/
























































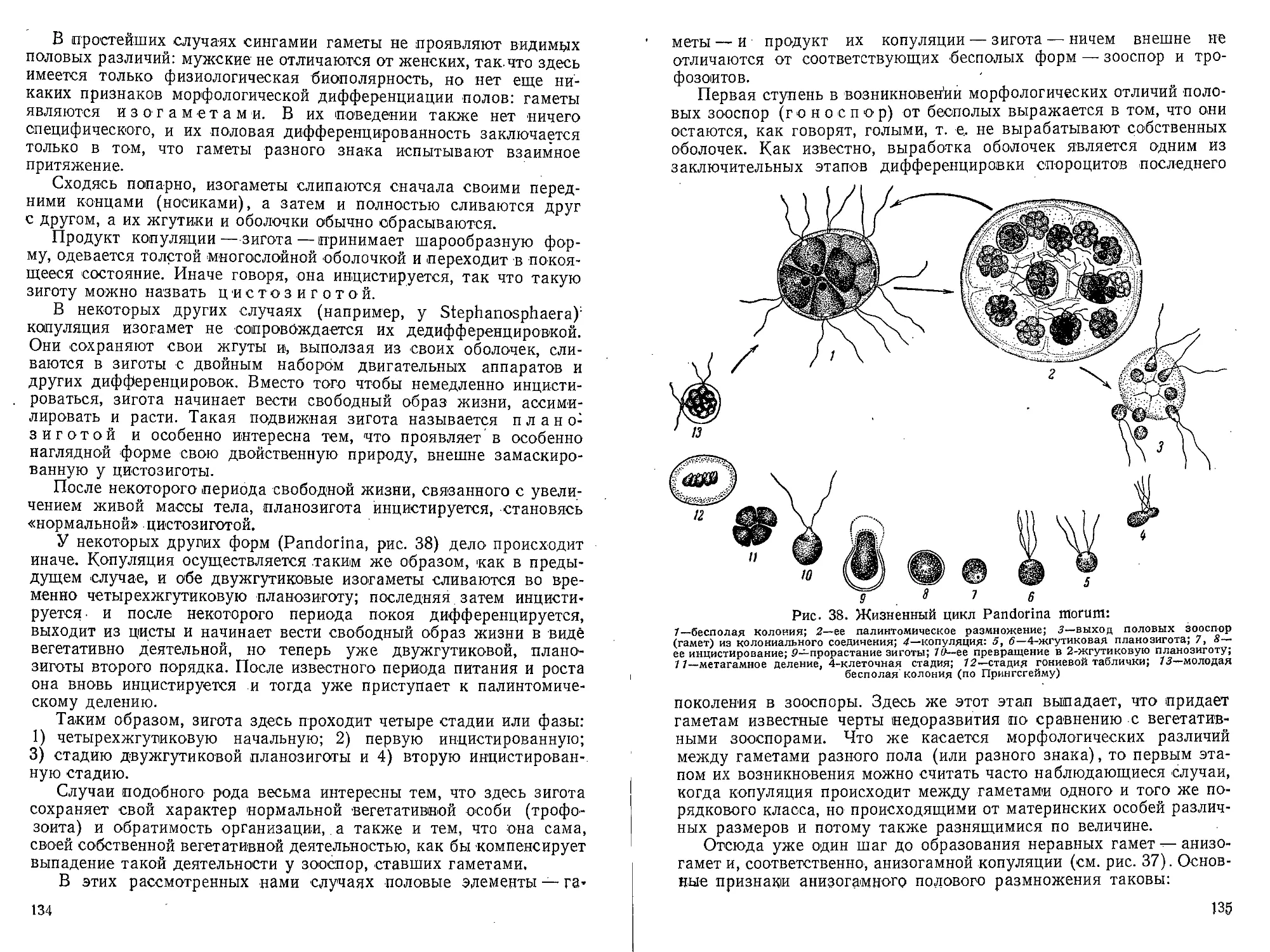
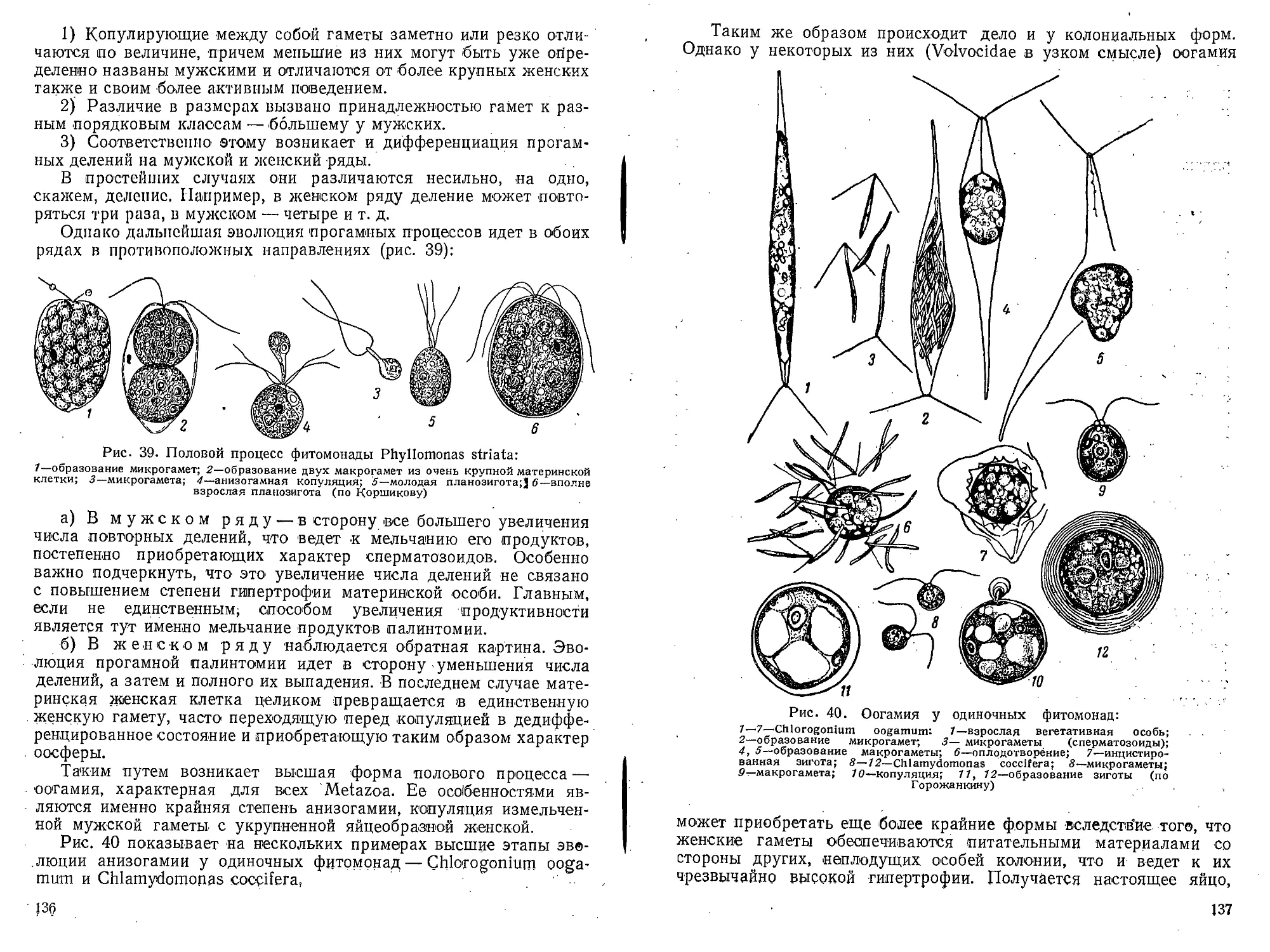





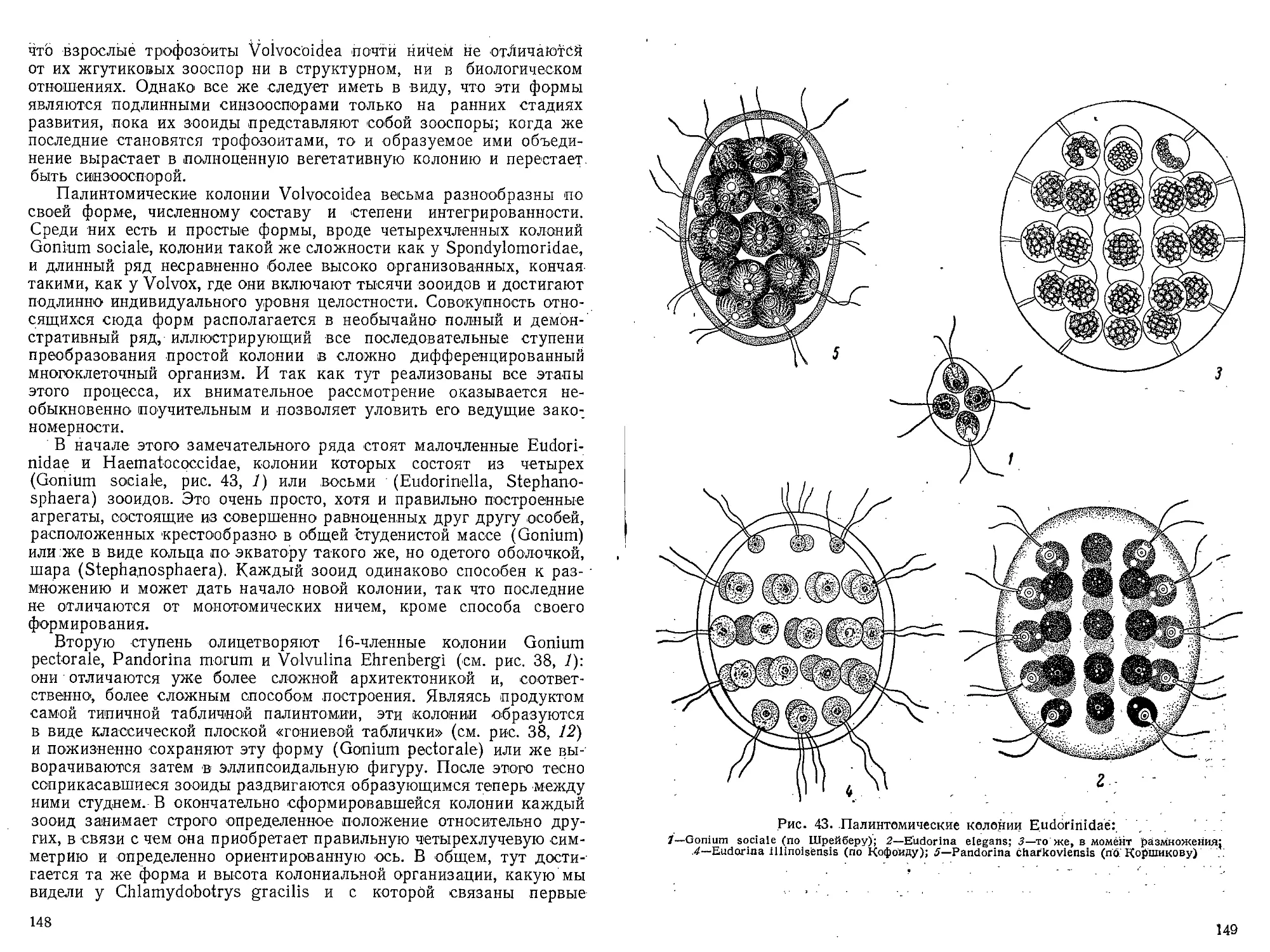
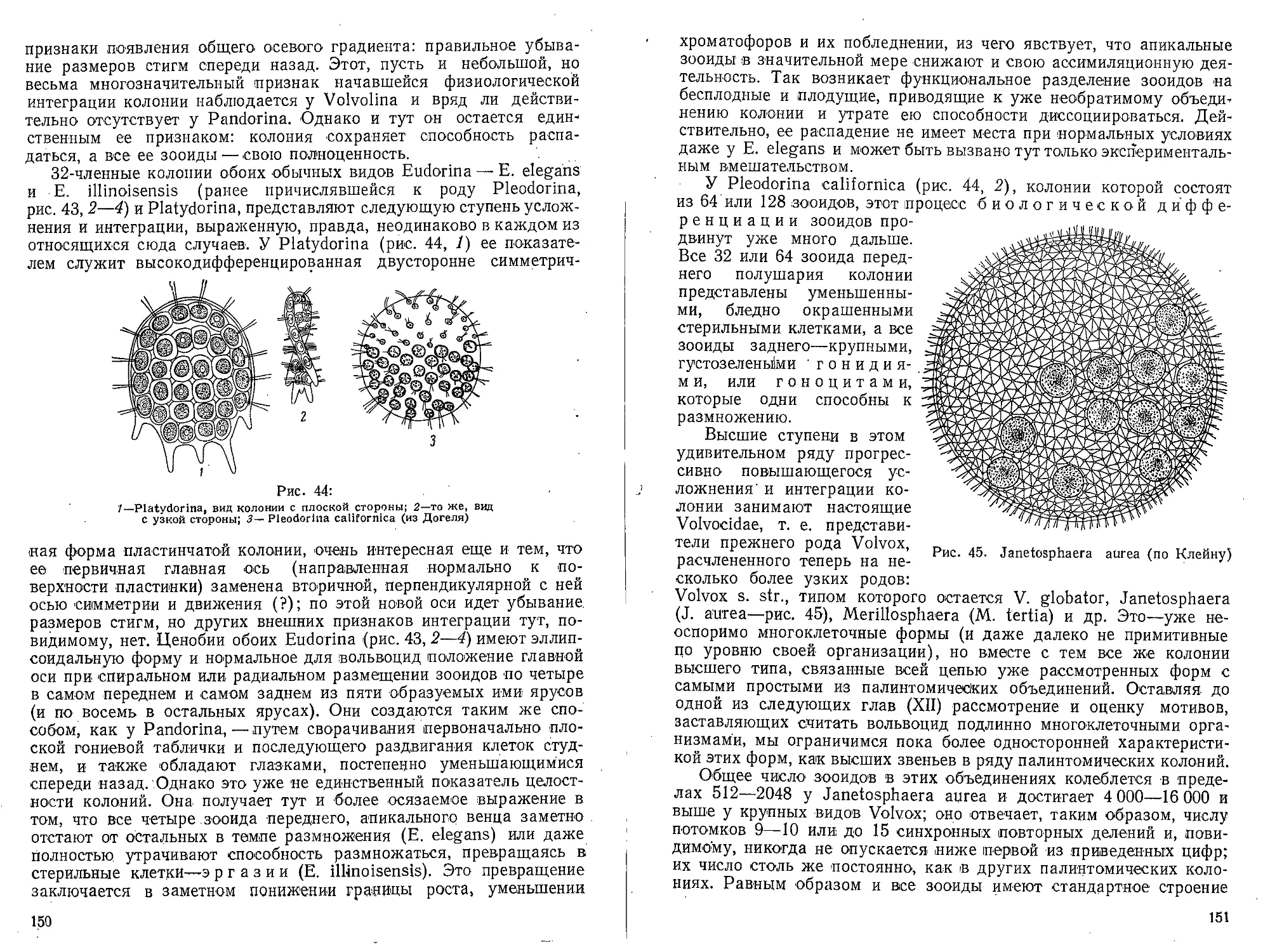























































































Author: Захваткин А.А.
Tags: биология зоология фауна эмбриология низшие беспозвоночные многоклеточные организмы
Year: 1949




Text
A.A.3AXBATKI/IH
СРАВНИТЕЛЬНАЯ
ЭМБРИОЛОГИЯ
НИЗШ ИХ
БЕСПОЗВО НОЧНЫХ
Проф. А. А. ЗАХВАТКИН
СРАВНИТЕЛЬНАЯ
ЭМБТРИОЛОГИЯ НИЗШИХ
БЕСПОЗВОНОЧНЬБХ
(Источники И пути формирования индивидуального
развития многоклеточных)
Допущено Министерством высшего обра-
зования СССР в качестве учебного пособия
для государственных „университетов
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО «СОВЕТСКАЯ НАУКА»
М о с к в а — 1 9 4 9
ПРЕДИСЛОВИЕ
Настоящая книга, представляющая собой переработку курса,
читаемого мною на Биологическом факультете Московского уни-
верситета, целиком посвящена изложению и разработке одной из
основных проблем эволюционной эмбриологии—проблемы проис-
хождения индивидуального развития Metaz-oa. _
В вопросе о том, как возник и офорлмился онтогенез многокле-
точных животных, каковы его источники и пути исторического
становления, сливаются воедино главнейшие из вопросов, стоящих
перед эволюционной эмбриологией беспозвоночных: вопросы проис-
хождения многоклет-очности Metazoa (или, что то же, их клеточного
строения) и свойственной им клеточной, тканевой и органоло-гич-е-
ской дифференцировки; вопросы генезиса половых клеток и .процес-
сов дробления, возникновения метаморфоза, личиночных форм И
характерных для Ме’са2оа особенностей всего их жизненного цикла.
Поэтому мы и считаем. эту проблему основной, краеугольной
проблемой сравнительной и общей эмбриологии беспозвоночных.
Развернутая ее постановка на современном-мичуринском——
этапе развития биологической науки представляется не только за-
конной, ьно и настоятельно необходимой. Действительно, поскольку
гмы «стоим сейчас перед задачей коренного пересмотра и переоценки
всех положенпий и концепций, нашей науки в свете всеобъемлющей
теории индивидуального развиггия организмов, созданнойИ. В. Ми-
чуринылм и Т- Д. Лысенко и составляющей основу советского твор-
ческого дарвинизмла,— разработка вопроса об источниках и путях
‘формирования самого онтогенетического процесса приобретает осо-
бое принципиальное зна-чение.
Между тем, эта важнейшая из общеэмбриологических проблем
доныне остается почти. не разработанной. Она, в сущности, и не
ставилась в наше время в полном своем объеме: обсуждаются толь-
ко отдельные ее аспекты, рассматриваемые раздельно, вне взаим-
ной своей связи, как вопросы, относящиеся к разным даже отрас-
лям биолопии-к эмбриологии, к сравнительной анатомии, гисто-
логии и к общей биологии. Поэтому и не приходится удивляться,
что все эти вопросы не получили еще удовлетворительного реше-
ния и что их теоретическая разработанность доныне остается на
уровне произвольных и мало обоснованных гипотез.
Попытка рассмотрения всего этого круга вопросов в их anoma-
ЦИОННОМ ЕДИНСТВЕ И СОСТЭВЛЯЕТ ОСНОВНОЕ СОДЕЕПКЭНИЕ И ближайшую
цель предлагаемой книги.
Коренное ее отличие от существующих сводок и руководств по
сравнительной эмбриологии беспозвоночных заключается прежде
всего в том, что в основу ее .п-о.ложено подробное рас-смотрение
организации, размножения и развития простейших, т. е. вопросов,
совершенно игнорируемых современной сравнительной эмбриолог
гией беспозвоночных, никогда не вовлекаемых теперь в орбиту ее
интересов, но‘ без которых она лишается своей самой существенной
основы. Второе главное отличие этой книги от других эмбриологи-
ческих руководств выражается в. том, что в ней рассматривается
развитие далеко не всех беспозвоночных, а- только самых низших,
наиболее примитивных из «них-низших стрекающих кишеч-но-
полостных (книдарийп), губок и немногих других, т. е. как раз тех
форм, развитие которых может дать ключ к пониманию интересую-
щих нас процессов.
Такое построение данной ›книги отнюдь не является произволь-
ным. Оно определяется целью, достижению которой посвя-
щена вся книга‚—приблизиться к пониманию источников и путей
происхождения онтогенеза многоклеточных, к историческому,
эволюционному освещению явлений, рассматриваемых нынешней
общей эмбриолопией метафиеически, без всяких даже попыток по-
нять их историческую обусловленн-ость. Вместе с тем, это кажу-
щееся нововведение не является в сущности таковым: оно опреде-
‘ляеттся тем, что направленность этой книги и, в значительной мере,
ЕЕ фактическое содержание подсказаны замечательным ТРУДОМ
'И_ И. Мечникова «Эмбри-отлогические этюды о медузах»—послед-
_ней‚ и самой блестящей из его эмбриологических работ.
"Таким образом, все мои усилия и свелись, в конечном счете,
к попытке внести свою лепту в важнейшее дело возрождения и
дальнейшей -разработки классического мечниковского направления
эволюционной эмбриологии.
Необходимость в восстановлении этого славного ‘направления
исключительно велика. Действительно, нельзя мириться дольше
с существующим в нашей эмбриологии положением, характеризую-
щимся, во-первых, полным ее отходом от эволюционной теории,
отсутствием как-ой-либо связи с советским творческим дарви-
низмом вообще и с биологией, развития в ‘частности; во-вторых,
забвением материалистических традиций классической русской эм-
бриологии, принесшей в свое время столько славы отечественной.
наужке;_и, наконец, непомерным и ничем ‚не оправданным развитием
методологически порочной механпики развития, фактически вытес-
нившей из эмбриологии все прогрессивные направления исследова-
_ния и, приведшей ее в состояние глубочайшего’ идейного, кризиса.
Если эта КНпИГа, являющаяся по существу прологом к еще не-
написанной никем эволюционной эмбриологии беспозвоночных ока-
жет хоть сколько-нибудь ощутимую роль в перестройке советской‘
4
эмбриологии на эволюционных, диалектико-материалистических
началах, ~_ моя Hem: ‘и мой долг будут казаться мне выполненными.
Конечно, я прекрасно отдаю ;себе отчет, что это—только первый
шаг на этом пути, что в_этой книге есть много существенных недо-
статков и что данное в ней только сравнительно морфологическое
освещение материала совершенно недостаточно для сколько-нибудь
полного достижения стоявших передо мною Целей,
При работе над курсом и этой выросшей из него книгой я ши-
роко пользовался критическими замечаниями и советами В. Н. Бек-
лемишева, В. А. Догеля‚ Л. А. Зенкевича, Л- Б. Левинсона,
Г. А. Шмидта и ряда друпих лиц, которым я рад выразить и здесь
свою самую глубокую признательность. Особенно много обязан я
Л. Левинсон_у‚ взявшему на себя нелегкий труд редактирования
этои книги, а также студентам Биологического факультета, слушав-
ши=м и критиковавщим мой курс и подсказавшим мне ряд ‚интерес-
ных мыслеи.
- А. Захваткин
Москва, Университет, август 1949.
ВВЕДЕНИЕ
ИСТОРИЯ И СОВРЕМЕННОЕ СОСТОЯНИЕ ВОПРОСА
О ПРОИСХОЖДЕНИИ ОНТОГЕНЕЗА И МНОГОКЛЕТОЧНОСТИ
METAZOA
Возможно полное выяснение пер-
вобытного состояния многоклеточных
необходимо, чтобы получить основу
для сравнительной морфологии.
И.А И. Мечников
эмбриологические этюды о медузах,
стр. 150 (1886),
1. Главные направления в разработке теории происхождения
многоклеточных и их онтогенеза
‘Вопрос о происхождении онтогенеза многоклеточных животных
не отделим от признания онтогенеза в той или иной мере адэкват-
ным отражением филогенетического прошлого вида, от признания
исторической обусловленности и значимости каждой стадии индич
ВпИДУаЛЬНОГО развития. С этой биогенетической точки зрения вопрос
о происхождении онтогенеза Metazoa совпадает с вопросом о проис-
хождении самих этих последних, их лмногоклеточности и других ха-
рактерных для них свойств организации. Поскольку основу инди-
видуального развития Metazoa составляет, вообще говоря, проце-сс
превращения одноклеточной исходной стадии онтогенеза в Целост-
ную многокл-еточную особь с той или иной дифференцировкой со-
ставляющего ее клеточного материала, вопрос о происхождении
этого онтогенетического процесса не отделим от вопроса о филоге-
нетическом становлении многоклеточности и дв-у- или трехслойной
организации Metazoa.
Намеченный в общих чертах Дарвином еще в 1844 г. и глубоко
разработанный им в ‹<Происхождении видов» би-огенетический закон
развивался далее в двух принципиально различных направлениях:
1) в сторону его упрощения и схематизации, что имело своим ре-
зультатом создание широко известной концепции Г еккеля‚ и
2) в» том, гораздо более широком, и плодотворном напра-влениИ‚
которое шло непосредственно от Дарвина, привело _к формулировке
7
„ад
т-еории филэмбриогенеза А. Н. Северцова и стало однойиз основ
советского творческого дарвинизма.
Не имея в виду подробнее касаться здесь проблемы соотношения
Индивидуального и филогенетического развития, достаточно полно
и всесторонне освещенной в нашей литературе, мы отмпетим только,
что первое из этих направлений рассматривало филогенез как «ме-
ханическую причину» онтогенеза и считало при этом субъектом
филогенетических изменений однли только взрослые, конечные стадии
развития. Таким образом, все следующие друг за другом стадии
онтогенеза Metazoa считались представляющими дефинитивные
состояния их гипотетических предков, сменившиеся в той же оче-
редности в течение филогенеза, а индивидуальное развитие в це-
лом—-своего рода архивом отживших взрослых состояний, на-
слоенных одно на другое в порядке их хронологической последо-
вательности.
По мнению Геккеля, прогрессивную роль в эволюции играют
только взрослые стадии развития. Новые признаки появляются
исключительно во взрослом со-стоянии животного, когда все его
органы. уже сформировались и функционируют. «С дальнейшим те-
чением эволюции эти приобретенные признаки сдвигаются на все
более ранние стадии, а во взрослом состоянии их место занимают
новые возникающие признаки, в свою очередь потом сдвигающиеся
на более ранние стадии, и т; д. Таким- путем обеспечивается повто-
рени-е в онтогенезе потомпков этапов эволюции взрослых предков
в их исторической последовательности» (Ежиков, 1940, стр. 24).
В ПРОТИВОПОЛОЖНОСТЬ ЭТОМУ, дарвиновское направление В раз- ч
витии биогенетической идеи основывалось на признании филогене-
тической изменяемости всех стадий онтогенеза, а соответственно,
и самого о-нтогенетического процесса, и считало его отражающим
в своем течении не только дефинитивные, .но и молодые стадии
предковых форм. «Интерес эмбриологии значительно повысится,
если мы будем вид-еть в зародыше «более или менее затемненный
добраз общего прародителя в-о взрослом или в личиноч-
_ном его состоян и и», -‹писал по этому поводу Дарвин
(«Происхождение видов», гл. XIV). C ЭТОЙ точки зрения филогенез
‘является не только «причиной» онтогенеза, -но и его продуктом, так
_что связь между ними оказывается обоюдной. Единство онтогенети-
ческого и филогенетического процессов вытражаетсяв том, что в лин-
дивидуальном развитии потомков находит свое отражение весь
„онтогенез предков, а отнюдь не только взрослые его стадии, что
первопричиной филогенетических изменений являются адаптивные
изменения хода онтогенеза, что эволюция онтогенеза составляет
основу всего филогенетического процесса.
‚ Обеим этим концепциям, базирующимся на признании историче-
ской значимости всех стадий индивидуального развития Metazoa;
противостоит господствующая теперь в эмбриологии механистиче-
ская теория развития. Отрицая возможность _видеть в ранних
‘эмбриональных стадиях Историческиесвидетельства прошлого, эта
теория рашсмтрилшет‘ Эмбриогенез как своегорода «онтогенети-
ческую ‘п-хнпку», обеспечивающую построение многоклеточного
ортннпнтмая ин яйца на основе простейших физических законов, на-
нрнмср. мсхнпьшеских правил деления целостных геометрических
фигур. ншшни Плато и т. д. Согласно этой концепции, ранние стади-и
чмтпппгсчнъни ничего не дают для понимания филогенеза Metazoa
н япрнвнэхшкдеггия их онтогенеза.
днолюциоггная трактовка онтогенеза послужила основой для
гпндинин значительного большинства эмбриологических теорий про-
нсхшндеъгиэг Metazoa, одновременно являющихся, благодаря этому,
Н 'l‘(!()[)l/ISIVMPI происхождения ИНДИВИДУНЛЬНОГО рЗЗВИТИЯ МНОГОКЛВ-
ччщнь1х. Как известно, все эти теории могут быть разделены на две
основные группы: теории колониального происхождения Metazoa,
выводящие многоклеточных от гипотетическлих колоний жгутико-
пых, и теории их н_еколониального происхождения, принимающие за
предков Metazoa многоядерных инфузориобразных простейших. Обе
ЭТИ ГруПШЫ теорий ПрОТОЗОЙНОГО ПРОИСХОЖДЕНИЯ МНОГОКЛЁТОЧНЫХ:
возникли почти одновременно и, развиваясь далее параллельно,
дошли в различных своих вариантах вплоть до настоящего момента.
Что же касается механистической интерпретации ранних стадий
развития, то она, естественно, исключает использование эмбриоло-
гических данных для построения теорий происхождения Metazoa.
Поэтому те из сторонников этого направления, которым пришлось
столкнуться с дан-ной проблемой, трактовали" ее исключительно
в -сравнительно анатомическом (отчасти и сравнительно физиоло-
гическом) аспекте, вне всякой связи с онтогенезохм. Рассматривая
последний как «новоприобретенную технику», они отказывались от
всяких попыток заглянуть в его и-сторию.
Борьбу этих противоположных тенденций можно проследить
в истории общей эмбриологии огг Дарвина до наших дней. Рас-
смъотрим, как она преломлялась в разработке интересующих нас
проблем, и каковы те ее результаты, с какими нам теп-ерь прихо-
дится считаться.
2. Первые гипотезы колониального происхождения Metazoa
Гипотеза Геккеля, сформулированная им еще в 1866 г. и ‚впослед-
ствии развернувшаяся в. общеизвестную «теорию гастреи», может
служить примером последовательного проведения палинэгенетиче-
ской точки зрения в оценке индивидуального развития, при-писы-
вантия каждой стадии последнего абсолютного рекапитуля-
ционного значения; В основу этой гипотезы положено представление
о том, что все Metazoa проходят в своем развитии пять последова-
тельных онтогенетических стадий, каждая из которых воспроизво-
дит, или рекапитулирует, определенну.ю стадию филогенеза их
общих предков. Первая из них —— стадия ц пи тул ы, представленная
одноклеточным, лишенным оболочки и часто амёбоидно подвижным
яйцом, олицетворяет собой прародительскую амёбу, цитею, от
9
которой берут свое ‘начало все аНИМаЛЬНО питающиеся организмы.
‘Вторая стадия онтогенеза, M o р ул а, представляющая собой плот-
`ное скопление равноценных голых клеток—бластомеров, рекапи-
тулирует древнейшую многоклеточную исходную форму всех жи-
вотных, называемую Г еккелем мореей, или $упагпоеЬ1ит——
простейшее колониальное соединение нескольких подвижных амёбо-
идных клеток. Следующая за морулой однослойная полая -стадия
б л а с т у л ы воспройзв-одит в онтоген-езе прарод-ительскую бластею.
Эта последняя развилась из шаровидной плотной мореи вследствие
накопления внутри нее жидкости или студня, вытолкнувших все ее
клетки на поверхность, где они и стали располагаться в виде ров-
ного клеточного -слоя. При этом псевдоподлии клеток начинали дви-
гаться правильнее и быстрее й постепенно превратились в постоян-
ные жгуты, что позволило колонии перейти от ползающего к ma-
вающему образу жизни. Превращение бластулы в двуслойную
гаструлу происходит путем инвагинации, начальные этапы которой
‘осуществляются стадкией депулы. Депула есть онтогенетическое
отображение предковой деп еи, а заключительная стадия прото-
типического онтогенеза —— г а с т р у л а олицетворяет фи-логенетиче-
скую стадию т а с т р е и. Последняя обладала совершенно простым
ТКРУГЛЫМ, ЯЙЦЕВИДНЫМ ИЛИ удлиненным телом, СОСТОЯЩИМ ИЗ ДВУХ
клеточных пластов, энтодермы и эктодеркмы, низ которых первая была
покрыта ресничками и выполняла двигательную функцию; широко
открытая на одном конце тела полость гаструлы ‘представляла собой]
первичный кишечник. -
Каждая из стадий этих двух параллельных рядов, за исключе-
нием депулы-депеи, иллюстрируется Геккелем определенными
современными формами. Такими живыми пМОДЕЛЯМИ цитей служат
амлёбы, морей—разлйч-ны›е колониальные корненожки й жгутико-
‘вые, бла-стеи—\/о1уох, а гастреи-гидра, олинтусь1 губок и ряд
других форм. - _
Всем известно, что теория гастреи Геккеля подверглась сокру-
шительной критике со стороны И. И. Мечникова, выступившего
против нее в целом ряде своих сочинений. Мечниковым было убеди-
тельнейшим образом показано, во-первых, что инвагинация, считав-
шаяся Геккелем первичным способом образования двуслойности
у Метаяоа, ймеет в действительности явно вторичный, производный
характер, й совершенно не наблюдается у самых примитивных из
многоклеточных, Во-вторых, Мечниковым было установлено господ-
ство-у этих последних внутриклеточного пищеварения, а не полост-
ного, требуемого теорией гастреи, что й находит свое отражение
в присущих йм способах формирования второго, внутреннего пласта
тела. Отсюда стала очевидной полная беспочвенность теории гастреи.
От всей схемы Геккеля уцелела лишь первая ее часть+‹<теория
бла-стей>›, касающаяся самых ранних этапов становления многокле-
точности, до стадии бластулы включительно. Она является в какой-
то мере рациональным зерном схемы Г еккеля и в известной степени
сохраняла до сих пор определенное научное значение. Правда,
10
гипотеза бластеи так пИ осталась сугубо умозрительным построением,
грешащим крайн-ей схематйзацйей, но положенная в ее основу
- есом-
мысль о происхождении Metazoa от колониальных предков Н
„(шло Имела и имеет большое прогрессивное значение.
Теория планулыР. Лэнкестера (1877) существенно Отличается OT “Тала
пых в том что онаг выводит дифференциацию первичных пластов тела перв
бптных Метаиоа из предшествующего этому расслоеиия их протоплазмы на
) I
физиологически отличные друг OT друга 3K'I‘0' И эидоииазмУ- Такое ее ра°`
слоение произошло еще на ОДИОКЛЭТОЧНОЙ Стадии филогенеза В СВЯЗИ с внутр?”
° ом эктоплаз- .
клеточным пищеварением этой гипотетической стадии, при котор
матическйй слой выполнял функцию 3flXBaTbIBflH1’F51 T‘BePl1“=1X пищевых Частиц’
. переваривающихся затем в эидоплазме. ПРИ ПОС-ЛЭДУЮЩЕМ превращении Эти
стадии в плотную колонию типа стерробластулы ее плазматииескии Материал
распределился Между образовавшимися теперь клетками таким образом, что
эндоплазма составила их Внутренние концы, а эктоплазма—периферическую
часть создавшаяся таким путем колония амёбоидных клеток продолжала пи-
таться тем же способом, как ПреДШеСТВОВЗВШаЯ е“ Одидииетоииая “Tamm-
B дальнейшем в центре колонии оформилась специальная пищеварительная
полость, в которой скапливались ферменты, выделяемые эндоплазматическими
кольцами, клеток; пищеварение превратилось из внутриклеточного в полостное.
Вслед за тем все клетки бластулообразнои КОЛОНИИ РЗЗдЕЛиЛиСЬ Параллельно
ее поверхности, ‚и колония стала двуслойпой. Клетки наружного слоя сохра-
нили за собой функцию движения и улавливания пищи’, а: внутренние-Пищева-
рительную. Еще позднее у этой замкнутой дВУСд°ии°и сТадии ПРОРВЗЛСЯ POT»
чему должна, была: предшествовать соответствующая локализация притока пищи
извне.
Эмбриологидеской моделью для Р. Лэнкестера’ послужили ранние стадии раз-
Emmi трахймедузы Geryonia, гаструляция которой совершается, или; Hgaercflrrfif,
путем первичной, или клеточной, деляминации. Как “видно из сказ ог о,
теза Ланкастера, выгодно отличается от геккелевскои уже тем, что в неи учтен
физиологический момент, совершенно итнорйруемый I‘ еккелеМ‚ И ЧТО Она ЗНЗЧИ-
тельно больше считается с действительными особенностями развития некоторыи
низших Metazoa. Однако оиа все же никого не удовлетворила, поскольку легший
в ее основу и якобы объясняемый ею способ образования первичных пластов не
только fie типичен для низших многоклеточных, но даже составляет у них очень
редкое исключение. Кроме тог-о, из него невозможно вывести какие-либо другие
способы гаструляций а принимаемое Лэнкестером раннее возникновение полост-
‚
ного пищеварения идет в разрез с присущим всем низшим формам сохранением
внутриклеточного пищеварения.
Не лучше обоснована и гипотера БалЬФУРа° (18_‚39)‚ Исходящая ИЗ ПРИЗНШИЯ
амфибластулы первобытной формой многоклеточнои организации и также пере.
двигающая таким ооразом, момент дифференциации первичных пластов тела на
‚
однослойную стадию бластулы. Как справедливо указывал Мечников, эта гипо-
теза совершенно не в состоянии объяснить обРа3°Ваиие Эитодермы ПУТеМ имми‘
грации и деляминации и к тому же опирается на неверную интерпретацию Баль-
фуром первичных пластов тела губок. _
K теории амфибластулы непосредственно примыкает теория плакулы Бючли
I88/1), которой он пытался придать в известном смысле синтетический характер.
‘гправной точкой в этой гипотезе служит однослойная клеточная пластинка, по-
добпня гониев-ой табличке вольвоцид. Деление всех ее клеток параллельно no-
хшрхпости пластинки привело K образованию двуслойнои пластинчатой стадии,
плакульх, оба слоя которой были вначале одинаковыми; последовавшее затем
rlulwcsmmc плакулы и ее сворачивание в трубку превратило ее в гаструлу. Каковы-
Moran быть причины расслоения пластинки и ее скручивания, Бючли указать не
мог. Современными моделями плакулы поспужилщс-олнои стороны, уплощенные
эмбриончальные стадии некоторых нематод, олигохет, асцидии и т. п., а с другой-
вегетативные стадии загадочного Trichoplax adherens. Преимущества своей гипо-
тезы перед остальными Бючли видел в том, что она дает возможность одновре-
11
Mel-II-IO ОбЪЯСНИТЬ И ДЕЛЯМИНЗЦЙЮ И ИНВаГХ/ГНЗЦИЮ, Их ЧТО СООТВЕТСТВУЮЩИЕ пре-
ОбРЗВОВЗНИЯ ПЛШКУЛЫ МОГЛИ ПргОИСХОДИТЬ «ПОСТСПЕННО, а1 не скачкообразно? И
тем СаМЫМ бЫТЬ «ДЭЙСТВИТЕЛЬНО ВЫГОЩНЫМИФ) ДЛЯ организма.
Очевидная ДЛЯ нас НЕСОСТОЯТЕЛЬНОСТЬ «теории плакуль1>> была ХОрОШО ПО- `
казана Мечниковым (1886), указавшим, что те плакулообразные зародышевые
стадии, которые имел в виду Бючли, в действительности представляют собой
вторично уплощенные бластулы, что наиболее распространенные способы га-
струляции не могут быть выведены из по-перечного (тангенциального) кле-
ТОЧНОГО ДЭЛЕНИЯ И 'П.‚Д.; С бЕСПОЧВЭННОСТЫО ЭТОЙ ГИПОТЕЗЫ связана И1 ее крайа
няя схематичность и отсутствие у нее какой-либо физиологической базы.
Таким образом, к моменту появления теории Мечникова концеп-
ция колониального происхождения Metazoa оформилась в. целый
ряд односторонних и противоречащих друг другу гипотез чисто умо-
зрительного характера, базирующихся на немнопих отрывочных и
произвольно подобранных фактах и лишенных реальной объясни-
тельной ценности. Поэтому нельзя не удивляться тому громадному
шагу вперед, который сделала филогенетическая эмбриология бла-
годаря исследованиям In обобщениям Мечникова.
' З. Теория фагоцителльц И. И. Мечникова
Теория Мечникова, к рассмотрению которой мы сейчас пере-
ходим, безусловно представляет собой самую серьезную, прорабо-
танную и стройную из всех гипотез происхождения Metazoa,
a вместе с тем и самую материалистичную из них. Ее п-ервый эскиз
был опубликован Мечниковым еще в 1877 г.‚ а законченное, развер-
нутое изложение составило предмет заключительной главы его
знаменитых «Эмбриологических этюдов о медузах>> (1886) ——по—
следней и самой, значительной из его эмбриологических.работ.
В основу теории фагоцителлы положено представление о коло-
ниальном происхождении многоклеточных, «позволяющее нам осве-
тить большин-ство и притом самых важных явлений>›,——как заяв-
ляет и убедительно доказывает это автор, и с позиций которого
«оказывается возможным сводить дробл-ение яйц.а и, особенно, более
примитивное тотальное, к делению жгутиковых, служащему обра-
зованию колоний, а также и объяснить жгутиконосную форму кле-
ток стольких бластул»... Однако, отдавая должное своим предшест-
венникам по разработке колониальной теории, Мечников не при-
соединяется к предложенным ими концепциям, а создает свою
собственную—— намног-о более обоснованную и продуманную.
Отмечая, что половое размножение Metazoa также служит одним
из веских аргументов в пользу их происхождения от колониальных
жгутиковых-«что явствует из отношений, наблюдаемых у воль-
воциней», Мечников прежде всего ставит вопрос, «нет ли возмож-
ности несколько ближе определить, с помощью имеющихся знаний,
каковы были свойства тех колоний жгутиковых, от которых произо-
шли Metazoa», и показывает весьма убедительным образом, что
такая возможность вполне реальна.
Мечников отвергает предположение Бючли о двояком происхо-
12
ждении многоклеточных (губок—4от колоний Choanoflagellata,
a остальных Метаиоа-от колоний настоящих Flagellata), считая,
что «для этого вообще крайне рискованного предположения нет
достаточных оснований», так как обе эти группы жгутиковых <<,ца-
леко не так резко отграничены друг от друга и вдобавок ворот-
ничок, составляющий главное их различие, в некоторых случаях
совершенно втягивается». «Независимо от того, обладали ли ворот-
ничком те жгутиковые, из которых образовались Metazoa, или же
нет, они несомненно имели способность воспринимать твердые пи-
щевые частицы, что явствует из распространенности у низших
Metazoa внутриклеточного пищеварения. Поэтому я не могу согла-
ситься с Бючли, когда он считает способ питания безразличным для
вопроса о происхождении многоклеточных на том основании, что
физиология питаниячасто меняется у жгутиковых безотносительно
к их -морфологии..._ Я скорее думаю, что дальнейшие дифференци-
ровки колоний жгутиковых, поведшие к образованию Metazoa, на-
ходились в отнюдь не безразличном отношении к способу питания.
В то время -как у некоторых из них образовывались растительные
пигменты, обеспечивающие ассимиляшию, у других жгутиковых,
сохранивших животный способ питания, происходило образование
особых индивидов, специально приспособленных к питанию И пище-
варению. Что это заключение не является только дедуктивным——
видно из сравнения таких колониальных жгутиковых, как вольво-
цинеи И Protospongia».
Отмечая далее существенное теоретическое значение возможно
большей конкретизации наших представлений об особенностях раз--
вития таких гипотетических ‹<метазоо--флагеллят>›‚ Мечников де-
тально анализирует „вопрос о присущих им формах деления. Обсуж-
дение этого вопроса он основывает на справедливом признании того
факта, что способ деления играет у колониальных простейших знаъ
чительную роль -в определении формы образуемого ими объедине-
ния, и на оценке с этой точки зрения различных форм деления совре-
менных жгутиковых. '<<Мы знаем‚—пишет Мечников-что
современные жгутиковые ведут себя п-о-разному в этом отношении.
У настоящих Flagellata господствует, в общем, продольное lie-T1eHP1e,
ХОТЯ среди них встречаются и виды, обладающие поперечным деле-
пием, как, например, Phalansterium consoclatum по Ценковскому,
и Ph. digitatulm по Штейну. УЧворотничковых жгутиковых также
наблюдаются оба вида деления, даже у близко родственных видов;
так, ‘по С. Кенту, Salpingoeca campanula делится продольно, тогда
mm другие ‚виды того же рода, например, S. mqmllata, S.,grac111s,
рпнмпожаются исключительно‘ поперечным делением. Одновремен-
поо существование продольного и поперечного деления у одного И
того же вида, повидимому, кон-статировано до сих -пор только у не-
которых хламидомонад». Напоминая далее подчерки-ваемо-е им
обстоятельство, что при дроблении Яйца «ТРИ ‘ПЕРВЫХ деления ‘we’
дуют в трех измерениях ‘пространства и осуществляются одинако-
вым образом у медуз с самым различным ходом ра-звити-я (как
1'3
у гипогенетиче-ских, так гИ -метагенетических, и независимо от укло-
нений в способе образования энтодермы и т. д.) и у различнейших
животных с тотальным дроблением», Мечников считает «вполне
оправданным предположить, что тем же способом деления обладали
и предки многокл-еточных». «Если мы вообще хотим использовать
процесс дробления для генеалогических построений, то указанное
предположение представляется неоспоримым», -—говорит далее
МЕЧНИКОВ, УКЗЗЫВЭЯ, ЧТО ЭТО предположение подтверждается Ta‘ ‘
КИМИ аналогиями, как существование у бактерий деления в трех
измеренпиях пространства. <<Н_аряду с бактериями, у которых наблю-
дается только один вид деления, имеются другие, у которых клетки
делятся в двух меридианальных плоскостях, как, например, микро-
кокки триппера и еще такие, как, например, Sarcina, деления кото-
рой идут в трех измерениях пространства, т. е. так же, как при
тотальном дроблении большинства Мета2оа и как это предполо-
жено нами в отношении гипотетических метазоо-флаъгеллят»
(стр. 135 — 136).
Решив таким образом никем не ставившийся ранее вопрос о свя-
занности процесса образования колоний с определенными геометри-
ческими формами деления, Мечников тут же высказывает мысль,
что сходная бластулообразная форма колонии может создаваться
разными средствами. Поскольку у Metazoa «(ПОЛОСТЬ дробления,
появляется в типичных случаях уже вслед за третьим делением и
зародыш ра-но превращается в бластулу, то очень вероятно, что и
предкчи Ме’па2оа плавали ввиде бластулообразных колоний», фор-
мирование которых, таким образом, происходит здесь в результате
трехмерного деления при ‘по-следующем раздвигании его продуктов,
дающем начало бластоцелю. Однако, наряду -с этим, возможно и
образование бластулы иным путем, «без посредства трех типичных-
первых делений». «Так, мы видим, что у Volvox все деления идут
в меридианальных плоскостях, что приводит к образованию пластин-
‘ЧЗТОГО зародыша, напоминающего СТОППШП; гПрИ ЭТОМ не o6pa3yeTcs1- Ф
ПОЛОСТИ дробления, И бластоцель создается ЛИШЬ ВПОСЛЕДСТВИИ,
благодаря разрастанию пластинки в направлении к одному из по-
люсов». Как отмечает в сноске Мечников (стр. 134), он сам наблюдал
этот процесс у Volvox minor, где «за образованием пластинки сле-
дует ее превращение в открытый шар».
«Если принять изложенные особенности метазоо-флагеллят
в основу дальнейшего рассмотрения, то мы будем, как мне кажется,
в состоянии внести известную ясность и в вопрос о происхождении
первичных органов». Напомнив установленное им ‘ разнообразие
способов формирования энтодермы .y МеДУ3‚ МеЧНИКОВ переходит
затем к развернутому обсуждению вопроса о том, какая из точек
зрения способна объединить все это разнообразие, «какая из
существующих теорий происхождения первичных органов может
объяснить наилучшим образом действительное положение вещей?»...
«Допущение происхождения многоклеточных от многоядерных про-
естейших бросает нас в этом смысле на произвол судьбы, так как оно
14
(“голь же Мало объясняет иммиграцию с поверхности иди и не а
"""‘”У!0 деЛЯМИНаЦИЮ или инвагинацию Эта гипотеза ‚олжна бь рь
поэтому отброшена, как совершенно бесплодная» (стрд 137) суёте
‘они и вссрма обоснованная критика, ‚которой Мечников подвергает
гул п-япоясзьт Г екке.пя, Р. Лэнкестера, Бальфура и ‚Бючди резюмш
или 7
PM мн ним в общем выводе, что все эти гипотезы совершенно не
" ‘ЩДШЯПИИ Объединить всю сумму известных нам эмбриологических
IIIHKIUU ‘ПОД ОДНИМ углом зрения И ‚страдают, кроме того Недоста-
тошной физиологической обоснованностью ’
U о ‚ ° о
б снование своих собственных представленирд на этот счет Меч.
пиков начинает с указания, что еще в 1877 г он Высказал «В Очечь
осторожной форме» некоторые замечания, которые хорошо согла-
‘УКЛСЯ Как ‘С результатами, полученными в отношении образования
чпто е м J
д Р Ы у низших Мета2оа, так и со ставшими тогда известными
Нпакеииями внутриклеточного пищеварения «Я полагал что это-
дерма впервые возникла не в виде трубчатой кишки с терминаль-
"f’”‘f‘ °TBePCTKeM» K3§Y*0 ‘M1>1uHaX0ILI/IM y гаструлы, а что это образо-
щшие ИМЕЮТ ‘За СОбОИ долгии путь исторического развития, докумен-
ШУеМЫИ формированием плотнои паренхимы с внутриклеточным
пищеварением. ‘Последняя также появилась не сразу, а постепенно,
"УМ-М ВЫСе-ПеНИЯ поверхностных клеток бластодермы в глубь бла-
иготцеля. В конце концов создалась двуслойная паренхимелла, кото-
Р"?! ПРИ сокращении эмбрионального процесса и прогрессирующей
дичдференцировтке пищеварительного аппарата ‘превратилась в га-
струлу». При невозможности для него тогда опереться на какую-
ллибо Ёзысоко дифференцированную форму жгутиковых с животным
пирсе ом питания, Мечников искал некоторой поддержки своим
т‘ ЛЯдаМ -В истории развития Volvox, высказав при этом Амнение,
по необходимо поискать такие низшие существа, у которых питаю-
щие клетки покидают свое место вскоре после принятия ими пищи
‘Г УХОДЯТ С Наружной поверхности «колонии» в ее внутреннюю по-
лость» (1879, стр. 382).
" Taé{:OeII<I/I'MeHH0 существо было вскоре ‚после этого открыто и опи-
(‚дно . ентом в качестве колониального воротничкового жгути-
крвого под многозначительным названием Protospongia Haeckeli
(‘ 880). Главная особенность Protos‘pon.g1a заключалась, по данным
(‚. Кента, в том, что отдельные особи колонии могут принимать
"Мубоиднуюцформу И Ухедить пр-и этом в глубь студенистой массы
(е ›разующеи_основу этои колонии), где они испытывают затем де-
пие и другие изменения, толкуемые С. Кентом, как споруляция.
дрходятся ли это изменение пформы клеток и их миграция в какой-
лп ›‹› связи с процессамли пита.ния,—С‚ Кснтом установлено не
(шило, И это осталось неизвестным.
дпи необычные особенности Р. Haeckeli сразу приковали к себе
внимание Мечникова. Он увидел в этом загадочном организме жи-
вую модель паренхимеллы И нашел в соответствующих данных
КеНта ряд показаний в пользу такой его трактовюи. Так, отмечая
недоказанность факта споруляции у Protospongia, Мечников вь1-
15
оказывает лрешноложениещ не являются многочисленные зерна,"
принятые С. Кентом за споры, либо резервными ‚включениями, либо '
непосредственно заглоч-енньпми пищевыми частицами? Самый факт‘
перехода воротничковых клеток в амёбоидное состояние на-поми-
нает Мечникову картины, наблюдаемые у двуслойных личинок неко-
торых губок, и он расценивает этот переход, как первый шаг к обра-
зованию энтодермы. <<'Bo всяком случае я думаю, что эти особен-
ности Protospongia гораздо лучше согласуются -с моей концепцией .
(обозначаемой некоторыми авторами, как теория паренхимеллы),
чем со всеми вышеизложенными теориями других исследователей».
Оценка различных опособов образования энтодермы с точки
зрения теории паренхимеллы приводит Мечникова к заключению
о возможности вывести все эти способы из одного и-сходного—-
смешанной деляминации. Посвященный этому вопросу раздел книги
Мечникова получил наибольшую известность, и иллюстрирующие
его рисунки часто воспроизводятся в эмбриологических ру-ководствах.
Однако в его изложении обычно опускаются многие интересные
детали, так что ‘нам кажется уместным остановиться на нем
подробнее.
«Из всеобщей распространенности трех ‚первых делений, идущих
по трем измерениям пространства (сагиттальному, фронтальному,
экваториальному) и представляющих, такимобразом, поперечные
И продольные деления, я вывел заключение, что предки многокле-
точных также делились в этих различных направлениях. Однако
направления деления постепенно фиксировались таким образом, что
одна форма размножалась преимущественно или исключительно
посредством продольных делений, тогда как другая удерживала и
поперечный способ деления. Что подобные деления действительно
возможны, показывают упомянутые виды $а1р1п9оеса. Итак, мы
можем принять, что у наших гипотетических метазоо-флагеллят
некоторые поверхностные клетки становились амебоидными и ухо-
ДИЛИ ВНуТрЬ КОЛОНИИ ‘СОВЕРШЕННО ТЭКИМ ЖЕ образом, как МЫ НЭХО- -
дим это ныне у Protospongia, НО что наряду с этим другие особи
делились в поперечном направлении, причем центральная дочерняя
особь уходила внутрь, а периферическая сохраняла свое поверх-
ностное положение. (Возможно, что, наряду с поперечными и про-
дольными делениями, и-мели место и деления в косом направлении,
как мы это видим теперь у некоторых простейших.)... В истории
развития этот двоякий способ образования энтодермальных клеток
реализован иммиграцией ‘целых клеток и отшнуровыванием цен-
тральных продуктов деления в таких случаях, как, напрИМеР‚
у Ро1ухеп1а 1еисозту1а‚ где мы имеем дело со смешаннои делями-
нацией. В то время как у одних форм поперечное деление выступило
на первый план, у друпих стало господствовать продольное, и, сле-
довательно, выселение. Целых поверхностных клеток. Танк от сме-
шанной деляцминации ответвилалсь, с одной стороны, первичная Ile-
.JIH‘MI/IHEIIII/IH, a с другой — мультиполярная иммиграЦИЯ. ВТОРРРЧНЗЯ
деляминация должна рассматриваться, как слабая МОДИФИКаЩИЯ
‘16
смешанной, так как она отличается от нее только несколько более
поздним «появлением различий между обоими видами кл-еток».
Согласно концепции Мечникова, все эти мультиполярные опо-
собы образования энтодермы должны быть признаны более первич-
ными, чем гипотропный «концентрированный» способ, тогда" как
обратное предположение сталкивается с большими трудностями.
«Переход от мультиполярной к гипотропной иммиграции, напротив,
л-егко себе представить, поскольку последняя преимущественно
встречается у очень подвижных личинок —-бластул, в то время как
первая наблюдается у неподвижных или слабо подвижных заро-
дышей. При описании образования энтодермы у метагенетических
медуз я подчеркивал, что продольное деление является тут прави-
лом, а поперечное, напротив, встречается как очень редкое
исключение».
«Если концентрированная иммиграция без труда выводится
из мультиполярной, то, с другой стороны, также не трудно вывести
из первой и инвагинацию. Первый шаг в направлении такого пере-
‚хода иллюстрируется примером Laodicea cruciata, y которой обра-
зование энтодермы отличается от наблюдаемого у других метагене-
тических гидромедуз в том, что оно происходит за счет особых
-прозра-чнь1х клеток, составляющих задний полюс тела бластулооб-
разной личинки. Путем постепенного вселения этих клеток в бласто-
цель образуется паренхимообразная энтодерма, в которой затем
появляется полость. Исходная стадия Laodicea с прозрачными клет-
ками обнаруживает бросающееся в глаза сходство со стадией бла-
стуль1 у N ausithoé, которая также характеризуется видимыми отли-
чиями гипотропного ряда клеток от остальных»... «Если мы допу-
стим дальнейшее сокращение хода развития,-то легко можем себе
представить, ‚что поверхностно дифференцированные энтодермаль-
ные клетки, вместо того чтобы выселяться по отдельности, впячи-
ваются сообща и ‚дают прямым путем .энтодермальную полость.
После того .как произошла инвагинация первично дифференциро-
‚ВЕЪННЫХ КЛЕТОК, ЭТОТ npoueoc распространяется на СОСЕДНИЕ ЭЛЕ-
‚МЕНТЫ ОЛЗСТОДЕРМЫ, ‘благодаря ЧЕМУ ВПЯЧИ-ВЭЮЩИЙСЯ_МЕШОК ПОСТЕ‘
_ПЕННО УВЕЛИЧИВЗЕТСЯ. МЫСЛИМОЕ ЗЗТЕМ дальнейшее СОКрЭЩЕНИЕ уЖЕ
‚НЕ ОСУЩЕСТВЛЕНО МЕДуЗ; ОНО МОЖЕТ ОЫТЬ представлено, как ЕЩЕ
_бОЛЕЕ ранняя ДИФфЕрЕНЦИРОВКЭ ЭНТОДЕрМЭЛЬНЫХ КЛЕТОК, ‘ПРИ КОТО-
`рОЙ ВСЕ ВПЯ-ЧИВЗЮЩИЕСЯ КЛЕТКИ РЯЗЛНИЧИМЫ УЖЕ у бла-стулы В ВИДЕ
особых элементов... Что дальнейший прогресс ранней дифференци-
ровки энтодермы приводит к ам-фибластуле, появляется затем еще
в течение процесса дробления, и в конце концов может стать выра-
жен.ной уже в структуре яйца,——'не требует пространного обсуж-
дения. И ‚если амфигаструла может образоваться из архиигаструлы
вследствие ранней эмбриональной дифференцировки, как это
является общепризнанным... то, с другой стороны, неоспоримо, что
подобная форма может возникнуть и путем неравномерного дром-
бления при смешанной деляминации, как мы это видели на при-
мере Ро1ухеп1а leucostyla. ‘ПОСЛЕДНИЙ случай учит нас тому, что
2 Эмбриология ‘беспозвоночных 17
амфигаетрульт могут создаваться двумя различными ‚путями из двух
различных исходных стадий, -и показывает › нам, следовательно,
полифилию названной зародышевой формы» (стр. 150).
Эти глубоко интересные и далеко еще н-е оцененные по досто-„
инству соображения Мечникова об источниках и о путях эволюции
процессов формирования энтодермы, а тем самым и эмбриогенеза
низших Metazoa, завершаются изложением заново созданной им
теории происхождения первичных пластов тела многоклеточных
животных. ‘Хотя эта теория неоднократно излагалась‘ в литературе
И ‘известна в общих чертах почти- каждомпу биологу, она все же не
заняла до сих ‘пор того места в нашей науке, которое она вправе
занять. И если ее значение в полной мере осознается теперь у нас
в области сравнительной анатомии (как об этом ясно свидетель-
ствует интересный труд Беклемишева, 1944) и отчасти в области
гистологии (3аварзин, 1945), то этого никак нельзя сказать о нашей
современной эмбриологии. Она доныне остается в большом долгу
перед Мечниковым и далеко еще не освоила его богатейшее идейное
наследие. Поэтому мы считаем нужным привести здесь полнее, ч-ем
это обычно делается, высказывания самого Мечникова по относя-
щимся сюда вопросачм. ` ‚ ‘
С защищаемой им точки зрения, <<энтодерма должна рассматри-
ваться, чкак сумма клеток, первоначально образованная в бластуле
путем иммиграции или обособления и сложившаяся в разнообразно
развитое скопление амёбоидных элементов»; Обособление этих
клеток от остальных отражает те отношения, «которые должны были
иметь вместо у колониальных предков чмногоклеточных, когда эти
клетки еще имели значение отдельных зооидов колонии, как это
ныне наблюдается у Protospongia»... «Между особями колонии жгу-
тиковь1х должно было возникнуть сначала неравенство, поведшее
затем пк дальнейшей дифференциации: в то время как одни_особи
оказывались более способными к питанию, другие были лучше при-
способлены к движению и к улавл-иванию пищи. Для подвижной
колонии должно было быть большим преимуществом, что более за-
груженные пищей и потому более тяжелые особи неоставались на
периферии, а сдвигались возможно ближе к центру. Другое преиму-
щество могло возникнуть тогда, когда болеепрожорливые особи
оказывались в состоянии возможно полнее выполнять свои обязан-
ности. Мы знаем, что многие жгутиковые переходят из монадообраз-
ного в амёбоидное состояние, в котором они как раз и принимают
пищу». Далее Мечников развивает мысль, что подобные амёбоид-
ные особи размножались более интенсивно, что и привело к уста_-
новлению известной связи между продуктивностью ‚питания и раз-
множения; Еще другое основание к возникновению иммиграции
можно видеть в том, что «расширение поверхности колонии невоз-
можно дальше известного предела, что‘ должно было иметь важное
значение именно для плавающих колоний». «Так как увеличение
числа особей в, колонии способствовало повышению ее деятельности,
для нее могло быть полезным, чтобы те клетки, которые не могли
18
найти места на поверхности, приспособлялитсь и существованию
внутри колонии. В течение длительного времени‘ особи одной коло
нии ве оят - ' ч ' '
р но различались между (юбок только В количественном
отношении. локомоторные клетки ‘п
пищевые частицы и отчасти заглатывали более ‚мелкие из НИХ
подобно Тому’ как Мы Видим Теперь У Некоторых кишечнополостных
факЁ/льтативное питание посредством клеток эктодермы Внутренние
а›ме оидные особи бы а ' '
крупные Объе’ . ЛИ‚ Напротив, способны пожирать пи ‘более
В » КТЫ, с которыми не ‘могли справиться локомоторные
клетки. озможно что
Девали находящимисяэчнун ОСОЁИ приближались K периферии И Оша‘
По Ы НеИ ЧаСТИЦами через ‘многочисленные
Р ПОВЕРХНОСТНОГО СЛОЯ. Как аналогичное этому явлениея могу
привести Заглатывание ЧУЖеРЮдНых тел амёбоидными клеткаии
„ - ‘.L
перезимовавших На115агса ропыса, утратившими ‘свои воротнички
и подобное же их ` ’
max ЖИВОТНЫХ Пзагёхёёёьтванлие клетками M'e30,ILep1MbI у многих НИЗ-
п а - НН0 дифференциация в наметившемся на-
р вленииделала большие успехи: локомоторные клетки в большей
или е " .
Ш EM HbH1e1’1._MePe ТЁРЯЛИ Свою функцию питания, сосредоточив-
у ся на амебоидных фагоцитах»... «При повышающейся индиви-
дуальности колонии (индивид
ностные особи +
. б ДН е внутренние особи объединились в фаго-
ЦИТо ласт (паренхиму или мезоэнтодерму) Когда клеткам после
неИ оказывалось не ‚под силу поглотить отдельные крупные пищевЁе
объекты они н в ° А `
Ныне В ЗНТОДеОЁ/ЛЁЁаЁЁЁЁЁВа-“И ‘ПЛЗЗМОДИИ, как последние образуются
Вокруг более Ё ОНОФОР ИЛИ В МЕЗОДер-Ме стольких животных
Жизнедеятельнору ных чужеродных тел. Дальнейшее повышение
Вело K em пе em“ Уже двугслоиного теперь многоклеточного при-
Р ду к питанию крупнычми организмами, сопровож-
ДЁВШЭМУСЯ образованием одного или нескольких более крупных
B'B°'j§:::0‘£I?§PCTKfi._ ЧТО И ‘могло привести к оформлению рта»,
ПРЕДПОЛЗГЗЁМОИ ИМ дифференциации первично одно.
Ё/рдных клеточных особеи на локомоторные клетки и фагоциты
еЧНИКОВ Видит в том, что у низших книдарий (и в том числе неко.
торых медуз, например, Oceania, Phfialidium, Cunina) все жгутш.
ЁЁЁЫ; KJ:[I;;I1fII:1IXaH(';rIc3l;iL(§eFE>r1\£I:I_ fixsélxilgllfiimw обе эти ФУНКЦИИ, в то время
В Энтодерме явственно Дифрфе ен, СИФОНОФОРЬЪ) И У гребневиков
альные клетки кото ь р Цированы амебоидные Эпител‘ _
‚ р 1е одни способны к захватыванию и перева-
риваниюапищи, и жгутиковые клетки, имеющие своей единственной
фУНкЦгиеи поддержание тока воды и не способные K заглатыванию
пищевых частиц. То же самое наблюдается и у губок у многих
представителей которых жгутиковые клетки (хоаноциты), являются
в то же время и фагоцитами, тогда как у некоторых кремневых
губок функция питания целиком переходит к амёбогцитам а- за во-
ротничковыми клетками остается только гидрокинетическая.
q)arI;1;:fn>élgH:§T§3%/Izggggiggogg ИМ представления об энтодерлме, как
‚р реименовывает паренхимеллу в фаго-
9* 19
ривлекали движением жгутов-
уальности второго порядка) поверх- .
V СТИЧНОЙ и обоснованной из всех теорий прои
Цйтеллу й дает ей следующую характеристику: «Можно гпредполо-
_ЖИТЬ, ЧТО фагоцителла обладала ДВУМЯ ПЭРВИЧНЫМИ‘ органами,
кинобластом и фагоцитобластом, которые, однако, не были столь
резко разграничены друг от друга, как у большинства Metazoa;
вероятно, пополнение фагоцитобласта вселяющимися в него клет-
ка-ми кинобласта длилось. продолжительное время. В отношении
истории развития можно сказать, что яйца фагошителлы (она уже
должна была обладать половым способом размножения)‘ подвер-
гались равномерному дроблению, при котором бластомеры дели-
лись в трех измерениях пространства и рано образовывался бласто-
цель, КОТОРЫЙ ЗЗТЕМ ‘ПОСТЗПЭН-НО ЗНПОЛНЯЛСЯ иммиграцией ЦЕЛЫХ-
клето-к и обособлением: обращенных к центру продуктов деления»
(стр. 154).
›В заключение Мечников останавливается еще на „вопросе о про-
исхождении мезодермы. ‹<В то время как часть фагоцитобласта
преобразовывалась в энтодер-му, в которой первично амёбоидные
клетки постепенно пр.иобретали эпителиальный характер,.другая
часть того же самого первичного органа дала начало мезодерме.
Последняя первоначально появилась в „форме отдельных блуждаю-
_ЩИХ KJIGTOK, l'IpO1LOJI}KaBII.III/IX, как раньше, ‘СЛУЖИТЬ ‘ФЗГОЦИТЗМИ.
Подобные ОТНОШЕНИЯ МЫ НЗХОДИМ у МНОГИХ КИШЕЧНОПОЛОСЪНЫХ (где,
_ ОДНЗКО, В некоторых ‘СЛУЧЗЯХ ОЛУЖДЗЮЩИЕ КЛЕТКИ ВЫХОДЯТ ИЗ ЭКТО-
дермы, например, у кораллов по Ковалевскому и Мариону), у игло-
кожих, многих червей `и т. д. „Возникнув в связи -с фагоцитобла-
стом, а отчасти и` с ки-нобластом, мезодерма раньше (например,
у греб-невиков) или позже эмансипируется как особый зародыше-
вый л.исток, играющий важную роль в эмбриональном развитии»
. `(стр. 158—159).
Итак, мечниковская теория фагоците.лль1 содержит эволюцион-
ное освещение всех основных моментов онтогенеза Metazoa. B ней
последовательно разбираются генезис дробления в его связи с обра:
зованием многоклеточности‚ причины и механизм начальнои
клеточной дифференцировки, ведущей к возникновению фагоцито-
бласта, дальнейшее расслоение этого последнего и все существую-
щее многообразие способов его образования. Каждый пункт гипо-
тезы обосновывается„Мечниковым "обильным ‘фактическим мате;
риалом, касающимся истории развития низших -.Metauzoa— с однои
стороны, и явлений размножения и развития простейших (‚и даж_е
бактерий) —с другой. Исключительное богатство эмбриологических
и протистологических фактов, используемых‘Мечниковымц для по-
строения его гипотезы, дополняется глубокой биологической обосно-
ванностью развиваемых ИМ представлений; бЛЗГОДИРЯ ЭТОМУ:
гипотетическая.гфагоцителла рисуется нам живым, полноценным
организмом. Таким образом, теория фагоцителлы представляет
полную противоположность надуманным и по существу безжизнен-
ным схемам Геккеля, Бючли и-друтих, и является самои материали-
схождения много-
клеточных.
20
Тем более приходится удивляться, что теория Мечникова не по-
лучила того распространения и не заняла того места в эмбриологии,
на которое она им-ела все права, и что опровергнутая ею теория
гастреи продолжала —и продолжает отчасти и теперь-занимать
ведущую роль B‘ филогенетической эмбриологии. Еще более стран-
ным кажется нам, однако, то обстоятельство, что взгляды Мечни-
кова, почти игнорируются и теперь и, в частности, совершенно
забыты нашей советской эмбриологией. В наше время их пропа.ган--
дировал и отчасти. развивал один только А. А. Заварзин, много-
кратно отмечавший их огромную теоретическую ценность.
Имеющаяся в литературе кр.итика теории фагоцителлы не смогла
поколебать ее основных положений. Сам Мечников легко опроверг
критические высказывания Баль-фура, Бючли И других по поводу
этой теории; нам кажется так же нетрудным возразить и его совре-
менным критикам. Так, мы не можем согласиться даже с Заварзи-
ным (горячо поддерживающим, в общем, теорию фагоцителлы)
в том; что слабым местом этой теории является признание ею коло-
ниального происхождения Metazoa, и нам кажутся совершенно
неоправданными выдвигаемые Н. Г. Хлопиным возражения против-
нее.
Вместе с тем приходится признать, что одно из существенных.
положений теории Мечникова лишилось к настоящему моменту-
той фактической базы, на которую оно -в свое время-опиралось. Это
касается предложенного им объяснения иммиграции, основанного
на использовании в качестве модели Prot-ospongi-a haeckeli, само
существование которой как самостоятельного организма не под-
твердилось последующими исследованиями. Это загадочное суще-
ство является, повидимому, не чем иным цкак реституционным телом.
или фрагментом» пресноводной’ губки, так что его -приходится. вь1-‚
черкнуть ИЗ‘ ЗООЛОГИЧЭСКОЙ ‘СИСТЕМЫ; ТаКИ|М образом, ВМЕСТЕ ‘С PTO-
tospongia исчезла та-кфактическая основа, на которой строилось
представление о первичностимультиполярной. иммиграции. Однако р
и сейчас основные положения теории фагоцителпы полностью, сохра- ,
нили свое значение, и она может попрежнему служить примером
правильного подхода к проблема-м филогенетической эмбриологии.
4. дальнейшее развитие теории колониального происхождения
Metazoa. Теория А. Н. Северцова
За периодом интенсивной разработки теории колониального про-
исхождения многоклеточных, завершившимся появлением мечни-
ковской теории фагоцителлы, последовал. длительный перерыв в ее
развитии, связанный- с наступившим теперь -охлаждением як основ-
ным вопросам филогенетической морфологии. Усилияподавляющего
большинства эмбриологов устремились в сторону поисков «каузаль-
ного» объяснения онтогенеза, и его историческая трактовка вре-
менно сошла со. сцены. Однако даже и в это время отдельные иссле-
дователи продолжали интересоваться проблемой происхождения
21
' Метаиба и пытались отчасти" модернйзи
созданные представления ‚на этот счет. ь _
`так› В 1911 Г- КОРШЁЛЪТ И Гейдер сделали‘ попытку вдохнуть
ЖИЗНЬ B_'I"eKK€JI€BCKy}O ТЕОрИЮ ГЭСТрЕИ, ВЫДВИНУВ HOBOC Объяснение"
возможных причин инвагинаци-онной Гаструляцирд по их мнению’
первым шагом в подготовке условий для ее осуществления явилось
приобретение одноосно гетерополярного строения гипотетическими
вольвоксообразными колония-ми предков Metazoa, вызванное уста-
новлением определенного предпочтительного направления „лава-
HM ЭТИХ КОЛОНИЙ- Наблюдая За движением жгутиконосных бластул
ГУбКИ ОЗСЗГЭПЭ В МОРСКОЙ Воде, ‘Содержащей взвешенные частицы
кармина, авторы заметили, что эти частицы относятся токами воды
назад вдоль тела и скопляются в мертвом пространстве у заднего
его конца, чем определяется, по яих мнению, локализация На этом
конце места предпочтительно-го заглатывания пищи. И хотя они
признают, что такое заглатывание оформленных частиц «совершенно
не имеет места у бластул Oscarella, вообще ‚не питающихся на этой
стадии, авторы делают отсюда вывод о возможном значении этого
гидродинамического момента для объяснения путей становления
гаструлы. Уже небольшое уплощение заднего конца тела могло бы
дать бластуле известные преимушествав обеспечении захвата при-
носимой сюда пищи, и эти преимущества становились все более
ощутимымли по мере углубления наметившейся тут впадины. Этой
гипотезе, конечно, нельзя отказать в остроумии, но этого явно недо-
статочно, чтобы сделать ее убедительной.
Намного более фантастический характер имела "известная по-
пытка Ланга (1912) придать конкретность геккелевским бластее и
гастрее путем чисто д-едуктивного воссоздания их организации во
всех ее самых частных подробностях. Поразительные гипотетические
организмы, составленные таким способом Лангом, могут служить
образцом бес-почвенных“ филогенетических реконструкц-ий. 4
Полную противоположность этому направлению в теоретической
разработке проблемы происхождения многоклеточных представляет
гипотеза, высказанная в 1910 г. B.’ A. Догелем. Основы-
ваясь на изучении открытого им своеобразного Нар1о2ооп (относя-
щегося к паразитическим представителям В1по11а9е11а1а), В. А. До-
гель высказывает предположение, что одним из способов происхо-
ждения многоклеточных организмов амогло быть гетерономное
деление одноклеточных без расхождения его продуктов, подобное
наблюдаемому у Haplozoon делению на трофическую и генератив-
ные клетки._ Большими достоинствами этой гипотезы являются ее
'IIpOTO300JIOI‘I/I'~I‘€CKaH O6OCHOBa'HHOCTIj; ‘трезвое ПРИЗНЗНИС СУЩЕСТВОВЗ- '
ния ряда различных путей к достижению многоклеточности an осто-
рожность в ее формулировке, столь "резко контрастирующаяс чисто _
спекулятивными построениями вышеупомянутых немецких авторов.
С тридцатых годов нашего столетия начинается заметное ожив-
ление интереса к занимающей нас-проблеме, явившееся очевидным
следствием и выражением наметившегося теперь возрождения эво-
22
ровать и оживить ранее
люцион-ного направления в биологии. Она вновь стала серьезно
обсуждаться в нашей и отчасти даже в зарубежной литературе;
были выдвинуты новые гипотезы происхождения Metazoa И модер-
низованы некоторые из прежних; и, что салмое главное-вновь
было осознано исключительное теоретическое значение этой про-
блемы и ее Непреходящая актуальность. Вместе с тем, в самой ее
постановке наметились крупные сдвиги, обусловленные как общим
прогрессом биологических знаний за время, отделяющее нас от поры
расцвета филогенетической морфологии, так и изменившимися
взглядами на соотношения между индивидуальным и филог-енети-
ческим развитием. Если раньше все гипотезы этого рода строились
на безоговорочном признании основного биогенетического закона
в его геккелевской трактовке, т-о ‚теперь их принципиальной основой
стало ограниченное его понимание в духе теории филэмлбриогенеза
А. Н. Северцова. Центр.тяжести данной проблемы оказался при
этом перенесенным с эмбриологического на сравнительн-о анатоми-
ческий ее аспект. Одним из- положительных результатов этого
явилось оформление проблемы происхождения онтогенеза Metazoa
B качестве самостоятельной эволюционной проблемы, лишь отчасти
связанной с вопросом о происхождении многоклеточности; отрица-
тельным результатом— ряд попыток решить последний вопрос
в одном только сравнительно анатомическом п.лан-е, вне- всякой
связи с эмбрио-логическими данными. ‚Особенно же характерной
чертой „нового этапа. теоретической разработки этого круга проблем
явилось то, что она осуществлялась теперь усилиями одних морфо-
логов‚ что эмбриологи фактически устранились от участия в ней- и
остались на прочно занятой ими позиции чисто механистической
трактовки эмбриогенеза. Таким образе-мн, современный период раз-
вития интересующей нас проблемы может быть охарактеризован,
как период ее реабилитации стараниями. морфологов-эволюцио-
нисгов. iv
Начало ее реабилитации положил глава новой эволюционной
морфолопии акад. А. Н. Северцов, впервые поставивший эту про-
блему в специально онтогенетической плоскости, в форме вопроса
о том, как возник и как развивался онтогенез многоклеточных’ жи-—
вотных. Этому вопросу А. ‚Н. Севергцов посвятил- особую статью,.
опубликованную в 1934 г.‚ и одну из заключительных глав своего
классического труда «Морфологические закономерности эволюции»
(1939). '
В- основу- развиваемых Северцовым - представлений - положена
мысль, что «у Protozoa нет эмбрионального развитияв том смысле,
в каком мы его находим‘ у многоклеточных, происшедших от них».
Определяя-онтогенез Metazoa, как «весь цикл деленияи гистогене-
тических изменений, происходящих от времени конъюгации яйца
и сперматозоида до взрослого состояния, т. е. до периода развития
новых половых продуктов (яиц или сперматозоидов)», СеверЦ0В
считает, чтоу Protozoa «этому циклу соответствует цикл делений
потомков одной особи после конъюгации до нового HePPI01Ia"K0HT=*0'
23
' гации», т. е. целый жизненный цикл, как он понимается протисто-
логами. ОТСЮДЭ СЛЕДУЕТ, ЧТО «ИЗМЕНЕНИЯ, ПрОИСХОДЯЩИЕ В ТЕЛЕ ОДНО-
КЛЕТОЧНЫХ ПрИ рЭЗВИТИИ В НЕМ ОРГЭНЕЛЛ‘ “ЭЕСНИЧЕК, сократимых
фибрилл и т. д.)‚ соответствуют не онтогенезу Metazoa, HO их гисто- '^
ГЭНЕЗУ, Когда в теле каждой данной клетки образуются сократимые
фибриллы, реснички и т. д.>›.
Это соответствие, правда‚'не является полным, поскольку гисто-
ген-езодноклеточных «происходит после каждого деления клеток,
так что между двумя дифференцировками органеллл нет поколения
индиферентных клеток, как это имеет место у Metazoa», y KOTOpbIX'
«типгичная специализация клеток любой ткани происходит не тог-
час после деления яйц-еклетки надвое, как у Protozoa, НО через боль-
шое число делений потомков яйцеклетки. У большинства многокле-
точных последовательные стадии развития зародыша (морула,
бластула, гаструла и т. д.) состоят из неспециализированных эм-брио-
нальных клеток, которые только в конце морфогенеза начинают спе-
циализоваться гистологически в разных направлениях».
«Итак, мы принимаем, что у одноклеточных нет эмбрионального
развития, что материнская клетка дает после деления начало не заро-
дышу, когоро-му надо проделать ряд сложных превращений, пр-ежде
чем он- превратится во взрослое‘ животное, а сразу двум взрослым
дочерним клеткам, начинающим самостоятельное существование».
Поэтому, если пМЫ примем, что многоклеточные животные про-
изошли от одноклеточных, и что у этих первичных одноклеточных
периода эмбрионального развития (онтогенеза) не было, то необхо-'
ДИМЫМ СЛЕДСТВИЕМ ИЗ ЭТОГО ОУДЕТ ПРЕДПОЛОЖЕНИЕ, ЧТО ОНТОГЕНЕЗ
МНОГОКЛЕТОЧНЫХ... рЭЗВИЛСЯ -В ТЕЧЕНИЕ ЭВОЛЮЦИИ МНОГОКЛ-ЕТОЧНЫХ.
форм, иперед гнами встает вопрос о том, как произошла эта эво-
люция онтогенеза» (1939, стр. 561-562).
Ответ на этот вопрос могут дать, ввиду отсутствия каких-либо
палеонтологических свидетельств, одни только сравнительно анато-
мические данные, ипрежде всего данные, касающиеся колониаль-
ных форм жгутиковых, «которые в своей эволюции остановились на
стадиях, близких к вымершим переходным форгмам между Protozoa‘
и Metazoa‘, что дает нам возможность восстановитьприблизительное
строение и развитие анцестральных форм первичных многокле-
точных». т ‘ |
Современной моделью одной из самых примитивных «среди таких
переходных форм Северцов считает колониальную ‘фитомонаду
Pandori'na' morum. «ЗДЕСЬ мы имеем нечто анало.гичн0е дроблению
яйца у многоклеточного’ животного, причем дочерние клетки не рас-
ходятся, а образуют новый многоклеточный (16-клеточный) взрос-
лый организм. Аналогия с эмбрионом Мета2оа, называемым мору-
лой, ясна: каждая клетка проходит определенный ряд стадий,
в данном случае четыре стадии, прежде чем-произвести новый мн-ого-
клеточный организм‘. Мы видим, что онтогенез здесь очень короток
и что он происходит путем‘ надставок стадий (анаболии): из одно-
клеточной стадии произошла двуклеточная (первая надставка), из
24‘
двух-клеточной — четырехклеточная (вторая надставка)
и т. д.>›. _ _
Следующий этап усложнения онтогенеза олицетворяется много-
клеточными бластулообразными колониями Volvox, y которых «мы
имеем начало разделения тела многоклеточного, организма на заро-
дь1шевь1е- клетки и сому», так что «здесь переход от Protozoa к Ме-
tazoa еще более ясен, чем у Pa_ndorina>>. Подробно разобрав особен:
ности строения и развития Volvox, Северцов формулирует общин
вывод, что «в колониях одноклеточных существует очень простои
онтогенез, происшедший путем последовательных надставок -стадии;
онтогенез этот очень короток у Pandorina, гораздо длиннее
у Volvox, НО в обоих случаях он протекает по одному и тому же
типу: сначала ряд надставок до стадии шарообразной многоклеточ-
ной колонии, затем стадия гистологической дифференцировки;
в общем это типичная анаболия».
СТЭДИЯ
ПЕрЕХОДЯ далее K ра-ссмотрению РЭЗВИТИЯ НИЗШИХ Metazoa, СЕ-
верцов констатирует, что оно удлинлилось, «т. е. к -стадии -шаровид-
ной пМНОГОКЛЕТОЧНОЙ колонии прибавился ряд новых стадии». Иллю-
стрируя это положение примером онтогенеза гидры, автор описывает
его ‚следующим образом: «Одноклеточная исходная стадия представ-
лена здесь оплодотворенной яицеклеткои. Эта клетка многокроатно
делится, и из нее получается сначала шарообразный клеточныи за-
родыш (морула) .а затем зародыш с однослоинои стенкои и по-
лостью внутри (бла-стула). Стадии дробления приблизительно
соответствуют стадиям развития колонии одноклеточных (Рапооппа,
Volvox), НО затем внутрь полости бластулы выееляются отдельные
клетки вбластодермьт, которые образуют второи, внутреннгиИ Од-Н?‘
клеточный слой. Зародыш из одно-слойного превратился в двуслои-
ный — в гаструлу с отверстием (первичнымртом, или бластопором).
Одновременно в этой гаструле появляются полые пальцевидные
выросты вокруг бластопора, и гаструла-превращается в нмаленькУю
гидру с эктодермой, энтодермой‘, гастральной полостью, бласто-
пором и щупальцами. Этим морфогенез гидры заканчивается
иск нему прибавляется только стадия роста и гистологической
дифференцировки; Мы видим, таким образом, что онтогене-з этои
примитивной формы многоклеточного животного УдЛИНИЛ'СЯ‚НеСК0ЛЬ'
КИМИ НОВЫМИ ‘СТНДКИЯМИ, ‘ПРИОЭВЛЕННЫМИ K ДЕфИНИТИВНОМу ‘СОСТОЯНИЮ’
колониальных форм Protozoa. Еще большее удлинение ‚онтогенеза
мы находим у более высокой формы‘ кишечнополостных, например,
у ктенофоры». ` '
Приводя еще ряд примеров усложнения онтогенеза Metazoa
путем анаболии, Северцов формулирует положение; «ЧТО НадСТаВКа
стадий (анаболия) есть действительно первичныи тип эволюции
многоклеточных животных, тот тип, благодаря которому у них мало-
помалу развился особый начальный период жизни — онтогенез, или
эмбриональное развитие. Этот период жизни не существовал у иХ
одноклеточных предков, был очень короток у первичных колониаль-
ных форм и постепенно удлинялся по гмере ОСЛОЖНЕНИЯ Организации
25
‘у высших многоклеточных». Отсюда следует важнейший вывод-т
о том, что «самое изменение этого онтогенеза пу-
тем надставок было способом эволюции_-мно-‘
г о к л е т о ч н ы х ж и в о т н ы х», и имело своим результатом ре-
капитуляцию признаков предков в онтогенезе потомков.
В дальнейшем ходе эволюции к этому первичному способу лпри-
бавил-ся ряд новых, вторичных способов — положительные и отрица-
тельные архаллак-сись1, девиации и гетерохронии. «Эти вторичные
изменения хода онтогенеза гменяют ход первичной эволюции и при-
водят к редукции последствий первичного способа, т. е. к редукции
рекапитуляций».
Общие выводы Северцова по интересующему нас вопросу резю-
мированы им в девяти тезисах, которые мы считаем нужным .при-
вести здесь полностью ввиду их выдающегося значения.
<<1. У одноклеточных предков Metazoa не было пе-рлиода эмбрио-
нального развития: после каждого размножения делением у дочер-
них клеток наступало взрослое состояние и новый период размно-
жения делением.
2. Начало эволюции многоклеточных состояло в образовании ко-
лоний ‚одноклеточных, происшедших дихотомлическим делением ама-
теринской клетки, причем дочерние клетки не расходились в разные
стороны, «а оставались" вместе, образуя двух-, четырехц восьми- и
шестнадцатиклеточную колонию (Pandorina), каждая из клеток ко-
торой была спо-собна таким же ДиИХОТОМИЧЕСКИМ делением дать
начало такой же нов-ой колонии. V
3. Первая дифференцировка в колониях, состоящих из большого
числа клеток,.заключалась в разделении всех, клеток колонии на
половые (с неопределенной потенциальной продолжительностью
жизни)» и на соматические. '
4. Первые стадии эволюции колоний одноклеточных состояли
в увеличении числа кл-еток колоний путем дихотомического деления
до образования многоклеточной колонлии. Pandorina и Vol-vox оста-
новились на этой стадии эволюции. в
5. У низших кишечнополостных прибавились еще две стадии
эмбрионального развития: к стадии морулы прибавилась стадия по-
лого шара с однорядной стенкой (гбластула) и= стадия двуслойного
бокала с щупальцами вокруг бластопора (гаструла).
6. У высших кишечнополостных (путем архаллаксиса) разви-
лась мезодерма и прибавился путем надставок ряд новых ‘при-знаков:
stomodaeum, билатериально симметричное строение тела, статоци-
сты и 1т._д., т. е. их эмбриональное развитие удлинилось еще на
несколько конечных стадий.
7. Таким образом, у потомков примитивных Metazoa. K концу
онтогенеза прибавляются все новые и новые конечные стадии и тем
самым удлиняется первичный онтогенез. У Protozoa онтогенеза
в собственном смысле этого. слова, т. е. онтогенеза путем деления
клеток, не было. Колониальные формы одноклеточных остановились
на стадии морулы. Coelemerata JIQIIIJH/1 до стадии: гаструлы- и т. д,
36
Мы видим, что онтогенез есть процесс, развившийся в течение (mino-
генеза путем1 надставок конечных стадий и прив-едший к усложне-
нию строения Metazoa. Этим путем развились первичные рекапиту-
ляции признаков предков. .
8. Этот первичный способ эволюжции в течение времени изменился
вторично, причем способов вторичного изменения хода онтогенеза
было несколько: арахаллаксицдевиация, гетерохрония и т. д. Эти
вторичные‘ способы эволюции начали действовать очень рано, с са-
мого начала образования колоний одноклеточных, ‘и прпивели
к весьма крупным и существенным изменениям строения органов
взрослых многоклеточных. Мы видели, что первые дифференцировки
в клетках произошли уже у колониальных форм (дифференцировка
колонии на половые и соматические клетки); этим же путем вторич-
ного изменения появились новые системы органов (мезодерма
и т. д.). .
9. Главный вывод, к которому мы приходим на основании изло-
женного ряда фактов и гипотез, тот, что у одноклеточных не ‘было
онтогенеза, но что у их многоклеточных потомков постепенно раз-
вился эмбриональный период жизни сомы. Р а з в и т и е о н т о г е-
неза есть средство, которым была достигнута
эволюцтия новых, более высоких форм жизни».
Исключительное значение этих работ А. Н. Северцова для эво-
люционной эмбриологии беспозвоночных явствует из самого факта,
что четко поставленный им вопрос-«как возник и как эволюи-
ровал онтогенез многоклеточных животных» -— никем ранее не" ста-
вился в качестве самостоятельной проблемы. И хотя при такои его
постановке он и в.мещает`в себе все те вопросы (о происхождении
многоклеточности‚ первичных пластов тела и т. д.)‚ пКакИМИ ИЗдаВНа
занималась филогенетическая эмбриология, принципиальная важ-
ность его по-становки в новой форме И На Н0В0М‚ СеВеРЦ0В'СК0М‚
этапе развития эволюционной морэфологии чрезвычайно ВеЛИКа- Не
=будет преувеличением сказать, ЧТО ПРИВЕДЕННЫЕ ВОЗЗРЕНИЯ Север-
цова знаменуют собой поворотный пункт в теоретической разработке
проблем исторической онтогенетикли животных и открыли весьма
перспективные пути ее дальнейшего развития, _o
Правда, конкретное содержание предложенной РеверЦОВЫМ
схемы мало чем отличается, как мы видели, о-т таковои теории га-
стреи. В этом—-ее самое "слабое место, И=ЭТИМ‚ ОЧЕВИДНО, ОбЪЯъС-
няется то обстоятельство, что она не получлила широкого признания.
Однако для нас очевидно, что все присущие этой схеме недостатки
не способны умалить ее громадное, на наш взгляд, принципиальное
значение, A
Общепризнанная роль А. Н. Северцова в возрождении интереса
к большим проблемам филогенетической ‘морфологижи ясно сказалась
в появлении у нас в последние годы ЦеЛ0Г0 ряда ЕПЕЦИЗЛЬНО ТЮСВЯ‘
щенных им работ капитального значения. Особыи интерес для нас
представляют среди них известные монографии А. А. Заварзмна
(1945) и Н. Г. Хлопина (1946) по эволюционной гистологии и книга
27
А. Ливанова_<<Пути эволюции животного мира» (1945), .в каждой
ИЗ которых-подробно рассматривается -про-блема ПрОИСХОЖДЕНИЯ И *
эволюции многоклеточных животных, причем в двух последних из
этих работ данная проблема трактуется с точки зренгия колониаль-
ного происхождения Metazoa. V
Представления Н. Г. Хлопина (1946) по этому вопросу являются
в своеи основе не чем иным пкак сочетанием гипотезы плакулы
О. Бючли с основными положениями теории гастреи. «Можно ду-`
мать, — говорит ХЛОПпИН, — что в связи с условиями существования
на дне водоемов предки многоклеточных животных должны были
приобрести более или менее резко сплющенную в вертикальном
направлении форму или форму плосковыпуклой чечевицы с упло-
щенной нижней поверхностью. Вместе с тем в теле таких организ-
мов должна была возникнуть ориентированная вертикально система
градиентов, на основе которой должны были образоваться два кле-
точных пласта с неодинаковыми свойствами. Один из пластов все
ООЛЕЕ И бОЛЕЕ специализировался В направлении ПИЩЕВЗрИТЕЛЬНОЙД
функлции, приобретая. свойства, характеризующие в дальнейшем
энтодерму, а другой пласт, соответствующий области высокого гра-
дие-нта, приобретал особенности наружного эктодермального по-
крова тела с присущей этому последнему более высокой чувстви-
тельностью. Свертывание двуслойной пластинки пищеварительным:
клеточным пластом внутрь или, что почти одно и то же, инвагина-
ция дифференцирующейся энтодермы привели в конце концов
к образованию двуслойных организмов типа геккелевской- гастреи,
являвшихся предками современных кишечнополостных»; Принимая
занисточник образования энтодермы нижний, обращенный як суб-ф
страту, клеточный слой-плакулы, Хлопин объясняет несоответствие
ее ориентации той, какую мы видим‘ у ныне живущих сидячих кни-А
дарий‚ допущением, что <<двуслойная плакула или гастрея перешла
затем к плавающему образу жизни. Такие плавающие организмы
при переходе к сидячим-формам кишечнополостных стали уже при-
крепляться не оральным, а аборальным концом. Это не могло не
ОЫТБ‘ СВЯЗЭНО С ИНВЕрСИЕЁГ СИСТЕМЫ. Гр-ЗДИЕНТОВ В ОТНОШЕНИИ ОСИ-
'I‘6JIa ЖИВОТНЫХ». Однако-не и-сключенатакже И ВОЗМОЖНОСТЬ, <<ЧТО
ОТВЕрСТИЕ на оральном КОНЦЕ ПОЛИПОВ образовалось ЗЭНОВО, а пер-
ВИЧНОЕ, обращенное ‚ВНИЗ, ОТВЕрСТИЕ ИЛИ заросло ‘COB’6pII.I6HHO,- ИЛИ
СОХрЁНИЛОСЬ В‘ ВИДЕ ‘НЕЗТНЭЧИТЕЛЬНОГО остатка, обнаруживающегося '
вподошве НЕКОТОрЫХ СОВРЕМЕННЫХ полипов».
‘Попутно отмечая далее, что «полая внутри Шарообразная бла-_
стула и морула как стадия онтогенеза современных форм должны
были возникнуть вторично в связи -с переходом к развитию в ‚пела-
гических у-словиях», Хлопин. подробно останавливается на вопросе
о возникновении клеточной и тканевой дифференцировки у, первич-
ных Metazoa--. Касаясь мечниковской теории паренхимеллы, Хлопин
считает ее необоснованной: «B KOHIL-6H‘I1HK.M61IH'HKOIBa мало понятным
и в‘ гистогенетическом отношении м-ало оправданным моментом
является образование энтодермы (ФЭГОЦИТОбЛЭСТа) В Виде ВНУТР®Н*
28
ней клеточной массы, оттраниченной от внешней среды пластом
кинобласта. Неясно, почему фагошитирующие элементы этого по-
следнего должнымигрироватв ‘внутрь тела для выполнения пищева-
рительной функции. Приэволюционно-м разви.тии тканей и органов...
мы нигде ‚не видяим с достаточной убедительностью превращения
элементов внутреннейтсреды организма в пограничные с внешней.
средой образования... Наоборот, весьма обычным и частым является
обратный процесс-процесс эволюционного превращения .покров-
ных и разграничительных образований в ткани внутренней среды
организма».
Собственные воззрения Хлопина на этот счет заключаются в сле-
дующем: «Как гИ в онтогенезе современных форм, клеточные эле-
‘МЕНТЫ ПЕРВИЧНЫХ МНОГОКЛЕТОЧНЫХ ДОЛЖНЫ бЫЛИ ПОСТЕПЕННО ‚приоб-
рести Морфофизиологическую-характеристику гетерополярных эпи-
телиальных клеток, тесно сомкнутых в первую очередь своими
апиикальными концами. Базальные концы клеток, возможно, образо-
вали цитоплазматич-еские отростки, соединявшиеся друг с, другом.
За счет этих базальных частей элементов экто- и энтодермывоз-
никли в дальнейшем образования типа опорной пластинки и мезо-
глеи, а также нервные и мышечные продукты дифференцировки
цитоплазмы. Неодинаковые свойства апикальной и базальной
-ча-стей ЭНТО- И ЭКТОДЕрМЫ развиваются, ОЧЕВИДНО, В СВЯЗИ С первич-
ной системой ‚градиентов, ориентированных нормально к поверхности
раздела между организмом и внешней средой». Подчеркивая затем
то обстоятельство, что «тело простейших современных кишечнопог
лостных уже содержит в своем составе покровные, железистые,
трофические, опорные, мышечные и нервные образованиядсоответ-
ствующие в функциональном. отношении элементам всех четырех
ОСНОВНЫХ групп тканей КЛЭССИЧЕСКОЙ 'ГИСТОЛОГИИ>>‚ автор приходит-
к выводу, что эктодерму и энтодерму низших кишечнополостных
следует рассматривать с точки зрения их морфофизиологической
характеристики «не. как эпителиальные ткани, а как своеобразные
мультифункциональные тканевые сгистемы, каждая из которых объ-
единяет B себе особенность трех или всех четырех общепринятых
груп.п тканей яболее высокоорганизованных животных».
"эволюционную дифференцировку примитивной энтодермы. гастрееподобньхх
гипотетических предков Можно себе представить, по Хлопину, следующим обра-
зом. «Клетки эктодермы первоначально в более или менее равной степени при-
обрели некоторые морфологические особенности нейроэпиггелиальных и миоэпи-
телиальных. чувствительных клеток, базальная часть _тела которых, возможно,
имела отросток или отростки, обладавшие известной степенью сократимости и:
вместе с тем способностью передавать получаемые апикальным концом клеточ-
ного тела раздражения отросткам других таких же элементов. Часть та-ких
примитивных «мионейроэпителиальных» элементов в дальнейшем стала равви-
‘ваться В направлении рецепторных нейроэпителиальньтх КЛЕТОК И утратилаг- СО-
кратимость, а другая их часть, наоборот, специализировалась в направлении
сократимых мир-эпителиальных элементов». Из рецепторных нейроэпителиаль-
ных. клеток затем дифференцировались первичные нервные элементы, а- мио-
эпителиальные развились ‘в мускулатуру соматического типа. При дальнейшем
УСЛОЖНЕНИИ организма H13 ЭКТОДЕрМЫ ЖЕ ВЫДЕЛИЛИСЬ ЭЛЕМЕНТЫ, дав-шие НЗЧЗЛО „
29"
тканям ‚внутре-ньней среды 1, в образовании которых, однако, кёдва/лй можно
исключить участие первичной энтодермы». «Таким образом, возможно себе:
представить, что элементы будущих тканевых систем должны были первона- _
чально появиться путем дивергентной дифференцировки клеток двух клеточных
пластов, эктодермы и энтодермы, из которых состояло тело. примитивных пред-
ков современных многоклеточных животных». -
Преемственная связь комплекса тканей высших Metazoa с обеими ‹<прими- .
тивными тканевыми системами» кишечнополостных не внушает никаких сомне-
ний. «Эктодерма и энтодерма сравнимы во всем животном мире»-—говори1?
Хл-опин. И так как они, по ето мнениюп, возникают одновременно в результате
свертывания или инвагинации плакулы‚-то все другие способы гаструляции дол-
жны считаться вторично видоизмененными. «Мне кажется, что паренхимуле в
развитии многих современных кишечнополостных едва ли возможно придавать
филогенетическое значение стадии, рекапитулирующей организацию предков
многоклеточных животных. Способ превращения паренхимулы в планулу путем
образования щелевидной поло.сти в несообщающейся с внешней средой энто-
дермально-й клеточной массе также не может рассматриваться, как сохраненный
в онтогенезе древнейший филогенетический момент. Г аструляция путем унипо-
лярной иммиграции, наблюдающаяся у некоторых Hydrozoa, является, по всей
вероятности, вторично измененным споообом образования энтодермы. То же са-
мое справедливо и ‘в отношении различных видов деляминации, а также муль-
ти.полярной иммиграции, которые возможно, следует трактовать, в противопо-
ложность взглядам Мечникова, как результат, с одной стороны, вероятного
сдвига момента обособления энтодермы и эктодермы наг более ранние стадии
онтогенеза, ас другой, накопления в яйцах питательного: желтка, но не как,
примитивные формы их развития». Это вытекает из того, что «филогенетиче-
ское развитие энтодермы вне непосредственной связи с окружающей средой...
малопонятно и необъяснимо на основе несомненно имеющего место взаимодей-
ртвия 7внутренних и внешних факторов в процессе организменной эволюции».
стр. 6). `
Как ясно вид-но из приведенных высказываний Хлопина, он
во многом разделяет взгляды Г еккеля, и развиваемые им пред-
ставления мало чем, в сущности, отличаются от теории гастреи.
Симптоматично, что все фактически наблюдаемые у низших Metazoa
способы образования энтодермы расцениваются им, как вторичные
и что вся его концепция на-правлена против теории Мечникова.’ Это
и понятно, так как все возражения против теории гастреи, сделан-
ные в свое время Мечниковым, попрежнему остаются в -силе.
На базе той же теории «гастреи построены и ‘представления
Н. А. Ливанова (1945) о происхождении Metazoa. Однако, если
Хлопин исходит |ИЗ первоначального, геккелевского, варианта этой
теории, то Ливанов целиком опирается на ее поздней-шее видоизме-
1 Важно отметить, что, по мнению автора, «промежуточную опорную пла-
стинку и студенистую мезоглею низших кишечнополостных нельзя рассматри-
вать, как самостоятельные тканевые образования, существующие наравне с
эктоделрмой «и энтодермой. Они являются производными одной из двух или обеих
первичных тканей кишечнополостных и должны рассматриваться, как продукт осо-
бой` дифференцировки базальных поверхностей пограничных клеточных пла-
стов. В состав основного вещества мезоглеи низших кишечнополостных могут
вторично выселяться в небольшом количестве свободные клеточные элементы,
не образующие тканевой структуры... Поэтому ее основная масса не может
безоговорочно. сопоставляться с межклеточным веществом мезенхимных сое-
динительных образований; с которыми она сравнима лишь в известной степени
по своейдморфофизиологической характеристике ' (положение внутри орга-
низма,‘ опорноезначение); но не`по опоообу сво.его развития».
30
пение — гипотезу Ланга. Единственное существенное измене-нив,
пносимое им в эту гипотезу, заключается в приписывании студени-
стому основному веществу колониальных предков Metazoa (и про-
изошедших от них бластеи, прогастреи итастреи) ведущей роли
в их эволюции, в признании за этим студнем исключительного инте-
грирующего значениядВозникая путем дегенерации экзоплазмы
клеток колонии, студень становится первоисточником основного
вещества внутренней среды вышестоящих в системе форм.
Таким образом, в гипотезе Ливанова находит свое отражение и
ранее высказанная им «теория галлерты» -(1913), как первично-
неклеточноиткани внутренней среды, дополняющая собой разви- .
ваемые им взгляды Ланга.
В ЗЗКЛЮЦЧЕНИЭ СЛЕДУЕТ упомянуть об относительно недавней ра-
боте Л. Хаимэн (1942), специально посвященной вопросу о проис-
хождении Metazoa. Интерес этой работы заключается в том, что она
представляет собой попытку обосновать новыми данными прини-
маемую автором теорию Мечникова и увязать ее с современными
представлениямио физиологических градиентах. Преобразование
вольвоксообразоной анцестральной колонии Flagellata ‘B целостный
многоклеточный организм определилось переходом от двухмерного
меридианального дробления к трехмерному, т. е. установлением
экваториальной плоскости деления. Ее появление явилось в свою
очередь результатом приобретения колонией тетерополярности,
и само способствовало дальнейшему усилению’ последней, вь1разив-
шемуся в постепенной физиологической дифференциации клеток
блцастулы согласно еепередне-заднему осевому градиенту. В даль-
неишем бластула превратилась в паренхимлеллу ‘путем иммиграции
части ее клеток внутри бластоцеля-«Мой собственный вывод из
рассмотрения всех фактовёпишет Хаймэн, — заключается в том,
чтомечниковская теория является самой правдоподобной».
5. Первые гипотезы неколониального происхождения
Metazoa
Сравнительно-морфологическую 0CHOBy I‘I/IIHOTC3 Неколониального
происхождения Metazoa составляет старинное представление о икон-
структивной равноценности олсобипростейшего целому многоклеточ-
ному организму, о единстве общего плана строения всех животных.
Это положение, казавшееся само-собой разуме-ющимся до возник-
новения клеточнои теории и нашедшее свое ярчайшее выражение
в «принципе равно совершенной организацию-Г. Эренберга (1838),
имеет и теперь многочисленных сторонников, опирающихся на непо-
средственное взаимное сравнение различных сложно расчлененных
«инфузорий с коловратками и турбелляриями. Обилие устанавливае-
мых при этом сходств (см. Заварзин, 1945, а также Гелеи, 1936)
естественно подсказывает мысль, что многоклеточное строение Ме-
tazoa могло получиться путемпростого распадения на клетки вы-
COKO дифференцированных многоядерных простейших, т. е. в ре-
31
зультате одновременного обособления протоплазмы вокруг каждого
из их ядер. A » .
правдоподобность ‚такого допущения подтверждалась И эмбрио-с
логическими данными, а именношраспространенностью у Metazoa
различных форм поверхностного дробления, которые и могли — при
желании — служить моделью подобного перехода из многоядерного
в многоклеточное состояние. л
Еще в 1877 г. Иеринг и за ним С. Кент высказывались за воз-
можность непосредственного превращения многоядерных, прибли-
зительно опалинообразных ин-фузорий в многоклеточных Metazoa,
приводя в качестве эмбриологических свидетельств в ‚пользу этой
возможности такие примеры распадения симпласта на клетки, как
образование клеток бластодермы на ранних стадиях развития тлей.
Одна-ко, как справедливо указывал Мечников (1886), сами эти сви-
детельства «ясно показывают, что такая гипотеза не может быть
поддержана. Игнорируя все особенности развития низших Metazoa,
она согласуется только с образованием бла-стодермы у насекомых,
т. е. у таких животных, которые вторично видоизменены во всех
отношениях. Однако даже среди последнихесть формы, противо-
речащие этой гипотезе, как, например, подуры, занимающие НИЗШЭЁ
место во всем классе...». у
Особое положение среди разбираемых нами представлений, зани-
Мает гипот-еза А. А. Тихомирова (1887), выгодно отличающаяся от
остальных своей большей эмбриологической обоснованностью.
Исходя из тщательного изучения .развития целого ряда. гипогенети-
ческих гидрополипов, Тихомиров приходит к заключению, что ти-
пичной формой образования зародышевых пластов у гидроидов
является простая, как он ее называет„ делям-ина-ция (т. е. тот ее
тип, который обозначался Мечниковым, как вторичная деляминация,
и обозначается теперь Беклемишевым, как ‚паренхимная). И, по-
скольку «вопрос о происхождении пластов у гидроидов есть, конечно,
одновременно и вопрос о происхождении пластов у всех Metazoa»,
нет, по «мнению автора, «никакой причины не допускать деляминации,
как основного способа образования пластов у всех Metazoa». Эти-м
определяется скептическое отношение Тихомирова к «весьма pac-
простране-нной теперь в эмбриологии» ‘гипотезе о происхождении
.Metaz0a OT KOJIOHI/IaJIbHbIX ЖГУТИКОВЫХ. .кажется «СОВЕРШЕННО
произвольными все гомологизирования разных стадий дробления
яйца с колониями простейших животных». «Я не думаю, например,
чтобы мы имели основания полагать, что онтогенетическим стадиям
деления на 2, 4, 8 „соответствовали непременно 2, 4,.8 ...n членные
колонии каких-то давних Protozoa —— предполагаемых родичей со-
временных Metazoa. Весьма возможно, например, что давними роди-
очами современных-низших M-etazoa были большие плазматические
существа, которые, ДОСПИГНУВ ИЗВЕСТНОЙ ВЕЛИЧИНЫ, дсразу преврати-
„лись в многоклеточные организмы; так ‚что, может быть, между
‚филогенетической стадией, соответствующей нераздробившемуся
„яйцу, И стадией, соответствующей моруле, не было «переходных
32
стадий. В том моем предположении нет ничего невозможного; быть
может, например, ближайшее знакомство с механизмом деления
клеток покажет нам, что, при известных условиях, деление на 2 вле-
чет за собой немедленное же деление на 4 и т. д.>›.
По мнению Тихомирова, шпоггеза колониального происхождения Metazoa
«трудно примирима с нашими представлениями об индивидуальности.‘ Гипотеза
ma требует допущения, что самостоятельный член колонии, т. е. самостоятель-
ное животное, нисходит до значения части животного, стоящего на высшей
ступени генетической лестницы. На каких аналогиях можем мыосновать это
допущение? Не проще ли себе представить, что плазматическое существо,
бывшее родичем той или другой ветви современных Metazoa (или всех их), в
известное время рас-п-алось на определенное количество составных частей
(клеток), которые никогда не были самостоятельными животными, никогда не
составляли никакой колонии, а всегда. были лишь частями животного орга-
низма. Впоследствии между этими составными частями началась определенная
дифференцировка, которая и привела к тому, что мы называем теперь обособле-
нием пластов. При этом деляминация представляется, конечно, простейшим спо-
собом этого обособления пластов» (цит. раб., стр. 613-641).
Очевидным преимуществом концепции Тихомирова по сравнению с остальными
ГИПОТЕЗЗМИ НЭКОЛОНИЗЛЬНОГО ПРОИСХОЖДЕНИЯ Metazoa ЯВЛЯЕТСЯ ее серьезная эмбри- ‹
ологическая мотивированность, опирающаяся к тому же на значительно более
подходящий для этой цели и гораздо более обширный фактический материал,
чем тот, которым пользовались Иеринг, иСеджвик и другие. Это — по сути дела
единственная попытка обосновать «неколониапьную» концепци-ю со строго эмбри-
ологичеокои точки зрения, попытка, несомненно, интересная и заслуживающая
внимания. Однако она осталась до сих пор почти незамеченной; работу Т.ихоми-
рова теперь мало кто знает, и ее странным образом протлядели все современные
сторонники кполиэнерпидной теории, для которых она явилась бы настоящим кла-
дом и откровением. ‚
ОДНЗКО ТСОРЁТИЧССКИЕ ВОЗЗрЕНИЯ Тихомирова ПО I/IHT€p'€Cy}OI.IJ,eMy
' нас вопросу должны быть признаны несравненно менее убедитель-
ными, чем оспариваемые им представления Мечникова. Слабым
местом концепции Тихомирова является самъо ее основное поло-
жение о первичности «простой» деляминации по сравнению -с дру-
гими способами образования зародышевых пластов, а, стало быть,
и о первичности -стадии морулы сравнительно с целобластулой.
В самом деле, доказать это совершенно недоказуемое положение
Тихомиров-у так и не удалось и его попытки сделать это могут быть
названы бе-спомощными. Заявляя, что он «готовсчитать» образо-
вание целобластулы с последующей иммиграцией клеток из стенки
внутрь ее полости <‹спо-собом вторичным, вызванным особыми усло-
вия-ми личиночной жизни», Тихомиров приводит в пользу этого
своего предположения только следующие соображения:
1) «По наблюдениям Ковалевско-го, Мережковского, особенно же Мечникова,
все личинки Craspedota, имеющих перемежающее размножение (т. е. метагене-
_ тические гидромедузы.——А. З.) в период окончательного дифференцирования пла-
стов, упорно возвращаются к стадии плотной морулы, столь характерной для гид-
роидов, не проходящих стадии бластулы». , '
2) Признавая, что из сравнения истории развития медуз с историей развития
изученных им гипогенетических гидроидов «можно было бы легко притти к мысли,
что развитие первых идет проще, а потому и ближе к первоначальному типу»,
Г ихомиров все же думает, что «простота развитиямедуз есть только кажущаяся,
и то обстоятельство, что мы видим здесь почти всегда бластулы с большими по-
д 3 Эмбриология беспозвоночных 33
‚ _ ‚ . — ч - › ‘ т том -личйнки,-
„ . - - - - об словливается быстрым РОС ‚
лбстямач (т. Has:-,1 ugngiifiifigygggfiqecrgom пластического материала, заключаюшб‘
с одН0и СТОРОН э 3 есь-иногда рост доста-
- _ ° чтобы видеть, как быстро Идет д ’ -
гося в яице, С дРУГОИ- V - t 305 аженную-Мечнико-
. бластулю) Осеашаъ arma а, и Р
точно взглянуть на личинку ( нам что Многие
исунки показывают ‚
вым на одном из его рисунков-п, другие Р K ТОМ. же Sm.‘
° быстро вырастают НО еще У
личинки по выходе из яица Не ТОЛЬКО б из“ а „д-
- . чи-нают вести сво одНУЮ Ж :
чительно изменяют свою :1aopl\1’\IagroFr:)c:<I:[;J;I;L;oI нчато не может не влиять на Задержку
‘TON?’ Koflelflllflll/(I)’H:)J,{a’1l"FJ:laq(EI‘}:)a}:O'li3BHI[y сказанного возможно ПРОЁПОЛОЖИТЬ’ ЧТ° кажу"
В О Ра3°Ва ' V МИ с авнительно с яйцами,
ц отложенных медуза ‚ Р
_щееся упрощение В Развитии “H ’ именно теми физиологиче-
их гонофорах‚ обусловленно
развивающимися в CHM” азал т е малым количеством пластического
скими моментами, на КОТОРЫЕ Я УК т - - „ E же МЫ Tamm-
° быстрым ростом-с дРУГОи- Оли ’ '
материала, с однои стороны, и н Явление а Вторич.
бластуля медуз ЕСТЬ Не ПОРВИЧ Ое ’
образом, примем, что цело ЯЧИМИ Гонофдрами
_почему все гидроиды с сид г .
H09: To ‘нам легче будет ПОНЯТЬ › о азвитие как бЫ
ак бы они: ни начинали св е Р э .
pr все МетагенетичеЪрРкЁеДЁ/гоеёггёеккаков бы НИ был Состав их желтка’ В конце
схо лило - = ' ' › „
ЁЁЁДЁЁИ приъодятук стадии СТЭРРОГЗОТРУЛИ (“”°TH°“ “"aHy”H)»‘
Легко Видеть, как недостаточны эти доводы, чтобы поколебать,
_ . что опровергнуть прекрасно обоснованные взгляды
а Не ТОЛЬКОИ я п едложенная Тихомировым точка зрения несо-
1T1e:1?1:do{?cBe;:).be3H::T остальных гипотез ‚неколониального ПРОИСХОЖДеТ
.ния Метаиоа, она только лишний раз СВИдеТеЛЬСТВУеТ О ‘He°‘0CT°H_
тельности этой концепции В ЦЕЛОМ‘ . _-
Ha работе Тихомирова фактически обрывается раёрёаэбзтиибарёёёё;
лониальной теориИ ПРОИСХОЖДЕНИЯ МНОГ°КлеТ`°’ЧНЬ”Е только сравню
гическом аспекте. Отныне она траКТУеТСЯ В Одноиблемами Эволюд
тельно-анатомической плоскости, вне связи с про
ционной онтогенетики.
' д ь ого
5_ последующее развитие гипотезы неколониал н
происхождения Metaloa
. H a
т
гипотеза неколониальногомсёр Игьельньхх И Недоказуемых предполд
nepezrlllf JIa3l'3ro0‘6Jc::lI(:1I;<o{ K(:1Te0 1')I,0MeI.EIaJIO ей иметь многих сторонников,
ЖЕН И- 9 ' _ _ д . _ И
да-же получить в работах А. А. 3аваР3И'На ( ущ ‘
и оригинальное „содержание-
° еи связанное с ИМЕНЗМИ.
Основное направление. развития этои ид ‚
„Х жи 1944 оказалось
П_Де.ляжа (1896)‚ с. в. Аверинцева и И ад ( )‚ Г п Инд
правда мало плодотворным. Оно, в сущности, ‚не внесло ниче о р
’ г . ента кп одолжая опи-
Ципиально нового в кОНЦепЦИРЁЁШЁЁЁЁЁГЁЛЁЕЁЮК JIe“’{OCpTb ‚выведения
раться’ как И прежде’ На ‘совённого многоклеточного из много-
«готового», уже диффер-ВНЦИР ' › М ‚е_
рного и инфузориеобразного «морфологического .пред1<а>>- Од
яде - ‘ _
- ИЛИСЬ: ЛЯ ДеЛЯЖа
лями такого первичного многоклеточного яв д S 1, leua
мифическая (или же явно ошибочно интерпретированёзёэе :1‘/in Ка];
с мат ива
Для Хаджи- реальные Typfienmzfpnu, ЁЁ сазомр ТУ) тРУдНо гово-
самые примитивные из Metazoa. аким р ‚
34
рить о каком-либо заметном п
ЭЪИХ НОВЫХ ‘вариаций на стару
ee аргументации.
Уже упомянутая работа Иована Хаджи
poI*peC*oe,.1»I ЕДИНСТВЕННЫМ КОЁЫрЕМ
Ю ТЕМУ ЯВИЛОСЬ НЕКОТОРОЕ ОСВЕЖЕНИЕ
(1944) Интересна для
нас В ТОМ ОТНОШЕНРИИ, ЧТО ЯСНО ПОДВОДИТ ТЕ ИТОГИ|‚ K КОТОРЫМ ТЕПЕрЬ.
пришла полиэнергидная (как ее называет автор) гипотеза проис-
Х0Ждения Metazoa. По мнению Хаджи, ее преимущества перед коло-
ниальной теорией Несомненны и заключаются в том, что она:
u1)U;Le.z1a~eT понятным укрупнение размеров тела еще на прото-
зоинои ступени развития;
2)цоблегча‚ет допущение непрерывной преемственности органи-
ческои целостности, или индивидуальности, при переходе в много-
клеточное состояние (возникновение которой заново оказывается
поэтому излишним);
3) объясняет непосредственное перенесение органелл простей-
ших, в качестве частей целого, в состав тела многоклеточных; и на-
конец, _
4) облегчает понимание происхождения половых фаз Metazoa
путем прямого преобразования половых энергид в половые же
клетки. -
Достоинством этой теории Хаджи склонен считать и то, что она
не только не вступает в «противоречие с господствующими в эмбрио-
логии взглядами на природу дробления, но и`даже как бы освобо-
ждает их от давления клеточно-колониальной теории. Очень пока-
зательно в этом смысле Высказываемое автором убеждение, что
_<<ДрО'бЛЕНИЕ.В ЦЕЛОМ И СВЯЗЭННЫЕ С НИМ МОргфОПЕНЕТИЧЕСКИЕ ПрОЦЕССЫ
_ ОТНОСЯТСЯ K ‘НОВОПРИООРЕТЕННОЙ ТЕХНИКЕ онтогенеза» И ЧТО «ПРОЦЕСС
целлюлации лишь впоследствии был перенесен на самые ранние
стадии онтогенеза». Отсюда становится понятным, что живучесть
полиэнергидной теории происхождения многоклеточности не -слу-
чайна, а в значительной мере объясняется возможностью ее мир-
ного сосуществования с самыми механистическими представлениями
современной общей эмбриоло_гии.
Переходя теперь к изложению и новейшей из гипотез неколо-
ниального происхождения Metazoa —— гипотезы А. А. Заварзина, мы
прежде всего должны отметить, что в основу ее положено то же,
в сущности, представление об организменной равноценности про-
стейших и Metazoa, на котором базируется и полиэнергидная теория.
Полагая, что «в группах одноклеточных Protozoa И многоклеточных
Metazoa органическая интеграция проделала очень сходную и
длинную эволюцию и что в отдельных случаях эта интеграция про-
.ЯВЛЯЛаСЬ ЕПОЧТИ ТОЖДЕСТВЕННО, НЕСМОТрЯ на ТО, ЧТО В ОДНОМ случае
интегрируются и дифференцируются части одноклеточного тела,
а в другом——ткани и органы многоклеточного организмам-За-
варзин высказывает предположение, что «разделение =на два эволю-
Ционнь1х ствола: протистов и Metazoa, произошло еще до возник-
повения клеточной организации. Относительно механизма этого
С разделения МЫ НИЧЕГО СКЗЗЭТЬ НЕ ПМОЖЕМ. МОЖЕМ ТОЛЬКО ПРЕДПО-
3* ч 35
-» . . с -- - - " оказалисбгр
лагать, что в одних случаях массы живого ВЁЁЁЁЁЁЁ СЛОЖНЯВ:
ИНТеГРИРОВаНЫ Таким Образов/Цчто Для осгачгёчной одноётлеточная"
ш-егося мета-болизма для НИХ Оказалась ДОС -
или ве нее (в смысле Добелла), неклеточная организация, а для-
fl y;"1z1xpcTa.ua возможна сразу МНОГОКЛЕТОЧНЗЯ Организация‘ Таким
оъбразом к моменту возникновения клеточной структуры в совре-
менном ее значении первые гетеротрофные организмы были уже
и одноклеточнылми и многоклеточными, и дальше параллельно pas-'
. bl.
BII/IBa.JII/ICb, увеличивая свою органическую дифференцированност ‚-
каждый в своем организационном типе». 04
В пользу этого недоказуемого, как говорит сам автор, ПреДПОЛЯШ
жения приводятся различные косвенные подтверЖдеНиЩ СВ°д_
щиеся в конечном счете к попыткам показать несостоятелырёст;
теории колониального происхождения MetaZ0a- HQ мнению аВ р ’
. у п оис-'
«колониальным способом весьма ПраВДОПОДОбНО ОбЪЯСНЯЁТСЯ Р ля
хождение многоклеточных растении», ПОСКОЛЬКУ Специфические д
, ° к х колониаль-
них черты интеграции носят явственныи отпечато и п ОЧ‘
ного прошлого. «Этот способ происхождения наложил очень р
U ' . ° е самых высших
ныи отпечаток на характер интеграции ЧаСТеИ Ёёёаётся Неизмеримо
из современных растении, организм которых ЁИЗМ ЖИВОТНЫХ Даже
большей независимостью частеи, не? оргЁТсЯ также у растений;
градиентная интеграция которая ‚На Люда б ЖИЫ
носит здесь несколько иной характер, так как градиенты о- нарУ
ваются на точках роста, т. е. связаны с явными различиямли м0Р'ф0'
ч U , , а а а к оме того
Логическои дифференцировки клеточного сМаТзЁчгаЧчиЁель-нойр автоном:
градиенты в различных частях отличают я Тныеше
ностью» В противоположность этому, многоклеточные живчо
обна живают никаких признаков колонпиальности в своеи интегра-
111/11/1>>p)i/IM свойствен совершенно иной, градиентный тип интеграции‚
' * U "ш х многоклеточных
проявляюЩиися сходным ОбРаЁОМ У ПРОСЁЁИВИЁНЬЁЙ|И Общий ЖИВЫМ
и представляющий собои «какой-то оченьоисрождения» Опираясь На
организмом Механизм очень Древнего пр фного питания по сравне-
Теорию Опарина О первичности Гетероагзрзвает едположение что
“mo С аВТОТРОфНЫМ’ Заварзин выси бр 1й эквивалент’,
<<г адиентная интеграция, иЛи‚ ВЕРНЩ ее Перво “TH” ’
].[O€I}KHa была возникнуть одновременно с появлением первых прич
ых живых с ЩЭСТВ И
Знаков ЖИЗНИ» Именно у первых ГеТЁРОТЁОЁЁестВе специцгического
в дальнейшем сохранилась только у Ёшентичность этого Типа ‘Инте;
для них интеграционного механизма. Д 3 З Нар
грации у простейших и Metazoa и составляет, ‘ПО МНеНИЮ amp И ’
. . осле них из колониаль-
непреодолимое препятствие к выведению п д Т ВОЗМОЖНОСТЬ
ных объединений простейших. Если предположить Э у 7
возникли‘ же после’
«то следует допустить, что многоклеточные У
низменн ю интеграцию
того, как одноклеточные приобрелиёёзёщёчиочных Заде ВЫВОДИТЬ
о ательно п оисхождение _
еЁакзих-то колонгхй одноклеточных ИНДИВИДУЭЛИЗРЁРОВШТНЫХ OP”
« . . ч отдельных ипдиВидУа-ЛЬ‘
низмов путем полного ПОДЭВЛЕНИЯ M
U v " остностью, Между те. ,
ностеи и замены их новои органическои цел
36
, с таким предположением стоят в резком противоречии приведенные
выше факты развития организменной диффер-енцировки у современ.
ных одноклеточных и многоклеточных существ, ‘а также гИ тот факт,
ЧТО даже у самых н-едифференцированных из современных много-
клеточных организмов, каковыми, несомненно, являются губки,
никаких признаков колониального прошлого ни в гистологических
структурах, ни в процессе эмбрионального развития не обнаружи-
гвается. Поэтому возможность одновременного происхождения И
одноклеточных и многоклеточных не только не исключена, но даже
представляется весьма вероятной с позиции наших современных
знании». ‘
Общие выводы Заварзина заключаются в предположении «об
одновременном с простейшимги и неколониальном происхождении
многоклеточных животных» и <<о более раннем, чем растения, проис-
хождении животных многоклеточных организмов, отнеся этот мо-
тмент к тому периоду, когда еще не определилась клеточная органи-
цзация. Мы имеем основания усматривать причину разнлицы и
JB строении клеток и в интеграционных механизмах растений и жи-
твотных в разном происхождении них многоклеточности: колониаль-
яном у растенийинеколониальном у животных». —
Излагая свою концепцию, Заварзин специально подчеркивает,
mo для ее построения не использовались эмбриологические данные.
«Мы не клали в основу ‘раннлих эмбриональных стадий,——пишет
вон, —— а сделали попытку подойти к оценке теорий (о происхождении
/Ме’па2оа.——А. 3.)‚ исходя из свойств существующих организмов,
лклеток и живого веще-ства вообще». Однако в следующей главе
своей книги (посвященной теории зародышевых листков, происхож-
‚дению и основам классификации тканей) автор приводит и свое
эмбриологическое вкредо. Совершенно правильно определяя эмбрио-
‚лоцию, как биологическую дисциплину, изучающую закономерности
онтогенеза ли его эволюции, Заварзин, однако ошибочно, считает, что
в «эмбриональном развитии имеются две стороны, два ряда биоло-
гических процессов. С одной стороны, при развитии яйца происхо-
-дкит ‚превращение его во взрослый организм. Этотпроцесс по своему
существу должен отличаться от филогенетического развития вида,
так как, вероятнее всего, появлениев филогенезе полового «процесса
в целом’ явилось результатом приспособительной эволюции уже
после того, как произошли многоклеточные животные, и нет ника-
‘max оснований придавать рекапитуляционное значение самым пер-
вым процессам эмбриогенеза. А между тем, эти первые процессы
в значительной мере определяют собой и последующие, на которых
уже вое больше и больше могут накладываться филог-енетически
обусловленные моменты. Наконец, обе эти стороны эмбриогенеза
в отдельных случаях могут то более, то кменее сильно нарушаться
различными n;eHoreHe3aM14, КОТОРЫЕ также ЯВЛЯЮТСЯ pe3yJIbTaTOM
приспособительной эволюции. Такимобразом, эмбриональный про-
цесс содержит два момента: момент становления многоклеточного
организма из единой яйцевой клетки и момент отражения в этом
з?
процессе филогенетического прошлого вида. Первый момент
является общим для всех животных; второй момент будет различен
для разных типов, классов си т. д.». При этом «весь эмбриональный
процесс в целом, очевидно, проделывает эволюцию, которая затра-
гивает обе стороны— и формообразовательные процессы и момент
отражения в них филогенетической обусловленности» (стр. 31).
Итак, общие всем животным формообразовательные процессы
эмбриогенеза-дробление, превращающее яйцо в многоклеточное
тело, гаструляция И т. д.‚—по самому своему существу должны
отличаться от филогенетического развития вида, не имеют никакого
рекапитуляционн-ого значения И, по мнению Заварзина, являются
не чем иным как процессами чисто онтогенетической механики или
даже техники.
Переходя, однако, далее к вопросу о происхождении зароды-
шевых листков и тканевой дифференцировки тела Metazoa, Заварзин
оказывается все же вынужденным строить свою точку зрения на
этот счет на основе мечниковской теории фагоцителлы и дать свое
филогенетическое толкование этих ранних эмбриональных процес-
сов и стадий. Учитывая большой интерес теоретических высказы-
ваний Заварзина по этому поводу, мы изложим их тут возможно
подробнее. '
«В отнош-ении первичного многоклеточного существа в его докле-
точном СОСТОЯНИИ для нас несомненным является только факт его
организменной интегрированности... При этом условии и у него
поверхностные И центральные ‘части находились с самого начала
в разных отношениях к внешнему миру. Весьма вероятно, что вна-
чале поверхностные и центральные части не были фиксированы и
могли меняться местами, но несомненно также, что живое вещество,
попадающее на поверхность, вступало в непосредственное взаим'о-
действие с внешним миром и -сразу же становилось в иные условия
по сравнению с теми, в каких оно находилось до выхода на поверх-
ность. Мы не знаем, как возникла в таком существе многоклеточная
структура, но можем с достаточным правом утверждать, что клетки
этого многоклеточного организма с самого начала оказались инте-
грированными. Мы можем также утверждать, что поверхностные и
центральные клетки оказались в разных условиях существования.
Весьма вероятно, что все вклетки обладали в одинаковой степени
способностью к амёбоидному движению, которую мы должны прти-
знать одним из первичных свойств клеток многоклеточного орга-
низма. Также весьма вероятно, что поверхностные клетки Momm-
меняться местами с центральными, но, ппопадая на поверхность, все
клетки неизбежно должны были попадать в такие. условия, в кото-
рых их метаболизм должен был обусловить полярную дифферен-
цировку. В самом деле, если организм представлял собой плотное
образование, тоодин ряд расположенных ‚поверхностно клеток
‘одним концом был обрачщен к внешней среде, а другим к внутреннеИ
массе» (стр. 44). ‘
«Основной функцией такого первобытного многоклеточного орга-
38
T""""”” П”
. низма ‚несомненно была функция питания... Восприятие пищи прежде
всего an поставило в разные условия поверхностные и внутренние
клетки: только поверхностные "клетки непосредственно соп.рикаса-
лись с пищей... Попадая на поверхность, клетки принимали форму
более или менее полярную, призматическую, а уходя внутрь, снова
становились аполярными, ацмёбоидными. Такой диморфизм вначале
должен был носить чисто функциональный характер, а .по суще-
ству все клетки такого многоклеточного интегрированного орга-
низма были одинаковы... Обусловленная взаимоотношениями с внеш-
ним миро-м лабильная функциональная дифференцировка на
поверхностные и внутренние клетки в процессе эволюции должна
была закрепиться. Такому закреплению, вероятно, много -способ-
ствовало то обстоятельство, что поверхностные клетки одновременно
с захватыванием пищи —— а, возможно, пи в связи с этой функцией ——
стали образовывать на «своей поверхности различные приспособле-
ния, превратившиеся в конце концов в жгутики... Появление жгу-
тиков Ha поверхностных клетках, им-евшвих вначале, возможно,
только трофическое значение, должно было привести к образованию
новой функции ——локомоторной, которая и закреплялась за поверх-
лностными клетками. В связи с тем, что тело многоклеточного орга-
низма с нашей точки зрения развивалось не как нколония, а как
интегрированное с самого начала существо, появившиеся жгутики
или реснички в своих движениях должны были подчиниться инте-
грирующему механизму, т. е. проявлять известную сочетанность,
известную ритмичность. Такой ресничный покров оказывался весьма
‘выгодным для организма, так как одновременно служил и органом
движения," и подгонял к поверхности пищевые частицы, и освежал
окружающую среду. Локомоторная функция поверхностного слоя
должна была послужить поводом к более резкой дифференцировке
поверхностных и внутренних клеток, которые сосредоточили.
в себе трофическую функцию... После наследственного закрепления
такой дифференцировки и появления отдельных камбиальностей И
у поверхностного локо.моторного слоя и у трофической внутренней .
клеточной массы в организме многоклеточного существа должна’
была осуществиться первая дифференцировка его тела на ткани.
При этом должны были появиться сразу две ткани; поверхностная,
или пограничная, отделяющая внутренние части, внутреннюю среду
такого примитивного организма от внешнего мира и одновременно
выполняющая локомоторную функцию и и-меющая эпителиальную
структуру, и ткань, состоящая из отдельных амёбоидных клеток,
несущих трофическую функцию, и рыхло выполняющая внутрен-
Ность организма, почему эту ткань можно назвать тканью внутрен-
ней среды. Таким образом, мы видим, что, если исходить из теории
неколониального происхождения многоклеточных животных и хо-
рошо обоснованной теории фагоцителлы, то вследствие взаимодей-
ствия‘ такой фагоцителлы с внешним миром ее недифференшнро-
ванное вначал-е на ткани тело должно-было неизбежно приобрести
тканевую ‘дифференцировку, выражающуюся в появлении погранич-
39.
НОЙ ТКЭНИ ЭПИТЕЛИЗЛЬНОГО СТРОЕНИЯ |И ТКЗНИ ВНУТРЕННЕЙ СРЕДЫ, СО-
ставленной из свободных амёбоидных клеток» (стр. 44——46).
Дальнейшая эволюция такой фагоцителлы заключалась в оформлении
кишечного, энтодерМального эпителия и соответствующем переходе от внутри-
клеточного пищеварения к полостному. Этот процесс рисуется Заварзину сле-
дующим образом: «Если допустить, что после дифференцировки организма на
поверхностный слой и внутренние клетки, последние сохраняют способность, _
Вклиниваясь между поверхностными клетками, выходить частично наружу, то, -
принимая ВО внимание ИНТЕГРИРОВЗННОСТЬ 0praHH3Ma, нетрудно ДОПУСТИТЬ, ЧТО
ЭТОТ ‘ВЫХОД МОГ ПРОИСХОДИТЬ на ОДНОМ КОНЦЕ ЕГО тела. Такое ДОПУЩЕНИЕ ВОЗ-
МОЖНО ‘СДЕЛШГЪ, ОЮНОВЫВНЯСЬ на эмбриональных ‘СТЗДИЯХ БНЕКОТОПРЫХ МЕДУЗ, у _
КОТОРЫХ ИМЕЕТСЯ ПОЛЯрНО-МИГраЦИОННаЯ гаструляция... ОТСЮДЗ УЖЕ МОЖНО переити
к таким формам, у которых на поверхности тела, наиболее часто сталкивающейся
с пищевыми частицами (а таковой, вероятно, был задний конец), образовалась
площадка наиболее частого выхода внутренних клеток. Весьма вероятно, что
в этом месте развился особый ресничный аппарат, который и начал загонять
сюда пищевые частицы. Вполне допустимо и предположение, что в этом месте
в массе внутренних клеток образовалось небольшое углубление, которое в даль-’
нейшем увеличивалось и образовало полость первичной кишки. Внутренние
клетки по отношению к внутренней поверхности этой полости оказались в том
же положении, в каком были все клетки тела более раннего предка по отноше-
нию к поверхности тела. Поверхностные ‘(ограничивающие кишечный просвет)
клетки заглатывали пищу и уходили внутрь переваривать ее, на их же место
становились новые и т. д. На этой почве возникла дифференцировка, аналогич-
ная первой дифференцировке на поверхностные и внутренние клетки. Поводом
к такой дифференцировке мог послужить постепенный переход от внутриклеточ-
ного пищеварения к полостному. Закрепившись наследственно, эта новая диффе-
ренцировка дала начало появлению кишечного эпителия, обеспеченного своей
камбиальностью, и выделению его из первичной единой ткани внутренней среды.
Таким образом, последняя, вступив вторично, притом в новых условиях, в непо-
ч
средственное соприкосновение с внешней средои, снова выделила из себя погра- Д
НИЧНУЮ ЭПИТЕЛИЗЛЬНУЮ ткань, Т. Е. снова ОСУЩЕСТВИЛИСЬ ОТНОШЕНИЯ, аналогичные
первичному происхождению кожной локомотор-ной поверхности и ткани внут-'
ренней среды».
«После образования кишки с полостньъм пищеварением и кишечного эпите-‘й
‘mm многоклеточные животные приобрели вторичную тканевую дифференцировку.
Пограничные ткани приобрели на этой почве различные функции. Кожный эпи-
телий, помимо ограничения внутренней среды, стал тканью локомоторной и вос-
принимающей внешние раздражения, а кишечный эпителий стал тканью, воспри-
нимающей пишу и передающей ее во внутреннюю среду. В связи с этим не могла
не измениться и функция ткани внутренней среды. Из ткани, непосредственн-0
воспринимающей пищу, она стала тканью-посредницей, через которую или, вер-
нее, посредством которой питательные вещества, уже переработанные rs КИШКЕ,‘
передаются в кожу, являющуюся органом движения... Кожный эпителий после-
образования кишки функционировал в трех направлениях: как пласт, отграничи-
вающий внутреннюю среду от внешних неблагоприятных воздействий, как орган;_
воспринимающий внешние раздражения, и как орган движения. На этой почве
возникла дальнейшая, уже третья тканевая дифференцировка тела мн-огоклеточ-
ного организма. В результате этой новой дифференцировки из кожного эпителия
возникли нервная система и соматическая мускулатура, в связи с-чем кожный
эпителий в значительной степени утратил свои локомоторные и чувствительные
(рецепторные) функции... Таким образом, после появления нервной системы
и мускулатуры в организме многоклеточного животного оказываются налицо все
четыре основных типа тканей, которые, несмотря на разновременное происхож-
дение, возникли в процессе эволюции настолько рано и имеют такое большое
историческое прошлое,-что у ныне живущих организмов почти совершенно не
сохранилосьпризна-ков, которые можно было бы истолковать как указания на
все те обстоятельства их развития, которые были нами приведены выше как бо-
лее или менее обоснованная гипотеза». (стр. 4б‚+51).
40
7. Вопрос о происхождении личиночных форм и метаморфоза
Metazoa
С господствующей теперь в эмбриологии точки" зрения всякая
вообще личинка представляет собой вторично видоизмененную ста-
дию развития —— «организм, вынужденный на более или Менее ранней '
стадии развития приспособиться к условиям свободного существо-
вания путем ранней дифференцировки необходимых для этого спе-
циально личиночных орга-нов» (Иванов, 1937, стр. 95), Hm, другими
словами,’ «зародыш -с преждевременно дифференцированной орга-
низациеи, достаточной для самостоятельной жизни» (Иванов, 1945,
стр. 55)... «личинка появляется в развитии животного потому, что
питающего материала, накопляюэщегося в ооците в период желтке.
образования, недостаточно для полного завершения зародышем
своего развития, и он выходит из яйца на ‘более или менее ранней’
эмбриональной стадии». Будучи -след-ствием недостаточного снаб-
женаия яица желтком, «развитие с личинкой является вторичным по
отношению к полному (т. е. завершающемуся, в основном" еще
В яице) развитию, так как разница между преждевременной
дифференгцировкой отдельных органов и образованием личинки
только количественная: известная стадия превращается в ли-
чинку потому, что значительная часть органов этой стадии
-H0JlB€]3‘I‘JIaCb преждевременной дифференцировке» (Иванов, 1937,
стр. 95, 735). -
В том же духе, хотя и на существенно иной базе, решается этот
вопрос и в некоторых, специально посвященных ему работах
(Гарстанг 1928, 1930), так что представление о вторичном харак-
тере личинки по сравнению с другими стадиями онтогенеза может
считаться господствующим в современной эмбриологии (сравните
также Де-Бер, 1940). При такой ‚постановке вопроса он рассматри-
вается, таким образом, вне -свя-зи с более общими проблемами онто-
генеза в целом. 1 _
Совсем в другом свете этот вопрос рисуется исследователями,
разрабатывающими общие вопросы эволюционной онтогенетики.
Так, И. И. Ежиков (1936, 1938, 1940) придерживается взгляда, что
«метаморфоз есть первичный для многоклеточных тип развития».
«Существо дличинки, —говорит он, — заключается не в наличии при-
способлении, отличающих ее от взрослой формы, а в эмбриональном
характер-е ее организации- Из изложенного ясно, сколь далеко
стоят от истины те авторы, которые склонны считать характерней-
шпм признаком личинки наличие у нее приспособительных upm-
знаков, а самый метаморфоз считают явлением, вторично ‚приобре-
тсппым» (1938, стр. 261, 263). По Ежикову, «наиболее примитивной
яйггеклоткой надо было бы считать такую, которая лишена оболочки,
обладает амёбоидным движением, самостоятельно питается и сво-
бодно живет в той же самой среде, что и` материнский организм
(например, морской воде);'если все последующее развитие имеет
примитивный характер, то каждая стадия развития ведет свободный
41
образ жизнпи и самостоятельно поддерживает свое существование, _' `
COBp€JM€HHbIe Metazoa не СОХРЗНИЛИ СТОЛЬ‘ ПрИМ|ИТИВН0Г0 типа раз-
вития, но некоторые отдельные его черты наблюдаются -среди низ- "Ё
щих групп многоклеточных; упомянем хотя бы смену бластуло-
образной личинки паренхимулой ‘и потом планулой у некоторых
Hydrozoa>>. Итак, здесь имеют место весьма разноречивые мнения.
представляющиеся почти в равной мере произвольны-ми. М-ежду тем’,
совершенно очевидно, что представление о вторичности развития
с личинкой (и узко приспособительном характере последней) не
может иметь того универсального значения, которое ему придается
эмбриологамэи. На самом деле, если оно частично и справедливо по
отношению к некоторым группам высших Мета2оа (на основе изу-
чения которых оно, кстати, и оформилось-как и большинство
других традиционных представлений эмбриологии), то его абсо-
лютно нельзя распространить на нпизшие группы, например, губок
или кишечнополостных. Последние характеризуются именно слож-
ностью присущего HIM «развития с личинкой>›, часто связанного
здесь с необычайно ‚громоздкими формаами метаморфоза и, наряду .
с этим, предельной примитивногстью своих личинок; особенно важно
подчеркнуть, что эти их онтогенетические особенности выражены
в своей крайней форме как раз у самых примитивных из предста-
вителей этих групп (метагенетических гидромедуз и известковых
губок). Следовательно, прнинятие этой эмбриологической версии
равнозначно признанию онтогенеза названных групп гораздо более
видоизмененным, чем у вышестоящих в системе форм, а здесь явно
вторичные проявления криптометаболиги или эмбри-онизации (напри-
мер, ведущие к образованжию актинулы) нам пришлось бы считать
более, примитивной формой развития, чем типичный для них мета-
морфоз. Отсюда ясно, насколько неприемлема данная концепция-Ё
в корне противоречащая, по существу, всем данным сравнительнои
эмбриологии. ' -
Таким образом, у нас нет пока достаточных оснований остановить
свой выбор ни .на какой-либо из предложенных до сих пор гипотез
происхождения личиночных форм и .метаморфоза многоклеточных.
Препятствием к этому является, прежде всего, их отвлеченность,
оторванность от конкретной действительности и, в конечном счете,
неуб-едительность. Они явно не отвечают фактическому положению‚
вещейщхотя бы в том отношении, что требуемой способностью
к самостоятельному питанию не‘ обладает нли одна из более нрими-
тивных форм личино-к— бластул, -паренхимул или пла-нул, так
что принятие этих концепций вынуждает нас считать вто-рично
видоизмененным онтогенез даже наиболее ‘примитивных губок и
Cnidaria.
Итак, вопрос о происхождении личиночных форм Метаиоа
остается, по сути дела, открытым——если ставить его в общей‘
фОрМ6— И, ВО В-СЯКОМ случае, КОНКРЕТНО Не решенным В ОТНОШЭНИИ
тех групп низших животных, где он имеет особую, Максимальную
остроту.
42
8. Некоторые итоги и перспективы
Попытаемся теперь дать общую характеристику современного
состояния комплекса проблем, связанных с происхождением онто-
генеза многоклеточных. V
Первое, что нужно констатировать, это то, что мы являемся сви-
детелями нов-ого подъема интереса к этим проблемам, обусловлен-
ного наступившим у нас расцветом эволюционного направления в
биологии. Все возрастающее внимание, уделяемое теперь этим про-
блемам морфологами и, в частности, представителями эволюцион-
ной гистологии, служит ярким свидетельством наступившего пере-
лома в отношении к этим .проблема.м. Можно сказать, что теперь,
после долгого перерыва, вновь стали осознаваться их актуальность
и непреходящее принципиальное значение, что они на наших глазах
завоевывают одну за другой ранее утраченные и‹ми позиции. ` '
Вместе с тем, "приходится признать, что возрождение интереса
биологов к основным вопросам эволюционной онтогенетики не нашло
еще нникакого от_клика со стороны дисциплины, главным предметом
которой должно быть их самое глубокое изучение. Действительно,
наша официальная эмбриология все -еще остается преимущественно
механикой развития, фактически безучастной к эволюционной
идее. Утратив свою некогда веду-щую роль в творческой раз-
работке учения Дарвина, эмбриология сейчас переживает глу-
бокий идейный кризис. Он настолько очевиден, что его откровенно
признают даже представители буржуазной «каузальной» эмбриолоа
гии, один из которых-Дж. НМДХЭМ (1947) — дает такую характе-
ристику ‘ее современного состояния: «Эмбриология, в особенности
теоретически, обветшала, если можно так выразиться, с тех пор как
эволюционная теораия потеряла свое значение метода объяснения
явлений». Констатируя, что строго эволюционное направление в эм-
бриологии не удержалось в ХХ веке, и не будучи склонным жалеть
об этом, Нидхэм. заявляет следующее: «Плохо, что мы до сих пор
нич-его не имеем взамен его. Экспериментальная, морфологическая,
физиологическая и химическая эмбриология не что иное, как сумма
фактических знаний, не связанных какой-либо объединяющей тео-
рией, так как мы не можем пока признать такой теорией учение об
осевых градиентах, теорию полей или гипотезы относительно гене-
тической роли энзимов» (1947). Однако, если трудно не согласиться
с такой оценкой нынешнего состояния эмбриологии, то, конечно, на.м
более чем чужды признание эволюционного объяснения явлений,
утратившим свое значение, и поиски выхода из создавшегося тупика
в тех «достижениях математического, даже математически логиче-
ского характера», на которые возлагает свои надежды Нидхэм.
Для нас, напротив, является аксиомой, что эмбриология безжиз-
ненна без эволюционной теории и -что единственный путь к подлин-
ному ее возрождению лежит в полном ее переходе на позиции со-
ветского творческогодарвиньизма. Только развиваясь по этому ‚пути,
эмбриология животных сможет Найти выход из переживаемого ею
43
кризиса _и занять подобающее ей место в нашей передовой биоло-
гической науке. _ в V
Как велика и ответствен-на должна быть ее роль на современном
этапе развития бИОЛОГИлИ, явствует из того, что в основе всего ми-
чуринского направления лежит о н т о г е н е-т и ч е с к о е понимание
природы живого, овладение закономерностями живой природы через
изучение индивидуального развития организ-.'
гм отв и сознательное управление им. Сформулированное с «предель-
ной четкостью акад. Т. Д. Лы-сенко в тезисе: <<о р г а н и з м ы, а о т-
сюда и их природа, создаются только в проце-ссе
р а-з в и т и я», — это‘ основное положение учения Мичурина —— Лы-
сенко выдвигает эмбриологию, как науку об индивидуальном раз-
витии организмов, на одно и=з первых мест среди всего комплекса
биологических дисциплин. Однако, еслина сегодняшний день это
|М6СТ0 ‚ужезавоевано эмбриологией растений, развившейся на на-
ших глазах в мичуринскую биологию развития растительных орга-
низмов‘ и ставшей краеугольным камнем советского творческого
дарвинизма, то этого никак еще нельзя сказать об эмбриологии
животных. Здесь все еще впереди, и почти все нынешние эмбрио-
логические концепции подлежат переоценке и глубокой творческой
перера-ботке.
Нам кажется несомненным, что как только наша эмбриология
вновь станет дарвинистической и будет развиваться в мичурин-ском
натправленищто иигнорируемые ею сей-час проблемы эволюционной
онтогенетики станут в ц-ентре ее внимания. Они вновь войдут в круг
интересов общей эмбриологии и бесспорно составят одну из самых
важных ее глав. При этом особое принципиаль-ное значение при-
обретает вопрос об источниках и путях исторического становления
индивидуального развития животных со в-семи входящпими в него
разделами, касающимися происхождения и эволюции процессов
гаметообразования, оплодотворениъя, дробления пи фор›мирования‹
многоклеточности, клеточной ‘и тканевой дифференцировки и т. д.
Очевидным наам представляется и то, что если разработка и ре-
manna этих вопросов являются одной из главных задач общеи эм-
бриологии, то-они пимеют не меньшее значение и для в-сех дРУгих
отраслей морфологии животных. Рассмотренные на-ми- работы
А. Н. Северцова, И. И. Ежикова, А. А. Заварзина и ряда других
авторов ясно показывают, что ни одна из больших проблем эволю-
ционной морфологии не может быть даже и поставлена вне связи
-с этими вопросами. И тут нельзя не отметить, что большая заслуга
‘наших морфологов заключается именно в том, что они сдВИНУЛИ
с имертвой точкивопросы эволюционной онтогенетики, почти поза-
(бытые со времен Мечникова, и поставили их на повестку дня.
"Правда, на-сдалеко ‚не могут удовлетворить гипотетические по-
гстроения, предложенные Сев-ерцовым, Заварзиным и ДРУГИМИ PIC‘.
‘следователями, ‘занимающимися этими вопросами. Все они, как мы
виделъи, крайне противоречивы, абстрактны и слабо обоснованы
"фактическими данными. Бросается в глазгц-чтоони либо вовсе не
`44
подкреплены эмбриологйческими фактами, либо проникнуты скеп-
сисом п-о отношению. к, последним, что существующие протистодоги-
ческие данные не находят в них никакого отражения, и что они
строятся, таким образом, на очень узкой ‚фактической основе. Этим,
естественно, и определяется крайняя абстрактность этих „гипотез, их
недоказуемость, ставящая их, в сущности, в один ряд с гипотезами
.гекк-елевского стиля. Отсюда невольно складывается впечатление,
что все построения подобного рода не могут, по самому своему су-
ществу, быть чем-то большим, чем простыми предположениями, и
‘что в этом сказывается объективная невозможность трактовать дан-
.ный круг вопросов в более конкретной и трезвой форпме, чем это
принято до сих пор. Иначе говоря, сугубо умозрительный характер,
придаваемыи этим ‚гипотезам, кажется единственно возможным ‹и
вытекающим из самой специфики тех вопросов, которые онипри-
званы осветить. И этим„в известной мере, оправдывается то обстоя-
тельство, что научная ценность ипродуктивность таких филогене-
тиче-ских построений ставятся многими под сомнение.
Это обескураживающее впечатление, однако, быстро рассеи-
вается, если-только при-смотреться к методологической стороне дела
и ‚попытаться оценить с этой точки зрения интересующие нас ги-по.-
тезы. Такая оценка ясно „показывает, что поскольку ни одна
из них не основывается на принциипах диалектического материа-
лизма, они не могут и ‚претендовать на правильное, объективное
освещение и истолкование рассматриваемых ими явлений природы,
такой, как она есть, без всяких посторонних прибавлений. Таким
_образом, становится ясным, что общей причиной имеющихся тут
неудач являются, прежде всего, моменты методологического по-
рядка.
Одним из частных выражений этого и -служит то обстоятельство,
что рассмотренные нами гипотезы Северцова, Заварзина, Хлопина
и других авторов мало счпитаются с реальной действительностью,
базируясь ‚на очень фрагментарном и произвольно подобранном
фактическом пматериале. Они оставляют почти совершенно неис-
пользованными ‚громадную мас-су данных, накопленных ax настоя-
щ-ему моменту эм-бриологией и сравнительной анатомией тех групп
организмов, на основе которых и очевидно должны бы строиться
все главные концепции филогенетической эмбриологии. Мы имеем
в виду, с одной стороны, всю огромную группу простейших и, с дру-
гой——наиболее примитивных из многоклеточных, к которым сле-
дует отнести, прежде в-сего, метагенетических книдарий и известко-
вых губок.
Плодотворность конкретного и всестороннего взаимного сравне-
‘Hm особенностей строения, размножения и развития разнообразных
представителей этих групп была так блестяще продемонстрирована
Мечниковым, что сомневаться в ней не приходится; и можно только
пожалеть, что никто -не решается следовать за ним „по столь ясно
указанному и-м пути. В самом деле, делатвшиеоя и до Мечникова
попытки сопоставления многоклеточных с простейшими и все ана-
45
Алогичные сравнения, проводившиеся после него, в корне отличаются
от соответствующих сопоставлений самого Мечникова, тем, что а
они строились: 1) на чисто произвольном и случайном подборе кон- A
кретнь1х объектов сравнения, на ничем не. оправданном предпочте-
нии в ‚выборе одних из них при полном игнорировании остальных;
2) .на использовании в таком плане, в качестве модельных объек-
‚тов, ряда высоко специализированных представителей простейших
и многоклеточных, обладающих тем самым минимальной степенью
обоюдной сравнимости; 3) на одной только анатомии простейших,
привлекаемых к сравнению с многоклеточными, без учета присущих
им особенностей размножения и развития. Таким образом, залог
успеха заключается тут в том, чтобы подобное сравнение было
действительно всесторонним и исчерпывающим, чтобы оно учиты-
вало всю совокупность относящихся сюда фактов B ИХ неразрывной
взаимной связи, а не было бы выборочным пи произвольным, и, что
самое главное, чтобы оно было насквозь проникнуто онто г е н е-
т и ч е с к и M пониманием явлений. `
Правда, этот путь постановки и рассмотрения проблем эволю-
шионной онтогенетики сталкивается со многими трудностями и вы-
зывает сейчас разнообразные, якобы принципиальные возражения,
сводящиеся в конечном счете к декларации объективной несравни-
мости друг с другом организмов различного конструктивного уровня.
Однако все эти трудности и возражения представляются скорее от-
ГОЛОСКОМ укоренившего-ся .представления Об оторванности «царства О
ПрОТИСТОВ» OT ОСТЗЛЬНОГО органического мира ИЛИ же следствием
МЕТОДОЛОГИЧЕСКИ неверной ПОСТЭНОВКИ ЭТОГО вопроса И В ЗНЭЧИТЕЛЬ-
НОМ СВОеМ ‘ООЛЬШИНСГВЭ ОКЭЗЬЕВЗЮТСЯ МНИМЫМИ, как ЭТО будет ВИДНО
ИЗ дальнейшего. ПОЭТОМУ МЫ Не В|ИДИМ реальных ПрВПЯТСТВИЙ K -ca-
MOMy ШИрОКОМу -применению И ТУТ МОГУЩЭСТВЗННОГО сравнительно-
ПМОРФОЛОГИЧВОКОГО метода.
Таки-м образом, перед нами стоит задача систематизации и обо-б-
щения огромной груды фактов, накопленных к настоящему времени‘
Апротистолопией и эмбриологией ‘низших беспозвоночных, с целью .
посильной разработки основных проблем общей эмбриологии жи-
ВОТНЫХ ОРГЕЪНИЗМОВ, приобретающих В НЭШИ ДНИ ИСКЛЮЧИТЕЛЬНОЕ
принципиальное ЗНЭЧЕНИЭ.
ЧАСТЬ ПЕР_ВАЯ
ОРГАНИЗАЦИЯ, РАЗМНОЖЕНИЕ и РАЗВИТИЕ
одноклеточных простейших
ВВОДНЫЕ ЗАМЕЧАНИЯ
Давний СПОР О ТОМ, следует ЛИ СЧИТЗТЬ простейших ОДНОКЛЭТОЧ-
НЫМИ организмами ИЛИ же ОрГЗНИЗМЁПМИ НЗДКЛЕТОЧНЫМИ (I/ILHI/I.HeK.He"
ТОЧНЫМИ, как ИХ ИНОГДЗ НЭЗЬХВЭЮТ), рЭЗрЕШЗЭТСЯ НЭИЛУЧШРЪМ И са-
' ‘МЫМ объективным ООРЭЗОМ «ПРОСТЫМ ПРИЗНЭНПИЁМ факта, ЧТО СрЭДИ
них есть и подлинно одноклеточные и действительно Неклеточные
формы. ' _
Истинно -одноклеточной организацией безусловно обладаютвсе
одноядерные простейшие. Однако основным критерием такой под-
линной одноклеточности‚ очевидно, должна служить не одна только
<<моноэнергидностЬ»—в том ее отвлеченном смысле, в каком она
понималась Сак-сом, а сочетание одноядерности с наличием полного,
НО ординарного комплекта постоянных клеточных органелл. Пере-
ход от понимаемой таким образом «истинной» одноклеточности к бо-
лее сложным ступеням надклеточ-ной (ил.и Неклеточной) организа-
ции осуществляется у простейших необычно широким фронтом, хотя
и довольно однообразным путем: путем умножения тех или иных
клеточных структур или даже всей их совокупности. Эта линия раз-
вития; до крайности характерная для простейших и наблюдаемая в
любой их группе, обозначается Догелем (1929‚ 1946), как .поли-
M е р и з а ц и я. Самым известным ее примером является умножение
чи-сла ядер, т. е. приобретение простейшим «полиэнертидного»
строения, но не менее распространено и умножение, например, дви-
‘гательных органелл или целых их комплексов (кинетид), трофиче-
ских структур (хроматофоров, пиреноидов) И т. д.; нередки также
случаи, ‘когда умножению подвергаются еще более сложные ком-
плексы, состоящие, например, из ядра и многожгутиковой кинетиды
‘(образованной в свою очередь, не только ими одними, но и соот-
ветствующими базальными зернами жгутов, ризопластом, пара-
базальным аппаратом и т. д.) и называемые тогда кариомастигон-
тами (у Polymastigina), или даже вся система органелл, входящих
в состав нормальной клетки (Diplozoa). Отсюда уже ясно, что в
47'
основе полимеризации лежит частичное или не доведенное до;
конца деление, И ПрИТОМ нередко многократное.
Полимеризация, НВСОМНСННО, ЯВЛЯЕТСЯ ОСНОВНЫМ СПОСОООМ «пе-
рерастания>› простейшими одноклеточной (моноэнергидной) основы
их организации. Очевидно, что полимерное простейшее — уже не
клетка, а организм более высокого, чем клетка, структурного уровня,
к которому гораздо лучше подходит морфологическое понятие плаз-
МОДИЙ— В тех СЛУЧЗЯХ, КОГДЭ речь ИДЕТ О м-ногоядерном ОРГЭНИЗМЕ-
амёбоидного склада, И сомателла—когда дело касается более
сложно организованных простейших с постоянной формой тела (на-
пример, инфузорий). Таким образом, полимерные (в том числе и
многоядерные, но не одни только они) простейшие представляют
СОбОЙ надклеточные ПЛЗЗМОДИИ ИъЛИ же сомателлы, а ОТНЮДЬ не ОД-
неклеточные «ЭЛЕЗМЕНТЗРНЫЗ организмы».
Дальнейшее усложнение организации, наблюдаемое «во многих‘
группах простейших, заключается, -главнь1м образом, в про-гресси-
рующей дифференциации первично однородных органов сомателлы
(например, множественного ядерного или двигательного аппарата),-
В ИХ СПЕЦИЭЛИЗЗГЦИИ |И объединении В СТРУКТУРЫ ВЫСШЕГО порядка
‘(хороши-м примером которых могут служить хотя бы мембранеллы
и-нфузорий); при этом часто возникают и оформляются образования’
принципиально нового характера, не имеющие подчас ничего об-
щего С органеллами КЛЕТОЧНЫХ Наконец, на ВЫСШИХ ступе-
^НЯХ организации простейших ПОЛИМеРИЗЗЦИЯ ОбЫЧНО уступает свое
место противоположному ей процессу о л и г о M е р -и з а Ц и и (т. е.
сокращению числа однородных частей). В отношении к простейшим,
достигшим такой высоты организации (излюбленным примером ко?
торь1х служат инфузории из семейства Ophryoscolecidae), не „может
быть и речи о какой бы ‚то ни было одноклеточности. От нее тут,
мало что остается; она оказывается п-очти полностью преодоленной,
и этим легко объединяется объективная несравнимость данных-
ф-орм с отдельными клетками Metazoa, которая так энергично ис-
пользуется противниками клеточной теории. -
Этими двумя основными уровнями организации—одноклеточ-
нь1м и надклеточным — еще не исчерпывается структурное разнооб-
разие Protozoa. B самом деле, среди них есть и немногие, правда,
формы, обладающие типично многоклеточны-м строением, полностью
отвечающим таковому Metazoa (высшие вольвоциды); наконец, есть,
такие, у которых многочисленные сложные сомателлы теснейшим
образом объединены в целостный организм высшего порядка (коло-
ниальные сувойки).
_ Итак, обиходное представление о простейших, как типично одно-
клеточных элементарных организмах, является справедливым и-с-
ключительно по отношению к тем из них, которых принято называть
моноэнергидными: к большинству свободно живущих жгутиковьтх,
и немногим и.з паразитических представителей этой группы, к части
одноядерных саркодовых и -споровиков, а также и к морфологически
равноценным им ранним стадиям развития (зооспоры, апланоспоры,
48
гаметы) большинства остальных Flagellata, Sarzcodina, Sporozoa и
Сп1с1о5рог1с11а.чВсе же остальные простейшие, а именно все без ис-
ключения пинфузории, основная масса саркодовых и споровиков и
очень многие паразитические жгутоносцы, безусловно должны‘ рас-
сматриваться, как организмы надклеточные, более или менее далеко
вышедшие из рамок одноклеточного плана строения и в той или иной
мере утратившие его специфпические черты. И так как эти неклеточ-
ные формы составляют среди простейших явное большинство, то
вряд ли будет преувеличением сказать, что для Protozoa B целом
характерно не столько пожизненное сохранение одноклеточности,
как отход от клеточной организации вообще, идущий здесь несрав-
ненно дальше, чем у более консервативных в этом отношении Ме-
tazoa. -A . , ,
Г л а в а 1
ОРГАНИЗАЦИЯ ОДНОКЛЕТОЧНЫХ PROTOZOA
1. Три главных типа одноклеточной организации
При всем разнообразии внешней формы, строения и образа жиз-
нъи одноклеточных простейших их организация „легко сводима к трем
основным типам, являющимся вместе с тем главными типами одно-
клеточной организации вообще: а м ё б о и д н о м у, т и п у с о л-
нечника и жгутиковому. .
А. Амёбоидньлй тип. Морфологически наиболее примитивным из
всех типов одноклеточной организации является амёбоидный,-ха-
рактеризующийся прежде всего отсутствием всякой симметрии
вообще, т. е. анаксонным строением при неопределенности и непо-
стоянстве общей формы тела, или же наличием самой низшей из
форм симметрии-неопределенно полиаксонной, отвечающей его
шарообразной форме (Беклемишев). Правда, у некоторых форм,
обладающих постоянной центросомой, неизменность ее положения
относительно ядра намечает наличпие зачаточной, но устойчивой глав-
-ной клеточной оси; в других- случаях о ее присутствии говорит отно-
сительно устойчивое положение’ постоянных вакуолей (имеющихся
у ряда форм) или же определенность ориентации животного при его
движении. Однако намечающаяся таким образом одноосность тела
остается»слабо выраженной и не приводит к определенной .диффе-
ренциации его полюсов. В связи с этим здесь нет M’ постоянных гра-
диентов свойств ни в одном направлении, кроме направления от
центра тела к его ‚периферии.
Органологическая (цитологическая) простота амёбоидных Рго-
tozoa выражается ярче всего в полном отсутствии хоть сколько-
нибудь устойчивых метаплазматических дифференциро-вок: могут
иметься только эуплазматиче-скиве, представленные иногда здесь
длительно существующими сократительныпми вазкуолями.» Однако
чаще всего однородная форма тела нарушается только чисто
4 Эмбриология беспозвоночных 49
метаболическими образованиями протоплазмы, возникающими в лю- '
бой точке тела и в любое время, а так же легко и быстро исчезаю-
щими. Их самым известным примером „служат различные- псевдо-
подии, но они могут дополняться другими, более сложными по своей
форме, хотя и столь же быстротечными образованиями: Трубча-
тыми цитостомами и глотками, -образуемыми в момент заглаты-
вания пиши, особыми опорными структурами в ложноножках, и т. д.
Не лучше представлены и дифференцировки ядерного происхо-
ждения: постоянная центросома, как правило, отсутствует, и= она
чаще появляется только вре-
менно, -в момент митоза (или
даже не при всех митозах,
tae) и не обнаруживается в
интеркинезе. Центросомы бы-
вают, повидимо-му, только вну-
триядерными: внеядерные наб-
людаются только у форм, имею-
щих жгутиковую фазу.
рис_ 1_ Amoeba Proteus с аппаратом мы, то она часто хорошо раз-
Гольджи, представленным шариками со вита в виде разнообразных И
светлым центроми черной осмиофиль- Иногда ‚весьма СЛОЖНЫХ раке;
“ой °б°"°‘“<°й (из дшел”) винок или домиков и цист как
7
ОРГЭНИЧЭСКОГО, так И H60-pI‘aHIrI-
'-IeCK-0I‘O составаВнутренний скелет, однако, отсутствует. _
Аппарат Гольджи обычно имеет диффузный характер и пред-
ставлен 601166 ИЛИ менее МНОГОЧИСЛЗННЫ-МИ ДИКТИОСОМШМИ, б6СП0рЯ-"
дочно разбросанными по всей плазме (рис. 1). Иногда в плазме
‚ИМЕЮТСЯ, сверх ТОАГО, И МНОГОЧИСЛЗННЫЗ 0СМИОф|ИЛЬНЫе ЗЗРНЫШКИ,
состоящие из того же материала как периферический -слой диктио-
сом; возможно, что из них воссоздаются диктиосомы, иногда исче-
зающие перед инцистированием (Endamoeba Ь1а’с’сае).+Таким обра-
зо-м, аппарат Г ольджи может и не быть здесь постоянным образо-
ванием. Заслуживает внимания трудность его разграничения от
вакуома, также свидетельствующая о его слабой оформленности.
Митохондрии и весьма различные непостоянные включения суще-
ствуют, вероятно, у всех представителей этого типа.
Переход чИЗ состояния активной жизнедеятельностив покоящееся
здесь не сопряжен с какими-либо особо ощутимыми структурными
изменениями. Происходит только округление тела и втягивание его
придатков, так что видимым выражением такого перехода служит
еще только выделение ‘цисты или преобразования трофических и
иных непостоянных включений цитоплазмы (например, появление
в этот момент зерен парагликогена, хроматоидных телец -1 рис; 2 а
и т. п.). -
Таким образом, организация простейших этого типа характери-
50
а то-льк-о предшествующих ин- ‚
цистированию: Endamoeba b1at--
Что же касается аллоплаз-
зуется относительным постоянством, определяемым, однако, только -
ее крайней простотой; она остается практически одинаковой inpa-
всех обстоятельствах-кроме, разумеется, тех, оче-нь частых, слу-.
чаев, когда амёбоидный организм превращается в жгутиковую’ или
надклеточную- форму. '
Б. Т an солнечника олицетворяет собой следующую, значительно
болеервысокую ступень одноклеточной организации простейших, но
организации еще определенно амёбоидного склада, связанной всеми
своими корнями с ее наиболее типичными и простыми формами.
Рис. 2.
A—En’camoeba histolyticaw: XPOMETOHIJHBIMH телами в плазме; Б-Епт. gingivalis; особь,
образовавшая трубчатую клеточную глотку при заглатывании добычи (по Элштейну)
Е-го можно рассматривать, соответственно, как непосредственную
МОДИФИКЗЦИЮ предыдущего типа, связанную с переходом к планкти-
ческому или прикрепленному образу жизни при неуклонно-м сохра-
нении анимального способа питания.
В основе. архитектоники одноклеточных этого типа (большин-
ство солнечников‚ ‚молодые вегетативные стадии радиолярий, осо-
бенно более примитивных групп, некоторые примыкающие сюда
хризомонады вроде Rhizochrysis ИЛИ Porochrysis) ЛеЖИТ многоос-
нь1и, гомополярный тип симметрии, создаваемый лучистым располо-
жением ложноножек, их опорных структур или скелета на базе
шароо-бразного, неопределенно полиаксонного клеточного тела. От-
сюда берут свое начало разнообразные направления прогрессивного
развития, ведущие, с одной стороны, к бесконечному проморфоло- .
гическому усложнению и дифференциации -скелета при сохранении
клеткой довольно примитивного строения (радиолярии), а с-дру-
гой —›к приобретению последней одноосно-гетерополярной -сим.мет-
рии радиального или даже пбилатерального типа (Centrohelidia),
4* 51
'V~
.7**..,.«.;$;;.§»2:@
317:2?“
Рис. 3. Образование аксоподии у солнечника Actz'nosphaerz'um:
1——na.nbueBH,11Hbm зачаток аксоподии; 2—ранняя стадия оформления 00950”? CT9P>*<H$I3
3 — аксоподия, обнаруживающая на месте сгиба дно-
4 — участок готовой аксоподии; (по Роскину)
рИЛЛЯрНОЁ строение осевого СТЕРЖНЯ}
без ощутимого гондолами-гения’
21pX-HT€K’I‘0HHKPI :CKeJIeTa. _.
Главную органологическую -
особенность «рассматриваемого
сейчас ти-па составляет наличие
поразительно разнообразного
внутреннего скелета, демонстри- '
рующего здесь все мыслимые
этапы своего формирования и
дальнейшей стабилизации. И-сход-
ной формой этого скелета (как
в морфологическом, так и в био-
химическом отношении), оче-
видно, следует считать весьма
лабильные аксоподии та-ких форм,
как Асйпошопаз или Ciliophrys,
y которых о-ни имеют характе-р
сокраггимых эуплазматич-еских
дифференцировок — легко созда-
ваемых в любой, невидимому,
точке ‘ПОВЕРХНОСТИ тела И так же.
легко расплывающихся; в этих
случаях формообразующие -опор-
ные структуры создаются путем
простой желатинизации прото-
плазмы.
оформлен-нести -скелета представ-
‚ляют солнечники с мезоплазмати-
ческими, значительно более устой-’
ЧИВЫМИ ЗКСОШОДИЯМИ, ИСЧЭЗЗЮ-
ЩИ‘МИ ТОЛЬКО ‘ПРИ инцистировании
И ИНОГДЗ ДЕЛЭНИИ.
Строение И образование аксо-‘
подий у солнечник-ов этого типа —
Auctinophrys И‘ многоядерного
Асйпозрйаегйпт — подробно изу-
чалось Роскиным (1918, 1937).
Аксоподии эти-х
имеют вид длинных лучей, в ко-
торых отчетливо видны осевые
стержни, .простирающиеся в виде
игл до самого их конца. Форми-
рование аксоп-одии начинается
с появления на поверхности тела
небольшого ‚пальцевидного прото-
плазматическо-го выступа, в толще
которого видны нити более, густой,
иначе преломляющей свет плазмы;
Следующую ступень А
СОЛНЭЧНИКОЕ
эти нити не являются ещё застывшими фибриллами, а скорее опре-
деленным образом ориентированными токами жидкой плазмы. По
Мере ВЫТЯГИВЗНИЯ И УТОНЧЗНИЯ 3aqaTKa ЭКООПОДИИ ЭТИ нитевид-
ные токи плазмы, занимавшие раньше большую его часть, сосредото-
чиваются по его оси, и каждая из нитей выпрямляется и становится
более резко видной (рис. 3). Затем отдельные нити сближаются все
болеери более и, наконец, сливаются, переходя теперь из жидкого
состоянеия в твердое или полутвердое. Сформировавшийся таким
образом стержень аксоподии представляет собой трубку, стенка
которой весьма отчетливо и иначе красится, чем одевающая его
плазма. Этот трубчатый стержень заполнен сильно красящейся
‘плазмой, имеющей очень вязкую консистенцию; его ст-енки состоят
из фибрилл, отличающихся от нитчатых структур начальных стадлий
развития аксоподий своей толщиной, массивностью и более ярк-ой
окрашиваемостью на препаратах. Исчезн-овение осевых стержней
при ликвидации аксоподий вероятно идет через стадию их пред-
варительного распадения на отдельные фибриллы.
У ряда солнечников стержнпи аксоподий пронизывают по радиу-
сажм все тело и сходятся к его центру, где они вплотную подходят
к ядру (Actinophrys) илилежащей здесь центросоме (все Centre-
helidia). У некоторых из представителей последней группы (Wagne-
rella, Cienkowskya), образовавшийся таким путем радиальный ске-
лет приобретает уже метаплазматиче-ский характер; он сохраняется
при инцистировании (целиком или без дистальной „гасли своих
лучей) и делится пополам при размножении делением (см. рис. 19).
Дальнейшее развитие такого скелета заключается в его минера-
лизации прли сохранении им делимо-сти. Этот этап его развития ха-
рактерен для ряда более примитивных радиолярий (из Acantharia)
И служит как бы отправной точкой для всех дальнейших его пре-
образований в этой группе. В общем, эти преобразования идут по
линии срастания всех лучей скелета в нерушимое целое › (путем
укрепления их соединения в центре отложением органического Be»
щества), присоединения к ним тангенциальных скелетных элемен-
тов (имеющих своим источником’ чехол из кремнез-емных игл, часто
наблюдаемый и у солнечников), а затем и разнообразнейших изме-
нений сложения все более минерализующегося теперь скелета.
Итак, радиальный скелет Heliozoa наглядно демонстрирует все
стадии нарастающей стабилизации, возникая в виде легко обрати-
мых эуплазматических дифференцировок и приобретая у высших
форм типично метаплазматическую природу. Дальнейшие этапы его
развития осуществлены у радиолярий, у которых оно связано с за-
мещенвием органического вещества минеральным.
, Другая линия специализации аксоподий солнечников заключает-
ся в их преобразовании в жгуты. Не ставя пока вопроса о генезисе
жгутов в его общей форме, можно ‘констатировать, во-первых, что
_они имеются у многих солнечников наряду с аксоподиямди и в этих
ЪСЛУЧЗЯХ ЯВНО ГОМОЛОГИЧНЫ ПОСЛЕДНИМ, представляя СОбОЙ ОТДЕЛЬНЫЕ
„лучи образуемойгими радиальной системьъ- Соответствие жгутов
sé
r
Вис. 14; Преобразование. аксоподий в жгуты у солнечников: ‚
J—_Cz‘li_ophrys marina в питаъощейся фазе солнечника; 2——НаЧа-'1_0 И ‹?—К_ОЁ*Э$НЫЙ Эта" ее "P9-'
вращения в расселительную жгутиковую фазу; 4—Actz'nomonas mzrabzl_zs ——сидячая форма _C
одним жгутом; прикрепленная як субстрату посредством одной из своих аксоподий; 5———Dz—
morpha титла, сидячая солнечниковая стадия с аксоподиями и двумя жгутамш ОДИН ИЗ K°T°'
b1;xX‘cny)x<HT осьюстуденистого прикрепительного стебёлька; б——свободная СОЛНЗЧНИКОВаЯ
„ _ . - . и 7——жгутик0вая фаза того же вида. Кана) . - ‘
5'4"”
a
_()C’I‘21JIbI-IBIIM ЗКСОПОДИЯМ выражается ЗДЗСЬ В ТОМ, ЧТО те И другие
представляются у Actinophrydia чисто поверхностными образова-
ниями, а у Centrohelidia отходят от центросомы и испытывают оди-
наковую судьбу при делении (рис. 4). Во-вторых, имеются дан-
ные, что у солнечников, способных превращаться в жгутиковую
форму, лишенную аксоподий (Ciliophrys, C1a’ch_ru1i'r1a), эта тран-с-
„формация происходит путем втягивания всех аксоподий, кроме од-
‚ной или двух, преобразуъощихся в жгуты. Далее, само строенлие
жгутов говорит здесь в пользу их признания функционально видо-
„ИЗМЭНЗННЫМИ ЕЪКСОПОДИЯМИ. Нак-онец‚ НЗЛЬЗЯ не обратить ВНИМЗНИЗ‘
и на то, что при переходе к сидячему образу жизни функции сте-
белька могут выполняться как обычными аксоподиями, так и жгу-
тами, чем еще подчеркивается отсутствие принципиальных различий
между ними.
В геометрическом центре радиальной системы Centrohelidia pac-
IIQJIEFEIBTCH крупное Центральное зерно, природа И ‘ФУНКЦИИ КОТОРОГО
служили предметом длительной дискуссии. Оставляя пока споры
в стороне, мы отмегтим следующие главные особенности этого
органоида .у солнечников: 1) его постоянство и генетическая непре-
рывиость в ряду поколений при размножении посредством деления;
2) неразрывность его связи с радиальной системой и обязательное
ведущее участие в делении последней; 3) факультативный характер
его участия в кариокинезе; он часто играет роль центра деления
‚(истинной центре-сомы) либо с самого его начала (Dimorpha), либо
приобщаясь ‚к нему уже после установления веретена, .но может и
оставаться в стороне от этого процесса; 4\ "явная морфологическая
и, как сказано, функциональная независимость отядра, сказываю-
щая.ся в их разобщенности в интеркинезе и в установлении непроч-
ной связи с ‘ним только с профазы деления; 5) крупные размеры
зерна и его полное морфологическое сходство с классическим типом
клеточного центра Metazoa; OH также состоит из сильно окрашеи-
вающегося тельца (Центриоли), непосредственно связанного с лучи-
_СТЬ1МИ структурами И окруженного участком СВВТЛОЙ протоплазмы
(жцентросферой). .
Всем этаим признакам постоянного и автономного клеточного
органоида противопоставляется тот факт, что центральное зерно
обычно не передается дочерним особям при почковании, так что
его преемственность оказывается тут прерванной. В этих случаях
описывалось его возникновение заново из ядра (у Acanthocysti.-3),
НО теперь кажется много вероятнее, что он образуется из цитоплаз-
матического материала.
_ С радиальным скелетом связан у Acantharia еще один вид диф-
ференцир-овок- мионемы, характеризующиеся своей поперечно no»
лосатой микроструктурой и играющие роль мышц своеобра-зного-_
гидростатического аппарата этих радиолярий. '
Ко всем описанным дифференцировкам присоединяется у радио-
лярий еще одна‘: мета-Плазматическая центральная капсула, состоя-
ЩЭЯ 1:13. .QP~I‘3HPI‘1€CKQI‘0 „вещества (Псевдохитина) и представляющая
55 _
собой ттипичное пограничное образование, отделяющее перифериче- _
ский слой эктоплазмы (представленный крупно ячеистой плазмой,
ЗЗПОЛНЗННОЙ ВЗКУОЛЯМИ студенистого вещества) ОТ ЦЗНТРЗЛЬНОЙ ча- ’
сти клеточного (или, у взрослых форм, плазмодиального) тела.
Итак, тип солнечника характеризуется богатством и разнообра- -
зием дифчференцировок, главные из которых (аксоподии, жгуты,
центросома) ВОЗШИКаЮТ тут, фигурально выражаясь, ‘на наших гла-‚р
зах. Примечательно, что они наблюдаются тут в одних случаях
in statu nascendi, B других-как еще ‘лабильные, но уже более
постоянные образо-ва.ния и, на-
конец, как вполне ста-билизи-
рованные структуры. На мгно-
гих морфологических рядах
шагом и переход некоторых
из дифференцировок -в заклю-
чительную стадию сложноорга-
низованной аллоплазмы, полу-
чающей необычайно пышное
развитие в причудливом ске-
лете радиолярий.
клеточной организации обна-
ружи-вает полное сходств-о с
амёбоидным, непосредствен-
ным дериватом которого он
несомненно и является. При
переходе в покоящееся состоя-
ние (обычно связанном с раз-
множением и наблюдаемом
‘Рис. 5. Организация одноклеточных
жгутиковых: ^
1—Polytoma имена: п. ш-сократительная
вакуоль; х — хондриосомы; я ~— ядро; на пра-
ВОЙ Пд-‘ЮВННВ Рисунка изображен только cer-
чатый лейх/Ёоплпст (rug) Blom<oHcKoMy); 2 —chat-
one a su за sa (no Бишле) МНОГИХ ‚соло > _
л новодных форм,
- вдроде Асттэдорппз) организа-
ЦИЯ ‘СОЛНЕЧНИКЗ теряет все СВОИ Специфические черты вследствие PIC-
чезновения всех имевшихся дифференцировок (из которых сохра-
няется иногда только центральное зерно — y Acanthoxcyastis) и тогда
ровно ничем не отличается от амебоиднойь Таким образом, этот тип
ОРГЗНИЁЗЦИИ приурочен исключительно к активной, вегетативно Hesi-
TeJT__1{H‘0H ‘P336 Определенных групп одноклеточных и сменяется
ЗМЭООИДНЫМ ‘при Их инпистированили. Он представляет собой,
CJ_I€lIOBaTeJIbH0, дИф-ференцированную фазу этих одноклеточных‚_рез-
кпие отличия которои от ИХ дедифференецированной, полн-остью амё-
боиднои, фазы составляют характерную’ для них черту, Эта диффед
ренцированная фазасолнечника удерживается в качестве единствен-
нои у всех высших представителеид этого типа, доетигающихфтолько
BI НеИ НадКЛеТоЧНой ступени организации. ' ‘A ° ~ `
' B.. Жгутиковььйгтип. одноклеточной организации олицетворяет, со
строго‘ пморфологическойхгочки зрения, ее еще-более-высокую cry»
56
может быть гпрослежен шаг за '
В остальном этот тип одно-
TO1JIbK0 y ‘ПРЭСНОВОДНЫХ И не:
-ггень——высшее выражение ОДНОКЛ-ЗТОЧПОЙ организации вообще
(рис. 5, 6). K ЭТОМу типу относятся большинство одноклеточных
Flagellata в их деятельном трофичес-ком состоянии, расселительнь1е
стадии некоторых амёб и солнечников, жгутиьковые зооспорьх и га-
меты всех остальных простейших, водорослей, гри-бов и высших
животных и растительных организмов. Самой общей биологической
особенностью представителей этого типа является их высокая и
всегда" определенно ориентированная подвижность, обеспечиваемая
наличием характерного для них жгутикового аппарата и оказываю-
Рис. 6. Представители Zooflagellataz
7—Cryptobia helicis; 2—Pro’ceromonas lacertae-viridis; 3—Trichomonas
duboscqi: видны аксостиль, парабазальный аппарат, митохондрии,
- вакуом, ризопласт (по Грассе)
щая глубокое формирующее воздействие на все стороны их орга-
низации. A '
B проморфологическом отношении одноклеточные этого типа
‘отличаются, прежде всего, ярко выраженной тетерополярн-остью.
Ею определяется наличие постоянной передне-задней главной (кле-
точной) оси тела, которую можно назвать также его первичной
"осью. У активно движущихся форм, каковыми является большин-
ство жтутиковых простейших, главная ось тела обычно совпадает
‘с предпочтительным направлением поступательного движения, яв-
ляясь, таким образом, ио-сью движения; у сидячих форм она соот-
ветственно-приобретает значение оси прикрепления, а если речь
57
идет О прикрепленных napaamax, —TO И ОСИ трофической ПОЛЯрИ‘
зации (см. нниже). У всех Euflagellata И отвечающих им по строению-
жгутиковых форм других групп главная ось имеет промор-фологи-
ческое значение оси симметрии; чаще всего она намечена положег
нием базального зерна жгута или центросомы по отношению к ядру, _
ПрОХОДЯ через ИХ центры, а нередко приобретает свое ВИДИМОе ВЫ-
ражение В наличии СТрОГО осевых ЦИТОПЛЭЗМЗТИЧЭСКИХ СТРУКТУР,
например, ритзопла-ста ИЛИ ЗКСОСТИЛЯ. В ЭТИХ наиболее ТИ‹П!И‘ЧНЫХ`
случаях очевидное совпадение физиологической OOH тела С ПрОМОр-
фОЛОГИЧеСКОИ еще подтверждается тем, ЧТО‘ С НИМИ совпадает И
направление ПЛОСКОСТИ деления.
ОСНОВНЫМ гСТруКТУрНЫМ ПрИЗНЗКОМ физиологически переднего.
полюса (т, е. полюса, неизменно- обращенного вперед при поступа-
ТеЛЬНОМ движении) тела СЛУ)КИТ отхождение отсюда ЖГУТЗ ХИЛИ‘
жгутов, так что этот -полю-с носит название жгутикового; кроме того,
здесь располагается место приема пищи, когда только оно локали-
зовано, отверстие часто имеющихся у флагеллят оформленных по-
лостей клеточного тела (например, сложно устроенной глотки
у криптомонад |ИЛИ резервуара эвглен) и т. д.; наконец, у этого по-
люса обычно группируются также сократительные вакуоли, глазок,
парабазальный аппарат или отвечающие ему образования. Проти-
воположный, задний, конец тела дополнительно характеризуется
тем, что он обычно служит полюсом, которым животное прикреп-
ЛЯЕТСЯ K субстрату; впрочем, имногие автотрофные формы, образую-
Mme неподвижные вегетативные колонии (Colacium, Hydrurus), при-
крепляются передним полюсом, теряющим при этом свои жгуты.
Кроме того, у заднего полюса тела нередко сосредоточивается ос-
новная масса резервных пара-плазматических включений. Таким
образом, разнополюсность выражена у жгутиковых Protozoa не
менее резко, чем у одноосных многоклеточных, и притом во вполне
аналогичных формах, что делает вполне применимыми здесь и поня-
ти-я об анимальном и вегетатив-ном полюсах. Однако, в противопо-
ложность большинству низших многоклеточных, функция заглаты-
вания пищи локализуется у жгутиковых (когда она вообще суще-
ствует, си ‚притом локализована) на их" анимальном полюсе, а при-
крепление к субстрату составляет тогда удел вегетативного.
Кажется очевидным, что такое взаимоотношение разноименных
полюсов укладь1ва_ется точно так же в представление о физиологи-
ческом доминировании анимального полюса над вегетативным, т. е.
говорит о наличиии здесь весьма универсального отношения домини-
„рованИя-—-подчинения и соответствующих осевых градиентов фи-
зиологических -свойств.
Разнообразие форм симметрииодноклеточных жгутиковь1х Рго-
fOZOa ОЧЕНЬ велико, ограниичиваясь В ТО же время ТОЛЬКО ТеМИ, какие V
вообще устанавливаются на одноосно-гетерополярной основе. Более
ПрИМИГГИВНЫе ее формы, характеризующиеся ОСЬЮ симметрии не-
определенно большого порядка, нередки у СИДЯЧИХ ЖГУТИКОВЫХ, ОСО-
бенно, Craspedomonadina И xp.H39M0_.H.a1L среди которых МЫ -.H.a>$0mrI.M
55
' !И ТИПИЧНО радиальные фОрМЫ, поразительно напоминающие ПОЛИ-
пов „по своему строению (и также обладающие, в частности, пра-
вильно расположенными щупальцами, представленными здесь аксо-
подиями-: Cyrtophora). Однако даже у этих форм обычно имеются„.
хотя бы незначительные, проявления симметрии билатеральной,
становящейся господствующим ее типом у многих свободноплаваю-
щих жгутиковых. Среди последних двубоковая симметрия иногда
является весьма полной, но нередко она совмещается с поступа-
тельно вращательной, или винтовой, проявляющейся преимуще-
ственно во внешнем строении таких форм и почти не затрагивающей
внутреннее (эвглены, особенноРпасцз). Гораздо чаще, однако,
правильность двубоковой симметрии нарушается асимметричным
положением ИЛИ строением тех ИЛИ ИНЫХ внутриклеточных CTpyK- ‘
тур (особенно часто хроматофоров и их пиреноидов, а также глазка,
вакуолей, парабазального аппарата и пр.), что делает ее в той или
иной мере неполной. Тем ‘не менее билатеральная основа организа-.
ции обычно остается тут вполне осязаемой.
Описанный план строения, характерный для жгутиковых Рго-
tozoa B целом, подвергается в ряде групп различным видоизмене-
ниям. Чаще других наблюдается смещение жгутикового полюса,
вместе со в-семи связанными с ним структурами, с переднего конца
тела ‘на его вентральную сторону. Он как бы соскальзывает вниз по
сагиттальной линии тела, что ведет к соответствующей переориен-
тировке главной оси относительно остальныхдэта ось приобретает
диагональное (см. рис. 17) или даже почти поперечное направление
(см. рис. 26) и более или менее сильно укорачивается, отклоняясь
таким образом на 45° ИЛИ больше ‚от длинной оси клеточного тела.
Последняя сохраняет свое значение оси движения (и, соответ-
ственно, оси прикрепления) и вследствие этого может принимать на
себя роль новой, вторичной, оси симметрии.
Подобное расхождение первично совпадающих осей тела харак-
терно для ряда криптомонадин (например, Protochrysis, некоторые
Cryptochrysis) И достигает высшего развития у генетически свя-
занных с ними Dinoflag-ellata, становясь у них ведущей чертой
организации. Таким образом, создается особый ти-п архитекто-
ники, который» можно обозначить как д в уос н ы й, противопостав-
- дПЯЯ его НОРМЭЛЬНОМУ ДЛЯ РХГУТИКОВЫХ ОДНООСНОМУ ТИПУ. В соответ-
ствии с этим можно говорить и о двух отвечающих им формах п-о-
ступательного движения: в первом случае, когда ось движения
не совпадает с главной осью тела—-о «вторичноосном дВИЖеНИИ»;
во втором —— о движении первичноосном. ' '
Крайнее своеобразие и сложи-ость архитектоники Dinoflagellata очевидио
определяется сочетанием у них двуосного плана строения с наличием очеиь слож-
ного панцыря. Ось движения здесь целиком проявляет свою организующую р-оль
оси симметрии именно в отэношении к наружному скелету, в „то времякак откло-
нившаяся от нее первичная ось сохраняет за собой ведущую роль во внутреннем
строении п-ротопласта и в определении положения плоскости его деления. Благо-
даря ЭТОМУ И ВОЗНИКЗЭТ та проморфологическая ДВОЙСТВВННОСТЬ, _KOT0pa$i Ta_K за-
трудняет понимание плана строения панцырных Динофлагеллят и об-условливаечт
59
в частности, разнообразие присущих им форм деления. Представители этой
группы легко могут б-ыть расположены в морфологические ряды, иллюстрирующие
самым наглядным образом все последовательнчыеэтапы формирования этих выс-
ших, проморфолотически наиболее сложных типов одноклеточной организации,
вообще. На: этих рядах, идущих от таких примитивных, одноосных форм, как Ргого- _
„септгйоае через различных Dinophysidae K самым уклоняющимся перидинеям
легко прослеживаются, во первых, все этапы описанного смещения жгутикового
полюса с его первоначально апикального положения на вентральное и далее
даже субтерминальное; во-вторых, можно убедиться в соответствующей перео-.
риентировке первичной главной оси и сагиттальной плоскости простейшего хотя
бы по тому признаку, что определяемая ею плоскость деления неизменно проходит
через точку прикрепления жгутов к телу, т. е. через жгутиковый полюс, незави-
симо от приобретаемого им положения. Далее можно видеть. как происходит
постепенно обособление оси движения; так, у динофизид она еще связана с пер-
вичной сагиттальной плоскостью тела (намеченной здесь даже внешне—сагит-
тальным «линочным шьвом»), а у высших, асимметричных, перидиней приобретает
полную независимость от нее; наконец, на этих рядах необычайно отчетливо
видно, что осью симметрии все более усложняющегося панцыря служит здесь
именно ось движения, а не первичная ось тела, далеко отклонившаяся от нее.
Кажущееся отклонение от общего плана архитектоники одно-
клеточных жгутиковых представляют трипанозомы, своеобразное
строение которых создает видимость обращения их морфологиче-
ской полярности. A `
В самом деле, со строго «морфологической точки зрения трипано-
зома отвечает, скажем, трихомонаде, перевернутой задом наперед.
Однако это мнимое обращение ориентации животного не отражается
на его физиологической полярности и характере деления и, вдоба-
вок, полностью исчезает при превращении трипанозомы в крити-
‚шиальную форму. Ход этого превращения ясно показывает, что дело
здесь сводится к простому смещению жгутикового-а.ппарата относи-
тельного клеточного тела—-смещению, не чсопровождаемому здесь
какими-либо более глубокими морфофизиологическими изменения-
ми. Столь свободное его перемещение может показаться странным,
если вспомнить все сказанное выше о проморфологическом значе-
нии жгутикового -полюса у других Flargellata И, в частности, крипто-
монад и Dinoflag-ellata. Однако оно легко объясняется тем, что
у Trypanosomidae (и Bodonidae) ЖГУТИЪСОВЫЙ аппарат мор:фологи-
чески и функционально не связан с ядерным и не принимает,
в частности, никакого участия в кари-окинезе, в ‘товремя как у
остальных жгутиковых они находятся в тесной связи, вы-ражаю-
щейся ярче всего в том, что базальные зерна-жгутов играют при
делении роль центросом. ' -
_ На этом именно признаке основывается недавно предложенное
объединение обоих только-что названных" семейств ‘в группу Мазй-
gosomea, „противопоставляемую совокупности остальных Zoofla—
gellata, образующих-группу Centrosomea. .
Архитектонической сложности жгутиковых (Protozoa соответ-
ствует их высокая органологичесгкая расчлененность. Онаесоздается
главным образом за счетобильных и`разнообрёазныхдифференци-
ровокрпышное развитие которых необычайно характерно дляпро-
стейших этого типа. ' _- - ~ ч ‹ . »
60
Первое место по своей типичности занимает, конечно, жгутико-
вый аппарат, состав И строение которого чрезвычайно разнообразны,
В самых простых случаях-——у жгутиконосных -солнечников и неко-
торых Rhizomastigina (Mastigella, Cercobodo), —_— OH состоит только
из одн-ого или нескольких жгутов, не связанных с ядром И не имею-
щих собственных базальных зерен. Одна-ко среди настоящих Fla-
gellata (включая и диморфных амёб) и генетически связанных с
ними групп жгутиковый аппарат никогда не бывает настолько про-
стым._ Он представляет собой тут гораздо более сложное и диффе-
ренцированное образование, киннетиду (Шаттон, 1924), нахо-
дящуюся в тесной связи с ядром и состоящую из ряда структур,
объединенных в одно функциональное целое:
1) самих жгутов, число которых в одной пкинетчиде никогда не
превышает четырех;
2) одного-двух базальных зерен, к которым прикрепляются жгу-
ты и из которых они образуются;
З) центросомы, представленной авнутри- ил-и внеядерным тепьцем,
могущим в ‚последнем случае служить базальным зерном для жгу-
тов, но нередко и отсутствующим;
4) соединительных тяжей, связывающих базальные зерна друг
с другом, с.ядром или с центросомой; _
5) побочных образований, какими являются парабазальный ап-
парат и аксостиль.
Разнообразие строения кинетиды очень велико, определяясь чис-
лом ее жгутов, наличием или отсутствием побочных ее частей,
а главным же образом тем, каковы соотношения базальных зерен
с центросомой или ее гомологами; как уже упоминалось, эти два
вида органоидов могут быть раздельными и тогда обозначаются,
соответственно, как M ac T И г о с о M ы, или базальные зерна соб-
ственно (раньше их обычно называли блефаропластами), И Цен-
т р осо м а; в дРУгих же случаях они как бы объединены в одной
структуре, получающей тогда названия к и н е т о с о M ы, центрома-
стигосомы или центробл-ефаропласта (ри-с. 7). Нитевидные тяжи,
связывающие друг с другом отдельные части кинетиды, также ‚обо-
значаются различными эпитетами, в зависимости от того, какие
именно из ее частей соединяются каждым из них: так, ризопластом
принято называть нить, тянущуюся между центросомой пи базаль-
ным зерном жгута, центродесмой— временный“ тяж, соединяю-
щий центросомы при их делении, мастигодесмои——аналогичный,
но чаще сохраняющийся тяж, получающийся при делении базаль-
ных зерен. У ряда Zooflagellata кинетида сохраняется и при инци-
стировании. С другой стороны, у ряда форм она вся может созда-
ваться заново и резорбироваться в деятельном состоянии организма.
Важнейший вопрос об источниках возникновения кинетиды и
первых этапах ее формирования принадлежит к числу самыхъзапу-
танных и неясных вопросов сравнительной морфологии простейших.
Имеющийся на этот счет большой, но весьма противоречивый фак-
тичес-кий материал не привел еще проти-стологов к хоть сколькоыни-
61
будь общепринятому и убедительному решению вопроса, так что,
существующее тут положение представляется совершенно обеску-
раживающим. -
Однако со стороны кажется очевидным, что имеющаяся тут не-‘
определенность объясняется главным образом тем, что все усилия
ПРОТМСТОЛОГОВ НЗПРЗВЛЯЮТСЯ ПО НЕПРЗВИЛЬНОМУ пути——на ПОИСКИ
ОбЩЕГО, ЕДИНОООРЗЗНОГО И абсолютного РЕШЕНИЯ ЭТОГО ВОПРОСЗ, Ир
что, вдобавок, им все же остается чуждой методология сравнитель-
ной, эволюционной, морфологии. Поэтому имеющиеся в их руках
обильные и разнообразные фактические данные по этому вопросу
служат в большей мере предметом дискуссии, чем сравнительной.
оц-енки, и энергично оспариваются каждым автором, если только
они не согласуются с резуль-
татами, полученными HM самим
на ином, подчас крайне отлич-
ном и систематически далеком,
объекте. Если же отрешиться
от так-ого ‘принципиально не-
оправданного скепсиса и по-
дойти к накопленным фактам
без предубеждения, и притом
с более общих зоологических
позиций, разрешение -вопроса
„о генезисе жгутикового аппа-
Рис. 7. Различные типы строения кине- рата перестает казаться безна-
тИдЫ= дежным.
1 -к и ‚ ки ето-
сомы "ЁЁЁЁЬ;СЁСТЁЯЁЦЁЁ‘кЁЁЁЁЁЁььНЁом-„Ёщие Первое? K Чему ПРИВОДИТ
ИЗ ЦЕНТРОСОМЫ, РИЗОПЛЗСТЗ, ОДНОЙ ИЛИ ДВУХ г
’3?1ZTc’i~2?°‘?s”_”£L‘§Z$3§;,4 cT;c‘§~%;’$;a"‘Ls6§§r‘;‘$3°é это» к допущению двоякого про-
мастигосомойипарабазального аппарата. Исхождения }KI~y»1-OB‘: путем
соответствующего преобразова-
ния аксоподий, т. е. дальнейшей дифференцировки образующей
их цитоплазмы, и 2) путем их создания из ядерного материала `и
его непосредственных производных. В первом случае можно гово-
рить о плазмогенном происхождении жгутов, во втором— о карио-
генном. ’
Плазмогенные жгуты характерны для солнечников и наблюда-
ются также. у некоторых амёбоидньтх форм, условно относимых к
Rhizomastigina (Mastizgella). У солн-ечников, обладающих одновре-
менно аксоподиями и жгутами, последние, как уже отмечалось, вхо-
дят вместе с первыми в общую радиальную систему в качестве рядо-
ВЫХ ee ЧЛЕНОВ И НЕ ОТЛИЧЗЮТСЯ ОТ ООТЭЛЬНЫХ НИЧЕМ, КРОМЕ СВОЕЙ,
тонкости, подвижности и специально выполняемых ими функций.
Очень показательным примером генетического единства этой си-
стемыипутей ее функциональной дифференциации может служить
примитивный м-орской солнечник Actinomonas mirabilis. Это-си-
дячая форма с ТОНКпИМИ аксоподиями, стержни которых не идут в
глубь тела,чи у которой, соответственно, нет и центрального зерна.
62
Одна из акс-оподий несколько толще и длиннее остальных и обра-
зует стебелек, посредством которого животное прикрепляется к
субстрату, а другая,‘ диаметрально противоположная -стебельку,
представлена длинным жгутом. ‘В результате движений жгута ча-
стицы пищи п-одгоняются к остальным аксоподиям, которыми они
улавливаются и втягиваются в цитоплазму; При делении жгуты
создаются заново каждой из дочерних особей, а материнский резор-
бируется. Таким образом, здесь налицо явственная функциональная
дифференциация радиальной системы на прикрепительный, подго-
няющий и улавливающий аппараты, что не мешает им сохранять
очень сходное строение и почти одинаковую степень обратимости.
Не менее наглядный пример того же рода составляет Dimorpha
mutans, также сочетающая в себе признаки. солнечника и жгутико-
вого. Все элементы ее радиальной‘ системы сходятся к связываю-
щему их центральному зерну, причем два соседних из них представ-
лены постоянными жгутиками, а остальные типичными аксопо-
диями. В сидячей стадпии солнечника один из жгутов используется
в качестве прикрепительного стерженька, обрастая при этом тол-
стым студенистым чехлом, а второй остается свободным; выполняет
ли он в это время какую-нибудь определенную функцию—неиз-
вестно. В этой же фазе происходит и деление, начинающееся с раз-
деления центросомы на две, из которых одна остается на месте
с материнским прикрепительным жгутом и половиной имевшихся
аксоподий, а вторая отходит вместе со свободным жгутом И другой
их половиной. При переходе в жгутиковую стадию «бродяжки» -со-
храняются только жгуты (один из них становится рулевым), в то
время как остальные элементы радиальной системы теряют свои
свободные, торчащие из тела концы, а гИх внутриклеточные отрезки
становятся едва видимыми (см. рис. 4, 5-7).
Сходным способом совершается превращение вегетативной фазы
солнечника у Ciliophrys и Clathrulina B чисто расселительную жгу-
тиковую; однако тут остающиеся аксоподии попутно превращаются
в жгуты. В этих случаях и у Dimorpha функциональное преобразо-
sample отдельных лучей в жгуты оформляется в связи с исчезнове-
нием остальных лучей, так что жгуты фактически оказываются более
устойчивыми, чем последние, и потому полностью заслуживают быть
признанными стабилизированными и видоизмененными аксоподиями.
Такую же природу аксоподий имеют и жгутьт у некоторых
Rhizomastigina. Их- образование изучалось у Mastigella Vi’trea—
одножгутиковой апмёбоидной формы, имеющей лопастевидные
псевдоподии (типа лобоподий), каждая из которых снабжена
СТЕРЕОПЛЭЗМЗТИЧЕСКИМ СТЕРЖНЕМ, ИДуЩИМ B I‘JIy‘6b ЭНДОПЛЗЗМЫ.‘
ЗЗКЛЗДКЗ ЭТИХ ОСЕВЫХ СТРУКТУР ПРОИСХОДИТ В ЭНДОПЛЗЗМЕ, OTKy.7lEl
ОНИ ВЫТЯГИВЗЮТСЯ B ЦЕНТРООЕЖНОМ НЭПРЗВЛЕНИИ; она ПРЕД‘
ШЕСТВУЕТ формированию ПСЕВДОПОДИИ, а РЗЗЖИЖЕНИЕ ЭТИХ ‘СТРУКТУР
ПРОИСХОДИТ ДО РЭСПЛЫВЗНШЯ ПОСЛЕДНИХ. СХОДНЫМ СПОСОбОМ обра-
зуется и жгут, отличающийся от этих дифференцировок только IBM,
ЧТО ОН ДЛИННЕЕ, ОДЕТ бОЛЕЕ ТОНКИМ ПЛ-ЭЗМЭТИЧЕСКИМ ЧЕХЛОМ И о-бла-
63
дает несколько большей, хотя и ограниченной, подвижностью-Пери
поступательном движении животного жгут обычно направлен впе-
ред, но не играет никакой роли в самом передВИЖеНИИ- .
Жгутами ядерного происхождения обладают ПОВИДИМОМУ, ВСЁ
настоящие Flagellata (включая и диморфных амеб) и очень многие
из жгути-ковых стадий различных других групп организмов. Такие
жгутынеизменно снабжены собственным базальным зерном, из ко-
торого они выра-стают. Крометого, он-и обычно объединеныс раз-
личными другими дифференцировками-в тот более или мценее слож-
ный моторный аппарат, который мы называем кинетидои. Другими
словами, такие жгуты характеризуются прежде всего тем, что они
Рис. 8. Образование кинетиды у Vahlkampfia gruberi при ее превращении
из состояния амёбы в жгугиковое состояние (по Ч. Вильсону)
являются не изолированными образованиями, а составной частью
Целой кинетиды, включающей, как минимум, и соответствующее ба-
зальное зерно. ‘
Формированиетакой кинетиды может идти весьма разнообразными путями,
что и объясняет отмеченное разнообразие ее состава, взаимоотнош-ении ее ком-
понентов и различия в степени и объеме генетической преемственности послед-
них. Имеющиеся данные иллюстрируют многие из этапов этого морфогенетиче-
ского процесса и позволяют уловить его главные направления, подчиняющиеся,
в целом, общему принципу повышающейся устойчивости дифференцировок.
Беря этот принцип в основу расположения соответствующего фактического
материала, мы, очевидно, должны идти от форм, вся кинетида КОТОРЫХ 05_Pa3Y€-T051
заново при превращении. в жгутиковую фазу и исчезает при инцистировании или
переходе в амёбоидное состояние.
Ход формирования такой временной кинетиды полнее всего прослежен
у Vahlkampfia (Naegleria) gruberl (рис. 8) и фитомоНадЫ P01Yt0m€1151'C1tT1, У 490‘
торых оно целиком происходит за счет материала ядрышка. Первым этапом этого
процесса является своего‘ рода неравномерное деление ядрышка (РИО 9: 2-3): ‘ПРИ
котором меньший из его продугктов выходит из ядра к будущему переднему
концу тела, оставаясь связанным с ядрышком посредством Все УДЛИНЯЮЩЭГОСЯ
тяжа. (рис_ 9, 4——7). Дойдя до переднего полюса, это зерно разделяется надвг,
каждое из которых продуцирует по одному (Naegleria) или по два (Ро1у’соп1е11а)
жгута; после этого соединительный тяж может исчезнуть, вследствие чего upe-
рывается связь между офо-рмившейся кинетидои и ядром. У Na-eglerla B жгути-
ковшу, фазе простейшее не способно ьк делению и оно осуществляется исключи-
тельно‘ в амёбоидн-ой, причем тут нет постоянной центросотш и ее роль выпол-
64
‘няетсхя самим ядрышком; у Polytornella, напротив, простейшее‘ делится только‘ и
активной жгутиковой фазе, деление кинетиды, идет независимо от, кариокинеза,
а ГЪОСЛЕДНИЙ связан С o6pavsoB_aH14eM ВРЕМЕННЫХ -внеядерных ЦЕНТРОСОМ И СОЭДИ"
няющей их центродесмы, исчезающих по его окончании. .
У другою представителя Vahlkampfiidae, Vahlkampfia bistadialis образование
временной кинетиды отличается от описанного тем, что общий (ядерный) зачаток
кинетиды делится в тангенциальном направлении еще на поверхности ядра, после
чего епопроизводные _сначала несколько расходятся, а затем делятся еще раз
радиально, давая таким образом начало сперва двум проксимальнымгранулам 11
порядка, а потом уже паре дистальных гранул III порядка, являющихся базаль-
ными зернами жгутов. Деление этой формы происходит также, как у V". (М) gru-
beri, только в фазе амёбы, и идет таким же образом, хотя тут иногда описыва-
лась временная центросома внутри ядрышка. A г '
.../
5 д "7
Рис. 9. Образование кинетиды у 'Po1ytomella citri
при возвращении в дифференцированное состояние:
7—инцистированная особь; 2, 3—первые стадии образования
зачатка кинетиды, отпочковыва иЩеГОСЯ от ядрышка; 4-
выход зачатка на поверхность ядра; 5—его разделение на
центросому и базальное зерно жгута, остающиеся соеди- .
ненными тяжем (ризопластом); б—разрастание ризопласта;
7—раздвоение базального зерна; «ЭК-появление жгутов и
выход животного из цисты (по катеру)
д
Еще сложнее дело обстоит у Euglena graci1is—-ecJ1IvI '1‘OJ]bK0 верить данным
Бейкера. (1926) и его интерпретации наблюдающихся тут картин 1. Согласно
этому автору, кинетида целиком обновляется при каждом делении (происходя-
щем во временно покоящемся, хотя и не инцистированном состоянии организма)
и создается каждый раз заново из такого же ядерного зачатка, как у V. gruberi
и Р. citri. Выходя на поверхность ядра, этот зачаток делится сначала тангенци-
ально, причем образовавшиеся дочерние гранулы (11 порядка) расходятся к про-
тивоположным полюсам ядра, но не достигают их, оставаясь связанными тяжем;
далее, каждая из них_ отделяет от себя по грануле 111 порядка (‹<блефаропласт>›
по Бейкеру), уходящей к самой поверхности резервуара (углубления, в котором
располагается основание кинетиды) и здесь делящейся еще раз; из получаю-
щейся теперь новой гранулы (IV порядка)—‹<ба3ального зерна» Бейкера выра:
стает дополнительный короткий жгут, тогда как главный вырастает из <<блефаро-
пласта». В дальнейшем десмозы, соединявшие гранулу II порядка с ядрышком
и с ее производными, распадаются, и эта гранула уходит в цитоплазму, где она
1 Эти данные подверглись суровой критике со стороны _Хэлла и Поуэлля
(1928) и не нашли подтверждения в позднейших (гораздо более поверхностных
кстати) исследованиях Крихенбауэра (1937) над Е. gracilis. Однако первые
авторы основывали свое суждение на изучении совсем другой эвглены. (Регапета)
и принятом у протистологов скептическом отношении к чужим данным и чужим
взглядам, а Е. gracilis Крихенбауэра, безусловно, не идентична с формой изучав-
шейся Бейкером и, в частности, делится в активном состоянии, чем полностью
могут быть объяснены имеющиеся расхождения (см. гл. П, 3). `
5 Эмбриология беспозвоночных '5-5
свободно лежит в виде остаточной «хроматоидной Массы» до следующего деле;
ния, при котором резорбируется. Что же касается кариокинеза, то он осущестд
вляется тут таким же путем Kax/Vahlkampfia (гл. 11, 2) без участия в нем,
центросом. - `
Итак, во всех этих случаях доказывается образование всей ки- Ё
НЕТИДЫ ИЗ кариосомного ЗЗЧЭТКЗ, НЕ принимающего участия В ДЕ?
ЛЕНИИ ядра И дающего начало (ЛИОО НЕПОСРЕДСТВЕННО, ЛИбО ЧЕрЕЗ
1-2 деления) базальным зернам жгутов. Эти последние имеют,‘
следовательно, значение мастигосом пи представляют собой типичные
дифференцировки временного характера. Они могут создаваться за-
ново либо при превращении активной амёбоидной фазы в жгути-
ковую (Vahlkampfiidae), либо при каждом акте деления (Euglena
gracilis), либо только при выходе из цисты (Polytomella citri), T. e.
обнаруживают ряд различных ступеней структурной устойчивости.
Такой же, сугубо временный, характер имеет и остаточное хрома-
Ruc.'_10. Образование кинетиды у зооспор Syndinium (no Шаттону)
тоидное тельце Е. gra-cilis И, разумеется, -сами жгуты. Стало быть,
кинетида не включает тут ни одн-ого постоянного элемента, и ее пря-
мая генетическая преемственность либо вовсе не имеет места, лиибо‘
ограничена интервалом между двумя последовательными инцисти-
рованиями. I „
Несколько особое положение занимают‘ процессы образования
кинетиды у жгутиковых зооспор некоторых паразитических» Dinof1a-
ge11ata.. Так, у Syndinium зачаток кинетиды внезапно появляется в
телофазе последнего спорогенетического деления на полюсе каж-
дой из половин веретена, т. е. Tam, где полагается быть центросоме,
поуже, если можно так. выразиться, по миновании надобности в ней.
Этот зачаток быстро вытягивается по длинной оси зооспоры и обра-
зует Ha своем дистальном конце базальное зерно, из которого вь1ра-
стают оба жгута; после этого он теряет свою связь с ядром
(рис. 10). При переходе зооспоры в плазмодиальную паразитиче-
скую стадию кинетида полностью и-счезает.
Все другие способы формирования кинетиды легко выводятся
из только что рассмотренного прототипа, причем выясняется,.что
все их разнообразие определяется исключительно тем, делится ли,
и сколько раз, общий ядерный зачаток кинетиды, сохраняется ли
связь лмежду его производными и какие из частей кинетиды стаби-
66
Щизируются или, напротив, исчезают; Однако во всех этих случаях
обычно уже не удается наблюдать образование самого зачатка: мы
теперь сталкиваемся с ним, как с постоянным уже клеточным орга-
ноидом |ИЛИ же совсем его не находим.
Одна из намечающихся тут линий развития кинетиды связана
с делением в покоящемся (преимущественно инци-стированном) со-
стоянии и демонстрируется лучше всего на примере различных
Volvocales. У большинства этих фитомонад имеется перманентный
внутри- или внеядерный зачаток кинетиды, играющий рольцентро-
сомы в делении ядра и представляющий собой ее единственный по-
стоянный элемент. Эта центросома, видимо, соответствует грануле 1
порядка Е. gracilis И других рассмотренных выше форм, но суще-
ственно отличается от нее своей активностью в кариокинезе, неслу-
чайно идущем здесь и в другой форме-в форме настоящего ми-
тоза (гл. 11, 2). B ходе митоза, материнская центросома (1) делится
на две дочерних (гранулы 11 порядка), а при переходе в активное
состояние каждая из них отделяет по мастигосоме (грануле 111 по-
рядка), могущей еще разделиться надве. Все эти зерна и соединяю-
пцие их тяжи, кроме самой центросомы, разрушаются при дедиф-
ференцировке (т. е. перед каждым делением), имея, таким образ-ом,
чисто «интеркинетический» хара-ктер.
Другая линия стабилизации жгутикового аппарата связана, на-
против, с делением в активном дифференцированном состоянии.
Здесь также имеется постоянный, но всегда внеядерный зачаток ки-
нетиды, выполняющий, как правжило, функции центросомы; однако
этот зачаток является вместе с тем и базальным зерном жгута (или
же представлен двумя с-ближенными зернами, каждый из которых
. несет жгуты), т. е. кинетосомой. Иначе говоря, в нем сосредоточены
ВСЕ фуНКЦИИ КИНЕТИЧЕСКОГО ar1.rIapaTa, распределяющиеся В ПрЕДЫ-
ДУЩЕМ СЛУЧЗЕ МЕЖДУ РЭЗЛИЧНЫМИ ЕГО 'I'IpOI/IBBOJIHBIMII/I, И ВСЯ КИНЕТИДЗ
В ЦЕЛОМ ОКЭЗЫВЗЕТСЯ ПОСТОЯННЫМ, стабильно-Дифференцированным
образованием. ТЗКОВО ЕЕ ‘СОСТОЯНИЕ у ОЧЕНЬ МНОГИХ ‘ПРОТОМОНЗД, '
сохраняющих свою кинетиду и в цисте или же вообще не способных
инцистироваться. Нельзя не обратить внимание на бросающееся
в глаза сходство подобной, высоко устойчив-ой, кинетиды с радиаль-
ным аппаратом солнечников из Centrohelidia; это сходство прости-
рается вплоть до одинаковой тенденции этих систем терять свою
связь с кариокинезом, или, лучше -сказать, проявлять свою незави-
симость or только что названного процесса. Высокая степень устой-
чивости кинетиды в целом сказывается и в такой характерной де-
тали, как длительная сохранность центродесмозы, обнаруживаемой
в .плаз-ме еще долгое время спустя ‚после разъединения ядер.„
Еще один, третий путь формирования кинетиды реализован у так
называемых Мазтйдоэотеа-трипаносом и Bodonidae И у хризо-
ЛМОНЗД; у ЭТИХ фОрМ ВСЕ НЗЛИЧНЬТЕ ЧЗСТИ ЖГУТИКОВОГО аппарата HB-
ЛЯЮТСЯ ПОСТОЯННЫМИ, НО ОН СОВЕРШЕННО НЕ ЗЗВИСИМ ‘ОТ ядра И
ВОСПРОИЗВОДИТСЯ ВНЕ ВСЯКОЙ СВЯЗИ С НИМ. В СОСТЗВ КИНЕТИДЫ ВХОДЯТ
только жгуты с их базальными зернами (мастигосомами), и при-
5* 67
соединенным иным, ‚парабазальнътм аппаратом, а Цёнтросома либо,
отсутствует, либо является внутриядерной (например, у C0110-. "
(Псйуоп) И, во всяком случае, не имеет касательства к кИнетиде.:
Правда, во время деления последняя часть сближается с полюсами
веретена, но это сближение происходит уже после установления ка-
рИокинетической фигуры и непосредственно с нжим не связано. Гене-
ЗИС такой оторвавщейся ОТ ядра КИНЕТИДЫ ТОЧНО не установлен, НО `
все. говорит за то, что в основе ееформирования лежит тот же ‘upma-
1I,Ir'IwH‘,,‘KaIQ у Polytomella или Euglena gracil__is, т. е. исчезновение свя-
зующихцдэлементов (десмоз) после образования мастигосом Из
внутриядерногозачатка. Однако этот процесс здесь непосредственно
нешнаблюдается, и мы имеем дело только с его уже закрепленными
результатами. _ A
Общими выводами из всех этих фактов, очевидно, должно быть
признание следующих_, принципиально важных положений:
1) все части кинетиды собственно образуются из единого за-
чатка, имеющего ядерное и чаще всего кариосомное происхо-
ждение; I A
2) этот зачаток имеет у примитивных форм все свойства обыч-
ной дифференцировки, создаваемой заново при формировании spe-
менного жгутикового аппарата (Vahlkampfiidae), mm при каждом .
делении (Euglena), или же после каждого инцистирования
(<I?o1-ytomella); т
3), стабилизация этого зачатка в форме постоянной кинетосомы
(центросомы) имеет явно вторичный характер и происходит далеко
не всегда; у У
.4) кинетосома может оставаться неразделенной _и тогда совме-
‚ьщает в себе функции центросомы и базального зерна жгутов, непо-_
средстве-пне вырабатывая эти последние, или же делиться на ряд
образований, получающих более узкое и специальное функциональ-
ноезначение; - '
5) „общий процесс стабилизации кинетиды, ведущий ее к превра-
щению в постоянный элемент организации, может идти различными
путями и касаться разных» ее частей; у
6) таким образом, генетическая непрерывность кинетиды в це-
лом или центросомы создается вторично, согласно общему правилу
дно-степенного повышения-устойчивости дифференцировок, и последо-
вательно ‚проходит через все ее ступени.
‘Что же касается самого механизма образования кариогенных
ткгутов, то он вряд ли принципиально отличен от того, каким созда-
‘ются плазмогенные, аксоподиальные жгуты; можно думать, что вся.
разница между ними сводится к тому, что киноплазматическая ось
первых состоит-из ядерного материала, а вторых- из цитоплазма-
тического; Во всяком случае, нет решительно никаких оснований
считать жгуты стабилизированными центродесмозами, как это пола.-
згал в-свое время Гартманн. I _ _
Побочные компоненты кинетиды образуются ;из разного исход-
ного материала и имеют, соответственно, неодинаковую „природу.
‘68
Наиболее распространенным Из ‘них является парабазальныйаппа-
рат, имеющийся у всех, повлидимому, Хоойадепата ‘и некоторых дру-
гих жгутиковых, (например, криптомонад). Это-высоко диффе-
ренцированный органоид весьма разнообразного и подчас сложного
строения, представляющий собой не что иное, как аппарат Гольджи
типа диктиосомы, вошедшей в тесный контакт с кинетидой, Еще не
так давно парапбазальньхй аппарат протомонад считался их вторым
двигательным ядром—<<кинетонуклеусом>›, и только в последнее
время была окончательно у-становлена его действительная пр.Ирода.
Однако до сих пор еще спорят о том, следует ли считать его со-став-
ной частью кинетиды, как это думает Шаттон, или же не зависимым
от нее образованием (Гра-с-се И другие). Исходя из совокупности
относящихся сюда фактов, следует признать, что оба эти воззрения
в равной мере справедливы, так -как, с одной стороны, парабазаль-
ный аппарат Zooflagellata безусловно гомологичен диктиосомам
других простейшиих и клеток Metazoa, T. e'.'.r1epBut1Ho не зависим от
кинетиды, а с другой — он прочно входит в ее состав у только что
названной группы жгутиковых. При этом он испытывает ту же
судьбу, как И другие дифференцировки, а именно постепенно стаби-
лизируется и переходит из разряда временных в постоянные эле-
менты органцизации. Очень поучительный этап данного процесса
реализован у Trichomonas caviae; При делении этой формы (Грассе,
1938) материнский парабазальный аппарат остается в одной из, цо-
черних особей, а в другой образуется заново; он возникает в плазме
вне всякой связи с мастигосомой И до ее деления, постепенно вь1ра-
стает до размеров материнского, и только послеэтого входит в кон-
такт с новой ма-стигосомой, отсылающей к нему соединительную
Н1ИТЬ (см. рис. 18). У более специализованных форм пара-базальный
аппарат становится постоянным органоидом (сохраняющимся И в
цисте), приобретает свойство делимости, а с ним И генетическую
непрерывность в ряду поколений; вместе с тем упрочивается и. его
связь -с кинетидой, находящей себе особенно яркое выражение в Их
полимеризации у многожгутиковых форм (гл. VIII, 3); однако эта
связь может прерываться; в цисте или в другие моменты жизне-
деятельности. ' ' ‘ 5 V
__ Побочными компонентами кинетиды’ являются также эндоске-
летные стержни И аксостиль трихомонад. Они образуются тут Из
кинетосомы или, во всяком случае, в тесной связи с нею. ц '
Изпдругих дифференцировок особенно характерны, стигмы (свето-
чувствительные аппараты, имеющиеся у "многих автотрофных форм)‚
более или менее сложный выделительный аппарат в ‘виде’ пульси-
рующих вакуолей, пиреноиды И многочисленные и разнообраз-
ные органоиды, связанные с анимальным’ способом питания: "unas-
матические воротнички (Craspedomonadina), ротовое отверстие,
глотка и т. п. Кроме того, большим распространением пользуются
здесь опорные И покровные дифференцировки. Примера-ми первых
могут служить, помимо аксостиля ряда Protomonad1na_, оригиналь-
ные7осевые структуры эвглен из семейства Per'anem1dae'-(‘HBJImo:-
69
ЩИССЯ СВОЕОбРЗЗНЫМ ВИДОИЗМВНЕНИЭМ ГЛОТКИ, нередко ИМЕЮЩЭЙ
опорное значение и в других группах, где она сильно развита—- ‚А
например, у Cryptomonadina), a также центральная ка-псула, обо-‘
собляющая внутреннюю часть цитоплазмы с ядром от ее периферпи- '
ческих слоев у некоторых перидиней ли Chloromonadina (Chatte-
пеПа). дифференцировки покровного характера особенно обычны
у эвглен, пелликула которых часто сильно уплотнена и имеет слож- ‘
ную скульптуру, и досгигап‹›'г своего крайнего развития в форме р
наружного скелета у нсръпдиггсй. Наряду с этим ‘нередки случаи,
когда поверхность эктоплазмы не более дифференцирована‚ чем у
амёбоишгтых форм, что обеспечивает не меньшую,
чем у них, мтетаиболичность формы тела.
цировками дополняется -еще одной характерной
для них чертой: большей сложностью И оформлен-
ностью постоянных клеточных включений, Наи-
лучшим примером этого может служить аппарат
Гольджи, часто приобретающий, как уже говори-
лось, высоко дифференцированную фо-рму пара-
базального аппарата, на в других случаях пред-
‘ставлениный диктиосомами. Н-ередко также услож-
Рис. 11. Дециффе-
ренцировка при
инцистировании у
Chlamydomonas sp;
округление клеточ-
ного тела и его
обособление от обо-
мжонадин, где он образует связную митохон-
дриальную сеть) и хромопластов, морфологиче-
ское разнообразие которых тут очень велико
(см рис. 5, б). г
Большим разнообразием обладают параплаз-
матические включения, в чем ярко отражается
общеизвестное физиологическое разнообразие
жгутиковых; при этом некоторые из них отличаются большой слож-
ностью строения (хотя бы у эвглен). ’
Что же касается алл-оплазмы, то она, напротив, не достигает
‚такого развития, как в некоторых группах амё-боидньтх Protozoa.
B ЧЗСТНОСТИ, здесь никогда не бывает минерального внутреннего
скелета, а особо сложные рак-ови.нь1 или домики имеются только
в одной группе ХрМЗОМОНЗД, у Silicoflagellata.
Таким образом, большая сложность строения одноклеточных
жгутикового типа проявляется в преобладании у них высших форм
симметрии, широчайшем развертывании разнообразнейших диф-
ференцировок и усложнении системы постоян-ных органоидов.
В полном соответствии с этими особенностями переход в инци-
стированное состояние связан здесь с крупными морфологическими
изменениями и ведет к очень сильному упрощению всей организации.
дифференцировки частью резор-бируются, частью отбрасываются,
постоянные органоиды нередко значительно упрощаются и от былой
сложности строения не остается почти ничего (ри-с. 11). Организа-
ция животного теперь сводится как бы к своей первооснове, прин-
ципиально мало чем отличающейся от таковой ачмёбоидных форм.
'70
почки и жгутико-
вого аппарата (по
Коршикову)
Богатство жгутиковых простейших дифферен-
няется строение хондриома (например, у фито-
Ь
ЭТО общее правило допускает ряд исключений в смысле пове-
дения отдельных дифференцировок B процессе ИНЦИСТИрОВаНИя
Так а '~
, y M shgosomea B цистах сохраняются мастигосомы, а нередко
и основание самих ‘жгутов, у Cerc-omonadidae__ атксдстидь’ у веко-
торых фитомонадин—то стигма, то пиреноиды и т. д. В этих с.лу-
чаях можно видеть первые проявления определенной тенденции
дИффеРеНЦИровок становиться постоянными органоидами, т. е. те-
о
Рис. 12. Polytomella citri, выход из цисты в полуамёбоидном состоянии;
стрелки показывают направление движения (по Катеру)
рять ‚при-сущую им обратимость. Эта тенденция получает свое
полное а в . °
ТИКОВЫХР гнёта?) ъёшдклеточньтх простейших (паразитических жгу-
-ВЫМИ ДЛ‚Я НееУЭЛЕМЁ И др.)Ви ведет K обогащению организации но-
НТаМИ- 0051119 РОВОРЖ Эта тенденция тем более
ВЫРЗЖЭНЗЬ ЧеМ сложнее строение простейшего.
Общее упрощение организации может зайти так Далеко что
"ИНЦИСТИРОВЗННОЗ ЖГУТИКОВОе теряет свои ‚самые основные промор-
Ёологические свойства-определенность оси тела и его полюсов
но еп о '
фолоёческиёя I; И 3 Уие Т С H. И Тогда уже лишается последних мор-
очень He I 0 ЛИЧИИ ОТ одноклеточных амебоидного типа, как это
Ш п но вирно на примере таких форм, как Polytomella, Pyrami-
п 0 а5« (РИС- 2)- Чаще, однако, этого не происходит, и покоя-
Щаяся особь сохраняет не только монаксонно-гетерополярное -строе-
‘Ёшёкёёобще’ H0 ‘И ТУ ЧаСТНУЮ ФОРМу симметрии, какую она имела
OTHO BHOM СОСЮЯНИИ» ПРЁВДЗ, ее ориентация нередко меняется
. сительно первоначальнои, что легко устанавливается по карти-
Нам деления (стр. 95).
Перед Выходом низ цисты все временно утраченные дифференци-
РОВКИ вырабатываются заново, и животное полностью восстанавли-
ае ‹ ‚ -
В Т Типичное для него строение (см. рис. 9). Таким образом, нспеци-
71
фИЧЗСКИЭ ОСОбЗННОСТИ ЖГУТИКОВОГО типа ОДНОКЛЕТОЧНОЙ Ор-ГЭНИЗЗЦИИ
„оказываются приуроченнымии к одной только активной, вегетативно-
;деятельной фазе существования, и в своем большинстве или даже
полностью исчезают у покоящейся, инцистированной фазы.` Иначе‘
говоря‚. эти две взаимообратимые физиологические фазы приобред
‚тают здесь крайне резкое морфологическое выражение. И так как
их различия целиком сводимы к явлениям дедифференцировки орга-
низ-ма при его инцистировании и его редифференцировки перед ‘
выходом из цисты, то будет справедливо рассматривать специфиче-
ские особенности этого типа одноклеточной организации, как выра-
жение ее дифференцированного состояния. Можно также сказать,
что самое глав-ное отличие простейших жгутико-
вого типа от амёбоидных заключается именно
в том, что ‘у первых активная жизнедеятель-
ность осуществляется в осо бом‚ морфологически
усложненном дифференцированном состоянии орга-
Низации, тогда «как у ‘вторых этого нет,иих активная
фаза морфологически мало чем отличается от покоящейся.
2. Взаимоотношения амёбоидного и жгутикового типов
организации
Вопрос о взаимоотношениях амёбоидного и жгутикового типов
одноклеточной организации имеет важнейшее принципиальное зна-
чение не только в том общем филогенетическом аспекте, в каком его
принято трактовать, но также чИ с конкретной морфологической
точки зрения. Речь идет о том, должно ли им приписываться пре-
имущественно систематическое значение, ‘как это обычно делается
зоологами, или же более общее значение двух первых «ступеней
морфологической дифференцировки» организмов из тех семи, ка-
кие различаются ботаниками, mm, может быть, еще более универ-
сальное значение взаимообратимых фаз клетки.
‘Первое из этих толкований решительно опровергается самим
фактом, что оба типа одноклеточной организации встречаются не
только во всех классах простейших, кроме инфузории, но также и
у три-бов, водорослей и подавляющего большинства высших, много-
клеточных, организмов. В одних группах тем или другим из них
представлены основные по своему биологическому значению веге-
тативные стадии трофозоитов; в других они оба сосуществуют, как
равноправные и взаимообратимые формы трофозоитов; В ТРЕТЬИХ
ими представлены только раоселительные стадии бродяжек или
‚зооспор; наконец, еще в других — половые стадии — гаметы и обра-
зуемые ими зиготы. Таким образом, каждый из этих типов может
олицетворять в одних случаях «взрослое», дефинитивное состояние
организации, а в других—— ран.ние онтогенетические стадии или же
определенные стадии жизненного Цикла: B‘M€CT€_C ТОМ ОНИ МОГУТ
закономерно или от случая к случаю чередоваться в последнем или
даже в онтогенезе. ` `
72
ОПРЕДЭЛЁННОЕ, фиксированное систематическое значение приоб-
РОТЗОТСЯ ЭМЭбОИ/Ёьным и жгутиковым типом только тогда «когда тот
или го” ’ . .
с дру И из них становится господствующим типом
тцроения вегетативных форм в какой-либо (естествен-
нои г ппе о г .
а Не? KaPK§;Ie~Jm61:) aHH;:IoB. Такая установка именно на вегетативные,
в основе всей бгёэру" е фазь} Жизненного Цикла’ Лежит’ По СУТИ дела’
U логическои систематики; она служит ее основ-ньхм
“r ПОЖЗЛУРЪ Наиболее специфическим принципом а если и не всегда
осознается ’
Этому МЫ Ьтгоопхочтаи неуёлонно применяется на практике. Благодаря
или ТИП Fla Пусч ем о ъективную возможность соединять в класс
‹ ‘ge а а Все „ТО ГРУППЬЪ У Которых эту главенствующую
роль играет жгутиковыи тип одноклеточной организации и его непо-
средственные производные; соединять в класс Sarcodina те-группы
Где Такая РОЛЬ ‘Принадлежит аМЁбОИДНОМУ типу и выделять в само-
ст ’
KO(;$(iTe:II)I:HbIe классы или типы все те организмы, вегетативные стадии
pg далеко отошли в своем строении от обоих этих типов,
прио ретая надклеточную или многоклеточную организацию.
аб-‘СЛЭДОВОТЕЛЬНО, Интересующие нас типы организации имеют не
сёлютное, а сугубо относительное систематическое значение.
толь же относительно и их значение в качестве ступеней или
УРОВНОЙ МОРФОЛОГИЧООКОИ дифференцировки- в том смысле, в ка-
глаза/г они понимаются альголо.гами. Дело в том, что они принципи-
_но отличаются от большинства других таких ступеней (а именно
протококковои, нитчатои, а отчасти и сифональной) двумя особен.
гостями, делёъющимиоих явно неравноценными остальным. Первой
з таких осо енностеи является их универсальность. В то время как
перечисленные ступени пользуются лишь относительно узким рас-
'“P0C’_{P3HeHH€M, ОГРЗНИЧЭННЫМ пределами низших растений, амё-бо-
идныи и флагеллятный типы организации не знают, в сущности ни-
7
Каких ЁИОТОМОТИЧОСКИХ границ. ВО-ВТОРЫХ, они обладают и несрав-
"WHO ' Одее ШИРОКИМ‚ изменчивым и разнообразным биологическим
3НаЧеНИеМ‚ Чем остальные ступени, представленные, одними только
вегетативными стадиями цикла, и никакими больше. Поэтому их
соотношения с лестницей этих ступеней в сущности таковы же, как
с таксономической системой: они приобретают значение единиц той
ИЛИ Другой из этих систем лишь в определенных конкретных слу-
чаях, но не сводимы к таким единицам и явно-не вмещаются в их
слишком тесные рамки.
С другой стороны, взаимообратимость этих типов представляет
настолько распространенное явление, что трудно не считать ее
весьма многозначительной. В самом деле, легкий переход вегетатив-
пых форм из жгутикового состояния в амёбоидное и "обратно, нз
амебоидного в жгутиковое, имеет место у громадного числа простей-
,ШИХ, ОТНОСЯЩИХСЯ K самым различным СИСТЕМЭТИЧЗСКИМ группам.
Классическими его примерами являются многочисленные хризомо-
надины, специально изучавшиеся в этой плоскости Пащером и уже
упоминавшиеся Vahlkampfiidae (рИС. 13); эти примеры можно до-
ПОЛНИТЬ длинным рядом других, касающихся Dirloflagellata (Dina-
73
moebidium, Syndinium), Crypt-omonadina (~Crypt0chfysis ашое-_
boidea), Chlor-omonadina, Phytomonadina (Pyramimonas и др.),
Pr-otomonadina, Polymastigina (Histiomonas, рис. 14) —из Flagel-~
lata, Dimastigamoeba и Trimasti,gamoeba— ИЗ голых амёб, многих-
солнечников и т. д. Еще чаще наблюдается превращение жгутико- д
вых начальных стадий развития в амёбоидные вегетативные. Оно.
характерно для радиолярий, фораминифер, параме-б, многих сол-
нечников, некоторых Dinoflagellata H групп, связывающих их со
Рис. 13. Vahlkampfia bistadialis:
1—питающаяся фаза амёбы; 2——начало ее превращения в жгутиковую; З——жгутиковая
фаза; 4—инцистированная особь
споровиками (Coccidiniidae), слизевиков и т. п. Наряду -с этим
известны случаи, когда амёбоидная стадия предшествует в онтоге-
незе жгутиковой (аимёбозиготы некоторых Chlamydomonas).
Прев ащение из жгуттиковой формы в амёбоидную сводится
к исчезновению жгутов и других дифференцировок и к деполяризации
тела, т. е. к тем же процессам, которые сопутствуют инцистирова-
нию И составляют содержание дедифференциировки. Вся разница
тут заключается в том, что деполяризация идет, как правило, зна-
чительно дальше и что эти процессы осуществляются в активном
состоянии организма, а «не при его превращении в цисту М, соот-
ветственно ЭТОМУ, не сопровождаются СПЗЦИЗЛЬНЫМИ ИЗМЕНЗНИЯМИ,‘
направленными к изоляции от внешней среды (округлением тела,
выработкой оболочек). Равным образом и превращение амебоиднои
стадии в жгутиковую происходит путем выработки кинетиды, уплот-
НЕНИЯ ПЕЛЛИКУЛЫ, СТЭбИЛИЗЗЪЦИИ фОрМЫ тела И его -П0ЛЯрИ3аЦИ!И‚‚
т, е., по сути дела, путем дифференцир-овки, но опять-таки проис-
ходящей Ha -базе активной, а не покоящейся стадии. Впрочем, быч
74
вают случаи, когда превращение из амёбоидного состояния в жгу-
тиковое происходит не непосредственно, а через инцистирование,
или же_когда за дедифференцировкой жгутиковой формы непосред-
CTBBHHO следует и.нци-стирование. ЭТИ случаи ОбЪЁШИНЯЮТ еще тесн-ее
ра-ссматриваемое явление С явлениями, СВЯЗЗННЫМИ С ИНЩИСТИрО-
_вание›м у ОДНОКЛЗТОЧНЫХ ЖГУТИКОВОГО ТИПЗ, ДЗДИффЗрЗНЦИРОВЗННЗЯ
фаза которых, как уже говоргилось, снижает свою организацию до
амёбоидного ее уровня.
Все эти факты, очевидно, свидетельствуют о том, что амёбоид-
ный и жгутиковый тип одноклеточной организации являются в ко-
нечном счете не чем иным, -
как ее альтернативными,
взаимообразными, фаза-
ми: первый олицетворяет
собой ее -недифференци-
р-ованное состояние, вт-о-
рой——состояние диффе-
ренцированное.
Большое внимани-е за-
служи-вает то обсто-ятель-
ство, что степень легко-
сти, с которой одна фаза
превращается в другую,
очень неодинакова в раз-
ных группах простейших
и находится в определен-
ной связи с высотой орга-
низации, достигаемой ими
в дифференцированном (жгутиковочм) состоянии. Так, непосред-
ственное превращение вегетативных стадий из одной фазы в другую
имеет мест-о, строго говоря, только тогда, когда жгутиковая харак-
теризуется значительной примитивностью—›протомонад.ным уров-
нем строения (Vahlkampfiidae) или же низшим из имеющихся в данн-
Рис. 14. Одножгутиковая, четырехжгутиковая
и амёбоидная фазы Histiomonas (по Уэнричу)
ной группе (например, уровнем организации Chromu1inidae——
у хризомо-над). При- более сложном же строении жгутиковой фазы
оно настолько стабилизируется, чт-о ее непосредственное -превра-
щение в, амёбоидную становится невозможным; оно может -осуЩе-
ствляться лишь в порядке реорганизации, сопряженной с ини_цисти-
рованием или же путем постепенных онтогенетических перестроек.
Другими словами, при слабом развитии дифференцировок их лик-
видация может осуществляться «походя», без коренного изменения
физиологического состояния организма, остающегося активным,
тогда как при сильном их развитии осуществление этих процессов
неизбежно требует перехода из вегетативно деятельного в покоя-
щееся, инцистированное состояние. Уместно напомнить, что в тех
случаях, когда дифференциро-вки приобретают особую сложность,
о-ни стабилизируются еще больше и тогда. сохраняются даже в цисте
(хотя бы частично).
75
Таким образом, мы не только можем говорить о налгичии у одно- п
клеточных двух фаз организации- недифференцированной и диф- `
ференцированной, но И различать у них три уровня Дифференци- '
ровки: 1) обратимый, при котором переход виз одной фазы
в другую осуществим без утраты подвижности пи без инцистирова-
ния; 2) л а б и л ьн ы й, при котором он требует инцистирования, И Ё
3) ст а б и л ь н ы й, характеризующийся тем, что раз он достигнут,
то уже сохраняется пожизненно, нс «снимаясь» инци-стированием.‘
Наличием трех различных уровней дифференцировки у одно-
клеточных устанавливается ее нарастающий характер у жгутиковой
фазы, являющейся в этом смысле прогрессивной по сравнению
с амёбоидной. Следовательно, создается ряд, идущий от постоянно
амёбоидных форм к таким, у которых амёбоидная фаза, оставаясь
господствующей, способна факультативно дифференцироваться
в несложную по своему строению и неустойчивую жгутиковую;
далее располагаются формы, жгутиковая фаза которых прогрес-
СИВНО УСЛОЖНЯЕТСЯ И тем самым стабилизируется, СТЗНОВЯСЬ ОСНОВ-п
ной формой вегетативного существования, в связпн с чем амёбоидная
теряет свои вегетативные функции и низвсдится до уровня пас-сив-
ной фазы жизнедеятельности; наконец, жгутиковая фаза становится
настолько сложной и устойчивой, что амёбоидная полностью исче-
зает. Иначе говоря, этим рядом иллюстрируется постепенное стано-
вление дифференцированной фазы существования и последующее
замещение ею более примитивной недифференцированной.
Если учесть, что с морфогенетической точки зрения дифферен-
цировки безусловно являются производными цитоплазмы или
постоянных органоидов клетки и сами эволюируют в направлении
от временных, эуплазматических образований к высоко устойчивым
и сложным метаплазматическим (как это было ясно показано в
‘отношении аксоподий солнечников и жгутов у Mastigella), то
трудно не придать упомянутому ряду вполне определенного эволю-
ционного значения.
Он показывает именно, что амёбоидная и жгутиковая фазы
одноклеточной организации не могут считаться одинаково первич-
ными; что вторая возникает и оформляется путем повышения спе-
циалиизации амёбоидной фазы, являющейся простейшей формой
одноклеточной организации вообще, и что существование в одной
только амёбоидной фазе отвечает более примитивному положению
вещей, чем наличие двух взаимообратимых фаз, а тем более, ко-
нечно, чем редукция или полное выпадение амёбоидной.
Это, разумеется, не значит, что амёбоидная организация не
может быть достигнута и вторично —- путем упрощения и деполяри-
зации жгутиковой. Примеров такого регрессивного развития можно
найти сколько угодно, особенно среди паразитических форм, но оно
вряд ли может иметь то универсальное значение, какое ему припи-
сывается теперь с легкой руки Пашера. Действительно, разрабо-
танная |ИМ общая теория происхождения амёбоидного типа орга-
низации от жгутикового (обычно трактуемая в одном только систе-
‘76
Матическом ее аспекте-как теория происхождения Sarcodina от
Flagellata) _убедительна лишь постольку, поскольку верно представ-
ление о первичности автотрофного питания, положенное в ее
основу. А так как оно оказывается теперь несостоятельным и вь1-
нуждено уступить свое вместо противоположному — об исторической
первичности гетеротрофного питания (Опарин), то рушится и вся
основывающаяся на нем концепция Пашера.
Рис. 15. Обратимость организации у Pseudospora:
P7“::I:é68O1iId0;(l)fgl B фазе солнечника; 2, 3—переход в состояние амёбы; 4, 5, б—переход
д 0 состояния в жгу/тиковое, 7—начало перехода из фазы солнечника
в жгутиков)! и, 8—Ps. VDIVDCISI фаза солнечника; 9—фаза амёбы; 7о—ее деление;
. 1, 72—переход из амебоидной_фазы_в жгутиковую (по Роскину)
Все сказанное о взаимоотношениях амёбоидного и жгутикового
типа одноклеточнои организации лможет быть резюмировано в сле-
дующих положениях: 1) у самых примитивных Protozoa имеется
TOJIBKUO 0lIHa— аМеб0ИдН_ая—форма существования; 2) жгути-
ковыи тип организации возникает и оформляется на основе амёбо-
идного путем выработки им все более специалиизованных и устой-
чивых СТРУКТУР (дифференцировок); З) возниикшее таким образом
дифференцированное состояние организации сосуществует -с nep-
ВИЧНЫМ ‘Недифференцированным на правах взаимообратимых,
альтернативных, фаз жизнедеятельности до той поры, когда даль-
77
нейшее усложнение дифференцпировок придаст им необратимый хат
рактер; 4) стабилизация жгутиковои фазы ведет к редукции, а
затем и исчезновению амёбоидной. Следовательно, и их взаимоотно-
ШЭНИЯ не ИМЕЮТ абсолютного, РЭЗ навсегда фИКСИрОВЭННОГО харак- у
тера.
Здесь уместно еще НаПОМНпИТЬ, что наличие двух взаимообратит
мых фаз наблюдается и у ряда солнечников, у которых одна из них‚ _
имеет характерную для них организацию, а другая является жгути-
ковой. Они тут обе дифференцированы, причем переход из солнеч-
никового состояния в жгутиковое совершается, как мы знаем,
путем ликвидации части имевшихся аксоподий и превращенир
оставшихся в жгуты, т. е. принципиально иным образом, чем у аме-
боидных форм.
Если добавить к этому, что у ряда одноклеточных имеются не
две, а, целых три взаимообратимых фазы, а именно солнечниковая,
жгутиковая и амёбонидная (хорошим примером чего могут служить
виды Pseudospora, паразитирующие в колониях вольвоцид, рис. 15)д
становится очевидным, что все три главных типа ОДНОКЛеТОЧНОИ
организации являются первично и по существу не чем иным, как,
различными биологическими фазами организмов ЭТОГО ТИПЭ, ПРИНИ‘
маемыми ими при разных у-словиях питания пи дВИЖеНИЯ- Иначе I9‘
воря, эти три типа олицетворяют разные приспособительные
состояния одноклеточной организаЦиИ‚ ЛеГКО СМеНЯЮЩИе-ДРУГ друга
у относительно низко организованных простеиших |И становяШдтеся
все более устойчивыми по мере усложнения их строения.
Глава Н
ПРОСТОЕ ДЕЛЕНИЕ (МОНОТОМИЯ)
1. Общее понятие о размножении одноклеточных
. . ных
Основным и первичным опособом размножения o,££HoJ1I<JIeITIo:H0_
Protozoa является п р ост о е дед е Н И е Н ад В 0614513 елении
томия (Шаттон и Львов, 1924), оно заключается в р дклетот
материнской особи на две, ~KaK ПРЭВИЛО, равноценные, ОДНО В
ные же дочерние особи, бЫСТРО ДОРаСТаЮЩИе Затем д° размеро
материнской. .
» V л Hono-
МОНОТОМИЯ равно-значна клеточному деЛеНИЮ‚ ИЛИ (1;/II:IrTeCI>VI3Yq¥1O она
клеточных организмов, качественно отличаясь от нег ‚
является делением надвое целого самостоятельного организма, Т- е-
я
"именно процессом его размножения. ДРУГИМИ CJ10BaM1’Iv MOHOTOMH
есть размножение одноклеточных посре/СЬСТВОМ 0IlH0KPaTH'0'T0 Митоза’
. ой мате-
результатом которого является образование из ОДНОКЛеТОЧН
инской особи двух таких же, Как Она: ддчерних-
р С биологической точки зрения, по своим биологическим резуль-
° . . ‚ вет-
‘татам, монотомия одноклеточных простеиших полностью соотмоцв
ствует делению надвое надклеточных и многоклеточных организ ‚
'78
например, инфузорий, ресничных червей или гидр, —— явлению, обо-
значаемому, как бесполое, или вегетативное, размножение (пара-
томия, архетомия). Поскольку во всех этих случаях материнская’
особь разделяется на ‚шве дочерние особи того же конструктивного
уровня, как она сама, и размножение происходит, "следовательно,
без изменения этого уровня, мо-нотомию можно с тем же правом
рассматривать, как вегетативное размножение одноклеточных, с ка-
ким деление надвое существ более высокого уровня организации
может считаться монотомией.
Однако, если придерживаться такого расширительного понима-
ния монотомии, как деление надвое организма любого уровня, мы
оставим в тени самые характерные ее сво-йства, а именно ее при-
роду размножения путем клеточного деления, митоза. Понятие мо-
- НОТОМИЯ ЛИШИТСЯ СВОЕЙ КОНКРЭТНОСТИ, а TQM СЭМЬЬМ И СВОЕЙ ОПИСИ‘
тельной ценности.
Поэтому мы считаем нецелесообразным ставить знак равенства
между монотомией и любым другим видом деления животного на-
двое; для нас монотомия есть частный случай такой делимости,
особая ее форма, строго связанная с одноэклеггочным уровнем орга-
низации и не имеющая места при других ее уровнях. Поэтому же
мы не можем согласиться с отнесением к монотомии «простого
деления» надклеточных Protozoa и рассматриваем это последнее,
какдругой частный случай вегетативного размножения (гл. 1Х).
У очень многих одноклеточных простое однократное деление
перерастает в повторное деление, или палинтомию.
‘Палинтомия представляет собой не что иное, как цепь быстро сле-
дующих одно за другим простых делений, т. е. быстрое повторение
‘подряд. двух или более монотомий, не преры-ваемых в интеркинезах
переходом дочерних особей в активное состояние и их дорастани-ем
до размеров материнской особи. Промежуточными продуктами
палинтомии являются с In о р о ц и т ы, а заключительными ——
зооспоры; чи-сло последних не бывает меньше четырех (‚по-
скольку деление повторно) и может достигать многих сотен и даже
тысяч, в зависимости от числа простых делений, объединенных в Ha:
ЛИНТОМИЧЭСКИЙ цикл. Таким образом, ‹палинтомия есть один из видов
множественного размножения посредством зооспор.
ВТОрЫМ ВИДОМ TaKOI‘0' MHO}Ke-CTBCHHOTO РЭЗМНОЖЭНИЯ ОДНОКЛЭТОЧ‘
ных Protozoa является в т о р и ч н а я с и н т о M и я, нередко обо-
‘значаемая, как делени-е распада; Этот тип деления представляет
собой вторичное видоизменение палинтомии, от которой он отли-
чает-ся тем, что рас-падение материнской особи на зооспо-ры (или
сп-оры) со-вершает-ся, вообще говоря, в одинжприем, минуя все ее
промежуточные деления на опороцито-в. `
Этими. тремя типами деления — монотомией, палинтомией и вто-
ричной синтомией- исчерпываются -существующи-е способы размно-
жения одноклеточных простейших. ’
' монотомия одноклеточных простейших чаще всего связана с бес-
полым размножением H‘ лишь как исключение-с половым. Моно-
79
ТОМИЯ СКЛЗДЫВаЕТСЯ ИЗ КОМ-ПЛЕКСЭ ВЗЭИМООВЯЗЭННЬЕХ ПРОЦЕССОВ ДЕЛЕ‘
НИЯ ядра И ЦИТОПЛЭЗМЫ, ВКЛЮЧЭЮЩИХ ДЕЛЕНИЕ И распределение
ДруГИХ КЛЕТОЧНЫХ органоидов И ВКЛЮЧЕНИИ, ПрИкВОДЯЩИХ K ДЕЛЕНИЮ
КЛЕТКИ.
2. кариокинез
Разнообразие форм кариокинеза, наблюдаемых у простейших;
как ИЗВЕСТНО, ОЧЕНЬ ВЕЛИКО. У ОДНОКЛЕТОЧНЫХ представлены ПОЧТИ
ВСЕ ЕГО ТИПЫ, ИМЕЮЩИЕСЯ у Protozoa B ЦЕЛОМ, а ИМЕННО ШЕСТЬ ИЗ ТЕХ"
семи, какие различаются Догелем (1937); многие из них о-суще-
ствляютося, as свою очередь, в нескольких вариантах. Постоянство
отдельных типов кариокинеза очень неодинаково в разных группах:
в то время как у одних из них доминирует лишь один, характерный
для нее, тип кариокинеза (наапример, у Dinoflagellata), B других
ИХ бывает НЕСКОЛЬКО, так ЧТО НЕрЕЩКИ СЛУЧЭИ, КОГДЭ СИСТЕМЭТИЧЕСКИ I
близкие друг к другу формы резко различаются между собой в этом
отношении. Более того, известны случаи, когда у одного и того же
вида кариокинез протекает в двух различных формах (например,
у Vahlkampfia gruberi).
T И П И Ч Н Ы Й М И Т О З, ЯВЛЯЮЩИЙСЯ чПОЧТИ ЕДИНСГВЕННЫМ em ВИ- `
дом у Metazoa, описан также у ряда одноклеточных: у голых амёб
(Hartmanella вытянет) и рако-винных (Euglypha) И целого ряда
солнечников из групгпы Centrohelidia (Dimorpha, Wagnerella И др.).
Он характеризуется внеядерным положением центросом, наличием
вокруг них более или менее оформленной лучистой Центросферы,
малозаметной центро-десмой, а также и тем, что ядрышко и ядерная
оболочка исч-езают при митозе. Вппрочем, последняя из этих особен-
ностей не всегда имеет место, и оболочка ядра нередко сохраняется
В ТЕЧЕНИЕ ВСЕГО rrpoue-oca ДЕЛЕНИЯ (ЧЕГО, как ИЗВЕСТНО, ООЫЧНО НЕ бы-
вает у многоклеточных).
I‘0pa31Lo бОЛЬШИМ распространением, ЧЕМ ТИПИЧНЫЙ МИТОЗ, ПОЛЬ-
зуется у одноклеточных тот тип кариокинеза, который обозначается
Догелем, как деление с внеядерным веретеном. Он
отличается от типичного митоза, в сущности, только сильным разви-
тием и устойчивостью центр-одесмы; она сохраняется в течение
всего деления (до окончательного разъединения дочерних ядер
на стадии телофазы) в виде мощного тяжа, соединяющего внеядер-
ныецентросомы и вплотную пприлегаю-щего снаружи к поверхности
ядра, а затем ахроматинового веретена. Кроме того, митозы этого
типа обычно лишены лучистого сияния. Что же касается ядерной
оболочки, то она либо сохраняется в течение всего деления (напри-
мер, у Ро1утогпе11а citri), либо исчезает, что происходит ‘иногда
только в поздней анафазе (Chlamyd-omonas nasuta; Parapolytoma),
НО в других случаях в метафазе (у Наетатососсцз) или даже еще
раньше (Eudorina, Polytoma), T. -e. B типичный для Метаиоа момент.
Известное разнообразие в кариокинез этого типа вносится тем, что
роль центра деления играют в одних случаях (у тех же фитомонад)
истинные Центросомы,‘морфологичеоки и функционально о-бособлен-
80
д ные от базальных зерен жгутов, а в других——ёдиные органоиды,
совмещающие обе эти функции. В последнем случае они могут быть-
значительно удаленными от ядра в период интеркинеза и прибли-
жаться к нему только перед делением; Это перемещение иногда
запаздывает, так что кариокинетическая фигура устанавливается
до того, как кинетосомы займут свои Места на полюсах веретена
(у Ochromonas granularis). А отсюда уже один шаг до следую-
щего типа кариокинеза, в котором центросомы не принимают
участия.
Этот третий тип——митоз при отсутствии центро-
с о M е является несомненно сборным. К н-ему относятся все те слу-
чаи, когда базальные зерна жгутов не принимают участия в делении
ядра; те случаи, когда имеющаяся центросома факультативно
- (у солнечника АсапШосузЫз) или регулярно лишается своих функ-
ций центра деления; и, Наконец, те многочисленные (о-собенно
у амёб) случаи, когда вообще нет оформленной центросомы.
Четвертый тип—-митоз с внутр иядерддыми центро-
сом ами, описанный у многих амёб, Vahlkampfiidae И жгутико-
вых, отличается от его второго типа только тем, что центросома
помещается под оболочкой ядра, а не на его поверхности.
Этим четырем типам кариокинеза, безусловно близким между
собой и представляющим, в конечном счете, различные варианты
типичного митоза, резко ‘противопоставляется пятый-м ито з
с с-охраненлиепм кариосомы. Он описан у эвглен, ряд?
Protomonadina, Oxyrrhis ИЗ Dinoflagellata, некоторых нкокцидии
и т. д., и принципиально отличается от остальных тем, что крупная
центральная кариосома ядра сохраняется в течение всего деления,
вытягиваясь сначала по оси ядра и принимая бисквитообраз-ную
форму и затем перешнуровываясь пополам. При этом хромо-сомь1
экваториальной, пластинки располагаются вокруг оси, образован-
ной делящейся кариосомой. К этому типу кариокинеза, очевидно,
примыкают те его формы, которые Беляр называет пкрпиптом и-
то з ом. Они создают видимость прям-ого, амитотического деления
ядра, осуществляются без образования хромосом и описаны у Це-
лого ряда амёб и некоторых Zooflagellata. U
СТОЛЬ же резко обособлен от остальных и последнии из типов
кариокинеза, различаемый Догелем, ~— т и п д е л е н и я я д р а
D i n о fl a g е1 1 а t a. Его специфические особенности определя-
ются своеобразным строением ядра (относящегося тут к особому,
хромосомному его типу), перманентно находящегося в состоянии
профазы и не имеющего стадии покоя. Во время митоза хромати-
новые зерна ядра, еще до этого расположенные в ряды, склады-
ваются в 200—300 длинных нитевидных хромосом; ХРОМОСОМЫ
сначала расщепляются вдоль, после чего обе половины каждои из
них выстраиваются в одну линию‘ и затем как бы HepeIlIHYP0Bb1'
ваются в области соединения обеих своих ПрОДОЛЬНЫХ ПОЛОВИН-
Ахроматиновая фигура при этом типе деления обычно отсут--
ствует. „ _‚
81
в Эмбриология беспозвоночных
3. Основные .типы монотомии
Размножение посредством монотомии может происходить как
в активном —кподвижном и вегетативно деятельном состоянии, так
И В ‘ПЭССИВНОМ, Н-ЭПОДВИЖНОМ: В СОСТОЯНИИ ЦИСТЫ, ПЭЛЬМЕЛЛЫ ИЛИ ЖЕ ‚
особом дедифференцированном состоянии, принимаемом трофо-
зоитом накануне деления. _ ‘
В зависимости от того, осуществляется ли деление в активной
фазе ЖИЗНЕДЕЯТЕЛЬНОСТИ ОДНОКЛЕТОЧНОГО ИЛИ ЖЕ ПЭОСИВНОЙ, ОНО СВО‘.
димо к двум основным типам, специфика которых целиком опреде-
ляется тем, является ли эта фаза дифференцированной или же нет.
Деление в дифференцированной фазе организа-
ции олицетворяет монотомию в той ее самой типичной форме, в
какой она всегда описывается и изображается в учебной литературе
в качестве примера простого деления у Protozoa. Оно в высшей сте-
ПЕНИ характерно ДЛЯ ПОСЛЕДНИХ И ВО МНОГОМ отличается ОТ ООЫЧ- I
HOI‘0 клеточного деления Metazoa, почему мы и будем называть эту"
форму монотомии монотомией п р о т о з о йт н о- г о т и п а.
Вторым основным типом монотомии является деление в дедиффе-
ренцированном, НЕПОДВИЖНОМ СОСТОЯНИИ; ОНО ПОЛНОСТЬЮ ОТВЕЧаЕТ.
общим нормам клеточного деления Metazoa, почему мы и называем
егофметазойным типом монотомии. -
К эти-м двум основным тиъпам монотомии сводится, в конечном
счете, все безграничное разнообразие частных ее вариантов, наблю-
даемое у одноклеточных. Поэтому кажется даже странным, что они
доны-не не привлекали к себе должного внимания протистологов,
что их обычно даже не различали и что они пока еще не ‘получили
хоть сколько-нибудь развернутой характеристики. .
1YpoTo3oLZHbLLZ Tun простого деления наблюдается у большин-
ства ОДНОКЛЕТОЧНЫХ, ТрОфОЗОИТЫ КОТОрЫХ обладают ЛЭОИЛЬНЫМ,
уровнем дифференцированности (за исключением, конечно, Tex,
которые размножаются B дедифференцированном состоянии); он‘
представляет собой, единственный вид размножения у форм, обла-_
дающих стабильным уровнем дифференцировки, но зато никогда
‘не имеет места у жгутиковых форм, временно образующихся из_
амёбоидных или из фазы с.олнечника (он не встречается, на-
пример, у Vahlkamphiidae, делящихся только в фазе амёбы, и у
диморфных Heliozoa, ЖГУТИКОВаЯ фаза которых вообще не спо-
собна к делению), Характерно, что у Zooflagellata этот тип деления
я-вляется основным видом монотомическото- размножения, тогда как
у Phytoflag-ellata очень распространены и остальные. Самые харак-
ТЕрНЫЕ ЧЕРТЫ ЭТОГО типа МОНОТОМИИ ТЭКОВЫЗ В, ДЕЛЕНИИ ТУТ уЧа-__
СТВуЮТ ВСЕ ПЛЭЗМЭТИЧЕСКИЕ ДИфтфЕрЕНЦИрОВКИ, КОТОРЫЕ, ОДНЗКО,
сами не делятся, а только распределяются между обеими.
дочерними особями; 2) их распределение между дочерними особями
обычно в той или, иной мере неравномерно; 3) каждая из последних
дополняет, доставшуюся ей часть материнских дифференцировок.
СВОИМИ СОбСТВЕННЫМИ, вырабатываемыми ЕЮ ЗЭНОВО В ПРОЦЕССЕ ДЕ-
82_
сления; 4) благодаря этому комплект плазматических образований
каждойособи оказывается двойственным по» своему происхожде-
нию и возра-стности, представляя собой сочетание материнских
дифференцировок с дополняющими. их собственными; 5) некоторые
Ё.
рис. 16, Продольное деление протозойного типа у жгутиковых:
7—4—Негре1:оп1опа$ faculum; 5, 6-Crithidia gerridis; 7—,71~—Trypanosoma brucei;
72—Leishmania donovani; 73—Copromonas pusilla (no Кюну)
из материнских дифференцировок обычно резорбируются или от-
брасываются в ходе деления‘ и вырабатываются заново обеими
` дочерними’ организациями; наконец, деление в дифференцир-ован-
ном-состоянии резко отличается от остальных его типов и своими
производными: результатом задержки плазмотомии является тут
образование сложных полимерных -сомат-елл‚ как временных,
так и постоянных (см. гл. 1Х).
5* ~ 83
Деление в дифференцированной фа-зе организации характери-
зуется также гораздо большим диапазоном изменчивости, чем ДРУ‘
гие типы монотомии, что является очевидным следствием исключи-
тельного структурного разнообразия этой фазы у одноклеточных.
Самые простые его формы наблюдаются у парножгутиковых
и несложно организованных форм, вроде Ръо-1утоте11а‚ ИЛИ ДВУЖГУ- I '
тиковых протомонадин. У этих форм (см. рис. 24, 25) каждой ИЗ
Рис. 17} Вторично поперечное деление протозойного типа
у Ceratium hirundinella:
7-‘MOMCHT разъединения дочерних особей; 2——апикальная и З-антапикальная _
дочерние особи; 4—вид панцыря сверху; 5—то же снизу (по Кмну)
дочерних особей достается по половине лматерпинской кинетидь1, т. е.
по одному из ее базальных зерен с относящимся к нему жгутом
„(или парой жгутов), после чего полученное MM базальное зерно
делится на два, одно из которых остается при материнском жгуте,
а другое вырабатывает новый жгут. В этих случаях распределение.
всех Материнских органоидов между дочерними особями ЯВЛЯЭТСЯ
полным и равномерным, а «дополняющие» морфогенезы, происхо-
дящие B каждой иИЗ них, невелики и вполне одина-ковы. Столь же
просто и деление одножгутиковых прото- и хризомонад (рис. 16).
отличающееся, однако, тем, что оно уже явно не равномерно: ма-
теринская кинетида остается Здесь ‘ПРИ ОДНОИ ИЗ ДОЧЭРНИХ КЛЭТОК»
84
а другой достается только один из продуктов деления базального
зерна, низ которого вырастает новый жгутик. Тут, следовательно,
одна из дочерних особей не вырабатывает никаких новых дифферень т
цировок, а другая, напротив, образует их заново, что является»
первым шагом к неравномерным формам деления. с
Такая -его неравномерность в смысле распределения материнских
дифференцировок, при их исчерпывающей передаче потомству в со-
вершенно неизмененном виде, находит свое крайнее выражение
в делении высоко дифференцированных панцырных Dinoflagellata.
Чрезвычайно показательным примером такого варианта монотомии
может служить деление общеизвестного Ceratium hirundinella
(рис. 17), при котором материнские дифференцировки распреде-
ляются между дочерними особями следующим образом:
Левая (апикальная) ос-обь по- Правой (антапикальной) ‘Ьсоби
лучает: достаются: ‚
а) все четыре апикальные пла- а) антапикальная пла-стинка‚
стинки панцыря, образую- образующая задний рог пан-
Щие его передний рог; цыря;
б) две из трех прецингулярных б) одна из прецингулярных
пластинок; пластинок;
в) две из трех постцингуляр- в) постцингулярная пластинка
ных его» пластинок; с боковым рогом;
г) одну из двух пластинок г) замыкающая пластинка;
кольцевой борозды (по- д) вторая поясковая пластинка;
Яска): е) пластинка продольной жгу-
Д) поперечный жгутик Ь с его
базальным зерном и ризо-
пластом.
тиковой борозды;
ж) рулевой жгут с его базаль-
ным зерно-м и ризопластом.
Стало быть, апикальной особи приходится здесь вырабатывать
заново все дифференцировки, перечисленные в правом столбце,
а антапикальной-перечисленные в левомдлюбопытно, что все
новообразующиеся пластинки панцыря д-олгое время легко отли—
чимы от «старых» по своей иной окрашиваемости иодом,`блародаря
чему двойственный характер пластинок приобретает, тут_ особую
осязаемость. . . i
He менее демонстративные примеры того же рода дают и дру-
гие панцырные Dinoflagellata, размножающиеся в активном состоя-
нгии, хотя бы продольно делящиеся Dinophysoidea. Так, у Pseudoi
phalavcroma nasuta каждой дочерней особи достается по одной из-
спворок панцыря, включающей половину всех его парных, двусто-
ронне симметричных частей, а непарные части панцыря распреде
ляются между ними так: правая особь получает значительно больд
шую часть склеритов, образующих борозду продольного жгута,
в том числе большой кусок окаймления поры жгута и оба крылод
видных киля гипотеки, тогда как левой передается еще только
пластинка, несущая пору поперечного жгута си ничтожная часть
скелета борозды рулевого жгута. ' ‘ В ` ' ` “ ' д
8.5.
Другие варианты деления в дифференцированном состоянии
отличаются от описанных тем, что при них происходит частичная
или полная ликвидация отдельных материнских дифференцировок,
не подвергающихся, таким образом, распределению между дочер-
ними особями и вое-создаваемых заново каждой из них.‘ Так,
у эвглен, делящихся таким способом (например, Е. gracilis), от-
бра-сывается непарная гдистальная часть жгута с ее утолщенным
Рис. 18. Деление Trichomonas caviae (по Грассе)›
основанием, а его, остающаяся парная базальная ча-сть -с обои-ми
базальными зернами поровну разделяется между дочерними осо-
бями. Так же равномерно распределяются и ‘остальные материн-
с-кие дифференцировки, например, резервуар и пиреноиды (подвер-
гающиеся, правда, определенной реконструкции), так что обе
дочерние особи испытывают здесь, -ка—к y~Polytome11a,- совершенно
тождественныеморфогенезы; они заключаются в разделении ба-
зального зерна на два и в- выработке недостающих частей жгути-
кового аппарата. У более сложно дифференцированной-эвглены
Регапедпа-деление уже менее-равномерно»- так как материнская
кинетида целиком переходит к одной, из дочерних 0c06eI?I,'a;lIpy1‘a7£
$6.-
образует ее заново обычным способогм—-— и сопровождается полной
рЭКОНСТрУКЦИЕЙ СЛОЖНОГО ГЛОТОЧНОГО скелета. ’
Еще сложнее протекает деление у таких сильно дифференциро-
ванных форм, как трихомонады. Так, у особенно детально изучен-
ной Trichomonas caviae (рис. 18) верхнее ‹И3 ‘базальных зерен
(несущее у трофозоита все четыре жгута) делится в эпрофазе на
два дочерних, из которых одно, достающееся правой особи, полу‘-
чает два из трех свободных жгутов и волнообразную перепонку,
а другое, переходящее к левой 5
особи, —только третий свобод- '
ный жгут; второе, ниж-нее ба-
зальное зерно, связанное у
тро-фозоита с боковым опор-
ным ребром, аксостилем и па-
рабазальным аппаратом, те-
ряет перед делением связь с
обоими этими последними И
также делится само, причем
материнск-ое ребро остается
при том из продуктов его де-
ления, который сохраняется
правой дочерней особью, а, ле-
вая образует новое; наконец
правой особи достается и весь
материнский пара-базальный
аппарат (подвергающийся, од-
нако, некоторым изменениям),
тогда как у левой он целиком
создается заново. Что же ка- _ _
(дается 3KC0'CT1’1-H71 И HHTH» СВ? Рис. 19. Деление протозойного типа
зывающей парабазальный ап- солнечника Actinophrys s01.(no Беляру)
парат с ниж-ним базальным `
зерно-м, то они целиком резо-рб-ируются при делении и создаются
затем вновь каждой из дочерних особей. Таким образом, распре-
деление материнских дифференцировок оказывается тут крайне
неравномерным: они почти все остаются у ‘правой дочерней особи,
в то время как левая получает только один из старых жгутов и вы-
рабатывает сама все остальные, * ‚д _ » -
Среди солнечников деление в дифференцированнойфазе орга-
низации, и именно в фазе солнечника, также является господствую-
щей формой монотомии. Оно осуществляется здесь в двух харак-
терных длязтой группы вариантах, которые можно обозначить
соответственно, как тип деления Actinophrys и тип деления -_C1athru-
lina. Первый из них характеризуется полной равномерностью
и приводит к простому распределению материнских дифференци-
рово-к (представленных тут одними только аксоподиями) между
совершенно тождественными ‚дочерними особями, дополняющими
их п-р-имерно таким _же числомновых (рис. 19)„_Полную дпротивог
. 8.7
положность ему Составляет тип деления Clathrulina (наблюдаемый.
также у Асйпоптопаз и других сидячих форм), который приводит
K образованию ДВУХ совершенно не СХОДНЫХ ДруГ ‘С ДРУГОМ дочер- '
них организмов, один из которых сохраняет организацию солнеч-
ника и остается на материнском стебельке, тогда как другой- диф-
ференцируется в жгутиковую форму и затем уплывает. Такое
неравноценное деление идет в начале абсолютно таким же образом,
как у Actinophrys (рис. 20), и также имеет своим непосредствен-
ным результатом образование двух одинаковых особей; вслед затем
1:
Рис. 20. Деление солнечника Clathrulina elegans:
1-1-xaqano разжижения и втягивания аксоподии одной из ДОЧЗРНИХ особей? 2--ВЫХ0д ЭТОЙ
особи, перешедшей в жгутиковое состояние, из материнской оболочки; оставшаяся на месте
особь образует новые аксоподии (по Беляру)
у одной из них тут же резорбируются аксоподии, образуется 1—
или 2—жгутиковая кинетида (повзидикмому, путем преобразования
соответствующих аксоподий в жгуты), и простейшее приобретает
удлиненную форму тела; вторая особь развивает-ся в типичного
солнечника. Таким образом, при формировании жгутиковой особи
происходит коренная реконструкция доставше-гося ей материнского
наследия, большая часть которого ликвидируется, а меньшая идет
на построение кинетиды. -
Итак, при делении рассматриваемого сейчас типа судьба мате-
ринских дифф-еренцировок может быть очень различной. В одних
случаях они передаются дочерним особям в неизмененном -состоя-
нии, в порядк-е прямой (‘или непосредственной) преемственности;
в других они вырабатываются ‚одной из них заново. но из доставше-
гося ей материнского материала‚ что ведет к установлению Hemm-
МОЙ или косвенной ттреем-ственности этих образований из одного
. .
88
ш
‚ поколения в другое; наконец, часть дифференцировок может уничто-
жаться при каждо-м делении.
Метазойньай тип простого деления характеризуется прежде всего
тем, что деление происходит в неподвижном состоянии и связано
с инцистированием трофоз-оита или его превращением в пальмеллу
или. же неподвижную "стадию размножения. Это превращение
выражается в округлении клеточного тела, освобождении цито-
плазмы от части своих пищевых включений и всех дифференци-
ровок,“измене-ний вязкости, окрашиваемости, а передко и видимого
строения цитоплазмы, в появлении в ней особых видов включений
Рис. И. Деление амёбы Amoeba proteus:
7—профаза; 2—метафаза; 3—ранняя анафаза; 4—ранняя тело-
фаза; 5 и б—закл.очительные моменты деления (по Лише)
и т. д. — короче говоря, в тех явлениях дедифференцировки, о кото-
рых уже говорилось в свое время. Объем этих перестроек столь же
изменчив, как и при обычном инцистировании, и зависит главным
образом от того, насколько развиты и устойчивы дифференцировки
трофозоита, подвергающиеся теперь ликвидации. Монотомия метан
зойного типа наблюдается у голых амёб, ряда солнечников и мно-
гих жгутико-вых, обладающих лабильно дифференцированной
активной фазой. »
Первый из различаемых нами ее подтипов —- р а з м н о ж е н и е
в состоянии шара деления-очень характерен для
свободноживущитх голых амёб, т. е. представителей сем. Amoebidae.
‘Приступающая к делению амёба останавливается, втягивает свои
ложно-ножки и принимает форму правильного шара, вся поверх:
ность которого равномерно одевается многочисленными мелкими
отростками (рис. 21); ее цитоплазма в значительной мере теряет
свою прозрачность и раздражимость и имевшаяся сократительная
вакуоль исчезает. K концу процесса деления (в его телофазе) его
89
продуктыпостепенно переходят в подвижное состояние: псевдо-
подии увеличиваются в размерах и сокращаются в числе, по-
ЯВЛЯЭТСЯ пульсирующая ВЭКУОЛЬ, ТЕЛО рапспластьтвается ПО суб-'
страту и теряет правильность очертаний. Таким образом, ‘явления
дедифференцировки имеют тут кратковременный, быстро преходя- .
щий характер, целиком приурочены к моменту деления и касаются _ =
одних только эуплазматических‚ едва оструктуренных образований.
Второй подтип метазойного деления——деление под защитой
материнского скелета—можно назвать и хламидомонадным, как
его часто им-еновали встарину. Эта, самая распространенная форма
монотомии у Phytomo-nadina И ряда других панцырных Phyt0f1a-
gellata, отличается от только что описанной лишь тем, что ей преды
шествует устранение многочисленных материнских дифференциро- ‘
вок‚ полностью восстанавливаемых затем заново каждой из дочер-
них особей. Вследствие этого делением охватывается здесь не весь
материнский материал, а только само клеточное тело со своими
ПОСТОЯННЫМИ органоидами И ЧЭСТЬЮ ВКЛЮЧЭНИЙ. ОНО так ИЛИ иначе '
обособляется от материнской оболочки и от кинетиды, освобо-
ждается от других своих дифференцировок и делится точно таким
же способом, «каким делятся голые амёбы. Подобное размежевание
неделящегося, остаточного материнского материала и его деля-
Щ-ейся массы нередко приводит к тому, что размножающая-ся особь
полностью сохраняет подвижность в течение всего процесса деле-
ния. Это замечательное явление находит свое объяснение в том,
ЧТО HCKOTOPEIH ча-сть МЗТЭРИНСКОЙ ПРОТОПЛЭЗМЫ, а ИМЕННО КИНО-А
плазматический конус, окружающий риз-оппласт и, может быть,
даже целый тонкий ее периферический слой, подстилающий обо-
лочку (перипласт)‚ остается неиопользованнымпри делении; она
не входит, таким образом, в состав тела дочерних ‚особей и про-
должает обеспечивать бесперебойную работу материнских жгутов
в течение всего периода деления. Поэтому и создается видимость
деленэия в активном состоянии, хотя в действительности оно проте-
кает здесь в утрированно метазойных формах. * ^
Эти особенности раэбираемого типа деления имеют своим
результатом то, что мезо- и метаплазматические образования
каждой особи приобретают полную однородность в смысле своей
возрастности и происхождения, становя-сь ее. «личной» принадлеж-
ностью и теряя свою преемственность о-т одного поколения в дру-
гое. Каждая молодая особь получает от матери только свою цито-
плазму и постоянные о-рганоиды и самостоятельно вырабатывает
все дифференцировки, в том чи-сл-е и собственный наружный скелет.
Конкретное представление о ходе этой формы деления может
дать пример Chlamydomonas nasuta (рис. 22). Как легко видеть
из рисунков, происходящие тут процессы дедифференцировки перед
делением идут совершенно так же, как при обычном инцистирова-
нии (ср. рис. 11), а дифференцировка дочерних особей ничем не
отличается от редифферендировки перед выходом из цисты (рис. -9,
стр, 65). Однако сочетаниеэтих процессов о размножением при-
90
ж {___.___4_ _____ _ _ _ _
'ВОДИТ K ТОМУ, ЧТО ОНИ -приобретают тут НОВОЕ ДЛЯ НИХ ЗНЭЧЭНИЭ.
Дедифференцировка получает характер специального про-цесса под-
готовки к делению, в результате которого вегетативная особь
-(трофозоит) превращается в особую генеративную -стадию. На-сту-
пающее затем деление даетдочерних особей, оказываюшихся сна-
чала в край-не характерном недифференцированном «эмбриональ-
ном» состоянии. Их дифференцировка становится тут процессом
особо-го послемитотического превращения, имеющего своим резуль-
татом преобразование обеих дочерних особей в дифференцирован-
ных и активных трофозоитов. Таким -о-бразом, этот тип нмонотомии
чрезвычайно резко отличается от первых двух, что еще подчерки-
вается его почти безудержной, можно сказать, тенденцией пере-
растать в повторное деление (палинтомию).
Третий подтип мета-
зойного деления — д е-
ление в инцисти-
pols анном состо-
я н и и —— отличается от
предыдущего только
тем, что он связан с об-
разовани-ем временных
оболочек самого разно-
образного характера
(начиная от студени-
стых и- кончая такими
сложными, как кремне-
земные цисты хризомо-
над). Однако он все же
заслуживает выделения, так как ведет во многих группах к иным
дериватам, чем хламидомонадное деление, и иначе комбинируется
с другими его типами. Так, у амёб он сосуществует с характерной
для них монотомией в форме шара деления, а у Polytomella agilis
И некоторых хрнизомонад- с делением протозойного типа; с другой
стороны, он никогда не сочетается с хламидомонадным делением
иначе, чем при половом размножении (гл. VI). '
Рис. 22. Деление метазойного типа и дифферен-
цировка дочерних особей у Chlamydomonas nasuta:
7 — еще не дифференцированные дочерние особи; 2 -- на-
чало их дифференцировки; 3 — дифференцированные мо-
лодые особи в материнской оболочке; 4 — взрослая особь.
(по катеру)
Своеобразным его вариантом является деление в стадии паль-
меллы, столь характерное для многих автотрофных жгутиковых. Оно отли-
чается от деления в цисте только тем, что осуществляется без выключения асси-
миляции и роста, т. е. в вегетативно деятельном, хотя и неподвижном состоянии
организма. Одним из проявлений „такой физиологической активности является и
то очень характерное обстоятельство, что каждая дочерняя особь немедленно
выделяет свою собственную оболочку. Таким образом, деление в стадии паль-
меллы, сочетает в себе признаки метазойното и протозойного деления, занимая
в известно-м смысле, промежуточное положение между ними.
ОТДЕЛЬНОГО УПОМИНЗ-НИЯ заслуживает‘ еще одна своеобразная
фо-рма МОНОТОМИИ —— так называемое деление почкования, ОЧЕНЬ
характерное для когрненожтетк. При этом типе деления ча-сть цито-
плазмы материнской особи выпячивается-из ее домика в виде
быстро увеличивающегося _выроста, . сразу же` начинающего
9'1
вырабатывать собственную метаплазматическую оболочку (рис. 23);
отпочковавшаяся таким -образо-м дочерняя осо-бь вначале лишена,
ядра, так как его разделение на дочерние ядра прои-сходит уже
после начала ее образования, и одно из них только позднее пере-
ХОДИТ В ее ТЕЛО‘. На ДЭЛЭНИИ ПОЧКОВЗНИЯ, HIO ТОЛЬКО не ДОВОДИМОМ ’›
ДО КОНЦЭ, ОСНОВЫВЗЮТСЯ СВОЭООрЗЗНЫЕ фОрМЫ роста ПЛЗЗМЭННОГО,"
тела многокамерных корненожек, приводящие к формир-ованию их‘
сложно построенных раковинок.
Рис. 23. Деление почкования у Euglypha alveolata (no Шевякову)
производными метазойного типа деления являются, с одной
стороны, палинтомия, представляющая собой не что иное, как ряд
быстро следующих друг за другом делений именно этого типа,
с ' другой стороны-переход к многоядерной, плазмодиальной
организации и связанному с ней синтомическому делению. Вообще
говоря, тенденция метазойного деления одноклеточных перерастать
в различные формы множественного размножения составляет одну
из его самых характерных черт. р '
4. Геометрические формы монотомии
Монотомия осуществляется у_одноклеточных в весьма разно-
образных геометрических форма . „-
Самые простые формы деления при-сущи‚ разумеется, анаксон-
92
ным и неполяризованным (амёбоидным) одноклеточнь1м. В этих
случаях нет фиксированной ‘плоскости деления, и ее положение
определяется таким непостоянным здесь моментом, как направле-
ние наибольшей массы или наибольшей протяженности чистой
протоплазмы, по отношению к которому эта плоскость ложится
нормально (перпендикулярно); к етим случаям может быть при-
ложимо второе ‚правило Г ертвига, с-огла-сно которому ось ядерного
веретена располагается в направлении наибольших протоплазмен-
ных масс, являющемся направлением их наименьшего сопротивле-
ния удлинению делящегося ядра. Поэтому при шаровидной форме
Рис‘; 24. Деление Bodo edax (IIO:KIOHy')
тела плоскость деления может располагаться по любому его диа-
метру, а если тело удлиняется в каком-либо направлении, то это
влечет за собой фиксирование этой плоскости. Такое деление,
согласующееся с правилом Гертвига, можно назвать п е рлв и ч но-
поперечным.
K этому, весьма примитивному, типу монотомии относится,
вероятно, и деление некоторых одноосно-гетерополярных, довольно
высоко дифференцированных Zooflagellata (рис. 24, 25), а имен-по
ряда протомонад M Polymastigina (например, Oikomonas, Bodo,
трихомонад и т. д.). Удлинение делящегося ядра идет здесь с са-
мого начала (или с ранней метафазы) по длинной (главной) оси
тела и сопровождается соответствующим смещением одной из до-
черних кинетид (и связанного с.нею парабазального аппарата),
‚ ПОСТЭПЭННО сдвигающейся K заднему КОНЦУ тела, В ТО время как
другая остается на ‘СВОЕМ МЕСТЕ. В результате ЭТОГО переднчим ПО-
люсом оказывается у одной из дочерних особей материнский,’
а у другой-противоположный ему конец тела, т. е. полюс, быв-
' 93
ший до деления вегетативным. Таким образом, в ходе митоза про-
исходит изменение полярности тела, вследствие которого продукты‘
деления противостоят друг ДРУГУ, соприкасаясь зад-ними‘ концами
ира-сходясь в противоположные стороны передними.
По Шаттону И ‘Вилльнёву (1936), деление этихьформ представляет собой"
уклонение от типичного для жгутиковых продольного деления (см. ниже);
объясняющееся отставанием плазменного деления от ядерного и от деления кле-
точных органоидов, или, другими словами, нарушением нормальной согласован-
ности и последовательности отдельных этапов монотомии. Таким образом, ему
` приписывается вторично ви-
Доизмененный характер. Од-
нако, если учесть, что такое
дел-ение характерно для при;
митивных Zooflagellata, тес-
но связанных с амёбоидными
формами (протомонады-че-
рез Vahlkampfiidae, трихо-
монадьь-через Dientamoeba
И Histiomonas), что оно co.-
пряжено с формами кариоки-
неза, особенно типичными
для амёб, и, наконец, то, что
разновременность отдельных
слагаемых деления тут не
более выражена, чем при
других формах деления, ——
кажется вероятным, что
французские авторы не пра-
вы.‚ Существо этого типа
деления, очевидно, заклю-
чается в том, что оно идет в
согласии со вторым прави-
лом Гертвита и как бы не
подчинено, таким образом,
п-роморфологической струк-
туре организма. А если так,
то наблюдающиеся его вари-
анты приобретают исключительный интерес, как переходные ступени от nep-
вично-поперечного к продольному делению. Окончательное решение этого воп-
роса, очевидно, принадлежит протистологам.
Рис. 25. Деление Trypanoplasma congeri (no
Мартину) -
У подавляющего большинства других одноосно-гетерополярных
одноклеточных положение плоскости деления уже не зависит от
общей формы тела и не сводимо к столь простым механическим
причинам, как направление наибольших масс чистой протоплазмы.
Плоскость деления располагается тут по главн-ой, органической
оси тела, становясь ее лучшим показателем. В соответствии с этим
плоскость деления имеет продольное, меридианальное направление
‚у одноосных форм и становится диагональной или даже поперечной
(экваториальной) у двуосных. Мстанавливающаяся тут зависи-
мость прекрасно иллюстрируется такими примерами, какделение‘
сильно вытянутых в длину жгутиковых (см. рис. 16), где оно
осуществляется, явно вопреки правилам Г ертвига, в продольном.
направлении; с другой стороны, она наглядно демонстрируется
различными Dinoflagellata, B ряду которых легко можнохпро-
94
следить постепенное отклонение плоскости деления от своего перво-
начально продольного положения, неотступно следующее за пере-
ориентировкой первичной «оси клеточного тела.‘
\' Таким именно образом возникают оба главных геометрических
ти-па монотомии, наблюдающиеся у полярно дифференцированных
(жгутиковых) одноклеточных: п р «о д о л ь н о е д е л е н и е, являю-
щееся здесь у основным непо типом, и «B т о р и ч н о-‘п о п е р е ч н о е,
характерное для ряда перидиней. Принципиальное различие между
ними “заключается ‘B
том, что при продоль-
ном делении оба его
продукта располагают-
ся симметрично отно-
сительно плоскости де-
ления, т. е. относятся
друг к другу, как ан-
тимеры,.тогда как при
истинном вторично-по-
перечном делении его
продукты уже не сим-
метричны, а только по-
добны друг другу и
оказываются во взаи-
моотношениях не анти-
мер, а метамер: ‚они
располагаются вдоль.
общей оси («оси пере-
носа», или оси метаме-
рии), а чне по ее сторо-
нам, чередуясь, таки-м . -
образом, ‚своими _ра3н0- Рис. 26. Деление Oxyrrhis marina (no Хэллю)
именными полюсами.
Строгое различение этих двух основных типов ‚монотомии ока-
зыва-ется во- многих случаях невозможным -без пользования только
что приведенными критериями. Дело в том, что очень нередки
случаи, когда продольное деление при-обретает видимость поперечд
ного. Эти случаи, которые можно объединить в понятие ложно-
п о п е р е ч н о г о деления, очень часты, например, у фитомонадин.
В этой группе деление обычно начинается, как правильное про-
дольное; затем делящееся яд-ро поворачивается на 9О°‚ так, что
ось веретена оказывается лежащей по длинной ‚оси материнского
тела и Наступающее затем деление разделяет его в определенно
поперечном направлении. Однако на ряде примеров нетрудно уста-
новить, что эта, лпоперечность является здесь только кажущейся:
так, у Haematococcus. (CM. pun-c. 28) отчетливо видно, что деление
в действительности осуществляется по всем правилам продольно-го,
но сопровождается ‘переориентировкой тела клетки относительно
ее. ОбОЛОЧКИЗ ПОСТЗПЭННО РЗЗДЭВЗЯСЬ. В ширину, ТеЛО ЦЕЛИКОМ 'ПОВО-..
95-
рачивается ‘на четверть круга, ОЛЗГОДЗРЯ чему его главная ОСЬ_'
(а соответственно и плоскость деления) расп-0-лагает-ся теперь под
прямым углом по ошоше1-1и1о к ее первоначальному положению.
У других фитомонадин (например, лучше в-сех изученной в этом '
смы-сле Chlaxnydomouas навыка) такая переориентировка тела про-
стейшег-о предшествует началу деления; интересно отметить, что в
этих случаях опа носит не полный, а частичный характер, выражаю-
щийся в том, что некоторые из дифференцировок не изменяют
своего первоначального положения. Так или иначе, можно считать
твердо установленным, что у Phytomonadma деленпие всегда про-
дольное, и это полностью подтверждается симметричным располо-
жением его продуктов.
По Шаттону (1931, 1936), ложно-поперечное деление имеет"
В СВОЭЙ ОСНОВЕ уже УПОМИНЕЪВШЭЭСЯ явление нарушения ОбЫЧНОИ
СОГЛЗСОВЗННОСТИ МеЖДУ РЭЗЛИЧНЫМИ слагаемыми деления. O,/I‘I-EEK9.
ЭТО ТОЛКОВЗНИЭ неприменимо И K кфИТОМОНЗДИНЭМ, ИЛИ, ПО краинеи
мере, K ООЛЬШИНСТВУ ИЗ НИХ, у КОТОРЫХ таких нарушении нет И все.
дело сводится к простому повороту всего клеточного тела в про-
цес-се деления. '
у всех двусторонне-симметричных ЖГУТИКОВЫХ (СОСТЗВЛЯЮЩИХ, как уже
ГОВОРИЛОСЬ, значительное ИХ ООЛЬШИНСТВО), ПЛОСКОСТЬ ПРОДОЛЬНОГО деления
совпадает С ПЛОСКОСТЬЮ симметрии, ЯВЛЯЯСЬ, таким образом, СНГИТТНЛЬНОИ-
ВОПРОС О ее ПОЛОЖЕНИИ у радиально-симметричных фОрМ неясен; ВОЗМОЖНО, ЧТО
она здесь не обладает ТЗКОИ определенностью, НО отчетливых данных Об ЭТОМ.
нет. Форм, делящихся в фронтальной плоскости среди одноклеточных Flagel-
lata неизвестно: такое положение ПЛОСКОСТИ деления характерно ТОЛЬКО ДЛЯ
двойственных, Морфологически двуклеточных Polymastigina, объединявшихся
в группу Diplozoa (см. ниже).
Деление тела простейшего всегда осуществляется путем обра-
зования постепенно углубляющейся борозды деления, характер
которой может быть различным. При поперечном делении эта
-борозда «всегда имеет -вид кольцевой перетяжки, появляю-
щейся сразу по всей окружности тела и затем углубляющейся
равномерно по всему ово-ему протяжению (при округлом сечении
тела) или же более быстрым темпом в одних направлениях, чем
в других (при уплощенной форме тела, когда она приобретает
скорее двусторонний, чем правильный кольцевой характер, быстро
продвигаясь IB направлении большого сечения тела и лишь очень
медленно—по меньшему). Постепенное углубление такой коль-
цевой борозды приводит пк перешнуровыванию тела особи, вслед-
ствие чего обе ее половины остаются дольше всего связанными по
середине (см. рис. 24). В тех случаях, когда ядро (а стало быть,
и веретено деления) расположено эксцентрично, борозда обычно
появляется сперва только на ближайшей к нему стороне тела и
оггсюда постепенно распространяется по всей ее окружности, пока
не достигнет противоположной стороны (рис. 26). 1акую борозду
можно назвать полукольцеив ой. Отсюда уже один „шаг до
целиком односторонне врезающейся борозды (Мечников,
1886), необыкновенно характерной для продольно "делящихся
96
жгутиковых рис. 16). ‚Врезающаяся борозда появляется, как
‘правило, в виде вдавления или желобка на жгутиковом полюсе тела
и затем постепенно углубляется отсюда в одном только направле-
нии ~— спереди назад, рассекая тело вдоль по всей его длине; в этих
случаях дочерние клетки остаются дольше всего «связанными сво-
ими задними концами, образуя к концу деления характернейшую
Ч-образную фигуру.
Эти два основных типа борозд деления, установленные Мечни-
ковымхв его классическом исследовании по эмбриологии медуз
(1886)_ и до сих пор не различавшиеся у ‘простейших, связаны
между собой рядом переходов. Следует также отметить, что у не-
. которых" фитомонадин борозда врезающегося типа появляется не
на жгутиково-м, а на противоположном ему конце тела и углуб-
ляется сзади наперед (Pyramimonas), T. e. B направлении, обратном
типичному; однако, дойдя до середины длины тела, бо-розда меняет
‚свой односторонний характер на кольцевой, что позволяет рассма-
тривать ее скорее, как односторонне кольцевую, чем как настоя-
щую вр-езающуюся. Наконец, заслуживает еще упоминания, что
к „борозды кольцевого типа иногда закладываются не как сплошной
желобок, а в виде многочисленных отдельных складочек enema-
кулы, потом только объединяющихся в общую <‹сборчатую>>
борозду.
- Появлению борозды деления н-еред.ко .предШествует образова-
ние экваториальной (относительно веретена) плазматической пла-
спинки, намечаюжщей будущую плоскость деления и более или менее
заметно отличающейся от остальной массы плазмы по своей струк-
туре, наличию пигментных включений и т. д.
Отдельного упоминания заслуживает еще то обстоятельство, что
взаимная ориентация продуктов. -симметричного деления оказы-
.вается .в разных случаях весьма различной. Типичным может счи-
таться их параллельное расположение при одинаковой ориентации
одноименных полюсов, но еще -чаще они направлены дРУг к другу
под большим или меньшим углом, доходящимдо 180°, что создает
‚видимость поперечного деления. Наконец, нередки и такие случаи,
A когда дочерние особи располагаются параллельно друг другу своими
.р главными осями, но так, что их одноименные полюса направлены
врозь. Чем обусловлены эти смещения и перео-риентировки ——‚ос-
‚таетчся, в сущности, неизвестным. Можно только сказать, что в не-
которых случаях (например, у трихомонад) окончательная ориен-
тация устанавливается еще в ходе деления и в связи с определен-
ными его особенностями, тогда как в других она является резуль-
татом видимых перемещ-ений уже разъединившихся или близких к
разъединению дочерних особей;
-В заключение надо еще отметить, что оба основных типа моно-
томии——протозойный и метазойный—могут осуществляться в лю-
бых геометрических формах.
7 Эмбриология беспозвоночных _ 97
Г л а в а I I1
ЙЁДИВИДУАЛЬНОЕ РАЗВИТИЕ монотомичвских
одноклвточньпх
1. Вводные замечания
д
До сих пор еще можно слышать, что простейшие не имеют
индивидуального развития, что- к ним неприменимо понятие онто-
генеза и что наблюдаемые у них возрастные изменения организации
принципиально несравнимы и не имеют ничего общего с онтогене-
зом многоклеточных. .
Это, конечно, недоразумение или, вернее, формальное рассмотре-
ние вопроса. В самом деле, наличи-е онтогенеза является н-еотъемле-
мым свойством всякого живого существа, любой физиологически и‘
экологически сам-остоятельной особи, сове_ршенно независимо от то-
го, каков ее конструктивный уровень.'Понятие «онтогенез» не имеет
иного смысла, чем ‹<развитие' особи» и не отделимо от понятия
«особь», поскольку невозможно себе пред-ставить живую особь
не развивающейся. Стало быть, это понятие применимо K любой
органической особи, и все виды онтогенеза в той же мере сравнимы
друг с другом, (B какой сравнимы между собой организмы любого
конструктивного уровня. ' ’
Такое широкое понимание онтогенеза, как универсального
свойства живых существ, ни кв какой мере не лишает его вполне
‘конкретного содержания. Действительно, „каков бы ни был ко-н-
структивный уровень развивающегося -организма, его онтогенез
неизменно включает, во-первых, процесс а к т и в н о г о р о с т а,
т. е. увеличение живой массы тела, и, во-вторых, те или иные
морфогенетические процессы. Далее, для любого онто-
генеза характерны строго закономерные изменения возрастного
состояния самого организма, выражающиеся в той или иной форме
этапности развития. _
Вместе с тем совершенно очевидно, что индивидуальное разви-
тие организмов разного конструктивного уровня обнаруживает
глубокие к а ч ес т в е н н ы е различия, что каждому из этих уров-
ней отвечает свой специфический т-ип онтогенеза. Поэтому индиви-
дуальное развитие одноклеточных Prot-0 zoa И имеет принци-
лиально иные формы и существенно ’ иное содержание, чем
у надклеточных простейших, с ‘одной стороны, и многоклеточных
организмов —=с другой. Таким образом, признавая наличие у одно-
клеточных само-го- подлинного онтогенетического развития, дмы
ни на минуту не должны забывать, что их онтогенез качественно
ОТЛИЧЭН ОТ ИНДИВИДУЗЛБНОТО развития ОрГЗНИЗМОВ НЗДКЛСТОЧНОЙ И _
мн-огоклеточной ступеней органаизации и в значительной мере от-
вечает по своему содержанию процеосам онтогении клеток Metazoa
B том смысле, в каком эта последняя понимается Б. П. Токиным
(1935, 1936).
98
2. Общее понятие об онтойенезе монотомических
одноклеточных
Индивидуальное развитие одно-клеточных Protozoa характери`-
зуется прежде всего тем, ‘что оно целиком осуществляется в рам`-
ках одного и того же—одноклеточного-уровня организаци-и и
не связано, таким образом, с его повышением. Организм оформ-
ляется B качестве самостоятельной особи, растет и развивается,
оставаясь одноклеточным, и все испытываемые им возрастные изме-
нения не ведут к перерастанию им его исходного конструктивного
уровня.
Вторая характерная особенность онтогенеза одноклеточных за-
ключается в том, что его началом является процесс деления надвое
материнской особи, а концом-профаза следующего митоза, т. е.
что он включает только один полный ядерный цикл. В соответствии
с этим онтогенетическое развитие одноклеточных может быть раз-
делено на два основных периода: '
1) период возникновения особи в ходе деления ма-
теринского организма, являющийся одновременно процессом рас-
щепления, -снятия материнской индивидуальности и зачатия
сменяющих ее дочерних; этот период развития протекает при мито-
. тическом состоянии ядра И ДРУГИХ клеточных органоидов и обни-
мает весь цикл их деления и распределения;
2) Период индивидуального—в прямом и точн-ом
смысле этого слова — p a 3 B и ти я (т. е. развития в н еде л и-
м о м с о сто я н и и), охватывающий все онтогенетические изме-
нения, испытываемые особью с момента ее полного обособления
до перехода к размножению путем деления; этот второй период
развития целиком протекает, стало быть, при интеркинетиче-ском
состоянии ядра.
Третьей характерной особенностью онтогенеза одноклеточных
Protozoa является та, что морфогенетические процессы осуще-
сгвляются у них, как правило, до начала активного роста, а по
краинеи мере частью—еще в ходе деления, т. е. фактически еще
До того, как произошло взаимное обособление обеих вновь обра-
зующихся особей; таким образом индивидуальный пери-од развития
часто протекает в форме «чистого» роста, т. е. дпри минимальном
изменении исходного строения организма.
Далее, еще одной особенностью, типично-й для одноклеточных,
можно считать характер испытываемого ими роста. Насколько
позволяют судить существующие на этот счет данные, рост одно-
клеточных Protozoa происходит с относительно постоянной удель-
нои скоростью.
Чго- же касается морфогенетических процессов, то они условно
могут быть разделены на две категории. K первой из этих катего-
PW‘ ОТНОСЯТСЯ МОРФОГЭНСЗЫ, которые зможно назвать `«м ит о ти ч е-
спким и», поскольку они осуществляются в порядке деления, рас-
пределения и реорганизации материнских органоидов в процессе
7* 99
д y ал Ь н ы е»,
самого деления. Сюда относится формирование дочерних ядер и‘
других органоидов, получаемыхдочерними особямив уже готовом
виде; при протозойном типе деления ‚в ету же категориювх-одят и
дифференцировки, образование которых происходит, как правило, _.
до разъединения дочерних особей, т. е. до начала их индивидуальд
Эного развития.
категорихо мо-рфогенез-ов составляют
осуществляемые дочерними
по-сле их взаимн0го- обособления, в течение их собственного инди-
видуального развития. Морфогенезы тако-го рода наблюдаются
преимущественно при метаз-ойном типе моното-мии и лишь как
исключение-при протозойном. у
Ведущая роль митоти-ческих морфогенезов в создании органи-
зации простейших составляет характернейшую их черту и особенно
Вторую
ярко выражена у одноклеточных представителей Protozoa, где oun-
морфогенетическими про-
часто оказываются
цессами.
Этими основными признаками исчерпывается, в сущности, все,
чем можно сейчас охарактеризовать, в общей форме, индивидуаль-
"н-ое развитие одноклеточных Protozoa. Общие онтогенетические раз-
личия между монотомическими и палинтомическими одноклеточ-
ЭДИНСТВЭННЫМИ
‘ными сводятся, в конечном счете, к тому, что у первых рост огра-
ничивается примерно удвоением исходной массы дочерних особей,
‚тогда как у вторых о-н приводит к многократному (по крайней
мере, Четырехкратному) увеличению начальной массы зооспоры.
К этому можно еще добавить, что онтогенез монотомиче-ских одно-
клеточных протекает в двух, резко различных формах, отвечающих
обоим главным типам монотомии, в то время -как у палинтомиче-
ских он развертывается на основе только одной из этих форм-
той, которая связана с Метазойным типом деления.
Прежде чем перейти к конкретной характеристике о-боих основ-
ных типов индивидуального развития монотомических, Protozoa,
уместно сказать еще несколько слов об общих особенностях роста
иморфогенетических процессов у одноклеточных. Первым делом
надо отметить, что, помимо активного, или истинного, роста, часто
‚наблюдается и пассивный рост, происходящий за счет накопления
запасных питательных включений, образования в цитоплазме
обширных вакуолей или же поглощения и связывания ею воды
‚(у Ceratium, по Энтцу). В этих случаях ‘можетпрои-сходить значи-
тельное увеличение общего объема организма без увеличения его
живой массы. Рост различных дифференцировок осуществляется,
вообще говоря, в течение всегодиндивидуального развития, так как
ооотношение их размеров -с размерами непрерывно увеличиваю-
щейся клетки остается, в общем, постоянным.‚То же можно -сказать
и -о метаплазматических образованиях. Так, клетчатковый пан-
‘цырь Ceratium непрерывно растет и усл-ожняет свой рельеф в
течение всей индивидуальной жизни, в результате чего общее
"увеличение его поверхности может составить, по „вычислениям
100
<<И Н д И В И- '
орга-низмами `
‘Энтца, до 360% к первоначально‚йдд(у только ‘что оформившейся
особи). Показано, что рост этих образований происходит преимуг
щественно путем интуссусепции, т. е. повсеместно и равномерно.
Помимо того, у Dinophysidae рост панцыря происходит также и
путем аппо-зиции (на-слоения новых порций вещества на ‘ранее
имевшиеся), что ведет к образованию у них временной зоны при-
роста между постепенно раздвигающимися створками панцыря.
Чисто аппозиционным способом происходит и увеличение массы
раковин или скелета, демонстрируем-ос нагляднее всего развитием
мн-огокамерных корненожек, и радиолярий группы Arcantharia,
Таким образ-ом, процессы роста обычно включают Не только актив-
ную, но и пассивные его формы.
3. ‘ОСНОВНЫЕ ТИПЫ онтогенеза МОНОТОМИЧЕСКИХ ОДНОКЛСТОЧНЫХ
- ‘ Самые простые формы индивидуально-го развития связаны с де-
лением в дифференцированном состоянии, т. е. с п р о т о з о й н ы М
типом монотомии.
Поскольку морфогенетические процессы совершаются тут В
основном (или даже полностью) в ходе деления, еще до разъеди-
нения дочерних особей, то они фактически оказываются закончен-
-нь1ми к началу индивидуального периода жизни каждой особи,
В «момент своего обособления о-бе дочерние -особи уже вполне офор-
мированы, так ЧТО ИХ ДЗЛЬНЕЙШЕЭ развгитие выражается главным
образом ‚в чистом росте, ведущем к удвоению и-сход-ной массы‘
особи. V
Исключительное своеобразие этих форм онтогенеза выражается
ИМОННО -В TOM, ЧТО не ТОЛЬКО ПОСТОЯННЫЕ ‘ОРГЗН-ОИДЫ, НО И ВСЗВОЗЧ,
можные дифференцировки оформляются тут еще в процессе
размножения, в порядке удгвноения материнских структур, и что это
УДВОЕНИЕ завершается, как правило, ДО ра-ЗЪЭДИНЭНИЯ ‘ПРОДУКТОВ’
деления. Таким образом, начальный морфогенетич-еский период
предшествует тут индивидуальному развитию: все морфогенезы
имеют характер митотичепских, если можно так выразиться, пред-
варительных морфогенезхов, в чем и заключается их протозойная
специфика. '
Правда, в ц-елом ряде случаев, пополнение материнских диф-
ференцировок собственными происходит ‚после обособления Upon
;Z[yK’I‘OB деления, в течение их индивидуального развития (например,
у Ceratium и других панцырных Dinoflagellata), HO это касается
лишь некоторой их части (в основном метаплазматических струк=
тур) и не меняет поэтому существа дела.
„Разделение ОНТОПГЭНЭЗОВ ЭТОГО ТИПЗ на ЭТЭПЫ ЗЗТРУДНИТЕЛЬНО,
поскольку они целиком протекают в дифференцированном состоя-
-нии организации. За достижением предела роста немедленно сле-
дует деление, так что размеры трофозоита не испытывают хоть
сколько-нибудь длительно-й стабилизации; поэтому «взрослое» -c-0-‘
стояние не м-ожетгбыть сатграничено; как ‘правилсдот периода‘ роста
101'
Принципиально иные формы индивидуального развития сопря-_'
жены с ‚Метазойн ым типом простого деления. Они характе-
РИЗУЮТСЯ прежде Bcero тем, ЧТО СКЛЭДЫВЭЮТСЯ ИЗ нескольких" очень
четко разграниченных периодов, а именно:
1) «Э M 6 р и-о н а л ь н о го» периода, проводимого молодыми
особями в недифферснцированном, вегетативно недеятельном со-
СТОЯНИИ, ПрИ ВЫКЛЮЧеППОМ ИЛИ, ПО крайней мере, замедленном‘
(у автотрофных форм) росте.
2) Периода дичпрсренцировки, ‘при котором у каж-
дой дочерней особи образуются все характерные для нее структуры,
в том числе все дичшчэерегпцировки, и она переходит из пассивного
в активное состояние. - I
3) «HOICTSM6131/IOIIZIJIBI-I‘OI"O>> вегетативного пе-
р и о д а, начинающегося с выхода молодых особей из материнской
оболочки и заканчивающегося их дедифференцировкой перед деле--
ние’м; в течение этого активного периода жизнедеятельности цели-
ком осуществляются питание и рост, и он весь проходит в диффе-
ренцированном состоянии. _
4) П е р и о д а з р е л -о-с ти, |при котором питание и рост прак-
тически прекращаются, особь дедифференцируется и переходит
в вегетативн-о недеятельное состояние, сохраняя до деления
относительно постоянные размеры (что может не соблюдаться
у автотрофных, окрашен-ных форм); следовательно, тут можно гово-
рИТЬ 0 НЭЛИЧИИ ВЗРОСЛОГО! СОСТОЯНИЯ, характеризующего-ся СТабИЛИт
зацией ранее достигнутой массы тела. _
Таким образо-м, онтогенезы этого типа резко отличаются от
остальных форм индивидуального развития одноклеточных прежде
всего в том отношении, что тут господствуют индивидуальные
морфогенезы, и что они могут быть разделены на все те этапы,
какие характерны для нормальной схемы онтогенетическог-о раз-
вития Metazoa.
Г л а в а IV
ПОВТОРНОЕ ДЕЛЕНИЕ (ПАЛИНТОМИЯ)
1. Общие определения
При простом делении надвое (мон-ото-мии) рост одноклеточного
организма ограничивается примерно двукратным увеличением его
исходного объема. Достигнув этой границы роста, особь делится
пополам, и получающиеся из нее две дочерние особи вновь прибли-
зительно удваивают свой объем. Таким образом, для этого вида
размножения характерно правильно-е чередование мито-зов с дора-
станием его продуктов до размеров материнской особи. Одним из
сл-едствий этого является то обстоятельство, что абсолютные раз-
меры монотомических одноклеточных, как общее правило, очень
невелики.
" При повторном же делении наблюдается совершенно иная Kap-
102
ТИНа. РОСТ ‘ОДНОКЛеТОЧНОЙ ОСОбИ далеко ВЫХОДИТ за пределы „удвоег
ния исходного объема, приводя як его увеличению -от 4 до многих
тысяч раз, т. е. к гипертрофии, или переразвитию, особи. Такая
укрупненная, подчас действительно гигантская, одноклеточная
особь не теряет своей способности к делению. Последнее наступает,
однако, только по окончании ее гипертро-фического роста, и уже
не в виде однократного простого деления, а в виде целой серии
таких делений, быстро следующих друг за другом, не прерываясь
периодами роста. Вследствие этого ‚материнская особь, при-ступив-
. щая к размножению, дробится сначала на 2, затем на 4, на 8 и т. д.
одноклеточных -особей, соответственно уменьшающихся в объеме
приблизительно в геометрической прогрессии.
Итак, для размножения путем повторного деления, или палин-
томии, характерно прежде’ всего то, что оно складывается из сле-
дующих закономерно чередующихся процессов:
1) гипертрофического роста материнской особи, приводящего
уже к многократному, а не только двукратному увеличению ее ис-
ходной массы;
2) наступающей затем серии сближенных во времени митозов,
имеющих своим результатом последовательное раздробление мате-
ринской особи на 2, 4, 8 и т. д. потомков все уменьшающихся
размеров.
Число повторных делений, имеющих место при палинтомии, на-
ХОДИТСЯ В тесной ЗЭВИЮИМОСТИ ОТ ТОГО, B‘0 СКОЛЬКО раз увеличилась-
материнская особь в ходе ее гипертрофического роста: так, если ее
исходный объем увеличился при этом вчетверо, то она обычно
делится дважды (т. е. на четыре потомка);'если он увеличился раз
в восемь, то особь делится трижды (т. е. на восемь) и т. д. Поэтому
можно сказать, что гипертрофированная материнская особь делится
подряд примерно столько ра-з, во сколько раз удвоился перед тем
ее объем, что соответствует числу как бы «пропущенных» ею
делений.
Отсюда ясно, что гипертрофический рост одноклеточной особи,
заканчивающийся ее повторным делением, имеет своей основой
временную задержъку, как бы выключение клеточного деления. Это
временное снятие деления, называемое г и п е р т р о ф и ч е с K о й
дистом ией (Шаттон, 1919, 1925), и позволяет растущей -особи
далеко выходить за <‹монотомическую›> границу роста (каковой яв-
ляется приблизительно удвоение ее исходного объема) и приобре-
тать относительно очень крупные раз-меры.
Г ипертрофическая дистомия связана с определенными биологи-
ческими условиями, главным из которых является избыточное пи-
тание. Поэтому она наблюдается чаще всего у паразитических
одноклеточных — в условиях неограниченного обилия пищи и непре-
рывного ее поступления, у колониальных форм, находящихся
в особо благоприятных трофических условиях, и лишь значительно
реже и в гораздо меньшем объеме у свободно живущих одиночных‘
‚видов, Помимо питания, определенную роль могут играть тут‚ п-о-
105
‚видимому, и другие факторы среды: осмотическое давление, гумо-1
нральное воздействие организма хозяина и ‘г. п.
ИТЭК, ГИПерТрОфИЧ-еСКИИ РОСТ материнскьои КЛЕТКИ, временная
‚задержка ее деления И последующая реализация «пропущенных».
‚ею делений в виде ускоренногогтп-овторного деления суть взаимно
обусловленные явления, образующие единый палинтомический
процесс.
‚Своими корнями палинто-мия тесно связана с метазойным типом
«простого деления. Она наблюдается среди простейших, только
‘у видов, размножающихся в дедифференцированнчом состоянии, и
лишь как исключение-у постоя-нно амёбоидных форм. Типичная
палинтомия особенно характерная для некоторых групп жгутико-
-вь1х‚ а именно для большинства фитомо-над (.в то-м числе всех кол-о-
ниальных), многих Dinoflagellata (особенно паразитических), неко-
торых Chrysomonadina, Chloromonadina и эвглен (Euglena gracilis,-.
'Ast'asia), встречаясь также „у единичных протомонад и- отдельных
форм неясного систематического положения, вроде Зрог-отопаз или
Gregarella. Случаи палинтомии описаны также у неко-торь1х кокци-
дий, с-олнечников (Vampyrellidium) и к-орненожек (Euglypha),
a производные, видоизмененные ее формы (OM. ниже) —обычны
ту радиолярий и целого ряда споровиков.
Особенно важно подчеркнуть, что наступление временно за-
держанного деления происходит исключительно в дедифферен-циро-
гванном состоянии организма. Это, очевидно, означа-ет, что ему
обязательно должна предшествовать реорганизация, до осуществле-
-НИЯ КОТОрОЙ O'H‘O не -Mom-e“r ВпСТуТПИТЬ В СИЛУ. И ‘РОЛБКО после нее
достигается то состояние готовности к (делению, при котором послед-
нее может наступать или немедленно, или же в результате особого
активирующего толчка извне.
Итак, непосредственными результатами гизпертрофической
дистомии являются перерастание особью ее нормальной границы
роста при сохранении ею типично одноклеточной организации, и
соответствующее перерастание нормального клеточного деления
в повторное, или палинтомию. -
2. Общие особенности палинтомии д
Все характерные особенности палинтомии Целиком опреде-
ляются тем, что она происходит при дедифференциров-анном состоя-
нии организ-ма и представляет сто-бой не что иное, ‘как серию простых
делений, ‘сильно сближенных во времени. В силу этого она обладает
всеми общими свойствами, характеризующими метазойный ‘тип
КЛеТОЧНОГО деления И, В ДО‘П0ЛНеН'Ие K НИМ, целым KO'MiI'IJI€KnCOM'
новых, специфических для нее особенностей,’ обусловленных самой
повторностью таких делений.
В самом деле, при палинтомии наблюдаются все те явления,
какие присущи метазойному типу моното-мии: дедифференцировка
особи перед размножением; устранение та_ким путем части ‚материи--
ДМ
' i
.CKOI‘O Материала о-т участия в процессе деления; исключительная
полнота и равномерность распределения остального материала
между обоими продуктами деления; их специфически «эмбриональ-
ное» состояние и т. д. Однако первым же повторением такого деле-
ния сюда вносится совершенно новый момент: выпадание процесса
дифференцировки дочерних особей и их перех-ода к активному
из пассивного состояния,
дочерние особи сразу же де-
лятся сами и дают -начало
либо продолжает делиться
дальше точно таким же спо-
собом, как первое. Так или
Рис. 27. Повторное деление фитомонады Нае- Рис. 28. Повторное -деление
matococcus pluvialisz
1—начало профазы первого деления; деление внеядер-
ной ценбросомы; 2, З-расхождение дочерних центро-
сом; 4-—метафаза; растворение ядерной оболочки;
5-—ранняя анафаза; б-завершение первого деления;
7—перекрест спороцитов в профазе второго деления;
8-—то же, ранняя анафаза; 9—начало превращения
спороцитов-продуктов второго деления в зооспо-
ры; 10—-3ооспоры, закончившие дифференцировку
(по Эллиоту)
Oicomonas tetraspora: »
1, 2—профа3а первого деления; 3—
анафаза; 4—телофаза; появление
врезающейся борозды на жгутико-
вом полюсе; 5--центробежные боро-
ры (по Сварчевскому)
иначе, дифференцировка составляет удел. одного только заключи-
тельного ‘ПОКОЛЭНИЯ ПРОДУКТОВ ПЭЛИНТОМИИ, а в-се промежуточные
так не ИСПЫТЫВЭЮТ ее, ПрОВпОДЯ ВСЮ непродолжительную ЖИЗНЬ
своего рода вкэмпбриональном» ‹С›0'СТО'ЯпНИИ.
Таким образом, создается наиболее своеобразная из ‘особенном
стей повторного деления: то, чт-о оно не прерывается -в интеркине-
зах переходом его продуктов -в подвижную вегетативную фазу
жизнедеятельности, а стало быть и их дорастанием до материнской
размерной нормы. И именно потому, что рост остается вь1ключен-
ным или, по крайней мере, сильно заторможенным в течение всего _;
этого процесса, размеры продуктов палинтомии убывают с каждым‘
‚делением —,в приблизительно геометрической прогрессии,
105'
существованию. ' ‘Не выходя -
второму их поколению, кото-'
рое либо дифференцируется,‘
зды на стадии 4', 6--l‘0’l‘0BbIe ЗООСПО- А
Следствием этого является в высшей степени замечательное
превращение одного или нескольких поколений дочерних особей
в сильно редуцированных споро-нитов, обладающих ничтож-
ной продолжительностью индивидуально-го существования, не выра- *
батывающих никакихсобственных дифференциро-вок. Это——в зна-
чительной мере «обезличенные» одноклеточные организмы, как бы
утративши-е свое знач-ение самостоятельных особей и болЬШинств-о
би-ологических проявлений своей индивидуальности; организмы,
превратившиеся в генеративные клетки, не имеющие другой функ-
ции, кроме функции размножеч-хия. Кроме того, палинтомия приво-
д_ит к превращению кон-ечъхьхх продуктов деления в жгутиковые,
как правило, зооспоры, отличающиеся, правда, от продуктов про- A
стого деления только тем, что они образуются в большем числе и
имеют более сложное индивидуальное развитие (ри-с. 27, 28).
Итак, в результате ги-пертрофической дистомии возникает тот
исключительно своеобразный процесс палинтомии, который мы на-
зываем у Metazoa дроблени-ем. Имея своей осново-й метазойный ‘
тип моно-томии, он ведет, во-первых, к переходу от ‚размножения
простым делением к размножению посредство-м более или мен-ее
многочисленных зооспор; во-вторых, к ‚замене ряда поколений нор-
мальных трофо-золитов (Max бы перешагиваемых при гипертрофиче-
ско-м рост-е) соответствующим числом генераций с-пороцито-в, обла-
дающих сугубо эмбриональным характер-ом бластом-еров и многими
их опеци-фическими свойствами; в-третьих, к возникающему вслед-
ствие этого первичному (кпалинтомическому) чередованию поколе-
ний, каждый цикл которого объединяет одно вегетативное одно-
клеточное поколение с несколыкими- генеративны-ми, и, наконец,
к созданию прямого пути к достижению много-клеточной орга-
низации.
З. факультативная палинтомия
Характерн-ейшую особенность метазойного тизпа деления состав-
ляет, как уже говорилось, его «неудержимая» склонность к неодно-
кратному повторению, т. е. к п-ерерастанию в палинтомию. Эта
тенденция огольнастойчиво проявляется во всех труппах, размно-
жающихся в дедиффер-енцированном состоянии, что монотомия
здесь лишь редко бывает устойчивой; гора-здо чаще она принимает
характ-ер ф а к у л ь т а т и в н -о й п а л и н т о-м и и, очевидно пред-
ставляющей собой самую примитивную форму последней.
Факультативная палинтомия очень характерна для свободно-
ЖИВУЩИХ ЖГУТИКОВЬКХ, обладающих метазой-ньгм ТИ'ПОМ ДеЛ-ЭНИЯГ
одиночных фитомонад, ряда хригзомо-над, .перидиней и немногих
протомонад. Общеизвестно, что у большинства представителей
пер-вой из перечисленных групп деление может быть то однократ-
ным, то дву- или даже трехкратным, а иногда даже повторяться
до 4—5 раз подряд; таким образом, число делений подвержено тут
значительно индивидуальной изменчивости. .
Эта изменчивость, однако, не имеет «случайного» характера.
106
ВСЕХ тех фОрМ, у КОТОрЫХ о-на СПЭЦИЭЛЬНО изучалась, всегда ока-
ЗЫВЭЛОЮЬ, ЧТО ЧИЮЛО делений, ИОПЫТЬПВЗЭМЫХ ОСОбЬЮ, НЗХОДИТЧСЯ
В ТЕСНОЙ 3aBI/PC-I/I‘M0'CTI/I OT наличных УСЛОВИЙ ПИТЭНИЯ, ОТ ‘СТЕПЕНИ
ее упитанности и достигнутых ею размеров. Так, у миксотрофнжой’
Haemato-coccus droebaki-ensis, одно-кратное деление наблюдается
только в средах, бедных питательными веществами, тогда как дву-
кратное является правил-ом при достаточной концентрации послед-
них, а трехкратное часто име-ет место при дальнейшем повышении
концентрации питательного раствора и, особенно, в первые недели
существования культур, когда их плотность еще нев-елика.
У Chlamydomo-nas nasuta легко заметить прямую связь между
дефинитивными размерами особи -и числом испытываемых ею
-делений, ‘ISM также y~CTaiHaBJII/I'Ba'€'I‘«CH 3a'BI/IICI/IIMOCTI3 ЭТОГО ЧИЮЛа OT
. CTe‘l'IeHI/I y‘l‘II/ITEIHHOICTI/1' материнской О'С0бИ‚ И Т. Д.
Таким образ-ом, наблюдающаяся здесь задержка деления на-
ступает при определенных условиях существования и не имеет
места пгри д-рутих, т. е. наследственно не закреплена.
4. Типичные формы палинтомии
В пределах тех же одиночных фитомонад очень легко прослежи-
вается переход от факультативной палинтомии к постоянной. Так,
у очень многих ‚форм нормой становится двукратное деление,
а однократное или трех-четырехкратное осуществляется лишь от
случая‘ к случаю; у других совершенно выпадает однократное и
число делений варьирует -в пределах двух-трех, редко четырех. П-ри
этом числ-о делений находится и здесь =в весьма наглядной зависи-
мости от степени гипертрофии, достигаемой материнской особью
к концу ее вегетативной деятельности. Правда, известные коррек-
тивы вносятся сюда тем фактом, что фот-осинтетичес-кая деятель-
ность опороцитов не является полностью выключенной; поэтому
_в ходе повторного деления происходит некоторое увеличение живой
массы дробящегося комплекса, своего рода эм-бриональный рост,
-в известной мере компенсирующий их прогрессивное м-ельчание.
У одиночных свободно живущих представителей других групп
простейших, размножающихся палинто-мией, число повторных де-
лен_ий колеблется в тех же пределах: чаще всего деление повторяется
от двух до трех раз, и лишь как исключение+до четырех.
Таким обра-зом, продуктивность палинтомическопо размножения
этих форм относительно невелика и выражается в образова-
нии 4—8‚ реже 16 зооопоэр; эти малопродуктивные формы палин-
томии весьма характерны именно для сво-бодноживущих и одиноч-
ных Protozoa.
V У колониальных фитомонад (Volvoacoidea); с одной стороны, и
Целого ряда паразитических форм из Вйпойаёепата (Oodinium И др.)
и других групп жгутиковых (Зрогогпопаз) —— с другой, число деле-
ний B ПаЛИНТОМИЧЭСКОМ цикле значительно повышается, доходя
до 8-10, а иногда даже (у некоторых видов Vo_1vo-x) до 14 или 15.
107
Столь высоко продуктивные формы типичной палинтомии наблю-
даются только в тех случаях, когда материнская о-со›бь оказывается
в особо благоприятных условиях избыточного питания, ‘что воз-
можно при ее паразитическом образ-е жизни или же'при использо-
Baum ею ассимиляционной деятельности других членов колонии.
В этих случаях особь подвергается крайне высокой гипертрофии,-
приобретая при этом многие характерные свойства яйца, а ее па-
линтомическое размножение может быть наввано M H o- г о эк р а т н о
повторным делением, или гиперпалинтомией.
‘У паразитических форм гиперпалинтомия ведет к массовой unpo-
дукции свободньпх-зооспор, а у кол-оттиалвных-к возникновению
многоклеточности, как это будет подробно показано ниже.
Необходим-о, однако, отметить, что и среди паразитических и
колониальных простейших имеется-немало форм (примером кого--
рых могут служить у первых такие роды, как Protouodiniurn, a y вто-
рых-все Eu_do-rinidae и Spondylo-moridae), палинтомический цикл, '
K'OTOpbIX ВКЛЮЧЭСТ ВСЕГО‘ O’I‘ ДВУХ ДО четырех ИЛИ ПЯТИ делени-й.
5. Морфология повторного деления
МОрфОЛОГИЧЭСКИЭ картины ШаЛ-ИНТОМИИ СОЗДЗЮТСЯ В pE‘:3yJIbTaTe'
B3211/IMOILCI‘/’I'C’IT‘BI/IS"-I ДВУХ МОтМЭНТОВ ПРИНЦИПИЗЛЬНО! РЗЗЛИЧНОГО ПОрЯДКЭГ
а) геометрических форм того типа просто-го деления, повторение
которого лежит в основе пали-нтомического дел-ения, и ‘б) перео-риен-
тировок спороцитов по ходу этих повторных делений. Первым из
них определяются истинные геометрические формы
этого „процесса, тогда как второй имеет своим результатом группи-
ровку продуктов деления в виде различных фигур и играет, таким’-
образом, главную роль в ооздании- в и д и м ы х к а р т и н повтор-
ного деления, часто совершенно не отражающих его действителы:
ный характер. ' '. ч
Разнообразие истинных форм палинтомии простейших очень_
невелико, сводясь здесь только-к трем ее типам: 1) л и н-ей н о й
палинтомии, в основе которой лежит повторение поперечного ‘де-
л-ения; 2) о д`н о р я д н о й, или строчечной, палинтомии, создаю-
щейся повторением продольных монотомических делений в одн-ой
и той же неизменной плоскости; 3) т алб л и ч н о й палинтомии,
возникающей в результате чередования продольных делений в двух,
взаимно перпендикулярных пл-оскостях. _
При л ин е й н ой ‘In a Л и н т о M ии все ее" продукты распола-д
гаются, в принципе, в виде цепочки, чередуясь своими разноимен-
ными полюсами, т. е. в. строго метамерном порядке. Однако,_ no-'
скольку такая цепочка образуется в результате повторного попереч-
ного деления каждого ее звена, и притом деления равномерного, —_
ee разрастание в длину идет одновременно в двух противоположных,
направлениях от плоскости. первого деления, а не только ‘в каком-
либо Одном, как-при образовании мета-мерных систем у различных‘
1085
. членистых Metaz/0a.. B соответствии с. порядок чередования
„плоскостей последовательных делений одинаков. по обе стороны
от плоскости первого поперечного деления; они располагаются
зеркально относительно этой плоскости. Линейная палинтомия
характерна для Dinoflagellata, вообще обладающих поперечным
делением‘, но чаще встречаетсяздесь не в типичной своей форме,
а в очень своеобразных формах повторного спорообразования
(см. ниже).
Одн орядн ая, или -строчечная, палинтомия имеет в своей
основе правильное повторение делений в одной и той же продоль-
ной плоскости, условно принимаемой за сатиттальную. Ее продукты
поэтому располагаются в один поперечный (по отношению к осям
составляющих его клеток) ряд, паралл-ельн-о друг другу своими
главными осями. Образующаяся таким путем «строчлкав обладает
теми же проморфолотическими особенностями, как цепочки,
создающиеся ПРИЛИНЭЙНОМ типе ПЭЛИНТОМИИ, НО С тем ТОЛЬКО раз-г
личием, что плоскости делений идут в направлении длинных осей
спороцитов, а не пересекают их под прямым углом. Этот тип палин-
томии встречается в чистом виде главным образом у зеленых водо-
рослей и олицетворен, например, известными 4-—8-клеточными
колониями Зсепеаезтцз. „
Несравн-енно большим распространением y-Protozoa пользуется
второй вариант продольной в своей основе палинтомии —— палинто-
мия т анб л и чан а я. При этом ее типе-плоскости всех составляю-
щих ее простых делений правильно ‚чередуются в двух взаимно
перпендикулярных направлениях (отвечающих -сагиттальной и фчрон-
тальной плоскостям-у двусторонне симметричных форм, деля-
щихся таким образом, и соответствующим им плоскостям симм-етрии
уформ, построенных по двулучевому плану). Благодаря такому
чередованию плоскостей всех следующих друг за ДРУГ-ом делений,
их пнродукты располагаются параллельно друг другу в „плоскую
пластинку, или табличку, растущую одновременно в двух измере-
ниях пространства. Иначе говоря, продукты табличн-ой палинтомии
располагаются эпителиальным строем; они поставлены не только
параллельно друг другу своими главными осями, но и их одноимен-
ные п-олюсы обращены в одну сторону, вследствие Чего этот тип
палинтомии можно было бы назвать еи э п и т е л и -о г е н н ы M ее
типом. ‘
Таблич-ная палинтомия особенно ‘характерна для фитомонад,
но встречается также у некоторых перидиней (Protoodirxium,
Dissodinium и др,), эвглен (Euglena 'gracilis),. хризомонад, Сгазре-
domonadina и ряд других форм, ‘обладающих, в типе, продольным
клеточным делением. Она также обычна у водорослей, особенной
при делении в стадии пальм-еллы. '
Этим трем основным типам палинтомии простейших теорети-
чески может быть противопоставлен четвертый тип повторного
клеточного деления -— к у лб и ч ес к а я п а л и н т о м и я, считаю-
щийся характерным для Metazpa. При_` кубической палинтомии
109
деления чередуются не в двух, а ‘во всех трех измерениях простран-
ства, -по трем взаимно перпендикулярпьхм (координатным) плоско-
ст-ям. Однако с-реди простейших 11ам не известно ни одно-го случая
подобного их правильного чсродовапия-чередования, фактически
означающего отход от основного принципа палинтомии, как серии
сближенных, ‘но точно пловторяюнщих дру-г друга простых клеточных
делений. Правда, у некоторых неридин-ей (Dissodinium, Blast0-di- Г
nium) действительно имеет место сочетание табличной палинтомии
(у Dissodiniurn) или нр‹›до.льтг‹›го простого деления (Blastodinium)
с линейной ‘паличтлтхжмией, что на деле равнозначно делению мате-
ринской осо-би и трех взаимно перпендикулярных плоскостях; ни
в одном из этих случаен, однако, нет их правильного чередования
по законам кубичсокогт ‘наличггомлии, так что тут приходится гово-
рить не об этой последней, а именно о последовательной смене.
0ILH'OKp21'I‘II0II‘() ИЛИ ПОВТОрцЪГОГО ‘ПрОДОЛЬНОГО деления ПОВТ-ОРНЫМ же
поперечным, О ПРОХОХЩОНИИ ДВХХ рЭЗЛИЧНЫХ И отчетливо ра3гра-.
ниченных между собой пали-нтомических Циклов, в одном из кото-
рых она иъме-ет табличный, а в другом-линейный характер
(см. следующий раздел данной главы). '
Более близкое касательство в кубической палинтомии имеет, возможно,
те очень немногочисленные случаи, когда правильное повторное деление
про-исходит у’ амёбоидных, неполятризованных одноклеточных. Мы имеем в виду
п им-еры факультативной палинтомии у краковинной корненожки Euglypha sp.
(- B3.-H‘Pl“{, 1935), И описанную этим же автором повторную палинтомию амёбоид-
ного солнечника Vampyrellidium. B этих случаях Можно допустить, что следую-
щие друкг за другом монотомии имеют характер первичночтоп-елречгного деления
и чередуются, таким образом, во всех измерениях пространства. Однако, чтобы
можно было это утверждать, требуется гораздо более тщательный анализ этих
случаев, чем это было сделано Иваничем.
Переходя теперь K рассмотрению вопроса O переорпиентировках
И смещениях C-H'0'p'OI.lPITOI3 В ходе КПЗЛШНТОМИЧеСКОГО деления, МЫ,
прежде всего, ДОЛЭКНЫ отметить, ЧТО механизм ЭТИХ перемещений
остается неисследоватгным И неизвестным. MOHGHO ТОЛЬКО СКЗЗЗТЬ,
ЧТО ЭТИ перемещения ИМеЮТ СТрОГОч определенный, закономерный ,
характер и связаны с неизученными явлениями взаимного притяже-
ния спороцитов.
Отчетлижвое представление о характере этих переориентиро-вок
может дать рис. 29, показывающий ход этого процесса у фитомо-
нады Haemat-ococcus пи перидинеи Blastodinium. Как видноиз этих
рисунков, они выражаются в том, что опороциты стадии 2 сначала
склоняются друг к другу своими анимальными полюсами, а затем
поворачиваются гво взаимно противоположных направлениях, ло-
жась крест-накрест под углом от 45 до 90°; при этом они тесно
прижимаются один к другому таким образом, что следующее, вто-
рое, дел-ение имеет своим результатом группировку спороцитов
стадии 4 в виде двух перекрещивающихся дуэтов. На этой стадии
противостоящие друг к другу спороциты входят в еще более тесны-й
контакт, приводя к образованию характер-нейшей фигуры—‹<‚п а-
л и н т о м -и ч -е с к о г о кр е с т а», слагаемые которой плотно смы-
110
‚ каются друг с Другом В двух различно ориентироватъньтх плоско-
стях, образуя широкие полярные спайки.
Сходные as принципе переориентировки спороцитов происходят
в течение всего дробления у высших’ Volvocidae (CM. рис. 81, 82),
где они создают иллюзию деления во всех измерениях простран-ства,
хотя оно и является тут в действительности строго табличным
- тетень-
›
5
9 т I2
Рис. 29. Повторное деление (палинтомия) жгутиковых:
7 —9 I-laematococcus pluviali-., изменения ориентации материн-
ского клеточного тела и спороцитов в ходе деления:
1—взрослая особь перед делением; 2—приближение ядра к
жгутиковому полюсу; 3—метафаза первого деления; появление
борозды На жгутиковом полюсе; 4—ранняя телофаза, поворот
клеточного тела; 5—конец первого делення; б——поворот обо-
их спороцитов перед вторым делением; 7—метафаза второго
деления; перекрест спороцитов и появление центробежных бо-
розд второго деления; 8—крестообразное положение споро-
цитов стадии 4 (по Рейхенову); 9—перекрест спороцитов
стадии 2 (по Эллиоту); 70—Parapolytoma satura; односторон-
няя врезакощаяся борозда втелофазе первого деления; 77-
телофаза второго деления; появление центробежных борозд;
72—перекрест спороцитов Btastodinium hyalinum; 73-75-
группировка спороцитов того же вида в форме «палинтомичес-
кого креста» (по Шаттону)
(т. е. продольным). Не исключено, что в основе всякой вообще
табличной палинтомии также лежит изменение взаимной ориента-
ции спороцитов (их поворот при каждом делении: на 90°)‚ а не чере-
дование деления в двух плоскостях, но имеющиеся на этот счет
данные недостаточны для решения этого вопроса.
Во всяком случае необходимо всячески подчеркнуть, что эти
‘явления Наблюдаются ‘ПОЧТИ ‘В ‘КЭЖДОМ случае I1'I‘a.TII/IH'I‘0MI/II/I, не ЗЗВАИ-
симы от ее истинных геометрических форм (как легко видеть из
сравнения соответствующих фигур у фитомонад, обладающих
111
табличной тталинтомией, и-у Blasfodinium, где она является линей-
ной), и играют огромную роль кв создании квидимых, кажущихся
ФОРМ ПЭЛИНТОМИЧЭСКОГО ДЕЛЭНИЯ. МОЖНО: -CKa8aTb, ЧТО ЭТИ ПОСЛЕД-
НИЕ 'Ha'CTOJIbKO ЗЭВИСЯТ ОТ ИЗМЕНЕНИЙ ‘ОУРИЭНТЗЦИЖИ СПОрОЦИТОВ, "ЧТО
ПО НИМ НЕВОЗМОЖНО СУДИТЬ Об ее ИСТРХННЫХ ГЭОМЕТрИЧЭСКИХ формах;
ТОЛЬКО ДЭТЗЛЬНЪШИ al-IZIJIPIB ПОЛОФКЭНИЯ КЛЭТОЧНЫХ ОСЁЙ И ПОЛЮСОВ
СПОрОЦИТОВ И ИСпПЫТЬПЗЕПЭМЬ1Х ИМИ ‘ПЭРЭМЫЦЭНИЙ МОЖЕТ СЛУЖИТЬ КрИ- '
терием истинных форм ‘нилидттомии, и это никогда не следует
упускать из ‚вида.
Все сказанное о геометрических формах палинтомии простей-
ших может быть 1)ез1‹›м1‹х1)о-вг111о в следующих двух положениях:
1) Все разнообразие исттн—н-1ь1х форм палинтомии сводимо, в ко-
нечном счете, к двум ее главным типам — л и н е й н о й, имеющей
в своей основе нонеречное (или диагональное) клеточное деление,
и табличной! налинтомии, базирующейся на продольном кле-
точном делении. _
2) Видимые формы пали-нтомического деления, ‚нередко кажу-
щегося трехмерным, обычно не отражают ее истинных геометрич-е-
ских форм и определяются главным образом изменениями взаим-
нпой ориентировки опо-роцитов в ходе этого проц-есса.
6. прерывистая палинтомия
Большой теоретический интерес представляют случаи прерыви-
стой палинтомии, описанные у некоторых паразитических перидиней
(Dissodinium 1_ur1u1a; Догель‚ 1910) и солнечнинков (Vampyrellidium;
И-ванич, 1936). ‘
У Van:-pyrellidiu-m амёбоидная материнская особь инцистируется
перед размножением и затем испытывает 2, 3 или 4 не вполне emi-
хронных последовательных делений, дающих соответственно 4, 7, 8
или до 15 амёбоспор, вскоре’ покидающих ци-сту. Выйдя из нее,
амёбоспоры тут же линцистируются вновь, превращаясь в «цисты
повторного размножения перво-го порядка», содержимое которых
опять делится два или ‘при раза. .Образующиеся таким путем
4-6 амёбоспор выходят из цисты и затем еще раз инцистируются,
давая начало «диетам повторного размножения второго порядка».
В этих последних происходит еще два быстро следующих д-руг за
другом деления, продуктыкоторых вновь расползаютчся. По Ива-
ничу, они могут инцистиръоваться еще раз, но таки-е «цисты повтор-
ного размножения третьего порядка» уже нежизнеспособны: ‘их
ядро, правда, может делитвспя еще два раза, но выхода из цисты
«уже не происходит. Таким образом, палинтомическое размножение
‘-Vampyrellidium складывается из трех циклов, разграниченнь1х
периодами инцистирования, и включает в о-бщей сложности до
девяти И(4д+ 3 + 2) делений, не прерываемых питанием и ростом.
В соответствии с этим, амёбоспоры и образуемые ими ци-сты повтор-
"ного размножения прогрессивно уменьшаются в объеме. Судя по
амёбоиднопму характеру Van:-pyrellidiu-m на всех стадиях развития,
112
8
Эмбриология беспозвоночных
3—-стадия 4 спороцитов в материнской оболочке; 4-
ех°де В З-КПЕТОЧНУЮ стадию; ьд-Ёрогатая-Циста», б, 7, 8—повторное
9—превращение содержимого этои цисты в единственную крупную
. Рис. 30. прерывистая палинтомия Dissodinium lunulat
7—материнская особь перед делением; 2——деление на 4;
Продольное деление спороцитов при пер
поперечное деление в «рогатой цисте»;
эооспору (по Догелю)
все испытываемые им повторные деления относятся к первично-
поперечному типу. „
'- Палинтомическое размножение Dissodinium складывается из
двух циклов, также разграниченных инцистированием. Первый из
них состоит из четырех последовательных ‚продольных делений
(рис. 30, 2-4), дающих 16 веретеновидных спороцитов, лежащих
параллельно друг другу внутри материнской оболочки. Вслед за’
этим каждый спороцит выделяет собственную оболочку, превра-
щаясь в характерную серповидную |ИЛИ рогатую цисту (рис. 30, 5).
Содержимое каждой из них делится еще один, два или три раза,
образуя таким путем 2, 4 или 8 округлы-х жгутиковых динос-пор
(рис. 30, 6—8), или же целиком преобразуется в единств-ен-ную, но
соответственно более крупную и более удлиненную диноспору
(т. е. зооспору типичного для Dinoflagellata строения; рис. 30, 9)._
Таким образом, второй палинтомическии Цикл (совершающийся
-в рогатых цистах) имеет, в отличие ‘отпервото, явно факультатив-_
ный характер. Самая замечательная его особенность заключается,
однако, не в этом, а в том, что составляющие его деления имеют
уже не ПРОДОЛЬНОЭ, а HJBCTBCHHO ПОПЕрЭЧчНОВ I-Ia‘l'Ipa'BJIeHI/Ie, как ЭТО
легко заметить по соответствующим картинам мито-за и располо-
ЖЭНМЮ продуктов деления в правильную цепочку (рис. 30, 8). Дру-
гими сл-овами,'табличная вначале палинтомия сменяется затем,
во втором ее туре, на линейную, что резко противопоставляет этот
случай прерывисто-й палинтомии описанному выше случаю Vampy-
rellidium. ' ' '
Стоит отметить, что столь необычайная смена направления
повторных делений находится в очевидно-й связи с отмеченной нами’
ранее (-стр. 59) двуосностью перидиней: она ярко подтверждает
привязанность плоскости деления этих форм к их первично-й глав-
ной оси, сохраняющей тут продольное ‘направление у с-пороцитов
четырех первых поколений и отклоняющейся на 90° у последующих
их генераций. у . ‚
7. Повторное спорообразование ’(палинспорогенез)
- Это. своеобразное видоизменение палинтомии характерно для
Многих паразитических перидиней, а именно представителей родов
Haplozoon, Blastodinium, Apodirxium, Atelodinium и др., и заклю-
чается в неоднократном периодически повторяющемся палинтоми-
ческом размножении одной и той же особи, остающейся при этом
в вегетативно деятельном состоянии. A
B основе этого удивительного явления лежит сочетание обыч-
ного у Dirxoflagellata повторного вторично-поперечного деления
с ярко выраженной трофической поляризацией простейшего, не
прекращающего питание» в течение всего периода размножения.
Взаимодействие этих трех моментов и ‚приводит к тому, что веге-
тативная особь периодически делится на две нералвно-ценные дочер-
ние особи-проксимальную, или трофоцит, остающуюся свя-
114
8*
Рис. 31. Повторное спорообразование Blastodiniumz
1_..молодая вегетативная особь; 2—ее диагональное деление на трофоцит (нижний) и гоноцит (верхний); З-образование второго.
r0Ho1_mTa;4—-Aanbfielfimafl стадия деления; 5—трофоцит (Т), окруженный продуктами деления первого гоноцита (г. 1); б-трофоцит-
' (T), второй гоноцит (г. II) и продукты деления первого гоноцита (г. 1) (По ШаТТОНУ)
115
116
Рис. 32. Продольный разрез Blastndinium pruvoti:
I-—npo11y1<1-bx палинтомического деления первого, 11-—второго и 111._треть.
его гоноцитов, ‘Г-трофоцит (по Шаттону)
ЗЗННОИ С ХОЗЯИНОМ И ПРОДОЛЖЭЮЩУЮ питаться И расти, И ДИСТаЛЬ-
.ную особь-гону-оцит, сразу же делящуюся дальше путем
линейной палинтомии. Таким образом, повторное спорообразование
складывается из чередования гетерономных простых делений мате-
ринской особи, при каждом из которых она отделяет от себя по
‚ОДНОМУ ГОНОЦИТУ, И нормального. ПЗЛИНТОМИЧЕСКОГО ДЭЛЭНИЯ ЭТИХ
последних, завершающегося образованием многочисленных жгути-
ковых зооспор (диноспор). Число гоноцитов, продуциру-емых каж-
дой особью, ‚может доходить до 6——7 (у некоторых Blastodinium).
a каждый из гоно-цитов дает до 64—128 и больше зооспор, так что
общая продуктивность ‚повёторного опороо-бразования может быть
чрезвычайно высока (рис. 31, 32).
Большего внимания заслуживают те обстоятельства, что повто-
"рение гетерономнь1х простых делений обычно наступает значительно
раньше ОКОНЧЭНИЯ ПЭЛИНТОМИЧЭОКОГО ДЕЛЭНИЯ первых ГОН0ЦИТОВ‚ не
ДОЖИДЗЯСЬ, так -сказать, ЕГО завершения, И ЧТО все ПРОДУКТЫ СПОрО-
“ГСНЭЗЗ остаются ДЛИТЭЛЬНО СВЯЗЗННЫМИ ДруГ С ДруГОМ И -C рОДИТЭЛЬ-
ской особью. Последнее чаще всегопроисходит в результате того,
что каждое гетерономное деление сопровождается «линькой», т. е.
выделениемвегетативной особью «новой метаплазматической обо-
лочки, внутри которой и происходят все дальнейшие деления соот-
_ветствующего. гоноцита; таким образом, размножающаяся особь
оказывается окруженной несколькими концентрическими,вложен-
нь1ми одна в другую оболочками, каждая из которых с-одержит в
себе всех потомков одного гоноцита. Однако в других случаях (на-
пример, у Нар1о2ооп) таких линек не происходит, «и продукты
повторного‘спорообразования остаются связанными друг с другом
пронизывающими их пучками фибрилл.‘
A Так или инач`е‚‚ при повторном спороо-бра-зовании создаются
своеобралзнейшие временные аг-регать1, состоящие из особи 5‹осно-
вательницы», продолжающей еще питаться И расти, и цело-го
комплекса последовательно образованных ею гоноцитов, находя_-
Таблица 1
Число г0_ Наличное число ‘потомков каждого гоноцита
. М! T%'$:g:’“_ 2:1’-;Jl’;Té3oB__z I _ Общее число
особи ртов танных 1-го `2-го З-го 4-го 5—г0 6-r'o 7-ro KOMWHEHTOB
особью
1 ‘1 Ё 32 8 - 4 ~. _‚ _ 4. .1 + 44
2 1 64 . 16 8 —~ -—- —- —- 1 -88
3 1 3 128 32 8 -—, т ~— — 1 i . 168
_4 1‘ 4 64 32 8 4 -- —— ——- 1~+ 108
5 1 4 128 64 32 8 -—- — —~ 7 1 + 232 `
6 ‘ 1 4 256 128 32, 8 --» —'~ ——- 1+ 424
7 . 1 5 128 4 64 32 16 8 — ~— н: 248
8 1, 5 256 128 64 32 8 —.—- т. 1+- 438__
9 1 5 512 256 128 32 8 ц 4 1+ 935-
- 10.. 1 6 512 .256 ‚128 64 32 8 в 1+1ооог'
- 11 1 7 512 265 128 64 32 ‚16 8 1’+ 16161
' 117
щихся на разных стадиях палинтомии (вследствие ‘чего каждый из
них и представлен разным числом опгороцитов соответственно pas-
личного размера). Конкретное представление о сложности состава-
ПОДООНЫХ МНОГОКЛВТОЧИЫХ агрегатов И ЧИСЛВННЫХ СООТНОШЕНИЯХ _
ВХОДЯЩИХ В НИХ КОМПОНЕНТОВ МОРКЕТ дать приведенная ВЫШЕ таб-
лица 1, касающаяся В1а51о‹11п1нпт contortum и составленная по соот-‚
ветствующим дат-иным Шаттопгх (1920). _'
Из этой таблички явствует‘ и еще одно примечательное обстоятельство, odo-
бенно характерное для ньюшнх, наиболее продуктивных форм повторного споро-
образования. Оно заключается в том, что повторные деления тоноцитов идут
строго ‘синхронно и в строгом согласии с последовательностью их обособления
от материнского трофицита, причем каждый последующий гоноцит отстает
только на 1-2 деления от предшествующего. Исключительно правильный ритм
всех делений-как гетерономных монотомическитх, так палинтомических-ил-
люстрнруется особенно наглядным образом особями N97 И N911 нашей таблич-ки,
у которых деление всех наличных опороцитов разнится всего на один такт.
В ЭТИХ случаях гетерономные ДЕЛВНИЯ вегетативной ОСОбИ ИДУТ В ТОМ же TEMIIB,‘
что и деление гоноцитов, так что каждое синхронное деление всей системы в це-'
лом включает отделение одного очередного трофоцита от родительской особи и
серию одновременных делений каждого из ранее образованных. Нельзя
мимоходом не заметить, что эта схема воспроизводит с поразительной точно-
стью классическую схему спирального дробления яйца Meta_zoa,'a именно после-
довательных делений производных одного из четырех его квадрантов.
Сложность наблюдаемых тут -картин иногда (у ряда видов
Blastodinium) еще усугубляется тем, что начальному (первому)
гетероно-мному делению материнской особи может предшествовать
ее продольное, или вернее, диагональное, деление надвое, дающее
двух вполне равноценных дРУг ДРУГУ дочерних особей, каждая из
которых может испытать еще одно такое же (гом-оно-мное) деление,
прежде чем приступить к опорообразованию. Таким путем создаются
Водном хозяине группы из двух, трех или четырех особей второго
или третьето порядка, которые затем уже сообща вступают в споро-
ген-етическое размножение. При одно-временно-м размножении такой
четверки его сходство с ходом опирального дробления особенно
бросается в глаза. - -
Завершающим актом палияопорогенеза является у части форм
(Ароойпйнш, Chytriodinium) отрыв материнской особи от хозяина,
после которого она сама подвергается палинтомическому делению
и- целиком расходуется на образование зооспор. Однако у B1astodi-
‘nium повторное спорообразование заканчивается таким образом
только -в- ‘тех случаях, когда материнская особь случайно, или
искусственно изгоняется из хозяина на относительно ранних ста-
диях размножения, задолго до ег-о. нормального к-онца. Еслиже
этого не происходит, то заключительным актом размножения яв-
ляется выработка материнской особью последнего гон-оцита (напри-
мер, 7-го), после чего она ‘подвергается дегенеративнымизменениям
и п-огибаетдСходным образом обстоит дело- и у Hapl-0«z0*0r1. Г-ОЛОВНаЯ
Клетка (трофоцит) которого также гибнет после образования по-
следнего гоноцита. - ь ' . . . Ц -
Необходимо еще разл подчеркнуть, что ‘ловт-орноет спорообразо-
118 h
»BaHIrI€ паразитич-еских ПЕрИДИНЕЙ ЗИЖДЕТСЯ на. характерном ДЛЯ НИХ
вторично-поперечном делении, представленном тут двумя вариан-
тами: гетерон-омными простыми делениями, имеющими своим ре-
зультато-м отделение гоноцитов от продолжающего‘ питаться трофо-
цита, и гомономной линейной палинтомией, ведущей к распаденню
гоноцито-в на более или менее многочисленные равноценные спо-ро-
циты. Однако у Blasto-dinium этим двум чередующимся видам ‚по-
перечного деления могут предшествовать, как мы видели и 1——2 про-
дольных го-миономных деления, приводящие к образованию
характерных для них «семей» из 2——4 особей. Таким образом, здесь
фактически имеет место то же самое, что у Diss-odinium, НО в еще
более усложненной форме, в форме повторного, а не одно-кратного,
образования рогаты-х цист (представленного здесь линьками) и
дифференциации происходящих ‚ь них поперечных делений на гете-
рономные и гомономные.
Итак, повторное с-порообразование представляет собой такое
видоизменение линейной палинтомии, при котором родительский
организм не прекращает, после дедиффсренцировки, своей вегета-
тивной деятельности, а продолжает питаться и расти. Поэтому он
и оказывается -в силах произвести на свет не один, а несколько
«пометов>> зооспор и достичь таким, необычайным для простейших,
способом исключительно высокой продуктивности размножения.
Принимая на себя роль с-воего рода ко-рмилки, материнская особь
постепенно доходит до высшего, неизменного, уровня дифференци-
ровки, _за которой уже неизбежно следует ее дегенерация и смерть.
8. Вторичная синтомия
Гораздо большим распространением, чем прерывистая палинто-
мия -и повторное спорообразование, пользуется другое видоизме-
нение повторного деления, которое можно назвчать вто р ичн о й
синтоми-е й. Оно. характеризуется тем, что задержка плазмен-
ного деления идет еще дальше, чем при нормальной палинтомии,
is результате чего многократное деление ядра значительно опер-е-
жает деление клетки, а не идет рука об руку с в порядке
ряда правильных, совершенно законченных, монотомий, как; это
происходит при правильном повторном делении. Благодаряэтому,
гипертрофированная особь переходит сначала во временное много--
ядерное СОСТОЯНИЕ, за КОТОРЫМ уже следует, В качестве заключи- -
тельного акта д-еления, одновременное распадение клеточного тела
на множество одноядерньтх отдельностей. Другими словами, деле-
ние принимает с и н т о M и ч е с к и й характер, присущий множе-
ственному делению всех многоядерных Ргофо2оа. Принципиальное .
отличие вторичной синтомии от первичной (см; гл. Х) заключается,
следовательно, только ‘в том, что она1разыгрывается после длитель-
ной задержки не только плазменных, но и ядерных деленийуна
основе предшествующей ей гипертрофической дисто-мии, тогда ка-к
пригпервичноймсинтомии умножение ядер „идет с- самого начала
119'
параллельно С РОСТОМ организма, В ПОРЯДКЕ ПОЛИМЕРИВаЦИИ. ИНЗЧЕ
*ГО‘ВОРЯ, ЭТИ РаЗЛИЧИЯ касаются ЛИШЬ ПОДГ-ОТОВИТЕЛЬНЫХ этапов РаЗ-
‘МНОЖЕНИЯ И В КОНЕЧНОМ ‘СЧЕТЕ СВОДЯТСН K ТОМУ, ЧТО ПРИ ПЕРВИЧНОЙ.
синто-мии простейшее вступает в разменожение‘ будучи уже в много-
ядерном (надклеточном) состоянии, в то время как при вторичной
синт-омии оно находится в этот Момент в одн-оклеточно-м состоянии,
СМЕНЯЮЩЕМПСЯ на МНОГОЯДЕРНОЕ ЛИШЬ ПОЗДНЕЕ, УЖЕ ХОДЕ pa8»‘
МНОЖЕНИЯ. — . _
Вторичная синтомия наблюдается уряда амёб, радиолярий и
РогапппНега и в высшей степени характерна для слп-ор-овиковз (точ-
нее, акнидоопоридий), обладающих гипертрофированными .одно-‘
клеточными стадиями; крайним ее вариантом является известное
«деление распада» таких форм, как Aggregata и некоторых ради-о-
лярий, при котором даже ядро делится 1не дихотомически, а одно-
временно путем многополюсного митоза, или же сразу распадаясь
на множество ‘вторичных ядер. Эти случаи, впрочем, уже граничат ‘
с синтомическим размножением надклеточных сомателл, поскольку’
-в ряде эиз них в ядре ясно различимы «генеративная» и «остаточ-
-ная>› части, последняя из которых остается не использованной при
делении. A - - - '
Переход. палинтомии во вторичную синтомию заключается,
таки›м образом, B» ПОЛНОМ нарушении ХРОНОЛОГИЧЕСКОЙ ‘СВЯЗИ МЕЖДУ
отдельными ядерными и плазменными делениями, правильно повто-
ряющимися при типичной палинтомии. Это ‘имеет своим результа-
том полноевы-падениег всехпромежгуточных продуктов «повторного
деления-спороцитотв, окончательное исчезновение всей олицетво-
ряемой ими цепи редуцированных клеточных п-ок-олений. Таким
образом, при крайней Модификации палинтомии она фактически
теряет целый ряд характерных для н-ее свойств и п-ереходит в св-ою
противоположгность——-в разМножени-е, полн-остью осуществляемое
Одним ‘единственным клеточным поколением: от былог-о о-бъедине-
НИЯ В ЭТО-М ПРОЦЕССЕ ЦЕЛОГО ЦИКЛЭ КЛЕТОЧНЫХ генераций НЕ OCTaeT‘CH _.
почти и следа;
п Г л а в.а V
_ ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ ПАЛИНТОМИЧЕСКИХ
' ОДНОКЛЕТОЧНЫХ
1. Вводные замечания
‚
Особенности индивидуального развития палинтомических‘ одно-
клеточных всецело ‘определяются спецификой этого цвида размно-„
ж-ения и заключаются, вообще говоря, в следующем:
_- 1) Молодая особь- начинает свое существование в виде недиф-
фЕРЕНЦИР‘0ВаННО'ГО' спороцита, T-. е. В ЖЕ «ЭМОРИОНЭЛЬШОМ» СО-7
СТОЯНИИ, котороМдна-чинаетлся ИНДИВИДУЭЗЪЬНОЕ развитие ПРОДУКТОВГ
всякого метазойного деления, ‘будь то однократное или повторное.
2)’ Этот первьтйдэмпбрионалэный, период онтогенеза завершается;
_1r20.-
"1II>Ic1)|c1)epeH.111»1p-oB1<3o1'31 спороцита в жгутиковую зооспоъру того или
‚иного типа или же ее переходом в активное агмёбоидное состояние
‘(в состояние амёбоспоры), при котором дифференцировка не имеет
места. ' ч
3) Преобразование зоосппоры ‚в трофозоита составляет следую-
щий, третий, этап онтогенеза палинтомических Protozoa; У подвиж-
ных автотрофных M амёбоидных форм оно не связано -с заметными
морфологическими изменениями, поскольку их зооспоры оказы-
ваются сразу же способными к вегетативной деятельности и по-
жизненно сохраняют свою исходную организацию. Напротив, у при-
крепленных и паразитических форм это преобразование может быть
очень значительным и иметь тогда своим результатом коренную
перестройку организации зооспоры. В подобных случаях стадия
зооспоры получает исключительно расселительное значение бро-
‘дяжки, а питание и рост молодо-й особи начинаются только после
ее оседания и превращения в вегетативную особь.
4) Четвертый период о-нтогенеза ‚заключается в гипертрофиче-
ском росте вегетаггивной особи. Пари относительно небольшой сте-
пен-и гипертрофии по-на не ведет к особо заметному изменению
организации особи, так что ее развитие имеет характер э пи мор-
ф-о з а. Однако в тех случаях. когда гипертрофический рост‘ очень
усилен, он связан с весьма значительными изменениями строения.
осо-би, что позволяет квалифицировать эти случаи, как г и п е р т р о-
фический метаморфоз. '
' 5) Заключительный этап-онтогенеза-период размножения-
обычно начинается с дедифференцировки организма, означающей
его переход в стадию размножения. Впрочем, у целого ряда _форм
дедиффе-ренцировнка может происходить задолго до окончания
гипертрофического развития особи, в порядке испытываемого ею
метаморфоза, и она теряет тогда свой заключительный характер.
Так или иначе, моментом‘ завершения"индивидуального развития
приходится считать начало первого „палинтомического деления.
Таким образом, «онтогенез ‚палинтомических одноклеточных пред-
ставляет собой не что иное, как клеточный онтогенез‘ метазойного
типа, дополненный явлениями гиперггргофического развития. Все
его специфические особенности сводятся именно- к вхождению
в‘ него этих я.влений,'к при-обретению онтогенезом гипертрофиче-
ско-го характера, которым он только и- отличается: от соответствую-
щихформ развития монотомических одноклеточных. И‘ хотя эта
гипертрофия нередко приобретает тут крайне своеобразноеи слож-
ное выражение, она никогда не заслоняет собой эту исходную
онтогенетическую схему. ' - -
2. Особенности организации зооспор
. „Подчеркиваемое “нами „отсутствие каких-либо
пиаль ных различий между з-ооспор-ами, с ‚одной стороны, и
продуктами‘метазойного‘простого.деления-ас другой, не означает,
.131
пи н цдие’
‘конечно, что,они всегда обладают тождественной организацией,
что зооспоры вообще ничем не отличаются от дочерних особей,
обра-зующихся при метазойном типе простого деления. Существо-
вание тех или иных различий между дними, напротив, далеко
не является редкостью, но все они не имеют настолько универсаль-
НОГО ЗНЗЧЕНИЯ, ЧТОбЫ МОЖНО бЫЛО 'II'p0'B€CTI/I ЧЕТКО-Е разграничение
между ними. -
Та‚кое их разграничение, в частности, совершенно невозможно
при факультативных формах палипто-мии, столь обычной у одиноч-'
ных фитомонад и многих других одноклеточных Protozoa. По-
CKOJIbKy B ЭТИХ случаях ЧИСЛО ДВЛВНИЙ‘ О-СОбИ ПОДВЕРЖВНО ИНДИЮ
видуальной изменчивости и зависит от достигнутых ею размеров
между продуктами однократного деления и двукратного, часто
нет даже и. ощутимых размерных отличий; достаточно взглянуть
еще раз на соответствующие рисунки (ом. рис. 22, 27), чтобы убе-
ДИТЬСЯ В ТОМ, ЧТО такие ЗООСПОРЫ практически НИЧЕМ не ОТЛИЧаЮТСЯ
от продуктов простого деления того же вида. _
Первые нам-еки на появление у зо-оопор некоторой морфологи-
ческой специфики замечаются при уж-е устойчивых формах палин-
томии, когда они образуются в числе 4——8. B этих случаях зооспоры
отличаются от молодых стадий близких к ним мон-отомических
видов нескольк-о меньшими абсолютными размерами, несколько
иными пропорция-ми (например, более стройными —при табличном
делении или более коренастыми ——при линейном; ом. ри-с. 30),
а ин-огда и такими признаками, как наличие одного только пире-
ноила вместо большего их числа, и т. д. Усиление тех же немногих
признаков составляет, в сущности, основную линию эволюции
‘зооспор при переходе к высшим, наиболее продуктивным- формам
палинтомии. Для этих последних характерны минимальные раз-
меры продуцируемых зо-ос\пор,'их округлая, или, напротив, значи-
тельно вытянутая в длину ‘форма и известная упрощенность орга-
низации, выражающаяся, «например, в спа-бомразпзитии или даже
отсутствии выделительного ап-парата, в недоразвитии метаплазма-
тическопго панцыря, хр-оматдофоров, в уменьшенном против нормы
‘числедиктиосом, отсутствии стигмы и т. д. . A
Большинство этих особенНо-стей зоосп-ор, становящихся все
более определенными по мере увеличения их общего числщслужат
явнымотражением особенностей самого палинтомическ-ого про-
ц-есса‚ а именно прогрессирующего мельчания его продуктов при
получении каждым из них все меньшего числа материнских по-
стоянных органоидов. Иначе говоря, эти признаки зооспор опреде-
ляются сами-м способом их образования, почему они и оказываются
наиболее выраженными при высших, самых продуктивных видах
палинтом.ии. . _ ‚ .
Однако самые характерные из особенностей организации зооспо-р
ЗЗКЛЮЧЭЮТСЯ не В ‚ЭТОМ; так сказать; (HaJII’IHT40‘MPIv1I‘6CKOM, KOMIIJIEKCB .
признаков, а связаньъ- с их сугубо раоселительным значением «бро-
lIH>KeK>>,_ В _c.a1M»o‘M деле, ‘все .жгутиковые. 3g0:0(;'I1Q.1l)bl.§.aMbIX различных,
122
простейших, водорослей и грибов характеризуются, прежде всего,
своей общей организацией свободно плавающего жгутиконо-сца.
Принадлежностью к этой жизненно-й форме определяется архитек--
тоника зооспор, их обтекаемая каплеобразная или «вытянутая
в длину форма тела, полное развитие кинетиды, а также и отсут-
ствие у них «всяких дифференцировок, связанных с сидячим обра-
з-ом жизни и приуроченньгми к пнему способами питания. Во всех
этих признаках жгутико-выезооспоры вполне схожи с трофозои-
тами автотрофных или осмотрофных свободноплавающих Flagel-
lata; при этом их строение ближе всего напоминает строение именно
более примитивных, пожизненно свободных форм, связанных систе-
матическими и филогенетическими узами с данн-ой группой палин-
том-ических Protozoa. .
Это последнее обстоятельство находит свое выражение в том,
что зооспоры всех, даже наиболее видоизмененных из паразитиче-
ских Dinoflagellata обладают единым —— гимнодиниевым — планом
строения, зооспоры‘ различных зеленых водорослей-хламидомпо-
надным, зооспоры вор-отничкочв-ы-х жгутиковых-стр-оением -св-о-
бодноплавающих Protomonadina и т. д. Таким образом, морфоло-
гическое разнообразие жгутиковых зо-оспор относительно невелико,
чт-о ясно- говорит о консерватиз-ме строения зооспор, связанном
с однообразием их образа жизни идвьпп-олняемых ими расселитель-
ных функций.
З. Гипертрофический эп_иморфоз
Наиболее простые формы- постэмлбрионалвного развития зооспор
могут быть обозначены, как г и п е р т р о ф и ч е с к и й э п и IM o р-
ф-о з. Известно, что под эппиморфозом принято‘ понимать развитие,
идущее при постепенных, плавных и в общем незначительных из-
менениях исходной организации особи—изменениях, не влекущих
за собой хоть сколько-нибудь ощутимой перестройки этой органи-
зации, устойчиво сохраняющейся, таким образом, в течение всего
индивидуального развития. В применении к онтогенезу палинтомиъ
ческих простейших это понятие годится для обозначения всех тех
его конкретных форм, при которых организация зооспоры удержи-
вается (хотя бы B главных своих чертах) М-олодой особью при ее
превращении в трофозоита и дальше в течение всего периода роста,
вплоть до момента перехода в дедифференцидрованно-е состояние.
Такое эпим-орфное развитие особенно типично для всех одиночных
и большинства колониальных ‚фитомонад (за исключением настоя-
щих Volvocidae) и, в сущности, для всех других палинтомических
одноклеточных, пожизненно остающихся свободноплавающими;
впрочем, и отдельные паразитические формы (вроде, например,
Protoodinium) „могут сохранять этот же тип развития. =
В самых простых слу-чаях эпиморкфозаизменения, испытываемые
развивающейся зооспо-рой, столь минимальны, что его можно с пол-
ным правом назвать п р я м ы м -р- а з в ит и е м. Исходная орга-
нивация зооспоры остается практическинеизменной, _И видимые
123
ОНТОГСПЁУГИЧЭСКИЕ ИЗМСНЭНИЯ исчерпываются ПОЧТИ ТОЛЬКО рОСТОМ
особи, связанным с очень небольшими- изменениями ее пропорции и ’
ничтожными по объему морфогенетическими процессами. Лучшие
примеры такого рода дают хламидомо-надь! И другие одиночные
Phytomonadina, ‘B'3pO'CJIbI.€-CT"a1UrIH которых недаро-м назывались-
встарину зооспорами, и, деистчзигелвп-о, мало чем отличаются от
них (см. рис. 27). Во всех ьугих случаях гипертрофический рост осу-
Ащоствляется в отно-сигелилио скромны-х масштабах, поскольку они
имеют место только гири МЯЛОМрОДуКТИВНЫХ формах повторного
деления. -
Повышение степень: гнакср'г1)‹о‹])ии, достигаемой к концу развития,
есть октнопппогт Цнпктор ус.п‹»кнсния онтовегтеза. Это ясно демонстри-
руется тн-кимгп lll])HMC1)ilMIIrI, как раз/питие плодущих особей ‚(гоно-
ниччнь) l3ml<n'ini(Im~., npn котором существенно изменяется- общая
форма и ирогнорция тела, намного увеличивается число пирен-оидов
и, особенно, диктиосо-м, еще усложняется строение хроматофзора
‚и т. д. (см. рис. 80, 1, З): Создающаяся таким путем шаровидная
взрослая стадия имеет уже многие признаки яйца, но остается
_дчикрференцированной до окончания своего гипертрофического роста.
Однако еще большее повышение гипертрофии оказывается уже не-
совместимым с пребыванием трофозоита в дифференцированной,
жгутизковой фазе:_ он удерживается в ней только до определенного
«момента (примерно отвечающего конечной стадии роста гоноцитов
у Епоогйпйоае), а затем переходит -в дедифференцированное состоя-
Hume, в котором уже протекают позднейшие этапы его роста. Так
возникает дополнительная онтогенетическая стадия гиперт‘ р о-
ф o 3 -o и т а, характеризующаяся сохранением вегетативных функ-
ций-в присущем_ейадедиф-ференцированном состоянии, достижением
сею крайней степени пипертрофии и своим ярко выраженнйям харак-
тером оосферы или яйца; ее образцом могут служить гоноциты
‚вольвоцид в заключительный период своего развития. Стадия гипер-
дтрофозоита особенно интересна тем, что она выполняет функции
вегетативно деятельной- особи, нах-одясь в структурно-упрощенном
состоянии; она олицетв-оряет, таким образом, край-не специфиче-
скую форму существования, связанную с избыточной гипертрофией
особи за счет ассимиляционной деятельности своих сочленов по
колонии или,- в других случаях, за счет ее паразитическ-огообраза
жизни. Прохождение этой -стадии в онтогенезе «фактически означает
перерастание гипертрофического эпиморфоза в метаморфоз и‘ слу-
жит, следовательно, соединительным звеном между обоими этими
типами развития. - -- V ~ - -
_ Как уже отмечалось выше, вполне типичные случаи гипертрофического arm-
морфоза наблюдаются, как исключение, и у паразитических одноклеточных. Это
иллюстрируется.примером‘ Рготооайпйитп спайопйчпанцырнойё перидинеи,’ жи-.
вущей в качестве _ эктопаразита на медузе Lizzia blondina и некоторых других.
зооспоры Protoodinium,. образующиеся обычно в числе 8. редко 16, имеют все
особенности ‘строения свободных Ркегйойпйаае, и _-в’ том числе д-вуосный харак-
тер, сотклонением- первичной главной оси от вторичной-на, примерно, 900; Про-
плавав, некоторое‘ время; они. прикрепляются к хоботку, или к нижней‘ поверхно-
124
- ётй зонтика медуз посредством выпускаемого ими" плазматического стебельке
внедряемого в ткани хозяинаиЭтот прикрепительный и сосательный стебелек вы-
растает из жгутикового полюса диноспоры, оказывающегося таким образом полю-
сом ее прикрепления. Далее следует гипертрофический рост молодой особи, при ‚
котором она полностью сохраняет свою организацию ‘свободноживущей перидинеи
со сложно расчлененным панцырем, пояском, жтутами и т. -д. дедифференци-
ровка происходит лишь по окончании питания и роста, непосредственно перед
размножением. Таким образом, развитие Protoodiniurn является по сути дела
прямым, и превращение зооспоры в сидячую вегетативную особь нисколько не
нарушает ее исходной организации; испытываемые ею морфологические измене-
ния исчерпываются образованием сосательного корешка и принятием на себя пер-
вичной осью нового тут для нее значения оси прикрепления и трофической поля-
ризации. . . v _ ‘
K гипертрофическому эпиморфозу должно быть также отнесено
постэмбриональное развитие всех ‘палинтомических простейших, .
‚размножающихся амёбоспорамаи и споро-зоитами, если только оно
не идет по пути полимеризации и соответствующего выхода трофо-
зоита за рамки одно-клеточного строения. Примерами онтогенеза
этого типа может служить развитие макрогамет кокцидий из меро-
зоитов (см. рис. 73, 76), развитие Eugregarina И множества дру-
гих простейших, размножающихся посредством вторичнойсинтомии.
Из всех относящихся сюда фактов может быть сделан общий
вывод, что гипертрофический эпиморфоз имеет место при люб-ом
типе исходного строения особи (т. е. при люб-ом типе строения
зооспоры), но ведет кдостижению «высших форм гипертрофии лишь
при строго определенных условиях. Такими условиями является
либо пож-изненное сохранение особью недиффер-енцированной (амё-
боидн-ой) или несложно дифференцированной организации, либо ее
специальный переход -из жгутикового в дедифференцированное со-
стояние гипертрофоз-оита (Уо-Ълосйоае). В последнем случае гипер-
трофический эпиморфо-з приобретает уже характер метаморфоза.
. 4. Гипертрофический метаморфоз
K этому типу онтогенеза мы относим весьма разн-о-образные
и разнородные его виды, общим признаком которых является только
наличие крупных перестроек организации в ходе ‚индивидуального
развития. Вообще говоря, такие онтогенетические изменения могут
создаваться тремя разными п-утями:
а) при превращении зооспоры в стадию вегетативно деятельного
трофозоита, пкогда оно -связан-о с изменением" образа жизни особи,
например, с ее переходом к сидячему или к паразитическому образу
жизни; метаморфоз‘ такого рода, предшествующий началу самостоя-
тельного питания молодой особи, мы будем называть н а ч _а л ь н ы M
M€‘T'aMO«p(i)013‘0M; “ . ' _ V
6) путем перехода дифференцированного трофозоита в уже оха-
рактеризованную выше стадию дедифференцированного гипертрофо-
з-оита; такой вариант метаморфоза можно назвать конечным
метамо'рфозом; - о A
в) путем сочетания -в индивидуальном развитии начального и
' 125
еверхгипертрофического метаморфоза, что ведет к приобретению
1»1MvxapaK'repa комбинированного .метаморфоза.
Самым распространенным из этих трех основных видов гипер-
трофического метаморфоза бесспорно является начальный метамор-
фоз, протекающий в разных случаях в самых различных конкретных
Рис‘. 33. Развитие Atelcdiniumr -
1—З—синтомическое размножение; 4——›кгутиковая зооспора; 5——
ее превращение в амёбоидную форму; б-молодои Вегетативный
плазмодий (по Шаттону)
формах. Чтобы дать ясное представление о его. разнообразии, мы
отметим следующие четыре прим-ера;
1) Превращение жгутиковой зооспо-ры .в неподвижную стадию
пальмеллы. При таком характере начального метаморфоза, очень
нередком -у автотрофных жгутиковых, он сводится к переходу жгути-
ковой зооспоры в недифференцированное неподвижное состояние,
в котором и осуществляется вегетативная деятельность особи.
2) Превращение так-ой же зооспоры в амёбоидную в-егетативную
стадию. Это——оч‹ень распространенное явление, примеро-мркоторого
может послужить начальный метаморфоз Atelodinium (рис. 33).
126
с
3) Превращение жгутиковой зооопо-ры В сидячую жгутиковую
вегетативную стадию, например, подгоняющего типа с плазматиче-
ским воротничком и другими резко выраженными отличиями от пред-
шествующей -стадии развития. ’ › A '
4) Превращение амёбоспоры в дифференцированную вегетатив-
ную -стадию. ‘
Наиболее сложные формы метаморфоза наблюдаются у парази-
тических перидиней и хорошо иллюстрируются примером онтогенеза
Oodinium. Жгутиковые зооспоры (диноспоры) (рис. 34, 1) обладают
вполне типичным для Peridinidae строение-м: одинаков-о развитыми-
верхним и нижним полущариями, оформленной кольцевой бороздой,
характерной двужгутиковой и _
кинетидой, сетчатым хромато-- - ‘
фором и ядром постоянно де-
лящегося типа, лишенным, ка-
риосомы и ядерно-й оболочки.
Зооопора прикрепляется к
хозяину "посредством выпус-
каемого ею стебелька, рас-
пластывчающегося -в широкую
присоску, и тут же линяет
(рис. 34, 2). Перейдя таким
образом в стадию безжгутико-
вою сидячего трофозоита, мо-
лодая особь приступает к пи- pm
танию и интенсивному росту,
постепенно перестраивая‘ при
этом‘ свою о-рганизацию. Ее
вое увеличивающееся в объеме
тело приобретает характерную
яйцевидную форму, что происходит путем н-епропорционального раз-
растания верхнего полушария; она тут еще более обгоняет в своем
развитии нижнее полушарие, чем у известного Poroceratium gra-
Идиш, поскольку ее эрост идет «не в одной только плоскости, как
там, а в трехизмерениях пространства. В течение всего периода
роста трофозоит сохраняет ‘свой сложный метаплазматический
34. Постэмбриональное развитие
Oodinium poucheti:
7—свободноплаваюцгая зооспора; 2—линька
зооспоры после ее прикрепления к хозяину;
З——8—последователъные стадии гипертрофиче-
ского метаморфоза (по Овассу)
панцырь; характерно, что‘ в области тела, окружающей основание _
стебелька, имеется коленчатая бороздка-очевидно, представляю-
щая с-обой общий рудимент кольцевой и продольной жгутиковых
бо-розд — от нижнего конца которой и отходит сам стебелек. Стало
быть, гипертрофированная сидячая (паразитическая) стадия 0061-
nium отнюдь не является дедифференцированн-о-й, как это кажется
с первого взгляда: она остается, напротив, полностью дифференци-
рованной, утрачивая только с-в-ои жгуты. — `
Параллельно с ростом трофозоита и изменением его внешнего
строения происходят и крупные изменения его внутренней органи-
зации. Они заключаются в распадении и последующем исчезновении
сетчатого хроматофора, в образовании системы фибрилл, связанных
127
сфприкрепительньтм аппаратом, и в полной перестройке ядра, сопут-
ствующей его гипертртофическому росту. У только что прикрепив-
шихся Oodinium ядро имеет еще типичное для Dino-flagellata
строение динокариона, т. е. элдра «хромосомного» типа. Затем «состав-
ляющие его хромосомы носколько- расходятся, и между образуемыми
ими скоплениями, в центре ядра, появляется ‚неправильно очерчен-
ная кариосома. Эта носллодпнтя носттснет-н-ю разрастается и образует
клубок, состоящий из нстнннтихсэт и частично анастомозирующих
петель; хромосомы (гггсснэлнутом нм =к периферии ядра (рис. 34, 3-4),
одевающегося тенсрп, янегнснтннт (шболоч-к-ой. К концу развития хр-о-
мосомы 11‹›.пн‹›ст‘ь1‹› шч|и3г1|‹›1*‚ а картсома распадается на, много-
численные изнит-ьке отде.нь11‹›ст‘т1, образующие .в своей совокупности
рыхлую норнфсрнчтетскупо сеть (рис. 34, 5-8), среди которой раз-
бросано несколько внутриядерных телец.
Таким образом, онтогенетиче-ское развитие ядра заключается,
по сути дела, в его переходе из нормального для Dino-flagellata
состояния перманентной профазы в состояние, отвечающее телоафазе,
за затем и в интеркинетическую фазу. ' ’
Индивидуальное развитие Oodinium завершается, как обычно,
дедифференцировткой, за которой непосредственно следует гпали-нто-
мическое размножение.
Глава VI
ПОЛОВОЙ ПРОЦЕСС И ЖИЗНЕННЫЙ ЦИКЛ ОДНОКЛЕТОЧНЫХ
ПРОСТЕЙШИХ
х
1. Вводные замечания
Половой процесс, или сипгамия, наблюдается лишь в немногих
группах ‚постоянно отдноклето-чных Pro-tozoa: В качестве редкого
явления у солнечников, эвглен, протомонад, хризомо-над, и может
быть, некоторых раковинных корненожек, несравненно чаще-
у фитомонад и как обязательный элемент жизненного цикла у не-
КОТОрЫХ ГруПП ~Cl'IOpOBI/IKOB. BO ВСЕХ ЭТИХ группах ПОЛОВОЕ размно-
жение сочетается с бесполым, факультативно или регулярно пре-
рывая однообразный ход этого последнего. Это имеет своим-
результатом установление тех или иных форм чередования беспо-
лых генераций с половыми, т. е. возникновение метагенетиче-ского
жизненного цикла. '
При, всем разнообразии полов-ого процесса у одноклеточных он
обычно совершается по той же -общей схеме, как у высших животных:
его основу составляет слияние воедино двух сексуально дифферен-
цированных гамет, дающее начало двойственной по своей природе
зиг-оте‚ и -он всегда сопровождается явлени-ями редукции (или
мейоза), „нередко охватывающей здесь и некоторые из дифференци-
ровок клетки, как, например, жгутиковый аппарат. Однако, .в т-о
время, как у Metazoa эта общая -схема реализуется только в одном,
очень постоянном у них варианте — в форме оогамии, половой-про-
128
цеес одноклеточных поражает своим разнообразием. Мы сталки-
ваемся тут уже не с одной, а с самыми различными формами
копуляции, начиная с аутогамии и ‘копуляции трофогамет и кончая
самой типичной оогамией.
2. Саркодовые
-Вряд ли можно считать случайным, что оформленный половой
процесс до сих пор достоверно не установлен ни у одной одноядерной
амёбы: среди Rhizopoda он составляет привилегию постоянно дву-
ядерных илимн-огоядерных форм и связан, таким образом, т-олькёо
c плазмодиальным типом организации. Среди одноклеточных сол-
нечников хорошо известен прнимер сингамии у Actinophrys so1.,
niocr-051-Hno используемый -в учебной литературе. У этого исолнечника
(рис. 35) половой процесс происходит исключительно в дедиффе-
ренцированном состоянии, под защитой специально выделяемой
двуслойной цисты, тогда как бес-полое размножение осуществляется
в'активном.„ состоянии, по протозойному типу ‚простого деления.
Гаметами являются оба продукта последнего, остающиеся амёбоид-
ными и внешне совершенно тождественными друг другу во всем,
кроме своего поведения при копуляции, когда один из них проявляет
свою мужскую природу однёосторонней активностью в этом акте.
Сразу же после своего образования, они оба испытывают подва
редукционных деления, воспроизводящих с поразительной точ-
н-остью все особенности созревания яйца у Metazoa.
.. ,При копуляции мужская гам-ета слипается с женскойсвоими
псевдоп-одиями (рис. 35, 5) и затем полностью сливается с нею; про-
нуклеусы образовавшейся теперь зиготы некоторое время остаются
обособленными, но затем сли.ваются ‹в синкарион, а зигота выделяет
новуюоболочку и превращается в цистозиготу. После известного
периода покоя она переходит -к свободному существованию в виде
амёбоидного вначале трофозогита, подвергающегося затем диффе-
ренцировке, и размножается дальше уже бесполым способом (деле-
нием в активном состоянии).
‘Итак, половой процесс Actinophrys представляет собой педога-
мию, поскольку гаметами тут оказываются продукты деления одной
итой: же родительской особи, и осуществляется в формах изогамии
(т. -е. мор-фологически тождественными гаметами), имеющей, однако,
и определенные признаки анизогамии (сказывающиеся в неодинако-
вом поведении -гамет вовремя копуляции). ‘Особого внимания заслу-
живают четыре его особенности: 1) приуроченность плолового про-
цесса к дедифференцированной, амёбоидн-ой фазе существования
игнаблюдающа-яся тут активность мужско-й гаметы; 2) paBHbIM обра-
зом амёбоидный и активный характер зиготы в момент ее выхода из
цисты и ее последующее превращение в нормального трофозоита;
3)] монотомический характер «прогамного деления, имеющего своим
следствием образование двух морфологи-ч-ески равноценных дочер-
них особей, преобразующихся в гаметы путем редукции; 4) порази-
9 Эмбриология беспозвоночных 129
. Тельное совпадение хода последней с самыми типичными формами
редукции при оогенезе Метаиоа. ‚ '
местно отметить, что половой процесс у многоядерного Acti-
nosphaerium eichhorni отличается лишь в деталя-х от таково-го
A-ctinophrys (в частности, тем, что «прогамн-ое деление является тут
синтомич-еским и дает начало не одной, а нескольким парам изо-
гамет)‚ что свидетельствует о ‚выраженности у солнечников одного,
и притом высоко оформленного, типа полового процесса. '
3. 3BI‘JIeHbl
Наиболее известный из примеров полового процесса .в этой
группе описан Добеллом (1911) у Copromonas pusilla. B этом
случае гаметы имеют строение и размеры нормальных вегетативных
ос-обей, копулируют в дифференцированном состоянии и вполне
::~«_
‘r-euvu...-nu.»a~=~>“"‘”"' м
'“ “дед.
(по БелЯрУ)
Рис. 36. Половой процесс у эвглены Copromonas pusilla: '
1—начало копуляции, редукция жгута у одного из партнеров; 2-—деления созревания; з-
подвижная зигота с жгутом и еще не вполне слившимися ядрами; 4, 5—образование цисто-
зиготы (по Добеллю)
° солнечника fActinophrys S01-2 - .
Рис. 35. Половои процеёвёниж 4 Второе Зеленую созревания; 5-—копуляция; б-оформление эиготьш
._ 2 дьмёйоз и первое деление cos 1 -‘ _
1-—aHa(Da3a прогамного деления, ‚
равноценны ‘друг другу -B морфологическом отношении и, видимо,
в смысле своего поведения (рис. 36). В начальной фазе популяции
обе гаметы срастаются своими передними концами и лишь затем
спаиваютс-я по всей длине; при этом жгут одно-й из них (Copromonas
представляет собой одножгутиковую форму) отбрасывается (друг-ой
остается при зиготе), а глоточный аппарат, «повидимому, ‘исчезает.
Вслед затем ядра обеих гамет испытывают по два деления созрева-
ния, ‚в результате которых образуется по два направительных тельца,
разрушающихся затем в плазме, жгутиковые затем копулируют
‘обычным порядком, а зигота ли-бо сохраняет свое строение трофе-
(хотя и представляет собой двойниковую подвижную зиготу——
оокинету), или же инцистирует-ся, превращаясь в цистозиготу.
9* 131
130
зоита И ТГРОДОЛЖЗЕТ активное существование В КЭЧЕЁСТВЕ TaK0-BOI‘O'
K сожалению, остается _'невь1ясненнь1м‚ в ‘каких генетических
отношениях находятся тут копулирующие осо-би и каковы детали
.мейотического процесса.- Этот случай, однако, интересен как пример
изогамной копуляции гамет„ являющихся продуктами- простого,
а не множественного. деления и бесспорно гаметной (заключитель-
ной) редукции.
4. Фитомонады
Исключительное разнообразие форм копуляции настолько харак-
тер-но для Phyt-omo-nadina, что оно может быть введено в диагноз
этого отряда. В его пределах наблгодаются и все мыслимые этапы
формирования полового процесса, начиная с самых зачаточных
и кончая -столь же совершенными, как у высших животных, так что
их сравнительное рассмотрение широко практикуется в литературе
и является .весьма поучительным. г
Бесконечное, разнообразие наблюдающихся тут вариантов поло-
вой дифференциации гамет, способов, копуляции и других слагаемых
полового процесса развертывается ›в этой группе на существенно
иной основе, чем у со-лнечников и эвглен, а именно на базе палинто-
мии, являющейся господствующим типом ка.к бесполого, та-к и по-ло-
вого размножения Phytomonadina. Поэтому роль гамет играют тут
уже зооспоры, образующиеся повторным делением.
Наиболее изменчивой стороной полового процесса фитомонад
является степень сходства или, наоборот, различия между икопули-
рующими друг с другом гаметами (рис. 37) или, другими словами,
степень изо- или: аниз-огамии, встречающейся тут, с одной стороны,
в самых крайних своих формах, а с другой—связанной много-
численными и самыми постепенными переходами.
При наиболее простых и примитивных фо-рмах изогамии гаметы
ничем не отличаются от зооспо-р, образуемых при бесполом размно-
жении, и формируются в том же, обычно очень небольшом, числе-
вколичестве 4 или 8, редко 16. Значительно чаще число повторных
делений в прогамной палинтомической с-ерии увеличивается по
сравнению с нормой (каковой можно считать их число при бесполом
размножении), что имеет своим результатом и соответствующее
увеличение числа половых зооспор против бесполых; с `
В результате такого обособления прогамного цикла делений
и увеличения его продуктивности возникают и размерные различия
между бесполыми и половыми зооспорами; их проще всего характед
ризовать порядковым классом зооспор 1, что позволило бы сказать,
‘что половые зооспоры часто превышают наодин ‘или два класса
бесполые з-ооспорь1 того же вида.
д Порядковые классызооспор отвечают may повторных делений, приво-
‘дяших к их образованию; Зооспора-ми первого порядкового класса являются
те, которые образуются в результате двух последовательных делений материя:
ской особи, т. е. в числе четырех; зооспорами второго класса-щпродуктьт трех
последовательных делений, т. е. образующиеся в числе восьми; третьего клас-
са-продукты четырех их делений из т. д.
132
73 - 74
Рис. 37. Половой процесс хламидомонад:
Chlamydomonas xjeinhardtiz 7_—B3pOCJ1b!e ‚особи; 2—изогаметы- .З—их о -
ция;4-—цистозигота; 5—9——СШ. ehrenbergi: 5—взрослые особи; бдё-ЕХЁЗ.
ЛЯЦНЯ; 8—4-жГУтиковая зигота; 9—ее инцистирование; 10—74—С|11. brauni.
70-—MaKporaMe'ra;_ 11—микрогаметы; 12-74 различные моменты копудяцид;
' (по Горожанкину)
~ I .133
В простейших случа-ях сингамии гаметы не проявляют видимых
половых различий: мужские‘ не отличаются от женских, так. что здесь
имеется только физиологическая ‘биополярность, но нет еще ни"-
каких признаков морфологической дифференциации полов: гаметы
являют-ся изо-гаметами. В их «поведении также нет -ничего
специфического, и их половая дифференцированность заключается
только в том, что гаметы -разного знака испытывают взаимное
притяжение. _
Сходя-сь попарно, изогаметы слипаются сначала своими перед-
ними концами (носиками), а затем и полностью сливаются друг
с другом, а их жгутики и оболочки обычно сбрасываются.
Продукт копуляции ——-зигота—принимает шарообразную фор-
My, одевается толстой «многослойной оболочкой и переходит -в по-коя-
щееся состояние. Иначе говоря, она инцистируется, так что такую
зиготу можно назвать ц-и с то з и г о т о й.
В некоторых других случаях (например, у Stephanosphaera)‘
КОПУЛЯЦИЯ изогамет не сопровождается их дедифференцировкой.
Они сохраняют свои жгуты и, выползая из своих оболочек, сли-
ваются в зиготы с двойным набором двигательных аппаратов и
других дифференцировок. Вместо тог-о чтобы немедленно инцисти-
роваться‚ зигота начинает вести свободный образ жизни, ассими-
лировать и расти. Такая подвижная зигота называется пл анод
зиготой и особенно интересна тем, что проявляет в особенно
наглядной форме свою двойственную природу, внешне замаскиро-
ванную у цистозиготы.
После некоторого ‚периода свободной жизни, связанного c УВЕЛИ-
чением живой массы тела, планозигота инцистируется, становясь
«нормальной» цистозиготой. `
У некоторых друпих форм (Pandorina, рис. 38) дело происходит
иначе. Копуляция осуществляется таким же образом, как в преды-
дущем случае, и обе двужгутиковые изогаметы -сливаются во вре-
менно четырехжгутиковую планозигоггу; последняязатем инцисти-
руется. и после некоторого периода покоя дифференцируется,
выходит из цписты и начинает вести свободный образ жизни в виде
вегетативно деятельной, но теперь уже двужгутиковой, плано-
зитоты второго п-орядка. После известного периода питания и роста
она вновь инцистируется .и тогда уже приступает к палинтомиче-
скому делению.
Таким образом, зигота здесь проходит четыре стадии или фазы:
1) четырехжгутиковую начальную; 2) первую ин-Цистированную;
З) стадию двужгутиковой планозиготы и 4) вторую инцистирован-
ную стадию.
Случаи подобного рода весьма интересны тем, что здесь зигота
сохраняет свой характер ‘нормальной вегетативной особи (трофо--
зоита) и обратимость организации‚_а также 1-1 тем, что гона сама,
своей собственной вегетативной деятельностью, как бы ‘компенсирует
выпадение такой деятельности у зооспор, ставших гаметами.
В этих рассмотренных -нами случаях половые элементы+га-
134
MET“- и- продукт их копуляции—зигота——ничем внешне не
отличаются от соответствУЮЩИХ ‘бесполых ФОРМ “ЗООСПОР И ТРО‘
фОЗОпИТОВ. у , ’ U п до
Первая ступень в возникновении МОрФОЛОГИЧёСКИХ ОТЛИЧИИ 0
ВЫХ зооспор (го н о с п op) от беополых выражается в TO1\6/I, что они
остаются, как говорят, голыми, т. е. не вырабатывают со ственных
о-болочек. Как известно, выработка оболочек является одним из
заключительных этапов дифференцировки )С‘П'0Р0ЦИТ0’В ‘H0CJ1'€l1H'€F0
д д 7
Рис. 38. Жизненный цикл Pandorina morum:
. _. эоосп-ор
_ . ° 2-—ее палинтомическое азмножение 3 выход половых _
ЁгаЁЁЁЁЁЁаЁолЁЁЁЁЬЁЁЪго соединения; 4"КОПУЛЯЁИЯ= 5: 5—4‘7"<"YT"“°Ba" "“a“°3"r°Ta’ 7’ 8'"
ин исти ование- 9-—'—11P0PaCTaH"° 3"|'°’"=13 70-ee Превращение в 2—жгутиковую планозиготу;
e7e1—-Is;-‘e1-araliaxqoe деление 4-клеточная стадия; 12-стадия ГОННВВОЙ Таблички; 73*-M0J'I0J2(ail
’ бесполаяколония (по Прингсгейму)
поколения в зооспоры. Здесь же этот Этап Въшадает, что зпридаеТ
гаметам известные черты недоразвития по сравнению с вегетатив:
ными зооспорами. Что же касается морфологических различии
между гаметами разного пола (или разного знака)‚ ТО ПЕРВЫМ Эта‘
пом их возникновения можно считать часто наблюдающиеся случаи,
когда к-опуляция происходит между гаметамии ОДНОГО И T0110 же по-
рядкового клаоса, но происходящими от материнских особеи различ-
ных размеров и потому также разнящимися по величине. -
Отсюда уже один шаг до образования неравных гамет— анизо-
гамет и, соответственно, анизогамной копуляции (см. рис. 37). О-снов-
ные признакли анизоганмного полового размножения Таковы‘
135
1) Копулирующие Между собой гаметы заметно или резко отлил `
чаются по величине, причем меньшие из них могут быть уже о-пре-
деленно названы мужскими и отличаются от более крупных женских
также и своим более avK'l'I’IBHHM поведением. _
2) Различие в размерах нькзвано принадлежностью гамет к раз-
ным порядковым классам —— большему у мужских. р .
3) Со-ответстветпно этому возникает и дифференциация прогам-
ных делений на мужской и женский ряды. . .
В простейших случаях они различаются несильно, на одно,
скажем, деление. Например, в женском ряду деление может повто-
ряться три раза, в мужском -— четыре и т. д.
Однако дальнейшая эволюция прогамных процессов идет в обоих
рядах в противоположных направлениях (рис. 39):
Рис. 39. Половой процесс фитомонады Phyllomonas striata:
1—образование микрогамет; 2—образование двух макротамет из очень крупной материнской
клетки; З-микрогамета; 4-—анизогамная Копуляция; 5-—-молодая планозиготщ] б—вполне
взрослая планозигота (по Коршикову)
а) В M у же к о м р я ду -—в сторону‘ все большего увеличения
числа повторных делений, что ведет к мельчатнию его продуктов,
постепенно приобретающих характер сперматозоидов. Особенно
важно подчеркнуть, что это увеличение числа делений не связано
с повышением степени гипертрофии материнской особи. Главным,
если не единственным, способом увеличения ‘продуктивности
ЯВЛЯЕТСЯ ТУТ ИМЗННО м-ельтчание ‘ПРОДУКТОВ палинтомии.
_ б) В ж е-н с-к о M ‘р я ду наблюдается обратная картина. Эво-
З-ЛЮЦИЯ прогамной ПЗЛИНТОМИИ ИДЕТ В 'C’I‘0pO'Hy~yM€HI:II_I€HI/ISI числа
ДЕЛЕНИЙ, а затем И ПОЛНОГО ИХ выпадения. В ПОСЛЕДНЕМ СЛУЧЭЕ мате-
рИНСКаЯ женская клетка ЦЕЛИКОМ превращается ‘В ЕДИНСТВЕННУЮ
‚женскую гамету‚ часто переходящую перед популяцией в деди-ффе-_
ренцированное состояние и приобретающую таким образом характер
. оосферы.
Таким путем возникает высшая фо-рма полового процесса-
„оогамия, характерная для всех ‘Metazoa. Ее особенностями яв-
~ ляются именно крайняя степень анизогамии, копуляция измельчен-
ной мужской гаметы с укрупненной яйцеобразной женской.
Рис. 40 показывает на нескольких примерах высшие этапы эво-
.люции анизогамии у одиночных q)14ToMoHa_L;—C.h1onrogonium ooga-
mum И Сыашубошопаз соссйега,
' 136
Таким же образом происходит дело и у колониальных ф-орм.
Однако у некоторых из них (V-olvocidae в узком смысле) оогамия
Рис. 40. Оогамия у одиночных фитомонад:
7—7—С111огоаоп1цп1 oogamum: 1—-взрослая вегетативная особь;
2—образование микрогамет; З- микрогаметы (сперматозоиды);
4, 5——образование макрогаметы; б-оплодотворение; 7—инцистиро—
ванная aurora; 8—72—Chlamydomonas coccifera; 8-—микрогаметы;
9.—MaKporaMe'ra; 10—копуляция; 17, 72—образование зиготы (по
Горожанкину) . . . ,
может приобретать еще более крайние формы ‘вследстви-етого, что
женские гам-еты обеспечиваются питательными материалами со
стороны других, неплодущих особей колонии, что и- в-едет к их
чрезвычайно высокой гипертрофии Получается настоящее яйцо,
137
с большими резервами питательных материалов, относительно очень
крупное и т. д. - '
Итак, в пределах отряда фитомо-надин мы видим в-сю цепь пере-
ходов от зачаточной «биполярной дифференциации гамет при изога-
мии до крайнего ее выражения при оогамии. При это-м половой
процесс возникает на основе палиптомии и ведет к ее изменению
в против-оположных направлениях в мужском и женском прогамных
_рядах.
5. Общее понятие о жизненном цикле
У тех простейших, у которых особи всех следующих друг за
другом одноклеточп-хьпх поколений относительно. равноценны и прак-
тически подобны друг другу в биологическом и морфологическом
отношениях, испытывают сходное индивидуальное развитие и
представляют собой «нормальных» трофозоитов,—нет соответ-
CTBEHHO И оформленного ЖИЗНЕННОГО ЦИКЛЗ (В ТОМ смь1сле,вкаком_
он понимается у простейших)‘. Последний возникает только-тогда,
когда однообразное воспроизведение себе подобных, характерное
для таких а ц и к л и ч е с к и х Protozoa, уступает свое место той или
ин-ой форме чередования такого вос-п роизведения
с воспроизведением себе неподобных, т. е. с обра-
зованием поколений, в той или иной мере неравноценных остальным,
Периодическое, закономерное появление такого рода клеточных
‘поколений, такили иначе отличающихся от других в биологическом
и структурном отношениях, и приводит к разбивке непрерывной
ранее цепи сменяющих друг ‘друга гене-раций на ряд повторяющихся
циклов; и поскольку каждый такой жизненный цикл начинается
и завершается оформле›нием‹одноклеточного поколения (или поколе-
ний), отличного от остальных, то он и п р и об р е т а е т с п е ц и ф и-
ч ес к и е св ой с т в а р а з в и т и я многоклеточных организмов,
т. е. процесса «воспроизведения с-ебе подобных не непосредственно,
а через длинную цепь превращений себе неподобных, п-ока не полу-
чится подобное начальному» (Лысенко, 1944; 398). `
С этой точки зрения ациклическое бесполое размножение одно-
клеточных Protozoa, заключающееся в непосредственном воспроиз-
ведении себе подобных каждойодноклеточной особью (путем моно-
томии)` во всем -ряду поколений, о то е ч а е т п р о ц ес с а M
роста многоклеточных
как известно, размножение и ‚развитие составляющих их клеток.
И только с появлением циклической дифференциации поколений
создается у простейших и качественно новая форма развития, со-
ответствующая индивидуальному развитию целых многоклеточных
организмов.
Соответствие жизненного цикла одноклеточных индивидуальному
развитию многоклеточных еще подчеркивается тем, что там и здесь
ведущим звеном цикла является п о л о в о й п р о ц е с с, которым
начинается и завершается этот циклический процесс развития. Та-
.ким образом, основной признак «развития», противопоставляющий
-138
о p г а н и з м о в, включающим, _
1
е ‹ _
агьдзз°‚:х::;‚„;а°322“:;2:®2“25? и
чится подобное начальномр EMo>:ecTe бе неподо‘ Ных’ пока не Homi-
смысле что Такое ИК У, ыть конкретизирован в том
‚ Ц лическое воспроизведение себе подобных осу-
ЩЕСТВЛЯЕТСЁЬ как ОбЩЕЕ ПРЗВИЛО, Путем полового размножения.
ЦИСЁЁЧЁЁЁЁГОЁЁЖЭТЁОЁЁЁЭЁГЗЁЕЁЁЁЁЁИХпЁедстгёвляюгг только жизненные
амёб)’ у которых нет ‘ПОЛОВОГО п а со 1па (например, кишечных
д рОЦе-сса и его биологическую роль
(обновления, усиления жизненности организма) играют повиди.
М°МУ› ОПРЕДЕЛЕННЫЕ бЕСЙОЛЫе Стадии Цикла (стр. 187). ’
6- ЖИЗНЭННЪПЗ Циклы одноклеточных
В основ r
дожить раЗЁИЁЁЁЁЕЁИФИКЗЦИИ ЖИЗНЕННЫХ ‘Циклов целесообразно по-
составляющих ИХ пЁТЕПеНИ PIE дифференциации и самом характере
следую Не ОСН * РИОДОВ. соответствии с этим мы различаем
I Щ рвные группы Циклов одноклеточных:
) K перво“ ИЗ НЕМЕЧЗТОЩИХШЗЯ Здесь групп относятся наименее
диф+ференцирова=нные И ОФОРМЛЕННЫе Циклы с неопределенным
периодом, которые можно назвать .н е р а .¢ q л е н ед н Ы М и Om
Xa’Pa'KT€P1’13Y10f1‘C5I прежде всего тем, что половой процесс имеет
ско . о о т
РЕЕ ФЗКУЛЪТЗТИВНЫИ, Чем ритмический характер, легко вызы-
ВЕЕТСЯ ОПРЕДеЛеННЫМИ внешними воздействиями и так же легко
‘поддаЕтся УЕТРЕРНЕНИЮ: И, что главное‚—что он не в-едет тут к отчет.
ливой и е -ен U
П 15’ (M) р Циации прогамных и метагамных поколении Цикла.
олов-ои процесс разыгрывается здесь на основе одного только поко
ления и практически не затрагивает остальных, что придает ему
известныи оттенок «внезапности» и позволяет ему смещаться в п
тич.ески неограниченных пределах. Роль тчамег B,,mo,,mHeTc;1p«.:KT
обычными вегетативными особями, приоб-ретающими половую диф
ФЕРЕНЦИРОВКУ, или же их прямыми и невидоизмененными потомками
так что з ес - ~ -‚ ’
Я д ь не приходится говорить 0.5 0¢,o,pM‘meHH,O,M _np,0FaM_HoM
p де и о наличии гамето-цитов. Равным образо-м нет и оформленного
метагамного ряда поколений: сама зигота непосредственно п ев а
щается в ‚вегетативную особь, внешне ничем не отличающухзсярот
трофоз-оитов. Такие неустойчивые циклы характерны для многих
фитомонад и являются единственным их типом, встречающимся
у эвглен идсолнтечников. Экспериментальные итосдеддваддия Беляра
над этими последними (Actinophrys и Act'in‘osph’aeriu'm) показали
что в оптим . ‚ . ’
б ' альных ‚УСЛОВИЯХ СРЕДЫ ОНИ размножаются только
есполым путем, тогда как половое размножение наступает при
перена-селении =и недостатке питания; у Actin-ophrys удается заста-
вить .пе в ю же о › - _ o
гаметы й Хове шенЁЁЁьраЗВИВШуЮСЯ из HHCTOBHPOTM’ образовывать
Лые генераци Однако гёеснять, таким образом, нормальные весло-
А 1 ОЖНО ‘С ТёКИжМ Же успехом Заставить этого
солнечника размножаться и исключительно беополым путем
2) Вторую группу сингамных жизненных Циклов образуют те
В КОТОРЫХ ‘прога-мная часть цикла более или менее явственно вы,
' 139
дедяется в ‚особый период г а М е то г е H e 3 a, протекающий сх-од- '
ным образом в МУЖСКОМ и женском рядах, т. е. имеющий своим ре-
зультатом образование изогамет. „Эти последние не выполняют
“нормальных вегетативных функции, копулируя сразу же ПОСЛЕ
своего оформления. Выпадение вегетативной функции гамет ком-
пенсируется в таких циклах активной ассимиляционной деятель-
ностью зиготы, имеюньсйя здесь, как правило, характер подвижной
пдаоозиготь1; она испытывает при этом гипертрофический рост, а
затем уже инпи-стирустся, после чего только и приступает кпали-н-
I TOM ИЧВСКОМУ pz13M11m1(e111110.
3) Циклы третьей группы отличаются от предЫдУЩИХ Тем, ЧТО
гамето-генсз идет здесь по-разн-ому в мужском и женском рядах:
в ‘первом он развивается по линии все большего повышения продук-
тивности ‚при соответствующем мельчании гам-ее, во втором—
в сторону полного выпадения прогамных делении И превраЩеНИЯ
самой материнской особи в единственную женскую макрогамету.
За копуляцией следует инЦи-стирование зигоггы и ее размножение,
сопряженное с редукцией зиготного типа.
„ ‘ Этими тремя типами и исчерпывается, в сущности, Известное
нам разнообразие сингамных циклов у свобо-дноживущих, ПОСТОЯННО
одноклеточных простейших. Легко заметить, что каждый из них отве-
чает определенной ступени эволюции ПОЛОВОГО процесса, ведущая
роль которого в формировании И дифференциации Цикла ТУТ РЕЗКО
бросается ‚В ‚Глаза действитедьъло, при самых примитивных формах
полового процесса __ копуляции разнополых вегетативных особей ——'
она не отражается хоть сколько-нибудь заметным образом на
дуотадьных поколениях цикла, остающихся относительно равноцен-
„ными. Сл-едующая ступень эволюции полового процесса, ‚олицетво-
ряемая изо-гамной копуляцией гоне-спор (т. е. половых зооспор,
"отличающихся от бесполых), приводит уже ‘к обособлению прогам-
Hope ‚периода цикла от вегетативного (-в качестве гаметогенеза)
‘и к определенным изменениям в его м-етагамн-ом периоде. Наконец,
при оогамии цикл оказывается "i€’PKO JIH*(13‘139P€HHHP0'1{aH‘Hb1M Над?
mpg правильно чередующихся периода: протамныи, сингамныи,
Ём-етагамный и Вегетативный, из которых 11e~p131:11»1U.tLH(13(13eP'eH1{1’IPY€'1‘C7i»
B‘ свою очередь, на раздельные ряды-—_мужскои и женский. Таким
образом, по мере эволюцииполового процесса в нег_о вовлекается
все большая часть Цикла, и цикл становится все более правильным
и четко расчлененным. у ‚ _. .1 .. о „х
7. Циклическая дифференциация И ИНТОГРЗЦИЯ“
оформление жизненного цикла у одноклеточныхРГ0ЁО2Оа‚‚ ВЫ-
званное возникновением полового процесса, приВОдИТд Как УЖЕ
отмечалось выше, к качественному преобразованию процессо, отвез
чаъощего росту многоклеточных; в процесс, соответствуЮЩИИ ИНДИ-
„видуальному развитию последних. Это ‘влечет за собой рЯд ВЕОЪМЗ
‘существенных последствий, первым из которых является ВОЗНИК“?
140
вение заметных морфофизиологических различий между особями
разных‘ поколений цикла, т. е. возникновение ц и кл и ч ес к о й
р а-з н о -к л-е т о ч н о с т и. Как известно, разноклеточность счи-
тает-ся до сих пор привилегией одних только многоклеточных и вь1-
ражением и следствием ‘разделения труда между клетками. На этом
представлении базируются как общепринятые взгляды ‚на происхо--
ждение клеточной дифференциации Metazoa, так и самое противо-
п-оставление многоклеточных организмов ——. в качеств-е разноклеточ-
ных — .про-стейшим. _ ‘ .
Однако из самого факта наличия циклической разноклето-чности
у одн-оклеточнь1х простейших явствует с полной очевидн-остью, что
общее противопоставление простейших и многоклеточных п-о этому
признаку неверно. Оно справедливо в гораздо более ограниченном
и конкретном смысле, а именно при условии четкого различения
двух видов разноклеточности — п о с л е д о в а т е л ь н о и, или ци-
клической, при которой она осуществляется во времени, в порядке
чередования клеточных генераций разного характера, и о д н о в р е-
м е н н о й, параллельной, или функциональной, разноклеточности,
заключающейся в одновременной разносторонней специализации
клеток в пределах одной их ген-ерации. Одновременная разноклеточ-
ность, действительно, почти совсем чужда простейшим, у которых,
однако, она может все же наблюдаться (например, у солнечников,
при дифференцировке одной дочерней особи в солнечнике, а дру-
той-в жгутиковую форму), _и весьма характерна, напротив, для мно-
‘гоклеточных. Тем не менее и у этих последних функциональная pas‘-
HOKJIETOIII-I0CTb не является единственной ее формой: наряду с ней
имеется и циклическая, находящая свое выражение в различиях
между половыми клетками разных генераций (гоноцитами, осгония-
ми и сперматогониями 1 и II порядка, гаметами)—с одной стороны,
'бластомерами—с другой, и разнообразными клетками тела — с тре-
"тьей. Таким образом, разноклеточность имеется не только у Metazoa,
‘НО и у простейших; единственная разница между ними -в это-м отно-
шении заключается в том, что у первых сосуществуют две ее
фор-мы- функциональная и «циклическая, а у вторых, как правило;
имеется только одна последняя.
С дифференциацией жизненного цикла неразрывно связана его
интеграция, составляющая друг-ое следствие эволюции полового про-
ц-есса. В самом деле, по мере перехода последнего -от низших степе-
ней к высшим, он оказывает все более ‘ощутимое организующее
влияние на весь жизненный цикл и придает ему все более целостный
и упорядоченный характер. Среди множества равнозначных и веге-
‘тативно полноценных клеточных генераций постепенно выделяются
узко специализованные, не способные к выполнению но-рмальных
вегетативных функций. Индивидуальность таких особей в значитель-
ной мере подавляется, как бы растворяясь в целостности цикла,
и они низводятся с уровня вполне самостоятельных особей до зна-
чения подчиненных компонентов того целого, каким теперь стано-
вится жизненный цикл. Циклическая интеграция выражается, таким
141
образом, в нарастающем и все более полном подчинении отдельных
клеточных генераций циклу, что является первым шагом в сторону
их объединения в индивидуальное развитие высшего по-ряд-ка, т. е.
в жИзненнь1й цикл, целик-ом осуществляемый одной единственной
много-клеточной особью.
Глава VII
ОБРАЗОВАНИЕ КОЛОНИЙ
1. Вводные замечания
Исклклпггелнлпс-е значение колониальности, как ступени к созда-
нию высших форм индивидуальности, и та роль, какая отводится ей
в особо интересующем нас вопросе -о происхождении ‘много-клеточ-
ности, —— общеизвестны. Однако мы до -сих пор не располагаем даже
хоть сколько-нибудь удовлетворительн-ой классификацией форм
колониальиости у Protozoa и, в-частности, их одноклето-чнпях пред-
ставителе-й.
Обычно коло-нии простейших разделяются на прикрепленные,
имеющие чаще всего древовидную форму, и свободноплавающие,
большинство кото-рь1х шарообразны. Такая чисто описательная
и статическая кла-ссификация колоний мало что дает; принимая ее,
мы ни на шаг не подвигаемся к пониманию их природы и не ‚полу-
чаем даже ОСНОВЫ ДЛЯ ИХ ICpaBHI/ITEJIBHOTO paCCMOTp€HI/151..
Такую основу, «однако, совсем нетрудно по-лучить, если только раз-
личать формы колониальности по способу их образования. С этой
точки зрения, колонии одноклеточных ьмогут быть распределены -в
две главные группы: 1) м о н о т о M и ч е с к и е ко л о н и и, обра-
зующиеся mar за шагом путем многократного размножения простым
делением, при бесперебойном выполнении зоо-идами своих вегета-
тивных функций и Непостоянном их числе; 2) п а л и н т о M и ч е-
ские колонии, возникающие в результате одновременного
объединения целой семьи зооопор, получившейся путем повторного
деления и уже не изменяющей своего численного состава.
Монотомические колонии одноклеточных представляют с-обой
оч-ень простые и не всегда ст-ойки-е объединения, состоящие здесь
из вполне равноценных зо-оидов. В противоположность им палинто-
‚мические колонии достигают очень большой сложности и среди _них
можно на-йти все ступени ‚возрастающей организменной целостности
колонии, ее превращения в многоклеточный орга-низм.
Этим двум главным типам колониальности, создающейся в связи
с размножением, должен быть противопоставлен еще третий, в зна-
чительн-о-й мере условный, ее тип, наблюдаемый только у асмёбоид-
ных организмов и солнечников: а г р е г а ц и о н н ы е -к ол он и и,
образующиеся путем слияния ряда первоначально независимых-
дособей. Обращает на себя внимание, что самый распространенный
_у Metazoa способ построения колонии почкованием не имеет места у
одноклеточных; равным образом мы не видим здесь и образования
‚142
Стодднов И ряда других форм бесполого размножения, встречаю-
щихся у колониальных многоклеточныхб олений т (‘ЮТ
. . С.
Очень большую роль в построении о оих типов KI P
студенистые или другие аллоплазматичес-кие выделения ее ‘IJIEHCB.
создающие конструктивную основу колонии и в значительной мере
определяющие принимаемую €10 ФОРМУ-
2. Монотомические колонии
На всех стадиях своею развития колонии этого типа СОСТОЯТ ИЗ
трофозоито-в, находящихся вдифференцированном состоянии и в нем
же размножающихся (рис. 41). ВВИДУ ЭТОГО ‘ОНИ ФОРЧИИРУЮТСЯ “О”
степенно И, В общем, ритмично, удваивая число своих ‘ЗООИЛЮВ’
с каждой ново-й их генерацией; число членов колонии, следова-
тедьнод неустойчиво и проявляет тенденцию к постояНдЮМУ Увеличе-
нию также ‚постепенноъ Шаг за Шаром, наращивается, когда она
есть, и аллоплазматическая основа колонии, так как каждыи вновь
образовавшийся зо-0-ид ВЫдЕЛЯеТ -C3010 ‘НОВУЮ ПОРЦИЮ a““‘°'“J1a3'MbI’
наслаивающуюся на ранее -имевшиеся.
У колоний довольно распространенного типа Phalansterium Hz:lp$I<())JI:4bI1I:z;sIIIpI~;I:1§t;l:a
6 аз ется как п авило в -проксима -
атУдениетой ‘;fe’:{‘:IIJ:I::M:I ngyinxy 5,1a.}o;1a»pa t1x)eMy она и растет преимущественно
НИИ и Ёораздо ‘каж ‚дм новым делением зооиды оттесняются к периферии коло-
центро ежно. с асЁтояние от основания последней Оказывается «пропорциональ-
Hm’ так что Ёхшрествовавших поколений зооидов, принимавших участие в ее по-
НЫМ ‘йъгллунпечэаёномерный рост аллоплазмы в сочетании с монотомическим спо-
СТРОе - томи-
. колоний этого типа и локализацию наличны
чеокое СТРОеНИЁб ая фо ма получающегося таким образом куста зависит глав-
ee развилках’ ‚ЁЦТО mg каким утлом друг ‘к дируъгу располагаются дочерние
HHM506pa:;(1):a;(Ene:HH3 если они остаются более или менее параллельными, коло-
осо и по - .
вее овидн ю или д 630
ния растет только в одном направлении и имеет узкую BE ная колония год?
видную форму; при большем расхождении их осеи древо д га me сильней
и г от е —
чается более раскидистои, а если они отклоняются дру „удру Ц у ОМ к ст
стет- по всем радиусам -и становится подобной ШарОВИДН У У Y:
колония pa всех нап авлениях Подавляющее боды-ПИНСТВО древшшдных
BeTB”m§MTyaC}:lOrl;OpOfla „Бляди сидячими прикрепленными к субстрату (:Pha1anste-
КОЛОНИИ ›
.. ' а бы ют свободно-
r1um, Spongomonas, Cladomonas и npyme), НО шаровидные Ba
. ижными Uro lena Uroglenopsis) или же хоть и сво-
ЁЛШЁШМЁ РЁюЁеЁМЁаЁсЁЁЁыми обёэазоёниями, медленно, перекатывающимися
одны и, н —
S haeroeca .
В фЁЩе и-‘латргпрмонотомйческих колоний, тип Dinobryon, представлен древовид-
Тороин ШИЯМИ тстроенными из жестких бокадовидных домиков и пассивно
'HZIM§a£(§f1f§ues воде’ Они созидаются несколько иначе, чем колонии первого типа,
п Ёгменно путем поочередного выселения одной из дочерних особеи из материн-
a a за которым следует ее прикрепление к дистальному краюпослед-
СКОРО домик ’ ние нового бокала Складывающаяся ТаКИМ ПУТЭМ‘ колония имеет
негоми lBeb(I!l'}'?3JeIfIHO дихотомическое СТРОШШЭ, НО ОНО совмещается ТУТ ° M°H°"°'
,I7I1I1?I:1.;IlbI3-$1-[M (когда на ‚краю материнского бокала сидит только один зооид) И
- но три). я
с ехветвистым (когда они располагаются тут о
Трследующий’ очень распространенный, тип колонии xapaKre1?uayD:':;;In:I::lI;4£4a';1;IC3
не образуют вовсе собственных оболочек, или же ТОЛЬКО
способные делиться, оболочки- В результате ЭТОГО е°°иды °°T"“°T°" труп‘
ёироваьными пучками ‚на конце общего аллоплазматического .стебелька .(выде-
143
' 1—Uroglenopsis amerlcana, общий вид; 2-—то же, оптический
ленного основателем колонии еще перед началом его размножения) или же
располагаются в виде шара, соприкасаясь в его центре своими задними концами.
Так или иначе, они остаются на том же месте, где находился материнский тро-
фозоит‚ и, умножаясь в
числе в каждом после-
дующем поколении, не
испытывают при этом
центробежного смеще-
ния. Такие колонии_ха-
ра-ктерны для воротнич-
ковых жгутиковых, где
они всегда являются си-
дячими, и обычны среди-
хризомонад, где имеются
как прикрепленные, сте-
бельчатыед, формы (Ant-
hophysa), так и многие
активно движущиеся пе-
лагические (Syncrypta,
Synura, Chrysosphaereb
Ia); впрочем, и сидячая
Anthophysa легко Impe-
ходит в свободноплаваю-
щее состояние‘. Ветвле-
ние колонии, наблюдае-
мое у ряда видов (АНЁ-
hophysa, Codonocla¢11-
um), достигается перио-
дическим выделением ал-
белька. Так уАСооопо-
cladium каждый из зоои-
дов первоначально обра-
зовавшегося пучка выде-
ляет по такому стебель-
Ky И, обособившись та-
дает начало уже новому‚
вторичному, пучку тро-
фозоитов. У Anthophysa
новые развилки общего
ст-ебля сложной колонии
. образуются после укаж-
A . цого деления пополам
' целых полушаровидНЫХ
_ групп зооидов и т. д.
К четвертому типу
монотомических КОЛОНИИ
3 б S I 5 Ch ОТНОСЯТСЯ известные Ile-
--TO же делящаяся осо ь‘ 4- yncrypta vq мох‘ —— ryso- - 05- аз .
зрпаегепа longispina; б-Ёупцга uvella, o6:.u1m’sm1; 7—то “ОЧКИ Ceratlum’ Mp та
же, депящаяся особь; 8-—-Sphacroeca volvox, общий вид; ЮЩИЁСЯ В РеЗУ-дьт е
.9-—один из членов колонии; 70-—одиночное воротничковое го’ что особи ряда после-
. „жгутиковое Codosiga (из Захваткина) доватшьных поколений
` ' остаются оклеенными
своими панцырями, и потому, что деление является тут поперечным.
` " так, все рассмотренные типы колоний всецело обязаны своим строением
`тому‚_ что онисоздаются в результате "монотомии протозоиного типа, и притом
преим’ щественно продольной, и что строящие их зооиды регулярно Выделяю?
алло лазматические массы. ‘
14;
Рис. 41. Шарообразные монотомические колонии
' жгутиковых:
разрез через части колонии (видно строение ее остова);
ЛОПЛ азматич BCKOFO CT‘e' '
КИМ ПУТЕМ ОТ ОСТЗЛЬНЫХ‚-
АЁо все нивах/тенты свое-го существования монотомичеокие колонии
состоят из вегетативно-полноценных особей —зооидов‚ практически
тождественных друг с друго-м в морфологическом и физиологиче-
ском отношениях и, в частности, одинаково способных к неограни-
ченному дальнейшему размножению." Число их в колонии может
быть очень большим (доходя, например, до нескольких тысяч
у Uroglen-opsis), ЧТО -не мешает им оставаться биологически равно-
ценными. Какого-либо полиморфизма и, соответственно, разделения
т-руда между зооидами не наблюдается здесь никогда.
Возобновление монотомических колоний происходит различными
способами. Чаще всего, оно осуществляется путем выселения отдель-
ных зооидов из колонии, их временного перехода к одиночному
существованию, что наблюдается чаще всего у прикрепленных форм.
В других случаях колония может целиком распадаться на состав-
ляющих ее зооидов, каждый из которых дает затем на-чало новой
колонии. Наконец, у некоторых хризомонад (Synura Anthophysa)
шарообразные колонии сп-особны делиться пополам или на несколько
фрагментов, что может совмещаться, однако, и с выселением отдель-
ных ее членов. Таким образом, каждый из зооидов обладает тут
возможностями основателя колонии, что, очевидно, свидетельствует
о примитивности этих форм колониального соединения.
Эта‘ примитивность ярко сказывается и в архитектонике шаро-
образных свободных колоний монотомическото происхождения. Все
такие коло-нии имеют многоосное, гомополярное строение, не обнару-
живая никаких морфологических признаков региональной, а тем
паче градиентной дифференциации своих членов, и не имея также
какого-либо предпочтительного направления при -плавании. Следо-
вательно, мы имеем здесь дело с колониями низшего, весьма прими-
тивног-о, характера, представляющими собой просты-е и относи-
тельно слабо спаянные объединения нормальных вегетативных
особей. Следует еще подчеркнуть, что зооиды этих колоний никогда
не связаны между собой протоплазматическими мостиками и обычно
разобщены студенистой массой или же собственными оболочками.
Таким образом, здесь нет и «общего тела» колонии, и ей =в целом
могут принадлежать только аллоплазматические образования.
Слабая степень интеграции этих колоний не означает, однако,
полного ее отсутствия. Определенная степень подчиненности зооидов
колониальному целому экспериментально до-казана у Зуппга, где
она проявляется различиями в норме реакции между зооидами,
искусственно выделенными из колонии и произвольно покинув-
шими ее.
3. naIIPIHTOMPI‘-ICCKPIC КОЛОНИИ
Колонии палинтомического происхождения встречаются толькоу
фитомонад (и генетически близких с ним трупп зеленых водорослей),
где они представлены свободноплавающими, чаще всего сфериче-
скими ‹<ценобиями>> вольвоцид и некоторых примыкающих к ним
‘семейств. Самую общую особенность этих ‚колоний составляет
10 Эмбриология беспозвоночных 145
именно то, что весь процесс их формирования имеет чистоэмбрио-
нальныйхарактер дробления; что вегетативн-о деятельными зоои-
дами СТЗНОВЯТСЯ ОСОбИ ОДНОГО ТОЛЬКО заключительного ‘ПОКОЛЕНИЯ’
Целой их палинто-мической серии, тогда как все остальные промежу-
ТОЧНЫЕ ПОКОЛЕНИЯ спороцитов никак не МОГУТ считаться таковыми;
ЧТО ЭТО заключительное поколение представлено зоосп-орами; короче
ГОВ-ОрЯ, ЧТО КОЛОНИИ ЭТОГО типа создаются как бы В ОДИН прием,"
путем одно-временного объединения целой семьи жгутиковых зооспо-р ~‘
И потому обладают постоянным численным составо-м.
Это объединение может совершаться двумя различными спо-
собами. Один из них заключается в том, что зооспоры активно
складываются в коло-нию уже после своего оформления, другой-
в том, ‘что их объединение происходит ‚пассивно и одновременно’
с их формированием.
Первый из этих способов очень характерен для сем. Spondy1o-
moridae и многих водорослей, ‘например, известной водяной сеточки,
(Hydrodictyon).
Зооспоры этих форм, образующихся в числе 8-16 (y Sp»0ndy-
lomoridae) внутри материнской оболочки, тут же разъединяются,
вырабатывают собственные (пектиновые) оболочки И становятся
подвижными. После этого зооспоры одновременно складываются
в очень правильную звездообразную фигуру, слипаясь носиками
своих оболочек. В восьмичленньтх колониях Chlamydobotrys st-ellata
ОНИ Л-ОЖаТСЯ ЧЕТЫРЬМЯ ярусами, |HO- ДВЕ ОС-ОбИ В каждом; ОСОбИ‘
верхней пары срастаются передними концами под очень тупым
углом, под ними крест-накрест располагается вторая пара особей,
уже не касаясь друг дрУга, =но прилегая боками к выше и ниже ле-
жащим; сходным образом, и так же чередуясь с остальными, рас-
полагаются и следующие два яруса- (Коршиков, 1924; рис. 42, 1—5).
В таком же порядке, но не по две, а по четыре в ярусе, соеди-
няются и зооспоры у Chl. gracilis и Зропс1у1оп1огпп1 quaternarium,
складываясь тут в не менее правильную 16-, редко 32-членную ко-
л-о-нию (ри-с. 42, 6-8). Дальнейшее развитие зооидов идет =по той же
схеме, как у других окрашенных фитомонад, имея своим р-езульта-
том их превращение из зооспор в тро-фо-зоитов и постепенное дора-
стание до Материнской нормы. у ` ’
Бо-льшого внимания заслуживает то обстоятельство, что колонии
Spondylo-mo-ridae одноосно-гетерополярны и обладают правильной
2—4-лучевой симметрией вращательного типа, что бесспорно гово-
рит об их значительной интегрированности. У 16—З2-з-оо-идных ко-
лоний Chlamydovbotrys gra-cilis это качество проявляется еще более
наглядным образом в правильном уменьшении размеров стигм зоо-
идов в ‘на-правлении спереди назад, т. е. в наличии тут определенного
осевого передне-заднего градиента и соответствующей зачаточной
дифференциации зооидов одной колонии. Однако дальше этого дел-о
тут не идет: все зооиды в одинаков-ой мере способны к размноже-
нию, а колония в целом способна распадаться. Следует также еще
раз подчеркнуть, что складывание зооспор в колонию происходит
146
ттутем их агрегации, наступающей после того, как они совершенно
оформились. . . ч - - . *
Второй способ объединения зооспо-р в палинтомическую колонию
наблюдается у всех представит-елей семейства Eudorinidae и
Уо1уос1оае и сводится .к нерасхождению продуктовпалинтомии.
Их объединение устанавливается еще в ходе повторного деления,
когда они находятся еще в стадии опороцитов, так что зооспоры А
оказываются тут с самого начала связанными друг сдругом в более
Рис. 42. Палиитомические колонии Spondylomoridae.
7—5—Ch1amydobotrys stellatat 1—общий вид молодой колонии; 2——колония среднего возра-
ста с начавшими округляться клетками; Змвзрослая колония с яйцевидными клетками;
4—расположение клеток при наблюдениис переднего полюса; 5—колония‚ приступившая
к размножению; 6—8—ChIamydobotrys gracilis: б-молодая 16—клеточная колония; 7—схема
расположения клеток при наблюдении с переднего полюса; 8—взрослая колония (по
Коршикову)
или мен-ее стойкое колониальное объединение. Такое объединение,
состоящее из многих зооспор (в данном случае целой их семьи) и
создаюше-еся в результате их нерасхождения, принято называть
сложнои зооспор-ой, или си-нзооспорой. Стало быть,
палинтомические колонии этого типа формируются в виде син-
зооспор; и так как их‘ компоненты остаются тут, как правило, в no-
жизненном соединении, то взрослые колонии этих фито-монад пред-
ставляют собой не что и-ное‚ как с т а б и л и з о в а н.н ы е
синзооспоры. `
Такая их стабилизация, сне встречающаяся в других группах
одноклеточных организмов, оказывается тут возможной потому,
10* ` 147
что взрослые трофозоиты Volvocoidea .nomm ничем не отличаются
от их жгутиковь1х зооспор ни в структурном, ни в биологическом
отношениях. Однако все же следует иметь в -виду, что эти формы
являются подлинными синзооспэорами то-лько на ранних стадиях
развития, пока их з-ооиды представляют собой зооспоры; когда же
последние становятся трофозоитами‚ то и образуемое ими объеди-
НЕНИЕ вырастает В пПОЛНОЦЕННУЮ ВЕГЕТЭТИВНУЮ КОЛОНИЮ И перестает.
быть синзоосп-орой.
Палинтомически-е колонии Уо1уосо1с1еа весьма разно-образны по
своей форм-е, численному составу и степени интегрированности.
Среди них есть и простые формы, вроде четырехчленных колоний
Gonium social-e, КОЛОНИИ такой же сложности как у Spondylomoridae,
И ДЛИННЫЙ рЯД НВСРЗВНЕННО I6-OJIEE ВЫСОКО ОРГЭНИЗОВа-ННЫХ, кончая-
‘ГЗКИМИ, КаК у VOIVOX, ГДЕ ОНИ включают ТЫСЯЧИ ‘ЗООИДОВ И ДОСТИГЗЮТ
п-одлинно индивидуального уровня целостности. Совокупность отно- _
сящихся сюда форм располагается в необычайно полный и демон-
стративный ряд-иллюстрирующий все последовательные ступени
преобразования простой колонии в сложно дифференцированный
многоклеточный организм. И так как тут реализованы все этапы
этого процесса, их внимательное рассмотрение оказывается не-
обыкновенно поучительным и позволяет ул-овить его ведущие закод
номерности.
‘В начале этого замечательного ряда стоят малочленные Eudori-_
nidae И Haematoco-ccidae, КОЛОНИИ которых состоят из четырех
(Gonium souciale, рис. 43, 1) или восьми '(Eud0rinve11a, Stephan-o-
sphaera) зооидов. Это очень просто, хотя и правильно построенные
агрегаты, состоящие из совершенно равноценных друг другу особей,
расположенных ‘крестообразно в общей студенистой мас-се (Gonium)
или:же в виде кольца по экватору та-кого же, но одетого оболочкой,
Шара (Stephanosphaera). Каждый зооид од.инаково способен к рази
множению и может дать начало новой колонии, так что последние
не отличаются от мо-нот-омических ничем, кроме способа своего
формирования.
Вторую ступень олицетворяют 16-членные колонии Gonium
pect-orale, Pandovrina m-orum и Volvulina Ehrenbergi (СМ. рис. 38, 1):
ониотличаются уже более сложной архитектоникой и, соответ-
ственно, более сложным способом ‚построения. Являясь продуктом
сам-ой типичной табличной палинтомии, эти колонии образуются
в виде классической плоской «гониев-ой таблички» (см. рис. 38, 12)
и пожизненно сохраняют эту форму (Оопшш pectorale) ИЛИ же вы-‘
ворачиваются затем ‘в эллипс-оидальную фигуру. После этого тесно
соприкасавщиеся зооиды раздвигаются образующимся теперь между
ними студнем.- В окончательно сформировавшейся колонии каждый
зоо-ид занимает строго определенное положение относительно дру-
гих, в связи с чем она приобретает правильную четырехлучевую -сим-
метрию и определенно ориентированную ось. В общем, тут дести--
гается та же ф-орма и высота колониальн-ой организации, какуюмы
видели у Chlamydo-botrys gracilis И с которой связаны первые-
148
Рис. 43. ‚Палиитомические колонии Еиоогйпйааё; ` ` ' . ‚.
7——Gonium so_ciale (no I.LIp_ei»’16epy)'; 2—E‘udorina elegans; З-тоже, в момент размножени '
‘4—-Eudorma illinoisensls (no Кофоиду); 5—Pandorina charkoviensis (n’6.‘I(op'u.u»u<oBy)'
149
признаки по-явления общего осевого градиента: правильное убыва-
ние размеров стигм спереди назад. Этот, пусть И небольшой, но
весьма многозначительный признак начавшейся физиологической
интеграции колонии наблюдается у Volvolina И вряд ли действи-
тельно отсутствует у Pandorina. Однако и тут о-н остается един-
ственным ее признаком: колония -сохра-няет способность распа-
даться, а все ее зооиды —свою полноценность. I __
32-членные колонии обоих обычных видов Eudorina ——- E. elegans
и „Е. illinoisensis (ранее причислявшейся к роду Pleodorina,
рис. 43, 2-4) и Platydorina, представляют следующую ступень услож-
нения и интеграции, выраженную, правда, неодинаково в каждом из
относящихся сюда случаев. У Platydorina (рис. 44, 1) ее п-о-казате-
лем служит высокодифференцированная двусторонне симметрич-
Рис. 44:
1—Platydorina, вид колонии с плоской стороны; 2—то же, вид
с узкой стороны; З-— Pleodorina callfornlca (из Догеля)
пая форма пластинчатой колонии, очень интересная еще и тем, что
ее первичная главная ось (направленная нормально к по-
верхности пластинки) зам-енена вторичной, перпендикулярной с ней
осью сикмм-етри-и и движения (Р); по этой новой ос-и идет убывание.
размеров стигм, но других внешних признаков интеграции тут, по-
видимому, нет. Ценобии обоих Eudorina (рис. 43, 2——4) ИМЕЮТ эллип-
соидальную форму и нормальное для вольвоцид положение главной
оси при спиральном или радиальном размещении зооидов по четыре
в само-м переднем и самом заднем из пяти образуемых ими ярусов
(и по_восемь в остальных ярусах). Они создаются таким же спо-
собом, как у Pandorina, —путем сворачивания первоначально пло-
ской го-ниевой табл-ички и последующего раздвигания клеток студ-
нем, и также обладают гла-з-ками, постепенно уменьшающимися
спереди назад. хОднако это уже не единственный показатель целост-
ности колоний. Она. получает тут и более осязаемое выражение в
ТОМ, ЧТО ВСЕ ЧЕТЫРВЗООИДЭ 'H€p€)1.H€I‘0, a‘l'II/IKaJIbHOI‘»OV венца ЗЗМЕТНО.
отстают огг остальных в темпе размножения (Е. elegans) ИЛИ даже
полностью утрачивают способность размножаться, превращаясь в
стерильные клеткит-эргазии (Е. illinoisensis). Это превращение
заключается в заметном пониж-ении границы роста, УМеНЬШеНИ-И
150
хроматофоров и их побледнении, из чего явствует, что апикальные
зооиды ев значительной мере снижают и свою ассимиляционную дея-
‘ёельность. Так возникает функциональное разделение зооидов на
ЁСПЛОДНЬЮ И ПЛОДУЩИЕ, ПРИВ-ОДЯЩИе к уже необратимому объеди-
нению колонии и утрате ею способности диссоциироваться. Дей-
СТВИТВЛЪНО, ее распадение не имеет м-еста при нормальных условиях
даже у Е. elegans и может быть вызван-о тут только эксперименталь-
ным вмешательством.
У4Р1еооог1па са11Тогп1са (рис. 44, 2), коло=нии которой состоят
из 64 или 128 зооидов, этот процесс б и о л о г и ч е с к-о- й д иф ф е-
ренциации зооидов про-
двинут уже много дальше. д; ,__. с
Все 32 или 64 зооида перед- ‘ 1-": .
него полушария колонии тишь: 9.
«ч 40“ .v~V~‘>
представлены уменьшенны-
ми, бледно окрашенными д%%гцщд$йшг
стерильными клетками, а все
зооиды заднего-крупными, ‘4 ‘.;'{§-rf,
rycroseneabflmu ‘ г о н и д и я- ,
ми, или гоноцитами,
которые одни способны к
размножению.
Высшие ступени в этом
удивительном ряду прогрес-
сивно повышающегося ус- _ о А _ _. ‚
ложнения' и интеграции ко- ' , д ' _ _ _
лонии занимают настоящие ‘ _."~“""~'
Volvocidae, T. e. представи-
тели прежнего рода Volvox,
расчлененного теперь на не-
сколько более узких родов:
Volvox s. str., THHOM которого остается V. globator, Janetosphaera
(J. аитеа-рис. 45), Merl-11ospha«era (M. tertia) и др. Этонуже Не-
оспоримо многоклеточные формы (и даже далеко не примитивные
по уровню своей организации), но вместе с тем все же колонии
высшего типа, связанные всей цепью уже рассмотренных форм с
самыми простыми из палинтомических объединений. Оставляя. до
0дН0И ИЗ следующих глав (ХП) -рассмотрение и оценку мотивов,
заставляющих считать вольвоцид подлинно много-клеточными орга-
низмами, мы ограничимся пока более односторонней характеристи-
кои этих форм, как высших звеньев в ряду палинтомических колоний.
ОЁще-е число зооидов в этих объединениях колеблется -в преде-
23:11 е 12—2048 y Janetosphaera aurea И достигает 4000—16 000 и
- у крупных видов Volvox; оно отвечает, таким образом, числу
потомков 9—10 или до 15 синхронных повторных делений и, пови-
дИМ'0‘МУ‚ НИКОГДа не опускается ниже первой из приведенных цифр;
их число столь же постоянно, как us других палинтомич-еских коло-
ниях. Равным образом и все зооиды имеют стандартное строение
М‘: V ›
Рис. 45. Janetosphaera aurea (no Клейну)
151
одиночных хламидомонад, и колония as цел-ом созда-ется совершенно
таким же способом, как у Eudorinidae. ” _
Однако дифференциация огромной массы вооидоав -на пл-одущие
и бесплодные заходит здесь несравненно дальше, чем у всех менее
богатых особями форм: она выражается в том, что подавляющее’
большинство зооидов представлено здесь стерильными, сильно
уменьшенными против` нормы эргазиями и лишь буквально еди-
ничные (6-12, редко до 30) -—сильно гипертрофированны-ми гони-
диями; все эти последние располагаются в заднем, вегетативном
полушарии колонии. Осевая дифференциация ее бесплодных з-ооидов
имеет ярко выраженный градиентный характер. Зооиды переднего
полюса мельче остальных, обладают более мелкими и бледными
хроматофорами, содержащими меньше хлорофилла, но зато более
крупными глазками; по направлению- кзади эти признаки очень
постепенно сменяются на об-ратные- зооиды становятся более
круп-ными, имеют более крупные и ярче окрашенные хроматофоры,“
ИХ глазки y'Mr€Hb1I_Ia}O’I‘vCH И, :HaKOH€‘Il, COrB'€pII.I-€H'HO исчезают. Эта
двоякая дифференциация наступает уже после оформления син-_'
зооспорьт и достижения ею окончательного численного состава;
в момент своего оформления колония состоит из совершенно одно-
родных жгутиковых з-ооопор, дальнейшее развитие которых идет,
однако, в противоположных направлениях. Основная их масса почти
не растет и лишь слабо изменяет свое строение (в порядке осевой
передне-задней дифференциации), тогда как будущие гонидии
дедифференцируются и испытывают после этого длительный гипер-
трофический рост. Таким образом, все зооиды взрослой колонии‘
сов-ершенно ‘утрачивают свои признаки нормальных трофозоитов:
подавляющее их- большинство остается недоразвитыми, . сохраняя
размеры и, отчасти, строение вооспор, а гонидии приобретают ‘все
свойства гипертрофозоитов.
Эта характернейшая особенность высших вольвоцид обуслов-
л-ена сохранением постоя-нной, или, во всяком случае, очень длитель-
‹ной круговой связи между всеми зооидами колонии: они все соеди-
нены друг с другом, протоплазжматическими мостиками (плазмод
десмами), образующими сплошную сеть и спаивающими всех
зооидо-в в синцитиальную систему. Эти мостики образуются при
дроблении частью за счет неполного разъединения его продуктов,
частью же, по-видимому, путем последующего: установления плазмаы
тических контактов между ними; -в дальнейшем, при образо-вании`
межклеточного студня, они сильно «вытягиваются, истончаются ‘и
могут к концу развития даже обрываться совсем; Наличие такой
синцитиально-й св-язи между всеми Членами колонии обеспечивает.
ее физиологическое единствододним из проявлений которого и
служит избыточный рост г-онидий за счет ассимиляционной деятель-
ности ‘всех остальных зооидов, почти вся ее продукция идет на фоГр-..
мирование немногих гонидий, поглощающих львиную долю матег.
риала, ‘СОЗИДЗЭМОГО УСИЛИЯМИ ЦеЛОЙ КОЛОНИИ.
K концу развития гонидий бесплодные члены колонии начинают”
152:
проявлять пер-вые признаки дегенерации, усиливающиеся при палин-
‚томическо-м размножении гонидий; -B ДаЛКЫНЕЙШ-ВМ ОФОРМИВШИЕСЯ _
молоды-е колонии прорывают стенки материнского ~op‘raIHI/I3Ma, K0’F0-'
рый при этом погибает. Таким образом, вся огромная колония,
кроме гонидий, оказывается чисто вегетативным образованием,
пластический обмен которого целиком поставлен -на службу немного-
численны-м гонидиям. Такое крайнее их доминирование приводит
к созданию своеобразного с-клада коло-ниальности, очень характер-
н-ого для вольвоцид; однако это доминирование касается тут только
одной из сторон жизнедеятельности колонии, а_ именно ее вегетатив-
ных функций; в отношении же анимальных функции колония пред-
ставляет собой высоко индивидуализованное и интегрированное
целое. u ’
Эта целостность сказывается в одинаковои мере в строении ко-
лонии и во всех проявлениях ее активности. _Все вольвоциды——
активно плавающие -существа, постоянно движущиеся анимальным
‚полюсом вперед, вращаясь при этом вокруг своеи оси в совершенно
определенном направлении (например, -в направл-ении часов-си
стрелки: V. globator); с этим связано и стр-ого согласованное дви-
жение жгутов двигательного аппарата.
Морфологические признаки целостности колонии остаются
у в-ольвоцид такими же, как у д-ругих колониальных фитомонадин,
но только ‘приобретают еще более отчетливое выражение. Так,
одноосно-гетерополярное строение колонии здесь подчеркнуто бро-
сающейся в глаза осевой дифференциацией зооидов (касающейся
уже не только размеров их глазков, но и самой их величины и omen-
пени раз-вития фотосинтетического аппарата), а также "и большей
сохранностью ‚во взрослой организации эмбриональной переднеи
розетки или креста и также эмбрионального фиалопора-отвер-
стия на заднем полюсе колонии. Равным образ-ом и признаки четы-
рехлуневой вращательной симпметрии и СОВМЕЩаЮ-ЩЕЙЮЯ с нею дву-
лучевой выражены у вольвоцид лишь "несколько определеннее, чем
у их ниже организованных сородичеи-; -в строении апикального
креста, спиральном расположении зоо-идов, крестообразной форме
фиалопора и заметном тут разграничении четырех квадрантов ко-
лонии нет ничего чуждого. эудоринидамили даже Зропоуюпюпоае;
и вся разница заключается в том, что все эти особенности получают
более развернутое и устойчивое выражение и более заметны _на
многотысячных колониях вольвоцид, чем на том, сравнительно
малочленном, материале, каким они являются у только что на-
званных семейств. _
Итак, рассматриваемый ряд колониальных Vo1voco1d~ea показы-
вает с исключительной наглядностью, каков может быть ход посте-
пенного преобразования простейших колоний в боле-е сложные,
а затем и -в высоко дифференцированные многоклеточные образо-
Baum. OH показывает также, что многоклеточность вольвоцид есть
конечный результат того же интегрирующего процессе КОТОРЫЙд
на более ранних своих этапах, приводит к формированию колонии
153
меньшей степени сложности. Наконец, на примере этого ряда
выявляются две главные особенности процесса интеграции’: ее пре-
рывистый, ступенчатый характер и связанность различных ее ступе-
ней с вполне определенным численным составом объединения; они
конкретно выражаются тут в том, что колонии разного численного
соста-ва обладают и разным уровнем целостности, что последняя
тем выше, чем больше чи-сло зооидов в колонии., и что здесь остаются
фактически ‘НЕЗЗПОЛНЭННЫМИ МНОГИЕ ШрОМЕХЦ/Т-ОЧНЫЕ УРОВНИ ЧИСЛЕН"
ного состава колоний. К этим, исключительно важным, моментам
мы еще вернемся в гл. XV. ‘
H
ЧАСТЬ ВТОРАЯ
от одноклвточности к высшим ступеням
НАДКЛЕТОЧНОЙ ОРГАНИЗАЦИИ
глава v111
ПУТИ ФОРМИРОВАНИЯ ПЛАЗМОДИЕВ и СОМАТЕЛЛ
1. Общее понятие о полимеризации
В предварительных зам-ечаниях к гл. 1 мы отм-ечали, что сравни-
тельно-морфологическое рассмотрение простейших показывает,
что ведущим направлением эволюции этих организмов является
«эперерастани-е» ими своей первично одноклеточной организации, что
эта линия раз-вития осуществляется посредство-м умножения, или
полимеризации, тех или иных клеточных структур (или даже всей
их совокупности) —без соответствующего разделения цитоплаз-
мы—— и что в основе полимеризации лежит, таким образом, частич-
ное или недоведенное до ко-нца клеточное деление.
Рассмотри-м теп-ерь более внимательно и подробно природу этих
явлений, их творческую роль -в морфогенезе различных груп.п про-
стейших и те исключительно разнообразные ‘последствия, к каким
они прив-одят.
Прежде «всего необходим-о еще раз подчеркнуть, что полимери-
зация ‘вс-егда осуществляется у простейших путем деления тех или
иных клеточных органоидов (ядер, кинетид и т. д.) или целых их
комплексов, идущего нормальным для .них образом, но не одновр-е-
менно с делением цитоплазмы, а в той или иной мере опережая его.
В самой наглядной форме полимеризация наблюдается в ходе
индивидуального развити-я многих паразитических са-ркод-овых,
споровиков и жгутико-вых и лишь значительно реже у свободно-
живущих видов. Она ведет тут к переходу одноклеточного молодого
организма -в надклеточное состояние взрослого плазмодия или
сомателлы; вновь раопадающегося ‘при размножении на -одноклеточ-
ные дочерние ос-оби. В этих случаях мы имеем дело с временными
плазмодиями или "сомателлами, образование которых путем поли-
.155.‘
а"
меризации, умножения ядер, кинетид и пр. целиком происходит на
наших глазах, в порядке онтогенетическом процесса.
Однако ав значительном большинстве случаев полимерным”
(многоядерным, многожгутикотвым) строением обладают все стадии
ИНДИВИДУЗЛЬНОТГО развития ‘ПрОСТЕИШЕГО, так ЧТ-О ОНО ‘ЦЕЛИКОМ 'Hp‘O--
текает ‘В НЗДКЛЕТОЧНОМ СОСТОЯНИИ организма. В ЭТИХ СЛУЧаЯХ МЫ
ИМЕЕМ ДЕЛО С 'HO'CTOv$I‘HHI:IMI/I ‘ПЛЭЗМОДИЯМИ И сомателлами, C УЖЕ <<1"O'- -
ТОВЬЪМИ» продуктами филе-генетически пройденн-ой полим-еризации.
Правда, мы знаем много таких примеров, когда в ходе онтогенеза
ПРОИСХОДИТ дальнейшая ППОЛИМЕрИЗЭЦИЯ УЖЕ надклеточной МОЛОДОЙ ‚
особи, продолжающей умножать свои ядра, кин-етиды, пара-базаль-
ные аппараты и пр. по мере роста, что существенно о-блегчает пони-
мание путей и опособов становления самых сложных типов -надкле-
точной органи-зации. Однако есть немало и таких примеров, когда все
индивидуальное развити-е протекает с самого начала ‘в: высоко поли-
мерном состоянии орга-низации, практически не изменяющемся
в онтогенезе. Таким образо-м, в ряду надклеточных простейших A
представлены ВСЕ ПОСЛЕДОВЗТЕЛЬНЫЕ этапы ВОЗНИКНОВЕНИЯ И стаби-
ЛИЗаЦИИ ПОЛИМЕРНОГО СТр0ЕНИЯ‚ НаГЛЯДНО ПОКЗЗЬЁВЭЁОЩИЕ НаМ ПУТИ
ЕГО И СТО-р И ЧЕС K0-I‘ O CT ан OB ЛЕН И›Я .
2. Плазмодии саркодовых и ‘споровиков
Мы: уже подчеркивали не раз, что одной из характерных о-собен-
ностей деления постоянно амебоидных простейших является значи-
тельная степень независимости’ плазмотомии от хода ядерного де-
ления. К этому можно еще добавить, что оба эти процесса реаги-
руют по-разно-му на внешние воздействия в том смысле, что плазмо-
т-о-мия значительно легче поддается торможению, чем деление ядра.
Поэтому не приходится удивляться, что ее подавлени-е может вызы-
ваться люпбыми, в сущности, условиями, что оно очень легко пере-
ходит из временного в постоянно-е и что, благ-одаря этому, тут та-к
часто реализуется переход из одноядерного в Многоядерное -—-Ё
плазмодиальное, или симпластическое состояние.
Действительно, даже свободноживущие амёбы отличаются край-
ним непостоянством числа своих ядер, чем совершенно стирается
грань между обоими этими сост-ояниями. Это приводит, например,
к реальной невозможности ‘пользоваться признаком числа ядер для
четкого различения преимущественно плазмодиальных Ре1оп1уха от
факультативно многоядерных видов Amoeba. Что же касается‘
паразитических амебоидных форм, то тут ‘прямо можно сказать,
что лишь очень немногие из них постоянно «удерживаются» в,
одноядерном состоянии; гораздо чаще оно обязательно - сменя-
ется много-ядерным при превращении в цисту, а’ у ряда форм
(Endamoeba) дву- или‘ многоядерность достигается, еще в активной,
трофической фазе существования. В последнем случае одноядер-
ность характеризует ранние стадии онтогенеза," а факультативная;
или устойчивая многоядерность-более поздние. Таким образом,
156
т
перерастание истинно одноклеточной организации ‚молодых стадий
в надклеточную плазмодиальную становится тут правилом и полу-
чаетзначение закономерно-го онтогенетич-ес-кого процесса.
Этот процесс становится одним из основных морфологических
процессов в онтогенезе у всех высших Sarcod1na (Forammlf-era, pa-
диолярий) и «плазмодиальных споров-иков, неизменно начинающих
свое развитие в виде типично одноклеточных амебоидных или вре-
менно жгутиковых стадий (зооспор) и затем постепен-но умножаю-
щих свопи ядра до наступления момента размножения. Таким обра-
зом, наблюдаюЩаЯСЯ ТУТ Задержка плазменного деления приобре-
тает выдающееся морфогенное значение: ею целиком обеспечивается
создание плазмодиальной организа-
ции у Protozoa при громадном Под-
_час увеличении жи-вой› массы их те-
ла (рис. 46).
Не менее существенна роль по-
лимеризации и в формировании мно-
жественного- размножения; ЕЮ ВЫ-
зь1вается переход от монотомии к
первичной синтомии, т. е.
более или менее одновременному
распадению материнского плазмодия
на одноядерные дочерние отдельно-
сти. Это——основная форМа размно-
жения плазмодиальных простеиших,
оказывающаяся, естественно, тем бо-
лее продуктивной, чем выше число
ядер плазмодия к моменту его раз-
множения.
Синтомия однако не ЯВЛЯЕТСЯ
все же единственным’ видом размножения плазМодИаЛЬНЫХ ПРО‘
стейших. У многих из них-— как свободноживУЩИХ (РеЁЁ/ЁХЁЁКЁЁ:
которые радиолярии типа Acanthoma), так и паразитиче еЛеНИЯ
доспоридий)——оно осуществляется еще путем простого д ИНЫХ
плазмодия надвое или же его распадения на несколько кружив-
многоядерных кусков. В этих случаях мы имеем дело си B14331 B me‘
НЫМ размножением, на котором мы специально Останов .
ДУКЁЩЕИ Гл-Ёхбеазом окончательное снятие плазмотомии ‘в ходе
ных делений" служит источником возникновения самых разно“ раз!
ных типов плазмОдИаЛЬН0Й '0P‘1‘aH1’13a11“Ha Высшие из КОТОРЫХ д“.
стигают не меньшей сложности и ,lIH¢‘13‘ePeHHHP°‘BaH“°°TH' Чем .Ta.{§
каков’; характеризуются некоторые многоклеточные. Можно га;
зать, следовательно, что задержка плазменного деления тйёбо.
здесь своим конечным результатом УСТОИЧИВОЁ СОХРЗНЁНИЁ да EKG
идного типа строения, в рамках. КОТОРОГО "ОСУЩЁСТВЛЯЕТСЯЗЁТЁМ И
идущая, но исключительно «ядерная» полимеризация, а A
- 157
Рис. 46. Индивидуальное разви-
тие плазмодия (микрогаметоцита)
споровика Eimeria magma:
7 —- молодой одноядерный микрога-
метоцит; 2 — дВУядбРНаЯ СТЗДИЯЭ 3"
4-ядерная; 4 — б-ЯДЭРНЗЯ? 5 ‘"3 "j "О"
следующие многоядерные стадии (ПО
Хейсину)
функциональная дифференциация ее первично однородных про-п
дуктов. Последняя же приводит к созданию у ряда симпластиче-
ских простейших (Myxosporidia, Actinomyxidia) высоко специалич
зованных форм размножения и развития,
совершенно парал-
лельных наблюдаемым у Metazoa.
3. Надклеточные жгутиковые
Образование «временных сомателл наблюдается среди ж-гутико-
ВЫХ ТОЛЬКО у НЕКОТОРЫХ ПЗРЗЗИТИЧЭСКИХ фОрМ, а I/IMGHVHO трихомонад
Рис. 47. Образование временных сомателл у паразитических жгутиковых’:
1 -— 3 — последовательные стадии образования сомателлы Trypanosoma lewisi; 4 — ее синто-
мическое деление (стадия розетки); 5 — сомателла Trichomonas (no Кюну)
и трипанозом, и также обусловлено здесь «временной задержкой
плазмотомии. Все остальные же фазы деления протекают вполне
нормально и повторяются н-есколько раз подря , благодаря чему
и образуются многочленные сомателлы, содержащие несколько
полных «индивидуальных наборов» ядер, двигательных и опорных
органоидов в одном нера-зделившемся плазменном теле; образовав-
шаяся таким путем временная сомателла имеет вид цельного шара
или эллипсоида (рис. 47). B дальн-ейшем она расщепляется на
158
столько одноклеточных особей, сколько в ней содержалось полных
комплектов органоидов, что происходит‘путем их одновременного
обособления (рис. 47, 4) или же поочередным отпочковыванием од-
ной особи за другой (трихомонады).
Гораздо большим распространением по-льзуются среди Flagellata
постоянно надклеточные формы, классическим примером которых
являются высоко дифференцированные тм-нотожгутиковые предста-
Рис. 48:
1 --молодая. особь Devescovina striata; 2 Цпоздняя телофгза деления
этого же вида, представляющая полное подобие двучленной сомателлы
Diplozoa; з - сомателла Octomitus intestinalis (no Яницкому)
вители отрядов: Polyrnastigina (ВКЛЮЧЕН! Diplozoa) и Hypermasti-
шла, паразитирующие в кишечнике различных животны-х.
С- сравнительно анато-мической точки зрения сомателлы этих
замечательных простейших должны расцениваться, как продукты
разнообразных форм полимеризации, имеющей в разных случаях
различный объем (в смысле числа охватываемых ею структур) и
различную, та-к сказать, интенсивность. Они могут быть располо-
жены В очень ‘поучительные морфологические ряды, показывающие,
что организация этих животных могла создатвсятолько в резуль-
тате снятия тех или иных фаз клеточного деления при нормальном
159
или же повторном осуществлен-ни остальных фаз этого процесса,
т. е. путем частичных, не доведенных до конца дел-ений,-продукть1
которых остались соединенными в виде целостной надклеточной
сомателлы. ‚ _ V .
Так, организация Diplozoa может быть объяснена снятием
одной только плазмотомии при однократном прОДОдЛЬНОМ деЛеНИИ‚
. ' так как их двоиственная сома-
телла. морфологически соответ-
ствует двум не разделившимся
до конца одноклеточнЬ1м осо-
бям типа Devescovina (рис. 48,‘
49), содержа два полных набо-
ра соответствующих органои-
дов. У таких форм, как Ste-
phanonympha, Ka-K бы «выклю-
томия, но другие фазы деления
повторены многократно, так что
сомателла содержит тут мно-
жество таких наборов, назы-
ваемых «кариомастигонтами_>›
(т. е. особями, состоящими из
ядра и жгутикового аппарата)
И располагающихся в ней оп-
ределенным образом (рис. 50,
Рис. 49. Двучленные постоянные сома- I—2). B случае Calonympha
_ _ _Tf“—IU!b1D_iP10_Z0a= _ (рис. 50, 3) дело усложняется
7“ C‘”"“”‘ аг"‘5{„ёдс{1зд‘ёёё)т“"з (“З д‘°б°' тем, что не все частичные деле-
ния вовлеченные в процесс
построения сомателльт, осуществлены в одинаковомобъеме: в то
время как одни изних охватили как ядро, так и кинетиду с пара-
базальным аппаратом, другие как бы свелись к умножению этих
последних. В результате этого к полносоставным кинетидам при-
соединяются у Calonympha И многочисленные «акариомастигонты»
(т. е. образования представленные одними жгутиковыми аппара-
тами, безядра), состоящие только из ки-нетиды с парабазальным
аппаратом (рис. 50; 4). Следующую ступень в этом морфологиче-
ском ряду представляют таткие формы,'как Trichonympha, y KOTO~
рЫХ деление ядра оказалось полностью «выключенным» и полиме-
ризации подверглись только кинетиды и „парабазальные аппараты.
_ Натконец, в заключение этопо ряда» можно поставить Par-ajoenia
(рис. 50, .5), y которой умножению подверглись только жгутиковые
аппараты и которая как бы олицетворяет, таким образом, резуль-
тат многократного повторения одних только делений кинетидьг.
1 дЭтогг же морфологический ряд показывает «с полной определен-
ностью, что усиление полимеризации неразрывно связано с увели-
чением размеров тела и общим повышением организации живог-
норо, а также и с повышением его целостности. Последнее выра-
160
чена» ТОЖЕ одна ЛИШЬ плазмо-ч
ж
Рис. 50. Взрослые многочленные сомателлы жгутиковых:
7 -— Stephanonympha silvestrii, общий вид; 2 -— разрез передней части сомателлы, показываю:
щий состав и расположение отдельных кариомастигонтов; 3 —Calonympha grassu, общин
вид; 4 — схема состава и расположения карио- и акариомастигонтов; 5 — Paramenia
grassu, общии вид (по Яницкому)
11 Эмбриология беспозвоночных 161
жается здесь в нарастающей дифференциации первично однородных с
множественных структур, в их упорядочении и превращении всей
их совокупности во все более связную систему (например, двига-
тельную). ‘ _
Еще ‘более наглядное представление -о ходе и результатах поли-
меризации, чем описашый срав-нителнно анатомический ряд, дает,
разумеется, непосредственное наблюдение этого процесса в ходе
ИНДИВИДУЗЛЬНОГО развития таких ВЫСОКО ПОъЛИМЕЭрНЫХ }КГУТИКОВЫХ,- р
как Spirotrichonympha И I-Iolomastigotes (Hypermastigina). У пер-
вого из этих родов самые молодые из известных нам стадий раз-
Рис. 51. Индивидуальное развитие Spirotrichonympha ветошь Объяснение
см. в тексте (по Дюбоску и Грассе)
вития еще очень малы (3——5р) и довольно похожи на представителей
Шр1о2оа, но имеют только одно ядро. Они обладают двумя сим-
м-етрично расположенными опереди кинетидами, состоящими каж-
дая из четырех жгутов и одного парабазального аппарата, тесно
приложенного к ядру; аксостиль на этой стадии (рис. 51,1), по-ви-
димому, еще отсутствует, появляясь лишь несколько позднее (2, 3).
На следующих стадиях онтогенеза продолжается повторное удвое-
ние числа кинетид и парабазальных аппаратов; последние распола-
гаются теперь венцом вокруг ядра (рис. 51, 4—II). После этого в раз-
витии молодой особи, наступает переломный момент, вн-ешн-е выра-
«жающийся в резком изменении характера ее морфогенезов. Все
умножающиеся кинетиды выстраиваются теперь в спиральные
ряды, сначала опоясывающие только переднюю часть тела, а зат-ем
162
распространяющиесянзади (рис. 51, 12-14); под въггкамгт спирали
располагаются также УМ-НОФКаЮЩИ-ЭСЯ, но теперь уже в более
медленном темпе, парабазальные аппараты; тело, имевшее до
этого форму волчка, становится веретен-овидньгм, его передний ко-
нец удлиняется в виде хоботка‚ Та-к создается спирально симметрич-
ная взрослая стадия Зрйготггспопушрпа, почти не уступающая по
Рис. 52. Индивидуальное развитие Holomastigotes elongatqm (по Дюбоску
и Грассе)
СЛОЖНОСТИ ‘СВОЕЙ ОрПГЕПНИ-ЗЗЦИИ ИНфУЗОРИЯМ, НО так И ОСТаЮ-ЩЭЯСЯ
- одноядерной.
Развитие другого представителя Hypermastigina, Holomastigotes, известно
начиная со стадии, изображенной на рис. 52, 1. Она имеетуже очень многочис-
ленные жгуты, «расположенные по спирали в два- завитка, но только один пара-
базальный аппарат. При дальнейшем развитии и росте тела винтовой ряд кине-
тид образует новые обороты; при этом полимеризация жгутиковых аппаратов
идет в гораздо более бу.рном темпе, ч-ем умножение паранбазальных телец, общее
число которых остается незначительным.
В обоих этих случаях мы видимфследовательно, очень сильное
усложнение начального строения простейшего в ходе его онтоге-
неза, идущее одновременно с его ростом и заключающееся в силь-
”* 163
нейшей полимеризации жгутикового и парабазального аппаратов.
У Spirotrichonympha ИХ умножение идет сначала почти синхронно,
н-о позднее кинетиды м-ножатся гораздо быстрее, чем парабазаль-.-.
ные тельца, а у I-Iolomastigotes полимеризация тех и других идет
с самого-начала с разной скоростью.
Приспособит-ельиый характер я-влений полимеризации прояв-
ляется ярче всего на примере двигательного аппарата. Усиление его
МОЩНОСТИ ПО Mepe роста, УВЕЛИЧЕНИЯ массы тела, ДОСТИГЗЕТСЯ"
именно этим способом. Благодаря полимеризации и совершается
замена движе-ния посредством жгутов, характерного для мелких
организмов, гораздо более выоокой- мерцательной формой пере-
движения, обеспечиваемой согласной работой громадного числа
жгутов.
4. Сомателлы опалин и ресничных инфузорий
Как бы ни решался вопрос о происхождении инфузорий, не-
сомненно, что они представляют собой продукт далеко заш-едшей
гипертрофии некогда одноклеточных жгутиковых форм, сопряжен-
ной с бурной полимеризацией жгутикового аппарата и большим
или меньшим умножением числа ядер, или, другими словами, что
их организация создавалась в филогенезе примерно тем же путем,
ка-к у Ро1у- и I-Iypermasti-gina.
Теперь стало также очевидным, что оба глав-ных подразделения
ресничных инфузорий (о сосущих мы будем «говорить дальше):
опалины (Protociliata), с одной стороны, и настоящие ресничные
(Ci1iata)——_c другой, не находятся хоть в сколько-нибудь тесной
генетической связи и обладают, в частности, фундаментально раз-
личной организацией. Однако в обеих этих группах очень ясно
проступает икх ж-гутиковая—хпоть и разная-первооснова и сх-од-
ное, в общем, направление ее видоизменения. - -
Имеющиеся сходства между Рготоспйата и Ciliata заключаются
прежде всего в- наличии у тех и других обильного ресничного по-
крова, состоящего из продольных, в тише, рядов тесно сближенных
друг c другом ресничек.
Пути формирования такого многочленного двигательного anna-
-рата, характернейшего из атрибутов «Ciliophora», долгое время
оставались неясными. Еще не так давно высказывались самые про-
тиворечивые мнения на этот счет, но теперь можно считать установ-
ленным, что каждая ресничка инфузорий представляет собой целую
кинетиду, состоящую из тех же частей, как кинетида одножгутико-
вь1х Zooflagellata (рис 53). Ее жгут представлен здесь самой рес-
ничкой‚ Мастигосома — ее базальным зерном, парабазальный ап-
парат — инфрацилиарным тельцем, также состоящим из дв-ух
. веществ —— хромофильного и хромофобного. Базальные зерна ресниц
. являются постоянными органоидами, обладающими непрерывной
ПрЕЕМСТВЕННОСТЬЮ ИЗ ОДНОГО ПОКОЛЕНИЯ В ДруГОЕ И СЛУЖЭТ ИСТОЧНИ-
КОМ образования ВСЕХ ОСТЗЛЬНЫХ ЭЛЕМЕНТОВ КИНЕТИДЫ; ОНИ умножа-
164
ются делением, при котором создается типичный парадесмоз. Сово-
купность базальных зерен образует постоянную и н ф р а ц и л и a-
Рис. 53. Инфрацилиатура ресничной инфузории Chi1odo-
nella uncinata И ее изменения в ходе деления:
1 -—базальное зерно (б) и парабазальный аппарат (п) отдельной рес-
нички; II—IV—. умножение кинетид в ходе деления, образование дочер-
них 6-го и 7-го ряда ресничек из материнского 5-го p>=ma;V—-VIII перест-
ройка инфрацилиатуры на последовательных стадиях деления; указаны
принятые цифровые обозначения отдельных Рядов ресничек (по Шат-
' тону и Львову)
туру, которая сохраняется при дедифференцировке инфузорий и
дает начало новым ресницам и инфрацилиарным тельцам при ее
возвращении в дифференцированное состояние. -
I65
Таким образом, двигательный аппарат инфузорий представляет
собой не‘ что иное, как систему кинетид, подвергшихся тут сильней-
шей полимеризации и упорядочению своих элементов.
ПРОИЗВОДНЫМИ КИНЕТИД ЯВЛЯЮФСЯ такж-е ТРИХОЦИСТЫ И ТрИХИТЫ .'
инфузорий, а также имеющи-еся иногда у них стрекательные
капсулы. При образовании первых базальное зерно кинетИдь1 де--
лится таким же образом, как при умножении ресниц, но иногда .
повторно; одно из дочерних зерен остается у инфрацилиарного
т-ельца, а другое (или дру-гие) отодвигается, и из него вырастает
трихит или трихоцист.
Мембранеллы, Цирры и другие сложные двигательные и осяза-
тельные органоиды настоящих инфузорий образуются также из
кинетид, но путем их склеивания в единицы более крупного
порядка. .
Итак, исключительное разнообразие двигательного и стрека-
тельного вооружения Ciliata имеет своим источником разносторон‹
ние видоизменения ЖГУТИКОВОГО аппарата: его безграничную ПОЛИ-
меризацию, преобразование части его элементов В ОРГЭНОИДЬ}
специального‘ назначения и далеко идущую дифференциацию
остальных. Что же касается Protociliata, то их двигательный аппа-
рат удерживается на более примитивной ступени развития, оста-
ва-ясь вполне однородным. '
Сомателлы опалин занимают эВ известной мер-е промежуточное,
положение между плазмодиями и истинными сомателлами. Они
характеризуются именно полной однородностью своего двойного
или, чаще, множественного ядерного аппарата, представленного
ядрами довольно своеобразного строения, и, как уже указывалось,
весьма однородным рес-ничным покровом, образующим однотипные
продольные ряды. Эти особенности строения опалин в сочетании
с присущими им особенностями размножения -и развития позволяют
считать, что гипертрофическое раз:витие Protociliata ШЛ0 от двуядер-
ных форм к многоядерным и сопровождалось интенсивным умно-
жением кинетид, не дошедшим, правда, до их дифференциации.
В общем, организация опалин все же ясно обнаруживает свою
жгутиковую основу, но основу, значительно более примитивную
(а тем самым и более амёбоидную), чем у исходных (одноклеточ-
ных) типов Polymastigina. Опалииы олицетворяют вместе с тем и
особое направление ядерно-жгутиковой полимеризации, довольно
близкое к наблюдаемому у только что названной группы жгути-
ковь1х‚ но отличающееся, во-первых, еще большей независимостью
умножения кинетид от ядерно-го деления, "а, вот-вторых, полной
однородностью ядер и ки-нетид.
Г ипертрофическое развитие настоящих ресничных инфузорий
также шло, по всем данным, от двуядерных, и притом ра-вноядер-
ных, сомателл, но уже ‘высоко дифференцированных и обладавших
анимальным способом питания. Вполне конкретное представление
о морфологическом прототипе Ciliata могут дать молодые вегета-
166
тивные особи замечательной морской равноресничной инфузории
Stephan-opongo-n mesnili.
Исключительная примитивность этой хищной формы выра-
жается ярче всего в том, что оба ее первичных ядра совершенно
идентичны, а также и в малых размерах ее ранних стадии разви-
тия, т. е. в фактическом отсутствии у них определенных признаков
гипертрофии. Последняя _целиком осуществляется в онтогенезе,
сопровождаясь простым умножением ядер по мере увеличения жи-
вой массы сомателлы. К концу индивидуального развития число
ядер доходит до 10—24, в зависимости от размеров, достигнутых
простейшим. По оконча-
нии питания и роста жи-
вотное инцистируется и
делится на 5—12 двуядер-
нь1х особей! (рис. 54).
Особо ‘следует ‘подчерк-
нуть, что все ядра Ste-
phanopogon mesnili — и
первичные, и образующие-
ся из них вторичные—
вполне равноценны друг
другу в морфологическом
и функциональном отно-
шении; единственным на-
меком на дифференциа-
цию ядерного аппарата Ч
является некоторое рас-- Рис. 54. Индивидуальное развитие равно-
Хождение в сроках (и, Мо- ядерной инфузории Stephanopogon mesniliz
ЖеТ быте темпах) деления Z.;;.,’i§¥.$’i§Z"E‘$§L$S“§£1’fm?;;°%‘L§1‘.?;‘;‘§§§§$’,3”.333;
первичных ядер, ‚из кото- почке материнской цисты (по Львову) -
рых заднее обычно _опере-
жает в делении переднее.
Как видно из этих данных, гипертрофич-еское развитие идет
у S. mesnili тем же, в сущности, «способом, как у амёбоидных про-
стейших и имеет своим результатом создание многоядерной вр-е-
мегн-ной сомателлы второго порядка, раьопадающуюся затем на
двуядерные «элементарные» сомателлы. Таким образом, двуядер-
ная (и притом равноядер-ная) основа организации Ciliata прояв-
ляется тут самым наглядным образом, и к тому же в сочетании
с наиболее Примитив-ной из форм частичной и временной дистомии.
Однако при всей своей примитивности в только что указанном отно-
шении S. mvesnili характеризуется и многими признаками высокой
специализации, как T02 неравномерным распределением реоничного
покрова, глоткой, усаженной трихитами, наличием -набора околоро-
товых мембранелл и т. д. Короче говоря, это-вполне типично
сложившаяся инфузория, хотя и самая примитивная из всех. '
Остальные Ciliata существенно отличаются от S. mesnili резко
выраженным ядерным диморфизмом при пожизненном сохранении"-
167
двуядерности И удержании ПрИ РЗЗМНОЖЭНИИ СВОИХ ги-пертрофирод
ванных размеров. У громадного большинспваСШаЪа ядерный аппа-
par ‚представлен, как известно, непропорционально маленьким:
микронуклеусом‚ относящимся к <<»CJI1~epMaJIbH0MY>> TPWY ядер, И_
сильно гипертрофированным макронуклеусом, принадлежащим-
к «массивному» их типу (Догель).
Рис. 55. Статомоторный аппа-
рат Boveria zenkevltshiz
7——базальнь1е зерна продольных
ресничных рядов; 2——продольные
фибриллы; 3—-тонкие продольные
фибриллы; 4—базальные зерна попе-
речного ряда ресничек; 5—базаль-
ные зерна околоротовых ресничек;
б-передняя околоротовая фиб-
рилла; 7—задняя околоротовая
фибрилла; 8—соединительные фиб-
риллы; 9——ротовое кольцо; 10-
Макронуклеус имеет самую разно--
об-разную форму, иногда очень причуд-—-
ливую (вплоть до сетчатой у некото-
рых эндопаразитических Foettingerii-
dae), делится амитотически и раство-
ряется при конъюгации и других фор-
мах реорганизации.
Новый] макронуклеус создается из
продуктов деления микронуклеуса‚
иногда (например, у Stentor) путем
СЛИЯНИЯ НЕСКОЛЬКИХ ИЗ НИХ В ОДНО це-
лое. Уклонения от этой нормы идут
чаще ВСЕГО Br >CTO‘pOHy УМНОЖЕНИЯ ЧИСЛЗ
микронуклеусов, заходящего особен-
но далеко у самых крупных разнорес-
ничнь1х, вроде трубача (Stentor) ИЛИ
Bursaria; увеличиваясь в числе, они
сохраняют, однако, характерные для
них мелкие размеры, если только не
превращаются в макронуклеусы (как
замыкающая фибрилла; 11—опорное
утолщение („о Левинсону) это происходит у Loxodes, где за счет
. постоянно делящихся микронуклеусов
возникают пузыревидньте макро-нуклеусы, лишенные здесь способ-
ности K делению). .
Итак, главн-ой особенностью ядерного аппарата ресничных
является необычная судьба м-акронуклеуса: подвергаясь далеко
идущей гипертрофии, он утрачивает способность к митозу (или
всякому делению вообще) и превращается в образование, Целиком
создающееся заново посл-е каждой конъюгации.
Для инфузорий характерно также сильное развитие опорных и
сократительных структур, образующих целостный статомоторный
аппарат (рис. 55).
5. COMaTeJlJIbl сосущих инфузорий
Анализ организации Suctoria не оставляет сомнений в том, ‘что
они представляют собой самых настоящих инфузорий, вегетативные
стадии которых подверглись ряду любопь1т-нь1х преобразований
в связи с переходом K особому способу питания и преимущественно
сидячему образу жизни. . .
Молодые овободноплавающие стадии э-тих инфузорий ~— бро-
дяжки или «эмбрионы» (рис. 56, 1) —мало ч-ем отличаются от беде
168
ротых форм Сшайа. Они обладают хорошо развитымресничным
покровом, то равномерно одевающим „всю поверхность тела- как
у равноресничных, то образующим поперечные плавательные пояса
и иногда «теменной» султан. Строение отдельных кинетид таково же,
как у Ciliata, HO OHH не образуют тут правильных продольных ря-
Рис. 56. Инфрацилиатура Ёосущей инфузории Рооорпгуа
V пха:
1 —свободноплавающий эмбрион; 2 —сидячая взрослая стадия;
3, 4 —- различные стадии образования, инфрацилиатуры эмбриона
при почковании (по Шаттону и Львову)
дов; однако, на ранних стадиях формирования инфрацилиатуры
мастигосомы имеют правильное рядовое расположение (рис. 56, 3).
‘Превращение «эмбриона» ив вегетативную особь сопряжено
С дедИфференЦировкой ресничного покров-а, от которого остается
одна инфрацилиатура (рис. 56, 4). Ha обнажившейся таким обрад
зом поверхности тела V, образуются радиальные сосательные щу-
пальцы, то равномерно разбросанные по всей его окружности, то
169’
собранные в пучки или венцы. Щупальцы Suctoria (рис. 57) обла-.
дают довольно сложным строением и состоят из: 1) слоя кортикаль:
ной плазмы, одетого снаружи тонкой пелликулой; 2) скелетной
нити, идущей по внутренней поверхности пелликулы; 3) ос-евого
плазматического столбика, заканчивающегося грушевидным взду-
тием (Роскин, 1937).
Таким образом, особи Suctoria ‘приобретают внешнюю форму и
‘НЕКОТОРЫЕ ИЗ ЧбрТ ОРГЭНИЗЗЦИИ, 'Ha|H'OMI_/IHEIIOLLLPIE ‘COJIH€‘«IHI/I‘KO'B.
Однако дифференциация щупальцев идет здесь много дальше, чем
у солнечников, особенно у сидячих форм, где они часто распола-
гаются в виде венца или собраны :в пуЧкИ. ДаЛЬШе Идет И дифферен‘
Циация общей формы тела, ведущая к созданию подлинно mowin-
ПОИДНЫХ форм, вроде
Ephelota ИЛИ колони-
ального Dendrosoma,
точно повторяющих в
Рис. 57. Сосательная трубочка Podophrya sandi миниатюре все архи-
(ПО РОСКИНУ) тектурные особенности
щолоний гидрополипов.
Характерным свойством Suctoria является также способность
полипоидной особи временно превращаться в свободноплавающую
мерцательную бродяжку совершенно такого же строения, ка-к
«эмбрион», и впоследствии вновь возвращаются к полипоидному
строению. Столь полная обратим-ость организации Sucto-ria при на-
личии у них дифференцированной ресничной фазы и’ частично
дедифференцированной «фазы солнечника» составляет в- высшей
степ-ени замечательную черту этой группы инфузорий. Особого вни-
мания заслуживает то обстоятельство, что переход в трофическую
вегетативную стадию осуществляется путем дедифференцировки
ресничной фазы и последующей редифференцировки организма,
в принципиально отличном направлении; или, другими словами,
что тут имеются не одна, а две глубоко различные дифференциро-
ванные фазы, реализуемые в зависимости от определенных внешних
условий. Замечательно также, что они обе развертываются на
основе двуядерной сомателлы с типичной для инфуз-орий постоян-
ной инфрацилиатурой, одинаково способной дифференцироваться и
нормальным способом, и путем новообразования целого комплекса
временных полипоидньтх или «псолнечнилковых» структур.
Г л а в а 1 Х
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ НАДКЛЕТОЧНЫХ PROTOZOA
1. Общие определения
Достижение надклеточной ступени организации неизбежно вле-
чет за собой переход от монотомии и ее непосредственных произ-
водных к иным, новым формам бесполого размножения. Если над-
клеточность имеет только временный, преходящий характер и.
170
сменяется при размножении возвращением к одноклеточному исход-
ному состоянию,—это происходит путем синтомии; если же она
становится постоянной, то размножение теряет тем самым свой
первичный характер клеточного и переходит ‚в разряд вегет а-
тивного " ' ’‚
_ В самом деле, все виды размножения ‹плазмодиев’и сомателл,
при которых они не делятся на одноклеточные (одноядерные)
отдельности—будь то их однократное или повторное «простое
деление» надвое или почкование, существенно отличаются от
любых форм клеточного деления уже тем, что их продукты имеют
не одноклеточное, а такое же надклеточное строение, как и соот-
ветствующие взрослые организмы. Мы видели, что, независимо от
того, двуядерна ли сомателла, как у Diplozoa И типичных инфузо-
рий‚ многоядерна —как у Calonymphidae И сверхгипертрофирован-
ных разноресничньгх, или же имеет одно, но гипертрофи-рованно-е
ядро, как у Hypermastigina, — она неизменно делится надвое, а не
каким-либо другим способом. Поэтому кажется даже странным, что
эти виды размножения до сих пор еще отождествляются с моното-
мией одноклеточных, а не с соответствующими им формами вегета-
тивного размножения Мета2оа. ‘ '
Основное разнообразие видов вегетативного размножения жи-
воггных может быть сведено к следующей схеме, одинаково .прило
жимой к Metazoa И большинству надклеточных простейших: ‘
1. Деление надвое.
А. Деление без предшествующей морфогенетической подготовки;
осуществляется простым перешнуровыванием или расщеплением
тела и обычно завершается регенерацией обеими особями недо-
стающих частей (а р х е т о м и я). ’
Б. Детте-ние после предшествующего удвоения частей, распреде-
ляемых между обоими его продуктами (п а р а т о м и я).
П. Повторное деление надвое.
А. Повторная архетомия.
Б. Повторная паратомия. —
111. Фрагментация.
А. Одновременное распадение материнского тела «на несколько
более или менее равноценных кусков, перестраивающихся затем
в целые небольшие особи (фрагментация в собственном смысле
слова). у
Б. Отщепление или автотомия отдельных кусочков от материн-
ского тела.
IV. Почкование.
А. Образование почек на самом теле материнской особи (на-
стоящее почкование).
Б. Образование столонов и почек на них (столонизация).
171
2. Саркодовьпе и плазмодиальные споровики
Вегетативное размножение этих форм осуществляется чаще;
всего путем деления надвое, обычно называемого плазмотомией,.
и различными видами фрагментации, частью близких к почкова-
НИЮ. ДЕЛЕНИЕ надвое часто СВОДИТСЯ K l'IpOCTOMy l'I€p‘€IHHypOBbI-_
I/IMSPOHLI/IXICH ядер между
обеими его половинами.
В этих случаях, нередких
у Myxosporidia, Foraminifera
H некоторых радиолярии,
-оно явно отвечает понятию
архетомии, особенно тогда,
когда вслед за делением
происходят _H регенератив-
ные процессы (например,
у Асапйюпйа), где они выра-
жаются B новообразовании
недостающих частей скелета.
В других случаях столь же
простой плазмотомии пред-
_ шествует удвоение ядер,
благодаря чему ее прих-одится уже считать паратомией (Ре1отуха‚
некоторые другие Amoebina). Архетомия может быть повторной
(Cal-cituba) H ин-огда переходить в синт-омию (приводя к образова-
нию одноядерных qa~cTe131)'HJIH в фрагментацию. Эта последняя
также нередка и часто приобретает формы множественного почко-
вания (например, у Ichtyosporidium, рис. 58). .
В общем, у саркодовых и пплазмодиальных опоров-иков осу-
ществляются почти все виды твег-етативного размножения, но они
слабо разграничены и ‘производят впечатление неустановившихся,
что вполне гармонирует с общей ‚бесформенностью плазмодиев.
ванию ПЛЭЗМОДИЯ И распределению
Рис. 58. Фрагментация
плазмодиального У
споровика Ichtyosporidium hertwigi (по
- Сварчевскому)
3. Полимерные жгутиковые
Полную противоположность сплазмодиям составляют и в этом
отношении некоторые надклеточные Flagellata, ‚размножающиеся,
как уже указывалось, исключительно продольным делением надвое.
Поскольку ему всегда предшествует удвоение частей, распределяеъ
мых __между обеими дочерними сомателлами (рис. 59, 60), оно,
безусловно, имеет характер паратомии, и притом продольной. Поло-
жение плоскости деления зависит от типа симметрии сомателлы.
Так, у билатеральных Diplozoa она располагается фронтально.
Таким образом, паратомия «полимерных Fla-gellata взначитель-
‚ной-мере сохраняет все особенности, характерные для монотомии
жгутиковых. Как происходитразмножение таких форм, ‘как Spiro-
trichonympha H Holomastigotes—HeH3BreQTHo, HQ ОНО, несомненно,
является множественным,
172
Рис. 59. Начальная стадия паратомического деления Stepha-
nonympha silvestrii; деление ядер и кинетид (по Яницкому)
Ч
«\ 1
I
c ‘
_ WI
Рис. 60. Продольная паратомия Calonympha grassii (no
Яницкбму) 7
I 3
4. Опалины `
БЭСПОЛОЭ pa3MHO}K€HI/I6 ОПЭЛИН ПРОИСХОДИТ ИСКЛЮЧИТЕЛЬНО ‘ПРО-р
дольным (вернее, диагональным) делением ‘надвое, совмещающим
в себе признаки паратомии и архетомии. Принципиальное отличие
деления опалин от такового ресничных инфузории заключается
в том, что его плоскость ложится п а р а л л е л ь н о рядам ресни-
чек, B одном из промежутков между ними, а не разрезает их, как
там, в поперечном направлении. В соответствии с этим материн-
ский двигательный аппарат разделяется примерно поровну между
обеими дочерними особями, и каждая из них пополняет достав-
шийся ей набор целых продольных рядов ресниц примерно таким же
числом новых, образующихся в промежутках между материнскими
рядами. Ист-очником формирования этих вставочных новых рядов
СЛУЖИТ рЗЗМНОЖЗЮЩИЭСЯ ОбИЛЬНЫе ‘МЁШСТИТОСОМЫ, располагающиеся '
по переднему ребру уплощенного тела животного. Таким образом
ряды ресниц, вырабатываемые каждой особью, образуют тут цели-
ком заново, вне всякой овязи с-о старыми, материнскими рядами
(Шатто-н и Брашон, 1936).
5. Ресничные инфузории (паратомия)
Господствующей формой бесполого размножения инфузорий
является, как известно, «простое» поперечное деление надвое,
осуществляемое в подвижном состоянии. У некоторых, ‚преимуще-
ственно сидячих, форм оно приобретает характер почкования,
а у ряда равноресничных—гла.вным образом паразитических-
происходит в цисте, и тогда повторяется неоднократно, подобно
настоящей палинтомии; наконец, у Stephanopogon наблюдается и
полное подобие синтомии. Однако при всех без исключения видах
размножения оно дает начало двуядерным (как минимум) сома-
теллам, чем убедительно показывается его характер вегетативного,
а не клеточного размножения.
В основе всех видов размножения Ciliata лежит самая типичная
поперечная паратомия, точно соответствующая ее классическим
обра-зцам у Metazoa. Одновременно с этим она сохраняет и все
свойства истинно поперечной мо-нотомии-как, впрочем, и у тур-
беллярий или аннелид.
Продуктами паратомического деления являются две подобные
друг другу, но не всегда одинаково крупные особидчередующиеся
своими полюсами и несколько разнящиеся между собой по способу
своего образования, почему их и обозначают особыми названия-ми:
переднюю называют п р и M И т о м, а заднюю — с а т е л л и т о м.
У всех разноядерных инфузорий паратомия связана с кариоки-
нетич-еским делением микронуклеуса и амитозом макронуклеуса, и
имеет в этом отношении строго равномерный характер; у видов
со многими ядрами последние также равномерно распределяются
между обеими дочерними особями. Гораздо менее равномерно рас-
174
пределяют-ся материнские дифференцировки и, ‚в частности, отдель-
ные составные части ДВИГаТ-ЭЛЬНОТО аппарата, имеющие B ЭТОМ
смысле весьма различную судьбу. V
B самых ПрОСТЫХ случаях ДЭЛЭПНИЯ В аКТИВНОМ СОСТОЯНИИ, K-OH-
кретным прим-ером КОТОРЫХ МОЖЕТ СЛУЖИТЬ равноресничная
Glaucoma, примит получает ОТ матери В -COBv€pU.I€HHO НЭИЗМЭНЭННОМ
‘виде рот со всеми окружающими его структурами и переднюю поло_-
вину всех 27 продольныьх рядов ресничек, тогда как задняя их
Рис. 61. Образование
зачатка рт_а у задней дочерней
особи (сателлита)
при делении инфузории Glaucoma
scintillans:
7—инфрацилиатура материнской особи; звездочкой отмечен ряд,
дающий начало инфрацилиатуре рта дочерней особи; 2 —б-—
последовательные стадии образования последней; 7—участок
одного из рядов при большем увеличении (по Шаттону,
А. и М Львовым и Моноду)
половина вместе с зачатком нового рта достается сателлиту. Этот
зачаток возникает здесь путем отщепления от инфрацилиатуры
одн-ого из рядов ресниц небольшойцеплочки новых базальных зерен,
начинающих затем усиленно делиться, образуя таким путем основу
вооружения нового рта (рис. 61). Таким образом, примиту прихо-
дится регенерировать только заднюю половину рядов ресниц (что
достигается соответствующим их удлинением при умножении со-
ставляющих их кинетид),тогда как сателлит регенерирует их п-еред-
ние половины и заново формирует свой ротовой аппарат.
Очень часто дело усложняется тем, что наиболее сложные из
частей двигательного аппарата дедифференцируются при делении,
причем не только исчезают реснички и инфрацилиарны-е тельца, но
175
часто происходят и более глубокие перестройки. Так, при делении’
паразитической Proboveria loripedis, инфрацилиатура околорото-
вых образований двигательного аппарата сильно раздается-в длину,‘
принимая диффузный характер, а затем распадается на несколько `
неравных частей, р-еорганизующихся совсем дно-разному у примита
и сателлита (рис. 62). Еще много сложнее протекают эти процессы
у брюхоресничных (Hypotri-cha), y которых обе дочерние особи реге-
нерируют весь реснич-ный аппар-ат, хотя каждая и получает свою
половину ресничного аппарата материнской особи: все материнские
реснички атрофируются и заменяются затем новыми.
Делению в цисте предшествует, как уже сказано, дедифферен-
цировка, выражающаяся в сведении ресничного аппарата к по-
стоянной части-инфрацилиатуре (т. е. в сбрасывании самих
ресниц, исчезновении и=нфрацилиарнь1х телец и, когда они несть, ри-
зопластов), соответствующей редукции рта и специальной подго-
товке других элементов двигательного аппарата к последующему
разделению и ра-спределению. Представление о ходе этих и сле-
дующих за ними реконструктивных процессов может дать- серия
рисунков, касающихся Chilodonella uncinata (CM. рис. 53). По ним
легко проследить, как происходит формирование всего сложного
двигательного аппарата сателлита из немногих частей материнского:
отщепление новых рядов ‹<7», а затем «6» от задн-ей половины
пятого материнского и создание‘инфрацилиатуры нового рта из
распадающейся на куски части ротового ряда (<<12» . _ *
Как видно из этих примеров, паратомия протекает тут почти
одинаковым образом в активном состоянии и в цист-е, сопрово-
ждаясь в обоих случаях той или и-ной степенью дедифференцировки
реоничного ап-парата и его значительной перестройкой. И хо-тя
дедифференцировнка- совпадает здесь по своему содержанию с ана-
логичными видоизменениями кинетиды у жгутиковых-паратоми-
ческое деление инфузорий все же имеет „больше общего с прото-
зойным типом Монотомии, ч-ем с метазойньхм. Это находит свое
выражение в постоянстве и генетической преемственности инфра-
цилиатуры и пелликулярной сети, как бы удерживающих сомателлу
в устойчивом, мало изменяющемся структурном состоянии даже
в момент дедифференцировки, и не допускающей, таким образом,
большого снижения ее организации.
Кроме того, высокая сложность инфрацилиатуры приводит
к тому, что она, являясь, по существу, у инфузорий системой «по-
стоянных» органоидов (мастигосом), сохраняет некоторые черты,
характерные для дифференцировок. Действительно, мы уже видели,
что ее преемственность из одного поколения в другое далеко
не всегда является прямой: наряду с прямой обычно имеет место
и непрямая преемственность -ее частей, а именно их новообразова-
ние из материнского зачаткового материала. Такое новообразо-
вание может осуществляться двумя путями: отщепл-ением новых
рядов мастигосом от материнской инфрацилиатуры ‘путем их одно-
временного деления (какг это «было показано у С1111ооопе11а) или же
176
rlrfivwi-coco.
’ ‘ i‘s":':3‘u
2
t
.
Х
а
‚ ....—-:.-._r.“
4%"
Рис. 62. Последовательные стадии паратомии инфузории Proboveria loripedis-
ХОД Перестройки и распределения инфрацилиатуры (поШаттону и Львову;
12 Эмбриология беспозвоночных
Через временный перех-од отдельных частей последней в диффузное
состояние, с ‚последующей перегруппировколй и умножением обра-
зующих их Мастигосом (как в случае околоротового в‹о›о'ружтения_
Probov-eria). Благодаря этим особенностям признаки протозойно-го
типа дел-ения и получают тут явный перевес над метазойными. ‘
6. Повторная паратомия ресничных инфузорий
Этот своеобразный вид размножения инфузорий обладает всеми
признаками тиупичнейшей линейной палинтомии, за исключением
Рис. 63. Повторная паратомия Gymnodinioides inkystans:
1 —— 3 — последовательные стадии подготовки материнской особи к делению; 4, 5 — первое
деление; б-стадия 16. На всех рисунках показана инфрацилиатура рядов ресниц (по Шаттону
и Львову) . -
ТОЛЬКО ОДНОГО, НО! зато капитально ВЗЖНОГО ПрИЗНЗКЗЗ ТОГО, ЧТО ОНО
совершается ПрИ УСТОЙЧИВОМ сохранении НЭДКЛЭТОЧНОГО УРОВНЯ
ОрГЗНХ/ЮЗЦИИ, так ЧТО ЭГО продуктами оказываются не одноядерные
КЛЕТКИ, а ПОЛИМЕРНЫЕ двуядерные ЮОМЭТЭЛЛЫ; ` _ ‚ _- ‘
178
Повторная паратомия встречается в факультативной форме
У ДВУХ родов свободноживущих равноресничныхт Co-lpoda. и
С1111ооопе11а, и в облигатной, ‘гораздо более продуктивной форме
"*-.........и
“Рис. 64. Индивидуальное развитие инфузорий сем. F0ettingeriid'ae: '
, 7‘*—'4 —Vampyrophrya ре1адйсаг1 — бродяжка (цилиоспора); 2 — сидячая покоящаясярстадия}
3 -— взрослая вегетативная особь; 4—дедифференцированная стадия накануне деления;
;-5‚ б — Spirophrya hypertrophicaz 5 — еще не’ дифференцированная цилиоспора; ‹б Ь-готовая
-; . 4 цилиоспора (по Шаттону и Львову)
у Целого ряда паразитических и.нфузорий: известного эктопаразита
рыб I»chty.o.phthirius всей обширной группы Арозйошщ Особым
вариантом повторной парат-омии, но уже скорее табличной; чем
12* fig
п 1 д- ЗЕ- Hyalospira
линейной, является и повторное делеттйе
колониальных еувоек;
о котором будет -ещ-е речь впереди.
7 б
Рис. 65. Морфологические картины повторной паратомии различных безротых
' инфузорий (Apostomata) . .
„ caridinae, заключительная стадия 2-го деления (1), стадия 4-(2),
З—е‚деление_ (З) (по Мияшита); 4—Sp1rophrya subparasitica, стадия 4; расположение
продуктов деления в виде «палинтомического креста»; 5—З-е деление у того же вида;
полная иллюзия деления в`трех взаимно перпендикулярных плоскостях; б, 7 ——Synophrya
hypertrophica; б — стадия 4; 7 — стадия 10; 8, 9—Polyspira delagei; часть цепочки стадии 16
(8) и_стадш»1 32 (9); 10—Chromidina elegans; цепочка продуктов повторной паратомии
(по Шаттону и Львову)
Подобно палинтомии, точны-м аналогом которойона является,
повторная паратомия представляет собой серию сближенных во
180
"I‘O>.TIbKO инфраЦилиатур-а не
- ной формой повторной пара-
‘времени «простых делений» в дедифференцированном состоянии
организации, не прерываемых «питанием и ростом и потому в-едущих
к мельчанию продуктов деления в геометрической прогрессии.
Равным образом ей предшествует гипертрофический рост материн-
ской особи и ее дедифференцировка перед размножением, выра-
жающаяся тут в сложном метаморфозе; так же, как при палинтомии
конечные продукты размножения имеют характер чисто рассели-
тельных стадий «бродяжек», а его промежуточные продукты —— ха-
_ рактер пассивных бластомеров. К этому можно еще добавить, что
у тех форм, у которых повторная паратомия имеет факультативный
характер и сочетается, стало быть. с простой паратомией, число де-
лений, испытываемых материнской особью, определяется в основ-
ном условиями ее питания. › ‘
Заслуживает особого упоминания, что геометрические формы
повторной ‘паратомии в- точности повторяют таковые настоящей
палинтомии; в частности, видимые или кажущиеся формы повтор-
ной паратомии" также не совпадают с истинными, так чт-о многие
наблюдаемые тут картины могли бы истолковываться, как трех-
мерное деление, если бы -
выдавала его неизменно по-
перечного характера. Кон-
кретное представление об
особенностях процесса по-
вторной парат-омии у неко-
торых представителей Foet-
tirlgeriidae могут дать
рис. 63 и 64, а об онтогенезе
Цилиоспор — ри-с. 65. ‘
7.`"Повторная паратомия
колониальных сувоек ‘
Рис. 66. Макрозоид колониальной сувойки
Zoothamnium arbuscula B свободноплаваю-
щем (7) и осевшем состоянии (2, 3, 4) при
различных стадиях формирования прикре-
пительного стебелька (по Фурсенко)
Поразительно своеобраз-
томии является тот особый
спос-об, которым создаются
сидячие древовидные или головчатые колонии Zoothamnium И не-
которых других колониальных су-воек. Развитие этих сложных
объединений, являющихся, по сути дела, скорее целостными много-
членными организмами, чем колониями в обычном смысле слова,
было подробнейшим образом изучено Фурсенко (1924, 1929)
у Z. arbuscula. а а
Основателем колонии служит здесь крупная, сильно гипертро-
фированная, почти шаровидная особь, — макрозоид‚ образующаяся
на материнской колонии и затем отрывающаяся от нее (рис. 66).
Проплавав некоторое время макрозоид прикрепляется задним Kon-
uoM к поверхности водного растения или какому-нибудь другому
I8!
67. Образование колонии
Zoothamnium arbuscula путем повторного продольного деления
_ 4 р макрозоида; _ у А
Т-сгадич 2; 2—сгадия 4; 3—стадия 8; 4——стадия 16; 5—стадия 28; б-стадия 42 (по Фурсенко)
субстрату, дедифференцируется и выделяет довольно длинный про-
стой стебелек. Сразу же после этого макрозоид приступает к много-
кратному продольному и в‘то же время неравномерному делению;
оно идет с такой правильностью и столь точ-но детерминировано, что
невольно напоминает дробление‘ «мозаичных» яиц некоторых много-
клгето-чных. '
Первое деление макрозоида дает две неодинаковые по величине особи,
отклоняющиеся друг от друга под определенным тупым углом (рис. 67, 1). При
втором делении (рис. 67, 2) большая особь делится слегка неравномерно
а—меньд11ая‚—на две-резко разнящиеся-по величине дочерние особи‘; каждая
Рис. 68. Образование макрозоидов у Ёоотпатпйит
arbusculaz
I — взрослый макрозоид, заканчивающий свой гипертрофи-
ческий рост; __11 — растущий молодой макрозоид следующей
«смены» (по Фурсенко)
из образовавшихся особей принимает’ строго определенное положение по отно-
шению к остальным. Третье, тоже синхронное, деление приводит к разделению
самой крупной из 4 особей на одну несколько более крупную и меньшую;
второй особи — на крупную и маленькую, третьей особ-и и четвертой, самой
маленькой, особи-на две еще меньшие. Таким же неравномерным и синхронным
является и четвертое деление, приводящее к оформлению стадии 16. При пятом
делении одновременно „делятся все особи, кроме четырех самых мелких, и ко-
лония становится 28-членной (рис. 67, 3-—б); B дальнейшем происходит еще
целый ряд частью синх’ онных делений, в результате которых число членов
колонии достигает неско ьких тысяч. Заслуживает внимания, что первые деле-
ния макрозоида совершаются в недифференцированном состоянии и не сопро-
вождаются заметным ростом их продуктов; это приводит к их мельчанию‚ чем
еще подчеркивается сходство этого процесса с дроблением. Делениям сопутст-
вует рост и постепенное ветвление стволнка колонии, идущие с большой пра-
вильностью, чем и определяется исключительная правильность строения колонии
этой замечательной сувойки. ’ . ‘ ' '
о
I На ранних стадиях развития КОЛОНИИ B06 СОСТЗВЛЯЮЩИе- ее’
ОСОбИ бИОЛОГИЧЕСКИ ВПОЛНЕ paBHOII'€HHbI И ПрОДОЛЖЗЮТ ДЭЛИТЬСЯ
183
дальше. В определенный момент некоторые из них (а именно,‘
сидящие у основания некоторых определенных ветвей колонии)
начинают УСИЛЭННО расти И раздуваться И мало-помалу превра-я
щаются в уже описанных макрозоидов (рис. 68), отрывающихся
от колонии по окончании сво-его развития. Размножение колонии
происходит исключительно посредством этих «бродяжек», образую-
ЩИХСЯ на ней В НЕСКОЛЬКО СМЭН, ‘С характерной ДЛЯ ЭТОГО ВИДЗ _
регулярностью.
Высокий уровень целостности колонии находит одно из своих
выражений в том, что гипертрофическое развитие макрозоидов
"Рис. 69. Макрозоид колониальной сувойки Systylis
hoffi и его повторное деление '
7 — макрозоид перед делением; 2, 3 —- первое деление; 4 -вре-
МЭННЗЯ дифференцировка его продуктов; 5 —-8 —- последователь-
НЫЕ ЭТЗПЪ! второго деления; 9 — 17 —- последовательные этапы
третьего деления; 72—начало четвертого деления (по Бресслау)
обеспечивается главным образом окружающими ее мелкими осо-
бями, а также и в том, что эти мелкие особи погибают к концу
существования колонии. _ _ `
И Еще большее, пожалуй, внешнее сходство одетерминирован-
ными формами дробления яй=ца „у Metazoa обнаруживает повтор-
ная паратомия другой колониальной сувойки- Systy1is., y которой
ЁРОДУЁЪТ деления остаются сближенными L виде плотной головки
РИС‚. _. . _ - ‚
184
Глава Х
ЦИТОГОНИЧЕСКОЕ РАЗМНОЖЕНИЕ И ЖИЗНЕННЫЙ ЦИКЛ
' НАДКЛЕТОЧНЫХ ПРОСТЕЙШИХ
1. Общее понятие о цитогонии
Ц И Т О Г О Н И Я, ИЛИ РЗЗМНОЖЭНИЭ ПОСРЕДСТВОМ СЁДНОКЛЭТОЧНЫХ
стадий жизненного цикла представляет собой настолько распростра-
ненное явление, что она часто обозначается как «основное явление»
размножения. Очевидно, что все виды размножения постоянно
одноклеточных Protozoa, как бесполые, так и половые, относятся
к этому его типу; Цитогония является тут единственным видом
размножения и ооуществляетсяна всем протяжении цикла в виде
простого или тех или иных видов повторного деления.
У больши-нства надклеточных простейших, а именно очень мно-
гих саркодовых, всех кокцидий, гемоспоридий, микроснпоридий и
некоторых жгутиковых, цитогония также остается ‘единственным
способом размножения (и бесполого и полового) на всех этапах
жизненного цикла; она осуществляется в форме монотомии и син-
томии, причем последняя приобретает здесь господствующее зна-
чение—по крайней мере при гаметогенезе и половом размножении.
Однако у ряда других саркодовых и многих споровиков к цитогонии
присоединяется ивегетативное размножение, часто становящееся
тут ведущим или даже единственным способом бесполого раз-
множения; в этих случаях за питогони-ей остается роль гамето-
генеза и метагамного размножения. Такую же роль она играет
и в жизненном ‹цикле грегарин‘ и некоторых других групп, размно-
жающихся, как известно, исключительно половым путем.
У опалин и большинства инфузорий Цитогония имеет место
только в прогамной части цикла, эприводя тут к образованию одно-
клеточных гамет, в то время как метага-мное (половое) идбесполое
размножение осуществляются исключительно -пут-ем паратомии.
У опалин одноклеточные гаметы образуются как в мужском, так
и в женском ряду путем продольного деления двуядерных гамето-
цитов и представлены вполне самостоятельными стадиями жизнен-
ного цикла, живущими и копулирующими вне материнского орга-
низма и обладающими соответственно высокой степенью морфоло-
гической дифференцировки. я
У настоящих же инфузорий одноклеточные гаметы имеются
только в мужском ряду. Они представлены теми подвижными клет-
ками, какими обмениваются конъюганты и которые обычно описьь
ваются, как «мигрирующие ядра». В действительности же— это на-
стоящие клетки амёбоидного типа (рис. 70) или же имеющие харак-н
тер сперматозоидов (рис. 70, 4) -I/I'CO‘CTO$II.IlI/Ie не только из ядра,
How: более или менее значительного количества цитоплазмы.
Что жекасается ‚женских гамет, представленных у инфузорий
«стационарными ядрами» то они, в „противоположность мужским,
18.5
с.
не обособляются от Материнской сомателлы, а остаются в ее со-—
став-е. Таким образом, конъюгация этих инфузорий заключается
в перекрестном слиянии одноклеточных Мужских гаМет, ВЫДЭЛЯЭ‘:
мых каждым из конъюгантов, с телом ег-о партнера, прио-бретаю-
щем теперь значение надклеточной женской гаметы. и д
Редукция Цитогонии, столь характерная для инфузории, завер-
ша-ется ее. полным исчезновением у сувоек и Suctoma, вообще не
сохранивших одноклеточных стадий в цикле: наблюдающаяся у них
Рис. 70. Цитогония инфузорий
1 —- 3 — конъ огация Uroleptus тошна; последовательные стадии
обмена мужскими гаметами между копъюгантами; на фиг. 2. они
проходят одна мимо другой; внизу видны женскае (стационар-
male) ядра L(no‘ калкинсу); 4 —- сперматозоид Cycloposthlum
' ' bipalmatum (no Догело)
копуляция заключается в слиянии двух разнополых сомателл, т. е.
происходит в том же- надклеточном состоянии, в котором‘ протекает‘
весь жизненный цикл этих замечательных животных,
Уместно отметить, что у подавляющего большинства много-
клеточных организмов цитогония (за исключением части Mesozoa)
неразрывно связана с половым размножением и является тут един-
ственным его способом. Характ-ерную особенность цитогонии Meta-‘
zoa составляет то, что она удерживает свое значение подлинного
размножения (т. е. процесса, ведущего к увеличениючисла. отделы
ных 'o.(5;o6e15'1) только ‘в прогамной части цикла, т. е. при гаметоге-
незе, тогда как метагамная цитогония Гтеряет здесьсвои исходный;
характер размножения и становится средством‘создагниямвогокле-г
точных-стадий Цикла (‘СМ- ГЛ- XI)-' . , ‚ ч ‘т’
186
Таким образом, цитогония действительно является главным
видом полового размножения как у простейших, так и у мног-окле--
точных, и во всяком случае служит основой самого полового про-
цесса. И только у на-стоящих инфузорий последний в той или иной
мере уклоняется от этой общей нормы, что позволяет противо-
поставить их всем остальным простейшим и всем многоклеточным.
2. Саркодовьпе
Цитогоническое размножение плазмодИальных голых амёб
происходит, как правило, только беополым путем, но обычно свя-
зано с реорганизацией и оформлением довольно сложного и устой-
чнвого жизненного цикла. Основным видом размножения является
здесь синтомия, нередко осуществляемая, правда, путем поочеред-
ного, а не одновременного обособления амёбоспор от материнского
плазмодия и при н-еполном его использовании: Характерно также,
что многоядерн-ость достигается чаще всего ‚в состоянии цисты или
непосредственно перед инцистированием, в особой предцис т-
ной ст ад и и развития, а синтомия имеет место уже после вы-
хода из Цисты, в подвижном в е г е т а т и в н о м состоянии.
Проще всего обстоит дело у свободноживущих амёб типа
А. proteus, факультативно становящихся многоядерными в состоя-
нии «шара деления»; в этих случаях образовавшийся 4—'6-ядерный .
плазмодий сначала возвращается в активное состояние и затем
уже распадается на соответствующее число одноядерных амёбоспор.
Однако такое чередование обычных одноядерных стадий с плазмо-
диальными происходит нерегулярно, и амёбы этого типа могут
неограниченно долго размножаться простым делением надвое.
Противоположную крайность, в смысле расчлененности и пра-
вильности цикла, представляет кишечная амёба таракана, Enda-
moeba-blattae. Исходным звеном цикла являются тут амёбоспоры,
образующиеся в большом числе (36—72) „в проглоченных тара-
каном Цистах; они интенсивно питаются и растут, превращаясь
со временем в кфупных (45—200p.), весьма активных амёб, харак-
теризующихся, м-ежду прочим, наличием многочисленных диктио-
сом и одного единственного, довольно крупного ядра..А-мёбы дли-
тельно размножаются монотомией, причем их деление идет все
ускоряющимся темпом, со все более короткими интервалами, что
ведет к прогрессирующему мельчанию особей из одного поколения
в другое. Когда амёбы достигнут таким путем определенного раз-
мера (25-35 р), монотомия прекращается, и они переходят
в предцистную стадию; на этой стадии число ядер увеличивается
до 8 (редко 16), no ОНИ заметно мельчают; диктиосомы умень-
шаются в числе, а иногда и вовсе исчезают, протоплазма
постепенно освобождается от имевшихся в ней пищевых включенийЕ
После стабилизации числа ядер на только-что указанном уровне
предцистные амёбы остаются еще долгое время активными, но уже
не‘ растут, Далее происходит своеобразная реорганизация ядер:
18‘?
ного аппарата, после чево амёбы инцистируются. В цисте ядра
делятся еще несколько раз И притом не в-сегда CI/IHXp*OHHO, так
что общее их число достигает 32—72; при этом они все рас-пола- I
ь
гаются В центральной ЧЕГСТИ шаровидного теперь ‘ПЛЭЗМОДИЯ. Зрелые 1
ЦИСТЫ ВЫВОДЯТСЯ наружу И рЭЗВИВЗЮТСЯ дальше уже ‚ПОСЛЁЗ ТОГО, _
как они попадут в кишечник нового хозяина. Перед эксцистирова-
нием ядра мигрируют
к периферии плазмо-
дия, располагаясь тут
подобно ядрам пери-
ностным типом дробле-
ния; оболочка Цисты
лопается, освобождая
плазмодий, от которого
шнуровывается столько
амёбоспор, сколько в
нем заключалось ядер;
тральная часть обра-
зует крупное остаточ-
ное тело. Так же, пови-
димому, протекает в
основном цикл некото-
рых из кишечных амёб
человека. Так, у дизен-
терийной амёбы (Enta-
«moeba histlolytica) ги-
гантские трофические
генерации, представлены
кр-упной и активной
патогенной «тканевой»
формой стойко сохра-
няющей однлоядерное
строение и делящейся
только монотоМически;
а более мелкие пред-
цистные — комменсаль-
ной «просветной» фор-
мои, , размножающейся
путем синтомии (рис.71).
‚я - - › - д В цикле у Е. coli также
имеются укрупненная и патогенная стадия (описан_ная под назва-
нием Councilmania. 1af1veuri)«H~ относительно более мелкие коммен-
сальные, Однако в обоих этих случаях господствующим и основным
звеном цикла являются скорее мелкие комменсальные, чемгипер-
трофированные паразитические стадии, появляющиеся факульта-
F333
Рис. 71. Жизненные циклы кишечных амёб
человека Entamoeba histolytica (вверху) и Ent.
coli (внизу) (по Гнездилову из Павловского)
последовательно от;
ПрИ ЭТОМ ВСЯ его цент.
бластулы яиц с поверх- —
тивно, при наличии особых условий. «Под воздействием разнооб-
разных факторов в организме хозяина, как в среде обитания пара-
-зита комменсала, может возникнуть комплекс, условий, при кото-
ром мелкая просветная форма (f. minuta), реализует эволюционно
выработавшуюся у нее способность к образованию крупных, ткане-
вых, патогенных эритроцито-фагоцитирующих форм (f. magma), ПО-
-селяющихся в тканях хозяина. Крупные тканевые формы, выпадая
Рис. 72. Жизненный цикл двуядерной амёбы Sappinia
diploideag
B просвет кишечника, выделяются с фекалиями во внешнюю среду
и гибнут, как и мелкие формы, или же они превращаютсяв исход-
ные мелкие, просветные формы, которые образуют цисты» (Пав-
ловский, 1947). С другой стороны, у ряда других кишечных амёб
(Е. dispar, E. hartmanni И видов Endolimax) укрупненные стадиисо-
вершенно отсутствуют ‚и имеются только мелкие одноядерные ком-
менсальные формы; наконец, у своеобразной Dientamoeba трофо-
‚ЗОИТЫ, как правило, двуядерны И очень MCJIKI/I, а встречающиеся
гигантские одноядерные формы носят скорее дегенеративный ха-
рактер, неспособны к размножению и не имеют Циклическогоха-
рактера. _ ‚ ' _
Та_ким образом, взаимоотношения между-мелкими и крупными
формами трофозоитов тут далеко не так ясны, как у Е. blattae, что
в 189
И допускает самые противоречивые и-столкоьания жизненного цикла
этих амёб.. ‚ а - ‘ъ
Так или иначе, предцистные стадии всех кишечных амёб чело-
века (за исключением Di-entamoeba, вообще не образующей Цист)
представлены мелкими и, как правило, одноядерными формами,
а увеличение числа ядер происходит уже в цисте, посредством двух
последовательных кариокинезов; только у Е. соП первое деление
ядра имеет обычно место еще в предцистном состоянии, а второе
(или, редко третье) в цисте, получающейся тут г8- (16-) ядерной.
После эксцистирования плазмодий либо одновременно распадается
на 4—8 амёбоспор, либо отделяет их от себя одну за другой.
Итак, жизненные циклы этих Amoebina характеризуются много-
членностью, чередованием многих одноядерных монотомических
поколений с периодически прерывающим их плазмодиальным,
цистическим поколением, размножающимся синтомией‚ а также
и тем, что тут не имеют места половые формы размножения.
Пол-овое размножение достоверно установлено среди надклеточ-
ных амёб, только у почвенной или копрозойной Sappinia diploidea,
где они осуществляются весьма своеобразно. Трофозоиты (рис. 72)
представляют собой постоянно дв-ойниковых особей, размножаю-
щихся простым делением при сопутствующем (<<конъюгированном>>)
делении их обоих, неизменно сближенных, ядер. При половом раз-
множении двуядерные амёбы соединяются попарно и затем покрь1-
ваются общей оболочкой. В цисте амёбы сливаются в общий плаз-
модий, затем сливаются попарно и ядра; после этого каждое из
п-оследних и-опытывает по два деления созревания, давая начало
трем абортивным ядрам (направительным тельцам) и одному пол-
ноценному. Эти ядра не сливаются, а только тесно сближаются
друг с другом в образовавшейся теперь зигот-е. Последняя затем
выходит из цисты и дает начало длинному ряду двуядерных бес-
полых п-околений.
З. Акнидоспоридии
Колоссальное разнообразие форм размножения и развития этих
споровиков не препятствует все же тому, чтобы его можно было
свести .к относительно немногим главным типам. Примечательно,
что каждый из этих типов оказывается характерным для какого-
либо одного определенно-го подотряда споровиков, что ясно говорит
об их естественности, тогда как общепринятые отрядные группи-
ровки Аспйоозрогйайа поддаются лишь с большим трудом такой
обобщающей характеристике. Поэтому рассмотрение материала
удобнее всего вести также по отдельным подоггрядам.
А. Haemosporidiidae. Главные особенности развития этих спо-
ровиков, общеизвестными ‚представителями которых являются
„малярийные плазмодий, сводятся, в сущности, K трем пунктам:
1) трофические поколения, развивающиеся в красных кровяных
ттельцах, представлены постоянно_ амёбоидными «шизонтами»;
‘ОНИ ВСЕ СГГЭНОВЯТСЯ многоядерными K КОНЦУ СВОЕГО ‘ИНДИВИ-
"190 I
Рис. 73. Жизненный цикл кокцидии Eimeria shubergi:
'1 — проникновение спорозоита (или мерозоита) в эпителиальную клетку кишечника хозяина; ‚
2—5—-его рост и превращение в многоядерный плазмоций,. делящийся синтомически (б) на
мерозоитов (7), которые вновь проникают в кишечные Клетки и повторяют тот же цикл или
превращаются там в половые особи — гаметоциты (8); 9—77--рост мужского гаметоцита и
‚его распадение на микрогаметы: 12, 13-—гиперт офический рост женской гаметы; 14, 15 —‘—
оплодотворение; 76 — 79 —деление зиготы и о разование четырех споробластов; 20_— пре-
вращение споробластов в споры; 21 —— выход спорозоитов из цисты (по Шаудинну)
191
дуального развития иразмножаются, соответственно, путем син-
томии;р2) гаметоциты оформляются еще в крови позвоночного
хозяина, сильно отличаются как от мерозоитов, так и от шизонтов;
в мужских гаметоцитах ядро делится на 5—7 дочерних ядер, кото-
рые окружаются тонким слоем протоплазмы, отрываются от оста-
точного тела и образуют подвижные нитевидные микрогаметы;
женские же гаметоциты целиком превращаются в
крупные макрогаметы; 3) таким образом, наступаю-
щая затем копуляпция имеет ярко выраженный анизо-
_I 0 гамный характер. Следует также отметить, что гамет-о-
циты образуются из мероз-о-итов различных по счету
поколений, например, начиная с четвертой или пятой
их генерации у Plasmodium Vivax ИЛИ третьей-четвер-
той —— у Р. falciparum.
о. Б. Кокцидии подотряда Eim-eriida. Развитие этих
животных идет по той же общей схеме, как у гемоспо-
ридий, но обладает и рядом отличий. в прогамном и,
особенно, метагамном отрезке жизненного цикла.
Кокцидии проникают в эпителиальную клетку хозяина
в стадии спорозоита (рис. 73) и превращаются здесь
в округлого амёбоидного трофозоита, долгое время
ш остающегося одноядерным; к концу своего развития
он _стан0вится многоядерным шизонгом и затем де-
лится синтомически на большое число мерозоитов;
' последние вновь внедряются в эпителий кишечника или
Э другого органа хозяина и проделывают те ж-е -онт-оге-
IY нетические превращения. Так повторяется от двух до
четырех ‘(Емпегйа separata И Е. magna, рис. 74) или‘
Рис. 74. Ме- даже пяти (Е. nieschultzi) pas, пока мерозоиты опре-
E)IIJ)30§I_TI;>I(I1I-)I7I деленного поколения .не начнут осуществлять прогам-
3_fiv(”I)“4_fi ную часть жизненного цикла. Начинается она с того,
(,V),eHepa_ что эти мерозоиты вырастают (в зараженных ими
ций Eimeria клетках) в гаметоцитов, испытывающих затем гипер-
танца (ПО трофический рост. В мужском ряду они впоследствии
Хеисину) становятся многоядерными; затем ядра микрогамето-
цита мигрируют к его периферии, отпочко-вываются от
него вместе с окружающим участком плазмы и дифференци-
руются B двужгутиковые микрогаметы. Как видно из рис. 75,
показывающего ход этого процесса у Aggregata, OH необыкно-
венно точно повторяет привычные картины спермиогенеза Metazoa:
развитие каждой почки проходит стадию, имеющую все признаки
двужгутиковой сперматиды очень обобщенного жгутикового типа
(рис. 75, 2), затем почка удлиняется, и, наконец, микрогамета при-
обретает типичный для нее облик сперматозоида (рис. 75_, 3—5).
"После образования микро-гамет остается большое остаточное
тело. В женском ряду делений не происходит, и микрогаметоцит
непосредственно превращается в объемистую, сильно гипер-
трофированную оосферу. Перед копуляцией оосфера образует
‘l 92
наодном ‘И-з своих полюсов выступ, вполне равнуоценньхй- ‘воспри-
нимающему холмику яйца многоклеточных и так же служащий .
местом проникновения микрогаметы. Сразу же после копуляции .
оосфера- одевается тонкой оболочкой оплодотворения, начинаю-
щей ‘ОТСЛЭИВЭТЬСЯ OT ее поверхности’ На МЭСТЭ внедрения МУЖСКОЙ ._
;-‘mum-: .1
Рис. 75. Спермиогенез кокцидий Aggregata schubergi:
1 — множественное почкование микрогаметоцита: 2 — 4 -— преобразование почек-микрогамет
в сперматозоиды; 5 —- готовый сперматозоид (по Добелл о)
гаметы. После этого пронукл-еусы обеих гамет сливаются в ‘синка-
рион, и дв зиготедпроисходит ряд ядерных делений, первое из кото-
рых является редукционным. Это деление и следующее з-а ним дают
четыре равноценных ядра, каждое из которых либо сразу же ста-
новится ядром споробласта (которых тогда образуется четыре),
либо делится перед этим еще раз. Формирование споробластов за-
ключается в распадении содержимого зигоггы на столько отдельино-
стей, сколько .в ней было ядер; после выработки каждым из них
своей собственной-оболочки они становятся спорами. В споре про-
исходит еще одно, два или до четырех-пяти делений, продуктом
которых является соответствующее число опорозоитов; жпри этих
делениях часть материала споры остается н-еизрасходованной в виде
более мили менее крупного остаточного тела. ч
13 Эмбриология беспозвоночных 193
Ё. A-delelda. У кокцидий— этого подотряда‘ бесполаэъ ТРОФЁЧЭ‘
ская часть цикла значительно более дифференцирована, чем у Elma-
riida; она всегда состоит из строго определенного числа поколении,
из которых каждое имеет свои особенности. Этих поколении бывает
два (Ad-elina deronis), четыре (Adelina cryptocercl), ПЯТЬ (К1о551а
100551) или Шесть (Adelina dimidiata).
;="’@@©@a@?'7
Рис. 76. Различныецгенерации цикла кокцидии Klossia loossi:
I —.1V— индивидуальное развитие четырех, следующих друг за другом тро- ‚
фических поколений; V—— развитие макрогаметоцита, его гипертрофический’
рост и превращение в оосферу; VI —— развитие микрогаметоцига и его деление
на четыре микрогаметы (по Наби)
Конкретные различия между особями разных по счету трофи-
ческих поколений К1о551а 1оо551 показаны на рис. 76 и заключаются
‚в данном случае в том, что их размеры прогрессивно увеличиваются.
Индивидуальное развитие этих трофических особей замет-но отли-
чается от такового‘ эймерий и Plasmodidae тем, что увеличение
числа ядер начинается с самых ранних ею этапов и сопутствует
194
росту, так что шизонты очень рано становятся многоядерными и их
[ЭЗЗМНОЖЭНИЭ ИМЕЕТ характер ПЕРВИЧНОЙ, а не ВТОрИЧНОЙ СИНТОМИИ.’
. ' I1
Рис. 77. Жизненный цикл кокцидии Adelea ovata:
1-—5—рост и бесполое размножение в стенках кишки хозяина;
б —мерозоиты заключительного бесполого поколения; 7—моло-
дой мужской гаметоцит; 8—молодой женский гаметоцит; 9-
соединение мужского и женского микрогаметоцитов в виде си-
эигии; 10—деление ядер микрогаметоцита; 71——копуляция;
одна из микрогамет проникла в макрогамету, покрывшуюся
оболочкой оплодотворения; 72—-74—зигота и ее превращение в
ооцисту; 15—деление ядра ооцисты; 76 —- образование в ооцисте
многочисленных спор, в каждой из которых содержится два
спорозоита и остаточное тело (из Догеля)
Половая дифференциация особей обычно начинается еще за
одно поколение до образования гаметоцитов и может быть заметной
уже у шизонто-в последней генерации, но она чаще проявляется
13* 195
только iéfroix/i,-%1TO особи, дающие макрогаметоцитов, обгоняют‘ в своем;
развитии тех, которые продуцируют микрогаметоцитов. В. резульд
тате этого последние появляются значительно позднее женских
гаметоцитов, успевающих к этому времени достичь значительных
размеров. Копуляции предшествует длительное объединение гамето-
цитов в сизигию (рис.. 77, 9—11)‚ оба. члена которой продолжают
расти и развиваться открыто или под выделяемой ими общей оболоч-
кой; макрогаметоцит становится оосферой, а приклеенный к ней в
виде колппачка мужской гаметоцит незначительно вырастает, испы-
тывает два ядерных деления и образует из небольшой части своег-о
материала четыре двужгутиковых сперматозоида, тогда как большая
его часть остается при этом неиспользованной. Непосредственно
перед копуляцией ядро оосферы смещается к периферии и несколько
выпячивается на полюсе, противоположном тому, к которому при-
креплен микрогаметоцит; после этого одна из четырех микрогамет
внедряется в ядерный воспринимающий бугорок, откуда сразу же
начинает отслаиваться оболочка оплодотворения; далее происходит
редукционное деление синкариона, отслаивание второй оболочки
зиготы и, обычно, дальнейш-ее умножение числа ее ядер. В зиготе
образуется от З (Chagasella hartmanni) до 32 споробластов
(K1. lvoossi);-" y последнего -вида они формируются точно таким же
образом, -как амёбооплоры Endamoeba blattae, T. e. B результате
миграции ядер K ПОВЕРХНОСТИ ПЛЭЗМОДИЯ И ИХ ОТПОЧКОВЫВЗНИЯ,‘
вместе с участком плазмы, от объемистого остаточного тела. Kam-
ДЫЙ споробласт проделывает по два ядерных деления, окружается
оболочкой и распадается затем на четыре опорозоита (рис. 77).
Итак, цикл Ad-eleida существенно отличается от такового Eimeriida
ранним наступлением прогамной половой дифференциации и по-
жизненным сизигиальным объединением растущих ‚гаметоцитов.
Г. Schizogregarina. Развитие грегариън этого подотряда отли-
чается, как известно, значительным разнообразием, выражающимся
в различиях в судьбе сиз-игиальных нар, формах копуляции, числе
образуемых спор и т. д. Примером наиболее развернутых вариантов
жизненного цикла схизогрегарии может служить цикл Lipocystis
polyspora, паразитирующей в клетках жирового тела скорпионовой
мухи (Рапогра). Трофическая часть гцикла состоит из нескольких
поколений многоядерных шиз-онтов, первые из которых характери-
зуются мелкими ядрами, а позднейшие-—более крупными; число
этих поколений точно не устан-овлено. Мерозоиты заключительного
поколения попарно объединяются в сизигии, причем оба партнера
не отличаются друг от друга и развиваются совершенно парал-
лельно, сильно увеличиваясь в размерах, причем увеличивается и
количество ядер. По окончании роста сизигии содержимое обеих
спаренных ‘особей распадается на множество округлых голых и3ога-
мет, интересных тем, что они снабжены недоразвитой, но хорошо
заметной кинетидой; гаметы копулируют тут же, в оболочке роди-
тельской сизигии, и дают множество зигот; последние’ испытывают
редукционные _И еще два спорогенетических деления ядра,.. оде-
196
ваются плотной оболочкой и распадаются затем на восемь споро-
зоитов.
Д. Настоящие epeeapuflbt (Eugregarzna). Главная особенность
Цикла этих животных заключается в том, что вся его Трофическая
и прогамная часть осуществляется одной особью, последовательно
проходящей онтогенетические стадии спорозоита, растущего и затем
взрослого трофозоита (что свя-
зано с колоссальным увеличением
начальнои массы каждого инди-
вида——в несколько тысяч или
даже миллионов раз); трофозоиты
затем объединяются попарно п
сизигию, ‘вначале часто свобод-
ную, а затем инчцистирующуюся
под общей оболочкой. В сизигии
происходит многократное деление
ядер обоих партнеров. Ядра сме-
шаются затем к периферии плаз-
м-одия и отпочковываются от
него вместе с участком прото-
плазмы в виде мелких одноядер-
ных гамет (ри-с. 78). Г аметы мо-
гут быть одинаковыми и тогда
амёбоидными (например, у Асы-
nocephalus), ИЛИ же мужские
дифференцируются в жгутиковые
сперматозоиды, иногда более
крупные, чем женские гаметы
(stylorhynchus, рис. 78, 5, б).
После происходящей затем копу-
ЛЯЦИИ Зигогга окружается двойной . A
плотной оболочкой И превра- Рис. 78. Половое размножение тре-
дщается В C1-IOpy_ Ядро Споры По_ 7 гарины Stylorhynchus longxcoluhsz
'CJ1€l10'B3T€JIbH'0 делится на не- „ЁЁЁИЁИЁ ЁЁЦЁСЁЁЁЗЁЁЁЁЁЁСЁ %§.§c?:?‘L3°3§I
‘°“°””°K° (Чаще Все“) ВОСЭМЬ) §’§é1?c‘ifL.Z “(yc’§§¥‘§‘£§e;‘“f§’~§‘é’§”5a_'ix§‘>‘§<¥f<‘;‘£.‘7.
ядер, а зат-ем все ее содержимо-е б——женская гаметы; 7—10-—копуляция и
Ёраспадается На C0*0TBePCTByrom-ee 3333323333‘: 3£‘.§3§‘,3’£. §.§$l,i“‘€fn"§§,$’§a’E‘2Z
число спорозоитов. Редукционное , догеля) . . ‚ ~
деление происходит у некоторых - . . _‚
‘грегарин (Рыусузйоае, Diplocystis, Zygosoma) B споре в порядке
гпервого деления ее син-кариона, тогда как у других (например,
MOHOCYSUS) ‘ОНО, ПОВИДИМОМУ, „предшествует оформлению гамет. I
Итак, конденсация жизненного цикла достигает у грегарин
,']CV1£Ce>%I;Z же высокого уровня, каким характеризуется большинство
~ 03- ‘B9013 UIIPIKJI осуществляется в онтогенезе одной особи, сома’-
теллацкоторои оказывается, таким образом, биологически равна-
ценнои многоклеточному телу Metazoa. Эта’ сомателла также закан-
чивает свою жизнь выработкой половых чпродуктов, ‚слияние mm-
"I97
рых в виде зиготы дает, однако, начало Не одной, а обычно восьми
особям следующего поколения.
У всех же остальных акнидоспоридий цикл сохраняет типично
протозойную многозвенность, принципиально отличаясь от сходных
метагенетич-еских циклов одноклеточных только/тем, что все входя-
.щие в них вегетативные стадии приобретают к концу своего онто-
генетического развития надклеточное (плазмодиальное) строение
шизонтов, а не остаются, как там, одноядерными.
4. Книдоспоридии
А. Миксоспоридии. Вегетативный отрезок цикла осуществляется
одним или Многими бес-полыми поколениями плазмодиев, все ядра
которых в это время вполне равноценны; -плазмодии размн-ожают-ся
вегетативным способом, путем архетомии, фрагментации и т. д.
(гл. IX)-. Прогамные явления ‘начинаются с образования в извест-
ных местах плазмодия особых генеративных ядер, отличающихся
от вегетативных (или трофических) меньшими размерами и некото-
рыми деталями -стро-ения. Вокруг каждого такого ядра обособляется
по островку протоплазмы. Эти эндогенные одноклеточные образо-
вания представляют собой гаметоцитов; они соединяются попарно
в -сизигии, п-осл-е чего ядра обоих партнеров делятся несколько раз
подряд, пока их не окажется 14, а их протоплазма слива-ется
в общую ма-ссу. Так образуются 14-ядерные п а нс п о-р о б л а с т ы,
каждый из которых дает по две споры; при формировании посл-ед-
них два из 14 ядер па1нспоробласта остаются неиспользованными,
а остальные 12 распределяются поровну между зачатками обеих
спор; в дальнейшем по два из них вместе с небольшой частью
плазмы идут на построение двух створок оболочки споры, -по два
других дают начало двум стрекательным кагпсулам — книдо-ци-стам,
располагающимся либо вместена одном из полюсов споры, либо
по одному на противоположных ее полюсах, а оставшиеся два ядра
с большей ча-стью плазмы зачатка образуют амёбоидный зародыш.
Образование споры завершается пикнотическим перерождением и
гибелью ядер обеих покровных и обеих стрекатель-ных ее клеток
с превращением всего их плазматического материала в метаплазму;
‚только после этого ядра амёбоидн-ого зародыша (представляющие
собой Мужской и женский пронуклеусы) -сливаются в синкарион,
и зародыш становится зиготой. При заглатывании спор рыбой
(в которых развиваются большинство миксоспоридий) их клеточ-
„ная оболочка разрушает-ся и освободившийся зародыш постепенно
растет, становится многоядерным и превращается таким образом
у В В-егетативную СТЭДИЮ ‘ПЛЗЗМОДИЯ.
,- Редукционные деления имеют место при формировании спор,
‚в порядке созревания гам-ет‚ и протекают по обычным нормам га-
‚метной редукции у Рт›о’со2оа—с образованием двух направитель-
ных телец. -Число многоклеточных спор, проду-цируемых "одним
‚плазмодием, ‚колеблется в очень широких пределах. Так, упр-едста-
к
вителей группы Dispor-ea каждый плазмодий продуцирует только по
одному панспоробласту (т. е. по две споры), а у Ро1узрогеа——до
нескольких сот у более мелких полостных форм и нескольких мил-
лионов у крупных тканевых видов, цисты которых могут достигать
размеров куриного яйца. В соответствии с этим число ядер в плаз-
м-одии и степень использования его материала при спорообразова-
хнии очень различны: плазмодии двуспоровых содержат всего лишь
один их комплект °(т. е. фактически равноценны одному панспоро-
бласту), и входящая в него пара вегетативных ядер остается в_теле
животного, гибнущего посл-е созревания спор, тогда как у много-
споровых видов плазмодий содержит огромное их количество и
нередко целиком расходуется на спорообразование.
Б. Acz‘z‘n0,myxL‘dia. Цикл этих своеобразных паразитов олигохет
замечателен прежде всего тем, что он не содержит обособленных
бесполых генераций и почти целиком осуществляется в многокле-
‚точном состоянии. Наиболее молодые из известных вегетативных
стадий имеют амхёбоидное строение и два одинаковых ядра. Далее
они становятся 4-ядертныпми и затем 4-клеточными (благодаря раз-
межеванию соответствующих околоядерных территорий), причем
две из оформивших-ся теперь клеток образуют оболочку тела, окру-
жающую обе другие. Эти две внутренние клетки, представляющие
собой гаметоцитов, делятся вновь, так чт-о их получается четыре;
две из этих четырех клеток некоторое время‘ не делятся дальше,
сохраняя свои крупные размеры, а дведРУРие быстро делятся по_
два раза, превращаясь -в восемь микрогамет; после этого делятся
и оба крупных макрогаметоцита, дающие также восемь гамет; на
этой стадии тело животного состоит из двух покровных клеток и
16 аниз-огамет, различающихся по величине ядер и окрашиваем-ости
плазмы. Затем гаметы копулируют и образуют восемь зигот, пре-
вращающихся в дальнейшем в споробласты. Процесс образования
спор _чрез-вы-чайно оригинален. Ядро зиготы (споробласта) сначала
делится на два, одно из которых сохраняет значение генеративного
ядра и идет на образование зародышей, а другое дает начало по=
бочным клеточным эл-ементам будущей споры. Из последнего полу-
чается шесть мелких ядер, группирующихся ‘в с п о р о г е н н у ю
M асс у, приобретающую при этом клеточное строение; три ее над
ружные клетки расходуются на построение трехлучевой метаплаз4
матической оболочки споры, а три внутренние служат источником
формирования -ее трех стрекательныхкапсул (того же типа‘ книдо-
цист, как у Myxo-sporidia). Одновременно с образованием и диффе-
ренциацией спорогенной массы начинает делиться и генеративное
ядро; его продукты образуют обособленную э M 6 ри о г-е н -н у ю‘
массу, состоящую из 8, 32 или большегочислаодноядерных амё-
боидных клеток, -или же представленную г многоядерным плазмтот
дием. Заключительным этапом спорогенеза является‘ акти-вное все-г
ленив эмбрио-генной массы вдгото-вуюгоболочку‘ споры, ‘где она‘
окончательно распадается на 84110 амёбоидных зародышей. ' ' *
. ЭТИ- „последние представляют: ;собой ; ‚единственную. одноклеточы
199‘
ную стадию развития, протекающую вне родительского организма.
Как видно из изложенного, они абсолютно не гомологичны ‘амёбоид-
ным же зародышам миксосп-оридий, как не гомологичны друг другу
и защищающие их многоклеточные образования, хотя и построен-
ные у тех и других по очень сходному общему плану. В самом
деле, в то время как у Myxosporidia они образуются из
потомков гаметоцитов, а зародыш является зиготой, у Actinomyxidia
дело обстоит совершенно иначе: как это образование, так и много- _
чттсленъньге амёбоидные зародыши являются потомками зиготы;
-иначе говоря, они только в последнем случае соответствуют спорам
и спорозоитам Acrlidosporidia, a B первом не имеют никакого отно-
шения к тем и другим, являясь производными эпрогамных клеточ-
ных п-околений. _ .
Все эти замечательные особенности развития плазмодиальных
книдоспоридий ставят их особняком -среди Protozoa H давно уже
служат поводом для дискуссии -об их действительно-м положении
в системе. Известно, что ряд авторов склонен относить их к Мезо-
zoa, с частью которых (Orthon-euctida) -oHH действительно имеют
много общего, тогда как другие энергично отстаивают их при-
надлежность к простейшим.
5. Опал ины
Капитальные различия’ между этими инфузориеобразными
формами и настоящими (ресничными и сосущими) и-нфуз-ориями,
столь ярко сказывающиеся в их вегетативном размножении
(гл. 1Х), получают еще более наглядное выражение в особенно-
стях полового процесса тех и других и самом построении их жиз-`
ненно-го цикла. ' ' И .
Вся бесполая часть цикла опалин осуществляется у Ор. ranarum,’
Ha примере которой мы его рассмотрим, во взрослых лягушках
летом, осенью и зимой ‘и заканчивается ранней весной, когда хо-
зяева уходят в воду и -сами приступают к размножению. Начинаю-
щаяся теперь прогамная ча-сть цикла заключается в распадении
крупных многоядерных вегетативных сомателл на небольшие 3—6.-
(редко до -12)‘. ядерные гаметоциты. Г аметоциты инцистируются
(что сопровождается их дедифференцировкой); образовавшиеся
цисты выходят наружу вместе с экскрементами и служат для зара--
ж-ения молодых головастиков. Проглоченные голова-стиками цисты
раскрываются ‘и ‘из них выходят реди-фференцированные- гамето-
циты, начинающиетут. же делиться дальше палинтомическим,
в сущности, путемг‘ по крайней мере последнее из этих поздних’
прогамных делений; имеющее своим? результатом расщепление дву-
ядерных гаметоцитов на одноядерные гаметы, осуществляется по
еамым строгим правилам продольного дел-ения Flagellataei 13pe3aIo4
щейся бороздой, постепеннотрассекающей т-ело гаметоцита? вдоль,-
по направлениюспереди назаддОфоТрми-вшиеся таким образом изо-
гаметы „копулируют, склеиваясь- сначала. своими дпередними-конттами
30.0
.01
H затем постепенносрастаясь по всей длин-е. Образовавшаяся зи-
гота дедифференцируется, округляется и одевается то-нкои оболоч-
кой. После некоторого периода покоя зигота одевается ресничками‚
выходит из своей цисты и начинает питаться и расти, причем ее
синкарион многократно делится по мере увеличения массы тела;
так создаются многоядерные сомателлы первого вегетативного по-
коления, размножающиеся дальше только парато-мией до наступле-
ния следующего полов-ого процесса. Следует еще отметить, что
редукция у опалин осуществляется в прогамн-ой части цикла, т. е.
является гаметной. '
6. Основные типы жизненных циклов надклеточных простейших
Сопоставляя -все рассмотренные здесь ЦИКЛЫ друг с другом и
с циклами постоянно одноклеточных простейших, легко заметить,
что их разнообразие все же не так велико, как это кажется с пер-°
в-ого взгляда; они все построены по одному и тому же общему плану,
складываясь из одних и тех же компонентов, чередующихся в со-
вершенно определенном порядке.
Центральным звеном всякого цикла является, естественно, по-
л-овой процесс, иногда только как бы замененный образованием
«реорганизационной цисты» (у паразитических амёб). За ним, чаще
всего непосредственно, следует синтомическое или палинтомическое
деление, обычно ведущее к образованию ампёбоидньтх или жгутико-
вых зооспор или же спорозоитов, т. е. спорообразование, или споро-
генез, являющееся, таким образом, первым этапом метагамного
или метацистического отрезка жизненного цикла. Далее идет наи-
более продолжительный отрезок цикла--троф.ический или вегета-
тивный, выполняемый более или менее длинным рядом бесполых
поколений и лишь в немногих группах—только одним. Наконец,
‚наступает прогамный (или предцистный) период раз-вития‚ реали-
зуемый чаще одним, но нередко и несколькими поколениями (число
которых -в этих случаях невелико и всегда строго фиксировано),
принимающими на себя подготовку к реорганизации; это—— заклю-
чительный период гамето-генеза или равноценных ему предцистных
преобразований. Таким образом, все эти циклы ‘являются посуше-
ству M е т а г е н ет и ч ес к и M H, T. e. выражаются в чередовании
биологически и морфологичпески неодинаковых поколений.‘
На этой стандартн-ой основе развертывается все наблюдаемое
многообразиецикла простейших. У с-вободноживущих форм оно
создается, как мы видели, преимущественно за счет различий
“В ПРОГЗМНОЙ ЧЭАСТИ цикла; ЭТО pa3Hoo-6pa3H-e ЗЭВИСИТ ЗДЕСЬ О’Г ТОГО,
на-сколько специализованы в разных случаях гаметы по сравнению
с бесполыми зооспорами (или нормальными ' вегетативными осо-
гбями), насколько различны между ‘собой мужские и женские га‘-
меты‘ и какова степень гипертрофии, достигаемой этими послед-
ними; Ведущим фактором, дифференциации жизненного‘ цикла оказ-
зывается тут половой процесс, испытывающий настолькобурную
1201
прогрессивную эволюцию, что он достигает св-о-их ‚высших форм
даже при слабой отрегулированности цикла в цело-м или же когда
он еще только факультативен. Так, оо-гамия со всеми ее атрибу-
тами—сперматогенетическим характером прогамной части цикла
В МУЖСКОМ ряду И ООГЭНСТИЧЕСКИМ В‘ женском—наблюдается,
у ТЭКИХ ОТНОСИТЕЛЬНО ПрИМИТИВНЫХ ФОРМ, как НЕКОТОРЫЕ ОДИНОЧ-
ные фито-монады. В этих случаях осо-бенно бросается в глаза, что
эти процессы м-огут Не затрагивать хода трофической части цикла
и во всяком случае не ведут к дифференциации образующих его
поколений. Их число может быть неограниченным, и сама циклич-
ность развития -в большей м-ере управляется внешними условиями,
чем собственным ритмом: фактическое осуществление полового
процесса смещается в очень широ-ких пределах и легко может быть
устранено экспериментальным вмешательством.
Совершенно другую картину представляют паразитические про-
стейшие. Биологический «центр» цикла смещается у них с половою
или равнозначн-ого ему процесса на момент обеспечения инвазии
хозяина, и весь цикл в той или иной Мере перестраивается приме-
нительно к -ее осуществлению. Не касая-сь здесь -специально пара-
зитологическог-о аспекта этого явления, мы должны то-лько отме-
тить, что создающееся таким путем вторичное, п а р а з и то г е н-
н ое че редов а н и e‘ п окол е н и й может заслонить собой их
первичное метагенетическое чередование. Это последнее может быть
полностью преодолено или же м-ожет приобрести особые, атипиче-
ские формы; Однако тако-е замещение одного из ‚видов смены поко-
лений другими -н-е только не является правилом, а, наоборот, скорее
исключением. ‘Гораздо чаще исходное (или, строже говоря, прото-
типическое) расчленение цикла сохраняется при всех его специаль-
ных приспособительных перестройках, но только получает новое
содержание, новый биологический смысл и нередко новое выраже-
ние. Так, спорогенетическая часть цикла обычно принимает на себя
у паразитов расселительные функции и обеспечение задачи зара-
жения нового хозяина; его прогамные этапы получают дополни-
т-ельную «нагрузку» в виде выработки тех или иных способов по-
выш-ения плодовитости или,- в некоторых случаях, осущест-вления
смены хозяев; наконец, на бесполы-е покол-ения цикла ложится
максимальное использование ресурсов, черпаемых ими из хозяина,
что и осуществляется при помощи интенсивного размнож-ения.
И именно в результате подобного разделения труда между отдель-
ными слагаемыми жизненного цикла‘ паразитов их трофические
стадии эволюируют в на-правл-ении специализации к особым усло-
виям существования в организме хозяина (и потому так часто ока-
зьпваются морфологически уяпростившимися вплоть до амёбоидного
уровня организации; см. гл. 1, IV), ‘расселительные метагамные‘
' стадии устойчиво сохраняют -0-рганизацию свободножи-вущих форм
{дополня-емую, "правда, специально инвазионными ‘приспособле-
ниями), а прогамны-е стадии женского ряда очень часто приобре-
тают‘ характер яйцеклетки. „ д . 1 - =
202
Итак, ЖИЗНЕННЫЕ ЦИКЛЫ’ ВСЕХ простейших ‘СХОДНЫ ‚В TOM, ЧТО
они основываются на единой метагенетической основе, в значи-
тельной мере сохраняющей свою силу и при их перестройке на
паразитический лад. Этим существенно -обл-егчается задачацкласси-
фикации жизненных циклов простейших-задача большои теоре-
тической важности, но не получившая «еще вполне удовлетворитель-
ного решения. В самом деле, имеющиеся классификации циклов
слишком отвлеченны или упрощенны, чтобы отвечать основным тре-
бованиям, предъявляемым ко всякой схеме такого рода: тре-бова-
ниям отразить в себе наличное разнообразие классифицируемых
явлений, передать в меру возможности их действительные соотно-
шения и служить достаточно продуктивным образом уяснению этих
явлений. Оценивая с это-й точки зрения рас-смотренные нами циклы
простейших, можно прежде всего констатировать, что они явственно
распределяются в две основные группы по самому характеру их
по-строения.
Первую, самую емкую, из этих групп образуют -п р а в и л ь н ы е
м е т а г е н е т и ч е с к и е ц и к л ы, удовлетворяющие следующим
основным признакам: 1) первым этапом мета-гамного (или м-етацИ-
стического) отрезка цикла является сп-орообразование, осущест-
вляемое синтомическим или палинтомическим делением зиготы
(цисты) и имеющее, как правило, я-вн-о расселительный характер;
2) образовавшиеся таким пут-ем споры (зооспоры, спорозоиты) вы-
растают в бесполых вегетативных особей; эти особи представляют
собой основателей целой серии трофических поколений (‚в много-
звенных циклах) или выполняют сами всю бесполую часть цикла
(в двух- или однозвенных); 3) прогамные этапы цикла имеют
в основном характер гаметогенеза (или предцистных преобразова-
ний), хотя могут и включать выполнение вспомогательных рассели-
тельных задач (например, связанных со сменой хозяев). A
‘K этой группе относятся циклы значительного «большинства про-
стейших. Среди них довольно ясно намечаются четыре типа, раз-
личающихся способом спорообразования и гаметоген-еза, а именно:
1. Тип, характеризующийся раздельным гаметогенезом и обра-
„зованием спор -в один прием путем однократной синтомии или
непрерывного палиинтомического шроцесса. Сюда относятся, .напри-
м-ер‚.циклы фитомонад. -
2. Тип, для которого характерно сочетание раздельного гамето--
генеза с образованием спор путем повторно-й синтомии, т. е. распа-
дания зиготы сначала -на споробласты, а затем уже каждого спо-ро-
„бласта на одноклеточные споры (спогрозоиты). Этот тип встре-
чается у Eimeriida. - - ~ - ‘ - A
3. Тип, отличающийся сизигиальнь1м гаметогенез-ом при та-
ком же, как в. перв-ом ти-пе, однократном спорообраз-овании. Он
представлен циклами-схизогрегарии и Eugregarina. ~ -
4._дТип, при котором сизигиальный гам-етоген-ез сочетается с фор-
мированием спор путем повторной двухступпенной синтомии. Приме-
ром этого типа могут служить жизненные циклы'А‹1-е1е1‹1а.
203
вторую главную ‘группу’ жизненных циклов простейших о-бра-
зуют те, которых можно назвать, за отсутствием лучшего термина,
неправильными метагенетическими циклами.
‘Они принципиально отличают-ся от таковых п-ерво-й группы с-овер-
шенно иным порядком чередования своих биологических фаз или,
другими словами, иными соотношениями между половым процес-
сом и остальными слагаемыми цикла. Они характеризуются, Именно,
тем, что: 1) зигота непосредственно превращается в бесполую веге-у
‘тативную о-собь, минуя м-етагамное спорообразование и связанное
с ним расселительное размножение; 2) эта о-со-бь или ее прямые
потомки, получающиеся в результате ее вегетативного размноже-
ния, вырабатывают педогамные или даже аутогамные гаметы или
гам-етоциты, ча-ст-о вступающие в сизигиальное соединение задолго
до копуляции или кариогамии; 3) прогам-ные этапы цикла или
сами гаметы принимают на- себя осуществление ра-сселитепьных
задач или же фактически не разграничены от трофических стадий
цикла.
Эти удивительно своеобразные циклы могут быть также разде-
лены на несколько типов, главные из которых таковы:
1. Тип, отличающийся тем, что гаметы образуются непосред-
ственно из бесполых трофических стадий путем простого деления
или синтомии; здесь нет оформленного прогамно-го ряда, так что
гаметы как бы совмещают в себе значение гаметоцитов с таковым:
настоящих гамет. Циклы этого типа олицетворяют, очевидно, низ-
шую ступень их биологической расчлененности; они характерны
для свободноживущих форм, например многоядерных солнечников
‘(Actir1osphaeriu'rr1, Trich~o»spha»eriurr1) и некоторых радиолярий.
2. Тип миксо-споридий, для которого характерна совершенно
необычная-роль, принимаемая прогамными КЛеТОЧНЫМИ HO-KOJI€'.
ниями цикла, а. именно их участи-е в построении многоклеточных
«спор», не имеющих здесь ничего общего с настоящими м-етагам-
ными спорами. Эти замечательные раоселительны-е образования
сравнимы, ‘B сущности, только с прогамным расс-елительным поко-
лением м-едуз у метагенетич-еских Hydro-zo-a. K этому можно еще
добавить, что вспомогательные прогамные образования (правда,
уже не рас-селительного характера) встречаются еще только у ‘губок,
где они представлены клетками сперматоц-ист. - -
' 3. Не менее своеобразный тип A-ctin-0-myxidia, замечател-ьный
прежде- всею размежеванием продуктов первого цикла ‚метагамных‘
делений зиготы на абортивные (расходуемые на построение шести-
клеточного тела кспоробласта) и генеративные (образующие -вну-
тр-еннюю эмбриогенную массу), а также и тем, что вегетативная
стадия цикла имеет необычное «малоклеточное» строение.
-- Оба последних типа неправильных метагамных циклов имеют
так_ много общего с циклом Mésozoa и вместе с тем столько специ-
фически метазойных черт, `что отнесение обеих этих труп-п к-пр-о-
стейшим очень трудно обосновать, с - V 1 ’ "
‚г‘
мы.
ЧАСТЬ ТРЕТЬЯ
ИСТОЧНИКИ И ПУТИ ФОРМИРОВАНИЯ
ПРИМИТИВНЫХ ВИДОВ МНОГОКЛЕТОЧНОСТИ
Вводные замечания
Рассмотрению и анализу истории раз-вития ‚низших Метаиоа
необходимо предпослать несколько самых общих замечании, ка-
сающихся мнотоклеточности вообще, путей ее онтогенетического-
становления и все еще дискуссионного вопроса о принципиальной
сравнимости многоклеточных с одноклеточными и надклеточными
Protozoa.
Первое, чточнам надо всячески еподч-еркнуть,—это то, что мы
не знаем ни одного „представителя Metazoa, который оставался бы
многоклеточным на протяжении всего своего жизненного цикла.
Все они размножаются цито-гонией и соответственно обладают
одноклеточными стадиями развития, представленными тут гамето-
цитами (оотониями, ооцитами, сп-ермато-гониями и сперматоцитами)
и самими мужскими и женскими гаметами. Таким образом, много-
клеточные -стадии цикла тут чередуются с одноклеточными, что
никогда нельзя упускать из вида.
Второе обстоятельство, требующее столь же настойчивого под-
черкивания, заключается. в том, что многоклеточное строение-
не является монополией одних только Metazoa (оставляя тут,
разумеется, в стороне все многоклеточные растения): будучи ха-’
рактернейшим для них признаком, оно, вместе с тем, наблюдается
и среди простейших, и притом в специфически, ‹<м-етазойном» виде
подвижной эпителиальной бластулы. Его олицетворяют тут высшие
колониальные Уо1мосо1с1еа, палинтомические ко-лонии которых обла-
дают всеми-свойствами подлинно многоклеточных организмов, в го-
раздо большей мер-е животного, `чем растительного характера.
Правда, этот разительный пример чи-сто «метазойной» многокле-
точности у простейших является, в сущности, единственным, по-
скольку состоящие из нескольких ‘клеток споры Асйп-отухйойа и’
своеобразные тела размножения Myxo-spo-ridia. в, не. меньшей мере
205’
уклоняются от этого ее типа, чем состоящие ‘из многих сомателл
колониальные су-войки. Однако уникальность этого п-римера
ни в коей мере не обесценивает его; она говорит только о том, что
м-ногокл-еточность встр-ечается у Protozoa лишь в виде исключения,
тогда как у Metazoa она является правилом.
Таким образом, признак многоклеточности не имеет того абсо-
лютного значения, которое ему обычно при-писы-вается. Можно
только сказать, что он гораздо более типичен для Metazoa, ЧЕМ
ДЛЯ ПРОСТ-ЕЙШИХ, ЧТО МНОГОКЛЕТОЧНЫЕ стадии несравн-енно чаще«
имеют у первых вегетативный характер и занимают, соответственно,
ведущее ‘место в их цикле. Это сущ-ественно важное, но часто не-
доучитываемое обстоятельство может б-ыть сформулировано и так,
что одно из ‘главных качественных различий между Protozoa,
с одной стороны, и Metazoa—c другой, -вь1ражается -в том, что
метагамная, трофич-еская и расселительные части жизненного цикла
осуществляются у первых-преимущественно одноклеточными или
надклеточными -стадиями и лишь как очень р-едкое исключение-
многоклеточными, тогда как у вторых эти стадии почти всегда
Многоклеточны и только иногда имеют плазмодиальное строение.
В отношении же прогамной части цикла и полового процесса между
ними н-ет даже и таких различий, а тем более различий абсолют-
НЫХ, так ЧТО СЭМО ‘ПРОТИВОПОСТЭВЛЕНИЕ ЭТИХ «ПОДЦЭРСТВ» ‘ОКЭЗЫВЭЕТСЯ С
еще более условным.
Не меньшую принципиальную важность имеет и тот факт, что
единственным существующим в природе способом достижения
многоклеточности является дробление яйц-еклетки, т. е. ее много-
кратное повторное деление. Этим, и только этим, путем совершается
на наших глазах превращение одноклеточной начальной стадии
развития всех без исключения Metazoa ‘B многоклеточное тело их
последующих онтогенетических стадий. Этим ж-е путем создается
многоклеточность и в ряду колониальных Volvocoidea. И поскольку
МЫ НЕ‘ знаем ОУКВЭЛЬНО НИ ОДНОГО СЛУЧЭЯ, КОГДЭ МНОГОКЛЕТ0ЧНОСТЬ›
образуется какими-либо другими средствами, чем дробление,
можно со всей уверенностью заявить, что таких средств и не суще-
ствует, что процесс дробления владеет монополией создания много-
клеточности в онтогенетическом развитии.
К этому следует еще добавить, что дроблению яйца (или любой
равноценной ему генеративной кл-етки) неизменно предшествует
его гипертрофический рост при сохранении им одноклеточного
строения. Следовательно, этот общеобязательный для многоклеточ-
ных процесс ооген-еза представляет собой не что иное, как процесс
гипертрофической дистомии, а следующее за ней многократное де-
ЛЕНИЕ ЯЙЦа НЕ МОЖЕТ РЭСЦЕНИВЭТЬСЯ ИНЭЧЕ, как ПРОЦЕСС ПЭЛИНТОМИ-
ЧЕСКОГО‘ характ-ера.
ЭТИМИ ТРЕМЯ ОСНОВНЫМИ мом-ентами ОПРЕДЕЛЯЕТСЯ наш ПОДХОД
K ВОПРОСУ О‘ ‘СОИОТНОШ-ЕНИЯХ МНОГОКЛ-ЕТОЧНОСТИ С ДРУГИМИ УРОВНЯМИ
ОРГЭНИЗЭЦИИ, а СТЭЛО бЫТЬ И K ТЕМ ПРИНЦИПЗМ, на КОТОРЫХ СЛЕДУЕТ
СТРОИТЬ ВЗЭИМНОЕ сравнени-е МНОГОКЛЕТОЧНЫХ И ПРОСТЕЙШИХ. МЫ СЧИ-
20б_
1
THEM, ВО-ПЕРВЫХ, ЧТО ‘ОНО ДОЛЖНОдб-ЭЗИРОВЭТЬСЯ на РЭЗДЕЛЬНОМ COHO-'
СТЭВЛЕНИИ ГОМОЛ-ОГИЧНЫХ ДРУГ ДРУГУ СТЭДИЙ ВСЕГО ЖИЗНЕННОГО цикла‘
в целом, а не только на какой-либо одной _из них, взятой в отдель-
НОСТИ; ВО-ВТОРЫХ, ЧТО ПРИ ТЗКОМ сравнении ОТДЕЛЬНЫХ стадий цикла
Над-О ПРЕЖДЕ ВСЕГО считаться С ПРОЦЕССЭМИ ИХ ВОЗНИКНОВЕНИЯ И раза
вития, а не брать их, как это обычно делается, только в «готов-ом»
состоянии. В-третьих, мы постоянно должны иметь в виду, что до-
стижению многоклеточногоуровня строения неизменно предше-
ству-ет про-хождение развивающимся организмом ряда качественно
отличных его уровней, начиная с одноклеточного, и что эти уровни,
следовательно, относятся друг к другу, как последовательные сту-
пени в едино-м восходящем ряду развития, а не как чуждые друг
другу и принципиально несравнимые состояния. Наконец, при этом
СРЭВНЕНИИ НЕООХОДИМО ЕЩЕ УЧЕСТЬ, ЧТО ДОСТИЖЕНИЮ МНОГОКЛЕТОЧНОГО,
состояния посредство-м повторного деления (дробл-ения) противо-
стоят все Te случаи, когда это последнее имеет своим конечным
результатом образование многих раздельных одноклеточных особей.
Это ясно говорит о наличии двух противоположных тенденций в раз-
витии процессов .палинтомического характ-ера, что и должно лечь
в основу их сравнительного анализа. ‚
С этих позиций кажется очевидным, что продолжающиеся еще
споры о самой возможности сравнения Protozoa И Metazoa ЯВ-
ляются, в сущности, беспочвенными и должны у-ступить свое место
трезвому, конкретному и всестороннему анализу особенностей орга-
низации, размножения и развития многоклеточных в свете тех полсо-
жений и фактов, с какими нас ознакомило соответствующее рас-
смотрени-е про-стейших.
II
Когда ГОВОРЯТ O МНОГОКЛ-ЕТОЧНОСТИ ЖИВОТНЫХ О-РГЭНИЗМОВ,‘ ТО
в это -поняти-е вкладывается, как пра-вило, гораздо более узкое,
конкретное, но-, вместе с тем, и более сложное морфологическое
СОДЕРЖЗНИЕ, ЧЕМ ЭТО СЛЕДУЕТ ИЗ СЗМОГО ЕГО СМЫСЛЭЗ МНОГОКЛЕТОЧ"
ность представляется в форме двуслойной, как минимум, организа-
ции, связанной с определенной тканевой и оргганологической
диф-ференцировкой и с полиморфизмом клеточного материала (при
наличии физиологического разделения труда между отдельными ка-
тегориями клеток); короче говоря, под мно-гоклеточностью подразу-
м-евается ТОЛЬКО та ЕЕ частная ФОРМЭ, КЭКЭЯ характерна ДЛЯ ВЗРОС‘.
лых вегетативных стадий относительно высоко организованных
Metazoa.
Ha основе такого понимания многоклеточной организации
строятся и все попытки понять ее происхождение, воплощенные
-в длинном ряде -сугу-бо умозрительных гипотез и не давшие пока
удовлетворительного результата; на _н-ем же базируется и обиход-
ное мнение о существовании глубокой и непреодолимой «пропасти».
между Protozoa И Metazoa, делающей не сравнимыми друг с дру-
гом представителей этих подцарств; и оно до сих пор служит пре‚
207
пятствием к трактовке сыбщей эмбриологии с эволюционной, исток
рической точки зрения.
Между тем, хорошо известно, что эта сложная, двуслойная
форма м-ногоклеточности далеко н-е является единственной ее фор-:
мой, реально существующей в .прир-оде. Наряду с нею имеют-ся И
намного более простые и, безусловно, более примитивные формы
подлинно многоклеточной организации: это—действительно много-
клеточные, в лпрямом и точномп смысле этого слова, жгутикон-осные
бластулоо-бразные организмы, гармонически сочетающие в своем
строении предельно. низкий для Metazoa уровень дифференцировки
с очень ярко выраженной организменной ц-елостно-стью.
В самом деле, бластулоо-бразные организмы характ-еризуются
прежде всего тем, что они обладают исключительно строго выдер-
:жанным клеточным строением, т. е. состоят только из типичных,
одноядерных клеток и не имеют в- своем составе каких-либо над-
клеточных элементов; далее, тем, что число составляющих их клеток
очень велико и тем самым удовлетворяет далеко не случайному\
конструктивному принципу м=ного-, а не мал-оклеточности, лежа-
ЩЭМУ В основе ОРГЭНИЗЗЦИИ Metazoa И TaK СПРЭВЭДЛИВО ПОДЧЭРКИ-
ва-емому Хлопиным (1946); далее, тем, что все эти клетки поляри-
зованы и образуют правильный эпителий, в чем, бесспорно, заклю-
чается одна из самых общих и характерных гистологических
особенностей Metazoa-; дал-ее, тем, что все эти клетки находятся
в очевидном «биологическом соединении», -со всеми вытекающими
отсюда последствиями; наконец, жгутиконосные бластулы характе-
ризуются тем, что имеют одн-оосно гетерополярное строение с теми
или иными признаками вращательной симметрии и наличия осевых
градиентов, что служит самым наглядным структурным выраже-
ни-ем высокого, и притом чисто «животного», уровня их организмен-
ной целостности, связанного с высокой степенью подвижности
бластул и определенными особенностями их движения в воде.
Не менее примечательны и другие особенности этих сво-еобраз-
ных ‘организмов. Многие из них, а именно все целобластулы, ха-
рактеризуются еще т-ем, что все составляющие их клетки имеют
совершенно одинаковое строение, или, другими словами, что они
построены из совершенно одинаковых и мо-рфологически равно-цен-
ных друг другу жгутиковых клеток. Это значит, что эти ‚примитив-
ные формы обладают р а в н о K Л е т о ч н ы м строением и состав-
ляют в этом смысле исключение среди Metazoa, которых мы -при-
выкли считать исключительно разноклеточными организмами.
Таким образом, большая гистологическая простота ‘бластул
не мешает им обладать относительно очень совершенной архитекто-
никой и достойно представлять многоклеточность, но только в одно-
слойной, а н-е двуслойной форме. А отсюда, очевидно, следует, что
эта последняя, равно как и сопутствующая ей тканевая и органо-
логическая дифференцировка, отнюдь н-е являются обязательными.
атрибутами многоклеточности, а тем более ‘ее главными при-
знаками. A . ‘ . . . . . .
208
Простая констатация этого факта показывает, что обычное
отождествление многоклеточности Metazoa с дву- или трехслойным _
и разноклет-очнь1м строением не отвечает действительности, что
между ними никак нельзя ставить знака равенства, и что их, на-
против, надо строго различать; или, иначе говоря, что самая подлин-
ная многоклеточная организация может быть представлена как
в дву- или трехслойных разновидностях, связанных с явлениями
гистологической дифференциации, так и в однослойном, эпители-
альном виде, при котором эти явления вовсе не имеют места.
Бластулоо-бразньхе организмы, воплощающие этот однослойный
тип многоклеточности, относятся к весьма различным категориям
естественной системы и имеют при этом неодинаковое онтогенети-
ческо-е значение. К ним относятся, с одно-й стороны, высшие коло-
ниальные вольвоциды, представляющие собой, как мы ‚видели,
палинтомические колонии Phytomonadina -и сохраняющие пожиз-
ненн-о свое строение целобластул; с другой стороны — разнообраз-
ные жгутиконосные личинки наиболее примитивных из Metazoa,
a именно целобластулы метагенетических медуз (из Leptolida), не-
которых низших сцифомедуз и ча-сти гомоцельных известковых и
кремневых губок, а также целобластулы морских ежей, голотурий
и морских звезд. Все эти весьма разнородные по своему характеру
организмы обладают вместе с тем настолько поразительным сход-
ст-вом друг с другом, что жгутиковые бластулы Metazoa вполне
заслуживают названия «животных вольво-ксов», данного им Г ексли
(1876). Особенно странным кажется то обстоятельство, что столь
определенный тип организации, как-этот, не связан с какой-либо
одной, пусть самой высокой, категорией естественной системы,
а встречается у очень далеких друг от друга гру.пп; это является
исключительным, в сущности, примером независимости -плана строе-
ния организма от его систематической принадлежности.
Это исключительное свойство жгутиковых бластул, наряду с еще
более шир-окой распространенностью безжгутиковых эмбриональ-
ных стадий этого же типа у представителей самых различных групп
Metazoa, издавна обеспечило им особую роль в филогенетических
построениях. Еще в 1828 г. К. M. Бэр пи-сал, что «мы не без основа-
ния можем предположить, что простая пузыревидная ф-орма есть
общая исходная ф-орма, из которой исторически развились все жи-
вотные», и эта мысль Бэра легла в основ-у общеизвестно-й гипотезы
Геккеля о происхождении Metazoa от вольвоксообразных предков,
‘а также и всех ее позднейших модификаций.
Установившееся в этой связи -палинген-етическо-е толкование
формыбластулы, как жив-ой модели «предкового метазоона», «про-
гтотипа гипотетического предка» всех Metazoua ИЛИ «прообраза анце-
стальной бластеи», настолько заслонило собой все иные возмож-
ности ее оценки и толкования, что организмы этого склада -п-о-чти
‘перестали-казаться, в конце концов, настоящими живыми сущест-
вами. Представление об отсутствии у бластул собственных биоло-
гических свойств так укоренилось в сознании биологов, что по мино-
14 Эмбриология беспозвочн-ьхх 209
sauna эпохи‘ филогенетических-увлечений бластулы были как бы
упразднены и вычеркнуты из списков реально существующих орга-
низмов. Дело дошлодо того, что н-еоспоримое и прославленное
в свое время сходство между <<жив-отнь1ми вольвоксами» и настоя-
щими было объявлено недействительным: стали считать, что это
сходство объясняется исключительно одинаковым действием зако-
нов поверхностного натяжения на инертный комплекс клеток,
не имеющих между собой внутренней связи, что оно не дает нам
права делать не только какие-ли-бо филогенетические, но даже и
морфологические выводы, и что даже нет никаких оснований видеть
в Volvox форму, отвечающую стадии бластулы.
Совершенно очевидно, однако, что если упрощенное палингене-
тич-еско-е толкование организации бластулы в духе геккелев-ской
гипотезы и не выдержало пробы времени, о-казавшись, по существу,
бесплодным и даже в известной мере дезо-риентирующим, то и про-
тивоположная, Механистическая ее интерпретация является еще
менее приемлемой; означая отказ от биологического и историче-
ского понимания этого типа животной организации, она, вдобавок,
находится и в вопиющем противоречии с фактами. -
Объективное сравнительно эмбриологическое объяснение бла-
стулы, как простейшей, действительно исходной (как в онтогене-
тическом смысле, так и, по всем данным, в филогенетическом),
формы многоклеточной организации, однако, вполне возможно.
Оно заключается именно в признании бластул сложными з-ооспо-
рами, или синзооспорами, обра ующимися путем объединения
в одно целое семьи зооспор, создающейся повторным делением
(Зах-ваткин, 1946). Это представление поддерживается, как мы
увидим, всей совокупностью ‚относящихся сюда фактов и сущест_-
„вен-но важно тем, что дает нам ключ к пониманию процесса станов-
ления многоклеточной организации. При этом выясняется, что срав-
нительный анализ особенностей формирования и строения бластул
позволяет уловить и некоторые основные закономерности, управ-
ляющие этим процесс-ом и при-водящие с железной необходимостью
к его осуществлению в одних и тех же формах эпителиальной бла-
стулы у всех подлинных «пионеров» многоклеточной организации.
/
Г а в а Х1
ДРОБЛЕНИЕ, КАК ПАЛИНТОМИЧЕСКОЕ ВОСПРОИЗВЕДЕНИЕ
1. Общие соотношения между спорообразованием и дроблением
Сра-внимо-сть дробления многоклеточных с процессами споро-
„образования у простейших, размножающихся повторным делением,
неизбежно вытекает из любого их сопоставления, в каком бы плане
оно ни про-водилось и какая бы цель им ни преследовалась. К та-
кому выводу приводит и „непосредственное взаимное сравнение
СЗМИХ ЭТИХ ПРОЦЕССОВ, И сравнительное РЭОСМОТРЕНИЭ" ЖИЗНЕННЫХ
210
циклов Protozoa и Metazoa, и сличение-хода полового процесса при
er‘-O”O»0I‘aMHOM характере; ПОЭТОМУ не ПрИХОДИТСЯ удивляться, что те
немногие исследователи, которые решались СОПОСТЗВЛЯТЬ многокле-
точных С простейшими‘ В TOM ИЛИ ИНОМ ИЗ’ ЭТИХ аспектов, неизменно
приходили к заключению о какой-то степени соответствия дробле-
НИЯ яица п-ервь1х спороген-езу вторых, О том, ЧТО ЭТИ ЯВЛЕНИЯ
"в какой-то мере гомо-логичны друг другу. Четкую постановку этого
важнейшего вопроса. в единственно правильном историческом,
эволюционном, аспекте мы ‘находим, однако, только у Мечни-
кова+в его классических «Эмлбриологических этюдах о меду-
зах» (1886). _
Как уже говорилось выше (стр. 12), Мечников считал дробле-
ние “яйца многоклеточных берущим свое начало от трехмерного
повторного деления тех гипотетических «метазоо-флагеллят>>, потом-
ками которых они являются. Напомним, что, по М-ечникову, только
такое деление могло обеспечить построение шаровидной колонии
типа бластулы у этих предковых «метазоо-флагеллят», что им
были приведены факты наличия подобного деления у некоторых
низших организмов и что он видел в закономерном чередовании
плоскостей первых делений яйца медуз в трех ‘измерениях про-стран-
`ства историческое свидетельствотого, что дробление яйца Metazoa
произошло от размножения их протозойных предков именно путем
трехмерного деления. Правда, во времена Мечникова различные
формы размножения простейших были еще так мало известны, что
он-не мог подробнее обосновать ‘и развить эту «лишь более или
менее вероятную гипотезу»; но самый факт, что Мечников первый
ясно высказал мысль -о филогенетической связи дробления Metazoa
’c делением, ведущим к образованию колоний у Protozoa, и пра-
вильно оценил роль этих процессов, как средств создания многокле-
точности,— является громадной его заслугой. с
После М-ечникова вопрос о взаимоотношениях спорообразова-
ния и дробления почти не обсуждался" в литературе: Только в ра-
' ботах Э. Шаттона и А. Львова (1919—1936) и Ова-сса (1936) вопрос
этот получил более подробное, хотя и одностороннее, освещение
в связи с изучением этими авторами палинтомического размноже-
ния простеиших. Шаттон и Львов считают дробление яйца Metazoa
›тИ‘ПИЧНЫМ примером палинто-мии, включающей, по их представле-
ния-м, любой“ вид многократного деления гипертрофированной
сисн-энергиднои клетки-независимо от того, связано ли оно или
нет с половыми_ формами размножения и представляет ли ообой
такая клетка яицо многоклеточного -или же отдельную особь про-
‘ стейшего.
Справедливость подобного сближения представляется совер-
шенно неоспоримо-и, если учесть все факты, приведенные нами
у в, IV, V, VI И Х главах. В самом деле, все, что было сказано там
‚ о палинтомии у Protozoa И, в частности, о метагамных ее формах,
полностью приложим-о и к дроблению яйца много-клеточных, так
что принадлежность дробления к кругу палинтомических процессов
14* - 211
не может быть поставлена под сомнение. А это .значит; что оно отно-
сится к явлениям спорогенетическо-го порядка, подобно любой дру-
гой форме палинтомии.
Вм-есте с тем нельзя закрывать глаза на факт, чт-о представле-
ния Мечникова и других авторов, связывавших дробление Metazoa
c процессами размножения простейших, не получили пока хоть
сколько-нибудь широкого признания. Это я-сно показывает, что ин-
тересующий нас ВОПрОС далеко не ‘ПРОСТ, ЧТО ‘ОАН все же не решен С‘
необходимой степенью доказательности и что громадным и неоспо-
римыми сходствам между дроблением и -палинтомическим спорооб-
разованием противостоят еще более явные различия между ними,
‚рЗЗЛИЧ-ИЯ, кажущиеся настолько пр-инципиальн-о ВЭЖНЫЁМИ, ЧТО ИМИ
как будто обесцениваются ‚все имеющиеся сходства ‚между этими
. процессами.
Самым главным и бросающимся в глаза из таких различий,
безусловно, является т-о-, что спорообразование есть, по самому
своему существу, процесс массовой продукции зародышей, т. -е.
высоко продуктивного р а з м н о ж е н и я з и г о т ы или б-есполой
одно-клеточной особи, в то время как дробление вообще не является
размножением: оно есть не что иное, как процесс во сп p o И з ве-
д е н и я з и го т ы (или агаметы), пр-оцеос -ее развития и пр-евраще-
ния из одноклеточного в многоклеточное состояние. Благодаря этому
и создается впечатление, что мы имеем здесь дело с явл-ениями раз-
личного порядка: в одном случае-с самым продуктивным из видов
подлинного размножения, каким является спорогония, а в- дру-
гом — с чисто онтогенетическим процессом.
Не менее существенным противопоказанием, ‘препятствующим
сближению дробления со сп-ороген-ез-ом, служит глубоко укоренив-
шееся мн-ение, что абсолютно характерным признаком последнего
является его обязательно б-есполый характер и то,.что он осуще-
ствляется только в гаплоидн-ой ядерной фазе, создающейся путем
зиготной или, чаще, особой споровой р-едукции; между тем дробле-
. ние Metazoa СТОЛЬ же неразрывно связано с половыми формами раз-
„ множения, диплофаз-ой и принципиально иным, гаметным, типом
_ редукции. Таким образом, и с этой точки зрения дробление попадает
в другую категорию явлений, чем спор-ообраз-ование.
К этим двум, самым капитальным, различиям между ними .при-
бавляется еще одно, касающееся геометрических форм деления при
сп-орообразовании и дроблении. Речь идет «о том, что дробле-
ние M-etazoa обычно считается идущим в трех измерениях
пр-остранства согласно правилам Г ертвига и не менее общим пра-
вилам деления целых геометрических фигур? В то же время споро-
. образование и, более конкретно, палинтомия Protozoa ооверша-ет-ся
тольк-о по линейному или табличному -ее типу и подчинена, как мы
видели, совершенно другим закономерностям. Итак, неминуемо
создается впечатление, что если дробление’ и относится, как мы это
считаем, к разряду палинтомических процессов, то все же занимает
среди них весьма обособленное место и должно быть противопо-
«212
ставлено всем остальным, поскольку оно осуществляется исключи-
тельно в диплофазе и в особых формах трехмерной, кубической
палинтомии и, что самое главное, н-е имеет характера размножения.
Этим, очевидно, и объясняется то предубеждение, с которым при-
ходится сталкиваться и считаться при гомологизации дробления
многоклеточных с процессами спорооб-разования у простейших.
Эти три основных различия, безусловно, им-еют место, если
сравнивать, как это обычно и де.лается, дробление «вообще», в его
кла-ссических формах, характерных для высших Metazoa, со столь
же каноническими видами спорообразования у низших растений.
Между тем стоит только перейти от такого общего (и потому неиз-
бежно абстрактного) сравнения к более конкретному, учитываяпри
этом все существующее разнообразие частных форм того и другого
процесса и ведя его, что главное, на максимально пригодном для
этого материале,-—как картина существенно меняется: становится
очевидным, что все эти различия далеко не являются абсолютными,‘
а частью даж-е и нереальны.
Так, первое, самое главное из них, значительно сглаживается,
прежде всего, тем обстоятельством, что фитомонады дают полную
, цепь переходов от самого подлинно-го спорообраз-ования, все много-
численные продукты которого разбегаются в виде зооспор и обра-.
зуют каждый отдельный одноклеточный организм, к таким его фор-
мам, где они все остаются в пожизненном соединении -в виде
многокл-еточн-ой палинтомич-еской к-олонии, т. е. дают начало одной
единственной особи. Далее, хорошо известна способность ряда низ-‚
шик Metazoa (например, гидром-едуз) к факультативной (а иногда
облигатной—у Mesozoa) полиэмбрионии, называемой тут тотипо-
тентностью бластожмеров и имеющей своэим результатом возмож-
ность образования нескольких особей из одного и того же яйца,
т. е. самоеподлинное его размнож-ение. Наконец, самые классиче-
ские примеры половото воспроизведения архег-ониат показывают
не менее наглядным образом, насколько условна грань между раз-
множением и воспроизведением. Все эти и многие другие при-
меры определенно свидетельствуют о том, что BIC я ко е в ос п р -о-
изведение представляет собой не что иное, как
дериват подлинного размножения и не может быть
противопоставлено ему, как нечто чуждое, как явление принци-
пиально иного порядка. Таким -образом, в самом факте, что дробле-
ние есть п-о существу и в норме процесс воспроизведения, а не раз-
множения, не содержится ничего такого, что заставило б-ы принци-
пиально противопоставить его спорогенезу; можно только -сказать,
что оно является па-линтомиче-ским воспроизведе-
н ием, возникшим и оформившимся здесь, как и во всех дРУгих
аналогичных случаях, на базе множественного размножения зиготы.
Не теряя- от этого своей характерности для Metazoa (a также.
и Volvocoidea), ‘ЭТО отличие дробления от типичного споро-генеза
является, стало быть, только относительным и ни в коей мере не
препятствует их самому тесному сближению. .
2 13
Не препятствует такому сближению и второй -из аргументов,
обычно используемых против него: противоположность ядерных .-
ф.аз, при которой совершаются эти процессы. Из т-ого, что уже гово- И
рИЛОСЬ О РЗЗЛИЧИЯХ В ЧЭРЁДОВЭНИИ ЭТИХ фаз В ЦИКЛЗХ РЭЗНЫХ ПрО-
стейших‚ явствует с полной очевидностью, что огульное распро- _
странение на них чисто ботанических правил соотношения ядерных
фаз не оправдыва-ется фактами; напомним известные примеры обра-
зования диплоидных спор у грегарин и стадий того же порядка7
(цилиоспор) у ряда паразитических инфузорий. Таким образом,-
это главное оружие противников всякой гомол-огизации дробления
со спорогенетическими процессами в действительности не имеет
силы.
Что же касается геометрических форм деления, то и здесь ока-
зыва-ется невозможным провести отчетливую грань между обоими
сравниваемыми процес-сами. В самом деле, типичнейшее по виду
дробление вольвоцид и, как будет показано дальше, ряда низших
Metazoa является в действительности не кубической, а табличной
палинтомией и подчиняется всем нормам этой последней, а не пра-
вилам Закса и Г-ертвига, как это кажется с первого взгляда.
Итак, основное общее различие между спорообразованием и
дроблением сводится, в конечном счете, к тому, что «в одном случае
споры разъединяются и образуют каждая отдельный организм,
тогда как в другом они остаются соединенными и дают начало
одному сложному организму» (Бальфу . И хотя это азличие.
Р Р
и не имеет абсолютного характера, оно все же настолько харак-
терно, что дробление Metazoa и вольвоцид правильнее «всего считать
палинт-омическим воспроизведением и видеть именно в это-м его
наиболее специфич-ескую ч-ерту.
Подробное обоснование этого положения на соответствующем
фактическом материале и выяснение путей и способов преобразо-
вания палинтомического размножения в эту, характерную для
многоклеточных, форму воспроизведения составят основную задачу
эт-ои и следующих за нею глав. ›
2. Переход от спорообразования к палинтомическому
воспроизведению у Volvocoidea
Процессы. формирования ‚шали-нтомических колоний Volvocoidea
посредством повторного деления служат превосходным и весьма
поучительным примером превращения размножения зооспорами,
характерного для всех фитом-онад, в воспроизведение посредством
сложных, многоклеточных синзооспор, образуемых -в единственном
числ-е каждой из плодущих особей коло-ний. Многие из особенно-
стей этих процессов были уже описаны в гл. VII, где они рассматри-
вались, однако, преимущественно с точки зрения способов сложения
колонии составляющими ее зооспорами. Теперь нам надлежит рас-
смотреть их в ином, сравнительно эмбриологическом освещении,
имея прежде всего в виду „открывающуюся здесь возможность про-
214
следить шаг за шаром ход замещения множественного палинтоми-
ческого размножения воспроизведением синзоосп-орами. Эта редкая
и заманчивая персп-ектива обеспечивается естественным располо-
жением представителей этой группы в чрезвычайно показательный
ряд, иллюстрирующий, как было уже описано, все ступени повы-
Шающейся- целостности колоний—от слабо интегрированных и
легко распадающихся объединений до подлинно многоклеточных
организмов. о
Просматривая этот ряд с интересующей нас сейчас точки зрения,
‘не трудно заметить, что у самых малочленных из его звеньев—
у 4——16-зооидных колоний Goniu-m, Pandorina И др. —палинтомия
вполне сохраняет свой исходный характер размн-ожения. Это выра-
жается B том, что все члены колонии обладают способностью к раз-
множ-ению, что означает, что каждый из продуктов повторно-го
деления становится физиологически независимым организмом, раз-
множающимся в свою очередь таким же порядком, как одиночные
фитомонадины. Правда», вся образуемая ими семья зооспор объеди-
няется при бес-п-олом размножении в более или менее п-остоянную
колонию; однако эта последняя сравнительно очень легко распа-
дается на один-очны-е особи, и это же происходит в обязательном
порядке при половых формах размножения. Учитывая, что при
столь низком уровне объединения колонии она «еще не имеет прав
считаться целостным организмом, здесь, очевидно, не приходится
еще и говорить о наличии воспроизведения: налицо самое подлин-
ное размножение, нисколько не умаляемое тем, что его продукты
соединены в колонию. Иначе говоря, каждая особь постоянно
сохраняет свое значение полноценного организма, и если и произ-
водит по одной только колонии, то колонии самого примитивн_ого
типа, являющейся простым собранием нескольких, относительно
независимых, особ-ей. ‘
Однако, как только интеграция такой колонии дойдет до уровня,
наблюдаемого уже у З2-зооидных колоний Eudorina, положение’
в корне меняется. Практически необратимая целостность этих
объединений приводит к тому, что они уже сами становятся орга-
'низмами‚ тогда как составляющие их одноклеточные особи теряют
это значение и соответственно низводятся до ранга клеток. Следо-
вательно, как только перейден определенный «порог интеграции»
колонии, или синзоосп-оры, палинтомическое размножение немед-
ленно уступает свое место самому подлинному воспроизведению,
воспроизведению, сводящемуся к превращению одно-клеточной
материнской о-соби в многоклеточную дочернюю, т. е. осуществляю-
щемуся таким же образом, как у всех других многоклеточных
организмов.
Такими одноклеточными материнскими особями являются
у Pleodorina И всех вышестоящих в системе Volvocoidea ИХ гонидии,
или гоноциты, общее число которых в составе тела прогрессивно
понижается, как‘ мы знаем, по мере усиления его целостности.
Однако. это ужен-е влияет на самый характер палинтомического
215
ПрОЦ-ЕССЭ! раз ПрИНЯВ ‘ФОРМУ ВОСПРОИЗВЕДЕНИЯ, ОН ОСТЭЕТСЯ таковым
И ПрИ ВСЕХ СЛЕДУЮЩИХ ‘СТУПЕНЯХ ИНТЕГРЭЦИИ, И ПОСЛЕДНЯЯ ВЕДЕТ
ТОЛЬКО K дальнейшему ‘СНИЖЕНИЮ ЧИСЛа генеративных ЭЛЕМЕНТОВ.
КОЛОНИИ И СООТВЕТСТВУЮ-ЩЕМУ падению ЕЕ ООЩЕИ, ‘СУММИрНОИ, ПЛО- '
ДОВИТОСТИ ПрИ ОДНОЗВРЕМЕННОМ ПОВЫШЕНИИ УрОВНЯ o»pra111»13a111»11r1
образуемых ею дочерних ‘колониальных -осо-бей.
Итак, размножение целостных, многоклеточных колоний Volvo-
cidae совершается исключительно путем (бесполого или полового)
палинтомического воспроизведения их гонидий, каждая из которых
продуцирует лишь по одной, зато многоклеточной, особи. Таким
образом, оно становится пподлиннным размноже-
нием только в силу того о-бстоятель-ства, что
в каждой колонии находится не одна, а по край--
н е й Me ре нес кол Ь ко -гон иди й; или, другими словами,
потому, что складывается из большего или меньшего числа отдель-
ных актов воспроизведения, осуществляемых одной и той же мно-го--
клеточной особью, и ведущих таким путем K действительному уве-
личению численности таких особей. В этом——характернейшая
особенность цитогонигческого размножения всех вообще многокле-
точных, противопоставляющая его любым другим видам цитогонии
и заключающаяся именно в том, что оно- совершается за счет многих,
а н-е одной генеративных клеток, каждая из которых дает начало
одной единственной доч-ерней особи.
З. Следы спорообразования в‘ дроблении низших Metazoa
СПОРОГОНИЧЕСКЗЯ основ-а ДрОбЛЕНИЯ ЯрКО проступает ‘ВО МНОГИХ
его признаках у низших Cnidaria,- ГУООИ и игл-окожих. Правда, мы1
уже не находим здесь столь эффектных ‘примеров перехода споро--
образования в спорогенетиче-ское воспроизведение, какие нам-
встретились в замечательном ряду Volvocoidea, и даже, более того,
НЕ находим ЗДЕСЬ НИ ОДНОГО случая ‘ПОДЛИННОГО СПОРОГОНИЧЕСКОГО:
размножения; ЭТО И ПОНЯТНО, ПОСКОЛЬКУ ВСЕ Metazoa (КрОМЕ, p2l3B1€,-
ДИЦИЕМИД) ЯВЛЯЮТСЯ ВПОЛНЕ СЛОЖИВШИМИСЯ МНОтГОКЛЕТОЧНЫМИ орга-
НИЗМЭМИ, УЖЕ ТВЕрДО ставшими на ПУТЬ ПЭЛИНТОМИЧЕСКОГО ВОС-
произведения. Однако у них все же легко обнаружить целый ряд
таких особенностей дробления, „в которых очень ясно сказывается-
CFO спорогонич-еская природа.
Прежде всего необходимо всячески ‘подчеркнуть, что дробление
у всех тех форм, где оно заканчивается образованием свободно-
плавающей жгутиконосной бла-стулы, имеет чисто цитотипический
характер, т. -е. не сопровождается какими-либо морфогенетическими
процессами и сводится, стало быть, к одному только повторному
делению яйца. При этом именно здесь находят свое место крайние
для Metazoa случаи недетерминированности дробления, т. е. макси--
мальной для них и предельно длительной равноценности бласто-
меров. Помимо того, о-но характеризуется еще тем, что включает
большое число ——„_чаще всего 9 или 10+повторных делений, так
gm.
что общее число образующихся кл-еток достигает 500-1000 и более,
Таким образом, здесь оказывается налицо такой характерный при-
знак споро-генеза, исчезающий у более высоко организованных
Metazoa, как очень большое количество равноценных продуктов
деления зиготы. Он еще дополняется, как будет показано в следую-
щей главе, и рядом очень специальных признако-в в морфологиче-
ских картинах дробления, выражающих собой значительно более
' высокий уровень автономности бластомер-ов, чем это типично для
Metazoa. B ЦЕЛОМ, все эти признаки придают дроблению этих низ-
ших "форм особый отпечаток «чего-то неустановившегося», как очень
правильно говорил Давыдов, отпечаток очень еще низкой и неусто-й-
чивой организменной целостности дробящейся системы.
Самым ярким ее выражением служит встречающаяся только
здесь способность дробящегося яйца к распадению на отдельные
бластомеры, сохраняющаяся в ряде случаев до относительно позд-
н_их стадий, а именно, до стадии 16 бластомеров включительно
у многих гидром-едуз. Это замечательное явление, впервые о-писан-
но-е Мечников-ым у Oceania и затем экспериментально изученное
Зойя (1895), Маас-ом (1905) и Т-ессье (1930) у Clytia, Laodi-cea'
и Amphisbetia, являет-ся, как отмечал-ось, своего рода факульта-
тивной полиэм-брионией, но полиэмбриони-ей, по всем данным, пер-
вичной, остаточной, свидетельствующей прежде всего об исключи-
тельно низком уровне объединенности этих ранних стадий
дробления. Однако самый факт, что это свойство встречается в та-
кой своей крайней форме как раз только у наиболее примитивных
метагенетических представителей низшего из отрядов Hydrozoa
(Leptolida) и ‘вдобавок сочетается: у них с рядом дРУгих, типично’
протозойных черт морфологии дробления, заставляет придать ему
и другое еще значение. Оно представляется именно отголоском
СПОрОГОНИЧЕСКОГО РЗЗМНОФКЕНИЯ ЯЙЦЭ, ‘СОВЕРШЕННО ЭНЭЛОГИЧНЫМ"
таким ЖЕ ПрОЯ-ВЛЕНИЯМ затухающей, ЕСЛИ МОЖНО ВЫРЭЗИТЬСЯ,
спорогонии у немн-о-гочленных колоний Eudo-rinida-e. Самое замеча-
. тельное в этой невольно напрашивающейся параллели заключается
в том, что свойство тотипотентности сохраняется там и тут до оди-
накового численного уровня системы — 16-клеточно-го, соответ-
ствующего в ряду Volvocoidea. определенному порогу интеграции.‘
Выше этого уровня о-но не простирается и здесь, у гидро-медуз,
исчезая уже на стадии 32..
Само собою разумеется, что сходные, но более ограниченные
проявления тотипотентно-сти бластомеров у морских ежей и дру-
ГИХ ФОРМ, ИЗУЧОННЫХ В ЭТОМ! ОТНОШЕНИИ, ТЭКЖЕ МОГУТ рассма-
триваться „как ‘СЛЕДЫ, ИЛИ ОТГОЛОСКИ, СПОРОГОНИЧЕСКОГО ‘раз-Т
МНОЖЕНИЯ.
В своей совокупности все эти особенности дробления у низших"
ГрУПП Metazoa ‘ПОЗВОЛЯЮТ, ВО ВСЯКОМ случае, ГОВОРИТЬ О НЕПОЛНОМ
И ЕЩЕ НЕУСТОИЧИВОМ ЗЭМЕЩЕНИИ спо-рообразования ‘ВОСПРОИЗВЕДЕ-
НИЕМ, О Т-ОМ, ЧТО ПОСЛЕДНЕЕ НЕ ДОСТИГЛО ЕЩЕ ЗДЕСЬ ТЕХ СВОИХ стабиль-
ныхформ, в каких мы привыкли его видеть у высших Metazoau
2f~7
4. Результаты И СЛЕДСТВИЯ перехода 0Т ПЗЛИНТОМИЧВСКОГО
cn0p005pa3oBaHmI К ПЗЛИНТОМИЧВСКОМУ воспроизведению
Ограничиваясь пока НВМНОГИМИ ПРИВЕДСННЫМИ данными В ПОЛЬЗУ
РЭЗВИВЭЭМО-ГО ЗДЕСЬ ПРЭДСТЭВЛЭНИЯ О ДрОбЛЭНИИ МНОГОКЛЁТ-ОЧНЫХ,
как ПЭЛИНТОМИЧЭСКОМ ВОСПРОИЗВЁДЕНИИ, ОТМЭТИМ y>K-e сеичас H€K-O-
торые примечательные общие моменты, выЯ-сняющиеся при такой '
его тракт-овке и служащие ей вместе с тем весьма немаловажной
поддержкой. Речь идет о тех прямых и косвенных следствиях заме-
щения -спорого-нии воспроизведением, которых мы обычно не заме-
чаем из-за их кажущейся обычности, а также и по той про-стой
причин-е, чтоони обнаруживаются и объясняются только при осве-
щении этих факто-в под определенным угло-м зрения.
Об одном из следствий такой замены было уже сказано несколько
слов на стр. 186. .Это—— замечательное в сущности замещение того
истинного цитогоническ-ого размножения, каким является споро-
образование (в любых его——›бесполых и половых—формах), тем
специфическим для многоклеточных‘способом размножения, кото-
рый мы ошибочно продолжаем еще считать равноценным и даже
тождественным истинн-о-й цитогонии простейших.
Во избежание каких-либо неясностей напомним, во-первых, что
размножением следует считать всякий процесс, ведущий к увели-
чению числа биологически независимых осо-бей. Во-вторых, по-вто-
рим еще раз, что гцитогоническое размножение простейших, которое
мы принимаем за типичное, заключается в образовании неско-льких
(по крайней мере двух) новых особей каждой материнской генера-
ТИВНОЙ КЛЕТКОЙ. ВО B-CHKOM случае, если РЭЗМНОЖСНИС ПРОИСХОДИТ ‘
половым путем, оно целиком осуществляется в метагамном отрезке
цикла, за счет подлинного размножения зиготы, являющегося тут
главным фактором во всем этом процессе и продуктивностью кото-
рого определяется (целиком или хотя бы в -главн-ой мер-е) общая
продуктивность размножения.
Совсем не то мы видим при <<цитогоническом>› размножении
многоклеточных. Здесь каждая генеративная клетка (яйцо, гони-
дия, агамета) дает начало- одной только особи, в чем именно и за-
ключается процесс воспроизведения. Эти клетки сами не размно-
жаются, и если бы они вырабатывались по одной каждым
материнским организмом, размножение не ‘им-ело бы здесь м-еста. ‚
Иначе говоря, з дес ь в ов с е о т с у т ст в у е т в типе (за исклю-
чением случаев полиэм-брионии) м е т а г а м н о е р а з м -н о ж е-
Hue, И ‘ЭТО ПОСЛЭДНСЭ ФЭКТИЧЕСКИ ‘ПЕРЕНОНСИТСЯ
на прогамную часть жизненного цикла. Действи-
тельно, единственным фактором, обеспечивающим наличи.е размно-
жения и определяющим его продуктивность, становится тут
производительность гаметогенеза в женском
е г о р я ду, от которой полностью зави-сит число образующихся
потомков. Поэтому и число метагамных (спорогенетических) деле-
ний зиготы совершенно лишается своего былого значения и -служит
ч
218 . °
только увеличению числа клеток, спаянных в один дочерний
организм.
Таким образо-м ' осуществляется второе важнейшее следствие
замещения спорогонии воспроизведением. Оно выражается в том,
что продуктивнейший п р о цесс с п о р оо б р а з-ов а н и я це-
ликом теряет свои прямые функции — ‘массовой выработки
одноклеточных спор — и пол н ос т ь ю об р а щ а-е TC я н а
построение своего единственного теперь про-
д у к т а. Так создается многоклеточная дочерняя особь, отражаю-
щая богатством своих клеток высокую интенсивность споро-генети-
ЧЕСКОГО npouecca. Эта НОВЭЯ рОЛЬ cnoporenesa, ВЫЗВЭННЭЯ K ЖИЗНИ‘
заменой одного способа размножения ДРУГИМ, заключается, следо-
вательно, в его превращении из деления, служащего размножению,
в деление, так сказать, ко н с т р у кт и в н о е, в тот самый процесс,
посредством которого одноклеточное яйцо становится многоклеточ-
ным зародышем. Нельзя не подчеркнуть в этой связи далеко не
всегда учитываемое обстоятельство, что образовавшееся таким пу-
тём дробление является единственным реальным, существующим в
природе способом построения целостных ‘мн-огоклеточных организ-
мов из одной исходной клетки. _
Третьим, не менее замечательным и важным последствием пре-
вращения спорогонии в спорогоническое воспроизведение является
оформление того процесса, который мы называем эмбриональным
развитием и с которого обычно начинается онтогенез всякого мно-
гоклеточного существа. Em первооснову образует дробление, состав-
ляющее вм-есте с тем Минимальное, а у многих низших Metazoa
и единственное, содержание эмбриогенеза; в ча-стности, им практи-
чески исчерпывается эмбриональное развитие большинства тех наи-
более примитивных форм, у которых последнее заканчивается
образованием свободноплавающей жгутиконосной бла-стуль1. Есте-
ственно, поэтому, что все характерные свойства таких простых
и примитивных форм эмбриогенеза „сводятся к самым общим при-
знакам палинтомического спорогенеза и не имеют еще здесь почти
ничего специфически «зародышевого», Действительно, основная
Масса этих свойств, квалифицируемых как эмбриональные, уже
имеется во вполне оформленном виде п.ри любых видах палин-
томического размножения простейших, и они сами сводятся,
в конечном счете, к еще более общим особенностям метазойного
типа деления. Иначе говоря, это — с в о- й с т в а, п е р в и ч н о- и
по существу чисто п алинтомически-е и приоб-
р-етающие значение эмбриональных лишь вто--
рично, с того ‘момента, как палинтомия -стано-
вится основой эмбриогенеза мн-огоклеточных.
П-оэтому нельзя отказать в остроумии замечанию Заварзина (1945),
что низшие представители Metazoa (речь шла конкретно о губках)
«не имеют, в сущности, эмбриологии». Эта мысль, правда, несколько
утриро-вана, но она правильно угадывает действительное положе-
ние в-ещей.
219
Еще одним р-езультатом занимающег-о нас сейчас явления ока:
зывается приобретение гипертрофическим развитием материнской
клетки зооспор нового для него значения; оно становится тенерь
оогенезом, сохраняя при этом ряд ранее ПРИСУЩИХ ЕМУ ‘Своими-
В сам-ом деле, развитие яйцеклетки. многих низших Metazoa проходит
те же этапы и стадии, какие чередуются В РаЗВиТии паразитических»
питающихся осмотическим ПУТЕМ, ИЛИ ДЭЖЕ ЭКТИВНО ХИЩНИЧЭЮЦЁИХ
трофозоитов простейших, и также завершается‘ превраЩеНИеМ В‘ A
НЕПОДВИЖНуЮ СТЭДИЮ, НЗЗЫВЗЕМУЮ ТУТ ЯЙЦОМ.
Таковы некоторые из любопытных сдвигов, к каким приводит.
замещение первичного спорогонического размножения выросшим
из него воспроизведением. Ими одновременно создается целыи
комплекс взаимосвязанных явлении, выливающихся в формирова-
нии многоклеточности со всеми ее наиболее характерными атрибу-
тами——оогенезом‚ дроблением, эмбриональным развитием и свое-
НОВЫМИ онтогенеза И РЗЗМНОЖЕНИЯ.
мы вправе сказать теперь же, без всяких колебании, что источником
всех видов многоклеточности у животных организмов служит про-
цесс замещения спорообразования палинтомическим ж-е воспроиз-
ведением. Как осуществляется это замещение и каковы его би-оло-
гические ‘причины — вопрос особый; его решение только намечается
некоторыми из приведенных здесь фактов и сможет быть достигнуто
только на основании использования еще множества дргугих.
Г л а в а ХП
ЭМБРИОГЕНЕЗ МНОГОКЛЕТОЧНЫХ ФИТОМОНАД
1. О многоклеточности Volvocoidea
O главных особенностях строения и развития многоклеточных
фитомонад уже говорилось в гл. VII, где они рассматривались в ка-п
честве палинтомических колоний и соответственно освещались
с этой точки зрения. Т-еперь мы рассмотрим их в другом плане-
в качестве целостных многоклеточных организмов, что позволит
дать этим особенностям и существенно ИН-Ое, сравнительно ЭМбРИО-ЛО-
гическое освеЩени-е. _
Представление о вольвоцидах как многоклеточных организмах
было выдвинуто и обосновано нашим известным альгол-огом
И. Горожанкиным еще в 1875 г. и получило дальнейшее -подтвер- `
ждени-е во всех решительно исследованиях, проведенных с того
времени над различными представителями этого замечательного‘
семейства. И если во времена Горожанкина оно могло базироваться‘
на лишь относительно немногих известных тогда признаках‘ их
строения и развития, то теперь иная их трактовка попросту исклюд
чена. Высокая организменная Целостность вольвоцид особенно‘
интересна тем, что она. выражается в таких специфических для
Metazoa свойствах, Как, ГРадИеНШНа51 (Осевая) диффеРеиЦиаииа
22.0
всех клтеток организма И Их строго ёпитеЛиаЛЬНое сложение, СПИ-
‚ральныйчхарактер дробления и т. д. (-см. "гл. ХШ).
Однако остается все же неясным, является ли многоклеточность
привилегией одних только настоящих вольвоксов (т. е. представи-
телей рода Volvox -в прежнем, широком его понимании), или же
ее делят с ними и какие-либо другие из членов того замечательного
‚ряда нарастающей организм-енной интеграции, в ‘который так
красиво располагаются все колониальные Volvocoidea. Иначе
РОВОРЯ, ПЕРЕД нами встает ВОПРОС О ТОМ, ГДЕ ЛЕРКИТ ГРЗНИЦЭ МЕЖДУ-
колониальностью и многоклеточностью «в этом удивительном
Ряд)’-
Мы уже видели, что в нем существует целый ряд уровней целост-
ности, настоящая их лестница, каждая ступень которой характери—
зуется различной степенью выраженности одних и тех ж-е признаков
морфофизиологической интеграции; что эти ее признаки ‚начинают
проявляться в ощутимой, хотя и зачаточной, форме у 16-членных
колоний, становятся уже оч-ень яркими у З2-членных колоний, еще
более наглядны у 64—128-членных и достигают максимума у на-стоя-
. щих, более чем 512-членных, Volvocidae s. str. Стало быть, только
самые бедные особями 4- или 8-членнь1е объединения практически
-ЛИШЕНЫ ЭТИХ ПРИЗНЭКОВ, а ВСЕ ОСТЭЛЬНЫЕ ПрО-ЯВЛЯЮТ ИХ В СТЕПЕНИ,
прямо пропорциональной числу своих одноклеточных компон-ентов.
Таким образом, здесь существует целый ряд градаций -организм-ен-
-НОЙ ЦЕЛОСТНОСТИ, ОНЛИЦ-ЕТВОРЯЮЩИХ РЭЗНЫЕ ЭТЭПЫ ЕДИНОГО npouecca
ИНТЕГРЭЦИИ, ПРОЦЕСС ПРЕООРЭЗОВЭНИЯ ПРОСТОГО КЛЕТОЧНОБО СОЕДИ-
НЕНИЯ В ЦЕЛОСТНУЮ МНОГОКЛЕТОЧНУЮ ОКСООЬ. `
С ЭТОЙ ТОЧКИ ЗРЕНИЯ МОЖНО ГОВОРИТЬ О СуЩ-ЕСТВОВЭНИИ ОПРЕДЕ-
_ЛЕННОГ0 nopora ИНТЕГРЭЦИИ, С ДОСТИЖЕНИЕМ КОТОРОГО ОФОРМЛЯЕТСЯ
первая, низшая и’-еще неустойчивая ступень многоклеточной орга-
низации, наблюдающаяся у 16-клеточных Зропоуютогйоае и Eudo-
rinidae; далее идетуже много более оформленная и устойчивая
32-клеточная ее ступень, при которой начинают проявляться первые,
но уже необратимые признаки дифференциации членов колонии
на немногие бесплодные и основные по своему числу полноценные
. плодущие клеткидна следующей, 64—12:8-чл'енной, ступени много-
клеточной организации эта дифференциация -зах-одит еще много
дальше и имеет своим результатом оформление особого типа амфи-
бластулы, наблюдаемого у Pl-eodorina; И только крайняя, самая
‚ВЫ-СОКЭЯ ИЗ ИМЕЮЩИХСЯ ЗДЕСЬ ‘СТУПЕНЕЙ ИНТЕГРЭЦИИ ПР-ЕДСТЭВЛЕНЭ
в форме эпителиальной бластулы вольвоксов, состоящей из гро-мад-
ного числа бесплодных и буквально нескольких плодущих клеток —
‚ г-он идий.
Отсюда очевидно следует, что многоклеточным-и организмами
_ должны быть признаны все члены этого рода, начиная с З2-кл-еточ-
_ных, тогда как 16-клеточные олицетворяют переходное, промежу-
_точное состояние между целостным организмом и простой мало-
_ клеточной колонией, а 4—8-членные агрегаты несомненно относятся-
K категории ЭТИХ ПОСЛЕДНИХ.
221
Неоубходимо отметить, что этот вьтвод, основывающийся’ на
современном уровне наших знаний о Volvocoidea, полностью со- .
‘гласуется с теми. заключениями, к которым пришел Г орожанкин
‘почти 75 лет тому назад. Сопоставляя особенности развития широ-
"Kore круга изученных им фитоМонад, Горожанкин писал (1875/ _
стр. 36): «У -Chlamydococcus плазма подвижной осо-би делится на
2-8 клеток, образующих зачаточную ткань; участки ткани здесь
скор-о ‘разъединяются и, получив жгуты, вых-одят на свободу.
У Зтернапоернаега ткань доходит до состояния 8-клеточной пла- у
стинки, у Gonium— 16-клеточной. В известную пору развития
клетки ткани также разъединяются, но разъединение не ведет
за собой вь1х-ода на свободу: у ЗЪерЬаН-озрначега они связаны общей
оболочкой (coerloubium) тканевой пластинки, у Схопшш—углами
своих оболочек. И в том и в другом случа-е образуются. коло-нии,
где клетки расположены в одной плоскости, и притом каждая от-
дельная клетка способна совершать те же процессы, какие ‘заме-
чаются в прочих клетках колонии. У Pan-dorina И Eudorina, где
клетки. расположены во взрослых колониях сфероидом, вначале
также образуются 16- и даже З2-клеточные тканевые пластинки, ко-
торые мы назвали гоничеокими состояниями этих колоний. Здесь, так
же, как и в предыдущих колониальных родах вольвоксовых, клетки
ткани в известный период разъединяются, и так же каждая кл-етка
колонии функционирует одинаково с прочими. Развитие и диффе-
ренцирование ткани у Volvox .g1-obator идет еще далееоСферо-ид
‚воль-вокса представляет постоянную ткань, клетки кого-рои никогда
не разъединяются и, кром-е того, как мы знаем, -одни из них остаются
‘вегетативными, ДРУГИЕ М-ЭТЭМОРФИЗУЮТСЯ В клетки бЭСПОЛОГО ИЛИ’
полового воспроизведения. Все это, вм-есте с неспособностью отдель-
‘ных клеток вегетировать самостоятельно, приводит нас к заключе-
‘нию, что Vo1vox.H-e есть колония особеи, как думают с-о времен
Эренберга, но единая особь».
2. Eudorinidae
Как было указано уже в другом. месте (гл. VII, 3), плодущие
зооиды и зиготы этих колониальных еще форм представляют собой
вполне нормальных особей, мал-о чем отличающихся от таковых
одиночных фитомонадин. Поэтому понятие яйца к ним еще мало
приложимо, и они могут быть сравнимы скорее с ооцитами, про-
‘шедшими только пер-в-ый этап своего гипертрофич-еского роста-
малый его период. Их проморфологические особенностиртаковы же,
как у других окрашенных Рнутотопафпа: эт0—-0дноосно-гет-еро-
полярны-е существа с симметрией преимущественно двулучево-го
‘типа, сохраняемой ими и ‘в Дедифф-ереНЦИрОВгЪННОМ состоянии.
тГлавным выражением двулучевой симметрии СЛУЖИТ, у дифферен-
ЦИРОВЭННОЙ СТЭДИИ, НЭЛИЧИВ ДВУХ РЭЗДЭЛЬНЫХ КИНеТИД, располагаю-
‘ 1щихся соответственно в о-дной из ее плоскостей, которую можно
НЭЗВЭТЬ ЖГуТИКОВ-ОЙ ПЛ-ОСКОЪСТЬЮ, ТОГДЭ как ВТОРЭЯ ПРОХОДИТ МЕЖДУ
п 2222
У-бесполых особей и на- ранних стадиях развития макрогам-ет
двулучевая симметрия дополняется двусторонней, намечаемой на-
личием глазка ‚ (стигмьт), лежащего в жгутиковой плоскости.
Отношение этих плоскостейк таковым; двух первых делений факти-
чески не установлено, но все говорит за то, что первая из них
ложится, как и удругих двужгутиковых Flagellata, между базаль-
ными зернами обоих жгутов.
Эмбриональное развитие Eudorinidae совершается -при бесполом
размножении в материнской колонии, а при половом иногда (на-
пример, у Pandorina) вне ее. Дробление является, как нам уже
известно (стр. 109), самой типичной табличной палинтомией, стоя-
щей еще только на грани превращения из спорообразования
в палинтомическое во-спроизведение. Такой ее переходный характер
выражается во многих признаках, и прежде всего именно в том,
что о-на сочетает в себе особенности простейших (спорогонических)
форм палинтомии с рядом специальных свойств истинного дробле-
ния. В целом Эмбриогенез может быть охарактеризован нижесле-
дующими особенностями. д
Все следующие друг -за другом деления (общее число которых
варьирует от 2 у Goniurn tso-cial-e до 4 или 5) строго продольны,
или, в переводе на язык эмбриологии, мжеридианальньх, и пправильно
чередуются в. двух взаимно перпендикулярных плоскостях, вероят-
нее всего соответствующих плоскостям двулучевой -симм-етрии мате-
ринской клетки з-ооспор. Характерно, что -в ходе первых З—4 деле-
ний бластомеры (спороциты) почти не меняют своей ориентациии
и остаются в очень тесном контакте; блачгодаря этому, видимые
геометрические формы этих делений довольно точно передают его
действительный табличный характер, и конечным результатом
дробления является типичная ‘гоническая стадия или <<го н и е в а я
п л а с т и н к а», состоящая из одного слоя клеток. Наименьшее
взаимное смещение бластомеров по ‘ходу деления наблюдается
как раз у видов Gonium И имеет своим следствием их группировку
в виде простого креста на стадий 4 и по-чти правильную квадратную
форму всей пластинки к моменту окончания дробления; небольшое
нарушение формы квадрата происходит уже позднее, в порядке
приобретения ‘готов-ой табличкой более устойчивой 2‹формы равно-
весия». У остальных представителей семей-ства бластомеры стадии 4
располагаются в виде палинтомического креста (тетраэдра), и го-
ниевая табличка имеет на всех стадиях более округлые очертания,
что свидетельствует ‚о том, что небольшие, но заметные переме-
щения бластомеров все же сопутствуют делениям (рис. 79). Все
этипоследние осуществляются посредством односторонних кольце--
вых борозд и при заметном митотическом растягивании делящихся
клеток. '
Хорошо известно,ечто гониевая табличка эудоринид практически
никогда не бывает плоской, а сначала в той или иной мере вогнутой
сверху, а затем в той же или большей степени выгнутой. Эти, очень
характерные, изменения носят название процессов и н к у р в а ц и и
223-
щи
и ‘э KC к у р в а п и и и замечательны своим далеко не случайным
сходством с «морфогенетическим» изгибанием эпителиальных пла-.
стов в эмбри-ональном развитии высших животных. У 4-клеточ-ного
Gonium sociale инкурвация в-овсе «He имеет места, а экскурваЦИЯ;
выражается только в отклонении верхних полюсов клеток кнаружи_
в момент ‚выделения ими -студенистой оболочки. У 16-клеточното
I0 ll _ I2
/5 д 9.
‚д
Рис. 79. Эмбриогенез различных Eudorinidae.
'1——4—Eudorina illinoisenslsz 1—-табличная 8—клеточная стадия, 2—стадия 16, 3——чашевидная
З2—клеточная стадия сверху и 4-—сбоку; 5—иНкУрВаЦИЯ‚ Порледовательньхе стадии экскур-
вации (6-17) И начало дифференцировки (72)6nac-rynb1Eudorma elegans; 73——77—дробление и
инкурвация Gonium pectorale (no Мертону и Гартману),
мм
17
G. p-ectorale инкурвация уже хорошо выражена и осуществляется
во время дробления, благодаря чему готовая табличка оказывается
вогнутой в виде блюдца или неглубокой чаши (рис. 79, I3—.—I7);
таким образом, все ееклетки склонены друг K другу своими ани-
лмальными полюсами (на которых вскоре ж-е появляются жгуты),
обращенными в этот; момент внутрь чаши. При вылуплении колоний
(синзооспоры) из материнской оболочки происходит ее выгибание
(экскурвация), в ‚результате которой его верхняя поверхность ста-
новится слегка выпуклой, а составляющие ее клетки радиально
3 224
расходятся жгутиковыми полюсами. У 16-клеточной Pandorina
и З2-к.лето-чных Eudorina оба этих процесса, и особенно экскурва-
ция‚ осуществляются в гораздо более явственных формах. Как видно
на примере обеих Eudorina (рис. 79, 2-5), инкурвация замет-на уже
с 8-клеточной стадии и приводит к концу дробления, на стадии 32,
к образованию глубокой чаши вчвиде полушария или даж-е почти
3/5 целого шара. Эту стадию можно назвать с т-о м о б л ас т у л о й,
как ее называют у губок, а. ее широко зияющее отверстие-
ф и а-ло п о р о м; главные оси всех клеток стомобластулы распо-
лагаются радиально, а их анимальные полю-сы обращены внутрь
ее -полости. При наступающей теперь экскурвации стомобластула
сначала выпрямляется, а затем все сильнее и сильнее выгибается,
постепенно вы-вор-ачиваясь наизнанку (рис. 79, д6—11); по мере ее
выворачивания края фиалопора все более сближаются и, наконец,
смыкаются; получается замкнутый полый шар — э к с к у р -B а-
Ц и о н н а я б л а с т у л а, вскоре вырабатывающая жгуты и при-
ступающая к самостоятельному существованию. В результате
выворачивания бластулы все ее клетки оказываются ориентирован-
ными в направлении, обратном первоначальному-жгутиковьгми
полюсами кнаружи, а бластула в целом-ориентированной Ta-
KI/IM же образом, как та материнская клетка, из которой она
возникла. - . I
Итак, конечным р-езультатом дробления является тут полушаро-
видная открытая стомобластула, превращающаяся путем вывора-
чивания, уже после окончания дробления, в истинную бл а-
стулу. Таким образом, формиров-ание последней складывается
из двух раздельных процессов: самого дробления, посредством
которого строится табличная, в принципе, основа будущей бластулы
и создается ве-сь ее клеточный материал, и действительно ‹<морфо-
генетического» процесса экскурвации, придающего этой «заготовке»
законченную сфероидальную форму бластулы.
Вторым немаловажным следствием экскурвации оказыва-ется
восстановление проморфологической преемственности между зо-
оидом—— родоначальником колонии и уже многоклеточным, в сущ-
ности, продуктом его воспроизведения, преемственности, как мы
видели, временно нарушаемой инкурваци-ей. Наконец, еще одним
прямым следствием процесса выворачивания является создание
вторичной (экскурваЦНонной)
принимающей роль бластоцеля, н-0 не имеющей никакого отношения
к первичной полости дробления. Эта последняя пр-едставлена тут
зияющей полостью стомобластулы, имеющей иное (инкурвацион-
ное) происхождение и впоследствии бесследно исч-езающей.
о Большого внимания заслуживает то обстоятельство, что инкур-
вация вносит очень существенные изменения в видимы-е геометри-
ческие формы табличного дробления. Эти изменения обусловли-
ваются самим фактом искривления гениев-ой таблички и заклю-
чаются в том, что по мере его усиления меридианальные плоскости
делений стан-о-вятся все более наклонными и в конце концов прини-
15 Эмбриология беспозвоночных 225
и
п ол ос т и бластулы,
мают то положение, каким характеризуются спиральные формы '
дробления; ОНИ ОКЭЗЬХВЗЮТСЯ частью ДЕКСИОТрОПНЫМИ, частью
` ЛЁОТРОПНЫМИ ПО ОТНОШЕНИЮ K ОСИ стомобластулы. ›
' Такое ИХ перемещение, очевидно, н-е означает изменения I/ICTI/IH-_
НЫХ форм дро-бления, остающегося двухмерным, но создает полное
впечатление ТОГО‘, ЧТО О-НО является трехмерным (СМ. ниже).
Рис. 80. Eudorina illinoisensisz
_ ва ная особь из очень молодой колонии’ 2 —— бесплодная особь;
Ё ggzifig ‘I{‘:>?-x1E:2:iq')eEfL;:%(t)1)a3: ее третьего деления; 5 — 011090‘-(“T ‚Стадии 16 В °°°Т°я“"“
—— у
предмитотического уширения; 6 —-MHKp0raMeT3- (ПО ОВЗССУ)
Необходимо еще отметить, что уже упоминавшийся «палинтомический крест»
и, ‚в частности, образуемая им характерная полярная спаика сохраняются в тече-
ние всего процесса формирования бластулы и разрушаются только в момент вы-
работки молодой особью (колонией) межклеточного студенистого вещества. Та:
кое постоянство этого образования делает его лучшим выразителем двулучевой
симметрии развивающейся бластулы и легко позволяет проследить ее устоичи-
вость с 4 до 32-клеточной конечнои стадии. Стоит, наконец, упомянуть, что все
повторные деления Eudorinidae осуществляются посредством односторонних,
врезающихся или полукольцевых борозд‚ при значительном митотическом удли-
нении бластомеров, и что им предшествует придвигание ядра к анимальному
полюсу этих последних.
Цитологические изменения, „происходящие B0 время дробления,
касаются главным образом аппарата Гольджи и пиреноидов. Они
226
заключаются в изменении формы и окрашиваемости диктиосом и
постепенном уменьшении их числа из одного поколения бластомеров
в другое, что происходит вследствие того, что их деление.не поспе-
вает за митозами; то же можно сказать о пиреноидах, получаемых
в единственном числе каждым из бла-стомеров заключительной гене-
рации (рис. 80).
Дифференцировка бла-сто-меров в жгутиковые зооспоры проис-
ходит совершенно таким же образом, как у одиночных фитомона-
дин. Окна происходит одновреме-нно во всей колонии и сходным
образом в каждой из его клеток, получающихся совершенно одина-
ковыми. Их осевая диффере-нциация осуществляется уже позднее,
в постэмбриональномя периоде развития.
3. Volvocidae
ГОНИДИИ пи зиготы вольвоцид удовлетворяют уже многим призна-
кам настоящих яиц, являясь специализованными генеративными
клетками, достигшими гипертрофического развития за счет обра-
зующей их материнской особи. Тут уже вполне уместно говорить
о прохождении ими полного, оогенетического цикла, включающего
в качестве второго, большого периода роста их увеличение за счет
использования ими ассимиляционной деятельности всех бесплодных
клеток колонии. Обладая на всех стадиях ярко выраженной гете-
рополярностью‚ яйца обнаруживают на ранних этапах развития и
те ж-е внешние признаки двулучевой симметрии, какие мы видели
у Euudorinidae, последняя, повидимому, остается проморфологиче-
скои основой строения и y зрелого яйца. '
В соответствии с высокой степенью гипертрофии яйца дробление
складывается из большого числа делений, равного 7 у одной только
Pleodorina, HO никогда не меньшего чем 9 у н.астояЩих Volvocidae,
т. е. представителей прежнего рода Volvox; уже упоминалось, что
оно варьирует тут в пределах 9—11 у Janetosphaera И 10——14—15
у Volvox собственно. Ввиду наличия существенных и чрезвычайно
Интересных различий в ходе дроблен-ия разных видов мы рассмотрим
по отдельности два лучше изученных примера. .
У Volvox globator ранние стадии дробления мало чем отличаются
от таковых Eudorina illinois-ensis. B результате двух первых, ме-
ридианальных, делений образуется типичный палинтомеический
крест (рис. 81, 3-5). Третье Деление осуществляется посредством
четырех совершенно разобщенных центробежных, врезающихся бо-
розд, однако уже не мериди-анальных, а заметно наклонных; при
этом вегетативные части всех бластомеров смещаются кверху и на-
право, в направлении движения часовой стрелки, т. е. дексиотропно.
Благодаря этому, образующаяся 8-клеточная гониевая табличка ока-
зывает-ся уже более вогнутой, чем у Eudorina. При четвертом де-
лении, реализуемом уже центростремительными полукольцевыми
бороздами, его спиральный характер становится еще более очевид-
ным. Он выражается тут в резком искривлении всех восьми деля-
15* 227
Щихся бластомеров, вегетативные части которых вновь сильно сме-
ЩаЮТСЯ вправо И кверху относительно анимальнь1х, ОСТаЮШИХСЯ
прочно фиксированнькмд-х пп своем Месте полярной и ДРУГИМИ спай- .
ками креста. В рСЗУ.П|›'|'П’Г0 OllIH'CElI-IHBIX ‘СМЭЩЕНИЙ 6.113-CTOMepOB-HO
восходящей спирали стадия I6 представляет собой уже стомобла- .
стулу и даже кмспсс широко открытую сверху, чем у Епаоппа на V
стадии 32; они уже стили скрсричлсской, и ее фиалопор приобрел
15 V6
Рис. 81. Дробление Volvox globatorz
7, 2—начало третьего деления; 3—стадия 8 с анимального и 4, 5-
с вегетативного полюсов; б, 7—переход от стадии 8 к стадии
16, вид с анимального (б) и вегетативного (7) полюсов; 8—стадия
16, вид с анимального полюса; 9, 70—пятое деление, вид с
анимального (9) И вегетативного (70) полюсов; 71, 72—ста-
дия 32, вид с анимального (7.7) и вегетативного (12) полю-
сов; 13, 74—шестое деление, вид с анимального (73) и вегета-
тивного (74) полюсов; 75 — 7б--схема искривления квадран-
тов шара дробления Volvox (76) в сравнении с нормой при
спиральном дроблении Metazoa (no дельсману из Захваткина)
характерную четырехлучевую форму (рис. 81, 7). Следующее, пятое,
деление идет таким же образомддкак и четвертое, но в о-братном,
л-еотропном направлении, оказывающемся фактически почти эквато-
риальным в связи с тем, что митотическое удлинение бластомеров
происходит в почти строго меридианальнлом направлении (рис. 81,
9—10). Шестое деление вновь является дексиотропным, а дальней-
шие, повидимому, чередуются .в этом «и леотропном направлении,
хотя и представляются скорее меридианальными и экваториальными;
они все (начиная, возможно, с пятого или шестого) осуществляются,
повидимому, посредством кольцевых, а не одно-сторонних борозд.
228 .
Происходящее увеличение числа клеток стомобластулы и достиже-
ние ею окончательного клеточного состава мало отражаются на ее
форме; только фиалопор постепенно сокращается в размерах, оста-
ваясь все же довольно широким. .
На этом заканчивается первый этап эмбриогенеза, содержание
которого сводится к дроблению, идущему рука обруку с инкурва-
цией; они тут находятся в столь тесной взаимной связи, что слива-
ются в единый морфогенетический процесс, в икотором инкурвашия
Рис. 82. Дробление Janetosphaera aurea:
1—4—стадии 4 (1), 8 (2)‚ 16 (3) И 32 (4) с вегетативного полюса; 5 —8 -стадии 8 (5), 16
(б), 32 (7) и 64 (8) с анимального полюса (по Жана)
кажется следствием смещений плоскостей деления, а это последнее-
результатом инкурвации.
Дальнейшее "объединение этих двух первоначально независимых
процессов в двуединое изнкурвационлное дробление можно видеть
у Janetosphaera. Здесь табличная группировка бластомеров оказы-
вается преодоленной с самого начала дробления, так что состояние
стомобластулы фактически устанавливается уже на стадии 4 И
получает вполне законченное выражение пна 8-клеточной (рис. 82').
IB результате этого уже третья борозда дробления ложится, как
правило, приблизительно экваториально и‘ приводит -к ‘созданию
совершенно такой группировки бластомеров стадии 8, как при са-
мых классических формах спирального дробления. Любопытно, что
степень наклона этой борозды по отношению к главной оси яйца
подвержена индивидуальной изменчивости, так что ‘она ибычвает то
.. ‹. . 220
почти меркидианальной, то явственно наклонной, то почти строго
экваториальной; к сожалению, остается неизвестным, является ли
она, в принципе, дексио- или леотропной. Такое кор-енное изменение
геометрических форм раннего дробления, приводящее к созданию
полной видимости чередования его трех первых плоскостей по трем
измерениям пространства, достигается тут энергичными переориен-
тировками самих делящихся бластомеров, т. е. таким же образом,
как у одиночных фитомонадин (ср. стр. 110). Заслуживает упоми-
нания, что борозды этих трех первых и по крайней мере еще четвер-
того PI пятого делений имеют явно односторонний характер, сменяю-
щийся кольцевым только на более поздних стадиях дробления.
Общий спиральный характер последнего у Janetosphaera выясняется
с полной определ-енностью из самого мозаичного расположения
бластомеров на всех стадиях. но порядок чередования его плоско-
стей остается неустановленным.
Итак, дробление вольвоцид нельзя назвать иначе, чем спираль-
ным. ‚Его сходство с наиболее равномерными формами гомоква-
дрантного спирального дробления п-ервичноротых не может не бро-
ситься в глаза, и оно еще подчеркивается особенностями геометри-
ческого плана, лежащего в егодоснове. Как видно из рис. 81 и 82, ТИП
симметрии дробящегося яйца Volvox и даже его конкретное морфо-
логическое выражение практически совпадают с таковыми гомоква-
дрантных аннелид или моллюсков. В том и другом случае мы прежде
всего отмечаем чрезвычайно правильную 4-лучевую симметрию вра-
щательного типа, проявляющуюся в самом наличии четырех иден-
тичных квадрантов шара; правда, у Volvox ОНИ ссильнее спирально
искривлены, чем у Металоа (и притом в дексиотропном направле-
нии, что легко объясняется тем, что дексиотропные деления тут не
столь регулярно чередуются с леотропнь1ми и потому не уравнове-
шиваются ими), но это, конечно, не меня-ет дела; далее мы видим
у тех пи других совмещение этого вида симметрии с правильной дву-
лучевой- в строении одинаковой у них апикальной розетки, пред-
ставленной у Volvox палинтомическим крестом; при этом одна из
плоскостей двулучевой симметрии определяется тем, что в ней лежит
характерная полярнаяспайка двух из клеток этого креста, а вто
рая—положением двух других, разъедлиняемых ею клеток. -
Нельзя также не отметить, что дробление вольвокса идет на-
столько закономерно, что позволяет точнейшим образом просле-
дить генеалогию и судьбу каждой из его клеток и дать ей соответ-
ствующее обозначение, и что ч-ередующееся, мозаичное расположе-
ние его клеток полностью отвечает норме спирального дробления.
Правда, этому большому сходству противостоит пи ряд различий,
самым главным из которых является то, что весь этот процесс раз-
вертывается, в сущности, в обратном направлении, буквально на-
изнанку, по сравнению с классическими его формами. Поэтому и
получается, что при рассмотрении дробящегося яйца с анимального
полюса мы видим смещения вегетативных, а не анимальных частей
бластомеров при их МИТОТИЧЕОКОМ удлинении; что весь процесс раз-
330
вертьпвается над этим полюсом, остающимся на дне инкурвацион-
ной бластулы, и что при таком положении объекта мы фактически
наблюдаем ход дробления с изнанки и вмлесте с тем-с вегета-
тивного полюса образующейся стомобластулы. Этого нельзя упус-
кать из вида, так что если мы будем смотреть на стомобластулу
с ее собственного анимального полю-са‚ вся картина дробления,
описанная у Volvox, потребует противоположной интерпретации:
дексиотропные деления окажутся леотропными, и наоборот.
Продуктом инкурвационного дробления Volvocidae является
удивительная стадия стомобластулы, существенно отличающаяся от
таковой Eudorinidae своей формой шатра, открывающегося На
Рис. 83. Последовательные стадии экскурвации _]anetosphaera_aurea (1'IO:_L[HM-
мерману) 4
одном из полюсов лишь очень небольшим отверстием фиалопора
(рис. 83). Все составляющие ее клетки обращены своими анималь-
ными (будущими жгутиковыми) полюсами внутрь просторного‘ бла-
стоцеля, являючщегося, по сути дела, полостью дробления, хотя она
пи создается, так сказать, обрастанием пространства в ходе инкур-
вационного дробления, а -не обычным способом раздви-гания бласто-
меров. Замечательнее всего, что прямым результатом этого процесса-
является полное обращение полярности организма: нетрудно видеть,
что стомобластула оказывается ориентированной в направлении,
обратном ориентации яйца. Ее передний, анимальный, полюс обра-
зован вегетативными полюсами клеток креста и направлен к Центру
материнского организма, а вегетативный смотрит кнаружи “и пред-
ставлен фиал-опором, в то время как первичный анимальныи полюс
дробящегося яйца оказывается лежащим внутри бла-стоцеля, на
самом его дне. Таким образом, стомобластула фактически вывер-
нута наизнанку, представляя собой своего рода «антибластулу».
231
Восстановление первоначальной ориентации организма происхо-
дит во время гпроцесса экскурвации, совершающегося путем вывора-
чивания стомобластулы через разверзающееся отверстие фиалопора.
Этот процесс, открытый впервые Кушакевичем (1919) у ряда
Volvocidae И обнаруженный затем у всех остальных Vo1vocoide_a,
показан на рис. 83, не нуждающемся в особых пояснениях. Так
создается истинная бластула‚ вскоре же дифференцирующаяся в,
жгутиковую таким же образом, как у Eudorina, HO с тем отличием,
что фиалопор остается здесь открытым.
Г л а в а XIII
ЭМБРИОГЕНЕЗ БЛАСТУЛООБРАЗУЮЩИХ МЕТА2ОА
1. Предварительные замечания
Общепринятое освещение ранних стадий онтогенеза самых при-
митивных из Metazoa —- известковых губок и метагенетических
Cnidaria — -c точки зрения тех представлений, на которых базируется
до сих пор об-щая эмбриология, приводит к поразительному, в сущ-
ности, результату: к тому, что эмбриогенез этих животных кажется
налм каким-то нагромождением эмбриологических парадоксов, укло-
нений от привычных норм и совершенно необъяснимых нарушений
самых эл-ементарных и общих законов всякого эмбрионального
развития.
Совершенно очевидно, однако, что раз эти «нарушения» имеют
тут настолько массовый характер, что становятся своего рода пра-
вилом, то они свидетельствуют только об одном: о неприложимости
к низшим беспозвоночным самих этих законов и норм, считающихся
общеобязательными для всех вообще Metazoa. Иначе говоря, дело
тут вовсе не в том, что развитие губок и книдарий и даже Mesozoa
атипично и_ уклоняется от общего для Metazoa стандарта, а в том,
‘что сам этот стандарт имеет гораздо более ограниченное и узкое зна-
чение, чем это ему обычно приписывается.
В самом де.ле, все эти бесконечные нарушения и беззакония сразу
же перестают казать-ся таковыми, если только оставить совершенно
безнадежную попытку подогнать развитие более примитивных групп
Ме’са2оа ик онтогенетическому шаблону, прочно установившемуся
у высших; если, вместо того чтобы ориентироваться на этих послед-
них, подойти к его анализу «снизу» —— с точки зрения тех фактов и
положений, которыми обогащает нас .знакомство с размножением и
развитием простейших. Однако, чтобы пможно было это сделать,
необходимо сначала рассмотреть конкретные особенности эмбрио-
генеза тех многоклеточных, у которых оно завершается образова-
нием жгутиконосной бла-стулы. Таковыми являются метагенетиче-
_ские гидромедузы и сшифомедузы и известковые губки, а также и
целый ряд групп иглокожих- морские ежи, морские звезды и го-
лотурии: ‘ - - '
2.32
V2. Метагенетические гидромедузы
Основным источником наших знаний о развитии метагенетиче-
ских гидромедуз является классическая‘ монография Мечникова
(1886), остающаяся непревзойденным‘ образцом сравнительно-эм-
бриологического исследования, но доныне почти неизвестная нашим
современным эмбриологам. Богатейший фактический материал, со-
держащийся в этом труде, касается онтогенеза целого ряда метаге-
нетических Leptolida (ИЛИ Craspedota, как их тогда называли),
а именно Rath_kea fasciculata, Tiara pileata, Oceania armata, 'C1y’cia
=flav1du1a, Clytla v1r1»dicans, Mitrocoma annae, Laodicea cruciata.
Этот материал, равно как иллюстрирующие его многочисленны-е и
Тщательно выполненные рисунки до сих пор «совершенно не исполь-
зованы в нашей эмбриологчической литературе, вследствие чего су-
шествовавшие вней представления о развитии этих медуз были
поразительно неточны. В настоящем очерке эти данные и-спользо-
ваны очень широко.
Монаксонно-гетерополярное строение яйца обычно имеет видимое
Выражение только в близости ядра «к одному из его полюсов, но
иногда ярко обнаруживается и в соответствующем распределении
пигмента-у” Amph1sbetia, где он сосредоточен в анимальном‘
полушарии яица (Тессье, 1930); в этом случае было показано, что
такое его строение устанавливается еще в гонаде, задолго до опло-
ДОТВОРеНИЖ-И ЧТО яйцо прикрепляется к фолликулярному эпителию
своим нео-кроашенным вегетативным ‘полюсом. Физиологическая
полярность яица выражена самым наглядным образом — в наличии
осевого анимально-вегетативного градиента восприимчивости и .ак-
тивности, показанного у той же Arnphisbetia и другой лептомедузы
РЪпаПаштп. Установлено и наличие у яйца двулучевой симметрии,
приобретающей, однако‚.свое видимое выражение только на более
поздних стадиях индивидуального развития.
Оплодотворение и дробление яйца происходят вне материнского
организма, в морской воде. Общее число делений в процессе дроб-
ления составляет 9 или 10. Первое деление яйца осуществляется
посредством о дн о с т о р о н н е й борозды, названной Мечниковым
«В Р е З а Ю Щ сои с я»; она появляется на его анимальном полюсе
в виде пологои сначала выемки, или желобка, затем постепенно
УГЛУбЛЯЮЩеЙСЯ Отсюда в на.правлении к ‚противоположному по-
люсу, ра-ссекая яйцо по ‘всей его длине (рис. 84-86); при делении
яицо обычно с-ильно вытягивается п-о своей поперечной оси (в направ-
лении веретена деления), а с появлением первых же признаков бо-
розды принимает характерную почковидную форму. Положение
плоскости первого деления в ее отношении к проморфологии самого
яица и дальнейших стадий развития толковалось по-разному раз-
личными авторами, считавшими ее то строго фиксированной, сагит-
1 Тессье придерживается обратного толкования ориентации яйца и счи-
тает его окрашенный полюс за: вегетативный; с этим, как будег показано,‘
НЕЛЬЗЯ СОГЛЗСИГГЬСЯ.
233 -
Рис. 84. Дробление метагенетических гидромедуз:
- . _ " о с выделенными им направительными тельЦёМИ;
;—8— Кап]??? ‚ЁЁЁГЁЁЁЗЁЁЁ; £en§33£’;°.’-"§‘?e4’i”'ia'Topoe деление; перекрест бПа0Ұ̰а и Центр?
—стадИя е - I _ - 6 7—pacrIon -
- 5- конец второго деления, ‚
бежный, врезающииСя Характер борозд деления’ и овка бластомеров cm-
4 крест-накрест; 8—— спиральная групп р
жение бластомеров стадии й а, ю_ телофаза Первого де-
. __ ' 'du1a: 9- анафаза первого деления я Ц з _ п
дии 8, 9-74 Clytla Паи й а о Носторонней врезающейся борозды, _.
Пения появление На анимальном полюсе я Ц д ° 12——т етье деление‘ 13- чет-
вертое и 1ЁБЁЯЁЁЁсЁЁЪ°ЁЁЁБ с одчосторонними центробежными бороздами ддюбления; т‘;
ДРУГ С др `. ._ ' ‘ : 17—с а ия 2‘ 18— расположени
картина анархического дробления, 771-2779_тёйаёа)рзё'ёгцсгъ_руппиръБди fiflsacmmepoa стадии 8;
бластомеров “pe°T'”aKp§§§;1'::. %1;l())1gJP':eH;l$l‘ 22 23-Octorchis gegenbauri: 22—-6-клеточная ста-
‘ ... ние
20, 21 более позд шт 23* Оптический разрез стадии („о Мечников”
234
тальной, то, наоборот, ложащейся произвольно, без всякой опреде-
ленной связи с архитектоникой позднейших стадий. Имеющиеся
Новые данные позволяют теперь считать установленным, что эта
плоскость меридианальна, т. е. продольна, и ложится по главной
оси яйца, и притом исключительно в той или иной из двух наме-
чающихся B HEM плоскостей двулучевой симметрии; однако она
может быть с одинаковым успехом как сагиттальной, так и фрон-
тальной, т. е. подвержена в этом смысле альтернативной изменчи-
вости. - .
Вследствие одностороннего (врезающегося) .характера борозды
первого деления разделяемые ею бластомеры остаются дольше всего
связанными своими задними полюсами посредством все утончаю-
щегося плазматического мостика, в конце концов обрывающегося
(рис. 84, 2; 85, 7-8). Однако по мере углубления. этой борозды бла-
стомеры обычно не раздвигаются, а наоборот, очень тесно прикла-
дываются друг к другу своими медиальными поверхностями; при
этом они обычно заметно поворачиваются в противоположных на-
правлениях, перекрещиваясь в виде буквы Х (рис. 84, 3——4), но
мпогут оставаться и строго параллельными. У Oceania, характери-
зующейся своим анархическим дроблением, первые два бластомера,
напротив, почти совершенно обособляются друг от друга, оставаясь
в соприкосновении только отдельными (и притом любыми) точ-
ками своей внутренней поверхности, очень легко теряя и эту
хрупкую взаимную связь (рис. 84, 15).
В профазе второго деле-ния яйца ядра обоих первых бластолмеров,
обычно несколько смешаются к Центру поверхности соприкоснове-
ния последних и располагаются одно против другого. В этом же
месте появляются затем борозды вггорого деления, закладываю-
Щиеся одновременно, но порознь у каждого бластомера и совер-
шенно таким же образомп, как первая -борозда. _
При своем появлении они также имеют вид пологих углублений,
придающих бласто-м-ерам такую же почковидную форму, и затем
тоже постепенно врезаются в их толщу и рассекают бластомеры по-
полам; стало быть, эти борозды являются тоже врезающимися,
односторонними бороздами. Возникая в центре дробящейся системы
и постепенно распространяясь отсюда кнаружи в диаметрально
противоположных направлениях, они тем самым приобретают тот
Центробежный характер, который так справедливо привлек
к себе внимание Мечникова. Особенно замечательно, что эти бо-
розды возникают раздельно, фактически независимо одна от другой,
в качестве, так сказать;«собственных» борозд деления каждого
бластомера в отдельности, и притом возникают не на анимальном
полюсе дробящегося яйца, а именно в его Центре (рис. 84, 3-4;-
85, 10-11). ‘ ‘
Иное направление этих двух борозд второго деления наблю-
дается лишь в немногих, повидимому, случаях. Так, среди метагене-
тических Leptolida оно описано у Tiara, где они также являются
раздельными ‚и врезающимися. но закладываются не на внутренней,
235
а на наружной поверхности бластомеров; благодаря этому они
имеют тут центростремительный, а не ‘Центробежный характер
(рис. 84, 7, 20).
‘Плоскость второго деленлия перпендикулярна к таковой первого
и равным образом имеридианальна; таким образом, она ложится,
вообще говоря, то во фронтальной, то -в сагиттальной плоскости
яйца—в зависимости от т’ого, по какой из них прошла борозда
первого деления. Однако такое ее те-оретическое положение далеко
не всегда отражается в действительных формах дробления. Дело
в т-ом, что обе борозды второго деления оказываются строго ориен-
тированными лишь относительно самих тех бластомеров, которым
они принадлежат, так что отмеченные обоюдные смещения послед-
них не остаются без влияния на фактическое положение этих борозд
в отношении яйца в целом. Так, у Rathkea ОНИ явно располагаются
в двух разных наклонных плоскостях, соответственно перекресту
бластомеров, и даже фактически разобщены, а в таких случаях, как
у Oceania, невозможно даже и говорить о наличжии одной общей
плоскости второго деления. Это——существенно важное обстоятель-
ство, наглядно показывающее высокую степень автономности деле-
ния отдельных бластомеров.
В ходе второго деления яйца бластомеры часто испытывают еще
более резкие переориентировки, чем наблюдающиеся при первом
делении, и также впервые отмеченные М-ечниковым. Бластомеры
поворачиваются теперь так, что перекрещиваются длинными осями
под углом 45—90°, оставаясь при этом склеенными своими проти-
воположными концами. Получается уже знакомая нам характер-
нейшая фигура «палинтомического креста», образуемая двумя на-
легающими друг на друга парами еще неразъединившихся бласто-
меров, тесно смыкающихся крест-накрест своими чередующимися
полюсами (рис. 84, 4-5). Подобное их скрепление оказывается
иногда устойчивым и тогда дает в дальнейшем начало двум харак-
терным структурам дробящейся системы: полярной опайке на ее
анимальном полюсе и аналогичному соединению на противопо-
ложьном; стоит отметить, что эти спайки располагаются одна над
другой, проецируясь под прямым углом, и направлены под 45° к
плоскостям двух первых делений. В других случаях, однако, такая
группировка бластомеров не отражается на последующих стадиях
дробления.
` Третье синхронное ДЕЛЕНИЕ ОСУЩЕСТВЛЯЕТСЯ, как ПРЭВИЛО, совер- ›
Шено таким же образом, как второе-посредством раздельно воз-
никающих и обычно центробежных (рис. '84, 12; 85, 13) борозд вре-
зающегося типа. ‘Плоскость этого деления раополагается, в прин-
ципе, экваториально по отношению к осям яйца, но фактически по-
вторяется то же самое, что и при втором делении: борозды всех
Четы-рех делящих-ся теперь бластомеров тне образуют единую пло-
скость, а перекрещиваются в разных направлениях, что особенно
хорошо видно у C1y’£ia (рис. 84, 12), где -они вс-е наклонны, лежат на Ц
разных уровнях и, вдобавок, Совсем разобЩеНЫ одна от другой. Сле-
взад,
›.о4
довательно, и здесь эти плоскости правильно ориентированы только
по отношеёию к самим бластомерам, а не ко Всей образуемой ИМИ
ЁЁстем-еб чень примечательно, что третье деление Amphisbetia
огда ывает не экваториальным, а меридианальным, как бы Me-
НЯЯСЬ Местом со вторым, борозда которого ложит-ся тогда эквато-
20
Рис. 85.-Дробление Amphisbetia operculata:
7- еще не отложенные яй а п и еплен в
‘НОМУ эпителию Гонады; 2-L-(7;1‘r'1u1()) REP M0Me::‘Ie0E:fK€§';raJ-filhlillfl;Kg}:-‘f-lr11§Bi?0pflg1x¥l<g;}1(;;g:
Hoe яйцо; 4-7- появление и постепенное углубление врезающейся борозды
первого деления; 8- момент максимального расхождения бластомеров стадии
2; вид с неокрашенного полюса; 9- последующее смыкание бластомеров. ‚д
77 — второе деление посредством центробежных борозд; 72 — стадия 4- 53 __’
начало третьего деления; 14- стадия 8; 15—начапо четвертого деления.
1б—- стадия 16 бластомеров; 17— стадия 32; 78—— пребластула в оптической
разрезе; 79, 20- пребластулы (по Тессье)
рииально; это лишний раз показывает, насколько мало фиксирован
тут порядок чередования последовательных плоскостей дробления.
Группировка бластомеров на стадии 8 быва-ет очень различной,
варьируя от совершенно беспорядочной у Oceania до правильной
опиральнои у Rathkea и Tiara. B последнем случае_(рис. 84, 8, I9) она
ИМееТ ВИД ДВУХ наложенных друг на другаквартетов, бластомеры
237
которых чередуются с такой же правильностью, как у спирально
дробящих-ся трохофорных первичноротых. - .
Четвертое деление идет таким же порядком, как предыдущее, и
так же осуществляется врезающимися бороздами. У Amphisbetia
оно идет обычно в двух вертикальных плоскостях (рис. 85, 15), паралч
лельных друг другу и одной из плоскостей симметрии яйца, нопро-
ходящих вдали от нее, по обеим ее сторонам. В результате такого
положения борозд создается характерная группировка 16 бласто-
меров в виде двухэтажной таблнички, сильно вытянутой в поперечном
направлении и. имеющей, таким образом, очень правильную двулуче-
вую симметрию; однако у этого же вида иногда наблюдается И
иное, тангенциальное, направление плоскостей четвертого деления,
что ведет, разумеется, и к иной группировке бластомеров на ста-
A дии 16. У других видов они располагаются как при спиральном
дроблении (ри-с. 84, 20), но у Clytia образуют, судя по рисунку Меч-
никова (рис. 84, 13, 14), такую же вытянутую в ширину фигуру,
как у Amphisbetia. ‘
Начиная с пятого деления дробление интересующих нас форм
начинает проявлять некоторые новые свойства, заключающиеся в
утере более или менее выдержъивавшейся до сих пор синхронности
деления отдельных бластомеров, в замене (в большинстве случаев)
центробежных -борозд дробления более обычными центростремитель-
ными, а также в усилении присущих ему признаков опиральности.
О наличии единой плоскости пятого деления обычно не может быть и
речи, поскольку представляющие ее плоскости деления отдельных
бластомеров располагаются самым различным образом: «частью
меридианально, частью Экваториально, частью "же в плоскости, про-
межуточной между ними» (Мечников, цит. раб. стр. 45). Некоторое
представление о наблюдаемых здесь картинах мьогут дать рис. 84 И
85, показывающие, между пр-очим‚. тот любопытный факт, что у
Clytia все раздельные борозды пятого деления продолжают быть
центробежными (ри-с. 85, 14) и что стадия 32 Amphisbetia также
вытянута поперек главной оси яйца, как 16-клеточная (рис. 85, 16).
Образование полости дробления происходит так или иначе путем
раздвигания бластомеров, т. е. по существу с х и з о ц е л ь н ы M
способом. Ее зачатком чаще всего оказываются небольшие и обычно
разобщенные между собой полости, возникающие при центробеж-
ных делениях между каждой новообразую-щейся парой бластомеров
и впоследствии сливающиеся между с-обой. Способ ее образования
лучше всего иллюстрируется примером Clytia, где очень хорошо
видна роль этих борозд, митотического удли-нения бластомеров и
изменения их взаимной ориентации в создании все увеличивающе-
гося бластоЦеля.
С момента оформления бласгоцеля совокупность делящихся
бластомеров приобретает характер бластулы или, точнее, п р е б л а-
стул ы. Ее форма становится теперь все более правильной, шаро-
образной, стенки утончаются пропорционально уменьшению разме-
ро.в клеток, из которых они складываются, и последние, все увели-
238
чиваясь в числе и располагаясь все более тесно, постепенно прини-
мают характерную форму эпителиальных клеток— шестигранных -и
вытянутых в продольном (радиальном по отношению к бластуле
в целом) направлении. Все происходящие теперь деления идут и-с-
КЛЮЧИТСЛЬНО В ЭТОМ же радиальном направлении, В ПОРЯДКЕ ПРО‘
дольного деления каждой отдельной клетки (см. рис. 95, 1). Таким
образом происходит постепенная эппителизация бластулы, и она при-
обретает свой окончательный ‚клеточный состав—-—порядка 500-
1000 и более клеток, уже став типичной э п и т е л и а л ь-н о й ц е-
л о -б л а с т у л о й. На этом заканчивается Эмбриогенез, заключи-
тельным актом которого является наступающая теперь дифференци-
ровка клеток бластулы в жгутиковые клетки и ее превращение та-
ким образом в жгутиконосную «мерцательную» личинку. ‘
C>QD<33@3g3
еёеее
Рис. 86. Дробление Stomatheca. Видны те же, характерные для
гидромедуз, осооенности, как на рис. 84 и 85 (по Риттенхоузу)
Итак, конечным результатом дробления непосредственно оказы-
вается сферическая, замкнутаяэпителиальная бластула с просторг
ным бластоцел-ем, состоящая из множества одинаковых, морфоло-
ГИЧЭСКИ РаВНОЦЭННЫХ ДРУГ ДРУГУ клеток. По способу своего форми-
рованния она одинаково сильно отличается и от инкурвационной сто-
мобластулы УоЬ/осочдеа, и от их экскурвационной бластулы, так
что можно говорить о ее принадлежности к особому (схизо-
цел ь но M у) типу бластул, отношение которого к остальным вы-
яснится из дальнейшего. .
Проморфол-огические соотношения бластулы с яйцом и промежу-
точными стадиями дробления кажутся с первого взгляда очень
простыми и ясными и обычно толкуются в том духе, что анимальный
полюс яица становится передним полюсом бластулы, а вегетатив-
ныи-задним. Однако Те-с-сье (1930, 1931) привел весьма веские
доводы против этои установившейся точки зрения, с которыми не-
возможно не -считаться. Они заключаются, во-первых, в’ том, что
естественная оранжевая окраска эмбриональных стадий Amphisbetia
занимает у ооцита его дистальный полюс, у яйца—-тот полюс, на
котором возникает первая борозда дроблениия и где ‘выделяются’
направительные тельца, а у бластулы и следующих за нею стадий —
239
задний полюс, на котором сосредоточено образование энтодермьх;
у паренхимулы и планулы в оранжевый цвет окрашена только эта
последняя, а все жгутиковые эктодермальные клетки- оказываются
бесцветными. Во-вторых, опыты этого автора с прижизненной ло-
кально-й окраской Amphisbetia ‚и других гидроидов (Dysanema,
I-Iydractinia) показали, что еслаи окрашивать полюс деления яйца,
то эта. окраска впослед-ствии
обнаруживается в задней поло-
вине бластулы и планулы и,
обратно, окрашивание нижнего
полюса яйца приводит к образо-
ванию личинок с цветным перед-
ним концом (рис. 87). Прослежи-
ваемая таким образом прямая
преемственность материала, ‘заро-
дыша прив-одит к заключению,
~ что полюс деления и выхода по-
. лярных телец яйца соответствует
полюсу гаструляции бластулы и
ротовому полюсу гидранта, т. е.
‘вегетативному, а не анимальному
полюсу. Иначе говоря, действи-
тельная ориентация ранних ста-
дий развития Нуаго2оа противо-
положна той, какая им до сих пор
приписывалась, и совершенно та-
кова же, как у гребневиков.
Дело, однако, еще услож-
_ ›- - _ няется тем, что «градиент физио-
5 8 логической активности этих ста-
_ дии не согласуется с такои интер-
Рис. 87. Amphisbetia operculata: претаЦией. Как бЫ-ПО ПОКЗЗаНО
7——3—определение полюса максимальной еще Чайльдом у Phialidium И
чувствительности к ядам у 2—клеточной
затем самим Тессье у Amphisbetia
положнь1м—задне-передним (вегетативно-анимальным’) —у более
поздних. Таким образом, полюс деления яйца ведет себя во всех
отношениях, как анимальный, и приобретает свойства вегетатив-
ного полюса только позднее, в результате произошедшего обраще-
ния физиологической полярности зародыша. _ .
Итак, приходится считать доказанным, что анимальный полюс
яйца становится вегетативным полюсом бластулы, ни наоборот, его
Вегетативный полюс дает на- „
чало анимальному полюсу ли- ^
чинки.
Эти более сложные, чем
казалось, отношения пред-
ставляют определенную анало-
гию с теми, какие существуют
между яйцом и стомобласту-
лой у вольвоцид и являются
там результатом инкурвацион-
но-го деления — результатом
чисто временным, переходя-
щим и «в дальнейшем своди-
мым на-нет той своеобразной
«перелицовкой» бластулы, ка-
кую представляет собой про-
цесс ее экскурвации.
Поэтому не было бы боль--
шой неожиданностью встре-
титься и у Leptolida с этими
или равноценными им процес-
сами действительного‘ обраще-
ния полярности зародыша в
ходе его формиярования——про-
цессами‚ о существовании ко-
торых и не подозревалось до‘ Рис. 88. Схема опытов по изоляции
Ётадии (1) бластулы (2) и планулы (З); 4-
——результаты прижизненной окраски оо- М . " В -
цита (4), стадии 2 (5) и бластулы (б); 7-— И других’ Макси альнои ос
их открытия в 1919 г. Кушаке-
вичем у вольвоксов и вслед за
бластомеров Amphisbetia operculata на
стадиях 2—16 и полученных результа-
тов. Пунктированы ОКрНШЭННЬТЁ бласто.
планула, получившаяся из яйца, окрашен-
ного нейтральной красной; 8— планула,
непосредственно окрашенная этой краской
(по Тессье)
приимчивостью к действию ядов
и наиболее энергичной окраши-
ваемо-стью витальными красками
обладают: у ооцита—-дисталь-
ный полюс, у яй-ца ——тот, с которого начинается деление, а у бла-
стулы и планулы-передний конец тела. Таким образом, полюсом
метаболической активности является на всех стадиях тот, который
мы привыкли считать анимальным и который в этом и проявляет
свое главное свойство физиол
противоречие в показаниях мор
огического доминирования. Такое
фологического и физиологического
критериев полярности приводит Т-е-ссье к выводу, что в течение раз-
-вития происходит обращение физиологической полярности орга-
низма, замена передне-заднею градиента ранних стадии противо-
240
НИМ В 1936 Г. Дю-боском И меры и получившиеся из них блас-
ТЮЗЭ у известковых губок тулы, оставлены белыми неокрашенные;
И хотя ОНИ до СИХ Пор фактш стрелками соединены бластомеры (или
бластомеров) и получившиеся из
чески не описаны и - Группы .
у Г дроидод них бластулы, контуры бластомеров, не
существуют все же некоторые использованных в каждом данном
факты, позволяющие п-одозре- опыте, намечены тонкой пунктирной
вать их наличие в эмбриогенезе линией (ПО Тессъе)
некоторых Leptolida.
dulell-IaI1r1O§Df)J1:6e: интересно с этой точки зрения развитие Clytia flavi-
‚ о р но описанное и изображенное Мечниковым в его Knac-
сическои монографии. Как видно из воспроизводимых"здесь рисун-
ков (см. риз. 84‚ 13-14), положение бластомеров при первых пяти
делен-иях яи›ца совершенно таково же, как у Janetosphaera (см.
16 Эмбрнология бйспозвоночных 241
рис. 83): они ьвсе обращены внутрь бластоцеля теми своими полю-
сами, у которых располагается ядр-о и где появляются Врезающиеся
борозды их (потому-то и центробежного) деления. Таким образом,
пребластула стадии 32 получает все специфически-е свойства стомо-
бластулы (кроме одного-наличия фиалопора). Однако уже при
следующем, шестом, делении такая характернейшая ориентация ее
клеток сменяется на обрат-ную. По Мечникову‚ это происходит в ре-
зультате перемещения ядер —их постепенной Миграции сначала к
центру бласт-омеров, а затем и к их наружной, периферической no-
верхности, где они и занимают свое окончательное положение.. Иначе
‚говоря, изменение первоначальной ориентации клеток опписьтвается
как процесс обращения их собственной полярности, т. е. точно таким
образом, как он описывался у вольвоцид и губок, пока не стало
известным, что оно д-остигается экскурвацией. „
Ту же, св сущности, картину мы видим у Amphisb-etia И других
метагенетических Dept-olida, обладающих центробежными бороздами
дробления. Достаточно еще раз взглянуть на соответствующие кар-
ти=нь1 дробления, чтобы убедиться в неизбежности такой пе-
реориэентировки бла-стомеров, чтобы их анимальные пол-осы ока-
зались направленными кнаружи и смогли стать жгутиковыми по-
люсаими клеток бластулы. Таким образом, вряд ли будет рискован-
ным утверждать, что у всех этих форм действительно существуют
процессы, равноценные по своим результатам экокурвации вольво-
цид и губок, но осуществляющиеся тут неизвестно еще каким_спо-
собом; абсолютно «не исключено, что им тоже окажется экскурвация,
так долго не замечавшая-ся у губок и Volvocoidea, несм-отря на их
исчерпывающую, как казалось, изученность.
Цитология дробления гидромедуз остается, ›к сожалению, почти
неизвестной. О дифференцировке мы знаем ‘только, что она заклю-
чается B более пили менее одновременной выработке жгутов всеми
клетками ‘бластулы и часто сопровождается вытягиванием ее клеток
в длину. Равным образом мы не располагаем удовлетворительными
данными о физиологии эмбриогенеза; можно только сказать, что
эмбриональный рост не показан ни для одного из метагенетических
пред-ставителей I-Iydrozoa, что и вполне естественно, так как их
эмбриональное развитие целиком протекает вне мтатеринского тела,
в морской -воде. ’
3. Низшие Scyphozoa
Из всех имеющихся сведений об эмбриогенезе сцифомедуз, про-
ходящих в своем развитии стадию жгутиконосной бластулы, ooo;
бенно интересны и важны данныеМечникова о дроблении пероме-
дузы Nausithoé ( 1886); существенны и исследования Тессье (1930),
- касаю-щпиеся эмбрионального роста-Сйгузаога. Дробление-Маизййюё,
ОТЛИЧЗЕТСЯ ТЕМ, ЧТО ЕГО первая бОРОЗДЗ ИМЕЕТ НЕОбЬТЧЗЙНО ЯРКО ВЫ‘
‘раженный врезающийся характер (рис. 89), а вторая-не менее
типично центробежный, тогда как уже третья является кольцевой e-
yrax же как и борозды всех последующих делений; таким образ-ом,‘
242
16*
Рис, 89. Дробление сцифомедузы Nausithoé:
- 1- зрелое яйцо; 2, З- конечные стадии первого деления; 4—— второе деление, осуществляющееся посредством центробежный борозд; _
5- четвертое деление; б—- молодая бластула в оптическом разрезе; 7—пребластула с поверхности (пБ Мечникову)
243
замена врезаюшихся борозд кольцевыми происходит тут гораздо
раньше, чем у метагенетических Leptolida. Эта замена одного типа
борозд другими связана тут с переходом к неравномерному деле-
нъию, так что третья, экваториальнаярборозда разделяет каждЫЙ
бластомер стадии 4 на две неравноценные половины-меньшую
Рис. 90. Дробление и рост бластулы сцифомедузы
Chrysaora hysocella:
1—7—стадии 4(7), 12(2), 30 (3), 60 (4), 120 (5), 160 (б) и 260 (7)
бластомеров; 8— желточные вклочения в яйце и 9- B растущей
бластуле; 10, 11- относительные размеры молодой (10) и вы-
росшей (71) бластул; клетки молодой (72, 13) и выросшей (74,
’ \ 15) бластул, с поверхности и в оптическом разрезе (по Тессье)
анимальную И 60.7186 объеми-стую вегетативную, ЧТО ПОЗВОЛЯЕТ y}K€
_говорить об их дифференциации на микро- и макромеры. Столь же
неравномерно и четвертое деление, нопри следующих эта неравно-
мерность постепенно сглаживается. Однако у оформившейся цело-
бластулы высота клеток бластодермы правильно возрастает в ее.
заднем полушарии спереди назад (см. рис. 93, 8). «B этом же ее «п-о-
лушарии сосредоточивается и большая часть пигментированнои
244
эндоплазмы, имев-шей у яйца диффузно-е, равномерное распределе-
ние, так_что разнополюсность целобластулы выражена тут самым
наглядным образом.
У Chrysaora (рис. 90) дробление осуществляется, повидимому,
одними только кольцевыми бороздами и с самого начала неравно-
мерно, но затем выравнивается даже в большей мере, чем у
Nausithoé, так что получающаяся бластула состоит из клеток одина-
кового размера. Самой интересной особенностью развития Chrysaora
является явно «преждевременная» дифференцировка бластулы в
жгутиконосную личинку и то, что эта личинка длительно остается
в материнском теле, где она питается осмотическим ппутем и очень
сильно растет, продолжая при этом дробиться. é
Дифференцировка происходит здесь на стадии 120 бластомеров,
отвечающей, следовательно, пребластуле других рассмотренных
нами форм; с этого момента и начинается усиленное осмотическое
питание шарообразной ЛИЧМНКИ и ее интенсивный рост, приводящий
к тысячекратному увеличению живой мас-сь1 бластулы к моменту ее
превращения в гаструлу. При этом размеры ее клеток и их ядер
остаются строго постоянными, так что это громадное увеличение
объема достигается исключительно умножением чи-сла клеток по-
средством пяти, быстро следующих друг за другом, делений. Таким
образом получается, что дробление, состоящее из 11 более или менее
синхронных делений, оказывается разбитым на два периода: период
нормального, эмбрионального, дробления, включающий семь первых
делений яйца, и период беспримерного, в сущности, постэмбрио-
нального дробления, осуществляемый в уже дифференцированном
состоянии зародыша и состоящий обычно из пяти последних де-
лений. `
Такое преобразование поздних делений яйца в процессе (вегета-
тивного уже) роста имеет своим следствием И коренное изменение
самого характе_ра этого последнего. Как видно низ соответствующих
данных Тессье, общий рост практически не имеет места в течение:
первого периода дробления: размеры бластомеров мельчают, как,
обычно, с каждым делением, и это же касается их ядер, общий‘
объем которых остается приблизительно постоянным до стадии 120
ВКЛЮЧИТЭЛЬНО; С НЭЧЗЛОМ же ЭКЗОТРОфНОГО питания ‘КЗЖДОЕ ДЕЛЕНИЕ.‘
полностью компенсируется ростом, что ‘и имеет своим результатом
постоянство клеточных размеров бластулы при непрерывном нара-
стании числа ее клеток. Иначе говоря, Постэмбриональное дробле-
ние теряет свой типичный характер палинтомии, не компенсируемой
ростом, и пидет уже по законам «нормального» вегетативного, т. е.
монотомического роста.
4. Известковые губки
Яйца этих губок (рис. 91) характеризуются своей уплощенной"
формой и одноосно-гетерополярным строением. Главная ось совпа-
дает с наименьшим, вертикальным, диаметром яйца, а анимальным
2.45
полюсом является тот, который обращен’ к стенке материнской жгу-
тиковой камперы впериод оплодотворения и созревания яйца. Однако
этот полюс обладает тут только одним из двух главных признаков
анимального -—- тем, что он служит местом внедрения сперматозоида,
тогда как выделение направительных телец происходит не здесь,
а в одной низ точек п-ериферии яйца, в направлении его большего, го-
ризонтального диаметра. Распределение желтка в яйце очень рав-
_ н-омерно, но имеющийся
в нем пигмент сосредото-
A ЧЭН, «по-видимому, ‘в одном
ф только вегетативном no-
д WW V W
- 1 2 3 Дробление осущест-
ВЛЯЭТСЯ ПОСРЕДСТВОМ‘ КОЛЬ-
ресекая-сь по главной оси
8 яйца, что имеет своим
результатом создание
п Ъ» очень характерной груп-
. пировки бласто-мпер-ов на
‚ стадии 8. У Sycon ОНИ‘
5 „ 7 ta:
располагаются в виде
плоской 8-лучевой розетки
или блюдца (рис. 91, 10),
Ьбращенной своей вогну-
той поверхностью кверху
(т. е. в сторону, в которую
был обращен анИМалЬнь1й
Рис. 91. Дробление известковых губок:
(1;—)—§——A3&etta prirrgorglialais, ScTaru»ug 2 (1). 4 (2% д? полюс яйца), и не compu-
П0 ЕЧНИКОВ ; '-‘ - can ra ста ии ‚
4 (5) и 6 (б) (из йванова); 7X-72- S’ycon nraphanus, касаются llpyl“ C ДРУГОМ
стадии 2 (7), 4 (8), 8 (10) и 16 (17), вид сверху; 9—- В центре образуемой HM“
стадия 8 и -12-— стадия 16, вид сбоку в оптическом
разрезе (по Шульцу)
фигуры; в других случаях
. (рис. 91, 3-6) (бласто-
меры тесн-ее смыкаются друг с другом и образуютфигуру, еще
более напоминающую гониевую табличку; в этих случаях группи-
ровка бластспиеров на этой стадии показывается не радиальной, как
в первом случае, а явственно двулучевой.
Четвертая борозда дроблен.ия ложится экваториально и четко
разграничивает пматериал обоих полушарий яйца,—так что это деле-
ние является неравноценным (гетерономным): бластомеры верх-
него круга оказываются несколько крупнее нижних, в которых те-
перь уже ясно локализуется весь имевшийся в яйце пигмент. На ста-
ди-и 16 бластомеры группируются в виде уплощенной стомобластулы,
обладающей довольно просторным бластоцелем -и зияющим фиалопо-
ром на своем верхнем полюсе (рис. 91, 11, 12). Она образуется из
блюдцеобразной стадии 8 в результате того, что бластомеры верх-него
246
цевь1х борозд. Плоскости
Фа первых трех делений ло-‚
, жатся меридианально, пе-
Ч круга, отсекаемые от нижних экваториальной бороздой, заметно
СКЛОНЯЮТСЯ друг K другу своими анимальными полюсами и создают
1 2 Рис. 92. эмбриональное развитие Sycon raphanus:
‚ —С’ГЗдИЯ стомобластулы‘ 3- стомобластула на стадии д
жгутиковые клетки; 4, 5- выворачивание стомобластулы (axcx‘§d§d§ZflZ‘§§‘;‘p8i“fio§%,1§§°§,‘;3§H‘i
бластула (по Дюбоску и Twas)
таким путем свод, в то ‚время как бластомеры нижнего круга обра-
зуют дно стомобластулы. Благодаря этому, края фиалопо-ра окайм-
лены анимальньтми полюсами восьми окружающих его бластомеров‚
247
и стом.областула в целом сохраняет ту организацию, какую имело
яйцо до дробления. Таким образом‘, формированные стомобластулы
происходит и здесь путем своего рода инкурвации, но осуществляе-
мой другим способом, чем у вольвоцид, и потому не влекущей за
собой переориентировки всей системы.
. Дальнейшие деления бластомеров идут в разном темпе пи в раз-
личных направлениях в каждом из полушарий стомобластулы.
В верхнем полушарии деления очень немногочисленны, идут в. раз-
личных плоскостях и имеют своим результатом образование неболь-
шого числа крупных макромеров, становящихся в дальнейшем зер-
ннистыми клетками заднего полушария амфибластулы; в нижнем же
полушарии деления быстро следуют одно за другим и приводят
к формированию очень многочисленных, но гораздо более мелких
микромеров, превращающихся впоследствии в жгутиковые клетки
амфибластулы; веретена всех этих делений лежат параллельно по”-
верхностчи бластулы.
В момент окончания дробления стомобластула имеет еще сильно
уплощенную, чечевицеобразную или плосковыпуклую форму
(рис. 92, 1, 2) И состоит приблизительно из 250 MI/IKpOvMep0B~ и 16—32
или более макромеров; последние могут быть расположены в один
слой (рис. 92, 2) или же образовать неправильный комок (рис. 92,3)
и в этом случае ориентированы без видимого порядка; микромеры
же образуют правильный эпителий, проявляют гораздо раньше и
яснее свою разнополюсность и все обращены своими анимальными
полюсами внутрь бластоцеля, временно замыкаемого теперь макро-
мерами. На этой стадии будущее жгутиковое полушарие занимает
до 3/4 всей поверхности бластулы, а макромеры оказываются стес-
ненными в узком пространстве вокруг верхнего полюса стомобла-
стулы.
Непосредственно по окончании дробления происходит одновре-
менная дифференцировка всех микромеров в одлинаковые жгутико-
вЫе клетки, жгуты которых оказываются направленными внутрь «no-
лости стомобластулы; только четыре из микромеров не вырабаты-
вают жгутов и начинают дифференцироваться в своеобразные чув-
ствительнь1е (зрительные) клетки. Вслед за тем наступает процесс
экскурвации, при котором стомо-бластула выворачивается наизнанку
через раскрывающийся теперь фиалопор точно таким же образом,
как у вольвоцид (ри-с. 92, 4—5).
Она превращается теперь в амфибла-стулу, все клетки которой
при этом сильно вытягиваются в длину и принимают форму узких
усеченных пирамид (рис. 92, б).
Та-ким образом, соотношения осей и полюсов стомобластулы и
экскурвационной амфибластулы тут совершенно такие же, как__
у вольвоцид. и
Однако их ориентация относительн-о первичнои оси и полюсов
‘яйца оказывается диаметрально . противоположной: анимальный
полюс яйца уступает свое место фиалопору стомобластулы и стано-
вится в к-онце концов задним, а не передним полюсом амфибластулы.
248
Это и понятно, поскольку стомобластула губок строится иным спо-
СО'б0М‚ ЧЕМ у ВОЛЬВОЦИД, ПИ сохраняет ИСХОДНУЮ ориентацию яйца,.
тогда как у последних она сменяется на обратную.
С-другой стороны, нельзя не обратить внимания на то, что гене-
тические соотношения полюсов яйца и готовой бластулы оказыва-
ются у губок такими же, как у гидроидов, что кладет конец всем
нередко высказывавшимся сомнениям о гомологии их одноименных
‘полюсов. Это——очень существенное обстоятельство,’ все значение
которого будет показано ниже, в конце этой главы. `
Переходя теперь к цитологической характеристике отдельных стадий эмбри-
огенеза Heterocoela, необходимо прежде всего дополнить несколькими указа-
ниями то, что уже говорилось об организации яйца. Надо отметить, именно, что
аппарат Гольджи представлен тут более или менее Многочисленными диктиосо-
мами, разбросанными в цитоплазме, что хондриом состоит из коротких, преиму-
щественно палочковидных или бисквитообразных митохондрий, что центросомы
незаметны, а ядро исключительно велико. Эти же особенности характерны и
для всех бластомеров первых поколений и остаются практически неизмен-
ными у макромеров до конца эмбрионального развития; -видоизменяется только
аппарат Г ольджи, принимающий форму колпачка, как бы надетого на ядро. Что
же касается микромеров, то испытываемые ими изменения заключаются в посте-
пенном приобретении признаков гетерополярности (удлинение клеточного тела,
смещение ядра к одному из:его полюсов), появлении около ядра одной чешуе-
видной (выпукло-вогнутой) диктиосомы, а затем и явственной центросомы. При
дифференцировке микромеров в жгутиковые клетки они еще вытягиваются в
длину; из центросомы вырастает длинный жгут, а диктиосома вступает с нею в
контакт и преобразуется в парабазальный аппарат. Превращение нескольких из
микромеров в зрительные клетки креста представляет еще более сложный про-
цесс; микромеры же так и не испытывают дифференцировки и только в самом
конце развития приобретают упорядоченное расположение и становятся поляри-
зованными.
5. Иглокожие. Морские ежи
Морские -ежи, голотурии пи морские звезды ‘могут служить при-
мером наличия (чтобы не сказать сохранения) стадии сво-боднопла-
вающей жгутиконосной бластулы в качестве первой личиночной
стадии у относительно очень высоко организованных беспозвоноч-
ных, систематически очень далеких от всех тех низших групп Meta-
zoa (а тем более от многоклеточных фитомонад), где личинки
этого типа пользуются наибольшим распространением. Особенно
любопытно то обстоятельство, что бластулообр-аз-ные ли-чинки
импеют, среди иглокожих, не более (примитивные Pelmato-zoa, a зна-
чительно выше организованные Eleutherozoa, И притом большин-
ство из них, а не отделвные их представители.
Это не мешает, однако, тому, чт-о жгутиковые бластулы иглоко-_
жих обладают тем же весьма специфическим типом организации,
ка4к и все остальные Целобластулы, и формируются в результате не
более сложного, чем у Leptolida, эмбриогенетического процесса,
также -сводящегося, по сути дела, к одному только дроблению. Та-
ким образом, и здесь эмбриональное развитие отличается большой
простотой и нримитивностью и, строго говоря, не заключает в себе
еще ничего специфически зародышевого.
249
ЗНЭЧИТЕЛЬНОЕ ОДНОООРЗЗЧИЕ ЭТОГО процесса у ВСЕХ ИГЛОКОЖИХ
ПОЗВОЛЯЕТ ОГРЗНИЧИТЬСЯ ЕГО РЗССМОТРЕНИЕМ на ОДНОМ ТИПИЧНОМ ПрИ-Е
мере, каковым нам послужит класоич-еский объект эксперименталь-
ной эмбриологии беопозвоночных —— морской еж Paracentr-otus
lividus. ОДНООСНО гетерополярное строение яйца Paracentrotus уста-
навливается, как известно, с самых ранних стадий развития ооцита.
Его анимальным полюсом становится тот, который обращен к стенке
яичника, так что этот п0лю.с вначале имеет значение трофическогр,
а намечаемая им главная осьяйца — значение оси трофическойпо-
ляризации. В дальнейшем на этом же полюсе выделяются направи-
тельные тельца ли он же служит ‘местом внедрения сперматозоида
при оплодотворении. Видимым выражением полярности зрелого
яйца служит наличие пигментированной зоны, опоясывающей его
нижнее полушарие непосредственно под экватором и появляющейся
сразу же после созревания; до этого образующий ее „красный пиг-
мент равномерно распределен в кортикальном слое всего яйца. Та-
ким образом, анимальный полю-с морских ежей совмещает в себе
признаки обоих полюсов яйца гидроидов, отличается от такового
губок большей определенностью своих анимальных свойств и тем,
что он проходит стадию трофического полюса.
Одноосно-гетерополярное строение яйца дополняется его дву-
сторонней или, «вернее, двулуневой симметрии, получающей видимое
выраженние в направлении плоскостей его двух первых делений._ Как
теперь хорошо известно (Гёрстадтиус, 19.28), одна из них является
строго сагиттальной, а другая —— фронтальной по отношению к позд-
нейшим билатеральным стадиям развития, что и -служит поводом
говорить о билатеральной симметрии сам-ого яйца; однако так же
хорошо известно, что борозда первого деления может лечь по любой
из этих плоскостей, т. е. быть либо сагиттальной, либо фронтальной,
что автоматически влечет за собой подобную альтернативную измен-
чивость положения второй борозды. А это, очевидно, говорит о рав-
ноц-енности обеих ‘плоскостей симметрии яйца, т. е. о двулучевом, а
не двустороннем ее характере.
Дробление яйца осуществляется посредством одних только коль-
цевых борозд и относится к радиальному ти-пу. Его третья
борозда экваториальна И РЗЗДЕЛЯЕТ ЯЙЦО на 4 неокрашенных ани-
м-альных и 4 вегетативных бластомера, по которым целиком рас-
пределяется его пигментированный материал; при четвертом деле-
нии бластомеры- анимально-го круга делятся равномерно и -в мяери-
дианальном направлении, а -вегетативные—— резко неравномерно
и в плоокости, параллельной экватору; каждый из них дает по
одному пигментированн-ому макромеру и лежащемгу под ним неокра-
шенному микромеру. Таким образом, анимлальное полушарие пред-
ставлено -на стадии 16 кольцом из восьми одинаковых бла-стомеров
средней величины, а вегетативное — Двумя лежащими ДРУГ на дРУге
квартетам1и макро- и микромеров. Борозды ппято-го деления ложатся
параллельно экватору в верхнем полушарии, в нижнем же макро-
мперы делятся меридианально и равномерно, а м-икромеры -—-
250
неравномерно и в горизонтальной ‚плоскости; стадия 32 СОСТОИТ,
соответственно, из двух аним-альных и одного вегетативного круга
по восемхь бластомеров в каждом из двух квартетов микромеров.
Следующими тремя делениями общая численность бластомеров
доводится до 250; к этому моменту их размеры совершенно вырав-
няли-сь‚ и они сложились в из-оморфный кубический эпителий, о-бра-
зующий стенки бластулы. Оформьпяющаяся бластула имеет пра-
вильную форму шара и ориентирована таким же образом, как яйцо,
что легко устанавливается по тождественному, субэква-ториальному
положению ее ппигментированного Il'lOHCKa; следовательно‚-и ее по-
. ЛЮЪСЫ СОХраНИЛИ ТУТ СВОЕ ПЕрВОНЗЧЭЛЬНОЕ ПОЛОЖЕНИЕ И значение.
Обширная полость бластулы образуется постепенным расхождением
бласт-омеров, начиная с 8-клеточной стадии, и имеет здесь характер
первичной схиз-оцельной полости дробления.
На стадии 250 начинается поклеточная дифференцир-овка клеток
бластулы в жгутиковые, постепенно охватывающая всю ее поверх-
ность; в это время в гбластуле ‘происходит еще два не вполне син-
хронных деления, доводящих чнисло ее клеток примерно до 808, при-
чем можно заметить, что дифференцировка неотступно следует за
заключительным делением и так растянута именно потому, что де-
ление происходит неодновремленно в разных точках бластулы. В про-
цессе дифференцировки клетки формируется одножгутиковая
полносоставная книнетида, образующаяся из имевшейся в ‘клетке
центросомы и в состав которой входит настоящий парабазальный
аппарат, образовавшийся из ранее разрозненных диктиосом (Овасс,
1935; см. рис. 94, 8); одновременно с этим клеточное тело сильно
вытягивается в длину, а кортикальный сло-й его ужплощенного жгу-
тиковог-о полюса ‘приобретает хара-ктер кутикулы. Такой дифферен-
цировке подвергаются все клетки бластулы, становящиеся одно-род-
ными и мо-рфологически тождественными жгутиковыми клетками,
независимо от занимаемого ими положения (в бластуле) и “PIX буду-
щего значения. Друпими словами, дифференцировка имеет здесь
такой же всеобщий и равномерный характер, как у всех других жгу-
ТИКОВЫХ ЦЕЛООЛЗСТУЛ. ПО ЕЕ ОКОНЧЭНИИ ЗЗРОДЫШ ПЕРЕХОДИТ K 'CaMO- ‚
стоятельному существованию в виде мерцательной личинки, в naus-
нейшем проходящей длительный и необычайно сложный путь
постэмбрионального развития.
6. Особенности примитивных форм дробления
Как ВИДНО ИЗ ТОЛЬКО ЧТО ПрИВЕДЕННЫХ данных, ОДНОЙ ИЗ характер- _
нейших особенностей эмбрионального развития низших Metazoa
приходится считать исключительное разнообразие присущих им
форм дробления. Оно варьирует здесь от внешне совершенного
аморфного «анархического» до строго радиального, часто имеет
все главные признаки спирального и иногда осуществляется в фор-
мах, очень близких к билатеральному._Наряду с этим, степень его
равномерности может быть очень различной, и это же можно ска- п
к
251‘
зать о согласованности составляющих его делений (всмысле их
большей или меньшей синхронности). Если еще добавить к этому,
‘ITO ЗМПЛИТУДЗ ИЗМ-ЭНЧИВОСТИ фИЗИОЛОГИЧЭОКИХ ТИПОВ дробления
идет здесь от предельно недетерминированного (у Ь-ертопоа) до бе-с-
спорно высоко детерминированного (у Heterocoela), легко понять
те трудности, с какими сталкивается всякая попытка свести этот
хаос к скудному набору типов и норм дробления, различаемых
общей эмбриологией.
Мы также могли убедиться, что большому разнообразию наблю-
даемых тут форм дробления сопутствует и не меньшее их свое-
образие. При этом до сих пор еще принято считать, что все-эти
проявления самобытности эмбриог-енеза низших кишечнополостных
и губок являются не более, как уклонениями от нормы, олицетво-
ряемой правильными формами развития вышестоящих в системе
групп. _
На этом представлении зиждется, между прочим, и обиходное
признание правильного радиального дробления самой примитивной
из в-сех его форм. Репутация первичной, «примордиальной» формы
дробления закрепилась за ним так прочно еще с геккелевских вре-
мен,-что его неизменно еще рассматривают ‘как прототип дробления
яйца Metazoa. Главным основанием для такой его трактовки
служит, очевидно, тот факт, что радиальное дробление подчиняется
много лучше других его типов правилам Закса и отвечает ближе
других идеальным формам последовательного деления ша-рообраз-
ной геометрической фигуры. Правда, приложимость этих законов
ограничивается и здесь порядком чередования плоскостей только
первых трех делений (сагиттального, фронтального и экваторяиаль-
ного) и н-е простирается на дальнейшие, идущие вопреки этим зако-
нам в антиклинальных (т. е. перпендикулярных ‚к поверхности яйца,
радиальных), а не в периклинальном (параллельном его поверх-
ности) направлении.
Однако в св-ете тех фактов и положений, о которых говорилось
в предыдущей главе, такая трактовка радиального дробления ока-
зывается явно несостоятельной. Ей в корне противоречат сами же
признаки, какими оно характеризуется, и она не находит себе под-
держки ни с одной, в сущности, стороны. Вместе с тем именно ‹<укло-
няющиеся» формы дробления обнаруживают самое большое сход-
ство с уже хорошо знакомыми нам картинами гпалинтомического
спорообразования у простейших и, как это специально уже подчер-
кивалось, проявляют наиболее ясным образом свою первичную
СПОрОГОНпИЧЗСКУЮ природу. Поэтому кажется очевидным, что самыми
примитивными формами дробления многоклеточных следует счи-
тать именно такие, которые, будучи приуроченными к развитию
наиболее примитив-ных из них, сохранили вместе с тем и наиболь-
шее число сходств с процессами палинтомического размножения.
Наиболее интересные дан.нь1е на этот счет дает дробление мета-
генетических гидромедуз. Оно. обычно описывается, как радиальное,
‘С нормальным ЧЪЬРЭДОВЗНИЭМ €_I‘0 ТРЕХ первых ПЛОСКОСТЭЙ COI‘JIaCHO_
252
правилу Мечникова, т. е. в трех- измерениях пространстваг первая
его борозда считается сагиттальной, с вторая—фронтальной,
а третья — экваториальной. Правда, при этом отмечается, что в гео-
метрических формах дробления отсутствует та правильность, опре-
деленность, которая свойственна высшим типам Метаяоа; и если
оно происходит по радиальномутип-у, <<-то все жедпроизводит впе-
чатление чего-то неустановившегося» (Давыдов), что’ заставляет
считать его не вполне правильным, а в некоторых случаях даже
беспорядочным. Таким образом, можно думать, чтодздесь имеет
место та самая кубическая палинтомия, которую мы тщетно искали
у Protozoa, H представляем себе неотъемлемым свойством дробле-
ния всех вообще многоклеточных животных.
Однако, если внимательнее отнестись «K соответствующим факти-
ческим данным и учесть все сказанное выше о независимости общих
геометрических форм палинтомии (а также и точно копирующей ее
повторной паратомии инфузорий) от действительного направления
плоскостей последовательного деления, принадлежность дробления
Leptolida к кубическому ее типу начинает внушать самые серьезные
сомнения из-за своих, крайне своеобразных, особенностей. Эти о-со-
бенности заключаются, как мы видели, в том, что дробление осуще-
ствляется тут не посредством обычных кольцевых борозд, идущих
от периферии яйца пк его центру, а посредством односторонних
<‹врезающихся»' или режущих борозд, становящихся к тому же
«центробежными», начиная -со второго деления; далее они выра-
жаются в том, что дроблению сопутствуют правильные повороты
и переориентировки бластомеров относительно друг друга, ведущие
к созданию характерных крестообразных или Х-образных группи-
-ровок на стадиях 2 и 4; наконец, эти специфические особенности
ранних стадий дробления дополняются на более поздних стадиях
многими другими, наиболее интересными из которых являются
типично спиральное расположение бласто-меров на стадии 8 и сле-
дующих, их «анархическое» поведение у Oceania И описанное у С1у-
tia перемещение ядер клеток бластулы из непосредственной бли-
зости к полюсу, о-бращенномпу внутрь благстоцеля в направле-нии
к противоположному, наружному их полюсу. `
Значение всех этих особенностей дробления Leptolida оставалось
до сих пор неясным и действительно не поддается объяснению на
основе того материала и в духе тех основных концепций, какими
оперирует до сих пор сравнительная эмбриология беспозвоночных.
Поэтому они «PI не были оценены по достоинству и даж-е стали посте-
ПЭННО забываться; XapaKT€pHO, ЧТО В COBPEMGHHBIX СВОДКЗХ И pyK0-|
водствах об этих замечательных открытиях Мечникова не упоми-
нается ни словом. '
Между тем, нельзя не заметить, что наблюдаемые тут картины
воспроизводят с совершенно удивительной точностью морфологи-
ческие картины пали.нтоми-и простейших и, в частности, все специ-
фические особенности ее табличных форм у различных фитомонад.
ВоспроизводИмь1-е здесь рисунки из классического-труда Мечникова,
253
-с одной стороны, M из ряда специальных работ по развитию Phyto-
m/onadina—~c другой, показывают самым наглядным образом, на-
сколько в-елики и специфичны имеющиеся тут сходства (см.
рис. 28 И 84). т =
Таким образом, не будет преувеличением сказать, что дробление -_
яйца Lept-0«1ida идет по тем же правилам, калк палинтомия Phyt-0mo- _
nadina, и в совершенно тождественных формах. Это делает более
чем вероятным, что к нему приложима и та ж-е интерпретация,
т. е. что оно относится в действительности не к кубической, а к таб-
личной палинтомии. Это значит, что наблюдаемое тут чередование
направления плоскостей дробления является в такой же мере мни-
мым, как у фитомонад, и достигается тем же путем ——переориенти--
ровкой спороцитов (бластомеров) в ходе дробления, при сохранении
каждым из них постоянного положения плоскости деления: эта
плоскость неизменно проходит по главной клеточной оси, а соот-
ветствующая -борозда деления всегда возникает на одном и том же —-—
анилмальном — полюсе клетки. Следовательно, все Непонятные иначе
особенности морфологии ранних стадий дробления Leptolida полу-
чают простое объяснение в том, что яйцо пи первые, по крайней мере,
поколения бластомеров обладают тут типично продольным спо-
собом деления. Иначе говоря, ранние стадии дробления этих медуз
еще не подчиняются тем общим правилам последовательного деле-
ния целостной геометрической фигуры, к которым принято сводить
дро-бление яйца всех Metazoa, а |ИДеТ в порядке самостоятельного
деления каждого из бластом-еров в отдельности. Каждый ‚из них
ведет себя тут как отдельная особь жгутикового с при-сущими elf
»npoMopqpoJ1on1»1I1e»cK14-MP1 особенностями и врезающейся продольном
"бороздой деления, т. е. обладает ровно такой же степенью индиви-
дуальности, как спороциты палинтомических Flagellata. Такое тол-
кование геометрических форм дробления Leptolida находит себе-
очень вескую поддержку со стороны экспериментальной физиологии
развития. Дело в том, что такое толкование означает признание
ранних стадий дробления этих медуз лишь слабо интегрированными
системами, состоящими из относительно равноценных друг другу
одноклеточных особей, подобных спороцитам жгутиковых. А если
так, то невысокая степень морфологической целостности дробя-
щейся системы, очевидно, должна иметь пи соответствующее физио-
логическое выражение. Известные опыты по изоляции бластомеров
Clytia viridicans, uaodicea cruciéta и Amphisbetia operculata (см.
рис. 88) показывают со всей очевидностью, что так оно и есть: что
именно эти, метагенетические, медузы обладают рекордной для
Metazoa степенью недетерминированности дробления. Она выра-
_жается здесь, во-первых, в том, что каждый изолированный бласто-р
мер стадии 2, 4, 8 и 16 может дать н_ачало нормальной, н-о только
соответственно уменьшеннои бластулообразн-ои личинке, и, во-вто-
‘РЫХ В ТОМ, ЧТО ИСКУССТВЕННОЕ ВЗЗИМНОЕ перем-ещение бласто-меров ‘В
дробящейся системе не нарушает нормального хода развития. Ко-
роче говоря, все бластомеры до стадии 16 включительно являются
254
тотипотентными и вместе с тем экв-ипотенциальными, что как нельзя
лучше согласуется с их ярко выраженной морфологической само-
стоятельностью.
Что же касается тех «уклонений от нормы», C какими мы встре-
чаемся на ранних стадиях дробления губок‚.,то главное из Них
заключается тут в том, что третья борозда дробления ложится не
экваториально, а в меридианальном направлении, гчто имеет своим
результатом табличную группировку бластомеров на стадии 8. При
этом их расположение бывает далеко не всегда радиальным —-OHO
с таким же успехом может проявлять и правильную щвулучевую
симметрию. В последнем случае борозды трех ‚первых делений
"ложатся ПОРОЗНЬ В КЗЖДОМ ДЕЛЯЩЕЪМСЯ бластомере И МОГУТ даже
появляться неодновременно, что может вести к образованию проме-
жуточных б-клеточных стадий с совершенно разобщенными плоско-
стями деления.
Таким образом, и здесь совершенно не соблюдены порядок чере-
дования и положения плоско-стей дробления, характерные для
радиального его типа. `
Вместе с тем легко заметить, что эти особенности раннего дро-
бления губок в точности повторяют картины образования гониевой
таблички у Volvocoidea, т. е. тех форм табличной палинтомии, при
которых не происходит заметной переорие-нтировки спороцитов.
Поэтому табличная группировка бластомеров Са1-сагеа на стадиях
4, 6 и 8 не может расцениваться иначе, как самое «откровенное»
проявление этими стадиями типичнейшей табличной палинтомии.
У Ногпосое1а ей подчиняются, повидимому, и последующие -стадии
дробления, насколько можно судить по немногим имеющимся лизо-
бражениям, во всяком случае не препятствующим такому их ист-ощ-
кованию. И только у I-Ieterocoela дробление идет дальше уже по дру-
гому пути, поскольку четвертое деление яйца является ‘тут неоспо-
римо эвкваториальным и приводит к рассечению яйца на две
неравноценные половины. Однако дальнейшие деления бластомеров
идут (сначала в каждом из полушарий, а затем только в будущем
жгутиковом) в порядке их продольного деления только в двух чере-
дующихся плоскостях, так что и оно оказывается сводимым к общим
принципам табличной палинтомии. Стало быть, отход от этих прин-
ципов заключается только в гетерон-омности четвертого деления
яйца и в том; что им не следуют бластомеры будущего зернистого
полушария. Поэтому можно сказать, что дробление Heter-ocoela есть
ВИДОИЗМЭНСННЗЯ табличная, НО никак не ТИПИЧН=О выраженная куби-
ческая палинтомия.
K великому сожалению, ранние стад.ии развития Ногпосое1а
не служили до сих пор объектом экспериментального исследования,
так что мы ровно ничего не можем сказать о существующей тут
степени детерминированности дробления. Тот факт, что оно резко
детерминировано у Heterocoela (Syc0n), He является, конечно, пре-
пятствием к допущению его недетерми.нирован.ного характера
у более ‘примитивных, чем они, Нотосое1а.
255
Таким образом, специфические для губок особенности дробления
также находят свое объяснение в том, что оно осуще-ствляется, по
крайней мере на ранних стадиях, по правилам табличной ‘палин-
томии, здесь совершенно не замаскированной. .
Особенности дробления иглокожих и, в ча-стности‚ аморских ежей
допускают ту же, в общем, интерпретацию, какая была дана в отно-
шении I-Iueteroc-0-ela. Тут определенно экваториальным и гетероном-
ным характером обладают, в сущности, только третье деление,
обособляющее анимальный квартет бластомеров от вегетативного,
и те борозды четвертого деления, которыми микромеры отделяются
от этих последних; все же остальные борозды дробления являются
определенно продольными для каждого бластомера в отдельности
п чередуются только в двух координатных пл-оскостях; в отношении
к яйцу в целом они получают любое, в том числе и диагональное,
положение, ч-ем и определяется спиральный, а не радиальный харак-
тер лклеточной мозаики бластулы. Таким образом, ‘и здесь можно
говорить о табличной в своей основе палинтомии, прерь1ваем0й дву-
кратным неравноценным делением дробящейся системы в целом.
Эти гетерономные и охватывающие всю систему деления служат
видимым показателем определенной степени детерминированности
дробления Их, как бы накладываясь на его табличную перпв-оосьнову,
придают ему, в конечном счете, характер правильного и типичного
радиального дробления.
Все сказанное позволяет придти к выводу, что «принмордиаль-
ными» типами дробления Metazoa являются такие, при которых оно
полнее всего проявляет признаки табличной палинтомии. Это как
раз и имеет место при якобы уклоняющихся, но в действительности
скорее «еще неустановившихся» формах дробления, наблюдаемых
у низших гидронидных и губок, что и заставляет нас считать именно
эти равномерные спиральные, табличные или же в той или иной .
«мере анархические ЕГО фОрМЫ за ДЕЙСТВИТЕЛЬНО самые ПРИМИТИВ-
ные из типов дробления Metazoa. Что же касается радиального, то
он представляется далеко не примитивным, а, напротив, сильно
видоизмененным типом той же табличной палинтомии.
Сводимость всех форм дробления бластулообразующих Metazoa
к табличной палинтомии находит се-бе очень существенное (можно
сказать, даже решающее) подтверждение в самом: факте, что про-
дукты дробления рас-полагаются обычно почти с самого его начала
в виде более или менее правильного однослойного эпителия. В самом
деле, наблюдаемое здесь первично-эпителиальное расположение
бластомеров означает не что иное, как их табличное, двухмерное
расположение по законам табличной палинтомии, и не может быть
достигнуто другими ее формами. Таким образом, истинные геомет-
рические формы дробления низших Метагоа ярко проявляются
в его эпителиогенном характере, в том, что конечным продуктом
дробления является однослойная -бластула, обладающая строго
эпителиальным строением и все клетки которой являются с самого
начала поляризованными.
256
7. Изменения полярности в ходе формирования бластулы
и их значение
Большой принципиальный интерес представляют описанные
выше явл-ения последовательной смены морфологической и физио-
логической полярности развивающегося многоклеточного организма.
Вообще говоря, ее изменения в онтогенезе проявляются втом, что
исходная морфо-физиолопическая ориентация яйца, выражающаяся
определенным набором свойств, уступает свое место в дальнейшем
иной, особой ориентации бластулы, характеризующейся другим
набором свойств и часто оказывающейся противоположной перво-
начальной.
Чтобы приблизиться к пониманию этих сложных отношений,
необходимо сначала внести возможно полную ясность .в вопрос
о том, каковы проявления полярности разных стадий развития и
чем характеризуются у каждой стадии и в разных труппах разно-
именные полюса. Дело -в том, что привычны-е представления на этот
счет, базирующиеся, как всегда, на особенностях эмбриогенеза
высших Metazoa, не оправдываются в отношении к низшим много-
клеточным, ведущим себя и в этом смысле несравненно более сво-
бодно, чем высшие. Поэтому-то и не приходится удивляться, что,
например, до сих пор нет твердой уверенности в правильности име-
нования переднего полюса амфибластулы губок анимальным полю-
сом или ж-е окрашенного полюса яйца Amphisbetia —— вегетативным,
И ЧТО МЫ ТУТ ЧЗСТО НЕ МОЖЕМ УСТЗНОВИТЬ И-СТИННУЮ ‘ПРЕЕМСТВЕННОСТЬ _
-П0ЛТОС0В В ОНТОГЕНЕЗЕ.
Первый вопрос, требующий уяснения, заключается в том, каковы,
в разных конкретных случаях, признаки разноименных полюсов
зрелого яйца или гонидий низших многоклеточных и. отвечают ли
они тому стандарту, к которому их принято сводить. Известно, что
самыми общими признаками а н и м а л ь н о г о п о л ю с а яиЦа
Metazoa считаются следующие ш-есть: '
1) приуроченность пк нему места выхода направительных
телец при созревании;
2) локализация на этом полюсе точки вхождения спермато-
зоида при оплодотворении;
3) появление на нем первых борозд дробления, когда только
они имеют хоть сколько-нибудь односторонний характер;
4) близость к этому полюсу центросомы, а часто и самого
ядра;
5) относительная бедность его плазмпьт желточными вклю-
чениями, если только они не распределены вполне равномерно;
6) он является полюсом максимальной физиологической
активности (и -в том числе восприимчивости) яйца.
В соответствии с этим, признаками в е г е т а т и в н о г о п о-
л ю с а являются почти только отрицательные, за исключением двух:
его большей загруженности желтком при неравномерном pampe-
делении последнего -и его пониженной физиологической активности.
17 Эмбриология беспозвоночных 257
Соотношения полюсов зрелого яйца с напра-влением его трофической '
поляризации на ра.нних стадиях развитчия н-е однозначны: в од-них
случаях полюс ооцита, которым он соприкасается с материнскими
питающими клетками, становится в дальнейшем анимальным, в дру-
гих же, на-против, вегетативным.
Просматривая под этим углом зрения материал, изложенный
в предыдущих разделах, можно констатировать, что эти стандарт-
ные отношения имеют
у морских ежей, низших сцифоидных и гидромедуз, тогда как губки
и Volvocoidea существенно уклоняются от шаблона. В саямом деле,
единственным признаком анимального полюса яйца губок является
близость к нему довольно изменчивой все же точки проникновения
спермоцисты, так как: а) полярные тельца выделяются тут в совер-
шенно другом месте; -б) борозды дробления являются с самого на-
чала кольцевыми; в) ядро дробления занимает в яйце строго цен-
тральное положение, а центросома не обнаруживается в интерки-
незе; г) желточные зерна равномерно рассеяны в ооплазме; на-
конец, д) мы не располагаем данными о наличии и направлении
физиологических градиентов яйца. '
Что же касается Volvocoidea, то тут первый признак анималь-
ного полюса вообще отпадает (в связи с равно-ценностью продуктов
редукционных делений), а полюсом оплодотворения является полюс,
противоположный тому, который удовлетворяет трем следующим
признакам‘ анимального. Таким образом, чтобы назвать один из
полюсов яйца губок или вольвоцид анимальным, приходится произ-
вольно предпочесть одни из его признаков другим, и притом в каж-
дом случае равные; а это ясно показывает значительную условность
их принятого обозначения. с
Оно становится еще более условным, если ввести в характеъри-
стику полюсов яйца будущее, презумптивное значение образующего
их материала. Нормой мы привыкли считать, что анимальное полу-
шарие яйца дает исключительно эктодерму‚ -а Вегетативный-
только энтодерму. Однако это общее правило полностью оправды-
вается, на нашем материале, только у низших сцифоидных, и даже
морские ежи не вполне подчиняются ему, так как материал вегета-
тивного полюса дает тут не энтодерму, а первичную .мезенхиму. Что
же касается Leptolida, то тут игмеют место как ‚раз обратные отно-
шения, а сказать что-либо о губках мы пока не можем ввиду неяс-
ности .в вопросе, что считать у них энтодермой и что — эктодермой.
Итак, строгой определенности характеристики полюсов яйца
"высших M-etazoa противостоит большая изменчивость их признаков
у низших.
Переходя теперь к характеристике полюсов сложившейся бла-
стулы и следующих за нею стадий развития, мы опять начнем с пере-
числения тех свойств а .н и м а л ь н о г о п о л ю с а, какие считаются
ДЛЯ НЕГО ТИПИЧНЫМИ. ЭТИ ПРИЗНЗКИ ТЗКОВЫ:
1) он ‚неизменно обращен вперед прьи поступальном движении
(плавании), т. е. является передним -полюс-ом оси движения;
258
IMECTO ДЗЛЕКО не всегда, а ИМЕННО ТОЛЬКО‘
Q) ОН служит полюс-ом прикрепления при оседании личинки
перед ее превращением в сидячую вегетативную стадию;
3) на этом полюсе сосредоточены, когда они есть, чувстви-
тельные и другие узко специализованные клетки «анимального»
‚значения;
4) материал всего анимального полушария имеет презумптив-
ное значение эктодермы; .
ё) этот полюс является полюсом: повышенной физиологиче-
скои активности.
Соответственно признаками в е г е т а т и в н о г о
бластулыпринято считать следующие четыре:
1) его направленность назад при плавании;
2) то, что он становится дистальным полюсом прикрепленной
стадии;
3) что этот полюс дает материал будущей энтодермы;
4) что он отвечает точке минимума физиологического гра-.
диента.
Сравнивая эти характеристики с те-ми, какие давались полюсам
яйца, нельзя не обратить внимание на то обстоятельство, что они
совпадают только в одном признаке—одинаковой направленности
физиологического градиента. Все же остальные признаки полюсов
яйца и бластулы не имеют фактически ничего общего между собою,
касаясь качественно различных явлений, не находящихся к тому же
и в хоть сколько-нибудь очевидной взаимной связи. Действительно,
ттрудно себе представить, какова может быть‘ внутренняя связь
между направлением поступательного движения личинки и, скажем,
траекторией, проходимой полярными тельцами или сперматозоидом
в яйце, и даже если допустить, что такая связь сущ-ествует,+
полюса
‚ПОЧЕМУ ‘МЫ ДОЛЖНЫ СЧИТЗТЬ ее ОбЯЗЕТЭЛЬНО однозначной.
Невольно создается впечатление, что мы ‚произвольно обозна-
чаем одними и теми же терминами—‹<анимальнь1й» или «вегета-
тивный» полюс — качественно различнь1-е свойства, сравнимые друг
с другом исключительно потому, что 0Н|И служат проявлением
полярной дифференциации соответствующих стадий онтогенеза.
Однако ничто, в сущности, не говорит об идентичности гетеропо-
лятрного строения яйца с таковь1.м пмногоклеточных личиночных ста-
дий: в ней ‘можно усомниться уже потому, что |ИХ «одноименные»
полюсы характеризуются принципиально различными признаками.
Это впечатление еще усиливается, если подойти с обычной
меркой к конкретны-м особенностям. бластул низших Metazo-a и
Volvocoidea. B самом деле, только у морских ежей ли, возможно,
Scyphozoa и, как это ни странно, у Volvocoidea передний полюс
бластулы удовлетворяет всем перечисленным общим признакам
стандартного анимального полюса; у гидроидов же он состоит из
материала, становящегося в дальнейшем энтодеярмой, а у губок его
материал дает не только последнюю, но и ряд других клеточных
систем. Стало быть, в одних случаях передний полюс оои движения
личинки является эктодермальным, а в других-энтодермальньхм,
17* 259
что свидетельствует о несовпадении двигательной полярности ли?
чйнки с той или иной локализацией будущих энто- и э-ктодермы.
Обращая-сь к вопросу об онтогенетической преемственности
одноименных полюсов яйца и личинки, мы о.пять сталкиваемся
с очень различными и противоречивыми отношениями Эта преем-
ственность является прямой у Echinoidea, Scyphozoa И УоЫосШае
и обратной у Leptolida и губок, у которых, как мы теперь знаем, A
анимальный полюс яйца становится задним полюсом жгутиковой
личинки.
Отсюда следует, во-первых, что мы должны не придавать этим
терминам абсолютного значения, а наоборот, ясно представлять себе
них относительность, зависимость их содержания от того, к какой
из стадий развития они применяются. Во-вторых, мы обязаны при-
знать, что первичный анимальный полюс яйца может стать в одних
случаях пер-едним—-<<анимальным>›, а в других задним-«вегета-
тивным» полюсом личинки, и что такое его «превращение» является,
по сути дела, мни мы M 1/1. не вызывается действительным обра-
щением полярности организма. Наконец, приходится констатиро-
вать, что у зани-мающих нас сейчас форм анимальный полюс яйца
‘СТЗНОВИТСЯ CO ВРЕМЕНЕМ задним ПОЛЮжСОМ ЛИЧИНКИ, а вегетатив-
ный- передним, тогда как в других группах могут иметь место и
иные отношения. Все дело в том, что одноклеточное яйцо и много-
клеточная‘ личинка—— организмы разного структурного уровня,
отличающиеся поэтому друг от друга и конкретными формами
своего одноосно-гетерополярного плана строения.
Как происходит в онтогенезе смена одного из них вторым-
другой вопрос, ответ на который можно, повидимому, найти в ряде
специальных особенностей эмбриогенеза Leptolida. МЫ имеем
в виду, во-первых, наблюдения и эксперименты Тессье, показавшие,
что все бластомеры яйца й любые фрагменты бластул являются в
равной мере ОгМНИПОТЭНТНЫМИ, независимо OT HpHCyII_l€1'O ИМ пре-
зумптивного значения, т. е. того, должны ли они дать экто- или энто-
дерму (см. рис. 88). Далее им было установлено, что промежу-
точные -стадии дробления, предшествующие оформлению бластулы,
‘не обнаруживают в экспериментах никаких признаков полярности
И никаких УЛОВИМЫХ ГРЗДИЭНТОВ ЖИ также лишены ‘ВНЕШНИХ ПрИЗНЗ-
ков симметрии. Далее, им описываются случаи, когда, п-осле виталь-
ного окрашивания, одного из полюсов яйца, окрашенные бласто-
меры оказываются беспорядочно перемешанными с белыми
в течение этого же промежуточного периода дробления и только
‚ПОЗДНЕЕ ЗЗНИМЭЮТ «ПОЛЁПЗЮЩЭЭСЯ ИМ МЕСТО. НЁЁКОНЭЦ, И ЭТО CEIMOE
главное, все, что говорилось выше о положении борозд дробления
и их своеобразном центробежном характере начиная со второго и
кончая четвертым или пятым делением, свидетельствует с полной
определенностью о т_ом, что все бластомеры этих стадий обладают
собственной „полярностью й ведут себя при делении как клетки,
относительно независимые друг от друга и от проморфологий дро-
бящегося организма как ‘целого.
260
Ёд
Сближа u
Нести И Г an эти факты с тем, что говорилось оструктддрнои подяр-
р диентах как руководящих проявлениях физиологической
целостности организма, нельзя не придти к выводу что сущность
- ' 9
промор.фологических изменений, происходящих в эмбриогенезе
Leptohda, заключается не в чем ином‚- как в посл едователь-
ной смене нескольких и
A различных овнеи .
лостности. ур He
B CEIMO}/II деле, начальные одноклеточные -стадии онтогене3а_
ооЦит и яйцо—обладают всеми проморфологическими и физиоло-
;I’:‘OeF°1fXH1‘gH СВОЙСТВЗМИ целостного одноклеточного организма и во
Н тнощениях эквивалентны отдельной особи простейшего
а QEIHHHX стадиях дробления этим-и же свойствами обладают
в равнои мере все потомки яйЦа-—-бластомеры‚ ярко проявляющие
СВОЮ ПРОМОРФОШОГРГЧЭСКУЮ ИНДИВИдуальность до стадии 16 или
даже 32; однако образуемая ими система еще лишена в это время
УСТОИЧИВОЙ организменной целостности, пи потому именно не про-
Ёвляет в полнои мере структурного и физиологического единства;
КЁа еще не поляризована, не имеет тои степени интеграции, при
торой последняя приобретает осязаемые черты. И лишь позднее,
На даЛЬНеИШИХ ‘СТадИЯХ дробления, его продукты объединяются
ЁаСТОЛЬКО ТеСН0‚ Что образуют целостный, но уже многоклеточный,
Шрганизм, вновь получающий все структурные и физиологические
HCC))£CTHBaafl «?]l1HOOCH(3-1‘eTepOl'lOJIflpH'OI‘O Зкивотного. Короче говоря,
Д ротоз-оиная›> полярность яица сменяется затем неполя-
P1’130'B3H'H1>1M1’1 1'IP0Me>KY’I‘0‘IHI>IMH СТаДИЯМИ дробления, сменяемыми,
В ДСВОЮ ОЧЕРЕДЬ, резко поляризованными стадиями бластом-ерной и,
в конце концов, эпителиальной бластулы.
Глава XIV
ОРГАНИЗАЦИЯ жгутиконосньпх влАстул
и их ЗНАЧЕНИЕ синзооспор
1. Проморфологйческие особенности целобластул
qac:Ide°;Ig:g:eH:fl'E11:I%*}?H£'i113AHP:1ILJ;e>:){H0cTb к самым различным и, от;
системы И больше А .ру т друга категориям естественной
е физиологическое разнообразие, „все жгутиковые
Цедобластудё‘ очень ‘СХОДНЫ Между собой по своей архитектонике
Ёзпхределёяемои одинаковым способом движения в водной среде. Все
НЁеМ ‘::OJ;;l1e5I1I*1C:1T}OIuP<I3I1f4LH11;Ia(I:<(:D6I3eI:I1I\;I -;I~3§§HoocHo-reTep0no.71s1pHbIM —— crpoe-_
Ной ёИММетрИИ Начиная с 2_ри 4 ки различных ивидов вращателдг.
с осью ВраЩеН‚ИЯ большого Н -лучевои и кончая спиральной.
, о всегда довольно определенного,
ПОВИДИМОМУ, порядка
‘Признаки 4-лучевой симметрии имеются, вероятно, у большин-
:3I:::1I,pe@)((3JII1;IfléITf30 :cHe:n I:1§:;1§$I1:-gig, выражаясь х-отя бы в положений
го креста или розетки, занимающих
261'
анимальный полюс тела у всех тех бластул, образование KOT0OpbIX
связано с более или менее спираЛЬНЫМИ ФОРМЁГМИ дробдения- C0‘ .
pm.__ 93, Жгутиконосньхе целобластулы. кнйадарий и иглокожих (в оптическом
. ' ‚ разрезе Z _ .
. ‚ . __ ' '- 3 _ м”; а ; 4 —~ C1 ‘ста flav1dula'
; "g3'fa§§?§§§t’cl,a,;ci§ta; R§t:K8%t§fi§}c£§u1:§:énbauri; r<'(?)(-:3nI1IaaLlaS§1tti"10eé; 9 — Mgpcxofi звезды
' Asterias; 70 —г0лотурии Synapta (9, 10 no Зеленке, остальное по Мечникову)
бенного развития достигает 4-лучевая симмеврия у вольвоксов, где
она ПРОЯВЛЯЕТСЯ еще ‘В достаточно ОТЧЕТЛИВОМ рНЗГрЗНИЧЕНЕ/{И ‘ЧЕТЫ-
рех квадрантов их шароооразного тела и в крестообразной форме
262
фиалопо_ра. -Наряду -с этим нередки и признаки двулучевой
симметрии, наиболее распространенным из которых можно считать
наличие полярной спаики в той же «теменной» розетке, устанавли-
вающей одну из ее плоскостей, в то время как другая определяется
положением двух других клеток розетки, разъединяемых этой
спайкой. Что же касается винтовой, спиральной, симметрии других
порядков, то она выражается преимущественно в соответствующей
клеточной мозаике и определяемом ею положениижгутов двига-
тельного аппарата. Правда, достаточно точными данными на этот
счет мы пока не располагаем‚'но вращательный характер поступа-
тельного движения бластул .при плавании наряду с эмбриологи-
ческими показания-ми (спиральный тип дробления) делает несо-
мненным наличие тех или иных проявлений такой симметрии
в строении всех жгутиконосных бластул.
Признаками анимального ‚полюса бластул служат, помимо уже
отмечавшейся розетки, только его направленность вперед при пла-
вании и его физиологические свойства. Вообще говоря, у бластул
не описано каких-либо отчетливых структурных проявлений
передн-е-задне-го осев-ого градиента, экспериментально устан-овлен-
ного у всех специально изучавшихся с этой точки зрения форм,
и это справедливо даже в отношении молодых бла-стул вольвоцид,
если брать их до на-чала соответствующей дифференциации их
клеток. Однако у Nausithoé, по Мечникову, высота клеток бласто-
дермы и их загруженность окрашенными включениями довольно
правильно возрастают в направлении спереди назад, так что клетки
ее заднего полюса обладают наибольшими размерами и наиболее
насыщены включения-ми. Сходное увеличение размеров клеток по
направлению спереди назад наблюдается -и у ряда других бластул
(рис. 93). д
Отдельного упоминания заслуживают (не- столь уже редкие
у целобластул) случаи противопоставления одного из полушарий
тела другому по какому-нибудь одному признаку, например, их
различной окраске: так, у бластул ряда морских ежей и гидроида
Amphisbetia пигментирована большая часть заднего, вегетативного,
полушария. Эти случаи интересны тем, что они -служат как бы зача-
точныим выражением той амфиполярности, какой характеризуются
"разно-клеточные бластулообразные личинки типа амфибластулы.
Уместно напомнить в этой связи, что у вс-ех‚ -в сущности, вольвоцид
эта же форма гетерополярности, но в другом конкретном выражении
(а именно, в отношении локализации всех гон.идий в одном только
вегетативном полушариибластулы), устанавливается на поздних
стадиях их развития, уже в его постэмбриональном периоде.
2. Гистологические особенности
С гистологической точки зрения подавляющее большинство
жгутиковых цело-бластул должно „быть признано типично эпителии
ЗЛЬНЫМ. ВСЯ стенка тела представлена ЗДЕСЬ ВПОЛНЕ ИЗОМОРФНЫМ,‘
263
как правило, однорядным столбчатым или куби-ческим жгутиковым
эпителием, шестигранные клетки которого находятся в тесном
взаимном соединении. Наличие связующих межклеточных мостиков‘
особенно хорошо заметно у вольвоцид при выделении их клетками
студенистого вещества, когда эти мостики сильно растягиваются
в длину, сохраняясь в виде радиально расходящихся плазмодесм;
однако на более ранних стадиях, обладающих сомкнутыми клет-
камжи, они так же мало заметны, как у бластул Metazoa.
Наружные, апикальные концы всех клеток стенки тела слага-
ются в правильную ровную поверхность или же равно-мерно выда-
ются в виде куполов с большей или меньшей кривизной; напротив,
их базальные концы часто растрепаны, не зависимы друг от друга
и вдаются на разную глубину в полость бластулы, не образуя тут
выровненной поверхности (рис. 93, 1). Как уже упоминалось
выше, размеры клеток бластулы часто обнаруживают известную
степень осевой дифференциации, для более подробной характери-
стики которой нехватает еще данных. В значительном большинстве
случаев ядра всех клеток бластул располагаются у их апикального
полюса, но у вольвоцид они занимают базальное положение, а у
своеобразной питающейся бла-стулы Спгузаога размещены без ви-
димого порядка, на самых различных уровнях (см. рис. 90, I2—I5).
Просторная полость целобластулы заполн-ена жидкой или сту-
денистой ма-с-сой, вырабатываемой ее клетками еще в ходе дробле-
ния или же в момент их дифференцировки (Volvocoidea).
3. Численность клеток
Отдельного рассмотрения заслуживает вопрос о числе клеток,
слагающих тело бластулы. Мы уже указывали в другом месте, что
у Volvocoidea оно строго определенно и достаточно по-стоянно
а
у каждого вида, варьируя только в пределах, устанавливаемь1х-
1-—2 синхронными клеточными делениями. При этом только у Р1ео-т
dorina californica оно равно числу потомков семи таких делений
(128), тогда как у всех настоящихвольвоцид в состав бластулы
входит от 512—1024——-2048 (Janetosphaera) ДО приблизительно‘
50 000 (крупные виды Volvox) жгутиковых клеток. _
У бластул различных Metazoa наблюдаются, примерно, те же
отношения, хотя точных подсчетов числа их‘ клеток производилось
очень мало. Известно только, что у иглокожих окончание форми-
РОВаНИЯ бластулы „ПРОИСХОДИТ ПОСЛЕ ДЭВЯТОГО! ИЛИ ДЭСЯТОГО не ВПОЛНЕ-
синхронного клеточного деления, т. е. при численности клеток
порядка 500-1000 (например, 808 у Paraucentrotus). B отношении
же кишечнополостных и губок данных такого рода почти нет.
Однако некоторое представление о численности клеток этих цело-
бластул может быть все же составлено на основании подсчета по
соответствующим рисункам, применяя для этой цели метод, пред-
ложенный Жанэ (1922). Основываясь на таких, неизбежно очень
приблизительных, расчетах‘, можно установить, например, по рисун-ц
264
3|
кам Мечникова, что у бластул метагенетических гидрометдуз
(Tiara pileata, Clytia viridicans, Octorchis, Mitrocoma annae, Laodi-
сеа cruciata) насчитывается примерно от 500 до 1000 клеток
у Nausithoé свыше 1000, а у Pelagia —-‘ITO-TO около 16000 клеток.
‘Подобный же подсчет клеток целобластулы губки Ascetta (C1athr'1-
na) дал цифру порядка 4000 и т. д. Таким образом, имеются все
основания думать, что диапазон изменчивости числа клеток «жи-
вотных вольвоксов» таков же, как у настоящих Vplvocidae ‘H что,
следовательно, общим признаком всех эпителиальных целобластул
является относительно очень высокая степень‘ многоклеточности,
отвечающая числу потомков девяти и более клеточных генераций.
Это весвм-а существенное обстоятельство приобретает особый инте-
рес, если учесть, что у большинства других типов мерцательных
личинок клеточный состав тела исчисляется много меньшими циф-
рами; достаточно сказать, что жгутиконосные морулы часто состоят
всего из нескольких десятков клеток.
Не менее характерна для целобластул и значительная устойчи-
вость их клеточного состава. Известно, что большинство бластул
Метаъоа н-е испытывает клеточных делений до начала превращения
в паренхимулу, а у Volvocoidea HCXOLLHOIE ЧИСЛО клеток не изме-
няется вообще; в последнем случае мы имеем, стало быть, налицо
самое подлинное постоянство числа клеток— явление, характерное
для некоторых очень высоко дифференцированных организмов
(коловратюи, апэпенди-кулярии) и с которым нам кайк-то странно
встретиться здесь.
Интересное уклонение от этих общих норм клеточного состава
тела целобластул наблюдается у не раз уже упоминавшейся
Chrysaora hys-ocella. У этой метагенетической сцифомедузы одно-
слойный полый зародыш одевается жгутами и становится подвиж-
ным на стадии около 120 бластомеров, т. е. после седьмого деления.
Однако„ став -жгутиконосной личинкой, бластула Chrysaora еще
долго остается жить в материнском теле, где она питается осмоти-
ческим путем и интенсивно растет, непрерывно увеличивая при этом
число своих клеток, доходящее только к концу ее развития до нор-
мального для других целобластул.
4. Цитологические данные
Строение жгутиковых клеток -бластул известно во всех деталях
только у вольвоцид и морского -ежа Paracentrotus lividus (Овасс,
1934, 1935); у других целобластул оно, к сожалению, остается еще
не изученным. Этот крайне досадный пробел в наших знаниях по
такому важному вопросу в какой-то мере восполняется, однако,
имеющимися цитологическими сведениями о жгутиковых клетках
амфибластул известковых губок Зусоп и Grantia (Дюбоск и Тюзэ,
1935, 1937). Таким о-бразом, при всей фрагментарности существую-
щих данных на этот счет, их все же оказывается достаточно, чтобы
получить определенное представление об организации жгутиковых
265
клеток бластул И уяснить ее необычайно интересные особенности.
А. Вольвоццдьь. Уже давно известно, что жгутиковые клетки
многоклеточных фитомонадин обладают той же организацией, что
и их одиночные, одноклеточные сородичи и, более конкретно, хла-
мидомонады. Это в полной мере подтверждается цитологическими
исследованиями Овасса (1936, 1937, 1938), охватившими довольно
широкий круг одиночных и колониальных Phyfomonadina, B '1‘-O-M
числе Eudorina illinoisensis, виды Volvox H T. д. Жгутиковые клетки
этих форм характеризуются, таким образом, своим устойчивым од-
ноосно-гетерополярным строением, связанным с двулучевдой и
частичной двусторонней симметрией; наличием двойной (двужгути-
ковой) кинетидыс раздельными базальными зернами и ризопла-
стами; обширным, очень сложно устроенным чашевидным хромато-
фором с различным числом пиреноидов; присутствием высоко диф-
ференецир-ованного глазка; аппаратом Гольдж-и, состоящим из не:
скольких чешуевидных или чполулунных диктиосомч, и тем же соста-
вом цитоплазматических включений, что у других окрашенных фито-
монад.
Такая ТИПКИЧНО ХЛЭМИДОМОНЗДНЭЯ ОрГЗНИЗЗЦИЯ характерна ДЛЯ
жгутиковых клеток Volvocoidea Ha всех стадиях их развития, на-
чиная с момента их начальной дифференцировки в виде однородных
жгутиковых зооспор, чИ удерживается ими пожизненно, как и у дру-
гих Phytomonadina. Однако молодые, еще нейтральные (т. -е. еще
не Дифференцированные на ьплодущие и бесплодные клетки»), жгу-
тиковые ‘клетки обладают и некоторыми специфическими для них
особенностями, отражающими ‚их природу зооопор. K числу таких
признаков относятся, во-пе-рвых, их часто бол-ее удлиненная и
стройная общая форма при малых абсолютных размерах; во-вто-
рых, наличие у них только одного пиреноида (их число позднее уве-
личивается) и, наконец, меньшее, чем у взрослых клеток, число
диктиосом. 5 к k _. .
_ Б. Бластулы Metazoa. Одноосно-г-етерополярное строение жгу-
тиковых клеток, наличие у них одного жгута и отсутствие аллоплаз-
матических оболочек (т. е. межклеточного вещества) составляют
самую общую характеристику этих клеток, и к этому же сводится °
все, что известно об их строении у Целобластул книдарлий и гу-бок.
Тем большую ценность представляют упомянутые выше цитологи-
че-ские данные о Paracentrotus H амфибластулах I-Ieterocoela, даю-
щие новое освещение этому, очень существенному, вопросу.
Жгутиковые клетки Para-centrotus (рис. 94, 3--6) интересны
прежде всего тем, что их жгутиковый аппарат представлен сложной,
высоко дифференцированной кинетидой, состоящей из жгута с ба-
зальным зерном, тонкого двойного ризопласта и парабазального ап-
парата, имеющего вполне типичное строение. Последний представлен
здесь парой удлиненных булавовидных образований, состоящих
каждое из хромофильно-го наружного слоя и слабо окрашивающе-
гося внутреннего; они сходятся кпереди под углом и соединены с
базальным зерном жгута посредством более или именее длинного
266
тяжа. У жгутиковых клеток Sycon H Grantia (рис. 94, 1——2) кинетида
построена по тому же плану, как у Paracentrotus, ‘И так же включает
типичнейший парабазальный аппарат, имеющий, впрочем, несколько
иное -строение. Он представлен чешуевиднымл образованием, соеди-
няющимся непосредственно с базальным зерном жгута; его осмио-
фильный корковый слой обращен кнаружи и одевает, таким образом,
только с одной стороны прикрываемое им хромофобное вещество.
От базального зерна к ядру тянется тонкий ризопласт, образующий
ось расположенного здесь киноплазматичеокого конуса. Самое ядро
расположено в апикальной части клетки и характеризуется сетчатым
‘распределением хроматина. В цитоплазме рассеяны палочковидные
а .
2 3
дРис. 94. Жгутиковые клетки (киноциты) бластулообразных личинок:
7,, 2 — губки Grantia compressa pennigera (no Дюбоску и Тюзэ); 3—б—мор-
ского ежа Paracentrotus lividus; 7-—деление парабазального - аппарата
и 8 — превращение диктиосом в парабазальъяьтй аппарат при дифференцировке.
бластулы Paracentrotus (no Овассу)
хондриосомь1 и многочисленные мелкие вакуоли, окрашиваемые ней-
тральным красным. Характерны сильно удлиненная форма клеточ-
ного_тела и отсутствие каких бы то ни было дифференцировок, свя-
занных с‘ анимальными способами питания; в частности, у жгутико-
вых клеток бластул нет и в помине плазматического воротничка,
столь характерного для трофических клеток губок-хоаноцитов,
с которыми их иногда ошибочно отождествляют. - ` ‘-
Итак, строеньие жгутиковых клеток иглокожих и губок‘ разли-
чается лишь в деталях; им свойствен одинаковый, в сущности, план
жгутиконосного строения——план, по всей вероятности общий для
‘жгутиковых клеток бластул M-etazoa. ' . _
Этот характерный для них план строения чрезвычайно интересен
тем, что он полностью отвечает общему плану строения одиночных
одноклеточных Zooflagellata H даже более конкретно — организации
267
одножгутиковых, осмотрофных :и свободноплавающих представите-
лей Protornonadina. Насколько велики и специфичны наблюдаю-
щиеся здесь сходства, явствует, прежде всего, из того, -что вполне
оформленным пара-базальным аппаратом обладают, среди простей-
ших, одни только Zooflagellata И что это образование считалось
присущим только им, пока оно не было найдено в жгутиковых клет-
ках бластул и в хоаноцитах низших губок (см. гл. XVIII). Таким
образом, сразу же устанавливается морфологическая сравнимость
жгутиковых клеток бластул не с жгутиковыми «вообще», а с одними
только Zooflagellata. Уточняя дальше это сравн-ение, мы, естест-вен-
но, должны отбросить полимерные и многожгутиковые формы, обра-
зующие отряды Polymastigina И Hypermastigina, что приводит
к дальнейшей конкретизации нашего сопоставления, а именно к его
ограничению рамками одних только Protomo-nadina. И если продол-
жить дальше сравнение жгутиковых клеток бластул с различными
протомонадами, то состав их одножгутиковой кинетиды, частные
особенности строения парабазального аппарата у Sycon И Grantia,
вытянутая форма клеточного тела |И отсутствие всяких дифферен-
цировок, допускающих анимальное питание, с такой же неизбеж-
ностью подводят нас к осмотрофным, свободноплаваюшижм пред-ста-
вителям Protomonadina, с какой подобное же сравнение хоаноцитов
приводит к их сближению с сидя-чими воротничк-овь1ми протомона-
дами (Craspedomonadina),
Особого внимания заслуживает при этом тот факт, что строение
парабазального а-ппарата жгутиковых клеток -бластул не только
удовлетворяет общей схеме его конструкции у Zooflagellata, НО и
отвечает во всех деталях некоторым из его конкретных типов, встре-
чающихся у этих жгутиковых. Так, у Sycon и Grantia OH построен
по типу Bodo, а у Paracentrotus обнаруживает и-сключительное
сходство с двойным парабазальным аппаратом некоторых Giardia
(см. рис. 49). Правда, тот и другой тип е-го строения связан у назван--
нь1х -Zooflagellata синой комбинацией прочих морфологических
признаков, что в известной степени ограничивает значимость этого
далеко идущего сходства; но все же остается фактом, что известные
нам типы парабазального аппарата жгутиковых клеток бла-стул
находят свое полное подобие у некоторых определенных зоофла-
геллят. _ ' I
Конечно, было бы ошибочным делать отсюда вывод, что клетки
жгутиконосных «бластул Ме’са2оа могут быть непосредственно ото-
ждествлены именно с этими представителями простейших, и сейчас
же делать отсюда соответствующие филогенетические заключения.
Одна-ко не менее ошибочным было бы и игнорирование этих бес-
спорных, неопровержимых -сходств, сводящихся, в конечном счете,
"К ТОМУ, ЧТО жгутиковые КЛЕТКИ «ЖИВОТНЫХ ВОЛЬВОКСОВ» ПОЧТИ В та-
кой же мере сходны по своему строению с некоторыми из осмотроф-
ных протомонад, как хоаноциты губок — с сидячими воротничко-
ВЫМИ пред-ставителя-ми ЭТОЙ же ГРУППЫ ЖГУЪИКОВЫХ, а КЛЕТКИ ВЫСШИХ
Volvocoidea - c ОДНОКЛВТОЧНЫМИ фитомонадами. *
268
5. Биологические особенности
С биологической точки зрения все жгутиковые целобластулы ха-
рактеризуются тем, что они ведут свободный, пелагический о-браз
жизни, активно плавая в воде при помощи своего жгутиков-ого man-
гательного аппарата; иначе говоря, что они являются типичными
«мерцательными» формами. При поступательном движении, идущем
по винтовой, спирали вследствие вращения бла-стул вокруг своеи
главной оси, передний полюс неизменно обращен вперед, ЧТО Не
мешает, впрочем, некоторым из них обладать способностью активно
двигаться, при случае, и в обратном направлении (соответственно
меняя при этом и предпочтительное направление вращения своего
тела „на обратное). ' О,
Таким образом, -все организмы этого типа имеют одиинаковыи
характер «бродяжек», сходную активность и совпадающий способ
движения. и
В противоположность этому, их способность к вегетативной nexu-
тельности оказывается резко различной у Vo-Ivocoidea, с одной
стороны, и -бластул Metazoa—c другой. В то время как первые
ведут себя в этом отношении как полноценные вегетативные формы,
интенсивно питаются и растут-бластулы многоклеточных жи-
вотных совершенно не выполняют вегетативных функции: они
фактически неспособны зк самостоятельному питанию, не растут И,
существуя только за счет расходования внутриклеточных резервных
включений, имеют исключительно расселительное значение.
Это общеизвестное, но далеко не всегда учитываемое физиоло-
гическое различние между ними естественн-0 связано с чисто расти-
тельной, автотрофной природой \/о1\госо1‹1еа и отсутствием у жи-
вотных бла-стул как фотосинтетического аппарата, так и всяких при-
способлений к анимальному способу питания. Являясь, по самому
‘СВОЕМУ СУЩОСТВУ, гетеротрофными ОРГЗНИЗМЗМИ, ЭТИ бЛаСТУЛЫ не
располагают вместе с тем средствами ни к поклеточному улавлива-
нию оформленных пищевых частиц, ни, тем более, к высшим ф%рмам
животного питания, связанным -с полостным пищеварением. аким
образом им фактически доступно одно только осмотическое -пита-
7
ние. Однако, поскольку это последнее вряд ли может иметь ощутимое
значение у свободно-живущих морских организмов (хотя оно
тут ‘полностью и не исключено), не приходится и› УДИВЛЯТЬСЯ Т0МУ‚
что бластулы не питаются вовсе, суЩеСТВУЯ ТОЛЬКО 33 ‘CWT Расход‘)
ванн-я внутриклеточных запасных включений (желтка). '
Способность к осмотическому питанию хорошо доказывается
тем фактом, что бластулы Chrysaora, развивающиеся в гонадах
этой сцифомедузы до достижения ими стадии «планулы (и находя-
щиеся) таким ‚образом, .3 особо бЛа-ГОПрИЯТНЫХ УСЛОВИЯХ ПИТаНИЯ,
обеспечиваемых материнским организмом), успешно используют
эту возмпожность осмотического питания и растут ЧреЗ-ВЫЧаРГНО
интенсивно, увели-чивая свою живую массу в тысяч)’ И бО-Пее Раз-
269
6. Общее сравнение вольвоцид с целобластулами Mefazoa
Сопоставляя только-что изложенные данные о вольвоцидах и
жгутиковых бластулах Metazoa, МОЖНО воочию убедиться в совер-
шенно исключительном масштабе и глубине взаимного сходства
«животных вольвоксов» с настоящими. Это сходство особенно ве-
лико, если брать тех идругих на взаимно гомологичной стадии раз-
вития, на стадии только-что закончившейся однородной «начальной»
ДИфФЭРЭНЦИРОВКИ KJIETOK, до п-ерехода к свободному образу жизни.
На этои стадии вольвоциды являются столь же типичными эпители-.
альными целобластулами, как таковые Metazoa. Они сходны с ними
как по своей архитектонике, так и в гистологическом отношении и
построены из сходного числа однородных клеток типично жгутико-
носного строения. И-нтегрированность вольвоцид не уступает на
этои стадии таковой жгутиковых бластул Metazoa, a в дальнейшем
развитии приобретает еще более яркое, чем у них, градиентное вы-
ражение. Не менее вели-ко и сходство в эмбриогенезе тех и других:
.в чисто цитотипиче-ском характере дробления, состоящего из одина-
„кового числа делений, в морфологических картинах дробления, в
характере завершающей его «начальной» дифференцировки, в про-
хождении стадии стомобластулы и в ее одинаковом вывора-
чивании.
это сходство так вели-ко, что можно было бы, в сущности, отнести
вольвоцид к подлинным Metazoa, если бы только этому не препят-
ствовала их типично растительная, автотрофная прпирода.
Морфологическим ее выражением на упомянутых ранних стадиях
развития вольвоцид является, по сути дела, только хламидомонад-
ный тип строения их клеток в отличие от ‚протомонадного пили иного
зоофлагеллятного типа организации клеток животных жгутиковых
бластул. Однако в дальнейшем развитии тех и других это основное
различие между НИМпИ приобретает капитальное значение, направляя
их онтогенез по диаметрально противоположным руслам. У воль-
воцид бластулы с самого начала способны к вегетативной деятель
ности и сразу же начинают интенсивно осуществлять эту деятелья
ность, не перестраивая для этого свою организацию. Их развитие
остается такимже «прямым», как у всех вообще фитомонадин, и
этим можно объяснить п-о-жизненное сохранение вольвоцидами их
исходной 0-рганизации бластулы.
Форма бластулы является у них полноценной формой вегетатив-
ного существования аи размножения. В этой форзме осуществляется
вся жизнедеятельность организма, его рост, развитие и гистологиче-
ская ‚дифференцировка, достижение его бесплодными клетками «не-
изменного» уровня последней и осуществление плодущими клеткам-и
оо- и сперматогенетических процессов. Выполнение всей суммы
жизненных задач бродячей вегетативной бластулой, очевидно, и
приводит к прохождению ею столь же полного жизненного Цикла,
как у высших Metazoa, HO B рамках одной |И той же жизненной '
.формь1. В этом отношении вольвоциды аналогичны не низшим мно-
270
ГОКЛЕТОЧНЫМ, а высоко специализированным постоянноклеточным
формам. И
В противоположность этому жгутиковые бластулы Metazoa
фактически не способны к выполнению вегетативных функций. Они
не питаются пи не растут, и всеих клетки неидут, как правило,
далее лабильного уровня дифференцировки. И только после пре-
вращения такой личинки в сидячуюформу (сопутству-смого пере-
дифференцировкой всего или части ее клеточного материала) ста-
новится возможной вегетативная деятельность организма, его даль-
нейшее развитие, рост и размножение.
Иначе говоря, все дело в том, что бластулы вольвоцид стано-
вятся вегетативно полноценными организмами, не изменяя своей
исходной организации, а бластулы Metazoa к этому не способны;
поэтому они и имеют биологическое значение лишь кратковремен-
ной, узко расселительной начальной стадии постэмбрионального
развития, сменяемой в дальнейшем его течении совершенно иначе
организованными и всегда сидячими вегетативными стадиями.
Этим, конечно, не обесценивается огромное сходство между выс-
шими .вольвоцидами и жгутиковыми бластулами Metazoa. Можно
утверждать, что высшие вольвоциды являются столь же подлинными
жгутиковыми целобластулами, как таковые Metazoa, и в не мень-
шей мере целостными, чем они, многоклеточными организмами,
организмами, сходно интегрированными, совершенно ПОДОбНЫМъИ им
по своей архитектонике и эпителиальному строению, но построен-
ными из ка-чественно иного материала: из клеток типичного фито-
монадного, а не зоофлагеллятного характера. -
K этому, в конечном счете, |И сводятся все главные различия
между ними.
7". Бластулы как синзооспоры 1
В предыдущей главе было показано, что наиболее примитивный
из типов многоклеточной организации—тип бластулы—создается
формами дробления, сохранившими полнее других первобытные, как
мы считаем, свойства ПЕЪШИНТОМИЧЕСКОГО размножения. Если это дей-
ствительно так‚ то естественно ожидать, что бластулы ‚проявляют
с наибольшей полнотой и морфофизиологические и биологические
признаки сложных зооспор (синзоос-пор) и что, в частности, их жгу-
тиковые клетки удовлетворяют наилучшим образом общим призна-
кам отдельных жгутиковых зооспор.
Оценивая под этим углом зрения соответствующий‚фактический
материал, нельзя не придти к выводу, что это ожидание оправды-
вается даже в большей мере, чем можно было бы пред-полагать: все
специфически-е признаки бластул, отличающие пИХ от всех других,
‚более СЛОЖНО ОРГЗНИЗОВЗННЫХ ТИжПОВ «мерцательных» ЛИЧИНОК Ме-
tazoa, сводятся как раз к самым типичным и общим признакам
синзооспор и находят в этом свое объяснение.
Как уже отмечалось в другом месте (гл. V, 3), оинзооепорами
следует считать любые постоянные и.ли временные объединения
271
многих или хотя бы нескольких зооспор, возникшие в результате‘
ИХ НЕРЗСХОЖДСНИЯ И СОХРЗНЯЮЩИВ ПрИ ЭТОМ СВОИ ‘ПРЯМЫЕ рассели-
тельные функции. Известно, что понятие <<синзооспора>› было выра- V
ботано еще в прошлом веке альгологами и применялось пока-что
только в этой области ботаники. Классическим примером синзооспор
водорослей служат удивительные симпластические «мерцательные
личинки» вошерий, но ими считаются также молодые стадии Volvo-
coidea и некоторых других зеленых и разножгутиковых «водорос-
лей» (см. Миллер, 1924, 1935), независимо от того, образованы ли
они всей семьей зооспор, получившихся от множественного деления
материнской особи, или же только ее многочленными фрагментами.
Таким образом, это понятие не имеет (или, по край-ней мере, не
должно иметь) другого значения, чем заключающееся в его бук-
вальном, этимологическом смысле, и потому может применяться
к любому агрегату, состоящему из зооспор. А отсюда, очевидно,
следует, что для признания жгутиконосных бластул сложными зоо-
спорами достаточно только показать, что составляющие их клетки
равноценны по своему строению и биологическому значению от-
дельным жгутиковым спорам и что организация самих бла-стул не
препятствует такому их пониманию.
Напомним, что самыми общими признаками жгутиковых зоо-
спор являются следующие:
1) их образование путем множественного, а не простого деле-
ния, в чем и заключается их единственное принципиальное отличие
от продуктов простой монотомии; . -
2) их минимальные, для данного вида, размеры, обусловленные
самым фачктом, что они создаются посредством делений, не компен-
сируемых общим ростом в процессе своего осуществления;
3) заключительный характер их дифференцировки, т. е. их пере-
ход из недифференцированного в дифференцированное состояние
уже по окончании спорообразования, неизменно базирующегося на
метазойных типах деления;
4) значительный объем м-орфогенезов, осуществляемых при диф-
ференцировке, главным из которых является выработка кинетиды
из материала центросомы или другого ее зачатка;
5) строго одноклеточная, одноядерная организация;
6) принадлежность готовых жгутиковых спор к жизненной
форме свободноплавающего жгутиконосца, характеризующейся,
между прочим, отсутствием всяких плазматических и иных диффе-
ренцировок, связанных с сидячим, прикрепленным образом жизни;
7) известная упрощенность организации по сравнению с позд-
нейшими вегетативными стадиями развития (в тех, разумеется, слу-
чаях, когда эти поздние стадии не подвергаются вторичному упро-
щению в результате, например, паразитического образа жизни),
объясняемая, с одной стороны, тем, что каждой зооспоре достается
только часть материнских постоянных органоидов (например, по
одному пиреноиду или небольшому числу диктиосом), а с другой ——-
тем, что многие дифференцировки создаются только позднее, уже
272
после перехода зооспоры -к активной жизнедеятельности (оболочки
и некоторые из параплазматических включений);
8) специально отм-ечавшаяся неспособность жгутиковых зооспор
к иным способ-ам питаниядкроме автотрофного и осмотического,
находящая свое структурное выражение в отсутствии у них клеточ-
ного рта и других особых дифференцировок, связанных с анимадь-
ным спос-обомпитания и, наконец,
9) их явно расселительный характер «бродяжек», получающий
особенно яркое выражение у форм с сидячими вегетативными ста-
диями,‘ зооспоры которых практически не питаются и не растут до
момента прикрепления к субстрату.
Сопоставляя эти основные признаки жгутиковых спор простей-
ших, водорослей и низших грибов со всем тем, что мы знаем об об-
разовании и особенностях организации жгутиковых клеток бластул
Metazoa, мы оказываемся вынужденными признать, что буквально
все эти признаки в такой же мере характерны и для последних, и
тем самым дают им недостававшее сравнительно морфологическое
объяснение. .
Это явствует само собой в отношении первых трех из перечис-
ленных признаков, поскольку сущность дробления заключается
именно в его палинтомическом характере, а достижение к концу
ДрООЛЕНИЯ МИНИМЗЛЬНЫХ КЛЭТОЧНЫХ pa3MepoB также ВХОДИТ В ЧИСЛО-
er-0 самых характерных и общих признаков и тем самым в общую
Характеристику всякой вообще бластулы, как конечного продукта
дробления. То же можно сказать о специально подчеркивавшейся
выдержанности клеточного строения бластул. Что же касается про-
цесса клеточной дифференциро-вки, то ее‘ всеобщий характер, одно-
временность, присущее ей: значение- акта, завершающего собой
дробление, и наконец, е-е однородность (в смысле однозначности ее
результатов) столь же характерны для бластул (точнее, для цело-
бластул и клеток жгутикового полушария аимфибластул), как и для
любого другого комплекса спороцитов, дифференцирую-щихся в зоо-
споры; особенно следует подчеркнуть‚ что жгутиковые клетки боль-
шинства целобластул совершенно идентичны друг другу в момент
своего оформления, и только позднее приобретают те или иные ре-
гиональные различия. По своему конкретному содержанию про-
цессы дифференцировки клеток бластулы также ничем не отлича-
ются, как мы видели, от процессов перехода спороцитов в диффе-
ренцированное состояние зооспор: там и здесь-они заключаются
в выработке кинетиды, нередко сопутствуемой преобразованием
диффузного аппарата Гольджи в зпарабазальноетел-о (у акнидоопо-
ридий), и установлении или усилении од-ноосно-гетерополярного
строения клетки. Далее, у жгутиковых клеток бластул обнаружи-
ваются и все те отличия от вегетативных клеток дальнейших стадий’
РЗЗВИТИЯ, ‘какие МЫ ОТМЭЧЗЛИ у ОДИНОЧНЫХ 3OOCl'lOp, ЧТО ДЕМОНСТРИ- _
рует-ся С ИСКЛЮЧИТЕЛЬНОЙ НЗГЛЯДНОСТЬЮ на KOHKp€THOM ПРИМЕРЕ
l"yI6OK ПрИ сравнении ЖГУТИКОВЫХ КЛЕТОК ИХ бластул С ХОЗНОЦИТЕЪМИ
ВЕГЭТЗТИВНЫХ стадии. ЭТО сравнение ПОКЗЗЫВЗЭТ, ЧТО ВСЕ характер.-
18 Эмбриология беспозвоночных ` 273
нейшие ПРИЗНЗКИ хоаноцитов -- наличие ПЛаЗМЗТИЧЕСКОГО ВОРОТНИЧ‘
ка, радиальный план строения и аппарат Гольджи в виде диктио-
сомы (см. рис. 115) —-—-совершенно чужды клеткам бластулы, у кото-
рых нет ни во-ротничка, ни других дифференцировок, связанных с
анимальными с-пособами питания, аппарат Гольджи представлен д
ТИПИЧНЕЙШШМ -Hap'a6a3aJIbHbIM аппаратом, а СИЛЬНО ‘BBITH-Hy'I‘OIe’ В
длину клеточное тело обладает отчетливо билатериальн-ой симМет-_
рией (намечаемой п-оложением парабазальнопо тела).
Таким образом, если воротничковые клетки губок обладают
ВСЕМИ ОСООЕННОЮТЯМИ СИДЯЧИХ, ЗНИМЭЛЬНО ПИТЗЮЩИХСЯ Protomona-'
dina (во-ротничковых жпутиковых—Сгазреоогпопаойаае), то жгу-
тиковые клетки бластул не в меньшей мере отвечают свободнопла-.
вающим, осмотрофным представителям этой группы Elagellata, как
легко в этом убедиться из их непосредственного сравнения
(см. рис. 117, 120). И поскольку эти особенности характерны также,
насколько мпы з-наем, и для жгутиковых клеток других бластул
(в том числе и морских ежей), то можем даже сказать, что они
все обладают не только жгутиковым типом одноклеточнои органи-
зации, но и еще более конкретно—типом строения свободнопла-
вающей осмотрофной протомонадины. '
Наконец, сугубо и специально рас-селительная биологическая
функция клеток жгутиковых бластул проявляется с предеЛЫЮИ ЯС‘
ностью в том факте, что жгутиковые бластулы являются по самому
своему существу расселительными стадиями развития, ти.пичнеи-
шими «бродяжкамит» и фактически не способны к чинынм видам пи-
тания, кроме автотрофного (Volvocoidea) H.mH осмотического (Chry-
saora), a чаще всего вообще не питаются.
Итак, вопрос о природе ‘бластулообразных личинок Metazoa H
‚многоклеточных Volvoucoidea решается наилучшим образом их при-
знанием сложными зооспорами, или синзооспорамли. С этой, и только
с этой то-чки зрения становятся легко объяснимыми все непонятые
иначе особенности их биологии и ОрГаНИЗаЦИпИ (и в частности, все
свойственные им цитологические особенности), а также и ттсключи-
тельное сходство жгутиковых целобластул Metazoa с вольвоЦиДамгИ‚
о котором уже столько говорилось. Оно оказывается Целиком сво-
димым к тому, что те и другие построены из жгутиковых спор, обра-
зующихся одинаковым способом (путем табличнои палинтомии),
что они в-едут сходный образ жизни в одинаковых условиях средЫ
и одинаковым образом интегрированы. Градиентнь1и‚ а не коло-
ниальный характер их интеграции свидетельствует, видимо, о том,
что достижение теМпИ и другими самой подлиннои многоклеточности
(и притом в идентичных формах——эпителиальнои Целобластулы)
есть проявление вполне определенных и специфичных законов инте-
ТРЗЦИИ ЗООСПОР, ОДИНЗКОВО ДЕЙСТВУЮЩИХ В самых различных группах
организмов при относительно сходных условиях их существования-
С этой точки зрения жгутиковые бластулы Metazoa H молодые ко-
дюнии Volvocoidae— вполне гомологичные ДРУГ ДРУГУ ОбРЗЗОВаНИЯ
и стадии, а именно синзооспоры, или многоклеточные «тела Pa3MH9'
274
жения», различающиеся главным образом своей различной система-
тической принадлежностью.
С этой же точки зрения становятся легко объяснимыми и сход-
ства этих многоклеточных форм с симплатическими синзооспорами
вошерий (Захватклин, 1946).
Глава XV
ЗАКОНОМЕРНОСТИ И УСЛОВИЯ ОБЪЕДИНЕНИЯ
ПРОДУКТОВ ПАЛИНТОМИИ В МНОГОКЛЕТОЧНЫЕ
СИНЗООСПОРЫ -
1. Постановка вопроса
Итак, морфологическая и биологическая примитивность жгутико-
носных бластул находит себе исчерпывающее объяснение в том, что
они представляют собой синзооспоры, образующиеся путем объеди-
нения целой семьи жгутиковых зооспор в одно многоклеточное целое.
Таким образом, замещение палинтомического размножения воспро-
-изведением конкретно выражается именно в том, что его многочис-
ленные продукты вступают в прочную связь друг с другом, вместо
того чтобы разбегаться, и дают сообща начало одному сложному
организму, не теряя, однако, от этого своего прямого раос-елитель-
ного значения. Такое соедин-ение зооспор в целостную синзооспору
представляет собой, вместе с тем, и процесс построения многокле-
точного бластулообразного -организма, имеющего у Metazoa зна-
чение чисто расселительной бластулообразной личнинки, а у Volvo-
со1‹1еа——значение колониального объединения высшего ‘типа, по-
жизненно сохраняющего свою организацию целобластулы. Стало
быть, процесс формирования синзооспоры типа бластулы есть в то’
же время простейшая форма эмбрионального развития Metazoa,
высшая форма ‚построения колонии у Protozoa H общее им средство
достижения многоклеточности в ее наиболее примитивном, одно-
слоином выражении. -
Простое признание этого факта открывает весьма перспективные
пути подхода к проблеме происхождения многоклеточности, позволяя
поставить ее в совершенно конкретной форме и использовать для ее
решения обширный и разнообразный фактический иматериал. В са-
›мом деле, при такой ее конкретизации эта проблема сводится,
в интересующем нас аспекте, к одному, но решающей важности,
вопросу: вопросу об условиях и причинах, ведущих к объединению
зооспор в целостную многоклеточную синзооспору. Еще конкретнее
этот вопрос может быть сформулирован так: почему в одних слу-
чаях палинтомипи ее продукты разъединяются в виде отдельных жгу-
тиковых зооспор, дающих столько же одноклеточных или надкле-
точных организмов, в то время как в других они остаются
соединенными в виде одной многоклеточной жгутиконосной бла-
стулы.
13* 275
2. Смена уровней интеграции в процессе формирования
синзооспоры
Чтобы ответить на этот вопрос, необходимо, прежде всего, при-
смотреться к особенностям самого процесса интеграции, к ходу этого . Е,
процес-са во всех тех случаях, когда он развертывается на наших
глазах.и находит свое осязаемое выраж-ение в сменах уровня целост-
ности развивающегося организма.
Такая их смена наблюдается в самой наглядной форме в эмбрио-
генезе тех низших Metazoa, дробление которых отличается мини-
мальной степенью детерминированности, т. е. у метагенетических
пидромедуз и морских ежей. Дей-ствительно, в обеих этих группах
очень отчетливо намечаются четыре последовательные ступени инте-
грации ‚дробящегося организма, закономерно сменяющие друг
друг друга по мере формирования целобластулы. -
У гидромедуз первая, начальная ступень интеграции дробяще-
гося яйца приуроченачк периоду его первых четырех делений,
т. е. характеризует собой его ранние стадии до 16-клеточной вклю-
чительно. На этих стадиях все бластомеры вполне равноценны друг
другу .по возможностям своего развития, т. е. обладают свойством
тотипотентности, способны нормально развиваться, будучи изолиро-
ванными друг от друга, и ведут себя при дроблении, как относи-
тельно независимые одноклеточные особи со своей собственной
плоскостью деления и т. д. Этот, явно еще зачаточный, л а-б иль-
H ы й у р о в е н ь и н те г р а u И и бластомеров сменяется при
пятом делении, т. е. со стадии 32, значительно более высоким ее
уровнем, -с достижением которого интеграция уже становится
необратимой. Отдельные бластомеры теряют свою способность
к образованию, при изоляции, целого организма, а вместе с нею
и имевшиеся протозойные черты в морфолопии деления; дробящийся
организм превращается в устойчиво многоклеточное образование,
уже не поддающееся диссоциации.
Однако на этой второй ступени интеграции, охватывающей у бла-
стулоо-бразующих Ьертопоа две следующие стадии дробления
(32-64), комплекс бластомеров еще не проявляет полного морфо-
физиологического единства: эти стадии характеризуются, как мы
видели, очень невысокой структурной упорядоченностью и отсут-
ствием оформленного градиента физиологической активности.
Поэтому здесь было бы уместно говорить о хотя устойчивой, но еще
далеко неполной интеграции. `
‘Переход к следующему.уровню интеграции бластулы выражается
в ПОЯВЛЕНИИ у нее передне-заднего физиологического градиента, что
осуществляется, насколько мы знаем, при VI:I—VIII синхронном де-
лении яйца. С этого момента бластула начинает обнаруживать все
признаки оформленной организменной целостности, в том числе и
проморфологические; она вскоре приобретает окончательный чис-
ленный состав (512-——1024 И более клеток), строго эпителиальное
строение и вслед затем дифференцируется в личинку. ‘
276
Те же ступени нарастающей интеграции наблюдаются и
в эмбриональном развитии ‚морских ежей‘, Общеизвестно, что тоти-
_потентность бластомеров проявляется и здесь до 16-клеточной ста-
дии включительно, но, в отличие от гидромедуз, не всеми бласто-
мерами этой стадии, а одними только макромерами. Далее следует
вспомнитьопыты комбинирования п трансплантации бластомеров
32- и 64-.K.71eT0I{H0131~cTa;LH1/1, показывающей со всей очевидностью
неполноту их интегращии в это время при ее все же более высоком
уровне, чем на стадии 16. Напомним далее, что до стадии 128 бла-
стомеры округляются после каждого деления ги только соприка-
саются друг с другом и лишь после восьмого деления плотнее при-
ЖИМаЮТСЯ друг к Другу, начиная складываться в эпителий; это —~
стадия преобразования пребластулы в бластомерную бластулу, за
‘КОТОРОЙ УЖЕ ‘СЛЕДУЕТ, ПрИ ДЕВЯТОМ-ДЕСЯТОМ ДЕЛЕНИИ, ЕЕ ОКОНЧЗТЕЛЬ-
ное оформление в виде эпителиальной бластулы. Следовательно,
и здесь процесс интеграции проходит через те же ступени, как у м-е-
_тагенетических ГИДрОМЕДуЗ‚ КЗЖДОЙ ИЗ КОТОРЫХ СООТВЕТСТВУЕТ СТрОГО
определенная численность -бластомеров.
Это позволяет предположить существование особых, строго
определенных закономерностей на р а-стающей инте-
г р а ц и и целобластул, действующих сходным образом на самом
‚РЗЗЛИЧНОМ ‘MaTep|I/13.116 И ПРИВОДЯЩИХ -ВО ВСЕХ случаях K СОВЕрШЕННО
СХОДНЫМ pe3yJ1bTa'raM_ ЭТИ ЗЗКОНОМЕРНОСТИ ИМЕЮТ, ВЕРОЯТНО, ВЕСЬМЗ
общее значение, так как низвестно, что тот же, в сущности, ряд по-
следовательных ступеней интеграции проходится, хотя и в более
‚ускоренном темпе, всеми животными, обладающими регуляцион-
‚ными яицами и оформленной стадлией бластулы. Однако такое раз-
вернутое их выражение составляет все же привилегию одних только
книдарии и иглокожих, т. е. именно тех Metazoa, бластулы которых
обнаруживают в наиболее чистом виде спешифические особенности
‚СИНЗОО-СПОРЗ- ПОЛНУЮ ОДНОРОДНОСТЬ СВОИХ ЖГУТИКОВЫХ КЛЕТОК "В М0°
мент их начальной дифференцировки при очень большом их числе
.и полном сохранении ими организации отдельных жгутиковых спор.
Если учесть, что неустойчивость интеграции ранних стадий дробле-
нияуэтих форм неможет быть расценена иначе, чем сохранение’ ими
признаков спорогониического. размножения;-—мы получим серьез-
зные основания .полагать, что описанный ход интеграционного upo-
Цеоса отражает, пло крайней мере в» приблизительной форме, основ-
.ные' этапы преобразования спорогонии в воспроизведение посред-
;ствоъм синзооспор. _ ’ *
3. ‘Различные Гуровни целостности‘ синзооспор у УоН/осойоеа
Эта бросающаяся "в глаза связь между уровнем интеграции дро-
-бЯЩЕЙСЯ СИСТЕМЫ И ЕЕ ЧМСЛЕННЫМ составом ПОКЗЖЕТСЯ ЕЩЕ ООЛЕЕ
примечательной, если вспомнить, что такая связь проявляется еще
более демонстративным образом в удивительном ряду колониаль-
ных Volvocoldea, . V -
277
В этом ряду, как мы видели (гл. VII, 3), существует несколько
различных ступеней организменной целостности колонии, распола-
гающихся в восходящем порядке. Каждая из этих ступеней харак-
теризуется здесь различной степенью выраженности одних и тех же
признаков морфофизиологической интеграции, а именно: а) в осе- "
вой, градиентной дифференциации клеток колонии, и б) в их диф-
ференциации на плодущие и бесплодные.
Первые признаки такой интеграции начинают проявляться здесь
в ощутимой, хотя и зачаточной, еще форме у 16-членных колоний
Eudorinidae (Pandorina, Volvulina И др.) и некоторых Spondy1omo-
ridae (Chlamydobotrys gracilis), где они выражаются только в убы-
вании размеров глазков в передне-заднем направлении; у этих форм
сохраняется способность к распадению колоний и нет еше и намека
на биологическое размежевание составляющих ее клеток. Следую-
щую ступень объединения представляют 32-членные колонии обеих
Eudorina, характеризующиеся начавшейся дифференциацией своих
клеток на плодущие и бесплодные при уже устойчивой, необрати-
мой, целостности колонии. На следующей, 64—128-клеточной сту-
пени организации (Р1еос1ог1па) такая дифференциация клеток
заходит уже имного дальше и приводит ›к превращению половины
из них в бесплодные. Наконец, на верху всего ‘этого ряда распола-
гаются настоящие вольвоксы, состоящие из 512 и более клеток, по-
давляющее большинство которых представлено бесплодными клет-
ками и только единичные—полноценными гонидиями. Стало-быть,
только самые бедные особями 4- или 8-членные колониальные
объединения лишены у Volvocoidea всяких внешних ‘признаков и-н-
теграции, тогда как все остальные, начиная с 16-клеточных, -прояв-
ляют их во все нарастающей степени, в степени, прямо пропорцио-
нальной члислу своих одноклеточных компонентов. Благодаря этомоу
здесь имеется налицо целый ряд градаций IIGJIOCTHOCTPI KOJIOHPIH.
совокупность которых иллюстрирует самым ярким образом после-
довательные этапы процесса органической интеграции, процесса
перерастания простого клеточного агрегата в =высоко интегрирован-
ную многоклеточную особь.
Замечательнее всего, что этих этапов намечается и здесь четыре,
и что они в точности отвечают, по соответствующей им численности
одноклеточных ‘компонентов колонии, указанным выше степеням
интеграции в эмбриогенезе -бластулообразующих Metazoa. Действи-
тельно, и здесь 16-клеточные объединения характеризуются лабиль-
ным, обратимым уровнем интеграции, а начиная С 32-KJI€T0LIHb1X
интегрированы уже необратимо: при этом различимы те же три
ступени такой стабильной интеграции——низшая З2-——б4-клеточная
(Eudorina, Platydorina), «бластомерная», отвечающая 128-KJPETOLI‘
HoMy составу объединения (Pleodorina), И наконец высшая, эпите-
лиальная‚ представленная тут действительно эпителиальными цело-
бластулами Volvox И примыкающих к нему родов, СОСТО-ЯЩИМИ ИЗ
512 и более клеток. '
Таким образом, тем ступеням интеграции, с какими мы встре-
278
чались в эмбриональном развитии гидромедуз, соответствуют опре-
деленные дефинитивные состояния палинтомхических колоний
Volvocoidea. Иначе говоря, намечающийся у первых онтогенети-
ческий ряд дополняется сравнительно анатомическим, PI ОНИ оба
дают совпадающие показания.
4. Связь уровня интеграции с численностью продуктов
палинтомии ' _
Итак, в обоих рассмотренных нами случаях интеграция про-
дуктов дробления идет одинаково нарастающим ходом, проходя
одни и те же ступени, одни и те же узловые точки, где количествен-
ное изменение состава дробящейся системы отвечает ее переходу
из одного качественного состояния .в другое. Эти качественные изме-
нения наступают тут не постепенно, а путем скачкообразного пере-
хода от одного уровня интеграции к следующему, более высокому
ее уровню, каждый из которых связан с точно определенной
численностью объединяющихся элементов.
Иначе говоря, этот процесс идет стр-ого закономерно, подчиняясь
известным общим правилам, действующим одинаково в двух,
столь удаленных друг от друга группах, .как низшие Cnidaria, с од-
ной стороны, и колониальные фитомонады -— с другой. В том и дру-
гом случае -в ходе этого процесса явственно намечаются три узло-
вых момента: 1) переход из состояния неустойчивой, лабильной ин-
теграции в состояние уже необратимой, но еще слабо выраженной
целостности, отвечающей 32-клеточному составу агрегата; 2) пере-
ход из этой‹ начальной фазы интеграции в следующую стадию бла-
ст-омерной бластулы, оформляющуюся на стадиях 64—128; 3) мо-
мент эпителизации ‘бластулы, отвечающий: достижению, стадии
512мз41024 и завершению объединения ее компонентов. Первый из
них имеет, таким образом, значение п о р о г а ус т о й ч и в ой
интеграции, а третий——пор«ога эпителиза ции бла-
стулы, с достижением которого ее целостность получает свое ярчай-
шее выражение.
Таковы первые выводы, непосредственно вытекающие из сопо-
ставления и анализапроцессов формирования -синзооспор у Volvo-
coidea и метагенетических Deptolida. Эти выводы. очень неожиданны
и могут показаться малоубедительными, поскольку они основыва-
.ЮТСЯ на ‘ОЧЕНЬ небольшом фактическом материале; ЁЙОЭТОМУ QCT_a__€_’_I‘C5I
еще неясным, каково их истинное значение и, даже более; того,
могут ли они ‚претендовать на приписываемую им степень _ досто-
верности. Для нас, однако, совершенно очевидно, что все отмечен-
‘НЫС совпадения В ХОДЕ! процесса интеграции ПРОДУКТОВ «ПЗЛМНТОМИ-
ческого деления невозможно объяснить простой случайностью, и
что они действительно отражают весьма общие и важ-ные закономер-
ности этого, столь загадочного еще, процесса. За это говорит мно-
жество фактов, к рассмотрению которых мы сейчас перейдем.
279
5. Условия интеграции продуктов палинтомии
Если эти закономерности действительно существуют в природе
и имеют при этом хоть сколько-нибудь общее значение, то есте-Т
ственно ждать многих, в достаточной мере убедительных, косвенныхг
подтверждений этому -со стороны обильного фактического мате-
риала, -каким мы теперь рас-полагаем в отношенили различных форм .
палинтомии. В самом деле, чтобы допустить наличие таких законо-
мерностей, необходимо было бы прежде всего показать, что инте-.
грация продуктов палинтомии никогда не имеет места при их
образовании в небольшом числ-е, уступающем конкретному цифро-
вому значению порога интеграции и, наоборот, обязательно насту-
пает в тех случаях, когда палинтомия включает более четырех син-
хронных делений, или, иными словами, когда она дает начало 32
и более конечным продуктам. '
Оцени-вая под этим углом зрения все известные нам случаи
истинной палинтомии у простейших (т. -е. не включая сюда повтор-
ное деление инфузорий), мы без труда можем убедиться, что первое
из этих требований полностью оправдывается фактами. Действи-
тельно, у всех форм, продуцирующих по 4-8-16 зооспор, а именно
громадного числа фитомонад, некоторых свободноживущих и пара-
зитических перидиней, у хризомонад, эвглен, размножающихся
палинтомией солнечников и корненожек (Euglypha), зооспоры либо
сразу же разбегаются, либо объединяются в простые колонии, eno-
собные к расхождению и не проявляющие большей целостности,
чем пмалочленные, со-впадающие с ниими по своей численности, коло-
ниальные объединения Volvocoidea. Этот факт, очевидно, не тре-
бует особых доказательств; и
Что же касается второго требования ——об обязательности обра-
зования синзооспор при более продуктивных формах палинтомии,
то первое впечатление таково, что оно совершенно не поддержи-
вается фактами. Тут сразу же приходится вспомнить, „во-первых,
‘что многие паразитические перидинеи, а именно виды Нар1о2ооп,
Blastodinium, Oodinium, Dissodinium ‘H дРУПие, продуцируют гро-
мадное число зооспор, доходящее до 128, 256, 512 и более, причем
последние свободно разбегаются; далее, очень большое число
зооспор образует малоизвестный еще паразит инфузорий-Зрого-
топаз, где они также разбегаются; наконец, они образуются,
также в очень значительном числе, при половом размножении
у Haematococcus, некоторых Chlorogonium, ряда Volvocoidea H не-
которых других жгутиковыхтв качестве апибо изогамет, либо
спермиообрпазных мужских гамет, и никогда в этих‘ слу-
чаях не объединяются пожизненно. Таким образом, тем пока-
заниям, которые дают нам Volvocoidea H метагенетические медузы,
противостоят более многочисленные факты, когда при равной сте-
пени продуктивности палинтомии мы н-е видим никакого намека на
интеграцию ее продуктов.
Если, однако, пристальнее присмотреться ко всем этим случаям
‘280
чисто расселительной палинтомии, при которой: она, следовательно,
полностью «сохраняет свой характер сп-орообразования, то оказы-
вается, что они все существенно отличаются от дробления Volvocoidea
-H гидромедуз и‘еЩе в других отношениях. Так, у Нар1о2ооп, Blaste-
dinium H Oodinium отличен самый характер палинтомии, относя-
щейся тутк линейному, а не табличномуее типу. В слу-
чаях Diss_odi'nium, Euglena авто H Vampyrellidium наблюдается
другое отличие: «палинтомический процесс, включающий тут
‘6-8 повторных ‘делений, оказывается подразделенным на два или
несколько циклов, включающих по 4 (Dissodinium) или 2-3 прос-
тых деления и разъединенных вклинивающейся между ними
паузой; в течение последней каждый спороцит окружает себя же-
сткой (у Dissodinium, где они превращаются в характерные ‹<рога-
ты-е цисты»)_ или студенистой оболочкой, внутри которой и проис-
ходят все дальнейшие его деления.
Что же касается гам-етогенеза Chlorogonium H дРУгих фито-мо-
над, то он здесь либо осуществляется путем своеобразного почко-
вания протопласта, имеющего скорее синто-мический, чем палинто-
-мический характер, либо ведет к образованию высоко специализи-
рованных, не способных к вегетативной деятельности сперматозои-
дов-, а не клеток нормального характера, как при всех других фор-
мах палинтомии. Стало быть, ни в одном из этих случаев не наблю-
дается той правильной табличной палинтомии, с которой мы встре-
чались у бластулообразующих форм и на основе которой там разыг-
рывается процесс нарастающей интеграции.
Отсюда, очевидно, может быть сделан вывод, что описанные
закономерности этого процесса действуют лишь в очень определен-
ных рамках, в пределах правильной табличной па-
л›инто м ии, и не распространяются на дРУгие, например, линей-
ные ее формы. В этом, в сущности, .нет НИЧЕГО удивительного,
поскольку одна только табличная «палинтомия имеетфсвоим р-езуль-
татом то однослойное эпителиообразное расположение клеток, с ко-
ТОрЫМ фактически СВЯЗЗНО ДОСТЧИЖЕНИС МНОГОКЛЕТОЧНОСТИ В ее пер-
вобытной форме эпителиальной синзооспоры. ~
Второй вывод из этихфактов заключается-в том, что объеди-
‘нение даже очень многочисленных продуктов табличной палинтомии
‘не пролисходитв тех случаях, когда они, в ходе своего образования,
‘распределяются. небольшими группаими,
изолируемыми друг дот
друга теми или иными оболочками, т. е. когда их непосредственный
взаимный контакт ‘фактически не имеет места. . I
~JIai.1Iee, можно констатировать, что такое объединение не про-
исходит и тогда,1когда зооспоры образуются в один прием путемтех
‘или иных видов сИНТОМвиИ или почкования, т. е. когда оформляю-
Тщиеся зооопоры находятся в лишь очень кратковременном кон-
такте.
Наконец, еще один вывод заключается в том, что устойчивое
объединение продуктов табличной палинтомии не пимеет места
‘PI В TEX случаях, КОГДЗ ЭТИМИ продуктами ЯВЛЯЮТСЯ СИЛЬНО ВИДОИЗ-
v 281
мененные МУЖСКИЕ ГЗМСТЫ, РЕЗКО ОТЛИЧНЫЕ ПО СВОИМ фИЗИОЛОГИЧЁ-
чским свойствам от обычных бесполых зооспор.
Итак, интеграция продуктов палинтомии вообще не имеетместа
при малом их числе, а при большом не наблюдается ни при линей-
-ном ее типе, но при каких-либо уклоняющихся ее формах; она
ОСУЩЕСТВЛЯЕТСЯ ТОЛЬКО при ПраВИЛЬНОИ ТЗОЛГИЧНОИ «ПЗЛИНТОМИИ, п‘
ведущей к образованию бес-полых зооспор.
В ИТОГЕ МОЖНО СКЭЗЗТЬ, ЧТО, ПОСКОЛЬКУ массовая ПрОДуКЦИЯ CBO-' ц
боднь1х зооспор посредством правильной табличной палинтомии
фактически не наблюдается в природе, э т о т т и п п а л и н т о м и и
ведет либо к образованию немногочисленных
свободных зооспор, либо к их объединению
в м н о г о к л е т о ч н о е ц е л о е; или, другими словами, что про-
дуцируемые таким способом зооспоры сохраняют способность рас-
ходиться лишь в пределах первых четырех делений, а дальше неот-
вратимо вступают в прочную взаимную связь. Это значит, что
продуктивность данного типа палинтомии ограничена теми узкими
рамками, в которых она удерживается у свободноживущих одиноч-
ных простейших в силу низкого уровня достигаемой ими пипер-
трофии. И стоит только материнскому организму, перешедшему
.в особо благоприятные условия питания перешагнуть известный
уровень гипертрофии и повысить вследствие этого продуктивность
своего раз-множения (как это и происходит у .вольвоцид и бластуло-
образующих Metazoa), как сразу же вступают в силу явления про-
грессирующей интеграции: все его многочисленные потомки объеди-
няются в одно многоклеточное целое.
6. Приспособления простейших к предотвращению интеграции”
продуктов палинтомии
ДОВОЛЬНО ВССКИС, как НЗМ КЗЖСТСЯ, ДОПОЛНИТЕЛЬНЫЕ СВИДСТЁЛЬ-
ства в пользу предполагаемой нами связанности интеграционного
‚процесса с правильными формами табличной палинтомии могут
быть почерпнуты из особенностей развития целого ряда паразити-
ческих ‘простейших. Наиболее интересны в этом отношении факты,
касающиеся размножения акнидоопориидий. Дело в том, что име-
_ются серьезные основания считать, что исходным типом размно-
жения в этой группе было продольное простое деление, сохранивч
шееся тут только у Piroplasmoidea (B трофической части их цикла)
и близких к кокцидиям Toxoplasma. Наряду с этим мы хорошо
.3,наем‚д что у подавляющего большинства представителей ‘этой
;Гр_уДЦЫ размножение осуществляется ;1'IyT€M различных ВИДОВ ВТО-
.ричной`синтомии, т. е. модифицированных, вторично видоизменен-
ных форм палинтомического спорообразования; правильная же
палинтомия представляет тут редчайшее исключение; она описана,
насколько мы знаем, только у двух видов кокцидий (Pseudoclossia
Patellae И Ps. chitonis; Дебезьё, 1920), у которых дает всего по
нескольку продуктов. Таким образом, колоссальная плодовитость
282
размножения споровиков достигается ими путем явного отхода от
тех форм спорообразования, какие должны были бы иметь тут
место в силу присущего им характера простого деления. Будучи
продольным оно должно бы вылиться, при гипертрофии материн-
ской особи, в формы табличной палинтомии, ч-его на самом деле тут
почти никогда не происходит. Этим подсказывается мысль, что
переход от более примитивной правильной палинтомии -к делению
распада явился в этой группе (а может быть и в других группах
паразитических простейших) своеобразным приспособлением к
«предотвращению» интеграции продуктов деления, способом сохра-
нения жизненно необходимой паразитам исключительно высокой
плодовитости при самых высоких степенях гипертрофии.
Возможно, что роль другого приспособления к ее .предотвраще-
нию играют уже упоминавшиеся видоизменения хода табличной
(продольной в своей основе) палинтомии, с какой мы сталкиваемся
у Dissodinium B ВИДЕ образования рогатых цист, и что ряд особен-
ностей размножения других простейших сможет найти себе объяс-
нение при оценке с этой несколько необычной точки зрения.
Во всяком случае, отсутствие у паразитических Protozoa высоко
продуктивных -форм правильной табличной палинтомии есть факт,
с которым невозможно не считаться. Взятый изолированно, он пред-
ставляется нам загадочным, но в сочетании со всеми другими при-
веденными фактами приобретает недостававшее ему биологическое
объяснение. '
В этом же смысле могут быть объяснены и такие факты, как
переход от табличной ‘палинтомии к внутреннему почкованию у ви-
дов Haematococcus при образовании ими многочисленных изогамет
(микрозооидов), своеобразное формирование последних путем
массового наружного почкования у Noctiluca И ряд других, пред-
ставляющихся. различными приспособлениями к сохранению высо-
кой плодовитости размножения зооспорами или спорами при удер-
‘ЖЭНИИ ИМИ СВОЕГО ПрЯМОГО расселительного ЗНЗЧСНИЯ.
7. Выводы
Итак, создается вполне определенное впечатление, что объеди-
нение продуктов правильной табличной палинтомии в многоклеточ-
ные синзооспоры отнюдь не является делом случая, а подчинено
строгим биологическим закономерностям, что оно обусловлено
специфическими особенностями этого вида размножения и есть
‘неизбежное его следствие. Интеграция этих продуктов наступает
всякий раз, когда число повторных‘ делений идет дальше строго
определенного предела, устанавлива-емого. существованием порога
интеграции, и последняя может быть избегнута исключительно
путем тех или иных видоизменений самого процесса массового
образования зооспор, путем перехода от табличной палинтомии
к другим способам спорообразования. Только таким образом и сни-
мается у-‚паразитических Protozoa противоречие, заключающееся
283
о
в ограничении продуктивности такой палинтомии очень тесными
ди жесткими рамками при прогрессирующей гипертрофии однокле-
-точного материнского организма и сохранении им способности
к массовому расселительному размножению, жизненно необходиг
мому для паразитов.
Иначе говоря, достижение многокл-еточности в форме эпители-
ЭЛЬНОЙ СИНЗОООПОрЫ ПРЕДСТЭВЛЯЕТСЯ бИОЛОГИЧЕСКИ НЕООХОДИМЫМ
СЛЕДСТВИЕМ ПОВЫШЕНИЯ ГИПЕрТрОфМЩ а ТЕМ самым И ПЛОДОВИТОСТИ‘
Материнской КЛЕТКИ ЗООСПОР, ГПрИ УСЛОВИИ образования ПОСЛЕДНИХ
путем правильной табличной палинтомии. Однако только вольво-
циды и бластулообразующие Metazoa ПОШЛИ ‚по пути подчинения
действующим тут законам нара-стающей интеграции, в то время как
паразитические простейшие либо уклонились от действия этих зако-
"HOB, сменив данный вид размноженпия на другие, либо оставались
и остаются вне сферы их влияния, поскольку интеграция не распро-
страняется на линейную палинтомию, а тем более на столь обыч-
ную у паразитов первичную синтомию. С этим представлением
хорошо согласуется тот факт, что образование много-клеточных слин-
зооспор не имеет места у настоящих паразитов, а наблюдается
только в тех случаях, когда избыточная гипертрофия материнской
клетки зооспор (яйца) обеспечивается ее питанием за счет много-
клеточного же материнского организма.
Общий вывод из всех сопоставленных на этчих страницах данных
заключается в том, что развиваемое тут представление о существо-
вании весьма определенных законов прогрессирующей интеграции
продуктов дробления получает косвенное подтверждение в ряде
‘особенностей размножения спорогонических простейших и позво-
ляет связать воедино мнопие из относящихся сюда фактов. Наме-
чающийся тут путь их систематизации и объяснения может ока-
заться плодотворным И, как нам кажется, заслуживающим
внимания. '
Что касается вопроса о движущих силах процесса интеграции
продуктов дробления, о его природе и тех способах, посредством
которых он реализуется, — сказатьхчто-либо определенное мы пока-
мест еще не можем. Стараясь показать, что он действительно суще-
ствует и, мболее того,_ развивается согласно вполне определенным
закономерностям и при некоторых очень строго выдержанных усло-
виях, мы, конечно, не можем даже и. пытаться дать ему готовое
объяснение. Напм кажется только очевидным, что любое его объяс-
пение, должно базироваться, во-первых, на глубоком анализе физио-
ялогического, взаимодействия одноклеточных компонентов развиваю-
"Щейся системы, хпри котором должна быть учтена-его связанность
.со строго определенной группировкой, численностью этих компонен-
тов и с определенным уровнем их обмена, пи с другой стороны, на
всех тех особенностях в ходе формирования зооспор, при которых
‚их интеграция оказывается исключе-нной.
Во-вторых, любое объяснение процесса интеграции, должно
тисходить из биологического понимания этого процесса, как про-
_284
дукта естественного ‘подбора, как явления приспособительного "Xa-
рактера, приведшего к приобретению палинтомическим размноже-
нием зооспорами качественно нового значения—-значения способа
создания многоклеточных тел размножения (синзооспор). Оно
должно исходить также из показанного здесь закономерного посту-
пательного восходящего характера интеграции, ее скачкообразного
перехода от одного уровня к другому, более высокому ее уровню,
из учета борьбы противоположных тенденций, составляющей внут-—
. реннее содержание процесса перерастания палинтомического раз-
множения в воспроизвед.ение многоклеточными сложными зоо-
спорами.
Мы твердо убеждены, что при этих условиях можно дбудет
вплотную подойти к правильному пониманию этого важнейшего
биологического .процесса, раз «нет в мире непознаваемых вещей;
а есть только вещи еще не познанные, которые будут раскрыты и
познаны силами науки и практики» (И. В. Сталин).
ЧАСТЬ ЧЕТВЕРТАЯ
ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ НИЗШИХ
КНИДАРИЙ И Г УБОК И ПУТИ ЕГО ЭМБРИОНИЗАЦИИ
Вводные замечания
Фактич-еские данные, изложенные в предыдущих главах, исклю-
чают всякую возможность сомневаться в реальности и значимости
СХОДСТВ, СУЩЭСТВУЮЩИХ Между всеми видами эпителиальных жгути-
ковых целобластул и, в частности, между «животными вольвоксами>>,
с одной стороны, и настоящими, автотрофными Volvocidae — с дру-
гой. Это поразительное сходство между теми и другими особенно
велико и осязаемо, как мы видели, на самых ранних стадиях их
развития‚›еще до перехода к свободному существованию, когда
только-что закончилась начальная дифференцировка клеток -бла-
стулы в однородные жгутиковые клетки. На этой стадии онизвсе
обладают совершенно тождественной архитектоникой, сходным
гистологическим строением и одинаковым чИсленнЬ1м составом и
проявляют сходную степень организменной целостности; при этом
и самый процесс их формирования идет во всех -случаях очень сход-
ным образом, так что одинаковый конечный результат достигается
тут и почти одинаковым, в сущности, путем. Поэтому-то мы и мо-
жем с полным правом относить всех молодых жгутиковых бластул
к одному и то-му же личиночному типу—синзооспор или жгути-
ковых бластул, независимо от их систематической принадлежности
И даже от того, обладают они животной пили же растительной
природой.
В самом деле, это капитальнейшее различие м-ежду жгутиковыми
бластулами Metazoa И таковыми Volvocoidea еще не сказывается
на данной стадии в хоть сколько-нибудь значительном объеме по
той простой причине, что онли еще не приступили к активному, само-
стоятельному существованию, при котором только и выявятся их
фундаментальные физиологически-е различия. Поэтому и не прихо-
дится удивляться, что только-что оформившиеся жгутиковые бла-
стулы вольвоцид внешне отличаются от молодых цело-бластул Meta-
zoa лишь B цитологическом отношении, по строению составляющих
286
их жгутиковых клеток: они обладают тут фитолмонадным типом
строения и оснащены готовым, работоспособным фото-синтетическим
аппаратом, вто время как клетки жгутиковых бластул Metazoa
имеют организацию зоофлагеллят и, более конкретно, осмотрофных
Protomonadina ‘И лишены каких-либо специальных приспособлений
к анимальному .питанию.
Однако как только синзооспора перейдет к активному существо-
ВаНпию в качестве «бродяжки» или мерцательной личинки, это раз-
личие развертывается в полном -своем объеме; оно сразу же полу-
чает ярчайшее физиологическое выражение и направляет даль;
нейшее ‘развитие бластул вольвоцид и Metazoa по диаметрально
противоположным руслам. _ -
Действительно, у Volvocidae все жгутиковые клетки бластулы
немедленно приступают к вегетативной деятельности, не перестраи-
вая для этого своей исходной организации. Их индивидуальное
развитие остается таким же прямым, как у всех вообще фитомо=
надин, и на этом, очевидно, основано пожизненное сохранение воль-
воцидами их исходной организации бластулы. Жизненная форма
однослойной жгутиковой целобластулы оказывается у них .полно-
ценной форм-ой вегетативного существования и последующего затем
размножения. В рамках этой, неизменнойздесь, жизненной формы
протекает вся жизнь многоклеточного организма, его рост, постэм-
бриональная дифференцировка на бесплодные и плодущие клетки
и осуществление последними оо- и сперматогенеза; в бластуле же
происходит эмбриональное развитие ее потомков до достижения ими
той же стадии бластулы.
В противоположность этому, жгутиковые бластулы Metazoa
фактически не способны к выполнению вегетативных функций. Как
общее правило, они не питаются и не растут, имея значение кратко-
временной и преходящей, чисто расселительной начальной стадии
постэмбрионального развития, -сменяемой в дальнейшем ходе онто-
генеза совершенно иначе организованными и почти всегда сидячими
вегетативными стадиями. Они становятся способными к вегетатив-
ной деятельности лишь после коренной перестройки всей своей орга-
низации, после глубочайшего метаморфоза, имеющего своим резуль-
татом их превращение в аНпИМаЛЬНО питающуюся сидячую форму.
Таким образом, коренное физиологическое различи-е между син-
зооспорами животной и растительной природы вступает в полную
силу лишь в постэмбриональном периоде |ИХ развития, при их пре-
вращении в трофические стадии и выполнении ими свояих прямых
функций-тпитанаия и роста.
С точки зрения развиваемого здесь представления о жгутиковых
бластулах, как синзооспорах, следует считать, что в основе этого
превращения лежит процесс преобразования расселительных, по
существу, зооспор, олицетворяемых жгутиковыми клетками бла-
стулы, во взрослых трофозоитов, представленных трофическими и
иными клетками позднейших вегетативных стадий развития. Иначе
говоря, первооснову этого превращения составляет тот самый upo-
287 -
цесс онтогенеза отдельных клеток бластулы,
ствляется превращение всякой зооспоры в одноклеточную же веге-
тативную особь и который является, вместе с тем, характерной ‚для
нее формой постэмбрионального развития. И так как этот процесс -
протекает у автотрофных простейших в формах прямого, эпиморф-
ного развития, а у анимально питающихся обязательно связан j‘
с метаморфозом, то у многоклеточных наблюдается такое же раз-
личие в характере их пост-эмбрионального развития; оно остается ~
эпиморфным у Volvocidae И ‚представлено сложнейшим п е рвич-
ным метаморфозом унизшихМетаиоа. „
Однако, рассматривая таким образом Постэмбриональное раз-
витие многоклеточных синзооспор, мы HM Ha минуту не должны упу- '
СКЗТЬ ИЗ ВИДЕ ИХ ГЛУЁОКОГО качественного СВОЭОбРЭЗИЯ |ПО -сравне-
НИЮ С морфологически рЭВНОЦЭННЫМИ ИМ СЭМЬЯМИ ОДИНОЧНЫХ РЯЗ-
бегающихся зооспор истинных простейших. Действительно, в то "А
время как у последних все зооопоры одной семьи развиваются
совершенно одинаковым образом и превращаются со временем
в равноценных друг другу особей, способных в дальнейшем к раз-
множению зооспорами,——развИтие многоклеточных син.зооспор
сильно уклоняется от этой схемы. Их специфические свойства це-
лостных многоклеточных организмов проявляются тут, во-первых,
в том, что развитие составляющих их клеток идет не в одном,
а в нескольких различных направлениях, что и имеет своим резуль-
татом создание р а з н очк л е т о ч н о г о строения, составляющего
характернейшую из особенностей всех их вегетативно деятельных
стадий. Во-вторьтх, способностью к нормальному для зооспор гипер-
трофическому росту обладают при этом не все клетки синзооспоры,
а лишь те из них, которые сохраняют значение генеративных
(Volvocoidea), ИЛИ же они все совершенно ее лишаются, и эта сп-о-
собность восстанавливается только их отдельными, более или менее
отдаленными, потомками (Metazoa).
Таким образом, в результате постэмбрионального превращения
синзооспорь1 в следующие за нею вегетативные стадии оформляется
характерный для них клеточный полиморфизм, приводящий в ко-
нечном счете у Метаиоа к созданию их столь специфичной трех-
слойной организации.
Глава XVI
ПЕРВИЧНЫЙ МЕТАМОРФОЗ НИЗШИХ КНИДАРИЙ И Г УБОК
1. Общее понятие о первичном метаморфозе
Превращени-е замкнутой однослойной синзооспоры, ведущей
пелагический образ жизни и не способной к активному питанию,
в совершенно иначе организованную сидячую ‚вегетативную стадию,
построенную по типичному для Metazoa трехслойному плану, и мо- .
гущую питаться анимальньтм способом-составляет содержание
288
которым осуще- `
сложнейшего онтогенетического процесса, называемого нами п е p-_
ВИЧНЫМ метаморфозом. `
Вообще говоря, первичный метаморфоз складывается из след
дующих четырех взаимосвязанных процессов, приводящих, в коа
Нечном счете, -к полной морфофизиологической перестройке разви-
вающегося организма: -
1) передифференцировки однородных жгутиковых клеток cun-
зооспоры в клетки самого разнообразного функционального значе-
ния, т. е. процессов цитологической дифференциа-
ц и и образующего ее клеточного материала;
2) оформления первичных пластов тела в виде двух тканей ‚по,
граничного характера и третьеи, создающейся между ними, ткани
. ВНУТРЕННЕЙ среды, Т. е. процесса Г И С Т О Г е Н е З а;
3) 0P1“aH0JI0I‘PILIe»CK0I"4, дифференцировки формирующейся теперь’
вегетативной стадии, т. е. процесса о р г а н о г е н е з а; - A
4) кореннои, как правило, перестройки архитектоники орга-
низма, связанной с его переходом от свободноплавающего к при-
крепленному образу жпизни и представляющей собой метаморфоз
в прямом морфологическом смысле этого слова. р
Характернейшей особенностью онтогенеза всех Metazoa, размно-
жающихся посредством п-елагических синзооспор, является то, что
все эти -сложНь1е морфогенетические процессы целиком совершаются
в постэмбриональножм периоде развития; поэтому
последнее и имеет здесь поразительно сложные формы, резко кон-
трастирующие с крайней простотой эмбриогенеза этих животных.‘
Так, у специально интересующих на-с сейчас низших книдарий
и губок первичный метаморфоз включает, как минимум, последо-
вательное прохождение личинкой следующих этапов развития:
а) ее преобразование из однослойной и равноклеточной бла-
СТУЛЫ В дВУСЛОИНуЮ паренхимулу, продолжающую еще вести своа
бОДНОП-ПЗВЗЮЩИЙ образ жизнги; б) оседание паренхимулы, ее .при-
крепление к- субстрату и дедифференцировку, при которой она
превращается в более или менее уплощенную клеточную массу,
являющуюся зачатком_сидячей вегетативной особи; в) превращение
этого, часто бесформенного, зачатка в радиально-симметричный,
гетцерополярньтйп полип или в губочку с оформленной пищеваритель-
нои полостью, сообщающейнся с внешней ср- дой одним ‘или многими
отверстиями, и со специальными приспо_соб ‘ениями к улавливанию
ПИЩИ.
Однако у многих из метагенетических книдарий эта, и без того
сложная, схема дополнительно усложняется прохождением личин-
койеще одной свободной. пелагическойр стадии——стадии планулы,
получающеися из паренхимулы путем гистологической дифферен-
цировки ее наружных клеток на чувствительные, железистые, стре-
кательные и т. д., и эпителизации ее внутренней клеточной ма-ссь1.
В. этих случаях, правда, соответственно несколько упрощается пр-
следующее развитие особи, поскольку некоторые из позднейших;
морфогенезов как бы «досрочно» выполняются ею на стадии пла- '
19 эмбриология беспозвоночных 389‘
нулы, до ее оседания и прикрепления к субстрату, но метаморфоз `
в целом становится еще более громоздким. ‚
Большая сложность первичного метаморфоза
Cnidaria И губок и вс
что он тут целиком осуществляется в постэмбриональном развитни‚._
составляют коренное отличие данных форм от их более высоко орга-’
низованных сородичей и всех вообще высших Metazoa. B самом
деле, у последних все процессы, составляющие первичный метамор-.
фоз, входят полностью (или, по крайней мере, большей своей
частью) в состав их эмбрионального развитчия в качестве его вто-'
рого, органотипического периода или даже отчасти осуществляются
попутно с дроблением, превращая его тогда из цитотипического
в органотипическое.
Таким образом, Постэмбриональный первичный
метаморфоз низших многоклеточных отвечает
содержанию И достигаемым `
ъримитивных
по своему
зультатам тем процессам гаструляции, обра-
зования зародышевых листков, их последую-
щей дифференцировки и органогенеза, которые
мы так привыкли считать «изначально» эмбриональными процес-
сами, процессами «зародышевыми» по самому своему существу, что
не «можем представить себе без них нормального эм-бриогенеза.
Между тем мы уже могли убедиться, что в наиболее примитив-
ных формах эмбриогенеза, имеющих своим результатом создание
Жгутиконосных синзооспор‚ ни один из этих процессов не находит
себе места. Тут нет ни гаструляции с образованием зародышевых
листков, ни каких-либо намеков на органогенез‚ т. е. нет еще ничего
специфически «эмбрионального», ни одного из проявлений того
органотипического развития, которое только и признается ‹<подлин-
ным» эмбриогенезом. Стало быть, мы уже сейчас можем сказать,
что приобретение последним его привычных сложных форм есть
явление вторичного порядка, результат его _ усложнения за счет
поглощения им первично и по существу постэмбриональных этапов
развития, за счет эмбрионизации первичного метаморфоза.
Как и почему происходит эта эмбрионизация, будет показано
в одной из следующих глав. Пока же мы займемся возможно более
подробным рассмотрением имеющихся данных о процессе первич-
ного метаморфоза низших книдарий и губок, являющемся, по сути
дела, процессом становления характерной для Metazoa трехслойной
организации.
2. Начальная дифференциация протобласта и формирование
паренхимулы
с Начальным этапом всех видов первичного метаморфоза является
приобретение синзооспорой двуслойного строения путем дифферен-
циации ее дотоле единой и единственной системы жгутиковыхукле-
ток на две системы клеток; одна из них сохраняет при этом свое
290"
ячески подчеркиваем-ее нами обстоятельство,-
Ием ре-П
“СЁОДНСЮ двигательное з т - - - .
положение, тогда как новЁЁЁЁЁЁЁю-Ётаёсояверхностн“ Эпителиалыюе
складывается в аморфную Клеточн Ю вторая система чаще всего
деда ЛИЧИНКРЪ Эта Н а Ч а л Ь-Н а HY KI»:1Ha§CTy(.) IE{3aHH;).I:1H511;Ul}(’bIOq)HOJIOC]\}¢
ИаЦия имее И е ен-
nepT;:T:::9o?;‘T:P€;y;‘:T;;°:w“-Hefiepaue
3 ЖГуТИКОВЫх к ‚ состоящей
т е л а, обычно обозначаемтйег ЁЁй эётгэ- п е р В И Ч Н Ы е п Л а с Т Ы
‚тельности не имеющие еще [Ёрав Н Т и энтодерма, но в действи.
топограёфические соотношения на эЁоЙШЁЁЁдЁЁОЁЁЁЁЕНИе’ так как их
жают д е ршенно не от а.
ё 3§ia§§’£;5’f§,y:?‘§§§B§°£Z ssssem их
Ё дает в дЁдЬнейшем преимущестрвеннс? ЁЁЁЁЁЁЁЁЁЁИ Ёлеточныййрлэй
лавным о разом энто е > ВНУТРЕННИЙ-
наоборот, а у метагенётёчуески: Zugfiii губок дело обстоит как раз
няя система клеток дает Начало ТОЛЬК дных и иглокожих рнутрен.
а 5KT0_ И энтодерма Целиком Образ юЁЯтканя-м вНУтреннеи среды,
Вот эпителия Уже двуслойной в этот’ MOM из оставшегося ЖГУТИКО`
дует еще Добавить, что у Некоторых Извес ент личинки. K ЭТОМУ one.
рЁсЁлоение протобласта на экто- и auTou::c1f:;1:HP;;§p§§HeBb‘X губок
› х дит на мест _ ловины п o-
TIOK И без перех‘); 5;:qPI;I:In::H:1:Lp]13a C15JaI30auHMHor0 расположения его Kge.
последняя Оформляется тут лицгь пЁзЁнЮ стадию пареНХИМУдЫ; Эта
чинтЁи уже третьей системы клеток — Kn§Te6xH§a:s:g§::a0BaHH" y ЛИ-
~ аким образом нача Ы‘
Личинки _ п P 0 т о’б л а cJ1I“b:a1:"1el;H§Jq):§:EuHaHHH первичной ткани
личные результаты, что Нет решителрно IX cJIy:Ia51x настолько раз-
вести ее, как это обычно делается по Никакои ВОЗМОЖНОСТИ mm"
ЭК“, И энтодермы И отождествлять, этоёъппонятие дифференциации
видами гаструляции действительно л Роцесс с теми или иными
не всех) вышестоящих в системе фо Ишь у HEKOTOPMX (но далеко
протобласта имеет своим Herlocpenc§1:aeHHHa1:;nb:::y£:2q3eP€HIlgIaLu»Is{
вание настоя " т том о азо-
СпециалИЗОВаЦЁЁЁЙГЗЭСДРЕЗ/Ёеьё 1;0gJ1:cTKoanqc>e1:oM, первичной кишкой ирузко
СЛОЯ у Всех же интересующих Нас сейёве 6ee второго, внУтреннего‚
дело обстоит Несравненно сложнее? ста ас олее примитивных форм
Ционально отвечающая гаструле -Omsfllfifl, морфологически и функ-
том длинной Цепи Морфогенетических пвается Конечным результа-
т 901190003, Обычно не имею-
ЩИХ ничего общего—ни по
- c
уществу» НИ “O ф0РМе—с классиче-
СКИВйИаВИДаМИ гаструляцирд
Т K. начальное асчленени
. е
системы клеток, ведУЕдее к преоержёёбласта На Две различные
синзооспоры в двуслойную личинк тип HMO первично однослойной
считаться гастРУЛяЦией П Р Я мзы M ар Еёренхимуль!’ Не ДОЛЖНО
процесса Является - Ультатом этого
‹ как мы Bu и
оформление 3lKTO_ H. 3HTOfleYpM11bxMB1I:£b1T1IHe, отнюдь не
лизированном состоянии “WHOM. специа-
кот
позднее и ’ . A ( Орое ОНИ ПРИОбРЕТУТ лишь много
совсем другими путями)‚ н тол ь ко деди ф ф е р ен
19’
291
ц ир отвк-аг-‘ча с ти 1<":'I1'黑*i*.'('3.t1 н о где ‘ми Tie-p-11-a'Jf'21""cuH«11’§ вед
спо р ы, при устойчивом пока сохранении другой его частью своего‘
исходного строения жгутиковых, зооспор; Биологическое и онтогед‘ .
нетическое значение этого своеобразного процесса- будет рассмотдЁ
рено. несколькодальш-е, после „того как мы сможем установить, ка-Ъ
ковадальнейшаясудьба: создаваемых им клеточных систем и какЁ
идет в разных случаях их последующая функциональная диффеьд
ренциация; „Однако, чтобы.‘ можно было описать соответствующий‘
фактический материал, необходимонайти уже сейчас такое обознад.
чение продуктов начальной дифференциации протобласта, которое"
было1бы применимо ко всем этим разнообразным продуктам и отра-°
жало бы, вместе с тем, их настоящее, а— не только будущее значенеие._
Эти-м требованиям вполне удовлетворяет одно только, введенное.
Мечниковым, понятие к и н о б л а с т а, оказывающееся подходящим
для _обознаЧения той части протобласта, которая сохраняет при его.
раздвоении свое первоначальное строение и функцию. Что же кад’
сается второй. его части, то к ней лучше подходит из имеющихся
терминов мечниковский термин ф а г о ц и т о б л а с т, хотя и имею-
щий более узкий, чем это требуется, смысл. Стало быть, р е з ул ь->
татом‘ начальной дифференциации протобласта-
является его рас-слоение на кинобласт пи фаго-'
цитобласт, имеющие в разных случаях самое‘
различное презумптивное значение. --‚
Посмотрим теперь, как совершается эта дифференциация у син-`
зооепор "различных систематических групп и к каким результатам
она ‚приводит. ь '
А. „Метагенетичеекие Leptolida. У всех метагенетических гидроч
медуз начальное. расслоение протобласта протекает в ‘форм-е гипо-д
тропной (т. е. локализованной в заднем конце тела) или; реже
мультиполярной иммиграции частчи его ‘клеток внутрь бластоцеля
j(pHc. 95). Как было установлено еще Мечниковым, этот процессё
заключается в дедифференцировке отдельных жгутиковых клеток
бластулы, сопровождающейся их округлением, утерей имевшихся
у них связей с соседнимпи клетками и выклиниванием этих клеток
в полость тела. Мечников особенно подчеркивал амёбоидную форму
и‘ подвижность, приобрета-емые данными клетками, пребывание их
ядра в покоящемся (т. е. интеркинетическом) состоянии и то харак—’
терное обстоятельство, что картины митоза «наблюдаются лишь‘
какмредкость» „в ‚образуемой ими -массе. Постепенное пополнение
этой внутренней клеточной массы происходит за счет вхождения в
нее все новых и новых иммигрантов, убыль которых в жгутиковом
эпителии постоянно "восполняется делением его остающихся на‘
месте клеток. Таким образом, происходящее теперь нарастание‘
численности гклеток личинки достигается преимущественно, если не‘
исключительно, делением клеток кинобла-ста, а не новообразуемой-
ею" паренхпимы. `
Эта последняя представляет ‘собой аморфную массу амёбоидНЬН;
‘клеток, выполняющую‘ весь1 бластоцель и Ёбезусловно удовлетворяю-ё
ч -1,, 2
292 у
‘ л
дн.
щ
чт-
.. ж,
чаш: .
=‘;.~:~=u
Ф \\\vg:;"\"‘:“.‘{£'.ii";ji§;a-mm и
ъ „е ‘M: e.%.éi!1:1;1:i:1s’.,
V V у Рисд. 95._ Первичный" метаморфоз С1у’с1а “пачяаьпаг ' - .
_ 1-3 -: последоватепьньте стадии образования (ban-<')uv1-ro6nacr'rai' путем- однопольной (гит)-
J тронной) HMM"FPaU""";'-11-5_,—, i1<f.J1H¢t1JepeHunpo13Ka._v1’131;1ce.r1e1-me КИНОЦИТОВ при большем yap-
Г ‚личении; 6_-8 -_-.>a‘I\fl§6<).1»l')11-1_1:_1e клеткифагоцитобласта; 9 —- деление такой тетки; 10 ——- паренхи-
‘- мула; '71, 72 —-ее превращение в ‘планулу; 73—+15°__.+_упдодщеьдиед и д-дедифференцировка
. дСЁВШЁЙ 1_maHynu;.16-.-J8 т последующие этапы ре лревращенияд радиалвнодсимметрйчнбго
. '~1'1oJ'11»1%r1a/ivecfi-osarream-?Ro31o1»11«11»1 (no-Me\1Hm<oay.);. .. .- ‚‚ ‚
5- "293
щую по своему строению всем признакам п е р в и ч н о й п а р е H:
х и M ы в смысле В. Н. Беклемишева («однородное соединение не-
определенно многих одинаковых клеток, расположенных друг отно-
сительно друга без какой-либо повторяющейся правильности, не"
слитых в синцитий и не разделенных промежуточным веществом»).
С ее оформлением бластула становится п а р е н хим у л о й (или,
как ее называл Мечников—-гпаренхимеллой) —второй свободно-
плавающей личиночной стадией, имеющейся у всех метагенетиче-
ских L-eptolida. __
Следовательно, в итоге начального расслоения первичного жгу-
тикового эпителия синзооспоры он частично преобразуется в амё-
боидные, дедифференцированные клетки, складывающиеся в ком-
пактную и вполне однородную первичную паренхиму———в ткань
внутренней среды, остающуюся пока не дифференцированной.
Б. Низшие Scyphozoa. У некоторых из сцифоидных, размножаю-
щихся посредством оинзооспор, их преобразование в паренхимулу
происходит точно таким же образом, как у метагенетических гидро-
медуз —путем преимущественно гипотропной иммиграции части их
клеток в бластоцель, где они складываются в сплошную массу.
Однако у ряда форм (Chrysaora, Aurelia) выселение клеток в no-
лость тела совершает-ся в гораздо более ограниченном масштабе,
так что они не образуют тут сплошной паренхимы, а остаются рас-
сеянными поодиночке в коллоидной среде, заполняющей эту полость.
Таки-м образом и создается диффузная ткань трофического типа,
действительно становящаяся таковой (а именно мезоглеей) при
дальнейшем развитии животного. Так или иначе, оформляющаяся
двуслойная личинка получает все основания считаться паренхиму-
лой, хоть и не компактного, а особого — мезоглеального —типа.
В. Морские ежи. Совершенно таким же образом, как в только
что описанном случае формируется и паренхимула у морских ежей.
Образование фагоцитобласта происходит тут исключительно путем
однополюсной иммиграции определенных клеток протобласта
(а и-менно потомков микромеров вегетативного, полюса бластулы),
носящих название первичной мезенхимы; уйдя в бластоцель, они
образуют комплекс свободных амёбоидных клеток, взвешенных в:
полостной жидкости личинки- и дифференцирующихся впоследствии
в ск-елетогенные клетки (склеробласты); следовательно, и здесн
паренхимула ОТНОСИТСЯ K мезоглеальному ТШПУ, И ее фагоцитобласт
целиком идет на построение ткани внутренней среды.
Г. Известковые губки. У части относящихся сюда форм фагоци-
тобласт образуется также путем ги-потропной (Ascetta reticulum,
А, primordialis; рис. 96) или же мультип-олярной иммиграции от-
дельных дедифференцирующихся клеток протобласта. Однако у ряда
других Нотосое1а клеточный диморфизм имеется налицо с самых
первых стадий существования «и обязан тогда своим происхождением
‘чрезвычайно ранней дедифференцировке ДВУХ (Clathflfla blaflca)
ИЛИ даже восьми (Ьепсоэо1еп1а) жгутиковых клеток ее заднего
полюса, «превращающихся тут же, на месте, вукрупненные первич-
, 294
ные зернистые клетки, или «личиночные археоциты». У Clathrina
дальнеишее развитие такой лпичинки заключается в мультиполярном
‚выселении клеток жгутикового эпителия, складывающихся и здесь
в диффузный комплекс амёбоидных клеток, плавающих в жидком
-содержимом полости тела личинки. В результате целобластула пре-
-вращается в паренхимулу, на заднем полюсе которой еще долго
видны обе первичные зернистые клетки (рис. 96, 1).
Совершенно иным способом развивается фагоцитобла-ст уже упо-
минавшейся Leucosolenia, первым зачатком которого являются во-
семь самых задних клеток протобласта личинки, превращающиеся
из жгутиковых в дедифференцированные з-ернистые. В дальнейшем,
Рис, 96. Личинки известковых губок:
1-— Clathrina"-'blanca, образование фаго
„ Цитобласта путем мультипо .
2—-,‚то же _y бластулы Clathrma reticulu лярной иммиграции’
m; 3 ——амфибластула Leucosolenia (из давыддва)
. Подобному же преобразованию постепенно‘ подвергаются и осталь-
‚ ные клетки заднего полушария личинки, превращающиеся На Месте
В крупные вторичные зернистые кл-етки‚ теряющие СВОИ ЖГУТЫ, НО
.0СТаЮЩиеся В э.пителиальном соединении (Минчин, 1896), ‘floore-
IT€H.H0 P3C1TP0CTP3H7f51Cb OT заднего полюса личинки к ее экватору,
„дедифференцировка охватывает один ряд клеток за другим пока
7
Ёичинка не превратится в типичнейшую амфибластулу (ppm 95., 3),
РОИСХОДИТ ли при этом и выселение отдельных клеток в полость
тела-неясно, но вполне вероятно, так как в этой, очень тесной
здесь, ПОЛОСТИ Находятся несколько плотно сомкнутых клеток‘
. I
_‘r1:;;T7=P$HIlPI‘M3€P€HILH-pyIoI11I«Ixc51 B примитивный зрительный аппарат;
иначе, начальное. расслоение протобласта осуществляется
_у Ьеисозо1еп1а принципиально отличным способом, чем во всех ‘pa-
gfge рассмотренных случаях, и соответственно приводит к суще-
ННО ИНОМУ Pe3YJII=TaTy: к превращению синзооспоры в амфибла-
СТУЛУ, а Не В -fIaP€HXPIMyJIy. Тем не менее, в основе этого превраще-
ния лежит и здесь процесс частичной дедифференцировки жгутика.
«BOW UP0T05J13CTa= ‘ЧТО делает амфибластулу Leucosolenia всеже
795 ‚
.
сравнимой-с зпаренхимулами других Са1сагеа и в известной мере
равноценной этим двуслойным личиночным фор-мам. '
Д. Низшие кремневые губки. У низших представителей кремне- _
двых губок, начинающих свое постэмбриональное развитие в виде
‘жгутиковых целобластул (ОзсагеПа, Placina), превращение послед-
них в разноклеточнь1е личинки идет по тому же, в сущности, прин-
ципу, как у Leucosolenia: путем передифференцировки на месте
клеток заднего полушария личинки. В результате этого процесса ли-
‘Чинка также перестраивается из равноклеточной вначале целобла-
5стуль1 в довольно типичную амфибластулу; ее переднее полушарие
„сохраняет исходное строение своих клеток, тогда как в заднем
клетки заметно видоизменяются, не теряя, однако, своих жгугов._
Как происходит их реконструкция ——точно не установлено. В nam-
нейш-ем часть этих видоизменившихся клеток заднего полушария
выклинивается поодиночке в полость тела личинки, в которой они
остаются взвешенными в виде рассеянных п-о ней амёбоцитов. Так
создается своеобразный тип замкнутой мерца ельной личинки, сов-
‚мешающей в себе признаки амфибластулы и паренхимуль1‚ но су-
ществ-енно отличающейся от тех и других наличием уже трех раз-
личных систем клеток вместо полагающихся двух. В этом отноше-
нии личинки данного типа стоят уже ближе к планулам и, в част-
ности, таковым сцифомедуз, как это будет видно низ дальнейшего
изложения. д - '
Подводя итог рассмотренным здесь фактам, пможно констатиро-
вать, что в результате первого этапа постэмбриональных превра-
щений, испытываемых синзооспорами самых различных беспозво-
ночных, они становятся’ двуслойнымаи’ личинками типа паренхимульп
Аили, у некоторых ‚губок, амфибластулами. В том и другом случае
создается особый тип многоклеточной и теперь уже разноклеточной
организации, характеризующейся тем, что ее ярко выраженный
клеточный диморфизм возникает путем перехода чансти жгути-
‘йковых клеток бластулы в дедифференцированное аМЁбОИДНФЭ
i состояние: О - - _.
Дальнейшее разви-тие оформившейся паренхпимулы (или амфи-
Ц-бластулы) может пойти по трем различным направлениям; в одних
случаях `она непосредственно превращается в прикрепленный saga-
-TOK будущей вегетативной осо-би, в других-продолжает passa-
ваться дальшев виде свободноплавающей мерцательной личинки,
' приобретающей усложненное строение планулы, и затем уже оседает
.на дно; наконец, она может еще длпительно развиваться дальшедвъ
{качестве пелагической личинки, но постепенно перестраивая свою-
ъиеход-ную.„организацию в гораздо более сложную, организацию
‘позднейших, "тоже пелагических" стадий развития. Случар=п_е‘рво15от
"рода наблюдаютсяу всех низших губок, последовательно проходя-
‘ЩИХ стадии бластульги паренхимулы, „а такжеиу’ некоторых дмета-
тенетическйх’ Léptolida; случаи второго рода обычны среди _L_§:pto1_id_a
‘H Hn3umx‘—ry6ox' и` "нередки у метагенетических сцифоидныхд-тогда
I как‘ превращен-иен паренхимул-ы.’ .131 пелагическую- же чмедузоидную
" 296
стадию [наблюдается лишь у немногих I‘PIIIO1"eHeTPl'1eCKPIX'C11I/Iq)OMe-
.дуз (например, Pelagia) И типично для иглокожих. 1
‘ Конкретное рассмотрение Этих типов дальнейшего развития па-
гренхимулы мы начнем, из соображений удобст-ва, с ее преобразо-
вания B планулу.
з. превращение двуслойной "личинки в’ трехслойную личинку
типа планулы .
А. Метагенетические гцдромедузьл. При преобразовании парен-
„химулы в следующую за ней стадию планулы (см. рис. 95, 11—12)
-- тело личинки обычно заметно вытягивается в длину, при-чем ее зад-
ний конец часто утончается по сравнению с передним, остающимся
‘широко закругленным. Иначе говоря, гетерополярность личинки
-приобретает теперь еще более наглядное выражение, чем раньше.
Среди жгутиковых клеток ктинобласта дифференцируются желези-
стые, чувствительные (нейро-эпителиальные) и сократительные
~ (нмышечно-эпителиальные) клетки и появляются интерстициальные,
.частью тут же превращающиеся в книдоцисты одного или несколь-
. ких. разных типов. Оформляется пограничная ‘пластинка, отделяю-
щая наружный эпителий личинки от ее внутренней клеточной массы.
. Периферические клетки этой последней выстраиваются в более пили.
менее правильный столбчатый эпителий и при этом поляризуются,
в то время как внутренние подвергаются. фагоцитозу. Освобождаю-
‘щееся таким образом пространство представляет собой зачаток
Пбудущей гастральной полости, постепенно расширяющейся затем
Зв направлении спереди назад. Особо следует подчеркнуть то об-
стоятельство‚ что эта полость возникает в компактной дотоле массе
- клеток" фагоцитобласта путем .фагоцитирования одних из них. дру-
Тгими (Мечников) и в связи с явно вторичной эпи-телизацией
"уцелевших. Это определенно свидетельствует, с одной стороны,
го невозможности подвести данный процесс под привычные нормы
тгаструляциии, с другой стороны, о начавшейся активизации амё-
‘боидных клето-к внутреннего пласта и проявлении ими свойст
настоящих энтодермальных жклеток. _ ‚г‘
: " Такимобразом, на стадии «планулы уже оформляется типично
трехслойная “организация.Ну‹1го2поа в характерной для нее ‚форме
Тдвух эпителиальных пластов, разграниченных тонкой гпластинкой
„промежуточного вещества; .При_этом кинобласт подвергается слож-
ной и разносторонней гистологической дифференцировке, в резуль-
дтате которой образуются"разнообразные высокоспециализованные
„т-"(нейро-эпителиальньхе, мышечные, хслэизистые),›или, напротив, кам-
с
I
к
г’: биальные клетки‘ (интерстициальные), из которыхчпоследние‚1втсвою-
дочередыгтгут, же" частично‘дифференцируются в характернейшую
для‘ Crridaria систему стрекательных иклеток. Все‘ эти- разнообразные
тсклеточные...‘ элементы ._ остаются; смешанными св виде разнородного: и
многорядного эпителия типично эпидермального‘Цха-рактера‘;‘полу-
Ьнатющего‘ дтеперьтгзначениет настоящей оформленнойэкагг.еде-др м ы.
297
к
Одновременно с этим в ранее аморфном иоднородном фагоцито-
бласте происходит разделени-е клеток на трофически активные,
складывающиеся со временем в полноценную эпителиоидную э н т о-
дер М у, И агборти-вные питательные КЛЕТКИ, рЭСХОДУЭМЫС, ПОВИДИ- '
мому, на поддержание жизнедеятельности личинки и обеспечение
ее морфогенеза. `
Общим результатом всех этих процессов является чрезвычайно
p€3KO€ повышение УРОВНЯ opranusaunu ЛИЧМНКИ, ПрИООРСТСНИС ЭЮ;
той цитологической и тканевой дифференцировки, в которых полнее
всего выражается качественная специфика планулы. Однако дости-
гаемая ею высота организации оказывается все же Недостаточной,
чтобы планула могла самостоятельно добывать себе пищу. Оста-
ваясь пока замкнутой, лишенной дееспособного пищеварительного
аппарата и типичной органологической дифференцировки, она
продолжает быть узко расселительной стадией развития подобно
предшествующим ей стадиям паренхимулЬ1 и бластулы. Сохраняя
таким образом свои биологические свойства синзооспоры, планула
сохраняет и все проморфологические особенности последней, при-
обретающие только еще более отчетливое структурное выражение.
Так, признаки ее переднего полюса обогащаются появлением здесь
чувствительных элементов в кинобласте и к этому добавляется ряд
друпих проявлений гетерополярности в гистологическом строении
личинки. Не изменяется и направление градиента физиологической
активности (-см. рис. 87), установившееся еще у бластулы. Таким
образом, весь личиночный период постэмбрионального развития
осуществляется здесь в рамках одной и той же жизненной формы
синзооспоры и, в соответствии с этим, без каких-либо перестроек
архитектоники личинки.
Б. Метагенетические Scyphozoa. Гистологическая дифференци-
ровка кинобла-ста протекает тут примерно так же, как у личинок
гидром-едуз и также приводит к его превращению в полиморфный
эпителий эпидермального типа, т. е. в подлинную эктодерму. В со-
став этого эпителия входят, помимо неизменжившихся жгутиковых
клеток, более мелкие нейро-эпителиальные, снабженные чрезвы-
‘чайно тонкими, нитевидными, ветвистыми и частью узловатыми от-
'pOCTKaMI/I, мышечно-эпителиальные, секреторные [И ДВЕ категории,
стрекательных клеток, более крупные из которых располагаются
только в заднем полушарии личинки. Других интерстициальных
клеток, кроме развивающихся в стрекательные, у планул Scyphozoa
не описано. I
У ЗЭДНСГО ПОЛЮСЗ ЛИЧИНХИ кгинобласт заметно утолщается всл-ед- -
ствие особенно интенсивного деления составляющих его клеток,
становящихся тут очень высокими и узкими. У Chrysaora общее
число эктодермальных клеток планулы намного выше, чем на
.CTa,71I/I'I/I паренхимулы‚ так как ОНИ все ПрОДСЛЫВЭЮТ еще ОДНО ИЛИ
два более или менее синхронных деления, и ‚их размеры достигают
теперь своего минимума. .
Образование энтодермы происходит, как правило, совершенно
-298
иным путем, чем у heptolida, a именно путем пипотропного враста-
ния в полость тела связного клеточного комплекса; такое врастание
обычно считается инвагинапией, но в действительности вряд ли оно
заслуживает этого Названия. Так, у наилучше изученной в этом ог-
ношении Chrysaora (рис. 97) врастание начинается с появления
в центре утолщенной обла-сти тела личинки (т. е. на ее заднем по-
люсе) плотного скопления клеток, вдающегося в бластоцель в виде
бугорка; вслед за тем внутри этого бугорка появляется небольшая
полость, ‘продолжающаяся
кнаружи в виде очень узкого
канала, а стенки образую-
щегося таким образом Me-
шочка приобретают эпите-
лиоидное строение. В даль- 0o
нейшем этот однослойный
мешочек все более и более
разрастается, оставаясь до-
вольно тонкостенным, при-
чем канал ‹<бластопора›> .
либо сохраняется в том же '
виде сквозного капилляра,
либо сжимается совсем, пре- о
вращая тогда мешковидный _
зачаток энтодермы в совер-
шенно замкнутую капсулу. '
Как видно из этого описания Рис. 97. «Гастрултция»
и соответствующих рисун-
ков, гаструляция Chrysaora
представляет собой скорее
врастание, чем инвагинацию, _
а образующийся зачаток гастральн-ой полости не имеет, пожалуй,
ничего общего с типичным, инвагинационным архэнтероном. Это-
полость схизоцельн-ого характера, мало чем отличающаяся по спо-
собу своего образования от таковой планул гидромедуз. Однако
сам материал, из которого строится будущий кишечник, создается
у Chrysaora совсем иначе, чем у гидромедуз,—— фактически заново
в результате бурной пролиферации клеток из одного узко л-окали-
зованного очага, так что его генетическая связь с протобластом
оказыва-ется очень отдаленной. Иначе говоря, зачаток кишечника
возникает тут не из ранее оформившегося фагоцитобласта, а как
особое новообразование.
У других метагенетических Scyphozoa, например изученной
Мечниковым Nausithoé, гаструляция уже больше напоминает, нови--
димому, инвагинацию, но и здесь далеко не типичную (рис. 98, 2-7).
По описанию Меч-никова, материал будущего кишечника представ-
лен уже у паренхимулы довольно многочисленными клетками, рав-
номерно заполненными окрашенными (желтыми) включениями и
тем отличающимися от всех прочих клеток кинобласта, у которых
Ё
спифомедузы
Chrysaora. Образовагие зачатка кишеч-
ника и его погледуюп ее развитие (по
Тессье)
299
риз gsgr: ч С ы. ‘ U ‘ . ,.«l.‘
Fm‘ f . . ePBV1‘1Hb1Pl метаморфоз метагенетическои сцифомедузы Naus1thoe:.
'Pic=1e3HoBer;)pI;leo‘::-xZ1‘a;zlx'1* 6n'acTy'na;' ' 5 н стадии Инвггингдииз .
'l3eHxnMYIIY‘“6 и 7‘ инационнчч п°л°°т".„(“"°Рв“Ч*?°й кишки
‘осевшаяпланула —r1n;:B;§:1]»1L::1: ас? астания бластопора" вид
-_ к м ~- I» g V _ Я Я- В, двтюляризованньхй зачаток сидя й -. _ .
. . .. . `\ ... .. . .. .-xwofipagggaflna-.n0n,hna .(nD_N1e.qH"K0B}sr)' .. „д“? .„_9Тад“'?›„9„ "3_“”,‘,r”,°
' "4; 5 -'замыкание' бластопора,
»)`и превращение личинки_ вла-
с заднего полюса‘ личинки; 8-
эти включениэтсосредоточены только B'»6a3aJ1bHo1i‘i ихполовине. Эти
клетки, располагающиеся на заднем. конце тела личинки, впослед-
ствиисообща впячиваются в бластоцель, увеличиваясь при этом’
в числе; возникающее таким образом вдавление на заднем: полюсе
личинки постепенно углубляется, суживается и в конечн-ом счете
закрывается -совсем. Получившийся замкнутый энтодермальный зам
чаток очень объемист (он практически заполняет всю полость тела,
плотн-0 прилегая по всей окружности к внутренней поверхности ки-д
нобласта) и массивен; его полость слабо развита, а в дальнейшем
совершенно исчезает: «Энтодермальные клетки так прочно‘ скреа
плены друг с другом, что их трудно разъединить; к тому же они ле-
жат так тесно, что гастроваскулярная полость не обнаруживается»
(Мечников, цит. раб.‚ стр. 87). Таким образом, к концу стадии пла-
нулы личинка МацзйЬо-ё приобретает строение компактной парен-
химулы, но приобретает его вторично, имея уже сильно дифферен-г
цированную эктодерму и другие специфические свой-ства планулы.
Так или ‚иначе, будущая энтодерма метагенетических сЦифоид-ё
ных являет-ся продуктом специального морфогенетического процесса,
происходящего уже после начального рассл-оения протобласта, по-
путно с преобразованием однородного кинобласта в сложно диффе-
ренцированную эктодерму. Этот процесс имеет все основания счи-
таться подлинной гаструляцией (хоть и не обязательно инвагина-
ционной)‚ а оформившаяся таким путем личинка—гаструлой, но
гаструлой трехслойной, еще замкнутой и остающейся пока не спо-
собной к анимальньлм формам питания.
4. Оседание личинки и ее превращение в зачаток сидячей
вегетативной стадии
Хорошо известно, что оседание мерцательных личинок метаг_е-'
нетиических книдарий и губок и их превращение в последующие, не-
изменно сидячие, стадии развития может происходить в разных
случаях либо еще на стадии паренхимулы или амфибластулы, либо
уже после преобразования личинки в трехслойную планулу. Вместе
с тем мы совершенно не знаем примеров непосредственного превра-.
щения бластулы (синзооспоры) ‚в прикрепленную вегетативную
форму. Это показывает, что достижение личинкой двуслойного
строения необходимо‘ предшествует ее оседанию, что перестройка
в сидячую фазу возможна лишь при условии предварительной диф- -
ференциации ее клеточного материала не пменее чем на две системы
клеток. ' '
с. А. Метагенетические Leptolida. H e II o с р е д ст в е н н о е п р е-
вращение паренхимулы В зачаток СИДЯЧЭЙ, Be-'
I‘ е т а т и в н о й с т а д И и наблюдается среди L,epto1ida как у не+
которых антомедуз (Oceania), так ни у ряда лептомедуз (L5aodic_ea',
Mitrocoma), протекая у тех и других приблизительно одинаковым
образом. «После более или менее продолжительного периода плаф-
вания, —фпишет Мечников, -.— они ‚оседают на дно, замедляя движе-
301
ние своих ресничек, но сохраняя при этом первоначальную форму‘
тела. Помещая в бокалы предметные стекла, мы находим на них
КОЛбЗСОВИДНЫХ, выпрямленных ИЛИ ИЗОГНУВШИХСЯ ЛИЧИНОК, еще-
обнаруживающих все ранее присущие им особенности строения, и в
том числе жгуты. Последние, однако, вскоре исчезают, и на всей
верхней поверхности появляется тончайшая кутикулярная оболочка,
которая постепенно утолщается при дальнейшем развитии и в не-
которых местах отслаивается от эктодермы». Црикрепившаяся
плашмя личинка превращается, таким образом, в зачаток гидро-
ризы, или «корневища» будущей колонии, нередко начинающий‘
затем ветвиться, прежде чем дать начало одному или сразу не-
скольким гидрантам. В других случаях личинка прикрепляется не
параллельно, а перпендикулярно к субстрату, причем ее передний
конец распластывается в виде диска, а задний остается свободным‘
и торчит тогда вверх в виде тонкого, отростка (рис. 99); любопытно,
что у Mitrocoma annae личинка может прикрепляться как тем, так
и другим способом. «Лишь долгое время спустя прикрепления обра-
зуются первые стрекательные капсулы, появляющиеся в месте, соот-
ветствующем заднему концу свободноплавающей личинки. Полость
в энтодерме появляется у личинок этого типа также долгое время
спустя после их прикрепления, причем мне не удалось установить,
возникает ли она тут также путем внутриклеточного пищеварения»
(Мечников, цит. раб.‚ стр. 75). Образование первого гидранта про-
исходит путем соответствующей перестройки свободного, заднего
конца гидроризы, а в случае ее ветвления они формируются либо
одновременно на двух ее противоположных концах, создавая харак-
тер-ные биполярные образования, либо на многих свободных концах
гидроризы, как это происходит, например, у Oceania.
Превращение планулы в сидячую вегетатив-
H у ю с т а д и ю протекает в разных случаях неодинаково в смысле
способа прикрепления личинки и объема испытываемых ею изменю-
ний, но всегда осуществляется путем ее далеко идущей дедифферен-
цировки, выражающейся в сбрасывании всего мерцательного по-
крова, в выделении кинобластом тонкой кутчикулярной оболочки—
пери-сарка и, разумеется, в переходе личинки в неподвижное состоя-
ние. Разнообразие конкретных форм такого метаморфоза может
быть сведено к следующим двум типам: .
1) Тип Stomatheca. Планула ложит-ся на субстрат плашмя, па-
раллельно его поверхности, и прикрепляется к нему по всей длине:
таким образом она н-епосредственно превращается в зачаток стеля-
щейся гидроризы будущего полипа. Первый гидрант образуется в
виде направленного вверх выроста в центре верхней поверхности
пидроризы, т. е. перпендикулярно к ее длинной оси; одновременно
с этим гидрориза дает ростки в различных направлениях, становясь
двух-, трех- или четырехлучевой (рис. 100).
2) Тип Clytia. Личинка прикрепляется к субстрату своим перед-
ним полюсом, распластываясь при этом в виде ди-ска, имеющего
вначале» неправильные очертания, но впоследствии могущего при-
302
в
таи
авт‘?
% и
%§§%g2“-€’«:‘§fi
Ёж
и ’
в,
гей?
.:?;%%
iliiliauszaiifiix
i
gig
«:3;
ilggii ‘
W
так
„из
W д Ё
ж? mg
.. „её“
питии
_.1„шишишазтинтииадаптации
ЁЁЁ`®"”‘ЙЁ g
liiififififiiza
V %§§§E?3‘35§i3EE:a:
Рис. 99. Первичный матаморфоз метагенетических гидромедуз:
1 2——'Г1ага pileata, Ha‘IaJ10, образования фагоцитобласта; Зтпаренхимула; 4-5 Octorchis
gégenbauri, превращение осевшей планулы в полипа. б—10-—М11:госота аппае, образование
фагоцитобласта (6,7); 8—l'laP8HX"MY-"3; 9'—0C€Il3HH€ TUIBHY-“bl H 7¢’—“P9BP3"4°H”9 “-“3“‘Y-“Ht
прикрепившейся плашмя к субстрату, в гидроризу основателя колонии (по Мечникову)
303
ц‘
а . . . . . b еи-
обрести довольно совершенную радиальную симметрию В дал н
шем центральная часть диска, отвечающая ЗаДНЭМУ ПОЛЮСУ Личинки’
начинает выпячиваться вверх и образует стебелек первоЁ^ЁМгиД5ЁНёЁ‚
натвершипе которого. ‚вскоре оформляется. его головка, . р т . ‚
13-18)-.- _ * _ › . A
отсюда" ‘видно, что проморфологически соотношения между “ma-
0 т __ ‘ И —
чинкои и образующимся из нее полипом основателем колрни
могут быть весьма различными. В ПерВОМ 'CJ1Y‘*3€. У Stomat eca’ Не
- "- ВЭННОСТИ
приходится даже и говорить о наличии какои либо преемст _
в архитектонике этих стадий, поскольку главная ось гидранта Рас
полагается перпендикулярно к первичной оси личинки, а эта послед-
няя фактически исчезает при pa3H0CT0P0HHeM разрастании Гидро‘
ризы. Происходит очевидная д е п о л я р и з а ц И я личиН0ЧН0Г0
- V_ тела (одним из первых
проявлений которой
служит исчезновение
у, осевшей личинки
‘имевшихся различий в
конфигурации ее кон-
цов), сменяемая затем
ее подчинением вновь
устанавливающейся ра-
диальной симметрии
гидранта. При разви-
тии по типу Clytia, при
„котором личинка сплю-
Рис. 100. Превращение планулы Stomatheca B
сидячую полипоидную стадиюг
1_пдануда‚ прикрепившаяся плашмя к субстрату:
2-—появление выроста на ее верхней стороне; 3—8;u-1; U_u,IBae-pcfl ‚В лепешку
па -— основат в
"штедующие °§3§‘.'3?:u§qi§I?;M3§¥¥'e’ixx§§:y") B. направлении СВОЭИ
главной оси и прини-
мает, при ЭТОМ неправильную форму, O*Ha несомненно ТОЖЕ ДЕПО-
ляризуется, и лишь затем перестраивается по новому. Pal1PIaJ1bH0My,
плану симметрии полипа. Однако главная ось ЦОЁЛЕДНЕГО ЁОЁГЁШЁЁЁ;
тут сосью прикрепления личинки, и его оральныи полюс о р у ая
из материала заднего конца личинки, так что временно утрачена)-
полярность восстанавливается гидрантом в ее прежнеи форме. М-
этому здесь можно говорить о наличии лишь н е п р я м о и п р е e-
Ствен н ост и, главной оси и ее полюсов. Таким образом, сохра-
нение первично-й личиночной оси в процессе дальнеишего развития
здесь не имеет места. Следовательно, обычное представление о кни-
дариях как первичноосных формах, пожизненно сохраняюЩИХ
Я
первичную ось в качестве главной оси на всех стадиях развития,
не оправдывается И в этом отношении.
_ Другие „изменения организации, происходящие при превращении
планулы в сидячую стадию полипа, в сильной мере зависят от того, '
no какому из типов осуществляется это превращение. Они относсрс;
тельно невелики у форм, сохранЯЮЩИХ ПРИ МЭТаМОРфОЗе СТР
эпителиальное строение и, естественно, гораздо более значительны
в тех случаях, когда планула совершенно Деполяризуется перед
3047
своим превращением в гидранта. Так, у С1у’с'1а фагоцитобласт т-еряет
В ЭТОТ МОМЕНТ СВОЕ ЭПИТЭЛТИЭЛЬНОС расположение, И НЗМЕЧЭВШНЯСЯ
в нем полость временно исчезает. Иначе говоря, деполяризация ве-
дет тут к возвращению к моноэпителиальному строению паренхи-
мулы, и гастральная полость полипа создается затем заново — путем
повторного упорядочения клеток внутреннего слоя распластав-
шеися личинки.-
Дальнейшие этапы метаморфоза включают дифференцировку
первоначально трубчатого зачатка гидранта в более сложно рас-
члененный полипоидный организм, формирование им щупалец и
появл-ение рта на его дистальном конце.
Б. Известковые губки. Превращение паренхимул Са1сагеа в си-
дячую вегетативную стадию во многом ‘напоминает ход этого про-
цесса у Heptolida. Личинка прикрепляется к субстрату своим перед-
ним концом, уплощается и полностью дедифференцируется, превра-
щаясь B аморфную, сплошную массу амёбоидных клеток. В первые
моменты после оседания они располагаются таким же образом, как
у личинки, но вскоре клетки первичной паренхимы начинают пооди-
ночке выходить на поверхность тела осевшей личинки и постепенно
складываются в сплошной -д е р M a Л ь н ы й с л о й, принимая при
этом характер покровных клеток—пинакоцитов. Таким образом
оформляется стадия «к у к о л к и», состоящая из однослойного по-
кровного эпителия, образованного производными клеток паренхимы,
и бесформенной внутренней клеточной массы, состоящей из бывших
жгутиковых клеток кинобласта. Меняясь своими местами, эти две
системы клеток и создают то «извращение зародышевых листков»,
которым так славятся губки. Однако подобное «извращение» вовсе
не имеет места при метаморфозе амфибластул—-будь то «вторич-
ные» амфибластулы типа Leucosolenia ИЛИ «первичны.е>>—типа
Syoon. B том и другом случае жгутиковое полушарие личинки впя-
чивается при ее оседании внутрь зернистого, которое, таким обра-
зом, сразу же становится наружным сводом «куколки» и тут же
преобразуется в ее дермальный -слой; последний обволакивает затем
со всех сторон содержащуюся в нем компактную массу дедифферен-
цир-овавшихся жгутиковых клеток. ‘Эти клетки располагаются вна-
чале совершенно беспорядочно, но затем наружные из них выстраи-
ваются в более или менее правильный эпителиоидный слой, под-
стилающий слой первичных пинакоцитов.
Дальнейшее развитие куколки (рис. 101) заключается в диффе-
ренциации первичных покровных клеток на собственно пинакоциты,
пороциты и склеробласты, вырабатывающие известковые спикулы,
в появлении полости в центре внутренней клеточной массы и в на-
чинающемся превращении ее элементов в жгутиковые клетки
(хоаноциты) ‚и различные типы амёбоидных (амёбоциты). На этой
стадии, называемой преолинтусом или клистолинту-
сом‚. молодая губочка остается еще замкнутой: ее воротничковые
клетки еще не оформились и элементы будущей мезенхимы не раз-
граничены от Энтодергмальных. Эти последние обычно представлены
20 Эмбриология беспозвоночных 305
пока однородными амёбоиднь1ми клетками с небольшим ядроми
довольно обильнымивключениями, среди которых рассеяны более
Мелкие клетки с пикнотическим ядром, очевидно дегенеративного
характера; многие из этих клеток поглощаются энтодермальньтми,
Рис. 101. Заключительные этапы первичного метаморфоза известковой губки
Clathrina:
1, 2.—- стадия «куколки»; 3 — появление в ней полости; 4 — дифференцировка Хоаноцитовз
а —пинакоциты; б —- пороцит; в-юкперобласты; г-—амебоциты мезенхимы; д— хоаиоциты,
е-— образуклциеся из них амёбоидные клетки; на всех рисунках дегенерируюцию клетки
зачернены (по Минчину)
обладающими ярко выраженной фагоцитарной способностью.
К концу стадии преолинтуса (а в Некоторых случаях даже и
раньше) среди клеток внутренней дмассы начинают появляться
отдельные амёбоциты, а некоторые из эпителизующихся теперь
энтодермальных клеток превращаются в зернистых эозинофилов и
мегацитов, уходящих затем отсюда в мезенхиму.
306
При переходе в следующую стадию-ю л и н т у с а—оформляется
ирригационная система губочкпи; на ее дистальном полюсе проры-
'Ba€TrC51'OCKyJIyM, 3HTOJ1€pMaJIbHbIe КЛЕТКИ выстраиваются В ПраВИЛЬг
ный эпителий, одеваю-ший всю полость тела (Ногпосое1а) или только
ее жгутиковые камеры (Heterocoela), и дифференцируются в хоа-
ноцитов; появляются поры с их прпиводящими каналами, а дермаль-
ный слой тела и мезенхима испытывают дальнейшую ди-фференциа- -
цию своего клеточного материала. Одновременно с этим постепенно
меняется и форма тела губки, становящаяся бокаловидной, и оно
приобретает характерные для него особенности архитектоники—
одноосно-гетерополярное строение с неопределенным числом пло-
скостеи симметрии.
Таким образом, архитектоника сидячей стадии олинтуса, явно
отвечающей полипу- основателю книдарпий, создается и здесь факл
тически заново, путем коренной перестройки деполяризованной ста-
дии «куколки»; стало быть, и здесь проморфологическая преем-
ственность между первичной вегетативной -стадией и личинкой фак-
тическпи не имеет места.
В. Scyphozoa. Преобразование осевшей планулы в поли-поидную
стадию, называемую тут сцифистомой, идет по тому типу, как у
Clytia (Mansithoe) ИЛИ же (у Сйгузаога), начинается с того, что
ее эллипсоидальное тело постепенно принимает характерную бока-
ловидную форму; его задние две трети превращаются при
этом в чашечку полипа, а передняя треть становится его ножкой, или
стебельком. Вскоре после этого на краях образующейся чашечки
одновременно появляются зачатки двух первичных щупалец, а в
центре ее .поверхности—округлое, несколько вдавленное светлое
пространство: зачаток будущего ротового отверстия. По мере роста
первичных щупалец зачаток рта становится все более заметным и
приобретает овальную форму, вытягиваясь в первой, щупальцевой,
плоскости симметрии тела. На первых стадиях образования рта экто-
дерма того полярного уча-стка тела, на котором он появляется,
сильно растягивается и истончается; образующий его эпителий пре-
вращается из столбчатого в плоский однорядный вследствие при-
остановки клеточных делений в этом участке эктодермы при интен-
сивном ее разрастании по всей его окруж_ности. Вслед за этим
энт-одерма‚ подстилаюжщая этот утонченный уча-сток покровного
эпителия, начинает выпячиваться горбом, все сильнее растягивает
его, вследствие чего он разрывается; обнажившаяся таки.м образом
Энтодерма в свою очередь лопается по середине, „после чего -края
разрыва обоих пластов срастаются друг с другом по всей своей
окружности. Формирование рта заканчивается к моменту достиже-
ния первичными щупальцами их окончательной длины. В это время
появляются зачатки второй пары щупалец, нампечающие собой
вторую плоскость симметрии сцифистомы, пересекающую первую
под прямым углом; таким образом, двулучевая симметрия двущу-
пальцевой стадии сменяется теперь правильной четырехлучевой, ста-
новящейся проморфологической основой строения всех следующих.
20* 307
стадий развития животного; эти две первые плоскости симметрии
обозначаются как перрадиальные. С появлением второй «пары щу-
палец овальное ротовое отверстие сцифистомы начинает раздаваться
в ширину и становится совершенно круглым, когда эти щупальЦЫ
достигнут того же размера, как первичные щупальцы; с этого мо-
мента молодая сцифистома начинает самостоятельно питаться. Вся
ее гастральная полость выстлана на этой стадии однородным жгу-
тиковым эпителием, состоящим из сильно вакуолизированных, бо-
гатых включениями клеток, среди которых рассеяны единичные
железистые и стрекательные клетки. Дальнейшая органолопическая
дифференцировка сцифистомы заключается в оформлении харак-
терной для нее системы тэниолей, появлении желудочных септ
и в о-бразовании ищупалец треть-ейи четвертой очереди. ТЭНИО-ТШ
возникаютв интеррадиусах ротовой поверхности чашечки, в непо-
средственном соседстве с ртом, в виде четырех симметричных впя-
чиваний эктодермы—септальнь1х воронок. Постепенно углубляясь
в полость тела, дно этих воронок дает начало плотным клеточным
тяжаам, врастающим между э-кто- и энтодермой и постепенно дости-
гающим внутренней поверхности подошвы полипа‚ С КОТОРОЙ ОНИ
срастаются.
Параллельно с органологической дифференцировкой СЦИФИСТОМЫ И ПОЁЛЁ‘
довательной сменой одной формы симметрии другои происходит И ее даль е
шая гистологическая дифференцировка. В эктодерме различаются, KEIK И У ГИД“
роидов, эпидермальныеклетки, мышечные, утрачивающие связР с эпителием и
располагающиеся под его базальной поверхностью, и неиро-эпителиальные
клетки, отростки которых складываются в субэпителиальное нервное сплетение.
Среди эпидермальных клеток различаются, в свою очередь, секреторные или
скелетогенные‚ вырабатывающие тонофибриллы и аморфныи хитиноидНЫИ Сек‘
рет, с помощью которых сцифистома прирастает к субстрату, и стрекательные
клетки трех различных типов. Таким образом, набор гистологических элементов
эктодермы, имевшийся уже у планулы, дополняется тут только однои новои
категорией стрекательных клеток. Однако их размещение и степень о развития
оказываются у сцифистомы существенно иными. Так, мышечные и неиро-эпите-
лиальные клетки сосредоточиваются,главным образом в области перистома и ЩУ-
палец и очень немногочисленны в ДРУГИХ ЧЗОТЯХ Теда- ТО Же, В Общем, МОЖНО
сказать о стрекательных, один из типов которых приурочен исключительно к
перичстому, а остальные появляются в изобилии, начиная с З-щупальцевои стадии,
сначала преимущественно на концах щупалец, а затем и по всеи их длине. На-
ряду с‘ этим первоначально однорядный столбчатый характер наружного эпите-
ЛИЯ ПОЧТИ ВСЮДУ ПОДВЭрГаЕТСЯ ТЕМ ИЛИ ИНЫМ изменениям-особенно В области
стебелька, где он становится многослойным.
Еще более существенны изменения, происходящие в энтодерме. Она диффе-
ренцируется на: 1) опорную энтодерму щупалец, имеющую характер однород-
ной паренхимы в их рукоятках и однорядного столбика-в жгутах; 2) очень
правильный столбчатый эпителий глотки, клетки которого отличаются отсутст;
вием жгутов и своей призматической формой; 3) общую выстилку гастральнои
полости, состоящую из слабо интегрированных жгутиковых клеток различного
характера; 4) «глубинную» энтодерму дна этой полости‘, состоящую преимуще-
ствен-но из очень крупных клеток, перегруженных резервными включениями;
наконец, 5) железистую энтодерму гастральных септ, образованную главным
образом высоко дифференцированными секреторными клетками.
’ ‘B процессе развития сцифистомы постепенно оформляется и мезоглея, выра-
батываемая обоими пластами тела и получающая все признаки диффузной_ткани
внутренней среды, образованной блуждающими амёбоцитами. Наконец, диффе-
308
‘тырехгранного хоботка.
ренцируется и мышечная ткань тэниолей, образуемая эктодермальными миобла-
стами дна септальных воронок. При этом наружные из этих клеток превраща-
ются в мышечные, а внутренние сливаются в синцитий, образующий стержень
тэниоли. Эти последние приобретают таким образом характер гладких продоль-
ных мышц, обеспечивающих высокую сократимость тела сцифистомы. Врастание
септальных воронок в первичную полость тела и формирование тэниолей имеют
своим результатом образование невысоких продольных выступов энтодермы—
септ, делящих гастральную полость на четыре сектора, и приобретение ртом его
дефинитивной крестообразной формы. Заключительный период органогенеза сци-
фистомы включает последовательное образование четырех интеррадиальнь1х и
восьми адрадиальных щупалец и окончательное оформление перистома в виде че-
Результатом всех этих превращений являются дальнейшие ви-
доизменения в характере симметрии сцифистомы. Став из двулучевой
чтетырехлучевой при появлении второй пары щупалец, она превра-
щается в восьмилучевую при появлении септальных воронок и ин-
террадиальных щупалец и дополняет-ся неполной 16-лучевой с мо-
мента образования адрадиальных щупалец. Таким образом, разви-
тие симметрии в онтогенезе сцифистомы идет в сторону последова-
тельного увеличения числа ее плоскостей, закономерно проходя все
этапы .от двулучевой до 16- (и более в других случаях) лучевой.
В целом можно сказать, что формирование полипоидной стадии
идет у Scyphozoa no тем же общим нормам, как у Нуагоиоа, но за-
ходит значительно дальше в смысле ее органологического расчле-
нения.
5. Дальнейшее развитие вегетативной стадии
Оформлением одиночной вегетативной особи типа олинтуса или
полипа завершается первый — э н д о т р 0- ф н ы й — период постэм-
брионального развития, обозначенный намти как ‚первичный «Mem-
морфоз. Теперь начинается следующий, обычно очень длительный,
период разв-ития‚ характеризующийся активным добыванием пищи
из внешней среды, т. е. уже экзотрофным питанием. Этот
‚период ЖИЗНЕННОГО ЦИКЛа ОСУЩЕСТВЛЯЕТСЯ ТРСХСЛОЙНЫМИ И МНО-
гоклеточными особямяи, сохраняющими или повторяющими строение
родоначальной одиночной сидячей стадии; характерной их чертой
служит присущая им неограниченная способность к -вегетативному
размножению путем почкования, столонизации, продольного или
поперечного деления и т. д., т. е. высокая степень делимости этих
стадий и исключительно высокая способность к регенерации. У ча-
сти форм получающиеся таким образом новые вегетативные особи
разъединяются и ведут дальше одиночный образ жизни; гораздо
чаще они остаются связанными в виде вегетативных колоний, члены
которых (зооиды) в той илни иной мере тесно объединены в физио-
логическом и структурном отношении. Характерным для губок яв-
ляется бесформенность создающихся таким образом группировок
при крайней обезличенности их компонентов, т. е. чрезвычайно низ-
кий уровень индивидуальности как колонили в целом, так и ее зоо-
идов. У низших Cnidaria, напротив, высоко индивидуализованы как
колония в целом, так и все располагающиеся на ней зооиды (пид-
309
ранты), так что здесь приходится говорить о сосуществовании и сов-
мещении двух уровней индивидуальности —- к о л о н и а л ь н ог о
и з о оидн ого. Так или иначе, Вегетативный период цикла боль-
шинства низших Metazoa характеризуется своей пмногочленностью,
тем, что он представлен целым комплексом ясно или слабо разгра-
ниченных генераций типа олинтуса или полипа.
А. Метагенетические Leptolida. Развитие сидячей! вегетативной
стадии метагенетических гидромедуз (рис. 102) ведет, как правило,
к построению сложно-и мно-
гочленной колонии, пред-
ставляющей единое морфо_-
физиологическое целое. Под-
ведение этого процесса под
понятие «бесполое размно-
жение» сталкивается с бо-ль-
шими трудностями, так как
он не ведет тут к увеличению
числа экологически иефизио-
логически самостоятельных
организмов, т. е. не является
подлинным размножением.
Равным -образом его невоз-
можно свести к не доведен-
ному до конца почкованию,
Рис. 102. Развитие колонии Amphisbetia ПОСКОЛЬКУ "почкование НОВЫХ
operculata: гидрант-ов составляет только
1, 2-—-образование первого гидрантщЗ-моло- одну из сторон этого Upo-
'§fi£‘p§3¥§§;"”§_‘3?i‘£.i,‘%‘§Z‘,’?ap?,?;'Z;%’l3”‘S§"éi“ ЁЁдБЁТЁ Цесса; другой его стороной
pi;:::;(::*m::;*;,::T::¢°6"*:'m;p:%:::*:z: являются реет И ветвл-ение
ного из ее гидрантов (по тессье) самого ствола колонии и си-
стемы столоно-в, образующих
цено с а р к, или общее тело колонии. Всл-едствие этого колония
гидроидов не разложима на отдельные зооиды (гидранты), н-е яв-
ляется простым их агрегатом, а представляет собой гораздо- более
сложную систему. Ее конструктивную основу образует разнообразно
в-етвящийся ценосарк, а выпочковываемьте им гидранты являются,
по существу, его вегетативными органами. Кратковременность инди-
видуального существования отдельных гидрантов (по сравнению
с продолжительностью жизни колонии в целом), обусловливающая
известную «текуче-сть>> их состава, в сочетании с отсутствием какои-
либо преемственности между ними (поскольку они чаще всего за-
кладываются совершенно независимо друг от друга, в различных
точках ценосарка) и целым рядом других признаков, определенно
свидетельствует о том, что гидранты являются скорее продуктами
общего тела колонии, чем наоборот, как это обычно принято считать.
.Фигурально выражаясь, можно сказать, что гидранты являются
детищем общего тела колонии, а не его создателями.
6
Таким образом, толкование колониальной сидячей стадии низ--
310
ших Нус1го2оа как едлиной многочленной особи гораздо лучше со-
гласуется с фактами, чем противоположное. Правда, сама ее поли-
мерность‘, способ, которым эта последняя создается, строго
локализованный характер роста и почкования ценосарка и его ти-
пично растительная архитектоника очень плохо вяжутся с тем пони-
мание-м индивидуальности, которое «принято в зоологии. Однако
если подойти к колонии гидроидов с другой — ботанической — Mep-
-кой, то ее значение особи покажется несомненным. Все ее, необыч-
‘Hue для животных, особенности окажутся типичными для Согпю-
phyta И найдут тогда свое объяснение в действительно и букваль-
но зоофитной природе сидячихвегетативных стадий низших. кни-
‘дарий. ‘ р
С этой точки зрения легко объяснимо и то обстоятельство, что
подлинное вегетативное размножение почками, отделяющимися от
материнского тела (т. е. путем доведенного до конца почкования),
имеет место лишь у немногих, более специализированных Hydrozoa
’(TaKHX, например, как гидры) и совершенно не встречается у более
примитивных. Оно является в этой группе не начальной, а конечной
СТЭДИСЙ ЭВОЛЮЦИИ вегетативного ПОЧКОВЗНИЯ, .принятием ИМ на себя
тех генеративных и расселительных функций, какие здесь нормально
и первично несут одни только половые почки-медузы. Таким
образом, перерастание почкования, столонизации и т. п. в настоящее
вегетативное размноженэие аналогично отделению разнообразных
вегетативных диаспо , наблюдаемом Соггпо h ta.
. У Р У
Б. Метагенетические Scyphozoa. Дальнейшее развитие метагене-
тических Scyphozoa идет по принципиально иному пути, чем
у большинства метагенетических гидромедуз, а именно как бы
в обход колониальности, путем сосредоточения вегетативных функ-
ций «в руках» самого полипа-основателя, называемого тут сцифи-
стомой, и превращения вегетативного размн-ожения из процесса
строительства ‘колоний в самое подлинное размножение. Столони-
"sauna, почкование и высокие регенеративные потенции полипа при-
водят к -созданию новых, совершенно независимых вегетативных
осо-бей, рано теряющих связь с материнским полипом и ведущих
‘дальше вполне самостоятельное существование, усаживаясь в близ-
ком соседстве с ним. Этот вид размножения в известной мере
ДОПОЛНЯСТСЯ рЭЗМНОЖЕНИСМ ПрИ ПОМОЩИ ТЭКИХ же ‚ПОЖНЫХ пла-
‘нул, или вегетативных личинок, какие наблюдаются у некоторых
-Leptolida.
Такое освобождение бесполого размножения от выполнения
‘функций построения колонии с очевидностью говорит о значитель-
ном повышении интенсивности вегетативной деятельности самого
‘l'I€pBOHOJII/I113, O ДОСТИЖЭНИИ И!М ТОГО ПОВЫШЭННОГО УРОВНЯ органи-
зации, .при котором колониальное существование уже не является
необходимостью. И действительно, сцифистома метагенетических
Scyphozoa олицетворяет значительно более высокую ступень поли-
поидной организации, чем наблюдаемая у первополипов—основа-
телей колонии метагенетических Нус1го2оа.
311
Глава XVII
ЭМБРИОНИЗАЦИЯ ПЕРВИЧНОГО МЕТАМОРФОЗА
У Г ИПОГ ЕНЕТИЧЕСКИХ Г ИДРОПОЛИПОВ
1. Строение яйца и первые стадии дробления
Оогенез часто протекает тут в весьма своеобразных формах,
существенно отличных от наблюдаемых у м-етагенетических гидро-
идных, например -в том, что второй период роста яица тут зачастую
осуществляется за счет активного хищничества амебоиднои «в это
время яйцеклетки (рис. 103) или пу-
тем слияния с нею многих других
клеток, плазма которых включается
в состав. такого плазмодия, а ядра
разрушаются. Организация зрелого
яйца мало чем отличается от тако-
вой яйца‘ метагенетических Ьерто-
lida. морфологическим выражением
одноосно-тетер-ополярного строения,
принимаемого яйцом к концу, по
крайней мере, его развития, служит
и здесь расположение ядра у ани-
мального его полюса, а иногда так-
же и вытянутая в длину‘ форма
яйца; физиологически это строение
проявляется в соответствующей ори-
ентации градиента восприимчивости
яйца к действию отрав-ляющих и
Рис. 103. Выросшее амёбоидное красящих веществ. Выделение на-
ЯЙПО Tubularia m€SembFYan’Ch9- правительных телец И плодотворе-
gggéfgggggfifigflfiteflfigfifiéafijflpg? ние приурочены к анимальному по-
ма яйца заполнена «ложноклет- “Юсу Яица? В ГОНОфОРаХ (В КОТОРЫХ
ками»—-ядрами ассимилирован- проходят не только заключительные
HEX ею I<fieT0K(110 ТИХОМИРОВУ) стадиих оогенеза, но «обычно и все
эмбриональное развитие) яйцо ори-
ентировано этим полюсом кнаружи, прикрепляясь к материнскому
телу своим вегетативным полюсом. Существующая в. яйчце двулу-
чевая симметрия проявляется только с начала дро-бления. Харак-
терны крупные размеры яиц: их диаметр лишь редко (напримлер,
у Hydractinia echinata) уступает 200 р и доходит до 350 д И
больше (пресноводные гидры), что ясн-о отражает интенсивность
их предшествующей вегетативной деятельности. Ее результатом ока-
зывается обильное, как правило, накопление в оо.плазме желточных
зерен, а часто также и еще не ассимилированных до конца пищевых
включений, наиболее характерными из которых являются так назы-
ваемые «ложноклетки», или желточные ядра, представляющие собой
ядра клеток, ранее заглоченных ооцитом (и находящихся на разных
312
стадиях распада). Размещение всех этих включений в плазме яйца
может быть довольно равномерным, но иногда они сосредоточива-
‚ются преимущественно в его центральной части. В последнем случае
„создается видимое разграничение эктоплазмы яйца от сильнее sa-
груженной желтком эндоплазмы; их разграничение отчетливее всего
у яиц Eudendrium capillare, y которых эктоплазма содержит лишь
мелкие темно оконтуренные зернышки, почти отсутствующие в эндо-
плазме, ‘заполненной зато более крупными оранжевыми или жел-
тыми желточными элементами. Яйца развиваются в гонофорах или
гонадах ‘в относительно очень небольшом числе, часто даже по
одному, что в одних случаях обусловлено малым числом ооцитов,
проникающих сюда извне (из обычно удаленных очагов их
образования или первоначального накопления), а в других-тем,
что из многих наличных ооцитов развивается только один (или во
всяком случае единичные), тогда как остальные идут ему на про-
корм.‘ Эмбриональное развитие почти всегда протекает тут же,
в гонофоре, под защитой и воздействием материнского тела, и лишь
как исключение — свободно, в морской воде (Hydractinia, яйца кото-
рой выбрасываются в воду сразу же после их созревания).
Первые стадии д-робления обычно не представляют каких-либо
специфических отличий по сравнению с соответствующими им ста--
диями развития м-етагенетических гидромедуз, по крайней мере
в отношении положения" и чередования его первых трех борозд и
самого характера этих боро-зд (рис. 104). Последние чаще всего бы-
вают односторонне врезающимися (Dynamena, Hydractinia, Go-
nothyrea, Cordylophora), HO иногда и полукольцевыми, постепенно
распространяющимися тогда по окружности яйца, начиная с одного
его полюса (Clava); так или иначе, первая из них неизменно закла-
дывается на анимальном полюсе яйца. «Углубление этой борозды
иногда идет весьма медленно, так что она долго остается очень
мелкой; при этом обыкновенно оба ее края скоро начинают за-
острять-ся, борозда же скоро значительно расширяется, так что на
разрезах она представляется более или менее широким углубле-
нием, ограниченным двумя резкими краями» (Tubularia; ТИХОМИрОВ,
1887; рис. 105).
Отклонения от этой нормальной схемы первых делений яйца
довольно обычны. Та-к, у Campanularia -calyculata И некоторых дру-
гих форм третье, экваториальное, деление явно неравномерно, имея
своим результатом разделение каждого из четырех бластомеров на
меньшую анимальную и значительно более крупную вегетативную
половину, т. е., в сущности, их разделение на микро- и макромеры;
в других случаях нарушается правильность группировки бласто-
меров уже со стадии 4, или же их размеры с этой же стадии неоди-
наковы (Dynamena); далее, известны случаи, когда борозда второго
деления появляется задолго до разъединения обоих первых бласто-
меров, пока первая борозда еще относительно неглубока.
Наконец, очень нередки случаи нарушения синхронности деле-
ния, вследствие чего число бластомеров возрастает почти ссамого
313
начала не в нормальной геом-етрической прогрессии, а, например,
согласно ряду 2——4—-6——8—10 и т. д.‚ включая и нечетные их числа
- ‘(17ubu1aria). Таким образом, прав Тихомиров, называядробление
"этих пидрополипов ‚полным неправильным ‘дробле-
' н, и е м. '
Неполнота последо-
вательного обособления
бластомеров с первых
же этапов дробления,
наблюдаемая у ту-бу-
лярии, может рассмат-
риваться как первый
шаг в направлении пе-
рехода такого, все же
V еще полного, дробле-
ния в неполное дробле-
ние, синцитального или
синтомическпо-го типа,
Рис. 104. Эмбриональное развитие гипогенети-
ЧЭСКИХ ГИЦРОПОЛИПОВЗ
'7-—6` -— Hydractinia echinata:7 -— первое деление яйца, осу-
ществляемое посредством врезавшейся борозды; 2— рис_ 105_ продольный раз. `
второеи 3 ——третье деления‘ хо ошо виден цент обеж- и ‚-
ный характер борозп; 4—- 1’2—кл1е’точная стадия; Ё, 6‘ — рез Яида ТУОУЛЯРИ“ В M0‘
две стадии превращения бластулыв псевдоморулу; 7 —— мент первого деления; на-
;1;.§;T;.$)uya'3a31 e::l1€-ia):I()1»l1l;?11i%3a': 770- '§f§S§§a-"139 ?T3E?f§°§5§§§§‘ верху ` вьщеленше "М на‘
ления эктодермы; 72—молодая, еще уплощенная Планула "разительные Тельца (по
(по Тессье) Тихомирову)
описанное у ряда форм, в частности некоторых видов Eudendriu-m.
B этих, перегруженных желтком, яйцах делению подвергается
только ядро с окружающим его скоплением плазмы, в то время как
‘желточная маоса яйца им не охватывается, и борозды дробления,
соответственно, не появляются вовсе.
2. Формирование морулы
Мы уже говорили выше, что дробление гипогенетических гидро-
_полипов принципиально отлично от дробления их метагенетических
сородичей в том, что оно имеет конечным результатом о-бразование
компактной морулы, а не бластулы. Этот результат достигается
(314
B разных случаях неодинаковыми путями: в одних — путем предва-
рительного формирования типичной пребластулы и последующего
заполнения ее полости продуктами дальнейших делений составляю?
щих ее клеток; в других —— вовсе минуя однослойную стадию в связи
с приобретением самым дроблением особых геометриче-ских форм
‘трехмерного, или кубического, дробления. Иначе говоря, морула
формируется в одних случаях тем же, в сущности, способом, как
паренхимула метагенетических видов, но только много раньше, за-
долго до завершения полного цикла повторных делений яйца и
дифференцировки их конечных продуктов, -в то время как в случаях
второго рода она получает с самого же начала дробления характер-
ные для нее свойства сплошного комка бластомеров.
Эти две крайние формы данного процесса связаны друг с другом
полной цепью промежуточных, показывающих весьма наглядным
образом, как совершается переход примитивного табличного дро-
_бления в трехмерное, как это его преобразование сдвигается, -в соот-
ветствующих рядах развития, на все более и более ранние онтоге-
нетические стадии и как постепенно сводится на-нет и исчезает хо-
рошо развитая до этого первичная однослойная стадия пребластулы.
Далеко ‘не случайным является то обстоятельство, что наиболее
примитивные, развернутые и сложные способы формирования мо-
руль1 наблюдаются у пресноводных Athecata — гидр и Cordylophora,
где они повторяют с замечательной полнотой начальные этапы nep-
BII/1'!-HOFO метаморфоза метагенетических Athecata. Tax, у гидр дроб-
ление идет ‚до седьмого деления включительно как правильное,
полное и равно-мер-ное, при устойчивом расположении. всех веретен
параллельно «поверхности зародыша, т. е. по всем правилам таблич-
ного дробления; гбластоцель появляется уже на 8-клеточной стадии
и «постепенно становится очень просторным. При восьмом оинхрон-
ном делении в различных точках окружности бластулы появляются
отдельные веретена, лежащие радиально или косо. Поэтому неко-
торые из клеток делятся теперь параллельно, а не перпендикулярно
к ее поверхности, давая таким образом нначало первым элементам
внутренней клеточной маосы зародыша. Одновременно с этим начи-
нается и выселение отдельных клеток «пребластулы в полость дроб-
ленпия, происходящее также в самых различных ‚местах стенки тела,
т. е. M у л ь т и ‘п о л я pm о или, точнее, аполярно, поскольку этот
вид иммиграции характеризуется именно тем, что он не приурочен
‘к каким-либо определенным точкам окружности. Сочетание этих
двух способов образования внутренней маосы клеток за счет пер-
вичной бластодермы, ‘называемое Мечниковым с М е ш а н н о й д е-
л я IM |И н а ц и е й, постепенно приводит к заполнению всего бласто-
целя. Пребластула превращается =в морулу, состоящую из множества
одинаковых, недифференцированных еще клеток, из которых по-
Зверхностные отличаются от остальных только более упорядочен-
ным расположением. ' =
Такая морула, образующаяся из предшествующей ей бластуло-
образной стадии путем заполнения полости этой последней, явно
315
„гомологична паренхимуле метагенетических Leptolida, отличаясь от
_нее главнымобразом тем, что она здесь формируется в чпорядке
‚эмбриогенеза, из еще недифференциро-ванного клеточного материала
и потому не проявляет “гитпичного для паречнхилмул клеточного ди-
морфизма. Иначе говоря, мы имеем здесь дело с эмбрионизажцией
стадии паренхимулы, с ее безусловно вторичным смещением на
несколько более ранний момент развития, при частичном сохранении
ею иммиграционного способа образования внутренней клеточной
массы. Однако в ее построении принимает участие и второй, "прин-
ципиально отличный морфогенетическгий механизм-деление _в
третьем измерении простран-
ства, или деляминация, при-
дающее такой эмбрионизи-
рованной паренхимуле, или
псевдоморуле (назы-
ваемой также стеррога-стру-
лой), преизнаки самой на-
стоящей морулы. '
Совершенно таким же
образом, как у гидр, со-
здается и псевдоморула у
Согс1у1ор11ога, где она начи-
нает формсироваться, однако,
Рис. 106. Дробление Gonoth r:a love 1‘ еще На ОДНО Деление раньпдё
1_8_mmHM mm” с umpoxumysnmomleim-_ Здесь уже на 4-клеточнои
ёгдггггечазгзгт стгггглттзгг; “д” ‘Юзникает Щеляевид‘
“WM; 4__24_KmoqHaH Momma (no Iglgnbggggg ная поло-сть дробления, по.
степенно расширяющаяся за-
тем до достижения пребла-
стулой приблизительно 64-клеточного состава (т. -е. до шестого де-
л-ения включительно). После этого начинают встречаться и ра-
диально расположенные веретена деления, а просторный бластоцель
постепенно заполняется клетками. Создается плотный многослой-
ный зародыш, все клетки которого представляются в это время
совершенно одинаковыми.
‘ Таким же образом, но начинаясь еще на одно или два "деления
раньше, идет формирование нсевдоморулы в некоторых из вариан-
тов эмбриогенеза у Gonothyrea И, возможно, Tubularia И в более
обязательном порядке——у Нуагасйпйа. У это-й последней (см.
рис 104, 2-6) полость дробления возникает на стадии 4 лили 8 бла-
стомеро-в, становится очень значительной при следующем делении
и достигает максимального объема у пребла-стул, состоящих из
20-30 бластомеров. В этой, очень неправильной по своим очерта-
‚ниям, полости уже со стадии 16 или немного позже появляются
„рассеянные «энтодермальные» клетки, обязанные своим происхо-
ждением как аполярной ииммиграции, так и периклинальным кле-
точным делениям. Постепенно пополняясь новыми пришельцами и
увеличиваясь в числе также за счет интенсивного деления ранее
.315
3 .
внедрившихся клеток, образуемая ими масса заполняет со време-
нем весь бласгоцель в виде плотной аморфной паренхимы.
b ~Hepexo11 OT такого окольнозго способа образования морулы путем
смешан-ной деляминации к ее непосредственному построению по-
средством одного только
трехмерного дробления
иллюстрируется нагляднее
всего примером таких '
форм, как Gonothyrea
loveni, y которой этот
процесс может идти как
тем, так и другим сп-осо-
бом, B зависимости от той
обстановки, в какой о-н
имеет м-есто (рис. 106).
Подмеч-ено, что когда за-
родыши G-onothyrea раз-
виваются в гон-офорах по
одному или в небольшом
числе, они дают начало
полой пребластуле, пре-
образующейся затем, на-
чиная с 64-клеточной ста-
дии, в такую же псевдо-
морулу, гкак у Cordy1opho-
ra. Если же число- заро-
дышей в гонофоре очень
велико, то периклиналь-Ф
ные деления появляются
очень рано, например со
стадии 8, так что. уже на
24-клеточной стад`ии заро-
дыш может быть пред-
ставлен плотной массой pm;
КЛЕТОК. V
Окончательное выпа-
дение однослойной стадии
дробления, а вместе с нею
и процессов поклеточной у
иммиграции, типично для эмбриогенеза подавляющего большинства
гидроидов. Как общее правило, деления периклинального характера
п-оявляются тут после 3 или 4 антиклинальных (радиальных), а за-
тем чередуются или комбинируются с ними в различных сочетаниях,
так что полость дробления не образуется вовсе, и бластомеры сса-
мого начала располагаются плотным комком. Так создается
истин н а я мо р ул а, сост-оящая из вполне однородных клеток
т
г ‘
ы
107. эмбриональное развитие Clava
squamata:
7—12-клеточная стадия; 2——32-клеточная морула; 3-
5—последовательные стадии обособления эктодермы;
б-—планула перед вылуплением с намечающейся
гастральной полостью (по Гарму)
и уже несравнимая по способу своего образования с-яморфологщ
чески равноценной ей паренхимулой (рис. 107),
317
V Итак, среди гипогенетических гидроидов фактически имеют
место все последовательные дступени «превращения дробления из
двухмерного, ведущего ‚к‘ образованию однослойной бластулы, в‘
трехмерное, дающее морулу, и это превращение имеет. своим ре-
зультатом 13106 более и более раннее оформление многослойной
стадии развития, достигаемой притом все более простым и прямым
путем, . "
Особого ‚внимания заслуживает то обстоятельство, что создаю-
щийся таким образом тип дробления подчиняется :МНОГО лугчше дру-
гих известным общим правилам деления целостных клеточных агре-
- 1 гатов, и именно правилам деления шара.
Эти правила требуют прежде всего, чтобы
первые три деления были антиклиналь-
ными и чередовались в трех координат-
ных плоскостях пространства, последова-
тельно деля шар на половины, ‘четверти
и осьмушки; во-вторых, чтобы ‘четвертое
деление было периклинальным, параллель-
ным поверхности шара и отделяло, таким
образом, внутренние половины о-ктантов
рез молодой морулы то- от наружных; и, в-третьих, чтобы плоско-
bularia mesembryanthemum сти дальнейших делений чередовались
(ПО ТИХОМИРОВУ) в антиклинальных и переклинальных
_ плоскостях.
‘При таком именно порядке чередования плоскостей дробления
и создается стадия истинной морулы у Gonothyrea, Clava И других
гидроидов, когда она оформляется сразу, непосредственно, как
первая же форма группировки немногочисленных еще бластомеров
(рис. 108). Поэтому можно считать, что э т а с т а д и я е с т ь
прямой результат подлинного трехмерного дроб-
ления и служит выражением той высокой степени
Целостности дробящейся системы, при которой
она только и подчиняется в известной мере зако-
нам деления целых геометрических фигур.
Рис. 108. Продольный раз-
3. Организация морулы
Итак,.стадия морулы представляет собой плотное скопление
бластомеров или эмбриональных клеток одинакового строения, рас-
положенных без определенной структурной правильности в виде
практически однородной первичной паренхимы. Правда, перифери-
ческий слой образующих ее клеток часто сохраняет несколько боль-
шую упорядоченность, чем облекаемая им внутренняя масса клеток
(особенно у псевдоморул, проходящих стадию бластулы), но не-
правильная форма этих поверхностных клеток и неполное их раз-
граничение от -внутренних не дают повода противопоставлять их
этим последним и видеть в поверхностных клетках особый клеточ-
ный пласъотличный от внутреннего. Поэтому нельзя не соглаоиться
3_18.
с.Беклемишевым, считающим морул типа Clava лучшим примером.
первично-паренхимной структуре у Metazo-a.
„_ ‚Общая форма морул довольно изменчива —— от почти строго-ша?
рообразной или эллипсоидальной до неправильной и неопределен-
ной (см, рис. 105-108); очень нередко она более или менее уплощена
в виде чечевицы. В проморфологическом_отношении морулы харак?
теризуются своей г о м о п о л я р н о с т ь ю при сохранении или же
временном исчезновении первичной главной оси тела, т. е. той или
иной степенью «структурной и физиологической деполяризации. Осе-
вой, градиент физиологической активности не обнаруживается у них
ни путем воздействия отравляющими веществами, ни прижизненной.
окраской (Тессье, 1931), восстанавливаясь лишь впоследствищпргг
преобразовании морулы в пла-нулу. Особенно любопытно, что у не-
которых видов, обладающих на стадии ранней морулы (по крайней
мере в период от 8-клеточной до приблизительно 100-к.леточной
стадии) довольно правильной неравнооснойч формой (Dynamena),
эта последняя не отражает действительной архитектоники заро-
ДЫШЗ; ЭТО ВИДНО ИЗ совершенно беспорядочного расположения окра-
шен-ных клеток ТЗКИХ иморул ОТНОСИТЕЛЬНО HeOKpaI.I.I€HHbIX, Ha6JIiO-'
даемого после маркировки одного из полюсов яйца, только-что на-
чавшего дробиться. Однако тв ряде других случаев меньшая ось
сходно уплощенной морулы определенно соответствует главной оси
яйца и планулы, что, очевидно, говорит о более ограниченном масг.
штабеее деполяризации. Так или иначе можно считать установлен-
ным, что у морул нет той отчетливой гетерополярности, какая имеется
у яйца и у следующих за нею стадий, что хорошо подтверждается
самым фактом формирования морулы путем нелокализованной, anno-
лярной иммиграции и также всесторонней клеточной деляминации.
Общим выводом из этих и имеющихся экспериментальных дан-
ных по изоляции бластомеров, показывающих, что она возможна
только на стадии 2, являетсяпризнание положения, что устойчивая
организменная целостность дробящегося яйца устанавливается у
гипогенетических гидроидов много раньше, чем у метагенетических,
хотя и получает более простое, чем у них, проморфологичес-кое вь1-
гражеъние.
4. дифференцировка эктодермы
Процеос формирования морулы в ходе дробления до сих пор
еще ‘ча-сто трактуется как гаструляция, а -сама морула -— ка.к стер-ро-
гаструла, т. е. компактная, сплошная ‚гаструла. Между тем к этому
нет решительно никаких оснований, поскольку на этой стадии еще
нет’ видимого разграничения между будущими пластами тела и оно
пока состоит из одинаковых недифференцированных клеток. И хотя
внутренние слои клеток морулы действительно дают в дальнейшем
энтодерму, а периферические—эктодер-Му (плюс систему интерсти-
циальных клеток), они пока представляют эти Ёпласты только в по-
тенции, в презумптивной форме, отражающей лишь размещение
соответствующего материала в яйце. Другими словами, здесь можно
319
говорить только о сохранении морулой той же предварительной
структуры, какая уже имелась в яйце. `
Что же касается действительной дифференцировки первичных
пластов тела, то она происходит много позднее, при преобразовании
морулы в планулу (или отвечающую ей эмпбриональную стадиию), и,
ка-к правило, в два ‹пргиеМа: первым ее этапом является обособление
и гистологическое оформление эктодермы, за которым уже много
позднее следует превращение внутренней клеточной массы заро-
дыша B энтодерму. Первый из этих двух, всегда раздельно проте-
кающих, процессов прослежен подробнее всего Тихомировым
у Tubularia и ряда других гидроидов‚ где он сводится к постепен-
ному «оспециализированию поверхностных клеток морулы и к обо-
соблению их от остальной массы, долгое время остающейся без
дальнейших изменений и представляющей собой зачаток эндо-
дермы» (цит. соч., стр. 45).
«Чтобы составить себе ясное представление о том, как совер-
шается здесь обособление эктодермы, всего лучше обратиться
ик сравнению м-оих трех рисунков: 19, 20 си 21 (рис. 109). На первом
из этих рисунков... мы видим самое чпервое начало той дифференци-
ровки клеток дробления, которая вконце концов должна привести
гк обособлению эктодермы, а именно`в этой еще крупноклеточной
моруле мы уже видим некотороеразличие между поверхностными
клетками, с одной стороны, и внутренними-с другой, выражаю-
щееся в том, что первые- представляются более плоскими. Легко,
однако, видеть, что это различпие не очень строго выражается в ка-
ждой из клеток этих двух родов. Мы видим также, что внизу раз-
реза =в одном месте большие внутренние клетки пне покрыты более
плоскими поверхностными кл-етками. Рис. 20 представляет нам даль-
нейшую стадию дифференцировки. Мы уваидим впоследст-вии, что
экто- и эндодерма в период своего полного обособления в значи-
тельной степени разнятся между со-бойпо своей окраске, а имен-но:
эктодерма, состоящая из более плазматических клеток, окраши-
вается сильнее; эндодерма, состоящая из более крупных, наполнен-
ных вакуолями клеток, красится слабее. Начало такого различия
мы видим и на рис. 20. Здесь нельзя не видеть, что корковый слой
яйца состоит из клеток более плазматических, более темных, между
тем как внутренность яйца наполнена более светлыми клетками.
ЛЕГКО «ВИДСТЬ, ОДНЗКО, ЧТО разделение ЭТО неполное В TOM OTHOI.LI€-_
НИИ, что в некоторых местах внутренние светлые клеткли доходят до
самой периферии яйца; с другой стороны, более темны-е клетки за-
мечаются также и в середине... С ходом дальнейшего развития обо-
собление обоих слоев идет все дальше и дальше. Корковый слой
плазматических, темнокрасящихся клеток все -более и более ровным
Слоем покрывает остальную массу более крупных светлых клеток,
со временем уже нлигде не доходящих до «периферии яйца, как это
мы видели приописании предыдущего рисунка.’ В конце концов,
получается стадия, которая представлена на моем рис. 21. Здесь
эктодерма уже обособилась. Правда, мы не видим еще, чтобы она
320
we-
Рис. 109. Три последовательные стадии дифферен-
цировки эктодермы морулообразного зародыша
тубулярии (по Тихомирову)
21
Эмбриология беспозвоночных
321
была отделена ровным контуром от эндодермы; скажем более: кон-
тура, отделяющего экто- от эндодермы, мы здесь вовсе не видим;
здесь еще каждая клетка эктодермы вдается своим закругленным
основанием в подлежащую эндодерму; некоторые же клетки еще
языком вдаются между клеток эндодермы. Всматриваясь ближе в
оо ооеодооо ’
00006, О Q5%g:é§g
O ‘о О
T 0%)
‘звёзд
Ego
Рис. 110. Процесс дифференцировки эктодермы у зародыша
' Aglaophenia tubulnfera (по Тихомирову)
строение эктодермы, ›мы замечаем, что -B O5Hl€M ОНа представляет
собой многослойный пласт, состоящий из неправильно многоуголь-
нь1х клеток; однако кое-где уже и теперь между этими клетками
начинают попадаться Цилиндрические клетки, пронизывающие всю
толщу эктодермы... В то время как эктодерма представляет уже
указанную степень развития, эндодерма ПРОДОЛЖЗеТ 'BC€ еще C0-
стоять из приблизительно одинаковых светлых клеток, заключаю-
щих B себе еще очень большое количество неассимилированных
322 '
ядер питательных клеток. Впрочем, эти последние встречаются и
в эктодерме, но, залегая в ней в меньшем количестве, попадаются
не на каждом разрезе» (там же, стр. 22—23). `
Вслед за этим происходит окончательное размежевание обоих
пластов, при котором между нимп появляется тонкая прослойка
основного вещества, а затем уже и эпителизация эктодермы. В ходе
этой последней значительное большинство клеток поляризуется и
выстраивается в правильный, однослойный, сначала многорядный
столбчатый эпителий, а часть остается неполяризованной и образует
залегающие в нем интерстициальные клетки. После этого уже про-
исходит функциональная дифференцировка клеток эктодермы на
эпителниально-мьтшечньге, чувствительные и железистые (рис. 110).
Описанная схема формирования эктодермы приложима к ho-
давляющему большинству гипогенетических гидроидов. Она, безу-
словно, являет-ся самой типичной и‘ господствующей у них формой
расслоения индиферентных до этого клеток морулы на два первич-
ных клеточных пласта, один из которых приобретает при этом ха-
рактер эпителизованной пограничной ткани, а второй сохраняет
пока ещеособенности эмбриональной первичной паренхимы. Как
известно, этот тип образования пластов тела был назван Мечнико-
вым вторичной деляминацией и противопоставлялся им
деляминации п е р в и ч н о й, которая осуществляется на базе бла-
СТУЛЫ, а не МОРУЛЫ, путемпериклинального деления ВСЕХ ее KJIBTOK-
Такая первичная, или клеточная (Беклемишев), деляминация,
уже знакомая нам .по ранним стадиям эмбриогенеза многих гидро-
полипов (в качестве одного из слагаемых смешанной деляминации
или же представленная у других четвертым или пятым делением дро-
бящегося яйца), наблюдается, однако, среди гидрополипов и в той
же классической своей форм-е, в "какой она- протекает у Geryonia,
Единственным, правда, примером такого рода может послужить
Eudendrium сарШаге, у которого, по Мечникову, `‹<равномерное де-
ление ведет к образованию крупноклеточного зародыша, в котором
рано появляется слабо развитая полость`дробления. Бластомеры
обнаруживают при этом явственное разделение на экто- и эндо-
ПЛЗЗМУ, ИЗ КОТОРЫХ перв-ая СОДЕРЖИТ МЕЛКИЕ темнооконтуренные
зернышки, в то время как последняя заполнена более крупными,
окрашенными в оранжевый цвет шариками желтка. После увеличе-
ния бластомеров в числе они делятся "таким образом, что экто
плазма остается в наружном клеточном слое, а_ эндоплазма концен-
трируется, наоборот, во внутренней клеточной массе, совершенно
заполняющей маленькую полость дробления. Таким образом, здесь
возникает такая же первичная деляминация, какую мы встречали
у одних только G-eryonidae, и хотя у Eudendrium вследствие богат-
ства желточным материалом ядерные веретена не могут быть за-
мечены, можно судить об их направлении по характеру образую-
щихся клеток» (Мечников, 1886, стр. 68).
Так или иначе, первым этапом в процессе образования первич-
ных пластов тела неизменноявляется обособление эктодермы и свя-
21* 323
ёанной с нею системы интерстициальных клеток ‘от пока еще аморф-
гной внутренней массы паренхимы, еще жне имеющей вэто время
значения подлинной энтодермы. По-этому подобное обособление
эктодермы может лишь с еще большей натяжкой считаться гастру-
ляцией, чем равноценное ей по своему конечному результату пре-
образование свободной бластулы в иммиграционную паренхимулу.
5. Формирование планулы
Значительное большинство пипогенетических гидрополипов на-
чинает свое свободное существование в виде планулы, являющейся,
та.ким образо-м, их первой и единственной личиночной стадией.
Однако у тубулярий, гидр и ряда других форм (Myriothela, A'ctino—
gonium и другие) стадия планулы про-ходит как эмбриональная, и
вылупление происходит много позднее, в виде уже полипоидной
актинуль1 или даже оформленного молодого полипа. Таким образом,
в пределах этой группы мы встречаемся с несколькими различными
ступенями прогрессирующей эъмбрионизации, заверШаЮЩеЙСЯ ЗДЭСЬ
включением в состав эмбрионального раз-вития всех этапов первич-
ного метаморфоза и соответствующим выпадением всего комплекса
мерцательных личиночных форм.
Начальным этапом преобразования морулы ‘в планулу является
только что описанное обособление эктодермы и ее эпителизация.
Еще до завершения этого процесса зародыш начинает постепенно
вытягиваться в дли-ну, в направлении своей главной оси, и вновь
приобретает гетерополярное строение. Вслед за тем начинается ‘И
э.пителизация его внутренней клеточной массы, при которой ее
глубокие слои подвергаются распаду и фагоципируются, а перифе-
рический слой располагается обычным порядком вокруг образую-
щегося схизоцеля. Наконец, «происходит и дифференцировка ЭКТО-
дермы; планула становится подвижной и приступает к самостоя-
тельному существованию.
Главная особенность процесса дифференцировки зародыша в
ли-чинку за-ключается, ‚во-первых, в том, что ею охватывается лишь
часть клеточного материала зародыша, а именно одни только экто-
дермальные и некоторые из интерстициальных его клеток, в то
время как энтодермальные и остальные интерстициальные сохра-
няют амёбоидное недифференцированное состояние. Во-вторых,
дифференцировка клеток идет сразу же ав разных направлениях,
в соответствии с их разносторонней специализацией, минуя таким
образом этап начальной однородной дифференцировки в однотип-
ные жкгутиковые клетки. В результате этого лишь часть эктодер-
мальных клеток получает характер обычных жгутиковых, тогда как
другая их часть непосредственно превращается в слизистые, пиг-
ментные и другие высоко специализированные категории клеток.
Короче говоря, многосторонняя гистологическая дифференцировка
эктодермы занимает место начальной, так что стадия однородного
кинобласта оказывается здесь пропущенной.
324
ЭТИ ОСОбЭНЧЮСТИ Хода дифференцировки планулы, имеющие
большое принципиальное зна=чение, не влекут, однако, за собой
особо приметных отличий в строении этих личинок по сравнению
с планулами метагенетических beptolida. Действительно, если не-
которые из них (Gonothyrea, Laomedea, некоторые сертулярии) пре-
восходят по-следних по степени гистологической расчлененности
эктодермы и уровню дифференцировки ее клеток, то в отношении
других (например Вупатепа) этого сказать нельзя; так, .в частности,
разделение жгутиковых клеток эктодермы на двигательные, эпите-
лиально-мышечные клетки и чувствительные (нейро-эпителиаль-
ные) далеко не всегда бывает за-Метнь1м, а наличие в составе тела
особых клеточных типов, вроде пигментного, являет-ся скорее исклю-
чением, чем правило-м. Стало быть, можно только сказать, что пла-
нулы гидрополипов только иногда достигают более высокой органи-
зации, чем планулы метагенетических форм. Более определенные
РЕЧЗЛИЧИЯ Между теми и другими могут быть, пожалуй, замечены
в том, что первые содержат в своих клетках более обильные резерв-
ные включения в виде остаточного желтка И других запасных пита-
тельных включений. Так, у планул Dyria-mena призматические
клетки всей эктодермы заполнены мелкими желточными гранулами,
увеличивающимися в размерах в их базальной части, в которой
содержатся и многочисленные капельки жира; в энтодерме эти
включения еще обильнее. В некоторых других случаях (Gor1othyrea_)
большое количество остаточного желтка сохраняется в обоих кле-
точных слоях задней поло-вины планулы и т. д.
Проморфологические соотношения между плануло-й и .предше-
ствующими ей стадиями развития таковы же, как у метагенетиче-
-ских Deptolidaz физиологически передний (агборальный) полюс пла-.
нулы отвечает вегетативному полюсу яйгца, а ее оральный полюс ——
анимальному. ‚Осевой градиент метаболической активности здесь
также ориентирован ‘в направлении, обратном его янаправле-нию
у яйца. ‚
6. Превращение планулы в сидячую полипоидную стадию
У всех, повидимому, гипогенетических Тпесарйога, обладающих
свободной планулой, ее превращение в полипа осуществляется по
типу Clytia: планула прикрепляется к субстрату (рис. 111) своим
передним КОНЦОМ И расплющивается В лепешку, ИЗ центра КОТОРОЙ-
вырастает затем стебелек основателя колонии; далее, из’ вершинной
части этого стебелька формируется головка первого гидранта. В не-
которых случаях, однако (например у Dynamena pumila), эта про-
стая схема развития значительно усложняется тем, что конец сте-
белька дает не одного радиально построенного гидранта‚’а сразу це-
лый членик колонии, состоящий из двух гидрантов и расположенно-
го между ними булавовидно-го выроста стебелька (рис. 112). Это об-
разование обладает яс-н-о выраженной двулучевой симметрией, осью
которой служит ось стебелька, а антимерами—- головки обоих гид-
рантов, отходящие от последнего под углом порядка 45° в диамет-
325
‘тально показано Тессье, эта глав-
"pore, занимая лишь в приблизи-
дрантов, то они обладают нор-
"er особенно обильными резервами
рально противоположных направлениях. Плоскость, в которой они
лежат, точно отвечает плоскости одного циз двух первых делении
яйца: как это было эксперимен- с
ная плоскость симметрии колонии
сов-падает в 40% случаев с та-
ковой первого из этих делений и
также в 40% — с плоскостью вто-
т-ельно 20% случаев промежуточ-
ное или неопределенное поьпоже-
ние. Что же касается самих ги-
мальной многолучевой симметри-
ей, ось которой оказывается та-
ким образом не зависим-ой от
первичной оси планулы, ставшей
осью колонии в целом.
Этот замечательный пример не-
обычно раннего почкования осно-
вателя колонии‚ предшествующего
здесь началу его самостоятель-
ново питания, находит свое физи-
ологическое обоснование в том,
что планула Dynamena рас-полага-
Рис. 112. Формирование сидячей
полипоидной стадии Dynamena
pumila:
1—осевшая планула с еще торчащим
и задним КОНЦОМ; 2—совершенно распла-
Рис. 111. Продольный разрез через осев- ставшаяся планула; 3—6—появление и
ш Ю п а А lao he - u u ч: рост стволика полипа; 7—15-—последо-
‚У л_ НУЛУ T§xoNI;?Hp(T)1;;)t b 11 era (ПО уательньте стадии оформления первой
пары гидрантов (по Тессье)
326
питательных И ЭНЕРГЕТИЧЕСКИХОМЗТЭрИЭЛОВ. ИХ хватает ЗДЕСЬ не
‘ТОЛЬКО на обеспечение ЭТОГО СЛОЖНОГО морфогенетического процесса,
но и на подготовку зачатка следующего двойного членика. Более
того, если даже удалить задний конец планулы, оставшейся части
оказывается достаточно для образования первичной пары гидрантов
нормального размера, а при расщеплениин задней части планулы из
нее могут образоваться даже две пары гидрантов; и только при
удалении всей задней ‘половины планулы образующийся стебелек
образует один единственный, непарный
гидрант. Важно отметить, что гид-
ранты, получаемые из фрагментов пла-
нул, сохраняют те же размеры, как
нормальные, что особенно резко про-
тивопоставляет Dynamena ТаКИМ фор-
мам, как Clytia, у которых размеры
основателя колонии всецело опреде-
ляются, в аналогичных опытах, вели-
чиной того фрагмента планулы, из ко-
торого он развивается. Все эти факты
определенно говорят о том, что в раз-
витии Dynamena имеет место особая
форма повышенной эмбрионизации,
выражающаяся в осуществлении за
счет желтка не только заключительных
этапов первичного метаморфоза, но и
первых этапов построения колонии.
У гипогенетических Athecata пре-
вращение планулы в гидранта совер-
ша-ется другими способами, чем у The-
caphora. Прикрепившаяся своим п-еред-
ним концом планула чаще всепоне _
становится плоской, а, сохраняя свою Рис. 113. Превращение осев-
вытянутую форму, непосредственно Шей ПЛ3НУЛЪ1 В полипа У
преобразуется -в целого полипа. Упло- Athecata’
о 7—3—Clava squamata (по Гарму), 4,
ЩаЁТСЯ ЛИШЬ ее абОРаЛЬНЫИ КОНеЩ 5—Cordylophora lacustris (no Мор-
образующий подошву полипа, в то “°“L“”P*‘V '
время как во-круг заднего появляются
щупальца, а само тело планулы превращается одной своей частью
в стебелек, а другой в головку гидранта. Впрочем, некоторое упло-
щение планулы после ее фиксации может ‚все ж-е иметь место
(Clava), НО в гораздо более ограниченном масштабе, чем это ти-
пично для Тпесарйога. Таким образом, устанавливается прямая
преемственность архитектоники от планулы к полипу, и метаморфоз
сводится к минимуму. Появление щупалец идет в различной после-
довательности; так, у Согоу1орйота их -сначала образуется два (на-
мечающих, таким образом, одну из плоскостей двулучевой симмет-
рии), затем еще два (в плоскости, перпендикулярно-й к первой,
щупальцевой) - и т. д., тогда как в других случаях они сразу закла-
327
дываются целым венчиком или даже одновременно двумя венцами .
(рис. 113). При высших формах эмбрионизации, наблюдаемых, на-
gang ‘
\ д;
г, д"{ё‘
о W *
Рис. 114. Tubularia mesembryanthemum: ‘ Po
- A—1'1P0J10J'1bHbII‘»’I разрез молодой актинулы, еще не вышедшей из гонофора; Б—продольный
Разрее актинульп, дифференцирующеися в полипа (по Тихомирову)
ПРИМЁБЪ У ТУбулярии (рис. 114), зародыш непосредственно разви-
вается в полипоидную -стадию—акт›инулу, минуя таким образом
„все мерцательные личиночные стадии. ‘
Глава XVIII
ОСОБЕННОСТИ КЛЕТОЧНОГО СТРОЕНИЯ СИДЯЧИХ
ВЕГЕТАТИВНЫХ СТАДИЙ
1. Предварительные замечания
Существование глубоких качественных отличий между клетками,
слагающими тело Metazoa, И одноклеточными особями простейших
не вызывает ни малейшего сомнения. Со времени крушения теории
клеточного государства каждому ясно, что клетки Metazoa, нахо-
328
дящиеся в специфическом для них‘ биологическом соединении и
представляющие собой, та-ким образом, тесно интегрированные ча-
сгги целого, далеко не пидентичны с одноклеточными же Protozoa,
являющимися самостоятельными целостными организмами. Это
признается даже самыми убежденными сторонниками клеточной
теории, для которых гклетка есть только элементарная структурная
единица живой материи, построенная по одному и тому же общему
плану у всех без исключения организмов. Таким образом, признание
‘реальности клеток, разложимости на клетки любого организма и
единства общего плана строения клетки, составляющее основу кле-
точной теории, как общей теории структуры живого вещества,
включает. в то же время и признание о р г а н и з IM е н н о й -неравно-
ценности жклеток простейших и Metazoa.
‘Однако остается, в сущности, неясным, насколько эта нера-вно-
ценность самостоятельных и интегрированных клеток отражается
на их организации, и можно ли ра-сценивать последнюю иначеднем
проявление теми и другими самых универсальных законов
строения живой материи. Вопрос заключается именно в том, исчер-
пывается ли сходство между ними единством общего им плана
строения как «клеток вообще», или же оно может иметь и более
конкретный характер, быть более содержательным и осязаемым.
Иначе говоря, у нас все же ‚нет ясного представления о том, в какой
мере отдельные клетки Metazoa отвечают по своей организации
одноклеточным простейшим, и каким именно из .них‚ и насколько
содержательно и плодотворно их непосредственное взаимное
сравнение. _
Что оно возможно пи законно —следует из самой сути клеточной
теории, из самого признания клетки элементарной структурной еди-
ницей всех вообще живых систем. О его плодотворности «говорят
с неменьшей очевидностью всем известные классические примеры
сравнения хоаноцитов губок с воротничковымпи жгутиковыми или
‘сперматозоида многоклеточных с те-ми или иными Flagellata. Тем
не менее такое сравнение не только не проводилось до сих пор в
хоть сколько-нибудь широком масштабе, но и встречается с очень
серьезными принципиальными возражениями. Так,
А. А. Заварзина (1945), «так называемые одноклеточные оргад
низмы никак нельзя сравнивать с отдельными клетками тканей гмно-
гоклеточных организмов; такое сравнение будет -но-сить всегда со-
вершенно формальный характер, будет крайне односторонним и
потому будет всегда вступать в решительные противоречия с кле-
точной теорией, что и лимело место -на протяжении всей истории
развития этой последней. Простейшие (Protozoa) являются прежде
всего организмами, а затем уже клетками. Их можно назвать
также клетками-организма‹ми...>>. «Огромный мир простейших,-
пишетцон (1945)‚— развился на почве одноклеточной организации,
но на эту организацию на_ложилась организменная интеграция;
клетка в этом случае развивалась -как органическое целое, и это
наложило такой отпечаток на клеточную структуру, что она стала
329
ПО МНЕНИЮ.
ПОХОЖЗ УЖЕ НЕ на КЛЕТОЧНЫЕ СТруКТурЫ ТКаНЕВЫХ КЛЕТОК МНОГО-
"КЛЕТОЧНЫХ организмов, а на МНО-ГОКЛЕТОЧНЫЕ ОрГЗНИЗМЫ BO ВСЕЙ
=ИХ СЛОЖНОСТИ И ЦЕЛОСТНОСТИ». ПОЭТОМУ «СЛЕДУЕТ ПРЕДПОЛОЖИТЬ, ЧТО
разделение на два эволюционных ствола: протистов и Me-Iazoa,
произошло еще до возникновения клеточной организации». В" еще
более категорической форме сходное мнение
Е. М. Вермель, согласно которому всякое вообще сравнение Между
простейшими и многоклеточными ‹<не только бессмысленно, но и
бесплодно и не может выйти из рамок поверхностных и ни к ‘чему
‘не обязывающих аналогий» (1940). Таким образ-ом, нам прихо-
ДИТСЯ считаться С ПрЕДСТаВЛЕНИЕМ‚ ЧТО СУЩЕСТВУЮЩИЕ МОрфОЛОГИ-
ЧЕСКИЕ СООТНОШЕНИЯ МЕЖДУ ‹<клетками-о-рганизмами» И «НаСТОЯ-
щими» тканевыми клетками Metazoa He превышают уровня ана-
логии (и может быть даже аналогии самой поверхностной) и что
имеющееся между ними сходство является, следовательно, чисто
конвергентным.
Этому представлению в корне противоречат, однако, вышеупо-
`МЯНУТЫЕ И ЦЕЛЫЙ рЯД ДрЬ/ГИХ, НЕ MeH-ee показательных, ЦИТОЛОГИЧЕ-
ских фактов, описание пи оценка которых составляют главную за-
дачу настоящей главы. Н-еверно оно и с принципиальной, чисто
теоретической точки зрения. Дело в том, что в основе этой концеп-
ции лежат хоть и различные, но одинаково произвольные допуще-
ния: о независимом, раздельном происхождении одноклеточных
организмов, с одной стороны, ли многоклеточных — с другой, о том,
что клеточное строение возникло исторически вторично, уже после
приобретения живыми существами присущей им всем органической
целостности (Заварзин); или же другое, не более вероятное, допу-
щенпие, что новое ‘качество многоклеточной целостности, создаю-
щееся при уста-новлении сложных взаимоотношений между клет-
ками тела Metazoa, полностью снимает собственные свойства этих
клеток, приводит к их полному «растворению» в лмнотоклеточном
комплексе, к полной утере ими своей индивидуальности; что, став
частями целого органгизма, они тем самым как бы перестали суще-
ствовать как отдельные клетки. .
О том, что в действительности это далеко не так, свидетель-
ствуют с полной очевидностью при-водимые ниже факты, касаю-
щиеся орга-низации клеток сидячих вегетативных стадий низших
губок пи книдарий.
2. Известковые губки. Состав клеточного ассортимента
и соотношения различных видов клеток
В состав тела взрослых известковых губок входит до десяти раз-
личных видов клеток, отличающихся по своему строению, происхо-
ждению, локализации в теле губки и функциональному значению,
а также и ряду других свойств структурного ‚и физиологического
характера. Эти десять видов клеток могут ‘быть разделены на сле-
дующие три основные группы:
330
высказывал И.
-—....._..„„ _.‚_—‚—___—_...‚..-.„_____..__ _
1) дифференцированные жгутиковые клетки,
каковыми являются характернейшие для всех губок вообще
воротничковые клетки, или хоаноциты;
2) п о к р о в н ы е к л е т.к и, представленные пинакоцитами
и пороцитами; `
3) а M ё б о`и д н ы е к л ет к и, или амёбоциты, к которым
относятся. оседлые звездчатые колленциты, вторичные склеро-
бласты, зернистые и две формы незернистых мелких амёбоци-
тов — мелкоядерные и крупноядерные, мегациты ни очень мелкие
клетки с признаками дегенеративных изменений ядра.
Клетки первой из этих групп образуют трофический аппарат
"губки, функционально отвечающий энтодерме остальных многокле-
ТОЧНЫХ И -СООТВЕТСТВЕННО ИМЕЮЩИЙ ЭШИТЕЛИОИДНОЕ СТрОЕНИЕ; КЛЕТКИ"
второй группы выполняют функции эктодермы, а все остальные
складываются в исключительно ‚пеструю здесь ткань внутренней
СрЕДЬ1‚ обладающую характером МЕЗЕНХИМЫ ИЛИ, ПО ТЕрМИНОЛОГИИ-
Беклемишева, разнородной паренхимы.
Обиходные представления о происхождении клеточного поли-
морфизма губок и генетических взаимоотношений между различ-
ными типами клеток, образующих их тело, как известно, зиждутся
на предположении, что жгутиковые клетки бластулы дают начало
одним только хоаноцитам, а зернистыеывсем остальным клеточ-
ным типам, и что родоначальником всех этих последних и их един-
ственным камбием являются незернисть1е амёбоциты (археоциты).
Однако теперь можно считать установленным, что действитель-
ное положение вещеи не отвечает этому, уже давно устоявшемуся,
взгляду |И что он совершенно не приложим, в частности, к образо-
ванию клеток у известковых губок. В самом деле, имеющиеся теперь
новые сведения по этому вопросу, полученные преимущественно
Дюбоском и Тюзэ (1934-1937), могут быть резюмированы сле-
дующим образом: ‹
1) Непосредственными производными зернистых клеток личинки
являются только первичные склеробласты, образующие покровный
клеточный слой аморфной «куколочной» стадии и в дальнейшем
дифференцирующиеся на наружные, кроющие элементь1-——пинако-
циты и пороциты и внутренние, соединительно-тканные клетки——
вторичные склеробласты и колле-нциты. ‘
2) Все эти эктобластические, по существу, элементы достигают
стабильного уровня дифференцировки и в нормальном развитии
не способны дать начало каким-либо другим клеточным типам. _
3) Непосредственными производными жгутиковых клеток am.-
CTyJIbI являются однородные сначала амёбоидные внутренние клетки
«куколки», большинство которых располагается затем в виде surn-
телия_ и дифференцируется в хоан-оцитов, а меньшинство частью пре-
вращается в незернистых амёбоцитов, частью же дегенерирует.
4) Хоаноциты не переходят лабильного уровня‘ дифференци-
ровки и могут на месте превращаться в эозинофилов или мегапитов,
уходящих затем в.мезоглею‚ или же. сами выходить из эпителиаль:
331
ного строя, погружаться в мезоглею и там дедифференцироваться
в мелкоядерные или другие‘ амёбоциты, в том числе и в первичные
половые клетки; стало быть, только хоаноцить1 и еще не дифферен-
цированные производные жгутиковых клеток служат родоначаль-
никами «археоцитов.» и действительно ом-нипотентны.
З. Трофические клетки губок
Хоаноциты характеризуются прежде всего своим общеизвестным,
поистине прославленным сходством с трофозоитами воротничковых
жгутиковых (Craspedomonadidae или Choanoflagellata) ——сход-
ством, выражающимся не только в наличии у них столь исключи-
26
Рис". 115. Хоаноциты известковых губок:
1-Clathrina coriacea; 2—Syc0n coronatum; 3—Leucandra johnstoni;
7a, 2a, и „га-парабазальный аппарат (по Волконскому);
4-группа хоаноцитов Sycon raphanus с чечевицеобразным параба-
зальным аппаратом (по Добоску и Tress); 5--01I(IrIHO'-IHOB воротничко-
вое жгутиковое Codonosiga с таким же парабазальным аппаратом (по
де-Седелеру)
’0 га
тельного образования, как, протоплазматический воротничок, не
встречающегося нигде, кроме как у этих клеток и у Craspedomor1a-
didae, HO И в сходстве буквально всех других особенностей их орга-
низации, coBnaJ1a}ou1ey°1';Lo мелочей с таковой названных жгутиковых.
Клеточное тело хоаноцитов Са1сагеа (рис. 115) обладает удли-
ненно-эллипсоидальной mm ‚ПОЧТИ цилиндрической, относительно
постоянной формой и увенчано бокаловидным или цилиндрическим
плазматическим воротничком, окружающим основание единствен-
ного Id здесь жгута. Этот последний отходит от базального зерна,
имеющего тут характер центромастигосомы, т. е. принимающего
на себя роль центросомы в процессе деления; в близком соседстве
с базальным зерном помещается типичнейший парабазальный аппа-
332
v рат, состоящий, как обычно, из наружного хромофиль-ного и внут-
реннего хромофобного вещества и отличающийся, у разных видов;
довольно разнообразной формой: от простой чечевчицеобразной
у Бус-оп raphanus до извитой колбасе-видной у Leucandra. Таким
образом, налицо полносоставная одножгутиковая кинетида того же
типа, как у многих Protomonadina, и даже более того-идентич-
ная до деталей с таковой некоторых .Craspedomo«r1adidae. Шаровид-
ное, довольно крупное ядро снабжено эксцентрично расположен-
ным ядрышком и имеет сетчатое распределение хроматина. В цито-
плазме содержатся не очень многочисленные гпалочковидные,
бисквитообразные или овальные хондриосомы, отдельные пищева-
рительные вакуоли, шаровидные элементы вакуома, окрашива1о-
щиеся нейтральной красной, и различные непостоянные включения,
например липоидного характера; кроме того, импрегнация клеток
тяжелыми металлами позволяет обнаружить между вакуолями об-
разование, очень напоминающее «внутренний сетчатый аппарат
Гольджи». Топографические соотношения элементов клеточной ор-
ганизации неодинаковы у разных видов, в соответствии с чем раз-
личают два ее типа: а) тип Grantia, характеризующийся дистальным
положением ядра и концентрацией всех включений цитоплазмы в
проксимальной половине клеток, в пространстве между ее основа-
ни-ем и ядром, и б) ТИъП Clathrina, отличающийся, напротив, прокси-
мальным положением ядра и локализацией всех плазменных вклю-
чений над ним, в дистальной половине клетки.
В пром-орфологическом отношении хоа-ноцниты могут быть охарак-
теризованы как одноосно-гетерогполярные клетки с нормальной для
сидячих жгутиковых ориентацией полюсов и радиальной основой
симметрии, преобразуемой в двустороннюю положением ли формой‘
пара.базального аппарата. Размеры хоаноцитов, повидимому, до-
вольно постоянны и в общем очень невелики.
Функцией хоаноцитов является, как известно, улавливание взве-
шенных в воде пищевых частчиц (главным образом бактерий‘) и их
переваривание. Она осуществляется точно таким же образом, как.
у воротничковых жгутиковых, путем подгонки таких частиц током
воды, вызываемым непрерывным винтообразным движением жгута,
к протоплазматическому воротничку, которым они улавливаются
и втягиваются затем внутрь цитоплазмы; поступающая в клетку
mama окружается пищеварительной вакуолью, в которой она и пер-е-
варивается. Характерная особенность известковых губок по срав-
н-ению с другими, вышестоящими в системе губками (кремневымид
роговыми) заключается в том, что как улавливание, так и перева-
ривание пищевых частиц совершается исключительно хоаноцитами;
стало быть, они являются тут единственными трофическими клет-
ками тела, а не делят эту функцию с амёбоцитами, как у более
высоко организованных формл. _ - _
Хоаноциты обладают неограниченной способностью K_ делению,
осуществляемому «ими без предшествующей дедифференцировки,
в необычайно характерных для них формах протозойного про-
333
дольного деления. Как видно из воспроизводимых здесь рисунков
(рис. 116), оно совершается тут таким же способом, ка.к у Сгазре-
domonadidae и многих других жгутиковых: оно начинается с деления
кинетосомы на две, из которых одна остается при еще имеющемся
в этот ямомент материнском жгуте, а другая отодвигается от нее,
растягивая соединяющий их тяж; далее они располагаются по сто-
ронам придчвигающегося к ним теперь ядра, причем этот тяж при-
кладывается к его, пока еще явственной, оболочке, а имевшийся
жгут сбрасывается. В наступающей метафазе кинетосомы играют
роль центров деления, располагаясь по концам веретена, в то время
как соединяющий их тяж ложится
на одной из сторон последнего,
в в качестве его внеядерной основы.
‘\ B анафазе из каждой кинетосомы
вырастает по новому жгуту, так
что при наступающем затем де-
ЛЕНИИ КЛЕТОЧНОГО тела ОбЕ ДОЧЕр-А
ние клетки уже снабжены офор-
МЛЕННЫМИ КИНЕТИДаМИ} ЭТО ДЕЛЕ-
ние совершается посредством
односторонней врезающейся бо-
РОЗДЫ, ВОЗНИКЗЮЩЕЙ на ПЕРЕДНЕМ
конце клетки, на дне материн-
Рис. 116. Деление протозойного ти- скот плазматёческого Ворот"
па'у хоаноцитов известковой губки НИЧКаЗ ПОСЛЁДНИИ Сохраняется до
С1а’с11г1па(поМинчинуиРобертсону) конца митоза, а затем исчезает,
после чего каждая из клеток
образует по новому воротничку. Удвоение парабазального аппа-
рата происходит .в профазе или несколько раньше, иногда даже
раньше деления базальных зерен; в ходе митоза парабазальные
аппараты располагаются у концов кариокинетической фигуры. Необ-
ходимо еще отметить, что карлиокинетическая фигура лишена лучи-.
стого сияния и что центродесмоза сохраняется очень долго и хорошо
заметна вплоть до момента разъединения обеих дочерних клеток.
Итак, беспримерное сходство хоаноцитов с воротничковымипро-
томонадинами находит поразительно яркое выражение и в харак-
тере их деления, относящегося к наиболее типичным формам
протозойного деления, точно к тому же типу, как и деление Сгазре-
domonadidae. '
Исключительно высокая степень «протозойности» хоаноцитов
Са1сагеа выражается еще в том, что они тут не спаяны друг с другом
(„как это бывает у более высоко организованных губок) и очень
легко выходят из эпителиоидного строя, переходя к самостоя-
тельному существованию в мезоглее в качестве амёбоцитов раз-
личного типа. Таким образом, они отличаются в этой группе и пол-.
ной обратимостью своей жгутиковой организации, т. е. лабильным"
уровнем дифференцировки, что также не имеет места у представи-
телей высших групп губок.
334
4. Трофические клетки книдарий
Трофические клетки книдарий, функционально соответствующие
хоаноцитам губок, имеют с ними много общего, будучи также
лабильно дифференцированными жгутик-оными клетками опреде-
ленно протомонадного типа организации. Они тоже характеризу-
ются.одноосно-гетерополярным строением, совмещающим в. себе
признаки радиальной ни двусторонней симметрии, наличием пре-
красно оформленной кинетиды и способностью легко переходить
в амёбоидное состояние; они также свободно выходят из эпители-
альн-ого строя и также обладают способностью к захватыванию
оформленных пищевых частиц и aux внутриклеточному ‚перевари-
ванию. Однако этим бесспорным чертам сходства с хоаноцитами
противостоят и многочисленные отличия, придающие трофоцитам
совершенно своеобразный характер.
Отличия от хоаноцитов выражаются прежде всего в соответствии
трофоцитов совершенно иной жизненной форме жгутиковых, чем та,
которой соответствуют хоаноциты, а именно—жизненной форме‘
амфимонады (сам. рис. 120).
Этот биологический тип сидячего, анимально питающегося жгу-
тоносца характеризуется прежде всего тем, что захватывание пище-
вь1х частиц происходит не посредством постоянного плазматиче-
ского воротничка, а путем их схватывания амёбоидными выростами
плазмы, окружающей‘ основание жгутов, т. е. при помощи хотя
и относительно локализованного, но структурно не оформленного
захватывающего аппарата. С таким, чисто амёбоидным типом
приема ‚пищи, по всей вероятности, связан другой характер строения
и функционирования жгутикового аппарата, чем наблюдающиися
у типичных <‹подгоняющих>> форм: он здесь чаще бывает двойным,
чем одинарным, и обладает, ПОВтИДИМОМу, существенно иными гидро-
кинетическими свойствами. Далее, этот тип жгутиковой организации
отличается от подгоняющего и тем, что жгутиковые могут перехо-
дить от сидячего к свободноплавающему образу жизни, без замет-
ного „изменения при этом строения клеточного тела. Наконец, отно-
сящиеся к „нему жгутиковые характеризуются, как правило, очень
слабым развитием пелликулы, легко позволяющим им временно
изменять свою форму, т. е. метаболировать. Таклим образоМ‚ ТИП
амфимонады существенно разнится от подгоняющего гораздо менее
выраженной специализацией к сидячему образу жизни.
Все эти общие особенности данной жизненной формь1 присущи
в полной мере и трофоцитам кишечнополостных. Всем_известна их
способность активно заглатывать пищу посредством псевдоподии,
образуемых в любой точке их свободного, жгутикового полюса,
и к внутриклеточному ее перевариванию, что дало возможносфь
Мечникову считать эти клетки фагоцитами, совмещающими в сеое
характерные свойства так-овых и первичных двигательных клеток.
Менее широко известна, повидимому, та легкость, с которой трофо-_
циты многих нкнидаргий могут выселяться ИЗ кишечного ЭПИТеЛИЯ и
335
вести затем in Vitro самостоятельное существование в активнопла-
вающей жгутиковой или в ползающей аМёбоиднои форме; при этом
они -либо сохраняют жгуты и поляризованное строение, mm же так
или иначе теряют их и совершенн-0 деполяризуются. Далее, для них
очень характерна и двужгутиковая, как правило, кинетчида, одина-
ково способная служить и подгоняющим“ и двигательным аппаратом.
Переходя теперь к более конкретнои морфологической характе-
ристике трофоЦитов‚ мы должны прежде всего отметить ту, обычно
сильно вытянутую в длину, цилнинДРИЧеСКУЮ ФОРМУ, какую ОНИ
Рис. 117. Различные типы жгутиковых клеток книдарий!
- ’ - 3——Tpoq)oum-
_ 2—-.желе3истая энтодермальная клетка гидры _(no Мак ДОННЭЛЛЮ),
1 трозътёЁнейро-эпителиальные клетки сцифистомы Chrysaora (no Чеу-Таи-ШЮИНУ)
Имеют, когда находятся в составе эпителия, а иногд; СОХРЗТРЯЮТ
и в подвижном, изолированном состоянии (рис. 117). та типичная
для них форма, постоянно вчидоизменяющаяся, впрочем, -в ХОДЁВРЁЁ
активного питания, нередко усложняется распластываниеь/Ёпзсгппа .
. 1 ' -
НИЯ тела клетки в виде подошвы, последняя является глацвн '
ратом, обеспечивающим соединение клеток между собои И, ИХ ПРИ‘
крепление к подстилающей жих базальнои пласЁинке. Х MH-
Жгутиковыи аппарат трофоцитов состоит о ычно из ДВУ Д V
ных одинаковых жгутов, имеющих раздельные базальные зерна,
э ’ .
типа масти.го-сом, т. е. не принимающих участия в кариоклинезёъ ОЧИ
способны самостоятельно делиться в интервалах между митозами,
что ведет к образованию у одной клетки трех, четырех или даже
пяти жгутов У некоторых форм (Chrysaora) в состав кинетидЫ
входит еще ризопласт и, возможно, округлый .napa6a3an1;IIeL;gIfl§IrTI:)II:l
рат (рис. 117, 3), строение которого, к сожалению, ост , .
336 '
еще детально не изученным. Ядро взрослых ‚клеток обыкновенно не
велико‚_ имеет обычное сетчатое распределение хром-атина и отно-
сительно крупное эксцентрическое ядрышко; его положение в клетке
может быть различным, проксимальным или дистальным и, кроме
того, меняться в различные моменты жизнедеятельности. Хондриом
обычно представлен очень '- ’ ‘
многочисленными зернисты-
Mn митохондриями, нор-
мально сосредоточенными
преимущественно у жгути-
кового полюса клетки. В с-о-
став вакуома входят весьма
разнообразные и многочис-
ленные включения, соотно-
шение которых .с диффузным
аппаратом Гольджи не мо-
жет считаться окончательно
выясн-енным; в целом он ха-
рактеризуется своей чрезвь1-
Рис. 118. Внутриядерньхй митоз энтодер-
чайной лабильностью. Сле- мальной клетки сцифистомы Chrysaora (по
дует еще отметить, что -в ЧеУ-ТаИ-ШЮРКНУ)
подошве у многих форм об-
наруживается сократительная нить, направленная поперечно к длин-
н-ой о-си трофоцита (рис. 117, 1).
‘Деление трофоцитов представляет совершенно исключительный
интерес в том отноше-нии, что оно часто совершается в очень свое-
образных формах «вну-
триядерного митоза»
(Вермель, 1926), харак-
теризующегося сохра-
нением ядерной обо-
ТЕЧЕНИЕ всего деления.
У Chrysaora, где митоз
- ЭТОГО типа является, ВО
Рис. 119. {Внутриядерный митоз энтодермальной всяко-м случае, господ-
‚клетки гидры (по Вермелю) ствующим у трофоци-
тов (рис. 118), он начи-
нается С ИСЧЭЗНОВСНИЯ ЯДЕРНОЙ СЕТИ И объединения ИМЕВЪПИХСЯ‘
в ней глыбок хроматина в одно округлое хроматиновое тельце;
вслед затем происходит сильная вакуолизация ядрышка, и оно
сливается с этим тельцем; образовавшаяся такимобразом округ-
лая масса н-ачинает удлиняться, перетягивается по середине и
затем разделяется на две дочерние отдельности, расходящиеся
к‘ противоположным концам постепенно вытягивающейся ядер-
ной оболочки; эта последняя теперь в свою очередь перешну-
ровыва-ется, завершая разъединение образовавшихся дочерних ядер.
При их реконструкции хроматин ядерной ‘сети вновь обособляется
22 Эмбриология беспозвоночных 337
ЛОЧКИ И КарИОСОМЫ Br
отдельно от деления ядер,
от карлиосомы и затем распадается на отдельные глыбки (Чеу-
Таи-Шюин, 1930).
У гидры, у которой этот тип митоза и был открыт Е. М. Вер-
мелем, он осуществляется несколько иначе (рис. 119).
Исключительное своеобразие этого типа кариокпинеза дало повод
некоторым авторам считать его дегенеративным уклонением нор-.
мального митоза и видеть в нем признак
старения клетки. Однако этому резко
противоречит хотя бы самый факт, что
внутриядерный митоз наблюдается имен-
но у трофоцитов (правда, не только
у них), т. е. наиболее омнип-отентных и
лабильных из клеток энтодермы и, кроме
того, пользуется, повидимому, очень ши-
роким распространением у Cnidaria. По-
этому ему должно «быть найдено иное,
более правдоподобное, объяснение, счи-
тающееся и с «его вероятной приурочен-
ностью к наиболее примитивным из кни-
дарий. Таким объяснением может, очевид-
но, послужить совершенно бесспорное
сходство внутрияд-ерного митоза кни-
дарий с различными вариантами уже
упоминавшегося нами (гл. П, 2) крипто-
митоза диморфных амёб (Vahlkamp-fi-
ida-e), ближайших к ним Polyb1ephari-
didae и некоторых постоянно амёбоидных
форм, врод-е Amoeba 'crysta11igera, a так-
ж-е и- настоящих протомонад из Bodonidae
или Атрпйшодпаойдае. Что оно не может
быть «случайным», — следует еще из, того,
что имеющееся тут сходство в ходе ядер-
ного д-еления дополняется и поддержи-
вается рядом других, ка-сающихся, напри-
мер,‘ поведения кинетиды при делении и
самого ее строения. В самом деле здесь
надо, во-первых, подчеркнуть еще раз,
что базальные зерна жгутов относятся
у гидры к в сущности очень редкому их
типу—к типу мастиг-осопм, очень харак-
терному как раз для тех из перечисленных форм, у которых имеется
жгутиковая фаза. Во-вторых, не легко подыскать примеры столь
полн-ого совпадения в строении жгутиковой кинетиды, как наблю-
даемое у Chrysaora, с одной стороны, и некоторых Vahulkampfia,
где каждый жгут также имеет два лежащих друг за другом зерна,
одно из которых является там и здесь мастигосомой,. а второе па-
рабазальным аппаратом.
Итак, трофоциты гидроидных и низших Scyphozoa обладают
338
Рис; 120. „Трофозоиты ко-
лониальной амфимонады
Spongomonas splendidum,
близкие по своей органи-
зации к трофоцитам гидр;
на рисунках видно, что де-'
ление кинетиды происходит
имеющего характер внутри-
ядерного митоза (по Гарт-
ману и Чагасу)
Ничуть не меньшей степенью «протозойности», чем прославившиеся
этим хоаноциты. Однако, в то время как последние точно отвечают
по своей организации и характеру деления представителям -семей-
ства Craspedomonadidae, трофоциты гидроидов явно тяготеют
к соседнему с ним семейству Amphimonadidae, а таковые сцифопо-
липов—скорее к двужгутиковым Vahlkampfiidae. TaKIdIM образом,
Существующее разнообразие их строения полностью укладывается
врамки систематического разнообразия низших Protomonadina.
5. Общие особенности дифференциации клеток тела низших Metazoa
Ввводных замечаниях к главе о первичном метаморфозе низших
Metazoa было указано, что в основе этого столь сложного процесса
лежит переход их клеточного материала из стадии киноцита, соот-
ветствующей стадии расселительной жгутиковой зооспоры простей-
ших, в стадию вегетативно деятельного трофоцита, отвечающей
стадии трофозоита у Protozoa; иначе говоря, первооснову этого
явления составляет‘ совокупность элементарных клеточных онтоге-
незов, прохождение каждой низ клеток тела определенного онто-
генетического цикла. _
А Основными доводами в пользу этой точки зрения служит, во-
первых, совершенно неоспоримый факт полного лморфофизиологи-
ческого соответствия киноцитов низших губок и кишечнополостных
_свободноплавающим протожмонадинам при СТОЛЬ же бесспорном И
детальном сходстве организации жгутиковых трофоцитов этих же
Metazoa c СИДЯЧИММ трофозоитами разных представителей назван-
ной группы Zooflagellata. Вторым доводом в ее пользу является то,
что киноциты низших губок непосредственно превращаются в хоано-
цитов. Хотя они проходят при этом ‘кратковременную стадию
дедифференцировки, усложняющую них онтогенез, остается фактом’,
ЧТО начальные ЕГО СТаДИИ ИМЕЮТ строение ЖГУТИКОВЫХ СПОр, а KO- '
He’-IHbI€—ICTpOeHIrIe трофозоитов. Таким образом, онтогенез эгих
клеток вполне сравним с онтогенезом любого одноклеточного, начи-
нающего свою жизнь в- качестве свободной расселительной жгути-
ковой споры или бродяжки и превращающегося затем в сидячего‘
«подгоняющего» трофозоита. Наконец, эта точка зрения поддержи-
вается всей совокупностью фактов, заставляющих считать дробление
частным случаем палиннтомического спорообразо.вания и =все виды‘
киноцитов —эквивалентами жгутиковых зооспор, что влечет за‘
собой и признание дальнейших превращений клеток бластулы отве-
чающим «постэмбриональному» развитию з-ооспор.’
Среди современных Metazoa мы не знаем, однако, ни одного
случая, когда все без исключения киноциты превращались бы при
оседании -синзооспорь1 в равноценные друг‘ другу и однотипные
вегетативные клетки характера хоаноцитов или других жгутиковых
трофоцитов. Этой участи подвергается здесь лишь известная часть
клеточного материала синзооспоры, дающая начало пищеваритель-
ному слою тела позднейших вегетативных стадий, в то время ‚как
22* 339
остальная дифференцируется в клетки иного стр-оения и иного био-
логическо.го значения. Следовательно, мы никогда не встречаемся
тут с одной этой формой клеточного онтогенеза, а одновременно
с несколькими его формами, с развитием однородных вначале кле-
ток в нескольких различных направлениях. И именно, благодаря
такой онтогенетической радиации киноцитов, а затем и их потомков,
создается клеточный полиморфизм, характеризующий собой веге-
тативные стадижи развития вс-ех без исключения Metazoa.
Таким образом, филогенетически исходный тип клеточного онто-
генеза, заключающийся в превращении из эквивалентов зооспор
в эквиваленты жгутиковь1х трофозоитов, сохраняется в развитии
Metazoa лишь частично как одна из нескольких (или многих) имею-
щихся у них форм клеточного онтогенеза. Что касается онтогенети-
ческой радиаципи других клеток, то ее характер и диапазон очень
не одинаковы в разных случаях. Так, у низших Са1сагеа она идет
всего в двух основных направлениях, одно из которых заключается
в пожизненном сохранении клеткойамёбоидной организации, при-
обретенной ею в порядке всеобщей дедифференцлировки клеток бла-
стулы, а другое—— в их более или менее далеко идущей специали-
зации в рамках той же амёбоидной организации. По первому
путиразвиваются тут все виды незернистых амёбоцитов, по вто-
рому—зернисть1е амёбоциты, колленциты, склеробласты и клетки
дермального слоя, достигающие крайнего уровня дифференцировки,
оставаясь, в сущности, амёбоидными. У книдарий дивергентное раз-
витие клеток идет по более разнообразным линиям. Одни из них
постоянно сохраняют и здесь свой характер амёбоидных клеток (те
из интерстициальных, которые получают значение камбия и служат
в дальнейшем источником образования самых различных клеточных
типов), другие переходят в такое же, как у губок, специализо-
ванное амёбоидное состояние (зернистые амёбоциты), третьи пре-
вращаются из амёбоидных в высоко дифференцированные клетки
совершенно ьиного строения (хотя бы книдотбласты); и, что самое
характерное, к этим трем направлениям развития клеток прибав-
ляется еще одно- преобразование уже дифференцированных жгу-
тиковых клеток в клетки самого различного специального наз-на-
ч-ения: половые, нейгро-эпителиальные, мышечные, железистые и дру-
me. Таким образом, первичный, как мы считаем, тип клеточного
онтогенеза видоизменяется либо путем отпадения его конечных
стадий, либо по модусу девиации, либо вследствие надставки новых
конечных‘ стадий.
Глава Х1Х
ГАМЕТОГЕНЕЗ И ПОЛОВОЙ ПРОЦЕСС НИЗШИХ
МНОГОКЛЕТОЧНЫХ
1. Вводные замечания
_ В огромной современной литературе, посвященной гаметогенезу
и половому процессу животных, -содержится лишь очень немного
данных, касающихся хода этих процессов у губок и низших кни-
340
дарий. Хотя именно эти примитивнейшие группы беспозвоночных
представляют совершенно особый интерес с данной точкпи зрения-
B силу их неограниченной способности к бесполому размножению
И отсутствия у них резкой грани между этим п-оследним и размно-
жением половым,——они далеко не пользуются тем вниманием, на
которое «былин бы вправе претендовать. Поразительное пренебре-
жение к ним, как к объектам генетического исследования, доп-ол-
няется еще тем обстоятельством, что имеющиеся теперь важнейшие
данные о гаметогенезе этих животных (в частности, известковых
губок) остаются не оцененными по достоинству и никем, в сущности,
He принимаются в расчет.
Создавшееся положение становится понятным, если учесть, что
подавляющее большинство исследований по гаметогенезу, созре-
ванию и оплодотворению животных базировалось на хромосомной
теории наследственности cu было направлено к ее обоснованию.
Между тем, развитие половых клеток низших Metazoa никак не
укладывается в формально генетические схемы, опровергает их
и потому естественно игнорируется их приверженцами. ‚
Весьма показательны в этом смысле те, поистине «героические
усилия, которые делались авторами, исследовавшими гаметогенез
известковых .губок—М. Иёргенсеном (1910), Дюбоском и Тюзэ
(1935—1937), чтобы как-то согласовать установленные ими факты
с положениями формально генетической теории мейоза. И хотя эти
усилия и не могли увенчаться успехом, они все же имели своим
результатом превратное истолкование соответствующих фактов и то,
что эти факты не получили до сих пор хоть сколько-нибудь широ-
кои известности.
Учитывая, что интересующий нас круг явлений достаточно хо-
рошо изучен в -настоящее время лишь у гетероцельных известковых
губок, мы ограничимся здесь рассмотрением фактического мате-
риала по одной только этой группе низших многоклеточных.
2. Источники образования первичных половых клеток
В полном соответствии с неустойчивостью клеточной дифферен-
ЦИрОВКИ ЭТИХ ОрГЗНИЗМОВ ИСТОЧНИКИ 0'6pa30BaHI/I51 ИХ ПЕрВИЧНЫХО
половых клеток очень разнообразны и непостоянны. Существовав-
шее раньше представление о происхождении этих клеток от «архео-
цитов» -—якобы обособляющихся от будущих соматических клеток
губки на самых ранних стадиях ее развития '-- теперь окончательно
опровергнуто, как и все строившееся -на этом мнение об обособлен-
ности «сомы» губок от их «зародышевого пути».
В -самом деле, теперь совершенно твердо установлено (Бриен,
1931—1932, Дюбоск и Тюзэ, 1935—1937), что первичными пол-овыми
клетками могут становлиться клетки различн-ого происхождения и раз-
ного функционального значения и в первую очередь сами хоаноцитьх
или их прямые потомки. Преобразование воротничковой клетки
в первичную половую может происходить на разных стадиях раз-
341
вития губки и заключается в выходе такой клетки из эпителиаль-
ного строя в Мезоглею и последующей ее дедифференцировкф В Ре-
‘зультате которой она становится амёбоидной, а затем и приобретает
‘специфические свойства гоноцита (см. рис. 124, 1-2). В первичные
половые клетк›и могут превращаться и крупноядерные незернистые
амёбоциты, входящие в состав ткани внутренней среды губки и,
повидимому, играющие тут роль фагоцитов (об их происхождении
см. стр. 331). Наконец, есть определенные указания (Иёргенсен),
что гоноциты могут образовываться из звездчатых колленцитов,
втягивающих свои отростки и становящихся не ОТЛИЧпИМЫМИ от дру-
гих незернистых амёбоцитов. Однако высоко дифференцированные
амёбоцить1—такие как зернисты-е или мегациты—не могут дать
начало первичным половым клеткам, и это же, разумеется, можно
сказать о пинакоцитах и склеробластах. Короче говоря, гоноцитами
могут стать только клетки, вообще обладающие высокой степенью
обратимости, т. -е. лабильно дифференцированные или же не диффе-
ренцированные вовсе. и
Превращение разнородных по своим исходным свойствам амё-бо-
цит-ов в первичные половые клетки выражается в их росте сверх
нормального предела и ряде ядерных изменений, в том числе в зна-
Читель-ном увеляичении массы ядра. У Grantia замечено, что взрослые
гоноциты бывают двух типов, но каково значение такого их димор-
физма —- неизвестно.
3. Общий очерк оогенеза
_ Как уже отмечалось выш-е, первичные половые клетки этих
губок располагаются в мезоглее, залегая чаще всего непосред-
ственно под жгутиковыми клетками. Здесь происходит них рост и
деление (идущее путем типичного митоза) на две о о г о н и и -`п е р-
вог-о порядка. По своему строению (рис. 121, 2-11) эти по-
следние мало чем отличаются от гоноцитов, и так же как и они,
питаются осмотически, вдвое увеличивая при этом свою исходную
массу,’ Тут же, в мезенхиме, выросшие оогонии первого порядка
вступают в "начальную фазу (профазу) своего деления; однако‘ для
его завершения они уходят отсюда в полость ирригационной си-
стемы губки, прорывая прпи ‘этом строй хоаноцитов, выстилающих
эту полость. Такой их переход извнутренней среды материнского
телав морскую воду. заполняющую его полость, является, повили-
мому,’ строго обязательным"; очевидно, он-о- свидетельствует о? том,
что деление оогонии первого. порядка требует" для своего осуще-
ствления совсежм иных условий существования, чем те, в которых
' протекает их рост.
Выйдя в воду, оогонии первого порядка заканчивают свое деле-
ние на две дочерние клетки—оогонии второго порядка,
сразу же делящниеся дальше каждая на два оо цита. Эти два
последовательных деления оогоний первого порядка совершаются
без всякого интервала, непосредственно переходя одно в другое’:
342
Рис. 121. Оогенез известковой губки Sycon raphanus:
. _ c «покоя имс » яд our 3-
;;{rzl1?1%BIgg;:§0:£?I:I2: a::,:1Hl;<>Je-21131-‘1;l1<;a:1fc-12:31:11:1/In,:4p‘e:r$:1I1:1}?0.;.—t:pz1!<§)nF:1;1§%l1f1il4e cr11»1peM1=1;m6, 7fl—o6p§3oB’a1me_:
тетрад; 8-—метафаза‚ 9—-70——анафаза и 77—тел0фаза деления оогонии первого порядуа,
12—оогония второго порядка метафаза и 1З—телофаза; 14—только что образовавшимся
ооцит из радиального канала губки; 15—ооцит из мезенхимьт, подготовка к образованию
спиремьх; 16-—лептотенная стадия; 17, 78, 79—стадия букета; 20-—пахитенная стадия, 21-
ооцит в начале периода большого роста (по ДюбоскУ И ТЮЗЭ)
343
телофаза первого из них как бы обрывается, не дойдя еще до
своего конца (заключительного этапа реконструкции дочерних
ядер), и непосредственно сменяется, таким образом, анафазой сле-
дующего деления (рис. 121, 12—13). Поскольку тут нет хотя бы
мимолетного интеркинетического состояния, нет, ‚конечно, и. роста
между делениями, так что масса каждого из четырех ооцитов‚ по-
лучающихся в результате этого «спаренного» деления, оказывается
вчетверо меньшей, чем масса оогонии первого порядка.
Только что образовавшиеся ооциты (рис. 121, I4) имеют вид мел-
ких (4‚5—7 р) амёбоидных клеток с небольшим ядром и одной
чешуевидной диктиосомой. Они вскоре же покидают водную среду,
в которой образовались, и внедряются в мезоглею мматеринской
губки, пробираясь при этом между хоаноцитами, под слоем которых
теперь оседают. Тут начинаются их питание и рост, проходящие три
последовательных этапа.
Первый из них, период M a Л о г о р о с т а, осуществляется, так
сказать, только собственными силами ооцита, за счет его осмоти-
ческого питания в мезоглее. K концу этого периода размеры ооцитов
достигают 15-18 р (Зусоп) или 20-22 р. (Grantia), и их ядро
становится относительно очень крупным (рис. 121, 15). Теперь -они
начинают стягиваться к основанию отводящих ирригационных кана-
лов, по-стоянно перестраивающихся в связи с продолжающимся
ростом материнского тела и расширением его атриальной полости.
Здесь ооцпиты находят необходимые им теперь клетки-кормушки,
соединяясь с которыми они переходят в следующий период своего
развития——в период большого роста.
Клетки-кормилки образуются из хоаноцитов, выстилавших рас-
падающиеся участкзи каналов: это—дедифференцированные ворот-
ничковь1е клетки, ставшие амёбоидными и испытавшие гипертрофи-
ческое развитие. Еще до своего объединения с яйцеклеткой они
захватывают по одной (редко две-три) дегенерирующей клетке,
которая постепенно высасывается, а впоследствии и поглощается
ими. Подошедшие сюда амёбоидные яйцеклетки в свою очередь
пристают к этому, уже двуклеточному комплексу, образуя вместе
с ним характернейшую фигуру (рис. 122, 1).
Вы-сасывая кл-етку-кормилку, а затем и поглощая.ее, яйцо зна-
чительно увеличивается в размерах и осуществляет таким образом
второй период своего роста. После этого оно возвращается к жгутик-
ковьгм камерам и вступает тут в третий, заключительный, период
своего развития. В течение этого периода ж е л т к о об р а з о в а-
н и я яйцо еще энергично растет, но уже без помощи клеток-корМи-
лок‚ и не фагоцитирует; источником его осмотического питания ony-
жит теперь только мезоглея, вероятно особо богатая питательными
веществами именно в соседстве с действующими хоаноцитами, чем,
очевидно, и объясняется соответствующее перемещение яйца.
Последнее прини-ма-ет теперь довольно правильную эллипсоидальную
форму и поляризуется, причем его главная ось устанавливается
перпендикулярно к стенкам жгутиковой камеры, а вегетативным
344
И!
полюсом становится тот, который обращен к этим стенкам, fr. e.
к основанию выстилающих ее хоаноцитов. Таким образом, можно
думать, что полярность яйца устанавливается в соответствии с на-
правлением преимущественного притока к нему питательных мате-
-o
Рис. 122. Поздние стадии оогенеза и оплодотворение яйца Sycon raphanus:
1—растущее яйцо (внизу), объединенное с питающей клеткой-кормилкой (в середине) и де-
генерировавшим хоаноцитом в характерное «трио»; 2-—-амебоидное яицо с кпеткой-кормил-
кой; 3——ЯЙцО‚ уже ассимилировавшее клетку-кормилку; 4-—яйцо с приставшеи к нему семя-
.носной клеткой, содержащей спермиоцисту; 5—выход спермиоцисты из семяносной клетки
и ее проникновение в яйцо; 6—яйцо с еще не ассимилированной спермиоцистой, заканчи-
ваёощее свой рост; 7—сближение мужского и женского ядер; на'правом конце яйца видны
выделенные направительные тельца (по Дюбоску и Тюзэ)
риалов (поставляемых хоаноцитами) и обусловливается именно им
и, "стало быть, имеет характер обычной трофической поляризации.
4. Явления созреванияв женском ряду
Эти явления необычайно интересны тем, что они п ов торя-
ются в оогенезе дважды: в первый раз—в порядке
равноценных делений оогониев, гидущих тут по нормам делении
созревания в мужском ряду, и в-о в т о р о й раз —— по обычнои схеме
345
созревания яйца с последовательным отделением двух редукцион-
ных телец. ’ - ' -
Этот неслыханный, можно сказать, факт, опрокидывающий все
устоявшиеся представления о редукции и ее сущности, дополняются
и рядом других не менее замечательных особенностей мейоза
Нетегосое1а, также не укладывающихся в привычные «нормы».
Поэтому не приходится удивляться, что исследователи, твердо
установившие эти факты ——М. Иёрген-сен (1910) и Дюбоск и Тюзэ
(1935, 1937), были не в силах согласовать их с ходячими представ-
лениями о мейозе и, считая эти теоретические представления незыб-
лемыми, не смогли осознать все з-начени-е сделанных ими открытий.
Это обстоятельство очень резко бросается в глаза при ознакомлении
с работами названных авторов—работами, могущими послужить
примером предвзятости в оценке и толковании фактов, добросо-
вестно и неопровержимо установленных ими самими в результате
очень тщательных исследований. И этим, конечно, следует объяс-
нить, что важнейшие выводы, неизбежно вытекающие из ‘этих
фактов, так и оставались до сих пор не сформулированными.
Таким образом, для того чтобы получить верное представление
о действительном положении вещей, о том, как происходит на самом
деле редукция в оогенезе НеЬегосое1а, надо исходить Не из явно
необъективных конечных выводов Иёргенсена, Дюбоска и Тюзэ, а из
сообщаемого ими фактического пматериала, к изложению которого
мы пи переидем.
Первый мейотический цикл в оогенезе Зусоп и
Grantia начинается с появления в ядре оогония первого порядка
призна-ков его перехода в стадию подготовки к образованию спи-
ремы (см. рис. 121, З). Далее следует стадия оформленной спиремы,
отвечающая по своим особенностям лептотенной стадии классиче-
_ского цикла и характеризующаяся значительной толщиной нити
спиремы; ее четко видным строением и отсутствием каких-либо сле-
дов двойственности этой ниити (см. рис. 121, 4). Ha следующей за-
тем стадии спирема распадается на нмногочисленные (выше 20)
палочковидные отдельности (см. рис. 121, 5), после чего происходит
вполне‘ типичный диакинез. На первых его стадиях (см. рис.121‚' б)
образовавшиеся биваленты представлены короткими палочковиднь1-
ми или гантелеобразными хромосомами, частью лежащими бок о бок,
Не СОПРИКаСЗЯСЬ друг с другом, а частью соединенными концами
в Ч-образную фигуру. На следующей стадии диакинеза (см?
рис. 121", 7) уже все биваленты соединены концами в такие фигуры,
а к моменту установления веретена они принимают форму кольце-
видных пили ромбических «тетрад». У Grantia‘ ромбические «тет-
рады» очень сходны с кольцевыми тетрадами'типа Arr.-phiu-ma И
частью явственно состоят из четырех хроматид, связанных своими
концами, .в то время как у Зусоп они имеют более округлые очерта-
ния (-см. рис. 121, 8) и не обнаруживают подразделения на отдель-
ные хроматиды; аи ‘в том и в другом случае нет никаких следов
продольного расщепления «тетрад» в плоскости кольца. Общее
346
число «тетрад», располагающихся в‘ Экваториальную пластинку,
‘равно, по одним данным, 8 (Иёргенсен и дрр), а по другим- 13.
В‘ анафазе первого деления созревания,‘ каковым оказывается
здесь деление оогония первого порядка на оогонии второго порядка,
биваленты разрезают-ся‚ как обычнодпоперек на соответствующее
число «диад». A A
- Эти последние сразу же после разъединения сильно изменяют
свою форму, превращаясь из У-образных в укороченные, лмассивные
хромосомы биоквитообразной или гантелевидной формы (см.
рис. 121, 11, 12).'Ка›к происходит это преобразованлие- в точно-сти
не известно; вероятнее всего, что оно заключается в выпрямлении
ранее Ч-образных диад и последующем их укорочении.
Второе деление созревания, представленное в оогенезе Hetero-
—coe1a' делением оогонпиев второго порядка на ооциты, осуще-
ствляется, как уже говорилось, немедленно после первого, без какой-
либо паузы между ними и без перех-ода ядра в интеркинетическое
состояние; телофазапервого деления непосредственно переходит в
метафазу второго, так что только что образовавшееся кольцо диад
сразу же оказывается расположенным п-о экватору вновь устанав-
ливающегося веретена. Как легко в-идеть из соответствующих ри-
сунков‘ (см. рис. 121, 12, 13), деление диад происходит в поперечном
направлении, в месте имевшейся у них перетяжки, чем объясняется и
округлая форма получающихся теперь хромосом. По окончании
этого деления ядра ооцитов переходят в интеркинетическое состоя-
ние, и на этом заканчивается первый мейотический цикл в женском
ряду гаметогенеза. -
Как видно из приведенных данных, этот цикл действительно
осуществляется по классической схеме процессов созревания в муж-
ском ряду, посредством двух сближенных во вр_емени делений. Этим
делениям равным образом предшествует образование спиремыци
диакинез, и оба они имеют ясно выраженный редукционный, меио-
тический характер. о
Однако наряду со всеми этими признаками типичного меиоза,
полностью исключающими возможность рассматривать данные
стадии оогенеза иначе, чепм как стадии созревания, в ‘самом ходе
этого процесса имеется немало существеннейших отличий от Tom
вейсманистской схемы мейоза, к которой пытаются подогнать все
наблюдаемые его формы. с '
‘Первое из ‘ниих заключается в‘ том, что тут никому не удалось
найти чего-либо напоминающего синапсис, видеть сближение и
конъюгацию нитей спиремы и обнаружить какие-либо намеки на
двойственность этих НпИТеЙ. Все имеющиеся фа-кты говорят о том,
что синаптические явления вообще не имеют места у оогонии пер-
вого порядка, чт-о тут нет никакого разделения фазы спиремы на
отдельные стадии. По всем данным, спирема формируется непосред-
ственно из распыленного хроматина покоящегося ядра, в виде-не-
прерывной одиночной нсити, еще сохраняющей четко видноеостроение
и‘ соответствующей, таким образом, лептонеме типичнои схемы,
. 347
а затем распадается на куски, подобно постсинаптической спиреме.
Другими словами, тут полностью отсутствуют как сама синаптиче-
ская стадия (с обеими ее подстадиями-синерезиса и букета), так
и следующие за ней стадии пахитены и диплотены; минуя эти стадии,
лептотенная -сразу переходит в диффузную, за которой наступает
следующая стадия — диакинеза. .
Процесс образования тетрад также не согласуется с традицион-
ными представлениями -на этот счет. Элементы, складывающиеся
в тетрады, соединяются друг с другом исключительно только- кон-
цами, тогда как параллельная их конъюгация никогда не имеет
места. .
Наконец, сам процесс расхождения и распределения хроматид_
в ходе обоих делений созревания совершенно не удовлетворяет-
требованиям морганистского канона. Если при первом из этих де-
лений‚ имеющем своим результатом поперечное разделение тетрад,
все идет примерно так, как полагается по канону, то при втором,
как мы видели, дело обстоит совсем иначе: вместо продольного рас-
щепления диад они делятся поперечно, чем окончательно доказы-
вается факт отсутствия синаптического соединения. Таким образом;
оогониальные «тетрады» известковых губок являются тетрадами
только в том смысле, что они состоят каждая из четырех элементов,
последовательно ра-спределяющихся посредством двух сближенных
делении, а отнюдь не «синаптическими партнерами», как их пыта-
лись изобразить (Иёргенсен). А это значит, что оба -эти деления
являются, по существу, редукционными и не имеют тут никакого
касательства к требуемым каноном, но фактичес-ки не существую-
щим здесь, явлениям синапсиса.
Таким образом, первый мейотический цикл в оотенезе Heterocoela
протекает в форме, внешне отвечающей общей схеме созревания
сперматоцитов, но по существу принципиально отличается от всех
общепринятых схем мейоза в том, что тут нет ни синапсиса, ни .па-
раллельной конъюгации хромосом и что оба деления созревания
являются редукционными.
Переходя теперь к описанию второго ‘цикла созревания в оо9енезе
известковых губок, необходимо преждевсего отметить, что он отде-
лен от первого сравнительно небольшим интервалом, а именно только
первыми этапами периода малого роста ооцита, протекающими, как
уж-е отмечалось, при «покоящемся» состоянии ядра. Так, у Sycon
первые признаки начавшегося мейоза наблюдаются у ооцитов раз-
мером 13-15 д . На этот раз подготовительные стадии процесса
созревания проходят в гораздо более развернутой, так сказать,
форме, значительно полнее, чем при созревании оогониев. Тут
удается легко проследить стадию появления «прохромосом», раз-
вертывания и лептотены, за которыми следуют сжатие петель спи-
ремы, а затем образование букета, пахитенная, стрепситенная и,
наконец, диффузная стадия, завершающаяся переходом ядра в сет-
чатое «покоящееся>> состояние. Таким -образом, тут имеются налицо
все последовательные стадии синаптического цикла, и единственное,
3218"
чего здесь нехвагает,—это попарного соединения нитей спиремы,
т. е. самого синапсиса. Тот факт, что ни в один из моментов описы-
ваемого чпроцесса нити спиремы не обнаруживают двойственности,
подчеркивается Дюбо-ском и Тюзэ в отношении Зусоп raphanus,
a о Grantia ОНИ пишут, что смогли найти в качестве лептотены
с двойной нитью только стадию, изображенную ‘на рис. 121, I7—I9,
y которой такая двойственность во всяком случае не видна. Так или
иначе, -попарное сближение нитей спиремы (или их отдельных ча-
стей) не было обнаружено искавшими его автора-ми. Отсутствие
здесь подлинного синап-сиса доказывается и изменчивым числом от-
д-ельностей на диффузной стадии, а также, как мы увидим, позд-
нейшими стадиями созревания. '
Эти поздние, заключительные этапы второго периода созревания
происходят в самом конце третьего периода роста яйцеклетки, зна-
чительное время спустя после проникновения в нее спермиоЦисты.
Наступающие теперь изменения в ядре заключаются в его пе-
реходе из пузыревидного состояния в стадию, отвечающую диффуз-
ной, но разнящуюся от нее намного большим (но также неопреде-
ленным) числом хроматиновых отдельностей. Эти последние пред-
ставлены многочисленными извитыми и как бы мохнатыми лентами,
перемешанными без всякого -порядка с мелкими‘ 4- или Б-угольными
хромомерами, заметно увеличивающимися в числе-на более поздних
стадиях с-озревания (см. рис. 122, б). «Стадий диакинеза обнаружить
не удается, равно как и оформленных тетрад; можно только сказать,
что многочисленные хроматиновые отдельности, имевшиеся в ядре,
дают начало 8 (по Иёргенсену) или 12-13 (по Дюбоску и Тюзэ)
коротким толстым хромосомам палочковидной или бисквитообраз-
ной формы, часть которых у Зусоп «иногда кажется состоящеи из
двух слипшихся хромосом. Однако никогда нет характерных бива-
лентов, как у оогония первого порядка» (Дюбоск и Тюзэ, 1937,
стр. 200). „ и
Перво-е деление созревания имеет определенно ‘гетеротипическии
характер. У Зусоп оно ведет к превращению очень массивных до
этого хромосом в намного более тонкие бисквитообразные, что дает
основание думать, что хромосомы тут делятся продольно. У Granha,
однако, наблюдается совершенно другая картина: разделившиеся
хромосомы представлены‘ в анафазе либо типичнейшими диадами
(состоящими из двух лежащих ‘бок о. бок хромосом), либо‘ простыми
округлыми тельцами. Результатом этого митоза. является вь1деле-
ние первого направительного тельца; оно долго остается приклеен-
ным к яйцу и часто испытывает тут деление, не доводимое, впрочем,
до конца. а
Телофаза первого деления созревания яица непосредственно
переходит в анафазу второго, имеющего своим результатом вь1де-
ление второго направительного тельца. В начале этого деления хро-
мосомы имеют округлую форму, -сменяемую в метафазе навытя-
нутую бисквитообраз-ную; как происходит их разделение- остается
пока неясным.
349
Вторым делением созревания яйца заканчивается его собствен-
ный мейотический Цикл, разорванный тут, как обычно, на два этапа,
разделенных длительным периодом роста. Приведенные фактиче-
ские данные убедительно свидетельствуют о том, что повторное
созревание безусловно является именно таковым и полн-остью ОТВе-т
чает типичным картинам созревания яйца у всех остальных много-
клеточных животных. Однако и здесь, как и в первом туре мейоза,
имеется ряд расхождений с прпинимавшейся до сих пор теоретиче-
ской схемой созревания.
Главное из них заключается в том, во-первых, ‘что тут нет синап-
тического процесса, требуемого этой схемой, и во-вторых, в том, что
здесь не приходится и говорить о существовании какой-либо преем-
ственности между элементами спиремы юного ооцита пи хромосо-
мами «выросшепо яйца, возникающими только накануне жего ре-
дукции.
Было бы, однако, большой ошибкой думать, что эти особенности
созревания яйца губок являются чем-то уклоняющимся от общих
норм: они только служат лишним доказательством того, что пипотеза
соединения «синаптических партнеров» при мейозе яйца и их пре-
образования в хромосомы делений созревания не оправдывается и
в этом случае, как и во множестве других. _
Итак, мейотические явления в оогенезе известковых губок ха-
рактеризуются тем, что они совершаются дважды-сперва по об-
разцу созревания сперматоцитов и затем уже в форме созревания
яйгца; что они оба раза обходятся без синапсиса; что оба деления,
входящие в первый из мейотических Циклов (оогониальный), явля-
ются определ-енно редукционнымжи и дополняются впоследствии еще
по крайней мере одним (если не двумя) редукционными делениями
яйца; и‘, наконец, что во втором из этих циклов не наблюдается
никакой реальной преемственности между элементами спиремы
ооцита и хромосомами созревающего яй-ца. ' ,
Совершенно очевидно, что все эти факты, в достоверности к-ото-
рых не. приходится сомневаться, в корне противоречат сущ_ествовав-
шим представлениям о ходе и существе мейоза и исключают, в ча-
стности, всякую возможность привычной интерпретации этого про-
цесса с позиций хромосомной теории наследственности. Нанося
этой последней еще один -сокрушающий удар, они могут служить
необычайно демонстративным конкретным примером несостоятель-
ности этой теории. '
5. Оплодотворение
Еще задолго до окончания роста яйца и его созревания к, нему
вновь присоединяется крупная амёбоидная клетка, напоминающая
по виду‘ клетку-кормилку и входящая в столь же тесный и дл-итель-
ный контакт с яйцом. Этоесемяносная -клетка, ил-и спермиофор,
также представляющая собой дедифференцированную воротничко-
вую лклетку, ушедшую из жгутикового эпителия в мезоглею, но
содержащую в себе сперматозоид. Образование таких семяносных
350
клеток из «обычных хоан-оцитов и их дальнейшая судьба, безусловно,
являются одной из самых любопытных особенностей процесса опло-
дотворения у губок —— процесса, целиком протекающего тут в весьма
необычных формах.
Первым этапом этого процесса является активное внедрение
спермия в тот из хоаноцитов (выстилающих жгутиковую камеру),
который оказывается ближе других к растущему под ни-м яйцу, или,
точнее-оказывается расположенным в продолжении главной осн
яйца, уже устанавливающейся в это время. Входя в хоаноцит через
его плазматический воротничок, спермий затем проникает в тело
этой клетки и постепенно смещается к ее о-снованию.
Спермий, находящийся в теле воротничковой клетки, помещается
в продолговатой ваку-оле, внутри которой он испытывает Целый ряд
характерных превращений. Хвост сперматозоида разжижается и_
исчезает, а его головка и митохондриальное тело значительно (раза
в три) увеличиваются в объеме. Одновременно с этим содержащий
его х_оаноц.ит теряет свой жгут и воротничок (т. е. дедифференци-
руется), округляется и затем погружается в мезоглею, становясь
уже амёбоидной семяносной клеткой. Теперь_спермий окружается
толстой оболочкой, превращаясь в характернейшую с п е р M И о,-
Цисту (см. рис. l22,'4, 5).
Оформившаяся семяносная клетка не сразу вступает в соедине-
ние с яйцом‚,а некоторф время блуждает в мезоглее. Затем она
прилипает к одной из сторон яйца и еще лишь позднее переме-
щается на его Вегетативный полюс. Складывающееся таким
образом «сизигиальчное» соединение яйца с -семяносной клеткой, со.-
держащей стпермиоцисту, поразительно напоминает истинные сизи-
гии некоторых кокцидий (например Adelea, CM. рис. 76, 9), причем
тут особенно бросается в глаза сходство семяносной клетки губок
C мужским гаметоцитом этих кокцидий, также являющимся, по су-
ществу, своего рода спермиоформ (см. рис. 122, 4). '
Процесс оплодотворения начинается сперехода спермиоцисты из
семяносной клетки в яйцо. При этом происходит еще одно необыч-
ное явление — образование в ооплазме ‚канала с резко очерченными,
двухконтурными стенками, по которому спермиоциста погружается
в глубь яй-ца (см. рис. 122, 5). Этот канал за-канчивается расширен-
ным ложем, в котором и помещается спермиоциста; его отверстие
еще долго остается прикрытым снаружи опустевшей семяносной
клеткой, лишь позднее отваливающейся от яйца. Инкапсулирован-
ная в яйце спермиоциста тоже долго не испытывает изменений;
яйцо -в это время продолжает расти и проделывает деления созре-
вания, при котором "направительные тельца выделяются у экватора
яйца. ‘
Только к моменту окончания делений созревания спермиоциста
выходит из; состояния покоя. Ее оболочка распадается, а ядро и ци-
топлазма освобождающегося -таким образом спермия обособ-
ляются от митохондриального тела, также подвергающегося ассими-
ляции. С этого момента начинается быстрый рост мужского прону-
351
клеуса в плазме яйца; он Многократно увеличивается в объеме и
часто обгоняет в росте женский пронуклеус, хотя последний и про-
должает расти достаточно интенсивным образом. После этого про-
исходит слияние обоих пронуклеусов в ядро дробления; вслед затем
сразу же устанавливается веретено перво-го метагамного деления.
Таким образом, процесс оплодотворения складывается у извест-
гковых губок из длинного ряда этапов, следующих друг за другом‘
в такой последовательности: 1) проникновение сперматозоида в ней-
тральную воротничковую клетку, в которой он тут же „испытывает
свои первые превращения; 2) преобразование этого хоаноцита в
блуждающую семяносную клетку, в к-оторой уже измененный и вы-
росший спермий превращается в спермиоцисту; 3) миграция семянос-
ной клетки, завершаемая ее объединением с еще растущим яйцом;
4) переход спермиоцисты в плазму яйца, где она еще долго остается
недеятельной; 5) разрушение спермиоцисты и освобождение из нее
сперматозоида; 6) интенсивный рост мужского ядра за счет ассими-
ляции ооплазмы; и, наконец, 7) слияние обоих пронуклеусов в ядро
дробления, сразу же вступающее в митоз. ~-
Начиболее примечательными особенностями этого на редкость
сложного и растянутого проце-сса безусловно являются, во-первых,
вовлечение в него хоаноцита в качестве вспомогат-ельной семяносной
клетки; во-вторых —энергичный рост с рмия как внутри этой
клетки, так и в яйце; и, в-третьих, очень аннее объединение семя-
носной клетки с яйцом, еще далеко не закончившим своего роста.
Все эти моменты, очевидно, могут рассматриваться как приспособ-
ления, необходимо связанные с той своеобразной формой внутрен-
него оплодотворения, ъкаким оно, в сущности, и является у известко-
вых и других губок.
6. Сперматогенез
Сперматогенез НеЬегосое1а был впервые изучен Н. ‘Полежаевым
(1882) у Зусоп raphanus, a затем немногими другими авторами
у того же Зусоп и у бгапйа, Сперматогончии первого порядка обра-
Рис. 123. Образование сперматоцисты у Sycon raphanus:
1, 2—начальные стадии объединения покровной клетки с семенной; З—-6—обрастание семен-
ной клетки покровной клеткой, образующей оболочку сперматоцисты
зуются в верхних ярусах губки из воротничковых клетокржгутико-
вых камер, подвергающихся при этом дедифференцировке и гипер-
трофическому развитию. В результате первых делении образуются
две амёбоидные клетки, содержащие зернистые митохондрии и по
одной чешуевидной диктиосоме. После полн-ого разъединения они
352
вновь тесно прикладываются друг к другу и одна из них {начинает
как бы обрастать или заглатывать другую, постепенно обхватывая
ее всю в виде все утончающейся оболочки (рис. 123, 1—2); эта по-
кровная клетка превращается, таким образом, в оболочку харак-
терной для губок сперматоцисты, внутри которой протекают в-се
последующие этапы сперматогенеза. Играя, по существу, роль кор-
милки для развивающихся в ней ысеменных клеток, покровная
клетка -сама сильно увеличивает свою массу, испытывая со време-
Рис. 124. Сперматогенез"кремневой губки Renieria simulans:
7, 2—хоаноцит, превращающийся в первичную половую клетку;
З-выросший гоноцит; 4—б—образование сперматоцисты и деление
обхватываемой ею семенной клетки на сперматогонии (5) и сперма-
’ тоциты (д); 7—готовый спермий (по Toss)
‘HQM дегенеративные ИЗМЕНЕНИЯ (В ЧаСТНОСТИ, пи-кнотичес-кое пере-
рождение ядра).
Семенная клетка, заключенная в сперматоцисте пи питаемая этой
посл-едней, делится два или три раза на подобные ей клетки, также
содержащие по одной диктиосоме.’ Получившиеся таким путем
сперматоциты первого порядка заметно увеличиваются в объеме, и
имевшаяся в каждом из них дликтио-сома делится на две. При насту-
пающем теперь редукционном делении имевшиеся в сперматоцитах
диктиосомы распределяются по одной между обеими дочерними
клетками-—сперматоцитами второго порядка. Иначе говоря, явле-
ние редукции охватывает тут не только ядерный материал, но и ап-
_парат Гольджи‚ и фактически сводится к тому, что при, делении
сперматоцитов первого порядка не происходит обычного удвоения
{клеточныхорганелл (в том числе и_хромосом), а имевшийся их
23 Эмбриология беспозвоночных 353
набор просто распределяется пополам. Происходящее затем деле-
ние сперматоцитоив второго порядка дает По две сперма-ЦИДЬЬ ВСКОРЕ
же переходящие в дифференцированное жгутиковое состояние.
едва’
рис 195 сперматогенез гидроида Tubularia mesembryanthemum:
. _. . W ‚
ТЫМИ
1 2_сперматоциты с двумя диктиосомами и многочисленными крупинча й
митохондриями; з._б._недифференцироВанные сперматиды с одной; дрётидозамаог‘
митохондриями, сливаощимся в четыре шаровидных тельца, л г „_
и н ировка спердлатидьп: 9—12——1'IP€BPaU-I61-I146 CTl9PM3'“’”1'=‘ В Сперми"! вид"
Ёзрептньобразование диктиосомы в акросому спермия и спаивание митохонд-
` риальных телец и их превращениев среднюю Часть СПЭРМИЯЗ 73"‘76‘r°'
ТОВЫЙ спермий (по Тюзэ)
ц
‚ Превращение сперматир ‘В Оформленные ‘СПЭРМИИ -YH:13;:C3TTK:I:
.ВЬ1Х губок спеЦИаЛЬНО не изучалосы Одна“ СУЩеЧТВУЪЁЩ 1932
Счет данные „о ‘другим представителям ‘губок (Remena, Тюзэ, )‚
354
имеющим спермии совершенно такого же строения, как у Heterocqelei,
позволяет в известной ‚мере ‘восполнитьэтот пробел. Для спермиоге-
неза губок характерны следующие главные особенности: ' ‘5
I 1) Рассеянные в плазмезернистые хондриосомы постепенно со-
бираются в четыре шаровидныхтельца, вначале лежащие разроз-
ненно,‘ но затем сосредоточивающиеся у жгутикового полюса ‘спер-
зматиды. В дальнейшем. они сливаются в одно массивное кольцо,
становящееся средней частью спермия- (рис. 124,‘ 7). _ ° А ‚
2) Диктиосома смещается к ‘полюсу ядра, противоположному
жгутиковому, и преобразуется здесь в акросому, полностью сохраэ
няющую свойства аппарата Гольджи (втом числе прижизненную
окрашиваемость янусом зеленым), ‘ '
3) пронизывающая ядро осевая нить образуется, повидимому,
за счет околоядерной (проксимальной) центросомы.
Готовый спермжий характеризуется удлиненной яйцевидной го-
ловкой с четко отграниченными от нее акросомой и соединительной
частью, и очень напоминает по своему‘ строению спермии гидроидов
(рис. 125) и некоторых моллюсков.
7. "Общие выводы
Подводя некоторые итоги фактам, изложенным в этой главе,
можно сформулировать ряд положений, имеющих существенное
принципиальное значение.
Первый из таких выводов заключается в том, что учение о не-
прерывн-ости зародышевого пути не только не находит себе под-
держки со стороны истории развития низших Metazoa, HO прямо
опровергается всей совокупностью относящихся сюда фактов. Пер-
вичные половые клетки интересующих нас форм неизменно образу-
ются -На -относительно поздних стадиях жизненногоцикла из кле-
точного материала, прошедшего через длинную Цепь преобразований
в ходе индивидуального развития той особи, в составе которой они
возникают. Половые клетки имеют ту же родословную, как и все
прочи-е клетки тела, из которых они выделяются только в опреде-
ленный жмомент Цикла после длительного пребывания предшествую-
ШИХ им поколений в состояняии чисто «соматических» клеток. Так,
у известковых губок в первичные половые клетки преобразуются
хоаноциты, функционировавшие до этого в качестве питающих кле-
ток тела и сами являющиеся потомками таких же воротничковых
клеток. У метазгенетических сцифоидных в «го-ноциты превращаются
ранее дифференцированные клетки сцифистомы, скорее всего энто-
дермальные, и это же можно сказать о метатенетических гидроме-
дузах‚ у которых они имеют-своим источником клетки энтодерМаль-
ного или же эктодермального происхождения.
Во всех этих случаях первичные половые клетки представляют
собой не что ино-е, как дедифференцированные клетки тела, как
клетки, перешедшие из дифференцированного жгутикового состоя-
ния в дедифференцированное амёбоидное.
. 93* 355
‚ Второй существенный вывод, непосредственно вытекающий из
рассмотренных здесь фактов, заключается в полной невозможности
подвести процессы созревания в оогенезе губок к основным поло-
жениям формально генетической теории мейоза. Уже самый факт,
mo созревание повторяется тут дважды-осуществляясь в -пер-
вый раз в форме, считавшейся строго специфичной для мужского
ряда "гаметогенеза, а во второй раз в виде вполне типичного созре-
вания яйца, —-показывает со всей очевидностью несостоятельность
этой теории. К такому же выводу неизбежно приводит все сказан-
ное вь1ше об отсутствии конъюгации хромосом при обоих процессах
созревания, об отсутствии какой-либопреемственности между спи-
ремой ооцита и хромосомами выросшего яйца и т. д. '
‘ ‘Наконец, нельзя н-е отметить, что явленлияредукции охватывают
в мужском ряду не только ядерные образования, но и чисто плазма-
тические (аппарат Гольджи), что еще раз подчеркивает невозмож-
ность объяснить этот процесс с точки зрения хромосомной, теории
наследственности.
ЗАКЛЮЧЕНИЕ
Попытаемся теперь подвести некоторые итоги всему тому Мате-
риалу, которого нам пришлось коснуться в этой книге, исформули-
ровать те общи-е выводы, к которым нас приводит их сравнительное
освещение с точки зрения -специально интересовавшей нас проб-
лемь1—об источниках и путях формирования индивидуального
развнития многоклеточных. _
I. Признание подлинного единства организации одноклеточных
простейших—с одной стороны, и клеток тела низших Metazoa—.-
c другой, есть первое по своей важности заключение, ‘к которому
обязывает весь рассмотренный нами материал.
Это единство проявляется особенно наглядным образом в отно-
шении «типичных» одноядерных клеток тела низших Metazoa и
находит свое выражение: '
1) в -самом факте обратимости клеточной организации, в способ-
ности клетки переходить из -недифференцированного состояния в
дифференцированное, и наоборот, т. е. в наличии у нее двух взаимо-
обратимых морфофизиологических фаз;
2) в том, что дифференцированная фаза клеточной организации
представлена ее жгутиковьгм состоянием. а недифференцированная
фаза-амёбоидным; действительно, жгутиковая фаза является
НИЗШИХ Metazoa HOpMa.TII>HbIM «рабочим СОСТОЯНИЕМ» КИНОЦИТОВ И
образующихся из них жгутиковых клеток других личиночных ста-
дий, хоаноцитов и доугих трофических жгутиковых клеток сидячих
вегетативных стадий, а также сперматид‚ а иногда и гоноцитов; что
касается амёбоидной фазы, то она тут наблюдается как времен-
ный этап при дедифференцировке жгутиковых клеток в ходе их раз-
вития (при образовании фагоцитобласта, при превращении личинки
в зачаток сидячей стадии и т. д.) и является «рабочим состоянием»
у всех видов амёбоцитов;
З) в наличии переходных, амёбоидно-жгутиковых состояний кле-
точной организации, наблюдаемых, например, у энтодермальных
клеток книдарий и губок; ‚ ' ‚
, 4) в совпадаюшем общем плане строения каждой из этих фаз;
жгутиковая неизменно характеризуется одноосно-гетерополярным
строением и комбинированными формами симметрии (например,
сочетанием радиальной симметрии с двубоковой у хоаноцитов губок,
357
или двулучевой с билатеральной и т. п.), обычно удлиненной и более
или Менее постоянной формой тела, наличием разнообразной по
своему составу кинетидь1, базальные зерна которой могут быть то
мастигосомами (книдарии), то центромастигосомами (губки) и су-
ществованием у вегетативных жгутиковых клеток т-ех или иных
специальных дифференцпировок (плазматические воротнички, цито-
стом, пульсирующие вакуоли); для амёбоидной фазы, наоборот,
характерны анаксонное или неопределенно многоосное строение,
непостоянная форма клеточного тела и отсутствие оформленных
дифференцировок; _
5) в совпадающем составе комплексаклеточных органоидов и
включений (ядерный аппарат, центросома, аппарат Гольджи в раз-
ных его вариантах, хондриом) ни постоянном наличии в клетке Tex"
или иных параплазматических включений. - _
_' _ Отсюда ясно, что единство организации отдельных клеток тела
„Мейаяоа и целых одноклеточных простейших является фактом, а не
предположением, и что оба основных ее типа у низших Метаяоа —
эпителиальный Жгутиковый и амёбоидный паренхимный-точно
соответствуют обоим главным типам одноклеточной организации
,у„_Рго’по2оа, типам, олицетворяющим, как мы видели, ее взаимооб-
ратимые морфофизиологические фазы. Таким образом, можно
с полным правом говорить, что -клетки тела низших Metazoa со-
‚храняют типично протозойное строение, а именно строение однокле-
точных, лабильно дифференцированных (диморфных) Flagellata.
п- Это „принципиально „важное положение может быть еще значи-
тельно конкретизировано в том смысле, что отсутствие у клеток
"МеЁа2оа_фотосинтетического аппарата и связанных с ним образова-
ний (пиреноидов) и оболочек, при способности многих из них к за-
{хватыванию оформленных пищевых частиц (как в амёбоидном, так
и`вжгутиковом их состоянии) и внутриклеточному их переваёрива-
нию, делает их подобнымине всяким вообще простейшим указан-
‘gore T‘1}1na,M'a одним толькоосмотрофным и анимально питающимся
‘его представителям, т. е. формам, относящимся к Zoof1agel1a_ta.
Стало быть, пможно еще уточнить приведенное положение, сказав,
(что „клетки низших Мета2оа сходны по своей ‚органгизации с одно-
‘клеточными, диморфными 2ооТ1а9еПата. ` ` " _ у
` Однако и _этим еще далеко" не достигнут предел возможной здесь
конкретизации; многие из наблюдаемых у низших Metazoa ‘КЛЕТОЧ-
ных типов и форм митоза нгичем не отличаются от некоторых кон-
кретных типов строения и деления различных одиночных Рготото-
пафпа. Наиболее поразительный исамый’ «парадный» из прим-еров
такого рода дают‘ нам хоаноциты губок`(особенно низших), практи-
чески не отлвичимые от трофозоитов Craspedomonadidae даже в та-
ких деталях, как строение парабазального аппарата, выполнение
базальными зернами жгутов роли центросом при делении, наличие
пульсирующей вакуоли ' (y" пресноводныхформ) и т: д. ‘Не менее
‘демонстративный другие приводпившиеся нами примеры, касаю-
щиеся энтодермальных жгутиковых клеток книдарий -- организация
‘$53
[которых крайне. близка к таковой ряда ёбезворотничковых ‘сидячпих
протомонад-и киноцитов различных бластул, имеющих строение
свободноплавающих осмотрофных Protomonadma. Вопиющии недо-
статок цитологических исследований по жгутиковым клеткам низ-
ших Cnidaria не позволяет пока продолжить этот список; но его,
‘ТЕМ НЕ МЕНЕЕ, ВПОЛНЕ ДОСТЗТОЧНО, ЧТОбЫ ПРИЗНЗТЬ, ЧТО ЭТИ ‘КЛЕТКИ
обладают организацией протомонад, и притом в целом ряде -ее ва-
риантов. Легкость .перехода жгутиковых клеток низших Метаяоа
(кроме, разумеетоя, оперматид) в амебоидное состояние (путем
дедифференцировки) очень хорошо совместима с их протомонад-
нь1м характером, и это же мгожно сказать о дедиффереНЦигЮВаННЫХ
и постоянно амёбоидных клетках этих жпивотных.
Правда, бедность любых амёбоцитов структурами и их морфо-
физиологическое однообразие таковы, что тут нельзя и рассчиты-
вать на хоть сколько-нибудь эффектные сравнения, особенно, если
учесть и ничтожную степень изученности цитологии амебоидньтх
Flagellata; однако в строении, способах деления и биологии этих
клеток нет и ничего чуждого амёбоидным протомонадам. Таким
образом, изменчивость организации клеток наиболее примитивных
›Ме’са2оа не выходит за ее пределы у одноклеточных Protomonadina.
Иначе говоря, основные клеточные типы низших книдарий и губок
олицетворяют различные морфофизиологические состояния, прини-
маемь1е при разных обстоятельствах одним и тем же -— протомонад-
нь1м—типом одноклеточной организации и представляют поэтому
полную параллель системе жизненных форм, существующей у самих
Protomonadina.
Устойчивость протом-онадного типа организации, характерного
для клеток низших M-etazoa, относительно очень велика. Так, у гу-
бок даже наиболее специализированные из клеток (такие, как
пинакоциты, пороциты и склеробласты из более оседлых клеточных
типов и гранулошиты-из блуждающих) не изменяют ему при иС-
‘пытываемой ими функциональной дифференциации, и ЭТО Же Спра‘
‘ВЕДЛИВО В ОТНОШЕНИИ ОСНОВНОЙ массыклеточных фОрМ НИЗШИХ КНИ-
дарий. При специализации в любом направлении клетки стоико
удерживают протомонадный план строения, дополняемыи только
теми или инынми специальными дифференцировками, и ‘Одии ЛИШЬ
клетки стрекательной системы далеко отходят, к концу развития, от
общего клеточного типа. Особо следует подчеркнуть и то обстоя-
тельство, что в тел-е низших M-etazoa практически отсутствуют мно-
гоядерные и другие надклеточные или неклеточные образования
(не говоря, конечно, о промежуточном веществе), так что оно Деи-
ствительно полностью разложимо на клетки одного- и того же струк-
турного уровня. Таким образом, мы имеем все основания утвер-
ждать, что все пили почти все клетки тела наиболее примитивных из
Metazoa обладают не зоофлагеллятным вообще, а типично протомо-у
НЗДНЫМ ТИПОМ ОрГаНИЗЗЦИИ. '
ЭТО ЗНаЧИТ, ВО-ПЕрВЬШХ, ЧТО уже НЕВОЗМОЖНО отрицать_ филогене-
тическую. связь низших многоклеточных с РЮЁФШОПЗСППЗ и ЧТФ
359
классическая теория происхождения Metazoa OT одноклеточных
зоофлагеллят не только остается в полной оиле но и поддержи-
вается всей совокупностью имеющихся, теп-ерь данных по цитологии
низших многоклеточных животных. Особенности их клеточного
строения совершенно необъяснимы с какой-либо иной точки зрения
и служат неопровержимым доказательством правильности этой
теории.
В°'ВТ°РЫХ› ИЗ СКЗЗЗННОГО следует, что клеточная теория есть не
только теория -существующего строения организмов, но имеет и
ГЛУбОКОЁ филогенетическое содержание, так часто игнорируемое
Даже ее сторонниками. Являясь, в общей своей форме, теорией един-
ства происхождения всего организованного мира, к л е т о ч н а я
теория имеет вместе с тем и’ различное конкретное
значение: теории протомонадного происхожде-
ния Metazoa, c одной стороны, и фитомонадного
ПРОИСХОЖДеНИя многоклеточных Volvocoidea "H,
вероятно, высших растений-мс другой,
Уместно отметить, что единство протомонадного происхождения
всех Metazoa (не исключающего, конечно, их происхождения от
дРЗЗНЫХ групп ОбШИрНОГО И теперь протомонадного комплекса) на-
ходит свое очевидное выражение и в клеточном строенили высших
животных. (Достаточно вспомнить о различных жгутиковых эпители-
альных клетках, сперматидах и амебоцитах‚ чтобы правильность
этого утверждения не ставилась под сомнение. ‚
П- Итакэ МЁЁЗЁОЗ H. особенно, ‘более примитивные из них выдаю
свое протомонадное происхождение всей спецификой своего кле-
точного строения и морфологически равноценны (на многих ста-
диях) организованному комплексу клеток типично протомонадного
характера. У низших Metazoa такими комплексами представлены
все стадпии, проходимые ими в онтогенезе с момента окончания
дробления и до мом-ента .полов-ого размножения, т. е. стадия еще
не дифференцированной бла-стулы, следующие затем жгутиконосные
личиноч-ные стадии, сидячие вегетативные и, наконец, бесполые или
половые расселительные стадии. Однако в ряде других стадий
цикла все Ме’са2оа имеют типично одноклеточное строение (молодые
гоноциты и ооциты, сперматиды и спермии) или же строение гипер-
трофированных клеток (взрослые гаметоциты и, в частности,
яйца) или же представлены комплексом гипертрофированных кле-
“ток-—бластомеров (стадии дробления). Возникает вопрос, являются
ли эти-стадии цикла и онтогенеза особыми клеточными типами,
‘выработавшимися у Metazoa B порядке дифференцировки некоего
‘гоно-бласта, как это теперь принимается многими, или же им следует
‘при-писать иное, принципиально отличное значение.
Из всего рассмотренного материала явствует с полной очевид-
ностью, что гаметогенетическая цепь одноклеточных поколений
Метаиоафотвечает прогамным поколениям жизненных циклов. upo-
“стейших и представляет эти ‘поколения в цикле Ме’са2оа, и что яйца
‘И спермии ПОСЛЕДНИХ действительно являются соответственно макро-
360
и микрогаметами. Это так же очевидно, как и то, что продуктом
копуляции последних является истинная зигота и что общая схема
полового процесса одинакова у всех организмов.
Равным образом невозможно отрицать, что избыточный рост
„яйцеклетки Metazoa есть типичнейший случай гипертрофической
дистомии; следовательно, нет решительно никаких оснований про-
должать сомневаться в палинтомической природе следующего затем
(метагамного) дробления и в соответствии бластомеров Metazoa
спороцитам палинтомических простейших.
Отсюда ясно, что все одноклеточные стадии Ме’са2оа и их стадии
дробления не могут считаться новоприобретением самих многокле-
точных, отсутствовавшимли у их предков. Наличие этих стадий «в
Цикле развития многоклеточных абсолютно необъяснимо иначе, чем
‘с точки зрения протозойной теории их происхождения, и служит еи
потому могущественной поддержкой.
Посмотрим теперь, что дают-одноклеточные ‚стадии цикла Ме-
tazoa (Ha особенностях самого их цикла мы остановимся далее)
для конкретизации этой теории в намеченном выше направлении".
Из того, что они все имеют амёбоидное (в прямом или, по крайней
‘смере, В ШИрОКОМ СМЫСЛЕ слова) ‘СТРОЕНИЕ, само СОбОЙ ЯВСТВУЕТ, ЧТО
от них нельзя ожидать столь же я-сных показаний на этот счет, как
те, какие дают нам высоко дифференцированные . жгутиковые
формы. Обнаружить у них жгутиковые, а тем более протомонадные
ЧЕРТЫ, кажется C I'IepBOF0 ВЗГЛЯДЗ даже МаЛОВЕрОЯТНЫМ‚ ХОТЯ МЫ ‘И
'ХОрОШО 3HaeM, ЧТО МНОГИЕ ИЗ НИХ 06pa3y1oTcs{ II/I3 ЖГуТИКОВЫХ КЛЕТОК
путем их дедифференцировки (гоноциты губок) или же сами пере-
ходят со временем B }KI‘yTI/IKOBOe состояние (сперматиды): ‘
При ближайшем рассмотрении, однако, такие жгутиковые черты
резко бросаются в глаза. Самым распространенным, хотя и далеко
не самым характерным из жгутиковых признаков одноклеточных
стадий Metazoa, СЛУЖИТ наличие у большинства из них постоянной
внеядерной центросомы, являющейся, как мы знаем, зачатком или
рудиментом кчинетиды. ?
Значительно более характерны в этом смысле явления гипер-
трофической дистомии яйцеклетки и ее последующее деление (по-
вторное), т. е. весь палинтомический процесс, который встречается
среди одноклеточных почти исключительно у диморфных Flagellata
ги достигает только у них форм, отвечающих дроблению Metazoa.
Таким образом, высокая ‘степень гипертрофии, достигаемая яйце-
"клеткой, палинтомический характер дробления и связанные с ним
особенности бластомеров являются, по самому своему существу,
"признаками жгутикового характера. Еще более типичны в этом
"отношении одноосно-гетерополярное строенпие зрелого яйца и бла-
стомеров и специфические особенности -их деления у наиболее при=
митивных Ну‹1го2оа—его продольный характер и осуществление
посредством односторонних врезающихся борозд.
Таким образом, жгутиковая организация половых клеток и бла-
стомеров многоклеточных вполнеосязаема и наглядна, иддопол-
361
няясь их гетеротрофной организацией, ясно указывает на связь
Metazoa с Zooflagellata-. ' ;
Вместе с тем приходится констатировать, что организация, раз-
множение и‘ образ жизни одноклеточных стадий Metazoa не Дают
оснований к дальнейшему уточнению этих связей. Об их половых `
клетках можно сказать то же, что о входящих в состав их тела
разнообразных амёбоцитах: ничто не препятствует их трактовке
‚как недифференцированных KJICTOK протомонадного ТИПЗ, НО МЗЛО‘
ЧТО И поддерживает ее. ЧТО же касается гипертрофированных ОДНО:
клеточных» стадий и явлений палинтомии‚ то они ничего не дают
в этом отношении, Дело в том, что палинтомия встречается у
Protomonadina, как редкое исключение (у некоторых Craspedomo-
nadidae, no старым данным С. Кента, у гигантских форм трипано-
[COM —- где она имеет характер вторичной синтомии, и у еще сомни-
тельной епо своему систематическому положению Sporomo-Has),
И, стало быть, не типична для них. '
Итак, если наличие одноклеточных стадий в жизненном цикле
‚МеЁаЪоа и организация этих стадий прочно поддерживают зоофла-
‘геллятную теорию происхождения"многоклеточных, они ничего не
говорят о филогенетических связях последних с какой-либо кон-
кретной группой Zooflagellata. Эти связи ярко проявляются только
в расселительном и трофическом отрезке ц-икла, „на материале диф-
ференцированных жгутиковых клеток——кианоцитов и трофошитов. A
v HI. МЫ ос-обо подчеркивали выше, что жизненный цикл низших
'MetaZoa развертывается по тому же общему плану, как у многих
Protozoa, a H=MeHHo no плану правильного расчлененного метагене-
тическогоцикла. Он также складывается из четырех периодов:
-1') м-етагамного спорообразования, представленного тут дроблением
ги оформлением синзооспоры; 2) вегетативного периода, включаю-
щего длинный ряд разнообразнейших, но всегда монотомических
клеточных поколений; 3) прогамного периода, состоящ-его из ряда
клеточных генераций (в женском ряду: гоноцнты — оогонии первого
порядка-оогонии второго порядка——ооциты-; в мужском: гоно-
циты —— C1'IepMaTOI‘OHHI»I~. первого порядка —— сперматогонии второго
порядка — сперматиды), из которых последняя подвергается гипер-
трофии B женском ряду и спермиогенезу- в мужском; и, наконец,
4) п-олового процесса, заключающегося в га-Метной редукции и
оогамии. ‘ ›
Постоянство этой схемы у Metazoa B целом хорошо известно.
В своем месте (гл. Х) было указано, что ее главной особенностью
по сравнению с цикла-ми большинства простейших является переход
метагамного спорогонич.еского размножения зооспорами в характер-
ный для многоклеточных процесс воспроизведения, ОбУСЛОВЛИВНЮ-
‚щий перенос размножения на прогамный период цикла. Это явле-
ние,— наблюдаемое также у Volvocoidea, есть прямой результат
объединения зооспор в одно многоклеточное целое ли самое заметное
отражение такого объединения во всем жизненном цикле много-
I КЛЕТОЧНЫХ. ЧТО же каса-ется ‘СООТВЕТСТВУЮЩЕГО УВЁЛИЧЕНИЯСПРОДУКГ
862
тивности гаметогенеза в женском ряду и дифференциации прогам-
НЬ1Х клеточных поколений, то их специфичность для Metazoa 3Ha1_11«I¢-
TeJIbHO более велика, хотя тут и следует вспомнить аналогичные
явления у кокцидий и предельно резкую дифференциацию данного
отрезка цикла у миксоспоридий. .
Если добавить к этому, что гаметный тип редукции не составляет
‘редкости у Protozoa (особенно гетеротрофных), то придется при-
‚знать фактом, что жизненный цикл Metazoa no уровню своей диф.-
ференциацпии и ее конкретным формам обнаруживает громадное
«сходство с некоторыми из циклов простейших. A
Сказанному не противоречит то обстоятельство, что жизненный
«цикл Metazoa осуществляется одной особью и составляет содержа-
ние ее онтогенеза, в то время как у простейших он складывается
из длинного ряда сменяющих друг друга поколений самостоятель-
ных особей и представляет совокупность их онтогенезов; стоит
только вспомнить, с одной стороны, множественность многоклеточ-
ных поколений у метагенетических книдарий, и с другой — особенно
подчеркиваемое нами обстоятельство, что нормальный жизненный
‘цикл всех Metazoa фактически включает не менее четырех одно-
клеточных генераций, из которых только одна приобретает затем
многоклеточное строение. ‚
Все это вместе взятое с определенностью говорит о том, что
сингамный характер жизненного -цикла Metazoa и его общая кон-
струкция-не могут считаться следствием их даногоклеточносттт, ре-
зультатом ее возникновения. Напротив, все говорит за то, что
оформление дифференцированного Цикла явилось и является необ-
ходимой предпосылкой возникновения многоклеточности, ‚что ци-
клическая пинтеграция и дифференциация клеточных поколений во
времени на протяжении жизненного цикла необходимо предшество-
вала их пространственному, физическому объединению. Только
с этой точки зрения объяснимы, во-первых, самый характер-инди-
видуального развития Metazoa, как «воспроизведения себе подоб-
ных не непосредственно, а через длинную цепь воспроизведения
себе не подобных»; во-вторых, то, что объединению в многоклеточг-
Hoe целое подверглись у Metazoa лишь клеточные поколения мета-
гамного и вегетативного отрезка цикла, в то время как прогамные
и половыеостались не интегрироваными; в-третьих, она делает по-
нятной множественность вегетативных поколений, характерную для
низших Metazoa; в-четвертых, ею объясняется поразительное одно-
образие жизненного цикла всех Metazoa; наконец; такая точка spe-
ния дает возможность понять, как будет показано дальше,многие
необъяснимые иначе особенжности онтогенеза ‚многоклеточных.
’ Противоположная трактовка цикла Metazoa, как выработанного
ими новообразования, не встречает никакой поддержки в относяг
щихсясюда фактах. Наоборот, она сталкивается c непреодолимыми
трудностями и лишена, вдобавок, какой-либо объяснительной цен-
ности. _ .. _ 4
Таким образом, мы должны признать сингамный жизненный
цикл фундаментом организации и онтогенеза Metazoa, a He над-
стройкой над ними, пи притом фундаментом протозойного характера.
Это положение, однако, нуждается в уточнении. Дело в том, что
если каждое, или, точнее, почти. каждое, из слагаемых жизненного
цикла Metazoa имеет свое подобие у различных свободноживущих
постоянно одноклеточных простейших (из сравнения с которыми
только и следует исходить в такого рода построениях), мы не знаем
среди них форм со столь сложно расчлененным, -как у Metazoa,
прогамным отрезком цикла. Иначе говоря, уровень расчлененности
цикла, существующий у Metazoa ДОСТИГНУТ среди простейших од-
ними только паразитическими формами. Поэтому, принимая нали-
чие метагенетического жизненного цикла с оогамией, гаметной ре-
дукцией и чередованием палинтомических и монотомических поко-
лений за унаследованное от простейших свойство Metazoa, за тот
фундамент, на котором они создались,— мы не можем считать та-
-ким свойством дальнейшую, более дробную дифференциацию про-
гамной части цикла, характерную для Metazoa. Тот факт, что сход-
ная и даже большая его дифференциация наблюдается и среди
высших Protozoa, говорит только об общности путей эволюции
цикла у них и у Metazoa, НО не дает этому прямого объяснения.
Однако стоит только учесть то, что говорилось о последствиях пере.-
хода размножения зооспорами в воспроизведение‚посредством мно-
гоклеточных синзооспор, чтобы это явление стало для нас понятным:
=оно оказывается именно тем способом п о в ы ш е н и я п р. о д у к-
т и в н о с т ли г а M е т ог е н е з а, которым биологически компенси-
руется выпадение метагамного размножения. Следовательно, эта
особенность цикла Metazoa есть необходимое следствие их много-
клеточно-сти, а ее сходство с аналогичными явлениями у споровиков
сводится к тому, что и у них гаметогенез испытывает подобное же
-.(но вызванное другими биологическими моментами) усиление про-
дуктивности. Иными словами, если расчлененность прогамной части
Цикла Metazoa биологически обусловлена самой их многоклеточ-
ностью, то сходная его дифференциация у споровиков вызвана
"K ЖИЗНИ совсем друпими причинами.
Итак, единство общего строя сингамного жизненного цикла про-
стейших и Metazoa, наличие у первых всех типичных компонентов
цикла M-etazoa и сами те конкретные особенности последнего, ка-
гкими он отличается от шиклов свободноживущих метагенетических
одноклеточных,‘ так же единогласно свидетельствуют о протозойном
происхождении многоклеточных животных, как свидетельствует
то нем организация клеток их тела и одноклеточных стадий цикла.
Конкретизировать филогенетические показания,‘даваемые жиз-
-неннь1м циклом Metazoa, оказывается пока затруднительным. Его
жгутиковые признаки усматриваются из сказанного в предыдущих
параграфах, но специальные признаки протомонадного характера
в нем практически отсутствуют, что и понятно потому, что у с-овре-
именных Protomonadirla не Известны оформленные Циклы метагене-
тического «типа. .
364
IV. Преобразование многочленного жизненного цикла, харак-
терного для простейших, в «нормальный» онтогенез, типичный для
высших Metazoa, проходит через ряд последовательных -ступеней.
Низшую ступень в этом р-яду повышающейся концентрации цикла
занимают среди книдарии метагенетические пидромедузы; их цикл,
как известно, осуществляется целой цепью сменяющих-ся генераций
и еще явно не отвечает, в целом, тому пониманию онтогенеза, кото-
РОЕ ПРИНЯТОЕ ЗООЛОГИИ. Характер ТИПИЧНОГО ИНДИВИДУЗЛЬНОГО раз-
вития имееттут только начальный этап цикла, включающий гипер-
трофический рост яйцеклетки и ее оплодотворение, дробление и
оформление жгутиконосной‘ бластулы, ее последующее преобразо-
вание B паренхимулу и затем в планулу, превращение последней
в зачаток сидячей стадии и, наконец, оформлени-е полипа — основа-
теля колонии. Далее следует длительный процесс формирования
многочленной гидроидной колонии, осуществляемый путем почкова-
ния H других процессов, могущих перерастать в подлинное вегета-
тивное размножение. В этой части цикла понятие об индивидуаль-
ности одинаково приложимо и к колонии в целом и к каждому виз
составляющих ее гидрантов; однако многочленность этой части
цикла все же несомненна и становится очевидной при вегетативном
размножении. Следующая часть цикла, заключающаяся в образо-
вании поколения медуз, ведущих самостоятельный образ жизни,
является истинным вегетативным размножением; каждый его про-
дукт испытывает св-оеобразный (в силу своего вегетативного проис-
хождения), но все же заслуживающий этого названия онтогенез и
обладает той же относительно высокой степенью метазойной инди-
видуальности, как и мерцательные стадии первой части цикла.
Впрочем, и медузы иногда способны к вегетативному размножению,
‚ |И ‘РОГДЗ ЭТа Ча-СТЬ ЦИКЛЗ ТОЖЕ ОК-ЗЗЫВЗЁТСЯ МНОГОЧЛЕННОЙ. ЧТО PKE
касается истинной его ‘прогамной части, т. е. самого гаметогенеза,
то он-осуществляется блуждающими клеточными генерациями- со-
матического происхождения, свободно перемещающимися ‘в преде-
лах материнского тела и только впоследствии оседающимпи ‚в гона-
дах медуз. 3 -
Таким образом, на этой низшей ступени концентрации цикла
лишь относительно небольшая его часть протекает в порядке офор-
Мленных онтогенезов: 1) начальная, включающая заключительные
этапы гаметогенеза, половой процесс, дробление и дифференцировку
его продуктов в киноцитов, а также монотомическое деление ‘no-
следних, т. е. первые этапы вегетативной части цикла; 2) ‚заключи-
тельная часть вегетативного периода, представленная образованием
медуз. Большая же Часть вегетативного отрезка цикла осущест-
вляется неопределенно многочисленными, но плохо разграниченными
поколениямли гидрантов, еще не выкристаллизовавшихся в само-
стоятепъньте особи. О-бразуемое ими колониальное целое обладает A
низким уровнем целостности и его развитие имеет мало общего с ти-
пичными формами онтогенеза M/etazoa (локализованный во многих
точках и почти неограниченный во времени рост, повторное умног-
365
жениед сходных многоклеточных зооидов и т. ‹д.). Что касаетсяуодно-
клеточных прогамных поколений, то относительно очень высокая
степень их автономности здесь особенно бросается в глаза.
Следующую, вторую, ступень объединения жизненного цикла
олицетворяют гип-огенетические гидрополипы, обладающие много-
ЧЛеННЫМпИ колониями. Они иллюстрируют самым наглядным об_ра-
зом все этапы постепенной редукции медузоидного поколения и его
превращения в гонофоры и споросаки, теряющие свое былоезна-
чение, самостоятельных организмов и становящиеся органами ко-
лонии, ее гонадами. Таким способом совершается первый шаг в на-
правлении преобразования громоздкого метагенетического развития
в гипогенетическое, характерное для всех вышестоящих Metazoa.
Дальнейший _шаг в этом направлении осуществлен одиночными
гипогенетическими гидрополипами. Почкование целиком обращается
тут в вегетативное размножение, и вся вегетативная часть жизнен-
ного цикла может осуществляться тут силами одной единственной
полипоидной особи. Ее развитие уже вполне укладывается в норму
типичного онтогенеза, и только неограниченная деЛИМОСТЬ особи и
ее способность к бесполому размножению делают жизненный цгикл
таких форм факультативно многочленным, На базе той же особи
развертывается и половое размножение; при этом прогамная часть
цикла остается не интегрированной, ярчайшим проявлением чего
служитвысокая степеньбиологической самостоятельности яйца,
ведущего весьма активный «хищный» образ жизни. '
Окончательная концентрация жизненного цикла характерна для
гипогенетических гидромедуз. Здесь обычно уже нет- вегетативного
раз-множения (сохраняющегося почти только у паразитических
форм), и степень интеграции единственной медузоидной особи много
ВЫШЕ, чем у ГИДрОПОЛИПОВ. ЕЮ ВЫПОЛНЯЕТСЯ В ПОРЯДКЕ 0HTOI‘eI-I633 ‘
полный жизненный цикл, и даже прогамная ча-сть последнего полу-
ЧЗЕТ. ‘ГО же значение-продукта ДЕЯТЕЛЬНОСТИ ПОЛОВЫХ ЖСЛЕЗ,‘
какое она имеет у высших Metazoa. Иначе говоря, тут оказывается
достигнутой полная концентрация жизненно-го цикла, его полное
преобразование в цикл индивидуальный.
‘ Сходные картины повышающейся концентрации жизненного
цикла дают различные губки, у которых, однако, дело -не доходит
до столь четкого‚:как у Hydroz-oa оформления единой индивидуаль-
ности. Равным образом и у Scyphozoa осуществлены различные
этапы превращения двучленного метагенетчического цикла в одно-
Членныйонтогенез (гип-огенетические сцифистомы и сцифомедузы).
Таким образом, преобразование неопределенно многозвенното
жизненного цикла протозойного‚ по существу, характера в типичное
для Metazoa. индивидуальное развитие, осуществляемое одной
особью, является более чем вероятным. Различные этапы этого
‘процесса наблюдаются у разных низших Metazoa (полнее всего
у Hydrozoa), ЧТО. позволяет ясно установить его особенности -и (как
об этом будет сказано еще дальше) его движущие силы. Другими
словами, нет не только необходимости, но даже и возможности при-
‘366
бегать к гипотезам для его реконструкции. И еспэи у нас нет теперы
перед глазами некоторых из самых первоначальных этапов этого
процесса, то мысленному восстановлению этих недостающих звеньев
п-одсказываются настолько определенные и тесные контуры, что оно
почти лишается гипотетического элемента.
Уместно отметить, что безусловная примитивность многозвенных
пметагенетических (в том смысле, в каком этот термлин применяется‘
к Metazoa) ЦИКЛОВ подтверждается также примером многоклеточ-
ных Volvocoidea. Их жизненный Цикл осуществляется, большим
числом поколений: бесполых многоклеточных особей (колонии),
заключительное из которых становится половым. Напомним, что
целостн-остьэтих особей несравненно более высока, чем у всех низ-
-ших Metazoa, так что цикл складывается тут кИЗ многих четко раз--
граниченных онтогенезов. и и
V. Ha протяжении многих глав первой и третьеи частеи настоя-
щей книги было подробно обоснован-о и развито положение о палине
томии, как источнике и. спос-обе создания многоклеточностли у Meta-
zoa И высших Volvocoidea, KaK о процессе, служаЩеМ rIepB0-
основой эмбриогенеза многоклеточных. Не возвращаясь тут к раз-
бору этой концепции, представляющейся нам скорее простым при-
знанием существующего положения вещей, чем гипотезои, напомним
только следующие моменты:
' 1) Палинтомический процесс простейших складывается из:
а) гипертрофического роста одноклеточной материнской особи;
б) повторного деления, все промежуточные продукты которого
инмеют своеобразный характер бластомеров (спороцитов), не подвер-
гающихся дифференцировке в иинтеркинезах и мельчаюЩЕ/ХХ ИЗ
одного поколения в другое в приблизительно геометрической про-
грессии; в) процесса дифференцировки заключительных продуктов
палинтомии в однородные жгутиковые зооспоры (лишь очень редко
остающиеся недифференцирова-нными, амёбоидными); и, наконец,
г) процесса преобразования зооспоры в вегетативно деятельную
одноклеточную особь—трофозоита‚ обычно, но не всегда, испыты-
вающего затем гипертрофический рост. Все эти взаимообусловлен-
ные компоненты палинтомического процесса связаны в одно нераз-
рывное целое. у ‘ у
2) Сравнение эмбриогенеза низших Metazoa c палинтомическим
процессом простейших вынуждает признать гипертрофический рост
ооцита-частным случаем гипертрофическои дИ-СТ0МИИ‚ бЛЗСТО‘
меры-гомологами спороцитов, начальную дифференцировку ПРО-
дуктов дробленлия в жгутиковые клетки- пр0цеСС0М‚ COOTBBTCTBYIO‘
ЩИМ оформлению жгутиковых зооспор, а состоящую из одних таких
клеток жгутиконосную бластулообразуюЩУЮСЯ ЛИЧИНКУ- СЛОЖНО“.
зооспорой‚ или синз-ооспорой. Равным о-бразом и последующее пре-
образование кин-оцито-в в сидячих трофоцтитов Не МОЖЭТ Раёценди‘
ваться иначе, чем процесс, отвечающии преобразованию сво ОДНО-
плавающей зооспоры в сидячего трофозоита. Такая трактовка HOLI-
держивается. всей совокупностью фактов, КаСаЮЩИХСЯ C00TB€'1"
‘367
ствующих стадий развития низших Metazoa, дает им недостававщее
ранее объяснение и является, по существу, простым называнием
вещей своими именами. '
3) То же самое можно -сказать о многоклеточных фитомонадах,
у которых, вдобавок, перерастание палинтомического размножения
зооспорами в воспроизведение посредством многоклеточных син-
зоо-спор иллюстрируется с совершенно идеальной наглядностью на-
личием полной цепи переходов м-ежду ними. , _
4) Как ясно видно из примера тех же Volvocoidea, объединение
целой семьи зооспор в одну синзооспору есть специальная форма
образования временной или постоянной колонии особого типа———
палинтомической колонии, качкотвыми, очевидно, являются также и-
синзооспоры Metazoa. Таким образом, формирование синзооспоры
есть в одно и то же время процесс формирования колонии однокле-
точных зооспор и процесс онтогенетический, имеющий своим резуль-
татом образование целостной многоклеточной особи,
Все это плюс многие другие данные, рассмотренные в соответ-
ствующих разделах текста, с неизбежностью приводят к выводу
о том, что многоклеточность низших Metazoa, Volvoc-oidea И, веро-
ятно, всякая вообще многоклеточность метазойного типа создается
на основе палинтомии, в результате объединения создаваемой ею
семьи зооспор в целостную многоклеточную синзооспору. Так <‹про-
тозойное» размножение зиготь1 зооспорами становится ее воспроиз-
ведением, характерным для всех многоклеточных организмов, а па-
линтомический процесс в целом приобретает новое для него
значение исходной базы их онтогенеза. О вызываемых этим особен-
ностях жизненного цикла Metazoa говорилось уже достаточно; по-
вторим только, что они согласносвидетельствуют в пользу указан-
ного пути происхождения многоклеточности.
VI. Как и почему происходит пи могло происходить. объединение
одноклеточных продуктов дробления в одну многоклеточную особь,
каковы движущие силы этого интегрирующего процесса-вопрос
нелегкий и, что главное, никого еще специально не занимавший
в интересующем нас аспекте. Мы так привыкли теперь считать лю-
бой организм изначально и неизменн-о целостным, что представляем
себе возникновение этой ц-елостности лежащим за пределами види-
мости науки. Между тем является фактом то, что она возникает на
наших глазах в развитии некоторых низших многоклеточных, что
ее зарождение можно тут воочию видеть и изучать, и что рано или
«поздно ее можно будет понять.
Напомним, прежде всего, что метагенетические гидромедузы
проходят на ранних стадиях развития ряд различных ступеней орга-
низменной целостности: 1) протозойную, представленную однокле-
точным яйцом, доказывающим свою целостность всем своим пов-е-
дением в ходе оогенеза, своим одноосно гетерополярным строением
и его физиологическим выражением-наличием передне-заднего
-осевого градиента; 2) ступень лабильной интеграции, охватываю-
Щую стадии от 2 бластомеров до 16 включительно, на которых они
368
все тотипотентньт и равноценны и легко поддаются разобщению; эти
стадии не обнаруживают еще градиентных признаков целостности;
3) первую ступень устойчивой, уже необратимой интеграции, при-
ходящуюся на стадии порядка 32-64 и характеризующуюся отсут—_
ствием общего градиента и структурной поляризации; составляющие
их бластомеры нередко образуют бесформенный комок; 4) следую-
щую — «бластомерную» ступень устойчивой интеграции, на которой
заново возникает видимая поляризация и осевой физиологический
градиент, но уже общие всей дробящейся системе (состава
100—250 бластомеров) и, что характерно, ориентированные в на-
правлении, обратном градиенту яйца; 5) заключительную ступень
эпителиальной интеграции, начинающуюся со стадии около512;
передне-задний градиент физиологической активности, установив-
шийся на предыдущей ступени, остается неизменным и у оформляю-
щейся теперь бластулообразной- личинки.
Сходный ряд нарастающей интеграции, но уже ряд сравни-
тельно ‘анатомический, дают нам Volvocoidea, y которых первая
ступень целостности представлена одноклеточными формами и ста-
диями (гонидиями), вторая——лабильно интегрированными коло-
ниями низших Endorinidae, a следующие три ступени стабильной
интеграции олицетворяются колониями Eudorina (третья), Р1еос1о-
rina (четвертая) и настоящих вольвоксов (пятая).
, В обоих этих рядах интеграция проходит одни и те же ступени
И приводит к одинаковым результатам, несмотря на "крайнюю систе-
матическую удаленность;сравниваемых групп. Этим ясно подсказы-
вается наличие очень общих законов палинтомической интеграции,
действующих сходным образом на весьма различном материале, но
пока еще напм неизвестных. Из рассмотренного материала явствует,
однако, чтонарастающая интеграция характеризуется прежде всего
тем, что ее узловые точки, означающие переход развивающейся си-
стемы из одного качественного состояния в другое, ‘соответствуют
совершенно определенным значениям ее численного состава; эти
качественные изменения _наступают тут не постепенно, а путем
скачкообразного перехода от одного’ уровня целостности к другому,’
более высокому ее уровню, каждый из которых связан с опреде-
ленной численностью объединяющихся‘элементов. Далее, из приве-
денных фактов явствует, что законы палинтомической интеграции
действуют‘ лишь при табличномее характере (наличие которого
у метагенетических Leptolida представляется несомненным) и не
распространяются на процессы линейной палинтомии, а также и на
различные, вторично видоизмененные формы палинтомиие преры-
вистые ее формы и вторичную синтомию.
Привлечение к сравнениюс интегрирующими формами палинто-
мии всего разнообразия вариантов этого процесса, наблюдаемого
у простейших, дает ряд косвенных показаний в пользу существо-_
вания определенных законов палинтомической интеграции. Таким
образом, создается вполне ‘определенное впечатление (но пока еще
именно впечатление,‘ а неуверенность), что объединение продуктов
24 Эмбриология беспозвоночных ‘ 369
правильной табличной палинтомии в много-клеточные синзооспоры-
отнюдь не является делом случая, а подчинено определенным СПЭЦИ"
фическим закономерностям. Необратимое ‘объединение этих про-т
дуктов наступает всякий раз, когда ЧИСЛО ПОВТОРНЫХ ПРОДОЛЬНЫХ
делений идет дальше известного предела, отвечаюшещ определен‘
ному порогу интеграции. Последняя может быть избегнута, при мае;
совом образовании зооспор (или спор), только путем перехода о
табличной палинтомии к другим ее формам, КЗКОВЫМИ K3K.P::H1’f
являются многие высоко продУКТИВНЫе ФОРМЫ P33MH°7KeH1’”* пар
тических простейших.
Учитывая, что число продуктов палинтомии, ПРедЕТаВЛЯЮЩе‘еЁЁ
столь важным для их объединения, находится в прямой зависимое
от степени гипертрофии, достигаемой материнскои клеткои зооспор,
H ‘ и
можно сформулировать еще положение, что необходимым урловиел
интеграции этих продуктов является высокая степень такои гипер-
трофии ——так сказать «сверхгипертрофия», действительно имеющая-
место у всех низших Metazoa H многоклеточных Volvocoidaea. ИНЗЧВ
говоря, достижение многоклеточности в форме синзооспоры пред-
ставляется в конечном счете, необходимым следствием перехода
. ’ ,_‚ ' г
гипертрофии яица (гонидии) за определенные предеЛЫ аззёгеёёё
делении табличным способом. Попутно отмечая подСК 1
ЭТИМ объяснение связанности многоклеточности с оогамными фор-
мами размножения, мы можем сказать, что только вольвоцидыиэи
Metazoa (насколько позволяют судить об этом их современные Н р.
шие представители) пошли Ho пути ПОДЧИНеНИЯ ДЕЙСТВУЮЩИМ ТУ?
законам нарастающей интеграции; вто же время паразитиче-скёте
.‚ ` ” . ли 0
простейшие, также достигающие избыточнои гдипертрёфёхёаёитель-
уклонялись от их деиствия, сменив в ходе своеи при б
ной эволюции табличную палинтомию на другие формы спороо ра-
зования, либо остались и остаются вне сферы деЙ-СТВИЁ ЁЁЁЁ ЁЁЪУЁ;
нов, поскольку они не распространяются ни на повторно бр ‚Ю
деление (линейную палинтомию)‚ НИ‚ K0He‘1H°, На СТОЛЬ ° ЫЧНУ‘
у них первичную синтомию.
Гипотетичность развитой тут .точки зрения очевидна, НО B09 “IE3/11:‘
женные в ее основу факты представляются твердо устан-овленн и
Таким фактом является, во всяком случае, прогрессирующии
скачкообразный характер интеграции продуктов ДРОбЛЕНИЯ яица
метагенетических Leptolida H Volvocoidea, связанность ее последо-
о’ . иями
вательных ступенеи с определенными количественными состоян Х
дробящейся системы и единство течения процесса интегрЁцИЁаЁаёЁй
И других. Причинное объяснение этого процесса являеойя от awn
ФИЗИОЛОГИИ’ развития, и мы должны его Ждать ОТ ЭТ P стер
эмбриологии; только на однои из стОРОН ИНТеГРаЦИИ МЫ еще Q
HoBuHMcH ниже (1Х). быть
VII. Все сказанное в двух предЫдуЩИХ ПУНКТ” может H СТЬ
резюмировано в следующих трех положениях: 1) 1v1H6<)roI1{<:I1eTo;;1B Дряю-
низших Metazoa H Volvocoldea создается путем дро He Я‚и их
щегося одновременно способом массовои ‘продукции КЛЕТОК, дУЩ
370
на построение их тела, и процессом, посредством кото-роггупроиёяоз
дит объединение этих клеток; 2) дробление является частным слу-
чаем- палинтомического размножения зооспорами и,‘ более конч
кретно, высоко продуктивной табличной (продольной всвоей. основе)»
палинтомии; 3) многоклеточное состояние первоначально сдо-стиз
гается в форме равноклеточной жгутиков-ой синзооспоры, с момента
установления устойчивого уровня её интеграции. .
‚ ' Этими обстоятельствами целиком определяются и объясняются
все специфические особенности организации и образа жизни жгутиё
коносных целобластул‚ давшие повод к стольким филогенетическим
догадкам, но так и остававшиеся непонятными. В самом деле, укаъ
занными обстоятельствами дается объективное, как нам думается;
И вместе с тем простое объяснение следующих характерныхособен-
ностей свободноплавающих бластул: ‚ ‘ „ ‘ I
1) "специфических свойств организации составляющих их жгутиа
ковь1х клеток—киноцитов, выражающих их природу жгутиковых
зооспор; - д 1 ;
_ 2) однородности и равноценности жгутиковых клеток, опр-едее
ляемой этим же; 2 .› ' " .-
3) большой численности жгутиковых клеток, составляющих тело
бластулы; ' . „ ' -_ д
_4)«строго эпителиального расположения киноцитов, отражаю-
щего табличный характер тех форм. дробления, которыми они
создаются; V .
_5) их предельно низкого, для многоклеточных, общего уровня
организации (а именно отсутствия какой-либо тканевой, а тем более
органологической дифференциации); . ‘ ‚ '
6) сугубо расселительного характера бродяжек, в котором атак
ярко проявляется их природа сложных зооспор;
7) неспособности животных бластул к‚другим видам питания,
кроме осмотического (у Chrysaora), обусловленной, как мьдстараьъ
лись показать, самой их организацией; ' ' ‚
_ 8) поразительного единства организации всех жгутиконосньхх
бластул, не считающегося, если можно так выразиться, нис какими
границами естественной системы организмов. V . :
Все это, вместе взятое, убедительно свидетельствует о том, что
первобытной формой многоклеточной организации Metazoa была пи
есть форма жгутиковой синзооспоры, и что ее точно олицетворяют
эпителиальные жгутиковые целобластулы современных низших
Metazoa. Существование впрошлом каких-либо иных, еще, более.
примитивных форм многоклеточности, чем форма жгутиковойбла-
стулы, маловероятно, так как подобное допущение идет вразрез со
всеми имеющимися фактами ине имеет под собой почвы. Едины
ственно, что можно, и, пожалуй, даже следует, допустить, исходя:
из существующих данных, это то, что палинтомическхте предки
Metazoa обладали K01“11a-.'r0 ‚менее продуктивными формами. повтора.
ного деления, чем их теперешние. потомки, и становились СЗМОС-ТОЯтЗ
тельными многоклеточными организмами на более ‘низких, чем:
24* ‘ 371
теперь, уровнях устойчивой интеграции-Легко можно себе upen-
ставить, что их синзооспоры последовательно прошли в своем эво-
люционном развитии те же, примерно, стадии оформления своей
целостности, на каких остановились различные с-овременные_\/`о1уо-
cioidea; однако пока трудно привести какие-либо конкретны-е факты,
свидетельствующие в пользу этой гипотезы.
Здесь уместно всячески подчеркнуть, что признавая бластуло-
образную синз-ооспору первобытным состоянием многоклеточности,
мы никак не можем считать ее единственной формой существования
первичных Metazoa. Она и у них могла быть только временной
личиночной стадией цикла, сменявшейся затем сидячими питаю-
щимися стадиями совершенно иного строения. ._
V VIII. B тесной связи с только что разобранными вопросами
стоит вопрос о происхожденииполяризованной структуры Metazoa
как в отношении цито- и гистологическом, так и в проморфологи-
ческом отношении. Известно, какие разногласия вызывают эти во-
просы, от решения которых в том или ином смысле ‘целиком зависит
соответствующая интерпретация генетических соотношений различ-
ных типов структуры многоклеточных. Поэтому пмы считаем себя
обязанными четко сформулировать те ‚выводы, которые диктуются
разобранными вышеданными. Эти выводы сводятся к следующему:
1) ПОЛЯрМЗОВЗННОСТЬ. клеток бластулы перви_чна, являясь. выра-
жением их природы жгутиковых зооспор; A
2) B равной мере первично и их эпителиальное расположение,
обусловленное табличным, эпителиогенным характером тех видов
дробления, "результатом которых является бластула; ъ
3) совокупность жгутиковыхг клеток, образующих тело бластулы,
есть одновременно система клеток одинакового характера, един-
ственный первичный‘ пласт тела и простейшая эпителиальная ткань,
имеющая в основном функциональное значение двигательного
пласта, кинобласта, -
Что же касается гетерополярности бластулы Metazo-a И прису-
щих ей форм-симметрии, то они ясно отражают расс-елительную
природу этой стадии, ее высокую подвижность, характерный для
нее способ поступательно-вращательного движения, в воде в одном_
предпочтительном направлении.
Поэтому и_ с чисто морфологической точки зрения одноосно-гете-.
рополярное строение бластулы первично, раз оно устанавливается
при ее оформлении и „выражает собой присущую ей степень и форму
целостности _и -ее ‘биологическую природу бродяжки. „
v IX. Возникновение многоклеточного организма путем объеди-
НЕНИЯ В ОДНО H;,eJIOE ‘МНОГИХ целостных же ОДНОКЛЗТОЧНЫХ ОРГаНИЗМОВ.
НЕИЗОЕЖНО.„ДОЛЖНО. сопровождаться ГЛУООКИМ качественным ИЗМЁНЭ-
HIHEM ЭТИХ ‘ПОСЛЕДНИХ, ИХ НИЗВЗДЗНИЭМ ДО ЗНЗЧЭНИЯ KJICTOK, ЯВЛЯЮ-
ЩИХСЯ ПОДЧИНЭННЫМИ частями ЦЕЛОГО, em ВЗЗИМОСВЯЗЭННЫМИ ЭЛЕ-
ментами. Трудность представить себе, как могла совершиться
подобная замена одной (протозойнои) формы организменнои
целостности другой, новой ее формой (метазойной)‚.является, как
372
известно, главным аргументом всех противников‘ колониальной
теории происхождения Metazoa, начиная с Тихомирова и кончая
3аварзинь1м. ‘
Однако именно такая замена первоначальной протозойной
целостности яйца скачкообразно повышающейся метазойной целост-
ностью позднейших (многоклеточных) стадий дробления отчетливо
наблюдается, как МЫ видели, в развитии метагенетических beptolida.
Таким образом, эта, еще оспариваемая, возможность оказывается
тут осуществленной, что должно положить конец всяким сомнениям
на этот счет.
Кажется более чем вероятным, что само осуществление этого
качественного скачка обеспечивается или, по крайней мере, облег-
чается. специфическими особенностями процесса дробления. Мы
имеем тут в виду явление палинтомической интеграции, составляю-
щее характернейшую черту этого вида спорообразования и Empa-
жающееся, как мы знаем, в том, что: а) все входящие в него одно-
клеточные поколения объединяются B Целостный палинтомический
цикл: б) эти поколения находятся в пассивном, вегетативно недея-
тельном состоянии спороцитов (бластомеров)‚ лишенных многих
свойств самостоятельных организмов. Инач-е говоря, биологическое
объединение продуктов палинтомии оказывается как бы подготов-
ленны-м их ярко выраженной циклической интегрированн-остью и,
если можно так выразиться, подавленностьюих собственных wanna:
видуальностей, выражающейся в сведении на-нет вегетативной дея-
тельности и подвижности спороцитов.
‘Этим легко объясняется, почему многоклеточность создается
исключительно на основе палинтомии и почему одни только палин-
томические колонии-синзооспоры, а не какие-либо другие коло-
нии, служат отправной точкой для ее оформления.
Нельзя не упомянуть в этой связи поразительно наглядной кар-
тчины низведения полноценных одноклеточных членов колонии до
значения узко специализированных клеток тела, наблюда-емой
в ряду колониальных Volvocoidea. Идя здесь 'ру-ка об руку с повы-
шением целостности колоний и служа ее наиболее осязаемым выра-
жением, это явление может служить, вместе с тем, прекрасной
сравнительно-анатомической иллюстрацией процесса снятия" одно-
клеточных индивидуальностей при их объединении в индивидуаль-
ность высшего порядка. И хотя она обусловлена тут другими причи-
нами, чем y_-Metazoa, сама’ ее возможность все же явно обеспечиё
вается и здесь палинтомической природой создающегося многоклеэ
точного целого. '
Х. Объединение продуктовпалинтомии в целостную многокле-
точную синзооспору является, таким образом, тем способом, каким’
создаются низшие, первобытные,’ формы многоклеточной органи-
зации метазойного типа. Оно осуществляетсяйна наших глазах’ у
наиболеепримитивных из современных Metaz"o.a ПИ у высших Чо1уо5
coidea, И. нет никаких серьезных оснований предполагатычто когдаэ
либо’. могли иметь место‘ ‘и ка`кие=нибудь 9 другие с способы дпрогтсхо-
т
зтз
Ждения такой многоклсточности. Тот факт, что последняя создается
исключительно путем, дробленияу абсолютно всех Metazoa В соче-
тании с вышеуказанными особенностями их жизненного Цикла,
исключает, как мы думаем, всякое иное толкование происхождения
многоклеточности ‹Ме’са2оа и, во всяком случае, делает его
ненужным.
Это значит, что колониальная или, лучше, «интеграционная»
теория происхождения многоклеточности Metazoa не только
остается в силе, но и должна окончательно восторжествовать, как
единственная объективная и методологически приемлемая из coor-
ветствующих теорий. Связывая воедино огромную массу фактов и
не вступая в противоречие ни с одним, в сущности, из них, она дает
этим фактам материалистическое эволюционное объяснение и обла-
дает несомпненной познавательной ценностью; - -
Конечно, на существующем_этапе наших знаний э'та теория
приобретает в значительной мере новые очертания по сравнению
с теми, какие она имела во времена Мечникова и Г-еккеля. Она ста-
новится скорее палинтомической, чем просто колониальной теорией;
однако самый факт, что многоклеточность первично достигается в
форме палинтомического объединения, имеющего значение синзоо-
споры, сохраняет за колониальностью видное место в этой ТЕОРИИЪ
„ XL Создание многоклеточной синзооспоры есть в то же время
и оформление первого этапа онтогенеза многоклеточных.» Оно пре-
творяет гипертрофический рост материнской клетки-зооспор в пред:
зародышевое развитие многоклеточного, ее повторное -деление—%
в дробление,-дифференцировку зооспор— в начальную дифферен-
цировку зародыша, и придает комплексу объединившихся зооспор
онтогенетическое значение» личинки. Так создается первооснова
онтогенеза нового, метазойного типа, в который объединяются,
таким образом, ‚элементарные онтогенезы целого ряда одноклеточ-
HEX‘ поколений цикла, а именно: а) индивидуальное развитие мате-
ринскойодноклеточной особи, сохраняющее тут значение исходного
и-ведущего .звена; б) редуцир-ованньте онтогенезывсех следующих
друщза другом „поколений спороцитов" «(число которых, -как уже
говорилось, обычно не у-ступает 9), -|И в) начальный этап онтогенеза
одноклеточных конечных‘ продуктов палинтомии-до момента их
дифференцировки в жгутиковые клетки. - ~- -
То обстоятельство, что «только незначительная часть- всего
неъгноро „цикла +' а именно заключительная; часть‘ его прогамного-
периодам период метагамдногоспорообразован-ият- идет на построе-
ние онтогенеза многоклеточной синзооспоры, а главное-игор-что
оформление последнейдзаканииваетоя ‚на-начальной стадии развития
зоосп-орд.(до‚их ‚перехода к вегетативной деятельности),показывает’
—со всей очевидностью, дЧТО-„Этац первооснова -~ онтогенеза «Met~az»o_a
никогдаднемогла ‚быть целым, законченным онтогенезом; ‚Она соста:
вилгаыпишьмего первую,„начфальнуюыхаеть, ‘фундамент эмбриогенеза,
за „неюддзсегда „должны; бьтли .‚ ‚следовать другие, Постэмбриональ-
Ндгйе „стадгщ -~ раёвидгид„ - тразиаэующдееи из; _ -Beierarnraaqm-.« .. отрезке
$1‘?
Цикла. Уже а priori можно сказать, что первооснову этих стадий
должно составить преобразование расселительных жгутиковых
клеток-зооспор в вегетативные клетки характера трофозоитов,
:а затем и длинный ряд трофических клеточных поколений. Это и
лимеет место у современных низших Ме’са2оа и не могло не иметь
места у их даже наиболее пипотетичных предков.
Х11. Первым этапом п-остэмбрионального развития всех низших
Metazoa, проходящих стадию целобластулы (синзооспоры) в своем
индивидуальном развитии, является преобразование последней
в двуслойную, разноклеточную паренхпимулу. Это происходит у го-
модельных известковых губок, метагенетических гидромедуз‚ мета-
генетических сцифоидных и бластулообразующих иглокожих,
и осуществляется у них всех одинаковым, в сущности, путем — пу-
тем перехода отдельных киноцитов в дедифференцированное (амё-
боидное) состояние и их выселения в полость Ёластулы; выселив-
шиеся клетки могут затем более или менее интенсивно размножаться
в этой полости, но этого на первых порах может и не быть. Известно,
что дальнейшая судьба иммигрировавших жгутиковых клеток (и их
ближайших прямых потомков) может быть крайне различной: так,
у Ношосое1а они дают начало поверхностным (дермальным) клет-
кам будущей сидячей стадии, у метагенетических гидромедуо-
фагоцитобласту (т. -е. мезо- и энтодерме), у 5сурЪ1о2оа—блуждаю-
щим клеткам мезоглеи, а у иглокожих-первой смене личиночной
мезенхимы. Известно также, что образуемый ими комплекс может
иметь характер связной первичной паренхимыили же быть пред-
ставленным разрозненными клетками, остающимися свободно взве-
шенными во внутренней среде личинки. Далее, мы хорошо знаем,
что высел-ение дедифференцировавшихся жгутиковых клеток бывает
гв одних случаях строго локализованным, тогда как в других оно
«совершается в любой точке стенки тела бластулы, практически со
[всей ее внутренней поверхности (мультиполярная иммиграция).
1Наконец, нам также известно, что происходящая убыль жгутико;
хвых клеток из стенок тела обычно восполняется продольным деле-
нием" остающихся на месте. '
Такова фактическая основа, которой мы располагаем в отно-
зшении начальных этапов формирования двуслойности у самых при-
митивных из Metazoa. OHa бЫЛа частично использована Мечнико-
‘вым для обоснования теории фагоцителлы, но фактически не
принималась в расчет большинством другихавторов, касавшихся проч
блемы происхождения двуслойной организации Метаиоа. Поэтому
и не приходится удивляться безуспешности всех попыток ее реше-
нлия как проблемы гаструляции-ос-обенно если учесть, что
‘последняя фактически имеет эместо среди бластулообразующих
Metazoa, только у некоторых сцифоидных `и у иглокожих, где она.
происходит, к тому же, уже после начальной иммиграции жгути-
ковых клеток (т. е. уже после превращения бластулы в парен-
химулу). ' ' ‘
Невозможность подведения этой начальной иммиграции под по-
375
НЯТИе гаструляЦии ИЛИ образования ЭНТОДерМЫ явствует _С ПОЛНОЙ
_очевидностью, во-пер-вых, из того, что дедифференцированные жгу-
тиковые клетки служат в разных группах источником самых раз-
лиичных пластов и тканей, и, во-вторых, что на стадии паренхимулы
они не дифференцируются, сохраняя в это время характер эмбрио-
нальных клеток. Хотя они и играют физиологическую роль клеток
внутреннеи среды личинки‚_это не отражается, как правило, на их
будущем (презумптивном) значении. Действительно, только в тех
_CJIY‘1a7iX, КОГДа За ИММИГраЦией вскоре же следует подлинная гаст-
‚РУЛЯЦИЯ (сцифоидные, иглокожие)‚, эта функция прочно закре-
пляется за ними, и они непосредственно преобразуются в настоящие
клетки внутренней среды.
Амёбоидное состояние, в котором находятся теперь жгутиковые
клеткпи, делает применимым к ним мечниковское понятие фагоцито-
бласта. Однако самый факт, что они на этой стадии развития лишь
редко проявляют свои потенции фагоцитов и в дальнейшем прояв-
ляют их далеко не всегда, заставляет нас делать главное ударение
не на потенциальную способность этих клеток к амёбоидному пита-
нию, а на то, что ончи находятся в дедифференцированном состоянии.
Особо следует подчеркнуть, что процесс образования фагоцито-
дбласта не имеет еще ничего общего с явлениями разносторонней
ФУНКЦИОНаЛЬНОЙ дифференциации клеток, с оформлением истинной
разн-оклеточности. В самом деле, он заключается только в том, что
часть жгутиковых клеток переходит в свою ажмёбоидную фазу, в чем
сказывается только лабильность уровня их дифференцировки, их`
«протомонадная» обратимость, проявляющаяся при изменении
(условии их существования.
Можно также констатировать, что частичная дедлифференцировка
жгутиковых клеток, ведущая к преобразованию ‘бластулы в дву-
слойную паренхимулу, является необходимой предпосылкой к пре-
вращению свободноплаваюшей личинки в сидячую вегетативную
стадию. Наконец, следует еще отметить, что связанное с иммигра-
циеи деление жгутиковых клеток означает вступление в монотоми-
ческий период Цикла, хотя и не имеющий пока характера вегета-
тивного. A
XIII. Непосредственное превращение паренхимулы в зачаток‘
‘сидячей вегетативной стадии наблюдается у гомоцельных известко-
выхгубок и у‘ относительно немногих метагенетических гидромедуз
(Oceania, Laodicea, Mitrocoma). Это — самый простой из существую-
щих вариантов первичного метаморфоза и, повидимому, самый прии-
митивныи. Начинаясь сходным образом у тех и других (‚оседание
{Хичинки и ее дедифференцлировка), метаморфоз- идет тут и-дальше
в-весьма сходном направлении. Напомним», что у Нотпосое1а первый‘
его era? заключается-в выходе клеток фагоцитобласта изнутри
осевшещличинки на ее поверхность, где они; постепенно склады-
вдюдт?‘ ‘Ё 5!-I!4T§=JI?I9K;ItH;>Ifi..9J_Iofi QIIHQPQZIHHX еще перв-ичньтх- дермалЬ-г
ЁЫХ КдеТ0К‚ В то время как дедифференцировавшиеся Ter1epb'>K1“yT1»1-.-
„одвые клетки роетаются.» расположеннылми в вице грыхлой- паренхимы:
$76
LII
последняя может быть практически однородной или же содержать
отдельные дегенерирующие клеточные элементы, нидущие на питание
остальных, а иногда и разрозненные клетки фагоцитобласта, еще
не вышедшие кнаружи. Далее начинается поклеточная дивергент-
ная дифференцировка поверхностных (первичных дермальных)
клеток в специализованные (пинакоциты, склеробласты и коллен-
циты), часть которых остается в составе дермального слоя
губки, а другая уходит в образующуюся теперь мезоглею; измене-
ний во внутренней клеточной'массе в это время не происходит. Сле-
дующий, третий, этап "метаморфоза характеризуется появлением
полости в толще паренхимы ‚и постепенной эпителизацией высти-
лающих ееэлементов. В заключение устанавливается сообщение
этой полости с внешней средой (через посредство пороцитов, а затем
‘и прорывающегося оскулума), а выстилающие ее клетки паренхимы
(бывшие жгутиковые клетки паренхимулы) поляреизуются и частью
превращаются на месте в хоаноЦитов, частью же выходят из эпите-
-лиального строя и, уходя в мезоглею, дают начало различным
‘амёбоцитам. - ‚
Ход метаморфоза упомянутых выше гидромедуз известен слиш-
ком недостаточно, чтобы можно было его подробно сопоставить
с только что сказанным о Ногпосое1а. Приходится ограничиться кон-
статацией, что он включает все те же этапы, ‘кроме первого
(поскольку жгутиковые клетки паренхимулы остаются тут при ее
дедифференцировке‚ в эпителиальной связи и сохраняют свое
поверхностное положение), а именно: 1) дивергентную функцио-
нальную дифференцировку клеток поверхностного слоя осевшей
личин-ки, -сопутствуемую образованием прослойки мезоглеи;
2) образование полости в паренхиме и упорядочение окружающих
ее клеток; 3) прорыв это-й полости наружу и дифференцировку вы-
стилающих ее клеток в жгутиковых трофоцитов. .
Достойно всяческого внимания, что у форм, обладающих стадией
планулы, она образуется из паренхимулы LB результате первых двух
из только что перечисленных процессов—-.дифференцировки ее
поверхностных клеток в эктодерму и следующего затем образова-
ния гастральной полости. Вся разница сводится к тому, что онти оба
осуществляются здесь-в свободноплавающем и дифференцирован-
ном состоянии организма, а не после его оседания и дедифферен-
цировки, так что жгутиковые клетки непосредственно преобразу-
ются частью ‘в различные эктодермальные клеточные типы, частью
же лв интерстициальные клетки. На долю осевшей стадии прихо-
дятсячтут, следовательно, только окончательное оформление пер-
вичной кишки и образование рта (о формировании щупалец мы
пока не говорлим)‘. А V - д
.Это сравнение особенно интересно тем, что оно позволяет
‘(в силу лучшей, хотя все же и далеко еще не достаточной изучен-
ности гистогенеза планул, чем отвечающих ей сидячих стадий pag-
ВИТИЯ) существенно дополнить имеющееся сходство между дермаль-
ным слоем губок пи „эктодермой книдарийд Мы имеем в виду образоё
3,77
‘ванне у последних системы амёбоидных интерстициальных клеток
и их дальнейшую судьбу, имеющую много общего с формированием
‘мезенхимы губок. Это общее выражается, во-первых, в самом факте.
образования при дифференцировке поверхностного клеточного слоя.
свободных амёбоцитов, скрывающихся затем в Мезоглею или, при
ее слабом развитии, в непосредственное соседство с нею-
‘ («и -клетки»); во-вторых-в том, что часть этих амёбоцитов non:
вергается затем очень сходному видолизменению и образует затем
либо-скелетные элементы (в спикулобластах или склеробластах
губок), либо сложный стрекательный аппарат (в книдобластах
Cnidaria). Наконец, следует еще учесть, что у губок все же не
исключена всеми признававшаяся раньше возможность преобразо-
вания части дермальных (по своему происхождению) амёбоцитов
в первичные половые клетки, подобно тому, что нормально имеет
место в отношении части эктодермальных клеток книдарий.
Таким образом, дермальный слой сидячих молодых стадий губок
полностью соответствует эктодерме тех же стадий и планул кни-
дарий: 1) по своему поверхностному положению; 2). по общему
характеру испытываемой ним функциональной дифференцировки на‘
покровные, скелетогенные и другие клетки специального назначе-
ния, остающиеся перемешанными; 3) по тому, что его дифференци-
ровка намного предшествует таковой внутренней клеточной ма-ссы
и, являясь первой по времени дивергентной дифференЦировкой‚
испытываем-ой развивающимся организмом, служит и первым источ-
ником его истинной—функциональной—разноклеточности; 4) по
овоему участию в образовании среднего пласта тела.
Этот средний пласт тела также полностью отождествим у губок
и низших книдарий в силу следующих его свойств: 1) самого его
‘промежуточного положения между Наружным——покров»ныМ›—
‘слоем тела и внутренним-трофическим, образующим выстилку
гастральной полости тела; 2) двойственности его состава в том
смысле, что он представлен промежуточным веществом и ком-
плексом клеток, либо погруженных вэто вещество, либо тяготеющих
к ‘нему; 3) участия в_этом клеточном комплексе скелетогенных
(в ‘широком смысле), камбиальных и генеративных клеточных эле-
ментов; 4) амёбоидного характера всех этих элементов; 5) преобла-
дания среди "них блуждающих форм; 6) постоянно выраженной
у губок и возможной у Cnidaria двойственности происхождения
амёбоцитов, образующихся и из поверхностного и из внутреннего
(у книдарий -—энтодермальные половые клетки) пластов тела.
Е-ще более наглядным является соответствие системы хоано-
Цитов губок-энтодерме кишечнополостных. За это говорит: 1) одно-
родность клеточного состава этого пласта, неизменная у всех губок;
а среди Дкнидарийд-характерная для одних только более прими-
тивных форм; 2) природа сидячих, анимально питающихся‘ жгути-
ковыхтрофозоитов, в ‘одинаковой ‘мере присущая этим клеткам
у губок и книдарий; 3)‘ то важнейшее обстоятельство, что диффе-
ренцировка этих клеток осуществляется отнюдь не в порядке раз-
$78
носторонней специализации клеточного материала животного (как
это происходит с его поверхностными клетками), а представляет
собой естественное завершение нормального клеточного онтогенеза
части этого материала, а именно достижение жгутиковыми клет-
ками (эквивалентами зооспор простейших) состояния взрослой
вегетативной клетки ‘(эквивалентной трофозоиту одноклеточных
Flagellata); 4) вторичный характер поляризации этих клеток и их
выстраивания в эпителий; 5) характерная для них непрочность
эпителиальной связи между ними—связи, как мы знаем, очень
легко утрачиваемой; 6) сп-особность этих клеток к фагоцитозу до
их превращения в жгутиковые (губки) или ‘как до, так и после
него (кныидарии); 7) в той или иной мере связанное с фагоцитозом
образование гастральной, полости, по окружности которой и вы-
страиваются эти клетки; и наконец, последнее—то‚ что на этих
клетках целиком лежит функция питания, осуществляемая ими
внутриклеточно. .
Этой массе точнейших соответствий между пластами тела низ-
ших губок, с одной стороны, и низших Hydrozoa —— с другой, проти-
востоит, в сущности, только одно различие: что поверхностный пласт
тела губок образуется из материала фагоцитобласта, а у книдарий,
наоборот, из оставшихся на месте жгутиковых _клеток. Этим уже
определяются выход клеток, фагоцитогбласта на поверхность тела
осевшей стадии и вторичность и неполнота эпителизации создаю-
щегося таким путем поверхностного клеточного‘ слоя. Однако даже
иэто различие не имеет абсолютного характера. В самом деле, у тех
Са1сагеа, личиночной стадией которых является амфибластула,
все происходит несравненно проще: при оседании личинки ее жгути-
ковое полушарие накрывается уже эпителизованньтм, как правило,
зернистым полушарием, так что их окончательное взаимное располо-
жение сразу ‘же оказывается ‚достигнутым. Не.происходит ника-
кого выползания внутренних клеток наружу, и эпителиоидность дер-
мального слоя получает значение «изначальнои». ' ' ‚
Выводы гистиогенетического и онтогенетического порядка, непое
-средственно вытекающие из всех этих данных, разнообразны и не=
маловажны. Наиболее существенными из первых представляются
следующие: ‘ ' ‘ ‘ а
‹ 1) Первым этапом становления трехслойности является преобра-
зование периферического, наружного слоя личинки (осевшей или
еще плавающей) в пограничную ткань эпидермального характера;
из‘ которой при этом попутно выделяются основные компоненты
ткани внутренней среды. Это преобразование заключается впере-
дифференцировке ранее однородных поверхностных‘ клеток в разд
нообразныеклетки специального назначения; оно происходит путем
ихчдивергентной функциональной дифференциации чисто приспосод
бительного характера. . . г
: „Так создаются специфические для Metazoa клеточные типы и
сама их разноклеточность, приуроченные пока кдодному только
дермтальному ‚слою тела: ‚у51сидячих стадий этот последний. „несет
3715
преимущественно защитные функции, у мерцательных-преиму-
Щественно локомоторные.
2) Первичные клеточные и Межклеточные компоненты среднего
пласта, выделяющиеся при такой специализации поверхностного
пласта, принципиально сходны у низших книдаркий и губок и удов-
летворяют всем признакам ткани внутренней среды. Важно под-_
черкнуть, что основа этой последней создается тут одновременно
с оформлением наружной пограничной ткани и за ее счет и что все
клеточные элементы среднего слоя сохраняют характер амёбоцитов.
-3) Третий, внутренний, слой тела, несущий исключительно
функции улавливания и переваривания пищи (как у губок, так
и у более примитивных Hydrozoa), создается путем эпителпизации
клеток внутренней массы тела вокруг возникающей в ней полости
и их перехода из амёбоидного в дифференцированное жгутиковое
состояние. Их дифференцировка не имеет диивергентного характера
и заключается именно в ничем не замаскированном и не нарушен-
н-ом здесь переходе этих клеток из одной приспособительной фазы
клеточной организации в другую. Следовательно, она принциппи-
ально отлична от разносторонней дифференцировки клеток других
пластов, заключающейся в перестройке их организации примени-
тельно к выполнению тех илпи иных узко специальных функций.
4) В процесс-е эпителизации внутреннего слоя тела некоторые из
его клеток уклоняются от общей линии развития остальных и, оста-
ваясь амёбоидными, уходят в глубь тела. Так возникает второй
компонент ткани внутренней среды, имеющей, стало быть, явно
двойственное происхождение.
5) Такой порядок и конкретный характер оформления трех пер-
вичных тканей тела—экто-, мезо- и энтобласта-на
основе двуслойной организации паренхимулы в одинаковой мере
типичны для низших книдарий и губок. Иначе говоря, это первичное’
тканеобразование совершается одинаковым образом на различном
МЗТЕРИЗЛВ, ОПрЕДЭЛЯЯСЬ В МЕНЬШЕЙ мере ЕГО ИСХОДНЫМИ СЗВОЙСТВЗМИ, ‘
чем положением, занимаемым им в этот момент относительно диф-
ференцирующих условий среды. Так, поверхностный слой тела
приобретает специфические свойства эктобласта или эктодермы,
независимо от того, состоял ли он из жгутиковых клеток (планулы),
дедифференцированньтх, но оставшихся на своем месте киноцитов
(осевшие стадии. кнъидарий), или же из клеток фагоцитобласта, вто-
рично вылезших наружу (те же стадии Homocoela), или же, нако-
нец, из еще ни разу не дифференцировавшихся бластомеров (Hete-
mcoela); решающим фактором является поверхностное, пограничное
положение всех этих клеток, а не |ИХ собственное «презумптивное»
значение. С превращения этого клеточного материала в эктобласт
потому и - начинается тканеобразо-вание, что он‘ находится под
‚прямым воздействием внешней среды и идет, так-им образом, на
построение первогооргана’ животного——его'кожного (в Широком
смысле) покрова. ' и г ч 1 .
Равным. ‚образом -и внутренний = слой _'reJ.1a._—- aHTo.6J1acT_.—.—. может,
380.
создаваться из различного ‘личиночногоматериала: фагоцитобласта
у книдарий и кинобласта у губок. Оформляясь во внутреннеи кле-
точной массе животного в порядке перехода части ее клеток в
дифференцированную фазу организации, этот слои лишь вторично
становится пограничным,_принимая на себя ведущую роль в выпол-
нении вегетативных функций организма. Характерным его отличием
от эктобласта служит то, что его мультифункциональность обеспе-
чивается у низших форм высокой биологической полноценностью
образующих его одинаковых клеток, в то время как в эктобласте
она достигается, наоборот, физиологическим разделением тр-уда
между отдельными его клетками. Наконец,_и мезобласт образуется
в обеих группах в два приема: путем выделения части клеточных
элементов сначала из эктобласта, а затем и из энтобласта.
Все это ясно говорит о том, что создание одинаковых трех пер-
вичных тканей у низших Metazoa отражает самые общие законо-
мерности первичной органологической и клеточной дифференциации
их многоклеточного тела, закономерности самого универсального
характера, разыгрътвающиеся сходным образом у любых многокле-
точных организмов при одинаковых условиях их существования.
6) Отсюдавытекает‚позиция, которую мы должны занять в от-
ношении вопроса о гомолопии первичных пластов тела у книдарии,
с одной стороны, и губок-с другой. Если вести генеалогию тканеит
от стадии паренхимулы и считать критерием гомологии пред-
шествующую судьбу жгутиковых клеток, то днам придется решать
его отрицательно, поскольку кинобласт этои личиночнои стадии’
дает у первых эктобласт последующих Стадии развитияу а гУ
Ногпосое1а—энтобласт (плюс, в обоих случаях, соответствующие
фракции мезобласта). Но тогда придется: также признать, что
эктобласт Heterocoela не гомологичен таковому Ноп1осое1а,траз его-
и-сточником служат не дедифференцировавшиеся жгутиковые клетки,
ушедшие внутрь личинки, а остановившиеся в евр-ем развитии
бластомерьт, даже не дошедшие до стадии жгутитговои клетки. Рав-
ным образом придется тогда признать эктобласт планулы flydrozoa
Her-oMo.11or1»1qHb1M этому пласту у форм, Не иМеЮЩиХ ЭТОИ стадии,
так как B П-ЕрВОМ случае ОН образуется НЕПОСРЕДСТВЕННО ИЗ KI/IH'O"
бласта, а во втором-после его дедифференцировки. Однако‘ Все
эти противоречия (и связанное с ними представление об извращении
пластов тела у губок) совершенно теряют свою силу, если не пы-
таться придавать обоим пластам тела паренхимул фиксированное
презумптивное значение и учесть все сказанное о тождественности
их приспособительных возможностей. .›
Короче говоря, разделение клеток паренхимульт на жгутиковые
клетки ли аМёбОЦИТЫ ЕЩЕ не ОПРЭДЕЛЯЕТ ИХ дальнеишеи СуДЬбЫ, так
что кинобласт и фагоцитобласт служат только субстратом для фор-
мирования: первичных пластов тела Metazoa И сами не являются
еще таковыми. ' -
Что же касается выводов более общего онтогенетического ха-
рактера, то они сводятся к следующему:
381
1) Протозоа-нуга первооснову начального тканеобраз-(твавив низа
ших Мета2о ‚ ,
клеток Очинаасоставляют Процеосыднтогенеза ОбРаЗУЮЩИХ ИХ
. . у . U . з ключаются в переходе этих.клеток из дифференци,
РОВЗННОИ РЁССеЛительнои стадии типа зооспоры в дедифферевциро-
ВаННУЮ 3Меб0ИдНУЮ стадию, способную к фагоцитироваиию и осмо
тическом питани - . J
второй сгадии в ста И в последующеи ' редифференцирдвке ЭТОИ,
2 ь о ДИЮ ВЗРОСЛЭГО ЖГуТИковто.гто‚ Трофоцитгр
) олныи онтогенетическии цикл такого типа испытывают
Только. клетки энтобласта; у всех остальных он либо Обрывается
на ста ии а " ‘
_ д Мебоцита, либо идет дальше в особом, специфическом
для клеток, Metazoa направлении, чем и создается их функциоиалв
НаЯ разноклеточность, _ д x V H ‘ „
3) Все клеточные онтогенезы, осуществляющиеся в ‚этом „е-
риодЁ первичного Метаморфоза, имеют характер монотомических
и Действительно заканчиваются (за (учет, редкими исключениями).
тем; ирти инымиформами М0-Н0тоМии_
; I м ГОВОРЯ 0 ГШЁТРУЛЯЦИИ У НИЗШИХ сцифоидных и иглокож
их
МЫ подчеркивали то очень важное обстоятельство что она воп е ’
ходячеМУ'дШЁНИЮыОсуществляется не бластулой а уже фактичЁеЁЁ
ц о г о » a
ДВУСЦЮИНОИ паренхимулои. _В самом деле, процессу гаструляции
ёРедШеСТВУеТ У ТЕХ И других образование диффузного фагоцито
лас у . o о `
, та, создающегося И здесь путем поклеточнои иммиграции кино;
ЦИТОВ В Полость бластуль1, и притом иммиграции вполне TH
:I;:1:c‘II‘/’;“:2LVI:’JJI1:p”}TIHgOJ1${pH0I3’I ё: Scyphozoa и локализованной у морёхиииё;
.. ~ .‚ е ша
MET Т очень п ИЗёСУДЬ а этих вь1селившихся элементов представ;
з; ) у у - р ЧаТеЁЬНУЮ Особенность, ЧТО она сходна, в обеих
Тих ГРУППад -C ~Ta‘K0Bo1»1 эктобластических цкомп-онентов‘ ‘среднего
пласта: с -и ои ‚ -~ ' ' '
3HaI1eHHey'6J1I:i>1(?,uaIlgfL:)1fI)2{< (:1?/1PI"60CTaIOTCH амебоидньтми и Приобретают
п ‘Зим - е °ЦИТ0В‚ а у морских ежеи являются
‚Р YCHIGCTBEHHQ спикулобластами и вырабатывают известковые
иглы. ле ователь- - „ I ' `.
соответствёющих ЁЁЁЁМЁСНЛИ у первых они Имеют Характер И СУдЬбУ
ственны части эктоблас тов Гидромедуз’ то у ВТОРЫХ ОНИ южде‘
Ч "тических производных губок.
„ то касается самои гаструляции, то она MWKET Происходить.
существенно различными способами: 1) путем образования. и раз-
аСТаНИЯ СВОСГО Рода НОЧКИ, Внутренняя полость которой возникает
вто ично и остается ° .
инвгагинации заве ШаЗюЛЁЁИНКИ замкнут“ (chfysaora), 2) путем
ным-исчезновениевг а хэЦТИСЯ ЗаРЁТаН-ЙЁМ" бластоспора и ВРЕМЕН"
ной инвагинаЦИи бысз он ‘ероъба ( au§1t 06-)» ИЛИ 3) ПУТЁМ ТИПИЧд
ОС Е Н _ ‚‚ р углу ляющеися в виде пальцевидного от.
р ma ( C 1“‘°'1d'ea)-J3 соответствии с этим материалом для построе ‘
ния кишки мог т сл "
ток интенсивно] азЁЁЁхЁЬ Либо ПОТОМКИ немногих жгутиковыхпкле‘
(в первом-случай) JII»I60a1f/Iomflecfl В Процессе формирования. КИШКИ
клеток теряющих ‚п и это аесы погрузившихся Вглубь ЖГУТИКОЕЬЁХ
-OM) г б - . Р_ "M правильность расположения: (во вто;
Р a ЛИ 0, HaKO~HeII, Определенная зона кинобла-ста сохраняющая‘-
строго эпителиальное строение -(в третьем случае) ‚Иначе Говоря
киш- V '
ечник может создаваться как из особого, локализованно’
3.82:
I
нарастающего зачатка, так и из части самого кинобласта погру-
Ежающейся в тело личинки. Если добавить к этому, что окончатель-
Hag гастральная полость интересующих нас сцифоидных образуется,
как у Leptolida, СХИЗОЦВЛЬНЫМ способом, т. е. путем раздвигания
и эпителизации плотно сомкнутых до этого клеток, а рот тоже про-
рывается лишь много позднее,_уже на сидячей стадии сци-фистомы,
то трудно не согласиться с Мечниковым в том, что атипичная гастру-
ляция этих форм может служить хорош-ей иллюстрацией того пути,
по которому шла эволюция процесса формирования «первичной»
‚кишки ц-елентерат. Нет нужды подробнее останавливаться на твердо
‚доказанном Мечниковым же положении о сугубо вторичном харак-
тере архигаструлы, архэнт-ерона и бластопора, но упо-мянуть о нем
необходимо, поскольку его иногда еще оспаривают и теперь.
Большой эмбриологический интерес резюмированных здесь яв-
лений заключается прежде всего в том, что о-ни подтверждают
единство хода и результатов процесса тканеоб-разования у самых
различных групп бластулообразующих Metazoa при отсутствии
однозначной связи между ним и предшествующими ему процессами
-начальной дифференциации прото-бласта. подтверждается, что эта
последняя не_имеет другого значения, чем переход части жгутико-
вых ‘клеток в .дедифференцирс.ванное состояние, и является только
предпосылкой для развертывающегося затем гистогенеза, а никак
не начальным этапом самого тканео-бразования. Эту мысль можно
выразить и так, что для оформления трех первичных тканей тре-
буется, в качестве субстрата,сосуществование в организме клеток
двоякого характера, разнящихся друг_ от друга либо только по
с-воему морфофизиологическому состоянию (дифференцированные
и де- или недифференцированные — у амфибластул), либо по своему
положению в организме (наружные и внутренние—у ранних си-
дячих стадий), либо—что бывает много чаще—-по обоим этим
признакам (у паренхимул); однако дальнейшая судьба, испытывае-
мая обоими сортами клеток при гистогенезе, может быть, как мы
видели, совершенно различной, так что им невозможно приписать
и значения двух первичных тканей.
Второй интересный момент, связанный с явлениями га-струля-
ции, заключается в том, что они отчетливо показывают редукцию
иммиграционного процесса при выработке новых, более совершен-
ных и экономных морфогенетических приспособлений. Поскольку
более примитивный способ «выкр-исталлизовывания» кишечника из
сплошной массы иммиграционной паренхимы уступает тут свое Me.-
сто все более H более простым приемам его непосредственного по-
строения из (еще эпителизованного) кинобласта, постольку сокра-
щается и роль первичной поклеточной иммиграции. Из ведущего
процесса подготовки к гистогенезу она превращается, в конечном
счете, в ор-удие формирования среднего пласта (точнее, одной только
эктобластической его фракции); и она могла бы считаться именно
таковым (т. е. процессом гистогенетическо-го характера), если бы
этому не препятствовали те обстоятельства, что: 1) выделение этих
383
клеток происходит еще на стадии бластулы, т. е. из образующего
ее протобласта, и 2) что оно может иметь характер подлинной
локализованной иммиграции. .
V Третье, на что необходимо указать,-—это то, что га-струляция
некоторых сцифоидных и иглокожих действительно представляет
собой первый среди низших Metazoa пример устойчивого удержа-
ния эпителиальности в ходе морфогенеза и ее использования для
этого в качестве «маневренного построения» (Беклемишев).
Наконец, следует подчеркнуть, что гаструляция имеет своим
прямым результатом длительное или даже пожизненное закрепле-
ние главно-й оси и полюсов тела личинки (т. е. установление хотя
бы относительно первичноосности) и другие проявления повыше-
ния целостности организма, ведущие к переходу от метагенетиче-
ских к гипогенетическим формам развития.
XV. Конечный этап первичного метаморфоза, заключающийся.
в образовании питающейся стадии типа полипа (книдарии) или
олинтуса (низши-е губки) из бесформенной замкнутой «куколки»,
представляет бо-льшой интерес с точки зрения генезиса общего
плана строения, характерного для сидячих радиально симметрич--
ных животных. Его экологическая обусловленность более чем оче-
видна, но пути становления толкуются по-разному. Между тем,
имеющиеся на это-т счет фактические данные столь определенны,
что м-еста догадкам не остается вовсе. :.
Первое, чего мы должны здесь коснуться,—это вопроса о сте-
пени постоянства главной оси тела wee полюсов у интересующих
нас групп низших Metazoa. Известно, что постоянство это обычно
признается абсолютным, что и послужило поводом называть эти
группы первичноосными (Protaxonia). Что в действительности это
не так-вытекает уже из факта отсутствия прямой преемственно-
сти м-ежду о-сьюи полюсами яйца, с одной стороны, и таковыми
позднейших одноосно-гетерополярных стадий эмбрионального раз-
вития——с другой. Однако и пр-оморфология бластулы не имеет де-
финитивного характера. Ее устойчиво сохраняют только мерцатель-
ные личиночные стадии (паренхимула, планула), а с. их оседанием
она вновь, как правило, исчезает. Мы специально отмечали то об-
стоятельство, что распластавшаяся по субстрату личинка обычно
полностью деполяризуется, что естественно увязывается с испыты-
ваемой ею при этом общ-ей дедифференцировкой. Мы видели также,
что главная ось образующегося теперь полипа (у книдарий эти
явления особенно показательны) может устанавливаться либо пер-
п-ендикулярно к оси личинки (тип Stomatheca), либо в том же
н.аправлении‚ какое она имела у личинки, но фактически ‘заново
(тип Clytia), либо, нак-онец, точно соответствовать о-си личинки
(тип Laodi-cea). Таким образом, возможны любые соотношения
' МЕЖДУ ОСЬЮ И ПОЛЮСаМИ ЛИЧИНКИ И таковыми ПОЛИПа-ОСНОВЗТСЛЯ,
вплоть до полного отсутствия какой-либо связи между ними. Если
вспомнить, что и низшие губки проходят аморфную стадию куколки,
прежде ЧЕМ превратиться В 0ЛИНТуСа (ВСЛЕДСТВИЕ qero СОВПаДеНИЕ
384
иходинаково ориентированных осей также не реально, как у гидро-
‘медуз типа Clytia), нельзя избежать выв-ода, что предп-олагавшееся
постоянство главной оси и ее полюсов «Рготахтопйа» в действитель-
ности не имеет места у их низших представителей.
Второй вопрос, на котором надо остановиться, касается порядка
установления радиальной симметрии у метагенетических гидроидов
и сцифистом. Наибольший интерес в этом отношении представляет
факт, что у многих Athecata И некоторых Scyphozoa (например,
Chrysaora) начальной формой симметрии полипа является Двулуче-
вая. Она устанавливается в момент появления единственной тут
пары первичных щупалец (намечающих положение одной из ее
плоскостей—- щупальцевой) и стменяется затем 4-лучевой (при по-
явлении второй пары щупалец), уступающей потом в свою очередь
место 8-лучевой и т, д. Короче говоря, порядок оси симметрии уве-
личивается в онтогенезе шаг за шагом по мере появления новых
щупалец.
Сопоставление этого факта с тем, что говорилось -о независи-
мости оси симметрии полипа от полярно дифференцированных осей
предшествующих ему стадий развития, а также и о вращательном
характере симметрии этих стадий, имеет не только сравнительно
анатомический, но и чисто эмбриологический инт-ерес. Действи-
тельно, совокупность этих фактов заставляет нас признать, что у
низших представителей Cnidaria архитектоника полипа-основателя
целиком создается заново, вне какой-либо определенной связи с
таковой предшествующих ему стадий. Это-факт большого зна-
ч-ения, к оценке которого мы еще вернемся.
При оформлении сидячей вегетативной стадии осуществляются
и первые морфогенетические процессы, заслуживающие названия
органогенеза. Исчерпываясь у низших губок образованием оску-
лума, они получают у книдарий гораздо более развернутое и типич-
ное выражение, заключаясь тут в появлении оформленного рта и
окружающей его системы щупалец и в расчленении тела полипа на
НЕСКОЛЬКО дифференцированных отделов.
Прорывом рта на дистальном конце тела полипа заканчивается
формирование его гастральной полости‘ (целэнтерона). О ней, сле-
довательно, можно сказать, что она создается схизоцельным спосо-
бом и открывается затем наружу посредством явно вторичного рта.
Вчобразовании щупалец интересно то обстоятельство, что у ме-
тагенетических Leptolida ОНИ лишь очень редко (у Зупсогупе) бы-
вают полыми; для них типичны щупальца массивные, с однорядным
энтодермальным стержнем. Опорное значение энтодермальной оси
щупалец, равно как и некоторых других участков энтодермы, на-
глядно подтверждает правильность тезиса Мечникова о наличии
в составе последней мезодермального компонента, имеющего опор-
НОЭ «Хордальное» значение. Ведущая роль энтобласта в -органо--
генезе вообще очень характерна для книдарий и губок; не будет
преувеличением -сказать, что прогрессивная эволюция обеих этих
rpynn ИДЕТ ПОД 3HaKOM ПОВЫШЗЮЩЕГОСЯ ОРГЗНОЛОГИЧЭСКОГО расчле-’
25 Эмбриология беспозвоночных 385
нения энтобласта, связанного с егодалеко идущей функциональной
дифференциацией. _ - ‚
XVI. Сидячие стадии о-линтуса (у губок) и полипа-основателя
(у книдарий) олицетворят тот минимум организации, при котором
современные низшие Ме’са2оа становятся способными к полноцен-
ному вегетативному существованию. Если предшествующие онто-
генетические стадии были чисто расселительными, не способными
к самостоятельному питанию, то теперь, достигнув такого строения,
они получают возможность самостоятельно питаться анимальным
способом, расти и нормально размножаться; только теперь они ста-
новятся, следовательно, настоящими животными организмами с
ЧЕТКО выраженными признаками ТОГО ИЛИ ИНОГО систематического д
ТИПа.
Этот минимум полноценной животной организации обнаружи-'
вает, однако, много общего даже‘у систематически столь отдален-
ных груптп, как низшие книдарии и известковые губки. Общее между
ними заключается, прежде всего и главнее всего, в сидячем и практи-
чески неподвижном характере соответ-ствующих вегетативных ста-
дий, в принципиально сходном тканевом и цитологическом расчле-
нении их тела, в сосредоточении функции питания на энтобласте и
в организации клеток последнего, отвечающей таковой сидячих
анимально питающихся жгутоносцев (протомонад).
Наряду -с этим, между начальными вегетативными стадиями
низших губок и книдарий имеются и капитальные физиологические
и структурные различия; самое главно-е из них заключается в том,
что захватывание и переваривани-е пищи совершается у первых по-
клеточно, в порядке улавливания пищевых частиц каждой из тро-
фических клеток в отдельности, в то время как у вторых захваты-
вание пищи является функцией организма, как целого, и питание
осуществляется не только посредством внутриклеточного пищеваре-
ния, но также и полостного. Это ведущее различие влечет за собой
и все остальнЫ-е. Н-ецентрализованным, диффузным характером
питания губок определяются: предельно низкий уровень индиви-
дуальной целостности, столь характерный для вегетативных стадий
губок; <‹профилирующее›> значение ирригационной системы, как
единственного у них орудия активного привлечения пищи; высокая
производительность гидрокинетической функции хоаноцитов; узко
покровное и опорное значение эктобласта и его производных и т. д.
Напротив, вся специфика организации книдарий находится в связи
с захватыванием этими животными крупных пищевых объектов и
их полостным перевариванием. Этому служат, во-первых,. сп-е-
циальные органы улавливания д0бь1чи——щупальца, снабженные
стрекательными батар-еями, биологически обусловленное радиальное
расположение которых устанавливает особый характер симметрии
животного; этим же определяется прогрессивное развитие подвиж-
ности щупалец и всего тела путем соответствующей приспособи-
тельной дифференциации мышечно- и нервно-эпителиальных кле-
ток, повышение его целостности (неизмеримо большей тут, чем у
386
губок), профилирующая роль кишечника, как фундамента органи-
зации, физиологическая дифференциация клеток энто-бласта на
трофические ‘и секреторные, обеспечивающие своим секретом по-
лостное пищеварение, и пр.
Таким образом, ранние вегетативные стадии низших губок и
книдарий служат олицетворением не только и не столько двух раз-
личных систематических типов, как двух различных жизненных
форм, или биологических типов сидячего многоклеточного живот-
ного, а именно: губки—-биологического типа подгоняющего и
фильтрующего животного, существующего только поклеточным пи-
танием и лишенного нервной системы, а книдарий- типа сидячего
хищника, приспособленного к схватыванию и перевариванию круп-
ной живой добычи и уже обладающего соответственно оформленной
н-ервной системой.
Ясно, что первый из этих типов несравненно ниже и примитивнее
второго и что они воплощают, таким образом, две различные сту-
пени эволюции многоклеточной организации; -в первой из них еще
явно доминируют протозойные черты, а во второй——специфически
метазойные. Иначе говоря, основное различие между вегетатив-
ными стадиями губок-с одной стороны, и книдарий-с другой,
сводится в итоге к различию в степени достигнутой ими <‹метазой-
ности», к различию в степени и форме интеграции их многоклеточ-
ного тела.
Что эволюцию анимального питания Metazoa невозможно себе
представить иначе, как шедшей от поклеточного улавливания мел-
ких пищевых частиц с их раздельным перевариванием каждой
клеткои в отдельности к питанию крупной добычей, при котором
внутриклеточное пищеварение постепенно уступало ведущую роль
полостному, а ирригационная система соответственно приобретала
значение пищеварительной,—явствует со всей очевидностью из
всего, что нам говорят об этом сами губки и ‘кишечнополостные.
А отсюда неизбежно следует, что вегетативные стадии книдарий не
могли придти к современной своей организации, минуя филогенеч
тическую стадию, во многом подобную той, на которой остановились
губки. Поэтому вегетативным стадиям губ-о-к можно в известной
мере! приписать значение «морфологических предков» прочих
Metazoa, знач-ение более или менее затемненного прообраза веге-
тативных стадий развития первобытных Metazoa. Это, разумеется,
не значит, что мы правомочны выводить всех Metazoa 1»13‘ry6oK.
Напротив, губки явно представляют собой слепой ‘Филогенетический
ствол многоклеточных, имеющий иные протом-онадные корни чем
7
книдарии и другие МеЪа2оа; однако все говорит за то, что этот
ствол возник и развивался параллельно стволу книдарий, под дей-
стви-ем тех же биологических закономерностей интеграции, но
только гораздо более односторонне и, в итоге, менее прогре-ссивно.
С другои стороны, представляется несомненным, что радиальная
тёолипоидная организация сидячих стадий книдарий создается на
азе аморфного, деполяризованного зачатка, получающегося из
?5* . 337
личинки при ее оседании и ничем принципиально не отличающегося
от соответствующей ему «куколки» (клистолинту-са) низших губок.
Создается она, как мы видели, совершенно заново в порядке само-
стоятельного морфогенетического процесса, а никак не в порядке
восстановления утраченной личиночной организации. Отсутствие
всякой преемственности между этой последней и организацией по-
липа, характерное для наиболее ‚примитивных из книдарий, наряду‘
с очевидной невозможностью выведения полипоидного плана строе-
ния из спирально симметрического личиночного, также служат
свидетельствами того, что эволюция сидячих вегетативных стадий
низших книдарий шла своим особым путем——от первично аморф-
ной жизненной формы типа губки к относительно высоко органи-
зованному биологическо-му типу полипа.
Таким образом, нет решительно никаких оснований рассмат--
ривать одиночную полипоидную стадию книдарий‚ как некую осев-
шую <<га-стрею>>. Все, что нам известно о ее развитии и строении,
противоречит такому допущению и заставляет нас считать, что
вегетативные -стадии низших Ме’са2оа являются искони сидячими.
То обстоятельство, что низшие губки стоят, по существу, на
грани между колониальными сообществами монотомических сидя-
чих жгутиковых и многоклеточными организмами, служит веским
аргументом в пользу этой точки зрения. Особенно характерно, что
палинтомическая колониальность, ответственная за создание много-
клеточности в виде высоко интегрированных личиночных форм
(синзо-оспор), сменяется тут, при переходе к сидяч-ему существ-о-
ванию, колониальностью монотомического, по сути дела, характ-ера,
одним из выражений чего является несравненно меньшая це-
л-остность вегетативных стадий ‚низших Metazoa по сравнению с
личиночными. .
XVII. Резюмируя все сказанное в предыдущих параграфах, мы
должны придти к заключению, что разделение постэмбрионального
развития на два резко противоположных ‚периода-личиночный,
протекающий в состоянии свободно-плавающей «бродяжки», не по-
nyqammefi ПИЩИ ИЗВНЕ, ТИ ВЕГЗТЗТИВНЪХЙ, ПрОВОДИМЫЙ В прикреплен-
НОМ СОСТОЯНИЩ-‘ЯВЛЯЕТСЯ ИЗНЗЧЗЛЬНЫМ ДЛЯ Metazoa. ОНО ПОЛНО-
СТЬЮ отражает ‚ПЕРВИЧНОЕ O.HTOI‘€H'€TI/I‘1€'CK'O’e РЗЗЛИЧИЕ [МЕЖДУ СТЗДПИЕЙ
плавающей зооспоры, представленной у них личиночными жгути-
ковыми клетками и стадией взрослого сидячего трофозоита, олице-
творяемой трофоцитами сидячих стадий. Иначе говоря, это разде-
ление есть прямое и неизбежное следствие того, что в осно-ве
онтогенеза лежит палинтомическое размножение зооспорами, пре-
вращающимися в дальнейшем в сидячих троф-оьзоитов.
Отсюда ясно, что трудно предположить возможность существо-
вания таких первобытных Metazoa, y которых не было подобного
различия между расселительной личиночно-й стадией и сидячей
вегетативной. Это различие целиком вытекает из их жгутикового
происхождения, подтверждает его (это происхождение) и входит
в число самых яркихиз протозойных особенностей низших много-
388
клеточных. Таким образом, первичным типом онтогенеза Metazoa
безусловно следует считать «развитие с-личинкой». а
Из это-го следует, во-первых, что предположение о пожизненном
сохранении формы бластулы гипотетическими предками Metazoa
является совершенно беспочвенным. Все говорит, напротив, за то,
что их вегетативные стадии развития были такими же сидячими,
как у их современных потомков, и что основу этих стадий -с-остав-
ляли сидячие жгутиковые трофоциты. Конечно, можно представить
себе эти стадии еще ниже организованными, чем губки, и не имев-
шими, в частности, функциональной клеточной дифференцировки;
однако вряд ли имеет смысл пытаться сказать о них что-либо еще
более конкретное. ‚
В-о-вторых, из сказанного следует, что теории вторичного про-
исхождения личинки и м-етаморфоза M-etazoa He верны. Это не зна-
чит, конечно, что вообще не бывает вторично образовавшихся ли-
чиночных форм, и что всякий метаморфоз ‘первичен: речь идет
только о первичном метаморфозе низших Metazoa, о происхождении
метаморфоза у многоклеточных. В частности, тут оказываются со-
вершенно неприменимыми как теория преждевременной, или ана-
хронической, дифференцировки, разработанная П. П. Ивановым
(справедливость которой в отношении к «развитию с несвободной
личинкой>› р смысле Г. А. Шмидта для нас несомненна), так и
представление о личинке, как свободноживущем зародыше, защи-
' щавшееся И. И. Ежиковым (икоторое так жесправ-едливо каса-
тельно многих форм развития высших Metazoa). Существо личинки
низших Metazoa в той же мере заключается в ее природе сложной
расселительной зооспоры, в какой существо вегетативных стадий
заключается в их относительно-й равноценности комплексу сидячих
трофозоитов. _
ХХ. Таким образом, мы приходим к общему выводу, что сложэ
ные и громоздкие формы онтогенеза, наблюдаемые у наиболее при-
митивных из современных Metazoa, B основном обусловлены их
происхождением от анимально ‘питающихся одноклеточных Flagel-
1а1:а типа протом-онад, обладавших лабильным характером диффе-
ренцировки и расчлененным жизненным циклом палинто-мическото
типа. В этом находят свое объяснение многие специфические для
них особенности организации и развития, являющиеся, по сути дела
пережитками их прото-зойного- прошлого и свидетельствующие о
‚глубоком единстве обоих «подцарств» животного мира. . с
XXI. Прогрессивная эволюция по-д-обных первобытных ‘форм
развития Metazoa, прив-едшая к приобретению им характера <<ти-
пичного» онтогенеза, шла и идет очень широким фронтом и в самых
различных направлениях. В-едущим из этих направлений, или, вер-
нее, их равнодействующей, является эмбрионизация развития. ‘
Под эмбрионизацией мы понимаем такое приспособительное
видоизменение первобытных форм развития, при котором -стадии
и процессы, прот-екавшие постэмбрионально, т. е. вне материнского
организма и при дифференцированном состоянии (хотя бы части)
.' 389
клеточного материала этих стадий, вторично приобретают значение
эмбриональных стадий и процессов, протекающих под защитой ма-
теринского тела (или сп-ециально выделяемых им оболочек) и до
дифференцировки соответствующего клеточного материала.
Ход процесса эм-брионизации развития прослеживается наибо-
лее наглядным о-бразом у различных гип-огенетических гидропо-
липов‚ иллюстрирующих все последовательные этапы преобразова-
ния первичного Метаморфоза во все более и более «нормальный»
эмбриогенетический процесс.
Первую ступень в намечающихся тут рядах прогрессирующей
эмбрионизации представляют те формы, у которых этой участи
подверглась стадия паренхимулы и осуществляемые ею процессы;
‘B этих случаях первой свободной личиночной стадией является
планула, оформление кот-орой тут происходит в порядке начальной
дифференцировки зародыша.
Вторую ступень в этих рядах занимают такие формы, у которых
в значительнои мере эмбрионизиро-вана и планула, и свободное раз-
витие во внешнеи среде начинается со стадии, соответствующей
Стадии уже осевшей и дедИффер-енцировавш-ейся планулы.
Третью, высшую, ступень заниимают тут формы, подобные тубу-
лярии, выходящие из материнского гонофора в виде полипоидной
актинулы, т. е. на стадии, отвечающей стадии полипа-основателя‘
9
в этих случаях полностью выпадают мерцательные" личиночные
стадии, И первичный метаморфоз в целом оказывается эмбриони-
зированным.
Таким образом, начальный период п-о-стэмбрионального разви.
тия тут, на наших, можно сказать, глазах, «уходит» в развитие
эмбриональное, как бы поглощается им; Эмбриогенез разрастается
и усложняется за счет постэмбрионального развития и приобретает
при этом все более и более типичные черты.
Однако было бы ошибочным считать, что дело сводится тут и
простому механическому переходу первично постэмбриоиальиьтх
этапов развития в эмбриональные, что эмбрионизируемые стадии
и процессы остаются при этом не измененными. Напротив, они под.
вергаются значительнои переработке, утрачивают ряд предкних
качеств и приобретают новые, уже специфически эмбриональные
качественные особенности. Главные из этих преобразований таковы‘
1) ПРОЦЕСС НаЧаЛЬНОЙ дифференцировки, т. е. перехода клеток
ЗЗРОДЫШЗ В дифференцированное состояние, сдвигается по мере
возрастания эмбрионизации на все более ‘поздние стадии развития‘
9
теряя свое исходное значение заключительного акта палиитомии
он становится в прямую зависимость от все Отодвигающегося МОЁ
мента перехода относительно изолированного от внешней среды и
инертного, зародыша в активную, свободноживущую личинку
2) Тем самым все эмбрионизированные стадии оказываются со-
СТОЯЩИМИ ИЗ еще не дифференцированных клеток в чем и состоит
, w
ИХ Первое (и главное) качественное отличие от гомологичных им
ЛИЧИНОЧНЫХ стадий, ИХ специфически эмбриональный Характер
390
3)} Начинающаяся с этого редукция эмбрионизируемых стадий
может идти далее, до их полного исчезновения; так исчезают стадии
иммиграционной паренхимулы,’ затем бластуль1 и, наконец, п-ребла-
стулы, замещаемые специфически эмбриональной стадией морулы.
4) Такое замещение связано с изменяющимся при эмбриони-
зации характером дробления: из двухмерного, эпители-огенного оно
становится трехмерным; продольные деления бластомер-ов уступают
место поперечным (паратангенциальным), что приводит к их груп-
пировке в виде плотного комка (морулы). Последовательные этапы
процесса замещения <<бластулогенного>› дробления «морулогенным»
хорошо иллюстрируется рядом’ Hydra—Cordy1ophora—Hydractinia—
Gonothyrea и особенно рядом изменчивости дробления у Gonothyrea
loveni.
5) Со стадией бластулы исчезает из эмбрио-генеза первично-
эпителиальная структура и поляризованность клеточных элементов.
Онтогенетически исходной становится первично-паренхимная струк-
тура, столь характерная для морулы.
6) Дифференциация первичных пластов тела становится состав-
н-ой частью эмбриогенеза, получая значение процесса образования
зародышевых листков. Первой фазой этого процесса остается обо-
собление эктобласта с частью элементов мезобласта от внутренней
клеточной массы (происходящее путем эпителизации поверхност-
ных клеточных слоев морулы, т. е. путем моруля-рной деляминации),
а второй-образование центральной полости в паренхиме с по-
следующей эпителизацией последней в виде энтобласта.
7) При полной эмбрионизации первичного метаморфоза форми-
ро-вание полипоидной стадии также осуществляется в порядке эм-
брионального проце-сса, в форме подлинного органогенеза.
8) По мере повышения эмбри-онизации развития повышается и
устойчивость проморфоло-гических свойств зародыша. Ориентация
главной оси и полюсов, установившаяся у поздней морулы, прочно
удерживается последующими стадиями до актинулы включительно.
Легко заметить, что все эти изменения, связанные с эмбриони-
зацией первичного метаморфоза, действительно приводят к созда-
нию вполне оформленногозародышевого развития, точно отвечаю-
щ-его эмбриологическому канону. Эмбриогенез получает теперь все
свои основные аттрибуты—дробление в трех координатных изме-
рениях пространства, процессы формирования зародышевых листков
и процессы органогенеза и, наконец, то индиферентное «эмбрио-
нальное» состояние клеточного субстрата, на котором разыгрыва-
ются эти процессы. Таким образом, свойства эмбрионального разви-
тия, которых мы привыкли считать изначальными и неотъемлемыми,
оказываются результатом сложных онтогенетических перестроек
явно вторичного характера.
ХХП. Эмбрионизация первичного метаморфоза обычно имеет
своим результатом переход от метагенетического развития к гипо-
генетическому, а при дальнейшем ее у-силении—переход от ко-
лониального развития к подлинно индивидуальному развитию,
391
Осуществляемому от начала до конца‘ одной единственной, высоко
индивидуализованной особью. Различные гипопенетически-е полипы
показывают всю цепь переходов между обеими этими крайностямрд
ЁЁЁЁОЁЁХЁЁЁИ (зжсь все мыслимпые этапы рудиментации медузоидного
р ущего по схеме. ‚прикрепленные медузоиды — крип-ю-
медузоиды — гонофоры —споросаки), ведущей к его низведению до
состояния гонад сидячей особи, а также и все переходы последней
от „первично-полимерного пи равночленного строения пк функциональ-
Н0и дифференциации ее гидрантов, с одной стороны, и .K Моно-
гидрантному состоянию — с другой.
В последнем случае почкование обращается из средства пестрое-
ния колоний в подлинное размножение, что ясно товорит о таком
л ~ Деятельности, при котором колониальная
форма существования перестает быть необходимостью,
- _Лишь в очень немногих случаях (например, у Corymorpha)
значительная степень эмбрио-низацгии развития уживается с чередо-
ванием поколений. Однако в таких случаях вегетативное тело жи-
.вотного представлено одной полипоидной особью, на которой и
отпочковываются половые медузки. ‚
cqJ;“’ff“‘é6§’JZ5é§Z%“§;u3“Efe‘i%“*;?§§““ развит “"“B°“"T В
‚ _ НЕННОГО Цикла в онтогенезе однои,
высоко индивидуализованной ' особи. Т а к м е т а г е н е т и ч е.
3'-K314” ЖИЗЪНЗННЫЙ Цикл Получает значение пан-диви-
дуального развития, прочно удерживаемое им у
всех Bb1cm1»1xMetazoa.
содержание
Предисловие
Введение
ИСТОрИЯ И современное СОСТОЯНИЕ вопроса О ПрОРЦЁХОЖЦЕНИИ OHTOI‘€H€3Z1 I-1
МПОГОКЛЕТОЧНОСТИ M€t.':1ZO2l - . . . . . - -
1.
mu mm дыр
Главные направления в разработке теории происхождения многокле-
точныхиихонтогенеза
Первые гипотезы колониального происхождения Ме а2оа.
.Теория фагоцителлы И. И. Мечникова . . . . . . . . .
. Дальнейшее развитие теории колониального происхождения ›е а2оа.
Теория А. Н. Северцова . - - -
. Первые гипотезы неколониального происхождения МеЁаШа - - -
. Последующее развитие гипотезы неколониального происхождения
Меъа2оа .
. Вопрос о происхождении личиночных форм °и метаморфоза Metazoa
. Некоторые итоги и перспективы . . . . . . . . . . . - -
Ч а с т ь п ер в а я. Организация, Размножение и развитие одноклеточных
простейших
Вводные замечания . . . . . - . . - - '- - - -
Глава 1. Организация одноклеточных Вгоъоиоа . ‹
1. Три главных типа одноклеточнои организации . -
Глава - - - -_
.. Общее понятие о размножении ОДНОКЛЗТОЧНЫХ -
. Кариокинез . . . . . . . . . . . . . . . ..
. ОСНОВНЫЕ ТИПЫ МОНОТОМИИ. .
2.
4:-L/J[\')*-‘
Глава
ma
>-«
1
'::l
SD
LON)’-‘
ва
4:-oa1\>—-3 сочтет-дымы
Взаимоотношения амёбоидного и жгугикового TH 03 ОрГаНИЗаХ-киг! -
П. Простое деление (монотомия) - - - - ° - - - - ' ' ' ‘
Геометрические формы монотомии . . . . . . . . . . .[.
111. Индивидуальное развитие мснотошкичезтпжх СДНОКЛЁТОТНЫ)‘ -
.Вводн.ыезамечания.......-.
. Общее понятие об онтогенезе монотомических ОДНОКЛЭТОЧНЫХ
. Основные типы онтогенеза монотомических одноклеточных .
IV. Повторное деление (палинтомия) - -
. Общие определения. .› . . . .
. Общие особенности палинтомип .
. факультативная палинтомия
. Типичные формы палиптомии .
. Морфология повторного деления
.Прерывистая палинтомия . . - - - - - - - - -
. Повторное спорообразованпе (палннспорогенаез)
. Вторичная синтомия . . . .
V. ИНДИВИДУЗЛЬНОЕ pasamue TIZUIHHTOMPI‘{€CI{I-IX ОДНОКЛСТОЧНЫХ .
. Вводные замечания . . . . . .
. Особенности организации зооспор .
. Гнпертрофический эпиморфоз.
. Гипертрофический "метаморфоз
Стр.
3
12
21
31
34
41
43
47
49
49
72
78
78
80
82
92
98
98
101
102
102
104
106
107
108
112
'. 114
119
120
120
121
123
125
393
Стр.
Глава VI. Половой процесс и жизненный цикл- одноклеточных простейших
ыошьрюы
Гла
CU
21
CJJl\J*—‘
. Вводные замечания .
. Саркодовые
Эвглены
.Фитомонады............
. Общее понятие о жизненном цикле .
. Жизненные циклы одноклеточных
Циклическая дифференциация и интеграция .
VII. Образование колоний . . . . . . . . .
. Вводные замечания . .
. Монотомические колонии.
. Палинтомические колонии .
128
128
129
131
132
138
139
140
142
142
143
"145
Ч а с т ь в т о р а я. От одноклеточности к высшим ступеням надклеточной
Глава
2.
3.
4.
5.
Глава
Глава
фшдшю—
Ч а с т ь т р е т ь я. Источники и пути формирования примитивных видов
oprannsaunn
VIII. Пути формирования плазмодиев и сомателл
Общее понятие о полимеризации . . . . .
Плазмодии саркодовых и споровиков
Надклеточные жгутиковые . . . . . . . .
Сомателлы опалин и ресничных инфузорий
Сомателлы сосущих инфузорий . . - . . - . . . . . .
IX. Вегетативное размножение надклеточных Protozoa
. Общие определения. . . . .
Саркодовые И ПЛЗЗМОДИЭЛЬНЫЕ СПОРОВИКИ
. Полимерные жгутиковые
„Опалины . . . .
Ресничные инфузории (паратомия). . .
. Повторная паратомия ресничных инфузорий .
Повторная паратомия колониальных сувоек .
с
Х. Цитогоническое размножение и жизненный Цикл иадклеточлаьхх
простейших . . . . . . . . .
. Общее понятие о цитогонии .
.Саркодовые. . . . . . .
. Акнидоспоридии
. Книдоспоридии
. Опалины. . . . .
ОСНОВНЫЕ ТИПЫ ЖИЗНЕННЫХ ЦИКЛОВ НЗДКЛВТОЧНЬХХ ПрОСТСЙШИХ
МНОГОКЛ ETD‘-IHOCTH
Вводные замечания
Глава
1.
2.
3.
4.
Глава
2.
3.
Глава
XI. Дробление, как палинтомическое воспроизведение .
Общие соотношения между спорообразованием и дроблением
Переход от спорообразования к палинтомическому воспроизведению
yV0lvocoidea.
Следы спорообразования в дроблении низших Metazoa . . . . .
Результаты и следствия перехода от палинтомического спорообразо-
вания к палинтомическому воспроизведению . . . . . . . . .
Х11. Эмбриогенез тиногоклеточных фитомонад
1.0 многоклеточности Volvocoidea .. . . . . .
Eudorinidae .
Volvocidae.
XIII. Эмбриогенез бластулообразуюших Metazoa
Предварительные замечания . . . . . . . .
Метагенетические гидромедуз
Низшие Scyphozoa . . . . . .
Известковые губки . .
Иглокожие. Морские ежи .
. Особенности примитивных форм дробления . . . .
Изменения полярности в ходе формирования бластулы и их значение
155
155
156
158
164
168
170
170
172
172
174
174
178
181
185
185
187
190
198
200
201
205
210
210
214
216
218
220
220
222
227
232
232
233
242
245
249
251
257
Стр.
Глава XIV. Организация жгутикоиосных бластул и их значение синзоо-
-doom:-oJm»—.
спор .
. Проморфологические особенности целобластул .
. Гистологические особенности . . . . . .
. Численность клеток . .
. Цитологические данные . .- .
. Биологические особенности . . . . . . . . . . . . . -
. Общее сравнение вольвоцид с целобластулами Ме’са2оа .
Глава
Часть четвертая.
Е’
N
BII2lCTy.TIbI, как синзооспоры. . . . . . . . . . . . . . . . . . .
XV. Закономерности и условия объединения продуктов палинтомии
в многоклеточные синзооспоры . . . . . . . . . . . . . . .
1‚Постановкавопроса........................
2. Смена уровней интеграции в процессе формирования синзооспоры .
З. Различные уровни целостности синзооспор у Volvocoidea .
4. Связь уровня интеграции с численностью продуктов палинтомии .
5. Условия интеграции продуктов палинтомии . . . . . . . . . . . .
6._ Приспособления простейших к предотвращению интеграции продуктов
ПЗЛИНТОМИИ. . . .
7.Выводы
Постэмбриональное развитие низших книдарий
и губок и пути его эмбрионизации
Вводныезамечания....................
Глава XVI. Первичный метаморфоз низших книдарий и губок
1. Общее понятие о первичном метаморфозе
2.
3.
4.
5.
Глава
чшшьшюызшьш юн Фшдшю—
Начальная дифференциация протобласта и формирование паренхимулы
Превращение двуслойной личинки в трехслойную личинку типа
планулы
Оседание личинки и ее превращение в зачаток сидячей вегетативной
Дальнейшее развитие вегетативной стадии». . . .~ . . . . . . . .
XVII. Эмбрионизация первичного метаморфоза у гипогенетических
гидрополипов._....
. Строение яйца и первые стадии дробления .
.Ф0рмир0вание морулы . . . . . - .
. Организация морулы.
. Дифференцировка эктодермы
‚Формирование планулы. ...................
Г . Превращение плапулы в сидячую полипоидную стадию . - . . - .
лава
XVIII. Особенности клеточного строения сидячих вегетативны
Предварительные замечания . . - . . - . . . . . - . . . . . .
Известковые губки. Состав клеточного ассортимента и соотношения
различных видов клеток .....----......-.
. Трофические клетки губок - . .
‚Трофические клетки книдарий . . . . . . . . . . . . . . . .
. Общие особенности дифференциации клеток тела низших Metazoa
XIX. Гаметогенез и половой процесс низших многоклеточных.
.Вводныезамечания.................
. Источники образования первичных половых клеток .
. Общий очерк оогенеза . . . . . . . . .
. Общие выводы .
Заключение.
. Явления созревания в женском ряду
Оплодотворение . . . .
Сперматогенез
261
261
263
264
265
269
270
271
275
275
276
277
279
280
282
283
286
288
288
290
297
301
309
312
312
314
318
319
324
325
328‘
328
330
332
335
339
340
340
341
342
345
350
352
355
I 357
Отв. редактор Левцнсон Л. Б.
_Технич. редактор Довольное В. В.
А08991. Подписано к печати 30.VIII. 49 r. Ти-
раж 10 000 экз. Объем 24“/4 п. л. 27,2 уч.-и3д л.
Формат 60><921/„. Цена 10 р. 50 к. Зак. 68.
З-я типография «Красный Пролетарий». Главпо-
лиграфиздата при Совете Министров СССР.
Мозква, Краснопролетарзкая, 16. Зак. 1711.