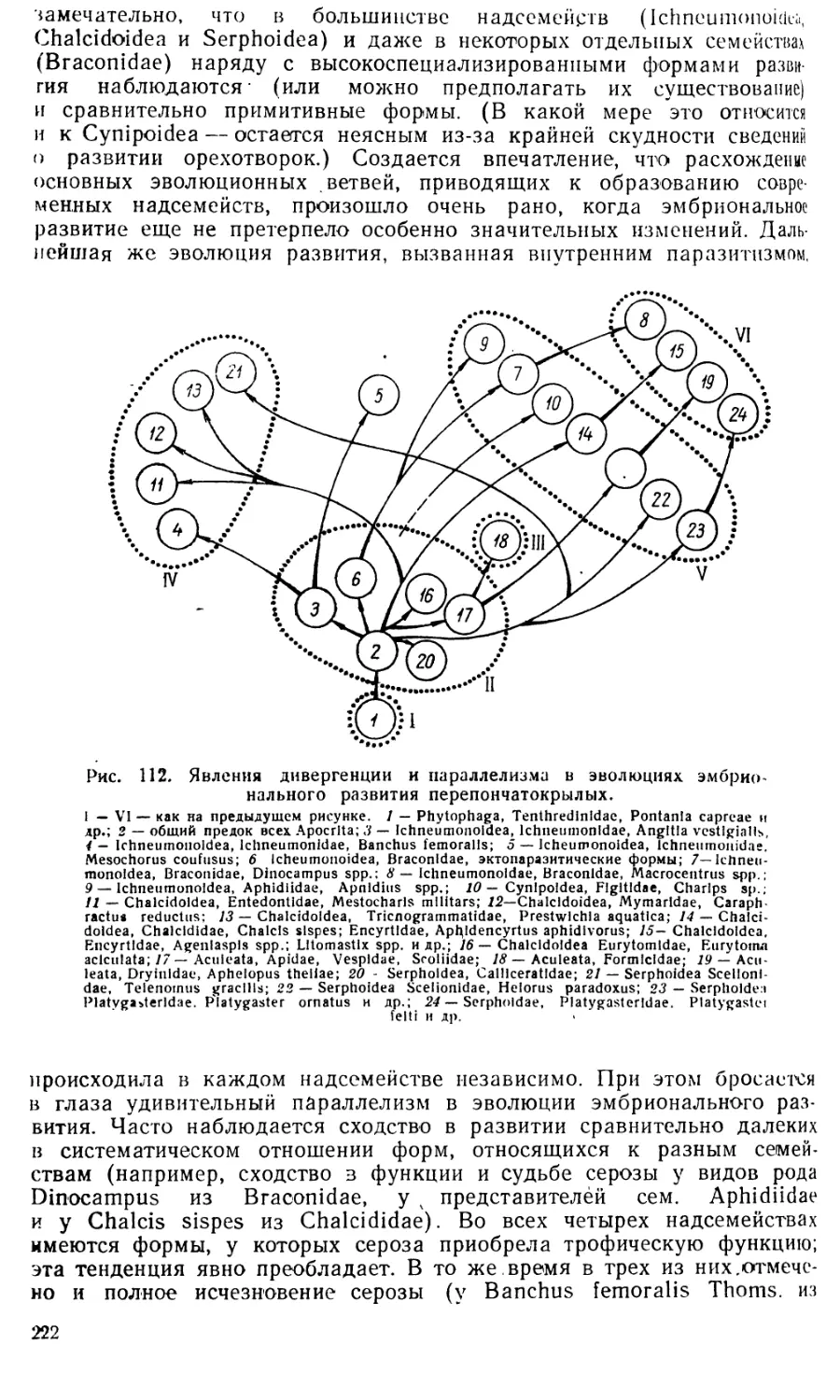
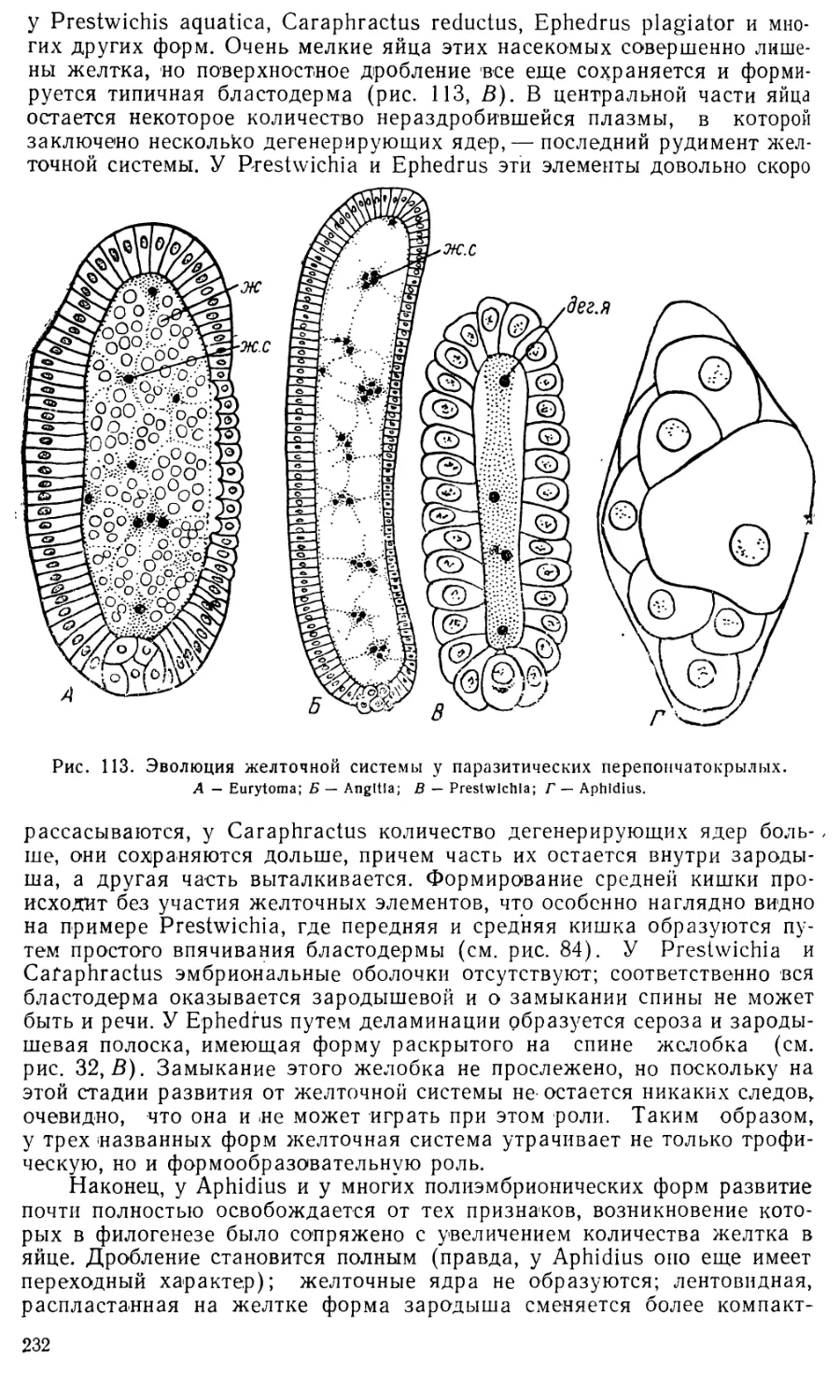
/


























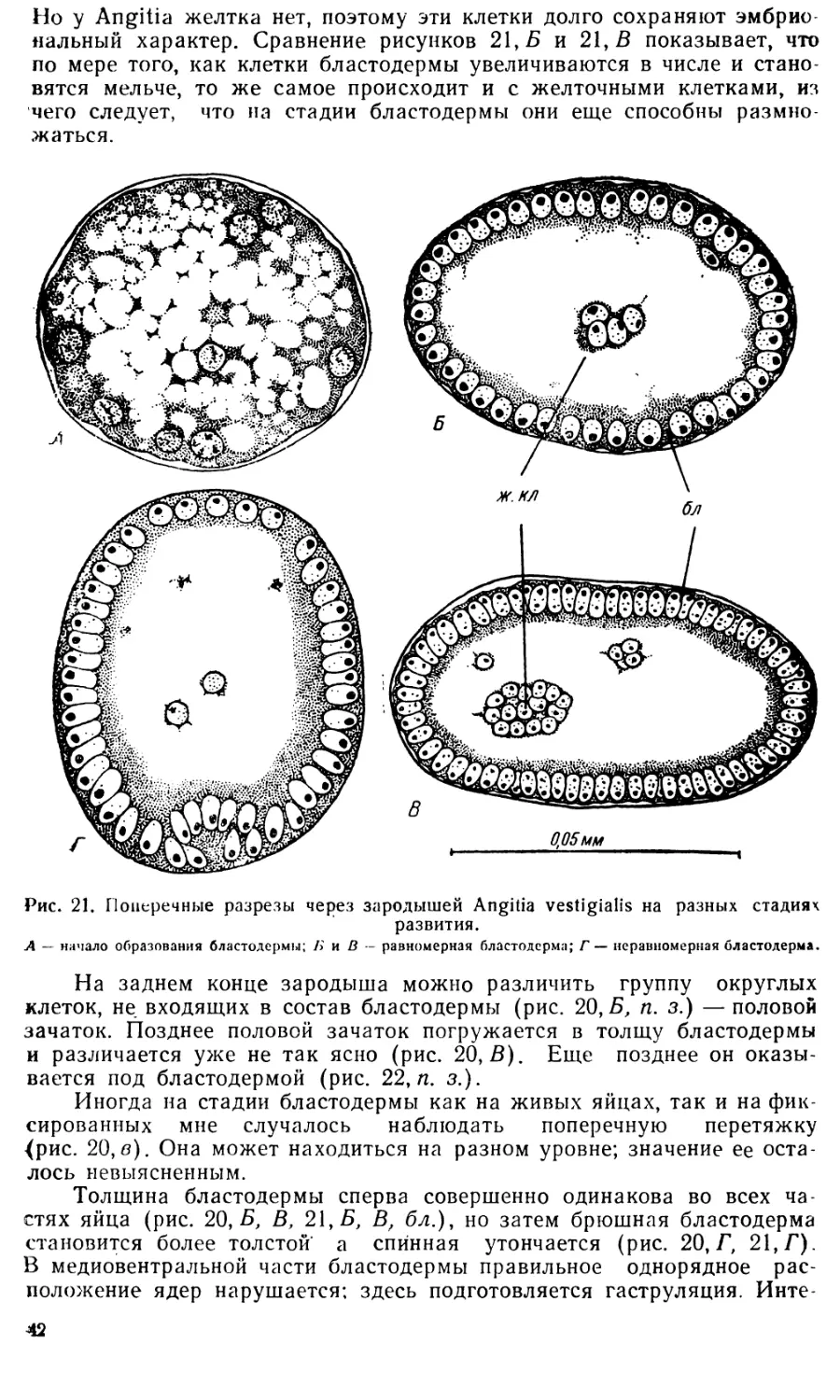

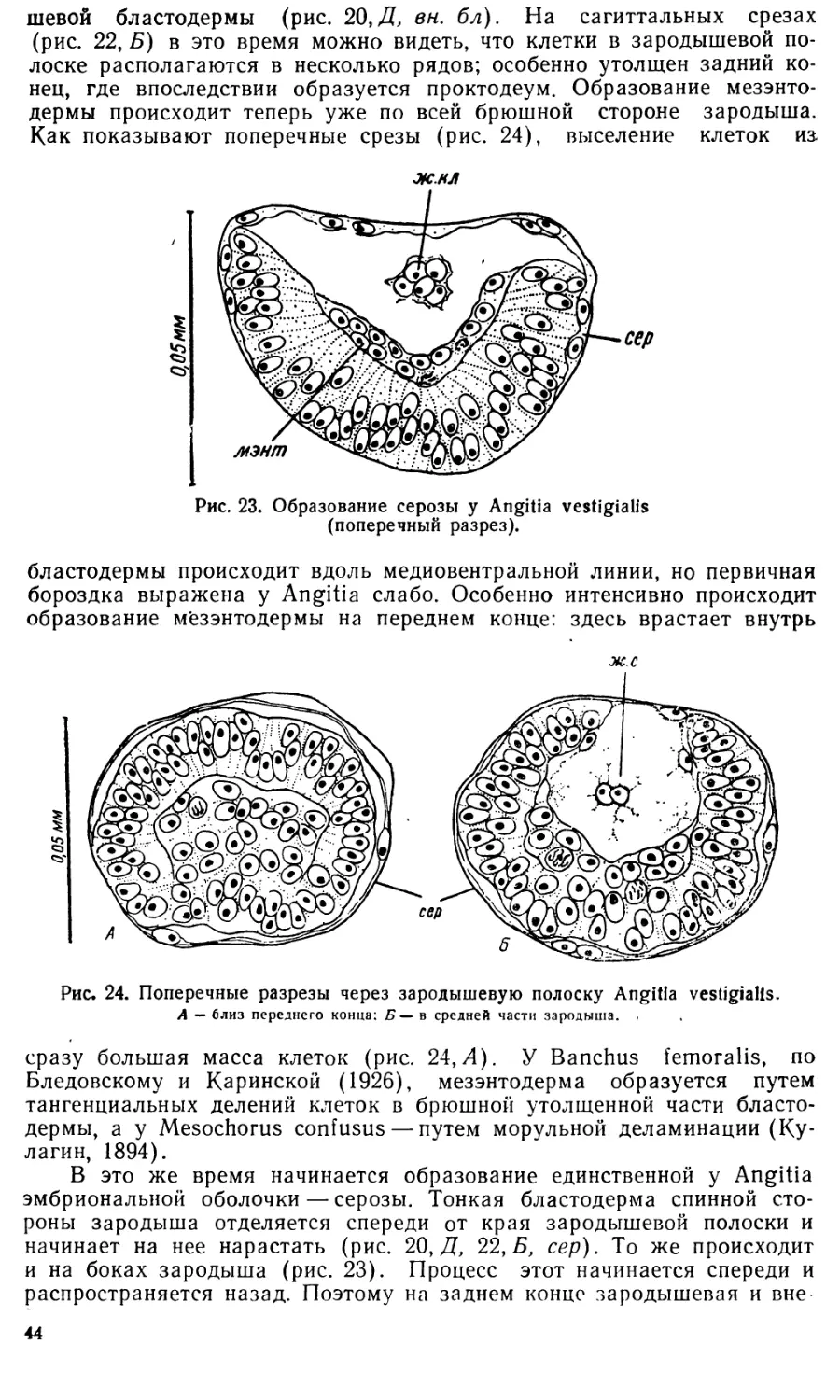






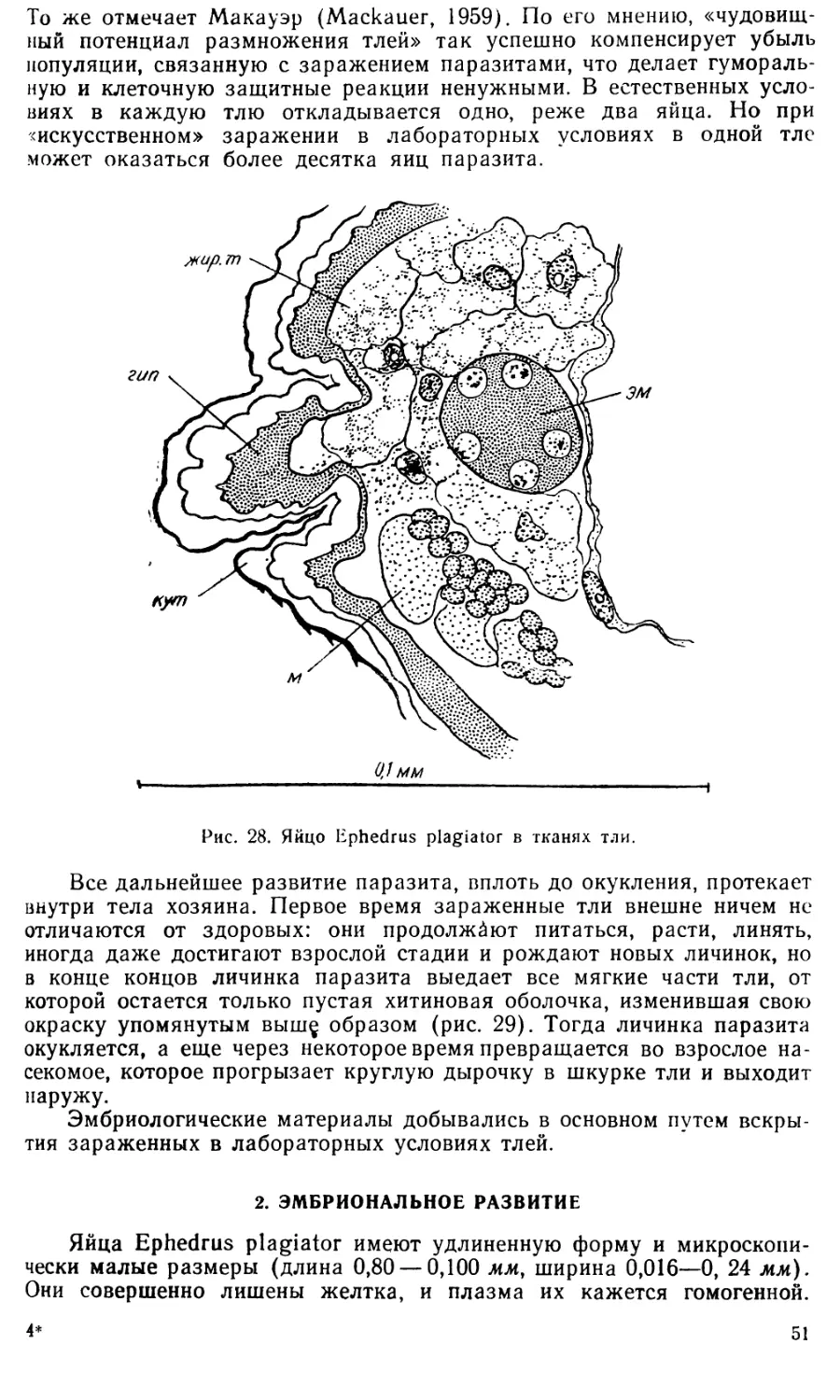
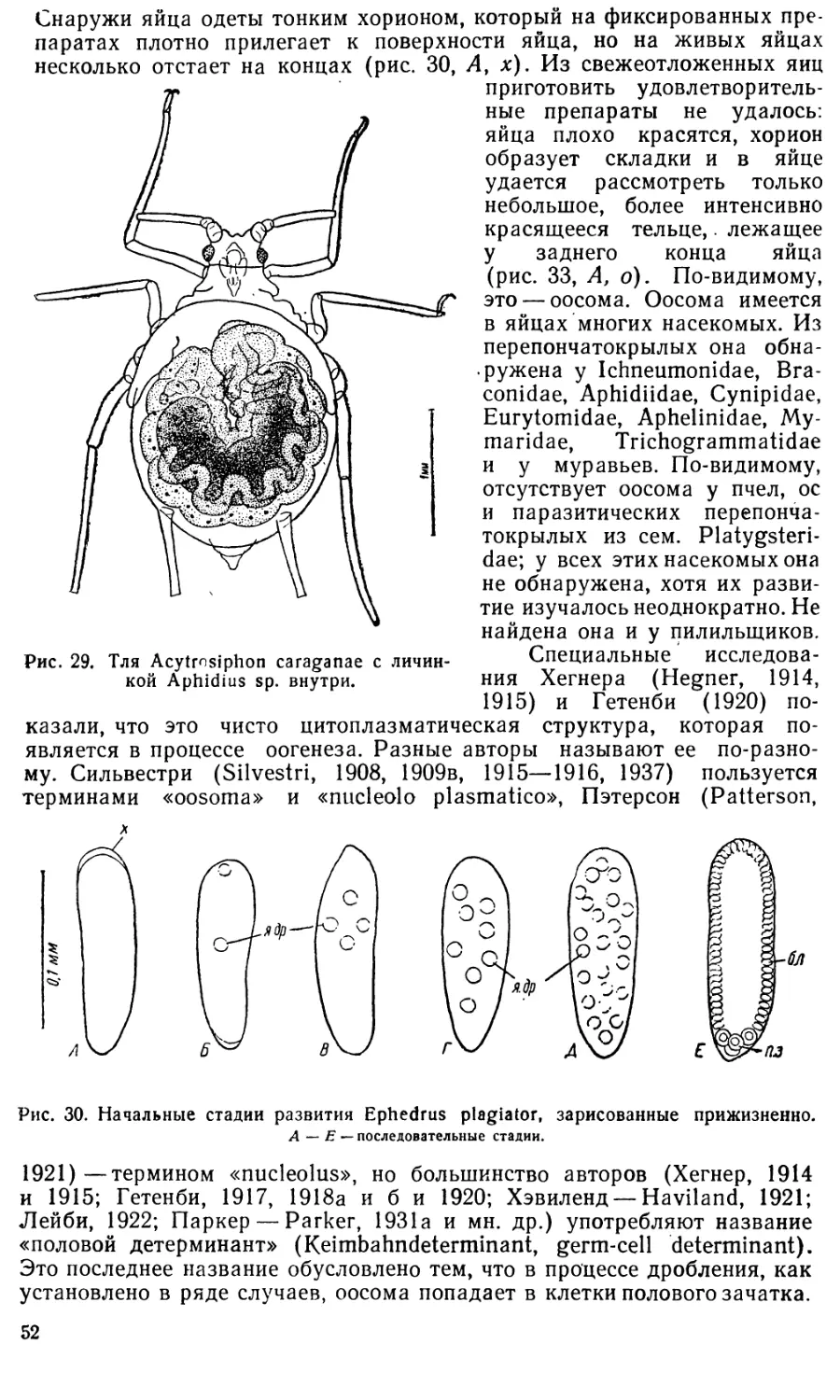
























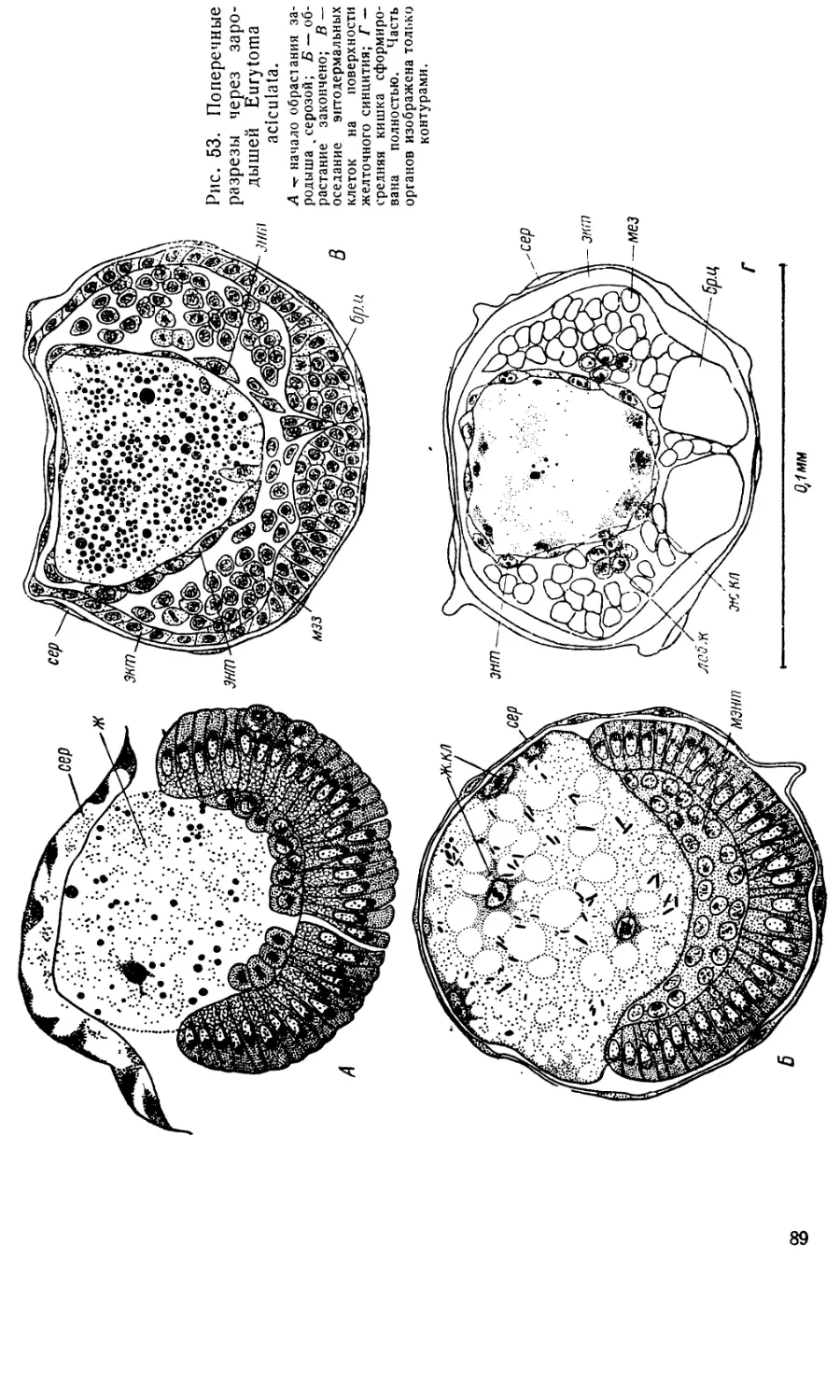

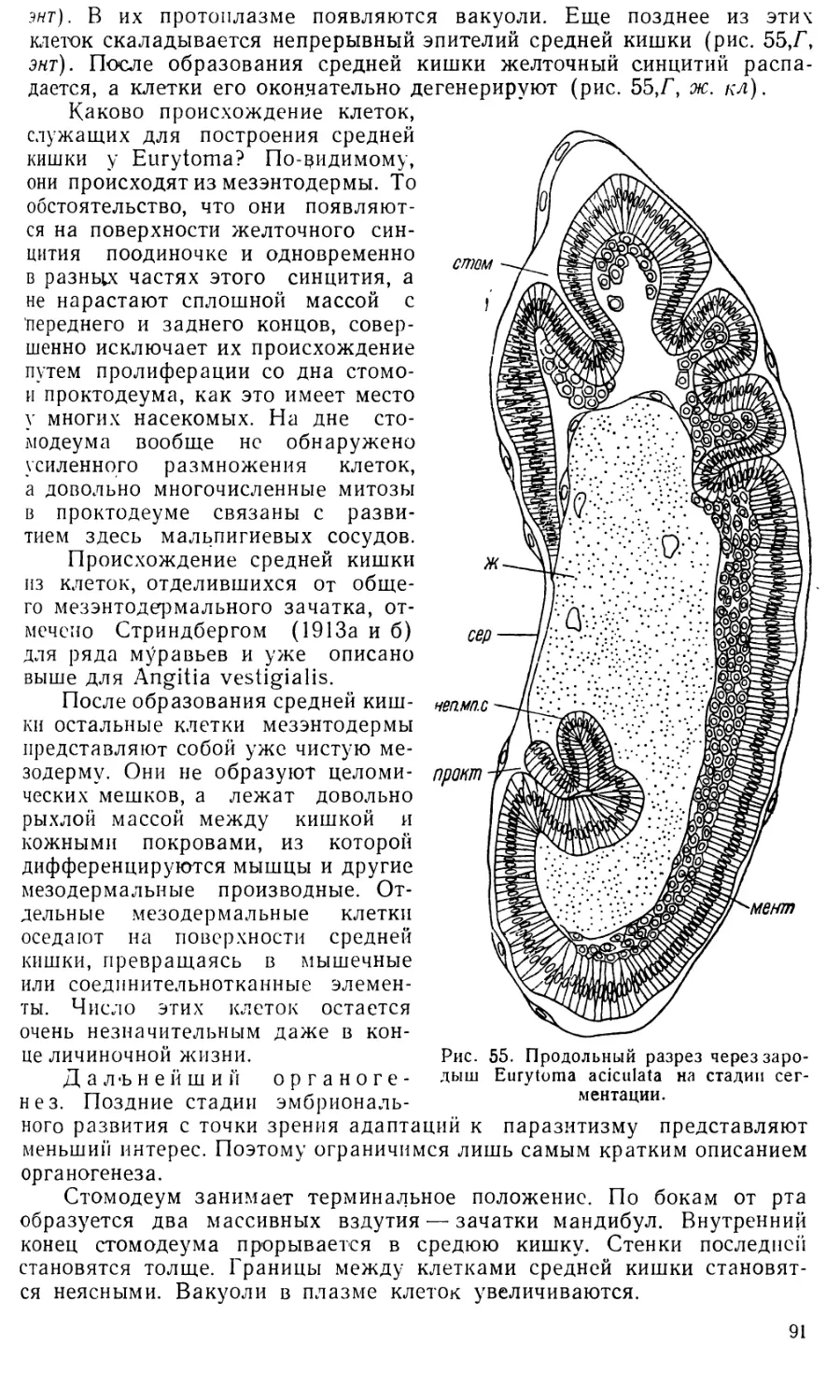







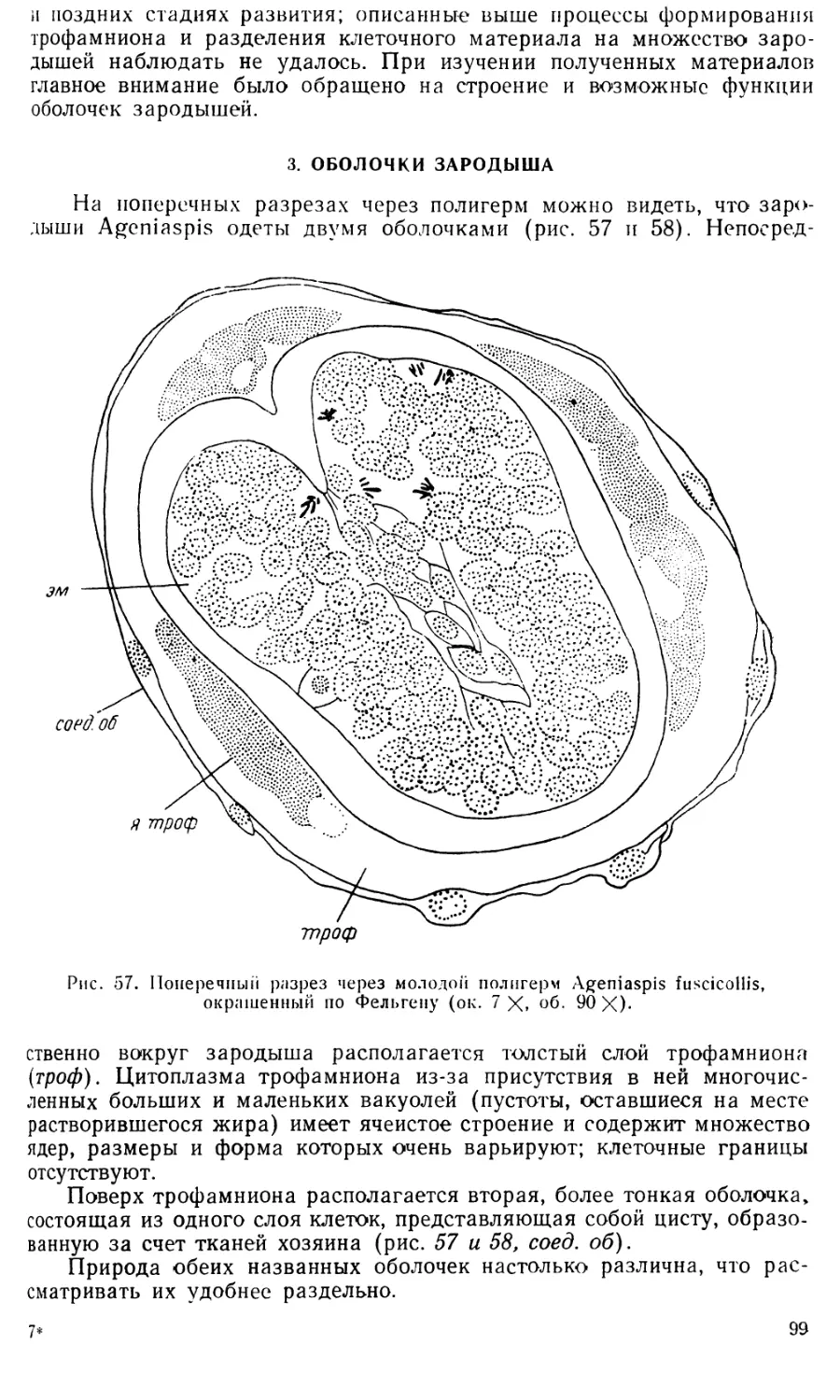
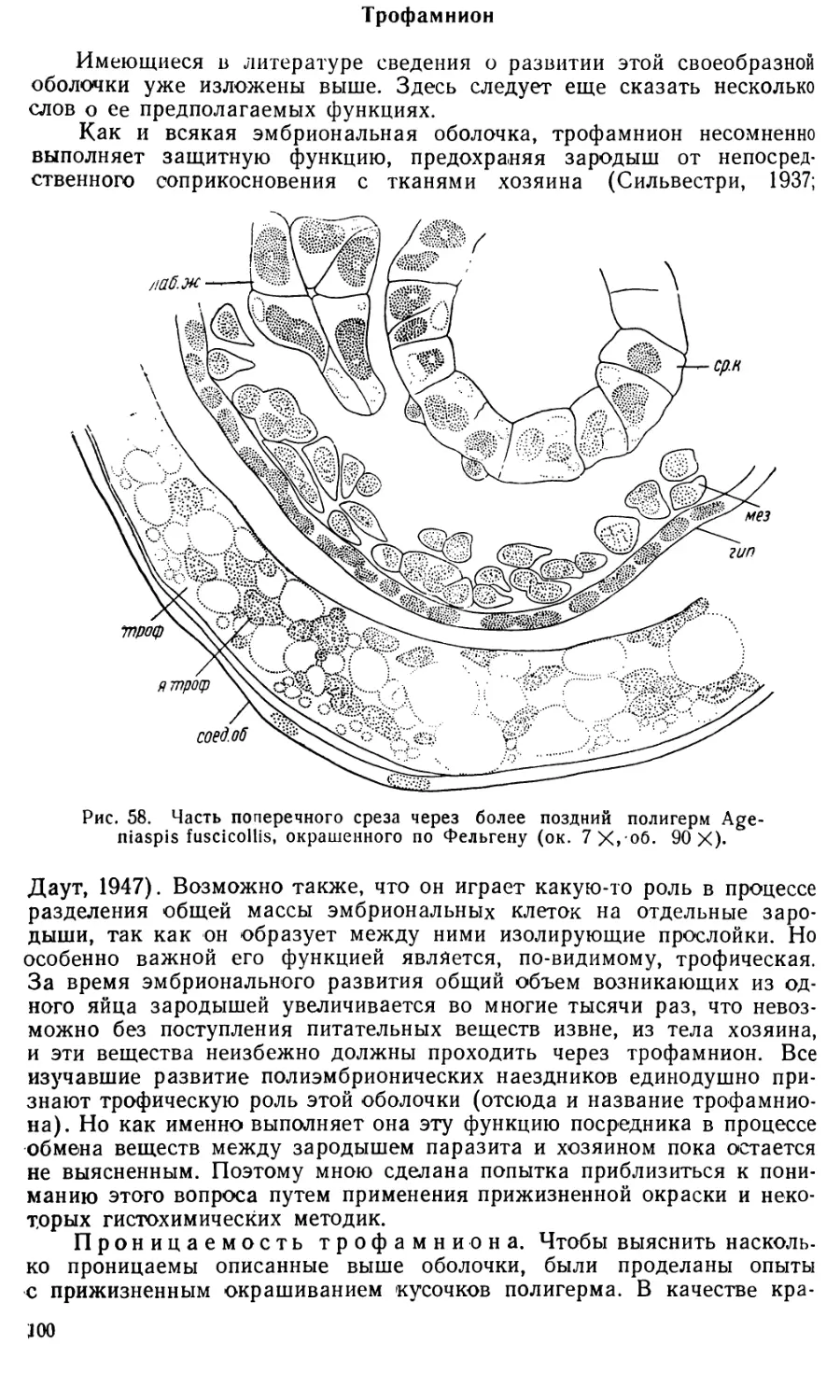














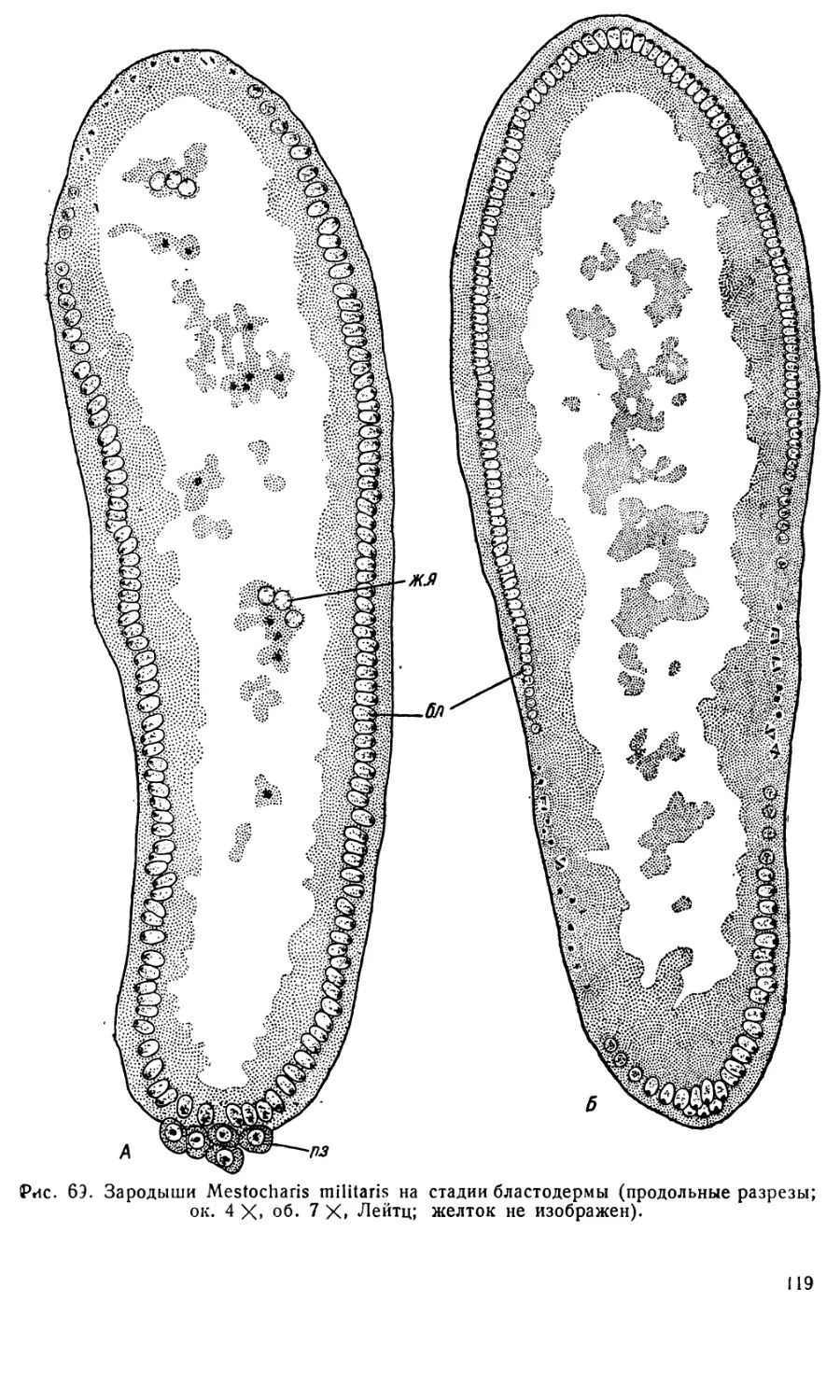
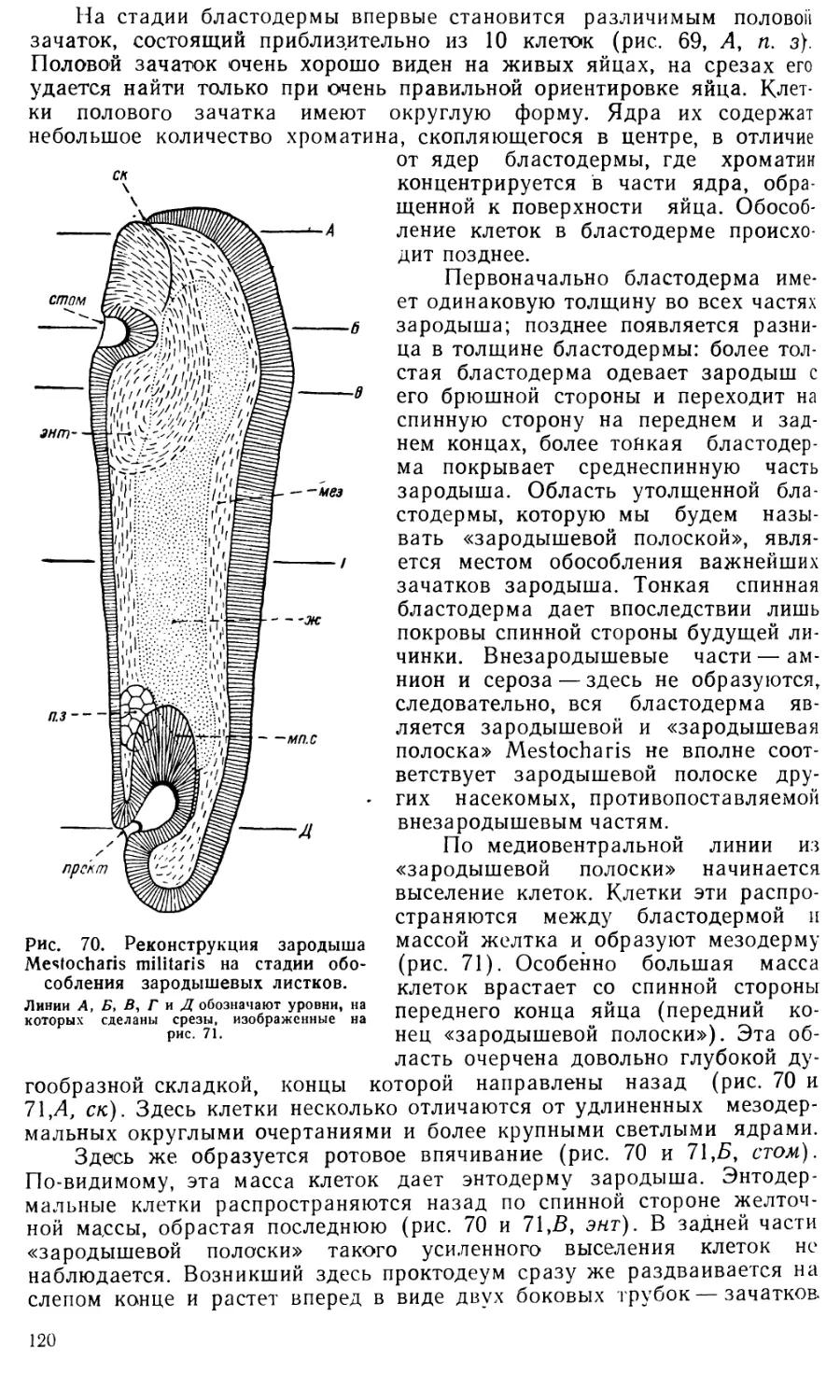
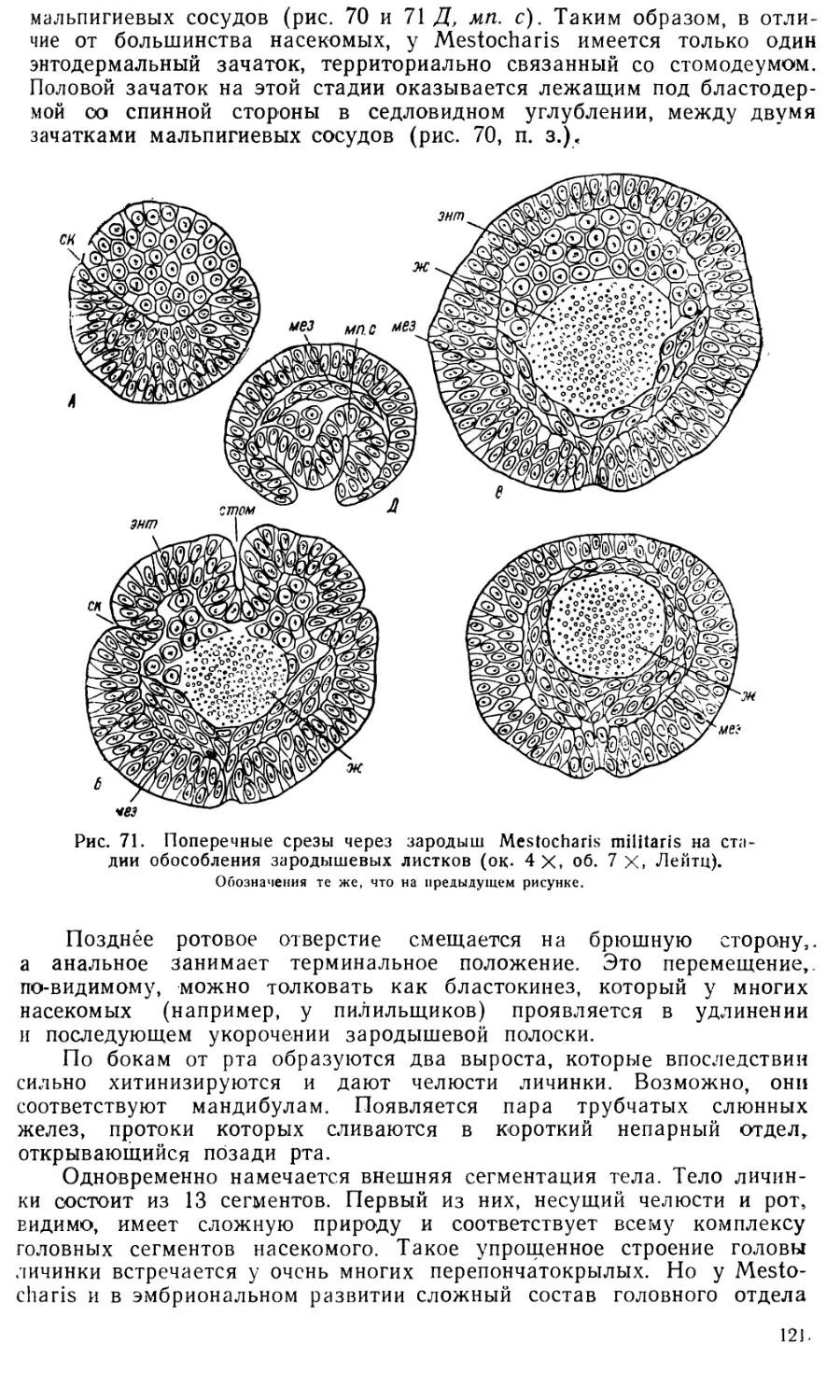



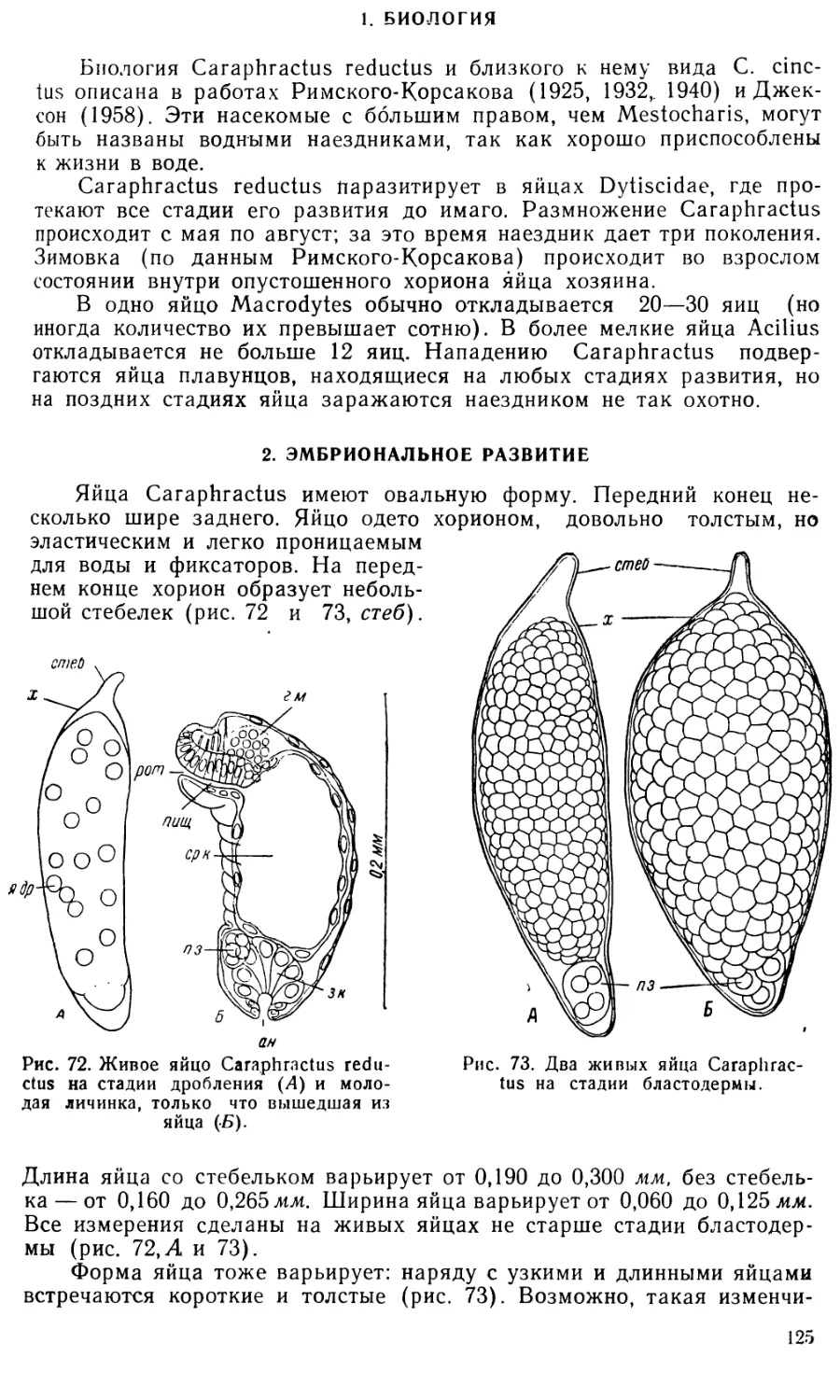
















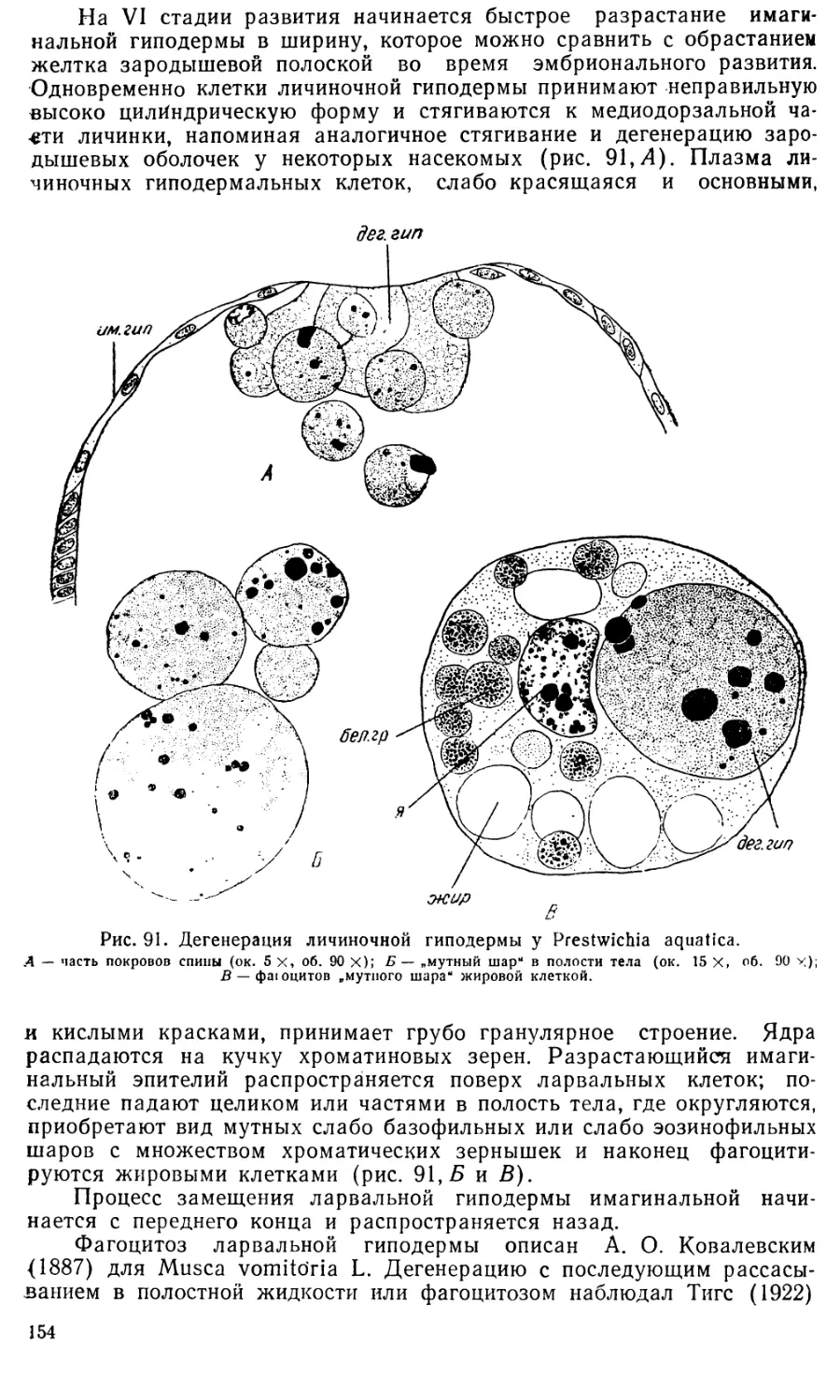
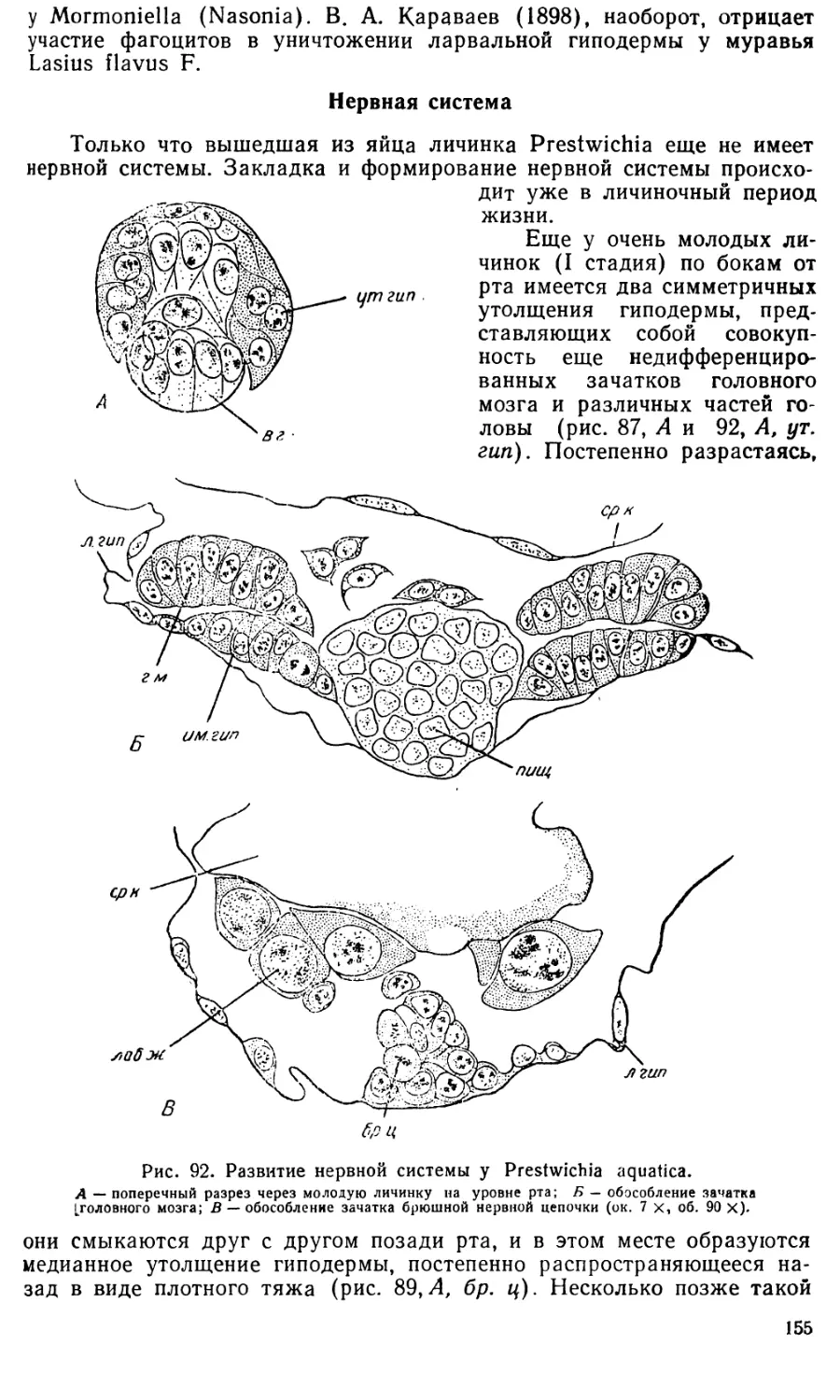
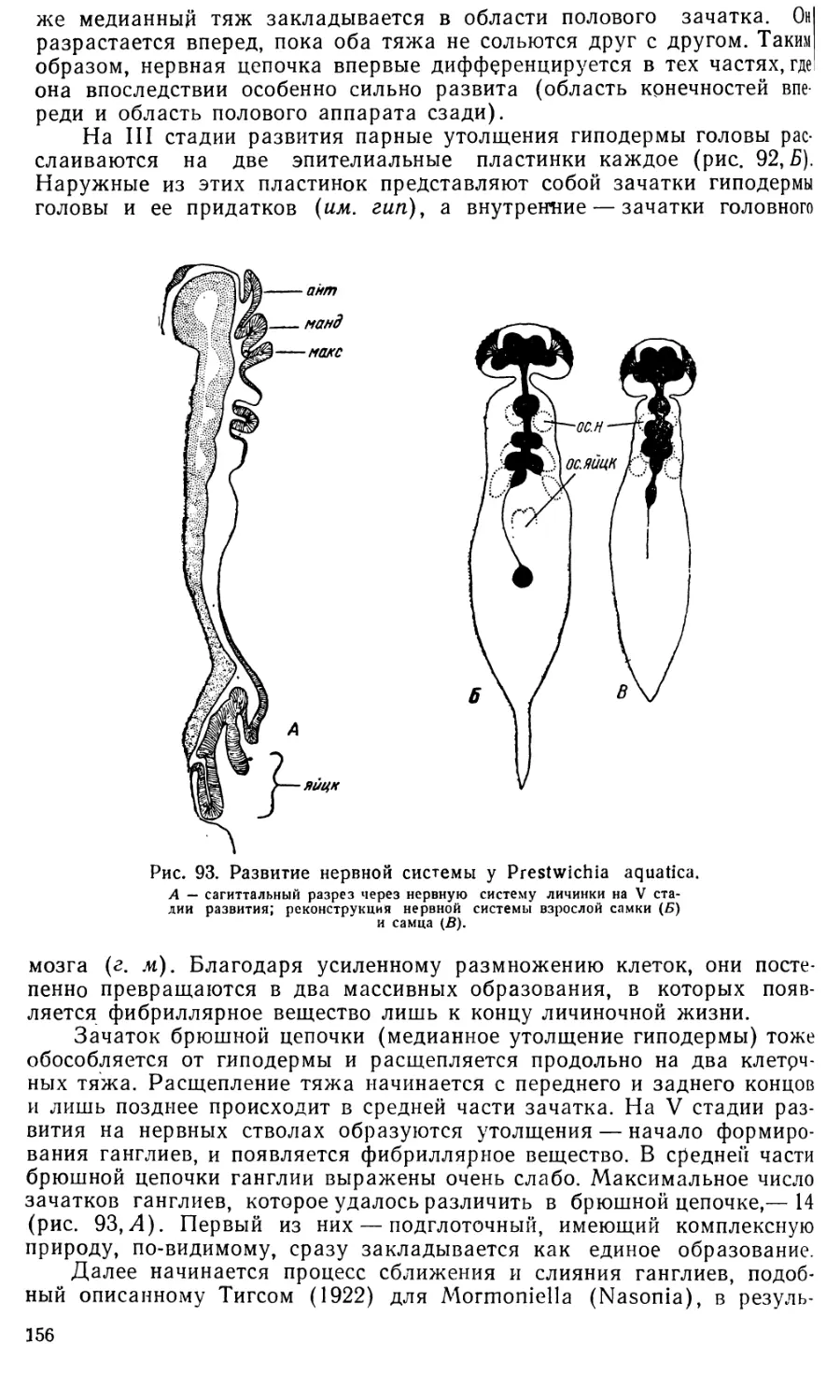

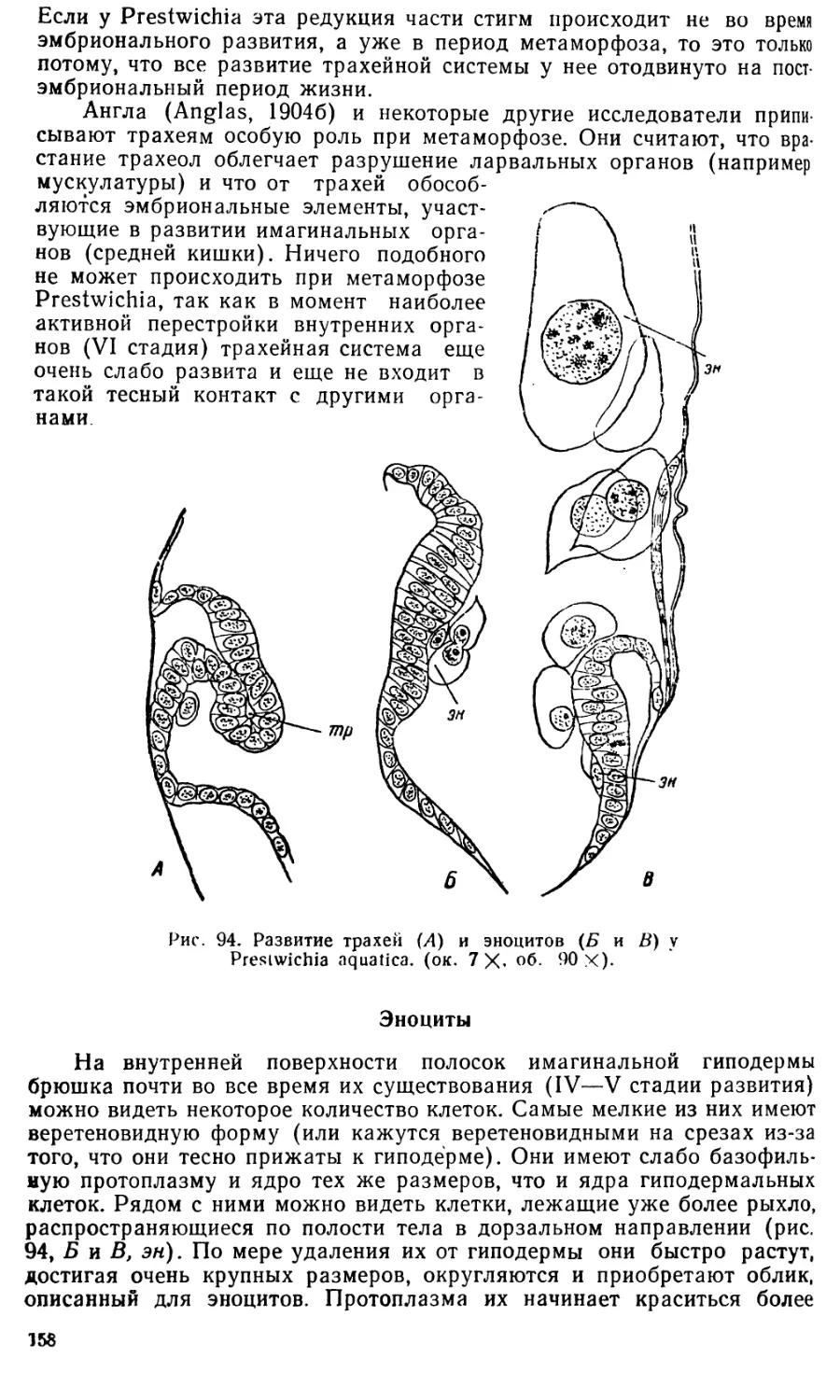



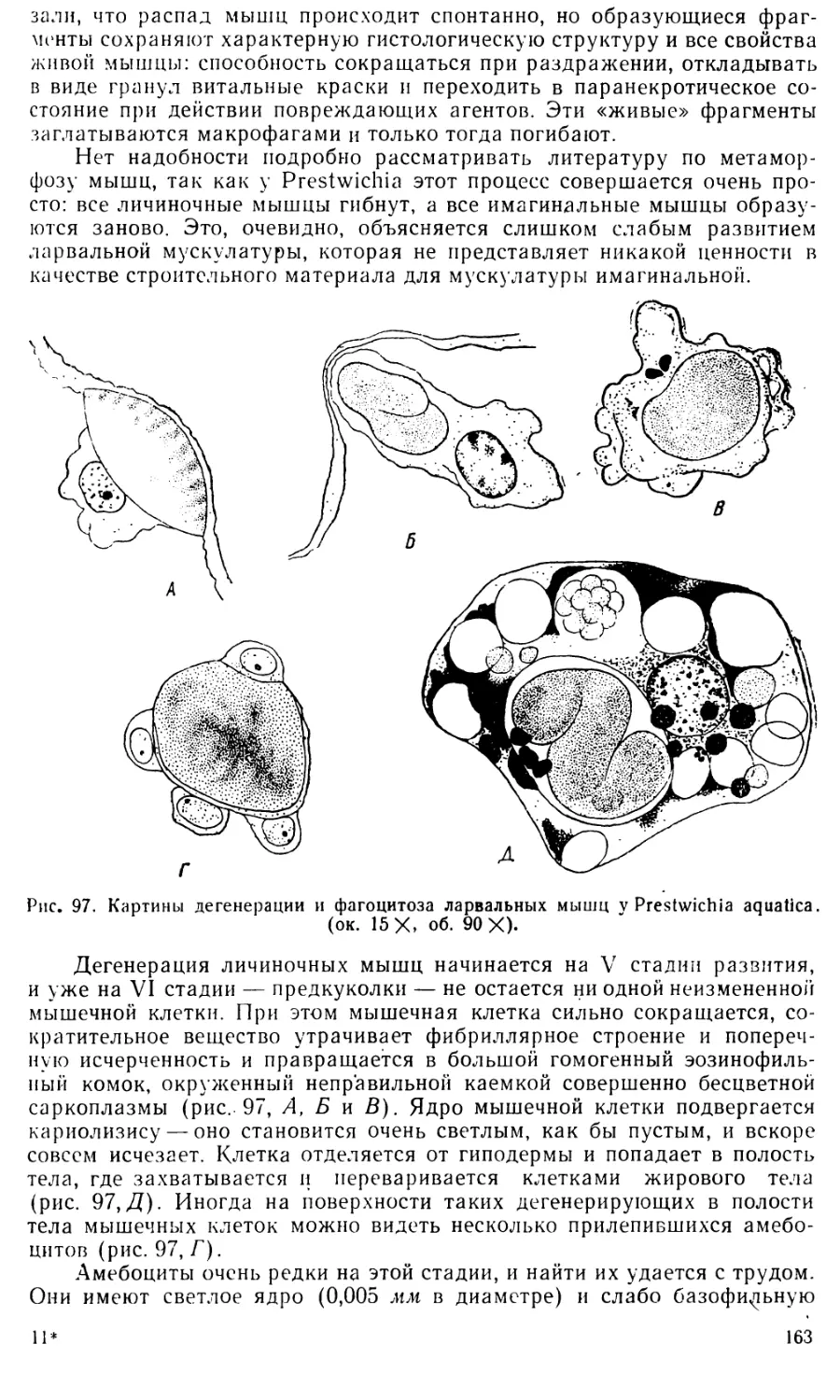




































































































Text
ЛЕНИНГРАДСКИЙ ОРДЕНА ЛЕНИНА ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ имени А. А. ЖДАНОВА
О. М. ИВАНОВА-КАЗАС
ОЧЕРКИ
ПО СРАВНИТЕЛЬНОЙ
ЭМБРИОЛОГИИ
ПЕРЕПОНЧАТОКРЫЛЫХ
ИЗДАТЕЛЬСТВО
ЛЕНИНГРАДСКОГО УНИВЕРСИТЕТА
1961
Печатается по постановлению
Редакционно-издательского совета
Ленинградского университета
Монография представляет собой обобщение много-
летних исследований автора, удостоенных в 1958 г. Ака-
демией наук СССР премии им. А. О. Ковалевского.
В книге приводится обширный оригинальный факти-
ческий материал по эмбриональному развитию ряда пе-
репончатокрылых, ' на основе которого выясняются во-
просы эволюции онтогенеза, рассматривается филогения
и обсуждаются адаптации к паразитизму и морфологи-
ческие закономерности эволюции.
Книга предназначена для научных работников —
эмбриологов, зоологов и энтомологов.
Посвящаю
светлой памяти моих учителей
В. А. Догеля, /7. П. Иванова
и А. П. Римского-Корсакова
ВВЕДЕНИЕ
Значение, которое имела сравнительная эмбриология для развития
эволюционного учения общеизвестно. Блестящие достижения эмбриоло-
гии 2-й половины XIX в., успешное разрешение ею многих спорных во-
просов систематики и филогении выдвинули эту науку на одно из
первых мест среди биологических дисциплин и обеспечили ей широкую
популярность.
В значительной мере благодаря успехам сравнительной эмбриоло-
гии удалось в общих чертах выяснить родственные отношения между
крупными группами животных, и трудно ожидать, что в этой области
могут быть сделаны какие-нибудь новые сенсационные открытия. Идея
эволюции восторжествовала и не нуждается в новых доказательствах.
Однако из этого не следует, что сравнительная эмбриология уже
сыграла свою историческую роль и должна сойти со сцены. Новое время
ставит перед наукой новые задачи.
Классическая сравнительная эмбриология играла в сущности под-
собную роль, способствуя разрешению вопросов общей зоологии, а соб-
ственных проблем не имела. Накопив огромный фактический материал
по развитию различных животных, она не пыталась раскрыть законо-
мерности, управляющие развитием, и мало внимания уделяла приспосо-
бительным изменениям онтогенеза, связанным с конкретными условия-
ми существования, эволюции самого процесса индивидуального разви-
тия. Первое и крупнейшее обобщение сравнительной эмбриологии —
теория зародышевых листков — было сразу же связано с общезоологи-
ческим вопросом о происхождении многоклеточных, а вопрос о проис-
хождении онтогенеза многоклеточных был поставлен лишь более чем
через полстолетия А. Н. Северцовым (1934, 1939) в его «Морфологиче-
ских закономерностях эволюции».
Вышедшая в конце XIX в. на сцену экспериментальная эмбриология
занялась каузальным анализом развития, но при этом сравнительный
и эволюционный подход к явлениям был в значительной степени утрачен.
Попытка Д. П. Филатова (1939, 1943) осмыслить с эволюционной точки
3
зрения данные «механики развития» не получила, к сожалению, даль-
нейшего развития. Это новое направление в эмбриологии, хотя и оттес-
нило сравнительную эмбриологию на второй план, но не смогло заменить
ее полностью.
Для формирования нового направления в сравнительной эмбриоло-
гии большое значение имели работы А. Н. Северцова (1934, 1939).
Л. Н. Северцов показал, что онтогенез не только продукт филогенеза,
но в значительной степени является его творцом, а эволюция есть ряд
постепенно изменяющихся онтогенезов. Если раньше эмбриолог-эволю-
ционист стремился путем сравнения развития далеких в систематиче-
ском отношении групп животных подметить черты сходства и тем дока-
зать общность их происхождения, то теперь гораздо больший интерес
представляет выявление различий в развитии близко родственных жи-
вотных, анализ причин, вызвавших эти различия, процесс адаптивного
изменения и эволюции самого онтогенеза. Поэтому в литературе все
чаще появляются работы, посвященные эволюции индивидуального
развития.
Конечно, проблема соотношения между индивидуальным и истори-
ческим развитием животных еще далека от полного разрешения, но
решить ее призвана именно сравнительная эмбриология, пользующаяся
как морфологическим, так и экспериментальным методом. Вообще раз-
деление эмбриологии на сравнительную и каузальную, исторически воз-
никшее в процессе развития этой науки, со временем должно исчезнуть.
Одно из крупнейших обобщений современной' эмбриологии — теория
первичной гетерономности П. П. Иванова (1944)—построено на основе
синтеза описательно-морфологических и экспериментальных данных.
Только после того, как мы всесторонне изучим развитие животных — с
морфологической, физиологической и экологической стороны,—только
тогда станет возможным познание подлинных закономерностей эволю-
ции онтогенеза. Но это еще дело будущего, а пока в наших суждениях
приходится опираться в основном на данные описательной эмбриологии
с учетом конкретных условий существования.
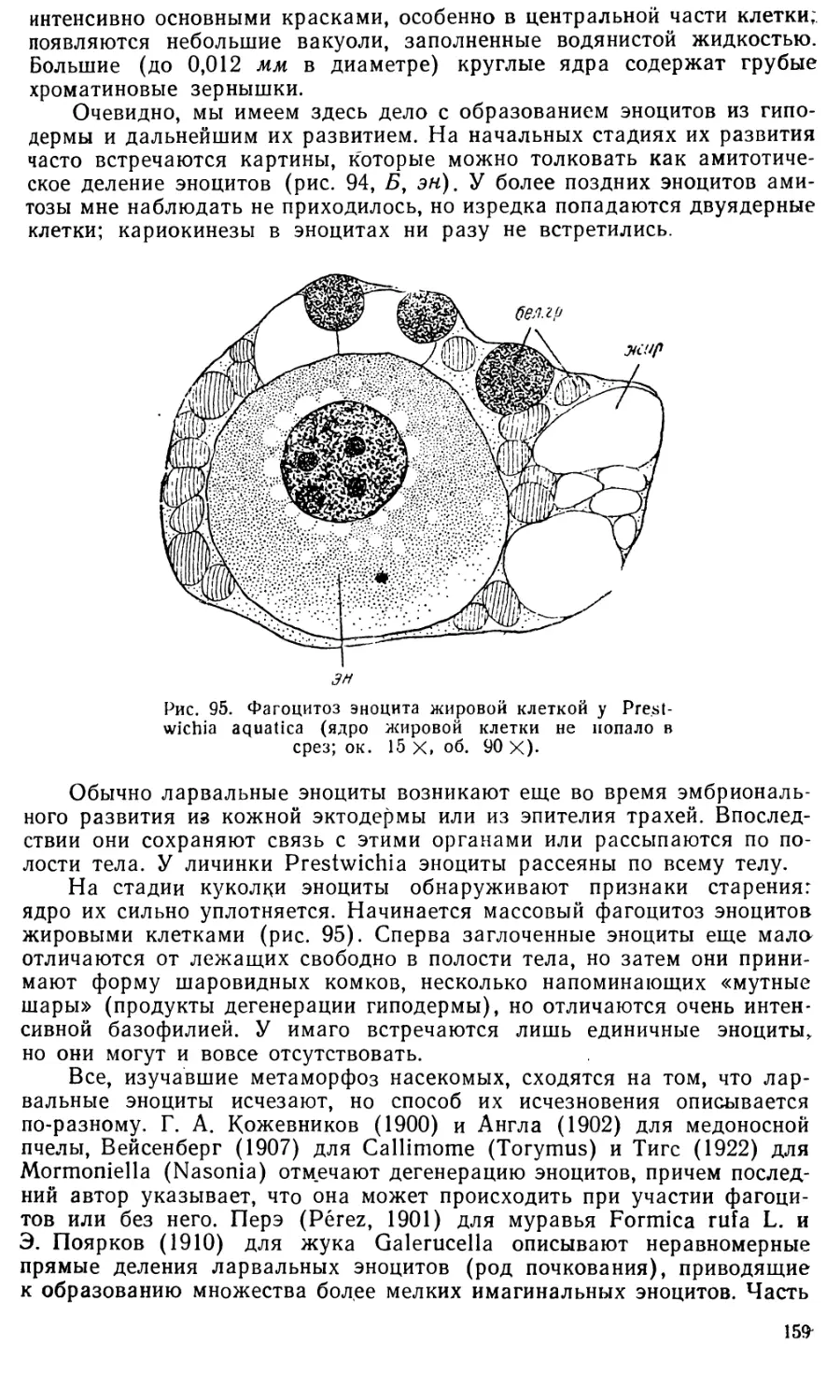
Еще И. И. Мечников (1886) в «Эмбриологических исследованиях
медуз» показал, как происходила эволюция процесса гаструляции у
кишечнополостных. Но классический труд Мечникова долгое время
оставался единственным^ Лишь недавно блестящую разработку его идеи
дал А. А. Захваткин (1949), показавший возможные пути возникновения
и начальных этапов эволюции онтогенеза Metazoa в плане теории фаго-
цителлы. Основные тенденции эволюции онтогенеза многоклеточных
намечены И. И. Шмальгаузеном (1938, 1946). В учебнике эмбриологии
Г. А. Шмидта (1951) делается попытка проследить эволюцию некоторых
процессов развития (дробления, гаструляции и т. д.). Эволюция внеза-
родышевых образований Sauropsida рассматривается в книге А. М. Сер-
геева (1943). Бластокинез и эмбриональные оболочки насекомых были
освещены с эволюционной точки зрения А. Г. Шаровым (1953, 1957)
и т. д.
4
Однако, пока речь идет об эволюции развития в пределах больших
групп животных (типов, классов), соображения авторов поневоле имеют
несколько отвлеченный спекулятивный характер. Чем меньше объем
изучаемой группы, чем более близки* рассматриваемые объекты, тем
более точный материал получает исследователь для суждения о причи-
нах и закономерностях изменения онтогенеза. Быть может, наилучшие
результаты дало бы изучение развития в пределах семейства или рада.
В настоящей работе, однако, речь будет идти об эмбриональном разви-
тии в отряде перепончатокрылых (Hymenoptera).
Изучение развития перепончатокрылых представляет не только
теоретический интерес. Многие паразитические представители этого от-
ряда имеют экономическое значение, так как используются для борьбы
с вредителями сельского хозяйства. Поэтому всестороннее изучение их
биологии может в конечном итоге принести и практическую пользу.
Перепончатокрылые дают благодарный материал для сравнительно-
эмбриологического исследования. Отряд этот интересен большим разно-
образием образа жизни относящихся.к нему насекомых: здесь встреча-
ются первичнорастительноядные формы, наружные и внутренние
паразиты, и насекомые, вторично вернувшиеся к растительноядности;
сюда относятся одиночные виды и «общественные», живущие большими
семьями со сложным разделением труда; в этом отряде наблюдаются
разнообразные и изощренные инстинкты «заботы о потомстве». Все это
не могло не наложить свой отпечаток на эмбриональное развитие
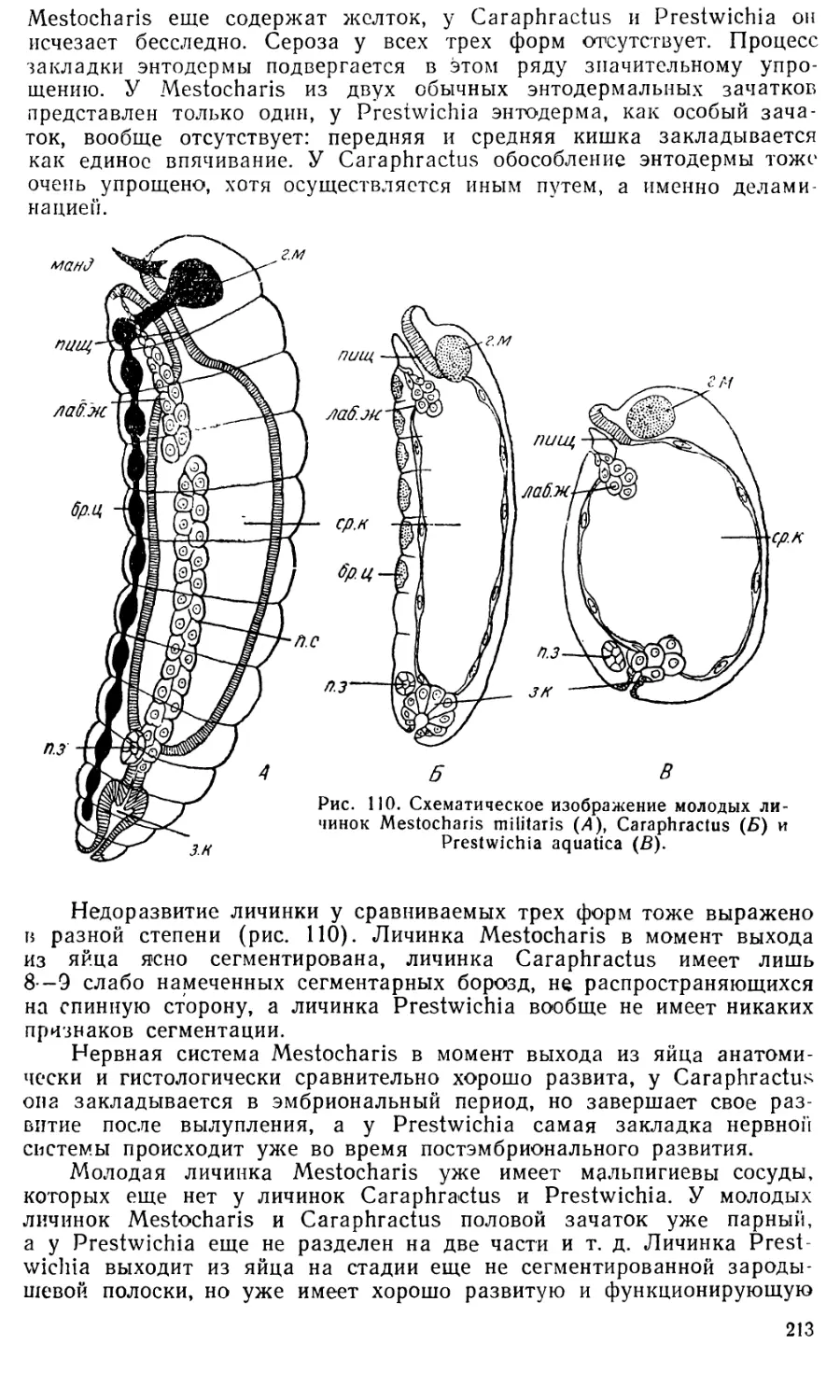
перепончатокрылых. Действительно, у них удается различить несколько
разных типов развития: примитивный—у пилильщиков, несколько
измененный — у низших наездников, ос и пчел, специализированный — у
муравьев и по крайней мере три разных типа развития у эндопаразнти-
ческих форм. Едва ли еще в каком-нибудь отряде насекомых можно
встретить такое разнообразие способов развития.
Настоящая работа не претендует на полный охват группы. Отряд
Hymenoptera включает около сотни семейств, и изучить эмбриональное
развитие хотя бы у одного представителя из каждого семейства не по
силам одному исследователю.
При выборе объектов особое внимание было уделено паразитиче-
ским видам, так как при паразитизме наблюдаются особенно сильные
адаптивные изменения, подчас охватывающие всю организацию живот-
ного. Яркие примеры морфологических адаптации к паразитизму
собраны в «Курсе общей паразитологии» В. А. Догеля (1947),,но влия-
ние паразитизма на эмбриональные стадии развития там не освещено,
так как данных такого рода во время написания «Курса» еще не было.
В то же время развитие паразитических форм представляет интерес
и с точки зрения разрабатываемой на кафедре эмбриологии Ленинград-
ского государственного университета проблемы иммунитета зародышей
(«Иммунитет зародышей». Токин, 1955), одну из сторон которой состав-
ляет вопрос об отношениях между развивающимся организмом и био-
5
тическими факторами среды. В случае паразитизма последние пред-
ставлены организмам хозяина.
Поэтому только один из изученных объектов, а именно: галлообра-
зующий пилильщик Pontania carpeae L., относится к подотряду
Phytophaga, все же остальные являются представителями подотряда
Parasitica: Eurytoma aciculata Ratz. ведет инквилинный образ жизни в
коконах Pontania, Angitia vestigialis Ratz:, Ageniaspis fuscicollis (Dalm.),
Eptiedrus plagiator Nees и виды родов Aphidiiis и Charips паразитируют
внутри различных насекомых, a Mestocharis militaris R.-Kors., Caraphrac-
tus reductus R.-Kors. и Prestwichia aquatica Lubb. относятся к так назы-
ваемым яйцеедам, т. е. развиваются в яйцах других насекомых.
Работа начата на кафедре зоологии беспозвоночных Ленинградско-
го университета, руководимой тогда профессором В. А. Догелем, а
закончена на кафедре эмбриологии (заведующий — профессор Б.П.То-
кин). Руководителям обеих кафедр и их коллективам я выражаю самую
глубокую благодарность за неизменно благожелательное и вниматель-
ное отношение к моей работе.
СПЕЦИАЛЬНАЯ ЧАСТЬ
ГЛАВА I
PONTANIA CAPREAE L.—ИВОВЫЙ ТОЛСТОСТЕННЫЙ
ПИЛИЛЬЩИК
Для сравнительного изучения развития перепончатокрылых очень
важно иметь представление о наиболее примитивных его формах, послу-
живших отправной точкой для последующей эволюции. Этот исходный
тип развития надо искать среди низших растительноядных перепончато-
крылых (подотряд Phytophaga), куда относятся пилильщики. Эмбрио-
нальное развитие некоторых пилильщиков освещено в работах Грабера
(Graber, 1890), Шафика (Shafiq, 1954) и Суня (Sun Sao-chen, 1959).
Ниже излагаются результаты наблюдений над развитием ивового толс-
тостенного пилильщика (Иванова-Казас, 1957, 1958а и 19596).
1. НЕКОТОРЫЕ ДАННЫЕ ПО БИОЛОГИИ
Pontania сартеае откладывает яйца в молодые листочки ивы
Salix fragilis L. и вызывает этим развитие характерных галлов, очень
многочисленных в окрестностях Ленинграда. Отложенное яйцо оказы-
вается в паренхиме листа, в узком щелевидном пространстве между
двумя пластинками эпидермиса, заполненном тканевой жидкостью.
Затем начинается формирование галла. Полость галла остается узкой,
но заполняется теперь уже воздухом. Таким образом, развитие яиц
Pontania сперва происходит в жидкой среде, а потом в воздушной, но
очень влажной.
Откладка яиц Pontania совершается на протяжении всего лета
(с июня по август). В июле наряду со свежеотложенными яйцами попа-
даются уже совершенно зрелые галлы. Личинка прогрызает в стенке
галла отверстие, через которое выбрасывает наружу экскременты.
В конце июля личинки начинают покидать галлы, они уходят в почву,
где плетут глянцевитые темно-коричневые коконы, в которых окукляются.
В августе наблюдается вылет первых пилильщиков. Личинки, вышедшие
из более поздних яиц, покидают галлы лишь в сентябре. Они зимуют
в галлах; линька на куколку и вылет имаго происходят только весной.
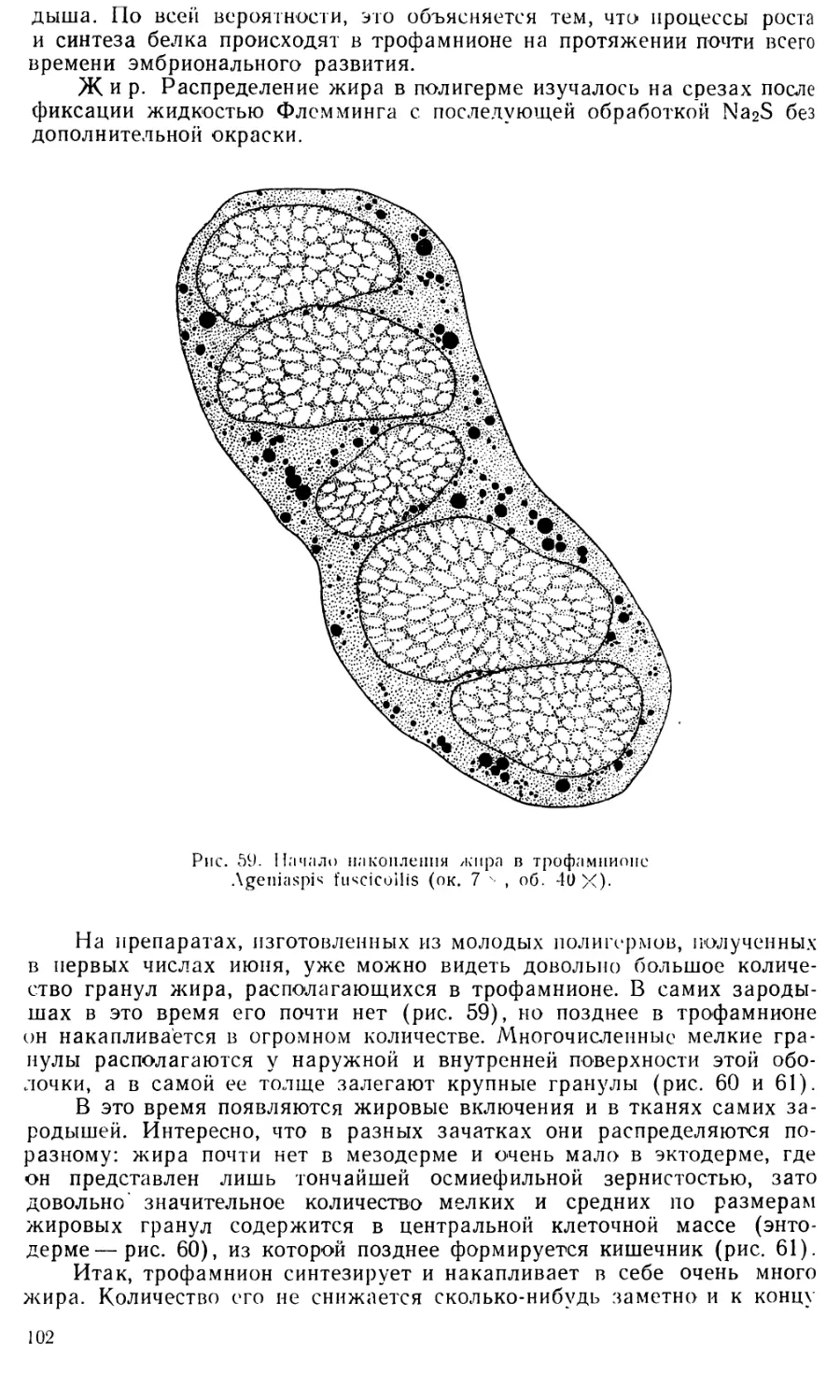
2. НАБУХАНИЕ ЯИЦ И ПРОНИЦАЕМОСТЬ ХОРИОНА
Яйца Pontania саргеае на ранних стадиях дряблые и нежные, но
к концу развития становятся упругими и плотными. При этом они не-
сколько увеличиваются в длину и в гораздо более значительной сте-
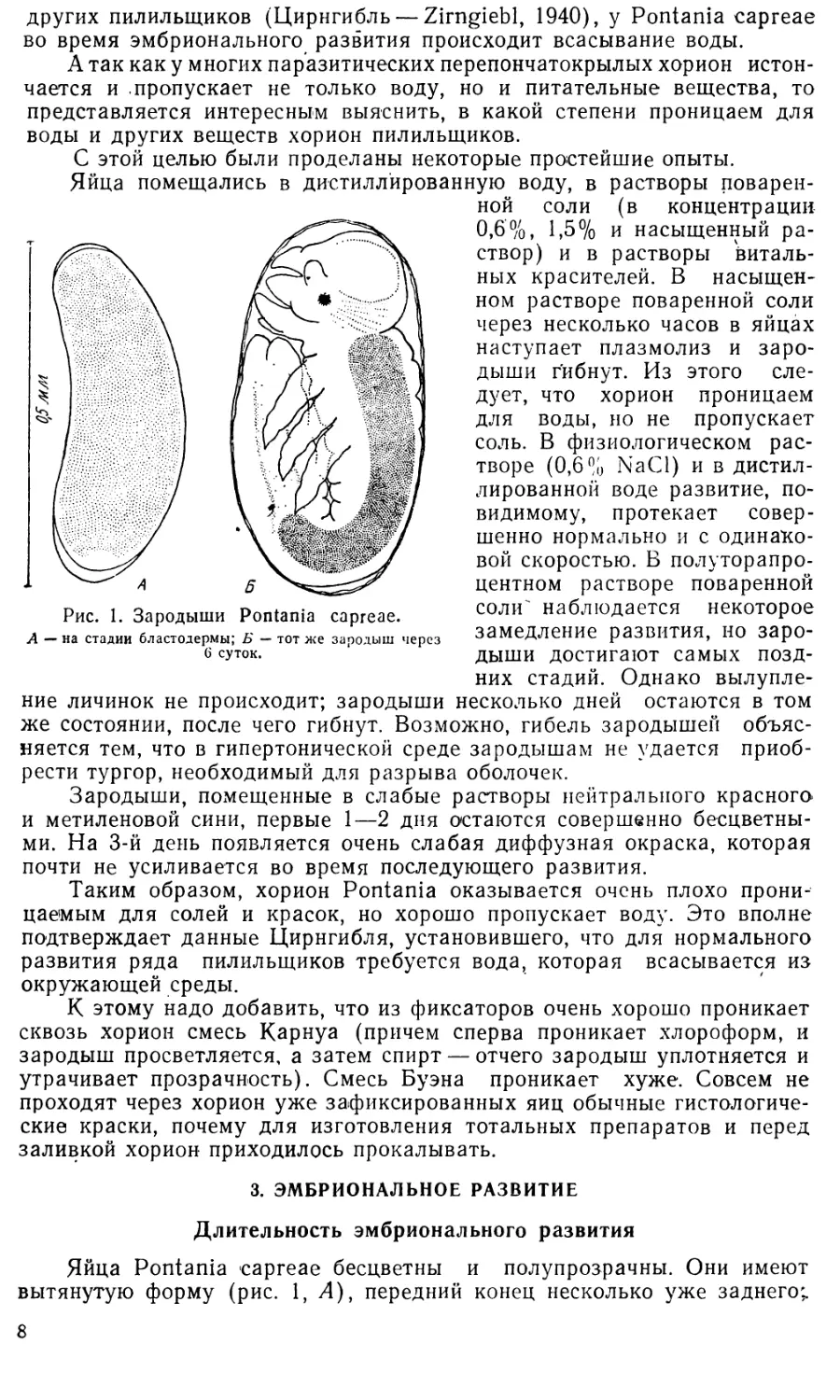
пени— в ширину (рис. 1, Л и £), так что объем яйца • возрастает в
среднем в 2,8 раза. Это заставляет предполагать, что, как и у многих
7
брюшная сторона выпукла, спинная — слабо вогнута. Длина ранних яиц
(не старше стадии бластодермы) составляет в среднем 0,49 мм, шири-
на—0,16 мм.
Как показала последующая гистологическая обработка, мне уда-
валось находить яйца Pontania, находящиеся на самых ранних стадиях
развития (стадия деления созревания). Такие зародыши (не старше
стадии бластодермы) помещались в воду или физиологический рас-
твор для прижизненных наблюдений. В лаборатории, где производились
наблюдения над скоростью развития Pontania capreae, температура
воздуха колебалась в разное время суток и в зависимости от погоды—'
от 20 до 24°. В этих условиях эмбриональное развитие Pontania дли-
лось 8—10 дней. Все развитие Pontania условно можно разделить на
4 больших периода, которые в свою очередь подразделяются на стадии:
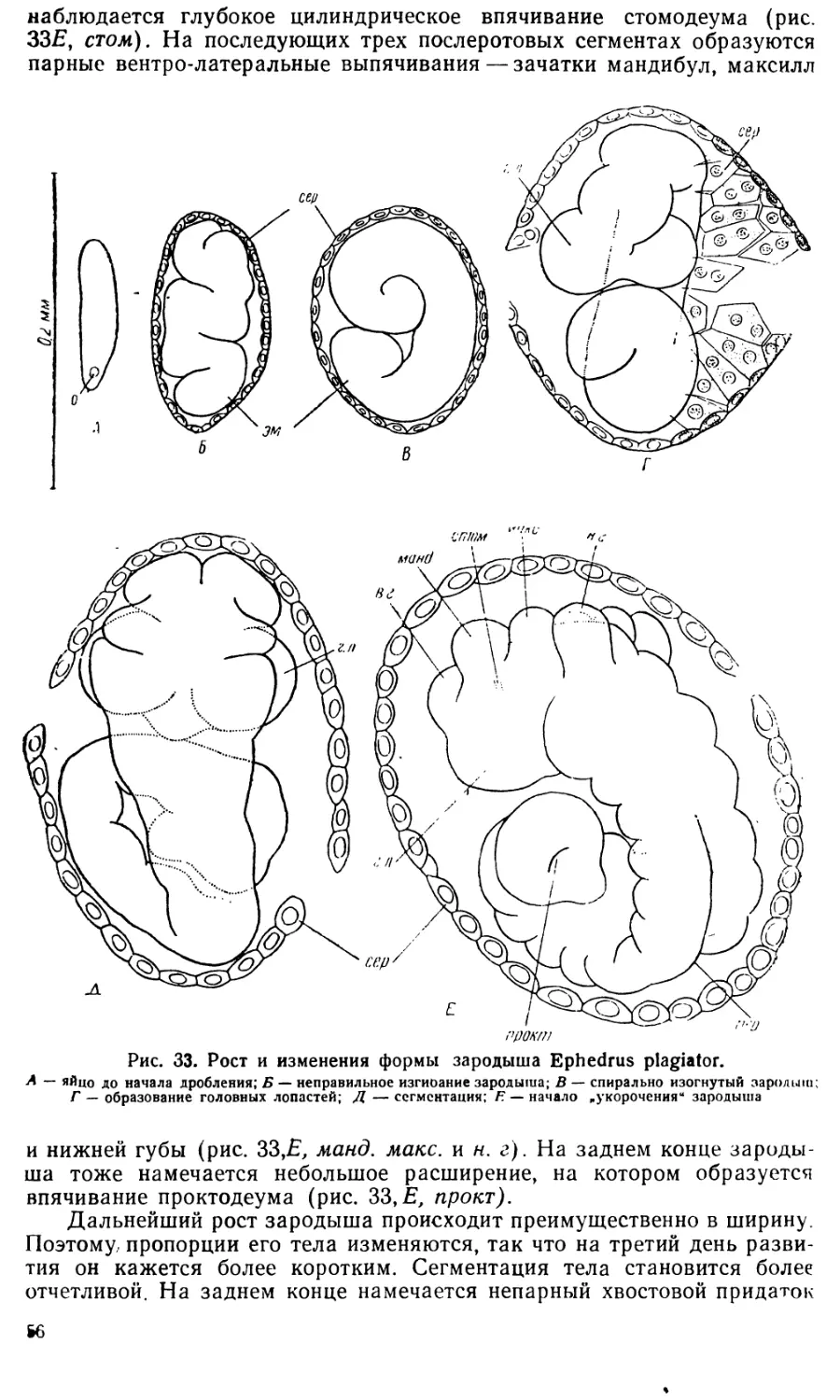
I. Период, охватывающий начальные стадии развития от откладки
яйца до образования зародышевой полоски:
1. Стадия яйца до начала дробления.
2. Стадия дробления.
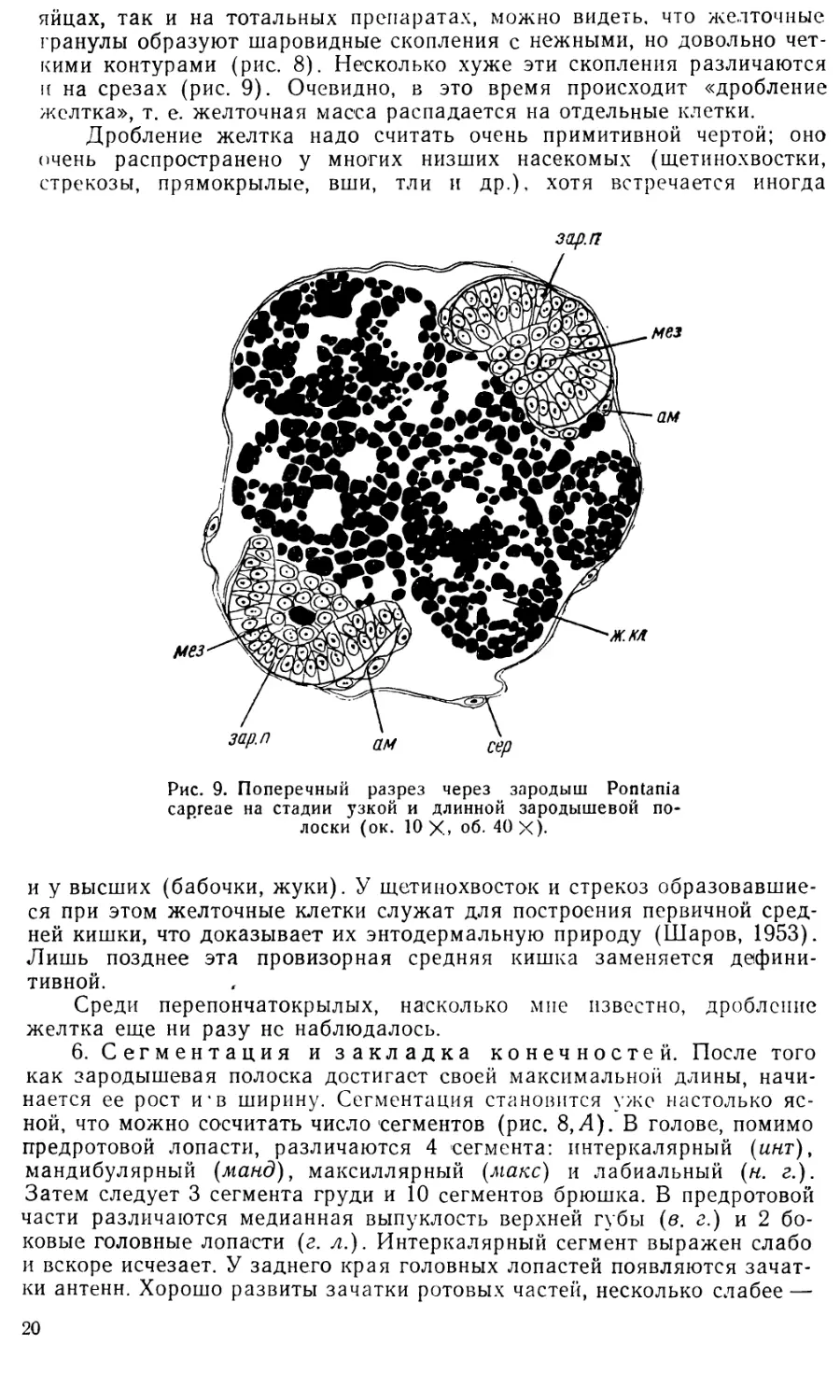
3. Стадия бластодермы (см. рис. 3 и 4, Л).
4. Стадия ранней (короткой) зародышевой полоски (см.
рис. 4, Б и 5).
Все эти стадии обычно проходили в течение первых суток на-
блюдения.
II. Период, удлинения зародышевой полоски.
5. Стадия узкой длинной зародышевой полоски (образование
мезодермы и эмбриональных оболочек; см. рис. 7 и 9; 2-й и 3-й день
наблюдений).
6. Стадия сегментации и закладки конечностей (см. рис 8 и 10;
4-й день наблюдений).
III. Период укорочения зародышевой полоски.
7. Стадия укорочения зародышевой полоски и роста ее в ши-
рину, когда закладывается большая часть внутренних органов (см.
рис. 11, 12, Л).
8. Стадия выпрямленного зародыша, задний конец которого
находится на заднем конце яйца, когда появляются зачатки брюш-
ных ног (см. рис. 12,5 и В; 5-й день наблюдений).
IV. Период формирования личинки.
9. Стадия роста зародыша в длину, сопровождающегося подги-
банием заднего конца на брюшную сторону (см. рис. 15). Эта ста-
дия, во время которой завершается органо- и гистогенез, обычно про- .
должается 2—3 дня. Обычно уже на 7-й или 8-й день зародыш начи-
нает поворачивать голову, двигать челюстями, слабо изгибает туло-
вище. Еще через день происходит вылупление личинки (см. рис. 18).
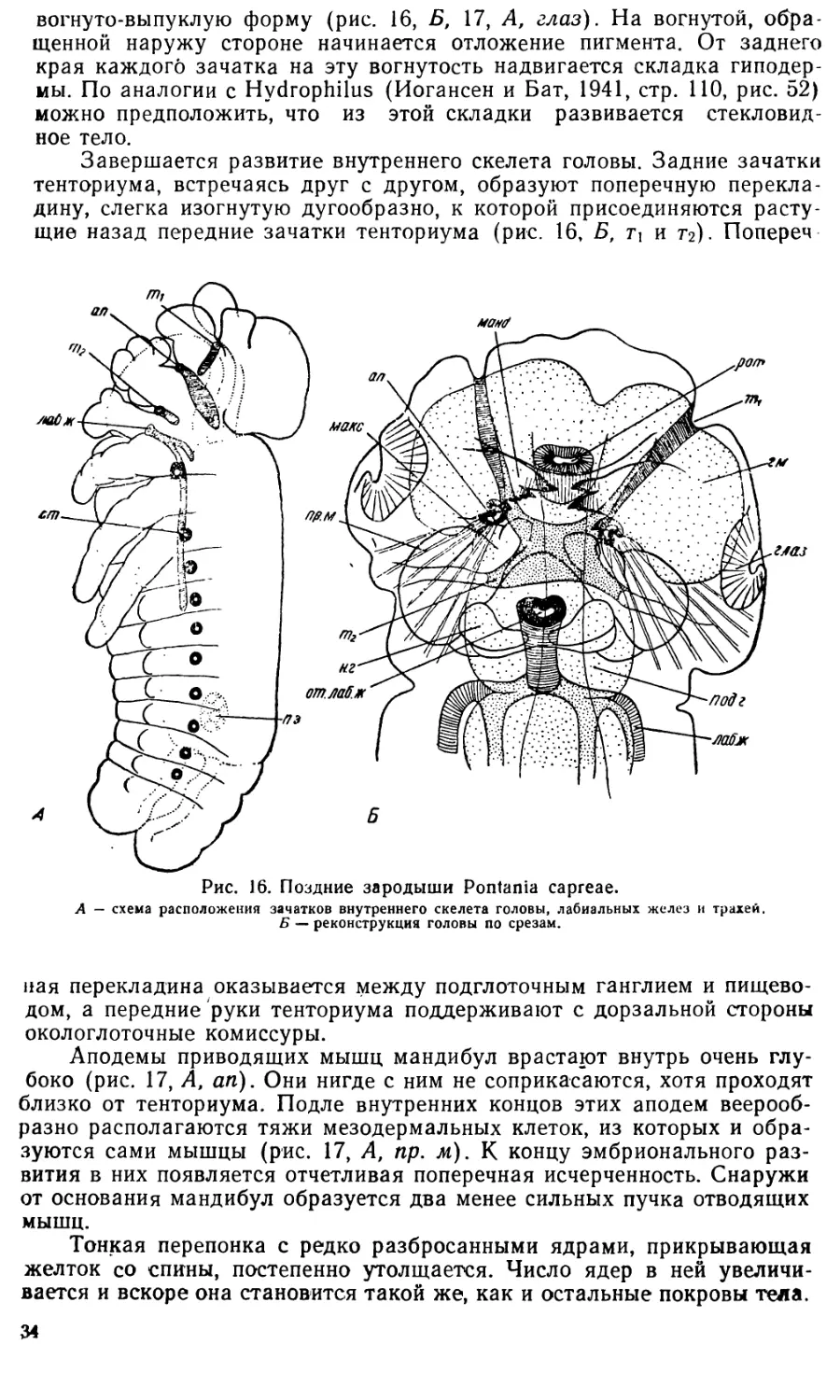
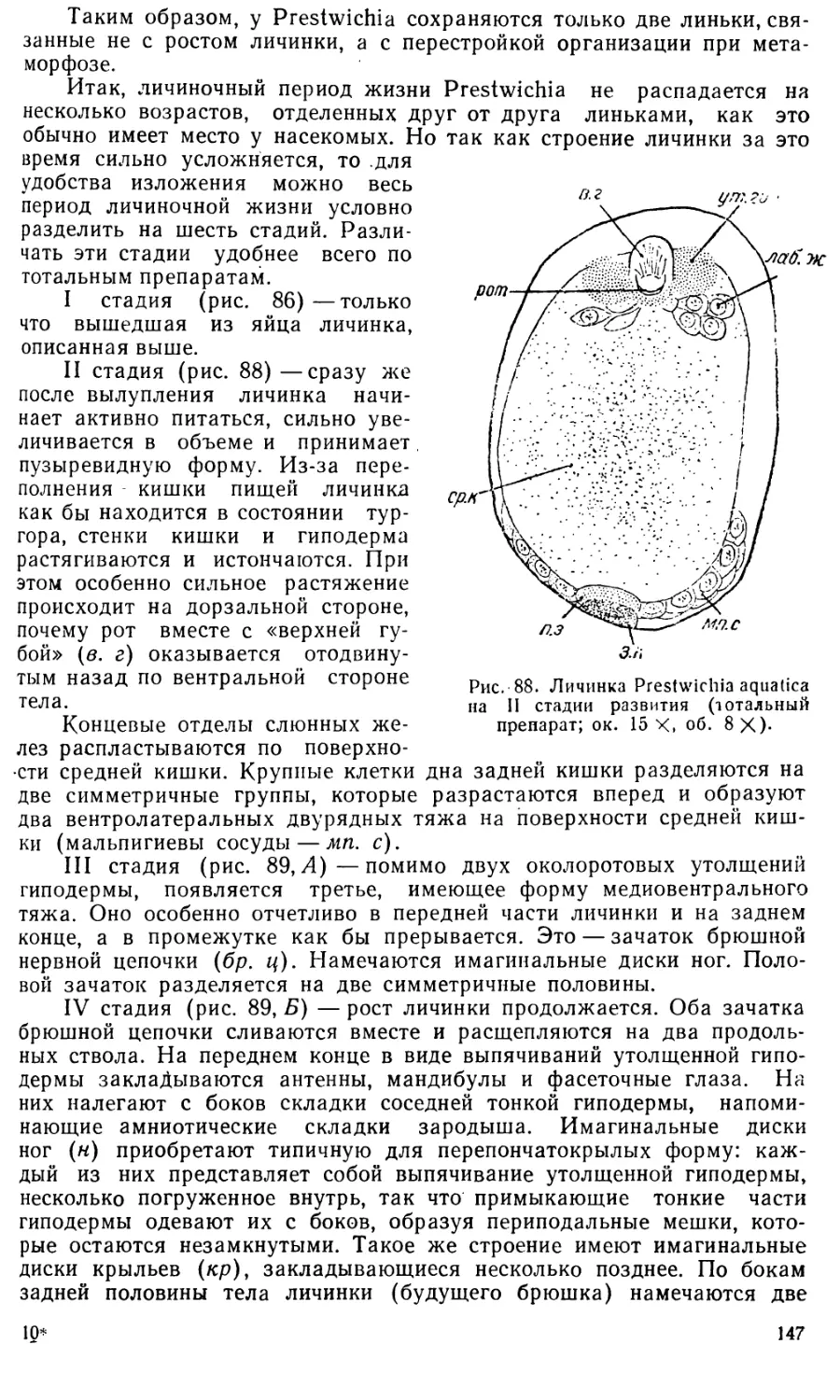
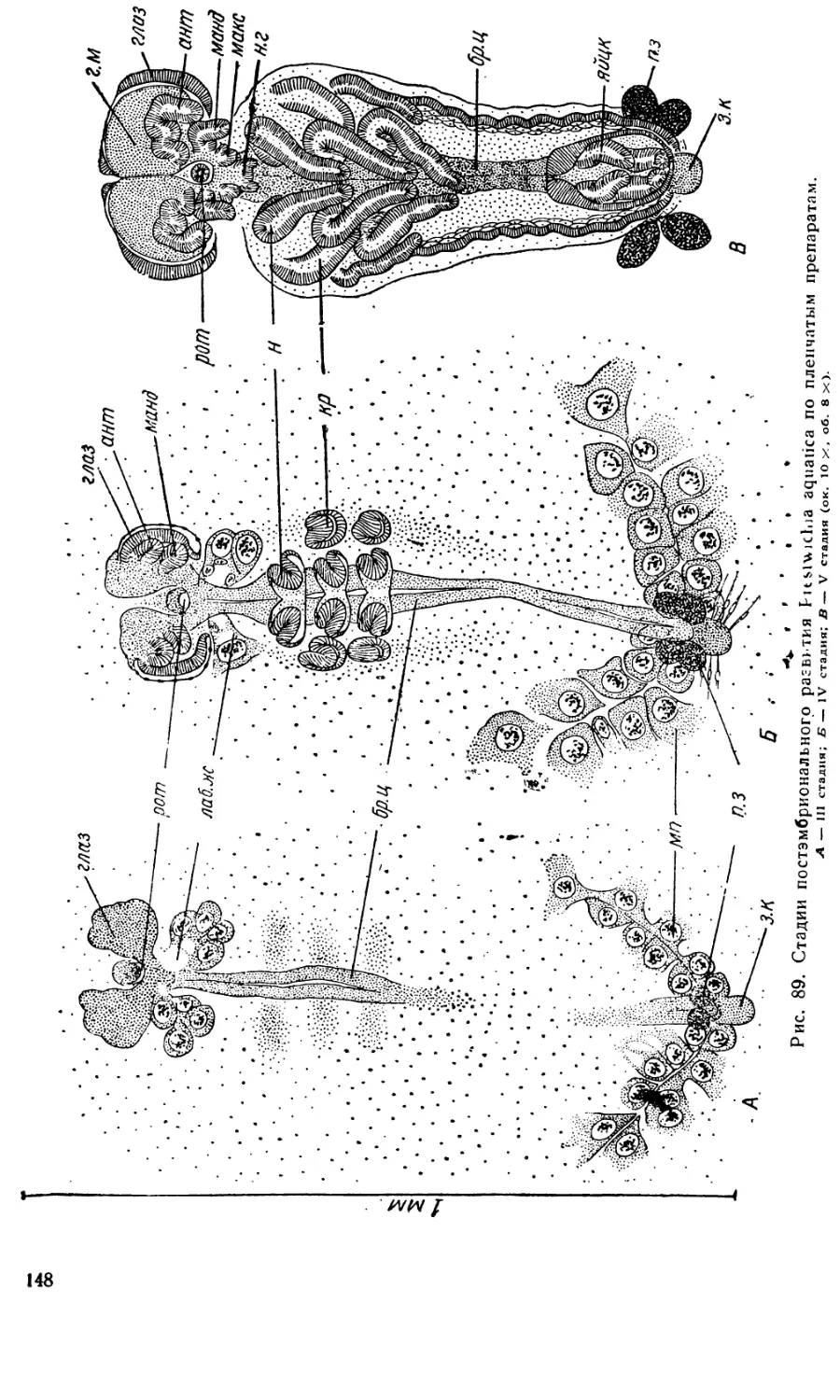
Эти периоды и стадии развития установлены в значительной мере
произвольно для удобства изучения и изложения материала по наиболее
заметным морфологическим признакам. Так как эмбриональное разви-
тие есть единый и непрерывный процесс, то само собой разумеется, что
резких граней между отдельными стадиями нет, и одна стадия перехо-
дит в другую постепенно.
Интересное сопоставление относительной длительности разных пе-
риодов развития у пилильщика Nematus (Pteronidea) ribesii Scop, и у
медоносной пчелы (Нельсон — Nelson, 1915) делает Шафик (1954).
Нельсон и Шафик делят все эмбриональное развитие на три периода:
I. От откладки яйца до образования оболочек и зародышевых лист-
ков (что соответствует первым пяти стадиям развития Pontania);
9
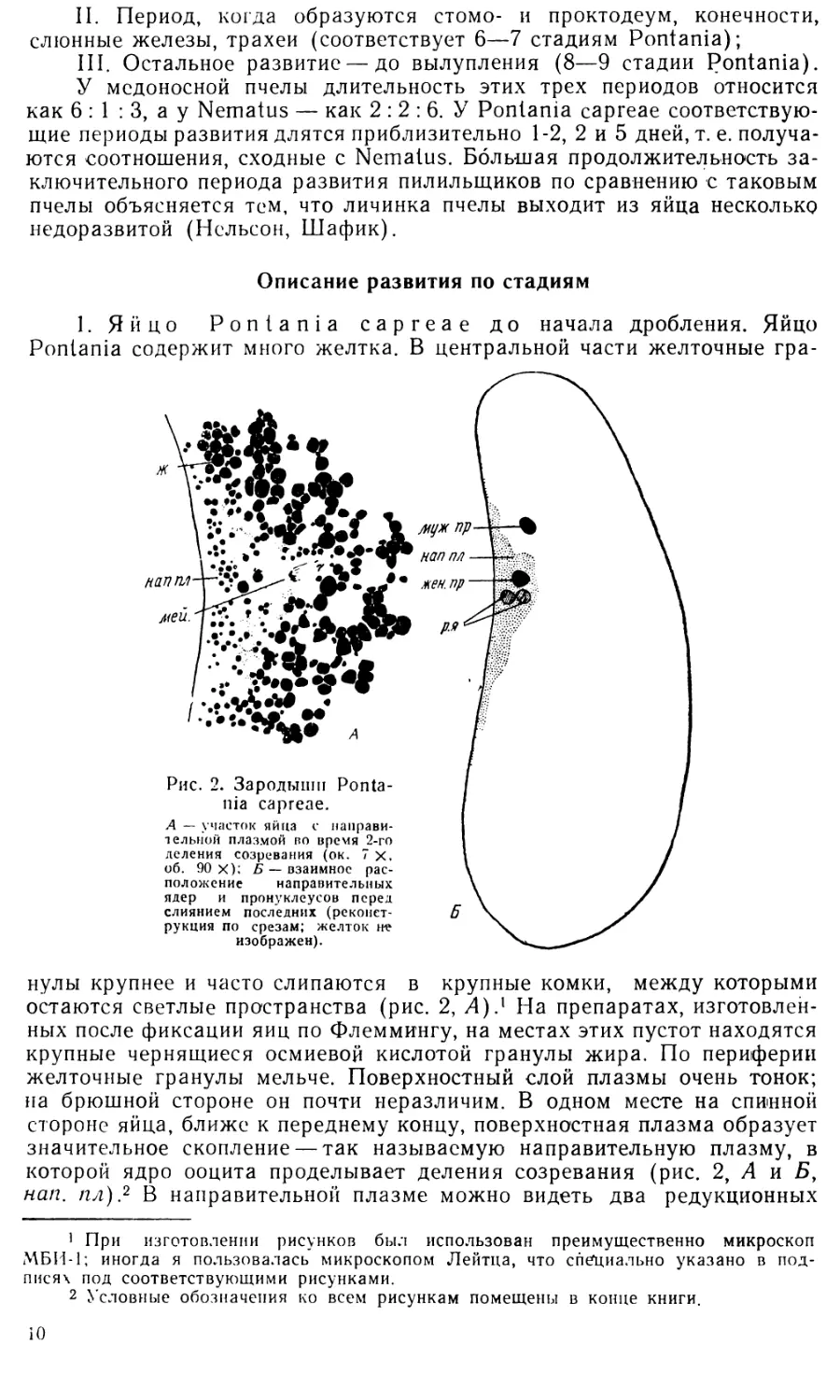
ядра (рис. 2, Б, р. я). Наблюдать три редукционных ядра мне ни разу
не случалось.
Мужской пронуклеус удалось обнаружить не на всех препаратах,
что может быть поставлено в связь с нередко наблюдающимся у пилиль-
щиков партеногенезом. Мужской пронуклеус, когда он есть, лежит в
желтке между направительной плазмой и передним концом яйца (рис. 2,
Б, муж. пр.).
На некоторых препаратах оба пронуклеуса лежат очень близко друг
к другу в более глубокой части яйца, готовые к слиянию.
Ознакомившись с внешней формой и распределением веществ в
яйце Pontania, уместно поставить вопрос о полярности яйца. Анимально-
вегетативная полярность яйца — одно из основных проморфологических
понятий эмбриологии, без которого невозможно сравнение яиц различ-
ных животных. Тем не менее, когда речь идет о насекомых, эти понятия
забываются. В учебниках эмбриологии К. Н. Давыдова (1914, 1928),
Мэк Брайд (Mac Bride, 1914), Коршельта и Гейдера (Korschelt и Heider,
1936), П. П. Иванова (1937, 1945) и Г. А. Шмидта (1951, 1953) в главе
о насекомых термины «анимальный» и «вегетативный» даже не упоми-
наются. То же относится к специально посвященным развитию насеко-
мых сводкам Ричардса и Миллера (Richards и Miller, 1937), Иогансена
и Бата (Johannsen и Butt, 1941), Пфлюгфельдера (Pflugfelder, 1952),
а также к «Общей энтомологии» Б. Н. Шванвича (1949) и к «Grundrifi
der Insektenkuride» Вебера (Weber, 1954), где эмбриология рассматри-
вается очень обстоятельно. Обычно просто подчеркивают, что яйца
многих насекомых имеют билатерально-симметричную форму, так что
в них с самого начала различаются концы — передний и задний, сторо-
ны— спинная, брюшная, правая и левая. Все эти обозначения прини-
маются в зависимости от того, какое положение занимает в яйце поздний
зародыш, незадолго до вылупления. Возможность различать еще до
начала эмбрионального развития передний и задний концы, правую и
левую сторону и т. д. представляет большой интерес с точки зрения де-
терминации в яйце частей будущего животного, но при этом надо пом-
нить, что действительное положение зародыша насекомого в яйце по
ходу развития иногда очень сильно изменяется в связи с бластокинезом.
Следует ли из этого, что в яйцах насекомых отсутствует полярность,
присущая яйцам большинства животных? А если анимально-вегетатив-
ная ось у насекомых все-таки существует, то каково ее отношение к
передне-задней оси зародыша?
Яйцо Pontania очень удобно для разрешения этих вопросов, так
как обладает отчетливой билатеральной симметрией, а положение за-
родыша в яйце по ходу развития почти не меняется (бластокинез здесь
сводится к удлинению и укорочению зародышевой полоски). Это очень
упрощает дело.
Как известно, полюсы яйца характеризуются следующими призна-
ками:
1) На анимальном полюсе происходят деления созревания и обра-
зуются направительные тельца.
2) При неравномерном распределении желтка он концентрируется
в вегетативном полушарии.
3) В процессе дальнейшего развития клетки анимального полуша-
рия бластулы дают эктодерму, а клетки вегетативного полушария —
энтодерму и мезодерму.
4) Процессы гаструляции (инвагинация или иммиграция) сосредо-
точены на вегетативном полюсе. При этом клетки анимального полуша-
рия в большей или меньшей степени перемещаются в сторону вегетатив-
11
ного полюса, становясь на место погрузившихся внутрь клеток. Это
перемещение клеточного материала особенно заметно и значительно
при эпиболической гаструляции, когда клетки энто- и мезодермы
кажутся относительно неподвижными, а клетки эктодермы активно об-
растают их с поверхности.
5) Полярность яиц проявляется еще в наличии физиологического
градиента, но так как никаких определенных данных, характеризующих
яйцо Pontania (и насекомых вообще) с этой точки зрения, нет, то к этому
признаку полярности мы больше возвращаться не будем.
Из приведенных выше описаний уже известно, что на спинной сто-
роне яйца, несколько ближе к переднему концу, находится скопление
активной цитоплазмы (направительная плазма), вокруг которого жел-
ток располагается не так плотно. В этой плазме происходят деления
созревания и остаются лежать редукционные ядра после того, как жен-
ский пронуклеус мигрирует в более глубокие части'яйца. Таким образом,
по первым двум признакам (положение редукционных телец и распре-
деление желтка) надо считать, что анимальному полюсу в яйце Ponta-
nia соответствует направительная плазма, лежащая на спинной стороне.
Что касается пунктов 3 и 4, приведенных выше, касающихся уже
начальных процессов развития, которые будут изложены более подроб-
но ниже, то, забегая несколько вперед, можно отметить только, что при
формировании зародышевой полоски у Pontania, происходит характер-
ное перемещение клеток от спинной стороны к брюшной, а процессы
гаструляции сосредоточены, как и у большинства насекомых вдоль
медио-вентральной линии. Все это говорит за положение анимального
полюса на-спинной стороне яйца, а вегетативного — на брюшной.
В пользу такой трактовки свидетельствует и судьба различных частей
бластодермы: большая спинная (анимальная) часть бластодермы
дает зародышевую и внезародышевую эктодерму, а брюшная (вегета-
тивная) часть—мезодерму и дефинитивную энтодерму.
Из этого можно сделать вывод, что анимально-вегетативная ось
в яйце Pontania соответствует спино-брюшной оси и проходит под пря-
мым углом к главной (передне-задней) оси тела будущего насекомого
(Иванова-Казас, 1957). Такое соотношение осей возникло из более при-
митивных отношений многощетинковых червей вследствие выпадения
стадии трохофоры. Аналогичные изменения в архитектонике яйца про-
изошли по тем же причинам и у некоторых других групп (олигохеты,.
пиявки).
Таким образом, на примере Pontania мы видим, что первичная по-
лярность яйца сохраняется у насекомых со всеми своими признаками,
но она в значительной степени замаскирована вторично приобретенной
билатеральной симметрией, которая возникла как результат «установки
развития» (термин П. П. Иванова, 1937, 1945) не на трохофору, а на
билатерально-симметричное животное.
2. Дробление. Дробящиеся яйца были представлены лишь
единичными препаратами; по-видимому, дробление протекает у Ponta-
nia очень быстро. Однако, можно было видеть, что дробление Pontania
типично поверхностное, что вполне соответствует крупным размерам
яиц и большому количеству в них желтка.
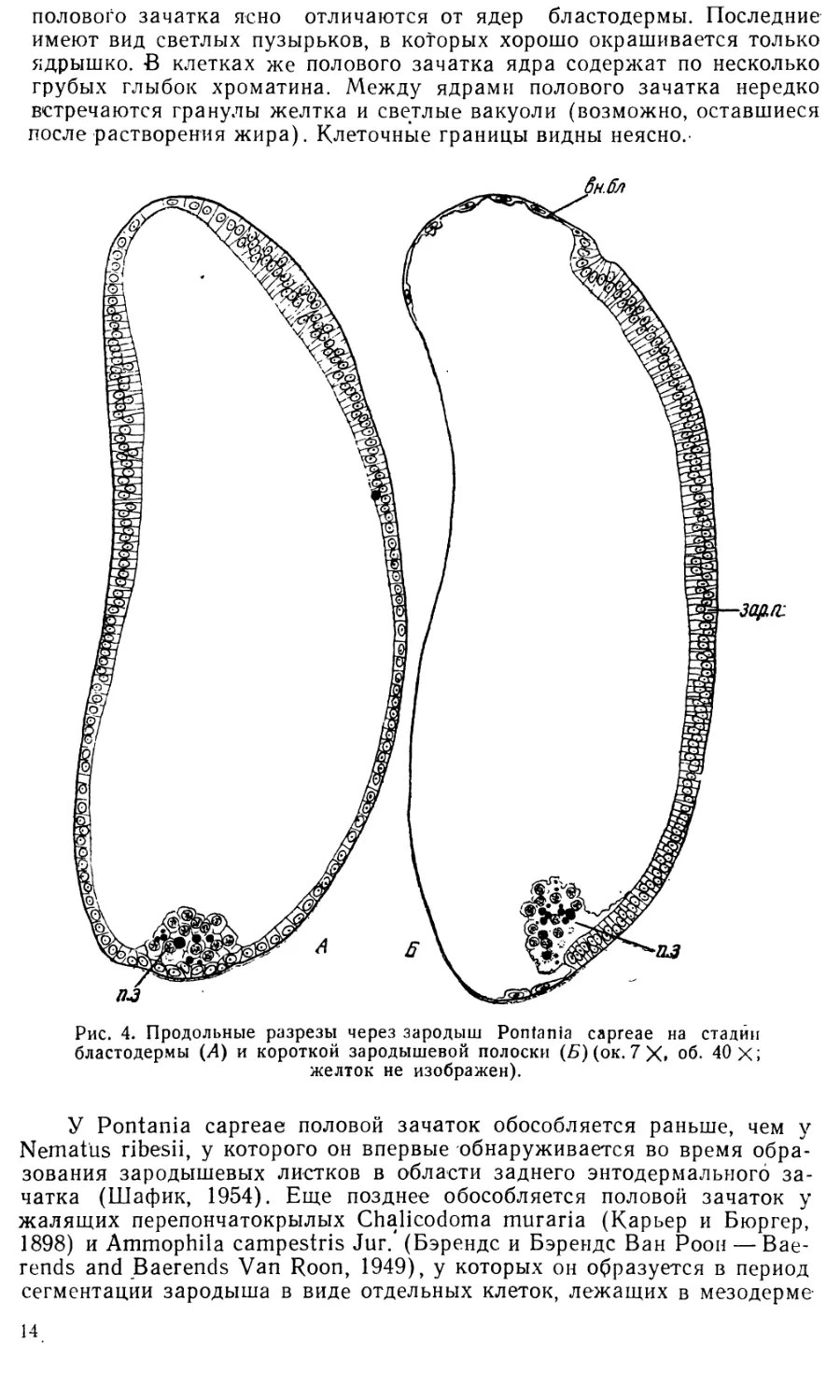
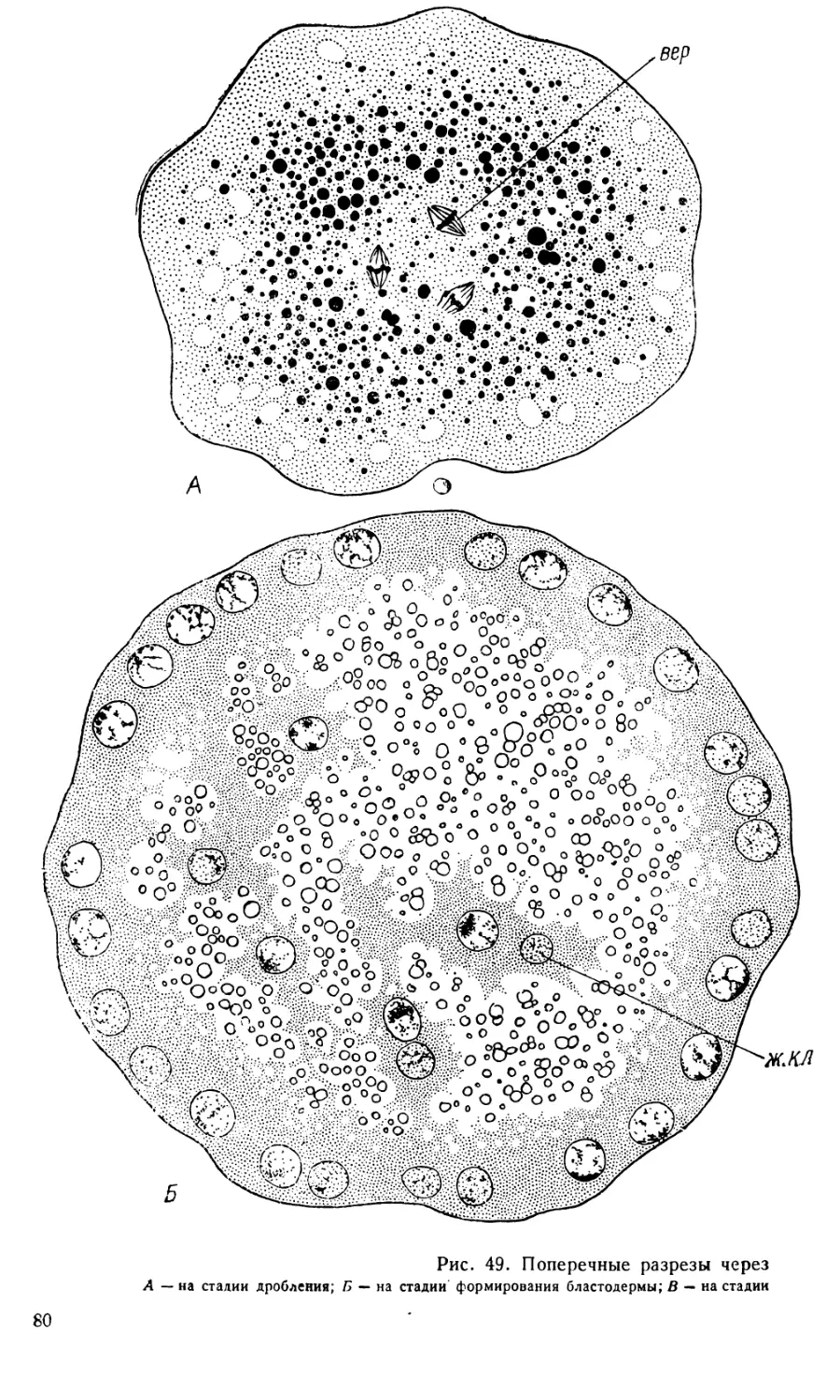
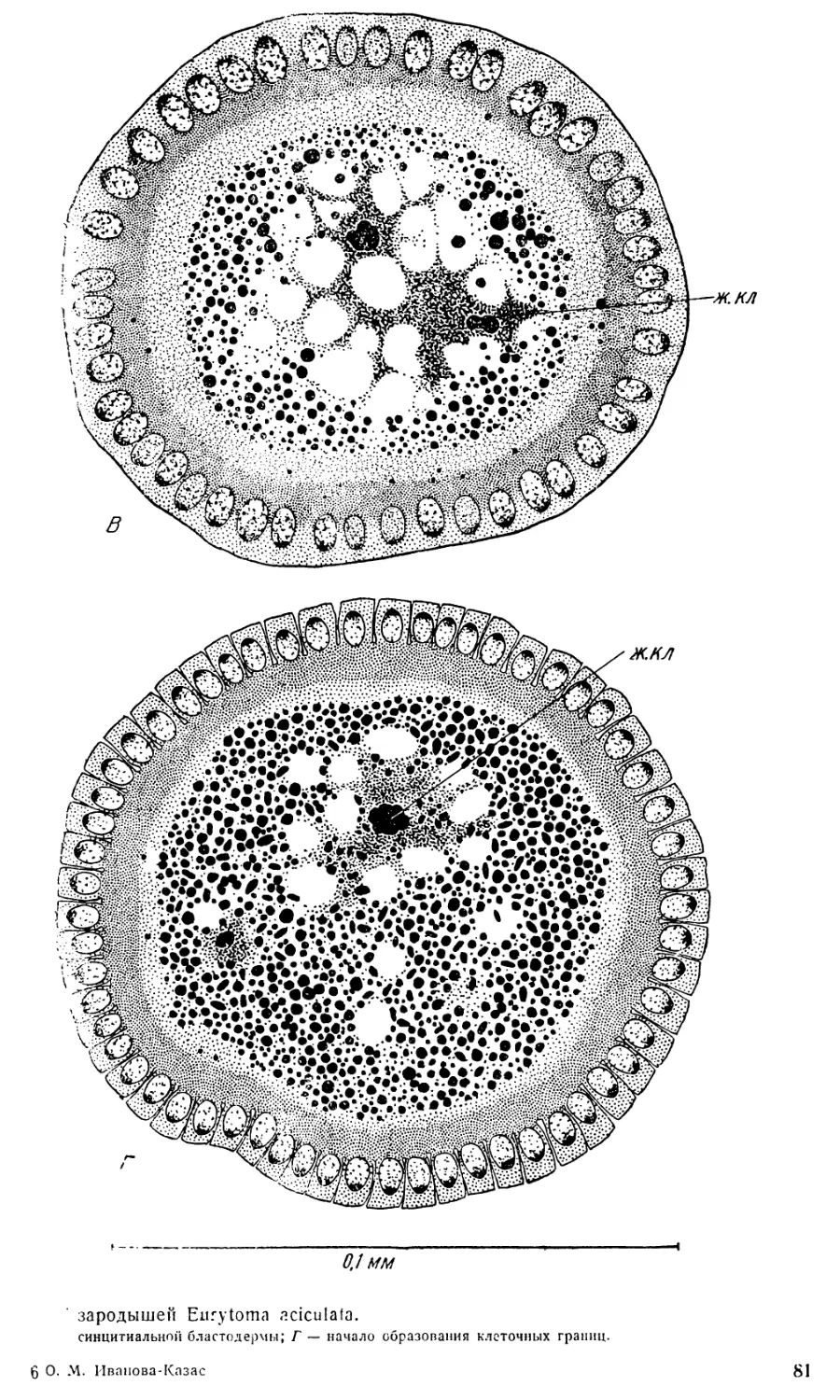
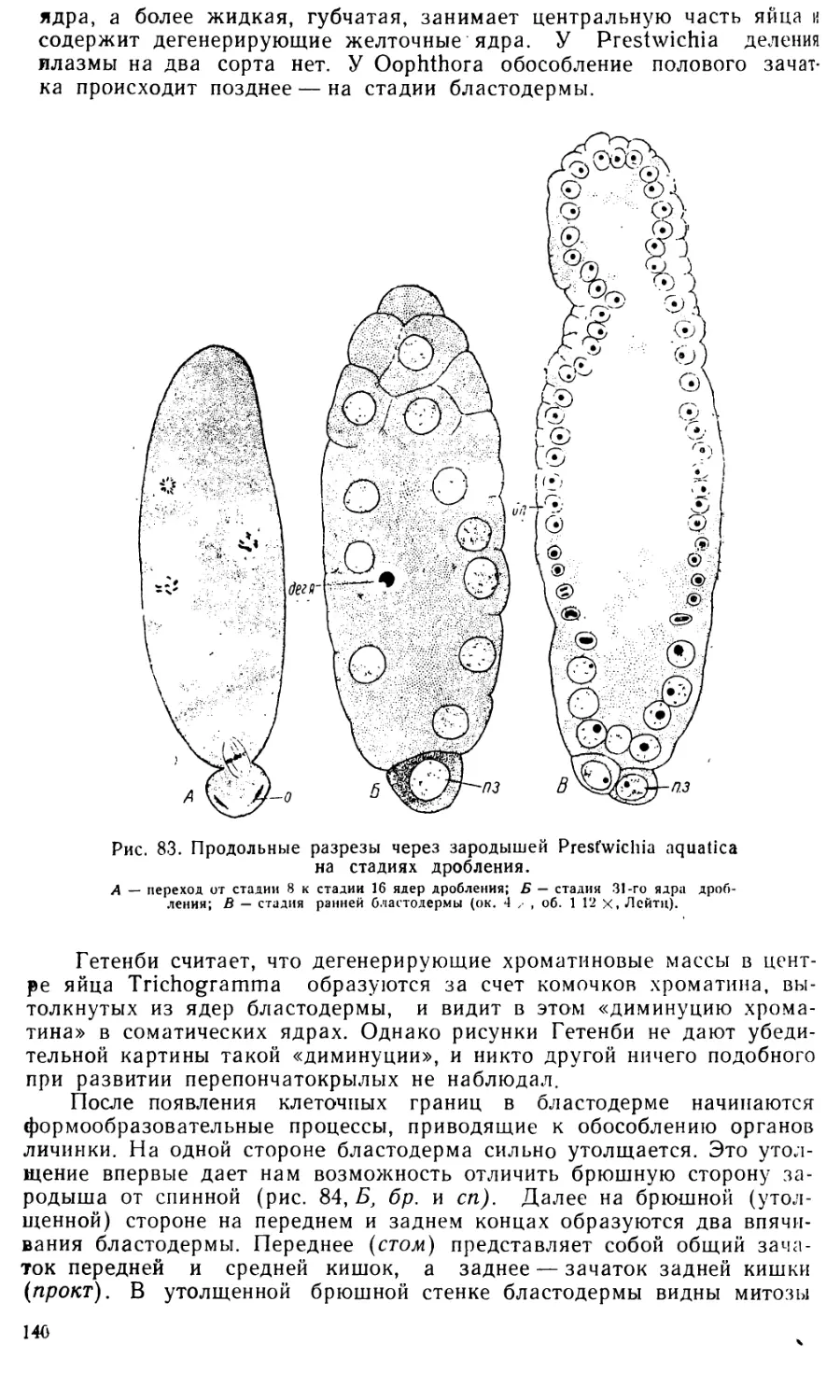
3. Стадия бластодермы. Сразу же после выхода ядер
в поверхностную бластему бластодерма представляет собой довольно
тонкий слой цитоплазмы, в которой разбросаны сравнительно крупные
светлые ядра. Клеточные границы еще не видны. Часть ядер дробления
остается в желтке, превращаясь в так называемые первичные желточные
ядра. Каждое такое ядро с примыкающим к нему участком плазмы обра-
12
3-го, 4-го, и 5-го сегментов брюшка. У паразитических перепончатокры-
лых, наоборот, известны случаи очень раннего обособления полового
зачатка еще на начальных стадиях дробления (см. ниже).
Вскоре становится заметным и перемещение клеточного материала
со спинной стороны на брюшную. Клетки внезародышевой бластодермы
начинают растягиваться и уплощаться, почему весь этот участок бласто-
дермы покрывает теперь гораздо большую поверхность яйца, чем вна-
чале. Зародышевая бластодерма, наоборот, начинает стягиваться к
брюшной стороне, так что обе слегка утолщенные боковые пластинки
яйца смыкаются в одну зародышевую полоску (рис. 4, Б и 5, зар. п.).
Более низкие медианные клетки теперь уже не различаются; по-видимо-
му, они уподобляются боковым.
Таким образом, зародышевая полоска закладывается у Pontania
в виде двух боковых утолщений бластодермы, которые лишь позднее
соединяются на брюшной стороне. Вебер (1954) и Кюн (Kuhn, 1955)
считают, что парная закладка зародышевой полоски у насекомых —
примитивный признак. Действительно, близкие явления наблюдаются у
ряда сравнительно низкоорганизованных насекомых: у Tachycines и
Gryllus (Orthopteroidea) сгущение клеток бластодермы происходит
сперва на спинной стороне, потом клеточный материал разделяется на
две группы, которые перемещаются на брюшную сторону, где и образуют
зародышевую полоску; у Plalycnemis (Odonata) и Notonecta (Hemiptera)
образование зародышевой полоски происходит сходно с Pontania из
двух «Vorkeimanlage» (Краузе — Krause, 1939).
4. Стадия короткой зародышевой полоски. Соеди-
нение двух утолщений бластодермы в одно непарное брюшное происхо-
дит сперва в задней части зародыша и несколько запаздывает спереди.
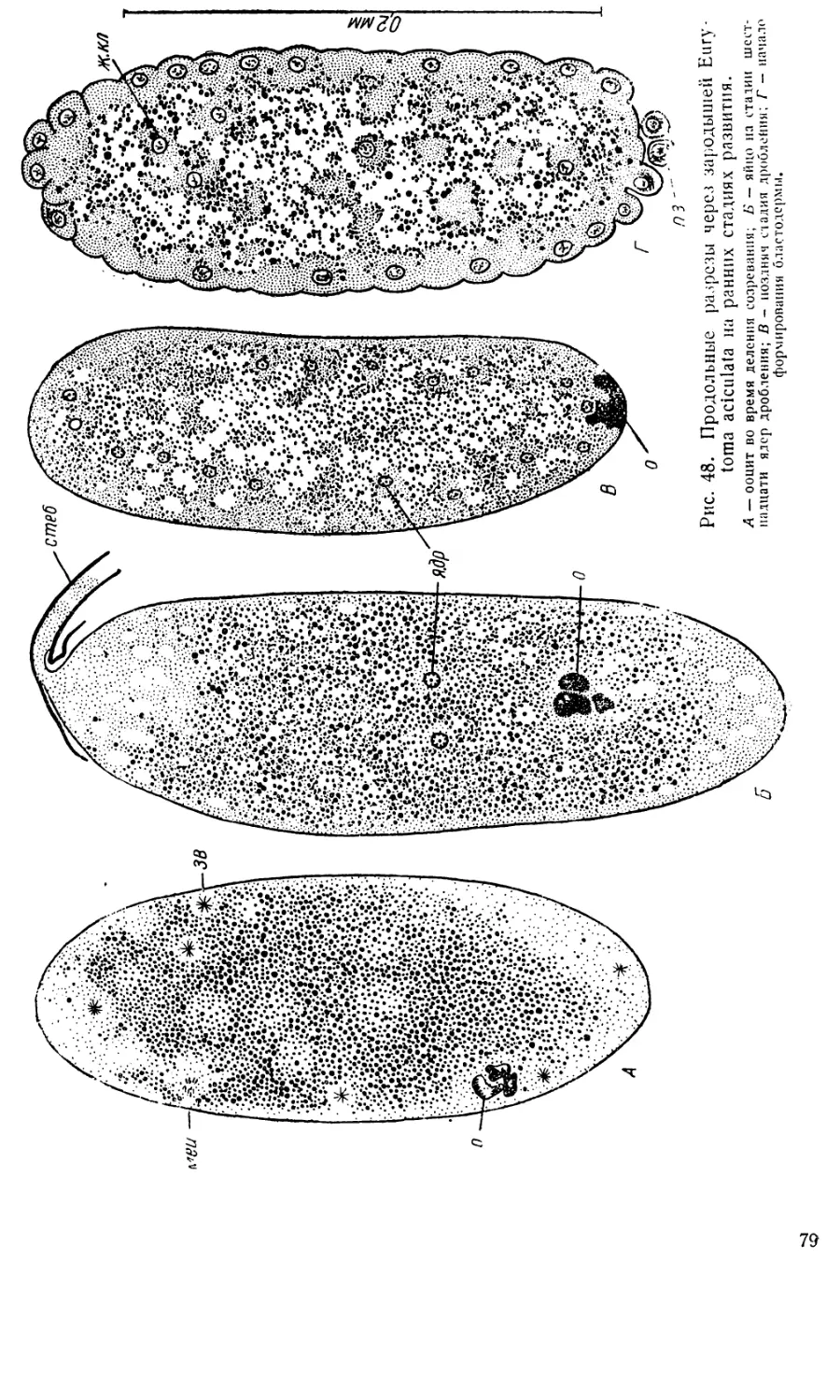
На рис. 5 представлено четыре поперечных разреза через зародыши
Pontania на стадии ранней зародышевой полоски. На первом срезе
(рис. 5, А) еще отчетливо различаются два утолщения зародышевой
бластодермы (возможно, это зачатки головных лопастей). Граница
между зародышевой и внезародышевой бластодермой здесь очень от-
четлива; в этом месте бластодерма образует небольшой излом, кото-
рый можно считать зачатком амниотической складки.
В средней части яйца (рис. 5, Б) зародышевая полоска имеет при-
близительно одинаковую толщину по всей ширине и лишь немного утон-
чается к краям.
Ближе к заднему концу (рис. 5, В) в медианной части зародышевой
полоски ясно намечается некоторое утолщение, словно две надвигаю-
щиеся с боков клеточные массы столкнулись здесь и устремились
внутрь. Это медианное утолщение означает начало образования мезо-
дермы.
На последнем срезе (рис. 5, Г) представлен задний конец зароды-
шевой полоски, подле которого лежит половой зачаток (п. з). В момент
своего обособления половой зачаток расположен терминально на зад-
нем конце яйца, но затем он смещается на брюшную сторону, что
хорошо видно на рис. 4. Это показывает, что в процессе формирования
зародышевой полоски происходит стягивание клеточного материала со
спинной стороны на брюшную не только через бока зародыша, но и
через его задний конец, хотя и в меньшей степени.
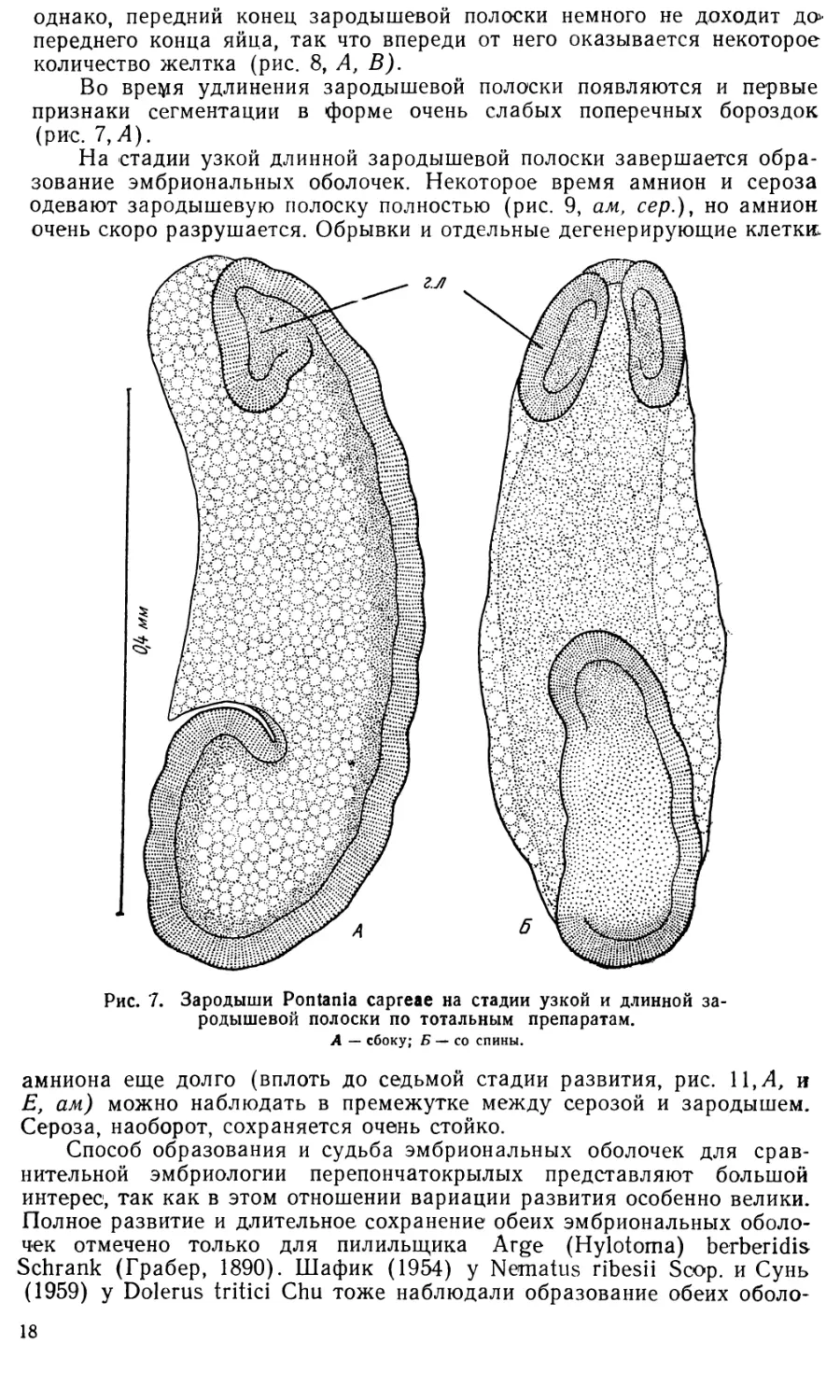
5. Стадия длинной узкой зародышевой полоски.
Вскоре после образования зародышевой полоски начинается и развитие
эмбриональных оболочек. Спереди внезаррдышевая бластодерма образу-
ет полулунную амниотическую складку, надвигающуюся на зародыше-
вую полоску (рис. 6, В, ам. ск.). Несколько позднее такая же складка
15
зачатки грудных ног (гр. н).Все эти зачатки конечностей довольно ши-
роко расставлены и обращены своими вершинами в стороны и назад.
Такой же сегментарный состав головы принимается Шафиком
(1954) для Nematus ribesii Scop., хотя у этого пилильщика интеркалярныи
сегмент не выражен. А у Chalicodoma muraria на переднем конце заро-
дышевой полоски наблюдаются многочисленные складки, которые Карь-
ер и Бюргер (1898) сочли за сегменты. Таким образом, в предчелю-
стной части они различали 4 сегмента: сегмент верхней губы, мозговой,
антеннальный и предчелюстной (интеркалярныи). Однако надо отме-
тить, что у Pontania саргеае ничего похожего не наблюдается, хотя
сегментация зародышевой полоски очень ясная.
На рассматриваемой стадии развития процесс образования мезо-
дермы уже завершен, но медианная бороздка еще сохраняется. По сто-
ронам от нее эктодерма утолщается и образует два нервных валика,
в которых происходит расслоение на более тонкий поверхностный дер-
матогенный слой и более толстый внутренний неврогенный слой, состоя-
щий из крупных невробластов с базофильной плазмой и светлым пузы-
ревидным ядром (рис. 10, невр).
В серозе {сер) все еще сохраняются гранулы желтка. Кнаружи
от нее появляется тонкая бесструктурная, плохо красящаяся перепонка,
сопровождающая все изгибы серозы. Это серозная кутикула (рис. 10,
сер. кут).
Амнион представлен небольшими обрывками и отдельными округ-
лившимися клетками, встречающимися кое-где между серозой и по-
верхностью тела зародыша (рис. 10, 11, А, Е, ам).
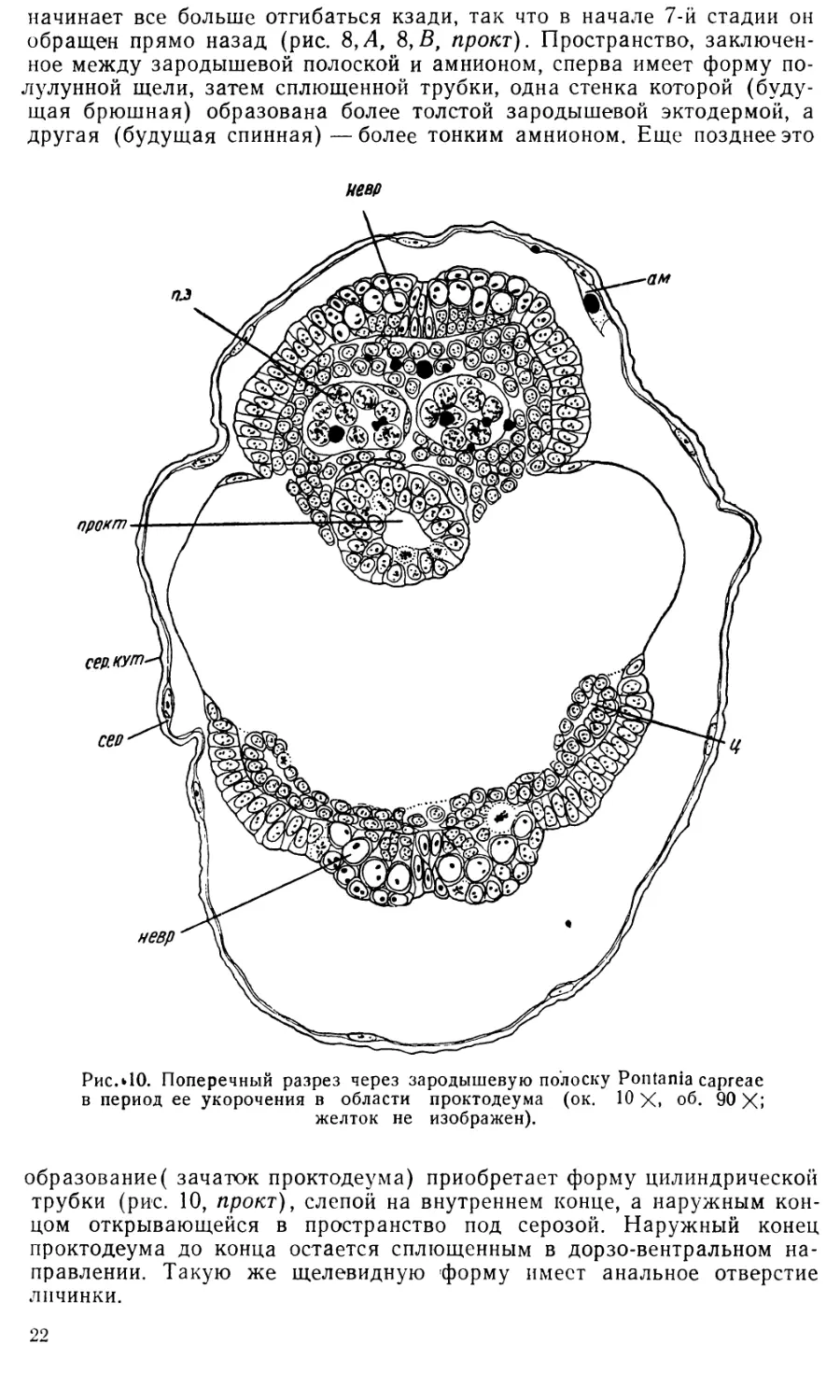
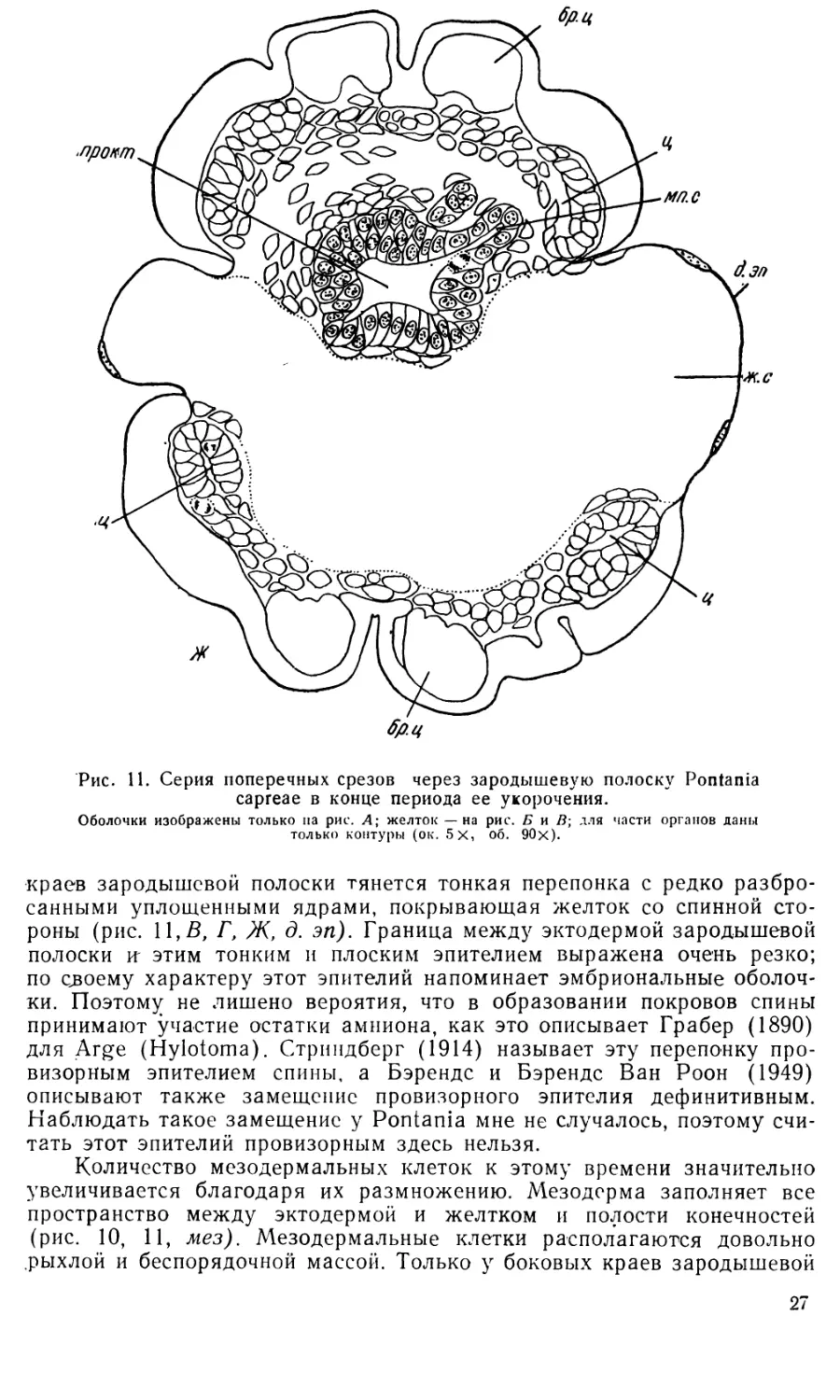
7. Укорочение зародышевой полоски. Эта стадия раз-
вития, внешне очень неясно отграниченная от предыдущей, характери-
зуется закладкой целого ряда новых систем органов и быстрым разви-
тием уже имеющихся.
Зачаток верхней губы приобретает более резкие очертания. На
некоторых препаратах он выглядит слегка раздвоенным (рис. 8,Б, в. г.).
У его основания между зачатками антенн образуется небольшое впячи-
вание стомодеума, которое вскоре превращается в цилиндрическую труб-
ку, растущую своим слепым концом назад (рис. 8, £, 11, Л, 12, стом).
Зачатки антенн остаются маленькими, а зачатки головных и груд-
ных конечностей значительно увеличиваются в размерах. Форма их
усложняется, а вершины их обращены теперь к медианной линии. Зачат-
ки грудных ног, которые становятся особенно длинными, отгибаются
назад (рис. 8, В, 12, Л).
По мере укорочения зародышевой полоски ее задний конец все
больше приближается к заднему концу яйца (рис. 12,Л). Удлинение
зародышевой полоски и последующее ее укорочение наблюдаются у мно-
гих насекомых и являются, вместе с образованием зародышевых оболо-
чек, проявлениями бластокинеза. Других явлений, сопровождающих
бластокинез (например, погружения зародышевой полоски в желток),
у Pontania и вообще у перепончатокрылых не наблюдается.
Одновременно с укорочением зародышевой полоски на ее заднем
конце происходит формирование проктодеума. После разрушения ам-
ниона у заднего конца зародышевой полоски сохраняется небольшой
его участок, довольно толстый в непосредственной близости от зароды-
шевой полоски и постепенно утончающийся по мере удаления от нее
(рис. 7, А, 8, А, 8, В). Складка, образованная эктодермой зародышевой
полоски и загибающимся на нее амнионом, на 5-й стадии развития на-
правлена своим гребнем внутрь яйца под прямым углом к его продоль-
ной оси (рис. 7,Л). Но уже на 6-й стадии развития гребень этой складки
21
Таким образом, небольшой участок амниона, сохраняющийся у зад-
него конца зародышевой полоски, несомненно принимает участие в по-
строении дорзальной стенки проктодеума.
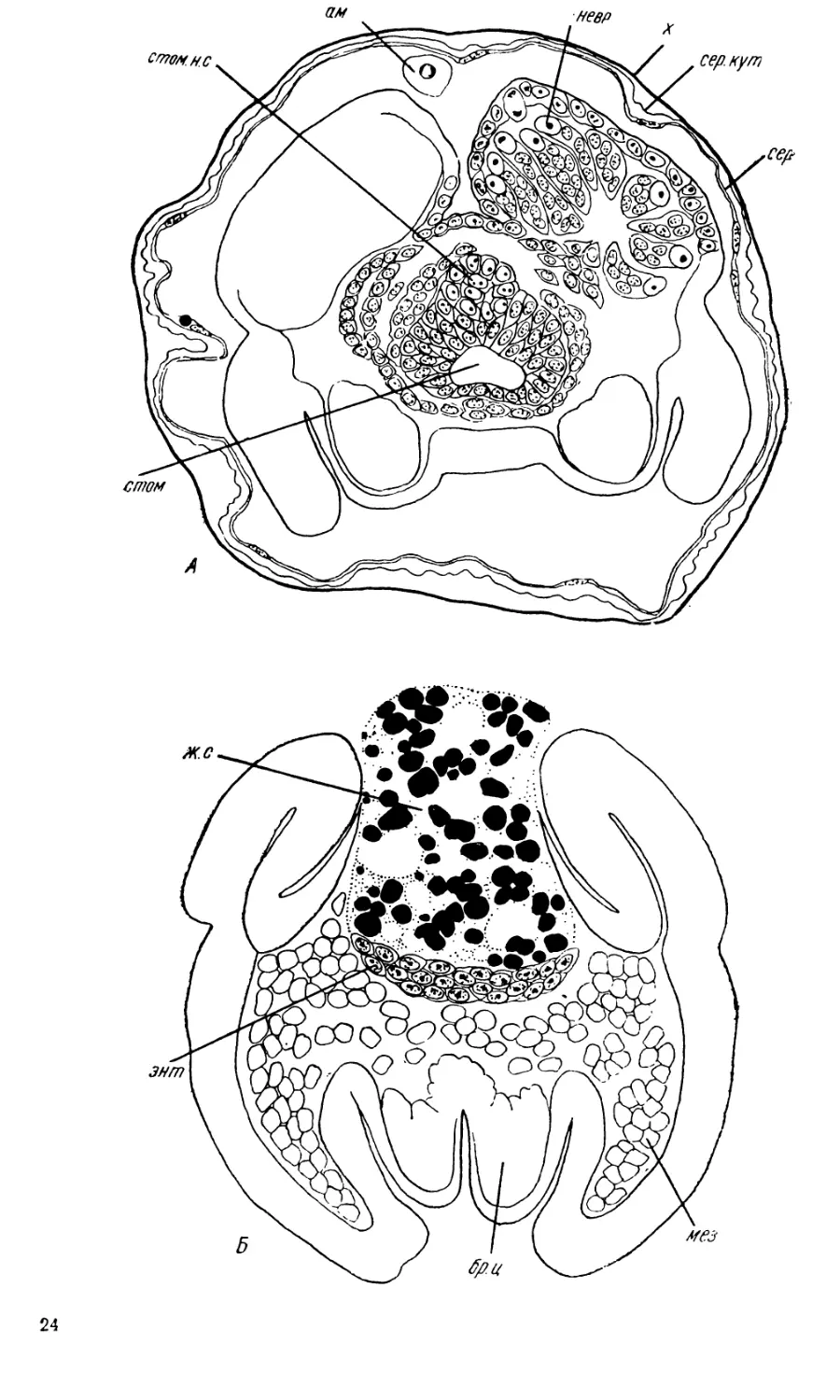
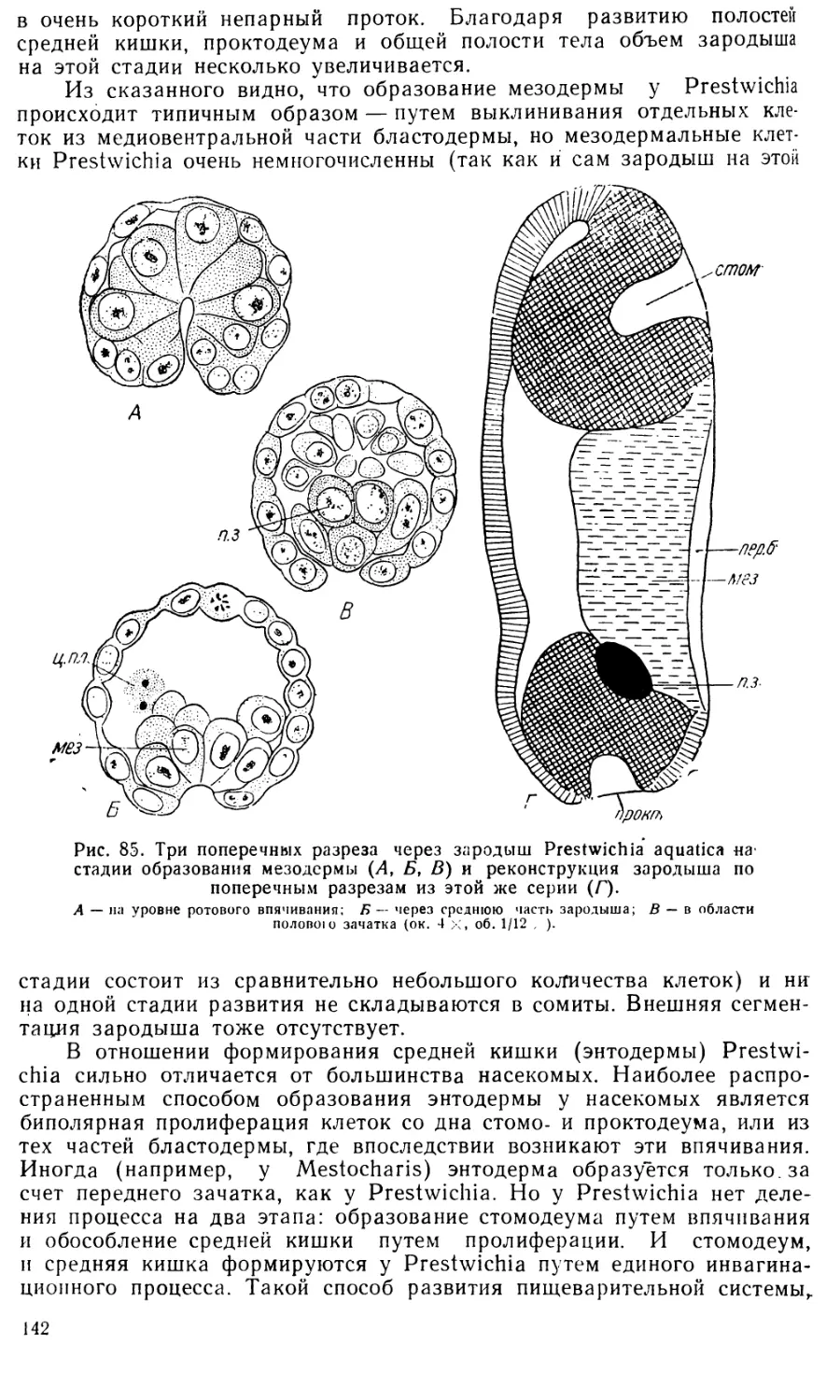
В стенках проктодеума происходит усиленное размножение клеток,
и проктодеум быстро удлиняется. На дне его появляются четыре пары
более тонких цилиндрических выпячиваний — зачатки мальпигиевых
сосудов (рис. 11,Ж, мп. с).
На этой стадии развития (довольно поздно) впервые появляются
зачатки средней кишки (дефинитивная энтодерма). Подле внутреннего
конца стомодеума лежит скопление клеток, налегающих на желточную
массу в виде дугообразного зачатка, концы которого тянутся по бокам
желтка назад (рис. 11,£, 11,В, энт). Боковые энтодермальные тяжи,
сперва довольно короткие, плотные и узкие, затем распластываются на
поверхности желточной массы в виде однослойных эпителиальных пла-
стинок, все дальше распространяющихся назад (рис. 11, В, энт). Такой
же дугообразный энтодермальный зачаток образуется и в области прок-
тодеума, но выражен он гораздо менее ясно. Позднее оба зачатка
смыкаются, образуя среднюю кишку. У Pontania capreae обособление
зародышевых листков происходит самым распространенным у насеко-
мых способом: мезодерма образуется путем выселения клеток со дна
первичной бороздки, а энтодерма, по-видимому, путем пролиферации
клеток на внутренних концах стомо- и проктодеума. Сходные простран-
ственные отношения между зародышевыми листками имеются у Nematus
ribesii, но Шафик (1954) склонен считать, что энтодерма и мезодерма
образуются путем разделения «внутреннего зародышевого листка», т. е.
из клеток, иммигрировавших из первичной бороздки.
На 7-й стадии развития объем желточной массы значительно сокра-
щается. Всякие следы раздробления желтка на клетки исчезают и снова
образуется желточный синцитий (рис. II, Б, В, ж. с). Количество гра-
нул желтка становится меньше, между ними появляются большие вакуоли
Из-за этих вакуолей плазма желточного синцития имеет губчатое строе-
ние; в перемычках между вакуолями и лежат ядра и гранулы желтка.
Снаружи желточный синцитий отчетливо кон гурирован. Он представляет
собой не просто скопление запасных питательных веществ и даже не
комплекс клеток, перегруженных этими веществами, а единую систему.
Желточный синцитий служит той основой, на которой происходит фор-
мирование средней кишки.
В литературе нередко встречаются указания на то, что желточные
клетки скопляются у поверхности желточной массы и образуют нечто
вроде провизорного эпителия средней кишки, поверх которого образуется
дефинитивный эпителий. Это отмечено для Chalicodoma muraria F.
Карьером и Бюргером(1898), для Vespa vulgaris L. Стриндбергом(1914),
и для Scolia quadripunctata F. Дондуа (1953). У Ammophila campestris
Jur., no наблюдениям Бэрендса и Бэрендс Ван Роон (1949), желточные
клетки концентрируются в непосредственной близости от мест, где про-
исходят усиленные формообразовательные процессы, т. е. под зароды-
шевой полоской и подле энтодермальных тяжей. Ничего подобного
у Pontania не наблюдается., rjo-видимому, образование «провизорного
эпителия средней кишки» у жалящих перепончатокрылых связано с уси-
лением функции вителлофагов, как клеток, которые перерабатывают
желток и через посредство которых происходит питание тканей зароды-
ша. Это скорее результат специализации желточных клеток, чем сохра-
нение ими первичной способности образовывать стенки кишечника, как
это имеет место у щетинохвосток и стрекоз.
Уже на этой стадии начинается обрастание желтка: от боковых
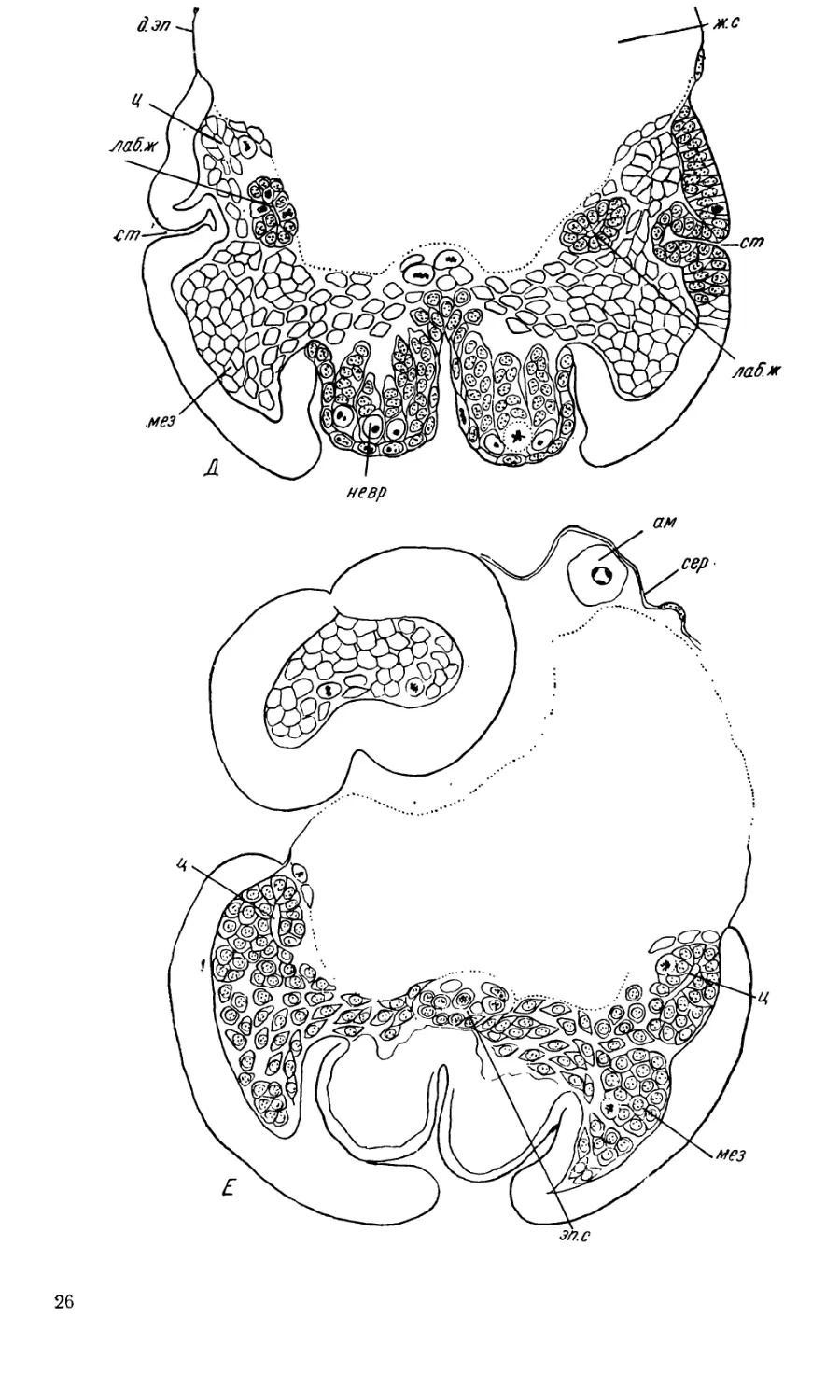
23
полоски мезодермальные клетки приобретают более правильное распо-
ложение. Здесь они складываются в пласт, латеральный край которого
заворачивается внутрь (рис. 10, 11,Г, Д, Е, Ж, 3, ц). В результате по-
бокам зародыша образуются две мезодермальные трубки, не вполне
замкнутые с медианной стороны, полость которых надо признать цело-
мом. Подобные же мезодермальные трубки описаны Карьером и Бюр-
гером (1898) для Chalicodoma, Нельсоном (1915)—для Apis, Бэренд-
сом и Бэрендс Ван Роон (1949) —для Ammophila, Шафиком (1954) —
для Nematus. Но разделения этих мезодермальных трубок на сегменты
в большинстве случаев (в том числе и у Pontania) не происходит.
В медианной части зародышевой полоски между желтком и зачат-
ком нервной системы обозначается трубковидное пространство, тяну-
щееся через все туловище (рис. 11, Е эп. с). Мезодермальные клетки,
лежащие в этом пространстве, отличаются округлыми очертаниями;
некоторые из них значительно крупнее остальных. Это эпиневральный
синус, внутри которого начинается дифференциация кровяных клеток.
Зачатки нервной системы на 7-й стадии значительно продвигаются
в своем развитии. Каждый невробласт (рис. 11, Л, Д, невр) отделяет от
себя внутрь тяж более мелких клеток, ядра которых отличаются от ядер
невробластов меньшими размерами, менее правильной формой и присут-
ствием нескольких хроматиновых зернышек. Границы клеток в этих
тяжах не видны; возможно их нет.
Все эти гистологические процессы происходят очень сходно в го-
ловных лопастях и нервных валиках. В последних невробласты распо-
лагаются сегментарными группами, вследствие чего образуется 16 пар
ганглиев брюшной цепочки. Кроме этих зачатков центральной нервной,
системы, на 7-й стадии развития появляется и зачаток стоматогастри-
ческой системы. Он представлен группой сходных с невробластами кле-
ток, отделяющейся от дорзальной стенки стомодеума (рис. 11, Л,.
стоя. н. с).
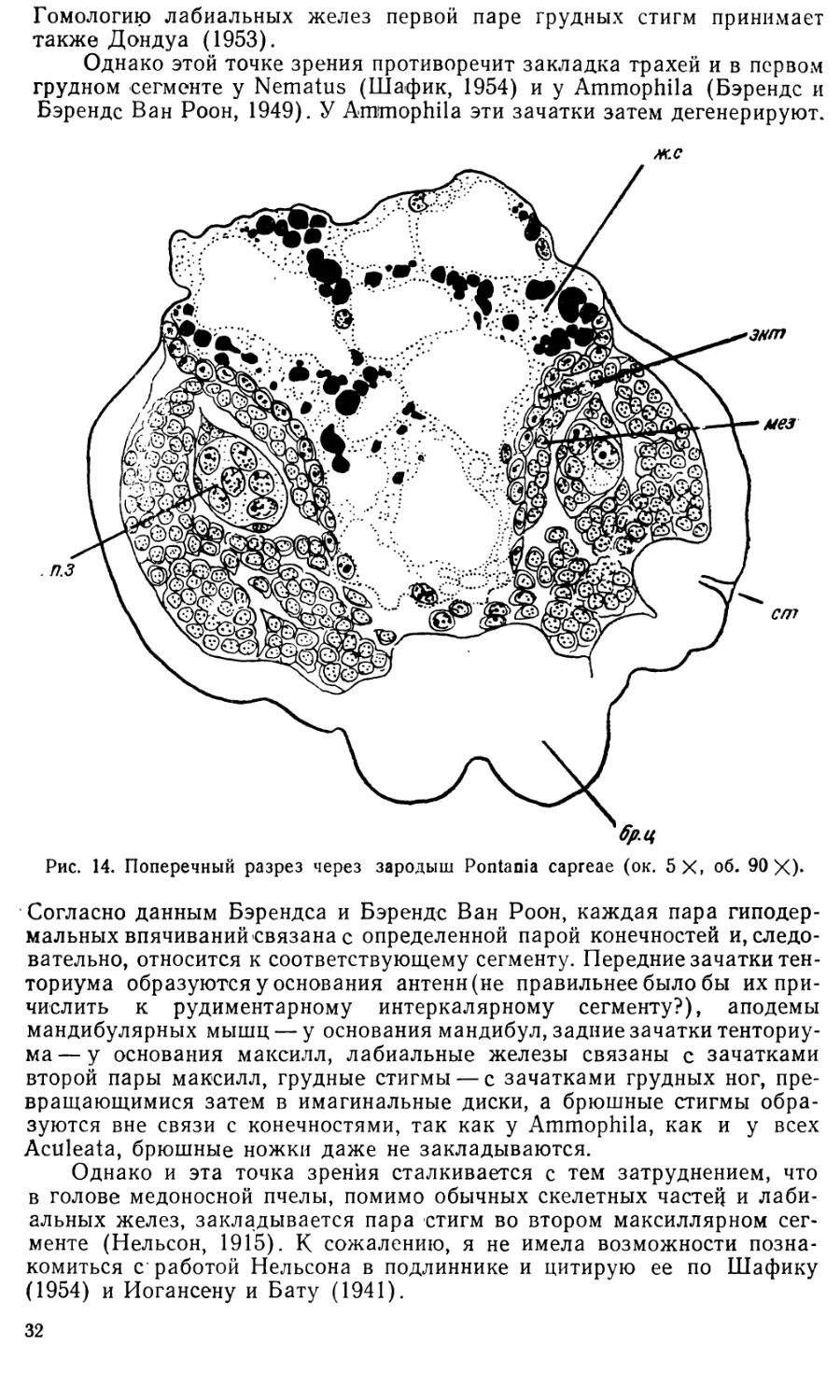
В то же время в эктодерме появляются зачатки трахей и лабиаль-
ных желез. Трахеи закладываются в количестве 10 пар в виде неглубо-
ких впячиваний эктодермы, со дна которых, сразу же, начинают расти
более тонкие трубочки (рис. 11, Г, тр, Д ст). На тотальных препаратах
на этой стадии зачатки трахей не видны, а по срезам точно установить
принадлежность каждого зачатка к определенному сегменту трудно.
Позднее (на 8-й и 9-й стадиях) становится видно, что первая пара стигм
лежит на границе первого и второго грудных сегментов, вторая пара —
в третьем грудном сегменте, а остальные—в восьми первых сегментах
брюшка (рис. 15,Л, ст). Впоследствии первая пара стигм целиком пере-
мещается в первый грудной сегмент, а вторая пара располагается между
вторым и третьим сегментом. Остальные стигмы сохраняют свое перво-
начальное положение (рис. 15,Б, ст).
Одновременно происходит закладка лабиальных желез. Относитель-
но эмбрионального материала, из которого развиваются лабиальные же-
лезы насекомых, существует два разных мнения. Подавляющее большин-
ство исследователей принимает чисто эктодермальное происхождение
этих желез (Карьер и Бюргер, 1898; Бэрендс и Бэрендс Ван Роон, 1949;
Дондуа, 1953; Шафик, 1954; Бронскилл — Bronskill, 1959). При этом
Карьер и Бюргер и Бэрендс и Бэрендс Ван Роон считают трахеи, лаби-
альные железы и скелетные части головы сериальными гомологами —
все они развиваются из сегментарно расположенных трубчатых впячи-
Еаний эктодермы.
Согласно другой точке зрения, только наружные отделы — вывод-
ные протоки лабиальных желез имеют эктодермальное происхождение,.
28
а железистые отделы развиваются из мезодермы. Так, Пфлюгфельдер
(1934), исследовавший развитие лабиальных желез у Pontania salicis
Christ.' описывает этот процесс следующим образом. Медианные части
целомических пузырьков обособляются в виде плотных тяжей, которые
позднее принимают форму длинных трубок и вступают в связь с пар-
ными эктодермальными зачатками выводных протоков. Часть целоми-
ческой стенки, из которой образуются лабиальные железы, соответ-
ствует половым валикам других насекомых; по всей длине зародыша
они превращаются в лабиальные железы и только в области будущих
половых желез идут на построение гонад.
Мезодермальное происхождение слюнных желез установлено также
Бледовским и Краинской (Bledowski и Krainska, 1926) для Banchus
l'emoralis Thoms. (Ichneumonidae).
Признание мезодермалыгого происхождения лабиальных желез
насекомых дало бы возможность гомологизировать их с таковыми мно-
гоножек и считать видоизмененными метанефридиями. Наличие в лите-
ратуре таких значительных разногласий по вопросу о происхождении
лабиальных желез в процессе индивидуального развития и вытекающие
из них различия в трактовке морфологической природы этих желез
заставляют относиться к решению этого вопроса с большой осторож-
ностью.
У Pontania capreae в период укорочения зародышевой полоски у ос-
нования каждого зачатка второй пары максилл образуется небольшое
впячивание, которое сразу же разделяется на две ветви (рис. 11, Г,
лаб. ж). Одна из ветвей лежит ближе к середине и брюшной стороне,
другая — более дорзо-латерально. Последняя сильно отстает в своем
росте, так что на всех стадиях развития она оказывается короче первой.
Внутренние ветви лабиальных желез лежат на значительном расстоянии
•от целомических полостей, поэтому развитие их из стенки целома исклю-
чается. Нередко можно видеть их одновременно с целомическими труб-
ками на одном поперечном срезе (рис. 11, Г). Наружные ветви лабиаль-
ных желез подходят довольно близко к мезодермальным трубкам, но
непосредственного контакта между ними не наблюдается. И на более
поздних стадиях, когда целомические трубки распадаются на отдельные
клетки, я не наблюдала, чтобы при этом формировались плотные кле-
точные тяжи, становящиеся затем полыми и присоединяющиеся к за-
чаткам лабиальных желез. Вообще рост этих зачатков происходит очень
постепенно, и в них постоянно наблюдается много делящихся клеток,
так что нет никаких оснований предполагать, что в какой-то момент
развития к этим зачаткам присоединяется клеточный материал извне.
Итак, несмотря на систематическую близость Pontania capreae L.
и Pontania salicis Christ, получить достоверные данные, подтверждаю-
щие наблюдения Пфлюгфельдера и указывающие на участие мезодер-
мы в построении лабиальных желез, мне не удалось. Общее впечатле-
ние таково, что лабиальные железы Pontania capreae развиваются це-
ликом из эктодермы.
Половой зачаток, оказавшийся после образования зародышевой
полоски под ее задним концом, проделывает вместе с ним все переме-
щения. На стадии длинной зародышевой полоски он лежит в самом
последнем сегменте подле формирующегося зачатка проктодеума. По-
ловой зачаток затем начинает продвигаться в сегменты, лежащие впе-
реди. В момент вылупления личинки он лежит в 5-м сегменте брюшка
и разделяется на две группы клеток (рис. 10, п. з.), которые к концу 7-й
стадии становятся компактными и одеваются тонкой оболочкой из рас-
пластанных на их поверхности мезодермальных клеток.
29
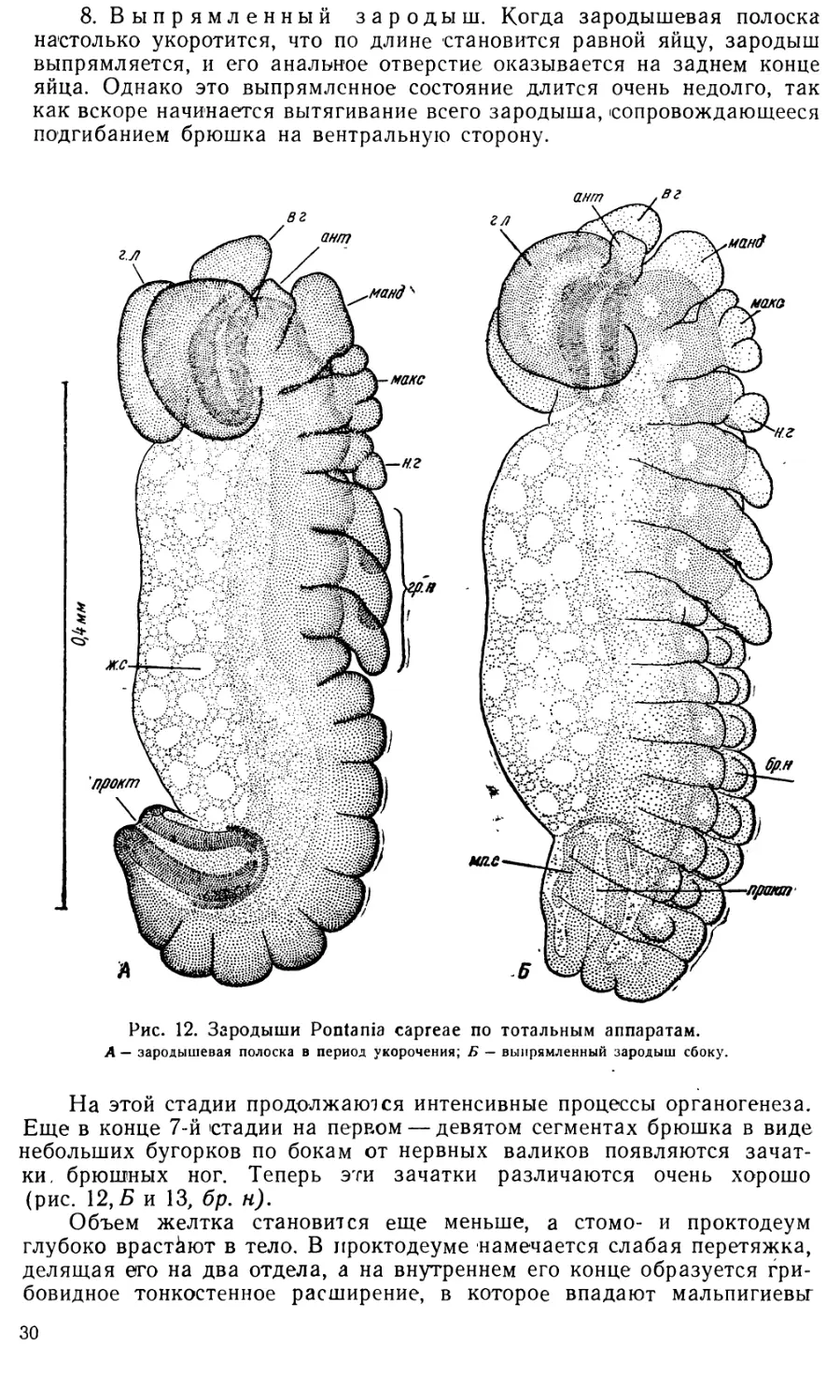
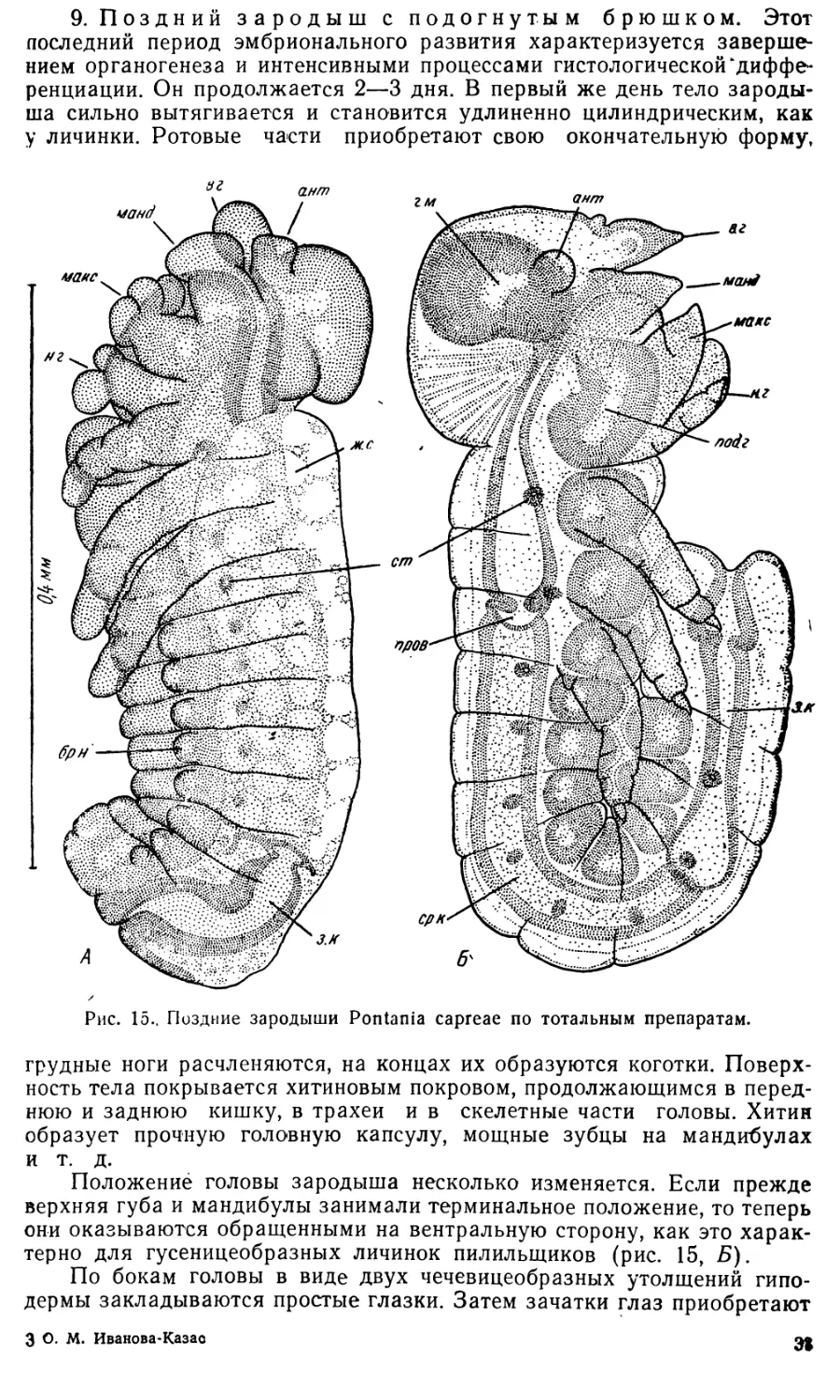
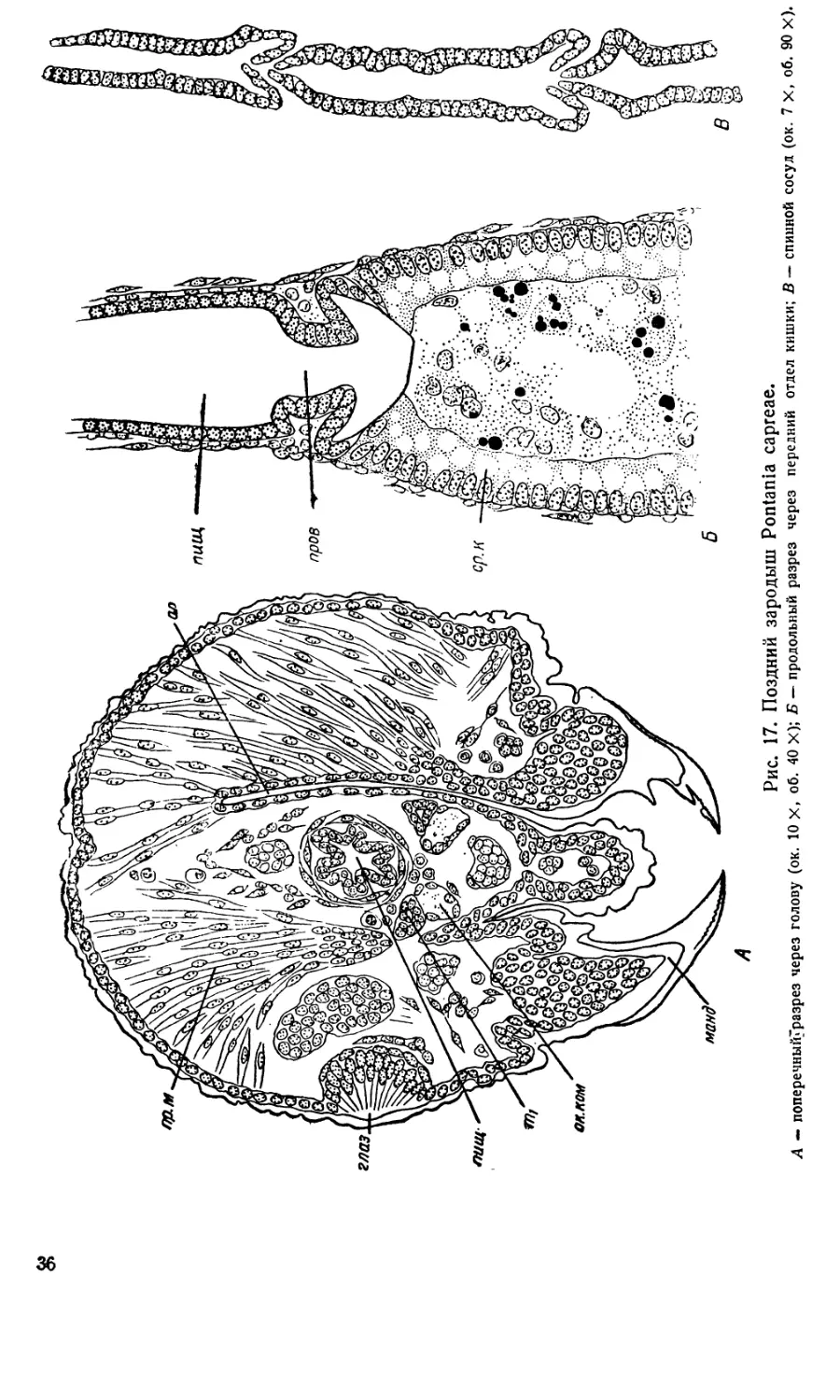
9. Поздний зародыш с подогнутым брюшком. Этот
последний период эмбрионального развития характеризуется заверше-
нием органогенеза и интенсивными процессами гистологической'диффе-
ренциации. Он продолжается 2—3 дня. В первый же день тело зароды-
ша сильно вытягивается и становится удлиненно цилиндрическим, как
у личинки. Ротовые части приобретают свою окончательную форму,
Рис. 15.. Поздние зародыши Pontania саргеае по тотальным препаратам.
грудные ноги расчленяются, на концах их образуются коготки. Поверх-
ность тела покрывается хитиновым покровом, продолжающимся в перед-
нюю и заднюю кишку, в трахеи и в скелетные части головы. Хитин
образует прочную головную капсулу, мощные зубцы на мандибулах
и т. д.
Положение головы зародыша несколько изменяется. Если прежде
верхняя губа и мандибулы занимали терминальное положение, то теперь
они оказываются обращенными на вентральную сторону, как это харак-
терно для гусеницеобразных личинок пилильщиков (рис. 15, Б).
По бокам головы в виде двух чечевицеобразных утолщений гипо-
дермы закладываются простые глазки. Затем зачатки глаз приобретают
3 О. М. Иванова-Казао
э*
При этом дегенерации одних клеток и замещения их другими не про-
исходит.
По мере того, как вторые максиллы сближаются и сливаются в
нижнюю губу (рис. 16, Г, н. г), происходит сближение и слияние в
непарный канал выводных протоков лабиальных желез. Этот непарный
канал открывается на вершине нижней губы и идет назад вентральнее
подглоточного ганглия. (Рис. 16, Бу от- лаб. ж). На границе головы и
груди он разделяется на правую и левую железы, которые уходят
в глубь тела, так что лежат уже ковнутри от нервной системы. Вскоре
каждая железа разделяется на две ветви тянущиеся вдоль всего тела,
но внутренняя при этом сильно изгибается, а наружная идет прямо, т. е.
все еще остается более короткой. Вокруг канальцев прядильных желез
образуются грозди железистых клеток, возникающих, по наблюдениям
Пфлюгфельдера (1934), путем выпячивания отдельных клеток из стенок
канальцев.
Нервная система полностью утрачивает свою связь с эктодермой.
Крупные эмбриональные невробласты исчезают. Образуются окологло-
точные комиссуры, нервные.пучки, связывающие между собой ганглии
брюшной цепочки, и периферические нервы. Три пары ганглиев челюст-
ных сегментов сливаются в подглоточный ганглий (рис. 15, Б, под. г).
Три последних ганглия брюшной цепочки тоже сливаются.
Энтодермальные пластинки, разрастаясь в ширину по поверхности
желтка, образуют, наконец, замкнутую трубку средней кишки.' Сперва
правая и левая Энтодермальные пластинки v смываются на брюшной
стороне, где они сближены, затем — на спинной. Клетки средней кишки
становятся цилиндрическими, а их плазма базофильной. Ядра лежат
у основания клеток, а во внутреннем конце каждой клетки содержится
крупная вакуоль (рис. 17, Б, ср. к). Заключенный в средней кишке
желточный синцитий превращается в пенистую массу с дегенерирующи-
ми желточными ядрами, которая затем полностью переваривается. Сто-
модеум становится пищеводом, начальный отдел которого имеет склад-
чатые стенки (рис. 17, £, пищ). Дальше назад пищевод несколько рас-
ширяется и складки разглаживаются. На границе пищевода и средней
кишки образуется провентрикулюс (рис. 17, Б, пров). Задняя кишка в
передней части имеет круглое сечение, а в задней — шестигранный или
звездообразный просвет из-за шести пучков растягивающих ее мышц.
Просветы передней и задней кишки даже у поздних зародышей отделе-
ны тонкой перепонкой от просвета средней кишки, но после вылупления
личинки полная проходимость кишечника устанавливается, по-видимому,
очень скоро. Это отличает низших растительноядных перепончатокры-
лых от жалящих и паразитических, у которых это происходит только
во время метаморфоза. Исключение составляет Ammophila campestris
Jur., у которой, по Бэрендсу и Бэрендс Ван Роон (1949), задняя кишка
прорывается в среднюю еще во время эмбрионального развития.
Мальпигиевы сосуды, сильно извиваясь, опутывают весь передний
отдел задней кишки.
После образования средней кишки последняя вместе с заключенным
в ней желтком несколько отодвигается от спинной стенки тела. В обра-
зовавшемся пространстве формируется спинной сосуд, образующий ме-
тамерные расширения (рис. 17, В). Передний конец его заходит в голову
до мандибулярного сегмента. Из мезодермы образуются мышцы туло-
вища, кровяные клетки и жировое тело. Последнее имеет вид тонкой
пленки, состоящей всего из одного слоя клеток, которая окружает ки-
шечник на некотором от него расстоянии. Ковнутри от жирового тела,
кроме кишечника, лежат еще лабиальные железы и мальпигиевы сосу-
з*
35
*тим нужно отметить, что зачатки брюшных ног появляются иногда и
при эмбриональном развитии паразитических перепончатокрылых, ли-
чинки которых их лишены, что имеет место, например, у Pimpla
turionellae (L.), по данным Бронскилл (1959). Во всяком случае, неза-
висимо от того, будем ли мы считать брюшные конечности пилильщиков
истинными или ложными, для большинства Hymenoptera это примитив-
ный признак, который был присущ растительноядным перепончатокры-
лым еще до того, как от них произошли стебельчатобрюхие, вторично
его утратившие.
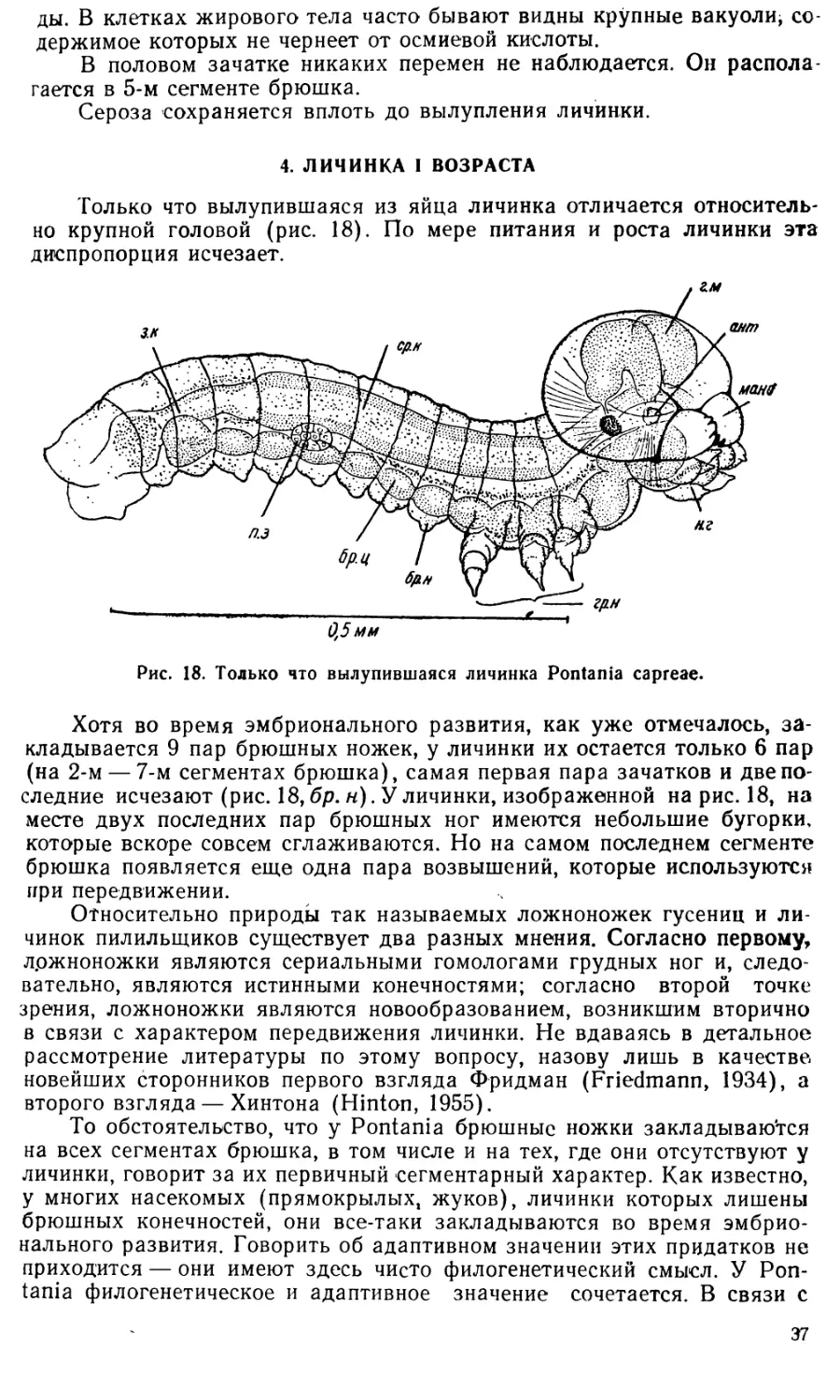
Сразу после вылупления личинки Pontania начинают питаться за
счет стенок галла, о чем свидетельствуют присутствие пищевых ком-
ков в кишечнике и появление в галле экскрементов.
Как это уже отмечалось Малышевым и Малышевой-Пузановой
(1952), личинки вялы и малоподвижны. Будучи извлечены из галла, они
не делают никаких попыток спастись бегством. Лишь в самом конце
личиночной жизни, накануне окукления, когда личинки покидают свой
галл, они становятся более активными. Такая слабая подвижность ли*
чинок, очевидно, является следствием их жизни в ограниченном про-
странстве внутри галла.
* *
*
Итак, для развития пилильщиков, с которыми мы познакомились
на примере Pontania capreae L., характерны следующие черты:
1. Довольно крупные яйца и большое количество желтка.
2. Поверхностное дробление; временное раздробление желтка и вто-
ричное его слияние в желточный синцитий.
3. Образование двух "эмбриональных оболочек, из которых амнион
довольно скоро редуцируется.
4. Очень полное развитие придатков тела: имеются антенны, ротовые
части, грудные ноги и так называемые «ложноножки» на брюшке.
5. Развитие органов зрения.
6. Установление сообщения между средней и задней кишкой к кон-
цу эмбрионального развития.
Почти все эти празнаки свойственны большинству насекомых, но их
приходится подчеркивать, имея в виду особенности развития высших
перепончатокрылых, о которых речь будет дальше.
Как особенно примитивные признаки следует отметить временное
раздробление желточной массы на клетки и парную закладку зароды-
шевой полоски.
ГЛАВА И
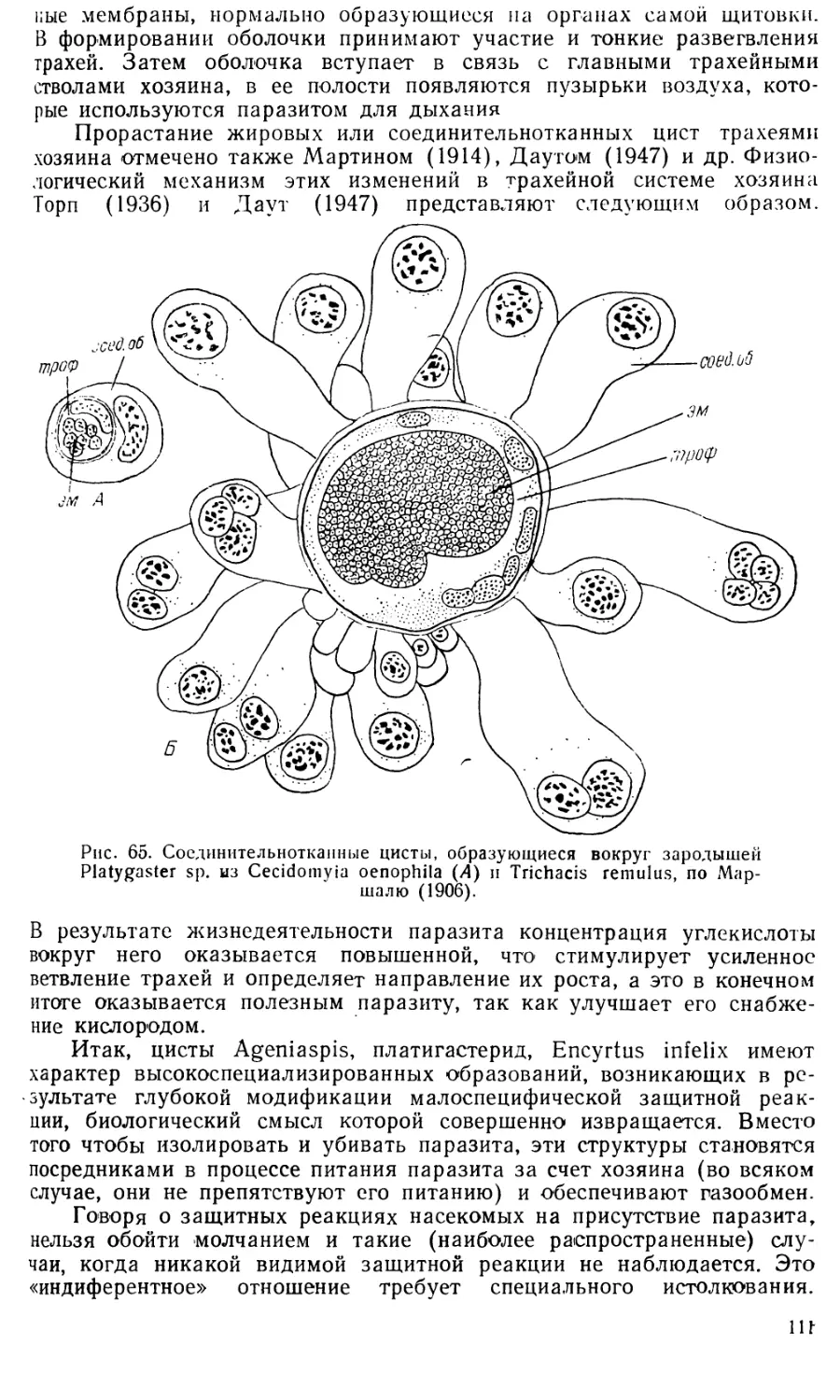
ANGITIA VESTIGIAL1S RATZ
Angitia vestigialis относится к подотряду Parasitica надсем.
lchneumonoidea сем. Ichneumonidae. Ихневмониды — одно из самых
больших семейств паразитических перепончатокрылых, представители
которого развиваются как наружные или внутренние паразиты за счет
насекомых из разных отрядов и пауков. Многие ихневмониды имеют
большое значение, так как ограничивают размножение вредных насе-
- комых. Однако эмбриональное развитие их освещено в литературе
очень слабо. Имеются лишь два обстоятельных описания развития:
Banchus femoralis Thorns. (Бледовский и Краинская, 1926) и Pimpla
turionellae (L.) (Бронскилл, 1959), а также довольно старые и непол-
ные сведения о развитии Mesochorus confusus Holmgr. (Кулагин, 1892
и 1894). Таким образом, Angitia vestigialis является четвертым пред-
ставителем сем. Ichneumonidae, ставшим объектом эмбриологического
исследования (Иванова-Казас, 19606).
1. БИОЛОГИЯ
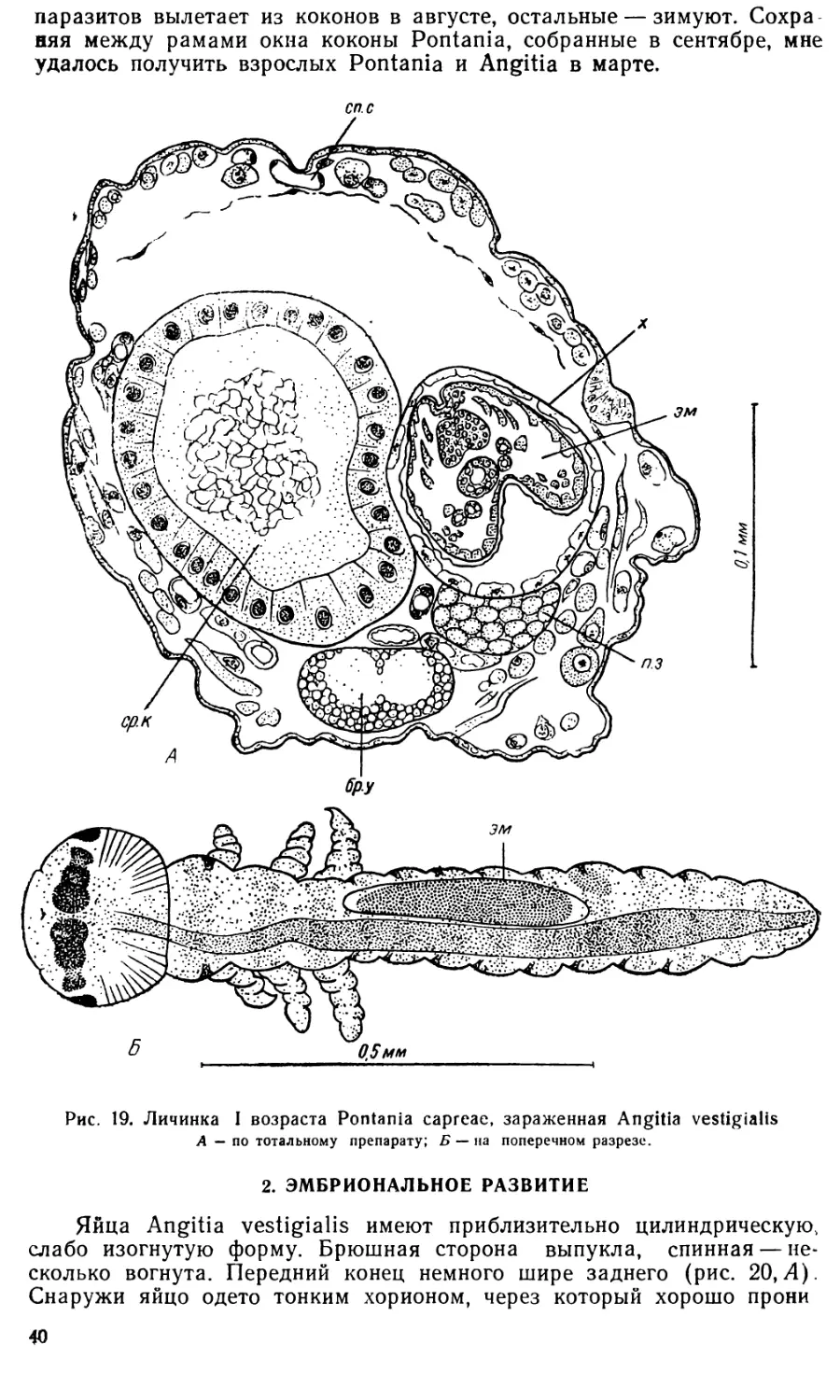
Angitia vestigialis откладывает яйца в полость тела личинок Роп-
tania capreae L. Личинки пилильщика подвергаются нападению наезд-
ника сразу же после выхода из яйца. Рис. 19, Б изображает личинку
Pontania I возраста, лишь недавно вылупившуюся, о чем свидетельст-
вует большая по сравнению с туловищем головная капсула. В полости
тела этой личинки справа от кишечника просвечивает яйцо Angitia
(эм). В личинках Pontania II возраста также встречаются яйца Angi-
tia, личинки же пилильщика более поздних возрастов, по-видимому,
уже не заражаются, в них встречаются только личинки Angitia.
Обычно в одну личинку Pontania откладывается только одно яйцо
Angitia, но как исключение в одной личинке оказывается 2, 3 или даже
4 яйца паразита. Личинки Angitia, как правило, встречаются пооди-
ночке. Очевидно, у Angitia vestigialis, как и у A. fenestralis Holmgr.
(Мейер, 1925а) и многих других наездников, имеет место личиночный
каннибализм.
Пораженные Angitia личинки Pontania внешне выглядят совер-
шенно здоровыми. На срезах, изготовленных из зараженных личинок,
никакой защитной реакции со стороны тканей хозяина на присутствие
паразита обнаружить не удается (рис. 19, Л). В конце лета заражен-
ные личинки пилильщика, так же как и здоровые, покидают свои гал-
лы и уходят в почву, где плетут свой кокон. Часть пилильщиков и их
Э9
зародышевая бластодерма дольше всего сохраняют связь друг с дру-
гом, и задний конец дольше остается не прикрытым серозой. Вполне
сформированная сероза имеет вид замкнутого со всех сторон мешка;
никаких следов амниона у Angitia нет. Такой способ образования се-
розы (без амниотических складок и амниона) путем отделения зачатка
серозы от зародышевой полоски и нарастания на нее довольно часто
встречается у жалящих перепончатокрылых: у Formica (Стриндберг,
1913а), у Vespa vulgaris L. (Стриндберг, 1914), у Apis mellifera L.
(Нельсон, 1915). У Pimpla turionellae (L.), по Бронскилл (1959), обра-
зуются рудиментарные амниотические складки, но амнион вскоре исче-
зает, и дальше развитие серозы идет сходно с таковым Angitia. У Ме-
sochorus confusus,'no H. M. Кулагину (1894), тоже образуется только
одна эмбриональная оболочка — путем деламинации бластодермы.
А у Banchus femoralis эмбриональные оболочки отсутствуют; вся бла-
стодерма здесь является «зародышевой», т. е. непосредственно участ-
вует в построении тела зародыша (Бледовский и Краинская, 1926).
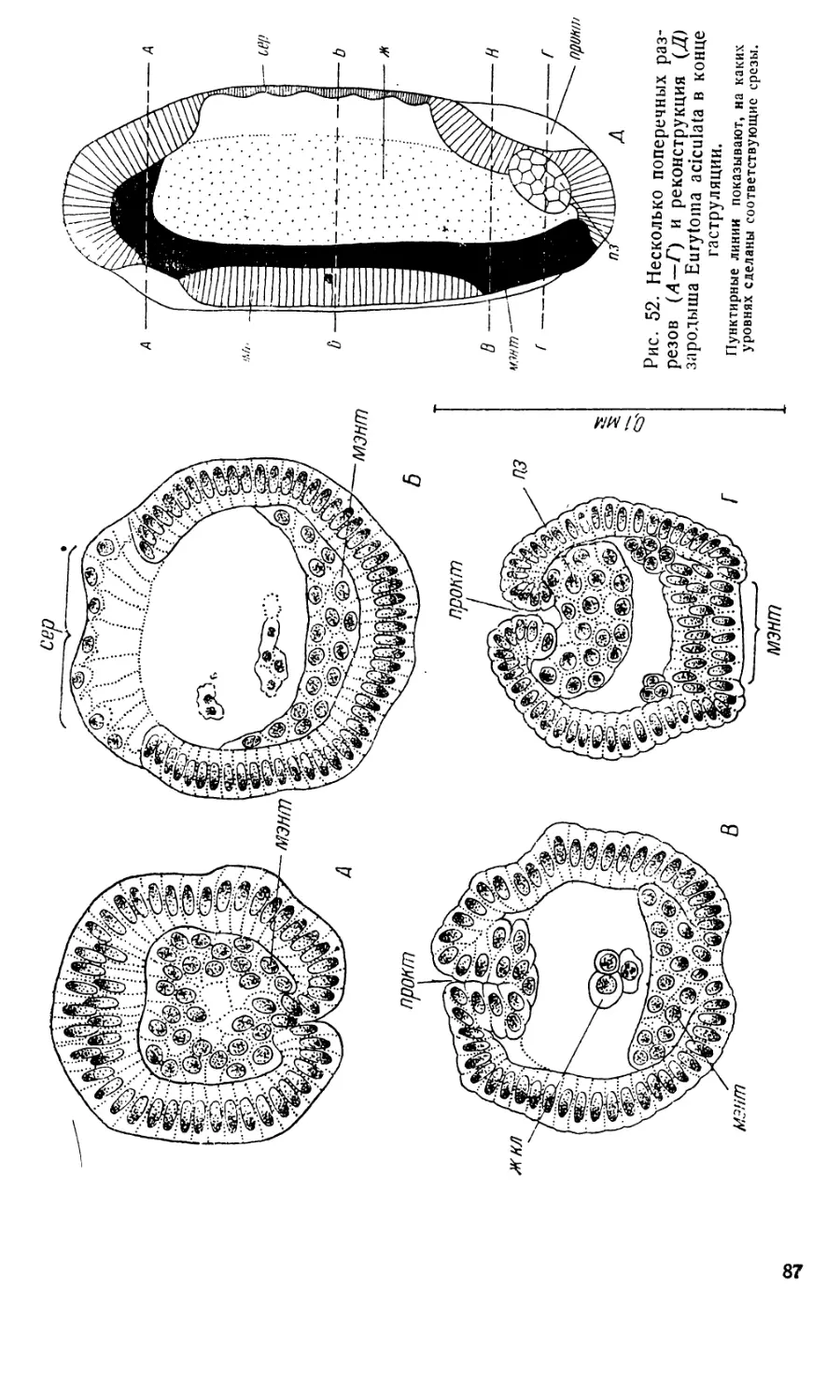
Затем в задней части зародышевой полоски, загибающейся на
спинную сторону, образуется небольшое впячивание проктодеума
(рис. 22, В, прокт). Вскоре после этого и на переднем конце, прибли-
зительно в том месте, где перед этим происходило массовое врастание
клеток, образуется стомодеум (рис. 22, В, стом).
Одновременно начинается сегментация зародыша. Первые меж-
сегментные борозды, по-видимому, очерчивают лабиальный и первый
грудной сегменты (рис. 22, В). Потом сегментация распространяется
на всю зародышевую полоску. У более поздних зародышей (рис. 20, Е)
на переднем конце различается большая непарная лопасть, нависаю-
щая над ротовым отверстием, соответствующая верхней губе (в. г), три
послеротовых сегмента головы с зачатками ротовых частей (манд,
макс, и. г) и ряд туловищных сегментЬв- На туловищных сегментах
зачатки конечностей не образуются.
К этому времени сероза уже одевает зародыш полностью. Сероза
и хорион растягиваются так, что между зародышем и этими оболоч-
ками возникает довольно значительное пространство (рис. 20, Е). Клет-
ки на свободных краях зародышевой полоски уплощаются и надви-
гаются на спинную сторону, ограничивая тело зародыша со спины.
Возникшая таким образом спинная стенка тела имеет сперва характер
очень плоского эпителия, затем клетки ее становятся кубическими.
Стомодеум и проктодеум врастают глубже. Начинается формиро-
вание средней кишки. Уже на стадии поздней зародышевой полоски
желточные клетки не образуют таких плотцых комков, как на стадии
бластодермы, а рассеиваются поодиночке в пространстве, ограничен-
ном с брюшной стороны и с боков зародышевой полоской, а со спины—
серозой. При этом -они сохраняют связь друг с другом посредством
плазматических перемычек, образуя сетевидную синцитиальную массу
(рис. 24, Б, ж. с). С поверхности этот синцитий довольно четко ограни-
чен тонким плазматическим слоем, к которому примыкает большинство
желточных ядер. Лишь единичные желточные ядра остаются в сере-
дине. Этот синцитий повторяет структуру, образуемую желточными
клетками в яйцах насекомых, богатых желтком, но желток, лежащий
обычно в петлях этой синцитиальной сети, у Angitia отсутствует.
У живых зародышей этот синцитий выглядит как пенистая масса
(рис. 20, Е, ж. с).
Вскоре на поверхности желточного синцития оседают отдельные
клетки, которые затем складываются в непрерывную стенку средней
кишки (рис. 25, энт). Таким образом, желточный синцитий Angitia
45
Сероза сохраняется у Angitia до конца эмбрионального развития.
Клетки серозы становятся более толстыми (рис. 27,Л). В их плазме
появляются светлые вакуоли и мелкие гранулы, выявляемые при фи-
ксации жидкостью Флемминга (рис. 27, В). Границы отдельных кле-
ток, неразличимые на более ранних стадиях, теперь становятся отчет-
ливыми. Связь между клетками становится очень слабой, так что меж-
ду ними появляются заметные щели (рис. 27, Б). Создается впечатле-
ние, что клетки серозы только потому сохраняют эпителиеобразное
расположение, что они тесно прилегают к хориону.
Поздний зародыш Angitia, все еще одетый серозой и хорионом,
способен энергично изгибаться вместе с одевающими его оболочками.
Однако Angitia vestigialis не относится к числу тех наездников, кото-
рые сохраняют яйцевые и эмбриональные оболочки на протяжении
всего I возраста. После разрыва оболочек вышедшая из них личинка
успевает значительно вырасти прежде, чем произойдет первая линька.
3. СРАВНЕНИЕ РАЗВИТИЯ ANGITIA VESTIGIALIS С ТАКОВЫМ
ДРУГИХ ICHNEUMONIDAE И ПИЛИЛЬЩИКОВ
По сравнению с пилильщиками, развитие Angitia отличается ря-
дом отрицательных признаков — отсутствием желтка, отсутствием ам-
ниона, отсутствием глаз, грудных и брюшных конечностей, отсутствием
сообщения между средней и задней кишкой. Последние из этих при-
знаков есть результат недоразвития ряда личиночных структур. Ли-
чинка Angitia выходит из яйца в морфологическом отношении менее
развитой, чем личинка Pontania. В такой же, а иногда еще большей
степени, дезэмбрионизация — термин А. А. Захваткина (1953), приме-
няемый им для обозначения таких эволюционных изменений онтогене-
за, при которых молодой организм покидает яйцевые оболочки на
более ранней стадии развития, — свойственна всем паразитическим и
жалящим перепончатокрылым. О том же говорят различия в длитель-
ности различных периодов эмбриогенеза у пилильщиков и медоносной
пчелы, о которых уже была речь выше.
По характеру развития другие ихневмониды могут в значительной
степени отличаться от Angitia. Так у Pimpla turioneilae. (L.) паразити-
рующей в Rhyaciona buoliana (Schiff.) и других бабочках, судя по
описанию Бронскилл (1959), развитие сохраняет больше примитивных
черт. Яйца Pimpla довольно крупны (1,615 X 0,265 мм) и содержат
много желтка. Дробление и образование бластодермы происходит
обычным способом. Формирование серозы сопровождается временным
появлением амниотических складок, но амнион не образуется. Зароды-
шевая полоска дифференцируется на срединную пластинку (мезодер-
ма) и две боковые пластинки (эктодерма). Срединная пластинка по-
гружается внутрь. Энтодерма образуется биполярно: на переднем и
заднем концах зародышевой полоски, где впоследствии происходит
впячивание стомо- и проктодеума. Появляются зачатки конечностей —
не только на голове, но и на груди и брюшке (очень примитивная чер-
та), но потом зачатки туловищных конечностей исчезают. К моменту
вылупления личинки уже имеются хорошо развитые нервная система,
трахеи, внутренний скелет головы, лабиальные железы, мальпигиевы
сосуды, мускулатура сердца.
У Banchus femoralis Thorns., откладывающего яйца в гусениц
Panolis flammea Schiff., по Бледовскому и Краинской (1926), яйца не
так велики (0,6X0,12 мм), но еще содержат желток. Эмбриональные
оболочки не развиваются. Мезэнтодерма образуется в результате тан-
48
генциальных делений клеток в брюшной бластодерме. Медианная
часть этого внутреннего листка дает зачаток средней кишки (энто-
дерму), а латеральные — мезодерму. Своеобразной особенностью Вап-
chus является участие внутреннего листка в построении глубоких частей
передней и задней кишки. Развитие Banhus нескблько упрощено: кроме
исчезновения эмбриональных оболочек, которые, видимо, утратили свое
функциональное значение, надо отметить и сокращение числа сегментов:
голова ни на какой стадии развития не обнаруживает своего сегмен-
тарного состава, а развивается сразу как единое образование. То же
относится и к подглоточному ганглию, который обычно у насекомых
получается слиянием трех пар челюстных ганглиев брюшной цепочки.
Дезэмбрионизация выражена у Banchus еще сильнее, чем у Angitia.
Развитие ротовых частей сильно запаздывает, мальпигиевы сосуды обра-
зуются только во время метаморфоза (функцию выделения у личинки,
по-видимому, выполняет задняя кишка, которая выпячивается наружу,
как «анальный пузырь»).
Mesochorus confusus Holmgr., по Н. М. Кулагину (1894), развивает-
ся в личинках пилильщика Nematus vallisnerii. Яйца Mesochorus очень
мелки (0,02 X 0,009 мм) и совершенно лишены желтка. Дробление,
однако, поверхностное. Образование оболочки и зародышевых листков
происходит сильно модифицированным способом («деламинация в мо-
руле»): периферический слой клеток зародыша образует оболочку;
клетки, непосредственно прилегающие к оболочке изнутри, дают экто-
дерму; еще глубже лежащие слои клеток составляют энтодерму и мезо-
дерму, а центр зародыша занимает полость, заполненная плазматиче-
ской жидкостью. Таким образом, начальные процессы эмбрионального
развития Mesochorus подверглись очень большим изменениям. Однако,
эмбриональная оболочка сохраняется, что дает основание думать, что
она выполняет какие-то функции. Впрочем, рудиментарные органы так
же возможны в эмбриональном состоянии организма, как и во взрослом.
Из вышеизложенного можно видеть, что даже в пределах одного
семейства (в данном случае — Ichneumonidae) возможны довольно
значительные вариации развития.
4 О. М. Иванова-Казас
ГЛАВА III
EPHEDRUS PLAGIATOR NEES
Ephedrus plagiator относится к сем. Aphidiidae (подотряд Parasitica,
Ichneumonoidea), представители которого специализировались как
внутренние паразиты тлей. Большинство афидиид — полифаги, т. е.
паразитируют во многих видах тлей. По мнению Н. А. Теленги (1950),
«развитию полифагии среди афидиид способствует морфологическое
однообразие их хозяев — тлей и большое однообразие условий обитания
тлей. С этим в известной степени связана и слабая морфологическая
дифференциация афидиид». Н. А. Теленга считает афидиид очень ак-
тивными паразитами, которые могут быть использованы для борьбы с
тлями, вредящими сельскохозяйственным культурам.
Биология и постэмбриональное развитие афидиид изучены рядом
авторов (Скрипчинский, 1930; Янишевская — Janiszewska, 1933; Спен-
сер— Spencer, 1926;'Бэйрн — Beirne, 1942, и др.), но относительно их
эмбрионального развития имеются лишь отрывочные сведения в рабо-
тах Бэйрн (1942), Спенсера (1926) и Гетенби (Gatenby, 1919).
Материалы по развитию Ephedrus и других Aphidiidae собраны
в парке Биологического института Ленинградского государственного
университета в Петродворце летом 1954 и 1955 гг. Изучены главным
образом афидииды, паразитирующие в тле Acyrtosiphon caraganae
Cholodk. с желтой акации, но использованы также материалы по раз-
витию паразитов других видов тлей Петродворца и некоторых афидиид
Туркмении, собранные в 1952 г. Таким образом, получены сведения по
развитию Ephedrus plagiator Nees из Acyrthosiphon caraganae и не-
скольких представителей рода Aphidius (Иванова-Казас, 1954в, 1956а, б
и 1961) биологию которых удобно рассмотреть вместе с таковой Ephed-
rus.
1. БИОЛОГИЯ
Пораженные афидиидами тли через некоторое время меняют свою
окраску и гибнут. Так например Acyrtosiphon caraganae, зараженные
Aphidius, из зеленых становятся соломенно-желтыми, а зараженные
Ephedrus — чернеют. Паразиты, вылетевшие из этих погибших тлей,
использовались для заражения новых тлей в лабараторных условиях.
Афидииды обычно заражают тлей, когда те находятся еще на ста-
дии личинки. Отложенное яйцо оказывается неглубоко под гиподермой
тли (рис. 28, эм). Оно лежит в жировом теле или свободно между раз-
личными внутренними органами. Никаких морфологически выраженных
защитных реакций со стороны тканей тли наблюдать не приходилось.
50
Сильвестри (1908, 1937) высказал предположение, что присутствие этого
включения необходимо для развития полового зачатка, и строит на этом
свое объяснение возникновения так называемых «бесполых» личинок
у некоторых полиэмбрионических видов (см. Общ. часть, глава III, 4),
но Гетенби (19186) и Пэтерсон (1918) подвергают его соображения
сомнению. Во всяком случае, прямых доказательств того, что оосома
детерминирует клетки, в которые она попадает, как половой зачаток,
нет.
Рейт (Reith, 1931, 1932), экспериментально изучавший развитие
муравья Camponotus, сделал попытку доказать, что оосома (которую он
называет «полярным тельцем») играет роль организатора, является
местом локализации «центра детерминации». Хотя попытка эта и не
имела успеха, но Рейт остался при мнении, что у муравьев присутствие
оосомы необходимо для осуществления цепи детерминационных про-
цессов при развитии.
Поскольку роль этого включения не может считаться выясненной,
мне кажется более осторожным пользоваться нейтральным термином
«оосома».
Проследить дальнейшую судьбу оосомы у Ephedrus не удалось.
В только что отложенном живом яйце ядро не различается; воз-
можно, что в этот момент оно находится в состоянии деления созревания.
После начала дробления ядра имеют вид округлых светлых пузырьков
(рис. 30, я. др.). Несмотря на отсутствие желтка и малые размеры яйца„
дробление у Ephedrus типичное неполное. Удалось наблюдать стадии
двух, четырех, восьми и многих ядер дробления (рис. 30, Б, В, Г и Д).
Уже на стадии четырех ядер дробления свободное пространство между
поверхностью яйца и хорионом исчезает, что можно объяснить набу-
ханием яйца. У зародыша, изображенного на рис. 30, Д, ядра уже очень
близко подходят к поверхности яйца, и начинается формирование-
бластодермы. Рис. 30, Е изображает яйцо Ephedrus на стадии бласто
дермы в оптическом разрезе. Число ядер здесь уже очень значительно.
Все они лежат одним слоем у поверхности яйца тесно прижимаясь
друг к другу. На заднем конце различается группа из четырех округ-
лых клеток — половой зачаток (рис. 30, Е, п. з). На тотальных препара
гах на этой стадии синцитиальной бластодермы в центральной части
яйца можно видеть несколько сильно красящихся комочков — дегенери
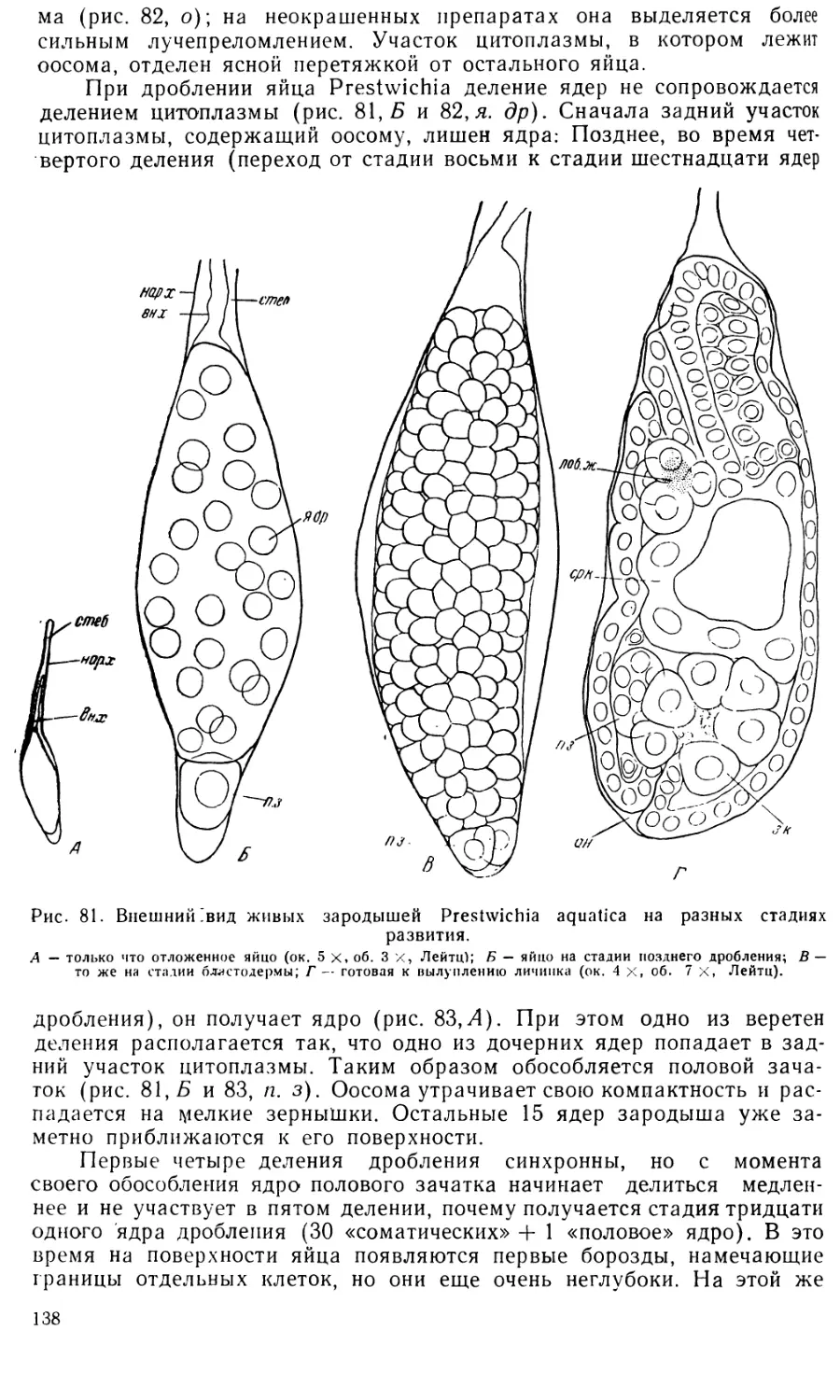
рующие ядра. (рис. 31, А, дег. я).
Подобную же дегенерацию части ядер на стадии бластодермы
наблюдали Маршаль (Marchal, 1906) у Synopeas rhanis Walk., Силь-
вестри (19096 и 1915—1916) у Aphidencyrtus (Encyrtus) aphidivorus
(Mayr), Trichogramma (Oophthora) semblidis (Aur.), Anaphoidea Luna
Masi) и Trichaporus (Encarsia) partenopeus (Masi) и Гетенби (1917)
у Trichogramma evanescens Westw. Сильвестри полагает, что дегенери-
рующие ядра соответствуют вителлофагам других насекомых. Гетенби
склонен толковать это явление как своеобразную диминуцию хрома-
тина, происходящую путем выталкивания части хроматина из ядер
бластодермы. Однако мнение Сильвестри кажется гораздо более прав-
доподобным.
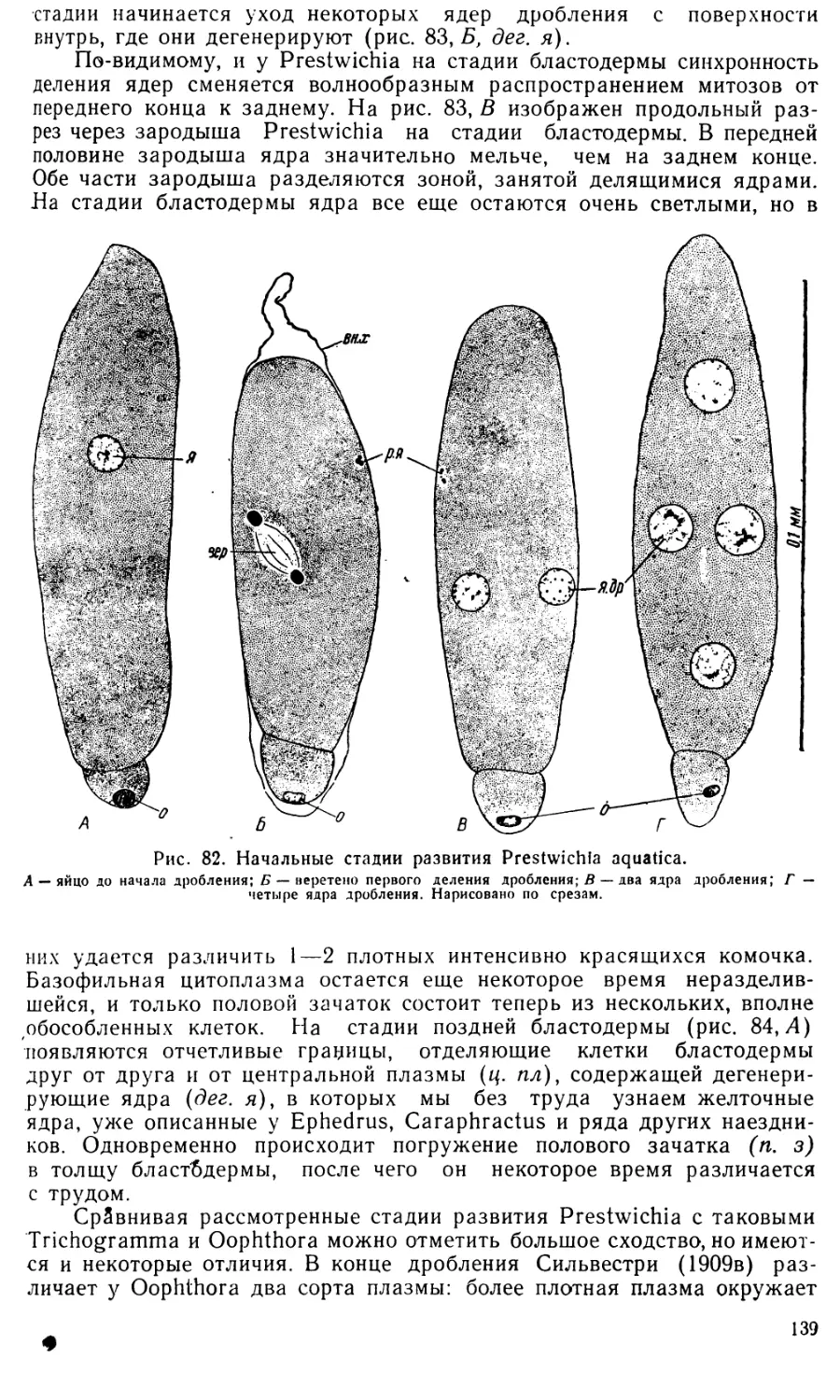
Затем появляется граница, отделяющая центральную плазму с
дегенерирующими ядрами от бластодермы. Границы отдельных клеток
бластодермы еще не видны, но, по-видимому, они уже имеются, так
как обычно боковые стенки клеток бластодермы образуются раньше
базальной. По мере увеличения числа клеток, бластодерма становится
неправильно двурядной. Затем бластодерма расслаивается на поверх-
ностный слой, характеризующийся более светлыми ядрами, лежащими
53
менее плотно, и внутренний слой, ядра которого красятся более интен-
сивно и лежат плотнее, а также вытянуты перпендикулярно поверх-
ности яйца (рис. 31,Б). Из наружного слоя клеток образуется оболочка
(сероза), из внутреннего — сам зародыш. Таким образом, у Ephedrus
plagiator сероза образуется путем деламинации бластодермы, т. е. еще-
более измененным способом, чем у Angitia.
Рис. 31. Зародыши Ephedrus plagiator.
Л — яйцо в конце дробления; Б — зародыш с дифференцированно»
бластодермой и серозой.
После образования эмбриональной оболочки клетки полового за-
чатка оказываются под серозой. Дальнейшая судьба их не прослежена.
Далее зародыш Ephedrus заметно вытягивается в длину. По осп
его образуется узкое трубковидное пространство, в котором лежат деге-
неративные остатки плазмы и желточных ядер (рис. 31, Б). На попереч-
ных разрезах через среднюю часть зародыша видно, что зародышевая
бластодерма не образует сплошного слоя, а прерывается на одной сто-
роне и представляет собой, таким образом, нечто вроде желоба (рис.
32, Б). В дальнейшем эмбриональная оболочка зародыша растягивается
и становится более просторной (рис. 32, В сер), а края желоба как бы раз-
двигаются (рис. 32, В, зар, п). В это время зародыш Ephedrus имеет
форму зародышевой полоски, но отличается тем, что не лежит на желтке
и края его не переходят в амнион так как ни желтка, ни амнион)
54
После вылупления личинки клетки серозы разъединяются и округ-
ляются. Они длительно сохраняются в полости тела тли, окружая ли-
чинку Ephedrus и превращаются в гигантские клетки, как это описано
для разных Aphidiidae Спенсером (1926), Янишевской (1933) и мною
(Иванова-Казас, 19566). Затем они съедаются личинкой (подробно об
этом см. Спец. часть, глава IV, 3).
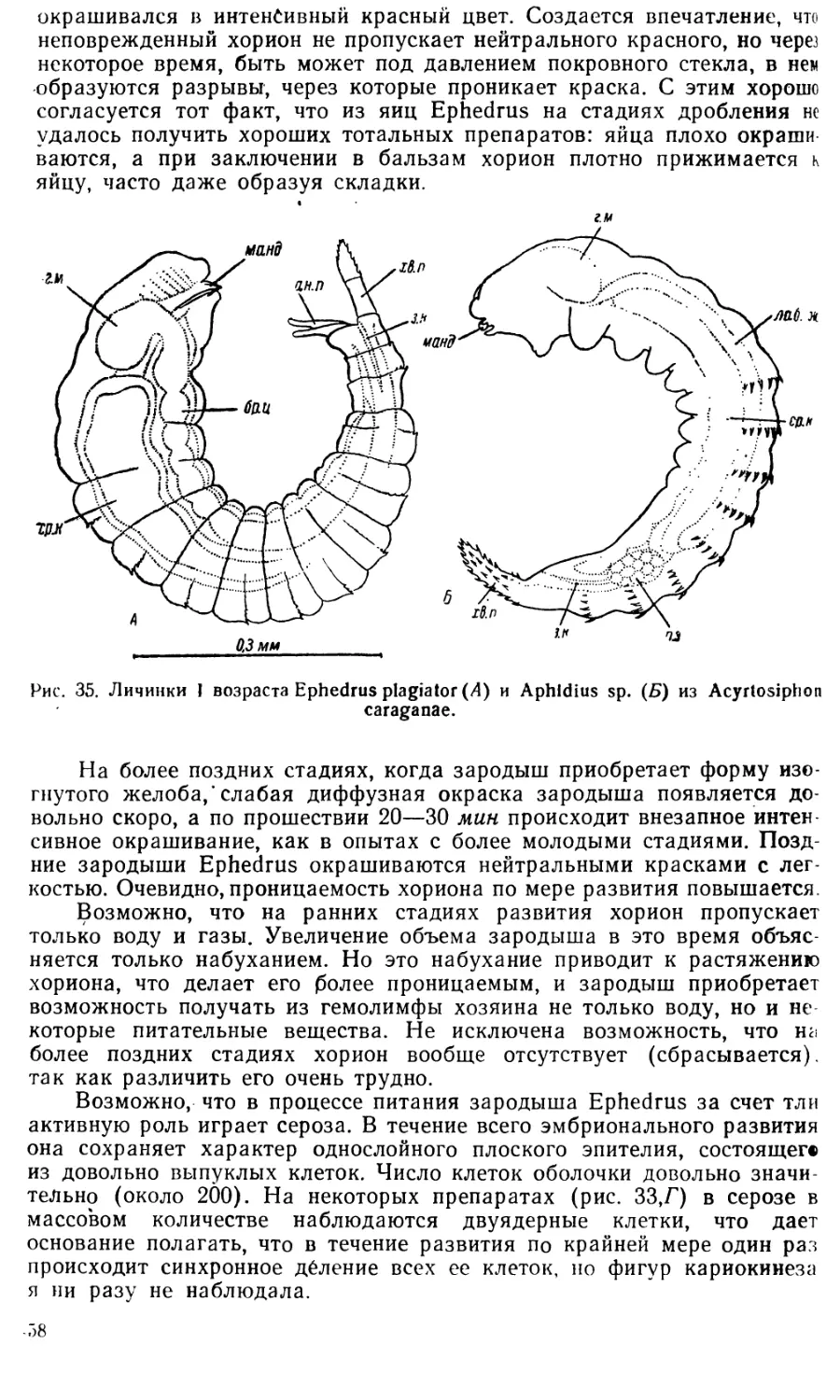
3. ПАЛИНГЕНЕТИЧЕСКИЕ ЧЕРТЫ В РАЗВИТИИ EPHEDRUS PLAGIATOR
В эмбриональном развитии Ephedrus наблюдается много черт па-
ли нгенетического характера, т. е. таких особенностей, которые утра-
тили свое первоначальное функциональное значение, но тем не менее
в силу наследственности все еще сохраняются. К числу таких особен-
ностей относятся следующие:
1) Дробление поверхностного типа, характерное для насекомых
и возникшее в связи с наличием в яйце большого количества желтка.
У Ephedrus желток отсутствует, но дробление остается поверхностным.
2) В процессе образования бластодермы некоторые ядра остаются
в центральной части яйца и там дегенерируют. Эти ядра гомологичны
вителлофагам насекомых, функция которых состоит в переработке
желтка. Здесь желтка нет, а вителлофаги все-таки образуются.
3) После разделения бластодермы на эмбриональную оболочку и
собственно зародыш, последний приобретает форму зародышевой по-
лоски. Обычно зародышевая полоска образуется в яйцах, богатых
желтком, на котором зародыш как бы распластывается. У Ephedrus
зародыш сохраняет форму полоски, несмотря на полную редукцию
желтка. Спинная сторона зародыша некоторое время остается откры-
той и лишь затем замыкается, что имитирует обрастание желтка при
развитии типичных яиц насекомых.
4) Наконец, значительное удлинение зародыша в начале развития
и последующее его относительное укорочение можно толковать как по-
следний след («рудимент») бластокинеза. Функциональное значение
бластокинеза насекомых до сих пор не может считаться точно установ-
ленным. По этому вопросу имеются только предположения. Можно ду-
мать, что бластокинез связан с уходом зародыша с поверхности яйца
и образованием эмбриональных оболочек, которым приписывается за-
щитное значение; в то же время — в процессе бластокинеза, особенно
когда он сопровождается удлинением зародыша, увеличивается кон-
такт зародыша с желтком.
У Ephedrus образование эмбриональной оболочки и последние
остаточные проявления бластокинеза происходят независимо друг от
друга, что, конечно, есть результат вторичного изменения процесса раз-
вития. Нет здесь и желтка, контакт с которым имеет такое важное
значение для питания зародыша. Впрочем, можно предполагать, что
удлинение зародыша Ephedrus тоже имеет трофическое значение, так
как при этом увеличивается вся поверхность зародыша, через которую,
очевидно, и происходит его питание.
Все эти особенности развития Ephedrus показывают, что течение
эмбриональных процессов, тип развития каждого вида живцтного не
являются простой функцией наличия или отсутствия желтка, а опреде-
ляются наследственными свойствами вида, изменения которых под
влиянием указанных факторов могут довольно сильно запаздывать.
Вышеуказанные черты развития Ephedrus не являются чем-то
исключительным. Аналогичные явления наблюдаются у дождевых чер-
вей (Светлов, 1928), у которых изгибание плакулы имитирует процесс
59
обрастания желтка, происходящий при развитии более примитивных
пресноводных олигохет. П. П. Иванов (1937) в «Общей и сравнитель-
ной эмбриологии» приводит еще ряд примеров того, что признаки, при
обретенные в связи с большим количеством желтка в яйцах, сохра-
няются и после его исчезновения при переходе зародыша к плацентар
ному питанию (Peripatus edwardsii, млекопитающие) или питанию
белковой жидкостью, окружающей яйцо (Lumbricidae, пиявки, Limax
и др.). Однако П. П. Иванов подчеркивает, что это явление не распро-
страняется на самые ранние стадии развития, «когда присутствие или
отсутствие желтка почти механически влечет за собой полное или не
полное дробление». Поэтому пример Ephedrus и некоторых других па-
разитически^ перепончатокрылых представляет особый интерес, так как
наследственная обусловленность процессов развития проявляется у них
на ранних стадиях так же отчетливо, как и на более поздних.
С другой стороны, нужно отметить, что во всех упомянутых
П. П. Ивановым случаях особенности развития, связанные с желтком,
после его исчезновения не утрачивают полностью свое функциональное
значение, а видоизменяют его. Зародыши дождевых червей и пиявок
рано начинают заглатывать белковую жидкость, которая, проникнув
внутрь зародыша, занимает там то место, которое у более примитив-
ных форм занимал желток. В результате части зародыша оказываются
расположенными в отношении источника их питания так же, как у их
предков, и функциональные отношейия остаются сходными.
Внезародышевые части Sauropsida, унаследованные млекопитаю-
щими, полностью сохраняют свое питательное, дыхательное и защит-
ное значение, но выполняют эти функции иначе, применительно к но-
вым условиям, претерпевая соответствующие перестройки.
Однако у Ephedrus большинство перечисленных «рудиментарных»
признаков в развитии не приобрело нового функционального значения
и потому, надо полагать, в процессе эволюции должны раньше или
позже исчезнуть. С этой точки зрения представляет интерес сравнение
развития Ephedrus с развитием близко родственного рода Aphidius
ГЛАВА IV
APHIDIUS FABARUM MARSH. И ДРУГИЕ ВИДЫ APHIDIUS
Биология наездников из рода Aphidius (сем. Aphidiidae) настоль-
ко близка к таковой Ephedrus, что нет надобности повторять сказанное
ранее. Мне удалось получить материалы по развитию Aphidius faba-
rum Marsh, из Chaitophorus sp., Aphidius cardui Marsh, из тлей на Zy-
gophyllum atriplicoides (Туркмения) и еще нескольких видов рода
Aphidius, точно определить которые оказалось затруднительным. Одна-
ко с эмбриологической точки зрения, это, по-видимому, не имеет боль-
шого значения, так как сколько-нибудь значительных различий в раз-
витии разных видов Aphidius не обнаружено.
!. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
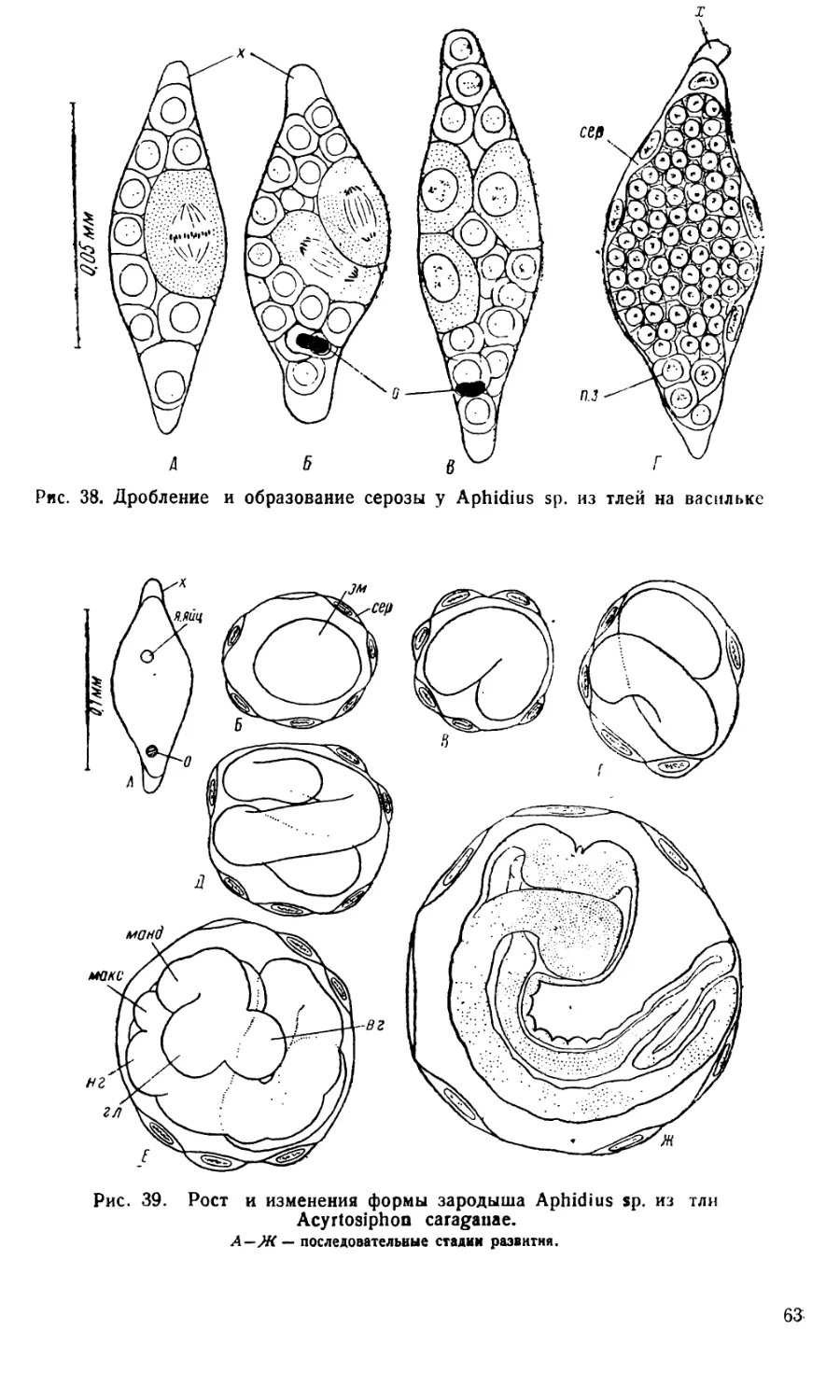
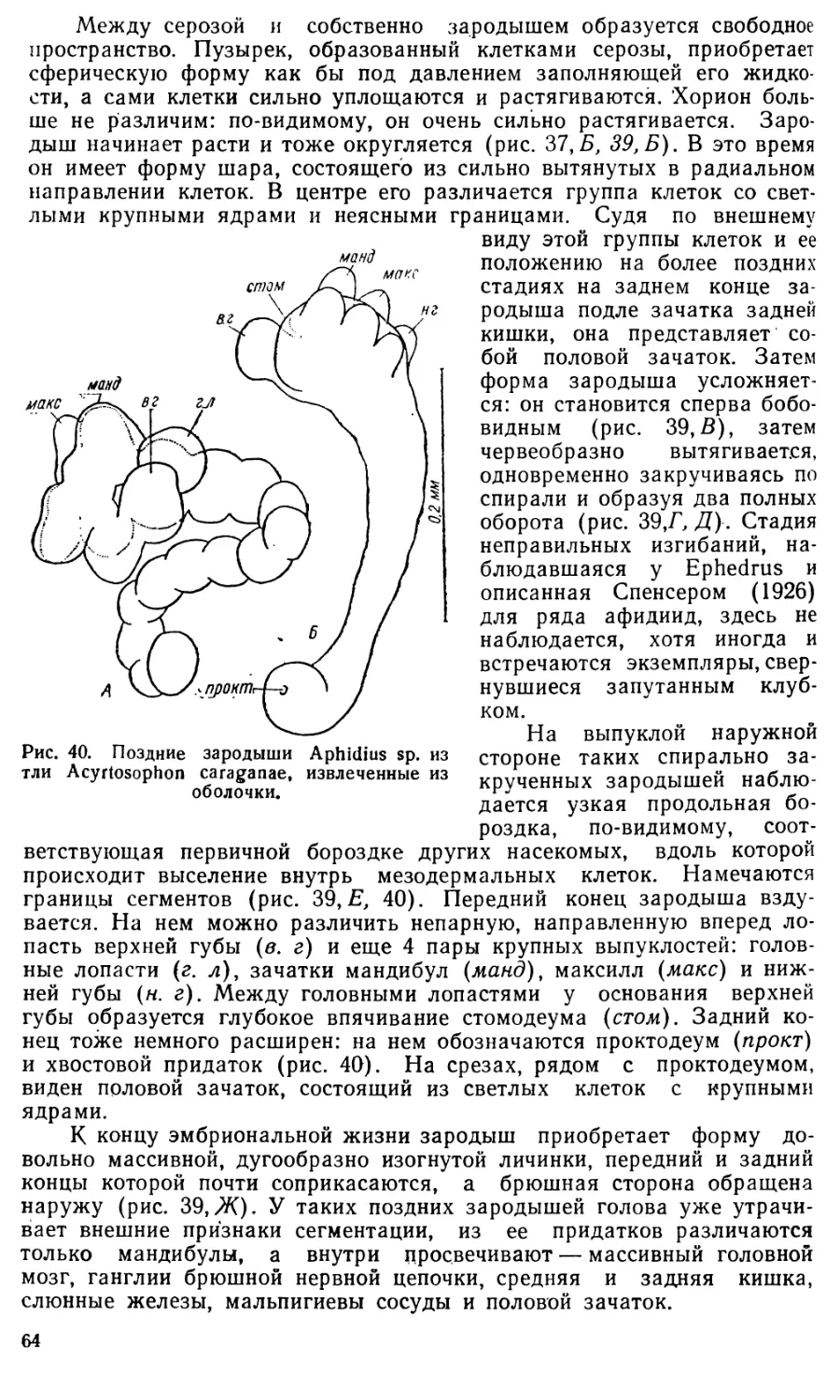
Яйца Aphidius имеют веретеновидную (рис. 36 и 38) форму.
Снаружи яйцо одето тонким хорионом (*), который обычно на концах
яйца несколько отстает от его поверхности. Плазма яйца кажется со-
вершенно гомогенной и не содержит желтка. В средней части яйца ле-
жит светлое пузыревидное ядро, на заднем конце — небольшое, более
темное тельце — оосома (о).
Размеры яиц варьируют довольно сильно, что, очевидно, связано
с видовыми отличиями. Так, яйца Aphidius fabarum Marsh, имеют раз-
меры 0,086X0,036 мм. Яйца Aphidius sp. из тлей на васильке (рис.38)
имеют длину 0,070—0,090 мм, а ширину 0,024—0,029 мм. Значительно
крупнее яйца Aphidius, паразитирующих в Acyrtosiphon caraganae
(длина 0,090—0,150 мм, ширина 0,030—0,060 мм).
Дробление яйца начинается как типичное неполное: ядро разде-
ляется дважды, а плазма остается неразделившейся (рис. 36, £, В).
Лишь после следующего деления яйцо сразу распадается на отдельные
клетки и дробление становится тотальным (рис. 36, Г, Д, Е). Характер-
ная для насекомых стадия бластодермы, на которой зародыш состоит
из поверхностного слоя клеток и центральной нераздробившейся части
яйца, у Aphidius отсутствует.
С самого момента образования клеток одна из них отличается бо-
лее крупными размерами и сильнее красится основными красками;
остальные клетки мельче и светлее. Эти различия удалось подметить
лри дроблении Aphidius sp. из тлей на васильке (рис. 38), у афидиусов
из Acyrtosiphon caraganae (рис. 36) и у Aphidius fabarum (Иванова-
Казас, 19546).
61
(Паио —за половые клетки, Безе—за эноциты). По недавним наблю-
дениям Макауера (1959), «гигантские клетки» образуются из клеток
жирового тела тли.
Однако специальное исследование развития «гигантских клеток»,
их происхождения, роста, накопления в них вакуолей и различных вклю-
чений показывает, что Безе и Пайо ошибались. Когда начинается раз-
витие «гигантских клеток», половая система тли еще сохраняет совер-
шенно нормальный вид, так что превращение половых клеток в гигант-
ские очень мало вероятно. Количество эноцитов в теле здоровой тли
невелико: размножение их ни в норме, ни при заражении не наблюда-
лось. Число же «гигантских клеток» бывает иногда (при заражении
тли сразу несколькими личинками Aphidius) очень значительным. Часто
рядом с «гигантскими клетками» можно видеть совершенно неизмененный
эноцит. Невозможность происхождения «гигантских клеток» от эноци-
тов или жировых клеток становится очевидной при изучении препара-
тов, окрашенных эозин-азуром. Молодые «гигантские клетки», как и
клетки еще не диссоциированной серозы, обладают резко азурофильной
плазмой. Эта азурофилия сохраняется в несколько более слабой сте-
пени и позднее, когда начинается образование вакуолей и накопление
различных включений. Эноциты же красятся в красновато-лиловый
цвет, эозинофилия у них явственно преобладает, а жировые клетки тли
имеют характер высоко дифференцированных элементов с цитоплазмой,
переполненной включениями, задолго до того, как начинается рост и
накопление включений у «гигантских клеток». Таким образом, эти на-
блюдения подтверждают правильность взглядов Спенсера и Янишев-
ской.
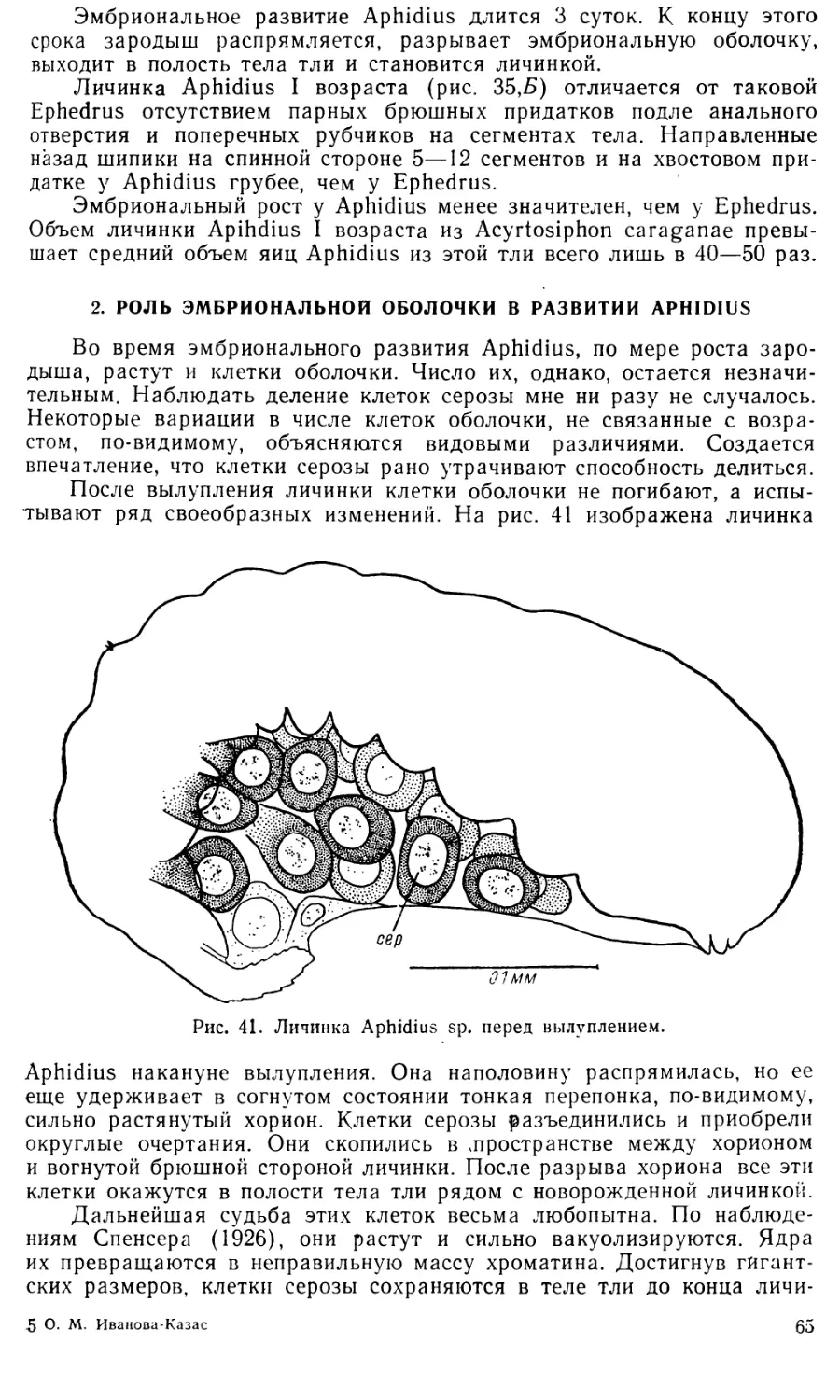
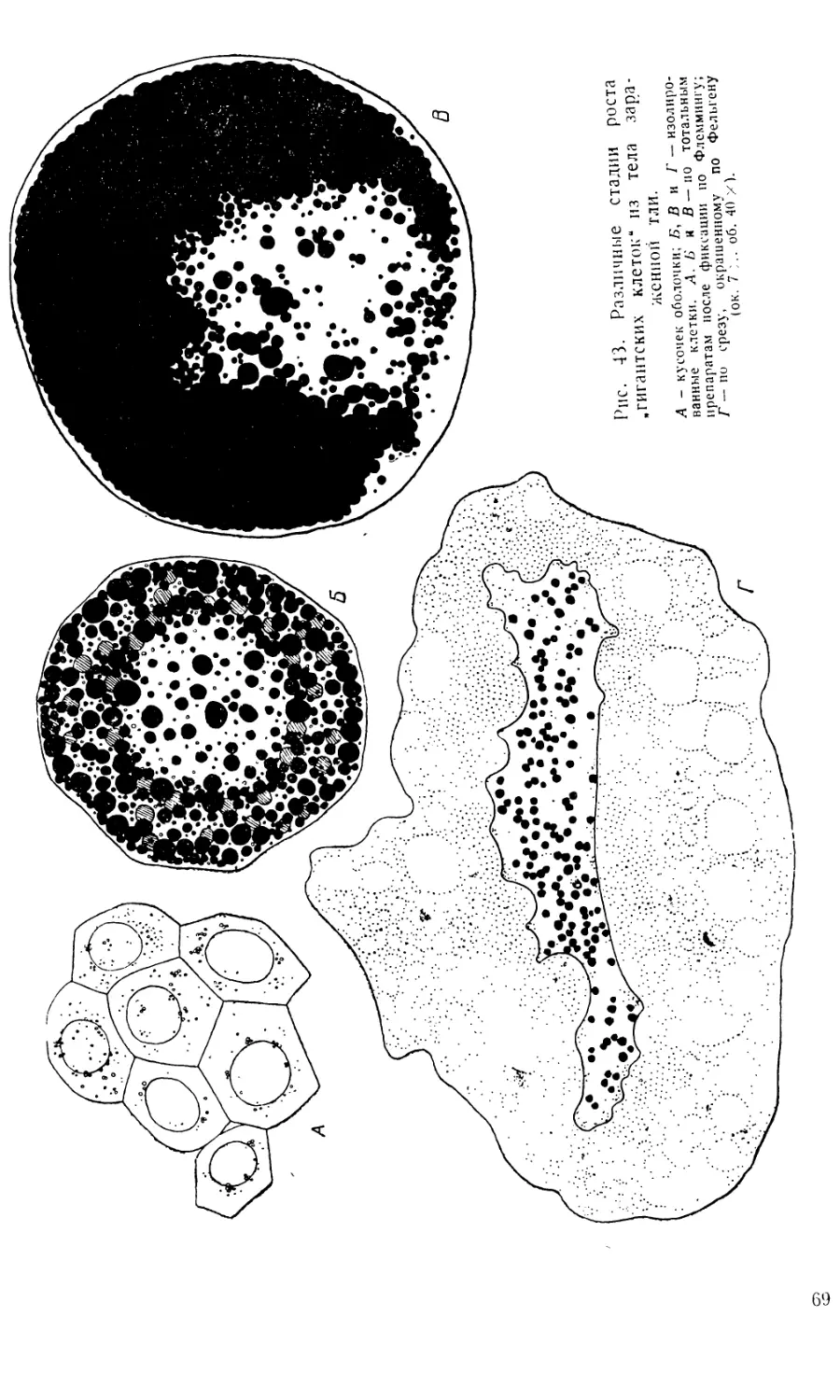
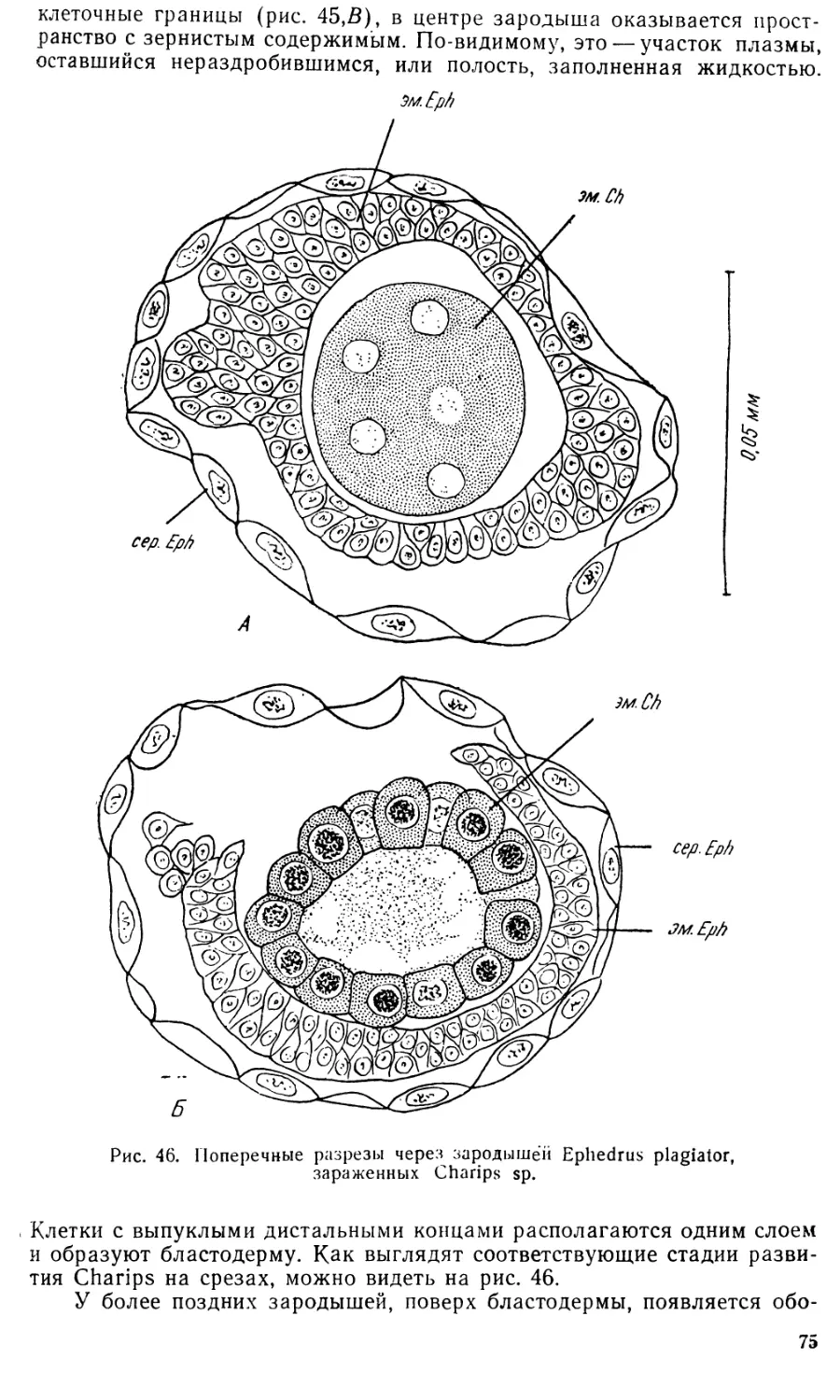
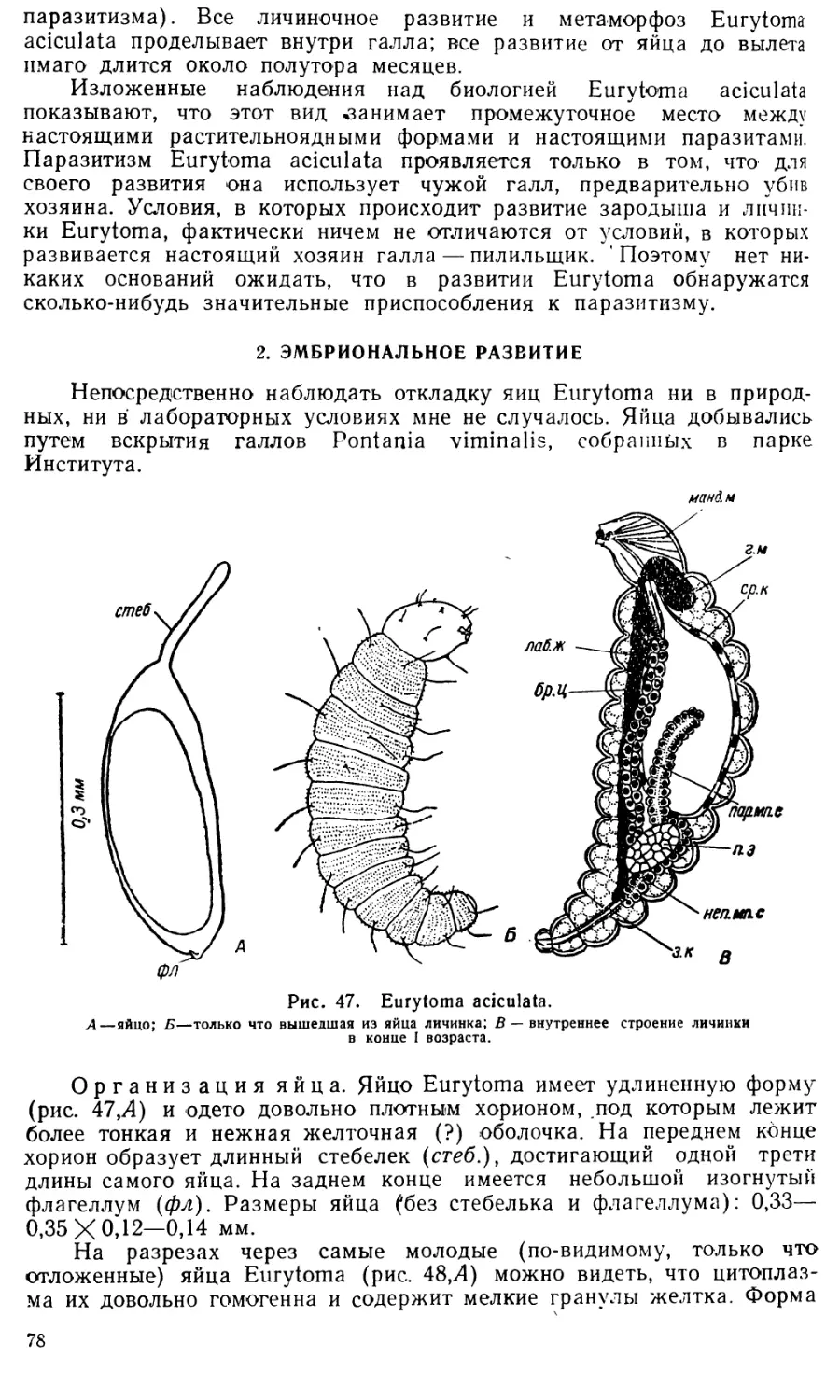
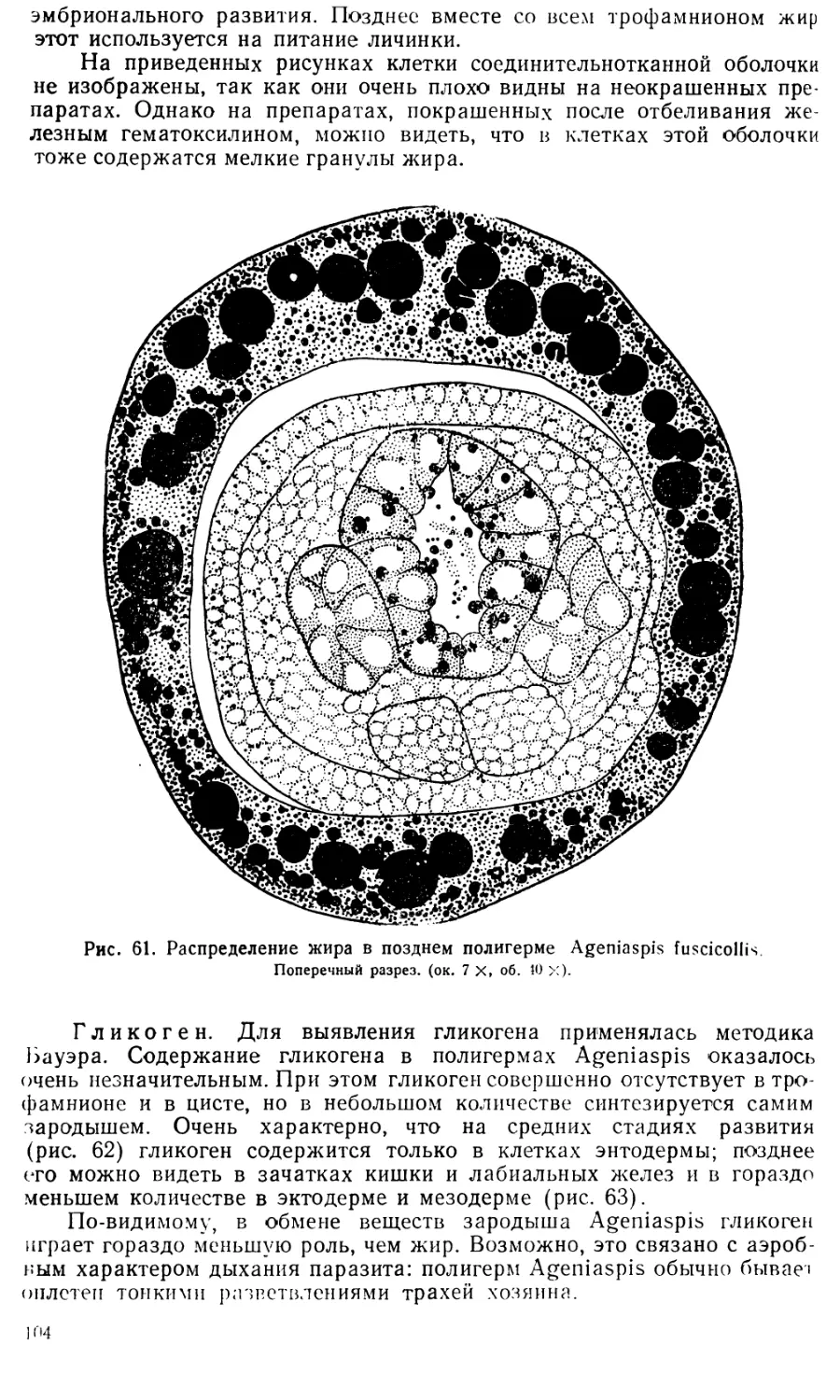
Увеличение размеров клеток серозы очень значительно: если в
момент их диссоциации диаметр их составляет 0,028—0,035 лш, то в
дальнейшем он достигает 0,26—0,29 мм и более (ср. рис. 43 Л и С).
По мере роста разъединившихся клеток оболочки в их плазме
накапливаются многочисленные, сперва мелкие, затем более крупные
блестящие гранулы. Эти гранулы окрашиваются Суданом III в оранже-
вый цвет и чернятся осмиевой кислотой (рис. 43Л, Б, В). Очевидно,
это капельки жира. Под действием люголя «гигантские клетки» окра-
шиваются в интенсивный коричневый цвет, а в проходящем свете ка-
жутся непроницаемо черными. Это дает основание предполагать присут-
ствие в них гликогена, но более точных гистохимических методов при-
менено не было. В периферической части клеток начинается накопление
«вакуолей, наполненных светлой жидкостью. К концу развития плазма
«гигантских клеток» вакуолизируется.
Накануне метаморфоза личинка Aphidius съедает все мягкие части
тли, от которой остается только хитиновый скелет (см. рис. 29). Такой
же участи, по-видимому, подвергаются и «гигантские клетки». Такое
же превращение после вылупления личинки испытывают и клетки эмб-
риональной оболочки Ephedrus.
Сходные явления наблюдал A. NA. Оглоблин (1913, 1924) при разви-
тии Dinocampus terminatus Nees. (Braconidae), паразитирующего в
жуках из сем. Coccinellidae. Эмбриональная оболочка образуется через
24 час после откладки яйца. «С этого момента, — пишет автор,-^-начи-
нается паразитическая жизнь Dinocampus, зародыш развивается и ра-
стет, за 8—10 дней (колебания зависят от температуры) объем его
увеличивается почти в 1000 раз». Оглоблин полагает, что сероза играет
во время эмбрионального развития двойную роль: снабжает зародыш
кислородом и питательными веществами, извлекаемыми из гемолимфы
хозяина, и наращивает хорион, который сильно растягивается, так что
5*
67
его поверхность увеличивается в 100 раз, а между тем он не становится
тоньше.
После вылупления личинки сероза распадается на отдельные клет-
ки, которые округляются и растут. Плазма их начинает краситься более
интенсивно, в ней появляются гранулы жира, гликогена, белка. Ядра
приобретают неправильную разветвленную форму, иногда фрагменти-
руются, что Оглоблин считает признаком интенсивного обмена между
ядром и плазмой, т. е. усиленного функционирования клеток. Сначала
клетки оболочки имеют диаметр 0,029—0,038 мм, к концу их существо-
вания он достигает 0,357 мм. Эти клетки служат единственной пищей
личинок Dinocampus. Таким образом, клетки эмбриональной оболочки
сохраняют здесь трофическое значение и в постэмбриональный период,
так как они извлекают питательные вещества из тела хозяина, накапли-
вают их в себе, после чего и поедаются личинкой паразита.
Совершенно аналогичные наблюдения сделаны Джексон (Jackson,
1928 и 1938) для Dinocampus rutilus Nees.: здесь оболочка тоже стано-
вится необходимым промежуточным звеном в процессе питания личин-
ки за счет хозяина. То же имеет место у Microctonus vittatae Muesb.
(Смит —Smith, 1952) и Meteorus (Zemiotes) nigricolHs Thomps. (Пар-
кер, 19316, цит. по Джексон, 1928).
Нечто подобное описано Маршалем (1906) для некоторых Platy-
gasteridae, у которых после вылупления личинки оболочка распадается
на куски, которые свободно плавают в полости тела хозяина (личинки
различных Cecidomyidae), растут, «размножаются» и превращаются
в многрядерные образования, имитирующие зародышей паразита («псев-
догермы»). Возможно, они тоже играют какую-то роль в питании ли-
чинок паразита.
Какова же роль эмбриональной оболочки при развитии Aphidius?.
Раннее образование оболочки Aphidius (еще во время дробления,
когда впервые возникают клеточные границы и число клеток не превы-
шает десятка) указывает на важную роль, которую сероза играет в
жизни зародыша, так как прогрессивные и важные для вида органы
обычно закладываются раньше и развиваются быстрее остальных (Ме-
нерт — Mehnert, 1897). Это положение особенно справедливо по отно-
шению к органам, специфическая функция которых связана с условиями
зародышевой жизни. Вспомним, что у мелкопитающих и особенно у
приматов внезародышевые части успевают достичь очень сложного стро-
ения — возникают синцитиальный трофобласт, внезародышевая мезен-
хима, амниотический пузырек и желточный мешок — и только тогда
появляется первый намек на образование самого зародыша—зароды-
шевая пластинка.
Среди паразитических перепончатокрылых, особенно полиэмбрио-
нических,. известен ряд случаев еще более раннего обособления клеток
эмбриональной оболочки, чем у Aphidius. Трофическая роль оболочки
часто не вызывает сомнений, почему и принято называть эту оболочку
трофамнионом, хотя правильнее было бы называть ее трофосерозой.
В какой мере можно приписывать трофическую функцию обо-
лочке афидиид?
Как известно, наступление гистологической дифференциации и вы-
полнение специальных функций обычно тормозят деление клеток. В этом
отношении клетки серозы Aphidius, рано перестающие делиться, ведут
себя как дифференцированные функционирующие элементы. Опыты с
прижизненным окрашиванием яиц Aphidius метиленовой синькой и ней-
тральным красным показывают, что хорион и сероза проницаемы для
этих красителей. По всей вероятности, не препятствуют они и проникно-
68
вению питательных веществ из тела тли к зародышу Aphidius. В то же
время можно предполагать, что сероза играет при этом активную роль:
проявляет избирательную проницаемость или как-то изменяет прохо-
дящие через нее вещества, почему под оболочкой создается специаль-
ная среда, необходимая для нормального развития зародыша. За это
говорят следующие наблюдения.
Хотя зародыш Aphidius имеет довольно сложные очертания, тонко-
стенный пузырек, образованный оболочкой, всегда сохраняет правиль-
ную сферическую форму, как бы под влиянием- внутреннего давления,
тургора, препятствующего спаданию его стенок. На это указывают и
сильно растянутые стенки серозы, несмотря на наличие довольно зна-
чительного пространства между ней и зародышем. При вскрытии зара-
женных тлей сероза часто разрывалась, при этом зародыш почти всегда
выталкивался из нее, а клетки серозы сокращались и становились более
выпуклыми.
Зародыши, выпавшие из серозного пузырька и попавшие в физио-
логический раствор (использовался 0,7% раствор NaCl и смесь Бидла
и Эфрусси), очень быстро утрачивали свои первоначальные очертания
и распластывались на дне солонки бесформенной массой. Уже через
1—2 мин появлялись признаки распада. Зародыши же с неповрежден-
ной оболочкой сохраняли нормальный вид в течение нескольких часов.
Очевидно использованные физиологические растворы являются не-
благоприятной средой для зародыша Aphidius, а эмбриональная оболоч-
ка способна в какой-то мере предохранить зародыш от их воздействия.
Возможно, что и полостная жидкость тли не является благоприятной
средой для развития зародыша Aphidius. На срезах через тлей Chai-
tophorus sp., зараженных Aphidius fabarum, несколько раз попались
зародыши с поврежденной серозой. Сам зародыш при этом был совер-
шенно дезинтегрирован: он представлял собой кучу клеток, не связан-
ных друг с другом.
Таким образом, целость эмбриональной оболочки есть необходимое
условие нормального развития зародыша Aphidius; оболочка изолирует
его от внешней среды и под ней создаются нужные для нормального
развития физико-химические условия. С другой стороны, оболочка
обладает известной независимостью от самого зародыша. Иногда, по
невыясненным причинам, зародыш Aphidius погибает внутри совершенно
неповрежденной оболочки. Однажды в теле тли Acyrtosiphon caraganae
была найдена группа «гигантских клеток», а личинка паразита отсут-
ствовала. По-видимому, это результат ранней гибели зародыша Aphi-
dius. Очевидно, присутствие нормального зародыша не необходимо для
дальнейшего развития клеток серозы и превращения их в «гигантские
клетки».
Сходные явления наблюдала Джексон (1923) у Dinocarnpus rutilis;
если зародышу Dinocarnpus случится погибнуть еще внутри хозяина,
клетки эмбриональной оболочки так же диссоциируются и растут вну-
три хориона, как при вылуплении личинки.
То же самое отмечает и Смит (Smith, 1952) для Microctonus vit-
tatae, и при этом делает любопытное наблюдение, что, когда личинка
Microctonus впадает в диапаузу, рост диссоциированных клеток серозы
приостанавливается. По-видимому, одни и те же причины задерживают
развитие личинки и рост «гигантских клеток».
Итак, можно полагать, что сероза афидиид служит для питания
зародыша и участвует в создании необходимой для его нормального
развития среды. Труднее решить каково значение клеток серозы после
вылупления личинки. Спенсер и Янишевская склонны были ви'деть в
70
гипертрофии «гигантских клеток» дегенеративные изменения, а
А. А. Оглоблин — функциональные. По Оглоблину (1924), личинки
Dinocampus не питаются непосредственно тканями хозяина: пищей им
служат эти гипертрофированные клетки.
У Aphidius «гигантские клетки» тоже в конце концов поедаются
личинкой, но личинка Aphidius не утрачивает способности питаться и
тканями хозяина. Поэтому развитие Dinocampus надо считать более
специализированным, а трофическая роль серозы в постэмбриональный
период у Dinocampus более значительна.
Итак, раннее обособление эмбриональной оболочки, прекращение в
ней клеточных делений, указывающее на дифференцированное состоя-
ние клеток, и немедленная гибель зародыша при ее повреждении, дают
основания считать эту оболочку важным органом зародыша, выполня-
ющим защитную и трофическую функции. Переживание клеток серозы
после вылупления личинки, их дальнейший рост и накопление запасных
питательных веществ, возможно, связаны с сохранением у них трофиче-
ских функций и в постэмбриональный период.
3. СРАВНЕНИЕ РАЗВИТИЯ EPHEDRUS И APHIDIUS
Интересно сопоставить развитие рассмотренных выше представи-
телей сем. Aphidiidae.
Несмотря на то, что Aphidius и Ephedrus представляют собой два
рода небольшого, слабо дифференцированного морфологически и био
логически семейства (Теленга, 1950), различия между ними проявля-
ются очень рано. На всех рассмотренных стадиях развития зародыши
Aphidius и Ephedrus различаются как по ряду чисто поверхностных,
бросающихся в глаза признаков, так и по течению внутренних процес-
сов. Некоторые из этих различий не имеют большого значения (форма
яиц), другие более или менее тесно связаны с существенными эмбрио-
логическими процессами. Так, образование эмбриональной оболочки
происходит у Ephedrus на более поздней стадии, чем у Aphidius. Вовремя
обособления зачатка оболочки зародыш Aphidius состоит только из
восьми клеток, а зародыш Ephedrus содержит уже множество ядер.
Это определяет и способ образования оболочки (дифференциация бла-
стомеров у Aphidius и деламинация у Ephedrus). Этим же объясняется,
что зачатки оболочки у Aphidius и Ephedrus с самого начала отлича-
ются по числу составляющих их клеток, а так как клерки оболочки
рано перестают делиться, это отличие сохраняется до конца.
Многоклеточность серозы у Ephedrus в свою очередь обусловливает
меньшую растянутость клеток, менее правильную форму пузырька, об-
разованного серозой, и дает возможность зародышу располагаться в
нем более свободно, не образуя такого крутого изгиба, как у Aphidius.
Но самое интересное состоит в том, что в развитии Aphidius наблю-
дается гораздо меньше палингенетических особенностей. Так, дробление
Aphidius рано становится тотальным; это влечет за собой выпадение
стадии бластодермы, никаких гомологов желточных ядер у Aphidius
не наблюдается, зародышевая полоска не формируется вовсе.
Способ образования эмбриональной оболочки у Aphidius более
изменен, чем у Ephedrus, и процесс этот сдвинут на более раннюю ста-
дию. Так как есть основания полагать, что сероза выполняет весьма
существенные функции, эти изменения имеют адаптивное значение.
Таким образом, Aphidius делает, по сравнению с Ephedrus, следующий
шаг эволюции в сторону адаптации к паразитическим условиям разви-
тия зародыша и еще больше уклоняется от исходного общего типа
7!
развития насекомых. По характеру эмбрионального развития Ephedrus
представляется более примитивным родом; по-видимому, этот вывод
можно распространить и на имагинальные стадии: жилкование крыла
у Ephedrus гораздо менее редуцировано, чем у Aphidius.
Однако из изложенных данных вытекает еще один, несколько неожи-
данный вывод: темпы морфологической эволюции могут не совпадать
с темпами физиологической адаптации. Одной из сторон физиологиче-
ской адаптации паразита к хозяину, интересующей нас в данном случае,
является способность зародыша паразита питаться за счет хозяина,
которая выражается количественно в эмбриональном росте. Хотя
Ephedrus морфологически отстал от Aphidius, физиологически он его
опередил, так как у него наблюдается гораздо более значительное
увеличение зародыша за время эмбрионального развития. Впрочем воз-
можно, что различное увеличение объема зародышей Aphidius и Ephed-
rus объясняется различной продолжительностью эмбрионального раз.-
вития: у Aphidius оно длится 3 суток, у Ephedrus — около 4.
ГЛАВА V
CHARIPS SP.
При сборе материалов по развитию афидиид было замечено, что
иногда личинки и даже зародыши Ephedrus бывают в свою очередь
поражены внутренним паразитом, который при ближайшем рассмотре-
нии оказался принадлежащим к сем. Figitidae (подотр. Parasitica,
Cynipoidea — орехотворки), к роду Charips. Большинство орехотворок
вторичнорастительноядны, представители некоторых семейств (в том
числе Figitidae) ведут паразитический образ жизни. Относительно
эмбрионального развития орехотворок почти ничего не известно. Хэви-
ленд (1921) описывает начальные стадии развития трех видов орехо-
творок, которые являются вторичными паразитами (гиперпаразитами)
тлей (Charips victrix Westw., Bothryoxysta curvata Kjeff и Alloxysta ery-
throthorax Hartig). Для них характерны очень мелкие яйца (0,0ЮХ
X 0,006 мм)у лишенные желтка, поверхностное дробление и наличие
одной эмбриональной оболочки. Джеймс (James,' 1928) тоже отмечает
наличие одной оболочки у Figites anthomyarum Bouche. Судя по его
рисункам, объем зародыша Figites увеличивается во время развития
приблизительно в 12 раз. Трофическая оболочка и довольно значитель-
ный эмбриональный рост (линейные размеры зародыша увеличиваются
в 3—8 раз) имеют место у Ibalia leucospoides Hoch., паразитирующей
в личинках Sirex (Кристал — Chrystal, 1930, цит. по Клоузен — Clausen,
1940). Об эмбриональном развитии растительноядных видов вообще
ничего не известно.
Слабая изученность орехотворок с эмбриологической точки зрения
послужила побудительной причиной для попытки собрать материалы
и по развитию Charips. Однако работа с гиперпаразитами представляет
значительные технические трудности и требует больше времени и вни-
мания, чем.те, которые можно было уделить этому объекту. Гиперпара-
зит стоит в конце четырехчленной пищевой цепи: 1 —кормовое растение
(в данном случае — карагана), 2 — насекомое-хозяин (тля), 3 — пара-
зит (Ephedrus) и 4 — гиперпаразит (Charips). Каждое из этих звеньев —
живой организм; достаточно неполадок в одном звене, и вся цепь будет
нарушена.
Поэтому по Charips удалось сделать немного: получена откладка
яиц in vitro, наблюдались прижизненно начальные стадии развития
if несколько зародышей Charips было обнаружено на препаратах, изго-
товленных из зародышей Ephedrus.
Обычно в литературе указывается, что паразитические орехотворки
заражают афидиид на стадии личинки III — IV возраста (Хэвиленд, 1921;
73-
лочка, состоящая из уплощенных линзообразных клеток (рис. 45,Д,
сер). Происхождение оболочки не прослежено. Можно лишь предпола-
гать, что она образуется, как у Ephedrus, путем деламинации. Эти
наблюдения подтверждают данные Хэвиленд, но, к сожалению, не идут
дальше.
Несмотря на всю неполноту сведений, которыми мы располагаем
относительно развития орехотворок, все же их можно суммировать
следующим образом. Эндопаразитические орехотворки утрачивают жел-
ток, но сохраняют поверхностное дробление. Имеется одна эмбриональ-
ная оболочка (сероза), способ образований которой, очевидно, сильно
изменен вторично. Несомненно имеет место эмбриональный рост, выра-
женный у разных представителей в различной степени. Общий характер
эмбрионального развития очень напоминает то, что мы уже видели у
Aphidiidae.
ГЛАВА VI
EURYTOMA ACICULATA RATZ.
Представители сем. Eurytomidae (подотряд Parasitica, Chal-
cidoidea) отличаются большим разнообразием образа жизни. Здесь
наряду с чисто растительноядными формами встречаются хищники-
яйцееды, настоящие паразиты и так называемые инквилины, отклады-
вающие яйца в галлы других перепончатокрылых, за счет которых
и питаются их личинки, предварительно убив (а иногда и высосав),
личинку хозяина галла (Никольская, 1952, 1956). Малышев (1947,
1949а, 1956, 1959), посвятивший ряд статей вопросам филогении и эво-
люции инстинктов у перепончатокрылых, предполагает, что инквили-
инзм был необходимой ступенью эволюции от растительноядности
к паразитизму, однако растительность современных Eurytomidae он
считает вторичной (Малышев, 1957, 1959). Того же мнения относи-
тельно фитофагии эвритомид придерживается Никольская (1956).
Большая биологическая пластичность эвритомид дала основание
полагать, что развитие представителей этого семейства еще не при-
обрело черт узкой приспособленности к какому-нибудь одному опреде-
ленному образу жизни. В полном соответствии с этим предположением
находится сравнительно мало специализированный характер развития,
обнаруженный у Eurytoma aciiculata (Иванова-Казас, 19586).
1. БИОЛОГИЯ
Eurytoma aciculata Ratz. развивается в галлах пилильщика Роп-
tania viminalis L. на иве и становится причиной гибели истинного
хозяина галла.
Обычно зародыш или личинка Pontania погибают еще до того, как
из яйца Eurytoma выйдет личинка. Очевидно, они погибают еще в мо-
мент откладки яйца самкой Eurytoma, но в очень редких случаях
(около .0,5%) яйца Eurytoma попадаются вместе с совершенно нор-
мальными личинками Pontania.
Обычно в галл Pontania откладывается только одно яйцо Eury-
toma, но иногда можно в одном галле встретить 2 и 3 яйца Eurytoma
и больше — результат многократного заражения. При вылуплении из
этих яиц нескольких личинок одна из них убивает остальных. По-види-
мому, личинка Eurytoma aciculata питается только растительной
пищей. Никаких признаков питания личинок Eurytoma за счет личинок
Pontania наблюдать не приходилось. Таким образом, Eurytoma acicu-
lata дает пример инквилинизма в чистом виде( без элементов экто-
77
гранул желтка у Eurytoma варьирует: чаще всего она шаровидная,
но иногда бывает слегка вытянутая или даже палочковидная
(рис. 53,£). Количество желтка и скорость его исчезновения тоже
непостоянны: иногда желток почти полностью рассасывается на ста-
дии зародышевой полоски, а иногда он сохраняется в довольно значи-
тельных кличествах даже после образования средней кишки. Но
в общем, в яйцах Eurytoma по сравнению с большинством насекомых
желтка не очень много.
Желток распределяется в яйце не срвсем равномерно: на обоих
концах имеются значительные скопления плазмы, свободные от желтка.
Тонкий периферический слой плазмы по всей поверхности яйца тоже
почти не содержит желтка. Переднее скопление плазмы продолжается
в стебелек яйца (рис. 48,В).
На рис. 48,Л недалеко' от переднего конца яйца, близ его поверх-
ности, виднеется группа хромосом. По-видимому, это фигура деления
созревания (мей). Ближе к заднему концу, на той же стороне яйца,
находится несколько крупных бесформенных комков, окрашивающих-
ся гематоксилином более интенсивно, чем окружающая плазма. Как
показыйает дальнейшее развитие, это — оосома (о). В нескольких ме-
стах в плазме яйца встречаются нежные звездообразные .фигуры
(зв\ на рисунке они изображены несколько утрированно). Нахтсгейм
(Nachtsheim, 1913) описывает, что у оплодотворяемых яиц медоносной
пчелы имеет место физиологическая полиспермия, и каждый спермий,
проникший в яйцо, образует вокруг себя плазматическую лучистость.
Может быть, зведчатые образования Eurytoma имеют тоже отношение
к полиспермии, но никаких структур, напоминающих ядро в состоянии
покоя, деления или дегенерации подле них обнаружить не удалось.
Дробление яйца Eurytoma происходит по обычному для насекомых
поверхностному типу. Рис. 49,Л изображает поперечный разрез через
дробящееся яйцо Eurytoma, на котором видны три кариокинетические
фигуры. Рис. 48,Б изображает продольный разрез через яйцо Eurytoma
на стадии шестнадцати ядер дробления. На обоих рисунках можно
видеть, что в протоплазме яйца появляются вакуоли, наполненные
водянистым содержимым. Ядра занимают центральное положение.
Оосома лежит уже не у поверхности яйца, а по его оси, приблизитель-
но на расстоянии 7з всей длины яйца от его заднего конца.
По мере дальнейшего дробления, число ядер увеличивается и они
приближаются к поверхности яйца (рис. 48,В). Количество вакуолей
в плазме возрастает, и они сливаются друг с другом в центре яйца
в общее пространство, в котором рассеяны гранулы желтка. Оосома
приобретает характер зернистой массы, лежащей на самом заднем
конце яйца.
Формирование бластодермы. Затем ядра выходят на
поверхность, и окружающие их плазматические островки сливаются
с периплазмой и образуют бластодерму, имеющую сперва синци-
тиальное строение. Формирование бластодермы происходит одновре-
менно по всей поверхности яйца (рис. 48,Г и 49,£). В середине яйца
остаются лишь отдельные ядра, окруженные некоторым количеством
плазмы — ядра желточных клеток (ж. кл). Желточные клетки не
имеют резких границ, они соединяются плазматическими перемычка-
ми друг с другом и с периферической плазмой.
В то время когда вся бластодерма представляет собой еще непре-
рывный плазматический слой без клеточных границ, на заднем конце
яйца полностью обособляется несколько крупных клеток, в плазму
которых попадает темная зернистость оосомы. Они образуют неболь-
£2
шую группу, лежащую вне бластодермы на заднем конце зародыша.
Это—половой зачаток (рис. 48,Г, п. з.).
Плазма бластодермы становится неоднородной (рис. 49,£).
У поверхности располагается слой довольно светлой гомогенной плаз-
мы, глубже —правильным рядом лежат ядра, за ними следуют более
темнокрасящийся слой, а еще глубже — слой, состоящий из светлой
тонко вакуолизированной плазмы, переходящей без резкой границы
в центральную плазму, содержащую желток, вакуоли и желточные
ядра.
Светлые пузыревидные ядра бластодермы имеют эллипсоидальную
форму. Хроматин в них концентрируется на том конце ядра, который
обращен к поверхности яйца. Такое расположение хроматина в ядрах
бластодермы наблюдается и у некоторых других перепончатокрылых
(см. рис. 68 и 69). Гетенби (1917, 1918) пытается объяснить это пред-
полагаемым питанием зародыша через бластодерму. В случае Euryto-
ma, яйца которой развиваются в воздушной среде внутри галла, о"та-
ком патании не может быть и речи. Очевидно, упомянутая полярность
клеток и их ядер не имеет отношения к питанию зародыша.
Ядра желточных клеток вскоре приобретают неправильную фор-
му. На многих препаратах они имеют вид плотных бесформенных ком-
ков; очевидно, в них рано начинаются дегенеративные изменения
(рис. 49,В, Г, ж. кл).
Итак, на этой стадии развития в зародыше уже отчетлива выра-
жена дифференциация частей: различаются бластодерма, желточные
клетки, половой зачаток. В самой бластодерме наблюдается зональ-
ная дифференциация плазмы. Затем по всей поверхности яйца по-
являются глубокие борозды, намечающие границы отдельных клеток
бластодермы (рис. 49,Г, 54,Л). Сперва эти борозды врезаются только
до глубины более темного плазматического слоя. Полное разделение
бластодермы на клетки происходит позднее (рис. 51,Л, 54£). По-
является и резкая граница, отделяющая бластодерму от центральной
части яйца. Теперь в бластодерме различается только два слоя плаз-
мы— более темный периферический и более светлый, слабо вакуоли-
зированный внутренний. Половой зачаток (рис. 54,5, п. з.) начинает
погружаться внутрь и оказывается в толще бластодермы.
Почти одновременно, с появлением клеточных границ, появляются
и различия в толщине бластодермы: на брюшной стороне она очень
толста и состоит из высоких узких цилиндрических клеток, а на спин-
ной стороне бластодерма значительно тоньше и образована короткими
широкими клетками (рис. 51,Л). Толстая и тонкая области бластодермы
соответствуют зародышевой и внезародышевой частям, без резких гра-
ниц переходящим друг в друга. »
На брюшной стороне зародыша обозначаются две слабые про-
дольные бороздки (на поперечном разрезе они выглядят как две не-
большие выемки), которые делят утолщенную бластодерму продольно
на три части: медианную пластинку (зачаток мезэнтодермы;
рис. 51,Л, мед. п) и. две боковые пластинки (зачаток эктодермы;
рис. 51А б. п). Эти изменения в бластодерме знаменуют собой начало
гаструляции.
Особенности деления ядер на стадии бластодер-
мы. Дробление Eurytoma происходит синхронно, (рис. 49,Л). Эта
синхронность сохраняется некоторое время и на стадии бластодермы.
Одновременно делятся не только все ядра бластодермы, но и желточ-
ные ядра и ядра полового зачатка.
Затем дифференциация частей начинает сказываться и на размно-
6*
83
мзнт
0J мм
Рис. 51. Поперечные разрезы через зародышей Eurytoma aciculata
на стадиях гаструляцни.
А — стадия неравномерной бластодермы; Б — погружение мезэнтодермы.
85
волны митозов у Eurytoma соответствует центру дифференциации
пчелы и других насекомых. На более поздних стадиях развития мне
уже не случалось наблюдать, массовых (синхронных или гетерохрон-
ных) митозов.
Гаструляция. Бороздки, разделяющие брюшную бластодерму
на срединную пластинку и боковые части, сперва намечаются на
переднем конце, а затем распространяются по всей длине зародыша.
Срединная пластинка начинает погружаться, а боковые части бласто-
дермы надвигаются на нее с боков (рис. 51,5). Смыкание боковых
пластинок сперва происходит спереди, но полное их слияние в непре-
рывный эпителиальный слой, полностью отграниченный от погрузив-
шегося клеточного материала, происходит раньше в средней части
зародыша, так как спереди долго сохраняется глубокая медианная
бороздка, и создается впечатление, что здесь продолжается уход
клеток с поверхности внутрь, -но не целым пластом, а поодиночке.
Поперечные срезы и реконструкция зародыша на описываемой стадии
изображены на рис. 52.
Впрочем надо отметить, что после полного погружения медианной
пластинки внутрь нередко можно наблюдать картины, говорящие
о выселении мезэнтодермальных клеток вдоль медианной линии по
всей длине зародышевой полоски.
Разделение у Eurytoma зародышевой полоски на медианную
пластинку и две боковые и образование мезэнтодермы из медианной
пластинки напоминают соответствующие процессы у Aculeata, для
которых они очень характерны. Такой способ гаструляции отмечен
для всех изученных с эмбриологической точки зрения жалящих пере-
пончатокрылых: для Chalicodoma muraria F. (Карьер и Бюргер,
1898), для ряда муравьев (Танквари — Tanquary, 1913; Стриндберг,
1913а и б), для Vespa vulgaris L. (Стриндберг, 1914), для Apis
mellifera L. (Снодграсс — Snodgrass, 1925), для Ammophila campestris
Jur. (Бэрендс и Бэрендс Ван Роон, 1949) и для Scolia quadripunctata F.
(Дондуа, 1953). В то же время этот способ не встречается у других
перепончатокрылых. Как мы видели, у пилильщиков мезодерма
образуется путем иммиграции клеток из первичной бороздки — спосо-
бом, наиболее распространенным у насекомых, который и надо считать
первичным (во всяком случае в пределах отряда Hymenoptera).
То же имеет место, у паразитических перепончатокрылых; Eurytoma
aciculata — исключение. Второе исключение представляет Pimpla
turionellae (L.), у которой, по наблюдениям Бронскилл, мезодерма
также образуется путем погружения медианной пластинки.
Любопытно, что у низших двукрылых, например у Anopheles
maculipennis Mg. (Иванова-Казас, 1949а), зародышевые листки обра-
зуются сходно с пилильщиками, а у высших, например у мух (по
Ковалевскому, 1886, и более поздним авторам), сходно с жалящими,
так что имеется известный параллелизм в эволюции гаструляции
у двукрылых и перепончатокрылых.
Образование зародышевой полоски и серозы.
Утолщенная область бластодермы, в которой происходит образование
мезэнтодермы, имеет форму зародышевой полоски, занимающей всю
брюшную поверхность яйца и загибающейся на переднем и заднем
концах на спинную сторону. Внезародышевая бластодерма (зачаток
серозы) занимает небольшую область в средне-спинной части зароды-
ша (рис. 52,£, Д, сер). На заднем конце зародышевой полоски имеет-
ся довольно глубокая медианная борозда, представляющая собой
«6
зачаток проктодеума (рис. 52,5, Г, прокт). Дно этой борозды в самой
глубокой ее части занимает половой зачаток (рис. 52,Г, п. з).
Интересные гистологические изменения происходят во внезароды-
шевой бластодерме. Здесь во внутренних концах клеток образуются
большие вакуоли, оттесняющие ядро и плазму клетки к ее наружному
концу. В результате внутренние концы клеток как бы запустевают
(рис. 52,£, 54,5 сер). Иногда в эти пустоты попадают гранулы
желтка. Границы клеток сохраняются довольно долго; особенно дли-
тельно различается базальная мембрана (рис. 53,Л, сер).
Передний край внезародышевой бластодермы \ отделяется от
зародышевой полоски и начинает на нее нарастать. Затем то же самое
происходит на боках зародыша (рис. 53,Л) и на заднем его конце.
Постепенно внезародышевая бластодерма одевает всего зародыша
непрерывной однослойной оболочкой (рис. 53,5, В, Г, 55, сер.).
Оболочка (сероза) принимает форму очень тонкой перепонки,
утолщающейся только в тех местах, где лежат ядра. Такой способ
формирования серозы (без образования амниона путем нарастания
внезародышевой бластодермы на зародышевую) мы уже видели
у Angitia vestigialis. Он довольно обычен у жалящих перепончато-
крылых.
Одновременно с образованием серозы зародышевая полоска
Eurytoma несколько удлиняется, а расстояние между ее передним
и задним концом на спинной стороне сокращается (рис. 55).
Образование стомодеума, проктодеума и начало
сегментации. Продольное щелевидное впячивание на заднем
конце зародышевой полоски принимает форму трубки с узким просве-
том, один конец которой кончается слепо», а другой—открывается
наружу (в пространство, ограниченное серозой). Слепой конец этой
трубки направлен внутрь зародыша и несколько вперед. Эта трубка
представляет собой проктодеум. Половой зачаток к этому времени
утрачивает связь с проктодеумом и лежит среди клеток мезэнтодермы.
Несколько позднее на переднем конце зародышевой полоски по-
является глубокое цилиндрическое впячивание — стомодеум (рис. 55,
стом),- а в области проктодеума образуются зачатки мальпигиевых
сосудов (рис. 55, неп. мп. с). Так как передний и задний концы за-
родышевой полоски загибаются на спинную сторону, то оба эти
эктодерм альные впячивания (стомо- и проктодеум) закладываются
на спинной стороне и лишь позднее, когда происходит укорочение
зародышевой полоски, занимают свои окончательные места.
Одновременно с укррочением зародышевой полоски происходит ее
рост в ширину, так что боковые края зародышевой полоски смыкают-
ся на спине. Вскоре после закладки стомодеума начинается сегмен-
тация зародыша (рис. 54, В).
Формирование средней кишки. Еще во время образова-
ния серозы становится заметным, что вакуолизированная центральная
плазма, содержащая желток и желточные ядра, ограничена тонкой
перепонкой. Изнутри к этой перепонке местами примыкают скопления
более плотной плазмы, содержащей ядра (рис. 53,5, ж. /ел). Эти
плазматические островке с ядрами ничем не отличаются от желточных
клеток, лежащих среди желтка. Как от поверхностных, так и от
центральных желточных клеток отходят тонкие отростки. Вся цен-
тральная плазма представляется непрерывным сетевидным синцитием,
в петлях которого лежат гранулы желтка и вакуоли.
Скопление желточных клеток у поверхности желточной массы
описано и для некоторых жалящих перепончатокрылых: для Chalico-
88
Связь между средней кишкой и проктодеумом не устанавливается.
От слепого конца проктодеума вырастают три трубковидных маль-
пигиевых сосуда. Два из них растут прямо вперед, а третий отгибает-
ся к спинной стороне и растет назад (рис. 55, неп. мп. с). Появляются
лабиальные железы, имеющие форму двух длинных трубок, тянущихся
по бокам тела. Происхождение их осталось невыясненным. Из утолще-
ний эктодермы образуются зачатки головного мозга и брюшной
цепочки.
Половой зачаток разделяется на две группы клеток, лежащие
справа и слева от проктодеума. Каждый зачаток одевается тонкой
соединительнотканной перепонкой, от ( которой назад тянется тяж
мезодермальных клеток—зачаток полового протока. Большая часть
мезодермальных клеток идет на построение мышц. К концу эмбрио-
нального развития образуется спинной сосуд, имеющий форму очень
тонкой трубки. На поперечном разрезе он состЪит только из двух кле-
ток. Гиподерма выделяет личиночную кутикулу с ее характерными
структурами.
3. ЛИЧИНКА I ВОЗРАСТА
Личинка I возраста Eurytoma aciculata (рис. 47,Б и В) имеет
веретеновидную форму. Тело личинки изогнуто на брюшную сторону
так, что она соприкасается с субстратом (стенкой галла) только лишь
в двух точках — головой и задним концом. Личинка способна пере-
двигаться наподобие пяденицы, но движения ее имеют беспорядочный
характер — то вперед, то назад, то в сторону.
Тело личинки состоит из небольшой головы и 13 сегментов тулови-
ща. На голове находятся маленькие антенны и мандибулы, причленяю-
щиеся к околоротовому кольцеобразному склериту. Сегменты тела
снабжены длинными щетинками: на каждом сегменте различаются две
спинные, две брюшные, а на первых четырех сегментах еще и две боко-
вые щетинки. На четвертом сегменте тела брюшные щетинки отсут-
ствуют. Кроме того, каждый сегмент опоясан широкой полосой мелких
шипиков, складывающихся в неправильные поперечные ряды.
Личинка I возраста снабжена парой длинных лабиальных желез
(рис. 47,£, лаб. ж). Пищевод ведет в объемистую мешковидную сред-
нюю кишку (ср. /с), которая не сообщается с задней (з. к). Последняя
имеет форму прямой трубки, от переднего конца которой отходят
три мальпигиевых сосуда; два из них идут вперед (пар. мп. с), а тре-
тий (спинной) направляется назад (неп. мп. с). По бокам от задней
кишки лежат два зачатка половых желез (п. з).
Нервная система представлена головным мозгом (г. м) и брюшной
цепочкой (бр. ц). Последняя неясно сегментирована, но содержит
хорошо развитое фибриллярное вещество. Клетки жирового тела, еще
немногочисленные и бедные включениями у только что вышедшей из
яйца личинки, к концу I возраста заполняют всю полость тела. Муску-
латура тела и мандибул развита хорошо. Трахейную систему удалось
обнаружить лишь у личинки U возраста.
4. АДАПТИВНЫЕ И СВОЕОБРАЗНЫЕ ЧЕРТЫ В РАЗВИТИИ
EURYTOMA ACICULATA
В эмбриональном развитии Eurytoma aciculata мы не встречаем
никаких я&ных признаков адаптации к паразитизму. В яйцах еще
сохраняется довольно много желтка. Процессы дробления, образова-
92
ния бластодермы, гаструляция, сегментация и т. д. протекают по
общей для всех насекомых схеме. Хотя из двух эмбриональных оболо-
чек, присущих насекомым, у Eurytoma развивается только одна —
сероза, редукция амниона у перепончатокрылых, по-видимому, не
имеет прямой связи с паразитизмом, так как начинается еще у пилиль-
щиков.
Что касается постэмбрионального развития, то для личинок
Eurytoma характерно полное отсутствие конечностей на груди и брюш-
ке и недоразвитие кишечника (отсутствие сообщения между средней
и задней кишкой, делающее невозможной дефекацию). Эти "особенно-
сти могли возникнуть, как приспособление к развитию в замкнутом
пространстве внутри галла. Правда, сам хозяин галла— личинка
пилильщика—не имеет таких адаптации: конечности и кишечник
хорошо развиты, а экскременты выделяются в полость галла, не
вызывая особенных осложнений в жизни личинки. Может быть,
в филогенезе Eurytoma период жизни в ограниченном пространстве
был более длительным, чем у пилильщиков.
Но вообще принято думать, что недоразвитие кишечника у пере-
пончатокрылых могло возникнуть только в условиях паразитизма.
Если мы примем, что жизнь внутри галла не является достаточной
причиной для недоразвития кишечника, то эктопаразитизм тоже не
может служить такой причиной, даже если он осуществляется внутри
галла, норки, ячейки сот или под куколочной кутикулой хозяина. Тогда
придется пойти дальше и признать, что недоразвитие кишечника яв-
ляется следствием эндопаразитизма, при котором выделение экскре-
ментов в тело хозяина действительно биологически очень невыгодно,
так как могло бы вызвать преждевременную гибель хозяина. Из этого
в свою очередь пришлось бы заключить, что не только Eurytomidae,
но и все жалоносные перепончатокрылые, у личинок которых средняя
и задняя кишка тоже разобщены, происходят от эндопаразитических
предков. Однако вторичный переход от эндопаразитизма к эктопарази-
тизму, и даже растительноядности, мало вероятен.
Особенности строения и функции кишечника можно (и даже
с большим основанием) поставить в связь с характером самой пищи.
Интересная мысль относительно причины возникновения разобщенности
кишечника у личинок всех стебельчатобрюхих перепончатокрылых
высказана Викторовым (1959). Этот автор сравнивает эктопаразити-
ческих личинок, питающихся гемолимфой, с кровососущими насеко-
мыми. Кишечник последних сильно раздувается от заглоченной кро-
ви, у них возникают специальные сфинктеры, препятствующие вытека-
нию* крови через анальное отверстие до конца пищеварения. Такой
же смысл, по мнению Викторова, имеет отсутствие сообщения между
средней и задней кишкой у личинок высших Hymenoptera. На этом
основании предполагается, что в филогенезе последних была экто-
паразитическая фаза.
По мнению Малышева (1959), недоразвитие кишечника высших
перепончатокрылых произошло в то время, когда их предки перешли
от инквилинизма к питанию яйцами других насекомых (хищная яйце-
едная или эктооофаговая фаза эволюции). Высокая питательность
такой пищи, усваивающейся почти полностью, сделала возможной
задержку дефекации до конца личиночной жизни. Итак, если разобщен-
ность отделов кишечника связана с паразитизмом или поеданием
яиц других насекомых (а других объяснений этого явления пока не
предложено), то наличие этой особенности у личинок Eurytoma гово-
рит за вторичный характер растительноядности этого насекомого.
93
В общем, развитие Eurytoma aciculata Ratz. имеет много общего
с развитием жалоносных перепончатокрылых. Оно несколько изменено
по сравнению с развитием первичнорастительноядных представителей
отряда (пилильщики), но сохраняет еще очень примитивный характер
по сравнению с развитием большинства паразитических форм. Эта
характеристика вполне соответствует примитивности эвритомид и их
слабой специализации как паразитов.
Однако одна особенность развития Eurytoma aciculata, сближаю-
щая его с таковым Aculeata— эпителизация мезодермального зачат-
ка,— не может считаться примитивной. В этом отношении развитие
Eurytoma и Aculeata обнаруживает вторичные изменения, которых нет
у Hymenoptera parasitica. Поэтому тип развития Eurytoma нельзя
считать исходным для эволюции развития паразитических перепончато-
крылых. В то же время это наводит на мысль о существовании генети-
ческой близости между Eurytomidae и Aculeata (Иванова-Казас, 1958).
Однако после обнаружения такого же способа закладки мезодермы
у Pimpla turionellae (L.) это последнее соображение кажется менее
убедительным.
ГЛАВА VII
AGENIASPIS FUSCICOLLIS (DALM.)
Ageniaspis (Encyrtus) fuscicollis (Dalm.) (Chalcidoidea, Encyrti-
дае) паразитирующий в гусеницах горностаевых молей из рода
Hyporiomeuta, знаменит тем, что это первое насекомое, у которого
было открыто' явление полиэмбрионии. Еще в 1891 г. Бюньон
(Bugnion, цит. по Martin, 1914, и др. авторам) обнаружил, что
в теле гусениц, зараженных Ageniaspis, содержатся длинные извитые
трубки, в которых заключены многочисленные зародыши паразита.
Маршаль (Marchal, 1898, 1904а, б) проследил развитие Ageniaspis
и доказал, что все эти зародыши возникают из одного яйца. Он назвал
это явление (развитие многих зародышей из одного яйца) полиэмбрио-
нией, после чего этот термин и вошел в зоологическую литературу.
Затем полиэмбриония была обнаружена и у ряда других наездников.
Развитие Ageniaspis fuscicollis изучалось также Сильвестри (1907)
и Мартином (Martin, 1914) и в общих чертах известно достаточно, хоро-
шо., но отдельные стороны этого процесса остаются еще не достаточно
ясными и нуждаются в дополнительном изучении. Поэтому, когда
в 1958 г. представилась возможность собрать кое-какие материалы по
Ageniaspis, я решила ею воспользоваться, чтобы лично ознакомиться
с развитием этого интересного насекомого.
1. ЛИТЕРАТУРНЫЕ ДАННЫЕ О РАЗВИТИИ AGENIASPIS
Ageniaspis fuscicollis имеет одногодичный цикл развития. В конце
лета самка Ageniaspis откладывает яйца в яйца моли. По наблюде-
ниям\Мартина (1914), яйца Ageniaspis имеют 0,025 мм в диаметре и со-
вершенно лишены желтка и других запасных питательных веществ.
Редукционные ядра лежат у переднего конца яйца, а оба пронуклеуса
располагаются ближе к заднему концу (рис. 56, Л). Образующийся
синкарион обособляется вместе с небольшим центральным участком
цитоплазмы, а вся периферическая цитоплазма с лежащими в ней ре-
дукционными ядрами превращается в оболочку — трофамнион
(рис. 56, Б, В, троф). Редукционные ядра распадаются па отдельные
зернышки, которые разбухают и снова сливаются в так называемый
парануклеус (я. троф). Заключенная в трофамнионе эмбриональная
клетка начинает дробиться.
Дробление Ageniaspis тотальное, т. е. разделяются и ядро, и плаз-
ма. Когда число бластомеров достигает 8—10, на поверхности зародыша
появляется циста, образующаяся за счет соединительнотканных клеток
хозяина (рис. 56, Б, соед. об)\ в стенке цисты разветвляются трахеи.
95
(так называемое «паразитическое тело» или «полигерм») быстро
растет, становится сперва колбасовидным, а затем приобретает харак-
тер разветвленных извитых тяжей. В июне паразитическое тело дости-
гает 5 см длины (не считая ответвлений) и 1/2—2 мм ширины (Мар-
шаль, 1904а). Таким образом, объем паразитического тела в конце
эмбриональною развития более чем в 4 миллиона раз превышает ис-
ходные размеры яйца. В это время в нем различаются уже вполне
сформированные личинки Ageniaspis.
Процесс разделения эмбрионального материала на отдельные
зародыши (полиэмбрионизация) в мае замедляется. В некоторых заро-
дышах число клеток достигает 60—80 и начинается прогрессивное разви-
тие, описанное Маршалем (1904с). Зародыш, представляющий собой
плотный многослойный комплекс клеток, приобретает бобовидную
форму. На спинной стороне появляется глубокое впячивание, на вы-
пуклой брюшной стороне — медианная бороздка, и намечается сег-
ментация.
Центральная масса клеток дает зачаток средней кишки. Перифе-
рическая многослойная масса клеток более толста на вентральной сто-
роне, тонка на боках и отсутствует в области дорзального впячивания.
По мнению Маршаля, этот утолщенный вентральный поверхностный
слой клеток соответствует зародышевой полоске других насекомых.
Передний и задний ее концы загибаются на спинную сторону и почти
соприкасаются друг с другом и разделены лишь дорзальной вогну-
тостью. Обособление мезодермы происходит вдоль медианной бороздки,
т. е. обычным образом.
Эмбриональное развитие завершается в июне. Молодая личинка
Ageniaspis достигает 1 мм длины. Она состоит из головы и 13 сегментов
и снабжена маленькими мандибулами. У нее есть хорошо развитые
слюнные железы и трахейная система. Первое время личинки питаются
трофамнионом (сходно с Dinocampus), затем выходят в полость тела
хозяина и начинают поедать ею ткани.
Полиэмбриония известна и у некоторых других представителей
сем. Encyrtidae, а также в сем. Braconidae, Platygasteridae и Dryinidae
(см. табл. 2). У большинства энциртид образование трофамниона про-
исходит несколько иначе: цитоплазма яйца разделяется на переднюю,
«полярную», часть, содержащую дериваты редукционных ядер, и зад-
нюю, «эмбриональную», часть, в которой лежит ядро яйца. Эмбрио-
нальная часть начинает дробиться, а полярная постепенно обрастает и
окружает ее со всех сторон, превращаясь в оболочку. Это имеет место
у Encyrtus mayri Masi (Сильвестри, 1915—1916), Copidosoma buyssoni
Мауг (Сильвестри, 1914), С. gelechiae How. (Лейби, 1922), С. koehleri
Blanch. (Даут —Doutt. 1947), Litomastix truncatellus (Dalm.) (Силь-
вестри, 1937) и L. floridanus (Ashm.) (Пэтерсон, 1921).
У полиэмбрионических платигастерид: Platygaster dryomyiae Silv.
(Сильвестри, 1916), PL hiemalis Forbes (Лейби и Хилл— Leiby and Hill,
1923), PI. vernalis Myers (Лейби и Хилл, 1924) трофамнион образуется
так же, как у Ageniaspis.
Ранние стадии формирования трофамниона у браконид и дриинид
пока не прослежены.
Обычно редукционные ядра перед тем, как образовать парануклеус,
утрачивают резкие контуры и распадаются на отдельные зернышка
(у Ageniaspis, по Мартину, 1914) или принимают «диффузный харак-
тер» (у Platygaster dryomyiae Silv.,) по Сильвестри, 1916). Затем пара-
нуклеус снова приобретает более четкие контуры; он растет и делится
амитотически или фрагментируется. Только у Litomastix truncatellus
7 О. М. Иванова-Казас
97
(Dalm.) (Сильвестри, 1937) три редукционных ядра сливаются в одно
триплоидное, которое затем размножается кариокинетически.
Многие авторы отмечают что поверхностный слой трофамниона
вступает в очень тесный контакт с тканями хозяина. Последние '-тасто
образуют вокруг паразитического тела так называемую цисту.
2. НЕКОТОРЫЕ НАБЛЮДЕНИЯ ПО БИОЛОГИИ
Материалы по развитию Ageniaspis fuscicollis (Dalm.) были собра-
ны в июне 1958 г. в учлесхозе «Лес на Ворскле» (Белгородская область).
В учлесхозе и его окрестностях Ageniaspis fuscicollis был обнаружен
в гусеницах Hyponomeuta malinellus Z. с яблони и Н. cognatellus Hb.
с бересклета. Гусеницы этих молей живут колониями и образуют на
своих кормовых растениях хорошо заметные паутинные гнезда. Колонии
Н. cognatellus гораздо чаще бывают поражены Ageniaspis: на европей-
ском бересклете было» поражено 58% колоний, причем зараженные гу-
сеницы составляли в этих колониях 48,5%; на бородавчатом бересклете
было поражено 18% колоний, а зараженные гусеницы составляли
в них 15%. Колонии Hyponomeuta с яблони были поражены лишь на
3%, но зараженные гусеницы составляли в них 54%. Возможно этот
более низкий процент заражения колоний моли на яблоне объясняется
тем, что наряду с Н. malinellus Z. здесь встречается Н. padellus L.,
гусеницы которых мрофологически плохо отличаются от первых. Гусе-
ницы Н. padellus, собранные на терне, ни разу не оказались заражен-
ными Ageniaspis.
Полигермы Ageniaspis добывались из гусениц путем вскрытия
последних в физиологическом растворе. Материал изучался прижиз-
ненно и в фиксированном состоянии. Иногда гусеницы из заведомо
зараженных колоний фиксировались целиком.
Как уже отмечалось, полигерм Ageniaspis имеет форму длинных
извитых и разветвленных шнуров, лежащих в полости тела гусеницы.
В начале июня зародыши располагаются внутри этих шнуров беспо-
рядочно, местами скопляясь неправильными комками. При этом от
взаимного давления зародыши сильно деформируются. Потом распо-
ложение зародышей упорядочивается: >все они ложатся в один ряд
с значительными интервалами и приобретают бобовидную форму. В это
время уже имеются мезодерма, стомо- и проктодеум и зачатки лабиаль-
ных желез, намечается сегментация. Тяжи полигерма становятся четко-
видными.
В «середине июня полигерм содержит уже вполне сформированных
личинок. Из-за их подвижности извлеченные из гусениц шнуры слабо
изгибаются. Еще до выхода личинок из оболочек в их кишечнике
появляется какая-то зернистая масса — по-видимому, личинки начи-
нают питаться трофамнионом. Полости, в которых лежат отдельные
личинки, сливаются, и шнуры превращаются в трубки, внутри которых
ползают личинки. Потом оболочки распадаются, и личинки Ageniaspis
оказываются непосредственно в полости тела гусеницы.
Развитие многочисленных зародышей Ageniaspis происходит не
вполне синхронно. Часто в разных частях полигерма оказываются заро-
дыши, находящиеся на разных стадиях развития. По всей вероятности,
отсутствие синхронности в развитии зародышей Ageniaspis зависит от
различных условий питания и газообмена в разных частях гусеницы
Hyponomeuta.
Так как время сбора материала ограничивалось лишь одним ме-
сяцем (июнь), то удалось получить зародышей лишь на средних
98
сителей были использованы метиленовый синий, нейтральный красный
и толуидиновый синий. Оказалось, что все эти красители очень
быстро проходят сквозь оболочки и окрашивают зародыш. При этом
обнаруживаются некоторые различия в окрашиваемости зародыша и
оболочек.
При помещении полигерма в метиленовую синь сначала происхо-
дит равномерное прокрашивание всех его частей. Однако уже через
20—30 мин. появляются некоторые различия: трофамнион красится
диффузно и более интенсивно, а зародыш и соединительнотканная
оболочка (циста) остаются более светлыми и накапливают краску
в виде гранул.
При использовании нейтрального красного первоначально тоже
получается равномерная диффузная окраска. Через 10 мин. в зародыше
и цисте начинается гранулообразованис, а через 40—50 мин. зародыш
становится вишнево-синим. При раздавливании его покровным стек-
лом видно, что окраска эта обусловлена наличием в клетках большого
количества гранул. Гранулы краски содержатся и в клетках соедини-
тельнотканной оболочки. К этому же времени трофамнион становится
совершенно бесцветным.
При использовании в качестве красителя толуидиновой сини тоже
проявляется способность тканей зародыша и цисты к гранулообразова-
нию, трофамнион окрашивается диффузно. При этом наблюдаются а
резкие отличия в оттенках. Гранулы, заполняющие клетки зародыша
и цисты, имеют чернильно-лиловый цвет, а трофамнион приобретает
грязный голубой тон с серым или зеленоватым оттенком.
Результаты этих опытов можно суммировать следующим образом:
1. Соединительнотканная оболочка и трофамнион хорошо пропу-
скают прижизненные красители. По всей вероятности, это относится
ко многим веществам, растворенным в гемолимфе хозяина.
2. Способность гранулообразования, хорошо выраженная у клеток
зародыша и цисты, отсутствует у трофамниона, что, по-видимому, следу-
ет считать признаком некоторой физиологической неполноценности
последнего.
3. Трофамнион отличается также от других составных частей по-
лигерма по интенсивности окраски и по оттенкам.
Нуклеиновые кислоты. Для изучения нуклеиновых кислот
в зародыше и трофамнионе были использованы срезы, окрашенные по
Фельгену и по Унна.
Ядра трофамниона обнаруживают прекрасно выраженную положи-
тельную реакцию Фельгена. В более молодых полигермах (рис. 57,
я. троф) эти ядра имеют гигантские размеры и очень неправильную
форму. Они не производят впечатления разбухших или водянистых,
наоборот, они красятся даже более интенсивно, чем ядра самого заро-
дыша. Мелкие и многочисленные комочки хроматина плотно и равно-
мерно заполняют все ядро. Вероятно, это высокополиплоидные ядра.
На более поздних стадиях развития (рис. 58, я. троф) ядра рас-
падаются на многочисленные более мелкие фрагменты. Они уже не
кажутся такими плотными, комочки хроматина распределяются в них
менее равномерно. Однако они не обнаруживают признаков дегенера-
ции. Лишь в самом конце эмбрионального развития, накануне распада
трофамниона, в его ядрах начинаются дегенеративные изменения.
При окраске по Унна плазма трофамниона принимает розовый
тон, не особенно интенсивный, но не уступающий по яркости окраске
цитоплазмы клеток зародыша. Таким образом, содержание рибонуклеи-
новой кислоты в трофамнионе ничуть не меньше, чем в тканях заро-
101
На основании изложенных материалов можно заключить, что
трофамнион Ageniaspis сохраняет жизнеспособность и участвует в об-
мене веществ до конца эмбрионального развития. Он растет вместе
с заключенными в нем зародышами, ядра его богаты дезоксирибону-
клеиновой кислотой, в цитоплазме содержится довольно значительное
количество рибонуклеиновой кислоты и происходит интенсивный синтез
жира. Опыты с прижизненным окрашиванием показали, что сквозь
трофамнион могут проходить витальные красители, а следовательно,
и многие другие растворенные в гемолимфе хозяина вещества. К сожа-
лению, примененные методики не дают возможности судить о том,
в какой мере трофамнион способен активно изменять химическую при-
роду проходящих через него веществ. Возможно, для разрешения
вопроса об участии трофамниона в процессе питания зародышей
Ageniaspis за счет хозяина могло бы оказаться более эффективным
применение метода радиоактивных индикаторов.
Заслуживает внимания еще одно обстоятельство. В то время, когда
почти все ткани зародыша лишь потребляют питательные вещества,
в клетках энтодермы происходит еще и накопление резервов (жира
и гликогена). Таким образом, энтодерма не только служит для построе-
ния органов пищеварения будущей личинки, но и сама играет какую-то
особую роль в процессе питания зародыша. К этому надо прибавить,
что в яйцах, богатых желтком, функцию его переработки обычно тоже
берут на себя элементы энтодермального происхождения. Энтодерма
как зародышевый листок очень рано начинает играть трофическую роль.
Представляло бы большой интерес выяснить, что собой представ-
ляет энтодерма Ageniaspis. По своему положению в теле зародыша на
стадии зародышевой полоски (рис. 60, энт) она очень напоминает
желточный синцитий других насекомых, а по своей дальнейшей судьбе
она соответствует кишечной энтодерме. К сожалению, способ ее обра-
зования настолько необычен (по Маршалю она образуется из цен-
тральных клеток морулообразного зародыша, т. е. путем вторичной
деламинации), что это не проливает света на ее морфологическую
природу.
Циста и ее роль в отношениях между паразитом и хозяином
Поверх трофамниона Ageniaspis лежит более тонкая оболочка,
состоящая из одного слоя клеток, имеющих сперва отчетливые границы.
Эта оболочка образуется за счет тканей хозяина и описана разными
авторами под названием цисты для многих полиэмбрионических
наездников, в том числе и для Ageniaspis fuscicollis.
На препаратах, изготовленных из более молодых полигермов, со-
бранных в первых числах июня, циста состоит из довольно крупных кле-
ток с резко очерченными контурами (рис. 64,Л, соед. об). Плоским
основанием они сидят на поверхности трофамниона, а наружные их
концы имеют неправильные очертания. Ядра клеток имеют округлую
или слегка вытянутую форму. Зернышки хроматина занимают в ядре
преимущественно периферическое положение. Различается одно или
два ядрышка. В цисте иногда можно наблюдать митозы, но- на более
поздних стадиях размножение клеток цисты, по-видимому, прекра-
щается.
На тангенциальных срезах, захвативших поверхностный слой поли-
герма ( рис. 64,5) можно видеть, что клетки цисты располагаются про-
дольными рядами. Такое упорядоченное расположение клеток цисты,
возможно, объясняется чисто механическими причинами и связано с не-
J06
равномерным ростом полигерма, который увеличивается преимуществен-
но в длину. Неравномерный рост паразитического тела обусловливает
и вытягивание осевших на его поверхности соединительнотканных* кле-
ток хозяина. Согласно правилу Гертвига, в вытянутых клетках веретено
Рис. 64. Ageniaspis fuscicollls.
Л — поперечный разрез через молодой полигерм (с00 X); Б — поперечный разрез через поздний
полигерм (600 X); В — тангенциальный разрез через полигерм, захвативший только соединитель-
нотканную оболочку; видны продольные ряды клеток (1000 х); Г — поперечный разрез через
полигерм; виден трахейный ствол (1000 X).
деления обычно располагается продольно, а сама клетка делится попе-
речно. Постоянное вытягивание клеток цисты в одном направлении
и многократно повторяющиеся поперечные деления должны были при-
вести к образованию подобных продольных рядов клеток.
107
На более поздних стадиях развития циста очень растягивается,
клеточные границы различаются плохо, ядра принимают уплощенную
форму. Очевидно рост цисты начинает отставать от роста полигерма.
К концу эмбрионального развития Ageniaspis циста превращается
в тонкую бесструктурную перепонку, в которой редко разбросаны пло-
ские плотные ядра (рис. 64, Б, соед. об). При этом, по-видимому, имеет
место не только растягивание цисты, но и дегенерация части ядер, как
это обычно происходит в различных соединительнотканных оболочках
насекомых (Заварзин, 1945).
Циста, одевающая полигерм Ageniaspis, совершенно обособлена от
остальных тканей хэзяина. Шнуры полигерма свободно лежат в полости
тела гусеницы. Лишь случайно некоторые участки этих шнуров оказы-
ваются впаянными в жировое тело. Единственная система органов
хозяина, с которой паразитическое тело оказывается в постоянном кон-
такте, это трахеи. Тончайшие разветвления трахей, а иногда и более
крупные стволики, оплетают петли полигерма, связывая их друг с дру-
гом и органами хозяина. Трахеи укрепляются на поверхности полигерма
через посредство клеток цисты (рис. 64,Л и Г, тр.).
Начальные стадии развития цисты мне, к сожалению, наблюдать
не удалось. По описанию Мартина (1914), формирование цисты вокруг
зародыша Ageniaspis начинается очень рано. Вскоре после откладки
яйца Ageniaspis в яйцо Hyponomeuta клетки жирового тела зародыша
Hyponomeuta оседают на поверхности трофамниона и вступают в такую
тесную связь с его «эктоплазматическим» слоем, что, по предположе-
нию Мартина, эта часть трофамниона тоже принимает участие в фор-
мировании цисты. После вылупления из яиц гусеничек Hyponomeuta
(осенью и зимой) расположение клеток цисты имеет беспорядочный
характер, весной же клетки равномерно распределяются по всей по-
верхности полигерма одним слоем, почему циста приобретает описанное
выше эпителиеподобное строение.
В приведенном описании развития цисты, данном Мартином, вызы-
вают сомнения два обстоятельства. Во-первых, образование ее из кле-
ток жирового тела хозяина мало вероятно, так как у зародышей Hypo-
nomeuta едва ли уже имеется настоящее жировое тело, и речь может
идти только о его зачатке. Кроме того, на изученных мною (правда
более поздних) стадиях развитие циста не обнаруживает сколько-
нибудь тесной и постоянной связи с жировым телом хозяина, а «*е клет-
ки не имеют никакого сходства с клетками последнего. Правда, после
фиксации жидкостью Флемминга в цитоплазме клеток цисты обнару-
живается некоторое количество мелких гранул жира, но в общем со-
держание жира в цисте по сравнению с таковым жирового тела или
трофамниона ничтожно. По всей вероятности, Мартин, говоря об уча-
стии клеток жирового тела в образовании цисты, имел в виду просто
слабо дифференцированные мезодермальныс клетки поздних заро-
дышей.
Тесная связь между поверхностным слоем трофамниона и цистой,
описанная Мартином, тоже кажется сомнительной. На изученных мною
срезах граница между обеими оболочками всегда хорошо видна; иногда
между ними имеется даже щелевидное пространство (рис. 58). Мало
г.ероятно и предположение, что такая тесная связь существует только
на ранних стадиях развития, а потом утрачивается: клетки молодой
цисты (рис. 57 и 64, соед. об) имеют слишком четкие границы, чтобы
предполагать, что в состав их тела вошли какие-то чужеродные части..
Возникает вопрос, какую роль играет эта соединительнотканная
оболочка в сложных отношениях между паразитом и хозяином. Опи-
108
санные Мартином начальные стадии формирования цисты во многом
напоминают защитную реакцию организма хозяина на присутствие
паразита. Образование соединительнотканных капсул, приводящих
к гибели паразита, отмечено для многих насекомых (Мейер, 1925 а, б,
1926а, б; Ларченко, 1933; Солт —Salt, 1955, 1956, 1957 и др.).
По наблюдениям К. И. Ларченко (1933) над гусеницами Loxostege
sticticalis L. и Pieris brassicae L., зараженными паразитами из родов
Eulimneria и Angitia, яйца паразитов вскоре после откладки окружают-
ся кровяными элементами хозяина, которые образуют концентрически
слоистую массу. Размножение клеток при этом не происходит, обра-
зующаяся капсула растет за счет присоединяющихся х ней новых кле-
ток гемолимфы. Потом во внутренних слоях капсулы границы клеток
исчезают и начинается их дегенерация. Этот внутренний слой все боль-
ше уплотняется, сжимает яйцо, деформирует его и тем обусловливает
гибель зародыша.
Такая реакция тканей насекомого-хозяина на присутствие паразита
имеет, по-видимому, малоспецифический характер, -гак как очень похожа
на реакцию соединительной ткани насекомых на введение индиффе-
рентного инородного тела.
Лазаренко (1925), вводивший целлоидиновую трубку в тело жука-
носорога на разных стадиях его развития, наблюдал скопление вокруг
нее веретенчатых клеток гемолимфы, располагающихся концентриче-
скими слоями. Затем, начиная с самых глубоких слоев капсулы, проис-
ходят: слияние клеток в широкопетлистый синцитий, дегенерация ядер
и превращение цитоплазмы в основное вещество.
Подобные же многослойные капсулы нередко встречаются и в
теле гусениц Hyponomeuta. Однако установить, к какому виду отно-
сились заключенные в капсуле паразиты (в гусеницах Hyponomeuta
встречается помимо Ageniaspis еще несколько видов паразитических
Hymenoptera и Diptera), не удалось, так как во всех обнаруженных
случаях они уже находились в состоянии распада.
Интересные эксперименты по изучению защитных реакций насеко-
мых были произведены Солтом (1955, 1956, 1957). Солт искусственно
заражал гусениц бабочек 8 разных видов Microlepidoptera (1955),
взрослых палочников Carausius morosus Br. (1956), личинок, куколок
и взрослых жуков Tenebrio molitor L. и личинок и взрослых мух
Calliphora erythrocephala Mg. (1957) яйцами заведомо несвойственного
им паразита Nemeritis canescens. Почти во всех случаях Солт наблюдал
хорошо выраженную защитную реакцию, завершающуюся гибелью
паразита еще в яйце или сразу после вылупления личинки. Эта реак-
ция проявлялась в двух формах: в фагоцитарной реакции, приводящей
к образованию многослойной капсулы, и в форме отложения меланина
на поверхности тела паразита и в его кишечнике. Такая блокировка
кишечника паразита пигментом тоже может служить причиной его
гибели. Первая (клеточная) реакция может быть вызвана и
инородным телом, вторая же вызывается химическими раздражителя-
ми, исходящими от паразита.
Хотя для всех использованных Солтом в качестве хозяев насекомых
Nemeritis был в равной мере случайным паразитом, однако в каждом
случае защитная реакция имела свои характерные особенности: раз-
ные виды насекомых-хозяев реагировали на яйца Nemeritis с разной
силой, иногда преобладала клеточная реакция, иногда — меланиновая.
Автору удалось подметить определенную корреляцию между характе-
ром защитной реакции и систематической принадлежностью хозяина,
из чего он сделал вывод, что родственные виды насекомых имеют
109
кровь более сходную химически, чем виды далекие, и что хозяин
реагирует на паразита прежде всего как на источник химических
стимулов. Таким образом, наблюдения Солта показывают, что даже
при заражении случайным, «неспецифическим» паразитом защитная
реакция может несколько варьировать в зависимости от видовых осо-
бенностей хозяина. Можно думать, что не меньшую роль играет и вид
паразита.
Однако отмеченные вариации имели лишь количественные отличия,
и описанную Солтом реакцию надо считать малоспецифической. Гораз-
до более специальное образование представляет собой циста Agenias-
pis. Как уже отмечалось, она состоит всего лишь из одного слоя кле-
ток и ее присутствие нисколько не вредит паразиту. Наоборот, есть
основания предполагать, что наличие цисты необходимо для нормаль-
ного развития паразита. Так, Мартин (1914) указывает, что яйца
Ageniaspis, попавшие при откладке в желток яйца Hyponomeuta, до-
стигают лишь стадии 2—3 бластомеров, после чего дегенерируют. Из
этого он делает вывод, что «яйца паразита нуждаются в посредниче-
стве эмбриональных клеток хозяина для питания и не способны сами
ассимилировать желток».
Аналогичные наблюдения сделаны над другими полиэмбриониче-
скимн наездниками. Необходимость для нормального развития пара-
зита тесного контакта с живыми тканями развивающегося хозяина и
особенно с жировым телом отмечена и для Litomastix truncatellus
(Dalm.) (Сильвестри, 1937), Copidosoma koehleri Blanch. (Даут, 1947),
Platygaster hiemalis Forbes (Лейби и Хилл, 1923) и др. Яйца Litoma-
stix floridanus (Ashm.), отло-женные в неоплодотворенное яйцо хозяи-
на (Autographa brassica) или попавшие при откладке в желток или
полость кишки хозяина, погибают; для нормального развития необхо-
димо, чтобы они оказались в тканях развивающегося зародыша (Пэтер-
сон, 1921).
По наблюдениям Лейби (1922), полигерм Copidosoma gelechiae How.
одевается цистой из жировой ткани хозяина (гусеницы Gnorimoschema
galleosolidaginis); если же циста не образуется, развитие задерживает-
ся. Яйца Amicroplis collaris Spin, обычно откладываются в жировое
тело гусениц Euxoa segetum Schiff.; если же яйцо попадает в полость
тела, оно вскоре прикрепляется к какому-нибудь органу, а со стороны
полости тела одевается амебоцитами, как бы для фагоцитоза, но не
фагоцитируется (Пайо—Paillot, 1940).
Образование соединительнотканных цист за счет хозяина описано
для ряда монэмбрионических платигастерид Маршалем (1906). Иногда
такая циста состоит только из одной клетки, так что создается впечат-
ление, что яйцо платигастра заглочено фагоцитом; однако оно не по-
гибает, а нормально развивается и так влияет на фагоцит, что он
испытывает гипертрофический рост (рис. 65,Л).
Своеобразную структуру имеет циста, развивающаяся вокруг
зародыша Trichacis remulus Walk. (Маршаль, 1906). Клетки ее при-
обретают высокоцилиндрическую форму и достигают гигантских раз-
меров. Многие из них содержат по 2—3 ядра (рис. 65,5 соед. об).
В цитоплазме накапливается большое количество жира. Но Маршаль
не склонен считать, что эти «внутренние животные галлы» имеют
какое-нибудь отношение к питанию зародышей Trichacis.
Иного рода соединительнотканная оболочка образуется вокруг личи-
нок IV возраста Encyrtus infelix (Embleton) за счет гемоцитов хозяина
(щитовки Saissetia), по данным Торпа (Thorpe, 1936). Эта оболочка
имеет вид прозрачной бесструктурной перепонки и напоминает базаль-
110
Существует мнение (Томпсон — Thompson, 1930), что активность
фагоцитов вовбуждается только больными и мертвыми паразитами,
а здоровые и жизнеспособные паразиты «не выделяют привлекающих
фагоциты веществ». Однако более распространено представление, что
инкапсуляция и гибель паразита происходит в тех случаях, когда он
еще не приспособился к данному хозяину (Кено — Cuenot, 1896;
Тимберлейк —Timberlake 1912; Коллери — Caullery, 1922, и др.).
Приспособившийся же паразит либо настолько уподобляется био-
химически своему хозяину, что продукты его жизнедеятельности уже не
производят раздражающего действия, либо выделяет какие-то веще-
ства, которые «иммобилизуют фагоцитарную активность». Фландерс
(Flanders, 1938), и Мелланби (Mellanby, 1939) делают шаг к при-
мирению этих двух точек зрения, предполагая, что, попав в непривыч-
ного хозяина, паразит начинает страдать от невозможности усвоить
питательные вещества, делается больным и поэтому становится добычей
фагоцитов.
По-видимому, можно принять, что случайный, неприспособленный
к данному хозяину паразит вызывает малоспецифическую защитную
реакцию, имеющую в большинстве случаев для него фатальный исход.
Если же паразит все-таки устоит перед этой реакцией, то отношения
между паразитом и хозяином эволюционируют в двух направлениях:
либо эта реакция ослабевает и сходит на нет, либо- она изменяется
таким образом, что становится для паразита безвредной или даже
полезной. Оба эти случая означают, что в антагонистических отноше-
ниях между двумя видами перевес находится на стороне паразита.
Описанные выше цисты и капсулы представляют интерес не
только с паразитологической точки зрения. Основным достижением
экспериментальной эмбриологии за последние десятилетия является
твердое установление того факта, что в процессе индивидуального
развития очень важную роль играет взаимодействие частей и зачат-
ков. В последнее время все большую популярность приобретает
представление, что в основе этих взаимодействий лежат явления
иммунологического характера (Токин, 1955). С этой точки зрения
особенно интересны для эмбриолога иммунологические реакции, имею-
щие своим результатом определенный формообразовательный эффект.
В данном случае речь идет именно о такой реакции, т. е. о взаимодей-
ствии двух организмов, приводящем к возникновению соединительно-
тканной цисты определенного строения.
Сходство между нормальными формообразовательными процесса-
ми и паталогическим процессом формирования цисты настолько ве-
лико, что при рассмотрении последнего можно пользоваться понятиями
механики развития. Д. П. Филатов пишет, что закладка органов
«возникает в системе формообразовательного аппарата, т. е. перед
обособлением закладки происходит морфогенное взаимодействие двух
частей зародыша, из которых одна представляет материал закладки,
а другая в закладку не входит, но оказывает на ее материал детерми-
нирующее действие, благодаря которому материал превращается
в закладку определенного органа (Шпеманновский организатор). Эти
две части временно, на срок, в течение которого происходит детерми-
нация закладки, я называю формообразовательным аппаратом»
(1943, стр. 28).
Сходный формообразовательный аппарат возникает на месте
соприкосновения двух биологических систем, представленных в нашем
конкретном примере гусеницей Hyponomeuta и зародышами Ageniaspis.
Полигерм Ageniaspis оказывает на соединительнотканные клетки
112
гусеницы детерминирующее действие, под влиянием которого образует-
ся циста. При желании здесь тоже можно говорить об организаторе
или индукторе и реагирующем материале. Реакция эта имеет очень
специфический характер — циста, одевающая полигерм Ageniaspis,
резко отличается от обычных защитных капсул и, по-видимому, не
вполне идентична соединительнотканным оболочкам, образующимся
на поверхности органов самой гусеницы. Этот специфический характер
цисты показывает, что отношения, существующие между орга-
низмами Hyponomeula и Ageniaspis, сложились уже давно. Но это не
существенно. Те же элементы формообразовательного аппарата могут
быть усмотрены и при менее специфических защитных реакциях.
Даже те случаи, когда образование капсулы вызвано мертвым инород-
ным телом, не умаляют правомерности сделанного сопоставления
между нормальными и патологическими формообразовательными про-
цессами. Они показывают только, что воздействия, оказываемые друг
на друга организмами или частями зародыша, имеют вполне материаль-
ный характер и в конечном счете могут быть сведены к механиче-
ским, химическим и физико-химическим стимулам.
8 О. М. Иванова Kairac
ГЛАВА VIII
MESTOCHARIS MILITARIS R.-KORS.
В окрестностях Ленинграда яйца плавунцов (Ma'Crodytes, Acilius,
Agabus), откладываемые в стебли водяных растений, часто бывают
заражены наездником Mestocharis militaris R.-Kors. (Римский-
Корсаков, 1916, 1932, 1940). Так как сем. Entedontidae (Parasitica,
Chalcidoidea), куда относится Mestocharis, с эмбриологической точки
зрения еще совершенно не изучено, этот наездник тоже был использо-
ван в качестве объекта исследования (Иванова-Казас, 19486, 19496,
19526 и 1954г).
1. БИОЛОГИЯ
Яйца плавунца окаймленного (Macrodytes marginalis L.) в окрест-
ностях г. Ленинграда (Озерки) в 1947—1949 гг. оказались заражен-
ными Mestocharis militaris на 16—17%. В одном яйце Macrodytes
можно встретить до 44 яиц Mestocharis, но чаще их бывает 8—9.
Нахождение большего количества паразитов объясняется, по-видимому,
двукратным и трехкратным заражением. В пользу этого говорит тот
факт, что нередко яйца Mestocharis, извлеченные из одного яйца
Macrodytes, можно разделить по стадиям их развития на 2—3 хорошо
разграниченные группы. .
Хотя личинки Mestocharis довольно активные «внутренние хищ-
ники» и охотно поедают личинок других наездников и своих собратьев,,
все-таки в одном яйце Macrodytes может благополучно достичь
зрелости несколько экземпляров Mestocharis.
Эмбриональное развитие Mestocharis длится очень недолго (по
данным Римскою-Корсакова 2—3 дня). Стадия личинки продолжается
около полутора месяцев, а стадия куколки—около 10 дней. Как долго
живет имаго — неизвестно. Яйца и личинки Mestocharis попадаются на
протяжении всего мая и июня. В конце июня яйца уже не встречаются,
но появляются куколки, из которых легко вывести взрослых наездни-
ков. Все стадии развития Mestocharis протекают внутри яиц хозяина.
Только взрослые наездники выходят во внешнюю среду.
Очень часто яйца и личинки Mestocharis встречаются в яйцах
Macrodytes одновременно с другими паразитами — Prestwichia aquatica
Lubb. и Caraphractus reductus R.-Kors.
Prestwichia и Caraphractus, в случае совместного заражения
одного яйца Macrodytes, по-видимому, нисколько не мешают друг
114
это тоже объясняется тем, что передний конец яйца жука доступнее
для наездников, чем задний.
Однако ни гармоничные карлики, ни уродливые зародыши никогда
не достигают стадии свободной личинки, паразиты уничтожают их
прежде, чем завершится их развитие. Если заражение происходит на
более поздних стадиях развития, когда зародыш жука уже заполняет
собой все яйцо, а весь желток заключен в его кишке, яйца паразитов
при откладке попадают внутрь зародыша, а вышедшие из них личин-
ки начинают уничтожать его изнутри. Подобный зародыш изображен
на рис. 66, В. Находящиеся внутри него личинки Prestwichia съели все
его ткани, оставив лишь хитиновые части, которые свидетельствуют
о том, что зародыш плавунца уже почти достиг стадии личинки
(форма ротовых частей, сильное развитие и пигментация трахей
и т. д.).
Таким образом, с чего бы ни начиналось вредное воздействие
паразитов, оно всегда приводит к уничтожению зародыша хозяина.
Мне ни разу не приходилось наблюдать какой-нибудь защитной реак-
ции со стороны зародыша жука на присутствие паразита.
2. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Яйца Mestocharis имеют удлиненную форму и одеты тонким
прозрачным хорионом. Передний конец несколько расширен. Длина
яиц достигает 0,35—0,48 лш„ наибольшая ширина — 0,08—0,11 мм.
Яйца содержат довольно много желтка. Цитоплазма образует сравни-
тельно толстый слой у поверхности яйца, а в центральной его части —
сеть с массивными перекладинками, между которыми лежит желток
(рис. 67,Л; на этом и на последующих рисунках место, занятое желт-
ком, оставлено пустым). В этой губчатой центральной плазме лежат
первые ядра дробления.
Как обычно при поверхностном дроблении, сперва размножение
ядер происходит в середине яйца и лишь позднее ядра входят в пери-
ферический слой плазмы, где начинается формирование бластодермы
(рис. 67, Б, 6л).
В процессе формирования бластодермы происходит все более
полное разделение желтка и плазмы. Почти вся цитоплазма стягивает-
ся к поверхности яйца. В- центре среди желтка остаются лишь не-
большие участки плазмы с желточными ядрами, которые довольно
скоро дегенерируют (рис. 68, 69, ж. я). Первоначально дробление
имеет синхронный характер (рис. 67,Л), но затем волны митозов
начинают зарождаться на переднем конце, после чего распространяют-
ся назад. На некоторых препаратах можно видеть, что на переднем
конце яйца все ядра находятся в состоянии деления, а на заднем --
в интеркинезе (рис. 67,£ и 69,Л).
В других случаях зародыши перепоясаны зоной митозов (рис. 68, Б
и 69,5), при этом отчетливо видна разница в размерах интеркинети-
ческих ядер в передней и задней половинах зородыша: передние
ядра значительно мельче задних, что объясняется тем, что они уже
прошли деление, к которому задние ядра еще не приступили. Наконец,
на рис. 68,Л изображен зародыш, у которого делящиеся ядра рас-
полагаются на обоих концах яйца, а интеркинетические — в средней
его части. Этот случай, по-видимому, можно объяснить, предположив,
что деления быстро следовали одно за другим и одновременно оказа-
лись представлены две волны — уходящая (сзади) и зарождающаяся
(спереди).
116
личинки выражен очень неясно. Однако изучение строения поздних
личинок показало, что все имагинальные части головы возникают за
счет этого первого «сегмента» личинки; второй, третий и четвертый сег-
менты содержат уже имагинальные диски ног и, следовательно, явля-
ются грудными. В этом случае не эмбриональное развитие, а начальные
стадии метаморфоза помогают нам понять морфологическую природу
головного отдела личинки.
Из внутренних зачатков зародыша сегментация ясно выражена
лишь в нервной системе. Последняя закладывается в виде парных
метамерных утолщений эктодермы, от которых обособляется мате-
риал соответствующих ганглиев. Таким образом образуется пара
надглоточных ганглиев и 12 пар ганглиев брюшной цепочки. Первая
пара брюшных ганглиев лежит в голове и может быть названа под-
глоточным ганглием. Он массивнее всех остальных ганглиев брюшной
цепочки, но его сложная природа в развитии не проявляется.
В момент своего образования и на средних стадиях эмбриональ-
ного развития мезодерма не обнаруживает сегментации и не образует
целомических пузырьков. Но позднее, когда начинается дифференциа-
ция мышц, последние приобретают сегментарное- расположение. Эта
запоздалая сегментация тесно связана с сегментацией гиподермы,
имеет функциональный характер и отнюдь не распространяется на ту
часть мезодермы, которая дает жировое тело.
К концу эмбрионального развития энтодермальный зачаток
окружает со всех сторон массу желтка и образует среднюю кишку,
в которую спереди открывается пищевод. Сообщение средней кишки
с задней не устанавливается. Оба отдела остаются разобщенными
и у личинки. После вылупления личинки средняя кишка переполняет-
ся заглоченной пищей и сильно увеличивается в объеме.
У молоденькой личинки мышцы и нервная система еще не дости-
гают полной гистологической дифференциации: мышцы еще лишены
поперечной исчерченности, в ганглиях нервной системы еще слабо
развито фибриллярное вещество. Такая личинка почти совсем не-
подвижна и совершает только глотательные движения. Но впослед-
ствии гистологическая дифференциация всех личиночных органов
завершается, и личинка приобретает способность к очень своеобраз-
ным, но энергичным сокращениям тела. Мускулатура личинки состоит
преимущественно из продольных мышц. Обычно в каждый данный
момент сокращаются мышцы на одной стороне тела, например —
слева. При этом личинка несколько изгибается на левую сторону
и ложится на левый бок. Затем сокращаются мышцы брюшной сто-
роны тела, потом правой, спинной, и так далее. Соответственно личин-
ка перекатывается вправо, вращаясь вокруг своей продольной оси.
По-видимому, это перекатывание может служить лишь для меха-
нического разрушения и перемешивания тканей хозяина; ни о каком
поступательном движении не может быть и речи.
*
Итак, мы видим, что первой своеобразной чертой развития
Mestocharis militaris является отсутствие эмбриональных оболочек —
амниона и серозы. Вследствие этого у Mestocharis отсутствует деление
бластодермы на зародышевую полоску и внезародышевую бласто-
дерму. Правда, в бластодерме Mestocharis тоже можно разли-
чить область утолщенной бластодермы, которая является местом
122
активных формообразовательных процессов (закладка мезо- и энто-
дермы, стомо- и проктодеума, нервной системы), почему я сохраняю
для нее название «зародышевой полоски» (в кавычках), но тонкая
бластодерма тоже участвует в построении тела зародыша и ни в коем
случае не может быть приравнена к внезародышевой бластодерме
других насекомых.
Далее следует отметить некоторое упрощение процесса развития:
сегментация мезодермального зачатка выражена слабо и сильно
запаздывает* энтодерма образуется только из одного зачатка, а не
из двух, упрощается не только строение головы личинки, но и ее
развитие. В этом отношении развитие Mestocharis militaris очень
напоминает развитие Banchus femoralis Thorns. (Tchneumonidae), no
данным Бледовского и Краинской (1926).
ГЛАВА IX
CARAPHRACTUS REDUCTUS R.-KORS.
Caraphractus reductus R.-Kors. относится к сем. Mymaridae (под-
отр. Parasitica, Chalcidoidea), большинство представителей которого
развивается в яйцах других насекомых. Первые сведения об эмбриоло-
гии Mymaridae мы находим у Ганина (1869а), описавшего развитие
ряда паразитических перепончатокрылых, и в том числе Polynema na-
tans из яиц Agrion. По мнению Римского-Корсакова (1940) Ганин
имел дело с Anagrus incarnatus Hal.
Несмотря на всю их неполноту, старые данные Ганина представ-
ляют большой интерес. Ганин сообщает, что яйца Anagrus' (= Poly-
nema) имеют длину 0,105 мм из которых 0,048 мм приходится на сте-
белек, и ширину 0,024 мм. В ооплазме почти нет включений. Хорион
рано сбрасывается, и зародыш остается одетым только тонкой кути:
кулой. Каково происхождение этой кутикулы, Ганин не пишет. Сам
зародыш в это время представляет собой массу недифференцирован-
ных клеток. Постепенно, в течение последующих 5—6 дней, путем
деламинации обособляется внутренняя масса клеток (энтодерма),
утолщенная брюшная стенка тела («зародышевая полоска») отделяет
от себя зачатки мышц и нервной системы; на переднем и зад-
нем конце формируются впячивания стомо- и проктодеума. Эмбрио-
нальные оболочки не образуются. Зародыш заметно растет. На одной
из более поздних стадий он имеет размеры 0,15Х0>06 мм; следова-
тельно, объем его увеличивается в 16,4 раза (если считать его исход-
ными размерами размеры яйца без стебелька). К моменту образования
личинки паразит еще несколько увеличивается. Остается неясным, как
понимать стадии развития после сбрасывания хориона: как продолже-
ние эмбриогенеза или как начало постэмбрионального развития. Сам
Ганин называет эту стадию «зародышем».
Кое-что известно и о другом представителе сем. Mymaridae —
Anaphoidea luna Girault, паразитирующем в яйцах Phytonomus varia-
bilis Herbst. Сильвестри (1915—1916) проследил начальные стадии
развития Anaphoidea — оплодотворение, поверхностное дробление и
формирование бластодермы. Более поздние стадии остались неизвест-
ными.
Таким образом, Caraphractus reductus R.-Kors. — третий предста-
витель этого семейства, ставший объектом эмбриологического исследо-
вания (Иванова-Казас, 19496 и 19546).
124
ного расположения и как бы проталкиваются внутрь зародыша^ обра-
зующимся вслед за ними стомодеумом; 2) либо зачаток средней киш-
ки формируется за счет основной массы внутренних клеток, обособив-
шихся при первоначальном расслоении бластодермы; в этом случае
надо принять, что все клетки, погружающиеся на переднем конце, идут
на построение стомодеума, а связь стомодеума со средней кишкой до-
стигается вторично. Первое предположение больше согласуется с тем,
что мы обычно видим у насекомых вообще и у Hymenoptera в частно-
сти: средняя кишка формируется из клеток, включенных в дно стомо-
деума. Но тогда надо допустить, что внутренняя масса клеток идет на
построение мезодермы, т. е., что образование мезодермы у Caraphractus
происходит за счет клеток, выселяющихся из всей бластодермы. Такой
способ образования мезодермы тоже отличается от того, что мы обыч-
но наблюдаем у насекомых. Итак, какова бы ни была природа вну-
тренних клеток, сочтем ли мы их энтодермальными или мезодермаль-
ными, — в любом случае несоответствие между типичными процессами
дифференциации зародышевых листков у насекомых и таковыми у
Caraphractus остается одинаковым.
Нечто подобное описано Н. М. Кулагиным (1894) для другого
паразитического перепончатокрылого — Platygaster instricator. У Pla-
tygaster в брюшной части бластодермы происходит усиленное размно-
жение клеток, почему брюшная бластодерма становится двух- и трех-
слойной. Эту утолщенную часть бластодермы Кулагин называет «заро-
дышевой полосой» и считает местом образования внутреннего слоя.
Далее он пишет: «Впрочем, клетки внутреннего слоя не только отде-
ляются от клеток зародышевой полосы, но отдельные клетки бласто-
дермы на спинной поверхности точно так же могут делиться и давать
начало внутреннему слою клеток. В конце концов, на всем протяжении
зародыша образуется внутренний слой, состоящий на брюшной поверх-
ности из трех или четырех слоев клеток, а на спинной — из двух слоев.
Следовательно, одновременно путем деления клеток бластодермы и
путем иммиграции их являются вместе энтодерма и мезодерма» (1894).
Какие именно элементы этого внутреннего слоя Кулагин считает энто-
дермальными, а какие мезодермальными, ни из его описания, ни из
рисунков не видно.
Мне кажется, что клеточный материал, получающийся путем им-
миграции или деламинации бластодермы у Caraphractus, надо считать
зачатком энтодермы, и вот по каким соображениям. Если бы вся вну-
тренняя масса клеток давала только мезодермальные элементы, то ко-
личество мезодермальных клеток на поздних стадиях развития должно
было бы быть очень значительным, тем более, что выселение мезодер-
мальных клеток еще некоторое время продолжается, а сравнительно
мелкие размеры мезодермальных клеток по сравнению с внутренними
клетками стадии многослойной бластодермы заставили бы предполо-
жить их размножение. На самом же деле число мезодермальных кле-
ток на поздних стадиях развития (рис. 79, м) невелико. Очень вероят-
но, что мезодерма зародыша образуется исключительно за счет клеток,
выселяющихся из медиовентральной части бластодермы. Поэтому
надо полагать, что большая часть внутренних клеток идет на построе-
ние средней кишки.
Такое понимание процессов гаструляции у Caraphractus согла-
суется и с наблюдениями Ганина (1869а) над Anagrus. Ганин тоже
описал образование энтодермы из группы крупных клеток в центре
зародыша и не отмечает никакой связи между этим зачатком и стомо-
деумом на ранних стадиях развития.
9 О. М. Ипанова-Казас
129
задней кишки со средней не устанавливается ни во время эмбриональ-
ного развития, ни на ранних личиночных стадиях. Нередко в полости
задней кишки можно наблюдать какое-то базофильное содержимое,
частично изливающееся наружу (рис. 79, Г). У молоденькой личинки
задняя кишка еще сохраняет эмбриональный характер.
Половой зачаток разделяется на две симметричные части, которые
лежат в задней части тела зародыша по бокам от задней кишки
(рис. 79, Б, п. з). На поверхности каждого полового зачатка распола-
гается .по нескольку соединительнотканных клеток, образующих нечто
вроде тонкой оболочки. Впоследствии, после вылупления личинки из
яйца, оба половых зачатка тесно прилегают к боковым стенкам тела
личинки (рис. 80, Б, п. з.).
Незадолго до вылупления личинки на брюшной стороне зародыша
эктодерма образует 7—8 поперечных складок, не распространяющихся,
однако, на бока и спину (рис. 79, В). В этой складчатости можно ви-
деть сильно редуцированную сегментацию. На мезодерму эта сегмен-
тация не распространяется. Мезодерма, как уже отмечалось, состоит
из очень немногочисленных клеток, редко рассеянных по полости тела
и уже частично вставших на путь гистологической дифференциации.
Так, под эктодермой можно видеть тяжи крупных веретеновидных
клеток с резко базофильной плазмой, которые, видимо, являются за-
чатком соматической мускулатуры. Часть мезодермальных клеток дает
мышцы пищевода, другая — соединительнотканные оболочки половых
зачатков. Количество недифференцированных мезодермальных клеток
на этой стадии едва ли превышает два десятка.
Единственная система внутренних органов, на которую распро-
страняется сегментация,— нервная система, развитие которой начи-
нается в самом конце эмбрионального развития. Головной мозг закла-
дывается как два симметричных линзовидных утолщения эктодермы
по сторонам от пищевода (рис. 79, Л, г. м). У молоденькой личинки
зачатки головного мозга уже отчетливо обособлены от эктодермы и
имеют вид двух эпителиальных пластинок, лежащих под кожей голов-
ного конца личинки (рис. 80, Л, г. м). Брюшная нервная цепочка за-
кладывается как ряд непарных утолщений эктодермы, имеющих сег-
ментарное расположение (рис. 79, £ и Г, бр. ц). Удается различить
6 зачаточных ганглиев брюшной цепочки. Каким именно ганглиям
взрослого насекомого соответствуют эти зачатки — можно было бы
сказать, лишь проследив метаморфоз Caraphractus. В момент вылуп-
ления личинки из яйца все эти части нервной системы находятся еще
в совершенно зачаточном состоянии.
Вскоре после вылупления личинка начинает заглатывать желток
яйца хозяина. Тело ее вытягивается и приобретает цилиндрическую
форму, слабо изогнутую на брюшную сторону (рис. 72, Б, 79, Г). По
мере наполнения кишечника пищей, внешняя сегментация почти совер-
шенно сглаживается.
Личинка способна совершать слабые сокращения и изгибания
тела, но ползать она не может. Такое слабое развитие двигательной
функции связано с паразитическим образом жизни.v Но и эти очень
ограниченные движения имеют на первых порах, по-видимому, автома-
тический характер, т. е. являются реакцией мышечных клеток на непо-
средственные раздражения со стороны внешней среды, так как нервная
система 'молодой личинки находится еще в эмбриональном состоянии.
Из всего вышеизложенного видно, что развитие Caraphractus об-
наруживает в еще большей степени, чем Mestocharis militaris, тенден-
цию к укорочению и упрощению.
132
Упрощение развития сказывается в отсутствии эмбриональных
оболочек, в слабом и неполном развитии сегментации. Укорочение раз-
вития проявляется в том, что личинка выходит из яйца, не достигнув
того морфологического уровня, на котором обычно находятся личинки
Holometabola: мускулатура ее развита слабо, нервная система нахо-
дится в состоянии раннего эмбрионального зачатка и неспособна функ-
ционировать, кровеносная, дыхательная и выделительная системы от-
сутствуют (Иванова-Казас, 19546).
Если считать стадии развития Anagrus после сбрасывания хориона
началом тюстэмбрионального развития, то этот наездник дает нам при-
мер вылупления из яйца в еще более недоразвитом состоянии.
ГЛАВА X
PRESTWICHIA AQUATICA LUBB.
Сем. Trichogrammatidae (подотр. Parasitica, Chalcidoidea), к ко-
торому относится Prestwichia aquatica Lubb., содержит около 40 родов
(Никольская, 1952), паразитирующих в яйцах насекомых. Некоторые
виды рода Trichogramma, например Tr. evanescens Westw., поражают
очень большой круг хозяев — по Клоузену (1940), более 200 — и имеют
хозяйственное значение. Литературные данные об эмбриональном раз-
витии трихограмматид относятся большей частью к этому же роду
Trichogramma. Сильвестри (1909в) описал развитие Trichogramma
(Oophthora) semblidis (Aur.) из яиц Mamestra brassichae L., а Гетенби
(1917, 1918a) —Tr. evanescens Westw., паразитирующей в яйцах Dona-
cia simplex Fab. Хенриксен (Henriksen, 1922) считает эти названия
синонимами. Описанный М. С. Ганиным (1869а) Ophioneurus, развиваю-
щийся в яйцах Pieris brassicae, по-видимому, тоже есть один из видов
Trichogramma (Шмидекнехт — Schmiedeknecht, 1930). М. Н. Никольская
(1932) отмечает, что Tr. evanescens может паразитировать и в яйцах
Pieris brassicae. Таким образом, очень вероятно, что Ганин, Сильвестри
и Гетенби имели дело с одним и тем же видом.
По всей вероятности, к сем. Trichogrammatidae относится и «Ptero-
malina» из яиц Chrysomela fastuosa, развитие которой (очень похожее
на таковое Prestwichia) описано Н. П. Вагнером (1868).
Кое-какие отрывочные данные, касающиеся эмбриологии Prestwi-
chia aquatica, можно найти у Хенриксена (1922), который дал изображе-
ния яйца Prestwichia на стадии бластодермы, позднего зародыша и мо-
лодой личинки.
Все названные формы развиваются в общем сходно. Об имеющихся
отличиях удобнее будет сказать по ходу изложения собственных мате-
риалов.
1. БИОЛОГИЯ
Образ жизни Prestwichia aquatica очень напоминает таковой Сага-
phractus (Римский-Корсаков, 1916, 1932, 1940; Иванова-Казас, 19496,
1950, 1952а, 1954г). Prestwichia паразитирует в яйцах Dytiscidae, где
проделывает все стадии развития. Даже копуляция взрослых насекомых
происходит под покровом хориона яйца хозяина, так что самкам по вы-
ходе во внешнюю среду остается только отложить яйца.
Эмбриональное развитие Prestwichia длится 2—4 дня, стадия ли-
чинки— 8—15 дней, стадия куколки — 12—15 дней, а стадия имаго —
136
Ю—15 дней. За лето Prestwichia успевает дать до четырех поколений,
используя, как Caraphractus, яйца разных видов плавунцов.
В одном яйце Macrodytes случалось находить до 178 экземпляров
Prestwichia, но чаще всего их было 10—20. В яйцах Acilius встречалось
до 45 экземпляров Prestwichia (обычно 7—8), а в яйцах одного из
видов Agabus — только до 13 (обычно 4—5). Эти различия в числе от-
кладываемых яиц находятся в соответствии с размерами яиц названных
видов жуков. Нападению Prestwichia подвергаются яйца плавунцов,
находящиеся на всех стадиях развития, но поздние стадии, по-видимому,
менее привлекательны для наездника.
Часто яйца Prestwichia откладываются целыми пачками, причем*
стебельки яиц бывают прочно склеены друг с другом. При этом яйца
могут быть отложены в любую часть зародыша жука. При заражении
в лабораторных условиях развивающиеся яйца Prestwichia можно найти
в желтке, в зачатке нервной системы, в полости крупных трахейных:
стволов и т. д.
Обычно в тканях хозяина не наблюдалось никаких реактивных из-
менений, связанных с присутствием яйца паразита. Но в яйцах Agabus
зародыши и ранние личинки Prestwichia часто погибали по неизвестным
причинам и выталкивались из зародыша, так что оказывались лежащи-
ми между ним и хорионом. Такие погибшие личинки имели сморщенный
высохший вид, а полости средней и задней кишки и лабиальных желез
были заполнены пигментом. Зародыши же Agabus благополучно завер-
шали свое развитие. По мнению Солта (1957), отложение меланина
в теле паразитов происходит под влиянием хозяина и может играть за-
щитную роль в тех случаях, когда оно мешает- жизненным отправле-
ниям паразита — его питанию, вылуплению из яйца и т. д.
2. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Яйца Prestwichia одеты прозрачным эластичным хорионом, под
которым лежит еще одна очень нежная, бесструктурная оболочка, про-
исхождение которой осталось невыясненным (рис. 81, Л, нар. х и вн. х).
Витальные красители через эти оболочки не проходят. Форма яйца
эллипсоидная. На переднем конце хорион образует длинный стебелек,
в который заходит и вторая оболочка (рис. 81, А, стеб). Длина яйца
вместе с хорионом и его стебельком варьирует от 0,2 до 0,5 мм. Размеры
самого зародыша значительно меньше: длина 0,055—0,253 мм, ширина
0,038—0,091 мм. (Эти измерения сделаны на живых яйцах, находящихся
на стадиях дробления). Такие значительные вариации в размерах яиц
зависят от вариации в размерах взрослых Prestwichia, которые в свою
очередь находятся в зависимости от условий питания на стадии личинки
(т. е. от того, сколько личинок развивалось в одном яйце жука, и от
размеров этого яйца). Вариации в размерах зародышей Prestwichia,
находящихся"на разных стадиях развития, почти не выходят за пределы
вариаций в исходных размерах яиц и не дают основания предполагать
питание зародышей Prestwichia за счет хозяина. Некоторое увеличение
объема, связанное с развитием различных полостей, конечно имеет
место, но «живая масса» едва ли увеличивается.
Цитоплазма яйца разко базофильна и выглядит совершенно гомо-
генной. Желток отсутствует полностью. В средней части яйца лежит
ядро, очень бедное хроматином (рис. 82, А, я). Недалеко от ядра, но
ближе к поверхности яйца, в цитоплазме лежит темный комочек —
дегенерирующие направительные ядра (рис. 82, £, р. я). У заднего конца
яйца имеется плотное тельце, окрашивающееся сильнее плазмы — оосо-
137
более прямой и короткий, при котором энтодерма как особый зароды-
шевый листок совсем не выражена, без сомнения, имеет адаптивный
характер и связан с паразитизмом.
По- описанию М. С. Ганина, средняя кишка Ophioneurus обра-
зуется путем деламинации в плотной, клеточной массе, каковой являет-
ся на этой стадии зародыш. Стомо- и проктодеум закладываются, как
обычно, из впячиваний эктодермы. Наблюдения Ганина в отношении
средней кишки Ophioneurus едва ли правильны, так как не согла-
суются с данными Сильвестри и Гетенби.
Trichogramma, развитие которой изучал Сильвестри (1909в), тоже
проходит стадию компактной клеточной массы. Такое строение дости-
гается благодаря выселению клеток из передней части бластодермы,
по Сильвестри, или из передне-спинной ее части, по Гетенби. Одновре-
менно происходит оттеснение центральной губчатой плазмы к спинной
стороне тела и выталкивание ее наружу (нечто подобное имеет место
и при развитии Caraphractus reductus). По Гетенби, зародыш все вре-
мя сохраняет синцитиальное строение (хотя на некоторых его рисун-
ках клеточные границы хорошо видны). Затем в общей синцитиальной
массе происходит дифференциация зачатков органов. Сильвестри, на-
оборот, везде изображает ясные клеточные границы. Вытолкнутая из
зародыша губчатая плазма с содержащимися в ней дегенерирующими
ядрами, растекается между зародышем и хорионом и имитирует обо-
лочку, которую Сильвестри назвал «псевдосерозой». По Гетенби, эта
плазма просто дегенерирует.
Данные М. С. Ганина, Сильвестри и Гетенби отличаются большой
неясностью и зачастую противоречат друг другу. Утверждение Гетенби,
что у Trichogramma зародыш долго сохраняет синцитиальное строение,
что врастание клеток внутрь происходит на спинной стороне и, нако-
нец, описанная им «диминуция хроматина», представляется очень со-
мнительными. Наблюдения Сильвестри, по-видимому, более точны,
если не считать описанной им эмбриональной оболочки.
3. ЛИЧИНКА
Молодая, только что вышедшая из яйца личинка Prestwichia по
размерам немного превышает яйцо и отличается от позднего зародыша
лишь большей гистологической дифференцированностью клеток
(рис. 86). На переднем конце субтерминально расположен рот {рот),
ведущий в пищевод, который совершает явственные глотательные дви-
жения. Пищевод сплющен в дорзо-вентральном направлении и на попе-
речных разрезах имеет полулунную форму (рис. 87, Л, пищ). Брюшная
стенка пищевода тонка и у самого ротового отверстия представлена
только складкой хитина, который тонким слоем одевает все тело личин-
ки и заходит в пищевод; спинная же стенка состоит из высоких эпите-
лиальных клеток, содержащих крупные ядра с грубо комковатым хро-
матином. Со стороны полости тела к пищеводу примыкает несколько
мезодермальных клеток, быть может, являющихся зачатком кольцевой
мускулатуры. Между пищеводом и спинной стенкой тела тянется не-
сколько удлиненных клеток, которые можно считать мышцами
(рис. 87, Л, ж), оттягивающими утолщенную дорзальную стенку пище-
вода и тем расширяющими его просвет. Никаких твердых хитиновых
частей около рта нет. Пищевод ведет в среднюю кишку, сильно разду-
вающуюся по мере наполнения ее пищей, так как у личинок Prestwi-
chia, как и у личинок других паразитических перепончатокрылых, сред-
няя кишка не сообщается с задней и дефекация не происходит
143
(рис. 86, ср. к). Клеточные границы в средней кишке исчезают
(рис. 87, В, ср. к). Имеются хорошо развитые лабиальные железы
(рис. 87,5, лаб. ж). Задняя кишка личинки сохраняет эмбриональный
характер (рис. 86 и 87, Г, з. /с).
Между брюшной стенкой тела и задней кишкой располагается
группа из 6—8 светлых клеток (рис. 86 и 87, Г. п. з) —половой зача-
ток. Несколько мезодермальных клеток распластываются на его по-
верхности, образуя соединительнотканную, оболочку.
Относительно нервной системы у личинок Prestwichia в литературе
имеются разногласия. М. Н. Римский-Корсаков (1916) отметил наличие
у личинок нервного ганглия, но Хенриксен (1922) обнаружить его не
смог.
У молодых личинок Prestwichia мне не удалось найти анатомиче-
ски выраженной нервной системы ни на живом материале, ни на то-
тальных препаратах, ни на срезах. Однако по бокам от ротового отвер-
стия эктодерма личинки образует два утолщения, состоящие из резко
базофильных округлых клеток эмбрионального типа (рис. 87, Л, ут.
гип). Они очень напоминают закладку головного мозга во время эм-
брионального развития других насекомых и, действительно, таковыми
и являются, только процесс формирования нервной системы сдвинут
у Prestwichia на постэмбриональные стадии развития.
Вследствие сильного растяжения средней кишки полость тела ли-
чинки низводится до состояния незначительных щелей, в которых ле-
жат немногие мезодермальные клетки. Вся личинка пузыревидно раз-
дувается и не обнаруживает никаких признаков сегментации (рис. 86).
Дыхательной, выделительной и кровеносной систем у только что вылу-
пившейся из яйца личинки Prestwichia еще нет. Личинка Trichogram-
ma, описанная Сильвестри (1909в), очень похожа на личинку Prestwi-
chia. Она имеет такую же пузыревидную форму и лишена всякой сег-
ментации. В отличие от Prestwichia она имеет по бокам от рта два
хитиновых зубца (мандибулы?), служащие для размельчения тканей
хозяина, но лишена слюнных желез. О нервной системе Сильвестри
ничего не пишет, а Гетенби считает головным ганглием массу недиффе-
ренцированных клеток над пищеводом.
Сходно устроена и личинка Ophioneurus. По описанию М. С. Та-
нина, она имеет мешковидную нерасчлененную форму, снабжена парой
хитиновых зубцов, но отличается от личинки Trichogramma наличием
пары лабиальных желез. Мускулатура личинки Ophioneurus пред-
ставлена только несколькими тяжами, втягивающими передний конец
тела. На брюшной стороне происходит усиленное размножение клеток,
в результате чего образуется утолщение, названное Ганиным «зароды-
шевой полоской», из "которого дифференцируются зачатки нервной
системы и мышц, реализующиеся только во время метаморфоза. Такую
же «зародышевую полоску» описывает Ганин у личинок Teleas и Р1а-
tygaster. Однако, поскольку речь идет об образовании, существующем
уже у личинки, из которого развиваются структуры взрослого насеко-
мого, мне кажется более правильным называть это утолщение гипо-
дермы не «зародышевой», а «имагинальной полоской».
Таким образом, упрощение и укорочение развития выражены у
Prestwichia еще сильнее, чем у Caraphractus. Упрощение проявляется
в выпадении из развития эмбриональных оболочек, по-видимому, утра-
тивших свое функциональное значение, в более слабом развитии жел-
точных ядер, которых у Prestwichia образуется меньше, чем у Caraph-
ractus, в способе формирования средней кишки. Укорочение развития
(дезэмбрионизация) доходит у Prestwichia так далеко, что из яйца вы-
144
ходит личинка, у которой отсутствуют выделительная, дыхательная,
кровеносная системы, мускулатура представлена только глотательны
ми мышцами, а нервная система — лишь зачатком головных ганглиев.
Эта личинка даже еще не сегментирована (Иванова-Казас, 1950).
Интересно, что, по данным Н. Г. Коломийца и А. В.Доваленок
(1958), такая же крайне упрощенная и несегментированная личинка
выходит из яйца Telenomus gracilis Mayr (Scelionidae), эмбриональное
развитие которого тоже сильно упрощено и не сопровождается образо-
ванием оболочек.
4. МЕТАМОРФОЗ
Из яйца Prestwichia aquatica выходит личинка так сильно отли-
чающаяся от типичных личинок насекомых, что невольно возникло
предположение, что и в течение последующих процессов развития у
Prestwichia должны быть значительные уклонения от обычного типа.
Это побудило меня предпринять изучение и постэмбрионального разви-
тия Prestwichia (Иванова-Казас, 1952а). Мне хотелось выявить своеоб-
разные черты метаморфоза Prestwichia, прямо или косвенно связанные
с паразитизмом. Поэтому были рассмотрены лишь самые общие про-
цессы, а вопросы, имеющие частный характер-и требующие примене-
ния специальных методик (например, гистогенез нервной системы и
сложных глаз, формирование наружных половых частей и т. д.), остав-
лены без рассмотрения. *-
Личиночные стадии и куколка Prestwichia изучались преимущест-
венно на срезах и тотальных препаратах. Последние изготовлялись в
форме пленчатых препаратов. У личинок средних возрастов гиподерма
и стенка кишки сильно растянуты из-за большого количества заглочен-
ной пищи. Содержимое средней кишки живой личинки выглядит совер-
шенно прозрачным и сильно преломляет свет; при проколе стенки тела
оно немедленно вытекает тягучими струями, которые от соприкоснове-
ния с водой мутнеют, а сама личинка при этом деформируется. При
фиксации содержимое кишки личинки превращается в плотное непро-
зрачное тело, которое мешает изготовлению и изучению обычных то-
тальных препаратов. Поэтому при изготовлении тотальных препаратов
затвердевшее содержимое средней кишки извлекалось через разрыв в
спинной стенке тела личинки, а оставшиеся неповрежденными брюшная
и боковые стенки тела разворачивались, как лоскут, окрашивались и
заключались в бальзам.
Стадии постэмбрионального развития
За время личиночной жизни, от выхода из яйца вплоть до окукле-
ния, Prestwichia на разу не линяет. Этот факт, отмеченный еще
М. Н. Римским-Корсаковым (1932), несомненно, связан с паразитиче-
ским образом жизни: хитиновый покров утрачивает свое защитное и
опорное значение, делается тонким и растяжимым и не препятствует
росту личинки. Сохраняются только куколочная и нмагинальная линь-
ки, что прекрасно согласуется с данными Э. Пояркова (1914), показав-
шего, что преобразование внутренних органов насекомого и особенно
мускулатуры требует двух линек: при первой линьке тело принимает
окончательную форму и начинается развитие имагинальных мышц, при
второй — осуществляется прикрепление мышц к кутикуле; это и послу-
жило, по его мнению, причиной появления в филогенезе стадии ку-
колки.
146
продольные полоски утолщенной гиподермы, представляющие собой
нечто вроде продолжения той активной области гиподермы груди, где
формируются диски ног и крыльев. У женских личинок каждый поло-
вой зачаток еще раз разделяется на два, так что получается четыре
зачатка яичников (п. з). У мужских личинок это второе разделение не
происходит. На этой стадии личинка уже совершает слабые перисталь-
тические сокращения тела, указывающие на дифференциацию сомати-
ческой мускулатуры.
V стадия (рис. 89, В) — на этой стадии личинка перестает питать-
ся. Объем кишечника значительно уменьшается, в нем начинают отла-
гаться экскреты, заметные благодаря своей непрозрачности. Форма
тела личинки из пузыревидно-овальной превращается в удлиненную.
Зачатки крыльев, ног и антенн заметно удлиняются. Ноги и антенны
образуют изгибы. Появляются зачатки мандибул, максилл, нижней
губы, а у самок — и яйцеклада (яйцк). Последний представлен двумя
парами пальцевидных выпячиваний гиподермы, первоначально очень
похожих на зачатки конечностей. Но окружающая их гиподерма тоже
утолщена и не образует тонкостенных периподальных мешков. Вся об-
ласть утолщенной гиподермы яйцеклада начинает погружаться внутрь,
одновременно смещаясь вперед.
Две латеральные гиподермальные полоски брюшка становятся
очень заметными. Они образуют симметричные волнообразные изгибы,
которые можно считать первыми проявлениями сегментации брюшка.
В брюшной цепочке личинки тоже появляются утолщения—зачатки
ганглиев.
Позднее (на VI стадии развития), в виде слабых изгибов гиподер-
мы, намечается сегментация груди и образуется более глубокая пере-
тяжка на границе груди и головы. Но в голове никаких признаков
сегментации, помимо конечностей, не появляется. С самого начала раз-
вития и до конца она имеет форму компактного образования. Подгло-
точный ганглий (см. ниже) тоже сразу закладывается как единое обра-
зование, ничем анатомически не проявляя свою трехмерную природу.
Таким образом, сегментация тела у Prestwichia, с момента своего
возникновения, имеет характер гетерономной имагинальной сегмента-
ции. Интересно отметить также, что сегментация выражена лишь в ги-
подерме и ее производных, но совершенно не затрагивает мезодерму.
На V стадии развития форма тела личинки уже сильно усложняет-
ся. Личиночная кутикула не следует за изменением рельефа тела; там,
где образуются различные внешние придатки, она просто, отслаивается,
и эти придатки лежат под ней свободно (рис. 90, кут). Но на спине, где
поверхность тела остается гладкой, кутикула продолжает прилегать к
гиподерме.
Следующая — VI стадия является переходной к стадии куколки и
может быть названа предкуколкой (praepupa, pronympha). Кутикула
отслаивается по всей поверхности тела и в виде просторного овального
пузыря, повторяющего форму личинки IV стадии, окружает личинку,
сильно уменьшившуюся в объеме и постепенно приближающуюся по
форме к куколке. Если до сих пор все имагинальные зачатки были
сконцентрированы на вентральной стороне тела, образуя так называе-
мую «имагинальную полоску», то теперь, вследствие сокращения силь-
но растянутой на предыдущих стадиях ларвальной гиподермы спины
и разрастания имагинальной гиподермы брюшной стороны, все зачатки
становятся на свое окончательное место. Так, например, зачатки
крыльев из первоначального вентролатерального положения переходят
в дорзолатеральное положение. Тот же процесс приводит к формиро-
149
ванию головы. Все внешние придатки еще больше удлиняются и приоб-
ретают свою дефинитивную форму.
На этой стадии происходит полное переваривание содержимого
средней кишки и заполнение ее экскрементами. Потом происходит де-
фекация, указывающая на установление связи между средней и зад-
ней кишкой.
Окончательное сбрасывание личиночной кутикулы знаменует пере-
ход к стадии куколки. Внешняя форма куколки в дальнейшем почти
не меняется. Постепенно происходит пигментация: сперва глаза де-
лаются красными, затем чернеют и наконец наступает потемнение всех
покровов. Вторая линька приводит к образованию имаго.
Более подробное рассмотрение преобразований, происходящих при
метаморфозе Prestwichia, удобнее производить для каждой системы
органов отдельно.
Гиподерма
У молоденькой личинки вся гиподерма кажется однородной и, по
всей вероятности, еще сохраняет свойства эмбриональной ткани. Уже
во время постэмбрионального развития из нее выделяются зачатки, го-
ловного мозга и брюшной нервной цепочки. Путем утолщения гиподер-
мы образуются имагинальные диски головы, дающие начало сложным
глазам, антеннам, ротовым частям и всем покровам головы, имагиналь-
ные диски ног и крыльев, которые, разрастаясь, дают всю имагиналь-
ную гиподерму груди, и диски половых придатков. На IV и V стадиях
развития в Ъентролатеральных частях будущего брюшка личинки на-
мечаются две продольные полоски утолщенной гиподермы, являющие-
ся прямым продолжением той области гиподермы груди, где образуют-
ся диски ног и крыльев. Наружные края полосок как бы погружены в
полость тела и переходят в тонкую личиночную гиподерму, которая
слегка нависает над ними (рис. 90,»Б, им. гип). В этих полосках впер-
вые проявляется сегментация. На почве этих двух полосок развивается
трахейная система и эноциты (см. ниже). Разрастаясь в ширину, эти
полоски дают всю имагинальную гиподерму брюшка. Очевидно, это
имагинальные диски покровов брюшка.
Специальные имагинальные диски, за счет которых происходит
образование гиподермы брюшка, наблюдаются у ряда двукрылых (Га-
нин, 1877; Ковалевский, 1887; Валь — Wahl, 1901; Робертсон — Robert-
son, 1936) и перепончатокрылых. Тигс (Tiegs, 1922) описал и изобра-
зил на рисунках парные имагинальные диски во всех сегментах тела
паразитического перепончатокрылого Mormoniella (=Nasonia) (сем.
Pteromalidae). Такие же парные диски имагинальной гиподермы най-
дены Вейсенбергом (Weissenberg, 1907) у Callimome abdominale
(Boh.) (=Torymus nigricornis; Callimomidae). У Prestwichia мы встре-
чаемся не со многими парами дисков, отвечающими каждому сегменту,
а с непрерывными полосками имагинальпых клеток. Очевидно, это свя-
зано с запаздыванием сегментации в развитии Prestwichia, происходя-
щей уже после образования полосок имагинальной гиподермы.
Помимо перечисленных производных гиподермы личинки, имею-
щих эктодермальную природу, в литературе встречаются указания на
то, что гиподерма личинки способна производить элементы, обычно
происходящие из мезодермы. Так, М. С. Ганин (1877) описал образо-
вание иг имагинального диска ноги мухи мезодермальных клеток, даю-
щих впоследствии мускулатуру ноги. В. П. Поспелов (1911) наблюдал
B'j время постэмбрионального развития бабочек новообразование жи-
152
рового тела за счет клеток, выселяющихся из гиподермы. Неоднократ-
но описывались особые «очаги кроветворения», тесно связанные с утол-
щениями трахей или с гиподермой имагинальных дисков крыльев
(Ларченко, 1937, 1946, 1956; Федотов, 1940; Штейнберг, 1948, и др.).
У Prestwichia мне не приходилось наблюдать ни очагов кроветворения,
ни новообразования жирового тела. Что же касается происхождение
мезодермальных элементов имагинальных дисков от гиподермы, то,
хотя я и не наблюдала вполне достоверных картин выклинивания кле-
ток из гиподермы диска, но все же эта возможность кажется очень
вероятной, так как на стадии личинки недифференцированные мезодер-
мальные клетки встречаются почти исключительно в полости имаги-
нальных дисков, в тесном соседстве с их утолщенной гиподермой.
Все это показывает, что формообразовательные возможности ги-
подермы очень велики и выходят за рамки наружного зародышевого
листка.
После выделения брюшных имагинальных дисков окончательно
очерчивается та область гиподермы, которая принимает участие в фор-
мировании имагинальных органов и которую можно назвать «имаги-
нальной полоской». Все имагинальные части гиподермы состоят из
мелких, интенсивно размножающихся клеток. Остальная (ларвальная)
гиподерма состоит из крупных, сильно уплощенных клеток. Размноже-
ние их прекращается на ранних стадиях постэмбрионального развития.
Ядра имеют сплюснутую форму дисков, округлых или несколько удли-
ненных (рис. 92, Б и В, л. гип). На V стадии развития диаметр ядер
равен 0,021—0,022 мм, толщина — приблизительно вдвое меньше.
Первоначально имагинальная полоска занимает лишь небольшую
поверхность на вентральной стороне личинки (приблизительно Уб всей
поверхности); всю остальную гиподерму мы вправе назвать личиноч-
ной. В какой степени утрачивает она способность давать новые произ-
водные, способность к регенерации и т. д. — может решить только
эксперимент, но несомненно, что при нормальном развитии она не при-
нимает никакого участия в построении имагинальных частей и дегене-
рирует во время метаморфоза. Д. М. Штейнберг (1938, 1948) экспери-
ментально показал, что у поздней личинки восковой огневки Galleria
melonella L. происходит регенерация имагинального диска крыла за
счет прилегающих частей гиподермы. Но у Galleria различия между
имагинальными и ларвальными частями гиподермы выражены не так
резко. Возможно, что гиподерма Galleria вполне однородна и имеет
двойственную природу, являясь одновременно и покровной тканью,
и источником образования имагинальных органов. Позднее способность
к регенерации исчезает, очевидно, в связи с наступающей гистологиче-
ской дифференциацией. У жуков большая часть кожных покровов
взрослого насекомого образуется за счет личиночной гиподермы, и ме-
таморфоз не сопровождается отмиранием сколько-нибудь значитель-
ных количеств гиподермальных клеток (Поярков, 1910; Мэррей и
Тигс —Murray a. Tiegs, 1935; Петей —Patay, 1939). По мнению Снод-
грасса (1954), резкое разграничение ларвальных и имагинальных уча-
стков в гиподерме личинки (как и в других органах), свойственное
высшим отрядам насекомых с полным превращением (Hymenoptera и
Diptera), есть признак специализации.
На V стадии развития Prestwichia имагинальная полоска занимает
уже половину всей поверхности личинки. Это объясняется, отчасти,
ростом имагинальной гиподермы, а также связано с уменьшением об-
щего объема личинки и сокращением ларвальной гиподермы, до того
чрезмерно растянутой.
153
тате которого нервная система имаго достигает высокой степени кон-
центрации. В нервной системе взрослых Prestwichia (рис. 93, Б и В)
можно различить головной мозг, который вместе с подглоточным ган-
глием образует компактную массу, пронизанную лишь тонкой трубкой
пищевода, три пары грудных ганглиев и два брюшных. Брюшные ган-
глии каждой пары сливаются друг с другом, так что распознать их
парную природу анатомически невозможно. У самцов оба брюшных
ганглия сближены, у самок коннективы между ними сильно растянуты
и огибают основание яйцеклада с левой стороны.
Таким образом, мы видим, что с момента своей закладки нервная
система Prestwichia все время находится в состоянии непрерывного
прогрессивного развития. Здесь совершенно невозможно различить
личиночные и имагинальные части, как это сделали Бауэр (Bauer,
1904) в нервной системе Culex, Тйгс(1922) —у Mormoniella (Nasonia),
Л. К. Титова (1940, 1949) —у Pieris brassicae L. Правда, на V стадии
развития в нервной системе Prestwichia, наряду с митозами, встре-
чаются и многочисленные случаи дегенерации клеток: клетки распа-
даются на базофильные гранулы, которые падают в полость тела и фа-
гоцитируются или рассасываются на месте. Быть может, дегенерируют,
не достигнув полного развития, как раз те клеточные элементы, кото-
рые гомологичны ларвальным элементам других насекомых, но это
только предположение, нуждающееся в доказательстве.
Такое же прямолинейное развитие нервной системы описал
В. А. Караваев (1898) для муравья Lasius flavus L.
Трахейная система
У молодых личинок Prestwichia aquatica никаких органов дыхания
нет. Трахеи закладываются очень поздно, на IV—V стадиях развития.
Зачатки трахей имеют вид парных врастаний гиподермы в форме плот-
ных клеточных тяжей, которые имеют просвет лишь у своего основания
(рис. 94, Л, тр). Таких зачатков образуется 6 пар: первая, очень неясно
выраженная пара, закладывается в области головы, три пары — в об-
ласти груди, одна — в первом сегменте брюшка и одна — на уровне
яйцеклада. Грудные зачатки трахей лежат на одной продольной линии
с крыльями, а брюшные — по ходу полосок имагинальной гиподермы.
Все они растут внутренними концами вперед, и только первая пара,
как кажется, направлена назад.
Накануне метаморфоза (VI стадия развития) все зачатки сливают-
ся в два продольных ствола, от которых начинают расти ответвления
к различным внутренним органам. Связь с гиподермой сохраняется
лишь в заднегруди, где у имаго имеется пара стигм. Все остальные
зачаточные стигмы исчезают. Наличие у Prestwichia только одной пары
стигм уже отмечено М. Н. Римским-Корсаковым (1916).
М. С. Ганин (1877), А. О. Ковалевский (1885) и Тигс (1922) опи-
сали значительные перестройки трахейной системы. Но у Prestwichia
развитие трахейной системы имеет прямой характерен она не подвер-
гается во время метаморфоза никаким перестройкам. Уменьшение числа
стигм нельзя считать проявлением метаморфоза. Это просто явление
олигомеризации (Догель, 1936, 1954), имевшее место в процессе фило-
генеза Prestwichia и рекапитулируемое в развитии. Редуцируются не
личиночные части трахейной системы, функционировавшие на ранних
стадиях, а эмбриональные зачатки. Подобный процесс олигомеризации
постоянно происходит во время эмбрионального развития, и никому не
придет в голову называть эту частичную редукцию метаморфозом.
157
ларвальных эноцитов фагоцитируется. Образование новой генерации
эноцитов из гиподермы описывают Г. А. Кожевников (1910) для медо-
носной пчелы, Вейсенберг (1907)—для Callimome (Torymus), Тигс
(1922)—для Mormoniella (Nasonia).
Сравнивая мои наблюдения над эноцитами Prestwichia с данными
других исследователей, можно сделать вывод, что образование ларваль-
ной. генерации эноцитов у Prestwichia сильно запаздывает, а имагиналь-
ные эноциты не образуются вовсе. Если двукратное образование эноци-
тов у всех исследованных насекомых объясняется теми же причинами,
что и двукратное формирование других органов, замещающихся новыми
при метаморфозе, то в наличии только одной генерации эноцитов у
Prestwichia можно видеть такой же пример «выпрямления» развития,
как в нервной системе и органах дыхания. Если функция эноцитов, как
полагает большинство авторов (Снодграсс, 1954), связана с линьками,
то позднее появление их у Prestwichia вполне понятно: ведь у Prest-
wichia личиночные линьки отсутствуют.
Мускулатура
У молодой личинки (I—II стадии) мускулатура представлена толь-
ко мышцами пищевода, обеспечивающими глотательные движения.
Мышцы эти состоят из отдельных, сильно вытянутых в длину клеток,
в которых лежат миофибриллы с неясно выраженной поперечной исчер-
ченностью. По положению они разделяются на две группы: часть мышеч-
ных клеток протягивается от дорзальной утолщенной стенки пищевода
до дорзальной стенки тела. Они расширяют просвет пищевода. Другая
часть мускульных клеток распластывается по поверхности пищевода в
поперечном направлении. Эти клетки суживают просвет пищевода и яв-
ляются антагонистами первых.
Число глотательных мышц сперва невелико, затем оно значительно
увеличивается. Так как мне ни разу не приходилось наблюдать размно-
жение мышечных клеток, то самое вероятное, что увеличение их числа
происходит за счет новообразования из недифференцированных мезо-
дермальных клеток.
На IV—V стадиях развития появ-ляются мышцы в области задней
кишки, растягивающие ее стенки в стороны, почему ее просвет имеет
форму поперечной щели.
В то же время возникает кожная (соматическая) мускулатура. Не-
многочисленные мезодермальные клетки, находящиеся в полости тела
только что вышедшей из яйца личинки, размножаются и постепенно диф-
ференцируются в трех направлениях: они дают личиночные мышцы
(рис. 96, м), жировые клетки (рис. 101, жир. кл) и некоторое количество
соединительнотканных клеток на поверхности кишечника и других вну-
тренних органов (рис. 101, соед. кл).
На IV стадии развития мускулатура личинки уже достигает полного
развития. Она представлена многочисленными отдельными клетками,
распластанными на внутренней поверхности гиподермы. Изучение их
особенно удобно не на срезах, а на препаратах, изготовленных из лоскут-
ков снятой с личинки гиподермы (рис. 96). Распределяются эти клетки
очень равномерно, и лишь на брюшной стороне, где сосредоточены има-
гинальные зачатки, мышечных клеток нет. Плазма мышечных клеток со-
вершенно бесцветна; она настолько истончается у периферии клетки, что
различить койтуры.клетки не удается. В плазме мускульных клеток рас-
полагаются пучки миофибрилл; направление их не строго параллельно:
они могут сходиться и расходиться под некоторым углом; на концах
160
стве, сколько содействует перемешиванию содержимого кишки, лишенной
собственных мышц, т. е. служит пищеварению. Такая ограниченность
движений и связанная с ней редукция мускулатуры безусловно являют-
ся прямым следствием паразитического образа жизни.
Дегенерация личиночных мышц. Превращение мышц
является одним из самых сложных и запутанных вопросов метаморфоза
насекомых. Даже наблюдения авторов, работавших с одними и теми же
видами насекомых, часто противоречат друг другу. Дело осложняется
тем, что разные мышцы во время метаморфоза имеют разную судьбу.
Так, можно различить мышцы, которые дегенерируют полностью, мыш-
цы, которые подвергаются сложной перестройке и, наконец, такие мыш-
цы, которые образуются заново из миобластов, сохранившихся у ли-
чинки.
Процесс дегенерации мышц обычно сопровождается распадом их на
содержащие ядро или безъядерные фрагменты, которые нередко описы-
вались под специальными названиями (саркоциты, кариолиты, сарко-
литы и т. д.) и смешивались с фагоцитами, эноцитами и другими полост-
ными элементами, не имеющими прямого отношения к мышцам.
По А. А. Коротневу (1892), у моли Tinea все имагинальные мышцы
образуются путем перестройки ларвальных мышц; фибриллярное ве-
щество дегенерирует, а ядра мышц с частью саркоплазмы складываются
в новые мышечные тяжи и выделяют новое сократительное вещество.
Но чаще в перестраивающихся мышцах различают ядра двух родов:
крупные ларвальные, погибающие во время метаморфоза, и мелкие
имагинальные, которые обособляются с небольшим участком плазмы,
размножаются и превращаются в миобласты, дающие новые мышцы.
Караваев (1898) наблюдал это у Lasius flavus L., Англа (1902) —у осы,
Поярков (1910) —у Galerucella, Перэ (1912) —у Polistes и т. д.
По Тигсу (1922), имагинальные миобласты Mormoniella (Nasonia)
образуются не внутри ларвальных мышц, а происходят от мезодер-
мальных клеток полости тела. То же имеет место у двукрылых — у Cal-
liphora (Перэ, 1910) и Drosophila (Робертсон, 1936). За счет миобластов
образуются и те имагинальные мышцы, которые развиваются совер-
шенно заново, — например, мышцы конечностей, отсутствующих у ли-
чинки. Иногда описывается перестройка и вторичное использование са-
мого фибриллярного вещества.
Много разногласий существует по вопросу о том, какую роль в деге-
нерации играет фагоцитоз. Одни исследователи утверждают, что дегене-
рация и рассасывание мышц происходят без непосредственного участия
фагоцитов (Англа, 1902, 1904 а, б — у медоносной пчелы и осы; Эртель —
Oertel, 1930 — у пчелы), другие же,— что фагоциты пожирают только
дегенеративные обломки мышц (Поярков, 1910 — у Galerucella; Перэ,
1912 — у Polistes; Тигс, 1922 — у Mormoniella, и др.), третьи,— что фа-
гоциты нападают на совершенно «здоровые» мышцы и служат причиной
их разрушения (Ковалевский, 1885, 1887 — для Musca). При этом роль
фагоцитов обычно выполняют кровяные элементы, но Э. Поярков, кро-
ме того, отмечает способность фагоцитировать обломки мы-
шечных клеток у клеток жирового тела. Имагинальные миобласты не-
редко ведут себя, как фагоциты: растворяют и всасывают вещества лар-
вальных мышц.
Интересно отметить, что совершенно такая же путаница существует
в литературе по вопросу о дегенерации мышц хвоста при метаморфозе
у амфибий. Я упомяну лишь работу В. Я. Александрова и 3. И. Крюковой
(1950), выполненную не только обычной гистологической методикой, но
с применением новейших методов прижизненного наблюдения. Они пока-
162
плазму, достигают приблизительно 0,007 мм. Очевидно, здесь происходит
объединение нескольких мелких клеток для фагоцитоза сравнительно
крупного объекта.
Процесс фагоцитоза, видимо, длится очень долго: уже на стадии
куколки с пигментированными глазами встречается множество жировых
клеток с заглоченными комками ларвальных мышц.
Многие авторы, изучавшие дегенерацию личиночных мышц, начи-
ная с А. О. Ковалевского (1885) и кончая новейшими наблюдениями
В. Я. Александрова и 3. И. Крюковой (1950), описывают ее совсем иначе:
сперва происходит расщепление мускульного волокна вдоль на более
тонкие пучки фибрилл, затем эти пучки разрываются поперечно; при
этом фибриллярная структура, поперечная исчерченность и жизненные
свойства фрагментов остаются без изменений. Окончательное уничтоже-
ние мышц совершается с помощью фагоцитов.
У. Prestwichia тоже иногда, хотя и редко, наблюдается фрагмента-
ция мышечных клеток. Но при этом фрагменты всегда имеют форму со-
вершенно неправильных гомогенных комков, не сохраняющих никаких
следов своей первоначальной структуры. Основным же способом дегене-
рации мышц у Prestwichia является описанный выше. Таким обра-
зом, у Prestwichia личиночные мышцы подвергаются глубоким деге-
неративным изменениям, прежде чем они становятся добычей фаго-
цитов. Нет никаких оснований думать, что к началу фагоцитоза они еще
сохраняют жизнеспособность.
Дегенерация мышц у Prestwichia, не сопровождающаяся фрагмен-
тацией, по-видимому, объясняется малыми размерами мышечных эле-
ментов; они представлены отдельными клетками, связанными друг с
другом посредством отростков. При желании можно считать фрагмента-
цией распадение всей мышечной сети на отдельные клетки. В таком виде
мышечные элементы уже могут быть заглочены фагоцитами, и дальше
фрагментация не идет.
Имагинальная мускулатура. Источником формирования
имагинальных мышц являются свободные многочисленные мезодермаль-
ные клетки, в изобилии встречающиеся в области имагинальных дисков.
Так как недифференцированные мезодермальные клетки нигде больше
не встречаются, то очень вероятно, что они тут же и возникают, выкли-
ниваясь из гиподермы, к которой они плотно прилегают, как это наблю-
дал М. С. Ганин (1877), хотя вполне достоверные картины такого вы-
клинивания не встречались. В конце личиночной жизни (V стадия) в
голове, подле конечностей и зачатка яйцеклада, эти клетки, которые
можно назвать миобластами, складываются в тяжи, постепенно преоб-
разующиеся в мышечные пучки. Мною прослежено несколько подробнее
лишь формирование продольных летательных мышц.
Первые зачатки летательных мышц можно видеть в небольших сгу-
щениях мезодермальных клеток впереди от первой пары крыльев на
IV стадии'развития. Гиподерма здесь утолщена, и, возможно, именно
она продуцирует эти клетки.
На V стадии развития зачатки крыловых мышц образуют уже ком-
пактные клеточные массы, которые разрастаются назад и могут быть вид-
ными на одном поперечном разрезе с дисками крыльев латеральнее их.
На VI стадии, когда зачатки крыльев из вентролатерального поло-
жения переходят,в дорзолатеральное, зачатки мышц оказываются лежа-
щими дорзальнее их (рис. 98,0, прод. м и верт. м). В это время зачатки
летательных мышц приобретают форму двух полосок, тянущихся вдоль
тела и соприкасающихся с гиподермой лишь своими концами. Благодаря
размножению клеток число их значительно увеличивается. Начинается ги-
364
стологическая дифференциация: часть миобластов каждого зачатка скла-
дывается в 4 продольных тяжа, в которых начинает формироваться со-
кратительное вещество (рис. 98, Б и Г, прод. м). Плазма слившихся мио-
бластов как бы вся истрачивается на построение сократительного веще-
ства, и их ядра, лежащие у поверхности тяжей, кажутся обнаженными.
Сократительное вещество имеет волокнистую структуру, но поперечная
исчерченность появляется значительно позднее.
Между четырьмя формирующимися мышечными волокнами каждой
стороны тела остается еще значительное количество свободных миобла-
ст.ов. Они имеют удлиненную или неправильную угловатую форму: боль-
шое светлее ядро содержит крупное ядрышко. Резко базофильная плазма
образует вокруг него тонкий неправильный ободок. Часть миобластов
присоединяется к волокнам впоследствии, другая же часть подвергается
дегенерации: они сливаются в сферическую массу, хроматин каждого
ядра собирается в плотный бесформенный комок, лежащий эксцентрично
у самой ядерной оболочки, почему ядро кажется наполовину пустым
(рис. 98,5, дег. м). Такие дегенеративные массы наблюдались мною
почти у всех личинок в обоих зачатках летательных мышц. Очевидно, де-
генерация части эмбрионального материала мышц явление закономерное.
Затем комки дегенерирующих миобластов фагоцитируются жировыми
клетками (рис. 98,Л). Первоначально зачатки продольных летательных
мышц тянутся лишь через 1-й и 2-й сегменты груди.
В области заднего конца каждой из продольных летательных мышц
гиподерма образует поперечную складку, направленную вершиной на-
встречу мышце. Здесь происходит прикрепление мышечного зачатка к
гиподерме. Затем зачатки мышц сильно удлиняются, задние их концы все
больше отодвигаются назад. Одновременно обе гиподермальные складки
становятся глубже и сливаются друг с другом в одну поперечную склад-
ку, пересекающую всю дорзальную часть груди. Эта складка представ-
ляет собой зачаток большой фрагмы. Внутри складки формируется хи-
тиновая пластинка, которая как бы прикрывает продольные мышцы свер-
ху и сзади. Вместе с мышцами она простирается далеко назад в брюшко
(рис. 99, фр). Гиподермальные клетки, выделившие хитиновую пластин-
ку фрагмы, исчезают, и мышца оказывается прикрепленной непосред-
ственно к хитину. К концу метаморфоза объем продольных летательных
мышц Prestwichia значительно увеличивается, но они по-прежнему со-
стоят всего из 4 мощных волокон каждая. «
Исчезновение летательных мышц у короткокры-
лых особей Prestwichia. Изучая метаморфозм Prestwichia,
я столкнулась с любопытным явлением редукции крыловой мускулатуры.
Как известно, Prestwichia aquatica ведет амфибиотический образ жизни
и совсем (или почти совсем) не пользуется крыльями для передвижения.
При передвижении по суше она бегает или прыгает (возможно, с по-
мощью крыльев), в воде она плавает, перебирая ногами, а крылья оста-
ются сложенными. В связи с этим, у части особей этого вида наблюдается
редукция крыльев. М. Н. Римский-Корсаков (1916, 1932) описывает ди-
морфизм самок Prestwichia: наряду с нормальными «длиннокрылыми»
самками встречаются самки с значительно укороченными крыльями.
У всех самцов крылья укорочены еще сильнее, чем у «короткокрылых»
самок. Короткокрылость самок передается по наследству.
Начальные стадии развития крыльев у всех личинок одинаковы.
Разница в размерах крыльев становится заметной лишь на стадии ку-
колки. По-видимому, при редукции крыльев имеет место не только их не-
доразвитие, но и дегенерация части клеточного материала. Но эти деге-
неративные явления происходят не в дистальнои части крыла, как это
166
с плотными ядрами, гиподермальнного происхождения. Интересно, что в
этом случае способ дегенерации зачатка мышц отличается от вышеопи-
санного (стр. 166). Эти различия, по-видимому, объясняются тем, что
в первом случае дегенерировали миобласты, а теперь речь идет о зачат-
ках мышц, уже содержащих сократительное вещество. Дальнейшая
судьба дегенерирующих летательных мышц не прослежена.
На стадии поздней куколки и имаго самцы и часть самок уже не
имеют никаких следов мышц. Дегенерации подвергаются не только про-
дольные, но и дорзо-вентральные мышцы.
Большая фрагма, к которой прикрепляются сзади продольные мыш-
цы, образуется у всех куколок, но у куколок с редуцированными крылья-
ми она имеет меньшие размеры. Очевидно, развитие ее приостанавли-
вается в момент дегенерации мышц. Несмотря на отсутствие мышц, она
сохраняется и во взрослом состоянии.
Редукция мышц и редукция крыльев у Prestwichia — явления, тесно
связанные друг с другом. Они имеют одну общую физиологическую при-
чину— отсутствие функционирования крыльев; роль, которую крылья
играют в жизни Prestwichia, незначительна.
Д. М. Федотов (1940), описывая редукцию крыльев у самок зимней
пяденицы, отмечает и параллельно идущую редукцию крыловых мышц:
мощность и количество мышечных пучков уменьшаются. У Prestwichia
редукция мышц значительно опережает редукцию крыльев. Это обстоя-
тельство можно было бы объяснить тем, что остановившийся в своем
развитии зачаток мышцы, состоящий только из клеточных элементов,
легко подвергается гистолизу и уничтожению; труднее уничтожить за-
чаток крыла, укрепленный хитиновыми частями. Но если вспомнить, чта
размеры и форма крыльев определяются еще на стадии куколки, отде-
ленной от стадии имаго линькой, когда хитиновые структуры сбрасыва-
ются, станет ясно, что частичное сохранение крыла у имаго связано и с
сохранением его гиподермальной основы.
Таким образом, в нашем случае мускулатура гораздо быстрее реа-
гирует на функциональные изменения, чем гиподерма крыла.
Кровеносная система
У личинки Prestwichia кровеносная система отсутствует, но у взрос-
лых Prestwichia она имеется, хотя сильно упрощена. Она состоит из длин-
ной тонкостенной трубки, которая проходит по медиодорзальной линии
от 5-го сегмента брюшка до шеи. Проследить эту трубку в голове не
удалось. Стенка трубки почти не окрашивается применявшимися гисто-
логическими красками. В ней редко разбросаны крупные ядра с грубо
зернистым хроматином (рис. 100,5). Сердечная трубка несколько напо-
минает по внешнему виду трахеи, но отличается от них меньшей упруго-
стью (часто она бывает смята или изломана) и отсутствием такого от-
четливого внутреннего контура, который в трахеях обусловлен наличием
тонкой хитиновой выстилки. Никаких специальных структур в стенках
трубки или подле нее обнаружить не удалось, лишь на заднем конце она
образует значительное расширение округлых очертаний. Это расширение
имеет губчатое строение — просвет его заполнен сетью неправильных пе-
рекладин соединительнотканной или сократительной природы, в которых
лежат ядра, сходные с ядрами узкой части трубки (рис. 100, В). Увидеть
остии мне не удалось. Возможно, задний булавовидно расширенный ко-
нец трубки снабжен порами и способен всасывать и проталкивать
вперед гемолимфу, но проверить это можно было бы лишь путем прижиз-
ненных наблюдений в эксперименте.
168
Жировое тело
Формирование жирового тела у Prestwichia начинается лишь у ли-
чинки. На плоскостных препаратах из стенки средней кишки на III ста-
дии развитии можно видеть сеть веретеновидных или неправильных кле-
ток, связанных друг с другом тонкими плазматическими отростками
(рис. 101, Л, жир. кл). Базофильная протоплазма этих клеток содержит
несколько светлых вакуолей. При фиксации жидкостью Флемминга эти
вакуоли чернятся — очевидно, у живых личинок они заполнены жиром.
Берлезе (Berlese, 1901), Англа (1902), Перэ (1912) и многие другие
■ описывают наличие в жировом теле насекомых специальных экскретор-
ных клеток, в плазме которых происходит накопление мочекислых солей.
Мне самой приходилось наблюдать такие клетки в жировом теле совсем
молодых личинок Anopheles (Иванова-Казас, 1949а). Э. Поярков (1910)
.и Тигс (1922) отмечают появление экскреторных гранул в обыкновенных
жировых клетках куколок Galerucella и Mormoniella.
Ничего похожего не встречается у Prestwichia aquatica. Все клетки
жирового тела этого наездника кажутся совершенно однородными. Да-
же временное появление в них экскретов никогда не наблюдалось. По-
этому участие жирового тела Prestwichia в процессе экскреции кажется
сомнительным.
По мере роста личинки размеры жировых клеток тоже увеличива-
ются. Количество и объем жировых включений значительно возрастают
(рис. 101,5, жир. кл). Э. Поярков (1910), В. П. Поспелов (1911) и
К. И. Ларченко (1937) указывают на размножение клеток жирового тела.
У Prestwichia, как и у осы Polistes (Перэ, 1912), размножение жировых
клеток не происходит. На V стадии развития ядра жировых клеток дости-
гают 0,011—0,012 мм в диаметре; поперечник самой жировой клетки в 3—
4 раза превышает диаметр ядра. Благодаря переполнению клеток вклю-
чениями, они приобретают округлую форму и утрачивают связь друг с
другом. Быть может, этот момент соответствует стадии распада жирово-
го тела на отдельные клетки, описываемой многими авторами: Г. А. Ко-
жевниковым (1900) — у медоносной пчелы; Э.Поярковым (1910) —у жу-
ка Galerucella; В. П. Поспеловым (1911) и К- И. Ларченко (1937) — для
ряда бабочек; Перэ (1912) —для Polistes. Начиная с V стадии жировые
клетки встречаются не только на поверхности кишки, но и з других участ-
ках полости тела.
В плазме жировых клеток происходит как бы отмешивание каких-то
ее компонентов. Большая часть плазмы становится бесцветной, но подле
ядра и особенно в периферических частях клетки встречаются комки
очень плотной, резко базофильной плазмы. Интенсивность окрашивания
этих участков плазмы можно сравнить лишь с таковой хроматиновых
компонентов ядра (рис. 102,Л).
Наряду с жировыми вакуолями, в плазме жировых клеток появля-
ются эозинофильные гранулы, по-видимому, белковой природы
(рис. 102, бел гр.). Накопление белков в жировом теле насекомых нака-
нуне или в начале метаморфозма отмечали Англа (1902), Э. Поярков
(1910), В. П. Поспелов (1911), Перэ (1912), Тигс (1922), Эртель (1930),
К. И. Ларченко (1937) и многие другие. Эозинофильное вещество нака-
пливается в таком большом количестве, что иногда вся клетка пропиты-
вается им и становится эозинофильной и лишь немногие, но крупные жи-
ровые вакуоли остаются неокрашенными (рис. 102, Б и В).
Затем внутри эозинофильных гранул появляются базофильные зер-
нышки, количество их постепенно возрастает и, наконец, вся эозинофиль-
ная гранула становится базофильной. Нередко рядом с совершенно неиз-
170
конец, и эта оболочка исчезает, и базофильные зернышки оказываются
лежащими прямо в полости тела. Очевидно, эозинофильное вещество по-
степенно растворяется в полостной жидкости.
Это явление можно толковать либо как начало дегенерации клеток,
либо как «клеточную автотомию», наблюдавшуюся Э. Поярковым (1910)
при метаморфозе жука Galerncella в очень многих его тканях (в гипо-
дерме, в эпителии передней и задней кишки, в мышцах, в трахеях, в нерв-
ных клетках, мальпигиевых сосудах, в клетках щетинок и в сухожиль-
ных клетках). Э. Поярков придает большое значение явлению клеточной
автотомии, считая, что таким путем клетка освобождается от своих спе-
циализированных частей и омолаживается.
Мне кажется, что в клетках жирового тела Prestwichia мы имеем
дело с дегенеративным явлением. Отбрасываемые части жировых кле-
ток ничем не отличаются от самих клеток, так что нет никаких оснований
считать их особенно специализированными. Кроме того, иногда встреча-
ются целые жировые клетки, ставшие совершенно бесцветными, прозрач-
ными, с неясными контурами, словно наполовину растворившиеся. Ча-
стичная или полная дегенерация жирового тела при метаморфозе отме-
чалась многими исследователями, начиная с М. С. Ганина (1877), при-
чем В. А. Караваев (1898), Э. Поярков (1910), Перэ (1912), Тигс (1922)
и К- И. Ларченко (1937) указывают на участие фагоцитов, а Эртель
(1930) для медоносной пчелы отрицает фагоцитоз. После окукливания
общий вид жировых клеток не меняется. Но к началу пигментации глаз
полостная жидкость приобретает способность интенсивно окрашиваться
эозином: В ней наряду с целыми жировыми клетками встречаются облом-
ки их и отдельные гранулы. Очевидно, у части клеток оболочки разру-
шаются и содержимое их изливается в полость тела, как это описали
Перэ (1901) —для муравья и Англа (1902) —для осы. Разрушение жи-
ровых клеток начинается, по-видимому, с заднего конца тела.
На этой стадии амебоциты становятся довольно многочисленными по
сравнению с амебоцитами на предыдущих стадиях. Они несколько умень-
шаются в размерах (0,006 мм в диаметре), имеют светлую плазму и бо-
лее плотное ядро (0,004 мм в диаметре). Возможно, они участвуют в пе-
реработке продуктов распада жировых клеток.
В результате разрушения жировых клеток количество их значитель-
но уменьшается. Базофильные гранулы исчезают совершенно, эозино-
фильные уменьшаются в размерах и числе. У молодых имаго Prestwichia
в жировых клетках мы встречаем почти исключительно жировые вакуо-
ли, которые несколько увеличиваются в размерах, возможно, путем слия-
ния друг с другом (рис. 102, Д). Размеры самих клеток уменьшаются,
ядра их уплотняются и нередко приобретают неправильную форму. Плаз-
ма делается бесцветной. Границы между клетками становятся неясными,
и скопления жировых клеток, приобретают вид бесцветной пенистой
©массы. Изредка встречаются двуядерные клетки.
У экземпляров, проживших несколько дней после выхода из куколки,
исчезают и жировые включения. Клетки становятся небольшими, округ-
лыми; они имеют бесцветную плазму и светлое ядро. Очевидно, они до
конца выполнили свою роль и находятся на пути к исчезновению. Ран-
нее использование основной массы запасных питательных веществ жи-
рового тела безусловно связано с ранним созреванием гонад у Prest-
wichia, как это показано В. П. Поспеловым (1911) и К. И. Ларченко
(1937) для бабочек.
Из всего сказанного выше следует, что при метаморфозе Prestwichia
aquatica значительная часть клеток жирового тела переходит к имаго. Это
вполне согласуется с данными большинства авторов; только В. Поспелов
173
(1911) для ряда бабочек описывает формирование совершенно нового
жирового тела из свободных полостных элементов, которые в свою оче-
редь происходят из гиподермы. «Пополнение» жирового тела за счет не-
дифференцированных элементов в течение почти всего постэмбриональ-
ного развития описывает и К. И. Ларченко (1937).
Начиная с VI стадии развития, в очень многих тканях личинки на-
блюдаются дегенеративные явления. Дегенерируют личиночные мышцы,
гиподерма, часть эноцитов, часть клеточного' материала летательных
мышц; массовые дегенерации наблюдаются в зачатке нервной системы
и т. д. Продукты дегенерации всех этих клеточных элементов заглатыва-
ются жировыми клетками и перевариваются (рис. 91, Б; 95; 97, Д;
98, Б и В). Хотя, по-видимому, в фагоцитозе отмерших тканей принимают
участие и амебоциты, но они.так мелки и немногочисленны, что главную
роль в уничтожении продуктов гистолиза играют жировые клетки. Спо-
собность жировых клеток к фагоцитозу отмечена также у жуков Э. По-
ярковым (1910") и Мэррей и Тигсом (1935).
Обычно фагоцитами являются подвижные клетки, активно блуждаю-
щие по телу и «отыскивающие» подлежащие уничтожению материалы, з
том числе и микроорганизмы. В данном случае фагоцитируют элементы
почти или вовсе неподвижные, перегруженные разными включениями.
Очевидно, они могут заглатывать только то, что случайно оказалось ря-
дом, и едва ли способны вступать в активную борьбу с паразитическими
организмами. Мне кажется, что жировые клетки Prestwichia и Galeru-
cella было бы правильнее называть не фагоцитами, а некрофагами..
Пищеварительная система
Пищеварительная система личинки Pretswichia состоит из передней,
средней и задней кишки. Передняя и средняя кишка закладывается во
время эмбрионального развития как единое впячивание бластодермы на
переднем конце зародыша. Задняя кишка образуется независимо как
впячивание бластодермы на заднем конце зародыша. Средняя и задняя
кишки соприкасаются друг с другом, но сообщения между ними, как это
обычно для личинок высших перепончатокрылых, еще нет.
Передняя кишка личинки. В период питания и роста пе-
редняя кишка личинки сравнительно коротка. Она имеет" форму слабо-
изогнутой трубки, которая от вентрально расположенного рта направ-
ляется к спинной стороне тела и назад. Пройдя сквозь нервное около-
глоточное кольцо, она сразу переходит в среднюю кишку.
Передняя кишка сплющена в дорзо-вентральном направлении и
имеет на поперечном разрезе полулунную форму (выпуклость обращена
к вентральной стороне). Дорзальная стенка пищевода утолщена и со-
стоит из крупных эпителиальных клеток, к которой в передней его части
прикрепляются мышцы, расширяющие просвет (см. выше). Вентральная
стенка состоит из более мелких клеток. В передней части пищевода
имеются также кольцевые мышечные волокна. В передней кишке личинки
Pretswichia хитиновая выстилка выражена очень слабо в начальной
части и совершенно незаметна в более глубоком отделе.
Средняя кишка личинки. Средняя кишка личинки Prest-
wichia имеет форму широкого просторного мешка и состоит из очень
крупных немногочисленных (около 30) клеток. Еще у только что вышед-
щей из яйца личинки клетки средней кишки (рис. 103, А л. ср. к) отли-
чаются своими размерами. В это время отдельные клетки еще хорошо
отграничены друг от друга и обладают базофильной протоплазмой. Свет-
лые шарообразные ядра имеют 0,011 мм в диаметре.
174
Когда личинка начинает питаться, объем кишечника увеличивается
очень значительно. Это сопровождается ростом клеток (но не размно-
жением их) и сильным растяжением стенок, так что на II и III стадиях
развития кишка кажется бесструктурной перепонкой, утолщающейся
только подле ядер, которые принимают расплющенную форму. Клеточ-
ные границы становятся невидимыми. Лишь на IV стадии, благодаря
росту всей личинки, стенка средней кишки снова приобретает ощутимую
толщину, особенно в вентральной части. К этому времени плазма клеток
утрачивает свою базофилию и становится почти бесцветной. На внутрен-
ней поверхности средней кишки удается различить палочковый слой, осо-
бенно отчетливый в вентральной части кишки, где стенка ее менее рас-
тянута (рис. 103,5, В. Г). Дисковидные ядра достигают к этому времени
0,040 мм в диаметре и около 0,020 мм толщины. В это время средняя
кишка занимает все тело личинки, простираясь даже кпереди от ротового
отверстия (рис. 88 и 90,Л, ср. к).
Когда личинка перестает питаться (V стадия), объем кишки умень-
шается, а стенки ее, спадаясь, становятся толще. В плазме клеток в это
время видны различные вакуоли, гранулы, и сама плазма образует ка-
кие-то местные уплотнения, красящиеся более интенсивно
(рис. 103, Д, л. ср< к).
В тесной связи со стенкой средней кишки стоят четыре группы ги-
гантских резко базофильных клеток (рис. 89,Л и Б). Две из этих групп
сравнительно невелики, состоят из 3—7 клеток каждая и лежат в перед-
ней части кишки (лаб. ж). Две другие группы лежат ближе к заднему
концу и имеют форму двух двурядных полос, состоящих из 8—15 клеток
каждая (мп. с). Сходство клеток, составляющих эти группы, настолько
велико, что на первый взгляд они кажутся совершенно однородными. Но
более внимательное изучение препаратов показало, что мы имеем дело
с лабиальными (слюнными) железами и мальпигиевыми сосудами, рас-
пластавшимися на поверхности кишки. Подробнее о них будет ^идти речь
ниже.
Кроме того, в стенку средней кишки вкраплены довольно крупные
клетки с огромной вакуолью, придающей им пузыревидную форму
(рис. 90, 103, Д; 104, Л; 105, им. ср. к). Вакуоль наполнена водянистым
содержимым, в котором имеется нежная зернистость, осаждающаяся в
силу тяжести у одной стороны вакуоли. Изредка в вакуоли могут по-
падаться небольшие базофильные гранулы. Вакуоль оттесняет ядро к
периферии клетки. Ядро имеет неправильную форму и размеры
0,012X0,020 мм. Диаметр вакуоли достигает 0,044 мм.
Благодаря наличию вакуоли эти клетки производят впечатление
функционирующих клеток, но, как покажет дальнейшее, именно они яв-
ляются имагинальными элементами, из которых развивается средняя
кишка взрослой Prestwichia. Подобные же вакуолизированные клетки
наблюдал Ван Левен (Van Leewen, 1908) у Harmolita hyaUpenne (Wlk.)
(=Isosoma graminicola). Средняя кишка личинки Prestwichia лишена
мышечной оболочки. На ее поверхности встречаются лишь редкие клетки
соединительнотканной природы (рис. 101, Б, coed. кл).
После прекращения питания вся заглоченная пища постепенно пе-
реваривается и всасывается, по-видимому, почти без остатка.. Но одно-
временно в средней кишке накапливается большое количество экскретов
в форме кристаллических конкреций (рис. 104,Л, экс). Они имеют сфе-
рическую форму и размеры до 0,008 мм. Кристаллы эти сильно прелом-
ляют свет и явственно радиально исчерчены, так что получается впечат-
ление, что в центре их пересекается множество тончайших иголочек. Не-
редко в просвете кишки можно встретить маленькие четырех-, пяти- и
175
внутриклеточных полостей к периферии клетки (попытка передать эту
структуру представлена на рис. 103, Д, мп. с).
В задних клетках капилляры становятся более объемистыми и от-
крываются в заднюю кишку, передняя часть которой образована немно-
гими клетками, более мелкими, чем клетки мальпигиевых сосудов, но
все же во много раз превышающими по размерам клетки заднего ее от-
дела. Последний имеет эпителиальные стенки и сплющен в дорзо-вен-
тральном направлении, благодаря наличию мышц, растягивающих его
в стороны. У передней границы заднего отдела, уже на V стадии разви-
тия, имеются три цилиндрических выпячивания, состоящих из таких же
мелких клеток. Это — зачатки имагинальных мальпигиевых сосудов
(рис. 105, Б, им. мп. с).
Во внутриклеточных канальцах ларвальных мальпигиевых сосу-
дов и в просвете задней кишки часто можно наблюдать эозинофильную
зернистость и более крупные эозинофильные гранулы. Очевидно, это
эозинофильное вещество является продуктом выделения ларвальных
мальпигиевых сосудов. В период интенсивного питания личинки в про-
свете ларвальных мальпигиевых сосудов я никогда не видела экскретор-
ных гранул, подобных тем, которые скопляются в средней кишке. Лишь
на стадии предкуколки, когда происходит слущивание и переваривание
ларвального кишечного эпителия, а вместе с ним и мальпигиевых сосу-
дов, внутри дегенерирующих клеток последних и между ними могут по-
падаться как упомянутые выше радиально-исчерченные гранулы, так и
конкреции иного рода, имеющие большие размеры (до 0,022 мм), непра-
вильную форму и концентрически слоистую структуру. Центральное ядро
их обычно красится очень интенсивно гематоксилином, но перифериче-
ские слои несколько светлее и иногда имеют слабую радиальную исчер-
ченность. Они очень немногочисленны, встречаются не всегда и, по-види-
мому, являются продуктом уже нарушенной функции.
Лабиальные (слюнные) железы личинки. В момент
вылупления личинки Prestwichia из яйца слюнные железы представляют
собой две группы пузыревидных клеток, как бы подвешенных на корот-
ких выводных протоках (рис. 81, Г, лаб. ж). Оба протока соединяются
вместе у самого выводного отверстия, лежащего позади рта. Протоки
состоят из небольшого числа клеток, постепенно уменьшающихся в раз-
мере по мере удаления железистого отдела.
При прижизненном изучении слюнные железы бывают хорошо вид-
ны у очень молодых личинок, но на более поздних стадиях становятся
неразличимыми, так как их клетки вступают в тесный контакт со средней
кишкой и распластываются по ее поверхности ( подобно мальпигиевым
сосудам). Это обстоятельство, по-видимому, и дало повод Хенриксену
(1922) утверждать, что слюнные железы Prestwichia функционируют
лишь в начале личиночной жизни, когда личинка еще настолько мала,
что неспособна заглатывать целые гранулы желтка, и служат для его
растворения. Позднее, когда личинка подрастает, слюнные железы, по
мнению Хенриксена, редуцируются. Изучение личинок Prestwichia на то-
тальных препаратах и на срезах показывает, что мнение Хенриксена
ошибочно.
Гистологически слюнные железы настолько напоминают собой маль-
пигиевы сосуды, что отличить их на срезах можно, только просмотрев
всю серию и установив их место в организме личинки. Имеются лишь
незначительные отличия: внутриклеточные капилляры развиты в слюн-
ных железах слабее, зато сохраняется межклеточный канал, отсутствую-
щий в мальпигиевых сосудах. В просвете слюнных желез в период пита-
ния тоже можно видеть эозинофильный секрет.
12*
179
Такое сходство гистологического строения невольно заставляет
предполагать и какое-то сходство в функции обоих органов. Какова же
может быть эта общая функция? Маловероятно, что слюнные железы
имеют какое-нибудь отношение к экскреции. Наоборот, скорее можно
усомниться в экскреторной функции мальпигиевых сосудов, так как эта
функция очень интенсивно выполняется средней кишкой. Следовательно,
мальпигиевы сосуды личинки Pretswichia если и сохраняют выделитель-
ную функцию, то лишь частично.
После прекращения питания, но еще до дефекации, тело личинки
значительно уменьшается в объеме. По-видимому, это связано с удале-
нием из организма большого количества воды. Удаление избыточной
воды, несомненно, происходит с помощью мальпигиевых сосудов, а, мо-
жет быть, и слюнных желез.
Прямой функцией слюнных желез Prestwichia является выделение
пищеварительных ферментов. Личинка Prestwichia не имеет твердых ро-
товых чдстей, которые содействовали бы откусыванию и размельчению
пищи. Поэтому личинка способна всасывать лишь полужидкую пищу.
Содержание кишки личинки всегда выглядит гомогенным, в ней встре-
чаются лишь небольшие количества отдельных клеток и желточных гра-
нул хозяина. Однако личинка Prestwichia способна уничтожить и очень
поздних зародышей Macrodytes, оставляя от них только хитиновые части.
Это возможно лишь благодаря тому, что ткани хозяина подвергаются
предварительной обработке, еще не будучи заглоченными. Можно пред-
положить, что продукты выделения мальпигиевых сосудов тоже прини-
мают участие в обработке тканей хозяина, тем более, что выделение пи-
щеварительных ферментов мальпигиевыми сосудами уже установлено
для некоторых насекомых (Шванвич, 1949; Кузнецов, 1953; Шовен,
1953).
Метаморфоз кишечника. На VI стадии развития (предку-
колка) клетки средней кишки и примыкающие к ней клетки крупнокле-
точного отдела задней кишки расходятся, благодаря чему устанавли-
вается связь между средней и задней кишкой, и масса экскретов, нако-
пившаяся в средней' кишке, и непереваренные остатки пищи извергаются
наружу. После дефекации объем средней кишки еще более уменьшается,
просвет ее иногда редуцируется вовсе. Клетки ларвальной средней киш-
ки, сменившие свою уплощенную форму на закругленно-кубическую,
образуют плотную массу, в которой снова становятся видными клеточные
границы (рис. 104,Л, л. ср. к). В эту массу вкраплены также клетки
мальпигиевых сосудов (мп. с) и железистых отделов слюнных желез. По
периферии этого клеточного комка располагаются пузыревидные клетки,
образующие почти непрерывный слой (им. ср. к). Напряжение жидкости
в вакуолях, по-видимому, несколько ослабевает, так как вакуоли теряют
свою сферическую форму, и вся клетка как бы растекается по поверх-
ности ларвальной средней кишки. Затем вакуоли лопаются, содержимое
их изливается на ларвальные клетки, а сами имагинальные клетки скла-
дываются в новый эпителий. Ван Левен, описавший такую же судьбу
вакуолярных клеток у Harmolita (= Isosoma), предполагает, что содер-
жимое вакуолей служит для перевариван-ия ларвальных элементов. Эта
мысль кажется вполне правдоподобной.
Иногда освобождение от экскретов совершается менее полно или
задерживается. В этом случае просвет кишки сохраняется, и можно на-
блюдать, как клетки ларвального эпителия освобождаются от взаимных
связей, округляются и падают в просвет кишки (рис. 104, Б, л. ср. к).
То же происходит и с клетками ларвальных мальпигиевых сосудов и
слюнных желез (рис. 104, Б, мп. с). Интересно отметить, что во всех этих
130
клеточных элементах признаки дегенерации появляются задолго до того
момента, когда они подвергаются действию пищеварительных соков. Так,
еще на V стадии развития плазма клеток приобретает грубо грануляр-
ное строение, клетки мальпигиевых сосудов и слюнных желез становятся
дольчатыми со стороны полости тела, словно начиная распадаться ил
куски (рис. 105, Л, лаб. о/с). В имагинальном эпителии очень скоро гра-
ницы клеток исчезают. Ядра становятся сплюснутыми и уплотняются,
почему размеры их уменьшаются до 0,015 мм в диаметре и 0,010 мм — в
толщину.
Слущивание и переваривание ларвального эпителия средней кишки
описано всеми, изучавшими метаморфоз насекомых. Относительно судь-
бы ларвальных мальпигиевых сосудов и слюнных желез, которая обыч-
но бывает сходной, в литературе имеются различные высказывания:
А. О. Ковалевский (1885, 1887) —для мухи и Перэ (1902) —для муравья
Formica описывают активное вторжение фагоцитов в слюнные железы,
а Перэ — и в мальпигиевы сосуды. В. А. Караваев (1898) —для муравья
Lasius, Англа (1902) и Эртель (1930)—для медоносной пчелы отри-
цают всякое участие фагоцитов в разрушении слюнных желез и мальпи-
гиевых сосудов. Ван Левен (1903) —для Harmolita (= Isosoma), Э. По-
ярков (1910) —для Galerucella описывают дегенерацию с последующим
фагоцитозом. Тигс (1922) указывает, что у Mormoniella (= Nasonia)
имеет место и растворение в полостной жидкости, и фагоцитоз дегенера-
тивных остатков слюнных желез. Только Берлезе (1901; цит. по Перэ,
1901) наблюдал у Cynips втягивание мальпигиевых сосудов в полость
средней кишки и переваривание их. То же наблюдается у Prestwichia —
слюнные железы и мальпигиевы сосуды подвергаются здесь общей
участи с ларвальным кишечным эпителием, что несомненно объясняется
той тесной связью, которая существует между этими органами у личинки.
Новые слюнные железы образуются, по А. О. Ковалевскому (1887)
и Ван Левену (1908), за счет сохраняющихся протоков желез, по Э. По-
яркову (1910)—путем впячивания гиподермы в области максилл, по
Тигсу (1922)—путем впячивания дна ротовой полости. Судьбу прото-
ков слюнных желез у Prestwichia проследить не удалось, а слюнные
железы у взрослых Prestwichia отсутствуют. Новые мальпигиевы сосуды,
и пр моим, и по литературным данным, образуются за счет имагиналь-
ных частей задней кишки (см. ниже).
Метаморфоз передней и задней кишки происходит у некоторых на-
секомых путем преобразования личиночных клеток, которые дедиффе-
ренцируются и снова приобретают способность делиться. «Омоложение»
личиночного эпителия передней кишки Galerucella, по Э. Пояркову
(1910), сопровождается отбрасыванием частей клеток («клеточной авто-
томией»). Подобная реконструкция этих отделов кишечника наблюдается
и у некоторых других жуков (Мэррей и Тигс, 1935; Петей, 1939), у ру-
чейников (Люббен — Lubben, 1907) и у медоносной пчелы (Добровский,
1951). Но во многих случаях, например у двукрылых (Ковалевский,
1885, 1887; Перэ, 1910) и перепончатокрылых (Ван Левен, 1908; Тигс,
1922; Эртель, 1930, и др.)» происходит частичное или полное разрушение
личиночных передней и задней кишки с последующей их регенерацией
за счет особых «имагинальных колец», лежащих на границе со средней
кишкой. У Drosophila, по Робертсону (1936), в образовании передней и
задней кишки имаго, помимо имагинальных колец, принимают участие
зачаток нижней губы и гиподермальный имагинальный диск последнего
сегмента брюшка.
В пищеводе у Prestwichia не наблюдается ни дегенерации, ни явле-
ния «автотомии». По-видимому, передняя кишка личинки Prestwichia
181
целиком сохраняет эмбриональные свойства. Сразу же после прекраще-
ния питания (V стадия) задний конец пищевода замыкается, и связь со
средней кишкой прерывается. В передней кишке начинается усиленное
размножение клеток, и она приобретает форму узкой цилиндрической
трубки, удлиняющейся по мере сокращения объема средней кишки
(рис. 105,А, пищ). У молодой куколки она уже проходит через всю
грудь. Затем в передней кишке начинается дифференциация на отделы:
образуется объемистый зоб, с тонкими перепончатыми стенками, очень
бедными ядрами, а небольшой отдел, примыкающий к средней кишке
приобретает строение провентрикулюса с трехгранным просветом. Впро-
вентрикулюсё хорошо видна куколочная кутикула, отслаивающаяся к
концу метаморфоза. Конец передней кишки в вияе кольцевой складки
вдается в просвет средней кишки, образуя нечто вроде клапана. Сообще-
ние между передней и средней кишкой восстанавливается.
Сходные изменения происходят в задней кишке (рис. 105,Б). Самый
начальный ее отдел, состоящий из крупных клеток, дегенерирует. Зад-
ний мелкоклеточный отдел начинает на него нарастать, почему крупные
дегенерирующие клетки оказываются внутри задней кишки куколки и
подвергаются той же участи, что и клетки ларвальной средней кишки и
ларвальные мальпигиевы сосуды (VI стадия). Благодаря усиленному
размножению клеток задняя кишка превращается в длинную узкую труб-
ку, тянущуюся до заднего конца брюшка, которое тоже вытягивается и
кончается заострением. Йа стадии куколки по ходу задней кишки обра-
зуется изгиб, в области которого формируется ректальный клапан
(рис*. 99,з. к). С этого момента в задней кишке можно различить два
отдела: тонкую кишку и rectum. Имагинальные мальпигиевы сосуды
значительно удлиняются, и диаметр их просвета увеличивается. Все ав-
торы описывают частичную дегенерацию задней кишки и восстановление
ее за счет сохранившихся частей. Лишь у Galerucella, по Э. Пояркову
(1910), клетки задней кишки омолаживаются путем «клеточной авто-
томии».
Происхождение имагинальных элементов сред-
ней кишки. Одним из наиболее трудных вопросов метаморфоза насе-
комых является вопрос о происхождении тех клеточных элементов, за
счет которых развивается имагинальная средняя кишка. М. С. Ганин
(1877), А. О. Ковалевский (1885, 1887), И. В. Насонов (1886), В. А. Ка-
раваев (1898), Англа (1904а), Ван Левен (1908), Э. Поярков (1910),
Перэ (1912), Тигс (1922), Эртель (1930) и многие другие описывают
наличие в ларвальном эпителии средней кишки особых замещающих
клеток, из которых строится новый эпителий. Однако происхождение
этих клеток толкуется по-разному. Обычно, убедившись, что замещаю-
щие клетки уже имеются у самых молодых личинок, исследователь за-
ключает из этого, что они происходят из энтодермы и являются как бы
«сестрами» ларвальных клеток, от которых они отличаются своим вре-
менно пассивным недеятельным состоянием, мелкими размерами и спо-
собностью размножаться. Эта мысль вполне правдоподобна. Однако ни
один исследователь, насколько мне известно, не проследил возникнове-
ния этих клеток до самого истока, никто не наблюдал, когда и как появ-
ляется дивергенция в судьбе ларвальных и имагинальных элементов эн-
тодермы. Эитодермальное происхождение замещающих клеток 'средней
кишки признают М. С. Ганин, А. О. Ковалевский, В. А. Караваев, Перэ
и др.
Несколько сложнее происходит метаморфоз средней кишки у неко-
торых жуков. У Galerucella, по Э, Пояркову (1910), еще в самом начале
метаморфоза начинается, усиленное размножение клеток пищеводного
182
клапана. Образующиеся клеточные массы обрастают личиночную сред-
нюю кишку, вклиниваясь между базальной мембраной и мышечной обо-
лочкой. Из этих клеток образуется куколочная средняя кишка, в которой
сразу же намечается дифференциация на более крупные клетки, состав-
ляющие непрерывный слой, и более мелкие, рассеянные у их основания.
Личиночная кишка, оказавшаяся внутри куколочной, переваривается.
При переходе к стадии имаго крупные клетки вытесняются мелкими, за
счет которых формируется окончательная имагинальная средняя кишка
жука.
Сходно протекает метаморфоз средней кишки у Leptinotarsa, по
Петей (1939).
Но существуют и другие точки зрения. Так, Англа (1904а) приписы-
вает очень большую роль в процессах метаморфоза трахейной системе.
По его мнению, у перепончатокрылых ймагинальные клетки проникают
в ларвальную среднюю кишку вместе с тончайшими разветвлениями тра-
хей, от которых они и происходят. Это наблюдение, не подтвержденное
никем из более поздних авторов, изучавших метаморфоз перепончато-
крылых, несомненно ошибочно. К Prestwichia оно не может иметь ника-
кого применения уже потому, что к моменту формирования имагиналь-
ного эпителия средней кишки трахейная система личинки находится са-
ма еще в эмбриональном состоянии и нигде не соприкасается с кишеч-
ником.
Наконец, по мнению Берлезе (1901) (цит. по Ван Левену) и Ван
Левена (1908), ймагинальные клетки средней кишки происходят из
блуждающих элементов полости тела, которые проникают в ларвальный
эпителий и становятся на его место, когда он гибнет. Нечто очень похо-
жее видел на своих препаратах В. А. Караваев (1898). У наружной по-
верхности личиночной средней кишки муравья Lasius, он встречал мел-
кие округлые недифференцированные мезодерадальные клетки, очень по-
хожие на ймагинальные клетки кишки, так что возникло предположение,
что' последние лроисходят из первых. Но на том только основании, что
ймагинальные клетки встречаются еще у очень молодых личинок 3 мм
длиной, он встал на точку зрения энтодермального происхождения има-
гинальных клеток.
У самых молодых, только что вылупившихся, личинок Prestwichia
(I стадия) средняя кишка состоит из довольно крупных и совершенно
однородных клеток (рис. 103, Л, л. ср. к). Ядра их еще не сплюснуты,
имеют сферическую форму и достигают 0,011 мм в диаметре, На II и
III стадиях развития, как уже упомянуто, кишечный эпителий сильно
растягивается, но остается однородным. Размножение его клеток не про-
исходит, а размеры ядер увеличиваются.
На IV стадии развития на внешней поверхности средней кишки мож-
но различить очень мелкие клетки, ничем не отличающиеся от тех мезен-
химатозных клеток, которые и впоследствии постоянно встречаются на
поверхности кишки (рис. 101,5, соед. кл), но интересных тем, что неко-
торые из них вступают в такую тесную связь с кишечным эпителием, что
кажутся погруженными в его толщу, лежащими под базальной мембра-
ной, а не над ней (рис. 103, Б, им. ср. к). Клетки эти имеют светлое ядро
с мелко распыленным хроматином и ядрышком, а также очень неболь-
шое количество базофильной плазмы. Форма клеток коротковеретено-
видная. Ядро имеет 0,004 мм в диаметре, т. е. очень близкие-размеры к
таковым ядер амебоцитов. Они могли бы произойти от сравнительно
крупных энтодермальных клеток ранних стадий развития лишь путем
неоднократных делений, однако делений клеток средней кишки я никогда
но наблюдала.
183
Клетки, проникшие в толщу кишечной стенки, быстро увеличиваются
в размерах: растет и ядро, и плазма. Форма клеток остается вытянутой
(рис. 103, Ву им. ср. к). Затем в плазме появляется одна или несколько
вакуолей (рис. 103,Г, им. ср. к). В это время описываемые клетки ста-
новятся очень похожими на жировые, так что далеко не всегда одни эле-
менты можно отличить от других. Но все же жировые клетки на этой
стадии обычно имеют гораздо больше вакуолей, более светлую плазму
и более разбросанно-отросчатую форму. Жировые клетки лежат всегда
на поверхности стенки кишки, а не в ней.
Проникновение новых клеток из полости тела, по-видимому, продол-
жается, так как наряду с уже значительно выросшими клетками продол-
жают встречаться еще совсем маленькие.
На V стадии развития вакуоли сильно увеличиваются, но число их
уменьшается. По-видимому, происходит слияние вакуолей. Обычно
остается только одна большая вакуоль, и клетки приобретают характер-
ную пузыревидную форму. Таким образом, не только строение и по-
следующая судьба этих клеток, но и их происхождение, очень на-
поминают таковые вакуолярных клеток Harmolita (= Isosoma) (Ван
Левен, 1909).
За внешнее происхождение имагинальных клеток Prestwichia aqua-
tica говорит еще то обстоятельство, что в тех областях средней кишки,
которые прикрыты клетками слюнных желез и мальпигиевых сосудов, эти
клетки с самого начала лежат над слюнными железами и мальпигиевы-
ми сосудами, а не под ними, как должно было бы быть, если бы они
возникли в самом кишечном эпителии. Впоследствии, когда в них разви-
вается вакуоль, эти клетки иногда как бы втискиваются между клетками
мальпигиевых сосудов, но никогда не лежат в промежутке между маль-
пигиевыми сосудами и кишкой.
Многократное и тщательное изучение препаратов показало, что ни
со стороны передней кишки, ни со стороны задней кишки, никогда не
происходит наползания клеток на среднюю кишку. Следовательно, воз-
можность такого происхождения средней кишки у Prestwichia, какое на-
блюдал Э. Поярков у Galerucella, отпадает. Исключается, как уже отме-
чено, и происхождение имагинальных клеток от храхей. Наиболее ве-
роятно происхождение имагинальных клеток от недифференцированных
полостных элементов, которое подтверждается изложенными выше на-
блюдениями, f
Мезодермальное происхождение имагинальных элементов средней
кишки (как и образование мезодермальных клеток из гиподермы имаги-
нальных дисков) противоречит обычным представлениям о зародыше-
вых листках. Известно немало и других фактов, когда при бесполом
размножении или в экспериментальных условиях при нарушении нор-
мальных коррелятивных связей в организме органы могут развиваться
не только из того зародышевого листка, от которого они происходят при
эмбриональном развитии, и многие клетки проявляют гораздо большие
потенции, чем обычно. Однако не следует понимать теорию зародышевых
листков слишком формально. Происхождение органа от того или иного
зародышевого листка не является непреложным законом развития, ко-
торый должен осуществиться во что бы то ни стало. Образование заро-
дышевых листков в ходе эмбрионального развития и их дальнейшая
дифференциация повторяют явления, имевшие место при возникновении
первичных органов многоклеточных животных. Естественно, поэтому, что,
чем примитивнее животное, чем ближе оно стоит к истокам Metazoa, тем
полнее рекапитулирует его развитие указанные исторические события.
И, наоборот, в развитии вышестоящих групп животных эти первичные
184
черты все более стираются. Не мудрено поэтому, что вопрос о зароды-
шевых листках даже при эмбриональном развитии насекомых оказался
очень запутанным; еще меньше основании удивляться тому, что и при
метаморфозе имеют место некоторые уклонения от наших привычных
представлений. Для этих уклонений имеются к тому же и определенные
физиологические причины. Почти все тело личинки Prestwichia состоит
из очень немногочисленных высокодифференцированных клеток, утра-
тивших способность делиться. В частности, очень невелико число клеток,
составляющих среднюю кишку личинки. Быть может, необходимость
интенсивного питания требует непосредственного участия в пищеваре-
нии 'всех этих клеток, и ни одна из них не может остаться в резерве для
построения имагинальной кишки.
Иные отношения наблюдаются в гиподерме имагинальных дисков,
состоящей,из недифференцированных, интенсивно размножающихся
клеток, которая служит источником образования не только соответствую-
щих наружных органов имаго, но и клеток, пополняющих мезодерму.
Таким образом, в полости тела личинки всегда имеется некоторое коли-
чество еще неспециализированных клеток. Возможно, что некоторые
именно из этих выселившихся из имагинальной гиподермы клеток ис-
пользуются и для построения имагинальной кишки. Будем ли мы назы-
вать эти клетки мезодермальными или нет, в сущности, не имеет принци-
пиального значения. Эти наблюдения находятся в соответствии с наблю-
дающейся в эволюции членистоногих тенденцией к ограничению морфо-
генетических потенций энтодермы и мезодермы и все более возрастаю-
щему значению гиподермы, «которая у высших представителей совме-
щает функции покровной, опорной и эмбриональной ткани» (Штейнберг,
1956).
Половая система
С точки зрения задач, которые имеет настоящая работа, развитие
половой системы не представляет особенного интереса, так как она
всегда имеет у насекомых прямолинейный прогрессивный характер. По-
ловая система функционирует только на стадии имаго и никаким пере-
стройкам во время метаморфоза не подвергается. Поэтому в отношении
половой системы сделаны лишь небольшие наблюдения.
У самых молодых личинок половой зачаток представлен компакт-
ной группой клеток, лежащей в полости тела вентральнее задней кишки
(рис. 87, Г, п. з). В это время зачаток уже одет тонкой соединительно-
тканной оболочкой.
На III—IV стадиях развития половой зачаток растяги-
вается в поперечном направлении и перешнуровывается на две симме-
тричные половины. Одновременно происходит утолщение гиподермы в
области полового зачатка; впоследствии здесь закладываются наруж-
ные половые части.
Зачатки семенников значительно увеличиваются в размерах бла-
годаря все продолжающемуся размножению клеток. Образуются за-
чатки выводных протоков. Строение их стенок очень похоже на строение
прилежащих частей гиподермы и нисколько не напоминают нежную со-
единительнотканную оболочку семенников. Поэтому можнск полагать,
что протоки семенников образуются путем впячивания гиподермы.
Уже на V стадии развития сперматогонии складываются в группы,
между которыми образуются тонкие перегородки, по-видимому, соедини-
тельнотканного происхождения. На стадии куколки начинается форми-
рование сперматозоидов: сперматиды сильно вытягиваются и приобре-
тают веретеновидную форму. Ядро располагается на одном конце спер-
185.
матиды. Все сперматиды одной сперматоцисты обращены ядерными кон-
цами в одну сторону. Между пучками развивающихся сперматозоидов
попадаются отдельные крупные клетки, по-видимому, версоновские
клетки. Созревание сперматозоидов происходит почти одновременно
по всему семеннику. Уже на этой стадии нет никаких следов зачатко-
вой зоны.
У поздних куколок мужская половая система достигает полного раз-
вития. Семенники становятся объемистыми органами и заполняются пуч-
ками длинных нитевидных сперматозоидов. В семепроводах дифферен-
цируется передний, по-видимому, железистый отдел, состоящий из круп-
ных вакуолизированных клеток. Эти железистые отделы идут от семен-
ников назад, затем они переходят в тонкостенные, более узкие отделы,
которые поворачивают вперед, и на уровне основания копулятивного
органа оба семепровода сливаются в короткий непарный отдел. Перед
слиянием оба семепровода образуют два довольно объемистых выпячи-
вания, направленных вперед — семенные пузырьки. Развитие и строение
копулятивного органа осталось неизученным.
Такое же строение имеет половая система взрослых самцов, извле-
ченных из яйца Macrodytes. По Римскому-Корсакову (1932), именно в
это время (до выхода из яйца хозяина) происходит спаривание Prest-
wichia. У самцов, которые прожили несколько дней после выхода во
внешнюю среду, сперматозоиды уже отсутствуют, а семенники имеют де-
генеративный вид и очень маленькие размеры.
У личинок самок на V стадии развития оба половых зачатка разде-
ляются еще раз, и в каждом из четырех получившихся половых зачатков
происходит дифференциация на вздутый концевой отдел, имеющий яйце-
видную форму (рис. 106,5, конц. отд), и суженный стебельчатый отдел
(рис. 106,5 стеб, отд.), отходящий от более широкого конца первого
отдела. Клетки первого отдела имеют приблизительно шарообразную
или многогранную форму и бедны протоплазмой. В стебельчатом отделе
клетки богаче протоплазмой, слабее красятся и имеют эпителиеобраз-
ное расположение, хотя настоящего просвета в этих узких отделах нет.
Оба стебельчатых отдела сходятся друг с другом. Соединительноткан-
ная оболочка (соед. об.)у одевающая гонаду, не всюду плотно к ней при-
легает. В области стебельков под ней оказывается довольно обширное
пространство, в котором встречаются отдельные клетки. На этой стадии
зачатки яичников Prestwichia необычайно похожи на таковые Habro-
bracon juglandis Ashm., описанные и изображенные на рисунке Генше-
ном (Henschen, 1928). Геншен описывает последующую дегенерацию
узкого отдела, и значение дифференциации зачатков яичников на взду-
тый концевой и суженный стебельчатый отдел в его работе остается
неясным.
У Prestwichia судьба суженного отдела яичника совсем иная. Не-
сколько забегая вперед, можно сказать, что вздутые концевые отделы
представляют собой зачатковую зону яичника (гермарий), а стебельча-
тые отделы — зачаток фолликулярных трубок (вктеллярия).
На этой же стадии происходит формирование яйцеводов. Две пары
зачатков яйцеклада вместе с прилегающей к ним утолщенной гиподер-
мой начинают погружаться внутрь тела, смещаясь одновременно вперед,
почему основания половых придатков оказываются впереди от зачатков
яичников. Со дна этого впячивания назад начинают расти две трубки,
достигающие в конце концов места соединения стебельчатых отделов
яичников каждой стороны. Таким образом, яйцеводы Prestwichia обра-
зуются из гиподермы. У молодых белоглазых куколок половые зачатки
начинают сильно удлиняться; особенно вытягиваются стебельчатые от-
186
делы, так что они приобретают вид двух пар трубок, тянущихся по бо-
кам брюшка. Концевые вздутия лежат теперь далеко впереди на границе
груди и брюшка. Стебельчатые отделы, благодаря легким пережимам,
приобретают четковидную форму (рис. 106,5 стеб. отд.). Во вздутиях
между пережимами эпителиальные стенки состоят из очень плоских кле-
ток, благодаря чему образуются довольно значительные полости, в ко-
торых лежат группы крупных клеток с базофильной протоплазмой и
светлым ядром, совершенно сходных с клетками концевых отделов.
В промежутках между вздутиями эпителиальные клетки стенок трубок
высоки и смыкаются друг с другом в центре, не оставляя ясного просве-
та. Очевидно, вздутия стебельчатых отделов представляют собой зача-
точные яйцевые камеры, а заполняющие их крупные клетки спускаются
сюда из большого концевого вздутия с зачатковыми клетками, но еще
недифференцированы на будущее яйцо и питающие клетки.
Однако, несмотря на начавшееся формирование яйцевых камер, на
стадии пигментации глаз каждый из четырех зачатков яичников распа-
дается на целый пучок яйцевых трубочек, которые постепенно приобре-
тают строение, типичное для яичников политрофического типа. Концевые
зачатковые отделы к этому времени уже утрачивают свою вздутую фор-
му, быть может потому, что основная масса содержащихся в них клеток
переходит в -нижние отделы яйцевых трубочек.
Какое значение имеет описанное выше предварительное формирова-
ние яйцевых камер в стебельчатых отделах, после которой следует раз-
деление их на несколько трубочек — сказать трудно. Геншен (1928) опи-
сывает превращение каждого из двух концевых отделов зачатков яични-
ков каждой стороны тела Habrobracon juglandis в одну яйцевую трубоч-
ку, так что во вполне развитой половой системе этого насекомого имеет-
ся лишь четыре яйцевых трубочки. Быть может, у Prestwichia произошло
вторичное увеличение числа яйцевых трубочек. Однако, чтобы утверж-
дать это положительным образом, нужно более подробное исследование
развития половых желез Prestwichia и близких перепончатокрылых
Основные черты метаморфоза Prestwichia
Результаты наблюдений, касающихся постэмбрионального развития
Prestwichia aquatica, можно резюмировать следующим образом.
1. В связи с паразитическим образом жизни (защищенностью ли-
чинки от внешних механических воздействий, редукцией личиночной мус-
кулатуры, для котррой хитиновый покров дает точки прикрепления
и т. д.) у Prestwichia происходит редукция хитинового покрова. Он ста-
новится тонким, растяжимым и не препятствует росту личинки. Поэтому
в развитии Prestwichia сохраняются только две линьки: куколочная и
имагинальная.
2. В момент выхода личинки из яйца гиподерма еще однородна, но
дифференцируется затем, образуя «имагинальную полоску» на брюшной
стороне и ларвальную гиподерму на спине и на боках. Ларвальная гипо-
дерма дегенерирует при окуклении и фагоцитируется жировыми клет-
ками. ,
3. Сегментация появляется впервые в конце личиночной жизни
в имагинальных частях гиподермы. Она с самого начала имеет характер
гетерономной имагинальной сегментации. Личиночные части гиподермы
остаются несегментированными.
4. Нервная система закладывается и обособляется от гиподермы уже
в период постэмбрионального развития. У личинки она еще находится и
эмбриональном состоянии и не функционирует. Развитие нервной систе
188
мы все время имеет прямой прогрессивный характер с «установкой» на
имаго.
5. Трахейная система закладывается впервые у поздней личинки: она
развивается прямо в имагинальную, не подвергается метаморфозу,
а испытывает лишь явление олигомеризации (из 6 пар зачатков стигм,
реализуется лишь одна).
6. Эноциты появляются в конце личиночной жизни и почти пол-
ностью дегенерируют перед выходом имаго. Таким образом, имеется
одна единственная генерация зноцитов, существование которых ограни-
чено довольно коротким периодом жизни Prestwichia.
7. Соматическая мускулатура развивается уже после выхода ли-
чинки из яйца и представлена редкой сетью анастомозирующих клеток*
лежащей под гиподермой. Пучки мышечных фибрилл имеют поперечное
направление. Эти мышцы не способны обеспечить поступательного дви-
жения и производят лишь перистальтические сокращения тела. Накануне
метаморфоза они дегенерируют и фагоцитируются.
Кроме того, имеются глотательные мышцы в области пищевода и
мышцы, растягивающие в стороны заднюю кишку. Во время метамор-
фоза они исчезают.
8. Имагинальная мускулатура развивается за счет миобластов, сво-
бодно лежащих в полости тела личинки. При этом часть миобластов под-
вергается дегенерации: слипается в комки, которые фагоцитируются
жировыми клетками,
В связи с редукцией крыльев у самцов и части самок происходит
недоразвитие и дегенерация крыловых мышц.
9. Кровеносная система развита у Prestwichia слабо и возникает
лишь во время метаморфоза.
10. Жироъое тело формируется у молодой личинки. Оно не участвует
в процессе экскреции и ограничивается трофической ролью. В клетках
жирового тела сперва накапливаются жировые включения, затем по-
являются и белковые. Во время метаморфоза часть жировых клеток
разрушается, остальные почти полностью утрачивают свои включения.
Новообразование жирового тела в конце личиночной жизни и во время
метаморфоза не происходит.
Клетки жирового тела играют роль некрофагов, заглатывая и пере-
варивая остатки дегенерирующих личиночных органов.
И. Передняя кишка личинки коротка и развита слабо. Во время ме-
таморфоза она не разрушается, а испытывает лищъ прогрессивные из-
менения— быстро растет и подразделяется на отделы.
12. Средняя кишка личинки >не сообщается с задней кишкой и со-
стоит из крупных клеток, которые во время метаморфоза слущиваются
в просвет кишки и перевариваются. Имагинальная средняя кишка фор-
мируется за счет особых пузыревидных клеток, содержащих большую
вакуоль и лежащих на внешней поверхности личиночной средней кишки.
Эти пузыревидные клетки образуются из недифференцированных эле-
ментов полости тела.
К концу личиночной жизни в средней кишке накапливаются ураты.
13. В задней кишке личинки Prestwichia можно различить два от-
дела: передний, состоящий из более крупных клеток, от которого отхо-
дят два личиночных мальпигиевых сосуда, и задний, состоящий из мел-
ких клеток, у переднего края которого к концу личиночной жизни фор-
мируется 3 имагинальных мальпигиевых сосуда. Во время метаморфоза
передний отдел задней кишки дегенерирует, а задний отдел нарастает
на передний и смыкается с имагинальной средней кишкой.
14. Личиночные мальпигиевы сосуды на всем своем протяжении
189
тесно прилегают к личиночной средней кишке. Вместе с ней они оказы-
ваются заключенными в имагинальной средней кишке и перевариваются.
15. Две слюнные железы личинки состоят из крупноклеточных же-
лезистых отделов и более узких протоков, которые открываются наружу
общим отверстием позади рта. Железистые отделы тесно связаны с личи-
ночной средней кишкой и вместе с нею перевариваются в имагинальном
кишечнике. Судьба протоков не прослежена. У имаго Prestwichia слюн-
ные железы отсутствуют.
16. Дегенерация личиночных органов у Prestwichia часто сопровож-
дается фагоцитозом. Наблюдался фагоцитоз личиночной гиподермы, ли-
чиночной мускулатуры, части эмбрионального материала имагинальной
мускулатуры и эноцитов. Роль фагоцитов выполняют преимущественно
жировые клетки и, отчасти, амебоциты. Фагоцитозу всегда предшествуют
дегенеративные изменения в ткани.
5. ВЛИЯНИЕ ПАРАЗИТИЗМА НА РАЗВИТИЕ PRESTWICHIA
Итак; мы видим, что весь ход развития Prestwichia aquatica (и эм-
бриональный и постэмбриональный его периоды) обнаруживают значи-
тельные отклонения от типичного развития насекомых, несомненно свя-
занные с паразитическим образом жизни. Очень мелкие яйца Prestwichia
лишены питательных запасов. Такая редукция желтка стала возможной
потому, что из яйца рано выходит личинка, стоящая по высоте организа-
ции на уровне ранних (еще несегментирова'нных!) зародышей других
насекомых, но уже способная активно питаться. У этой личинки вполне
развиты и могут функционировать только органы пищеварения: глотка
с ее мускулатурой, средняя кишка и лабиальные (слюнные) железы. Все
остальные системы органов появляются в постэмбриональный период.
Для метаморфоза насекомых характерно двукратное функциониро-
вание органов: на стадии личинки и на стадии имаго. В промежуток
между этими стадиями вклинивается период дедифференцации или глу-
бокой перестройки органа, сопровождаемой замещением ларвальных
элементов имагинальными. У Prestwichia фактически подвергаются ме-
таморфозу только органы, связанные с функциями пищеварения и вы-
деления: слюнные железы, пищеварительный канал, мальпигиевы сосу-
ды. Кожная мускулатура-с ее перистальтическим характером движения
тоже служит скорее для перемешивания содержимого слепо замкнутой
средней кишки, чем для целей перемещения в пространстве. Все осталь-
ные органы Prestwichia имеют прямое развитие и функционируют одно-
кратно: либо у имаго (трахеи, сердце, нервная система), либо на стадии
куколки (эноциты). У жирового тела период функционирования более
длителен, но остается единым, и разделение жирового тела на ларваль-
ное и имагинальное отсутствует.
Далее, постэмбриональное развитие Prestwichia существенно отли-
чается от такового других насекомых отсутствием личиночных линек и
неравномерным характером роста. Обычно у Holometabola благодаря
действию ювенильного гормона рост личинки имеет равномерный'харак-
тер, и на протяжении ряда возрастов сохраняется почти без изменений
то строение, которое личинка имела в момент вылупления из яйца (Но-
вак — Novak, 1959). А у Prestwichia, как уже было показано, организа-
ция личинки значительно усложняется уже после вылупления: возникает
ряд чисто личиночных органов (мальпигиевы сосуды, мускулатура
и т. д.) и одновременно происходит прогрессивное развитие имагиналь-
ных органов (нервная система, трахеи, сердце). Развитие инкреторных
органов у Prestwichia осталось, к сожалению, неизученным, но выше-
190
указанные особенности ее метаморфоза дают основание полагать, что иг
железы внутренней секреции у личинки сильно 'недоразвиты и вступают
в действие с большим опозданием.
Таким образом, простота строения личинки в момент ее выхода из.
яйца привела к некоторому упрощению ее метаморфоза. Процессы про-
грессивного развития явственно преобладают над процессами разруше-
ния. Это положение находится в некотором противоречии с известной
теорией Берлезе—Ежикова о происхождении полного превращения у на-
секомых. По этой теории, причиной возникновения метаморфоза у Но-
lometabola послужило обеднение яиц желтком и овязанное с этим раннее
вылупление личинки, сохраняющей многие эмбриональные черты и тем:
самым резко отличающейся от взрослого насекомого. Берлезе (1913)
(цит. по Ежикову, 1939, и Шванвичу, 1949) различает несколько стадий
эмбрионального развития насекомых, на которых может происходить вы-
лупление личинки из яйца: протоподиальную олигомерную стадию, на
которой сегментация и зачатки конечностей имеются лишь в голове и
груди, полиподиальную стадию, когда сегментация распространяется
на брюшко и закладываются брюшные ножки, и, наконец, олигоподиаль-
ную стадию, когда брюшные конечности вторично редуцируются. Этим
трем стадиям развития соответствуют различные типы личинок Holome-
tabola. Наиболее ярко эмбриональные черты выражены, по Берлезе, у ли-
чинок паразитических перепончатокрылых Platygasteridae, относящихся
к протоподиально-олигомерному типу (что, несомненно, как и у Prest-
wichia, связано с паразитизмом).
Сходные взгляды как применительно к насекомым, так и по отноше-
нию к личинкам всех Metazoa развивал Ежиков (1929, 1939, 1940): «Су-
щество личинки заключается не в наличии приспособлений, отличающих
се от взрослой формы, а в эмбриональном характере ее организации»
(1939, стр. 261).
У Prestwichia эмбриональный характер организации выражен резче,
чем у какого бы то ни было другого насекомого. Для личинки Prestwichia
нет места в классификации Берлезе. Она соответствует еще,более ранней
«амерно-аподиальной» стадии. Тем не менее, метаморфоз Prestwichia не
более глубок, чем у других Holometabola, и даже 'несколько сглажен в
отношении ряда систем органов. Объяснить это можно только бедностью
личинки Prestwichia специальными личиночными приспособлениями,
что в свою очередь зависит от крайне благоприятных условий паразити-
ческой жизни. Таким образом, формулировка Ежикова страдает одно-
сторонностью, так как не придает должного значения условиям суще-
ствования личинки и связанным с ними приспособлениям.
Гораздо более полным; а потому и верным, является определение
личинки, данное Ивановым: «.. .личинка может быть охарактеризована,
как организм, вынужденный на более или менее ранних стадиях разви-
тия приспособиться к условиям свободного существования путем ранней
дифференцировки необходимых для этого специальных личиночных ор-
ганов» (1937, стр. 95). Где нет этой ранней дифференцировки, там нет и
метаморфоза. Это мы со всей наглядностью видим на примере Prest-
wichia: перестройка затрагивает только те системы органов, которые
функционируют у личинки. А те системы органов, которые у личинки без-
действуют— дыхательная, нервная, кровеносная, — слабое развитие ко-
торых особенно подчеркивает эмбриональный характер личинки, разви-
ваются прямолинейно и без метаморфоза.
ОБЩАЯ ЧАСТЬ
ГЛАВА I
ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ ЭМБРИОНАЛЬНОГО
РАЗВИТИЯ ПЕРЕПОНЧАТОКРЫЛЫХ
Из рассмотренного выше фактического материала видно, что про-
цесс эмбрионального развития у перепончатокрылых подвержен значи-
тельным вариациям. Варьируют размеры яиц, содержание в них
желтка и характер дробления; варьируют время и способ образова-
ния эмбриональных оболочек, их функция, и даже самое их наличие
не обязательно; варьирует степень морфологического развития в мо-
мент вылупления личинки из яйца; варьирует течение эмбриональных
процессов и т. д. Если мы привлечем к рассмотрению литературные
данные по развитию Hymenoptera, то убедимся, что эти вариации
еще более значительны.
На основании имеющихся сведений по эмбриологии перепончато-
крылых можно установить наличие в этом отряде нескольких основ-
ных типов развития: наиболее примитивный, свойственный пилильщи-
кам, сравнительно мало измененный у большинства Aculeata и низших
Parasitica, несколько более измененный у муравьев и три различных
типа развития у более специализированных форм. Один из них
характеризуется крайней дезэмбрионизацией, другой — наличием тро-
фической оболочки, а третий — полиэмбрионией.
К более детальному рассмотрению этих типов и преемственности
между ними мы сейчас и перейдем.
1. ПРИМИТИВНЫЙ ТИП РАЗВИТИЯ ПИЛИЛЬЩИКОВ
Так как развитие пилильщиков уже было подробно рассмотрено
ранее, отметим здесь лишь наиболее существенные его черты;
1) довольно крупные яйца, богатые желтком;
2) поверхностное дробление;
3) наличие типичной синцитиальной «желточной системы», испы-
тывающей лишь временное разделение на клетки;
4) образование двух эмбриональных оболочек, из которых одна
(амнион) может довольно рано исчезать;
5) полное развитие сегментации и всех систем внутренних орга-
нов, присущих личинкам насекомых (в частности установление сооб-
щения между средней .и задней кишкой);
6) наличие у личинок органов зрения;
7) наличие у личинок максимального количества конечностей —
помимо обычных головных и грудных придатков, закладывается еще
по паре придатков на всех сегментах брюшка, большая часть которых
сохраняется и функционирует у личинки.
J 92
Развитие пилильщиков мало отличается от такового других насе-
комых и, по-видимому, является исходным в эволюции эмбриональ-
ного развития перепончатокрылых.
2. ТИП РАЗВИТИЯ НИЗШИХ PARASITICA И БОЛЬШИНСТВА ACULEATA
Так как развитие жалящих перепончатокрылых в предыдущих
главах рассмотрено не было, то теперь следует на нем несколько
заде]ржаться.
Эмбриональное развитие ос и пчел изучено довольно хорошо.
Развитие медоносной пчелы (Apis mellifera L.) исследовалось неодно-
кратно (Бючли — Butschli, 1870; Диккель — Dickel, 1904; Нахтсгейм,
1913; Нельсон, 1915; Снодграсс, 1925; Шнеттер, 1934а и б, 1936;
Мюллер, 1957; Рейнхардт — Renchardt, 1960, и др.). Подробно изучено
развитие Chalicodoma muraria F. (Карьер и Бюргер, 1898), несколько
хуже — Vespa vulgaris L. (Стриндберг, 1914), Ammophila campes
tris Jur. (Берендс и Берендс-Ван Роон, 1949) и Scoila quadripuncta-
ta F. (Дондуа, 1953); известны ранние стадии развития Polistes
pallipes Lepel. (Маршал и Дернел — Marchall and Dernehl, 1905).
Развитие всех перечисленных форм очень похоже.
Яйца, как правило, довольно крупны: у Ammophila — 2,5 X 0,75 мм,
у Chalicodoma — 4Х 1 мм, у Apis — около 1,6 мм. У Xylocopa valga
Gerst., по Б. Н. Шванвичу (1949), яйца достигают. 15 мм длины.
Типичное поверхностное дробление завершается образованием бласто-
дермы, толщина которой часто в разных частях яйца с самого
начала различна. У всех перечисленных форм имеется только одна
эмбриональная оболочка, которую Стриндберг называет серозой,
а Нельсон амнионом. У Chalicodoma, Ammophila и Scolia сперва
образуются настоящие амниотические складки, но серозный и амнио-
тический листки рано отделяются друг от друга по наружному краю
складки. Рост амниотического листка вскоре приостанавливается,
амнион не образуется и только сероза получает полное развитие. Эти
наблюдения показывают, что единственная оболочка ос и пчел, (а также
паразитических перепончатокрылых) есть сероза, а не амнион.
Интересно, что в одной из первых работ, посвященных эмбрио-
нальному развитию медоносной пчелы, А. О. Ковалевский (1871)
описывает две эмбриональные оболочки. Как описание, так и рисунки
Ковалевского не дают никаких оснований сомневаться в точности его
наблюдений. Образуются эти оболочки так: возникают две амниотиче-
ские складки (передняя и задняя). Задняя складка появляется
позднее и растет медленнее, так что оболочки формируются преиму-
щественно за счет передней складки, и замыкание амниона происходит
не в средней части зародыша, а на заднем его конце. Другая особен-
ность формирования оболочек пчелы, отмеченная Ковалевским, состоит
в том, что передняя амниотическая складка растет не только своим
наружным выпуклым краем, но и внутренним вогнутым, так что одно-
временно происходит формирование покровов спины. Амнион вскоре
исчезает, и остается только сероза. Грасси (1884) тоже наблюдал
образование у пчелы, как исключение, двух эмбриональных оболочек.
Все эти данные показывают, что редукция амниона у пчел не
полная, и амнион может сохраняться как атавистический признак
у отдельных рас или семей-. По мнению Стриндберга, редукция амнио-
на у рассматриваемых форм объясняется тем, что материал, обычно
идущий на построение этой оболочки, здесь расходуется на раннее
провизорное замыкание спины зародыша.
13 О. М. Иванова-Казас
193
Дальше, на зародышевой полоске появляются две продольные
бороздки, разделяющие ее на три области: медианную и две лате-
ральные. Медианная область бластодермы почти на всем своем протя-
жении представляет собой мезодерму. Она погружается внутрь
иногда путем настоящей инвагинаций, так что формируется мезодер-
'мальная трубка (Chalicodoma). У Apis мезодерма погружается в фор-
ме пластинки, но позднее эта пластинка расслаивается на два листка,
и в результате возникает временная целомическая полость. Энтодер-
ма образуется путем обильного врастания клеток из бластодермы на
концах медианной пластинки. Позднее на этих местах возникают
впячивания стомо- и проктодеума.
Далее, зародыш испытывает ясную сегментацию, распространяю-
щуюся и на мезодерму. На голове у зародыша Apis появляются
зачатки антенн и челюстей, на груди зачатки ног. Затем зачатки антенн
и грудных ног превращаются в имагинальные диски, развитие кото-
рых задерживается и возобновляется только во время метаморфоза.
У Chalicodoma, кроме тою, появляются зачатки конечностей на пер-
вых двух сегментах брюшка, но они вскоре исчезают.
Нервная система закладывается посегментно. Головной мозг
Chalicodoma складывается из трех пар зачатков (по Карьеру и Бюр-
геру, которые различают в голове Chalicodoma 7 сегментов, они соот-
ветствуют мозговому, антенн ал ьному и предчелюстному сегментам).
Брюшная цепочка Chalicodoma состоит первоначально из 17 пар
зачатков ганглиев, но затем передние три пары и задние три пары
сливаются так, что окончательное число парных ганглиев в брюшной
цепочке равно четырнадцати. В виде впячиваний гиподермы заклады-
ваются зачатки скелетных частей головы, лабиальных желез и трахей
(об этом уже была речь выше). Две энтодермальные клеточные массы
(передняя и задняя) постепенно обрастают желток и образуют сред-
нюю кишку.
Из мезодермы развивается мускулатура, жировое тело, спинной
сосуд, кровяные элементы. Стомодеум разделяется на ряд отделов
и прорывается в среднюю кишку, но связь между средней и задней
кишкой не устанавливается. Половые клетки впервые становятся
заметными уже довольно поздно. Они образуются из мезодермы
3—5-го сегментов брюшка.
Итак, в развитии ос и пчел, по сравнению с пилильщиками,
можно отметить следующие особенности:
1) полную редукцию амниона;
2) отсутствие брюшных ног и задержку развития грудных ног,
превращающихся в имагинальные диски;
3) отсутствие глаз;
4) отсутствие сообщения между средней и задней кишкой.
О возможных причинах редукции амниона уже было сказано
раньше. Все же остальные особенности развития пчел и ос есть про-
явления некоторого общего недоразвития личинки, характеризующего
всех высших перепончатокрылых. Такая дезэмбрионизация развития
в данном случае ни в коей мере не может быть объяснена недостат-
ком питательных материалов в яйце. Как мы уже видели, яйца ос
и пчел крупны и содержат много желтка. Недоразвитие ряда личи-
ночных органов объясняется экологическими причинами и, в частности,
сильным развитием инстинкта «заботы о потомстве»: обычно самки
откладывают яйца в специальные норки или ячейки и снабжают личи-
нок необходимой пищей. Поэтому личинкам не приходится отыскивать
пищу или охотиться за добычей, они в какой-то мере защищены от
194
врагов и неблагоприятных условий. Они малоподвижны и беспомощны
и лишены не только конечностей, но и органов зрения.
Причины, которые могли вызвать разобщенность частей кишечни-
ка, уже рассматривались применительно к паразитическим перепончато-
крылым. Эти соображения могут быть применены и к жалящим, тем
более, что многие' авторы предполагают происхождение Aculeata от
низших Parasitica или, по крайней мере, от каких-то общих предков
(Гандлирш — Handlirsch, 1908; Малышев, 19496, 1959; Теленга, 1952;
Викторов, 1955, 1956, 1959, и др.). Это тем более вероятно, что раз-
витие некоторых низших представителей подотряда Parasitica (напри-
мер, Eurytoma aciculata Ratz. из Eurytomidae или Angitia vestigialis
Ratz. из Ichneumonidae) тоже происходит по этому же типу.
3. ТИП РАЗВИТИЯ МУРАВЬЕВ
Эмбриональное развитие муравьев впервые было изучено Гани-
ным (18696). Несмотря на то, что микроскопическая техника тогда
была еще слабо развита, и исследование велось почти исключительно
путем прижизненных наблюдений, Ганину все-таки удалось сделать
ряд очень верных наблюдений. Так, он отмечает, что у муравьев воз-
никает только одна эмбриональная оболочка (которую он называет
амнионом), образующаяся путем отслоения бластодермы от желтка.
Далее он описывает и изображает сегментацию зародышевой полос-
ки, формирование ротовых частей, нервной системы, лабиальных желез,
передней и задней кишки и замыкание спины.
Позднее эмбриональное развитие муравьев изучали Танквари
(1913), Стриндберг (1913а, б, 1915, 1916а, б, 1918, 1919) и Гехт
(Hecht, 1924).
* Эмбриональное развитие муравьев обнаруживает следующие осо-
бенности:
1) тенденцию к тотальному дроблению;
2) раннюю дифференциацию бла-стодермы;
3\ дальнейшую (по сравнению с осами и пчелами) редукцию
эмбриональных оболочек.
Тенденция к тотальному дроблению у муравьев выражается в том,
что, несмотря на значительное количество желтка, борозды дробления
при формировании бластодермы врезаются в яйцо очень глубоко.
Вследствие этого клетки некоторых внезародышевых частей бласто-
дермы приобретают высокую цилиндрическую форму, и их базальные
концы включают в себя большие количества желтка. Крайнее свое
выражение эта тенденция находит в развитии Azteca sp., яйцо кото-
рого целиком распадается на клетки (Стриндберг, 19166).
Дифференциация бластодермы проявляется прежде всего в том,
что настоящая клеточная бластодерма покрывает лишь брюшную
и боковые поверхности яйца, а на дорзальной его стороне остается
область, покрытая лишь тонким слоем плазмы, в котором разбросаны
редкие ядра — так называемый «дорзальный синцитий» (рис. 107,А).
В самой бластодерме очень ясно различаются зародышевая и внезаро-
дышевая области. Последняя тоже не вполне однородна. Так, у Сатро-
notus ligniperda Latr., по данным Стриндберга (19136) и Гехта
(1924), в бластодерме различаются 4 поперечных зоны (рис. 107, Л):
/—Передний полюс яйца занят клетками, богатыми вакуолями
и желтком; связь между ними непро«чная. Вскоре часть клеток
разъединяется и дает неправильную, постепенно дегенерирующую мас-
су. Другая часть их идет на построение серозы.
13*
195
В образовании серозы принимает участие только передняя часть
бластодермы. Наиболее полное развитие серозы происходит у Сатро-
notus и Formica. Так, у Formica (рис. 107, Б, В, и Г), растущим
зародышевый диск отделяется по всему краю от внезародышевых ча-
стей. Передний конец зародышевого диска распространяется вперед
по поверхности желтка таким образом, что внезародышевые клетки
оказываются над лим. Задний конец зародышевого диска, наоборот,
сохрняет поверхностное положение и как бы подминает под себя вне-
зародышевые клетки, которые оказываются внутри зародыша и там
дегенерируют. Клетки передней части бластодермы, составляющие
зачаток серозы, образуют эпителиальный слой,. постепенно обрастаю-
щий все яйцо. Замыкание серозы происходит на заднем конце. В том
обстоятельстве, что сероза формируются за счет переднего внезароды-
шевого материала, можно усмотреть черту сходства с развитием
эмбриональных оболочек у медоносной пчелы (по Ковалевскому).
У других муравьев такого полного развития серозы не происхо-
дит. Внезародышевые клетки Tapinoma и Pseudomyrma (рис. 108, Л, Б,
и В) образуют две бесформенные массы на полюсах яйца и дегене-
рируют.
Такая же редукция серозы наблюдается у форм, характеризующих-
ся задним положением зародышевого диска: Leptothorax (рис. 108,Г
и Д)у Laskis, Myrmica (рис. 108, £, Ж и 3), Tetramorium (рис. 108,,//
и /С). У Leptothorax появляется нечто вроде амниотической складки,
но все же оболочки не развиваются.
Образованию серозы, как эпителиального слоя, по мнению Стринд-
берга, препятствует переполнение внезародышевых клеток желтком,
почему они становятся крупными, неуклюжими, утрачивают связь друг
с другом. Возможно, внезародышевая бластодерма у муравьев прини-
мает на себя функцию виталлофагов.
Дальнейшее развитие муравьев в общем мало отличается от тако-
вого ос и пчел, ко изучено оно недостаточно, и потому остаются не-
которые неясности. Так, не вполне выяснен вопрос о происхождении
энтодермы. По Стриндбергу (1913а), зародышевый диск Formica
и Camponotus разделяются на три продольные полосы, из которых
медианная погружается внутрь и дает мезодерму и энтодерму. Послед-
няя обособляется от мезодермы по всей длине зачатка в виде отдель-
ных клеток (как у Angitia и Eurytoma). Биполярную закладку энто-
дермы Стриндберг отрицает. На соответствующих местах он находил
лишь скопления погрузившихся внутрь дегенерирующих внезародыше-
вых клеток.
Иначе описывает образование средней кишки Танквари (1913).
По его наблюдениям, у Camponotus от заднего конца зародышевой
полоски в промежутки между крупными клетками заднего конца
(соответствующим внезародышевым клеткам Стриндберга) врастают
тонкие пальцевидные тяжи клеток. За счет этих пальцевидных отрост-
ков образуется тонкая сеть, оплетающая крупные клетки. Часть кле-
ток зародышевой полоски растет поверх крупных клеток, и происходит
замыкание спины. Крупные клетки, вместе с оплетающими их мел-
кими, оказываются внутри зародыша, распространяются вокруг желтка
и образуют среднюю кишку. В построении средней кишки, по Танквари,
принимает участие и «внутренний периферический слой протоплазмы».
Под этим названием автор описывает, по-видимому, тот непрерывный
слой желточных клеток, который нередко образуется на поверхности
желтка. Следовательно, по описанию Танквари, средняя кишка му-
равьев образуется из трех разнородных элементов.
198
Другая неясность касается полового зачатка. Некоторые картины,
описанные Стриндбергом и Гехтом, очень напоминают раннее обособле-
ние полового зачатка — явление, которое вполне естественно можно
было бы ожидать у муравьев, поскольку здесь дифференциация частей
бластодермы вообще наступает очень рано. Сам Стриндберг (1919)
предполагает раннее обособление полового зачатка только у Pseudo-
myrma и решительно отвергает эту возможность для Camponotus.
Гехт (1924) изображает на заднем конце яйца Camponotus «полярное
тельце», сильно красящееся гистологическими красками (оосому).
Вскоре «полярное тельце» исчезает, на его месте в бластодерме
оказывается несколько мелких, резко базофильных клеток, некоторые
очень напоминают половой зачаток, так что Гехт допускает, что рас-
пыленное вещество «полярного тельца» попадает в эти клетки. Однако,
проследив дальнейшую судьбу этих клеток, он показал, что они не
имеют отношения к половой системе, а являются носителями симбио-
тических бактерий — мицетоцитами. Развитие половой системы Гехт
не описывает и лишь вскользь замечает, что она развивается из мезо-
дермы. Быть может половой зачаток муравьев утратил свою перво-
начальную функцию и стал носителем симбионтов, а, может быть, обе
эти функции совмещаются одними и теми же клетками (что было бы
довольно естественно). Все эти вопросы-очень интересны и безусловно
заслуживают специального изучения.
Дальнейший органогенез протекает также, как у ос и пчел.
Личинки муравьев также беспомощны и неспособны к самостоятель-
ному существованию.
Итак, в развитии муравьев имеется ряд своеобразных черт. Ран-
няя дифференциация бластодермы, редукция эмбриональных оболо-
чек, особенности развития, связанные с трансовариальной передачей
симбионтов, изученные, пока, недостаточно, — все это заставляет
призанть развитие муравьев гораздо более специализированным, чем
у ос и пчел.
4. РАЗВИТИЕ МОНЭМБРИОНИЧЕСКИХ НАЕЗДНИКОВ
С ТРОФИЧЕСКОЙ ОБОЛОЧКОЙ
Более существенные изменения развития наблюдаются у эндо-
паразитических представителей подотряда Parasitica. Развитие экто-
паразитических наездников в литературе не освещено, однако, можно
думать, что оно мало отличается от такового ос и пчел, тем более, что
даже некоторые эндопаразитические формы (например, Angitia vesti-
gialis Ratz.) развиваются по тому же типу.
Основное преимущество паразитического образа жизни у пере-
пончатокрылых состоит в том, что их зародыши и личинки развивают-
ся в среде, насыщенной питательными материалами; но, чтобы полнее
использовать это преимущество, нужно как можно раньше начать
питаться за счет хозяина. Эта биологическая задача разрешается
двумя различными способами: 1) путем раннего вылупления из яйца
сильно упрощенной личинки, которая сразу же начинает питаться
соками и тканями хозяина, и 2) путем появления у зародыша спо-
собности извлекать необходимые для развития питательные вещества
из тела хозяина еще во время эмбрионального развития. По-видимому,
подавляющее большинство эндопаразитических перепончатокрылых
в своей адаптации к паразитизму пошло по второму пути. При этом
роль органа, через посредство которого происходит питание зародыша,
берет на себя эмбриональная оболочка.
199
»*ч
а
5
з
.^
V©
й
X
крылы
о
н
онча
с
а
еских п<
рых паразитич
о
н
т неко
о
о.
«
2
X
алы
X
s
Эмбр
1 о.
Авто]
Увеличение
объема
родыша
Размеры за
Вид
1 °°
1 22
1 °^
1 "о
1 *-■
1 °
row
1 и
1 см
X
ы
1 о
1 о
X ~
со
о* -*
§ X
0,3
0,52
гнное яйцо:
ся личинка:
0ТЛ0Ж(
1 мм
ившая<
8©"§
и со
.
ress
и 1
ел 1
СО
uteli
rufisc
lypta
О |
Ichneumonidae |
со
ел
с
СО
3*
'£
ел
ledow
1926
00
ОО
X
-od
СО
со
2
Он
зме
СО
Он
CD CD
звития линейны
[ичиваются вдво
со >=;
О, О)
m
оо 2
СО*
°°
ел
о
■в
ел
orali
s fem
anchu
CQ
vo
о
CO
05
1 T"~l
^
ео
СП
со
^Г
со
я
о
ван
• s
оо
X
о
н
N
"5
ел
giali
1 vesti
ngitia
< 1
со
СО
to
o>
С.Г
.-o
CO
со
jp
CO
CQ
О
ван
s
°
LO
|
X
us sp.
phidii
< |
Aphidiidae
CO
CO
Ю
O)
w
CO
CO
* CO
1 ^
1
со
X
! о
ван
5
-400
о
о
СО
X
ел
си
Z
о
igiat
us р1г
phedr
tu 1
со
ел
<u
anc<
>
CM
X
s
ST
К
вел
2oo
о ю
о.
а за]
рина
звития длин
раза, а ши
СО СО
лга
а*
г: с-»
и со
со ffl
ГО
yle
с
°
ел 1
omp:
les tb«
pante
< |
Braconidae
ю
S
«_
owe
H
CO
CO
X
5>*
KOI
0 70
рина
ЪС 1 к
.соЭ
ч» о о
^ M H
on S £
OO ~ CO
-0,02
стиг;
ИЧИЕ
0,10X0,025-
го длина до
твенно увел
| V О
1 н
ОЭ '05 CD
о к со
° S О
.. га о
* °-s
ее;
Isch.
а
^
ел
litar
les mi
pante
< |
1 °>
О
[11Ш1
оо
*
ю
со
X
ел
ел
си
о
ris (
vulga
racon
OQ 1
сч
О
X
X
ч
1 VO
ГЛО
о
*
1 о
OlX
ел
си
CD
2
atus
termin
mpus
inoca
а
N
1
1
>»
1
0
^)
о
§
1 с,
1 о
1 н
1 °Q
<
1 ^
1 s
1 х та
1 CU S
1 Р* 0)
1 *» О
си о
>>
Размеры зародыша
Вид
о
<-> 1
э5
CD
0>
U
1 °°
1 СМ
1 CJ>
1 о
1 О
1 о
с->
та
*~*
"**
о
X
2 s
со ex,
сл S
ия длин;
раз, а ш
время эмбрионального развит
одыша увеличивается в 5—7
а в 12—14 раз
таЛК
СО
ees
Z
СЛ
til
со 1
73 1
Dinocamp
си
та
!2
"3
о
О
та
<—
СО
см
195
.п
mit
1 ^
*
°
СЛ
X
uesb.
8
tae
та
s vit
=s
Microctom
3
Cla
CO
Я?'
1 о
"S3
та
S а
.У си
«- сл
СЛ
СО
О
О
»о
X
* о
си
2 a
•*«
о
^ н
О S
О «
яйцо: 0.14Х1
зародыш станс
тром 1,5 мм
^жеотложенное
онцу развития
идным с диаме
м * м
CJ
та
imidi
•а
^s
tes
о
(Zem
mps.
Meteorus
tus Tho
1
1 CM
О
oo
1 f—1
^
3
b£
1 ^
1 С
£
CM
/00
CM
1 x
еет размеры
X0,42 мм
я зародыш им
а в конце—0,6
начале развити
0,15X0,050 мм,
CQ
та
Smic
11
щ>т0'
j
ispes
Chalcis s
clavipes
о
та
Г2
3
ale
X
и
CO
ст>
»**»
о
:ian
1 u
CO
^
CO
X
1 §
• со н
О Я
яйцо имеет длину 0,
у эмбрионального разв]
достигает 0,25 мм
вежеотложенное
0,04 мм; к конц
длина зародыша
и
Timb
со
3
S0I
"о
О.
«о '
3 1
Tetracnem
CU
та
rti
>>
п
щ
сч
192
>>
eib
см
•-^
СО
см
X
1 к си
со s- 2
О К со
-0,05X0,
ервых л
нгает ра
>жеотложенное яйцо: 0,04-
,035 мм; к моменту выхода п
ок паразитическое тело дост]
ов: 1,37X1,02X0,57 мм
Sosa
(J
How.
CD
та
<->
—
*cu
en
Copidosom;
CM
en
1 ~"T
к
•о
c
та
>*
eib
-j
00
CO
CM
X95
cu cu
tr 5
1 u
008 лш;
10 ДОСТИ
яйцо: 0,021X0,
зитическое тел
0,4 мм
:жеотложенное
ез месяц пара
азмеров: 0,88х
ё сиси
U 1
*-« 1
г) ve
си
(О
та
"та
CU
со >-, 1
Polygnotu
па lis M
dae |
си
"со
та
04
£»
Plal
со
1
1
1 ю
сл
lives
1 ^
сл
оо
Х313
3 та
сип
ет разме
ПОЗДНИЙ
дке яйцо име
07X0,008 мм;
-0,75 мм
овое к откла,
,015-0,017x0,0
одыш: 0,7x0,5-
g О Си
'
Silv.
cu
та
omyi
dry
V*
Platygaste
Способность всасывать воду из окружающей среды присуща
яйцам очень многих насекомых, вовсе не паразитических (например,
пилильщиков). Вопрос о том, в какой мере яйца каждого изучаемого
вида способны усваивать из окружающей среды и органические веще-
ства и какие именно, требует специального экспериментального
исследования. Такого рода исследования пока отсутствуют. О наличии
способности зародыша питаться за счет хозяина и о большей или
меньшей интенсивности питания мы можем судить лишь по увеличе-
нию размеров зародыша во время эмбрионального развития. Некото-
рые данные, характеризующие эмбриональный рост различных пере-
пончатокрылых, почерпнутые, главным образом из литературы, пред-
ставлены .на табл. 1. В ней приведены сведения о размерах зароды-
шей, а также показано- во сколько раз увеличился объем зародыша
за время развития. Лишь в немногих случаях авторы сами называют
эту цифру (отмечено звездочкой), чаще же этот расчет произведен
мною. При этом была использована формула объема эллипсоида:
V = 4/зЯ а2Ь,
где а — короткий радиус (половина поперечника яйца); b — длинный
радиус (половина длины яйца).
Как видно из таблицы во многих случаях объем зародыша уве-
личивается в несколько тысяч раз. При этом многие авторы приписы-
вают эмбриональной оболочке трофическую роль (Маршаль, 1906;
Оглоблин, 1913, 1924; Лейби и Хилл, 1924; Джексон, 1928; Симмондс —
Simmonds, 1947, и др.). Однако, несомненно, эмбриональная оболочка
наездников выполняет, наряду с трофической и защитную функцию,
изолируя зародыш от тканей хозяина, его фагоцитов, гормонов и т. д.
О значении эмбриональной оболочки уже была речь в связи с разви-
тием Aphidiidae (см. спец. часть, гл. III и IV); ниже мы еще вернемся
к этому вопросу.
Примером монэмбрионического типа развития с трофической
оболочкой может служить развитие афидиид. Другой хороший пример
этого типа представляет Synopeas rhanis Walk. (Platygasteridae), no
данным Маршаля (1906). Очень маленькое, лишенное желтка яйцо
Synopeas сперва дробится неполно (делится только ядро). Но уже на
стадии четырех ядер дробления одно из ядер, а именно «эмбриональ-
ное», вместе с небольшим участком плазмы отделяется от остальных —
«амниотических» (рис. 109,Л). В результате дробления эмбриональ-
ного ядра образуется «бластула», полость которой заполнена «вител-
лоидной массой» (рис. 109, В). Последняя представляет собой, по-
видимому, участок нераздробившейся плазмы, к которому присоеди-
няются продукты выделения бластулы. Иногда в вителлоидной массе
встречается некоторое количество ядер, которые Маршаль приравни-
вает к парацитам (вителлофагам). Уже на стадии бластулы размеры
зародыша значительно увеличиваются.
Затем бластодерма становится многослойной, и зародыш приобре-
тает бобовидную форму. Лишь на вогнутой (спинной) стороне заро-
дыша остается участок однослойной бластодермы, которую Маршаль
сравнивает с внезародышевой бластодермой других насекомых. Этот
однослойный участок разделяет головной и хвостовой концы зародыша,
которые почти соприкасаются. По брюшной стороне зародыша от перед-
него конца до заднего проходит медианная борозда, вдоль которой
происходит усиленное размножение клеток и разделение эктодермы
и мезодермы. Одновременно с мезодермой на заднем конце обособляет-
ся группа довольно крупных клеток—половой зачаток. Энтодерма
.202
этом типе развития личинки обычно не испытывают сильного недо-
развития и упрощения. Приток питательных веществ, поступающих из
тела хозяина через посредство оболочки, которую Маршаль называет
трофамнионом, компенсирует недостаток их в яйце и делает возмож-
ным развитие -личинки, не уступающей по сложности организации
личинкам Aculeata, яйца которых обильно снабжены желтком.
5. ПОЛИЭМБРИОНИЧЕСКИЙ ТИП РАЗВИТИЯ
Благодаря тому, что некоторые паразитические перепончатокрылые
приобрели новое физиологическое качество — способность питаться
за счет хозяина еще в эмбриональный период их развития, зародыш
оказываемся в условиях избыточного питания и перед ним открывают-
ся новые эволюционные возможности — возникает полиэмбриония.
Сущность полиэмбрионии состоит в том, что из одного яйца раз-
вивается не один зародыш, а несколько. Это явление было известно
еще Клейненбергу (Kleinenberg, 1879), описавшему развитие двух
зародышей из яйца Lurnbricus trapezoides Duges, и Мечникову (1886),
который наблюдал распадение зародышей медузы Oceania armata
Kolliker на отдельные фрагменты, превращавшиеся затем каждый в
нормальную жизнеспособную планулу.
Среди насекомых полиэмбриония была открыта Маршалем (1898).
У Ageniaspis (Encyrtus) fuscicollis Dalm. (сем. Encyrtidae), с разви-
тием которого мы уже познакомились раньше. В 1903 г. Маршаль
обнаружил полиэмбрионию также у Platygaster zosinae Walk.
(= Polygnotus minutus Lind.)—паразита гессенской мушки. После
этого в литературе стали появляться сообщения о полиэмбриониче-
ском способе развития у целого ряда паразитических перепончато-
крылых (подробную историю открытия и изучения полиэмбрионии
у Hymenoptera можно найти у Сильвестри, 1937, и Яхонтова, 1940).
Развитие разных полиэмбрионических наездников может несколь-
ко различаться в отдельных деталях. Так, у полиэмбрионической
энциртиды — Litomastix (Paracopidosomopsis) floridanus (Ashmead)
из гусениц Autographa brassicae (Пэтерсон, 1921) трофамнион обра-
зуется во время первого дробления: яйцо разделяется сразу на три
части, из которых две являются бластомерами, а третья, большая, со-
держит редукционные ядра. Эта «полярная» клетка постепенно облекает
весь зародыш и превращается в оболочку. Дробление Litomastix тоталь-
ное. Первые признаки полиэмбрионии появляются на стадии 220—225
бластомеров. Часть эмбриональных клеток приобретает веретеновид-
ную форму и образует перегордки, разделяющие всю совокупность
клеток на 15—20 отдельных групп. Вокруг каждой группы появляется
оболочка из этих веретеновидных клеток. Первичные группы эмбрио-
нальных клеток могут подразделяться вторично и общее количество
таких групп может превысить тысячу.
Среди Encyrtidae полиэмбриония отмечена также для представи-
телей родов Copidosoma и Berecyntes (список полиэмбрионических
видов приведен в табл. 2).
Целый ряд полиэмбрионических представителей имеется и в сем.
Platygasteridae. Процессы обособления эмбриональной оболочки (обо-
собление периферического слоя ооплазмы, в которой остаются редук-
ционные ядра) и подразделение общей эмбриональной массы на груп-
пы, дающие начало отдельным зародышам, протекают сходно с тем,
что описано для Ageniaspis. Но в отличие от Encyrtidae число зароды-
шей, развивающихся из одного яйца, в этом семействе никогда не
204
^
<2
3
Табл
рылых
пончаток
пере
х 1
S
-CJ
а>
т
S
X
о
к
сх
2
полиэ
о
X
с
CJ
га
Я
эГ
чество паразитов,
[ЦИХСЯ ИЗ ОДНОГО Я
Коли
звиваю]
га
СХ
Автор
хозяина
ние
га
азв
X
аразита
вание п
со
га
X
Семейство
10
нем
:ред
со
1931а
Parker,
lis Hiibn.
bilai
3
с
га
yraust;
сх
s
Л
uensis As
us gifi
*■*
С
CD
<->
Mac
1
Braconidae
1932
Daniel,
CJ
lesta Bus
о
В
га
lith
irapho
°
•c
о
ylivorus R
us anc
u.
cu
О ui
u. CD
1 *
-25
нем
ред
OQ
land,
Ber
1946,
Gadd,
1951
us homonae
i-i
0)
<->
v-
Mac
о
50
Около
1940
Paillot,
Schiff.
В
3
Id
cu
uxoa !
w
"■"
is Spin.
collai
CO
3
*CX
О
<->
S
1 <
180
о
904в
oT
1, 1898a-
n, 1914
Marcha
Marti
ex
ex
CO
.ra
"з
cu
в
yponc
i J-
-P
СЛ
yrtus) fu
(Enc
aim.)
.2Q
cx^
СЛ
.2 ~
c1^
CD О
<
Encyrtidae
^
-10-15
нем
ред
CQ
, 1908
Silvestr
CO
3
lee
rays о
a
ra
<_
dIHs var. p
fuscici
ilv.
со С/Э
■8\a
«3 О
*— CJ
с а
CD •-
1 ttf"
1 **
1
1
cn
, 1904
Marcha
ttl
imerella
те
u
CO
llet
ithoco!
-j
N
eipes Rat
testae
1 .M
'cu
1 M
1 .2
1 "c
cu
c^O
1 <
CN
CN
°
usen,
Cla
1932,
Jancke,
1940
niella L.
pru
ra
S
rgyres
<
His Dalm.
atrico
1 M
CO
Л
a
CD
1 ьс
<
О cf^r
X cd ra
О СХ со
CD « .
гусеницы G. Galla
Riley вылетает е
паразита, а из G
sck.—191
СЛ __ ГЗ
o.S2m
3 txO ел
^ ra s "C
о -a cd ra
S
t^
Oi
n, 1915,
Pattersoi
•dds
ema
JS
osc
norim
° •
chiae How
i gele
Ё
со
°
•a
'ex
о
u
^N
? табл.
4J
5
Продолже
тов,
го яйца
1 5 О
1 со х
1 ^ cf
1 О, о
1 га
Количество п
звивающихся из
1 та
1 ^
Автор
1
зяина
о
X
звание
га
X
ита
го 1
ие пара
ван
ГО 1
га 1
X 1
Семейство
1 °^
41 до
1 н
О
о
ri, 191
ilvest
СЛ
с
га
о
:=
с
с
со
эга stef
•с
Он
oleo
CJ
Цн
га
£
buyssoni
га
Е
о
сл
о
32
Он
° 1
и
Encyrtidae
[-38
реднем
DQ
"•f
ri, 192
ilvest
СЛ
d
s
J
CO
с
CO
oeflingi
X
ortri
H
*- 1
ra
tortricis
ra
E
о
CO
°
"H
"Oh
о
и
1 Of-
чено 68:
^
ITO
с
гусеницы
аразита
одной
80с? п
« s
S
1918
агга,
СЛ
iibn
X
га
с
га
04
си
tes vari
со
leth
О
Он
СЛ
га ;
Е
о
СЛ
°
*°
'Он
О 1
и
3 до 1
*-
О
СЛ
со"
ri, 192
ilvest
1940
СЛ
ibn
X
га
~
<v
с
га
С
.2
'С
га
ecur
еж
>
Sil
cu
nanellai
.га
Е
°
СЛ
°
•а
'о.
° '
_^ 1
00
т
реднем
о
са
ompson,
t х:
1 Н
and
агкег
1928
*о"
tau
СЛ
JS
OJ
senticet
.-
othr
z
cet.
1ч
CU
:>
с
thompso
га '
S
о
СЛ I
о
*о
Он
о 1
_^ 1
-38
реднем
и
CQ
ompson,
ь*
and
агкег
1928
Он
СЛ
ал
Tri<
SB
си
с
pingui
га
!с
elec;
О
atz. I
05*
urn
с
bouchea
га
Е
о
со
°
тэ
'Он
о
и 1
, гз га
[летает
паразит
J о
1 а ^
i т—*
3 со
гусениц
21с? или
3 ОДНОЙ
среднем
s
1947
►outt,
n
j2
ulel
•ere
Он
о
schema
о «-
Е —
nori
(Zel
О
nch.
Bla
.
koehler
га 1
S 1
о
СЛ
о
*а
'а.
о
U |
ЗООО
©
с*
Leiby,
07,
ri, 19
ilvest
1929
СЛ
га
Uh
met
yto
x:
Он
3
. >>
mma L.
:ae Rile
ra <-»
lusia g
brassi
Oh
В
'та
Q
СЛ
3
truncetell
.и
га
E
5
"^- 1
Li \
to
7
реднем
о
CC
и 1927
^
on, 19
atters
Q-
Ы
ra
CJ
tra brass
cu
Й
hyto
(X
юр-
E
О y—s
о Е
•ox;
Paracopi
inus (As
X 'C
"h3_0
ra **"*
Е1Г
<2"й 1
'Л
120
21 до
н
О
е, 1926
errier
Du
геу
tin
га
ell
с
'So
ria alpi
га
СЛ
СЛ
ерге
Сн-
егу
Е
bau
Хн
kriec
х
2 >>
Era
S%
~
М 1
-I
.^
^
-
s
ние
Продолже
го
Г>Е
- к
о о
s о
го Яй
го f=l
OiO I
Количество па
звивающихся из
го
О.
Он
Авто
яина
со
о 1
ние х
ее
Назв
се
н
парази
ние
то
03
со
ее
X
ейство
s
о
о
сч
, 1951
•о
X
CD
CQ
trei
dela
.-
mast
о
J
cyrtidae |
i
:реднем—28
C-»
CQ
915
~*
rra,
ее
C/3
Ul
CD
ell
N
ella
cd
CD
С
.2
"К
ее
С
<
ees.
mis N
о
iriia
TO
>
3
>►*
С
щ
реднем—1289
00
1925
ow,
с
СЛ
rote
а
СЛ
ее
**
3
се
ее
о
X
3
щ
=
CU
bak
СЛ
3
erecynl
1 CQ
10 до 12
н
О
Х>
, 1904
*сё
irch
ТО
ее
СУЭ
tor
а
3
estr
*а
ее
ол
pha
о
>*
XJ
а
alk. |
ind.)|
ел
3
sinae
minuti
О ел
N 3
О
аЗ &
«-• >»
СЛ —
се о
laty
а
asteridae
Ь<)
Platy
00
о
н
о
.927
3
>п, 1921
о
ел
tter
се
а
hopa-
<х
3
го
С
texa
пае
^2
2 «*
>,"
E.S
2 ^
сл С
■3©
Fouts
*НЗ
4-1
<-*
0>
gas
>>
lat
о-
8 до 12
н
О
со
CN
Оэ
z£
nd Hi
се
iby
CD
J
ее
СЛ
tor
и
3
estr
ТЭ
се
СХО
pha
о
>>
x:
a
yers
alis M
J с
ven
i *—
CD
gas
I >>
lat
1 a
"S c-[ CU
P О го
§ 2 S
2 s ч
° H CQ
TO = ^
я « 2
~ ГО Я
« c^s
•дну личинку хоз:
ается 4—8 яиц па
ой личинки хозя
о 15 паразитов
«ияч
CQ
929
i~*
926 и
"~'
iby,
CD
-1
- ТО
С
саго
то
lomy
се
О
-С
°*
outs
ibilis F
го
vari
*-•
CD
gas
>»
lat
Oh
к I
CO
CO
то
Q-
н
=
ro S
a-«
одного яйца па
ается 1—2 личин
со m
S
СО
од
О
•—;
nd Hi
се
iby
CD
-1
се
СУЗ
tor
a
3
estr
1
то
1 ЬО
pha
2
"►»
•c
CU
.SJ
x:
aj
tygasti
1 я
(PI
rb.)
i *£
з tu
olygno
malis
1 ^
-
5
to
ro
Q-
ro —
н с
а к
со S
TO CC
Q-s
каждого яйца па
ается 40—60 инд:
со m
£
OJ
ст>
iser, 1
го
rnh
о
^Г
^
хз
се
U-.
lata
3
(J
ее
Е
5
lia
си
.с
Н
ihan
ТО
о
ее
thel
СЛ
3
lop
си
CJ-
<
inidae
>%
а
бывает значительным. На ряде представителей Platygasteridae можно
проследить возникновение и развитие полиэмбрионии чиаг за шагом.
Здесь встречаются следующие варианты развития:
1) типичное монэмбрионическое развитие, например у Synopeas
rhanis Walk. (Маршаль, 1906);
2) случайная полиэмбриония: у Allotropa burelli в 3% случаев
развиваются двойники (Кланси, 1944);
3) факультативная полиэмбриония: Platygaster hiemalis Forbes,
у которого, по предположению Лейби и Хилла (1923), из оплодотворен-
ных яиц, дающих впоследствии самок, развиваются двойники, а не-
оплодотворенные яйца, дающие впоследствии самцов, развиваются
монэмбрионически; '
4) прочно установившаяся, хотя и не очень «продуктивная»
полиэмбриония: две личинки из одного яйца у Platygaster variabilis
Fouts.; 8 личинок — у Polygnotus (Platygaster) vernalis Myer (Лейби,
1929); 10—12 личинок —у Platygaster zosinae Walk. ( = Polygnotus
minutus Lind.), по Маршалю (1903 и 1904в); 11 — 18 личинок — у Platy-
gaster felti Fouts, no Пэтерсону (1927).
Среди сем. Braconidae полиэмбриония встречается у представите-
лей родов Macrocentrus и Amicroplus (см. табл. 4). К сожалению,
эмбриональное развитие этих форм изучено так слабо, что сопоставле-
ние его с развитием Encyrtidae и Platygasteridae пока невозможно.
Наконец, имеются сведения, тоже очень скудные, о полиэмбрионии
у Aphelopus theliae Gahan паразита цикады Thelia bimaculata (Fabr.)
(Корнхаузер, 1919).
По-видимому, полиэмбриония представляет собой высшую форму
адаптации эмбрионального развития к паразитизму у перепончато-
крылых. В защиту этого положения можно привести следующие
соображения.
1. При полиэмбрионическом развитии преимущество паразитиче-
ского образа жизни (избыток питательных материалов) используется
наиболее полно. Об этом свидетельствуют цифры, характеризующие
эмбриональный рост у разных наездников (табл. 1).
2. Известно, что в жизненном цикле многих паразитов имеются
такие критические стадии развития, когда большая часть зародышей
гибнет, и лишь немногие из них попадают в благоприятные условия.
Поэтому паразиты особенно нуждаются в повышении плодовитости,
что чаще всего выражается в так называемом «законе большого
числа яиц», в появлении партеногенетических поколений и разных
форм бесполого размножения, увеличивающих интенсивность зараже-
ния хозяина. К числу последних надо отнести и полиэмбрионию.
В. А. Догель в своем «Курсе общей паразитологии» (1947) опре-
деляет полиэмбрионию как «одно из приспособлений к умножению
потомства, которые столь разнообразны у паразитов» (стр. 175).
«Легко видеть, — пишет он, — каким мощным средством размноже-
ния является полиэмбриония и насколько она экономит силы отклады-
вающей яйца самки паразита. При небольших размерах вышеназван-
ных наездников каждый из них может продуцировать за свой соб-
ственный счет лишь небольшое количество яиц. Но, попадая в среду,
богатую питательными веществами, яйцо наездника дает начало мно-
жеству зародышей, увеличивая интенсивность продукции каждой
особи наездника в 100—1000 раз» (стр. 176).
Хорошей иллюстрацией этого положения могут служить цифры,
приведенные Клоузеном (1940) в его сводке по насекомым-энтомо-
фагам. Клоузен подсчитал потенциальные возможности размножения
208
у двух близких видов Platygaster hiemalis Forbes и PI. zosinae
Walk., паразитирующих в одном хозяине — гессенской мушке.
Яичник зрелой самки PI. hiemalis содержит в среднем 3300 яиц, из
которых каждое дает 1—2 зародыша. Возможное число потомков
PI. hiemalis составляет 5000. Самки PL zosinae производят в среднем
только 228 яиц, но каждое из них дает 2—12 зародышей (в среднем.—
7—9). Общее потомство должно составить 18 000.
Далее Клоузен замечает, что полиэмбриония перепончатокрылых
не столько содействует увеличению численности вида, сколько служит
средством поддержания этой численности на необходимом для сохра-
нения вида уровне. Полиэмбриония, по мнению этого автора, пред-
ставляет собой попытку со стороны вида, далеко не всегда успешную,
преодолеть особые, лимитирующие его размножение факторы.
3. Полиэмбриония. встречается у видов, наиболее глубоко приспо-
собившихся к паразитизму. В литературе можно найти ряд указаний
на то, что полиэмбрионические виды откладывают яйца в строго опре-
деленные части тела хозяина, причем между тканями хозяина и пара-
зита устанавливаются очень тесные отношения (Мартин, 1914; Пэтер-
сон, 1921; Лейби и Хилл, 1923; Лейби, 1922 и 1929; Пайо, 1940; Даут,
1947, и др.). На присутствие паразита хозяин реагирует образованием
соединительнотканной капсулы, которая, однако, не только не приводит
паразита к гибели, но становится необходимым условием его нормаль-
ного развития.
Таким образом, полиэмбрионические перепончатокрылые настолько
специализированы как паразиты, что в процессе эволюции преодолели
защитную реакцию хозяина, извратили ее, обратили себе на пользу.
Между тканями паразита и хозяина устанавливаются такие интимные
трофические отношения, какие встречаются лишь у некоторых живо-
родящих животных между тканями матери и зародыша.
4. Наконец, замечательно, что полиэмбриония наблюдается в четы-
рех разных семействах перепончатокрылых: Braconidae, Encyrtidae,
Platygasteridae и Dryinidae, причем первые три относятся к подотряду
Parasitica, а последние — к Aculeata. В каждом из этих семейств, на-
ряду с полиэмбрионическими видами, есть и типичные монэмбриониче-
ские. Все это говорит о четырехкратном и независимом возникновении
полиэмбрионии у перепончатокрылых (см. следующую главу). Очевид-
но, полиэмбриония возникла здесь не случайно, а представляет собой
одну из основных тенденций эволюции паразитических перепончато-
крылых. ч
Все это заставляет признать, что полиэмбриония перепончатокры-
лых тесно связана с их паразитическим образом жизни.
Интересно, что бесполое размножение, одной из форм которого
является полиэмбриония, встречается у членистоногих крайне редко.
Помимо полиэмбрионии насекомых, оно наблюдается только у корне-
головых раков, где возникло тоже на почве паразитизма (Догель, 1947).
Однако следует отметить, что не все зародыши, возникшие в ре-
зультате диссоциации первичной массы эмбриональных клеток, успеш-
но завершают свое развитие. Очень часто образуются неполноценные
зародыши, получившие названия «псевдогермов», «псевдоморул»,
«псевдоэмбрионов», «псевдоличинок», «абортивных зародышей», «бес-
полых личинок», «тератоидных личинок» и т. п. При этом в большин-
стве случаев имеет место ранняя дегенерация зародышей, вызванная
или слишком далеко зашедшим разделением полигерма, вследствие
чего некоторые зародыши получают слишком мало эмбриональных
клеток (иногда только одну), или тем, что они оказываются непол-
14 О. М. Иванова-Казас
209
жилью одетыми трофамнионом и тканями хозяина, либо лежащими
слишком далеко от жирового тела хозяина, и испытывают недостаток
в пище. Такое объяснение принимается для «абортивных зародышей»
у Copidosoma gelechiae How. (Пэтерсон, 1915 и 1918; Лейби, 1922,
1929) и Platygaster hiemalis Forbes (Лейби и Хилл, 1923; Лейби, 1929)
Иногда под названием псевдогермов описывают обособившиеся
участки трофамниона, не содержащие в себе эмбриональных клеток.
Они длительно сохраняются в теле хозяина и растут, а затем поедаются
личинками паразита. Такие псевдогермы наблюдаются у Macrocentrus
gifuensis Ashm. (Паркер, 1931а), М. ancylivorus Roh. (Даниель, 1952)
и Amicroplus coljaris Spin. (Пайо, 1940). У некоторых платигастерид
сходные псевдогермы образуются #з трофамниона после выхода личинкм
(Маршаль, 1906). Эти образования очень напоминают гигантские клет-
ки, образующиеся из серозы у Braconidae и Aphidiidae.
Гораздо, более сложное явление представляют собой бесполые
личинки, обнаруженные Сильвестри (1906) наряду с нормальными при
развитии Litommastix trucatellus Dalm. В многоклеточном зародыше
Litomastix еще до проявления полиэмбрионии можно различить
две области: задний отдел, состоящий из мелких клеток, и передний
отдел, состоящий из мелких и крупных клеток. Задний отдел Силь-
вестри назвал «massa monembrionale» — из него получается одна бес-
полая личинка. Передний отдел — «massa germinigena» — дает в даль
нейшем некоторое количество бесполых личинок и много нормальных.
В одном случае из одного яйца Litomastix развилось 1700 нормальных
личинок и 220 бесполых. Бесполые личинки выходят из оболочек
несколько раньше нормальных и некоторое время (до 10 дней) оста-
ются живыми, но затем все же погибают. У бесполых личинок отсут-
ствуют половая, дыхательная, кровеносная и выделительная системы,
но они более мускулисты и подвижны, чем нормальные. Уилер (1910)
отмечает, что бесполые личинки больше похожи (внешне) на личинок
ихневмонид, чем на таковых хальцид. Предположение Уилера, что эти
личинки принадлежат не Litomastix, а какому- то другому насекомому,
было отвергнуто Пэтерсоном (1918) на основании специально пред-
принятых экспериментов.
Подобные же бесполые личинки были найдены еще у Litomastix
(Paracopidosomopsis) floridanus Ashm. (Пэтерсон, 1915), Copidosoma
thompsoni Mercet (Паркер и Томпсон, 1928; Лейби, 1929) и С. bouche^-
num Ratz. (Лейби, 1929).
Попытки уяснить причины образования бесполых личинок вызвали
в литературе целую дискуссию. Сильвестри (1906) высказал предполо-
жение, что бесполые личинки образуются из тех зародышей, в которые
не попала часть, полового детерминанта. В то же время он сравнивает
диморфизм личинок Litomastix с полиморфизмом общественных насеко-
мых и приписывает бесполым личинкам функцию предварительного
разрушения и размельчения тканей хозяина, облегчающую эту работу
для нормальных личинок.
Эта гипотеза Сильвестри вызвала ряд возражений со стороны
Пэтерсона (1915, 1917, 1918, 1921), наблюдавшего бесполых личинок
при развитии Litomastix (Paracopidosomopsis) florilanus (Ashm.).
Пэтерсон отмечает, что у изученного им вида никакой правильности
в месте образования бесполых личинок, подобных указанным Силь-
вестри, не наблюдается. Не проявляют здесь бесполые личинки и ни-
какой «полезной деятельности» по разрушению тканей хозяина. Труд-
но представить себе, пишет автор, чтобы в полигерме Litomastix был
бы какой-то механизм, обеспечивающий попадание первичных половык
21Q
клеток, снабженных веществом полового детерминанта, в нескольк<>
сотен нормальных личинок. Но если даже допустить, что такой меха-
низм существует и бесполые личинки образуются лишь вследствие
его несовершенства, все же отсутствие у них систем выделительной,
дыхательной и кровеносной остается непонятным, так как эксперимен-
тально доказано, что даже развитие вторичных половых признаков не
зависит у насекомых от присутствия полового зачатка. Пэтерсон выска-
зывает предположение, что nucleolus, унаследованный полиэмбриони-
ческими наездниками от монэмбрионических, утратил здесь функцию
полового детерминанта. Собственная гипотеза Пэтерсона состоит
в том, что при дроблении из-за случайного нерасхождения хромосом
могут образоваться бластомеры, лишенные х-хромосомы; из них-то
и развиваются бесполые личинки. Но никаких цитологических наблю-
дений, подтверждающих эту мысль, Пэтерсон не приводит.
В 1937 г. Сильвестри еще раз возвращается к этому вопросу;
полемизируя с Пэтерсоном, он продолжает развивать свои прежние
взгляды. Чтобы объяснить отсутствие ряда соматических систем орга-
нов у бесполых личинок, он делает допущение, что «вещество полового
детерминанта» имеет организующее значение для развития этих систем
и что соматические клетки насекомых, при отсутствии половых, не
могут дать полноценный организм.
Далее Сильвестри отмечает, что у многих паразитических перепон-
чатокрылых первая личиночная стадия значительно отличается от по-
следующих и высказывает новое предположение, что бесполые личинки
атавистически воспроизводят такую стадию, бывшую у предков Lito-
mastix, но затем утраченную в связи с их специализацией как внут-
ренних паразитов. Эта выпавшая из современного нормального разви-
тия стадия сохранилась, однако, в скрытом виде в генотипе и прояв-
ляется только при отсутствии половых клеток. Но отсутствие половых
клеток и ряда других органов у бесполых личинок делает невозможным
завершение их развития, поэтому они обречены на гибель. Спекуля-
тивный характер всех этих сложных построений Сильвестри очевиден.
Последние высказывания по этому вопросу принадлежат Дауту
(1947, 1952, 1959), который наблюдал образование бесполых личинок
у Copidosoma koehleri Blanchard, паразитирующей в гусеницах Gnori-
moschema operculata (Zeller). Как отмечает Даут, в каждой заражен-
ной гусенице хозяина встречается 20—30 нормальных личинок и 1—2
бесполых. Даут сомневается, что рассматриваемые личинки являются
бесполыми, так как изучение их производилось лишь самыми грубыми
методами, с помощью которых обнаружить маленький половой зачаток
на этой очень молодой стадии трудна. Поэтому он сам пользуется
термином тератоидные личинки. Присоединяясь к мнению Паркера
н Томпсона (1928), Даут считает этих личинок просто уродством. Он
заметил, что тератоидные личинки Copidosoma koehleri образуются
только при развитии оплодотворенных яиц, из чего делает заключе-
ние, что вхождение в яйцо сперматозоида имеет какое-то отношение
к их возникновению. Он допускает, что тератоидные личинки возни-
кают путем андрогенеза, или в результате слияния спермия с поляр-
ным тельцем (Даут, 1959).
Из изложенного видно, что возникновение бесполых или тера-
тоидных личинок остается загадочным. Если бы причиной их развития
были бы просто неблагоприятные условия, в которые попадают не-
которые из зародышей, то дефекты этих личинок были бы очень разно-
образными, как в количественном, так и в качественном отношении.
Однако на самом деле (судя по литературным данным) бесполые
14*
211
личинки каждого вида имеют вполне определенную морфологическую
характеристику, почему и можно говорить о диморфизме личинок.
Очевидно имеются какие-то постоянно действующие причины, законо-
мерным образом вызывающие в известном проценте случаев определен-
ные нарушения развития Litomastix, Copidosoma и др. Но что это за
причины и каков механизм их действия — пока остается тайной.
Наконец, заслуживает упоминания парадоксальный факт, обнару-
женный Даниелем (1932) при изучении развития Macrocentrus
ancylivorus Roh., паразита Grapholita molesta Busk. Первая личинка
этого паразита, вышедшая из оболочек, оказывает подавляющее дей-
ствие на развитие остальных зародышей. Даниелю иногда случалось
находить в одной гусенице хозяина одновременно двух личинок
Macrocentrus I возраста, но он никогда не находил больше одной
личинки II возраста. Таким образом, несмотря на полиэмбрионический
тип развития, зрелости достигает только одна личинка Macrocentrus.
Для объяснения этого несоответствия выдвинуто предположение, что
некогда Macrocentrus ancylivorus паразитировал в более крупном
насекомом, в котором успешно завершало свое развитие несколько
личинок, а затем, в силу неизвестных причин, он был вынужден
перейти на более мелкого хозяина, который оказался в состоянии
«прокормить» только один индивид Macrocentrus.
6. КРАЙНЕ ДЕЗЭМБРИОНИЗИРОВАННЫЙ ТИП РАЗВИТИЯ
НЕКОТОРЫХ ЭНДОПАРАЗИТИЧЕСКИХ НАЕЗДНИКОВ
Тенденция к дезэмбрионизации, проявляющаяся в развитии
Aculeata и у наиболее примитивных Parasitica получает дальнейшее
развитие у некоторых эндопаразитических форм. С этим типом раз-
вития мы уже познакомились на примере Mestocharis militaris R.-Kors.,
Caraphractus reductus R.-Kors. и Prestwichia aquatica Lubb.
Литературные данные показывают,, что этот же тип развития
присущ Banchus femoralis Thorns, (по Бледовскому и Краинской,
.1926), Anagrus incarnatus Hal. (= Polynema natans, по Ганину,
1869a), Trichogramma (Ганин, 1869а; Сильвестри, 1909в; Гетенби,
1917, 1918a) и Telenomus gracilis Mayer (Коломиец и Коваленок,
1958). Быть может, не случайно, что все названные виды (кроме
Banchus femoralis Thorns.) являются яйцеедами.
Для развития всех этих форм характерны следующие черты:
1. Паразит начинает питаться за счет хозяина только после вы-
хода личинки из яйца. Зародыш не получает питательных веществ
извне и, следовательно, не является по отношению к хозяину парази-
том, а скорее напоминает инородное тело.
2. Размеры яиц и содержание в них желтка уменьшается. Из
яйца выходит сильно недоразвитая морфологически личинка.
3. Намечается тенденция к упрощению эмбрионального развития,
выражающаяся в упрощении отдельных процессов или даже в выпа-
дении их: например, выпадение из развития эмбриональной оболочки,
видимо утратившей свое функциональное значение, или выпадение
сегментированной стадии в развитии головы у Banchus и Mestocharis,
или упрощение процесса развития средней кишки у Prestwichia и т. д.
Три последовательные стадии развития наиболее характерных
особенностей этого типа можно видеть при сравнении Mestocharis,
Caraphractus и Prestwichia.
Средние размеры яиц у Mestocharis составляют 0,415 X 0,095 мм}
у Caraphractus—0,213X0,093 мм, Prestwichia— 0,154 X 0,065. Яйца
«12
ГЛАВА II
СРАВНИТЕЛЬНАЯ ЭМБРИОЛОГИЯ И ФИЛОГЕНИЯ
У ПЕРЕПОНЧАТОКРЫЛЫХ
Схема, изображенная на рис. 111 имеет сравнительно-эмбриологи-
ческий характер и показывает только основные направления эволюции
эмбрионального развития в отряде Hymenoptera и преемственность
между отдельными типами развития, не претендуя на отображение
филогенетических отношений. Однако представляло бы большой инте-
рес выяснить, в какой мере данные сравнительной эмбриологии соот-
ветствуют современным представлениям о филогении внутри отряда
перепончатокрылых. Ближайшее знакомство с литературой, к сожале-
нию, показало, что родственные отношения даже между большими
группами семейств перепончатокрылых до сих пор hq могут считаться
выясненными и являются предметом спора между специалистами.
Ввиду этого пришлось поставить вопрос несколько иначе: в какой
мере различные типы развития перепончатокрылых являются харак-
терными для определенных систематических групп, и не может ли
сравнительная эмбриология и в этом случае (как это не раз бывало)
содействовать решению вопросов филогении.
Попытаемся рассмотреть эмбриональное развитие по отдельным
систематическим группам. Однако сразу же надо сделать оговорку —
развитие многих семейств все еще остается 'неизвестным, поэтому
высказываемые ниже соображения имеют лишь предварительный
характер.
1. ПОДОТРЯД PHYTOPHAGA
Из подотряда Phytophaga пока известно только развитие пилиль-
щиков, которое, как мы видели, сохраняет очень примитивный харак-
тер, следуя общей схеме развития насекомых почти без отклонений,
что вполне соответствует представлению о примитивности первично-
растительноядных перепончатокрылых. Поэтому есть все основания
считать тип развития пилильщиков исходным для последующей эво-
люции развития перепончатокрылых (рис. 111, 1).
Эмбриональное развитие других Phytophaga не изучено. Интерес-
но, что у личинок многих эндофитных форм произошла редукция
брюшных (а иногда и грудных) конечностей, т. е. они уже несколько
недоразвиты. Именно эти эндофитные формы—цефоидные пилиль-
щики (Росс —Ross, 1955; А^алышев, 1947, 1949а, 1957; Викторов,
1955, 1956) или рогохвосты (Гандлирш, 1908, Теленга, 1952) обычно
считаются близкими к предкам всех стебельчатобрюхих перепончато-
215
крылых (Apocrita). У личинок последних, как правило, грудь и брюш-
ко лишены конечностей, но иногда имеется различное число парных
придатков, морфологическое значение которых остается пока неясным.
2. ПОДОТРЯД PARASITICA
Надсемейство Ichneumonoidea
Из семейств, объединяемых в группу Jchneumonoidea, в эмбрио-
логическом отношении изучались лишь Ichneumonidae, Braconidae и
Aphidiidae.
Ichneumonidae. Развитие ихневмонид нам известно по
Mesochorus confusus Holmgr. (Кулагин, 1892, 1894), Banchus femora-
lis Thorns. (Бледовский и Краинская, 1926); Pimpla turionellae (L.)
(Бронскилл, 1959) и Angitia vestigialis Ratz. (Иванова-Казас, 19606).
Эти данные были довольно подробно изложены во II главе Специальной
части. Здесь же отметим только, что наиболее примитивный характер
сохраняет развитие Pimpla turionellae (много желтка, зачаточные
амниотические складки, закладка туловищных конечностей). Об отсут-
ствии значительных физиологических приспособлений к эндопаразити-
ческим условиям развития у зародышей Pimpla, Banchus и Anomalon
свидетельствует неоднократно отмечавшийся в литературе факт, что
при содержании в неволе самки этих наездников иногда откладывают
яйца в разные щели и что эти яйца способны нормально развиваться,
если их поместить в воду (Шевырев, 1912; Мейер, 1937).
Яйца Angitia vestigialis отличаются отсутствием желтка. Увеличе-
ние объема зародыша настолько незначительно, что не дает оснований
предполагать восприятие питательных веществ извне. Таким образом,
развитие Pimpla и Angitia, еще очень мало модифицировано
(рис. 111, II).
У Banchus femoralis еще сохраняется желток, но в других отно-
шениях развитие изменено сильнее, чем у Angitia. Полное отсутствие
эмбриональных оболочек, запаздывание в закладке ротовых частей
и мальпигиевых сосудов, неполнота сегментации — все это признаки
дезэмбрионизации и упрощения развития, зашедших гораздо дальше,
чем у Pimpla и Angitia. В этом отношении Banchus очень напоминает
Mestoeharis militaris R.-Kors (рис. Ill, IV).
У Mesochorus confusus желтка нет, а образование серозы и заро-
дышевых листков сильно модифицировано («деламинация в моруле»).
То обстоятельство, что сероза сохраняется, несмотря на очень сильную
редукцию размеров яйца и отсутствие желтка, наводит на мысль, что
она уже выполняет трофическую функцию. Если это верно, то по
характеру развития Mesochorus приближается к типу развития, обозна-
ченному на рис. 111 цифрой V.
Итак, у всех изученных с эмбриологической точки зрения ихневмо-
нид наблюдаются лишь начальные этапы приспособления к парази-
тизму, что хорошо согласуется с мнением о примитивности этого
семейства среди Parasitica (Гандлирш, 1908; Теленга, 1952, и др.)
В то же время в пределах этого семейства уже намечается диверген-
ция, характеризующая эволюцию эмбрионального развития паразити-
ческих перепончатокрылых.
Braconidae. Имеется очень мало работ, специально посвящен-
ных эмбриональному развитию Braconidae, и нет ни одной; дающей
подробное описание, однако отрывочные сведения нередко встречаются
в работах биологического характера.
216
Для зародышей всех браконид описано наличие одной оболочки
и для очень многих отмечен значительный эмбриональный рост
(см. табл. 1), что дает основание предполагать у них трофическую
роль серозы (рис. 111, V). У представителей родов Dinocampus,
Microctonus, Meteorus сероза сохраняет свое трофическое значение
и после вылупления личинки, превращаясь в «гигантские клетки»
(Оглоблин, 1913, 1924; Джексон, 1928, и др.).
Наконец,.у ряда браконид из родов Macrocentrus и Amicroplus
имеет место полиэмбриония (Паркер, 1931а; Даниель, 1932; Пайо,
1940) (рис. 111, VI).
Таким образом, в сем. Braconidae основное направление эволюции
развития ведет к возникновению и усилению трофической функции
серозы (линия дальнейшей дезэмбрионизации и упрощения здесь не
выражена).
Генетические отношения между ихневмонидами и браконидами
толкуются по-разному. Гандлирш (1908) выводит Braconidae из
Ichneumonidae, Брэдли (1908, цит. по Теленге, 1952) предполагает
обратное, а Н. А. Теленга (1952) считает, что оба семейства произошли
независимо. В эмбриологическом отношении ихневмониды кажутся
более примитивными, чем бракониды. Однако надо учесть, что все-
упомянутые выше представители сем. Braconidae — эндопаразиты, т. е.
уже довольно специализированы как паразиты. Но среди браконид
есть и эктопаразитические виды, у которых, надо полагать, развитие
едва ли уступает по примитивности таковому ихневмонид. По-види-
мому, дивергенция обоих семейств произошла очень рано—еще на
эктопаразитической ступени эволюции, когда эмбриональное развитие
претерпело только те изменения, которые характеризуют всех стебель-
чатобрюхих перепончатокрылых. Определить точнее характер отно-
шений между обоими семействами на основании эмбриологических
данных пока не представляется возможным.
Сем. А р h i d i i d a e близкородственно с Braconidae, и многие
авторы даже не считают афидиид самостоятельным семейством. Это
находит свое отражение и в развитии, которое очень напоминает раз-
витие некоторых монэмбрионических браконид с трофической оболоч-
кой вроде Dinocampus (рис. 111, V). По-видимому, афидииды отчлени-
лись от браконид, уже достигших какого-то среднего уровня эволюции
развития.
Надсемейство Cynipoidea
У Cynipoidea неизвестны ни примитивные формы развития ни
высокоспециализированные. Однако придавать этому обстоятельству
большое значение нельзя, так как эмбриологические данные относи-
тельно орехотворок вообще крайне скудны. Те формы, о которых все-
таки кое-что известно (напр., Charips sp. или Ibalia leucospoides Hoch.)
(Кристалл, 1930; Клоузен, 1940), достигли приблизительно того же
уровня эволюции, что и Aphidiidae (рис. 111, V).
Надсемейство Chalcidoidea
Из 24 семейств, объединяемых в это надсемейство (Никольская,
1952), довольно подробно и по многим представителям освещено разви-
тие Encyrtidae; еще о 6 семействах можно судить по развитию 1—3 видов:
остальные же 17 семейств в эмбриологическом отношении совершенно
не изучены.
217
Leucospidae и Chalcididae. Наиболее примитивными се-
мействами Chalcidoidea надо считать Leucospidae и Chalcididae (Ни-
кольская, 1958). О развитии Leucospidae никаких данных нет. Очень
краткие сведения о развитии Chalcis sispes L. (= Smicra clavipes F.,)--
внутреннего паразита личинок Stratiomyia — дает Эннеги (Henneguy,
1892, 1904). Автор описывает тотальное дробление и образование обо-
лочки («псевдоамниона») путем деламинации (возможно, ошибочно, так
как самые ранние стадии он не наблюдал) и увеличение объема заро
дыша более чем в 200 раз. К концу эмбрионального развития оболочка
диссоциируется на отдельные клетки, которые испытывают жировое пере-
рождение и, как полагает Эннеги, поедаются личинкой (сходство
с Dinocampus и афидиидами). В развитии Chalcis sispes отчетливо
выражены черты адаптации к эндопаразитическому образу жизни
(наличие трофической оболочки и эмбриональный рост; рис. 111, V).
Eurytomidae. Гораздо более примитивный характер сохраняет
развитие Eurytomidae, с которыми мы познакомились на примере
Eurytoma aciculata Ratz. — инквилина из галлов Pontania viminalis L.
В общем оно представляется довольно мало измененным и находится
приблизительно на той же ступени эволюции, что и развитие Angitia;
степень дезэмбрионизации в обоих случаях приблизительно одинаковая,
способ образования серозы сходен (рис. 111, II).
В отношении желтка Eurytoma даже более примитивна. Единствен-
ный признак, возникновение которого, очевидно, связано с паразитиз-
мом — разобщенность средней и задней кишки, — говорит в пользу
представления о вторичном характере растительноядности у Euryto-
midae. Сравнительная примитивность развития Eurytoma aciculata,
по всей вероятности, присуща и другим Eurytomidae: трудно ожидать
ярких приспособлений к паразитизму в семействе с такой неустойчивой
вариабильной биологией. В этом отношении Chalcididae представляют
группу, гораздо более однородную, что и находит свое выражение
в большей специализации развития.
В то же время в развитии Eurytoma есть одна своеобразная черта:
мезэнтодерма образуется не путем выселения клеток из первичной
бороздки, а путем погружения внутрь медианной эпителиальной пла-
стинки. Такое изменение способа закладки внутреннего зародышевого
листка несомненно имееет характер усовершенствования, «рационали-
зации» самого эмбрионального процесса (подобно тому, как инвагина-
ция высших кишечнополостных пришла на смену иммиграции у низших
представителей этого типа) й не имеет адаптивного значения. У всех
остальных Chalcidoidea сохраняется более примитивный способ обособ-
ления мезодермы. Поэтому тип развития Eurytoma нельзя считать
исходным для развития всех остальных хальцид.
Encyrtidae. В литературе имеются сведения о развитии
15 представителей сем. Encyrtidae. У Aphidencyrtus (Encyrtus) aphidi-
vorus (Мауг) (Сильвестри, 1909а) и Pseudaphycus sp. (Иофф, 1948)
имеет место поверхностное дробление (правда, Иофф называет дроб-
ление Pseudaphycus тотальным, но, судя по его же рисункам, он
ошибается) и эмбриональная оболочка образуется, по-видимому,
путем деламинации. У обеих этих форм из каждого яйца раз-
вивается одна личинка (рис. 111, V). Большинство же изученных
видов Encyrtidae, относящихся к родам Ageniaspis, Copidosoma
и Litomastix, с большей или меньшей достоверностью считается поли-
эмбрионическими (Маршаль, 1898; Мартин, 1914; Пэтерсон, 1921;
Сильвестри, 1937 и др.) (рис. 111, VI). Дробление очень мелких, не
содержащих желток яиц, становится тотальным, что облегчает разде-
21 н
лонис клеточного материала на многочисленные группы, дающие нача
ло множеству личинок. Трофическое значение оболочки очень сильно
возрастает, косвенным подтверждением чего служит ее очень раннее
обособление — еще до начала дробления, причем роль ядер оболочки
выполняют дериваты редукционных ядер.
Некое промежуточное место между монэмбрионическими и поли-
эмбрионическими энциртидами занимает Encyrtus mayri Masi (Силь-
вестри, 1915—1916), у которого дробление и формирование трофам-
ниона происходит, как у полиэмбрионических форм, а развитие остается
монэмбрионическим.
В семействах Entedontidae, Trichogrammatidac и Му-
m а г i d a e эволюция развития направлена в сторону прогрессирующей
дезэмбрионизации (рис. 111, IV), что уже было показано & специаль-
ной части и в предыдущей главе. Представляют ли эти три семейства
единую филогенетическую ветвь или возникли независимо, на основа-
нии чисто эмбриологических материалов, — сказать нельзя. Пожалуй,
Entedontidae и Trichogrammatidae сближаются тем, что в обоих слу-
чаях энтодерма образуется на биполярно, как у большинства насеко-
мых, а только на переднем конце (у Mestocharis — на том месте, где
впоследствие образуется стомодеум, у Prestwichia — вместе со стомо-
деумом).
Mymaridae же в этом отношении стоят особняком — материал
средней кишки обособляется у них путем деламинации. Впрочем, спо-
соб образования энтодермы у насекомых вообще так сильно варьирует,
что на этот признак опираться нельзя, гораздо более устойчивым
признаком представляется образование мезодермы, которое у всех
Phytophaga и Parasitica (кроме Eurytomidae) происходит однотипно.
А р h е 1 i n i d а е. Чтобы закончить обзор эмбрионального разви-
тия Chalcidoidea, надо еще сказать несколько слов об Aphelinidae.
Судя по данным Сильвестри (1909а, 1915—1916), 'относительно
Encarsia partenopea Masi, Encarsia tricolor Foerst. ( = Prospalta conju-
i^ata Masi) и Prospaltella berlesei (How.), очень мелкие яйца афелинид
лишены желтка, но дробление их остается неполным и приводит
к образованию типичной бластодермы и желточных ядер. Более поздние
стадии развития не описаны, так что остается неясным, в каком напра-
влении осуществляется у афелинид адаптация к паразитизму.
Все высказанное о развитии Chalcidoidea можно резюмировать
следующим образом. Хотя мы располагаем сведениями только для
одной трети семейств этой группы, но даже то, что известно, показы-
вает, что у хальцид представлены почти все основные типы развития
Hymenoptera. Хальциды могли бы быть выведены из ихневмонид
с наименее измененным типом развития, но не лишено вероятия также,
что Chalcidoida представляют совершенно самостоятельную, независи-
мую группу среди Parasitica.
- Надсемейство Serphoidea
Р 1 a t у ga s ter i d а е. Из надсемейства Serphoidea лучше всего
изучено развитие Platygasteridae (имеются эмбриологические дан-
ные для 14 представителей этого семейства). Как уже отмечалось,
в этом семействе наблюдаются как моно- так и полиэмбрионический
типы развития (рис. 111, V и VI), и переход от одного типа к другому
можно проследить шаг за шагом. Несмотря на то, что полиэмбриония
у Platygasteridae возникает совершенно независимо от такой Encyrti-
dae, начальные стадии развития в обоих случаях поражают своим
219
сходством — у Platygasteridae трофамнион тоже образуется до начала
дробления из значительной части цитоплазмы яйца, и ядра его тоже
происходят от редукционных ядер.
Sceilonidae. Другое направление эволюции онтогенеза в не менее
яркой форме представлено у Telenomus gracilis, Мауг (Scelionidae)
(Коломиец и Коваленок, 1958). Чрезвычайно мелкие яйца этого наезд-
ника 0,130—0,193 мм длины, включая стебелек, и 0,022—0,031 мм ширины)
совершенно лишены желтка, однако дробление остается поверхностным.1
После образования бластодермы в центре яйца начинает скапливать-
ся жидкость, так что возникают отношения, напоминающие таковые
у Angitia. Зародыш значительно увеличивается в объеме и принимает
форму полого шара. Эмбриональные оболочки не образуются.
Затем зародыш становится удлиненным и изогнутым на спинную
сторону. В передне-брюшной его части образуется впячивание стомо-
деума, а в передне-спинной происходит пролиферация клеток, дающих
начало средней кишке. На заднем конце закладывается проктодеум.
В самом конце эмбрионального развития путем миграции клеток
вдоль медиовентральной линии образуется мезодерма, а на переднем
конце путем впячивания стенки тела появляются зачатки слюнных
желез. Из яйца выходит крайне упрощенная, пузыревидная, еще не
сегментированная личинка, лишенная каких бы то ни было придатков,
даже ротовых. Нервная система представлена у нее лишь двумя
зачатками головного мозга, мускулатура — мышечными клетками,
обеспечивающими ритмические глотательные движения.
Стадия личинки продолжается у Telenomus трое суток, лишь
к концу первых суток появляются сегментация и трахейная система.
Мальпигиевы сосуды и брюшная цепочка развиваются на вторые
сутки. У трехсуточной личинки уже есть мускулатура тела и имаги-
нальные диски. Общий ход развития Telenomus сильно напоминает
таковой Prestwichia, отличаясь лишь в деталях (рис. 111, IV). Остается
неясным только, чем объясняется отмеченное авторами довольно значи-
тельное увеличение объема зародыша.
Относительно развития других Scelionidae известно мало. Имеют-
ся очень старые данные о развитии Teleas sp., откладывающего яйца
в яйца водомерок Gerris (Мечников, 1866; Ганин, 1869а). По мнению
Клоузена (1940), Ганин имел дело с представителем рода Tiphodytes.
И. Й. Мечников и М. С. Ганин предполагают тотальное дробление,
описывают стадию бластодермы, наличие одной эмбриональной оболоч-
ки, срединную бороздку и т. д. Эмбриональную оболочку можно видеть
и у зародыша Helorus paradoxus Prov. (Heloridae), развивающегося
в яйцах Chrysopa majescula Banks (см. рис. 122 у Клоузена, 1940).
В последнем случае размеры зародыша значительно увеличиваются.
У обеих названных форм развитие монэмбрионическое.
Еще более примитивные формы развития можно предполагать
у эктопаразитических Calliceratidae. Таким образам, и в пределах
надсемейства Serphoidea, по-видимому, наблюдаются различные степени
специализации развития.
3. ПОДОТРЯД ACULEATA
Наиболее высокоорганизованными перепончатокрылыми считаются
Aculeata. Однако эмбриональное развитие ос и пчел мало изменено по
1 В цитируемой работе даются иные размеры яиц. Из переписки с авторами выяс-
нилось, что приведенные там цифры ошибочны.
220
сравнению с таковым пилильщиков. Оно характеризуется отсутствием
амниона и недоразвитием личинки — отсутствием у нее глаз, грудных
и брюшных конечностей и сообщения между средней и задней кишкой.
Эти проявления дезэмбрионизации развития в данном случае, очевидно,
не связаны с недостатком питательных материалов в яйцах, а зависят
скорее от причин экологического или филогенетического характера
(развитие в тесном помещении, ограничивающем подвижность, обеспе-
ченность личинок питанием, происхождением от форм с уже дезэмбрио-
низированным развитием). Самое вероятное, что экологические и фило-
генетические причины действовали одновременно. Сероза чаще всего
образуется, как у Angitia и Euryloma, без образования амниотических
складок, путем нарастания внезародышевой бластодермы на зароды-
шевую, а внутренний зародышевый листок образуется путем погруже-
ния медианной пластинки, как у Eurytoma (рис. 111, II).
Развитие муравьев, с которым мы уже познакомились ранее,
близко к таковому ос и пчел, но обнаруживает признаки специализа-
ции: тенденцию к тотальному дроблению, раннюю дифференциацию
бластодермы и дальнейшую редукцию оболочек, которые у многих форм
представлены лишь группой крупных клеток, перегруженных желтком
и рано дегенерирующих (рис. 111, III). Причины всех этих изменений
в развитии муравьев пока остаются неясными.
Резко обособленное положение среди Aculeata занимает Aphelopus
theliae Gahan (Dryinidae) — внутренний паразит цикады Thelia
bimaculata F. (Корнхаузер, 1919), у которого яйца мелки, как у наезд-
ников (0,145 X 0>060 мм) у и лишены желтка. У Aphelopus развивается
«плацентарная оболочка», имеет место- эмбриональный рост и из
одного яйца развивается до 60 личинок (рис. 111, VI). Полное дробле-
ние, трофическая оболочками полиэмбриония возникают у Aphelopus,
независимо от наездников, на почве эндопаразитизма.
Большинство специалистов считает, что жалящие произошли от
каких-то очень примитивных наездников. Строя свои филогенетические
представления на изучении биологии и инстинктов перепончатокрылых,
С. И. Малышев (1949, 1950), полагает, что жалящие произошли от
наездников, которые находились в «эктопаразитической экзофитной
(панискоидной) фазе» эволюции. Гандлирш (1908) выводит Aculeata
из Ichneumonidae. H. А. Теленга (1952) принимает бифилетическое
происхождение Aculeata: большую часть жалящих он производит от
«ихневмоноидного комплекса», а меньшую (куда входит и Dryinidae) —
от «проктотрупоидного комплекса». Г. А. Викторов (1959) считает,
что между примитивными представителями обоих подотрядов нет
принципиальных различий и что «в основе эволюции всех высших
перепончатокрылых лежит один и тот же образ жизни — парализация -
добычи самкой при помощи яйцеклада и развитие на ней безногой
личинки, не способной к самостоятельному добыванию пищи».
Близость Aculeata с примитивными Parasitica подтверждается
и данными эмбриологии: развитие ос и пчел очень напоминает раз-
витие некоторых наездников, таких как Eurytoma aciculata (Иванова-
Казас, 19586) и Pimpla turionellae (Бронскилл, 1959).
4. ЗАКЛЮЧЕНИЕ
Итак, наиболее примитивные формы развития бесспорно наблю-
даются у Phytophaga. Сравнительно мало изменено и развитие
Aculeata. Наиболее значительные изменения развития встречаются
в подотряде Parasitica и связаны с внутренним паразитизмом. При этом
221
Ichneumonoidea, в сем. Entedontidae, Trichogrammatidae и Mymaridac
из сем. Chalcidoidea и у Telenomus gracilis Мауг из Serphoidca).
Переход от монэмбрионического развития к полиэмбрионическому
совершился параллельно и независимо четыре раза: у Braoonidaer
Encyrtidae, Platygasteridae и Drynidae.
Итак, для эволюции эмбрионального развития Hymenoptera
крайне характерно явление параллелизма. Дивергенция проявилась
в основном лишь в эволюции серозы (трофическая функция и прогрес-
сивное развитие в одном случае и полная редукция — в другом), но
и эта дивергенция дублируется тремя надсемействами. Соотношение-
явлений дивергенции и параллелизма в эволюции эмбрионального
развития перепончатокрылых показано на рис. 112.
Из всего сказанного видно, что сравнительная эмбриология очень
мало может дать для понимания филогенетических отношений между
крупными группами семейств Hymenoptera, так как значительные
изменения развития начали совершаться только после образования этих
групп. Однако при выяснении отношений,между отдельными семейства-
ми внутри надсемейств большая или меньшая специализация разви-
тия должны приниматься во внимание, как одна из существенных
черт, входящих в общую характеристику каждого семейства. С сожа-
лением надо еще раз отметить, что многие семейства с эмбриологиче-
ской точки зрения мало или вовсе не изучены.
ГЛАВА III
ПРИСПОСОБЛЕНИЯ К ПАРАЗИТИЗМУ В ЭМБРИОНАЛЬНОМ
РАЗВИТИИ ПЕРЕПОНЧАТОКРЫЛЫХ
Как мы видели, наибольшие изменения развития наблюдаются у эн-
допаразитических перепончатокрылых. И не удивительно. В. А. Догель
писал: «Нигде в животном царстве не сказываются с такой силой при-
способительные потенции организмов, как у паразитов» (1947, стр. 85).
Хозяин становится для паразита фактором внешней среды первостепен-
ной важности, он служит для него источником пищи и средой обитания,
определяет физико-химические, температурные и прочие условия его су-
ществования. У паразитов возникают разнообразные морфологические
и физиологические адаптации, обеспечивающие проникновение в хозяи-
на и осуществление основных функций организма в условиях парази-
тизма; вырабатываются сложные жизненные циклы со сменой ряда ли-
чиночных стадий и хозяев, нередко охватывающие несколько поколений
с различными формами размножения. Особенно сильные адаптивные
изменения наблюдаются именно на тех стадиях развития, на которых
осуществляется паразитизм. Один из наиболее ярких и известных при-
меров морфологических изменений под влиянием паразитического образа
жизни дают корнеголовые раки (Sacculina, Peltogaster и др.)» парази-
тирующие во взрослом состоянии в крабах и подвергающиеся такой
глубокой деградации, что утрачивают все характерные черты типа чле-
нистоногих. В то же время ранние стадии развития саккулины вплоть
до циприсовидной личинки ни биологически, ни морфологически не от-
личаются от таковых свободноживущих усоногих раков. Лишь после
прикрепления личинки к кожным покровам хозяина начинается своеоб-
разный некробиотичеокий метаморфоз, сопровождающийся отбрасыва-
нием большей части тела (Делаж — Delage, 1884). Сходные и не менее
значительные изменения испытывает и брюхоногий моллюск Parentero-
xenos dogieli A. Ivanov, паразитирующий в голотуриях (А. Иванов, 1949).
Паразитизм только на личиночных стадиях тоже вызывает некото-
рое упрощение или появление различных провизорных структур, но та-
кая глубокая перестройка всей организации, как при имагинальном
паразитизме, обычно не наступает. Глубокая деградация личинки сде-
лала бы невозможным возврат к свободному образу жизни и более
сложной организации взрослого животного.
Что же касается эмбрионального паразитизма, то это явление встре-
чается крайне редко. Яйца довольно часто используются как средство
заражения хозяина, но обычно остаются совершенно пассивными — чем-
то вроде инородного тела, вплоть до вылупления личинки. Такая ^пассив-
ность зародышей обусловлена, по-видимому, тем, что обычно яйца бы-
вают снабжены необходимыми для развития питательными материалами
224
и в большей или меньшей степени изолированы от окружающей среды
яйцевыми оболочками. Лишь у самых примитивных многоклеточных
(губки, гидроиды) яйца еще не имеют оболочек. Дальнейшая эволюция
яиц направлена в сторону эмансипации зародыша от окружающей сре-
ды, которая достигает наибольшей степени у наземных членистоногих и
Sauropsida (Нидхэм — Needham, 1931; Токин, 1958). Лишь с переходом
к плацентарному живорождению, представленному в наиболее совер-
шенной форме у млекопитающих, между зародышем и внешней средой
(т. е. матерью) устанавливаются новые отношения; запасы питательных
веществ в яйце становятся ненужными, а яйцевые оболочки даже вред-
ными, так как они препятствуют обмену между матерью и плодом. По-
этому желток и яйцевые оболочки подвергаются редукции. Такие же
резкие изменения в отношениях с внешней средой должно претерпеть
яйцо и при переходе к паразитизму.
По-видимому, такие перемены происходят легче, когда роль внеш-
ней среды выполняет материнский организм, а не организм другого вида.
Во всяком случае живорождение с питанием зародыша за счет матери
распространено очень широко. Помимо млекопитающих, оно встречается
у гидроидов, актиний, скорпионов, первичнотрахейных, насекомых, асци-
дий, сальп, костистых рыб и рептилий. v
Иначе обстоит дело с эмбриональным паразитизмом. Пожалуй,
единственным примером такого рода являются наездники. Поэтому рас-
смотрение эмбрионального развития наездников, с точки зрения адапта-
ции к паразитизму, представляет особый интерес.
Первые эмбриологические исследования, посвященные паразитиче-
ским перепончатокрылым, принадлежат И. И. Мечникову (1866),
Н. П. Вагнеру (1868) и М. С. Ганину (1869а). В работе Ганина, описав-
шего развитие Polynema natans из стрекозы Agrion virgo L. (=Anagrus
incarnatus Hal), Platygaster sp. из личинок Cecidomyidae, Ophioneurus
sp.,n3 яиц Pieris brassicae L. и Teleas sp., мы уже находим ряд интерес-
ных обобщений. Так, для всех изученных им форм М. С. Ганин отмечает
малые размеры яиц и отсутствие желтка, что связывает с паразитиче-
ским образом жизни. Отсутствие желтка приводит к тотальному дробле-
нию. Сроки эмбрионального развития сокращаются, вследствие чего за-
родыш покидает эмбриональные оболочки, будучи еще слаборазвитым.
Многие органы, которые у других членистоногих развиваются в эмбрио-
нальный период, здесь закладываются позднее. Ганин описывает у Pla-
tygaster и Teleas одну эмбриональную оболочку, которую не решается
гомологизировать ни с амнионом, ни с серозой. Так как она имеется не
у всех изученных Ганиным видов, он полагает, что значение оболочки
невелико. Таковы основные выводы, сделанные Ганиным.
После опубликования наблюдений Ганина прошло около 100 лет,
но вопрос о влиянии паразитизма на развитие наездников никем больше
не обсуждался. Однако накопленный за это время фактический мате-
риал нуждается в обобщении.
Мы уже видели, что по характеру отношений между зародышем
паразита и хозяином, наездников можно разделить на две группы.
В первом случае зародыш не заимствует из тела хозяина ничего, кроме
воды и кислорода, и остается пассивным до вылупления из яйца личинки.
Но эмбриональный период сокращается, из яйца выходит сильно недо-
развитая морфологически, но способная к активному питанию личинка
(дезэмбрионизация); течение отдельных процессов развития упрощает-
ся, исчезает сероза. Это делает ненужным большое количество запасных
питательных веществ в яйце. Количество желтка и размеры яиц редуци-
руются.
15 О. М. Иванова-Казас
225
У наездников второй группы, -наоборот, между зародышем и хозяи-
ном рано устанавливается интенсивный обмен веществ. Соответственно
эмбриональное развитие испытывает более значительные изменения. Так
же, как в первом случае, размеры яиц уменьшаются, а желток исчезает,
поверхностное дробление .заменяется тотальным. Но, кроме того, хорион
становится проницаемым или исчезает, эмбриональная оболочка приоб-
ретает трофическую функцию. За время эмбрионального развития заро-
дыш значительно вырастает, питаясь за счет хозяина. Наконец, в ряде
случаев возникает полиэмбриония.
Некоторые из этих изменений в развитии, заслуживают специального
рассмотрения.
!. УМЕНЬШЕНИЕ РАЗМЕРОВ ЯИЦ
В числе следствий паразитического образа жизни выше были указа-
ны уменьшение размеров яиц и содержание в них желтка. Основанием
для такого утверждения послужили следующие соображения:
1. Размеры яиц в основном определяются содержанием в них желт-
ка. Накопление в яйцах большого количества желтка биологически
очень выгодно и, по мнению И. И. Шмальгаузена (1938, 1946), является
основной тенденцией эволюции онтогенеза Metazoa, так как дает воз-
можность зародышу дольше оставаться под защитой яйцевых оболочек,
причем из яйца выходит уже довольно сложный организм, способный к
активной борьбе за существование. Но в условиях паразитизма большое
количество желтка становится излишним — зародыш оказывается в
среде, богатой питательными веществами и благоприятной в других отно-
шениях. В такой среде возможно существование крайне упрощенной
личинки, развитие которой не требует большого количества питательных
материалов. Желток становится еще менее нужным в тех случаях, когда
зародыш паразита приобретает способность питаться с помощью трофи-
ческой оболочки. Таким образом, полное исчезновение желтка в яйцах
многих наездников — прямое следствие паразитизма.
2. Одной из причин уменьшения размеров яиц у наездников несо-
мненно является уменьшение размеров тела взрослых насекомых. Это,
в свою очередь, непосредственно связано с их образом жизни: насеко-
мые, развивающиеся внутри других насекомых и даже внутри их яиц
(иногда сразу по нескольку штук),'не могут быть очень крупными.
3. Наконец, существует мнение, что мелкие яйца дают наездникам
известные преимущества: они легче проскальзывают по яйцекладу, что
облегчает их откладку в подвижную, иногда сопротивляющуюся жертву
(Клоузен, 1940). ' .
Итак, все возможные причины уменьшения размеров яиц у наездни-
ков прямо или косвенно связаны с паразитизмом.
2. ПЕРЕХОД ОТ ПОВЕРХНОСТНОГО ДРОБЛЕНИЯ К ТОТАЛЬНОМУ
Некоторые признаки, характеризующие различные типы дробления
яиц животных (полнота дробления, его равномерность), обычно ставят в
зависимость от количества и распределения желтка, который, как пола-
гают, механически препятствует делению цитоплазмы. Если в яйце желт-
ка мало, то дробление бывает полным; равномерность или неравномер-
ность дробления зависят от равномерности или неравномерности в рас-
пределении желтка. Если же в яйце оказываются значительные
количества желтка, дробление становится неполным: поверхностным —
при равномерном распределении желтка или дискоидальным — при кон-
226
Название вида
Chalicodoma muraria
Camponotus herculeanus
Pontania capreae
Mestocharis militaris
Eurytoma aciculata
Angitia vestigialis
Caraphractus reductus
Prestwichia aquatica
Aphelopus theliae
Macrocentrus gifuensis
Aphidius fabarum
Copidosoma gelechiae
Автор
Carriere u. Burger
(1898)
Tanquary (1913)
Иванова-Казас (1958а)
Иванова-Казас(19526)
Иванова-Казас (19586)
Иванова-Казас (19606)
Иванова-Казас (19546)
Иванова-Казас (1950)
Kornhauser (1919)
Parker, (1931a)
Иванова-Казас (1954в)
Leiby (1922)
Длина
яйца
(в мм)
3,5-4
1,4
0,49
Ширина
яйца
(в мм)
0,75-1
0,5
0.16
0,415 !-0,095
0,33—0,350,13—0,14
0,3 L 0,05
0,213 0,093
0,154 1 0,065
0,145 ! 0,060
0,08—0,12 0,028—
0,086
0,04-0,05
о!оз'б
0,03 .
0,035
Коли-
чество
желтка
Много
»
п
Мало
„
Желтка
нет
.
»
»
Тип
дробле-
ния
Поверх-
ностный
*
я
ж
„
-
V
„
Тоталь-
ный
Переход-
ный
Тоталь-
ный
вершается со значительным опозданием. Очевидно поверхностный тип
дробления насекомых, 'возникший в связи с накоплением в яйцах желт-
ка, закреплен наследственно, и этот «наследственный механизм дробле-
ния», природа которого нам пока неизвестна, продолжает действовать
некоторое время и после исчезновения желтка, бывшего его первопри-
чиной.
Любопытно, что почти все виды с тотальным дроблением являются
в то же время и полиэмбрионическими. Это дало основание Сильвестри
(1937) предполагать, что тотальное дробление" перепончатокрылых воз-
никло как приспособление к полиэмбрионии. Таким образом, полиэм-
бриония явилась как бы дополнительным биологическим стимулом, спо-
собствующим появлению тотального дробления.
Однако тотальное дробление имеет место у мо'нэмбрионического
Encyrtus mayri Masi, приближается к тотальному и дробление предста-
вителей рода Aphidius. Поэтому можно предполагать и обратное: что
тотальное дробление облегчило возникновение полиэмбрионии (см.
ниже).
Интересно отметить, что такой же, не сразу преодолеваемый консер-
ватизм, обнаруживает дробление и в тех случаях, 'когда эволюция яйца
происходит в противоположном направлении, т. е. в сторожу накопления
желтка, например у низших позвоночных. При этом перемены в харак-
тере дробления тоже отстают от изменений в организации яйца. Яйцо
травяной лягушки Rana temporaria, имеющее в диаметре 2—3 мм, по
объему в 24 000 раз превышает яйцо Prestwichia и все же сохраняет хотя
и неравномерное, но типичное тотальное дробление. Переходный тип
дробления, приближающийся к дискоидальному, 'наблюдается у Sala-
mandra maculosa Laur., яйца которой имеют диаметр 3,8—5 мм (Грен-
рооз—Gronroos, 1895). Борозды дробления, появляющиеся на анималь-
ном полюсе, достигают вегетативного полюса с таким опозданием, что
здесь оказывается неразделенная масса желтка с лежащими в ней ядра-
ми. Однако это состояние — временное; позднее яйцо разделяется пол-
ностью; у Gymnophiona яйца достигают еще более, крупных размеров
(у Ichthyophis glutinosa L. — 6X9 мм), и'дробление становится диско и-
дальным. Самые крупные яйца и прочно установившееся дискоидальное
дробление характеризуют рептилий и птиц.
Таким образом, унаследованный от предков характер дробления и
в этом случае преодолевается не сразу. Отсюда следует, что желток
хотя и оказывает большое влияние на процесс дробления, как это обычно
и принимается, но это влияние нельзя понимать как чисто механическое
воздействие.
3. ЭВОЛЮЦИЯ ЖЕЛТОЧНОЙ СИСТЕМЫ
Переход от поверхностного дробления к тотальному есть лишь одно
из следствий, вытекающих из исчезновения желтка. Влияние, которое
оказывает на последующее развитие это изменение исходной структуры
яйца, гораздо значительнее. Морфологические особенности развития на-
секомых, связанных с большим количеством желтка, сводятся к следую-
щему:
1. вырабатывается поверхностное дробление;
2. зародыш распластывается на поверхности желтка в форме диска
или зародышевой полоски;
3. меняется характер гаструляции и органогенеза: инвагинация за-
меняется иммиграцией;
- 4. возникают эмбриональные оболочки, косвенно тоже связанные с
большим количеством желтка: существует предположение, что первич-
228
ной причиной возникновения оболочек было погружение зародыша в
толщу желтка (бластокинез), биологический смысл которого состоит в
достижении контакта с более глубокими его слоями (Иванов, 1937; Ша-
ров, 1953). Согласно этой точке зрения защитная функция эмбриональ-
ных оболочек у насекомых возникла вторично;
5. появляются специальные клеточные элементы, служащие для пе-
реработки желтка (вителлофаги) и образующие вместе с ним так назы-
ваемую желточную систему;
6. большое количество желтка делает возможным развитие в яйце
высокоорганизованной личинки.
Почти все эти особенности достаточно широко известны. Более под-
робного рассмотрения заслуживает только желточная система, функция
которой отнюдь не исчерпывается резорбцией желтка.'
Как известно, в результате поверхностного дробления богатых
желтком яиц насекомых образуется бластодерма и центральная масса
желтка, в котором остаются отдельные разрозненные ядра, одетые каж-
дый своим участком цитоплазмы,— желточные клетки, или вителлофаги.
В зависимости от времени их возникновения (во время формирования
бластодермы, после образования бластодермы путем выселения из нее
или еще позднее) различают первичные, вторичные и третичные желточ-
ные клетки. Образовавшись тем или иным путем, они не имеют резких
границ, а образуют синцитий, в петлях которого лежат гранулы желтка.
Иногда желточный синцитий распадается на совершенно обособленные
перегруженные желтком клетки (так называемое «вторичное дробле-
ние» или «дробление желтка»). Существование обособленных желточных
клеток чаще всего бывает непродолжительно, вскоре они снова сли-
ваются в синцитий. У большинства насекомых желточный синцитий
к концу эмбрионального развития дегенерирует, но у некоторых сравни-
тельно примитивных форм — Lepisma saccharinum L. (Геймопс — Неу-
mons, 1896, 1897; Шаров, 1953), стрекоз (Чупрова, 1903; Иогансен и Бат,
1941), Melanoplus differentialis (Thomas), (Стюарт—Stuart, 1935)
и др. — желточные клетки, или часть их, могут сохранять свою индиви-
дуальность и участвовать в 'построении провизорной средней кишки. Эти
наблюдения убедительным образом показывают, что желточные клетки
насекомых не стоят вне зародышевых листков, а представляют собой
рано обособившуюся часть энтодермы, специализировавшуюся для пере-
работки желтка.
Однако, несмотря на то, что желточные клетки у большинства насе-
комых не принимают непосредственного участия* в построении зароды-
ша, имеется ряд экспериментальных данных, показывающих, что формо-
образовательное значение желточной системы очень велико.
Первые наблюдения такого рода принадлежат Зейделю (Seidel,
1931, 1934, 1936), экспериментировавшему с яйцами стрекозы Platycne-
mis. З'ейдель показал, что действие так называемого «центра дифферен-
циации» имеет динамический характер и проявляется \в волнах сокраще-
ния желточной системы, распространяющихся от «центра»— вперед и
назад. В пространство, возникшее между сократившимся участком жел-
точной системы и хорионом, устремляются клетки бластодермы и возни-
кает то утолщение бластодермы, которое потом становится зародышевой
полоской. Искусственно вызывая местные сокращения желточной систе-
мы путем точечного ожога, наложением лигатуры и последующим ее
расслаблением и т. д., можно изменить время закладки, место, размеры
и форму зародышевой полоски.
В более поздней работе, совместной с Боком и Краузе (Seidel, Bock
и Krause, 1940), Зейдель еще раз повторяет те же положения и сравни-
229
вает желточную систему с формой для отливки (Pragungsstoek) зароды-
шевой «полоски. Взгляды Зейделя нашли свое подтверждение и в ряде
позднейших работ. Так, Зауэр (Sauer, 1954), экспериментально изучая
образование мезодермы и дефинитивной энтодермы у медоносной пчелы,
пришла к выводу, что эти процессы тоже зависят от активности желточ-
ной системы, которая, сокращаясь, представляет место для погружаю-
щихся клеток.
Краузе и Краузе (Kjrause и Kjause, 1957) показали, что желточная
система играет важную роль в процессе бластокинеза у сверчка Tachy-
cines. Сперва в желтке происходят некоторые структурные, изменения
(намечается «путь бластокинеза»), после чего; благодаря движениям
желточной системы зародыш ,как бы втягивается внутрь. Нарушение
желточной системы *без повреждения самого зародыша часто делает
дальнейшее нормальное развитие .невозможным.
Особенно всесторонне рассматривает функции желточной системы
Смречинский (Smreczynski, 1939), изучавший развитие жука Agelastica.
Он тоже отмечает способность желточной системы реагировать на укол
термокаутером содроганием, волнообразно распространяющимся от ме-
ста укола. Удаление части желточной системы не мешает нормальному
функционированию оставшейся части и приводит к развитию гармони-
ческих зародышей уменьшенных размеров. Но умерщвление части жел-
точной системы без ее удаления стесняет подвижность оставшейся части,
и нормальное развитие становится невозможным. Смречинский прихо-
дит к выводу, что, помимо питательной функции, желточная система
играет в развитии зародыша жука активную и пассивную формообразо-
вательную роль. Активная роль проявляется при формировании зароды-
шевой полоски и ее погружении в желток и в окончательном сокращении
желтка после его разделения на обособленные клетки. Последнее делает
возможным обрастание желтка эпителием средней кишки. После этого
желточная система выполняет уже пассивную роль каркаса, на который
«натягиваются» стенки тела.
Приведенное выше объяснение механизма формирования зароды-
шевой полоски и мезэнтодермы кажется несколько упрощенно-механи-
стическим. Возможно, что активная формообразовательная роль жел-
точного синцития несколько преувеличена. Но приведенные выше дан-
ные не оставляют сомнения в том, что желточный синцитий действительно
представляет собой живую систему, реагирующую на раздражение,
участвующую в процессе взаимодействия частей зародыша и в формо-
образовании.
Можно думать, что все сказанное о желточной системе насекомых
вообще относится и к перепончатокрылым (во всяком случае к тем и?,
них, яйца которых содержат много желтка).
В яйцах пилильщиков желтка так много, что активные элементы
желточной системы (желточные клетки) различаются с большим трудом.
В период удлинения зародышевой полоски происходит «дробление желт-
ка» (черта, по-видимому, примитивная), но затем желточные клетки
снова сливаются в синцитий. Объем желточной массы относительно объ-
ема всего зародыша становится к этому времени уже значительно мень-
шим. Это, несомненно, связано с потреблением желтка, так как желточ-
ные гранулы располагаются теперь уже <не так плотно, между ними
появляются крупные вакуоли, клеточные элементы в желтке различают-
ся гораздо лучше (gm. рис. 11). В какой мере уменьшение объема жел-
точ<ной системы обусловлено ее активным, сокращением — сказать
трудно, так как развитие пилильщиков изучалось только описательно-
морфологическим методом. Обрастание желточной массы энтодермаль-
230
ными зачатками и боковыми стенками тела происходит у пилильщиков
обычным образом. После образования средней кишки последние остатки
желтка исчезают, а желточный синцитий дегенерирует.
Таким образом, трофическая и пассивная формообразовательная
функция желточной системы выражены у пилильщиков с полной ясно-
стью. Можно предполагать и наличие активной формообразовательной
функции.
У жалящих перепончатокрылых (судя по литературным данным),
развитие которых подверглось сравнительно незначительным эволю-
ционным изменениям, желтка в яйце много и желточные клетки хорошо
выражены. Многие авторы (Карьер и Бюргер, 1898; Стриндберг, 1914;
Дондуа, 1953) отмечают, что желточные клетки нередко скопляются в
периферических частях желточной массы, образуя нечто вроде прови-
зорного эпителия средней кишки, исчезающего к концу эмбрионального
развития. Обрастание желтка дефинитивной энтодермой и замыкание
спины происходят по общей схеме. Как уже упоминалось, у пчелы
имеется хорошо выраженный центр дифференциации (Шнеттер, 1934)
и отмечена динамическая роль желточной системы (Шнеттер, 1936, и
Зауэр, 1954).
У Hymenoptera parasitica желточная система претерпевает далеко
идущую редукцию, различные степени которой можно наблюдать у раз-
ных представителей подотряда. '
Так, у Eurytoma aciculata, ведущей инквилинный образ жизни-в гал-
лах ивового пилильщика, желтка уже не так много, почему составные
части желточной системы различаются с большой ясностью. Желточные
клетки богаты плазмой и соединяются своими отростками в синцитий,
резко отграниченный от остальных тканей зародыша тонкой плазмати-
ческой перепонкой (уплотненный периферический слой синцития). Часть
желточных клеток непосредственно примыкает к этой перепонке
(рис. 113, А). Формирование средней кишки происходит у Eurytoma не-
сколько иначе, чем у большинства насекомых, хотя эти отличия и не
имеют принципиального значения. Из общей мезэнтодермальной кле-
точной массы 'выселяются отдельные клетки, которые оседают на поверх-
ности желточного, синцития, распластываются на нем и постепенно скла-
дываются в кишечный эпителий (см. рис. 53). Формообразовательное
значение желточной системы выступает здесь особенно отчетливо, так
как на ней объединяются разрозненные до того эндотермальные клетки.
Замыкание спины происходит обычным путем.
В яйцах Angitia vestigialis — внутреннего паразита личинок Ponta-
nia — желтка уже нет вовсе. Однако дробление остается поверхностным.
Уже на начальных стадиях развития в центральной части яйца появ-
ляются и накапливаются многочисленные вакуоли, наполненные водяни-
стой жидкостью. На стадии бластодермы все эти вакуоли сливаются в
единую центральную полость (рис. 113, Б), занимающую место желтка.
В этой полости рассеяны небольшие комочки цитоплазмы, содержащие
по нескольку ядер, связанные друг с другом и бластодермой тонкими
плазматическими мостиками. В этих структурах без труда можно узнать
гнезда желточных клеток, которые образовались, несмотря на отсутст-
вие желтка. Формирование средней кишки и обрастание желточного
синцития (в котором желток отсутствует) боковыми краями зародыше-
вой полоски происходит у Angitia совершенно так же, как и у Eurytoma
(см. рис. 25). Таким образом, желточная система Angitia, полностью
утратившая свою трофическую функцию из-за редукции желтка, все еще
сохраняет первоначальное формообразовательное значение.*
Следующую стадию редукции желточной системы можно наблюдать
' 231
ной. Пожалуй, немногими признаками, свидетельствующими о проис-
хождении этих форм от насекомых с большим количеством желтка, яв-
ляются пролиферационный способ образования мезодермы (способ обо-
собления энтодермы так варьирует, что на него опираться нельзя) и на-
личие серозы, которая не только сохраняется, но испытывает прогрес-
сивное развитие в связи с ее трофической функцией. Последовательные
стадии редукции желточной системы схематически изображены на
рис. ИЗ.
Изложенные выше наблюдения касаются изменений морфологиче-
ской картины развития. Но надо полагать, что не менее значительными
должны быть и физиологические изменения и в частности изменения
формообразовательного аппарата. Если основу действия центра диффе-
ренциации составляют сокращения желточной системы, то исчезновение
последней означает коренное изменение, если не разрушение, того «меха-
низма», с помощью которого осуществляется развитие большинства на-
секомых. За это же говорит и наличие полиэмбрионии, достигающей у
некоторых паразитических перепончатокрылых необычайной продук-
тивности.
4. ПОЛИЭМБРИОНИЯ
Выше уже было отмечено, что полиэмбриония у наездников пред-
ставляет собой наивысшую форму адаптации развития к паразитизму,
достигнутую у перепончатокрылых. Однако, вопрос о непосредственных
причинах и условиях, вызвавших или хотя бы способствовавших такому
коренному изменению всего хода развития, остался вне рассмотрения.
Теперь, после того как было показано, насколько глубоки изменения,
происшедшие в структуре и физиологии развития яиц наездников, умест-
но обратиться и к этому вопросу.
Первые соображения по этому поводу высказаны Маршалем
;(1904а и б), которому принадлежит и честь открытия самого факта по-
лиэмбрионии у насекомых. Маршаль различает три формы полиэмбрио:
нии: 1) экспериментальную, 2) случайную или тератологическую (близ-
нецы и двойниковые уродства, встречающиеся у всех видов животных)
и 3) специфическую, являющуюся нормальным способом развития неко-
торых видов. Необходимыми условиями полиэмбрионии Маршаль счи-
тает избыток питательных веществ и возможность их использования.
Однако первое условие имеется у всех паразитических Hymenoptera вто-
рое — у многих, а полиэмбриония наблюдается лишь в исключительных
случаях. Поэтому непосредственную причину «полиэмбрионизации» раз-
вития Маршаль пытается искать в каких-то внешних воздействиях, ко-
торым подвергается яйцо изученных им полиэмбрионических видов,
подобных воздействиям, применяемым экспериментаторами для искус-
ственного разделения бластомеров.
Так, Platygaster zosinae Walk. (= Polygnotus minutus Lind.) откла-
дывает яйца в кишечник поздних зародышей Cecidomyia. Когда личин-
ка Cecidomyia выходит из яйца и начинает активно питаться, в ее ки-
шечнике резко изменяются осмотические условия, что и вызывает, по
мнению Маршаля, разъединение бластомеров. Этому содействует также
механическое встряхивание зародышей Platygaster, обусловленное пе-
ристальтикой кишечника хозяина. Сходные (в основном осмотические)
причины диссоциации зародыша принимает Маршаль и для Ageniaspis
(Encyrtus) fuscicollis (Dalm.), развитие которого протекает в полости
тела гусениц Hyponomeuta; во время зимовки происходит сильная деги-
дратация тканей гусеницы, а весной, с началом ее активного питания,
осмотиуеские условия резко меняются.
233
Другой исследователь, лично изучивший развитие ряда полиэмбрио-
нических видав, — Сильвестри (1937)—непосредственную причину раз-
деления эмбриональной массы усматривает в исключительно благопри-
ятных условиях питания зародыша в его «переедании» (hypernutrition).
После исчезновения хориона и начала функционирования трофамниона
у Litomastix начинается усиленное питание и размножение эмбриональ-
ных клеток, которые начинают вести себя, как в культуре ткани. Они
«пытаются расползаться», оказывают изнутри давление на эмбриональ-
ную оболочку, которая, в противовес им, образует перетяжки и1 разделяет
всю массу эмбриональных клеток на отдельные группы. Это любопытное
объяснение оставляет, однако, неосвещенным вопрос — почему такой, ка-
залось бы, благоприятный фактор, как избыток питания, действует так
дезорганизующе на зародыш, что он уподобляется культуре ткани.
Стремление найти причину полиэмбрионии в каких-то чисто внеш-
них обстоятельствах (обычно неблагоприятных) характерно дл-я боль-
шинства авторов, подходящих к этому вопросу с позиций эксперимен-
тальной эмбриологии. Так, Гексли и де Бер (1936) полагают, что причи-
ны появления близнецов и полиэмбрионии надо искать в разъединении
бластомеров и нарушении полярности у одних животных, в задержке
плацентации у броненосцев, охлаждении яиц перед гаструляцией у птиц,
гибели части зародышей и их гниении у Oligochaeta и т. д.
Причину возникновения однояйцевых близнецов у человека Канаев
(1959) видит в нарушении нормальных физиологических корреляций, в
возникновении физиологической изоляции между частями зародыша. По
мнению Токина (1959), в основе полиэмбрионии наездников лежит рез-
кое падение уровня интеграции зародыша, обусловленное неблагоприят-
ными хим-ическими влияниями со стороны хозяина.
Однако, как справедливо отмечено Пэтерсоном (1927), такие чисто
внешние факторы могут служить лишь причиной спорадической поли-
эмбрионии, наблюдающейся как исключение, но не как правило. Поэто-
му гораздо правильнее пытаться объяснить полиэмбрионическое разви-
тие, исходя из внутренней структуры яйца. Попытка такого рода сделана
Даутом (1959), который связывает полиэмбрионию перепончатокрылых
с особенностями ядерно-плазменного отношения в яйце. Он указывает
на то, что у полиэмбрионических форм значительная часть ооплазмы
истрачивается на образование трофамниона; тем самым нарушается рав-
новесие между хроматином и цитоплазмой, что и является фактором, ин-
дуцирующим полиэмбрионию. В тех случаях, когда имеет место оплодо-
творение, эта диспропорция еще больше увеличивается, вследствие при-
соединения ядра спермия. Соответственно, оплодотворенные ядра «более
полиэмбрионичны». Лейби и Хилл (1923) указывают, что неоплодотво-
ренные яйца Platygaster hiemalis Forbes развиваются монэмбриони-
чееки, а оплодотворенные образуют двойников. Флиндерс (Flanders,
1942, 1946) отмечает, что женское потомство Copidosoma koehleri Blanch,
(развивающееся из оплодотворенных яиц) обычно бывает более много-
численным, чем мужское (развивающееся из неоплодотворенных яиц).
По данным самого Даута (1947), среднее число индивидов в чисто муж-
ском потомстве Copidosoma koehleri Blanch, составляет 21, а в чисто-
женском— 31,4 и т. д.
Приведенные факты, однако, еще недостаточны, чтобы принять ги-
потезу Даута. Кроме того, остается неясным, в чем сущность этой пред-
полагаемой связи между количественным преобладанием ядерного ве-
щества и разделением зародыша на отдельные клеточные комплексы,
способные к самостоятельному развитию.
Хотя соображения Даута выглядят и не особенно убедительными, но
234
он прав в том' отношении, что причина полиэмбрионии или хотя бы ка-
кие-то способствующие ей условия должны заключаться в самой орга-
низации яйца. Под организацией яйца следует понимать не только его
видимые структурные особенности, но и те формообразовательные аппа-
раты, с помощью которых осуществляется развитие, судить о которых
мы можем лишь на основании экспериментальных данных и косвенных
соображений. Полиэмбриония возможна только в том случае, если орга-
низация зародыша на какой-то стадии развития способствует, или, по
крайней мере, не препятствует разделению его на части.
Возрос о том-, в какой мере полиэмбрионическое развитие насекомых
согласуется с имеющимися представлениями об организации их яйца,
впервые был поставлен Гетенби в 1918 г. В то время уже было известно,
что в яйце насекомого в момент его откладки уже предопределены его
передний конец и задний, спинная сторона и брюшная. Опыты Хегнера
(1911) с умерщвлением частей зародыша жука Leptinotarsa обнаружи-
ли высокую степень детерминации частей, при которой оставшиеся части
зародыша образуют только зачатки, возникающие из них и при нормаль-
ном развитии, а утраченные части не восстанавливаются. Исходя из этих
сведений, Гетенби указывает, что большая часть протоплазмы яйца по-
лиэмбрионических наездников, из которой должны были бы развиться
(голова, мозг и т. д., идет на образование трофамниона. «Казалось бы
нет никаких оснований думать, — пишет Гетенби, — что полиэмбриони-
ческое яйцо иначе организовано, чем монэмбрионическое, а оказывается
как раз обратное. Мы не различаем никакого специального плана в дроб-
лении, даже половой детерминант после того, как он входит в ту часть
яйца, крторая у моноэмбрионических наездников дает половые клетки,
исчезает, и, принимая во внимание случайный способ формирования
внутренних эмбриональных масс (т. е. зачатков отдельных зародышей),
кажется очень маловероятным, чтобы какое-'нибудь специальное приспо-
собление обеспечивало бы попадание полового детерминанта в гонады
будущих эмбрионов» (1919, стр. 190). Всё это заставило Гетенби принять,
что у полиэмбрионических перепончатокрылых нет такой ранней детер-
минации, частей, и судьба каждой клетки зависит от ее положения в теле
зародыша.
Вся сложность организации яйца насекомых, еще неизвестная Ге-
тенби, стала выясняться лишь в тридцатых годах. В результате ряда ис-
следований Зейдель (1931, 1934, 1936) пришел к выводу, что эмбрио-
нальное развитие стрекозы Platycnemis осуществляется с помощью не-
скольких «центров», которые вступают в действие последовательно один
за другим: центра дробления, формативного центра (Bildungs-
Zenirum) и центра дифференциации. Центр дробления представляет со-
бой то место, где первоначально находится ядро яйца и начинаются его
деления. По мере увеличения числа ядер они расходятся к периферии.
Некоторые из них достигают заднего конца яйца, где находится форма-
тивный центр и активируют его. От последнего начинают исходить ве-
щества, распространяющиеся вперед и* необходимые для формирования
бластодермы. Эти вещества возбуждают деятельность центра дифферен-
циации, под контролем которого находятся процессы образования за-
родышевой полоски, сегментации, обособления мезодермы и т. д. Как
уже указывалось выше, действие центра дифференциации имеет дина-
мический характер и осуществляется путем сокращений желточной си-
стемы. Так как цепь процессов детерминации начинается у Platycnemis
только в конце дробления, то на более ранних стадиях яйцо обладает
довольно значительными регулятивными возможностями, которые про-
являются в образовании карликовых зародышей и удвоении частей в ре-
235
зультате некоторых опытов с перетяжками и прижиганием яйца. В одном
случае удвоение зашло так далеко, что было получено два почти полных
зародыша, из которых один был заключен внутри другого. Образование
сросшихся «янусовидных» двойников наблюдал и Краузе (1953) в его
опытах со сверчком Tachycines.
В настоящее время имеются данные по физиологии развития целого
ряда насекомых, относящихся к разным отрядам (Ричарде и Миллер,
1937; Пфлюгфельдер, 1952). Оказалось, что изложенная схема развития
приложима далеко не ко всем насекомым. Остается неясным, имеет ли
«центр дробления» какое-нибудь специальное физиологическое значение,
кроме того, что здесь лежит ядро яйца до начала дробления. Форматив-
ный центр найден лишь у немногих насекомых: у цикад Зандером (San-
der, I960) и у муравьев Рейтом (1931, 1932).
Наиболее постоянным и важным звеном в этой цепи формообразо-
вательных реакций, является, по-видимому, центр дифференциации, не
найденный только у мух, в яйцах которых детерминация частей будущей
личинки завершается к моменту откладки яйца (Пфлюгфельдер, 1952).
По времени детерминации основных отделов тела личинки все изу-
ченные насекомые могут быть расположены в ряд, на одном конце кото-
рого находятся стрекозы, а на другом — двукрылые (Ричарде и Миллер,
1937; Пфлюгфельдер, 1952). Перепончатокрылые занимают в этом ряду
промежуточное положение. Опыты Шнеттера (1936) с перешнуровкой
яиц медоносной пчелы показывают, что потенции частей яйца значитель-
но превышают их проспективное значение, но для полного развития ор-
ганов необходимо присутствие центра дифференциации.
У пчелы и муравьев удавалось получить зародышей уменьшенного
размера, но не двойников. Впрочем, изученные представители перепон-
чатокрылых (пчелы и муравьи) относятся к высшему подотряду Aculea-
ta. Быть может, низшие перепончатокрылые (пилильщики) и наездники
обладают большими регулятивными возможностями.
Однако даже наиболее гибкий формообразовательный аппарат, при-
писываемый яйцам стрекоз, едва ли совместим с образованием из одного
яйца многих десятков и сотен зародышей. Несовместимость эта настоль-
ко очевидна, что заставила Краузе (1939)—известного исследователя
физиологии развития насекомых — высказать малообоснованное пред-
положение, что у паразитических перепончатокрылых имеет место не по-
лиэмбриония, а раннее партеногенетическое развитие половых клеток:
«В яйцах этих паразитов полностью отсутствуют все параплазматические
компоненты. На заднем конце имеется вещество зародышевого пути
(Keimbahnsubstanz). Внезародышевая часть яйца содержит не ядра
дробления, а ядра направительных телец и функционирует, как трофам-
нион. Соматическая часть бластомеров, по-видимому, очень рано деге-
нерирует, а зародышевые клетки начинают свое развитие прежде, чем
дифференцируется „соматический зародыш". 'Специальная разработка
этого нового толкования полиэмбрионии еще отсутствует» (стр. 527).
Характерно, что Краузе даже не допускает мысли о возможности
эволюционного изменения самого «механизма» развития. В этом отно-
шении гораздо более плодотворным представляются взгляды Филатова
(1939, 1943), который не только допускал возможность изменений фор-
мообразовательных аппаратов, но даже утверждал, что эти аппараты
более подвержены изменениям, чем тот морфологический результат, к
которому они приводят.
Относительно физиологии развития наездников можно сделать два
предположения. Можно допустить, что формообразовательный аппарат
у них не подвергся сколько-нибудь значительным изменениям, но всту-
236
паст в действие позднее, в каждом из многих зародышей, происшедших
из одного яйца, особо. Но при этом надо иметь в виду, что первоначаль-
но все многочисленные зародыши располагаются совершенно беспоря-
дочно; даже преемственность между их главными осями и первичной
полярностью яйца отсутствует. Таким'образом, между предварительной
структурой яйца и формообразовательными аппаратами заро-
дышей связь утрачивается; в каждом зародыше этот аппарат должен
возникнуть заново, и очень маловероятно, что он вполне идентичен тако-
вому непаразитичееких перепончатокрылых.
Но можно сделать и другое предположение, что упомянутый слож-
ный механизм развития у наездников просто исчезает и заменяется более
простым. Формообразовательный аппарат насекомых предполагает до-
вольно крупные размеры яиц с сравнительно большой протяженностью
продольной оси, вдоль которой распространяются влияния «центров», и
наличием проводящей системы в форме желточного синцития. У парази-
тических перепончатокрылых яйца становятся очень маленькими, жел-
точная система редуцируется, а в результате дробления получается ма-
ленькая кучка клеток, в которой и происходит дифференциация. Едва ли
в этом случае нужны такие сложные аппараты, координирующие разви-
тие, во всяком случае на ранних стадиях, на которых как раз и происхо-
дит разделение клеточного материала на отдельные «морулы». Особенно
большое 'значение в упрощении механизма развития могло иметь исчез-
новение желточной системы, которая служит основой действия центра
дифференциации, наиболее важного и достоверного из всех упомянутых
«центров». Переход к тотальному дроблению, тоже связанный с редук-
цией желтка, несомненно также облегчает разделение эмбрионального
материала на независимые части. Очень может быть, что ведущую роль
в этом процессе играет трофамнион, изолирующий отдельные группы
клеток. Выделение из числа эмбриональных клеток Litomastix специаль-
ных веретеновидных клеток, играющих такую же изоляционную функ-
цию, надо считать уже более поздним приобретением, специальным при-
способлением, способствующим разделению зародышей.
Подводя итог всему вышеизложенному, можно сказать, что парази-
тические условия развития настолько Меняют структуру яйца, что есть
все основания предполагать и значительные изменения (упрощение)
формообразовательных аппаратов, так что на начальных стадиях раз-
вития эмбриональный клеточный материал очень слабо интегрирован.
Эта слабая интегрированность подготовила почву для возникновения
полиэмбрионии. Очень вероятно, что специфическая полиэмбриония раз-
вилась из случайной, причиной которой являются какие-то внешние по
отношению к зародышу обстоятельства, нарушающие нормальное тече-
ние развития. Но так как образование двух, трех и более зародышей из
одного яйца оказалось очень выгодным, то полиэмбрионический тип раз-
вития стал для некоторых видов наследственно закрепленным призна-
ком. Подобная замена в процессе эволюции внешних факторов развития
внутренними уже известна на примере развития легких амфибий (Маш-
ковцев, 1935, 1936).
5. ЭВОЛЮЦИЯ ЭМБРИОНАЛЬНЫХ ОБОЛОЧЕК
Чтобы лучше оценить те морфологические, функциональные и он-
тогенетические изменения, которым подвергаются эмбриональные обо-
лочки паразитических перепончатокрылых, надо сперва уяснить, что
вообще представляют собой эмбриональные#оболочки насекомых.
237
Первое.правильное описание строения и развития эмбриональных
оболочек мы находим у Мечникова (1866), который наблюдал их у Si-
mulium, Corixa, Aspidiotus, тлей и у паразитического перепончатокрыло-
го Teleas, развитие которого происходит в яйцах водомерок. Мечников
описал дифференциацию бластодермы на утолщенную зародышевую по-
лоску и более тонкую внезародышевую часть и образование двух оболо-
чек либо путем нарастания складок тонкой бластодермы на зародыше-
вую полоску, либо' в связи с погружением зародыша в желток; внутрен-
нюю оболочку, края которой связаны непосредственно с зародышевой
полоской, Мечников назвал складчатым или покровным листом, а на-
ружную— амнионом. У Teleas Мечников обнаружил только амнион.
В заключительной части своей статьи Мечников сравнивает эмбриональ-
ные оболочки насекомых с таковыми позвоночных и иногда называет
«амнион насекомых» серозным листом.
Наблюдения Мечникова настолько точны, что не нуждаются ни в
каких поправках; позднейшие работы их только подтвердили; но терми-
нология установилась иная: теперь мы называем складчатый или
покровный лист Мечникова амнионом, а амнион Мечникова — се-
розой.
В настоящее время развитие эмбриональных оболочек изучено у
очень многих насекомых. Способы' их возникновения до(ВОльно разнооб-
разны, но из них можно выделить три основных (Шаров, 1953). Наибо-
лее простой случай имеет место у Thysanura — здесь сравнительно ко-
роткая зародышевая полоска целиком погружается внутрь желтка, пере-
гибаясь посередине и увлекая за собой прилегающие части внезароды-
шевой бластодермы. У стрекоз, полужесткокрылых и некоторых других
насекомых, образование зародышевых оболочек начинается с погруже-
ния заднего конца зародышевой полоски и сопряжено с переворачива-
нием ее на 180°. Наконец, в третьем случае (жуки, бабочки и другие)
зародышевая полоска остается относительно неподвижной и сохраняет
поверхностное положение, а внезародышевая бластодерма нарастает на
нее двумя складками. Именно этот последний тип формирования эмбрио-
нальных оболочек наблюдается у наиболее примитивных Hymenopiera
(пилильщиков).
Вопрос о функциональном значении зародышевых оболочек насеко-
мых освещен в литературе недостаточно. П. П. Иванов, излагая данные
по развитию скорпионов, пишет: «Образование амниона и серозной обо-
лочки является специальным приспособлением-яиц, развивающихся вне
воды, так как, с одной стороны, защищает зародыш от непосредственного
соприкосновения с хорионом, который мог бы при подсыхании при-
клеиться к зародышу, с другой — жидкость, заполняющая амнион, соз-
дает благоприятную среду для тканей зародыша и вместе с тем, задер-
живает образование хитина, который мог бы препятствовать росту и из-
менениям формы зародышевой полоски, происходящим на этих стадиях
особенно интенсивно» (1937, стр. 318).
Говоря о насекомых, П. П. Иванов отмечает, что процесс бласто-
кинеза, в результате которого возникают эмбриональные оболочки, вы-
зван необходимостью использовать более глубокие слои желтка, а обра-
зующаяся при этом амниотическая полость заполнена амниотической
жидкостью, которая предотвращает выделение эмбриональной ку-
тикулы.
Соображения П. П. Иванова о значении амниотической жидкости,
Как препятствия для образования хитина, очень интересны, но требуют
экспериментальной проверки. Более раннее или бо!лее позднее образова-
ние кутикулы может зависеть просто от времени наступления гистологи-
238
ческой дифференциации в гиподерме. Кроме того, известны случаи,
когда амнион и амниотическая жидкость отсутствуют, а между тем хи-
тинизация покрова происходит не раньше, чем у других насекомых.
Большинство авторов считает эмбриональные оболочки защитными
структурами, предохраняющими зародыш прежде всего от /высыхания,
и связывает их 'возникновение с переходом к наземному образу жизни.
Эта точка зрения, наиболее подробно изложенная Шаровым (1953), ка-
жется очень убедительной, тем более, что совершенно сходные образова-
ния возникают и в эволюции наземных позвоночных.
Однако во многих случаях эмбриональные оболочки насекомых про-
ницаемы для воды. Зародыши некоторых насекомых поглощают воду из
окружающей среды. Всасывание воды яйцами пилильщиков уже было
отмечено выше. Ряд примеров такого рода-приводят Рунвол (Roonwal,
1936) и Слайфер (Slifer, 1938). По данным Слайфер (1938, 1946, 1948),
у Melanoplus differentialis (Thomas) сероза выделяет два кутикулярных
слоя, из которых наружный слой (желтая кутикула) очень слабо прони-
цаем для воды, а внутренний (белая кутикула) служит защитой от ме-
ханических повреждений. Но в небольшой области на заднем конце яйца
желтая кутикула отсутствует; в этом месте, названном «гидропиле», вода
может входить в яйцо и выходить из него. Подле гидропиле сероза
устроена иначе, чем в других местах. Перед наступлением диашаузы
сероза выделяет восковой слой, закрывающий гидропиле, и доступ воды
прекращается. Удаление этого слоя ксилолом и другими растворите-
лями может предотвратить диапаузу или прервать ее. Через эмбриональ-
ные оболочки происходит также газообмен.
Все эти очень интересные данные показывают, что эмбриональные
оболочки насекомых не только служат для изоляции зародыша от внеш-
ней среды, но в значительной степени являются и посредником между
{зародышем и средой. К сожалению, пока еще трудно сказать, в какой
мере результаты изучения оболочек Melanoplus могут быть распростра-
нены на других насекомых. По всей вероятности, дальнейшие работы в
этом направлении обнаружат значительные различия в тонком строении
и функции эмбриональных оболочек различных насекомых.
Среди перепончатокрылых наиболее примитивные отношения обна-
руживают эмбриональные оболочки пилильщиков. Образование оболо-
чек происходит путем нарастания на зародышевую полоску двух амнио-
тических складок. Сначала имеются две эмбриональные оболочки, но
затем амнион редуцируется. Клеточный материал амниона частично де-
генерирует, а частично используется для построения задней кишки и
кожных покровов спины. Однако если верить старым данным Грабера
(1890), то и среди пилильщиков есть виды, у которых амнион длительно
сохраняется.
Относительно функции эмбриональных оболочек у пилильщиков, по-
видимому, можно принять то же, что принимается для оболочек боль-
шинства насекомых, т. е. что основная их функция — защитная. В то же
время надо отметить, что сероза пилильщиков хорошо пропускает воду,
на что указывают наблюдения, результаты которых изложены в I главе
Специальной части, и данные Цирнгибля (1940).
У жалящих перепончатокрылых имеется только одна эмбриональная
оболочка, которая чаще всего образуется путем отделения внезароды-
шевой бластодермы от зародышевой полоски и нарастания первой на
вторую. Эту единственную оболочку Aculeata разные авторы называют
по-разному. Шнеттер (1934) пришел к выводу, что зародышевая оболоч-
ка медоносной пчелы является амнионом, а серозе соответствует часть
внезародышевой бластодермы, известная под названием «дорзальной
239
■полоски». Вслед за Шнеттером называют амнионом эмбриональную
оболочку пчелы Краузе (1939) и Зауэр (1954).
Другие авторы, например Танквари (1913)—в отношении му-
равьев,— считают эту оболочку серозой.
Для выяснения этого вопроса решающее значение имеют те случаи,
когда образование эмбриональной оболочки сопровождается появлением
зачаточных амниотических складок. Это имеет место у Chalicodoma mu-
raria Р. (Карьер и Бюргер, 1898), у Ammophila campestris Jur. (Бэрендс
и Бэрендс Ван Роон, 1949) и у Scolia quadripunctata F. (Дондуа, 1953).
В амниотических складках первоначально представлены оба листка:
внутренний (амниотический) и наружный (серозный), но затем внутрен-
ний листок перестает расти, а наружный отделяется от него, надвигается
на зародыш и образует оболочку — серозу.
Более существенные изменения испытывают эмбриональные оболоч-
ки у муравьев. Здесь внезародышевая бластодерма состоит из крупных
перегруженных желтком клеток, которые в ряде случаев оказываются
неспособными образовать даже одну эмбриональную оболочку (Стринд-
берг, 1913—1918). Редукция эмбриональных оболочек у муравьев, воз-
можно, связана с утратой ими защитной функции, что в свою очередь
может быть следствием развития инстинкта «заботы о потомству у этих
насекомых (как известно, муравьи заботливо охраняют свои яйца, очи-
щают их от. прорастающих спор грибов и в случае надобности переносят
их из одной части муравейника в другую в поисках оптимальных усло-
вий температуры и влажности).
У паразитических перепончатокрылых обычно есть одна эмбриональ-
ная оболочка, строение и способ образования которой у разных видов
очень разнообразен. Относительно функции этой оболочки высказаны
следующие предположения. М. С. Ганйн (1869)—один из первых эм-
бриологов, познакомившийся с развитием наездников, — обнаружил на-
личие одной эмбриональной оболочки у Anagrus (= Polynema) и
Ophioneurus (= Trichogramma) и отсутствие ее у Teleas, из чего заклю-
чил, что эта оболочка не играет большой роли в развитии.
Большинство современных исследователей приписывает зародыше-
вой оболочке наездников трофическую функцию, хотя, конечно, это от-
носится лишь к тем видам, у которых имеет место значительный эмбрио-
нальный рост. Некоторые авторы, наряду с питательной ролью оболочки,
предполагают и наличие у нее защитной роли, так как во многих случаях
(особенна у полиэмбрионических форм, у которых хорион развит слабо
и рано сбрасывается) она предохраняет зародыш от непосредственного
соприкосновения с тканями хозяина (Сильшестри, 1937; Даут, 1947). Не-
которые факты, говорящие в пользу этого мнения, были обнаружены и
при развитии афидиид.
Таким образом, предполагается, что зародышевая оболочка парази-
тических перепончатокрылых выполняет те же функции, что и оболочки
других насекомых — изоляции от внешней среды и посредничества при
получении некоторых веществ из этой среды, но выполняет эти функции
по-новому, применительно к условиям жизни внутри другого организма
и к паразитическому способу питания.
Но, кроме того, можно думать, что оболочка (трофамнион) играет
особенно важную роль при полиэмбрионическом типе развития, так как
именно она образует прослойки, изолирующие друг от друга «вторичные
эмбриональные массы». Эта функция разделения единого зародыша на
«вторичные эмбриональные массы», по-видимому, становится тоже очень
существенной. По всей вероятности не случайно, что у Litomastix flori-
danus (Ashm.), у которого полиэмбриония достигает наибольшей про-
240
дуктивности, к выполнению этой функции привлекается также часть
«эмбриональных» клеток (Пэтерсон, 1921).
Итак, у большинства паразитических перепончатокрылых сероза
выполняет очень важные функции. Лишь у сравнительно немногих пред-
ставителей этой группы она не приобрела трофического значения и, по-
видимому, каким-то образом утратила свою защитную роль, что послу-
жило причиной ее редукции у некоторых видов, исследованных М. С. Га-
ниным (1869), у Banchus femoralis Thorns. (Бледавский и Краинская,
1926), у Prestwichia aquatica Lubb., Mestocharis militaris R.-Kors., Ca-
raphractus reductus R.-Kors. (Иванова-Казас, 1950, 19526 и 19546) и
Telenomus gracilis Ma у г (Коломиец и Коваленок, 1958).
Вопрос о морфологическом значении единственной эмбриональной
оболочки разных представителей Hymenoptera parasitica и гомологии их
друг другу и оболочкам других насекомых нигде в литературе не об-
суждается. Имеются лишь разрозненные высказывания по поводу оболо-
чек отдельных видов. Отношение автора к описываемой им оболочке
часто выражается только в применяемом им для ее обозначения назва-
нии. Так, Кулагин (1894), Тоуэр (Tower, 1915) и Гетенби (1919) счи-
тают эту оболочку гомологичной и амниону и серозе других насекомых.
Маршаль (1906) называет ее амнионом, а Оглоблин (1924) и Ване (Van-
ce, 1931) —серозой. Ряд авторов не решается признать эту оболочку ни
амнионом, ни серозой и обозначает ее каким-нибудь новым термином.
Джексон (1928) и Кроуфорд (Crawford, 1933) называют ее «клеточной
оболочкой», Симмондс (1947)—«трофической оболочкой», Корнхаузер
(1919)—«плацентарной оболочкой», Сильвестри (1909) и Хэвиленд
(1921) —«псевдосерозой» и т. д. А. А. Захваткин (1953) высказал пред-
положение, что эмбриональная оболочка паразитических перепончато-
крылых гомологична желточным клеткам4 других насекомых. Основанием
для этого суждения послужили, вероятно, трофическая функция, выпол-
няемая обоими сближаемыми образованиями, и превращение вителло-
фагов в трофическую оболочку у некоторых Strepsiptera. Оболочку по-
лиэмбрионических наездников принято называть «трофамнионом» и счи-
тать продуктом слияния и гипертрофии направительных телец (Мар-
шаль, 1904в; Мартин, 1914; Рунвол, 1935; Сильвестри, 1937; Шванвич,
'1949, и др.). Никто из перечисленных выше авторов не пытался при раз-
решении вопроса о морфологическом значении зародышевой оболочки
наездников привлекать широкий сравнительно-эмбриологический мате-
риал. Поэтому каждому казалось, что он имеет дело с чем-то совершенно
исключительным, с образованием sui generis.
Полагаю, что вопрос о гомодогии эмбриональных оболочек различ-
ных перепончатокрылых может быть не только поставлен, но и разрешен
в положительном смысле (Иванова-Казас, 1954д). Единственная эмбрио-
нальная оболочка наездников соответствует единственной оболочке
Aculeata, т. е. серозе. Различия в способах ее образования объясняются
тем, что по мере того, как она приобретала все большее значение как
орган питания зародыша, ее обособление сдвигалось все на более ранние
стадии. С этой точки зрения можно наметить следующие этапы эволюции
эмбриональных оболочек у перепончатокрылых.
I. Исходная стадия эволюции — 2 оболочки, образующиеся из амнио-
тических складок. Такие отношения сохранились у пилильщиков
(рис. 114, Л).
II. Амнион исчезает, и остается только сероза, которая образуется
путем нарастания внезародышевой бластодермы на зародышевую. Это
имеет место у многих Aculeata и некоторых Parasitica (Eurytoma, Angi-
tia) (рис. 114, Б).
16 О. М. Иванова-Казас
241
gasteridac, представляющийся гипертрофированным направительным
тельцем, оказывается возможным вывести из типичных оболочек насе-
комых.
Несомненно, что сложный эволюционный >путь, пройденный серозой
паразитических перепончатокрылых, тесно связан с их образом жизни.
Недаром раньше всего и наиболее измененным способом происходит
образование эмбриональной оболочки у наиболее специализированных
как паразиты полиэмбрионических видов.
6. НЕКОТОРЫЕ АНАЛОГИИ МЕЖДУ ЖИВОРОЖДЕНИЕМ
И ЭМБРИОНАЛЬНЫМ ПАРАЗИТИЗМОМ
Выше уже было отмечено, что условия эмбрионального развития при
паразитизме и живорождении сходны в том отношении, что в обоих слу-
чаях роль внешней среды выполняет другой организм (хозяина или ма-
тери), от которого зародыш получает все необходимые питательные ве-
щества, а также кислород и которому он отдает продукты своего обмена.
Такое сходство в условиях существования не могло не вызвать более
или менее сходных измененией в характере развития, появления ряда
аналогичных морфологических адаптации.
Первое сопоставление живорождения и паразитизма было сделано
Фаусеком (1913), который определял живорождение как «частный слу-
чай паразитизма, временный паразитизм в пределах одного вида каж-
дого последующего поколения на предыдущем» (стр. 37).
Конечно, между живорождением и эмбриональным паразитизмом
имеются и принципиальные различия. В первом случае устанавливается
тесный контакт между организмами, относящимися к одному виду, кон-
такт полезный для этого вида, хотя- интересы матери часто приносятся в
жертву зародышу. При паразитизме не менее тесные отношения уста-
навливаются между представителями разных видов, и пользу из них из-
влекает только вид паразита — в ущерб виду хозяина. Но при этом, как
уже было отмечено, в случае успешного паразитизма защитная реакция
хозяина может быть ослаблена или извращена и зародыш паразита ока-
зывается почти в таком же привилегированном положении, как и заро-
дыши живородящих животных.
После ознакомления с особенностями развития паразитических на-
секомых, представляется интересным сравнить их с таковыми у живоро-
дящих форм. Подобные же сопоставления делали и авторы, имевшие
дело с живородящими насекомыми, например Носкевич и Полюжинский
(Noskievvicz und Poluszynski, 1927), изучавшие развитие веерокрылых.
Среди насекомых известно много случаев живорождения, возникше-
го независимо у представителей разных отрядов и осуществляемого раз-
личными способами. По характеру отношений между матерью и зароды-
шем Хаган (Hagan, 1951) различает у них следующие формы живорож-
дения:
1) Яйцеживорождение, при котором развитие происходит S3a счет
желтка, накопленного во время овогенеза. Никаких трофических отно-
шений между матерью и зародышем в период эмбрионального развития
нет. Хорион хорошо выражен. Развитие протекает обычным для насеко-
мых образом.
Эта форма живорождения представляет интерес лишь в том отно-
шении, что, по-видимому, она явилась подготовительной ступенью для
возникновения истинного живорождения с питанием зародыша за счет
матери. Аналогичные отношения имеют место у менее специализирован-
ных наездников, яйцо которых служит лишь средством заражения и еще
16*
243
не обладает способностью извлекать питательные вещества на теле хо-
зяина. Но на почве этих простых отношений возник настоящий эмбрио-
нальный паразитизм.
2) Аденотрофическое живорождение, свойственное куклородным
мухам (Pupipara) и некоторым другим двукрылым. Эта форма живо-
рождения отличается от предыдущей только тем, что по окончании эм-
брионального развития личинка не покидает тело матери, а задержи-
вается в ее половых путях, питаясь выделениями специальных желез.
Рождение молоди происходит лишь после завершения личиночного раз-
вития, причем новорожденные личинки сразу же окукливаются.
У таких вынашиваемых в теле матери личинок возникают некоторые
черты .сходства с личинками Стебельчатобрюхих' перепончатокрылых:
средняя кишка утрачивает сообщение с задней кишкой, которая служит
в это время лишь для накопления экскретов. Эта особенность строения
кишечника личинок куклородных мух может быть также связана с пи-
танием жидкой пищей.
3) Псевдоплацентарное живорождение, при котором в процессе эм-
брионального развития, зародыш получает дополнительное питание от
матери. При этом возникают специальные структуры, образующиеся из
тканей матери или зародыша, или обоих организмов, через посредство
которых и осуществляется передача питательных веществ. Этот питаю-
щий аппарат (плацента или псевдопланцента— по разным авторам) в
разных случаях устроен очень различно.
Так, у живородящих щитовок из родов Stictococcus и Apiomorpha
питание зародыша происходит с помощью серозы и фолликулярного эпи-
телия (Бухнер — Buchner, 1954, 1955 и 1957).
Своеобразные структуры осуществляют питание зародыша за счет
матери у Hesperoctenes fumarius Westw. (Hemiptera, Polyctenidae) —
эктопаразита летучих мышей. Здесь тоже трофическую функцию сперва
выполняет сероза; потом ей на смену приходит новая оболочка, возни-
кающая путем разрастания зачатков первой пары брюшных ног (плей-
роподий). Плейроподии становятся органами птитания и у живородяще-
го таракана Diploptera dytiscoides (Хаган, 1951).
Сложная плацента, состоящая из зародышевой части (амнион и се-
роза) и материнской части (разрастание фолликулярного эпителия),
образуется при развитии Hemimerus talpoides Walk. (Dermaptera) no
данным Геймонса (1912).
4) Последняя форма живорождения представлена гемоцельным жи-
ворождением цецидомиид и веерокрылых и характеризуется тем, что
развитие зародышей происходит не в половых путях самки, а непосред-
ственно в полости тела (гемоцеле), как при эндопаразитизме наездников.
Физиология питания зародышей изучена недостаточно, но у Cecido-
myidae, по-видимому, главную роль играет при этом сероза (Kahle,
1908), а у веерокрылого Helictoxenos simplics образуется трофическая
оболочка из желточных клеток. Эмбриональные клетки группируются на
внутренней стороне этой оболочки таким образом, что дают начало мно-
гим (до 50) зародышам (Носкевич и Полюжинский, 1935). В результате
у веерокрылых возникает типичная полиэмбриония.
Во всех тех случаях, когда питание зародыша совершается за счет
матери, в эмбриональном развитии проявляются те же эволюционные
тенденции, что и при паразитизме:
1. Исчезновение хориона наблюдается у тлей, некоторых щитовок,
веерокрылых, Hesperoctenes, Hemimerus, и Cecidomyidae.
2. Количество желтка резко снижается (щитовки, веерокрылые,
Hesperoctenes, Hemimerus).
244
3. Дробление становится полным или приближается к таковому
(Stylops из веерокрылых и Stictococcus).
4. Возникают различные структуры, обеспечивающие питание заро-
дыша, которые чаще всего имеют строение оболочки. Такими питающими
оболочками могут стать амнион или сероза (Apiomorpha, Stictococcus,
Hesperoctenes, Hemimerus), видоизмененные конечности (Hesperoetenes,
Diploptera), желточные клетки (Helictoxenos) и, наконец, фолликуляр-
ный эпителий 'или стенки яйцевода матери (Apiomorpha, Stictococcus,
Hemimerus, Strepsiptera), которые обнаруживают ряд морфологических
изменений, свидетельствующих о выполнении ими трофической функции.
В некоторых случаях (Stylops из веерокрылых) Оболочка из материнских
тка'ней является единственным приспособлением для питания зародыша.
Активное участие тканей матери в питании зародыша, пожалуй, наибо-
лее характерная черта, отличающая развитие живородящих форм от
такового паразитических. Хотя некоторые авторы и полагают, что соеди-
нительнотканные капсулы хозяина вокруг яиц паразита становятся не-
обходимым условием их развития, но питающая их роль не доказана и
требует специального изучения. Во всяком случае неизвестно ни одного
достоверного случая, когда питание зародышей паразитического пере-
пончатокрылого происходит без помощи собственной трофической обо-
лочки.
5. В некоторых случаях (Strepsiptera) развитие становится полиэм-
брионическим, но полиэмбриония не имеет при живорождении большого
распространения, а биологические преимущества такого способа раз-
множения в этом случае гораздо менее очевидны, чем при паразитизме.
Несомненно лишь одно, что, как и у перепончатокрылых, она стала воз-
можной лишь благодаря тому, что количество питательных материалов,
которые могут быть использованы при развитии, не лимитируется за-
пасами, имеющимися с самого начала в яйце.
Мы рассмотрели сейчас особенности развития живородящих насе-
комых, но в сущности такие же черты приобретает и развитие других
живородящих животных, например млекопитающих, у которых они вы-
ражены еще более ярко.
[ДАВА IV
РАЗВИТИЕ ПЕРЕПОНЧАТОКРЫЛЫХ И МОРФОЛОГИЧЕСКИЕ
ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
В одной из предшествующих глав были намечены основные направ-
ления эволюции эмбрионального развития у перепончатокрылых. Теперь
посмотрим, какие частные закономерности эволюции могут быть особен-
но ярко продемонстрированы на нашем материале.
Ив. Ив. Шмальгаузен (1946) указывает, что эволюция онтогенеза
складывается из процессов совершенно различного характера: а) из из-
менений развития, связанных с эволюцией взрослого организма, т. е.
филэмбрногенезов, б) из приспособлений зародыша или личинки к усло-
виям окружающей среды и в) из усовершенствования (рационализации)
самого онтогенеза с его факторами развития. Попытаемся раздельно
рассмотреть эти три группы явлений в развитии перепончатокрылых.
Теория филэмбрногенезов, разработанная А. Н. Северцовым (1934,
1939), рассматривает те изменения в развитии, в результате которых из-
меняется строение органов взрослого животного. Эволюция взрослых
перепончатокрылых и формирование признаков, характеризующих их
принадлежность к определенным видам, родам, семействам и т. д., про-
исходящее уже во время метаморфоза, не были предметом сравнитель-
ного изучения в настоящей работе, а потому задерживаться на них мы
не будем. Главное внимание обращено на эмбриональные стадии разви-
тия. На этих стадиях, осуществляются самые первые и наиболее общие
процессы развития: дробление и гаструляция, и происходит формирова-
ние личиночных органов, которые хотя и являются необходимой подго-
товительной ступенью для развития имагинальных органов, но не опре-
деляют непосредственно особенностей их окончательного строения. Но
как раз на этих стадиях возникают различные эмбриональные и личи-
ночные провизорные органы, через посредство которых и происходит
адаптация зародыша и личинки к условиям развития. Именно эти про-
визорные структуры и испытывают наиболее значительные изменения в
разных условиях развития, и прослеженная выше эволюция развития
перепончатокрылых в значительной степени сводится к эволюции этих
структур.
Хотя учение А. И. Северцова о значении функции для эволюции ор-
ганов и о способах их эволюционных изменений разработано в основном
для органов взрослого животного, но основные его положения в полной
мере приложимы и для органов, провизорных, функционирующих у за-
родыша и личинки.
Одним из таких чисто эмбриональных образований является жел-
точная система насекомых, представляющая собой часть первичной эн-
246
тодермы зародыша (а по некоторым представлениям — всю первичную
энтодерму), которая специализировалась на переработке желтка, не
участвует в построении средней кишки личинки и имаго и образование
которой сдвинуто на ранние стадий развития (образование вителлофа-
гов происходит в значительной степени еще во время дбрбления, а от-
части и на более поздних стадиях). Дифференциация единого энтодер-
мального зачатка на две части, одна из которых служит в основном для
усвоения питательных материалов желтка, а другая — для формирования
средней кишки и осуществления пищеварительной функции на активно
питающихся стадиях, связанная с накоплением в яйцах большого коли-
чества желтка, имеет место у очень многих животных: Polyclada, некото-
рых Polychaeta и Mollusca, пиявок, первичнотрахейных, многоножек,
огнетелок, осетровых рыб и т. д. Так же можно расценивать желточную
и кишечную энтодерму костистых рыб и Sauropsida, хотя в этих группах
оба зачатка так сильно разошлись по времени, месту и способу обособ-
ления, что единство их филогенетического происхождения оспаривается
(Светлов, 1959). Когда желтка не так много, одни и те же энтотермаль-
ные клетки осуществляют и эмбриональное и дефинитивное питание.
К'этому случаю вполне применим принцип филогенетического разделе-
ния функций и органов Се/верцова, согласно которому, орган, выполняю-
щий несколько функций, разделяется на части, превращающиеся в са-
мостоятельные органы и выполняющие разные функции. Особенностью
рассматриваемого примера является то, что единый зачаток разделяется
на две части, функционирующие не одновременно.
Согласно другой точке зрения на энтодерму насекомых (Геймонс,
1896; Шаров, 1953, 1959; Светлов, 1959), в подьзу которой тоже имеется
немало веских доводов и рассмотрение которых увело бы нас далеко от
основной темы, вся первичная энтодерма насекомых низводится до роли
вителлофагов, т. е. здесь имеет место уменьшение числа функций, при-
чем сохраняется отнюдь не первичная и не главная функция эндотермы,
так что можно говорить даже о смене функции. Роль же дефинитивной
энтодермы, из которой образуется средняя кишка, берет на себя совсем
новый зачаток — эктодерма дна стомо- и проктодеума. Такую подмену
одного зачатка другим, выходящую даже за пределы зародышевых
листков, авторы толкуют как один из примеров принципа меторизиса
В.М. Шимкевича (1908).
Наряду со специфической функцией переработки желтка желточная
система насекомых выполняет и важные морфогенетические функции:
желточный синцитий с заключенными в нём массами желтка служит
основой для формирования средней кишки. Ряд экспериментальных дан-
ных (см. Общ. часть, гл. И 1,3) свидетельствует об активной роли, которую
играет желточная система при образования зародышевой полоски, обо-
соблении зародышевых листков, бластокинезе и т. д.
Такова желточная система большинства насекомых, а из перепонча-
токрылых— первичнорастительноядных и жалящих форм. Но у парази-
тических перепончатокрылых в связи с редукцией желтка в силу функ-
циональной корреляции начинается и процесс редукции желточной
системы. Эта редукция происходит очень постепенно: у Angitia желточ-
ная система развивается полностью, хотя желтка уже нет. Можно ду-
мать, что и формообразовательное ее значение сохраняется здесь в пол-
ном объеме. Но у ряда других форм (Prestwichia, Ephedrus и др.) жел-
точные клетки хотя и образуются, но рано дегенерируют. Сохраняем ли
здесь желточная система свое морфогенетическое значение? В полной
мере едва ли. Однако трудно себе представить существование зачатка,
.даже находящегося на пути к полному уничтожению, который бы не
247
участвовал во взаимодействии частей зародыша, играющем такую важ-
ную роль в процессе индивидуального развития. Согласно И. И. Шмаль-
гаузену (1938, 1946), явления рекапитуляции и зародышевого сходства,,
сохранение в развитии древних признаков объясняются «трудностью
перестройки системы взаимозависимостей между морфогенетическими
процессами». Однако и эти взаимозависимости'в конце концов тоже
изменяются: у Aphidius и некоторых других наездников желточная си-
стема редуцируется полностью, даже зачаток ее выпадает из развития
(отрицательной архаллаксис). Таким образом, редукция желточной си-
стемы у паразитических перепончатокрылых происходит по схеме, наме-
ченной И. И. Шмальгаузеном (1938): причиной ее служит нарушение
эргонтических (функциональных) связей, затем разрушаются морфоге-
нетические 'корреляции, после чего становится возможным полное исчез-
новение органа.
Другим важным эмбриональным органом насекомых являются за-
родышевые оболочки: амнион и сероза. У перепончатокрылых амнион
подвергается постепенной редукции: у пилильщиков он получает полное
развитие, -но рано дегенерирует, у некоторых жалящих наблюдаются
лишь начальные стадии формирования амниона, у других жалящих и
большинства паразитических амнион не образуется вовсе. Функция
амниона и причины его редукции не вполне ясны. Морфологическая же
картина редукции амниона создает впечатление, что здесь имеет место
ряд отрицательных филэмбриогенезов, спускающихся все на более ран-
ний стадии' (то, что Северцов назвал вторичным архаллаксисом).
Так же неясны причины редукции серозы у Prestwichia, Caraphrac-
tus и др. В этом случае нам неизвестны никакие переходные состояния
между видами,- имеющими серозу, и (видами, лишенными ее. Возможно,
что редукция серозы произошла здесь путем первичного отрицательного
архаллаксиса.
Зато прогрессивные изменения испытывает сероза у тех паразити-
ческих видов, которые приобрели способность питаться за счет хозяина
на эмбриональных стадиях. Прежде всего надо отметить, что у этих
форм произошла субституция функций: функция питания зародыша за
счет желтка, осуществляемая с помощью желточной системы, заменилась
у них функцией питания за счет веществ окружающей среды, утилизи-
руемых с помощью серозы. Для самой этой эмбриональной оболочки
трофическая функция является в значительной степени новой. Эмбрио-
нальные оболочки насекомых по своей природе мультифункциональны
(как и всякий другой орган — Северцов, 1939),. Хотя сероза насекомых
допускает обмен газов и воды, но основной функцией серозы является
защита зародыша. У серозы рассматриваемых наездников функция об-
мена активизируется (или по крайней мере интенсифицируется) и выхо-
дит на первый 'план. Происходит типичная смена функций по принципу,
установленному Дорном и разработанному А. Н. Северцовым. Северцов
отмечает, что смена функций чаще всего происходит при перемене сре-
ды обитания. В данном случае она связана с переходом зародыша к ак-
тивному паразитированию.
Таким образом сероза превращается в очень важный провизорный
орган питания; чем раньше начнет она функционировать, тем полнее
сможет зародыш использовать преимущества паразитического образа
•жизни. Поэтому появляется тенденция к более раннему обособлению се-
розы (гетерохрония). На ряде представителей паразитических перепон-
чатокрылых можно наблюдать разные степени развития этой гетерохро-
нии. У крайних членов этого рода (шолиэмбрионические Encyrtidae и
Platygasteridae) обособление серозы («трофамниона») происходит еще
248
до начала дробления. Параллельно, изменяется и самый способ образо-
вания серозы. Эти изменения зависят от различий в строении зародыша
на тех стадиях, когда образуется сероза (так, например, возникновение
типичных амниотических складок просто немыслимо на стадии, когда
зародыш состоит всего лишь из нескольких бластомеров). Любопытно,
что на самых последних ступенях эволюции при формировании трофам-
ниона полиэмбрио'нических форм ядрами этой оболочки становятся не
ядра дробления (которых еще нет, так как дробление еще не началось),
а ядра направительных телец. Здесь имеет место своеобразная субсти-
туция, происходящая на таких ранних стадиях развития (нераздробив-
шееся яйцо), что речь идет о замене одних клеточных элементов дру-
гими.
Что касается эволюции личиночных органов, то им было уделено
гораздо меньше внимания. Однако бросается в глаза более низкий урр-
вень организации личинок у всех высших Hymenoptera (Apocrita) no
сравнению с таковым Phytophaga. Это упрощение организации прояв-
ляется у жалящих и менее специализированных наездников в редукции
глаз и конечностей и некотором недоразвитии кишечника и связано с
сильным развитием инстинкта «заботы о потомстве». Жизнь личинок про-
текает в тесных норках или ячейках, где они более или менее защищены
от неблагоприятных внешних условий, питание их обеспечено заранее
заготовленной матерью пищей в виде убитой или парализованной добы-
чи или растительных продуктов, поэтому личинкам не приходится актив-
но отыскивать пропитание и укрываться от врагов, они становятся вя-
лыми и малоподвижными. Последнее дает основание предполагать, что
и нервно-мышечный аппарат развит у них гораздо слабее, чем у более
примитивных сидячебрюхих, личинки которых ведут свободный экзо-
фитный образ жизни. Недоразвитие кишечника, состоящее в отсутствии
у личинок сообщения между средней и задней кишкой тоже связано с
экологией личинок и имеет свои биологические преимущества.
Итак, мы видим, что личинки высших перепончатокрылых испыты-
вают некоторый морфо-физиологический регресс, выражающийся в ис-
чезновении и ослаблении ряда личиночных органов. Эти регрессивные
изменения могут осуществляться по-разному. Личиночные глаза даже не
закладываются, так что можно говорить об отрицательном архаллаксисе.
То же относится и к конечностям брюшка — из всех Apocrita временное
существование зачатков брюшных ног описано только для Pimpla tu-
rionellae Thomps. (Бронскилл, 1959). Появляющиеся во время эмбрио-
нального развития у жалящих зачатки грудных ног и антенн вскоре
прекращают свое развитие и превращаются в скрытые под гиподермой
имагинальные диски соответствующих органов. Таким образом, редук-
ция грудных конечностей личинки осуществляется путем их недоразви-
тия (отрицательная анаболия). Так же можно расценивать и неполное
развитие кишечника.
Особенно далеко заходит упрощение личинки у некоторых эндопара-
зитических наездников. Ярким примером такого рода является личинка.
Prestwichia aquatica Lubb., которая в момент выхода из яйца имеет пу-
зыревидную форму, не сегментирована и совершенно лишена конечностей
(даже ротовых); мускулатура представлена только мышцами пищевода,
дыхательная и кровеносная системы еще отсутствуют, достаточно хо-
рошо развит и функционирует только кишечник. Существование такой
крайне упрощенной личинки возможно только благодаря исключительно
благоприятным условиям паразитической жизни.
Общее впечатление таково, что личинки Apocrita выходят из яйца
на гораздо более ранней стадии морфологического развитая, чем личин-
249
ки более примитивных представителей отряда, например пилильщиков.
Пользуясь терминологией Берлезе (1901), можно сказать, что личинки
жалящих вылупляются из яйца на протоподиальной стадии развития, а
личинки некоторых эндопаразитов, как Prestwichia. — на еще более ран-
ней стадии, 'когда еще нет ни зачатков конечностей, ни сегментации.
Существование различных соотношений между морфологическими
и биологическими стадиями развития (возможность перехода от эмбрио-
нального состояния к личиночному на разных стадиях морфологического
развития) может расцениваться как своеобразная гетерохрония. Одной
из основных тенденций эволюции онтогенеза многоклеточных животных,
по И. И. Шмальгаузену (1938, 1946), является удлинение эмбрионально-
го развития за счет включения в'него начальных стадий постэмбриональ-
ного развития. Такая «эмбрионализация» имеет то преимущество, что
развивающийся организм дольше остается под защитой яйцевых оболо-
чек, а ткани зародыша дольше сохраняют пластичность, так как гисто-
логическая дифференциация наступает в них позднее. А. А. Захваткин
называет это явление «эмбрионизацией» и тоже придает ему большое
значение в выработке характерных форм онтогенеза Metazoa (1949) и
в эволюции развития членистоногих (1953). При этом отмечается, что
удлинение эмбрионального развития и усложнение организации выходя-
щего из яйца животного возможны лишь при увеличении запасов желт-
ка в яйцах или при наличии постоянного притока питательных веществ
извне, что имеет место при живорождении. Но в некоторых случаях про-
исходит и • обратный процесс укорочения эмбрионального развития и
упрощения выходящего из яйца организма. Это явление было названо
Захваткиным (1953) «дезэмбрионизацией». Именно такое «преждевре-
менное» вылупление личинки, обусловленное уменьшением'запасов желт-
ка, послужило, по мнению Берлезе (1901) и И. И. Ежикова (1929, 1939,
1940), причиной возникновения метаморфоза Holometabola. Захваткин
придерживался иных взглядов на происхождение полного превращения
насекомых и усматривал явления дезэмбрионизации лишь в развитии
паразитических перепончатокрылых.
Однако не совсем правильно было бы понимать дезэмбрионизацию
перепончатокрылых как простое сдвигание момента вылупления на более
раннюю стадию и связывать ее с недостатком питательных материалов
в яйце. Как известно, у Aculeala яйца крупны и содержат много желтка.
Кроме того, органы, «не успевшие» развиться в эмбриональный период,
не закладываются и не завершают своего развития и в постэмбриональ-
ный период, они просто выпадают из развития, так как становятся не-
нужными в тех условиях, в которых протекает жизнь личинки.
У наездников упрощение новорожденной личинки сочетается с ре-
дукцией желтка в яйцах, и многие процессы органогенеза действительно
перемещаются из эмбрионального периода в постэмбриональный. Но и в
этом случае причины изменения характера развития кроются не в умень-
шении количества желтка. Скорее можно предположить обратное и
искать эти причины в условиях личиночной жизни: возможность суще-
ствования сильно упрощенной личинки сделала излишними запасы пи-
тательных материалов яйца и привела к их исчезновению.
Итак, мы рассмотрели основные изменения в развитии перепончато-
крылых, непосредственно связанные с условиями их существования. Те-
перь остается сказать несколько слов о третьей категории эволюцион-
ных изменений, состоящих в усовершенствовании («рационализации»)
самого онтогенеза.
Как внешняя морфологическая картина различных эмбриональных
преобразований, так и скрытый за ней формообразовательный аппарат
250
могут испытывать наряду с изменениями адаптивного характера (напри-
мер, изменение типа дробления в связи с изменением содержания в яйце
желтка или же неизвестные нам, но несомненно происходящие преобра-
зования формообразовательных механизмов при редукции желточной
системы у насекомых) такие изменения, которые не связаны непосред-
ственно с конкретными внешними условиями. Преимущества этих изме-
нений проявляются в любых условиях и совершенствуют, делают более
простым и экономным самый процесс развития. Хорошим примером та-
кого рода может служить эволюция процессов гаструляции у, кишечно-
полостных, прослеженная И. И. Мечниковым (1886). Окончательная
элиминация из развития различных рудиментарных органов тоже мо-
жет рассматриваться как «рационализация» развития, так как при этом
упрощаются процессы и экономится клеточный материал.
На примере перепончатокрылых удается подметить лишь немногие
шаги эволюции в этом направлении, касающиеся образования зароды-
шевых листков. У первичнорастительноядных форм и у большинства Pa-
rasitica мезодерма, или мезэнтодерма, образуется наиболее распростра-
ненным у насекомых способом — путем иммиграции клеток со дна пер-
вичной бороздки внутрь. Для Aculeata и некоторых Parasitica (Euryto-
ma aciculata Ratz., Pimpla turionellae Thomps.) характерен другой спо-
соб образования внутреннего листка — путем погружения внутрь целого
эпителиального пласта—так называемой срединной пластинки. Этот
второй способ надо считать более простым и рациональным, подобно то-
му как инвагинация высших кишечнополостных есть более совершенный
способ гаструляции, чем иммиграция низших представителей этого типа
(Мечников, 1886).
В отношении дефинитивной энтодермы перепончатокрылые представ-
ляют очень пеструю картину: средняя кишка может развиваться за счет
разрастаний дна стомо- и проктодеума (Pontania), за счет пролиферации
клеток на переднем и заднем концах зародышевой полоски (Chalicodb-
ma) или только на переднем ее конце (Mestocharis), за счет отделения ча-
сти клеток от общего мезэнтодермального зачатка (Eurytoma, Angita, му-
равьи), за счет деламинации бластодермы (Caraphractus, Ageniaspis и
Platygaster ornatus, по Маршалю, 1904с, 1906) и, если верить Маршалю
(1906), даже за счет тонкой дорзальной бластодермы (Synopeas, Tri-
chacis, Inostemma). Независимо от стомо- и проктодеума путем впячи-
вания дорзальной бластодермы образуется энтодерма и у Telenomus
gracilis Мауг. (Коломиец и Коваленок, 1948).
Такое многообразие процесса вызывает недоумение. Даже если при-
знать дефинитивную энтодерму насекомых метористическим новообра-
зованием, это не разъясняет вопроса, так как трудно себе представить,
что замещение первичной энтодермы вторичной происходило у разных
представителей одного отряда независимо друг от друга и разными спо-
собами.
Какие внешние или внутренние обстоятельства могут служить при-
чиной такой значительной изменчивости одного из самых основных про-
цессов развития — образования энтодермы, — пока остается неясным.
Более понятным представляется лишь пример Prestwichia aquatica, у ко-
торой передняя и средняя кишка образуется как единое влячивание бла-
стодермы в переднебрюшной части зародыша. Этот наиболее простой и
экономный (а 'потому и наиболее совершенный) способ закладки средней
кишки _ продиктован в этом случае необходимостью в кратчайший срок
завершить эмбриональное развитие и перейти в личиночную фазу, когда
становится возможным питание извне. Рационализация процесса форми-
рования кишечника имеет здесь явно адаптивный характер.
251
Говоря об эволюционных изменениях, которым подвергается эм-
бриональное развитие перепончатокрылых, трудно обойти молчанием
такое поразительное явление, как полиэмбриония. Что означает появле-
ние полиэмбрионии Hymenoptera с точки зрения общих закономерностей
эволюции? Есть ли это изменение хода развития, связанное с изменением
взрослого насекомого, или приспособление зародыша к тем условиям, в
которых протекает его жизнь, или, наконец, «рационализация» самого
способа развития?
Очевидно, что ни одно из этих определений сюда .не подходит. По-
лиэмбриония несомненно адаптивна, но ее биологический смысл состоит
не в удовлетворении потребностей зародыша или личинки на одной ка-
кой-то стадии развития, а в увеличений численности вида. Возникнове-
ние полиэмбрионии у перепончатокрылых, пожалуй, надо рассматривать
как своеобразный филэмбриогенез, и в частности архаллаксис; ведь это
резкое нарушение обычного хода развития, которое отражается на его
конечном результате. А так как это нарушение происходит на очень ран-
ней стадии развития (во время дробления), то и эффект получается
особенно значительный: происходит не изменение в строении какого-ни-
будь органа или числа составляющих его метамеров (как это имеет
место в наиболее наглядных примерах архаллаксиса, приводимых
А. Н. Северцовым) и даже не изменение числа однозначных органов,
а изменение (увеличение) числа развивающихся из яйца индивидов.
Чтобы обзор явлений, имеющих место при эволюции онтогенеза пе-
репончатокрылых, был полным, следует еще раз отметить, что наряду с
дивергенцией большую роль играют при этом параллелизмы. Адаптации
к паразитизму и возникновение полиэмбрионии совершались независимо
друг от друга в нескольких филогенетических ветвях. Заслуживают вни-
мания также и параллелизмы в возникновении различных эмбриональ-
ных приспособлений и изменении характера развития у живородящих и
паразитических насекомых. А сходство развития и внезародышевых
структур у паразитических и живородящих насекомых, с одной стороны,
и у живородящих животных из других групп (например, млекопитаю-
щих) представляет собой пример конвергенции. Дивергенция, параллель-
ное развитие и конвергенция в такой же мере характерны для эволюции
онтогенеза, как и для эволюции взрослых животных.
СПИСОК УСЛОВНЫХ ОБОЗНАЧЕНИЙ НА РИСУНКАХ
ам
ам. с к
ан
ан. п
ант
an
б. п
бел. гр
бл
бр
бр. н
Ор. ц
в. г
вер
вер т. м
вн. бл
вн. кл
вн. х
г. л
г. м
гиг. к л
гип
глаз
глик
гр. н
д. с
д. эп
дег. гип
дег. м
дег, я
ж
ок. кл
ок. с
ок. я
жен. пр.
жир
жир. к л
жир. т
з. вн. к
3. К
зар. д
— амнион
— амниотическая складка
— анальное отверстие
— анальные придатки
— антенна (или ее зачаток)
— аподема приводящей мышцы
мандибулы
— боковая пластинка
— белковые гранулы
— бластодерма
— брюшная сторона
— брюшные ножки (или их за-
чатки)
— брюшная цепочка (или ее за-
чаток)
— верхняя губа
— веретено дробления
— вертикальные мышцы
— внезародышевая бластодерма
— внутренние клетки
— внутренний слой хориона
— головные лопасти
— головной мозг (или его зача-
ток)
—„гигантскле" клетки
— гиподерма
— зачаток глаза
— гликоген
— грудные ножки (или их за-
чатки)
— дорзальный синцитий
— дорзальный эпителий
— дегенерирующая гиподерма
— дегенерирующие миобласты
— дегенерирующие ядра
— желток
— желточные клетки
— желточный синцитий
— желточные ядра
— женский пронуклеус
— жировые включения
—жировая клетка
— жировое тело
— задние внезародышевые клет-
ки
— задняя кишка
— зародышевый диск
зар. п
зв
им. гип
им. мп. с
им. ср. к
инт
к. кут
кр
конц. отд
кут
л
л. гип
л. кут
Л. €/?. К ,
лаб. ж
м
макс
манд
манд, м
мед. п
мез
мей
миц
мп. с
муж. пр
мэнт
н.
н. г
н. мп. с
н. пл
нар. х
невр
о
ок. ком
ос. н
ос. яйцк
от. лаб. ж
п. з
пар. мп.с
пер. б
пер.вн. кл
пищ
— зародышевая полоска
— звездчатые структуры
— имагинальная гиподерма
— имагинальные мальпигиевы
сосуды
— имагинальная средняя кишка
— интеркалярный сегмент
— куколочная кутикула
— имагинальный диск крыла
— концевой отдел яичника
— кутикула
— личинка паразита
— личиночная гиподерма
— личиночная кутикула
, — личиночная средняя кишка
— лабиальная железа (или ее
зачаток)
— мышцы (или их зачатки)
— максилла (или ее зачаток)
— мандибула (или ее зачаток)
— мандибулярная мышца
— медианная пластинка
— мезодерма
—фигура деления созревания
— мицетоцит
— мальпигиевы сосуды (или их
зачатки)
— мужской пронуклеус
— мезэнтодерма
— зачаток ноги
— нижняя губа (или ее зачаток)
— непарный мальпигиев сосуд
— направительная плазма
— наружный слой хориона
— невробласты
— оосома
— окологлоточная коммиссура
— основание ноги
— основание яйцеклада
— отверстие лабиальной же-
лезы
— половой зачаток
— парный мальпигиев сосуд
— первичная бороздка
— передние внезародышевые
клетки
— пищевод
253
под. г
пол. т
пр. м -
проз
проб, м
прокт
р. я
рот
сер
сер. кут
ск
соед. кл
соед. об
СП
сп. с
ср. к
cm
стеб
стеб, отд
с том
стом. н. с
— подглоточный ганглий
— полость тела
— приводящие мышцы манди-
булы
— провентрикулюс
— продольные мышцы
— проктодеум
— редукционное ядро
— ротовое отверстие
— сероза
— серозная кутикула
— складка, очерчивающая
ласть образования энтс
мы
— соединительнотканная
клетка
— соединительнотканная
л очка
— спинная сторона
— спинной сосуд
— средняя кишка
— стигма
— стебелек яйца
об-
>дер-ч
"
обо-
— стебельчатый отдел яичника
— стомодеум
— стоматогастрическая нерв-
ная система
тх
т2
тр
троф
ут. гип
фл
ФР '
X
хв. п
Ц
ц. пл
экс
эм
эм. к л
эн
энт
эп
я. др
я. гип
я. яиц
я. ср. к
я. троф
яйц
яйце
яйцк
— передний зачаток тенториума
— задний зачаток тенториума
— зачаток трахеи
— трофамнион
— утолщение гиподермы
— флагеллум
— фрагма
— хорион
— хвостовой придаток
— целом
— центральная плазма
— экскреторные гранулы
— зародыш паразита
— эмбриональная клетка
— эноцит
— энтодерма
— эпиневральный синус
— ядро дробления
— ядро гиподермальной клетки
— ядро яйца
— ядро клетки личиночной сред-
ней КИН1КИ
— ядро трофамниона
— яйцо
— яйцевод
— яйцеклад
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Александров В. Я. и 3. И. Крюкова. 1950. Цитофизиологический анализ гибе-
ли мышц хвоста головастика при метаморфозе. Изв. АН СССР, сер. биол., № 2.
Вагин В. Л. 1947. О вторичном кожно-мускульном мешке и филогенетическом зна-
чении этого явления. ДАН СССР, LVIII, 3.
Вагнер Н. П. 1868. О развитии одной из4 форм Hymenoptera из сем. Pteromalinae.
Тр. 1-го съезда русских естествоисп. Протокол 3-го засед. Отд. зоол.
Викторов Г А. 1955. Некоторые вопросы эволюции паразитизма наездников (Hy-
menoptera). ДАН СССР, 104, 1.
Викторов Г. А. 1956. Особенности паразитизма некоторых наездников (Hymenoptera,.
Ichneumonidae, Braconidae). Энтомол. обозр., XXXV, 1.
Викторов Г. А. 1959. О происхождении паразитизма наездников. Тр. Ин-та морфол.
животных, вып. 27.'
Ганин М. С. (Ganin M.). 1869a. Beitrage zur Erkenntnis der Entwicklungsgeschichte
bei den Insekten. Zs. wiss. Zool., XIX, 3.
Ганин M. C. 18696. Ober die Embryonalhulle der I Iymenopteren und Lepidopterenem-
bryonen. Mem. de Г Acad. Imp. Sci. St. Petergsbourg, ser. 7, 14.
Ганин M. C. 1877. Материалы к познанию постэмбрионального развития насекомых.
Тр. 5-го съезда русских естествоисп. и врачей в Варшаве.
Гексли Дж. и Г. де Бэр. 1936. Экспериментальная эмбриология. Биомедгиз. М.—Л.
Давыдов К. Н. 1914. Курс эмбриологии беспозвоночных. Пг.
Давыдов К. Н. (Dawydoff С). 1928. Traite d'embryologie comparee des inver-
tebres. Paris.
Догель. 1936. Олигомеризация гомологичных органов как один из процессов эволю-
ции животных организмов. Арх. анат., гист. и эмбр., XV, 3.
Догель В. А. 1947. Курс общей паразитологии. Л., Учпедгиз.
Догель В. А. 1954. Олигомеризация гомологичных органов. Л.
Дондуа А. К. 1953. К эмбриональному развитию Scolia quadripunctata F. Зоол. журн.,
XXXII, 4.
Ежиков И. И. 1929. Метаморфоз насекомых. М.
Ежиков И. И. 1939. О типах развития многоклеточных из яйца. Сб. памяти акад.
А. Н. Северцова. I. M.—Л.
Ежиков И. И. 1940. О ранних эмбриональных стадиях и их связи с типами постэм-
брионального развития у насекомых. ДАН СССР, XXVIII, 6.
Заварзин А. А. 1945,- Рчерки эволюционной гистологии крови и соединительной тка-
• ни. Вып. 1. М., Медгиз.-
Захваткин А. А. 1949. Сравнительная эмбриология низших беспозвоночных. М.,
, изд. «Сов. наука».
Захваткин А. А. 1953. Конспект курса «Эмбриология членистоногих». Сб. научн.
работ.
Иванов А. В. 1949. Строение и развитие эндопаразитического брюхоногого моллюска
Parenteroxonos dogieli A. Ivanov (семейство Entoconchidae). Сообщ. II. Орга-
низация личинки и метаморфоз. Изв. АН СССР, сер. биол., № 2.
Иванов П. П. 1937. Общая и сравнительная эмбриология. М.—Л.
Иванов П. П. 1945. Руководство по общей и сравнительной эмбриологии. Л.
Иванова-Казас О. М. 1948а. Особенности эмбрионального развития наездников,
связанные с паразитизмом. «Усп. соврем, биол.», XXV, 1.
Иванова-Казас О. М. 19486. Волнообразное распространение митотической ак-
255
тивности при дроблении Mestocharis militaris (Hymenoptera). ДАН СССР,
LXI, 4.
Иванова-КазасО. М. 1949а. Эмбриональное развитие Anopheles maculipennis Mg.
Изв. АН СССР, сер. биол., № 2.
Иванова-Казас О. М. 19496. Карликовые зародыши Macrodytes marginatus L.
как результат заражения его яиц наездниками. ДАН СССР, LXVII, 1.
Иванова-К азас О. М. 1950. Приспособление к паразитизму в эмбриональном раз-
литии наездника Prestwichia aquatica (Hymenoptera) Зоол. ж., XXIX, 6.
И в а н о в а - К а з а с О. М. 1952а. Постэмбриональное развитие Prestwichia aquatica
(Hymenoptera). Тр. Ленингр. о-ва естествоисп., LXXI, 4.
Иванова-КазасО. М. 19526. Эмбриональное развитие наездника Mestocharis
militaris R.-Kors. (Hymenoptera Chalcididae). Энтом. обозр., XXXII.
Иванова-К азас О. М. 1954а. Об эволюции эмбрионального развития у перепон-
чатокрылых. ДАН СССР, XCVI, 6.
Иванова-К азас О. М. 19546. Влияние паразитизма на эмбриональное развитие
Caraphractus reductus R.-Kors. (Hymenoptera). Тр. Ленингр. о-ва естествоисп.,
LXXII, 4.
И в а н о в а - К а з а с О. М. 1954в. Ранние стадии развития Aphidius fabarum March.
(Hymenoptera). ДАН СССР, XCVIII, 1.
Иванова-Казас О. М. 1954 г. Зараженность яиц плавунцов наездниками в неко-
торых водоемах г. Ленинграда. Уч. зап. ЛГУ, 172, 35.
Иванова-Казас О. М. 1954д. Вопросы эволюции эмбрионального развития пере-
пончатокрылых. Тр. Всес. энтомол. о-ва, 44.
Иванова-Казас О. М. 1955. О зависимости форм дробления от содержания' в
яйце желтка. ДАН СССР, 104, 3.
Иванова-Казас О. М. 1956а. Сравнительное изучение эмбрионального развития
афидиид (Aphidius и Ephedrus). Энтомол. обозр., XXXV, 2.
Иванова-Казас О. М. 19566. О роли эмбриональной оболочки в развитии наезд-
ников из рода Aphidius (Hymenoptera Aphidiidae). Сб. «Проблемы современной
эмбриологии». Л.
Иванова-Казас О. М. 1957. Об архитектонике яйца у насекомых. Тр. Ленингр.
о-ва естествоисп., LXXIII, 4.
Иванова-Казас О. М. 1958а. Об эмбриональном развитии пилильщика Pontania
proxima Lepel. Научн. докл. высш. школы, биол., 1.
Иванова-Казас О. М. 19586. Биология и эмбриональное развитие Eurytoma
aciculata Rstzeb. (Hymenoptera, Eurytomidae). Энтом. обозр., XXXIII, 1.
Иванова-Казас О. М. 1959а. Сравнительная эмбриология и филогения у пере-
пончатокрылых. Тр. ин-та морфол. животн. АН СССР, вып. 27.
Иванова-Казас О. М. 19596. Die embryonale Entwicklung der Blatwespe Pontania
capreae L. (Hymenoptera, Tenthredinidae). Zool. Jb., Abt. Anat. und Ontog., 77, 2.
Иванова-Казас О. М. 1959в. Желточная система насекомых и ее эволюция у пе-
репончатокрылых. Вестник Ленингр. ун-та, 21.
Иванова-Казас О.М. 1960а. Развитие перепончатокрылых (Hymenoptera) и мор-
фологические закономерности эволюции. Вестник Ленингр. ун-та, 9.
Иванова-Казас О. М. 19606. Эмбриональное развитие Angitia vestigialis Ratz.
(Ichneumonidae) внутреннего паразита пилильщика Pontania capreae L. Энтом.
обозр., XXXIX, 2.
Иванова-Казас О. М. 1961. К вопросу об эмбриональном развитии Ephedrus
plagiator Nees. Научн. докл. высш. школы, биол.
Иофф И. А. 1948. К вопросу об эмбриональном развитии наездников. ДАН СССР,
IX, 8.
Канаев И. И. 1959. Близнецы. М.—Л., Изд. АН СССР.
Караваев В. A. (Karawaiew W.). 1898. Die nachembryonale Entwicklung' von
Lasius flavus. Zs. wiss. Zool., LXXV.
Ковалевский А. О. (Kowalevsky A.). 1871. Embryologische Studien an Wur<
mem und Arthropoden. Зап. Петерб. акад. наук, сер. 7, XVI, 12.
Ковалевский А. О. 1885. Beitrage zur nachembryonalen Entwicklung der Musciden.
Zool. Anz., 8.
Ковалевский А. О. 1886. Embryonalentwicklung der Musciden. Biol. Zbl., 6.
Ковалевский А. О. 1887. Beitrage zur Kenntnis der nachembryonalen Entwic-
klung der Musciden. Zs. wiss. Zool., XLV.
Кожевников Г. A. (Koschevnikov). 1900. Ober den Fettkorper und die Oeno-
cyten der Honigbiene (Apis mellifera L.). Zool. Anz., XXIII.
Коломиец Н. Г. и А. В. Ковале но к. 1958. Биология яйцееда Telenomus gracilis
Мауг (Hymenoptera,. Scelionidae). Изв. Сибирск. отд. АН СССР, № 10.
Коротнев A. (Korotnef f А.). 1892. Histolyse und Histogenese des Muskelgewe-
bes bei Metamorphose der Insekten. Biol. Zbl., XII.
Кузнецов Н. Я. 1953. Основы физиологии насекомых. М.— Л.
256
Кулагин Н. М. (Kul a gin). 1892. Zur Entwicklungsgeschichte der parasitischen
Hautflugler. Zool. Anz., 15.
Кулагин Н M. 1894. Материалы по естественной истории паразитических перепон-
чатокрылых- (Platygaster, Mesochorus, Microgaster). Изв. Ими. о-ва любит, есте-
ствознания, антроп. и этнографии, LXXXV.
Лазаре н ко Ф. (Lazarenko F.). 1925. Beitrage zur vergleichenden Histologie
des Biutes und des Bindegewebes. II. Die Morphologische Bedeutung der Blut-
und Bindegewebeelemente der Insekten (Experimentelle Untersuchung). Zs. mikr.
anat. Forsch., 3, 4.
Ларченко К. И. (L а г t s с h e n k о). 1933. Die Unempfanglichkeit der Raupen von
Loxostege sticticalis und Pieris brassicae gegen Parasiten. Zs. Parasitenk., 5, 4.
Ларченко К. И. 1937. Цикл развития жирового тела лугового мотылька и озимой
совки и его связь с созреванием и плодовитостью. Энтом. обозр., XXVII, 1—2.
Ларченко К. И. 1946. Роль клеток крови и жирового тела в цикле развития вино-
градного червеца (Pseudococcus citri R.). Изв. АН СССР, сер. биол., 2—3.
Ларченко К. И. 1956. Закономерности онтогенеза насекомых. Тр. Зоол. ин-та АН
СССР, XXIII.
Малышев С. И. 1947. Пути и условия эволюции низших перепончатокрылых в связи
с вопросом о происхождении наездников (Hymenoptera). ДАН СССР, VIII, 6.
Ма л ышев С. И. 1949а. Пути и условия эволюции инстинктов низших перепончато-
крылых (Symphyta и Terebrantia). Ж. общ. биол., X, 1.
Малышев С. И. 19496. Пути и условия эволюции осообразных перепончатокрылых
(Vespoidea и Sphecoidea). ДАН СССР, XV, 4.
Малышев С. И. 1950. Пути и условия происхождения пчелиных (Hymenoptera Apoi-
dea). ДАН СССР, XXII, 5.
Малышев С. И. 1956. Пути и условия происхождения паразитоидпых перепончато-
крылых (Hymenoptera Terebrantia s. parasitica). ДАН СССР, 109, 5.
Малышев С. И. 1957. Происхождение вторичной фитофагии у наездников (Hyme-
noptera terebrantia s. parasitica). Тр. Ленингр. о-ва естествоисп., ХХШ, 4.
Малышев С. И. 1959. Перепончатокрылые, их происхождение и эволюция. М.
Малышев С. И. и Е. В. Пузанова-Малышев а. 1952. Инквилинизм у пилиль-
щиков (Hymenoptera, Tenthredinidae) в условиях эксперимента. Тр. Ленингр.
о-ва естествоисп., LXXI, 4.
Машков пев А. А. 1935. Роль функции в развитии морфологических структур у по-
звоночных животных. К проблеме механики индивидуального развития. Усп. совр.
биол., IV, 3.
Машковцев А. А. 1936. Смена эндогенных и экзогенных факторов эмбрионального
развития в онтогенезе и филогенезе. Изв. ОМЕН АН СССР, сер. биол., 5.
Me и ер Н. Ф. 1925а. Несколько слов об Angitia fenestraiis Holmgr. (Ichneumonidae) и
об иммунитете у насекомых. «Защ. раст.», 3.
Мей ер Н. Ф. 19256. Об иммунитете у некоторых гусениц по отношению к их пара-
зитам — наездникам. Изв. Гос. ин-та опыт, агр., 3*
Мей ер Н. Ф. (Meyer N.). 1926a. Biologie von Angitia fenestraiis Holmgr., des Pa-
rasiten von Plutella maculipennis Curt, und einige Worte uber Immunitat der In-
sekten. Zs. angew. Entom., 12, I.
Мейер Н. Ф. (Meyer N. F.). 19266. Ober die Immunitat einigen Raupen ihren Para-
siten, den Schlupfwespen, gegenuber. Zs. angew. Entom., 12, 2.
Мейер Н. Ф. 1937. Биологический метод борьбы с вредными насекомыми. Л.
Мечников И. И. (MecznikowE.). 1866. Embryologische Studien an Insekten.-
Zs. wiss. Zool., 16, 4; To же в русском переводе в Академич. собр. соч. Изд.
АМН, 1955.
Насонов Н. В. 1886. К постэмбриональному развитию муравья Lasius flavus. Прото-
колы Зоол. отд. о-ва любителей естеств., 1.
Никольская М. Н. 1952. Хальциды фауны СССР. Л., Изд. АН СССР.
Никольская М. Н. 1956. Хальциды-семееды фavны СССР и роль фитофагии в эво-
люции группы (Hymenoptera, Chalcidoidea)."Энтом. обозр., XXXV, 3.
Никольская (Nikolskaja M. N.). 1958. Geographical distribution and evoluti-
onary affinities of Chalcididae and Leucospidae (Hym., Chalcidoidea), XV Intemat.
Congr. Zool., papers reed in title, 5. London.
Оглоблин A. A. 1913. К биологии божьих коровок (Coleoptera, Coccinellidae). Русск.
энтом. обозр., 14.
Оглоблин (Ogloblin А.). 1924. Le role du blastoderme extraembryonnaire du Di-
nocampus terminatus Nees, pendent l'etat larvaire. Vestnik Krai. Ces. Spolec
Nauk. Tf. II. У
Поспелов В. 1911. Постэмбриональное развитие и имагинальная диапауза чешуе-
крылых. Зап. Киевск. о-ва естеств., XXI, 3.
Поярков Э. (Poyarkoff). 1910. Recherches histologiques sur la metamorphose
d'un Coleoptere (la Galeruque de l'Orme). Arch. anat. microsc, XII.
17 О. М. Иванова-Казне осу
Поярков Э. 1914. Опыт теории куколки насекомых с полным превращением. Тр.
Русск. энтомол. о-ва, X, 1. _
Римский-Корсаков М. Н. 1916. К фауне русских водных наездников. Русск.
энтом. обозр., XVI, 3—4.
Римский-Корсаков М. Н. 1925. О водных наездниках из рода Caraphractus Ha-
liday. Тр. Ленингр. о-ва естествоисп., IV/ 2.
Римский-Корсаков М. Н. (Rimsky-Korsakow). 1932. Methoden zur Unter-
suchung von Wasserhymenopteren. Handbuch biolog. Arbeitsmethoden v. Abder-
halden, IX, 7. т v w
Римский-Корсаков M. H. 1940. Перепончатокрылые (Hymenoptera). «Жизнь
пресных вод СССР», 1.
Светлов П. Г. 1928. Исследование над развитием дождевых червей. Тр. Особой
зоол. лабор. и Севастоп. биол. станции, сер. II, 11—13.
Светлов П. Г. 1959. Субституции при образовании зародышевых листков. Тр. Ин-
та морфологии животных АН СССР, вып. 27.
Северцов А. Н. 1934. Морфологические закономерности эволюционного процесса.
N M., Изд. Сорена, вып. 3.
Северцов А. Н. 1939. Морфологические закономерности эволюции. М. —Л.
•Сергеев А. М. 1943. Эволюция эмбриональных приспособлений у рептилий. М., «Сов.
наука». ^
С к р и п ч и н с к и й Г. 1930. К биологии Aphidius granarius March, и Ephedrus plagiator
Nees (Braconidae), паразитов черемухово-овсяной тли, Aphis padi L. Изв.
ГИОА, IV.
Теленга Н. А. 1950. К вопросу об использовании паразитов сем. Aphidiidae в борьбе
с мигрирующими тлями. Научн. труды Ин-та энтом. и фитопатол., № 2.
Теленга Н. А. 1952. Происхождение и эволюция паразитизма у насекомых-наездни-
ков и формирование их фауны в СССР. Киев, Изд. АН УССР.
Титова Л. К. 1940. Особенности роста ларвальных и эмбриональных имагинальных
элементов у Pieris brassicae. ДАН СССР, XXVII, 7.
Титова Л. К. 1949. Особенности роста ларвальных и имагинальных элементов в личи-
ночной стадии развития Pieris brassicae. Уч. зап. ЛГУ, № 113, сер. биол., 20.
Токин Б. П. 1955. Иммунитет зародышей. Л.
Токин Б. П. 1958. Эмбриональное развитие и среда. Вестник Ленингр. ун-та, 21.
Токин Б. П. 1959. Регенерация и соматический эмбриогенез. Изд. Ленингр. ун-та.
Фаусек В. А. 1913. Биологические этюды. Живорождение <и паразитизм. СПб.
Федотов Д. М. 1940. Явления редукции органов в онтогенезе зимней пяденицы
(Operopthera brumaia L.). Сб. памяти академика А. Н. Северцова, II.
Филатов-Д. П. 1939. Сравнительно-морфологическое направление в механике раз-
вития, его объект, цели и пути. М.—Л., Изд. АН СССР.
Филатов Д. П. 1943. Механика развития как метод изучения некоторых вопросов
эволюции. Ж. общ. биол., IV, 1.
Ч v n р о в а Е. (Tschfiproff W.). 1903. Ober die Entwicklung der Keimblatter bei den
Libcllen. Zoo!. Anz., 27.
Шаров А. V. 1953. Развитие щетинохвосток (Thysanura, Aplerygota) в связи с про-
блемой филогенеза насекомых. Тр. Ин-та. морфол. живот:, 8.
Ш.аров А. Г. 1957. Сравнительно-онтогенетический метод и его применение в си-
стематике и филогении (на примере насекомых). Зоол. ж., XXXVI, 1,
Шаров А. Г. 1959. Смена зачатков в эмбриональном развитии и ее связь с измене-
ниями условий существования. Журн. общ. биол., XX, 2.
Ш в а н в и ч Б. Н. 1949. Курс общей энтомологии. М., «Сов. наука».
Шевырев И. 1912. Паразиты и сверхпаразиты из мира насекомых. СПб.
Шимкевич В. М. 1908. Меторизис как эмбриологический принцип. Изв. Акад. паук
Шмальгаузен И. И. 1938. Организм как целое в индивидуальном и историче-
ском развитии. М.—Л., Изд. АН СССР.
Шмальгаузен И. И. 1946. Проблемы дарвинизма. М., «Сов. наука».
Шмидт Г. А. 1951—1953. Эмбриология животных. М., «Сов. па\ка».'
Шовен Р. 1953. Физиология насекомых. М.
Штейн бе р г Д. М. 1938а. Регуляционные процессы при метаморфозе насекомых.
I. Экспериментальные исследования над регенерацией крыльев v бабочек. Биол
ж.. VII, 2.
Штейпберг Д. М. 19386. Регуляционные процессы при метаморфозе у насекомых. II.
Эмбриональная территория крыла и ноги в гиподерме гусениц. Биол. ж., VII, 5— 6.
Штейпберг Д. М. 1948. Формообразовательные возможности личиночной гиподер-
мы насекомых. Докт. дисс. Л., Зоол. ин-т, АН СССР.
Штейпберг Д. М. 1956. Пути эволюции метаморфоза членистоногих. Сб. «Пробле-
мы современной эмбриологии». •
Яхонтов В. В. 1940. Полиэмбриония (гермипогонин) у насекомых «Yen соврем
биол.». 13. 3. '
258
A n g 1 a s J. 1902. Nouvelles observations sur les metamorphoses internes. Arch. anat.
microsc, V.
A n gl a s J. 1904a. De Г origin des cellules de remplacement de l'intestin chez les Hyme-
nopteres. С r. Soc. bioK Paris, LVI.
A n g 1 a s J. 19046. Du role des trachees dans le metamorphose des insectes. С. г. Soc.
biol. Paris, 56.
A n g 1 a s J. 1904b. Rapports du developpement de Г appareil trachees et des metamorpho-
ses chez les insectes. С. г. Acad, sci., Paris, CXXXVII.
Baerends С P. and J. M. BaerendsVan Roon. 1949. Embryological and eco-
logical investigations on the development of the egg of Ammophila campestris.
Jur. Tijdschr. Ent., 92.
Bauer V. 1904. Zur inneren Metamorphose des Centralnervensystems der Insekten.
Zool. Jb., Abt. Anat., 20.
Beirne B. P. 1942. Observations on the life-history of Praon volucre Haliday, a para-
site of the mealy plum aphis (Hyalopterus arundinis Fab.). Proc. Roy. ent. soc.
London (A), 17.
Be г land L. 1951. Order des Hymenopteres, reproduction. Traite de Zool., X, fasc. I.
Berlese A. 1901. Vorgange, welche wahrend der Nymphosis der metabolischen Insek-
ten vorkommen. Zool. Anz., XXIV.
Bledowski R. und M. К. К г a i n s к a. 1926. Entwicklung von Banchus
femoralis Thorns (Hymenoptera, Ichneumonidae). Bibl. Univ. Liberae Poloniae,
fasc. 16.
Boese G. 1936. Der Einfluss tierischer Parasiten auf den Organismus der Insekten.
Parasitenkunde, 8.
Bronskill J. F. 1959. Embryology of Pimpla turionellae (L.) (Hymenoptera, Ichneu-
monidae). Can. j. zool., 37.
Buchner P. 1954. Endosymbiosestudien an Schildlausen. I. Stictococcus sjoestedti. Zs.
Morph. u. Okol. d. Tiere, 43, 3.
Buchner P. 1955. Endosymbiosestudien an Schildlausen. II. Stictococcus diversiseta.
Zs. Morph. u. Okol. d. Tiere. 43, 5.
Buchner P. 1957. Endosymbiosestudien an Schildlausen. VI. Die nicht in Symbiose
lebende Gattung Apiomorpha und ihre ungewohnliche Embryonalentwicklung. Zs.
Morph. u. Okol. d. Tiere, 46, 5.
Butschli O. 1870. Zur Entwicklungsgeschichte der Bijene. Zs. wiss. Zool., 20.
Carriere J. and O. Burger. 1898. Die Entwicklungsgeschichte der Mauerbiene (Cha-
licodoma muraria Fabr.) im Ei. Nova Acta Abhand. Kaiserl. Leop.-Carol. Deutsch.
Acad. Naturf., LXIX, 2.
С a u I 1 e г у М. 1922. Lc parasitisme et la symbiose. Paris.
С h г у s t a 1 E. W. 1930. Studies of the Sirex parasites. Oxford. Forestry mem., II.
Clancy D. W. 1934. The biology of Tetracnemus pretiosus Timberlake. Calif. Univ.
Publ. Ent., 6.
С 1 a n с у D. W. 1944. The biology of Allotropa burrelli, a gregarious parasite of Pseudo-
coccus comstocki. J. agric. res., 69, 4.
Clausen C. 1940. Entomophagous insects. N. Y.
Crawford A. W. 1933. Glypta rufiscutellaris, an ichneumonid larval parasite of the
• oriental fruit moth. N. Y., Agr. Exp. Sta. Tech. Bull., 217.
Cue not L. 1896. Etudes physiologique» sur les Orthopteres. Arch, biol., 14.
Daniel D. M. 1932. Macrocentrus ancylivorus Rohwer, a polyembryonic braconid pa-
rasite of the oriental fruit moth. N. Y., Agr. Sta., Tech. Bull., 187.
D e I a g e J. 1884. Evolution de la Sacculine. Arch. zool. exp. gen. (2), 2.
Dickel O. 1904. Entwieklungsgeschichtliche Studien am Beienenei. Zs. wiss.
Zool., 77.
D о b г о v s к у Т. W. 1951. Postembryonic changes in digestive tract of the worker honey
bee, (Apis meilifera L.). Cornell Univ. Agr. Exp. Sta., Mem., 301.
Doutt R. L. 1947. Polyembryony in Copidosoma koehleri Blanchard. Amer. nat., 81.
Doutt L. 1952. The teratoid larva of polyembryonic encyrtidae (Hymenoptera). Canad.
ent., 84 (8).
Doutt R. L. 1959. The biology of parasitic Hymenoptera. Ann. review entom., 4.
E d e D. A. and S. J. С о u n с e. 1956. A cinematographic study of the embryology of Dro-
sophila melanogaster. Roux' Arch. Entw.-Mech., 148.
F e r r i e r e Ch. 1926. Note sur un chalcidien a developpement polyembryonique. Rev.
Suisse Zool. Geneve, 33.
Flanders S.E. 1938. Starvation of developing parasites as an explanation of immunity.
J. econ. entom., 30.
Flanders S. E. 1942. Sex differentiation in the polyembryonic proclivity of the Hyme-
noptera. J. econ. entom., 35.
Flanders S. E. 1946. Control of sex and sex-limited polymorphism in the Hymenopte-
ra. Quart, rev. biol., 21, 2.
Fridmann H. 1934. Ein Beitrag zur Kenntnis der embryonalen Entwicklung der
17*
259
Abdominalfiisse der Schmetterlingsraupen. Soc. Sci. Fennica, Commentationes
biol., 4 (10).
G a d d С H. 1946. Macrocentrus homonae, a polyembryonic parasite of the tea tortrlx
(Homona coffearia). Ceylon j, sci. (В), 23-, 2.
Gatenby J. B. 1917. The embryonic development of Trichogramma evanescens, mo-
nembryonic egg parasite of Donacia simplex. Quart, j. micr. sci., 62, 2.
Gatenby J. B. 1918a. The segregation of the germ-cells in Trichogramma evanescens.
Quart, j. micr. sci., 63, 2.
Gatenby J. B. 1918b. Polyembryony in parasitic Hymenoptera. Quart, j. micr. sci., 63.
Gatenby J. B. 1919. Notes on the bionomics, embryology and anatomy of certain Hy-
menoptera parasites. J. Linn, soc, XXXIII.
Gatenby J. B. 1920. The cytoplasmic inclusions of the germ-cells IV. On the origin
and probable constitution of the germ-cell determinant of Apanteles glomeratus
with a note on the secondary nuclei. Quart j.'micr. sci., 64, 2.
G г a b e г V. 1890 Vergleichende Studien am Keimstreifen der Insekten. Denkschr. Kai-
serl. Akad. Wiss. Wien, Math.-Nat. CI., 57.
Grassi B. 1884. Intorno alio sviluppo delle api nell'uovo. Atti Acad. Gioenia Sc. Nat.,
Catania, 18.
Gronroos H. 1895. Zur Entwicklugsgeschichte des Erdesalamanders (Salamandra ma-
culosa Laur.) I. Fortpflanzung, Ovarialei, Furchung, Blastula. Anat. Hefte, I Abt., 18.
Hagan H. R. 1951. Embryology of the viviparous insects. N. Y.
Hand lirsch A. 1908. Die fossilen Insekten und die Phylogenie der rezenten Formen.
Leipzig.
H a v i 1 a n d M. S. 1921. On the bionomics and postembryonic development of certain
Cynipid hyperparasites of Aphids. Quart, j. micr. sci., 65, 3.
Hecht O. 1924. Die Embryonalentwicklung von Camponotus. Zs. wiss. Zool., 122.
Hegner R. W. 1911. Experiments with chrysomelid beetles. III. The effects of Killing
parts of the e^ of Leptinotarsa decemlineata. Biol, bull., 20.
Hegner R. W. 1914. Studies on" germ cells. J. morph., 25.
H e g n e г R. W. 1915. Studies on germ cells. IV. Protoplasmic differentiation in the oocy-
tes of certain Hymenoptera. J. morph., 26.
HenneguyL F. 1892. Contribution a Pembryogenie des Chalcidiers. С. г. Acad, sci.,
CXIV.
Henneguy L. F. 1904. Les insectes. Paris.
Henri к sen K. 1922. Notes upon some aquatic Hymenoptera. Ann. biol. lacustre, II,
fasc. I.
Henschen W. 1928. Ober die Entwicklung der Geschlechtsdrusen von Habrobracon
juglandis Ashm. Zs. Morph. u. Okol. d. Tiere, 13, 1/2.
Heymons R. 1896. Entwicklungsgeschichtliche Untersuchungen an Lepisma sacchan-
na. L. Zs. wiss. Zool., 62.
Heymons R. 1897. Ober die Bildung und den Bau des Darmkanals bei niederen Insek-
ten. Sitzb Ges. Naturf. Freunde, 111 — 119.
Heymons R. 1912. Ober den Genitalapparat und die Entwicklung von Hemimerus
talpoides Walker. Zool. Jb., Suppl., 15, 2.
H in ton H. E. 1955. On the structure, function and distribution of the prolegs of tiie
Panorpoidea, with a criticism of the Berlese-Imms theory. Trans. Roy. ent. soc.
Loridon, 106.
Jackson X) J. 1928. The biology of Dinocampus (Perilitus) rutilus, a braconid parasi-
te of Sitona lineata. Proc. Zool. soc. London. 597—630.
Jackson D. J. 1938. Giant cells in insects parasitized by Hymenoptera larvae. Na-
ture, 135.
Jackson D. J. 1958. Observations on the biology of Caraphractus cinctus Walker (Hy-
menoptera, Mymaridae), a parasitoid of the eggs of Dytiscidae. Trans. Roy. ent.
soc. London, 110, 17.
J a m es H. C. 1928. On the life-history and economic status of certain cynipid parasites of
dipterous larvae with description of some new larvae forms. Ann. appl. biol., 15, 2.
J an eke O. 1932. Die Kirschblutenmotte (Argyresthia prinella) und ihr Parasit (Ageni-
aspis atricollis Dalm.). Gartenbauwissenschaft, 6.
Janischewska J. 1933. Untersuchungen tiber die Hymenoptere Aphidius sp.,-Para-
siten der Blattlaus Hyalopterus pruni Fabr. Bull. Intern. Acad. Polonaise, Class,
Math, et Nat., Ser. B.
Johannsen O. and F. Butt. 1941. Embryology of insects and myriapods. N. Y.
К a hie W. 1908. Die Paedogenesis der Cecidomyiden. Zoologica (Stuttgart), XXI, 5—6.
Kleinenberg N. 1879. The development of the earth-worm Lumbricus trapezoides
Duges. Quart, j. micr. sci., 19.
Kornhauser S. J. 1919. The sexual characteristics of the membracid Thelia bimacu-
lata (Fabr.). I. External changes induced by Aphelopus theliae (Gahan). J. morph.,
32, 3.
260
Korschelt E. and K. H e i d e г 1936. Vergleichende Entwicklungsgeschiehte der
Tierc. Iena.
Кг a use G. 1939. Die Eitypen der Insekten. Biol. Zentralblatt, 59, 9—10.
К га use G. 1953. Die Aktionsfolge zur Gestaltung des Keimstreifens von Tachycines
(Saltatoira), insbesondere das morphogenetische Konstruktionsbild bei duplicitas
parallela. Roux'Arch. Entew.-Mech., 146, 3.
Кг a use G. und J. Krause, 1957. Die Regulation der Embryonalanalage von Tachy-
cines (Saltatoria) im Schnitt versuch. Zool. Jb., Abt. Anat., 75, 4.
Kuhn A. 1955. Vorlesungen iiber Entwicklungsphysiologie. Berlin.
Leiby R. W. 1922. The polyembryonic development of Copidosoma gelechiae, with notes
on its biology. J. morph., 37.
L e i b у E. W. 1926. The origin of mixed broods in polyembryonic Hymenoptera. Ann. Eat.
soc. Amer., 19.
Leiby R. W. 1929. Polyembryony in insects. IV. Intern. Congr. Entom., Inthaca, II.
Leiby R. W. and V. C. Hill. 1923. The twinning and monembryonic development of
Platygaster hiemalis, a parasite of the hessian fly. J. Agr. res.. XXV, 8.
Leiby R. W. and С. С. Hill. 1924. The polyembryonic development of Platygaster
vernalis, J. Agr. res., XXVIII, 8.
L и b b e n H. 1907. Uber die innere Metamorphose der Trichopteren. Zool. Jb., Abt.
Anat., 24.
Mac Bride E. W. 1914. Textboox of embryology. London.
M а с к a u e r M. 1959. Histologische Untersuchungen an parasitierten Blattlausen. Zs.
Parasitenkunde, 19, 4.
Marchal P. 1898a. La dissociation de l'oeuf d'Encyrtus fuscicollis. Ann. nat. hist..
ser., 7, II.
Marchal P. 1898b. A new method of asexual reproduction in hymenopterous insects.
Nat. sci., 12.
Marchal P. 1898c. Un exemple de dissociation de l'oeuf. Le cycle de l'Encyrtus fuscicol-
lis. С. г. soc. biol., 5.
Marchal P. 1898d. Le cycle evolutif de l'Encyrtus fuscicollis. Bull. Soc. entom. France,
109—111.
Marchal P. 1898e. La dissociation de Pceuf en un grand nombre d'indivi-
dus distinctes et le cycle evolutif chez l'Encyrtus fuscicollis. C. r. Acad, sci., 126.
Marchal P. 1899. Comparaison entre de developpement des hymenopteres parasites
a developpement polyembryonnaire et ceux a developpement monoembryonnaire.
C. r. soc. biol., I.
Marchal P. 1903.Le cycle evolutif du Polygnotus minutus. Bull. Soc. entom. France, 72.
Marchal P. 1904a. La determinisme de la polyembryonie specifique et la determinisme
du sexe chez les hymenopteres a developpement polyembryonnaire. С. г. soc. biol., 56
Marchal P. 1904b Recherches sur la biologie et le developpement des hymenopteres
parasites. I. La polyembryonie specifique ou germinogonie. Arch. Zool. exper. et
gen., 4 ser., 2.
Marchal P. 1904c. Sur la formation de Tintestin moyen chez les Platygasters. C. r.
soc. biol., 56.
Marchal P. 1906. Recherches sur la biologie et le developpement des Hymenopteres.
Arch. Zool. exper. et gen., 4 ser., 4.
Marshall W. S. and P. H. Dernell. 1905. Contributions toward the embryology and
anatomy of Polistes pallipes. Zs. wiss. Zool., 80.
Martin F. 1914. Zur Entwicklungsgeschiehte des polyembryonalen Chalcidiers Ageni-
aspis (Encyrtus) fuscicollis Dalm. Zs. wiss. Zool., 110.
M e 1 1 a n b у К. 1939. The function of insect blood. Biol. rev. Cambridge Phil, soc, 14, №13.
Miller M. 1957. Entwicklung und Bedeutung der Vitellophagen in der Embryonalen-
twicklung der Honigbiene. Zool. Jb., Abt. allg. Zool., 67, I.
Murray F. and O. W. T i e g s. 1953. The metamorphosis of Calandra oryzae. Quart.
j. micr. sci., 77.
Nachtsheim H. 1913. Cytologische Studien iiber die Geschlechtsbestimmung bei der
Honigbiene (Apis mellifera). Arch. Zellforsch., II.
Needham J. 1931. Chemical embryology. Cambridge.
Nelson J. A. 1915. The embryology of the honey-bee. Princeton Univ. Press.
Noskiewicz J. und G. Polyszynski. 1927. Embryologische Untersuchungen an
Strepsipteren. I. Embryogenesis der Gattung Stylops Kirby. Bull, inter. Г Acad. Po-
lonaise, Kl. Math, et Nat., ser. В., 3.
Noskiewicz J. und G. Poluszynski. 1935. Embryologische Untersuchungen an Strep-
sipteren. II. Polyembryonie. Zool. Polon., I.
Novak V. J. A. 1959. Insektenhormone. Praha.
О е г t e 1 E. 1930. Metamorphosis in the honey-bee. J. morph., 50.
P ai 1 1 ot A. 1934. Modifications cytologiques et organiques engendrees chez les pucerons
par les Hymenopteres. С. г. Acad, sci., 199.
261
P a i 1 1 ot A. 1940. Contributions a l'etude du developpement polyembryonnaire d'Amicro-
plus collaris Spin., braconide parasite d'Euxoa segetum Schiff. Ann. epiphytes, 6.
Parker H L. 1931a. Macrocentrus gifuensis. Aschmead., a polyembryonic braconid pa-
rasite' in the European corn borer. Univ. St. Dept. Agr., Tech. Bull., 230.
Parker H. L. Notes on Meteorus (Zemiotes) nigricollis Thompson, an occasional para-
site of the European corn borer. Proc. Ent. Soc. Wach., 33.
Parker H. L. et W. R.Thompson. 1928. Contribution a la biologie des Chalcidiens
entomophages. Ann. Soc. Ent. France, 97.
P a t а у R. 1939. Contributions a l'etude d'un Coleoptere (Leptinotarsa decimlineata
(Say.) Evolution des organes au course du developpement. Rennes.
Patterson J. T. 1915. Observation on the development of Copidosoma gelechiae. BioL
bull., 29.
Patterson J. T. 1917. Studies on the biology of Paracopidosomopsis. I. Data on sexes.
Biol, bull., 32.
Patterson J. T. 1918. Studies on the biology of Paracopidosomopsis. IV. Asexual
larvae. Biol, bull., 35.
Patterson J. T. 1921. The development of Paracopidosomopsis. J. morph., 36.
Patterson J. T. 1927. Polyembryony in animals. Quart, rev. biol., 2, 3.
Perez Ch. 1901. Sur quelques points de la metamorphose de fourmie. Bull. Soc.
& entom., 22.
Perez Ch. 1901. Histolyse des tubes de Malpighi et des glandes sericigenes chez la
fourmi rousse. Bull. Soc. entom., 307.
Perez Ch. 1901. Sur les oenocytes de la fourmi rousse. Bull. Soc. entom., 351.
Perez Ch. 1910. Recherches histologiques sur la metamorphose des Muscides (Callipho-
ra). Arch. biol. exper. et gen., ser., 5, 4.
Perez Ch. 1912. Observations sur l'histolyse et I'histogenese dans le metamorphose des
Vespides (Polistes gallica L.). Mem. Acad. Roy. Belg., 2 ser., 111.
Pfliigfelder O. 1934. Bau und Entwicklung der Spinnendrusen der Blattwespen. Zs.
wiss. Zool., 145.
Pfliigfelder O. 1952. Entwicklungsphysiologie der Insekten. Leipzig.
Reinhardt E. 1960. Kernverhaltnisse, Eisystem und Entwicklungsweise von Drohnen-
und Arbeiterinneneiern der Honigbiene (Apis mellifera). Zool. Jb., Abt. Anat. u..
Ontog., 78, 2.
Reith F. 1931. Versuche uber die Determination der Keimesanlage bei Camponotus lig-
niperda. Zs. wiss. Zool., 139.
Reith F. 1932. Uber Lokalisation der Entwicklungsfaktoren im Insektenkeim. I. Zentri-
fugierungsversuche an Ameiseneiern. Roux'Arch. Entw.-Mech., 127.
Richards Av G. and A. Miller. 1937. Insect development analysed by experimen-
tal methods: a review. J. New York Entom. soc, XLV.
Robertson С W. 1936. The metamorphosis of Drosophila melanogaster, including an
accurately timed account of the principal morphogenetic changes. J. morph., 59.
Roonwal M. L. 1935. Fate of embryonic membranes in insects. Current sci., IV, 5.
R о on w a 1 M. L. 1936. Growth changes and structure of the egg of the African migra-
tory locust, Locusta migratoria migratorioides. Bull. Ent. res. Lond., 27.
Ross H. 1955. The evolution of the insect orders. Entom. news, 66, No 8.
Salt G. 1955. Experimental studies in insect parasitism. VIII. Host reactions following
artificial parasitization. Proc. Roy. soc, В., 144, No 916.
Salt G. 1956. Experimental studies in insect parasitism. IX. The reactions of a stict in-
sect to an alien parasitie. Proc Roy. soc. B, 146, No ,922.
S a 1 t G. 1957. Experimental studies in insect parasitism. X. The reactions of some endop-
terygote insects to an alien parasitie. Proc. Roy. soc, В., 147, No 927.
Sander K. 1960. Analyse des ooplasmatischen Reaktionsystems von Euscelis plebejus
Fall (Cicadina) durch Isolierung und Kombinierung von Keimtheilen. Roux'Arch.
Entw.-Mech., 151, 5.
Sarra R. 1915. Observazioni biologiche sulTAnarsia lineata Z. dannosa al frutto del
mandorlo. Boll. Lab. Zool. R. Scuola Agric. Portici, 10.
Sarra R. 1918. La Variegana ed i suoi parassiti. Boll. Lab. Zool. R. Scuola Agric
Portici, 12.
Sauer E. 1954. Keimblatterbildung und Differenzierungsleistungen in isolierten Eiteilen
der Honigbiene. Roux'Arch. Entw.-Mech., 147.
Schmiedeknecht O. 1930. Die Hymenopteren Nord- und Mitteleuropas. Jena.
Schnetter M. 1934a. Morphologische Untersuchungen uber das Differenzierungszen-
trum in der Embryonalentwicklung der Honigbiene. Zs. Morph. u. Okol. Tiere, 29,
Schnetter M. 1934b. Physiologische Untersuchungen uber das Differenzierungszen-
trum in der Embryonalentwicklung der Honigbiene. Roulx'Arch. Entw.-Mech., 131.
Schnetter M. 1936. Die Entwicklung von Zwerglarven im geschniirten Bienenei-
Verhandl. Deutsch. Zool. Ges., 38.
262
Shafiq S. A. 1954. A study of the embryonic development of the goosberry sawfly,
Ptcronidea ribesii. Quart, j. micr. sci., 95, 1.
Seidel F. 1931. Die Reaktionsfolge im Determinationsgeschehen des Libelienkeimes.
Verhandl. Deutsch. Zool. Ges., 34.
Seidel F-1934. Das Differenzierungszentrum im Libellenkeim. I. Die dynamischen Vo-
raussetzungen der Determination und Regulation. Roux'Arch. Entw.-Mech., 131, 2.
Seidel F. 1936. Entwicklungsphysiologie des Jnsektenkeimes. Verhandl. Deutsch. Zool.
Ges., 38.
Seidel F., E. Bock und. G. Krause. 1940. Die Organisation des Insekteneies. Na-
turwiss., 28.
Silvestri F. 1906. Sviluppo dell' Ageniaspis fuscicollis. Atti reale accad. Lincei, 15, 1.
Silvestri F. 1907. Contribuzioni alia conescenza biologica degli Imenotteri parassiti.
I. Biologia del Litomastix truncatellus (Dalm.). Boll. Lab. Zool Scuola Agr. Por-
tici, 1.
Silvestri F. 1908. Contribuzioni alia conoscenza degli Imenotteri parassiti. II. Svi-
luppo dell'Ageniaspis fuscicoll.is. Ann. Regia Scuola Super. Agr. Portici, ser. 2, 8.
Silvestri F. 1909a. Appunti della Prospalta berlesei. How. Boll. Lab. Zool. Scuola.
Super. Agric. Portici, III.
Silvestri 1909b.. Contribuzioni alia, conoscenza degli Imenoteri parassiti. Boll. Lab.
Zool. Scuola Super. Agric. Portici, III.
Silvestri F. 1910. Notizie preliminari sullo sviluppo del Copidosoma buyssoni. Moni-
tore Zool. Ital., 21.
Silvestri F. 1914. Prime fasi di sviluppo del Copidosoma buyssoni (Mayr), Imenotte-
ro chalcididae. Anat. Anz., 47.
Silvestri F. 1916. Contribuzioni alia conoscenza biologica degli Imenotteri parassiti.
V. Sviluppo del Platygaster Dryomyiae Silv. Boll. Lab. Zool. Gen. Agr. R. Scuoia.
Sup. Agr. Portici, 11.
Silvestri F. 1915—1916. Struttura dell'ovo e prime fasi di sviluppo di alcuni Imenotte-
ri parassiti. Boll. Lab. Zool. Scuola Super. Agr. Portici, X.
Silvestri F. 1923. La Tignola nanella (Recurvaria nanella Hiibn.) Bull. Lab. Zool.
Scula Super. Arg. Portici, XVI.
Silvestri F. 1924. Contribuzioni alia conoscenza dei Torlricidi delle Querce. Boll.
Lab. Zool. Scuola Super. Agr. Portici, XVII.
Silvestri F. 1937. Insect polyembryony and its general biological aspects. Bull. Mus.
Compar. zool. at Harvard College, 81.
.S i m m о n d s F. J, 1947. The biology of the parasites of Loxostege sticticalis L. in North
America—Bracon vulgaris (Cress,) (Braconidae, Agathinael. Bull, eritom. res.,
38, 1. \ i
S 1 i f er E. IT. 1938. The formation and structure of a special waterabsorbing area in the
membranes covering the grasshopper egg. Quart, j. micr. sci., 80, 3.
S .1 i f e r E. H. 1946. The effects of xylol and other solvents on diapause in the grasshop-
per e^, together with a possible explanation for the action of these agents. J.
exper. zool., 102, 3.
S lifer E. H. 1948. Isolation of a wax-like material from the shell of the grasshopper
egg. Discussions of the Faraday soc, No 3.
Smith O. J. 1952. Biology and behavior of Microctonus vittatae Muesbeck (Braconidae)
with description of its immature stages. Univ., Calif. Public, in Entom., 5..
Smreczynski St. 1939. Entwicklungsmechanische Untersuchungen am Ei dcs Kafers
Agelastica alni L. Zool. Jb., Abt. allg. Zool., 59.
S n о d g r a s s R. E. 1925. Anatomy and physiology of the honey-bee. N. Y.
Snod grass R. E. 1954. Insect metamorphosis. Smithson. miscell. collections, 122, 3.
Snow S. J. 1925. Observations on the cutworm Euxoa auxiliaris Grote, and its princi-
pal parasites. Л. ecqn. entom., 18.
Speicher B. R.1936. Oogenesis, fertilization and early cleavage in Habrobracon. J.
morph., 59.
Spencer H. 1926. Biology of the parasite and hyperparasite of aphids. Ann. Ent. soc.
Amer., XIX, 2.
Strickland E. H. 1923. Biological notes on parasites of prairie cutworms (Meteorus
dimidiatus—Braciniclae). Canada. Dept. Agr. ent. branch, bull., 22.
Strindberg H. 1913a. Embryologische Studien an Insekten. Zs. wiss. Zool., 106.
Strindb-erg H. 1913b. Einige Studien der Embryonalentwicklung bei Myrmica rubra
unter besonderer Berucksichtigung der sogenannten Entodermfrage. Zool.
Anz., 41.
Strindberg H. 1914. Zur Kenntnis des Hymenopterenentwicklung. Zs. wiss. Zool., 112.
Strindberg H. 1915. Zur Entwicklung der Hymenopteren nebst einigen clamit zu-
sammenhangenden Fragen. Zool. Anz., 45.
Strindberg IT. 1916a. Noch eine Ameise ohne Serosa (Tertramorium caespitum).
Zool. Anz., 46.
Strindberg II. 1916b. Azteca sp.,. eine Ameise mit totaler Furchung. Zool. Anz., 48.
263
Strindberg H. 1918. Neue Studien uber Ameisenembryologie. Zool. Anz., 49.
Strindberg H. 1919. Die Eifurchung von Tapinoma erraticum. Zool. Anz., 50.
Stuart R. R. 1935. The development of midintestine in Melanoplus differentialis. J.
morph., 58.
Sun Saochen 1959. The embryonic development of Delorus tritici Chu. Acta entom.
sinica, IX, No 1.
Tanquary N. С 1913. Biological and embryological studies of Formicidae. Bull. Illi-
nois State Lab. Nat. Hist., 9.
Thompson W. R. 1930a. Entomophagous parasites and phagocytes. Nature,125, No 3144.
Thompson W. R. 1930b. Reaction of the phagocytes of Arthropodes to their internal
insect parasites. Nature, 125, No 3154.
T h о г p e W. H. 1936. On a new type of respiratory interrelation between an insect (Chal-
cid) parasite and its host (Coccidae). Parasitology, 28. No 4.
Tiegs O. W. 1922 Researches on the insect metamorphosis. Trans. Roy. soc. S. Austra-
lia, 46.
Timber lake P. H. 1912: Experimental parasitism. A study of the biologie Limnerium
validum (Cresson). U. S. Dept. Agr. Bur. Entom. Plant Quarantine, Entom. Tech.,
ser. 19.
Tower D. G. 1915. Biology of Apanteles militaris. J. agr. res., V, No 12.
Van Leewen W. 1908. Beitrage zur Kenntnis der Metamorphosen. Die mikroskopische
Anatomie des Darmkanals, und dessen Drtisen von Isosoma graminicola Giraud.
Tijdschr. Nederl. Dierk. Vereen, 2 ser., XI.
Vance A. M. 1931. Apanteles thompsoni Lyle, a braconid parasite of the European corn
borer. U. S. Dept. Agr. Tech. Bull., No 233.
W a hi B. 1901. Ober die Entwicklung der hypodermalen Imaginalscheiben in Thorax und
Abdomen von Eristalis Latr. Zs. wiss. Zool., 70.
Weber H. 1954. Grundriss der Insektenkunde. Jena.
Weissenberg R. 1907. Ober die Oenocyten von Torymus nigricornis Roh., mit beson-
derer Beriicksichtigung der Metamorphose. Zool. Jb., Abt. Anat., XXIII.
Z i rn gi e b 1 L. 1940. Beitrage iiber die Entwicklung der Blattwespeneieren (Hvm. Tenth-
red.). Deutsche Entom. Zs., 181 — 189.
ОГЛАВЛЕНИЕ
Стр.
Введение 3
СПЕЦИАЛЬНАЯ ЧАСТЬ
Глава I. Pontania capreae L. — ивовый толстостенный пилильщик 7
1. Некоторые данные по биологии —
2. Набухание яиц и проницаемость хориона
3. Эмбриональное развитие ... S
Длительность эмбрионального развития
Описание развития по стадиям 10
4. Личинка I возраста 37
Глава II. Angitia vestigialis Ratz. 39
1. Биология . .
2 Эмбриональное развитие 40
3. Сравнение развития Angitia vcsligialis с таковым других Ichneumonidae
и пилильщиков . . . - .48
Г.шва III. Ephedrus plag'iator Nees . 50
1. Биология
2 Эмбриональное развитие -г>1
3. Палингеиетические черты в развитии Ephedrus plagiator 59
Глава IV. Aphidius fabarum Marsh, и другие виды Aphidius 61
1. Эмбриональное развитие —
2. Роль эмбриональной оболочки в развитии Aphidius 6.г>
3. Сравнение развития Ephedrus и Aphidius ... 71
Глава V. Charips sp. . 73
Глава VI. Eurytoma aciculata Ratz. . - 77
1. Биология .... . . .. —
2 Эмбриональное развитие . . 78
3. Личинка I возраста 92
4. Адаптивные и своеобразные черты в развитии Eurytoma aciculata
Глава VII. Ageniaspis luscicollis (Dalm.) 95
1. Литературные данные о развитии Ageniaspis —
2. Некоторые наблюдения по биологии 98
3. Оболочки зародыша 99
Трофамиион 100
Циста и ее роль в отношениях между паразитом и хозяином 106
Глава VIII. Mestocharis militaris R.-Kors 114
1. Биология .... —
2 Эмбриональное развитие 116
265
Глава IX. Caraphractus reductus R.-Kors 124
1. Биология .... 125
2. Эмбриональное развитие
Глава X. Prestwichia aquatica Lubb. 136
1. Биология ... —
2. Эмбриональное развитие 137
3. Личинка .... 143
4. Метаморфоз 146
Стадии постзмбрионального развития —
Гиподерма 152
Нервная система ... . . 155
Трахейная система 157
Эноциты ... 158
Мускулатура ... 160
Кровеносная система 168
Жировое тело ... . . 171
Пищеварительная система . 174
Половая система 185
Основные черты метаморфоза Prestwichia . 188
5. Влияние паразитизма на развитие Prestwichia .' 190
ОБЩАЯ ЧАСТЬ
Глава I. Основные направления эволюции эмбрионального развития перепон-
чатокрылых ... .192
1. Примитивный тип развития пилильщиков .... . —
2. Тип развития низших Parasitica и большинства Aculeata 193
3. Тип развития муравьев 195
4. Развитие монэмбрионических наездников с трофической оболочкой 199
5. Полиэмбрионический тип развития 204
6. Крайне дезэмбрионизированнын тип развития некоторых эндопаразитиче-
ских наездников 212
7. Преемственность между основными типами перепончатокрылых . . 214
Глава II. Сравнительная эмбриология, и филогения у перепончатокрылых 215
1. Подотряд Phytophaga
2. Подотряд Parasitica 216
Надсемейство Ichneumonoidea —
Надсемейство Cynipoidea 217
Надсемейство Chalcidoidea —
Надсемейство Serphoidea . 219
3. Подотряд Aculeata : . 220
4. Заключение . 221
Глава III. Приспособления к паразитизму в эмбриональном развитии перепон-
чатокрылых 224
1. Уменьшение размеров яиц . 226
2. Переход от поверхностного дробления к тотальному . —
3. Эволюция желточной системы ... ... 228
4. Полиэмбриония .... 233
5. Эволюция эмбриональных оболочек 237
6. Некоторые аналогии между живорождением и эмбриональным парази-
тизмом ... . 243
Глава IV. Развитие перепончатокрылых и морфологические закономерности
эволюции 246
Список условных обозначений на рисунках 253
Цитированная литература .... 2Г>Г>
Иванова-Казас* Ольга Михайловна
•Омерки по сравнительной эмбриологии
перепончатокрылых
Редактор О. Л. Петровичсва
Техн. редактор С. Д. Водолагина
Корректоры И. Т. Земсково
и М. В. Унковская
Сдано и набор 28 IV 1961 г. М-06320
Подписано к печати 28 X 1961 г.
Уч.-изд. л. 21,82. Печ. л. 16,75 (условных
22,95). Бум. л. 8,38.
Формат бум. 70X108Vi<i.
Тираж 1300 экз. Заказ 468.
Типография ЛОЛ ГУ. Ленинград,
Университетская наб., 7/9.