зимовках, а при насильственном перемещении
всегда стремятся возвратиться в условия соответствующе-
го положения солнца. Если в период перелета содержать
птиц в железных или медных клетках, исключая якобы
тем самым действие электрических и магнетических сил,
то птицы не проявляют типичного беспокойства, или пе-
релетного поведения. Предполагалось, что органом вос-
приятия направляющих излучений служит оперение.
Однако во многих случаях предположения братьев Штим-
мельмайр не соответствовали фактам. Кроме того, Бессе-
рер и Дрост (1935) не смогли подтвердить результаты
опытов с «отгораживающими» клетками '.
Предположения о влиянии на птиц электрических
волн нередко вызывают беспокойство и озабоченность,
особенно в кругах голубеводов, которые нередко объяс-
няют неудачи состязаний голубей в полете влиянием
мощных радиостанций. В 1921 г. проф. Пфунгст устано-
вил, что такие высокочастотные колебания, какими яв-
ляются электрические волны, не могут проникнуть в те-
ло голубя или другой птицы. Опыт, приобретенный во
время второй мировой войны, противоречит этим дан-
ным. Дрост и другие авторы (1949), бесспорно, доказа-
ли, что сверхкороткие волны, применявшиеся в радар-
ных установках, оказывали влияние на пролетающих
птиц.
Как уже отмечалось, предположения о том, что пти-
цы, возможно, реагируют на действие магнитного поля
Земли, впервые были высказаны Миддендорфом в 1855 г.
Но за пределами России его данные не были подтвер-
ждены. 15 лет назад этим вопросом, основываясь на ста-
1 В более поздних сообщениях А. Штиммельмайра и некоторых
его единомышленников о космических и астрономических причинах
перелетов птиц не приведено никаких новых фактов, и поэтому эти
работы не способствовали обогащению наших знаний.
128
Глава III
рой теории Вигье, занимался Штреземан. Вигье предпо-
лагал, что птица обладает высокоразвитым магнитным
чувством, т. е. способностью определять магнитное на-
клонение и склонение. В результате птицы из любого
места в состоянии возвратиться к цели прямым путем.
Это предположение поддержали в 1923 и 1927 гг. физики
Морен и Казамайор. Орган предполагаемого магнитного
чувства Вигье искал в полукружных каналах внутренне-
го уха. Штреземан обратил внимание на статолиты, на-
ходящиеся в улитке, круглом и овальном окошке. Опыты
с применением сильных магнитных полей, при которых
присутствовал и автор, не дали удовлетворительного ре-
зультата. Не был успешным и опыт Водзицкого и др.
(1939) с прикреплением намагниченных железных пало-
чек к голове подопытных птиц с целью исключения
влияния магнитного поля Земли. Данье (1936) не без
основания подвергал сомнению теорию Вигье и показал,
что магнитное поле Земли, возможно, действует на птиц,
но что с его помощью птицы могут определить лишь
географическую широту, но не долготу своего местооби-
тания. В более поздней работе (1941) он подчеркнул
важность содержания подопытных птиц в клетках без
железных деталей и высказал мнение о том, что воспри-
нимающий раздражение орган находится не во внутрен-
нем ухе. В широко разрекламированных сообщениях
американских журналов, которые частично нашли отра-
жение и в немецкой печати, утверждалось, будто физик
йегли (1948) в Пенсильванском университете после мно-
гочисленных опытов с почтовыми голубями, наконец,
объяснил их чувство «местности». Согласно его мнению,
воздействия от точек пересечения одинаковых магнитных
силовых линий с параллелями якобы воспринимаются
определенным органом «ориентации» в теле птицы. Пред-
полагают, что таким органом являются веерообразные
образования, окружающие глаза птицы. Во всех случаях,
когда к крыльям птиц прикрепляли небольшие «мешаю-
щие» магниты, ориентация нарушалась. Не обсуждая
эти сообщения, отметим только, что, согласно данным
Хейнрота и бесчисленным известным фактам, ориентацию
у голубей можно объяснить и без влияния магнитного
поля Земли и что нарушение ориентации при прикреп-
Результаты изучения перелетов птиц
129
лении к крыльям магнитов, повидимому, связано с изме-
нением нормального состояния подопытных птиц. Такие
предположения уже высказывались в отношении опытов
с аистами, проведенных Водзицким и др. (1939). Крамер
(1948) также отклонил гипотезу Йегли, причем в основ-
ном по физическим соображениям. После первоначально
положительной оценки со стороны авторитетных физиков
она подверглась критике даже в Америке, так что в на-
стоящее время объяснения йегли надо считать, по мень-
шей мере, сомнительными.
Таким образом, наши знания о влиянии внешних фак-
торов на ориентацию птиц, о специальных органах, вос-
принимающих раздражение извне, и о возможностях
определения направления перелета при помощи этого
«компаса» ничтожны. Неоднократно указывалось, что при
разрешении этих вопросов необходимо сотрудничество
физиков, анатомов и физиологов с орнитологами, изучаю-
щими перелеты птиц. В противном случае можно легко
ошибиться, опереться на несостоятельные предпосылки
или пойти по ложному пути, который заранее может
быть отвергнут специалистом в этой области. В резуль-
тате страдают и наука о перелетах птиц, и смежные ис-
следования.
Наблюдающееся в последние годы сотрудничество
американских физиков и физиологов уже дало опреде-
ленные результаты, хотя удо(влетворительных данных по-
ка не получено.
Закончим наш обзор по изучению ориентации птиц
меткими словами Кёлера (1942): «Таким образом, мы
пока не видим пути, который приблизил бы нас к разре-
шению загадки, поставленной перед физиологией органов
чувств фенологическим изучением перелетов птиц. Пока
нам остается лишь не очень приятная обязанность от-
вергнуть фантастические, надуманные гипотезы и убрать
их с дороги как строительный мусор. Для начала будет
правильным продолжить критическое изучение перелетов
птиц, стараясь при этом накопить по возможности боль-
ше данных; эти данные дадут нам возможность судить о
том, чего мы можем достигнуть, основываясь только на
знакомых нам психических способностях и управляемых
ими известных механизмах ориентации. Если при этом
130
Глава III
окажется (что уже сейчас представляется в какой-то ме-
ре вероятным), что одних этих данных недостаточно, то
указания к раскрытию этой физиологической загадки
«компаса» мы, возможно, найдем при изучении отрица-
тельных случаев, т. е. при неудачах. Поэтому в дальней-
шем последним следует уделять не меньшее внимание,
чем положительным результатам».
Стимулы перелетов
При изложении наших знаний о причинах перелетов
птиц мы можем начать с того, что уже говорилось в
гл. II. Тем самым мы установим связь между предпосыл-
ками всей проблемы и частным вопросом о происхожде-
нии «перелетного» инстинкта.
Известно, что решающее значение для всех жизнен-
ных функций организма имеют железы внутренней секре-
ции и их продукты — гормоны. Выделяемые в кровь в
ответ на внутренние или внешние раздражения, они влия-
ют на нервную систему и функции различных органов.
Следовательно, от них зависит общее состояние орга-
низма. Иногда достаточно очень незначительного стиму-
ла, чтобы нарушить гормональное равновесие. Такое же
воздействие могут оказать и психические факторы. В ре-
зультате происходит перестройка всего обмена веществ и
изменение инстинктивного поведения животных.
Нам известно, что многие птицы приступают к раз-
множению непосредственно после весеннего перелета, да-
же при неблагоприятных внешних условиях. Мы знаем
также, что смена оперения у перелетных птиц часто свя-
зана с началом осенних миграций (рано отлетающие пти-
цы линяют на зимовках, поздно отлетающие — на роди-
не) . Отмечено, что птицы задерживаются на гнездовьях,
если ко времени отлета молодые птицы еще не научились
летать. В аналогичных случаях другие птицы (в частно-
сти ласточки) покидали свой выводок. Все эти факты
указывают на внутреннюю связь между перелетным
инстинктом, инстинктом заботы о потомстве и линькой.
Эти процессы, регулярно сменяя друг друга, определяют
повадки птиц.
Насколько сильна может быть зависимость одного
Результаты изучения перелетов птиц
131
процесса от другого, особенно наглядно видно' на при-
мере жулана: старые птицы отлетают раньше и линяют
на зимовке, тогда как молодые меняют мелкие перья до
отлета и поэтому позже приступают к миграции. Линька
требует больших затрат энергии, и птицы в этот период
очень истощены; для перелета они должны вновь нако-
пить известные энергетические запасы. У тех птиц, у ко-
торых перелетов нет, например у поганки, согласно Кра-
аку и Хоогерхайде (1942), возможны перелеты, непосред-
ственно связанные с линькой («Mauserzug»). Подобные’
миграции наблюдаются у взрослых уток, в большом чис-
ле отлетающих в начале августа от берегов Северного
моря и Южной Швеции в Гельголандской бухте, где они
меняют свои маховые перья; затем эти птицы вновь воз-
вращаются в гнездовую область. Такую линьку, вероятно,
следует рассматривать как одну из форм «промежуточ-
ного перелета», с той только разницей, что за ней не сле-
дует отлета птиц. Даанье и другие авторы доказали, что
некоторые виды птиц, например чибисы, свиристели и
ряд хищных птиц, могут линять во время осеннего пере-
лета. Линька совершается во время миграции и у буре-
вестников Oceanites oceanicus, которые уже упоминались
как летующие птицы в наших широтах. Приведенные
данные еще раз показывают, насколько разнообразна
действительность, вновь и вновь нарушающая схему
общего «правила».
Исходя из связи между возвращением птиц весной с
начинающимся вслед за этим размножением, американ-
ский исследователь Роуэн (1926, 1929) пришел к выво-
ду, что между перелетным инстинктом и состоянием
половых желез, невидимому, существует тесная зависи-
мость; последние во время перелетов развиты очень сла-
бо и, напротив, в период размножения имеют максималь-
ное развитие. В опытах над серой овсянкой (Junco hiema-
lis) Роуэну удалось доказать влияние солнечной энергии
на рост половых желез. Длительное облучение искус-
ственным светом вызвало развитие, а затемнение — ин-
волюцию этих органов. Когда половые железы достиг-
ли известного уровня развития, у птицы пробудился
перелетный инстинкт, выразившийся в том, что она поки-
нула место эксперимента. Поэтому Роуэн предположил,
132
Глава III
что причиной проявления перелетного инстинкта являет-
ся секреция половых желез. Но так как на последнюю
влияет количество ежедневно поступающего света, он
был склонен считать увеличение или уменьшение свето-
вого дня решающим фактором в возникновении перелет-
ного инстинкта. Другой американский исследователь
Биссоннетт (1933), работая со скворцами в том же плане,
что и Роуэн, также получил аналогичные результаты.
Вскоре после этого (1933) Шильдмахер на орнитологи-
ческой станции Гельголанд продолжил работу Роуэна. Он
пытался доказать зависимость перелетного инстинкта от
гормонов половых желез, инъицируя самкам горихвосток
во время осеннего перелета препарат «прогинон». При-
меняя определенные дозы этого препарата, ему удалось
ослабить или даже погасить проявляемое птицами по но-
чам перелетное беспокойство. Последнее постоянно реги-
стрировалось при помощи особого прибора. Полученные
результаты позволили Шильдмахеру сделать вывод, что
осенью проявление перелетного инстинкта является след-
ствием уменьшения количества гормонов, выделяемых
половыми железами. Гирсберг и Штади (1934), напротив,
пытались возбудить перелетный инстинкт гормональным
воздействием. Они воздействовали на пребывающих зи-
мой в спокойном состоянии чижей и щеглов фолликули-
ном. В результате у птиц появилось ясно выраженное
«перелетное беспокойство», которое авторы объяснили
стимуляцией фолликулином выделения гормонов половой
железой, в свою очередь вызвавших проявление перелет-
ного инстинкта.
Здесь мы должны дать краткое объяснение понятия
«перелетное беспокойство» и описание аппаратуры, при
помощи которой оно регистрируется. Ранее уже упоми-
налось о том, что содержащиеся в неволе перелетные
птицы, когда другие особи их вида приступают к мигра-
ции, начинают проявлять беспокойство даже при наилуч-
шем уходе. Они машут крыльями и носятся по клетке,
как бы имитируя миграцию, т. е. удовлетворяют свой
перелетный инстинкт. Такое явление наблюдается у
большинства мелких певчих птиц (насекомоядных) пре-
имущественно ночью, так как на воле их перелет совер-
шается ночью. В опытах это перелетное беспокойство
Результаты изучения перелетов птиц 133
фиксируется при помощи так называемых регистрацион-
ных клеток. Каждый прыжок и каждое движение птицы
от одной жердочки к другой замыкает электрическую
цепь, которая соединена с часовым механизмом, регистри-
рующим каждое замыкание и размыкание электрической
Рис. 42. Прибор для регистрации перелетного беспокойства
птиц на орнитологической станции Гельголанд (в дальней-
шем он был усовершенствован).
цепи на передвигающейся бумажной ленте (как в ап-
парате Морзе). По этим записям можно затем судить о
длительности и силе прыжков и беспокойства (рис. 42).
Однако теории Роуэна и Шильдмахера вскоре под-
верглись критике. Прежде всего, они относятся только к
тем видам птиц, которые не перелетают из северного
полушария в южное, где укорочение светового дня сме-
няется его удлинением, но тем не менее половые железы
зимующих там птиц не развиваются. Кроме того, этим
взглядам противоречит поведение уток, половые железы
которых сильно развиты зимой, следовательно, задолго до
начала весеннего перелета. Указанные теории не могут
объяснить перелеты молодых птиц с еще совершенно не-
развитыми половыми железами. Наконец, подобные ши-
10-176
134
Глава III
рокие обобщения нельзя делать из-за небольшого числа
положительных результатов по сравнению с количеством
отрицательных данных. Итог этим соображениям подвел
сам Роуэн (1932) опытами над воронами. У одной груп-
пы подопытных птиц он удалил половые железы, другую
облучал светом, а третьей инъицировал гормон. В резуль-
тате оказалось, что поведение всех этих птиц по существу
мало чем отличалось от поведения контрольных птиц, ко-
торые не подвергались никакому воздействию. Ряд даль-
нейших опытов, проведенных Шильдмахером, а затем и
Путцигом (1937—1939) на орнитологической станции Рос-
ситтен, показал, что освещение многочисленных мелких
птиц зимой хотя и вызвало развитие половых желез, но не
привело к преждевременному проявлению перелетного
беспокойства. И наоборот, летом птицы в регистрацион-
ных клетках часто находились в состоянии перелетного
беспокойства до тех пор, пока оно не прерывалось линь-
кой. В соответствии с результатами Роуэна Путциг уста-
новил, что как весной, так и осенью выпущенные на волю
кастрированные перелетные птицы (клуши и обыкновен-
ные чайки) вели себя точно так же, как и физиологи-
чески вполне нормальные представители того же вида.
Сходное поведение наблюдалось также у ворон и галок,
которым был введен половой гормон самок. Ган (1939)
проводил аналогичные опыты с североамериканскими
певчими птицами и получил такой же результат.
Таким образом, было установлено, что в противопо-
ложность первоначально распространенным взглядам ин-
стинкты спаривания и насиживания, обусловленные воз-
действием гормонов половых желез, никоим образом не
следует приравнивать к инстинкту весеннего перелета.
Первые могут рассматриваться только как дополни-
тельный фактор, способный ускорить или прекратить пе-
релет. Что касается инстинкта осеннего перелета, то он
не находится в прямой зависимости от гормона половых
желез.
После первых поразительных результатов в этой обла-
сти исследователи стали предостерегать против возмож-
ной переоценки единичных данных. Учитывая видовые
различия в поведении подопытных птиц, необходимо из-
бегать всякого рода обобщений. Тем не менее весьма
Результаты изучения перелетов птиц
135
вероятно, что приведенные выше данные о зависимости
между развитием половых желез и миграцией относятся
ко всем перелетным птицам.
При обсуждении связи между перелетами птиц и по-
годой мы указывали на отличия птиц, перелет которых
определяется внешними метеорологическими условиями,
от птиц, миграции которых зависят от инстинкта (см.
стр. 50). При этом мы отмечали, что в противополож-
ность первоначальному пониманию этих терминов даже
типичные «инстинктивные» птицы в известных условиях
ведут себя как «погодные» птицы, так как у представи-
телей обеих групп миграции могут вызываться воздейст-
виями внешней среды.
Влияние таких факторов на птиц первой группы ска-
зывается и в том, что у птиц, залетевших в Германию из
чужих стран, часто в связи с приспособлением к местным
климатическим условиям сдвигаются периоды размноже-
ния. Германские птицы, перевезенные в тропические об-
ласти или южное полушарие, меняют время перелета или
же вообще перестают мигрировать. Кроме того, отмечено,
что содержавшиеся в течение многих лет в клетках пере-
летные птицы постепенно переставали проявлять беспо-
койство в период перелета. Характерно, что это чаще все-
го наблюдалось у таких видов, которые на воле склонны
к оседлости. Выше мы уже неоднократно упоминали эти
виды в связи с другими вопросами.
В этом отношении представляет интерес установлен-
ный Томасом (1934) и Хиксом (1938) факт, что европей-
ские скворцы вели себя в США в течение нескольких
десятилетий, как оседлые птицы, но в последнее время
стали совершать перелеты, направления которых соот-
ветствовали направлениям пролетов европейских сквор-
цов. Переведенные в Шотландию канадские казарки —
типичные перелетные птицы — превратились в оседлых;
птицы, выращенные Бенгт Бергом в окрестностях Каль-
мара (Швеция), улетели вместе с серыми гусями, следуя
вдоль берегов Центральной и Западной Европы. Пересе-
ленные из Англии в Финляндию кряквы приспособились
к поведению местных птиц этого вида (см. стр. 112).
Но если «погодные» птицы всегда отвечают на внеш-
нее раздражение началом миграции, то, как уже упоми-
ю*
136
Глава III
Рис. 43. Зависимость между перелетным беспокойством и весом, тела
у серой славки (Sylvia с; communis Lath.) (Из Меркеля, 1940.)
----------------—перелетное беспокойство;---— вес тела.
далось, «инстинктивные» птицы приступают к перелету
только при наличии особых предпосылок. В некоторых
случаях внешнее влияние оказывается неэффективным
и непосредственной причиной отлета являются сдвиги
гормонального равновесия, влияющие на обмен веществ.
Таким образом, мы переходим к физиологически обосно-
ванным различиям между этими группами птиц и к оцен-
ке значения физиологии обмена веществ для изучения
перелетов вообще, на что уже указывалось вкратце на
стр. 102.
Результаты изучения перелетов птиц
137
Первым ученым, положившим начало такого рода ис-
следованиям, был Греббельс. В своем большом труде
«Птица» (1932) он изучал зависимость между питанием
птицы и ее перелетом и исследовал значение усиленного
отложения жира в период перелета, которое он рассмат-
ривал как физиологический эффект кастрации. В связи
Рис. 44, Зависимость между температурой и перелетным беспокой-
ством у серой славки {Sylvia с. communis Lath.). (Из Меркеля,
1940.)
О—О температура; -------перелетное беспокойство.
с этим он собирал данные о весе птиц, определял состоя-
ние питания мигрирующих птиц, их пищеварение и тем-
пературу тела, чтобы таким путем получить представле-
ние о связях между обменом веществ и перелетом. При-
близительно в то же самое время Вагнер (1930) изучал
зависимость суточного ритма перелетных птиц от внеш-
них факторов. В дальнейшем (1937) он установил, что у
типичных перелетных птиц количество пищи оказывало
небольшое влияние на возникновение перелетного бес-
покойства, а у менее выраженных перелетных птиц это
влияние было очень значительным. Высокая температура
оказывала задерживающее действие, низкая — способст-
138
Глава III
вовала проявлению ночного беспокойства. Меркель
(1937) принял терминологию Греббельса о «предрасполо-
жении» и «готовности» к перелету и на основании ряда
собственных опытов пришел к выводу, что предрасполо-
жение к перелету, по существу, идентично способности
накапливать жир, а перелетное настроение равнозначно
началу перелетного беспокойства и вызывается неболь-
шим повышением секреции щитовидной железы, вследст-
вие чего мобилизуется резервный обмен веществ (см.
гл. III). Следовательно, при наличии готовности к полету
небольшое понижение температуры явилось бы (через по-
вышение секреции щитовидной железы) непосредственной
причиной отлета.
Американские исследователи в опытах над голубями,
домовыми воробьями и крапивниками получили иные ре-
зультаты. Они обнаружили видовые отклонения, явившие-
ся следствием различий в характере обмена веществ у
разных видов. В то время как гуси и утки оставались
совершенно спокойными в течение всего периода голода-
ния и тем самым понижали свой обмен веществ до ми-
нимума, волнистые попугайчики и канарейки пытались
поддержать температуру тела при помощи движений, за
счет запасов жира и гликогена. Согласно Путцигу
(1938а), этот факт указывает на значительные различия
в обмене веществ и в связи с этим также и в механизме
терморегуляции «погодных» и «инстинктивных» птиц.
Последние — к ним относятся в первую очередь насекомо-
ядные птицы — «должны при более быстром переварива-
нии и значительном расходе кислорода получать гораздо
больше пищи и много двигаться, чтобы поддержать свою
в норме высокую температуру тела». В условиях укора-
чивающегося дня в конце лета эти птицы не могут добы-
вать необходимое для поддержания процессов окисления
количество пищи, особенно когда первые холодные ночи
ускоряют сгорание жира и гликогена. В результате по-
является «психическое» беспокойство, птицы начинают
усиленно двигаться, что соответствует началу истинного
перелета.
Однако последние работы Путцига (1939) показы-
вают, насколько мало разработаны эти вопросы и сколь
ошибочно было бы переносить выводы о причинах пове-
Результаты изучения перелетов птиц 139
дения одного вида птиц на другой. В этих работах Пут-
циг в противоположность своим прежним предположе-
ниям и данным других авторов пришел к выводу, что,
несмотря на то, что накопление жира способствует пере-
лету, а его отсутствие задерживает миграции, птицы
иногда мигрируют и с минимальными энергетическими
запасами. Во всяком случае, у скворцов, зарянок и гори-
хвосток проявление перелетного инстинкта не зависит от
их физиологической конституции. Возможно, это отно-
сится и к другим видам, однако точных данных об указан-
ной закономерности пока нет. Вне зависимости от того,
будут ли получены положительные или отрицательные
результаты, мы приближаемся к разрешению очень важ-
ного вопроса о взаимосвязи перелетов и процессов обме-
на веществ. Путциг предложил заменить старые понятия
«Wettervbgel» и «Instinktvogel», которые он считает
слишком односторонними и в некоторых отношениях
вводящими в заблуждение, более нейтральными определе-
ниями: «птицы, перелеты которых обусловлены внешней
средой, и птицы, перелеты которых обусловлены внутрен-
ней средой» («Aussenwelt- und innenweltbedingte Zug-
vogel»). Различие между обеими группами он определил
следующим образом: для того чтобы представители пер-
вой группы приступили к миграции, необходим внешний
толчок, заключающийся в неприятном самочувствии
вследствие низких ночных температур в сочетании с дли-
тельной темнотой или же вследствие недостатка корма.
«В новых условиях у таких птиц обнаруживается не-
посредственное воздействие пережитого на выполняемые
ими движения. У птиц, перелеты которых обусловлены
внутренней средой, также возможен подобный толчок к
началу перелета и его усиление в результате воздействия
внешних раздражений. Последние передаются рефлек-
торно или же посредством эндокринных желез и обмена ве-
ществ. Однако гораздо чаще «пусковой механизм» пере-
летов этих птиц обусловлен автоматизмом нервной си-
стемы. У обеих групп целесообразное поведение наблю-
дается и при отсутствии опыта, что составляет основную
проблему перелетов птиц».
Говоря об обмене веществ и температурном режиме у
перелетных птиц, мы неоднократно указывали на роль
140
Глава III
щитовидной железы, а также коснулись ее значения для
начала перелетов. При низкой температуре происходит
усиление выделения гормона щитовидной железы (тирок-
сина) , который путем повышения интенсивности всех про-
цессов жизнедеятельности организма вызывает расходо-
вание запасных веществ. Установлено, что щитовидная
железа перелетных птиц выделяет большое количество
гормона. Зимой, однако, ее секреция значительно пони-
жена. Исходя их этих ритмичных изменений функции
железы, был сделан вывод, что тироксин играет решаю-
щую роль для начала перелета. Вагнер (1930) отметил у
подопытных птиц признаки перелетного беспокойства пос-
ле кормления их щитовидной железой. Путем инъекции
тироксина Меркель (1937) также вызвал усиление актив-
ности у птиц, перелет которых уже был закончен. Про-
веденные им гистологические исследования показали,
однако (в отличие от данных других исследователей,
например Кюхлера, 1934), что в период перелетов тиро-
ксин выделяется в небольшом количестве. Его содержа-
ние особенно велико в период линьки. После опытов Ваг-
нера Греббельс отметил, что повышение функции щито-
видной железы во время перелетов невозможно согла-
совать с понижением основного обмена веществ.
Гистологическими исследованиями (Путциг, 1937) уста-
новлено, что состояние щитовидной железы у разных
видов залетных птиц (клёст, пестрый дятел, свиристель,
снегирь, а также чайки, аисты и некоторые хищные пти-
цы) очень изменчиво, и поэтому на основании ее функ-
ции нельзя вывести никаких общих заключений относи-
тельно перелетов птиц. Ритмичность функции щитовидной
железы (как и половой железы и других желез внутрен-
ней секреции) иногда находится в противоречии с ее
реакцией на раздражения окружающей среды, результат
которой в данном случае зависит от общего состояния
организма.
Путциг (1938) обращает внимание на то, что усиле-
ние функции щитовидной железы отражается не только
на обмене веществ, но и на нервной системе, что
приводит к повышению чувствительности на раздражения
извне. И, наоборот, воздействие центральной нервной си-
стемы может вызвать изменения в органах. Этих взгля-
Результаты изучения перелетов птиц 141
дов придерживается и Штади (1938); он особенно подчер-
кивает возможность ошибок из-за непосредственного
сравнения поведения подопытных птиц, содержащихся в
клетках, с поведением птиц, живущих на воле. Содержа-
ние птиц в клетках может через центральную нервную
систему вызвать усиление функции щитовидной железы.
Нормальному протеканию процессов обмена мешает так-
же ограничение подвижности подопытных птиц, что не мо-
жет не отразиться на функции щитовидной железы. Подоб-
ные влияния, разумеется, трудно учесть в эксперименте.
Однако возможно, что именно они и являются ключом к
пониманию огромного разнообразия явлений, на основе
которых возникли обобщения, которые всегда оказыва-
лись лишь индивидуальной или же, в лучшем случае,
свойственной определенному виду реакцией.
В нескольких работах Путциг указывал на особую
роль гипофиза во всей системе желез внутренней секре-
ции, а следовательно, и в процессе перелета птиц. Он от-
мечает необходимость изучения гонадотропного гормона
передней доли гипофиза, влияющего на секрецию поло-
вых желез. В дальнейшем разработкой этого вопроса за-
нялись американские и английские исследователи, кото-
рые за последние годы достигли в этой области значи-
тельных результатов. Некоторые из них мы здесь вкратце
приведем.
Рилей, Гарднер и Уитши (1938) нашли, что гипофиз
самок очень слабо реагирует на световые раздражения и
что для стимуляции функции гипофиза, а следовательно,
и для созревания яичника очень большое значение имеют
«психические» факторы. Бергер, Биссоннетт и Дулиттл
(1942) в опытах над скворцами пришли к выводу, что
половые железы активирует не свет, а длительное бодр-
ствование. Вольфсон (1941) исследовал роль гипоталами-
ческой области промежуточного мозга, которую следует
рассматривать как центр регуляции сна. Если гипоталамус
не возбуждается зрительными раздражениями, то умень-
шается его активность, а следовательно, и активность
мозга, так же как и всего организма. Но, поскольку
гипоталамус воздействует на гипофиз, длительное свето-
вое раздражение (при длинном дне или искусственном
освещении в эксперименте) усиливает функцию гипофиза
142
Глава III
и тем самым — развитие половых желез. Хотя последова-
тельность биологических циклов у птиц — размножения,
линьки и перелета — как будто наследственно закрепле-
на, однако проявление каждого из них зависит от внеш-
них раздражений, различных у разных видов и нередко
даже у разных особей. Вольфсон (1942) исследовал во-
прос о происхождении готовности к перелету. Изучив
перелетных и оседлых представителей одного из видов
овсянок (Junco oregonus), он установил, что на готов-
ность к перелету влияет непосредственно гипофиз. Его
действие заключается в создании определенного физиоло-
гического состояния, которое обеспечивает птице необхо-
димую для миграции энергию. В отличие от перелетных
птиц у оседлых Вольфсон не обнаружил накопления жира
весной. У последних рано активируется функция семен-
ников. Вольфсон особенно подчеркивал важность для ре-
гулирования всех видов внутренней секреции «психиче-
ских» факторов (наряду с такими, как длина дня, темпе-
ратура и пища). На это, как известно, уже указывали
Путциг и 'другие авторы. В итоговой работе Вольфсон
(1945) пытался точнее определить, какой гормон влияет
на готовность к перелету, однако он не смог получить
исчерпывающих результатов. Изучение составных частей
комплекса внутренних раздражителей выявило огромное
влияние на все функции организма гипофиза, который,
собственно, управляет общим физиологическим состояни-
ем, а следовательно, и поведением животного.
Приведенные данные указывают, что вопрос о значе-
нии гипофиза в общем гормональном балансе организма
не решен. Подробное изложение влияния гипофиза на
перелет птиц выходит за рамки этой книги, и, поскольку
указанные вопросы еще только разрабатываются, это
может отрицательно сказаться на общем обзоре, который
мы предлагаем читателю.
Если мы после анализа физиологических исследова-
ний и сравнения полученных результатов с фаунистиче-
скими данными хотим еще раз охарактеризовать совре-
менный взгляд на сущность перелетов птиц и вызывающе-
го их перелетного инстинкта, то мы должны признать,
что привычное мнение о перелете, как о строго опреде-
ленном, доминирующем и неизменном явлении, может
Результаты изучения перелетов птиц 143
существенно измениться. В настоящее время (во всяком
случае, в отношении целого ряда видов, о которых
имеется достаточно данных) установлено, что перелеты
птиц не определяются факторами наследственности, что
находится в известном противоречии с данными, приво-
дившимися при характеристике методов изучения переле-
тов птиц. Но тогда речь шла только об основном правиле,
которое и в настоящее время в известной степени имеет
силу. То обстоятельство, что это правило в ряде отноше-
ний кажется нарушенным и что в результате развития
наших знаний такие нарушения, возможно, будут обнару-
живаться и в дальнейшем, служит лишь доказательством
изменчивости всех процессов в природе, среди которых и
перелеты птиц не являются исключением. «Перелет
птиц — это не статика, а динамика»,— говорил Путциг в
одной из своих последних работ. Он дал следующее опре-
деление понятия «перелетного инстинкта»: «Перелетный
инстинкт — это обусловленный историей вида и физиоло-
гическими реакциями организма приобретенный автома-
тизм, основанный на преобразовании сил внешней среды
во внутренние, осуществляющийся непосредственно или
же через систему желез внутренней секреции, результат
действия которого гарантирует сохранение вида».
На этом мы заканчиваем главу об изучении переле-
тов птиц, слишком краткую и сжатую по сравнению с ее
значением. По решению частных вопросов и по выводам
эта глава наименее зрелая, поскольку работы в этой
области еще не закончены и окончательных результатов
не получено. Это лишь первые попытки проникновения в
неизведанные края, куда наука получила доступ лишь
недавно. Но за это короткое время сделано так много,
что мы вправе надеяться со временем получить ответ и
на самый трудный, основной вопрос науки о перелетах
птиц: «почему?» Путь к этой цели известен — это методы
физиологического исследования, а достижение ее — во-
прос времени.
ПРИЛОЖЕНИЕ
В этой части картины перелетов нескольких видов
птиц излагаются несколько подробнее, чем в основной
части книги. В каждом случае мы дадим краткий обзор
особенностей перелета данной формы, сопровождая опи-
сание одной или несколькими иллюстрациями. Это позво-
лит создать своего рода монографическое описание соот-
ветствующего явления.
Выбор примеров производился с точки зрения «особо-
го отношения к перелету» и известных особенностей, от-
личающих перелеты этих птиц от миграций большинства
родственных им форм.
Этот обзор даст возможность точнее пояснить ряд по-
нятий и явлений, которые в основной части книги только
упоминались или о которых кратко говорилось в связи с
более общими вопросами.
Таким образом, это приложение служит для лучшего
понимания основной части книги, в которой приводились
соответствующие ссылки на помещаемый ниже материал.
Перелеты скворца
Скворец в большей части области своего распростра-
нения является одновременно и перелетной, и кочующей,
и оседлой птицей. Только наиболее северные и восточные
популяции зимой отлетают все без исключения, и лишь
на Британских островах скворцы ведут себя преимущест-
венно как оседлые птицы. По направлению к западу и
югу отмечается прогрессирующая склонность к зимовке
на месте (от исключительных случаев зимовок отдельных
особей до регулярных зимовок большинства птиц данной
популяции).
Перелеты скворца могут служить примером распреде-
ления различных популяций как во время перелета, так и
Приложение 145
на зимовках. Это выражается в следующем: скворцы из
Голландии, Бельгии, Северной Германии, Дании и При-
балтики, следовательно из всего района Северного и Бал-
тийского морей, отлетают в западном направлении, пре-
имущественно к Британским островам, где зимуют и скан-
динавские популяции. Скворцы из Средней Германии
(Тюрингия, Саксония) отклоняются на юго-запад; их зи-
мовки находятся в Средней и Южной Франции, Испании
и Марокко. По мере углубления внутрь европейского мате-
рика направление перелета популяции, например из Швей-
царии или Силезии, все отчетливее отклоняется на юго-
запад и юг. Эти популяции зимуют в Южной Франции,
восточной части Пиренейского полуострова, в Марокко,
Алжире и Тунисе, а также в Италии (рис. 45).
Указанные основные особенности перелетов скворцов
можно всегда выявить, несмотря на наличие многочислен-
ных переходов. Отдельные популяции не только мигри-
руют в совершенно определенном направлении, но и зи-
муют в совершенно определенных областях, которые неред-
ко строго разграничены. Расположение зимовок зависит от
распределения температур в зимнее время в западном по-
лушарии, так что направления пролетов различных популя-
ций приблизительно перпендикулярны январским изо-
термам.
Картины перелетов скворца усложняются явлением
«промежуточного перелета», наблюдающегося ранним
летом почти у всех среднеевропейских популяций часто в
очень изменчивых формах. В Швейцарии молодые сквор-
цы кочуют вскоре после вылета из гнезда вместе со ста-
рыми птицами, которые не размножались. Особенно
выделяются очень частые кочевки далеко на север
(до 550 км). В Средней и Восточной Германии подобные
кочевки проходят совершенно ненаправленно и не на
такие большие расстояния (в Силезии приблизительно до
ЛОО км). В районе Северного и Балтийского морей кочев-
ки молодых скворцов превратились в направленный пере-
лет, предшествующий истинному осеннему перелету, во
время которого покрывается приблизительно половина
пути к зимовкам (из б. Восточной Пруссии, например,
в Померанию, Мекленбург или Голштинию). В районе
«промежуточной цели» птицы в течение нескольких недель
146
Приложение
Рис. 45. Ареалы скворцов. (Из Нитхаммера, 1937.)
А — швейцарские; Б — из восточной части Прибалтики; В — саксонские; Г-си-
лезские. Гнездовая область не заштрихована и обведена жирной линией, область
зимовки заштрихована в косом направлении, область раннелетнего перелета —
в вертикальном. Стрелками обозначены редкие находки птиц.
отдыхают, а затем снова мигрируют. Этот особый харак-
тер перелетов у скворцов в районе Северного и Балтий-
ского морей напоминает «преждевременный отлет» чиби-
сов и других болотных птиц. Но у последних этот перелет
совершают не только молодые птицы, и его направление
почти всегда соответствует направлению осеннего пере-
лета (см. стр. 93).
Приложение
147
Рис. 46. Ареал жулана. (Из Нитхаммера, 1937.)
Черным обозначены гнездовая область и место зимовок только германских
жуланов; горизонтально заштрихована область зимовки всех европейских жу-
ланов; пунктиром обозначены пролетные пути.
Перелеты жулана
Жулан — типичная перелетная птица, которая очень
рано (в начале августа) покидает свою гнездовую об-
ласть и поздно возвращается (обычно лишь в мае).
148
Приложение
Он зимует в Восточной и Южной Африке (на западе —
приблизительно до Камеруна).
Пролет жулана проходит из Центральной и Северной
Европы в юго-восточном направлении. Он пересекает
Балканские страны и Средиземное море от острова Кри-
та до восточного побережья Эгейского моря в юго-восточ-
ном направлении. Согласно Гейру, преобладающие в
восточном районе Средиземного моря северные ветры от-
клоняют перелет на этом участке к югу. Начиная от дель-
ты Нила, перелет после отчетливого отклонения проходит
в южном направлении вглубь Египта до Судана и даль-
ше в область зимовки. Весной перелет, невидимому, под
влиянием проникающих глубоко в Египет сильных север-
ных ветров, начиная с Судана, проходит на север с лег-
ким отклонением к востоку, восточнее дельты Нила, над
Красным морем, Синайским полуостровом и Палестиной.
В Сирии вновь происходит характерное для миграции
этого вида отклонение. Пролетая дальше над Кип-
ром и центральными районами Малой Азии, над
Черным и Мраморным морями, птицы вновь достигают
области осеннего пролета над Балканским полуостровом
(рис. 46).
Таким образом, птицы летят осенью и весной различ-
ными пролетными путями над областями, расположен-
ными приблизительно между 26 и 44° с. ш. Это пример
«петлеобразного перелета», который в данном случае
имеет форму равнобедренного треугольника.
Еще более резко выраженный петлеобразный перелет
мы встречаем, например, у иволги и сизоворонки. У этих
птиц весенний и осенний пролетные пути расположены на
расстоянии сотен километров друг от друга.
Перелеты белой трясогузки
Белая трясогузка в южной части своей гнездовой об-
ласти — оседлая птица, а дальше к северу — кочующая
или перелетная. Но некоторые особи зимуют даже в
Скандинавии и сравнительно часто —в Западной Герма-
нии. Регулярная область зимовок — Южная Европа и
Южная Африка до Сенегамбии или, соответственно, Ке-
ния и Уганда, а также Палестина и Месопотамия.
Приложение 149
Германские белые трясогузки отлетают в юго-запад-
‘ ном направлении, рассеиваясь при этом широким фронтом
от юга — юго-запада до запада — юго-запада. Очень
сходно ведут себя популяции, населяющие Голландию,
Бельгию и Великобританию. Последние в соответствии с
положением их родины по отношению к материку отле-
тают в более южном направлении. Особенно много околь-
цованных птиц было найдено в бассейне р. Гаронны,
где1 в большом числе встречаются птицы из Англии, Гол-
ландии и Западной Германии, преимущественно еще на
пролете. Очень много данных получено также из Испании
и Португалии, а о силезских птицах — из Северной Ита-
лии. Менее часто окольцованных птиц находили на се-
верном побережье Африки (Марокко, Тунис).
Заслуживают внимания перелеты белых трясогузок из
восточной Прибалтики и Швеции (рис. 47). Эти птицы
мигрируют иным путем — в юго-восточном направлении.
Немногие данные указывают на то, что перелеты прохо-
дят через Малую Азию, Кипр, Палестину, Египет и Су-
дан. Эта форма миграции, следовательно, резко отличает-
ся (без переходов) от перелетов популяций, населяющих
области западнее Вислы. Где проходит пролетный
рубеж, должны показать данные кольцевания из б. За-
падной и б. Восточной Пруссии, которых, к сожалению,
! еще нет.
’ Таким образом, перелеты белой трясогузки в извест-
ной мере аналогичны перелетам белого аиста, хотя, ко-
нечно, их нельзя сравнивать, так как предпосылки этого
сходства, очевидно, совершенно различны. В последнее
' время стало известно, что такие различия в направлениях
• пролетных путей свойственны еще ряду птиц (пеночка-
j кузнечик, горихвостка-чернушка, удод, серая мухоловка).
О причинах этих особенностей перелета у различных по-
; пуляций пока существуют лишь предположения.
( Перелеты серебристой чайки
Серебристая чайка — преимущественно перелетная
птица (в большей степени, чем все другие виды чаек).
Лишь небольшая часть птиц зимует в районе Северного
пс
га
Рис. 48. Карта перелетов серебристых чаек из Западной и Северной
Европы.
Различия в перелетах у разных популяций: у западных пролет проходит только
над побережьем моря, у восточных —в основном над внутренними районами
суши. Соответственно распределяются места их зимовок в Африке.
152
Приложение
и Балтийского морей. Большинство чаек мигрирует зна-
чительно дальше на юг, вплоть до Тропической Африки.
При этом у отдельных рас и популяций отмечаются сле-
дующие различия. Птицы, гнездящиеся на западе, летят
вдоль берегов Франции, Испании и Португалии к запад-
ной части Средиземного моря, а также к западноафри-
канским водоемам вплоть до Нигерии. Исключение со-
ставляет небольшое число птиц, которые остаются в рай-
оне Северного моря. Из числа птиц, гнездящихся на
востоке — в Швеции, Финляндии и на оеверозападе
СССР, —только небольшая часть придерживается берегов
Балтийского и Северного морей. Подавляющее большин-
ство мигрирует на юг и пересекает при этом материк без
заметной связи с водоемами. Пролетев над Черным мо-
рем, они попадают в восточную часть Средиземного
моря, в Египет, в долину Нила, к Красному морю и даль-
ше, в Центральную Африку, до озера Виктория и сред-
него течения Конго.
В области гнездовья, расположенной между этими
районами, поведение птиц различно: из Норвегии, Дании
и некоторых районов Швеции они пролетают как над
внутренними районами суши, так и вдоль западных бе-
регов Европы и Африки, вплоть до устья Конго. С во-
стока они, очевидно, долетают даже до озера Виктория,
если не попадают туда по течению Конго с запада
(рис. 48).
Особенно часто над внутренними районами суши про-
летают старые птицы (при этом они развивают иногда
большую скорость: за 15 дней они долетают от Росситтена
до Каспийского моря), Молодые птицы придерживаются
побережья и вначале кочуют без определенного направ-
ления.
Перелеты серебристых чаек — характерный пример
того, как экологические связи обусловливают пролеты
птиц вдоль берегов морей. Однако перелеты этой чайки
указывают также на возможность освобождения от этих
связей у сильных летунов и водяных птиц. Поразитель-
ным остается резкое различие в отношениях к перелету
у западных и восточных популяций одного и того же
вида, что, несомненно, способствовало развитию различ-
ных рас серебристых чаек.
П риложение
153
Перелеты журавля
Журавль — типичная перелетная птица, лишь очень
редко зимующая в области гнездовья или пролета (за
последние годы подобные случаи известны в Централь-
ной Швеции, Лужице, Восточной Франции, Голландии и
Венгрии). Зимовки журавля находятся в западной части
Северной Африки. В эти места летят птицы, гнездовья
которых находятся в Центральной и Северной Европе, а
также часть популяций из Прибалтики и западной части
СССР. Но особенно много восточноевропейских журавлей
зимует в районе Белого Нила. Предполагается, что неко-
торые птицы остаются в Ливии.
Перелеты журавля проходят узким фронтом и по строго
ограниченным пролетным путям.
Но границы весеннего и осеннего перелетов могут не
вполне совпадать (например, северная граница пролета
журавлей в Ганновере проходит осенью южнее, чем вес-
ной, в то время как в Саксонии на юг сдвигается южная
граница). Пролетные пути, по которым следует узким
фронтом основная масса птиц, проходят следующим об-
разом: из Восточной Европы в направлении с севера на
юг над Литвой, Румынией, по западному берегу Малой
Азии или Греции, через Ливийскую пустыню — к Судану
и Белому Нилу; из Прибалтики и с запада СССР над
Венгрией с отклонением на запад, над Южной Италией
и Сицилией — в Тунис; из Северной и Центральной Евро-
пы в юго-западном направлении над Рейнской областью,
Юго-восточной и Юго-западной Францией — в Испанию и
Марокко. Из центральных районов и с востока СССР
журавли летят вдоль больших рек через Черное и Кас-
пийское моря (некоторые птицы остаются в Крыму) к
северному побережью Малой Азии, Кипру и Египту. Уда-
лось установить наличие перелетов над Кавказским хреб-
том, а также над Альпами. Наблюдающиеся колебания в
направлениях перелетов иногда приводят журавлей в та-
кие страны, которые не лежат на пути их пролетов, напри-
мер Швейцария, Британские острова (рис. 49).
Все эти данные, основывающиеся не на кольцевании
птиц, а на наблюдениях, доказывают, что журавли, в отли-
чие от белых и черных аистов во время своих миграций
154
Приложение
Рис. 49. Карта перелетов журавля в Европе и Африке. (ИзЛиббер-
та, 1936,'с дополнениями.)
Узкий фронт обозначен жирными черными линиями, зимовки — черным цветом,
области гнездования — пунктиром.
перелетают моря, горы и пустыми. Аисты больше ле-
тят скользящим или парящим, полетом, журавли — пре-
имущественно гребным.
Для миграции журавлей характерны места сборов
птиц, расположенные на пути пролетов, в которых осенью
в течение нескольких недель скапливаются тысячи птиц.
Приложение
155
(Основное время пролета над Германией: 11—20 ок-
тября.) Птицы, которые не высиживали, держатся от-
дельно уже начиная с августа.
Перелеты чибиса
Чибис в Средней и Северной Европе — перелетная
птица, дальше на западе и юге — оседлая или кочующая.
Но иногда некоторые птицы зимуют даже в Швеции или
Дании, нередко в большом числе — в Германии и многие—
в Голландии и Бельгии, а также южнее (Швейцария,
Венгрия). Среди последних очень редко встречаются
птицы, гнездящиеся в указанных местах. Обычно это пти-
цы, прилетевшие с севера.
Области зимовок чибисов почти всех европейских по-
пуляций находятся на Британских островах, северном и
западном побережье Франции, в Испании и Португалии,
Северной и Средней Италии, а также на северо-западном
побережье Африки. Приблизительно половина британских
чибисов остается в Англии, многие отлетают в Ирлан-
дию, другие распространяются на юг. В Германии и Гол-
ландии эти птицы встречаются редко (рис. 50); один раз
чибис был обнаружен в Болгарии. Из Дании и Южной
Швеции, а также из Финляндии и прибалтийских стран
чибисы летят в Англию и Ирландию, а при сильном мо-
розе — на юг, охватывая всю обширную область между
Португалией и Италией. Представляет интерес находка
окольцованной птицы, летевшей от Эланда до Мадеры.
Среди зимующих на Британских островах птиц пока не
удалось обнаружить только венгерскую популяцию, пред-
ставители которой нередко переселяются в новые области
гнездования: в СССР, Западную Сибирь и Вестфалию.
Наряду с этим данный вид отличается также особенно
сильной приверженностью к гнездовой области.
О больших летных способностях чибиса говорят на-
ходки этих птиц в Исландии, Гренландии и даже Север-
ной Америке, куда в декабре 1927 г. занесло большую
стаю английских чибисов.
Сроки отлета чибисов с родины не установлены; по
этому поводу приводились разные данные; наиболее пра-
вильно, повидимому, мнение Гейра (1942): чибисы поки-
156
П риложение
дают гнездовые области в начале мая и совершают ти-
пичный ранний пролет — промежуточный пролет, — кото-
рый не приводит птиц к местам зимовки. У небольшой
части птиц этот промежуточный пролет вначале не на-
правлен, но чаще он совершается в одном направлении
к «промежуточным целям», расположенным в направле-
нии зимовок. От промежуточных целей птицы часто от-
летают очень поздно. Среди чибисов, гнездовые области
которых расположены в Англии, промежуточный перелет
совершают лишь немногие особи; перелет на зимовки про-
исходит поздно, и птицы летят без остановок.
ЛИТЕРАТУРА
Besserer I., Drost R., 1935. Ein Beitrag zum Kapitel „Vogelzug
und Elektrizitat", Vogelzug, 6.
Bi ssonnette T. H., 1933. Light and sexual cycles in starling and
ferrets, Quart. Rev. Biol., 8.
Bretscher K., 1923. Der Vogelzug in Suddeutschland, Mitt. Vogelw.
Bretscher K., 1926. Der Vogelzug in der Schweiz auf Grund von
Beobachtungen, Tierwelt.
Brouwer G. A., 1936. De invasie van de Steltkluut in 1935,
Ardea, 25.
Burckhardt D., 1948. Sammelbericht, Orn. Beob., 45.
Burger I. W., Bi ssonnette T. H., Doolittle H. D., 1942.
Some effects of flashing light on testicular activation in the
male starling, J. Exp. Zool., 90.
С о о к e W. W., 1915. Bird migration, U. S. Dept. Agriculture Bull., 185.
С r e u t z G., 1941. Zum Zug des grauen Fliegenschnappers, Vogelzug, 12.
C r e u t z G„ 1942. Ergebnisse der Verfrachtung von Grunfinken,
Vogelzug, 13.
Daanje A., 1936. Haben die Vogel einen Sinn fiir den Erdmagnetis-
mus, wie Deklination, Inklination und Intensitat, Ardea, 25.
Daanje A., 1941. Heimfindeversuche und Erdmagnetismus, Vogel-
zug, 12.
D а у A. M., 1949. North American Waterfowl, New York.
Dircksen R., 1932. Die Biologie des Austernfischers, der Brand-
seeschwalbe und der Kiistenseeschwalbe nach Beobachtungen
und Untersuchungen auf Norderoog, J. Orn., 80.
Dobben van W. H., Makkink G. F., 1934. Vogeltrek over Neder-
land, Org. Cl. nederl. Vogelk., 6.
Drost R., 1931. Zug gegen den Wind bei Finkenvogeln, Vogelzug, 2.
Drost R., 1934. Uber die Ursachen der Herbstzugrichtungen, Ber.
Ver. schles. Orn.
Drost R., 1938. Uber den Einfluss von Verfrachtungen zur Herbst-
zugzeit auf den Sperber, С. г. IX. Intern. Orn. Congr.
Drost R., 1941. Uber den Vogelzug auf Helgoland wahrend des
Krieges, Vogelzug, 12.
158
Литература
Drost R., 1949. Zugvogel perzipieren Ultrakurzwellen, Vogelwarte.
Drost R., Schiiz E., 1940. Uber den Zug der europaischen Bach-
stelzen, Vogelzug, 11.
Geyr von Schweppenburg H. Frhr., 1930. Zur Terminologie
des Vogelzuges, Proc. VII. Internal. ’Orn. Congr. Amsterdam.
Geyr von Schweppenburg H. Frhr., 1931. Zug bei starkem
Gegenwind, Vogelzug, 2.
Geyr von Schweppenburg H. Frhr., 1933. Zwischenzug,
Vogelzug, 4.
Geyr von Schweppenburg H. Frhr., 1933. Zur Theorie der
Leitlinie, Ardea, 22.
Geyr von Schweppenburg H. Frhr., 1937. Storchzug, Zugwin-
kel, Normalrichtung, Vogelzug, 8.
Geyr von Schweppenburg H. Frhr., 1939. Stuwbaan-Leitlinie,
Vogelzug, 10.
Geyr von Schweppenburg H. Frhr., 1941. ,,Zuglinien“, Vogel-
zug, 12.
Geyr von Schweppenburg H. Frhr., 1942. Zur Terminologie des
Kiebitzzuges, Vogelzug, 13.
Geyr von Schweppenburg H. Frhr., 1949. Zuggeselligkeit.
Ornithologie als biologische Wissenschaft, Heidelberg, C. Winter
Verlag.
Giers berg H„ Stadie R., 1934. Uber experimentelle Auslosung
des Zugtriebes durch weibliches Sexualhormon, Vogelzug, 5.
Goethe F., 1937. Beobachtungen und Untersuchungen zur Biologie
der Silbermowe auf der Vogelinsel Memmertsand, J. Orn., 85.
G r i f f i n D., 1940. Homing experiments with Leach’s Petrels, Auk., 57.
Griffin D., 1943. Homing experiments with Herring Gulls and Com-
mon Terns, Bird Banding, 14.
Groebbels F., 1932. Der Vogel, Band 1. Atmungswelt und Nah-
rungswelt, Berlin, Verlag Borntrager.
G г о t e H., 1930. Wanderungen und Winterquartiere der palaarktischcn
Zugvogel in Afrika, Mitt. Zool. Mus., Berlin, 16.
Grote H., 1937. Neue Beitrage zur Kenntnis der palaarktischen Zug-
vogel in Afrika, Mitt. Zool. Mus., Berlin, 22.
Hann H. W., 1939. The relation of castration to migration in birds,
Bird Banding, 10.
Harrison T. H., 1931. On the normal flight speed of birds, Brit.
Birds, 25.
Heinroth 0. und K., 1941. Das Heimfindevermogen der Brieftaube,
J. Orn., 89.
Литература
159
Hicks L. E„ 1938. Population studies of the european Starling in
America, С. г. IX, Congr. Orn. Internat., Rouen.
H i 1 p r e c h t A., 1935. Heimfindeversuche mit Wintervogel, Vogel-
zug, 6.
HoogerheideJ., KraakW. K-, 1942. Voorkommen en trek van de
Bergeend naar aanleiding van veldobservaties aan de Gooische
kust, Ardea, 31.
К a 1 e 1 a 0., 1942. Die Ausbreij'ing der kulturbedingten Vogelfauna
als Glied der spatquartaren Faunengeschichte Europas, Ornis
fenn., 19.
Kipp F., 1936. Studien fiber den Vogelzug im Zusammenhang mit
Fliigelbau und Mauserzyklus, Mitt. Vogelw.
Kipp F., 1942. Uber Fliigelbau und Wanderung der Vogel, Biol.
Zentralbl., 62.
Kluyver H. N., 1935. Ergebnisse eines Versuchs fiber das Heim-
findevermogen von Staren, Ardea, 24.
Koch I. C„ 1934. Vogelzug unter Einfluss von Leitlinie und Wind-
richtung, Vogelzug, 5.
Koehler 0., 1942. Zum Heimfinden der Tiere, Z. Tierpsychol., 5.
Kramer G., 1948. Neue Beitrage zur Frage der Fernorientierung der
Vogel, Orn. Ber.
Kramer G., 1949. Uber Richtungstendenzen bei der nachtlichen
Zugunruhe gekafigter Vogel. Ornithologie als biologische Wis-
senschaft, Heidelberg, C. Winter Verlag.
К r a t z i g H., S c h fi z E., 1936. Ergenbis der Versetzung ostbaltischer
Stare ins Binnenland, Vogelzug, 7.
Lack D., Lockley R. M., 1938. Stokholm bird observatory homing
experiments. I. Puffins, Storm-Petrels and Manx Shearwaters,
Brit. Birds, 31.
Lincoln F. C., 1935. The migration of the North American Birds,
US Dept. Agricult. circ. 363.
L i p p e n s L., 1943. Migration au-dessous des nuages, Gerfaut, 33.
Mayr E., Meise W., 1930. Theoretisches zur Geschichte des Vogel-
zuges, Vogelzug, 1.
M e i n e r t z h a g e n R., 1921. Some preliminary remarks on the velo-
city of migratory flight among birds, with special reference to
the palaearctic region, Ibis, XI, ser. 3.
Meise W., 1933. Kinasthetisches Gedachtnis und Fernorientierung der
Vogel, Vogelzug, 4.
Mennig S., 1940. A propos de 1’engourdissement des hirondelles.
Gerfaut, 29,
160
Литература
Merkel F. W., 1937. Zur Physiologie des Vogelzugtriebes, Zool.
Anz., 117.
Merkel F. W., 1940. Neuere Untersuchungen liber die Ursachen des
Vogelzug-Triebes, Natur u. Volk, 70.
Moreau R. E., 1927. Migration as seen in Egypt, Ibis, XII, 3.
Moreau R. E., 1938. Bird migration over the Nordwestern part of
the Indian Ocean, the Red Sea and the Mediterranean, Proc.
Zool. Soc., 108.
О о r d t G. J. van, 1943. Vogeltrek, Leiden, Verlag E. J. Brill.
Posingis M„ Schuz E., 1938. Massen-Winterflucht von Rebhiih-
nern am Kurischen Haff Dez. 1937, Vogelzug, 9.
Put zig P., 1936. Das Wunder des Vogelzuges, Bd. 5 in: Die Welt
im Fortschritt, Berlin, Verlag F. A. Herbig.
Put zig P., 1937. Von der Beziehung des Zugablaufs zum Inkret-
driisen system, Vogelzug, 8.
Put zig P., 1938. Beobachtungen liber Zugunruhe beim Rotkehlchen,
Vogelzug, 9.
P u t z i g P„ 1938. Weitere Versuche liber die Beziehungen der Keim-
drlisen zum Zugverhalten, Vogelzug, 9.
Putzig P., 1938. Die Triebkrafte des Vogelzugs, Die Umschau, 42.
Put zig P., 1938. Der Friihwegzug des Kiebitzes, J. Orn., 86.
Putzig P., 1939. Beitrage zur Stoffwechselphysiologie des Zugvo-
gels. I., Vogelzug, 10.
P u t z i g P., 1939. Problematik und Stand der Vogelzugforschung, Gef.
Welt, 68.
Rens ch B., 1938. Einwirkung des Klimas bei der Auspragung von
Vogelrassen, mit besonderer Beriicksichtigung der Fliigelform
und Eizahl, Proc. VIII. Intern. Orn. Congr., Oxford.
Riley-Gardner M., Wits chi E„ 1938. Comparative effects of
light stimulation and administration of gonadotropic hormones
on female Sparrows, Endocrinology, 23.
Rowan W., 1926. On photoperiodism, reproductive periodicity, and
the annual migration of birds and certain fishes, Proc. Boston
Soc. nat. Hist., 35.
Rowan W., 1929. Experiments in bird migration. I„ Proc. Boston
Soc. nat. Hist., 38.
Rowan W., 1932. Experiments in bird migration. III. Proc. nat. akad.
sci., 18.
R u p p e 11 W., 1934. Versuche zur Ortstreue und Fernorientierung der
Vogel. II. Verfrachtungsversuche am Star u. a. Arten von
W. Schein, Vogelzug, 5.
Литература
161
R u р р е 11 W., 1934. Versuche zur Ortstreue und Fernorientierung
der Vogel. III. Heimfindeversuche mit Rauchschwalben und
Mehlschwalben von H. Warnat, Vogelzug, 5.
Rflppell W., 1935. Heimfindeversuche mit Staren 1934, J. Orn., 83.
Rfippell W„ 1936. Heimfindeversuche mit Staren und Schwalben, J.
Orn., 84.
Rflppell W., 1937. Heimfindeversuche mit Staren, Rauchschwalben,
Wendehalsen, Rotruckenwiirgern und Habichten 1936, J. Orn., 85.
R fl p p e 11 W., 1938. Ergebnis eines Heimfindeversuchs mit aufgezoge-
nen Staren von Wilhelm Schein, Winsen, Vogelzug, 9.
Rflppell W., 1940. Neue Ergebnisse fiber Heimfinden beim Habicht,
Vogelzug, 11.
Rflppell W., 1942. Heimfinden ostbaltischer Nebelkrahen nach Ver-
frachtung wahrend des Frflhjahrsdurchzuges am Kurischen Haff,
J. Orn., 90.
Rflppell W., 1942a. Versuch einer neuen Storch-Karte, Vogelzug, 13
R fl p p e 11 W., S c h i f f e r 1 i A., 1939. Versuche fiber Winter-Ortstreue
an Larus ridibundus und Fulica atra. 1935, J. Orn., 87.
Rflppell W., Schein W., 1941. Uber das Heimfinden freilebender
Stare bei Verfrachtung nach einjahriger Freiheitsentziehung,
Vogelzug, 12.
Schiff er И A., 1942. Verfrachtungsversuche mit Alpenseglern.
Solothurn-Lissabon, Orn. Beob., 39.
Schildmacher H., 1933. Zur Physiologic des Zugtriebes, Vogel-
zug, 4.
Schiiz E., 1929. Der Vogelzug, Aus der Heimat, 42.
Schiiz E., 19*30. Ergebnisse der Vogelberingung, Proc. VII Intern.
Orn. Congr., Amsterdam.
Schiiz E., 1931. Alte und neue Versuche fiber das Sich-Zurechtfinden
der Vogel, Vogelzug, 2.
Schiiz E., 1937. Ringfunde europaischer Rauchschwalben, Orn. Mber.,
45.
Schiiz E., Wei go Id H., 1931. Atlas des Vogelzuges, nach den
Beringungsergebnissen bei palaarktischen Vogeln, Berlin.
Siivonen L., 1943. Artenstatistische Daten fiber die Veranderungen
in der Vogelfauna Finnlands wahrend der letzten Jahrzehnte,
Ornis fennica, 20.
Sliwinsky U,, 1938. Isopiptesen einiger Vogelarten in Europa,
Zool. Polon., 2.
Southern H. N., 1938. The spring migration of the Swallow over
Europe, Brit. Birds, 32.
162
Литература
Southern Н. N., 1938. The spring migration of the Willow Warbler
over Europe, Brit. Birds, 32.
S t a d i e R., 1938. Licht- und Schattenseiten des zugphysiologischen
Experiments, С. г. IX, Congr. Orn. Intern. Rouen.
Stimmelmayr A., 1930—193Й. Neue'Wege zur Erforschung des
Vogelzuges, Verh. Orn. Ges. Bayern, 19.
Stresemann E., 1927/1934. Aves. Handbuch der Zoologie, VII, 2.
Halfte.
Stresemann E., 1935. Haben die Vogel einen Ortssinn, Ardea, 24.
Stresemann E., 1944. Der Fruhjahrsdurchzug einiger Vogelarten
durch die Mittelmeerlander, Orn. Mber., 52.
Stresemann E., 1944. Die Wanderungen der Blauracke, Orn. Mber.,
52.
Stresemann E., 1944. Der zeitliche Ablauf des Friihjahrszuges
beim Neuntoter, Orn. Mber., 52.
Stresemann E., 1948. Die Wanderungen des Pirols, Orn. Ber.
Thienemann J., 1926. Rossitten, Neudamm, Verlag J. Neumann.
Thienemann J., 1931. Vom Vogelzug in Rossitten, Neudamm, Ver-
lag J. Neumann.
T h о m a s E. S., 1934. A study of starlings banded at Columbus, Ohio.
Bird Banding, 10.
T h о m s о n A. L., 1926. Problems of bird migration, London.
Tri eb H. E., 1939. Das Rotfilter im Vogelauge, Gebrauchsphotogra-
phie, 46.
Valikangas I., 1933. Finnische Zugvogel aus englischen Vogel-
eiern, Vogelzug, 4.
Wachs H., 1926. Die Wanderungen der Vogel, Ergebn. Biol., 1.
W e i g о 1 d H., 1924. Das Wetter und der Herbstzug der Waldschnepfe,
J. Orn., 72.
Wei go Id H., 1930. Der Vogelzug auf Helgoland, graphisch darge-
stellt, Berlin.
Wodzicki K-, Puchalski W., Liche H., 1939. Untersuchungen
fiber die Orientation und Geschwindigkeit des Fluges bei Vo-
geln, J. Orn., 87.
Wolfson A., 1941. Light versus activity in the regulation of the
sexual cycles of birds: the role of the Hypothalamus, Condor, 43.
Wolfson A., 1942. Regulation of spring migration in Juncos,
Condor, 44.
W о 1 f s о n A., 1945. The role of the pituitary, fat disposition and body
weight in bird migration, Condor, 47.
Wuczeticz W., Tugarinov A., 1941. International Wildfowl.
Inquiry, 1, Cambridge, University Press.
СОДЕРЖАНИЕ
Стр.
Предисловие к русскому изданию ............................ 5
Глава I. Введение.......................................... 9
Что понимается пйд перелетом птиц? .................. 9
Как узнают перелеты птиц? ........................ 11
Где происходят перелеты птиц? ...................... 15
Как возник перелет птиц? ........................... 17
К истории изучения перелетов птиц . . .............. 20
Глава II. Методы изучения перелетов птиц ................. 24
Полевые наблюдения.................................. 25
Кольцевание......................................... 39
Изучение ориентации птиц............................ 46
Изучение причин перелетов .......................... 49
Глава III. Результаты изучения перелетов птиц............. 52
Цели перелетов (места зимовок)..................... 53
Направления перелетов............................... 65
Пути перелетов ..................................... 72
Формы перелетов ................................... 86
Время перелетов..................................... 89
Скорость перелета .................................. 94
Высота пролета...................................... 99
Перелет как физиологическая проблема............... 102
Перелет и погода................................... 105
Ориентация при перелете.............................. Ш
Стимулы перелетов ................................. 130
Приложение .............................................. 144
Перелеты скворца................................... 144
Перелеты жулана ................................... 147
Перелеты белой трясогузки ......................... 148
Перелеты серебристой чайки......................... 149
Перелеты журавля................................... 153
Перелеты чибиса.................................... 155
Литература .............................................. 157
И. Штейнба хер
ПЕРЕЛЕТЫ ПТИЦ И ИХ ИЗУЧЕНИЕ
Редактор Н. В. КРЕНИЧ
Художник Б. Н. Гладков-
Технический редактор С. В. Клименко
Корректор И. Г, Янова
Сдано в производство 29,'II 1956 г.
Подписано к печати 13/VI 1956 г.
Т05416. Бумага 84Х1081/,,-=2, 6 бум. л.
8,4 печ. л. Уч.-изт. л. 8,3. Изд. № 4,273 3
Цена 7 р. 80 к. Зак, 176.
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва, Ново-Алексеевская, 52
20-я типография Главполиграфпрома
Министерства культуры СССР
Москва, Ново-Алексеевская, 52