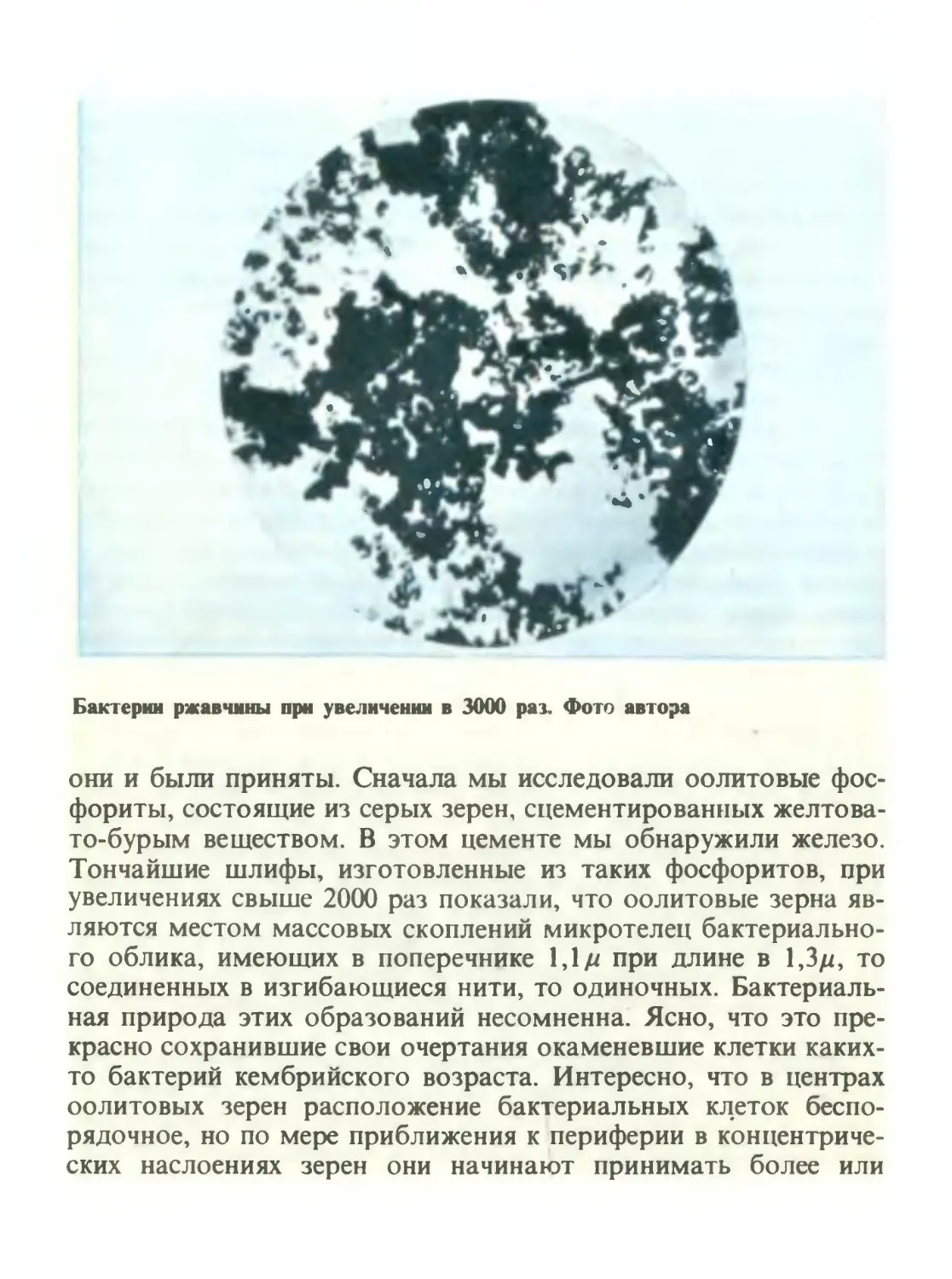
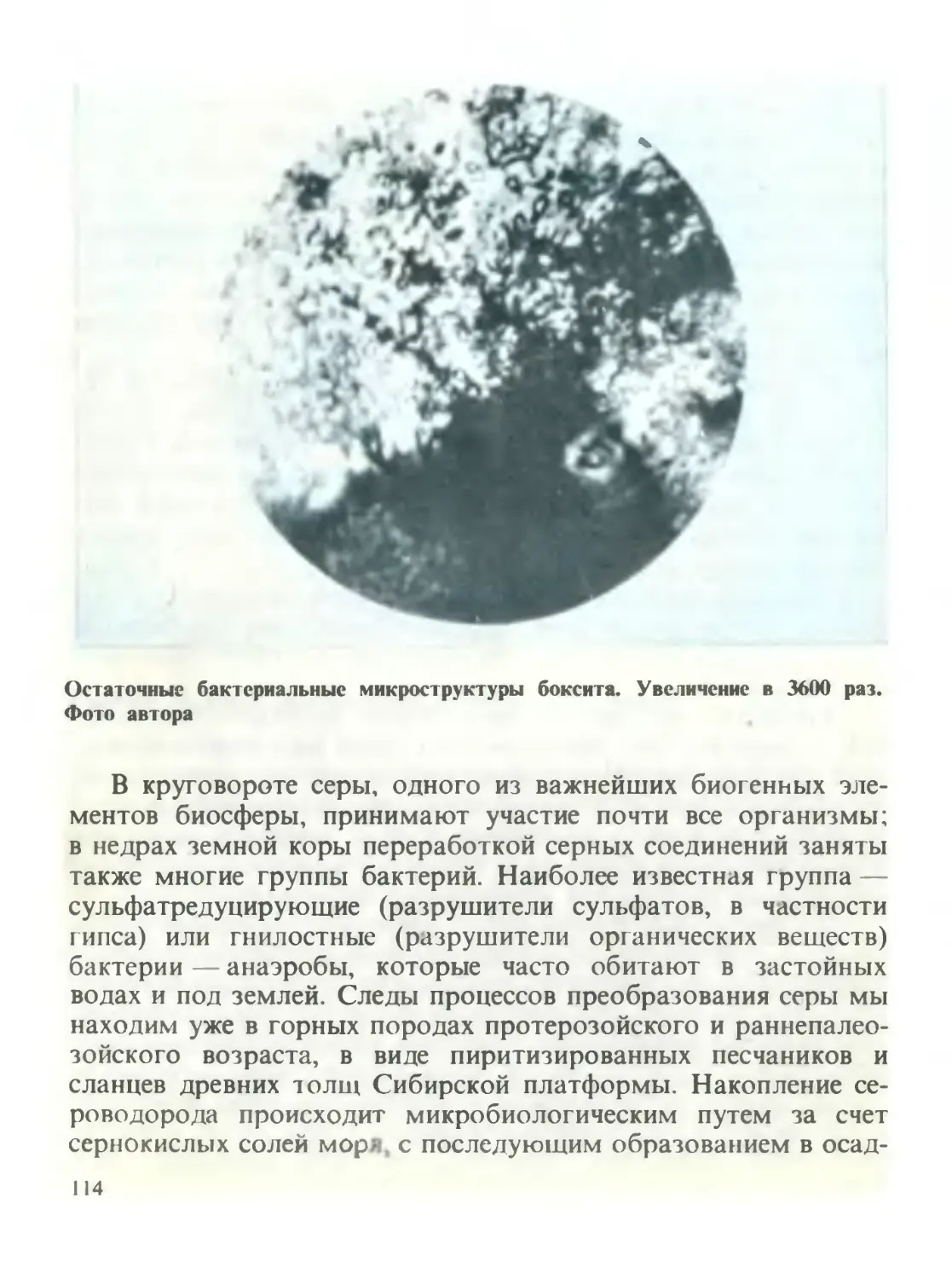
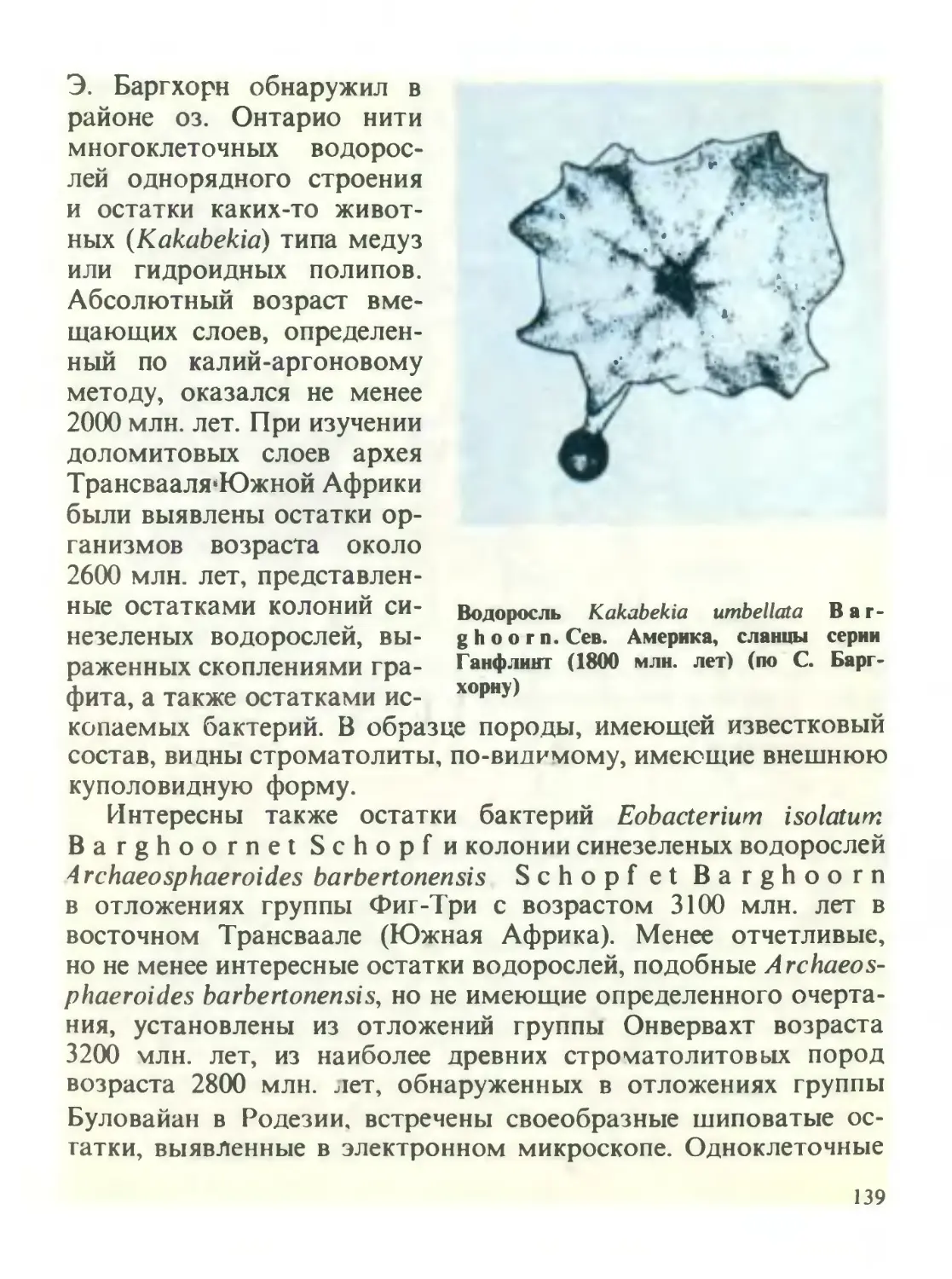
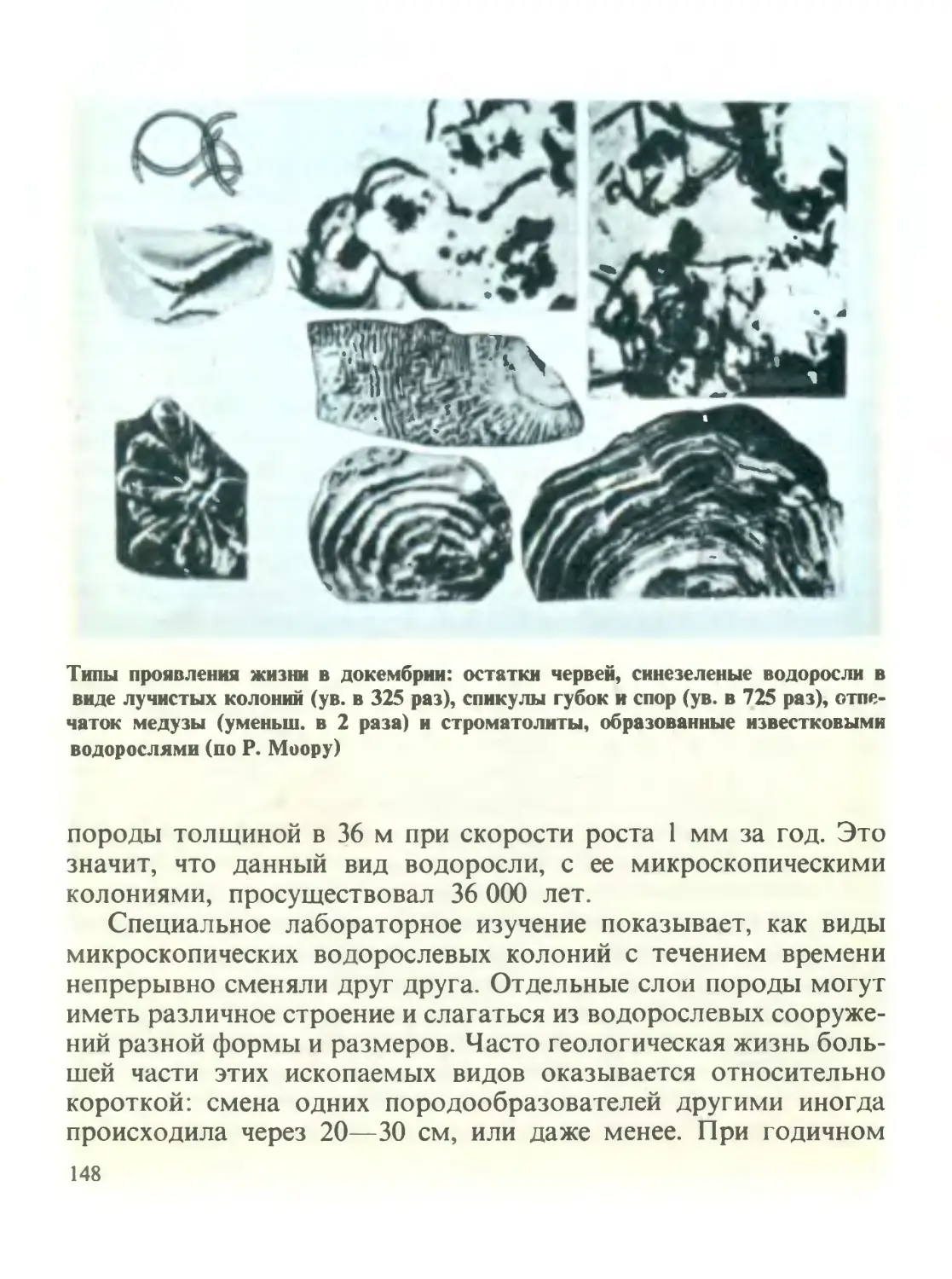
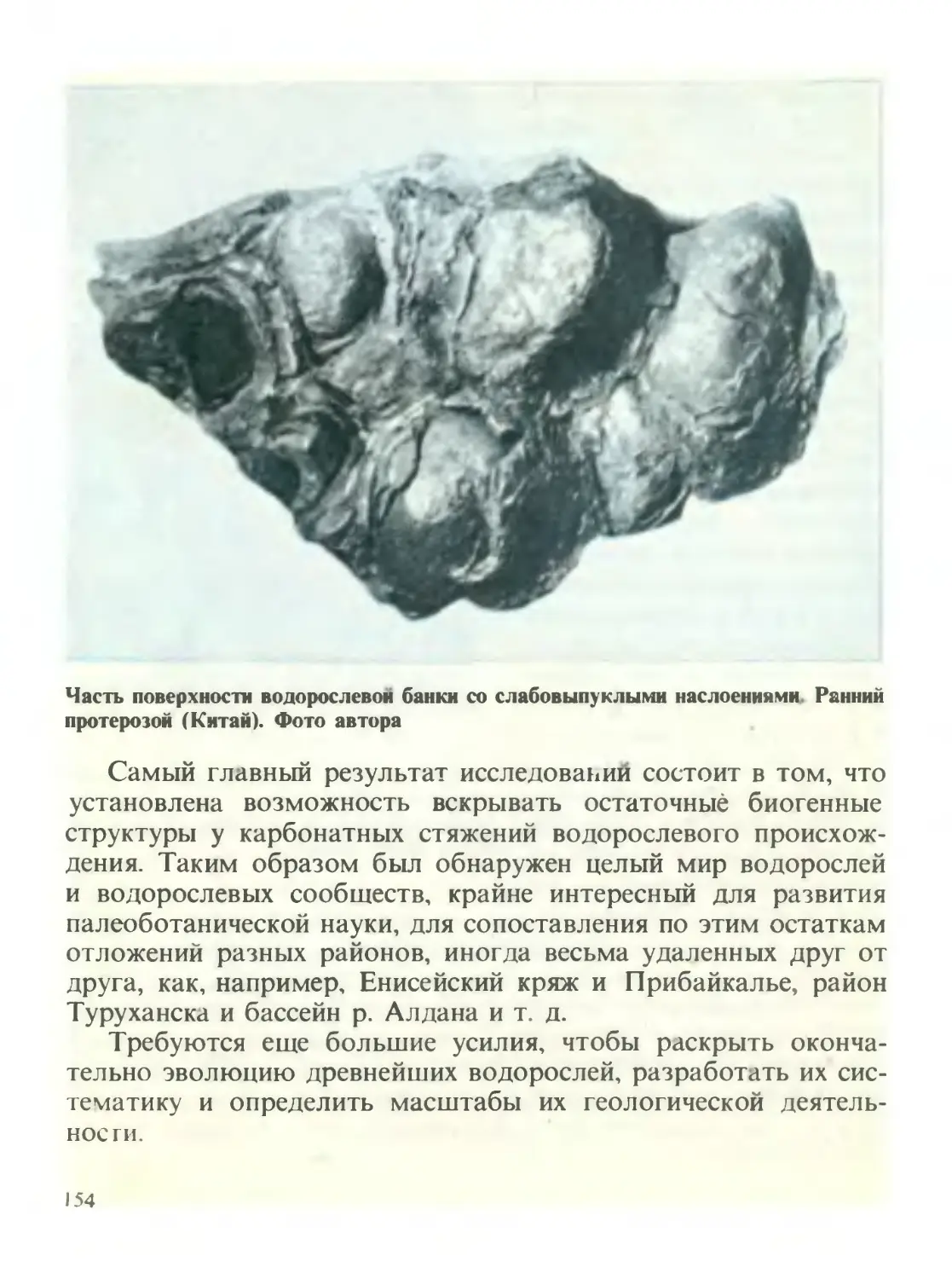
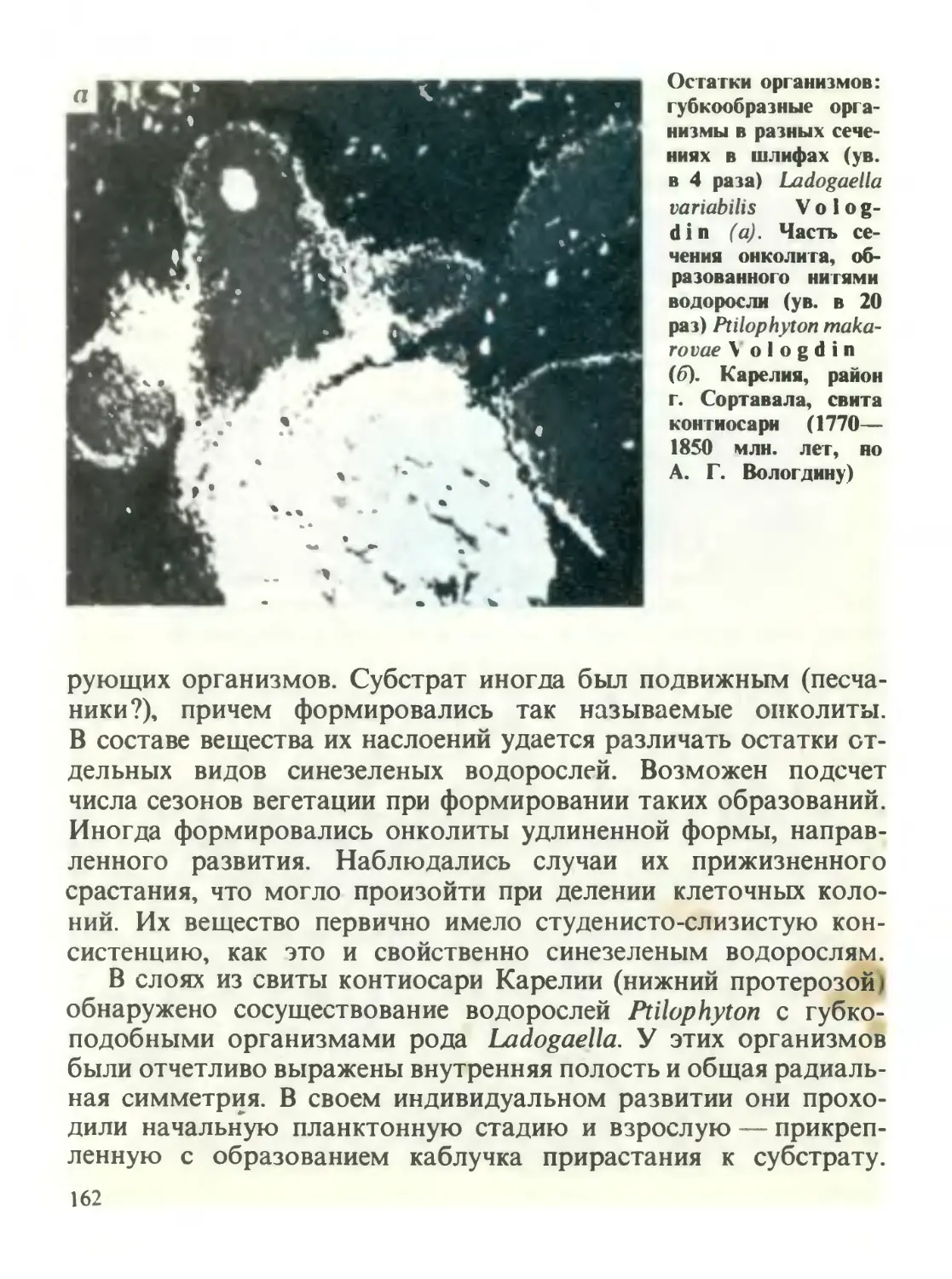
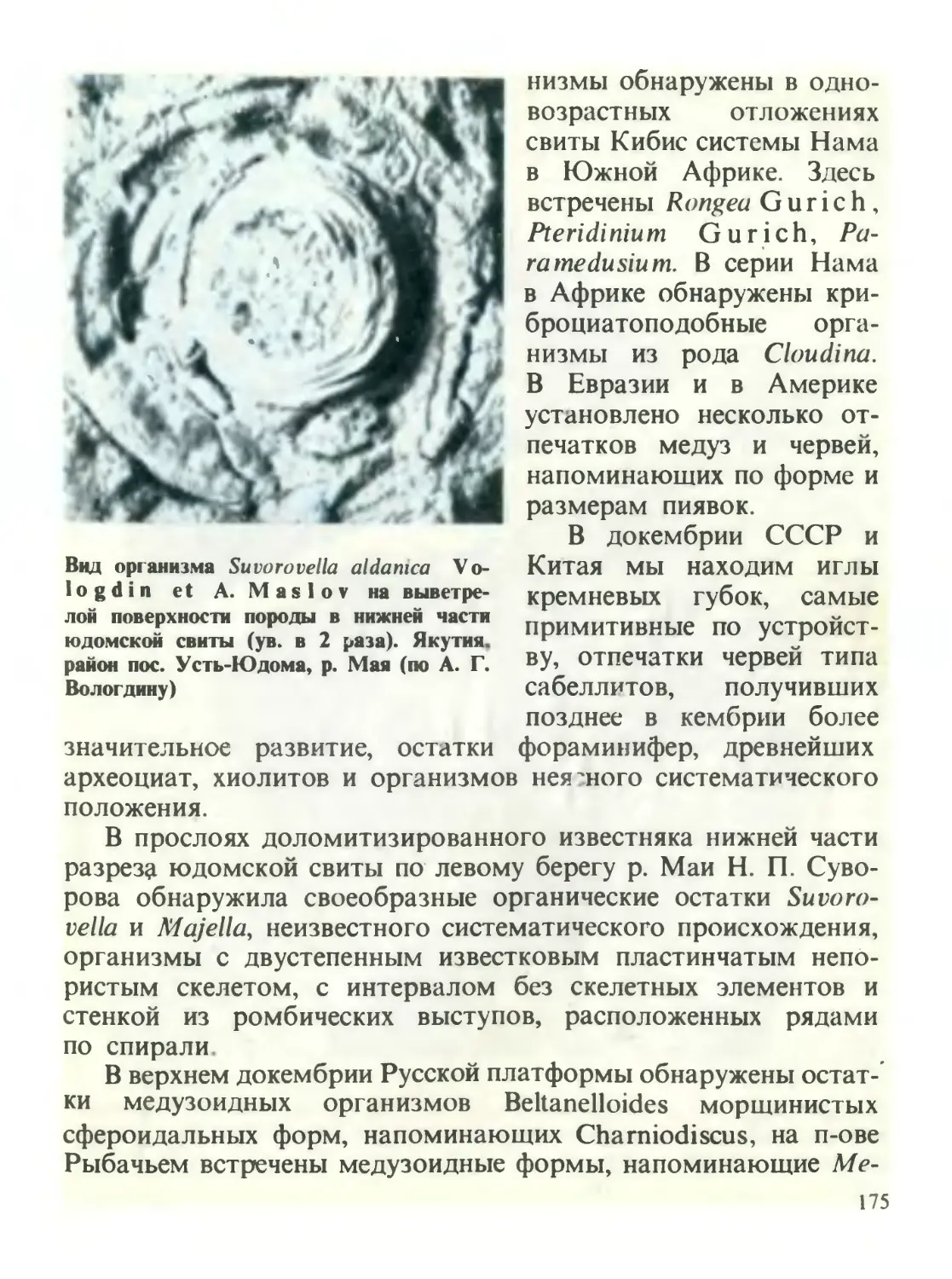
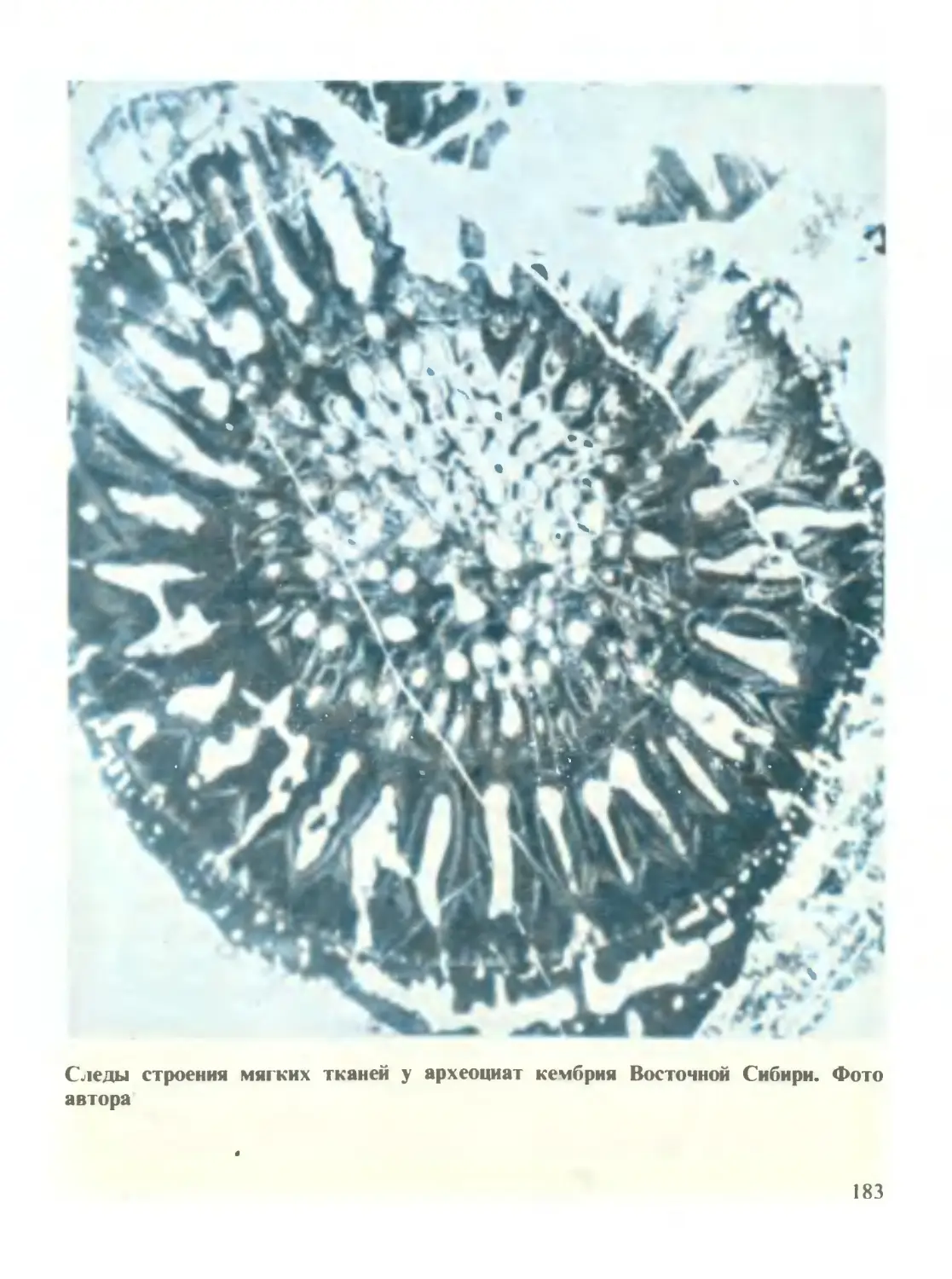
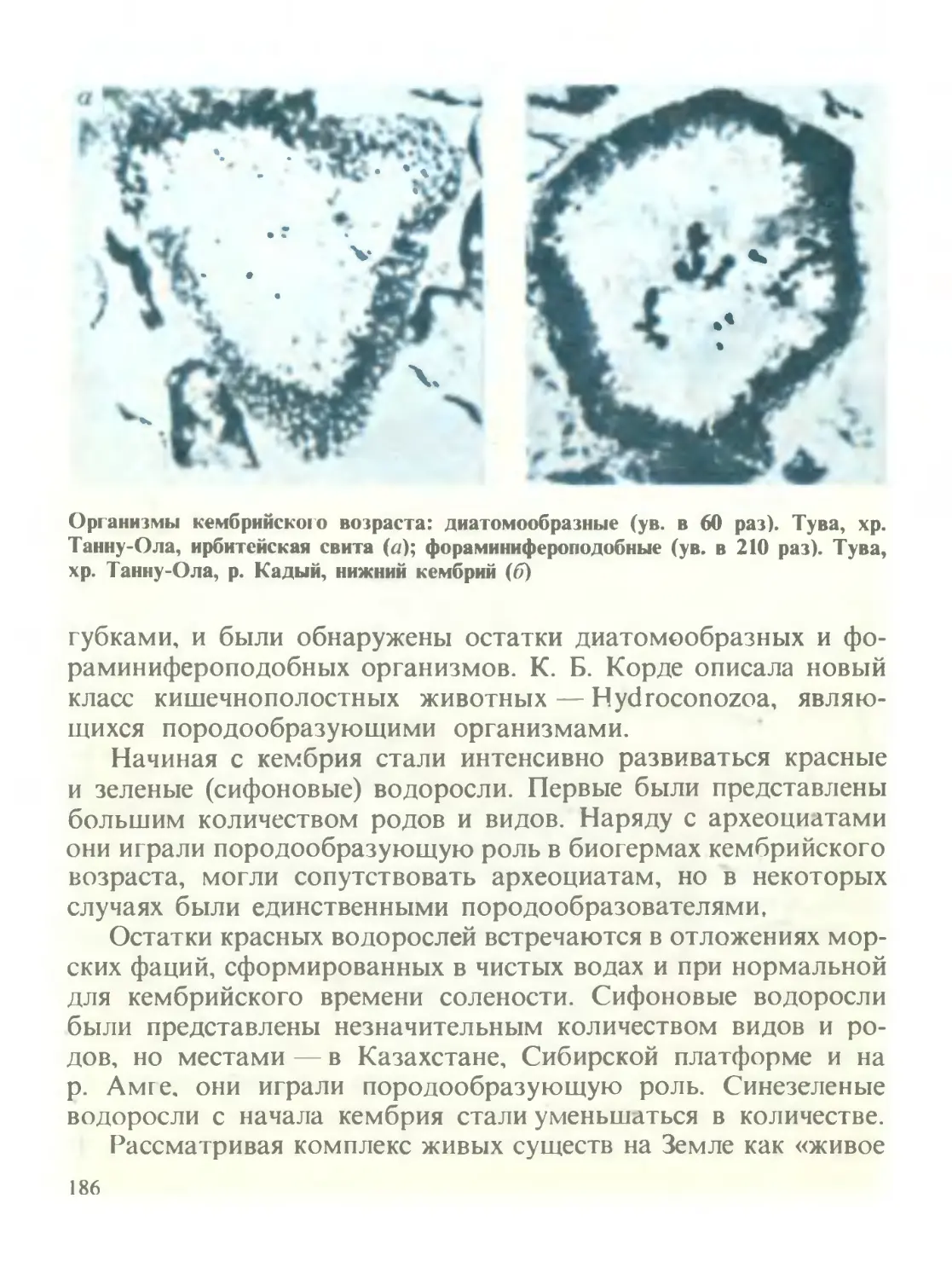
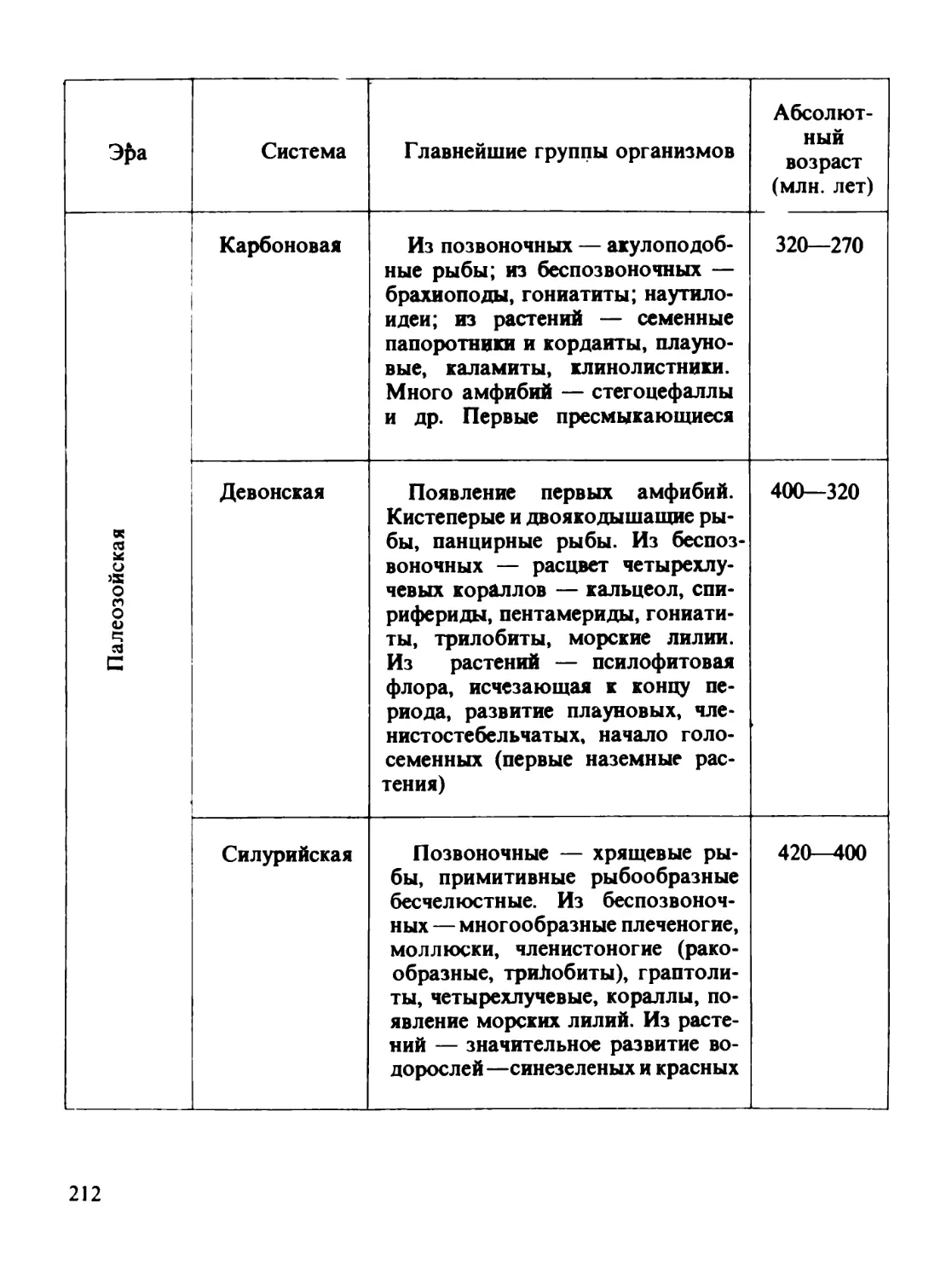
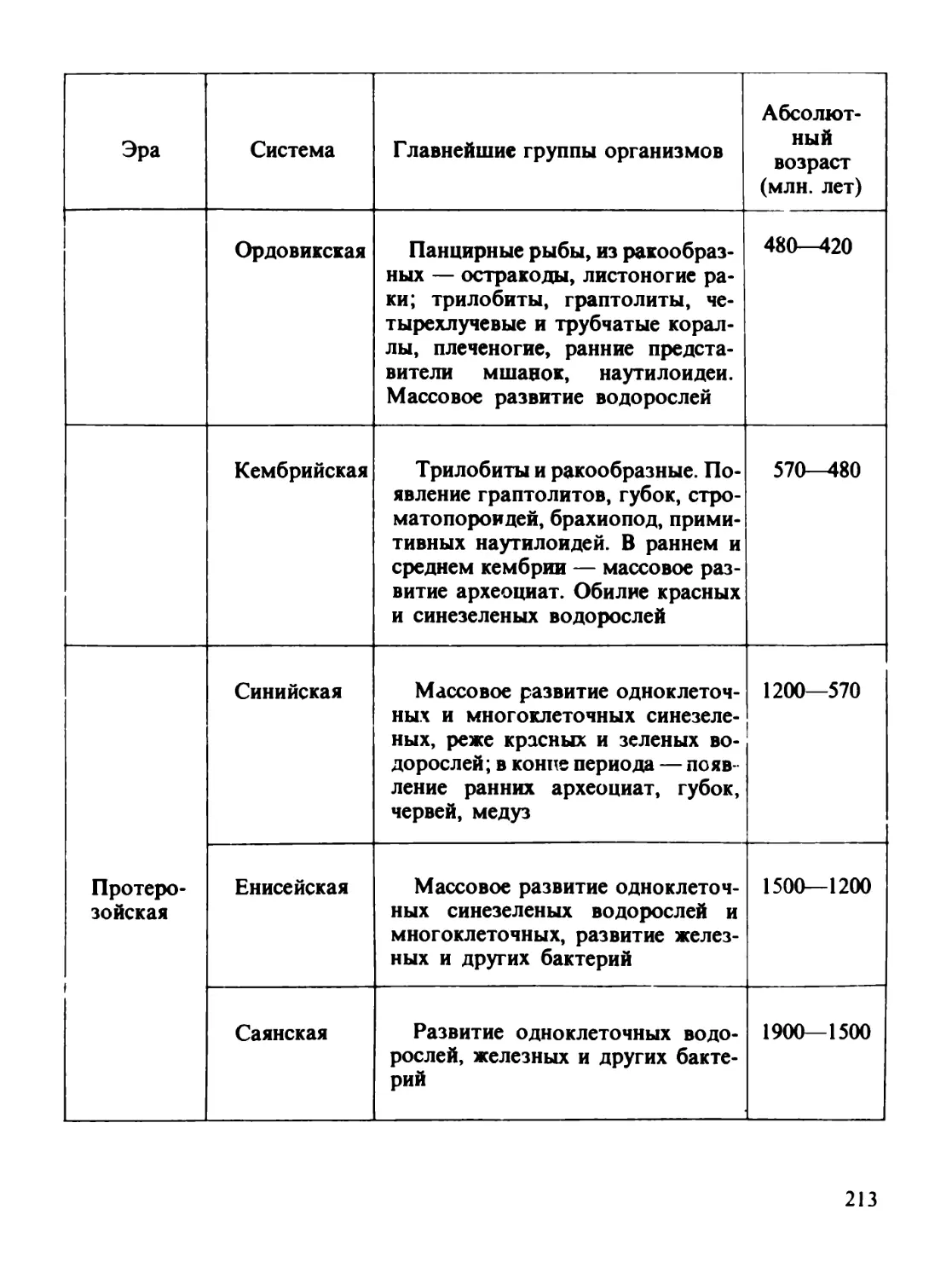
/
































































































































































































































Author: Вологдин А.Г.
Tags: науки о земле геологические науки эволюция история земли жизнь древние организмы
Year: 1976




Text
Skaning, Djvuing Lykas
А. Г. ВОЛОГДИН
ЗЕМЛЯ И ЖИЗНЬ
Издание второе, переработанное
и дополненное
МОСКВА . «НЕДРА» . 1976
УДК 55(20)
20801—195
В----------73—76
043(01)—76
© Издательство «Недра», 1976
От редактора
Книга А. Г Вологдина посвящена одной из интереснейших проблем науки —
изучению древнейших этапов истории Земли и жизни. В ней изложены современ-
ные данные по этой проблеме, особенно о древних организмах и вероятных
путях их эволюции в связи с изменением среды обитания, химического состава
и лика первобытной Земли.
К настоящему времени сравнительно хорошо изучен и широко известен
лишь послекембрнйский этап истории Земли и жизни — фанерозой, включаю-
щий палеозойскую, мезозойскую и кайнозойскую эры продолжительностью
около 600 млн. лет. Древнейший и значительно более продолжительный этап
существования нашей планеты, так называемый докембрий (криптозой), в те-
чение которого протекала ранняя эволюция жизни, был почти неизвестен.
И хотя в настоящее время в проблеме изучения этого древнейшего из этапов
истории Земли и жизни еще много неразгаданного, неясного - быстрое разви-
тие науки: накопление новых данных в области космогонии, биогеохимии,
палеонтологии, абсолютной геохронологии и в различных областях геологии
и биологии — позволяет более глубоко заглянуть в это волнующее и во многом
загадочное прошлое нашей планеты.
Палеонтологам и геологам удалось проникнуть в глубь веков по меньшей
мере на 2,5 - 3,0 млрд, лет назад. Возникла новая отрасль знания палеонто-
логия докембрия, — изучающая древнейшие проявления жизни на нашей пла-
нете, которая позволяет на основе историко-геологических документов «рестав-
рировать», изучать докембрийские этапы эволюции жизни на Земле. Об этом
в популярной форме в основном и рассказывает книга А. Г Вологдина, раскры-
вая древнейшие этапы истории Земли и жизни.
Александр Григорьевич Вологдин, крупный советский палеонтолог и гео-
лог, всю свою полувековую научную деятельность посвятил изучению водо-
рослей, примитивных многоклеточных животных, - археоциат, а также древ-
нейших отложений кембрия и докембрия СССР и других стран. Среди печатных
трудов А. Г Вологдина есть работы по региональной геологии и полезным ис-
копаемым, палеонтологии и стратиграфии. Однако подавляющее число тру-
дов по палеонтологии посвящено археоциатам, водорослям. А. Г Вологдину
принадлежит большая роль в распространении палеонтологического метода
5
в изучении древнейшей истории Земли, который он широко и горячо пропаган-
дировал.
А. Г Вологдин родился 11 марта 1896 г в селе Рождественском б. Пермской
губернии в семье учителя. В 1920 г он студент Петроградского Горного инсти-
тута и одновременно работает коллектором в Геологическом комитете* Окон-
чив в 1925 г. Горный институт, он возглавил работу Сибирской секции Всесоюз-
ного геологического института, где работал до 1941 г. В этот период А. Г Во-
логдиным были сделаны важнейшие палеонтологические открытия в области
изучения древних водорослей, археоциат и стратиграфические выводы, которые
легли в основу современной биостратиграфии и изучения геологии кембрия
Сибири, а также Монголии, Китая и других районов Евразии. Начиная с 1939 г
особое внимание А. Г Вологдин уделял органическим остаткам докембрия,
а также внедрению палеонтологического метода в изучение древнейшей исто-
рии Земли.
С 1941 г. работы по изучению археоциат и водорослей велись А. Г Волог-
диным в Палеонтологическом институте Академии наук СССР, куда он был
приглашен академиком А. А. Борисяком. В 1943 г. А. Г Вологдин организовал
в этом институте лабораторию по изучению древнейших организмов, которой
руководил до конца своей жизни.
За плодотворную научную деятельность А. Г Вологдину была присуждена
ученая степень доктора геолого-минералогических наук (1937 г ), и он бы;
избран член-корреспондентом Академии наук СССР (1939 г.). Правительство
высоко оценило научные заслуги А. Г Вологдина он был награжден Орле-
ном Трудового Красного Знамени и медалями.
Большое количество научных работ в области палеонтологии древних орга-
низмов принесли А. Г Вологдину мировую известность. По решению между-
народного жюри палеонтологов в 1945 г. А. Г Вологдин был удостоен Между-
народной палеонтологической премии и медали Чарлза Уолкотта «За изучение
организмов докембрия».
А. Г Вологдин был талантливым и неутомимым популяризатором науки
о Земле и жизни. Обладая живым умом и прекрасной памятью, он был замеча-
тельным лектором и увлекательным рассказчиком. Долгое время он являлся
заместителем председателя Методического совета по пропаганде наук о Земле
и членом правления общества «Знание» РСФСР.
А. Г Вологдин прожил долгую, насыщенную деятельностью жизнь уче-
ного, путешественника, популяризатора науки.
6
Второе издание книги Александра Григорьевича Вологдина переработано
и дополнено по сравнению с первым изданием (1963 г.) в значительной степени
самим автором.
Новые сведения по отдельным разделам работы были внесены сотрудни-
ками Палеонтологического института АН СССР — одной из ближайших уче-
ниц А. Г Вологдина доктором геолого-минералогических наук К. Б. Кордэ
и кандидатом биологических наук Б. А. Трофимовым, а также сотрудником
Института Физики Земли АН СССР кандидатом физико-математических наук
С. В. Маевой. При подготовке рукописи к изданию оказала помощь Г А. Во-
логдина.
Книга рассчитана на широкий круг читателей и, в известной мере, воспол-
нит пробел, который существует в научно-популярной литературе по этой
увлекательной проблеме.
Б. А. Трофимов
Светлой памяти
Владимира Ивановича
Вернадского
посвящает свой труд
автор
ПРЕДИСЛОВИЕ
Развитие человеческого знания, быстрый рост науки, теснейшим
образом связанный с великими победами социализма в Совет-
ской стране, ежедневно и ежечасно ускоряют научно-техниче-
ский прогресс. Научные идеи, рожденные практикой, и могучая
современная техника создают немыслимые ранее возможности
познания огромных пространств Вселенной, куда отправляются
космические корабли и где побывал уже человек. В глубинах
морей работают научно-исследовательские станции, которые
изучают сложную и богатую жизнь морского дна. Развитие
атомной физики ведет нас к познанию типов и свойств мельчай-
ших частиц и проявлений огромных энергий, которым в будущем
надлежит обеспечить человечество самыми совершенными ви-
дами топлива, неисчерпаемыми источниками энергии. Бурно
развивается химия и химическая промышленность, создающие
новые вещества с новыми свойствами. Больших успехов достиг-
ла и биологическая наука, способствующая подъему сельского
хозяйства — земледелия и животноводства, развитию совет-
ской и мировой медицины.
В этой небольшой книге мы попробуем обратить внимание
читателя в глубь геологических времен, на те этапы развития
нашей планеты и жизни на ней, которые еще недавно рисовались
очень смутно.
(Палеонтологическая наука, занимающаяся изучением орга-
низмов геологического прошлого Земли, историей их появления,
развития и расселения в морях и на суше, раскрывает перед нами
картину жизни планеты в основном только за последние пол-
миллиарда лет, а именно с начала кембрийского периода,
длившегося по современным данным от 440 до 570 млн. лет.
Уже в начале этого периода существовали морские водоросли,
некоторые простейшие брахиоподы, черви, древнейшие рако-
образные — трилобиты, губки, ранние головоногие и т. д.
Но еще раньше, во время так называемого докембрия — этого
9
огромного по продолжительности этапа развития нашей пла-
неты, закончившегося 570 млн. лет назад, — возникли архео-
циаты, беспозвоночные животные, замечательные строители
древнейших рифов. Особенно пышное развитие они получили
в ранне- и среднекембрийское время.
Что же представляла собой органическая жизнь на Земле
570 млн. лет назад? Как и в каких условиях зародилась и разви-
валась она задолго до этого рубежа?
I Для объяснения происхождения жизни на нашей планете
мы имеем весьма интересную гипотезу, предложенную акаде-
миком А. И. Опариным. Эта гипотеза, использующая резуль-
таты исследований геологов, геохимиков и астрономов, раскры-
вает первые страницы истории нашей планеты — истории, на-
считывающей по меньшей мере 6—7 млрд. лет. Ультрафиоле-
товое Излучение Солнца и грозовые явления в первичной атмо-
сфере и парах Земли, обильно содержавших метан (СН4), угле-
кислоту, аммиак (NH3), водород и другие элементы, при опре-
деленных температурах и давлениях должны были приводить
к массовому образованию сложных углеродных соединений,
в том числе белков. Поэтому момент возникновения жизни на
Земле ученые и относят к самым ранним этапам формирования
земной коры, примерно около 3 — 3,5 млрд, лет назад или даже
ранее.
Проблема происхождения жизни на Земле и ее эволюции —
это прежде всего проблема происхождения веществ определен-
ного состава, в первую очередь белков, и их изменения в ходе
геологического времени. С другой стороны, жизнь — это особая
сфера выявления и использования энергии, поступающей извне
или возникающей внутри организмов, и, таким образом, жизнь
есть форма энергетического состояния, зависящего от энерге-
тического и вещественного состава окружающей среды.
Происхождение и развитие жизни на Земле — это одна из
важнейших проблем для целого ряда наук, каждая из которых
помогает выяснить ту или иную сторону проблемы. Это прежде
всего биогеохимия, основоположником которой был академик
В. И. Вернадский (1863 — 1945). Она раскрывает связи между
ю
Владимир Иванович Вернадский (1863—1945)
живым и минеральным — косным веществом Земли в биосфере,
что важно для разработки проблемы возникновения и развития
жизни.
Геохимия, создателями которой были также академики
В. И. Вернадский и А. Е. Ферсман (1883 — 1946), позволяет по-
нять роль химических элементов и их изотопов в образовании
минеральных и органических соединений, а это раскрывает
также и особенности геохимических процессов далекого геоло-
гического прошлого. Кроме того, геохимия дает замечательный
метод определения абсолютного геологического возраста ми-
неральных и органических образований, а стало быть, и меру
геологического времени для различных процессов, имевших
место в земной коре или на ее поверхности.
Палеонтология дает нам конкретные указания о составе
организмов отдаленных геологических периодов, эволюция
которых привела к появлению бактерий и современного мира
растений и животных на Земле.
Микробиология (бактериология, вирусология) дает замеча-
тельные материалы по морфологии и физиологии мельчайших
живых существ, среди которых насчитывается немало родст-
венных, давно вымерших форм.
Астрономия и астрофизика, изучая состав и строение косми-
ческих тел, раскрывают особенности протекающих физико-
химических процессов, исследуют состав и строение Солнца
и других планет.
Биохимия позволяет составить представление о том, как
первичные химические компоненты Земли преобразовались в
органические, сначала неживые, а затем в живые вещества,
а также поясняет сущность процессов, протекающих внутри
организма. Современная экспериментальная биохимия уже мо-
жет из простых углеродных соединений создавать сложные,
в том числе и основные составные части белка — аминокисло-
ты, а из комбинаций аминокислот — и белок (правда, еще не-
живой).
Так, сотрудничество целого ряда наук позволяет проследить
развитие организмов на Земле в процессе ее эволюции под влия-
12
нием эволюции среды. Благодаря достижениям этих наук, изла-
гаемые здесь взгляды постепенно превращались из гипотез в
научные теории.
Предлагаемая вниманию читателя книга посвящена рас-
смотрению великой и прочной связи жизни с Землей, со средой
ее развития, связи жизни и минерального, косного вещества
земной коры. Мы попытаемся проследить, как изменялись
формы жизни в результате изменения среды ее обитания, пока,
наконец, после длительной эволюции жизнь не приобрела те
формы и связи, которые мы можем наблюдать ныне.
Наука, как и жизнь, раз зародившись, развивается и непре-
рывно решает одну задачу за другой. Открывая новые и новые
факты, она все полнее и точнее раскрывает тайны природы.
Не было большей загадки, чем происхождение жизни на Земле.
Поэтому этой проблеме посвящаются многочисленные труды
различных специалистов. Попыткой обобщить эти труды и явля-
ется предлагаемая книга.
Автор выражает глубокую признательность академику
В. Г Фесенкову, члену-корреспонденту АН СССР С. И. Кузне-
цову и профессору А. А. Яковлеву за ценные указания при под-
готовке книги.
ЗЕМЛЯ В СОЛНЕЧНОЙ системе
Благодаря успехам астрономии, астрофизики и ядерной физики,
развитию техники и приборостроения пытливый ум человека
раскрывает все новые и новые детали строения и состава Вселен-
ной. Развитие наук наряду с успехами техники, развитием мето-
дики познания окружающего нас мира продвинулось весьма
значительно. Решаются многие вопросы, прежде признававшие-
ся очень сложными или даже неразрешимыми.
Мы знаем сейчас, что наша Солнечная система изолиррвана
от других групп космических тел. Если представить, что удален-
нейшая от Солнца планета Плутон отстоит от него на расстоя-
нии 40 см, тогда радиус орбиты Земли составил бы всего 4 см,
а ближайшая к нам звезда Альфа Центавра оказалась бы на
расстоянии 2,7 км. Астрономическая наука раскрыла свойства
огромного количества космических тел, и все это солнца-звезды.
Открытие даже крайних планет Солнечной системы было труд-
ным делом, поскольку они светятся лишь отраженным светом.
Поэтому можно полагать, что отстоящие друг от друга на ог-
ромных расстояниях другие системы космических тел содержат
не только звезды с их разнотипными ядерными реакциями, но и
планеты — относительно мелкие тела, подобные планетам Сол-
нечной системы. Из них астрономы могут обнаружить лишь
тела, обладающие собственным светом.
Солнечная система, включающая 9 больших планет, 32 их
спутника, более 1500 астероидов, или так называемых «малых
планет», около 1000 комет и бесчисленное множество мелких
космических тел, — летит в космическом пространстве со ско-
ростью 19 км/с. Все тела, входящие в Солнечную систему, дви-
жутся с различными скоростями по своим орбитам, подчиняясь
влиянию Солнца и друг друга.
Солнечная система к настоящему времени исследована до-
статочно хорошо. Изучение ее продолжается и приносит все
новые результаты.
14
Краткие сведения о Солнце и планетах приведены в следую-
щей таблице.
Небесное тело Среднее расстояние от Солнца в едини-1 цах расстояния от Солнца до Земли Масса (масса Земли -1) ч 1 Средний диаметр, । км Плот- ность, г/см3
Солнце 333 000 1392 000 1,41
Меркурий 0,39 0,0553 4870 5,45
Венера 0,72 0,815 12400 5,25
Земля 1,00 1,00 12 700 5,52
Марс 1,52 0,107 6780 3,94
Юпитер 5,20 317,8 139400 1,34
Сатурн 9,54 95,27 115 800 0,70
Уран 19,19 14,59 49080 1,41
Нептун 30,07 17,29 50 000 1,58
Плутон 39,52 ? ? ?
Масса Солнца в 744 раза больше массы всех планет вместе
взятых. Солнце более чем на 70% по массе состоит из водорода
и на 28% — из гелия. На все остальные элементы приходится
менее 2%. Все планеты Солнечной системы по физическим свой-
ствам и химическому составу делятся на 2 группы. Планеты
земной группы — Меркурий, Венера, Земля и Марс облада-
ют значительной плотностью и состоят в основном из силика-
тов и металлов. Планеты-гиганты — Юпитер, Сатурн, Уран
и Нептун — обладают малой средней плотностью, имеют мощ-
ные атмосферы и состоят в основном из легких летучих элемен-
тов. Самая далекая от Солнца планета Плутон, открытая в
1930 г., еще мало изучена.
ПРОИСХОЖДЕНИЕ ЗЕМЛИ
Происхождение Солнечной системы долгое время было предм*
том научных дискуссий. Было выдвинуто несколько гипоте
по-разному объяснявших ее возникновение. Очень долго в кос
могонии господствовало представление о конденсации плане
из раскаленных сгустков солнечных газов. Однако сейчас тверд
установлено, что наша планетная система образовалась и
холодного газово-пылевого облака, которое некогда существо
вало вокруг Солнца. Вопрос о происхождении этого допланет]
ного облака пока еще не решен, но наиболее вероятно, что оно
образовалось совместно с Солнцем. В настоящее время уста|
новлено, что атомы вещества, из которого «построены» СолнЛ
и планеты, образовались при взрывах сверхновых звезд, котор Л
происходили в нашей Галактике в течение, по крайней мерЙ
10 млрд. лет. Последняя доля из продуктов такого взрыва
была получена веществом Солнечной системы от 30 до 300 млн^|
лет назад.
Протопланетное облако, из которого образовалась Земля и
другие планеты, первоначально, к тому времени, когда Солнце
уже сформировалось, имело температуру около 2000 С и нахо-
дилось в газообразном состоянии. На 98% оно состояло из
водорода и гелия, а 2% приходилось на долю всех остальных
элементов, которые при охлаждении облака сконденсировались
в пылевые частицы. При температурах 1800- 1900° С сгущались
самые нелетучие вещества. При температуре 1600°С конденси-
ровалось железо. Благодаря присутствию водорода оно выде-
лялось в металлическом виде. При остывании облака до 1400° С
затвердевали силикаты. При температуре 600° С образовыва-
лось сернистое железо.
Процесс объединения образовавшихся при охлаждении
облака пылевых частиц в планеты был рассмотрен акад.
О. Ю. Шмидтом. При этом О. Ю. Шмидт дал объяснение основ-
ных закономерностей планетной системы. Космогонист
16
пиральная туманность нз созвездия Гончих Псов
В. С. Сафронов, продолживший разработку теории О. Ю. Щмид-
ia изучив законы объединения и дробления в протопланетном
облаке, вычислил время роста Земли. Земля приобрела 98% сво-
ей массы за 100 млн. лет. В различных частях протопланетного
облака температура была неодинаковой. В областях далеких
от Солнца она была очень низкой, й все вещества, кроме водо-
рода и гелия, конденсировались в частицы. Среди них преоб-
ладали ледяные частицы метана, аммиака, воды, которые
вошли в состав планет-гигантов. В области планет земной груп-
пы температура была значительно выше, и все летучие вещества
(в том числе метан и аммиак) остались в газовом состоянии,
а образовавшиеся здесь небольшие плотные планеты состоят
в основном из силикатов и металлов. Незамерзавшие щхдорвя
иI гелий рассеялись в пртягтртнстве под--деистВйеКГ^
W'8 « / дСГ.ОПЕРЕ ........., 17
«солнечного ветра» — потока быстрых частиц, извергавшихся
Солнцем на заключительной стадии его формирования. Эти
газы были потеряны на раннем этапе образования планет, когда
массы зародышей почти всех будущих планет были еще малы.
Только зародыши Юпитера и Сатурна были достаточно велики
для того, чтобы своим тяготением удерживать значительные
количества газов на поверхности. Весь газ, оставшийся в облаке
в области Юпитера и Сатурна, был поглощен их зародышами,
и потому их масса оказалась много больше масс других планет,
а химический состав мало отличается от солнечного состава.
При изучении процессов роста планет в протопланетном об-
лаке В. С. Сафроновым была выяснена значительная роль круп-
ных тел, поперечником до 1000 км. Падая, такие тела сильно
нагревали растущую Землю. Их тепло проникало на значи-
тельную глубину. Наибольшая температура 1500 К достигала
на глубинах 300 — 500 км. Средняя же начальная температура
всей Земли лишь немного превышала 1000 К, т. е. области паде-
ния крупных тел на Землю дополнительно нагревались на сотни
градусов. Эти большие температурные неоднородности явились
основным фактором дальнейшей эволюции Земли. После обра-
зования Земли ее недра разогревались в результате накопления
тепла, выделяющегося при распаде радиоактивных элемен-
тов (U, Th, К), вошедших в ее состав. Дополнительно подогре-
тые ударами, области первыми достигали температуры плав-
ления, и в них раньше всего начались процессы, связанные с
расплавлением вещества и с его последующей гравитационной
дифференциацией. Более тяжелые вещества опускались вниз,
а более легкие поднимались вверх и образовывали земную кору
Весьма вероятно, что именно над сильно нагретыми областями
началось образование континентов. В Земле, разогреваемой
радиоактивными элементами, гравитационная дифференциация
происходила очень медленно и продолжается до сих пор. С ней
связана современная внутренняя активность Земли — горооб-
разование, сейсмичность и т. п.
Сейсмическими наблюдениями давно установлено, что в
Земле существует плотное ядро. Однако природа его до сих
18
пор не выяснена. По одним предположениям, ядро состоит или
из железа с примесью более легких элементов или из смеси
окислов железа, по другим — из силикатов, перешедших в
металлизованное состояние под действием высокого давления,
господствующего в глубине Земли.
Земля, непосредственно после своего образования, не имела
атмосферы. Это было связано с тем, что газы из протопланет-
ного облака были потеряны раньше, чем растущая Земля стала
настолько массивной, чтобы своим тяготением удерживать
заметное количество газа на поверхности.
Длительный этап формирования нашей планеты шел в со-
провождении бурно и сложно проявлявшихся химических реак-
ций, что, несомненно, осуществлялось с поглощением свобод-
ного кислорода при образовании кремнекислоты и ряда других
компонентов магматических пород. При разогревании недр
Земли газовые компоненты планеты, вовлеченные во внутрен-
ние зоны, выделялись на ее поверхности и создали, таким обра-
зом, вторичную, собственно земную атмосферу.
Достигнув к этому времени значительных размеров в резуль-
тате накопления падавших на поверхность Земли частиц веще-
ства туманности, планета стала способной удерживать эти газы,
в основном углеводородные газы, аммиак, углекислоту, свобод-
ный водород, при отсутствии свободного кислорода и азота.
Все запасы кислорода, которые могли быть вовлечены в про-
цесс формирования вследствие присутствия неокисленных со-
единений, несомненно, полностью шли на их окисление. Вода в
парообразном состоянии образовывала тогда пелену облаков.
Взаимодействие между углекислотой и водородом должно было
приводить к образованию метана и водяного пара. В связи с
этим появилась вода в атмосфере и на поверхности Земли.
Возник круговорот воды, возникли моря и океаны.
Первичный кислород мог образоваться в результате воздей-
ствия ультрафиолетовой радиации Солнца, а также и других
космических излучений на молекулы углекислого газа.
Поверхность земной коры, покрытая газовой оболочкой —
атмосферой,, является ареной проявления многих разнообраз-
19
ных геологических процессов, и естественно предполагать, что
именно здесь сначала образовались сложные углеродные соеди-
нения, а затем на их основе возникла и жизнь. Вторичная атмо-
сфера Земли состояла из водорода, метана, аммиака, окиси
углерода и сероводорода с парами воды в неясных количествен-
ных соотношениях.
С самого образования атмосферы из ее состава весьма интен-
сивно ускользал молекулярный водород. Позднее, с появлением
молекулярного кислорода, этот процесс замедлился, хотя не
прекратился и к настоящему времени. Как известно, Земля
отдает в космос значительные количества молекулярного водо-
рода, порядка 107 атомов в секунду с 1 см2 Это соответствует
расходу 20 г воды с 1 см2 в течение 4,5 млрд. лет.
Первоначально Земля не имела водных бассейнов, и первич-
ные запасы воды в виде пара входили в состав Древнейшей атмо-
сферы. Впоследствии с понижением температуры ее поверх-
ности метан и аммиак, соединяясь с парами воды, переходили
в раствор.
Так образовались первичные водные бассейны. Некоторые
летучие соединения водорода — СН4, С2Н2, НС1 и другие —
оставаясь в составе атмосферы, подвергались воздействию
ультрафиолетовых лучей и разлагались с выделением молеку-
лярного водорода. Ультрафиолетовые лучи разлагали и воду,
причем кислород быстро вступал в новые связи, а выделявший-
ся водород тоже частью ускользал с поверхности Земли.
Из недр планеты поднимался углекислый газ. Он входил в
состав атмосферы, а также интенсивно растворялся в воде,
входя при этом в разнообразные химические соединения с ме-
таном и аммиаком. Количества углекислого газа, попадавшие
в атмосферу, были огромны. Об этом говорят, с одной стороны,
его извечные выбросы в биосферу при вулканических явлениях,
с другой стороны, об этом свидетельствуют огромные массы
образовавшихся в разные периоды истории Земли отложений
карбонатных пород, представляющих собой соединения угле-
кислого газа с окисью кальция и магния.
Имеются основания полагать, что образование карбонатов,
20
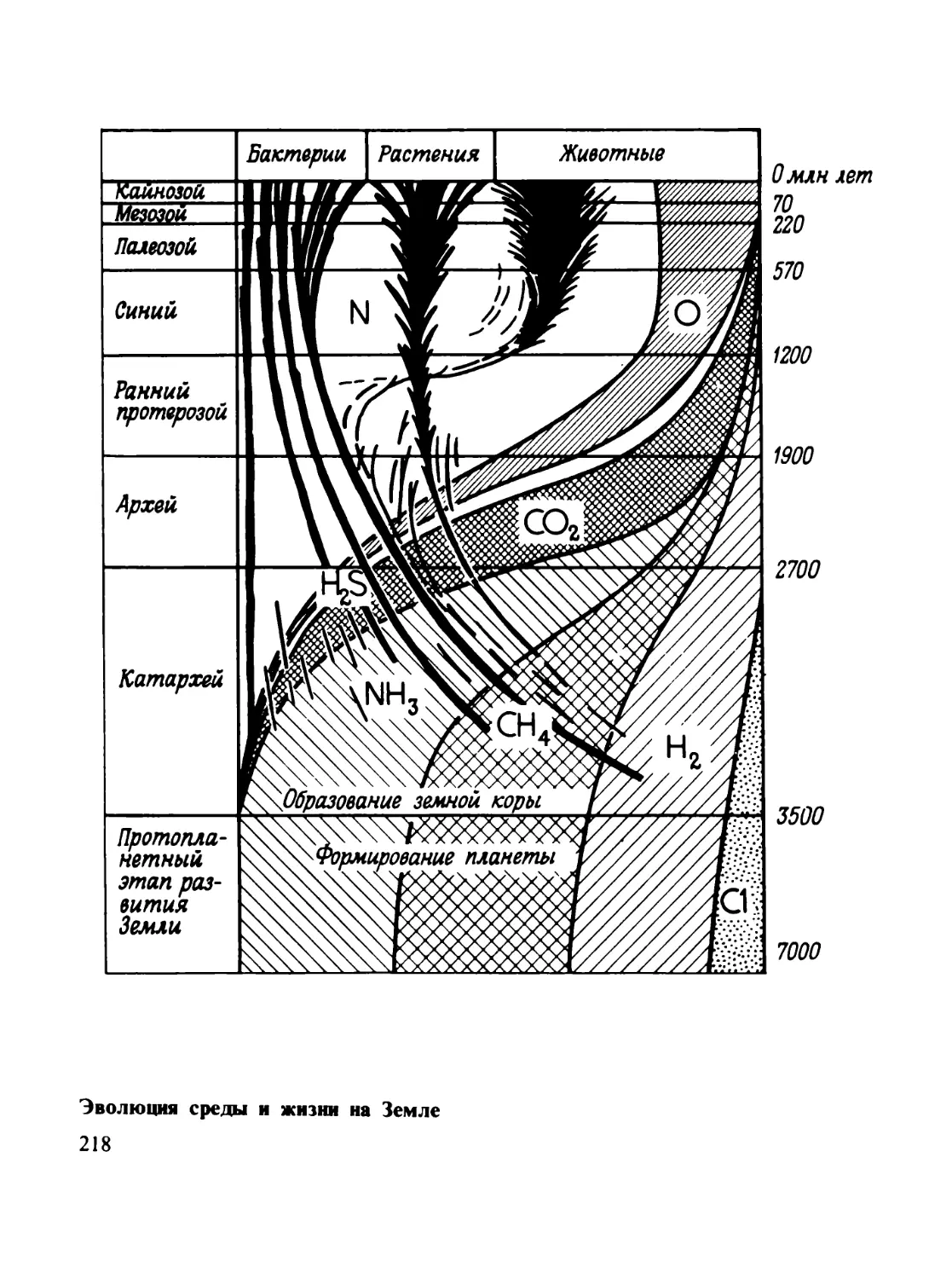
Миллиарды лет назад на Земле не было жизни и свободного кислорода
(по А. И. Перельману)
как и последовавшее позже возникновение наземной раститель-
ности и утлеобразованис, были подвержены определенному
ритму, который находит разгадку в ритме вулканической дея-
тельности на Земле.
На конференции в Москве в 1957 г по происхождению жизни
на Земле приводились интересные мнения относительно состава
вторичной атмосферы, собственной атмосферы нашей планеты.
Большинство ученых считают ее^ главными компонентами Н2,
СО2, N2, H2S, в меньшей степени — СН4, NH3. Обращалось
внимание на избыток в верхней зоне атмосферы Н2О, СО2.
СО2 и других компонентов; все эти газы рассматривались как
«равновесная смесь». Предполагалось, что, если не учитывать
паров воды, то основными компонентами вторичной атмосферы
Земли быдй СО2 (около 91%) и N2 (около .6,4%). В первую
фазу существования вторичной атмосферы в ней находилось
много водорода, в частности образовавшегося при разложении
21
воды под влиянием космических излучений. Но большая часть
его улетучивалась в космическое пространство. Вследствие
активности газов Н2, СО, СО2, SO2, SO3, NO, N2O, NO2 они
входили в присутствии воды в соединения, задерживаясь в по-
родах и в водах, тогда как СО2 и СО в значительной части по-
ступали в атмосферу.
Таким образом, складывается представление, что собствен-
ная атмосфера Земли состояла тогда из Н2, NH3, СН4, СО2,
H2S при малом содержании других компонентов, в том числе
и О2, что в общем создавало восстановительную обстановку.
В новой атмосфере, быстро разросшейся под влиянием актив-
ных радиохимических и вулканических процессов, стала преоб-
ладать углекислота с парами воды. Такая атмосфера, вероятно,
хорошо удерживала тепло геохимических реакций. Огромная
мощность и обилие паров воды делало ее малопроницаемой для
солнечных и отчасти космических лучей.
Возникали и угасали- разнообразные воздушные потоки.
Движение воздушных масс, вызываемое разностью давления
за счет притяжения Солнца и Луны, рождало ветры и бури.
Сильные ураганы и сейчас приносят огромные непоправимые
бедствия, особенно в приморских странах. Но, вероятно, они —
ничто перед бурями и ураганами далекой геологической древ-
ности.
При конденсации паров воды в атмосфере образуются ка-
пельки с электрическими зарядами. Каждая капелька имеет
положительный заряд в центре и отрицательный - на поверх-
ности. Потоки воздуха дробят эти капельки на мелкие части,
которые соответственно знаку своих зарядов способны группи-
роваться. Отрицательно заряженные капельки поднимаются
вверх, а более крупные положительно заряженные опускаются
вниз. При этом верхняя часть облака или тучи получает более
или менее мощный заряд отрицательного электричества, а ниж-
няя — положительного.
Так возникает электрический заряд между облаками или
же между облаком и землей.
22
Большая интенсивность атмосферных процессов в период
образования Земли приводила к тому, что над поверхностью
земной коры непрерывно сверкали и грохотали мощные элект-
рические разряды. Электрические разряды в атмосфере имели,
вероятно, большое значение для природного образования слож-
ных углеродных соединений. На то, как происходило связыва-
ние всех весьма активных химических компонентов вторичной
атмосферы Земли в разнообразные соединения, в том числе и
в сложные органические, проливают свет замечательные опыты
Стэнли Г Миллера.
Миллер пропускал электрический разряд через смесь ме-
тана, аммиака, воды и водорода. В результате действительно
удалось получить аминокислоты — исходный материал для
создания белков — и другие сложные органические вещества.
Тогда же Т. Е. Равловской и А. Г Пасынским были получены
аминокислоты при действии ультрафиолетовых лучей на рас-
твор смеси формальдегида и хлористого или азотнокислого
аммония.
Эти эксперименты подтверждают положение теории
А. И. Опарина о том, что в первородном океане ультрафиоле-
товое излучение Солнца и электрические разряды атмосферы
могли приводить к накоплению аминокислот, необходимых для
образования белковых соединений.
Интересны сравнительные данные о составе атмосфер планет
Солнечной системы. На Венере атмосфера в основном состоит
из углекислого газа (97%). Есть также азот, кислород не обнару-
жен. Можно полагать, что современное состояние атмосферы
Венеры соответствует состоянию атмосферы Земли в один из
ранних этапов ее добиосферного периода, когда в результате
накопления гравитационного тепла, ядерных и электронных
процессов, происходящих в толще атмосферы, богатой угле-
кислым газом, температура литосферы была в то время осо-
бенно высокой.
Марс больше других планет похож на Землю. О нем много
пишут ученые и не меньше — писатели-фантасты. Его атмосфе-
ра на 90% состоит из углекислого газа. Есть вода и кислород.
23
Атмосферы планет-гигантов состоят в основном из легких
газов. В атмосферах Юпитера и Сатурна преобладают водород
и гелий, углекислый газ отсутствует. Газообразный СО2 при
наличии водорода полностью превращается в метан и воду.
В верхних слоях атмосферы вода переходит в лед. Облака на
Юпитере состоят из аммиачного льда. В атмосферах Урана и
Нептуна преобладает метан, аммиак и вода.
В наше время исследования планет, которые проводятся
не только наземными методами, но и при помощи космических
аппаратов, приносят множество новых фактических данных о
Солнечной системе. Сравнительное изучение химического со-
става планет очень важно для понимания химической эволюции
допланетного облака, а следовательно, и для выработки пра-
вильных представлений о строении и эволюции Земли.
ЗЕМНАЯ КОРА
И ПАЛЕОНТОЛОГИЧЕСКАЯ ЛЕТОПИСЬ
Геология - наука о Земле - изучает происхождение нашей пла-
неты и ее развитие, в особенности развитие ее твердой оболоч-
ки — земной коры и проявлений на ней жизни с ее геологиче-
ской деятельностью.
Геология - наука историческая. Геологическая наука в на-
стоящее время сильно разветвилась, превратившись в комплекс
наук. Это: историческая геология, изучающая историю Земли;
физическая геология, изучающая геологические процессы на
Земле и их влияние на строение земной коры; палеонтология —
учение об ископаемых организмах, об их морфологии и разви-
тии; биостратиграфия — наука о руководящих ископаемых
организмах, как вехах геологического времени, помогающих
расчленять вмещающие отложения на относительно маломощ-
ные серии, свиты и горизонты, а также сопоставлять отложения
разных районов; учение о фациях — наука об условиях формиро-
вания осадочных горных пород и полезных ископаемых; петро-
графия - учение о минеральном составе, структуре и сложении
горных пород изверженного происхождения; литология — уче-
ние об осадочных горных породах, их составе и свойствах;
наука о рудных и нерудных полезных ископаемых', гидрогеология —
учение о подземных водах, их составе, движении и геологической
деятельности; инженерная геология и гидрогеология — наука о
грунтах, горных породах и подземных водах, связанная с зада-
чами строительства; минералогия — наука о минералах, их со-
ставе, свойствах и практическом применении; кристаллогра-
фия — наука о кристаллах; геофизика — учение о физических
явлениях Земли, о методах поисков полезных ископаемых и
изучении геологических структур; геотектоника — учение о
движениях земной коры и их результатах; геохимия — учение
о химическом составе геологических образований, о поведе-
нии химических элементов и их изотопов в земной коре во вре-
25
мени и пространстве; абсолютная геохронология — наука, имею-
щая целью определение абсолютного возраста горных пород
и минералов на основе изучения продуктов распада радиоактив-
ных элементов, включенных в минералы и горные породы при
их образовании.
Все эти науки характеризуются своими особыми объектами
и методами исследования. При этом геологический цикл наук
теснейшим образом связан, с одной стороны, с циклом наук
химических, изучающих химические элементы, их изотопы и их
поведение в различных соединениях и процессах, с другой сто-
роны — с циклом наук физических, изучающих явления внутри
Земли, на ее поверхности и в окружающем космическом прост-
ранстве; и с третьей — с циклом биологических наук благода-
ря палеонтологическим материалам, часто находимым в слоях
осадочных пород.
Для успеха палеонтологических исследований оказываются
часто очень важными следующие науки: физиология животных
и растений, микробиология (бактериология), биохимия, биогео-
химия (учение о геохимической роли организмов), сравнитель-
ная остеология и т. д. Можно добавить, что и сама палеонтоло-
гическая наука разветвляется на эволюционную теоретическую
и прикладную. Кроме того, она делится соответственно группам
ископаемых организмов на палеоботанику и палеозоологию
(или более подробно — археоциатологию), палеоконхиологию
(учение об ископаемых моллюсках), палеоэнтомологию (учение
об ископаемых насекомых) и т. д. При этом выделилась специаль-
ная отрасль палеонтологической науки — палеоэкология, изу-
чающая условия обитания ископаемых организмов.
Земная кора имеет весьма сложное строение. Она образована
породами различного происхождения, различного минерально-
го состава и различного геологического возраста в самых раз-
нообразных соотношениях составляющих их химических эле-
ментов. Распространение же различных типов пород обычно
подчиняется определенным закономерностям.
Основу земной коры составляют породы магматические,
т. е. некогда образовавшиеся из расплавленных масс, застывших
26
на глубинах близ земной поверхности или излившихся на нее.
В последних случаях образовывались лавовые потоки и отла-
гались иногда большие массы пепла, превращавшегося в вул-
канические туфы. Выветривание пород на суше приводило к об-
разованию огромных масс мелкозема — измельченного камен-
ного материала, который под действием воды, ветра и силы
тяжести перемещался и отлагался в виде слоев так называемых
осадочных пород, к которым добавлялись в тех или иных коли-
чествах осадки самих водных бассейнов. В одних случаях это
были известковые илы бактериального или химического проис-
хождения, превращавшиеся со временем в твердые известняки.
В других случаях, если воды бассейнов были сильно минерали-
зованы или если испарение вод превышало их приток, выпадали
различные соли кальция, магния, натрия, калия и других метал-
лов, иногда с образованием соляных залежей. В определенных
условиях при смешивании вод, стекавших с континентов, с во-
дами древних морей создавались местные скопления глин
(монтмориллониты, бентониты), фосфороносных и алюминие-
носных минералов (фосфориты и бокситы), к которым в том
или ином количестве подмешивался обломочный материал,
приносимый реками с суши.
Мощная водная оболочка, покрывающая более двух третей
поверхности Земли, является местообитанием огромного коли-
чества разнообразных живых организмов, которые тоже участ-
вуют в процессе образования донных осадков и осадочных
пород. Они обладают способностью накапливать минеральные
вещества, играющие роль опоры для их мягких тканей. Многие
организмы в процессе жизнедеятельности непрерывно выделя-
ют твердые минеральные массы, в частности углекислый каль-
ций, фосфорнокислый кальций, кремнезем и другие вещества.
В наше время созидающая геологическая деятельность свой-
ственна бесконечному множеству морских животных и большим
группам водорослей, живущих как в верхних слоях воды, так
и на глубинах. Организмы в истории Земли издавна были актив-
ными геологическими агентами и выполняли огромную работу
по формированию осадочных пород.
27
Создававшиеся в водных бассейнах геологического прошло-
го разнообразные минеральные, минерально-органические и
органические осадки уплотнялись и преобразовывались в раз-
личные типы осадочных пород. В сложной геологической жизни
земной коры с ее магматическими и вулканическими явлениями,
с опусканиями и поднятиями, с образованием впадин и выступов
и формированием складчатых горных сооружений, первичные
«нормальные» горные породы претерпевают те или иные вто-
ричные изменения. Подвергаясь давлению, они превращались
в слоистые сланцеватые породы. При воздействии на исходное
вещество пород, поднимавшихся из глубин нагретых минерали-
зованных вод и паров, преобразовывался их минеральный состав,
создавались новые минералы. Такие породы называют метамор-
фическими — измененными. Чем глубже залегали породы в
земной коре, тем сильнее они изменялись под воздействием
поднимавшихся из земных недр эманаций, тем больше они
отличались от первичных нормальных пород.
Исследователи природы давно заметили, что один слой оса-
дочной породы лежит на другом. Это привело к появлению поня-
тия более «молодой» и более «древней» породы. Постепенно
накапливались описания выходов пород на дневную поверх-
ность (по берегам рек, на склонах гор или в искусственных вы-
работках — каналах, ямах, колодцах, каменных, песчаных или
глинистых карьерах. Были выделены серии слоев — свиты, обра-
зование которых связывалось с более или менее длительными
этапами прошлого геологического времени. Формирование
каждого слоя, несомненно, было теснейшим образом связано
со средой (вода или воздух суши), с окружающей частью земной
поверхности (ландшафт), с климатическими особенностями.
Каждая смена слоев обусловливалась изменениями этих или
каких-либо других факторов. При этом могли проявляться и
деятельность ветра и течений, та или иная роль живых организ-
мов. Сама последовательность слоев, отличающихся друг от
друга, отражала историю водного бассейна или впадины на
поверхности континента за соответствующий отрезок геологи-
ческого времени.
28
Наблюдение свит помогало ученым представить себе этапы
геологического прошлого, но|не давало понятия общей меры
времени в каких-либо единицах. Впервые вопрос о времени
образования осадочных пород был решен, когда геологи стали
использовать для этого остатки животных и растений или их
отпечатки, находимые на поверхности наслоения древних осад-
ков, а также следы жизнедеятельности организмов. Постепенно
опыт изучения осадочных пород в разных районах мира и на
разных этапах геологического времени привел геологов к пред-
ставлению, что каждому пласту осадочной породы соответст-
вует как свидетель определенный состав остатков живых орга-
низмов. Наблюдавшееся при этом различие состава «окамене-
лостей» в отдельных слоях, последовательно налегающих друг
на друга, первоначально объяснялось «катастрофами» на Земле.
Но со времени работ Ж. Б. Ламарка (1744—1829) и Ч. Дарвина
(1809—1882) это стало бесспорным доказательством эволюции
животных и растений на нашей планете.
Палеонтологическая наука, выросшая на основе изучения
органических остатков в горных породах и специальных иссле-
дований геологических разрезов, раскрыла грандиозную кар-
тину непрерывной смены в ходе эволюции жизни на Земле.
Уже на ранних этапах развития палеонтологии в отдельных
слоях были обнаружены скопления остатков каких-либо опре-
деленных видов организмов, групп видов, родов, которые,
несомненно, были особенно приспособлены к существовавшей
тогда обстановке. Поскольку в соседних слоях наблюдались
остатки уже других организмов, то отсюда возникло представ-
ление о «руководящих» формах — об организмах, указываю-
щих на определенное время образования данного слоя и, таким
образом, служащих «вехами» геологического времени, своего
рода «часами».
Палеонтологические «часы» верно служат всему геологиче-
скому циклу наук. Они дали возможность построить схему
истории геологического развития земной коры с делением ее
на отдельные этапы. При этом геологические единицы времени
непосредственно связывались с соответствующими геологиче-
29
Смятые в крупные и мелкие складки слои доломитов и известняков докембрия
(Китай). Фото автора
скими образованиями. Так возникло представление об эрах,
периодах, эпохах и геологических «веках», а также о группах,
геологических системах, их отделах, ярусах и горизонтах.
Геологический возраст пород иного, не осадочного проис-
хождения — магматических и метаморфических, лишенных ор-
ганических остатков, тоже можно точно определить именно
по соотношению с осадочными породами, содержащими орга-
нические остатки. Так, если какое-либо гранитное тело или
жила изверженной породы «прорывает» осадочную серию опре-
деленного геологического возраста, то, следовательно, рас-
плавленная магма внедрилась сюда после формирования этой
серии. Если на поверхности гранитного тела или выше верхней
части жилы изверженной породы залегает свита осадочных
пород — значит это гранитное тело или жила является более
древней, чем осадочные породы.
Совершенно аналогично палеонтологические материалы по-
могают определить геологический возраст движений земной
коры. Земная кора никогда и нигде не находится в состоянии
абсолютного покоя — отдельные ее часги движутся в различ-
ных направлениях. Временами, когда эти движения приводили
к погружению части поверхности земной коры под уровень моря
или, наоборот, к появлению целых участков дна на поверхности,
характер геологических процессов коренным образом менялся.
Образованные в глубинах моря породы начинали в условиях
суши или мелководья выветриваться и размываться, а на по-
груженных участках формировались новые осадочные слои,
в которых появлялись соответствующие комплексы организ-
мов. Таким образом, возраст некоторых фаз движений земной
коры и районы их проявления могут быть также определены
палеонтологическими методами. Поэтому тектоника — учение
о движениях земной коры—стала наукой исторической только
с помощью палеонтологии.
Палеонтология довольно хорошо раскрывает историю жиз-
ни на Земле за последние 570 млн. лет. История живых существ
записана, как говорят, на камне. Она выражена остатками твер-
дых частей тела древних организмов, следами их ползания,
проявлениями жизнедеятельности (сверление раковин, отложе-
ние минерального или органического вещества). Но по отпечат-
кам, а еще лучше по раковинам и костям можно воссоздавать
и сами организмы. Интересно, что. вещества, из которых форми-
ровались и формируются твердые части тела организмов, —
это соли кальция, углекислая соль у большинства беспозвоноч-
ных животных и фосфорная у животных позвоночных. Причина
этого несомненно лежит в энергетических особенностях перево-
да солей кальция из растворенного состояния в нерастворимое,
что осуществляется с выделением свободной энергии. С этим
обстоятельством необходимо связывать неизбежное для орга-
низмов освоение солей кальция, преобразующихся при этом в
раковину* у моллюска или в скелет у позвоночного животного.
31
В процессе бесчисленных смен поколений организмов, кото-
рый выразился в видимом проявлении эволюции бесчисленных
линий развития с их временными и местными этапами формо-
образования новые виды, образуясь в условиях изменения
среды, были то удачными, то неудачными, приобретали одни
признаки, утрачивали другие. Вследствие этого, как оказывается,
в каждом геологическом периоде, в каждом этапе геологического
времени наибольшую жизнедеятельность проявляли разные
группы организмов. Наряду с проявлением и развитием форм
более сложных, приспособившихся к существованию в опреде-
ленных условиях, всегда существовали и более примитивные
формы, эволюция которых, по-видимому, замедлялась вслед-
ствие относительной устойчивости условий их существования
в древних средах, или же такие условия изменялись очень мед-
ленно в ходе геологического времени. Например, обстановка для
бактерий в недрах Земли, куда они проникают на глубины в не-
сколько километров, остается длительно относительно неиз-
менной, часто вполне обеспечивающей осуществление ими
обмена веществ со средой.
Исследования остатков организмов имеют двойную цель.
Во-первых, они помогают различать осадочные отложения раз-
личного геологического возраста, т. е. принадлежащие к раз-
личным стратиграфическим подразделениям их, образованным
в ходе геологического развития земной коры в водных бассей-
нах или на суше. Во-вторых, остатки организмов, находимые
в ископаемом состоянии, показывают ход и характер их эволю-
ционного развития. Они указывают на эволюцию жизни, в раз-
ные моменты оставлявшую свой след во вмещающих породах,
показывают общий ход эволюции за длительные этапы и пояс-
няют происхождение современного мира животных и растений
на Земле.
Чем древнее осадочные породы, тем менее они оказываются
похожими на первичные рыхлые осадки древних морских бас-
сейнов или пресноводных бассейнов суши. Известно, что некото-
рые организмы вообще не могли оставлять следы своей жизне-
деятельности в осадочных горных породах. Остатки других
32
иногда изменялись уже в момент их захоронения настолько
сильно, что оказывались для палеонтолога весьма трудно рас-
познаваемыми. Все же в прежде известковых илах, известково-
глинистых и иных осадках, ставших метаморфизованными по-
родами — кристаллическими известняками и мраморами, крем-
нистыми сланцами и кварцитами (сливными песчаниками), —
иногда сохраняются древние ископаемые остатки. Бережное
отношение к этому материалу, внимательное его изучение в
специально приготовленных препаратах (прозрачных шлифах
и в пришлифовках) позволяет различать эти следы древней
жизни.
Одним из важнейших факторов, способствующих сохране-
нию органических остатков в ископаемом состоянии в древних
породах, является содержавшееся в них первоначально остаточ-
ное органическое вещество. Пока оно н$ удалено из породы
геологическими процессами, органические остатки и структуры
не могут быть совершенно уничтожены. Их видно на выветрелой
поверхности породы и в прозрачных шлифах. Они придают
породе часто более или менее темную окраску. При этом при-
менение биологического микроскопа с увеличениями до 2000 раз
часто позволяет выявлять очень интересные явления. Оказыва-
ется, что остаточное органическое вещество как бесструктурное,
рассеянное в породе, так и в остатках организмов при наблю-
дении его в прозрачных шлифах под микроскопом выражено
скоплениями черного, непрозрачного углистого вещества в виде
сплетений тонких нитей, имеющих форму бактерий. Создается
впечатление, что все органическое вещество при переходе его в
ископаемое состояние перед завершением процесса окаменения
осадка (литификация, диагенез) становилось объектом освоения
бактериями и в виде отмерших клеток попадало затем в твердую
породу. Это свойство установлено в массе образцов древних
и поздних осадочных пород, включая и древнейшие, так назы-
ваемые докембрийские, т. е. древнее 570 млн. лет.
Совершенно то же явление, но в несколько иных формах и
других размерах микроскопических телец наблюдается в захо-
роненном в осадочных породах железе. Всюду, в самых разно-
образных осадочных породах железо представлено сгустками
окислов в виде тонких округлого сечения палочек или нитей,
напоминающих строение современных железобактерий.
Возможно ли сохранение в ископаемом состоянии остатков
бактерий? Сущность явления их сохранности несомненно впол-
не объяснима. Иногда сгустки органического вещества отмер-
ших клеток проходили стадию минерализации. Чаще они про-
питывались известью. Некоторые виды бактерий, способные к
использованию закисного железа, создавали чехлики из его
гидроокислов, хорошо сохраняющиеся в осадке в породе.
Б. Л. Исаченко считает, что отложение углекислого кальция в
морских осадках происходит под влиянием бактерий. Установ-
лено также, что некоторые культуры бактерий способны выде-
лять из морской воды также фосфорнокислую известь, фосфор-
но-аммонийно-магнезиальные соли. В то же время изучение
древних осадочных пород морского происхождения с примене-
нием биологического и электронного микроскопов позволяет
видеть и остаточные клеточные структуры бактериального ха-
рактера. Поэтому или минерализация остатков, или их отпе-
чатки в структуре цементирующего минерального материала,
иногда являвшегося минеральным коллоидом (например, крем-
ниевым гелем), и другие условия захоронения бактериальных
клеток несомненно имели широкое распространение в средах
геологического прошлого. При этом часто остатки бактерий
получали в породах и минералах прочную упаковку.
Большое значение имеют палеонтологические методы для
изучения изменений в геологическом прошлом Земли, ее климата
и земной поверхности, т. е. для палеогеографии. Материалы
палеофитогеографии (истории расселения растений) и палео-
зоогеографии (истории распространения животных) поэтому
являются основой палеогеографического исследования отдель-
ных районов нашей планеты или ее поверхности в целом.
Данные палеонтологии, как мы говорили выше, помогают
ученым представить себе последовательность отдельных этапов
геологического прошлого Земли. Однако они не могут помочь
нам определить абсолютную продолжительность этих этапов,
34
точный возраст тех или иных пород. Поэтому уже давно иссле-
дователи искали способ, который дал бы возможность устанав-
ливать геологический возраст отложений в абсолютных едини-
цах. В частности, для этой цели пытались использовать изуче-
ние скорости накопления на дне бассейнов различного типа
осадков. Этот метод, если его применять к отложениям, образо-
вавшимся из материала, принесенного с суши в моря, дает
интересные результаты, например, помогает подсчитать дли-
тельность времени отложений, образовавшихся из материала,
принесенного с областей бывшего оледенения в морские и внут-
риконтинентальные водные бассейны.
Принципиально новые способы определения абсолютного
возраста горных пород и минералов были получены геологи-
ческой наукой в результате изучения радиоактивных элементов.
Суть этих способов такова.
Ядра атомов всех химических элементов состоят из мель-
чайших частиц — положительно заряженных протонов и не
несущих заряда нейтронов. Масса протона почти равна едини-
це — массе атома водорода (ядро которого и состоит из одного
протона). Нейтрон же имеет массу чуть большую, чем протон.
Как оказалось, так называемый атомный номер элемента,
установленный Д. И. Менделеевым в периодической системе
элементов, соответствует числу протонов, или положительному
заряду' ядра. Нейтроны же присутствуют в составе ядра неза-
висимо от атомного номера. Число их в ядре варьирует у раз-
личных химических элементов — от атома водорода, не имею-
щего нейтронов, до атомов плутония и америция, содержащих
по 149 нейтронов. Химические элементы, ядра атомов которых
имеют одно и то же число протонов и, следовательно, один и
тот же положительный заряд, но различное число нейтронов,
называются изотопами. В периодической системе элементов
они занимают одно и то же место, но вследствие различий в
числе нейтронов имеют различные свойства.
Ядра бывают прочными (стабильными), не проявляющими
никаких изменений во времени, или нестабильными, неустой-
чивыми, распадающимися с измеримой скоростью. Радиоак-
35
тивность — это и есть распад ядер, который сопровождается
различным типом излучения.
Так называемое альфа-излучение представляет собой поток
положительно заряженных ядер гелия со значительной кинети-
ческой энергией; бета-излучение — это поток отрицательно
заряженных электронов; гамма-излучение — аналогично по
свойствам лучам Рентгена.
Все встречающиеся в природе химические элементы, имею-
щие атомный номер более 80, являются радиоактивными. Они
образуют четыре радиоактивных ряда. Иногда эти ряды назы-
ваются «семействами». Распад атомов исходных элементов —
урана-233, урана-235, урана-238 и тория-232 (U233, U235, U238,
Th232) — приводит к образованию неустойчивых ядер различ-
ной продолжительности существования, в свою очередь рас-
падающихся. Сам процесс распада не зависит ни от каких внеш-
них условий. Он идет с неизменной скоростью различной у
различных элементов. Половина всех имевшихся в наличии
ядер данного элемента распадается за строго определенный
промежуток времени — так называемый период полураспада.
Каждый радиоактивный ряд заканчивается образованием од-
ного из трех изотопов свинца с выделением гелия. Эти про-
цессы, и оказались теми «часами» истории Земли, которые по-
зволяют исчислять возраст геологических образований в виде
так называемой абсолютной геохронологии.
Что же такое абсолютная геохронология и что она дает
к настоящему времени?
Интерес геологов к природным процессам распада радиоак-
тивных элементов связан с тем, что скорость этих процессов,
во-первых, постоянна, во-вторых, очень мала. Лабораторные
испытания, в которых радиоактивные вещества подвергались
воздействию температур от —270 до +7000°С, давлений свыше
200 атм, сильных магнитных полей, бомбардировки космиче-
скими лучами не могли изменить нормального хода распада
радиоактивных элементов.
Периоды полураспада основных естественных радиоактив-
ных изотопов следующие.
36
Изотоп Период полурас- пада, лет Изотоп „ i Период полурас- пада, лет
Rb87 6,0 • 10*° и235 7,13 108
Th232 1,39 Ю10 с14 5568
К40 1,1 Ю10 Ra226 1622
и238 4,59 10’ Ас227 22
На ранних этапах по определению аосолютного возраста
геологических пород наибольшее применение получил метод,
основанный на определении количеств свинца и гелия как про-
дуктов распада урана-238, урана-235 и тория-232. Оказалось,
что радиоактивные химические элементы, входящие в состав-
минералов земной коры, способны давать указание на время
их образования и поэтому могут служить для установления
времени геологических событий, иногда весьма древних. По-
скольку урано- и ториеносные минералы встречаются в природе
реже, чем калиеносные, то преимущество калиевого метода
оказывается очевидным особенно для осадочных пород.
В применении к изучению абсолютного возраста минералов
осадочного происхождения наиболее интересным показал себя
стронциевый метод, основанный на распаде рубидия-87 с обра-
зованием стронция. Для этого необходимо, чтобы испытуемый
материал содержал рубидий, например, в виде хлористой соли
или в виде калийсодержащих глинистых минералов группы
иллита, или в виде глауконита, а иЗ магматических минералов —
микроклина, флогопита, мусковита и др.
Особое место в абсолютной геохронологии занимает угле-
родный м^тод, основанный на измерении содержания углеро-
да-14, период полураспада которого составляет 5568 лет. Этот
метод применим для сравнительно самых молодых геологиче-
ских образований — четвертичных. Исходным материалом для
него являются углеродсодержащие горные породы, лигниты,
торф и т. д.
37
Углерод в природе представлен двумя стабильными изото-
пами С12 и С13, соотношение количеств которых в некоторых
известняках составляет 98,892 и 1,108%. При этом интересно,
что С12 является биогенным изотопом углерода, поскольку
его содержание в организмах и в органогенных геологических
образованиях всегда относительно выше по отношению к С13
Принято различать углерод небиогенного происхождения
(С12/С13 =88,0-90,2) и биогенного (С12/С13 =90-92,9).
Методы определения абсолютного возраста прошли доволь-
но длительную проверку. Выяснено, что причиной ошибок при
вычислении могут быть утечка изучаемого элемента при вы-
ветривании породы, утечка гелия, образовавшегося при распаде,
примеси нерадиогенного свинца и т. д. Поэтому различные оп-
ределения одного и того же объекта способны давать более или
менее значительные отклонения. Но все же эти отклонения не
столь значительны, как можно было думать ранее, и цифры
возрастов тех или иных минералов и горных пород постепенно
уточняются.
Одним из важнейших достижений современной науки явля-
ется определение возраста Земли, точнее возраста слоев земной
коры.
Так, возраст ряда весьма древних геологических образова-
ний составлял для гранитов Беломорья 1800 млн. лет, габбро-
норитов Беломорья 2060 млн. лет, гранитов Балтийского щита
2030 млн. лет, пегматитов Карелии 2450 млн. лет. Аналогичные
цифры получены для древних пород Канады, Бразилии, Индии,
Южной и Центральной Африки и т. д. Возраст этих пород со-
ставляет от 2500 до 3350 млн. лет. Последняя цифра некоторыми
исследователями принималась за возраст земной коры, другие
принимали цифру 3,6 млрд, лет, полагая, что формирование
каменной оболочки — коры нашей планеты началось 4,5 млрд,
лет назад и длилось многие сотни миллионов лет в условиях
уже относительно медленного накопления дополнительного
космического вещества. Итак, наилучший способ проследить
процессы и события истории земной коры — это сочетание
палеонтологического и геохронологического методов. На осно-
38
Эра Период Эпоха Границы периодов, эпох и т. д. (млн. лет назад)
Третичный 1—0
Кайнозойская Неоген Плиоцен Миоцен 10-1 25—10
Палеоген Олигоцен Эоцен Палеоцен 40—25 70-40
Мезозойская Меловой Юрский Триасовый 100—70 140—100 185—140 225—185
Пермский Карбоновый 270—225 320-270
Палеозойская Девонский Силурийский Ордовикский Кембрийский 400—320 420-400 480-420 570-480
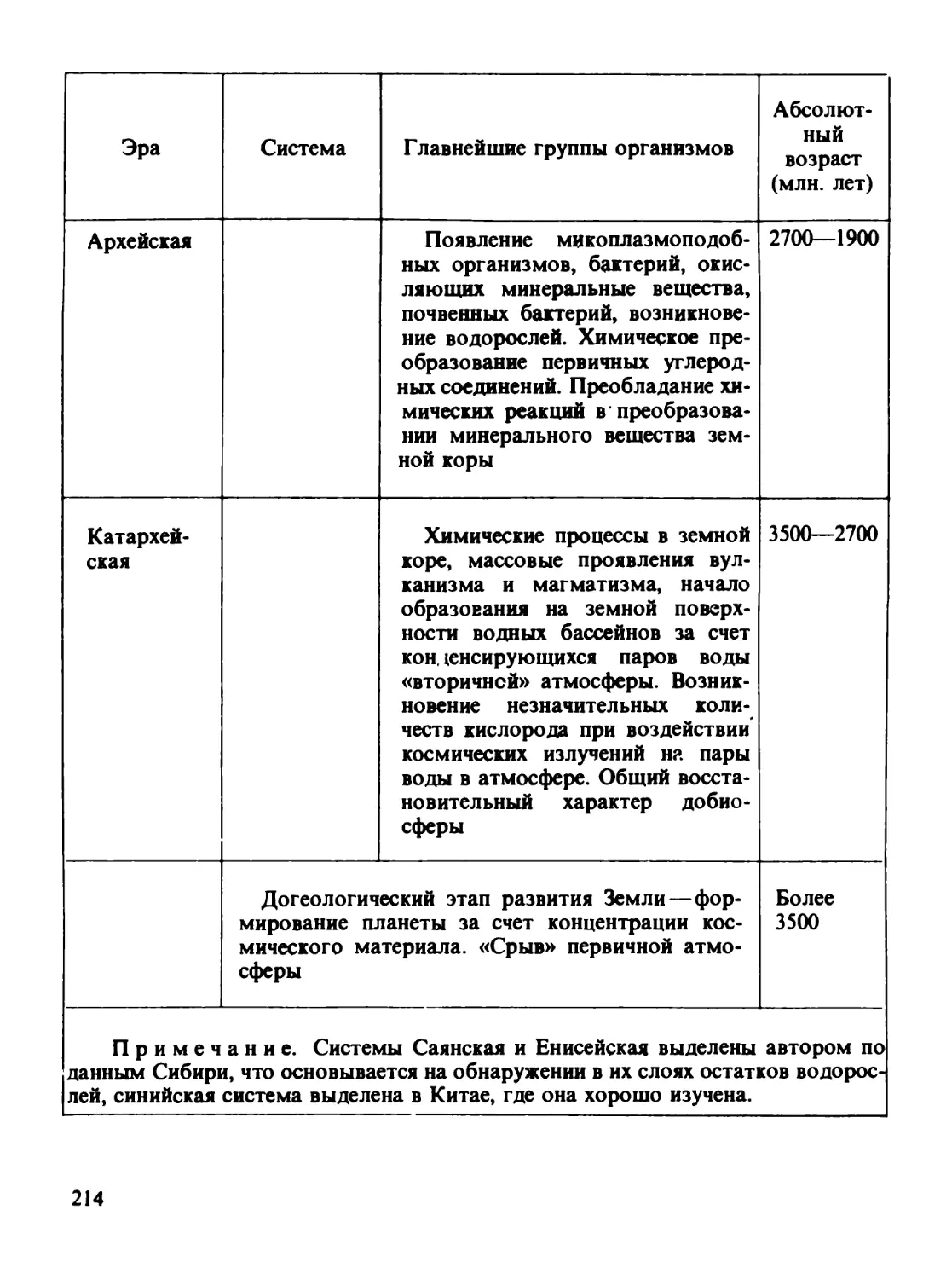
Докембрий IV (синий* — рифсй) Докембрий III (Протерозой нижний и средний) Докембрий II (Архей) Докембрий I (катархей) Формирование Земли и земной коры * 1200-570 1900—^1200 1700—1900 3500-2700 7000-3500
* Чаще геологи считают синий верхним протерозой. — Прим, автора.
39
ве этих данных выделены определенные этапы геологического
развития земной коры и была принята геохронологическая
шкала по данным определения абсолютного возраста (по
Д. И. Щербакову, 1961 г.). Эта схема сильно раздвинула воз-
растные границы эр и периодов, в особенности в отношении
более ранней части истории земной коры. Докембрий на терри-
тории СССР был разбит на четыре крупных подразделения,
имеющих огромную длительность. Весьма велика и мощность
образований, соответствующих этим подразделениям. Все че-
тыре подразделения докембрия можно рассматривать как эры.
Обращают на себя внимание различия в длительности выде-
ленных в схеме периодов. Чем ближе к нашему времени, тем
они становились как бы короче и короче. Это объясняется тем,
что основой для схемы были полученные геохимиками от гео-
логов и палеонтологов палеонтологические данные. Эти данные
отражали эволюцию органического мира Земли в ходе геоло-
гического времени, которая осуществлялась как бы убыстряю-
щимися темпами. От слоя к слою, от одного геологического
века к следующему изменения в составе организмов происхо-
дили все быстрее, что позволяло выделять менее мощные серии
отложений, образовавшиеся за сравнительно короткие проме-
жутки времени.
Заменяет ли метод абсолютной геохронологии другие ме-
тоды определения геологического возраста, в частности палеон-
тологический? Совсем наоборот! Именно сочетание палеон-
тологического метода, раскрывающего вместе с геологически-
ми данными последовательность формирования осадочных от-
ложений целыми свитами и сериями свит, с методом геохроно-
логии способствует наилучшему раскрытию истории развития
нашей планеты.
СРЕДА ЖИЗНИ
Средою жизни издавна являются моря и океаны. Растворяющие
свойства в жидком состоянии, способность в парообразном
состоянии соединяться с различными химическими элементами
и их соединениями делают воду важнейшим фактором и ком-
понентом в химических и биологических процессах в природе.
В воде растворяются газы атмосферы и газы, поднимающиеся
из земных недр. Поэтому воды суши, морей и океанов до самого
их дна являются продолжением атмосферы, обеспечивая тем
самым существование организмов — растений, животных и
бактерий. Поэтому влажному почвенному покрову суши, с его
богатыми проявлениями бактериально-водорослево-грибной
жизни, вполне соответствует верхний слой рыхлых донных
осадков. Ниже, на суше и в водах, органическая жизнь просле-
живается, но в количественном отношении оказывается более
скудной.
Важно отметить, что воздух, растворенный в воде, содержит
кислорода относительно больше, чем воздух газообразный,
так как растворимость кислорода в воде лучше, чем раствори-
мость азота. Высокую растворимость в воде имеет и углекислота,
запасы которой в водах гидросферы во много раз превосходят
ее запасы в атмосфере, обеспечивая тем самым существование
и развитие водных растений.
Водная масса Земли всегда находится в непосредственном,
теснейшем контакте с твердым .веществом земной коры, с гор-
ными породами и минералами и продуктами их выветривания
на суше и на дне моря. Поэтому вбды гидросферы обычно на-
сыщены минеральными веществами или содержат их в том или
ином количестве.
Вода основной компонент живого вещества прото-
плазмы клетки, крови, лимфы. Она совершенно необходима для
жизнедеятельности — процессы питания и выделения немысли-
мы без нее. Особо важную роль вода играет для водных орга-
41
низмов, являясь не только средой их жизни и эволюции, но и
происхождения.
Вода — минерал, как считают геологи и минералоги. Хими-
ки говорят, что вода — самое распространенное на Земле ве-
щество, имеющее определенный химический состав и физиче-
ские свойства (текучесть, отсутствие цвета и запаха, определен-
ную плотность и т. д.) и являющееся активным растворителем
газов и твердых веществ.
Процессы растворения в воде твердых веществ связаны с
разрушением их структуры, с образованием связей между их
молекулами и молекулами воды. Оказывается, абсолютно
нерастворимых веществ в природе не существует, но раствори-
мость разных элементов и их соединений проявляется в разной
степени в зависимости от их свойств. В результате высокой
растворяющей способности воды в природе она всегда оказы-
вается обогащенной веществами, с которыми входит в соеди-
нения, чаще в так называемые гидраты (сольваты). Образование
гидратов вызывается наличием у молекул растворенных веществ
электрического заряда и притяжением их к молекулам раство-
рителя обычно с выделением при этом тепловой энергии.
В природных водах, таким образом, оказываются в взаимо-
связи и во взаимовлиянии самые разнообразные по свойствам
химические элементы и соединения, на взаимоотношениях ко-
торых сказываются температура, ее изменение, изменение дав-
ления. Между растворенными в воде веществами возможны
самые разнообразные химические связи. При пересыщении
раствора каким-нибудь компонентом выпадает осадок, качест-
венно отличный от первично растворенных веществ, — то в
виде гидратов окислов элементов, то в виде их фосфатов, суль-
фатов, галоидных солей, с тем или иным количеством молекул
воды на молекулу осажденного из раствора соединения.
Таким образом, вода в природе — это сложнейшая химиче-
ская лаборатория, где встречаются и входят во взаимодействие
разнообразные химические элементы земной коры и атмосферы.
Поэтому в природе не было более благоприятной обстановки,
чем водная среда, для предбиологического этапа химического
42
развития углеродных соединении и для возникновения первич-
ных организмов.
Роль воды в природе оценивалась человеком различно в
странах с различным климатом. В условиях жаркого климата
южных стран вода — благо, без которого не может обойтись
ни одно живое существо. Например, древние индийские сказа-
ния гласили, что вода — основа всей жизни мира, из которой
произошло все остальное. Древний Египет с его своеобразной
историей и культурой, существовавшей благодаря Нилу, был
всем обязан этой реке, ее периодическим разливам, увлажняв-
шим и улучшавшим почву долины, и, естественно, представлял
ее божеством. Аналогичное отношение к воде других древних
народов Северной Африки, Ближнего Востока и Южной Азии
являлось основой представлений о мироздании. Возникали
учения древних мыслителей, по которым даже звезды, Луна и
Солнце признавались вызванными к жизни испарением вод.
В странах же с влажным климатом, с периодическими ливнями,
разливами рек, затопляющих обширные участки суши, вода
представлялась злым началом, олицетворяла злое божество,
приносящее людям иногда большие несчастья. Началом мира
здесь признавалось Солнце и его свет, породившие Землю.
В них видели и начало жизни.
Некоторые мыслители (Ш —II в. до н. э.) полагали, что мир
состоит из пяти элементов: воды, огня, дерева, металлов и зем-
ли, что изменения в природе происходят под влиянием взаим-
ного уменьшения или увеличения этих пяти материальных сил.
Позднее возникло учение, по которому все вещи произошли от
первоначального жизненного «эфира».
Философские обобщения XVII —XVIII вв. в странах Запад-
ной Европы были первыми попытками создать представление
о космосе. В XVIII в. французский естествоиспытатель Ж. Бюф-
фон говорил о воде, что она может уходить с поверхности в
глубины и вновь появляться на свет, что воды неба, падая на
землю и стекая в океаны, разрушают ее поверхность. Тогда же
высказывались и мысли о расплавленной массе Земли, о разру-
шительной силе небесной воды и созидающей силе вод океанов.
43
Постепенно, побеждая религиозную трактовку вопросов
устройства Вселенной, научная мысль приближалась к совре-
менному мировоззрению. Уже в XIX в. стало общепризнанной
истиной суждение о круговороте воды в природе, складываю-
щемся из двух циклических процессов. Один из них, «внешний»
цикл выражается в испарении воды под действием солнечных
лучей с поверхности суши, морей и океанов, затем вода осажда-
ется, стекая в ручьи и реки, пока не собирается вновь в водные
бассейны земной поверхности. Второй, «внутренний» цикл
состоит в просачивании воды в землю, где она проникает в глу-
бины земной коры, в область высоких температур. Там, близ
очагов расплавленной магмы, вода переходит в пар с высокой
температурой и давлением, в результате чего создаются мощные
запасы энергии, проявляющиеся затем при вулканических из-
вержениях и при образовании гейзеров. Пары воды способст-
вуют поднятию лавы на поверхность и выбросу пепла в атмо-
сферу. При этом они поднимаются над жерлами вулканов на
большие высоты, образуя облака. Вулканизм тогда связывался
именно с деятельностью воды в этом «внутреннем» цикле ее
движения и объяснялся проникновением морских вод в глубь
земной коры по трещинам.
В XX в. новая наука уже по-новому трактует происхождение
вод. Появилось представление о существовании вод земной по-
верхности, так называемых вадозных, и вод глубинных, под-
нимающихся из недр, где они были связаны с веществом земной
коры, — вод ювенильных, или молодых. На эту мысль Э. Зюсса
натолкнуло исследование поднятия горячей трещинной воды
до высоты в 12 — 15 м известного гейзера Шпруделя в Карловых
Варах. Воды ювенильные, подмешиваясь к вадозным, таким
образом увеличивают количество воды в биосфере. Отсюда
пришла идея о том, что гидросфера Земли создана глубинными
процессами земной коры, которые, протекая в течение долгих
геологических периодов, и создают на Земле запас вод атмо-
сферы, тчорей и океанов, непрерывно пополняемый и в настоя-
щее время. Вместе с парами ювенильной воды в биосферу тем
же путем поступали и поступают разнообразные ювенильные
44
газы, так называемые «благородные» - гелий, аргон, а также
и «неблагородные» — сернистый газ, окись углерода, закись
углерода, водород и др. В соответствии с этой идеей, ныне
прочно вошедшей в представления геологов, вся вода земной
поверхности является наследием догеологической и геологиче-
ской истории Зе^ли.
А. П. Павлов следующим образом рисует историю образо-
вания морей и океанов. «Суша, неравномерно нагретая, с мно-
гочисленными вулканами несколько иного типа, чем теперь,
и с горячими источниками владела почти нераздельно поверх-
ностью планеты. Это и была древнейшая на земле пустыня...
Первобытные бури потрясали атмосферу Земли могучими
страшными концертами. По временам разражавшиеся ливни
выметали из диких скалистых горных долин в необозримые
безжизненные и голые равнины разнообразные продукты дроб-
ления... Солнце льет свой жар сверху в тех местах, где холодные
верхи гор не сгущают паров в тучи. Море еще не родилось или
только еще нарождалось в наиболее глубоких впадинах... Снизу,
еще близко к поверхности, недавно заключенный в каменную
оболочку отрывок солнечной массы — раскаленная магма Зем-
ли. Местами она льется по Земле могучими потоками, достав-
ляя свежий каменный материал для грядущих процессов раз-
рушения, или выбрасывает из глубин новые массы паров —
созидателей будущего моря».
Первичная гидросфера и атмосфера Земли еще не заслужи-
вали название биосферы, но уже были средой ее зарождения.
Что же представляла собой древняя среда жизни?
Изучение ферментативных процессов обмена веществ у орга-
низмов приводит к выводу, что они у разных групп сходны по
своему характеру, хотя и отличаются по структуре, и что они
всегда связаны с восстановительной обстановкой. Следова-
тельно, первичная среда, создавшая их, тоже должна была иметь
восстановительный характер. Значит первичный обмен у орга-
низмов был бескислородным.
Жизнь на Земле связана с большим количеством так называе-
мых «биофильных» химических элементов, среди которых вид-
45
нейшее место занимают углерод, кислород, водород, азот и
сера.
Появлению жизни на Земле предшествовала длительная
и сложная эволюция веществ, состоящих из этих элементов, —
от простых соединений к сложным, с их новыми свойствами и
поведением в «добиосфере».
Уже давно возникло представление о «главном слое жизни»,
за который принимается в гидросфере Земли самый верхний
«слой» водной среды с многообразным комплексом организмов
планктона, среди которых основное место занимают микроско-
пические одноклеточные водоросли - фитопланктон. На суше
роль главного слоя жизни играют поверхность и верхний слой
почвы. В прибрежных зонах, по краям выступов суши, у границы
их с водными бассейнами эти два типа главного слоя жизни
совмещаются.
В геологической жизни Земли такие районы оказывались,
в частности, основными производителями больших запасов
углерода и, вообще, органического вещества. Исследователи
отводят побережьям древнейших морей и океанов особое место,
поскольку здесь, в условиях больших колебаний температуры,
взаимодействовали самые разнообразные компоненты состава
биосферы: горные породы, вода, солнечная и космическая ра-
диация, углеводородные и иные жидкие и газообразные соеди-
нения.
Таким образом, древнейшая среда на Земле, в частности
водная, представляла собой концентрированный раствор слож-
ных органических соединений, механически и химически тесно
связанный с твердым веществом земной коры и находившийся
под воздействием мощных излучений. Эта среда — колыбель
первичных организмов — была восстановительной, бедной кис-
лородом. Это была арена разнообразнейших химических и гео-
химических реакций, из которых многие шли с выделением теп-
ловой энергии.
В такой обстановке Должны были успешно развиваться
многие микробиологические процессы, в частности, окисление
водорода кислородом углекислоты с одновременным образо-
46
ванием метана и многие аналогичные процессы с участием
других, исходных веществ древнейшей природы.
Создается впечатление, что древнейшая среда на поверх-
ности нашей планеты не была гибельной для многих групп
микроорганизмов. Наоборот, можно полагать, что все главней-
шие компоненты первичного состава биосферы, каждый в от-
дельности, могли быть основой, жизненным субстратом соот-
ветствующих проявлений жизни. В отношении типов сред,
способствовавших возникновению жизни и ее развитию, между
учеными существуют значительные расхождения во взглядах.
Известны весьма решительные высказывания в пользу почвен-
ных растворов в качестве предполагаемой среды возникнове-
ния жизни, притом в условиях высыхания мелких материковых
водоемов.
Имеются высказывания в пользу прибрежных зон мелко-
водных бассейнов, где этому могло благоприятствовать соче-
тание природных факторов: увеличения запасов углекислоты,
влияния химически действующих солнечных лучей, одновре-
менного присутствия в растворах усложненных углеродных
соединений и минеральных веществ земной крры. Под защи-
щающим от гибельных лучей слоем почвы или на малых глуби-
нах водоемов средой жизни в основном была, конечно, вода.
Итак, средой жизни является вода. Она же является обяза-
тельным компонентом в веществе живой клетки — внутрикле-
точной средой при процессах обмена, а также материалом в
этих процессах.
Зародыши жизни — коацерваты — представляли собой по-
лужидкие коллоидные вещества, проницаемые для некоторых
растворенных в водной среде солей, газов и самой воды. Как
известно, коллоидным веществом весьма свойственна способ-
ность к поглощению и связыванию различных веществ из окру-
жающей среды. К сожалению, природные коллоиды, в частно-
сти минеральные, в специальньдх исследованиях, касающихся
иногда явно органогенных образований, как железо-марганце-
вые конкреции на дне современных морей и океанов, вещества
фосфоритового состава, кремниевый гель (силикогель) и дру-
47
гие, обычно пока трактуются авторами вне связи с биологиче-
скими явлениями.
Между тем в вопросе первичной среды жизни и ее возник-
новения Н. Г Холодный придавал большое значение коллоид-
ным органическим соединениям. Поэтому, подытоживая из-
ложенное выше, можно сказать, что средой возникновения
жизни на Земле были природные водные растворы сложных
органических и минеральных соединений с соответствующими
температурами и давлениями, создававшиеся в результате
сложного химического взаимодействия вещества земной коры,
гидросферы и атмосферы.
В связи с этим можно признать, что средой и местом массо-
вого возникновения жизни были влажные почвы, водная среда
прибрежной зоны морских бассейнов, лагун и внутриконтинен-
тальных водоемов, их зоны прибоя, в особенности зона при-
ливов и отливов.
жизнь и изотопы
и
Изменение температуры химических процессов способно отра-
жаться на изотопном составе углерода, благодаря чему откры-
вается возможность определения температур древних сред,
в частности температур, при которых образовались каменно-
угольные пласты.
По содержанию С13 и О18 Г Юри измерил в 1951 г. темпера-
туру отложения карбоната кальция в юрском белемните. Вы-
раженная слоистость раковины — проявление сезонности раз-
вития животного. Установление повышенного содержания С12
в биогенных карбонатных образованиях было проверено на
архейской окаменелости возраста 1,4 млрд. лет.
Интересно, что метаморфизм и, вообще, любой нагрев,
даже искусственный, различных графитов не изменяет у них
отношение С12 к С13 Углерод животного происхождения имеет
отношение изотопов углерода в пределах от 90,1 до 92,5, углерод
растительного происхождения в пределах от 90,6 до 93,1, иско-
паемая древесина - от 90,1 до 92,2, торф до 92,8, лигнит -
от 90,87 до 91,10, битуминозные угли - от 90,53 до 92,1 как
и антрациты и полуантрациты.
Выяснено, что неорганические процессы, включая вулканизм,
способствуют увеличению запаса С13, тогда как органические
обогащают вещество изотопом С12. Полагают, что современная
атмосфера Земли почти полностью была биогенного происхож-
дения. В этой атмосфере первичной двуокиси углерода присут-
ствует немного.
С14 — радиоактивный изотоп углерода — образуется под
действием космической радиации на азот атмосферы, поскольку
его ядро способно поглощать нейтроны космического излуче-
ния. Возникающий таким путем изотоп С14 реагирует с кисло-
родом атмосферы, образуя С14О2 или С14О, которая окисля-
ется в озоновом слое до окиси углерода. Опускаясь, С14 погло-
щается наземными и водными растениями.
49
Элементарный углерод не играет роли в процессах на земной
поверхности в ее водах и на суше. Он представлен здесь в виде
относительно прочного соединения — углекислоты СО2.
Углекислота происходит из трех источников: а) углекислота
атмосферы; б) углекислота болотная, торфяная и получающаяся
при преобразовании вещества бурых и каменных углей в природе
и благодаря деятельности человека; в) углекислота из карбонат-
ных пород, главным образом из известняков, а также и из кар-
бонатных руд железа и марганца, образовавшаяся при их вовле-
чении в процессы выветривания.
Поступая в атмосферу, углекислота в той или иной степени
поглощается дождевыми водами, проникает в почву и в корен-
ные породы, где становится сильным геологическим деятелем
вследствие ее способности образовывать более сложные соеди-
нения с встречающимися веществами земной коры. Воздейст-
вуя на них в качестве угольной кислоты, она образует карбо-
натные и бикарбонатные соединения. Первые чаще являются
твердыми минеральными образованиями, вторые — образу-
ются в растворах и могут в таком виде, поднимаясь с водами
источников на поверхность Земли, входить в химические связи
с различными другими растворенными или твердыми мине-
ральными или органическими соединениями.
Несомненно, значительные количества окиси углерода об-
разуются при естественном преобразовании в земной коре ве-
щества каменных и бурых углей и лигнитов, которые извечно
проявляют общую тенденцию к потере так называемых лету-
чих компонентов. В числе этих компонентов углекислота обыч-
но занимает видное место с примесями закиси углерода (угар-
ного газа) и часто метана. Процессы выделения газов из уголь-
ного вещества связываются с его окислением при участии кисло-
рода, заносимого туда проникающими с поверхности водами.
Общеизвестно, что скопления газов в угольных пластах иногда
вызывают их активные выбросы, наподобие взрывов, представ-
ляющих опасность для шахтеров. Подземные воды, насыщае-
мые этой углекислотой, становятся сильными растворителями
и потому оказываются способными выносить на поверхность
50
из углей многие химические элементы, в числе которых кремний,
барий, олово, железо, марганец, бор, никель и др.
Известен и другой путь образования окиси углерода в уголь-
ных пластах. Содержащиеся часто в них в виде конкреццй пи-
рит и марказит, окисляясь при участии кислорода из притекаю-
щих поверхностных вод, что, как мы знаем, обычно происходит
при участии бактерий, дает с водой серную кислоту. Серная
кислота, воздействуя на известняки, залегающие там же, обра-
зует сульфат кальция (гипс) с выделением углекислоты. Таким
образом, в выделении свободной углекислоты в биосферу участ-
вуют различные процессы в недрах Земли и в связи с осадочны-
ми отложениями.
Между тем углекислота в указанных условиях способна обра-
зовывать ряд органических кислот, таких, как муравьиная,
уксусная и другие, которые могли подниматься к древней по-
верхности насыщенными соответствующими металлическими
основаниями или не насыщенными, т. е. способными к реаги-
рованию с другими веществами в других условиях. Известны
многочисленные разнообразные минеральные углекислые источ-
ники, часто используемые для лечебных целей, воды которых
бывают насыщены различными солями (карбонаты, хлористые
и сернокислые соли щелочных металлов, кальция и магния
и т. д.). Примеры того — источники Кисловодска, Карловых
Вар.
На относительно больших глубинах в недрах Земли свобод-
ная углекислота образуется при взаимодействии известняков с
кремнеземом в условиях высоких температур, с образованием
кремнистых пород, где к поверхности поднимаются горячие
соленые источники, богатые углекислотой. Поэтому полагают,
что влияние высокой температуры недр и вулканических образо-
ваний на породы осадочных толщ состоит в выделении из по-
следних углекислоты.
Наибольшее количество углекислоты биосфера получает в
процессе вулканической деятельности. Эта углекислота — юве-
нильная, выделяется при остывании лавовых масс и в последние
фазы вспышек вулканической деятельности. Это явление было
51
изучено в районах действующих вулканов Италии и древних
вулканов Чехии и Оверни.
В вулканических областях известны струи газов с содержа-
нием СО2 до 59%, например на о-ве Мартиника у Мон-Пеле,
а также около Неаполя. В области Тихоокеанского вулканиче-
ского пояса, в Северной Америке, в «Долине десяти тысяч ды-
мов» сухие газовые струи углекислоты, вырываясь в атмосферу,
сильно охлаждаются и газ выпадает у самых источников в виде
твердой углекислоты.
Громадные запасы углекислоты в земной коре находятся в
связанном состоянии в разнообразных изверженных породах,
куда она попала в процессе их первичного формирования.
Азот, количественно основной компонент атмосферы Земли,
представлен в ней двумя стабильными изотопами N14 и N15 в
соотношении: N14 — 99,635%, N15 — 0,365%. Отношение
N14/N15=273. Изотоп N14 под влиянием космического излуче-
ния превращается в верхних слоях атмосферы в радиоактивный
углерод С14 и в радиоактивный тритий — Н3 При искусствен-
ной бомбардировке альфа-частицами изотоп N14 превращается
в кислород О . На больших высотах отношение между изото-
пами N14 и N15 возрастает, в частности, на высоте 58,3 км до
282,6. Полагают, что в разделении изотопов проявляется сила
тяжести.
Важно отметить, что в радиоактивных минералах различного
геологического возраста отношение изотопов азота иное. Так,
отношение N14/N^ в урановой смолке различного геологиче-
ского возраста следующее:
Возраст, млн. лет Местонахождение N14/N15
1251 Большое Медвежье озеро, Канада 170
625 Катанга, Заир 173
625 Шинколобве, Заир 173
57 Вуд Колорадо, США 187
52
Из этих данных отчетливо видно, что у определенного вида
минералов отношение изотопов отражает геологический воз-
раст, проявляя тенденцию со временем к относительному умень-
шению изотопа N15. По мнению некоторых ученых, отношение
изотопов в минералах, содержащих азот, не зависит от химиче-
ского их состава, хотя наблюдаются и исключения.
Откуда появился азот в атмосфере Земли и ее минеральных
образованиях? Один из возможных путей — это распад какого-то
азотного соединения под влиянием излучений радиоактивных
элементов на азот и гелий. Таким исходным веществом мог
быть скорее всего аммиак, известный в составе атмосферы ряда
планет Солнечной системы. При распаде U238 также образу-
ется азот. Наличие азота в составе многих минералов должно
указывать на особую геологическую, или точнее догеологиче-
скую, древность этого процесса, относящегося к стадии форми-
рования нашей планеты и ее коры.
Между тем есть и другие точки зрения на происхождение азо-
та в атмосфете. Так, В. И. Вернадский считал, что азот постоян-
но приходит в атмосферу из глубоких зон земной коры в составе
газов, выделяющихся при деятельности вулканов и минераль-
ных. источников. На Земле во многих местах известны газовые
струи, почти нацело состоящие из азота с примесью благород-
ных газов (гелий, аргон). Под влиянием электрических разрядов
азот входит в соединение с водородом и кислородом, образуя
NH3, NO2 и NO3, азотистокислые и азотнокислые соли аммония,
а также и более сложные азотные органические соединения,
«строящие тела организмов».
В природе имеет место грандиозный процесс извлечения из
биосферы свободного азота бактериями почвы и вод, а также
некоторыми водорослями. Азот при этом поступает в тела ор-
ганизмов в виде разнообразных соединений, а потом иногда
накапливается в виде азотсодержащих минералов, например,
селитры, азотных соединений нефтей, каменных углей и т. д.
В то же время в природе идет не менее грандиозный процесс
выделения свободного азота биохимическим путем, тоже при
участии организмов, в основном бактерий, принадлежащих
53
к группе так называемых денитрифицирующих. Таким образом,
часть азота из состава живых тел и захваченных ими аммиачных
и кислородных соединений возвращается назад в атмосферу,
часть откладывается в составе ряда минералов или снова за-
хватывается живым веществом биосферы (селитра, аммиачные
соединения) или захороняется в осадочных отложениях.
В. И. Вернадский полагал, что количество свободного азота
в атмосфере, по-видимому, не меняется и что его приток из бо-
лее глубоких слоев Земли возмещает его потребление организ-
мами и образование азотсодержащих минералов. Решающая
роль в этом процессе отводится живым организмам.
Если учесть приведенные выше данные, что биофильный
изотоп азота N14 в истории Земли проявляет тенденцию к на-
коплению в биосфере, по отношению к более тяжелому изо-
топу, то относительное обогащение некоторых минералов изо-
топом N15 позволяет считать его в большей степени связанным
с химическими процессами. Между тем в первичной атмосфере,
где преобладали лишь химические реакции преобразования ее
вещества под влиянием электрических разрядов, трудно допу-
стить большое содержание свободного азота. Скорее всего,
он был преимущественно связан в соединения с водородом и
углеродом в виде аммиака и углеводородных газов.
Водород представлен в природе тремя изотопами: Н1 —
протий, Н2 — дейтерий и радиоактивный Н3 — тритий.
Присутствие свободного водорода в составе атмосферы и
гидросферы нашей планеты не вполне еще исследовано., В окру-
жающей среде мы не видим его заметного содержания, но име-
ется достаточно данных о его присутствии в верхних слоях
атмосферы, что установлено исследованиями с помощью ра-
кет. В то же время известно, что свободный водород присутству-
ет в глубинах земной коры и выделяется при вулканических
процессах. По данным Резерфорда, весьма вероятен синтез
водорода в процессе разрушения атомов других элементов.
Значение водорода определяется его свойствами, в частности
исключительной легкостью, вследствие чего Земля может об-
мениваться им с космосом.
54
В земной коре водород наряду с другими газами захваты-
вается магматическими и лавовыми массами. Крайне интерес-
но, что в комплексе этих газов оказываются Н2, СН4, С2Н4,
СО, СО2, N2 и другие, поскольку их ассоциация, несомненно,
отражает исконное свойство земной материи и ее первичный
газовый состав вообще.
Содержание водорода в твердых породах достигает 36 — 38%.
Водород входит в состав вод минеральных источников, нефтей,
часто выделяется мощными струями из залежей морских солей,
в частности калийных. В газовой фракции выбрасываемых вул-
канами веществ он составляет до 3% у Везувия, до 22,3% у
Мон-Пеле, а в Исландии вулканические источники выбрасы-
вают струи газа, содержащие 54% водорода. Кроме того, во-
дород высвобождается при гниении органических (белковых)
веществ вследствие жизнедеятельности некоторых видов бак-
терий, что должно давать большой эффект в ходе геологиче-
ского времени.
На высоте около 100 км водород является господствующим
газом. Предполагается, что он частично улетучивается из зем-
ной атмосферы в космическое пространство. Под влиянием
азона верхних слоев атмосферы, под влиянием почвенных
бактерий и другими путями водород окисляется в воду.
Изотопы водорода дейтерий и протий обычно присутствуют
в соотношении 1 :6409. В связи с различием в массе протия и
дейтерия органические и неорганические процессы с их участием
идут различно. Обычно протиевые соединения вступают в ре-
акцию энергичнее и быстрее, чем дейтериевые, примерно в
6 раз. Тритий радиоактивен; его Содержание в веществах опре-
деляется путем измерений радиоактивности. Полагают, что из
изотопов водорода именно протий проявляет тенденцию к
утечке в космическое пространство, тогда как дейтерий в ходе
геологического времени накапливается. Окись дейтерия
так называемая «тяжелая» вода — имеет особые свойства, в
частности является менее активным химическим веществом по
сравнению с водой. Тяжелая вода вредна для организмов, а
чистая окись дейтерия даже ядовита, способна задерживать био-
55
логические процессы. Между тем организмы, усваивая воду,
способны разделять изотопы в соотношении обычно около
4500 — 6000.
Водород — активный биофильный элемент. Многие бакте-
рии, способные к автотрофному существованию, т. е. не тре-
бующие для себя готовых органических соединений, могут по-
лучать жизненную энергию при окислении водорода: Н2 +
+ 1/2О2 =Н2О + 56 больших калорий энергии. Правда, эти ми-
кроорганизмы не являются обязательными автотрофами, так
как могут существовать и на обычных органических средах.
Имеется группа водородных бактерий, которые усваивают во-
дород, окисляя его. в присутствии кислорода среды (аэроб-
ные бактерии). Некоторые бактерии развиваются автотрофно
на минеральной среде в присутствии окиси углерода и кислорода.
Существование автотрофных бактерий, добывающих энер-
гию за счет окисления водорода в присутствии углекислоты,
представляется фактом исключительного значения, к которому
далее мы вернемся.
В природе существует три изотопа кислорода: О16, О17 и
О18 При этом в атмосферном кислороде изотопы составляют
соответственно 99,760, 0,042 и 0,198%. Изучение их распростра-
нения в природе позволила прийти к выводу, что тяжелый кис-
лород О18 преимущественно концентрируется в кислороде дву-
окиси углерода, в частности в кислороде карбонатов и образо-
ванных из них кислородных соединений. Установлено, также,
что О16 скорее усваивается организмами, в частности почвен-
ными бактериями и в процессе фотосинтеза растений. Этот
изотоп в основном составляет кислород воды. Именно О16,
легкий изотоп кислорода, преобладает в процессе жизнедея-
тельности морских организмов, поглощающих кислород.
Поскольку тяжелый изотоп кислорода в большей степени
оказывается связанным с минеральным веществом земной коры,
а легкий О16 — с жизнедеятельностью организмов и водой
гидросферы, то геологическая история их, несомненно, была
различной. Малые запасы тяжелых изотопов О17 и О18 вероят-
но, представляли собой кислород древнейшей атмосферы Земли,
56
тогда как легкий изотоп создавался организмами, вероятно,
за счет соответствующих соединений, в связи с его более высо-
кой химической активностью.
Ранее полагали, что кислород атмосферы происходит из
кислорода двуокиси углерода при фотосинтезе. Между тем ока-
залось, что кислород водных растений образуется при фотосин-
тезе из воды, что подтверждено данными изучения фотосинтеза
у ряда видов современных водорослей (хлорелла, гелиатус и др.).
В выделяемом водорослями кислороде изотопа О18 оказыва-
ется меньше, чем в составе атмосферы. Изучение А. П. Вино-
градовым, Р В. Тейс и другими учеными процесса фотосинтеза
у элодеи показало, что состав фотосинтетического кислорода
совпадает с изотопным составом кислорода воды, но сущест-
венно отличается от общего изотопного состава кислорода
атмосферы и кислорода углекислоты. Изотопный состав фото-
синтетического кислорода оказывается средним по отношению
к составу кислорода атмосферы и углекислоты. Это обстоятель-
ство породило гипотезу о том, что в растениях содержатся
атомы кислорода, происшедшего как из воды (две трети), так
и из двуокиси углерода (одна треть). Путем определения изо-
топного состава кислорода ряда древних изверженных пород
был установлен первоначальный изотопный состав земного
кислорода. Отношение между изотопами О16/О18 для него по-
лучилось в пределах 495,6—495,3. Выяснено, что содержание
О18 в карбонатных осадках способно указывать температуру
процесса их осаждения в геологическом прошлом — палеотем-
пературу. Г Юри на основе тщательного измерения содержа-
ния О18 в породах разработал метод измерения температуры
воды в момент образования отложения древних карбонатных
осадков.
В. И. Вернадский, отмечая наибольшую роль водорода и
кислорода в образовании живого вещества, считал кислород,
в виде свободного кислорода биосферы, «продуктом жизни».
Он доказывал, что для большинства элементов их геохимия
обусловливается живым веществом и вообще до 99% (по весу)
вещества земной коры считал «обусловленным жизнью», из
57
чего вытекало его представление о теснейшей вещественной
связи между живой и косной (мертвой) природой.
Общее содержание кислорода в современной атмосфере —
2,8-1014 т, между тем все растения земного шара в процессе
фотосинтеза за год выделяют его 1,2 1011 т. Таким образом,
весь запас кислорода современной атмосферы Земли мог бы
образоваться всего за 2 — 3 тыс. лет, если бы он не расходовался
на разнообразные окислительные процессы химического и био-
логического порядка. Правда, имеются высказывания о неорга-
ническом пути образования свободного молекулярного кисло-
рода вследствие термической диссоциации воды, но они не по-
лучают поддержки со стороны геологов и химиков.
Возможно, что диссоциация паров воды и происходит в верх-
них слоях атмосферы при участии солнечной и космической ра-
диации. Приводимые подсчеты количеств кислорода, который
мог бы образоваться таким путем за три миллиарда лет сущест-
вования Земли оказывались в десятки раз больше, чем современ-
ные запасы молекулярного кислорода в атмосфере. Однако
количество паров воды в верхних слоях атмосферы весьма не-
значительно и потому эта гипотеза не получила признания
специалистов.
Сера — активный биофильный элемент. На Земле она в
основном находится в зоне биосферы, где входит в органиче-
ские и минеральные соединения, отличающиеся огромным
разнообразием. Последнее объясняется существованием у серы
способности образовывать полимеры в результате различной
валентности (S8, S6, S4, S2 и SJ. Известно четыре устойчивых
изотопа серы (S32, S33, S34, S36) и получено искусственным путем
несколько радиоактивных ее изотопов. Так, например, период
полураспада S35 равен 87,1 дня, что делает его пригодным для
применения в опытах с мечеными атомами. Изотопный состав
серы зависит от того, с минеральными или с биологическими
процессами она связана. Общее отношение S32/SM варьирует
в пределах от 21,4 до 23,2. Природная биогенная сера и биоген-
ный сероводород имеют отношение этих изотопов в пределах
от 22,4 до 22,6, тогда как сульфатные минералы химического
58
происхождения — 21,6 — 22,0. Природная вулканическая сера
имеет отношение изотопов чуть выше 22,2, тогда как сера суль-
фидных минералов изверженного происхождения — чуть ниже
22,2. Считают, что деятельность серных бактерии моует при-
вести к более или менее полному разделению изотопов серы.
Интересно отметить, что количественное отношение изото;
пов серы S32/SM, наблюдаемое в сульфидных минералах, в част-
ности в пирите, не зависит от геологического возраста мине-
рала, но дает указание на температуру среды кристаллизации.
При этом выяснено: что при высоких температурах в начале
процесса осаждается изотоп S3*, а при понижении температур
S32 в виде мелко- и затем крупнокристаллического пирита.
В первом случае отмечались температуры 235 — 210°, а во вто-
ром 185 — 135° Открытие такого «серного термометра» геохи-
миками Тодом, Макнамара и Коллинсом очень ценно для изу-
чения состояния сред «ископаемой» биосферы. Правда, этот
«термометр» при его практическом использовании требует
очень внимательного изучения условии осаждения минерала и
последующих процессов, которые могли бы изменить соотно-
шение между изотопами.
Таким образом, доказано, что в природе происходит повы-
шение концентрации S3* при неорганических процессах и обо-
гащение S32 при биологических, что может служить ключом
для понимания условий отложения серы в далеком геологиче-
ском прошлом. Принято считать, что отношение S32/SM боль-
ше 22,3 указывает на биогенное происхождение материала,
а меньше 22,18 — на минеральное. Установлено, что изотопное
разделение серы в докембрии выражалось слабо и началось
лишь 900 млн. лет назад, когда оно стало более заметным и
доступным для исследования. Объяснение явления, данное
Г Юри, может оказаться очень важным, если оно будет под-
тверждено фактическими данными. Г Юри полагает, что около
800 млн. лет назад восстановительные (бескислородные) среды
сменились окислительными (кислородными).
Известны группы нефотосинтезирующих автотрофных сер-
ных бактерий, способных окислять сероводород, с выделением
59
свободной серы (процесс формирования серы!), воды и с выделе-
нием при этом 172 кДж тепловой энергии на 1 грамм-молекулу
вещества. Существуют окислители серы в присутствии воды,
приводящие к образованию серной кислоты и к получению при
этом 496 кДж энергии.
Открытие автотрофных бактерий — окислителей серы в
природе, сделанное С. Ваксманом и Дж. Иоффе в 1921 г., при-
вело к возможности дальнейшего успешного исследования про-
цессов выделения серы в истории Земли. Впоследствии было
установлено, что сера выделяется и рядом других организмов,
не вполне автотрофных или гетеротрофных. Были выделены
группы организмов, жизнедеятельность которых протекает в
кислородной среде, а т#кже и такие, которые пользуются кисло-
родом, связанным в соединениях различного типа. Такие ана-
эробные микроорганизмы должны быть признаны древнейшей
из групп серных бактерий. Они берут из нитратов кислород, за
счет которого окисляют молекулярную серу.
Таким образом, биохимическое выделение самородной серы
или образование сероводорода в геологическом прошлом то
проявлялось, то исчезало в зависимости от условий, причем
восстановительная обстановка, преобладавшая в ранней био-
сфере Земли, для этого, несомненно, не служила препятствием.
В биосфере существуют факторы, регулирующие состав
атмосферы, — это водная оболочка Земли и живое вещество
природы.
Вода растворяет газы воздуха. Огромное их количество на-
ходится в растворе во всех типах природных вод: морских, соло-
новатых и пресных. Поскольку кислород растворяется лучше,
то воды содержат его в относительно больших количествах по
сравнению с азотом, что делает водные среды в химическом и
биологическом отношениях более активными. Этим путем
газы доходят до самых больших глубин морей и океанов, делая
их доступными для организмов. Море то выделяет кислород
в атмосферу, то поглощает его вновь.
Как отмечал В. И. Вернадский, в природе известны тысячи
реакций, связанных с поглощением, связыванием кислорода.
60
Между тем свободный кислород в истории Земли мог быть
образован только при жизненных процессах, при выделении его
зелеными хлорофильными организмами под влиянием энергии
света. Поэтому процесс выделения кислорода приурочен исклю-
чительно к так называемой кислородной поверхности — земной
коре, суше и водам. С глубиной выделение кислорода прекра-
щается и там уже господствует восстановительная обстановка,
В. И. Вернадский не допускал мысли, что в природе мог сущест-
вовать какой-либо иной источник свободного кислорода, кроме
биологического, биохимического, что это Является основной
чертой*истории кислорода Земли. Таким образом, весь кислород
атмосферы проходит через живое вещество. При этом подсчиты-
валось его количество (1,2 Ю15 т), что указывает на масштаб
проявлений жизнедеятельности фотосинтезирующих организ-
мов в ходе геологической истории Земли.
Естественно, возникает вопрос о форме существования кисло-
рода в догеологический период истории Земли. Ответом на него
может служить, по-видимому, мнение В. И. Вернадского, что
кислород был тогда полностью в связанном виде в многообраз-
ных соединениях типа окислов.
ПРОИСХОЖДЕНИЕ ОРГАНИЗМОВ
Бесчисленные виды растений, животных и микроорганизмов,
обитающие ныне на Земле в различных условиях, поражают
разнообразием своего строения и жизнедеятельности.^Организ-
мы обитают на поверхности суши, в почвах, в пресных и соленых
водных бассейнах, в глубинах земной коры и под дном морей
и океанов. Они прослежены до глубин, которых достигают бу-
ровые инструменты (до 7 км). Организмы можно обнаружить в
воздухе, который переносит их на значительные расстояния,
иногда на высоте в несколько километров. Споры и пыльца
растений, огромное разнообразие микроорганизмов, переноси-
мые вместе с земной и космической пылью, захватывают не
только тропические, теплые и умеренные пояса, но и области
высоких широт, вплоть до полярных. Эти зародыши жизни,
микроорганизмы и их споры проявляют жизнедеятельность
всюду, где они только находят для этого благоприятную об-
становку.
С тех пор, как наука решительно победила веками бытовав-
шие представления о «чудесном» происхождении всего мира,
мысль человека постоянно обращается к вопросу: как возникло
это огромное разнообразие организмор на нашей планете?
Микробиологи, ботаники и зоологи часто рассматривают
современные организмы с точки зрения уровня эволюционного
развития, располагая их в системы и ряды от наиболее простых
к сложным по жизненным функциям и строению. Точно так же
биогеохимическая наука создала и разрабатывает убедительную
историческую картину связи организмов с минеральным ве-
ществом Земли. Чем ниже уровень организации живого су-
щества, тем большую роль играют в его жизнедеятельности
(например, бактерии — геологические деятели) элементы мерт-
вой природы. Поэтому даже в современном живом населении
Земли можно подобрать такой ряд организмов, который как
бы отражает ход общей эволюции жизни.
62
Настоящее млн лет 500 1000 1500 1000 500 млн лет Настоящее
еммя время
Развитие организмов на Земле (но Суиннертону)
Однако все современные организмы являются продуктом
длительного хода эволюции жизни на Земле. Они приобрели
через бесчисленные смены поколений разнообразные особен-
ности и свойства, которых, несомненно, не имели их далекие
предки, поэтому палеонтологическая наука, изучающая остатки
прежде живших организмов, способна внести в данную проблему
верные решения. Идеи Ж. Б. Ламарка и Ч. Дарвина — стройное
учение о естественном отборе форм и происхождении видов —
дарвинизм, — успешно дополняемые данными мичуринского
учения о связях между организмом и средой, о неразрывности
этой связи, вскрывают общее направление эволюции жизни на
Земле. Этапы развития определенных комплексов организмов
63
геологического прошлого признаны этапами геологического
развития самой планеты, разделенного на эры, периоды и гео-
логические века. При этом, чем древнее подразделение осадоч-
ных пород с остатками организмов, тем примитивнее по построе-
нию и часто мельче по размерам оказываются представленные
там организмы, а чем позднее образовался слой породы, тем
остатки организмов в нем оказываются более сложно устроен-
ными и более разнообразными.
Обращая внимание на уровень организации ископаемых
форм жизни и основываясь на еще относительно скудных па-
леонтологических материалах того времени, Ф. Энгельс призна-
вал эволюцию жизни как движение материи, как переходы от
простого к сложному, как процессы обмена белковых соедине-
ний со средой, с основной ролью в них белков как носителей
жизни. Ф. Энгельс писал, что остается добиться еще только од-
ного: объяснить возникновение жизни из неорганической при-
роды.
Белковые тела представляют собой цепи связанных между
собой аминокислот (полипептидные цепи), уложенные в моле-
кулах таким образом, что в точках соприкосновения между
частями этих цепей возникают более или менее прочные химиче-
ские связи. Процесс образования белковых молекул требует
поэтому затраты энергии. Кроме того, в белковых телах могут
самопроизвольно образовываться вторичные связи, требующие
меньшей затраты энергии. Особенностью живой ткани организ-
ма, условием его существования является образование новых
аминокислот и распад части старых.
Практика лабораторной работы по синтезу более сложных
соединений из более простых при помощи соответствующих
энергетических воздействий добилась больших успехов. Обычно
для этого применяются высокие давления, поскольку при син-
тезе объем получаемого вещества меньше, чем сумма объемов
исходных веществ.
Опираясь на идеи А. И. Опарина и Г Юри о первичном
составе атмосферы Земли из метана, аммиака, воды и водорода,
профессор Колумбийского университета Стэнли Г. Миллер в
64
1955 г. провел очень интересный опыт синтеза органических
соединений в искровом электрическом разряде. Он подводил
к смеси метана, аммиака и водорода нагретые пары воды под
давлением в пределах 100 — 200 мм рт. ст. Эта смесь подвергалась
действию искровых разрядов с максимумом напряжения в
60 тыс. вольт. Было обнаружено что, кроме исходных газов,
в приборе присутствует целый ряд новых углеродных соедине-
ний. Получившаяся сложная смесь соединений была разделена
различными методами.
Результаты этого замечательного опыта подтвердили воз-
можность природного образования сложных органических сое-
динений из немногих простых компонентов, составляющих
первичную атмосферу Земли С. Миллер получил глицин, ала-
нин, аспарагиновую, глютаминовую кислоты и другие соеди-
нения, которые входят в состав белков или участвуют в биохи-
мических процессах обмена веществ организмов. Многие из
этих веществ могут быть единственными источниками углерода
и энергии организмов. Таким образом, опыт С. Миллера по-
казал, что органические соединения можно получить из простых
неорганических соединений.
Аминокислоты и полипептиды еще далеки от живого орга-
низма. Нужно найти механизмы синтеза еще более сложных
органических соединений — пуринов, пиримидинов, жир’ов, ко-
ферментов и нуклеиновых кислот, в особенности ферментов —
полипептидов, способных содействовать обмену веществ.
С. Миллер отмечает, что роль катализаторов в реакциях, про-
исходящих в организмах, могут играть и некоторые металлы
и простые органические соединения, но вряд ли их деятельность
могла быть достаточной для обеспечения жизнедеятельности.
Поэтому он приходит к заключению, что именно полипептиды
были катализаторами почти с самого начала возникновения
обмена веществ, еще до того, как развился воспроизводящий
себя организм. Таким образом, чтобы существующая гипотеза
о происхождении организмов стала вполне доказанной научной
теорией, экспериментальному направлению химии и биологии
предстоит еще получить искусственно эти полипептиды.
65
Ч С. Миллер считает возможным два пути образования пер-
вичных живых организмов. Первый из них — это появление,
согласно представлениям А. И. Опарина, коацерватов — разно-
роднейших по структуре и свойствам первичных сгустков белко-
вых веществ. Попадая в виде микрокапель в сферы развития
геохимических реакций с выделением тепла, эти сгустки на мо-
мент или на более длительные сроки «оживали», становясь сами
средой таких реакций. Продолжительность «жизни» соответ-
ствующих систем зависела от того, насколько успешно налажи-
валась их связь со средой, их породившей. Например, закисное
соединение, проникнув внутрь коацервата путем осмоса через
его граничный слой, превращалось в окись с выделением неко-
торого количества энергии. Поскольку закись исчезает, то сюда
из среды путем осмоса поступает новая ее порция, чем поддер-
живается процесс и, следовательно, жизнь. Коацерваты могли
накапливать еще больше белков и, таким образом, в течение
длительного времени вырабатывать механизм для синтеза бел-
ка и для генетического размножения, что составляет признаки
организма.
Вторую возможность развития органического вещества
С. Миллер видит в возникновении так называемой дезоксири-
бонуклеиновой кислоты — сокращенно ДНК, свойственной ве-
ществу современных организмов. Молекулы этой кислоты мог-
ли содержать «биологическую информацию», образовывать
мутации (скачкообразные изменения), в результате чего мета-
болические (созидательные) способности первичных форм жиз-
ни могли увеличиваться, мог расшириться обмен их веществ со
средой. Полезные изменения — это уже признаки живого орга-
низма. Естественный отбор должен был обеспечить дальнейший
ход эволюции организмов.
Какой из этих двух путей проявился в процессе возникнове-
ния жизни, пока еще не ясно. С. Миллер связывает решение этого
вопроса с изучением путей синтеза ферментов и воспроизводства
генов в современном организме.
Обширные геологические материалы говорят об определен-
ном этапе истории нашей планеты, когда возникли процессы
66
почвообразования на выступах суши, когда отлагались массы
мелкозема в древнейших бассейнах, когда начали отлагаться
микробиологическим и химическим путями известковые илы,
гидрозакиси железа, сульфиды железа и других металлов в
условиях восстановительных, т. е. при отсутствии в атмосфере
и водах запасов свободного кислорода.
Общеизвестно, что перевод бикарбонатов в карбонаты, за-
кисных соединений и элементов в окислы и закиси всегда сопро-
вождается выделением свободной энергии. Поэтому естественно
представить себе, что древнейшие формы жизни использовали
для своей жизнедеятельности ту или другую геохимическую ре-
акцию с выделением энергии. Таким образом, создается впечат-
ление, что древнейшими формами жизни являлись автотрофные
организмы, т. е. такие, которые не требуют готовых органиче-
ских соединений и способны довольствоваться лишь углекисло-
той и перерабатываемыми ими минеральными веществами зем-
ной коры.
Известно, что наиболее распространенные в природе авто-
трофы — это зеленые растения. Для их жизнедеятельности
нужна углекислота при источнике энергии в виде света, источ-
ник азота, серы и других элементов. На самой низкой ступени
по уровню организации стоят одноклеточные микроскопиче-
ские водоросли, которые иногда придают зеленую окраску стоя-
чим пресным водам, верхнему слою морских и океанских вод,
которые получают нужные им питательные вещества в виде
углекислоты и минеральных солей из воды через клеточную
оболочку. Среди бактерий есть формы пигментированные
окрашенные и бесцветные. Для их развития тоже необходим
свет, и многие из них также принадлежат к автотрофным орга-
низмам. Есть и бактерии, жизнедеятельность которых подавля-
ется светом и которые могут жить только в темноте. Но и они
способны производить белки, углеводы, жиры и другие сложные
соединения из соединений углерода типа углекислоты, исполь-
зуя энергию окисления некоторых минеральных веществ.
Весьма важно, что химический анализ живого вещества авто-
трофных бактерий показал в них наличие того же набора амино-
67
кислот, углеводов, жиров и биологически активных веществ —
ферментов, который свойствен и гетеротрофным организмам.
Из этого следует, что в широком смысле все формы жизни по-
строены по одному образцу.
Возникновение жизни произошло, несомненно, сразу же,
как только температура в водных бассейнах или на смежных
выступах суши снизилась до пределов, допускающих сохран-
ность белковых веществ. Идеи А. И. Опарина в отношении абио-
генной эволюции углеродных соединений на основе синтеза из
первичных газовых компонентов атмосферы при наличии паров
воды или воды в жидкой фазе не встречают возражений ни со
стороны биохимиков, ни со стороны геологов. Однако могут
быть некоторые расхождения в представлениях, какими были
первичные организмы — гетеротрофами или автотрофами. Не-
сомненно при этом, что те или другие или те и другие обязатель-
но должны были обладать способностью существования в ана-
эробных, т. е. в бескислородных условиях.
когда исследователи задумываются над первичной формой
и первичным носителем жизни, первым комочком органическо-
го вещества, приобретшего способность к размножению, то
возникают представления или о бактериях, или о иных орга-
низмах, с той или иной поправкой на «древность» таких существ.
Естественно, какую бы группу современных микроорганизмов
мы не взяли под наблюдения как модель первых организмов,
мы не можем упускать из виду их длительной эволюции, дли-
тельного приспосабливания к средам обитания с совершенство-
ванием в процессе отбора функций клеточного вещества.
Микробиологией изучено огромное разнообразие бактерий,
которые еще не так давно считались самыми мелкими предста-
вителями жизни на Земле. Несколько десятилетий назад были
обнаружены еще более мелкие элементы, невидимые в самые
лучшие микроскопы, которые были названы вирусами. Свойст-
ва и химический состав многих вирусов в настоящее время
раскрыты и хорошо изучены. По-видимому, все вирусы способ-
ны развиваться только в живых клетках организмов раститель-
ных и животных или в культурах их тканей.
68
Еще одна группа мельчайших организмов, которая по раз-
мерам укладывается между бактериями и вирусами, а по свой-
ствам близка к бактериям — это так называемые микоплазмы,
микроорганизмы, способные жить и развиваться как в организ-
мах, так и вне их. Способность некоторых из них жить в сточных
водах и почвах на продуктах разложения растений и животных
показывает, что среди них есть и сапрофиты, т. е. организмы,
использующие для своей жизнедеятельности готовый углерод
из органических остатков.
Вероятно, исследователям предстоит еще много поработать
над современными микоплазмами. То, что известно о них те-
перь, указывает на сильное морфологическое непостоянство —
пластичность, а также большую эластичность их клеток, ли-
шенных ригидной клеточной оболочки. Микоплазмы оказы-
ваются «голыми», тогда как бактерии имеют хорошо развитую
клеточную оболочку. Клетки микоплазм это нити и шаровид-
ные сгустки протоплазмы, которые способны менять форму в
процессе развития микоплазм. Нить может затем переходить
в цепочку округлых телец. Клеточное ядро имеется. Размноже-
ние осуществляется делением, или материнская клетка распа-
дается на массу мелких дочерних телец.
Основное питание современных микоплазм — белковые ве-
щества кровяной сыворотки, холестерин. Сами они, оказыва-
ется, неспособны синтезировать многие органические вещества,
в которых нуждаются. Но ведь микоплазмы — продукт дли-
тельной эволюции. Какими они были в геологическом прош-
лом? Н. Г Холодный дал им название «пробионтов», т. е. еще
«не вполне организмов» и приписывал им проявление высокой
приспособляемости, «пластичности» при изменениях условий
обитания. «Толчковые» изменения условий среды, уничтожая
одних пробионтов, способствовали подъему на более высокий
уровень эволюции других с превращением их в подлинно живые
существа — «археобионты».
Жизненные проявления у клетки организма зависят от физи-
ко-химического состояния ее веществ, характеризуемых в осо-
бенности концентрацией в них свободных водородных ионов —
69
так называемой величиной pH. От pH зависят вязкость белков,
осмотическое давление, способность набухания, поверхностное
натяжение и т. д. Поэтому считают, что соответствующий pH
является и необходимым условием и причиной жизнедеятель-
ности клеток. Различия в величине pH, несомненно, объясняются
составом веществ клетки, так как по существу химический со-
став клетки или организма в целом является признаком устой-
чивым, признаком систематического значения. Можно сделать
вывод, что осуществление жизненных процессов в первичном
сгустке белкового вещества связано с pH, обусловившим воз-
никновение процесса обмена вещества со средой, т. е. жизни.
Первичные организмы, естественно, могли не иметь оболоч-
ки, могли быть пластичными в виде сгустка протоплазмы и
при размножении делиться на две или на большее число дочер-
них частиц. Они могли питаться рассеянным в природе органи-
ческим веществом, вследствие чего должны были быть сходны-
ми с современными микоплазмами. Именно отсутствие оболоч-
ки и пластичность тел организмов типа микоплазм могли быть
основой бурной эволюции, которую прошла жизнь в ходе геоло-
гической истории Земли. От микоплазм могли произойти бак-
териальная клетка и одноклеточный животный или раститель-
ный организм, питанием для которых могли служить минераль-
но-органические среды.
Отметим, что понятия «окисление» и «восстановление» свя-
зываются с получением элементов положительного заряда (от-
дача отрицательного электрона) или с получением отрицатель-
ного заряда (получение отрицательного электрона). Ранее окис-
лением называли только присоединение кислорода. Действи-
тельно, в природе веществом, которое дает положительный
заряд (отнимает электрон), часто является кислород, тогда как
водород — восстановитель, так как он дает отрицательный
электрон. Но в современном понятии окислением является не
только присоединение кислорода, но и отдача водорода, отда-
ча электрона без участия водорода или кислорода. При этом
водород, потерявший электрон, является восстановленным,
получивший электрон — окисленным.
70
Вещественная основа жизнедеятельности организмов со-
здается производством и воспроизводством ими различных
белковых соединений, в основном представляющих собой так
называемые ферменты, белковые тела, которые вызывают про-
цессы обмена веществ, характеризующие жизнь. Необходимо
постоянное образование новых запасов таких тел в организме,
чтобы он мог существовать и развиваться. Изменение в составе
ферментов способно изменять свойства организмов, придавать
им новые признаки.
Химия белковых тел в настоящее время располагает возмож-
ностями анализа белковых тел клеток организмов, которые
представляют собой смесь весьма многочисленных различно
построенных белковых соединений, выполняющих определен-
ные функции. Белковые тела, это сложные, гигантские по срав-
нению с молекулами, известными неорганической химии, мо-
лекулы, в образовании которых принимают участие сотни
более простых органических веществ - аминокислот. Предпо-
лагается, что из 25 типов различных аминокислот в разных ка-
чественных и количественных сочетаниях друг с другом можно
получать бесчисленное множество самых разнообразных бел-
ковых тел с разнообразными свойствами. Установлена последо-
вательность, с которой образуются такие вещества.
Всюду и везде, где на Земле проявляется жизнь, идет преоб-
разование веществ под влиянием энергии, выделяемой внутри
организмов или получаемой ими извне. В первом случае про-
исходит превращение одних минеральных или органических
веществ в другие, причем выделяется тепловая или химическая
энергия, используемая организмом для его роста и размноже-
ния. Во втором случае организм должен располагать каким-то
органом, который является «ловушкой» для энергии, поступаю-
щей извне. Это имеет место у некоторых бактерий и у всех рас-
тений, от самых низших до самых высших представителей.
Этими «ловушками» у них являются вещества хлорофильного
ряда — так называемые хлоропласты.
Итак, химический состав вещества земной коры — твердого
и измельченного, неподвижного и перемещаемого бурями,
71
ветрами, дождями и волнами первичных бассейнов — неизбеж-
но и обязательно приводил к возникновению на самом раннем
этапе истории Земли микроскопических примитивных организ-
мов в тех районах, где для этого только-только сложились бла-
гоприятные условия температурного режима. На других участ-
ках поверхности планеты, преобладавших по площади, массо-
вое проявление магматических и вулканических процессов,
выбросы из недр остаточного тепла гравитационного происхож-
дения и тепла от распада радиоактивных элементов поддержи-
вали высокую температуру. Здесь падавшие на землю дождевые
воды обращались в пар и снова поднимались в атмосферу.
Бури и ураганы проносились над земной поверхностью. Волны
первичных водных бассейнов разрушали выступы суши, пре-
вращая камень в мелкозем — гальку, песок и тонкий ил. Возни-
кали химические связи между ними и некоторыми другими раст-
воренными веществами земной коры.
Космические, преимущественно солнечные, излучения, про-
биваясь через почти сплошную облачность тяжелой вторичной
атмосферы, то дробили атомы химических элементов, то пере-
страивали их соединения, вызывали ^аеоад паров воды на во-
дород и кислород, причем*первый**частично улетал в космиче-
ское пространство, а второй активно вступал в химическую связь
с веществом земной коры, твердым и растворенным.
Огромное разнообразие веществ древнейшей литосферы
(каменной оболочки Земли), находившихся в резулыате общего
восстановительного характера химических процессов в закисном
состоянии, представляло собой своего рода «топливо», к кото-
рому нужно было только поднести зажженную спичку. Этой
«спичкой» послужили окислительные процессы при участии
слабо связанного кислорода и его масс, возникавших при рас-
паде паров воды. Процессы такого порядка всегда сопровожда-
ются выделением свободной энергии. При химическом ходе
реакций окисления эта энергия в виде тепла разными путями
растрачивалась. Например, выбрасываемые на поверхности
Земли при современных вулканических извержениях лавовые
массы, всегда богатые закисями, приобретают еще более высо-
72
кую температуру, чем имели в недрах. Встречаясь с кислородом
атмосферы, они как бы загораются, начинают светиться, при-
обретают еще большую подвижность и иногда текут, подобно
рекам, далеко по поверхности Земли. При недостатке же кисло-
рода вулканические массы становились вязкими, как становятся
вязкими и лавы, бедные закисями и богатые кремнеземом, —
так называемые «кислые». Лавы древнейших фаз вулканизма
образовывали огромные скопления каменных масс, наращивав-
ших земную кору на суше и в водах, а пепел разносился вихрями
древнего углекисло-метано-аммиачного воздуха молодой пла-
неты с парами воды и осаждался на обширных площадях с теми
или иными химическими изменениями его компонентов.
Угольная кислота была первым химическим агентом доор-
ганического этапа истории Земли, имевшим биологическое зна-
чение. Она активно влияла на многие вещества, в частности об-
разовывала разнообразные карбонаты, в числе которых преобла-
дающее развитие получили карбонаты кальция и магния. В при-
сутствии молекул вода эти карбонаты превращались в раство-
римые бикарбонаты, которыми обогащались водные массы
древнейших бассейнов. Поэтому древнейшие воды гидросферы
Земли содержали много углекислых и, вероятно, азотнокислых
солей и очень мало хлористых. Другими словами, древние моря
были почти не солеными. Их известковые воды по типу были
близки к многим водам современной суши, связанным с карбо-
натными породами. Содержавшиеся в воде бикарбонаты были
тоже «топливом», поскольку они способны переходить в твер-
дые кальцит и доломит с выделением энергии.
Таким образом, вещество земной коры и геохимические про-
цессы, включая химическую эволюцию углеродных соединений,
неуклонно и необратимо вели к возникновению жизни и ее тор-
жеству в последующие геологические времена.
Протекание и развитие микробиологических процессов в
почвах представляет исключительный интерес, так как вообще
почвенные условия являются издавна одной из самых распро-
страненных сред питания микроорганизмов на земной поверх-
ности. Как известно, новообразование, т. е. разрушение корен-
ных скальных пород с превращением их в равномерно зернистый
мелкозем в присутствии бактериального населения, протекает
в 20 раз быстрее, чем в результате деятельности только агентов
физического выветривания пород (колебаний температуры и
растрескивания пород в присутствии воды при ее замерзании).
В первом случае мелкозем образуется в сложной биогеохимиче-
ской среде, где к деятельности микроорганизмов (бактерии,
грибы, водоросли) добавляется геологическая деятельность
выделяемых ими органических кислот.
Процесс отсортировки обломочного минерального материа-
ла в почвах обусловлен отделением кварцевых зерен извержен-
ных пород от кристаллов полевых шпатов, которые под влия-
нием микроорганизмов (диатомовые водоросли) превращаются
в глинозем, с образованием почвенных минеральных коллоидов;
это может приводить и к переотложению этих коллоидов в виде
«вторичных» глин на дне водных бассейнов. За счет этого ма-
териала в водах могут отлагаться глинистые минералы — бен-
тониты, монтмориллониты. Железо- и марганецсодержащие
минералы разрушаются в результате жизнедеятельности железо-
бактерий.
Какие условия в природе наиболее подходят для развития
сложных химических процессов при участии веществ, находя-
щихся в трех фазах - твердой, жидкой и газообразной - под
воздействием различных физических явлений, в частности излу-
чений? Как уже отмечалось выше, таким местом оказываются
почвы суши влажных районов, образованные за счет коренных
пород, содержавших соответствующие химические элементы.
Как известно, наиболее богаты биофильными химическими
элементами некоторые типы изверженных пород. Наиболее
бедными являются продукты переноса и переотложения почвен-
ного материала, из которых активные химические элементы
и их соединения удалены, но это имеет место в отношении пе-
сков и песчаников кварцевого состава.
Важнейшим агентом химических процессов является вода,
которая служит и важнейшим компонентом живого вещества
биосферы. Граница соприкосновения скальной горной породы
74
с водой, содержащей в растворе углекислоту, кислород воздуха
и азот, является местом взаимодействия между важнейшими
компонентами биосферы. Здесь развиваются соответствующие
реакции. Включение сюда же органических кислот или кислот
минерального происхождения усиливает воздействие таких сред
на вещество скальной горной породы, и она более или менее
быстро разрушается. При этом продукты разрушения деятель-
ностью силы тяжести, вода или ветра могут быть частью вы-
несены, частью вовлечены в раствор и унесены с водой. Здесь
за счет минеральных веществ разрушаемой горной порода об-
разуются новые минералы, часто содержащие воду.
Естественно, что почвы обычно являются местообитанием
огромного многообразия микроорганизмов, среди которых
много различных бактерий, грибов, водорослей и простейших
животных, создающих почву и использующих ее как среду своей
жизнедеятельности. О присутствии почвенного покрова на вы-
ступах суши в геологическом прошлом, если не сохранились
сами почвы или вообще кора выветривания горных пород в зоне
контакта с атмосферой, могут говорить массы обломочного
материала наземного происхождения, снесенные водами в бас-
сейны и превращенные затем в те или иные скальные породы
(песчаники — в кварциты, глины — в глинистые или кристалли-
ческие парасланцы, галька — в конгломераты). Чистота их
сортировки, однородность формы частиц и характерный мине-
ральный состав могут служить признаками активного хода про-
цессов почвообразования на суше. Поэтому очень интересно и
важно, что эти признаки характерны для древнейших осадочных
пород. Песчаники или образовавшиеся из них кварциты обнару-
жены даже в составе пород архея, так же как и мраморизованные
известняки и превращенные в кристаллические сланцы песчано-
глинистые породы.
Поскольку преобразование многих минеральных веществ
в биосфере происходит с выделением свободной энергии, то в
почвенных условиях эта энергия поддерживает жизнедеятель-
ность организмов. В далеком геологическом прошлом в особых
физико-химических условиях именно эта обстановка могла
75
быть наиболее подходящей для возникновения примитивных
форм жизни и их развития.
Таким образом, почвы — это микробиологическая лабора-
тория природы. По продуктам процесса породообразования
мы можем судить и о наличии почвенного населения в опреде-
ленные геологические эпохи. Имеются данные о существовании
почвенных бактерий и, возможно, почвенных водорослей уже
в позднем архее, не говоря уже о протерозое, когда почвенные
процессы, несомненно, захватили обширные выступы суши.
Наличие в составе древнейших обломочных пород сероцветов
или зеленоцветов указывает на преимущественный восстанови-
тельный характер как на суше, так и в бассейнах. Как известно,
продукты перемыва некоторых вулканогенных пород докембрия
часто бывают выражены так называемыми серыми вакками,
в которых встречаются и мало измененные первичные материа-
лы материнских пород (базальтов). Такие породы указывают на
времена и области преобладания физических агентов почвооб-
разовательного процесса.
Что касается железосодержащих и марганецсодержащих
минералов коренных пород суши, то их разрушение из химиче-
ских агентов идет наиболее активно под действием углекислоты
с образованием растворимых в водах бикарбонатов, которые
легко разрушаются и в почвах и в бассейнах биосферы деятель-
ностью разнообразных железобактерий. Поэтому состав и
сложение осадочных пород, сформировавшихся в водных усло-
виях, могут указать на характер обстановки на смежных высту-
пах суши и на деятельность тех или иных организмов в геологи-
ческом прошлом.
Возникновение первичного живого вещества на Земле на
основе запасов разнообразных углеродных соединений биосфе-
ры сопровождалось вовлечением в круговорот жизни косного
вещества земной коры и атмосферы с использованием энергии
его превращений. Энергетический процесс, который возникал в
минерало-органических веществах, оформленных в мельчайшие
каплеобразные тела, проявлялся сначала как вспышка, как
взрыв. Позднее он стал принимать более спокойное течение.
76
Жизнь, загоревшаяся, как искры света во тьме, то вспыхивала,
то угасала, то, получив достаточный приток «пищи», снова
становилась устойчивой и «горела» ровным маленьким пла-
менем.
Энергетические процессы, происходящие внутри живого ор-
ганизма, идут на более высоком качественном уровне, чем явле-
ния неживой природы. Обмен веществ, являющийся источником
энергии, если она не поступает извне в форме световой или теп-
ловой, приводит к получению энергии при окислительных про-
цессах, с превращением ее в химическую, механическую и элек-
трическую. По мнению Н. Г Холодного, основным источником
энергии в обмене веществ служит водород. Углерод окисляется
косвенно в процессе последовательной отдачи водорода, при-
соединения частицы воды, новой отдачи водорода и т. д. Ха-
рактерно, что отнимаемый от окисляемого вещества водо-
род соединяется с кислородом и освобождает весь свой запас
химической энергии не сразу. Он проходит через систему про-
межуточных переносчиков, от одного соединения к другому.
Отдавая электрон, водород переходит в ионную форму и, на-
конец, соединяясь с кислородом, образует воду.
Поскольку многие виды окислительных процессов, происхо-
дящих в природе, действительно проявляют свечение (светящие-
ся бактерии, светящаяся в темноте гниющая древесина), возник-
новение жизни в первичных сгустках - «коацерватных каплях»
А. И. Опарина — действительно можно уподобить искрам све-
та, бесчисленным огонькам, загорающимся в природе, то быст-
ро угасающим, то постепенно разгорающимся все сильнее и
сильнее.
Нельзя не отметить роли тепловой энергии, возникающей
при процессах обмена. Нередко эта энергия бывает избыточной,
излишней для биохимических процессов организма. Она выде-
ляется в среду обитания, которая при этом претерпевает нагре-
вание (окисление серных руд в штабелях, пирита и марказита в
углях и в штабелях, гниение сена, «созревание» торфа в торфя-
никах и т. д.). Это явление ведет к обособлению деятельности
так называемых «термофилов» (теплолюбивых организмов),
77
хотя изменение обстановки может в действительности идти
лишь в сторону уже чисто химических процессов во вред упомя-
нутым организмам. Так, организмы, возникшие при соответ-
ствующем составе и особенностях среды, влияют на нее и под-
вергаются при этом изменениям под влиянием ими же вызван-
ных изменений последней.
Раз появившись и приобретя способность к воспроизводству
новых поколений, жизнь стала быстро завоевывать среды всех
типов. Используя запасы углеродных соединений земной коры,
живое вещество — новый геологический фактор — быстро по-
глощало их. Поэтому на Земле могла быть только одна эпоха
возникновения жизни, так как это был определенный этап в
развитии вещества планеты, больше никогда уже не повторяв-
шийся.
В сферу жизнедеятельности организмов, возникших на осно-
ве минерало-органических соединений, с качественно новыми
отношениями их со средой, были вовлечены элементы, которые
до того были недоступными; поле деятельности организмов
быстро расширилось. Расселяясь, организмы стали осваивать
все новые и новые места обитания: все более глубокие зоны
морей, всю поверхность суши, от низин до высот, а также и
недра Земли. Таким образом, первый и главный слой жизни,
несомненно, расширяясь, быстро охватил выше и ниже лежа-
щие зоны и превратился в биосферу, размер которой по верти-
кали стал измеряться километрами.
Так жизнь как энергетический процесс стала на Земле неот-
делимой от минерального вещества, стала неотъемлемой частью
природы, планетарным явлением. Подчиняясь до известного
предела условиям среды своего развития, она вносила, как вно-
сит и теперь, часто значительные изменения в состав и свойства
среды. Жизнь расширила круг химических процессов на Земле,
добавив к нему огромное разнообразие биохимических реакций.
Жизнь с самого начала стала осуществлять разнообразную
огромную и все увеличивающуюся породообразующую геоло-
гическую работу. Она стала играть все большую роль в форми-
ровании пород, ландшафтов и самой среды.
78
По мнению Г Лиса, первичные организмы, обитавшие в
условиях атмосферы, содержавшей углекислоту и лишенной
кислорода, в средах, бедных органическими веществами, могли
иметь источником жизненной энергии только свет. Поэтому
этот исследователь полагал, что первыми живыми существами
на Земле были фотосинтезирующие организмы, которые жили
в анаэробных условиях, пока в результате того же фотосинтеза
не создался в биосфере и запас кислорода и органических ве-
ществ. Следует отметить, что хлорофилл, зеленый компонент
живого вещества фотосинтезирующих организмов, по своей
конструкции стоит на уровне аминокислот и потому мог быть
действительно одним из древнейших белковых соединений в
природе.
Фотосинтезирующие бактерии имеют в своем составе кра-
сящие вещества — пигменты, близкие к хлорофиллу растений.
У бактерий, выращенных в темноте, эти пигменты не образу-
ются. Бактериальный хлорофилл представляет собой различ-
ные близкие по составу пигментирующие вещества, каждое из
которых, свойственно соответствующим группам бактерий —
пурпурным, серным и несерным, зеленым серобактериям и др.
Схема процесса фотосинтеза у таких бактерий имеет следую-
щий вид:
Световая энергия Н (восстановление СО2)
Н2О ----------------
Пигменты, ферменты ОН
Фотосинтезирующие бактерии выделяют в составе среды не
молекулярный кислород, а группы ОН. Это приводит к окисле-
нию в среде их обитания некоторых восстановительных ве-
ществ — сероводорода, серы водорода. Как мы отмечали, во-
дородом древняя биосфера была богата.
Как пишет Г Лис: «...автотрофные бактерии могут быть
охарактеризованы как организмы, которые могут жить, расти
и размножаться в среде, где единственным источником угле-
рода служит углекислота, и для конструктивного обмена кото-
рых не требуется готового органического вещества... Гетеро-
79
трофные организмы к этому не способны, поскольку для их
развития требуется присутствие по крайней мере одного органи-
ческого вещества, например какого-либо сахара или аминокис-
лоты. Гетеротрофные организмы нуждаются в органических
веществах не только как в материале для роста, но и как в ис-
точнике энергии. Рост и развитие этих организмов регулируются
количеством энергии, получаемой при усвоении нужного им
органического вещества. Между тем автотрофные бактерии
проявляют свою жизнедеятельность, используя световую энер-
гию или энергию, получаемую ими при преобразовании неорга-
нических веществ».
Вот некоторые пути получения энергии (по Г Лису).
1. Окисление сероводорода до элементарной серы в присут-
ствии кислорода:
H2S + О = Н2О + S + 172 кДж
2. Окисление элементарной серы в присутствии кислорода:
Н2О + S + 30 = H2SO4 = 496 кДж
3? хОкислейие азотистой кислоты в азотную:
HNO2 + О = HNO3 + 71 кДж
4. Окисление водорода:
Н2 + О = Н2О + 235 кДж.
5. Световая энергия —6 • 1023 квантов красного света дает
автотрофу 40 ккал энергии.
Для сравнения мы можем привести энергетический эффект
усвоения гетеротрофными бактериями глюкозы:
С2Н,2О6 + 60 2 = 6СО2 + 6Н2О + 2900 кДж.
Из этого сравнения отчетливо виден значительно более
высокий уровень химического развития «пищевого» вещества
гетеротрофов сравнительно с автотрофами. Получаемый в по-
следнем случае огромный энергетический эффект не мог иметь
80
места на самом раннем этапе существования организмов и мог
развиться лишь в процессе длительной эволюции живого ве-
щества в биосфере при последовательном усложнении химиче-
ской структуры веществ, включаемых в сферу биологических
процессов. Способность вырабатывать эфиры, нектар, масла,
сахара растениями могла возникнуть и развиться лишь в ходе
длительной эволюции растительного живого вещества природы.
Отсюда естественно представлять развитие жизни от первичных
преимущественно автотрофных организмов к преимущественно
гетеротрофным, от анаэробных форм жизни, не нуждающихся
в свободном кислороде, к более высокоорганизованным ана-
эробным же и к аэробным, требующим кислорода для своей
жизнедеятельности.
Форма и абсолютные размеры тела живого организма игра-
ют рещдющую роль в его отношениях со средой. При этом опре-
деляющим фактором является поверхность тела, осуществляю-
щая обмен веществ.
Произведем некоторый расчет. Представим себе кубик с
ребром в 1 см. Будем дробить его на кубики меньшего размера
и при соответственно большем их количестве плпробуем^под-
считать их суммарную поверхность:
1 кубик с ребром в 1 см — 600 мм2;
1 тыс. кубиков с ребром в 1 мм — 6 тыс. мм2;
1 млн. кубиков с ребром в 0,1 мм -- 600 тыс. мм2;
1 млрд, кубиков с ребром в 0,01 мм — 60 млн. мм2;
1000 млрд, кубиков с ребром в 0,001 мм — 6 млрд, мм2, или
600 м2
Таким образом, если мы имеем дело, например, с бактериями
размером около 0,001 мм (1 микрон!) в поперечнике, то оказы-
вается, что 1,см3 такого бактериального вещества имеет суммар-
ную поверхность связи со средой 600 м2! Таким образом, у бак-
терий, если выразиться техническим языком, коэффициент по-
лезного действия оболочки и самого вещества оказывается
весьма низким. Живые существа с более высокой организацией
имеют иное отношение к среде, выражающееся, в частности,
большими размерами и иной конфигурацией тел.
81
Форма живого тела, его размер и конфигурация в простран-
стве теснейшим образом связаны с содержанием, со свойствами
и количеством органического вещества, осуществляющего жиз-
ненные функции особи — единицы жизни. Поэтому совершенно
ясно, что первичной формой при возникновении жизни на Земле
была каплеобразная сфероидальная, имевшая сверхмикроско-
пические размеры, до 1 д Малые размеры тела — результат
естественного отбора — составляли преимущество этих пер-
вичных организмов, унаследованное затем всем миром бакте-
рий и зародышевой стадией более высокоорганизованных су-
ществ. Таким образом, первичная сфероидальная клетка стала
основной первичной стадией индивидуального развития всех
живых существ Земли.
Важное свойство любого природного тела — его электриче-
ский заряд, возникающий на границе сред. Он есть у падающей
капли дождя, у пылинки, плавающей в воздухе и в воде, он есть
у бактерии, находящейся в жидкой среде. У тел неживой при-
роды заряд способен возникать и исчезать, но знак его не ме-
няется. У живых же организмов, от самых простейших их пред-
ставителей до высших, электрический заряд создается ими
самими и их определяет; в момент гибели особи заряд теряется.
Говоря об организмах как древнейших, так и современных,
мы не можем не коснуться связи между содержанием и формой.
При этом в понятие содержания входят представления о внут-
ренних источниках единства, целостности и развития части ма-
терии с совокупностью свойственных ей процессов. В сгустке
живой материи содержание — это все биологические, физиче-
ские и химические процессы, а форма — та организация, кото-
рая обеспечивает взаимодействие этих внутренних процессов
и связь их с внешними условиями среды. Содержание обладает
собственным движением, тогда как форма имеет относительную
самостоятельность. Содержание заключает в себе возможность
беспредельного развития, тогда как форма его ограничивает.
Развитие организмов выражается и обусловливается повто-
ряющимися в определенном порядке формообразовательными
процессами в виде бесконечных циклов. Наиболее простые фор-
82
мы развиваются в более сложные или приобретают при тех же
формах и размерах более сложные функции (современные
бактерии).
Геологические материалы, в частности образование осадоч-
ных горных пород, способны давать существенные указания на
характеристику водных сред геологического прошлого. Мине-
ралы, образовавшиеся одновременно с отложением осадков
на дне древних бассейнов, способны давать исчерпывающие
указания на условия среды осаждения. Чаще всего это были
щелочные и восстановительные среды; в придонной зоне, в ее
«бактериальной» пленке, кислород обычно был израсходован
в результате деятельности аэробных (потребляющих кислород)
бактерий. Особенно важно, что геологи находят под позднее
отложившимися осадками «коры выветривания» продукты раз-
рушения горных пород, относящиеся к разным моментам гео-
логической истории Земли. Оказывается, что древнейшие такие
«коры» не несут признаков развития окислительных процессов.
Отложение осадочных железных руд при участии железобак-
терий, происходящее в современности только на площадях
суши, в условиях кислых аэробных сред в течение всего докемб-
рия происходило в бассейнах морского типа. Это указывает на
их тогда еще низкую засоленность и близость по свойствам к
водам суши. Для бассейнов докембрия было свойственно также
огромное развитие водорослей, которые их исследователям —
палеоальгологами признаются за пресноводные. Таким обра-
зом, мы имеем теперь много «ключей» для разгадки особенно-
стей древней биосферы Земли, которые использовали пока еще
недостаточно.
Считаясь с признанием большинством исследователей пер-
вичной среды жизни как среды восстановительной, первые про-
явления жизни следует представлять подчиненными этому
фактору. Так, некоторые микробиологи, задумывающиеся над
путями эволюции обмена веществ, обращают внимание на тип
бактерий, способных фотоассимилировать угольную кислоту
в условиях анаэробной среды, как это осуществляется, в част-
ности, современными пурпурными и зелеными серобактериями.
83
Эта форма обмена к тому же является самой примитивной. Она
оказывается типично автотрофной. В. Н. Шапошников полага-
ет, что от такого типа обмена легко перекинуть мостик к типич-
ному гетеротрофному питанию, через гетеротрофную ассими-
ляцию несеропурпурных микроорганизмов, которые развива-
ются на свету за счет органических веществ.
Из углеродных соединений первичная среда жизни имела
углекислоту и углеводороды, в основном метан, и продукты их
соединения с водой в условиях почти полного отсутствия сво-
бодного кислорода.
Теория А. И. Опарина прекрасно поясняет ход процесса
уплотнения молекул органического вещества химическим путем
и механизм деятельности коацерватных капель, которые долж-
ны были слагаться из органических веществ типа белковых.
Причиной возникновения жизни в первичных сгустках белко-
вых веществ могло быть только одно — осуществление в них
химических реакций за счет вещества среды, поступавшего
через граничную пленку, с выделением при этом энергии, если
она не поступала извне. Оба эти пути равно могли иметь место
с самого начала проявления жизни на Земле.
Первыми возникли скорее всего автотрофы, так как их струк-
тура проще, а энергетический эффект, даже у современных их
представителей, значительно ниже, чем у гетеротрофов. Формы
типа гнилостных бактерий тогда отсутствовали, что прекрасно
доказывается относительно высокой сохранностью даже следов
клеточного строения у многих ископаемых представителей
древнейших водорослей и других организмов. Только появле-
ние древнейших животных могло стимулировать развитие групп
гетеротрофных — болезнетворных микроорганизмов паразити-
ческого, или сапрофитного, характера.
Различные автотрофные микроорганизмы, как известно, спо-
собны в природе образовывать разнообразные минеральные
вещества. Из аммиака нитрофицирующие бактерии в природе
образуют нитраты — соль азотной кислоты. Тионовые (серные)
бактерии обладают способностью из элементарной серы про-
изводить серную кислоту.
84
В анаэробных — бескислородных — условиях некоторые ав-
тотрофные бактерии способны восстанавливать сульфаты до
сульфидов, используя при этом молекулярный водород. В озе-
рах и реках автотрофные фотосинтезирующие бактерии в ана-
эробной обстановке могут окислять сероводород до серы, а
затем до серной кислоты, при этом могут создаваться местные
скопления серы.
Например, в одном небольшом озере в Киренаике ежегодно
накапливается около 100 т элементарной соли. Очень интересно,
что микроорганизмы предпочтительно накапливают один из
изотопов серы — серу-32. Изотопный анализ древних залежей
серы, начиная с докембрия, показал повышенное содержание
в них этого изотопа. Значит в их образовании сыграли роль
микроорганизмы.
Современные автотрофные микроорганизмы осуществляют
широкий круг природных процессов, где их «работа» оказыва-
ется то полезной для человека (образование месторождений
серы тионовыми и серными бактериями), то вредной (разруше-
ние бетона и кирпича при участии тионовых бактерий). Иногда
они закупоривают железные трубы отложениями гидрата окиси
железа, усиливают или осуществляют целиком коррозию ме-
таллических конструкций.
Современные автотрофные микроорганизмы, по данным
микробиологов, характеризуются весьма сложным составом
ферментов.
Первичные автотрофы не могли походить в* этом отноше-
нии на их потомков, хотя в общем характер их биогеохимиче-
ской деятельности остался тем же. Между теми и другими
прошел длительный этап биохимического развития. Коренным
образом изменилась и сама среда их существования. Следует
полагать, что деятельности первичных форм жизни существенно
помогала та среда, которая их породила и что ход эволюции
организмов является проявлением саморазвития их вещества,
с развитием свойств новых, с изменением при этом и требований
к среде обитания. Поэтому древнейшие автотрофы должны быть
сходны с современными лишь отчасти.
85
Мы остановились на вопросе об автотрофных микроорга-
низмах потому, что эти формы жизни наименее требовательны
к обстановке. Для них нужна в основном углекислота как росто-
вый материал и различные минеральные вещества при наличии
слабо связанного или свободного кислорода или без него.
Древняя среда жизни содержала огромное многообразие необ-
ходимых для них закисных минеральных соединений и являлась,
таким образом, средоточием доступнейших для организмов
источником энергии. Химический процесс преобразования этих
закисей поэтому легко и естественно стал биологическим.
Энергия геохимических реакций стала энергетической базой
жизни.
Данные геологии показывают наличие в древнейших оса-
дочных толщах докембрия скоплений сульфидов в виде пирита,
марказита, свинцового блеска и т. д. Нами установлено обиль-
ное развитие бактериальных микроструктур у многих кремнис-
то-железистых сланцев и железных руд, в частности докембрий-
ских. Напрашивается единственно возможный вывод, что древ-
нейшими формами жизни на Земле были по преимуществу
именно группы автотрофных бактерий и одноклеточные водо-
росли. Они играли важную роль не только в преобразовании
минеральных веществ, но и в накоплении высокомолекулярных
углеродных соединений, затем ставших питательной средой
для развития гетеротрофных бактерий и предков других групп
организмов.
Геохимические реакции в первичной биосфере Земли шли
при несомненном участии фотосинтезирующих бактерий и
простейших представителей растительного мира. Клетки этих
организмов, развиваясь с помощью поступавшей извне лучис-
той энергии, должны были располагать зеленым веществом
типа хлорофилла и рядом ферментов для удаления продуктов
фотосинтеза.
Возникновение хлорофилла в ходе эволюции было заклад-
кой нового типа живого вещества биосферы — всего мира рас-
тений-.
Мир растений оказался мощным геологическим факто-
86
ром, способным высвобождать кислород, выделять его в атмо-
сферу и воду, а также накапливать в биосфере органическое Веще-
ство, частью захоронявшееся в осадочных отложениях. Искус-
ственное получение в 1961 г. путем синтеза вещества феофорби-
да, почти соответствующего хлорофиллу, явилось новым до-
стижением в области изучения строения органических веществ
и истории их развития.
Наиболее вероятным временем возникновения на Земле про-
цессов синтеза органических веществ с помощью ультрафиоле-
товых лучей Солнца и электрических разрядов атмосферы был
этап перехода от катархея к архейской эре истории нашей пла-
неты. Это было временем возникновения массы первичного
живого вещества, чему способствовали вода в жидкой фазе и
запас сложных углеродных соединений.
ЖИВОЕ И МИНЕРАЛЬНОЕ ВЕЩЕСТВО
В течение тысячелетий передовые умы человечества бьются
над вопросами, что такое материя, каково ее строение. Появле-
ние атомистического представления о строении материи, горя-
чим сторонником которого был М. В. Ломоносов, стало важным
этапом в развитии знаний. Так, М. В. Ломоносов видел проявле-
ние атомного строения вещества в их свойстве нагреваться.
Он считал, что теплота — это не что иное, как движение мель-
чайших частиц тела, а степень нагрева зависит от скорости дви-
жения частиц. Открытие и изучение химических элементов пока-
зало, что вещества бывают простые и сложные и что имеются
возможности разложить одни вещества, получая при этом
другие, более простые. Постепенно были открыты химические
законы, в частности закон постоянства состава химического
соединения, открыто молекулярное строение веществ. При этом
доказано, что свойства элементов находятся в периодической
зависимости от зарядов ядер их атомов, от величины атомных
весов элементов.
Развитие идей Д. И. Менделеева, открытие ряда новых эле-
ментов с предсказанными свойствами, открытие электронной
структуры атомов Э. Резерфордом и Н. Бором привели к уста-
новлению единой природы всех химических элементов. Затем
были вычислены размеры электронов, определен состав ядер,
и новая физика научилась превращать одни элементы в другие.
Развитие материалистической мысли и создание К. Марксом
и Ф. Энгельсом диалектического материализма привело, к идее
развития, к идее непрерывного обновления всего существующе-
го. Диалектический материализм считает способность к разви-
тию неотъемлемым свойством материи. Материалистическое
миропонимание приводит нас к идее вечной и неизменной перво-
основы всех вещей, к идее существования предметов и явлений
объективно, т. е. независимо от нашего сознания. В. И. Ленин
писал, что материя есть объективная реальность, данная нам
88
в ощущении. Основной причиной того, что материя действует
на чувства, является то, что она находится в движении.
Открытие радия и радиоактивности необычайно обогатило
физику и химию, вплотную приблизив человечество к понима-
нию вещества Земли. Ядерная физика еще развивается на этой
основе и не сказала своего последнего слова, тем не менее она
привела к учению об изотопах в природе и позволила в содруже-
стве с химией, биохимией и геохимией подойти к пониманию
сущности связей между элементами мертвой и живой природы,
а также внутри этих двух категорий веществ.
В свете новых взглядов на живое вещество природы основ-
ным моментом оказывается понятие об энергетической стороне
явлений жизни.
В. И. Вернадский считал, что вся биосфера, во всех ее формах
находится в непрерывном движении. Температура свободной
тропосферы (нижней, охватываемой жизнью зоны атмосферы
Земли) кверху понижается, температура гидросферы понижается
книзу, температура литосферы (верхнего слоя земной коры)
постепенно повышается с глубиной. В тропосфере азот и кисло-
род находятся в определенном соотношении. Под землей быст-
ро исчезает кислород и существует «кислородная поверхность»,
ниже которой располагается азотная подземная атмосфера.
Вещество биосферы В. И. Вернадский делил прежде всего
на две крупные категории: «живое» вещество (совокупность всех
живых организмов), богатое действенной энергией, и «косное»,
действенная радиоактивная и химическая энергия которого в
подавляющей ее массе, в ходе исторического времени, ничтож-
на. Живое вещество охватывает и перестраивает все химические
процессы биосферы. В. И. Вернадский поэтому считал, что «жи-
вое вещество есть самая мощная геологическая сила биосферы,
растущая с ходом времени (разрядка моя — А. В.).
Оно не случайно и независимо от нее, в ней живет, она есть про-
явление физико-химической организованности биосферы».
Обратимся к рассмотрению тех химических элементов, ко-
торые являются общими как для живого вещества природы,
так и для косного. Эти общие элементы, поскольку они являются
89
основными компонентами веществ — носителей жизни, называ-
ются биофильными. Их не так много, хотя вообще в состав жи-
вых тел входит много десятков других элементов, но обычно
в ничтожных количествах.
Биофильные элементы следующие: углерод, азот, водород,
кислород и сера. Как выяснено, «биофильные» свойства этих
элементов нужно понимать и рассматривать с оговорками.
Суть в том, что почти все известные химические элементы ока-
зались смесями изотопов, почти не отличающихся друг от
друга по своим химическим и физическим свойствам. При этом
не все изотопы элементов, упомянутых выше, имеют равное
отношение к процессам жизни.
Стремление понять вещественный состав Земли, прежде
всего той ее части, которую называют биосферой, привело
исследователей — химиков и минералогов, а также и геоло-
гов — к изучению распространения химических элементов в
природе, в земной коре, к поискам закономерностей на основе
геологической карты.
Геологическая карта призвана показать распространение тех
или иных горных пород по их выходам на дневную поверхность
и искусственным выработкам и буровым скважинам, с указа-
ниями на их геологический возраст и вещественный состав,
на различные взаимоотношения этих пород.
Для пород земной коры было очень важно учесть распро-
странение в ней химических элементов. В результате трудов
В. И. Вернадского, Ф. У Кларка, А. Е. Ферсмана, их учеников
и последователей возникла и непрерывно расширяется таблица
количественной распространенности химических элементов,
или, как говорят геохимики, «кларков» элементов. Отсюда воз-
никло представление о среднем составе земной коры, на фоне
которого местные более высокие или более низкие концентра-
ции представляли практический или научный интерес. Высокие
концентрации практического значения — это месторождения
полезных ископаемых. Поэтому геохимия оказывает помощь
практической геологии, на материалах которой она развивалась
и развивается, дополняя ее специальными исследованиями.
90
Крайне важно, что в то же время родилась идея об эволюции
вещества земной коры. В. И. Вернадский высоко расценивал
работы в этом направлении, имея в виду возможность глубоко-
го изучения вещества Земли и полагая, чго химический атомный
состав его не есть случайное явление, что он связан с многооб-
разными свойствами атомов в физическом поле, прежде всего
в энергетическом — термодинамическом.
Количественные отношения атомов Земли были отражены
в так называемой таблице Филипса — Кларка — Фохта. Она
показала, что атомный состав Земли тождествен составам по-
верхностных слоев звезд и Солнца.
Живые организмы В. И. Вернадский не считал в основном
углеродными. Он полагал, что с геохимической точки зрения
живое вещество есть кислородное вещество, богатое углеродом,
и лишь иногда оно является углеродистым и содержит более
10% углерода по весу. Значение углерода в живом веществе,
в организмах, не объясняется его количеством, но есть функция
его химических свойств.
Замечательную особенность взглядов В. И. Вернадского со-
ставляет единство его подхода к представлениям об элементар-
ном составе живой и косной природы. Выяснилось, что элемен-
ты, слагающие тела организмов, свойственны природе вообще,
не только живому, но и косному ее веществу, в частности мине-
ральному. Отсюда естественным было категорическое призна-
ние постоянства химического облика земной коры в течение
геологического времени, признание того, что «средние количе-
ства и состав живого вещества оставались приблизительно оди-
наковыми в течение всего этого непостижимого по длительности
времени». Поэтому он признавал, что количество живого ве-
щества, по-видимому, является планетной константой (постоян-
ной величиной) со времени архейской эры, т. е. за все течение
геологического времени.
Следует отметить, что и ранее некоторые мыслители, в част-
ности в XVII в. Ж. Бюффон, высказывались за идею об относи-
тельной неизменности количества живого вещества. Между
тем и в настоящее время нет точных данных об изменениях коли-
91
чества живого вещества во времени и потому решение этогб
вопроса пока оказывается невозможным.
Рассматривая элементарный состав живого вещества,
В. И. Вернадский разбил элементы в зависимости от их распро-
странения на декады:
Элементы Весовые проценты Декады
О, Н
С, N, Са
S Р Si К
Mg, Те,’ Na, Cl, Al, Zn
более 10 I
1—10 II
0,1—1 III
0,01—0,1 IV
В составе тел всех ныне живущих организмов мы находим
кислород, водород, серу, азот и в особенности углерод. Эти
элементы и ряд других, присутствующих в меньших количест-
вах, железо, марганец, медь, образуют простые и сложные
химические соединения, главнейшими из которых являются
белковые. Из простых соединений следует отметить воду, угле-
кислоту. метан, угарный газ. закись азота и другие, свойствен-
ные. оказывается, не только живым организмам, ко и так назы-
ваемой «мерз вой природе».
Упомяну । ые лементы и их простые и сложные соединения
слагаю! !ела организмов, но все они также свойственны и окру-
жающей их среде. Элементы и соединения извлекаются орга-
низмами из среды с образованием нужных им веществ для по-
строения соответствующих клеток и тканей, а затем в процессе
обмена со средой и после смерти организма они возвращаются
снова в состав среды.
Таким образом, органическая жизнь базируется на химиче-
ских элементах и соединениях, являющихся общими и для нее
и для окружающей «мертвой природы». Отсюда возникает
один из важнейших законов биологии — «единство среды и
жизни», прежде всего в вещественном понимании.
Основной признак каждого живого существа — способность
усвоения соответствующих элементов и веществ природы
способность к ассимиляции. Ф. Энгельс считал ассимиляцию
существенной функцией жизни.
92
Элементарное вещество, обусловливающее жизнь, — белок,
или комплекс белковых соединений. По Ф. Энгельсу, белок —
совершенно бесструктурное вещество, обнаруживает все основ-
ные свойства жизни, осуществляемой путем ассимиляции: пище-
варение, выделение, сокращение (раздражимость, реакцию на
раздражение), размножение. Великий мыслитель считал, что
должны были пройти тысячелетия, пока не создались условия,
необходимые для следующего шага вперед, когда из этого бес-
форменного белка благодаря образованию ядра и оболочки
возникла клетка. Он полагал, что сначала возникли бесконечные
количества бесскелетных бесклеточных и клеточных протистов
(простейших), о которых говорит наука палеонтология и кото-
рые преобразовались постепенно в первые растения, в первых
животных. В настоящее время на первом месте в этом перечне
мы ставим микроорганизмы — бактерии и близкие к ним фор-
мы жизни.
Сущность связи организма со средой состоит в энергетике.
Всем организмам свойственно постоянное извлечение из среды
энергии, которая затем используется организмом в его много-
образных функциях, прежде всего на построение его тела, на
движение. Питание в общем смысле и есть тот путь, который
позволяет организму постоянно и непрерывно пополнять запас
энергии, и как только приток энергии по каким-либо причинам
прекращается — организм погибает. В древнейших водных бас-
сейнах или во влаге древнейших почв на суше, где возникали и
развивались наидревнейшие формы жизни, она могла развить-
ся путем усвоения в основном неорганических веществ, которые
оказывались способными к окислению их несвободным или
свободным кислородом. Это могли быть углекислота, метан,
соли, растворенные в воде, которые, попав в организм, способ-
ны были окисляться, причем выделялась свободная энергия
(тепловая и химическая), а продукты окисления или шли на
построение тела и его оболочки, или извергались обратно в со-
став среды, где они чаще шли в осадок, на дно. Накопление орга-
нических веществ, развитие органической жизни обусловило и
другой путь извлечения свободной энергии — окисление орга-
93
нических веществ, относительно более эффективное в энерге-
тическом отношении, чем окисление неорганических соединений.
Как известно, академик А. И. Опарин создал гипотезу про-
исхождения жизни на Земле именно на основе представлений
о первоначальном уплотнении органических соединений на
Земле, возникших химическим путем, до образования белка.
Белок сгустился в капли — коацерваты, которые, усваивая тоже
белковые соединения среды, стали первыми живыми клетками.
Не исключается, что жизнь, массово и беспорядочно, удачно и
неудачно, возникавшая и отмиравшая, шла и тем и другим
путем, а также и промежуточным, т. е. с одновременным исполь-
зованием и неорганической и органической пищи (миксотрофы).
Растение, получая энергию солнечных лучей извне, исполь-
зует ее на разложение воды, на ассимиляцию углекислоты из
атмосферы, на построение из синтетически создаваемых угле-
родных соединений нужных веществ в процессе своего роста
и размножения. Клетка железобактерии, ассимилируя из среды
растворенные в ней закисные соединения железа или марганца,
окисляет их в окисные соединения, выделяющиеся в твердом
виде или в состав среды, или на поверхности клетки в виде
чехла. Ржавая железистая вода есть результат растворения и
окисления железа бактериями «ржавчины» (в технике это назы-
вается коррозией металлов). Такие организмы, использующие
неорганические соединения для своего существования, называ-
ются автотрофами. Организмы, питающиеся органическими
соединениями, как, например, гнилостные бактерии, а также
животные называются гетеротрофами.
В. И. Вернадский выделял также промежуточный тип су-
ществ — миксотрофы — организмы, способные питаться за счет
неорганических и органических соединений. Можно полагать,
что морской моллюск — устрица, способная при весе тела около
40—50 г накапливать за несколько лет оболочку из углекислого
кальция (раковину) весом до 3 кг, принадлежит именно к миксо-
трофам. У всех организмов важная роль принадлежит оболоч-
ке. Сквозь оболочку внутрь организма проникают вещества из
среды, если они внутри него отсутствуют. Из организма выво-
94
дятся через оболочку вещества, содержание которых в среде
более низкое.
Первичный белковый комочек мог стать живым организмом
лишь во взаимодействии с энергией, поступавшей извне или об-
разовывавшейся внутри него. Первое имеет место у зеленых
автотрофов — всех растений Земли, второе—у других орга-
низмов. Существование первичных сгустков только из белко-
вых соединений, ставших организмами, в мировом древнейшем
океане или в древнейших почвах на суше маловероятно. Несом-
ненно, имелась, как и теперь, переходная — вирусоподобная,
«ультрамикробная» стадия с весьма менявшимися свойствами,
сходная то с организмами, то с мертвой материей. Ведь извест-
ны вирусы кристаллизующиеся, способные переносить и крайне
низкие и высокие температуры и т. д. Для белка же 60°С — пре-
дел, при котором он свертывается и ни к какой жизнедеятель-
ности уже не способен. Веществом первичных «вирусов» —
пробионтов, зародышей первичной жизни в геологическом
прошлом, могли быть различные металлоорганические соеди-
нения, каких в природе существует бесчисленное разнообразие.
Как известно, они образованы при участии ряда металлов из
группы «биофильных». При этом роль атомов металлов в слож-
ных металлоорганических соединениях — служить мостиками,
связями между частицами органических соединений, вследствие
чего молекулы металлоорганических соединений могут быть
весьма сложными и обладать разнообразными свойствами.
Известно, что бактерии, в частности «железные», способны
к жизнедеятельности в определенном температурном диапазоне,
в котором они действительно процветают, интенсивно размно-
жаются. Следовательно, температура среды в какой-то момент
«поджигает» жизненный процесс. Вещества, извне поступающие
в клетку, развязывают внутренние силы, как бы служа для них
запалом, стимулятором. Так, бактерия может из недеятельного
состояния перейти в деятельное. Так могли возникать и много-
образные первичные организмы. Удачно создавшиеся — суще-
ствовали, неудачные — исчезали в средах далекого прошлого
Земли, способствуя накоплению органического и минерального
95
вещества в водных бассейнах и в осадках. Наладившийся обмен
веществ означал жизнь, нарушившийся — гибель.
Естественный отбор организмов, приспособленных к среде
и условиям обитания в ней, далее уже моделировал первичный
сгусток жизни — живого вещества, обусловливая дальнейшую
эволюцию жизни. Возьмем пример хотя бы из истории техники.
Создателям бензинового мотора далеко не сразу, конечно,
удалось наладить режим его работы, чтобы превращать внут-
реннюю химическую энергию горючей смеси (пары бензина +
-I- воздух) в тепловую, способную производить работу. Но
когда мотор, засасывая регулярно «питание», ритмично зарабо-
тал, он как бы «ожил». Подобно этому возникла и жизнь.
Живое и минеральное (косное) вещества находятся в природ-
ных условиях во взаимосвязи и взаимодействии в процессах
жизнедеятельности автотрофных, полуавтотрофных, миксо-
трофных и гетеротрофных организмов. В. И. Вернадский, ко-
торый был первым исследователем, обратившим серьезное
внимание на геологическое значение явлений жизни, предложил
понятие о живом веществе, о биосфере — области развития
этого вещества с соответствующими энергетическими процес-
сами, сопровождающими его превращения. Поверхностную
оболочку Земли он предложил рассматривать не только как
область распространения вещества биосферы, но и как область
энергии, причем он учитывал влияние на развитие живого веще-
ства геологических и геохимических процессов, солнечных и
космических излучений. В. И. Вернадский говорил о роли лу-
чистой солнечной энергии для существования жизни и для
геологических явлений — движения воздушных масс, морских
водных масс (течения), морской волны, берегового прибоя;
отмечал теплорегулирующую способность водных масс морей
и океанов, играющую oi ромную роль в явлениях погоды и кли-
мата и связанных с ними процессах жизни и выветривания;
в живом веществе, совокупности живых организмов Земли,
видел преобразователя лучистой энергии Солнца в химическую
энергию биосферы, где в реакциях фотосинтеза создается беско-
нечное число новых химических соединений.
96
А. И. Опарин в своем замечательном труде «Возникновение
жизни на Земле» нарисовал очень убедительную картину хими-
ческой эволюции органических веществ, предшественников «жи-
вого вещества» В. И. Вернадского, путем трех основных типов
реакций: конденсации первичных органических веществ химиче-
ского происхождения, полимеризации — гидролиза, и окисли-
тельно-восстановительной реакции. А. И. Опарин создал много
новых интересных представлений о возникновении жизни, но
он недостаточно учитывает роли в этом процессе солнечных
и космических излучений, чему В. И. Вернадский придавал боль-
шое, почти ведущее значение.
Нижнюю границу области жизни — биосферы — В. И. Вер-
надский определял на глубине в три и более километра от днев-
ной поверхности. Он делал это на основании планетного значе-
ния микробного живого вещества, главным образом анаэробно-
го (живущего в бескислородной среде), достигающего таких
глубин. И на всю мощность биосферы, от этой границы в недрах
Земли до нижних слоев атмосферы он распространял проявле-
ния биогенной миграции химических элементов, биогеохимиче-
ской энергии организмов, которые, являясь «закономерной
функцией биосферы», распределены в ней в строгой связи с ее
структурой.
По состоянию знаний того времени о биохимических процес-
сах В. И. Вернадскому о многом приходилось говорить в общей
форме. Большей частью он не конкретизировал свои представ-
ления о деятельности организмов в круговороте веществ. Этой
области вопросов посвятил большие труды академик А. П. Ви-
ноградов, уделивший серьезное внимание химизму организмов
моря. Ему принадлежит и первая, единственная в своем роде,
замечательная схема взаимосвязи между живым и минеральным
(косным) веществом для условного эволюционного ряда совре-
менных организмов, приведшая его к идее «пирамиды» мине-
рального субстрата жизни, к идее постепенной химической эво-
люции живого вещества, освобождения его от минерагенной
части химических элементов, постепенного отхода организма
от использования биогеохимической энергии и перехода к ис-
97
пользованию энергии только биохимических превращений. Чем
выше уровень химического развития живого однородного ве-
щества, тем меньшее участие в его составе принимают минёро-
генные химические элементы.
Взаимосвязь жизни и среды обитания В. И. Вернадский ино-
гда определял понятием «биокосное тело», как, например, почва,
вода пресного водоема и т. д. Это области, зоны и участки
биосферы, где живое вещество перерабатывает геологическую
оболочку биосферы, меняет ее физические, химические и меха-
нические свойства. В связи с этим «косным» — неживым —
В. И. Вернадский называл главную массу вещества биосферы,
не входящую в живое вещество, — ее горные породы и минера-
лы, химические элементы которых в процессе биогенной мигра-
ции атомов могут «входить в естественные живые тела и уходить
обратно». Это проявляется в процессах дыхания, питания, раз-
множения, выделений, свойственных живому веществу, которое
он считал «закономерной функцией» биосферы, подчеркивая
постоянную теснейшую связь между организмом и средой, что,
как он отмечал, «не всегда правильно учитывалось» исследова-
телями. В. И. Вернадский отмечал, что биокосные тела харак-
терны для биосферы. К ним он относит, в частности, и зону
выветривания горных пород, которое считал биокосным про-
цессом, подчеркивая биогеохимическую природу разрушения
горных пород в коре выветривания.
Мы поставили перед собой задачу проследить влияние живого
вещества на минеральное — косное, на основании прежних и
новых данных о породо- и рудообразующей деятельности иско-
паемых автотрофных организмов. Новые материалы повлекли
за собой новые выводы, частью очень интересные, касающиеся
ряда смежных научных дисциплин. В связи с этим возникла и
неотложная необходимость внесения существенных поправок
в некоторые взгляды, а именно по вопросу происхождения не-
которых горных пород и минералов. Но следует при этом отме-
тить, что наши представления о связи живого и минерального
вещества в природе не выходят за общие рамки, гениально очер-
ченные В. И. Вернадским. Мы можем предложить лишь ряд
98
конкретных представлений о некоторых биокосных телах, телах,
сложенных живым и минеральным веществом.
Коснемся теперь более детально организмов — концентрато-
ров минерального вещества, минерогенных элементов, косного
вещества, расположенных в основании «пирамиды» А. П. Ви-
ноградова. Их существовало и существует в природе множество.
Есть концентраторы специализированные — автотрофы и рабо-
тающие биогеохимически, так сказать, «по совместительству» —
миксотрофы.
Алюминий химически осаждается в слизистой оболочке бак-
териями, например рода кренотрикс, содержащими его до 17,7%.
Кремний усваивается диатомеями, кремневыми губками и др.
Кальций осаждается разнообразными водорослями (синезеле-
ные, красные, харовые и др.). Водоросли осаждают карбонат
кальция, поглощая СО2 и Са(НСО3)2. Известен вид бактерий,
осаждающих СаСО3 внутри клетки. У других карбонат кальция
выделяется в виде кристаллов при распаде их клеточного веще-
ства. Кальций в огромных количествах, как это хорошо извест-
но, потребляют и позвоночные животные, строящие из фосфатов
кальция и кальцийорганических соединений свой скелет и раз-
личные части организма.
Магний накапливается литотамниями, некоторыми мшан-
ками и др.
Железо, кроме представителей железобактерий, усваивается
многими представителями водорослей. По существу оно исполь-
зуется всем миром животных.
Марганец усваивается целым рядом марганцевых организ-
мов, такими как лептотрикс, некоторыми диатомовыми.
Фосфор, в виде фосфатов, отмечался В. И. Вернадским в
составе раковин лигнул — брахиопод с роговой раковиной.
Роль бактерий в его накоплении ранее исследователями не учи-
тывалась.
Сера является источником энергии для колоссального коли-
чества организмов различного систематического положения,
от бактерий (тионовые, пурпурные и др.) до высших организмов,
поскольку сера непременная составная часть белка.
99
Из упомянутых организмов для нас особо интересны авто-
трофные, к которым В. И. Вернадский относил такие организмы,
которые «берут все нужные им для жизни химические элементы,
в современной биосфере, из окружающей их косной материи и
не требуют для построения своего тела готовых органических
соединений другого организма». Среди автотрофов он различал
две группы: зеленые хлорофильные организмы — «главный ме-
ханизм биосферы, создающий фотосинтезом органические тела»,
«источник действенной химической энергии биосферы» — и мир
автотрофных бактерий.
Массу зеленых автотрофов В. И. Вернадский считал количе-
ственно близкой к половине животного живого вещества. Важно
отметить, что количество живого вещества, собранное в форме
автотрофных бактерий, он считал гораздо меньшим. Об этих
бактериях он говорил, что они рассеяны всюду: в почвах, иле
водных бассейнов, в морской воде, — но что «нигде нет тех
их количеств, которые были бы сравнимы с количеством авто-
трофной зелени суши, не говоря о зеленом планктоне океана».
Он признавал при этом, что «геохимическая энергия бакте-
рий гораздо выше той же энергии зеленых масс... является мак-
симальной для живых существ». Причину же «малого скопления»
живой материи в этой форме он считал связанной с недостаточ-
ными количествами находящихся в биосфере не вполне окислен-
ных и неокисленных соединений азота, серы, железа, марганца,
углерода. По его мнению, автотрофные бактерии находятся
ныне в состоянии непрерывного недостатка пищи, «недоедания».
Он отмечал своеобразное «вторичное равновесие между бакте-
риями, восстанавливающими сульфаты и их окисляющими, как
это, например, наблюдается в озерных грязях, сырых почвах,
морской воде. Такое же равновесие он отмечал между бактерия-
ми, окисляющими азот, и гетеротрофами, раскисляющими
нитраты. Поэтому главную массу живого вещества В. И. Вер-
надский считал сосредоточенной в освещенной Солнцем части
нашей планеты, где собраны гетеротрофные организмы и авто-
трофные бактерии, тесно связанные с продуктами жизни зеле-
ных организмов и свободным кислородом. Из этой зоны авто-
юо
трофы проникают и в темные области биосферы, где нижнюю
границу для анаэробов он определял до глубины в 500 м и где
размах их жизнедеятельности считал ограниченным.
В наших исследованиях мы начали подбирать материалы по
рассеянному, законсервированному в горных породах органи-
ческому веществу, ранее вообще не изучавшемуся, которое
должно было иметь местами весьма сложный состав. Его изуче-
ние позволило бы нам подойти к представлениям о составе
исходного первичного органического вещества, к химическому
составу самих древних организмов. Это органическое вещество
осадочных горных пород во многих случаях не связано с соста-
вом видимых остатков организмов, обычных объектов палеон-
тологического исследования. Поэтому нам пришлось в ряде
случаев принять его отчасти за остаточное органическое веще-
ство бактериального происхождения. Местами оно оказалось
четко связанным с веществом ископаемых синезеленых водо-
рослей.
Зародившаяся мысль о значительной роли ископаемых древ-
них автотрофных и иных бактерий в породо- и минералообразо-
вании привела нас в некоторые районы Тянь-Шаня, в горные
районы Сибири — к разрезам древнейших осадочных отложе-
ний. где мы знали или предполагали аномальные концентрации
ряда интересных минеральных веществ. Это были залежи фос-
фатов в кембрии Каратау (Казахстан), а также слди с осадоч-
ными рудами железа и марганца, различные железисто-крем-
нистые породы, известняки различных типов. Как оказалось,
очень интересные материалы имеются в районах известных
контактовых месторождений железа и месторождений марган-
ца неясного происхождения. Интересные материалы дало и
изучение процесса цементации некоторых древних обломочных
пород, а также процессов выветривания горных пород, в том
числе и интрузивных.
Эти материалы привели к необходимости пересмотреть
взгляды на породообразующую роль микроорганизмов, осо-
бенно ископаемых. Первое место среди них теперь приходится
приписывать бактериям.
101
Изучая деятельность морских денитрифицирующих бакте-
рий в Атлантическом океане, Г Дрю открыл формы, осаждаю-
щие из морской воды карбонат кальция. Эти бактерии оказа-
лись денитрифицирующими и декальцинирующими. Они как бы
снижают содержание нитратов в морской среде, ограничивая
тем самым масштаб жизнедеятельности фитопланктона. Бак-
терии проявляют свою «деятельность» в районах тропических и
субтропических морей и океанов. Оптимальная температура
для них — около 29,5 ° С. Рост их отмечается при температуре
выше 10°, при 33 ° С он начинает замедляться. Образуемый ими
карбонатный осадок в виде белого облака — мути — медленно
оседает (в лабораторных условиях) или остается в течение дли-
тельного времени во взвешенном состоянии. Такое явление мож-
но наблюдать и в СССР на оз. Балхаш. Осадок получается или
иловидный, или в виде мелких шариков типа оолитов, если
в воду добавлять мелко раздробленный гипс. Шарики эти,
в частности, со дна Большого Соленого озера в Северной
Америке, состоят из арагонита с постепенными переходами в
кальцит. Кальцит иногда образовывался непосредственно из
известковой мути при отстаивании пробы морской воды, при-
чем отмечалось выделение сероводорода, которое происходило
при участии бактерий.
Советские микробиологи полагают, что выделение карбона-
та кальция бактериями из морской воды и других минерализо-
ванных сред происходит за счет сульфата кальция (гипса) в про-
цессе его разрушения, иногда с получением конечного продукта
в виде серы. Ход реакций представляется таким:
CaSO4- CaS
CaS + СО2 - СаСО3 + S
Таким образом, выпадение карбоната кальция, а иногда и
магния из природных водных сред приходится считать связан-
ным в той или иной степени с жизнедеятельностью бактерий.
Процесс этот в морях геологического прошлого некоторые
литологи и геологи иногда недооценивают, иногда совсем не
принимают во внимание как массовое явление.
102
Как известно, карбонатные породы начали входить в состав
осадочных отложений непосредственно с архейского времени,
с момента начала формирования древнейших нормальноосадоч-
ных пород. Если биогенное происхождение многих известных
слоев и горизонтов известняков, например: рифовых, археоциа-
товых, коралловых, брахиоподовых, фораминиферовых и про-
чих, не вызывало никаких сомнений, то в отношении многих
толщ и свит таких «плотных» пород вопрос о генезисе не всегда
был ясен. Чем древнее карбонатные породы, тем чаще мы встре-
чаем так называемые «немые» слои и толщи, лишенные види-
мых остатков ископаемых организмов. Подобные породы мы
находим, в сущности, во всех геологических системах. Слои
с органическими остатками в природе все же редкость, но там,
где они имеются, мы уделяем им преимущественное внимание,
в результате чего наши коллекции относительно богаче остат-
ками ископаемых макроорганизмов, чем отображаемые ими
естественные разрезы соответствующих геологических свит и
серий.
Между тем тщательные поиски органических остатков часто
позволяют охарактеризовать палеонтологически многие толщи
древних морских отложений, в которых обнаруживаются остат-
ки породообразующих синезеленых водорослей в виде строма-
толитов, онколитов и даже иногда плотных внешне пород.
Тем не менее еще бытует мнение о распространенности «немых
толщ» и вследствие этого часто некоторые морские осадки при-
знаются имеющими химическое происхождение, что не всегда
правильно.
Известно, что пересыщенный раствор карбоната кальция в
морской воде мог бы получаться лишь при выпаривании послед-
ней на 75%, что и осуществляется в природе, но не в нормально
морских, а в лагунных условиях. Выпадение карбоната кальция
при этом может сопровождаться осаждением сернокислого
кальция в виде гипса. Некоторые исследователи, подсчитывая
растворимость карбоната кальция в морских водах, полагают,
что и там могут возникать обстановки насыщения среды этой
солью, т. е. условия для ее химического выпадения. Но нужно
103
учитывать запасы в морских водах свободной углекислоты,
способной растворять твердый карбонат кальция. Таким обра-
зом, от идеи массового химического осаждения морских извест-
няковых осадков приходится решительно отказаться. Главную
массу их приходится определенно считать микробиогенной.
Следовательно, мы должны поддерживать ранее высказанные
взгляды Мэррея, В. И. Вернадского, Б. Л. Исаченко, А. П. Ви-
ноградова и других.
Изготовляя тончайшие прозрачные шлифы из различных
известняков древнейших толщ, мы смогли в ряде случаев, окра-
шивая препараты, различать палочковидные, кокко-бацилло-
видные тельца, в которых нельзя не узнавать фоссилизирован-
ных, пропитанных карбонатом кальция клеток каких-то породо-
образующих бактерий. Открытые нами палочковые бактерии
в кембрийском известняке имеют толщину около 1 д при длине
в 2,0—2,5 д.
Нам думается, что выпадение карбоната кальция из морской
воды происходит не при косвенном участии бактерий, как это
представляли себе многие исследователи ранее, а в теле бакте-
рий, в процессе превращения бикарбоната кальция морской
воды в кальцийорганическое соединение, близкое к кальциту.
Поэтому образование карбонатной мути, известкового илового
осадка в теплых морях, вероятно, есть процесс осаждения от-
мерших клеток бактерий, пересыщенных твердым минеральным
веществом в виде кальцийорганических соединений. При этом
органическая оболочка этих сгустков на пути ко дну бассейна,
в осадке, а иногда даже после превращения его в известняк,
защищает их более или менее длительно от перераспределения
вещества. Последнее происходит, несомненно, лишь после пол-
ного распада органических веществ клетки с выпадением в оса-
док карбоната кальция в виде мелких кристалликов. Вследствие
этого нам и удается наблюдать иногда подобные образования
в очень древних породах.
По данным исследований академика Б. Л. Исаченко, изучав-
шего условия образования известкового ила в озере Севан
(Армения), а также известковых образований в горячих источ-
им
никах, существует ряд видов бактерий, способных выделять
углекислый кальций сначала в виде кальцийорганических соеди-
нений, потом в виде бесформенных скоплений карбоната каль-
ция, затем в кристаллической форме кальцита или арагонита.
Иногда образуются округлые тела — сферолиты, или оолиты,
радиально-лучистые или концентрически наслоенные. Внутри
последних исследователь обнаруживал клетки бактерий. Поэто-
му Б. Л. Исаченко полагал, что таким путем и в древних пресных
и соленых водоемах могли образовываться известковые отло-
жения.
Принимая такой взгляд, мы должны допустить, что группа
денитрифицирующих (высвобождающих азот) бактерий явля-
ется одновременно и накопителем в биосфере органического
вещества. Этого нельзя хотя бы отчасти не связать с одновре-
менным выделением сероводорода, т. е. с деятельностью серо-
водородных бактерий, разрушающих при этом белковые веще-
ства отмерших клеток «кальциевых» бактерий.
Таким образом «немые», плотные, массивные и слоистые
известняки, часто перекристаллизованные, лишенные, следова-
тельно видимых остатков животных и растений, мы иногда
имеем основание считать микробиогенными образованиями,
биолитами, биокосными телами, что указывает на колоссаль-
ную роль кальциевых бактерий в формировании живого вещест-
ва — вообще органического вещества в биосфере. Если учесть,
что эти организмы усваивают большую часть приносимого в
моря и океаны с суши кальция (по Кларку, 577 670 000 т кальция
ежегодно!), то живому веществу кальциевых бактерий будет
принадлежать видное место среди породообразователей кон-
центраторов минерального вещества.
К «кальциевым» бактериям, по их роли в породообразова-
нии, приближаются лишь карбонатные водоросли группы си-
незеленых, в ископаемом состоянии представленные изучае-
мыми нами строматолитами, скопления которых, особенно в
древнейших отложениях, часто наблюдаются в виде слоев и
толщ мощностью во много сотен метров. В протерозое и кемб-
рии эти образования формировались в изобилии, местами до
105
Микроскопические синезеленые водоросли образовывали в сивинское время
часто мощные толщи, состоящие из ветвящихся слоистых известковых колонок.
Мощность слоя, часть коюрого показана на рисунке, составляла 36 м. Фото
автора
40 50°о по отношению к микробиогенным карбонатным по-
родам. Синезеленые водоросли живут в тесном общении с раз-
личными бактериями, чаще с железобактериями. Формирование
живого вещества здесь завершалось почти полной фоссилиза-
цией остаточного материала в виде упомянутых строматоли-
тов продуктов их многолетней жизнедеятельности. Следует
думать, что экзотермический характер реакции выделения твер-
дого карбоната кальция в организме играл также свою роль и в
энергетическом балансе некоторых древнейших водорослей,
как это, несомненно, имело место в слоевищах мезозойских и
более поздних кораллинацей.
106
В связи с отмеченным выше мы можем позволить себе по-но-
вому взглянуть и на процесс накопления карбоната кальция
многими беспозвоночными, например в виде раковин моллюс-
ков, брахиопод, кораллитов кораллов, скелета археоциат, спи-
кул известковых губок и т. д. Клетки мантий и соответствующих
других тканей этих организмов, живших и живущих в той же
морской среде, восстанавливая тот же бикарбонат кальция,
должны получать и освобождающуюся при этом энергию.
Мы можем вспомнить при этом о безобразно толстой, неуклю-
жей многослойной раковине некоторых современных и ископае-
мых устриц, о чем выше мы уже говорили. Такие раковины изу-
чать очень интересно, можно видеть, как она развивается, как
непрерывно утолщается корка за коркой, иногда даже без пря-
мой связи с ростом мягкого тела животного. Чем вызвано это
явление? Поскольку этот процесс непрерывен, хотя и идет в
течение года с неодинаковой скоростью, он больше похож на
процесс питания, чем на тенденцию к защите от механических
воздействий извне. Некоторые ученые думают, что такой уст-
рице нужно увеличивать свой вес, чтобы ее «не беспокоила»
волна или течение. Естественный отбор мог бы создавать и та-
кие организмы, но он бессилен создать упомянутую функцию
мантии, если она не возникла в ходе эволюции данной группы
для выполнения определенной физиологической задачи. А эта
задача для карбонатотлагающих организмов — превращение
бикарбоната кальция в карбонат (экзотермическая реакция!).
Поэтому в жизненном цикле устриц предполагаемый нами источ-
ник энергии (вероятно, исторически первичный) занимает, по-ви-
димому, видное место. В энергетическом балансе других живот-
ных, из числа вышеупомянутых, кроме кораллов, тоже постоян-
но отлагающих карбонат кальция, этот источник жизненной
энергии, по-видимому, играет относительно скромную роль
в виде «дара предков», в ходе эволюции иногда постепенно
утрачиваемого.
Палеонтология дает нам немало таких примеров, до перехо-
да к «бесскелетным» формам, как это намечается у некоторых
археоциат к началу позднею кембрия, а также у кораллов.
107
Таким образом, для нас представляется весьма соблазни-
тельным предположить, что все карбонатотлагающие много-
клеточные организмы, не исключая и высших представителей
животного мира (с фосфатами кальция в скелете), относятся
к числу участвующих в процессе энергетического использования
восстанавливаемого ими растворенного карбонатного вещества,
т. е. к числу полуавтотрофных, в той или иной степени несвобод-
ных от этого примитивного древнейшего источника химиче-
ской энергии. Исходя из этого, мы получаем новое представле-
ние о размахе взаимосвязи между живым и минеральным ве-
ществом природы, отличное от наших прежних представлений.
При этом мы признаем, конечно, обломочное, а местами хими-
ческое происхождение многих известняков. ( идериты, магне-
зиты, доломиты и другие карбонаты имеют, по-видимому,
лишь косвенную связь с деятельностью живого вещества.
В существующих учениях о рудных месторождениях и мине-
ралах миграция железа и марганца в земной коре и на ее поверх-
ности часто трактуется в свете чисто химических процессов.
Микробиологические отложения окислов железа и марган-
ца в наземных — дерновых, болотных и озерных условиях
известны давно. В изучении этих образований принимали уча-
стие многие исследователи.
Поиски морских железобактерий, начавшиеся с экспедиций
«Челленджера», вначале были безуспешными. Открытие мор-
ских железо-марганцевых бактерий было у нас осуществлено
лишь В. С. Буткевичем. Тем не менее многие ученые не сомне-
вались в том, что величайшие скопления железа в земной коре
образовались биохимическим путем при незначительном уча-
стии в этом процессе водорослей. Так, В. И. Вернадский считал
мезозойские железные руды Эльзас-Лотарингии и керченские
третичные руды биогенными, вероятно, бактериальными, как
и марганцевые чиатурские руды. Большинство исследователей
железобактерий склонялось к взгляду, что наряду с биогенными
скоплениями железа в осадке бассейнов отлагаются и хемоген-
ные. Железобактерии проявляют жизнедеятельность в трубах
водопровода, образуют конкреции на морском и озерном дне.
108
Биогенный путь состоит в окислении гидратов закиси же-
леза и марганца, а также их бикарбонатов, с переводом, с по-
мощью соответствующего фермента бактерий, двухвалентного
железа в трехвалентное, при переработке колоссальных коли-
честв закиси железа, выносимой из недр Земли подземными
водами.
Мы предприняли поиски микробиогенных структур в же-
лезных и марганцевых рудах осадочных толщ различного геоло-
гического возраста, начиная с раннепротерозойского, а также
в районах подобных руд первично магматогенного происхож-
дения, в обломочных терригенных породах и в зоне выветрива-
ния массивных пород. Результаты оказались замечательными.
Изучая под микроскопом шлиф из образца железистого
кварцита бедной руды Кривого Рога (докембрий), мы обнару-
жили массовые скопления чехликов железобактерий, заключен-
ных в коллоидную кремнеземную массу, по-видимому, и сохра-
нившую эту микробиогенную структуру сгустков окислов же-
леза. Первичные гидроокислы железа в результате метаморфиз-
ма представлены теперь магнетитом. Такие же скопления
ожелезненных клеток железобактерий в ассоциации с коллоид-
ным кремнеземом мы открыли и в железистых сланцах гребен-
ской свиты протерозоя р. Ангары, нижнего палеозоя хребта
Каратау.
Бурые железняки, образование которых В. Линдгрен счи-
тал химическим, представляли для нас большой интерес. Мы
изучали их по материалам Кокчетавского района в Казахстане,
в отложениях, относимых к докембрию. Для всех изученных
образцов руд происхождение оказалось общим. В них удалось
ясно различить чехлики древнейших представителей железо-
бактерий, из которых слагается вся масса руд. Размеры ожелез-
ненных клеток близки к современным. Изучение бурых железня-
ков и лимонитов многих других месторождений, до так назы-
ваемых «железных шляп», формирующихся в поверхностных
условиях при выветривании сульфидных руд железа, показало
решительно то же самое. Все бурые железняки и лимониты пре-
красно сохраняют свою микробиогенную структуру. При этом
109
стало ясно, что жизнедеятельность железобактерий, типич-
ных аэробов, осуществлялась и осуществляется и в недрах
Земли, в зоне циркуляции кислородных вод. Обычные в магне-
титовых месторождениях магматогенного происхождения бу-
рые железняки показали ту же микробиогенную структуру и,
следовательно, огромный размах этого процесса.
Мы изучали природные скопления марганца в месторожде-
ниях Казахстана, Сибири, относящиеся к нижнему и среднему
палеозою, и получили такие же результаты. Руды, за исключе-
нием части их, подвергшейся перекристаллизации, оказались
сложенными сплошь микротельцами фоссилизированных кле-
ток бактерий, имеющих около 1,33 д в поперечнике и до 2,0—3,6д
в длину, соединенных в длинные изогнутые, плотно сплетенные
нити. Последние различимы тем лучше, чем «беднее» изучае-
мая руда, в которой рудная масса перемежается с нерудными,
обычно прозрачными, минералами. Общим признаком для от-
меченных марганцевых биолитов, как и для железных, по ка-
захстанским и сибирским материалам, является плотное кол-
лоидоподобное их сложение, или волокнисто-лучистое (псило-
мелан, лимониты), тогда как в поверхностных, «осадочных»
условиях чаще распространены конкрепионно-желваковые и
гороховые — пизолитовые структуры. Природа конкреционных
образований, по-видимому, связана с разложением органиче-
ского вещества бактериальных клеток и потерей микробиоген-
ной структуры таких руд.
Микробиогенными образованиями являются и так называе-
мые «дендриты» — налеты окислов марганца на стенках тре-
щин горных пород, вблизи от скоплений марганцевых руд и
минералов. Природа рисунка дендритов, вероятно, связана со
строением колоний марганцевых и железобактерий. Такого же
происхождения, по-видимому, и «пустынный загар» горных
пород, нами пока ближе не изучавшийся. Железистый цемент
красноцветных песчаников, например, пермского возраста с
р. Шугор (бассейн Печоры), оказался массовым скоплением
между песчинками породы фоссилизированных телец железо-
бактерий, пропитанных окислами железа. Такое же происхож-
по
дение имеют различные охры, ржавчины, распространенные
в природе в трещинах горных пород, в карстовых полостях,
образовавшихся в процессах выветривания пород, где бакте-
риям, несомненно, принадлежит ведущая роль.
Постоянное присутствие органических веществ, часто в виде
углистой массы, в фосфоритовых слоях Каратау давно уже при-
влекало к себе внимание, но выяснить его природу и происхож-
дение самих фосфоритов мы смогли тоже лишь в последние
годы.
Накопление фосфатов в осадках до сих пор объяснялось уче-
ными с помощью двух теорий. Первая, так называемая «биолит-
ная», теория рассматривала образование фосфоритов как след-
ствие массовой гибели организмов и перехода их фосфора в
фосфатную горную породу. Согласно второй теории, происхо-
дило непосредственное осаждение фосфорных солей из морской
воды под влиянием изменения их растворимости. По первой
теории, причиной гибели организмов считались изменения
уровня моря, направления течений, степени солености в эпохи
«перерыва», крупных тектонических движений. Химическая тео-
рия родилась в свете представлений о повышенном содержании
фосфора в водах морских глубин (до 300 мг/м3). Поднимаясь
с гипотетическими восходящими течениями до глубин в
50—150 м, эти воды должны переносить содержащийся в них
фосфор и отлагать его вслед за несколько ранее выпадающим
в осадок карбонатом кальция. Но ведь здесь фосфор снова
становится объектом использования его фитопланктоном, т. е.
субстратом для морских водорослей. Верхние слои морских
вод всегда поэтому бедны фосфором. Слабое место этой тео-
рии — отсутствие в морях массовых «восходящих течений» и
отсутствие в нормально морских условиях возможности хими-
ческого осаждения карбоната кальция, о чем выше мы уже го-
ворили.
Каратауские фосфориты развиты в слоях среднего кембрия
на обширной площади. Они представлены здесь плотными мас-
сивными («плита») и оолитовыми разновидностями. Последние
очень напоминают внешне бокситы, за которые первоначально
Бактерии ржавчины при увеличении в 3000 раз. Фото автора
они и были приняты. Сначала мы исследовали оолитовые фос-
фориты, состоящие из серых зерен, сцементированных желтова-
то-бурым веществом. В этом цементе мы обнаружили железо.
Тончайшие шлифы, изготовленные из таких фосфоритов, при
увеличениях свыше 2000 раз показали, что оолитовые зерна яв-
ляются местом массовых скоплений микротелец бактериально-
го облика, имеющих в поперечнике 1,1 р при длине в 1,3/z, то
соединенных в изгибающиеся нити, то одиночных. Бактериаль-
ная природа этих образований несомненна. Ясно, что это пре-
красно сохранившие свои очертания окаменевшие клетки каких-
то бактерий кембрийского возраста. Интересно, что в центрах
оолитовых зерен расположение бактериальных клеток беспо-
рядочное, но по мере приближения к периферии в концентриче-
ских наслоениях зерен они начинают принимать более или
менее радиальное расположение, указывающее на формирова-
ние колоний «фосфатных бактерий» в условиях сезонно измен-
чивой среды. В составе концентрических наслоений, часто пере-
межаясь с ними, наблюдаются тонкие пленки, сложенные тель-
цами железистого состава в виде обогащенных железом отмер-
ших клеток железобактерий. Цемент природы, скрепляющий
оолитовые зерна, целиком состоит из ожелезненных клеток от-
мерших железобактерий знакомого уже для нас облика. Послед-
ние развивались позже, в условиях несколько измененного тем-
пературного режима.
Изучение немногих образцов плотных разновидностей ка-
ратауских фосфоритов показало, что они или сплошь состоят
из клеток «фосфатных» бактерий и бесструктурной массы карбо-
ната кальция, или же к минерализованным клеткам «фосфатных
бактерий» примешиваются тельца клеток железобактерий. По-
следние в таких случаях местами группируются в виде тонких
пленок, линзочек.
Раскрытие природы каратауских фосфоритов позволяет нам
думать, что и другие известные месторождения фосфоритов
имеют, возможно, такое же органическое, бактериальное про-
исхождение.
Интересно, что при изучении разреза фосфоритовой свиты
мы наблюдали смену фосфоритовых слоев марганцево-желези-
стой породой с прослоем карбонатных синезеленых водорослей.
Разрез показывает на близость экологических условий для авто-
трофных бактерий, усваивавших фосфор, железо и марганец,
и для синезеленых автотрофов. Известна местная засоренность
каратауских фосфоритов терригенным материалом.
О современных бактериях — накопителях фосфатов почти
ничего неизвестно. Известны лишь почвенные бактерии, раство-
ряющие фосфаты, фосфатредуцирующие, способные образовы-
вать растворимые соли фосфорной кислоты. Между тем концент-
рирующие фосфор бактерии в кембрии, по-видимому, существо-
вали. Может быть, подобные организмы формируют и редкие
в наше время скопления, фосфоритов на дне моря, как, например,
в районе южной оконечности Африки.
Остаточные бактериальные микроструктуры боксита. Увеличение в 3600 раз.
Фото автора
В круговороте серы, одного из важнейших биогенных эле-
ментов биосферы, принимают участие почти все организмы;
в недрах земной коры переработкой серных соединений заняты
также многие группы бактерий. Наиболее известная группа —
сульфатредуцирующие (разрушители сульфатов, в частности
гипса) или гнилостные (разрушители органических веществ)
бактерии — анаэробы, которые часто обитают в застойных
водах и под землей. Следы процессов преобразования серы мы
находим уже в горных породах протерозойского и раннепалео-
зойского возраста, в виде пиритизированных песчаников и
сланцев древних толщ Сибирской платформы. Накопление се-
роводорода происходит микробиологическим путем за счет
сернокислых солей моря, с последующим образованием в осад-
114
ке скоплений сернистого железа (пирит, марказит) в восстанови-
тельной обстановке. Существуют различные аэробы, которые
с успехом окисляют сульфиды как в зоне земной поверхности —
«выветривания» горных пород, так и в зоне окисления земной
коры. Часто бактериальная сероводородная зона развивается
на разлагающихся органических веществах, при разрушении
белковых и иных соединений.
«Обохривание» пиритовых залежей, по нашим наблюдениям,
как и окисление магнетитовых, происходит и в недрах (зона
окисления) и в субаэральной зоне (в воздушной среде) благодаря
жизнедеятельности одновременно железных и тионовых бак-
терий, с выносом сернокислых солей и с преобразованием ве-
щества пирита и магнетита в лимонит, что хорошо видно в
препаратах под мироскопом. Поэтому массовое гипсообразо-
вание в ряде случаев мы можем считать связанным с продукта-
ми жизнедеятельности тионовых бактерий.
Естественно, мы здесь не смогли перечислить еще многих
других процессов связи между живым веществом природы и
минеральным субстратом, играющим косвенно роль источ-
ника энергии.
Таким образом, органическая жизнь в природе в ее наибо-
лее примитивных бактериальных формах оказывается тесней-
шим образом связанной с веществом земной коры. Поэтому
было бы совершенно правильно трактовать все подобные ма-
териалы, как относящиеся к новой науке — геологической ми-
кробиологии. Советские ученые, начиная с В. С. Буткевича,
А. И. Самойлова, В. И. Вернадского и Б. Л. Исаченко, уже
внесли большой вклад в создание основ этой науки. Интересные
и практически важные работы по геологической деятельности
микроорганизмов ведутся в Институте микробиологии АН СССР
С. И. Кузнецовым и группой его сотрудников, которым удалось
значительно продвинуть решение многих вопросов геологиче-
ской микробиологии.
Множество интересных вопросов, связанных с древней при-
родой, уже разрешено. В данном случае мы могли бы обратить
внимание на.роль древних организмов как создателей тех или
115
других веществ, в водных средах прошлого, захороненных в
горных породах или же принявших характер горных пород.
Так, мы знаем, что из вещества наземных и водных растении в
геологическом прошлом Земли образовывались различные ти-
пы каменных и бурых углей, минерального топлива. Простей-
шие формы жизни — бактерии и простейшие животные —
фораминиферы создавали на дне водных бассейнов отложения
извести, из которых потом получались твердые горные породы
известняки и мраморы.
Долго казалось загадочным происхождение нефти и горю-
чих газов в недрах. Одни ученые считали, что эти вещества
первично в виде простого газа метана (углеводородный газ СН4)
являются остаточным компонентом древнейшей атмосферы
Земли, еще космической атмосферы, как остаток той атмосфе-
ры, которая создалась вокруг нашей планеты во время ее обра-
зования, другие — что нефть и газ есть продукты преобразова-
ния органического вещества, остатков организмов, в основном
различных групп водорослей, в результате его разложения и
преобразования при участии соответствующих групп бактерий.
Выяснено, что в древнейших водных бассейнах нашей пла-
неты жизнь на ранних этапах ее развития особенно обильно и
разнообразно была представлена бактериями и водорослями.
Первые перерабатывали местами и временами огромные массы
железа, марганца, серы, азотных соединений, образовывали
«осадочные» железные и марганцевые руды, серные руды,
свободный азот и т. д. Вторые благодаря энергии солнечных
лучей накапливали огромные массы органического вещества
за счет углекислоты и воды. При этом за счет воды выделялся
в водную среду и атмосферу кислород. Так и возникла современ-
ная атмосфера Земли, как известно состоящая на четыре пятых
из азота и на одну пятую из кислорода.
Отмиравшее водорослевое вещество в водной среде или уже
в осадках разрушалось бактериями. В условиях отсутствия
кислорода это разрушение не было полным. При этом образо-
вывались углеводородные газы (соединения углерода с водоро-
дом в разных пропорциях). Эти газы частично могли уходить
116
в древнюю атмосферу, растворяться в древних водах. Остатки
водорослей в толщах рыхлых осадков на дне бассейнов и углево-
дородные вещества захоронялись то скученно, то рассеянно и,
таким образом, оказывались при погружениях участков земйой
коры на разной глубине. Будучи более легкими, чем вода, эти
вещества всегда были склонными к стремлению перемещения
вверх, к земной поверхности в сторону меньших давлений.
Местами они встречали препятствия в виде более плотных по-
род и потому там накапливались. При этом происходило часто
их преобразование в более тяжелые углеводородные соединения,
в тот сложный комплекс веществ, который мы знаем под назва-
нием «нефть» — «кровь» Земли.
Ранее считалось, что чем древнее какая-нибудь нефтеобра-
зующая, нефтеносная толща осадочных пород, тем благоприят-
нее ее скопления (месторождения) в отношении количества.
Так, более ста лет назад разрабатывались на территории Евро-
пы нефтяные месторождения Карпат и Кавказа, имеющие воз-
раст в несколько десятков миллионов лет (меловые и третичные
отложения). Потом с конца 20-х годов на территории СССР
были открыты месторождения нефти и газа, гораздо более древ-
ние, — Волго-Уральская нефтяная провинция — «Второе Баку»
с возрастом нефтематеринских отложений около трехсот и
более миллионов лет (девонские отложения). Несколько лет
назад были открыты в Восточной Сибири месторождения нефти
и газа в осадочных породах морского происхождения возраста
около пятисот пятидесяти миллионов лет (нижнекембрийские
отложения). При этом последние оказались не связанными един-
ством происхождения с вмещающими породами (песчаниками
и доломитами).
Выяснено, что в наиболее древние геологические времена на
территории Восточной Сибири располагались обширные моря,
в которых особенно обильно развивались морские водоросли.
Остатки этих водорослей часто облекались корками извести и
сильно известковистой воды бассейнов. Органическое вещество
такого происхождения за сотни миллионов лег суммарно на-
капливалось, (условно подсчитывая) в толщи в сотни метров.
Последующее погружение древних (докембрийских) толщ в
недра Земли, где господствуют высокие давления и повышенная
температура, способствовали движению углеводородов вверх.
Так, в ряде районов так называемой Сибирской платформы,
между Енисеем и Леной, к северу от Восточного Саяна и Бай-
кальского нагорья в кембрийских отложениях сформировались
месторождения нефти и горючего газа.
Итак, органические вещества не могли не образовываться
химическим путем при участии физических (электрические раз-
ряды атмосферы, ультрафиолетовые лучи Солнца) и химиче-
ских (уплотнение, синтез молекул) явлений в более сложные
соединения (в белковые, в аминовые и нуклеопротеиновые кис-
лоты) А это при участии химической или лучистой энергии
должно было привести к возникновению жизни. При этом роль
жизни в геологических процессах оказывается по материалам
геологов и палеонтологов очень значительной.
ПЕРВИЧНЫЕ ФОРМЫ ЖИЗНИ
И ИХ ЭВОЛЮЦИЯ
л
Для первичных организмов Земли среда была столь же благо-
приятной, как и для существующих сейчас, — современная
среда их жизни. В жидких средах биосферы — в почвенной влаге
и в прибрежной водной среде — возникали организмы, условия
питания и жизни которых были различны. Состав первичной
биосферы, несомненно, определял и состав организмов. Это бы-
ли потребители углеводородных соединений и аммиака: серные
бактерии — окислители сероводорода вулканического происхож-
дения, водородные бактерии — окислители молекулярного во-
дорода и т. д. Такие группы организмов прекрасно представ-
лены и в современном бактериальном мире. Появились же их
предки в то время, когда они были лучше всего обеспечены
питанием. Вообще можно полагать, что эти группы живых су-
ществ во все геологические времена на Земле были в достаточ-
ной мере обеспечены пищей и что естественный отбор быстро
привел к выработке устойчивых биохимических форм. Многие
из этих групп расселились потом шире и глубже. В частности,
распространились они и в недра Земли, где стабильные условия
не давали им особых стимулов для проявлений изменчивости.
Отмеченную группу микроорганизмов можно считать древ-
нейшей группой живых существ на Земле, размах жизнедеятель-
ности которой уже на ранних этапах ее развития, вероятно,
быстро приобрел планетарный характер. Эти организмы спо-
собствовали сокращению в биосфере исходных запасов аммиа-
ка, водорода, метана, сероводорода. Они потребляли и углекис-
лоту, хотя запасы последней в общем все же увеличивались за
счет выделения из недр Земли.
Зарождение фотосинтезирующих бактерий и одноклеточных
микроскопических водорослей, подобных тем, которые состав-
ляют в основном морской и пресноводный фитопланктон, было
событием большого геологического значения. Имеющиеся в
119
настоящее время находки остатков бактериальных организмов,
начиная с отложений возраста 3100 млн. лет, на разных стра-
тиграфических уровнях докембрия датируют время их возник-
новения и начала интенсивного развития. Бактерии содейство-
вали накоплению в биосфере высокоорганизованных углерод-
ных соединений, водоросли приводили к обогащению биосфе-
ры свободным кислородом с одновременным обеднением ее
углекислотой.
Появление свободного кислорода, возможно еще в малых
дозах, стимулировало массовое развитие микроорганизмов,
окислявших железо самородное, закисное, вообще недоокислен-
ное. Продукты жизнедеятельности этих организмов в виде мощ-
ных залежей железных руд или рассеянных масс окислов железа
указывают на большое их распространение в бассейнах и почвах
древней биосферы. Геологическое время проявлений этого про-
цесса в основном известно. Это в большей степени поздний до-
кембрий — гуронские слои в Северной Америке, раннепроте-
розойские отложения в Европе и Азии.
Замечательно, что осадочные горные породы и многие’руд-
ные образования осадочного происхождения, сформированные
именно на дне водных бассейнов далекого геологического про-
шлого, часто способны сохранять следы бактериальных струк-
тур. По-видимому, это связано со стойкостью некоторых орга-
нических соединений остаточного вещества бактерий, а отчасти
с условиями захоронения.
Многие осадочные породы, если из них изготовить тонкие
прозрачные шлифы, позволяют видеть под микроскопом обиль-
ные остатки телец по форме и размерам близких к бактериям.
Осадочные железные и марганцевые руды часто представляют
собой сплошную массу оруденелых оболочек — «чехликов»
клеток железобактерий. Бокситы, фосфориты, глинистые мор-
ские и пресноводные отложения часто содержат большое раз-
нообразие таких микроскопических бактериеподобных телец.
Таким образом, появившись около 3000 млн. лет назад, мир
микроорганизмов быстро овладел биосферой нашей планеты.
Его история исследователями-микропалеонтологами еще рас-
120
крыта слабо, но имеются достаточные основания надеяться, что
она будет раскрыта. Она может быть исследована по форме и
размерам окаменевших остатков бактерий и их скоплений,
а также по геологическим продуктам их жизнедеятельности,
как это имеет место в отношении железных и марганцевых пла-
стовых руд, отложений пирита, глубинных залежей бактериаль-
ного азота и т. д.
Удивительное, бесспорно доказанное сохранение в ископае-
мом состоянии остатков спор древнейших растений, обнаруже-
ние множества осадочных пород с явными остатками бактери-
альных структур позволяют надеяться, что этот раздел палеон-
тологии получит свое развитйе.
Естественно, современные процессы не могут помогать вы-
явлению процессов далекого геологического прошлого, но прин-
цип актуализма помогает понять их, если мы правильно учи-
тываем при этом ход и характер явлений, процессов эволюции
среды и жизни за геологическое время.
Микробы, которые способны получать жизненную энергию
путем окисления различных веществ, являются аэробами. Ми-
кробы, способные получать жизненную энергию в отсутствие
свободного молекулярного кислорода в среде их обитания,
являются анаэробами. Какая же из этих групп микроорганиз-
мов является на Земле древнейшей0
Мнение о большей древности автотрофных бактерий по
сравнению с гетеротрофными поддерживается и специалистами,
занимающимися проблемой происхождения нефти, причем при-
нято считать, что в общем случае существование гетеротрофных
организмов немыслимо без существования автотрофов. Но в
общем миры автотрофов и гетеротрофов образуют в биосфере
единую неразделимую систему, которая геологически возникла,
несомненно, очень рано.
Обратимся к фактам. Изучение под микроскопом в тончай-
ших шлифах при значительных увеличениях древнейших осадоч-
ных горных пород, часто даже измененных последующими про-
цессами, указывает на местные концентрации остаточного орга-
нического вещества, которое почти никогда не бывает полностью
121
бесструктурным. Оно не бывает ни коллоидным, ни кристалли-
ческим, если не представлено чистым графитом. Обычно это
скопления микроскопических бактериеподобных образований,
размещенных с различной степенью густоты. Чаще это ните-
видные образования, прихотливо изогнутые, иногда сфероидаль-
ные, как бы смотавшиеся в маленькие клубки, прежде чем заглох-
ла их жизнь. При этом все следы древнейшей жизни ясно указы-
вают на водную среду.
Прослеживая состав организмов в различные геологические
этапы жизни нашей планеты, исследователи обычно переходят
от более поздних к более ранним — в направлении, противопо-
ложном ходу эволюционного процесса.
Используя материалы палеонтологии, мы можем убедиться,
что следы человека прослеживаются в глубь веков не далее чем
на 3 млн. лет. Следы млекопитающих исчезают 60 млн. лет назад.
Следы пресмыкающихся отсутствуют в слоях, возраст которых
более 185 млн. лет. Земноводные (амфибии) и костистые рыбы
не существовали во времена, удаленные от современности более
чем на 340 млн. лет. Остатки примитивных рыбообразных —
бесчелюстных и хрящевых рыб — не встречаются в слоях,
образовавшихся более 420 млн. лет назад. Археоциаты — свое-
образные древнейшие строители морских рифов с изящнейшим
известковым скелетом, видимо, начали свое развитие 600 млн.
лет назад. В более древних отложениях, образовавшихся до
1800—1900 млн. лет назад, мы находим лишь остатки водорослей
и следы жизнедеятельности бактерий.
Если же рассматривать развитие жизни по группам организ-
мов в нормальном ходе их эволюции, то мы увидим, что каждая
из групп проходила свои этапы развития. Появлялись новые
формы, вымирала часть старых. Некоторые группы существо-
вали ограниченное время, потомки других живут даже в наше
время. Таким образом, «корни жизни» на Земле оказываются
весьма многочисленными, представленными действительно наи-
более просто построенными, притом преимущественно мель-
чайшими по размерам организмами. Несомненно, жизнь по-
явилась более 2700 млн. лет назад.
122
ПАЛЕОНТОЛОГИЧЕСКИЙ МЕТОД В ИЗУЧЕНИИ
ДОКЕМБРИЯ
Граница между докембрием и кембрием обычно резко выраже-
на не только по степени вторичных изменений осадочных пород,
но и по кажущемуся коренному отличию пород кембрия, отло-
жения которого почти всюду охарактеризованы обильными
разнородными остатками беспозвоночных, водорослей и др.
Поэтому исследователи всего мира занимаются изучением орга-
нических остатков из палеозойских и более поздних отложений.
Некоторые исследователи уделяют внимание изучению докем-
брия, его литологии, петрологии, в еще меньшей степени па-
леонтологии, несмотря на огромное распространение и мощ-
ность его осадочных отложений.
В трансваальской серии докембрия Южной Африки остатки
водорослей известны в виде мелких строматолитов и онколитов.
Абсолютный возраст, определявшийся Ф. Абельсоном и Э. Барг-
хорном в институте Карнеги в Вашингтоне, дал цифру около
2600 млн. лет. В отложениях кремнистых сланцев гуронского
возраста (2000 млн. лет), в районе оз. Онтарио в Северной Аме-
рике Э. Баргхорн нашел хорошо сохранившиеся остатки нит-
чатых синезеленых водорослей и одноклеточных животных.
Таким образом, жизнь на Земле является очень древней,
имеющей возраст порядка 3000 млн. лет или более. Следы древ-
нейшей жизни можно наблюдать в некоторых геологических
образованиях докембрия, в виде непосредственных морфологи-
ческих материалов по конкретным группам организмов и в виде
косвенных данных.
Существование жизни на Земле можно проследить в ее про-
явлениях во времени и в пространстве, т. е. в данном случае в
разрезах разновозрастных осадочных горных пород, в их до-
вольно большом разнообразии.
Общеизвестно большое распространение в составе отложе-
ний архея и протерозоя нормальноосадочных пород терриген-
ного состава. Учитывая степень дифференциации мелкозема,
выделение достаточно хорошо отсортированного материала
123
обломочных пород, в частности песков, превращенных в квар-
циты, приходится допускать на выступах древнейшей суши
развитие почвенного покрова.
Как известно, почвообразование идет интенсивно с диффе-
ренциацией минерального вещества лишь при участии почвен-
ных микроорганизмов, роль которых в создании почвенного
мелкозема обычно в 20 раз превосходит роль почвенных коллои-
дов химического происхождения. Поэтому почвообразование,
связанное с деятельностью микроорганизмов, является, несом-
ненно, процессом очень древним. В связи с этим наблюдение
в составе беломорской серии архея Карелии магнетитовых слан-
цев и кварцитов, первично представляющих собой отложения
осадочных железных руд и песчаные отложения, показывает,
что они, несомненно, были продуктом развития на смежном
выступе суши почвообразовательных процессов с участием
почвенных микроорганизмов и железобактерий.
Многими ^следователями происхождение тонких морских
карбонатных илов рассматривается в связи с жизнедеятель-
ностью некоторых специфических морских бактерий, в частности
из группы денитрифицирующих.
Процесс денитрификации водной среды при участии бакте-
рий обусловлен в природе наличием нитратов, усвояемого
органического вещества, анаэробных (бескислородных) усло-
вий и нейтральной реакции среды. Представление многих
авторов о составе древней атмосферы и гидросферы Земли
сводится к признанию большой роли там свободного водорода,
метана, углекислоты и аммиака. Благодаря высокой раствори-
мости аммиак должен был легко переходить в раствор, чтобы
обеспечить развитие денитрифицирующих микроорганизмов,
продуктами жизнедеятельности которых могли .быть иловые
карбонатные отложения, с превращением их в известняки, до-
ломитизированные известняки (в порядке замещения кальция
осадка магнием раствора) и доломиты. Лишь образование
магнезитов можно отнести к процессам химического характера,
происходившим и происходящим в условиях выпаривания
морских вод полузамкнутых лагун.
124
Наличие кристаллических известняков в беломорской серии
архея Карелии Беломорского побережья позволяет предпола-
гать развитие в древнем бассейне соответствующих бактериаль-
ных процессов, частичным выражением которых могут быть
такие породы.
Имеющиеся цифры абсолютного возраста в 2000—2300 млн.
лет для архея Беломорья некоторые исследователи склонны
считать возрастом метаморфизма архейских пород. Таким об-
разом, процессы древнейшего карбонатного осадконакопления
при участии бактерий должны быть признаны здесь весьма
древними. В некоторых мраморах архея Кольского полуостро-
ва, по наблюдениям автора, между зернами кальцита установ-
лено содержание обуглероженных остатков микротел бакте-
риального размера.
Известно широкое распространение в докембрийских отло-
жениях всего мира пластовых железных руд, горизонтов пири-
тизированных пород, которые можно связать по условиям их
происхождения только с жизнедеятельностью ряда групп микро-
организмов.
Прямые морфологические данные мы можем получить,
изучая под микроскопом препараты из железных руд осадоч-
ного происхождения. Так, автором остатки железобактерий бы-
ли установлены в ряде образцов докембрийских железных руд,
в частности Кривого Рога, некоторых месторождений докемб-
рия Казахстана и Сибири.
На территории Карелии, характеризующейся широким рас-
пространением мощных отложений докембрия, был открыт ряд
ценных палеонтологических материалов в трех горизонтах
ятулия (среднего протерозоя). Р. В. Бутиным и автором описано
много видов водорослей, которые сохранились в виде остатков
строматолитов и других карбонатных образований. Это сви-
детельствует о большом разнообразии ценозов водорослей
в бассейнах протерозоя Северо-Запада СССР С этими откры-
тиями перекликаются открытия остатков строматолитов в
слоях серии Хяохе Северного Китая, которые по возрасту от-
носятся также, к среднему протерозою. Наряду с местной и
125
временной жизнедеятельностью синезеленых водорослей,
в меньшей степени красных водорослей, сыгравших местами
роль крупных геологических деятелей, имело место отложение
карбонатных илов, что осуществлялось, по-видимому, при учас-
тии денитрифицирующих микроорганизмов.
В ятулии Карелии открыто большое разнообразие проблема-
тических образований, предположительно отнесенных к водо-
рослям или остаткам ряда групп беспозвоночных животных.
Не подлежит сомнению, что теперь весь протерозой становится
объектом биостратиграфических и палеонтологических иссле-
дований. Выделена группа образований, которые можно услов-
но принять за остатки водорослей из группы багрянок — ко-
раллинацей. Некоторые из них отличаются необычайно круп-
ными размерами клеток (?). Возможно, подобные водоросли,
описанные из мезозоя, не являются самыми древними на Земле,
как это предполагалось. Известны также находки одноосных
кремневых спикул губок, остатки радиолярий и др.
Более поздние докембрийские отложения (верхний протеро-
зой — синий, иотний и гиперборей, овруч, бавлы, альгонкий,
рифей и др.) богаче остатками организмов, особенно синезеле-
ных и красных водорослей. Так, автором описано более 200 ви-
дов водорослей среднепротерозойского и позднепротерозой-
ского (синийского) возраста из отложений Карелии, Средней
и Восточной Сибири и Дальнего Востока, а также из отложений
КНР, которые он отнес к 80 родам и 20 семействам.
Морфология клеточных колоний, общий, местами хорошо
сохранившийся вид талломов, скопления остаточного органи-
ческого вещества в породе нередко оказываются близкими к
современным. Между тем преобладающая часть форм очень
своеобразна и потому является хорошим опорным материалом
для корреляции вмещающих отложений. Из сотен горизонтов
с остатками водорослей пока выявлено в верхнем протерозое
до 40, характеризующихся наиболее хорошей сохранностью
форм. В среднем протерозое (ятулии) Карелии Р В. Бутин
выявил три водорослевых горизонта.
Установлены случаи дальней миграции некоторых водорос-
126
лей древнее протерозойского возраста, например: северная
часть КНР — Оленекское поднятие, северная часть Сибирской
платформы — Кольский полуостров (о-в Кильдин), Енисейский
кряж — Восточное Верхоянье и т. д. Таким образом, выявлены
возможности открытия остатков древнейших водорослей в от-
ложениях докембрия как нормального палеонтологического ма-
териала. Впрочем, поиски его часто еще не являются система-
тическими. Открытие их чаще носит случайный характер.
Одним из интересных выводов, полученных при изучении
конкретного палеонтологического материала по древнейшим
водорослям, было прямое указание на незначительность или
отсутствие гнилостных процессов в докембрийских бассейнах.
Вследствие этого колонии микроводорослей, выделяя карбонат
калышя на поверхности, в условиях среды, богатой бикарбона-
тами, переходили в ископаемое состояние почти без изменения
их морфологии. Минерализацией охватывались сгустки студе-
нисто-слизистого вещества синезеленых водорослей, оболочки
их трихомов и клеточных колоний. Остаточное органическое
вещество, в той или иной степени обуглероженное переходило
в толщу осадков и в некоторых случаях за счет его могли обра-
зовываться скопления нефти и горючего газа. Так образовались
нефть и газ в нижнем кембрии Сибирской платформы, такого
же происхождения нефть и газ в бассейне верхнего течения
р. Лены в Иркутской области. Поэтому проблема шунгита
Карелии также может быть решена в связи с водорослевыми
ценозами докембрия и соответствующими геологическими усло-
виями.
В случае метаморфизма, как известно, битумы часто бывают
выражены скоплениями графита, и потому вкрапленность гра-
фита, его тонкие пленки в породах докембрия или более или
менее значительные скопления должны указывать на распреде-
ление продуктов преобразования исходного органического ве-
щества растительного происхождения в виде одноклеточного
или многоклеточного фитопланктона. Поэтому битумы, шун-
гиты, графиты, скопления обуглероженного вещества следует
признать заслуживающими специального изучения с целью
127
выяснения их связи с организмами докембрия. Осадочные же-
лезные и марганцевые руды пластового типа, пиритизирован-
ные породы и продукты гранулометрической дифференциации
материалов сноса с древних выступов суши должны быть при-
знаны связанными с соответствующими жизнепроявлениями в
докембрии, как следы этой жизнедеятельности. Специально
подготовленные препараты могут позволить обнаружить в
этих веществах следы остатков микроорганизмов.
Фании и их осадки составляют единство. Единство состав-
ляют также среда и жизнь. Среды прошлого проходили стадии
непрерывного и направленного изменения под влиянием об-
щих причин и отчасти при участии самих организмов. Организ-
мы выводили из гидросферы большие запасы углекислоты и
бикарбонатов. Они обогащали ее свободным азотом и свобод-
ным кислородом. Измененная среда становилась не пригодной
для одних организмов и пригодной для других. Поэтому иско-
паемые организмы можно рассматривать как индикаторы древ-
нейших сред, как свидетели тех или иных геологических про-
цессов, а иногда даже материалов астрономического значения.
Так, по проявлениям фототропизма и гелиотропизма древней-
ших синезеленых водорослей, отраженных в построении специ-
фических строматолитов (параллелофи гонов), можно устано-
вить положение палеомеридианов. По проявлениям циклич-
ности в строении слоистой карбонатной массы строматолитов
можно устанавливать 11-летние циклы в развитии солнечной
радиации. Таким образом были получены данные об абсолют-
ном приросте строматолитов за сезон, о масштабе накопления
органического вещества в осадочных толщах докембрия, о
длительности существования видов водорослей, о длительности
формирования горизонтов и свит.
Остатки докембрийских организмов могут оказать сущест-
венную помощь в раскрытии палеоклимата для соответствую-
щих этапов геологического времени, а также палеогеографиче-
ской эволюции лика Земли в докембрии.
Особое значение имеет в породах докембрия поведение изо-
топов углерода, кислорода, азота, серы, водорода и их отноше-
128
ний, поскольку известно, что легкие изотопы преимущественно
оказываются связанными с жизнедеятельностью организмов,
тогда как более тяжелые — лишь с минеральными — химиче-
скими процессами.
Остатки организмов в осадочных породах докембрия из-
вестны во многих районах мира. Можно отметить находку
Дж. Лаусона (1865 г.) Eozoon canadense в районе Оттавы. Л. Кайо
(1894 г.) открыл остатки радиолярий, фораминифер, спикул
кремневых губок в углистых сланцах докембрия Бретани.
Чарльз Д. Уолкотт в докембрии Большого Каньона и в серии
Белт в США выявил интереснейшие остатки многощетинковых
червей и ряд других организмов.
Известны находки остатков радиолярий в докембрии юга
Австралии. Правда, некоторые исследователи не признали
этого открытия, другие склонны относить его к образованиям
бактериального происхождения. И. Седерхольм открыл в ка-
рельском шунгите остатки проблематического животного Go-
ridum, которого, по-видимому, следует отнести к проблема-
тическим образованиям.
В среднепротерозойских отложениях Енисейского кряжа
Ю. А. Кузнецов обнаружил остатки одноосных спикул кремне-
вых губок. Между тем в бакальской свите позднего докембрия
Урала автором были обнаружены предполагаемые отпечатки
медуз (?), отчасти подобные найденным в докембрии Боль-
шого Каньона в слоях Нанкоуин. В Северо-Западной Африке
(в Анти-Атласе) установлены остатки членистоногих в виде
разрозненных сегментов. Во многих районах мира известны,
правда, еще редкие и случайные находки остатков простейших
животных, примитивных целетерат, брахиопод, червей, члени-
стоногих, заслуживающих специального изучения.
Больше всего в мире открыто в докембрийских отложениях
остатков фотосинтезирующих организмов — морских водорос-
лей, являвшихся часто активными породообразователями. Изу-
чение таких остатков особенно успешно идет в Советском Сою-
зе. Этот нормальный палеонтологический материал ныне часто
обнаруживается в отложениях докембрия и нижнего палеозоя.
129
Он представлен строматолитами, а иногда тонкослоистыми
карбонатными породами с «курчавой» текстурой.
Что такое строматолиты и онколиты? Строматолиты и
онколиты — это микрослоистые сгустки карбонатного, иногда
вторично измененного вещества, находимые в составе преиму-
щественно древнейших морских и пресноводных отложений,
формировавшихся в мелководьях в основном синезелеными,
реже красными водорослями на твердом субстрате (первые)
или на подвижном (вторые) в условиях сильно карбонатной
(богатой бикарбонатами кальция и магния) водной среды за
длительные этапы времени, измеряющиеся часто сотнями и
тысячами лет. В их построении обычно участвовало несколько
видов микроскопических водорослей, проявлявших жизнедея-
тельность одновременно (сосуществование) или в порядке по-
следовательной смены одного вида другим, причем часто через
малые промежутки времени (естественный отбор, быстрота
темпов эволюции).
Поскольку строматолиты и онколиты представляют собой
продукты сложных и длительных процессов и не являются
остатками единого конкретного организма, то они не заслужи-
вают применения к ним латинизированной номенклатуры, как
это делается некоторыми исследователями. Они являются з
геологических разрезах лишь частью того каменного материала,
который несет в себе (если этому не мешают вторичные явления)
остатки микроскопических породообразующих фотосинтези-
рующих организмов (синезеленых, реже красных, водорослей).
Поскольку между формами и размерами конкретных видов
породообразующих водорослей и формами и размерами вклю-
чающих их остатки строматолитов непосредственная связь
отсутствует, то эти сгустки карбонатного материала, обычно
характеризующиеся бесконечным разнообразием по виду, мо-
гут знаменовать лишь местную временную фацию, сочетание
экологической обстановки с тектоническим режимом области
бассейна и не более.
Строматолиты могут указывать на результаты взаимосвязи
процессов нарастания элементарных наслоений и накопления
130
биогенного (фитопланктонного) или обломочного материала
между ними.
Эти процессы протекали с относительно равными скорос-
тями (нависания и наросты на боковой поверхности стромато-
литов), с опережением первого процесса (гладкие боковые
поверхности или с пережимами экологического происхождения)
или отставанием его. Это создавало условия для прекращения
развития строматолитов (их сужения, приобретения неправиль-
ной общей формы и т. д.). Так шло их местное отмирание.
Естественно, что внешние признаки как-то отражают особен-
ности их конкретных образователей — водорослей (строение
и отчасти форма элементарных наслоений микрослоев, соответ-
ствующих накоплению карбонатного материала за сезоны ве-
гетации). Форма наслоений, степень их выпуклости отражают
чувствительность водорослевых колоний к свету, степень про-
явления фототропизма, иногда выражавшуюся в виде прямого
гелиотропизма (параллелофитоны).
При условии благоприятной сохранности микроструктур
строматолиты и онколиты, находимые в отложениях различ-
ного геологического возраста (от нижнего протерозоя и почти
до наших дней), являются ценнейшим материалом для изучения
истории развития мира микроскопических водорослей (синезе-
леных и красных), их эволюции и породообразующей деятель-
ности. В них мы видим свидетелей обстановок далекого геоло-
гического прошлого с указанием на палеоклимат, палеотемпера-
туру среды, палеотечения, батиметрические данные, скорости
вертикальных движений литосферы и гидрохимию бассейна.
Это дает представление о связях между бассейнами и палеогео-
графических перестройках лика Земли во времени.
Успех исследования строматолитов и онколитов, водоросле-
вых пород вообще как фитогенных образований, требует от
исследователей палеоботанического, особенно альгологическо-
го изучения и глубоких знаний современных представителей
названных типов микроскопических водорослей, их морфоло-
гии, экологии, физиологии, биохимии и природного распро-
странения (геоботаники водорослей).
Результаты изучения остатков водорослей из докембрийских
отложений СССР показали возможность решения важных во-
просов стратиграфии. В частности, отложения протерозоя
ряда районов Сибири, часто неправильно сопоставлявшиеся
друг с другом, ныне получили определенное положение на осно-
вании данных эволюции выявленных видов и родов синезеле-
ных и красных водорослей. Так, морской протерозой Енисей-
ского кряжа оказался более древним по отношению к карбонат-
ным отложениям Туруханского поднятия, древние же карбо-
натные отложения р. Нижней Тунгуски — более древними по
отношению к улунтуйской свите Прибайкалья. Между тем
водоросли нижнего кембрия Иркутского амфитеатра оказались
генетически связанными с формами, выявленными в улунтуй-
ской свите верхнего синия.
Необходимо отметить полную бесперспективность исследо-
ваний строматолитов как таковых, без изучения их микроско-
пических структур. При таком методе изучения строматолитов,
связанным с массовым распиливанием каменного материала,
уничтожается большое количество материала, который был бы
полезен для палеонтологического метода.
Организмы — индикаторы среды. Знание их раскрывает
особенности среды, ее обстановку. Поэтому ископаемые орга-
низмы как свидетели могут помогать раскрытию природных
обстановок геологического прошлого. В связи с исключитель-
ной древностью жизни на Земле перед исследователями может
раскрыться картина ее развития. По литофациям и органиче-
ским остаткам докембрия исследователи несомненно могут
вскрыть многие важные детали истории земной поверхности,
биосферы и состава и строения земной коры в ее геологическом
развитии. Ископаемые водные организмы могут указывать на
степень солености среды, температурный режим, глубинность,
характер и направления течений, взаимосвязь между древними
бассейнами и т. д.
Поскольку древность жизни на Земле доказана, то экологи-
ческие и другие данные могут быть исключительно полезными
для геологии, если учесть, что в дальнейшем на ее основе может
разрабатываться стратиграфия осадочных отложений, палеогео-
графия, могут вестись поисково-разведочные работы. Организ-
мы участвовали и участвуют в накоплении различных минераль-
ных веществ и даже некоторых видов руд.
Палеонтологи и палеоэкологи, занимающиеся изучением
палеозойского и более молодого палеонтологического материа-
ла, успешно получают соответствующие данные о древней среде
и условиях обитания ископаемых организмов. Необходимо при-
менить палеонтологический метод к докембрийским осадочным
отложениям. Этому должны способствовать специальные ра-
боты литологов по изучению проявлений метаморфизма, вос-
становлению первичных типов осадков и выявлению биогеохи-
мической роли древнейших организмов.
В настоящее время находки остатков докембрийских живот-
ных организмов пока эпизодичны. Наблюдаются огромные
пробелы в палеонтологической летописи докембрия. Тем более
следует внимательнее относиться к каждой находке и пытаться
найти для нее место в общей системе организмов.
Раскрытие физико-географических условий на границе до-
кембрий — кембрий раньше встречало действительные и кажу-
щиеся препятствия: следы крупных перерывов в осадконакопле-
нии, местная изменчивость состава горных пород докембрия,
особенно из-за обилия вулканогенно-туфогенных образований
и проявлений различной степени метаморфизма первичных
осадочных пород.
Открытие органических остатков дает новый материал для
изучения истории жизни и ее эволюции в докембрии, что в целом
для отдельных районов распространения докембрийских отло-
жений раскрывает перспективы разработки биостратиграфии и
палеогеографии.
Органические остатки докембрия помогают выяснить проис-
хождение известных типов животных, растений и бактерий, так
как корни их затеряны среди отложений глубокой геологической
древности. Изучение особенностей древних фаций по литофа-
циям и организмам способствует выяснению причин изменчи-
вости организмов во времени и пространстве и темпов этой
эволюции во времени в конкретных физико-географических,
физико-химических и других обстановках. Таким образом, изу-
чение древнейших организмов помогает решить целый ряд важ-
ных вопросов биологии и геологии.
Палеонтологическая наука в содружестве с геологической
дает обильные материалы к истории жизни на Земле в ходе гео-
логического времени, по эрам, периодам и эпохам. Палеонтоло-
ги и художники многих стран мира пытаются воссоздать кар-
тины среды и жизни на отдельных этапах геологического прош-
лого. Мы попробуем воспроизвести некоторые из этих картин.
КАТАРХЕЙ
Мрачной, еще безжизненной, мертвой была земная поверхность
в раннем докембрии — в катархее. Не было еще ни свободного
кислорода, ни жизни. Запасов воды было меньше, чем сейчас.
Атмосфера, мощная и тяжелая, состояла из обильных паров
воды, углекислоты, аммиака, метана, водорода и других газов.
Этот выделенный советскими геологами ранний этап раз-
вития Земли, в диапазоне геологического времени 3500—
2700 млн. лет назад, был временем бурней перестройки молодой
земной коры, интенсивных проявлений вулканизма и магмати-
ческой деятельности. Первичные осадочные породы образовы-
вали складки, преобразовывались под влиянием давления и
повышенных температур.
Во «вторичной» атмосфере, которой оделась Земля, став пла-
нетой, и на ее поверхности, как и в зоне коры, развивались про-
цессы только химические, которые привели к образованию пер-
вичных белковых соединений. В парах и водных массах первич-
ных бассейнов интенсивно растворялись и накапливались аммиак
и углеводородные газы, которые и преобразовывались в более
сложные углеродные соединения. Этому тогда способствовали
более высокое давление атмосферы, температурный режим и
жесткие космические излучения, которые в большей степени
участвовали в преобразовании вещества атмосферы, гидро-
сферы и земной коры, чем позднее, когда на Земле накопились
134
запасы свободного кислорода, выделенного фотосинтезирую-
щими организмами. Углекислый газ, поднимавшийся из недр
Земли при вулканических явлениях расходовался и на образо-
вание сложных углеродных соединений и известняков.
Именно в этих особых термодинамических условиях из ис-
ходных сложных органических соединений возникла простей-
шая и примитивнейшая жизнь, промежуточным этапом пути к
которой были подобные вирусам или микробам организмы.
АРХЕЙ
Длительность архея, по данным абсолютной геохронологии,
составляет 800 млн. лет — от 2700 до 1900 млн. лет назад.
В толщах пород архейского возраста, обычно сильно изме-
ненных более поздними процессами, известны осадочные вод-
ные отложения — мраморы, песчаники, глинистые сланцы, в ко-
торые превратились известняковые илы, пески и глины. Песчин-
ки и глинистые частицы, как известно, образуются преиму-
щественно на суше, откуда они могут быть смыты в моря.
Мелкозем на суше образуется под влиянием факторов почво-
образования, и это говорит о большой древности на Земле
почвенных процессов.
Измельчение пород в почвенных условиях, распад их облом-
ков на мельчайшие частицы, на отдельные минеральные зерна
или их части вызывают обычно не только агенты физического
выветривания — вода, ветер, температура. Значительно боль-
шую роль играют многообразные почвенные бактерии и орга-
нические кислоты, образующиеся за счет разложения вещества
почвенных организмов, в основном растений. Их активность
в почве примерно в 20 раз сильнее физических агентов. Столь
совершенное измельчение материала, из которого часто выне-
сены в закисном виде железо, марганец, алюминий, с образова-
нием в конечном счете светлоокрашенных песков (кварцитов)
явно указывает на то, что почвы на сушах архея уже существова-
ли и интенсивно развивались и что в этом процессе принимали
участие почвенные бактерии. В водных бассейнах бактерии от-
135
лагали железо, образуя местами крупные месторождения. Бак-
терии могут проявлять жизнедеятельность в весьма широком
диапазоне температур, от отрицательных до + 170 С.
Жизнеспособные бактерии можно обнаружить даже в Арк-
тике — в снегу, в погребенных льдах. Б. Л. Исаченко полагал,
что развитие бактерий началось еще в докембрии, причем до-
пускал, что первые периоды докембрия были более благоприят-
ными для развития автотрофных организмов, восстанавливав-
ших углекислоту.
Как уже отмечалось, некоторые ученые полагают, что угле-
водороды и нефть являются компонентами состава протопла-
нетного облака и что на поверхности Меркурия они быстро
рассеялись и разложились под влиянием высокой температуры.
На поверхности Земли обстановка была для них крайне неустой-
чивой из-за действия ультрафиолетового излучения Солнца
и окислительных процессов, и они тоже разложились и исчезли.
Длительное время считалось, что живые существа, населяв-
шие кембрийские моря (570—440 млн. лет), были самыми древ-
ними на нашей планете, хотя они нередко и удивляли исследова-
телей разнообразием представленных тогда групп организмов,
большой местами густотой населения ископаемых биотопов,
их обильными популяциями, в числе которых отмечалось при-
сутствие многих видов археоциат, трилобитов, хиолитов, реже
губок, криноидей, червей, в сопровождении разных типов во-
дорослей. Во всем мире, за малыми исключениями, палеон-
тологи не изучали и не изучают систематически организмы бо-
лее древнего возраста. Данные региональной геологии, каза-
лось, предрешали неудачу геологов и палеонтологов в поисках
органических остатков в более древних, чем кембрий, осадочных
горных породах.
Осадочные горные породы докембрийского возраста, исклю-
чительно широко представленные на выступах суши нашей пла-
неты, обычно более или менее сильно изменены вторичными
процессами по отношению к их первичному облику. Часто это
бывает выражено перекристаллизацией вещества пород, с по-
терей ими частично или полностью первичных признаков.
136
Поэтому у исследователей возникло представление о полном
исчезновении в таких образованиях признаков прошлой орга-
нической жизни. Поэтому палеонтологи почти не занимались
такими породами, проявляя лишь некоторый интерес к строма-
толитам, своеобразным сгусткам углекислого кальция и магния,
природа которых длительно оставалась неразгаданной.
Мощность докембрийских осадочно-метаморфических, оса-
дочно-вулканогенных, туфогенных толщ обычно огромна. Ме-
стами она достигает многих десятков километров, но точному
ее установлению как раз мешает недостаточность или отсут-
ствие критериев для расчленения и корреляции этих толщ,
особенно в условиях их интенсивной складчатости. При этом
известно, что серии таких образований часто вмещают место-
рождения весьма разнообразных полезных ископаемых, напри-
мер руд черных и цветных металлов, редких и рассеянных эле-
ментов, золота, фосфоритов, алюминиевого сырья и т. д.
Следы древнейшей органической жизни на Земле ученые
устанавливают по присутствию в древнейших горных породах
органических веществ и продуктов их изменения, как, например,
в виде графита, шунгита и других или же в виде непосредствен-
ных остатков организмов, по их отпечаткам на поверхности
пород или с помощью того же остаточного органического ве-
щества, обычно сильно обуглероженного. При этом иногда
открываются в совместном развитии остатки морских водорос-
лей и простейших животных.
Параллельно с изучением в СССР обильных материалов
палеонтологии кембрия автор более 30 лет вел поиски органи-
ческих остатков и в породах докембрия, в особенности его позд-
ней части — протерозоя (2700—570 млн. лет), широко представ-
ленной во всех горных районах нашей страны. Исследовались
некоторые осадочного происхождения руды. Изучению подвер-
гались метаморфические породы, обычно внешне лишенные
каких-либо признаков связи с ними остатков организмов. Еще
в 40-е годы автору удалось собрать большой материал по иско-
паемым бактериям, оказавшимся и в далеком прошлом важны-
ми геологическими «деятелями». Эти работы подтвердили за-
мечательные идеи В. И. Вернадского о роли организмов в геоло-
гических процессах Земли, поскольку раскрылась особо важная
роль ископаемых микроорганизмов в создании рудных веществ,
накоплении различных соединений и химических элементов
(железо, марганец, сера, фосфор, алюминий и др.).
Теперь можно сказать, что в породах докембрия, а именно
всего протерозоя и частью архея, открыты обильные новые
палеонтологические объекты, притом весьма различного уров-
ня развития — от бактериальных и бактериеподобных до слож-
ных многоклеточных, принадлежащих к целому ряду групп
беспозвоночных животных. При этом господствующими в ко-
личественном отношении оказались представители водорослей
различных типов. Так впервые появились данные к биострати-
графии докембрия, в частности для районов хребта Малого
Хингана, Удокана, Восточного Саяна, Байкальского нагорья,
Кузнецкого Алатау, Западно-Сибирской низменности (по сква-
жинам), Урала, Тимана, Карелии, Кольского полуострова и
докембрия Украины и Молдавии. При участии автора подоб-
ные материалы были выявлены на территории Китая, Монго-
лии, Польши и Чехословакии.
Таким образом, метаморфический осадочный докембрий
оказался способным сохранять остатки организмов в различ-
ных его морских и терригенных породах, особенно в кремнистых.
Вся северная Евразия стала благодарным объектом для поисков
остатков организмов докембрия вплоть до пород, возраст ко-
торых достигает 3 млрд. лет. Иначе говоря, выявилась возмож-
ность крупного «приращения» (термин М. В. Ломоносова)
палеонтологической науки, ранее в данном направлении не раз-
вивавшейся. Практическая же геология получила необходимый
ключ для расшифровки истории формирования, расчленения и
корреляции древнейших осадочных серий. Одновременно от-
крылась возможность прослеживать историю жизни на Земле
до самых ее «корней», где-то около 3,5 млрд, лет до н. э.
Особого внимания заслуживают работы Эльсо С. Баргхор-
на, П. Клода и В. Шопфа, открывших в породах архейского
возраста очень интересный комплекс органических остатков.
138
Э. Баргхорн обнаружил в
районе оз. Онтарио нити
многоклеточных водорос-
лей однорядного строения
и остатки каких-то живот-
ных (Kakabekia) типа медуз
или гидроидных полипов.
Абсолютный возраст вме-
щающих слоев, определен-
ный по калий-аргоновому
методу, оказался не менее
2000 млн. лет. При изучении
доломитовых слоев архея
Трансвааля*Южной Африки
были выявлены остатки ор-
ганизмов возраста около
2600 млн. лет, представлен-
ные остатками колоний си-
незеленых водорослей, вы-
раженных скоплениями гра-
фита, а также остатками ис-
Водоросль Kakabekia umbellata В ar-
gh оо г п. Сев. Америка, сланцы серии
Ганфлинт (1800 млн. лет) (по С. Барг-
хорну)
копаемых бактерий. В образце породы, имеющей известковый
состав, видны строматолиты, по-вилимому, имеющие внешнюю
куполовидную форму.
Интересны также остатки бактерий Eobacterium iso latum
Barghoornet Schopf и колонии синезеленых водорослей
Archaeosphaeroides barbertonensis Schopf et Barghoorn
в отложениях группы Фиг-Три с возрастом 3100 млн. лет в
восточном Трансваале (Южная Африка). Менее отчетливые,
но не менее интересные остатки водорослей, подобные Archaeos-
phaeroides barbertonensis, но не имеющие определенного очерта-
ния, установлены из отложений группы Онвервахт возраста
3200 млн. лет, из наиболее древних строматолитовых пород
возраста 2800 млн. лет, обнаруженных в отложениях группы
Буловайан в Родезии, встречены своеобразные шиповатые ос-
татки, выявленные в электронном микроскопе. Одноклеточные
139
Остатки бактерий (а), колонии Eobacterium isolatum Barghoorn et Schopf
и синезеленых водорослей Archaeosphaeroides barbertonensis Schopf et
Barghoo rn (б). Африка, Восточный Трансвааль, отложения группы Фиг-Три
(3100 млн. лет. по В. Шопфу), шиповатые остатки из строматолитов (в), Африка,
Восточная Родезия, отложения группы Буловайан (2800 млн. лет, по Ф. Оберлис
и А. Пресновскому)
синезеленые водоросли описаны профессором П. Клодом и
Н. Лайкери из отложений формации Соуден-Айрен, возраст
которой определяется в 2700 млн. лет.
Таким образом, уже в архее, даже в раннем архее, на Земле
существовали и многоклеточные водоросли и даже организмы
типа медуз или гидроидных полипов, т. е. многоклеточные жи-
вотные, жизнедеятельность которых при этом, вероятно, обеспе-
чивалась тесным соседством с водорослями, выделявшими сво-
бодный кислород.
В общем за последнюю четверть века, а особенно за послед-
нее десятилетие, в поисках следов древнейшей жизни получены
замечательные результаты. На территории северной Евразии
140
на разных уровнях стратиграфии докембрия выявлены местдмц
массовые скопления остатков различных организмов состав
которых менялся иногда при мощностях слоев всего в 20— £5 см.
На основе каменного материала лабораторным путем можно
изучать осадочно-метаморфическую серию пород докембрия
вплоть до магматогенных образований катархея и архея.
Полученный материал не всегда отличается достаточной
четкостью, поскольку он действительно местами бывает сильно
поврежден вторичными процессами. Главным образом прихо-
дится сталкиваться с остаточным органическим веществом
в его бесструктурном проявлении, но в большом числе случаев
это остатки конкретных организмов с признаками живого ве-
щества (способность к росту особей, к тем или иным способам
размножения, проявления биологических связей с сосущество-
вавшими организмами, срастания, прирастания, проявления
наследственности на популяциях и т. д.).
Размеры ископаемых организмов в основном микроскопи-
ческие. Самые древние в СССР остатки организмов пока обна-
ружены в слоях беломорской серии архея Кольского п-ова и в
докембрии Украинского кристаллического массива (криворож-
ская серия пород). Наиболее древние органические остатки в
настоящее время установлены в докембрии Африканского мас-
сива в отложениях группы Онвервахт возраста 3200 млн. лет,
группы Фиг-Три, возраста 3100 млн. лет. Они представлены
одноклеточными и колониальными водорослями и бактериями.
Не менее интересна природа тонкозернистых морских карбо-
натных илов, получивших название дрюитов, которым приписы-
вается минеральное происхождение и коллоидная структура,
в действительности, в условиях открытого моря и ненасыщен-
ности вод солями кальция, также являются продуктом жизне-
деятельности денитрифицирующих бактерий. Жизнедеятель-
ность этой группы микроорганизмов основана на переводе раст-
воренного в воде бикарбоната кальция в карбонат кальция.
При этой реакции выделяется значительное количество энергии.
Следует отметить, что и в настоящее время далеко не все
литологи и геологи правильно учитывают вышеприведенные
141
данные, проявляя склонность объяснять происхождение тра-
вертинов и известняков с позиций коллоидной или общей хи-
мии, что следует считать для условий биосферы ошибочным
представлением.
Отметим, что биологический путь осаждения карбоната каль-
ция из водных растворов проявляется в более широких преде-
лах изменения среды (pH от 6 до 9), тогда как химическая садка
кальцита идет лишь при pH около 9,5, т. е. в условиях сильного
испарения и садки всех солей вообще.
Известно, что отложения карбоната кальция — кристалли-
ческие известняки — имеются в составе самых древних образо-
ваний архея Карелии, Кольского полуострова и в других местах
(беломорская серия) среди слюдяных, гранатовых, полевошпа-
товых и иных гнейсов, которые тоже следует признать первично
представленными нормальными осадочными отложениями гли-
нистого или песчано-глинистого характера с примесью вулка-
нических пород. Для этих отложений первые определения аб-
солютного возраста дали цифры от 2000 до 2300 млн. лет.
Установление в геологическом прошлом массового отложения
карбоната кальция в открытых бассейнах гидросферы Земли
заставляет нас полагать, что начало массовой жизнедеятель-
ности денитрифицирующих бактерий относится уже к этому
раннему этапу формирования земной коры.
На Кольском полуострове выше отложений беломорской
серии залегает Кольская серия нормальноосадочных и вулкано-
генных образований, подвергшихся значительному вторичному
изменению. В их составе обнаружены песчаники, превращенные
в кварциты, и железистые отложения, превращенные в железные
руды (магнетитовые сланцы). Поэтому приходится делать вы-
вод, что на суше тогда уже шли процессы почвообразования,
вернее всего при участии микроорганизмов, а в водных условиях
временами очень интенсивно проявлялась жизнедеятельность
железобактерий.
Известняки и мраморы установлены в обилии в составе
архея горных систем Тянь-Шаня и Памира. Первичнокарбонат-
ные (известняковые) отложения геологи открыли в составе
142
архея Анабарского кристаллического массива на севере Сибири.
Карбонатные породы были первично, вероятно, илами бакте-
риального происхождения.
В пределах Енисейского кряжа, в Ангаро-Питском районе,
автор наблюдал очень интересные осадочные железные руды,
имеющие нередко лепешковидное строение, что прямо указы-
вало на их бактериальное происхождение. С помощью остатков
водорослей из слоев того же разреза автор установил докем-
брийский возраст этих руд.
Таким образом, древнейшие известные нам остатки орга-
низмов относятся еще к архейскому времени и ясно указывают
на то, что жизнь на Земле существует не меньше 3000 млн. лет.
Архейская эра представляется важнейшим этапом в истории
Земли, когда возникли условия для неизбежного и обязатель-
ного происхождения жизни. Вещественный состав нашей пла-
неты, в особенности ее твердой коры, жидкой и газообразной
оболочек, и природные энергетические проявления должны
были этому способствовать. Именно в это время среда была
наиболее обогащена сложными углеродными соединениями.
Уже первичная жизнь, используя запасы сложных углеродных
соединений, создавшиеся на более раннем химическом этапе
истории планеты, израсходовала их настолько, что в последую-
щие времена жизнь уже не могла больше зарождаться.
Жизнь возникла в мельчайшей микроскопической форме,
в огромном многообразии биохимических, биогеохимических
проявлений. Пластичность живого вещества и естественный
отбор обусловили быстрое повышение интенсивности жизне-
проявлений, пока они не приобрели планетарного значения.
ПРОТЕРОЗОЙСКАЯ ЭРА
Огромный этап геологического развития, выделенный под на-
званием «протерозой», соответствует времени 1900—570 млн.
лет. Он представлен в пределах всех выступов суши разнообраз-
ными горными породами огромной мощности, в числе которых
видное место занимают породы биогенные.
143
Бактериальная и водорослевая жизнь в протерозое достигла
исключительного размаха и представлена многообразными гео-
логическими факторами — породо- и рудообразователями. Про-
дуктами этой жизнедеятельности были карбонатные осадки
открытых водных бассейнов, железные и марганцевые руды,
осадочные сульфидные минералы, отложения кремнезема в виде
кремнистых сланцев, силицилитов и т. д.
Осадочное отложение железа происходит на Земле во все
геологические времена, от архея до современности. Его сущ-
ность давно уже разгадана. Это бактериальный процесс, при ко-
тором бактерии используют в качестве энергетической базы
растворенные закисные соединения железа. Оказывается, при
переводе железа в нерастворимое состояние происходит выделе-
ние тепла.
Исследования Н. Г Холодного показали, что железобакте-
рии необычайно распространены в природе. Они проявляют
жизнедеятельность обычно в условиях относительно низких
температур (0°—16°С), при наличии СО2 и карбонатных или
иных закисных соединений железа в слабокислых или нейтраль-
ных средах (pH от 5,8 до 7,6) в присутствии любой концентрации
кислорода. В бедных кислородом средах железобактерии разви-
ваются около колоний водорослей. В органических веществах
эти бактерии или не нуждаются совершенно, или относятся к
ним безразлично, так же как и к наличию света. Железобактерии
превращают бикарбонаты железа в гидрат его окиси, причем
углекислота является строительным материалом для вещества
их клеток.
Биологическое формирование осадочных месторождений
железа в огромных масштабах происходило в особенности в
протерозое, т. е. в позднем докембрии. Крупнейшие железные
месторождения мира принадлежат к группе отложений этого
возраста.
Н. Г Холодный считал, что железобактерии имеют прямое
отношение к поведению железа в биосфере, где они выполняют
значительную биохимическую работу по окислению закисей
железа и превращению их в нерастворимую гидроокись — в же-
144
лезные руды различных типов, по условиям юс образования и
последующим химическим изменениям, под влиянием различ-
ных геологических обстановок (лимониты, гематиты, гетиты,
а также силикаты, фосфаты и сульфиды железа). Эти руды очень
часто содержат остаточное органическое, по-видимому, бакте-
риальное вещество. Докембрийские руды Кирунавары (Швеция),
как правило, содержат до 5% этого органического вещества.
Богатейшие месторождения Северной Америки в районе Верх-
него озера тоже отложились в раннем протерозое.
К отложениям нижнего протерозоя относятся и криворож-
ские железные руды, а также железные руды Курской и Воронеж-
ской областей, Прибалтики. Руды кремнистые. В рудах Кривого
Рога и в аналогичных кремнистых рудах Кольского полуострова
автор наблюдал настоящие бактериальные структуры, лучше
различимые именно в рудах, бедных железом, на фоне светлого
кремнезема.
Начало отложений курской железорудной серии датируется
2060 млн. лет, а конец ее формирования — 1500 млн. лет.
И почвы на суше, и водные бассейны протерозоя были широ-
кой ареной деятельности целого ряда групп микроорганизмов,
из которых уже тогда эволюционно обособились группы авто-
трофов, приспособившиеся к освоению реакций распада и пре-
образования ряда минеральных веществ, связанного с выделе-
нием свободной энергии. В породах архея мы не находим зерен
пирита, но уже в протерозое пирит отлагался в осадочных по-
родах (пиритизация пород). Значит, в это время появились в
массовом количестве окислители серы, а затем и сульфатреду-
цирующие (разрушающие сернокислые соли) бактерии. Веро-
ятно, существовали и денитрифицирующие бактерии, выделяв-
шие элементарный азот в состав древней атмосферы за счет
первичного аммиака и соединений азота, возникших на его
основе.
Таким образом, органическая жизнь в виде железобактерий
была представлена уже более 2000 млн. лет назад. Для сущест-
вования этих бактерий был нужен, хотя бы в очень малых коли-
чествах, элементарный кислород. Часто они его получали от
145
водорослей, вместе с которыми иногда создавали концентрации
гидроокислов железа и карбоната кальция. Иногда деятельность
бактерий чередовалась с деятельностью водорослей: лето было
временем оптимального развития железобактерий. Часто в до-
кембрии так образовывались строматолиты.
Мир водных растений, настоящих фотосинтезирующих, уже
в позднем архее был представлен одноклеточными формами.
В следующую эру — в протерозое — во всех водных бассейнах
Земли развились разнообразные виды многоклеточных, начав-
ших играть важную роль в породообразовании. Лучистая энер-
гия Солнца, особенно красная часть спектра, и значительные
запасы углекислоты в биосфере, пополнявшиеся при вулкани-
ческих явлениях, способствовали развитию водной раститель-
ности не только плавающей, но и донной. Воды морей, еще
слабо соленые, но богатые бикарбонатами кальция и магния,
легко осаждали их химически. Подщелачивая воду, водоросли
играли важную роль в осаждении таких карбонатов, образуя
строматолиты.
Развитие многоклеточных водорослей произошло более или
менее одновременно с появлением в осадках протерозойской
эры слоев плотных известняков. Интересная зависимость суще-
ствует между вспышками вулканизма и количеством раститель-
ного водорослевого вещества. Создается впечатление, что вре-
менами развитие водных растений на Земле ограничивалось
содержанием углекислоты в биосфере, что подтверждается и
сокращением отложения карбонатных отложений вообще.
^Протерозой — это эра водорослей и бактерий. Лишь к концу
ее возникли самые ранние представители многоклеточных жи-
вотных — черви, губки и археоциаты. Это была также эра одно-
клеточных простейших животных, пока слабо вскрываемых при
исследовательских работах. Но главнейшими видимыми про-
явлениями жизни в протерозое были водоросли типа пресновод-
ных. Последние, вероятно, участвовали наряду с бактериями
и в развитии на суше процессов почвообразования. Перемыв
и смыв рыхлых масс почвенного мелкозема уже играл сущест-
венную роль в осадочном породообразовании во внутренних
146
и внешних морях того времени. Жизнь по преимуществу захва-
тывала в то время области морских мелководий. Многоклеточ-
ные растения еще не выходили на сушу.
В протерозойскую эру на выступах суши, не имеющих рас-
тительного покрова, интенсивно шло выветривание горных
пород при участии физико-химических и микробиологических
процессов; в морских и пресноводных мелководьях развивались
водорослевые банки, часто причудливого облика. Водные рас-
тения создавали очень своеобразные накопления известняка и
даже древнейшие рифы и банки. «В поле» исследователь далеко
не всегда отличит небольшие водорослевые скопления карбо-
натной породы от вмещающих отложений. Но повторяемость
рисунка поверхности породы иногда подсказывает, что найдены
остатки ископаемых древнейших водорослей. Порода местами
оказывается кривослоистой, и это уже частый признак ее водо-
рослевого происхождения; иногда видны известковые тела,
которые при разрушении, при выветривании как бы расслаива-
ются на отдельные пластинки. Это водорослевые известковые
образования — строматолиты, иногда достигающие значитель-
ных размеров. Они бывают построены или неправильно, или в
виде относительно правильных куполов, состоящих из последо-
вательно наросших друг на друга куполовидных годичных
наслоений. В некоторых слоях поперечные сечения водоросле-
вых образований кажутся округлыми, концентрически слоис-
тыми, тогда как в действительности это вытянутые вверх тела,
чаще конической формы, до метра в высоту и до полуметра в
поперечнике у основания. Иногда это крупные тела с округлой
поверхностью, тоже сложенные как будто из отдельных корок.
В изломе видно, что первоначально маленькое известковое тело
постепенно, слой за слоем, обрастало все больше сверху и с бо-
ков, сохраняя почти шаровидную форму. Кое-где на скалах как
будто нарисованы колонки, состоящие тоже из наслоений, тя-
нущиеся в одном направлении и время от времени ветвящиеся.
Лишь при более внимательном рассмотрении видно, что места-
ми они срастаются друг с другом, отклоняются в ту или другую
сторону. В одном случае колонии водоросли построили слой
147
Типы проявления жизни в докембрии: остатки червей, синезеленые водоросли в
виде лучистых колоний (ув. в 325 раз), спикулы губок и спор (ув. в 725 раз), отпе-
чаток медузы (уменып. в 2 раза) и строматолиты, образованные известковыми
водорослями (по Р. Моору)
породы толщиной в 36 м при скорости роста 1 мм за год. Это
значит, что данный вид водоросли, с ее микроскопическими
колониями, просуществовал 36 000 лет.
Специальное лабораторное изучение показывает, как виды
микроскопических водорослевых колоний с течением времени
непрерывно сменяли друг друга. Отдельные слои породы могут
иметь различное строение и слагаться из водорослевых сооруже-
ний разной формы и размеров. Часто геологическая жизнь боль-
шей части этих ископаемых видов оказывается относительно
короткой: смена одних породообразователей другими иногда
происходила через 20—30 см, или даже менее. При годичном
148
Колонковые строматолиты из отложении докембрия (Северный Ки1ай). Фото
автора
приросте, составлявшем около 0,1—0,5 мм, длительность жиз-
ни видов этих водорослей измеряется всего сотнями лет.
Для изучения строматолитов из них выпиливаются тонкие
прозрачные шлифы, определенным образом ориентированные
по отношению к направлению нарастания строматолитов. Если
сохранность первичных структур водорослевых колоний доста-
точная, в шлифе можно видеть особенности строения остатков
водорослей. При зарисовках с увеличениями в 10—400 раз
видны округлые клеточные колонии из окаменевших сгустков
слизистой массы в виде тяжей разной формы, или же удлиненные
многорядные нити, тоже с остаточными структурами слизистых
масс колоний синезеленых водорослей. Специальными иссле-
дованиями шлифов, зарисовок с них и микрофотографий рас-
149
Сосуществование множества видов водорослей привело к построению многообраз-
ных строматолитов в одном слое породы. Поздний гинии (Китай). Фото автора
крыто огромное разнообразие водорослевых структур из отло-
жений докембрия. Это позволило описать массу видов, принад-
лежащих к нескольким десяткам родов синезеленых и красных
водорослей.
Таким образом, непонятные прежде известковые стяжения —
строматолиты, ныне во все большем количестве обнаруживае-
мые в слоях древних морских и пресноводных отложений и часто
составляющие мощные слои породы, оказываются продуктами
жизнедеятельности древнейших фотосинтезирующих организ-
мов.
У некоторых водорослей при быстрой фоссилизации газовые
пузырьки, оставшиеся в слизи, входили в структуру своеобраз-
ных лентовидных строматолитов, обладающих временной спо-
собностью подниматься над субстратом. В результате образо-
150
Остатки одноклеточных водорослей из строматолитового известняка верхнего
докембрия на о-ве Кильдин. (Ув. в 4С тыс. раз). Фото В. В. Любиова
вывались своеобразные заросли из жестких известковых лент,
которые, по мере рассасывания пузырьков кислорода и запол-
нения их объемов вторичным кальцитом, опускались на дно и
накапливались иногда слоем в несколько метров мощностью.
Часто водорослевые банки протерозойского времени приобре-
тали общее куполовидное очертание, по-видимому, выгодное
в условиях обитания в зоне морского волнения (до глубины
в 20 м). Интересно отметить, что лишь в конце кембрийского
периода, т. е. много позднее, эта способность некоторых водо-
рослей, обитавших в условиях рифа или мелководной банки,
получила очень изящное выражение. Колонии водорослей сезон
за сезоном образовывали сотни и тысячи колонок, куполовид-
ные завершения которых вверху составляли в свою очередь
более крупице купола, что, несомненно, возникало в процессе
151
естественного отбора под влиянием волн. Рифы менее правиль-
ной формы под ударами волн часто разрушались. Между тем
куполовидная поверхность групп водорослевых колонок ока-
зывалась защищенной от такого разрушения. Интересно, что
в построении колонок иногда участвовали последовательно
многие виды водорослей, принадлежащие даже к разным родам.
Ниже волновой зоны моря часто многие виды водорослей
существовали совместно, по соседству, и образовывали извест-
ковые тела, строматолиты разной формы.
Интересный тип остроконических строматолитов образовы-
вали разные виды водорослей, одинаково подвижные колонии
которых скользили по субстрату, прежде Чем частично от него
оторваться. В таких случаях водорослевые банки имели вид
остроконечного частокола, с предельными поперечниками до
10—15 см.
Удалось также выяснить, почему часто строматолиты имеют
выпуклые вверх, иногда даже куполовидные наслоения. Оказы-
вается, в слизистой массе древнейших синезеленых водорослей
выделяемый ими кислород часто застревал в виде пузырьков.
Эти пузырьки, стремясь подняться вверх, способствовали сколь-
жению колонии по субстрату к более высоким точкам его по-
верхности. Образовывались вздутия колоний с тенденцией к
образованию конических форм, если пузырьки скапливались в
осевой зоне в большем количестве, чем по краям дерновинки
водоросли. Часто в последующих наслоениях колонии водорос-
ли скользили вверх по этим вздутиям, а иногда даже частично
срывались с субстрата и всплывали в верхние слои воды.
Так за столетия и тысячелетия формировались конические,
иногда даже остроконечные, строматолиты.
Это свойство проявлялось у многих видов водорослей в
разное геологическое время. Конические строматолиты образо-
вывались во все времена докембрия, в разные эпохи кембрия и
даже ордовика, исчезнув вообще лишь при общем спаде геологи-
ческой деятельности водорослей, когда появились древнейшие
группы морских беспозвоночных животных.
Таким образом, как конические, так и куполовидные
152
формы строматолитов не
имеют прямого отношения
к систематике породообра-
зующих водорослей
Следует учесть, что пре-
обладающее количество ви-
дов древнейших водорос-
лей не образовывало чет-
ких тел постоянной формы
типа строматолитов, а на-
слаивало осадок, превращая
его в плотную породу, где
лишь частично сохранялась
морфология колоний и
продукты их жизнедея-
тельности, то в виде при-
хотливо изогнутых столби-
ков. то в виде неправильных
стяжений мелких известко-
вых комочков, подобных
цветной капусте. Изучен-
ность этих древнейших во-
дорослей еще очень низка.
Однако за последние годы
в слоях морских отложений,
образовавшихся от 1200 до
600 млн. лез назад (синий-
Микроструктура строматолита из отло-
жений докембрия.
Видна быстрая смена породообразующих
форм. Фото автора
ский период), выявлено большое разнообразие родов и видов
синезеленых и красных водорослей, одновременно раскрывших
перед нами и абсолютные скорости роста водорослевых коло-
ний и отражение на их жизнедеятельности влияния 11-летних
циклов солнечной радиации. По тому, как ориентированы в
пространстве скопления карбонатного материала, созданные
жизнедеятельностью водорослевых колоний за длительные про-
межутки времени, можно определить общее направление свето-
вого потока. .
Часть поверхности водорослевой банки со слабовыпуклыми наслоениями. Ранний
протерозой (Китай). Фото автора
Самый главный результат исследований состоит в том, что
установлена возможность вскрывать остаточные биогенные
структуры у карбонатных стяжений водорослевого происхож-
дения. Таким образом был обнаружен целый мир водорослей
и водорослевых сообществ, крайне интересный для развития
палеоботанической науки, для сопоставления по этим остаткам
отложений разных районов, иногда весьма удаленных друг от
друга, как, например, Енисейский кряж и Прибайкалье, район
Туруханска и бассейн р. Алдана и т. д.
Требуются еще большие усилия, чтобы раскрыть оконча-
тельно эволюцию древнейших водорослей, разработать их сис-
тематику и определить масштабы их геологической деятель-
ности.
154
Мелководье раннесинииского моря. Строматолитовая банка, образованная сине-
зелеными водорослями. Реконструкция автора
Можно сказать, что водоросли, и только они, примерно за
два миллиарда лет извлекли из биосферы колоссальные коли-
чества углекислоты, образовав на ее основе столь же колоссаль-
ные количества органических веществ. При отмирании колоний
эти вещества послужили жизненной средой для развития других
организмов (бактерий и простейших) и в итоге — исходным
материалом для образования биохимическим путем углеводо-
родных соединений — нефти и горючего газа. Водоросли спо-
собствовали отложению на дне древнейших морей огромных
количеств углекислого кальция и отчасти магния. Можно счи-
тать, что в пределах современных континентов водоросли от-
ложили за время протерозоя карбонатные породы обшей мощ-
ностью более 1000 м. Эта их геологическая деятельность сопро-
155
Поперечное сечение конического строма-
толита. Видны годичные наслоения и
11-летние циклы. Средний синий. Фото
автора
вождалась часто отложени-
ем известковых илов при
участии «кальциевых» (де-
нитрифицирующих) бакте-
рий. Синезеленым водо-
рослям часто сопутствова-
ли также железобактерии,
иногда господствовавшие в
межвегетационные сезоны и
образовывавшие в строма-
толитах железистые пленки.
Систематически очищая
биосферу от углекислого га-
за и бикарбонатов (раство-
римых карбонатов кальция
и магния), водоросли выде-
лили за счет кислорода во-
ды огромные количества
свободного кислорода.
Этим они существенно из-
менили свойства среды жиз-
ни на Земле, сделав ее при-
годной для появления и раз-
вития аэробов в широком
смысле, т. е. для новых групп микроорганизмов и для живот-
ных. Восстановление аммиака и углеводородных соединений
деятельностью многих групп бактерий при одновременном на-
коплении в биосфере больших масс органического вещества
привело к концу протерозоя (синия) к накоплению на Земле
свободного азота.
Конечно, свободный азот moi накапливаться в атмосфере
не только в результате жизнедеятельности денитрифицирующих
бактерий, но и при разложении органического вещества бел-
кового состава в бескислородной среде биосферы того времени.
В круговороте веществ в природе такой путь также мог играть
важную роль.
156
Плитообразные строматолиты верхнего сииия с однородной ориентировкой в про-
странстве, соответствующей древнему меридиану. Фото автора
Совместная деятельность всех отмеченных групп организ-
мов, обитавших лишь в водных средах Земли, таким образом,
в корне изменила состав атмосферы и гидросферы, значительно
приблизив его в конце протерозойской эры (570 млн. лет назад)
к современному.
Остатки животных в отложениях протерозоя очень редки,
но нет сомнения в том, что основы животного мира были зало-
жены одновременно с возникновением мира бактерий и фото-
синтезирующих растений. Животные представлены в протеро-
зое мелкими формами, не получившими массового развития
и не принимавшими участия в породообразовании. Они пока
теряются среди обильно проявившейся водорослевой и бакте-
риальной жизни Однако все же находки остатков организмов
157
Отпечатки червей (?) на выветрелой поверхности известняка. Нижний синии.
Фото автора
в метаморфических толщах докембрия известны уже в ряде
стран мира, в особенности они участились в пределах некоторых
горных районов Дальнего Востока, Сибири и на севере европей-
ской части СССР. К таким районам можно причислить и Украи-
ну, в частности докембрий Украинского кристаллического мас-
сива. В 1958 г. В. В. Бесе в породах криворожской серии наблю-
дал некоторые «проблематические» остатки организмов, остав-
шиеся ближе не изученными.
В 1965 г. при микроскопическом изучении конгломератов
верхней свиты криворожской серии из района рудника им.
М. В. Фрунзе были обнаружены уже хорошо сохранившиеся
остатки организмов. В геологическом строении района этого
рудника участвуют породы метабазитовой и криворожской
158
Кораллиты Aseptalia ukrainika Vologdin, обнаруженные в нижнем проте-
розое криворожской серии. Украина. Кривой Рог (около 2000 млн. лег,
по А. Г. Вологдину)
серий глубокого докембрия, образующие синклинальную струк-
туру субмеридионального простирания. Верхняя из этих серий
сложена различными сланцами, мраморами, песчаниками и
конгломератами. Характерной особенностью некоторых слан-
цев, реже песчаников и мраморизованных известняков, является
местами содержание углеродистого материала с переходами
даже в углистые сланцы Конгломераты состоят из окатанных
обломков кварцитов, песчаников, гранитов и очень редко из
известняков, по-видимому, принадлежащих к более древней
серии отложений докембрия. В штуфе керна с глубины в 1344—
1345 м, представленном именно конгломератом, была обна-
ружена галька такого состава с одиночной галькой карбонатной
породы с явными остатками организмов Цемент песчанистый
159
Спикулы губок. Дальний Восток, бассейн р. У чур. Отложения гонамской свиты
(1500 млн. лет, по А. Г. Вологдину и Н. А. Дроздовой)
перекристаллизованный, показывающий зеленосланцевую фа-
цию метаморфизма на переходе к амфиболитовой. Абсолютный
возраст конгломерата, точнее время его метаморфизма, по дан-
ным Ф. И. Котловской (1961), А. П. Виноградова, Л. В. Комле-
ва и А. И. Тугаринова (1965), указывается около 2000 млн. лет.
С помощью прозрачного шлифа в гальке мраморизованного
известняка установлены несомненные остатки синезеленых во-
дорослей, в сопровождении нескольких экземпляров остатков
кораллообразных представителей беспозвоночных животных,
что, конечно, не только для докембрия Украины, но и вообще
является большой сенсацией. Остатки водорослей представлены
в виде местных скоплений одноклеточных форм, а также водо-
рослевым трихомом, свернутым в спираль, со следами его окру-
160
жавшей студенисто-слизистой массы (?). Подобный тип сохран-
ности ранее многократно наблюдался. Остатки кораллообраз-
ных организмов вскрыты в косых и продольных сечениях, при-
чем их внутренняя полость заполнена или остатками однокле-
точных водорослей, или густой массой метаморфизованного
органического вещества (битумами?). Кораллиты Aseptalia были
узкоконической или роговидной формы с двуслойной стенкой
и без каких-либо перегородок.
Не мецее древние остатки фауны червей, трубкожилов —
Udocania problematica обнаружены А. М. Лейтесом в отложе-
ниях битунской свиты удоканской серии Забайкалья, возраст
которых определен в 2000—1600 млн. лет. На этом же страти-
графическом уровне в туломозерской свите сегозерской серии
ятулия Карелии В. А. Перевозчиковой обнаружены створки
раковины.
В отложениях гонамской свиты в бассейне р. У чур (Дальний
Восток) автором установлены остатки спикул губок и двух типов
проблематик. Фрагменты спикул губок представлены одноос-
ными спикулами — рабды и спикулами крючковидной формы.
В этой же свите в Аяно-Майском районе установлены колониаль-
ные водоросли Gonamophyton, широко распространенные в му-
рандавской свите хребта Малый Хинган Приамурья и щекурьин-
ской свиты восточного склона Приполярного Урала, где были
распространены также колониальные водоросли Murandavia.
Обильный материал по древнейшей органике получен из
слоев нижнего и среднего протерозоя Южной и Центральной
Карелии. Он представлен здесь как исключительно мелкими,
так и относительно крупными формами, в числе которых глав-
нейшая роль принадлежит синезеленым водорослям, обитавшим
в составе планктона и бентоса. Они представлены как однокле-
точными, так и, чаще, колониальными формами, частью в виде
сложных биоценозов. Представители бентоса часто развивались
последовательно друг на друге в общем процессе, приводившем
к образованию соответствующего осадка, охватываемого лито-
генезом. Отчетливо выражены микрослои, вероятно, обуслов-
ленные сезонными изменениями условий жизни фотосинтези-
11-818
161
Остатки организмов:
губкообразные орга-
низмы в разных сече-
ниях в шлифах (ув.
в 4 раза) Ladogaella
variabilis Volog-
din (а). Часть се-
чения онколита, об-
разованного нитями
водоросли (ув. в 20
раз) Ptilophyton така-
rovae Vologdin
(б). Карелия, район
г. Сортавала, свита
контиосари (1770—
1850 млн. лет, но
А. Г. Вологдину)
рующих организмов. Субстрат иногда был подвижным (песча-
ники?), причем формировались так называемые онколиты.
В составе вещества их наслоений удается различать остатки от-
дельных видов синезеленых водорослей. Возможен подсчет
числа сезонов вегетации при формировании таких образований.
Иногда формировались онколиты удлиненной формы, направ-
ленного развития. Наблюдались случаи их прижизненного
срастания, что могло произойти при делении клеточных коло-
ний. Их вещество первично имело студенисто-слизистую кон-
систенцию, как это и свойственно синезеленым водорослям.
В слоях из свиты контиосари Карелии (нижний протерозой |
обнаружено сосуществование водорослей Ptilophyton с губко-
подобными организмами рода Ladogaella. У этих организмов
были отчетливо выражены внутренняя полость и общая радиаль-
ная симметрия. В своем индивидуальном развитии они прохо-
дили начальную планктонную стадию и взрослую — прикреп-
ленную с образованием каблучка прирастания к субстрату.
162
Иногда губкообразные организмы прикреплялись к колониям
водорослей, oi раничивая их жизнедеятельность. Случалось, что
стадия свободного плавания задерживалась, причем соседние
организмы иногда срастались. У сидячих особей верхняя часть
оформлялась в виде воронкообразного углубления, чем они
как бы предрешали внешнюю форму поздних кораллов и других
кораллообразных организмов. В поле прозрачного шлифа под
микроскопом удается наблюдать сложные картины таких древ-
нейших жизнепроявлений. Морской бассейн охватывал тогда,
по-видимому, всю Фенноскандию, поскольку автору удалось
в той же свите сделать вторую в мире находку Corycium enigma-
ticum Sederholm. При изучении данной формы оказалось,
что ей свойственна трубчато-коническая форма тела с радиаль-
ной симметрией. Оболочка (стенка) утолщенная, подчеркнутая
остаточным органическим веществом При этом были обнару-
жены внутренние перегородки на разных стадиях индивидуаль-
ного развития, обычно пористые, плоские. Этот новый признак,
и* 163
возможно, указывает на принадлежность формы к более высо-
коорганизованной группе животных. Не исключено, что разви-
тие шло в сторону цефалопод (?). У совместно находимых губ-
кообразных (ладогелл) на взрослой стадии проявлялась способ-
ность к почкообразованию, очевидно, прогрессивная по отно-
шению к простому делению. Как корициумы, так и ладогеллы,
по-видимому, имели студенисто-слизистую консистенцию тел,
что способствовало длительному свободному развитию особей.
Последующая фоссилизация, вероятно, была проявлением ста-
рения особей, что приводило к их отмиранию и погружению в
илистый осадок бассейна.
Обнаружено много остатков организмов с радиально-осевой
симметрией тела и внутренней полостью или с системой полос-
тей и каналов. Поражает, что они совсем не деформированы в
породе; это можно объяснить своеобразием проявлений фосси-
лизации. Вероятно, эти организмы были близкими к гидроид-
ным полипам, к гидромедузам.
Очень интересна сплошная черная окраска пород (кристал-
лических сланцев) свиты контиосари. Она оказалась обусловлен-
ной скоплением черной (вторичной) слюды — биотита и сильно
измененных битумов, частью вполне поддающихся экстракции
сильными растворителями. Поиски особо мелких остатков с
помощью шлифов и порошковых проб привели к открытию
множества своеобразных мелких скелетных образований, частью
целых, частью ломаных, среди которых присутствуют фосси-
лизированные остатки водорослей типа золотистых (?), периди-
неевых (?), диатомовых по построению очень еще примитивных,
чаще сложенных кремнеземом, что может быть следствием как
первичных явлений, так и вторичных. Данное местонахождение
в возрастном отношении близко к установленному в слоях
свиты ганфлинт на берегах оз. Онтарио, но в отношении богат-
ства органическими остатками оно значительно интереснее.
В Восточной Сибири — в Забайкалье, в Удоканском хребте
представлены отложения протерозоя большой мощности. Здесь
в слоях бутунской свиты тоже обнаружено много органических
остатков, в основном водорослевых. Местами породы свиты,
164
явно имеющей морское происхождение, целиком состоят из
подвергшихся фоссилизации органических остатков, местами
встречается бесструктурное остаточное органическое вещество.
Водоросли по типу принадлежат к красным. Следы слизеобра-
зования у их остатков выражены незначительно. Некоторые
формы с нитчатым талломом образуют сплетения, свивающиеся
в две пряди, подобно шпагату, что можно различить в сечениях
породы соответствующего направления. При хорошей сохран-
ности у однорядных нитей можно отчетливо различить последо-
вательность наращивания клеток или клеточных колоний. Не-
которые участки породы сложены однородно ориентированны-
ми тесно сомкнутыми фоссилизированными нитями с ясной
полярностью развития водорослевой дерновинки. Фоссилиза-
ция превосходно закрепила морфологические черты ряда видов
таких водорослей. Открыты виды, вполне сопоставимые с изу-
ченными автором из свиты цзинлин китайского синия. Удалось
также обнаружить остатки своеобразных двухстенных скелет-
ных образований, имеющих пластинчатое строение и порис-
тость, благодаря чему они несколько напоминают археоциаты
кембрия. Примерный возраст вмещающих пород — средний
протерозой.
В Юго-Западном Прибайкалье слои улунтуйской свиты верх-
него протерозоя оказались преимущественно фитогенными,
но органические остатки в них сохранились по большей части
плохо. Все же по р. Сарме удалось обнаружить разновидность
улунтуйского водорослевого известняка, почти целиком состоя-
щего из фоссилизированных остатков водорослей с округлыми
и нитевидными клеточными колониями. В результате был
описан ряд видов и рядов микроскопических водорослей, очень
четких. В их составе присутствуют одноклеточные с округлыми
клетками двух размеров, отнесенные к двум видам одного рода,
и интересная нитчатая форма. Сходная форма затем была от-
крыта в мурандавской свите протерозоя Приамурья.
Из учурской серии верхнего протерозоя на юго-востоке Си-
бирской платформы в районе пос. Нелькан Аяно-Майского
района был описан ряд интересных родов и видов водорослей
165
группы хроококковых и ревуляриевых. Первые (гонамофитоны)
были представлены в породе отдельными клеточными колония-
ми, окруженными прижизненно толстой оболочкой студенисто-
слизистой массы. В шлифах это видно отчетливо. В последней
видны радиальные каналы, по которым клетки могли выбрасы-
ваться наружу, подобно тому, как это имеет место у современ-
ного рода воронихиния (Voronichinia Elenkin). Интересно,
что очень сходную водоросль удалось обнаружить в докембрий-
ской породе в забое одной нефтяной скважины в Тюменской
области, что позволяет коррелировать вмещающие слои. Во-
доросли группы ревуляриевых — нельканеллы с колониями ра-
диально-лучистого строения развивались от центра колонии
или же, как казалось, только в зоне периферии с образованием
дерновинки сферического очертания в пространстве. Важно
отметить, что микроводоросли с радиально-лучистым расхож-
дением нитей и свободной полостью внутри найдены также в
протерозое восточного склона Кузнецкого Алатау. Накоплен-
ный опыт позволяет относить такие формы предположительно
к зеленым водорослям типа обнаруженных автором на Тимане.
Докембрийские отложения Батеневского кряжа в Кузнецком
Алатау также оказались изобилующими остатками водорослей
из мартюхинской свиты. Автором были выявлены и описаны
колониальные водоросли из порядка Chroococales G е i 11 е г
роды Vesiculophyton и Echaninia. Остатки своеобразной водорос-
ли Pustularia были встречены в верхнепротерозойских отложе-
ниях (джурская свита) низовьев р. Амбары восточного склона
Енисейского кряжа, среди строматолитового материала. Ее
остатки представлены известковыми лентовидными образова-
ниями весьма своеобразного строения, характеризующимися
отчетливо выраженными пузырчатыми микроструктурами.
Колония водоросли из микроскопических прямостоячих
нитей обладала способностью последовательного линейного
разрастания с образованием лентовидных пластин впоследст-
вии обызвествлявшихся. Причем скопления пузырьков воздуха,
находящихся в слизи колонии, мешавшие ее сплошному обыз-
вествлению, обусловливали наличие в дальнейшем ее пузырча-
166
той структуры. Обызвествленные колонии водоросли залегают
в породе одна над другой через промежуток 0,3 до 2,0 мм.
На западном склоне Тиманского кряжа, по р. Мезенская
Пижма, в слоях протерозойской серии, выраженной алевроли-
товыми породами, выявлен крупный биогерм, образованный
при участии зеленых водорослей большого размера. В резуль-
тате этого открытия автором была описана форма рода тима-
нелла, имеющая таллом в поперечнике до 10—12 см, при отчет-
ливо выраженной гигантской осевой части и постепенно утол-
щающихся массовых нитевидных ответвлениях в стороны и
вверх. Водоросль имеет черты, сближающие ее с типичными
палеозойскими сифоновыми, но по размерам таллома она в де-
сятки раз крупнее. Упомянутый биогерм развивался на грунте
илистого состава. Особи водоросли нарастали друг на друга,
образуя сплошную массу, отмершее вещество которой пропиты-
валось илистым материалом. Первичная консистенция этого
вещества была несомненно студенисто-слизистой. Материал
можно различать только на увлажненных пришлифовках и в
прозрачных шлифах благодаря сохранившемуся остаточному
органическому веществу.
Выше уже упоминались строматолиты — своеобразные из-
вестковые сгустки слоистого строения, встречающиеся в древ-
них толщах от архея до современных отложений. Некоторые
исследователи изучают такой материал на основе лишь внешних
признаков, учитывая их форму и размеры, не раскрывая его
микроскопического строения, неправильно придавая им трой-
ную латинизированную номенклатуру, как будто это нормаль-
ный палеонтологический материал. Между тем внутри таких
образований, по существу являющихся микробиогермами, не-
редко присутствуют остатки конкретных форм микроскопиче-
ских водорослей. Они вполне различимы для опытного глаза
и обычно заслуживают специального палеонтологического изу-
чения. Так, в строматолитах протерозоя Приамурья (хребет
Малый Хинган, левый берег р. Амура) в слоях мурандавской
свиты автору удалось обнаружить в веществе крупных, измен-
чивой формы-строматолитов множество четких видов и родов
167
синезеленых водорослей, среди которых видное место занимают
развивавшиеся округлыми колониями мурандавии. Чтобы
различить их и понять, необходимо было только при изучении
препаратов перейти от малых увеличений к большим, до не-
скольких сотен раз. Их остатки отчетливо видны в препаратах,
нередко напоминая картину живой колонии водоросли на пред-
метном стекле. Видны также фоссилизированные следы наруж-
ной студенисто-слизистой оболочки колоний с особенностями
их микростроения. Выделенные из препаратов таксоны разли-
чаются то размерами клеточных колоний, то толщиной и внут-
ренними деталями строения их оболочек, размерами клеток
и т. д. Благодаря этому значительно обогатился список руково-
дящих форм для мурандавской свиты, позволивший определить
и ее возраст как низы верхнего протерозоя, что заставило при-
знать докембрийским и возраст железорудной свиты Малого
Хингана. Как оказалось, выделенные мурандавии распростра-
нялись отсюда на запад, достигая Карелии, а весьма возможно
и дальше. Наряду с округлыми колониями здесь же были от-
крыты нитчатые формы водорослей и колонии с особо круп-
ными клетками. Одна из нитчатых форм особо примечательна.
Ее можно рассматривать как развитие двух-трех спирально
свернутых однорядных нитей в сгустке студенисто-слизистого
вещества. Она имеет сходство с Thisanaplanta filamentosa V.et J.,
выделенной автором и Т. Н. Титоренко в слоях улунтуйской
свиты протерозоя Юго-Западного Прибайкалья.
Водоросль нитчатого строения со спирально свернутой
нитью была обнаружена автором во внутриформационном
конгломерате в низах криворожской серии на Украине. К сожа-
лению, метаморфизм сильно изменил ее остатки. Нить водо-
росли тоже, по-видимому, находилась внутри слизевого чехла.
Сравнивать эти водоросли преждевременно; обе они относятся,
вероятно, к одной группе, к одному семейству (?). Вместе с
этой «спирогироподобной» водорослью в гальке того же кон-
гломерата были открыты интереснейшие остатки организмов
Aseptalia, тяготеющих к явным целентератам, о которых гово-
рилось выше.
168
Много нового, интересного материала по организмам до-
кембрия дал автору Чешский массив (Западная Чехословакия).
В районе к югу от г. Пльзень, близ г. Вотице, в слоях докембрия,
серия молданубика, была обнаружена большая группа ископае-
мых форм, в числе которых оказалось много водорослей с коло-
ниями сферической формы. У них варьируют размеры, очерта-
ние в пространстве и проявления следов клеточной структуры.
Они существенно отличаются от подобных сибирских микрово-
дорослей докембрия. Наряду с этим обнаружены фоссилизиро-
ванные дерновинки, которые тоже состоят из округлых, но раз-
новеликих клеточных колоний, имеющих форму неправильных
тяжей — нитей. Обычно им сопутствуют там крупные местные
скопления обуглероженного битума, иногда размером с голову
ребенка. Эта группа водорослей, кажущаяся эндемичной, дала
возможность автору выделить и описать в специальной моно-
графии ряд довольно четких видов и родов, которые характерны
именно для слоев пестрой свиты молданубика Южной Чехии.
Интересно, что в ассоциации с этими водорослями были обна-
ружены остатки беспозвоночных животных, которые очень на-
поминают скелеты археоциат, но отличаются от типичных кемб-
рийских представителей этой группы исключительно малыми
размерами. Возможно, что это предковые формы, требующие
дальнейшего исследования.
В составе докембрия Чешского массива, в его спилитовой
серии, автором также были открыты четкие нитчатые водоросли
в ассоциации с одноклеточными планктонными формами. Если
они удачно вскрыты при распиловке образца, именно продольно,
то в препаратах местами кажутся образующими целые заросли,
в частности водоросль Bystraia boucekt Там же, в районе
бассейна р. Бероунки, в лидитах альгонкия выявлена нитчатая
водоросль Berounkia, несколько сходная с соответствующей
находкой в докембрии района оз. Онтарио.
В чешском материале обнаружены следы сгустков бесструк-
турного органического вещества, иногда хорошо зафиксирован-
ных процессами окаменения, притом наблюдаемых среди мас-
сового развития одноклеточных водорослей. По-видимому,
169
Pustularia taeniata Vologdin. Уменью, в 10 раз. Реконструкция
Timanella gigas Vologdin. Внешний вид водоросли. Уменью, в 2 раза. Ре-
конструкция
Timarietta gigas Vologdin: a — внутреннее строение слоевища; б — боковые
ответвления. Нат. вел. Реконструкция
это тоже остатки водоросли, способной образовывать округлые
клеточные колонии с обильным слизеотделением (?). Там же
были обнаружены остатки конических или трубчатой формы
тел с довольно толстой стенкой. К водорослевым остаткам их
причислить трудно.
В докембрии Чешского массива при неясном стратиграфиче-
ском положении были найдены остатки почти настоящих архео-
циат с двухстенными скелетами, у которых стенки соединены
между собой пористыми радиальными перегородками. Таким
образом, накапливается достаточно данных, чтобы считать
археоциаты характерными и для протерозоя. В слоях пестрой
свиты района г. Вотице автором выявлено несколько групп
беспозвоночных животных, имевших пористо- пластинчатый
известковый скелет. Некоторые организмы отличались двух-
камерным скелетом. У других он был многокамерным, по сво-
ему строению напоминающим дольки апельсина. Возможно,
это тоже археоциатоподобные или приближающиеся к прос-
тейшим, к фораминиферам, организмы. Вопрос этот пока
остается открытым. В слоях чешского моравика в районе г. Гру-
дима были выявлены остатки многокамерных организмов со
следами пузырчатой ткани во внутренней полости. Еще важнее,
что там же были обнаружены сечения маленьких раковинок,
лишенных замка, скорее всего принадлежавших лингулоподоб-
ным брахиоподам. Раковины лежали на грунте вогнутой сто-
роной вверх и на них видны фоссилизированные скопления одно-
клеточных планктонных водорослей.
Большую группу горных пород составляют породы, образо-
ванные за счет остатков органического вещества и самих расти-
тельных организмов, такие, как горючие сланцы (кукерситы
Эстонии), каменные и бурые угли, вся группа ископаемых биту-
мов (нефть и горючий газ, асфальт и озокерит). Породы, сложен-
ные растительными остатками — водорослями, имеются и в
протерозое.
В составе пород докембрия Карелии издавна известны шун-
гиты, своеобразные горные породы осадочного происхождения,
встречающиеся в ряде районов Заонежья, в частности в районах
172
Кондопоги, Великой Губы, пос. Шунги. Шунгиты образуют осо-
бую свиту в верхах ятулия (среднего протерозоя). Имеются шун-
гитовые сланцы — пластообразные шунгиты, залегающие жи-
лообразно вблизи от покровных диабазов. Внешне шунгиты
напоминают каменные угли и антрацит. В особых условиях им
свойственна способность к сгоранию, чаще при большой золь-
ности, объясняемой высоким содержанием углерода. Эти шун-
гиты содержат до 98% углерода. Их плотные кремнистые разно-
видности используются как поделочный материал или как
пробирный камень. В порошке шунгиты пригодны для получе-
ния черной краски. Почвы на шунгитах отличаются повышенной
урожайностью сельскохозяйственных культур вследствие на-
личия в них большого комплекса фитофильных микроэлементов:
молибдена, меди, ванадия, мышьяка, калия, магния и т. д.
Некоторые разновидности шунгитов при температуре до
1090—1105 ° С вспучиваются, превращаясь в легкую пористую
массу, пригодную для использования ее в качестве теплоизоля-
ционного материала — шунгизита. Вспученная масса шунги-
та — пенокералит — хорошо пилится, имеет прочность на сжа-
тие до 40 кг/см2, что делает его тоже ценным строительным
материалом.
Происхождение шунгитов многие десятилетия представля-
лось загадкой. Более близко к объяснению природы шунгитов
подошел Б. А. Борисов, считавший, что они созданы за счет
метаморфизма пород сапропелевого типа, образованных в
стоячих морских мелководных бассейнах — бухтах и лиманах,
где интенсивно развивалась жизнь того времени. В 1968 г. в
шунгитовых породах из Приладожья В. И. Горлов действитель-
но обнаружил остатки организмов, представленных микроско-
пическими формами. При исследовании этой находки, а также
изучении других материалов из шунгитовой свиты Прионежья
было установлено в шлифах массовое скопление остатков мик-
роскопических водорослей типа синезеленых. По-видимому,
их периодическое развитие в прошлом обусловливало «цвете-
ние» воды в периоды годичных сезонов вегетации. Это оказа-
лись планктонные водоросли, по своим биологическим свойст-
173
вам действительно сходные с водорослями, образующими
сапропелевые осадки в современных стоячих пресноводных и
солоноватоводных водоемах.
Описанная значительная группа родов и видов микроскопи-
ческих водорослей, по их остаткам из карельских шунгитовых
пород характеризует особенности жизнепроявлений в водных
бассейнах глубокого геологического прошлого в пределах Се-
веро-запада СССР и, вероятно, Финляндии. Кроме того, можно
считать, что одновременно разгадывается происхождение шун-
гитоподобных образований из ряда других районов СССР,
в частности Восточной Сибири, Дальнего Востока и Украины.
Интересно, что процессом окаменения осадков в древнейших
бассейнах указанного выше типа закреплены, зафиксированы
вполне ясно стадии индивидуального развития отдельных кле-
ток водорослей, этапы их размножения посредством простого
деления, что указывает на их особую примитивность, как это
и можно было ожидать у таких древних форм жизни. Кроме
того, в средних и более поздних слоях протерозоя, в основном
в синии, известны замечательные находки остатков радиолярий,
фораминифер и кремневых губок в углистых сланцах докембрия
Бретани. Чарльз Д. Уолкотт открыл в докембрии Большого
Каньона Северной Америки остатки многощетинков.ых червей
и ряд других организмов. На юге Австралии обнаружены в
докембрии остатки радиолярий. В слоях докембрия Швеции
открыт отпечаток членистоногого животного — «ксенусиона»,
которого можно признать представителем ветви животных,
из которой в кембрии развился класс трилобитов. Своеобразные
остатки животных неясного систематического положения, отне-
сенного к проблематическому роду «чарний», были открыты в
докембрии Англии и Австралии.
Чрезвычайно разнообразна в систематическом составе фауна
Эдиакарского месторождения на юге Австралии близ г. Аделаи-
ды. Здесь в кварцитовых песчаниках Паунд возраста около
600 млн. лет установлены остатки бесскелетной фауны, представ-
ленной медузоидными формами, остатками червей, организмов
неизвестного систематического положения. Подобные же орга-
174
Вид организма Suvorovella aldanica Vo-
logdin et A. Maslov на выветре-
лон поверхности породы в нижней части
юдомской свиты (ув. в 2 раза). Якутия,
район пос. Усть-Юдома, р. Мая (по А. Г.
Вологдину)
значительное развитие, остатки
низмы обнаружены в одно-
возрастных отложениях
свиты Кибис системы Нама
в Южной Африке. Здесь
встречены Rongea G и г i с h ,
Pteridinium G urich, Pa-
ra tnedusium. В серии Нама
в Африке обнаружены кри-
броциатоподобные орга-
низмы из рода Cloudina.
В Евразии и в Америке
установлено несколько от-
печатков медуз и червей,
напоминающих по форме и
размерам пиявок.
В докембрии СССР и
Китая мы находим иглы
кремневых губок, самые
примитивные по устройст-
ву, отпечатки червей типа
сабеллитов, получивших
позднее в кембрии более
фораминифер, древнейших
археоциат, хиолитов и организмов неявного систематического
положения.
В прослоях доломитизированного известняка нижней части
разрезу юдомской свиты по левому берегу р. Маи Н. П. Суво-
рова обнаружила своеобразные органические остатки Suvoro-
vella и Majella, неизвестного систематического происхождения,
организмы с двустепенным известковым пластинчатым непо-
ристым скелетом, с интервалом без скелетных элементов и
стенкой из ромбических выступов, расположенных рядами
по спирали.
В верхнем докембрии Русской платформы обнаружены остат-
ки медузоидных организмов Beltanelloides морщинистых
сфероидальных форм, напоминающих Charniodiscus, на п-ове
Рыбачьем встречены медузоидные формы, напоминающие Ме-
175
Отпечатки протомедуз Sajanella arshanica Vologdin на поверхности глинис-
того песчаника в низах карагасской свиты (нат. вел.). Восточный Саян, Иркут-
ская обл., район пос. Аршан
dusina и Ediacaria, в керне скважины у г. Яренска из ляминари-
товых глин установлена Vendia, в ляминоритовых глинах на-
блюдаются пиритизированные следы червеобразных организ-
мов Vendo ver nites. В валдайских отложениях Прибалтики встре-
чены формы, подобные Beltanella, и отпечатки оболочек типа
Sahellidites.
Небезынтересно отметить находки в слоях карагасской сви-
ты Восточного Саяна отпечатки медуз Sajanella. Они были от-
несены к отряду брукселлид, выделенному по материалам ниж-
него палеозоя. Почти в тех же слоях обнаружены остатки червей
сабеллидитов, а также крупных ракообразных, несколько напо-
минающих палеозойских эвриптерид. Они были описаны авто-
ром как карагассии и отнесены к хелицеровым из-за их клино-
видных тельсонов и других особенностей панциря. Карагассии
достигали в длину полметра и более. Пока что это самые круп-
ные организмы из беспозвоночных позднего докембрия. Впро-
чем, некоторые исследователи склонны считать их «скрючен-
ными корками высыхания глинистых прослойков на поверхно-
сти песка». Карагассии являются, по-видимому, вымершей
группой членистоногих, живших в некоторых пресноводных
бассейнах конца докембрийского времени.
Таким образом, в мощнейших толщах осадочно-метаморфи-
ческого докембрия северной Евразии выявлен принципиально
новый обильный палеонтологический материал, охватываю-
щий огромный стратиграфический диапазон — от верхов архея
до начала кембрия. Он характеризует многие горизонты пород
и этапы геологического времени и начинает укладываться в
основу новой единой, по крайней мере для Евразии, схемы стра-
тиграфического расчленения докембрия.
Границу между докембрием и кембрием пытались устано-
вить многие исследователи. В настоящее время этот вопрос
в корне проясняется, поскольку оказывается, что переход от
протерозоя к палеозою (570 млн. лет) был отчетливо выражен
соответствующей сменой представителей растительной и живот-
ной жизни в общем ходе ее эволюции. Преобладание синезеле-
ных водорослей сменилось преобладанием водорослей крас-
12-818
ных. Сам протерозой и даже поздний архей оказались не только
потенциальными, но и фактическими вместителями остатков
организмов за огромный этап геологического времени, а именно
более чем на два миллиарда лет древнее той границы, ниже кото-
рой палеонтологи в сущности не заглядывали. Можно отчет-
ливо отделить начало кембрия от верхов докембрия по остаткам
специфических форм водорослей, археоциат, трилобитов и дру-
гих групп организмов, поскольку история формирования пород
земной коры отчетливо записывалась в них, одновременно с
общим ходом истории жизни. Поэтому вырисовывается, притом
во многом по-новому, общий ход развития организмов Земли
в палеофациях ее биосферы, почти с момента возникновения
жизни — около 3500 млн. лет до н. э. В результате этого имеется
возможность широко внедрить в практику стратиграфических
и геологосъемочных работ, связанных с древнейшими осадоч-
ными толщами, палеонтологический метод и выделить с его
помощью руководящие формы ископаемых организмов как
меру геологического времени одновременно с прослеживанием
развития самой жизни на ранних этапах.
В общих чертах выясняется древность известных науке орга-
низмов, устанавливаются новые, не известные ранее, группы
и конкретные предковые формы всех типов бактёрий (?), расте-
ний и животных, сформировавшихся в процессе общего измене-
ния жизненной среды в биосфере Земли, в процессе биохимиче-
ской эволюции живого вещества, сопровождавшейся неразрыв-
но эволюцией морфологической. Следует отметить, что органи-
ческое вещество в виде продукции организмов и их конкретных
остатков во вмещающих породах почти никогда полностью не
исчезает. Будучи в той или иной степени измененным — редуци-
рованным, обуглероженным, фоссилизированным, оно часто
способно сохранять во вмещающей горной породе исходные
морфологические черты, достаточные для палеонтологического
изучения. В то же время это вещество представляет немалый
интерес и для специальных биохимических исследований, по-
скольку местами в породах докембрия уже установлено присут-
ствие многих химических компонентов из состава живого веще-
178
ства. Важно лишь, чтобы палеонтолог и биохимик работали
совместно. Геолог при этом получит новый опорный материал
для своих стратиграфических, палеогеографических, фациаль-
ных и иных построений, а также данные о палеоклимате, иско-
паемых фациальных обстановках породо- и рудообразования
в осадочных условиях, именно от живых свидетелей геологиче-
ского прошлого. Биогеохимик и биохимик могут получить важ-
ный документальный материал и данные о биохимической эво-
люции жизни на нашей планете.
Таким образом, протерозойская эра истории нашей плане-
ты была в основном временем исключительного господства
бактерий и водорослей в водных средах. За этот этап времени,
длившийся, по данным абсолютной геохронологии, около
1200 млн. лет, упомянутые группы организмов выполнили
огромную геологическую работу по образованию ряда типов
осадочных пород и руд, а также по переработке вещественного
состава самой биосферы и атмосферы Земли.
ПАЛЕОЗОЙСКАЯ ЭРА
Начало палеозойской эры (кембрий — 570—480 млн. лет) отме-
чено появлением целого ряда новых групп организмов, в числе
которых виднейшую геологическую роль сыграли археоциаты,
интересные морские беспозвоночные животные с внутренним
известковым скелетом очень изящного и часто сложного строе-
ния, по преимуществу обитавшие на дне мелких морей, где они
строили своеобразные барьерные и береговые известковые ри-
фы. Это был особый тип морских животных, быстро распростра-
нившихся в морях Земли в пределах всех широт, за исключением
лишь самых приполярных областей. Нужно отметить, полю-
сы Земли тогда располагались в зоне современных шестидеся-
тых параллелей. Об этом «говорят» и водоросли, й археоциаты.
Захватывая средние и небольшие глубины морей, археоциа-
ты сильно потеснили обитавшие там водоросли. Вероятно, они
питались микроскопическим фитопланктоном, спорами водо-
рослей и бактериями. Их личинки, садясь на дно, прирастали к
12* 179
Известняк со скелетными остатками археоциат. Средний кембрий Западного
Саяна (ув. в 2 раза)
разным твердым предметам, чаще к скелетам отмерших особей
тех же археоциат. Они образовывали то малые, то крупные
скопления извести, возвышавшиеся над соседними участками
дна. Такие ископаемые рифы были новым явлением в истории
Земли, в истории ее морских бассейнов.
180
Так выглядели археоциаты — первые животные-ci роите ли морских известковых
рифоз. Реконструкция автора
В морских отложениях кембрия продолжают обнаруживать-
ся остатки археоциат в виде трубчатых или дискообразных
п !астинчатопористых скелетов кальцийорганического состава.
Их лабораторное изучение позволило выделить и описать до
1000 видов из более чем 60 семейств и 25 отрядов, принадлежа-
щих к четырем классам Установлено, что в пределах Советского
Союза кембрийские отложения особенно богаты остатками этой
фауны, долго считавшейся загадочной, пока она не получила
признания как особый самостоятельный тип животных, не свя-
занных ни с кораллами, ни с губками
В Сибири и Средней Азии выявлено много остатков архео-
циат. которые удается выделять из вмещающей породы. В про-
тивоположность правильному, изящному строению элементов
181
скелета их внешняя форма оказывается часто не столь правиль-
ной. Это, видимо, связано с влиянием на них среды, на что ука-
зывают поперечные пережимы и расширения кубков, встречаю-
щиеся у ряда развивавшихся по соседству особей, принадлежа-
щих не только к разным видам, но даже и к разным родам.
Археоциаты могли достигать значительных размеров: до 40—
80 см в поперечнике и до 100—150 см в высоту. В обломках
пород часто наблюдаются следы нарастания одной особи на
боковой поверхности или на дистальном конце скелета другой.
Наиболее интенсивно жизнедеятельность археоциат прояв-
лялась в ранне- и среднекембрийские эпохи. В более древних и
более поздних отложениях их остатки пока редки и еще недоста-
точно изучены. На обширных пространствах мелководных мо-
рей на востоке СССР, на Китайской платформе, в относительно
глубоких морях — на юге Сибири и в Монголии в начале кемб-
рийского времени археоциаты строили подводные банки, созда-
вая огромные массы рифогенных известняков в виде береговых
или барьерных рифов. Аналогичные материалы по археоциатам
известны на территориях Северной Америки, Австралии, За-
падной Европы и Северо-Западной Африки. Их широкое рас-
селение, связанное со способностью личинок перемещаться с
морскими течениями на дальние расстояния, приближает нас к
возможности осуществления палеогеографических построений
для отдельных моментов геологической истории Земли в интер-
вале раннего и среднего кембрия.
Что же представляли собой археоциаты как живые орга-
низмы далекого геологического прошлого? В 1948 г., на основа-
нии следов строения мягких тканей археоциат из кембрия р. Кии
в Кузнецком Алатау, удалось доказать, что археоциаты имели
не наружный скелет, а внутренний. Губчатое строение централь-
ной полости в целом было принято за внутренний орган ассими-
ляционного назначения. Эта находка позволила выяснить назна-
чение центральной полости, помогла установить роль пор сте-
нок, перегородок и днищ и, что особенно важно, — внутренний
скелет у этой группы организмов. Наружная мягкая оболочка
прекрасно объясняла, почему археоциаты обладали способно-
182
Следы строения мягких тканей у археоциат кембрия Восточной Сибири. Фото
автора
183
Эпифитоны — микроскопические красные водоросли кембрия. Фото автора
стью прикрепляться к субстрату, образуя те или иные дополни-
тельные элементы скелета до отложения иногда сплошной мас-
сы твердого скелетного вещества.
Находки археоциат с остаточными структурами мягких тка-
ней внутри кубка и на его поверхности в СССР уже перестали
быть редкостью. В южной части Сибири, Восточном Забайкалье
среди остатков скелетов археоциат попадаются и такие, у кото-
рых в разной степени сохранились следы строения мягких тка-
ней. В некоторых шлифах можно проследить замечательные
картины биологических взаимоотношений форм, принадлежа-
щих разным видам и являющихся самостоятельными организ-
мами. Своеобразно выглядело дно кембрийского моря, где,
кроме водорослей, появились археоциаты, губки, копошились
184
и плавали разнообразные древнейшие ракообразные — трило-
биты.
Самое значительное, что принесло изучение археоциат,
это обнаружение их остатков со следами сгроения мягких тка-
ней. Причина этого, по-видимому, в отсутствии в водах кемб-
рийских морей гнилостных бактерий. У многих экземпляров
археоциат во внутренней полости сохранились следы сложно
построенной системы взаимосвязанных каналов, сопряженных
с порами внутренней стенки. Сюда поступала морская вода с
пищевыми веществами. Продукты ассимиляции перемещались
затем по капиллярным сосудам в межстенное пространство,
где имелись многочисленные камеры со стенками, покрытыми
слоем мягкой ткани. Сквозь поры мягкой наружной оболочки
проходили наружу тонкие капиллярные сосуды.
Полученные микрофотографии со следами строения мягкого
тела археоциат имеют исключительную ценность для дальней-
шего изучения этого интересного типа животных, поздние пред-
ставители которого к девону совершенно вымерли.
Наряду с археоциатами в кембрии появилась большая группа
древнейших членистоногих — трилобиты, эволюция которых
растянулась на несколько геологических периодов — до триаса
включительно (570—185 млн лет назад). Появились древнейшие
панцирные рыбы, строматопороидеи, брахиоподы и т. д. С на-
чала кембрийского периода мир животных начал развиваться
бурно, сложно, с проявлением многих этапов формообразова-
ния, с вымиранием одних групп и появлением новых.
За последнее время в отложениях кембрия обнаружены пред-
ставители многих новых классов. Автором были установлены
криброциаты, сопутствующие почти повсеместно археоциатам,
остатки фораминифер в образцах из нижнего кембрия р. Ирби-
тей на южном склоне хребта Восточного Танну-Оли, встречен-
ные ранее Е. А. Рейтлингер в синской свите р. Синий, левого
притока р. Лены. В образцах своеобразного пятнистого извест-
няка, формировавшегося явно в условиях прибрежно-морского
мелководья, собранных в отложениях ирбитейской свиты на
южном склоне хребта Танну-Ола совместно с археоциатами и
185
Организмы кембрийского возраста: диатомообразные (ув. в 60 раз). Тува, хр.
Танну-Ола, ирбитейская свита (а); фораминифероподобные (ув. в 210 раз). Тува,
хр. Танну-Ола, р. Кадый, нижний кембрий (б)
губками, и были обнаружены остатки диатомообразных и фо-
раминифероподобных организмов. К. Б. Корде описала новый
класс кишечнополостных животных—Hydroconozoa, являю-
щихся породообразующими организмами.
Начиная с кембрия стали интенсивно развиваться красные
и зеленые (сифоновые) водоросли. Первые были представлены
большим количеством родов и видов. Наряду с археоциатами
они играли породообразующую роль в биогермах кембрийского
возраста, могли сопутствовать археоциатам, но в некоторых
случаях были единственными породообразователями,
Остатки красных водорослей встречаются в отложениях мор-
ских фаций, сформированных в чистых водах и при нормальной
для кембрийского времени солености. Сифоновые водоросли
были представлены незначительным количеством видов и ро-
дов, но местами — в Казахстане, Сибирской платформе и на
р. Амге. они играли породообразующую роль. Синезеленые
водоросли с начала кембрия стали уменьшаться в количестве.
Рассматривая комплекс живых существ на Земле как «живое
186
б
вещество биосферы», следует отметить, что это вещество вы-
полнило в течение палеозоя грандиозную работу. Непрерывно
и бурно развиваясь качественно, давая новые и новые формы,
оно уже с начала эры стало решительно распространяться и на
области суши. Первыми, освоив прибрежные зоны бассейнов,
вышли на сушу микроскопические синезеленые водоросли, ко-
торые затем довольно быстро распространялись на площадях
увлажнения почв. Водоросли с более высокой организацией,
красные и зеленые, располагавшие корневой системой и специа-
лизацией клеточных тканей, смогли при этом выйти в субаэраль-
ную среду. Несомненно, все присущие наземной растительно-
сти особенности —анатомическое строение, смена поколений
в цикле индивидуального развития и другие — все это было за-
ложено еще в предшествующей, водной, стадии развития.
Выход растений на сушу заставил их приспособиться к самым
разнообразным условиям обитания. Поскольку ни свет, ни вла-
187
Часть поверхности водорослевого рифа — биогерма. Нижний ордовик. Реконст-
рукция автора
га, ни углекислота не лимитировали развития растений, то они
быстро расселились на обширных пространствах, одновременно
приобретя многие новые специфические черты строения. Бота-
ники считают, что при этом, кроме ранее господствовавшего
полового размножения растений, проявилось и стало развивать-
ся бесполое — вегетативное. Переход на сушу вызывал возник-
новение у растений способности к защите от потери влаги и
высыхания, развитие тканей, которые цолжны были играть
роль опорного каркаса и т. д.
В истории палеозоя были этапы массового развития расте-
ний. В условиях влажного климата это приводило местами к
накоплению растительного вещества в составе осадочных отло-
жений с последующим превращением его в результате вторичных
188
Верхняя поверхность водорослевой банки. Сииур. Фото К. К. Во.длосовича
процессов в каменный уголь. В водных условиях усилива-
лась жизнедеятельность водорослей, что также приводило к
накоплению органических веществ с частичным их преобразо-
ванием потом, в толще накапливаемых минеральных осадков,
в нефть и горючий газ. По-видимому, стимулом для подобных
усилений жизнедеятельности фотосинтезирующих организмов
были выбросы в биосферу, уже значительно обедненную угле-
кислотой, новых ее запасов.
В'ордовике (450 420 млн. лет) на дне морей развивались
колонии кораллов, ползали крупные головоногие животные —
ортоцератиты, также трилобиты и т. д. Местами развивались
водорослевые рифы — биогермы с участием синезеленых водо-
рослей, то в виде неправильных скоплений массы карбонатного
вещества, то в виде довольно изящно оформленных рифов, в мо-
189
Придонная зона позднесилурийского моря с гигантскими ракообразными —
эуриптерами и с зарослями водорослей. Реконструкция Р. Моора
делировании которых, несомненно, сказалась приспособлен-
ность к разрушающей деятельности морских волн. Их строение
часто показывает быстрые темпы эволюции.
На дне силурийских морей (420—400 млн. лет) жизнь была
уже совсем иной. Красные водоросли местами преобладали над
синезелеными. Из животных крупнейшими были ракообраз-
ные — эуриптеры, пышно развивались моллюскообразные —
брахиоподы, птероподы. Кораллы строили рифы или распола-
гались малыми скоплениями на различных участках дна. В силу-
ре заметно распространилась наземная флора, представленная
споровыми растениями.
В девоне (400—320 млн. лет) развились псилофиты — древ-
нейшие стеблевые, древнейшие древовидные и кустарниковые
190
Наземная флора девонского периода. Реконструкция Р. Моора
папоротники, появились первые голосеменные — кордаиты,
гингко.и другие, также примитивные ранние хвойные. Местами
началось углеобразование. На дне мелких морей при участии
различных видов синезеленых водорослей формировались свое-
образные строматолиты.
В девонское время коралловые фауны развились очень пыш-
но. Водоросли делали уже решительные попытки выхода на
сушу, постепенно приспособившись в береговой зоне бассейнов.
Из животных крупнейшими были акулоподобные и панцирные
рыбы. Появились также кистеперые рыбы, у которых развитие
скелета шло к выработке конечностей, пригодных для хождения
по твердой земле. Эта ветвь жизни поэтому дала позднее амфи-
бий — двоякодышащих, вышедших на сушу. На низких высту-
пах суши девонского времени уже возникали леса
191
Поjднекарбоновая древесная растительность — лепидодендроны и другие расте-
ния. Реконструкция Р. Моора
В карбоне (320 —270 млн. лет) и перми (270—225 млн лет)
сравнительное однообразие флоры девона сменилось массой
более высоорганизованных форм, в свою очередь быстро рас-
пространившихся по всему миру. Впервые наметились особые
палеоботанические и палеогеографические области. Хотя корне-
вые системы новых растений все еще хранили черты водоросле-
вых, а также папоротникообразный характер верхнего строения,
новые кордаиты, каламиты и другие формы образовали уже
леса, часто типа тропических. При этом наземная флора поздне-
го палеозоя оставила в строении земной коры крупнейший след
в виде пластов каменных углей в ряде районов мира на самых
разных широтах (Шпицберген, Гренландия, Южная Африка
и т. д.). Эволюция водорослей также продолжалась Морские
192
Позднекарбоновыи ландшафт с пресмыкающимися петролакозаврами, скорпио-
нами, жуками, стрекозами и представителями наземной флоры. Реконструкция
Р. Моора
фауны беспозвоночных проявляли в карбоне большое разнооб-
разие и развитие характерных форм
К началу палеозойской эры фотосинтезирующие организмы
биосферы, а также химические процессы почти полностью исчер-
пали запасы углекислоты. С этого времени жизнедеятельность
фотосинтезирующих, в особенности наземных организмов, ока-
залась подчиненной периодическим выбросам ювенильной угле-
кислоты из недр Земли при вспышках вулканической деятель-
ности. В связи с этим общая масса живого вещества растений
была в разные моменты истории Земли различной. Это прояви-
лось в колебаниях количества органических и минеральных ве-
ществ, накапливавшихся в результате жизнедеятельности водо-
13-818
193
Раннепермские пресмыкающиеся — эдафозавры. Реконструкция Р. Моора
рослевых сообществ в бассейнах типа пресноводных и в морских
мелководьях. Особенно это выразилось в возникновении ритма
в развитии наземных растений. Некоторые исследователи про-
цессов угленакопления на Земле пришли к выводу, что такие
процессы должны быть причинно связаны с фазами складчато-
сти земной коры. Между тем дело не в «фазах складчатости»,
а в выбросах углекислоты из недр при вулканических явлениях,
хотя они часто были связаны с горообразовательными движе-
ниями.
Истории развития высших организмов на нашей планете —
позвоночных животных — посвящено много прекрасных спе-
циальных и научно-популярных исследований во всем мире.
В частности, следует особо отметить книги Б. А. Трофимова
(1957) и Ю. А. Орлова (1961), богато иллюстрированные худож-
194
Пресмыкающиеся пермскси о периода — сеймурии, сохраняющие сходство с
амфибиями. Реконструкция Р. Моора
195
Хищный пермский ящер иностранцевия зерзаез травоядную рептилию парейазав-
ра. Реконструкция А. П. Быстрова
196
ником — палеобиологом К. К. Флеровым. В связи с этим в
данной книге мы коснемся истории развития позвоночных
животных предельно кратко.
Высшие представители мира животных вышли на сушу тоже
в девоне. Это были кистеперые рыбы, превратившиеся в амфи-
бий — двоякодышащих, а затем, ко времени карбона, и в реп-
тилий — пресмыкающихся. В пермское время на суше обитали
уже крупные представители древних четвероногих, поедавших
водоросли, наземные растения или себе подобных. Появились
и многообразные насекомые, достигавшие огромных размеров,
до 1 м в размахе крыльев. В морях появились новые группы
беспозвоночных животных.
МЕЗОЗОЙСКАЯ ЭРА
Мезозойская эра началась около 200 млн. лет назад. Она ха-
рактеризуется расцветом пресмыкающихся, возникновением
высших представителей органического мира — покрытосемен-
ных растений, двукрылых и перепончатокрылых насекомых,
костистых рыб, птиц, млекопитающих.
В начале мезозоя, в триасовой период суша имела максималь-
ное за всю историю Земли распространение, климат был теп-
лый. В океанах и морях пышно развиваются новые формы мор-
ских лилий и ежей, образуют мощные рифы шестилучевые ко-
раллы. Плеченогие уменьшаются в числе и на смену им прихо-
дят двустворчатые моллюски. Трилобиты и ракоскорпионы
исчезли, появились длиннохвостые высшие раки. Особенно
характерными для мезозойской эры являются головоногие
моллюски — закрученные аммониты и белемниты, остатки
которых известны часто в народе под названием «чертовых
пальцев».
На суше также происходит энергичная замена низших форм
жизни высшими. Начинается пышный расцвет мезозойских
голосеменных растений, покрывших землю новыми лесами.
Эти леса и вся наземная растительность состояла в основном
из древних хвойных и их родичей — гингковых и беннеттитовых
197
Для наземных позвоночных триаса более, чем для пермских,
характерно следующее контрастное развитие, по-видимому,
связанное с резко континентальным климатом этого периода.
У одних наблюдается «тяготение» к водным условиям, у дру-
гих — к сухопутным. Многие пресмыкающиеся приспосабли-
ваются к жизни в море. В болотах и низменных лесах широко
распространились двуногие архозавры — предки юрских и ме-
ловых динозавров. Передвижение архозавров и динозавров на
двух конечностях было результатом приспособления их к жизни
среди высокой растительности. Они быстро передвигались на
задних конечностях и могли хорошо ориентироваться среди вы-
соких деревьев.
В триасе, вероятно, возникли первые млекопитающие, ко-
торые, однако, не играли заметной роли в органическом мире в
течение почти всей мезозойской эры.
Несмотря на то, что у млекопитающих есть признаки, общие
с земноводными и отсутствующие у пресмыкающихся, в част-
ности, обилие кожных желез, происхождение их от пермских и
триасовых териодонтов не вызывает сомнения. У териодонтов
постепенно появляются признаки, свойственные исключитель-
но млекопитающим. Скелет их мало чем отличается от скелета
древних млекопитающих и по ряду физиологических особен-
ностей они, вероятно, также были им близки. Хорошо развитое
вторичное костное нёбо и сложные зубы давали возможность
териодонтам непрерывно дышать и в то же время хорошо пере-
жевывать пищу. Они так же, как и млекопитающие, уже высоко
держались на ногах и были весьма активными животными.
Можно указать на следующие основные вехи прогрессивного
развития млекопитающих: преобразование рогового покрова
в волосяной, защищающий тело от потери тепла; преобразо-
вание черепа, связанное с питанием и развитием сложного уха:
развитие органов дыхания, кровообращения; прогрессивное
развитие головного мозга, особенно коры больших полушарий:
живорождение и вскармливание детенышей молоком. Комплекс
этих признаков обусловливал развитие теплокровности. В те-
чение всей мезозойской эры млекопитающие существовали в
198
Амфибия (риасового периода — мае го шнозавр. Реконструкция Р. Моора
199
виде мелких форм, от которых нам известны остатки обычно
в виде черепов, челюстей, зубов.
В юрский и меловой периоды моря были обширны и, напри-
мер, Европа представляла тогда архипелаг островов. Климат
был ровный, мягкий. В морях были широко распространены
простейшие фораминиферы, губки, шестилучевые кораллы,
морские лилии и ежи, двустворчатые моллюски, десятиногие
раки, крабы, но особенно многочисленны аммониты, белемни-
ты и различные рыбы. Акуловые рыбы были близки к современ-
ным, а костные имели строение, промежуточное между осетро-
выми и настоящими костистыми, развившимися в меловом
периоде.
На суше, в условиях средней влажности и довольно высоких
температур были распространены голосеменные растения. Тра-
вянистый покров составляли мелкие папоротники, мхи, хвощи,
плауны.
Пресмыкающиеся достигли огромной численности и разно-
образия. Они заселяют всю сушу, моря, поднимаются в воздух.
На суше появились речные крокодилы, черепахи, ящерицы,
но полными хозяевами ее были динозавры.
В юрском* периоде динозавры уже поедставлены самыми
гигантскими наземными животными — бронтозаврами, дипло-
доками и др. с очень длинными хвостом и щеей, маленькой го-
ловой и громадным туловищем. Эти гиганты, достигавшие
30 м длины, жили в прибрежных зонах больших водоемов,
питались мягкой растительной пищей. Другие динозавры имели
четырехлучевой таз птичьего типа. К ним относятся панцир-
ные ящеры — стегозавры, четвероногие с маленькой головкой.
Их спина была усажена длинным рядом костных вертикальных
пластин. Появились также хищные карнозавры, передвигавшие-
ся на задних конечностях.
В морях обитали прекрасно плавающие дельфинообразпыс
рыбоящеры — ихтиозавры. Они имели веретеновидное чело,
ласты, хорошо развитые спинной и хвостовой плавники. Kopoi-
кие с боченкообразным туловищем и длинной шеей плезиозав-
ры плавали при помощи ласт в более мелких, чем ихпюзавры,
200
1андшафт юрского периода с летающими пресмыкающимися — птерозаврами
вверху), первыми птицами — зубастыми археоптериксами (в центре) и мелкими
инозаврами (внизу). Реконструкция Р. -Моора
20)
зонах моря. Эти водные живородящие пресмыкающиеся были
хищниками и нередко достигали в длину 15 м.
Летающие пресмыкающиеся — птерозавры — были двух ти-
пов. Рамфоринхи с длинными узкими крыльями и длинным
хвостом — рулем, имели планирующий полет, ширококрылые
и короткохвостые птеродактили — порхающий. Крыло у пте-
розавров было образовано кожной складкой, отходившей от
боков тела и поддерживаемой длинным четвертым пальцем
передней конечности.
В юрском периоде появились птицы. Птицы имеют много
общих черт с пресмыкающимися и, несмотря на ряд существен-
ных новоприобретений и разнообразие форм, представляют
группу пресмыкающихся, приспособившихся подобно птеро-
заврам к полету. На основании палеонтологических данных
можно не сомневаться, что птицы произошли от лазающих
псевдозухий — мелких ящерицевидных хищных пресмыкаю-
щихся триасового периода, живших на деревьях, где они были
хорошо защищены от врагов и питались насекомыми, ягодами
и пр.
Особенно ясно это родство показывают юрские перво-
птипы—археоптериксы. Эти длиннохвостные величиной с го-
лубя животные имели на теле, хвосте и трехпалых, передних
лапах перья, причем пальцы лап были свободны и вооружены
когтями. Несмотря на наличие перьев, задних конечностей и
таза птичьего типа, в их строении еще много черт, свойственных
предкам — слабая грудина, наличие брюшных ребер и длин-
ного хвоста (18—20 позвонков), присутствие зубов и др. Но эти
в основном лазающие по деревьям птицы уже могли совершать
планирующий прыжок, что было переходной ступенью к полету.
Меловой период длительностью около 70 млн. лет является
переходным к эре новой жизни — кайнозою. В конце мелового
периода моря и суша приблизились по очертанию к современ-
ным. На побережьях океанов возвышались грандиозные горные
цепи. Влажный и теплый климат становится более прохладным,
континентальным, резче намечается разница между климати-
ческими поясами и ландшафтными зонами.
202
Рогатый динозавр — стиракозавр мелового времени. Реконструкция К. К. Фле-
рова
203
Органический мир меловых морей сходен по общему облику
с юрским: по-прежнему обильны аммониты, белемниты и осо-
бенно костистые рыбы. К концу мелового периода морские
ящеры, а также аммониты и белемниты вымирают.
На суше прежде всего изменяется растительный покров.
Уже в раннем мелу появляются покрытосеменные, или цветко-
вые, растения, а из голосеменных лишь некоторые новые хвой-
ные продолжали играть значительную роль в растительном
покрове Земли.
Мир наземных позвоночных в меловой период резко изме-
няется. Правда, до конца его продолжают жить различные ди-
нозавры, достигая апогея в своем развитии. Стройные, с длин-
ной шеей и маленькой головой двуногие бегуны — струтиоми-
мусы, похожие на страусов; величайшие из когда-либо живущих
хищников — тиранозавры, двуногие гиганты весом в несколь-
ко тонн и достигавшие 9 м в высоту при длине тела 14 м. Много-
численны птицетазовые утконосые динозавры — с вытянутым
вроде утиного черепом, с многочисленными зубами, передви-
гаются на двух ногах, опираясь на массивный хвост. Млекопи-
тающие жили в теплых влажных лесах. Они были величиной с
мышь и крысу, что помогало им вести скрытый образ жизни и
защищало от громадных ящеров. В воздухе вместе с появив-
шимися зубастыми птицами парили птеродактили и беззубые
гигантские птеронодонты, достигавшие 8 м в размахе крыльев.
Итак, на протяжении истории органического мира мы всегда
наблюдаем вымирание одних и процветание других организмов.
КАЙНОЗОЙСКАЯ ЭРА
Кайнозойская эра, длившаяся около 70 млн. лет, — время рас-
цвета высших форм жизниТВ эту эру происходило энергичное
поднятие больших участков материков, формировались высо-
чайшие горы Земли — Гималаи, Памир, Альпы, Анды, — исче-
зали или уменьшались в размерах огромные внутриконтинен-
тальные водоемы — океан Тетис окончательно распался на
ряд морей — Средиземное, Черное, Каспийское и др. Усили-
204
вается континентальность климата и происходит быстрая сме-
на растительных покровов.
Появление и развитие цветковых растений — один из круп-
нейших шагов в истории жизни на Земле — они увеличили об-
щую продукцию пищи, создали новые, лучшие условия (разно-
образную кормовую пищу) для многих животных.
С появлением покрытосеменных растений однообразные
леса из голосеменных и папортниковых стали исчезать. Приспо-
сабливаясь к изменяющимся и усложняющимся условиям среды,
цветковые растения создали новый разнообразный раститель-
ный покров Земли из деревьев, кустарников, травянистых од-
нолетних и многолетних растений.
Наземная растительность и млекопитающие первой поло-
вины третичного периода (палеогена) — еще мало похожи на
-современные. Тропическая растительность широко распростра-
нена в Европе, Азии, Северной Америке. В средней и южной
полосе Европы, например, в южной части Русской равнины
наряду с пальмами, араукариями, лаврами, магнолиями, ки-
парисами, каштанами, эвкалиптами росли дубы, липы, ивы,
ольха, тополь, клен, кустарниковые — бересклет, туя, мирта,
а в более влажных местах — разнообразные папоротники и
мхи.
Мир наземных позвоночных в кайнозойской эре резко от-
личается от мезозойской. В кайнозое млекопитающие быстро
заняли господствующее положение, приспособившись к раз-
личным условиям суши, к жизни в воздушной и водной среде,
и как бы заменили мезозойских пресмыкающихся, из которых
сохранились только крокодилы, черепахи, ящерицы, змеи.
Уже в мезозое появились низшие млекопитающие — сумчатые,
плацентарные (насекомоядные), которые были предками хищ-
ных, копытных и других млекопитающих.
Млекопитающие, найденные во многих местах Азии, Евро-
пы, Северной Америки, еще сочетали особенности, свойствен-
ные представителям различных отрядов или семейств. Пред-
ками современных копытных были первичные копытные —
кондиляртры.. Их непосредственными потомками были также
205
разнообразные копытные нередко причудливого строения и
гигантских размеров. Из них можно назвать громадных рога-
тых, похожих на носорогов, титанотериев, безрогих носорогов,
мелких, с зайца, безрогих парнокопытных. Последние были
предками оленей, жираф, полорогих. Самым крупным назем-
ным млекопитающим был носорог индрикотерий. Он дости-
гал 5 м в высоту.
Во второй половине третичного периода — неогене — очер-
тания морей и континентов, рельеф земной поверхности уже
близки к современным, климат стал холоднее, в горных райо-
нах были оледенения. Происходят большие изменения назем-
ной флоры и фауны, в умеренных поясах тропические и субтро-
пические растения сменяются растительностью с опадающей
листвой, возникают поросшие злаками и другими травами степ-
ные и лесостепные пространства, и позднее, в конце третичного
и в четвертичном периоде появляется растительность тайги,
тундр. На юге растительность, а также и животный мир изме-
няются меньше и сохраняют более древний облик.
Изменение климата и особенно появление нового раститель-
ного покрова — широкое распространение сухих лугов и лесо-
степей, — как показал еще в 70-х годах прошлого века основа-
тель современной палеонтологии В. О. Ковалевский,- резко ска-
залось на развитии млекопитающих и в первую очередь травояд-
ных копытных.
Несколько миллионов лет назад появились уже все семейст-
ва и многие роды ныне существующих млекопитающих: лоша-
диные, свиньи, олени, жирафы, полорогие (быки, антилопы),
гиены, кошки, ластоногие, обезьяны, в том числе человекообраз-
ные, хоботные — мастодонты и древние слоны. У многих ко-
пытных (лошадиных, оленей, полорогих) выработались хоро-
шие приспособления к быстрому бегу и питанию травянистыми
растениями. Ноги их удлинились, коренные зубы стали по-
стоянно растущими с высокой коронкой. Впервые эти основные
изменения в истории копытных млекопитающих в зависимости
от изменения условий жизни были установлены В. О. Кова-
левским.
206
Индрикотерии — крупнейшие млекопи i ающие позднетретично! о времени. Ре-
конструкция К. К. Флерова
207
В конце миоцена и в плиоцене в Европе, Азии, Африке была
широко распространена так называемая гиппарионовая фауна,
в состав которой входили трехпалые лошади — гиппарионы,
свиньи, олени, жирафы, разнообразные антилопы, мастодонты,
гиены, барсуки, виверры, медведи, кошки, в том числе гигант-
ские тигры — саблезубы, грызуны. Из птиц характерны страусы.
Во второй половине плиоцена эта фауна исчезла в Европе, боль-
шей части Азии, Средней Америке. В Африке она просущест-
вовала дольше и превратилась в современную фауну саванн^
Огромные скопления остатков гиппарионовой фауны обнару-
жены и раскопаны на территории Молдавии, Украины, Кавка-
за, Средней Азии, Казахстана.
Четвертичный период, или антропоген, начавшийся более
1 млн. лет назад, богат событиями. В это время образовались
многие виды существующих высших растений и животных,
формировались современные флора и фауна. В начале антро-
погена даже на севере было тепло, растительный и животный
мир отличался от современного. В Евразии, например, были
широко распространены гигантские древние слоны, крупные,
высотой до 2-х метров и с громадным рогом на лбу носороги —
эласмотерии, дикие однопалые лошади, верблюды, антилопы,
олени, различные грызуны, в том числе похожие на-гигантских
бобров, трогонтерии, страусы. Несколько позже появились
пещерные гиены и медведи, крупные длиннорогие бизоны,
большерогие олени, трогонтериевые слоны, олени и мало от-
личавшиеся от современных волки, лисицы, зайцы, кунипы и др.
Во второй половине антропогена в связи с изменением релье-
фа, морских течений, образованием холодных морей имело
место резкое похолодание, приведшее к обилию снегового по-
крова и возникновению оледенений больших пространств Ев-
ропы, Азии, Северной Америки. Одни млекопитающие не при-
способились к новым условиям и вымерли, другие изменились.
Наиболее распространенными животными были мамонты, во-
лосатые носороги, овцебыки и живущие теперь на крайнем
севере песпы, лемминги, северные олени и др. К этому времени
вымерли мамонты, волосатые носороги, гигантские олени,
208
Древнейшим безр<м ми носорог — хилотерии. Реконструкция К. К. Флерова
14-818
209
возник современный животный и растительный мир. Таким об-
разам, лишь во второй половине антропогена возникли отчет-
ливые умеренная и арктическая ландшафтные зоны, их флора
и фауна.
Антропоген — период развития человека, создателя новей-
ших форм органического мира — культурных растений и до-
машних животных.
Итак, развитие жизни на Земле от начала архея до современ-
ности можно проиллюстрировать следующей таблицей.
Абсолют-
Эра ~ ный Система Главнейшие группы организмов возраст
(млн. лет)
Антропоге- Человек, млекопитающие, мно- 1—0
новая гие группы морских и пресновод- ных моллюсков, кораллы, морские ежи и лилии, губкЕ, форамини- феры
Кайнозой- Неогеновая Разнообразные млекопитаю- 1—25
ская щие, в том числе трехпалые ло- шади, человекообразные обезьяны
Палеогено- Млекопитающие (появление 70—25
вая низших обезьян), пресмыкающие- ся; из беспозвоночных — пелеци- поды, гастроподы, нуммулиты, орбитоиды, из водорослей — диатомовые
210
Эра Система Главнейшие группы организмов Абсолют- ный возраст (млн. лет)
Меловая Млекопитающие, массовое раз- витие пресмыкающихся, костис- тых рыб; птицы; из беспозвоноч- ных — устрицы, белемниты, ко- раллы, губки, мелообразующие глобигерины, роталии и орбито- лины. Покрытосемянные растения 140—70
Мезозой- ская Юрская Массовое развитие пресмыкаю- щихся и амфибий (двоякодыша- щие), млекопитающие (мало), кос- тистые рыбы (мало), рифообра- зующие кораллы, аммониты; уст- рицы; насекомые 185—140
Триасовая Появление первых млекопитаю- щих, костистых рыб, много прес- мыкающихся — наземных, вод- ных, летающих; из беспозвоноч- ных — морские лилии, цератиты, аммониты, белемниты, беллеро- фоны 225—185
Палеозой- ская Пермская Амфибии, примитивные прес- мыкающиеся; из беспозвоноч- ных — брахиоподы, гониатиты, аммониты, пелециподы, мшанки, наземные растения — папоротни- ки, голосеменные в виде первич- ных хвойных, гингковые, кордаи- ты 270—225
14*
211
3f>a Система Главнейшие группы организмов Абсолют- ный возраст (млн. лет)
Палеозойская Карбоновая Из позвоночных — акулоподоб- ные рыбы; из беспозвоночных — брахиоподы, гониатиты; наутило- идеи; из растений — семенные папоротники и кордаиты, плауно- вые, каламиты, клинолистники. Много амфибий — стегоцефаллы и др. Первые пресмыкающиеся 320—270
Девонская Появление первых амфибий. Кистеперые и двоякодышащие ры- бы, панцирные рыбы. Из беспоз- воночных — расцвет четырехлу- чевых кораллов — кальцеол, спи- рифериды, пентамериды, гониати- ты, трилобиты, морские лилии. Из растений — псилофитовая флора, исчезающая к концу пе- риода, развитие плауновых, чле- нистостебельчатых, начало голо- семенных (первые наземные рас- тения) 400—320
Силурийская Позвоночные — хрящевые ры- бы, примитивные рыбообразные бесчелюстные. Из беспозвоноч- ных — многообразные плеченогие, моллюски, членистоногие (рако- образные, трилобиты), граптоли- ты, четырехлучевые, кораллы, по- явление морских лилий. Из расте- ний — значительное развитие во- дорослей—синезеленых и красных 420—400
212
Эра Система Главнейшие группы организмов Абсолют- ный возраст (млн. лет)
Ордовикская Панцирные рыбы, из ракообраз- ных — остракоды, листоногие ра- ки; трилобиты, граптолиты, че- тырехлучевые и трубчатые корал- лы, плеченогие, ранние предста- вители мшанок, наутилоидеи. Массовое развитие водорослей 480—420
Кембрийская Трилобиты и ракообразные. По- явление граптолитов, губок, стро- матопороидей, брахиопод, прими- тивных наутилоидей. В раннем и среднем кембрии — массовое раз- витие археоциат. Обилие красных и синезеленых водорослей 570—480
Протеро- зойская Синийская Массовое развитие одноклеточ- ных и многоклеточных синезеле- ных, реже красных и зеленых во- дорослей; в конпе периода — появ- ление ранних археоциат, губок, червей, медуз 1200—570 1
Енисейская Массовое развитие одноклеточ- ных синезеленых водорослей и многоклеточных, развитие желез- ных и других бактерий 1500—1200
Саянская Развитие одноклеточных водо- рослей, железных и других бакте- рий 1900—1500
213
Эра Система Главнейшие группы организмов Абсолют- ный возраст (млн. лет)
Архейская Появление микоплазмоподоб- ных организмов, бактерий, окис- ляющих минеральные вещества, почвенных бактерий, возникнове- ние водорослей. Химическое пре- образование первичных углерод- ных соединений. Преобладание хи- мических реакций в преобразова- нии минерального вещества зем- ной коры 2700—1900
Катархей- ская Химические процессы в земной коре, массовые проявления вул- канизма и магматизма, начало образования на земной поверх- ности водных бассейнов за счет кон. денсирующихся паров воды «вторичной» атмосферы. Возник- новение незначительных коли- честв кислорода при воздействии космических излучений на пары воды в атмосфере. Общий восста- новительный характер добио- сферы 3500—2700
Догеологический этап развития Земли—фор- мирование планеты за счет концентрации кос- мического материала. «Срыв» первичной атмо- сферы Более 3500
Примечание. Системы Саянская и Енисейская выделены автором по данным Сибири, что основывается на обнаружении в их слоях остатков водорос- лей, синийская система выделена в Китае, где она хорошо изучена.
214
ЭВОЛЮЦИЯ СРЕДЫ и жизнь
с
Изучение истории Земли и жизни позволяет видеть, что жизнь —
важнейший геологический фактор, что она издавна вносила и
вносит коренные изменения в состав геологических образова-
ний, в состав среды, вследствие чего изменялась среда и изме-
нялась сама жизнь.
Древнейшая бескислородная или бедная кислородом среда
узнавалась нами по ее геологический образованиям; она оста-
вила свой след в виде отложений в осадках водных бассейнов
закисных, недоокисленных соединений. Это способствовало
накоплению в природе органического вещества вообще и осо-
бенно вещества отмерших организмов, в частности, в виде би-
тумов жидких и газообразных, отчасти минерализованных, в
виде шунгитов докембрия Карелии. Восстановительный харак-
тер сред докембрия виден и по присутствию в его составе пири-
тоносных слоев, свидетельствующих о сероводородных фациях.
Эпоха возникновения жизни приходится на границу с археем,
когда водные бассейны уже образовались, когда углеродные
соединения еще естественно синтезировались химическим путем
при участии воды и внешних факторов и когда энергия геохими-
ческих реакций с выделением тепла могла стать основой оживле-
ния белковых микрокапель — коацерватов, став энергией био-
химических процессов.
Ареной зарождения жизни была восстановительная среда.
В древнейшей атмосфере углекислоты содержалось, по-види-
мому, в 100 000 раз больше, чем в наше время, причем около
40% ее запасов биологическим и в меньшей степени геохимиче-
ским путем было вовлечено в земную кору в виде твердых кар-
бонатов кальция и магния, карбонатных железных и других
руд. Древнейшие фотосинтезирующие организмы тоже извле-
кали огромные количества углекислоты из биосферы, высво-
бождая за счет воды большие запасы свободного кислорода.
Таким образом, органическая жизнь изменила первичные вос-
215
становительные среды на окислительные, в условиях которых,
естественно, углеродные соединения уже не могли создаваться,
а могли лишь разрушаться. Поэтому возникновение организмов
в дальнейшем уже не происходило.
Протерозой был временем проявления массовой жизнедея-
тельности водорослей и бактерий, в том числе железобактерий.
Водные среды были сильно карбонатными из-за обилйя угле-
кислоты в биосфере и мало солеными. Но к концу этой эры
расход углекислоты уже недостаточно восполнялся ее выбро-
сами при вулканических явлениях. Конечно, мы не располагаем
количественной характеристикой компонентов, которые вхо-
дили в протерозое в гидро- и атмосферу Земли. Но общий со-
став их, по-видимому, разгадан исследователями правильно,
что подтверждается как минеральными образованиями, так и
организмами, остатки которых мы находим в отложениях про-
терозоя. Высокая растворимость углекислоты, поступавшей в
биосферу при вулканических явлениях, способствовала быстро-
му захвату ее дождевыми и затем морскими водами. В морях
ее приток должен был стимулировать развитие фотосинтези-
рующих организмов, одноклеточного фитопланктона и донных
форм водорослей в мелководьях.
Изучение водорослей протерозоя, в основном синезеленых,
показывает на огромный размах их развития. Извлечение угле-
кислоты из среды, насыщенной бикарбонатами и нитратами
кальция и магния, вело к интенсивному выделению карбонатов
кальция непосредственно на тканях водорослей, поэтому уче-
ные нередко наблюдают в прозрачных шлифах пород прекрас-
ное сохрание их морфологии в ископаемом состоянии. Известь,
садившаяся на пленки и колонии микроскопических водорослей,
облекала их со всех сторон, что приводило за длительные про-
межутки времени к образованию известковых тел различной
формы и размеров (строматолитов), которые указывают часто,
какими были эти водоросли, и свидетельствуют об относитель-
ной устойчивости их жизнедеятельности. При этом, если водо-
рослевые строматолиты не подвергались вместе с вмещающей
породой существенным преобразованиям в процессе метамор-
216
физма, в них можно обнаружить остаточные микроструктуры,
которые обычно говорят о быстром ходе эволюции этой группы
организмов. Подсчитывая продолжительность жизни и дея-
тельности водорослей в постройках строматолитового типа,
мы можем говорить о длительности жизни их видов, что дает
цифры от сотен до нескольких тысяч, редко до двух-трех десят-
ков тысяч лет.
Все три первые эры существования Земли отличались от по-
следовавших за ними высоким содержанием в биосфере угле-
кислоты, запасы которой начали иссякать лишь к концу проте-
розоя в результате огромного размаха жизнедеятельности
«морских» придонных водорослей и, несомненно, фитопланк-
тона. Одновременно создавались запасы молекулярного кисло-
рода. Моря, освобождаясь от высокого содержания в водах
бикарбонатов, постепенно засолонялись. Таким образом, водная
среда, как и атмосфера, постепенно, но неуклонно изменялась,
влияя тем самым на развитие в биосфере живого вещества.
Из потребителей свободного кислорода около остатков
водорослей можно отметить, как видно по остаточным микро-
структурам, только железобактерии. Таким образом, наличие
железистых пленок в строматолитах все же скорее говорит о
бедности биосферы кислородом. По-видимому, он жадно по-
глощался тогда при химическом окислении минеральных ве-
ществ на выступах суши. Во всяком случае несомненно, что
появление запасов кислорода в биосфере еще не стимулировало
заметно развитие животных. Вплоть до начала палеозоя они
были представлены очень и очень бедно.
Вероятно, что к началу палеозоя сформировалась атмосфера
почти современного состава. Возможно, что это и так, но ос-
новной фактор, который при этом проявил себя как планетар-
ное явление, — это появление признаков повышенной соле-
ности некоторых бассейнов, приводившей к развитию лагун с
садкой морских солей, и снижение магнезиальности вод. В про-
терозое таких явлений не отмечено. Начало палеозоя ознаме-
новалось значительным развитием красных водорослей, так
называемой эпифитоновой флоры, и именно около мелких
217
Кайнозой
№SM&—
Палеозой
Синий
Ранний
протерозой
Археи
Катархей
Протопла-
нетный
этап раз-
вития
Земли
О млн лет
70
220
570
1200
1900
2700
3500
7000
Эволюция среды и жизни на Земле
218
«зарослей» таких водорослей мы находим и остатки археопиат,
трилобитов и других беспозвоночных животных кембрия. Па-
леоэкологические наблюдения позволяют думать, что стиму-
лом для развития многих ранних групп животных были условия
питания.
Вещество синезеленых водорослей (пектины) отличается
значительной химической стойкостью и вообще мало пригод-
но в качестве пищевого материала для каких-либо животных или
бактерий. По-видимому, развившийся фито-зоопланктон, состав
которого мы пока раскрыли лишь отчасти, послужил пищевым
субстратом, который определил возникновение в пресных и
морских водах синийского периода жизненного никла в виде
пепи организмов, биологически связанных между собой, что
обусловило появление животных более сложной организации.
Если для зоопланктона фитопланктон является базой, то для
более высокоорганизованных животных нужны были вещества,
вырабатываемые тем и другим, в большей степени — вторым.
Поэтому необратимое общее изменение вещественного состава
сред биосферы во все геологические времена с момента возник-
новения жизни влияло на микрожизнь, на бактерии, на микро-
водоросли, на грибы и другие организмы, приспособительные
свойства которых в общем вели ко все большему охвату разно-
образных сред. На базе этого микромира как пищевого субстра-
та в водных бассейнах развивались разнообразные организмы
в виде цепей, в которых представители одного звена поедались
другим, более сильным, хотя и не всегда более крупным.
Длительное отсутствие в протерозое животных в заметных
количествах представляло собой загадку. Но эта загадка как
будто разрешима.
Между вещественным составом фито- и зоопланктона име-
ется, конечно, большая разница. Она, несомненно, была и в то
далекое время, когда ход эволюции жизни только-только при-
вел к развитию зоопланктона — той живой «мелочи», имев-
шей огромное значение в жизненном цикле, без которой живот-
ные более высокой организации еше не могли существовать.
Потребовалось, по-видимому, длительное время для развития
219
разнообразного зоопланктона — простейших животных, остат-
ки которых мы находим теперь. Так, в слоях позднего синия
местами в составе отложений морских карбонатных пород при-
сутствуют мощные слои, сложенные мельчайшими округлыми
известковыми тонкостенными образованиями, внутри которых
мы находим или только вторичный кальцит, или кальцит и
битумоподобное остаточное органическое вещество. Их нельзя
принять за минеральные образования и трудно еще принять за
остатки организмов, хотя ничем иным они не могли быть.
Особи, составлявшие жизнь докембрия, были очень мелки.
Они имели микроскопические размеры, и их остатки нам удается
различать по большим скоплениям, закрепленным в ископае-
мом состоянии процессами минерализации, окаменением. Стро-
матолиты, часто огромные известковые тонкослоистые тела,
которые многими исследователями принимались, да и теперь
принимаются за остатки крупных живых существ не то расти-
тельного, не то животного происхождения, в действительности
оказались именно скоплениями огромных количеств окамене-
лых микротел — клеток и тел микроскопических водорослей,
развивавшихся тончайшими пленками жизни на поверхностях
субстратов слой за слоем, год за годом, тысячелетиями. Между
тем микроскопическое изучение строматолитов показало ог-
ромное разнообразие их строения, и, что очень и очень важно,
быструю смену одних породообразующих видов водорослей
другими. Эволюция и в протерозое шла быстрыми темпами,
с сильным проявлением естественного отбора в условиях не-
прерывно менявшей свои свойства среды.
Далек был путь от первичных комочков жизни — археобион-
тов до закрепившихся в водных средах биосферы видов бакте-
рий и бактериеподобных, до первых одноклеточных представи-
телей фитопланктона! Не менее длинным был путь развития
из тех же первичных комочков белковых веществ первых пред-
ставителей микроскопического зооплактона; на это по существу
ушло в истории жизни на Земле более двух миллиардов лет.
Появление в позднем докембрии червей, хиолитов и других
ранних представителей более высокоорганизованных существ
220
заставляет считать, что зоопланктон только к этому времени
развился достаточно и стал разнообразным и в морфологиче-
ском и в биохимическом отношениях, чему способствовала его
пищевая база — микроскопический фитопланктон и фито-бентос
водных бассейнов протерозоя. Так удлинялась биологическая
цепь жизни, удлинялись циклы жизни в древнейших средах,
дававшие в отдельные моменты геологического времени более
крупных и анатомически и биохимически сложных представи-
телей жизни, которые лишь в ничтожной доле попадают в виде
окаменелых остатков к палеонтологу.
Ничтожно малая частица «смеси» исходных белковых веществ
превратилась в бесконечное многообразие таких же малых час-
тичек жизни с колоссальным многообразием их свойств, мор-
фологии и жизнестойкости. Несомненно, огромные запасы
исходных сложных углеродных соединений осваивались жизнью
и были вобраны ею в круговорот полностью. Они стали живым
веществом Земли. Поэтому понятно, что в более поздние вре-
мена новые формы жизни уже не возникали.
Развитие углеродных соединений химическим путем в сто-
рону усложнения до стадии белковых должно было иметь свои
скорости и занять большой этап геологического времени.
Развитие микроскопических форм разного строения могло про-
изойти только в результате эволюции белковых веществ и физио-
логических и морфологических изменений организмов. При
этом в бесконечной смене особей шла не только смена поколе-
ний, но и формирование видов, родов, семейств, соответственно
их приспосабливаемое™ к условиям среды и к сообществу друг
с другом. Выявлен медленный темп повышения размеров тел
организмов, наиболее совершенных для соответствующих эта-
пов геологического времени.
Остаточное органическое вещество, часто захороненное в
горных породах вместе со скелетными образованиями форм
древней жизни, возможно, содержит осколки молекул первич-
ного его состава. Можно выразить надежду, что развитие био-
химической науки поможет нам распознавать исчезнувшие в
ходе эволюции жизни на Земле формы по вещественному их
221
составу, по комплексам микроэлементов, содержащихся в ис-
копаемых остатках. Важнейшую роль при этом играет закон
единства организма с условиями его существования, а также
фактор количественных соотношений между компонентами
среды. Поэтому по организму мы уже смело говорим теперь о
свойствах среды, а по признакам «ископаемой» древней среды —
^ возможных проявлениях жизни в ней.
По существу все организмы сложны. Их жизнепроявления
состоят в образовании вещества, построенного из «тяжелых»
сложных молекул с огромным молекулярным весом по срав-
нению с веществами косной мертвой природы. Ничтожное из-
менение в свойствах среды влияет на организм, вызывая у поко-
ления за поколением перестройку каких-то молекул, каких-то
органов или тканей, исполняющих их функции, и это приводит
к изменению затем и внешних морфологических черт. Когда
мы говорим о «направленности» таких изменений, то мы, ко-
нечно, имеем в виду, что естественный процесс, пока к нему не
прикасалась рука человека, шел и идет стихийно, с одновремен-
ным существованием и низших и высших, а главное в комплек-
сах, во взаимосвязи со средой и между собой, тогда как мы,
в наших интересах, прослеживаем, например, иногда только
развитие высших.
В основе существования жизни лежит зародышевая клетка
и индивидуальное развитие особей в их поколениях. Простей-
шие формы жизни и во взрослом состоянии особей неотличимы
от зародышевой клетки. Сложные многоклеточные организмы
проходят в своем индивидуальном развитии часто много про-
межуточных по уровню жизни этапов, повторяя при этом в
ускоренном темпе морфологическое развитие данной ветви
жизни, пока не появятся признаки вида. Это так называемый
закон рекапитуляции, закон повторения признаков предков в
развитии каждой особи, много давший зоологам и ботаникам
и ставший важнейшим теоретическим положением при изуче-
нии палеонтологами ископаемых организмов. Этот закон
позволяет воссоздавать ход эволюции жизни и понять, в конеч-
ном счете, как отдельные этапы ее развития в прошлом, так и
222
возникновение того мира живых существ, среди которого мы
живем. Поэтому он обычно и именуется основным биогенетиче-
ским законом.
Материалы по ископаемым организмам от начала кембрия
давно стали основой выделения этапов геологического времени;
для действительной меры времени и длительности этих этапов
абсолютная геохронология дала цифровую характеристику и
продолжает давать к ней поправки и дополнения.
Глядя на схему развития жизни на Земле, можно прийти к
мысли об усилении геологических процессов на Земле в ходе вре-
мени, о постепенном ускорении ее развития вплоть до современ-
ности: для четвертичного периода (антропогена) эта длитель-
ность определена около 1 млн. лет. Но это ложное впечатление.
В действительности земная кора в ходе геологического времени
постепенно как бы успокаивается, процессы складкообразова-
ния, фазы и площади вулканизма сокращаются. Прежние склад-
чатые области земной коры «захватываются» режимами плат-
форменного типа, со спокойными, только вертикальными дви-
жениями и т. д.
Сокращение этапов геологического времени обусловлено
способом выделения их при помощи палеонтологических ма-
териалов. Оно отражает действительное убыстрение хода эво-
люции живого населения Земли. Причина этого убыстрения
достаточно ясна. Поскольку в эволюции жизни сказывается
эволюция многих факторов — среды в геологическом ее пони-
мании, условий жизни организмов, пищевого субстрата, биохи-
мических процессов, живого вещества организмов и их органов,
то суммарное действие этих факторов приводило и приводит к
ускорению изменений формы организмов, их размеров, способ-
ности к миграции и т. д.
Органическая жизнь на Земле активно вмещалась в ход гео-
логических процессов. Она проявила себя как фактор изменения
самой среды, образовав атмосферу за счет элементов, до того
находившихся в связанном состоянии. Она стала мощным гео-
логическим фактором, поскольку активно участвовала и участ-
вует в породообразовании, в отложении вещества многих оса-
223
дочных горных пород и руд, нефти и газов, поскольку она влр<
ла на изменение глубины бассейнов и т. д. В общем, раз появй!*
шись в биосфере, бактериальный мир менялся морфоло! ическ|:
очень мало, совершенствуясь в основном биохимически и фи-
зиологически. Только с появлением высших организмов расти-
тельного и животного мира в массовом развитии он дал и даеГ
множество новых форм сапрофитного и паразитического рядов,
в том числе болезнетворных.
Мир растений, существующий с архея, был длительно пред-
ставлен только водными и отчасти почвенными формами.
Размах водорослевой жизни в протерозое был неповторимым
по своим проявлениям и по породообразующей роли этой груп-
пы организмов благодаря огромным запасам углекислоты.
Эволюция вещества спор, приобретение ими относительно проч-
ной оболочки позволило растениям выйти на сушу, где в разно-
образных условиях жизни их эволюция ускорялась обилием сол-
нечной энергии, развитием почвенных процессов и т. д.
К началу палеозоя живое зеленое вещество Земли удалило
из палеоатмосферы все ее запасы углекислоты, которая частич-
но сохранилась в растворенном состоянии лишь в глубинах
вод гидросферы. Для наземных растений палеозоя и более позд-
них эпох это обстоятельство временами ставило предел их ко-
личественному развитию. Поэтому наземные флоры в палеозое
и позднее оказывались подчиненными некоторому ритму, от-
части установленному геологами по эпохам углеобразования
на Земле. Сущность этого ритма, по-видимому, состоит в тесной
связи с выбросами углекислоты в атмосферу в эпохи вулканизма
и массовым развитием живого вещества фотосинтезирующих
организмов в соответствующий этап геологического времени.
Мир животных, возникший в тот же отдаленный момент,
как и другие типы живого вещества, долго «прозябал», не играя
геологической роли. Но эволюция вещества фитопланктона,
микроскопического зоопланктона и накопление в биосфере за-
пасов кислорода создали мощный стимул для его массового
развития к концу синия и началу кембрия. С этого момента на
Земле развилось много типов животных — от простейших
224
Эволюция позвоночных животных (по Л. Ill. Давиташвили)
(инфузорий и др.) к фораминиферам, от червей к примитивным
хордовым, к рыбам, от рыб к амфибиям, от амфибий к репти-
лиям, от рептилий к плацентарным млекопитающим. Эта выс-
шая по строению тел группа организмов на ранней стадий раз-
вития была представлена животными весьма небольшого раз-
мера, в основном насекомоядными. Вероятно, бурное развитие
насекомых с конца палеозойского времени во многом стимули-
ровало развитие этих животных с последующей их специализа-
цией как растениеядных, червеядных и др. Выделялись хищные
формы, которые приобрели соответственно оформленные зуб-
ные аппараты, форму тела, когти. Другие из растениеядных
приспосабливались к быстрому бегу, в связи с чем пятипалые
конечности древних форм у них становились одно-двухпалыми,
как у лошадиных и парнокопытных. Многие группы млекопитаю-
щих ушли снова в воду, став морскими организмами, как киты,
белухи, тюлени, сохранив при этом способность дышать с по-
мощью легких. Многие группы приспособились к лазанию по
деревьям, например обезьяны, и т. д.
Поэтому первая половина третичного времени была ознаме-
нована широкой специализацией млекопитающих, которые
освоили при этом все природные обстановки — сушу, воды и
воздух и пришли на смену пресмыкающимся, в большинстве
исчезнувшим на грани мелового и третичного периодов, 70 млн.
лет назад. Весь этот богатый формами мир млекопитающих
быстро достиг массового развития, чему, вероятно, способ-
ствовала благоприятная для него эволюция пиши — развитие
цветковых растений, плоды, семена, листья, кора и корни ко-
торых содержали многие новые в природе вещества, полезные
и важные для животных.
Ранние киты были небольшими животными, имевшими круп-
ные зубы. Позднее естественный отбор привел к питанию китов
мелкими рачками с помощью «китового уса». От древних на-
земных хищников развились роды кошек, собак, куниц, медве-
дей и барсуков. Наибольшего разнообразия в третичное время
на суше достигли растениеядные формы наземных и водных
животных.
226
[инотерии — древние хоботные. Реконструкция К. К. Флерова
15*
227
Широко были представлены хоботные животные, предки сов-
ременных слонов — индийского и африканского. Ископаемых
хоботных известно около 400 видов. Ранние их формы были
не крупнее лошади. У них развились передние зубы — резцы,
превратившиеся в бивни различного строения. Развился хобот —
с помощью которого животные могли подносить пищу ко рту,
пить воду, срывать плоды с дерева и т. д. Меритерии, мастодон-
ты, динотерии были распространены широко, как и целый ряд
других групп млекопитающих. Несмотря на их бивни — резцы,
которые, может быть, в большей степени служили копательны-
ми орудиями, чем средством защиты или нападения, эти живот-
ные часто становились жертвами хищников.
При специализации мелких насекомоядных животных в на-
чале мезозойского времени обособилась группа таких, у которых
не оказалось острых зубов и когтей, не развились копыта на
конечностях, не возникли крылья. Представители этой группы,
обитая в лесных условиях, находили там и приют и пищу в виде
плодов, ягод и насекомых. Они ходили по земле, но научились
лазить по деревьям. Из этой группы вышли в ходе времени
обезьяны, полуобезьяны и человек. Это так называемые при-
маты, т. е. «первенствующие», поскольку они составляют груп-
пу самых высокоорганизованных существ на Земле.. Общей их
чертой явилось значительное увеличение объема черепной ко-
робки по отношению к величине всего тела, пятипалое строение
конечностей. Особенно развилась у них хватательная способ-
ность передних конечностей. В ряду этих животных были полу-
обезьяны, лемуры и другие формы. Ответвились обезьяны,
перешедшие целиком к лазающему способу существования в
лесах. Появились бесхвостые человекообразные, среди которых
учеными выделены, по данным раскопок, остатки так называе-
мых дриопитеков (древесные обезьяны); среди них возникали
иногда гиганты, достигавшие 3-х метров в высоту (гигантопи-
теки). Остатки человекообразных обезьян найдены и на терри-
тории СССР, близ селения Удабно в Грузии. Наибольшее коли-
чество материала по древним предкам человека обнаружено
в Африке. Это были австралопитеки (южные обезьяны), сушест-
228
Меловой
Юрский
Пермский
Геологические,
периоды
Четвертичный'
Неогеновый
Палеогеновый
Триасовый
Карбоновый
Девонский
Силурийский
Ордовикский
Кембрийский
Эволюция человекообразных и человека
ва, которые ходили на двух ногах вертикально или слегка на-
клоняясь вперед и, несомненно, уже имели возможность срав-
нительно далеко уходить от лесной зоны. Их череп уже походил
на человеческий, мозг весил около 650—700 г. Челюсти их
менее выступали вперед, чем у обезьян. Зверообразный вид их
предков у них сменился чертами, приближающимися к облику
человека. Поэтому австралопитеки и гигантопитеки признаются
за прямых предков человека.
Особенно интересные данные о прямых предках человека
недавно были открыты в пределах Восточной Африки. В резуль-
тате многолетних работ английского археолога Луиса С. Б. Ли-
ки в ущелье Олдовай, к северу от оз. Паси, в северной части
Танганьики, были открыты остатки древнего человека — зинд-
жантропа, который уже умел изготавливать каменные орудия,
но еще не знал огня.
Калий-api оновым методом абсолютного летоисчисления
была получена цифра от 1 570 000 до 1 890 000 лет. Была При-
нята средняя цифра в 1 750 000 лет, что не означает, конечно,
даты появления этого человека (по последним сообщениям,
возраст олдовайских находок Л. С. Б. Лики сейчас пересматри-
вается в сторону увеличения). Судя по строению зубов, очень
крупных с гладкой жевательной поверхностью, зинджантроп
был еще вегетарианцем. Он умел охотиться на зебру, антилопу,
крокодила и даже на крупнейшего хоботного динотерия. Его
современниками были саблезубый тигр и ряд других, позднее
вымерших, животных. Зинджантроп питался растительной пи-
щей, но мог ловить птиц, черепах, грызунов, змей, варанов и
крокодилов.
Известны остатки китайского человека — синантропа, еше
более близкого к современному человеку. Синантроп, живший
в период времени 700—600 тыс. лет до современности, уке умел
поддерживать огонь.
Его каменные орудия были уже лучше и, по-видимому,
он одевался в шкуры и занимался охотой. Остатки синантропа
обнаружены в пещере Чжоукоудянь, недалеко от Пекина, где
установлены следы его длительного обитания.
230
Неандертальский человек. Реконструкция Р. Моора
В центральной Европе в го время обитал неандертальский
человек, который владел огнем и примитивными каменными
орудиями.
Человек по своему происхождению связан с миром животных,
из которого уже миллионы лет назад выделились и обособились
его предки, начав путь самостоятельного развития. Блаюдаря
благоприятному для развития анатомическому строению, тру-
ду, приспособляемости и развившейся способности мыслить
человек постепенно овладевал окружающей природой и одно-
временно изменялся сам. Поэтому, находясь в отдаленном
родстве с остальными позвоночными, человек представляет
собой в непрерывной цепи развития самое совершенное звено.
Эго помогло ему расселиться по всем континентам нашей пла-
неты.
233
Таким образом, благодаря физико-химическим процессам^
химико-биологическому возникновению живого вещества мерт-
вая, безжизненная планета Земля за миллиарды лет эволюции
организмов естественным ходом развития стала местообита-
нием сложнейшего мира бактерий, растений, животных и людей.
Осваивая многообразные жизненные ресурсы природы, человек
создает для себя наиболее благоприятную среду (жилье, одежда,
пищевая база), изучает ее и в наше время проявляет интерес уже
к другим планетам, к космосу.
ЗАКЛЮЧЕНИЕ
Несмотря на гипотетичность, предположительность наших пред-
ставлений о непосредственном механизме возникновения жиз-
ни из неживых углеродных соединений с достаточной ясностью
установлено, как возникла жизнь на Земле и как она затем раз-
вивалась с проявлением биохимической эволюции ее живого
вещества в условиях непрерывного изменения среды. Всеми
своими корнями органическая жизнь на Земле связана с косным,
мертвым, минеральным веществом. Эта связь составляет осно-
ву самого процесса жизни, энергетическую вещественную базу
жизни, от самых ее примитивных форм до самых высокооргани-
зованных.
Химические элементы и их соединения, вовлеченные в про-
цессы жизнедеятельности организмов, претерпели отбор с вы-
делением тех изотопов и их соотношений, которые в наиболь-
шей степени обусловливали ход биохимических и биогеохими-
ческих реакций со средой и внутри организмов. Выясняется
особое значение для проявлений жизни изотопа углерода — 12,
изотопа кислорода — 16, определенных соотношений изотопов
серы (S3i/S34) и т. д. Это раскрывает для нас возможности и
далее прослеживать проявления жизни в глубинах геологиче-
ских времен и их последующий ход методами геохимии. Наме-
тившееся плодотворное сотрудничество между геологией, гео-
химией, биогеохимией, физиологией, астрофизикой, экспери-
ментальной биохимией способно приносить и далее новые
успехи в разработке проблемы жизни и ее развития.
Очень важно развивать поиски и изучение конкретных ос-
татков всех жизнепроявлений прошлого, а также их следов по
вещественному (элементарному и изотопному) составу, посколь-
ку к сохранению в «ископаемом» состоянии оказываются спо-
собными даже многие органические вещества. Несомненно,
особое значение имеют прямые поиски органических остатков
во всех толщах осадочных пород, где они до сих пор еше не об-
235
наружены. Действительно «немых» отложений с момента воз-
никновения жизни на Земле практически ныне уже не оказы-
вается.
Особо следует считать важным и увлекательным прослежи-
вание проявлений жизни организмов в древнейших осадочных
отложениях. Почти во всех горных районах нашей страны го-
рообразовательными процессами подняты на огромных пло-
щадях складчатые серии древнейших отложений самого различ-
ного состава. По своей мощности, измеряющейся многими де-
сятками километров, эти докембрийские отложения, в той или
иной степени обогащенные различными вулканогенными и
туфогенными образованиями, превосходят в несколько раз мощ-
ность всех более поздних отложений. Их понимание, расчленение
и сопоставление встречают еще большие трудности. Между
тем в составе серий древних пород всюду имеются минеральные
богатства, часто огромного практического значения, для поис-
ков и выявления которых нужны надежные методы и средства.
Поэтому очень важно, что мы имеем возможность распро-
странять палеонтологический метод на отложения докембрия
и выявлять в его слоях остатки разнообразных организмов,
в основном пока в виде бактерий и водорослей. Развитие таких
работ весьма перспективно.
Единство жизни и среды, единство Земли и жизни и великая
роль жизни в развитии биосферы доказаны, но существует еще
большое количество вопросов научного и практического зна-
чения, которые ждут своего разрешения с помощью плодотвор-
ного сотрудничества ученых различных профилей.
СПИСОК ЛИТЕРАТУРЫ
Я
Вернадский В. И. Избранные сочинения, М., Изд-во АН СССР
1, 1954; т. IV, 1959.
Вернадский В. И. Химическое строение биосферы Земли и ее окру-
жение. М., «Наука», 1971.
Виноградов А. П. Химический состав морских организмов. — «Тру-
ды Биогеохим. лабор. АН СССР», 1935—1936.
Виноградов А. П. Геохимическая история кислорода и фотосин-
тез. — «Изв. АН СССР, серия биол.», 1947, № 3.
Вологдин А. Г Геологическая деятельность микроорганизмов.—
«Йзв. АН СССР, серия геол.», 1947, № 2.
Вологдин А. Г Разгадка происхождения строматолитов. — «При-
рода», 1955, № 9.
Вологдин А. Г Древнейшие строители рифов. — «Природа», 1955,
№ 11.
Вологдин А. Г Палеонтология и поиски полезных ископаемых.
Изд-во «Знание», 1960.
Вологдин А. Г Свидетели миграции полюсов. — «Природа», 1961,
№ 5.
Вологдин А. Г Древнейшие водоросли СССР М., Изд-во АН СССР,
1962.
ВойткевичГ В. Происхождение и химическая эволюция Земли. М.,
«Наука», 1973.
Криштофович А. Н. Палеоботаника. Изд. 4-е, М. Гостопгехиздат,
1957.
Крылов И. А. На заре жизни. Органический мир докембрия. М.
«Наука», 1972.
Л и с Г Биохимия автотрофных бактерий. М. Изд-во иностр, лит-ры,
1958.
Ломоносов М. R О слоях Земных. М., Госгеолиздат, 1949.
Миллер Стэнли Л. О синтезе органических веществ на первичной
Земле. — «Природа», 1958, № 5.
Некрасов А. Д. Чарлз Дарвин. М., Изд-во АН СССР, 1957.
Опарин А. И. Возникновение жизни на Земле. М. Изд-во АН СССР,
1957.
Орлов Ю. А. О мире древних животных. М., Изд-во АН СССР, 1961.
Перельман А. И. Геохимия древних ландшафтов. — «Природа»,
1961, № 1.
Ранкама К. Изотопы в геологии. М., Изд-во иностр, лит-ры, 1956.
Рассел Г Н. Солнечная система и ее происхождение. М., Гостехиздат,
1944.
237
наружены. Действительно «немых» отложений с момента воз-
никновения жизни на Земле практически ныне уже не оказы-
вается.
Особо следует считать важным и увлекательным прослежи-
вание проявлений жизни организмов в древнейших осадочных
отложениях. Почти во всех горных районах нашей страны го-
рообразовательными процессами подняты на огромных пло-
щадях складчатые серии древнейших отложений самого различ-
ного состава. По своей мощности, измеряющейся многими де-
сятками километров, эти докембрийские отложения, в той или
иной степени обогащенные различными вулканогенными и
туфогенными образованиями, превосходят в несколько раз мощ-
ность всех более поздних отложений. Их понимание, расчленение
и сопоставление встречают еще большие трудности. Между
тем в составе серий древних пород всюду имеются минеральные
богатства, часто огромного практического значения, для поис-
ков и выявления которых нужны надежные методы и средства.
Поэтому очень важно, что мы имеем возможность распро-
странять палеонтологический метод на отложения докембрия
и выявлять в его слоях остатки разнообразных организмов,
в основном пока в виде бактерий и водорослей. Развитие таких
работ весьма перспективно.
Единство жизни и среды, единство Земли и жизни и великая
роль жизни в развитии биосферы доказаны, но существует еще
большое количество вопросов научного и практического зна-
чения, которые ждут своего разрешения с помощью плодотвор-
ного сотрудничества ученых различных профилей.
СПИСОК ЛИТЕРАТУРЫ
Вернадский В. И. Избранные сочинения, М., Изд-во АН СССР,
1, 1954; т. IV, 1959.
Вернадский В. И. Химическое строение биосферы Земли и ее окру-
жение. М., «Наука», 1971.
Виноградов А. П. Химический состав морских организмов. — «Тру-
ды Биогеохим. лабор. АН СССР», 1935—1936.
Виноградов А. П. Геохимическая история кислорода и фотосин-
тез.— «Изв. АН СССР, серия биол.», 1947, № 3.
Вологдин А. Г Геологическая деятельность микроорганизмов.—
«Йзв. АН СССР, серия геол.», 1947, № 2.
Вологдин А. Г Разгадка происхождения строматолитов. — «При-
рода», 1955, № 9.
Вологдин А. Г Древнейшие строители рифов. — «Природа», 1955,
№ 11.
Вологдин А. Г Палеонтология и поиски полезных ископаемых.
Изд-во «Знание», 1960.
Вологдин А. Г Свидетели миграции полюсов. — «Природа», 1961,
№ 5.
Вологдин А. Г Древнейшие водоросли СССР М., Изд-во АН СССР,
1962.
ВойткевичГ В. Происхождение и химическая эволюция Земли. М.
«Наука», 1973.
Криштофович А. Н. Палеоботаника. Изд. 4-е, М. Гостоптехиздат,
1957.
К р ы л о в И. А. На заре жизни. Органический мир докембрия. М.
«Наука», 1972.
Л и с Г Биохимия автотрофных бактерий. М., Изд-во иностр, лит-ры,
1958.
Ломоносов М. R О слоях Земных. М., Госгеолиздат, 1949.
Миллер Стэнли Л. О синтезе органических веществ на первичной
Земле. —«Природа», 1958, № 5.
Некрасов А. Д. Чарлз Дарвин. М., Изд-во АН СССР, 1957.
Опарин А. И. Возникновение жизни на Земле. М. Изд-во АН СССР,
1957.
Орлов Ю. А. О мире древних животных. М., Изд-во АН СССР, 1961.
Перельман А. И. Геохимия древних ландшафтов. — «Природа»,
1961, № 1.
Ранкама К. Изотопы в геологии. М., Изд-во иностр, лит-ры, 1956.
Рассел Г Н. Солнечная система и ее происхождение. М., Гостехиздат,
1944.
237
Рыжков В. Л. Ультрамикробы. — «Природа», 1961, № 2.
С а у к о в А. А. Эволюция геохимических условий в истории Земли.
«Природа», 1958, № 2.
Пешков М. А. Первые шаги жизни на Земле. — «Природа», 1970, № 1
Трофимов Б. А. Жизнь в глубинах веков. М., Госкультпросветиздг
1957.
Тугаринов А. И., ВойткевичГ В. Докембрийская геохроч*
логия материков. М., «Недра», 1970.
Фасенков В. Г Происхождение и развитие небесных тел по совр<
менным данным. М., Изд-во АН СССР, 1953.
Холодный Н. Г Железобактерии. М., Изд-во АН СССР, 1953.
Шмидт О. Ю. Роль твердых частиц в планетной космогонии. — «Прг
рода», 1956, № 11.
Шмидт О. Ю. Четыре лекции о теории происхождения Земли и плане!
М. Изд-во АН СССР, 1950.
Штрауб Ф. Б. О синтезе белковых тел. — «Природа», 1956, № 2.
Яковлев Н. Н. Организмы и среда. М., Изд-во АН СССР, 1956.