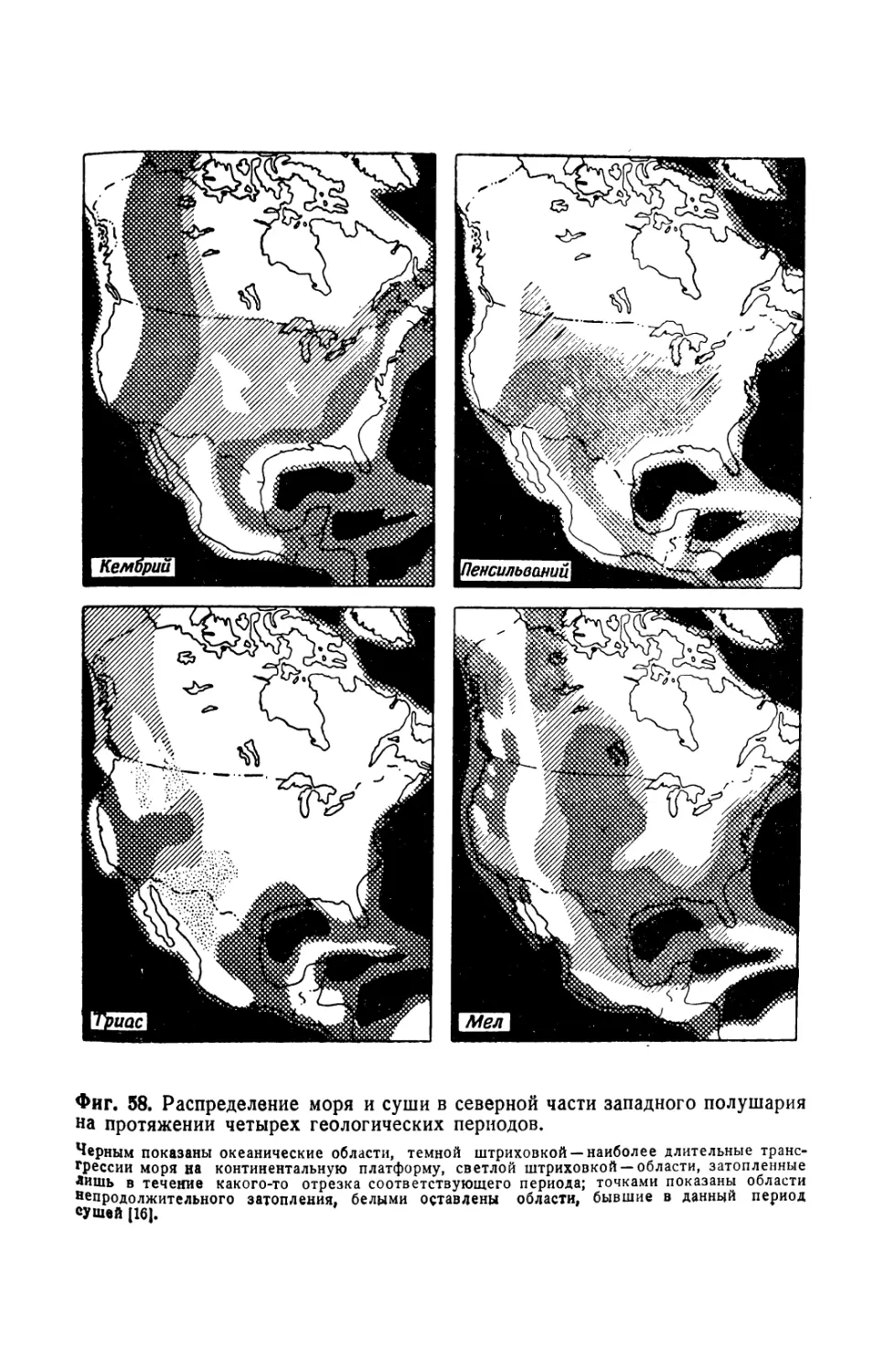
/









































































































































































































































































































































Text
IL&pwwx
P.XojVMf
Процесс
эволюции
ИЗДАТЕЛЬСТВО
«М И Р»
THE PROCESS OF EVOLUTION
Paul R. Ehrlich and Richard W, Holm
Department of Biological Sciences
Stanford University
19 6 3
McGRAW-HILL BOOK COMPANY, INC.
NEW YORK, SAN FRANCISCO, TORONTO, LONDON
\ II. Эрлилс
Р. Холм
ПРОЦЕСС ЭВОЛЮЦИИ
ПЕРЕВОД С АНГЛИЙСКОГО
10, И, Л а ши е в и ч а
ПОД РЕДАКЦИЕЙ П С ПРЕДИСЛОВИЕМ
просф, В, В, Алпатова
и
д-ра биол, наук В, Н, Сидорова
ИЗДАТЕЛЬСТВО „N И Р«
МОСКВА 19 3 6
УДК 575 + 576.1
Книга посвящена важнейшей проблеме биологии — эволюци-
онному процессу. В ней изложены современные представления о
происхождении жизни, об эволюции животных и растений, о фак-
торах и механизмах, управляющих этим процессом, и особенно
о его генетических основах. Большое внимание уделено общим
вопросам экологии и генетики популяций, которые за последнее
время мало освещались в нашей литературе. Книга написана яс-
ным, простым языком и хорошо иллюстрирована. Кроме того, к
ней приложен толковый словарь основных терминов, что облег-
чает ее чтение и позволяет пользоваться ею в качестве справоч-
ника.
Книга может служить учебным пособием при изучении об-
щей биологии и генетики.
Предназначена для студентов, аспирантов и преподавателей
биологических факультетов университетов, сельскохозяйственных,
педагогических и медицинских институтов, для преподавателей
средней школы, а также для биологов разных специальностей.
Редакция биологической литературы
ПРЕДИСЛОВИЕ К РУССКОМУ ИЗДАНИЮ
После выхода в свет «Происхождения видов» Чарлза Дар-
вина эволюционная идея стала стержневой идеей не только био-
логии, но и других наук, соприкасающихся с теоретическими и
практическими аспектами биологии. Представление, согласно
которому все разнообразие живых существ на Земле возникло
в результате действия законов природы, и знание движущих
факторов и механизмов эволюционного процесса важны в двух
отношениях. Во-первых, представление об эволюции чрезвычай-
но сильно укрепляет мировоззрение, основанное на диалектиче-
ском материализме. Во-вторых, знакомство с закономерностями
эволюции позволяет понять очень важные явления окружающей
нас живой природы и помогает разрабатывать практические
приемы для управления этой природой. Не все отдают себе от-
чет в том, что эволюция живых существ отнюдь не является
процессом, характерным только для далеких доисторических
времен. Это процесс, непрерывно идущий и в наши дни. Приве-
дем некоторые примеры эволюции, протекающей буквально на
наших глазах. Человеку постоянно приходится сталкиваться, с
одной стороны, с вредными для него организмами, и с другой —
с полезными. К вредным организмам в первую очередь отно-
сятся многочисленные насекомые — вредители сельского хозяй-
ства, паразиты человека и сельскохозяйственных животных. Хи-
мические, физические и механические меры борьбы с вредителя-
ми на фоне быстрой смены поколений у насекомых приводят к
образованию новых, устойчивых к разнообразным воздействиям
рас. Это вызывает необходимость в создании все новых и но-
вых препаратов и методов, которые позволяли бы бороться с
возникающими устойчивыми формами (примером таких форм
могут служить ДДТ-устойчивые расы насекомых-вредителей).
Человеку с давних времен приходится заниматься усовершен-
ствованием полезных организмов — создавать новые породы
сельскохозяйственных животных и сорта растений. По существу
племенное дело и селекЦия растений представляют собой не что
иное, как биотехническую эволюцию, управляемую человеком.
Особенно наглядны результаты такой эволюции у микроорганиз-
мов. Так, под влиянием химиотерапевтических препаратов на
наших глазах создаются устойчивые расы патогенных микробов,
6 | Предислозие к русскому изданию
Приходится поэтому все время создавать новые и новые препа-
раты. Иногда говорят, что медицина в настоящее время пере-
живает эпоху антибиотиков. В самом деле, по статистическим
данным, собранным, например, в Англии, чаще любых других
лекарств врачи выписывают антибиотики. Успех фармацевтиче-
ской промышленности, производящей антибиотики, в послевоен-
ные годы объясняется исключительно успехами селекционной
работы с организмами, которые являются продуцентами анти-
биотиков. Использование культурных штаммов продуцентов
вместо исходных штаммов в сотни раз повышает производи-
тельность заводов.
В отечественной литературе давно не появлялось книг, по-
священных проблемам эволюции с позиций генетики, цитологии
и экологии. Предлагаемая читателю книга написана двумя пре-
подавателями биологического отделения Стэнфордского универ-
ситета в Калифорнии, П. Эрлихом и Р. Холмом, авторами мно-
гих работ по экологической генетике, изменчивости и эволюции.
Оригинальные рисунки изготовлены Анной Эрлих. В книге ши-
роко освещены многообразные факторы и механизмы эволюци-
онного процесса и дан их глубокий анализ.
Авторам книги можно сделать один упрек. Они уделили мало
внимания количественным закономерностям борьбы за существо-
вание и явлениям отбора в экспериментальных и природных ус-
ловиях. Инициаторами изучения этих проблем были итальян-
ский математик Вито Вольтерра и американский статистик Аль-
фред Лотка. Для ознакомления с этим направлением можно
рекомендовать три монографии; две из них написаны советским
ученым проф. Г. Ф. Гаузе1.
При переводе из книги выпущены две главы, посвященные
эволюции человека. Вопрос об эволюции человека и его куль-
туры столь сложен, что 35 страниц книги, которые уделены это-
му вопросу и написаны к тому же несколько поверхностно, дают
читателю чрезвычайно мало.
В. В. Алпатов
Б. Н. Сидоров
1 Гаузе Г. Ф., The struggle for existence, Baltimore, 1934; Гаузе Г. Ф.,
Verifications experimentales de la theorie mathematique de la lutte pour la
vie, Paris, 1935; Umberto D’Ancona, The struggle for existence, Leiden,
1954.
ПРЕДИСЛОВИЕ
Современная эволюционная теория — это великая объеди-
няющая концепция биологии. Она представляет собой триумф
теоретической мысли в области биологических наук, всеобъем-
лющую теорию, которая стремится объяснить все многообразие
и сложность биологических явлений. Биохимик, пытающийся
расшифровать генетический код, нейрофизиолог, исследующий
сложные механизмы, лежащие в основе сознания, эмбриолог,
старающийся понять, каким образом одна ткань влияет на раз-
витие другой, — в сущности все биологи работают над пробле-
мами, теоретическое значение которых измеряется только их
вкладом в понимание эволюционных явлений. Биохимик сможет
в конце концов найти путь к излечению рака, нейрофизиолог —
понять природу психических расстройств, эмбриолог — раскрыть
процесс построения организма на основе генетического кода. Но
без теории, связывающей все эти явления между собой, их ра-
бота имела бы только прикладное значение.
Центральное положение теории эволюции в биологии давно
признано. Тем не менее большинство неспециалистов и многие
биологи плохо представляют себе современную эволюционную
теорию. В этой книге сделана попытка по возможности кратко
рассмотреть проблемы эволюции органического мира. Книга на-
писана для читателя, которого больше интересует процесс эво-
люции, чем ее результаты как таковые. Он не найдет в ней изо-
бражений динозавров, таксономических описаний разнообразных
представителей органического мира и экскурсов в историю эво-
люционной мысли.
Мы предполагаем, что наши читатели достигли по крайней
мере того уровня биологической подготовки, который дает пол-
ный университетский курс биологии. Мы надеемся, что эта книга
послужит дополнительным материалом для студентов, изучаю-
щих эволюционную теорию, основным курсом для аспирантов и
просто полезной книгой для биологов других специальностей,
которым может понадобиться краткий обзор того, что известно
о процессе эволюции. Большая часть излагаемого материала
была использована в курсе лекций по эволюционной теории для
студентов и в лекциях по избранным проблемам эволюции, ко-
торые читались в Стэнфордском университете.
8 | Предисловие
Мы сделали попытку представить эволюционную теорию как
единое целое. При этом мы исходили из допущения, что чита-
тель имеет хотя бы поверхностное представление о таких явле-
ниях, как, например, отбор или митоз; поэтому мы считали воз-
можным упоминать о них еще до того, как они рассматриваются
подробно. Жизнь, мейоз, генетические системы и т. п. не прини-
маются в книге как нечто само собой разумеющееся. Напротив,
мы пытаемся показать, что сами эти явления возникли в резуль-
тате эволюционного процесса. Это неизбежно влечет за собой
построение гипотез, что, как нам кажется, небесполезно и сти-
мулирует мысль. Важно, однако, не забывать, что это всего лишь
гипотезы. Многие эволюционисты, несомненно, найдут, что в не-
которых вопросах мы отходим от общепринятых представлений.
В частности, мы не придаем слишком большого значения кон-
цепциям вида и подвида, которые, по нашему мнению, сужают
пути мышления биологов в области эволюционных проблем.
Адаптации мы отводим сравнительно второстепенную роль, как
она того и заслуживает по нашему мнению.
Мы старались сделать наши описания и рассуждения как
можно более строгими. Обычно мы старались избегать таксоно-
мических концепций или употребления таких слов, как отбор
или адаптация, кроме тех случаев, когда это было совершенно
необходимо. Встречающиеся иногда выражения, которые могут
показаться телеологическими, следует понимать в телеономиче-
ском смысле. Как нам кажется (и мы надеемся, что читатель
согласится с нами), можно дать более цельную и логичную
интерпретацию эволюционных явлений, если не ограничивать
себя строгими рамками таксономии. Научные названия, упо-
требляемые в этой книге, указывают лишь на определенные
организмы и не связываются с какими-либо генетическими атри-
бутами или представлениями о филогении.
В конце каждой главы приводится список литературных
источников, преимущественно тех, которые легко достать и ко-
торые содержат обширную библиографию. Авторы, с именами
которых связаны определенные концепции или эксперименты,
часто упоминаются в тексте без прямых ссылок на источник;
точные ссылки можно найти в обзорных работах, приведенных
в списке. В конце книги дан довольно обширный словарь -тер-
минов.
77. Эрлих
Р, Холм
Часть первая
ОРГАНИЗМЫ,
ИХ ПРОИСХОЖДЕНИЕ
И ФУНКЦИИ
Книга «Процесс эволюции» делится на три части:
1) происхождение и функции организмов; 2) свойства
популяций; 3) пути дифференциации популяций и свя-
занные с нею типы филогенетических процессов.
Первая часть посвящена главным образом таким
вещам, которые часто при обсуждении проблем эво-
люции принимаются как нечто само собой разумею-
щееся. Между тем основные свойства жизни сами яв-
ляются продуктом эволюции. В первых главах дается
очерк некоторых свойств живых систем, имеющих кар-
динальное значение для исследования эволюции. Осо-
бое внимание уделяется способам поддержания не-
прерывности информации в циклическом потоке жиз-
ни и способам усовершенствования этой информации.
Там, где это возможно, приводятся более или менее
обоснованные гипотезы о путях возникновения меха-
низмов, общих для всех форм жизни, и о ранних эта-
пах их эволюции. Подобные гипотезы, какими бы не-
точными они ни оказались впоследствии, должны
напоминать нам о том, что, например, фотосинтез,
ДНК, мейоз, явление доминантности генов и клеточ-
ная дифференцировка не всегда существовали в их
современной форме. Мы отнюдь не стремились дать
энциклопедический обзор этих важнейших проблем
биологии; скорее мы пытались создать некоторую ос-
нову, которая позволила бы исследовать, каким обра-
зом в процессе эволюции возникли те или иные орга-
низмы в их современном виде.
Глава 1
ПРОИСХОЖДЕНИЕ ЖИЗНИ
Мы настолько привыкли к аксиоме, согласно которой все
живое происходит от живого, что редко задумываемся над тем,
каким образом жизнь возникла впервые. Древние решали эту
проблему, основываясь на представлении о самопроизвольном
зарождении из неживой материи даже таких сложных организ-
мов, как мухи и мыши. Однако эти идеи, так же как и более
хитроумные представления подобного рода, были опровергнуты
экспериментами Реди и Пастера. Но при этом на первый план
выступил основной вопрос: как же возникла жизнь? Каким об-
разом, совершенно исключив самопроизвольное зарождение, мо-
жно объяснить появление бесчисленного множества существ,
которых мы называем «живыми»? Эту проблему часто запуты-
вала тенденция отождествлять жизнь со свойствами сложных
организмов. Контраст между птицей и камнем или между бак-
териями и железными опилками очевиден; он столь велик, что
различие между живым и неживым можно было бы ошибочно
истолковать как качественное, а не количественное.
Огромное большинство биологов полагает, что между живым
и неживым нет резкой границы, хотя эти биологи могут при-
держиваться разных точек зрения относительно того или иного
определения «жизни» или даже относительно «свойств» живого.
Можно обойти многие трудности, рассматривая жизнь просто
как особое свойство материи, возникающее на определенном
уровне сложности, и не пытаясь дать ей строгое определение.
Во всяком случае можно сказать, что живые системы регули-
руют обмен энергии таким образом, чтобы создать энергетиче-
ский потенциал между организмом и окружающей его средой;
несомненно, одно из самых основных свойств живого состоит в
непрерывном и направленном перемещении электронов между
сложными молекулами, из которых построены живые организмы.
Важно отметить, что эти превращения энергии строго контро-
лируются. Регулируемое освобождение энергии в таком количе-
стве, которое при отсутствии контроля вызвало бы небольшой
взрыв, дает то, что мы понимаем под словом «жизнь». Кроме
того, живые системы обладают способностью воспроизводить са-
мих себя. Таким образом, исследуя проблему происхождения
жизни, нужно искать ответы на вопрос о том, как могли воз-
никнуть системы, извлекающие и использующие энергию
12 | Глава 1
окружающей среды, и как они могли воспроизводить себя. По-
видимому, существует своего рода переходная зона между жи-
вым и неживым — область, в которой эти понятия могут ока-
заться неприменимыми. В этой-то области, куда относятся ви-
русы, нуклеиновые кислоты и специализированные формы кол-
лоидов, возможно, и следует искать ответы на некоторые из
интересующих нас вопросов.
Кажется маловероятным, чтобы спонтанное возникновение
жизни можно было наблюдать в настоящее время. Если бы но-
вая жизнь и зародилась самопроизвольно, она, вероятно, была
бы быстро уничтожена современными гетеротрофными организ-
мами, даже если бы среда оказалась благоприятной для нее.
Совершенно иначе обстояло дело в условиях, которые, видимо,
существовали на Земле миллиарды лет назад.
РАННИЕ ЭТАПЫ ЭВОЛЮЦИИ
Необходимым условием возникновения той жизни, которую
мы знаем, является образование органических веществ — соеди-
нений, построенных на основе углерода, в которых атомы этого
элемента связаны в различных конфигурациях с атомами азота,
кислорода, водорода, фосфора и серы. На ранних этапах хими-
ческой эволюции земной поверхности, вероятно, существовали
гораздо более простые, неорганические молекулы. В связи с
этим сразу же возникает вопрос: как из этих простых веществ
синтезировались более сложные соединения, характерные для
живых систем, и что могло служить источником энергии для та-
ких превращений?
Имеется довольно много фактических данных, подтверждаю-
щих тезис Опарина о том, что примитивная атмосфера Земли
носила восстановительный характер и состояла главным обра-
зом из метана, водяных паров, аммиака и водорода. Было изу-
чено поведение этих веществ при разнообразных условиях. Мил-
лер помещал смесь этих газов в специальный аппарат (фиг. 1),
в котором на них можно было воздействовать электрическими
разрядами. В одной части этого аппарата воду доводили до ки-
пения, а в другой части пары конденсировались; таким образом
создавалась циркуляция газов. С помощью хроматографического
анализа, проведенного в конце эксперимента, удалось обнару-
жить присутствие в реакционной смеси аминокислот, оксикислот
и алифатических кислот, т. е. трех основных типов органических
молекул, в том числе и структурных компонентов белка. Из
аминокислот в реакционной смеси были обнаружены глицин и
аланин (аминокислоты, наиболее распространенные в белках),
аспарагиновая кислота и глутаминовая кислота. Интересно отме-
Происхождение жизни | 13
тить, что а-аланин преобладал в этих смесях над р-аланином;
белки современных организмов содержат только а-аминокисло-
ты (в а-аминокислотах группы NH2 и СООН присоединены к
одному и тому же атому углерода). Миллер полагает, что эти
же типы соединений должны были синтезироваться под влия-
нием ультрафиолетовых лучей и электрических разрядов, если
примитивная атмосфера Земли действительно носила восстано-
вительный характер. Он утверждает также, что в атмосфере
окислительного типа образование органических веществ было бы
невозможно, и указывает, что если аминокислоты (и другие ор-
ганические соединения) необходимы для жизни, то наличие
жизни на Земле является свидетельством восстановительного
характера примитивной атмосферы.
Идея о восстановительном характере примитивной атмо-
сферы получила сейчас широкое признание. Свободный кисло-
род, обусловливающий окислительные свойства современной ат-
мосферы, появился примерно 800 млн. — 2 млрд, лет назад или
даже раньше; источником его были фотохимические реакции
(фотолиз воды в верхних слоях атмосферы; образовывавшийся
при этом водород улетучивался в межпланетное пространство)
и фотосинтез в клетках живых организмов (в настоящее время
это главный источник свободного кислорода на Земле).
Фиг. 1. Аппарат для воздействия
искровыми разрядами на смеси
газов [7].
14 | Глава 1
Помимо обрисованного выше пути абиогенного синтеза орга-
нических веществ, простые органические соединения могут воз-
никать также при взаимодействии водяного пара с карбидами
в магме, выносимой на поверхность в результате вулканической
деятельности (ЗРетСп + 4тН2О = тРезО4-ЬС3пН8т). Как показа-
ли Кальвин и другие исследователи, проводившие эксперименты
с ионизирующими излучениями такого рода, источником кото-
рых могли бы быть радиоактивные вещества или космические
лучи, в присутствии молекулярного водорода может происхо-
дить частичное восстановление углекислоты. Дальнейшее облу-
чение водных растворов получаемых при этом веществ (муравь-
иной кислоты, формальдегида) ведет к образованию таких
соединений, как щавелевая и уксусная кислоты. Затем молеку-
лы с 2 углеродными атомами (уксусная кислота) могут соеди-
няться друг с другом, образуя вещество с 4 углеродными ато-
мами (янтарную кислоту). В этих экспериментах происходит
также образование аминокислот.
Фокс показал, что при нагревании смесей сухих аминокислот
образуются синтетические полипептиды (протеиноиды). В воде
протеиноиды склонны образовывать сферические тела, так назы-
ваемые микросферы, имеющие различные размеры и форму в
зависимости от характера взаимодействия этих полипептидов с
другими веществами, присутствующими в реакционной смеси.
Эти микросферы в некоторых отношениях напоминают коацер-
ваты и другие модели клетки. Между прочим, протеиноидные
микросферы способны сохраняться в течение довольно долгого
времени и не разрушаются при высокоскоростном центрифуги-
ровании.
ВОЗНИКНОВЕНИЕ САМОВОСНРОНЗВОДЯЩИХСЯ СИСТЕМ
Таким образом, в ранний период истории Земли могло суще-
ствовать несколько различных путей образования органических
соединений. Поэтому есть основания предполагать, что первич-
ный океан представлял собой нечто вроде разбавленного «буль-
она» из органических веществ. Не существует единого мнения
о том, каким образом в этом «бульоне» возникли первые само-
воспроизводящиеся системы. Очевидно, первым условием для
этого должно было быть избирательное построение определен-
ных молекул. Кальвин указал на то, что автокатализ носит ха-
рактер избирательного процесса. Автокатализом называют яв-
ление, заключающееся в том, что продукт химической реакции
обладает свойством каталитически влиять на скорость своего
собственного образования. В результате происходит постепенное
накопление все более и более сложных веществ, образующихся
Происхождение жизни | 15
из веществ более простых. Отбор такого типа (на сложность),
вероятно, и происходил в «органическом бульоне».
Автокаталитические реакции можно уподобить самовоспро-
изводящейся живой системе лишь отчасти. Ни одно из извест-
ных ныне веществ, будучи изолировано, не сможет воспроизво-
дить себя. Такой способностью обладают только системы ве-
ществ. Знакомые нам живые системы построены из белков и
нуклеиновых кислот, а также содержат некоторые средства для
мобилизации энергии. Полипептидные цепи — основа структуры
белковых молекул — строятся путем соединения аминокислот в
линейной последовательности. Образование связей между ами-
нокислотами в процессе удлинения цепи сопровождается выде-
лением воды. Связи между аминокислотными мономерами носят
название пептидных связей. Спонтанный синтез даже неболь-
ших белковых молекул в растворе аминокислот требует затраты
внешней энергии и представляет собой весьма маловероятное
событие. Однако в отсутствие свободного кислорода и организ-
мов-хищников аминокислотный «бульон» мог бы существовать
чрезвычайно долгое время — достаточно долго для того, чтобы
невероятное стало вероятным. (Шансы быть пораженным мол-
нией за 70 лет жизни весьма незначительны, но если бы человек
жил 7000 млн. лет, это случилось бы с ним почти наверняка.)
Однако, как указывал Уолд, тенденции к спонтанному обра-
зованию белковых молекул противостоит тенденция к самопро-
извольному их распаду. Действительно, равновесие обратимой
спонтанной реакции образования белка сдвинуто в сторону рас-
пада, а не синтеза. В то же время Уолд высказывает мысль, что
молекулы, по-видимому, способны противостоять тенденции к
распаду как за счет увеличения своих размеров, так и за счет
агрегирования с другими молекулами. Можно предположить,
что белки как бы находятся в положении неустойчивого равно-
весия, т. е. могут либо расщепляться на составляющие амино-
кислоты, либо образовывать более стойкие агрегаты. Вполне
возможно, что первые «организмы» возникли именно в резуль-
тате образования все более и более крупных агрегатов.
ИСТОЧНИКИ ЭНЕРГИИ
Обычно считают, что главным источником энергии для про-
цессов синтеза, протекавших на примитивной Земле, служили
ультрафиолетовые лучи. Простые молекулы поглощают лишь
очень короткие волны в этой области спектра, но по мере того,
как появлялись более сложные молекулы, они начинали погло-
щать более длинные волны. После того как вокруг Земли обра-
зовались плотные слои атмосферы, коротковолновые лучи
16 | Глава 1
ультрафиолетовой области спектра не могли более достигать
земной поверхности и использоваться в качестве источника энер-
гии. С возникновением пигментов (например, порфиринов, о ко-
торых будет сказано ниже) появилась возможность реализовать
энергию видимой области спектра. Однако каким бы ни был
источник энергии, от поглощения кванта энергии еще довольно
далеко до его использования в биологических процессах.
Особенно сложны проблемы использования энергии для бел-
кового синтеза и вопрос об условиях, при которых такое исполь-
зование было бы возможным. В современных биологических си-
стемах ферменты, ответственные за мобилизацию энергии и за
самый синтез, являются белками. Таким образом, если мы по-
стулируем участие таких систем в образовании первых белков,
то вновь окажемся перед вечной проблемой: что раньше — ку-
рица или яйцо? Было высказано предположение, что в отсут-
ствие белков катализаторами могли служить иные вещества
(например, глиноземы), ибо многие из известных нам фермента-
тивных процессов в своей основе представляют собой реакции
на поверхности молекул. В связи с этим возникает вопрос, ка-
ким образом эта функция впоследствии перешла к белкам. Как
бы то ни было, мы знаем, что явления, имеющие место на по-
верхности глин, в некоторых отношениях сходны с явлениями,
наблюдаемыми на поверхности белков.
В современных биологических системах химическая энергия
для синтезов запасается в фосфатных связях органических со-
единений; при расщеплении таких связей освобождаются очень
большие количества энергии. Энергия, выделяемая при разрыве
и переносе этих связей, регулируется сложной системой катали-
заторов (ферментов с их коферментами). Характеристики этих
комплексов реакций являются уникальными для живых систем.
Скорость этих реакций способна изменяться в ответ на измене-
ния концентрации продукта, а их зависимость от физических
условий (температуры, давления) носит особый характер, не
свойственный реакциям, протекающим в неживых системах; на-
конец, они отличаются высокой степенью сохранения химиче-
ской энергии, нечасто наблюдаемой в неорганических процессах.
Подобные реакции в живых системах в совокупности называют
биологическим окислением. Энергия, необходимая гетеротроф-
ному организму, почти во всех случаях записывается в фосфор-
ных связях макроэргического (богатого энергией) соединения,
называемого аденозинтрифосфатом (АТФ). При освобождении
энергии в ходе биологической реакции от АТФ отщепляется одна
фосфатная группа, и он превращается в аденозиндифосфат
(АДФ). В АДФ остается еще одна макроэргическая фосфатная
связь, и он очень легко подвергается рефосфорилированию в
Происхождение жизни | 17
АТФ; это фосфорилирование АДФ (называемое окислительным
фосфорилированием) осуществляется при участии системы фер-
ментов, о которых говорилось выше. Эти ферменты в свою оче-
редь нуждаются в энергии расщепления связей АТФ, но суще-
ственно здесь то, что на каждую связь АТФ, отдавшую свою
энергию для работы этих ферментов, образуется более одной
такой же связи. Добавочная энергия извлекается из молекул
глюкозы или других соединений, на которые ферменты дейст-
вуют непосредственно. В результате функционирования этой
цепи взаимосвязанных реакций химическая энергия, получаемая
организмом с молекулами углеводов, липидов или белков пищи
в форме, недоступной для непосредственного использования,
преобразуется в энергию связей АТФ.
В неживых системах при экзотермических реакциях расщеп-
ления образуются молекулы меньшего размера и освобождается
значительное количество тепла. В живых системах образуются
макроэргические фосфатные связи, молекулы меньшего размера
и выделяется поразительно мало тепла. В самом деле, одна из
важнейших особенностей энергетических механизмов в живых
системах — тесное сопряжение реакций, связанных с освобожде-
нием и запасанием энергии, благодаря чему энергия может быть
сохранена для последующего использования. Разумеется, пер-
вичным источником энергии для всех современных организмов
(за исключением хемосинтезирующих бактерий) служит солнеч-
ный свет, энергия которого улавливается фотосинтезирующими
организмами и в конечном счете запасается в форме фосфатных
связей в результате фотофосфорилирования — процесса, сход-
ного с описанным выше процессом, — или же в молекулах угле-
водов, синтезируемых также при участии процессов фосфорили-
рования. Многие важные ферменты фотосинтеза и биологиче-
ского окисления представляют собой окрашенные комплексы,
содержащие ионы металлов (Fe, Mg) и органические соедине-
ния, называемые порфиринами. Кальвин построил схему
(фиг. 2), показывающую, как могли бы возникнуть такого рода
важные для жизни вещества в процессе химического отбора,
связанного с автокатализом. В этом ряду постепенно усложняю-
щихся молекул образующиеся продукты служат катализатора-
ми для последующих этапов синтеза. Поскольку порфирины ис-
пользуются многими нефотосинтезирующими организмами,
Кальвин предполагает, что небольшие изменения случайного
характера в порфиринах могли привести в конце концов к воз-
никновению хлорофилла и к «изобретению» фотосинтеза. Гра-
ник, кроме того, полагает, что все окрашенные соединения в
ряду, ведущем к хлорофиллу, возможно, выполняли ту же
Функцию, что и хлорофилл. На ранних этапах химической эво-
2 Зак. 152
18 | Глава 1
люции ионы металлов, входящие в состав минералов, могли
катализировать те же реакции, которые они катализируют сей-
час, входя в состав металлсодержащих ферментов.
Фотосинтез представляет собой сложную последовательность
реакций. Некоторые из них могут протекать в темноте, тогда
как другие возможны только на свету. Большинство тех много-
численных реакций, которые обычно объединяют под названием
фотосинтеза в широком смысле, относятся в действительности
к «темновым» реакциям, связанным с присоединением СО2 к це-
пям С—С—С—. Такие темновые реакции могут осуществляться
большинством клеток. Кажется вероятным, что эти реакции воз-
никли независимо и, быть может, даже раньше, чем реакции
фотосинтеза в строгом смысле. При световой реакции происхо-
дит образование восстанавливающего агента (обычно это во-
дород, но в некоторых случаях — фосфорное соединение) и ка-
кого-либо макроэргического фосфата, вероятно адепозинтрифос-
фата (АТФ). Затем при участии этих веществ осуществляется
цикл восстановления углерода и синтезируются молекулы саха-
ров — гексоз. Энергия каким-то еще не вполне выясненным спо-
собом мобилизуется таким образом, что возбужденный хлоро-
филл трансформирует другие молекулы, в результате чего обра-
Янтарная кислота
СО2Н
СН2 СО2Н
СН2 сн2
СО2н СН2
у U
СН2-СО2Н /СН—СО2Н
nh2 nh2
Глицин a-NHz-/3Kemo-
адипиновая кислота
со2н
I
СО2Н сн2
сн2 сн2
». Н2С-О=С п стади^
.С, -СИ,
ын2—СН2 II
° Н2
д- Дминолевули-
новая кислота
Протопорфирин 9
Fe-
Порфирины
Фиг. 2. Возможные этапы синтеза порфириновых соединений из молекул,
образующихся случайным образом под действием излучения.
В пунктах, отмеченных стрелками, катализатором могло служить железо. В соединении
с порфирином железо является гораздо более эффективным катал иатором. Таким образом,
синтез протопорфирина 9 является автокаталитическим процессом [1].
Происхождение жизни | 19
зуются восстанавливающий агент и АТФ. Итак, фотосинтез в
типичном случае включает фотофосфорилирование, т. е. преоб-
разование света в «энергетическую валюту» фосфатных связей.
Вероятно, никогда нельзя будет сказать, на каком именно
этапе в процессах биосинтеза стали принимать участие окрашен-
ные соединения — до или после появления систем, которые мы
сегодня назвали бы живыми. Кальвин полагает, что конечный
этап в развитии современного фотосинтеза — выработка способ-
ности к выделению кислорода—возник сравнительно недавно.
Таким образом, реакции вроде тех, какие протекают у некото-
рых современных фотосинтезирующих организмов, неспособных
выделять кислород, появились раньше.
ВОЗНИКНОВЕНИЕ СТРУКТУРЫ
В свете сказанного выше не так уж трудно представить себе,
каким образом могли возникнуть вещества, необходимые для
протекания процессов, совокупность которых мы определяем как
«метаболизм». Однако живые системы не являются жидкими
бесструктурными образованиями. Обычно они обладают специ-
фической и сложной организацией и содержат механизм преоб-
разования энергии. Поэтому необходимо рассмотреть факторы,
связанные с эволюцией не только функции, но и структуры.
В первичном океане примитивные молекулы, вероятно, были
рассеяны в виде довольно однородной коллоидной суспензии.
Однако в коллоидных растворах различных веществ образуются
полужидкие коллоидные гели, или коацерваты, и можно предпо-
лагать, что они действительно возникали по мере того, как «ор-
ганический бульон» становился все более сложным. Из физиче-
ской химии мы многое знаем о свойствах коацерватов. Часто
они не образуют сплошного слоя, а выделяются из равновесной
жидкой системы (которая при этом обедняется коллоидами) в
форме обособленных капелек. Эти капельки не только способны
концентрировать в себе органические частицы высокого молеку-
лярного веса, но и обладают определенной внутренней структу-
рой, а также хорошо развитой поверхностью, отделяющей их от
равновесной жидкости. Каплю коацервата можно рассматривать
как первое отчетливое обособление структурного комплекса ор-
ганического вещества от окружающей его среды.
А. И. Опарин предполагает, что коацерватные капли в изве-
стном смысле конкурировали между собой за «питательные» ве-
щества, причем некоторые из этих капель, случайно обладавшие
благоприятным химическим составом или внутренней структу-
рой, росли быстрее других. Эти «более приспособленные» капли
Первыми достигали размеров, при которых они становились
2*
20 | Глава 1
неустойчивыми и распадались на более мелкие частицы. Послед-
ние затем росли, снова делились, и таким образом процесс про-
должался. Можно также представить себе, что в результате
случайного слияния капель друг с другом их размеры начинали
превышать предел, при котором сохраняется устойчивость, что
приводило к их распаду на капельки меньшей величины. Если
сливающиеся капли имели разный химический состав, то про-
исходил своего рода первичный процесс половой рекомбинации.
Таким образом, у коацерватов имеется целый ряд свойств, ко-
торые позволяют рассматривать их как звенья цепи, ведущей
к структуре живого вещества в том виде, в каком она нам изве-
стна, а поскольку структура связана с функцией, то и к живым
системам знакомого нам типа. Они отграничены от окружаю-
щей среды, обладают внутренней структурой, поглощают извне
«питательные» вещества и в достаточной мере способны к раз-
множению и «рекомбинации», чтобы служить материалом для
естественного отбора.
Кажется очевидным, что механизмы, подобные описанным
выше (а также другие, пока еще не открытые механизмы), дей-
ствуя на протяжении геологического времени, привели к появле-
нию предков современных живых -систем. В самом деле, если
представить себе безбрежные океаны, озера и горячие источ-
ники, богатые органическими веществами и создающие огром-
ное разнообразие условий температуры, освещенности, концен-
трации солей, а также физических субстратов (кристаллы,
глины), то трудно будет не поверить, что живые системы воз-
никали, и притом не однажды. Вполне возможно, что совре-
менные организмы являются потомками организма, вышедшего
победителем в жестокой борьбе за энергию, которая шла ме-
жду первичными организмами.
ВОЗНИКНОВЕНИЕ ГЕНЕТИЧЕСКОГО КОДА
Уровень сложности гипотетического предкового организма,
о котором шла речь до сих пор, не предполагает наличия систе-
мы, при помощи которой он мог бы воспроизводиться как целое.
Такой организм мог разделиться на равные или неравные части;
легко себе представить, что одна из частей оказывалась при
этом лишенной какого-нибудь компонента, необходимого для
поддержания жизни. На этой стадии не существовало никакой
системы наследственности, никаких генетических механизмов,
которые обеспечивали бы непрерывное воспроизведение функ-
циональных единиц. Основной принцип генетики состоит в том,
что «подобное порождает подобное». Этот принцип не следует
из какого-то великого и неизменного «закона природы», а яв-
Происхождение жизни | 21
ляется результатом функционирования сложной системы пере-
дачи генетической «информации», т. е. информации, необходи-
мой для построения нового организма. Кажется несомненным,
что без такой системы эволюция не пошла бы дальше уровня
коацерватных капель. Эффективность этого механизма передачи
информации была одним из важнейших факторов, определявших
пределы сложности живых систем.
По-видимому, за 2 или 3 млрд, лет химической эволюции,
предшествовавшей эволюции жизни, было испытано и забрако-
вано в процессе отбора много различных систем передачи ин-
формации. Ясно, что генетическая система должна быть сопря-
жена с системой синтеза, а также с системой преобразования
энергии; неудивительно поэтому, что вещества, из которых стро-
ится генетическая система, — нуклеиновые кислоты — содержат
в качестве структурного элемента аденозинтрифосфат. У боль-
шинства клеточных организмов в основе генетической системы
лежит кодирование информации в молекулах двух нуклеиновых
кислот — рибонуклеиновой (РНК) и дезоксирибонуклеиновой
(ДНК).
Из этого обычно заключают, что гигантские упорядоченные
молекулы, из которых построены живые системы, являются ко-
нечным результатом какого-то процесса агрегации более мелких
молекул и что этот процесс регулировался факторами отбора.
В результате случайного спонтанного образования упорядочен-
ной последо1вательности молекулярных субъединиц (примером
такой последовательности служит нуклеиновая кислота) могла
возникнуть матрица, на которой синтезировались новые копии
этой последовательности. Но это не дает и не может дать ответа
на вопрос о том, как возникла первоначальная, исходная после-
довательность.
Однако, как отметил Патти, это не единственная возможная
гипотеза. Патти высказывает мысль, что предшественниками
биологических макромолекул были не случайные последователь-
ности, а естественным образом упорядоченные кристаллические
структуры, свойства которых обусловлены ограничениями, свя-
занными с природой любого процесса кристаллизации. Возник-
шая на одном этапе конфигурация может определять, какие мо-
номеры будут присоединяться на следующих этапах роста поли-
мера. Таким образом, вполне возможно, что рост макромолекул
регулируется посредством обратной связи. Используя вычисли-
тельное устройство для моделирования этого процесса (фиг. 3),
Патти показал, что при наличии обратной связи из простых кон-
фигураций могут создаваться сложные, периодические и весьма
упорядоченные последовательности. Вовсе нет необходимости
постулировать (как это делало большинство авторов) предсу-
22 | Глаза 1
шествование статистически крайне маловероятной последова-
тельности, которая должна затем копироваться. По мнению
Патти, современные генетические механизмы сами представляют
собой результат эволюции возникновения в природе упорядочен-
ных макромолекулярных последовательностей.
Функционирование генетических механизмов выяснено еще
далеко не полностью, но в общих чертах оно сводится к следую-
щему. Единицы наследственной информации, называемые ге-
нами, «встроены» в структуру гигантских са'мовоспроизводящих-
ся молекул ДНК. Эти молекулы, которые воспроизводятся и пе-
редаются из поколения в поколение, служат как бы рабочими
чертежами, по которым строятся все живые организмы; они под-
ВВАВААВАВВАААВВАВВВВАВВАВАВ ВАВВААВААВ
АААВВВААААВАВВВВВААВАВАВВВА АВВАВАААВА
АВВВВАААВАВААААВВАААААВАААА ААВВВВВВВА
ВАВАВААВВААВВВАВВВАВААВАВВА ААВВАВВВВА
ВВАВАВВАВВААВААВАААВВВААААВ АВВВВВААВА
ВАВВВААВВАВАААВААВВВВАААВАВ ААААВВАААА
АВААААААВВВВВВВАВАВАВААВВАА ВВВАВВВАВА
Фиг. 3. Механическая модель для получения упорядоченных последователь-
ностей (с помощью обратной связи).
Коромысло весов управляет устройством, пропускающим шар А, когда чашки весов уравно-
вешены, и шар В, когда они не уравновешены’ Шары А тяжелее шаров В [8].
Происхождение жизни | 23
держивают непрерывность жизни. Небольшие изменения в этих
«чертежах» также воспроизводятся — они-то и обусловливают
изменчивость, благодаря которой возможна эволюция. Молеку-
лы ДНК представляют собой цепи, составленные из нуклеоти-
дов четырех типов: дезоксигуаниловой, дезоксицитидиловой, дез-
оксиадениловой и дезокситимидиловой кислот. Точно так же и
молекулы РНК составлены из четырех видов нуклеотидов — из
гуаниловой, цитидиловой, адениловой и уридиловой кислот.
Каждая нуклеотидная группировка состоит из остатка сахара —
пентозы — и присоединенных к нему основания и фосфатной
группы. Осевой скелет молекул ДНК и РНК образован остат-
ками сахара (пентозы), связанными между собой фосфатными
группами. В этой регулярной цепи к каждому остатку сахара
присоединено одно из пяти азотистых оснований: аденин, гуа-
нин (пуриновые основания), цитозин, тимин или урацил (пири-
мидиновые основания; урацил входит только в состав РНК).
Конфигурация молекулы ДНК, по-видимому, имеет вид двойной
спирали из пентозо-фосфатных осевых цепей, соединенных по-
перечными мостиками из спаренных оснований (фиг. 4). Хими-
ческие свойства оснований обусловливают их способность обра-
зовывать лишь следующие пары: аденин — тимин и цитозин —
гуанин. Спиральная структура наилучшим образом объясняет
результаты рентгеноструктурного анализа и другие данные о
физических свойствах молекулы ДНК.
Когда при подготовке к клеточному делению происходит
удвоение хромосом, комплементарные спирали, вероятно, «рас-
кручиваются» и каждая из них служит химической матрицей,
к которой присоединяются предшественники ДНК; происходит
формирование комплементарной цепи и тем самым воссоздание
двойной спирали. Это и есть тот механизм, благодаря которому
генетический код передается от клетки к клетке и (через га-
меты) от поколения к поколению. Показано, что генетический
код представлен последовательностью нуклеотидов в цепи ДНК.
СЧИТЫВАНИЕ КОДА
Синтез белка происходит главным образом в рибосомах
(микросомах) — цитоплазматических структурах, пространст-
венно обособленных от ядер ной ДНК. В рибосомах содержится
основная масса РНК цитоплазмы. ДНК служит матрицей, на
которой может строиться другая, комплементарная цепь ДНК
или же цепь РНК. Таким способом код может быть перенесен
на молекулы информационной РНК (РНК-посредника), которые,
как полагают, переносят его на рибосомы, где происходит син-
тез белка. (Остается неизвестным, каким образом клетка «сооб-
24 | Глава 1
щает» своей ДНК, что именно последняя должна синтезиро-
вать— ДНК или РНК.)
Возможны самые различные гипотезы, объясняющие, ка-
ким образом ДНК и информационные РНК управляют по-
строением разнообразных белков. Необходимо, чтобы код, пере-
несенный с ДНК на РНК, был способен однозначно определять
последовательность 20 обычных аминокислот, которые входят
в состав белков. Одна гипотеза была основана на математиче-
ском расчете, показавшем, что из четырех типов нуклеотидов
можно составить 20 (и только 20) -различных триплетов (после-
Основание
( Тимин )
Основание
(Аденин)
Основание
( Гу ан ин)
Основание
- Сахар
- Сахар
- Сахар
- Сахар
( Цитозин)
Фиг. 4. Структура ДНК.
Вверху слева —комплементарные цепи нуклеотидов. Буквами обозначены основания, пяти-
угольниками—пентозные .остатки, кружками —остатки фосфата, двойными линиями —водо-
родные связи. Внизу слева —схема участка одной из цепей. Справа —модель двойной спи-
рали, которую образуют нуклеотидные цепи. Спиральные ленты соответствуют пентозо-
фэсфатныхм осевым цепям, а поперечные перекладины— парам оснований, связывающим
эти цепи. Эта конфигурация^ виде двойной спирали известна как модель Уотсона —Крика [9].
Происхождение жизни | 25
довательностей из 3 нуклеотидов), которые не создавали бы
путаницы в результате «перекрывания». Обозначим, например,
нуклеотиды цифрами 1, 2, 3 и 4; триплеты 131 и 312 будут пере-
крывающимися; если бы они стояли рядом, скажем, в последо-
вательности 141131312, то получилось бы два триплета 131, на-
кладывающихся друг на друга. Неперекрывающимися трипле-
тами в случае такого кода будут триплеты 112, 212, 131, 132,
133, 231, 232, 233, 141, 142, 143, 144, 241, 242, 243, 244, 341, 342,
343, 344.
Недавно нескольким группам исследователей удалось пока-
зать, что код РНК состоит из неперекрывающихся триплетов,
каждый из которых определяет положение одной аминокислоты.
К моменту написания этих строк были частично расшифрованы
коды для 19 из 20 аминокислот. Например, аминокислота ала-
нин кодируется триплетом, состоящим из урацила, цитозина и
гуанина (последовательность оснований в этом триплете не
установлена), а аминокислота серин — триплетом, содержащим
два урацила и один цитозин.
В настоящее время кажется вероятным, что неперекрываю-
щийся характер кода определяется не структурой самих три-
плетов, а существованием какого-то приспособления для фикса-
ции начальных точек «считывания». Например, в приведенном
выше случае (141131312) никакой неоднозначности не будет,
если начальной точкой служит левый конец и если код пред-
ставлен только триплетами; ясно, что в этом случае он читается
как 141 —131—312. Если все это верно, то мы имеем 43 = 64 воз-
можных триплета; этого с избытком достаточно для кодирова-
ния двадцати аминокислот. Возможно, что каждая аминокислота
может кодироваться более, чем одним триплетом и что некото-
рые сочетания служат «заглавными» триплетами (с которых
начинается отдельная последовательность). Работа по расшиф-
ровке кода продвигается сейчас настолько быстро, что некото-
рые из этих вопросов почти несомненно будут вскоре решены.
Каким бы ни оказалось решение, оно наверняка будет содер-
жать интереснейшие указания относительно эволюции самого
кода.
Но каким же образом аминокислоты «считывают» последова-
тельность кодонов информационной РНК и складываются в бел-
ковые молекулы с надлежащей последовательностью амино-
кислотных остатков? Еще до своей конденсации в белковую мо-
лекулу аминокислоты присоединяются к сравнительно неболь-
шим молекулам растворимой (транспортной) РНК. Полученные
данные с большой вероятностью указывают на следующий ме-
ханизм построения белков. Для каждой аминокислоты суще-
ствует особый вид молекул транспортной РНК. В одной части
П рои с хождение жизни | 27
молекулы транспортной РНК имеется последовательность нукле-
отидов, определяющая, с какой аминокислотой она должна вза-
имодействовать, а в другой части — последовательность (три-
плет?), определяющая положение на РНК-матрице, которое
должна запять эта молекула-переносчик. Комплексы транспорт-
ной РНК с аминокислотами образуются за счет энергии, достав-
ляемой АТФ; эта реакция, катализируемая ферментом, схема-
тически представлена на фиг. 5. Затем образовавшиеся комп-
лексы находят свои места на длинной цепи РНК-матрицы — как
полагают, путем спаривания с комплементарными последова-
тельностями в этой матрице. Аминокислоты при этом продол-
жают сохранять активированное состояние; оказавшись в не-
посредственной близости друг к другу, они конденсируются
(вероятно, при посредстве фермента), образуя белок с нужной
последовательностью аминокислотных остатков.
Понятно, что эта система очень далека от первичной системы
передачи генетической информации. Ее совершенство выражает-
ся, в частности, в том, что ДНК во всех случаях служит исход-
ной (основной) матрицей; тем самым уменьшается возможность
ошибок, неизбежных в системе, в которой копии изготовляются
с копий. Далее, короткие молекулы транспортной РНК в высо-
кой степени специализированы для выполнения функции надле-
жащей расстановки предшественников белка. Многообразные
взаимодействия ДНК и РНК в организме не ограничиваются
процессами, связанными с передачей генетической информации
и синтезом белков. Существует гипотеза, согласно которой РНК
функционирует и в тех системах высших организмов, которые
ответственны за научение и память. Возможно, что психические
способности приматов в конце концов будут описаны в понятиях
фундаментальных химических свойств, присущих также простей-
шим клеткам и организмам.
Фиг. 5. Схема процессов синтеза белка под контролем информационной
РНК, согласно современным представлениям.
1 — молекула АТФ; 2—аминокислота (триптофан); 3 — фермент, катализирующий образование
макроэргической связи между остатком триптофана и адениловой кислотой (АМФ); 4-*фос-
фатные _группы, которые были ранее связаны с АМФ, теперь отщепились от субстрата фер-
мента; 5 —тот же фермент, катализирующий перенос остатка триптофана и макроэргической
связи с АМФ на .молекулу соответствующей транспортной РНК; 6 — молекула АМФ, отде-
лившаяся от фермента; 7 — молекула транспортной РНК с присоединенным к ней глицином
отделяется от фермента, активирующего глицин; ^ — информационная РНК, служащая матри-
цей; 9 — начало синтеза белковой цепи (молекула транспортной РНК с присоединенным
к не! «головным» метионином пристроилась к «головному» метиониновому триплету (УАГ) на
РНК-матрице); 10 — молекула транспортной РНК с присоединенным к ней «промежуточным»
метианином приближается к «промежуточному» метиониновому триплету (УГА) на матрице;
11 — фермент, действующий наподобие застежки-молнии; он соединяет аминокислотные
остатки в цепь белка; 12 — вновь синтезированный белок, который отделился от матрицы;
при этом произошло освобождение молекул транспортной РНК, участвовавших в его син-
тезе. На схеме представлена лишь небольшая часть многочисленных реакций, протекающих
В клетке одновременно.
28 | Глава 1
Может показаться невероятным, что сложнейшая функцио-
нальная организация, характерная для клеток современных ор-
ганизмов с их высоко'специализированными органеллами, эффек-
тивными системами для использования макроэргических фос-
фатных связей, точными механизмами клеточного деления и
распределения генетической информации, могла возникнуть из
примитивной структуры описанных выше коацерватных капель.
Это все равно, что, глядя на одноклеточный организм и на че-
ловека, пытаться представить себе первый в качестве предка
второго, ничего не зная о промежуточных ступенях. Следует
помнить, что время, которым располагала природа для эволю-
ции от коацервата до сложной клетки, было почти таким же
длительным, что и время для эволюции от простейших до че-
ловека.
ВЫВОДЫ
Жизнь представляет собой особую, очень сложную форму
взаимосвязи материи и энергии, происхождение которой можно
логически объяснить исходя из самых общих понятий. Важными
событиями в истории возникновения жизни, несомненно, явля-
лись: образование и усложнение органических соединений, обо-
собление их в структурные отдельности, возникновение циклов
мобилизации энергии и развитие механизмов самовоспроизведе-
ния. Предполагается, что эти события должны были совершать-
ся отчасти параллельно и регулировались своего рода прими-
тивным отбором (протоселекцией). Современный механизм са-
мовоспроизведения использует информацию, закодированную в
макромолекулах нуклеиновых кислот, управляющих синтезом
белков.
Жизнь можво рассматривать как некое проявление матери-
ально-энергетического континуума, характеризующееся непре-
рывной репликацией. Идеальная репликация невозможна, по-
этому неизбежен естественный отбор.
ЛИТЕРАТУРА
1. Calvin М., Round trip from space, Evolution, 13, 362—377
(1959).
2. Miller S. L., The formation of organic compounds on the primitive earth,
Ann. N. Y. Acad. Sci., 59, 260—275 (1951).
Проис хождение жизни | 29
3. Needham А. Е., The origination of life, Quart. Rev. Biol., 34, 189—209
(1959).
4. Опарин А. И., Браун штейн A. E., Пасы некий А. Г., Павлов-
ская T. Е. (редакторы), The Origin of Life on the Earth, Pergamon Press,
New York, 1959.
5. Опарин А. И., Life: Its Nature, Origin and Development, Academic Press,
New York, 1961.
6. S a g a n C., On the origin and planetary distribution of life, Radiations Res.,
15, 174—192 (1961).
7. M i 11 e r S. L, Ann. N. Y. Acad. Sci., 69 (1957) .
8. PatteeH. H., Biophysical Journal, 1 (1961)
9. Sinnott E. W., Dunn L. C., D о b z h a n s k у T., Principles of Geneticst
McGraw-Hill, 1958.
Глава 2
единицы РЕПЛИКАЦИИ
Одним из самых важных результатов развития современных
методов научных исследований и появления электронного микро-
скопа было возрождение интереса к цитологии. Высокая разре-
шающая способность электронного микроскопа позволила вы-
явить структуры поразительной сложности там, где раньше не
находили вообще никакой структуры, что по существу подвело
нас к идее о единстве формы и функции на уровне макромоле-
кул и их агрегатов. С эволюционной точки зрения эти результаты
особенно интересны; они позволяют предположить, что мембран-
ные системы, из которых главным образом и построены клетки,
возникли как неизбежный продукт смешивания крупных слож-
ных молекул, таких, как белки и липиды, еще до появления са-
мой жизни (см. гл. 1). Эти случайно возникающие макромоле-
кулярные структуры, вероятно, сходны с мембранами, разнооб-
разные сочетания которых можно видеть в клетках. Главные
клеточные компоненты являются общими для растений и живот-
ных, чем и определяется единство основного плана строения
всего живого, за исключением наиболее специализированных
клеток или организмов. Ниже мы даем краткий обзор строения
клетки, который послужит основой для обсуждения эволюции
генетических механизмов и систем.
Каково бы ни было происхождение клеток, сходство в строе-
нии растительных и животных клеток столь разительно, что -на-
водит на предположение либо о некоем общем предковом тиле,
либо о том, что с жизненными функциями — если говорить о той
жизни, которую мы знаем, — совместимы структуры лишь од-
ного типа (фиг. 6). Определить химический состав клеток срав-
нительно легко; многие физические свойства клеток и их состав-
ных частей могут быть измерены. Однако то, что является уни-
кальным свойством жизни — клеточное строение, возникает
лишь при определенной организации этих химических компонен-
тов. Главными структурными элементами клеток являются круп-
ные молекулы белков, углеводов и липидов. В промежутках ме-
жду элементами этого физического остова, образованного путем
агрегации крупных молекул, а иногда и в соединении с этими
Единицы репликации | 31
Фиг. 6. Обобщенная схе-
ма строения клетки и ее
компонентов, как они вы-
глядят в электронном
микроскопе (некоторые
компоненты увеличены
или представлены в упро-
щенном виде).
элементами, находятся бесчисленные виды более мелких моле-
кул: растворимые белки, аминокислоты, витамины, минеральные
вещества и т. д.
СТРОЕНИЕ КЛЕТОК
Как растительные, так и животные клетки, по-видимому,
имеют наружную оболочку, называемую плазматической мем-
браной, которая обладает важным свойством избирательной
проницаемости. Как показывают данные физических, химиче-
ских и биологических исследований, эта мембрана представляет
собой сложную структуру, состоящую из белковых и липидных
молекул, расположенных слоями. Биполярные молекулы липида
располагаются в два слоя, причем их гидрофобные «хвосты» на-
правлены к поверхности раздела между этими слоями, а гидро-
фильные «головки» — к гидратированным белковым тяжам,
длинные оси которых располагаются перпендикулярно к осям
липидных молекул. На многих клетках, особенно свободноживу-
щих и на клетках, выращиваемых в культуре, можно наблю-
дать явление пиноцитоза: быстро и непрерывно выпуская и
втягивая на своей поверхности мельчайшие выступы наподо-
бие псевдоподий, клетки могут поглощать воду и другие моле-
кулы.
У одиночных клеток и клеток, входящих в состав тканей, на
плазматической мембране, по-видимому, всегда имеются доба-
вочные слои молекул. В случае животных клеток эти слои по-
строены в основном из молекул белков и сахаров, и их проч-
32 | Глава 2
ность зависит от баланса кальция в окружающей клетку среде.
Что касается растительных клеток, то у них имеется два суще-
ственных отличия. Во-первых, наружные слои, или стенка, расти-
тельной клетки состоят главным образом из молекул углеводов.
Наиболее важный их компонент — целлюлоза, образованная из
остатков глюкозы, соединенных в длинные цепи. В состав кле-
точной стенки входят и другие углеводы, а также вещества, со-
держащие жирные кислоты (суберин, кутин и др.). Смежные
клетки в тканях сцементированы между собой материалом сре-
динной пластинки, которая состоит из' пектиновых веществ и
прочность которой также определяется присутствием кальция.
Второе важное различие между растительными и животными
клетками заключается в том, что в тканях растений клетки ор-
ганически соединены друг с другом цитоплазматическими тя-
жами — плазмодесмами. В то же время в клетках животных,
как показали электронные микрофотографии, так называемые
плазмодесмы в большинстве случаев не создают непрерывности
протоплазмы: поперек тяжей, внешне похожих на плазмодесмы,
проходит двойная плазматическая мембрана. Можно сказать,
что ткани Metazoa составлены из клеток, как бы склеенных ме-
жду собой, тогда как растительные ткани образуются .в резуль-
тате более или менее неполного разделения «протоплазмы».
У животных границы между клетками могут иметь чрезвычайно
сложную конфигурацию. Структурные и химические свойства
межклеточных областей позволяют предполагать, что специали-
зация периферии клетки играет важную роль в клеточной диф-
ференцировке.
Самым заметным образованием внутри большинства клеток
является, конечно, ядро. Новейшие электронные микрофотогра-
фии показывают, что ядро отделено от цитоплазмы двойной
мембраной, которую можно называть ядерной оболочкой.
Внутренняя мембрана, по-видимому, окружает содержимое
ядра наподобие мешка. Однако наружная мембрана без пере-
рыва переходит в цитоплазматическую мембранную систему, ко-
торая может быть выражена в большей или меньшей степени.
Клетка как бы пронизана системой трубочек, канальцев, пу-
зырьков и цистерн (обилие и тип которых зависят от природы
клетки и от состояния ее активности); эта система носит назва-
ние эндоплазматического ретикулума. Содержимое ядра в изве-
стном смысле находится вне клетки, ибо цитоплазматические
мембраны без перерыва переходят также и в наружную плаз-
матическую мембрану. В го же время наружная и внутренняя
мембраны ядерной оболочки соединены между собой, так как
обе они пронизаны порами, довольно правильно распределен-
Единицы репликации | 33
ними по поверхности. Эти поры ведут в цитоплазму, окружаю-
щую эндоплазматический ретикулум.
Судя по электронным микрофотографиям, эндоплазматиче-
ский ретикулум далеко не однороден. В активно синтезирующих
клетках с элементами, образующими ретикулум, связаны мел-
кие плотные гранулы, отсутствующие в тех участках, где эти
элементы переходят в плазматическую мембрану. Эти гранулы,
богатые нуклеопротеидом, могут также лежать свободно в цито-
плазме. Они известны под названием рибосом; полагают (см.
гл. 1), что они связаны с синтезом белков. Частицы, названные
физиологами микросомами, являются, видимо, артефактом: это
скопления рибосом и фрагментов эндоплазматического ретику-
лума, образующиеся при фракционировании клеток. Рибосомы
не встречаются там, где эндоплазматический ретикулум перехо-
дит в более или менее уплощенные, концентрически располо-
женные цистерны, образующие аппарат- Гольджи как в расти-
тельных, так и в животных клетках. Комплекс Гольджи трудно
отделить от других клеточных органелл, и его функция пока
еще точно не установлена.
В растительных и животных клетках между канальцами и
пузырьками эндоплазматического ретикулума разбросаны мито-
хондрии. Эти сферические и трубковидные образования также
имеют двойную пограничную мембрану, внутренний слой кото-
рой образует ряд складок в виде поперечных перегородок, или
крист. В митохондриях протекает большинство реакций, связан-
ных с клеточным дыханием, в том числе образование аденозин-
трифосфата (АТФ). Имеются также некоторые данные о том,
что митохондрии, возможно, играют роль в цитоплазматической
наследственности — в том смысле, что они способны к самовос-
произведению.
В структурном отношении к митохондриям очень близки
пластиды, ио встречаются они только в клетках растений. По-
добно митохондриям, они служат местом протекания важных
реакций, за счет которых клетка (а фактически почти все живое
на Земле) получает энергию. Пластиды имеют ламеллярную
структуру; в них к чередующимся слоям липидных и белковых
молекул прилегают слои особых пигментов — хлорофиллов раз-
личного типа, каротиноидов и других (в зависимости от группы
растений). Энергия света, поглощенного пластидой, преобра-
зуется в химическую энергию; в результате ряда реакций энер-
гия окисления используется для фосфорилирования АДФ в АТФ.
Это фотофосфорилирование родственно фосфорилированию,
протекающему в митохондриях. Функциональными единица-
ми хлоропластов высших растений служат мельчайшие ча-
стицы, называемые гранами. У фотосинтезирующих бактерий
3 Зак. 152
34 | Глава 2
и сине-зеленых водорослей организация гран в пластиды отсут-
ствует.
В цитоплазме большинства животных и многих раститель-
ных клеток вблизи ядра имеется образование, называемое цен-
тросомой. В этом относительно прозрачном участке цитоплазмы
находятся одна или две гранулы, — центриоли. Эти органеллы
играют важную роль в образовании и функционировании жгу-
тиков и ресничек, а также в делении ядра и всей клетки. Девять
нитей, образующих наружную часть жгутика или реснички, яв-
ляются продолжением девяти трубчатых или нитевидных ком-
понентов центриоли или базального зерна. Это поразительное
единообразие структуры центриолей, базальных зерен и ресни-
чек наблюдается во всем животном царстве и даже в таких вы-
сокоспециализированных клетках, как фоторецепторы сетчатки.
Центриоль и явно родственные ей структуры (базальные гра-
нулы и т. п.), видимо, обладают свойствами, обусловливающими
построение из фибриллярных белковых молекул особых струк-
тур, например канальцев (нитей) веретена во время митоза.
Генетическая информация клетки содержится в основном в
хромосомах ядра. Многие детали организации хромосом не вы-
яснены. В так называемой интерфазе, или метаболической ста-
дии, хромосомный материал обыкновенно мало доступен для
наблюдения. Иногда обнаруживаются части хромосом, не под-
вергшиеся характерным изменениям, сопровождающим митоз.
Часто один или несколько таких участков хромосом бывают свя-
заны с ядрышком, обычно хорошо видимым в метаболически
активном ядре. Как полагают, во время метаболической стадии
происходит обмен материалом между ядром и цитоплазмой;
в этот период информационная и транспортная РНК осуществ-
ляют свои функции по переводу кода ДНК в структуру белка.
Однако механизм этого обмена еще не выяснен. Полученные
данные ясно указывают на передвижение нуклеиновой кислоты
из ядра в цитоплазму. С помощью электронного микроскопа в
порах ядерной оболочки обычно не удавалось обнаружить ча-
стицы. По сообщениям цитологов, участки ядерной оболочки
могут «отшнуровываться» или «отпочковываться» в цитоплазму,
где они приобретают вид элементов эндоплазматического рети-
кулума или митохондрий.
ДЕЛЕНИЕ КЛЕТКИ —МИТОЗ
Обычно при делении клетки (фиг. 7) в первую очередь
удается наблюдать изменение внешнего вида ядра. В ядре как
живых, так и окрашенных клеток становятся видимыми хромосо-
мы, что часто сопровождается уменьшением размеров ядрышка.
Фиг. 7. Митоз.
Л — ранняя профаза; Б —поздняя профаза; В —прометафаза; Г —метафаза; Д — ранняя ана-
фаза; Б—поздняя анафаза; Ж—поздняя телофаза; 3—дочерние ядра [3].
3*
36 | Глава 2
Первая из произвольно выделяемых стадий митоза называется
профазой. К концу этой стадии в клетках многих организмов
хромосомы удваиваются, так что каждая из них состоит уже
из двух половинок (хроматид). У большинства организмов ис-
чезновение ядерной оболочки отмечает наступление промета-
фазы. Во время прометафазы или несколько раньше в цито-
плазме появляется веретенообразный пучок нитей (представляю-
щих собой, как теперь выяснено, тончайшие трубочки). К концу
прометафазы хромосомы скапливаются в экваториальной пло-
скости этой структуры.
Веретено (у тех организмов, у которых его можно выделить
и подвергнуть химическому исследованию) состоит из фибрил-
лярных белковых молекул, отличающихся высоким содержанием
сульфгидрильных связей; по-видимому, ориентация этих моле-
кул определяется центриолями, находящимися на обоих его
концах. У животных (а также у некоторых растений) центриоли
окружены венчиком из волокон, образующим фигуру звезды.
В соматических клетках большинства растений не видно ни
звезд, ни центриолей, но они могут быть хорошо видны в по-
ловых клетках. Во время короткой стадии, называемой ме-
тафазой, хромосомы расположены около экватора веретена;
при этом по крайней мере точки их прикрепления к нитям ве-
ретена — центромеры — лежат в самой экваториальной плоско-
сти или совсем близко к ней. Очень скоро после этого можно
видеть, что центромеры делятся (в действительности они, воз-
можно, расщепились на более ранней стадии), а затем хрома-
тиды — теперь это дочерние хромосомы — расходятся к полю-
сам.
Стадия расхождения хромосом называется анафазой] меха-
низм ее до сих пор не понят. Ни одна из существующих теорий
не позволяет удовлетворительно объяснить поведение хромосом
у всех организмов. У некоторых животных, например, опреде-
ленные хромосомы обладают большой автономией. Половые
хромосомы могут «забегать вперед» или «отставать» по сравне-
нию с аутосомами. Особые специализированные хромосомы
могут содержаться только в клетках зародышевого пути и эли-
минироваться при последующих делениях. В клетках грибного
комарика Sciara при одном делении образуется однополюсное
веретено, причем одна группа хромосом движется к его «непо-
лярному» концу. В отдельных случаях у животных ядерная
оболочка не исчезает и деление хромосом происходит внутри
мембраны, которая в конце концов перешнуровывается на две.
По мере дальнейшего исследования малоизученных представи-
телей беспозвоночных, водорослей и грибов несомненно будут
выявлены и другие случаи необычного протекания митоза. Не
Единицы репликации | 37
исключено, что, когда в результате систематических исследова-
ний все эти явления предстанут в надлежащей перспективе,
механизмы, найденные у высших растений и позвоночных, пока-
жутся необычными.
После того как хромосомы разошлись к полюсам веретена,
вокруг каждой группы дочерних хромосом образуется новая
ядерная оболочка, которая, возможно, строится из материала
одной из мембранных систем цитоплазмы. На этой стадии, нося-
щей название телофазы, животные клетки обычно делятся путем
образования перетяжки, а растительные — путем образования
клеточной пластинки (новой клеточной стенки, разделяющей
старую клетку). На это-м, т. е. на формировании двух дочерних
клеток, процесс клеточного деления заканчивается. Таким об-
разом, в результате эквационного деления и деления цитоплаз-
мы, при котором цитоплазматические органеллы распределяются
между дочерними клетками примерно поровну, образуются две
клетки с одинаковым генотипом.
В процессе митоза хромосомы претерпевают ряд интересных
и важных изменений (фиг. 8). Если сравнить профазную хромо-
сому с анафазной, то обнаружатся резкие различия. В анафазе
хромосома не только хорошо видна и легко окрашивается, но
она также толще и значительно короче. С помощью надлежа-
щей обработки можно показать, что анафазная хромосома за-
кручена в плотную спираль, витки которой выглядят так, словно
они окутаны окрашивающейся субстанцией, обычно называемой
матриксом. Часто бывает видно, что основная нить, или хромо-
нема, анафазной хромосомы образует мельчайшие витки вто-
рого порядка. Хорошо заметные витки первого порядка как бы
накладываются на них за счет какого-то процесса, вызывающего
укорочение хромосом. Если число хроматид больше одной (на-
пример, в метафазе), то они могут быть закручены одна вокруг
Другой.
Митоз обеспечивает эквационное деление хромосом, в ре-
зультате которого обе дочерние клетки (если исключить появ-
ление новой мутации) получают одинаковую генетическую
информацию. Во многих случаях, когда клетки уже специализи-
рованы в отношении формы и функции, деление хромосом мо-
жет и не сопровождаться делением ядра или клетки в целом.
В результате возникают либо клетки с двумя или большим чис-
лом ядер, либо ядра с большим по сравнению с зиготой числом
хромосом. Последнее, по-видимому, встречается чаще; это явле-
ние называют эндополиплоидией. У некоторых организмов (на-
пример, у насекомых) каждая ткань характеризуется своей соб-
ственной степенью плоидности. Высокоспециализироватщые клет-
ки слюнных желез водомерки (Gerris) могут быть 2048-плоид-
Фиг. 8. Цикл спирализации хромосом.
А — интерфаза; Б, В, Г—изменения, происходящие в профазе; появление матрикса, Д — про-
метафаза: хроматиды представляются двойными; Е— метафаза; Ж— анафаза; 3—телофазные
изменения, начинающиеся в дочерних хромосомах; исчезновение матрикса [3].
Единицы репликации | 39
ними. Ткани других организмов могут состоять из клеток раз-
личной плоидности (обычно нормальный уровень — выше дипло-
идного, с преобладанием, например, тетраплоидных или окто-
плоидных клеток). Такая картина наблюдается, в частности, в
печени человека или в тканях корня и стебля у цветковых ра-
стений.
Значение феномена эндо-полиплоидии не вполне ясно. Не-
сомненно, это явление коррелирует с секреторной активностью
клетки и, возможно, играет роль в развитии, а также в диффе-
ренцировке. Следует, однако, подчеркнуть, что эндополиплоидия
не сопровождается, по-видимому, какими-либо качественными
изменениями в генетическом материале. Клетки зародышевого
пути у животных и спорогенные клетки у растений не бывают
эндополиплоидными. Таким образом, репродуктивные клетки
сохраняют зиготическое или гаметическое число хромосом; в то
же время соматические клетки могут претерпевать последова-
тельное увеличение плоидности. При размножении растений
вегетативным путем, с помощью окулировки или отпрысками,
т. е. с помощью соматических тканей, может возникать потом-
ство с увеличенным числом хромосом. Такие случаи будут рас-
смотрены в гл. 9.
ДЕЛЕНИЕ КЛЕТКИ —МЕЙОЗ
Кроме этих механизмов сохранения или увеличения суще-
ствующего числа хромосом, у организмов, очевидно, должны
существовать механизмы для его уменьшения. Механизмы
уменьшения большого числа хромосом, наблюдающегося при
эндополиплоидии, мало изучены, но есть данные о существова-
нии таких механизмов у насекомых (Culex) и у растений (Al-
lium). У огромного большинства организмов имеется общий
механизм уменьшения зиготического числа хромосом до гаме-
тического; этот механизм называется мейозом (фиг. 9). Мейоз
происходит в тканях, не подвергшихся эндополиплоидизации, —
в клетках зародышевого пути у животных и в спорогенной тка-
ни у растений. Так же, как и митоз, он протекает в общих чер-
тах одинаково у всех организмов, но у животных он приводит к
образованию гамет, а у большинства растений — к образованию
спор. Клетку, подготовленную к мейозу, называют ауксоцитом.
В результате мейоза почти всегда образуются 4 дочерние клетки
с половинным по сравнению с ауксоцитом числом хромосом.
При образовании яйцеклеток у животных и мегаспор у растений
три из этих четырех клеток часто имеют значительно меньшую
величину и в конце концов исчезают.
Фиг. 9. Мейоз.
Л —стадия одной хроматиды, 5 —конъюгация, приводящая к образованию бивалентов; В —ста-
дия четырех хроматид; /' — расщепление бивалентов: видны хиазмы; Д — первая анафаза,
расхождение хромосом, образующих биваленты, без деления центромер; В—дочерние ядра
с кроссоверными хроматидами; Д< -вторая анафаза, центромеры делятся; 3— четыре гаплоид-
ные дочерние клетки [3]. *
Единицы репликации | 41
Редукция числа хромосом в процессе мейоза обусловлена
тем, что при этом происходит два клеточных деления и лишь
одно деление хромосом. Ниже дается описание типичной кар-
тины поведения хромосом, обнаруживаемой при исследовании
с помощью обычного микроскопа. (Используя специальные ме-
тоды исследования, у некоторых организмов удалось обнару-
жить отклонения и исключения.) Ауксоцит имеет обычно не-
сколько больший по сравнению с нормальной клеткой размер,
и его ядро окрашивается слабее. Часто бывает видно, что в про-
фазе мейоза хромосомы представляют собой одиночные, а не
двойные нити, как при митозе. Их дальнейшее поведение на-
столько сложно, что профаза первого деления мейоза растянута
во времени и разделяется на ряд подстадий, названия которых
нет нужды здесь приводить. Сначала происходит синапсис, т. е.
конъюгация гомологичных хромосом друг с другом; при этом
хромосомы сближаются по всей своей длине. После того как
произошло образование пар, или бивалентов, хромосомы при-
обретают вид двойных нитей. (Вспомните, что в митозе хромо-
сомы уже в начале профазы состоят из двух нитей.) Таким об-
разом, каждый бивалент содержит 4 хроматиды; две из них
принадлежат хромосоме, полученной от материнской гаметы, и
две — от отцовской. Хромосомы, не имеющие гомологов, напри-
мер половые хромосомы, остаются унивалентами, но удваи-
ваются примерно в то же время, что и остальные хромосомы.
По-видимому, на стадии удвоения хромосом тонкие хрома-
тиды в бивалентах разрываются и вновь соединяются концами
в местах разрывов. Будучи тесно сближены и перекручены, они
часто воссоединяются в несестринских комбинациях, т. е. вместо
воссоединения частей сестринских хроматид после разрыва мо-
гут соединяться части материнской и отцовской хроматид. Это
явление известно под названием кроссинговера (перекреста).
У некоторых организмов кроссинговер происходит только у осо-
бей одного из полов; так, например, у самцов Drosophila и Cal-
limantis он никогда не наблюдался. После удвоения хромосомы
ведут себя так, словно что-то заставляет их отталкиваться
друг от друга. Биваленты располагаются теперь в ядре обосо-
бленно, а составляющие их парные хромосомы остаются соеди-
ненными только в тех случаях, когда произошел перекрест
(если же перекреста не произошло, то они на этой стадии часто
отделяются друг от друга). В результате такого отталкивания
плечей хромосомы друг от друга (об «отталкивании» мы гово-
рим здесь в чисто описательном смысле) бивалент принимает
форму, которая зависит от числа и расположения перекрестов;
последние становятся теперь видимыми как крестообразные
конфигурации, или хиазмы.
42 | Глаза 2
На этой стадии мейоза присутствие хиазмы указывает на
кроссинговер. Позднее, когда хромосомы спирализуются, уко-
рачиваются и приобретают способность более интенсивно окра-
шиваться, хиазмы (но не точки перекреста) сдвигаются к кон-
цам хромосом. В результате этого процесса, называемого тер-
минализацией (фиг. 10), биваленты приобретают характерную
форму: соседние петли лежат теперь перпендикулярно друг
другу.
В конце первой профазы мейоза ядро содержит гаметическое
число бивалентов (плюс унивалелты, когда они имеются). В пер-
вой метафазе образуется веретено — по-видимому, в точности
так же, как и при митозе, — и биваленты располагаются на его
экваторе. Во время анафазы первого деления мейоза к противо-
положным полюсам расходятся не половинки разделившейся
центромеры, как при митозе, а две центромеры бивалента. Таким
образом, хромосомы не делятся, а расходятся-, в результате это-
го расхождения образуются два дочерних ядра, которые пре-
терпевают затем изменения, характерные для телофазы (послед-
няя может быть сильно укорочена). Центромеры унивалентов
также не делятся, и унивалент отходит к тому или другому
полюсу. Материнские и отцовские хромосомы распределяются
совершенно случайным образом.
Во втором делении мейоза хромосомы ведут себя так же,
как в митозе, с той лишь разницей, что в результате происшед-
шего кроссинговера хроматиды, прикрепленные к центромере,
оказываются не идентичными. Второе деление, при котором про-
исходит деление центромер, ведет к образованию из одного
ауксоцита четырех клеток. Каждая из них содержит гаметиче-
ское число хромосом, но вероятность того, что одна из них ге-
нетически тождественна какой-либо другой, чрезвычайно мала,
поскольку материнские и отцовские хромосомы, а также хрома-
тиды распределились случайным образом и, наконец, в резуль-
тате кроссинговера произошел частичный обмен генетическим
материалом между родительскими геномами.
В этом именно и заключается огромное значение мейоза и
кроссинговера. Эти процессы обеспечивают непрерывную пере-
тасовку генетического материала при воспроизведении. Все
время образуются новые комбинации генов, и поскольку соеди-
нение гамет в основном носит случайный характер, появление
двух особей с одинаковым генотипом чрезвычайно маловероятно.
Этот цитологический механизм составляет часть генетической
системы — ту ее часть, которая определяет число рекомбинаций,
возникающих в популяции и создающих материал для дейст-
вия отбора. Наиболее знакомые нам организмы, как, напри-
мер, кошки и собаки, дубы и сосны, диплоидны и размножаются
Единицы репликации | 43
Фиг. 10. Терминализация хиазм.
Слева направо— последовательные стадии этого процесса: хиазмы сдвигаются к концам
хромосом. (Обратите внимание на то, что точки кроссинговера нс перемещаются). На край-
нем правом рисунке— произошел поворот хромосом [3].
половым способом, путем перекрестного оплодотворения. В по-
следующих главах (гл. 8 и 9) будут рассмотрены иные генети-
ческие системы в качестве примеров механизмов, посредством
которых число рекомбинаций, возникающих при функциониро-
вании этой обычной генетической системы, может быть изме-
нено (обыкновенно снижено, возможно, до нуля).
ВЫВОДЫ
Клетка представляет собой единицу, в которой происходит
обмен веществ и которая состоит из крупных и мелких молекул,
объединенных специфическим образом, часто в форме мембран-
ных систем, в субъединицы, или органеллы, со специализирован-
ными функциями. При делении соматических клеток их орга-
неллы распределяются между дочерними клетками примерно
поровну. При помощи митоза достигается эквационное деление
хромосом между двумя новыми клетками. В процессе мейоза
число хромосом в клетках, из которых образуются гаметы,
уменьшается. При первом делении мейоза гомологичные хромо-
сомы образуют пары (синапсис), а затем расходятся, причем
центромеры не разделяются. Во время первого деления про-
исходит кроесинговер, и после того как центромеры во втором
Делении разделяются, возникают 4 дочерние клетки с рекомби-
44 | Глава 2
ни-рованными хромосомами и сегментами хромосом. Мейоз обе-
спечивает рекомбинацию, что обусловливает изменчивость, со-
ставляющую основу действия отбора.
ЛИТЕРАТУРА
1. Swanson С. Р., Cytology and Cytogenetics, Prentice-Hall, Englewood
Cliffs, N. Y., 1957.
2. Wilson G. B., Morrison J. H., Cytology, Reinhold, New York, 1961.
3. De Robertis E., Nowinski W., Saez F., General Cytology, 2d ed.,
W. B. Saunders Company, 1954 (Де Робертис E., Новинский В.,
Саэс Ф., Общая цитология, ИЛ, М., 1962).
Глава 3
ГЕНЕТИКА
Кажется разумным предположить, что факторы отбора, дей-
ствовавшие в процессе эволюции генетического аппарата у пер-
вичных организмов, вероятно, были направлены на стабилиза-
цию того, что вначале представляло собой почти безгранично
изменчивую систему. Механизмы репликации генетического ма-
териала, как правило, обеспечивают его точное удвоение, так
что у потомков синтезируются белки, тождественные белкам
родителей. У более сложных многоклеточных организмов само-
регулирующиеся механизмы развития связаны с ядерным и вне-
ядерным генетическим материалом; в совокупности они обра-
зуют систему, которая обычно обеспечивает построение нормаль-
ного, функционирующего организма дикого типа. Как уже гово-
рилось, сущность явления наследственности сводится к основ-
ной формуле: «подобное рождает подобное».
ИЗМЕНЧИВОСТЬ И НЕ ИДЕЛЕВСКАЯ ГЕНЕТИКА
В процессе репликации тем не менее происходят ошибки; в
результате создается изменчивость, обеспечивающая материал
для действия отбора. Вообще говоря, анализ природы изменчи-
вости и передачи измененных признаков из поколения в поко-
ление служит единственным средством изучения механизма
наследственности. При исследовании характера изменчивости у
живых организмов оказывается, что одни организмы, по-види-
мому, более изменчивы, чем другие. Кроме того, типы изменчи-
вости различны у разных организмов и для разных признаков.
В тех случаях, когда наблюдаются дискретные различия, измен-
чивость можно назвать прерывистой. В других случаях измен-
чивость представляется непрерывной', отдельные особи при этом
нельзя отнести к четко выраженным классам, которые легко
охарактеризовать. Гальтон в своих классических работах по изу-
чению наследования умственных способностей и других призна-
ков у человека пытался изучать непрерывную изменчивость.
Другие ученые, в том числе Линней и его предшественники,
изучали непрерывную изменчивость путем скрещивания расте-
ний или животных, различающихся по тем или иным призна-
кам. Однако генетика как наука по существу родилась только
тогда, когда начали изучать наследование признаков, для
46 | Глава 3
которых характерна прерывистая изменчивость. Организмы, об-
ладавшие такими признаками, можно отнести к тому или иному
из очень немногих четко разграниченных классов. Наблюдая
распределение по таким классам потомства, полученного от
экспериментального скрещивания родителей, обладающих раз-
личными признаками, Мендель смог описать основные правила
поведения ядерных единиц наследственности.
Работы Менделя не получили общего признания; некоторые
ученые, чувствовавшие, что Мендель далеко отошел от прото-
ренных путей научного исследования, даже убеждали его отка-
заться от публикации своих результатов. В 1900 г. статьи Мен-
деля были обнаружены тремя биологами, которые осознали всю
их важность. Почти тотчас же генетика превратилась в важную
и быстро развивающуюся отрасль биологии. Однако многие
ученые полагали, что выводы Менделя лишь в небольшой сте-
пени приложимы к эволюции в природных популяциях или к
тому типу непрерывной изменчивости, который преобладает в
популяциях диких и одомашненных растений и животных. Только
в результате многолетней работы было установлено эволюцион-
ное значение законов Менделя.
ЕДИНИЦЫ НАСЛЕДСТВЕННОСТИ
Единицы наследственности, постулированные Менделем и
впоследствии названные генами, были идентифицированы как
специфические участки хромосомы; это отрезки между двумя
ближайшими точками кроссинговера L Более поздние исследо-
вания, особенно в области биохимической генетики микроорга-
низмов, привели к другим определениям гена. Например, обще-
признано, что специфические гены контролируют образование
специфических ферментов. Выращивая микроорганизмы на сре-
дах известного химического состава, можно показать, что мута-
ция гена приводит к утрате способности осуществлять ту или
иную внутриклеточную реакцию. В результате мутации бакте-
рия может, например, утратить способность к синтезу какого-
либо определенного вещества, скажем триптофана. Было обна-
ружено, что число мутантных аллелей у генов обычно очень
1 Автор в соответствии с большинством современных руководств и книг
по генетике (в особенности по молекулярной генетике) категорически прини-
мает в качестве общепризнанного положения классической генетики отсут-
ствие кроссинговера внутри гена. Между тем это совсем не так. Еще в 1926 г.
в работе А. С. Серебровского (Журнал экспериментальной биологии, сер. А.,
т. II, вып. 1—2) была сделана интересная попытка измерить ген, именно ис-
ходя из представления о том, что кроссинговер возможен как между генами,
так и внутри отдельных генов. — Прим. ред.
Генетика | 47
велико. Как показали эти исследования, ген следует представ-
лять себе как участок хромосомы, состоящий из многих неза-
висимо мутирующих единиц. Бензер назвал эти места возможных
мутаций внутри гена мутонами. Полученные данные позволяют
предполагать, что отдельный мутон состоит всего из одного или
из очень немногих нуклеотидов.
Эксперименты показали, что рекомбинация может происхо-
дить в пределах единичного гена, т. е. внутри функциональной
единицы. Наименьшую единицу, способную при этом подвер-
гаться обмену, называют реконом. Как показывают исследова-
ния рекомбинации, рекон также близок по размерам к нуклео-
тиду, а мутоны в функциональной единице расположены в ли-
нейном порядке. Для фенотипа, по-видимому, безразлично, как
расположены гены у организма, гетерозиготного по двум фак-
4- “Ь а ~h
торам, т. е. имеет ли место расположение & или тр-*»
интересно, так ли обстоит дело для частей одного гена? Будет
ли синтез фермента идти одинаково успешно в том случае, если
мутации, затрагивающие один и тот же ген, распределены ме-
жду двумя хромосомами (так называемое транс-положение), и
тогда, когда они находятся в одной и той же хромосоме (цис-
положение)? Ясного ответа на этот вопрос пока не получено;
известно, однако, что иногда части гена могут быть разделены
между гомологичными хромосомами, хотя наименьшая группа
мутаций, которая должна находиться в ^we-положении, довольно
велика. Однако случаи комплементации (взаимного дополне-
ния) между мутантами еще не поддаются однозначной интерпре-
тации. Цис-транс-тес,т можно использовать и для определения
функциональных генных единиц, получивших название цистро-
нов L
При изучении популяций — так же как по существу и во
всех областях биологии — единице наследственности следует
придавать строго операциональное значение в соответствии с
контекстом данного исследования. В эволюционных исследова-
ниях эта единица не может быть той же, что и в биохимической
генетике. В этом нужно отдавать себе ясный отчет, когда при-
ходится сопоставлять рассуждения о генах в эволюционном
аспекте с выводами, базирующимися на изучении микроорга-
низмов в лаборатории. Когда изучают наследственность в при-
родных популяциях, единица наследственности в большинстве
случаев приобретает статистический характер, ибо факторы,
1 Автор преувеличивает противоречивость данных по эффекту взаимо-
действия мутонов в цис- и Tpawc-положениях. Как правило, цис-т ранс-чесу
служит хорошим критерием для выделения функциональных единиц — цист-
ронов. — Прим. ред.
48 | Глава 3
контролирующие количественные признаки, многочисленны и
находятся между собой в сложном взаимодействии. В настоящее
время только методы статистики позволяют идентифицировать
взаимодействия наследственных единиц (которые, как предпо-
лагается, сходны с единицами, определяющими дискретную из-
менчивость), онтогенетические механизмы, с помощью которых
они проявляются, и воздействие среды на обе эти системы.
Окружающие организм условия в каждый данный момент и в
каждом данном месте уникальны — они не повторяются и не
могут быть воспроизведены в точности. Это означает, что в экс-
периментальных исследованиях важно многократно дублиро-
вать опыты в пространстве и во времени, что, к сожалению, по-
глощает массу времени и средств. На последующих страницах
мы дадим сводку основных фактов, касающихся наследования
количественных и качественных изменений, и рассмотрим те
аспекты поведения генов, которые особенно важны при иссле-
довании эволюции.
ЗАКОНЫ МЕНДЕЛЯ
Основные правила наследственности, выведенные Менделем,
знакомы каждо-му, кто прослушал элементарный курс общей
биологии. Скрещивая разные сорта гороха, Мендель нашел, что
если родительские формы различаются по тому или иному при-
знаку, потомство первого поколения (Fi) похоже либо на одну,
либо на другую из них. Признак, проявившийся в Fi, он назвал
доминантным по отношению к тому, который не проявился, т. е
по отношению к рецессивному признаку. При скрещивании в
основном однотипных растений Fi возникло разнотипное потом-
ство F2, в котором появлялись особи с рецессивным признаком;
следовательно, факторы, ответственные за появление этих при-
знаков, не утрачивались, а только находились в скрытом со-
стоянии. Изучив типы потомства в Fi и F2, Мендель заключил,
что каждый потомок содержит два гомологичных фактора — по
одному от каждого из родителей, — влияющих на проявление
изучаемого признака. Потомки Fb полученные от скрещивания
различающихся по данному признаку родительских форм, долж-
ны содержать два различных, но гомологичных фактора: один
для доминантного признака и другой — для рецессивного. (Ины-
ми словами, Fi гетерозиготно.) При формировании F2 эти фак-
торы распределяются среди потомства приблизительно в отно-
шении 3 : 1 (на каждые три особи с доминантным признаком
одна особь с рецессивным признаком). Особь, у которой обна-
руживается рецессивный признак, является гомозиготной по
этим факторам. Дальнейшие скрещивания (в том числе воз-
Генетика | 49
вратные скрещивания с одной из родительских форм) показы-
вают, что из трех потомков, имеющих доминантный признак,
один будет содержать идентичные, а два других (подобно осо-
бям поколения Fi)—разные факторы.
Скрещивая формы, различающиеся по нескольким призна-
кам и гомозиготные по факторам, определяющим эти признаки,
Мендель нашел, что факторы различных исследованных им при-
знаков передаются независимо друг от друга. У потомков Fi
наблюдались только доминантные признаки, а в поколении F2
происходило расщепление по каждому признаку в отношении
3:1. Если, например, гомозиготные родители различались по
двум признакам, то количественное соотношение между типами
потомства в поколении F2 соответствовало как бы (3 : I)2, т. е.
9:3:3: 1. Производя возвратные скрещивания потомков с ро-
дительскими формами, можно получить подтверждение выводов
о числе факторов и об их независимой передаче.
Ясно, что поведение наследственных факторов соответствует
поведению хромосом, являющихся, как теперь известно, их но-
сителями. Факторы, влияющие на изученные Менделем признаки
у гороха, находились в разных, негомологичных хромосомах.
Проведенные позже исследования показали, что факторы, лежа-
щие в одной и той же хромосоме, сцеплены между собой (т. е.
появляются совместно чаще, чем следовало бы ожидать, если бы
распределение было независимым). Исследование большого
числа признаков показало, что соответствующие факторы обра-
зуют столько групп сцепления, сколько имеется хромосом в
гаплоидном наборе. В пределах одной группы сцепления процент
рекомбинаций варьирует от очень низких величин для генов,
близко лежащих друг к другу, до 50% —для генов, разделен-
ных большим расстоянием (практически этот случай нельзя
дифференцировать от независимой передачи, имеющей место
при локализации факторов в разных хромосомах).
РЕКОМБИНАЦИЯ
Причиной рекомбинации сцепленных генов является кроссин-
говер, происходящий в клетке при мейозе (см. гл. 2). При изу-
чении наследственности «на макроуровне» фактор, ответствен-
ный за тот или иной признак, отождествляют с минимальным
промежутком между двумя точками кроссинговера. (Это и будет
операциональным определением гена для данного случая.) Крос-
синговер наблюдается у всех организмов, у которых установлено
наличие мейоза и половой рекомбинации. Основной механизм
его, по-видимому, во всех случаях одинаков, и некоторые иссле-
дователи высказывали мнение, что мейоз не может протекать
4 Зак. 152
50 | Глава 3
надлежащим образом без кроссинговера (или его специализиро-
ванного заменителя).
Цитологический механизм кроссинговера с точностью не из-
вестен. Предполагается, что в профазе первого деления мейоза
во время синапсиса, когда хромосомы обвивают друг друга,
хроматиды разрываются и затем концы их фрагментов вновь
соединяются. Если соединятся несестринские хроматиды, про-
изойдет кроссинговер. Кроссинговер происходит с различной ча-
стотой в разных участках хромосомы. Вблизи от центромеры
и у самых концов хромосом кроссинговера не бывает или он
наблюдается редко. Поблизости от блоков гетерохроматинового
материала частота кроссинговера также снижена. У некоторых
организмов кроссинговер обычно происходит только .в опреде-
ленных участках хромосом; у других он, по-видимому, встре-
чается довольно равномерно по всей длине плеча хромосомы.
Вероятно, такие различия, по крайней мере отчасти, зависят от
количества гетерохроматина и его распределения. В некоторых
случаях кроссинговер бывает полностью подавлен; это наблю-
дается, например, у самцов дрозофилы и у самок тутового
шелкопряда (Bombyx mori). Кроме того, перекрест в одном
участке хромосомы снижает вероятность другого перекреста в
смежном участке. Степень этой так называемой интерференции
можно оценить, изучая сцепление; можно показать, что она раз-
лична для разных участков одной хромосомы и для разных
хромосом. Вероятно, интерференция также обусловлена струк-
турными причинами.
Если два фактора расположены в хромосоме на достаточном
расстоянии друг от друга, между ними может произойти более
одного перекреста. При двойном кроссинговере рекомбинацион-
ный эффект зависит от того, какие хроматиды в нем участвуют
(фиг. 11): если в обоих перекрестах участвуют одни и те же
две хроматиды из четырех, составляющих бивалент, то это со-
бытие нельзя обнаружить (оно выявляется только тогда, когда
удается наблюдать также поведение третьего фактора, распо-
ложенного между двумя первыми). Если же второй кроссинго-
вер происходит между двумя хроматидами, не участвовавшими
в первом перекресте, то в каждой хроматиде обнаруживается
только один перекрест. Эти два случая можно назвать двуххро-
матидным обменом и четыреххроматидным обменом. Треххро-
матидный обмен приводит к образованию одной некроссоверной
хроматиды и трех хроматид с одиночными перекрестами. Вообще
хромосомы с четным числом перекрестов или вовсе без перекре-
стов, образовавшиеся при множественном кроссинговере между
факторами, будут проявляться как хромосомы родительского
типа. Хромосомы с нечетным числом перекрестов между изучае-
Фиг. И. Различные результаты двойного кроссинговера.
/ — двуххроматидный обмен; 2 —четыреххроматидный обмен; 3, 4 —треххроматидный
обм ж [4, 5].
4*
52 | Глава 3
мыми факторами будут рекомбинантными. А так как число хро-
мосом с рекомбинациями равно числу хромосом без перекрестов
или с четным числом перекрестов, доля рекомбинаций не может
превышать 50%.
Кроосинговер подвержен влиянию факторов среды, напри-
мер температуры, и находится также под генетическим контро-
лем. Механизм генетического контроля довольно сложен и еще
не вполне ясен. По-видимому, гены определяют возможную дли-
тельность синапсиса (которая влияет на кроосинговер) и лока-
лизацию перекрестов; существуют гены, действие которых
состоит в полном исключении синапсиса. Следовало бы ожидать,
что и другие генетические аномалии, возникающие в процессе
мейоза, повлияют на кроосинговер. С эволюционной точки зре-
ния эти факторы, непосредственно влияющие на кроссинговер,
сами по себе, видимо, не имели большого значения в контроле
рекомбинаций. В тех случаях, когда частота рекомбинаций у
определенных организмов понижена, — как полагают, в резуль-
тате естественного отбора — это обычно достигается иными пу-
тями, например за счет инверсий, транслокаций или полного
исключения мейоза (у апомиктических форм — см. гл. 9).
ДЕЙСТВИЕ ГЕНОВ
Действие генов у отдельных особей оказалось чрезвычайно
изменчивым. Гены в популяции редко встречаются всего лишь
в двух альтернативных состояниях, или аллелях. По-видимому,
большинство признаков, например группы крови у животных,
системы несовместимости у растений, окраска меха у млекопи-
тающих, окраска цветков у растений и многие другие, чаще всего
контролируются системами множественных аллелей. Взаимодей-
ствие между генами в генетическом материале или взаимодей-
ствия на уровне механизмов развития, реализующих фенотип,
приводят к сложным формам расщепления. Например в случае
систем пигментов, обусловливающих определенную окраску,
разные этапы в синтезе пигмента могут контролироваться раз-
личными генами. При отсутствии любого из этих генов окраска
не проявляется. Такие гены называют комплементарными.
В то же время во многих случаях проявление одного гена
маскируется действием другого, находящегося в другом локусе.
Это явление носит название эпистаза; эпистатический ген маски-
рует или предотвращает выражение гипостатического гена. На-
пример, ген белой окраски у леггорнов является эпистатическим
по отношению к многим генам, определяющим любую другую
окраску и ее распределение. При наличии соответствующего
фактора никакая окраска, кроме белой, проявиться не может.
Генетика | 53
Очевидно, что эффекты комплементарное™ и эпистаз наряду
с другими видами модифицированного проявления генов обусло-
влены аналогичными механизмами. Сходное явление наблю-
дается, когда имеется один ген или группа генов, которые сами
по себе обладают сравнительно слабым действием, но способ-
ны совместно изменять эффект какого-либо гена, обладаю-
щего сильно выраженным действием. Такие «второстепенные»
гены носят название модифицирующих факторов (модификато-
ров).
Часто бывает необходимо охарактеризовать тип проявления
того или иного гена, ибо часто оно бывает изменчивым. При
наличии определенных факторов у всех или почти всех особей
с одинаковым генотипом развивается характерный фенотип,
отличающий их от особей с иными генотипами в известном диа-
пазоне внешних условий. В таких случаях говорят, что гены об-
ладают высокой пенетрантностью, так как данный признак про-
является у большинства носителей данного гена. Существуют
гены, далеко не всегда вызывающие в данных условиях замет-
ный фенотипический эффект у своих носителей. Это гены с низ-
кой пенетрантностью. Бывает также, что фенотипическое прояв-
ление гена может быть изменчивым даже в случае его полной
пенетрантности. Если оно относительно единообразно в более
или менее «нормальной» среде, то говорят, что ген обладает
постоянной экспрессивностью (или постоянной степенью выра-
жения); если же в проявлении данного признака наблюдаются
индивидуальные различия, то говорят, что экспрессивность гена
изменчива.
Изучение множественного, или плейотропного, действия генов,
а также систем генов, контролирующих выражение данных при-
знаков, позволило сделать вывод, что, по-видимому, все свой-
ства организма определяются более чем одним геном. И наобо-
рот, каждый ген влияет на большое число (если не на все)
свойств, составляющих в совокупности фенотип организма.
В то же время кажется вероятным, что последовательность
аминокислот в данном специфическом полипептиде всегда опре-
деляется одним геном.
МУТАЦИИ
В контролируемых скрещиваниях часто удается идентифици-
ровать специфические гены, ответственные за определенные при-
знаки, проявляющие дискретную изменчивость. Наиболее оче-
видная особенность генов с таким четким действием состоит в
том, что они изменяются, или мутируют; в самом деле, ведь
54 | Глава 3
только это и позволяет обнаружить их существование. Мутации
происходят спонтанно, с различной частотой; их можно также
вызывать, воздействуя на организмы ионизирующим излучением,
ультрафиолетовым светом и разнообразными химическими ве-
ществами. По-видимому, эти агенты оказывают более или менее
специфическое действие на ДНК. Обсуждение вопроса о частоте
спонтанных мутаций оказывается делом весьма сложным, ибо
частоту можно выражать различными способами. Насколько из-
вестно, мутация является случайным событием; определить,
какой именно ген окажется измененным, приписать появление
данной мутации действию какого-либо определенного мутагена
не представляется возможным. Обычно желательно бывает знать
вероятность возникновения одной мутации в расчете на одну
клетку на одно деление. Эта величина выражает зависимость
мутационного процесса от времени; она сравнительно легко под-
дается определению только у микроорганизмов. Даже у бакте-
рий при этом учитываются, по существу, фенотипические изме-
нения, а они не всегда зависят только от одного гена или от
одной мутирующей единицы. Частоты мутаций, найденные для
микроорганизмов, составляют от 10-6 до J0-9.
Для многоклеточных организмов частоту мутаций обычно
приходится измерять иными способами (кроме тех случаев,
когда возможна культура ткани), ибо доступным критерием
здесь служит число гамет, дающих мутантные особи, в расчете
на одно поколение. Таким образом, подсчитываются гаметы, а
не особи. Мутация, возникшая в зачатковой клетке половой
железы, может привести к появлению большого или ма-
лого числа мутантных гамет в зависимости от того, на какой
стадии гаметогенеза произошла мутация. Частота мутаций на
одно поколение у разных генов различна, но в среднем она
составляет 10-5. Например, частота мутаций на одно клеточ-
ное деление в культурах клеток костного мозга в среднем рав-
на ЮЛ
Частота мутаций, очевидно, находится под генетическим кон-
тролем и может поэтому изменяться в процессе эволюции. Из-
вестны гены, действие которых, по-видимому, в основном состоит
в изменении частоты мутаций других генов. Следует ожидать,
что у большинства организмов отбор будет приводить к возник-
новению генотипа, у которого поддерживается оптимальный уро-
вень мутаций в популяции. Этот вопрос труден для изучения, и
относящихся к нему данных немного. Он тесно связан с вопро-
сом об эффектах гетерозиготности в онтогенезе («канализация»
развития) и о генетическом гомеостазе и будет обсуждаться в
последующих главах.
Генетика | 55
ЭВОЛЮЦИЯ ДОМИНАНТНОСТИ
Проблема мутаций имеет ряд других интересных аспектов,
о которых мало что известно. У Bicex хорошо изученных орга-
низмов большая часть мутантных аллелей рецессивна по отно-
шению к гену дикого типа. В связи с этим возникает вопрос
о происхождении доминантности и рецессивности. Ясно, что
большая часть возникающих мутаций должна быть вредношдля
организма, так как они изменяют ту или иную функциональную
систему. Если они вызывают сильно выраженный эффект, то
почти наверное сложная взаимосвязь механизмов развития бу-
дет резко нарушена и организм погибнет. Но интеграция гено-
типа так совершенна, что даже в тех случаях, когда ген обла-
дает лишь сравнительно слабым действием, мутантный организм
окажется менее приспо1собленным, чем его родители, если только
не произойдет изменения в окружающей среде. (Шансы улуч-
шить работу радиоприемника, внося в его схему небольшие слу-
чайные изменения, пойти ничтожны.) Рецессивные мутации,
возникающие у диплоидного организма, пополняют резервы его
изменчивости. Если рецессивные мутации встретятся в гомози-
готном состоянии (что бывает редко), они почти наверняка
будут элимированы при условии, что среда (в самом широком
смысле этого слова) не изменится настолько, что они приобре-
тут положительную селективную ценность.
Каким же образом возникает рецессивность? На этот счет
существует несколько гипотез, из которых в настоящее время
трудно сделать выбор, хотя все они, вероятно, содержат элемен-
ты истины. Фишер высказал предположение, что мутации не
обязательно должны быть полностью рецессивными при своем
первом появлении. Поэтому они, как правило, будут неблаго-
приятны для организма, если только другие гены-модификаторы,
расположенные в иных локусах, не уменьшат вредных послед-
ствий их проявления. Поскольку в гомозиготном состоянии му-
тации встречаются редко, отбор будет воздействовать на чаще
встречающиеся гетерозиготы, вырабатывая системы модифика-
торов, действие которых приблизит фенотип гетерозиготы к фе-
нотипу доминантной гомозиготы.
Райт и Холдейн обсуждали проблему происхождения доми-
нантности с точки зрения взаимосвязи между ферментом, обра-
зующимся под контролем гена, и субстратом этого фермента.
По их представлениям, рецессивны те гены, которые в сравне-
нии с соответствующим геном дикого типа менее активны как
факторы образования определенного фермента. Отбор, по-види-
мому, должен был обусловить создание некоторого функциональ-
ного резерва, благодаря чему обеспечивается избыток фермента
56 | Глава 3
над субстратом; поэтому мутация, уменьшающая количество
фермента, не окажет заметного действия в гетерозиготном со-
стоянии. Биохимическая генетика уже позволяет получить ответы
на вопросы, касающиеся количественной стороны функции генов
в процессе синтеза ферментов.
Райт и Холдейн высказали также предположение, согласно
которому селективная ценность генов-модификаторов настолько
мала, что доминантность вырабатывалась бы слишком медленно,
для того чтобы иметь большие шансы на развитие, если учиты-
вать и другие факторы. Однако судя по данным, полученным на
некоторых объектах, коэффициенты отбора для генов-модифи-
каторов могут быть значительно выше, чем предполагали Райт
и Фишер. Как бы то ни было, отбор играл важную роль в эво-
люции фенотипического проявления генов. Опыты по гибриди-
зации, проведенные на таких различных организмах, как хлоп-
чатник и бабочки, ясно показали, что функционирование гена
может изменяться при переносе его из одного генетического
окружения в другое. Таким образом отбор, изменяя генетический
фон (например, модификаторы), может влиять на выражение
данного гена.
Существует достаточно данных в пользу того, что именно
так обстояло дело при развитии индустриального меланизма у
бабочки Biston betularia (см. гл. 7). В первых исследованиях,
посвященных этому вопросу, было обнаружено, что гетерозиготы
по аллелю, вызывающему меланизм, резко отличаются от гомо-
зиготных меланических особей. К середине XX века гетерозигот-
ные особи В. betularia почти перестали отличаться от гомозигот-
ных. Совершенно очевидно, что в этом случае выработалась
доминантность.
ХРОМОСОМНЫЕ МЕХАНИЗМЫ
Открытие методов искусственного получения мутаций у легко
выращиваемых организмов с относительно коротким жизненным
циклом (таких, как дрозофила, кукуруза и нейроспора) позво-
лило тщательно изучить явление сцепления и группы сцепления,
т. е. хромосомы. Если принять, что частота кроссинговера между
двумя факторами пропорциональна расстоянию между ними, то
можно определять расстояния и порядок расположения генов
вдоль хромосомы. Можно построить для хромосомы карту сцеп-
ления, основанную на данных о рекомбинациях. При этом сле-
дует помнить, что возможна интерференция между соседними
точками перекреста, что между расположенными на большом
расстоянии друг от друга факторами может произойти множе-
ственный кроссинговер и, наконец, что карты можно строить
Генетика | 57
только для тех участков хромосомы, в которых расположены
гены, обладающие выраженным фенотипическим эффектом. Су-
ществуют, однако, иные способы построения хромосомных карт;
с их помощью можно проверять карты, построенные на основе
изучения сцепления. Карты составляют, например, изучая эф-
фект индуцированных делеций небольших участков хромосомы
или эффект таких изменений, как инверсии и транслокации, а
также другими методами. С помощью всех этих методов был
подтвержден тот порядок линейного расположения генов, кото-
рый был установлен на основании данных по кроссинговеру.
В то же время полученные карты отличались (нередко весьма
существенно) от карт сцепления в отношении расстояний и дру-
гих характеристик. Гетерохроматиновые участки хромосом, в
которых, по-видимому, нет генов с сильным действием, трудно
исследовать путем анализа рекомбинаций, а эти-то участки в
основном и ответственны за различия. Большие успехи достиг-
нуты в установлении локализации специфических генетических
локусов в определенных участках хромосом у дрозофилы (ра-
боты проводились на гигантских политенных хромосомах слюн-
ных желез) и у кукурузы (у которой хромосомы обладают ха-
рактерными хромомерами, видимыми в микроскоп).
ПОЛОВЫЕ ХРОМОСОМЫ
У раздельнополых животных и растений наблюдается особый
тип сцепления. У этих организмов, имеющих специальные поло-
вые хромосомы, отличные от остальных хромосом (так назы-
ваемых аутосом), один пол обычно имеет две гомологичные
половые хромосомы. У другого пола имеется лишь одна хромо-
сома, гомологичная этим двум, вторая же либо отсутствует,
либо отличается от первой и лишь отчасти гомологична ей.
У дрозофилы и человека женский пол является гомогаметным
(в каждой гамете содержится по одной Х-хромосоме), а муж-
ской пол — гетерогаметным (образуются гаметы двух типов, со-
держащие либо Х-хромосому, либо ее частичный гомолог, Y-xpo-
мосому). Ясно, что передача генов, находящихся в половых
хромосомах, будет отличаться от передачи аутосомных генов.
Кроме того, определяемые этими генами признаки будут обна-
руживать генетическое сцепление с полом. Половые хромосомы
у таких организмов отличаются от аутосом тем, что они диффе-
ренцированы на два различных участка. Определенная часть
двух различных половых хромосом гетерогаметного пола спо-
собна вступать в синапсис, что обусловливает возможность крос-
синговера. В отличие от этих синаптирующих сегментов суще-
ствуют дифференциальные сегменты X- и Y-хромосомы, которые
58 | Глава 3
не конъюгируют. Дифференциальный сегмент Y-хромосомы
обычно не содержит или почти не содержит генов, обладающих
сильным действием, состоит из гетерохроматина и имеет мень-
шую величину, чем дифференциальный сегмент Х-хромосомы Г
В том случае, когда он все же содержит такие гены, они пере-
даются от отца только к сыну — соответствующие признаки
никогда не проявляются у особей женского пола. Гетерохрома-
тиновый участок Y-хромосомы необходим для плодовитости;
значит, он не может быть полностью неактивным.
У некоторых групп организмов гетерогаметным бывает жен-
ский пол, а мужской пол является гомогаметным. У многи/Х
видов Y-хромосома отсутствует; тогда самец имеет формулу ХО.
Встречаются и более сложные системы половых хромосом; они
будут рассмотрены в гл. 9. Например, у некоторых организмов
имеется много половых хромосом. Следует подчеркнуть, что де-
тали механизма определения пола варьируют у разных групп
животных, хотя хромосомный аппарат и может казаться у них
одинаковым. Эволюция полового процесса как средства, обусло-
вливающего изменчивость и обеспечивающего ее реализацию
(путем генетической рекомбинации), рассматривается в гл. 8.
ХРОМОСОМНЫЕ ПЕРЕСТРОЙКИ
Помимо мутаций, рассмотренных выше (так называемых ген-
ных, или точковых, мутаций), наблюдаются изменения в строе-
нии хромосом, возникающие спонтанно или вызываемые теми же
агентами, что и генные мутации. Хотя эти хромосомные пере-
стройки также иногда называют мутациями, будет меньше пу-
таницы, если относить термин «мутация» только к изменениям
отдельных генов. Перестройки называют также хромосомными
аберрациями или аномалиями, но только потому, что их сравни-
вают с произвольно выбранным стандартным фенотипом хромо-
сом (обычно с диким типом); их не следует принимать за какое-
то необычное или особо вредоносное явление, во всех случаях
невыгодное для организма. Подобно генным мутациям, хромо-
сомные перестройки при своем возникновении обыкновенно
имеют низкую или отрицательную селективную ценность, но они
вполне могут закрепиться в популяции или даже вытеснить
стандартный тип, если их селективная ценность повысится
вследствие изменений окружающей среды при условии, что ядро
и гены также рассматриваются как часть среды.
1 У человека и, по-видимому, у всех млекопитающих дифференциальный
сегмент Y-хромосомы содержит факторы, определяющие развитие мужского
пола. — Прим. ред.
Генетика | 59
В сущности все, что теоретически могло бы случиться с хро-
мосомами во время жизненного цикла и деления клетки, было
действительно обнаружено как у лабораторных, так и у диких
животных. Часто можно показать, что то или иное изменение
закрепилось в качестве постоянной особенности данного орга-
низма. К основным типам хромосомных перестроек относятся
делеции (утрата участка хромосомы), дупликации (повторение
какого-либо участка в той же или другой хромосоме), инверсии
(при которых участок хромосомы оказывается перевернутым
на 180°) и транслокации (перенос участка хромосомы на дру-
гую, негомологичную хромосому; обычно при этом происходит
взаимный обмен участками). Изменения числа хромосом, часто
называемые «хромосомными мутациями», будут рассмотрены
в гл. 9.
Делеции и дупликации. Роль в эволюции утраты или доба-
вления хромосомного материала еще очень неясна. В гомози-
готном состоянии делеции обычно детальны. Делеции служат
полезным инструментом при построении хромосомных карт; их
эволюционное значение в настоящее время неизвестно. Значение
дупликаций состоит в том, что они составляют возможный цито-
генетический механизм увеличения общего количества генети-
ческого материала. Как изменялось это количество в ходе эво-
люции — мы не знаем. В настоящее время невозможно также
сказать, обязательно ли увеличение сложности организмов
должно сопровождаться увеличением количества генетического
материала.
Инверсии и транслокации. Инверсии и взаимные транслока-
ции представляют собой более существенные перестройки; они
лучше изучены, и их эффект хорошо известен. Организмы, у ко-
торых эти изменения стали постоянной особенностью генетиче-
ской системы, будут рассмотрены в гл. 9. Здесь мы обсудим
только цитогенетические аспекты таких изменений. Если не ис-
следовать конъюгацию хромосом, то перевернутый участок
обычно нельзя обнаружить визуально; исключение составляют
организмы, имеющие политенные хромосомы или хромосомы с
хорошо видимыми хромомерами. Во время профазы первого де-
ления мейоза, когда гомологичные хромосомы образуют пары,
гетерозиготность по инверсии выявляется благодаря тому, что
для точного спаривания одной из конъюгирующих хромосом
приходится образовать петлю (фиг. 12). Эту характерную инвер-
сионную петлю можно также наблюдать в хромосомах слюнных
желез дрозофилы и других двукрылых, у которых политенные
хромосомы конъюгируют и в соматических клетках. Если ин-
версия короткая, кроссинговер в инверсионной петле может*не
происходить. Если инверсия достаточно длинная и находится в
60 [ Глава 3
определенных пространственных отношениях с центромерой, то
кроссинговер возможен и результатом его бывает образование
дицентрической хромосомы и ацентрического фрагмента. В ана-
фазе ацентрический фрагмент не отходит к полюсу веретена, а
дицентрический фрагмент разрывается. Среди образующихся га-
мет, помимо сбалансированных, будут гаметы с нехватками
больших участков хромосом, и соответствующие зиготы ока-
жутся нежизнеспособными.
Генетически все это выглядит так, как если бы кроссинговер
был подавлен. Однако на самом деле происходит только эли-
минация его продуктов. Последствия зависят от числа перекре-
стов в инверсионной петле и от их распределения между хрома-
тидами бивалента. Так как у гетерозигот гены, лежащие в
инверсионной петле, при известных условиях оказываются эф-
фективно сцепленными, инверсии будут подробнее рассмотрены
в связи с рекомбинацией (см. гл. 9). Формы, гомозиготные по
хромосомной инверсии, могут быть распознаны только путем
скрещивания их с другими формами и исследования синапсиса
хромосом у полученного гибрида (исключение составляют орга-
низмы с политенными хромосомами, у которых такие формы
могут быть выявлены путем тщательного изучения хромосом-
ных дисков). Поскольку у гомозиготы инверсия не повлияет на
синапсис, число рекомбинаций не будет снижено, но линейная
последовательность генов и сцепление между ними, конечно, из-
менятся.
Во взаимных транслокациях участвуют негомологичные хро-
мосомы. При этом часть одной хромосомы присоединяется к
другой и наоборот, так что группы сцепления изменяются. Ве-
личина обмениваемых участков может быть одинаковой или
разной, большой или малой. Иногда гетерохроматиновый уча-
сток хромосомы (с незначительной специфической генной актив-
ностью) обменивается на эухроматиновый участок (обладающий
типичными генетическими свойствами). Гетерозиготность по
крупным взаимным транслокациям визуально выявляется в мей-
озе, а также у организмов со спаренными политенными хромосо-
мами в соматических клетках. При синапсисе происходит соеди-
нение четырех хромосом — двух стандартных и двух с транс-
локациями. Форма такого квадривалента в метафазе зависит,
помимо всего прочего, от распределения хиазм. Обычно хромо-
сомы разделяются, образуя кольцо. Концы хромосом удержи-
ваются вместе терминализованными хиазмами. Расхождение мо-
жет происходить таким образом, что к одному и тому же по-
люсу отойдут смежные центромеры. Существует два возмож-
ных способа расхождения такого типа (фиг. 13), но оба они
ведут к образованию несбалансированных гамет (т. е. гамет с
Фиг. 12. Результаты кроссинговера в петле
парацентрической инверсии.
бивалента, гетерозиготного по
Фиг. 13. Результаты кроссинговера на дистальных концах хромосом и рас-
хождения хромосом у организма, гетерозиготного по взаимной транслокации.
Хромосомы или их участки, являющиеся гомологичными, показаны линиями
одного типа (сплошными или прерывистыми).
62 | Глава 3
дупликациями и нехватками). Сбалансированная гамета может
возникнуть только в том случае, если к одному полюсу отойдут
несмежные центромеры. При слиянии таких гамет в случайных
комбинациях образуются стандартные гомозиготы, гетерозиготы
с транслокацией и гомозиготы с транслокацией в отношении
1 : 2 : 1. У организмов, гомозиготных по взаимной транслокации,
в мейозе не обнаруживается ничего необычного, так как синап-
сис хромосом не нарушен. Группы сцепления, однако, оказы-
ваются в этом случае измененными, что можно установить пу-
тем генетического анализа.
В ядре и в хромосоме может быть несколько инверсий или
транслокаций. Инверсии в хромосоме могут целиком лежать в
одном плече, но могут захватывать и центромеру. Если их не-
сколько, то они могут быть обособленными или частично пере-
крывающимися; кроме того, возможно включение одной или не-
скольких инверсий в другие. Детальные исследования этих со-
стояний проводились на дрозофиле и будут рассмотрены в гл. 9.
При наличии двух или нескольких транслокаций результат за-
висит от того, какие хромосомы в них участвуют. Если после
первой транслокации обменивается участками другая пара хро-
мосом, то в метафазе мейоза образуются два квадривалента.
Если же одна из хромосом, участвовавших в первой транслока-
ции, обменивается участком с третьей хромосомой, то мы обна-
руживаем кольцо из трех хромосом. Наконец, у некоторых орга-
низмов все хромосомы обмениваются плечами, и в мейозе все
они соединены в кольцо. Соответствующие примеры мы рас-
смотрим в гл. 9.
НЕПРЕРЫВНАЯ ИЗМЕНЧИВОСТЬ
Переходя к изучению наследования признаков, подвержен-
ных непрерывной изменчивости, мы сталкиваемся с еще более
трудными проблемами. Обычно невозможно бывает выявить спе-
цифические гены, контролирующие, определенные признаки. Как
уже упоминалось, существует, по-видимому, непрерывный спектр
изменчивости от типично качественной до чисто количественной.
Предполагается, что нет коренного различия между генами с
легко выявляемым, резко выраженным действием (которые ча-
сто называют олигогенами или генами-переключателями) и та-
кими генами, которые в отдельности оказывают лишь слабое
действие, но функционируют как часть системы, состоящей из
бесчисленного множества факторов (эти гены часто называют
полигенами, хотя этот термин употреблялся и в более узком
значении). В операциональном смысле можно сказать, что раз-
личие между этими двумя типами признаков больше зависит
Генетика | 63
от относительного значения генетического материала и среды
в определении фенотипа, чем от величины эффекта отдельных
генов. Однако существуют признаки, для проявления которых
среда относительно несущественна, — таково, например, число
абдоминальных щетинок у дрозофилы.
При изучении какого-либо количественного признака произ-
водится анализ возможно большего числа различающихся ме-
жду собой особей. Все эти особи уникальны, но полученные для
них величины могут быть сгруппированы в ряд размерных клас-
сов. Представив эти величины в виде графика распределения
частот, можно исследовать характер изменчивости. Можно, на-
пример, рассчитать среднее арифметическое для всех особей
данной выборки. Степень изменчивости в выборке можно оха-
рактеризовать стандартным отклонением (S) или его квадра-
том— дисперсией (S2).
Обычно бывает трудно разделить генетические и средовые
компоненты изменчивости и исследовать отдельно ту ее часть,
которая обусловлена генетически. Для изучения и разделения
этих компонентов были разработаны различные эксперименталь-
ные методы и использован довольно сложный математический
аппарат. Например, один из способов оценки роли этих двух
компонентов основан на уменьшении генетического компонента
до пренебрежимо малой величины. Определяют дисперсию для
данного признака в популяции, находящейся в «нормальной»
среде. Затем дисперсию того же признака определяют для осо-
бей высокоинбредной линии, выращенных в тех же условиях.
Поскольку эти особи можно считать по существу генетически
тождественными, полученную дисперсию можно всецело припи-
сать влиянию среды. Тогда различие между двумя полученны-
ми величинами дисперсии служит характеристикой генетической
изменчивости, так как общая дисперсия (фенотипическая дис-
персия Sj) в этом случае равна сумме генетической и средо-
вой дисперсий, Таким образом, Sp — S1e = S2q- Этот ме*
тод оценки двух компонентов изменчивости основан на предпо-
ложении, что средовая изменчивость не зависит от генотипа, а
это часто оказывается неверным.
Даже если удается количественно охарактеризовать генети-
ческую изменчивость, остаются еще другие сложности. Не для
всех признаков можно принять, что эффекты генов аддитивны,
т. е. просто суммируются. Генетическая изменчивость сама мо-
жет быть разделена на ряд компонентов. Существует аддитив-
ный компонент, представляющий различия между гомозиготами
и гетерозиготой (или гетерозиготами) по каждому локусу. Не-
обходимо также учитывать компонент, обусловленный взаимо-
64 | Глава 3
действием аллелей, т. е. явлением доминирования, и компонент,
связанный с взаимодействием неаллельных генов (эпистатиче-
ский компонент). Во многих случаях аддитивный генетический
компонент является единственным компонентом, величину кото-
рого нетрудно оценить. Таким образом, фенотипическая измен-
чивость оказывается подразделенной на аддитивную генетиче-
скую и «остаточную» изменчивость. К последней относятся не-
аддитивные компоненты генетической изменчивости, средовая
изменчивость и взаимодействия генов со средой. Доля феноти-
пической изменчивости, которую можно приписать аддитивным
генетическим эффектам, служит показателем наследуемости
Ii-Sq/Sp) Иногда наследуемость определяют в б9лее узком
смысле как Sa/Sp, где Sa — аддитивная генетическая дисперсия.
Наследуемость является хорошим показателем степени сходства
между потомством и родителями и поэтому имеет большое зна-
чение при селекции растений и животных.
Специалисту, изучающему эволюцию, приходится иметь дело
с этими сложностями потому, что огромное большинство измен-
чивых признаков у организмов изменяется количественно и на-
ходится под контролем множества факторов. В тех случаях, ко-
гда оказываются возможными скрещивания между расами, ви-
дами и даже родами, потомство в Fi обычно бывает более или
менее промежуточным по своим признакам, а в Р2 обнаруживает
непрерывную изменчивость, характерную для полигенной на-
следственности. Это слишком обобщенная картина сложной си-
туации; более детальные сведения читатель найдет в книге Фол-
конера [1], особенно в разделах, посвященных инбредной депрес-
сии и гетерозису [1, гл. 14].
Определение признака многими генами обусловливает такую
стабильность фенотипического выражения, какой может и не
быть, если признак определяется лишь одним геном. Например,
единичная мутация вряд ли серьезно нарушит проявление при-
знака, зависящего, скажем, от аддитивных эффектов 35 локусов.
Напротив, единичная мутация в гене, подавляющем окраску
у лука, приведет к образованию белой луковицы, а не красной
или желтой. Если учесть возможность резко выраженных эф-
фектов, обусловленных изменениями в «главных» генах, не по-
кажется удивительным, что значительное число признаков кон-
тролируется многими генами. Отбор должен был благоприят-
ствовать выработке подобных систем, так как они в общем
уменьшают вредные последствия таких незначительных собы-
тий, как замена одной пары оснований в ДНК.
Полигенные системы, характеризующиеся сравнительно ма-
лой изменчивостью, могут обладать безгранично большой по-
Генетика | 65
тенциальной способностью к изменчивости: они способны под
действием отбора создавать такие генотипы, какие при отсут-
ствии отбора никогда не возникли бы. Представим себе, что
какой-то признак контролируется 40 локусами, в каждом из ко-
торых имеются два аллеля, + и —, и что эффекты этих генов
аддитивны (самые крайние фенотипы соответствуют гомозигот-
ности + + или гомозиготности---по всем локусам). Если бы
частота аллелей в каждом локусе была 0,50+ и 0,50—, то при
отсутствии отбора вероятность появления одной диплоидной осо-
би с крайним (> +)-фенотипом (т. е. особи, гомозиготной + +
по всем локусам) составляла бы (72)80, что неизмеримо меньше
единицы, деленной па число электронов во всей вселенной, т. е.
для всех практических целей эта вероятность равна нулю. Од-
нако при наличии отбора, благоприятствующего особям с наи-
большим числом аллелей +, эта потенциальная возможность
могла бы реализоваться за какие-нибудь 8 или 10 поколений.
Таким образом, полифакториальные системы наследственности
могут быть важным механизмом поддержания равновесия ме-
жду приспособленностью к условиям данного момента и гибкой
приспособляемостью к медленным, длительным изменениям
среды.
выводы
У большинства организмов генетический материал (ДНК)
связан с длинными белковыми тяжами, образуя хромосомы.
Хромосомы в продольном направлении дифференцированы на
функциональные единицы, называемые генами, которые могут
быть представлены многочисленными аллелями и контролируют
образование специфических ферментов. Мутирование генов к
различным аллеломорфным состояниям происходит спонтанно с
частотой от 10"5 до 10-9 на одно поколение. Мейоз и кроссинго-
вер приводят к рекомбинациям, причем наименьшие рекомбини-
руемые единицы обычно эквивалентны функциональным генам.
Если исключить сцепление в хромосомах, то можно сказать, что
гены распределяются между гаметами и комбинируются в зиго-
тах независимо друг от друга. Наблюдается взаимодействие
между аллелями (доминирование) и между локусами (эпистаз).
Некоторые признаки зависят от генов, обладающих сильным,
хорошо выраженным действием, но могут существовать и мо-
дифицирующие факторы. Большинство признаков контроли-
руется очень большим числом негомологичных генов, каждый
из которых обладает сравнительно слабым действием. Изуче-
ние возникающей при этом количественной изменчивости ослож-
няется трудностью отделения различных «фракций» генетиче-
5 Зак. 152
66 | Глава 3
ского компонента изменчивости друг от друга и от средового
компонента. Основным источником изменчивости являются ген-
ные мутации. В популяциях высших организмов, размножаю-
щихся половым путем, более важным источником быстрых из-
менений оказывается процесс рекомбинации.
ЛИТЕРАТУРА
1. Falconer D. S., Introduction to Quantitative Genetics, Ronald, New York,
1960.
2. Sager R., Ryan F. J., Cell Heredity, Wiley, New York, 1961.
3. Sinnott E. W., Dunn L. C., D о b z h a n s k у T., Principles of Genetics,
McGraw-Hill, New York, 1958.
4. De Robertis E., Nowinski W., Saez F., General Cytology, 2d ed.,
W. B. Saunders Company, 1954. (Де Робертис E., Новинский В.,
Саэс Ф., Общая цитология, ИЛ, М., 1962.)
5. White V. S., Animal Cytology and Evolution, 2d ed., Cambridge University
Press, 1954.
Глава 4
РАЗВИТИЕ
Генетические механизмы, описанные в гл. 3, очевидно, выра-
ботались потому, что они закрепляли выгодные комбинации ге-
нетических структур. Возможно, что некоторые первичные орга-
низмы просто росли, до тех пор пока какие-то случайные об-
стоятельства не приводили к их дезинтеграции. Многие из них,
быть может, погибали вследствие того, что менялось отношение
поверхности к объему; это нарушало их несовершенную внутрен-
нюю организацию. Некоторые могли распадаться на более мел-
кие единицы, причем один лишь случай определял, будут ли
дочерние фрагменты обладать организацией, обеспечивающей
возможность дальнейшего роста. Любые случайно возникающие
механизмы, способствующие сохранению у продуктов фрагмен-
тации способности к росту (и к дальнейшей успешной фрагмен-
тации), должны были автоматически закрепляться. Так вырабо-
тались механизмы, которые привели к существенному ограниче-
нию изменчивости, вероятно имевшей место при делении и раз-
витии первичных организмов. Возникновение этих механизмов
составляет в известном смысле основную проблему в происхо-
ждении живых систем, как уже говорилось в гл. 1. В шутку
была высказана мысль, что человеческие существа — это всего
лишь приспособления, которые выработала ДНК для синтеза
все новых количеств ДНК; можно также сказать, что ДНК
представляет собой просто средство, используемое человеком
для того, чтобы избежать появления инородного потомства. Ге-
нетический материал неспособен к репликации в отсутствие дру-
гих компонентов живых систем. В ходе эволюции вырабатыва-
лись все более и более сложные системы, в том числе и генети-
ческая система.
РОСТ И ГОМЕОСТАЗ
Как полагают, самые первые организмы были одноклеточ-
ными (или неклеточными). У таких организмов всего лишь одно
или два клеточных деления (и, возможно, одно слияние клеток)
ведут к образованию отдельных функционирующих единиц.
Здесь нередко трудно бывает провести различие между наслед-
ственностью и развитием, или дифференцировкой, что, напротив,
5*
68 | Глава 4
совсем нетрудно сделать, имея дело с многоклеточными орга-
низмами. Каждый клеточный компонент представляет собой на-
следственную единицу, которая воспроизводится более или ме-
нее точно в процессе размножения клетки.
С развитием многоклеточности и увеличением сложности воз-
никают другие проблемы. В конце концов происходит обособле-
ние зародышевых и соматических клеток. Функции ядерных и
внеядерных самовоспроизводящихся компонентов, имевшихся у
одноклеточных организмов, теперь, по-видимому, до некоторой
степени дифференцируются. Для ядерного материала, с которым
мы связываем генетическую информацию (генотип), характерны
большая стабилизация и более строгая регуляция. Внеядерный
материал играет важную роль в развитии и дифференцировке,
изменяя свои свойства с течением времени и взаимодействуя
с ядерным материалом и внешней средой (в том числе с дру-
гими клетками). Одноклеточный организм в известном смысле
бессмертен. «Концом» существования клетки обычно бывает ее
случайное разрушение (в частности, использование в пищу дру-
гим организмом) или разделение одной клетки на две. Сомати-
ческие клетки многоклеточных существ рано или поздно отми-
рают, что можно рассматривать как элемент генетической си-
стемы.
Непрерывное воспроизведение предполагает увеличение раз-
меров, или рост. Под развитием понимают изменения, которым
подвергается организм на протяжении своей жизни. От простых
изменений соотношения между поверхностью и объемом, к ко-
торым, возможно, сводилось развитие первичного организма,
еще очень далеко до жизненного цикла бабочки-данаиды (яйцо—
гусеница — куколка — имаго), но разница здесь количественная,
а не качественная. В процессе роста организмы изменяют свою
величину; но структура, эффективная при одних размерах, мо-
жет оказаться совершенно неэффективной при других. Если
учесть физически обусловленные пределы возможной величины
клетки млекопитающего (определяемые величиной белковых мо-
лекул, скоростями диффузии и многими другими факторами),
то легко понять, что нормально устроенный миниатюрный чело-
вечек величиной с яйцеклетку или сперматозоид длиной 180 см
просто не могли бы существовать. Генетический механизм обес-
печивает не построение копий родительского многоклеточного
организма, а образование систем, которые в известных преде-
лах изменчивости способны развиваться в организмы родитель-
ского типа.
Такого рода регуляция может быть названа онтогенетиче-
ским гомеостазом и тесно связана с понятием «дикого типа».
В большинстве групп организмов значительной изменчивости
Развитие | 69
подвержены, по-видимому, только более или менее второстепен-
ные признаки. Дрозофила дикого типа изучена генетически
вплоть до мельчайших подробностей. У человека в норме не
бывает глаз разного цвета или шести пальцев на каждой руке.
Красное дерево может быть разной высоты и с разным числом
ветвей, но оно всегда имеет характерные зеленые листья, рас-
положенные типичным образом, и шероховатую красную кору.
У крупных организмов изменчивость больше бросается в глаза,
чем у мелких, но это не обязательно означает, что мелкие орга-
низмы менее изменчивы. По-видимому, у всех организмов жиз-
ненно важные механизмы развития относительно мало подвер-
жены наследственным изменениям. Организмам выгодно
избегать большого количества «брака» среди потомства путем
развития оптимальных фенотипов из целого ряда несущественно
варьирующих генотипов. У многих организмов процессы разви-
тия специфической формы сделались «канализованными», что
приводит к фенотипической однородности особей в данной
популяции, несмотря на генетические различия между ними.
Благодаря этому механизму генетическая изменчивость (важ-
ная в отдаленной перспективе) может вызывать лишь мини-
мальное снижение приспособленности.
Как достигается этот «буферный» эффект? Используя модель
Лернера [4], предположим, что ген А продуцирует вещество а,
которое под действием вещества b (продукта гена В) превра-
щается в г, а г, взаимодействуя с с (продуктом гена С), дает
вещество t. .
А —
Если теперь ген В исчезнет в результате мутации или же во
внешней среде не окажется субстрата, из которого может быть
синтезировано вещество Ь, то будет накапливаться вещество а.
Система, в которой а в высокой концентрации взаимодействует
с d (продуктом гена О), образуя вещество s, способное превра-
щаться под действием с в t, представляет собой буферную си-
стему.
70 | Глава 4
Это настоящая система с обратной связью, так как конкретный
механизм, ведущий к нормальному проявлению признака, опре-
деляется получаемой организмом информацией о концентрации
вещества а. Это, однако, не значит, что буферный эффект всегда
имеет генную природу.
Обычно принято проводить резкую границу между генотипом
и фенотипом. Как педагогический прием это полезно, поскольку
при этом подчеркивается постоянство и непрерывность передачи
генетической информации; такое разграничение, хотя оно и упро-
щает действительность, привело к значительному прогрессу в
генетической науке. Однако в то же время оно создавало впе-
чатление, что генотип — это в некотором смысле основная сущ-
ность, а фенотип — всего лишь грубое отражение генотипа (об-
раз которого искажен влиянием среды). Можно было бы удив-
ляться, почему отбор не разделался с фенотипом начисто,
позволив генотипу развиваться без «посторонней примеси». От-
вет, разумеется, ясен: то, что можно разделить в учебниках или
в теории, нельзя разделить в живых организмах. Отпрепариро-
вав генетический материал плодовой мушки, мы получили бы
длинную, ничего не значащую цепочку нуклеотидов, которая
сама является компонентом «фенотипа». Ясно, что на этом уров-
не исследования различие между генотипом и фенотипом теряет
смысл. Генетическая информация становится биологически ос-
мысленной только тогда, когда она «расшифровывается» в ре-
зультате контакта с окружающей средой. В самом деле, о цен-
ности информации судят только по ее расшифрованной форме,
а не по оригиналу. Естественный отбор воздействует на фено-
тип, а не прямо на генотип, который лишь определяет реакции
развивающегося организма на внешнюю среду.
Только в последние годы специалисты, изучающие эволюцию,
обратили должное внимание на процессы развития, приводящие
к построению взрослого функционирующего организма из опло-
дотворенного яйца, или зиготы. Эти процессы связаны между
собой в систему, которую Уоддингтон назвал «эпигенотипом».
Последний можно представлять себе как разветвленную систему
путей развития, каждый из которых ведет к одному из компо-
нентов взрослой формы. Поскольку биохимические реакции,
определяющие различные пути, тесно связаны друг с другом
(о чем говорилось выше), наблюдается сильная тенденция к до-
стижению нормального конечного результата даже тогда, КОГДа
на ранних этапах произошли значительные нарушения. Таким
образом, эти пути «канализованы», или стабилизированы, объ-
единяющими их механизмами — обратными связями.
Эта эпигенетическая система, вероятно, явилась продуктом
естественного отбора, который действовал на гены, оказываю-
Развитие | 71
щие более или менее прямое влияние на проявление определен-
ных признаков у организмов. Однако отбор должен был затра-
гивать также многие гены, единственный явный фенотипический
эффект которых состоит в видоизменении проявления других ге-
нов. Уоддингтон указал на то, что в популяции организмов, жи-
вущих в определенной среде, каждая особь должна иметь свой
собственный генотип, а значит, и собственный эпигенотип, кото-
рый должен в конечном счете привести к развитию фенотипа
взрослой особи. Отбор, направленный на сохранение приспособ-
ленности в данной специфической среде, может приводить к
элиминации генотипов, обусловливающих отклонения в фено-
типе. Он может также устранять особей, недостаточно «стаби-
лизированных», для того чтобы противостоять влияниям среды.
В этом случае происходил бы отбор на лучшую стабилизацию
(канализацию) эпигенетической системы.
Можно ожидать, что в случае изменения среды у некоторых
хорошо стабилизированных особей будут развиваться приспособ-
ленные фенотипы даже без немедленного генотипического изме-
нения. Однако в новых условиях по прошествии некоторого вре-
мени генотипическое изменение неизбежно, и следует ожидать,
что отбор приведет к стабилизации новых путей развития. Если
затем такие организмы вернутся в свою первоначальную среду,
то окажется, что в результате изменения в генотипе у них уже
не разовьется прежний фенотип. Таким образом то, что вна-
чале было фенотипической (по существу эпигенетической) ре-
акцией на изменение внешних условий, оказывается включен-
ным в генотип вследствие отбора на хорошо стабилизирован-
ную систему механизмов развития в новой среде. То, что каза-
лось «приобретенным признаком», становится наследственной
особенностью в результате действия естественного отбора. Этот
процесс, известный под названием генетической ассимиляции,
будет рассмотрен в гл. 7.
ЖИЗНЕННЫЕ ЦИКЛЫ
Цикличность роста характерна для всех организмов. Дрож-
жевые клетки проходят циклы слияния и деления, включающие
как гаплоидную, так и диплоидную фазы (гл. 3). У большин-
ства высших растений развитие совершается и в гаплофазе, и
в диплофазе, хотя гаплофаза (мужской и женский гаметофиты)
гораздо короче и менее заметна, чем диплофаза (спорофит).
У большей части животных в гаплофазе, которая обычно пред-
ставлена только гаметами, фактически не происходит никакого
развития. (Самцы перепончатокрылых, являющиеся гаплоид-
ными, составляют разительное исключение — см. гл. 9.)
72 | Глава 4
Простой цикл «рост — фрагментация — рост», постулирован-
ный в качестве наиболее примитивной формы развития, претер-
пел в результате отбора разнообразные изменения. Примером
сложного цикла развития может служить жизненный цикл про-
стейшего Plasmodium — возбудителя малярии. Спорозоиты
Plasmodium в организме комара мигрируют в слюнные железы,
а затем, попадая при укусе в кровяное русло человека, способны
проникать в клетки определенного типа. Здесь они могут раз-
множаться бесполым путем, порождая мерозоиты, которые ин-
фицируют другие клетки или проникают в эритроциты. Мерозо-
иты, попавшие в эритроциты, могут либо вновь размножаться
бесполым способом, образуя новые мерозоиты, заражающие
другие эритроциты, либо развиваться в гаметоциты и в конце
концов образовать гаметы, которые сливаются в кишке другого
комара. Образующаяся при этом подвижная зигота мигрирует
к стенке кишки и превращается в спороцист. В спороцисте пу-
тем клеточного деления образуются спорозоиты.
Генотип, ответственный за построение той эффективной «ма-
шины» для накопления питательных веществ, которую мы назы-
ваем гусеницей, содержит также и информацию, необходимую
для создания совершенно несходной формы, осуществляющей
размножение и расселение вида, — так называемой бабочки. Зи-
гота, из которой разовьется гигантская секвойя, содержит также
информацию, нужную для образования ее крохотного пыльце-
вого зерна (мужского гаметофита, состоящего всего из несколь-
.ких клеток). Из зиготы человека путем деления в конце концов
образуются такие несходные между собой потомки, как эритро-
циты, мышечные волокна и нервные клетки. Эти отклонения от
простейшего цикла развития возникли в результате отбора, воз-
действовавшего на весь жизненный цикл организма от образо-
вания зиготы до смерти.
Каким образом действует отбор и каким образом он выра-
ботал механизмы развития, будет ясно в деталях только тогда,
когда будут выяснены сами эти механизмы. Поэтому нам необ-
ходимо вкратце рассмотреть их.
ДИФФЕРЕНЦИРОВКА И МОРФОГЕНЕЗ
Митоз был охарактеризован как механизм, обеспечивающий
равное распределение генетической информации между дочер-
ними клетками в ходе клеточного деления. То, что митоз может
достигать этого результата, легко доказывается целым рядом
способов. Если, например, зиготическому ядру стрекозы Platyc-
nemis дать возможность пройти через 7 делений (до стадии
128 клеток), а затем все дочерние клетки, кроме одной, убить
Развитие | 73
с помощью узкого пучка ультрафиолетовых лучей, то из этой
клетки все-таки разовьется нормальный эмбрион. Очевидно, ис-
ходное ядро передало каждому из своих потомков всю необхо-
димую генетическую информацию. Ввиду наличия сложного ме-
ханизма для обеспечения передачи всей необходимой генетиче-
ской информации (и учитывая эффективность этой системы, ко-
которую легко продемонстрировать) уместно поставить вопрос,
каким образом клетки и ткани дифференцируются и организу-
ются в жизнеспособный организм. Почему нервная клетка столь
отлична от эритроцита, если это потомки одной и той же зи-
готы?
Один из возможных ответов состоит в том, что эти две клетки
развивались в различных условиях. Даже на очень ранних ста-
диях дробления, когда клеток еще немного, окружающая их сре-
да может быть резко различной. У животных на дифференци-
ровку клеток могут влиять такие факторы, как положение отно-
сительно анимального и вегетативного полюсов, нахождение на
поверхности или внутри бластулы, близость к бластопору в га-
струле. От положения клетки может зависеть количество полу-
чаемых ею важных питательных веществ, количество доступного
кислорода, скорость накопления продуктов обмена и т. п. Раз
начавшись, дифференцировка углубляется, и ее последствия
растут в геометрической прогрессии. Различные сочетания диф-
ференцированных элементов еще больше увеличивают гетероген-
ность клеточной среды, так что сложные взаимодействия могли
бы создать основу для развития целого организма. (Степень
сложности механизмов развития, конечно, весьма различна
у разных организмов.) Таким образом, есть все основания ска-
зать, что развитие организма всецело определяется взаимодей-
ствиями внутри конгломерата делящихся и растущих клеток.
Все клетки обладают одной и той же информацией, но исполь-
зуют ее по-разному, так как действуют в различной физиологи-
ческой среде.
Такое представление о процессе развития подкрепляется ог-
ромным множеством данных экспериментальной эмбриологии.
Взаимодействия клеток можно наблюдать в культурах микроор-
ганизмов, в которых плотность популяции способна влиять на
скорость роста (и даже определять самую возможность его).
Литература об индукции (при контакте или на расстоянии) и
об организаторах свидетельствует о мощном влиянии клеточного
окружения и о сложности механизмов, выработанных эволюцией.
Однако эти данные не содержат никаких доказательств того,
что генетическая информация играет в развитии лишь пассив-
ную роль. Эксперименты вроде упомянутых выше опытов на
74 | Глава 4
Platycnemis доказали, что у большинства организмов клетки, по
крайней мере на первых стадиях дробления, омнипотентны, т. е.
сохраняют всю информацию, необходимую для развития целого
организма. Кроме того, клетки, взятые из различных участков
тела и исследованные под микроскопом, не обнаруживают не-
хватки хромосомного материала, которой следовало бы ожи-
дать в случае неравного разделения его между клетками. Далее,
у тех насекомых, у которых политенные хромосомы с ясно ви-
димыми поперечными дисками имеются в разных тканях тела,
характер расположения этих дисков, как установлено, во всёх
тканях одинаков.
При изучении этих гигантских хромосом получены и другие
решающие данные в пользу изложенной интерпретации разви-
тия. В ходе онтогенеза определенные диски сильно увеличива-
ются, образуя так называемые пуфы. Это явление обратимо. На
той же стадии развития в других тканях в состоянии пуфов на-
ходятся другие диски. Ясно, что различные участки хромосом-
ного материала ведут себя по-разному, но механизм, контроли-
рующий эти различия, неизвестен. Полагают, что наличие пуфов
указывает на активность генов. Число пуфов, которые могут
образоваться на хромосоме, гораздо меньше числа дисков; зна-
чит, активность других частей генетического кода должна быть
«невидимой».
Если генетический материал участвует в дифференцировке
более активно, чем следует из сказанного выше, он должен дей-
ствовать каким-то очень тонким способом. Интересные экспери-
менты, проведенные недавно Бриггсом и Кингом, показали, что
ядра в клетках некоторых эмбрионов лягушки каким-то образом
изменяются в ходе развития. Когда из яйца лягушки удаляли
ядро и заменяли его ядром из бластулы, развитие протекало
нормально. Когда же его заменяли ядром, взятым из гаструлы
или нейрулы, получались деформированные эмбрионы: единст-
венными нормальными тканями у них были ткани, развиваю-
щиеся из того зародышевого листка, из которого было взято
пересаженное ядро. Эти данные не обязательно говорят о том,
что генетическая информация в процессе развития изменилась,
но в то же время, несомненно, не противоречат такому предпо-
ложению. Разумеется, могли измениться другие компоненты
ядра, что могло привести к изменению в процессах расшифров-
ки кода, а не в первоначальной информации.
Быть может, величайшей задачей, стоящей в наши дни перед
эмбриологами, является полное выяснение механизма, управ-
ляющего дифференцировкой. Этот вопрос не случайно интере-
сует исследователей. В том случае, если части генотипа каким-
Развитие | 75
то образом могут включаться и выключаться (на что, по-види-
мому, указывает образование пуфов), действие отбора может
быть совершенно иным, нежели в случае, если всегда активен
весь генотип. Если часть информации, контролирующая, скажем,
окраску тела у гусеницы, каким-то образом инактивируется в
период дифференцировки тканей имаго, то отбор мог бы изме-
нять окраску гусеницы, не оказывая никакого влияния на взрос-
лую бабочку. Точно так же, если бы гены, действующие на во-
лосы, были неактивны в эктодермальных тканях, форма и цвет
волос могли бы изменяться без каких-либо последствий для ки-
шечника. Если же, напротив, такой избирательной активации и
инактивации не существует, тогда все изменения в генетической
информации должны в большей или меньшей степени сказы-
ваться на всех этапах жизни организма, на всех стадиях его
жизненного цикла. Во многих случаях этот эффект мог бы быть
настолько незначительным, что он терялся бы на фоне «шума»,
сопровождающего нормальное развитие. Если учесть данные о
рассеянной локализации генов, влияющих на один и тот же при-
знак (например, гены, определяющие особенности крыла у Dro-
sophila melanogasier, не сосредоточены в какой-то одной хромо-
соме), а также широкое распространение плейотропии и отсут-
ствие механизма «включения» и «выключения» больших частей
генотипа, то кажется весьма вероятным, что генетический мате-
риал в большинстве клеток организма в основном одинаков
(исключая количественные отличия в эндополиплоидных тка-
нях).
Известно, что клетки в различных тканях одного и того же
организма, содержащие, как предполагают, идентичную генети-
ческую информацию) обладают неодинаковым набором белков.
Для белкового синтеза, следовательно, должен существовать
специфический управляющий механизм, регулирующий количе-
ства различных генных продуктов.
Интересную модель такого механизма предложили Жакоб и
Моно. Эта модель касается транскрипции кода ДНК (см. гл. 1).
Предполагается, что наследственные факторы можно подразде-
лить на структурные гены и гены-регуляторы. Первичным про-
дуктом структурных генов является информационная РНК,
синтез которой представляет собой последовательный процесс,
начинающийся с определенных участков цепи ДНК. Эти уча-
стки получили название операторов. Один оператор можег
контролировать транскрипцию информации, заключенной в
нескольких структурных генах. Соседние гены, контролируе-
мые одним оператором, образуют единицу транскрипции —
оперон.
16 | Глава 4
Продуктом генов-регуляторов является цитоплазматическое
вещество — репрессор, быть может РНК, воспроизводящая код
данного гена. Предполагается, что репрессор обратимо связы-
вается определенным оператором и что присоединение репрес-
сора к оператору препятствует транскрипции всего оперона.
Синтез белка таким образом блокируется. Кроме того, согласно
этой модели, репрессор способен к обратимому взаимодействию
не только с оператором, но и с небольшими цитоплазматически-
ми молекулами, так называемыми эффекторами. В некоторых
системах только неизмененный репрессор может связываться
с оператором и блокировать оперой; в таком случае присутствие
эффектора снимает действие репрессора и освобождает оперон
от репрессии. В других системах с оператором может связывать-
ся только продукт соединения репрессора с эффектором; при
этом эффектор препятствует функционированию структурного
гена. Упрощенная схема этой модели представлена на фиг. 14;
ее следует сопоставить с фиг. 5.
Мы не будем подробно описывать модель Жакоба и Моно.
Отметим лишь, что в генетике микроорганизмов получено много
данных, подтверждающих существование ряда постулированных
| Ген-регулятор |
Оперон
| Оператор Структурный ген 1 Структурный ген 2 |
Белки, синтезирован-
ные рибосомами
Белки,синтезирован -
ные рибосомами
Фиг. 14. Модель функции генов.
У? —цитоплазматическое вещество-репрессор; Е— вещество-эффектор; —репрессор, видо-
измененный в результате соединения с эффектором; ^ — эффектор, видоизмененный в ре-
зультате соединения с репрессором [1].
Развитие | 77
в этой модели процессов и систем !. Кажется несомненным, что
какой-то аналогичный механизм с обратной связью действует на
уровне транскрипции кода ДНК и синтеза белка; точно так же
существуют, по-видимому, сходные механизмы, ответственные за
гомеостаз процессов развития на более поздних этапах.
ЭВОЛЮЦИОННОЕ ИЗМЕНЕНИЕ ПРОЦЕССОВ ОНТОГЕНЕЗА
Ни одно из явлений морфогенеза не привлекало большего
внимания, чем так называемая рекапитуляция. Эмбриологи
очень скоро заметили, что ранние стадии развития разных позво-
ночных сходны друг с другом (по крайней мере поверхностно)
в гораздо большей степени, чем взрослые формы. Некоторые
исследователи заключили из этого, что каждый организм в про-
цессе своего развития повторяет в сокращенном виде свою фи-
логенетическую историю, что человек, например, проходит ста-
дию одной клетки (зигота), стадию рыбы (когда появляются
жаберные мешки), стадию млекопитающего и т. д. Первоначаль-
но Геккель назвал это обобщение биогенетическим законом, и
его часто формулируют словами «онтогенез повторяет филоге-
нез». Однако эта прямолинейная интерпретация последователь-
ных стадий эмбрионального развития при ближайшем рассмот-
рении оказывается неверной. На ее недостатки указывают почти
все современные авторы, но эта идея все еще занимает видное
место в биологической мифологии.
Сходство ранних стадий эмбриогенеза позвоночных легко
объясняется и без привлечения таинственных сил, заставляющих
каждый индивидуум снова «взбираться на свое филогенетиче-
ское древо». Прежде всего следует подчеркнуть, что зародыш
млекопитающего на ранних стадиях похож на зародыш рыбы,
а не на взрослую рыбу. Фактически все организмы начинают
свое развитие с одной-единственной клетки. Огромное разнооб-
разие форм жизни обусловлено различием путей развития, в
значительной мере определяемых наборами генетической инфор-
мации, которые вызывают изменения в процессах морфогенеза.
Однако не каждое изменение означает перестройку в системе
механизмов развития. Эти необычайно сложные интегрирован-
ные системы могут быть успешно видоизменены только путем
накопления мелких изменений, что должно сопровождаться
1 Данные, указывающие на существование у высших организмов систем,
подобных устанавливаемым Жакобом и Моно для микроорганизмов, име-
лись уже давно в классических работах Б. Мак-Клинток, проведенных на
кукурузе (см. В. McClintock, 1961, Some parallels between gene control
systems in maize and in bacteria. American Naturalist, 95, 884; 265—277),—
Прим. ped.
78 | Глава 4
«перенастройкой» сбалансированных взаимодействий разнооб-
разных факторов. Тщательное исследование живых и ископае-
мых организмов позволяет нам восстановить эти постепенные
изменения путей развития. Хорошим примером служит медлен-
ное изменение строения нижней челюсти у позвоночных: сочле-
новная и квадратная кости, входившие в состав челюсти у реп-
тилий— предков млекопитающих, используются в качестве слу-
ховых косточек у млекопитающих. Все такие изменения были
связаны с модификацией существовавшей ранее последователь-
ности процессов развития и были возможны только в тех слу-
чаях, когда эта последовательность могла быть видоизменена
без гибельного расстройства сбалансированных взаимоотноше-
ний. Например, жаберные мешки (эмбриональные предшествен-
ники жаберных щелей у рыб) превратились у высших позво-
ночных в иные образования — евстахиевы трубы и зобную же-
лезу. Такой ход эволюции позволил избегнуть осложнений, ко-
торые могли возникнуть при изменении всего комплекса про-
цессов, в том числе процессов, приводящих к формированию са-
мих жаберных мешков. Подобное изменение, вероятно, могло
бы сильно расстроить индуктивные системы, ответственные,
скажем, за развитие дуг аорты.
Идея рекапитуляции предполагает сходство промежуточных
стадий онтогенеза с предковыми формами. Однако есть случаи,
когда взрослые формы, по-видимому, сходны с эмбриональными
стадиями предполагаемых предков. Например, самки некоторых
жуков и бабочек похожи на личинок; некоторые амбистомы не
превращаются во взрослых особей, а размножаются на стадии
личинки. У взрослых людей многие признаки (слабое развитие
волосяного покрова, большая голова и др.) напоминают соот-
ветствующие признаки детенышей человекообразных обезьян.
У австралопитека — самого древнего из известных ископаемых
людей — молочные зубы сходны с постоянными зубами Ното
sapiens, а постоянные зубы австралопитека сходны с постоян-
ными зубами человекообразных обезьян. В этих и во множе-
стве других подобных случаев эволюция как будто бы изме-
нила процесс развития таким образом, что промежуточная ста-
дия предка сделалась окончательной формой у потомка. Это яв-
ление известно под названием неотении.
Последовательные стадии индивидуального развития орга-
низма часто рассматривают просто как ступени, ведущие к ко-
нечной цели — взрослой форме. Биологически более реалистич-
ным, несомненно, было бы представление об онтогенезе как о
непрерывно изменяющейся реакции данного комплекса генети-
ческого материала па данную внешнюю среду. Разнообразные
процессы эпигенотипа регулируют в различной степени проявле-
Развитие | 79
ние инициирующего их генотипа. Эволюционное изменение мо-
жет затрагивать любую из произвольно выделяемых стадий раз-
вития.
выводы
Эволюционный ряд предков и потомков следует рассматри-
вать не как непрерывную последовательность особей, а как се-
рию циклических фаз. Каждый полный цикл представляет собой
ряд стадий развития, причем за конец одного цикла и начало
следующего по традиции принимают начало диплофазы (обра-
зование зиготы). Изменения в генетической информации ведут
к отклонениям в процессе развития, и накопление этих генети-
чески обусловленных отклонений составляет эволюцию. Важно
помнить, что эволюционирует весь жизненный цикл и что все
стадии любого данного цикла существенны для выживания и,
таким образом, одинаково важны с точки зрения эволюции.
Многие исследователи, которые рассматривали процесс начи-
ная с конца диплофазы, были склонны игнорировать этот био-
логический факт.
ЛИТЕРАТУРА
1. Cellular Regulatory Mechanisms, Cold Spring Harbor Symposia on Quanti-
tative Biology, vol. 26 (1961). (Регуляторные механизмы клетки, ИЛ, М.,
1964.)
2. Darlington С. D., The Evolution of Genetic Systems, 2d ed. Basic Books,
Inc., New York, 1958.
3. Waddington С. H., The Strategy of the Genes, G. Allen, London, 1957.
4. Lerner I. M., Genetic Homeostasis, Wiley, New York, 1954.
5. Jacob F., Monod J., Cold Spring Harbor Symp. Quant. Biol., 26 (1961).
(Регуляторные механизмы клетки, ИЛ, iM., стр. 477, 1964.)
Часть вторая
СВОЙСТВА
ПОПУЛЯЦИЙ
Биологи, изучая популяционный уровень организа-
ции, в значительной степени руководствуются особен-
ностями изучаемых организмов. Например, цитологи-
ческие аспекты генетических систем легче изучать на
Drosophila и Oenothera, чем на Papitio или Sequoia.
В силу ряда обстоятельств действие естественного от-
бора часто удобнее изучать на определенных организ-
мах, которые в цитологическом отношении совершенно
или почти совершенно не изучены. Весьма значитель-
ная доля наших знаний накоплена в процессе работы
с организмами, имеющими хозяйственное значение,
например с полевыми культурами, домашними живот-
ными и вредителями. Таким образом, по ряду причин
комплексное описание всех аспектов эволюции в пре-
делах популяций оказалось невозможным.
Теория популяционной генетики была разработана
главным образом применительно к диплоидным орга-
низмам с перекрестным оплодотворением. Поэтому
разумно будет сначала изложить эту теорию и относя-
щиеся к ней примеры, взятые из природы, а потом
уже рассмотреть во всей их сложности системы, конт-
ролирующие рекомбинацию у различных типов орга-
низмов. Есть надежда, что в конце концов будет соз-
дана теория, охватывающая взаимосвязи между гене-
тической системой организма и действующими на не?о
эволюционными силами. А пока не следует забывать о
предостережении Норберта Винера: очень трудно изу-
чать взаимодействия двух систем, сильно различаю-
щихся по своим временным масштабам. Это относится
как к попыткам понять историю на основе каждоднев-
ного поведения людей, так и к попыткам понять фило-
генез путем исследования изменений отдельных генов
U современных организмов.
Глава 5
ПОПУЛЯЦИИ
В известном смысл? каждое явление уникально. Никакие два
объекта не могут занимать одно и то же место в пространстве
и во времени. Комплексы энергетических взаимодействий, если
даже они повторяются с абсолютной точностью, различаются по
крайней мере по времени. Однако воспринимаемой нами вселен-
ной свойственна упорядоченность уникальных явлений. Челове-
ческий ум — это аппарат, функционирующий путем установле-
ния связей между уникальными событиями. Набор объектов, об-
ладающих общими свойствами, объединяется в некоторый класс
(например, стол, раса, бабочка), и подобные групповые понятия
оказываются полезными при передаче сообщений. Фактически
само существование любого организма зиждется на его спо-
собности в известном смысле производить обобщения, исхо-
дя из наборов уникальных событий. Совершенно уникальное
событие, для которого нельзя было бы установить никакой свя-
зи с каким-либо другим событием, было бы полностью лишено
смысла.
Понимание человеком действительности всегда основано на
изучении «популяций» объектов и событий, а также характера
предполагаемых взаимоотношений между ними. Для того чтобы
понять клеточные функции, биохимик исследует «популяции»
химических компонентов и процессов внутри клетки. Чтобы по-
стигнуть организацию живых систем, физиологи и эмбриологи
изучают популяции клеток и тканей и взаимодействий между
ними. На высшем уровне биологической организации биолог ис-
следует популяции организмов и отношения внутри них и ме-
жду ними. В этой книге термин «популяция» будет означать
только совокупности индивидуальных организмов; иначе говоря,
этот термин применяется здесь в том смысле, в каком его обыч-
но употребляют биологи, изучающие эволюцию. Таким образом,
популяционная биология имеет дело со свойствами совокупно-
стей организмов, особенно с эмерджентными1 свойствами, не
присущими составным элементам популяции. Популяции редко
поддаются сплошному обследованию: обычно приходится брать
1 Качественно новые свойства системы, отсутствующие у ее компонен-
тов. — Прим. ред>
G*
84 | Глава 5
выборки из них в одном или нескольких пунктах. К сожалению,
из одной и той же популяции невозможно взять две идентичные
выборки.
ОСОБИ И КОЛОНИИ
Первая проблема возникает в связи с определением «инди-
видуального организма». На первый взгляд кажется, что дать
такое определение нетрудно, так как знакомые нам растения
и животные существуют в виде дискретных единиц. Однако по-
ложение осложняется при рассмотрении таких форм, как лишай-
ники. Эти растения состоят из гриба и водоросли: как теперь
известно, гриб паразитирует на клетках водоросли, включенных
в его таллом. Разные лишайники имеют различные морфологи-
ческие и биохимические особенности, но последние развиваются
только тогда, когда образуется надлежащая комбинация гриба
и водоросли. Водоросль и гриб размножаются независимо друг
от друга, но размножается и лишайник в целом — с помощью
пропагул, состоящих как из водоросли, так и из гриба. Часто
водоросль удается выращивать без гриба, но гриб без своего
хозяина — водоросли — не выживает.
Сложные колониальные организмы также представляют
трудности в этом отношении. Колонии общественных насекомых
в некоторых отношениях аналогичны организмам; обычно такие
колонии называют квазиорганизмами. Португальский кораб-
лик— колониальный представитель Hydrozoa — можно было бы
«разложить» на составляющие его полипы, которые обнаружи-
вают удивительное разделение функций. Среди водорослей и
простейших есть менее специализированные агрегаты особей,
состоящие из единиц, которые могут существовать как самостоя-
тельно, так и в составе колонии. Что касается дрожжей Saccha-
romyces, то они ведут себя по-разному в зависимости от условий
среды. В жидкой культуре дрожжевые клетки (растеньица?)
представляют собой мелкие яйцевидные тельца, чрезвычайно
быстро размножающиеся почкованием. Могут встречаться и ко-
роткие цепочки клеток. Однако при выращивании на твердой
среде дрожжи образуют гигантские колонии — уплощенные об-
разования диаметром в несколько сантиметров, часто с харак-
терной окраской, с характерной структурой поверхности и опре-
деленными биохимическими свойствами. Клетки из наружного
слоя, из толщи колонии и из слоя, прилегающего к поверхности
среды, весьма различны как по форме, так, вероятно, и по функ-
циям. Несмотря на такие различия, для того чтобы получить но-
вую типичную колонию или жидкую культуру дрожжей, можно
взять клетки из любого участка колонии.
Популяции | 85
Не удается избежать трудностей и при определении границ
отдельных особей у высших организмов. Рассмотрим, к примеру,
растения, размножающиеся вегетативным путем (см. гл. 9);
если «потомки» остаются прикрепленными к родительскому рас-
тению, то все это в целом можно рассматривать как индивиду-
ум. Но если они окажутся отделенными, то каждое дочернее
растение также обычно считают отдельной особью, хотя в гене-
тическом отношении оно идентично своему «родителю». Само-
стерильный триплоидный лилейник Hemerocallis fulva является
в генетическом отношении одним индивидуумом по всей области
своего распространения, охватывающей значительную часть во-
сточных районов США. Популяции гидры, возникшие из одной
почкующейся особи, также в генетическом отношении представ-
ляют собой индивидуум, хотя в экологическом и функциональ-
ном отношениях они состоят из многих особей.
Встречаются также комплексы особей, принадлежащих к
разным видам. Многие червецы образуют поразительные комп-
лексные колонии, в которых в качестве симбионта присутствует
гриб Septobasidium. У лесных деревьев очень часто происходит
«самопрививка», когда их корни соприкасаются в процессе ро-
ста. Как выяснилось, если одно из таких деревьев срубить, пень
может многие годы оставаться живым. Лишенный собственной
фотосинтезирующей ткани, он тем не менее способен образовы-
вать новую кору из камбия, так что может полностью покрыть-
ся корой. Индивидуальные организмы в подобных случаях гене-
тически различны, но слиты в одну экологически значимую еди-
ницу. Сходным образом клон растений «живородящего» лука
(см. гл. 9), которые генетически идентичны, составляет один
экологически значимый комплекс, так как образует элемент
среды для других организмов.
Индивидуум можно представить себе как «машину», заранее
запрограммированную для выполнения определенных операций.
В организмах эта программа определена кодированной генети-
ческой информацией. Группа генетически тождественных особей
с точки зрения процесса воспроизведения представляет собой
один индивидуум. Экологически они образуют популяцию особей
с различными эпигенотипами. Если бы исторически первичным
в биологии был экологический, а не таксономический подход, то
теперь мы, вероятно, рассматривали бы биологические «факты»
совсем по-иному.
Очевидно, что понятию индивидуум (или особь), как и дру-
гим понятиям в биологии, можно придавать только операцио-
нальное значение. Чтобы сделать определение ясным, мы дол-
жны указать, применительно к какой области оно дается — к
таксономии, генетике или экологии. В дальнейшем мы будем
86 | Глава 5
пользоваться генетическим -определением индивидуума. В основ-
ном работа по изучению эволюции популяций проводилась на
организмах, отдельные особи которых являются продуктом поло-
вого размножения и поэтому в генетическом отношении обычно
разнородны. У организмов, размножающихся половым путем,
обширными популяциями обычно считаются совокупности осо-
бей, достаточно сходных для того, чтобы при подходящих об-
стоятельствах они могли успешно размножаться путем перекре-
стного оплодотворения. Точные критерии для суждения о том,
какая совокупность особей может быть названа популяцией,
установить трудно, и о степени соответствия этих критериев той
или ипой природной совокупности особей обычно приходится
лишь догадываться.
В настоящей главе мы, как правило, будем заменять опреде-
ления примерами. Бабочка и бизон явно принадлежат к разным
популяциям; пара малиновок, выводящих птенцов в саду, не-
сомненно, состоит из представителей одной популяции. Это край-
ние примеры; однако во многих других случаях вопрос решить
труднее. Можно ли считать, что европейский бурый медведь и
американский гризли или восточный и западный платан входили
в одну популяцию? Последние, хотя они и были географически
изолированы со времен миоцена, дают гибрид — мощное дерево,
широко используемое для озеленения улиц. Поскольку нас инте-
ресует в первую очередь процесс эволюции, а не поиски опреде-
лений, которые всегда в значительной степени произвольны, мы
не будем искать ответа па эти вопросы.
РАСПРЕДЕЛЕНИЕ В ПРОСТРАНСТВЕ
Одно из свойств популяции, которым (если понимать его
в том же смысле) не обладают входящие в нее организмы, —
это распределение в пространстве. В любой данный момент вре-
мени можно наблюдать распределение бабочек Euphydryas
editha по обнажению серпентиновых пород на биологическом
экспериментальном участке Стэнфордского университета в Джа-
спер-Ридже. На фиг. 15 показано распределение взрослых осо-
бей на этом участке в 1960 и 1961 гг. В 1961 г. особи распре-
делялись несколько иначе, чем в 1960 г. Подобные колонии
Euphydryas editha встречаются во всем районе бухты Сан-Фран-
циско; их можно найти на всем протяжении Западного побе-
режья — от Нижней Калифорнии до Британской Колумбии.
Трудно указать границы той наиболее обширной популяции,
к которой можно было бы отнести особей из Джаспер-Риджа.
Большинство биологов отнесло бы к этой совокупности даже
особей из такого отдаленного штата, как Монтана.
Фиг. 15. Распределение особей бабочки Euphydryas editha в районе Джа-
спер-Ридж в 1960 и 1961 гг.
Колония находится на островке луга, окруженном чаппаралем. Каждая точка указывает
место первой поимки одной особи. Буквами обозначены участки, на которые колония быда
произвольно разделена для ее изучения [6].
Фиг. 16. Иерархия скоплений Clematis fremontii var. riehlii.
А —ареал в целом; Б —отдельный участок; В —группа полян; Г — поляна; Д — скопление
особей (7].
Популяции | 89
У растений также наблюдается группировка особей в сово-
купности различной величины. Clematis fremontii var riehlii, ра-
стущий на известняковых полянах в штатах Среднего Запада,
представляет собой многолетнее растение, довольно детально
изученное Эриксоном. Отдельные особи группируются в скопле-
ния из нескольких сотен особей, причем много таких скоплений
занимает одну поляну. Выходы известняка также распределены
группами, приуроченными к горным системам и рекам. На Сред-
нем Западе Clematis fremontii var. riehlii имеет гораздо более
широкое распространение, составляя здесь наиболее обширную
популяцию (фиг. 16).
В самом общем смысле областью распространения называют
наименьшую географическую область, включающую все терри-
тории, в обычных условиях заселенные данными организмами,
На мелкомасштабной карте мира область распространения Но-
то sapiens захватила бы всю территорию США, так что неза-
штрихованными остались бы лишь моря и большая часть ледя-
ной шапки Гренландии. Но если бы мы отмечали распростра-
нение человека на крупномасштабной карте Калифорнии, то
незаштрихованными остались бы многие высокие горные пики
и некоторые другие районы. Трудности составления таких карт
очевидны. Организмы на определенных этапах своего жизнен-
ного цикла подвижны, и их распределение подвержено непре-
рывным изменениям. К тому же ни один из известных организ-
мов не распределен на обширных территориях равномерно.
Таким образом, чем большей детализации мы стремимся до-
стигнуть при описании распространения организмов, тем труд-
нее наша задача.
ЭКОЛОГИЧЕСКОЕ РАСПРЕДЕЛЕНИЕ
Неравномерность географического распространения особей
обычно удается объяснить взаимоотношениями организмов с
живыми и неживыми компонентами окружающей их среды. Бо-
лее общие аспекты зависимости распространения организмов от
экологических факторов понять легко; факторы, определяющие
более тонкие особенности распространения того или иного орга-
низма, по существу, никогда не бывают поняты до конца. В рай-
оне бухты Сан-Франциско гусеницы Euphydryas editha питают-
ся на Plantago erecta — небольшом местном подорожнике, осо-
бенно обильно произрастающем на обнажениях серпентиновых
пород. В этом районе Е. editha встречается только в тех местах,
где растет Р. erecta и распространены серпентины, но наличие
обоих этих факторов еще не является гарантией того, что мы
обязательно найдем здесь бабочку. В районе Джаспер-Ридж
90 | Глава 5
участки с выходом серпентиновых пород, изобилующие подоро-
жником, иногда остаются незаселенными, даже если находятся
в непосредственном соседстве с колонией Euphydryas. Для того
чтобы данное местообитание было занято, мало того, чтобы все
условия были благоприятными: нужно еще, чтобы организму
представился случай попасть на подходящий участок. Так, в не-
которых пригодных для Е. editha участках эта бабочка отсут-
ствует— просто потому, что ни одна оплодотворенная самка не
залетала сюда. Человек способствовал проникновению многих
организмов в ранее незаселенные, хотя и подходящие для этой
цели районы; примером могут служить скворец, домовый воро-
бей, капустница, медоносная пчела, одуванчик и горчица.
СТРУКТУРА ПОПУЛЯЦИЙ
Под структурой1 популяции мы понимаем здесь совокуп-
ность всех факторов, определяющих характер комбинаций, в ко-
торых гаметы различных особей соединяются друг с другом.
Структура может быть весьма разной — начиная от таких си-
туаций, в которых эти сочетания могут казаться в основном слу-
чайными (как, например, у некоторых морских животных, выде-
ляющих гаметы в воду, и у некоторых ветроопыляемых расте-
ний), до таких, где вероятность одних комбинаций значительно
больше, чем других. Последний случай, несомненно, является
правилом, хотя бы потому, что близкие соседи обычно имеют
больше шансов на спаривание, чем особи, живущие дальше друг
от друга.
Большое значение имеют и такие факторы, как продолжи-
тельность жизни поколения и размеры особей. Если нас интере-
сует число новых генных комбинаций, возникающих в данном
районе в единицу времени, то для мелких организмов эта пе-
ременная окажется больше, чем для крупных. В любом месте
крупных организмов меньше, чем мелких, для которых харак-
терна большая частота рекомбинаций. У организмов с коротким
жизненным циклом, возможно, больше генных комбинаций, чем
у организмов с большой продолжительностью жизни поколения,
и частота мутаций у тех и других форм также различна.
У животных, особенно высших, выработалось много форм
поведения, оказывающих глубокое влияние на структуру попу-
ляции. Многие животные в популяционном отношении близки
к малоподвижным формам, несмотря на большие потенциальные
возможности расселения. Птицы многих видов возвращаются из
дальних перелетов к тем же местам гнездовий, которые они за-
1 Автор дает здесь определение структуры популяции, отличное от опре-
деления, принятого в биоценологии. — Прим. ред<
Популяции | 91
нимали в предшествующие годы. Твитти показал, что тритоны
Калифорнии проявляют невероятное упорство и поразительную
способность к ориентировке, возвращаясь для размножения в
совершенно определенный участок ручья. В специально прове-
денных опытах было показано, что перемещенные особи возвра-
щались к своему родному водоему, преодолевая по несколько
километров в горной местности. Приверженность к определен-
ному участку в пределах одного ручья у тритонов показана на
фиг. 17. Бабочки часто используют свою способность к полету
только для патрулирования весьма ограниченного участка. В ко-
лонии Euphydryas editha в Джаспер-Ридже 625 из 647 (98,6%)
помеченных взрослых особей были вторично пойманы в районе
первичной поимки. Такого рода ограничение физической способ-
ности к расселению, обусловленное особенностями поведения,
по-видимому, является у немигрирующих бабочек скорее прави-
лом, чем исключением.
Вероятно, лишь немногие животные ведут истинно кочевой
образ жизни. В большинстве случаев животные (так же как
и люди) оседают поблизости от места своего рождения, занимая
территорию, которая была достаточно хороша для их родителей.
Многие животные защищают всю свою территорию или часть ее
от вторжения других особей того же вида — это хорошо изве-
стное явление территориальности. Такое поведение весьма обыч-
но у птиц, млекопитающих, рептилий, рыб и некоторых беспо-
звоночных и приводит к неслучайному распределению особей в
популяции, которое оказывается гораздо более равномерным,
чем можно было бы ожидать при чисто случайном рассеянии.
Одно из следствий этого состоит в том, что величина популяции
удерживается на уровне, при котором ресурсы окружающей
среды (пища, места для гнезд и т. п.) не могут оскудеть или
оказаться полностью исчерпанными. Те особи (часто это моло-
дые половозрелые члены популяции), которым не удается за-
нять и защитить территорию, должны отыскать себе лучшие
места — иначе они погибнут от голода; так в популяции возни-
кает стремление к рассеянию.
Со статистической точки зрения территориальности мы про-
тивопоставляем стадное поведение, которое обусловливает боль-
шую «скученность», чем при случайном распределении особей.
У животных, обнаруживающих такое поведение, может, насколь-
ко мы знаем, и не быть никакой (или почти никакой) общест-
венной организации; именно так обстоит дело, к примеру, в
скоплениях, которые образуют божьи коровки или змеи перед
зимовкой. Противоположную крайность представляют общест-
венные насекомые, у которых имеются морфологически диффе-
ренцированные «касты», причем в колонии может быть всего
8°/о поймано вновь в 1957 г.
Ьы!-........................
] ю 20 30 40 50
।--, — ______f
15% поймано вновь в 1958 г.
И fl I I I I I I I I I I I I I I I TI I I I I I Г I I I I I Г1 I М I I -I i '1 т
10 20 30 40 50
21% поймано вновь в 1959г.
I Г1 I I I I I I I I I 1ТТГТ1Т1ТГ I I I 1-1-Г Г I Г I I I I I I.
20 30 40 50
А
Номер станции
10
3% поймано вновь в 1957г
10
11 % поймано вновь в 1959 г
Г1 I fl Г ГГТ'А fl П-1 I I-1 I Я I I I I I 1 Т-| Г I Г Ат1 1'1 fl I I I I 1'1
1 10 20 30 40 50
Г2 Номер станции
Фиг. 17. Инстинкт дома у тритонов.
А —вторичные поимки особей, перемещенных в 1956 г. примерно на 1 км вниз по течению
ручья от места первоначальной поимки. Б —вторичные поимки особей, перемещенных
в 1956 г. примерно на 2 км вверх по течению от места первоначальной поимки. Район пер-
воначальной поимки и место выпуска отмечены стрелками. „Станции" — участки, на которые
произвольно разделено течение ручья. Все особи покидают ручей после каждого сезона
размножения [8].
Популяции | 93
одна пара размножающихся особей. В этом случае действие
отбора проявляется главным образом в форме дифференциаль-
ного размножения колоний, а не особей.
У многих позвоночных устанавливается общественная иерар-
хия; одни особи доминируют над другими и пользуются различ-
ными привилегиями — от права на выбор партнера для спарива-
ния до первого места в колонне, проходящей через калитку за-
гона. Доминирующие самцы иногда располагают большим
«гаремом» и в отличие от своих менее агрессивных собратьев
вносят крупный вклад в фонд генетической информации, пере-
даваемой последующим поколениям. Общественным группам
(например, колониям общественных насекомых или стадам
обезьян-ревунов) часто свойственна территориальность.
ЧИСЛО ОСОБЕЙ
Одним из самых очевидных атрибутов всякой популяции яв-
ляется число элементов, событий или индивидуумов, которое ха-
рактеризует ее в данное время. Число особей в биологических
популяциях имеет огромное значение, но, к сожалению, с точ-
ностью установить размеры какой-либо природной популяции
обычно бывает трудно или вовсе невозможно. Наиболее обшир-
ные популяции могут состоять из миллиардов особей (например,
популяции человека, комнатной мухи, микроорганизмов, различ-
ных водорослей) или менее чем из сотни особей (популяции
журавлей или некоторых редких эндемичных растений, как, на-
пример, Pedicularis dudleyi и Tetracoccus ilicifolius). Строго го-
воря, всякий раз, когда изменяется число особей в популяции,
изменяется и ее распределение; изменение распределения также
часто сопровождается изменением численности. Анализ резуль-
татов, приведенных на фиг. 15, на которой указаны места пер-
вой поимки взрослых Е. editha в 1960 и 1961 гг., дает известное
представление о проблемах, возникающих при попытках рас-
сматривать распределение и численность популяции по отдель-
ности. В области С число особей к 1961 г. почти не увеличи-
лось, но произошло некоторое изменение в их распределении.
В области G популяция уменьшилась и одновременно несколько
сузилась занимаемая ею территория. В области TI число особей
резко возросло и популяция захватила территорию, на которой
в предыдущем году ее особей фактически не было. Важно от-
метить, что такие схемы, подобно любым отображениям распре-
деления особей, по существу представляют собой статическое
отображение динамической ситуации. Численность и распределе-
ние особей в больших популяциях непрерывно изменяются, и
быстрота этих изменений у разных организмов весьма различна,
94 | Глава 5
Динамика популяций, изучение изменений их величины—
предмет очень сложный, который мы не можем здесь рассматри-
вать. Он представляет значительный интерес для исследователя
эволюции, ибо, как мы увидим, изменения размеров популяции
оказывают многообразное влияние на ее эволюцию. Это можно
понять интуитивно: поскольку каждая особь составляет элемент
среды для каждой другой особи, любое изменение величины по-
пуляции автоматически ведет к изменению ее собственной сре-
ды, а ведь в сущности именно изменения среды и заставляют
популяцию эволюционировать. Факторы, от которых зависит
величина популяции, разнообразны и во многих случаях неиз-
вестны. Вряд ли можно сомневаться, однако, что обычно раз-
меры популяции регулируются механизмами типа обратной
связи; иначе говоря, сама величина популяции влияет на ско-
рость ее роста. По этому вопросу имеется обширная литера-
тура; основные работы указаны в конце этой главы.
ВНЕШНЯЯ СРЕДА
Индивидуальный организм, когда таковой может быть выде-
лен, в известном смысле является морфологическим результатом
физиологических процессов, из которых он слагается. Каждый
из этих процессов связан с другими функциями организма, и
весь комплекс процессов лишь искусственно можно отделить от
окружающей среды. Функционирование организма возможно
только на основе взаимосвязи протекающих в нем процессов
с факторами среды. Каждый процесс характеризуется диапазо-
ном толерантности к факторам среды, интенсивность которых
не должна выходить за его пределы. Организм функционирует
только при условии, что интенсивность всех факторов среды не
выходит за пределы устойчивости всех жизненных процессов.
Эти диапазоны толерантности у каждого организма определя-
ются, конечно, генотипом, создавшимся в зиготе, и развиваю-
щимся эпигенотипом (см. гл. 4). Важно отметить, что пока не
наступает период размножения, между особями возможны самое
большее физико-химические, но не биологические взаимодей-
ствия. До тех пор пока организмы не вступают во взаимоотно-
шения, связанные с размножением, их .можно рассматривать как
часть физической среды для других организмов.
В настоящее время нет удовлетворительного подхода к изу-
чению сложных взаимодействий организма со средой. Обычно
пользуются какой-нибудь весьма упрощенной классификацией
факторов среды. Например, Андреварта и Бёрч расчленили
окружающую организм среду на четыре компонента: климати-
ческие условия, доступные источники пищи, другие организмы
Популяции | 95
и «место», где организм мог бы жить. Эти компоненты могут
быть в свою очередь разбиты на другие подразделения. Все они
непрерывно изменяются в той или иной степени. Как в ходе
развития организма изменяются границы устойчивости физиоло-
гических процессов (устойчивость к обезвоживанию, чувстви-
тельность к теплу или холоду), точно так же изменяется и ин-
тенсивность внешних факторов — циклически или же каким-либо
сложным, малоизученным образом. К примеру, в почве, окру-
жающей корни дуба, может произойти выщелачивание мине-
ральных элементов, а затем потеря их восполняется в резуль-
тате разложения опавших листьев. Растение, которое служит
пищей для данного насекомого, может исчезнуть в засушливый
год. Условия, необходимые для укоренения проростков и для
опыления цветков, могут существовать лишь в течение корот-
кого времени один раз в году; поэтому биологические особен-
ности растения должны быть хорошо согласованы с временем
появления этих условий.
В природных популяциях встречаются взаимодействия пора-
зительной сложности. Например, «другие организмы», поскольку
они являются частью среды, также можно подразделить на
«источники пищи» (организм-хозяин, добыча, кормовое растение
травоядных животных) и «местообитания» (организм-хозяин, де-
рево, на котором поселяется данный организм, и т. п.). Молодая
ондатра может столкнуться с тем, что все подходящие места
для устройства норы («место для поселения») окажутся заня-
тыми более старыми, более сильными особями («другие орга-
низмы»), и ей придется мигрировать. Во время миграции она
может замерзнуть («условия погоды»), погибнуть от голода
раньше, чем найдет подходящий корм («источники пищи»), или
быть съеденной койотом (опять-таки «другие организмы»).
Составляя часть среды, организмы могут изменять микро-
климат (например, в области затенения, в зоне листопада, в
корневой зоне дерева). Они могут служить переносчиками гене-
тического материала при заражении или размножении, а также
переносчиками пропагул. У цветков, плодов и семян покрытосе-
менных растений наблюдается огромное разнообразие приспо-
соблений, благодаря которым обеспечивается успешное опыле-
ние и распространение семян животными-переносчиками. Ка-
ждому знакомы насекомые-опылители, переносящие с цветка на
цветок пыльцевые зерна (мужские гаметофиты). Менее извест-
ны виды орхидей, цветок которых настолько похож на брюшко
самки мухи или осы, что самцы соответствующих видов делают
попытки спариться с цветком. Пыльца переносится с одного
цветка па другой в крошечных мешочках, прикрепляющихся на
Конце брюшка самцов, — так называемых поллиниях.
96 | Глава 5
Сходные явления встречаются и у животных. Интересно в
этом отношении двукрылое Dermatobia hominis. D. hominis ло-
вит комаров и прикрепляет к ним свои яйца, а затем отпускает
их. Когда комар садится на теплую кожу человека, из яиц вы-
лупляются личинки; они вбуравливаются в кожу и начинают
свое развитие. Это двукрылое паразитирует и на ряде других
млекопитающих.
Оставив в стороне обычные для умеренной зоны взаимодей-
ствия между растениями и животными, можно привести менее
известные примеры крайне сложных взаимоотношений, наблю-
даемых во влажных тропических лесах. На ветвях гигантских
деревьев поселяются различного рода растения, накапливающие
среди своих листьев воду и почву. Здесь, в этой специализиро-
ванной нише, выводятся, растут и претерпевают метаморфоз
личинки комаров и лягушек. Фауна комаров распадается на ряд
вертикальных зон, зависящих отчасти от распределения эпифи-
тов, на которых комары выводятся. В этих же лесах живет трех-
палый ленивец, шерсть которого окрашена в зеленоватый цвет
симбиотическими водорослями. Весь жизненный цикл бабочки
Bradypodicola hahneli проходит на ленивцах: гусеницы ее, по-
видимому, питаются этими водорослями. Объяснение истории
возникновения связей такого рода, а также других, быть может
еще более причудливых, связей представляет нелегкую задачу
для исследователя, интересующегося структурой экосистем.
Изменения климатических условий в большой степени опре-
деляют состав фауны в данной области. Находки ископаемых
тропических растений и амфибий в Арктике свидетельствуют
о более теплых временах в прошлом, а остатки древних городов
в пустынях — об изменениях в количестве атмосферных осадков
и в плодородии почвы. Многолетнее использование пастбищ,
когда не происходит возврата важных элементов в почву с раз-
лагающимися остатками растений и животных, изменило пита-
тельные свойства почв во многих частях Великих равнин США,
что привело в итоге к изменению флоры и фауны. Популяции
домового воробья в наших городах значительно уменьшились
после исчезновения лошадей, в навозе которых содержится мно-
го семян. Площади лугов в некоторых областях сокращались по
мере развития сети дорог и появления других факторов, облег-
чающих борьбу с лесными пожарами.
Много ярких примеров изменения влияния «других организ-
мов» возникло в результате деятельности человека, способство-
вавшей переселению растений и животных в новые для них
области. Завезенная в Северную Америку бабочка-репница,
Pieris гарае, размножилась здесь и вытеснила местный вид
Р. protodice, как полагают, победив его в конкуренции за пищу.
Популяции | 97
В ряде районов Калифорнии местные растения были вытеснены
аистником (Erodium), горчицей (Brassica) и диким овсом
(Avena). Завезенные на судах крысы фактически уничтожили
местную фауну мелких позвоночных на многих островах.
Постоянно происходят также изменения в имеющихся место-
обитаниях. Заиление ручьев и замедление их течения делает их
непригодными для личинок и куколок мошек и других организ-
мов, которым нужна быстротекущая, богатая кислородом вода.
Создание лесных полос в Великих равнинах привело к расшире-
нию ареалов лесных птиц, гнездящихся на деревьях. В резуль-
тате медленного накопления гумуса и разрушения горных пород
в процессе почвообразования создаются условия жизни для гри-
бов, клещей-орибатид, многоножек и других любителей влаж-
ных, темных мест. Обитатели областей, подверженных частым
катастрофическим изменениям, нередко обладают специализирр-
х ванными генетическими системами, определяющими особенности
их микроэволюции (см. гл. 9).
Все эти примеры касались сравнительно резко выраженных
изменений среды. Однако существенны и менее резкие, но более
частые изменения: колебания температуры в разные дни, вли-
яющие на популяцию планктонных организмов в мелком пруду,
или колебания количества осадков в разные годы, определяю-
щие состояние кормового растения какой-нибудь бабочки. Ни
одно из этих изменений нельзя рассматривать как изолирован-
ное событие. В результате повышенной влажности могут воз-
никнуть условия, благоприятные для роста кормового растения
гусениц, что приведет к появлению большой популяции бабочек.
Помимо всего прочего, это может означать, что будет больше
пищи для птенцов певчих птиц, а в дальнейшем — больше пищи
для ястребов. Обилие бабочек может привести к большей чис-
ленности гусениц в следующем году, возможно засушливом и
поэтому неурожайном для данного кормового растения. Тогда
лишь немногие гусеницы смогут превратиться в бабочек, оста-
нется мало кормовых растений и верхний слой почвы будет сне-
сен в результате эрозии. Может оказаться, что многие из пев-
чих птиц, появившихся в предыдущем году, не найдут другой
подходящей пищи и будут гибнуть от голода, эмигрировать и
плохо размножаться. Окажутся обреченными на голод и
ястребы.
В этой несколько утрированной схеме показаны лишь неко-
торые из множества возможных последствий простого измене-
' ния количества осадков. В действительности столь выраженные
эффекты сравнительно редки, ибо большинство экологических
систем включает огромное множество элементов и является
чем-то. исторически сложившимся. Сложность системы ведет к
7 Зак. 153
98 | Глава 5
тому, что такая строгая однозначная зависимость не соблю-
дается. Так, если в результате засухи уменьшается популяция
бабочек, то при этом смогут более обильно размножаться прямо-
крылые; тогда певчие птицы легко изменят свой пищевой ра-
цион. Длительно существовавшие ценозы должны были уже не
раз испытывать действие многих обычных климатических изме-
нений, и члены сообщества, вероятнее всего, уже выработали
способность к соответствующей адаптации. Например, кормовое
растение бабочек, вероятно, способно пережить засуху (быть
может, в виде непроросших семян, если это однолетник) и снова
пышно разрастется, как только вновь появится влага. К тому
же, если эрозия не зашла слишком далеко, она может прекра-
титься. Несомненно все же, что для многих организмов условия
могут необратимо ухудшиться.
СООБЩЕСТВА
Если даже единая скрещивающаяся в себе популяция ока-
зывается в области, отдельные части которой различаются по
условиям среды, следует ожидать, что со временем генетические
процессы приведут к ее дифференциации (см. гл. 6). Процессы,
участвующие в образовании сложных сообществ организмов по
существу неизвестны. Ясно, что какой-либо опустошенный уча-
сток будет вновь заселен. На ранних этапах такого вторичного
заселения группировки растений и животных недолговечны.
Несколько различных группировок сменяют друг друга, пока,
наконец, не сформируется относительно стабильное сообщество.
Совокупность этих последовательных стадий в экологии носит
название сукцессии, а конечную стадию часто называют кли-
максным или терминальным сообществом.
Экологи расходятся во мнениях относительно того, какой из
методов изучения сукцессий и терминальных сообществ считать
лучшим. Современные исследователи полагают, что сообщества
составляют часть некоего континуума и что выделять их можно
только искусственно. В самом деле, поскольку нет никаких био-
логических функций, которые осуществлялись бы между изоли-
рованными в репродуктивном отношении популяциями, сообще-
ства вряд ли могут быть дискретными единицами. Каждая
скрещивающаяся в себе популяция ведет себя в соответствии с
характерными для нее цитогенетическими процессами, порождая
особей, генотипы которых определяют диапазон условий, при-
годных для их жизнедеятельности. Каждая группа организмов
имела свою эволюционную историю, зависевшую, помимо всего
прочего, от присущих ей генетических процессов. Ввиду таких
различий трудно ожидать, что какие-нибудь две популяции
Популяции | 99
хотя бы в течение короткого времени будут развиваться одина-
ково. Сообщество, как бы мы его ни определяли, есть ревультат
наложения друг па друга диапазонов толерантности отдельных
организмов по отношению к разнообразным факторам среды,
действующим в данном месте.
Сообщества, завершающие сукцессию, при изучении их в
течение сравнительно недолгого времени кажутся стабильными
или находящимися в состоянии равновесия. В таких сообще-
ствах круговорот энергии и веществ остается постоянным и ре-
гулируется механизмами обратной связи. Много энергии сохра-
няется в органическом материале (растения, животные, гумус
и т. п.). Такое сообщество лишь с трудом выводится из равно-
весия внешними влияниями. Полагают, что организмам из дру-
гих местообитаний трудно проникнуть в сложившееся сообще-
ство. Энергетические отношения представляются четко устано-
вленными, так что можно различать первичных продуцентов
(зеленые растения), первичных потребителей (травоядные жи-
вотные), вторичных потребителей (хищники) и разрушителей
органического вещества (микроорганизмы). Естественный отбор
привел к созданию экологической единицы огромной сложности.
Организмы эволюционировали в соответствии со своим положе-
нием в этом комплексе (или в последовательных стадиях его
развития), по мере того как условия земной среды изменялись
во времени.
Очевидно, что различные популяции, существующие в дан-
ном месте, не являются случайной выборкой из всего многооб-
разия организмов. Канадский северный олень встречается в тех
же районах, где произрастает олений мох (Cladonia), а волки
встречаются совместно с северным оленем. Если собиратель
бабочек ищет в пустыне Аризона гусениц Battus philenor, он
обязательно найдет там и стелющееся растение Aristolochia
watsonii. Он быстро обнаружит, что A. watsonii растет в тени
других растений пустыни, особенно по краям впадин. Из пове-
дения взрослых бабочек-самок становится ясно, что в поисках
места для откладки яиц они используют подобные же ассоциа-
ции. Дубы и гикори нередко встречаются вместе, и леса из дуба
и гикори служат хорошим местом для охоты на виргинского
оленя. Пшеница, собака, комнатная муха, вошь и Treponema pal-
lidum наряду со многими другими организмами часто сопут-
ствуют человеку.
Большая часть проведенных экологических работ носит опи-
сательный характер. Результаты в целом дают мало интерес-
ного, особенно для исследователя, изучающего эволюцию.
Недавно оформившаяся школа экологов пытается дать матема-
тическое описание структуры, наблюдаемой в сообществах. Пока
7*
100 | Глава 5
еще, по-видимому, слишком рано решать, приведут ли эти уси-
лия к успеху, но полученные до сих пор результаты обнадежи-
вают. Как бы то ни было, кажется несомненным, что для объ-
яснения «эволюции сообществ» не требуется постулировать уча-
стие каких-то специальных механизмов. Чрезвычайно сложные
взаимодействия процессов, описываемых в этой книге, объяс-
няют все наблюдаемые в сообществах явления.
выводы
Хотя применительно к некоторым организмам понятие инди-
видуума определить трудно, у большинства организмов известны
группировки индивидуумов, называемые популяциями. Эти груп-
пировки или их комплексы могут образовать скрещивающиеся
в себе популяции; последние различным образом комбинируют-
ся, составляя таксономические группы. Жизнедеятельность осо-
бей — явление физиологическое и определяется генотипом и
эпигенотипом, от которых зависит диапазон толерантности орга-
низма к факторам внешней среды, подверженным многообраз-
ным изменениям. Популяция несет генетические функции и опре-
деляет генетическую конституцию зигот. Сообщества представ-
ляют собой группировки различных популяций, образующих
часть среды друг для друга; в них возникает определенная си-
стема энергетических отношений. Составляющие сообщество
популяции не связаны между собой никакими биологическими
функциями. Сообщества обязаны своим существованием только
взаимному соответствию диапазонов толерантности входящих
в них организмов в данный период времени.
ЛИТЕРАТУРА
1. Andrew а г th а Н. G., Birch L. С., The Distribution and Abundance of
Animals, Univ, of Chicago Press, Chicago, 1954.
2. Erlich P. R., Holm R. W., Patterns and populations, Science, 137, 652—
657 (1962).
3. Slob od kin L. B., Growth and Regulation of Animal Populations, Holt,
Rinehart and Winston, New York, 1962.
4. Stebbins G. L., Variation and Evolution in Plants, Columbia Univ. Press,
New York, 1950.
5. Wynne-Edwards V. C., Animal Dispersion in Relation to Social Beha-
vior, Hafner, New York, 1962.
6. Erlich P. R., Science, 134 (1961).
7. Erickson, Ann. Missouri Bot. Gard., 32 (1945).
8. T w i 11 y, Science, 130 (1959),
Глава 6
ТЕОРИЯ ПОПУЛЯЦИОННОЙ ГЕНЕТИКИ
В первой части этой книги мы рассмотрели происхождение
жизни, кодирование и передачу генетической информации, раз-
витие организмов и некоторые особенности популяций. Теперь
мы перейдем к центральному предмету эволюционной теории —
к изменениям уже не отдельных особей, а популяций организ-
мов. В настоящей главе будут рассматриваться теоретические
аспекты генетики менделевских популяций. Знание основных
концепций популяционной генетики абсолютно необходимо для
понимания того, каким образом наследственная информация,
содержащаяся в популяции, изменяется от поколения к поколе-
нию. Знакомство с изложенными здесь простыми математиче-
скими представлениями позволит читателю разобраться в более
сложных случаях, рассмотренных в последующих главах. Хотя
различные алгебраические примеры будут сопровождаться и не-
математическими описаниями, для твердого усвоения материала
необходимо разобраться в этих несложных алгебраических пре-
образованиях.
Приводимые в этой главе примеры представляют собой гру-
бые упрощения. Интегративные аспекты генотипа, множествен-
ные аллели, одновременное действие различных эволюционных
факторов и иные осложняющие явления большей частью не будут
учитываться. Мы на некоторое время допустим, что можно вы-
делить одиночный локус из его окружения и поставить его в
любые условия по нашему выбору; сложные взаимодействия
будут рассмотрены позднее.
МЕНДЕЛЕВСКИЕ ПОПУЛЯЦИИ
Только организмы, размножающиеся половым путем, обра-
зуют менделевские популяции, которые можно приближенно
определить как совокупности скрещивающихся между собой
особей. Более точное определение невозможно, да и нежела-
тельно, так как слова «скрещивающиеся между собой» могут
относиться к любой ситуации — от панмиксиса до почти полной
изоляции. Колорадских жуков на одном кусте картофеля уже
можно рассматривать как менделевскую популяцию, но можно
и расширить определение, охватив им всех колорадских жуков
102 | Глава 6
на одном картофельном поле, на группе смежных полей, в це-
лом округе или даже на большей территории. Поэтому важно
указывать масштабы рассматриваемой популяции и отмечать
все, что известно о ее структуре.
ПАНМИКСИС
Популяция является панмиктической, если особи внутри нее
скрещиваются случайным образом. В пределах такой популяции
каждая особь имеет одинаковые шансы на спаривание с любой
особью противоположного пола. Ожидаемая частота скрещива-
ний каждого данного рода равна произведению частот самцов
и самок соответствующих типов. Возьмем в качестве примера
гипотетическое животное, которое представлено черной и белой
формами и образует панмиктическуЛ популяцию, состоящую из
100 самцов (90 черных, 10 белых) и 100 самок (70 черных,
30 белых). Ожидаемые частоты спаривания в различных ком-
бинациях приведены в табл. 1.
ТАБЛИЦА 1
Частота спариваний
Комбинации родительских форм Произведение частот родительских форм Ожидаемая частота
самец самка
Черный Черная 0,90 X 0,70 0,63
Черный Белая 0,90 X 0,30 0,27
Белый Черная 0,10X0,70 0,07
Белый Белая 0,10 X 0,30 0,03
1,00
С помощью статистического исследования частот различных
скрещиваний можно было бы показать, что наблюдаемые от-
клонения от этих ожидаемых частот удовлетворительно объяс-
няются ошибкой выборки (т. е. случайностью), а это означает,
что популяция, по крайней мере в отношении признака окраски
тела, является панмиктической. Рассматривая панмиксис в дру-
гом аспекте, можно сказать, что он имеет место в том случае,
когда генотипы участников любого акта спаривания представ-
ляют случайную выборку из генотипов, имеющихся в популяции.
По-видимому, полный панмиксис в природе не существует или
встречается редко, хотя бы по той причине, что родственные
особи часто живут поблизости друг от друга и поэтому чаще
Теория популяционной генетики | 103
спариваются между собой. В таких случаях пары не являются
случайной выборкой из популяции и в популяционно-генетиче-
скую картину входит элемент инбридинга.
ГЕНОФОНД И ЧАСТОТА ГЕНОВ
Совокупную генетическую информацию, которой обладает
популяция, можно назвать генофондом этой популяции. Если бы
генофонд можно было описать полностью, мы знали бы не
только то, какие виды информации он содержит, но частоты
различных видов информации. В настоящей главе речь будет
идти главным образом о распределении внутри генофонда ин-
формации, относящейся к одиночному локусу.
Одно из основных понятий популяционной генетики — это
понятие частоты генов. Если мы предположим, что исследуемый
локус представлен только двумя аллелями (Л и а) и что имеется
N диплоидных особей, то D может означать число гомозигот по
одному аллелю данного локуса (ЛЛ), Н — число гетерозигот
(Ла) и R— число гомозигот по другому аллелю (аа). Тогда
D + H-\-R~N, и существуют три типа особей, в совокупности
несущих гены двух типов. N особей содержат в этом локусе
2;V генов. Поскольку каждый индивидуум ЛЛ имеет два гена Л,
а каждый индивидуум Аа~ по одному гену Л, общее число ге-
нов Л в популяции составит 2D + H. Доля р генов Л в популя-
ции равна
2D^H
р 2N 2V
Величина р носит название частоты гена А. Частоту другого ал-
леля (а) принято обозначать q. Поскольку в данном локусе
только два аллеля, /7 + ^ч=1 и, следовательно, q=\—р.
ЗАКОН ХАРДИ — ВЕЙНБЕРГА
Если спаривание в популяции происходит случайным обра-
зом и если родительские гаметы комбинируются случайным
образом, то будет иметь место совершенно случайное комбини-
рование всех гамет, образующихся в популяции. Поскольку ка-
ждая гамета содержит только один из аллелей, частота двух
различных типов гамет (Л и а) и частота соответствующих ге-
нов одинаковы. Случайное соединение гамет в зиготы даст нам
следующие количественные соотношения:
[р (спермии Д) + <7 (спермии а)] X [р (яйцеклетки Л) -Е
Д q (яйцеклетки а)] = (р + <?)2 = р1 (особи ЛЛ)4~
(особи Ла) + #2 (особи аа).
104 | Глава 6
Популяции с таким распределением частот генотипов находятся
в состоянии равновесия.
Это равновесие описывается законом Харди — Вейнберга,
который можно сформулировать следующим образом:
Если в большой панмиктической популяции имеются альтер-
нативные формы какого-либо аутосомного гена, то при отсут-
ствии мутирования, отбора или дифференциальной миграции
первоначальные численные соотношения (частоты) этих аллелей
(Pi, Рг, Рз, •.Рп) будут сохраняться из поколения в поколение,
и после одной смены поколений частоты генотипов также до-
стигнут равновесия. Равновесные частоты генотипов* соответ-
ствуют членам выражения, получаемого после возведения в
квадрат в формуле (/?1 + р2+Рз • • - + рп)2.
В дальнейшем обсуждении вопросов популяционной генетики
мы постоянно будем опираться на этот закон, составляющий
одну из фундаментальных концепций биологии. Алгебраическое
доказательство поддержания равновесия Харди — Вейнберга
приводится в табл. 2.
ТАБЛИЦА 2
Типы спаривания и состав потомства в популяции, находящейся
в равновесии Харди —Вейнберга
Тип спаривания ’ Частота спаривания Соотношение генотипов в потомстве
АА Аа aa
АА X АА (р2 X Р2) р4 р4
АА X Аа {р2 X 2pq) 2p3q p3q p3q
Аа X АА (2pq X р2) 2p3q p3q p3q
Аа X Аа (2pq X 2pq) 4p2q2 Р2Ч2 2p2q2 p2q2
АА X аа (р2 X ?2) p2q2 p2q2
аа X АА (р2 X q2) P2Q2 p2q2
Аа X аа (2pq X q2) 2pq3 pq3 pq3
аа X Аа (2pq X q2) 2pq3 pq3 pq3
аа X аа (q2 X q2) q4 q4
1,00 ** р2 *** 2pq**** q2 *****
* Частота типов спаривания как для самцов, так и для самок соответствует членам
выражения p2-\-^PQ + q2; поэтому при случайном спаривании частоты различных спариваний
будут (Й+2Р<7 + <72) (Р2 + 2/><7+<72) = ^ + 4р8^ + 6р242 + 4р(78 + (74.
** Сумма в этом столбце равна р< + 4р37 + 6р2<72 + 4р<73 + (74 = (р2 + 2р7 + <72)2 = Кр + <7)2]2--
«= [(I)2]2 = 1,00.
*** Сумма в этом столбце равна p* + 2p3q + P2q2=Р2 (р2 + 2pg + (?2) = р2 (1) = р2.
**** Сумма в этом столбце равна 2p*qA-4p2q2A-2pq3 = 2pq (p2 + 2pq-[- q2) = 2pq ‘KV) = 2pq.
***** Сумма в этом столбце равна p2q2 + 2pqt-\-q* — q2 (р2 +2p^4-g2) = <72 (1)==</2.
Если в произвольно выбранной исходной популяции частота
некоторого гена р=0,2 (^ = 0,8) и частоты генотипов ДА = 0,10,
Теория популяционной генетики | 105
Л<2 = 0,20 и rm = 0,70, то популяция за одно поколение достигает
равновесия и затем остается в этом состоянии (табл. 3).
ТАБЛИЦА 3
Поколе- ние Частота генотипов Частота генов
АА Аа аа
1-е 0,10 0,20 0,70 р = 0,2 7 = 0,8
2-е 0,04 0,32 0,64 р = 0,2 7 = 0,8
3-е 0,04 0,32 0,64 Р = 0,2 7 = 0,8
4-е 0,04 0,32 0,64 р = 0,2 7 = 0,8
У-е Р2 2р? Q2 Р Q
Обратите внимание на то, что никакого изменения частоты
генов не происходит; ее можно определить уже после 1-го поко-
ления, извлекая квадратный корень из частоты соответствующей
гомозиготной формы. При описанных условиях менделевским
популяциям свойственна генетическая инерция. Если равнове-
сие не нарушается в результате мутаций, отбора, дифференци-
альной миграции, определенных изменений в выборе родитель-
ских пар или уменьшения размеров популяции, то генетическая
структура последней остается без изменений. В очень большой
степени эту инерцию преодолевает (в частности, изменяя часто-
ту генов) тот процесс, который мы называем эволюцией.
Представления, связанные с законом Харди — Вейнберга,
служат основой любого рассмотрения эволюционных процессов,
и важно, чтобы читатель хорошо освоился с ними. О следствиях
закона Харди — Вейнберга и о свойствах равновесных популя-
ций вы можете узнать из книги Ли [2], посвященной генетике
популяций.
Хотя природные популяции разве только в редких случаях
находятся в равновесии, этот закон чрезвычайно полезен при
описании ситуаций, в которых не происходит никакой эволю-
ции, так как он указывает «нулевой» уровень для измерения
эволюционных изменений. Рассмотрим теперь некоторые откло-
нения от равновесия Харди — Вейнберга.
ВЕЛИЧИНА ПОПУЛЯЦИИ
Во всех популяциях наблюдаются небольшие случайные ко-
лебания частоты генов. Вследствие этого частота генов в лю-
бой выборке гамет, взятой из родительской популяции, будет
106 | Глава 6
отличаться от частоты генов в популяции в целом. Кроме того,
популяция зигот, образующихся по закону случая при соедине-
нии гамет, никогда не будет характеризоваться точно такой же
частотой генов, что и популяция гамет. Наконец, даже если ги-
бель созревающих особей распределяется совершенно случайным
образом, «ошибка выборки» приведет к тому, что в размножаю-
щейся популяции второго поколения частоты генов и генотипов
опять-таки будут отклоняться от соответствующих частот в
исходной популяции зигот. В обширных популяциях эти случай-
ные отклонения, обусловленные ошибкой выборки, более или
менее уравновешивают друг друга, поскольку они различны в
разных частях территории и на разных этапах процесса воспро-
изведения. Поэтому изменениями в частоте генов, обусловлен-
ными ошибкой выборки, обычно можно пренебречь. Однако в
небольших популяциях эти случайные отклонения приобретают
существенное значение.
Таким образом, размеры популяции могут оказывать значи-
тельное влияние на ее генетическую структуру. Биологические
популяции всегда имеют конечную величину. Общая величина
популяции — это просто число осебей, входящих в ее состав
в данный момент времени. Гораздо важнее для характеристики
популяции величина размножающейся части популяции (репро-
дуктивная величина) N, определяемая как число родителей, от-
ветственных за генетический состав следующего поколения. То,
что эта часть бывает значительно меньше популяции в целом,
видно на примере человека, у которого индивидуумы моложе 10
и старше 60 лет, как правило, не входят в размножающуюся
часть популяции.
ЭФФЕКТИВНАЯ РЕПРОДУКТИВНАЯ ВЕЛИЧИНА ПОПУЛЯЦИИ
Для эволюции чрезвычайно существенна так называемая
эффективная репродуктивная величина популяции, Ne. Она экви-
валентна репродуктивной величине W только в случае идеальной
популяции, все время сохраняющей большие размеры, в которой
оба пола имеют равную численность, спарива-ние происходит
случайным образом и так же случайна комбинация гамет роди-
тельских особей. Такая идеальная популяция представляет собой
абстракцию; практически же эффективная репродуктивная ве-
личина всегда меньше репродуктивной величины. В некоторых
случаях, однако, разница бывает совсем незначительной. Нерав-
ное соотношение полов, инбридинг, периодическое уменьшение
репродуктивной величины и неслучайная выборка гамет — все
эти факторы уменьшают эффективную репродуктивную величину
популяции. Рассмотрим в качестве примера популяцию из по-
Теория популяционной генетики [ 107
стоянного числа размножающихся особей, N, в которой спари-
вание происходит случайным образом и так же случайна вы-
борка гамет родителей. Результаты, приведенные в табл. 4,
иллюстрируют влияние численного соотношения полов на эффек-
тивную репродуктивную величину Ne для случая, когда
JVc/1 + Ar$=:400; Ne = 4/V N$/(N+ N ?) (вывод форму-
лы дается в книге Ли).
ТАБЛИЦА 4
Влияние кисленного соотношения полов на эффективную
репродуктивную величину популяции *
Число самцов Число самок Ne Число самцов Число самок Ne
200 200 400,00 25 375 93,75
100 300 300,00 5 395 19,75
50 350 175,00 1 399 3,99
* Репродуктивная величина популяции /V=400.
Если эффективная репродуктивная величина для четырех
последовательных поколений составляет 10, 100, 10 000 и 100,
то средняя эффективная величина (которая представляет собой
среднее гармоническое, т. е. величину, обратную среднему ариф-
метическому величин, обратных величинам для каждого поко-
ления) для всех этих поколений равна приблизительно 33. Сле-
дует отметить, что эта величина гораздо ближе к минимальной
величине (10), чем к максимальной (10 000). Эффективная
репродуктивная величина соответствует величине идеальной по-
пуляции, генетическое поведение которой было бы таким же,
как и у рассматриваемой популяции. Это позволяет получать
сравнимые характеристики величин различных популяций. При-
равнять друг другу две популяции из 400 размножающихся
особей, в одной из которых 200 самцов и 200 самок, а в дру-
гой — 5 самцов и 395 самок, значило бы игнорировать генети-
ческие последствия неравной численности полов во второй из
них (например, при прочих равных условиях у второй популя-
ции степень изменчивости снижалась бы гораздо быстрее, чем
У первой, так как самцы по существу оказались бы «узким ме-
стом» в процессе передачи информации из поколения в поко-
ление). Иногда можно указать особи, принадлежащие к раз-
множающейся популяции, но невозможно выделить какую-то
группу особей и назвать ее «эффективной популяцией».
108 | Глава в
В дальнейшем всюду (кроме оговоренных случаев) термином
«величина популяции» мы будем обозначать эффективную ве-
личину и будем обозначать ее просто N.
ДРЕЙФ ГЕНОВ 1
Представьте себе урну, содержащую 10 000 черных и 10 000
белых шариков, соответствующих гаметам некоторой популяции
с частотой генов p = q = 0,50 (в некотором локусе). 2000 шари-
ков, случайным образом взятых из этой урны, будут соответ-
ствовать 1000 диплоидных особей, которые составят репродук-
тивную популяцию следующего поколения. Допустим, что в
первой выборке оказалось 979 белых шариков и 1021 черный
шарик (р = 0,49). Тогда изменившийся генофонд можно будет
представить 9790 белыми и 10 210 черными шариками. Затем
вновь берут выборку в 2000 шариков. Предположим, что на этот
раз частота генов в репродуктивной популяции следующего по-
коления будет соответствовать 1033 белым и 967 черным шари-
кам (/7 = 0,52). Еще раз наполняем урну шариками, среди кото-
рых теперь должно быть 10 330 белых и 9670 черных. Продол-
жение этого процесса в большинстве случаев приводило бы лишь
к очень небольшим колебаниям частоты генов около первона-
чальной величины (р = 0,50). Отметим, что эта модель построена
таким образом, чтобы величина популяции оставалась посто-
янной.
Теперь представим себе другую урну, содержащую 500 бе-
лых и 500 черных шариков, которые соответствуют гаметиче-
скому генофонду меньшей популяции с частотой генов /7 = 0,50.
Предположим, что из нее наугад будут вынуты 10 шариков
(соответствующие 5 особям)—из них 6 черных и 4 белых
(/7 = 0,60). Представим новую популяцию гамет 600 черными и
400 белыми шариками и повторим прежнюю процедуру. На этот
раз мы вынули, допустим, 8 черных и 2 белых шарика (/7 = 0,80)
и для следующего опыта соответственно кладем в урну 800 чер-
ных и 200 белых шариков. В третьей выборке может оказаться
7 черных и 3 белых шарика (/7 = 0,70), и мы кладем в урну
700 черных и 300 белых шариков. Легко видеть, что в ходе этой
процедуры у нас возникали гораздо большие колебания частоты
генов, чем в случае более обширной популяции. Эти случайные
колебания частоты генов известны как дрейф генов. Термин
1 В советской литературе явление дрейфа генов принято называть вслед
за его первооткрывателями генетико-автоматическими процессами (см. Д у-
б и н и н Н. П., Р о м а ш е в Д. Д., Биологический журнал, т. V, вып. 5—6,
1932). — Прим, ред.
Теория популяционной генетики [ 109
«дрейф» очень хорошо отражает суть дела, так как частота р
как бы блуждает, не приближаясь ни к какой определенной ве-
личине, в отличие от направленных смещений, обусловленных
так называемым систематическим давлением мутационного про-
цесса, отбора и дифференциальной миграции.
На фиг. 18 показано распределение вероятностей частот р
в группах из 50 и 5000 потомков родительской популяции, в ко-
торой частота р составляла 0,50. В этом и в последующих при-
мерах будет предполагаться наличие всех условий для поддер-
жания равновесия Харди — Вейнберга, за исключением изучае-
мого фактора. Другими словами, мы будем рассматривать
факторы по отдельности для того, чтобы показать, как каждый
из них может влиять на изучаемое равновесие.
Возможные величины р в двух различных группах потомков
представлены на фиг. 18 двумя гистограммами. Ордината соот-
ветствует приблизительной вероятности р для каждого интер-
вала частот. Как и следовало ожидать, для меньшей группы
потомков вероятность крупных отклонений, обусловленных
ошибкой выборки, оказывается гораздо большей.
Графики, приведенные на фиг. 18, характеризуют также
судьбу большого числа локусов, для каждого из которых часто-
та аллелей в родительской популяции составляла 0,50. Когда
М = 50, следует ожидать, что только для 3 локусов из 100 000 ча-
стота отклонится больше чем до 0,70; в то же время при М = 5000
только для 3 локусов из 100 000 отклонение частоты будет пре-
вышать 0,52. Нас может интересовать распределение частот ал-
лелей того или иного локуса в большом числе популяций, в
каждой из которых первоначальная частота составляла 0,50.
Согласно фиг. 18, при М = 5000 следует ожидать, что в 99,994%
0,40 г-
1
0,300 0,360 0,400 0,450 0^00 0,550 0.600 0650 0 700
Частота гена
Фиг. 18. Распределение вероятностей р для популяций из 50 и 5000 потом-
ков родительской популяции, в которой частота генов р равна 0,50.
Подробности см. в тексте.
ПО | Глава 6
особей частота аллелей данного локуса будет лежать в преде-
лах от 0,48 до 0,52, тогда как при М = 50 в том же проценте осо-
бей мы найдем величины от 0,30 до 0,70.
ЗАТУХАНИЕ ИЗМЕНЧИВОСТИ
Возможные последствия дрейфа генов в небольшой популя-
ции схематически представлены на фиг. 19. Частота генов ведет
себя аналогично упругому шарику, который, скатываясь по
наклонной доске, отскакивает от вбитых в нее штифтиков, пока
не дойдет до одного из краев доски. С течением времени часто-
та переходит от одной величины к другой, пока не достигнет 0
или 1,0. При отсутствии мутирования и миграции величины 0
(утрата данного гена) и 1,0 (фиксация гена, т. е. полное вытес-
нение им других аллелей) не могут более изменяться. Если ша-
рик попадает в один из краевых желобков, он остается там;
аналогичным образом ген А либо фиксируется (все особи АА),
либо утрачивается (все особи аа). Итак, частота гена А может
переходить от любой промежуточной величины к конечным точ-
кам 0 и 1, но не обратно; в конце концов она достигнет 0 или 1,
если популяция достаточно долго не будет испытывать никаких
«возмущений». Так как по достижении одной из этих величин
(0 или 1) образование гетерозигот по данному локусу стано-
Теория популяционной генетики | 111
вится невозможным, чем больше генов будет утрачено или фик-
сировано, тем меньше гетерозигот останется в популяции. Про-
цесс снижения гетерозиготности в результате утраты или
фиксации аллелей в различных локусах получил название зату*
хания изменчивости.
Скорость этого затухания находится в тесной зависимости
от величины популяции. Гены утрачиваются со скоростью V4W
на одно поколение и фиксируются также со скоростью на
одно поколение. Таким образом, затухание изменчивости про-
исходит со скоростью V2W генов на одно поколение. Рассмотрим
некоторые крайние случаи. В репродуктивной популяции, со-
стоящей всего лишь из 4 особей и имеющей величину р = 0,125,
единственный ген, характеризующийся этой частотой, будет
присутствовать у одной особи и окажется переданным V8 всех
гамет. Если эта единственная особь не сможет принять участие
в размножении, ген будет утрачен. Если же эта особь даст по-
томство, то вероятность утраты гена будет равна !/2 в случае
одного потомка, V4—при 2 потомках, У8— при 3 потомках и
т. д. Однако в популяции из 4000 размножающихся особей при
частоте некоторого гена 0,125 минимальное число содержащих
этот ген особей составляет 500. В этом случае все они были бы
гомозиготными по данному гену, и для того чтобы ген оказался
утраченным, ни одна из них не должна была бы оставить потом-
ства. Дрейф генов (ошибка выборки) есть математический факт.
Однако вопрос о значении дрейфа как фактора, действующего
независимо от отбора, вызвал много споров. Кажется несомнен-
ным, что дрейф генов имеет очень небольшое значение в боль-
ших популяциях (скажем, при AZ>500), но в небольших попу-
ляциях он может быть активным фактором эволюции.
ПОТЕРЯ МУТАЦИЙ
Еще один аспект изменчивости в результате ошибки выборки
связан с вероятностью утраты мутантного гена. Представим
себе менделевскую популяцию, у которой М — величина посто-
янная (пара взрослых особей дает в среднем двух потомков) и
в которой определенный локус фиксирован (все особи по этому
локусу гомозиготны, АА). Если у какой-то одной особи А му-
тирует в а, так что возникает гетерозигота Аа, то в случае раз-
множения этой гетерозиготы происходит возвратное скрещива-
ние с особью АА (никаких других особей в популяции нет).
Среди потомков от этого скрещивания в среднем 50% особей
будут иметь генотип А А и 50% особей — генотип Аа. В зависи-
мости от случая число потомков может быть 0, 1, 2, 3, . . ., г,
причем вероятности этих чисел распределены по закону
112 | Глава 6
Пуассона со средним 2 (среднее число потомков). Если не по-
явится ни одного потомка, то ген будет утрачен. Если будет 1
потомок, вероятность утраты гена составит 0,50, при двух по-
томках эта вероятность равна 0,25, а при г потомках она рав-
на 2-г. Используя параметры распределения Пуассона, можно
вычислить, что предел совокупной вероятности утраты гена бу-
дет равен 0,3679. Фишер вычислил вероятность потери мута-
ции, возникшей у одной особи, при условии, что эта мутация
не имеет никакой селективной ценности, а также при условии,
что селективное преимущество мутантного аллеля перед алле-
лем дикого типа составляет 1%. Эти вероятности приведены
ТАБЛИЦА 5
Вероятность исчезновения мутации,
возникшей у одной особи
Поколение Вероятность исчезновения мутации
при отсутствии селективного преимущества при селективном преимуществе, равном 1%
1-е 0,3679 0,3642
3-е 0,6259 0,6197
7-е 0,7905 0,7825
15-е 0,8873 0,8783
31-е 0,9411 0,9313
63-е 0,9698 0,9591
127-е 0,9847 0,9729
Предел 1,0000 0,9803
в табл. 5, из которой явствует, что новая мутация практически
не имеет шансов сохраниться в популяции, если потере измен-
чивости не противодействует отбор.
МУТАЦИОННЫЙ ПРОЦЕСС
Мутации — это изменения в генетической информации, и в
этом аспекте они рассматривались в гл. 3. Теперь мы раесмо*
трим мутационный процесс как один из факторов, стремящихся
вызвать отклонение от равновесия Харди — Вейнберга. Если Л—
«типовой ген», а — мутантный ген, и — частота мутаций А-^а и
v — частота обратных мутаций А <-а, то такая система имеет
точку равновесия, как видно из нижеследующего:
Kq = up (прирост) — vq (утрата)
Д^Ов точке равновесия (/?, q).
Теория популяционной генетики | 113
Таким образом,
&q = 0 = и (1 — q) — vq
Q = u — uq — vq
— u = q(— и — <v)
~ и
q = —i—
и -|- у
Z 1 Z 1 U, U-X-V— U V
p=i — q = 1-----j— = ——------= —i—,
r 4 U-\-V «4“V u,-\-v ’
где p—частота гена A; q — частота гена a; p + ?=l; <7=1—p\
&q— изменение q\ q — равновесная величина.
Это графически представлено на фиг. 20. Обратите внимание,
что А*? — это результирующее изменение частоты гена а за 1 по-
коление и что Д</ положительно, когда q < q, и отрицательно,
когда <7 > <?. Изменение в точке 1 (все а) вдвое меньше, чем
изменение в точке 0 (все Л), так как Л—>a = u = 0,00004, а
а—>Л = v = 0,00002. Таким образом, мы можем видеть, что в по-
пуляции, удовлетворяющей всем требованиям равновесия Хар-
ди — Вейнберга, кроме требования отсутствия мутаций, равно-
весная величина для частоты гена определяется частотой пря-
мого и обратного мутирования в данном локусе.
ОТБОР
Отбор — это неслучайное (дифференциальное) воспроизве-
дение генотипов. Поток жизни популяции можно было бы рас-
сматривать как состоящий из непрерывно изменяющихся, деля-
щихся, сливающихся и исчезающих частиц, текущий из прошлого
в настоящее через ряд необычайно сложных отсеивающих сит,
§ Зак. 152
114 | Глава 6
Можно сказать, что произошел отбор, если различие в составе
этого потока в двух последовательных сечениях настолько ве-
лико, что его нельзя отнести за счет ошибки выборки (дрейфа)
или мутаций. При попытках выявить действие отбора необхо-
дима уверенность в том, что поток на исследуемом отрезке не
разветвляется (нет эмиграции) и не принимает в себя «при-
токов» (нет иммиграции).
Если популяция генетически неоднородна, то вероятность
успешного воспроизведения одних генотипов будет больше (за
возможными редкими исключениями), чем других. Таким обра-
зом, определенные типы генетической информации будут встре-
чаться в генофонде популяции все чаще и чаще, а другие типы
все реже и реже. Частоты генов, р и q, будут изменяться во
времени, а не сохранять постоянную величину, как следовало бы
ожидать при условиях равновесия Харди — Вейнберга.
Очень часто об отборе говорят как о великой «созидательной
силе», как о причине, определяющей направление наблюдаемых
процессов эволюции. На самом деле это феномен, который мож-
но наблюдать только a posteriori — при описании прошедших
событий. Когда некоторая неслучайная выборка генотипов остав-
ляет больше потомства, чем другие типы, мы говорим, что про-
исходит отбор. В самом широком плане — отбор уменьшает
разнообразие живых организмов; организмы, содержащие опре-
деленные типы и сочетания генетической информации, нежизне-
способны или не оставляют потомства. Этот процесс и «создает»
то, что мы осознаем как некоторый порядок в природе, наподо-
бие того как порядок, который мы видим в команде из тяжело-
весных, мускулистых игроков, «создается» игрой в футбол.
Только в этом ограничительном смысле — в смысле противо-
действия тенденции к возрастанию энтропии — отбор является
творческим фактором.
Критики теории естественного отбора утверждали, что отбор
никак не может быть творческой силой, так как он действует
только путем элиминации определенных форм. Они указывали
на то, что если через сито, с которым обычно сравнивают есте-
ственный отбор, просеивать крупные и мелкие камни, конечным
результатом будет куча из более мелких камней, отобранных из
первоначальной кучи; ничего не будет создано заново. Однако
критики в данном случае пользуются слишком упрощенной ана-
логией, ибо в ней не учитывается потенциальная мутационная
и рекомбинационная изменчивость биологических единиц. Ана-
логия была бы более точной, если бы два мелких камешка
могли скрещиваться и в их потомстве встречались камешки еще
меньше, чем каждый из родителей. -Тогда при использовании
достаточно мелкого сита образовалась бы куча камешков
Теория популяционной генетики [ 115
более мелких, чем те, которые содержались в первоначальной
куче.
Существует два типа отбора: естественный отбор, при кото-
ром окружающая среда определяет, какиё генотипы являются
наиболее приспособленными, и искусственный отбор, при кото-
ром это определяет человек. Важно отметить, что если есте-
ственному отбору всегда приходится действовать на генотип
через фенотип, то искусственный о'Лбор может иногда воздей-
ствовать непосредственно на генотип. Предположим, например,
что имеется меланическая форма бабочки, обусловленная доми-
нантным геном М. В случае полного доминирования воздействию
среды будут подвергаться только два различных фенотипа —
меланический и немеланический (М— и mm). В отношении
действия естественного отбора доминантная гомозигота (Л1М)
и гетерозигота (Мт) неразличимы. В лаборатории дело обсто-
яло бы совсем по-иному: надлежащие анализирующие скрещи-
вания позволили бы отобрать любой из этих генотипов.
ПРИСПОСОБЛЕННОСТЬ, ИЛИ АДАПТИВНАЯ ЦЕННОСТЬ
Для каждой данной среды произведение относительной цен-
ности данного генотипа для выживания на его относительную
репродуктивную способность есть приспособленность, или адап-
тивная ценность этого генотипа. Приспособленность включает
много компонентов, примерами которых могут служить отно-
сительная плодовитость, длительность репродуктивного периода,
способность находить партнера для спаривания (у 5кивотных),
эффективность механизма опыления (у растений) и общая вы-
носливость особей с данным генотипом. Адаптивную ценность
условно принято выражать величинами от 0 (адаптивная цен-
ность зиготы, гомозиготной по летальному гену) до 1 (адап-
тивная ценность генотипа, дающего максимальное число гамет,
участвующих в создании особей следующего поколения). Эго
максимальное число зависит, конечно, от того, с каким орга-
низмом мы имеем дело. Таким образом, в нашей книге термин
«приспособленность» означает эффективность генотипа в отно-
шении передачи генетической информации следующему поколе-
нию, причем для сравнения эффективностей генотипов исполь-
зуют шкалу, в которой за 0 принимается отсутствие передачи
информации, а за 1 — передача всей информации, заключенной
в генотипе. Некоторые авторы включают в понятие приспосо-
бленности такие вещи, как «перспективная» приспособленность,
т. е. способность популяции адаптироваться к предполагаемым
будущим изменениям среды. Не может быть сомнения в том,
что некоторые типы популяций (как будет отмечено в главах,
8*
116 I Глава 6
посвященных рассмотрению генетических систем, и ниже в на-
стоящей главе) более способны адаптироваться к изменениям
среды, чем другие типы. Однако ввиду трудностей анализа этого
аспекта приспособленности лучше, по-видимому, употреблять
это понятие применительно к ограниченному времени, как оно
было определено выше.
Следует отметить, что эти определения отбора очень далеки
от той картины «кровавой борьбы», которую обычно рисуют
себе несведущие люди. Творческая сторона этого процесса почти
целиком связана с воздействием среды, которая через посред-
ство отбора влияет на генетическую структуру популяций таким
образом, что в них возникают наиболее приспособленные фено-
типы.
ТИПЫ ОТБОРА
Существует три основных типа* отбора в популяциях: на-
правленный, стабилизирующий идисруптивный отбор. О направ-
ленном отборе говорят тогда, когда в популяции происходит
сдвиг средней величины рассматриваемого признака. Стабили-
зирующий отбор выражается в пониженной приспособленности
крайних вариантов с одновременным уменьшением дисперсии
данного признака, что ведет к большей однородности популя-
ции. О дисруптивном отборе говорят тогда, когда условия бла-
гоприятствуют двум или нескольким различным типам, но не
благоприятствуют промежуточным типам. Такой отбор обычно
увеличивает дисперсию и при некоторых условиях может вести
к дроблению популяции на части. Примеры всех трех типов от-
бора читатель найдет в следующей главе.
Проведена обширная работа по математическому описанию
различных типов отбора. Большая часть ее не может быть рас-
смотрена в этой книге, и интересующегося ею читателя мы снова
отсылаем к книге Ли. Однако мы все же приведем несколько
примеров этого количественного подхода как для иллюстра-
ции применяемых принципов, так и для того, чтобы читателю
не пришлось принимать на веру результаты некоторых селек-
тивных процессов.
СЛУЧАЙ ПОЛНОЙ НЕЖИЗНЕСПОСОБНОСТИ ГОМОЗИГОТНЫХ РЕЦЕССПВОВ
Если мы имеем ряд генотипов (например, АА, Аа, аа), мы
можем, приписывая каждому из них некоторую адаптивную
ценность W, количественно оценить их способность передавать
свои гены в генофонд следующего поколения. Как уже говори-
лось, для наиболее приспособленного генотипа 1F=1, тогда как
Теория популяционной генетики | 117
менее приспособленным комбинациям соответствуют меньшие
величины W7. Таким образом, возможна следующая ситуация:
Генотип А А Аа аа
Адаптивная ценность W 1 11 — $
В этом случае гомозиготные рецессивы уступают по своей при-
способленности как гетерозиготам, так и доминантным гомози-
готам. Степень невыгодности этого генотипа измеряется коэф-
фициентом отбора s, а его приспособленность, или адаптивная
ценность, выражается как 1—s. Коэффициент отбора мог бы
* здесь варьировать от 0 (отрицательного отбора нет, что делает
сравнение излишним) до 1 (летальность рецессивных гомо-
зигот).
Последствия полной элиминации рецессивов из популяции
(рецессивные гомозиготы нежизнеспособны, s=l) показаны в
табл. 6.
Отношение между частотами генов в любых двух последо-
вательных поколениях определяется по формуле
Чп+Х~ 1_|_Ул .
где индексы указывают порядковые номера поколений. Таким
образом,
<7о = <7о
^+i = ?i = t^7
п =п ЧоНУ 4~ ?о) ________?о/(1 + ?о)_________
ЧХ+Х 42 1+<7о/(1+<7о) (1+<7о)/(1+<7о) + <7о/(1+<7о) ~
?о/(1 4~ Чо) Чо
(1+2<70)/(1+9о) 1+2<70
Эти последовательные величины q (частоты генов) образуют
гармонический ряд, т. е. такой ряд, члены которого являются
величинами, обратными членам арифметической прогрессии.
Если известна первоначальная частота генов, то частоту их для
любого из последующих поколений можно найти по формуле
?п = ?о/(1+^?о). Изменение частоты генов за 1 поколение опять-
таки обозначается \q и определяется по следующей формуле:
Обратите внимание на то, что скорость изменения частоты генов
сама является функцией частоты генов. Когда частота гена
ТАБЛИЦА 6
Полная элиминация рецессивов
Поко- ление До или после отбора Частота генотипов Частота гена а
АА Аа аа
0 До отбора р2 2pq я2 Q
После отбора * р2 р2Л-^ря ^РЯ Р2 + ^РЯ 0 Я 1+4
1 До отбора ** 1 (1 + <7)2 (1+<7)2 я2 (1 + <7)2 Q 1+4’
После отбора *** 1 1 + 2? 2? 1+2? 0 Q 1+2?
...
* p2-\-2pq — это то, что остается после элиминации генотипов аа (72). Для того чтобы
найти частоты двух оставшихся генотипов, их нужно представить как доли общей суммы.
Эти две величины определяются просто следующим образом:
Р2 = Р (Р) = 1~7 = 1~7
Р2 + 2р7 Р(Р + Ц) 1 —7 + 27 1 + 7 ’
и аналогичным образом
2^7 27
Р2 + 2/?7 1 + 7 ’
___ 1 — 7 , 2q
Поскольку - , + -гп—
1 + 7 1+7
7
. ч 1 27 7
(частота гена л) = — х г * = -г-2—
2. 1 + 7 1 7
и
р=1—7=1
7
** Частоты генотипов
вая скобки в выражении
до отбора находят, используя новые величины р и q и раскры-
7
/ 1 \2 1
Например, частота зигот А А = р2 = (^ - -^ J (здесь используются новые значения
Р и 7).
*** Частоты после отбора вычисляются так же, как и для предыдущего поколения после
отбора, например:
1/(1+ 7)2 ; 1
1/(1+ 7)2 +27/(1+ 7)2 1 + 27 *
Теория популяционной генетики | 119
высока, ген элиминируется из популяции быстро. Несколько чи-
словых примеров приводится ниже.
Частота гена Снижение частоты за 1 поколение
0,9 0,426
0,5 0,167
0,1 0,009
0,05 0,0024
0,01 0,000099
Причина такого изменения скорости элиминации состоит в.
том, что с увеличением частоты данного гена доля рецессивных
генов, находящихся в гетерозиготном состоянии, быстро возра-
стает. Если q — частота гена а, то доля генов а, находящихся
в гетерозиготном состоянии, изменяется определенным образом
(табл. 7).
ТАБЛИЦА 7
Частота генотипов Гены я, находящиеся в гетерозиготном состоянии, %
АА Аа аа
0,9 0,01 0,18 0,81 10
ОД 0,81 0,18 0,01 90
0,01 0,9801 0,0198 0,0001 99
В гетерозиготах рецессивные гены «укрыты» от действия от-
бора, так как они детальны только в гомозиготном состоянии.
Чем ниже частота гена, тем меньше доля рецессивных генов,
подверженных элиминации в гомозиготах, и соответственно тем
сильнее замедляется процесс элиминации гена из популяции.
Этот результат представляет особый интерес для исследовате-
лей в области евгеники. Если частота q какого-либо нежела-
тельного гена (а) в популяции равна (У,01, так что q2 (доля
индивидуумов аа) составляет 0,0001 (один дефект на 10 000
нормальных индивидуумов), то пришлось бы производить сте-
рилизацию обладателей этого гена на протяжении 100 поколе-
ний (около 2500 лет), чтобы снизить частоту гена вдвое и
уменьшить число гомозигот до 1 на 40 000. Ввиду трудности
осуществления такой программы без ошибок в течение столь
длительного периода кажется несомненным, что столь ничтож-
ные результаты вряд ли могут оправдать необходимые для их
120 | Глава 6
достижения усилия. В то же время отбор, направленный против
доминантных генов, относительно эффективен. Например, если
бы доминантный ген стал летальным, он был бы элиминирован
из популяции за одно поколение.
СЛУЧАЙ МЕНЬШЕЙ ПРИСПОСОБЛЕННОСТИ ГОМОЗИГОТНЫХ РЕЦЕССИВОВ
Ситуация, в которой доминантные особи имеют преимуще-
ство перед рецессивными, но гомозиготные рецессивы все же
вносят известный вклад в генофонд последующего поколения,
встречается чаще, чем полная летальность гомозиготных рецес-
сивов.
Если мы припишем обеим доминантным формам (гомози-
готной и гетерозиготной) приспособленность 1, а гомозиготным
рецессивам — приспособленность 1—s, то при действии отбора
в течение одного поколения мы будем иметь р2 особей АА, 2pq
особей Аа и q2—sq2 особей аа из общего числа p2 + 2pq + q2—
—sq2=\—sq2 особей. Пользуясь той же процедурой, что и в пре-
дыдущем примере, мы получим изменение частоты генов за
1 поколение:
кп_ РЯ + ч2(У — s) п _ — s?2(l — q)
\ — sq2 Ч~ \—sq2 ’
Таким образом, изменение частоты генов при этих условиях
будет малым, когда q очень велико или очень мало, и сравни-
тельно большим при промежуточных величинах q. Когда q ве-
лико, процесс будет медленным из-за относительно большого
вклада рецессивных гомозигот (в противоположность доминант-
ным формам) в репродукцию генов. Когда q становится малым,
защитное действие доминантного фенотипа гетерозигот замед-
ляет процесс — так же как и в случае летальности гомозиготных
рецессивов. Несколько примерных цифр приведено ниже (коэф-
фициент отбора, направленного против гомозиготных рецесси-
вов, s, принят равным, 0,5).
7 Д q Д q
0,99 —0,00961 0,30 —0,03298
0,90 -0,06807 0,05 —0,00119
0,60 —0,07143 0,01 —0,00005
Теория популяционной генетики | 121
СЛУЧАЙ МЕНЬШЕЙ ПРИСПОСОБЛЕННОСТИ ГОМОЗИГОТ ПО СРАВНЕНИЮ
С ГЕТЕРОЗИГОТАМИ
В качестве последнего детального примера моделей отбора
рассмотрим случай, когда оба гомозиготных генотипа уступают
по своей приспособленности гетерозиготам (т. е. когда данный
локус проявляет сверхдоминантность в отношении адаптивной
ценности):
Генотип А А Аа аа
Адаптивная ценность 1 — s „ 1 1 — sa.
Тогда после действия отбора в одном поколении получаем
р2(1—5а) особей АА, 2pq особей Аа и ^2(1—sa) особей аа из
общего числа 1—sAp2—saq2 особей. Последнее число получаем
путем сложения соответствующих величин для всех трех гено-
типов и последующего упрощения:
р2(1 - 5д)4-2^ + ^2(1 - 5а) =
= Р2 + Zpq + q2 — sAp2 — saq2 = 1 — sAp2 — saq2.
Поскольку начальная частота генов qo равна pq + q2, частота
qt генов а после действия отбора в одном поколении будет равна
P? + ?2 (l —sfl) _ PQ + q2 — saq2 Q — s^q2
1 “ SaP2 Sa^ 1 SaP2 ~ Sa^ 1 “ SAp2 ~~ Saq2 ’
Тогда изменение q за 1 поколение находим следующим образом:
А _ Я — saq2 _q — saq2 — q + sAp2q + saq2(l—p)_
l — sAP2 — saq2 q~ \ — sAp2 — saq2 ~
_ — saq2 + sAp2q + saq2 — sapq2 pq (sAp — saq)
1'~SAP2~~Saq2 ~ i—sAP2~~Saq2
Следовательно, если sAp>saq, то \q положительно и частота
генов а возрастает, а если sAp<saq, то Ад отрицательно и ча-
стота генов а понижается. Когда sAp = saq, &q = 0 и частота ге-
нов а находится в равновесии. Таким образом, в этой ситуации
равновесная величина q(q) определяется только величиной ко-
эффициентов отбора:
sAP=saq sA(\—q) — saq = sA — sAq—saq = O
~ ~ S
SA = sAq + saq = q(sA + sa) q = .
A 1 a
Аналогичным образом
122 | Глава 6
Если адаптивные ценности всех трех генотипов остаются
постоянными, то равновесная величина частот генов также по-
стоянна. Если же какое-нибудь обстоятельство, например при-
бытие группы мигрирующих особей, сдвинет частоту генов с
равновесной точки, силы отбора восстановят равновесие. По-
этому мы называем это равновесие устойчивым.
СБАЛАНСИРОВАННЫЙ ПОЛИМОРФИЗМ И СОХРАНЕНИЕ ИЗМЕНЧИВОСТИ
Ситуация, при которой гетерозиготы имеют преимущество
перед обеими гомозиготными формами, приводит к сохранению
в популяции обоих аллелей, а не способствует фиксации того
или другого из них. Состояние популяции, при котором в ней
сохраняются две или более форм данного организма, причем
частота даже наиболее редкой формы слишком высока, чтобы
ее можно было объяснить одним лишь мутированием, носит на-
звание полиморфизма. Когда гетерозиготы приспособлены луч-
ше гомозигот, установление равновесной частоты генов ведет
к сбалансированному полиморфизму. Этот тип полиморфизма
важен для эволюции отчасти потому, что он позволяет сохра-
нять известный уровень изменчивости в популяции. Это озна-
чает, что популяция может быть способна к очень быстрой ре-
акции на изменение среды и таким образом может избежать
вымирания. Предположим, например, что у какого-то вида са-
ранчи, обитающего в полузасушливых условиях, адаптивные
ценности фенотипов для некоторого локуса будут следующими:
В В — 0,50; ВЬ—1,00, bb— 0,40. Гетерозиготы имеют физиоло-
гическое преимущество перед обеими формами гомозигот, а
нимфы ВВ несколько более устойчивы к обезвоживанию, чем
нимфы bb. В такой популяции частота генов достигла бы устой-
чивого равновесия при
В = —— = °’6°п аг = 0,545.
sb + SB 0,60 + 0,50
(Нужно учесть, что при адаптивной ценности особей bb, равной
0,40, коэффициент отбора равен 0,60, так как адаптивная цен-
ность равна 1—s.) Поддержание равновесия при В = 0,545 и
6 = 0,455 для этой ситуации показано в табл. 8.
Предположим, что изменение климата в области, где обитает
наша гипотетическая саранча, внезапно приведет к увеличению
осадков в виде дождя, что будет способствовать росту плесени,
которая смертельна для яиц ВВ и ВЬ, но к которой яйца bb
относительно устойчивы. Следовательно, адаптивная ценность W
равна 0 для особей ВВ, 0 для особей ВЬ и 1 для особей bb. Вы-
живание популяции всецело зависит теперь от яиц bb. Допустим,
Теория популяционной генетики | 123
что до повышения влажности существовала полиморфная попу-
ляция из 100 пар взрослых особей и что самка откладывает
в среднем 100 яиц; в таком случае воздействию плесени под-
верглось бы 10 000 яиц. Из них 2070 (0,207 • 10 000) яиц были бы
резистентного типа, что, по-видимому, дало бы популяции ре-
альные шансы на выживание.
Между тем, если бы до изменения климата популяция была
мономорфной (все особи имели бы генотип ВВ, например вслед-
ствие сильного отбора, направленного против особей ВЬ и bb),
исход почти наверное был бы иным. Если принять, что частота
мутаций В->Ь равна 10-5, то лишь 1 из 10 млрд, яиц будет
иметь генотип, обеспечивающий выживание. (Вероятность му-
тантного состояния обоих членов какой-либо пары аллелей у од-
ной особи равна произведению соответствующих вероятностей
для каждого члена пары: 10-5 • 10-5 = Ю-10 = 1/10 000 000 000.)
Преимущество сбалансированного полиморфизма для популя-
ции очевидно.
Сколь бы упрощенным и утрированным ни был этот пример,
он показывает, каким образом сбалансированный полиморфизм
может быть полезным для популяции: он предотвращает потерю
ТАБЛИЦА 8
Сбалансированный полиморфизм
Поко- ление До или после отбора Частота генотипов Частота генов
ВВ вь ьь В ъ
0 До отбора * 0,297 0,496 0,207 1,000 0,545 0,455
После отбора ** 0,148 0,496 0,083 0,727 0,545 *** 0,455
1 До отбопа 0,297 0,496 0,207 1,000 ,0,545 0,455
После отбора 0,148 0,496 0,083 0,727 0,545 0,455
В последующих поколениях эти соотношения будут повторяться до тех пор, пока
остаются в силе исходные предположения.
* При вычислении предполагается случайное комбинирование при спаривании; тогда
мы получаем следующие частоты генотипов: В2 = 0,5452 = р,297; 2236 = 2 (0,545) (0,455) = 0,496;
62 = 0,4552 = 0,207.
** Давление отбора учитывается путем умножения частоты каждого генотипа на его
адаптивную ценность: 0,297 (0,50) = 0,148; 0,496 (1,00) = 0,ч96; 0,207 (0,40) = 0,083.
*** Частота гена В определяется как (D + y2H}IN. Половина от 0,496 равна 0,248; таким
образом (0,1484-0,248)/0,727 = 0,545. Частота гена Ь равна 1—0,545 = 0.455,
124 | Глава 6
изменчивости в результате фиксации локуса. Другим механиз-
мом, замедляющим потерю изменчивости в том или ином ло-
кусе, служит доминантность, благодаря которой нежелательные
по своим проявлениям мутации могут быть «укрыты» в гетеро-
зиготах. Однако с другой точки зрения можно было бы оказать,
что оба эти явления помогают сохранению нежелательных ге-
нов в популяции. Чтобы понять эту точку зрения, нужно рас-
смотреть вопрос о «генетическом грузе».
ГЕНЕТИЧЕСКИЙ ГРУЗ
Если один генотип в популяции в данное время обладает
преимуществом перед всеми остальными, то можно принять, что
популяция, состоящая только из особей с этим генотипом, обла-
дала бы наивысшей возможной приспособленностью. Таким об-
разом, если этому наиболее приспособленному генотипу мы при-
пишем адаптивную ценность W = 1,00, то приспособленность по-
пуляции W также будет равна 1,00, поскольку U7=2 Wigt
(где Wi — адаптивная ценность f-ro генотипа, gi — частота f-ro
генотипа, i — любое число от 1 до п и п — число различных ге-
нотипов). Согласно этой модели любая популяция, состоящая
из смеси генотипов, будет менее приспособленной, чем идеаль-
ная мономорфная популяция. Величина, на которую приспособ-
ленность данной популяции отличается от этого идеала, представ-
ляет собой генетический груз L, который можно приближенно 1
рассматривать как дополнение W до единицы, т. е. Л=1— W.
Исходя из этой модели можно представить себе два крайних
случая. В одном случае идеальный генотип гомозиготен по всем
или почти по всем своим локусам. В другом крайнем случае все
локусы могут быть сверхдоминантными в отношении приспособ-
ленности, и идеальным генотипом будет тогда множественная
гетерозигота. Оживленную дискуссию вызывал вопрос о том, ко-
торая из этих двух крайних возможностей ближе к действитель-
ности. Данные о реакции популяций на инбридинг пока еще не
позволяют решить этот вопрос. Имеются веские данные, говоря-
щие о большом значении сверхдоминантности для приспособ-
ленности у животных; в связи с этим было высказано предпо-
_ 1 В действительности L определяется как —In W. Таким образом,
№=e"L, так что L — это среднее число потенциальных смертей на 1 особь,
a W — вероятность «генетического выживания», т. е. вероятность того, что
особь не погибнет из-за свойств своего генома. (Разумеется, особь, содер-
жащая в геноме эквивалент пяти летальных генов, погибнет лишь однажды,
но при исчислении среднего ее гибель учитывается как 5 смертей.) Читатель,
знакомый со статистикой, узнает в выражении первый член распре-
деления Пуассона.
Теория популяционной генетики | 125
ложение, что у многих организмов крайние фенотипические
уклонения могут быть результатом выщепления множественных
гомозигот в популяциях, в которых нормальными генотипами
являются множественные гетерозиготы. В то же время многие
растения обладают генетическими системами, обеспечивающими,
видимо, почти полную гомозиготность, однако они как будто бы
нисколько не страдают от этого.
Как говорилось в предыдущем разделе, сбалансированный
полиморфизм позволяет популяции накапливать изменчивость
для дальнейшей эволюции. Таким образом, межпопуляционный
отбор, возможно, благоприятствовал популяциям со сбаланси-
рованными полиморфными системами во многих локусах, и в та-
ком случае сопутствующее увеличение генетического груза мо-
жно было бы рассматривать как расплату за повышенную
эволюционную гибкость. Нужно отметить, однако, что генети-
ческий груз вычисляется для определенного момента времени и
что вопрос о будущих эволюционных потенциях не имеет отно-
шения к вопросу о том, какого рода генотип является идеаль-
ным в указанном выше смысле.
Серьезную проблему представляют два из тех предположе-
ний, с которыми в значительной мере связана дискуссия о гене-
тическом грузе. Первое предположение сводится к тому, что
идеальная популяция должна быть мономорфной (т. е. пред-
ставленной только одним генотипом). Оно было бы неверным,
если бы в занимаемой популяцией области существовали раз-
личные ниши, причем разные генотипы внутри популяции имели
бы высокую адаптивную ценность в одной (или одних) из этих
ниш и низкую — в других. Оно не оправдывалось бы и в случае
существования какого-то экологического синергизма, при кото-
ром наличие нескольких различных генотипов повышает адап-
тивную ценность каждого из них.
Второе исходное предположение состоит в том, что W слу-
жит мерой какой-то биологически важной величины и что могут
быть основания считать одну популяцию «лучше приспособлен-
ной» по сравнению с другой. Однако можно с уверенностью ут-
верждать, что любая поддерживающая себя популяция приспо-
соблена ничуть не хуже всякой другой и что любой стандарт
для сравнения приспособленности является произвольным. На-
пример существует одна популяция Drosophila tropicalis, в ко-
торой все выживающие особи гетерозиготны по инверсии, а обе
гомозиготные формы нежизнеспособны. Несмотря на такое
огромное возрастание «груза», связанное с расщеплением, труд-
но найти какое-либо основание считать эту популяцию «плохо
приспособленной».
126 | Глава 6
В заключение следует отметить, что характер генетической
груза у человека имеет некоторое практическое значение. HaN
необходимо знать, какая доля этого груза связана с расщепле
нием (т. е. с выщеплением нежелательных гомозигот по сбалан
сированным полиморфным локусам) и какая доля обусловлена
мутированием (т. е. возникновением вредных мутаций в локу-
сах, уже гомозиготных по «благоприятным» аллелям). Эти све-
дения помогли бы нам оценить отдаленные последствия мута-
ций, вызванных ионизирующим излучением. Хотя эта проблема
чрезвычайно сложна, можно думать, что если генетический груз
в основном мутационного происхождения, то добавочные мута-
ции увеличат его в относительно большей степени, чем если бы
он был связан главным образом с расщеплением. (Упрощенное
объяснение этого состоит в том, что в локусах со сверхдоми-
нантностью в отношении приспособленности новые мутации да-
вали бы положительный эффект при появлении их у гетерозигот,
тогда как в локусах, не обнаруживающих такой сверхдоминант-
ности, все они были бы «плохими».)
СЛУЧАЙ МЕНЬШЕЙ ПРИСПОСОБЛЕННОСТИ ГЕТЕРОЗИГОТ ПО СРАВНЕНИЮ
С ГОМОЗИГОТАМИ
Когда отбор направлен против гетерозигот, существует точка
неустойчивого равновесия при p = ^ = 0,50. Любое отклонение от
этой (величины ведет к элиминации аллеля, частота которого
снизилась в результате этого отклонения. Как видно из цифр,
приведенных при рассмотрении отрицательного отбора рецессив-
ных гомозигот (стр. 119), чем ниже частота аллеля, тем боль-
шая доля его находится в гетерозиготах. Поэтому, когда отбор
направлен против гетерозигот, более редкий аллель находится
в невыгодном положении, которое ухудшается по мере умень-
шения его частоты. Это значит, что в случае отклонения от точ-
ки равновесия не возникает никакой тенденции возвращения
к ней; отклонение будет возрастать, пока тот или другой аллель
не окажется закрепленным,
МИГРАЦИЯ И СТРУКТУРА ПОПУЛЯЦИЙ
Когда популяция не полностью изолирована от других попу-
ляций, частота генов в ней подвержена изменению вследствие
включения групп иммигрантов, отличающихся по частотам
генов от принимающей их популяции. Для любого локуса из-
менение частоты генов за одно поколение определяется по фор-
муле
М --= - tn (q --qm) ~ — mq-\- mqm,
Теория популяционной генетики | 127
где tn — число иммигрирующих особей, деленное на величину
принимающей их популяции; q — частота генов в этой популя-
ции и qm— частота генов в иммигрирующей группе. Преобразуя
правую сторону уравнения путем прибавления и вычитания ве-
личины tnqmq, получаем
— mq + mqmq + mqm — rnqmq — qm) + mqm (1 — q) -
- mq(pm) + rnp(qm).
Это окончательное уравнение имеет ту же форму, что и выраже-
ние для Л7, полученное при вычислении последствий прямого и
обратного мутирования (стр. 112); и действительно, эти две си-
туации аналогичны.
На фиг. 21 показано, как изменяется распределение частот
генов при изменении пг или Af, когда частота генов у иммигран-
тов равна 0,60. Кривые вроде представленных на фиг. 21 изве-
стны как стацицнарные распределения частот. Каждая кривая
здесь представляет собой функцию плотности вероятностей сле-
дующего вида:
где С — нормирующий множитель, а значения остальных сим-
волов те же, что и выше. Подобная функция может представ-
Фиг. 21. Распределение частот гена между различными подразделениями
популяции при частоте генов у иммигрантов р = q — 0,50.
Объяснение графика этого типа см. в тексте [5].
128 | Глава в
лять характер распределения вероятностей возможных событий.
Площадь под каждой кривой равна 1, а площадь между любым
участком кривой и соответствующим участком оси абсцисс
(оси q) соответствует вероятности того, что частота гена будет
лежать в пределах этого участка оси q. Так, например, из
фиг. 21 сразу видно, что при данных условиях вероятность того,
что q будет лежать между 0,4 и 0,6, при tn = xUN гораздо мень-
ше, чем при Стационарные распределения частот —
очень удобный способ иллюстрировать влияние различных эво-
люционных факторов на популяции разного типа, широко исполь-
зуемый для этой цели. Читателей, желающих получить бо-
лее полные сведения о функциях плотности вероятностей и о
других проблемах, связанных с математической теорией вероят-
ностей, мы отсылаем к любому вводному курсу теории вероят-
ностей.
Стационарными распределениями частот можно представ-
лять распределение частоты изучаемого гена в большом числе
популяций при одних и тех же условиях, распределение частот
генов во многих локусах при одинаковом давлении отбора в од-
ной популяции или распределение вероятностей частоты данного
гена по отдельным поколениям. Например, кривая Nm = 4 (т. е.
кривая m = ^IN) на фиг. 21 может быть истолкована следую-
щими способами. Можно оказать, что при данных постоянных
условиях частоты генов в большом числе популяций (или локу-
сов в одной популяции) имели бы тенденцию довольно плотно
группироваться около величины 0,50 или что внутри популяции
для любого из локусов, находящихся в одних и тех же условиях,
вероятность частоты гена в пределах от 0,35 до 0,65 высока. На-
конец, эта кривая выражает вероятность того, что частота гена
в одном определенном локусе будет иметь в том или ином поко-
лении данную величину. Она показывает, например, что вероят-
ность величины 7=1 для любого поколения исчезающе мала.
Тип и объем перемещающейся генетической информации
(«потока генов») внутри популяции или между популяциями
является важным фактором в определении их эволюции. Оче-
видно, что ситуация, в которой между членами группы полуизо-
лированных субпопуляций происходит случайный обмен генети-
ческой информацией («островная модель»), совершенно отлична
от такой ситуации, в которой имеется односторонний поток ге-
нов вдоль линейной последовательности субпопуляций («поточ-
ная модель»). В свою очередь обе они отличаются от третьей
ситуации, когда группа организмов непрерывно распределена на
большой площади. Хотя в последнем случае может и не быть
полудискретных группировок особей, вероятность скрещивания
между двумя отдаленными друг от друга особями может быть
Теория популяционной генетики | 129
очень мала из-за одной их взаимной отдаленности. Последствия
такой изоляции, обусловленной расстоянием, были математи-
чески исследованы Райтом, который показал, что степень ло-
кальной дифференциации внутри популяции зависит главным
образом от величины составляющих ее панмиктических скопле-
ний, лежащих по соседству друг с другом. Когда популяция раз-
делена на полуизолированные субпопуляции и когда популяции
в целом свойственна некоторая степень инбридинга, общим ре-
зультатом будет снижение частоты гетерозигот. Один важный
аспект этого изменения в частоте генотипов иллюстрируется на-
блюдаемыми в таких популяциях последствиями отбора, напра-
вленного против гомозиготных рецессивных форм. Такой отбор
1. оказался бы гораздо более эффективным в подразделенной по-
пуляции и в популяции с инбридингом, так как при пониженном
- числе гетерозигот было бы «укрыто» меньше рецессивных генов.
СОВМЕСТНОЕ ДЕЙСТВИЙ ФАКТОРОВ ЭВОЛЮЦИИ
До сих пор мы рассматривали действие на изолированные
локусы отдельных факторов эволюции. Однако по существу во
всех изучаемых случаях частота генов в том или ином локусе
подвергается совместному воздействию двух или нескольких
факторов. Кроме того, генные частоты в различных локусах на-
ходятся в зависимости друг от друга и частота в одном локусе
может оказывать глубокое влияние на частоту в другом локусе.
Чтобы понять это, достаточно вспомнить о явлении сцепления.
В математическом описании последствий различного рода
взаимодействий в менделевских популяциях достигнуты извест-
ные успехи. Еще неясно, станет ли когда-нибудь возможным
вполне удовлетворительное математическое описание одновре-
менного действия всех эволюционных сил (варьирующих в за-
висимости от внешних условий) на целостный генотип. Развитие
вычислительной техники позволяет надеяться на это, однако
ввиду чрезвычайной сложности ситуации, подлежащей анализу,
здесь понадобилась бы машина также невообразимой слож-
ности. В настоящее время мы вынуждены довольствоваться ком-
бинированием моделей, являющихся лишь грубым приближе-
нием к оригиналу. Отчасти нас утешает то, что эти простые
модели, по-видимому, приближаются к некоторым ситуациям,
< наблюдаемым в природе, и оказались весьма полезными при их
описании.
Для краткого ознакомления с более сложными ситуациями
рассмотрим фиг. 22 и 23. Фиг. 22 иллюстрирует действие раз-
личных давлений отбора в популяциях разной величины. На
графиках Л, Б и В частоты прямых и обратных мутаций
9 Зак. 152
130 | Глава 6
приняты постоянными и равными друг другу (г/ = у).На фиг. 22, А
величина популяции ЛА=1/40г>; на фиг. 22, Б N =10/40у; на
фиг. 22, В М= 100/40V. На всех трех графиках сплошной линией
представлен случай минимального отбора (s =—у/100), преры-
вистой линией — случай в 10 раз более сильного отбора (не
представленный на фиг. 22, А, так как здесь он практически не-
отличим от предыдущего) и пунктирной линией—случай в 100 раз
более сильного отбора. Обратите внимание, что в очень ма-
лых популяциях (фиг. 22, А) отбор лишь незначительно влияет
на симметричность распределения частот генов: вероятность по-
тери или фиксации гена остается большой. С увеличением попу-
ляции (фиг. 22, Б и В) эффекты отбора становятся гораздо бо-
лее выраженными. Фиг. 22, Г иллюстрирует распределение ча-
стот в случае, когда отбор благоприятствует гетерозиготам и
различия в селективной ценности между двумя формами гомо-
Фиг. 22. Распределение частот генов при различном давлении отббра и
в популяциях различной величины. i
Подробности см. в тексте [7].
Фиг. 23. Распределение частот генов в популяциях разной величины и раз-
личной степени подразделенности при различных давлениях отбора и мута-
ций.
Подробности см. в тексте [5].
9*
132 | Глава б
зигот отсутствуют. Здесь снова w = u, 7V= l/40z? и s=100a. (На
этих графиках и на фиг. 23 коэффициент отбора берется не в
том виде, как мы его определили ранее, а принимает как поло-
жительные, так и отрицательные значения. Таким образом, ра-
венство s=100y служит показателем преимущества гетерозигот,
тогда как равенство s = —а/100 было бы показателем меньшей
приспособленности данного аллеля.)
На фиг. 23 показано распределение частот генов в популя-
циях разных размеров и различной степени «подразделенности»
при разных величинах давления отбора и мутаций. На фиг. 23, Л
представлена небольшая популяция, в которой мутационный
процесс и отбор практически отсутствуют. Большинство аллелей
фиксируется или утрачивается по закону случая. Популяция
промежуточной величины при противоположных друг другу на-
правлениях мутирования и отбора представлена на фиг. 23, Б.
Здесь имеются случайные колебания около модальных величин,
установившихся в результате противоположных давлений. Слу-
чай большой популяции с равновесными частотами генов, опре-
деляемыми величинами противоположно направленных давлений
мутирования и отбора, представлен на фиг. 23, В. Наконец,
фиг. 23, Г показывает частоты генов в отдельных частях боль-
шой популяции, колеблющиеся около модальных величин, кото-
рые установились под действием противоположно направленных
сил миграции и отбора.
АДАПТАЦИЯ И ГЕННЫЕ СОЧЕТАНИЯ
Как указал Сьюэлл Райт, у организмов, размножающихся
половым путем, возможно огромное число различных комбина-
ций генов. У вида, имеющего всего лишь 1000 локусов, каждый
из которых представлен серией из 10 аллелей, путем рекомби-
нации .могло бы создаться Ю1000 различных генотипов (это число
неизмеримо больше числа электронов во всей вселенной). Хотя
многие из теоретически возможных комбинаций были бы не-
жизнеспособны или давали бы идентичные генотипы, ясно, что
такой вид обладал бы очень большим объемом возможной гено-
типической изменчивости. Среди этого необозримого множества
возможных комбинаций должно быть большое число высоко-
адаптивных сочетаний, а также много не столь благоприятных
или даже летальных сочетаний. Райт представлял это множе-
ство в виде двумерной контурной карты с «адаптивными пика-
ми» и «неадаптивными впадинами» и говорил что «проблема
эволюции... это проблема механизма, с помощью которого вид
мог бы непрерывно находить в таком поле путь от менее высо-
ких пиков к более высоким». Популяцию комаров, у которых
Теория популяционной генетики | 133
отбор выработал способность избегать инсектицидов — не са-
диться на отравленные поверхности, — можно было бы рассмат-
ривать как занимающую адаптивный пик. Переходом к более
высокому адаптивному пику могла быть выработка какого-либо
механизма физиологической устойчивости (например, механизма,
обеспечивающего лучший доступ в жилища). Для приобретения
более эффективной физиологической устойчивости потребова-
лась бы значительная реорганизация генотипа; эта реорганиза-
ция могла бы привести к утрате устойчивости, обусловленной
поведением. В таком случае, для того чтобы достигнуть более
высокого пика, виду пришлось бы пересечь неадаптивную впа-
дину (где устойчивости нет или почти нет).
На фиг. 24,(7 — е показано, что могло бы произойти с попу-
ляциями определенных типов в адаптивном поле при различных
условиях. Это поле представлено в виде топографической карты,
на которой изолинии соответствуют различным уровням адапта-
ции. Жирной пунктирной линией указано исходное положение
популяции, а стрелками — направление последующего измене-
ния. На фиг. 24, а можно видеть результат повышения частоты
мутаций или снижения давления отбора — общее увеличение из-
менчивости и снижение средней приспособленности популяции.
Фиг. 24. Поле комбинаций генов, занимаемое популяциями различного типа
при различных условиях.
Объяснение в тексте |4].
134 | Глава 6
При достаточном расширении диапазона изменчивости часть по-
пуляции может запять нижние склоны другого адаптивного
пика, который выше первоначального; если это случится, вся
популяция начнет перемещаться и займет новый пик.. Послед-
ствия усиления отбора и снижения мутационного давления по-
казаны на фиг. 24, б. Здесь средний уровень адаптации возра-
стает за счет уменьшения эволюционной пластичности. Шансы
на захват соседнего пика понизились.
Фиг. 24,в иллюстрирует последствия глубоких изменений во
внешних условиях (адаптивные пики превращаются во впадины,
и наоборот). Результат зависит здесь от интенсивности отбора и
от скорости изменения среды. Вид, занимающий небольшое поле,
при сильном давлении отбора может оказаться недостаточно
изменчивым, для того чтобы перейти на вновь образующиеся
пики; он может остаться, таким образом, во впадине и вымереть.
В условиях менее интенсивного отбора популяция просто будет
перемещаться по мере изменения среды. На фиг. 24, г показан
эффект сильного сокращения численности популяции и тесного
инбридинга. В результате случайной фиксации и утраты генов
популяция спускается по извилистому пути со своего адаптив-
ного пика, а инбридинг (повышающий однородность особей)
уменьшает величину адаптивного поля, занимаемого популяцией.
Этот процесс (если ему ничто не противодействует) ведет в кон-
це концов к вымиранию. На фиг. 24, д показаны следствия про-
межуточного соотношения между величиной популяции и тем-
пом мутирования. Популяция сходит с пика и начинает блу-
ждать, оставаясь, однако, поблизости от него. Это ситуация
проб и ошибок, которая может вести к захвату все более высо-
ких пиков, но при указанных условиях этот процесс протекал
бы крайне медленно.
На фиг. 24, е вид с обширным ареалом подразделен на мно-
гочисленные полуизолированные популяции. Участок поля, заня-
тый каждой из субпопуляций, быстро перемещается в основном
неадаптивным образом и гораздо быстрее, чем в предыдущем
случае (так как это перемещение определяется не частотой му-
таций, а интенсивностью межпопуляционной миграции). При
быстром перемещении в окрестностях одного пика какая-нибудь
субпопуляция попадет на нижние склоны другого, более высо-
кого пика и взберется на него. Тогда эта субпопуляция увели-
чит свою численность; в результате миграции особей ее гены
будут вливаться в другие субпопуляции, и весь вид переместится
в зону влияния нового пика. Эта ситуация, выдвигающая на пер-
вый план межгрупповой отбор, позволяет испытывать новце
комбинации генов с меньшим риском для вида, чем при наличии
одного лишь внутригруппового отбора. Такая подразделенная
Теория популяционной генетики | 135
популяция создает поэтому наилучшие возможности для эволю-
ционного изменения, связанного со сравнительно небольшим
риском.
ВЫВОДЫ
В этой главе была сделана попытка дать краткое вводное
описание эволюционных процессов с точки зрения теории попу-
ляционной генетики. Одной из фундаментальных концепций био-
логии является закон Харди — Вейнберга, согласно которому в
идеальной популяции при отсутствии эволюционных сил частоты
аллелей в аутосомных локусах не изменяются и количественные
отношения генотипов за одно поколение достигают равновесия.
Типы возможных отклонений от этого равновесия в результате
мутирования, миграции, дрейфа и отбора описаны в математи-
ческой форме. Влияние этих факторов зависит не только от вза-
имодействий между ними, но также от структуры популяции и
от ее обратных воздействий на сами эти факторы. Были сде-
ланы попытки теоретически рассмотреть возможные реакции
различных популяций на разнообразные сочетания факторов.
Рассмотрение популяций как изменяющихся и взаимодействую-
щих систем, характеризуемых частотами генов, дало эволюцио-
нисту средства (хотя и весьма грубые), позволяющие анали-
зировать фактические данные с известной строгостью и точ-
ностью. Популяционно-генетическая теория чрезвычайно полезна
при описании того, что может происходить в природных попу-
ляциях, и при истолковании имеющихся данных о таких попу-
ляциях.
ЛИТЕРАТУРА
1. Grow J. F., Some possibilities for measuring selection intensities in man.
In J. N. Spuhler (ed.), Natural Selection in Man, Wayne State Univ. Press,
Detroit, 1958.
2. L i С. C., Population Genetics, Univ, of Chicago Press, Chicago, 1955.
3. Population Genetics: The Nature and Causes of Genetic Variability in Popu-
lations. Cold Spring Harbor Symposia on Quantitative Biology, vol. 20, 1955.
4. Wright S., The roles of mutation, inbreeding, crossbreeding and selection
in evolution, Proc. Sixth Intern. Congr. Genet., 1, 356—366 (1932).
5. Wright S., Systems of Mating and Other Papers, Iowa State College Press,
Ames, Iowa, 1958 (в книгу включена статья, напечатанная в журнале Ge-
netics, 16, 97—159, 1931).
6. Wright S., The New Systematics, Oxford University Press, 1940,
7. WrightS., Proc. Nat. Acad. Sci., 23 (1937),
Глава 7
ИЗМЕНЕНИЯ В ПОПУЛЯЦИЯХ
Теперь, когда мы рассмотрели некоторые теоретические ас-
пекты популяционной генетики, уместно поставить вопрос, всег-
да ли при исследовании эволюционных механизмов приходится
полагаться на косвенные заключения или возможно прямое
изучение. В последние годы был исследован ряд примеров эво-
люции в природных популяциях, и некоторые из них будут рас-
смотрены в первой части этой главы. Остальная часть главы
будет посвящена более общему обсуждению некоторых сторон
эволюционных изменений в популяциях.
ПРИМЕРЫ ИЗ ПРИРОДЫ
ДИФФЕРЕНЦИАЛЬНАЯ СМЕРТНОСТЬ У ВОРОБЬЕВ
Предпринимались многочисленные попытки продемонстриро-
вать действие естественного отбора путем сравнения признаков
выживающих и невыживающих особей. В качестве примера опи-
шем кратко одну из самых ранних работ такого рода — работу
Бампуса на воробьях. Зимой 1898 г., после сильного дождя и
снегопада, X. К. Бампус принес в лабораторию (Университет
Брауна) 136 оглушенных домовых воробьев. Из них 72 выжили
и 64 погибли. Бампус измерил у всех особей общую длину тела,
размах крыльев, вес тела, длину клюва и головы, длину плече-
вой кости, бедра и тибиотарзуса, ширину черепа и длину киля.
Его измерения показали, что у выживших птиц все эти различ-
ные признаки в большей степени приближаются к средним ве-
личинам, чем у погибших.
Эта повышенная смертность воробьев с крайними морфоме-
трическими показателями служит примером стабилизирующего
отбора. Во многих других исследованиях также было подтвер-
ждено распространенное представление о том, что отбор часто
приводит к элиминации отклоняющихся особей. Как уже упоми-
налось (гл. 6), это только один аспект естественного отбора.
В дальнейших примерах мы нередко будем встречаться со слож-
ными взаимодействиями так называемых основных типов отбора.
Следует ожидать, что стабилизирующий отбор в форме элими-
нации особей с крайними отклонениями действует, по существу,
во всех естественных популяциях.
Изменения в популяциях [ 137
ИНДУСТРИАЛЬНЫЙ МЕЛАНИЗМ
За последние 120 лет в некоторых областях северной Европы
и Северной Америки у многих бабочек, обладающих защитной
окраской, появились темные формы. Во многих из этих обла-
стей такие меланические формы стали преобладать, полностью
или частично вытеснив «типичные» формы, имеющие защитную
окраску. Эти изменения отмечались главным образом в районах
с сильно развитой промышленностью и были особенно ярко вы-
ражены в Англии, где они подвергались интенсивному изучению.
По произведенным оценкам, в районе Манчестера в 1848 г. тем-
ная форма бабочки Biston betularia составляла не более 1% по-
пуляции, а в 1898 г. в том же районе ее относительная числен-
ность уже превышала 99%. В большинстве известных случаев
меланизм вызывается одним доминантным геном.
Для объяснения явления индустриального меланизма была
выдвинута следующая гипотеза. Распространение меланических
форм, по-видимому, тесно связано с загрязнением лесов копотью
Фиг. 25. Две особи Biston betularia — типичная и меланическая— на не-
загрязненном стволе, покрытом лишайниками [15].
138 [ Глава 7
в промышленных районах. В незагрязненных областях темные
формы, хорошо заметные, когда они сидят на стволах деревьев,
покрытых лишайниками (фиг. 25), очевидно, истребляются хищ-
никами, находящими добычу с помощью зрения. Возможно, в
данном случае это неблагоприятное обстоятельство превышает
селективное преимущество гусениц меланических особей, кото-
рые, по-видимому, лучше приспособлены в физиологическом отно-
шении. В загрязненных районах возникает противоположная ситу-
ация: типичные формы (светлые, с мелкими пятнышками), почти
невидимые на незакопченной, покрытой лишайниками коре, рез-
ко выделяются на деревьях, покрытых копотью, вызвавшей ги-
бель лишайников (фиг. 26). Таким образом, в загрязненном рай-
оне меланическая окраска оказывается защитной; кроме того,
если темные особи обладают к тому же каким-то физиологиче-
ским преимуществом, то оно приобретает большее значение в
неблагоприятных условиях, связанных с потреблением загряз-
Фиг. 26. Две особи Biston betularia — типичная и меланическая — на стволе
дерева, покрытом копотью [15].
Изменения в популяциях [ 139
Фиг. 27. Распространение Bistort betularia.
Относительная численность типичных и меланических особей представлена белым и черным
секторами соответственно (16].
ненной пищи. Поэтому отбор сильно благоприятствует мелани-
ческим формам в промышленных районах и типичным — в не-
загрязненных местностях. В загрязненных районах направлен-
ный отбор изменял частоту меланических особей, приближая ее
к 100%.
Какие фактические данные подкрепляют эту гипотезу? Одно
из наиболее убедительных свидетельств — сильная корреляция
(как в пространстве, так и во времени) между индустриализа-
цией и появлением меланических форм. Эта корреляция так
четко выражена, что делает почти несомненным наличие извест-
ной связи между обоими явлениями. Состав различных англий-
ских популяций Biston betularia показан на фиг. 27. Популяции
промышленных центральных графств отличаются высоким про-
центом меланпстов, так же как и популяции восточной Англии,
140 | Глава 7
живущие с подветренной стороны от промышленных районов,
где выпадение копоти было наиболее интенсивным.
Один фактор, ответственный за распространение индустри-
ального меланизма, был наиболее изящно продемонстрирован
в наблюдениях и экспериментах Кетлуэлла и Тинбергена, касав-
шихся дифференциального уничтожения бабочек врагами. В не-
загрязненном лесу в Дорсете были выпущены в одинаковом
числе меланические и типичные особи; уничтожение их птицами
наблюдалось и фотографировалось из укрытия. Мухоловки Mus-
cicapa striata, поползни Sitta еигораеа, овсянки Emberiza citri-
nella, малиновки Erithacus rubecula и дрозды Turdus ericetorum
съели 164 меланические и только 26типичных особей (Р<сО,О1)
В загрязненном лесу около Бирмингема были получены проти-
воположные результаты: здесь горихвостками (Phoenicurus
phoenicurus) было съедено всего лишь 15 меланических особей
и 43 типичные особи (Р<^0,01). Эти эксперименты запечатлены
в кинофильмах, в которых очень убедительно показано, как
птицы обследуют стволы деревьев и поедают хорошо видимых
бабочек, не замечая находящихся рядом особей с защитной
окраской.
Гипотезу покровительственной окраски подкрепляют и ре-
зультаты экспериментов с вторичным отловом выпущенных ба-
бочек. В той или иной местности выпускали известное число ме-
ченых особей обоих типов и для каждого типа подсчитывали
процент особей, впоследствии снова пойманных на свет. Про-
цент повторно выловленных особей среди бабочек, выделявших-
ся на окружающем фоне (меланических форм в незагрязненных
районах и типичных — в загрязненных районах), был значи-
тельно ниже, чем среди бабочек с защитной окраской. Напри-
мер, близ Бирмингема (где меланическая форма составляет
85—87% популяции) было помечено и выпущено 154 особи тем-
ной окраски и 73 типичные особи. Впоследствии было вновь пой-
мано 98 меченых бабочек, из них 82 темные (53% от 154 выпу-
щенных) и 16 типичных (25% от64 выпущенных). Если игнориро-
вать ошибку эксперимента и принять адаптивную ценность более
выгодной меланической окраски за 1, то адаптивная ценность
(1—$) типичного фенотипа составит 0,25/0,53 = 0,47. Отсюда
1 Вероятность значительно меньшая чем 1 : 100. Это означает, что от-
клонение от ожидаемой частоты, обусловленное чисто случайными причи-
нами, должно было бы наблюдаться значительно реже, чем в 1 % случаев.
Выражение Р < 0,02 означает, что вероятность меньше чем 2:100. Многие
биологи условно считают «достоверным» любой результат, для которого ве-
роятность чисто случайного осуществления меньше 5%. Элементарное изло-
жение статистических методов в применении к биологии можно найти в
книге Симпсона и сотр. [17].
Изменения в популяциях ।
5 = 0,53 (что случайно совпадает с процентом вновь пойманных
темных особей). Крайне маловероятно, чтобы эти результаты
были обусловлены чистой случайностью (Р<0,01). Аналогичные
эксперименты в незагрязненных районах дали обратные резуль-
таты. Связь между экспериментами с повторным отловом и на-
блюдениями за истреблением выпущенных бабочек птицами та-
кова: эксперименты показывают, что происходит отбор, а на-
блюдения раскрывают природу факторов отбора.
В многочисленных экспериментах была исследована жизне-
способность гусениц меланической формы. Форд, работавший с
бабочкой Cleora repandata, получал выводки от возвратного
скрещивания меланических гетерозигот с типичными рецессив-
ными гомозиготами. Как во всех анализирующих скрещиваниях,
здесь ожидалось отношение 1:1, и при хорошем питании гусе-
ниц в Fi достоверного отклонения от этого отношения не наблю-
далось. Однако при ежедневно повторяющемся голодании гу-
сениц (т. е. при неблагоприятных физиологических условиях)
отношение числа меланических особей к числу типичных состав-
ляло 51:31 (Р<0,02), что существенно отличалось от ожидае-
мой цифры 41,5 для каждого типа. Кетлуэлл подвергал 6 вы-
водков от возвратных скрещиваний воздействию неблагоприятных
условий и насчитал среди выживших особей 108 меланических
и только 65 типичных (Р = 0,01).
Вряд ли можно сомневаться в том, что при известных небла-
гоприятных условиях гусеницы меланических бабочек более
жизнеспособны, однако, как показывают новейшие исследования,
дело обстоит сложнее, чем предполагали раньше. В ряде прове-
денных недавно экспериментов ожидавшейся пониженной жиз-
неспособности типичных особей обнаружено не было. В других
экспериментах выявилась неоднородность различных выводков:
в потомстве от одних скрещиваний процент меланических осо-
бей был, несомненно, повышен, в других скрещиваниях этого не
наблюдалось. Кроме того, при изучении выводков от возвратных
скрещиваний Biston betularia, проведенных в период между
1900 и 1905 гг., не обнаруживалось никакого избытка меланиче-
ских особей — скорее процент их был несколько снижен (хотя
это снижение нельзя считать статистически значимым). Воз-
можно, что в ранний период возникновения индустриального ме-
ланизма меланические гусеницы не обладали физиологическим
преимуществом и что это преимущество (в тех случаях, когда
оно существует в настоящее время) выработалось сравнительно
недавно. Быть может, только после ослабления сурового отбора,
направленного против взрослых меланических особей, числен-
ность таких особей в популяциях достаточно возросла, так что
стала возможной избирательная реорганизация меланического
142 | Глава 7
генотипа, приведшая к появлению физиологического преимуще-
ства. Если отбор генов-модификаторов увеличил доминантность
факторов меланизма (гл. 3), то он мог точно так же усилить
влияние этих факторов на жизнеспособность.
Кетлуэлл пробовал экспериментально выяснить, не садятся
ли меланические особи преимущественно на темные поверхно-
сти, а типичные особи — на светлые поверхности. Он окрашивал
внутренность бочонка так, чтобы в нем чередовались черные и
белые участки, а затем выпускал туда бабочек обоих типов и
наблюдал, какие участки они выберут. 77 бабочек выбрали не-
контрастирующий фон, а 41 — контрастирующий фон. Было сде-
лано -предположение, что такой выбор возможен благодаря спо-
собности бабочки определять степень контраста между чешуй-
ками, окружающими ее глаза, и той поверхностью, на которой
она сидит.
В некоторых районах меланическая форма начинает преобла-
дать, несмотря на то что местность кажется незагрязненной. Это
можно объяснить двумя причинами. Во-первых, загрязненность
часто бывает больше, чем это представляется глазу. Облака
смога нередко относятся ветром на большие расстояния, и ме-
стность, несмотря на ее общий зеленый колорит, в действитель-
ности может быть покрыта значительным слоем сажи и хими-
ческих продуктов. Во-вторых, на сельскую местность влияют и
другие виды человеческой деятельности, и возникающие измене-
ния (например, исчезновение несекомоядных птиц) могут ока-
заться достаточными для сдвига равновесия в пользу меланиче-
ских особей.
Хотя известны и рецессивные меланические формы, распро-
странение индустриального меланизма обусловлено рассеянием
доминантных генов. Возможные причины этого обсуждались в
гл. 3 в связи с вопросом о происхождении доминантности.
Индустриальный меланизм служит примером переходного
полиморфизма. Вспомним, что полиморфизм — это существова-
ние в одном и том же местообитании двух или нескольких раз-
личных форм какого-либо вида в таком количественном соотно-
шении, что даже наиболее редкая из них не могла бы поддер-
живаться мутированием. В случае переходного полиморфизма
одновременно существуют две формы, одна из которых посте^
пенно вытесняет другую. Еще один вероятный случай переход-
ного полиморфизма в связи с меланизмом у бабочек был иссле-
дован в одном старом лесном массиве Шотландии. В этих почти
незагрязненных лесах популяция одного вида бабочек (Cleora
repandata) содержала 10% меланических особей. Типичные осо-
би, неподвижно сидящие на покрытых лишайниками стволах
(светлый фон), были незаметны. На темных стволах незаметны
Изменения в популяциях | 143
были меланические особи, но их защитная окраска была не так
эффективна (для человеческого глаза), как у светлой формы.
Во время полета в темном лесу гораздо заметнее оказывались
светлые формы (за период наблюдения птицами были схвачены
на лету 3 светлые бабочки и ни одной темной). Если бы как
в покое, так и во время полета преимущества были одинако-
выми, процесс фиксации гена происходил бы быстрее, чем в опи-
санной ситуации. Тем не менее фиксация все же произойдет,
если только условия не изменятся.
МИКРОЭВОЛЮЦИЯ БРИТАНСКИХ ЧЕШУЕКРЫЛЫХ
Британские специалисты по бабочкам первыми начали изу-
чать другие типы эволюционных изменений в популяциях чешуе-,
крылых. Фишер, Форд и Шепард провели длительное исследо-
вание частоты генов в одном локусе у бабочки-медведицы Ра-
naxia dominula. В единственной колонии (близ Оксфорда), в ко-
торой данный ген был обнаружен, начиная с 1939 г. ежегодно
производили оценку его частоты; приблизительную величину по-
пуляции определяли каждый год, начиная с 1941 г. В этой ко-
лонии встречаются 3 фенотипические формы: dominula (гомо-
зиготная по обычному аллелю), medionigra (гетерозиготная) и
bimacula (гомозиготная по редкому аллелю). Частота редкого
гена составляла 0,012 до 1928 г., 0,092 — в 1939 г. и 0,111 — в
1940 г. После 1940 г. она быстро упала, около 1947 г. выровня-
лась и с тех пор колеблется между 0,011 и 0,037. Кажется поч-
ти несомненным, что эти изменения обусловлены главным об-
разом колеблющимся давлением отбора. Известно, что данный
ген, помимо прочего, влияет на рисунок, поведение при спари-
вании, плодовитость самцов и жизнеспособность гусениц. Су-
ществовали некоторые разногласия относительно возможной
роли дрейфа генов в этом случае; степень взаимодействия слу-
чайных изменений с отбором не была определена.
Даудсвелл, Фишер, Форд и их сотрудники провели длитель-
ные исследования, в которых изучали распределение по числу
пятен на нижней поверхности задних крыльев Maniola jurtina
(Lepidoptera, Satyrinae). В большей части южной Англии это
распределение поразительно однотипно, несмотря на огром-
ное разнообразие внешних условий. Однако у самок обнаружи-
лось большое различие по этому признаку между популяциями
Девона и Корнуэлла. К тому же переход оказался чрезвычайно
резким; действительно, в 1956 г. границей между популяциями
с различным распределением по числу пятен была, как выясни-
лось, живая изгородь, которая не служила препятствием для
передвижения отдельных особей. Не было никаких признаков
144 | Глава 7
наличия градиента между двумя типами; пожалуй, различие
было наибольшим в точке их соприкосновения. В 1957 г. гра-
ница между двумя формами оказалась перемещенной на 5 км
к востоку по сравнению с 1956 г., и переход стал более посте-
пенным. Сама граница представляла собой полосу шириной око-
ло 135 м, занятую промежуточной популяцией. Распределение
по числу пятен было также основательно изучено у бабочек не-
большого архипелага Силли, у западной оконечности полуостро-
ва Корнуэлл. Были найдены многочисленные различия между
популяциями отдельных островов, но в общем каждый остров
имел характерную популяцию, которая не менялась из года
в год.
Число пятен, несомненно, находится под сильным генетиче-
ским контролем, хотя кажется очевидным, что селективная цен-
ность соответствующих генов определяется не малозаметными
изменениями в пятнах, а другими эффектами. Наличие резкой
и подвижной границы между девонским и корнуэллским типами
трудно объяснить чем-либо иным, кроме перехода от одного
в высокой степени интегрированного генного комплекса к дру-
гому. Оба типа кажутся весьма приспособленными, так как они
распространены на значительном расстоянии в ту и другую сто-
рону от границы (особенно к востоку), и на них сравнительно
мало сказывается неоднородность занимаемых местообитаний.
Вряд ли внешние факторы, определяющие различие в коэффи-
циентах отбора, изменялись здесь так же внезапно, как распре-
деление по числу пятен; причина резкой границы почти навер-
ное связана с генетическими особенностями Maniola.
ПОЛИМОРФНЫЕ НАЗЕМНЫЕ УЛИТКИ
Была детально изучена микроэволюция полиморфных на-
земных улиток рода Сераеа, главным образом очень изменчи-
вого вида С. nemoralis. Раковина этой улитки может быть жел-
той, коричневой или любого оттенка от бледного желтовато-ко-
ричневого до красного, включая розовый и оранжевый. Губа
раковины может быть черной или темно-коричневой (обычный
тип), а также розовой или белой (редкий тип); иногда раковину
украшают до пяти черных или темно-коричневых (редко про-
зрачных) продольных полос. Генетическая основа многих йз
этих признаков довольно хорошо изучена, а ископаемые наход-
ки показывают, что этот полиморфизм существовал со времени,
предшествовавшего неолиту.
Эти улитки были основательно исследованы в Европе, где
частота различных форм сильно варьирует от одной колонии
к другой. Вопрос о роли отбора и дрейфа генов в возникновении
Изменения в популяциях | 145
этих различий между колониями оставался спорным. Ламотт
(Франция) вначале утверждал, что наблюдаемое многообразие
объясняется дрейфом генов. Однако Кейн и Шеппард показали,
что по крайней мере в некоторых колониях в Англии действо-
вали факторы отбора. Они нашли, что близ Оксфорда частоты
различных типов раковины коррелируют с характером местооби-
тания улиток (фиг. 28). Материал собирали в местообитаниях
шести основных типов: в низинных буковых лесах, дубовых ле-
сах, смешанных лиственных лесах, на живых изгородях, на от-
крытых участках с высокой грубой травой и на открытых участ-
ках с очень коротким дерном. Анализ встречаемости различных
Фиг. 28. Карта района Оксфорда, на которой показана корреляция между
типом раковины у Сераеа и условиями среды.
На каждой гистограмме левым столбиком представлен процент желтых раковин, а правым
столбиком— процент раковин с хорошо заметными полосами. Местности, занятые лесом,
обозначены па карте точками; вне лесов все колонии обитают в живых изгородях или жест-
кой травянистой растительности (18].
10 Зак. 152
146 | Глава 7
типов окраски и рисунка раковины показал наличие высокой
корреляции с цветом фона и степенью его однородности. На-
пример, процент раковин без полос в пяти местностях с наи-
более однородным фоном и пяти — с наименее однородным
фонохм варьировал следующим образом (в каждом ряду одно-
родность убывает слева направо):
Наиболее однородные фоны 100 100 100 93 79
Наименее однородные фоны 39 35 34 25 4
Процент желтых раковин в пяти самых зеленых и пяти наи-
менее зеленых участках составлял (в каждом ряду интенсив-
ность зеленого цвета убывает слева направо):
Наиболее зеленые участки 76 64 45 43 41
Наименее зеленые участки 17 8 4 0 0
Эта связь между частотой различных типов раковин и харак-
тером местообитания наводила на мысль, что фактором отбора,
вызывающим дифференциацию колоний, могли быть враги, по-
едающие наиболее хорошо заметных улиток. Одним таким вра-
гом является певчий дрозд (Turdus ericetorum). Летом 1951 г.
Кейн и Шеппард изучали колонию улиток на небольшом болоте
на склоне холма в Уитхэмском лесу близ Оксфорда. Дрозды
хватали улиток из колонии, разбивали раковины о камни у
протекавшей поблизости реки и съедали мягкие части. Таким
образом, можно было получить выборку из уничтожаемой части
популяции, собирая разбитые раковины около используемых
дроздами камней, и выборку из всей популяции, собирая улиток
на болоте. Из 560 особей, взятых с болота, 296 (52,8%) не имели
полос на раковине, тогда как из 863 раковин, собранных вокруг
камней, без полос были только 377 (43,7%). Это статистически
значимое различие (Р<0,02) показывает, что в этой колонии
особи без полос имели меньше шансов быть съеденными, чем
«полосатые» особи.
Сходные методы Шеппард использовал при изучении двух
других колоний в 1950 г. Он нашел, что весной процент съеден-
ных улиток с желтыми раковинами постепенно снижается, при-
мерно с одинаковой скоростью в обеих местностях. Не было
никаких указаний на то, что это обусловлено изменением мест
охоты дроздов или изменением процента желтых улиток во всей
популяции. По-видимому, селективная ценность желтого фено-
типа по крайней мере отчасти зависела от фона, на котором
находились улитки. Ранней весной, когда поверхность почвы в
лесу была преимущественно бурой, желтые улитки были сравни-
тельно заметны и отбор не благоприятствовал им. В дальней-
шем фон становился все более зеленым и влияние этого небла-
Изменения в популяциях | 147
гоприятного обстоятельства (фактора) уменьшалось. К концу
апреля или началу мая желтые раковины были в отношении
отбора нейтральным признаком; к середине мая они давали уже
селективное преимущество.
Эти данные говорят о довольно сильном давлении отбора.
Поэтому следовало бы ожидать, что популяции в условиях
однородного фона будут целиком состоять из особей без полос,
а в условиях неоднородных, сильно расчлененных местообита-
ний — только из полосатых особей. Чем же тогда поддержи-
вается полиморфизм? Сезонные изменения в селективной цен-
ности фенотипов замедляют, но не предотвращают элиминацию
менее приспособленных форм. Отчасти изменчивость можно
было бы объяснить обменом особями между колониями из раз-
ных местообитаний, однако расстояния, на которые способны
передвигаться улитки, слишком малы, так что эту гипотезу
нельзя применить к более изолированным колониям.
Ответ состоит в том, что существуют физиологические фак-
торы, генетически коррелированные с типом окраски. Так, экспе-
рименты с Сераеа nemoralis показали, что особи без полос на
раковине (особенно желтые) более теплоустойчивы, чем поло-
сатые. Желтые улитки лучше переносят холод, чем розовые,
а улитки без полос — лучше, чем полосатые. Эти и другие сход-
ные наблюдения указывают на то, что окраска и полосы на
раковине подвержены сильному отбору, не связанному с их
броскостью вследствие того, что они коррелируют с какими-то
важными физиологическими преимуществами. Можно ожидать,
что в некоторых случаях гетерозиготы окажутся жизнеспособнее
обеих гомозигот, так что физиологический отбор будет прибли-
жать популяцию к стабильному полиморфизму. Эти селективные
факторы, взаимодействуя с отбором по признаку заметности для
врагов, по-видимому, и обусловливают в значительной степени
наблюдаемую у Сераеа картину изменчивости.
В некоторых ситуациях может иметь значение ряд других
факторов. Один из них — наличие хищников, при котором да-
вление отбора определяется относительной частотой особей типа,
подверженного истреблению. У некоторых хищников могут вы-
рабатываться такие формы поиска жертвы, которые приводят
к отбору, направленному против наиболее обычного в популяции
типа. Результатом такого поведения хищника мог бы быть ста-
бильный полиморфизм популяции жертвы. Стохастические про-
цессы, некогда считавшиеся главным фактором дифференциации
популяций Сераеа, возможно, имеют относительно второстепен-
ное значение. Несомненно, дрейф генов играет известную роль
в популяциях меньшего размера и, быть может, лежит в основе
форм изменчивости, отмеченных Ламоттом во Франции. Недавно
10*
148 | Глава 7
образовавшиеся колонии могли еще не достичь равновесия со
средой, и на их составе может сильно сказываться генетическая
информация одной или нескольких улиток, основавших данную
колонию. Это влияние генетического вклада особей, участвовав-
ших в формировании новой колонии, известно как «принцип
родоначальника» и будет подробнее рассмотрено в гл. 10.
Один из приводившихся доводов против роли отбора на за-
щитную окраску в определении характеристик отдельных коло-
ний Сераеа состоял в том, что в смешанных колониях из пред-
ставителей двух близкородственных видов С. tiemoralls и С. hor-
tensis в частоте соответствующих генотипов не обнаруживалось
корреляции. Кларк удовлетворительно объяснил это, показав,
что на давление такого отбора реагируют оба вида, но неодина-
ковым образом. Весьма вероятно, что селективная ценность раз-
личных генов, определяющих рисунок и окраску, в разных гено-
типах этих двух видов различна. Предположим, например, что
сильно выраженная полосатость в определенном сухом место-
обитании невыгодна. Предположим также, что у одного вида
гены сильно выраженной полосатости участвуют в создании
особей, устойчивых к недостатку влаги, а у другого вида этого
может и не быть. Тогда в одном и том же местообитании в
популяции первого вида значительная часть улиток будет иметь
раковины с резко выраженными полосами (в результате суро-
вого отбора на устойчивость к недостатку влаги), а в популяции
второго вида полосатые улитки могут полностью отсутствовать
(вследствие отбора против полосатости). Хотя гены, контроли-
рующие различные признаки, представляются в этом случае
гомологичными, они не могут действовать независимо от своего
генетического окружения. Напоминаем, что в гл. 6 гены рас-
сматривались так, как если бы они были независимыми едини-
цами; результаты, подобные тем, которые были получены при
исследовании Сераеа, постоянно напоминают нам о том, что
подобное представление слишком упрощено.
ПОПУЛЯЦИИ УЖЕЙ НА ОСТРОВАХ
Кэмин и Эрлих изучали микроэволюцию в популяциях ужей
(Natrix sipedon) на островах западной части озера Эри. Ужи на
этих островах различаются рисунком поперечных полос. Они
были разделены на 4 класса: А, Б, В и Г (у змей типа А по-
лосы отсутствуют, у змей типа Г полосы наиболее выражены;
фиг. 29). За исключением ужей, встречающихся в западной
части озера Эри (и одной местности в Теннесси), все известные
популяции АЛ sipedon состоят из особей типа Г. Фактически все
ужи, найденные в области, окружающей это озеро, относятся
Новорожден-
ные особи
Дифференциальная
смертность
Размножение
Случайные
иммигранты
Взрослые особи
местного
происхождения
Фиг. 29.
Эри.
Естественный отбор среди ужей, обитающих на островах озера
На наличие естественного отбора указывают
У взрослых особей и у потомства.
различия в частоте разных типов рисунка
150 | Глава 7
к типу Г. На островах же значительная часть ужей относится
к другим типам, в частности встречается много особей совер-
шенно без полос. На островах очень мало внутренних водоемов,
так что ужи встречаются главным образом в береговой зоне.
Единственными подходящими местообитаниями для них яв-
ляются плоские известняковые скалы, известняковые береговые
обрывы и галечные отмели.
С островов брали большие выборки взрослых ужей, и опло-
дотворенных самок содержали живыми до появления потомства.
Распределение различных типов рисунка в полученных вывод-
ках сравнивали с их распределением среди взрослых змей. Из-
менчивость взрослых особей и их потомства для одной группы
островов показана на фиг. 30. Статистический анализ получен-
ных данных сопряжен с известными трудностями, поскольку
приходится сравнивать популяцию взрослых особей, предста-
вленную случайной выборкой, и популяцию потомства, представ-
ленную неслучайными группами. Несмотря на это, удалось
показать наличие статистически значимого различия в харак-
тере полосатости между обеими популяциями. Процент особей
с относительно слабо выраженной полосатостью (типы А и Б)
в популяции взрослых особей был выше, чем в потомстве.
Наблюдаемые достоверные различия между популяциями
взрослых особей и детенышей можно объяснить только изби-
рательной элиминацией или возрастной изменчивостью окраски
тела у отдельных особей. Фактические данные почти с полной
определенностью говорят в пользу первой из этих гипотез, так
как у змей, выращенных в лаборатории, не наблюдается ника-
ких признаков возрастного изменения окраски. Кроме того, как
у взрослых особей, так и у детенышей были найдены все четыре
типа рисунка; различными оказались только их частоты.
Роль эволюционных факторов, не связанных с отбором, в
данном случае легко исключить. Высокий процент особей, ли-
шенных полос, можно было бы объяснить миграцией ужей из
других «неполосатых» популяций, но ближайшая популяция
такого типа находится в Теннесси. Для поддержания «неполо-
сатого» генотипа за счет одних лишь мутаций требовалась бы
частота мутаций, далеко превосходящая частоту мутаций в лю-
бом локусе, когда-либо изученном у какого-либо организма,
даже если предположить, что окраска тела — монофакториаль-
ный признак (каковым она в действительности не является).
Наблюдаемые различия нельзя объяснить и дрейфом генов. Нет
никаких указаний на то, что популяции этих островов когда-
либо уменьшаются до размеров, при которых дрейф мог бы
играть существенную роль. Один раз 7 человек поймали на
одном острове 400 ужей за 5 часов, а в другой раз — 234 ужа
Изменения в популяциях | 151
Фиг. 30. Сравнение потомства у взрослых особей Natrix sipedon с группы
островов Басс на озере Эри.
На оси ординат —процент особей данного типа, на оси абсцисс —тип рисунка (к типу А отно-
сятся особи с наименее выраженной полосатостью, а к типу Г —с максимально выраженной
полосатостыо |19].
за 4 часа. На всех островах возрастной сдвиг направлен в сто-
рону ослабления полосатости, что указывает на систематическое
давление отбора, а не на случайный дрейф. Таким образом,
изменение частот различных типов скорее всего обусловлено
отбором — процессОлМ избирательной элиминации.
Чтобы доказать совершившийся факт отбора, нет необходи-
мости вскрывать механизм его действия. Интересно, однако,
представить себе возможные факторы, создающие описанную
ситуацию. Для человеческого глаза змеи без полос плохо раз-
личимы на фоне плоских известняковых скал в прибрежных
частях островов, тогда как полосатые особи очень заметны.
В то же время полосы, вероятно, разбивают контуры змеи и
делают ее менее заметной в более типичном для ужей, зрительно
менее однородном болотистом местообитании. У змей есть вра-
ги, находящие их с помощью зрения (чайки, цапли, ястребы и
др.); много змей убивает человек из огнестрельного оружия.
Очевидно, что интенсивность отбора в данном случае очень
высока. Если весь спектр вариантов окраски произвольно раз-
делить на две части (с полосами и без полос), то вероятность
152 | Глава 7
выживания для особей «полосатой» половины будет примерно
в 4 раза меньше вероятности выживания особей без полос (s для
«полосатого» фенотипа равно приблизительно 0,75). В связи с
этим возникает вопрос, почему при столь суровом отборе в по-
пуляции вообще сохраняются полосатые особи. Ответ, по-види-
мому, состоит в том, что в результате миграции создается посто-
янный приток генов полосатости в генофонды островных попу-
ляций. Многократно видели ужей, плавающих вдалеке от суши,
а расстояние от берегов озера до островов не настолько велико,
чтобы его не могли преодолеть мигрирующие особи. Таким обра-
зом, материковые популяции представляют собой резервуар по-
лосатых особей, часть которых время от времени попадает на
острова. Возникающее при этом взаимодействие между отбором
и миграцией создало ситуацию, которую чрезвычайно удобно
анализировать.
ХРОМОСОМНЫЙ ПОЛИМОРФИЗМ У ДРОЗОФИЛЫ
Классические примеры микроэволюции мы находим в иссле-
дованиях, начатых впервые Добжанским, который изучил ча-
стоты инверсий примерно у 30 видов дрозофилы, обнаруживаю-
щих полиморфизм структуры хромосом. Такого рода исследова-
ния на дрозофилах возможны благодаря тому, что в слюнных
железах личинок этих мушек имеются гигантские политенные
хромосомы. Эти хромосомы находятся в состоянии тесной
конъюгации, наблюдаемой обычно лишь в зигонеме мейоза.
Их размеры и упомянутая соматическая конъюгация делают
их весьма полезными для исследователя. Гетерозиготность по
инверсиям у дрозофилы можно выявлять по наличию харак-
терных инверсионных петель на хромосомах слюнных желез
(фиг. 12).
Различным последовательностям поперечных полос на очень
изменчивой 3-й хромосоме D. pseudoobscura были даны особые
названия. В литературе по эволюции больше всего обсуждаются
типы инверсии «стандартный» (ST); «Эрроухед» (AR) и «Чи-
рикахуа» (СН). Если в одной популяции встречаются два раз-
ных типа хромосом, то она будет содержать особей трех типов:
две гомозиготы по инверсии и одну гетерозиготу. Например,
когда имеются только хромосомы ST и СН, этими тремя типами
особей будут две гомозиготы ST/ST и СН/СН и гетерозигота
ST/СН. Гомозиготы можно различить путем исследования по-
томства или тщательного изучения характера расположения
дисков, тогда как гетерозиготы легко распознаются по инвер-
сионной петле.
Изменения в популяциях | 153
Различные типы хромосом у D. pseudoobscura имеют разное
географическое распространение — очевидно, вследствие их не-
одинаковой адаптивной ценности в различных местообитаниях.
На фиг. 31 показана географическая изменчивость частот трех
названных типов хромосом в ряде местностей, образующих
трансекту через юго-западную часть США. Частоты хромосом
изменяются также с высотой над уровнем моря, как это явствует
из фиг. 32, где представлены количественные соотношения трех
типов на разных высотах в горах Сьерра-Невада в Калифорнии.
На эту географическую изменчивость частот накладывается
сезонная изменчивость. Например, в горах Сьерра-Невада
Фиг. 31. Частота (в процентах) различных типов хромосом в популяциях
Drosophila pseudoobscura (юго-запад США).
Черные столбики —тип ST, белые столбики —тип AR, заштрихованные столбики тип РР |20].
154 | Глава 7
стандартная последовательность генов чаще всего встречается на
средних высотах, с увеличением же высоты частота ее постепенно
убывает. В то же время частота типа AR обнаруживает поло-
жительную корреляцию с высотой: она достигает максимума в
субальпийской зоне (около 3000 ж) и оказывается наименьшей
у подножия гор (250 ж). Однако в общем частота хромосом ST
имеет тенденцию повышаться в течение сезона, тогда как ча-
стота типа AR обычно снижается. Таким образом, по частотам
этих хромосом позднелетние популяции на больших высотах
имеют тенденцию приближаться к весенним популяциям на
меньших высотах.
Эти изменения легче всего объяснить довольно сильным да-
влением отбора. Добжанский проверял эту гипотезу в серии
экспериментов, в которых он выращивал D. pseudoobscura в по-
пуляционных садках. Начальные частоты различных типов хро-
мосом в его экспериментальных популяциях были известны, а
затем он повторно брал из них пробы, чтобы установить, не
произошли ли в них какие-либо изменения. Он нашел, что в по-
пуляциях, поддерживаемых при температуре 16,5°, никаких из-
менений не происходит. Однако при 20° частоты изменяются и
обычно достигают точки равновесия, при которой все первона-
чальные типы сохраняются, но в совершенно иных соотношениях
по сравнению с исходными.
Фиг. 32. Частота различных типов хромосом в популяциях Drosophila
pseudoobscura на разных высотах в горах Сьерра-Невада.
Черные квадраты —тип ST, белые квадраты —тип AR, заштрихованные квадраты —тип РР.
Площадь квадратов пропорциональна частоте различных типов хромосом. На оси ординат —
высота в метрах; на оси абсцисс — горизонтальное расстояние в километрах. Высота мест-
ности соответствует уровню основания самого нижнего квадрата [21].
Изменения в популяциях [ 155
Фиг. 33. Частота хромосом стандартного типа (в процентах) у мух из одного
популяционного садка в разные месяцы [20].
Результаты одного эксперимента с лабораторной популяцией
представлены на фиг. 33. Колония в садке была заложена
1 марта 19*46 г., причем особи подобраны так, чтобы в популя-
ции было 10,7% хромосом ST и 89,3% хромосом СН. В течение
всей остальной части года доля хромосом ST неуклонно повы-
шалась, вначале быстро, а затем медленнее, и к концу года
(примерно через 15 поколений) достигла стабильного уровня
(около 70%). Такой характер изменения и установление равно-
весия давали веские основания предполагать наличие селектив-
ного преимущества структурных гетерозигот (ST/СН) по сравне-
нию с обеими формами гомозигот. С точки зрения популяционной
генетики это сравнимо со сверхдоминантностью по приспо-
собленности в единичном локусе. Такие же результаты могли бы
получиться, однако, и при отрицательном ассортативном скре-
щивании (спаривание несходных особей). Эта последняя воз-
можность была исключена путем взятия из популяционного
садка выборки яиц и* выращивания личинок в оптимальных
условиях, при которых почти все они выживали. Оказалось, что
частоты различных генотипов соответствуют закону Харди —
Вейнберга; это доказывало, что спаривание было случайным и
что причиной наблюдаемых изменений не могли быть различия
в плодовитости или в интенсивности размножения. Однако взя-
156 | Глава 7
тые из популяции выборки взрослых мух показали следующие
отклонения от частот Харди — Вейнберга:
ST/ST ST/CH сн/сн
Наблюдаемое число 57 169 29
Ожидаемое число 78,5 126,0 50,5
Отклонение —21,5 +43,0 —21,5
Эти результаты «наряду с установлением равновесия на уров-
не около 70% ясно указывают на дифференциальную элимина-
цию двух типов гомозигот между стадиями яйца и имаго. Было
установлено, что в случае яиц, отложенных дикими мухами, ста-
тистически значимое отклонение от ожидаемых частот Харди —
Вейнберга не обнаруживается, но среди взрослых самцов ге-
терозиготы встречаются значительно чаще, чем следовало бы
ожидать при наличии равновесия Харди — Вейнберга в популя-
ции.
Крайний случай селективного преимущества наблюдается в
одной популяции Drosophila tropicalis, где существует два рас-
пространенных типа хромосом. Структурные гомозиготы обоих
типов погибают на ранней стадии развития, и только гетерози-
готы выживают и размножаются. Хотя жизнеспособна лишь по-
ловина образующихся зигот, эта популяция процветает.
Были описаны и долговременные изменения в частотах ин-
версий. Среди 20 000 исследованных до 1941 г. в Калифорнии
хромосом Drosophila было обнаружено всего 4 хромосомы типа
«Пайке Пик» (РР). В 1957 г. хромосомы РР были найдены во
всех 10 местностях этого штата, где были взяты пробы из по-
пуляций, причем средняя частота составляла 0,08 (в 400 раз
больше прежней). Это увеличение произошло за счет хромо-
сом СН. Чем оно было вызвано, неизвестно, но почти наверное
причиной был отбор. С 1941 г. хромосомы РР неизменно пре-
обладали в популяциях восточных склонов Скалистых гор и
Техаса. Повышение частоты хромосом типа СН в Калифорнии
не могло объясняться миграцией из этого резервуара хромосом
РР, так как географические популяции в штатах Аризона и
Юта в период между 1941 и 1957 гг. не изменились сколько-
нибудь существенным образом.. Действовавший при этом селек-
тивный фактор неизвестен, но к числу наиболее очевидных фак-
торов, участие которых можно было бы предположить, следует
отнести сильную засуху и возрастающее задымление атмосферы.
Мы сделали лишь краткий обзор некоторых результатов
исследований по генетике природных популяций дрозофилы.
Вряд ли можно сомневаться в том, что различные группировки
генов или супергены в хромосомах с инверсиями (образующие
довольно устойчивые блоки вследствие элиминации продуктов
Изменения в популяциях | 157
кроссинговера при наличии инверсий) имеют разную адаптив-
ную ценность при разных условиях. Было обнаружено много
случаев изменения частоты тех или иных типов хромосом, кор-
релирующего с изменениями среды. Менее успешными были по-
пытки определить, в какой период жизненного цикла прояв-
ляются преимущества или невыгоды изменений. Выяснение
конкретной природы этих адаптивных изменений также оказа-
лось трудным делом. Воспроизвести природные условия в ла-
боратории невозможно, а о биологии и экологии различны,х
видов дрозофилы известно очень мало. При изучении D. pseudo-
obscura и D. persimilis, личинки которых развиваются в «нате-
ках слизи» (зараженный дрожжевыми грибами и бактериями
сок на стволах деревьев) в горах Сьерра-Невада, а также при
изучении многих видов Drosophila в лаборатории были получены
ценные сведения. Необходима еще большая работа в этом на-
правлении. Представление о сложности проблемы можно полу-
чить на примере факторов, определяющих место окукливания
личинок D. melanogaster; оно имеет, как известно, существенное
селективное значение в лабораторных популяциях. Место, кото-
рое личинка выбирает для окукливания, зависит, помимо про-
чего, от температуры, от влажности воздуха и питательной
среды, от плотности популяции личинок и продолжительности
периода личиночного развития. Экологическая генетика дрозо-
филы еще долго будет оставаться плодотворной областью иссле-
дования.
ПРИМЕРЫ ИЗ ГЕНЕТИКИ ЧЕЛОВЕКА
Как можно было ожидать, естественный отбор играет актив-
ную роль и в формировании генетического состава популяций
человека. Необычайно ярким примером может служить влияние
отбора на частоту гена так называемой «серповидноклеточно-
сти». У индивидуумов, гомозиготных по этому рецессивному
гену, наблюдается изменение формы эритроцитов, сопровождаю-
щееся резко выраженной анемией и тяжелым общим состоянием.
Заболевание обычно приводит к смерти. Гетерозиготных инди-
видуумов можно выявить по деформации эритроцитов, происхо-
дящей при снижении содержания кислорода в крови (при этом
эритроциты становятся серповидными). Однако гетерозиготы,
очевидно, в известной степени устойчивы к малярии, и поэтому
в географических районах, эндемичных по малярии, отбор бла-
гоприятствует им. Аллисон установил, что частота гетерозигот
по гену серповидноклеточности среди взрослых выше, чем среди
маленьких детей, что указывает на селективное преимущество
этого генотипа. Это преимущество гетерозигот имеет примерно
такую величину, какая теоретически необходима для поддержа-
158 | Глава 7
ния наблюдаемой частоты гена серповидноклеточности (около
0,20). Оно, по-видимому, и обусловливает поддержание сбалан-
сированного полиморфизма по локусу серповидноклеточности на
значительной части территории Африки, несмотря на очень ма-
лую жизнеспособность гомозиготных рецессивов.
Гласс и его сотрудники исследовали частоту генов, опреде-
ляющих некоторые признаки (группы крови, типы оволосения
на средних фалангах пальцев и др.), в религиозной общине дан-
керов в Пенсильвании (вся популяция насчитывала менее
100 человек). В ряде локусов частоты изученных генов сильно
отличались от соответствующих частот у окружающего населе-
ния и у населения тех мест, откуда ведет свое происхождение
эта группа (западная часть Германии). Предполагают, что эти
отклонения обусловлены дрейфом генов. Интересно, что по дру-
гим локусам, о которых известно, что они подвержены довольно
сильному отбору (например, локус Rh), никакого или почти ни-
какого отклонения обнаружено не было.
Один известный пример отбора, направленного против гете-
розигот, касается локуса Rh у человека. После оплодотворения
яйцеклетки /^-отрицательной матери (гомозиготной по рецес-
сивному аллелю) /?Л-положительным сперматозоидом образую-
щийся плод может погибнуть вследствие антигенной несовмести-
мости с матерью. Этот отбор, видимо, в известной степени ком-
пенсируется склонностью ^-несовместимых родителей к повтор-
ному деторождению. Однако само по себе это еще недостаточно,
для того чтобы объяснить существование постоянного полимор-
физма в человеческой популяции. Равновесие частот генов, хотя
оно уже не соответствует частоте 0,5 (см. гл. 6, разделы, посвя-
щенные отбору, направленному против гетерозигот), остается
неустойчивым. Возможно, что фиксации локуса Rh во всей по-
пуляции человека препятствовала миграция: одни субпопуля-
ции приближались к фиксации гена /?А+, другие — к фиксации
гена Rh~, но в результате последующего смещения этих попу-
ляций вновь восстанавливался полиморфизм.
РАСТЕНИЯ ПАСТБИЩ
К сожалению, ботаники нашли очень мало природных си-
туаций, доступных для такого анализа, какой был возможен в
рассмотренных выше случаях. Кемп изучал в южном Мэриленде
пастбище, засеянное бобово-злаковой смесью и впоследствии
разделенное на участки. Одна половина его охранялась от скота,
а другая использовалась для пастьбы. Спустя 3 года с обеих
половин пастбища были выкопаны растения мятлика (Роа рга-
tensis), ежи сборной (Dactylis glomerata) и белого клевера (Tri-
Изменения в популяциях | 159
folium repens) и пересажены на опытный участок. Оказалось,
что на использовавшейся под выпас половине пастбища процент
генотипов, обусловливающих низкий, стелющийся рост, был
выше. Растения с другой половины были прямостоячими, без
склонности к полеганию. Это ясно показывает, что в популя-
циях происходил интенсивный отбор адаптивных форм роста.
МПМИКРИЯ У СОРНЯКОВ
Чрезвычайно сложная система мимикрии была обнаружена
у растений рода Camelina (рыжик) из сем. Cruciferae. Различ-
ные типы рыжика растут как сорняки на полях культурного
льна (Linum, сем. Linaceae). Было высказано предположение,
что по мере того, как культура льна становилась более интен-
сивной, рыжик испытывал все более сильное давление отбора.
Для семян рыжика селективное преимущество состояло, напри-
мер, в том, чтобы при отвеивании мякины они оставались вме-
сте с семенами льна и затем высевались вместе с ними. Расте-
ния вырастали только из семян с надлежащими аэродинамиче-
скими свойствами; остальные семена не попадали в почву. Чем
тщательнее производится отвеивание, тем сильнее давление от-
бора. В результате этого отбора у рыжика появились семена,
сходные с семенами льна — не по виду, а по тому, на какое рас-
стояние они выдуваются струей воздуха определенной силы.
Кроме того, отбор благоприятствовал формам с высоким тон-
ким стеблем; такие формы не погибают от недостатка света в
густых посевах льна. В результате отбора формы, имитирующие
лен, появились не только в роде Camelina, но и среди других
растений, в частности в родах Spergula и Silene (Caryophyl-
laceae). Следует отметить, что свидетельства в пользу отбора
здесь более косвенные, чем в предыдущих примерах.
ДПСРУПТИВНЫЙ ОТБОР У МИМЕТИЧЕСКИХ ФОРМ БАБОЧЕК
Когда отбор благоприятствует сохранению двух или большего
числа модальных фенотипов, <рго называют дисруптивным. Экс-
перименты с отбором на число щетинок у дрозофилы показали,
что тип отбора, больше благоприятствующий крайним формам,
нежели промежуточным, может создать бимодальную популя-
цию с повышенной изменчивостью. Очевидно, случаи дисруп-
тивного отбора в природных популяциях можно найти у миме-
тических бабочек. Например, у широко распространенного и
служившего объектом многих исследований африканского па-
русника Papilio dardanus (фиг. 34) имеется целый ряд различ-
ных миметических самок, хотя у самцов никогда не наблюдается
Фиг. 34. Мимикрия у африканского парусника Papilio dardanus.
Вверху самец Р. dardanus\ справа три бабочки подсем. Danainae (Danais chryslppus,
Arnauris niavlus и Amaurls echerlay, слева формы самок P. dardanus. имитирующие этих
бабочек [22j.
Изменения в популяциях | 161
мимикрии. По-видимому, это объясняется тем, что без «нормаль-
ной» окраски и формы крыльев самцы утратили бы привлека-
тельность для самок. В одной и той же местности постоянно
встречается несколько различных форм самок, каждая из ко-
торых точно имитирует какой-либо вид бабочек, имеющих
неприятный вкус.
Имеются данные (резюмированные Шеппардом [12]), говоря-
щие о том, что определенные сочетания признаков обеспечивают
наилучшую имитацию той или иной модели и дают селективное
преимущество. Другие формы не сходны ни с какой моделью,
и отбор им не благоприятствует. По крайней мере в некоторых
случаях отбор, по-видимому, уменьшил возможность появления
плохо защищенных сочетаний признаков, увеличив сцепление
между локусами, участвующими в создании защитного комплек-
са. Благодаря этому возможен отбор групп сцепленных локусов
(супергенов) как единых комплексов и сохранение лучших со-
четаний. (Это явление вполне сравнимо с закреплением генных
комбинаций в популяциях дрозофилы благодаря инверсиям.)
У некоторых форм бабочек дисруптивный отбор может также
действовать путем накопления генов-модификаторов, еще более
совершенствующих сходство с моделью. Это предположение
подкрепляется экспериментами с гибридизацией, в которых су-
пергены оказываются перенесенными в новое генетическое окру-
жение. Здесь они не обеспечивают такого точного фенотипиче-
ского подражания модели, как на прежнем генетическом
фоне.
УСТОЙЧИВОСТЬ К АНТИБИОТИКАМ И ИНСЕКТИЦИДАМ
Рассмотрение эволюции не было бы полным, если бы мы не
упомянули о реакции некоторых организмов на попытки чело-
века сократить их численность или истребить их. Важнейшие и
наиболее яркие примеры реакции природных популяций на вме-
шательство человека связаны с явлением резистентности.
Сплошная обработка инсектицидами обширных территорий со-
здала весьма мощный фактор отбора. В огромнохМ большинстве
случаев высокая численность популяций насекомых-вредителей
обеспечивает резерв изменчивости, достаточный для того, чтобы
у этих насекомых могли выработаться линии, устойчивые прак-
тически к любым веществам, которые может создать изобрета-
тельность химиков-органиков. Конечно, возможности химиков
сильно ограничиваются требованием безвредности применяемых
веществ для организмов, не подлежащих уничтожению, напри-
мер для человека. И все же имеется немало данных о том, что
человек и используемые им растения и домашние животные не
избегли вредного действия мощных ядохимикатов,
11 Зак. 152
162 | Глава 7
Интересно отметить, что насекомые ответили на брошенный
им вызов самым различным образом. Описаны примеры пове-
денческих механизмов устойчивости, когда насекомое перестает
садиться на опрысканные поверхности, и много видов физиоло-
гической устойчивости, связанной с выработкой разнообразных
механизмов, препятствующих проникновению или действию ин-
сектицида.
Аналогичным выработке устойчивости к инсектицидам было
появление штаммов микроорганизмов, в высокой степени устой-
чивых к антибиотикам. Это вызвано тем, что врачи из самых
лучших побуждений злоупотребляют антибиотиками, применяя
их в тех случаях, когда другие средства могли бы быть доста-
точными или даже более эффективными. До сих пор с размно-
жением этих устойчивых организмов борются главным образом
путем поспешного изыскания новых видов химического оружия,
а не с помощью методов, которые были бы более утонченными
с биологической точки зрения и несравненно более благотвор-
ными в отношении отдаленных последствий.
Между тем такие подходы хорошо известны. Некоторые
устойчивые формы насекомых в среде без инсектицида оказа-
лись менее жизнеспособными, чем их нерезистентные сородичи.
В этих случаях достаточно длительный перерыв в применении
инсектицида позволил бы силам естественного отбора вернуть
популяцию к прежнему восприимчивому состоянию. Путем ра-
зумного использования инсектицидов в критические моменты
можно было бы достаточно успешно бороться с вредителями,
сводя к минимуму опасность возникновения устойчивых линий
(достигающую максимума при «массовом» применении ядохи-
микатов). В случаях когда перерывы неосуществимы, много-
кратное применение большого числа различных ядов, быть мо-
жет, исключило бы одновременную выработку в популяциях
устойчивости ко всем этим ядам. Поскольку, однако, невозмож-
но воздействовать на одного члена сообщества, не влияя на всю
экосистему, этот подход таит в себе непредвиденные опасности.
Хорошо известно, что химические яды могут накапливаться в
различных звеньях пищевых цепей. И если подлежащие истре-
блению организмы не погибнут, то могут сильно пострадать их
враги, а также враги их врагов.
К тому же со многими вредными насекомыми легче и эко-
номичнее бороться путем нехимического вмешательства в их
биологию, нежели с помощью инсектицидов. Например, эффек-
тивными оказались такие меры, как осушение болот, в которых
выводятся комары, интродукция организмов-врагов для борьбы
с завезенными вредителями, выпуск множества стерильных сам-
цов, с тем чтобы они конкурировали с дикими самцами за са-
Изменения в популяциях | 163
мок, и распространение выращенных в лаборатории патогенных
организмов. Точно так же и против высокоустойчивых микро-
организмов классические методы антисептики часто могут ока-
заться гораздо более эффективными, чем дорогостоящие новые
антибиотики.
ЛАБОРАТОРНЫЕ ПОПУЛЯЦИИ
Понятие искусственный отбор означает воздействие человека
на величину вклада различных генотипов в генофонд последую-
щих поколений. Искусственный отбор применяется в селекции
растений и животных, а также при экспериментальном изучении
последствий отбора в лаборатории. Он может быть сознательно
направлен на один признак — в этом отношении он также отли-
чается от естественного отбора, который воздействует на все
фенотипические признаки, влияющие на приспособленность, и
не имеет какой-либо «цели». Тем не менее даже при самом тща-
тельном регулировании лабораторных условий наряду с искус-
ственным отбором продолжает действовать и естественный
отбор. Множество исследований в области искусственного от-
бора касалось так называемых количественных признаков. Коли-
чественные признаки — это те признаки, на которые влияет
много пар аллелей во многих различных локусах (например,
рост и умственные способности у человека, число щетинок у
дрозофилы, окраска тела у ужей и т. п.).
Довольно типичным примером искусственного отбора служит
работа Мазера и Харрисона. Здесь отбор был направлен на
изменение числа абдоминальных щетинок у Drosophila tnelano-
gaster. На фиг. 35 графически представлены результаты отбора
на увеличенное число щетинок, проводившегося в течение более
чем 100 поколений. На протяжении 20 поколений отбор был
чрезвычайно эффективен. На 20-м поколении снижение плодо-
витости и интенсивности размножения заставило исследователей
прервать отбор, чтобы сохранить линию от вымирания. При
отсутствии отбора в течение нескольких поколений популяция
достигла равновесия в отношении числа щетинок; равновесный
уровень был выше исходного, но значительно ниже пика, достиг-
нутого в 20-м поколении. На 24-м поколении из линии, в кото-
рой отбор был прерван, была выделена новая линия для даль-
нейшего отбора, в ходе которого число щетинок быстро повы-
силось до уровня, близкого к достигнутому в исходной линии.
Когда отбор в этой новой линий был ослаблен, произошло лишь
незначительное уменьшение числа щетинок. Кроме того, в ре-
зультате непрерывного отбора еще в течение примерно 85 по-
колений был получен дальнейший сдвиг.
11*
164 | Глава 7
Искусственный отбор часто вначале дает быстрый эффект;
затем достигается плато, после чего дальнейший прогресс
затруднителен или невозможен; кроме того, жизнеспособность
линии настолько падает, что приходится прекращать (или
ослаблять) отбор, иначе линия будет потеряна. Обычно если
отбор прекращают, когда плато еще не достигнуто, линия ре-
грессирует в направлении к исходному уровню. Если же отбор
заканчивают после того, как популяция достигла плато, регрес-
сии может не быть или она оказывается очень незначительной.
Продолжение отбора в популяции, достигшей плато, часто не
дает заметных результатов в течение долгого времени. Однако
при достаточно длительном отборе снова может быть получен
сдвиг.
Одной из причин описанных явлений, как полагают, может
быть равновесие между искусственным и естественным отбором.
Хотя детали здесь неясны и не всегда одинаковы, естественный
отбор должен, по-видимому, создать сбалансированную систему,
в которой достигнуты наилучшие возможные соотношения при-
знаков, определяющих приспособленность. Иными словами,
Фиг. 35. Искусственный отбор по числу абдоминальных щетинок у Droso-
phila melan)gaster.
На оси ординат — среднее число щетинок; на jock абсцисс — число поколений. Линии дрозо-
филы, подвергающиеся отбору, представлены сплошными линиями, линии, не подвергаю-
щиеся отбору, — прерывистыми линиями. Буква а указывает на то, что эксперимент с дан-
ной линией прекращен; буква б —указывает на то, что линия вымерла вследствие стериль-
ности. Цифра справа—номера линий. В линии 1 проводился отбор на малое число щетинок,
кроме случая, отмеченного буквой в,— здесь отбор проводили на увеличенное число щети-
нок [23].
Изменения в популяциях | 165
приспособленность должна быть доведена до максимума. Имею-
щиеся данные как будто бы говорят о том, что высокая степень
гетерозиготности генотипа, особенно у животных, создает высо-
кий уровень физиологической приспособленности. Кажется так-
же вероятным, что крайние величины количественных признаков
часто бывают результатом высокой степени гомозиготности по
соответствующим локусам. Поэтому вполне возможно, что искус-
ственному отбору на большое или малое число щетинок проти-
водействует естественный отбор на приспособленность, если
крайние отклонения числа щетинок обусловлены гомозиготно-
стью по ряду локусов.
Здесь возможна аналогия (хотя и довольно грубая) с кон-
струированием самолета. Можно было бы попытаться улучшить
самолет, сделав мотор более мощным, но это принесло бы мало
пользы, если бы в результате возросшей скорости отламывались
крылья. Проблему можно было бы разрешить, укрепив фюзе-
ляж или структурные элементы крыльев, но и это оказалось бы
бесполезным, если бы привело к утяжелению самолета, так
что он не смог бы оторваться от земли. У организма, как и у
самолета, должно быть достигнуто необходимое равновесие всех
факторов, обеспечивающих успешное функционирование систе-
мы. Каждый отдельный фактор можно изменять лишь до из-
вестного предела, иначе их «работоспособная комбинация»
будет серьезно нарушена.
Лернер [8] создал математическую модель, позволяющую
объяснить появление плато ниже максимального уровня выра-
жения признака, по которому ведется отбор. Он основывает эту
модель на системе, в которой существует некоторый обязатель-
ный уровень гетерозиготности, определяющий приспособлен-
ность. В результате кроссинговера потенциальная генетическая
изменчивость может превращаться в «свободную» генетическую
изменчивость, обеспечивая возможность дальнейшего отбора
без потери приспособленности. Тогда всю ситуацию можно рас-
сматривать с точки зрения сдвигов равновесия. Интенсивный
отбор по одному или нескольким локусам деформирует сбалан-
сированный генотип. После того как сдвиг выражения признака
превысит определенную величину, эта деформация приведет
к утрате приспособленности, за которой последует либо выми-
рание, либо достижение нового состояния равновесия. В случае
прекращения отбора до наступления одного из этих событий
линия проявляет тенденцию регрессировать к исходному уровню.
По вопросам искусственного отбора накопилась огромная
литература, так как в процессе работы над экономически важ-
ными проблемами (улучшение пород домашних животных и
сортов растений, резистентность вредителей к инсектицидам и
166 | Глава 7
т. п.) получены чрезвычайно ценные сведения. Значительная
часть наших знаний по таким разноббразным проблемам, как
происхождение доминантности, интегративные свойства гено-
типов и эффективность отбора при различных условиях, явилась
прямым или побочным результатом изучения таких прозаических
вопросов, как яйценоскость кур, вес свиней, урожайность хлопка
и кукурузы, устойчивость пшеницы к ржавчине и продуктивность
молочных желез у коров. Читателя, желающего получить хоро-
шо систематизированные вводные сведения по этому обширному
и сложному предмету, мы отсылаем к книге Лернера [9].
ГЕНЕТИЧЕСКИЙ ГОМЕОСТАЗ
Здесь уместно будет упомянуть об одном важном свойстве
менделевских популяций — о часто наблюдаемой тенденции по-
пуляций, подвергающихся действию направленного отбора,
регрессировать в сторону первоначальной средней величины
соответствующего признака. Лернер назвал это явление гене-
тическим гомеостазом. Менделевская популяция обладает ха-
рактеристиками, которые нельзя отнести к составляющим ее осо-
бям. Например, бессмысленным было бы утверждение, что не-
которая особь находится в равновесии Харди — Вейнберга.
Популяции стремятся сохранять такой генетический состав, при
котором высокой степенью приспособленности обладает макси-
мальное число особей. Короче говоря, имеет место отбор, бла-
гоприятствующий поддержанию сбалансированного «макси-
мально приспособленного» генотипа. Это в основном стабилизи-
рующий отбор, направленный против отклоняющихся особей.
Генотип, обнаруживающий высокую степень приспособленности,
адаптирован не только к среде в классическом смысле этого
слова, но и к своей генетической среде, т. е. генофонду, на фоне
которого он существует. Иными словами, частота и распреде-
ление генов в популяции содействует повышению приспосо-
бленности любого генотипа в данной популяции. Представление
о генетическом гомеостазе можно резюмировать, сказав, что
отбор организует генофонд популяции таким образом, чтобы
многочисленные компоненты этого фонда были адаптированы
друг к другу и создавали максимальное число высокоприспо-
собленных генотипов. Хорошо интегрированные генотипы пред-
ставляют собой «выигрышные комбинации», и, как показали
эксперименты Мазера и Харрисона, отбору, направленному на
их изменение для адаптации к какому-либо новому требованию
внешней среды, противодействует отбор на сохранение эффек-
тивного интегрированного комплекса. Возможно, что необычно
резкий переход между двумя типами популяций Maniola jurtina,
Изменения в популяциях [ 167
0 котором упоминалось выше, соответствует границе между
двумя такими высокоинтегрированными комплексами. Стойкий
прогресс достигается направленным отбором только тогда, когда
факторы, благоприятствующие изменению, могут получить пере-
вес над факторами, способствующими сохранению эффективной
комбинации. Многое указывает на то, что явление сверхдоми-
нантности в отношении адаптивной ценности (селективное пре-
имущество гетерозиготы перед гомозиготами) служит одним из
основных механизмов, играющих роль в возникновении генети-
ческого гомеостаза у большинства животных и многих растений.
Однако термин «генетический гомеостаз» объединяет все методы
генетического саморегулирования популяций.
ГЕНЕТИЧЕСКАЯ АССИМИЛЯЦИЯ
При переносе растений Achillea lanulosa (сем. Compositae)
из местностей, расположенных на различной высоте в горах
Сьерра-Невада, на экспериментальный участок на уровне моря
не все они вырастали до одинаковых размеров (фиг. 36).
В ставших ныне классическими экспериментах Клаузена, Кекка
и Хизи растения с больших высот достигали значительно мень-
ших размеров, чем растения с меньших высот. Поскольку все
они выращивались в одинаковых условиях, было ясно, что кар-
ликовый рост растений из более высоких местностей обусловлен
генетически; иными словами, их генотипы определяли в общем
меньшую высоту растений независимо от того, в какой среде
они развивались. Однако если растения с меньших высот раз-
деляли на части (так что получались генетически идентичные
линии) и выращивали их на уровне моря и на средних высотах,
го в последнем случае они были ниже, чем их идентичные «близ-
нецы» на уровне моря. Другими словами, при взаимодействии
генотипа малых высот с внешними условиями средних высот
получались растения, похожие на растения с генотипом средних
высот. Такие формы, у которых фенотипическое изменение ими-
тирует генотипическое, получили название фенокопий.
Такого рода фенокопирование, по-видимому, широко рас-
пространено в природе, хотя строгие доказательства в большин-
стве случаев отсутствуют. Например, у многих бабочек особи
весенних генераций мельче и темнее, чем о-соби летних генера-
ций, и это различие, как полагают, обусловлено сезонными изме-
нениями среды. Однако в более северных частях ареала у этих
бабочек имеется только одно летнее поколение, в котором особи
Мельче, темнее и напоминают особей весенней генерации южных
Местностей. Предполагается, что в северных популяциях особи
имеют генотипы, обусловливающие меньшую величину и темную
CM
Фиг. 36. Рост растений Achillea lanulosa (п — 18), взятых с разных высот в горах Сьерра-Невада, после перенесе-
ния на опытный участок в Стэнфорде.
Приводятся данные для популяции, состоящей примерно из 60 особей. Полигоны частот показывают изменчивость высоты растений в каждой
из этих популяций. Горизонтальными линиями разделены интервалы в 5 см, соответствующие шкале на оси ординат, а расстояние между вер-
тикальными линиями соответствует 2 особям. Справа от каждого полигона частот указано число нецветущих растений. Стрелки указывают
среднюю величину в каждом распределении {24].
Изменения в популяциях | 169
окраску. Хотя решающие эксперименты с переносом бабочек не
производились, большее постоянство северных форм при изме-
нениях среды подкрепляет это предположение.
Как можно объяснить формирование высокогорных рас ра-
стений, северных рас бабочек и другие сходные явления? Вы-
званные средой изменения не могут быть непосредственно пере-
даны последующим поколениям; ламаркистское представление
о наследовании приобретенных признаков давно отвергнуто.
Уоддингтон постулировал процесс, в результате которого такие
приобретенные признаки могли бы при посредстве отбора асси-
милироваться в генотипе, и это предположение подкрепляется
рядом экспериментов самого Уоддингтона и других исследова-
телей. Сущность этой генетической ассимиляции лучше всего
можно пояснить на следующем примере.
Лабораторную популяцию одной линии Drosophila melanoga-
ster дикого типа подвергали тепловому шоку на стадии куколки.
У отдельных мух, выводившихся из таких куколок, обнаружи-
вался аномальный разрыв жилок на крыльях (фенотип cross-
veinless). В качестве родителей для получения следующей гене-
рации использовали только особей, обладавших приобретенным
фенотипом crossveinless. На втором десятке поколений частота
особей crossveinless, выводившихся из подвергнутых тепловому
шоку куколок, превышала 90%, но, кроме того, отдельные мухи
crossveinless начали появляться из необработанных куколок ли-
нии, в которой шел отбор. Скрещивания между такими мухами
дали линии с высокой частотой фенотипов crossveinless при
отсутствии теплового шока. Могло бы показаться, что приобре-
тенный признак стал наследственным. На самом деле в этом
эксперименте отбор, по-видимому, благоприятствовал тем гено-
типам, которые обусловливали низкий порог индукции жела-
тельного (с точки зрения отбора) фенотипа. В конце концов этот
порог снизился настолько, что уже не требовалось никакого
теплового шока для того, чтобы развитие шло в направлении
фенотипа crossveinless. Аналогичные результаты были получе-
ны при изучении других признаков дрозофилы.
Очень важно помнить о том, что диапазон возможных жизне-
способных фенотипов определяется наследственностью и что
отбор может изменить этот диапазон таким образом, что фено-
типы, развитие которых раньше могло быть индуцировано внеш-
ними условиями, наследственно фиксируются в качестве един-
ственно возможного результата взаимодействия генов со средой.
Когда последовательные поколения бабочек встречаются с раз-
личными условиями среды, отбор может благоприятствовать
фенотипической пластичности. Но он может вырабатывать и
В высокой степени «канализованное» развитие, приводящее
170 | Глава 7
к оптимальному фенотипу, — в тех случаях, когда существенные
факторы среды почти не изменяются из поколения в поколение.
Онтогенетические аспекты генетической ассимиляции были рас-
смотрены в гл. 4.
ПРИСПОСОБЛЕНИЕ К СРЕДЕ
Разнообразные и остроумные способы, при помощи которых
организмы решают проблемы выживания и размножения, слу-
жат косвенным свидетельством высокой эффективности есте-
ственного отбора. Это приспособление к среде обычно называют
адаптацией, но по ряду причин мы по возможности избегаем
этого неопределенного термина. Нам нет надобности входить в
детали эволюции крыла птицы, шеи жирафа, глаза позвоночных,
инстинкта постройки гнезда у некоторых рыб и т. д., так как
можно предполагать, что выработка этих и других органов или
форм поведения обусловлена, по существу, такими же процес-
сами отбора, как и те изменения, которые мы рассматривали
выше (индустриальный меланизм и т. п.). Даже незначительные
преимущества или невыгоды, связанные с теми или иными ге-
нетическими изменениями, создают дестаточную основу для
дифференциального действия естественного отбора. Например,
чувствительность к свету у одноклеточных организмов служит
отправным пунктом для развития на основе отбора чрезвы-
чайно сложных глаз позвоночных, насекомых и некоторых мол-
люсков.
Старый аргумент антиэволюционистов, утверждавших, что
глаз позвоночного был бесполезен, пока он не достиг современ-
ной сложности, совершенно неубедителен. Многие организмы
с более простыми и менее специализированными фоторецепто-
рами отлично используют их, и не вызывает сомнений, что даже
человек лучше ориентировался бы, обладая фоторецептором, не
создающим изображений (т. е. таким, который дает информацию
только о количестве света), нежели совсем без всякого фото-
рецептора. Точно так же любое безвредное изменение у съедоб-
ной для птиц бабочки, увеличивающее ее сходство с симпатри-
ческим видом бабочек, имеющих неприятный вкус, дает изме-
ненной форме селективное преимущество.
То, что явления мимикрии и защитной окраски могут быть
выражены в самой различной степени, служит веским доводом
против предположения, что такое сходство неэффективно, если
оно не достигает совершенства. Имеющиеся экспериментальные
данные показывают, что и недостаточно совершенное подража-
ние некоторым неприятным на вкус бабочкам обеспечивает
известную защиту, хотя, быть может, и меньшую, чем в случае
л Изменения в популяциях | 171
близкого сходства с моделью. Интересно отметить, что мимети-
ческие формы различных бабочек обычно не встречаются в тех
областях, где отсутствует модель; это указывает на то, что для
предотвращения регрессии к «дикому» типу необходимо давле-
ние отбора; благоприятствующее мимикрии. Даже такие трудно
объяснимые явления, как эволюция общественного поведения
у пчел, начинают уже поддаваться анализу (см., например, ра-
боту Миченера [10]). Эта проблема очень сложна потому, что
эволюционирующей единицей является здесь колония, а не особь
(большинство пчел в улье неспособно к размножению). Отбор
у медоносной пчелы выражается главным образом в дифферен-
циальном воспроизведении колоний, а не отдельных генотипов.
Одно из самых распространенных эволюционных явлений —
это утрата тех или иных признаков, когда они уже не дают се-
лективного преимущества. По-видимому, нейтральные в отно-
шении отбора признаки редки. Глаз, весьма полезный большин-
ству животных, у пещерной рыбы может стать легко повреждае-
мым, склонным к инфекции органом, который служит лишь
помехой. Волосы на теле, некогда защищавшие человека от хо-
лода и повреждений, с изобретением одежды утратили эту свою
функцию и превратились просто в весьма удобное прибежище
для вшей.
ВЫВОДЫ
В этой главе был приведен ряд примеров исследования эво-
люционных изменений в популяциях—примеров, отражающих
разнообразие подходов и материала. Кроме того, были рассмо-
трены косвенные данные относительно эффективности процессов
отбора. Становится очевидным, что если продемонстрировать
наличие изменений сравнительно просто, то связать их с теми
или иными эволюционными факторами гораздо труднее. Эту за-
дачу особенно затрудняют сложные взаимодействия внутри гене-
тических механизмов и систем, реализующих онтогенез. Труднее
всего документировать роль дрейфа генов. Почти невозможно
«доказать» эффективность дрейфа в природных популяциях, так
как мы всегда можем предполагать существование какого-то
еще не открытого фактора отбора, способного вызвать наблю-
даемые явления. И действительно, известен ряд случаев (в част-
ности, среди улиток), когда было показано, что изменения, при-
писывавшиеся ранее действию дрейфа генов, на самом деле вы-
зываются отбором. Однако неизбежность дрейфа генов, а также
результаты изучения частот генов в небольших популяциях
Дрозофилы (как в природе, так и в лаборатории) заставляют ду-
мать, что дрейф, взаимодействуя с другими факторами, может
172 | Глава 7
быть важной эволюционной силой. Необходимо провести
обширные исследования на самых разнообразных организмах,
прежде чем можно будет с полной уверенностью делать широкие
обобщения по поводу относительной роли различных факторов.
ЛИТЕРАТУРА
1. Allison А. С., Metabolic polymorphisms in mammals and their bearing
on problems of biochemical genetics, Am. Naturalist, 93, 5—16 (1959).
2. Camin J. H., Ehrlich P. R., Natural selection in water snakes (Natrix
sipedon L.) on islands in Lake Erie, Evolution, 12, 504—511 (1958).
3. Clarke B., Divergent effects of natural selection on two closely related
polymorphic snails, Heredity, 14, 423—443 (1960).
4. Dobzhansky T., Genetics and the Origin of Species, Columbia Univ.
Press, New York, 1951.
5. Dobzhansky T., Variation and evolution, Proc. Am. Phil. Soc., 103,
252—263 (1959).
6. Dobzhansky T., Evolution and environment. In Sol Tax (ed.), Evolu-
tion After Darwin, Univ. Chicago Press, Chicago, vol. 1, pp. 403—428 (1960).
7. Genetics and 20th Century Darwinism, Cold Spring Harbor Symposia on
Quantitative Biology, vol. 24, 1959.
8. Lerner I. M., Genetic Homeostasis, Wiley, New York, 1954.
9. Lerner I. M., The Genetic Basis of Selection, Wiley, New York, 1958.
10. Mich en er C. D., Distinctive type of primitive social behavior among
bees, Science, 127, 1046—1047 (1958).
11. Sheppard P. M., Natural Selection and Heredity, Hutchinson, London,
1959.
12. Sheppard P. M., Some contributions to population genetics resulting
from the study of the Lepidoptera, Advances in Genet., 10, 165—216 (1961).
13. Stebbins G. L., Variation and Evolution in Plants, Columbia Univ.
Press, New York, 1950.
14. Waddington С. H., Genetic assimilation, Advances in Genet., 10, 257—
293 (1961).
15. К e 11 1 e w e 11, Proc. X Int. Cong. Ent., 2, 1958.
16. К e 111 e w e 11, Heredity, 12, 1958.
17. Simpson G., et al., Quantitative Zoology, 2d ed., Harcourt, Brace and
Co., New York.
18. C a i n, S h e p p a r d P., Genetics, 39 (1954).
19. C a m i n, E h r 1 i c h P., Evolution, 14 (1958).
20. D о b z h a n s k у T., Evolution, 1 (1947).
21. Dobzhansky T., Genetics, 33 (1948).
22. P u n n e 11, Mimicry in Butterflies, Cambridge University Press, 1915.
23. M a t h e r K. et al., Heredity, 3 (1949).
24. Clausen J., Keck D., Hiesey W., Carnegie Inst. Wash. Publ. № 581
(1948).
Глава 8
ГЕНЕТИЧЕСКИЕ СИСТЕМЫ I
Рассмотренные в предшествующих главах цитогенетические
механизмы обеспечивают воспроизведение потомства, сходство
которого с родительскими типами достаточно для выживания
данной группы, если только среда не изменяется чересчур
быстро. Они обеспечивают также изменчивость, достаточную для
того, чтобы при постепенном изменении среды организмы тоже
изменялись и таким образом выживали. Для сохранения вида
или группы необходимо надлежащее равновесие между двумя
факторами: приспособленностью к условиям среды, существую-
щим в данное время, и так называемой перспективной приспо-
собленностью, т. е. подготовленностью к всевозможным измене-
ниям, происходящим в среде с течением времени. Как мы уже
указывали, у большинства организмов генетический аппарат
позволяет сохранять изменчивость в невыявленной форме, а так-
же регулировать выявление этой изменчивости. Разнообразие
средств для сохранения и внешнего проявления изменчивости,
которые, подобно другим признакам, должны находиться под
генетическим контролем, означает, что эти средства с течением
времени изменяются; иными словами, они подвержены эволю-
ции. Совокупность механизмов, контролирующих количество и
характер новых сочетаний генов, может быть названа генетиче-
ской системой. Таким образом, можно говорить об эволюции
генетических систем, понимая под этим эволюцию механизмов,
создающих и регулирующих изменчивость.
В этой главе будут описаны различные типы генетических
систем, встречающиеся у растений и животных. В следующей
главе мы более подробно рассмотрим ряд специализированных
генетических систем, которые приобрели особое значение у неко-
торых организмов. Выяснение взаимоотношений и эволюцион-
ного значения этих систем и включение их в эволюционную тео-
рию — одна из важнейших задач, стоящих перед современным
исследователем эволюции.
Хотя действительный ход эволюции генетических систем не-
известен, выдвигалось немало гипотез об основных направле-
ниях, в которых она должна была идти. Здесь мы рассмотрим
эти системы, начиная с самых простых и кончая наиболее
174 | Глава 8
сложными. Не следует отождествлять эту последовательность
с последовательностью развития рассматриваемых систем в про-
цессе эволюции. Нередко простейшие биологические явления
представляют собой, возможно, результат упрощения чего-то
более сложного.
ГЕНЕТИЧЕСКИЕ СИСТЕМЫ У МИКРООРГАНИЗМОВ
ТРАНСФОРМАЦИЯ
Трансформация — это особая форма наследственного изме-
нения, наблюдаемая у некоторых бактерий. Исследования на
штаммах пневмококка (Diplococcus) показали, что ДНК, вы-
деленная из штамма, обладающего определенным признаком,
способна «наделять» этим признаком штамм, ранее его лишен-
ный. Вирулентность пневмококков зависит от наличия у них
полисахаридной капсулы. Если бескапсульный, авирулентный
штамм выращивать на среде с очищенной ДНК вирулентного
штамма, то в культуре образуются вирулентные клетки с кап-
сулой. Таким образом, ДНК определяет образование полисаха-
ридной оболочки. Трансформация была получена в отношении
большого числа признаков, причем оказалось, что существует
равная вероятность трансформации в обоих направлениях. Об-
наружено также сцепление факторов в материале ДНК: в экс-
периментах с различными признаками двойные трансформации
происходят чаще, чем можно было бы ожидать в случае отсут-
ствия сцепления.
РЕКОМБИНАЦИЯ У ВИРУСОВ
Довольно сложные взаимоотношения существуют между
некоторыми бактериями и их вирусами, известными под назва-
нием бактериофагов. Каждая фаговая частица состоит из двух
главных структурных компонентов — хвоста и расширенной
головки (часто шестигранной формы). Белковая оболочка го-
ловки заключает в себе внутреннее «ядро» из ДНК. При зара-
жении бактериальной клетки фаговая частица прикрепляется
хвостом к специфическому рецепторному участку клеточной
стенки. В последующие несколько минут ДНК выходит из обо-
лочки фага и проникает в клетку. Внутри этой клетки фаг раз-
множается, и клетка в конце концов разрушается (подвергается
лизису), высвобождая новые частицы фага. В период размно-
жения синтезируются новая фаговая ДНК и новый фаговый
белок, и в результате образуется около сотни новых частиц того
типа, что и частица, вызвавшая заражение. Оказывается,
первоначальной частице удалось с помощью своей ДНК завла-
Генетические системы I | 175
деть синтетическими механизмами бактерии и использовать их
для собственных целей — для воспроизведения самой себя.
Были найдены фаги с самыми различными признаками.
Можно заразить культуру бактерий смесью фагов, отличаю-
щихся друг от друга какими-либо определенными признаками.
При этом у дочерних фаговых частиц наблюдается рекомбина-
ция. При некоторых сочетаниях фагов рекомбинанты появляются
с равной частотой; при других сочетаниях число рекомбинантов
оказывается сниженным, что указывает на сцепление генетиче-
ских факторов, влияющих на данные признаки. Таким образом,
мы имеем дело с рекомбинацией генетического материала бакте-
риофага внутри клетки бактерии-хозяина. Как показали иссле-
дования с радиоактивными изотопами, этот генетический мате-
риал представляет собой ДНК. Эти данные вместе с результат
тами экспериментов по трансформации бактерий, в которых
трансформирующим агентом также служит ДНК, свидетель-
ствуют о том, что и у этих форм генетическим материалом
является ДНК. Они показывают также, что при отсутствии по
лового размножения рекомбинация генетического материала
может происходить во время процессов синтеза, обеспечивающих
репликацию ДНК.
ТРАНСДУКЦИЯ
Рассмотренные выше бактериофаги вирулентны — они уби-
вают бактерию-хозяина, вызывая лизис ее клетки. Однако суще-
ствуют и «умеренные» фаги, способные вступать с бактерией в
своего рода симбиотические отношения, которые не приводят
к лизису клетки-хозяина. Бактерии, содержащие такой фаг, на-
зывают лизогенными (такова например, Salmonella) Неинфек-
ционную стадию умеренного фага называют профагом. Процессы
воспроизведения профага и бактерии так отрегулированы и со-
пряжены между собой, что присутствие профага не причиняет
клетке никакого вреда. Однако время от времени, с частотой
10~2—10-5 на одну клеточную генерацию, происходит лизис бак-
териальной клетки и высвобождающиеся фаговые частицы мо-
гут адсорбироваться на других клетках.
В процессе синтеза ДНК в профаг может включаться часть
генетического материала бактерии-хозяина. Если случится так,
что профаг перейдет в стадию фага, клетка-хозяин подвергнется
лизису. Когда освобожденная фаговая частица заражает новую
бактериальную клетку, она переносит с собой фрагмент генети-
ческого материала своего прежнего хозяина. Этот фрагмент
может быть включен в генотип бактерии-реципиента — явление,
известное под названием трансдукции. Переносимый генетиче-
ский материал обычно содержит не один наследственный
176 | Глава 8
фактор, а целую группу их. Таким образом, трансдукция — это
особый вид генетической рекомбинации, который опять-таки свя-
зан не с половым процессом, а с процессом инфекции. Фаг дей-
ствует как переносчик, осуществляющий инфекционную пере-
дачу бактериальных генов. Известны вирусы (в том числе и
фаги), которые вместо ДНК содержат РНК, но их изучение в
сущности еще только начинается.
ПОЛОВАЯ РЕКОМБИНАЦИЯ У БАКТЕРИЙ
У бактерий встречается и половая рекомбинация. Татум, Ле-
дерберг и другие исследователи показали, что у Escherichia coli
может происходить конъюгация клеток. Генетический материал
переходит из одной клетки в другую, причем одна из конъюги-
рующих клеток играет роль реципиента, а другая — роль до-
нора. Рекомбинация признаков, свойственных разным штаммам,
была изучена довольно детально. Процесс конъюгации можно
прерывать, разделяя клетки путем встряхивания; таким образом
удалось показать, что количество перенесенного генетического
материала пропорционально продолжительности конъюгации.
Генетический материал бактерии ведет себя так, как если бы он
весь находился в одной хромосоме. Для этой хромосомы были
построены карты сцепления. Может происходить спонтанный
разрыв хромосомы во время ее перехода в другую клетку. Были
исследованы также гибриды Е. coli и Shigella dysenieriae, а
также гибриды Е. coli и Salmonella typhimurium.
ГЕНЕТИКА И ЭВОЛЮЦИЯ МИКРООРГАНИЗМОВ
Значение и само существование таких явлений, как транс-
формация, трансдукция и половая рекомбинация, у многих ми-
кроорганизмов в естественных условиях не выяснены. Распро-
страненность этих процессов у различных микроорганизмов
также мало изучена. Совершенно очевидно, что популяционная
генетика этих организмов должна отличаться от генетики более
крупных форм. Вопрос о возможности какого-то «генетического»
родства между очень мелкими самовоспроизводящимися систе-
мами и более крупными организмами еще только начинают ис-
следовать. Большая часть известных явлений, относящихся к
этой области, описывается как заражение. На этом уровне раз-
граничение между генетическими процессами, заражением и
развитием становится, конечно, невозможным. Многие вирусы,
особенно содержащие РНК вирусы растений (которые могут
передаваться сосущими насекомыми), вызывают морфологиче-
ские эффекты, сходные с теми, которые контролируются хромо-
сомным генетическим материалом.
Генетические системы I [ 177
Некоторые штаммы парамеций (Paramecium) получили на-
звание «убийц», так как они выделяют в культуральную среду
вещество, токсичное для «чувствительных» штаммов парамеций.
Оказалось, что особи штаммов-«убийц» содержат особые ча-
стицы ДНК, называемые каппа-частицами. Для длительного
сохранения каппа-частиц в клетке необходимо, чтобы генотип
содержал доминантный ядерный фактор К. В процессе бесполого
размножения (деления) клеток каппа-частицы воспроизводятся
таким образом, что их получают все клетки образующегося
клона. В том случае, если клетки будут размножаться быстрее
каппа-частиц, могут появиться особи АТС, лишенные этих частиц.
Такие «чувствительные» особи — уже не «убийцы»; они могут
вновь приобрести каппа-частицы только извне, если будут по-
мещены в бесклеточную суспензию, приготовленную из клеток
«убийц». Они неспособны к новообразованию каппа-частиц, не-
смотря на то что их генотип допускает существование цитоплаз-
матического фактора каппа.
Половой процесс у парамеций связан с конъюгацией клеток
и с обменом цитоплазмой, при котором может происходить и
перенос каппа-частиц. При скрещивании особей КК и kk обра-
зующиеся парамеции Kk будут «убийцами», если конъюгация
длилась достаточно долго для того, чтобы каппа-частицы могли
перейти из клетки в клетку. Парамеции, гомозиготные по ре-
цессивному фактору k, могут получить каппа-частицы с цито-
плазмой, по при дальнейших клеточных делениях эти частицы
утрачиваются.
Известное сходство между этой ситуацией у Paramecium и
более разнообразными явлениями у бактерий и их вирусов оче-
видно. От наследования каппа-частиц до наследования разли-
чий в хлоропластах у растений всего один шаг. Хлоропласты —
это обладающие генетической преемственностью цитоплазмати-
ческие тельца, способные мутировать, что сопровождается изме-
нением формы и окраски. Мутантный хлоропласт может продол-
жать воспроизводиться в измененной форме. Отдельные участки
растения оказываются измененными, так что образуется мо-
заика из желтоватых или белых участков, чередующихся с обыч-
ными зелеными. Хлоропласты передаются через цитоплазму
яйцеклетки, но не через пыльцу. Некоторые мутации возникают
и поддерживаются в результате изменений в хромосомных ге-
нах; другие возникают вследствие какого-то изменения в ядре,
но сохраняются и при отсутствии хромосомного фактора; нако-
нец, есть такие пластидные мутации, которые, по-видимому, не
зависят от ядерного контроля. Если бы другие клеточные ком-
поненты, например митохондрии, было так же легко изучать,
как пластиды, то у них, возможно, обнаружились бы такие же
12 Зак. 15?
178 | Глава 8
явления. Следует отметить, что при вирусных инфекциях у ра-
стений иногда наблюдаются такого же рода изменения пластид,
какие возникают спонтанно или вызываются воздействием рент-
геновских или ультрафиолетовых лучей.
Этот экскурс в область явлений, обычно называемых цито-
плазматической наследственностью, показывает, насколько труд-
но провести границу между ядерной и цитоплазматической
наследственностью или между генетической рекомбинацией на
клеточном уровне и инфекцией у микроорганизмов. Нужно еще
раз повторить, что эволюционное значение таких явлений, как
цитоплазматическая и инфекционная наследственность, мало
изучено, но важность их вряд ли подлежит сомнению. Резуль-
таты ряда работ, проведенных как на растениях, так и на жи-
вотных, указывают на то, что некоторые признаки, считавшиеся
наследственными, на самом деле обусловлены вирусной инфек-
цией, или на то, что генетически детерминированные признаки
могут проявляться благодаря присутствию вирусов. Если рас-
сматривать такие процессы в связи с ролью микроорганизмов в
экосистеме или сообществе, можно обнаружить очень важные
взаимоотношения. Бактерии и другие микроорганизмы могут не
только играть роль разрушителей, но и различными способами
участвовать в передаче генетической информации в экосистеме.
ГЕНЕТИЧЕСКИЕ СИСТЕМЫ ДРУГИХ ОРГАНИЗМОВ
Основные черты генетических систем у огромного большин-
ства организмов одинаковы. У них имеет место чередование
гаплоидной и диплоидной фаз. У тех немногих вирусов и бак-
терий, которые были изучены, генетическая система проста и
более изменчива. Рекомбинация неизвестна только у сине-зеле-
ных водорослей (Cyanophyceae). В своих весьма специализиро-
ванных местообитаниях сине-зеленые водоросли, по-видимому,
представляют собой хорошо приспособленную и широко распро-
страненную группу. Кажется вероятным, что у них существует
какой-то пока еще не обнаруженный способ генетической реком-
бинации. Возможно, что он близок к механизму рекомбинации у
бактерий, а может быть, в нем участвуют и вирусы.
пол и диплоидпость
Явление полового размножения настолько широко распро-
странено и его эволюционное значение настолько очевидно, что
большинство биологов относят его возникновение к очень ран-
нему этапу эволюции жизни. Во многих группах растений и
животных каждый индивидуум образует как женские, так и
Генетические системы I | 179
мужские гаметы. Но даже и в этих случаях перекрестное опло-
дотворение часто является правилом. У раздельнополых орга-
низмов на тип полового развития могут влиять такие внешние
факторы, как интенсивность света, длительность фотопериода,
температура, химический состав среды и т. д. По-видимому, в
ходе эволюции этот довольно лабильный способ определения
пола заменяется, или по крайней мере дополняется, более ста-
бильными механизмами.
Первый вопрос, который мы должны рассмотреть, — это во-
прос о происхождении диплоидности. Обычно считают, что
диплоидность имеет большую селективную ценность, так как
дает возможность сохранять рецессивные гены, а тем самым и
изменчивость. Кажется несомненным, что диплоидное состояние
обеспечивает также необходимую стабильность развития и, сле-
довательно, большую независимость от влияний среды. По-ви-
димому, значение этих факторов должно было бы возрастать по
мере усложнения организмов. В тканевых культурах и в куль-
турах одноклеточных организмов настолько часто наблюдается
слияние свободных клеток, что нетрудно представить себе про-
исхождение сингамии. Возможно, что большая стабильность
генетических функций у возникающих при этом диплоидных кле-
ток могла бы сразу же иметь селективное преимущество.
Во многих случаях слияние клеток ведет к неустойчивости,
которая устраняется делением. В гл. 1 было высказано предпо-
ложение, что у первых протоорганизмов могла происходить
своего рода первичная рекомбинация. Поскольку многие низшие
водоросли и грибы в течение большей части своего жизненного
цикла находятся в гаплоидном состоянии (единственной дипло-
идной клеткой является зигота, которая сразу же делится),
можно было бы заключить, что рекомбинация, происходящая
при делении непосредственно после сингамии, имела большую
селективную ценность.
Половое размножение — это сложный процесс, происходящий
с участием многих компонентов, которые должны быть функцио-
нально интегрированы. Этапы его эволюции неизвестны. У про-
тоорганизмов с их нестабильной генетической системой каждый
«индивидуум», по-видимому, должен был отличаться от всех
остальных. Слияние «особей» в отдельных случаях могло приво-
дить к соединению дополняющих друг друга «генотипов», и
возможно, что именно в этом и состояла основа селективного пре-
имущества такого слияния. Исходя из соображений, приведен-
ных в гл. 1, можно предположить, что стабилизация генетиче-
ского механизма у ранних протоорганизмов была связана с
утратой излишнего генетического материала (или с переключе-
нием его на новые функции), с образованием связей между
12*
180 | Глава 8
нуклеотидами ДНК и белковыми молекулами, так что образовы-
вались хромосомы, и, наконец, с ограничением функций отдель-
ных генов, в результате которого выражение их стало более по-
стоянным, а контроль над признаками — более точным. В разно-
образных генетических системах растений и животных можно
встретить много причудливых и необычных явлений. Общие осо-
бенности этих систем указывают, по-видимому, на то, что почти
во всех случаях в них сочетаются преимущества, даваемые дип-
лоидностыо и рекомбинацией. Мейоз (редукция ядерного генети-
ческого материала) и слияние клеток сочетаются в жизненных
циклах различной степени сложности. Такое сочетание, видимо,
возникло у растений и животных независимо, несколькими раз-
личными путями.
Стеббинс пришел к выводу, что широкое распространение
гаплоидности у жгутиковых и у нитчатых водорослей обуслов-
лено их коротким и простым развитием и высокой скоростью
размножения; у таких организмов выработка сложных механиз-
мов, обеспечивающих генетический контроль над процессами
онтогенеза, не столь важна. Селективная ценность стабилизации
фенотипа и длительного сохранения рецессивных факторов тоже
должна быть меньшей. С усложнением организма продолжи-
тельность периода развития увеличивалась, что сопровожда-
лось удлинением жизненного цикла. В связи с этим оказалось
полезным образование интегрированных генных комплексов,
обеспечивающих стабилизацию онтогенеза и генетический гомео-
стаз, и диплоидное состояние приобрело высокую селективную
ценность.
ДППЛОЛДПЫЕ ЖИЗНЕННЫЕ ЦИКЛЫ И ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Некоторые группы Protozoa преимущественно диплоидны, и
им свойственны сложные процессы конъюгации, ведущие к ре-
комбинации генов. У многих групп водорослей, особенно у неко-
торых бурых водорослей (Phaeophyceae), диатомовых (Diato-
meae) и некоторых зеленых водорослей (Siphonales), большая
часть жизненного цикла также представлена диплоид-
ной фазой. В результате мейоза образуются гаплоидные
гаметы. Metazoa обладают генетической системой такого
же рода.
Как полагают, этот диплоидный жизненный цикл возникал
независимо в нескольких группах организмов с преимущественно
гаплоидным циклом. Кроме того, у растений и грибов вырабо-
тались жизненные циклы с чередованием поколений. У этих
форм наблюдается регулярная смена поколений гаплоидных
особей, производящих гаметы, и поколений, состоящих из дипло-
Генетические системы I | 181
идных особей, производящих бесполые споры. Гаметогенез про-
исходит путем митоза. Спорогенез связан с мейозом, который
сопровождается рекомбинацией. У некоторых водорослей и гри-
бов чередующиеся поколения изоморфны (морфологически не-
различимы) .
По мере усложнения организации наблюдается, по-видимому,
тенденция к редукции гаплоидного поколения — гаметофита —
и к преобладанию диплоидного спорофита. Например, у боль-
шинства папоротников гаметофит представлен небольшим заро-
стком, обычно не более сантиметра в поперечнике, в то время
как спорофит может достигать внушительных размеров. У голо-
семенных и цветковых растений гаметофит редуцирован до
сравнительно небольшого числа клеток (пыльцевое зерно — это
мужской гаметофит), а то растение, которое мы видим, пред-
ставляет собой спорофит. Напротив, у мхов привычное для нас
растение является гаметофитом. Быть может, правильнее было
бы рассматривать тип Bryophyta (мхи и печеночники) как
специализированную ветвь древних наземных растений, а не как
ответвление главного филогенетического ствола сосудистых ра-
стений.
У животных, за возможным исключением некоторых Sporo-
zoa, неизвестен процесс, сравнимый с чередованием поколений,
и гаплоидная фаза представлена у них только гаметами. У ки-
шечнополостных существует так называемое чередование поко-
лений медуз и полипов, но эти стадии различаются лишь мор-
фологически, а не цитологически.
Грибы как группа обладают рядом своеобразных генетиче-
ских систем, интересных для нас постольку, поскольку они про-
ливают свет на факторы отбора, действующие в эволюции гене-
тических систем. Эти высокоспециализированные организмы
мало изучены в цитогенетическом отношении. Наличие у них со-
матического кроссинговера и систем множественных «половых
типов» свидетельствует о селективном преимуществе рекомбина-
ции. У фикомицетов — водных плесневых грибов с развитым
мицелием, лишенным поперечных перегородок, — многочислен-
ные гаплоидные ядра лежат в общей цитоплазме нитей, или гиф.
У этих грибов в одном и том же мицелии могут возникать гене-
тически различные популяции ядер, создающих так называемое
гетерокариотическое состояние.
Базидиальные грибы (Basidiomycetes) — наиболее сложные
из грибов — и некоторые сумчатые грибы (Ascomycetes) пред-
ставляют интересную параллель с эволюцией диплоидности
У растений и животных. Их гаплоидные мицелии, часто пред-
ставленные различными «половыми типами скрещиваемости»,
182 | Глава 8
развиваются в почве. Клеточные гифы разных мицелиев, при-
ходя в соприкосновение, могут сливаться между собой. В конце
концов образуется мицелий, в котором каждая клетка содержит
два гаплоидных ядра (по одному от каждого из гаплоидных ми-
целиев), не сливающихся до тех пор, пока не сформируются
органы размножения. Этот особый вид диплоидности носит на-
звание дикариоза. Как диплоидность связана со сложностью
строения и развития у растений и животных, так дикариоз, по-
видимому, необходим для достижения большой морфологической
сложности у грибов.
РЕКОМБИНАЦИЯ И ГЕНЕТИЧЕСКИЕ СИСТЕМЫ
Итак, при сравнении многочисленных типов генетических си-
стем, выработавшихся в процессе эволюции, ясно обнаружи-
вается ряд общих особенностей. Во всех группах организмов,
кроме простых, быстро размножающихся одноклеточных форм,
диплоидность, по-видимому, имела селективную ценность. Про-
стые, быстро размножающиеся организмы благодаря самим
этим свойствам обладают большой гибкостью, и главным источ-
ником изменчивости у них служит мутационный процесс. Такие
организмы, как дрожжи и Paramecium, составляют исключение.
По мере усложнения организации создаются интегрированные
комплексы генов, управляющие процессами развития. Диплоид-
ное состояние (или дикариоз) обеспечивает необходимую ста-
бильность. Оно дает также возможность сохранять изменчивость
в форме рецессивных генов и полигенов в сбалансированных
системах. Мейоз и сингамия приводят к выявлению изменчиво-
сти в виде новых сочетаний генов.
Обычно считают, что при отсутствии изменений среды боль-
шая часть новых генных комбинаций будет обладать меньшей
селективной ценностью, чем существовавшие прежде комбина-
ции. В этом случае при половом размножении к потерям гамет
добавляется еще некоторая потеря зигот. Однако эта потеря
у диплоидных организмов с перекрестным оплодотворением, не-
сомненно, ниже, чем у гаплоидных. Потери в виде дефектных
рекомбинантов в системах из бактерий и бактериофагов, вероят-
но, еще значительно выше. Это позволяет предполагать, что про-
исходил также отбор в пользу таких генетических систем, кото-
рые не только обеспечивают стабилизацию, образование генных
комплексов, сохранение и постепенное выявление изменчивости,
но и снижают потери биологического материала, а тем самым и
энергии в процессе эволюции. (Однако не всякую «потерю» сле-
дует считать неблагоприятным событием.)
Генетические системы I | 183
СНИЖЕНИЕ ЧАСТОТЫ РЕКОМБИНАЦИЙ
Почти во всех генетических системах возникали изменения,
снижающие количество рекомбинаций. Результат этого — сни-
жение потерь, так как приспособленность к условиям данного
момента повышается. По общему мнению, наиболее примитив-
ные организмы были бесполыми; поэтому эти изменения обычно
рассматривают как проявления возврата к бесполому состоянию.
Поскольку механизмы рекомбинации были найдены почти у всех
организмов и поскольку способность к рекомбинации, по-види-
мому, является одним из фундаментальных свойств ДНК,
такое истолкование, пожалуй, неудачно. Половой процесс и дип-
лоидность, вероятно, выработались сравнительно рано. Сниже-
ние рекомбинации принимает много различных форм. У одно-
клеточных организмов может отсутствовать половое размноже-
ние. У более сложных многоклеточных организмов количество
рекомбинаций может снижаться в результате частичного или
полного подавления кроссинговера, выработки специализирован-
ных систем спаривания, а также в результате инбридинга, са-
мооплодотворения или утраты полового процесса. Как указывает
Стеббинс, такие видоизменения чаще всего встречаются у орга-
низмов-пионеров, численность популяций которых испытывает
значительные колебания. В условиях изменения окружающей
среды такие формы могут использовать новые для них место-
обитания путем быстрого воспроизведения немногих очень сход-
ных генотипов. При этом наряду с ускорением темпов размно-
жения часто уменьшаются размеры тела и упрощается развитие.
Цитогенетические механизмы, влияющие на рекомбинацию, бу-
дут рассматриваться в гл. 9; системы скрещивания рассмотрены
в настоящей главе.
Так как количество рекомбинаций подвержено влиянию от-
бора, генетическая система почти всякого организма все время
находится в состоянии изменения. Постоянное существование
изменчивых механизмов, повышающих или снижающих реком-
бинацию, создает систему, стабилизированную по отношению к
кратковременным изменениям среды, но способную реагировать
на ее долговременные изменения. Большинство систем скрещи-
вания в этом отношении «бивалентны» или сочетаются с дру-
гими механизмами, обеспечивающими эту бивалентность.
СИСТЕМЫ СКРЕЩИВАНИЯ И РЕКОМБИНАЦИЯ
Основным типом внутрипопуляционного скрещивания, с ко-
торым можно сравнивать другие типы, является случайное спа-
ривание. Если сходные особи скрещиваются между собой чаще,
Чем можно ожидать исходя из представления о случайности этого
184 | Глава 8
процесса, то систему называют положительно ассортативной.
Если имеет место отклонение от случайности в пользу спарива-
ния несходных особей, то систему называют отрицательно ассор-
тативной. Учитывая природу механизмов размножения у расте-
ний и животных, вряд ли можно думать, что когда-либо
происходит действительно случайное скрещивание. Трудно с точ-
ностью определить степень отклонения от случайности. Системы
скрещивания, разумеется, не распадаются на дискретные клас-
сы, и в любой природной популяции может встречаться целый
ряд произвольно выделяемых типов. Кроме того, на тип системы
скрещивания влияют такие факторы, как селективное преиму-
щество определенных признаков и размеры популяции.
Как указывалось в гл. 6, случайное скрещивание приводит
к постоянству частот генов и постоянству уровня изменчивости.
При наличии отбора частота генов изменяется, но изменчивость
популяции и генетическая корреляция между родственниками
остаются почти неизменными. Когда численность популяции до-
стигает нижних пределов, ошибка выборки приводит к генети-
ческому дрейфу. Малые размеры популяции ведут также к ин-
бридингу и к отклонению от скрещивания по закону случая,
если рассматривать вид в целом.
СИСТЕМЫ С ИНБРИДИНГОМ
Инбридинг можно рассматривать как положительное ассор-
тативное скрещивание, так как он увеличивает шансы на спари-
вание организмов, имеющих сходный генотип. Системы скрещи-
вания, ведущие к инбридингу, обусловливают распад популяции
па более мелкие группы, которые лишь редко обмениваются ге-
нетической информацией. Гетерозиготность популяции умень-
шается по мере фиксации генов в этих подгруппах. Генетическая
изменчивость популяции в целом возрастает, если только нет
сильного отбора в пользу определенных гомозигот. ЛАорфологи-
ческие и физиологические механизмы, ведущие к инбридингу,
весьма обычны у растений. Степень ограничения перекрестного
опыления может быть различной — от факультативного само-
опыления до облигатного самооплодотворения у форм с клей-
стогамными цветками (см. гл. 9).
Детальные исследования Стеббинса, Гранта и других пока-
зали, что степень ограничения рекомбинации у растений тесно
коррелирует с характером их роста и местообитанием. Как мы
увидим в гл. 9, в популяциях нередко сочетаются генетические
системы, влияющие на рекомбинацию противоположным обра-
зом. Травянистые растения, у которых продолжительность гене-
рации невелика и поэтому рекомбинация идет более интенсивно,
Генетические системы I | 185
обладают малым числом хромосом. У многолетних растений
с более длительным временем генерации число хромосом бывает
больше. У растений, входящих в экологические сообщества, ко-
торые обычно считают замкнутыми (в таких сообществах боль-
шая часть особей не доживает до созревания), более обычны
генетические системы, способствующие рекомбинации. Приме-
ром могут служить дубы. С другой стороны, организмы-пионеры,
где бы они ни оказались, являются членами открытых сооб-
ществ. Для того чтобы их зиготы выживали, у них должны быть
надлежащие генотипы: у таких организмов нет времени экспе-
риментировать с рекомбинантами. Неудивительно поэтому, что
растения пустынных областей, лугов и расчищенных от леса уча-
стков в тропиках обычно имеют генетические системы, обуслов-
ливающие пониженный уровень рекомбинации.
При вынужденном инбридинге в популяциях, которым обыч-
но свойствен аутбридинг, наблюдается снижение приспособлен-
ности, называемое инбредной депрессией. Отношения между
приспособленностью, гетерозиготностью и аутбридингом пока
еще не вполне ясны. Во многих группах растений инбридинг
успешно использовался в качестве основной системы скрещива-
ния в течение длительных периодов, сопровождаясь лишь спора-
дическим аутбридингом. Нельзя утверждать, что у всех орга-
низмов онтогенетический и генетический гомеостаз поддержи-
вается только с помощью аутбридинга и гетерозиготности.
СИСТЕМЫ, ОСНОВАННЫЕ НА АУТВРПДППГЕ
Отрицательное ассортативное скрещивание характеризуется
тем, что несходные особи скрещиваются чаще, чем этого следо-
вало бы ожидать исходя из представления о случайности про-
цесса. Дифференциация полов у животных обычно приводит к
невозможности самооплодотворения. Кроме того, у большинства
животных выработались системы различной сложности, которые
влияют на степень аутбридинга. Они были кратко рассмотрены
в гл. 5. Есть данные о том, что у насекомых на аутбридинг и
распространение генов влияют генетически контролируемые ком-
поненты процесса расселения особей. Поведенческие механизмы
как у беспозвоночных, так и у позвоночных нередко снижают
частоту скрещивания ближайших соседей. У Homo sapiens та-
кого рода этологические механизмы достигают крайней степени
развития.
У микроорганизмов на рекомбинацию влияет разнообразие
типов скрещиваемости. У растений, как и у животных, меха-
низмы расселения делают перекрестное оплодотворение между
ближайшими соседями более редким, чем следовало бы ожидать
186 | Глава s
при чисто случайном комбинировании. У цветковых растений
имеются механизмы опыления, которые также определяют коли-
чество и тип рекомбинаций. Обычно считают, что в эволюции
имела место тенденция к переходу от открытых цветков с много-
численными тычинками и пестиками, которые опыляются более
или менее случайным образом, к цветкам с немногими тычин-
ками и рыльцами, расположенными так, что пыльца с тычинок
попадает прямо на тело опылителя, а принесенная им пыльца —
прямо на рыльца.
У других растений имеются системы физиологической несов-
местимости, которые и обеспечивают аутбридинг. Поскольку для
размножения высших растений требуется не только опыление,
но и оплодотворение, этот процесс может быть прерван на мно-
гих этапах. Одна из обычных систем основана на взаимодей-
ствии множественных аллелей гена несовместимости S. Разно-
образные возможные генотипы влияют на процесс опыления
таким образом, что пыльцевая трубка растет очень медленно,
если пестик содержит тот же аллель S, который находится и
в мужском гаметофите. У цветковых растений изучены и дру-
гие, более сложные системы несовместимости.
ВЫВОДЫ
Способность к рекомбинации — одно из основных свойств
нуклеиновых кислот. Вирусы и бактерии обладают простыми,
изменчивыми генетическими системами, в которых рекомбинация
дает большое число нежизнеспособных продуктов. У жгутико-
вых и у нитчатых водорослей жизненный цикл оказывается пре-
имущественно гаплоидным с диплоидной зиготой, которая тот-
час же после образования делится путем мейоза. У растений и
животных — по-видимому, независимо в нескольких различных
группах — произошел переход от такого цикла к диплоидным
жизненным циклам и циклам с чередованием поколений. У гри-
бов выработались специализированные механизмы, приводящие
к рекомбинации и диплоидизации. Селективное преимущество
диплоидности и половой дифференциации, видимо, состоит в
том, что они обеспечивают стабилизацию онтогенетического раз-
вития, сохранение и выявление изменчивости и уменьшение бес-
полезной потери гамет и зигот. У многоклеточных организмов
процесс рекомбинации может видоизменяться благодаря систе-
мам спаривания, обусловливающим либо преимущественный ин-
бридинг, либо преимущественный аутбридинг.
Глава 9
ГЕНЕТИЧЕСКИЕ СИСТЕМЫ II
Как мы видели при рассмотрении эволюционных изменений
количественных признаков, рекомбинация является более суще-
ственным источником изменчивости, чем мутирование. Так как
генотип организма эволюционирует в условиях, определяемых,
помимо прочего, конституцией и организацией его хромосом, ис-
следование хромосомного механизма особенно важно при изуче-
нии эволюции. Основа изменчивости заложена в генетическом
коде, но изменения в структуре и числе хромосом, в которых
заключен этот код, могут прямо или косвенно влиять на степень
рекомбинации.
Гены мы определяем здесь как участки хромосомы между
ближайшими точками кроссинговера. Поэтому у высших орга-
низмов они представляют собой наименьшие единицы рекомби-
нации. Более крупными единицами рекомбинации могут быть
определенные части хромосомы, в которых кроссинговер или
его последствия ограниченны; их можно называть супергенами.
Существуют целые хромосомы (например, половые хро-
мосомы), которые ведут себя как единицы рекомбинации.
У организмов, размножающихся бесполым путем (некото-
рые категории апомиктических организмов), единицей пере-
дачи наследственной информации и рекомбинации служит ядро
в целом.
Цитологические механизмы, частично определяющие количе-
ство рекомбинаций в популяции, составляют важный элемент
генетической системы организма. Часто они очевидны из фено-
типической картины хромосом, т. е. кариотипа. Однако струк-
турные и генные изменения могут и не сопровождаться какими-
либо явными изменениями в кариотипе. Цитологические явления,
которые нам предстоит рассмотреть, обычно называют аберра-
циями или хромосомными мутациями. Поскольку они, по-види-
мому, происходят с измеримой частотой у большинства организ-
мов и становятся характерными признаками целых популяций
или таксономических групп животных и растений, их лучше все-
го рассматривать как аспект «цитологического репертуара» орга-
низмов. Подобно любым признакам, они возникают спонтанно
и поддерживаются отбором до тех пор, пока дают какое-либо
преимущество.
188 | Глава 9
МЕЙОТИЧЕСКИЙ ДРЕЙФ
У очень многих организмов различные аллели одного локуса
имеют неодинаковые шансы на включение в гаметы. Это явле-
ние было названо мейотическим дрейфом. Этот процесс является
элементом генетической системы, так как он снижает количество
рекомбинаций. Цитогенетические механизмы, влияющие на рас-
пределение аллелей, довольно подробно исследованы у Droso-
phila melanogaster. Очевидно, что при наличии мейотического
дрейфа частота какого-то аллеля в популяции могла бы возра-
стать, даже если бы он оказывал вредное действие. В этом слу-
чае отбор, по-видимому, приводил бы к накоплению факторов,
ослабляющих действие гена, ответственного за мейотический
дрейф. Не исключено, что мейотический дрейф мог бы привести
и к распространению благоприятного гена. Насколько часто это
происходило в природе, неизвестно.
ИЗМЕНЕНИЯ В СТРУКТУРЕ ХРОМОСОМ
Обычные цитологические механизмы, характерные для гапло-
идных и диплоидных организмов, уже были описаны. Прежде
чем идти дальше, читателю следует просмотреть приведенный
в гл. 3 материал о поведении хромосом, так как мы переходим
к обсуждению «производных» генетических систем — таких си-
стем, в которых цитологические изменения повышают или (ча-
ще) снижают количество рекомбинаций по сравнению с уров-
нем, который обычно принимают за «стандартный».
ИНВЕРСИИ
Инверсии — это такие структурные изменения хромосом, ко-
торые могут вести к образованию супергенов. Здесь участок хро-
мосомы оказывается перевернутым по сравнению со стандарт-
ной формой. У организма, гетерозиготного по инверсии, при
синапсисе в мейозе хромосомы одного из бивалентов закручива-
ются, образуя характерную петлю. Как мы уже видели, это ока-
зывает глубокое влияние на кроссинговер и рекомбинацию. Оди-
ночный перекрест внутри петли парацентрической инверсии (ин-
версии, не включающей центромеру) ведет к образованию ди-
центрической хроматиды и хромосомного фрагмента, лишенного
центромеры. В анафазе I эти продукты выявляются в виде мо-
стика, соединяющего дочерние ядра, и фрагмента, который не
движется к полюсу (фиг. 12). В зависимости от числа перекре-
стов и от того, какие хроматиды в них участвуют, все или часть
гамет оказываются генетически несбалансированными и поэтому
Генетические системы II ] 189
нежизнеспособными. Таким образом, гены всего инвертирован-
ного участка остаются сцепленными в суперген соответственной
длины. Гаметы, содержащие рекомбинации в этом супергене,
либо не выживают сами, либо дают нежизнеспособную зиготу.
В том и в другом случае рекомбинанты не образуются.
В длинной инверсии может произойти не один перекрест; ре-
зультаты будут зависеть от того, какие хроматиды участвуют во
втором перекресте — те же, что в первом, или другие. Если
это те же хроматиды, то второй перекрест компенсирует первый
и никакого видимого цитологического эффекта не обнаружится.
Если же в перекресте примет участие новая пара хроматид, то
в анафазе I появятся двойной мостик и два фрагмента. Возмож-
ны и различные другие конфигурации в зависимости от числа,
и локализации перекрестов. В случае перицентрической инвер-
сии (включающей центромеру) образуется суперген, но инвер-
сию нельзя обнаружить по наличию мостика и фрагментов, как
в случае парацентрических инверсий.
Эволюционный результат инверсий зависит от их относитель-
ной длины. Короткие или не очень длинные инверсии нередко
встречаются спорадически в популяциях растений и животных,
где они возникают спонтанно. Они получают широкое распро-
странение тогда, когда в них случайно окажется благоприятная
комбинация генов. Многочисленные примеры этого известны в
роде Drosophila (см. гл. 7). Оказалось, что одновременно с регу-
лярным появлением инверсий в определенных хромосомах может
наблюдаться заметное повышение частоты перекрестов в дру-
гих хромосомах. Таким образом, в результате инверсий не про-
сто снижается количество рекомбинаций: существенно то, что
они приводят к образованию супергеиов. Ведь если бы отбор
был направлен па общее подавление процесса рекомбинации, то
повышения частоты перекрестов в других хромосомах, вероят-
но, не наблюдалось бы.
При скрещивании в лаборатории особей из географически
изолированных популяций или особей разных видов часто ока-
зывается, что они различаются между собой очень длинными
инверсиями (случается, что инвертированным оказывается це-
лое плечо хромосомы). Наличие нескольких таких инверсий пре-
пятствует появлению плодовитого потомства; поэтому их можно
рассматривать как механизмы физиологической изоляции. Их
значение в природе не столь ясно, как значение коротких инвер-
сий. Кажется несомненным, что спонтанные инверсии могли бы
Довольно быстро вести к дифференциации кариотипов популя-
ций, оказавшихся изолированными друг от друга. Когда они воз-
никают регулярно, как у Drosophila и Sciara, они, видимо,
играют какую-то особую роль.
190 | Глава 9
ВЗАИМНЫЕ ТРАНСЛОКАЦИИ
Взаимные транслокации тоже могут приводить к образова-
нию супергенов в популяциях, гетерозиготных по этим пере-
стройкам. Если поменялись местами короткие участки, то у гете-
розиготного организма обычно могут образоваться только две
сбалансированные комбинации. В том случае, если между цен-
тромерой и местом транслокации произошел кроссинговер, толь-
ко две из четырех хроматид, сгруппированных в любом данном
участке, могут при своем расхождении образовать сбалансиро-
ванные комбинации (см. гл. 3). При двух или большем числе
обменных транслокаций между двумя парами хромосом в мей-
озе образуются кольца, состоящие более чем из четырех хромо-
сом. У некоторых организмов, например у Rhoeo и ряда видов
Oenothera, все хромосомы обмениваются участками и все оказы-
ваются соединенными, образуя одно общее кольцо в мейозе
(фиг. 37). В этих специализированных системах может происхо-
дить регулярное «поочередное» расхождение хромосом. Так как
синапсис бывает очень точным, отношения гомологичности при-
водят к тому, что каждая хромосома занимает в кольце опреде-
ленное место, которое может измениться только в результате
дальнейших структурных изменений.
Если происходит более одного обмена, то обособляется хро-
мосомный участок нового типа. Гомологичные плечи хромосом
называют парными сегментами, а часть хромосомы между ме-
стом обмена и центромерой — интерстициальным сегментом. Но-
вая область, связанная с центральной частью хромосомы, с кон-
цевыми участками которой она не конъюгирует, представляет
собой дифференциальный сегмент, который может содержать
или не содержать центромеру. В этой области кроссинговер на-
блюдается редко; если он происходит, то картина обменов видо-
изменяется, причем может измениться расположение хромосом
или число колец. Исследование взаимных транслокаций связано
с тщательным и трудоемким анализом скрещиваний между
большим числом различных особей, что позволяет определить
гомологию парных сегментов.
Спорадические транслокации встречаются у многих дипло-
идных растений (например, у Campanula, Paeonia), а также
у скорпионов и тараканов. У многих беспозвоночных трансло-
кации участвуют в системе половых хромосом. В тех случаях,
когда самка имеет формулу XX, а самец — Х0, одна из аутосом
может быть транслоцирована на Х-хромосому. В результате
одна из аутосом со своим гомологом становится частью видоиз-
мененной половой хромосомы (которую можно назвать «нео-Х»).
В дальнейшем эта аутосома ведет себя подобно Y-хромосомам
или гетерохроматиновым участкам хромосом вообще. Говорят,
ине
Oenothera hookeri
Oenothera biennis
Фиг. 37. Поведение хромосом у двух видов Oenothera,
и~Ое. hookeri, 7 бивалентов; б —кольцо из 14 хромосом в профазе мейоза у Ое. biennis;
в —кольцо из 14 хромосом в момент их „поочередного" расхождения в первой анафазе
мейоза у Ое. biennis [9, 10].
192 | Глава 9
что она подверглась гетерохроматизации, и называют ее «нео-Y».
Видимо, такой процесс произошел у некоторых прямокрылых и
богомолов, а также у ряда других беспозвоночных. Полагают,
что это сопровождается компенсаторным изменением активности
генов. Гены аутосомы, транслоцированной на Х-хромосому,
у самцов присутствуют теперь в половинном числе, так как ге-
терохроматизация, по-видимому, приводит к потере их актив-
ности в гомологичной аутосоме (нео-Y). Возможно, что системы
множественных половых хромосом (с несколькими X- и Y-хромо-
сомами) образуются именно таким путем, как это, очевидно,
произошло у Drosophila miranda. Эволюция кариотипа в группе
видов, близких к Drosophila virilis, также включает транслока-
ции между аутосомами и половыми хромосомами. Крайний слу-
чай множественности половых хромосом мы находим у палестин-
ского жука Blaps polychresta, самцы которого имеют 12 Х-хро-
мосом и 6 Y-хромосом, а самки — 24 Х-хромосомы.
Генетические системы, основанные на регулярно встречаю-
щейся структурной гибридности по взаимным транслокациям,
характерны для нескольких групп растений. Поведение транс-
локационных систем в природных популяциях растений в боль-
шинстве случаев мало изучено, хотя транслокационные кольца
описаны у представителей многих родов (например, Phoeo, Рае-
onia, Datura, Hypericum) и у многих Onagraceae. Исследования
Льюиса, Рейвена и их сотрудников приближают нас к понима-
нию роли таких механизмов в природных популяциях Clarkia и
Oenothera. Clarkia довольно детально изучена цитологически. Ее
виды, как правило, отличаются друг от друга по хромосомным
перестройкам, и межвидовые гибриды часто в высокой степени
стерильны. Как выяснилось, у многих видов Clarkia в природе
более 20% растений имеют хромосомные кольца, что указывает
на наличие транслокаций. Такие перестройки составляют часть
генетической системы и могут характеризовать целые популяции.
Интересный пример эволюции с участием хромосомных пере-
строек мы находим у Clarkia franciscana, изученной Льюисом и
Рейвеном. Это в основном самоопыляющееся растение встре-
чается лишь на одном участке (с выходом серпентинов) в окре-
стностях Сан-Франциско, который входит в ареал близкород-
ственного вида С. rubicunda. По-видимому, С. franciscana род-
ственна также северному виду С. атоепа. Исследование мейоза
у гибридов показывает, что С. franciscana отличается от С. ато-
епа по крайней мере двумя транслокациями и двумя парацент-
рическими инверсиями. От С. rubicunda она отличается по мень-
шей мере тремя транслокациями и четырьмя инверсиями
(фиг. 38). (С. атоепа и С. rubicunda различаются по крайней
мере тремя транслокациями и двумя инверсиями.) Льюис и
13 Зак. 152
Фиг. 38. Эволюция, связанная с хромосомными перестройками у Clarkia,
а —С. franciscana, 7 бивалентов; б —анафаза 1, гибрид С. rubicunda X С. franciscana', видны два мостика и два фрагмента; в —анафаза 1.
гибрид С. rubicunda X С. franciscana', видны цепь из 5 хромосом, цепь из 3 хромосом и 3 бивалента; г —анафаза I, гибрид С. атоепау^С. ru-
bicunda-, видны два мостика, два фрагмента и одна отставшая хромосома. На карте показано распространение этих трех видов Clarkia в районе
бухты Сан-Франциско |5].
194 | Глава 9
Рейвен заключили, что вид С. franciscana сравнительно недавно
произошел от С. rubicunda и что это, возможно, было резуль-
татом быстрой перестройки хромосомного комплекса, при кото-
рой возникло много различий в относительно короткое время.
Кажется также вероятным, что С. rubicunda произошла от
С. атоепа в более ранний период.
У некоторых растений, например у представителей рода
Oenothera (подрод Euoenotherd), выработались системы транс-
локаций поразительной сложности. Развитие этих систем, по-ви-
димому, было связано с отбором на гетерозиготность. Эта груп-
па растений хорошо изучена благодаря работам Клилэнда и
других исследователей. «Мутационная гипотеза» Де Фриза фак-
тически была основана на наблюдениях над Oenothera erythro-
sepala (Ое. lamarckiana). У некоторых видов подрода Euoenothe-
га, например у Ое. hookeri, имеются 7 пар хромосом (фиг. 37)
и небольшие кольца. У других, например у Ое. biennis, все
14 хромосом образуют в мейозе одно кольцо (фиг. 37). Кроме
взаимных транслокаций, в которых участвуют все хромосомы,
у них имеются сбалансированные группы летальных генов —
комплексы Реннера, которые препятствуют выживанию гомози-
готного потомства, так как ведут к образованию нежизнеспособ-
ных гамет или зигот. (Как уже говорилось выше, здесь наблю-
дается регулярное «поочередное» расхождение хромосом.) Виды,
у которых образуются транслокационные кольца, имеют мелкие
цветки и являются преимущественно самоопыляемыми (в проти-
воположность Ое. hookeri, например). Поэтому у них возникает
множество в высокой степени гетерозиготных, чаще всего нерас-
щепляющихся, частично изолированных рас. Происходящее ино-
гда перекрестное опыление между расами ведет к возникнове-
нию новых кольцевых систем и рас нового типа. Большое число
таких рас, представляющих собой первичные эволюционные еди-
ницы в группе «кольцеобразующих» форм Oenothera, сильно за-
трудняет изучение систематики этой группы.
Как указывает Клилэнд, на примере этих «кольцеобразую-
щих» форм Oenothera видно, что различные типы «мутаций», ко-
торые по отдельности можно было бы считать вредными, в соче-
тании могут создавать эффективную генетическую систему. Эти
формы составляют широко распространенную группу; среди них
бывают даже сорняки. Взаимные транслокации (приводящие
обычно к стерильности), летальные гены и самоопыление совме-
стно образуют систему, в которой поддерживается гетерозигот-
ность по всем хромосомам, и растения сохраняют высокую пло-
довитость. Подобная генетическая система обладает свойствами,
которые кажутся невыгодными. Рекомбинация здесь снижена,
возможна бесполезная потеря гамет и зигот. Нарушения в лю-
Генетические системы II | 195
бой части системы выводят из строя всю систему. Полиплоидия,
по-видимому, исключена, так как она несовместима с системой
единого кольца и комплексами летальных генов.
ИЗМЕНЕНИЯ ВЕЛИЧИНЫ И ФОРМЫ ХРОМОСОМ
Инверсии и транслокации по самой своей природе ведут к
изменению величины и внешнего вида хромосом. Перицентри-
ческие инверсии могут изменять положение центромеры, а тем
самым и относительную длину плечей хромосомы. Неравные
взаимные транслокации также могут изменять длину хромо-
сомы. Поскольку число перекрестов пропорционально длине пле-
ча, количество рекомбинаций может изменяться просто вслед-
ствие изменений этой длины. Как мы увидим дальше, если хро-
мосома очень мала (например, в результате транслокации) и
конъюгация ее нарушена, то она может быть утеряна, что при-
ведет к изменению основного числа хромосом.
ИЗМЕНЕНИЯ ЧИСЛА ХРОМОСОМ
Одним из показателей, характеризующих число рекомбина-
ций в той или иной генетической системе, может служить ре-
комбинационный индекс Дарлингтона. Этот показатель равен
сумме гаплоидного числа хромосом и среднего числа хиазм. Чем
больше рекомбинационный индекс (R.I.), тем больше число но-
вых сочетаний, возникающих в результате рекомбинации и рас-
щепления. Этот индекс не учитывает существования супергенов,
образующихся вследствие структурных изменений или апомик-
сиса, но он позволяет делать интересные сопоставления.
Увеличение основного числа хромосом повышает R.I.; по этой
причине изменения в числе хромосом можно рассматривать как
аспект генетической системы, имеющий отношение к балансу
между непосредственной приспособленностью и приспособляе-
1 мостью к медленным изменениям среды. Стеббинс произвел ана-
лиз распределения R.I. (по данным о числе хромосом) у цветко-
вых растений. У деревянистых растений (деревьев и кустарни-
ков) основное гаметическое число хромосом значительно боль-
ше, чем у травянистых растений. Это наводит на мысль, что
У долгоживущих растений отбор благоприятствовал высокому
рекомбинационному индексу. У организмов с короткой продол-
жительностью жизни, которые быстро размножаются, генети-
ческая система, по-видимому, в большей степени обеспечивает
непосредственную приспособленность ценой гибкости. Однако
известная гибкость обусловлена самой краткостью жизненного
Цикла с его быстрым «оборотом» генов. Стеббинс нашел, что
13*
196 | Глава 9
среди строго перекрестноопыляющихся травянистых цветковых
растений однолетние виды имеют в общем более низкие реком-
бинационные индексы, нежели многолетние. Большинство этих
однолетних видов — либо перекрестноопыляющиеся с низким
R.I., либо преимущественно самоопыляющиеся с высоким R.I.
Они, по-видимому, специально приспособлены для быстрого за'
селения однотипных местообитаний.
ПОЛИПЛОИДИЯ
Основное число хромосом, по-видимому, явно имело значе-
ние для отбора. Как происходят изменения этого числа? Мини-
мальное число различных хромосом, функционирующих как еди-
ное гармоничное целое, образует геном. Для целей нашего об-
суждения это число хромосом в геноме можно обозначить че-
рез х. Общее число хромосом в любом ядре называется его хро-
мосомным набором. Легко видеть, что такой набор может вклю-
чать один или несколько геномов или их частей. Рассматривая
регулярное чередование гаметического и зиготического числа
хромосом, удобно пользоваться символом п. Гаплоидное и ди-
плоидное число у всех организмов обозначают соответственно п
и 2п. Организмы, у которых произошло увеличение числа хро-
мосом, называют полиплоидными. Здесь п соответствует двум
или более х или долей х. У полиплоидных организмов п и 2п
обычно называют гаплоидным и диплоидным числами, хотя у
тетраплоида, например, п = 2х и 2гг = 4х. Самым распространен-
ным типом изменений является эуполиплоидия — необратимое
добавление целых геномов. Наиболее старые члены эуполипло-
идного ряда те, у которых числа хромосом наименьшие. Напри-
мер, если найдено, что у растения 2п = 22 (х=11) хромосом, то
близкородственная ему тетраплоидная форма должна иметь
2лг = 44 (х= 11) хромосом. Число хромосом в одном геноме тоже
может изменяться, что ведет к анэуплоидии; анэуплоидия обра-
тима и может возникать различными способами.
АНЭУПЛОИДИЯ
Имеются цитогенетические данные о том, что у организмов
в природных популяциях центромеры не возникают заново. Не-
возможно также добавление или исключение из генома генети-
чески активных хромосом. Однако условия, благоприятствующие
потере или приобретению хромосомы, могут возникнуть в ре-
зультате неравных взаимных транслокаций. У многих организ-
мов участок хромосомы, прилегающий к центромере, является
генетически инертным. Дарлингтон высказал предположение,
Генетические системы // | 197
что в случае транслокации активных эухроматических плечей на
другие хромосомы оставшийся гетерохроматиновый фрагмент с
центромерой обычно будет неспособен конъюгировать со своим
гомологом и может быть утрачен. При этом в геноме будет од-
ной хромосомой меньше, но количество генетического материала
останется прежним (фиг. 39). Таким образом, потомство ока-
жется цитогенетически изолированным от родительской формы;
если оно образует новую популяцию, можно ожидать дальней-
шей дифференциации. С другой стороны, в результате второй
транслокации к «центрическому» фрагменту с центромерой сно-
ва могла бы присоединиться часть активного генетического ма-
териала какой-нибудь другой хромосомы. Тогда конъюгация
опять будет нарушена и получатся гаметы разнообразных типов.
Может образоваться зигота, к генотипу которой добавлена
Потомство от сходных гамет
п+3 пар
Фиг. 39. Схема, показывающая, каким образом в результате взаимной транс-
локации неравных участков хромосом может произойти увеличение или
уменьшение основного числа хромосом.
Негомологичные хромосомы изображены белыми и заштрихованными; черным цвето.м обо-
Значецы участки хромосомы, которые, по-видимому, являются инертными [6, 7J.
198 | Глава 9
лишняя хромосома. При этом должно сохраняться исходное ко-
личество генетического материала, иначе новая комбинация бу-
дет нежизнеспособной.
Анэуплоидные изменения числа хромосом широко изучались
у растений. Обычно наблюдается уменьшение основного числа
хромосом. У скерды Crepis (сем. сложноцветных) можно
распознать индивидуальные хромосомы. У видов, которые, как
считают, остались примитивными и по другим фенотипическим
признакам, х=7, 6 или 5. У наиболее специализированных видов
х = 4 или 3. Правильность описанных выше представлений Дар-
лингтона подтвердил Тобги, который показал, что вид Crepis
fuliginosa (п = 3) произошел от С. neglecta или его предка
(гг = 4). У С. neglecta одно плечо С-хромосомы, по-видимому,
инертно, а другое перемещено на А-хромосому. Это видно по
конъюгации хромосом у гибрида Fi между обоими видами. Ана-
логичным образом вид С. kotschyana (п = 4) произошел от фор-
мы с /1 = 5, близкой к С. foetida.
Существуют многочисленные примеры, в которых можно
предполагать прогрессивное увеличение числа хромосом, но экс-
периментальных данных, к сожалению, очень мало. В роде
Clarkia, по-видимому, произошло добавление хромосом к геному.
Это увеличение числа хромосом может быть связано с образо-
ванием добавочных хромосом (см. ниже). Когда за потерей или
приобретением одной хромосомы следует амфидиплоидизация
(удвоение числа хромосом в результате гибридизации двух ди-
плоидов), может возникнуть анэуплоидия, имитирующая про-
грессивное увеличение числа хромосом. Часть такого ряда воз-
никла в роде Brassica (горчица), где числа х=8, 9 и 10, воз-
можно, представляют восходящий филогенетический ряд. Изве-
стны у природных форм или получены экспериментально числа п,
равные 17, 18, 19, 27 и 29. Род Carex (осока) содержит са-
мый обширный из известных анэуплоидных рядов. Здесь были
найдены гаплоидные числа от /1 = 6 до /г = 56, причем все числа
в пределах от 12 до 43 представлены одним или несколькими
видами. Как полагают, в этом ряду некоторые из чисел возник-
ли в результате структурных изменений или полиплоидии.
У дрозофилы, у некоторых прямокрылых и у рептилий не-
редки структурные перестройки, при которых из двух акроцен-
трических хромосом образуются большая метацентрическая хро-
мосома и мельчайший фрагмент, впоследствии исчезающий. Этот
процесс известен как центрическое слияние и представляет собой
особый вид взаимных транслокаций. Во многих семействах и
родах число длинных плечей постоянно, тогда как относитель-
ное число акроцентриков и метацентриков колеблется. Можно
привести много таких случаев. Интересным примером служит
Генетические системы II | 199
род сверчков Nemobius. У N. fasciatus имеется метацентрическая
Х-хромосома и 7 акроцентрических аутосом. У других видов
имеются добавочные метацентрики и меньшее число акроцентри-
ков — цо-видимому, это результат структурных перестроек.
Помимо основного набора хромосом, образующих геном, как
у растений, так и у животных могут быть лишние хромосомы,
называемые добавочными. Обычно эти добавочные хромосомы
не закрепляются, ибо они вызывают нарушения в генном ба-
лансе, а также в процессе мейоза, как это бывает у эксперимен-
тально получаемых трисомиков, тетрасомиков и т. д. Именно
поэтому, как правило, изменение основного числа может проис-
ходить только в результате взаимных транслокаций. Генетиче-
ский материал остается тем же самым: изменяется лишь его
распределение по отношению к центромерам. Очевидно, что до-
бавочные хромосомы должны быть в известном смысле ней-
тральными или иметь специальную функцию. Часто число их
неодинаково в различных клетках и у разных особей. Кажется
все же маловероятным, чтобы добавочные хромосомы былй пол-
ностью инертными, так как в некоторых случаях они могут со-
храняться в популяции.
У растений, у которых эти добавочные хромосомы обычно
называют В-хромосомами, они встречаются поразительно часто.
В-хромосомы меньше других хромосом и конъюгируют только
между собой. В большинстве случаев это гетерохроматиновые
структуры. Число их у разных особей варьирует, оно может быть
четным или нечетным, их может не быть вовсе и присутствие их
нельзя обнаружить по фенотипу растения. Возможно, В-хромо-
сомы вызывают какие-то физиологические эффекты, эволюцион-
ное значение которых, однако, неизвестно. У Clarkia добавоч-
ные хромосомы, вероятно, возникли в результате структурных
изменений в хромосомном наборе. У этих растений при гетеро-
зиготности по транслокациям часто наблюдаются кольца или
цепи из хромосом, и неравное расхождение может вести к об-
разованию трисомиков. У представителей рода Clarkia добавоч-
ные хромосомы по какой-то причине не нарушают генетический
баланс и размножение.
Добавочные хромосомы встречаются у многих беспозвоноч-
ных. По-видимому, они в основном представляют собой гетеро-
хроматиновые структуры. Некоторые из них в процессе митоза
стабильны; другим свойственно нерасхождение. Может быть,
этим добавочным хромосомам родственны так называемые
«ограниченные» («limited») хромосомы некоторых двукрылых.
Эти хромосомы имеются только в клетках зародышевого пути,
где они часто отличаются весьма своеобразным цитологическим
поведением. Например, у Sciara coprophila в соматических
Элиминируются
Отцовские
а
а
Зародыше- v\V
7
а
путь
„ чч & 1 s'
Яйцеклетка »L'
u** / Раннее
дробление
ххх
Зигота
t
1-я элиминация
I
I
2,-я элиминация
Сперматогонии
* 2-е деление
неравное
1-е деление,
избирательное
расщепление у.
!
Л
а 1
М 4-
а" й
X X
Спермий
Соматические_
клетки
xW
Сома
Соматические^
клетки ------
m
Л
I „Ограниченные” хромосомы
1 Материнская X- хромосома
• Отцовская Х-хромосома
а Аутосомы
Фиг. 40. Изменения хромосомного комплекса в жизненном цикле Sciara со prophila.
Объяснение в тексте [4].
? Сома
Генетические системы II | 201
клетках самцов имеется 7 хромосом, а в соматических клетках
самок — 8 (фиг. 40). Клетки зародышевого пути, из которых об-
разуются гаметы, содержат 10 (иногда 9 или 11) хромосом;
этот набор состоит из 4 пар хромосом, общих с соматическими
клетками, и еще одной пары крупных хромосом, никогда не
встречающихся нигде, кроме как в клетках зародышевого пути.
Это и есть «ограниченные» (L) хромосомы. При мейозе у самок
образуются яйца с полным гаплоидным набором, в который вхо-
дит и L-хромосома. У самцов мейоз необычен в том отношении,
что отсутствует синапсис и в первом делении образуется одно-
полюсное веретено. L-хромосома и по одной хромосоме каждой
из остальных пар отходят к полюсу веретена, а остальные хро-
мосомы (по генетическим данным, это отцовские хромосомы)
элиминируются, отходя в противоположном направлении и От-
шнуровываясь от клетки внутри небольшой почки. Во время вто-
рого деления образуется двухполюсное веретено, и хромосомы
ведут себя обычным образом, кроме Х-хромосомы; хотя она и
делится, но обе дочерние хромосомы отходят к одному полюсу.
Созревает только та сперматида, которая получает эти Х-хромо-
сомы; поэтому при каждом мейозе образуется лишь один спер-
мий. Он содержит 2L- и 2Х-хромосомы и по одной из аутосом
каждой пары; за исключением L-хромосом, все эти хромосомы
материнские.
После оплодотворения, приводящего к образованию зиготы
с тремя парами аутосом, тремя Х-хромосомами и (обычно) тре-
мя L-хромосомами, происходит ряд дроблений, при которых L-
и Х-хромосомы избирательно элиминируются из клеток. В конце
концов восстанавливается то число и распределение хромосом,
которое было в начале цикла. У эмбрионов будущих самцов
Х-хромосомы отцовского происхождения элиминированы, а у
эмбрионов будущих самок одна из отцовских Х-хромосом также
элиминируется и дегенерирует в цитоплазме. Здесь есть много
удивительных деталей, не упомянутых в нашем описании, и мно-
го нерешенных проблем, представляющих интерес для генетиков,
эмбриологов и исследователей эволюции. Поразительно, сколь
многие из наблюдаемых цитогенетиками явлений остаются не-
известными переменными в общей картине эволюционного про-
цесса.
ЭУПОЛППЛОИДИЯ
Эуполиплоидия как генетическая система очень часто встре-
чается у растений, но, по-видимому, крайне редка у животных.
Было высказано предположение, что увеличение числа хромо-
сом у животных нарушало бы механизм определения пола и по-
этому отбор препятствовал такому процессу. Только у партено-
202 | Глава 9
генетических форм встречаются полиплоидные ряды, сравнимые
с такими же рядами у растений. Однако число хро'мосом опре-
делено у сравнительно немногих организмов, и делать оконча-
тельные выводы о распространенности полиплоидии было бы
рискованно. Судя по имеющимся данным, полиплоидия, по-види-
мому, наиболее обычна у сосудистых растений. Поэтому в после-
дующем обсуждении мы будем касаться только этой группы.
Исследование числа и поведения хромосом у диких форм и
у гибридов, полученных в лаборатории, позволяет обычно иден-
тифицировать полиплоидные организмы. Иногда удается даже
синтезировать в лаборатории вид, известный в природе. Если
удается получить диплоидный гибрид, то можно вызвать удвое-
ние числа хромосом с помощью веществ, разрушающих меха-
низм веретена (например, колхицином). Если данный вид имеет
большое число хромосом и не скрещивается с другими формами,
то можно сделать лишь косвенное заключение о его полиплоид-
ной природе. Если синапсис у этого растения совершается без-
упречно и плодовитость высока, то следует считать, что оно сде-
лалось функционально диплоидным. Только методы сравнитель-
ной морфологии (включая сравнение белков) дают надежду на
выяснение происхождения этого растительного вида.
Рассматривая такого рода генетическую систему, обычно
проводят различие между теми полиплоидами, у которых все
геномы одинаковы, или автополиплоидами, и теми, у которых
геномы различны, — аллополиплоидами. На практике часто бы-
вает трудно провести различие между ними. Очевидно, что орга-
низмы являются автополиплоидными, если они возникли в ре-
зультате соматического удвоения числа хромосом (если только
родительская форма не была диплоидным гибридом). Растения,
полученные в результате слияния гаплоидных или полиплоидных
гамет одного и того же вида, обычно считают автополиплоида-
ми. Предполагается, что если не гены, то геномы у них в основ-
ном одинаковы. Трудность возникает тогда, когда речь идет
о скрещиваниях менее родственных форм и когда у гибридов
происходит удвоение хромосом, ведущее к сложным сочетаниям
автополиплоидии и аллополиплоидии.
Среди высших растений формы с гаплоидным спорофитом
с гаметическим числом хромосом (п = х) в соматических тканях
в природных популяциях неизвестны. Они, вероятно, могут быть
получены в культуре и представляют большой интерес для ци-
тогенетики, но не имеют прямого эволюционного значения.
Автополиплоиды в строгом смысле также редки в природе.
Наличие у автотриплоидного (Зх) или автотетраплоидного (4х)
организма трех или четырех одинаковых геномов приводит к за-
труднениям р мейозе: в каждой точке могут конъюгировать
Генетические системы // | 203
только две гомологичные хромосомы. Вместо обычных бивален-
тов могут возникать мультиваленты и (или) униваленты, слу-
чайно или в зависимости от отношения длины хромосом к числу
хиазм. Хотя многочисленные автополиплоидные организмы мо-
гут быть более «мощными» или обладать другими физиологиче-
скими свойствами, ценными с точки зрения селекционера, они
обычно неспособны размножаться половым путем и в природе
элиминируются отбором, если им не удастся выработать эффек-
тивный способ бесполого размножения (см. ниже). Понижение
плодовитости может не доходить до полного бесплодия (оно мо-
жет быть весьма незначительным); к тому же большинство выс-
ших растений обладает одним или несколькими способами
вегетативного размножения. Многие из наиболее важных деко-
ративных и плодовых растений являются триплоидами (большин-
ство сортов банана, многие сорта яблони, некоторые вишни,
японский ирис, тигровые лилии, тюльпаны). Кроме того, авто-
триплоиды и автотетраплоиды изредка спонтанно возникают
у животных (например, у некоторых саламандр). Однако в при-
роде автополиплоиды редко закрепляются в виде популяций.
В отличие от этого аллополиплоидия известна по существу
во всех основных группах растений. Хорошо известными и часто
обсуждаемыми примерами служат аллотетраплоидный гибрид
редьки с капустой — Raphanobrassica — и такой же гибрид двух
разводимых в культуре видов примулы — Primula kewensis.
Первый является результатом межродового, а второй — межви-
дового скрещивания; поэтому два объединенных в гибриде ге-
нома почти наверное должны быть различными.
У диплоидных межвидовых и межродовых гибридов хромо-
сомы могут вести себя по-разному. Например, у Raphanobras-
sica между хромосомами родительских видов имеется, по-види-
мому, так много небольших различий, что конъюгации хромосом
не происходит и в профазе мейоза оказываются только унива-
ленты. Так же обстоит дело при скрещивании Allium fistulosum
с А. сера. Униваленты расходятся беспорядочно, и лишь в ред-
ких случаях возникают клетки со сбалансированными наборами
хромосом. В противоположных крайних случаях, у гибрида
Primula и у гибрида Festuca pratensis (овсяница) X Lolium ре-
геппе (плевел многолетний), у диплоидов образуются бивален-
ты. Это позволяет предполагать, что число различий между ге-
номами меньше. Однако тем не менее диплоидные гибриды
стерильны.
Если число хромосом удваивается, хромосомы каждого ге-
нома получают в мейозе подходящих гомологов; образуются
биваленты и плодовитость восстанавливается. Получающиеся
при этом аллотетраплоиды иногда называют амфидиплоидами,
Фиг. 41. Полиплоидный
комплекс видов в роде
Asplenium (папоротники).
Объяснение в тексте [81.
rhlzophyllum
206 | Глава 9
так как по существу они диплоидны в отношении каждого из
родительских наборов: 2п = 2%i4-2%2. Аллотетраплоиды этого
типа встречаются в природе. Наблюдения над хромосомами ис-
кусственно полученных гибридов между предполагаемым тетра-
плоидом и его возможными «родителями» могут дать указания
относительно его происхождения. После возвратного скрещива-
ния с истинным родителем возникает смесь бивалентов и унива-
лентов. Очевидно, однако, что структуре строгого аллотетрапло-
ида присуща одна невыгодная в эволюционном отношении
особенность. Геномы могут оказаться настолько дифференциро-
ванными, что при соединении их в плодовитом тетраплоидном
гибриде между ними не будет происходить рекомбинации.
В каждом из родительских геномов рекомбинации, видимо, бу-
дут возникать в прежнем количестве, как это было у диплоида.
Поскольку конъюгация происходит совершенно нормально, гены
различных геномов не смогут рекомбинироваться и расщеп-
ляться.
В природе, вероятно, более обычны тетраплоиды, у которых
геномы частично дифференцированы, но все-таки еще настолько
сходны между собой, что образуются мультиваленты, у которых
возможен кроссинговер между хромосомами обоих родителей,
ведущий к генетической рекомбинации. Стеббинс называет это
сегментарной аллополиплоидией, так как здесь различия затра-
гивают лишь некоторые участки хромосом. Хотя сегментарные
аллополиплоиды имеют наибольшее значение для эволюции, их
трудно распознавать в природе. Они могут возникать в резуль-
тате гибридизации морфологически сходных форм; вследствие
образования мультивалентов они симулируют автополиплоидные
формы. Кроме того, из-за своей частичной стерильности они не-
стабильны. Отбор, действуя в направлении повышения плодови-
тости, может способствовать дальнейшей дифференциации хро-
мосом. Может произойти образование бивалентов, и тогда по-
лучится строгий аллополиплоид.
Есть много групп растений, в которых эволюционная диффе-
ренциация была связана с аллополиплоидией. Одним из лучших
примеров служит род папоротников Asplenium, изученный Ваг-
нером и его сотрудниками. Как показано на фиг. 41, существует
три первичных вида, у которых п = 36. Мейоз у них протекае!
нормально, при этом образуется 36 бивалентов. Изучение мор-
фологии и анализ поведения хромосом у других видов показали,
что A. bradleyi, A. ebenoides и A. pinnatifidum являются алло-
тетраплоидами, происшедшими от различных пар первичных
видов. Интересно, что известен стерильный диплоидный гибрид,
возникший из тех же исходных форм, что и A. ebenoides; у него
образуется 72 унивалента вместо 72 бивалентов. Asplenium iru-
Генетические системы // | 207
dellii произошел от возвратного скрещивания A. pinnatifidum
с одной из родительских форм, так как у него образуется 36 би-
валентов (геном montanani) и 36 унивалентов (геном rhizophyl-
lum). С другой стороны, A. gravesii — гибрид двух разных алло-
тетраплоидов, имеющих в качестве общей исходной формы
A. montanum. Это видно опять-таки из поведения хромосом:
хромосомы двух геномов montanum образуют биваленты, тогда
как в геномах platyneuron и rhizophyllum они остаются неспа-
ренными (36 бивалентов + 72 унивалента).
В других группах известны гораздо более сложные поли-
плоидные комплексы. Возможные комбинации геномов и сочета-
ния автополиплоидии и аллополиплоидии показаны на схеме
Стеббинса (фиг. 42) (хотя были найдены и комбинации более
высокого порядка). В некоторых группах аллотетраплоиды
способны давать частично плодовитые гибриды с автополипло-
идными формами каждого из своих родителей. У растений,
Дутоаллооктоплоид
A A A A BjBtBjBj
2-74 IV
24 0 П
Дутоаллогексаплоид
A A Bj Bj Вг Bj
1-7 IV
13-7 ц
Аллогексаплоид
A A BjBj С С
21 П
Сегментарный
аллотетроплоид
Дутотетраплоид
АААД
1-7 IV
12-0 II
Аллотетроплоид
A A BjBj
14 U
Bj Bj В2 В2
0-7 IV
14-011
A Bj Bj
711 71
Bud A Fj
A A A Bj
711 -------- 141
Триплоид Ft
A BjC
21 I
Bud Bj P} Bud B2 Bud C
Bj Bj Bj B2 B2 B2 CC
7 П -*-141 711 7П
ДиплоиЪныи вид n = 7
Фиг. 42. Схема, показывающая различные уровни и типы полиплоидии, со-
став геномов полиплоидов и пути их возникновения.
Буквами обозначены геномы; римские цифры указывают на поведение хромосом при конъю-
гации в мейозе: / — униваленты; // — биваленты и т. д. (например, 24-0// означает; от 24 до Q
бивалентов) (6).
208 | Глава 9
особенно у тех, которые в какой-то мере могут размножаться
вегетативно, результатом этого может быть «размывание» или
полное стирание морфологических границ между исходными
таксонами. Может возникнуть группа организмов с разными
степенями плоидности, разнообразной морфологией и различ-
ными требованиями к условиям среды. Такие комплексы можно
разделить только путем цитологического анализа, да и то иногда
лишь частично. Ситуация может оказаться еще сложнее, если
растения размножаются апомиктически, так что поддерживается
популяция особей, в половом отношении абсолютно стерильных
(см. ниже).
Трудно определить общее значение полиплоидии как генети-
ческой системы. По произведенной оценке, до половины всех
цветковых растений имеют полиплоидное происхождение. Рав-
ным образом полиплоидия распространена у папоротников и
родственных им форм, а также у водорослей и мхов. Только
грибы составляют исключение, однако лишь очень небольшая
часть их изучена цитологически. К сожалению, мало что из-
вестно о тропических видах, так что исследованные растения
далеко не составляют представительной выборки. Для целых
семейств или подсемейств (например, Pomoideae — яблони и
родственные формы — из сем. Rosaceae), а также родов харак-
терны числа хромосом, позволяющие предполагать полиплоид-
ное происхождение. Хотя у животных также происходит кратное
увеличение числа хромосом, его обычно не объясняют полиплои-
дизацией. Как мы уже говорили, причиной его считают центриче-
ское слияние.
Следствия полиплоидии как генетической системы много-
образны и подчас противоположны друг другу. Например, авто-
полиплоидия может резко ограничить или устранить генетиче-
скую рекомбинацию у данного организма, и тогда она служит
связующим звеном между аллополиплоидом и одним из его ро-
дителей. У строгого амфидиплоида рекомбинационные потенции
ограничены теми возможностями, которые были у исходных
форм, но при сегментарной аллополиплоидии может происходить
и рекомбинация родительских признаков, в том числе признаков,
касающихся «экологических предпочтений». В целом можно бы-
ло бы сказать, что полиплоидия ведет к увеличению размеров
эволюционных единиц, так как способствует повышению частоты
случайных скрещиваний между группами организмов. Эта си-
туация сходна с постулированными Райтом условиями наиболее
быстрой эволюции: частично изолированные в пространстве
субъединицы время от времени обмениваются генетическим ма-
териалом. Это делает возможным дальнейшее использование
преимуществ гибридизации на диплоидном уровне,
Генетические системы II | 209
АПОМИКСИС
Предельным случаем ограничения рекомбинации является
полное исключение полового размножения. В крайних случаях
рекомбинация может быть совершенно исключена. Видимо, это
бывает сравнительно редко; существует много типов бесполого
размножения, при которых рекомбинация в той или иной мере
сохраняется. Термин апомиксис обозначает все формы бесполого
размножения, т. е. те способы воспроизведения, которые частично
или полностью заменяют половое размножение. Классификация
типов апомиксиса чрезвычайно сложна, так как могут встре-
чаться разнообразные случаи, промежуточные между типичным
половым размножением и облигатным апомиксисом. У боль-
шинства растений в апомиксисе участвует либо гаметофит, либо
спорофит, либо обе эти фазы. Терминология, относящаяся к апо-
миксису, также стала чрезмерно сложной. В нижеследующем
обзоре она будет по возможности упрощена.
Апомиксис может быть факультативным или облигатным.
Некоторые организмы размножаются иногда половым, а иногда
бесполым способом, обычно в зависимости от внешних условий.
Однако у животных регулируемое чередование полового и бес-
полого размножения не является, строго говоря, факультатив-
ным апомиксисом, так как половая часть цикла облигатна.
Простейшая форма апомиксиса — вегетативное размножение.
Оно может быть единственным способом размножения. Из-
вестно, например, что некоторые водные растения размножаются
только бесполым путем в северной Европе (такова Elodea, у ко-
торой здесь встречаются только женские растения) и половым
путем — в других частях своего ареала. Самостерильный трипло-
идный лилейник (Hemerocallis fulva) в Америке размножается
бесполым путем. Такого рода вегетативное размножение — пу-
тем укоренения и роста отломанных побегов или путем обра-
зования специализированных пропагул (видоизмененных побегов
или почек) — ведет к созданию популяции генетически тожде-
ственных организмов. Такую популяцию называют клоном.
Клоны могут образовать животные, размножающиеся делением
(Protozoa) или почкованием (Coelenterata, Annelides и др.).
С точки зрения генетики клон представляет собой один инди-
видуум. Так как члены такой популяции обычно в конце концов
отделяются друг от друга, физически они являются особями и
в экологическом отношении также играют роль особей.
Как у растений, так и у животных иногда происходит вегета-
тивное размножение особей, способных также и к половому
Размножению и обладающих, таким образом, двумя способами
воспроизведения. Такое вегетативное размножение обычно не
Называют апомиксисом, кроме тех случаев, когда оно служит
14 Зак. 152
210 | Глава 9
Фиг. 43. Растение фиалки (Viola) с типичными цветками, клейстогамными
цветками и органами вегетативного размножения (ползучими подземными
побегами).
единственным способом воспроизведения на протяжении какой-
либо части жизненного цикла. Обыкновенно, как, скажем, у не-
которых видов Viola (фиалка), половое и бесполое размножение
происходят одновременно (фиг. 43). Например, у основания
отдельных растений Viola возникают отпрыски или столоны; из
них в конце концов вырастают новые розетки листьев, которые
могут отделиться от родительского растения. У фиалки обра-
зуются и типичные цветки; они опыляются насекомыми и дают
гетерозиготные семена. В то же время могут образоваться и
клейстогамные цветки. Они расположены среди листьев у самой
земли, никогда не раскрываются и самоопыляются в бутоне.
Генетические системы II | 211
Фиг. 44. Растение лука (Allium), у которого
некоторые цветки в соцветии замещены луко-
вичками.
Семена от этих цветков, попадая прямо на землю после растре-
скивания плода, дают растения, генетически сходные с роди-
тельскими. Таким образом, здесь у одного и того же растения
одновременно осуществляется три способа размножения, обусло-
вливающих различную степень генетической рекомбинации.
Известны также несколько менее явные формы вегетативно-
то размножения, без образования семян. Сюда относится ряд
примеров так называемого «живорождения» у растений. Пропа-
гУлами здесь служат миниатюрные луковички, которые обра-
зуются в соцветиях и могут замещать цветки. Таким способом
Размножаются, например, некоторые виды лука (Allium, фиг- 44)
14*
212 | Глава 9
и мятлика (Роа). С точки зрения развития можно сказать, что
отдельные цветки, представляющие собой видоизмененные спо-
роносные побеги, превращаются в специализированные вегета-
тивные побеги, которые отпадают и впоследствии укореняются
и начинают расти. В результате возникает клон.
Кроме этих сравнительно простых способов апомиктического
размножения, существуют еще способы размножения, связанные
с образованием семян; эти формы апомиксиса поразительно
сложны, а их последствия очень трудны для понимания. В сово-
купности эти типы размножения называют агамоспермией. Рас-
сматривая их на примере одних только высших растений, сле-
дует прежде всего вспомнить о том, что в жизненном цикле
последних имеется фаза гаметофита, которая отделяет мейоз от
оплодотворения и чередуется с поколением спорофита. Гамето-
фит может состоять всего лишь из нескольких клеток (напри-
мер, из 8 клеток — у многих цветковых растений), но может
быть представлен и относительно массивным многоклеточным
растением, физиологически независимым от спорофита (как у
большинства папоротников). В обоих случаях в этом поколении
возможен апомиксис, если яйцеклетка развивается без оплодо-
творения. Четыре основные формы апомиксиса, связанные с по-
колением гаметофита, известны под общим названием гамето-
фитного апомиксиса; сюда относятся и такие формы, при кото-
рых новый спорофит развивается не из яйцеклетки, а из других
клеток. Развитие яйцеклетки без оплодотворения называется
партеногенезом, тогда как апогамией называют образование
спорофита из других клеток гаметофита. Продукты апомиксиса
обоих типов бывают обычно диплоидными; гаплоидный парте-
ногенез и гаплоидная апогамия не имеют эволюционного значе-
ния у сосудистых растений.
Иногда в одном из спорофитных поколений происходит
какое-нибудь нарушение мейоза или мейоз может полно-стью
выпасть. В этом случае споры и развивающиеся из них гамето-
фиты будут диплоидными. Здесь возможны два варианта. Те
клетки спорофита, которые обычно делятся путем мейоза (на-
пример, в семяпочке), могут подвергнуться митотическому или
частично мейотическому делению; это называется диплоспорией.
У некоторых растений гаметофит может развиваться из сомати-
ческих клеток репродуктивных структур путем митотических
делений. Этот процесс — апоспория — также ведет к образова-
нию диплоидного гаметофита. В ходе дальнейшего развития
такого гаметофита возможны партеногенез и апогамия, о кото-
рых говорилось выше.
Для развития может требоваться воздействие пыльцевой
трубки или образование питательной ткани, называемой эндо
Генетические системы II | 213
спермой (это тоже продукт оплодотворения), которая окружает
возникший в результате апомиксиса зародыш. Таким образом,
опыление необходимо, но оплодотворения яйцеклетки не проис-
ходит. Эта так называемая псевдогамия — одна из самых труд-
ных для выявления форм апомиксиса.
Апомиксис у высших растений может доходить до полной
редукции гаметофита; при этом наблюдается так называемая
адвентивная эмбриония, разновидность агамоспермии, когда из
клеток интегумента или нуцеллуса семяпочки путем митотиче-
ских делений образуется новый спорофит. Таким образом, ткань
спорофита непосредственно дает начало новому спорофиту, ко-
торый заключен в обычные покровы семени и внешне неотличим
от «нормального». Адвентивная эмбриония может быть также
псевдогамной.
Из сказанного ясно, что при апоспории и адвентивной эм-
брионии возникают растения, генетически тождественные роди-
тельскому растению. Поскольку диплоспория может быть свя-
зана с неполным мейозом, существует возможность генетической
рекомбинации. Природа и возможные результаты частичной
рекомбинации этого рода рассматриваются ниже в связи с те-
литокией у животных.
Если у высших растений апомиксис усложняется из-за чере-
дования поколений, то у высших животных он усложняется из-за
половой дифференцировки. В некоторых группах животных сам-
цы развиваются из неоплодотворенных яиц и бывают гаплоид-
ными. Самки образуются из оплодотворенных яиц и поэтому
диплоидны. Очевидно, что апомиксис этого типа, называемый
еаплодиплоидией, гаплоидным партеногенезом или арреното-
кией, может служить механизмом определения пола. По-види-
мому, гаплодиплоидия возникла независимо в семи (или около
того) группах Metazoa — у некоторых насекомых, паукообраз-
ных и коловраток. Среди насекомых эта форма апомиксиса и
определения пола характерна для всего отряда Hymenoptera и
известна также у Homoptera, Coleoptera и Thysanoptera.
При гаплодиплоидии у самца не происходит истинного мейо-
за, сперма образуется путем митоза или ложного мейоза и
поэтому генотип ее (если исключить мутации) идентичен роди-
тельскому. В эволюционной генетике гаплодиплоидных организ-
мов еще много неясного. Очевидно, например, что у видов с
такой генетической системой рецессивные гены должны тотчас
проявляться у самцов, если этому не препятствуют какие-то осо-
бые механизмы. Высказывалось предположение, что благодаря
Какому-то неизвестному механизму дупликации генов эти самцы
Могут быть функционально диплоидными. Хорошо известно, что
большинство высокодифференцированных тканей организма
214 | Глава 9
бывает эндополиплоидным (часто в высокой степени); это могло
бы защищать сому от вредных мутаций. С другой стороны,
гаплоидные самцы могли бы служить своего рода фильтрами
против рецессивных леталей.
Несколько иной вид апомиксиса у животных называют тела-
токией; это разновидность партеногенеза, при котором из не-
оплодотворенных яиц развиваются самки. По-видимому, он воз-
ник независимо в большинстве основных групп животных, в том
числе у позвоночных, в естественных условиях. У некоторых
животных наблюдается полная телитокия: самцы редки или не-
известны, все особи развиваются из неоплодотворенных яиц.
Более обычна циклическая телитокия, когда имеет место чередо-
вание (обыкновенно годичный цикл) половых и бесполых поко-
лений. Этот вид телитокии встречается у тлей, орехотворок, вет-
вистоусых раков, многих паразитических червей и коловраток.
Как и у растений, редукционное деление в случае телитокии
может полностью отсутствовать, и яйцеклетка образуется в ре-
зультате митотических делений. В этом случае она диплоидная.
Возможен и более или менее нормальный мейоз, но тогда позже
происходит удвоение хромосом, восстанавливающее диплоидное
состояние (фиг. 45). Это сравнимо с результатами диплоспории
у растений. Обычными продуктами мейоза являются 4 гаплоид-
ные клетки. У высших растений это мегаспоры, а у животных —
яйцеклетка и 2 (или 3) полярных тельца. Если совершается пер-
вое деление мейоза, то может происходить кроссинговер, что
создает основу для рекомбинации. Если же дочерние клетки
после второго деления мейоза попарно сливаются, то образуются
только две клетки — в таком случае мейоз и рекомбинация ча-
стично подавлены. У животных это может быть результатом
слияния ядра яйцеклетки с ядром второго полярного тельца.
Получаются диплоидные клетки, в которых могла произойти
ограниченная рекомбинация.
Уайт указал на то, что поскольку при амейотической тели-
токии не может происходить никакого расщепления, рецессии
ные мутации и перестройки хромосом имеют тенденцию нака-
пливаться. Дочерние особи будут похожи на своих матерей, но
линия будет становиться все более и более гетерозиготной. Воз-
можно, что этим объясняется мощность некоторых апомиктиче-
ских организмов. В то же время при телитокии с частичным
мейозом, когда возможно расщепление, существующая гетерози-
готность будет со временем уменьшаться, так как она не будет
восстанавливаться в результате слияния гамет.
Влияние апомиксиса на организмы многообразно и сложно.
И у растений, и у животных апомиксису обычно сопутствуют
гибридность и полиплоидия, однако нет оснований усматривать
Генетические системы II | 215
причинную связь между этими тремя явлениями. Если главной
причиной сравнительной редкости полиплоидии у животных
является их механизм определения пола, то ясно, что у живот-
ных, которым свойственна телитокия, нет препятствий для поли-
плоидизации. Кроме того, разнообразные элементы цитологиче-
ского механизма апомиксиса контролируются большим числом
генов, которые по отдельности чаще всего имеют отрицательную
селективную ценность. Например, мутации, нарушающие про-
цесс формирования веретена, должны быть вредными, если они
не сочетаются с мутациями, ведущими к образованию реститу-
ционных ядер и к развитию яйца без оплодотворения. В резуль-
тате гибридизации эти гены могут оказаться соединенными в
Родительские
хромосомы
Диакинез
Первое
деление
мейоза
Второе
деление
мейоза
ь- ь- ь-
2-е полярное 2-е полярное
Единствен^
ное поляр*
-а'ное тельце
тельце тельце
а | b aib' а
O-J I O-L—, I
О-Т—г- От-- I О-Г-
а b а Ь' а’
Хромосомы
потомства
Мейотический
тип 1а
Меиотический
тип 1 b
Амейотический
тип
Фиг. 45. Схема, показывающая три типа партеногенеза и их генетические
следствия.
^®йотический тип 1а —соматическое число хромосом восстанавливается в результате слия-
С«со ЯДРа ВТ0Р0Г0 полярного тельца с ядром яйцеклетки; мейотический тип 16 — соматиче-
Них Число хромосом восстанавливается после дробления яйца в результате слияния дочер-
* ядер; амейотический тип —яйцеклетка образуется путем митотических делений [4].
216 | Глава 9
общей функциональной системе. Было также высказано предпо-
ложение, что полиплоидный организм обладает некоторого рода
«буферностью».
У тех растений, у которых апомиксис сделался постоянной
особенностью генетической системы, очень часто возникают так
называемые агамные комплексы — сущее наказание как для си-
стематика, так и для цитолога. Роды Rubus (малина и ежевика)
и Роа (мятлик) —хорошо известные примеры групп, внутри
которых родственные отношения настолько запутаны, что сколь-
ко-нибудь ясное понимание филогении группы в целом стано-
вится невозможным.
Американские виды Crepis — единственный крупный агамный
комплекс, который во всем своем объеме изучен как в си-
стематическом, так и в цитологическом отношении. Интересно,
что из 196 видов Crepis (сгруппированных в 27 секций) апомик-
сис и полиплоидия известны только у 5 видов секции Pyrimaches
в юго-восточной Азии и у 9 видов секции Psilochaena в Америке.
10 североамериканских видов секции Psilochaena составляют
наиболее гетерогенную группу. Свойственное им основное число
хромосом (х=11) не встречается нигде в других секциях и воз-
никло оно, возможно, в результате межвидовой гибридизации
азиатских видов с п = 7 и п = 4 или с п = Ъ и /г = 6. Crepis runci-
nata (2п = 22) —единственный представитель секции, у которого
нет ни апомиксиса, ни полиплоидии. Из остальных видов семь
включают одну 22-хромосомную диплоидную форму, к которой
примыкают многочисленные апомиктические полиплоиды. Два
других вида содержат только полиплоидные апомиктические
типы, возникшие путем гибридизации двух или большего числа
видов из упомянутых выше семи.
Таким образом, полиплоидные апомиктические формы про-
исходят от семи первичных диплоидных типов. Пять из них
имеют весьма ограниченное географическое распространение и
сильно различаются по своим экологическим «наклонностям».
Неизвестно ни одной области, где два диплоида встречались бы
совместно и где не было бы найдено диплоидных гибридов.
У полиплоидов, имеющих 33, 44, 55, 77 или 88 хромосом, соче-
таются признаки двух или большего числа диплоидов; все это
апомиктические формы (у некоторых апомиксис факультатив-
ный). Как правило, ареалы у апомиктов шире, чем у их дипло-
идных предков.
Анализ этого агамного комплекса может служить примером
решения таких проблем вообще. Для раскрытия взаимоотноше-
ний и построения полезной классификации необходимо было рас
познать первичные сингамные (размножающиеся половым пу
тем) диплоидные формы. Вокруг них можно было сгруппировать
Генетические системы II | 217
Ц. производные автополиплоидные и аллополиплоидные апомикти-
•; ческие формы. Хотя изменчивость вначале кажется совершенно
J не поддающейся анализу, в ней все-таки можно разобраться, и
\ при этом выясняются интересные факты. Например, разнообра-
зие оказывается наибольшим в областях распространения син-
гамных видов и меньшим — в периферических участках ареала,
где встречаются только апомикты. Последние, по-видимому, рас-
селились (после своего образования путем гибридизации и по-
липлоидизации) за пределы ареала своих сингамных предков.
Агамные комплексы у растений, не относящихся к роду Сге-
pis, менее изучены. Среди них есть как совсем небольшие, так
и чрезвычайно обширные и сложные комплексы. Число хромо-
сом может сильно возрастать, а набор их становится несбалан-
сированным. Изучение других родов оказалось крайне трудным
в тех случаях, где предковые сингамные диплоиды вымерли
(Rubus) или где может быть несколько агамных комплексов
внутри одного вида (Роа).
У животных крупные агамные комплексы встречаются реже.
Хорошо известен живущий в соленых озерах и лиманах рачок
Artemia salina — вид, представленный сингамными диплоидами
и размножающийся путем образования телитокных триплоидов,
тетраплоидов, пентаплоидов, октоплоидов и декаплоидов. У Сиг-
culionidae (жуки-долгоносики) и Lumbricidae (дождевые черви)
обнаружены более обширные комплексы; в них могут входить ди-
плоиды, триплоиды, тетраплоиды и пентаплоиды, а иногда даже
гексаплоиды и декаплоиды. Есть основание полагать, что у долго-
носиков изредка происходит оплодотворение партеногенетически
развивающихся яиц спермой от двуполых рас и видов.
Тринадцать видов дождевых червей, размножающихся путем
телитокии, были исследованы цитологически; все они оказались
полиплоидами — от триплоидов до возможного декаплоида. Не-
которые сингамные виды также являются полиплоидными. Так
как Lumbricidae обычно гермафродиты, телитокия связана у них
с видоизменением или утратой мужских половых органов. Ооге-
нез сложен, образование хиазм и бивалентов происходит даже у
«нечетных» полиплоидов. Наблюдаемую приспособленность этих
форм (если судить по их широкому географическому распро-
странению) можно было бы объяснить гетерозисом, обусловлен-
ным аллополиплоидией.
Апомиксис был описан и у позвоночных, но его распростра-
ненность и эволюционное значение неизвестны. Некоторые под-
ниды европейской ящерицы Lacerta saxicola размножаются пар-
теногепетически. Ряд видов американской ящерицы Cnemido-
Phorus, возможно, представлен только самками, так как самцов
никогда еще не находили; это позволяет предполагать партено-
218 [ Глава 9
генез. Интересная ситуация была довольно подробно изучена
у рыб рода Mollienesia. Было исследовано несколько популяций
этих рыб в северном Техасе, где они живут в ручьях и дренаж-
ных канавах. Их выращивали также в лаборатории, где опре-
деляли степень их генетического сходства с помощью пересадки
тканей. Трансплантаты донорской ткани у рыбы-реципиента
отторгаются (из-за наличия тканевых антигенов, образующихся
под контролем генов реципиента) по прошествии некоторого
времени, приблизительно пропорционального степени генетиче-
ского сходства между реципиентом и донором.
Mollienesia formosa— партеногенетический вид, но яйца ее
не развиваются без стимуляции спермой. Поскольку самцы
М. formosa в природе чрезвычайно редки, для стимуляции разви-
тия требуется сперма родственных видов (в данном случае —
М. latipinna). Этот способ размножения, при котором генетиче-
ская информация спермы не включается в зиготу, носит назва-
ние гиногенеза. Как и в других случаях апомиксиса, генетически
идентичное потомство одной самки образует клон. Исследования
с трансплантацией показали, что около 80% всей популяции
М. formosa в одной дренажной канаве близ Ольмито в долине
Рио-Гранде составляют два клона.
Остальные особи принадлежат к третьему клону или не мо-
гут быть идентифицированы. В 1961 г. были найдены те же
клоны, что и в 1960 г.
В 1954 г. несколько десятков особей М. formosa было взято
из канавы у Ольмито и выпущено в реку Сан-Маркос, примерно
в 400 км к северу. Этот вид прижился здесь, так же как и
М. latipinna, интродуцированная в эту область на много лет
раньше. Таким образом, можно было сравнить популяцию, обра-
зовавшуюся в реке Сан-Маркос, с рыбами района Ольмито; эту
популяцию также исследовали в 1960 и 1961 гг. Было найдено
только два клона, и это были клоны, обычные для Ольмито.
Значит, эти клоны весьма мало изменились (насколько позво-
ляют судить результаты пересадки ткани) по меньшей мере в
течение 10 лет.
Нам осталось рассмотреть только циклический партеноге-
нез— своеобразную генетическую систему, встречающуюся у
тлей, орехотворок, ветвистоусых раков и коловраток. У тли
Tetraneura ulnii, вызывающей образование галлов на листьях
ильма, наблюдается последовательность поколений, которые по-
лучили названия, характеризующие поведение особей: fundatri-
ces (основательницы колоний), emigrantes (самки-расселитель-
ницы), exules, sexuparae (полоноски) и sexuales (половые осо-
би). Весной самки из поколения основательниц достигают поло-
возрелой стадии внутри галлов на листьях ильма. Здесь же
Генетические системы II | 219
каждая из них партеногенетически производит второе поколение
самок, которые позже, после развития у них крыльев, улетают
питаться на корнях трав. Это самки-расселительницы (emigran-
ts), которые также партеногенетически производят несколько
поколений exules (тоже самки). В конце концов эти последние
порождают sexuparae — крылатых самок, которые улетают об-
ратно на ильм и там партеногенетически производят самцов и
самок, называемых sexuales. Последние спариваются, и из опло-
дотворенных яиц вновь выводятся самки-основательницы —
галлообразующее поколение.
Цитологическое исследование показывает, что у самок sexua-
les 2п=14, тогда как у самцов 2п= 13; очевидно, имеется систе-
ма половых хромосом XX : ХО. Размножающиеся путем телито-
кии самки типов fundatrices, emigrantes и exules имеют диплоид-
ный набор из 14 хромосом. При оогенезе происходит лишь одно
деление созревания. Поскольку это деление нередукционное, об-
разуются диплоидные яйца и из них развиваются генетически
идентичные (если не учитывать мутаций) самки. Sexuparae
откладывают яйца двух типов. В тех яйцах, из которых разо-
вьются самки, все хромосомы расщепляются, как в митозе.
В яйцах, предназначенных для развития самцов, хромосомы ве-
дут себя так же, за исключением Х-хромосом, которые конъю-
гируют и подвергаются редукции, как в мейозе. Одна хромосома
остается в яйцеклетке, а другая отходит в полярное тельце.
При сперматогенезе у самцов sexuales Х-хромосома обычным
образом переходит в один из сперматоцитов второго порядка.
Однако клетки, не получившие Х-хромосому, в конце концов
дегенерируют, и только из содержащих Х-хромосому клеток
образуются спермин. Таким образом, самцы sexuales могут да-
вать только дочерей, которые и завершают цикл как основа-
тельницы колоний.
У других тлей наблюдаются сходные циклы, но число поко-
лений может быть иным: так, возможны два типа sexuparae —
производящие самцов и производящие самок — и т. п.
Внутри комплексов, подобных описанным у животных и ра-
стений, очень трудно применить обычное понятие вида. Размно-
жающиеся половым способом диплоиды могут быть сравнимы
с видами других организмов. Но авто- и аллополиплоиды, соче-
тающие в себе признаки двух или нескольких диплоидов, у раз-
множающихся бесполым путем и потому очень плодовитых орга-
низмов обесценивают критерии, основанные на постепенных
Морфологических переходах, обмене генами и географическом
Распространении. И все же сочетание классических методов
таксономии с методами цитогенетики может позволить биологу
Дифференцировать основные эволюционные единицы внутри-
220 | Глава 9
такого комплекса. Эти единицы он по традиции возводит в ранг
видов, тогда как множество апомиктических форм можно опи-
сывать или с помощью формальных таксономических катего-
рий, или без них, в зависимости от того, что именно кажется
более целесообразным.
Каковы следствия апомиксиса как генетической системы, по-
мимо того, что они сильно затрудняют работу биолога? Очевид-
но, что апомиксис делает возможным выживание многих гено-
типов, обеспечивающих мощность и хорошую адаптацию, но по
той или иной причине обусловливающих половую стерильность,
как, например, у несбалансированных полиплоидов. Апомиксис
позволяет также во множестве производить генетически сходные
особи для быстрой колонизации новых местообитаний. Апомик-
тическая система часто встречается у сорняков и у организмов,
впервые осваивающих необжитые местообитания, а также у оби-
тателей мест, часто или регулярно подвергающихся катастрофи-
ческим изменениям (песчаных дюн, газонов и т. п.).
Но в то же время апомиксис ограничивает генетическую из-
менчивость организмов, у которых он стал единственным спосо-
бом размножения. Поэтому он чаще всего служит альтернатив-
ной или второстепенной генетической системой. Апомиксис обычно
не сочетается с другими системами, снижающими потенциальную
эволюционную гибкость организма ради большей приспособлен-
ности к условиям данного момента (например, с самооплодотво-
рением). Интересно, что даже в таких группах, как род Роа, где
использование апомиксиса и высокая полиплоидность доведены,
казалось бы, до крайней степени, создавшаяся ситуация не яв-
ляется, как ее обычно описывают, «тупиком». Пыльца облигат-
ных апомиктов бывает способна функционировать, и опыление
апомикта может случайно привести к такому расщеплению, ко-
торое позволит организму вернуться к половому размножению.
ВЫВОДЫ
Популяции растений и животных часто обладают цитогене-
тическими механизмами, контролирующими количество и при-
роду генетических рекомбинаций. Эти механизмы наряду с дру-
гими, упомянутыми выше, составляют генетическую систему
популяции, определяющую число новых генных комбинаций, об-
разующихся в единицу времени. Сюда относятся, в частности,
инверсии и транслокации, создающие сравнительно небольшие
группы сцепленных генов, полиплоидия с ее разнообразными и
изменчивыми следствиями и, наконец, апомиксис, при котором
рекомбинация исключена. Такие механизмы нередко считают
невыгодными с точки зрения отдаленной перспективы. Однако
Генетические системы П | 221
они чрезвычайно распространены как у растений, так и у жи-
вотных и должны давать какое-то селективное преимущество.
Рассмотренные нами причудливые и сложные генетические си-
стемы некоторых организмов еще мало изучены и не связаны
удовлетворительным образом с теорией эволюции.
ЛИТЕРАТУРА
1. Darlington С. D., The Evolution of Genetic Systems. 2d ed. Basic Books,
Inc., New York, 1958.
2. S a g e г R., R у a n F. J., Cell Heredity, Wiley, New York, 1961.
3. S t e b b i n s G. L., The comparative evolution of genetic systems. In Sol
Tax (ed.) Evolution After Darwin, Univ. Chicago Press, Chicago, vol. I. The
Evolution of Life, pp. 197—226, 1960.
4. White M. J. D., Animal Cytology and Evolution. 2d ed., Cambridge Univ.
Press, New York, 1954.
5. Lewis, Raven, Evolution, 12 (1958).
6. Stebbins G. L., Variation and Evolution in Plants, Columbia University
Press, 1950.
7. Darlington C. D., Recent Advances in Cytology, Blakiston, 1937.
8. Wagner, Evolution, 8, (1954).
9. Cleland, Bot. Rev., 2 (1936).
10. Abrams, Illustrated Flora of the Pacific States, Stanford University Press,
1951.
Часть третья
ДИФФЕРЕНЦИАЦИЯ
ПОПУЛЯЦИЙ
Эволюционный процесс иногда подразделяют на
микроэволюцию (возникновение изменений внутри по-
пуляций) и макроэволюцию (возникновение различий
крупного масштаба). Границу между этими двумя
типами эволюции можно провести лишь произвольно,
и мы предпочитаем вовсе не делать этого. До сих
пор мы рассматривали главным образом изменения
внутри популяций. В третьей части книги будет рас-
смотрен вопрос о том, как в результате изменения и
взаимодействия эволюционирующих популяций воз-
никают разнообразные формы жизни.
В гл. 10 речь будет идти об основном процессе
расщепления групп организмов — о том, каким обра-
зом группа, эволюционирующая как единое целое,
разделяется на две или несколько групп. Сначала мы
сравним различные наблюдавшиеся типы дифферен-
циации, а затем попытаемся выяснить, как они могли
осуществляться.
В гл. 11 будут рассмотрены результаты длитель-
ных процессов изменения, расщепления и вымирания
популяций. Мы не постулируем никаких особых фак-
торов для объяснения данных, полученных при изу-
чении палеонтологической летописи, которая носит
фрагментарный характер и во многом искажает дей-
ствительную картину. Мы полагаем, что за последо-
вательную эволюцию популяций, протекающую во
времени, ответственны те же процессы, которые при-
водят к созданию различных популяций в разных ча-
стях дифференцированного местообитания. Мнимую
проблему возникновения «высших» таксономических
категорий мы рассматриваем как артефакт, создан-
ный применением таксономического метода в усло-
виях нехватки фактических данных из-за бесследного
исчезновения многих организмов.
Глава 10
ДИФФЕРЕНЦИАЦИЯ ПОПУЛЯЦИЙ
Каждому, кто наблюдает изменчивость живых организмов в
природе, должно быть очевидно, что эта изменчивость не всегда
бывает непрерывной. Внутри какой-либо группы однородных
организмов варианты могут образовать непрерывный ряд, но ме-
жду такими рядами существуют разрывы. При наших обычных
методах исследования растения и животные представляются
нам разделенными на дискретные или почти дискретные группы,
обыкновенно называемые видами. Конечно, ученый может систе-
матизировать объекты живой природы многими способами, от-
личными от традиционной таксономии; некоторые из этих спо-
собов представляют значительный интерес для исследователя.
Тем не менее, поскольку возможность распознавать таксономи-
ческие единицы и классифицировать их все-таки существует,
хотя это и сопряжено часто с большими трудностями, биологи
поставлены перед необходимостью попытаться понять происхо-
ждение таких единиц в природе. При этом прежде всего обычно
возникает вопрос о том, каким образом единая популяция, вну-
три которой предполагается свободное скрещивание, может диф-
ференцироваться на обособленные группы организмов. Связан-
ные с этим процессы в совокупности составляют то, что в биоло-
гии называют видообразованием.
Выяснение механизмов видообразования нередко считали
центральной проблемой эволюции. Классический труд Дарвина
был озаглавлен «Происхождение видов...», а во многих вышед-
ших в последние годы монографиях по зоологии и ботанике осо-
бое внимание уделяется так называемому видовому уровню на-
блюдаемых биологических различий. Возможно, что именно по-
этому оказался в тени ряд чрезвычайно важных и интересных
проблем, которые обычно относят к области экологии (напри-
мер, вопрос о природе и эволюции растительных и животных
сообществ). Однако при обсуждении вопроса о том, как возни-
кают в природе различимые группировки сходных организмов,
мы будем пока придерживаться общепринятой интерпретации и
общепринятых терминов.
Даже при поверхностном знакомстве с живыми организмами
нетрудно заметить, что степень сходства между ними может
быть самой различной. Одни формы кажутся весьма отличными
от всех остальных; другие как будто бы почти незаметно пе-
15 Зак. 152
226 | Глава 10
реходят в иные, очень сходные с ними формы. Приступая к ис-
следованию вопроса о том, как происходит дифференциация по-
пуляций, полезно рассмотреть природу и величину «разрывов»
между группами сходных организмов. В этой главе будут рас-
сматриваться случаи, когда степень дифференциации относи-
тельно невелика. Другими словами, мы будем иметь дело с
ситуациями, которые кажутся очень близкими к точкам развет-
вления эволюционного древа, т. е. с такими организмами, кото-
рые, по-видимому, находятся на пороге расщепления на несколь-
ко форм, и с такими группами форм, которые возникли, очевидно,
сравнительно недавно. Снач-ала мы приведем ряд примеров
с тем, чтобы дать читателю некоторое представление о суще-
ствующих типах дифференциации, а затем обсудим причины,
ведущие к возникновению того или иного типа. Формы с резко
выраженными отличиями будут рассмотрены в гл. 11.
В последующем изложении термином признак будет обозна-
чаться любая особенность, варьирующая у обсуждаемой группы
организмов в целом. Так, наличие или отсутствие пластид есть
признак, когда речь идет обо всех вообще живых организмах.
Для каждого данного организма можно установить, обладает
он пластидами или нет. Но их наличие или отсутствие не будет
признаком, например, для роз, так как у роз пластиды имеются
всегда. Длина бедренной косги — это признак для человека, так
как она варьирует у этого вида и может быть измерена у каж-
дого индивидуума. Длину бедра также можно считать призна-
ком в том случае, если мы сравниваем, к примеру, взрослых
мышей и взрослых людей, но диапазоны наблюдаемой изменчи-
вости этого признака не перекрываются. Такого рода прерыви-
стость в изменчивости (по отдельным признакам или по сочета-
ниям признаков) мы будем называть разрывами, или хиатусами.
ПРИМЕРЫ ДИФФЕРЕНЦИАЦИИ
НЕПРЕРЫВНАЯ ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Во многих случаях изменчивость бывает в достаточной сте-
пени непрерывной, так что между группами нельзя провести
разграничительных линий. Изменение некоторых признаков мо-
жет носить характер градиентов. Такие градиенты по отдель-
ным признакам называют клинами, а изменчивость этого типа
называют клинальной.
Изменчивость окраски, рисунка и размеров тела у животных.
Географическая изменчивость окраски, рисунка и размеров те-
ла— это одно из наиболее широко изучаемых биологических
Дифференциация популяций | 227
явлений. Такая изменчивость часто принадлежит к типу, уже
описанному в гл. 7, при котором популяции отличаются друг от
друга главным образом частотой различных форм — например,
популяции Biston, Сераеа, Natrix. Еще одним примером может
служить североамериканская бабочка-парусник Papilio glaucus\
некоторые ее популяции состоят из желтых и «тигровых» (с чер-
ными полосками) самцов и самок, тогда как другие популяции —
из «тигровых» самцов, «тигровых» самок и одноцветных темно-
коричневых самок. В южной Канаде и на крайнем севере США
популяции Р. glaucus состоят только из особей тигровой рас-
цветки. На юге центральной части Флориды доля темно-корич-
невых самок в популяциях очень невелика (6—8%), а на са-
мом юге Флориды темные самки иногда совершенно отсутству-
ют. Однако в большинстве южных штатов популяции характе-
ризуются высокой частотой (до 50%) темно-коричневых самок.
Во многих случаях различия между популяциями могут не
касаться частоты тех или иных форм (как при внутрипопуляци-
онной изменчивости). В пределах популяции все особи могут
быть очень сходными (низкая внутрипопуляционная изменчи-
вость), но окраска и рисунок могут изменяться от популяции к
популяции в соответствии с направлением изменения основных
условий среды в пределах их географического ареала. У млеко-
питающих и птиц тенденция к более светлой окраске в более
холодных и засушливых частях ареала (по сравнению с более
теплыми и влажными областями) настолько обычна, что полу-
чила название «правила Глогера». Другие, так называемые «эко-
логические правила» касаются изменчивости (не обязательно
непрерывной) размеров тела и формы его частей. Одно из них,
правило Бергмана, гласит, что гомойотермные позвоночные в хо-
лодных областях в общем крупнее, чем в теплых. Согласно дру-
гому правилу — правилу Аллена, все выступающие части тела —
крылья, ноги, нос и т. д. — в теплых областях обычно длиннее,
чем в холодных.
Экотипическая изменчивость у растений. Ботаники придава-
ли локальной популяции как основной единице большее значе-
ние, чем зоологи, — быть может, потому, что менее подвижные
растения легче изучать. Клаузен, Кекк и Хизи в течение многих
лет исследовали изменчивость внутри популяций и между по-
пуляциями растений, широко распространенных в Калифорнии.
Выращивая растения в полевых условиях на опытных станциях
в Стэнфорде (на уровне моря), Мэтере (высота 1380 м) и
Тимберлайне (высота 3000 л/), они смогли в значительной
степени разделить средовые и генетические компоненты измен-
чивости. Многолетние растения, способные к вегетативному раз-
множению, можно выращивать во всех этих местностях и таким
15*
228 | Глава 10
образом изучать их физиологические реакции на факторы внеш-
ней среды. По существу, одну и ту же генетическую особь мож-
но исследовать одновременно в трех различных экологических
ситуациях. Такого рода исследования привели к открытию у
растений экологических рас, или экотипов.
Мы уже упоминали род Achillea (тысячелистник) из семей-
ства сложноцветных. Методом пересадок Клаузен, Кекк и Хизи
довольно детально изучили комплекс A. millefolium. Эти расте-
ния встречаются во всем северном полушарии, где они растут
в зоне от уровня моря до верхней границы леса. Для них ха-
рактерна непрерывная морфологическая изменчивость. Так, на-
пример, растения A. millefolium бывают высотой около 180 см
(в долине Сан-Хоакин) или всего несколько сантиметров (на
альпийских лугах). Другие морфологические признаки также
постепенно изменяются от популяции к популяции, так что
таксономические подразделения установить в данном случае
очень трудно.
В основе приспособления растений Achillea к условиям среды
лежит надлежащая интеграция многих физиологических показа-
телей, таких, как интенсивность фотосинтеза и дыхания, холо-
достойкость, длительность зимнего покоя и других периодиче-
ских явлений. Каждая местная популяция содержит много раз-
личных генотипов. В зависимости от исследуемого уровня эти
генотипы можно объединять в группы различной величины. На
основании опытов, проводившихся на 320-километровой трансек-
те в Калифорнии, Клаузен, Кекк и Хизи заключили, что гено-
типы и локальные популяции группируются по меньшей мере
в 11 физиологических рас. На протяжении этой трансекты пред-
ставлены два таксономических вида, встречающиеся в различ-
ных местообитаниях. Вид Achillea lanulosa, распространенный
в более возвышенных местностях^ типичен для континентальных
местообитаний, тогда как A. borealis — в основном приморский
вид, растущий на меньших высотах. Интересно отметить, что
у A. lanulosa в северной части ареала, где он доходит до побе-
режья, выработались приморские экотипы, сходные с экотипами
A. borealis.
Клинальная изменчивость у растений. Asclepias tuberosa так-
же обнаруживает географическую изменчивость, но изучали ее
совсем по-иному. Подвиды, встречающиеся в восточной и сред-
ней частях США, весьма детально изучил Вудсон. Распростра-
нение этих подвидов показано на фиг. 46. Здесь мы рассмотрим
только A. tuberosa tuberosa и A. t. interior. На большей части
их ареалов эти подвиды можно различить по форме листьев.
К счастью, два существенных элемента формы листа можно
охарактеризовать количественно, что облегчает изучение геогра-
Дифференциация популяций | 229
фической изменчивости: это угол А, который служит показате-
лем заостренности верхушки листа, и угол В, характеризующий
форму основания листа. Два указанных подвида встречаются
друг с другом на широком фронте в восточной части США, и,
как явствует из фиг. 46, здесь имеется зона постепенного заме-
щения одного подвида другим. Вудсон исследовал географиче-
скую изменчивость, разделив карту страны на равные по пло-
щади квадраты и проведя измерения гербарных экземпляров,
собранных на этих квадратах. Он изучал также выборки из ло-
кальных популяций и определял изменчивость в пределах одной
особи и между особями и колониями.
Сравнивая результаты измерений экземпляров, собранных в
1948 г. на 1930-километровой трансекте от Канзаса до Вирги-
нии, и имеющихся гербарных экземпляров из соответствующих
квадратов, Вудсон смог оценить изменения во времени. Гербар-
Фиг. 46. Карта, показывающая распространение Asclepias tuberosa.
Каждый знак соответствует округу, в котором найдена данная форма. Крупные черные
г?/Жки~-А* interior', мелкие черные кружки — A. t. tuberosa', белые кружки —предпоЛа-
гибриды между подвидами tuberosa и interior, значки в форме полукруга —Д. t.
230 | Глава 10
ный материал, собранный за много лет, представляет собой
выборку, относящуюся в среднем к более раннему периоду, чем
трансекта 1946 г. Было ясно, что признаки A. t. interior распро-
странялись в восточном направлении, а признаки A. t. tuberosa
в западном, но гораздо медленнее. Когда в 1960 г. вновь были
собраны экземпляры со всей трансекты, оказалось возможным
дать точную количественную оценку изменений, происшедших
за 14-летний промежуток времени. Очевидно, происходило вза-
имное проникновение восточных и западных генотипов, опреде-
ляющих два названных признака (углы А и В). По мнению
Вудсона, восточный подвид находится в процессе генетического
вытеснения западным подвидом, так как он сравнительно мень-
ше распространился на запад. Тем не менее можно ясно видеть
его влияние на форму листа у западного подвида.
Клинальная изменчивость у животных. Экспериментальные
данные указывают на то, что, несмотря на непрерывный харак-
тер географической изменчивости, имеет место значительная
дифференциация. Изменчивость лягушки Rana pipiens широка
и дискордантна. В табл. 9 сведены данные об изменчивости по
12 признакам. Изменчивость здесь не обнаруживает какой-либо
определенной направленности; в самом деле, многие признаки
варьируют, казалось бы, совершенно независимо. Проведенное
Муром детальное исследование изменчивости процессов развития
дало обильный материал, позволяющий сделать определенные
ТАБЛИЦА 9
Формулы популяций у лягушек из восточной части
Северной Америки [17] *
Район Формула
Квебек ABC DEFGHIJKL
Мэн ABC D Е F g HIJk L
Верхмонт ABC DEFGHIJKL
Нью-Йорк, северная ABCDeFGHIJKL
часть штата
Массачусетс A bC DEFGHIJKL
Род-Айленд ABC DEFGHIJKL
Нью-Йорк, южная часть Abe dEfghiJk 1
штата
Нью-Джерси Abe defghiJk I
Мэриленд a Be DeFghiJk I
Продолжение табл. 9
Район Формула
Северная Каролина Abe DeFGhiJk 1
Южная Каролина — b — Def g hiJ — —
Джорджия a be defghiJk I
Флорида a be defghijKl
Онтарио — BC DEFGHIJK —
Мичиган — BC DeFGHIJ — —
Висконсин A BC DEFg HiJK L
Миннесота — BC De Fg HiJK L
Южная Дакота ABC De Fg HiJK L
Небраска — BCDeFghiJk L
Индиана ABCDeFgHiJk 1
Кентукки a Be DeFgHiJ — —
Иллинойс a BCDeFgHiJk 1
Миссури ABCDeFgHiJk L
Канзас ABCDeFgHiJk L
Арканзас ABc DeFgHiJk 1
Оклахома aBCDeFgHiJk 1
Миссисипи abc DeFGhiJk 1
Луизиана aBCdeFGhiJk 1
Техас aBCDeFgHiJKL
♦Значения символов:
А—отношение ширины головы к ее длине
составляет 0,92 или более
В —50% или более особей имеют тибиальные
полосы
С—среднее число тибиальных полос (когда
они есть) 1,4 или более
D—50% или более особей без бедренных
полос
Е —50% или более особей без тимпанального
пятна
F —50% или более особей со светлым сет-
чатым рисунком
G—число пятен на спине меньше 13
К—число пятен по бокам 12 или больше
* —число боковых пятен больше числа
спинных пятен
J —50% или более особей без сетчатого
к рисунка на боках
£ —50% или более самцов с яйцеводами
ъ "Наружные голосовые мешки отсутствуют
или слабо развиты у 50% или более
самцов
а—отношение ширины головы к ее длине
менее 0,92
Ь —менее 50% особей имеют тибиальные
полосы
с —среднее число тибиальных полос (когда
они есть) менее 1,4
d —менее 50% особей без бедренных полос
е—менее 50% особей без тимпанального
пятна
f — менее 50% особей со светлым сетчатым
рисунком
g —число пятен на спине 13 или больше
h —число пятен на боках меньше 12
I — число боковых пятен равно или меньше
числа спинных пятен
j —менее 50% особей без сетчатого рисунка
на боках
к—менее 50% самцов с яйцеводами
1—наружные голосовые мешки отсутствуют
или слабо развиты менее чем у 50% сам-
цов
232 | Глава 10
заключения. Например, у северных и южных популяций
лягушек обнаружены различия в диапазоне температур, допу-
скающих нормальное эмбриональное развитие (фиг. 47). Эти
различия коррелируют с различиями между южными и север-
ными лягушками, принадлежащими к четко обособленным
группам. (Так, северный вид R. sylvatica, ареал которого про-
стирается от субарктической зоны до центральных районов
США, может нормально развиваться при температурах от 2,5 до
24°, тогда как R. catesbeiana, живущая в области от южной Ка-
нады до Мексики, — при температурах от 15 до 32°; в совпа-
дающей части их ареалов R. sylvatica размножается ранней вес-
ной, a R. catesbeiana — в середине лета.)
При скрещиваниях особей из разных популяций R. pipiens
образуется нормальное потомство, если родители взяты из гео-
графически смежных популяций (например, из центральной и
южной Флориды) или с примерно одинаковых широт (скажем,
из Техаса и центральной Флориды). Однако чем больше широт-
ный разрыв между популяциями, к которым принадлежат роди-
тели, тем большей оказывается доля нежизнеспособного потом-
ства. Яйца от самок из Вермонта, оплодотворенные самцами из
Нью-Джерси или Висконсина, развиваются так же, как яйца,
оплодотворенные самцами из Вермонта. Если для оплодотворе-
ния взять самца из Луизианы, то возникают некоторые анома-
лии развития, но смертность повышается лишь незначительно.
У техасско-вермонтских гибридов развитие уже во многих отно-
Широта Местоположение
46° N ------------------------------------ Квебек
45 --------------------------------- Вермонт
44---------------------------------------- Висконсин
40 --------------------------------- Нью-Джерси
30 --------------------------------------- Луизиана
2 9 ----------------------------------- Окала (Флорида)
2 7 ------------- 1 Ингленд (Флорида)
32 —-------------------------------- Техас
22 ------------------------------- Мексика
О 5~" 10 15 20 25 30 35 40
Температура, °C
Фиг. 47. Пределы температур для нормального эмбрионального развития
Rana piplens из различных местностей.
Нижний предел для лягушек из Квебека и Висконсина не установлен, но полагают, что он
такой же, как и для лягушек из Вермонта [2].
Дифференциация популяций | 233
щениях нарушено, и смертность может достигать 100%. После
одного скрещивания самца из восточномексиканской популяции
с самкой из Вермонта большинство гибридных эмбрионов погиб-
ло на стадии гаструлы или нейрулы. Таким образом, можно
было бы сказать, что с лягушками R. pipiens дело обстоит при-
мерно так же, как с саламандрами Ensatina (см. ниже). Однако
ареал Rana не образует кольца, так что популяции крайних ти-
пов в природе нигде не существуют совместно.
Сходный характер носит дифференциация популяций бабоч-
ки Coenonympha tullia (Satyrinae) в Англии. При скрещивании
особей из отдаленных популяций в некоторых выводках встре-
чались интерсексуальные «самки», что указывает на известную
степень генетической несовместимости. (Это находится в соот-
ветствии с правилом Холдейна, согласно которому нежизнеспо-
собность или стерильность среди гибридов наблюдается обычно
у гетерогаметного пола, в данном случае у самок.) При скре-
щиваниях между менее отдаленными популяциями никаких ано-
малий обнаружено не было. Однако эта бабочка изучена менее
детально, чем непарный шелкопряд (Lymantria dispar), у кото-
рого Гольдшмидт описал много степеней интерсексуальности в
потомстве от скрещиваний между популяциями, дивергировав-
шими в разной степени. Интересующиеся найдут хорошее изло-
жение этой работы в книге Добжанского.
Первые этапы генетической изоляции. Популяции некоторых
животных, связанные длинным рядом промежуточных форм,
могут существовать совместно, сохраняя обособленность. Инте-
ресная картина изменчивости была, например, описана у сала-
мандр рода Ensatina (сем. Plethodontidae). Эти животные рас-
пространены вдоль западного побережья Северной Америки от
юго-западной части Британской Колумбии до южной Калифор-
нии. В Калифорнии они встречаются в прибрежных районах, в
горах Сьерра-Невада и на юге внутренних горных районов стра-
ны. Для них характерна значительная географическая изменчи-
вость окраски и в меньшей степени — размеров тела (фиг. 48).
У представителей прибрежных популяций верхняя сторона тела
бурая или красновато-бурая, тогда как у представителей попу-
ляции из других областей на теле появляются желтые, кремовые
или оранжевые пятна, тем более выраженные, чем южнее мест-
ность. В горах Сьерра-Невада, на широте бухты Сан-Франциско,
имеется «островок» популяций, сходных с прибрежными, и
встречаются также особи, промежуточные между прибрежным
типом и типом, характерным для Сьерра-Невады. В отношении
изученных признаков наблюдается, по-видимому, довольно по-
степенное изменение в направлении с севера на юг (за исключе-
нием упомянутого «островка»), хотя систематики и разбили этот
Фиг. 48. копуляции Ensatina на западном побережье Северной Америки
Объяснение в тексте 113].
Дифференциация популяций | 235
непрерывный ряд форм на несколько «подвидов» и выделили
«зоны интерградации». Однако к югу от Центральной долины,
где ареалы южного берегового и внутриматерикового типов со-
прикасаются, имеется довольно резкий разрыв непрерывности.
Разительно несходные формы — одноцветные и пятнистые —
были найдены на расстоянии 0,32 км друг от друга на юго-во-
сточном склоне горы Паломар.
В каньоне Милл, выше Баннинга (Калифорния, примерно на
80 км по прямой линии севернее горы Паломар) Стеббинс и
Браун в 1962 г. обнаружили обе эти формы, живущие совместно,
а также одну, по всей видимости, гибридную особь и несколько
особей, произошедших, возможно, от возвратного скрещивания.
Быть может, гибриды со временем будут найдены и в месте
сближения обеих форм на склонах горы Паломар.
Сходная ситуация недавно была описана у неотропической
плодовой мушки Drosophila paulistorum. В этом случае картина
оказалась сложнее, чем у Ensatina: здесь были обнаружены три
района, в которых совместно обитают две группы мушек, не
скрещиваясь между собой. При этом не только не было обна-
ружено гибридизации в природе, но и в лабораторных опытах,
в которых была исключена возможность спаривания с себе по-
добными, не наблюдалось даже перекрестного осеменения (не
говоря уже о развитии жизнеспособных гибридов). Однако в
эксперименте можно было осуществить обмен генетической
информацией между этими формами, используя ряд промежуточ-
ных, «связующих» лабораторных популяций, полученных от осо-
бей из других географических областей. В какой степени осуще-
ствляется такой обмен через посредство промежуточных попу-
ляций в природе, неизвестно. По мере проведения все более
тщательных исследований сложные ситуации такого рода обна-
руживаются у все большего числа организмов.
БЛИЗКОРОДСТВЕННЫЕ ИЗОЛИТЫ
Скопления близких видов («рои» видов, species swarms) у
рыб. В восточноафриканских озерах Виктория, Танганьика и
Ньяса встречается довольно значительное число близкород-
ственных видов рыб сем. Cichlidae. Например, в озере Вик-
тория найдено около 70 эндемичных и 6 неэндемичных видов ро-
да Haplochromis, обитающих в трех различных экологических
зонах. Одна группа состоит из форм с высоким телом, коротким
рылом, горизонтальным ртом, одинаковыми по длине челюстями
и двувершинными хватательными зубами (фиг. 49). Эти формы
Живут около берегов и являются бентофагами. Цихлиды второй
группы имеют более стройное тело, рыло у них длиннее, рот
Фиг. 49. Рыбы Haplochromls (сем. Cichlidae) из озера Виктория (Африка).
Сверху вниз Н. macrops, И. bayonl, Н, cavlfrons. Длина тела составляет около 100, 180
и 145 мм соответственно.
Дифференциация популяций | 237
слегка скошен вверх, нижняя челюсть выступает, хватательные
зубы конические, клыковидные (фиг. 49). Представители этой
группы питаются рыбой, за которой охотятся в толще воды как
у берегов, так и вдали от них. Третья группа видов Haplochro-
mis отличается стройным, удлиненным телом и длинным ры-
лом. Рот сильно скошен вверх (у двух форм — почти вертикаль-
ный), нижняя челюсть очень сильно выдается, хватательные зу-
бы клыковидные (фиг. 49). Это хищные рыбы, питающиеся у
поверхности воды главным образом другими рыбами и насеко-
мыми.
В этом обширном комплексе близкородственных, но четко
разграниченных групп наблюдается лишь умеренное разнообра-
зие форм. Хотя некоторые формы практически неразличимы по
своей морфологии, оказалось, что они дифференцированы в эко-
логическом отношении и по брачной окраске. Морфологически
наиболее изменчивы зубы и структуры головы, что вполне по-
нятно ввиду различий в биологии питания.
Виды-двойники альпийских бабочек. В некоторых случаях
под поверхностным сходством может скрываться значительное
разнообразие. Лоркович показал, что голарктические бабочки из
группы Erebia tyndarus, очень сходные между собой по внеш-
нему виду, тем не менее сильно различаются по числу хромосом
(и = 8, 10, 11, 15, 24, 25, 51 и, возможно, 52) и в меньшей сте-
пени — по морфологии мужских и женских копулятивных орга-
нов. Такие внешне сходные формы часто называют видами-двой-
никами (sibling species). В западных Альпах (фиг. 50) две фор-
мы, Erebia cassioides (п=10) и Е. nivalis (п= 11), занимают две
почти неперекрывающиеся экологические области — первая в
субальпийской (1400—2400 м), а вторая в альпийской зоне
(2300—2700 м и более). Хотя существует узкая полоса, где ареа-
лы Е. cassioides и Е. nivalis перекрываются, данных, свидетель-
ствующих об обмене генами, почти нет. Из 400 исследованных
экземпляров только 2 нельзя было с уверенностью отнести к ка-
кому-либо из двух названных видов. Эти две формы резко раз-
личаются по своему жизненному циклу: Е. cassioides завершает
развитие в один год, а для Е. nivalis требуется 2 года. Резуль-
таты лабораторных скрещиваний указывают на сильно выра-
женную поведенческую изоляцию (самки не реагируют на самцов
другого вида; копуляция, если ее вызвать, быстро прекра-
щается); искусственно полученные гибриды совершенно сте-
рильны и не похожи ни на каких особей, встречающихся в при-
роде. Короче говоря, эти две формы, видимо, генетически
полностью изолированы друг от друга.
Erebia cassioides имеет более широкое географическое рас-
пространение, чем Е. nivalis, и в районах, где последний вид не
238 | Глава 10
встречается, ареал ее захватывает те же высоты, на которых жи-
вет Е. nivalis в районах, где имеются оба вида. Напротив, в не-
которых областях, где горы не так высоки, Е. nivalis живет на
меньших высотах, а Е. cassioides соответственно обитает ниже
или отсутствует.
Erebia tyndarus (/т=10) обитает, по существу, в той же эко-
логической зоне, что и Е. cassioides, но, как можно видеть из
фиг. 50, это не симпатрические виды: Е. tyndarus встречается
в центральных Альпах, а Е. cassioides — восточнее и западнее.
В экспериментальных скрещиваниях (£. tyndarus $Х Е. cassioi-
des </) обнаружилась слабо выраженная поведенческая изоля-
ция; примерно из 15% яиц от этого скрещивания вывелись гусе-
ницы, из которых на ранних стадиях выжило лишь 25%. Хотя
ареалы Е. tyndarus и Е. cassioides очень тесно соприкасаются
как на востоке, так и на западе, они, по-видимому, нигде в сколь-
ко-нибудь значительной степени не перекрываются. Действитель-
но, все три вида — Е. tyndarus, Е. cassioides и Е. nivalis — обна-
руживают поразительное отсутствие склонности к сосуществова-
нию. Четвертый альпийский вид, Е. calcarius (п = 8), встречается
в Юлийских Альпах, но его точные географические взаимоотно-
шения с Е. cassioides в настоящее время неизвестны.
Вьюрки Галапагосских островов. Большой комплекс близко-
родственных, но обособленных групп имеется в подсемействе
Фиг. 50. Карта распространения бабочек группы Erebia tyndarus в Централь-
ной Европе.
ТАБЛИЦА 10
Виды дарвиновых вьюрков (Geospizinae) [1]
Название вида Характеристика Число островов, на кото- рых данный вид по- стоянно обитает
Geospiza tnagni- Крупный; кормится на земле, в кустарниках и
rostris на деревьях. Пища: очень твердые, обычно крупные семена сравнительно немногих растений. 14
Geospiza fortis Средней величины. Пищей служат разнообраз-
ные семена умеренной твердости, как мел- кие, так и крупные. 12
Geospiza fuligi- Небольшой; кормится на земле в большей
nosa степени, чем G. fortis или G. magnirostris. Пищей служат разнообразные мягкие, обычно
мелкие семена. 14
Geospiza diffi- Средней величины; ищет корм на земле. Как
cilis полагают, выбирает мягкие семена. 7
Geospiza scandens Средней величины; чаще всего сидит на как-
тусах Opuntia. Питается умеренно твердыми семенами сравнительно немногих растений, а также мягкими растительными тканями и нектаром. 11
Geospiza coniros- Крупный; образ жизни мало изучен, но сходен
tris с таковым G. scandens. 3
Platyspiza eras- Крупный; живет главным образом в густом
sirostris кустарнике и на высоких деревьях. Питается преимущественно мясистыми плодами, мяг- кими и умеренно твердыми семенами, моло- дыми листьями и цветками. 10
(^amarhynchus Средней величины; кормится на деревьях,
psittacula кустарниках и иногда на земле. Преимуще- ственно насекомоядный вид, довольно глу- боко долбит деревянистые ткани, обычно
на крупных ветвях. 11
Camarhynchus Подобен предыдущему, которого он замещает
pauper на о. Чарлз. 1
240 J Глава 10
Продолжение табл. 10
Название вида Характеристика Число островов, на которых данный вид по- стоянно обитает
Camarhynchus Небольшой; кормится на деревьях и кустар-
parvulus никах, на кактусах и на земле. Преимуще- ственно насекомоядный вид, долбит менее глубоко, чем С. psittacula, обычно на более мелких ветвях и в лишайниках. 12
Cactospiza pal- Средней величины; по образу жизни сходен
lida с танагрой. Зондирует выдолбленное отвер- стие веточкой или шипом от кактуса; пре- имущественно насекомоядный вид 7
Cactospiza helio- Средней величины; образ жизни мало изучен,
bates преимущественно насекомоядный вид. Встре- чается только в прибрежных мангровых
зарослях. 2
Certhidea oliva- Небольшой, похож на славку. Кормится па
cea деревьях (на всех уровнях), изредка и на земле. Питается только животной пищей, в особенности мелкими личинками. 16
Pinaroloxias Средней величины; отмечено питание на земле
inornata и на деревьях. Как полагают, использует в пищу насекомых, нектар и некоторые
плоды. 1
Geospizinae семейства вьюрков (Fringillidae). Эти птицы, извест-
ные под названием «дарвиновы вьюрки», встречаются, за одним-
единственным исключением, только на Галапагосских островах.
Один представитель этой группы живет на острове Кокос. Изу-
ченные впервые Дарвином, они послужили темой блестящих
монографий Лэка и Боумэна [1]. Существует около 14 ясно раа-
личимых форм, которые орнитологи распределяют между 6 ро-
дами. Эти виды перечислены в табл. 10.
Ареалы этих 14 видов в пределах архипелага Галапагос весь-
ма различны; на отдельных островах встречается от 3 до 10 ви-
дов (фиг. 51). Они различаются между собой главным образом
размерами тела, формой и размерами клюва и некоторыми дру-
гими признаками, связанными с биологией питания (фиг. 52).
По степени дифференциации они образуют почти полный копти-
Дифференциация популяций | 241
нуум. Группа форм, известная как вид Platyspiza crassirostris,
встречается на 8 островах, причем между популяциями отдель-
ных островов почти нет различий в отношении изученных при-
знаков. Другой вид, Certhidea olivacea, распространен на всех
Галапагосских островах и обнаруживает значительные различия
в окраске на разных островах. Например, окраска оперения спи-
ны у обоих полов варьирует от серо-коричневой (о. Джемс) до
бледно-серой (о. Баррингтон). На эту изменчивость наклады-
90°
92°
91°
0°
0°
92°
90°
фиг. 51. Карта архипелага Галапагос; показаны главные острова.
Цифры указывают число различных форм Geosplzlnae, найденных на каждом из островов [1],
За^. J5?
242 | Глава 10
Фиг. 52. Схема корреляции между строением клюва и биологией питания
у 10 видов Geospizinae, обитающих на острове Индефатигэбль [1].
ваются различия в примеси оливкового оттенка. Окраска брюш-
ной стороны варьирует от светлой желто-оливковой (о. Джемс)
и светло-коричневой (о-ва Кулпеппер, Венмен, Чарлз) до белой
(о. Баррингтон).
Распространенный на периферии архипелага вид Geospiza
conirostris во всем, кроме размеров тела, сходен с видом G. scan-
dens, обитающим в центре архипелага, причем это полностью ал-
лопатрические формы. Три вида земляных вьюрков, G. magni-
rostris, G. fortis и G. juliginosa, являются в значительной мере
симпатрическими и различаются лишь общими размерами тела
и относительной величиной клюва. В областях совместного оби-
тания этих трех форм морфометрические данные почти не пере-
Дифференциация популяций | 243
крываются, и наблюдения показывают, что особи способны рас-
познавать представителей своего вида и спариваются только с
ними. Иногда все три формы потребляют одну и ту же пищу, но,
как видно из табл. 10, особенности их питания совпадают лишь
частично.
Два представителя рода Cactospiza — С. pallida и С. helioba-
tes — очень сходны между собой. Они питаются главным обра-
Фиг. 53. Крайние формы дифференциации черепа у дарвиновых вьюрков.
®верху — Qeosplza magnirostris, внизу — Certlilclea olivacea (1].
16*
поползней.
Фиг. 54. „Сдвиг" признаков у азиатских
в^те”3районах^ гдТони^вс^аютТя оТЛковы^ SX™
Д-Лу^с?ан°^БПах?иа?иВ“-Яф^сеаТ: ^е₽байджа» и «верныйИран, /-КермаГш^?
(S. Anota)-. З-Ир^кий а₽Аал°в
Е.г,« ж-йг : x=-?:,u"r »г~™
Дифференциация популяций | 245
зом насекомыми, причем С. pallida пользуется единственным в
своем роде приемом: в поисках жуков и других насекомых пти-
ца долбит кору деревьев клювом, а затем зондирует проделан-
ные отверстия шипом от кактуса или веточкой. Эта замечатель-
ная форма поведения компенсирует отсутствие длинного языка
и является одним из немногих примеров использования птицами
«орудий». На единственном острове (о. Альбемарль), где со-
вместно обитают С. pallida и С. heliobates, они изолированы
друг от друга благодаря своим экологическим различиям (С. he-
liobates живет в прибрежных мангровых зарослях, а С. palli-
da—во внутренних районах). Таким образом, это аллопатри-
ческие виды, хотя в данном случае они встречаются на одном и
том же острове. О диапазоне дифференциации в группе в це-
лом можно судить, сравнивая черепа двух в значительной части
симпатрических видов Geospiza magnirostris и Certhidea olivacea
(фиг. 53).
Характер дифференциации у дарвиновых вьюрков противо-
положен наблюдаемому у большинства других птиц. Наиболее
близкие формы этих вьюрков различаются главным образом ве-
личиной и формой клюва, тогда как у других птиц близкие фор-
мы обычно в наибольшей степени дифференцированы по окра-
ске оперения. Очень близкие азиатские виды поползней Sitla
tephronota и S. пеитауег явственно дифференцированы как по
оперению, так и по длине клюва, но только там, где эти виды
встречаются совместно (фиг. 54). Там же, где их ареалы не сов-
падают, они почти неразличимы. Это усиление различий в об-
ласти совместного обитания служит примером «сдвига призна-
ков» — явления, обычного у птиц. Другие примеры описаны у
муравьев, жуков, ракообразных, рыб и лягушек. Есть данные о
двух формах термитов, которые роятся в одно и то же время су-
ток в несовпадающих частях своих ареалов и в разное время —
в области совместного распространения. Возможные причины
«сдвига признаков» будут рассмотрены во второй части этой
главы.
ПРЕДПОЧТЕНИЕ ХОЗЯЕВ У ПАРАЗИТИЧЕСКИХ ОРГАНИЗМОВ
Среди паразитов широко распространена дифференциация в
отношении предпочитаемых хозяев. Это явление изучалось у та-
ких различных организмов, как кукушки, вши и нематоды. У ку-
кушек выработалась необычная форма специализации — то, что
называют «гнездовым паразитизмом»: они откладывают яйца в
гнезда других видов птиц. В большинстве случаев яйцо кукуш-
ки высиживают приемные родители, и прожорливый кукушонок
ньцесняет своих мнимых братьев из гнезда, начиная со временем
Перехватывать всю пищу, приносимую «родителями». Обычно
246 | Глава 10
Фиг. 55. Миметические формы яиц.
В верхнем ряду яйца азиатских воронов Corvus coronoldes, С. splendens insolens и С. s. pro-
tegatus-, в нижнем ряду —яйца кукушки Eudynamys scolopacea, откладываемые в гнезда
соответствующих видов вброна.
отложенное кукушкой яйцо поразительно сходно с яйцами
птицы-хозяина (фиг. 55). Виды кукушек, по-видимому, подраз-
делены на ряд форм, представители которых обыкновенно от-
кладывают яйца в гнезда лишь одного определенного вида птиц.
Эти подразделения, так называемые фратрии (gentes), не изо-
лированы друг от друга географически: особи одной из них
встречаются в непосредственной близости к особям из другой
или других фратрий.
Существуют две четко обособленные формы человеческих
вшей (Pediculus humanus') — головная и платяная, которые,
сильно различаясь в экологическом отношении, морфологически
почти идентичны. Головная вошь, как указывает ее название,
живет главным образом в волосяном покрове головы; яйца ее
приклеиваются к волосам. В отличие от головной платяная вошь
живет в складках одежды и сосет кровь в тех местах, где оде-
жда соприкасается с телом. Платяная вошь прикрепляет свои
яйца к одежде.
Дифференциация популяций | 247
Распространенный паразит человека круглый червь Ascaris
lutnbricoides морфологически и серологически неотличим от па-
разита свиньи A. suum, но в большинстве случаев яйца одного
вида не могут нормально развиваться в хозяине другого вида.
ОБСУЖДЕНИЕ НАБЛЮДАЕМЫХ ТИПОВ ДИФФЕРЕНЦИАЦИИ
В основе разнообразных типов дифференциации, описанных
в предыдущем разделе, лежит неоднородность физической сре-
ды. Эта неоднородность привела к тому, что эволюционные фак-
торы, особенно отбор, действовали неодинаковым образом. А это
в свою очередь создавало гетерогенность биотической среды и
вызывало дальнейшую дифференциацию сил, действующих в.
любой части этой среды.
Очень легко впасть в ошибку, представив себе дифференциа-
цию популяций на генетически изолированные формы как некую
«цель» эволюции. Дифференциация происходит очень часто, но
обычно те случаи, когда зарождающиеся виды вновь «раство-
ряются», остаются незарегистрированными. Когда морфологиче-
ская дивергенция двух популяций достигает определенного уров-
ня, мы часто уже не говорим, что они просто скрещиваются,—
мы говорим, что между ними происходит гибридизация. Это
значит, что мы рассматриваем такое скрещивание как проис-
шедшую по какой-то причине «ошибку». Мы забываем о том,
что спорадический обмен генами обогащает изменчивость обеих
популяций. Это просто одно из многих событий, происходящих
с популяциями в природе.
ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ДАВЛЕНИЕ ОТБОРА
Географическую изменчивость в большинстве случаев можно
объяснить различными направлениями отбора в разных местно-
стях. Например, растения Achillea на больших высотах испыты-
вают давление отбора, благоприятствующего карликовым фор-
мам, физиологически приспособленным к суровым условиям гор-
ных вершин. В низинах они подвергаются действию не тех же
самых, а иных факторов отбора. Популяции Natrix sipedon при-
способились как к «нормальным» для них болотистым местооби-
таниям, так и к особым условиям, существующим на островах
озера Эри. Популяции Сераеа подвергаются действию факторов
отбора, отчасти определяемых типом местной растительности, а
Популяции Biston — факторов, зависящих от степени промыш-
ленного загрязнения местности. Географическую изменчивость
обнаруживает и частота гена так называемой серповидноклеточ-
Ности в популяциях человека; она высока в областях, где
248 | Глава 10
встречается «злокачественная» тропическая малярия, и низка
в других местностях.
Хорошо знакомые с биологией читатели смогут припомнить
множество случаев, в которых различия можно истолковать
как результат разного направления отбора (в разных условиях
среды). Для студента мы добавим еще несколько примеров.
У песцов (в соответствии с правилом Аллена) уши и морды
короткие, тогда как у видов лисиц, обитающих в тропиках, уши
и нос длинные — видимо, потому, что в первом случае низкое
отношение поверхности к объему помогает сохранять теплоту те-
ла, а во втором — более высокое отношение этих величин обус-
ловливает ускорение теплоотдачи. Действительно, как правило
Аллена, так и правило Бергмана, по-видимому, просто отра-
жают значение определенного соотношения между поверхностью
и объемом для сохранения или отдачи тепла. У огромного мно-
жества гомойотермных организмов наблюдается клинальная из-
менчивость, согласующаяся с этими правилами. Однако физио-
логических данных, которые подкрепляли бы такую их интер-
претацию, в большинстве случаев не имеется.
В некоторых областях популяциям Papilio glaucus свойствен-
на высокая частота темных самок — по-видимому, в связи с тем,
что эти темно-коричневые самки похожи на парусников Battus
philenor, также встречающихся в этих районах. Было показано,
что бабочки В. philenor неприятны на вкус для птиц; в то же
время отмечено, что птицы в природе поедают взрослых Р. glau-
cus. Очевидно, в этих областях отбор способствовал выработке и
сохранению темной формы Р. glaucus.
Высоко в горах Сьерра-Невады в Калифорнии встречаются
две формы бабочки Oeneis chryxust светлая форма в районах
распространения гранитов и темная — в районах распростра-
нения базальтов. Ответственный за это фактор отбора не выяв-
лен, но наличие такой корреляции позволяет предполагать,
что у этих бабочек имеется какой-то неизвестный нам враг,
использующий при охоте на них зрение. Аналогичные при-
меры географической изменчивости окраски тела и типа место-
обитания описаны для многих групп животных, и их можно
было бы умножать до бесконечности — географическая измен-
чивость наблюдается повсюду, и для большинства тщательно
изученных популяций показано, что отбор играет важную роль
в дифференциации. Утверждение, что популяции организмов в
разных местах почти при всех обстоятельствах будут генетиче-
ски различны, можно считать трюизмом.
Изменчивость, описанную у Asclepias, по-видимому, можно
объяснить совместным действием различных факторов отбора
и явления дрейфа генов. Asclepias tuberosa — долгоживущий
Дифференциация популяций | 249
многолетник, образующий колонии разной величины. Эффектив-
ная величина популяции, по произведенной оценке, меньше ме-
дианы (равной 11) распределения по числу растений в одной ко-
лонии. Можно было ожидать, что в таких небольших популя-
циях генетический дрейф окажется важным фактором эволюции,
и фактические данные как будто бы подтверждают это. Тем не
менее в изменениях, происшедших на изученной трансекте за
14 лет, существенную роль играл, по-видимому, и отбор. Мы не
можем здесь подробно рассматривать эти изменения, однако яс-
но, что западные генотипы для признаков ZA и ZB обнаружи-
вают большую приспособленность, чем аналогичные восточные
генотипы. У A. tuberosa interior селективная ценность генотипа
для ZB приблизительно втрое выше, чем для ZA. За 14 лет из-
менчивость обоих признаков (ZA и ZB) снизилась примерно
на 30%.
Как в 1946 г., так и в 1960 г. в средней части разреза суще-
ствовала, по-видимому, зона гетерозиготности, что выражалось в
большей изменчивости, а также большей величине и мощности
растений. Возможно, что гибридизация этих подвидов, которые
были разделены в период плейстоценового оледенения и потом
вновь пришли в соприкосновение, привела к увеличению разме-
ров растений и расширению их ареала. Это одно из немногих ис-
следований, в которых изучение популяций охватывает период,
измеряемый годами. К сожалению, однако, Asclepias tuberosa не
является хорошим объектом для генетических исследований. Ци-
тогенетическая основа явлений, открытых методом биометрии,
неизвестна. Недостаточно ясен целый ряд других интересных ас-
пектов изменчивости этого вида Asclepias, но их сейчас изучает
Вудсон. К ним относятся «центробежная» изменчивость у A. tu-
berosa interior, которая привела к образованию концентрически
распространенных подвидов в западной и северной частях ареа-
ла, и интересная картина изменчивости окраски его цветка (ко-
торую можно исследовать биохимически).
ОБМЕН ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИЕЙ
У организмов с относительно непрерывным ареалом обмен
* генетическим материалом между популяциями ограничивает воз-
можную степень дифференциации. «Выравнивающий» эффект
рекомбинации сильнее всего проявляется в смежных популя-
циях, которые из-за относительно сходных условий среды менее
Подвержены дифференцирующему действию отбора. Иначе об-
стоит дело с удаленными друг от друга популяциями, соединен-
ными между собой цепью промежуточных популяций. Некоторое
количество генетической информации передается по этой цепи
250 | Глава 10
Фиг. 56. Схема, показывающая относительное число растений с голубой
(черные секторы) и белой (белые секторы) окраской венчика в выборках из
популяций Linanthus раггуае из одного района южной Калифорнии [14, 15].
в том и в другом направлении, но способность такого ослаблен-
ного генного потока выравнивать наследственные различия весь-
ма незначительна. В большинстве случаев более отдаленные
популяции подвергались совершенно различным воздействиям от-
бора, что повышало тенденцию к дифференциации. Это хорошо
видно на примере Rana pipiens и Coenonympha tullia, у которых
отдаленные друг от друга популяции сильно дифференцирова-
ны, тогда как между соседними популяциями почти не обнару-
живается различий1. Классический случай пространственной
изоляции мы находим в популяциях одного вида растений, встре-
чающихся в пустынях Калифорнии. На юге и западе пустыни
Мохаве у небольшого однолетника Linanthus раггуае (сем. Ро-
lemoniaceae) обнаруживается интересная картина изменчивости.
При обильных дождях растения образуют почти сплошной ко-
вер на обширных территориях. В 1941 г. L. раггуае интенсивно
изучали в области между Палмдейлом и Лыосерн-Валли, где
это растение образовало по существу одну непрерывную попу-
ляцию на площади около 1820 кв. км. В большинстве исследо-
ванных участков выборки растений состояли из форм с белыми
1 Возможно, что в некоторых из таких цепей обмен генами между от-
даленными популяциями идет настолько медленно, что эффект миграции как
источник изменчивости может быть практически неотличим от мутирования.
Дифференциация популяций | 251
цветками, но в трех изолированных районах были собраны вы-
борки, содержавшие в различном количестве растения с голу-
быми цветками. Состав выборок в материале, собранном в са-
мом западном из этих районов — между Палмдейлом и Льяно,—
показан на фиг. 56. Центральная область изменчивости отде-
лена от западной промежутком в 40 км, а от восточной — про-
межутком в 13 км.
Частота растений с голубыми цветками в выборках из этих
районов изменчивости представлена на фиг. 57. Это распреде-
ление частот фенотипов напоминает теоретическое распределе-
ние частот генов в небольших популяциях, подвергающихся да-
влению отбора (см. фиг. 23, Л). График распределения, приве-
денный на фиг. 57, не удается полностью интерпретировать, так
как генетические основы наблюдаемой изменчивости неизвестны.
Однако его U-образная форма делает весьма правдоподобным
фиг. 57. Частота распределения выборок Linanthus parryae (ордината) по
^Держанию растений с голубыми цветками (абсцисса) [14].
252 | Глава 10
предположение о существовании популяций, достаточно малых
для того, чтобы в них довольно часто могла происходить слу-
чайная фиксация и потеря генов, определяющих голубую окра-
ску цветка. Кажется вероятным, что наблюдаемая у Linanthus
картина изменчивости — это результат взаимодействия отбора,
дрейфа и пространственной изоляции. По-видимому, в районах
изменчивости какой-то неизвестный фактор отбора создает не-
большое преимущество для растений с голубыми цветками и в
то же время малая эффективная величина популяции (по рас-
четам Райта — от 14 до 27 растений) делает возможным значи-
тельный дрейф. Кроме того, в обычные для пустыни годы (го-
раздо более засушливые, чем 1941 г.) возникали бы лишь от-
дельные разбросанные популяции Linanthus. В сочетании с воз-
можным низким уровнем обмена генами внутри вида (если
опылители переносят пыльцу большей частью на ближайшие
цветки), это способствовало бы поддержанию установившейся
ситуации.
ПРЕКРАЩЕНИЕ ОБМЕНА ГЕНАМИ
Если между частями ареала того или иного организма возни-
кают широкие разрывы, то обмен генетической информацией
между популяциями (или группами популяций) может пол-
ностью прекратиться. Такое положение может создаться вслед-
ствие многих причин. Поднявшаяся суша может разделить море
на части. Известно, например, что Панамский перешеек на про-
тяжении геологического времени неоднократно затоплялся во-
дами океана. При каждом его новом поднятии некоторые попу-
ляции морских организмов, ареалы которых были до того
более или менее непрерывными, обособлялись. Имеется до-
вольно обширный фактический материал, говорящий о том,
что вследствие наступления ледников в плейстоцене ареалы
многих организмов неоднократно дробились. В результате
повторных трансгрессий моря (фиг. 58) происходила изоляция и
воссоединение участков материков, что создавало многообраз-
ные изменения в непрерывности ареалов различных организмов.
На распространение растений и животных глубокое влияние
оказывают изменения климата. Образование степей и пустынь
вследствие уменьшения осадков создает барьеры для миграции
организмов, нуждающихся в высокой влажности или густом ра-
стительном покрове. По причине засухи крупные озера могут
разделиться на многочисленные мелкие озера, а реки — превра-
титься в ряд обособленных водоемов. Напротив, увеличение ко-
личества осадков часто ведет к воссоединению изолированных
водоемов, что способствует обмену генами у водных организ-
мов и в то же время создает барьеры для наземных организмов.
Фиг. 58. Распределение моря и суши в северной части западного полушария
на протяжении четырех геологических периодов.
Черным показаны океанические области, темной штриховкой —наиболее длительные транс-
грессии моря на континентальную платформу, светлой штриховкой —области, затопленные
Лишь в течение какого-то отрезка соответствующего периода; точками показаны области
непродолжительного затопления, белыми оставлены области, бывшие в данный период
сушей [16J.
254 | Глава 10
Зоны высокой влажности создают барьеры для обитателей пу-
стынь и степей.
Интересно отметить, что разрывы в ареалах животных не
обязательно обусловлены барьерами, которые особи неспособны
преодолеть. Во многих случаях особенности поведения живот-
ного не позволяют ему переходить участки, которые оно легко
могло бы пересечь, если бы попыталось. Например, реки могут
изолировать популяции птиц на противоположных берегах, а уз-
кая полоска леса, делящая луг на две части, может служить эф-
фективной преградой между популяциями бабочек на этих двух
лугах.
изоляция
Физическая изоляция, чем бы она ни была обусловлена, соз-
дает возможность дифференциации популяций организмов, раз-
множающихся половым способом. Часто изоляцию описывают
как причину дифференциации. Это, конечно, не так; все дело
в том, что рекомбинация между изолятами невозможна, так что
каждый изолят реагирует только на селективные факторы своей
собственной среды. Если какой-то вид имеет непрерывное рас-
пространение вдоль градиента влажности, то образованию по-
пуляций, приспособленных к высокой и низкой влажности, на
обоих концах градиента будет мешать перенос генетической ин-
формации в обоих направлениях. Если середину этой цепи раз-
рушить, то дифференциация сможет продолжаться без такого
рода помех. По определению, условия среды, в которых ока-
зываются два только что изолированных фрагмента популяции,
не могут быть идентичными. Таким образом, эти два изолята
подвергаются действию различных давлений отбора. Вследствие
ошибки выборки два новых изолята будут иметь равную вели-
чину и нетождественные генофонды, так что эволюционные си-
лы будут действовать в неодинаковой генетической обстановке.
Поскольку мутирование носит случайный характер, нельзя ожи-
дать, что в обоих изолятах возникнут одни и те же мутации.
Таким образом, в результате физической изоляции одна эволю-
ционирующая единица может превратиться в две или большее
число эволюционирующих единиц. До тех пор пока такие еди-
ницы остаются изолированными, они будут независимо реаги-
ровать на воздействие эволюционных факторов. Но если об-
стоятельства позволят им восстановить контакт, открываются
многочисленные возможности.
Геологическая история Земли сопровождалась бесчисленны-
ми изменениями среды, которые могли приводить к встрече изо-
лированных ранее популяций. Последствия древних катаклизмов
дошли до нас в сильно измененном виде или не дошли совсем.
Дифференциация популяций | 255
Но два сравнительно недавних, частично совпадающих по вре-
мени события, несомненно, привели к изменениям, которые сей-
час трудно не заметить. Первое из этих событий — плейстоце-
новые оледенения, настолько изменившие климат, что растения
и животные должны были мигрировать или погибнуть. Ледники
к тому же «перепахивали» землю, смешивая почвы разных ти-
пов, создавая озера и перестраивая системы стока вод.
Сходные по масштабу последствия имело и появление чело-
века. Трудно переоценить значение человека как фактора эволю-
ции. В результате использования им огня, а позже — домашних
животных и культурных растений среда видоизменялась. Многие
случаи скрещивания ранее изолированных друг от друга форм
явились следствием преднамеренного или непреднамеренного
вмешательства человека. Получив доступ к огромным энергети-
ческим резервам климаксных экологических сообществ, он стал
отбирать часть этой энергии для своих собственных целей и рез-
ко изменил то, что мы называем «естественной» (существовав-
шей до появления человека) средой.
Слияние популяций. Существуют крайние случаи, когда по-
пуляции оказываются изолированными столь непродолжитель-
ное время, а условия их жизни могут быть настолько сходными,
что происходит лишь минимальная дивергенция. После воссо-
единения таких популяций особи спариваются случайным об-
разом, и при скрещивании представителей разных популяций
потомство оказывается столь же приспособленным, как и при скре-
щивании членов одного и того же изолята. Тогда ранее изоли-
рованные популяции сливаются в одну популяцию. Так всегда
бывает при объединении линий Drosophila, изолированных в ла-
боратории на протяжении нескольких поколений, или при скре-
щивании рыбок гуппи, содержавшихся в течение нескольких лет
в- разных аквариумах. Подобная ситуация, несомненно, весьма
обычна и в природе, но зоологи не уделяли ей особого внимания.
Смешение популяций без обмена генами. Другой крайний
случай — соприкосновение популяций, уже в значительной сте-
пени дивергировавших, так что взаимная привлекательность осо-
бей разных популяций полностью утрачена и межпопуляционные
скрещивания отсутствуют. В большинстве случаев это означает,
что обмен генами между эволюционными единицами прекратил-
ся, хотя (как, например, у Drosophila paulistorum) генетическая
информация может еще передаваться от одного такого изолята
к другому по «кругу рас». Случай с популяциями Ensatina
представляет большой интерес, так как еще не известно, сольют-
ся ли дифференцированные «концевые» популяции или же они
будут продолжать дифференцироваться, так что в конце концов
образуется типичный «круг рас». Такие ситуации, где две или
256 | Глава 10
большее число сходных форм живут в тесной близости друг к
другу без заметной гибридизации, многочисленны, но нет, по-
видимому, документированных случаев полной взаимной ин-
дифферентности между особями воссоединившихся сегрегатов.
Это не значит, что такие ситуации никогда не возникают — мо-
жет быть, они просто остались незамеченными.
Если возможен случай, когда две близкие формы вновь вос-
соединяются, причем для закрепления изоляции не требуется от-
бор, направленный против гибридов, то наличие такой ситуации
наиболее вероятно в роде Empidonax (птицы сем. мухоловок).
Недавно Джонсон исследовал область в восточной Калифорнии,
где совместно встречаются два вида: Е. wrightii и Е. oberholseri.
Возможно, что эта симпатрия очень недавнего происхождения;
она, по-видимому, возникла в результате изменения местообита-
ний вследствие расчистки местности от лесов, проводившейся в
середине прошлого столетия. Е. wrightii вьет гнезда среди по-
лыни или на низкорослых деревьях и обычно кормится на от-
крытых местах, тогда как Е. oberholseri живет в лесах и чаппа-
рале. В местностях смешанного типа, там, где эти виды живут
бок о бок, они сохраняют раздельность своих местообитаний, и
особи каждого вида защищают свою территорию от особей дру-
гого вида. Эти две формы настолько похожи друг на друга, что
их можно с уверенностью различить только путем тщательного
измерения крыльев и определения отношения длины крыла к
длине хвоста. Джонсону удалось обнаружить ясные различия
в особенностях брачных песен, однако сигналы, означающие вы-
зов, и внешний вид птиц настолько сходны, что особи Е. oberhol-
seri защищают свою территорию от Е. wrightii, и наоборот. Не-
смотря на это сходство, все несколько десятков пар, собранных
Джонсоном, состояли из особей одного вида (среди них не было
пар oberholseri!wrightii)\ следовательно, для птиц выбор надле-
жащего партнера не составлял трудности. Возможно, что отли-
чительные признаки (в чем бы они ни состояли) первоначально
были выражены слабее, а позже были усилены отбором, но в
таком случае этот процесс должен был завершиться очень бы-
стро.
Ограниченный обмен генами. Известен другой аналогичный
случай, также связанный с изменением среды человеком: два
мексиканских вида тауи, Pipilo erythrophthalmus и Р. ocai, при-
шли в соприкосновение друг с другом, и сейчас происходит их
гибридизация. В этом случае предшествовавшая экологическая
изоляция (представители вида Р. erythrophthalmus жили глав-
ным образом на дубах и в кустарниковом подлеске, тогда как
Р. ocai преимущественно в хвойных лесах и их подлеске), по-
видимому, нарушилась, когда сельское хозяйство и лесоразра-
Дифференциация популяций | 257
ботки создали местообитания, пригодные для обеих форм. Дей-
ствительно, гибридизация на том или ином уровне широко рас-
пространена среди птиц (например, чаек, уток, трупиалов, дубо-
носов, медососов, райских птиц), и обычно считают, что она
происходит в зонах вторичного контакта. Но как бы мы ее ни
истолковывали, ясно, что дифференциация в популяциях птиц —
процесс гораздо более сложный, чем можно было бы предста-
вить себе, просматривая аккуратные перечни «видов» и «под-
видов» птиц в фаунистических списках.
Точно так же довольно ясно показано, что во многих груп-
пах растений значительный обмен генами возможен и после до-
стижения так называемого видового уровня дифференциации.
Он может совершаться полностью на диплоидном уровне, но
может быть также связан с полиплоидией и апомиксисом (см.
гл. 9). Большинство таких случаев относится к категории явле-
ний, получивших название интрогрессивной гибридизации. Шан-
сы гибрида Fi на скрещивание с другими такими же гибридами
на ранних этапах гибридизации значительно ниже, чем вероят-
ность скрещивания с той или другой из родительских форм. Гиб-
риды почти всегда оказываются промежуточными формами как
в отношении требований к условиям среды, так и в отношении
морфологических признаков. Поэтому у потомства от возврат-
ных скрещиваний гибрида с тем или иным из его родителей
больше шансов найти подходящую экологическую нишу, чем
у особей Fi или F2. Ввиду этого генетическое «растворение» двух
вступающих в гибридизацию форм маловероятно. Скорее сле-
дует ожидать инфильтрации генотипа одного вида компонента-
ми зародышевой плазмы другого. Изменчивость каждого из ро-
дительских типов будет расширяться в направлении другого
типа; таким образом, вид или подвид может оказаться в состоя-
нии увеличить свой ареал и проникнуть в новые для него ме-
стообитания.
Интересный пример этого, изученный Хейзером, мы находим
у рода Helianthus (подсолнечник) в Калифорнии. Н. annuus —
обычный сорняк, встречающийся в большей части США; целый
ряд данных говорит о том, что он был интродуцирован в Кали-
форнию индейцами относительно недавно. Н. bolanderi пред-
ставлен в Калифорнии двумя расами. Одна из них распростра-
нена почти исключительно на серпентиновых почвах — экологи-
ческая особенность, свойственная лишь сравнительно немногим
специализированным растениям. Другой подвид является сор-
няком. Хейзер показал, что сорная раса Н. bolanderi, вероятно,
возникла в результате интрогрессии генетического материала
широко распространенного сорняка Н. annuus в расу Н. bolan-
deri, растущую на серпентиновых почвах. Мы не будем входить
17 Зак. 162
258 | Глава 10
в детали этого процесса, но отметим, что путем надлежащих
скрещиваний получившийся при этом подвид может быть «син-
тезирован» в лаборатории. Интересно, что интрогрессия была
двусторонней; она привела к созданию не только более крупного
и более выраженного сорняка Н. bolanderi, но и более мелкой
формы Н. annuus, обладающей, очевидно, более широкой эко-
логической амплитудой.
Распространение большинства видов растений, по-видимому,
сильно ограничено экологическими факторами. Там, где эколо-
гические барьеры эффективны, генетический обмен происходит
не часто, а гибриды с их промежуточными требованиями к усло-
виям среды не выживают. Но когда экологические барьеры раз-
рушаются, как, например, в местообитаниях, подверженных
эрозии или изменяющихся под воздействием ледников или дея-
тельности человека, промежуточные типы могут неожиданно
найти подходящие ниши и размножиться. Когда происходит
скрещивание двух видов, оно обычно ведет не к «растворению»
исходных видов, а к обогащению их изменчивости. По-видимому,
это действительно очень обычное явление для многих родов
многолетних растений, исследованных в США.
Отбор, направленный против гибридов. Кажется вероятным,
что при воссоединении сильно дифференцировавшихся популя-
ций отбор, направленный против гибридных особей, усиливает
факторы, противодействующие гибридизации. Обмен генетиче-
ской информацией между изолятами часто становится ничтожно
малым. Вполне возможно, что когда процессы дифференциации
будут лучше изучены, выяснится, что гибридизация между осо-
бями воссоединяющихся изолятов — явление почти всеобщее;
это означало бы, что механизмы, предотвращающие генетиче-
ский обмен между дифференцированными формами, обычно воз-
никают только в результате относительно неэффективной гибри-
дизации после восстановления симпатрии.
Интересное экспериментальное доказательство существова-
ния такого механизма получил Купмен; он синтезировал искус-
ственные смешанные популяции Drosophila pseudoobscura и
D. persimilis и выдерживал их при низких температурах (16°),
при которых половая изоляция между ними незначительна (гиб-
риды легче образуются при низких температурах, чем при высо-
ких). В экспериментальных условиях гибриды оказались чрез-
вычайно неприспособленными, но Купмен полностью исключал
возможность гибридизации, удаляя все гибридные особи раньше,
чем они могли бы приступить к размножению (гибриды рас-
познавались по генетическим маркерам).
За несколько поколений доля гибридов в потомстве заметно
снизилась, что указывало на усиление факторов, противодейст-
Дифференциация популяций | 259
вующих гибридизации (какова бы ни была их природа). Куп-
мену удалось показать, что изолирующий механизм, по крайней
мере отчасти, был связан с взаимоотношениями полов; оказа-
лось, что самцы «предпочитали» спариваться с самками своего
вида.
Хотя в природе D. pseudoobscura и D. persimilis встречаются
в одних и тех же географических районах, они не скрещиваются
между собой, что, видимо, объясняется двумя причинами. Пре-
жде всего в данном случае имеется значительная экологическая
изоляция, так как D. persimilis обычно обитает выше в горах
и предпочитает более прохладные, тенистые места, чем D, pseudo-
obscura. Некоторую роль должна играть и половая изоляция,
так как при обычных, не пониженных, температурах недавно
пойманные мухи обнаруживают мало склонности к гибридиза-
ции. Предполагают, что изоляции обеих форм в природе спо-
собствуют и другие, еще не выявленные факторы. В только
что описанных экспериментах эти два известных фактора изо-
ляции устраняли путем содержания плотной смешанной попу-
ляции при низких температурах. В очень короткое время есте-
ственный отбор создавал не известный ранее барьер, по крайней
мере отчасти связанный с выбором партнеров для спаривания.
К сожалению, мы очень мало знаем о том, при каких соот-
ношениях двух факторов миграции генов и нежизнеспособности
гибридов произойдет слияние воссоединяющихся сегрегатов и
при каких — окончательная дифференциация. Здесь перед нами
широкое поле для исследований.
РАЗЛИЧНЫЕ ТИПЫ ДИФФЕРЕНЦИАЦИИ
Вьюрки Галапагосских островов и африканские цихлиды. Не
следует недооценивать сложность процессов дифференциации.
Характерная картина, наблюдаемая ныне в группе дарвиновых
вьюрков, возникла, по-видимому, в результате взаимодействия
множества факторов. Весьма возможно, что все Geospizinae
произошли от небольшой стайки (быть может, одной пары)
птиц из сем. вьюрковых, случайно попавших на острова с мате-
рика Южной Америки. Крайне маловероятно, чтобы мигранты
составляли в данном случае большую и случайную выборку из
материковой популяции; напротив, скорее всего это была не-
большая выборка с определенным «уклоном», представлявшая
лишь ограниченный сегрегат генофонда соответствующего вида.
Эта ошибка выборки означает, что силы отбора, даже если
они были такими же, как на континенте, должны были влиять
На островную популяцию птиц по-иному, так как они действова-
ли здесь на совершенно иной генетический материал. Вспомните
17*
260 | Глава 10
о том, что при анализе действия эволюционных факторов от-
дельный локус нельзя рассматривать изолированно; давление
отбора на какой-либо локус будет частично зависеть от генотипа
в целом. Часто наблюдаемое явление, когда при основании но-
вых колоний ошибка выборки ведет к изменению генетической
среды, было описано как принцип родоначальника. Это, конечно,
особый случай генетического дрейфа.
После того как вьюрки закрепились на одном острове, рас-
пространение мигрантов на другие острова архипелага было
только вопросом времени. Популяции на разных островах, испы-
тывая действие различных факторов отбора и последствия гене-
тических различий между своими родоначальниками, вероятно,
быстро дифференцировались. Когда в результате миграций (в
том числе обратных миграций) происходило соприкосновение
дифференцированных форм, отбор должен был во многих слу-
чаях противодействовать наклонности к гибридизации, как опи-
сано выше. Величина и форма клюва у галапагосских вьюрков
связаны с характером потребляемой пищи, но, кроме того, слу-
жат для распознавания особей другого пола. Обилие различных
видов пищи на разных островах неодинаково, и весьма воз-
можно, что в условиях изоляции отбор привел к некоторой диф-
ференциации в отношении клюва и связанных с ним структур.
Когда же контакт восстанавливался, отбор, вероятно, вызывал
еще большую дифференциацию по этим признакам, так как они
важны для распознавания. Подобная дивергенция, по-видимому,
создает добавочное преимущество, уменьшая число форм, по-
требляющих пищу одного и того же типа. Очевидно, что такой
отбор на «пониженную конкуренцию» происходит, ио конкрет-
ный механизм его неясен.
История «роя» видов Haplochromis, вероятно, сходна с исто-
рией галапагосских вьюрков. Мигранты из речных систем коло-
низовали древнее озеро Виктория. Многократная колонизация
через надлежащие промежутки времени сама по себе позволяла
бы объяснить наблюдаемое разнообразие форм. К моменту при-
бытия новых иммигрантов прежние уже успевали частично диф-
ференцироваться; отбор, направленный против гибридов, дово-
дил дело до конца. Кроме того, повторные циклы пересыхания
и затопления могли поочередно приводить к дроблению и вос-
соединению изолятов, что позволяло действовать механизмам
дифференциации. Возможно также, что различия в характере
дна, береговой линии, глубины и т. п. играли (и продолжают
играть) роль внутренних барьеров для расселения различных
форм в пределах озера.
В иды-двойники у бабочек. Группа Erebia tyndarus — еще
один превосходный пример тонких взаимодействий, возможных
Дифференциация популяций | 261
между дифференцированными формами. Например, несмотря на
различия в адаптации жизненных циклов Е. cassioides и Е. ni-
valis к высокогорным условиям, присутствие или отсутствие
каждой из этих форм, по-видимому, сильно влияет на другую
форму: только там, где обе встречаются вместе, наблюдается
ограничение их распространения определенными высотами. Кар-
тина их современного распространения, видимо, обусловлена
взаимодействием дифференцированных популяций. Как и у дар-
виновых вьюрков, конкретную природу этих взаимодействий ука-
зать трудно. Отбор, направленный на устранение конкуренции,
сам по себе не может служить объяснением. Необходимо точно
знать, чем обусловливалось дифференциальное воспроизведение
генотипов.
Должно быть, эти формы несколько дифференцировались
еще до своего соприкосновения. Возможно, например, что перед
тем, как популяции предков современных Е. nivalis и Е. cassioi-
des встретились, Е. nivalis обитала ближе к верхней границе их
общего ареала, а Е. cassioides — в среднем ближе к нижней гра-
нице. Тогда генотипы Е. nivalis, «предпочитавшие» высокогор-
ную зону, и генотипы Е. cassioides, «склонные» оставаться на
меньших высотах, должны были получить известное преимуще-
ство. Быть может, относительная редкость другой формы сни-
жала возможность потери гамет вследствие неэффективной гиб-
ридизации. К тому же несходные гусеницы могли иметь склон-
ность поедать друг друга. Не исключено также, что экскреты
гусениц Е. nivalis тормозили рост Е. cassioides, и (или) наобо-
рот. Пищевые потребности различных изолятов могли быть
слишком сходными или число доступных ниш слишком ограни-
ченным, для того чтобы могла возникнуть специализация в от-
ношении местообитаний вроде той, которая наблюдается у дар-
виновых вьюрков.
В результате каждая форма занимает ныне те области, к ко-
торым она лучше всего приспособлена. Мы, вероятно, никогда
не узнаем всех деталей этого процесса. Какие-то формы — быть
может, не обладавшие достаточной генетической изменчивостью
для дальнейшей дифференциации — вымерли. Это могло про-
изойти потому, что из-за суровых условий в определенной обла-
сти сильно уменьшились пищевые ресурсы, и гусеницы какого-то
другого вида Erebia оказались способными гораздо лучше ис-
пользовать ограниченные возможности. Некоторые формы (на-
пример, Е. calcarius и западная Е. cassioides), видимо, никогда
не сталкивались между собой. Интересно отметить, что в группе
Е. tyndarus наблюдается стерильность гибридов между более от-
даленными формами, такими, как Е. iranica (/г=51) и Е. calca-
rius (/г = 8), при отсутствии поведенческой изоляции. Эти формы,
18 Зак. 152
262 | Глава 10
очевидно, не соприкасались друг с другом; следовательно, се-
лективной основы для развития изолирующих механизмов ни-
когда не существовало.
Дифференциация паразитов. Типы дифференциации, связан-
ные с выработкой линий, предпочитающих разные виды пищи
или разных хозяев, еще во многом неясны. Часто отмечалось,
что в лаборатории линии паразитических организмов можно с
успехом переключать с обычного хозяина на другого путем пере-
носа большого числа особей. Например, человеческая вошь,
Pediculus humanus, может быть превращена в кроличью. Для
этого переносят большое число вшей, но лишь сравнительно не-
многие из них, способные питаться на кролике, выживают и
размножаются. Процесс отбора в конце концов приводит к вы-
работке линии, благоденствующей на кроликах и не особенно
склонной переходить на человека. (Отдельные особи могут пи-
таться на человеке, но колония не будет процветать.) Вполне
возможно, что аналогичный процесс отбора ответствен за пре-
вращение головной вши в платяную в случаях, когда первая
попадает в условия, обычные для второй. Высказывалось пред-
положение, что генетическая структура популяции вшей способ-
ствует пластичности, благодаря которой превращение в обоих
направлениях совершается сравнительно просто. Нет надобности
говорить о том, что сама эта структура, несомненно, есть резуль-
тат отбора. Различие между Ascaris и Pediculus состоит, воз-
можно, главным образом в том, что аскариды генетически не
столь изменчивы.
Вряд ли можно сомневаться в наличии селективного преиму-
щества у тех кукушек, яйца которых очень похожи на яйца
птицы-хозяина. Яйца, отложенные в гнезда обычного хозяина,
гораздо реже остаются невысиженными, чем отложенные в гнез-
да необычных хозяев. Однако конкретный механизм образова-
ния и сохранения фратрий до сих пор остается несколько зага-
дочным. Кажется маловероятным, чтобы фратрии были генети-
чески изолированы друг от друга. Никаких иных признаков
дифференциации между ними, кроме внешнего вида яиц, обна-
ружено не было; между тем, если бы фратрия представляла со-
бой изолированную эволюционную единицу, следовало бы
ожидать дифференциации и в других отношениях. Поскольку, од-
нако, во многих орнитологических работах исследуются только
внешние признаки, возможно, что такая дифференциация суще-
ствует, но еще не выявлена. По-видимому, нет также никакого
механизма, который препятствовал бы обмену генетической ин-
формацией в местах, где несколько фратрий существуют
совместно, особенно ввиду значительной величины территории,
занимаемой самцами.
Дифференциация популяций | 263
Сходство между миметическими формами яиц у кукушки и
серией миметических форм самок у РарШо dardanus является
довольно поверхностным. Главное отличие состоит в том, что
самкам кукушки необходимо откладывать яйца в гнездо хозя-
ина надлежащего вида. (Выбор соответствующих гнезд кукуш-
ками можно было бы объяснить явлением импринтинга.) Здесь
недостаточно того, чтобы генетическая структура популяции
обеспечивала образование ряда различных форм; эта структура
должна быть такой, чтобы птица, выбирающая своим птенцам
приемных родителей определенного вида, откладывала яйца
надлежащего типа. Например, если самка кукушки, выросшая в
гнезде тростниковой камышовки, будет осеменена самцом из
фратрии, паразитирующей в гнездах черной трясогузки, то ее
яйца могут оказаться больше похожими на яйца трясогузки, че^м
на яйца камышовки. Но она будет откладывать яйца «не в те
гнезда» и, по всей вероятности, у ее потомства окажется срав-
нительно мало шансов на выживание. К сожалению, мы ничего
не знаем о генетике окраски яиц у кукушки, так что о роли от-
цовской наследственности можно лишь строить догадки Г
Кажется вероятным, что дифференциация фратрий у кукуш-
ки и миметических форм у Papilio dardanus происходила в усло-
виях изоляции. Когда два дифференцированных изолята Р. dar-
danus приходили в соприкосновение в областях, где имелись обе
модели, в результате дисруптивного отбора возникала поли-
морфная система (см. гл. 7). Но если две разные фратрии куку-
шек встречались в районе, где обитают оба вида хозяев, к этому
добавлялся, вероятно, еще один фактор. Помимо дисруптивного
отбора (менее успешное воспроизведение генотипов, дающих
яйца промежуточного типа), возможно, действовал также отбор,
благоприятствовавший тем формам поведения, которые способ-
ствовали положительному ассортативному скрещиванию (спари-
ванию особей, воспитанных приемными родителями одного
вида). По-видимому, такой отбор был наиболее успешным в тех
случаях, когда хозяева несколько различались по своим эколо-
гическим потребностям: тогда кукушки-самцы оставались в при-
вычном местообитании, в котором они выросли, и встречали по-
лучивших такое же «воспитание» самок.
Там, где местообитания разных хозяев частично совпадают,
кукушки часто откладывают яйца промежуточного типа и ми-
микрия нарушается. По-видимому, в данном случае фратрии у
кукушек представляют собой нечто промежуточное между поли-
морфными формами и тем, что можно было бы назвать расами,
1 Окраска яиц у птиц, например у кур, определяется исключительно ге-
нотипом матери. — Прим. ред.
18*
264 | Глава 10
специфичными по отношению к определенному хозяину. Степень
совершенства мимикрии, очевидно, зависит от степени экологи-
ческой изоляции данной фратрии. Там, где разные фратрии со-
существуют в одном географическом районе, нет никаких при-
знаков того, что такая степень изоляции уже может обеспечить
генетическую дифференциацию, достаточную для развития ре-
продуктивной несовместимости. Судя по всем данным, такие
фратрии не являются «зарождающимися видами», дивергенция
же на видовом уровне происходит в условиях географической
изоляции. Несомненно, для точного определения статуса фрат-
рий у кукушек требуются дальнейшие тщательные исследова-
ния.
АЛЛОПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
Дифференциацию физически изолированных популяций, ко-
гда она достигает степени, при которой в случае восстановления
контакта воссоединения популяций уже не происходит, назы-
вают аллопатрическим видообразованием. Множество данных
позволяет предполагать, что аллопатрическое видообразование
является основной причиной многообразия органического мира.
Именно механизм «расщепления» в сочетании с вымиранием
необходим для объяснения того, как возникло огромное число
сравнительно обособленных форм растений, животных и микро-
организмов, населяющих землю.
О времени, необходимом для дифференциации популяций,
мало что известно. В каждом данном случае на это время вли-
яли бы многие переменные, в том числе размер популяций, ве-
личина давления отбора, степень изоляции и генетическая си-
стема организма. Видимо, в большинстве случаев видообразова-
ние — процесс, слишком растянутый во времени, чтобы его легко
было наблюдать. Однако результаты многих недавно проведен-
ных исследований (например, данные об индустриальном мела-
низме у бабочек и о популяциях Сераеа и Natrix) указывают
на то, что, возможно, интенсивность отбора в природе в общем
выше, чем полагали ранее; если это так, то и видообразование,
быть может, идет быстрее, чем предполагалось в прошлом. Как
бы то ни было, представление об аллопатрическом видообразо-
вании как о главном механизме дивергентной эволюции основы-
вается не на результатах прямого наблюдения, а на существо-
вании таких картин изменчивости, которые, по-видимому, соот-
ветствуют каждому из всех мыслимых этапов постулированного
процесса. Некоторые из таких ситуаций были рассмотрены в на-
стоящей главе; в биологической литературе описано множество
других случаев.
Дифференциация популяций | 265
СИМПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
Могут ли новые, ясно отличимые от прежних формы орга-
низмов возникать при отсутствии физической изоляции популя-
ций? Можно с уверенностью ответить «да» — в случае аллопо-
липлоидии (см. гл. 9), а также организмов, не имеющих поло-
вого процесса. У организмов, размножающихся только бесполым
путем (например, у некоторых коловраток), каждая особь
представляет собой генетический изолят, а виды — это группы
клонов, сходство которых обусловлено межклоновым отбо-
ром.
Одна из длительных дискуссий между теоретиками эволюции
касалась вопроса о возможности симпатрического видообразо-
вания у организмов, размножающихся половым путем. Как уже
говорилось, огромное большинство данных указывает на то, что
аллопатрическое видообразование является правилом, и многие
случаи, якобы свидетельствовавшие о симпатрическом видооб-
разовании (например, скопления близких видов у цихлид), легко
могут быть объяснены по-иному. Один из доводов против сим-
патрического видообразования состоит, однако, в том, что обмен
генами должен вести к выравниванию любых различий, возни-
кающих под действием дисруптивного отбора. Недавно Тодей
с сотрудниками и Стримс и Пайментел провели исследования,
неожиданно показавшие, что это не обязательно должно быть
так. Эти исследователи в лабораторных экспериментах с дрозо-
филой показали, что дисруптивный отбор может привести к ди-
вергенции и при отсутствии изоляции. Например, Тодей и Гиб-
сон подвергали популяции дикого типа дисруптивному отбору
на число щетинок, причем отбираемые особи как с большим, так
и с малым числом щетинок помещались для скрещивания в об-
щий сосуд. По прошествии 12 поколений исходная популяция
разделилась на две, причем гибриды между ними возникали
редко. Таким образом, теоретически симпатрическое видообра-
зование, видимо, не исключено, но его значение в природе пока
еще не установлено. Слободкин удачно сформулировал ситуацию
следующими словами:
«В некотором смысле различие между теоретиком, экспери-
ментатором и исследователем, работающим в полевых условиях,
состоит в том, что теоретик рассматривает все мыслимые миры,
тогда как экспериментатор имеет дело со всеми возможными
мирами, а полевик ограничен реальным миром. Эколог-экспери-
ментатор должен спрашивать теоретика о том, представляет ли
сто возможный мир какой-либо интерес, а исследователь-поле-
вик — о том, имеет ли этот мир вообще отношение к реальному
Миру».
19 Зак. 152
266 | Глава 10
В споре о симпатрическом видообразовании слово, по-види-
мому, за исследователями, ведущими наблюдения в полевых
условиях.
ВЫВОДЫ
Имеющиеся данные указывают на то, что дифференциация
в условиях изоляции служит главным источником многообразия
органического мира. Когда популяции физически обособлены
друг от друга (так что обмен генами сведен к минимуму или
отсутствует), их «эволюционная история» различна и поэтому
они генетически дифференцируются. Дойдя в этом процессе до
определенной точки, популяции уже не могут воссоединиться в
случае возобновления контакта между ними. Исследование этой
генетической «точки необратимого разделения» и формулировка
касающихся ее обобщений — одна из самых трудных проблем
в изучении эволюции. Популяции, дивергировавшие в такой сте-
пени, что слияние их в будущем исключено, все еще могут об-
мениваться генетической информацией, взаимно обогащая свою
изменчивость, что стимулирует их дальнейшую эволюцию. Не
следует, однако, рассматривать создание разнообразия как
«цель» эволюции и считать происходящий изредка генетический
обмен между почти изолированными группами в каком-то смыс-
ле вредным или аномальным явлением.
ЛИТЕРАТУРА
1. Bowman R. I., Morphological differentiation and adaptation in the Gala-
pagos finches, Univ. Calif. Publ. Zool., 58 (1961).
2. Brown W. L., Wilson E. O., Character displacement, Systematic Zool.,
5, 49—65 (1956).
3. С 1 a u s e n J., Stages in the Evolution of Plant Species, Cornell Univ. Press,
Ithaca, N. Y., 1951.
4. Dobzhansky T., Genetics and the Origin of Species, Columbia Univ.
Press, New York, 1951.
5. Keck D. D., Trends in systematic botany. In Survey of Biological Prog-
ress, vol. 3, Academic, New York, 1957.
6. Ma yr E., Animal Species and Evolution, Harvard Univ. Press, Cambridge,
1963.
7. Ma yr E., (ed.), The species problem, Am. Assoc. Advance. Sci. Symposium
50 (1957).
8. S t e b b i n s G. L.. Variation and Evolution in Plants, Columbia Univ.
Press, New York, 1950.
9. T h о d a у J. M., Gibson J. B., Isolation by disruptive selection, Nature,
193, 1164—1166 (1962).
Дифференциация популяций | 267
10. Woodson R. Е., Butterflyweed revisited, Evolution, 16, 168—185 (1962).
11. Woodson R. E., Ann. Missouri Bot. Gard., 34 (1947).
12. Moore J., Evolution, 3, 1949.
13. Stebbins G. L., University of California Publications in Zoology, 48
(1949).
14. Eplin g, Dobzhansky T., Genetics, 27, 1942.
15. Abrams, Illustrated Flora of the Pacific States, Stanford University Press,
1951.
16. Moore J., Introduction to Historical Geology, McGraw-Hill, 1949.
17. Moore J., Bull. Am. Mus. Hist., 82 (1944),
19’
Глава 11
ВЫСШИЕ УРОВНИ ИЗМЕНЧИВОСТИ
Можно ли объяснить все огромное разнообразие жизни теми
же процессами, которые ответственны за дифференциацию попу-
ляций? В предыдущей главе мы показали эффективность мута-
ций, отбора, миграции (в генетическом смысле) и дрейфа генов
как факторов, обусловивших возникновение различной окраски
цветков у Linanthus, географических вариантов у бабочек или
новых видов у птиц. Теперь можно задать себе вопрос: не от-
ветственны ли эти же комплексы факторов и за различия ме-
жду цветковыми растениями, бабочками и птицами? Может
быть, именно этим факторам обязаны своим возникновением не
только сравнительно близкие группы организмов, но и совер-
шенно несходные формы жизни? Преимущественно таксономи-
ческая ориентация ранних эволюционных исследований послу-
жила причиной того, что этот вопрос часто рассматривают как
проблему происхождения высших таксономических единиц. Не-
которым палеонтологам и генетикам казалось, что высшие ка-
тегории — такие, как роды, семейства и отряды, — могли воз-
никнуть в результате эволюционных процессов (макроэволю-
ция), отличных от тех, которые действуют на уровне вида
(микроэволюция). Данные, полученные во многих областях био-
логии, по-видимому, ясно показывают, что ввиду огромной дли-
тельности прошлой эволюции нет необходимости постулировать
действие каких-то иных процессов, помимо тех, которые были
рассмотрены выше.
ВЫМИРАНИЕ И БИОГЕОГРАФИЧЕСКАЯ ОБОСОБЛЕННОСТЬ
Существование крайне обособленных групп можно объяснить
вымиранием промежуточных форм или же недостаточной изу-
ченностью флоры и фауны многих стран. Конечно, за возмож-
ными редчайшими исключениями (к ним относятся, например,
некоторые виды рыб, встречающиеся, по-видимому, лишь в од-
ном небольшом ручье), даже наиболее обособленные группы со-
стоят из подгрупп, в той или иной степени различающихся ме-
жду собой.
Высшие уровни изменчивости | 269
ВЫМИРАНИЕ
Единственный ныне живущий представитель рептилий под-
класса Rhynchocephalia — гаттерия (Sphenodon punctatus) —
встречается лишь на десятке с небольшим мелких островов у по-
бережья Новой Зеландии. Несомненно, группы особей, обитаю-
щие на разных островах, принадлежат к различным менделев-
ским популяциям, но степень генетической дивергенции между
этими популяциями (и обмена между ними) неизвестна. В жи-
вотном мире известно много примеров подобного рода изолятов.
Необычная перуанская бабочка Styx infernalis принадлежит к
сем. Lycaenidae (она родственна нашим обычным голубянкам),
но целый ряд своеобразных анатомических черт ставит ее особ-
няком от других лиценид, так что ее относят к особому подсе-
мейству. В данном случае о степени дифференциации внутри
этой группы опять-таки ничего не известно.
Если игнорировать ископаемые находки, то человек пока-
жется весьма обособленной формой. Между Homo sapiens и его
ближайшими ныне живущими родственниками (человекообраз-
ными обезьянами) имеет место полная репродуктивная изоля-
ция. Насколько нам известно, дифференциация внутри вида
Homo sapiens не достигла той степени, при которой представи-
тели внутривидовых подразделений дают бесплодное потомство.
Однако в отличие от Sphenodon и Styx вид Homo sapiens харак-
теризуется весьма значительной географической изменчивостью.
Наши представления о «расах» основаны главным образом на
изменчивости нескольких очень заметных внешних признаков
(цвет кожи, тип волос, форма черепа), но существуют и разли-
чия по менее явным признакам, в частности по группам крови
и типам гемоглобина (см. гл. 7). Таким образом, группу форм
Homo sapiens нелегко разбить на обособленные группы. У чело-
века обмен генами в той или иной степени возможен практи-
чески между всеми подгруппами, поэтому картина изменчивости
оказывается чрезвычайно сложной. Действительно, здесь очень
обычна дискордантная изменчивость, при которой различные
исследуемые признаки ведут себя весьма различным образом.
Хороший пример такой дискордантной изменчивости дает срав-
нение распределения групп крови в популяциях человека
(фиг. 59). Распределение генов А и В, определяющих группы
крови, оказывается совершенно различным.
Примеры сильно обособленных форм нередки и в царстве
растений. Гинкго (Ginkgo biloba), подобно гаттерии, является
единственным живым представителем своей группы — весьма
обособленного порядка, принадлежащего к типу голосеменных
Растений. Филогенетическую линию, к которой принадлежит
этот вид, можно проследить по ископаемым находкам далеко
Фиг. 59. Карты, показывающие дискордантный характер изменчивости в от-
ношении генов групп крови (вверху—ген Л, внизу — ген В), проявляющийся
в их распространении в аборигенных популяциях человека.
Одинаковой штриховкой показаны области, в которых популяции характеризуются примерно
одинаковой частотой данного гена [4].
Высшие уровни изменчивости | 271
в глубь времен. В мезозойскую эру существовало много родов
и видов порядка Ginkgoales (фиг. 60). По какой-то неизвестной
нам причине все представители этого порядка, за исключением
Ginkgo biloba, вымерли в третичном периоде. Естественное ме-
стообитание гинкго нам также с точностью неизвестно, но ки-
тайцы на протяжении столетий культивировали этот вид ради
его съедобных семян. В Китае гинкго выращивали главным об-
разом в садах при храмах. Так как гинкго практически не пора-
жается грибами и насекомыми и так как его листья осенью при-
обретают красивую ярко-золотистую окраску, его стали широко
использовать для озеленения улиц во многих странах умеренной
зоны. Гинкго, по-видимому, выживает при наихудших условиях
загрязнения среды, какие только может создать человек. Вид
G. biloba, подобно многим другим изолированным группам голо-
семенных, в генетическом отношении очень многообразен и
представлен большим числом декоративных вариантов.
Metasequoia glyptostroboides тоже является, видимо, послед-
ним сохранившимся представителем рода голосеменных, близко-
го к Sequoia и Sequoidendron. В отличие от Ginkgo Metasequoia
впервые была описана как ископаемое растение. Лишь спустя
примерно 5 лет после ее описания по отпечаткам листьев в пли-
оценовых отложениях Японии в Китае были обнаружены живые
экземпляры. И это дерево, очевидно, сохранилось потому, что
его выращивали в Китае.
Наряду с такими полностью изолированными видами суще-
ствуют также группы из двух или нескольких близких между
собой форм, ясно обособленные’ от своих ближайших ныне жи-
вущих родственников. В роде Neophasia (бабочки сем. Pieridae)
обнаружены две формы, которые все специалисты по чешуекры-
лым считают отдельными видами; оказалось, что промежуточ-
ные формы отсутствуют, а внутривидовая изменчивость выра-
жена лишь незначительно. Один вид, N. menapia, широко рас-
пространен в западной части Северной Америки, где он питается
на различных хвойных деревьях. Оба пола имеют белую окрас-
ку с черными пятнышками главным образом у верхушки и пе-
реднего края первой пары крыльев. N. terlooti встречается в се-
верной Мексике и Аризоне; этот вид также питается на хвойных.
Самцы N. terlooti похожи на самцов N. menapia, но у первых
темные пятнышки на передних крыльях крупнее. Самки сходны
с самцами, только основная окраска у них не белая, а оранже-
во-красная. У /V. menapia красные самки неизвестны, а у N. ter-
looti не встречаются белые самки.
Ныне живущие слоны представляют интересный случай, ко-
гда в многообразной группе сохранились только два сильно раз-
личающихся вида. Африканский слон (Loxodonta africana),
Фиг. 60. Ginkgo biloba и вымершие родственные формы.
В середине — Ginkgo biloba; вверху слева — Sphenobaiera f areata (верхний триас); вверху справа — Ginkgoites innzensis сгриас); внизу
слева — Baiera longl folia ^юра); внизу справа-Ginkgo leplda (юра) (5J.
Высшие уровни изменчивости | 273
несомненно, является ближайшим живым родственником индий-
ского слона (Elephas maximus), и наоборот. Однако оба вида
имеют еще более близких родственников среди известных вы-
мерших форм, которые не были их прямыми предками.
Сходная ситуация была выявлена при детальном изучении
видов сосны на западе Северной Америки. Вид Pinus masoni
известен только по плиоценовым остаткам. На основании иссле-
дования многочисленных ископаемых находок было высказано
предположение, что этот вид вымер в плейстоцене, после того
как дал начало двум обособленным линиям эволюции. Одна из
этих линий привела к виду Р. linguiformis, также вымершему
в плейстоцене. От Р. linguiformis произошла сосна Р. attenuata,
палеонтологическая история которой уходит довольно далеко
в глубь плейстоцена и которая является ныне самым распро-
страненным видом сосны в Калифорнии. Вторая эволюционная
ветвь, происходящая от Р. masoni, дала Р. muricata. Этот вид
существует поныне и весьма обычен на тихоокеанском побе-
режье, так же как и произошедшие от него Р. radiaia и Р. гето-
rata. Если рассматривать только современные виды, то Р. atte-
nuata и Р. muricata покажутся морфологически очень близкими
друг другу. Однако оба вида имеют близких родственников
среди ископаемых форм, которые предшествовали им, некоторое
время существовали вместе с ними и вымерли в плейстоцене.
Обе эти формы являются в настоящее время процветающими
видами, расширяющими свои ареалы и обладающими значи-
тельной наследственной изменчивостью. Ареал Р. radiata в не-
давние времена сократился, и этому виду, возможно, грозило
вымирание, но его спасло то, что его стали широко культивиро-
вать и интродуцировали во многие части света, где он выращи-
вается для изготовления бумажной массы.
Все только что описанные ситуации, когда вид или группа
видов кажутся сильно обособленными от других таких же групп,
связаны с вымиранием. Группа оказалась обособленной потому,
что наиболее близкие формы не выжили. Когда рассматривают
и ископаемые формы (если они имеются), эта обособленность
исчезает. Неверные представления возникают также в тех слу-
чаях, когда формы, населяющие достаточно обширную геогра-
фическую область, изучают лишь в какой-то части их ареала.
При рассмотрении изменчивости в ограниченной области ка-
жется, что существуют обособленные группы, однако при изуче-
нии всех встречающихся на Земле форм такого впечатления уже
не создается. Как и в предшествующих примерах, все дело в
степени репрезентативности выборки, причем в данном случае
Речь идет о недостаточной репрезентативности выборки в про-
странстве, а не во времени.
274 | Глава 11
БИОГЕОГРАФИЧЕСКАЯ ОБОСОБЛЕННОСТЬ
В Северной Америке обнаружены четыре четко разграничен-
ные формы бабочек, принадлежащих к роду Nymphalis (N. ап-
tiopa, N. milberti, N. calif arnica и N- vau-album). Любую особь
можно с уверенностью отнести к одной из этих четырех форм,
так как никаких промежуточных форм не найдено. Но если
включить в рассмотрение палеарктических представителей рода
Nymphalis, впечатление четкой подразделенности исчезает. От-
ношение группы, которую мы называем N. vau-album, к одно-
именной евразийской форме не вполне ясно. По немногим изу-
ченным признакам (величина, рисунок, строение гениталий) из-
менчивость в выборках из Северной Америки и из Евразии в
значительной степени взаимно перекрывается. Как и в большин-
стве других случаев, для оценки действительной степени диффе-
ренциации нужны тщательные исследования, охватывающие
много признаков. Дальнейшего изучения требует и вопрос о сте-
пени дивергенции между N. milberti и палеарктическими пред-
Фиг. 61. Близкие формы бабочек рода Nymphalis.
Вверху слева?/, milberti (Вайоминг); вверху справа N. caschm Irens Is (Непал); внизу слева
Аг. urticae (Сахалин); внизу справа N. urticae (Венгрия).
Высшие уровни изменчивости [
ставителями комплекса N. urticae (фиг. 61), а также между
N. californica и обитателями Старого Света N. polychloros и
N. xanihotnelas. Таких ситуаций бесчисленное множество.
В Йеллоустонском национальном парке живут два вида мед-
ведей: черные медведи (Ursus atnericanus) и гризли (U. horri-
bilis). Если бы весь ареал гризли ограничивался этим парком,
то гризли составляли бы обособленную однородную группу того'
же типа, что и совокупность особей гаттерии (Sphenodon). Од-
нако ареал гризли простирается и дальше к северу, до Аляски,
где медведи представлены очень крупными формами (аляскин-
ские бурые медведи U. middendorffi), а еще дальше, в Евразии,
медведи представлены видом U. arctos. Кажется вероятным, что
все виды образуют одну цепь групп, связанных переходами. Бе-
лый медведь (t/. maritimus)—очень близкая группа, приспо-
собленная главным образом к жизни у моря, в условиях Аркти-
ки. Ареалы бурого и белого медведей почти не перекрываются,
но отдельные особи встречаются друг с другом у арктического
побережья. Насколько мы знаем, промежуточные особи в при-
роде не обнаружены (в то же время хорошо известны гибриды,
полученные в зоопарках). Таким образом, хотя таксономия мед-
ведей в пределах йеллоустонского парка могла бы показаться
несложной, при рассмотрении картины в масштабе земного шара
возникает ряд проблем. Относятся ли европейские медведи к
«тому же виду», что и медведи Йеллоустонского парка? Дей-
ствительно ли белые медведи относятся к «иному виду»? И важ-
ны ли эти вопросы с точки зрения биологии?
С этой проблемой часто приходится встречаться и в ботани-
ке. Как и в зоологии, затруднения особенно очевидны в отноше-
нии форм, распространенных циркумполярно или циркумборе-
ально. Нередко один и тот же вид может скрываться под
разными названиями в Канаде, Европе и России, пока моногра-
фическое исследование, в котором впервые будут сопоставлены
экземпляры из всех районов, не выявит его истинного распро-
странения. Столь подробное рассмотрение здесь этого вопроса
может показаться излишним. Удивительно, однако, то, что био-
географы часто используют результаты локальных флористиче-
ских и фаунистических исследований* без учета этого существен-
ного источника искажений.
СЕТЕВПДНАЯ ИЗМЕНЧИВОСТЬ
В различных группах организмов встречается и совсем иная
ситуация, сущность которой еще не вполне ясна. Эта ситуация
характеризуется тем, что вид хорошо выражен и легко отличим
от других видов. Морфологической иитерградации, по-види-
мому, нет или почти нет, так что каждая группа ясно очерчена.
276 | Глава 11
В семействе ласточниковых (Asclepiadaceae), например, легко
выделить ряд неперекрывающихся таксонов (видов), стоит толь-
ко понять довольно сложную морфологию цветка. Известно, что
в этом семействе цветковых растений очень трудно выделять
группы этих первичных таксонов. Характер изменчивости таков,
что определить границы родов, по существу, невозможно. Явле-
ние, когда важные с точки зрения специалиста-систематика
признаки встречаются у большого числа видов в разных сочета-
ниях, называют сетевидной изменчивостью. Быть может, в дан-
ной группе растений это результат сложных механизмов обли-
гатного опыления насекомыми, однако такая изменчивость,
по-видимому, обычна на уровне вида в самых различных груп-
пах растений и животных.
Сетевидную изменчивость следует отличать от сетевидной
эволюции. Сетевидная эволюция — это слияние филетических
линий (в результате скрещивания); такая эволюция была ясно
продемонстрирована у многих растений, у которых она обычно
связана с удвоением числа хромосом у гибридов. Примеры этого
как у растений, так и у животных рассмотрены в гл. 9. В случае
Asclepiadaceae нет оснований подозревать соединение ранее обо-
собленных филетических линий и нет никаких указаний на то,
что в видообразовании сыграла какую-то роль полиплоидия.
АЛЕОНТОЛОГИЧЕСКАЯ ЛЕТОПИСЬ
Время — источник многих трудностей в проблеме «макроэво-
люции» в отличие от «М1икроэволюции». Прошлую историю жиз-
ни можно изучать только путем исследования ископаемых и
сравнения их с существующими формами. Благоприятные слу-
чаи для проверки таких наблюдений и сделанных на их основе
выводов поистине редки. Открытие таких «живых ископаемых»,
как Latimeria и Metasequoia, дает возможность подобной про-
верки. Latimeria — живой представитель кистеперых рыб —
группы, которую считали вымершей в меловой период. Metase-
quoia была впервые описана по ископаемым остаткам из плио-
ценовых отложений, но несколько лет спустя ее нашли в Китае.
Еще одной проверкой наших косвенных заключений, основан-
ных на сравнении современных и ископаемых форм, служит от-
крытие постулированных ранее «недостающих звеньев». Самой
известной из таких находок было, пожалуй, сделанное Дюбуа
открытие остатков Homo erectus erectus (Pithecanthropus) —
ископаемого обезьяночеловека.
Ископаемые находки следует рассматривать как далеко не
адекватную выборку остатков прошлой жизни. Изменения зем-
ной коры едва не стерли ключевые главы палеонтологической
Высшие уровни изменчивости | 277
летописи, причем самый интересный и важный раздел ее ока-
зался искаженным в наибольшей степени. Определенные части
некоторых организмов, например кости позвоночных, раковины
моллюсков и фораминифер, деревянистые части растений и ку-
тикула листьев, легче превращаются в окаменелости, чем другие
компоненты. Организмы, в меньшей мере снабженные твердыми
и устойчивыми к различным воздействиям частями (например,
черви, личинки насекомых, большинство водорослей и грибов),
сохраняются реже. Местообитание организма также в большой
степени определяет вероятность его превращения в окамене-
лость. Растения и животные- степей имели гораздо меньше шан-
сов сохраниться, чем обитатели болот. Факторы, благоприятные
для сохранения остатков растений, часто способствуют разру-
шению остатков животных (например, в кислой среде листья
могли бы сохраняться, но раковины и кости растворились бы
в ней). Наконец, некоторые специализированные участки среды,
такие, как асфальтовые ямы, болота с минерализованной водой
и склоны вулканов, по той или иной причине иногда сохраня-
ются почти целиком, со всеми организмами, которые оказались
в них.
И все же ископаемый материал достаточно полон и разнооб-
разен для того, чтобы палеонтологи могли распознать много ти-
пичных эволюционных ситуаций и описать предположительно
ответственные за них процессы. Многие проблемы специфичны
для палеонтологии. Например, в дополнение к таксономическим
трудностям, связанным с дифференциацией организмов в разных
участках неоднородной среды (горизонтальное видообразова-
ние), палеонтологу приходится иметь дело еще и с дифферен-
циацией во времени (вертикальное видообразование).
ТИПЫ ЭВОЛЮЦИОННЫХ ИЗМЕНЕНИЙ
Симпсон и другие авторы суммировали то, что известно об
основных формах эволюционных изменений, особенно в отноше-
нии животных. Симпсон делит эти изменения на три типа. Мно-
гие случаи, относящиеся к первому типу, — расщепление филе-
тических линий — могут быть прослежены на палеонтологиче-
ском материале. Здесь возможны процессы самого различного
масштаба — от разделения ствола рептилий-архозавров на
наземную и «воздушную» ветви (представленные ныне кроко-
дилами и птицами) до образования отдельных видов млеко-
питающих. С другой стороны, некоторые линии, очевидно, не
расщепляются в течение длительных периодов времени; в них
происходят настолько постепенные морфологические изменения,
что проведение таксономических границ на протяжении этого
278 | Глава 11
непрерывного ряда по существу невозможно нигде, кроме как в
его начале и конце. Этот тип эволюции называют филетической
эволюцией. Симпсон выделил также третий тип — квантовую
эволюцию, т. е. быстрое изменение, связанное с приспособле-
нием к совершенно иному образу жизни. Частичное или полное
слияние филетических линий трудно распознать по палеонтоло-
гическим данным, хотя в прошлом оно, по-видимому, играло
такую же важную роль, как и в наши дни.
Одна из самых интересных проблем, которые удается иссле-
довать на палеонтологическом материале, — это вопрос о ско-
рости, с которой протекала эволюция. Есть много различных
способов измерения темпов эволюции. В настоящее время ни
один из них не является вполне удовлетворительным, но тем не
менее они представляют огромный интерес. Во многих хорошо
изученных филогенетических линиях позвоночных и моллюсков
скорость изменения некоторых признаков можно определить,
пользуясь абсолютной шкалой времени. Возможно также изме-
рять скорость «формирования» таксономических категорий,
исходя из предположения, что она пропорциональна времени до-
стижения определенной степени морфологической дифференциа-
ции или генетической изоляции. Скорость вымирания таксоно-
мических групп, выраженная в виде кривых выживания, также
может представлять интерес. Следует помнить, конечно, что
сравнения, основанные на таксономических различиях между
группами, необходимо интерпретировать с большой осторожно-
стью. Изучение современных и ископаемых форм дает много
свидетельств в пользу того, что, например, «род» в одной группе
может не быть в биологическом отношении эквивалентным одно-
именной единице в другой группе.
Метод множественной корреляции, применяемый в настоя-
щее время в количественной таксономии, по-видимому, откры-
вает реальные возможности для разработки более удовлетво-
рительных методов измерения темпов эволюции. Изучая изме-
нения в большой группе признаков, происходящие во времени,
можно получить объективные количественные оценки общей
структурной дивергенции. Чем меньше степень корреляции ме-
жду последовательными выборками, тем больше происшедший
эволюционный сдвиг. Подобного рода анализ позволяет избе-
жать произвольных таксономических выводов.
Измерение времени также сопряжено с трудностями. Хотя
существуют современные методы датирования, дающие, как по-
лагают, надежные результаты, нет общего согласия относитель-
но начала и длительности основных геологических периодов.
Определение геологического возраста слоев связано с теми же
проблемами взятия выборок, что и исследование окаменелостей,
Высшие уровни изменчивости | 279
и результаты должны быть соотнесены с наличием известных
ископаемых форм. Ошибка может быть относительно большой.
Вообще установление абсолютной шкалы времени наиболее
эффективно в отношении более или менее длительных периодов.
Для более коротких отрезков геологического времени часто при-
ходится пользоваться относительной шкалой, располагая орга-
низмы во времени соответственно их относительному положе-
нию в осадочных слоях, уровню прослоек их пыльцы в ленточ-
ных глинах (varves) и т. п. В большинстве случаев при
исследовании эволюции приходится довольствоваться приблизи-
тельными данными о возрасте ископаемых остатков, хотя в том,
что касается последовательности появления форм данной серии,
можно быть почти уверенным.
ТЕМПЫ эволюции
Скорость филетической эволюции и расщепления линий очень
сильно варьирует. Темпы эволюции являются сложными функ-
циями многих факторов, находящихся в среде и в организме, и
взаимодействий между ними. Изменения в генетических систе-
мах, появление и исчезновение хищников или паразитов, изме-
нения температуры, повышение солености морской воды, подъем
уровня моря, горообразование и изменения интенсивности сол-
нечной радиации — вот несколько выбранных наудачу факторов,
которые по отдельности или в разных сочетаниях могут влиять
на скорость эволюции.
Если скорости эволюции для различных представителей ка-
кой-нибудь крупной таксономической группы отобразить в виде
диаграмм частот, то получается картина асимметричного рас-
пределения, в котором модальная скорость будет ближе к мак-
симальной. Симпсон назвал такое распределение горотеличс-
ским, а образующие его скорости можно назвать горотелически-
ми скоростями. Для большинства групп окажется, что некоторые
организмы, по-видимому, эволюционировали со скоростями не-
сколько меньшими или большими, чем те, которые характерны
для горотелической эволюции. Эти скорости были названы соот-
ветственно брадителическими и тахителическшш.
Для иллюстрации горотелических скоростей можно сравнить
наземных хищников (Carnivora) и двустворчатых моллюсков
(Pelecypoda). Симпсон построил кривые выживания для этих
групп организмов (фиг. 62). Тот факт, что средняя длитель-
ность существования родов моллюсков составляет 78 млн. лет,
а одного рода млекопитающих — 8 млн. лет, дает основание
предполагать, что хищники эволюционировали, возможно, в
10 раз быстрее двустворчатых моллюсков (исходя из продол-
время,млн лет
Фиг. 62. Кривые выживания для родов двустворчатых моллюсков Pele-
cypoda (Л) и наземных хищников Carnivora (Б).
Кривая / —длительность существования вымерших родов; кривая // — длительность суще-
ствования ныне живущих родов, известных и в ископаемом состоянии. На оси ординат —
процент выживающих родов; на оси абсцисс —длительность существования вымерших родов
по данным палеонтологии (для кривой /) или время, истекшее с момента появления ископа-
емых форм ныне живущего рода (для кривой //) [2].
Высшие уровни изменчивости | 281
Фиг. 63. Частота распределения родов двустворчатых моллюсков Pelecy-
poda (Л) и наземных хищников Carnivora (Б) по скоростям эволюции.
Гистограммы построены на основе кривых выживания вымерших родов (см. фиг. 62) и на
предположении, что существует полная отрицательная корреляция между длительностью вы-
живания и скоростью эволюции. На оси ординат —процент родов каждого класса; на оси
абсцисс —скорость эволюции, выраженная в условных единицах (децилях), что позволяет срав-
нивать гистограммы, несмотря на различие абсолютных скоростей [2].
Для сравнения на каждой гистограмме приведена кривая нормального распределения, пло-
щадь под которой равна площади этой гистограммы.
жительности существования произвольно выделенных родов).
Если допустить, что корреляция между временем существования
рода и скоростью эволюции является полной и положительной,
то можно построить график распределения частот скоростей
(фиг. 63). Такие графики для обеих групп совершенно одина-
ковы (хотя абсолютные скорости весьма различны) и соответ-
ствуют горотелическому распределению как для Pelecypoda, так
и для Carnivora.
Если исследовать Pelecypoda более детально, сравнив время
существования ископаемых родов и родов, существующих по-
ныне, то обнаружится явление брадителии. Исходя из вычисле-
ний Симпсона и из графика, приведенного на фиг. 62, можно
видеть, что действительное время выживания родов больше ожи-
даемого. Некоторые из ныне живущих родов существовали зна-
чительно дольше, чем вымершие роды. Самые долговечные из
вымерших родов существовали 275 млн. лет, но среди ныне жи-
вущих родов известны роды, существующие 400 млн. лет.
20 Зак. 152
282 | Глава 11
Таким образом, согласно Симпсону, все ныне живущие роды,
возраст которых превышает примерно 250 млн. лет, являются
брадителическими. Скорость их эволюции, измеренная этим по-
казателем, лежит вне горотелического распределения, ближе к
«медленному» концу шкалы.
Нетрудно назвать много групп растений и животных, в ко-
торых темп эволюции кажется очень медленным. Не следует
забывать, однако, что брадителия — статистический эффект,
выявляемый специальными средствами анализа. Поэтому не сов-
сем правильно было бы называть брадителическими современ-
ные организмы, мало изменявшиеся в течение длительных пе-
риодов. Представители многих групп организмов, например
опоссумы, гигантские секвойи, крокодилы и плауны, эволюцио-
нировали, по-видимому, очень медленно. Но можно ли отнести
их к брадителическим формам — это другой вопрос. При совре-
менном состоянии наших знаний об этих группах или о таких
организмах, как двустворчатые моллюски, невозможно также
установить, почему эволюция здесь как будто бы приостанови-
лась (по крайней мере, частично). Нет оснований полагать, что
такие формы более «примитивны», отличаются низкой частотой
мутаций или обедненностью генофонда и меньшей изменчи-
востью.
Метод исследования с помощью кривых выживания, выяв-
ляющий брадителию, не позволяет обнаружить наличие скоро-
Время
Фиг. 64. Схема, иллюстрирующая горотелию, брадителию и тахителию.
Объяснение в тексте.
Высшие уровни изменчивости | 283
стей, больших, чем те, которые укладываются в горотелическое
распределение. У организмов с очень быстрой эволюцией, полу-
чившей название тахителии, эта особенность проявляется, ви-
димо, в течение коротких периодов, а затем скорость их эволю-
ции снижается до горотелической или брадителической, если
только они не вымирают. Большая скорость эволюции, или та-
хителия, часто бывает связана с новым образом жизни — с тем,
что Симпсон называет новой адаптивной зоной, — и характерна
для квантовой эволюции. Под адаптивной зоной здесь имеется
в виду не просто место, где живет организм, а также и его образ
жизни в этом месте. Было высказано предположение, что тахи-
телия является одной из разнообразных возможных причин воз-
никновения пробелов в палеонтологической летописи, которые
часто встречаются там, где, по-видимому, начинается новая
филетическая линия. Если эволюция протекала в необычно
быстром темпе и особенно если предки новой линии были
рассредоточены в частично изолированных небольших популя-
циях, шансы на сохранение их в виде окаменелостей должны
быть очень невелики. Представления Симпсона о возможных
отношениях между горотелией, брадителией и тахителией по-
казаны на фиг. 64. Здесь от брадителической линии отходит
тахителическая ветвь, которая вскоре становится горотелической.
ОСНОВНЫЕ пути эволюции
АДАПТИВНАЯ РАДИАЦИЯ
Птицы, вероятно, представляют собой группу, испытывав-
шую вначале период ускоренной эволюции. Они быстро отде-
лились в мезозое от рептилий-архозавров — по-видимому, в
результате совершенствования полета в процессе отбора на даль-
ность и быстроту перелетов, который происходил в группе реп-
тилий, взбиравшихся на деревья и планировавших оттуда на
землю. Как только выработался настоящий полет, его преиму-
щества (при спасении от врагов, расселении, поисках пищи и
т. п.) привели к быстрой пролиферации группы. Такую пролифе-
рацию иногда называют адаптивной радиацией. Очевидно, адап-
тивные зоны летающих и планирующих форм в достаточной
степени перекрывали друг друга, так что оказавшиеся более
приспособленными птицы вытеснили группу, от которой они
произошли. Интересно отметить, что «зона планирующих форм»
До сих пор в значительной части остается незанятой (если не
считать нескольких рыб, ящериц, змей и млекопитающих), хотя
; Млекопитающие успешно проникли в воздушную зону (илиееспе-
^Циализированную подобласть), что было достигнуто, по крайней
Опышю мшми
(РЫш&тиш шги1ет)
Фиг. 65. Схема, иллюстрирующая разнообразие способов
опыления у представителей семейства Polemoniaceae.
Отдельными цветками представлены структурные типы, связанные
с различными механизмами опыления; стрелками показаны возможные
направления их эволюции.
Клеистогамные цватки
(Pvtemmtsfri mcranthuns)
286 | Глава 11
мере отчасти, путем приспособления к ночному полету и ловли
добычи с помощью звукового локатора.
Особенно ярким примером адаптивной радиации у млеко-
питающих может служить необычайное разнообразие сумчатых
в Австралии, где не было плацентарных млекопитающих. Здесь
среди сумчатых появились травоядные формы (некоторые кен-
гуру), роющие формы (сумчатые кроты), формы, напоминающие
белок и летяг (кускусы, сумчатые летяги) и кроликов (некото-
рые валлаби), хищники, напоминающие волков (сумчатый волк)
и барсуков (сумчатый дьявол), насекомоядные (сумчатый му-
равьед), а также древесные формы, не имеющие очевидных ана-
логов среди плацентарных млекопитающих (коала, древесные
кенгуру). Специалисты по сравнительной психологии, лингвисты
и социологи могли бы пожалеть о том, что в группе сумчатых
не возникло форм, аналогичных приматам. В Австралии адап-
тивная радиация наблюдается также у ядовитых змей сем.
Elapidae (кобры и родственные формы). Среди них появились
наряду с другими формы, внешне сходные с гадюками или с
ямкоголовыми змеями (сем. Viperidae и Crotalidae). Существует,
конечно, много других случаев адаптивной радиации. Хорошим
примером может служить эволюция вьюрков на островах Гала-
пагос и рыб сем. Cichlidae в озерах Африки, хотя масштабы
явления здесь не столь внушительны, как в случае автралий-
ских млекопитающих и рептилий.
Примеры адаптивной радиации встречаются и у растений.
В большинстве крупных семейств имела место дифференциация
на древесные формы, кустарники, лианы и различные типы тра-
вянистых форм, в том числе водные растения. Кроме того, в
ряде семейств независимо возникли сапрофитные и насекомояд-
ные формы. В эволюции отдельных биологических механизмов
у растений наблюдаются специализированные типы адаптивной
радиации. Как показал Грант, в сем. Polemoniaceae (куда отно-
сятся флоксы) существует замечательное разнообразие систем
опыления; они схематически представлены на фиг. 65. Основной
тип цветка с 5 лепестками, 5 тычинками и верхней завязью
(обычно с трехлопастным рыльцем) подвергся таким видоизме-
нениям, что получились формы, приспособленные к опылению
пчелами, колибри, дневными бабочками, бражниками, мухами-
жужжалами, жуками и даже летучими мышами. Существуют
также цветки, которые регулярно самоопыляются (автогамные),
в том числе цветки, самоопыляющиеся в закрытом бутоне
(клейстогамные). Кажется вероятным, что это разнообразие,
благодаря которому используются чуть ли не все типы возмож-
ных опылителей, явилось результатом видоизменения процессов
развития вследствие отбора. Специализация у Polemoniaceae
Высшие уровни изменчивости | 287
произошла на уровне родов и внутриродовых подразделений.
У других покрытосеменных растений иногда почти все семейство
может быть приспособлено к опылению одним агентом, обычно
ветром, как, например, у злаков (сем. Gramineae)- В почти
исключительно ветроопыляемом семействе осоковых (сем. Суре-
гасеае) существует одна или несколько насекомоопыляемых
форм (например, Dichromena), у которых группы редуцирован-
ных цветков с примыкающими к ним листьями похожи на цветки.
Эти примеры показывают, что понятие адаптивной радиации
применялось в отношении изменений, охватывавших как сравни-
тельно длительные, так и сравнительно короткие периоды вре-
мени. Оно может относиться и к уровню, который систематики
назвали бы видовым, и к разветвлению крупных стволов фило-
генетического древа в очень широких адаптивных зонах. Его
можно даже применять для описания эволюции специальных
взаимоотношений, например между цветковыми растениями и
их опылителями. Ясно, что значение термина «адаптивная ра-
диация» должно в существенной степени определяться контек-
стом, в котором его применяют.
РАЗЛИЧИЯ В ТЕМПАХ ЭВОЛЮЦИИ И АДАПТИВНЫЕ ЗОНЫ
Было бы неразумно выдвигать слишком конкретные гипотезы
о причинах различий в темпах эволюции. Возможно, что бради-
телия свойственна организмам в таких экологических ситуациях,
которые лишь медленно изменяются на протяжении геологиче-
ского времени. Было высказано предположение, что именно так
обстоит дело в океанах, но по существу о морских местообита-
ниях мы знаем гораздо меньше, чем о наземных. В одном и
том же местообитании одни организмы могут эволюционировать
медленно, а другие — быстро. Обычно так же невозможно уста-
новить «причину» того или иного случая тахителии. Очевидно,
она может быть связана с проникновением в новую адаптивную
зону. Почти невозможно избежать упрощенного представления
об адаптивных зонах и разрывах между ними (неадаптивпых
зонах), хотя они образуют некое абстрактное многомерное про-
странство, измерениями которого служат факторы среды, в том
числе и другие организмы. В прошлом, как и сейчас, ситуация
должна была быть чрезвычайно запутанной; вероятно, происхо-
дили сложные (нередко циклические) перемещения градиентов.
По-видимому, некоторые филогенетические ответвления ка-
ким-то образом получают «доступ» в новую адаптивную зону,
которая непрерывно изменяется. Эта новая зона должна быть
«смежной» с предыдущей зоной во всех многочисленных изме-
рениях. Организмы должны находиться физически поблизости
288 | Глава 11
от нее, у них должны оказаться (в результате действия генети-
ческих механизмов на уровне популяций) генотипы, способные
сохраняться и функционировать в ней, и, наконец, они должны
быть способны сосуществовать с организмами, уже занимаю-
щими эту'зону, или вытеснить их. Успешно «проникнув» в адап-
тивную зону, группа может затем дифференцироваться, с тем
чтобы занять различные участки этой зоны и распространяться
дальше в другие, смежные зоны. Количество новых адаптивных
подтипов, по-видимому, зависит главным образом от обширности
(внутреннего разнообразия) адаптивной зоны. Веерокрылые
(Strepsiptera) — интересные, сильно видоизмененные насекомые,
эндопаразиты пчел, ос и других насекомых — как будто бы во-
шли в довольно узкую адаптивную зону. Птицы, очевидно, на-
шли себе относительно широкую зону. Важно также помнить о
том, что область, которая кажется сейчас сравнительно неустой-
чивой (и сравнительно мало занятой) промежуточной зоной,
могла в свое время быть вполне стабильной. Зона планирующего
полета, вполне возможно, «дестабилизировалась» в результате
успешного проникновения потомков рептилий в истинно воздуш-
ную зону.
Как мы уже подчеркивали в гл. 5, биологи очень склонны
делить внешнюю среду на части в соответствии со своими ана-
литическими целями. Хотя понятие адаптивной зоны относится
к образу жизни, а не к участку физического пространства, оно
тем не менее искусственно вычленяет некоторую часть простран-
ственно-временного континуума, в котором находятся организ-
мы. В конечном счете эволюционирует вся ситуация в целом,
хотя нам, может быть, удобнее отделять органическую эволюцию
от изменений окружающей среды. Результат нашего исследова-
ния в очень большой степени зависит от выбора искусственных
разграничений. Наука продвигается вперед путем сравнения
результатов изучения сходных ситуаций с разных точек зрения,
пока не будет получен относительно однозначный ответ. Кажется
очевидным, что изучение эволюции организмов вместе с измене-
ниями их «местообитания» еще не достигло того этапа, на кото-
ром ответы получаются однозначными.
КОНКУРЕНЦИЯ
Те же проблемы возникают в отношении последствий «столк-
новения» двух организмов водной адаптивной зоне. Обыкновенно
полагают, или по крайней мере утверждают, что неизбежным
результатом должна быть какого-то рода «конкуренция».
В понятие конкуренции вкладывалось так много различных
значений, что его использование без каких-либо уточнений
Высшие уровни изменчивости | 289
смысла скорее затемняет, чем разъясняет дело. Кроме того, в
применении к растениям этот термин должен иметь значение,
несколько отличное от его возможного значения в применении
к животным. В расширенном смысле слово «конкуренция» при-
менялось и к таким случаям, когда одна крупная группировка
организмов вытесняла другие в чрезвычайно обширных адаптив-
ных зонах. Например, часто говорят, что плацентарные млеко-
питающие в результате конкуренции вытеснили сумчатых и одно-
проходных на большей части землой поверхности или что млеко-
питающие как группа полностью «уничтожили» группу сходных
с ними рептилий. Покрытосеменные растения, по-видимому,
также вытеснили голосеменные в большинстве районов земного
шара; это тоже рассматривают как результат конкуренции, ка-
ким-то образом распространившей свое действие до уровня
высших таксономических категорий. Ретроспективно мы можем
взглянуть на любую из этих ситуаций и сказать, что по самому
определению выжившие организмы были «более приспособлен-
ными», чем те, которые вымерли. Однако вымирание одной груп-
пы — не единственный возможный исход «конкурентной оккупа-
ции» одной и той же адаптивной зоны. У одного или двух
«оккупантов» может выработаться большая специализация, что
ограничит или устранит взаимодействие между двумя типами.
Тараканы, некогда доминирующая группа насекомых, занимав-
шая, вероятно, зону неспециализированной травоядности или
всеядности, в настоящее время являются в основном некрофа-
гами. По-видимому, их прежняя зона оказалась разделенной
между множеством специализированных групп насекомых, осо-
бенно из отрядов Hemiptera, Homoptera, Orthoptera, Coleoptera,
Lepidoptera, Diptera и Hymenoptera, в большинстве своем отно-
сительно поздно появившихся на эволюционной сцене.
КОНВЕРГЕНЦИЯ
Иногда организмы, не находящиеся в близком родстве, по-
падают в сходные адаптивные зоны и в результате отбора при-
обретают внешнее сходство друг с другом. Классическими
примерами служат киты и рыбы, а также суккулентные расте-
ния пустынь, принадлежащие к различным семействам. В вод-
ной среде очевидно селективное преимущество генотипов,
обусловливающих тот или иной тип обтекаемости формы тела.
Многие растения пустынь (фиг. 66) отличаются толстой кутику-
лой и низким отношением поверхности к объему (то и другое
уменьшает потерю воды) и часто бывают снабжены колючками,
что в известной мере предохраняет их от поедания обитающими
в пустыне животными. Растения, изображенные на фиг. 66,
Eunhorbtaceae
(Африка)
Asclepiadaceae
(Африка)
Cactaceae
(Новый свет)
Фиг. 66. Конвергенция у растений пустынь.
Слева — Euphorbia (Euphorbiaceae, Африка), в центре — Huernla (Asclepladaceae, Африка),
справа — Cereus (Cactaceae, Латинская Америка).
Фиг. 67. Конвергенция, по-видимому, обусловленная мимикрией.
Вверху: слева Alcidis agathyrsus, справа Cyclosia hestinioides\ внизу: слева парусник
Papilio laglalzel, справа данаида Ideopsis daos. Все формы из Новой Гвинеи [6].
Высшие уровни изменчивости | 291
относятся к трем разным семействам — Euphorbiaceae, Asclepia-
daceae (Старый свет) и Cactaceae (Новый свет). Шипы в этих
трех случаях являются производными различных анатомических
структур.
Более сложны разнообразные формы мимикрии, при которых
отбор благоприятствует генотипам, имеющим наименьшие шан-
сы на уничтожение хищниками. Один пример миметической кон-
вергенции показан на фиг. 67. Полагают, что в этом случае ба-
бочки Alcidis agathyrsus и Ideopsis daos неприятны на вкус для
своих врагов, a Papilio laglaizei и Cyclosia hestinioides охотно
поедаются врагами. Отбор в популяциях двух последних форм,
по-видимому, благоприятствовал генотипам, обусловливавшим
ту или иную степень фенотипического сходства с защищенными
формами.
Конвергенцию среди форм, находящихся в относительно
близком родстве между собой, называют параллелизмом. На-
пример, в нескольких линиях рептилий, сходных с млекопитаю-
щими, независимо возникли признаки, по которым мы отличаем
млекопитающих. Между конвергенцией в строгом смысле слова
и параллелизмом существует непрерывный переход, и различие
между этими двумя явлениями несущественно.
ВЫСШИЕ КАТЕГОРИИ
В литературе много внимания уделяется вопросу, довольно
нечетко определяемому как «проблема происхождения высших
таксономических категорий». Строго говоря, это проблема для
психологов, ибо высшие категории (Reptilia, Lepidoptera, Rosa-
ceae, Basidiomycetes и т. п.), так же как и виды, представляют
собой понятия, а не объекты. К формированию этих понятий
причастны многие факторы, которые нередко могут показаться
неожиданными: ранее сложившиеся подходы к анализу, струк-
тура языка, особенности словоупотребления и т. д. Тем не ме-
нее эти понятия соответствуют чему-то реальному, и уместно бу-
дет поставить вопрос о причинах возникновения «естественных»
группировок, обычно наблюдаемых в природе.
Кажется вероятным, что крупные разрывы в палеонтологи-
ческой летописи обусловлены случайными факторами. Если
какая-либо группа вступает в новую адаптивную зону в период
тахителии, то шансы на образование окаменелостей в этот корот-
кий переходный период снижены просто по причине ее кратко-
сти. В массе листвы дерева могут быть просветы, но даже
филогенетическое древо имеет непрерывные ветви. Система
категорий линнеевской таксономии несколько увеличила раз-
меры кажущихся разрывов.. Редкие организмы, занимающие по
292 | Глава 11
своим признакам более или менее промежуточное положение
между основными группами, обычно ради удобства относили
к той или иной крупной группировке. Так, однопроходных объ-
единили с млекопитающими, Micropterygidae — с бабочками,
грибы — с растениями. На относительно низких таксономиче-
ских уровнях возникали сложности в связи с сетевидной эволю-
цией. Если новый вид растений — амфидиплоидный гибрид,
произошел от двух других видов, относящихся к разным под-
родам, то его часто включают в один из существующих под-
родов, а не описывают как единственного представителя третьего
подрода.
Как мы уже отмечали, высшие таксономические категории
отчасти представляют собой результат вымирания, приводящего
к появлению морфологических хиатусов. Это проблема, к реше-
нию которой не так легко подойти, и биологи уделяли ей лишь
поверхностное внимание. Она имеет отношение к рассмотренно-
му выше вопросу о конкуренции. Специализированные группы,
взаимодействуя с примитивными группами, часто оказывались
более приспособленными, так что примитивные типы в конце
концов исчезали. Сравнение примитивных и специализированных
форм не связывается здесь с каким-либо представлением о «ка-
честве»; две группы сравниваются в отношении того, насколько
они предположительно удалились от исходного пункта — от сво-
его наиболее позднего общего предка. Например, крокодилы в
этом смысле более примитивны, чем птицы. Известны и такие
случаи, когда примитивные типы сохранились, а специализиро-
ванные исчезли. То, что эти случаи можно «объяснить» как вы-
живание менее специализированных групп в условиях конкурен-
ции с чрезмерно специализированными типами, служит лишь
иллюстрацией трудностей, с которыми связано понимание та-
кого рода экологических ситуаций.
Многие из наблюдаемых нами сейчас разрывов, по-видимому,
возникли еще до того, как начала создаваться удовлетворитель-
ная палеонтологическая летопись. Даже в тех случаях, когда
существует адекватный ископаемый материал по промежуточ-
ным типам, нельзя с уверенностью указать точные причины
наблюдаемых в настоящее время разрывов. Распространенная
гипотеза о том, что отсутствие организмов типа Archaeopteryx
обусловлено конкуренцией со стороны птиц, может быть совер-
шенно неверной. Возможно, что эти ранние типы специализиро-
вались на питании какими-то организмами, вымершими из-за
климатических перемен. Можно было бы, конечно, построить
множество гипотез, большинство которых имели бы примерно
равные шансы оказаться ошибочными.
Высшие уровни изменчивости | 293
ЭВОЛЮЦИОННЫЕ ТЕНДЕНЦИИ
Ретроспективно мы можем наблюдать эволюционные тенден-
ции фактически любого масштаба — от повышения частоты ме-
ланизма у одного вида британской бабочки до увеличения слож-
ности во всей биосфере. К сожалению, такие тенденции, наблю-
даемые a posteriori, слишком часто истолковывались так, как
если бы эволюционный процесс был средством для достижения
заранее поставленной цели. Тех, кто интересуется такого рода
тенденциями, прослеженными по ископаемым находкам, мы от-
сылаем к работам Ренша [1] и Симпсона [2]. Этим авторам
удается весьма успешно интерпретировать такие тенденции, ис-
ходя из представлений об основных механизмах мутирования,
рекомбинации, отбора и дрейфа генов.
УВЕЛИЧЕНИЕ РАЗМЕРОВ ТЕЛА
В этом и следующем разделах будут кратко рассмотрены две
очень общие тенденции. Во многих филогенетических линиях
животных наблюдается тенденция к увеличению размеров тела
(получившая название правила Копа). Правомерность этого об-
общения в его самом широком плане может оспаривать только
тот, кто склонен предполагать, что величина первичных живых
организмов лежала выше или около середины современного диа-
пазона различий по этому признаку. Из обсуждения вопроса о
происхождении жизни (гл. 1) легко видеть, что это маловероят-
но. Большие размеры связаны с потенциальным селективным
преимуществом в отношении чуть ли не любой физиологической
функции: так, увеличение размеров делает возможным макси-
мальное использование листьями света для фотосинтеза (у де-
ревьев) и максимальное сохранение тепла (у китов), создает
достаточное вместилище для сложного мозга (у высших живот-
ных), определяет устойчивость к обезвоживанию, обусловлен-
ную низким отношением поверхности к объему (у некоторых
кактусов).
Ископаемые находки свидетельствуют о многочисленных эво-
люционных линиях, характеризующихся увеличением размеров
тела. Хорошо документированным примером служит филогения
лошадей: от Eohippus (эоцен) величиной с собаку произошли,
помимо прочих форм, современные рабочие лошади. В палеон-
тологии млекопитающих подобных историй множество, хотя
известны и исключения (например, современные сумчатые и ле-
нивцы мельче соответствующих плейстоценовых форм). Необхо-
димо, хотя подчас весьма трудно, различать два процесса — об-
ратный переход к уменьшению размеров тела и вымирание
294 | Глава 11
более крупных организмов, ведущее к тому, что группа оказывает-
ся представленной лишь более мелкими родственными формами.
У некоторых животных отбор на большую величину тела, по-
видимому, был связан с тем, что при этом вследствие алломе-
трической корреляции происходил отбор по каким-либо другим
признакам, дававшим явное преимущество. Аллометрией назы-
вают дифференциальный рост частей тела. Рогам жвачных,
например, свойственна сильно выраженная положительная алло-
метрия. С увеличением размеров тела они становятся значитель-
но крупнее и по относительной величине. (У человека в постна-
тальный период голова растет относительно медленнее, чем
остальные части тела в целом; ей свойственна отрицательная
аллометрия.) Рога быков и оленей важны как при защите от
врагов, так и в битвах между самцами из-за самок. Поэтому не-
безосновательным кажется предположение, что крупные разме-
ры тела у многих жвачных объясняются отбором на большую
мощность вооружения. Это один из случаев, когда отбор на
большую относительную величину придатков тела, возможно,
привел благодаря положительной аллометрии к одновремен-
ному увеличению размеров тела в целом.
Возможно также, что в некоторых случаях отбор на большую
величину тела — например, в связи с требованиями лучшего раз-
мещения внутренних органов — мог привести к чрезмерному с
точки зрения предполагаемой функции росту придатков. Быть
может, так обстояло дело с эволюцией Megaceros — гигантского
ирландского оленя (плейстоцен), обладавшего самыми крупными
среди всех известных оленей рогами. Такое направление эволю-
ции продолжалось бы до тех пор, пока отрицательные стороны
гигантских рогов не перевесили преимущество, связанное с боль-
шими размерами тела. Наши рассуждения, быть может, слиш-
ком умозрительны, но они показывают, что нет никаких основа-
ний рассматривать большую величину этих рогов (или гигант-
ских мандибул жуков-оленей, а также многие другие случаи
подобной гипертрофии) как результат своего рода «эффекта мо-
мента», доводящего некогда адаптивную тенденцию до неадап-
тивной крайности. Как должно быть очевидно из рассмотрения
генетики популяций (гл. 6), эта аналогия лишена смысла. От-
бор, направленный против крайних форм, сразу же пресечет
дальнейшее изменение, как только его отрицательные послед-
ствия уравновесят преимущества. Как отметил Симпсон, орга-
низмы с какими-либо признаками, выраженными в крайней сте-
пени, могут казаться нам очень странными, но это отнюдь не
означает, что эти признаки неадаптивны. Действительно, процве-
тающие обладатели многих причудливых особенностей (павли-
ны, некоторые долгоносики, нарвалы, растения с цветками не-
Высшие уровни изменчивости | 295
обычной формы или такими сложными способами опыления,
как псевдокопуляция) вряд ли служат свидетельством в пользу
представления о невыгодности таких признаков.
УВЕЛИЧЕНИЕ СЛОЖНОСТИ
Пожалуй, наиболее общей и всеобъемлющей из постулиро-
ванных тенденций является тенденция к увеличению сложности.
Как ни трудно с точностью оценить уровень организации раз-
личных групп организмов, вряд ли кто-нибудь из биологов стал
бы отрицать, что трубкозуб имеет более высокую и сложную ор-
ганизацию, чем коацерватная капля. Бабочка-данаида — явно
более сложная система, чем амеба или даже папоротник. Хотя
в некоторых адаптивных зонах все еще существуют относительно
простые организмы (быть может, это более заметно в водных
местообитаниях), видимое преобладание более сложно органи-
зованных типов как будто бы свидетельствует об их превосход-
стве. Однако доминирование в той или иной экологической си-
туации чрезвычайно трудно поддается количественной оценке.
Можно ли сказать, что сосны в хвойном лесу «доминируют» над
грибом, который обитает в их корнях и без которого они не мог-
ли бы существовать? Доминируют ли млекопитающие над бак-
териями или другими представителями своей кишечной флоры,
вырабатывающими необходимые для их жизни витамины?
В действительности тщательное изучение любой экологической
ситуации вскрывает такие взаимодействия между организмами,
которые крайне затрудняют сравнение по признаку «превосход-
ства» или «доминирования».
Но тем не менее нельзя получить трубкозуба путем сложе-
ния множества коацерватных капель. Природе нужно было раз-
решить много сложных проблем структуры и функции, прежде
чем ДНК смогла использовать тело трубкозуба или человека
для воспроизведения себя самой. Эти проблемы, очевидно, ре-
шались медленно, по одной или по нескольку одновременно, на
протяжении длительных промежутков времени, причем их реше-
ние нередко обрекало на гибель менее эффективные типы ор-
ганизмов, так что перед нами предстает летопись жизни, свиде-
тельствующая о неуклонном повышении сложности или уровня
организации. Некоторые более простые организмы также сохра-
нились и, по-видимому, объединились с другими высокооргани-
зованными формами в сложные экологические системы. Основ-
ной вопрос остается без ответа: почему в ходе эволюции ДНК
создала для своего собственного воспроизведения трубкозубов
и людей, тогда как бактерии и другие простые организмы, ка-
залось бы, могут не хуже служить для этой цели? Вероятно, все
296 | Глава 11
дело в сложности экосистем, кибернетические механизмы кото-
рых обусловливают их высокую стабильность. Но эту проблему
трудно изучать на палеонтологическом материале. Одна из глав-
ных эволюционных тенденций, очень тесно связанная с увели-
чением сложности, состоит в повышении степени гомеостаза у
особи, в популяции и в экосистеме.
ВЫВОДЫ
Нет никаких данных, оправдывающих представление о нали-
чии существенной разницы между теми механизмами, которые
вызывают изменения частот генов в популяциях, и теми, которые
создали различия между человеком и микробами. Ввиду дли-
тельности времени, на протяжении которого шел эволюцион-
ный процесс, разнообразие форм жизни вполне можно объяс-
нить мутированием, рекомбинацией, отбором и дрейфом генов.
Разрывы непрерывности в картине изменчивости, видимо, об-
условлены главным образом вымиранием. Разрывы в таксоно-
мическом описании форм изменчивости отражают как вымира-
ние, так и многочисленные ошибки выборки. В результате иссле-
дования палеонтологического материала были выделены основ-
ные типы филогенетических изменений — расщепление линий,
филетическая эволюция и квантовая эволюция. Темпы эволю-
ции, по-видимому, сильно варьировали по времени. В каждой
данной группе наряду с типичными для нее скоростями часто
имели место необычно малые и необычно высокие скорости. Так
называемую проблему происхождения высших таксономических
категорий мы рассматриваем как комплекс лингвистических,
методологических и эволюционных проблем.
ЛИТЕРАТУРА
1. Re ns ch В., Evolution Above the Species Level, Columbia Univ. Press, New
York, 1960.
2. Simpson G. G., The Major Features of Evolution, Columbia Univ. Press,
New York, 1953.
3. Stebbins G. L., Variation and Evolution in Plants, Columbia Univ. Press,
New York, 1950.
4. Mourant, The Distribution of Human Blood Groups, Blackwell Scientific
Publications, Oxford, 1954.
5. Andrews, Studies in Paleobotany, Comstock Publishing Company, 1947.
6. P u n n e 11, Mimicry in Butterflies, Cambridge University Press, 1915.
СЛОВАРЬ ТЕРМИНОВ
Абиогенез — возникновение жизни из неживых систем.
Автополиплоид — полиплоидный организм, образовавшийся путем увели-
чения числа идентичных геномов у одного организма или в результате слия-
ния диплоидных или полиплоидных гамет от организмов с практически
идентичными геномами.
Автотетраплоид — автополиплоид с 4 одинаковыми геномами.
Автотриплоид — автополиплоид с 3 одинаковыми геномами.
Автокатализ — ускорение реакции под влиянием ее собственных конеч-
ных продуктов.
Агамные комплексы — комплексы организмов, в которых все особи раз-
множаются бесполым путем.
Агамоспермия — образование семян без оплодотворения; если мужские га-
меты и участвуют в этом процессе, то только в качестве стимуляторов
деления зиготы.
Адаптивная зона — «образ жизни» какой-либо таксономической группы
организмов в широком смысле; может подразделяться на адаптивные под-
зоны.
Адаптивная радиация — эволюционное образование в какой-либо группе
организмов разнообразных форм, часто за сравнительно короткий проме-
жуток времени, — как полагают, вследствие проникновения этой группы в
новую адаптивную зону; говорят также об адаптивной радиации в эво-
люции отдельных органов.
Адаптивная ценность — относительная способность данного генотипа к
выживанию и воспроизведению по сравнению с другими генотипами в по-
пуляции.
Адвентивная эмбриония — образование зародыша путем митотических
делений из тканей спорофита, таким образом, здесь отсутствует чередова-
ние поколений.
Аденин — одно из двух пуриновых оснований (6-аминопурин), входящих в
состав ДНК и РНК.
Аденозиндифосфат (АДФ) — соединение, состоящее из аденина, рибозы
(сахар с 5 атомами углерода) и двух фосфатных групп; участвует в мо-
билизации энергии для клеточного метаболизма.
Аденозинтрифосфат (АТФ) — АДФ с добавочной, третьей фосфатной
группой, присоединенной с помощью макроэргической связи; при расще-
плении АТФ до АДФ освобождается энергия, используемая для других
реакций.
298 | Словарь терминов
Акроцентрическая хромосома — хромосома с почти терминальной цен-
тромерой.
Аллель — одно из нескольких альтернативных состояний гена как функцио-
нальной единицы.
Аллометрия — разная скорость роста различных частей организма.
Аллопатрические формы — организмы, ареалы которых не перекры-
ваются.
Аллопатрическое видообразование — дифференциация популяций в ус-
ловиях географической изоляции, приводящая к образованию форм, кото-
рые систематики признают отдельными видами.
Аллополиплоид — полиплоидный организм, в клетках которого объеди-
нены геномы двух (или большего числа) различных типов.
Аллотетраплоид — полиплоид, образующийся в результате удвоения
числа хромосом у диплоидного гибрида, полученного скрещиванием двух
форм с различными геномами, или в результате слияния диплоидных га-
мет таких форм.
Аминокислоты — структурные компоненты белков; при гидролизе белки
распадаются на аминокислоты.
Амфидиплоид — тетраплоидный организм, который является диплоидом
по двум геномам, обычно происходящим от разных видов.
Анеуплоидия — такое изменение числа хромосом, при котором добавлен-
ные или утраченные хромосомы не составляют целых геномов.
Апогамия — род агамоспермии, при которой зародыш образуется не из
яйцеклетки, а из каких-либо иных клеток гаметофита.
Апомиксис— все формы размножения, при которых мейоз и сингамия ча-
стично или полностью исключены.
Апоспория— род агамоспермии, при котором диплоидный гаметофит об-
разуется путем митотических делений из соматических клеток споро-
фита.
Арренотокия — партеногенез, приводящий к образованию гаплоидного
мужского потомства, тогда как самки являются диплоидными.
Ауксоцит — клетка, ядро которой делится путем мейоза.
Аутосома — хромосома (обычно эухроматиновая), не являющаяся половой
хромосомой.
Ацентрический фрагмент — фрагмент хромосомы, не содержащий цен-
тромеры.
Базальные зерна (гранулы) — цитоплазматические органеллы, имеющие
характерную тонкую структуру; к этим органеллам прикрепляются рес-
нички и жгутики.
Бактериофаг— вирус, поражающий бактериальные клетки и часто вызы-
вающий их лизис.
Белки — класс высокомолекулярных веществ, образующихся в результате
соединения большого числа аминокислот; в состав молекулы белка могут
входить и другие компоненты.
Словарь терманов | 299
Бесполое размножение — то же, что апомиксис (см.). Часто имеется
в виду вегетативное размножение.
Бивалент — пара хромосом, конъюгирующих друг с другом в профазе
мейоза.
Бимодальная популяция — популяция, в которой величины, характери-
зующие данный признак, распределяются бимодально (т. е. распределение
имеет два различных максимума частот).
Бластопор — отверстие, образующееся в результате впячивания поверхно-
сти эмбриона на стадии гаструлы.
Бластула — ранняя стадия эмбрионального развития, на которой клетки
в типичном случае образуют полый шар.
Брадителическая эволюция — эволюция, протекающая значительно мед-
леннее, чем гороте личе ская эволюция (см.).
Буферность — способность системы противостоять действию внешних сил,
стремящихся изменить ее.
Веретено — структура, состоящая из белковых волокон, видимая во время
деления клетки; полагают, что эти волокна играют роль в передвижении
хромосом и делении цитоплазмы.
Взаимная транслокация — обмен участками между негомологичными
хромосомами.
Видообразование — процесс эволюционного расщепления форм, в резуль-
тате которого возникают организмы, признаваемые систематиками за раз-
личные виды (см.)'.
Возвратное скрещивание — скрещивание между гибридом и одной из
родительских форм или особью, относящейся к одному из родительских
типов.
3-Хромосомы — добавочные, гетерохроматиновые хромосомы, имеющиеся в
различном числе у некоторых растений (не обязательно во всех клет-
ках).
Выборка —некоторое подмножество, входящее в популяцию (см.); группа
отдельных объектов, которую извлекают каким-либо способом из популя-
ции и по которой исследователь рассчитывает узнать что-либо об этой по-
пуляции.
Высшие таксономические категории — подразделения выше уровня
рода в принятой таксономической иерархии.
Гаметофит — половое поколение у многих растений; на этой стадии жиз-
ненного цикла образуются гаметы, обычно путем митоза.
Гаметоцит — клетка, из продуктов деления которой образуются га-
меты.
Гаплодиплоидия — генетическая система некоторых животных, у которых
самцы развиваются из неоплодотворенных яиц и бывают гаплоидными,
тогда как самки являются диплоидными.
300 | Словарь терминов
Гаплоид — организм, соматические клетки которого содержат гаплоидное
число хромосом (соответственно говорят также: гаплоидные клетки, ткани
и т. п.).
Гаплофаза — часть жизненного цикла, в которой репродуктивные клетки
содержат гаметическое число хромосом.
Гаструла — стадия развития, следующая за бластулой; гаструла форми-
руется путем впячивания поверхности бластулы внутрь; при этом обра-
зуется полость первичной кишки, открывающаяся наружу отверстием —
так называемым бластопором (см.).
Ген — участок хромосомы между двумя ближайшими точками кроссинго-
вера; единица наследственности, которая может находиться в различных
состояниях, что ведет к различиям в фенотипе; участок генетического ма-
териала — носитель информации, определяющей структуру одного белка
или одной полипептидной цепи 1 (см. также Цистрон, рекон).
Ген-«переключатель» — см. Олигоген.
Генетическая ассимиляция — включение в генотип (в результате от-
бора) какого-либо признака, возникшего в онтогенезе под действием опре-
деленных условий среды.
Генетическая дисперсия — часть фенотипической дисперсии, обусловлен-
ная различиями генотипов особей в популяции.
Генетическая система — совокупность всех факторов, внутренних и внеш-
них, влияющих на рекомбинацию наследственных детерминантов у дан-
ного организма.
Генетический гомеостаз — свойственная популяциям тенденция регрес-
сировать под действием отбора к исходному среднему состоянию (в отно-
шении того или иного признака).
Генетический груз (genetic load) популяции — среднее число потенциаль-
ных смертей, обусловленных генетическими причинами (деталями, полуле-
талями и т. п.), в расчете на одну особь.
Генный поток (gene flow) — передвижение генетической информации вну-
три популяции и между популяциями.
Генбм — минимальный набор негомологичных хромосом, необходимый для
надлежащего функционирования клетки.
Генотип — совокупность генетического материала клетки (обычно имеется
в виду только ядерный материал); наследственность особи, взятая в це-
лом; генетическая структура особи в отношении отдельного локуса.
Генофонд^епе pool) — совокупность генетической информации данной по-
пуляции.
Гермафродит—особь, обладающая фенотипическими признаками обоих
полов, — у раздельнополых организмов.
1 Автор дает, в сущности, три разных определения гена, из которых мож-
но признать правильным только третье, говорящее о гене как о функциональ-
ной единице. Первое определение соответствует современному понятию рекон,
второе — понятию мутон или трансген X. С. Серебровского. — Прим. ред.
Словарь терминов | зо1
Гетерогаметный пол—пол, образующий гаметы, различающиеся в отно-
шении набора хромосом.
Гетерокариоз — наличие в одной клетке (или ценоците) ядер с различ-
ным генотипом.
Гетеротрофный организм — организм, для питания которого необхо-
димы органические вещества.
Гетерохроматиновая хромосома (или участок)—хромосома (или ее
часть), не претерпевающая типичных изменений при клеточном делении и,
по-видимому, не содержащая генов с заметным фенотипическим проявле-
нием.
Гибридизация — обмен генами между популяциями; термин применяют
обычно только в отношении популяций, относимых систематиками к от-
дельным видам.
Гидратированный белок — белковая молекула, удерживающая около
себя молекулы воды.
Гиногенез — род партеногенеза, при котором для развития яйцеклетки не-
обходима стимуляция ее сперматозоидом.
Голо метаболическое развитие — развитие, ясно расчлененное на ста-
дии яйца, личинки, куколки и взрослой формы (у насекомых).
Гольджи комплекс — клеточная органелла с неизвестной функцией, при-
сутствующая в большинстве растительных и животных клеток; нередко на-
ходится вблизи ядра; состоит из ряда концентрически расположенных ци-
стерн с гладкими контурами (без рибосом).
Гомолог — одна из двух или большего числа хромосом, идентичных по
содержащимся в них локусам.
Граны — содержащие пигмент структуры в пластиде; обычно состоят из
многочисленных сложенных в стопки дисков.
Гуанин —одно из двух пуриновых оснований (2-амино-6-оксипурин), вхо-
дящих в состав как ДНК, так и РНК.
Дезоксирибонуклеиновая кислота (ДНК)—высокомолекулярное соеди-
нение, гигантские молекулы которого у большинства организмов служат
носителями ядерной генетической информации, обладают способностью
к самовоспроизведению и распределяются поровну между дочерними яд-
рами (см. также нуклеиновые кислоты).
Деления — утрата участка хромосомы.
Денатурированный белок — структурно измененный белок, характери-
зующийся пониженной растворимостью и изменением биологической ак-
тивности.
Дикариоз — наличие двух ядер в каждой клетке.
Дикий тип — наиболее часто встречающийся фенотип, который чисто ус-
ловно принимают за «нормальный».
21 Зак. 152.
302 | Словарь терминов
Диплоид — организм, в соматических клетках которого содержится зиготи-
ческое (диплоидное) число хромосом, т. е. два генома. (Соответственно
говорят также; диплоидные клетки, ткани и т. п.)
Диплоспория — вид апомиксиса у растений, при котором диплоидный га-
метофит образуется путем митотических или неполных мейотическпх де-
лений спорообразующих клеток.
Диплофаза — часть жизненного цикла, в которой клетки содержат зиго-
тическое число хромосом.
Дисперсия —мера изменчивости, определяемая как сумма квадратов инди-
видуальных отклонений, деленная на число особей; квадрат стандартного
отклонения.
Дисруптивный отбор — отбор, благоприятствующий двум или несколь-
ким различным генотипам и элиминирующий промежуточные формы.
Дисульфидные связи — ковалентные связи между остатками цистеина
(S—S) в белковых молекулах или между ними; полагают, что эти связи
ответственны за образование веретена при делении клетки.
Дифференциальные сегменты — участки хромосом, не конъюгирующие
в мейозе.
Дифференцировка — происходящие в ходе эмбрионального развития из-
менения, в результате которых возникают различные типы клеток, тканей
и органов.
Дицентрический— имеющий две центромеры.
ДНК—см. Дезоксирибонуклеиновая кислота.
Добавочные хромосомы — хромосомы, наблюдающиеся (часто в непо-
стоянном числе) в дополнение к хромосомам типичного, относительно не-
изменного, набора.
Доминирование — явление, при котором гетерозигота по данному локусу
фенотипически более сходна с одной из гомозигот, чем с другой; аллель,
проявляющийся в большей степени, называют доминантным аллелем;
при полном доминировании гетерозигота не отличается по фенотипу от
доминантной гомозиготы.
Дрейф генов (genetic drift) — случайные колебания частот генов (обус-
ловленные обычно ошибкой выборки, неизбежной при функционировании
генетического механизма); имеет место в любой популяции, но послед-
ствия дрейфа наиболее заметны в очень малочисленных популяциях.
Дробление — деление оплодотворенной яйцеклетки, ведущее к образова-
нию эмбриона у животных.
Дупликация — наличие или возникновение в геноме повторений некоторого
участка хромосомы.
Естественный отбор — неслучайное (дифференциальное) воспроизведе-
ние генотипов без намеренного вмешательства человека.
Словарь терминов | 303
Живорождение. у растений — вегетативное размножение, при котором на
месте цветков в соцветиях образуются пропагулы; у животных — рожде-
ние живых детенышей (в отличие от откладки яиц).
Жизнеспособность (viability)—способность выживать или продолжать
развитие; этот термин часто неправильно употребляют как синоним При-
способленности (fitness) (см.).
Зигота — продукт соединения гамет (Сингамии, см.).
Импринтинг — возникновение устойчивой формы поведения в результате
воздействия специфического раздражителя на определенном этапе раз-
вития.
Инверсия — перестройка, в результате которой в хромосоме возникает «пе-
ревернутый» участок с обратной последовательностью локусов.
Индукция — взаимодействие эмбриональных тканей, при котором одна
масса клеток определяет направление развития другой.
Интерстициальный сегмент — участок хромосомы между транслоциро-
ванным сегментом и центромерой (см.).
Интрогрессия — включение генов одной популяции в генофонд (см.) дру-
гой популяции в результате возвратных скрещиваний гибридов; термин
применяется обычно только в отношении популяций, относимых система-
тиками к обособленным формам.
Искусственный отбор — любая процедура, с помощью которой человек
определяет выбор генотипов, включаемых в генофонд (см.) последующих
поколений.
Канализация развития — стабилизация путей развития, которую обычно
представляют себе как результат действия каких-то «буферных» сис-
тем.
Каппа-частицы — содержащие ДНК частицы в цитоплазме отдельных
особей некоторых видов парамеций; эти частицы обусловливают образо-
вание особого вещества, которое, выделяясь в окружающую среду, вызы-
вает гибель других парамеций с определенным генотипом.
Кариотип — характерный фенотип хромосом организма, который обычно
удается наблюдать в метафазе митоза.
Каротиноиды — жирорастворимые пигменты, цвет которых варьирует от
, желтого до красного; большинство каротиноидов представляют собой те-
тратерпены.
Квадратная кость — одна из челюстных костей многих позвоночных, из
которой, как полагают, образовалась наковальня среднего уха млекопи-
тающих.
Квантовая эволюция (quantum evolution) — термин Симпсона — быстрый
' эволюционный процесс, приводящий за сравнительно короткое время к об-
* разованию группы, которую систематик мог бы счесть новой крупной таксо
комической единицей,
21*
304 | Словарь терминов
Клейстогамные формы — растения с малозаметными цветками, котор1
совсем не раскрываются и поэтому подвергаются самоопылению.
Климаксное (терминальное) сообщество — относительно стабильно
сообщество, завершающее сукцессию (см.); обычно характерно для т<
или иной климатической зоны.
Клин— градиент изменчивости количественно варьирующего признака hj
комплекса признаков популяции. Часто крутизна этого градиента быва
различной на его протяжении, причем возможны и разрывы непреры
ности.
Клон — популяция, полученная от одной особи путем бесполого размн
жения.
Коацерваты—агрегаты различной сложности, образующиеся при взаим
действии двух или нескольких коллоидов (см.).
Коллоид—дисперсная система, обладающая некоторыми свойствами Ki
раствора, так и суспензии.
Комплекс Гольджи — см. Гольджи комплекс.
Конвергенция— поверхностное сходство организмов, обусловленное схо
ством занимаемых ими адаптивных зон(см.).
Конкуренция — использование одних и тех же ограниченных ресурсе
двумя или большим числом организмов.
Конспецифичные формы — формы, которые систематики относят к о,
ному виду.
Конъюгация — соединение клеток или организмов, во время которого вс
генетическая информация или часть ее переходит из одной особи (и;
клетки) в другую.
Конъюгация хромосом (синапсис) — спаривание гомологичных хромосс
в мейозе.
Конъюгирующие сегменты — эухроматиновые участки хромосом, кон'
югирующие со своими гомологами в первой профазе мейоза.
Коферменты — вещества, присоединение которых к специфическим белка
ведет к образованию каталитически активных комплексов, ферме!
тов (см.).
Кроссинговер — обмен участками (обычно соответственными) между хр<
матидами гомологичных хромосом; приводит к образованию хиазм (см
и к рекомбинации генов (см.).
Лизогенная бактерия — бактериальная клетка, содержащая одну ш
несколько фаговых частиц в форме профага.
Локус — участок хромосомы, соответствующий одному гену; участок, зан
маемый эквивалентными генами (аллелями) на гомологичных хромосома
Ъ-Хромосомы—см. «ограниченные» хромосомы.
Макроэволюция — эволюционные события, рассматриваемые обычно в пе
спективе геологического времени, например эволюция лошадей от млек
питающего величиной с собаку до Equus caballus.
Словарь терминов | 305
Мегаспоры — споры (у сосудистых растений), в результате деления кото-
рых образуется женский гаметофит.
Мезозой — одна из геологических эр, характеризовалась расцветом рептилий;
началась около 200 млн. лет назад, закончилась примерно 70 млн. лет
назад.
Мейотический дрейф (meiotic drive) — большая вероятность включения в
гаметы одного из аллелей (по сравнению с другими аллелями данного
локуса).
Меланическая форма — черная, темно-коричневая или вообще темноокра-
шенная форма какого-либо организма.
Меловой период—последний период мезозойской эры; начался около
130 млн. лет назад.
Менделевская популяция — группа размножающихся особей, имеющая
общий генофонд.
Метацентрическая хромосома—хромосома, в которой центромера
расположена примерно посредине, так что оба плеча примерно одина-
ковы.
Миграция — перенос генетической информации из одной популяции в дру-
гую; расселение организмов за пределы мест их выплода; периодические
передвижения особей.
Микросомы — цитоплазматическая фракция, выделяемая при центрифуги-
ровании гомогенизированных клеток; полагают, что она состоит из рибо-
сом и фрагментов эндоплазматического ретикулума (сейчас микросомную
фракцию чаще отождествляют с фракцией эндоплазматического рети-
кулума, см.).
Микроэволюция — эволюционные события, обычно наблюдаемые в тече-
ние короткого периода времени, например изменение частоты генов в по-
пуляции за несколько поколений.
Мимикрия — внешнее сходство одного организма с другим, имеющее, по-
видимому, защитное значение.
Миоцен — средняя эпоха третичного периода; началась около 26 млн. лет
назад, закончилась примерно 11 млн. лет назад.
Митохондрия — цитоплазматическая органелла нитевидной или сфериче-
ской формы, состоящая из двух мембран, из которых внутренняя обра-
зует складки; в митохондриях сосредоточены многие реакции клеточного
дыхания.
Множественные аллели^ аллели локуса, для которого известно более
двух альтернативных состояний.
Модификаторы —гены, фенотипическое действие которых состоит в изме-
нении проявления других генов.
Морфогенез — процесс, ведущий к формированию зрелого организма с ха-
рактерной для пего организацией.
Мультиваленты — объединение в мейозе более чем двух хромосом, го-
мологичные участки которых конъюгируют попарно.
306 | Словарь терминов
Мутагенный агент — фактор, вызывающий повышение частоты мутаций
по сравнению с уровнем спонтанных мутаций.
Мутон — отдельная мутирующая единица внутри гена.
Направленное поочередное расхождение — регулярное поочередное от-
хождение центромер к одному и тому же полюсу веретена в ядрах, гете-
розиготных по одной или нескольким взаимным транслокациям.
Направленный отбор—отбор, приводящий к сдвигу средней величины
данного признака в популяции.
Наследуемость — генетическая дисперсия, деленная на фенотипическую
дисперсию; показатель степени сходства между родителями и потомством
по какому-либо признаку.
Нейрула —стадия эмбрионального развития позвоночных, следующая за
гастулой и характеризующаяся быстрой дифференцировкой.
Нерасхождение — отхождение хромосом бивалента или мультивалента к
одному полюсу в анафазе первого деления мейоза.
Нита (экологическая) — специфический образ жизни определенного орга-
низма в сообществе, отношение его к физическому субстрату и т. п.; поня-
тие ниши родственно понятию адаптивной зоны (см.).
Нуклеиновые кислоты— макромолекулярные соединения, полимеры, по-
строенные из мономолекулярных единиц, нуклеотидов (см.).
Нуклеотид — фосфорный эфир N-гликозида азотистого основания, стоук-
турный элемент (мономер) нуклеиновых кислот.
Обратная связь — влияние результатов какого-либо процесса на ход са-
мого этого процесса.
«Ограниченные* хромосомы (limited chromosomes)—гетерохроматино-
вые хромосомы, содержащиеся только в клетках зародышевого пути у не-
которых видов грибного комарика Sclara.
Окисление — реакция, в которой вещество присоединяет кислород или от-
дает водород или в которой увеличивается валентность металла.
Олигогены—гены с ясно выраженным фенотипическим проявлением; гены-
переключатели.
Онтогенез — индивидуальное развитие особи.
Оператор—предполагаемый генетический элемент, который служит прием-
ником специфических сигналов, приходящих из цитоплазмы (в виде моле-
кул репрессора), и контролирует функцию оперона.
Оперон — группа цистронов (см.) (обычно сцепленных), выражение кото-
рых контролируется одним оператором (см.); генетическая единица транс-
крипции кода ДНК.
Органелла — структурный компонент клетки, обладающий характерной
морфологией и функцией; по отношению к клетке играет ту же роль, что
и орган в многоклеточном организме.
Организатор — часть эмбриона, определяющая направление развития масс
клеток, с которыми он приходит в соприкосновение.
Словарь терминов | 307
Ориентированное мейотическое деление — меойз (в оогенезе), в ко-
тором веретено расположено таким образом, что определенная группа
хромосом всегда попадает в полярное тельце, а не в будущую яйце-
клетку.
Отбор — дифференциальное воспроизведение генотипов.
Ошибка выборки—отклонение, обусловленное элементами случайности в
процессе получения выборки.
Панмиксис — случайный подбор пар родительских особей в популяции.
Параллелизм — конвергенция между близкородственными формами.
Парацентрическая инверсия — инверсия, не включающая центромеру.
Партеногенез — развитие особи из неоплодотворенной гаметы (яйце-
клетки) .
Пенетрантность — относительная частота проявления гена в гомозигот-
ном состоянии (доля гомозиготных особей, у которых данный ген прояв-
ляется в фенотипе).
Перекрест — см. Кроссинговер.
Перицентрическая инверсия—инверсия, включающая центромеру.
Пиримидин — органическое азотистое основание, имеющее структуру ше-
стичленного кольца, в котором атомы азота стоят в положениях 1 и 3.
Плазматическая мембрана — наружная пограничная мембрана цитопла-
ста; без перерыва переходит в мембраны эндоплазматического ре-
тикулума (см.); снаружи к ней может примыкать пелликула или клеточ-
ная стенка.
Планктон — популяция организмов, находящихся во взвешенном состоя-
нии в воде.
Пластиды— органеллы растительной клетки, в которых обычно содер-
жатся пигменты и происходит синтез растворимых и нерастворимых угле-
водов.
Плейотропия— влияние одного или нескольких аллелей на два или боль-
шее число признаков, не находящихся в очевидной связи между собой.
Плейстоцен — эпоха, составляющая большую часть четвертичного пе-
риода; началась примерно 1 млн. лет назад, закончилась около 10 000 лет
назад.
Плиоцен —последняя эпоха третичного периода; началась около 11 млн.
лет назад, закончилась примерно 1 млн. лет назад.
Плоидность— число геномов в зиготе данного организма.
Полигены — гены, по отдельности не вызывающие заметного фенотипиче-
ского эффекта; многочисленные факторы, влияющие на какой-либо при-
знак.
Полиморфизм—существование в популяции (на одной и той же стадии
развития организмов) двух или нескольких форм в таких количественных
соотношениях, что наличие даже наиболее редкой из них нельзя объяснить
одним лишь мутированием.
Полип — прикрепленная форма кишечнополостного животного.
308 | Словарь терминов
Полипептид — продукт соединения аминокислот в цепь; при образовании
полипептидной цепи кислотная группа одной молекулы аминокислоты со-
единяется с основной группой другой молекулы (при этом выделяется
одна молекула воды).
Полиплоид— организм, в соматических клетках которого содержится бо-
лее двух гаплоидных наборов хромосом или более двух геномов.
Полисахариды — макромолекулярные углеводы, образующиеся путем со-
единения большого числа молекул простых углеводов (а иногда, кроме
того, и других молекул).
Политенная хромосома — гигантская хромосома, состоящая из множе-
ства хромосомных нитей, тесно соединенных по всей длине.
Половая хромосома — хромосома, по крайней мере частично принимаю-
щая участие в определении пола (обычно в значительной части состоит из
гетерохроматина).
Половое размножение — способ размножения, при котором происходит
слияние гамет, образовавшихся в результате мейоза.
Полярные тельца — две или три клетки, образующиеся в результате
мейоза (в оогенезе), но не превращающиеся в яйцеклетки.
Популяционная биология— отдел биологии, исследующий взаимоотно-
шения организмов в пространстве и времени и включающий такие дисци-
плины, как экология, систематика, этология, популяционная генетика и дру-
. гие, в которых рассматриваются главным образом взаимодействия целых
организмов.
Популяция — набор исследуемых объектов; совокупность особей, находя-
щихся в данное время в данной области; при обсуждении биологических
вопросов под этим термином часто подразумевается менделевская попу-
ляция (см.).
Порфирины—класс соединений (производные пиррола), которые в форме
хелатных комплексов с металлами образуют пигменты, играющие важную
роль во многих биологических процессах.
Признак — любая особенность, варьирующая внутри изучаемой группы
объектов.
Примитивный (организм или признак)—тот, который в процессе эволю-
ции в меньшей степени отклонился от предполагаемого предкового типа
(по сравнению с каким-либо другим организмом или признаком).
Принцип основателя (founder principle)—принцип, состоящий в том, что
при образовании новой изолированной популяции ее генофонд (см.)
вследствие ошибки выборки (см.) несколько отличается от генофонда ро-
дительской популяции; это различие в дальнейшем увеличивается, так как
неодинаковые эволюционные факторы в районах, занимаемых двумя по-
пуляциями, будут к тому же действовать в различных популяционно-гене-
тических условиях; результатом будет возрастание дивергенции.
Протеиноид — сложный полипептид.
Протист — одноклеточный организм.
Словарь терминов | 309
Приспособленность (fitness) — относительная способность данного гено-
типа к выживанию и воспроизведению по сравнению с другими геноти-
пами в популяции.
Псевдогамия — партеногенетическое развитие яйца после стимуляции его
S: (но не оплодотворения) мужской гаметой или гаметофитом.
Псевдо копуляция — способ опыления у некоторых орхидей, у которых
цветки внешне сходны с самкой насекомого, так что самцы делают по-
5 пытки совершить копуляцию и таким образом переносят пыльцу.
Пурин — органическое основание, молекула которого состоит ив двух кон-
v. денсировапных гетероциклов — пиримидинового и имидазольного колец.
? Расхождение — передвижение центромер бивалента к противоположным
- полюсам веретена в анафезе первого деления мейоза (и вообще расхожде-
ние хромосом к разным полюсам при делении клетки).
Регулятор (ген-регулятор)—геп, контролирующий функцию оператора
в соответствии с поступающими из цитоплазмы сигналами в виде молекул
С репрессора.
Рекапитуляции теория — представление о том, что в процессе индиви-
дуального развития организм проходит стадии, сходные со взрослыми фор-
мами его предполагаемых предков; эту теорию часто формулируют сло-
вами «онтогенез повторяет филогенез».
Рекомбинационный индекс (RI)—показатель количества новых генных
сочетаний, которые могут возникнуть в течение данного времени. По Дар-
лингтону, определяется как среднее число хиазм плюс гаметическое число
хромосом.
.. Рекомбинация — образование таких сочетаний генов, которых не было
< у родительских форм.
Рекон — наименьшая единица, обмен которой в процессе рекомбинации ме-
жду хромосомами можно выявить с помощью методов, применяемых при
изучении тонкой генетической структуры.
Репликация — многократное последовательное удвоение.
Репрессоры — молекулы, способные соединяться с определенным метабо-
литом (который либо активирует, либо инактивирует их) и с определен-
ным оператором, действие которого при этом блокируется.
Репродуктивная величина популяции — число особей данного поколе-
ния (в популяции), фактически участвующих в размножении.
Репродуктивные клетки—гаметы и их непосредственные предшествен-
ники, из которых они образуются в результате клеточного деления.
Реснички — короткие, обычно многочисленные, подвижные протоплазмати-
ческие выросты на поверхности клетки; им свойственны характерная тон-
кая структура и своеобразное координированное движение (биение).
Рецессивность — свойство, противоположное доминантности (см.); пол-
‘ ностью рецессивный аллель проявляется только в гомозиготном со-
стоянии.
310 | Словарь терминов
Рибонуклеиновые кислоты (РНК) — макромолекулярные соединения, мо-
лекулы которых у большинства организмов обладают разнообразными
функциями; одна разновидность РНК копирует генетический код ДНК и
переносит его к местам синтеза белков (информационная РНК), а другие
участвуют в организации этого синтеза (транспортная РНК и рибосомная
РНК) (см. Нуклеиновые кислоты).
Рибосомы — гранулы, связанные с мембраной эндоплазматического ретику-
лума; предполагаемые места синтеза белков (см. также Микросомы).
РНК — см. Рибонуклеиновая кислота.
Самозарождение жизни — см. Абиогенез.
Сбалансированный полиморфизм—полиморфизм, который поддержи-
вается в популяции обычно благодаря тому, что гетерозиготы по данному
локусу имеют более высокую адаптивную ценность по сравнению с любой
из гомозигот.
Сверхдоминантность — большая степень выражения признака у гетеро-
зиготы по сравнению с обеими гомозиготными формами.
Сегментарный аллополиплоид — аллополиплоид, геномы которого го-
мологичны по многим небольшим участкам (сегментам) различных хромо-
сом хромосомного набора; в этом случае при кроссинговере возможна ре-
комбинация материала между различными геномами.
Сетевидная эволюция (reticulate evolution) —тип эволюции, при котором
линии не только разветвляются, но и сливаются друг с другом.
Симпатрические формы—организмы, области распространения которых
перекрываются или совпадают.
Синапсис — соединение гомологичных хромосом в профазе мейоза.
Сингамия — соединение гамет с образованием зиготы.
Систематическое давление — действие неслучайного эволюционного
фактора: отбора, мутирования или миграции.
Случайная выборка — подмножество объектов, отобранных из некоторого
множества таким образом, что все члены множества имеют равную ве-
роятность быть включенными в это подмножество.
Сдвиг признаков — явление, состоящее в том, что симпатрические попу-
ляции двух видов больше различаются между собой, чем аллопатрические
популяции тех же видов.
Сообщество — группа организмов, существующих в определенном место-
обитании.
Сочленовная кость — одна из челюстных костей многих позвоночных, из
которой, по-видимому, образовался молоточек среднего уха млекопитающих.
Специализированный (организм или признак) —тот, который в процессе
эволюции дальше отошел от предполагаемого общего предкового типа (по
сравнению с каким-либо другим организмом или признаком).
Средовая дисперсия—часть фенотипической дисперсии, обусловленная раз-
личиями во внешних условиях, в которых находились особи данной попу-
ляции.
Словарь терминов | 311
Стабилизирующий отбор — отбор, благоприятствующий тем генотипам,
которые обусловливают меньшее отклонение признака от средней величины 1.
Стационарное распределение частот — представление функции плот-
ности вероятностей, показывающее, как распределяются вероятности воз-
можных событий в данной ситуации.
Структурная гибридность—гетерозиготность по хромосомной пере-
стройке.
Структурный ген—функциональная единица генетического материала,
которая через свой первичный продукт — информационную РНК — контро-
лирует синтез определенного белка или полипептида.
Сукцессия — последовательность сообществ, сменяющих друг друга до тех
пор, пока в данном участке не сформируется климаксное (терминаль-
ное) сообщество (см.).
Суперген— группа генов, наследуемая как единое целое.
Сцепление — совместная передача генов, обусловленная их локализацией в
одной и той же хромосоме.
Таллом — тело растения, не вполне четко дифференцированное на корень
и стебель.
Тахителическая эволюция — эволюция, протекающая значительно быст-
рее го роте личе ской эволюции (см.).
Телитокия — форма апомиксиса у животных, при которой диплоидные
самки развиваются из неоплодотворенпых яиц.
Терминализация — перемещение хиазм к концам хромосом на поздних
этапах профазы первого деления мейоза.
Территориальность — защита особью определенного участка среды от
вторжения других подобных ей особей.
Тетраплоид — организм, клетки которого содержат 4 генома.
Тетрасомик — организм, у которого одна из хромосом представлена че-
тырьмя гомологами.
Тимин — пиримидин (2,4-диокси-5-метилпиримидин), входящий в состав
ДНК, но отсутствующий в РНК.
Тране гидрогенирование — последовательный перенос электронов в клеточ-
ных циклах окислительных процессов.
Трансдукция — механизм переноса генов у бактерий, при котором ге-
нетический материал переносится бактериофагом из одной клетки в
другую.
Транслокация - см. Взаимная транслокация.
1 Не следует смешивать с явлением стабилизирующего отбора в понима-
нии И. И. Шмальгаузена: это явление скорее соответствует тому, что автор
называет генетической ассимиляцией или «канализацией» (стабилизацией)
развития, — Прим. ред.
312 | Словарь терминов
Тран с-положение — такое расположение двух мутационных изменений (в
одном или двух близких цистронах), когда они находятся в разных гомо-
логичных хромосомах.
Трансформация—изменение генетической информации бактерии в резуль-
тате соприкосновения с экстрактом ДНК из бактерий, обладающих иным
генотипом.
Трисомик — организм, у которого одна хромосома представлена тремя го-
мологами.
Унивалент — хромосома, не имеющая гомолога и поэтому не вступающая
в профазе первого деления мейоза в конъюгацию.
Урацил — пиримидин (2,4-диоксипиримидин), входящий в состав РНК, по
отсутствующий в ДНК.
Фактор — см. Ген.
Фенокопия — фенотипическое изменение, имитирующее проявление какой-
либо мутации.
Фенотип — результат взаимодействия генетической информации со средой;
все признаки организма, за исключением его генетической структуры.
Фенотипическая дисперсия — полная дисперсия данного признака.
Ферменты — белковые катализаторы биохимических реакций.
Филетическая эволюция — любые изменения, происходящие последова-
тельно в одном филогенетическом ряду, термин Симпсона.
Филогенетический — имеющий отношение к филогенезу, эволюционной
истории организма.
Фосфатная связь (макроэргическая) — обычно ангидридная связь между
остатками фосфата, которая может служить важным источником энергии
(для биохимических реакций).
Фосфорилирование — присоединение фосфата к молекуле органического
вещества.
Фотолиз — расщепление воды за счет энергии света.
Фотосинтез — совокупность процессов, связанных с синтезом органических
соединений из неорганических с участием света в качестве источника энер-
гии.
Фотофосфорилирование — присоединение фосфата к органическому ве-
ществу за счет энергии света.
Фратрия (gens, множ, gentes) у кукушек — внутривидовая группировка
особей, откладывающих яйца преимущественно в гнезда одного определен-
ного вида птиц; такие группировки не обязательно изолированы друг от
друга географически.
Функция плотности вероятностей— функция, описывающая распре-
деление вероятностей возможных событий, т. е. ожидаемые относительные
частоты этих событий.
Харди — Вейнберга закон — закон, согласно которому в панмиктической
популяции при отсутствии систематических давлений и дрейфа генов ча-
Словарь терминов | 313
стота генов в данном аутосомном локусе остается постоянной и частота
генотипов по этому локусу достигает равновесия за одну смену поколений.
Хемо синтезирующее организмы—организмы, способные синтезировать
сравнительно сложные углеводы, используя в качестве источников энергии
только неорганические вещества (например, серу или железо).
Хиазма — крестообразная конфигурация хромосом в биваленте (в профазе
первого деления мейоза); обычно это результат происшедшего ранее крос-
синговера (см.).
Хиатус (gap) —разрыв непрерывности в картине изменчивости.
Хлорофиллы — магнийпорфириновые комплексы, представляющие собой
зеленые пигменты фотосинтезирующих организмов; функция этих пигментов
состоит в поглощении световой энергии.
Хроматида — видимое продольное подразделение хромосомы, имеющее
лишь одну центромеру (или ведущее себя так, как если бы центромера не
разделилась); продольная половинка хромосомы.
Хроматография — метод разделения компонентов какой-либо смеси, осно-
ванный на их дифференциальной миграции в одном или нескольких рас-
творителях; полученное распределение этих компонентов (выявляемых
обычно с помощью цветных реакций) называют хроматограммой.
Хромонема — тончайший из видимых продольных морфологических элемен-
тов хромосомы.
Хромосомный набор — полный набор находящихся в ядре хромосом, со-
держащий один или более геномов в зависимости от типа клетки и ее
состояния.
Хромосомы — клеточные органеллы, с которыми связана большая часть
ядерной генетической информации; хромосома содержит центромеру
(см.), которая служит местом прикрепления веретена (см.).
Центриоль — цитоплазматическая органелла (обладающая характерной
тонкой структурой), с которой соединены волокна веретена и звезды во
время цитокинеза и кариокинеза.
Центрический фрагмент — фрагмент хромосомы, содержащий центро-
меру.
Центромера — участок хромосомы, обычно очень короткий, к которому
при клеточном делении прикрепляются нити веретена; то же, что кине-
тохор.
Центросома— область, окружающая центриоль; полагают, что она имеет
отношение к организации белковых волокон.
Цис-положение — такое расположение двух мутационных изменений (в
одном или двух близких цистронах), когда они находятся в одной из двух
гомологичных хромосом.
Цистерны — полости в эндоплазматическом ретикулуме, представляющие
собой (судя по их контурам на электронных микрофотографиях) уплощен-
ные пузырьки.
314 | Словарь терминов
Настрой — функциональная единица, эквивалентная участку генетического
материала, контролирующему синтез специфического белка (точнее, одной
из его полипептидных цепей).
Цитозин —пиримидин (2-окси-4-аминопиримидин), входящий в состав ДНК
и РНК.
Цитоплазматическая наследственность — наследование признаков,
детерминанты которых находятся не в хромосомах, а в цитоплазме; то же,
что нехромосомная наследственность.
Частота гена (gene frequency) — число локусов в популяции, занятых
данным аллелем, деленное на общее число локусов, в которых этот ал-
лель мог бы находиться; то же, что концентрация гена.
Экосистема — комплекс организмов, занимающих определенное местооби-
тание, взаимоотношения между которыми можно описать термодинами-
чески.
Экспрессивность — мера единообразия фенотипического проявления гена
в определенных условиях среды.
Эндоплазматический ретикулум — система канальцев и пузырьков
(цистерн, см.) в цитоплазме растительных и животных клеток; ограничи-
вающие их мембраны без перерыва переходят в плазматическую мембрану
и в наружную мембрану ядерной оболочки; с мембранами ретикулума
связаны рибосомы (см.).
Эндополиплоидия — полиплоидия, обусловленная делением хромосом, не
сопровождающимся делением ядра.
Эпигенотип — ряд связанных между собой процессов развития, в резуль-
тате которых формируется взрослый организм.
Эпистаз — взаимодействие неаллеломорфных генов.
Эуполиплоидия — увеличение числа хромосом в результате добавления
целых геномов.
Эухроматиновые участки — участки хромосом, претерпевающие типич-
ные изменения при делении клетки и содержащие гены с заметным фено-
типическим эффектом.
Эффективная репродуктивная величина популяции — величина по-
пуляции, к которой можно математически «приводить» величины популя-
ций с различным отношением полов, различной степенью инбридинга
и т. п., что позволяет сравнивать их между собой.
Эффекторы — небольшие молекулы (метаболиты), которые соединяются
с молекулами репрессора, активируя или инактивируя их в отношении
способности связываться с оператором.
Ядерная оболочка — отделяет ядро от цитоплазмы; состоит из двух эле-
ментарных мембран с порами; наружная мембрана без перерыва перехо-
дит в эндоплазматический ретикулум.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Аберрации 187
Автокатализ 14, 15, 17
Автополиплоиды 202, 203, 208, 216,
219
Автотетраплоиды 202, 207
Автотриплоидия 202
Агамный комплекс 216, 217
Агамоспермия 212
Адаптация 170
— друг к другу 166
— и генные сочетания 132
Адаптивная зона 283
----- доступ 287
— радиация 283, 286, 287
-----в растениях 286
— ценность 115
Адаптивные пики 132—134
Адвентивная эмбриония 213
Аденин 23, 24
Аденозиндифосфат (АДФ) 16, 17,
33, 34
Аденозинтрифосфат (АТФ) 17, 18,
19, 33
Акроцентрические хромосомы 198
Аллели 46, 52, 103, 123, 132
— множественные 53
— частота 65, 103—105, 108, 109,
112, 122, 126, 128
Аллена правило 227, 248
Аллометрия 294
Аллопатрическое видообразование
264
Аллополиплоидия 202—208, 216, 217,
219, 265
— сегментарная 206, 208
Аллотетраплоидия 203, 206, 207
Аляскинские бурые медведи 275
Амейотическая телитокия 214, 215
Аминокислоты 12—15, 23, 25
Амфидиплоидизация 198, 203
Анафаза 33—38
Анеуплоидия 196—201
Антибиотики 161, 162
Апогамия 212
Апомиксис 195, 209—220, 257
Апомиксис у позвоночных 217,
218
Апоспория 212, 213
Арренотокия 213
Ассортативная система спаривания
184, 185
Атмосфера, восстановительные свой-
ства 12, 13
— окислительные свойства 13
Ауксоцит 39, 41
Аутбридинг 185, 186
Аутосомы 36, 57, 190
Африканский парусник 159, 160
Ацентрический фрагмент 60
Бабочка (и) 56, 68, 72, 89, 90, 96, 97,
99, 137, 142, 167, 233, 268, 291
— виды-двойники 237, 260
— индустриальный меланизм 137,
159
Бабочка-данаида 68
Бабочка-парусник 227
Бабочка-репница 96
Базальная гранула 24
Базальное зерно 34
Базидиальные грибы 181
Бактерии 54, 174—178
— лизогенные 175
— хемосинтезирующие 17
Бактериофаги 174, 175, 182
Бананы 203
Белки 12, 13, 30, 31, 33
— синтез 15, 16, 75
Белый медведь 275
Бергмана правило 227, 248
Биваленты 41, 50
Биогенетический закон 77
Биогеографическая обособленность
274
Богомолы 192
Брадителические скорости 279, 282,
283
316 | Предметный указатель
Бурые водоросли 180
Бурый медведь, европейский 86
«Буферный» эффект 69
Валлаби 286
Ваточниковые 276
Вегетативное размножение 209
Вертикальное видообразование 277
Вещество-репрессор 76
Взаимодействие гена со средой 64
Видообразование 225, 247
— аллопатрическое 264
— вертикальное 277
— горизонтальное 277
— симпатрическое 265
Виды 225, 247, 292
Виды-двойники бабочек 237, 260
Вирусы 174, 176, 177
Вишни 203
Внешняя среда 94—98
Внутригрупповой отбор 134
Внутрипопуляционная изменчивость
227
Водоросли 84, 178, 179, 208, см.
также Сине-зеленые водоросли,
Бурые водоросли, Зеленые водо-
росли
Возраст геологический, определение
278
Волк 99
Воробьи 96, 136
Вошь кроличья 262
— человеческая 245, 246, 262
Вьюрки 239, 245
— Галапагосских островов 238, 259,
286
Вымирание 268, 269, 271—273, 278—
283, 289, 292
Высшие категории 268, 291, 292
Гаметическое число хромосом 196
Гаметогенез 181
Гаметофит(ы), апомиксис 212
— мужской 181, 186
— поколение 71, 181, 212
Гаметы 39, 41, 54, 60, 90, 103,
180
— потеря 182, 194
— случайное соединение 42
Гапло-диплоидия 213
Гаплоидия 178 — 181, 196
Гаплоидная апогамия 212
Гаплоидный партеногенез 212, 213
Гаплоиды 178
Гаплофаза 71
Гаттерия 269
Ген(ы) 22, 46, 52—55, 187
— адаптация 132
— гипостатические 52
Ген(ы), дрейф 108—110, 114, 145, 158,
184, 260
— комбинации и адаптация 132
— комплементарные 52
— летальные 194
— определяющий окраску цветка
252
— потеря 130
---и фиксация 108—111, 134, 252
— скорость изменения частоты 117
— случайная утрата 134, 252
---фиксация 134, 252
— структурные, транскрипция 75,
76
— частота 103—105, 134
---мутаций 65, 103—105, 108,
109, 112, 114, 121, 126, 128
— эпистатические 52
Гены-модификаторы 53, 55, 56, 142,
161
Гены-операторы, транскрипция 75,76
Гены-переключатели 62, см. также
Супергены
Гены-регуляторы, транскрипция 75,76
Генетическая ассимиляция 71, 167
— изоляция 233
— инерция 105
— информация 70, 77
Генетические системы 43, 59, 68, 97,
173, 174, 182, 195, 221
Генетический гомеостаз 166, 167
— груз 124, 125
— код 20—27, 74, 75, см. также
Генетическая информация
— материал 42, 45, 59, 67, 68, см.
также Генетическая информация
Генетическое сцепление с полом 57
Генные сочетания 132
Генный поток 250—252
Геном 196
— число в хромосомном наборе 196
Генотип 68, 70, 74, 94
Генофонд 103
Географическая изменчивость 226—
235, 247
Географическое распределение 86,87,
89
Геологический возраст, определение
278
Гетерогаметный пол 57, 58
Гетерозиготность 48, 54, 122—126,
165, 184, 214
Гетерозис 64
Гетерокариотическое состояние 181
Гетерохроматиновые участки 50, 57,
60, 190, 197, 199
Гетерохроматиновый фрагмент с цен-
тромерой 197
Гибриды, отбор, направленный про-
тив гибридов 258
— экология 257
Гидра 85
Гиногенез 218
Глаз, эволюция 170
Глиноземы, реакции на поверхности
молекул 16
Глогера правило 227
Гнездовой паразитизм 245
Голосеменные 181, 289
Гольджи аппарат 33
Гомеостаз 67—71, 180
— генетический 166, 167
Гомогаметный пол 57, 58
Гомозиготность 48, 55
Гомойотермные организмы 248
Гомологичные хромосомы 41, 43, 47
Горизонтальное видообразование 277
Горихвостки 140
Горотелическое распределение 279,
281, 282
Горчица 96
Грана 33, 34
Грибы 84, 97, 179—181, 208
Гризли 275
— американский 86
Груз, связанный с расщеплением 126
Группы крови, определение 269
Гуанин 23, 24
Гуппи 255
Гусеницы 72, 75, 141
Дарвиновы вьюрки 239—245
Двукрылые 59, 199
Двустворчатые моллюски 279—281
Двуххроматидный обмен 50, 51
Дезоксирибонуклеиновая кислота
(ДНК) 21—24, 27, 34, 54, 67, 75,
174—177, 180, 183
Деление клетки 34—39
Делении хромосом 57—59
Диатомовые 180
Дикариоз 182
«Дикий тип» 68
Предметный указатель | 317
Диплоидия 213
Диплоиды 55, 178—183, 196
Диплоспория 212—214
Диплофаза 71
Дисруптивный отбор 116, 159, 161,
265
Дицентрическая хроматида 60, 188
Дицентрический фрагмент 60
ДНК см. Дезоксирибонуклеиновая
кислота
Добавочные хромосомы 198
Дождевой червь 217
Доминантность 48, 55, 115, 142
— экологическая 2, 295
Достоверный результат 140
Древесные кенгуру 286
Дрейф генов 108—ПО, 114, 145, 158,
184, 260
Дрожжевые клетки 71, 84, 182
Дрозды 140, 146
Дрозофила 50, 56, 57, 60, 69, 152—
157, 161, 165, 171, 198, 265
Дубоносы 257
Дубы дикие 96, 185
Дупликация 59
Евгеника 119
Единичная мутация 64
Ежа сборная 158
Естественный отбор 20, 52, 70, 99,
114, 115, 162
Жаберные мешки 78
Жгутики 34
Жгутиковые 180, 186
Живорождение у растений 211
Живые ископаемые 276
Жизненные циклы 71
---- гаплоидов 180
---- диплоидов 180
Жуки 192
Жуки-долгоносики 217, 294
Жуки-олени 294
Загрязненность местности 142
Зародышевая клетка 39, 68, 199
Защитная окраска 170
Звезда 36
Зеленые водоросли 180
Зигота 179
Зиготическое число хромосом 196
Злаки 287
Змеи 286
22 Зак. 15?
318 | Предметный указатель
Избирательная илиминация 151
Изменчивость 45
— генетическая 63, 64
— дискордантная 269
— затухание ПО—112
— клинальная 248
---у животных 231
------- растений 228
Изменчивость количественная 63
— непрерывная 62
Изоляция 254
— обусловленная расстоянием 129
— половая 258
Иммиграция 114
Инбредная депрессия 64
Инбридинг 129, 183, 184
Инверсия (и) 188, 189
— Арроухед (AR) 152—157
— парацентрическая 188, 192, 194
— перицентрическая 189
— «стандартная» (ST) у дрозофи-
лы 152—157
— хромосомная 57, 59, 60, 152, 188,
189, 192
— «Чирикахуа» (СН) у дрозофилы
152—157
Идивидуум 85, 86
Индукция 73
Индустриальный меланизм 56, 137—
142
--- у бабочек 264
Инсектициды 161, 162
Интерсексуальность 233
Интерстициальный сегмент 190
Интерфаза 34
Интерференция 50, 56
Интерогрессивная гибридизация 257,
см. также Гибридизация
Инфекционная наследственность 178
Инфекция вирусная 178
Информационная РНК 23, 25, 34, 75,
76
Информация генетическая см. Гене-
тическая информация
Ионизирующее излучение 126
Ирландский олень 294
Ископаемые люди 78, 269
— находки 276
Искусственный отбор 115, 162
Капуста 203
Кариотип 187, 192
Катализаторы 16
Квадриваленты 60
Квазиорганизмы 84
Квантовая эволюция 278, 283
Кенгуру 286
Киты 289, 293
Клевер белый 158
Клейстогамные цветки 210
Клетка 27, 30
— дифференцировка 72—76
Клеточное деление 34
Климаксное сообщество 98
Клинальная изменчивость 248
— — у животных 231
------- растений 228
Клины 226, 228—232
Клон 85, 209, 218, 265
Коала 286
Коацерваты 14, 19, 20, 28
Кобра 286
Коллоидные гели 19
Коловратки 213, 214, 218, 265
Колониальные организмы 84
Колонии 84, 85, 93, 171
Колхицин 202
Кольцо хромосом 60, 194
Комары 96, 162
Комплементация 47
Компонент аддитивный генетический
63
Компоненты изменчивости, аддитив-
ные 64
Конвергенция 289, 290
Конкуренция 288, 289
Конъюгация хромосом 59
Копа правило 293
Коферменты 16
Кривые выживания 278, 280, 282
Крокодилы 282
Кроссинговер 41, 42, 46, 49, 50, 56,
59, 60, 187
— соматический 181
Крысы 97
Кукуруза 57
Кукушки 245, 262—264
Кускусы 286
Кутин 32
Кактус 239
«Канализованные» процессы 69, 70
Канальцы нитей 34 '
Каппа-частицы 177
Ленивец 96
Леса из дуба и гикори 99
Летальные гены 194
Лизогенные бактерии 175
Лилейник 85, 209
Липиды 30, 31, 33
Локус Rh 158
Лошади 293
Луга 96
Лук 211
Лягушка 74, 96, 230, см. также
Rana
Макромолекулы 21, 30
Макроэволюция 268, 276
Малиновка 140
Мангровые заросли 241
Матрикс 37
Матрица 27
Медведи 86, 275
Межгрупповой отбор 134
Мейоз 39-43, 49, 50, 180, 181
Мейотический дрейф 188
Менделевская генетика 45
Менделевские популяции 101
Менделя законы 48, 49
Мерозоиты 72
Местообитание, изменение 256
— улиток 145
Металлсодержащие ферменты 18
Метафаза 33, 34, 38
Метацентрическая хромосома 198
Механизм с обратной связью 77
Миграция 90, 126—132, 158
Микросомы 23, 33
Микроэволюция 268, 276
Мимикрия 159, 160, 170, 263
— у сорняков 159
Митоз 34-39, 72, 73
Митохондрия 33, 177
Млекопитающие плацентарные 286
Многоклеточные организмы 68
Многолетние растения, рекомбинация
185, 195
Многоножки 97
Морфогенез 77
Морфологическая изменчивость 228
Мошки 97
Мультиваленты 203
Муравьед сумчатый 286
Мутаген 54
Мутагенные агенты 54
Мутант 47
Мутация 47, 53, 56, 64, 69, 104, 112—
114, 129—132, 187, 252—290
— частота 54, 90, 282
Мутоны 47
Мухоловки 140
Предметныйь указатель [ 319
Мхи 181, 208
Мятлик 158, 212, 216
Нарвалы 294
Насекомоядные 286
Насекомые 213, 288
— общественные, «касты» 91, 93
Наследственность, единицы 46—48
Наследуемость 64
Неадаптивные впадины 132, 133
Неотения 78
Непарный шелкопряд 233
Непрерывная изменчивость 62
Нерасхождение 199
Нитчатые водоросли 180, 186
Нуклеиновые кислоты 21, см. также
Дезоксирибонуклеиновая кислота,
Рибонуклеиновая кислота
Нуклеотиды 23—25, 47
Обратное мутирование 113, 127
Общественная иерархия 93
Общественные насекомые 84, 91, 171
Овсяница 203
Овсянки 140
Одноклеточные организмы 67
Однопроходные 289, 292
Окисление биологическое 16
Олений мох 99
Олень виргинский 99
— северный 99
Олигогены 62, см. также Супергены
Ондатра 95
Онтогенез 77
Онтогенетический гомеостаз 68
Оператор, транскрипция 75
Оперон 75
Опоссумы 282
Опыление 95, 184—186, 213, 276, 286,
287, 294
Организаторы 73
Организмы, взаимодействующие со
средой 94, 95
— колониальные 84
— одноклеточные 67
Организмы-пионеры 185
Органический бульон 15
Орехотворки 214, 218
Орхидеи 95
Особь (и) 84—86, 93, 94
— размер и рекомбинация 90
— число 93
22*
320 | Предметный указатель
Осоковые 287
«Остаточная» изменчивость 64
Островная модель 128
Отбор 113
— дисруптивный 116, 159, 161, 265
— естественный 20, 52, 70, 99, 114,
115, 162
— искусственный 115, 162
— коэффициент 117
— направленный 116, 139
— ослабление 163—165
— плато 164, 165
— созидательная сила 114, 115
Ошибки выборки 106, 111, 254, 260
Павлины 294
Палеонтологическая летопись 276,
291
Память 27
Панмиксис 101, 102
Папоротник 206
Паразитизм 245, 262
Параллелизм 291
Партеногенез 201, 202, 212, 213,
217—219
— циклический и последователь-
ность поколений 218
Парусник 248
— африканский 159, 160
Паукообразные 213
Пектиновые вещества 32
Пенетрантность 53
Перекрест 41, 42, см. также Крос-
синговер
Перекрестноопыляющиеся растения,
рекомбинации 196
Перестройки хромосом 58—62
Переходный полиморфизм 142
Перспективная приспособленность
173
Песцы 248
Печеночники 181
Пиноцитоз 31
Пиримидиновые основания 23
Плазматическая мембрана 31, 32
Плазмодесмы 32
Планирующие формы 283
Пластиды 33, 177
Платан 86
Плато в естественном отборе 164,
165
Плауны 282
Плацентарные млекопитающие 289
Плевел многолетний 203
Плейотропное действие генов 53,
75
Плейстоценовые оледенения 255
Плесневые грибы водные 181
Плодовая мушка 70, см. также Дро-
зофила
Плоидность 37, 39
Пневмококки 174
Поведение 90—91, 99, 185, 245, 254
— стадное 91
Поведенческая изоляция 237, 238,
261
Подсолнечник 257
Поколение, продолжительность жиз-
ни и рекомбинации 90
Покрытосеменные 95, 289
Пол 178
— дифференциация 57, 185
— определение 213, 214
Полет 283
Полигенная наследственность 64
Полигенные системы 64, 182
Полигены 62, 182
Полиморфизм сбалансированный
122—125, 158
Полипептиды 14, 53
Полиплоидия 195—208, 214—216, 257
Полиплоиды 202
Полипы 84
Политенные хромосомы 57, 60
Половая изоляция 258
Половое размножение 179, 209
Половые типы 181
— хромосомы 36, 41, 57, 58, 187
---множественные 192
---системы 190
---тип ХО 58
Поползни 140, 245
Популяция (и) 83—86, 91
— величина 91, 93, 105, 106
— журавлей 93
— менделевская 101
— мономорфная 125
— приспособленность 124
— слияние 255
— структура 90, 126
— эффективная величина 106, 249,
252
Португальский кораблик 84
Порфирины 16, 17
Потеря гамет 182, 194
— зигот 182, 194
Поточная модель 128
Предпочтение хозяев 245
Примула 203
Принцип родоначальника 148, 260
Приспособленность 115, 124, 125, 164,
165, 173, 185, 195
Прометафаза 35, 36, 38
Простейшие 84
Пространственная изоляция 250
Протеиноиды 14
Профаг 175
Профаза 35, 36
Прямокрылые 192, 198
Псевдогамия 213
Псевдокопуляция 96, 295
Птицы 238—245, 257, 259—263. 277,
283
— особенности пения 256
— райские 257
Пуриновые основания 23
Пуфы 74
Пчелы, эволюция общественного по-
ведения 171
Пыльцевое зерно 72, 181
Развитие 53, 67, 179, 180
Размеры тела, увеличение 293, 294
Размножение бесполое 203, 209, 265,
см. также Апомиксис
— вегетативное 39, 85, 203, 209
Раки ветвистоусые 214
Распределение в пространстве 86, 87,
89
— случайное 42
— экологическое 89, 96
Распространение особей 91
— семян 95
Расселение 90, 91, 185
Рассеяние 90, 91
Растения многолетние, рекомбинация
185, 195
Расы 269
Регулярное поочередное расхожде-
ние хромосом 190, 194
Редис 203
Рекапитуляция 77, 78
Рекомбинация 20, 42, 44, 47, 49, 56,
90, 174—176, 249
— геномов 206
— и окружающие условия 183, 184,
195, 219
----продолжительность жизни по-
коления 90
----размеры особей 90
— снижение частоты 182
— у микроорганизмов 176
----растений деревянистых 195
—------многолетних 185, 195
—------однолетних 196
— — — травянистых 184, 195, 196
Предметный указатель | 321
Рекомбинационный индекс 195
Рекон 47
Реннера комплексы 194
Рептилии 198, 269, 283, 288
Рептилии-архозавры 277, 283
Реснички 34
Рецессивность 55
Рецессивный признак 48
Рибонуклеиновая кислота 21, 23—25,
27, 76
----информационная 23, 25, 34,
75, 76
Рибосомы 23, 33
РНК см. Рибонуклеиновая кислота
— вируса 176
— транспортная 23, 27, 34
Рост 67
Рыба(ы) 218, 289
— скопления близких видов 23
Саламандры 203, 233
Самооплодотворение 183, 184, 196
Самопроизвольное зарождение 11
Сверхдоминантность 167
— в отношении адаптивной ценно-
сти 121, 126
Сверчки 198
«Сдвиг признаков» 245
Сегментарная аллополиплоидия 206,
208
Секвойя 72, 282
Селективная ценность 55, 58, 59, 130,
254
Серповидноклеточность 157, 247
Сетевидная изменчивость 275, 276
Симбиоз 85, 96
Симпатрическое видообразование 265
Симпатрия 256
Синапсис хромосом 41, 43, 50, 60,
188, 201
Синаптирующие сегменты хромосом
57, 190
Сине-зеленые водоросли 34, 178
Система с обратной связью 70, 71
Системы спаривания 183
— физиологической несовместимо-
сти 186
Скерда 198
Скопления близких видов у рыб 235
Скорпионы 190
Скрещивание возвратное 49
— случайное 102, 183
Слияние филетических линий 278
Сложность, увеличение 295
322 | Предметный указатель
Сложноцветные 198
Слон 271
— африканский 271
— индийский 273
Смещение популяций без обмена ге-
нами 255
Совокупность организмов 83
Сообщество (а) 98
— замкнутое 185
— климаксное 98
— открытое 185
— состояние равновесия 99
— эволюция 100
Соотношение полов 106
Сорняки, мимикрия 159
Сосна 273
Спирализация хромосом 36, 38
Спорадические транслокации 190
Спорогенез 181
Спорофит 71, 181, 212
Средовые компоненты, изменчивость
63, 64
Стабилизирующий отбор 116, 136,
166
Стандартное отклонение 63
Стационарные распределения частот
127
Стелющееся растение 99
Стенка растительной клетки 32
Стрекоза 72
Суберин 32
Суккуленты, конвергенция 289
Сукцессия 98, 99
Сумчатые 286, 289
— грибы 181
— кроты 286
Сумчатый волк 286
— дьявол 286
Супергены 156, 161, 187—189, 195
Сцепление 49, 50, 56, 60, 129, 161,
175, 176
Территориальность 91
Тигровые лилии 203
Тимин 23, 24
Типы отбора 116
Тля 214, 218, 219
— последовательность поколений
218
Толерантность 94, 99
Травянистые растения, рекомбинации
184, 195, 196, 286
Трансдукция 175, 176
Транслокации взаимные 57, 59, 190,
192, 194, 195
— реципрокные 57, 59
Трянс-положение 47
Транспортная РНК 23, 27, 34
Трансформация 174, 176
Треххроматидный обмен 51
Триплеты 24, 25, 27
Триплоидный лилейник 85
Триплоиды 203
Трисомики 199
Тритоны 91, 92
Тропические леса 96
Трупиалы 257
Тутовый шелкопряд 50
Тюльпаны 203
Углеводы 30, 32
Ужи водяные 148, 150, 151, 163
Улитки, полиморфизм 144—148, см.
также Сераеа
Ультрафиолетовый свет 16
«Умеренные» фаги 175
Униваленты 41, 203
Уникальные события 83
Уотсона — Крика модель 24
Упорядоченность уникальных явле-
ний 83
Урацил 23, 25
Таксономические группы 278, 279
Таксономия 278
Тараканы 190, 289
Тауя 256
Тахителические скорости 279, 282,
283, 287
Телигокия 213, 214, 219
— полная 214
— циклическая 214
Телофаза 37—42
Терминализация 42, 43
Термиты 245
Фаги 174, 175
— «умеренные» 175
Факторы среды, классификация 95
— эволюции, совместное действие
129
Фенокопии 167
Фенотип 69, 70
Фенотипическая дисперсия 63
Ферменты 16, 55, 56
Фиалка 210
Фикомицеты 181
Предметный указатель | 323
Фиксация гена 108—111, 130—132,
143
— случайная и потеря гена 108—
111, 134, 252
Филетическая эволюция 278, 279
Филетические линии, расщепление
277, 278
Фильтры против рецессивных лета-
лей 214
Фосфатные связи 16, 17, 19
Фоторецепторы сетчатки 34
Фотосинтезирующие бактерии 33
Фотофосфорилирование 17, 19, 33
— окислительное 17
Фратрии кукушек 246, 262—264
Функция плотности вероятностей 127
Харди-Вейнберга закон 103—105, 112,
113, 155, 156
Хиазмы 41, 42, 195
Хищники 279, 280, 286
Хлоропласты 177
Хлорофилл 17, 33
Холдейна правило 233
Хроматида 36, 40—42, 50
— дицентрическая 60, 188
Хромонема 37
Хромосомное кольцо 60, 194
Хромосомные мостики 188, 189
— мутации 187
Хромосомы 23, 34—43, 49, 206, 207
— акроцентрики 198
— дифференциальный сегмент 57,
58, 190
— добавочные 198
— интерстициальный сегмент 190
— конъюгация 59
— метацентрические 198
— нео-Х 190
— нео-Y 190, 192
— «ограниченные» 199
— парные сегменты 190
— перестройки 58—62
— половые 36, 41, 57, 58, 187
— расхождение 42, 61
— сегменты 57, 190
— синапсис 41, 43, 50, 60, 188, 201
— слюнных желез двукрылых 57,
59, 152
— карты сцепления 56, 57
— типа «Пайке Пик» (РР) 156, 157
— типы по инверсии 152—157
— цикл спирализации 36, 38
число в гамете 42, 195, 196
В-хромосома 199
L-хромосома 201
Х-хромосома 57, 58, 190, 192, 201
Y-хромосома 57, 58, 190, 192
Цветки клейстогамные 210
Цветковые растения 181, 208, см.
также Голосеменные
Целлюлоза 32
Центриоли 34, 36
Центрическое слияние 198
Центромеры 36, 42, 60, 196
Центросома 34
Т^цс-положение 47
Цистроны 47
Цитозин 23—25
Цитоплазма 32—34, 37
Цитоплазматическая наследствен-
ность 33, 178
Частота генов 103—105, 134
--- изменение 117
Человек 57, 72, 96, 158, 163, 246, 247,
255
Челюсть позвоночных, эволюция 78
Червецы 85
Черви 214
Чередование поколений 180, 181
Черный медведь 275
Четыреххроматидный обмен 50, 51
Штаммы-«убийцы» 177
Эволюционные изменения, типы 277,
278, 286
— тенденции 293—296
Эволюция квантовая 278
— основные пути 283
— скорость 278—283, 287
— сообществ 100
— филетическая 278, 279
— эффект момента 294
Эквационное деление 37, 43
Экологическая доминантность 2, 295
— сукцессия 98, 99
Экологическое распределение 89, 96
Экосистемы 96, 178, 296
Экотипическая изменчивость 227
Экспрессивность 53
Эмбриология экспериментальная 73
Эмерджентные свойства 83
Эмиграция 114
324 | Предметный указатель
Эндоплазматический ретикулум 32--
34
Эндополиплоидия 39
Эндосперм 212, 213
Энергия 11, 15—19
Эпигенетическая система 70, см. так-
же Система с обратной связью
Эпигенотип 70, 78, 85, 94
Эпистаз 52
Эпистатический компонент 64
Эпифиты 96
Эуполиплоидия 196, 201—209
Эухроматин 60, 197
«Эффект момента» 294
Эффектор 76
Яблони 203, 208
Ядерная оболочка 34
Ядро 32—37, 187
Ядрышко 34
Яйцеклетка, оплодотворение 213
Ямкоголовые змеи 286
Японский ирис 203
Ящерицы 217
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Achillea 228, 247
— borealis 228
— lanulosa 168, 228
— millefolium 228
Alcidis agathyrsus 290, 291
Allium 39, 211
— сера 203
— fistulosum 203
Annelides 209
Archaeopteryx 292
Aristolochia watsonii 99
Artemia salina 217
Ascaris 262
— lumbircoides 247
— suum 247
Asclepiadaceae 276, 290
Asclepias 248
— tuberosa 228, 229, 231, 248, 249
Ascomycetes 181
Asplenium 204—206
— ebenoides 204, 206
— montanum 207
— pinnatifidum 205—207
— platyneuron 204, 207
— rhizophyllum 204, 205, 207
— trudellii 205
A vena 97
Basidiomycetes 181, 291
Battus philenor 99, 248
Biston 227, 247
— betularia 56, 137—139, 141
Blaps polychresta 192
Bomb у x mori 50
Bradypodicola hahneli 96
Brassica 97
Bryophyta 181
Camarhynchus parvulus 240
— pauper 239
— pasittacula 239
Camelina 159
Campanula 190
Carex 198
Cariophyllaceae 159
Ccpaea 144, 145, 147, 148, 227, 247,
264
— hortensis 148
— nemoralis 144, 147, 148
Certhidea olivacea 240, 241, 243, 245
Cichlidae 235, 286
Cladonia 99
Clarkia 192—194, 198, 199
— атоепа 192—194
— franciscana 192—194
— rubicunda 192—194
Clematis fremontii 88, 89
Cleora repandata 141, 142
Cnemidophorus 217
Coelenterata 209
Coenonympho tullia 233, 250
Coleoptera 213, 289
Crepis 198, 216, 217
— foetida 198
— fuliginosa 198
— kotschyana 198
— neglecta 198
— runcinata 216
Crotalidae 286
Cruciferae 159
Culex 39
Curculionidae 217
Cyanophyceae 178
Cyclosia hestinioides 290, 291
Cyperaceae 287
Cactaceae 291
Cactospiza heliobates 240, 243, 245
— pallida 240, 243, 245
Callimantis 41
Dactylis glomerata 158
Datura 192
Dermatobia hominis 96
Diatomeae 180
Dtchromena 287
326 | Указатель латинских названий
Diplococcus 174
Diptera 289
Drosophila 41, 189, 255
— melanogaster 75, 157, 163, 169,
188
— miranda 192
— paulistorum 235, 255
— persimilis 157, 258
— pseudoobscura 152—154
— tropicalis 125, 156
— virilis 192
Hemerocallis fulva 85, 209
Hemiptera 289
Homo erectus 276
— sapiens 78, 89, 185, 186, 269
Homoplera 213, 289
Hydrozoa 84
Hymenoptera 289
Hypericum 192
Ideopsis daos 290, 291
Elapidae 286
Elephas maximus 273
Elodea 209
Emberiza citrinella 140
Empidonax 256
— oberholseri 256
— wrightii 256
Ensatina 233, 234, 235
Erebia calcarius 238, 261
— cassioides 237, 238, 261
— iranica 262
— nivalis 237, 238, 261
— tyndarus 237, 238, 260
Erithacus rubecula 140
Erodium 97
Escherichia coli 176
Euphorbiaceae 291
Euphydryas editha 86, 87, 89, 90, 91,
93
Festuca pratensis 203
Fringillidae 240
Lacerta saxicola 217
Latimeria 276
Lepidoptera 143, 289, 291
Linaceae 159
Linanthus 252, 268
— parryae 250—252
Linum 159
Lolium perenne 203
Loxodonta africana 271
Lumbricidae 217
Lymantria dispar 233
Maniola jurtina 143, 166
Megaceros 294
Metasequoia 276
Metasequoia glyptostroboides 271
Metazoa 180, 213
Micropterygidae 292
Mollienesia 218
— formosa 218, 219
— latipinna 218
Muscicapa striata 140
Geospiza conirostris 239, 242
— difficilis 239, 242
— fortis 239, 242
— fuliginosa 239, 240, 242
— magnirosiris 239, 242, 243, 245
— scandens 239, 242
Geospizinae 239
Gerris 37
Ginkgo 269, 272
— biloba 269, 271, 272
Ginkgoales 271
Gramineae 287
Haplochromis 235—237, 260
Helianthus 257
— annuus 257
«— bolanderi 257
Natrix 227, 264
— sipedon 148, 247
Nemobius fasciatus 199
Heophasia 271
— menapia 271
— terlooti 271
Nymphalis 274
— antiopa 274
— californica 275
— milberti 274
— polychloros 275
— urtica'e 275
— van-album 274
— xanthomelas 275
Oeneis chryxus 248
Oenothera 190—192, 194
Указатель латинских названий | 327
Oenothera biennis 191, 194
— erythrosepala 194
— hooker i 191, 194
— lamarckiana 194
Onagraceae 192
Opuntia 239
Orthoptera 289
Rana pipiens 231—233, 250
— sylvatica 232
Raphanobrassica 203
Rhoeo 190, 192
Rhynchocephalia 269
Rosaceae 208, 291
Rubus 216, 217
Paeonia 190, 192
Panaxia domintda 143
Papilio dardanus 159, 160, 263
— glaucus 227, 248
— laglaizei 291
Paramecium 177, 182
Pedicular is dudleyi 93
Pediculus 262
— humanis 246, 262
Pelecypoda 279—281
Phaeopliyceae 180
Phoenicurus phoenicurus 140
Pieridae 271
Pieris protodice 97
— rapae 97
Pinaroloxias inornata 240
Pinus attenuata 273
— linguiformis 273
— masoni 273
— muricata 273
— radiata 273
— remorata 273
Pipilo erythrophthalmus 256
— ocai 256
Pithecanthropus 276
Plantago erecta 89
Plasmodium 72
Platycnemis 72, 74
Platyspiza crassirostris 239, 241
Poa 212, 216, 217, 220
— pratensis 158
Polemoniaceae 284, 286
Pomoideae 208
Primula kewensis 203
Protozoa 180, 209
Psilochaena 216
Saccharomyces 84
Salmonella 175
— typhimurium 176
Satyrinae 143
Sciara 36, 189
— co prophila 199, 200
Septobasidium 85
Sequoia 271
Sequoiadendron 271
Shigella dysenteriae 176
Silene 159
Siphonales 180
Sitta europaea 140
— neumayer 244
— tephronota 245
Spergula 159
Sphenodon 269, 275
— punctatus 269
Sporozoa 181
Styx 269
— infernalis 269
Tetracoccus ilicifolius 93
Tetraneura ulnii 218
Thysanoptera 213
Treponema pallidum 99
Trifolium repens 158
Turd us ericetorum 140, 146
Ursus ameriqanus 275
— arctos 275
— horribilis 275
— maritimus 275
— middendorffi 275
Rana 233
— catesbeiana 232
Viola 210
Viperidae 286
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию......................................5
Предисловие ............................................... . . . 7
Часть первая
ОРГАНИЗМЫ, ИХ ПРОИСХОЖДЕНИЕ
И ФУНКЦИИ
Глава 1. Происхождение жизни......................................11
Ранние этапы эволюции.........................................12
Возникновение самовоспроизводящихся систем....................14
Источники энергии .... I5
Возникновение структуры ... 19
Возникновение генетического кода..............................20
Считывание кода...............................................23
Выводы .......................................................28
Литература .................................................28
Глава 2. Единицы репликации.......................................30
Строение клеток............................................. 31
Деление клетки — митоз...................................... 34
Деление клетки — мейоз...................................... 39
Выводы .......................................................43
Литература .................................................44
Глава 3. Генетика ................................................45
Изменчивость и менделевская генетика .........................45
Единицы наследственности......................................46
Законы Менделя................................................48
Рекомбинация ............................................... 49
Действие генов........................................... .... 52
Мутации .................................................... 53
Эволюция доминантности........................................55
Хромосомные механизмы.................................... .... 56
Непрерывная изменчивость.................................... 62
Выводы .... .............................. . . 65
Литература .................................................66
Оглавление | 329
Г лава 4. Развитие...............................................67
Рост и гомеостаз..............................................67
Жизненные циклы...............................................71
Дифференцировка и морфогенез.................................72
Эволюционное изменение процессов онтогенеза...................77
Выводы ..................................................... 79
Литература . , » ...................................... 79
Часть вторая
СВОЙСТВА ПОПУЛЯЦИЙ
Глава 5. Популяции.................................................83
Особи и колонии................................................84
Распределение в пространстве ................................. 86
Экологическое распределение .................................. 89
Структура популяций .......................................... 90
Число особей...................................................93
Внешняя среда..................................................94
Сообщества ....................................................98
Выводы .......................................................100
Литература .................................................101
Глава 6. Теория популяционной генетики...........................101
Менделевские популяции .................;....................100
Величина популяции ......................................... 105
Дотационный процесс..........................................112
Отбор .......................................................113
Миграция и структура популяций . . ..........................126
Совместное действие факторов эволюции' ‘.....................129
Адаптация и генные сочетания.................................132
Выводы ......................................................135
Литература ................................................135
Глава 7. Изменения в популяциях..................................138
Примеры из природы...........................................138
Лабораторные популяции ..................................... 163
Генетический гомеостаз . 166
Генетическая ассимиляция.....................................167
Приспособление к среде ......................................170
Выводы ......................................................171
Литература ................................................172
330 | Оглавление
Глава 8. Генетические системы I................................. 173
Генетические системы у микроорганизмов........................174
Генетические системы других организмов........................178
Выводы ... .... .................... 186
Г лава 9. Генетические системы II.................................187
Мейотический дрейф............................................188
Изменения в структуре хромосом................................188
Полиплоидия ..................................................196
Апомиксис.....................................................209
Выводы .......................................................220
Литература ................................................ . 221
Часть третья
ДИФФЕРЕНЦИАЦИЯ ПОПУЛЯЦИЙ
Г лава 10. Дифференциация популяций............................ 226
Примеры дифференциации......................................225
Обсуждение наблюдаемых типов дифференциации.................246
Выводы ................................................... 266
Литература ................................................268
Г лава 11. Высшие уровни изменчивости...........................267
Вымирание и биогеографическая обособленность................268
Палеонтологическая летопись . . 276
Основные пути эволюции ............... . .................283
Высшие категории............................................291
Эволюционные тенденции......................................293
Выводы .....................................................296
Литература ................................................296
Словарь терминов ...............................................297
Предметный указатель........................... . 315
Указатель латинских названий ...................................325
ИЗДАТЕЛЬСТВО „МИР"
Выходят в свет в 1967 г.
Мюнтцинг А. Генетика. Общая и прикладная (2-е издание). Перевод
с английского, 35 изд. л.
Учебник генетики, написанный одним из крупнейших современных
генетиков — шведским ученым А. Мюнтципгом, широко известным сво-
ими исследованиями в области теоретической и прикладной генетики.
Книга эта хорошо знакома советскому читателю по предыдущему
изданию, выпущенному на русском языке Издательством иностранной
литературы в 1963 г. под названием «Генетические исследования».
В ней систематически изложены все разделы генетики: материальные
основы наследственности, законы Менделя, наследование количествен-
ных признаков, кроссинговер, мутационный процесс и различные типы
мутаций, физиологическая и биохимическая генетика, основы молеку-
лярной генетики, природа гена, различные типы гибридов, полиплоидия,
селекция животных и растений, некоторые сведения по генетике чело-
века. К книге приложен словарь основных генетических терминов.
В новом издании значительно дополнены главы по молекулярной и
биохимической генетике; внесены также последние данные науки и
в некоторые другие главы.
Предназначена для студентов и преподавателей биологических фа-
культетов университетов, педагогических, медицинских и сельскохозяй-
ственных институтов, для преподавателей средней школы и техникумов,
для научных работников различных специальностей (биологов, меди-
ков и т. д.).
Кастлер Г. Возникновение биологической организации. Перевод
с английского, 5 изд. л.
Имя Г. Кастлера хорошо известно всем ученым, интересующимся
вопросами применения теории информации в биологии. В этой книге
с исключительной четкостью рассмотрены весьма тонкие вопросы об
информационном содержании биологических систем, о путях спонтан-
ного возникновения сложных организаций, способных к созданию, на-
коплению и передаче информации, о конкретных механизмах функцио-
нирования ферментов и генетического аппарата с информационной
точки зрения. Хотя изложение всегда строго и книга не может быть
отнесена к числу научно-популярных, чтение ее требует знания мате-
матики в объеме, не превышающем курса средней школы (если не
считать самых элементарных сведений по теории информации). Моно-
графия Кастлера, безусловно, привлечет внимание широких кругов био-
логов, математиков и физиков. Нет сомнения, что она найдет чита-
телей и среди неспециалистов, учащихся старших классов, студентов и
просто лиц, интересующихся одним из самых увлекательных вопросов
современной науки.
П» Эрлих Р. Холм
ПРОЦЕСС ЭВОЛЮЦИИ
Редактор Е. Э. Казакевич
Художник А. Д. Смеляков
Художественный редактор Ю. Л. Максимов
Технический редактор А. Г. Резоухова
Корректор В. П. Горячева
Сдано в производство 6/1V 1966 г.
Подписано к печати 9/IX 1966 г.
Бумага 60x901/1G= 10,38 бум. л.
20,75 печ. л.
Уч.-изд. л. 20,09. Изд. № 4/3359
Цена 1 р. 63 к. Зак. 152
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2
Ленинградская типография № 2
имени Евгении Соколовой Главполиграфпрома
Комитета по печати при Совете
Министров СССР. Измайловский проспект, 29.