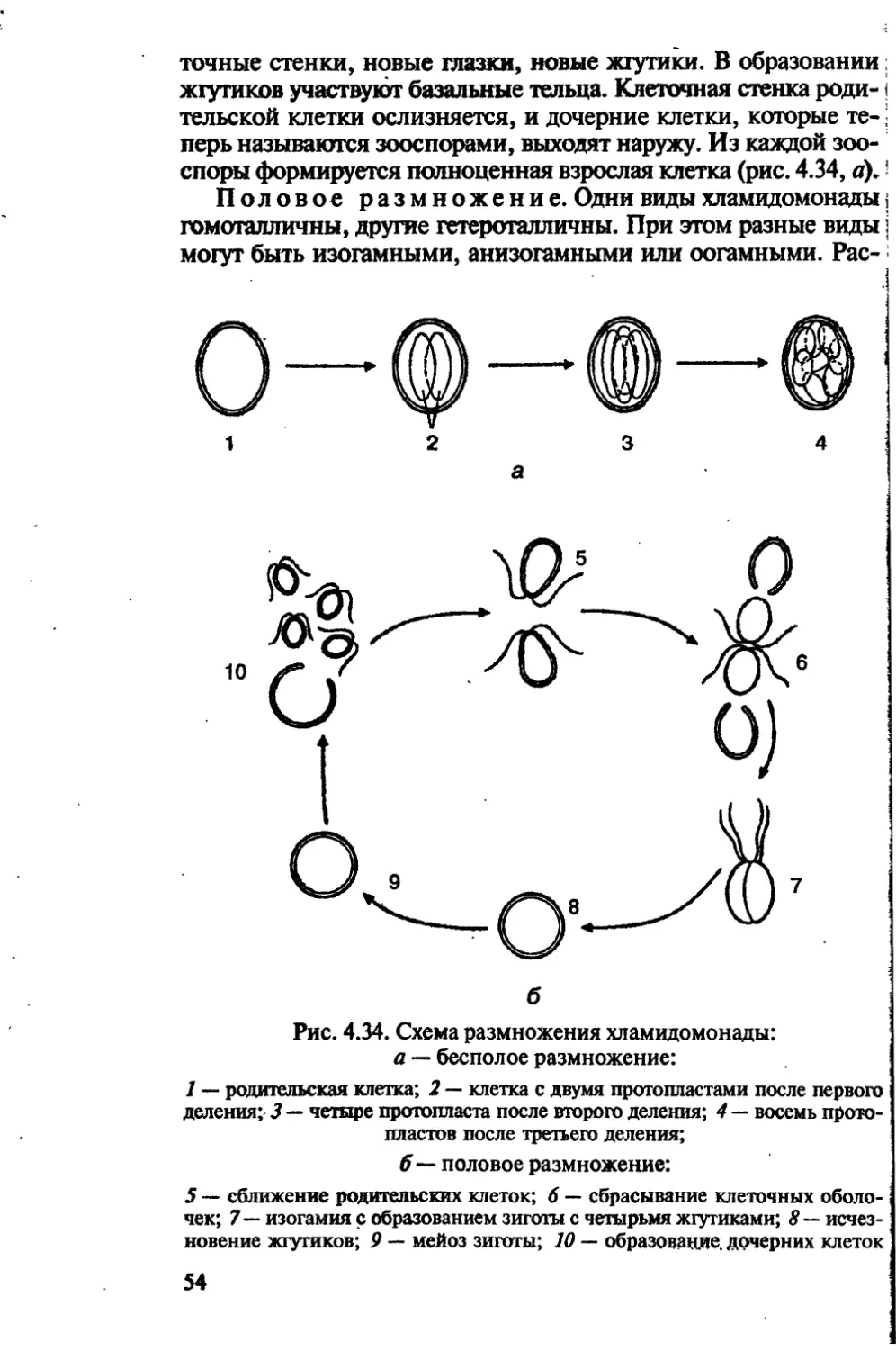
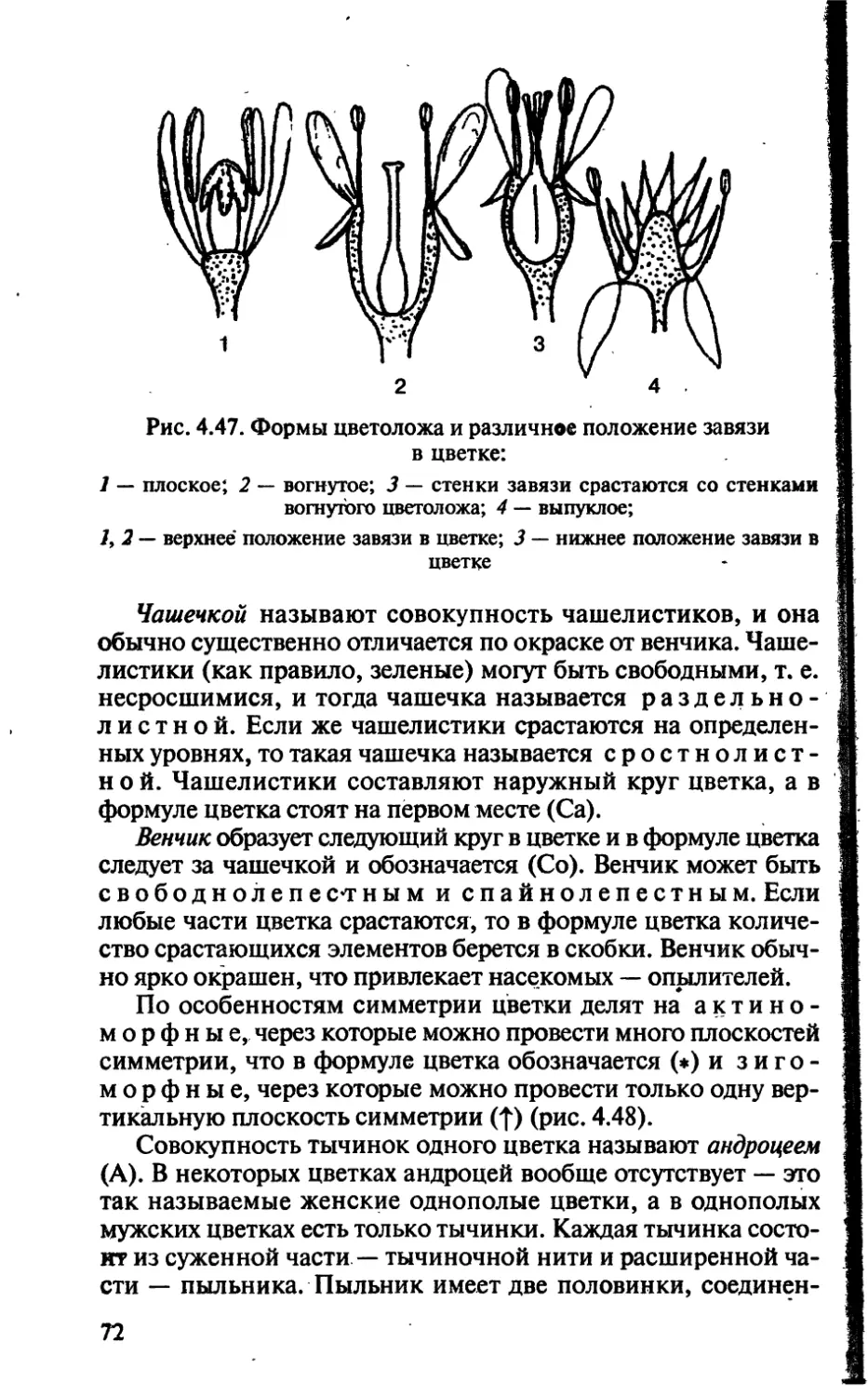
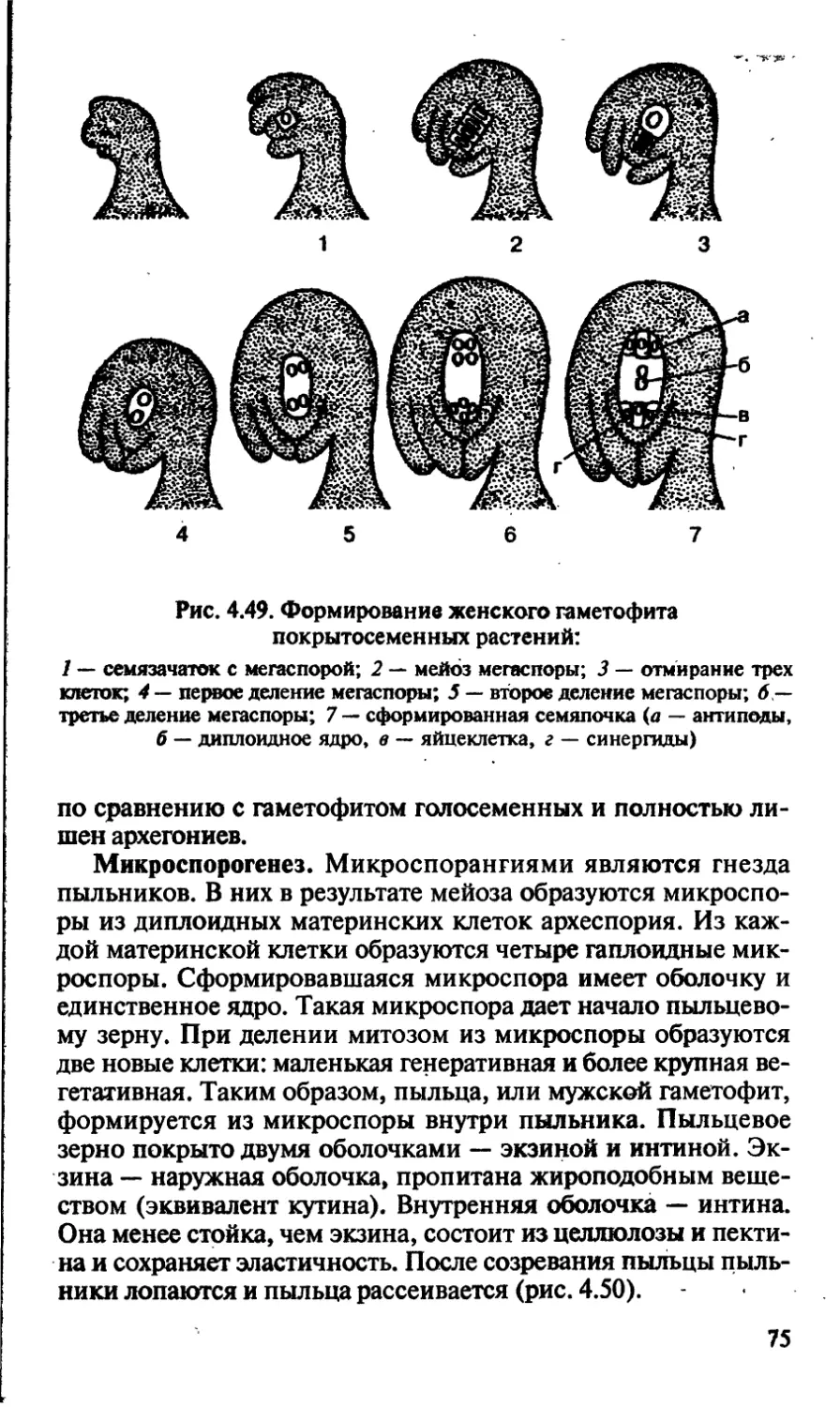
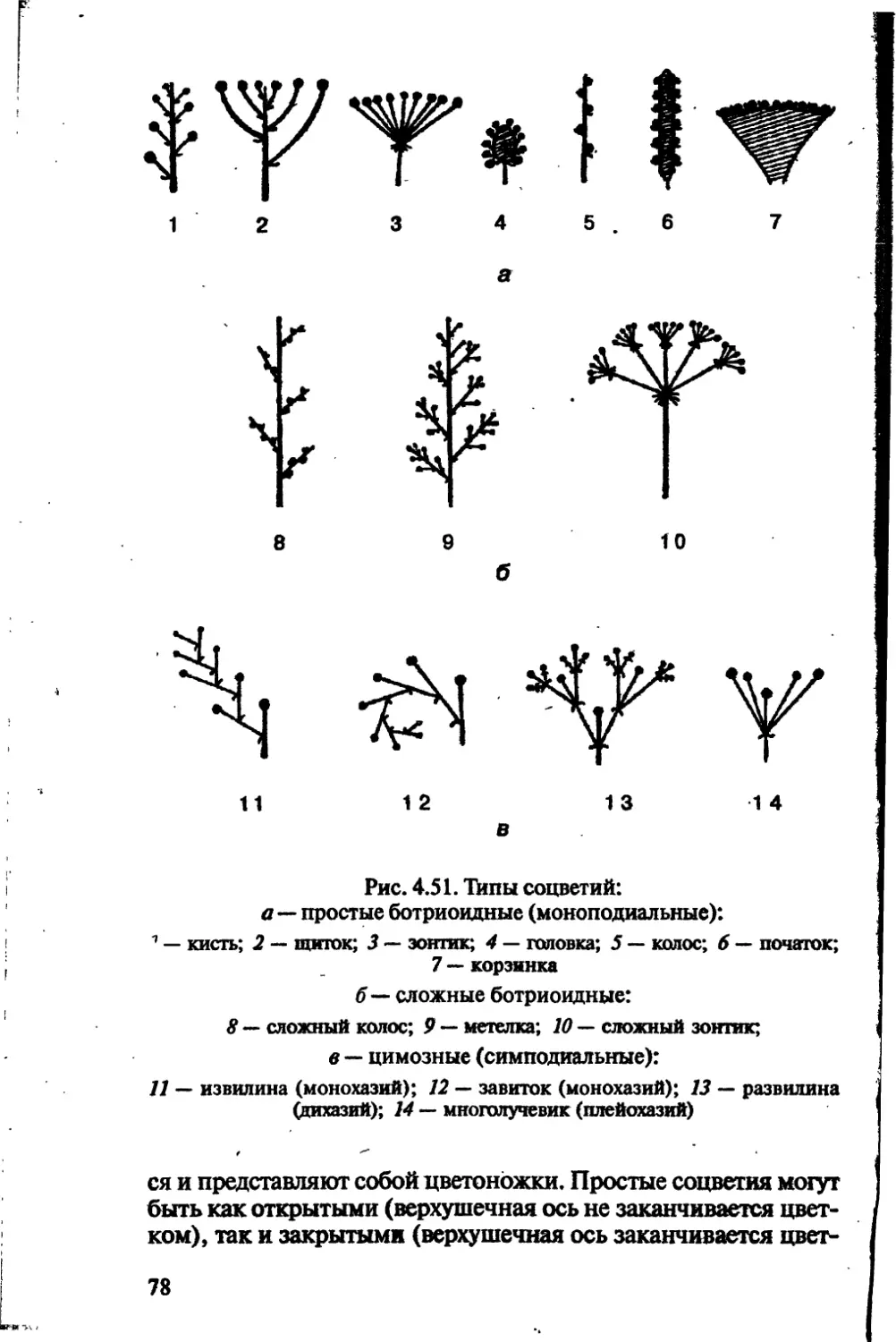
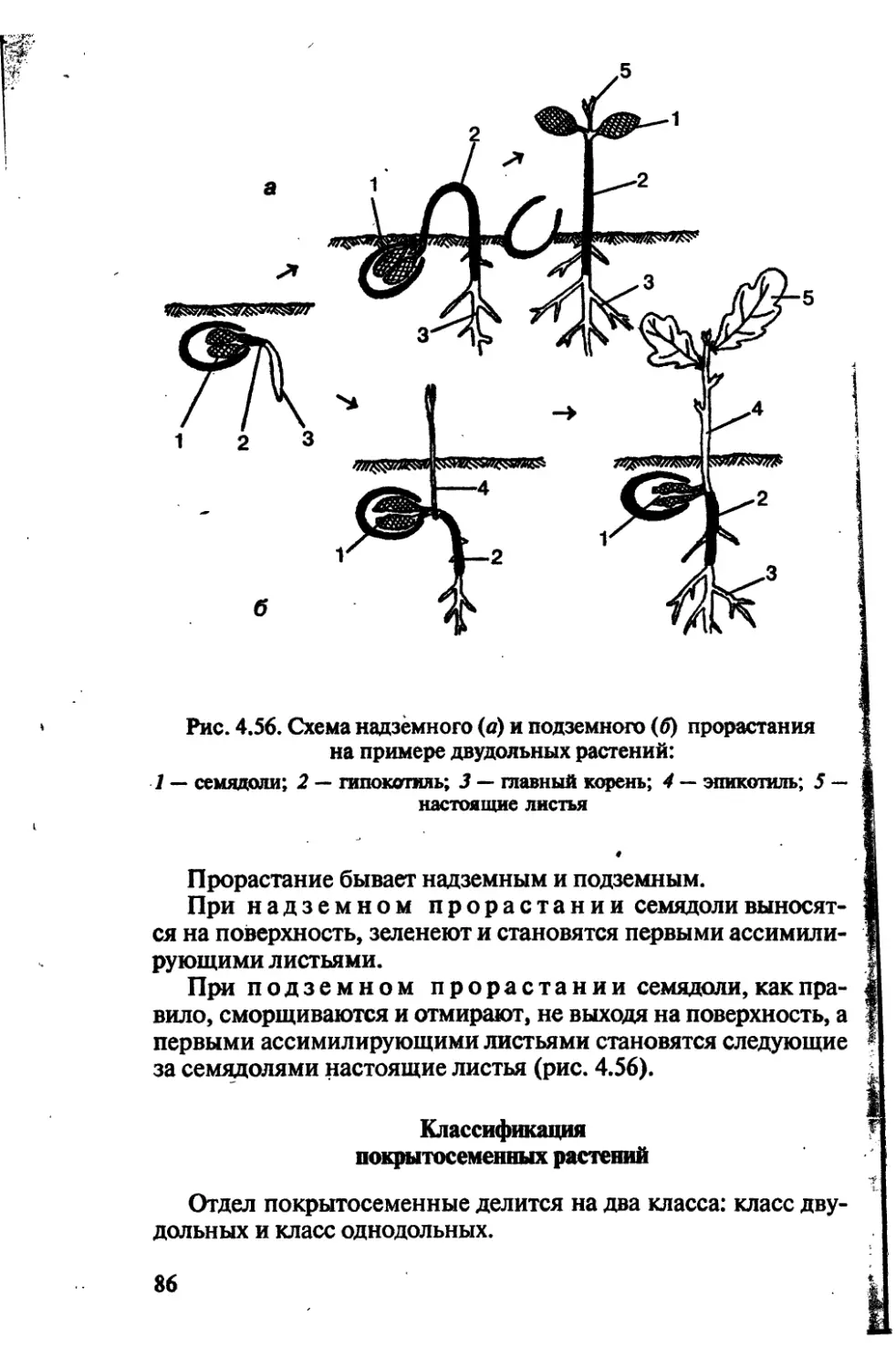
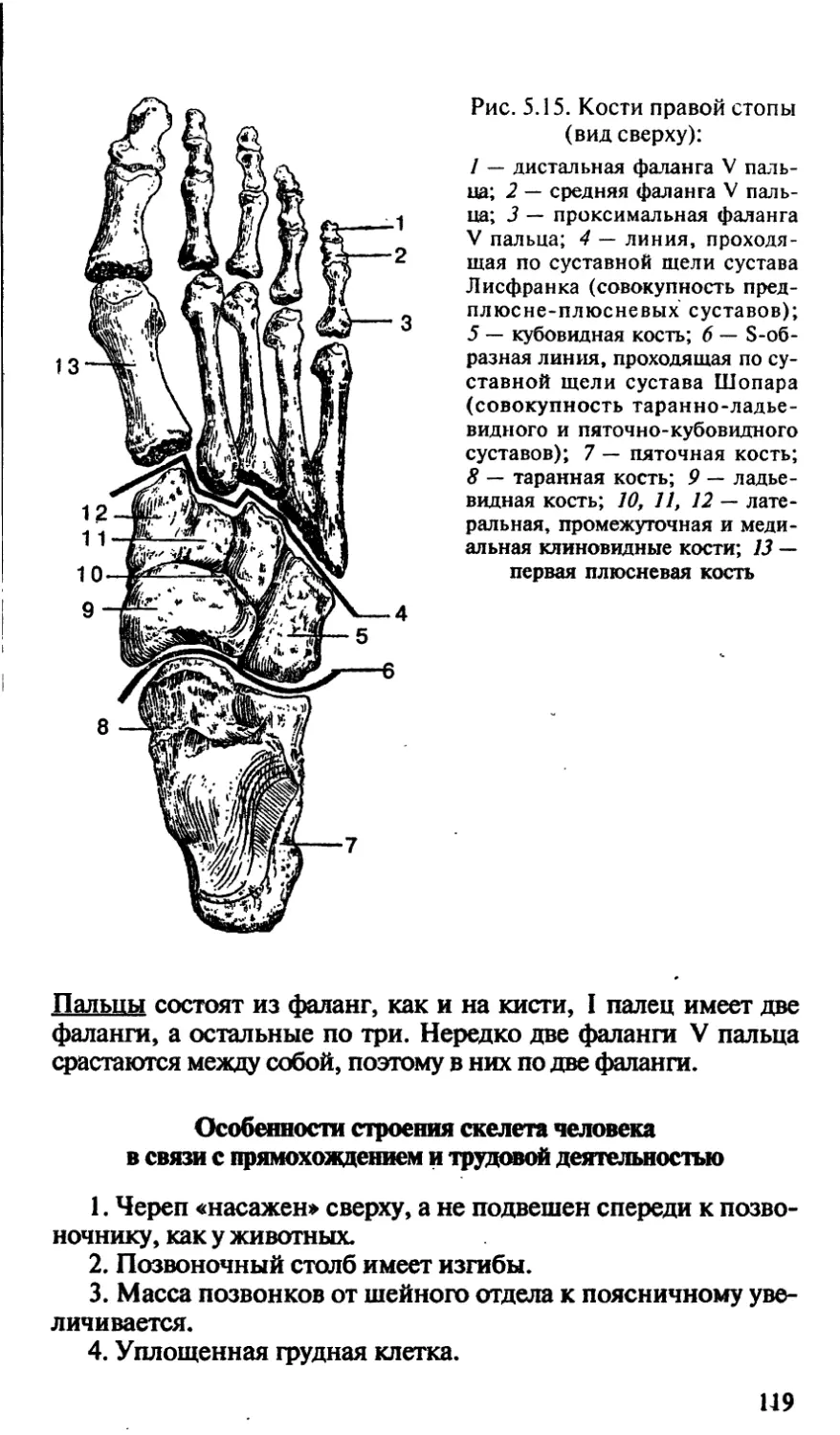
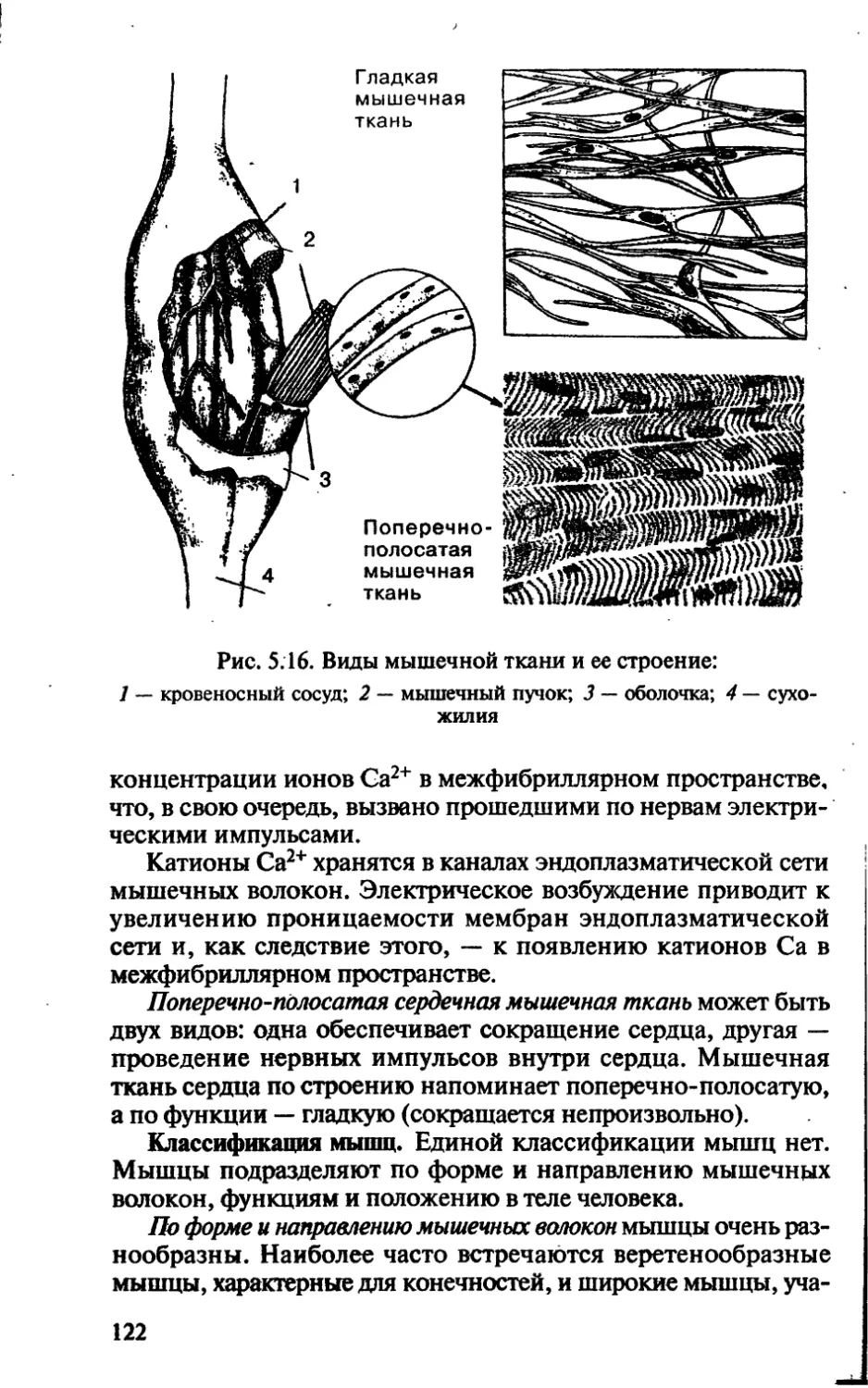
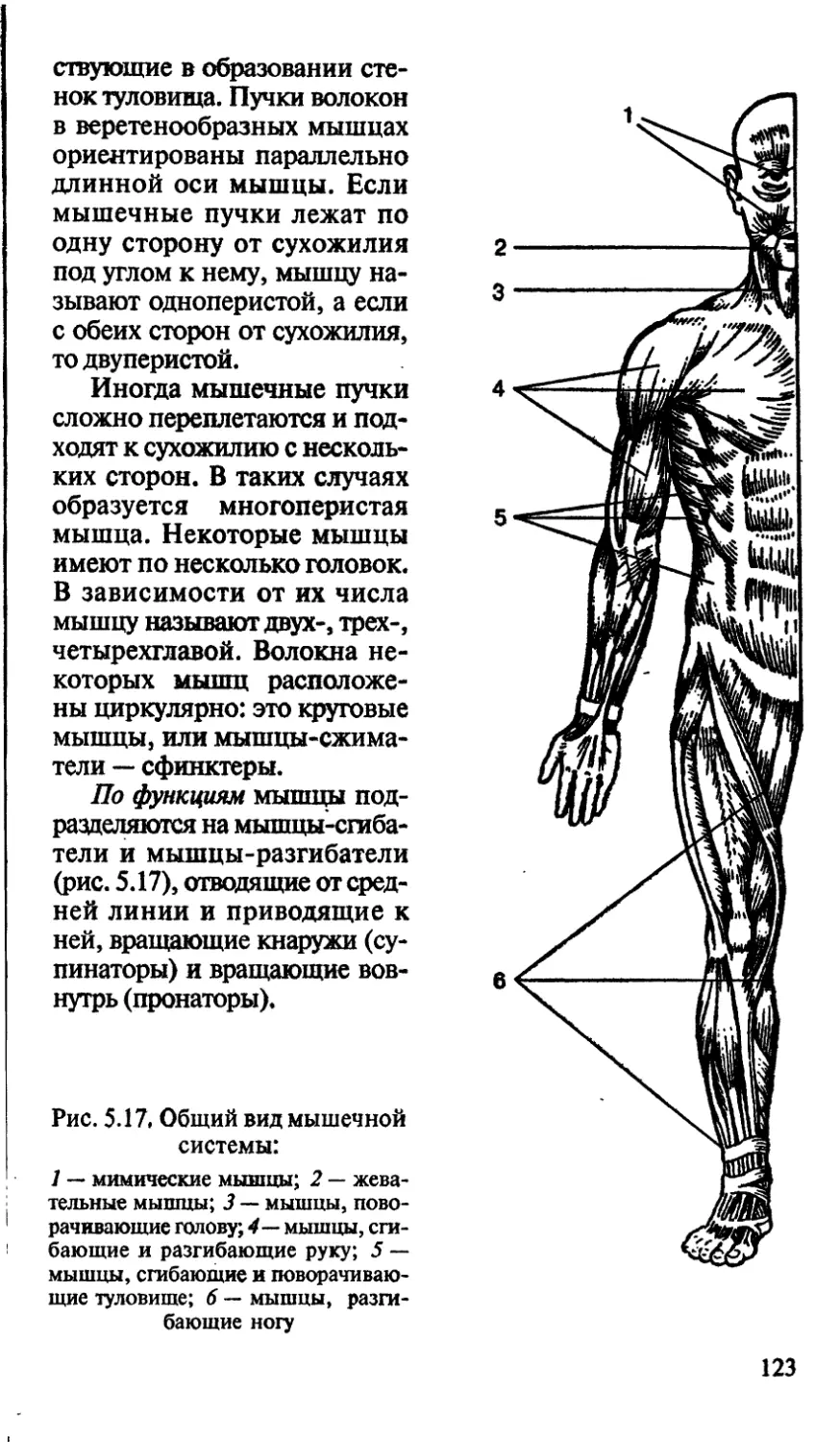
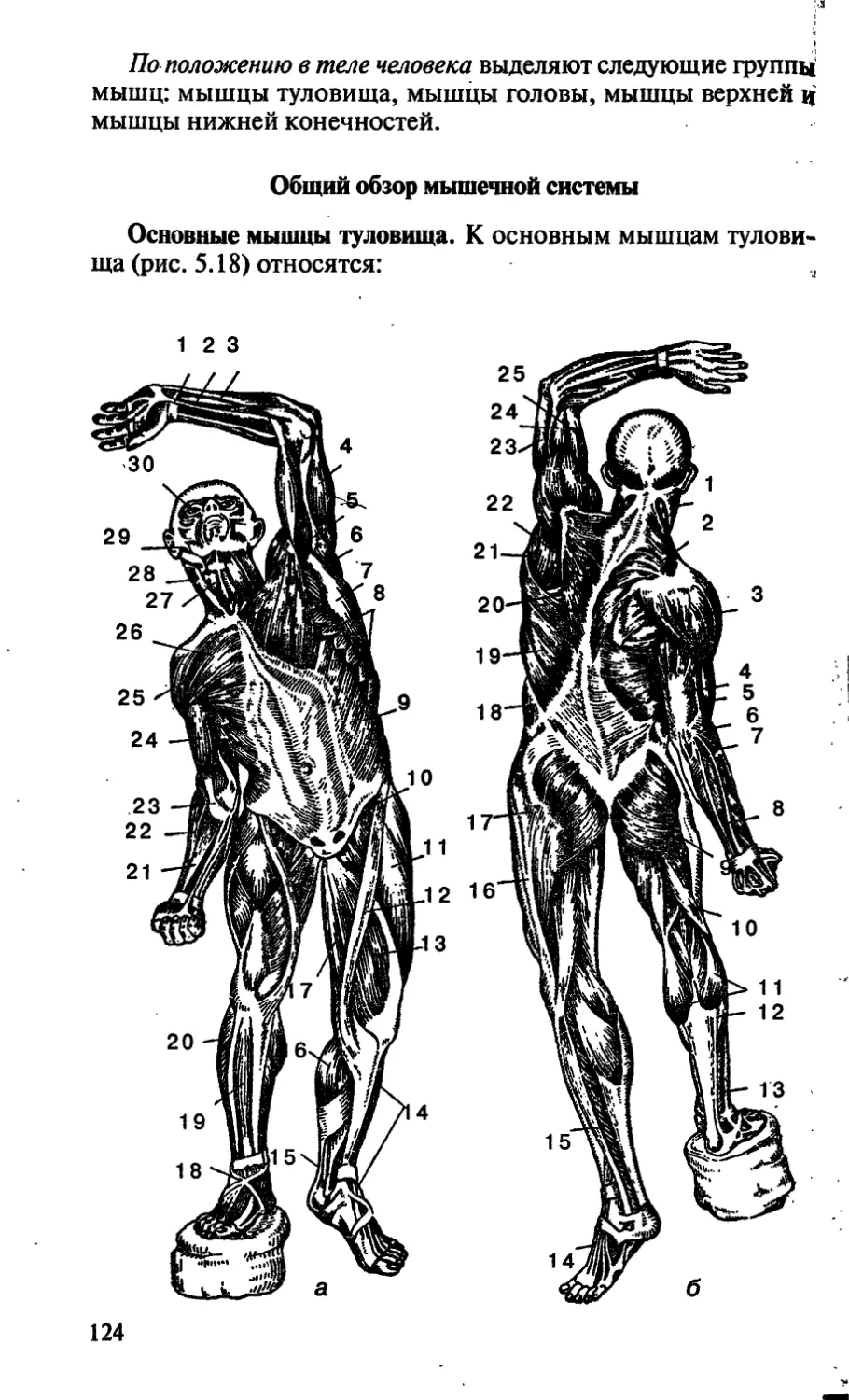
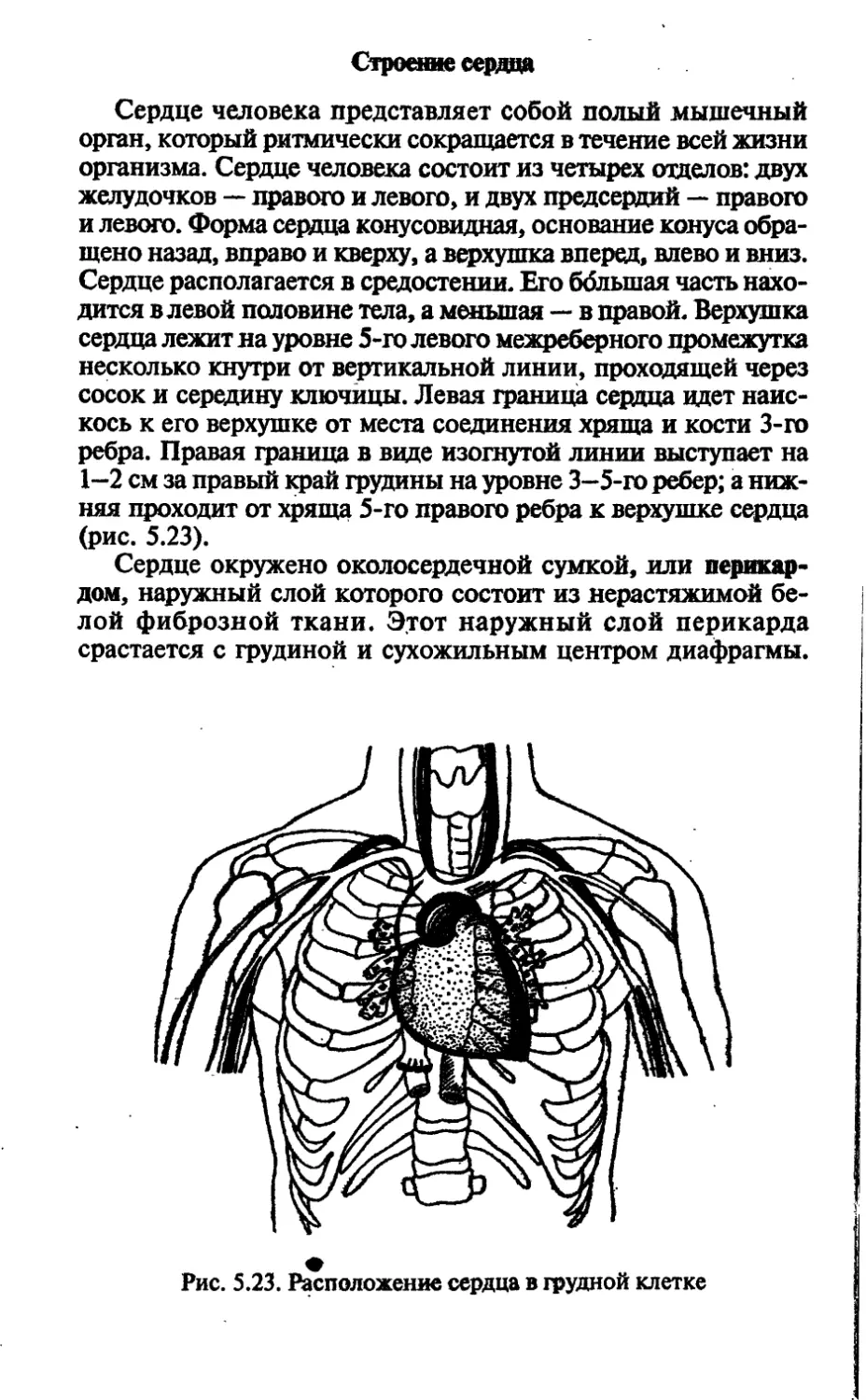
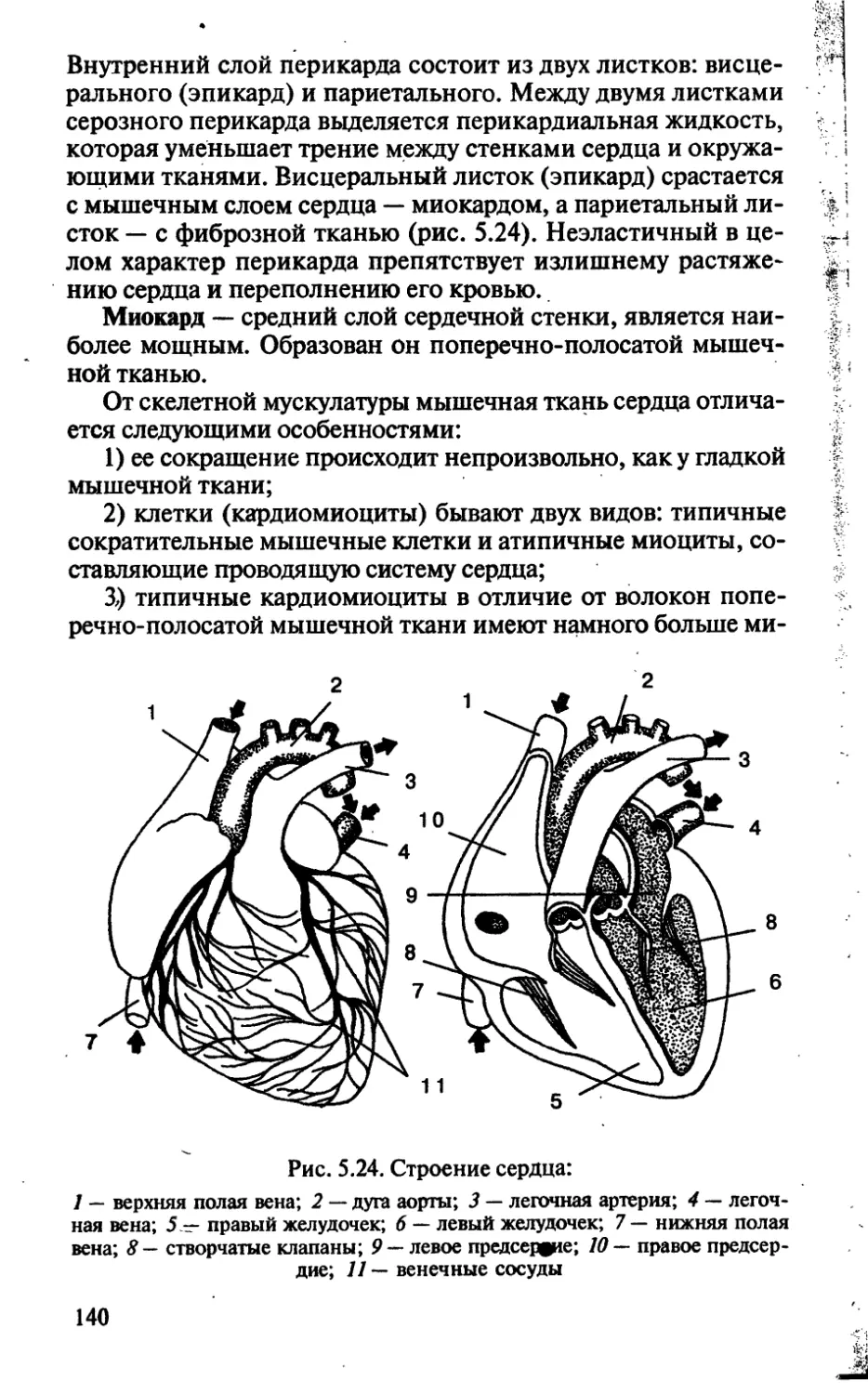
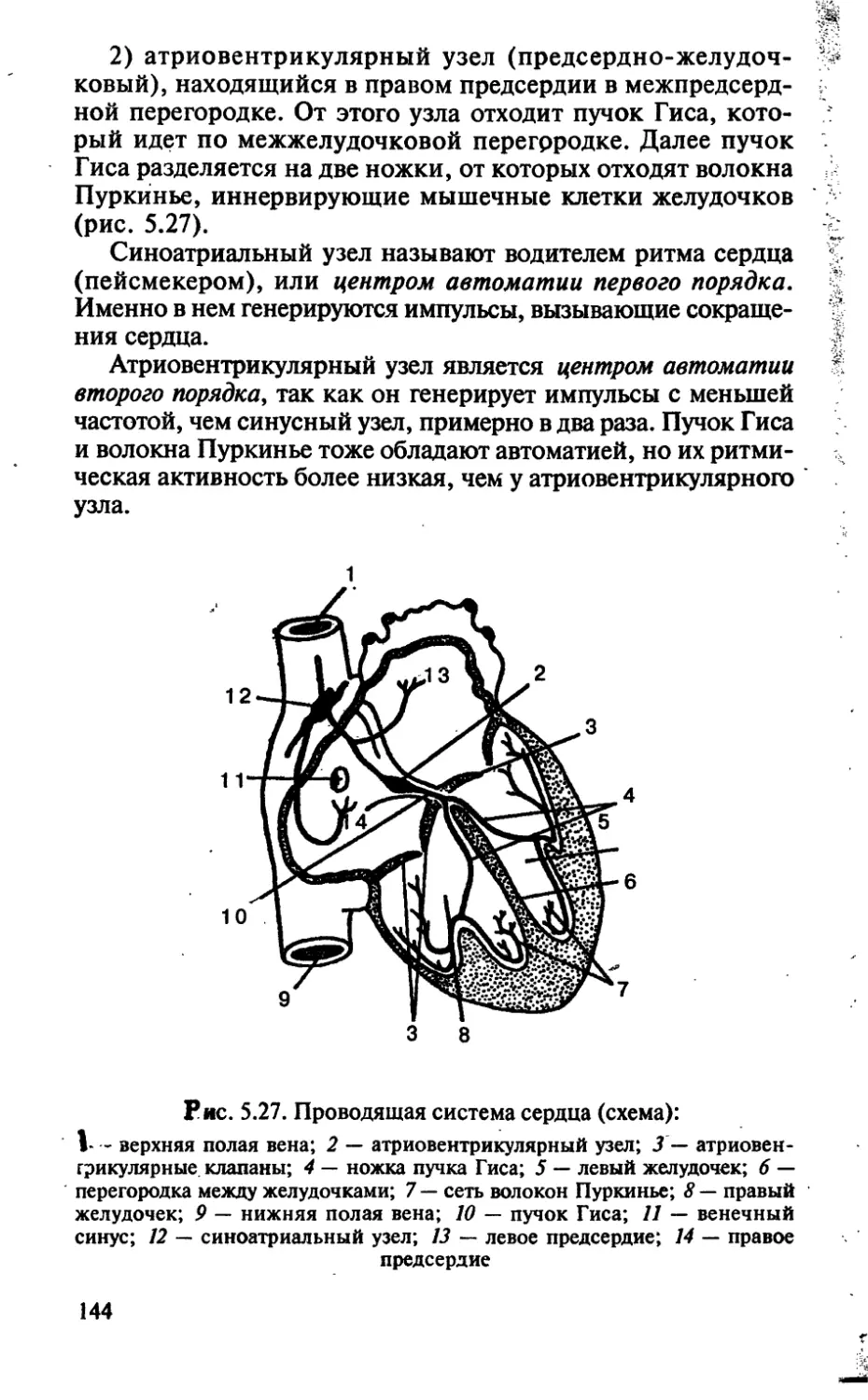
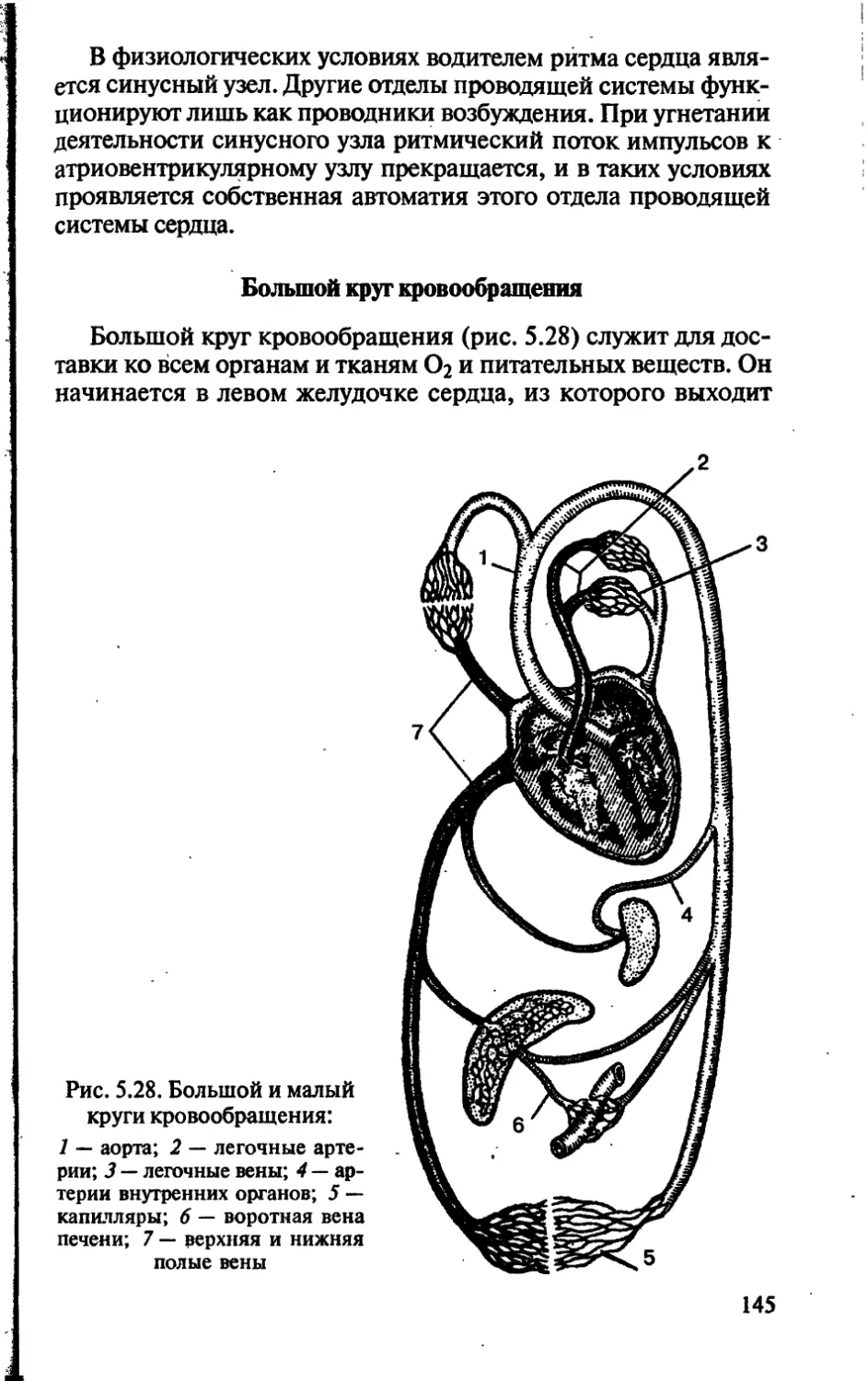
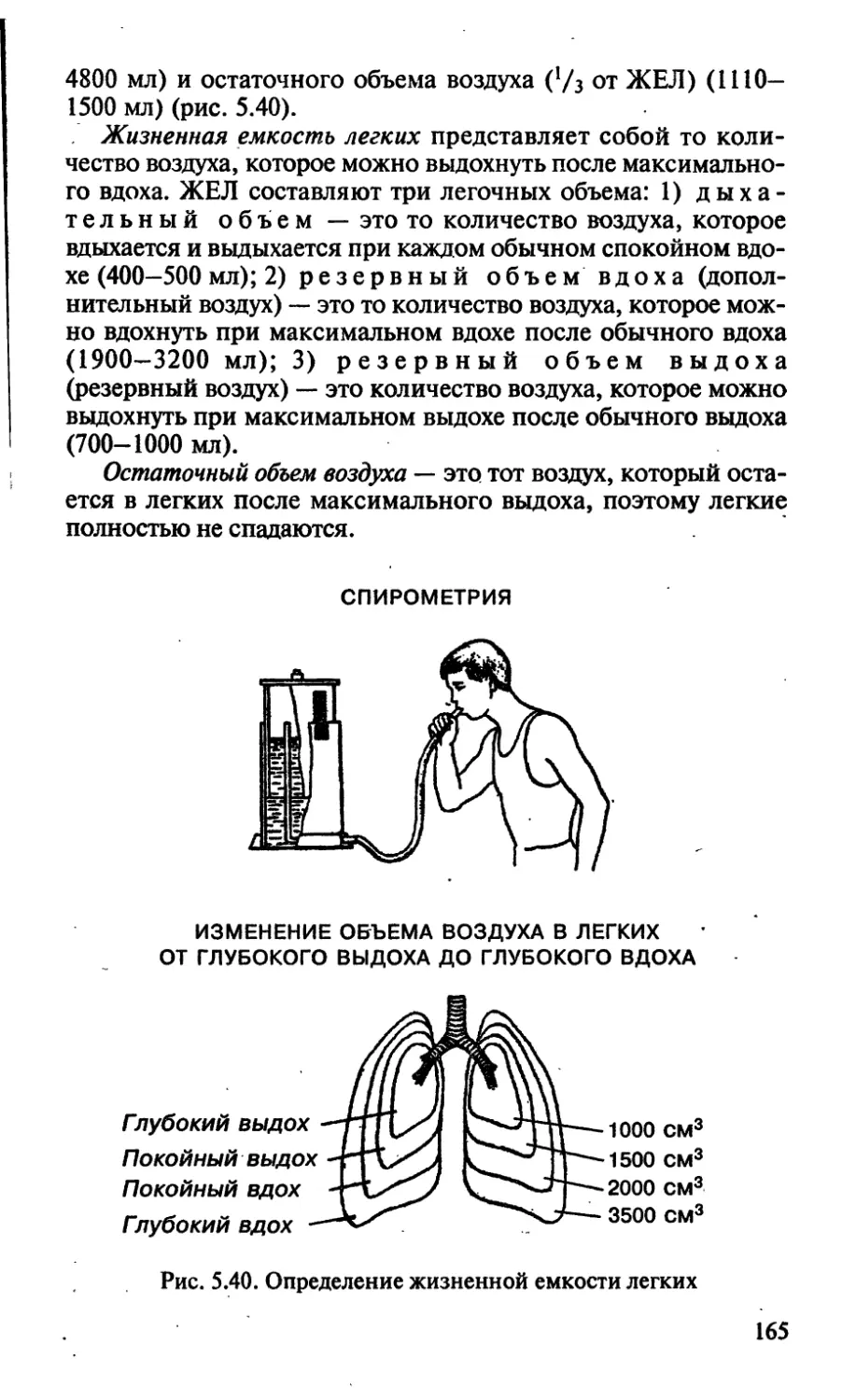
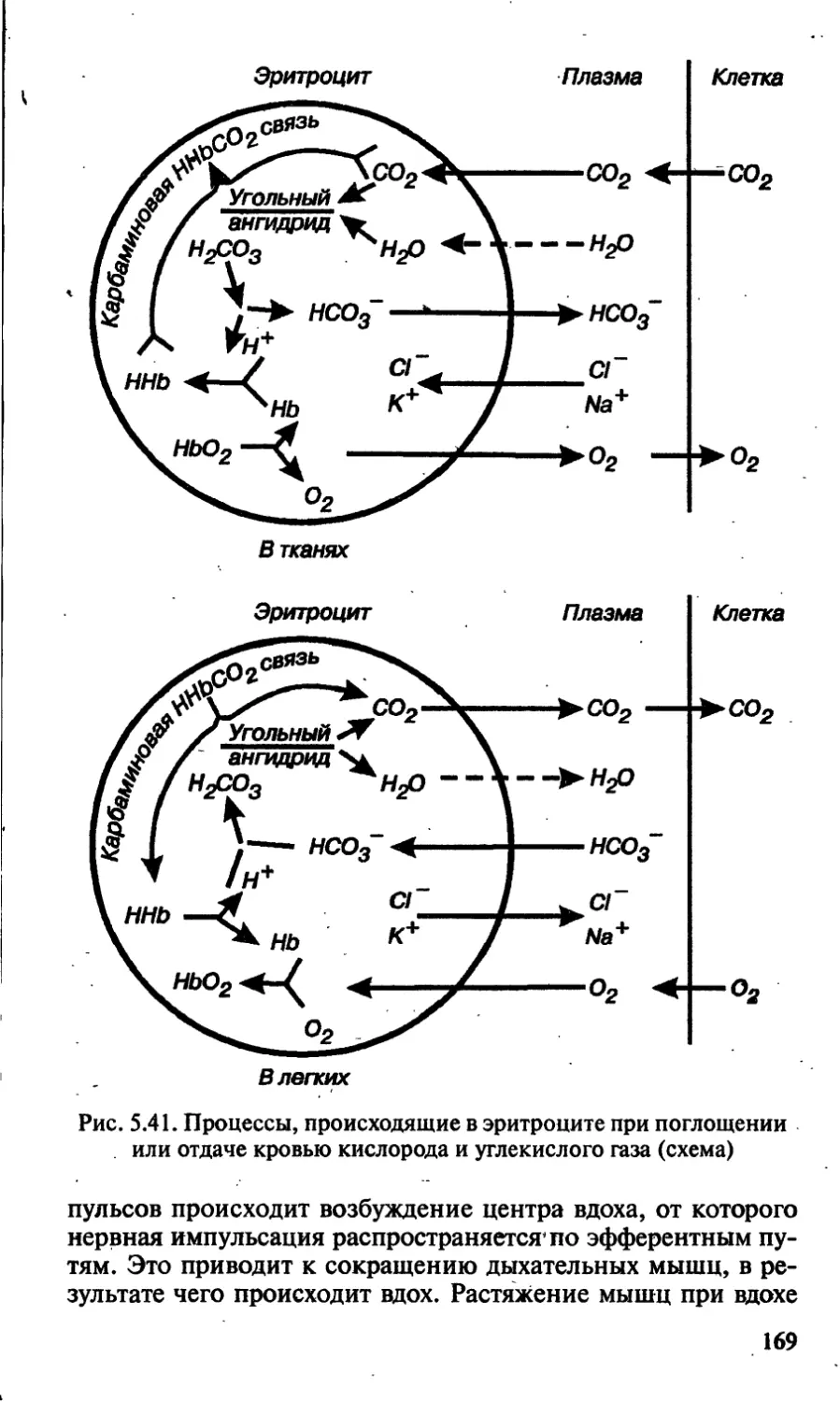
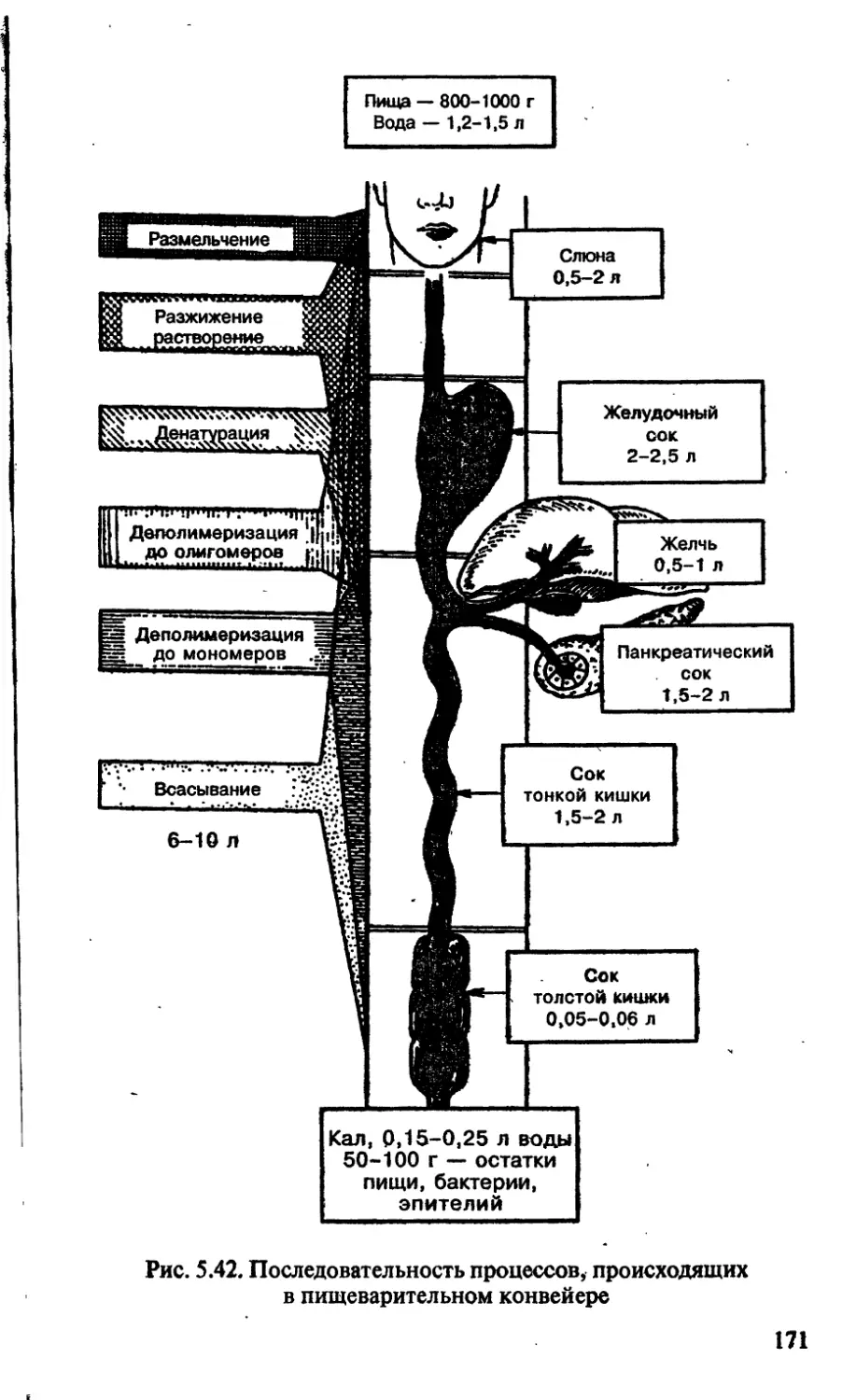
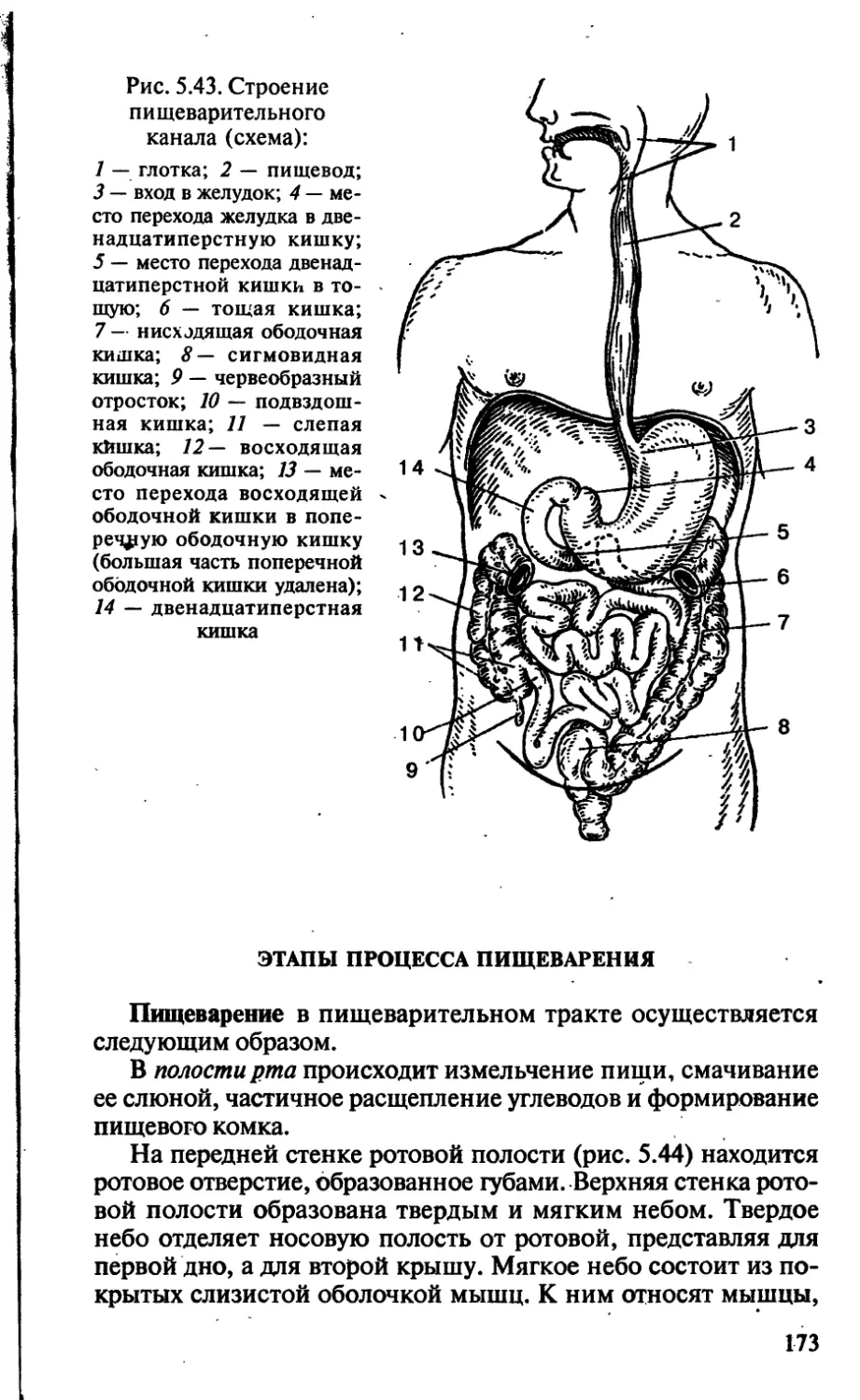
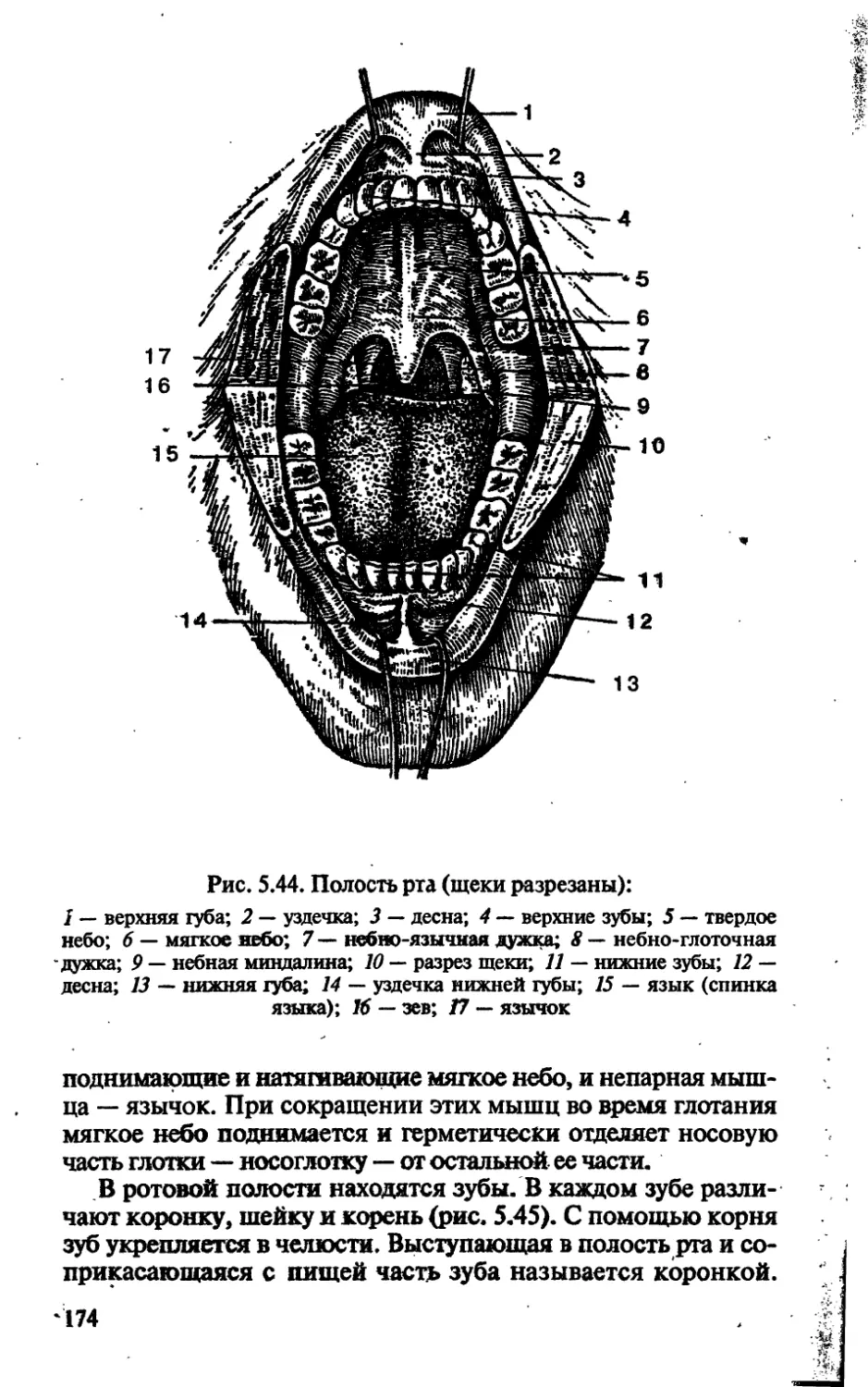
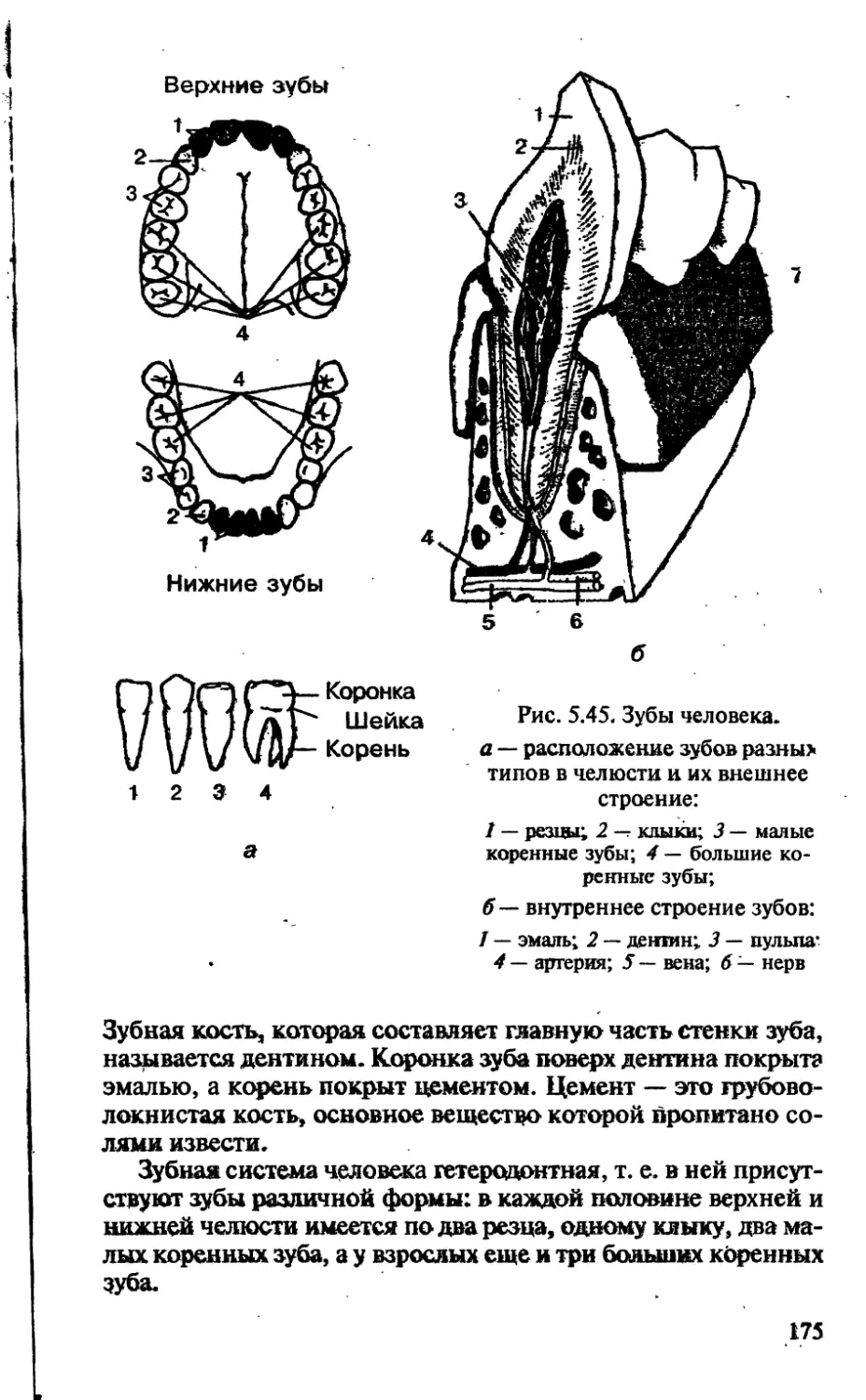
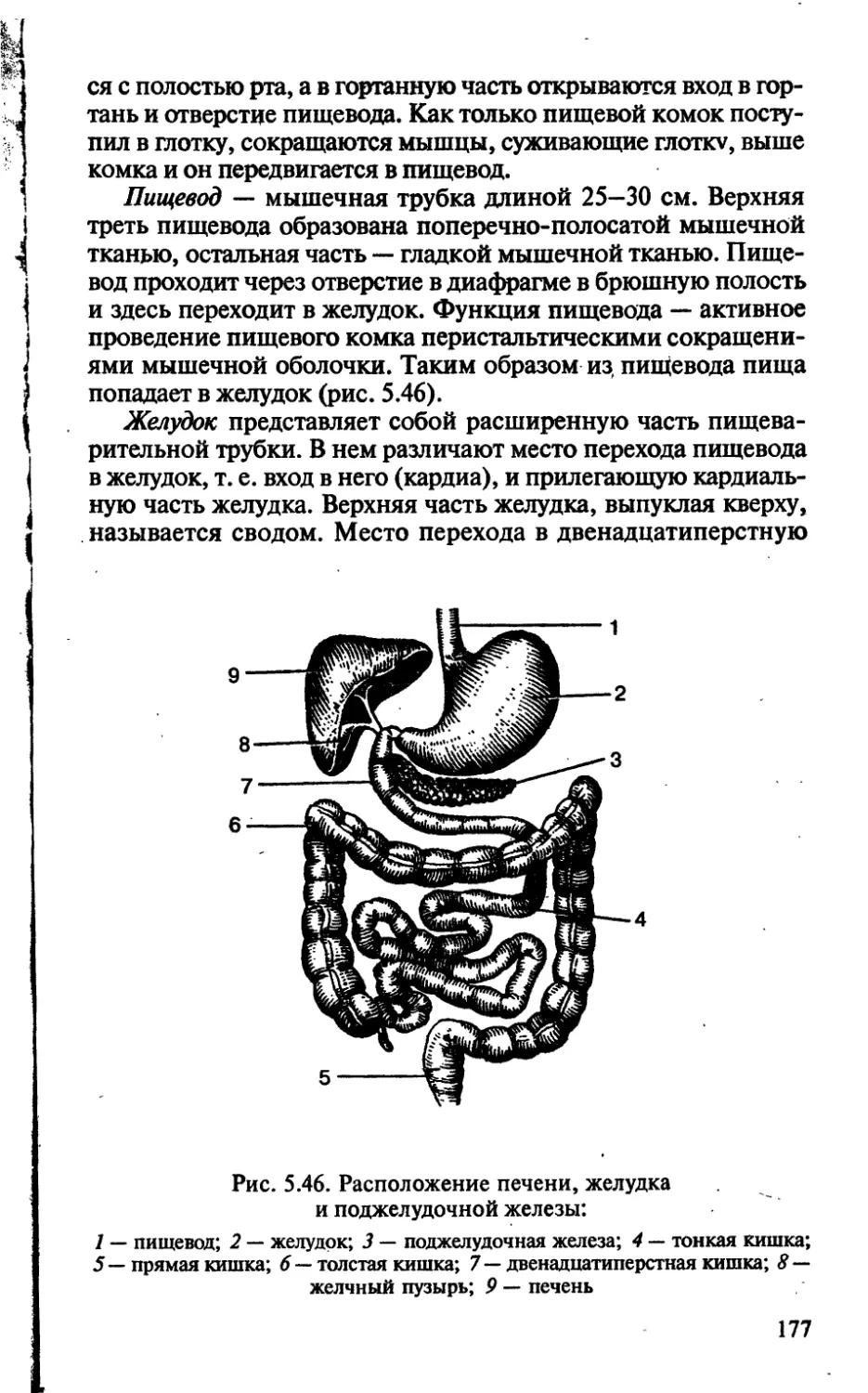
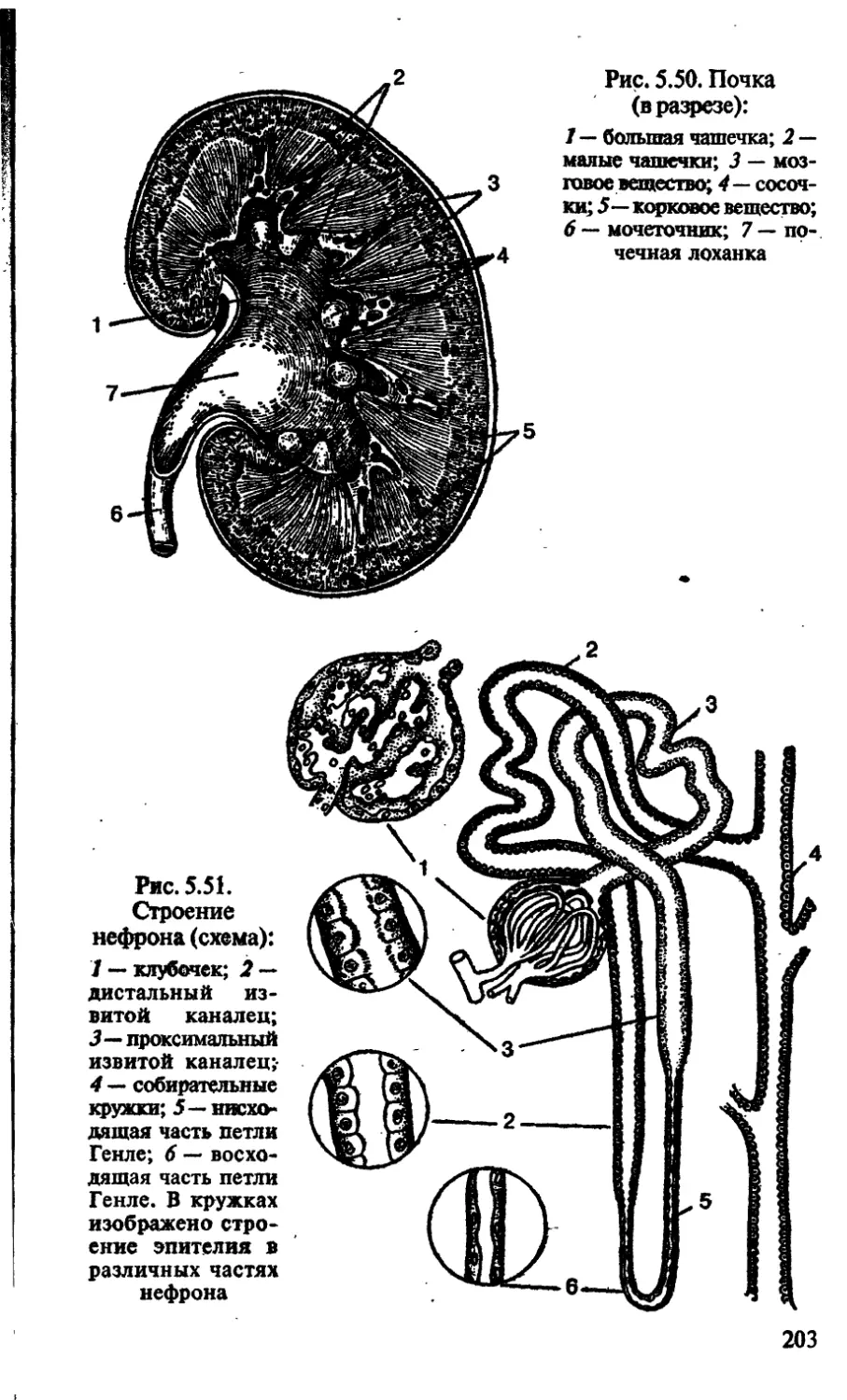
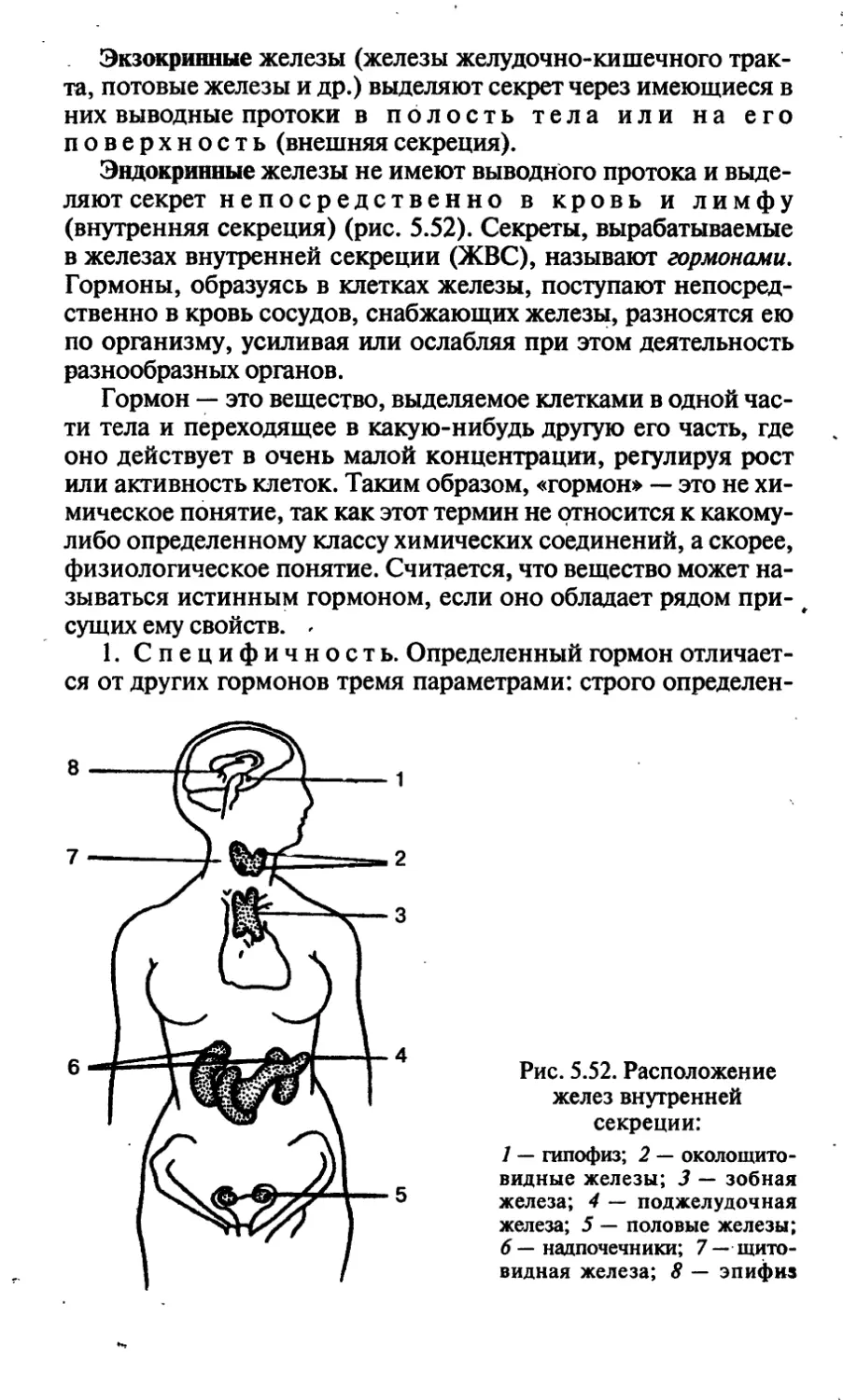
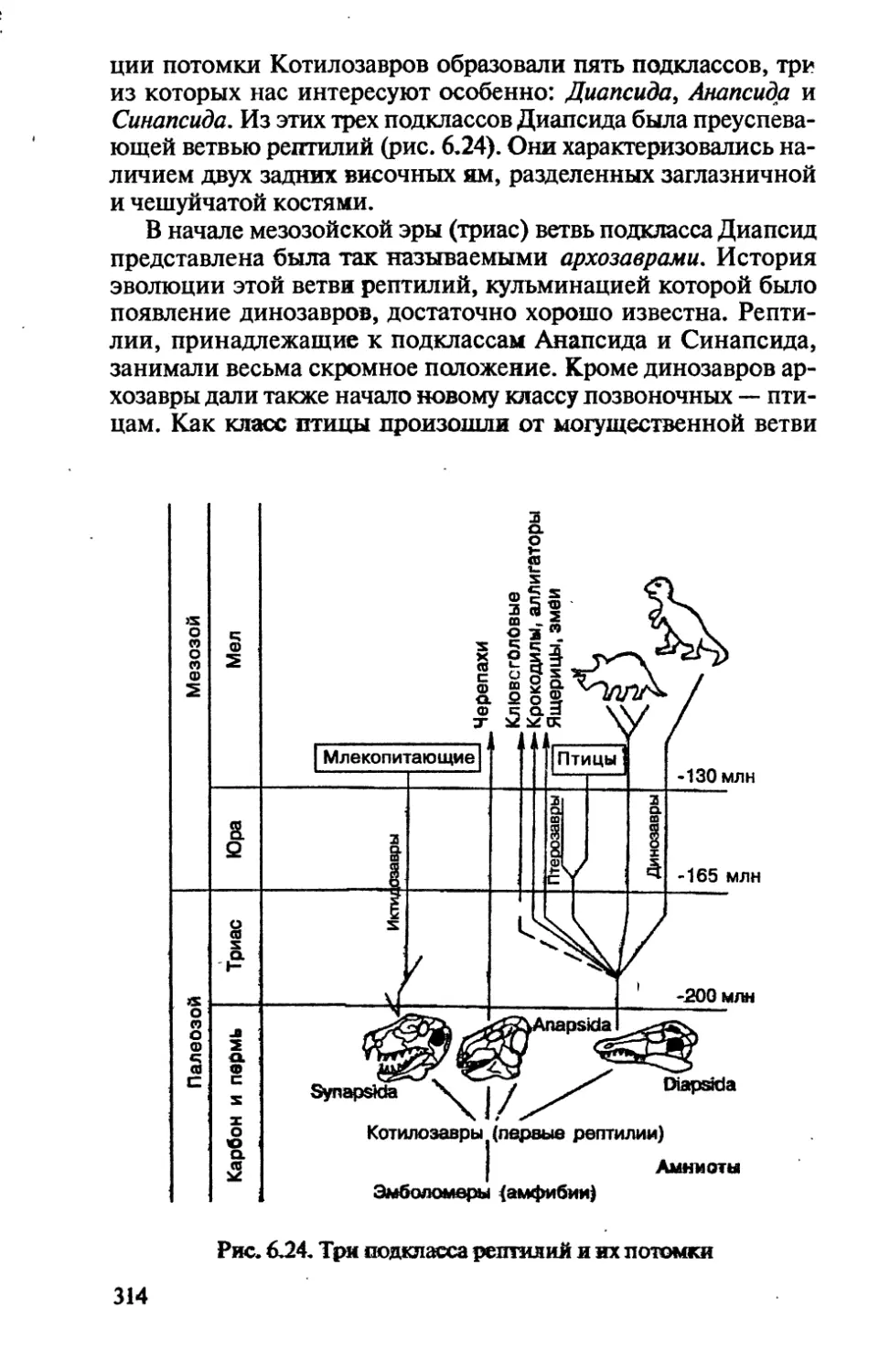
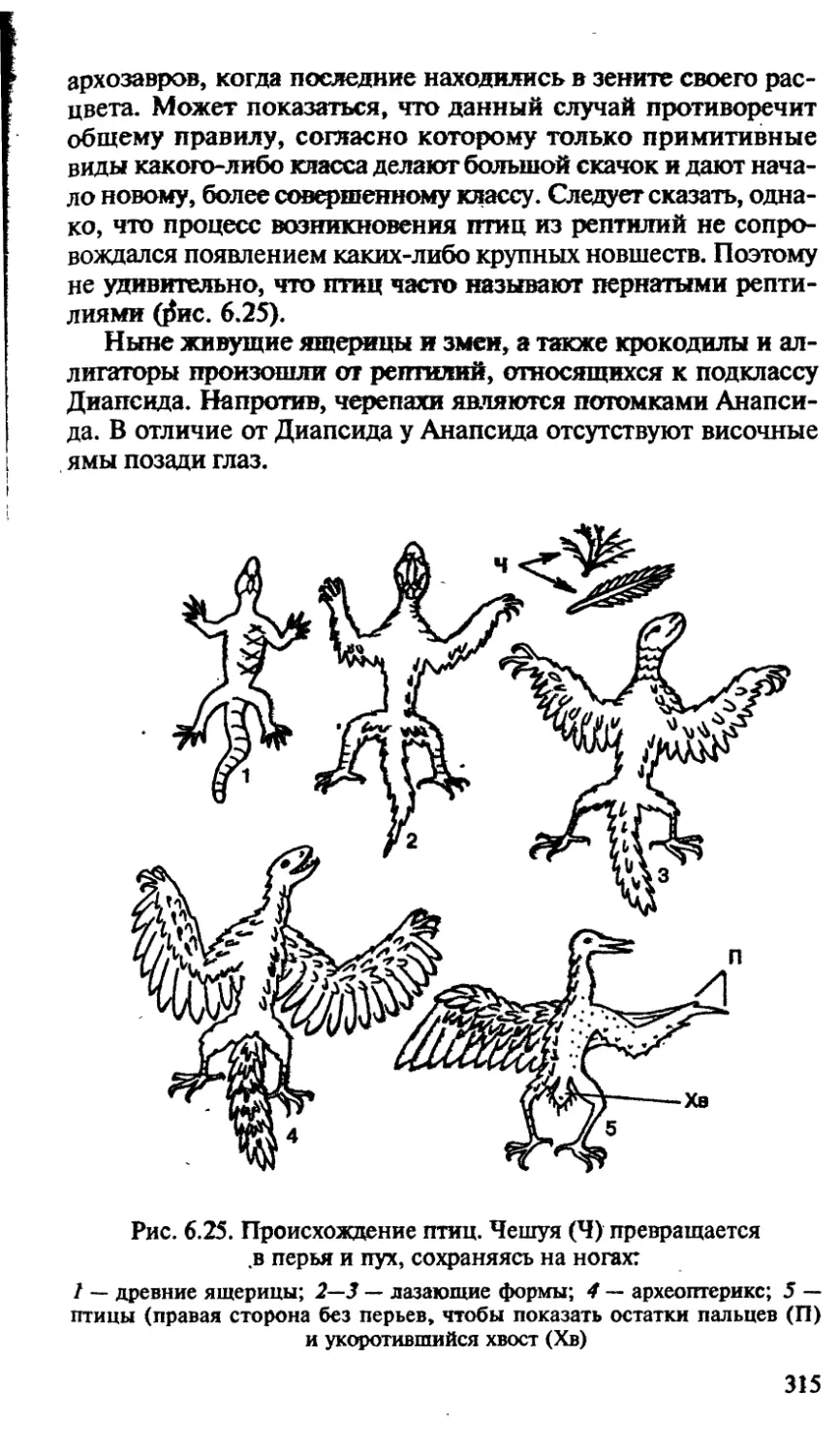
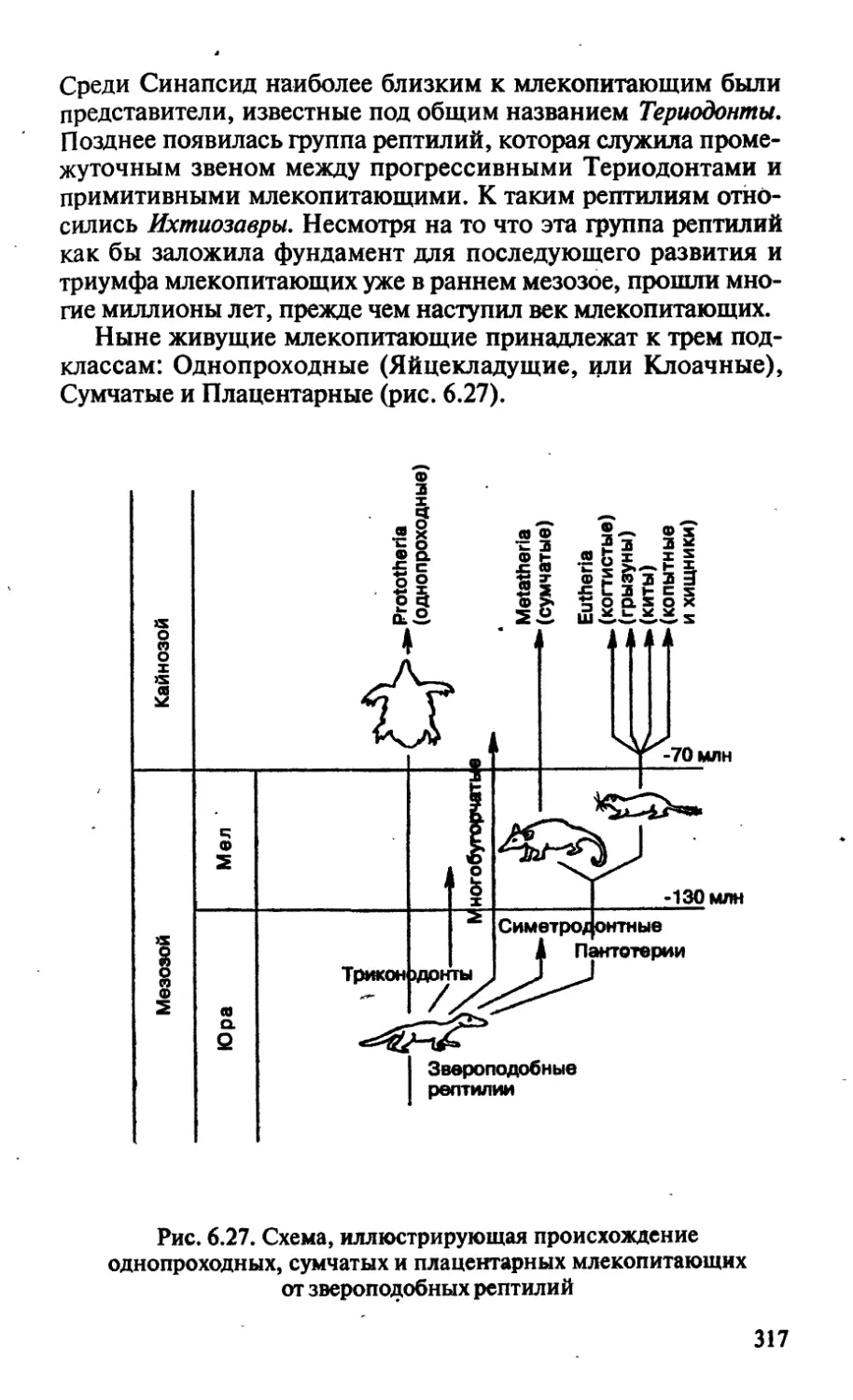
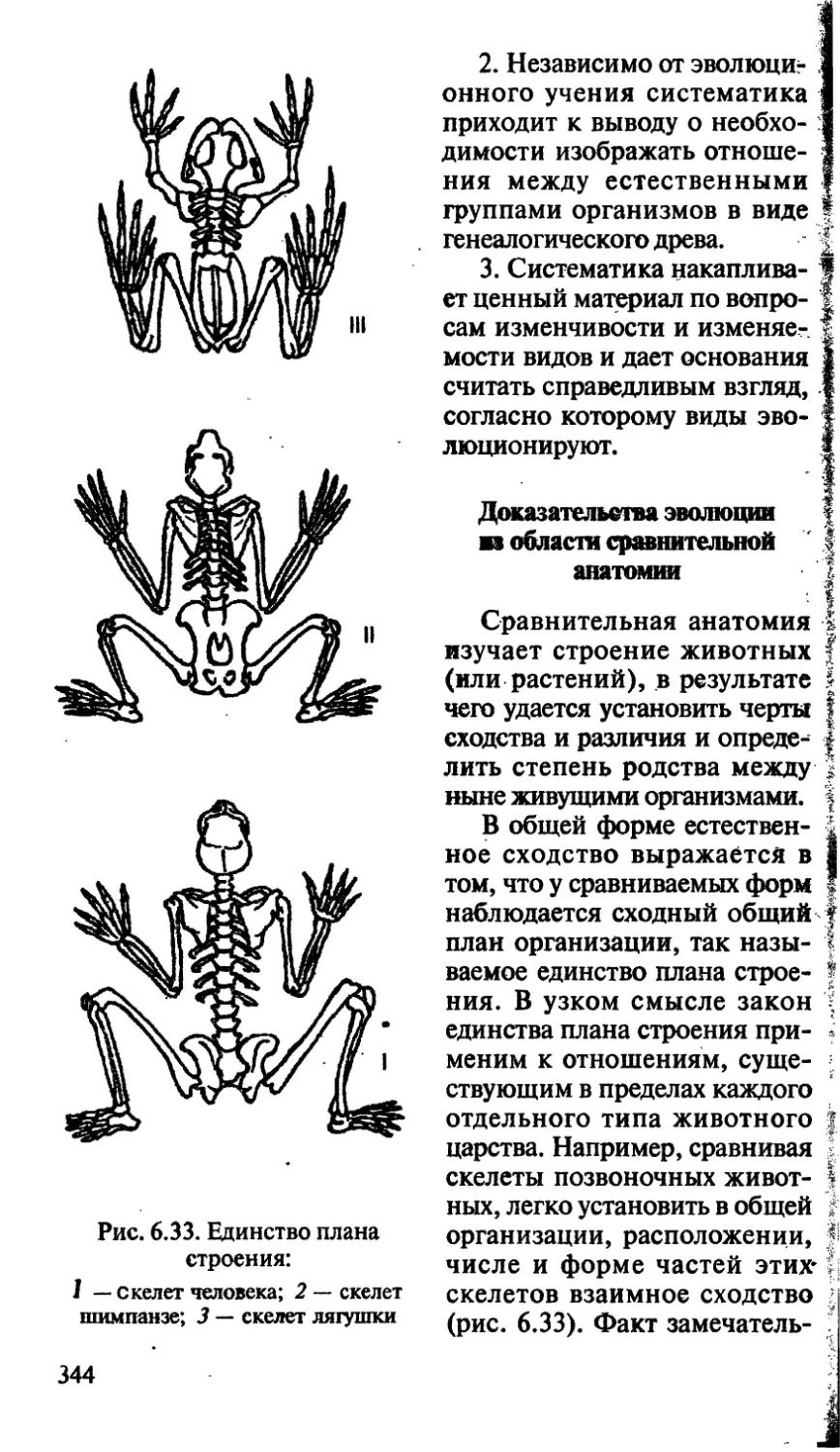
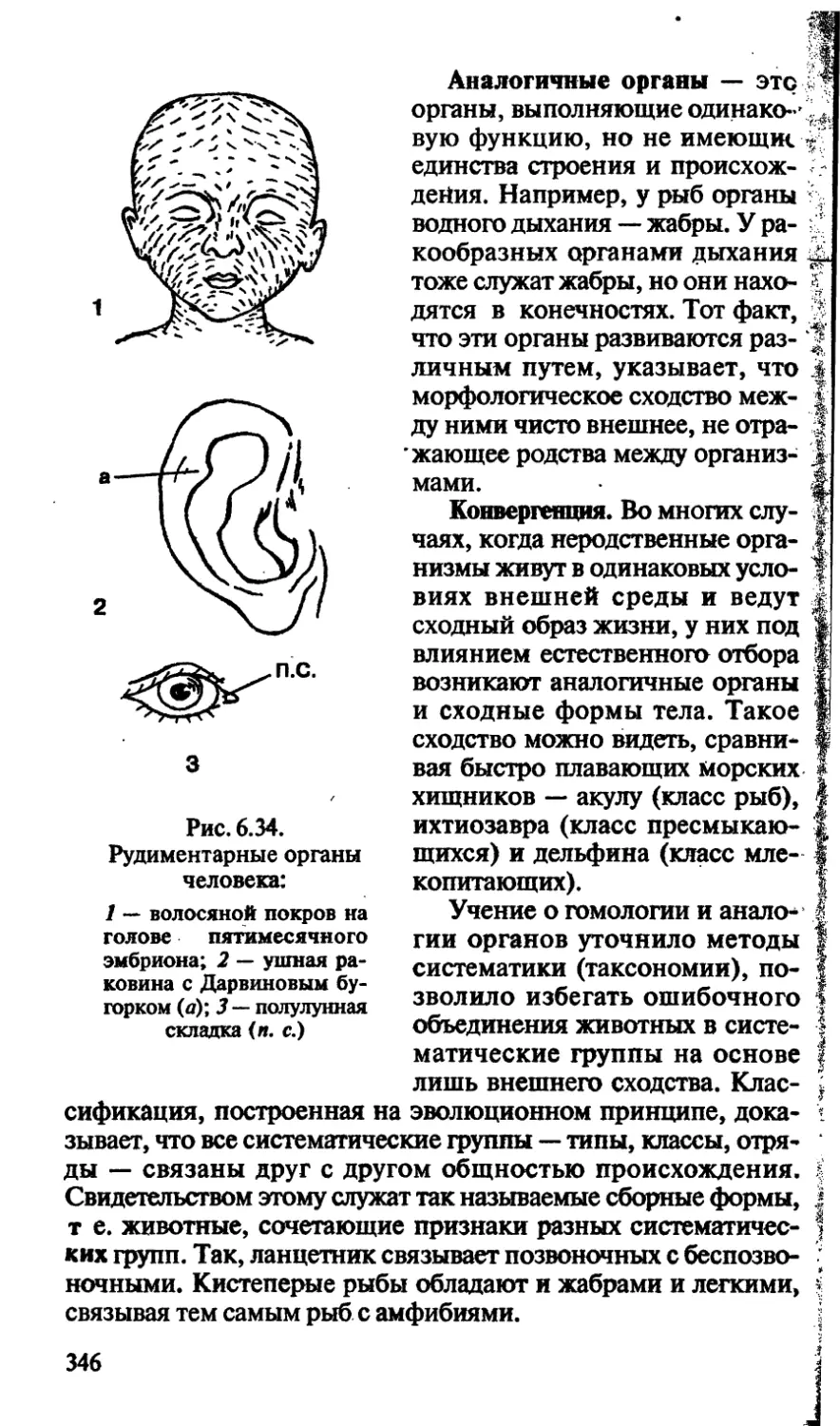
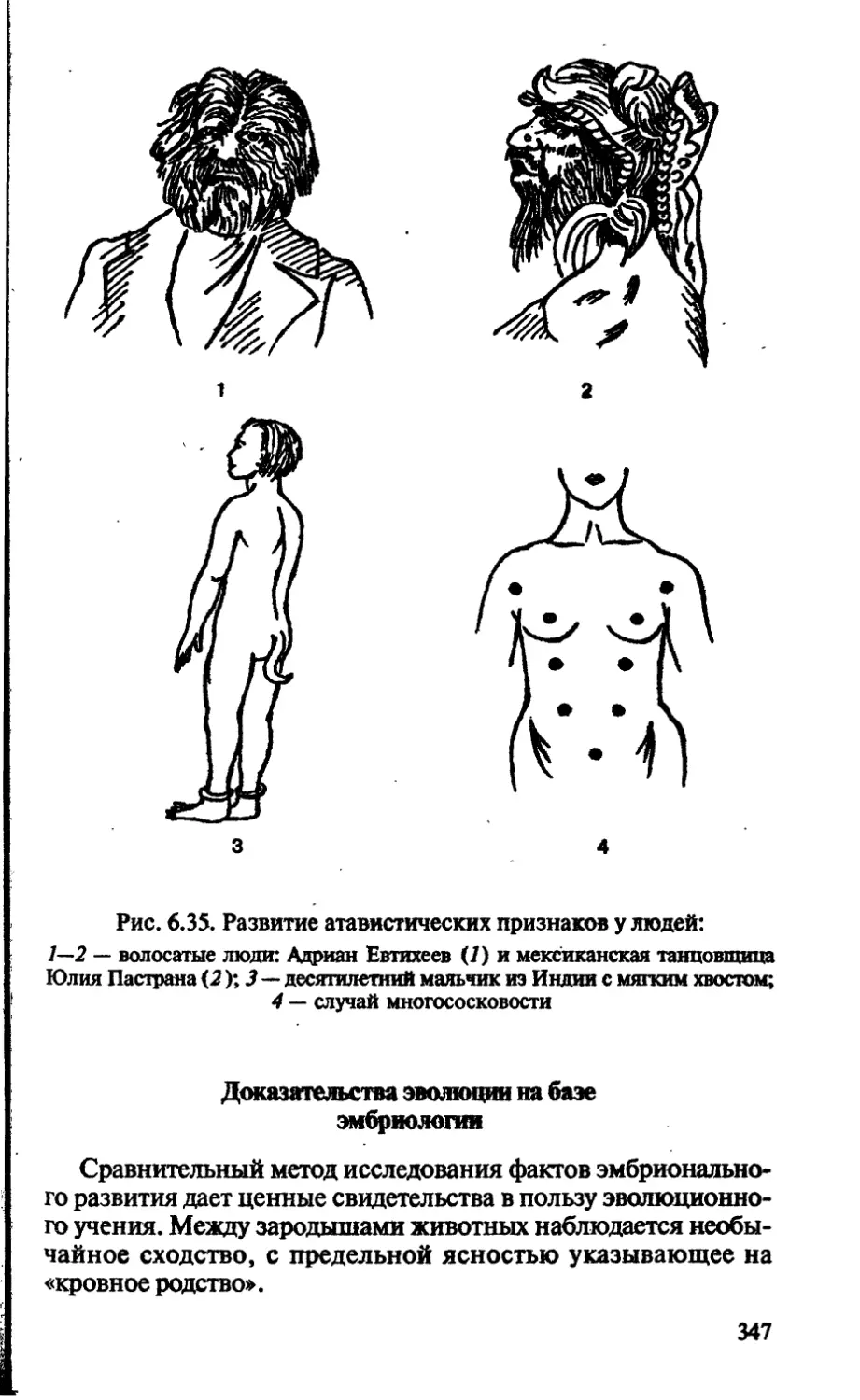
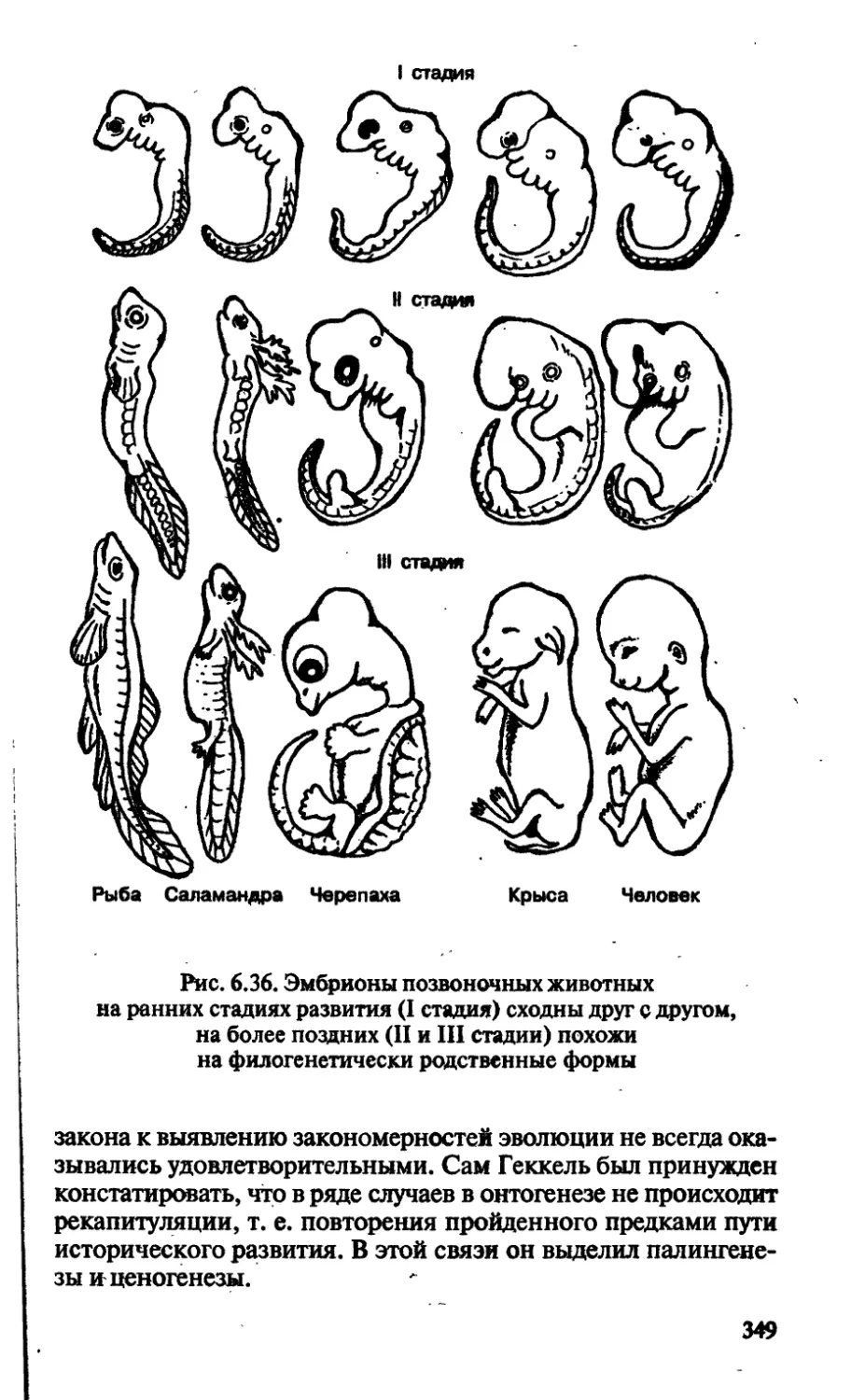
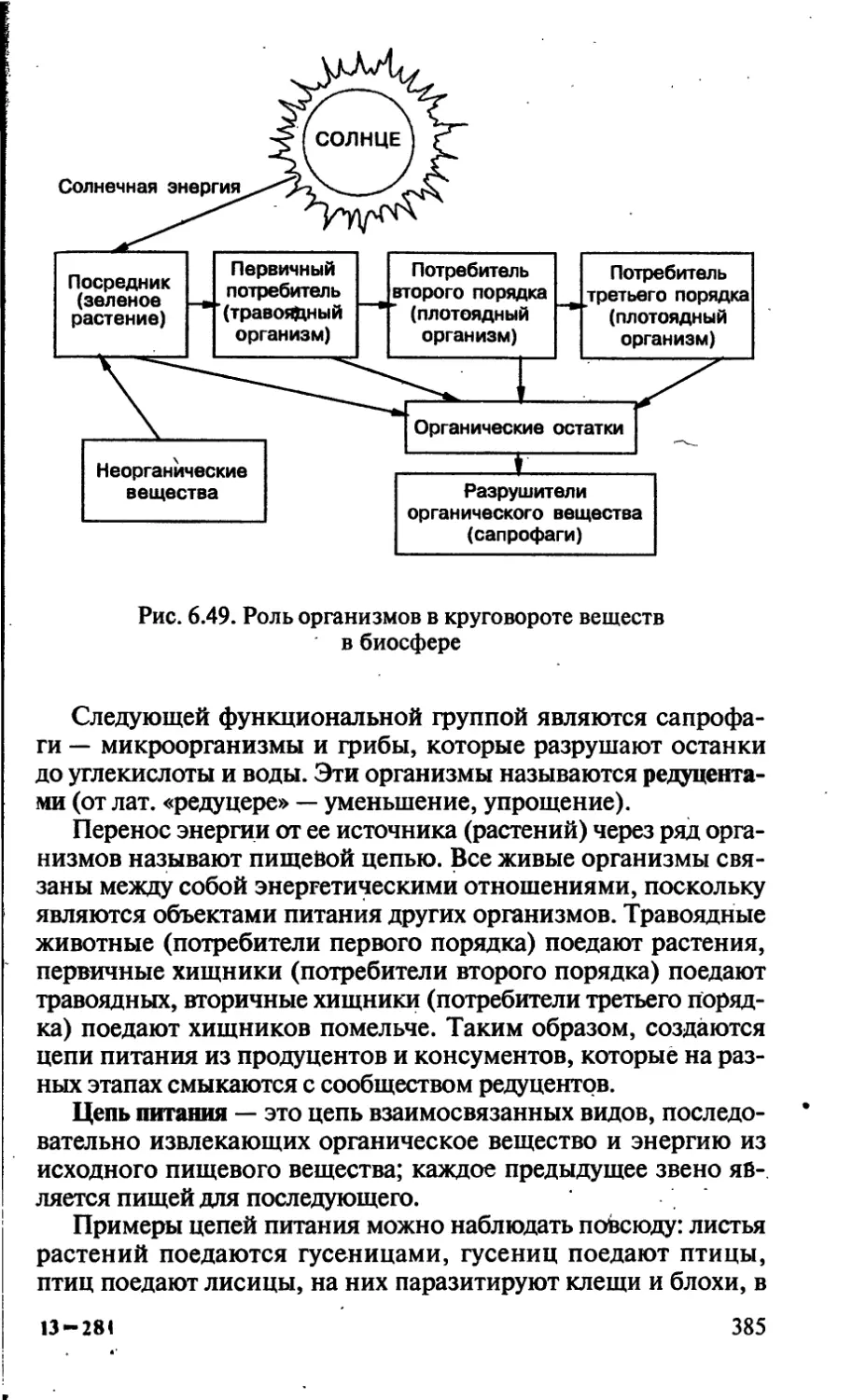
/



















































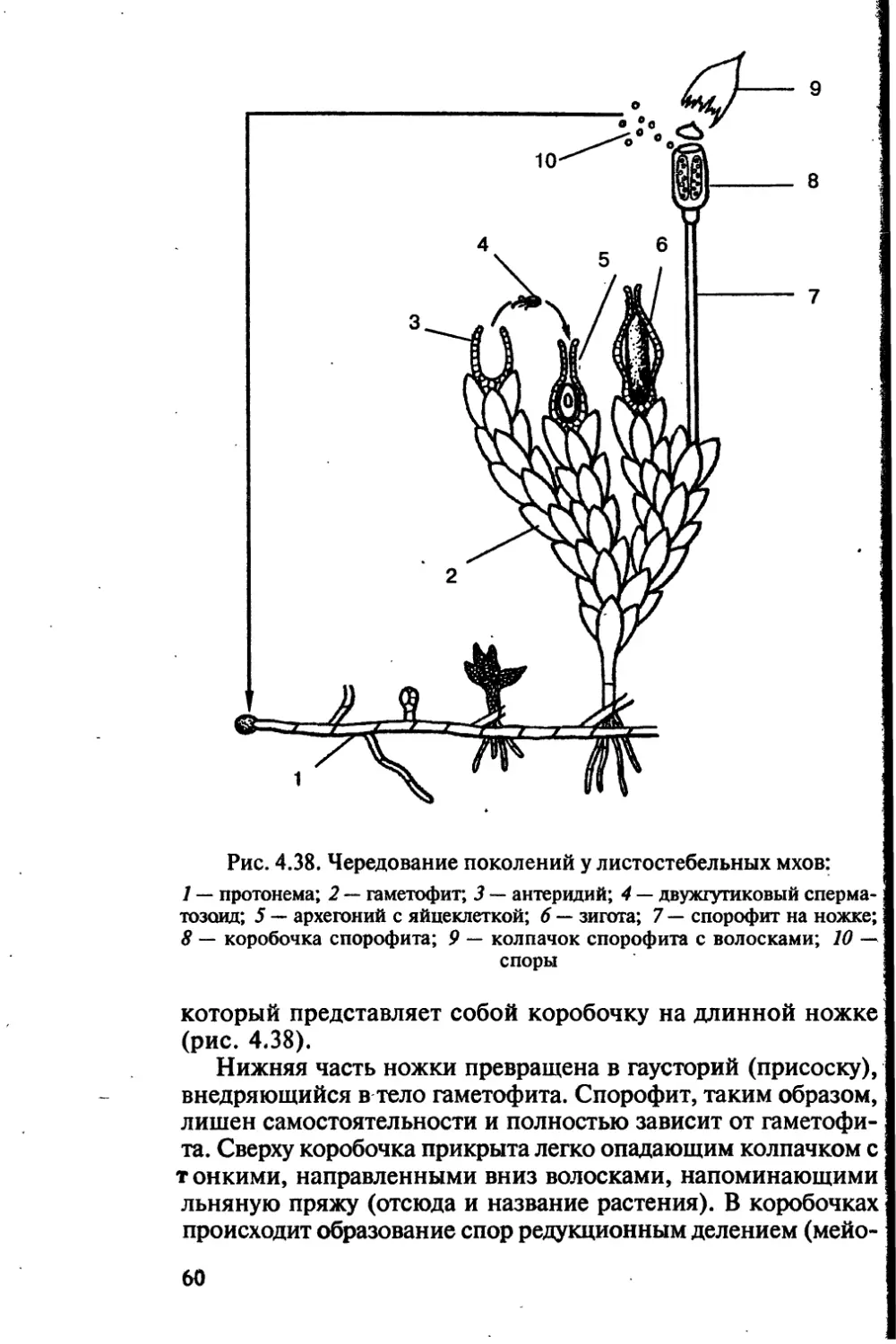




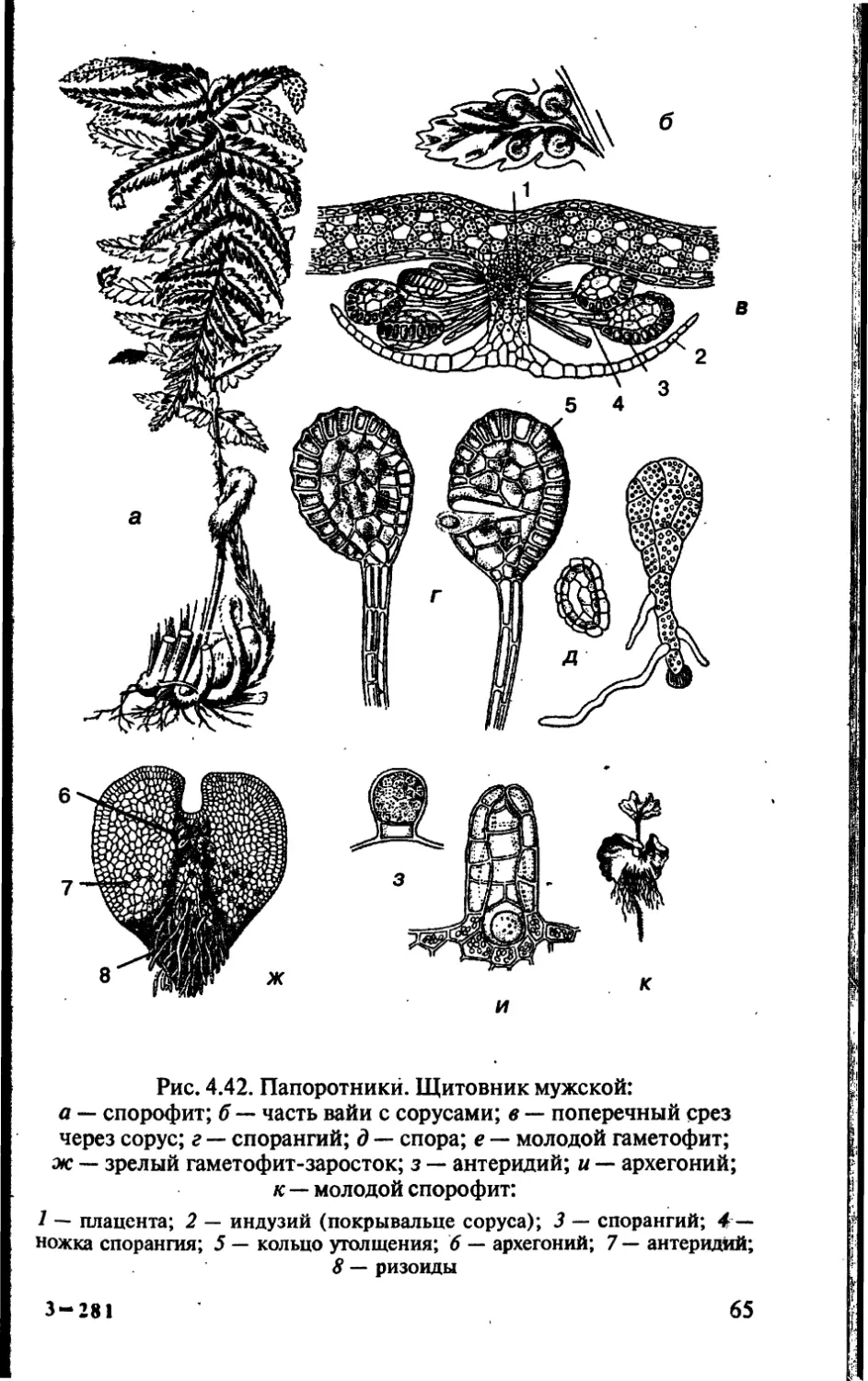





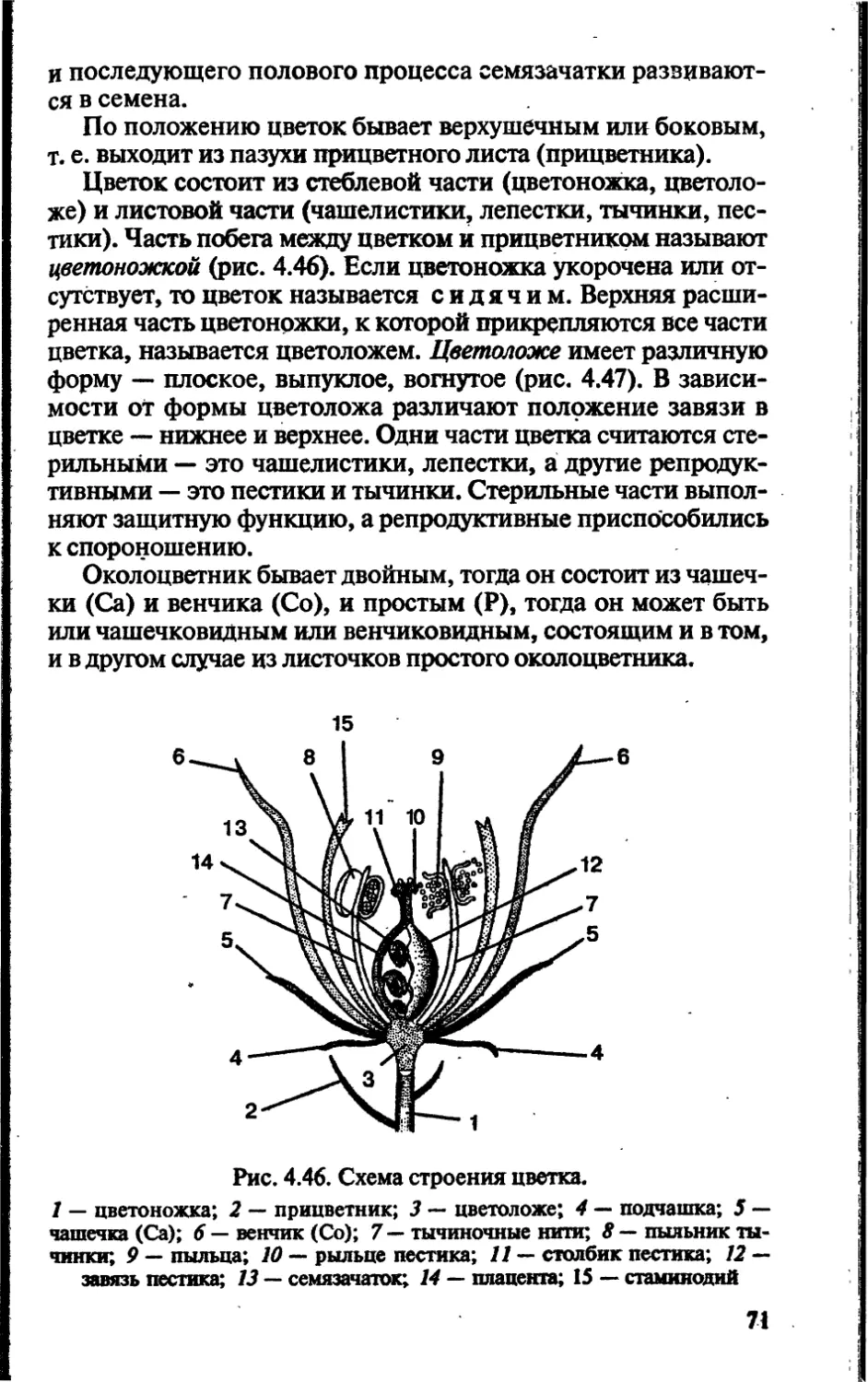


















































































































































































































































































Author: Чебышев Н.В.
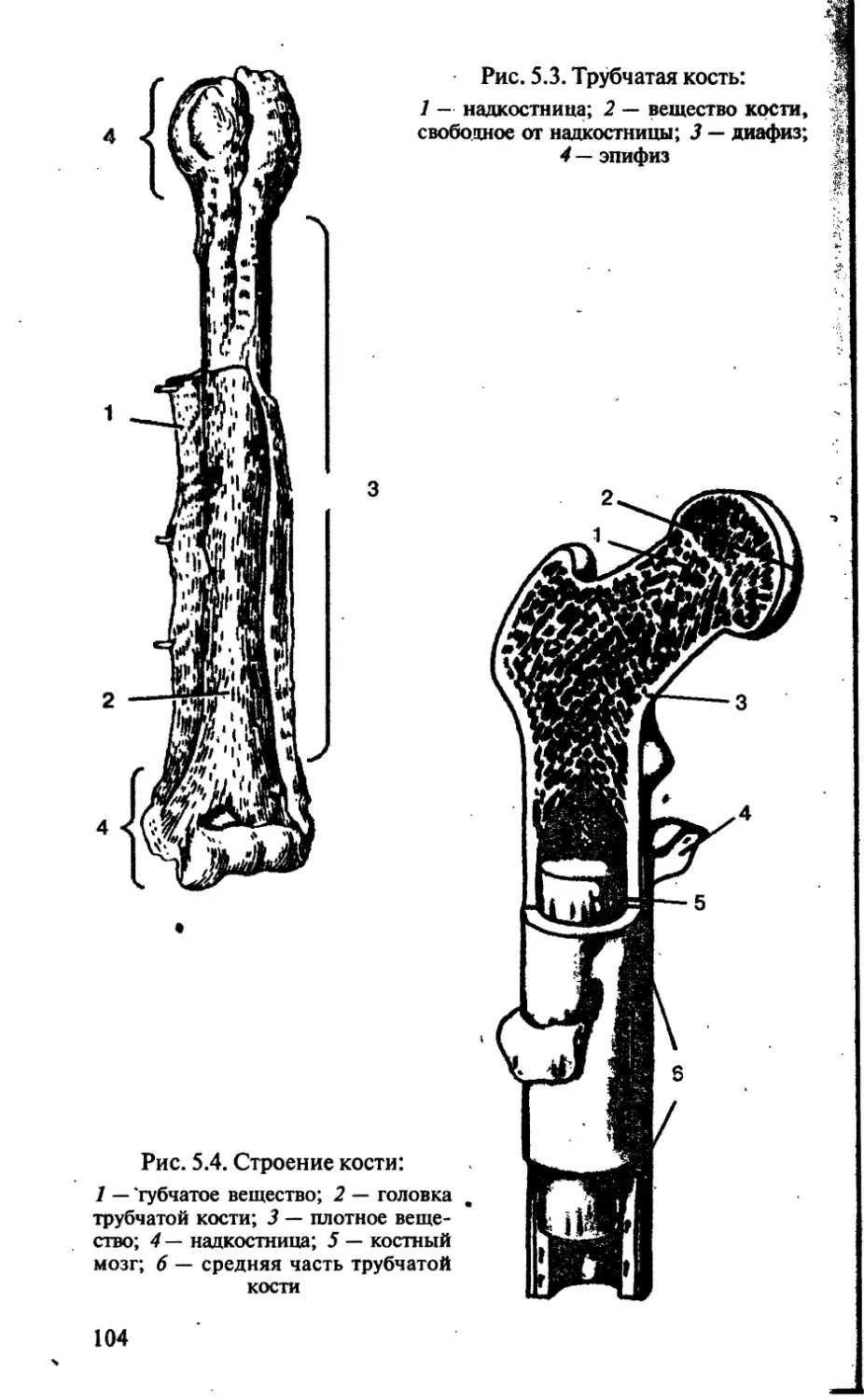
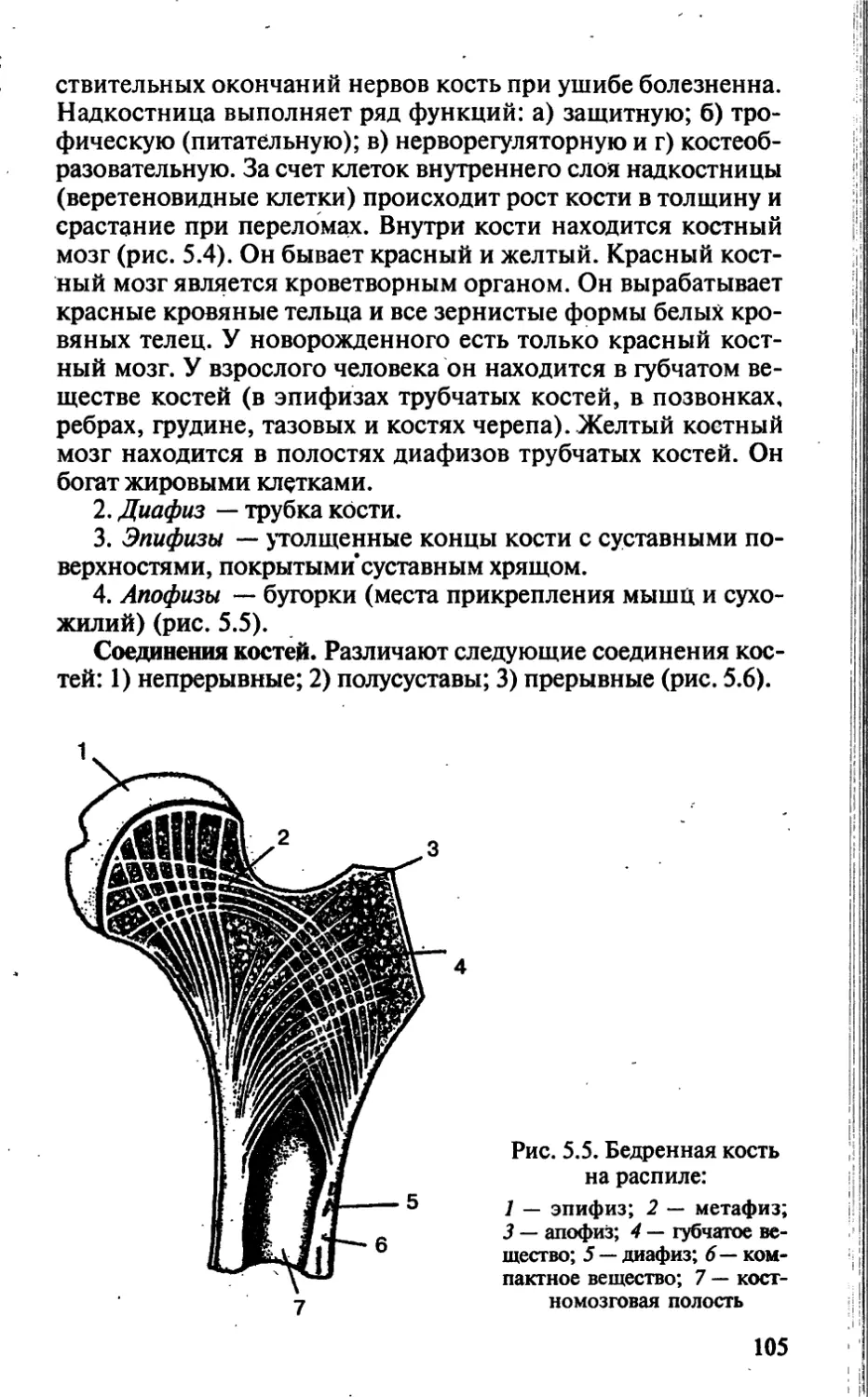
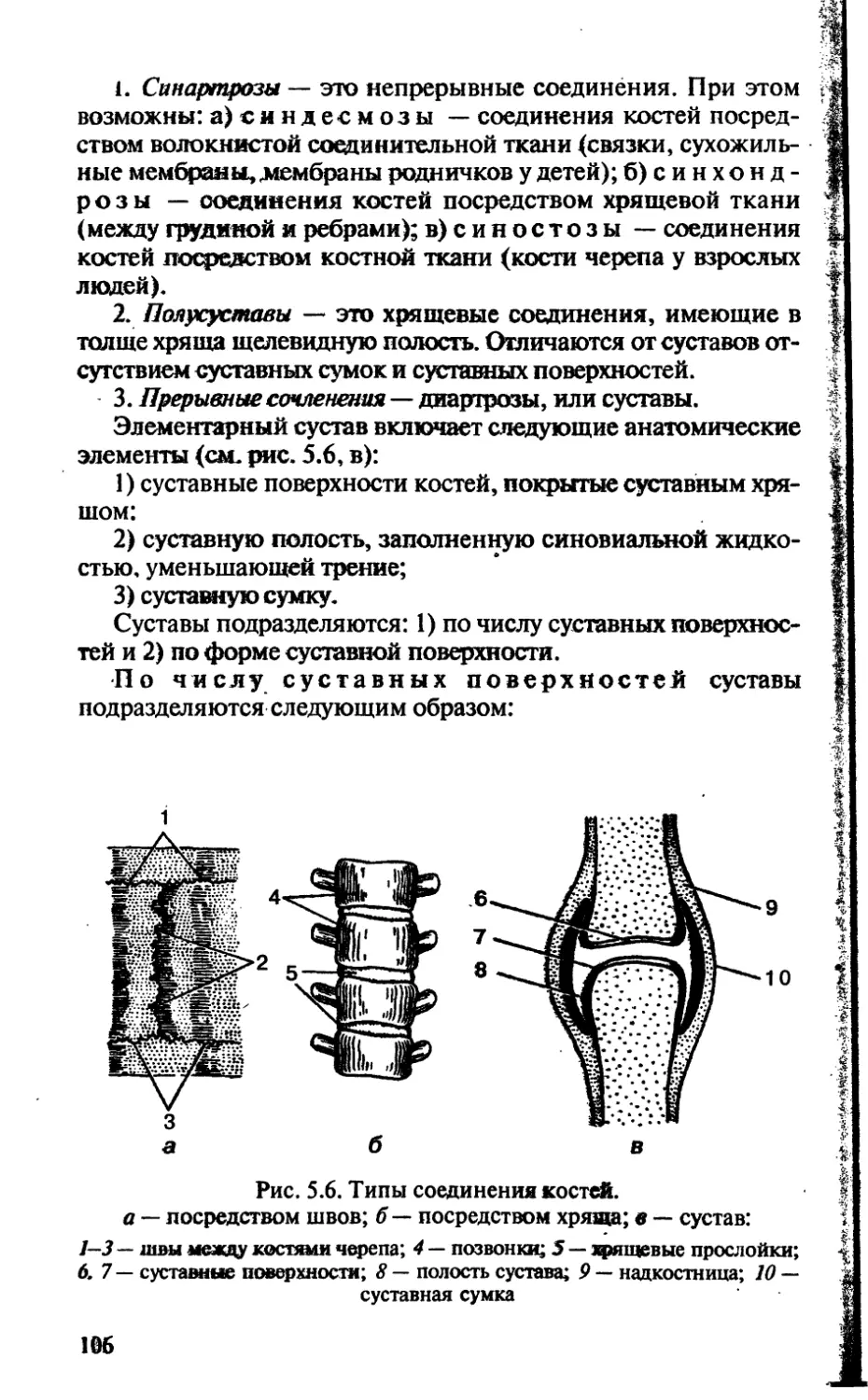
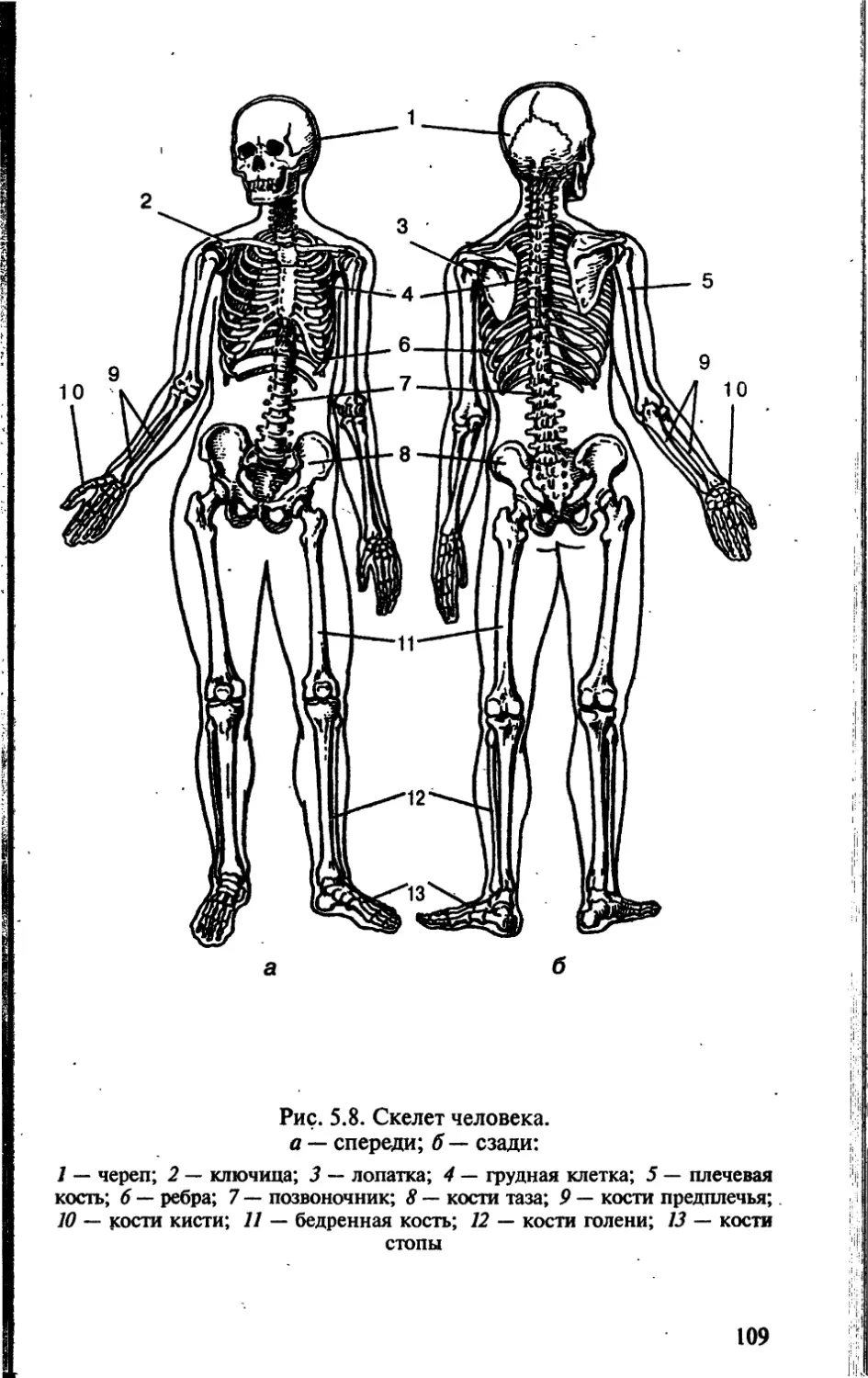
Tags: биологические науки в целом биологические науки биология экология физиология анатомия ботаника эволюция
ISBN: 5-7864-0112-X
Year: 2002




Text
Новая волна
аника
анатомия
эволюция
и экология
ИОЛОГИЯ
Н. В. Чебышев, С. В. Кузнецов
Зайчикова
4^ ____________________
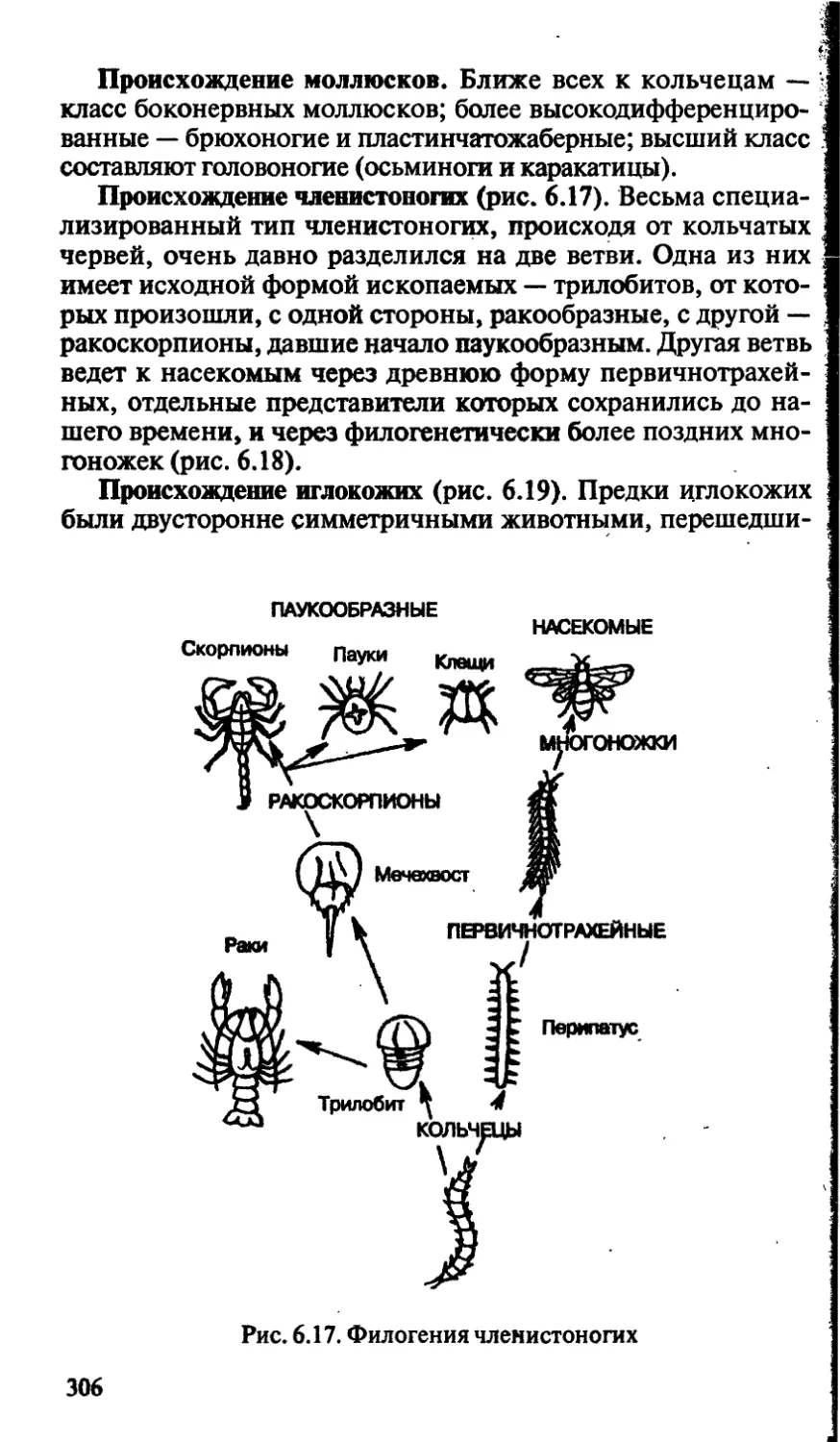
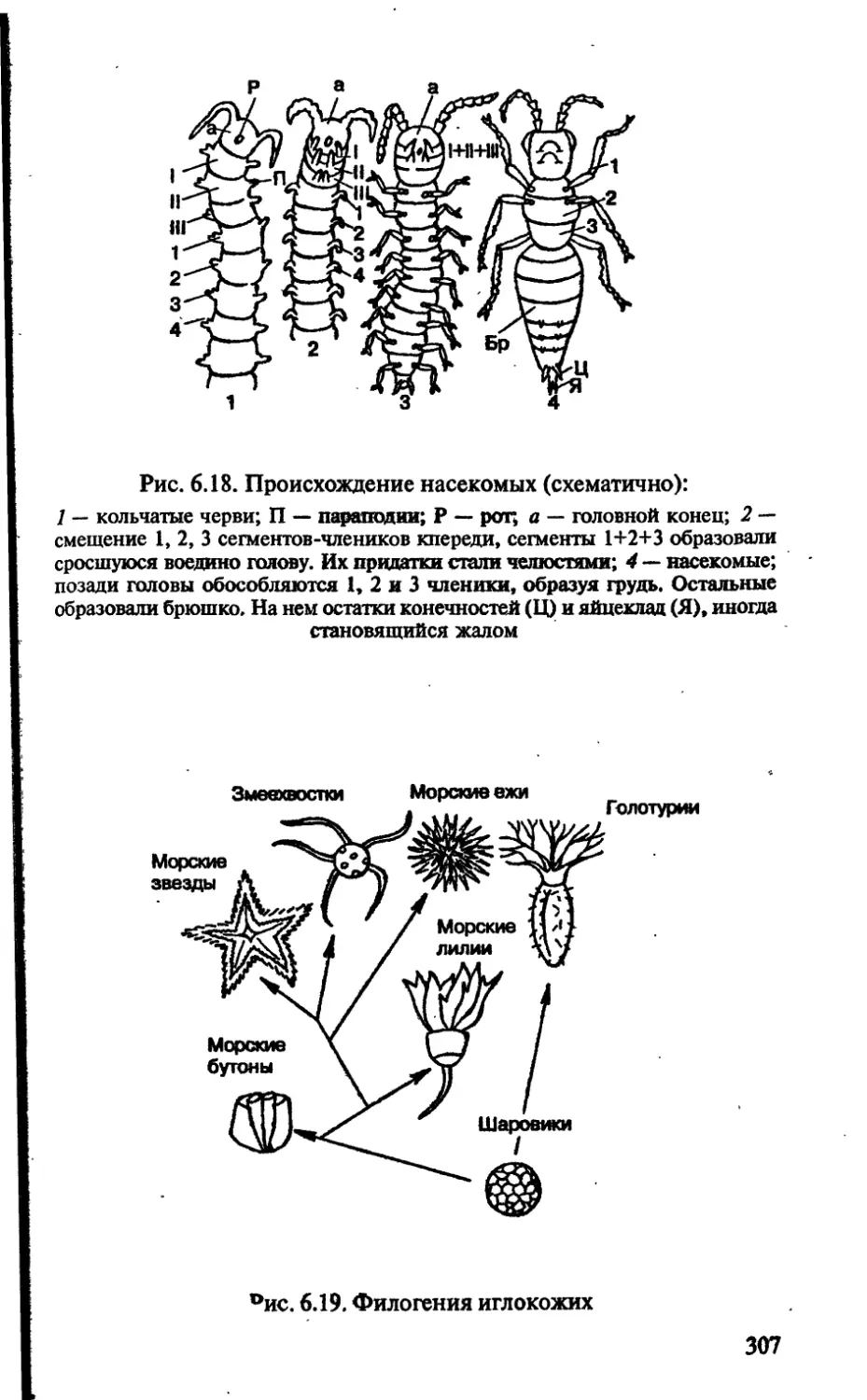
пособие
для поступаю!.
& 2У
биология
пособие
для поступающих
в вузы
под редакцией академика РАО Н. В. Чебышева
| ботаника
* анатомия и физиология
ЭВОЛЮЦИЯ и экология
Москва
«Новая Волна»
2002
УДК 57(075.3)
ББК 28я729
Б63
Авторы:
д-р биол. наук, профессор Н. В. Чебышев,
канд. биол. наук, доцент С. В. Кузнецов,
канд. фармац. наук, доцент С. Г. Зайчикова,
канд. биол. наук С. И. Гуленков
Биология: Пособие для поступающих в вузы. Том 2.—
Б63 М.: ОСЮ «Издательство Новая Волна»: Издатель Умерен-
ное, 2002.—413 с.
ISBN 5-7864-0112-Х (Изд. Новая Волна)
ISBN 5-94368-019-5 (Изд. Умеренков)
Данный курс биологии написан с учетом современных научных
достижений и уровня знаний, необходимого для успешной сдачи всту-
пительных экзаменов в вуз. В первый том вошли разделы «Биология
клетки», «Генетика и онтогенез», «Зоология», во второй том — «Бо-
таника», «Анатомия и физиология» и «Эволюция и экология». Обра-
щено внимание на органичную связь всех биологических дисциплин.
Четкость и доступность изложения делают возможным самостоятель-
ное изучение предлагаемого материала. В книге имеется большое ко-
личество иллюстраций, схем и таблиц.
Издание предназначено для подготовки абитуриентов к вступи-
тельным экзаменам в вузы, а также может быть использовано учащи-
мися лицеев с медицинским и биологическим уклоном в качестве ба-
зового учебника.
УДК 57(075.3)
ББК 28я729
ISBN 5-7864-0112-Х (т. 2) (Изд. Новая Волна)
ISBN 5-94368-019-5 (т. 2) (Изд. Умеренков) 1
ISBN 5-7864-0110-3 (Изд. Новая Волна) I
© Коллектив авторов, 1998
© Оформление. ООО «Издательство Новая Волна», 1999
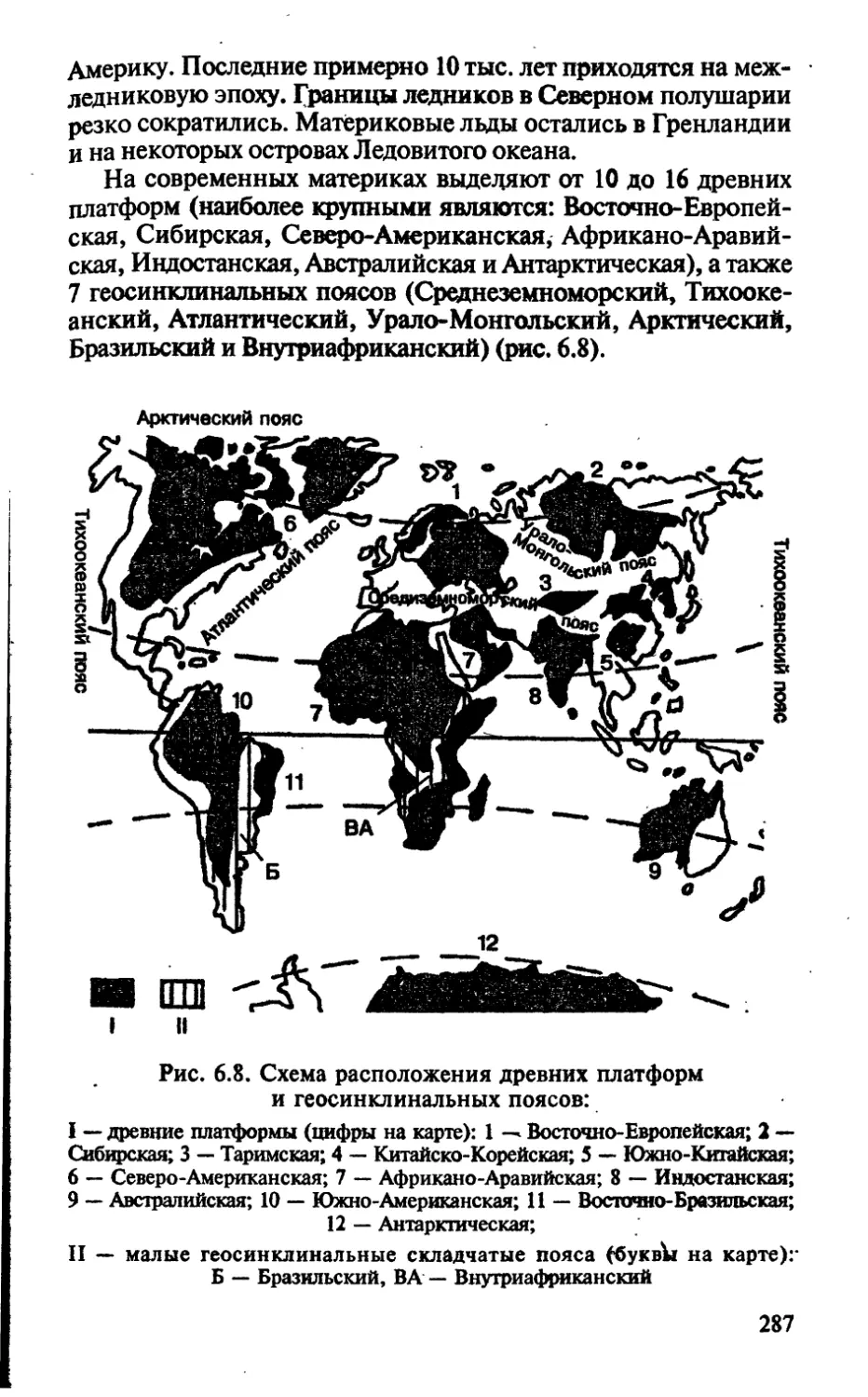
раздел 4
РАСТИТЕЛЬНЫЕ ТКАНИ
Понятие о ткани. Царство растений состоит из простейших
(одноклеточных) и многоклеточных организмов. Если это од-
ноклеточный организм, то в его единственной клетке проис-
ходят все необходимые процессы жизнедеятельности; если
многоклеточный, то различные клетки объединяются в груп-
пы одинаково функционирующих клеток, в так называемые
ткани.
Ткани — это устойчивые, закономерно повторяющиеся
комплексы клеток, сходные по происхождению, строению и
приспособленные к выполнению одной или нескольких функ-
ций. Ткань называется простой, если все ее клетки одинаковы
по форме и функциям (паренхима, склеренхима, колленхима)
Сложные ткани (потраничные, проводящие) состоят из кле-
ток, неодинаковых по форме, внутреннему строению и функ-
циям, но связанных общим происхождением (например, кси-
лема, образованная камбием).
Классификация тканей. Растительные ткани делят на не-
сколько групп в зависимости от основной функции.
1. Образовательные ткани, или меристемы, обладают
способностью к делению и формированию всех прочих тка-
ней.
•>2. Основные ткани составляют большую часть тела расте-
ния. Они бывают:
1) ассимиляционные (хлорофиллоносные);
2) запасающие;
3) дыхательные — аэренхима;
4) водоносные.
3. Пограничные ткани:
1) наружные пограничные ткани (покровные), с преоблада-
нием функций газообмена и транспирации (испарение воды),
выполняют также функцию механической защиты от влияния
внешней среды;
2) внутренние пограничные ткани (эндодерма, экзо-
дерма, обкладочные клетки сосудисто-волокнистых пучков)
с преобладанием функций регуляции продвижения веществ
(барьерная функция);
3)ризодерма — наружная ткань с преобладанием функ-
ций всасывания (покровно-всасывающая ткань корня).
4. Выделительные ткани:.
1) наружные: а) железистые волоски; б) г и д а т о р ыво-
дяные устьица; в) нектарники;
4
2) внутренние: а) выделительные клетки с эфирными мас-
лами, смолами, дубильными веществами; б) многоклеточные
вместилища выделений, млечники.
5. Механические ткани (опорные, скелетные):
1) колл енхима;
2) склеренхим а.
6. Проводящие ткани:
1)ксилема (древесина) — ткань восходящего тока;
2) ф л о э м а (луб) — ткань нисходящего тока.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ
Образовательные ткани благодаря постоянному митотичес-
кому делению их клеток обеспечивают не только рост, но и об-
разование всех тканей растения, т. е. фактически формируют
его тело. Часть дочерних клеток дифференцируется, превра-
щаясь в клетки различных тканей. Другие, сохраняя свои ме-
ристематические свойства, продолжают делиться и образуют
все новые и новые клетки. Различают первичные и вторичные
меристемы.
Первичные меристемы:
1) апикальные (верхушечные) — это такие меристемы, кото-
рые закладываются в теле зародыша (промеристема), сохраня-
ются на верхушках стеблей и кончиках корней?
У стеблей в конусе нарастания выделяют два меристемати-
ческих слоя: тунику, из которой образуется покровная
ткань и периферическая часть первичной коры, и корпус,
из которого образуется внутренняя часть первичной коры и
центральный осевой цилиндр (рис. 4.1).
В кончике корня различают три слоя — дерматоген, из
которого в зоне всасывания образуется первичная покровно-
всасывающая ткань ризодерма; п е р и б л е м а, из которой'об-
разуются ткани первичной коры, и плером, образующий
ткани центрального осевого цилиндра (рис. 4.2);
2) прокамбий — меристема, из которой формируется кам-
бий, перицикл и первичные элементы сосудисто-волокнистых
пучков;
3) вставочные меристемы возникают в виде отдельных уча-
стков в зонах активного роста в различных частях растения
(например, в основании черешков листьев, у оснований меж-
доузлий). В основании междоузлий у злаков деятельность
этой меристемы ведет к удлинению междоузлий, что обеспе-
чивает рост стебля злака в длину.
5
Рис. 4.1. Верхушечная
меристема побега элодеи:
1 — конус нарастания; 2 —
зачаток листа; 3 — сформи-
ровавшийся лист; 4 — буго-
рок пазушной почки
5
Рис. 4.2. Кончик корня:
1 — чехлик; 2 — эона деле-
ния; 3 — зона растяжения;
4 — зона всасывания; 5—
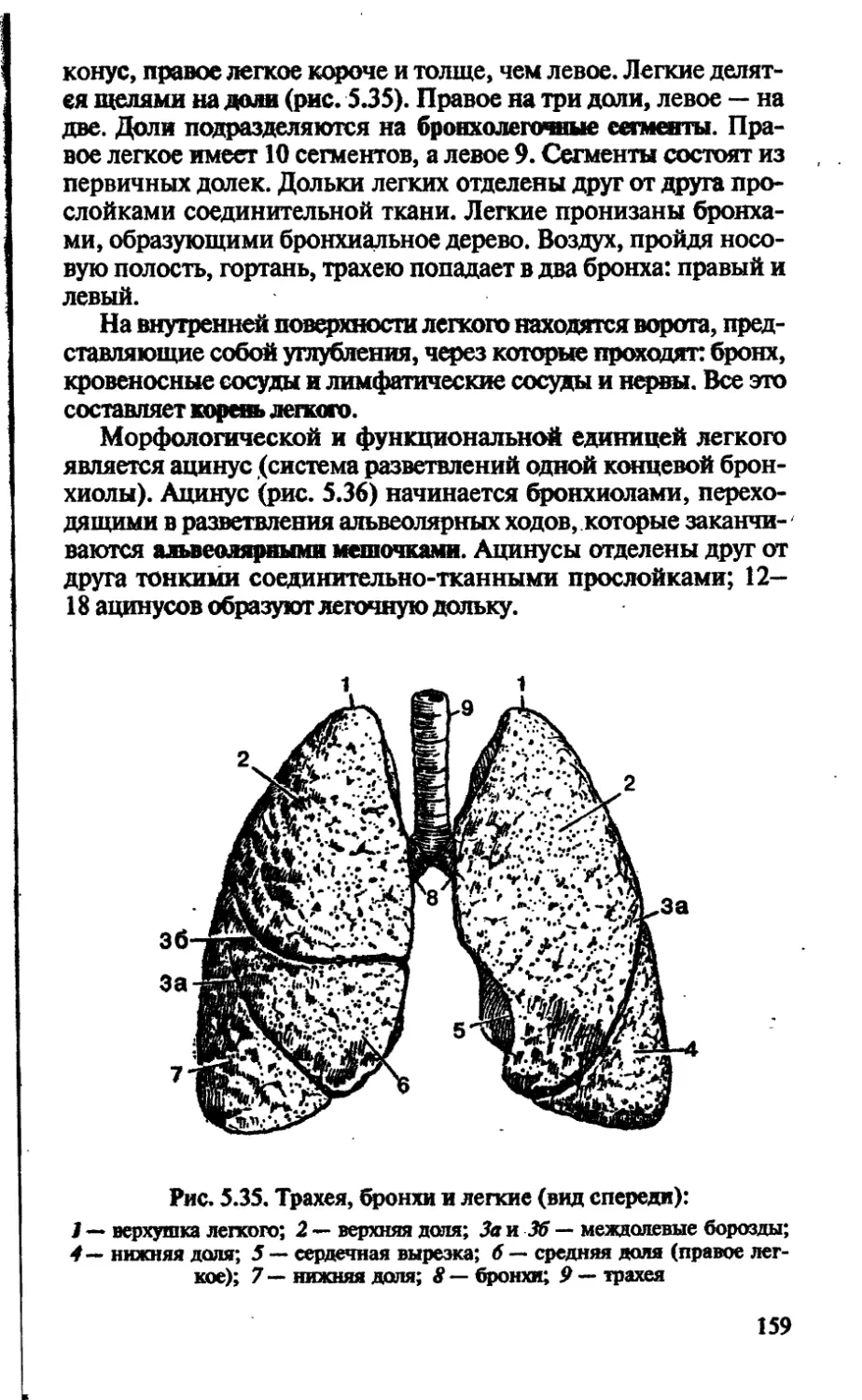
зона проведения
Вторичные меристемы — камбий я фелюген (пробковый кам-
бий) — образуются из клеток постоянных тканей. Они
занимают боковое положение по отношению к оси органа и
обеспечивают его рост в толщину. Среди вторичных меристем
выделяют так называемые раневые, которые дают начало осо-
бой защитной ткани в местах повреждения растения.
ОСНОВНЫЕ ТКАНИ
Основные ткани (паренхимы) составляют бблыпую часть
всех органов растений. Они заполняют промежутки между
проводящими и механическими тканями и присутствуют во
всех вегетативных и генеративных, органах. Эти ткани обра-
зуются за счет дифференцировки апикальных меристем и со-
стоят из живых паренхиматозных клеток, разнообразных по
строению и функциям. Различают ассимиляционную, запаса-
ющую, воздухоносную и водоносную паренхимы.
В ассимиляционной, или хлорофиллоносной, паренхиме осу-
ществляется фотосинтез. Она встречается в надземных органах
растений (листьях, молодых зеленых стеблях). »
б
Запасающая паренхима преобладает в стебле, корне корне-
вище. В клетках этой ткани откладываются запасающие веще
ства — белки, жиры, углеводы.
Воздухоносная паренхима, или аэренхима, состоит из возду-
хоносных полостей (межклетников), представляющих собой
резервуары для запаса газообразных веществ. Эти полости ок-
ружены клетками основной паренхимы (хлорофиллоносной
или запасающей). Аэренхима хорошо развита у водных расте-
ний в различных органах и может встречаться у сухопутных ви-
дов. Главное назначение аэренхимы—участие в газообмене, а
также в обеспечении плавучести растений.
Клетки водоносной паренхимы содержат в вакуолях слизис-
тые вещества, способствующие удержанию влаги. Преимуще-
ственно эти клетки бывают у суккулентов(кактусы, алоэ, агава).
ПОГРАНИЧНЫЕ ТКАНИ
Пограничные ткани бывают наружными и внутренними.
Наружные пограничные ткани, называемые покровными, по-
крывают тело растения и предохраняют внутренние ткани от
различных воздействий внешней среды — перегрева, охлажде-
ния, проникновения различных бактерий и грибов. Различают:
1) первичную покровную ткань — эпидерму;
2) вторичную покровную ткань—перидерму;
3) третичную покровную ткань — корку, или ритидом.
Эпидерма — первичная покровная ткань, образованная из
апикальных (верхушечных) меристем — является сложной и
многофункциональной тканью, так как в нее входит ряд морфо-
логически различных клеток: а) собственноэпидермальные
клетки; б) замыкающие клетки устьиц; в) околоустьичные клет-
ки; г) трихомы—различные выросты эпидермальных клеток.
Собственноэпидермальные клетки плотно сомк-
нуты между собой. Эти клетки у листа двудольных растений
имеют извилистую форму, у однодольных — прямоугольную
форму и располагаются параллельными рядами.
Снаружи эпидерма покрыта слоем кутикулы, состоящей из
липоподобного вещества кутина и полисахарида пектина.
Иногда эпидермальный слой покрывается восковым налетом
различной толщины (например, эпидерма листа фикуса).
В собственноэпидермальных клетках имеются лейкопласты,
а хлоропласты обычно отсутствуют.
Замыкающие клетки устьиц имеют бобовидную
форму, между которыми образуется устьичная щель — устьи-
це. Устьице это межклетник между двумя замыкающими
клетками бобовидной формы. Устьица также можно назвать
порами эпидермы. Они имеются главным образом в листьях,
но есть и на стебле (рис; 4.3).
Стенки замыкающих клеток утолщены неравномерно: та
часть клеточной стенки, которая формирует устьичную щель, /
значительно утолщена («брюшная» стенка) по сравнению с ос- ,;
тальной клеточной стенкой («спинная» стенка). Щель может j
расширяться и сужаться, регулируя транспирацию и газооб- 1
мен. Под щелью располагается крупная полость (межклет- I
ник), называемая дыхательной, которая окружена клетками <
мезофилла листа. "
Замыкающие клетки окружены околоустьичными
клетками, которые совместно с замыкающими клетками
образуют устьичный комплекс. Устьица обычно рас-
полагаются на нижней стороне листа. Однако у водных расте-
ний, обладающих плавающими листьями, они находятся толь- .
ко на верхней стороне листа.
Механизм работы устьиц обусловлен осмотическими свой-
ствами клеток.
При освещении поверхности листа солнцем в хлоропластах
замыкающих клеток активно происходит процесс фотосинте- |
за. Насыщение клеток продуктами фотосинтеза — сахарами и
крахмалом — влечет за собой активное поступление в клетки
ионов калия, вследствие чего концентрация клеточного сока в
замыкающих клетках возрастает. Возникает разность концент-
рации клеточного сока околоустьичных и замыкающих клеток.
Рис. 4.3. Строение устьица (схема):
а — вид сверху; б — поперечный срез:
1 - • эпидермальные клетки; 2 — побочные клетки; 3 — замыкающая клет-
ка; 4 — устьичная щель; 5 — «дыхательная» полость; 6 — кутикула;
7 — хлоропласты клеток мезофилла листа
8
В силу осмотических свойств клеток вода из околоустьичных
клеток поступает в замыкающие клетки, что ведет к увеличе-
нию их объема и резкому возрастанию тургора. Утолщение
«брюшных» стенок замыкающих клеток, обращенных к усть-
ичной щели, обеспечивает неравномерное растяжение клеточ-
ной стенки и замыкающие клетки приобретают выраженную
бобовидную форму, вызывая открытие устьичной щели. При
падении интенсивности фотосинтеза вечером снижается обра-
зование сахаров в замыкающих клетках. Приток ионов калия
прекращается. Концентрация клеточного сока падает в замы-
кающих клетках по сравнению с околоустьичными. Вода пу-
тем осмоса уходит из замыкающих клеток и тургор этих клеток
понижается, что ведет к закрытию устьичной щели ночью.
Трихомы — это различные по форме, строению и. функ-
циям выросты клеток эпидермы — волоски, чешуйки, щетин-
ки и т. п. Размеры трихом варьируют в значительных преде-
лах. Наиболее длинные трихомы (до 5—6 см) покрывают
семена хлопчатника.
Перидерма — это сложная покровная ткань стеблей, корней
и корневищ многолетних растений (реже однолетних). Она
сменяет эпидерму осевых органов, которая постепенно отми-
рает и слущивается. Перидерма образуется, из. ф е л л о ге н а
(вторичная меристема). Феллоген закладывается в основной
паренхиМе, лежащей под эпидермой. Клетки феллогена делят-
ся: наружу откладываются клетки пробки, а внутрь — живые
паренхимные клетки феллодермы. В клетках феллодермы со-
держатся хлоропласты (рис. 4.4).
Рис. 4.4, Перидерма стебля бузины:
а — чечевичка; б — участок перидермы:
1 — выполняющая ткань; 2 — остатки-эпидермы; 3 — пробка (феллема);
4 — феллоген; 5 — феллодерма
9
Пробка состоит из мертвых клеток, у которых клеточная
стенка пропитана жироподобным суберином. Клетки распола-
гаются ровными рядами, имеют прямоугольную форму (на по-
перечном срезе), плотно прилегают друг к другу, формируя
многослойный футляр. Пробка охраняет внутренние живые
ткани от потери влаги, резких температурных колебаний и
проникновения микроорганизмов. Живые ткани, лежащие
под пробкой, нуждаются в газообмене и удалении избытка
влаги. Поэтому на месте воздухоносной полости под устьицем
феллоген откладывает живые, рыхло расположенные, со мно-
жеством межклетников паренхимные клетки, называемые вы-
полняющей тканью, которая разрывает эпидерму и создает
возможность газообмена и транспирации со внешней средой.
Это структурное образование называется ч е ч е в и ч ко й.
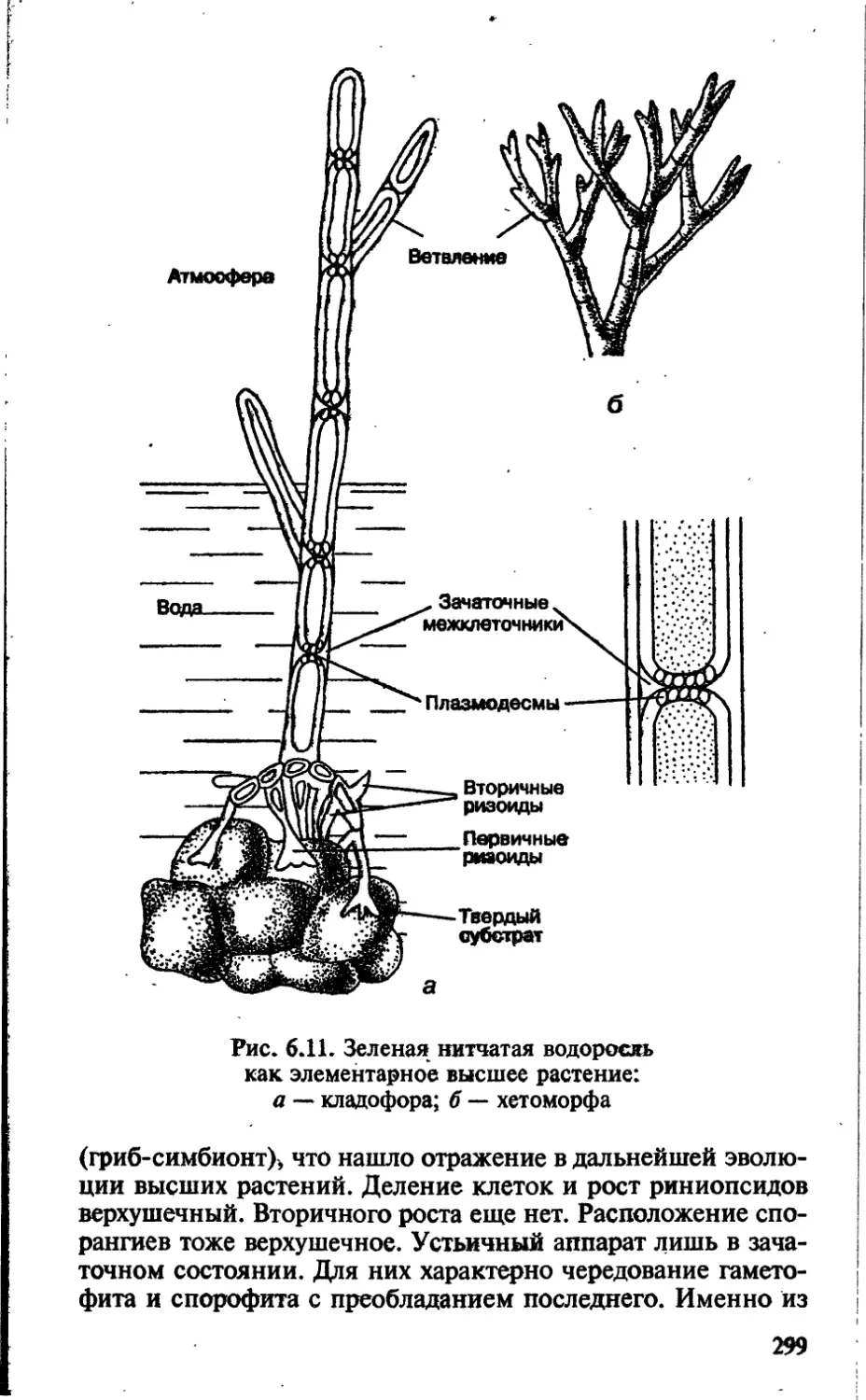
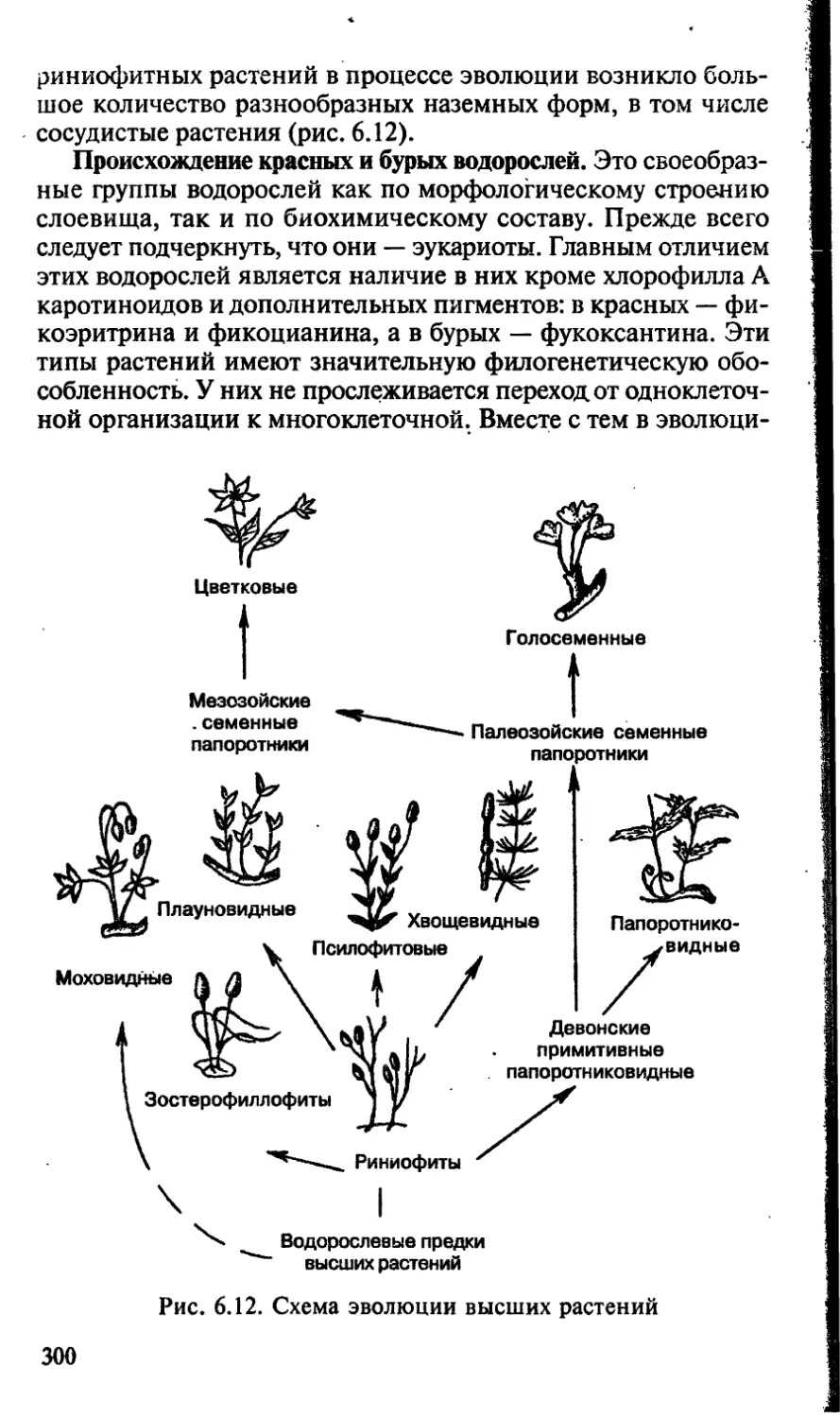
Чечевички, имеющие вид небольших бугорков, отчетливо
видны на поверхности побегов деревьев и кустарников. На
стволах березы их остатки наблюдаются в виде характерных
поперечных черных полосок и «черточек».
Ab/жа {ритидом) является третичной покровной тканью, ко-
торая образуется у многолетних растений в корне, стебле, корне-
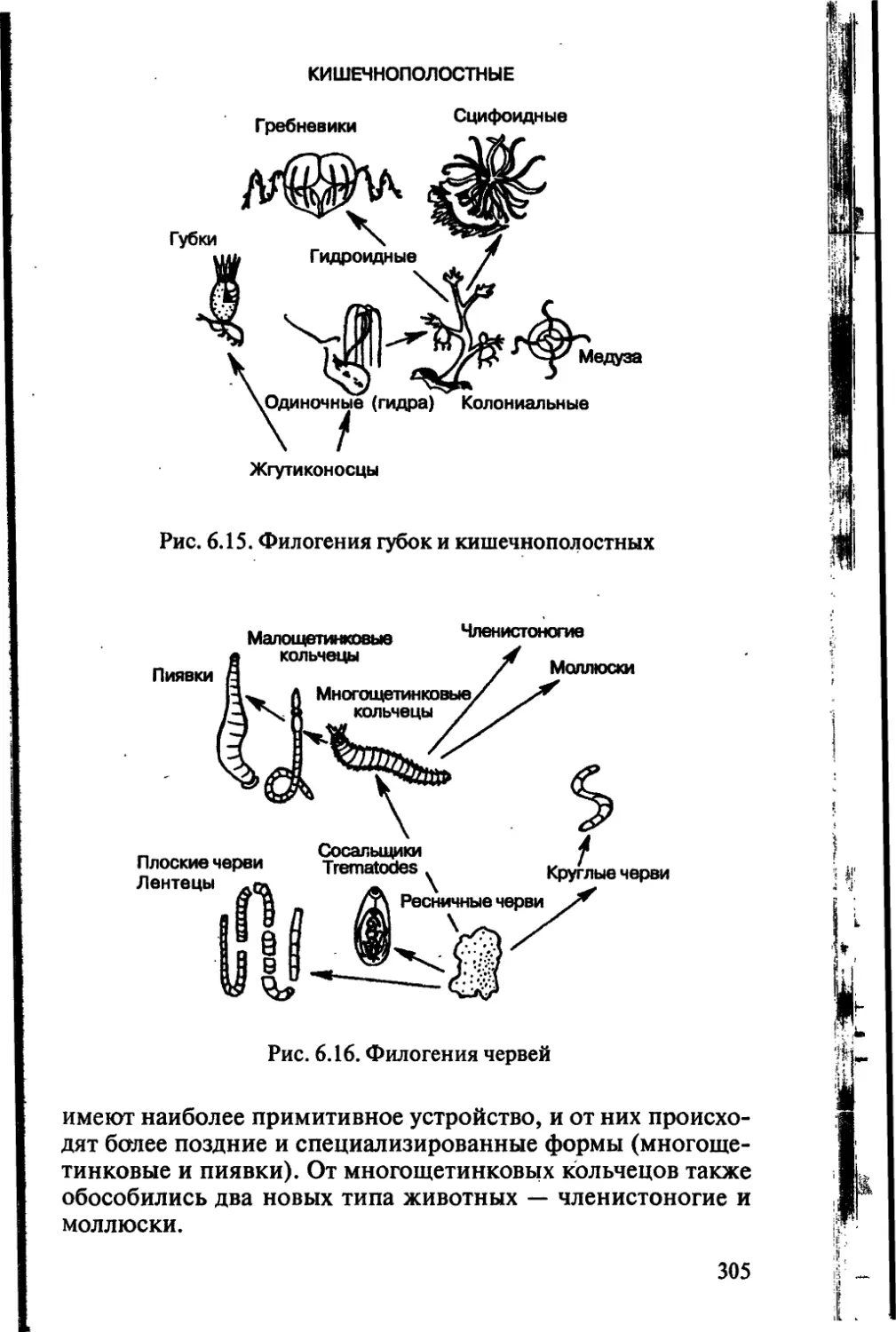
вище. Каждой год в более глубоких слоях первичной корызакла-
довается новый слой перидермы. Наружный слой перидермы—
пробка — изолирует все вышележащие ткани, в результате чего
они отмирают. Таким образом, совокупность многочисленных
перидерм с отмершими между ними тканями и является коркой..
О внутреишк пограничных тканях (эндодерма, экзодеома) и
ризодерме подробнее см. в разд. Корень.
ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
Выделительные ткани представлены различными образова-
ниями (чаще многоклеточными, реже одноклеточными), вы-
деляющими из растения или изолирующими в его тканях про-
дукты обмена веществ либо воду.
У растений различают следующие выделительные ткани:
Вместилища выделений — образуются из межклетников и
классифицируются по происхождению:
1) лизигенные вместилища, образующиеся за счет растворе-
ния клеточных стенок (лимон);
2) схизогенные вместилища, образующиеся за счет расхож-
дения клеток (у хвои сосны).
Идиобласты — состоят из отдельных клеток, содержащих
минеральные продукты выделения.
10
Млечники — представляют собой систему полостей, содер-
жащих млечный сок.
Кроме того, к выделительным тканям относят железки и
железистые волоски — многоклеточные образования, встреча-
ющиеся в эпидерме, где происходит накопление эфирных ма-
сел, смол и дубильных веществ.
МЕХАНИЧЕСКИЕ ТКАНИ
Механические ткани — это опорные (арматурные) ткани,
образующие скелет растения и обеспечивающие его проч-
ность, вследствие чего растение способно противостоять на-
грузкам на растяжение, сжатие и изгиб. Различают механи-
ческие ткани с равномерно и неравномерно утолщенными
клеточными стенками.
Колленхима. Ткань, клетки которой имеют неравномерно
утолщенные клеточные стенки, называются колленхимой. Раз-
личают уголковую и пластинчатую колленхиму. Стенка клеток
колленхимы состоит из целлюлозы, гемицеллюлозы и пектино-
вых веществ. Клетки являются хлорофиллоносными, поэтому в
подземных органах колленхима не встречается (рис. 4.5).
Клетка уголковой колленхимы имеет форму шестиугольного
многогранника, у которого утолщение целлюлозной оболочки
идет вдоль ребер, а на поперечном срезе утолщения клеточной
стенки заметны по углам этого многогранника. Уголковая кол-
ленхима встречается у стеблей двудольных растений (в основ-
ном травянистых), в черешках листьев и по обеим сторонам
1 — полость клетки; 2 — утолщенная стенка;
— объемное изображение
11
крупных жилок листа. Колленхима не препятствует росту
органа в длину, в котором она расположена.
Клетка, пластинчатой колленхимы имеет форму параллелепи-
педа, у которой утолщается только пара граней (стенок), заметных
на поперечном срезе с тангентальных сторон, т. е. находящихся
параллельно поверхности стебля. Пластинчатая колленхима
встречается, как правило, в стеблях древесных растений.
Склеренхима. Механическая ткань, состоящая из клеток с
одревесневшими и равномерно утолщенными клеточными
стенками, называется склеренхимой. Ее клеточная стенка од-
ревесневает, т. е. пропитывается лигнином, а ядро и цитоплаз-
ма клетки разрушаются. Существуют две разновидности скле-
ренхимы — склеренхимные волокна и склереиды.
Склеренхимные волокна образуют ткань, состоящую из кле-
ток вытянутой формы с заостренными концами и поровыми
каналами в клеточных стенках. Эти клетки плотно примыкают
друг к другу и их оболочки обладают высокой прочностью. На
поперечном срезе клетки многогранны (рис. 4.6).
Если склеренхимные волокна встречаются в древесине
(ксилеме), то они называются древесинными волок-
нами (либриформ). Они защищают сосуды от давления дру-
гих тканей, являясь механической частью ксилемы.
Если склеренхимные волокна встречаются в лубе (флоэме),
то они называются лубяными волокнами (камби-
форм). Лубяные волокна могут быть и неодревесневшими, об-
ладая при этом большой прочностью и эластичностью, что на-
ходит большое применение в текстильной промышленности
(например, волокна льна).
Если волокна возникают на месте перицикла, то тогда они
называются перициклическими волокнами. *
Рис. 4.6. Древесинные
волокна листа герани
луговой:
а — на поперечном срезе;
б — на продольном срезе:
1 — простая пора; 2 — полость
клетки; 3 — стенка клетки;
12
Склереиды обычно возникают из клеток основной паренхи-
мы в результате утолщения и лигнификации их клеточных сте-
нок. Они имеют различную форму и встречаются во многих
органах растения. Склереиды более или менее изодиаметрич-
ной формы (с одинаковым диаметром клетки) называются
брахисклереидам и, или каменистыми клетка-
ми (в плодах груши). Склереиды, имеющие расширение на
обоих концах клетки — остеосклереиды — встречаются в
листьях чая. Склереиды, форма которых напоминает звезду,
называются астросклереидами (в листьях камелии).
Удлиненные палочковидные клетки склереид находятся в се-
менах бобовых.
ПРОВОДЯЩИЕ ТКАНИ
Проводящие ткани обеспечивают восходящий и нисходя-
щий ток растения. Восходящий ток — это ток минеральных со-
лей, растворенных в воде, идущих от корней по стеблю к лис-
тьям. Восходящий ток осуществляется по сосудам и трахеидам
ксилемы (древесины).
Нисходящий ток — это ток органических веществ, направ-
ляющийся от листьев к корням по ситовидным элементам
флоэмы (луба).
Ксилема и флоэма — это сложные ткани, состоящие из
трех основных элементов (табл. 4.1).
Таблица 4.1
Основные элементы ксилемы и флоэмы
Проводящая ткань Основные элементы
проводящие механические запасающие
Ксилема Флоэма Сосуды и трахеиды Ситовидные трубки и клетки-спутницы Древесинные волокна Лубяные волокна Древесинная паренхима Лубяная паренхима ~
Проводящие элементы ксилемы. Наиболее древними прово-
дящими элементами ксилемы являются трахеиды — эго
вытянутые клетки с заостренными концами. Они дали начало
древесинным волокнам. Трахеиды имеют одревесневшую кле-
точную стенку с различной степенью утолщения, кольчатую,
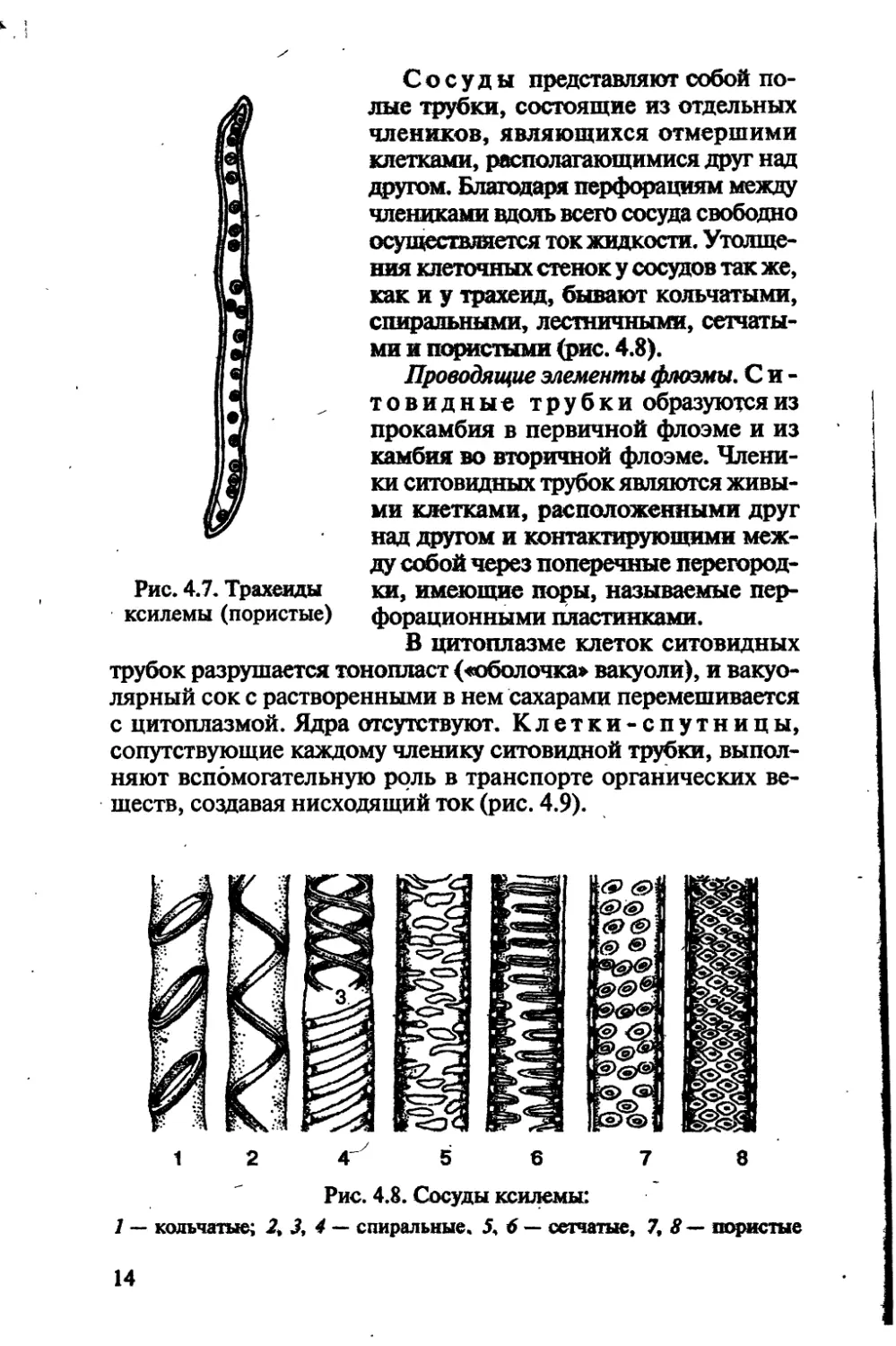
спиральную, точечную, пористую и т. д. форму (рис. 4.7).
13
Сосуды представляют собой по-
(лые трубки, состоящие из отдельных
члеников, являющихся отмершими
клетками, располагающимися друг над
другом. Благодаря перфорациям между
члениками вдоль всего сосуда свободно
осуществляется ток жидкости. Утолще-
ния клеточных стенок у сосудов так же,
как и у трахеид, бывают кольчатыми,
спиральными, лестничными, сетчаты-
ми и пористыми (рис. 4.8).
Проводящие элементы флоэмы. С и -
„ товидные трубки образуютсяиз
прокамбия в первичной флоэме и из
камбия во вторичной флоэме. Члени-
ки ситовидных трубок являются живы-
ми клетками, расположенными друг
над другом и контактирующими меж-
ду собой через поперечные перегород-
Рис. 4.7. Трахеиды км, имеющие поры, называемые пер-
ксилемы (пористые) форационными пластинками.
В цитоплазме клеток ситовидных
трубок разрушается тонопласт («оболочка» вакуоли), и вакуо-
лярный сок с растворенными в нем сахарами перемешивается
с цитоплазмой. Ядра отсутствуют. Клетки-спутницы,
сопутствующие каждому членику ситовидной трубки, выпол-
няют вспомогательную роль в транспорте органических ве-
ществ, создавая нисходящий ток (рис. 4.9).
Рис. 4.8. Сосуды ксилемы:
1 — кольчатые; 2, 3, 4 — спиральные. 5, 6 — сетчатые, 7,8 — пористые
14
a
Рис. 4.9. Ситовидные элементы флоэмы тыквы:
а — продольный срез; б — поперечный срез:
1 — незакупоренная ситовидная пластинка; 2 — клетка-спутница; 3 — лу-
бяная (флоэмная) паренхима; 4 — закупоренная ситовидная пластинка;
5 — функционирующий членик ситовидной трубки; 6 — членик ситовид-
ной трубки с закупоренной ситовидной пластинкой (образовалось мозоли-
стое тело)
Сосудисто-волокнистые пучки. Флоэмаи ксилема образуют
сосудисто-волокнистые пучки. Пучки располагаются в цент-
ральном осевом цилиндре и бывают открытыми и закрытыми.
Закрытые пучки состоят из ксилемы и флоэмы, между кото-
рыми отсутствует камбий, и, таким образом, образования но-
вых элементов флоэмы и ксилемы не происходит. Закрытые
сосудисто-волокнистые пучки встречаются в стеблях и корне-
вищах однодольных растений.
Открытые пучки имеют камбий между флоэмой и ксиле-
мой. В результате деятельности камбия пучок разрастается и
происходит утолщение органа. Открытые сосудисто-волокни-
стые пучки встречаются во всех осевых органах двудольных и
голосеменных растений.
В зависимости от взаимного расположения флоэмы и кси-
лемы различают четыре типа сосудисто-волокнистых пучков
(рис. 4.10).
Чаще всего флоэма лежит по одну сторону от ксилемы, т. е.
флоэма примыкает к ксилеме и обращена к периферии органа.
Такие пучки называют коллатеральными. Они могут быть как
15
1 2 3 4 5 6
Рис. 4.10. Схемы разных типов сосудисто-волокнистых пучков
(поперечный срез):
1 — открытый коллатеральный; 2 — биколлатеральный; 3 — закрытый
коллатеральный; 4, 5 — концентрические (4 — центрофлоэмный; 5 — цен-
троксилемный); 6 — радиальный. Ксилема зачернена, флоэма показана
точками, камбий — штрихами
закрытыми, так и открытыми. Закрытые коллатеральные пучки
встречаются у стеблей и корневищ однодольных растений, а от-
крытые — у стеблей, корневищ и корней двудольных растений.
Биколлатеральные пучки — это такие пучки, в которых фло-
эма примыкает к ксилеме с двух сторон (сверху и снизу). Верх-
ний, более мощный участок флоэмы, обращен к периферии
органа и отделен от ксилемы слоями камбия. Нижний учас-
ток — внутренняя флоэма — развит слабо и примыкает к кси-
леме без камбия. Биколлатеральные пучки встречаются у стеб-
лей, корней и корневищ двудольных растений.
Радиальные пучки составлены в основном ксилемой, кото-
рая располагается по радиусам. Между лучами ксилемы нахо-
дятся участки флоэмы. Эти пучки не имеют камбия и встреча-
ются у корней в первичном строении.
Концентрические пучки представляют собой круг. Если в
центре круга располагается флоэма, окруженная кольцом кси-
лемы, то такой пучок называется центрофлоэмным (у
большинства корневищ однодольных); если в центре круга
расположена ксилема, окруженная флоэмой, то такой пучок
называется концентрическим центроксилем-
ным (у корневищ папоротников).
КОРЕНЬ
Корень — осевой орган, имеющий цилиндрическую форму
и обладающий радиальной симметрией. Он способен к росту
до тех пор, пока сохраняется апикальная меристема. Морфо-
логически корень отличается от побега тем, что на нем никог-
16
да не возникают листья, а апикальная меристема прикрыта
корневым чехлом.
Главный, или первый, корень семенного растения развивает-
ся из зародышевого корешка семени. У двудольных и голосе-
менных растений от главного корня отходят боковые корни
первого порядка, которые закладываются в перицикле. Боко-
вые корни первого порядка дают начало боковым корням вто-
рого и третьего порядков. Корневая система, образованная
системой главного и боковых корней, называется стержне-
вой, а с развитой системой боковых корней — ветвистой.
Существуют еще и придаточные корни, берущие начало от лис-
тьев, стеблей и старых корней. Корневая система, образован-
ная придаточными корнями, называется мочковатой. Раз-
росшийся главный корень образует корнеплод (морковь,
свекла). Разросшиеся придаточные корни образуют корнеклуб-
ни (георгин).
Функции корня.
1. Минеральное и водное питание (поглощение воды и ми-
неральных веществ).
2. Закрепление растения в почве (заякоревание).
3. Синтез органических веществ.
4. Накопление запасных веществ.
5. Вегетативное размножение.
6. Симбиоз с бактериями. На корнях бобовых и других рас-
тений возникают особые образования — клубеньки, в которых
поселяются бактерии. Эти бактерии способны фиксировать
атмосферный молекулярный азот, переводя его в связанное
состояние. Часть азотистых соединений, образованных таким
путем, усваивает высшее растение — хозяин, отдавая при этом
бактериям углеводы.
Зоны корня. В корне различают четыре зоны: деления, растя-
жения, всасывания, проведения (илиукрепления) (см. рис. 4.2).
К зоне деления относят верхушку конуса нарастания, где
происходит активное деление клеток. Эта зона состоит из тон-
костенных паренхимных клеток первичной меристемы, кото-
рые прикрыты корневым чехликом, выполняющим защитную
функцию при продвижении корня между частичками почвы.
От соприкосновения с почвой клетки чехлика постоянно раз-
рушаются, образуя слизистый чехол. У большинства растений
корневой чехлик восстанавливается за счет первичной мерис-
темы, а у злаков 3£ счет особой меристемы — калиптрогена.
В зоне растяжения клетки меристемы увеличиваются в раз-
мерах, вытягиваются в длину и деление клеток постепенно
прекращается. Вследствие вытягивания клеток в продольном
17
направлении осуществляется рост корня в длину и продвиже-
ние его в почве. Эти две зоны можно объединить в одну —
зону роста. Ее протяженность составляет несколько милли-
метров.
В зоне всасывания происходит дифференциация тканей
корня. По происхождению эти ткани первичные, так как они
образуются из первичной меристемы конуса нарастания. По-
кровной тканью является ризодерма, или эпиблема. Клетки
ризодермы живые, с тонкой целлюлозной стенкой. Из некото-
рых клеток ризодермы формируются корневые волоски. Каж-
дый корневой волосок представляет собой длинный вырост
одной из клеток ризодермы, ядро клетки обычно находится в
кончике выроста. Корневой волосок содержит тонкий при-
стенный слой цитоплазмы, более плотный на верхушке волос-
ка, а в центре — крупную вакуоль. Корневые волоски недолго-
вечны, и в зоне укрепления они отмирают. Зона всасывания
представляет собой физиологически очень важную часть кор-
ня. Клетки ризодермы поглощают водные растворы всей по-
верхностью наружных стенок. Развитие корневых волосков во
много раз увеличивает поверхность поглощения. Зона всасыва-
ния имеет протяженность от 1 до 1,5 см.
Зона проведения. Зона всасывания постепенно переход ит в
зону проведения (или укрепления). Она тянется вплоть до
корневой шейки и составляет большую часть протяженности
корня. В этой зоне идет интенсивное ветвление главного кор-
ня и появление боковых корней.
Первичное строение корня. Микроскопическое строение кор-
ня в зоне всасывания называется первичным потому, что диф-
ференциация тканей в этой зоне происходит из первичной ме-
ристемы конуса нарастания. Первичное строение корня в зоне
всасывания можно наблюдать у двудольных растений, а у одно-
дольных — еще и в зоне проведения. На поперечном срезе кор-
ня первичного строения выделяют три основные части: покров-
но-всасывающая ткань, первичная кора и центральный осевой
цилиндр.
Покровно-всасывающая ткань — ризодерма (эпиблема) —
выполняет как покровную функцию, так и функцию интен-
сивного всасывания воды и минеральных веществ из почвы.
Первичная кора корня развита более мощно, чем централь-
ный осевой цилиндр. Она состоит из трех слоев тканей: экзо-
дермы, мезодермы (паренхимы первичной коры) и эндодер-
мы. Клетки э к з о д е р м ы многоугольные, они плотно
сомкнуты и расположены в несколько рядов. Клеточные стен-
ки пропитаны суберином (опробковение) и лигнином (одре-
18
веснение). Опробковение обеспечивает непроницаемость кле-
ток для воды и газов. Одревеснение делает ее более прочной.
В экзодерме, обычно под корневыми волосками, сохраняются
клетки с тонкими целлюлозными стенками — пропускные
клетки, через которые проходит вода и минеральные вещества,
поглощенные риэодермой. Экзодерма может выполнять по-
кровную функцию по мере отмирания клеток ризодермы.
Под экзодермой находятся живые паренхимные клетки
мезодермы. Это наиболее широкая часть первичной коры.
Клетки мезодермы выполняют запасающую функцию, а также
функцию проведения воды и растворенных в ней солей от кор-
невых волосков в центральный осевой цилиндр.
Внутренний однорядный слой первичной коры представ-
лен эндодермой. В зависимости от степени утолщения
клеточной стенки различают два типа эндодермы — эндодер-
му с поясками Каспари и эндодерму с подковообразными
утолщениями.
Эндодерма с поясками Каспари — это начальный этап фор-
мирования эндодермы, при котором утолщению подвергаются
лишь радиальные стенки ее клеток. В утолщениях можно об-
наружить лигнин и суберин.
Эндодерма с подковообразными утолщениями развивается
у однодольных растений, когда в клетках эндодермы происхо-
дит дальнейшее утолщение клеточных стенок и пропитка их
суберином. Неутолщенной остается только наружная клеточ-
ная стенка. Однако среди толстостенных клеток в эндодерме
встречаются клетки с тонкими целлюлозными оболочками.
Это пропускные клетки. Они обычно располагаются напротив
лучей ксилемы радиального типа пучка. Считается, что эндо-
дерма выполняет роль гидравлического барьера, способствуя
продвижению минеральных веществ и воды из первичной
коры в центральный осевой цилиндр и препятствует их выходу
обратно.
Центральный осевой цилиндр начинается с клеток перицик-
ла, который обычно в молодых корнях состоит из живых тон-
костенных паренхимных клеток, расположенных в один ряд.
Клетки перицикла дольше других тканей корня сохраняют
свойства меристемы и способность к новообразованиям.
Из перицикла образуются в зоне укрепления боковые корни,
поэтому его называют корнеродным слоем. Проводя-
щая система корня представлена одним радиальным сосудис-
то-волокнистым лучком, в котором группы элементов пер-
вичной флоэмы чередуются с лучами первичной ксилемы.
У двудольных растений в радиальном пучке встречается от од-
19
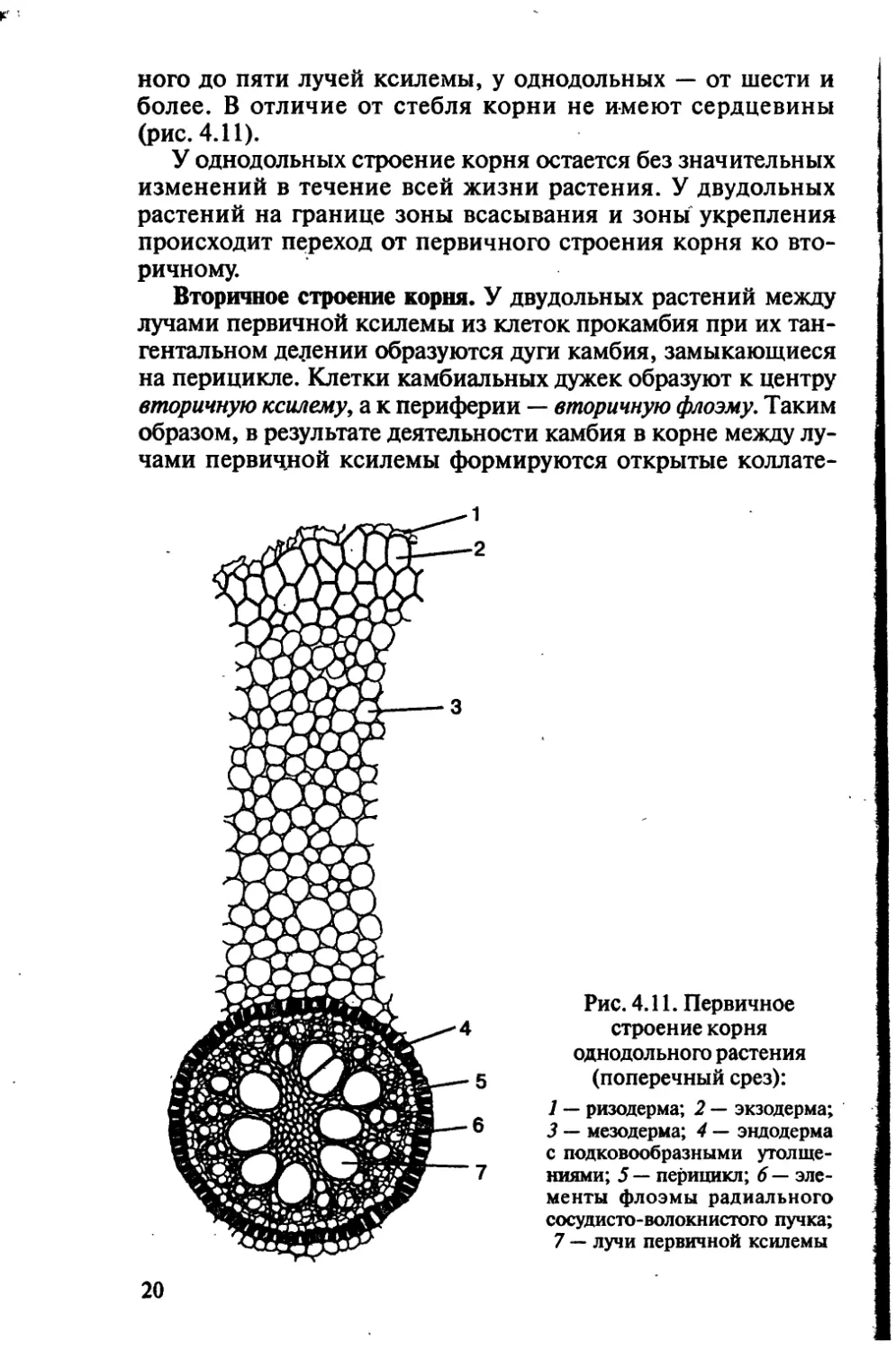
ного до пяти лучей ксилемы, у однодольных — от шести и
более. В отличие от стебля корни не имеют сердцевины
(рис. 4.11).
У однодольных строение корня остается без значительных
изменений в течение всей жизни растения. У двудольных
растений на границе зоны всасывания и зоны укрепления
происходит переход от первичного строения корня ко вто-
ричному.
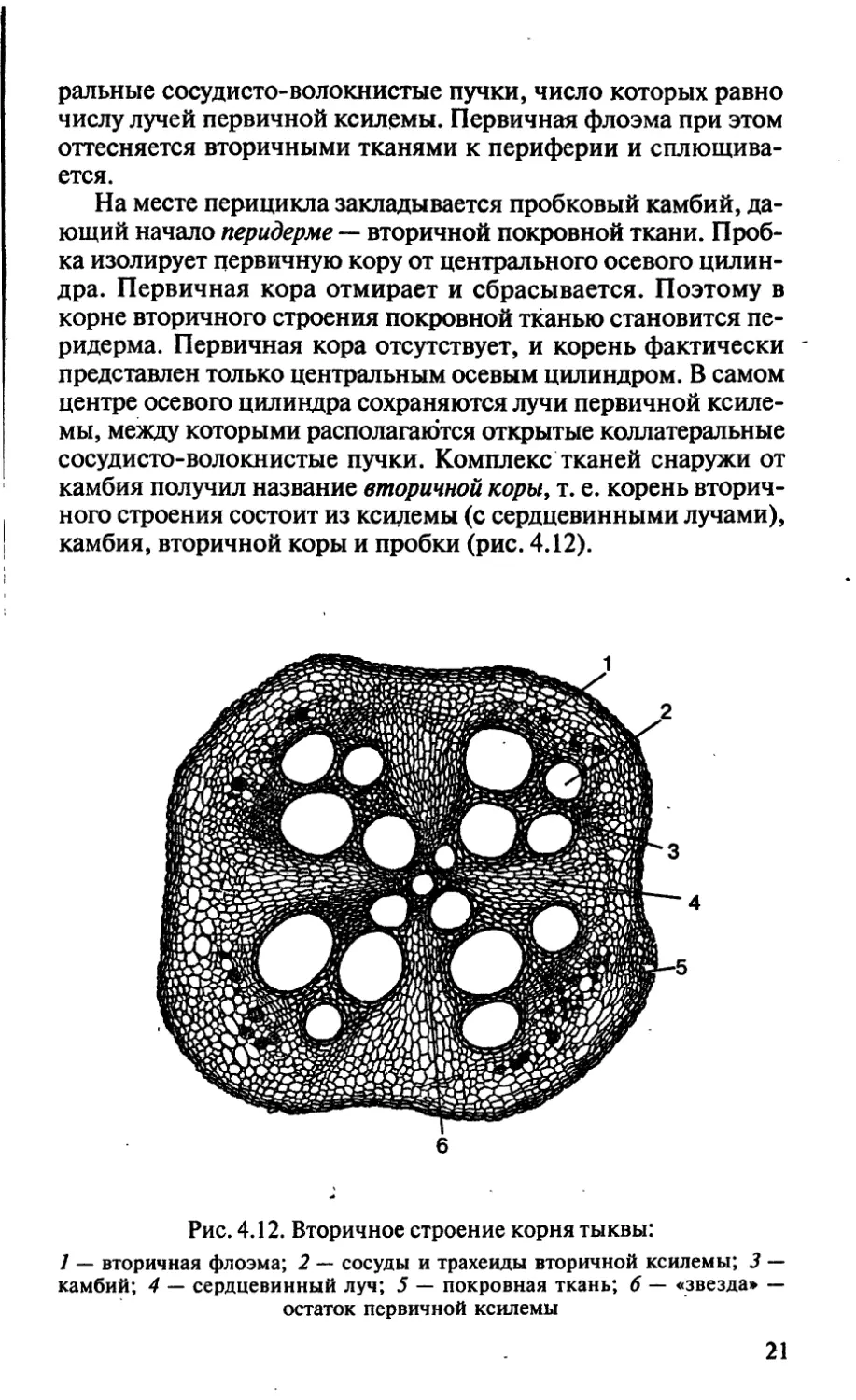
Вторичное строение корня. У двудольных растений между
лучами первичной ксилемы из клеток прокамбия при их тан-
гентальном делении образуются дуги камбия, замыкающиеся
на перицикле. Клетки камбиальных дужек образуют к центру
вторичную ксилему, а к периферии — вторичную флоэму. Таким
образом, в результате деятельности камбия в корне между лу-
чами первичной ксилемы формируются открытые коллате-
Рис. 4.11. Первичное
строение корня
однодольного растения
(поперечный срез):
1 — ризодерма; 2 — экзодерма;
3 — мезодерма; 4 — эндодерма
с подковообразными утолще-
ниями; 5 — перицикл; 6 — эле-
менты флоэмы радиального
сосудисто-волокнистого пучка;
7 — лучи первичной ксилемы
20
ральные сосудисто-волокнистые пучки, число которых равно
числу лучей первичной ксилемы. Первичная флоэма при этом
оттесняется вторичными тканями к периферии и сплющива-
ется.
На месте перицикла закладывается пробковый камбий, да-
ющий начало перидерме ~ вторичной покровной ткани. Проб-
ка изолирует первичную кору от центрального осевого цилин-
дра. Первичная кора отмирает и сбрасывается. Поэтому в
корне вторичного строения покровной тканью становится пе-
ридерма. Первичная кора отсутствует, и корень фактически -
представлен только центральным осевым цилиндром. В самом
центре осевого цилиндра сохраняются лучи первичной ксиле-
мы, между которыми располагаются открытые коллатеральные
сосудисто-волокнистые пучки. Комплекс тканей снаружи от
камбия получил название вторичной коры. т. е. корень вторич-
ного строения состоит из ксилемы (с сердцевинными лучами),
камбия, вторичной коры и пробки (рис. 4.12).
Рис. 4.12. Вторичное строение корня тыквы:
1 — вторичная флоэма; 2 — сосуды и трахеиды вторичной ксилемы; 3 —
камбий; 4 — сердцевинный луч; 5 — покровная ткань; 6 — «звезда» —
остаток первичной ксилемы
21
Минеральное питание растения. Растение поглощает воду
из почвы с помощью корневых волосков. В их клеточной обо-
лочке имеется значительное количество пектиновых ве-
ществ, обеспечивающих ослизнение корня. К корню при-
липают комочки земли, что позволяет ему легче проникать
в поры почвы. Поступление воды в корневой волосок обус-
ловлено осмотическими свойствами клетки. В случае силь-
но засоленных почв осмотические свойства клеток поддер-
живаются особыми биоколлоидами цитоплазмы.
Продвижение воды по клеткам первичной коры до сосудов
происходит за счет корневого давления по свободным про-
странствам между клеточными стенками.
СТЕБЕЛЬ
Стеблем называют растительный орган, представляющий
ось побега и несущий на себе листья, почки и цветки.
Основные функции стебля. Стебель выполняет опорную,
проводящую, запасающую функции и, кроме того, является
органом вегетативного размножения. Через стебель осуществ-
ляется связь между корнями и листьями. У некоторых расте-
ний стебель выполняет функцию фотосинтеза (хвощ, алоэ).
Побег. Побегом называется стебель с листьями и почками.
В более конкретном смысле под побегом нужно понимать од-
нолетний неразветвленный стебель с листьями и почками,
развившийся из почки или семени.
За счет верхушечной почки осуществляется рост основного
побега; за счет боковых почек — ветвление побега. Главная
внешняя черта, отличающая побег от корня — наличие листь-
ев. Участок стебля, от которого отходит лист (или листья), на-
зывают узлом, а расстояние между узлами — междоузлием.
Каждый повторяемый узел с междоузлием называется метаме-
рам.
Почка — это зачаточный, еще не развернувшийся побег.
Почки бывают вегетативными, вегетативно-генеративными и
генеративными.
Вегетативная почка состоит из конуса нарастания
стебля, зачатков листьев, зачатков почек и почечных чешуй.
В вегетативно-генеративных почках заложен
ряд метамеров, а конус нарастания трансформируется в зача-
точный цветок или соцветие (у травянистых растений).
22
Генеративные,или цветочные, почки заключа-
ю-тл себе-тОлько зачаток соцветия (вишня) или одиночный
цветок (рис. 4.13).
Если верхушечная почка отмирает, то тогда трогается в
рост боковая почка. Генеративная верхушечная почка после
развертывания верхушечного цветка или соцветия больше не
способна к верхушечному росту.
Развертывание побега из почки. Превращение почки в побег
начинается с разрастания листовых зачатков и роста междоуз-
лий. Наружные чешуи почек быстро подсыхают и отпадают
при начале развертывания почки. От них у основания побега
часто остаются рубцы, так называемое почечное кольцо, кото-
рое хорошо заметно у многих деревьев и кустарников. По чис-
лу почечных колец можно подсчитать возраст ветви. Побеги,
вырастающие из почек за один вегетационный период, раз в
год, называют годичными побегами или годичным приростом.
В нарастании побега в длину и ширину участвует ряд мерис-
тем. Рост в длину происходит за счет верхушечной и вставоч-
ной меристем, а в толщину — за счет боковых меристем (кам-
бия и феллогена).
На начальных этапах развития побега формируется первич-
ная анатомическая структура стебля, сохраняющаяся у одно-
дольных в течение всей их жизни. У двудольных древесных и
голосеменных растений в результате деятельности вторичных
Рис. 4.13. Строение различных типов почек:
а — вегетативная почка дуба;
б — вегетативно-генеративная почка бузины;
в — генеративная почка вишни:
1 — почечные чешуи; 2 — зачатки листьев; 3 — зачаток цветка
23
образовательных тканей довольно быстро из первичной струк-
туры формируется вторичное строение стебля.
Листорасположение — порядок размещения листьев на оси
побега (рис. 4.14).
1. Очередное, или спиральное — от каждого узла
стебля отходит один лист.
2. Супротивное — на каждом узле сидят друг против
друга два листа.
3. Мутовчатое — из каждого узла выходят три и более
листьев.
Характер ветвления побега (рис. 4.15). Ветвление побега у
растений необходимо для увеличения площади соприкоснове-
ния со средой — водной, воздушной и почвенной. Различают
моноподиальное, симподиальное, ложнодихотомическое и
дихотомическое ветвление побега.
1. Моноподиальное — длительное время сохраняет-
ся рост верхушечной почки (у ели).
2. Симподиальное — ежегодно верхушечная почка
отмирает, а рост идет за счет боковой почки (у березы).
3. Ложнодихотомическое (при супротивном лис-
торасположении, вариант симподиального) — верхушечная
почка отмирает, а рост идет за счет двух боковых почек (у idre-
на).
Рис. 4.14. Листорасположение:
а — очередное у персика обыкновенного; б — супротивное у бирючины
овальной; в — мутовчатое у олеандра
24
4. Дихотомическое - конус нарастания верхушеч-
ной почки делится надвое (плаун).
Характер расположения стебля в пространстве (рис. 4.16).
1. Прямостоячий стебель — все время растет
вверх.
2. Приподнимающийся стебель — в гипоко-
тильной части развивается в горизонтальном направлении, а в
дальнейшем растет вверх как прямостоячий (гипокотилем,
или подсемядольным коленом, называют участок от основа-
ния корня до семядолей).
3. Стелющийся с т е б е л ь — растет в горизонтальном
направлении, параллельно поверхности земли. Если стелю-
щийся стебель укореняется, он называется ползучим.
Рис. 4.15. Типы ветвления
побега:
1 — моноподиальное; 2 — сим-
подиальное; 3 — ложнодихото-
мическое, 4 — дихотомическое
Рис. 4.16. Характер расположения стебля в пространстве:
1 — прямостоячий; 2 — приподнимающийся; 3 — стелющийся; 4 — вью-
щийся; 5 — цепляющийся
25
4. Вьющийся стебель —растет вверх, но ему нужна
дополнительная опора, так как в нем плохо развиты механи-
ческие ткани.
• 5. Цепляющийся стебель —растет так л®, каки
вьющийся, вокруг дополнительной опоры, но с помощью спе-
циальных приспособлений — усиков.
Видоизменения побегов. Видоизменение побегов произош-
ло в процессе длительной эволюции, как следствие приспо-
собления к выполнению специальных функций. Например,
корневища, клубни и луковицы, являясь запасающими по-
бегами, часто выполняют функцию вегетативного размноже-
ния. Кроме того, видоизменения побега способны служить
органом прикрепления (усики) и средством защиты (ко-
лючки).
1. Подземные побеги (рис.4.17):
а) корневище (папоротник, ландыш) — многолетний под-
земный побег, имеющий редуцированные листья в виде бес-
цветных или бурых мелких чешуек, в основании которых лежат
почки;
б) клубень (картофель) — метаморфоз побега с ярко выра-
женной запасающей функцией стебля, наличием чешуевид-
ных листьев, которые быстро сшелушиваются, и почек, фор-
мирующихся в пазухах листьев и называемых глазками.
У клубня также имеются столоны — подземные недолговеч-
ные корневища, верхушечная почка которых разрастается и
превращается в клубни;
в) клубнелуковица (гладиолус) — это лиственный клубень,
т. е. клубень, покрытый чешуевидными сухими пленчатыми
листьями и внешне напоминающий луковицу;
г) луковица — это укороченный побег, стеблевая часть ко-
торого называется донцем. В луковице различают два типа
видоизмененных листьев: листья с чешуевидными сочными
основаниями, запасающими воду с растворенными в ней пи-
тательными веществами, главным образом сахарами, и листья
сухие, покрывающие луковицу снаружи, выполняющие защит-
ную функцию. Из верхушечной и пазушных почек вырастают
фотосинтезирующие надземные побеги, а на донце образуют-
ся придаточные корни.
2. Надземные побеги (рис.4.18).
а) колючки (боярышник);
б) усики (виноград).
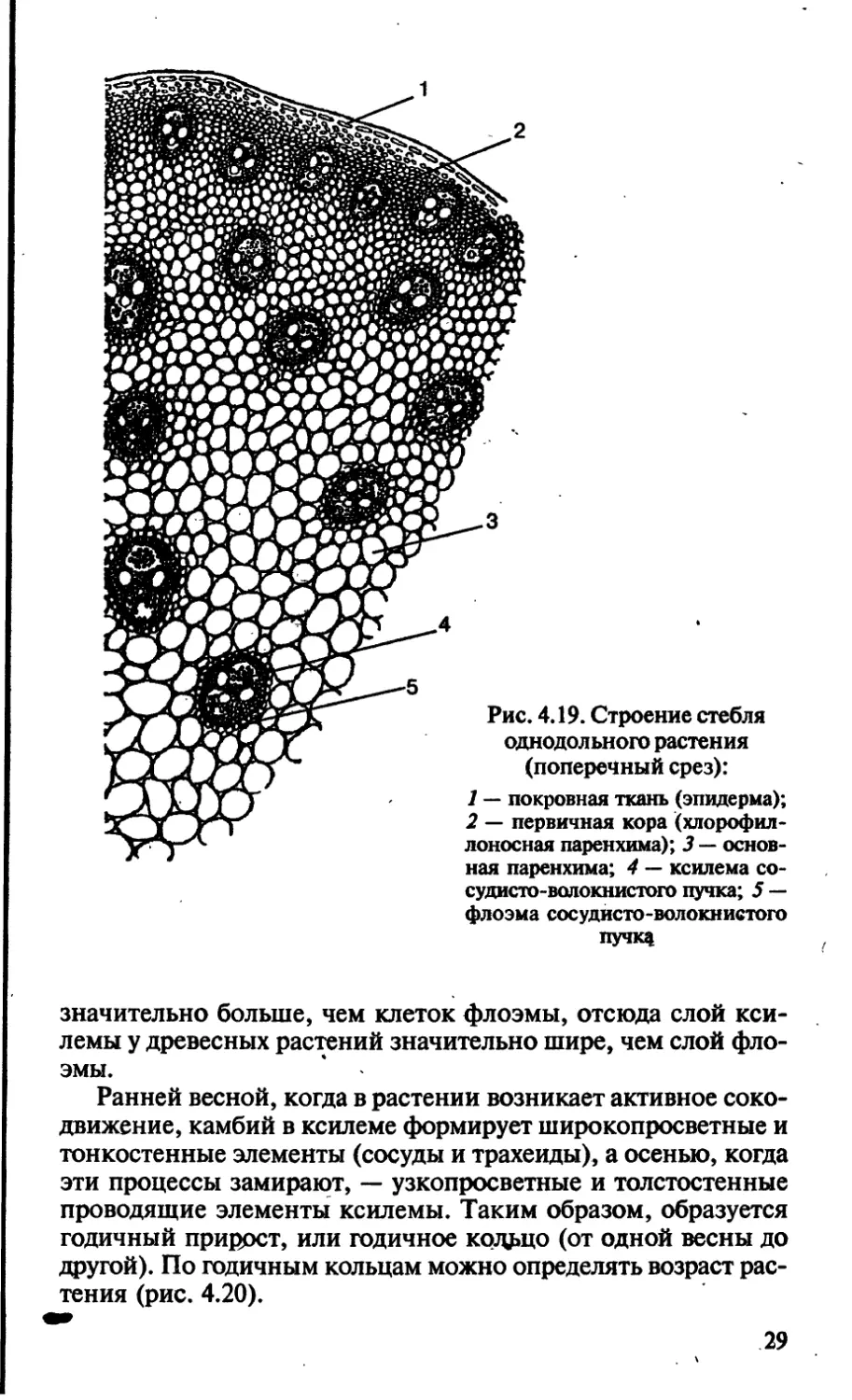
Строение стебля однодольных растений. Развитие стебля
осуществляется за счет разрастания туники и корпуса — пер-
вичных меристем. Из них образуется первичная покровная
26
Рис. 4.17. Подземные видоизменения побегов:
а — корневище; б — клубень; в — клубнелуковица
(продольный разрез); г—луковица (продольный разрез):
1 — отмершие чешуи; 2 — зачаток цветоносного побега; 3 — листья будуще-
го вегетативного периода; 4 — почки; 5 —• укороченный стебель (у луко-
виц — донце); 6 — придаточные корни
Рис. 4.18. Надземные
видоизменения
побегов:
а — колючки; б — усики
27
ткань — эпидерма, первичная кора и центральный осевой ци-
линдр.
Первичная кора представлена однородными клетками
хлорофиллоносной паренхимы, граничащей с перицикли-
ческой склеренхимой центрального осевого цилиндра.
Иногда хлорофиллоносная паренхима отсутствует, и тогда
перициклическая склеренхима располагается сразу под эпи-
дермой.
Весь центральный осевой цилиндр пронизан изолированны-
ми сосудисто-волокнистыми пучками, которые образовались
за счет прокамбиальных клеток конуса нарастания. Сформи-
рЬвавшиеся сосудисто-волокнистые пучки у стеблей одно-
дольных всегда закрытые, поэтому стебель не способен к даль-
нейшему утолщению. От периферии к центру происходит
увеличение размеров пучков. Для однодольных, в отличие от
двудольных растений, не характерно наличие сердцевины в
центре стебля, хотя может быть развита центральная воздуш-
ная полость (например, у стеблей злаков — соломина). Про-
странство между пучками занято запасающей или основной
паренхимой (рис. 4.19).
Строение стебля двудольных растений (на примере древесно-
го стебля). У древесных стеблей в результате деятельности
камбия формируется непучковое строение проводящих тка-
ней, т. е. флоэма и ксилема, сливаясь, образуют кольцо флоэ-
мы и кольцо ксилемы (кольцевое расположение проводящих
тканей). Покровной тканью в результате деятельности фелло-
гена становится перидерма — вторичная покровная ткань.
Ежегодно закладывающиеся перидермы меняют структуру
первичной коры, и поэтому у древесных растений принято вы-
делять кору, камбий, древесину и сердцевину. Граница коры
проходит по камбию.
За перидермой располагаются пластинчатая колленхима,
затем хлорофиллоносная паренхима и однорядный крахмало-
носный слой, называемый «крахмалоносное влагалище».
В этом слое находятся зерна «оберегаемого» крахмала, которые
растение не расходует. Считается, что этот крахмал играет
роль в поддержании растением равновесия. Далее располагает-
ся слой флоэмы (луба), в котором, как правило, имеются лубя-
ные волокна, составляющие твердый луб, а мягкий луб состав-
ляют ситовидные трубки с клетками-спутницами и лубяная
паренхима.
Камбий, располагающийся между ксилемой и флоэмой, к
периферии стебля откладывает вторичную флоэму, а внутрь —
вторичную ксилему. Причем клеток ксилемы он откладывает
28
Рис. 4.19. Строение стебля
однодольного растения
(поперечный срез):
1 — покровная ткань (эпидерма);
2 — первичная кора (хлорофил-
лоносная паренхима); 3 — основ-
ная паренхима; 4 — ксилема со-
судисто-волокнистого пучка; 5 —
флоэма сосудисто-волокнистого
пучк$
значительно больше, чем клеток флоэмы, отсюда слой кси-
лемы у древесных растений значительно шире, чем слой фло-
эмы.
Ранней весной, когда в растении возникает активное соко-
движение, камбий в ксилеме формирует широкопросветные и
тонкостенные элементы (сосуды и трахеиды), а осенью, когда
эти процессы замирают, — узкопросветные и толстостенные
проводящие элементы ксилемы. Таким образом, образуется
годичный прирост, или годичное кодьцо (от одной весны до
другой). По годичным кольцам можно определять возраст рас-
тения (рис. 4.20).
29
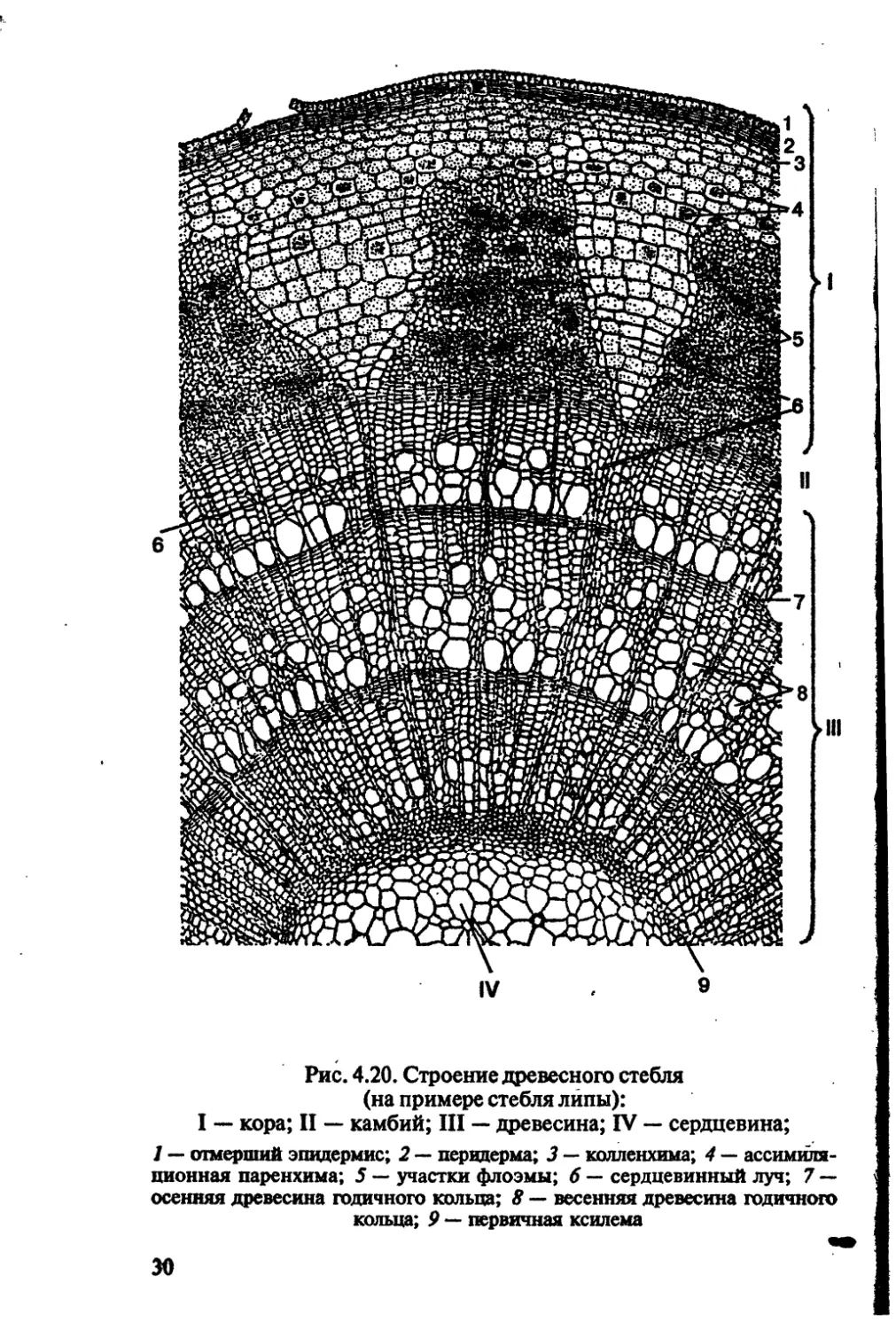
Рис. 4.20. Строение древесного стебля
(на примере стебля липы):
I — кора; II — камбий; III — древесина; IV — сердцевина;
1 — отмерший эпидермис; 2 — перидерма; 3 — колленхима; 4 — ассимиля-
ционная паренхима; 5 — участки флоэмы; 6 — сердцевинный луч; 7 —
осенняя древесина годичного кольца; 8 — весенняя древесина годичного
кольца; 9 — первичная ксилема
30
лист
Лист — боковой орган побега. Нарастает за счет основания
и имеет ограниченный верхушечный рост. Основными функци-
ями листа являются фотосинтез, транспирация и газообмен.
Первые листья семенных растений являются семядолями
зародыша. Следующие листья формируются в виде меристе-
матических бугорков — примордиев, возникающих из верху-
шечной меристемы побега.
Основные части листа:
1) листовая пластинка;
2) черешок;
3) основание листа;
4) прилистники — выросты из основания листа.
Прилистники прикрывают боковые почки и предохраняют
их от различных повреждений. В почке прилистники обяза-
тельно закладываются вместе с листьями, однако у многих рас-
тений быстро опадают или пребывают в зачаточном состоя-
нии. Если прилистники срастаются, то образуется раструб
(семейство гречишные); если же разрастается основание лис-
та, то образуется листовое влагалище (семействозон-
тичные) (рис. 4.21).
Черешки ориентируют листовые пластинки по отношению
к источнику света, создавая листовую мозаику, т. е. такое раз-
Рис. 4.21. Основные части листа:
1 — листовая пластинка; 2 — черешок; 3 — прилистник; 4 — основание
листа; 5 — раструб; 6 — листовое влагалище
мещение листьев на побеге, при котором они не затеняют друг
друга. Это достигается: а) различной длиной и изогнутостью
черешка; б) различной величиной и формой листовой плас-
тинки; в) светочувствительностью листьев.
Лист называется сидячим, если черешок отсутствует, и
тогда лист прикрепляется основанием листовой пластинки к
стеблю.
Жилкование листа бывает:
1) дуговое (лист ландыша и других однодольных растений);
2) параллельное (лист злака и других однодольных расте-
ний);
3) пальчатое — все жилки сходятся в одной точке у основа-
ния листовой пластинки (характерно для двудольных расте-
ний, например, клен татарский);
4) перистое — выражена центральная жилка (характерно
для двудольных растений, например, лист черемухи обыкно-
венной).
Форма листа. Лист, состоящий из одной листовой пластин-
ки, называется простым. Когда на одном черешке с общим ос-
нованием располагается несколько листовых пластинок, то
лист называется сложным. Их общий черешок называется ра-
хисом.
Простые листья делятся на листья с цельной и расчленен-
ной листовой пластинкой,.
Простые листья с цельной листовой пластин-
кой получают название в зависимости от соотношения дли-
ны и ширины листовой пластинки, например: округлый, про-
долговатый, линейный (рис. 4.22).
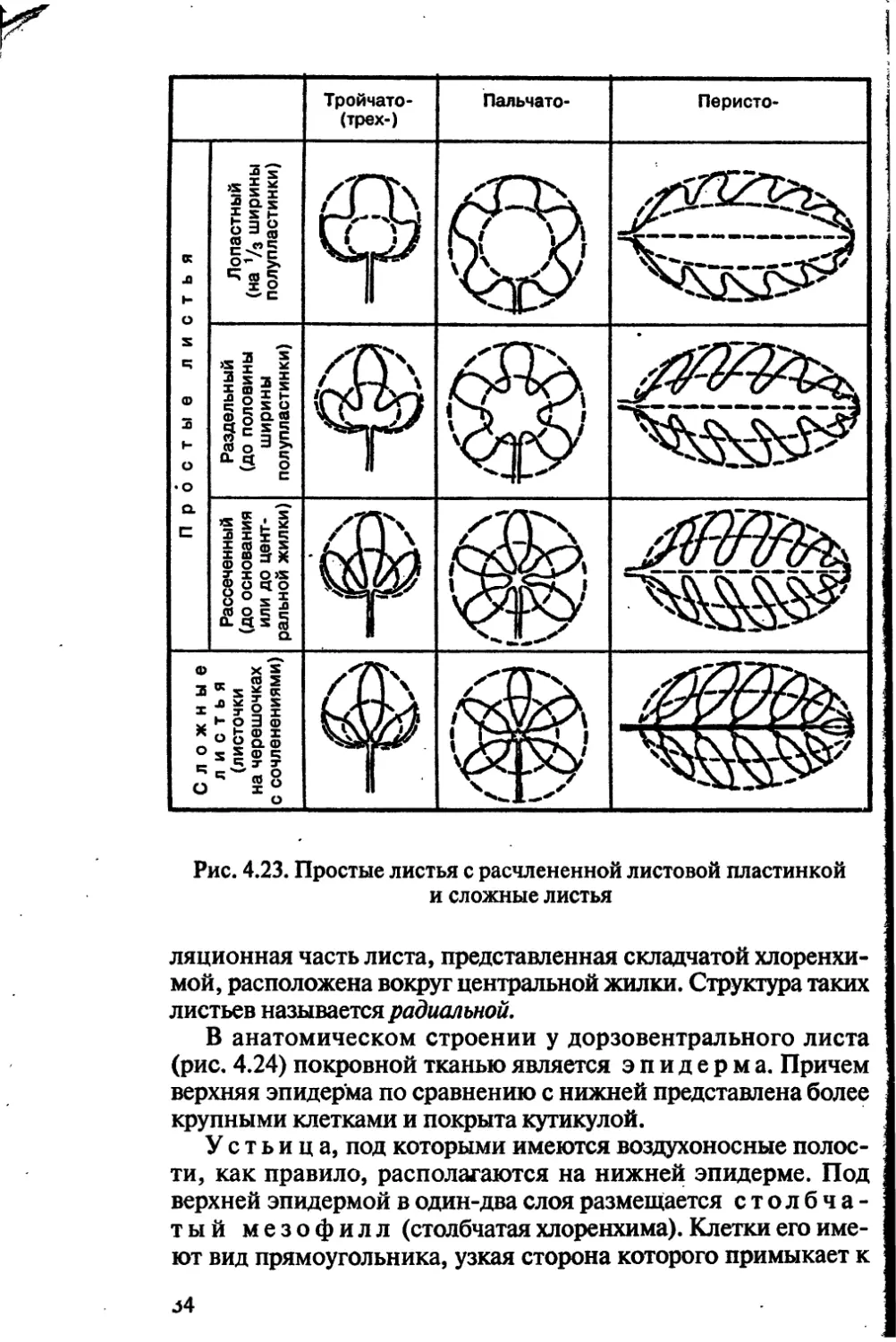
Различают следующие простые листья с расчленен-
ной листовой пластинкой:
а) пальчатолопастной, или перистолопастной, если рас-
членение листовой пластинки доходит до */з всей ее поверх-
ности;
б) пальчатораздельный, или перистораздельный, если рас-
членение листовой пластинки доходит до ‘/г всей ее поверхно-
сти;
в) пальчаторассеченный, или перисторассеченный, если
степень расчленения листовой пластинки доходит до ее осно-
вания или центральной жилки (рис. 4.23).
Сложные листья бывают тройчатосложные, состоящие из
трех листочков (земляника), и пальчатосложные, состоящие
из множества листочков (каштан). У этих типов сложных лис-
тьев все листочки прикрепляются к верхушке рахиса.
Кроме того, есть сложные листья, листочки которых распола-
32
Рис. 4.22. Простые листья с цельной листовой
пластинкой
гаются по всей длине рахиса. Среди них различают
парноперистосложные, если они заканчиваются на верхушке
листовой пластины парой листочков (горох посевной), и не-
парноперистосложные (рябина обыкновенная), заканчиваю-
щиеся одним листочком (см. рис. 4.23).
Анатомическое строение листа. По анатомическому строе-
нию выделяют дорзовентральные, изолатеральные и радиаль-
ные листья.
Если ассимиляционная часть листа (столбчатая хлоренхи-
ма) размерена только на верхней стороне листовой пластинки,
а губчатая — на нижней, то листья с такой структурой называ-
ются дорзовентральными (свекла сахарная). Если столбчатая па-
ренхима располагается с двух сторон листа, то лист называется
изолатеральным, что свойственно листьям, размещающимся
под острым углом к стеблю (гладиолус). У хвои сосньгассими-
2“ 281
33
Рис. 4.23. Простые листья с расчлененной листовой пластинкой
и сложные листья
ляционная часть листа, представленная складчатой хлоренхи-
мой, расположена вокруг центральной жилки. Структура таких
листьев называется радиальной.
В анатомическом строении у дорзовентрального листа
(рис. 4.24) покровной тканью является эпидерма. Причем
верхняя эпидерма по сравнению с нижней представлена более
крупными клетками и покрыта кутикулой.
Устьица, под которыми имеются воздухоносные полос-
ти, как правило, располагаются на нижней эпидерме. Под
верхней эпидермой в один-два слоя размещается столбча-
тый мезофилл (столбчатая хлоренхима). Клетки его име-
ют вид прямоугольника, узкая сторона которого примыкает к
J4
Рис. 4.24. Строение дорзовентрального листа:
1 — верхняя эпидерма; 2 — столбчатая хлоренхима; 3 — склеренхима; 4 —
сердцевинные лучи ксилемы; 5 — сосуды ксилемы; 6 — флоэма; 7 — губча-
тая хлоренхима; 8 — воздухоносная полость; 9 — устьице; 10 — колленхима;
11 — нижняя эпидерма
эпидерме. Прямоугольная форма клеток обеспечивает сохран-
ность хлорофилла, содержащегося в хлоропластах. Находясь
большую часть времени на вытянутых радиальных стенках, че-
чевицеобразные хлоропласты не подвергаются воздействию
прямых солнечных лучей. Солнечные лучи скользят вдоль
них, равномерно освещая хлоропласты, но не разрушая при
этом хлорофилла. Все это способствует активному протеканию
фотосинтеза.
Ниже лежит губчатый мезофилл, характеризую-
щийся рыхло расположенными округлыми клетками с боль-
шими межклетниками. Как столбчатый, так и губчатый мезо-
филл содержат хлоропласты. В центре листа находится
центральная жилка, которая представлена сосудисто-
волокнистым пучком открытого типа. Вверху сосудисто-во-
локнистого пучка располагается ксилема, а снизу флоэма.
Вокруг пучка кольцом лежит склеренхима, оберегающая пу-
чок от давления разрастающихся клеток мезофилла листа.
Над пучком и под ним, сразу после эпидермы, располагается
уголковая колленхима, также выполняющая опорную функ-
цию.
2»
г
РАЗДРАЖИМОСТЬ И ДВИЖЕНИЕ
РАСТИТЕЛЬНЫХ ОРГАНИЗМОВ
Раздражимость — это общебиологическая способность кле-
ток и организмов реагировать (отвечать) на влияние факторов
внешней среды. Важнейшим элементом в процессе раздражи-
мости являются рецепторы. Рецепторные клетки называют
биологическими датчиками или преобразователями, так как
они превращают энергию давления, света, химические и дру-
гие факторы в электрические импульсы. У растений рецепто-
ры не такие дифференцированные, как у животных. Ими явля-
ются эктодесмы, крахмальные статолиты, чувствительные
волоски и др. ,
Основными формами проявления раздражимости организ-
мов являются разного типа двигательные реакции, которые
осуществляются целым организмом или отдельными его час-
тями. Наиболее распространенными двигательными реакция-
ми живых организмов на изменение условий среды являются
таксисы, а у растений (кроме таксисов) — тропизмы, настии,
нутации и автономные движения.
Таксисы — это движение организма, проявляющееся в про-
странственном перемещении его Относительно раздражителя
(амеба, инфузория). Если движение организма осуществляет-
ся в направлении действующего фактора, то такой таксис на-
зывается положительным; а отрицательным, когда движение
происходит в противоположном направлении.
Классифицируют таксисы в зависимости от вида раздражи-
теля. Реакция на действие: света — фототаксис, химических
соединений — хемотаксис, температуры — термотаксис.
Примером положительного фототаксиса является ориентиро-
ванное движение жгутиковых одноклеточных водорослей (хла-
мидомонада) к зоне оптимального освещения в аквариуме или
водоеме, целесообразная ориентация хлоропластов в клетках
мезофилла листа; хемотаксиса — скопление бактериальных
клеток возле отмершей клетки инфузории, движение лейко-
цитов к бактерии и т. д.
Тропизмы — это двигательная реакция органов и частей
растений на одностороннее влияние фактора окружающей
среды (света, силы притяжения, воды, химических веществ
и т. д.).
В зависимости от растительного организма тропизмы могут
быть положительными, когда из-за неравномерного роста
орган или часть растения изгибается в сторону действующего
36
фактора и отрицательным, когда процессы роста вызывают от-
клонения органа в противоположную сторону. У растений
лучше всего выражен геотропизм — реакция отдельных его ор-
ганов на одностороннее воздействие силы земного притяже-
ния.
Различают три типа геотропизма: положительный — когда
орган растет вертикально вниз, отрицательный — когда на-
правление движения противоположное, и поперечный, или
диагеотропизм, когда орган старается занять горизонтальное
положение. Главный стержневые корни имеют, как правило,
положительный геотропизм; ветви первого порядка древесных
растений, черешки многих листьев — отрицательный; многие
корневища, боковые корни — поперечный.
Фототропизмы — это ростовые движения растений в ответ
на одностороннее воздействие света. При одностороннем воз-
действии света (на поляне, около строений, в комнате и т. п.)
фототропизм отдельных побегов или даже всей надземной ча-
сти проявляется особенно четко. Растения как бы тянутся к
свету (растения на подоконнике, соцветия подсолнуха, листья
на побегах).
Одностороннее влияние на растущие органы могут оказы-
вать и другие физические и химические факторы. Соответ-
ственно этому различают еще хемотропизмы, гидротропизмы,
термотропизмы, магнитотропизмы (т. е. классификация тро-
пизмов зависит от источника раздражения).
Настин. К настическим принадлежат движения, которые
являются ответной реакцией органов или частей растений на
действие раздражителей, которые не имеют определенного на-
правления, а влияют диффузно и равномерно с разных сторон.
Именно поэтому установить какой-либо односторонний фак-
тор двигательной реакции невозможно.
Эпинастии — когда изгиб органа (чаще листа) происходит
вниз. Это может быть связано с ускорением роста или тургор-
ного растяжения верхней стороны черешка (опускание листоч-
ков мимозы, вики, акации белой).
Гипонастии — изгиб органа за счет ускоренного роста или
растяжения клеток нижней стороны черешка и центральной
жилки (поднятие листовых пластинок на ночь вверх у лебеды,
табака).
Никтинастии — двигательные реакции, вызванные наступ-
лением темноты, так называемый сон у растений (закрывание
цветков, опускание на ночь соцветий у моркови).
Фотонастии — раскрывание лепестков цветков при усиле-
нии освещения (соцветия цикория, одуванчика, картофеля).
37
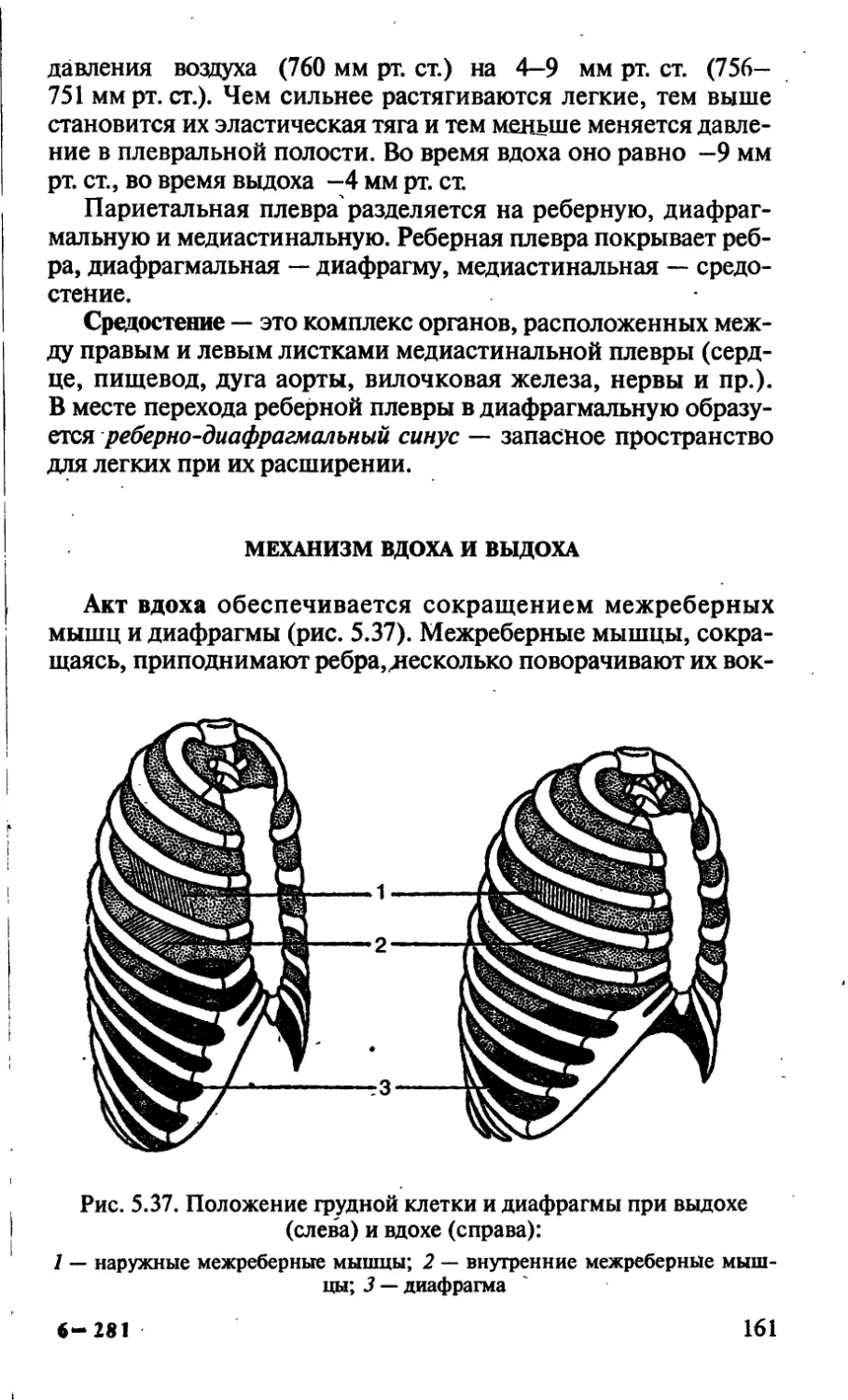
Термонастии — раскрывание лепестков при повышении
температуры (тюльпан, мать-и-мачеха, мак огородный).
Сейсмонастии — движение органов растений, которые яв-
ляются ответом на удар или сотрясение (мимоза, кисличка,
портулак).
Нутации. Под нутациями понимают способность растений
к круговым или маятниковым движениям за счет периодичес-
ки повторяющихся изменений величин тургорного давления и ;
интенсивности роста противоположных сторон определенно- !
го органа. Лучше всего это выражено у верхушек и усиков вью-
щихся растений. У вьющихся растений верхушка во время рос-
та делает равномерные нутационные движения и при контакте
с опорой начинает обвиваться вокруг нее (хмель, тыква, горох,
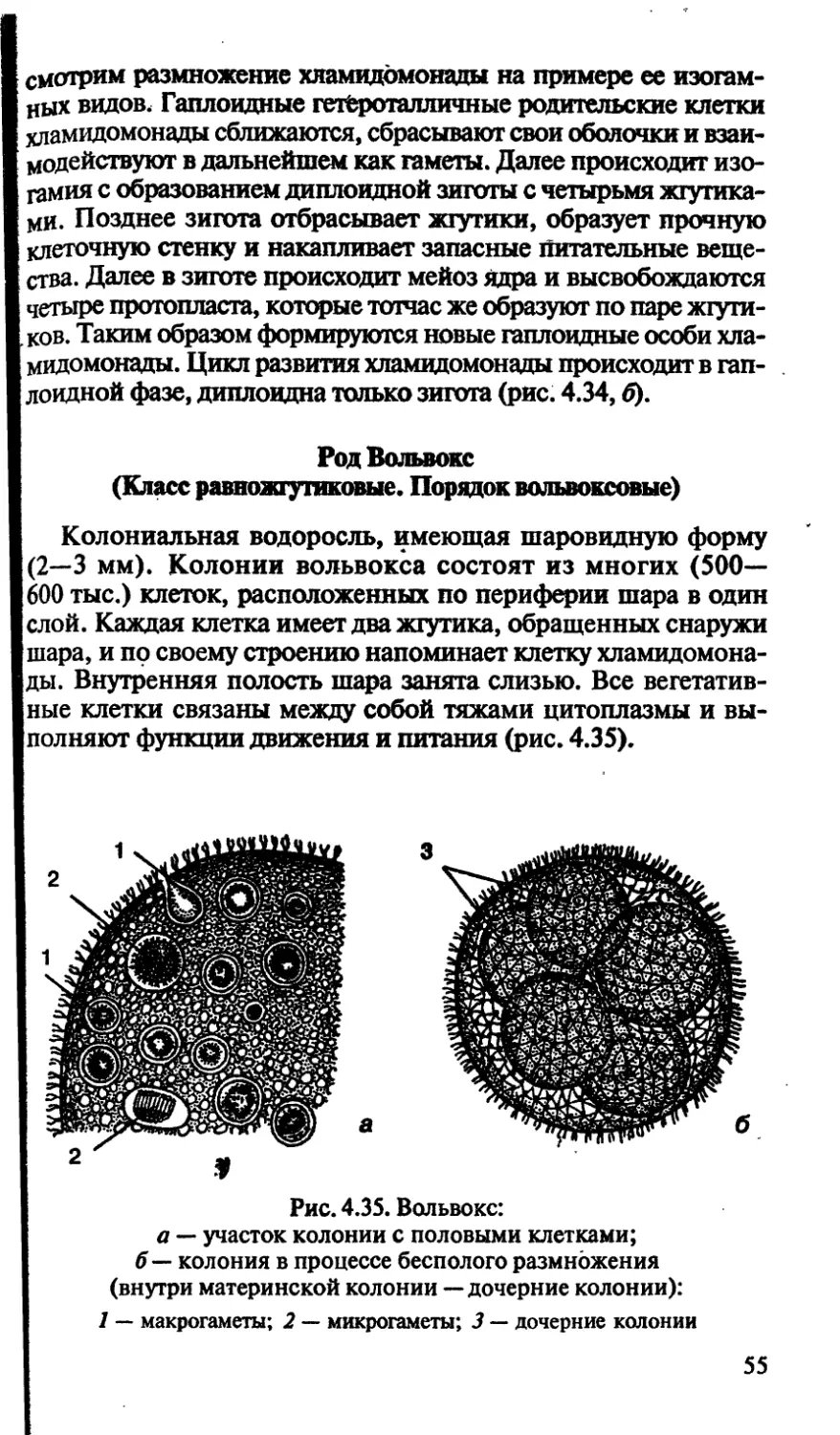
фасоль).
ТАКСОНОМИЧЕСКИЕ КАТЕГОРИИ И ТАКСОНЫ.
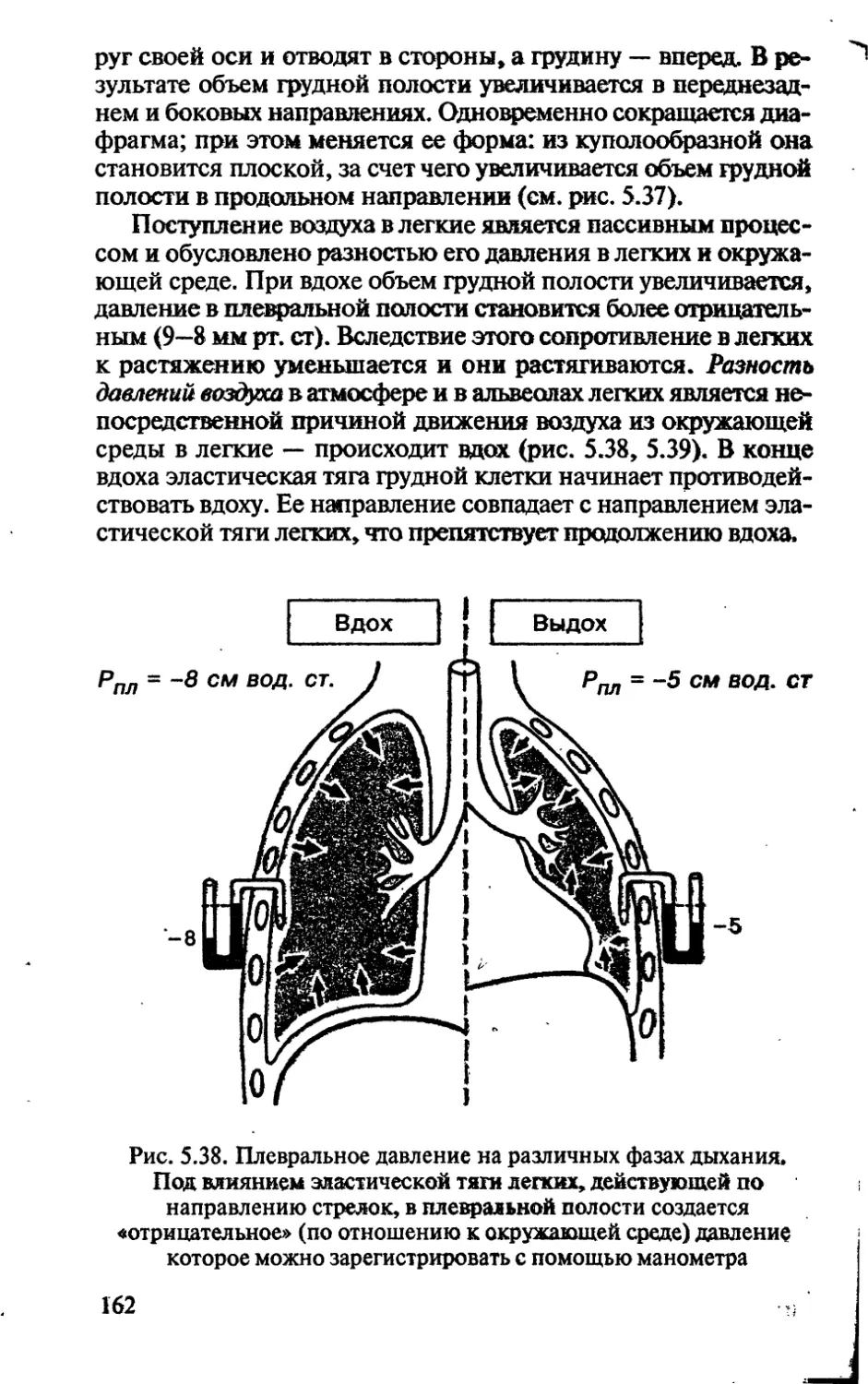
БИНАРНАЯ НОМЕНКЛАТУРА
Согласно правилам ботанической номенклатуры основны-
ми таксономическими категориями считаются: вид, род, се-
мейство, порядок, класс, отдел (тип в животном мире), цар-
ство.
При необходимости могут использоваться и промежуточ-
ные таксономические категории, например подвид, подрод,
подсемейство, надпорядок, надцарство.
Таксонами принято называть реально существующие или
существовавшие группы организмов, которые в процессе
классификации отнесены к определенным таксономическим
категориям. Например, ранги рода или вида являются таксо-
номическими категориями, а род крапива и вид крапива дву-
домная — два конкретных таксона. Первый таксон охватыва-
ет все существующие виды рода крапива, второй — все особи,
относимые к виду крапива двудомная.
Научные названия всех таксонов выше вида состоят из од-
ного латинского слова, т. е. униноминальны. Для видов, начи-
ная с 1753 г. — дата выхода в свет книги К. Линнея «Виды рас-
тений», — приняты биноминальные-названия, состоящие из
двух латинских слов. Первое обозначает род, к которому отно-
сится данный вид, второе — видовой эпитет, например, лан-
дыш майский. Введение такой бинарной номенклатуры —
одна из заслуг Карла Линнея.
38
НАДЦАРСТВО ДОЯДЕРНЫЕ ОРГАНИЗМЫ
(ПРОКАРИОТЫ)
ЦАРСТВО ДРОБЯНКИ
ПОДЦАРСТВО НАСТОЯЩИЕ БАКТЕРИИ
Бактерии — это в основном одноклеточные и колониаль-
ные организмы. Условно их относят к царству растений, одна-
ко связь бактерий с высшими растениями или водорослями не
доказана.
Существуют следующие морфологические типы бактерий.
1. Кокки (сферические формы):
а) диплококки (располагаются попарно);
б) стрептококки (образуют цепочки);
в) стафилококки (скопление в виде грозди винограда);
г) сарцины (образуют скопления кубической формы).
2. Бациллы (палочковидные формы):
а) диплобациллы (располагаются попарно);
б) стрептобациллы (образуют цепочки).
3. Спириллы (спиральные формы — негнущиеся и состоя-
щие из одного или большого числа витков спирали).
4. Вибрионы (негнущиеся, представляющие собой лишь
часть витка спирали).
5. Спирохеты (спиральные формы — изгибающиеся и со-
стоящие из нескольких витков) (рис. 4.25).
Строение. Совокупность всех веществ, составляющих со-
держимое клетки, называется протопластом. К первичным
производным протопласта относится клеточная оболочка и
вакуоль. Снаружи протопласт одет цитоплазматической мем-
браной. Цитоплазма бактерий имеет зернистый вид. В хими-
ческом отношении цитоплазма представляет собой сложную
смесь белков, жиров, углеводов, многочисленных других орга-
нических соединений, минеральных веществ и воды. Значи-
тельная часть органических веществ находится в коллоидном
состоянии, причем они всегда тесно перемешаны.
Ядерный аппарат бактерий обычно называют нуклеоидом.
Генетический материал представлен одной гигантской моле-
кулой ДНК. Нить ДНК у бактерий прикрепляется какой-либо
частью к цитоплазматической мембране или ее выростам с по-
мощью специфических белков. Таким образом, цитоплазма-
39
Рис. 4.25. Различные формы бактерий:
а — кокки:
1 — кокки; 2 — стрептококки (многие виды вызывают инфекционные за-
болевания верхних дыхательных путей); 3 — стафилококки (разные штам-
мы вызывают фурункулез, воспаление легких, пищевые отравления); 4 —
диплококки (две клетки в одной капсуле, например, пневмококк — возбу-
дитель пневмонии);
'б — бациллы (палочковидные):
1 — одиночные палочки (возбудитель брюшного тифа); 2 — бациллы с
эндоспорами (например, возбудитель столбняка); 3 — палочки, образую-
щие цепочки клеток (возбудитель сибирской язвы);
в — спириллы (спиралевидные) — спиральная палочка с одним
жгутиком. По форме клеток на спириллы похожи спирохеты,
но есть различия по способу передвижения, например,
возбудитель сифилиса;
г — вибрионы — короткие палочки, всегда изогнутые в виде
запятой (возбудитель холеры)
тическая мембрана у бактерий принимает участие в делении
нуклеоида. Кроме рибосом в цитоплазме бактерий можно об-
наружить сложные мембранные структуры в виде самостоя-
тельных телец — мезосом. Мезосомы выполняют функцию ап-
парата Гольджи.
Плазматическая мембрана бактерий сверху покрыта клеточ-
ной оболочкой. В составе клеточной стенки бактерий отсутствует
хитин и целлюлоза, характерные для грибных и растительных
клеток. Опорный каркас стенок образован гликопептидом (му-
копротеидом) муреином, Поверх клеточной оболочки распола-
40
гается капсула или слизистый слой. Эта часть клеточной стек
ки, являясь дополнительной защитой бактерий, также служит
для формирования колоний из отдельных клеток.
Многие бактерии подвижны. Органами движения у них слу-
жат жгутики и тонкие палочковидные белковые выросты —
пили, или фимбрии. Жгутики берут свое начало под цитоплазма-
тической мембраной, закрепляясь там с помощью пары дисков
(рис. 4.26).
Колонии большинства бактерий, растущих на неокрашен-
ных средах, бывают сероватыми или беловатыми; иногда они
полупрозрачны, а иногда совсем непрозрачны. Однако для
некоторых видов характерно образование самых разнообраз-
ных пигментов (бактериохлорофилл, цитохромы, липохромы
и др.).
По типу питания бактерии подразделяются на автотрофы
(хемосинтетики) и гетеротрофы (сапрофиты, симбионты, па-
разиты).
По типу дыхания бактерии бывают анаэробами (столбняч-
ная палочка) и аэробами (туберкулезная палочка).
Бесполое размножение бактерий. После достижения опреде-
ленных (критических) размеров клетка подвергается делению,
Для подавляющего большинства прокариот характерно равно-
великое бинарное поперечное деление, приводящее к образова-
нию двух одинаковых дочерних клеток. Весь цикл деления
прокариот можно разделить на три стадии:
Рис. 4.26. Обобщенная
схема строения
палочковидной бактерии:
1 — жгутик; 2 — капсула; 3
клеточная стенка; 4 — плазма-
тическая мембрана; 5 — фото-
синтетические мембраны; 6 —
рибосомы; 7 — запасные пита-
тельные вещества; 8— пили, или
фимбрии; 9 — цитоплазма; 10 —
кольцевая молекула ДНК; 11 —
мезосома
41
1. Редупликация (начинается в точке прикрепления
кольцевой хромосомы к цитоплазматической мембране, кото-
рая определяет начало и конец ее репликации).
2. Синтез мембраны в области контакта ДНК с ци-
топлазматической мембраной. Это приводит к разделению
(растаскиванию) дочерних молекул ДНК и оформлению обо-
собленных хромосом.
3. Образование поперечной перегородки.
Синтез поперечной перегородки идет от периферии к центру.
Она разделяет две дочерние хромосомы, каждая из которых
прикреплена к цитоплазматической мембране (рис. 4.27).
Простое деление клетки в благоприятных условиях насту-
пает каждые 15—20 мин.
Половое размножение бактерий. При равновеликом бинар-
ном делении материнская клетка, делясь, дает начало двум до-
черним клеткам, и сама, таким образом, исчезает.
У некоторых бактерий известен половой процесс, при кото-
ром происходит липп» генетический обмен между клетками, но
не происходит образования новых клеток. Он состоит в прямом
контакте двух клеток, при этом формируется (клеткой-донором, -
выполняющей мужские функции) специальный вырост — копу-
ляционный канал, по которому генетический материал (ДНК)
передается в клетку-реципиент (имеющую женскую потенцию).
Такой процесс называется конъюгацией. Очень часто наблюд ается
передача не всей молекулы ДНК, а только ее фрагментов.
У бактерий имеются и другие способы передачи наследствен-
ного материала. Это трансформация и трансдукция. Первая осу-
ществляется путем внесения ДНК разрушенных клеток одной
Рис. 4.27. Схема деления
бактериальной клетки
при бесполом размножении:
1 — материнская клетка; 2 — ре-
дупликация; 3 — синтез мембра-
ны; 4 — образование поперечной
перегородки
культуры в живую культуру даугой бактерии. Трансдукция прояв-
ляется в переносе генетического материала от одной культуры к
другой с помощью бактериофагов. Эти способы передачи наслед-
ственного материала расцениваются как хромосомные мутации.
Значение бактчпй.
1. Участвуют в экосистемах в разрушении мертвого органи-
ческого материала и тем самым принимают непосредственное
участие в круговороте углерода, азота, фосфора, серы, железа и
других элементов.
2. С активностью бактерий связаны такие важнейшие про-
цессы в природе, как симбиотическая (клубеньковые бакте-
рии) и несимбиотическая (азотобактерии) фиксации молеку-
лярного азота.
3. Многие виды бактерий человек использует в народном
хозяйстве: получение органических продуктов в результате !
брожения (уксуснокислые, лактобактерии). ;
4. Служат источником для получения антибиотиков (стреп-
томицин, грамицидин). »
5. Бактерии используются для создания новых способов |
получения важнейших для промышленности веществ, в. том ;
числе спиртов, органических кислот, сахаров, полимеров, .
аминокислот и ряда ферментов. ।
6. Симбиотические бактерии кишечника млекопитающих 1
(микрофлора) участвуют в синтезе ряда витаминов группы В и
витамина К, а также расщепляют клетчатку. |
7. На основе знаний генетики и молекулярной биологии
работы на микроорганизмах проводятся с помощью манипу-
ляций с индивидуальными генами (генетическая инженерия). '
В настоящее время удалось успешно перенести гены челове-
ческого инсулина в геном кишечной палочки, и уже началось
промышленное получение этого гормона.
8. Многие виды бактерий служат причиной болезни расте-
ний и животных, в том числе человека.
ПОДЦАРСТВО ОКСИФОТОБАКТЕРИИ I
ОТДЕЛ ЦИАНОБАКТЕРИИ (СИНЕ-ЗЕЛЕНЫЕ ВОДОРОСЛИ)
5
Общая характеристика. Сине-зеленые водоросли — очень F
древние организмы, возникшие свыше 3 млрд лет назад. Их '
клетки — по форме округлые, эллиптические, цилиндричес- :
кие, бочонковидные — могут оставаться одиночными, обведи- j
няться в колонии или образовывать многоклеточные нити. р
43
Они часто выделяют слизь в виде толстого чехла, окруженного
у определенных форм плотной оболочкой. Клеточная стенка
довольно толстая, содержит некоторое количество целлюлозы,
но главными ее компонентами являются иные полисахариды
и пектиновые вещества. Подобно многим прокариотам, кле-
точная стенка сине-зеленых водорослей содержит муреин
(гликопептид). Настоящие вакуоли с клеточным соком редки,
но в цитоплазме многих видов этих водорослей часто встреча-
ются вакуоли, наполненные азотом. Считается, что эти вакуо-
ли регулируют плавучесть клетки и позволяют ей парить в тол-
ще воды. У цианобактерий найдены пигменты: хлорофилл А,
несколько каротинов и ксантофиллов, а также фикобиллины,
модифицирующие их окраску от сине-зеленой, фиолетовой и
красноватой до почти черной. Фотосинтезирующий аппарат
цианобактерий имеет фотосистемы I и II, поэтому способен к
аэробному фотосинтезу с выделением кислорода. Продукты
•фотосинтеза могут накапливаться, хотя и в небольших количе-
ствах. Чаще всего это гликопротеид, похожий по химическому
составу на гликоген. Большинство цианобактерий, будучи ав-
тотрофными организмами, могут синтезировать все вещества
клетки за счет энергии света. Однако они способны и к сме-
шанному типу питания (как гетеротрофы). Кроме того, в циа-
нобактериях возможно фиксирование атмосферного азота.
Нитчатые формы водорослей (осциллятория, рис. 4.28) по-
мимо обычных клеток имеют более крупные клетки с утол-
щенными стенками — гетероцисты. Гетероцисты способны
фиксировать азот, снабжая таким образом азотистыми веще-
ствами прочие клетки. В отличие от настоящих бактерий жгу-
тиков никогда не имеют. Размножаются
Рис. 4.28.
Общий вид нити
осцилляторий
путем деления клетки надвое. Полового
процесса нет.
Сине-зеленые водоросли встречаются
в воде и на суше. Цветение воды в водо-
емах часто вызывается цианобактериями,
которые как гетеротрофы питаются остат-
ками органических веществ, что отрица-
тельно сказывается на жизни обитателей
водоема. На суше образуют налеты на
камнях и коре деревьев. Среди относи-
тельно немногих случаев полезного ис-
пользования человеком цианобактерий
— искусственное разведение видов анабе-
на на рисовых полях в тропиках с целью
обогащения почвы соединениями азота.
44
НАДЦАРСТВО ЯДЕРНЫЕ ОРГАНИЗМЫ
(ЭУКАРИОТЫ)
ЦАРСТВО ГРИБЫ
ОТДЕЛ НАСТОЯЩИЕ ГРИБЫ
Грибы — это гетеротрофы. Могут быть паразитами, сапро-
фитами и симбионтами. Клетки грибов одно- или много-
ядерные. Жесткая клеточная стенка белковая и содержит хи-
тин.
Тело состоит из длинных тонких нитей, называемых гифа-
ми. Совокупность гиф образует мицелий. Клетки мицелия за-
пасают углеводы в виде гликогена. Размножаются спорами.
Грибы обособлены по своей морфофизиологической орга-
низации от остального мира живых существ. Их нельзя отнес-
ти ни к растениям, ни к животным. Тело грибов состоит не из
типичных клеток, а из нитей — гиф. С миром животных их
связывает наличие мочевины в обмене веществ, запасного гли-
когена, а также хитина в стенках гиф.
Теория растительного происхождения грибов предполагает
их происхождение от зеленых водорослей, из чего следует, что
грибы прежде всего явно регрессивная группа растений, утра-
тивших хлоропласты.
Теория животного происхождения основывается на том,
что грибы изначально являются бесхлорофильными организ-
мами, т. е. происходят от простейших гетеротрофных организ-
мов, а не от водорослей. Эта теория предпочтительней, по-
скольку бесхлорофильные водоросли, относимые к зеленым, в
качестве запасного продукта накапливают крахмал. У грибов
же крахмала нет.
Отдел настоящие грибы делится на шесть классов. К низ-
шим грибам относятся хитридиомицеты, оомицеты, зигоми-
цеты, а к высшим грибам — сумчатые, базидиомицеты, несо-
вершенные. Низшие грибы имеют неклеточное строение
мицелия, так как его гифы не разделены на перегородки. Рас-
смотрим некоторых представителей плесневых грибов, относя-
щихся к зигомицетам и аскомицетам, паразитов, относящихся
к аскомицетам, и шляпочных грибов, относящихся к базидио-
мицетам.
45
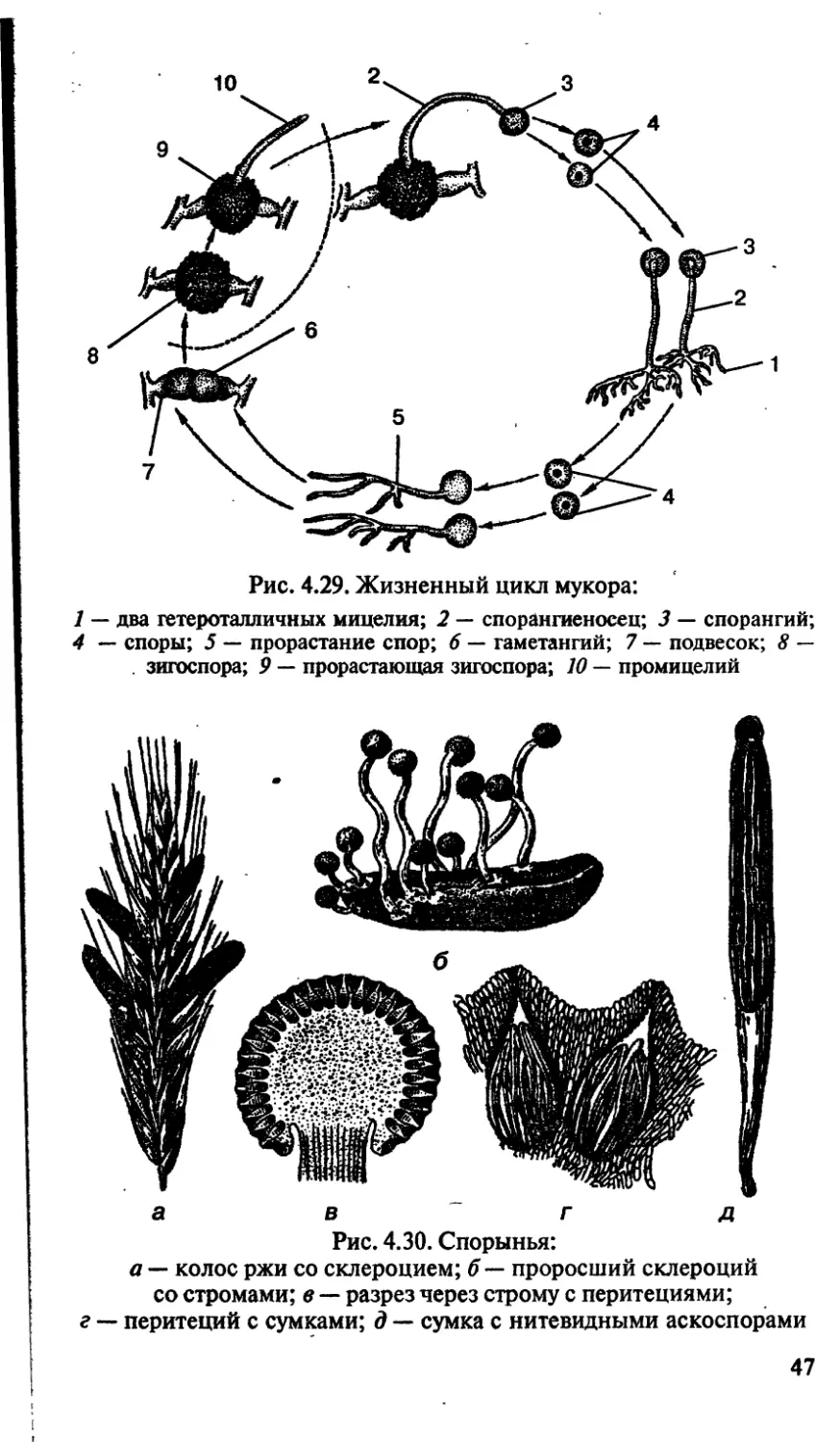
Класс зигомицеты
К данному классу относятся мукоровые грибы, а также гри-
бы, паразитирующие на насекомых. Мукор широко распростра-
нен в природе под названием белой плесени. Сапрофит по спо-
собу питания, развивается на почве, на пищевых продуктах.
Гифы мицелия представляют собой вытянутую разросшуюся
гигантскую клетку со множеством ядер (неклеточное строение).
Ядра имеют гаплоидный набор хромосом (п). На мицелии раз-
виваются многочисленные вертикальные спороангиеносцы со
спорангиями. В результате митоза содержимое спорангия рас-
падается на множество спор беспалого размножения. После со-
зревания оболочка спорангия лопается и споры рассеиваются,
прорастая в новые особи. Размножение может быть беспалым
(спорами), вегетативным (частями мицелия), редко половым
(зигогамия).
Зигогамия делится на несколько стадий:
I) образование гиф;
2) образование утолщенных и обособленных участков на
концах гиф (гаметангии);
3) слияние обособленных участков с последующим слияни-
ем ядер и образование зиготы (2п);
4) прорастание зиготы.
После периода покоя зигота делится мейозом и прорастает
в гифу, увенчанную спорангием (рис. 4.29).
Класс сумчатые (аскомицеты)
Аскомицеты — класс высших грибов с многоклеточным ми-
целием и органами спороношения — сумками (асками). Рас-
смотрим особенности данного класса грибов на примере спо-
рыньи. Спорынья — паразит злаков (рис. 4.30). Осенью на
злаковых растениях образуются склероции — темно-фиолето-
вые рожки, представляющие собой мицелий гриба в стадии
покоя. На зиму склероции выпадают из злаков на почву и зи-
муют в ней. Весной на почве склероции прорастают, образуя
нитевидные выросты, увенчанные головками — стромами.
В этих стромах в результате полового процесса образуются
плодовые тела — перитеции, заполненные длинными цилин-
дрическими сумками (асками), содержащими нитевидные ас-
коспоры. Созревание спор идет во время цветения злака в ре-
зультате мейоза — это споры полового размножения. Споры
активно выбрасываются с помощью ветра, попадают на рыльце
злаков и прорастают. Образующийся мицелий проникает в за-
46
10
2
3
Рис. 4.29. Жизненный цикл мукора:
1 — два гетероталличных мицелия; 2 — спорангиеносец; 3 — спорангий;
4 — споры; 5 — прорастание спор; 6 — гаметангий; 7 — подвесок; 8 —
. зигоспора; 9 — прорастающая зигоспора; 10 — промицелий
а в гд
Рис. 4.30. Спорынья:
a — колос ржи со склероцием; б — проросший склероций
со стромами; в — разрез через строму с перитециями;
г — перитеций с сумками; д — сумка с нитевидными аскоспорами
47
Рис. 4.31. Пенициллиум:
1 — членистый мицелий;
2 — конидиеносец; 3 — сте-
ригмы; 4 — конидии
вязь пестика и разрушает ее. На
концах гиф мицелия в результате
митоза отшнуровываются кони-
дии — споры бесполого размно-
жения, т. е. наступает кониди-
альное спороношение. При этом
гифы гриба выделяют капельки
сладкой жидкости — медвяную
росу. Насекомые переносят ко-
нидии на цветки соседних расте-
ний и заражают их. Склероции
содержат алкалоиды, вызываю-
щие отравления.
Пенициллиум гриб сапрофит
(рис. 4.31). Имеет членистый ми-
целий, состоящий из члеников.
Вверх отходят грифы конидие-
носцы, ветвящиеся на верхнем
конце — стеригмы. Каждый чле-
ник одноядерный. Стеригмы за-
канчиваются цепочкой наружных
спор — конидий. В результате полового процесса образуются
сумки с восемью аскоспорами, которые помещены внутри за-
крытого плодового тела — клейстотеции. Из мицелия пени-
циллиума Флемингом впервые был выделен антибиотик пени-
циллин. Применяется также в пищевой промышленности для
приготовления особых сортов сыра.
Класс базвдиомицеты
В этом классе объединены почти все группы шляпочных гри-
бов. Вегетативное тело представлено членистым мицелием, со-
стоящим из членистых гиф. Специальных органов полового
размножения не образуется. Половое размножение осуществ-
ляется следующим образом. Из прорастающей гаплоидной ба-
зидиоспоры развивается первичный мицелий (рис. 4.32). За-
тем этот мицелий превращается в членистый. Каждый членик
одноядерный. Вскоре происходит гологамия — слияние ко-
нечных клеток гиф. Однако слияние содержимого члеников не
сопровождается слиянием ядер. Образуются дикарионы, кото-
рые затем синхронно делятся. Так формируется вторичный
дикарионный мицелий.
На дикарионном мицелии образуется плодовое тело, которое
состоит из пенька и шляпки. Гимениальный слой шляпки может
48
Рис. 4.32. Цикл развития базидиомицетов:
/ — базидия; 2 — базидиоспора; 3 — первичный мицелий; 4 — дикарион-
ный мицелий; 5 — плодовое тело
быть пластинковым или трубчатым. В гимениальном слое на
концах дикарионных гиф из двух ядерных клеток образуются ба-
зидии. По своему развитию базидии напоминают сумку и гомо-
логичны ей. В базидии завершается половой процесс, т. е. слива-
ются ядра дикариона и образуется диплоидное ядро. Это ядро
делится мейозом с образованием четырех базидиоспор.
У высших грибов в цикле развития чередуются три фазы:
гаплоидная, дикарионная (длится основную часть жизни) и
диплоидная (кратковременна).
Велико значение грибов в деятельности человека. Они играют
роль в круговороте веществ в природе. Грибы, как и бактерии,
минерализуют органические вещества и принимают участие в
образовании гумуса. Их используют в пищевой промышленно-
сти для производства спирта, вина, пива, кваса, в хлебопечении,
в получении белков и витаминов. Грибы образуют органически
активные вещества — антибиотики, ферменты, органические
кислоты и др. Многие грибы наносят существенный вред челове-
ку, животным и растениям: вызывают ряд заболеваний (микозы,
стригущий лишай, парша), коррозию металлов, разрушают кожу,
бумагу, ткани, производят порчу пищевых продуктов, могут слу-
жить причиной отравлений.
ОТДЕЛ ЛИШАЙНИКИ
Это группа симбиотропных растений, состоящих из двух
компонентов — автотрофных водорослей и гетеротрофных
грибов. Грибная основа лишайников формируется сумчатыми
49
грибами. Водорослевый компонент состоит из видов, относи-
мых в большинстве случаев к представителям отделов зеленые
и сине-зеленые водоросли. Выделенные из лишайника водо-
росли не отличаются от свободно живущих форм. Физиологи-
чески этот тип симбиоза основан на межклеточном обмене
между водорослями и грибами. Гриб питается углеводами во-
доросли, а водоросли получают от грибов минеральные веще-
ства. Однако симбиоз с грибами приводит к появлению ново-
го биологического качества, которое выражается у лишайника
в его способности размножаться как единый организм.
Вегетативное тело лишайников представлено слоевищем,
имеющим различную окраску (серую, зеленоватую, буро-корич-
невую, желтую или почти черную). Морфологически различают
три основных типа слоевища лишайников: накипной (корко-
вый), листоватый и кустистый, между которыми существуют и
переходные формы. Наиболее низкоорганизованные — накип-
ные, или корковые, слоевища, имеющие вид порошковатых,
зернистых, бугорчатых налетов, плотно срастающихся с суб-
стратом и не отделяющихся от него без значительных повреж-
дений.
Более высокоорганизованные лишайники имеют листова-
тое слоевище в форме пластинок, чешуек или розеток, при-
крепленных к почве или деревьям с помощью ризин — анало-
гов ризоидов, состоящих из пучков-грибных гиф.
Наивысшей организации в своем строении достигают ли-
шайники с кустистым типом слоевища, имеющие вид разветв-
ленного кустика 12—15 см в высоту и срастающиеся с субстра-
том только основанием.
По анатомическому строению лишайники бывают гомео-
мерными и гетеромерными. У гомеомерных, более прими-
тивных, по всей толще слоевища равномерно расположены
грибные гифы и водоросли. При гетеромерном строении на
поперечном разрезе лишайника сверху можно видеть так на-
зываемую верхнюю кору. Она образована переплетающимися
и тесно смыкающимися грибными гифами. Под корой гриб-
ные гифы лежат более рыхло и между ними находятся клетки
водоросли. Внутри слоевища можно выделить сердцевину,
состоящую из рыхлых грибных гиф и больших пустот, запол-
ненных воздухом. Под ней размещается нижняя кора, кото-
рая по строению сходна с верхней. Через нее из сердцевины
проходят отдельные гифы, закрепляя лишайник в субстрате.
Большинство лишайников легко переносят высыхание. Фо-
тосинтез и питание у них в это время прекращаются, чем и
объясняется их незначительный ежегодный прирост.
5к
Разююжаются лишайники преимущественно вегетативно.
Вегетативное размножение основано на способности лишай-
ников регенерировать из отдельных участков. Оно осуществля-
ется путем фрагментации (отделения участков слоевища) или
с помощью обособленных групп клеток водорослей, окружен-
ных гифами гриба и различных по своей форме — соредий,
изидий и лобул. Соредии — мельчайшие образования округлой
формы, включающие одну или несколько клеток водоросли и
окруженные грибными гифами. Изидии — бугорчатые палочко-
видные выросты на верхней поверхности слоевища. Лобулы
имеют вид маленьких чешуек, расположенных вертикально на
поверхности слоевища или по его краям.
Кроме того, наблюдается бесполое размножение с помо-
щью спор, самостоятельно образующихся и у водорослей и у
грибов.
Половое размножение достаточно не изучено, но в общих
чертах протекает как таковое у свободно живущих грибов.
Значение лишайников велико. Они разлагают и минерали-
зуют органические вещества почвы. Одними из первых заселяя
скалы, разрушают их поверхностный слой, и, отмирая, образу-
ют гумус, на котором расселяются другие растения. Лишайни-
ки являются индикаторами чистоты воздуха, так как не выно-
сят даже малейших примесей сернистых газов. Из некоторых
их видов получают краску и особое вещество — лакмус — для
химической промышленности. В тундре и лесотундре лишай-
ники (ягель) являются основным кормом для оленей. Встреча-
ются съедобные лишайники и в полупустынных и пустынных
районах Киргизии и Туркмении.
ЦАРСТВО РАСТЕНИЯ
Царство растений принято делить на три подцарства: баг-
рянки, настоящие водоросли и высшие растения.
К настоящим водорослям относятся отделы пирофитовые,
золотистые, диатомовые, желто-зеленые, зеленые и бурые во-
доросли.
Водоросли (низшие растения) являются самыми прими-
тивными растениями, и у них нет разделения на стебель, ко-
рень и листья. Особая роль растений в жизни нашей планеты
состоит в том, что без них невозможно существование живот-
ных и человека. Только содержащие хлорофилл зеленые расте-
51
ния способны аккумулировать энергию солнца, создавая орга-
нические вещества из неорганических. Благодаря фотосинтезу
поддерживается уровень кислорода в атмосфере, при этом по (
меньшей мере половину всего кислорода выделяют водоросли,
и их вклад в этот процесс намного больше, чем вклад назем-
ных лесов.
ПОДЦАРСТВО БАГРЯНКИ (КРАСНЫЕ ВОДОРОСЛИ)
Красные водоросли представляют собой древнюю группу
многоклеточных организмов. Таллом (вегетативное тело водо-
росли) у багрянок имеет вид разветвленных многоклеточных
нитей ярко-красного цвета, прикрепленных к субстрату с по-
мощью ризоидов.
В хроматофорах красных водорослей содержатся пигменты:
хлорофилл, каротиноиды и фикобиллины (красные фикоэрит- ,
рины и синие фикобиллины, дающие различную окраску в за-
висимости от преобладания тех или иных пигментов).
Продуктами ассимиляции является «багрянковый крах- i
мал», откладывающийся вне связи с хлоропластами. Этот по-
лисахарид более близок к амилопектину и гликогену, чем к >
крахмалу. j
Размножение может быть вегетативным (частями таллома),
и половым (оогамия с чередованием ядерных фаз).
Среди багрянок ценны виды рода Родимения. Некоторые.
багрянки из рода Анфельция служат сырьем для получения j
агар-агара, основой которого являются фикоколлоиды, а также
используются в пищевой промышленности.
ПОДЦАРСТВО НАСТОЯЩИЕ ВОДОРОСЛИ
ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ
Это самый обширный отдел среди всех водорослей, насчиты-
вающий свыше 13 тыс. видов. Включает в себя несколько классов,
наиболее распространенными из которых являются равножгути-
ковые, сцеплянки И харовые. Зеленые водоросли разнообразны
по внешнему виду: одноклеточные, колониальные, многоклеточ-
ные, нитчатые и т. д. Представители отдела большей частью оби-
тают в пресных водоемах, имеют зеленую окраску из-за преобла-
дания хлорофилла над другими пигментами.
52
Род хламидомонада
(Класс равножгутиковые. Порядок вольвоксовые)
Включает около 320 видов одноклеточных организмов. Эти
виды обитают в пресных водоемах, лужах, канавах. Хламидо-
монада совсем не похожа на растение, так как она активно
движется, л у нее имеются пульсирующие вакуоли. Характер-
ны типичные для эукариот органеллы: аппарат Гольджи, ми-
тохондрии, рибосомы и мелкие вакуоли. В хроматофорах мно-
гих водорослей выявлена особая структура — пиреноид. Это
белковое образование, которое осуществляет фиксацию дву-
окиси углерода. Пиреноид участвует в запасании углеводов,
например, крахмала.
Красный глазок (стигма) воспринимает изменения в интен-
сивности освещения, и клетка либо перемещается туда, где
интенсивность света оптимальна для фотосинтеза, либо оста-
ется на месте, если освещенность достаточна (рис. 4.33). Такая
ответная реакция называется фототаксисом.
Взрослая особь хламидомонады гаплоидна. Имеет два типа
размножения — бесполое и половое.
Бесполое размножение осуществляется с помощью зооспор.
Родительская клетка теряет жгутики, стенки ее ослизняются
(обычно это происходит при подсыхании водоема), и прото-
пласт клетки делится на два дочерних протопласта. В это время
происходит митотическое деление ядра; кроме того, делится и
хроматофор. У дочерних протопластов образуются новые кле-
Рис. 4.33. Строение клетки
хламидомонады:
7 — жгутики; 2 — клеточная стен-
ка; 3 — базальные тельца; 4 —
ядро с ядрышком; 5 — цито-
плазма; 6 — чашеобразный
хроматофор; 7 — пиреноид; 8 —
зерна крахмала; 9 — красный
глазок
53
точные стенки, новые глазки, новые жгутики. В образовании;
жгутиков участвуют базальные тельца. Клеточная стенка роди- j
тельской клетки ослизняется, и дочерние клетки, которые те-
перь называются зооспорами, выходят наружу. Из каждой зоо-
споры формируется полноценная взрослая клетка (рис. 4.34, а),!
Половое размножение. Одни виды хламидомонады i
гомоталличны, другие гетероталличны. При этом разные виды
могут быть изогамными, анизогамными или оогамными. Рас-
б
Рис. 4.34. Схема размножения хламидомонады:
а — бесполое размножение:
1 — родительская клетка; 2 — клетка с двумя протопластами после первого
деления;' 3 — четыре протопласта после второго деления; 4 — восемь прото-
пластов после третьего деления;
б — половое размножение:
5 — сближение родительских клеток; 6 — сбрасывание клеточных оболо-
чек; 7 — изогамия с образованием зиготы с четырьмя жгутиками; 8 — исчез-
новение жгутиков; 9 — мейоз зиготы; 10 — образование, дочерних клеток
54
смотрим размножение хламидомонады на примере ее изогам-
ных видов. Гаплоидные гетЬроталличные родительские клетки
хламидомонады сближаются, сбрасывают свои оболочки и взаи-
модействуют в дальнейшем как гаметы. Далее происходит изо-
гамия с образованием диплоидной зиготы с четырьмя жгутика-
ми. Позднее зигота отбрасывает жгутики, образует прочную
клеточную стенку и накапливает запасные питательные веще-
ства. Далее в зиготе происходит мейоз ядра и высвобождаются
четыре протопласта, которые тотчас же образуют по паре жгути-
ков. Таким образом формируются новые гаплоидные особи хла-
мидомонады. Цикл развития хламидомонады происходит в гап-
лоидной фазе, диплоидна только зигота (рис. 4.34, б).
Род Вольвокс
(Класс равножгутиковые. Порядок вольвоксовые)
Колониальная водоросль, имеющая шаровидную форму
(2—3 мм). Колонии вольвокса состоят из многих (500—
600 тыс.) клеток, расположенных по периферии шара в один
слой. Каждая клетка имеет два жгутика, обращенных снаружи
шара, и пр своему строению напоминает клетку хламидомона-
ды. Внутренняя полость шара занята слизью. Все вегетатив-
ные клетки связаны между собой тяжами цитоплазмы и вы-
полняют функции движения и питания (рис. 4.35).
Рис. 4.35. Вольвокс:
а — участок колонии с половыми клетками;
б — колония в процессе бесполого размножения
(внутри материнской колонии—дочерние колонии):
1 — макрогаметы; 2 — микрогаметы; 3 — дочерние колонии
55
Функцию полового размножения выполняют другие специ-
ализированные клетки, отличающиеся более крупными разме-
рами и отсутствием жгутиков.
Вегетативное размножение осуществляется с помощью
дочерних колоний, образующихся в материнских колониях
путем последовательных продольных делений их протопла-
стов. Затем дочерние колонии выпадают внутрь материн-
ского шара и освобождаются лишь после ее разрушения.
При половом размножении часть специализированных кле-
ток преобразуется в оогонии с яйцеклетками. В других спе-
циализированных клетках развиваются двужгутиковые спер-
матозоиды. Сперматозоиды одного материнского шара
оплодотворяют яйцеклетки другого. Диплоидная зигота на-
зывается ооспорой, которая обычно прорастает весной. При
этом сначала происходит мейоз, а затем многократное деле-
ние. В результате возникает пластинка гаплоидных клеток,
после чего постепенно формируется новое шаровидное тело
вольвокса. Основная форма существования простейших в
колонии происходит в гаплоидной фазе, диплоидна только
зигота.
. Род Хлорелла
(Класс равножгутиковые. Порядок хлорококковые
или протококковые водоросли)
Объединяет одноклеточные водоросли, широко распрост-
раненные в пресных водоемах, почве. Клетки одеты оболоч-
кой, содержат чашевидный хроматофор, одно ядро. Половой
процесс отсутствует. При бесполом размножении материн-
ская клетка делится на 4—10 частей. Образуются неподвиж-
ные, т. е. без жгутиков, автоспоры, которые еще в материнской
клетке одеваются собственными оболочками и выходят из нее
после разрыва материнской оболочки.
Хлорелла характеризуется очень быстрыми темпами ра. -
множения. Каждая клетка хлореллы в течение суток может об-
разовывать до 10 дочерних клеток. Она способна использовать
от 10 до 12% световой энергии (против 1—2% у наземных рас-
тений). В сухом веществе содержит: 50% белков, 22% жиров,
12% углеводов, 10% минеральных солей, витамины В, К, С
(больше, чем в лимонном соке). Свойство хлореллы оыстр»
накапливать биомассу используется для получения дешевого
корма. Получила применение при полетах космонавтов как
источник пищи и кислорода.
56
Класс сцеплянки
У представителей этого клас-
са наблюдается особый половой
процесс (конъюгация) и обра-
зование зооспор отсутствует.
Представителем этого клас-
са является спирогира — много-
клеточная нитчатая водоросль,
свободно плавающая на поверх-
ности рек и прудов. В центре
каждой клетки находится ядро,
«подвешенное» на тяжах ци-
топлазмы. Имеется спирально
закрученный хроматофор с пи-
реноидами. Вегетативное раз-
множение происходит частями
таллома, т. е. путем разрыва
нитей на отдельные участки
или даже отдельные клетки.
Половой процесс — конъюга-
ция. При конъюгации у двух
гетеротал;шчных)параллельно
Уасполагающй£ся нитей водо-
росли у супротивных клеток напротив друг друга образуются
выросты. В месте соприкосновения этих двух выростов оболоч-
ки растворяются, и образуется популяционный канал. По это-
му каналу происходит слияние содержимого двух клеток с об-
разованием зиготы. После периода покоя зигота редукционно
делится, и образуются четыре новые клетки, три из которых
отмирают, а одна прорастает в новую особь (рис. 4.36).
Рис. 4.36.
Конъюгация спирогиры:
1 — образование копуляцион-
ного канала; 2 — начало копу-
ляции; 3 — зигота
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ
Бурые водоросли — многоклеточные организмы, обитаю-
щие в морях. Это самые крупные из известных водорослей.
Рассмотрим этот отдел на примере ламинарии сахарной, отно-
сящейся к порядку ламинариевых.
Ламинария сахарная — представитель Северных морей и Даль-
него Востока. Особи достигают от 10 до 60 м в длину. Все тело
водоросли разделено на части: листовая пластина, стволик и ри-
зоиды (рис. 4.37). Клетки содержат одно ядро, несколько вакуо-
лей и имеют сильно ослизняющиеся стенки. Хроматофор округ-
лый, в виде зерен, содержащий пигменты: хлорофилл, каротин,
57
Рис. 4.37. Внешний вид
ламинарии сахарной:
1 — листовая пластина; 2 — стволик;
3 — ризоиды
ксантофилл и фукоксантин. Избыток фукоксантина придает во-
доросли бурую окраску. Запасной полисахарид—ламинарии. Он
откладывается вне хлоропласта — в цитоплазме.
Размножение половое — оогомия с чередованием диплоид-
ной й гаплоидной ядерных фаз.
Взрослое растение ламинарии является спорофитом (беспо-
лая фаза). В результате мейоза в зооспорангиях, находящихся
на листовой пластине, образуются зоосцоры. Зооспоры попада-
ют в почву и прорастают в заростки (гаметофиты) — микроско-
пические образования, несущие гаметы. На одних гаплоидных
заростках образуются женские половые органы — оогонии с яй-
цеклетками, на других заростках образуются мужские половые
органы — антеридии со сперматозоидами. После оплодотворе-
ния яйцеклеток сперматозоидами образуется диплоидная зиго-
та, которая сразу же прорастает во взрослую особь.
Бурые водоросли находят различное применение в хозяй-
ственной деятельности человека. Из них получают агар, альги-
наты — вещества, широко используемые при приготовлении
консервов, красящих и клеющих веществ, а также кровезаме-
нитель и маннит. Бурые водоросли, кроме того, служат сырьем
для получения йода и брома.
ПОДЦАРСТВО ВЫСШИЕ РАСТЕНИЯ,
ИЛИ ВЫСШИЕ СПОРОВЫЕ РАСТЕНИЯ
К высшим споровым растениям относятся моховидные,
плауновидные, хвощевидные и папоротниковидные.
58
ОТДЕЛ МОХОВИДНЫЕ
Моховидные включают в себя класс листостебельных мхов
и класс печеночников.
Среди высших растений мхи образуют обособленную груп-
пу. Это — единственная в истории растительного мира линия
эволюции, связанная с регрессивным развитием спорофита.
Они представляют собой тупиковую, или слепую, ветвь разви-
тия растений, а по общей организации и по экологии еще близ-
ки к водорослям.
Характерными признаками мхов являются:
1) отсутствие настоящих корней. У некоторых представите-
лей подземная часть представлена ризоидами;
2) полное преобладание гаметофазы в общем цикле разви-
тия. Половое и бесполое поколение представляют собой одно
растение, при этом спорофит не может существовать без гаме-
тофита.
Класс листостебельные мхи делится на два подкласса: под-
класс зеленые мхи и подкласс белые, или торфяные, мхи.
Подкласс зеленые мхи.‘Рассмотрим подкласс зеленых мхов
на примере кукушкиного льна.
Кукушкин лен — один из наиболее высокорослых мхов,
его стебель достигает 50 см в высоту. Растет крупными по-
душкообразными дернинами. Стебель мха густо покрыт ли-
стьями и имеет ризоиды. Относится к двудомным растени-
ям. На женском растении (гаметофите) между верхними
междоузлиями образуется архегоний — женский половой
орган. Архегоний является многоклеточным образованием
бутыльчатой формы. Суженная часть — шейка, а расширен-
ная часть — брюшко, в котором помещается крупная яйце-
клетка. На мужском растении (гаметофите) среди верхних
красных листьев развиваются антеридии —мужские поло-
вые органы, в которых образуются двужгутиковые спермато-
зоиды. Антеридии имеют вид продолговатых или округлых
мешочков на ножке. При созревании архегония шейковые
или брюшные клетки ослизняются и на их месте формиру-
ется узкий канал, по которому сперматозоид может проник-
нуть в яйцеклетку. В период обильных дождей сперматозои-
ды подплывают к архегониям. Как предполагают, они
обладают хемотаксисом к содержимому слизи архегония.
Один из сперматозоидов проникает в архегоний и продол-
жает движение к яйцеклетке. Слияние гамет и дальнейшее
развитие зиготы происходит внутри архегоний. Из зиготы
через несколько месяцев прорастает спорофит (спорогон),
59
9
Рис. 4.38. Чередование поколений у листостебельных мхов:
1 — протонема; 2 — гаметофит; 3 — антеридий; 4 — двужгутиковый сперма-
тозоид; 5 — архегоний с яйцеклеткой; 6 — зигота; 7 — спорофит на ножке;
8 — коробочка спорофита; 9 — колпачок спорофита с волосками; 10 —
споры
который представляет собой коробочку на длинной ножке
(рис. 4.38).
Нижняя часть ножки превращена в гаусторий (присоску),
внедряющийся в тело гаметофита. Спорофит, таким образом,
лишен самостоятельности и полностью зависит от гаметофи-
та. Сверху коробочка прикрыта легко опадающим колпачком с
т онкими, направленными вниз волосками, напоминающими
льняную пряжу (отсюда и название растения). В коробочках
происходит образование спор редукционным делением (мейо-
60
зом). При этом происходит редукция хромосом вдвое, и, сле-
довательно, споры имеют гаплоидный набор хромосом. Все
споры одинаковы (изоспоры). Спора попадает в почву и про-
растает в протонему (типа нитчатой водоросли), на которой
формируется гаметофит (взрослое растение), имеющий гапло-
идный набор. На этом цикл замыкается, и все повторяется
снова.
Подкласс белые, или торфяные, мхи. Подкласс белых, или
торфяных, мхов рассмотрим на примере мха сфагнума. Торфя-
ные мхи беловато-зеленого цвета не имеют ризоидов. От стеб-
ля растения кверху отходят боковые веточки, собранные в го-
ловку. Мхи растут верхушкой стебля, а его нижняя часть
отмирает («отторфовывается»). Так в течение многих лет обра-
зуются огромные залежи торфа. Процесс торфообразования
происходит благодаря застойному переувлажнению, отсут-
ствию кислорода и созданию мхами кислой среды. Эти усло-
вия в совокупности оказываются неблагоприятными для про-
цессов гниения, т. е. для развития грибов и бактерий, что
препятствует разложению сфагнумов.
Листья сфагнума обладают гигроскопическим свойством
за счет того, что в своем строении имеют мертвые гиалино-
вые клетки, способные удерживать воду. Фотосинтез проте-
кает в живых ассимиляционных клетках, содержащих хло-
ропласты.
ОТДЕЛ ПЛАУНОВИДНЫЕ
Плауновидные — один из самых древних отделов высших
растений; их ископаемые остатки известны из силурийского
периода палеозойской эры. В настоящее время они представ-
лены только порядком плауновые, единственные представите-
ли которого сохранились до наших дней. Плауны — вечнозе-
леные травянистые растения. Это жители хвойных лесов.
Рассмотрим характеристику плаунов на примере плауна бу-
лавовидного. Взрослое растение плауна является спорофитом,
т. е. имеет диплоидный набор хромосом (бесполое поколе-
ние). Стебель плауна стелется по земле, дает дихотомически
ветвящиеся побеги и имеет дихотомически ветвящиеся прида-'
точные корни. Стебель покрыт мелкими узкими листьями.
Верхушечные побеги заканчиваются спороносными колоска-
ми. Спороносный колосок состоит из оси, покрытой тонкими
листочками — спорофиллами, на которых располагаются ок-
руглые спорангии (рис. 4.39).
В спорангиях происходит мейоз и образуются гаплоидные
споры. Споры желтого цвета, мелкие, округло-тетраэдричес-
кой формы. Морфологически и физиологически все споры
одинаковы (изоспоры). Споры высыпаются из спорангия и в
благоприятных условиях прорастают в маленький 2—3-мм за-
росток — обоеполый гаметофит, лишенный хлорофилла (по-
ловое поколение). В клетки заростка внедряются гифы грибов.
В присутствии гиф почвенного гриба гаметофит, питаясь сап-
рофитно, медленно растет и развивается в течение 12 лет. На
верхней стороне гаметофита образуются многочисленные ан-
теридии и архегоний. Оплодотворение яйцеклетки, находя-
щейся в архегоний, происходит в капельно-жидкой среде.
После оплодотворения из зиготы образуется зародыш, а из
него — взрослое растение (рис. 4.40).
Некоторые виды плауна содержат парализующий яд, сход-
ный по действию с ядом кураре. Очевидно поэтому позвоноч-
ные животные не употребляют плауны в пищу. Споры плауна
булавовидного использовали в медицине в качестве детской
присыпки, а также для обсыпки пилюль.
Рис. 4.39. Продольный
разрез через спороносный
колосок плауна:
1 — ось колоска; 2 — микро-
спорофиллы; 3 — спорангий
со спорами
62
Рис. 4.40. Внешний вид плауна
булавовидного:
7 — придаточные корни; 2 — основной
стебель; 3—вертикальные побеги; 4—спо-
роносный колосок; 5 — спорофилл со спо-
рангием; 6 — спора
ОТДЕЛ ХВОЩЕВИДНЫЕ
В геологическом прошлом хвощевидные были весьма раз-
нообразны. Ископаемые хвощевидные, например, древовид-
ные каламиты, наряду с древними плауновидными образовы-
вали леса каменноугольного периода. Современные хвощи —
травянистые растения, представленные в растительном мире
единственным родом хвощ.
Охарактеризуем хвощевидные на примере хвоща полевого.
Это многолетнее травянистое растение, встречающееся в усло-
виях избыточного увлажнения. Ранней весной из глубоко зале-
гающих корневищ вырастают спороносные побеги до 20 см.
На узлах этих побегов сидят листья, срастающиеся своими ос-
нованиями в листовое влагалище. Заканчиваются эти побега
спороносными колосками. Каждый из них состоит из оси, пер-
пендикулярно которой крепятся щитки — спорангиефоры, под
которыми находятся 6—10 спорангиев, содержащих споры, об-
разующиеся в результате мейоза. В начале щитки сидят плотно,
без зазоров, но позднее, к моменту созревания спор, стержень
колоска удлиняется. Между щитками образуются промежутки,
через которые и высыпаются споры из созревших спорангиев.
Шаровидная спора обмотана четырьмя пружинами — эла-
терами, с помощью которых споры лучше разносятся токами
воздуха. Попадая во влажную среду, споры прорастают. Разви-
вающиеся из спор гаметофиты разнополые. На одних созрева-
ют антеридии со сперматозоидами, на других — архегоний с
яйцеклеткой. Во влажной среде происходит оплодотворение.
Из образовавшейся зиготы развивается зародыш, а из него —
взрослый спорофит. У хвоща полевого после рассеивания спор
спороносные побеги отмирают и на смену им вырастают зеле-
ные ассимилирующие побеги с мутовчатыми боковыми побе-
гами, выполняющими вместо пленчатых листьев ассимилиру-
ющую функцию (рис. 4.41).
Хвощ полевой применяют как кровеостанавливающее и мо-
чегонное средство. В основном же, быстро размножаясь, хво-
щи становятся сорняками пастбищ, так как являются несъе-
добными растениями для животных. Вследствие содержания
сапонинов и алкалоидов могут вызывать отравления.
ОТДЕЛ ПАПОРОТНИКОВИДНЫЕ
Папоротники относятся к числу наиболее древних групп выс-
ших споровых растений. По своему возрасту они уступают только
63
Рис. 4.41. Хвощевидные.
Хвощ полевой:
а — вегетативный побег;
б — спороносный побег:
1 — корневище с клубеньками;
2 — спороносный стробил
плауновидным и имеют при-
близительно один геологи-
ческий возраст с хвощевыми.
Папоротники имеют самые
разные местообитания. Наи-
большее их разнообразие ха-
рактерно для влажных тропи-
ческих лесов.
Рассмотрим характерные
особенности папоротников
на примере щитовника муж-
ского (рис. 4.42). Внешне па-
поротник представляет со-
бой пучок листьев — вай,
отходящих от хорошо разви-
того корневища. Вайями ли-
стья папоротника называют
из-за того, что по своему
происхождению они возник-
ли в результате уплощения
крупных ветвей предковых
растений. Это доказывается
тем, что вайи папоротника
долго сохраняют верхушеч-
ный рост, образуя при этом
характерную разворачиваю-
щуюся улитку, что не свой-
ственно листьям. Листья
простые, дважды-трижды пе-
ристо-рассеченные. Взрослое
растение папоротника явля-
ется спорофитом (бесполое
поколение). На нижней сто-
роне вайи образуются сору-
сы — собрания спорангиев.
В спорангиях после мейоза
созревают гаплоидные споры.
При созревании спорангии вскрываются и споры высыпа-
ются. Из гаплоидных спор прорастают гаплоидные заростки,
или гаметофиты (половое поколение). Заросток представляет j
собой зеленую сердцевидную пластинку, которая с помощью
ризоидов прикрепляется к почве. На заростке формируются
архегонии и антеридии. После оплодотворения во влажной
среде на заростке образуется зигота, из которой далее развива-
64
Рис. 4.42. Папоротники. Щитовник мужской:
а — спорофит; б — часть вайи с сорусами; в — поперечный срез
через сорус; г — спорангий; д — спора; е — молодой гаметофит;
ж — зрелый гаметофит-заросток; з — антеридий; и — архегоний;
к — молодой спорофит:
1 — плацента; 2 — индузий (покрывальце соруса); 3 — спорангий; 4 —
ножка спорангия; 5 — кольцо утолщения; 6 — архегоний; 7 — антеридий;
8 — ризоиды
3-281
65
ется зародыш. Зародыш состоит из гаустории — ножки, кото-
рой он внедряется в ткани заростка и которой поглощает пита-
тельные вещества, зародышевого корня, почки, первого листа
зародыша — семядоли. Зародыш после его формирования на-
чинает расти. Вскоре зародышевый корешок редуцируется, и
на смену ему возникают придаточные корни.
Значение папоротников велико. Они выступают в роли
важнейшего компонента многих лесных сообществ. Щитовник
мужской обладает антигельминтным действием, молодые по-
беги орляка в некоторых странах употребляют в пищу.
J
СЕМЕННЫЕ РАСТЕНИЯ
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Семенные растения подразделяют на два отдела: голосемен-
ные, размножающиеся семенами, но не образующие плодов, и
покрытосеменные, имеющие семена, заключенные в плоды.
Голосеменные растения подразд еляют на следующие классы:
1) семенные папоротники;
2) саговники;
3) беннетитовые;
4) гнетовые; .
5) гинкговые;
6) хвойные.
Голосеменные — древняя группа растений, появившаяся
еще в девоне. По сравнению с высшими споровыми голосе-
менные представляют собой более высокий уровень организа-
ции растений, так как средством для расселения вида является
семя. Половой процесс у них не связан с капельно-жидкой
средой. Благодаря такой независимости семенные растения
получили возможность расселиться по всей Земле и стать гос-
подствующей труппой растений.
В отличие от высших споровых растений семенные произво-
дят семена, являющиеся многоклеточными образованиями, в
то время как споры одноклеточны. Большинство систематиков
считают труппу голосеменных естественной группой. По их
мнению, все голосеменные произошли от одной из боковых
ветвей разноспоровых древнейших папоротниковых растений.
Рассмотрим характерные особенности класса хвойных (со-
сновых) на примере сосны обыкновенной.
Взрослое растение сосны является спорофитом. В мае у ос-
нования ее молодых побегов образуются пучки желтоватых
66
мужских шишек. Каждая шишка имеет ось, к которой спи-
рально прикрепляются микроспорофиллы (гомологи тычи-
нок), имеющие с нижних сторон по два пыльцевых мешка —
микроспорангия. Вначале в них имеются диплоидные мате-
ринские клетки, из которых весной в результате мейоза обра-
зуются гаплоидные микроспоры. При этом из одной диплоид-
ной материнской клетки образуются -четыре гаплоидные
микроспоры. Микроспора одноядерна, покрыта двумя обо-
лочками — интиной (внутренней) и экзиной (наружной), и
несет два воздухоносных мешка. Они возникают в результате
отслоения экзины от интины. Далее микроспора начинает де-
литься, т. е. преобразуется в заросток, но при этом не покида-
ет микроспорангия. Ядро микроспоры делится, и образуются
две клетки. Одна клетка превращается в две проталлиальные
клетки, которые вскоре исчезают (их роль пока еще не выясне-
на). Из другой клетки в результате митоза образуются антери-
дальная клетка, формирующая в дальнейшем мужские поло-
вые гаметы — спермин, и более крупная вегетативная клетка,
из которой развивается пыльцевая трубка.
Таким образом, у голосеменных растений образуется пыль-
ца, которая и является мужским гаметофитом (рис. 4.43).
При вскрытии пыльцевого мешка пыльца высыпается нару-
жу и летит к женской шишке. Воздушные мешки облегчают
перенос пыльцы ветром. Дальнейшее развитие мужского гаме-
тофита происходит после опыления, т. е. на женских шишках
i внутри семязачатка.
Рис. 4.43. Образование пыльцы (мужского гаметофита)
у голосеменных:
1 — мужская шишка; 2 — микроспорофилл с микроспорангиями; 3 — пыль-
ца с двумя воздушными мешками
3
67
Женские шишки образуются на верхушках молодых побе- ;
гов. Женская шишка — это группа укороченных видоиз'ме- 1
ненных боковых побегов, состоящих из наружных кроющих,
под которыми располагаются семенные чешуи. У основания
семенной чешуи находятся два семязачатка. Молодой семя-
зачаток состоит из нуцеллуса и интегумента. Нуцеллус явля-
ется мегаспорангием. Он срастается с интегументом. Лишь
около вершины, обращенной к оси шишки, интегумент име-
ет отверстие, называемое микропиле, или пыльцевход. В на-
чале развития семязачатка нуцеллус состоит из однородных
диплоидных клеток. Затем в средней части обособляется
одна материнская археспориальная клетка. Она мейотически
делится с образованием четырех гаплоидных мегаспор. Далее
три из них отмирают, а одна остается способной к развитию.
Эта мегаспора многократно делится митозом, в результате
чего образуется новая ткань — эндосперм, являющийся жен-
ским гаметофитом. Из двух наружных клеток эндосперма,
ориентированных к микропиле, образуются два архегония —
женские органы полового размножения (рис. 4.44).
С помощью ветра и воздушных мешков пыльца из мужских
шишек переносится на семязачатки. Улавливается пыльца
каплями густой жидкости, заполняющей пространство между
нуцеллусом и интегументом и выступающей через микропиле.
Эта жидкость, подсыхая, затягивает пыльцу внутрь семязачат-
ка на нуцеллус (в пыльцевую камеру). После опыления микро-
пиле зарастает. Чешуйки женской шишки смыкаются, и муж-
Рис. 4.44. Развитие женского гаметофита сосны обыкновенной:
1 — археспориальная клетка; 2 — тетрада мегаспор;
3 — деление мегаспоры; 4 — женский гаметофит (эндосперм),
развившийся из мегаспоры
68
ской гаметофит продолжает свое развитие уже на мегаспоран-
гии. Экзина лопается, и вегетативная клетка, окруженная ин-
тиной, развивается в пыльцевую трубку, которая внедряется в
ткань нуцеллуса и растет в направлении архегония. Антери-
дальная клетка деДится и образует две клетки — клетку-ножку
и генеративную (спермагенную) клетку. Функция клетки-нож-
ки до сих пор не выяснена. Генеративная клетка перемещается
в пыльцевую трубку, по которой и достигает архегония. Обра-
зование из спермагенной клетки двух спермиев, лишенных
жгутиков, происходит непосредственно перед оплодотворени-
ем. Один из спермиев сливается с ядром яйцеклетки, а второй
отмирает. Также отмирает и второй архегоний. От опыления
до оплодотворения проходит около 13 месяцев. Из образовав-
шейся зиготы тотчас развивается зародыш, и развитие его идет
за счет запасных продуктов эндосперма.
Сформированное семя сосны соединяет в себе гаметофит и
спорофит. Одна часть семени — первичный эндосперм —
представляет собой женский гаметофит. Другая часть семе-
ни — зародыш — зачаток дочернего спорофита. Кожура семе-
ни — бывший интегумент семязачатка и прилегающий к нему
слой перисперма (бывшая ткань нуцеллуса), являются обособ-
ленной частью материнского спорофита (рис. 4.45).
Семена созревают осенью, на второй год после опыления.
Шишки к этому времени достигают длины 4—6 см, чешуи
одревесневают, из зеленых становятся серыми. Следующей
весной чешуйки расходятся и се-
мена высыпаются.
Хвойные леса определяют ланд-
шафт огромных территорий, служат
основой многих биоценозов, итра-
ют значительную водоохранную
роль. Наибольшую часть таежных
лесов России занимают лиственни-
ца, сосна, ель, пихта. Древесину
хвойных деревьев используют как
строительный материал, топливо;
из хвои пихты получают эфирное
масло, идущее на производство ла-
ков. В молодых ветвях пихты содер-
жится борнеол — исходный про-
дукт для получения камфоры.
Из сосны сибирской получают ски-
пидар и канифоль, из кедровых
орешков — масло.
Рис. 4.45. Строение
семени сосны сибирской
(кедровый орешек):
1 — кожура семени (бывший
интегумент); 2 — перисперм
(бывший нуцеллус); 3 — за-
родыш с зачатками листьев,
стебля и корня; 4 — эндо-
сперм
69
ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ
В отличие от других высших растений покрытосеменные
имеют особый орган — плод, развивающийся из цветка. Важ-
нейшие функции плода: защита семязачатка (семени) и рас-
пространение семян.
Голосеменные
1. Семязачаток лежит от-
крыто на семенной чешуе
(мегаспорофилл).
2. Женским гаметофитом
является эндосперм с двумя
архегониями.
3. Развитие семязачатков
и образование семени проис-
ходит очень медленно — око-
ло 18 мес.
4. Оплодотворение оди-
нарное, в результате форми-
руется зародыш, который
формируется за счет первич-
ного эндосперма.
Покрытосеменные
1. Семязачаток находится
под покровом мегаспоролис-
тиков.
2. Женским гаметофитом
является зародышевый ме-
шок с восемью ядрами.
3. Развитие семязачатка и
образование семени происхо-
дит сравнительно быстро,
особенно у трав — 3—4 неде-
ли — вегетационный период.
4. Двойное оплодотворе-
ние, в результате формирует-
ся диплоидный зародыш, ко-
торый развивается за счет
триплоидного вторичного
эндосперма.
Покрытосеменные — самая совершенная и многочислен-
ная группа высших растений, насчитывающая не менее 250
тыс. видов. Эти растения наиболее приспособлены к совре-
менным условиям жизни и распространены в самых различ-
ных местообитаниях во всех климатических зонах и на всех
континентах.
Отдел покрытосеменные растения делится на два класса:
класс двудольных и класс однодольных растений.
Высшим этапом эволюции полового размножения в мире
растений является цветок и его производные — семя и плод.
Цветок
Цветок — орган семенного размножения покрытосемен-
ных растений. Цветок представляет собой видоизмененный
укороченный спороносный побег ограниченного роста,
приспособленный для образования микро- и мегаспор, га-
мет и для перекрестного опыления, В результате опыления
70
и последующего полового процесса семязачатки развивают-
ся в семена.
По положению цветок бывает верхушечным или боковым,
т. е. выходит из пазухи прицветного листа (прицветника).
Цветок состоит из стеблевой части (цветоножка, цветоло-
же) и листовой части (чашелистики, лепестки, тычинки, пес-
тики). Часть побега между цветком и прицветником называют
цветоножкой (рис. 4.46). Если цветоножка укорочена или от-
сутствует, то цветок называется сидячим. Верхняя расши-
ренная часть цветоножки, к которой прикрепляются все части
цветка, называется цветоложем. Цветоложе имеет различную
форму — плоское, выпуклое, вогнутое (рис. 4.47). В зависи-
мости от формы цветоложа различают положение завязи в
цветке — нижнее и верхнее. Одни части цветка считаются сте-
рильными — это чашелистики, лепестки, а другие репродук-
тивными — это пестики и тычинки. Стерильные части выпол-
няют защитную функцию, а репродуктивные приспособились
к спороношению.
Околоцветник бывает двойным, тогда он состоит из чашеч-
ки (Са) и венчика (Со), и простым (Р), тогда он может быть
или чашечковидным или венчиковидным, состоящим и в том,
и в другом случае из листочков простого околоцветника.
Рис. 4.46. Схема строения цветка.
I — цветоножка; 2 — прицветник; 3 — цветоложе; 4 — подчашка; 5 —
чашечка (Са); 6 — венчик (Со); 7— тычиночные нити; 8 — пыльник ты-
чинки; 9 — пыльца; 10 — рыльце пестика; 11 — столбик пестика; 12 —
завязь пестика; 13 — семязачаток; 14 — плацента; 15 — стаминодий
71
Рис. 4.47. Формы цветоложа и различное положение завязи
в цветке:
1 — плоское; 2 — вогнутое; 3 — стенки завязи срастаются со стенками
вогнутого цветоложа; 4 — выпуклое;
1,2 — верхнее* положение завязи в цветке; 3 — нижнее положение завязи в
цветке
Чашечкой называют совокупность чашелистиков, и она
обычно существенно отличается по окраске от венчика. Чаше-
листики (как правило, зеленые) могут быть свободными, т. е.
несросшимися, и тогда чашечка называется раздельно-
листной. Если же чашелистики срастаются на определен-
ных уровнях, то такая чашечка называется сростнолист-
ной. Чашелистики составляют наружный круг цветка, а в
формуле цветка стоят на первом месте (Са).
Венчик образует следующий круг в цветке и в формуле цветка
следует за чашечкой и обозначается (Со). Венчик может быть
свободнолепестным и спайнолепестным. Если
любые части цветка срастаются, то в формуле цветка количе-
ство срастающихся элементов берется в скобки. Венчик обыч-
но ярко окрашен, что привлекает насекомых — опылителей.
По особенностям симметрии цветки делят на актино-
морфные, через которые можно провести много плоскостей
симметрии, что в формуле цветка обозначается (♦) и зиго-
морфные, через которые можно провести только одну вер-
тикальную плоскость симметрии (f) (рис. 4.48).
Совокупность тычинок одного цветка называют андроцеем
(А). В некоторых цветках андроцей вообще отсутствует — это
так называемые женские однополые цветки, а в однополых
мужских цветках есть только тычинки. Каждая тычинка состо-
ит из суженной части — тычиночной нити и расширенной ча-
сти — пыльника. Пыльник имеет две половинки, соединен-
72
Рис. 4.48. Актиноморфные (1) и зигоморфные (2) цветки
ные связником, т. е. продолжением Тычиночной нити. Каж-
дая половинка несет два гнезда — микроспорангия. Гнезда
пыльников называют пыльцевыми мешками. В зрелом пыль-
нике перегородки между гнездами часто не выявляются. Сна-
ружи пыльник покрыт эпидермой. Под ней располагается
слой клеток, за счет которых при подсыхании пыльника
вскрываются гнезда. Самый внутренний слой, выстилающий
полость пыльников, называется тапетумом. Содержимое кле-
ток тапетума идёт на питание микроспор.
Элемент листовой части цветка, несущий один или не-
сколько семязачатков, а после оплодотворения семя (семена),
называется плодолистиком. Совокупность плодолистиков од-
ного цветка, образующих один или несколько пестиков, назы-
вают гинецеем (от греч. «гине» — женщина). Плодолистик яв-
ляется структурной единицей гинецея. Если плодолистик
образует единственный пестик в цветке, то такой гинецей на-
зывается монокарпным. Если множество (два и более)
плодолистиков образуют множество самостоятельных пести-
ков, то такой гинецей называется апокарпным. Гинецей,
формирующийся из нескольких сросшихся плодолистиков,
образующих единый пестик, называется ценокарпным.
Каждый пестик состоит из разросшейся завязи, столбика и
рыльца. Завязь пестика выполняет функцию влажной каме-
ры, предохраняющей семязачатки от высыхания. Пестик на-
дежно укрывает семязачатки от поедания их насекомыми и
отчасти от резких колебаний температуры. Структура пести-
ка идеально приспособлена к опылению и оплодотворению.
В семязачатках происходит процесс образования мегаспор и
гамет.
Место прикрепления семязачатков в завязи называется
плацентой. Плацента обычно имеет вид вздутия, выроста или
выступа, образованного тканями завязи. Семязачатки распо-
73
лагаются на плацентах завязи. Они сообщаются с плацентой с
помощью семяножки. В семязачатках у семенных растений
- происходит: 1) мегаспорогенез — образование мегаспор;
2) формирование женского гаметофита; 3) процесс оплодотво-
рения.
Семязачаток состоит из центральной части — нуцеллуса.
Нуцеллус является мегаспорангием. Его окружают один или
два покрова — интегумента, которые над верхушкой нуцеллуса
образуют канал — микропиле. В семязачатке различают апи-
кальную (верхушечную) — микропилярную часть, и базаль-
ную, ей противоположную — халазальную часть, которую час-
то называют просто халазой.
Мегаспорогенез. У цветковых растений в нуцеллусе (мега-
спорангии) из клеток археспория обособляется одна мате-
ринская клетка (мегаспора), имеющая, как и все растение,
диплоидный набор хромосом. После мейотического деления
этой клетки образуются четыре гаплоидные мегаспоры. Далее
три клетки из них отмирают, а четвертая, обращенная ближе
к халазе, сохраняется. Из нее и развивается женский гамето-
фит — зародышевый мешок. Далее мегаспора разрастается,
оттесняя ткань нуцеллуса к интегументам. Ядро мегаспоры
претерпевает три последовательных митотических деления
(эндомитоз), в результате чего образуются восемь монопло-
идных ядер. После первого деления мегаспоры образуются
два ядра, которые расходятся к полюсам удлиняющейся ме-
гаспоры. Далее каждое из этих ядер синхронно делится еще
два раза; в результате у каждого полюса возникают по четыре
ядра. На этой восьмиядерной стадии процесс деления ядра
женского гаметофита заканчивается. От каждой из двух чет-
верок ядер в центральную часть зародышевого мешка отходят
по одному ядру. Эти так называемые полярные ядра сближа-
ются в центральной части и, сливаясь, образуют диплоидное
вторичное ядро. В дальнейшем семь ядер зародышевого меш-
ка претерпевают фазу цитоплазматического деления, т. е.
каждое ядро с участком цитоплазмы окружается плазмати-
ческой мембраной, следовательно, образуются семь отдель-
ных клеток (рис. 4.49).
У микропилярного полюса гаметофита три клетки форми-
руют яйцевой аппарат, одна из которых, наиболее крупная,
является яйцеклеткой, а две другие вспомогательными клетка-
ми — синергидами. Три клетки халазального полюса получи-
ли название антипода. В центральной части зародышевого
мешка располагается клетка с диплоидным ядром. Так возни-
кает женский гаметофит, который сильно редуцирован даже
Рис. 4.49. Формирование женского гаметофита
покрытосеменных растений:
1 — семязачаток с мегаспорой; 2 — мейоз мегаспоры; 3 — отмирание трех
клеток; 4 — первое деление мегаспоры; 5 ~ второе деление мегаспоры; 6 —
третье деление мегаспоры; 7 — сформированная семяпочка (а — антиподы,
б — диплоидное ядро, в — яйцеклетка, г — синергиды)
по сравнению с гаметофитом голосеменных и полностью ли-
шен архегониев.
Микроспорогенез. Микроспорангиями являются гнезда
пыльников. В них в результате мейоза образуются микроспо-
ры из диплоидных материнских клеток археспория. Из каж-
дой материнской клетки образуются четыре гаплоидные мик-
роспоры. Сформировавшаяся микроспора имеет оболочку и
единственное ядро. Такая микроспора дает начало пыльцево-
му зерну. При делении митозом из микроспоры образуются
две новые клетки: маленькая генеративная и более крупная ве-
гетативная. Таким образом, пыльца, или мужской гаметофит,
формируется из микроспоры внутри пыльника. Пыльцевое
зерно покрыто двумя оболочками — экзиной и инти ной. Эк-
зина — наружная оболочка, пропитана жироподобным веще-
ством (эквивалент кутина). Внутренняя оболочка — интина.
Она менее стойка, чем экзина, состоит из целлюлозы и пекти-
на и сохраняет эластичность. После созревания пыльцы пыль-
ники лопаются и пыльца рассеивается (рис. 4.50).
75
Опыление. Для того чтобы образовался зародыш, должны ;
произойти опыление и оплодотворение. Опыление — это про-, j
цесс переноса пыльцы с тычинки на рыльце пестика. Опыле- 1
ние впервые появляется у голосеменных, но наибольшего со-
вершенства оно достигает у покрытосеменных.
Различают два типа опыления: самоопыление и перекрест-
ное опыление.
При самоопылении на пестик цветка попадает пыльца того
же растения.
Если перенос пыльцы осуществляется между цветками
разных особей, то происходит перекрестное опыление. Счи-
тается, что оно свойственно 90% растений. Перекрестное
опыление обусловливает высокий уровень гетерозиготности
популяций. Это создает большие возможности для есте-
ственного отбора. Строгое самоопыление встречается отно-
сительно редко (например, у гороха) и может вести к рас-
щеплению вида на ряд чистых линий, т. е. делает популяции
гомозиготными.
Для эволюционного процесса оптимально сочетание са-
моопыления и перекрестного опыления, что чаще всего и
имеет место в природе. Одна из форм, ограничивающих са-
моопыление, — двудомность, т. е. на одних растениях разви-
ваются только мужские, или тычиночные, цветки, а на других
женские, имеющие только гинецей. Однодомные растения
имеют цветки, содержащие и андроцей и гинецей. Другая
форма, ограничивающая самоопыление, — полная и физио-
логическая несовместимость. Она выражается в подавлении
при самоопылении прорастания пыльцы на рыльце пестика
той же особи.
Рис. 4.50. Строение пыльцы
покрытосеменных растений:
а — микроспора;
б — пыльцевое зерно:
1 — вегетативная клетка; 2 — экзина;
3 — интина; 4 — генеративная клетка
76
Выделяют два типа перекрестного опыления — биотичес-
кое и абиотическое. Биотическое опыление осуществляется жи-
вотными, абиотическое — с помощью неживых факторов
внешней среды (ветер, вода у водных растений).
Оплодотворение. Пыльца, попав на рыльце пестика, начина-
ет прорастать. Из вегетативной клетки образуется пыльцевая
трубка, а из генератйвной — два спермия. Пыльцевая трубка
проникает в зародышевый мешок, достигнув яйцеклетки ло-
пается и обеспечивает проникновение в него спермиев. В это
время вегетативная клетка дегенерирует. Один из спермиев ко
пулирует с яйцеклеткой, образуя зиготу, дающую начало заро-
дышу. Второй спермий сливается со вторичным диплоидным
ядром. В результате получается триплоидная клетка, развиваю-
щаяся в специальную питательную ткань — эндосперм. Таким
образом, происходит двойное оплодотворение, свойственное
только покрытосеменным растениям.
У покрытосеменных растений эндосперм называется «вто-
ричным», или «белком». Только у покрытосеменных расте-
ний зародыш (спорофаза) начинает свое развитие самостоя-
тельно за счет триплоидной фазы. У всех предшествующих
групп (голосеменные и др.) зародыш развивается за счет га-
метофазы.
Таким образом, из составных частей цветка образуются:
1) из оплодотворенной яйцеклетки — зародыш;
2) из диплоидного ядра — эндосперм;
3) из интегументов семязачатка — семенная кожура семени;
4) из нуцеллуса — перисперм семени;
5) из стенок завязи и часто с участием других элементов
цветка (чашечки, цветоложа) — стенка плода (перикарпий).
Перикарпий имеет три слоя: наружный — экзокарпий, сред-
ний — мезокарпий и внутренний — эндокарпий.
У некоторых видов покрытосеменных, принадлежащих к
разным семействам, зародыши и семена развиваются апомик-
тически, т. е. без оплодотворения по типу партеногенеза (на-
пример, у одуванчика).
Соцветия. Соцветия — это побег или система специализи-
рованных побегов, несущих цветки.
Классификация соцветий может осуществляться на основе
особенностей ветвления побегов. В соответствии с этим соцве-
тия подразделяются на моноподиальные, или ботриоидныс
(от греч. «ботрион» — кисть), и симподиальные, или цимоз-
ные (рис. 4.51).
Моноподиальные соцветия бывают простые и сложные. Бо-
ковые оси простых, в отличие от сложных соцветий, не ветвят-
Рис. 4.51.Типы соцветий:
а—простые ботриоидные (моноподиальные):
7 — кисть; 2 — щиток; 3 — зонтик; 4 — головка; 5 — колос; 6 — початок;
7 — корзинка
б — сложные ботриоидные:
8 — сложный колос; 9 — метелка; 10 — сложный зонтик;
в — цимозные (симподиальные):
11 — извилина (монохазий); 12 — завиток (монохазий); 13 — развилина
(дихазий); 14 — многолучевик (плейохазий)
ся и представляют собой цветоножки. Простые соцветия могут
быть как открытыми (верхушечная ось не заканчивается цвет-
ком), так и закрытыми (верхушечная ось заканчивается цвет-
78
ком). Роспуск цветков происходит снизу вверх, если соцветие
вертикальное (кисть, колос), и от периферии к центру, если
цветки в соцветии расположены горизонтально (зонтик, кор-
зинка).
Кисть — все цветки сидят на цветоножках, которые
поочередно прикрепляются к главной оси соцветия (чере-
муха).
Щиток — сходен с кистью, но у щитка нижние цветонож-
ки длиннее верхних, и поэтому цветки его расположены на од-
ном уровне в одной плоскости (рябина, боярышник).
Зонтик — все цветоножки с прицветниками выходят от
верхушечной точки укороченной оси соцветия (вишня).
Головка — видоизмененный зонтик, у которого разрас-
тается главная ось в виде головки, а на ней располагаются
цветки на укороченных цветоножках (клевер).
Колос — производное кисти, отличающийся от нее от-
сутствием цветоножек (подорожник).
Початок — отличается от колоска разросшейся главной
осью соцветия (кукуруза).
Корзинка — главная ось соцветия разрастается в виде
чаши или ложа, на котором располагаются сидячие цветки,
окруженные со всех сторон (часто в два ряда) видоизменен-,
ными листьями — листовой оберткой (подсолнечник, ро-
машка).
Сложные моноподиалъные соцветия. Метелка (или слож-
ная кисть) — главная ось ветвится, т. е. образует боковые побе-
ги первого или второго порядка, на которых располагаются
цветки на цветоножках (сирень).
Сложный колос — на главной оси от каждого узла
вместо цветков отходят боковые побеги с сидячими на них
цветками, т. е. простые колоски (пшеница, рожь).
Сложный зонтик — на верхушке главной оси вместо
цветков отходят простые зонтики (морковь).
Симподиальные соцветия. Соцветия этой группы имеют
симподиальное (мрнохазии) и ложнодихотомическое (диха-
зии) ветвление. Главная ось заканчивается цветком. Другие
цветки возникают в пазухах прицветных листьев, т. е. распус-
кание цветков происходит центробежно, а не снизу вверх, как
у моноподиальных. Различают три типа симподиальных со-
цветий — монохазий, дихазий и плейохазий.
Кмонохазиям относятся: извилина и завиток.
Извилина — от главной оси отходит ось второго порядка,
от второго — третьего порядка и т. д., то с одной, то с другой
стороны (лютик едкий).
79
Завиток — от главной оси отходит ось второго порядка, от
второго — третьего порядка и т. д., но при этом оси отходят
только в одном направлении, как бы закручиваясь в улитку
или завиток (незабудка и другие представители семейства Бу-
рачниковые).
К дихазиям относятся: развилина и двойной завиток.
Развилина — под цветком главной оси образуются супро-
' тивно расположенные ветви (оси), заканчивающиеся цветком.
В дальнейшем каждая из этих осей также образует две супро-
тивно расположенные оси (гвоздика).
Двойной завиток — под цветком главной оси супротивно
образуются два простых завитка (картофель).
Плейохазий — ниже цветка, которым заканчивается
главная ось, развивается несколько боковых осей (с цветками
или соцветиями), перерастающих главную ось и расположен-
ных мутовкой (молочай).
Плод
Плод — орган размножения покрытосеменных растений,
заключающий семена и развивающийся из одного цветка.
Функции плода: формирование, защита и распространение
семян. Максимальное число семян в плоде равно числу семя-
зачатков в завязи, но обычно меньше, так как не все семяза-
чатки достигают Зрелости. Все многообразие плодов в соответ-
ствии с типом гинецея делят на четыре типа: монокарпии,
апокарпии, ценокарпии и псевдомонокарпии.
Стенка плода или отдельных плодиков называется пери-
карпием (рис. 4.52) и имеет три слоя: экзокарпий; мезокар-
пий, эндокарпий.
Плоды монокарпии (рис. 4.53, а) — возникают из цветков,
чмеющих монокарпный гинецей и всегда верхнюю завязь.
Однолистовка — многосеменной монокарпий, вскры-
вающийся только по брюшному шву (живокость полевая).
Рис. 4.52. Строение плода (однокостянки)
на примере персика обыкновенного:
1, 2, 3 — околоплодник, или перикарпий (1 — эк-
зокарпий, 2 — мезокарпий, 3 — эндокарпий); 4 —
семя; 5 — след плодоножки; 6 — плодоножка
80
Рис. 4.53. Схема классификации плодов по типу гинецея:
а —монокарпии:
1 — однолистовка; 2 — боб; 3 — однокостянка;
б — апокарпии:
1 — многолистовка; 2 — многоорешек; 3 — фрага; 4 —• многокостянка;
в — ценокарпии:
1 — яблоко; 2 — коробочка; 3 — ягода; 4 — стручок;
г — псевдомонокарпии;
1 — зерновка; 2 —• орех; 3 — желудь
81
Однокостянка — односеменной невскрывающийся^
монокарпий, эндокарпий которого (косточка) твердый, скле-1
рефицированный. Мезокарпий может быть сочным, как в пло-j
дах вишни, черемухи, или сухим кожистым (миндаль).
Б о б — одно-, дву-, многосеменной монокарпий, раскрыт
вающийся по двум швам — спинному и брюшному — от вер-’
шины к основанию. Семена, располагаются непосредственно^
на створках (донник, арахис, акация).
Плоды апокарпии (рис. 4.53, б) — возникают из цветков с;
верхней завязью. I
Многолистовка — многосеменной сборный плод, со»;
стоящий из простых листовок (от двух и более), например, ку-
пальница, калужница болотная.
Многоорешек—сборный многосеменной плод, состо-
ящий из отдельных односеменных невскрываклцихся плоди-
ков — орешков. Многоорешки характерны для видов лютиков,
лапчаток, горицветов.
Многокостянка — сборный плод, состоящий из двух
или многих плодиков — костянок. Мезокарпий каждого тако-
го плодика сочный, а эндокарпий — твердый, склерефициро-
ванный.
Плоды ценокарпии (рис. 4.53, в) — самая многочисленная
группа плодов. Возникают из цветков как с верхней, так и с
нижней завязью; при этом, хотя завязь в цветке одна, она об-
разована двумя и более сросшимися плодолистиками.
Ягода — многосеменной невскрывающийся плод с соч-
ным перикарпием, в который погружены семена. Например,
плод помидора, картофеля, возникающий из гинецея, образо-
ванного двумя сросшимися плодолистиками.
Тыквина — плод, близкий к ягоде, но отличающийся
кожистым экзокарпием (тыква, арбуз, огурец).
Коробочка — многосеменной плод с сухим вскрываю-
щимся перикарпием. Возникает из гинецея, образованного
двумя и более плодолистиками. Коробочки очень варьируют
по форме и по способам вскрывания (мак, наперстянка, дур-
ман).
Стручок — сухой многосеменной плод, возникает из ги-
нецея, образованного двумя сросшимися плодолистиками.
По краям срастания плодолистиков на рамке располагаются
семена. Стручки бывают вскрывающиеся и невскрывающиеся.
Стручочки — это стручки, у которых длина приблизительно
равна ширине.
Я б л о к о — многосеменной невскрывающийся плод, обра-
зованный из цветка с нижней завязью. Завязь в цветке образу-
82
ет от трех до пяти сросшихся плодолистиков (яблоня, рябина,
боярышник).
Плоды псевдомонокарпии (рис. 4.53, г) — образуются из
цветков с псевдомонокарпным гинецеем, в котором заклады-
ваются два или более плодолистиков, но развивается только
один, а остальные редуцируются.
Семянка — односеменной плод с кожистым перикарпи-
ем, несрастающимся с семенем. Характерен для всех предста-
вителей семейства сложноцветных (подсолнечник, василек,
ромашка).
Зерновка — односеменной плодх пленчатым перикар-
пием, срастающимся с семенем. Характерен для всех злаков
(пшеница, рожь, тимофеевка).
Орех — односеменной, редко двусеменной плод с сильно
склерефицированным, почти деревянистым перикарпием (ле-
щина, ольха, береза).
Соплодия — это совокупность зрелых плодов, возникающих
путем срастания завязей цветков, в пределах одного соцветия.
Примеры соплодия: плод ананаса, шелковицы, свеклы.
Семя — орган полового размножения и расселения расте-
ний, развивающихся в основном из оплодотворенного семяза-
чатка.
Семя состоит из зародыша, эндосперма, семенной кожуры.
Зародыш является основной частью семени, и в нем различа-
ют три зародышевые органа: зародышевой корешок, зароды-
шевой стебелек с зародышевой почкой и зародышевые листья
(семядоли).
Семенная кожура обычно многослойна и присутствует у се-
мени всегда. Ее главная функция — защита зародыша от чрез-
мерного высыхания; она также предохраняет зародыш от
преждевременного прорастания. При прорастании первые
порции воды проникают внутрь семени через отверстие в се-
менной кожуре — микропиле.
Эндосперм обычно состоит из округлых клеток запасающей
ткани. Это могут быть зерна крахмала или капли жирного мас-
ла, нередко в сочетании с запасными белками. Вещества эн-
досперма гидролизуются при набухании семян под действием
ферментов и поглощаются зародышем в процессе прораста-
ния; после этого клетки его разрушаются.
Зародыш — это миниатюрный спорофит. Зародышевый по-
бег представлен осью (зародышевым стебельком) и семядоль-
ными листьями, или семядолями: двумя у двудольных и одной
у однодольных растений. У зародыша однодольных намечает-
ся так же, как и у двудольных, два бугорка—зачатки двух семя-
83
долей, но у однодольных эта симметрия быстро исчезает, так |
как одна семядоля не получает дальнейшего развития. Учас- |
ток стебелька в зародыше выше семядолей называется эпико- 1
тилем, ниже семядолей — гипокотилем, или подсемядольным |
коленом. 1
Виды семян. Семена двудольных бывают с эндоспермом, без |
эндосперма и с периспермом. Семена однодольных бывают с i
эндоспермом и без эндосперма. I
Семена двудольных без эндосперма. Кэтой f
категории относят семена бобовых, тыквенных, сложноцвет- I
ных, крестоцветных и др. (рис. 4.54, а). |
Семена двудольных с эндоспермом. Между |
семядолями находится конус нарастания побега; почечка еще |
не сформирована (у семени клещевины) (рис. 4.54, б). >
Семена двудольных с периспермом. В неко- |
торых случаях, помимо эндосперма, в семенах развивается за- |
пасающая ткань иного происхождения — перисперм, лежа- |
щий под кожурой и возникающий из нуцеллуса семязачатка. |
Функционально эндосперм и перисперм равноценны, хотя |
морфологически имеют разное происхождение: они аналоги, |
но не гомологи (у семени свеклы) (рис. 4.54, в). I
Семена однодольных с эндоспермом.Кэтой
категории принадлежит подавляющее большинство семян од-
нодольных. Большую часть объема семени занимает эндо-
сперм. В него погружен палочковидный зародыш. Семядоля ’
цилиндрическая; ее нижняя часть представляет собой влагали-
ще, охватывающее конус нарастания со всех сторон и прикры-
вающее его. Функция влагалища семядоли — защита точки ро-
ста (рис. 4.55, а).
Рис. 4.54. Строение семян двудольных:
а — без эндосперма, б — с эндоспермом, в — с периспермом:
1 — корешок; 2 — эпикотиль; 3 — семенная кожура; 4 — гипокотиль, 5 —
семядоля; 6 — эндосперм; 7 — перисперм
84
Рис. 4.55. Строение семян однодольных:
а — с эндоспермом; б — злака; в — без эндосперма:
1 — воздухоносная полость; 2 — семенная кожура; 3 — эндосперм; 4 —
семядоля; 5 — почечка; 6 — щиток; 7 — корень; 8 — колеоптиль; 9 —
колеориза
Семена злаков. Строение семени злаков-довольно
своеобразно. Зародыш в зерновке соприкасается с эндоспер-
мом одной стороной, а не окружен его тканью, как у большин-
ства других однодольных. Вследствие такого расположения се-
мядоля злаков имеет форму плоского щитка, прижатого к
эндосперму. Всасывающая функция щитка обеспечивается
сильно специализированными клетками его поверхностного
слоя. В отличие от большинства однодольных, почечка заро-
дыша злаков обычно довольно сильно развита, имеет несколь-
ко листовых зачатков. Наружный колпачковидный листок
почечки называется колеоптилем. Гипокотиль у злаков недо-
развит; зародышевый корешок окружен специальным много-
слойным чехлом — колеоризой, которая при прорастании на-
бухает и развивает на поверхности всасывающие волоски,
корень пробивает ткань колеоризы, чтобы выйти наружу, в
почву (рис74.55, б).
Семена однодольных без эндосперма.Такой
тип семян характерен для болотниковых (частуха болотная,
рдест плавающий). Семя имеет форму подковы, под тонкой
кожурой находится зародыш, сосредоточивший в семядоле все
запасы, поглощенные им в ходе созревания семени; эндо-
сперм им уже «съеден» (рис. 4.55, в).
Прорастание семян. Для прорастания семени требуются оп-
ределенные условия.
1. Увлажнение семени.
2. Холодная стратификация, т. е. длительное выдержива-
ние их при пониженной температуре.
3. Скарификация (искусственное нарушение целостности
кожуры).
85
Рис. 4.56. Схема надземного (а) и подземного (б) прорастания |
на примере двудольных растений: |
1 — семядоли; 2 — гипокотиль; 3 — главный корень; 4 — эпикотиль; 5—1
настоящие листья 1
Прорастание бывает надземным и подземным. I
При надземном прорастании семядоли выносят- 1
ся на поверхность, зеленеют и становятся первыми ассимили- 1
рующими листьями. |
При подземном прорастании семядоли,какпра- |
вило, сморщиваются и отмирают, не выходя на поверхность, а 1
первыми ассимилирующими листьями становятся следующие I
за семядолями настоящие листья (рис. 4.56). <
I
Классификация г
покрытосеменных растений
Отдел покрытосеменные делится на два класса: класс дву-
дольных и класс однодольных.
86
Двудольные
1. Зародыш обычно имеет
две семядоли.
2. Листья простые и слож-
ные, жилкование перистое и
пальчатое..
3. Характерен вторичный
рост осевых органов за счет
камбия.
4. Первичный зародыше-
вый, корешок развивается в
главный корень, от которого
отходят боковые корни.
5. Корневая система стер-
жневая или ветвистая.
6. Цветки главным обра-
зом пяти-, реже четырех-
членные. Чашечка и венчик
хорошо обособлены.
Однодольные
1. Зародыш с одной семя-
долей.
2. Листья всегда простые,
жилкование параллельное и
дуговое.
3. Вторичный рост за счет
камбия отсутствует. Пучки
закрытые.
4. Первичный корешок
рано отмирает, главный и бо-
ковые корни не развиваются,
заменяясь придаточными кор-
нями.
5. Корневая система моч-
коватая.
6. Цветки обычно трех-
членные, четырехчленные, но
никогда не бывают пятичлен-
ными. Околоцветник обычно
простой.
Класс двудольные
Семейство розоцветные. Розоцветные включают свыше
3000 тыс. видов. Семейство подразделяется на четыре под-
семейства: спирейные, розовые, яблоневые и сливовые
(рис. 4.57, табл. 4.2).
Таблица 4.2
Характерные признаки представителей Семейства
розоцветных
Признак Подсемейство
Спирейные Розовые Яблоневые Сливовые
Гинецей Апокарпный Апокарпный Ценокарп- ный Монокарп- ный
Число пло- долистиков 8—5(1) 5 (редко 1) (3—5) 1
Завязь Верхняя Верхняя Нижняя Верхняя
Тип плода Многодис- товка Многоорешек Многокостянка Яблоко Однокостянка
Подчашие Нет Имеется у трав Нет Нет
Предста- вители Спирея Малина» земля- ника Яблоня» ря- бина Слива, мин- даль, вишня
Формулы •CajCosA^G* •CasCosAooGoo ^^es+sCojAeJGoo •CajCojA^ *Ca5Co5A*Gj
П
Рис. 4.57. Розоцветные. Цветки представителей
различных подсемейств:
а — лабазник вязолистный (подсемейство спирейные);
б — малина (подсемейство розовые); в — шиповник майский
(подсемейство розовые); г — миндаль степной (подсемейство
сливовые); д—яблоня дикая (подсемейство яблоневые)
Среди розоцветных известны вечнозеленые и листопадные
деревья и кустарники, полукустарники и травы. Листья оче-i
редные, простые и сложные, с прилистниками. Соцветия’
ботрические (кисть, зонтик, щиток) и цимозные (развилина,
извилина). Цветки правильные, обоеполые, с двойным около-
цветником. Чашечка из пяти, реже из четырех чашелистиков,
иногда снабженная подчашием (в основном у травянистых ви-
дов), которое образовано прирастающими к ней прицветника-
ми. Лепестки в числе пяти или четырех, свободные, прикреп-
ленные подобно тычинкам к краю гипантия — образования,
возникающего из-за срастания оснований чашелистиков, ле-
пестков и тычинок с тканями цветоложа. Форма гипантия мо- ’
жет быть.различной (блюдцевидная, колокольчатая и т. д.).!
Особенно характерен гипантий для шиповника, где он участ-
вует в образовании плода. У подсемейства яблоневых проис-
ходит срастание гипантия с завязью, и плод при созревании
становится сочным. Андроцей состоит из тычинок, располо-
женных в несколько рядов (их более 12). Гинецей апокарпный
или ценокарпный. Число плодолистиков может быть различ-
ным — от одного (редко) до множества. Каждый плодолистик
содержит, как правило, два семязачатка.
Розоцветные имеют большое практическое значение как
плодовые и ягодные культуры (яблоня, груша, слива, малина,
земляника). Из розы добывают розовое масло, используемое в
88
парфюмерии и медицине. Лекарственными являются корни
кровохлебки, корневища лапчатки, плоды черемухи.
Семейство бобовые. Огромное семейство, объединяющее до
18 000 видов. Среди бобовых известны почти все жизненные
формы. От древесных растений высотой 60—80 м до однолет-
них трав в несколько сантиметров. На корнях значительной
части бобовых имеются клубеньки, которые возникают из-за
разрастания паренхимной ткани корней, вследствие внедре-
ния туда азотфиксирующих бактерий.
Листья всегда сложные — пальчато- и перистосложные,
с прилистниками. Соцветия ботрические — кисть, метелка,
реже головка. Формула цветка: TCa^Coi^AsMOl (рис. 4.58).
Чашечка состоит из пяти сросшихся чашелистиков, венчик —
из пяти лепестков. Самый крупный лепесток — парус — спо-
собствует привлечению насекомых (опылителей). Два боковых
лепестка называются веслами и используются насекомыми в
качестве посадочной площадки. Два сросшихся лепестка назы-
ваются лодочкой, которая представляет собой защитный фут-
ляр, предохраняющий тычинки и пестик от посещения мел-
кими насекомыми, поедающими пыльцу цветка. Тычинок
может быть десять сросшихся,
десять свободных или девять
сросшихся и одна свободная,
лежащая в этом же круге. Плод
монокарпий — боб, одно-, дву-
или многосеменной. Опыле-
ние перекрестное, в основном
крупными насекомыми — пче-
лами и шмелями, но есть и са-
моопыляющиеся виды (горох).
После злаков бобовые —
важнейшая по практической
значимости для человека груп-
па растений. В семенах бобо-
вых содержится много запасно-
го белка, крахмала и жирного
масла, что определяет их боль-
шую пищевую и кормовую
ценность (соя, фасоль, горох,
арахис). Лучшие кормовые
растения — это виды люцер-
ны, клеверов. Лекарственные
виды — кассия (александрийс-
кий лист, употребляющийся
Рис. 4.58. Строение
венчика у бобовых:
1 — парус; 2 — весло;
3 — лодочка
89
как слабительное средство), софора японская (источник для
получения рутина — флавоноида, обладающего Р-витаминной
активностью).
Семейство крестоцветные (капустные). Семейство, насчиты-
вающее около 300 видов. Распространены на всех континентах
мира, но преимущественно в умеренных и холодных областях,
вплоть до Арктики. Крестоцветные — преимущественно одно-
летние и многолетние травы, реже кустарники и кустарнички.
Листья обычно очередные, цельные и расчлененные с перис-
тонервным жилкованием. Правильные обоеполые цветки со-
браны в моноподиальные соцветия, чаще это верхушечная
кисть или метелка. У разных представителей цветки имеют
одинаковое строение. Околоцветник двойной, чашечка состо-
ит из четырех свободных чашелистиков, а венчик — из четы-
рех свободных лепестков, располагающихся крестообразно
(отсюда и такое название семейства). Формула цветка:
*Ca4Co4A2+4Gi2). Тычинок шесть: две, более короткие, отно-
сятся к внешнему кругу, четыре, более длинные, — к внутрен-
нему кругу. Гинецей ценокарпный, сросшийся из двух плодо-
листиков. Завязь верхняя двухгнездная, с многочисленными
семязачатками (рис. 4.59). Плод ценокарпий — стручок или
стручочек.
Для крестоцветных характерно накопление гликозидов, ко-
торые при расщеплении дают ядовитое горчичное масло.
Опыляются насекомыми.
По своему значению для человека крестоцветные уступают
лишь злакам, бобовым и пасленовым. Некоторые представите-
Рис.4.59.
Строение цветка
крестоцветных:
1 — цветоложе; 2 —
нектарник; 3 — ты-
чинка; 4 — пестик
ли относятся к числу важнейших овощ-
ных культур (капуста огородная, репа,
редька, редис). В качестве лекарствен-
ного и пищевого растения используют-
ся горчица, желтушник. Большое коли-
чество видов являются злостными
сорняками.
Семейство пасленовые. Семейство
насчитывает 2900 видов. Пасленовые в
основном многолетние травы или полу-
кустарники, реже кустарники или даже
деревья (в тропиках). Листья простые,
цельные или разнообразно расчленен-
ные, как правило, очередные, всегда без
прилистников. Соцветия цимозные —
завиток, двойной завиток. Плод цено-
карпный — коробочка, ягода. Цветки
90
Рис. 4.60. Строение
цветка пасленовых
правильные, с двойным околоцветни-
ком. Чашелистики срастаются в пяти-
зубчатую чашечку, а срастающиеся
лепестки образуют трубчатый, коло-
кольчатый или колесовидный венчик.
Андроцей состоит из пяти тычинок,
прикрепленных к трубке венчика. Ги-
нецей ценокарпный, образован двумя
плодолистиками. Завязь верхняя, двух-
гнездная. Семязачатки многочислен-
ные. Формула цветка “Ca^Co^^jG^,
(рис. 4.60).
Пасленовые — перекрестноопыля-
ющиеся насекомыми растения, иногда
(у картофеля) наблюдается самоопыле-
ние.
Пасленовые — растения исключительной практической
ценности. Среди них первое место принадлежит картофелю.
Важное место занимают томаты, баклажан и др. Из лекар-
ственных видов белладонна является источником атропина;
также в медицине широко применяются препараты из дурма-
на, белены, скополин, табака, богатые алкалоидами.
Семейство сложноцветные. Огромное семейство, насчиты-
вающее около 20 000 видов. Встречаются везде, редки только
во влажных и тропических лесах. Большинство сложноцвет-
ных — многолетние или однолетние травы, но в тропиках
есть лианы, кустарники, небольшие деревья. Листья без при-
листников, простые, цельные или различным образом рас-
члененные. Листорасположение очередное, реже супротив-
ное. Цветки собраны в соцветия корзинки. В подавляющем
большинстве корзинки являются составной частью сложных
соцветий — сложных метелок, колосьев, щитков и др.
Различают четыре типа цветков, составляющих корзинки
(рис. 4.61):
трубчатые: *Ca0_K,Co(5)A(5)G(2), например, у пижмы;
язычковые. tCao_00Co(5)A(5)G(5), например, у одуванчика; -
ложноязычковые. tCao_wCO(j)AoG(5), например, краевые
цветки у ромашки;
воронковидные (бесполые): TCao_«,Co(6.9)AoG(o3)» например,
краевые цветки у василька.
Некоторые корзинки могут быть образованы только из
трубчатых (пижма) или только из язычковых цветков (одуван-
чик). Такие цветки, как ложноязычковые или воронковидные,
являясь недоразвитыми, располагаются по краям корзинки,
91
Рис. 4.61. Сложноцветные. Цветки различных типов:
а — трубчатые цветки василька синего (Z) и полыни горькой (2); б — г'
язычковые цветки одуванчика лекарственного (3) и цикория обыкновен- '?
кого (4 ); в — краевые ложноязычковые цветки подсолнечника однолетне-
го (5) и тысячелистника обыкновенного (6); г — краевые воронковидные .
цветки василька синего (7) и василька лугового (8)
привлекая своим видом насекомых, а в центре находятся труб-
чатые цветки, которые и дают плоды.
Андроцей обоеполых трубчатых и язычковых цветков состо-
ит из пяти тычинок. Нити тычинок, несросшиеся между собой, ’
прикреплены к трубке венчика, а пыльники срастаются своими
боковыми сторонами и образуют пыльниковую трубку, через
которую проходит столбик пестика. Гинецей псевдомонокарп-
ный, сросшийся из двух плодолистиков, образующих нижнюю
одногнездную завязь с единственным семязачатком, прикреп- ;
ленным ко дну завязи. Плоды псевдомонокарпии — сухие не- i
вскрывающиеся семянки, односеменные. Очень часто снабже- . '
ны хохолками из волосков чашечки. Семена без эндосперма. >
92 .
Большинство растений насе-
комоопыляемые. У одуванчика
наблюдается апомиксис.
Из пищевых растений можно
отметить подсолнечник, цико-
рий. Лекарственными видами
являются ромашка лекарствен-
ная, тысячелистник, календула
и др.
Класс однодольные
Семейство лилейные. Семей- Рис. 4.62. Строение
ство лилейные насчитывает цветка лилейных
около 470 видов. Лилейные —
многолетние травянистые луковичные растения. Цветоносный
стебель может быть с листьями и без них. Цветки правильные,
обоеполые, одиночные или собраны в соцветие кисть (рис. 4.62).
Листья простые с дуговым или параллельным жилкованием.
Околоцветник простой, венчиковидный, из шести сросшихся
или свободных листочков, расположенных в два круга. Тычинок
шесть, они также расположены в два круга. Гинецей ценокарп-
ный, из трех сросшихся плодолистиков. Завязь верхняя, с мно-
гочисленными семязачатками: *Р3+3Аз+зОхз). Плод — вскрыва-
ющаяся коробочка. Цветки'опыляются насекомыми. Среди
лилейных много декоративных растений, таких, как тюльпан,
лилия, гиацинт, аспарагус. Многие виды являются лекарствен-
ными растениями — ландыш майский, купена лекарственная.
В пищу употребляются лук (репчатый, батун, порей), чеснок,
спаржа и др.
Семейство злаки. К этому семейству принад лежит 11 000 ви-
дов. Среди злаков много однолетников, но преобладают много-
летние растения. Ветвление осуществляется близ основания,
где находится зона кущения. Стебель почти всех злаков — соло-
мина, членистая в узлах и полая в междоузлиях. Листья очеред-
ные, с листовым влагалищем, с параллельным жилкованием.
Листовые влагалища служат защитой междоузлий, сохраняю-
щих довольно долго способность к вставочному или интерка-
лярному росту. Цветки злаков обоеполые (рис. 4.63).
Раздельнополые цветки, например, у кукурузы, встречаются
редко. Соцветия — метелки, сложные колосья. Основой всех со-
цветий являются элементарные колоски. Типичный многоцвет-
ковый колосок состоит из оси, в основании которой находятся
верхняя и нижняя колосковые чешуи. В их пазухах нет цветков.
93
,5
Рис. 4.63. Строение
цветка злаков:
а — цветок;
б — диаграмма цветка:
1 — ось колоска; 2 — верхняя
цветковая чешуя; 3 — андроцей;
4—лодикулы; 5— верхняя цвет-
ковая чешуя; б — гинецей
Колосковые чешуи — это видоизмененные листья. Выше колос-
ковых чешуй на оси располагаются цветки. При основании цвет-
ка располагается верхняя и нижняя цветоносные чешуи. Выше
верхней цветковой чешуи, сросшейся из двух листочков, на оси
цветка располагаются две маленькие бесцветные чешуйки, назы-
ваемые лодикулами. Полагают, что это остатки околоцветника.
- Большинство злаков имеют три свободные тычинки. Гинецей
псевдомонокарпный, состоит из двух сросшихся плодолистиков
(один редуцируется по сравнению с лилейными). Формула цвет-
ка: t Рф+гАзСф. Плод псевдомонокарпий — зерновка, у которой
пленчатый околоплодник плотно прилегает к семени и иногда
срастается с семенной кожурой. Большую часть семени составля-
ет эндосперм. Большинство злаков ветроопыляемые растения.
Опыление перекрестное. Характерно вегетативное размножение
с помощью ползучих корневищ или укореняющихся побегов.
Основными представителями культурных злаков являются
рис, кукуруза, пшеница, ячмень. Кроме них широко возделы-
ваются также овес, рожь, просо, сорго, сахарный тростник.
К злаковым относится и много кормовых трав (тимофеевка,
лисохвост и др.). В субтропических и тропических странах как
строительный и поделочный материал широко применяются
бамбуки — своеобразные злаки с одревесневающим стеблем.
раздел 5
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК
РАЗВИТИЯ АНАТОМИИ
Начальный период развития анатомии своими корнями
уходит в далекое прошлое человечества. Среди наскальных
изображений встречаются рисунки, сделанные рукой перво-
бытного человека и представляющие животных, бывших пред-
метом его охоты. По этим рисункам можно судить о том, что
наши далекие предки хорошо знали, например, где располага-
ется сердце у мамонта или у бизона.
Изучение анатомии в древних культурах Востока началось с
незапамятных времен. В первую очередь это относится к Ки-
таю. Еще в третьем тысячелетии до н. э. в Древнем Китае была
написана книга под названием «Канон медицины». Судя по со-
держанию этой книги, в то далекое от нас время китайцам было
известно наличие в человеческом организме кровеносных сосу-
дов и внутренних органов. Сведения, изложенные в «Каноне
медицины», основывались на фактах, полученных при вскры-
тии трупов. В более поздней книге «Нейцзин» (XI—VIII вв.
до н. э.) имеются уже конкретные упоминания о сердце, пече-
ни, легких и других органах. В индусской книге «Аюр-Веда»
(«Знание жизни», VI в. до н. э.) описываются кости, мышцы,
связки, сосуды, нервы и другие анатомические структуры.
Позднее, в эпоху рабовладельческого строя, строение чело- ,
веческого тела стало предметом специального изучения, по-
видимому, в связи с тем, что некоторые из этих знаний были
необходимы при бальзамировании трупов, как это имело мес-
то в Древнем Египте. Об этом можно судить по тем мумиям,
которые сохранились до наших дней. Особенно интересен в
этом отношении «папирус Эберса», в котором изложены ана-
томические и медицинские знания древних египтян.
Наибольшие успехи в изучении анатомии в Древнем мире
были достигнуты в Античной Греции.
Среди первых известных истории науки ученых-анатомов
следует назвать Алкмеона Кротонского, который жил в первой
половине V в. до н. э. Он первым начал вскрывать трупы жи-
вотных для изучения строения их тела. Выдающимся является
его утверждение, что органы чувств непосредственно связаны с
мозгом и от него же зависят восприятия ощущений.
Гиппократ (ок. 460—377 до н. э.), знаменитый древнегре-
ческий врач, «отец медицины», оставил после себя многочис-
ленные сочинения по анатомии, физиологии и практической
медицине. Как анатом Гиппократ известен тем, что описал
некоторые кости черепа, указал на наличие швов между от-
96
дельными его костями и на наличие мышц. Он занимался так
же вопросами эмбриологии. Гиппократ, по-видимому, произ-
водил анатомические наблюдения только на животных, а по-
лученные результаты переносил на человека.
Аристотель (384—322 до н. э.), величайший философ и уче-
ный Древней Греции, писал, что все артерии идут от сердца, а
вены — к сердцу. Он отличал нервы от сухожилий, однако счи-
тал сердце, а не мозг центром умственной деятельности.
Герофил (род. в 304 до н. э.), живший в Александрии, про-
изводил анатомические исследования на трупах людей и жи-
вотных, применяя метод рассечения. Ему принадлежит целый
ряд открытий в области анатомии. Он различал двигательные
и чувствительные нервы, считал мозг органом мышления,
описал его оболочки и исследовал строение его отдельных об-
разований. Ему принадлежит первое описание продолговатого
мозга. Герофил указал на различие в строении артерий и вен.
Все это вместе взятое позволяет считать Герофила основате-
лем анатомии как самостоятельной науки.
Эразистрат (ок. 300 — ок. 240 до н. э.) — древнегреческий
врач, был анатомом и физиологом. Он исследовал поверхность
полушарий мозга и указал на наличие извилин и борозд, изу-
чал сокращение мышц и механику движения человека. Кроме
того, ему принадлежит описание строения двустворчатого и
трехстворчатого клапанов. Эразистрат открыл, что артерии и
вены соединяются между собой с помощью невидимых глазу
тонких кровеносных сосудов.
Клавдий Гален (ок. 130—201) был врачом школы гладиато-
ров в Риме. Оставил большое число сочинений. Анатомичес-
кие исследования Гален проводил на трупах животных. Он
описал многие мышцы и считал, что «без нерва нет ни одного
движения, называемого произвольным», Он различал отдель-
ные кости по их форме. Доказал, что при жизни животного в
его артериях течет не воздух, а кровь.
После Галена и до эпохи Возрождения анатомия и физио-
логия человека как наука практически не развивалась. В те-
чение целого тысячелетия врачи и анатомы эпохи Средне-
вековья занимались главным образом изучением сочинений
Гиппократа, Галена, Аристотеля, Герофила и их комментари-
ями, не прибавляя к ним почти ничего нового.
Абу Али-ибн Сина (Авиценна, 980—1037) написал «Канон
врачебной науки» — итог взглядов и опыта греческих, рим-
ских, индийских и среднеазиатских врачей, который в течение
нескольких веков был обязательным руководством, в том чис-
ле и в средневековой Европе. Авиценне принадлежит первое
4-281
97
описание мышц глаза. Он указал на то, что сетчатка глаза яв-
ляется его световоспринимающим образованием.
Леонардо да Вин*» (1452-1519), выдающийся итальянский г
ученый и художник эпохи Возрождения, сделал многочислен- '
ные зарисовки костей, мышц, сердца и других органов с пись- ;
менными пояснениями к ним. Он составил классификацию
мышц, объяснил их работу с точки зрения законов механики.
Андрей Везалий (1514—1564), профессор Падунского уни-
верситета, на основании собственных наблюдений, сделанных
при вскрытии трупов, написал труд «О строении человеческо-
го тела» (1543), где были обстоятельно описаны многие органы
человеческого тела. Его считают основоположником описа-
тельной анатомии.
Вильям Гарвей (1578—1657) открыл кровообращение и навсег-
да остался знаменит в истории науки благодаря своему сочине-
нию «Анатомическое изыскание о движении сердца и крови у
животных» (1628). Он положил начало физиологии как самосто-
ятельной науке, отделившейся в дальнейшем от анатомии.
Отечественная анатомия развивалась под воздействием
положений и взглядов Галена. В древнерусских рукописях X—
XIII вв. имеются некоторые анатомические сведения о строе-
нии органов («Церковный устав», X в., «Изборник Святосла-
ва», XI в., «Русская правда», XI—XII вв.).
В XV—XVI вв. в России в книгах «Врата Аристотелевы» и
«Тайна Тайных» были изложены наряду с теоретическими воп-
росами медицинские представления античных авторов об ана-
томии, их обсуждение и краткое описание телосложения чело-
века. В 1658 г. Епифаний Славинецкий перевел с латинского
языка труды Везалия задолго до их перевода на другие языки
(на немецкий — лишь в 1781 г.).
М. И. Шеин (1712—1762) известен тем, что издал первый
анатомический атлас на русском языке (1744).
С. Г. Зыбелин (1735—1802), профессор Московского уни-
верситета и академик, известен своими трудами «Слово о дей-
ствии воздуха в человеке» и «Слово о сложении тела человечес-
кого и о способах, как оные предохраняют от'болезней».
П. А. Загорский (1764—1846) создал новый оригинальный
учебник анатомии «Руководство к познанию человеческого
тела», выдержавший пять изданий. Он является основателем
первой крупной русской научной анатомической школы.
И. В. Буяг&ский (1789-1866) — профессор анатомии Меди-
ко-хирургической Академии, ученик П. А. Загорского, автор
анатомического атласа «Таблицы хирургической анатомии».
Предложил метод бальзамирования трупов.
98
Е. О. Мухин (1766—1850), профессор Московского универ-
ситета, известен как автор оригинального учебника по анато-
мии на русском языке. В числе учеников Е. О. Мухина был
Н. И. Пирогов.
И. М. Сеченов (1816-1872) — профессор анатомии и хирур-
гии Московского университета, автор «Анатомо-хирургичес-
ких таблиц». Значительно обогатил коллекцию анатомических
препаратов Московского университета и первым начал вести
преподавание анатомии на русском языке.
Н. И. Пирогов (1810-1881) высоко поднял уровень анатоми-
ческих знаний в России и за границей. Он явился основополож-
ником топографической анатомии, развившейся в самостоятель-
ную науку. Н. И. Пирогов известен также как замечательный
хирург, много сделавший в области военно-полевой хирургии,
первый применивший гипсовую повязку и эфирный наркоз во
время войны на Кавказе и Крымской кампании.
Д. Н. Зернов (1843—1917) — профессор Московского уни-
верситета, автор многих исследований по анатомии органов
чувств, центральной нервной системы и внутренних органов.
По его книге «Руководство описательной анатомии человека»
изучали анатомию многие поколения русских врачей.
В. М. Бехтерев (1857—1927), выдающийся невропатолог,
психиатр и анатом. Своими замечательными сочинениями
«Проводящие пути спинного и головного мозга», «Учение о
функциях мозга», «Рефлексология» и др. много сделал для рас-
ширения знаний по анатомии и физиологии центральной не-
рвной системы.
И. М. Сеченов (1829—1905) является «отцом русской физи-
ологии», развившей научные представления о физиологии не-
рвной системы, дыхания, утомления и др. В работе «Рефлексы
головного мозга» (1866) изложил свои взгляды на природу
произвольных движений и психических явлений.
И. П. Павлов (1849—1936) — гениальный физиолог, кото-
рый своими трудами способствовал развитию не только фи-
зиологии, но и всех биологических и медицинских наук. Он
создал учение об условных рефлексах и современную физиоло-
гию высшей нервной деятельности.
Н. Е. Введенский (1852—1922) — крупнейший представи-
тель отечественной физиологии. Первым обнаружил в мышцах
явления оптимума и пессимума и частые колебания биотоков.
Он разработал оригинальную теорию о физиологической при-
роде возбуждения и торможения, сформулировал понятие о
лабильности нервов и мышц, развил представление о пара-
биозе.
4*
99
В. П. Воробьев (1876-1937)— известный анатом, исследо-
ватель вегетативной нервной системы. Он создал пятитомный
«Атлас анатомии человека».
В. Н. Тонков (1872—1954) — профессор Военно-медицин-
ской академии им. С. М. Кирова, автор известного руковод-
ства по анатомии. В своих исследованиях сочетал методы ана-
томического и макромикроскопического изучения органов.
А. А. Ухтомский (1875—1942) — крупнейший физиолог, от-
крывший закон доминанты в деятельности нервной системы.
В своей книге «Физиология двигательного аппарата» привел
обширный материал, непосредственно относящийся к функ-
циональной анатомии скелета и мышц человеческого тела и к
изучению его движений.
Л. А. Орбели (1882—1958) — ученик И. П. Павлова, один из
создателей нового направления в физиологической науке —
эволюционной физиологии. Он внес важный вклад в физиоло-
гию мышечной деятельности, симпатической нервной систе-
мы, мозжечка и органов чувств.
Д. А. Жданов (1908-1971) — профессор кафедры анатомии
1-го Московского медицинского института (с 1956). Известен
своими работами по анатомии лимфатической нервной систе-
мы. В своих исследованиях сочетал методы анатомического;
микроскопического и электронно-микроскопического изуче-
ния органов.
П. К. Анохин (1898-1974) — ученик И. П. Павлова, профес-
сор кафедры физиологии 1-го Московского медицинского ин-
ститута. Разрабатывал многие вопросы нейрофизиологии и
высшей нервной деятельности. Он выдвинул концепцию о си-
стемной деятельности нервной системы.
Во второй половине XX в. в отечественную анатомию вне-
дряется ряд новых современных методов. Успешно использу-
ется микроскопическая и ультромикроскопическая техника,
авторадиография, рентгеновский и биометрический методы.
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Опорно-двигательный аппарат человека состоит из двух
компонентов — костей и мышц. Скелет и его соединения яв-
ляются пассивной частью аппарата, а мышцы с их вспомога-
тельными образованиями (фасциями, влагалищами сухожи-
лий и др.) — активной частью.
ПАССИВНАЯ ЧАСТЬ
ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
(УЧЕНИЕ О КОСТЯХ - ОСТЕОЛОГИЯ)
Строение и классификация костей
Значение скелета. Скелет человека имеет следующие функ-
ции:
1) механическую, связанную со способностью костной сис-
темы осуществлять: а) опорное; б) двигательное; в) защитное
действие.
Опорное действие осуществляется прикреплением мягких
тканей и органов к различным частям скелета, двигательное —
за счет строения костей в виде д линных и коротких рычагов, со-
единенных подвижными сочленениями и перемещающихся в
результате сокращения мышц, защитное — образованием от-
дельными костями костного канала (позвоночного), предохра-
няющего спинной мозг, костной коробки (черепа), защищаю-
щей головной мозг и т. д.;
2) метаболическую (биологическую), связанную с участием
костей в обмене веществ, особенно в минеральном обмене (это
депо фосфора, кальция, железа и др.);
3) кроветворную, осуществляемую в костях.
Химический состав кости. Кость состоит из оссеинами
неорганических веществ, главным образом фосфорнокис-
лого кальция. Эластичность зависит от оссеина, а твер-
дость — от минеральных солей.
Строение костной ткани. Кость построена из остеонов (рис. 5.1).
Они являются структурными элементами кости, видимыми под
лупой или под малым увеличением микроскопа. Из остеонов
формируются более крупные образования кости, видимые уже
невооруженным глазом — перекладины костного веществами
балки. Клетки, вырабатывающие костное вещество, называют
остеобластами. Клетки костной ткани — остеоциты,
находятся в полостях межклеточного вещества. Остеоклас-
ты — пожиратели костного вещества — костные макрофаги.
Вещество кости. 1. Компактное вещество образуют плотно
прилегающие друг к другу перекладины и балки. 2. Губчатое
вещество представляет собой перекладины и балки, между ко-
торыми имеется пространство, заполненное костным мозгом.
Классификация костей. Различают три вида костей:
1) трубчатые’, а) длинные трубчатые кости (плечо, кости
предплечья, бедра); б) короткие трубчатые кости (фаланги
пальцев, кости плюсны и пясти);
101
a
Рис. 5.1. Костная ткань:
а — строение
под микроскопом
(7 — система пластинок остео-
на; 2 — канал остеона; 3 — си-
стема вставочных пластинок);
б — схема
(в левой половине изображены
костные полости и канальцы, ч
правой — направление волокон
в отдельных пластинках)
102
2) губчатые: а) длинные губчатые кости (ребра;; б; корот
кие губчатые кости (позвонки, кости запястья и предплюс-
ны):
3) плоские (рис. 5.2): а) кости черепа; б) кости поясов ко-
нечностей.
Строение кости. Рассмотрим строение кости на примере
трубчатой (рис. 5.3).
1. Надкостница покрывает всю кость (кроме суставных по-
верхностей). Она представляет собой соединительно-ткан-
ную оболочку, состоящую из двух слоев — наружного (волок-
нистого) и внутреннего (костеобразующего). Надкостница
богата кровеносными сосудами и нервами, дающими ответ-
вления внутрь кости. Вследствие наличия в надкостнице чув-
Рис. 5.2. Череп (вид сбоку):
А — венечный шов; Б — чешуйчатый шов; В — ламбдовидный шов; 1 —
височная линия (лобной кости); 2 — лобная кость; 3 — верхняя височная
линия; 4 — нижняя височная линия; 5 — теменная кость; 6 — затылочная
кость; 7 — чешуя височной кости? 8 — сосцевидный отросток; 9 — наруж-
ный слуховой проход; 10 — скуловая дуга; 11 — нижняя челюсть; 12 —
подбородочное отверстие; 13 — верхняя челюсть; 14 — скуловая кость;
75— слезная кость; М— носовая кость; 77— большое крыло клиновид-
ной кости
103
Рис. 5.3. Трубчатая кость:
1 — надкостница; 2 — вещество кости,
свободное от надкостницы; 3 — диафиз;
4 — эпифиз
Рис. 5.4. Строение кости:
1 — губчатое вещество; 2 — головка е
трубчатой кости; 3 — плотное веще-
ство; 4 — надкостница; 5 — костный
мозг; 6 — средняя часть трубчатой
кости
104
ствительных окончаний нервов кость при ушибе болезненна.
Надкостница выполняет ряд функций: а) защитную; б) тро-
фическую (питательную); в) нерворегуляторную и г) костеоб-
разовательную. За счет клеток внутреннего слоя надкостницы
(веретеновидные клетки) происходит рост кости в толщину и
срастание при переломах. Внутри кости находится костный
мозг (рис. 5.4). Он бывает красный и желтый. Красный кост-
ный мозг является кроветворным органом. Он вырабатывает
красные кровяные тельца и все зернистые формы белых кро-
вяных телец. У новорожденного есть только красный кост-
ный мозг. У взрослого человека он находится в губчатом ве-
ществе костей (в эпифизах трубчатых костей, в позвонках,
ребрах, грудине, тазовых и костях черепа). Желтый костный
мозг находится в полостях диафизов трубчатых костей. Он
богат жировыми клетками.
2. Диафиз — трубка кости.
3. Эпифизы — утолщенные концы кости с суставными по-
верхностями, покрытыми*суставным хрящом.
4. Апофизы — бугорки (места прикрепления мышц и сухо-
жилий) (рис. 5.5).
Соединения костей. Различают следующие соединения кос-
тей: 1) непрерывные; 2) полусуставы; 3) прерывные (рис. 5.6).
Рис. 5.5. Бедренная кость
на распиле:
1 — эпифиз; 2 — метафиз;
3 — апофиз; 4 — губчатое ве-
щество; 5 — диафиз; 6 — ком-
пактное вещество; 7 — кост-
номозговая полость
105
1. Синартрозы это непрерывные соединения. При этом г
возможны: а) синдесмозы — соединения костей посред-
ством волокнистой соединительной ткани (связки, сухожиль-
ные мембраны, .мембраны родничков у детей); б) синхонд-
розы — соединения костей посредством хрящевой ткани
(между грудиной и ребрами); в)синостозы — соединения
костей посредством костной ткани (кости черепа у взрослых
людей).
2. Полусуставы — это хрящевые соединения, имеющие в
толще хряща щелевидную полость. Отличаются от суставов от-
сутствием суставных сумок и суставных поверхностей.
3. Прерывные сочленения — диартрозы, или суставы.
Элементарный сустав включает следующие анатомические
элементы (см. рис. 5.6, в):
1) суставные поверхности костей, покрытые суставным хря-
щом:
2) суставную полость, заполненную синовиальной жидко-
стью, уменьшающей трение;
3) суставную сумку.
Суставы подразделяются: 1) по числу суставных поверхнос-
тей и 2) по форме суставной поверхности.
По числу суставных поверхностей суставы
подразделяются следующим образом:
Рис. 5.6. Типы соединения костей.
а — посредством швов; б — посредством хряща; в — сустав:
1—3 — швы между костями черепа; 4 — позвонки; 5 — эзрящевые прослойки;
6. 7— суставные поверхности; 8 — полость сустава; 9 — надкостница; 10 —
суставная сумка
106
простой сустав имеет только две суставные поверхности
(например, межфаланговый сустав);
сложный сустав имеет три и более суставные поверхности
(например, локтевой сустав);
комбинированный сустав представляет собой два и более
анатомически самостоятельных сустава, функционирующих
одновременно (например, височно-нижнечелюстные суста-
вы);
комплексный сустав — это сустав, внутри которого между
сочленяющимися поверхностями имеется диск или мениски,
которые делят сустав на две камеры (например, коленный сус-
тав).
По форме суставной поверхности суставы
подразделяются следующим образом (рис. 5.7):
одноосные суставы: а) цилиндрический, или колесовид-
ный, сустав (между 1-м и 2-м шейными позвонками); б) бло-
ковидный сустав (межфаланговые сочленения пальцев);
лвуосные суставы: а) эллипсоидный, или яйцевидный, сустав
(лучезапястный); б) седловидный сустав (запястно-пястный сус-
тав первого пальца); в) мыщелковый сустав (каленный и атланто-
затылочный);
многоосные суставы: а) шаровидный (плечевой); б) плос-
кий (межпозвонковый).
Рис. 5.7. Различные формы суставов (схема):
а — шаровидный; S — эллипсоидный, или яйцевидный;
в — седловидный; г — плоский; д — блоковидный;
е— цилиндрический, или колесовйдный
107
Строение скелета
Скелет человека состоит из скелета туловища, скелета верх-
них и нижних конечностей и черепа.
К скелету туловища (рис. 5.8) относятся позвоночник и
грудная клетка.
Позвоночник делится на пять отделов (рис. 5.у):
1) шейный (7 позвонков);
2) грудной (12 позвонков);
3) поясничный (5 позвонков);
4) крестцовый (крестец из 5 слившихся позвонков);
5) копчиковый (копчик из 4—5 сросшихся позвонков).
Общее число позвонков 33—34.
Позвонки относят к коротким губчатым костям. Каждый
позвонок имеет тело (рис. 5.10), обращенное вперед, и
дугу, расположенную сзади от тела. Дуга и тело позвонка
ограничивают позвоночное отверстие.
Тело позвонка с наружной поверхности покрыто плотным
костным веществом, а внутри содержит губчатое вещество, со-
стоящее из пластинок, вертикально и горизонтально направ-
ленных и расположенных почти под прямым углом друг к дру-
гу. Своей верхней и нижней поверхностями тела позвонков с
помощью межпозвоночных дисков соединяются с выше- и ни-
жележащими позвонками. Передняя поверхность тела позвон-
ка так же, как боковые и задняя, несколько вогнутая.
Позвоночные отверстия, располагаясь одно над другим, в
совокупности образуют позвоночный канал, в котором
находится спинной мозг и связанные с ним образования. Дуга
позвонка у места прикрепления к телу имеет снизу и сверху
вырезки. Вырезки ниже и выше лежащего позвонка образу-
югмежпозвоночное отверстие, через которое про-
ходят нервы, кровеносные и лимфатические сосуды. От дуги
позвонка отходят семь отростков. Непарный, обращенный на-
зад отросток называется остистым. Он служит для при-
крепления связок и мышц. Остальные отростки — парные.
К ним относятся поперечные отростки, направлен-
ные в стороны от позвонков и лежащие приблизительно во
фронтальной плоскости, а также верхние и нижние
суставные отростки. Поперечные отростки служат для
прикрепления мышц и связок, а суставные — для соединения
с такими же отростками выше- и нижележащих позвонков.
Размеры позвонков неодинаковы. Чем больше нагрузка, тем
больше размеры позвонков. Максимальные размеры имеют
поясничные и первые крестцовые, которые воспринимают ос-
108
1
б
а
Рис. 5.8. Скелет человека.
a — спереди; б — сзади:
1 — череп; 2 — ключица; 3 — лопатка; 4 — грудная клетка; 5 — плечевая
кость; 6 — ребра; 7 — позвоночник; 8 — кости таза; 9 — кости предплечья;
10 — кости кисти; 11 — бедренная кость; 12 — кости голени; 13 — кости
стопы
109
Шейные позвонки
Грудные позвонки
Поясничные позвонки
Сросшиеся крестцовые
.позвонки
Копчиковые позвонки
Рис. 5.9. Строеьжетозвоночника (я) и отдельного позвонка (5)
Рис. 5.10. Строение позвонков.
а — шейного; б — грудного; в — поясничного:
1 — дуга; 2 — отросток; 3 — позвоночный канал; 4 — тело
новную нагрузку. Самые небольшие позвонки — копчиковые»
что связано с уменьшением приходящейся на них нагрузки и
редукцией хвостовой мускулатуры. Позвонки разных отделов
позвоночного столба имеют свои особенности, позволяющие
отличать их друг от друга.
Грудная клетка. Скелет грудной клетки образуется сзади из
грудных позвонков с их соединениями и связками, а также
НО
задних частей ребер. С боков грудная клетка представлена реб-
рами. Спереди ома состоит из передних концов ребер, ребер-
ных хрящей и грудины.
У человека имеется 12 пар ребер, из которых первые
семь, называемые истинными ребрами, переходят в реберные
хрящи, соединенные с грудиной. Следующие три пары ребер
(8-, 9-, 10-я) имеют хрящи, недостигающие грудины и соеди-
няющиеся с хрящом вышерасположенного ребра (рис. 5.11).
Эти ребра называют ложными. Две последние пары (11-я и
12-я) не имеют реберных хрящей и не соединяются передни-
ми свободными концами ни с грудиной, ни с другими ребра-
ми, вследствие чего оказываются наиболее подвижными. Эти
ребра называют блуждающими, или колеблющимися.
Грудина является непарной костью, напоминающей по
форме кинжал, в котором различают: рукоятку, тело и мече-
видный отросток.
16
Рис. 5.11. Строение грудной клетки:
1-7— истинные ребра; 8-10 — ложные ребра; 11, 12 — колеблющиеся
ребра; 13 — рукоятка грудины; 14 — тело грудины; 15 — мечевидный отро-
сток; М — Т грумюй позвонок; 17 — хрящевые част* ребер
111
Череп. Скелет головы (рис. 5.12) состоит из двух отделов:
мозгового и лицевого черепа. Лицевой отдел черепа образует
костную основу лица, а также дает опору начальным отделам
дыхательной и пищеварительной систем. Мозговой череп об-
г разует вместилище для головного мозга.
К лицевому отделу черепа относятся:
1) верхнечелюстная кость — парная кость, кото-
рая составляет большую часть лицевого черепа, участвует в об-
разовании глазницы, носовой и ротовой полостей, а также
подвисочной и крылонебной ямок;
2) скуловая кость — парная, наиболее крепкая
кость, которая имеет четырехугольную форму и определяет
форму средней части лица;
3) небная кость — парная кость из двух пластинок,
составляющая задний отдел лицевого черепа и участвующая в
образовании костного неба, боковой стенки носовой полости
и глазницы;
4) носовая кость — парная кость в виде желоба. Она
образует скелет спинки носа;
5) слезная кость — парная кость, располагающаяся в
области переднего отдела внутренней стенки глазницы.
Сверху она соприкасается с лобной костью, а снизу — с телом
верхней челюсти. Принимает участие в образовании носослез-
ного канала;
6) нижняя носовая раковина — парная кость,
расположенная на боковой стенке носовой полости;
7) сошник — непарная кость, образующая нижнюю
костную часть перегородки носовой полости;
8) нижняя челюсть — непарная крепкая толстая
' кость, сочлененная справа и слева с височной костью;
9) подъязычная к о с т ь — непарная кость, по внеш-
нему виду напоминающая подкову. Она находится сзади и не-
сколько ниже нижней челюсти, в передней поверхности шеи.
Мозговой отдел черепа состоит из крыши и основания. Ко-
сти плоские, внутренняя их поверхность имеет углубления и
выступы, соответствующие извилинам и бороздам наружной
поверхности головного мозга. Кроме того, на внутренней по-
верхности костей черепа есть борозды, в которых проходят ар-
терии твердой мозговой оболочки.
К мозговому отделу черепа относятся:
1) затылочная кость (непарная), принимающая уча-
стие в образовании основания черепа и заднего отдела его кры-
ши. В затылочной кости имеется большое отверстие, через ко-
торое проходит продолговатый мозг и позвоночные артерии;
112
1
2
8
Рис. 5.12. Скелет головы:
1 — теменная кость; 2 — кост-
ный шов; 3 — лобная кость; 4 —
носовая кость; 5 — верхне-
челюстная кость; 6 — нижнече-
люстная кость; 7 — скуловая
кость; 8 — височная кость; 9 —
затылочная кость
113
2) клиновидная кость (непарная), тело которой я
имеет кубовидную форму. Она содержит воздухоносную пазу- *
ху, сообщающуюся спереди с носовой полостью. На верхней
поверхности пазухи есть углубление — турецкое седло, где рас- |
полагается гипофиз. Турецкое седло с боков имеет борозды, в #
которых залегают внутренние сонные артерии. Через зритель- i
ный канал клиновидной кости проходят зрительный нерв и
глазная артерия. Через многочисленные отверстия проходят |
ветви тройничного нерва, а также глазодвигательный, боковой |
и глазничные нервы; |
3) лобная (непарная), участвующая в образовании кры- |
ши и основания черепа. Эта кость имеет лобную часть, глаз- |
ничные и носовую частя; |
4) решетчатая кость (непарная), легкая и хрупкая, |
состоящая из двух пластинок: решетчатой пластинки и решет- |
чатого лабиринта. Решетчатая пластинка располагается гори- |
зонтально в решетчатой вырезке лобной кости. Она имеет |
множество отверстий, а в средней плоскости от нее отходит 1
обращенный кверху костный выступ — петушиный гребень, к |
которому прикрепляется отросток твердой мозговой оболоч- |
ки. Через отверстие решетчатой пластинки проходят из носо- |
вой полости в полость черепа обонятельные нервы; |
5) теменная кость (парная), составляющая цент- |
ральную часть свода черепа. На ее выпуклой поверхности есть 1
возвышение — теменной бугор, легко прощупываемый под ко- ;
жей. Внутренняя (вогнутая) поверхность теменной кости, об- "j
ращенная в полость черепа, имеет артериальные и синусои- *
дальние борозды, являющиеся местом кровеносного синуса ;
твердой мозговой оболочки; I
6) височная кость (парная), принимающая участие в i
образовании основания черепа и отчасти его крыши. Кость со-
стоит из трех частей: каменистой (или пирамиды), чешуйча-
той и барабанной. В каменистой части располагается костный
лабиринт и формируется внутреннее ухо. Через каменистую
часть височной кости проходят лицевой и преддверно-улитко-
вые нервы; здесь внутренняя сонная артерия входит в полость
черепа. Чешуйчатая часть височной кости принимает участие
в образовании крыши черепа. От нее отходит скуловой отрос-
ток, образующий вместе с височным отростком скуловой кос-
ти легко прощупываемую скуловую дугу. У основания этого от-
ростка находится суставной бугор для сочленения с головкой
нижней челюсти. Барабанная часть височной кости представ-
ляет собой изогнутую пластинку, которая снизу и спереди ог-
раничивает наружный слуховой проход.
114
Скелет верхней конечности подразделяется на две частя:
1) скелет пояса верхней конечности и 2) скелет свободной
верхней конечности.
К скелету пояса верхней конечности относятся лопатки и
ключицы.
Лопатка представляет собой плоскую треугольной фор-
мы кость, расположенную на задней поверхности туловища.
Боковой край лопатки утолщен и имеет суставную впадину,
которая служит для соединения с головкой плечевой кости.
Ключица представляет собой S-образно изогнутую по
длинной оси кость. Она располагается горизонтально спереди
и сверху трудной клетки на границе шеи и легко прощупывает-
ся на всем своем протяжении. Ключица имеет д ва конца —
грудинный и акромиальный. Первый утолщен, имеет сустав-
ную поверхность с грудиной. Второй также утолщен и соеди-
няется с акромионом (акромион — выступающая кнаружи бо-
ковая часть лопатки). Ее используют при измерении ширины
плеч (портные). Функция ключицы заключается в том, что
она способствует укреплению положения лопатки, удерживая
плечевой сустав в некотором отдалении от грудной клетки.
Пояс верхних конечностей соединяет свобод ную верхнюю ко-
нечность с туловищем, служит местом прикрепления мышц и
увеличивает амплитуду движения верхних конечностей.
Скелет свободней верхней конечности состоит из трех отделов.
1. Плечевая кость является типичной длинной труб-
чатой костью. На проксимальном (верхнем) конце плечевой кос-
ти различают головку, которая обращена к лопатке и имеет сус-
тавную поверхность. Головка кости отд елена от остальной части
анатомической шейкой, по краю которой прикрепляется сумка
плечевого сустава. Наиболее суженное место плечевой кости
ниже анатомической шейки называется хирургической шейкой,
так как здесь часто происходят переломы. Дистальный (нижний)
отдел плечевой кости образует мыщелок, суставная поверхность
которого служит для соединения с костями предплечья.
2. Скелет предплечья образуют локтевая и лучевая
кости.
Локтевая кость — трубчатая, трехгранной формы. Прокси-
мальный конец ее утолщен и имеет блоковидную вырезку для
соединения с плечевой костью. Дистальный конец представ-
ляет головку локтевой кости. С боковой стороны головка обра-
зует суставную поверхность для сочленения с лучевой костью,
а снизу — с треугольным хрящом.
Лучевая кость является длинной трубчатой костью. В отли-
чие от локтевой кости у лучевой утолщен дистальный конец.
115
Проксимальный конец имеет головку, на верхней поверхности
которой находится ямка для соединения с головкой мыщелка
плечевой кости. А по краю располагается суставная поверх-
ность для соединения с локтевой костью.
3. Скелет к и с т и делят на три отдела:
а) запястье — восемь мелких костей неправильной формы,
расположенных в два ряда (рис. 5.13). Они относятся к корот-
ким губчатым костям;
б) пясть состоит из пяти коротких трубчатых костей. Пяст-
ная кость I пальца короче остальных, но массивнее. Каждая
пястная кость имеет основание, тело и головку;
в) кости пальцев — фаланги.'Это короткие трубчатые кос-
ти. Каждый палец состоит из трех фаланг: проксимальной,
Рис. 5.13. Кости правой кисти (вид с тыльной стороны):
1 — головчатая кость; 2 — ладьевидная кость; 3 — трапециевидная кость;
4 — кость трапеции; 5 — пястная кость (II); 6 — проксимальная фаланга;
7— средняя фаланга; 8 — дистальная фаланга пальца; 9 — головка V пяст-
ной кости; 10 — тело; 11 — основание; 12 — крючковатая кость; 13 —
гороховидная кость; 14 — трехгранная кость; 15 — полулунная кость
116
средней и дистальной. Исключение составляет первый палец,
имеющий только две фаланги: проксимальную и дистальную.
Суставные поверхности дистальных и средних фаланг, а также
суставная поверхность головки проксимальной фаланги имеют
блоковидную форму. Суставная поверхность основания про-
ксимальных фаланг соединяется с головкой пястной кости су-
ставом шаровидной кости.
Кроме указанных костей кисть еще имеет сесамовидные ко-
сточки. Они находятся в толще сухожилий мышц (чаще всего
идущих к большому и указательному пальцам со стороны ла-
донной поверхности кисти) и увеличивают плечо силы тех
мышц, которые к ним прикрепляются.
Скелет нижней конечности подразделяется на две части:
1) скелет пояса нижней конечности и 2) скелет свободной
нижней конечности.
Скелет пояса нижней конечности образуют кости тазового
пояса и крестец.
Тазовый пояс состоит из двух тазовых костей, кото-
рые образуются за счет срастания подвздошной, седалищной
и лобковой костей. Эти кости участвуют в образовании верт-
лужной впадины, которая служит для соединения таза с го-
ловкой бедра. Каждая из этих костей закладывается как само-
стоятельная, но в 14—16 лет они срастаются между собой.
Тазовые кости относят к плоским костям. Сзади они соединя-
ются с крестцом, а спереди обе тазовые кости соединяются
друг с другом лобковым симфизом (полусустав), образуя таз
(рис. 5.14).
Крестец состоит из пяти сросшихся позвонков; участву-
ет в образовании таза.
К костям свободной нижней конечности относят бедренную,
кости голени и кости стопы.
Бедренная кость — наиболее длинная трубчатая
кость, имеющая цилиндрическую форму и несколько изогну-
тая впереди. На проксимальном конце бедренной кости голов-
ка несет суставную поверхность для сочленения с вертлужной
впадиной. Головка соединяется с телом кости шейкой, ось ко-
торой по отношению к продольной оси тела бедренной кости
располагается приблизительно под углом 130°. Дистальный
конец бедренной кости расширен в два мыщелка. Мыщелки
бедра имеют суставные поверхности для сочленения с боль-
шой берцовой костью и надколенником.
Надколенник находится спереди дистального эпифиза
бедренной кости. По форме это двояковыпуклая линза. Надко-
ленник является наиболее крупной сесамовидной костью.
117
1
Рис. 5.14. Мужской таз (вид сверху):
1 — подвздошно-поясничная связка; 2 — место расположения крестцово-
подвздошного сустава; 3 — большое седалищное отверстие; 4 — малое седа-
лищное отверстие; 5 — лобковый сустав (лонное сращение); 6 — погранич-
ная линия; 7 — крестцово-бугорная связка; 8 — крестцово-остная связка
Большая берцовая ко ст ь.расположена с внутрен-
ней стороны голени (со стороны большого пальца стопы).
Проксимальный ее конец расширен и образует два мыщелка.
На мыщелках сверху находятся суставные поверхности, служа-
щие для соединения с мыщелками бедра, а между ними распо-
лагается межмыщелковое возвышение, к которому фиксиру-
ются крестообразные связки коленного сустава. Тело большой
берцовой кости трехгранной формы. Дистальный отдел боль-
шой берцовой кости книзу имеет выступ — медиальную ло-
дыжку; кнаружи — вырезку для соединения с малой берцовой
костью. Снизу большая берцовая кость образует суставную по-
верхность для соединения со стопой.
Малая берцовая кость — длинная трубчатая
кость, расположенная снаружи голени. Проксимальный конец
заканчивается головкой, которая соединяется с большой бер-
цовой костью, а дистальный конец образует латеральную ло-
дыжку. Дистальная голень переходит в стопу.
Стопа (рис. 5.15) состоит из трех частей: а) предплюсны;
б) плюсны и в) пальцев. К костям предплюсны относят: таран-
ную, пяточную, ладьевидную, кубовидную и три клиновидные.
Все они являются короткими губчатыми костями. К костям
плюсны относят пять коротких трубчатых плюсневых костей.
118
Рис. 5.15. Кости правой стопы
(вид сверху):
/ — дистальная фаланга V паль-
ца; 2 — средняя фаланга V паль-
ца; 3 — проксимальная фаланга
V пальца; 4 — линия, проходя-
щая по суставной щели сустава
Лисфранка (совокупность пред-
плюсне-плюсневых суставов);
5 — кубовидная кость; 6 — S-об-
разная линия, проходящая по су-
ставной щели сустава Шопара
(совокупность таранно-ладье-
видного и пяточно-кубовидного
суставов); 7 — пяточная кость;
8 — таранная кость; 9 — ладье-
видная кость; 10, 11, 12 — лате-
ральная, промежуточная и меди-
альная клиновидные кости; 13 —
первая плюсневая кость
Пальцы состоят из фаланг, как и на кисти, I палец имеет две
фаланги, а остальные по три. Нередко две фаланги V пальца
срастаются между собой, поэтому в них по две фаланги.
Особенности строения скелета человека
в связи с прямохождением и трудовой деятельностью
1. Череп «насажен» сверху, а не подвешен спереди к позво-
ночнику, как у животных.
2. Позвоночный столб имеет изгибы.
3. Масса позвонков от шейного отдела к поясничному уве-
личивается.
4. Уплощенная грудная клетка.
119
5. Чашеобразный таз.
6. Массивные нижние конечности.
7. Сводчатая стопа.
8. Преобладание мозгового отдела черепа над лицевым.
9. Появление подбородочного выступа.
10. Совершенное противопоставление большого пальца х
всем остальным.
а
АКТИВНАЯ ЧАСТЬ ОПОРНО-ДВИГАТЕЛЬНОГО АППАРАТА
(УЧЕНИЕ О МЫШЦАХ - МИОЛОГИЯ)
Функции, строение и классификация мышц
Активной частью опорно-двигательного аппарата являются <
мышцы с их вспомогательными образованиями (фасциями, :
влагалищами, сухожилиями). <
Строение мышечной ткани. По строению мышечную ткань ’
делят на гладкую, поперечно-полосатую (скелетную) и сердеч-
ную поперечно-полосатую.
Гладкая мышечная ткань состоит из вытянутых в длину ве-
ретенообразных клеток — миоцитов. В ее цитоплазме нахо-
дится ядро и сократительные нити — миофибриллы. Гладкая
мышечная ткань является основной в составе гладких мышц,
которые осуществляют движение внутренних органов, крове-
носных И лимфатических сосудов. Гладкие мышцы совершают ”
длительные тонические сокращения.
Скелетная мышца не имеет клеточного строения и состоит
из пучков поперечно-полосатых мышечных волокон, которые
имеют соединительно-тканную оболочку. Пучки волокон от-
делены друг от друга соединительно-тканными прослойками.
Мышцу покрывает соединительно-тканная оболочка, которая
вместе с внутренними соединительно-тканными оболочками
переходит в сухожилие. Мышечные пучки образуют мясистую
часть органа — брюшко. Проксимальное сухожилие (находит-
ся ближе к срединной оси тела) носит название «головки», ди-
стальное сухожилие обозначают термином «хвост». Артерии и
нервы входят в мышцу с ее внутренней стороны. Здесь же из
мышцы выходят вены и лимфатические сосуды. Артерии вет-
вятся до капилляров, к каждому мышечному волокну приле-
жит не менее одного кровеносного капилляра. Лимфатические
капилляры начинаются между пучками мышечных волокон.
На мышечных волокнах имеются двигательные' (нервные)
бляшки. Ими заканчиваются нервные волокна, несущие им-
120
пульсы к мышце. В мышцах и сухожилиях располагаются чув-
ствительные нервные окончания, которыми начинается реф-
лекторный путь. Эти окончания воспринимают «мышечное
чувство» — информацию о тонусе мышц и степени их сокра-
щения и передают ее по нервам в мозг.
Скелетные мышцы выполняют следующие функции:
1) являются органами движения;
2) участвуют в образовании стенок полостей тела (ротовая,
брюшная), выполняя при этом защитную функцию;
3) входят в состав стенок внутренних органов (глотки, гор-
тани и др.);
4) их активность на ранних стадиях онтогенеза обеспечива-
ет условия для нормального развития ЦНС.
Поперечно-полосатое мышечное волокно образуется в ре-
зультате слияния большого числа одноядерных клеток в про-
цессе гистогенеза. Поперечно-полосатое мышечное волокно
имеет цилиндрическую форму с диаметром до 0,1 мм и длиной
до нескольких десятков миллиметров. В нем имеется много
ядер, расположенных по периферии вблизи плазматической
мембраны волокна — сарколеммы, и большое число митохонд-
рий, лежащих между миофибриллами.
Миофибриллы — это сократительные элементы мышечных
волокон, составляющие до 80% их объема.
Поперечно-полосатая исчерченность волокон связана с че-
редованием в миофибриллах светлых и темных участков, име-
ющих различные оптические свойства. Стопки толстых нитей
(рис. 5.16), образованных белком миозином, обладают двой-
ным лучепреломлением и кажутся более темными в световом
микроскопе. Миозиновые нити образуют темный участок 1 в
составе миофибриллы. Чередующиеся с ними светлые участ-
ки 2 образованы стопками тонких нитей белка актина.
Тонкие актиновые нити (длина 1000 нм, диаметр 8 нм) час-
тично заходят между толстыми миозиновыми волокнами
(длина которых 1500 нм, диаметр 15 нм).
Тонкие белковые нити прикрепляются к так называемой
полоске Z — сложному переплетению белковых нитей. Учас-
ток миофибриллы между двумя полосками называют саркоме-
ром 3. Центральная зона темного участка 7, лишенная тонких
нитей, носит название субдиска Н.
При мышечном сокращении тонкие нити вдвигаются меж-
ду толстыми и светлые участки Н укорачиваются или^переста-
ют быть видимыми. Миофибриллы укорачиваются, а вместе с
ними — и мышечные волокна. «Скольжение» нитей относи-
тельно друг друга — результат скачкообразного увеличения
121
Рис. 5.16. Виды мышечной ткани и ее строение:
7 — кровеносный сосуд; 2 — мышечный пучок; 3 — оболочка; 4 — сухо-
жилия
концентрации ионов Са2+ в межфибриллярном пространстве,
что, в свою очередь, вызвано прошедшими по нервам электри-
ческими импульсами.
Катионы Са2+ хранятся в каналах эндоплазматической сети
мышечных волокон. Электрическое возбуждение приводит к
увеличению проницаемости мембран эндоплазматической
сети и, как следствие этого, — к появлению катионов Са в
межфибриллярном пространстве.
Поперечно-полосатая сердечная мышечная ткань может быть
двух видов: одна обеспечивает сокращение сердца, другая —
проведение нервных импульсов внутри сердца. Мышечная
ткань сердца по строению напоминает поперечно-полосатую,
а по функции — гладкую (сокращается непроизвольно).
Классификация мышц. Единой классификации мышц нет.
Мышцы подразделяют по форме и направлению мышечных
волокон, функциям и положению в теле человека.
По форме и направлению мышечных волокон мышцы очень раз-
нообразны. Наиболее часто встречаются веретенообразные
мышцы, характерные для конечностей, и широкие мышцы, уча-
122
ствующие в образовании сте-
нок туловища. Пучки волокон
в веретенообразных мышцах
ориентированы параллельно
длинной оси мышцы. Если
мышечные пучки лежат по
одну сторону от сухожилия
под углом к нему, мышцу на-
зывают одноперистой, а если
с обеих сторон от сухожилия,
то двуперистой.
Иногда мышечные пучки
сложно переплетаются и под-
ходят к сухожилию с несколь-
ких сторон. В таких случаях
образуется многоперистая
мышца. Некоторые мышцы
имеют по несколько головок.
В зависимости от их числа
мышцу называют двух-, трех-,
четырехглавой. Волокна не-
которых мышц расположе-
ны циркулярно: это круговые
мышцы, или мышцы-сжима-
тели — сфинктеры.
По функциям мышцы под-
разделяются на мышцы-сгиба-
тели и мышцы-разгибатели
(рис. 5.17), отводящие от сред-
ней линии и приводящие к
ней, вращающие кнаружи (су-
пинаторы) и вращающие вов-
нутрь (пронаторы).
Рис. 5.17. Общий вид мышечной
системы:
1 — мимические мышцы; 2 — жева-
тельные мышцы; 3 — мышцы, пово-
рачивающие голову; 4— мышцы, сги-
бающие и разгибающие руку; 5 —
мышцы, сгибающие и поворачиваю-
щие туловище; 6 — мышцы, разги-
бающие ногу
123
Па положению в теле человека выделяют следующие группы
мышц: мышцы туловища, мышцы головы, мышцы верхней ц
мышцы нижней конечностей.
Общий обзор мышечной системы
Основные мышцы туловища. К основным мышцам тулови-
ща (рис. 5.18) относятся:
124
1. Трапециевидная мышца — плоская, треугольной формы,
занимающая верхнюю часть спины и заднюю область шеи.
2. Широчайшая мышца спины — треугольной формы, зани-
мающая нижнюю половину спины на соответствующей стороне.
3. Большая грудная мышца — веерообразная, занимающая
значительную часть передней стенки грудной клетки.
4. Малая грудная мышца — треугольной формы, располага-
ющаяся непосредственно под большой трудной мышцей.
5. Наружные межреберные мышцы, располагающиеся меж-
ду ребрами, при сокращении поднимающие их, обеспечивая
расширение грудной клетки.
6. Внутренние межреберные мышцы, находящиеся под на-
ружными межреберными почти под прямым углом к ним.
7. Диафрагма — подвижная куполообразная мышечно-сухо-
жильная перегородка между грудной и брюшной полостями. Вы-
пуклой стороной диафрагма направлена в грудную полость, име-
ет сухожильный центр и мышечные пучки, расположенные по
периферии. Форма купола обусловлена положением внутренних
органов и разностью давлений в трудной и брюшной полостях.
Рис. 5.18. Мышцы тела человека.
а — спереди:
1 — длинная ладонная мышца; 2 — поверхностный сгибатель пальцев; 3 —
локтевой сгибатель кисти; 4 — трехглавая мышца плеча; 5 — клювоплечевая
мышца; 6 — большая круглая мышца; 7 — широкая мышца спины; 8 —
передняя зубчатая мышца; 9 — наружная косая мышца живота; 10 — под-
вздошно-поясничная мышца; 11, 13 — четырехглавая мышца бедра; 72 —
портняжная мышца; 14 — передняя большеберцовая мышца; 75 — пяточ-
ное сухожилие; 16 — икроножная мышца; 17 — стройная мышца; 18—
крестообразная связка; 19 — передняя большеберцовая мышца; 20 — ма-
лоберцовая мышца; 21 — лучевой сгибатель кисти; 22 — плечелучевая мыш-
ца; 23 — фиброзная пластинка; 24 — двуглавая мышца плеча; 25 — дельто-
видная мышца; 26 — большая грудная мышца; 27 — грудино-поясничная
мышца; 28 — грудино-ключично-сосцевидная мышца; 29 — жевательная
мышца; 30 — круговая мышца глаза
б — сзади:
7 — грудино-ключично-сосцевидная мышца; 2 — трапециевидная мышца;
3 — дельтовидная мышца; 4 — трехглавая мышца плеча; 5 — двуглавая
мышца; 6 — плечелучевая мышца; 7 — длинный лучевой разгибатель кисти;
8 — общий разгибатель пальцев; 9 — большая ягодичная мышца; 10 —
двуглавая мышца; 77 — икроножная мышца; 72 — камбаловидная мышца;
13, 15— длинная малоберцовая мышца; 14 — длинный разгибатель пальцев
(сухожилие); 16 — часть широкой фасции бедра; 77 — мышца, напрягающая
широкую фасцию бедра; 18 — наружная косая мышца живота; 19 — широ-
чайшая-мышца спины; 20 — широкая мышца спины; 27 — ромбовидная
мышца; 22 — подостная мышца; 23 — трехглавая мышца; 24 — плечевая
мышца; 25 — двуглавая мышца плеча
125
8. Наружная косая мышца живота — самая поверхностная и
обширная из мышц живота.
9. Прямая мышца живота — плоская, длинная, лентовид-
ная. Мышечные пучки прерываются тремя или четырьмя пос-
ледовательно ориентированными сухожильными перемычка-
ми. Входит в стенку брюшной полости.
10. Подкожная мышца шеи — тонкая, плоская, занимаю-
щая место непосредственно под кожей.
11. Грудинно-ключично-сосцевидная мышца — наиболее
развита у человека в связи с прямохождением. Форма этой
мышцы определяет форму шеи с переднебоковой стороны.
Мышцы шеи также относятся к мышцам туловища.
Мышцы верхней конечности. Мышцы верхней конечности
делят на мышцы плечевого пояса и мышцы свободной верхней ко-
нечности.
1. Дельтовидная мышца —одна из наиболее круп- .
ных и поверхностно расположенных мышц плечевого пояса.
Вся мышца при сокращении отводит руку от туловища, пере-
дняя часть сгибает плечо, задняя разгибает.
Рука как орган труда выполняет многочисленные и разно-
образные движения, которые осуществляет большое число
мышц. Наиболее крупные мышцы свободной верхней конеч-
ности это двуглавая и трехглавая мышцы плеча.
2. Двуглавая мышца плеча имеет две головки.
При сокращении она сгибает и супинирует предплечье в лок-
тевом суставе, сгибает плечо в плечевом суставе.
3. Трехглавая мышца плеча разгибает и приво-
дит плечо в плечевом суставе (одна из трех головок), разгибает
предплечье в локтевом суставе.
Мышцы нижней конечности. На долю мускулатуры нижних
конечностей приходится более 50% всей массы мышц. Из
мышц таза (пояса нижней конечности) наиболее развита
большая ягодичная мышца, а из мышц свободной нижней ко-
нечности — четырехглавая мышца бедра и трехглавая мышца
голени.
1. Большая ягодичная мышца разгибает бедро в
тазобедренном суставе, поддерживает равновесие туловища.
2. Средняя ягодичная мышца отводит бедро.
3. Малая ягодичная мышца отводит бедро.
4. Четырехглавая мышца бедра сгибаетногув
тазобедренном суставе и разгибает в коленном суставе.
5. Портняжная мышца начинается от подвздош-
ной кости таза и прикрепляется к большеберцовой кости,
являясь самой длинной мышцей человеческого тела. При
126
сокращении сгибает ногу в тазобедренном и коленном сус-
тавах.
6. Трехглавая мышца голени состоит из двух
мышц: икроножная мышца сгибает голень и стопу, а камбало-
видная мышца, расположенная под брюшками икроножной
мышцы, сгибает стопу.
Мышцы головы. Мышцы головы делят на две группы: ми-
мические и жевательные. В ряде случаев мышцы этих групп
функционируют совместно (членораздельная речь, жевание,
глотание). Мимические мышцы располагаются под кожей. Они
начинаются от костей черепа и заканчиваются в коже. При
своем сокращении они сдвигают кожу, изменяют ее рельеф,
формируют мимику.
1. Надчерепная мышца имеет затылочное брюш-
ко, сокращение которого тянет конец волосистой части голо-
вы назад, и лобное брюшко, при сокращении которого обра-
зуются поперечные складки кожи лба и брови поднимаются
кверху.
2. Мышца гордецов начинается от носовых костей и
прикрепляется к коже между бровями. При ее сокращении
над переносьем образуются поперечные морщины.
3. Круговая мышца глаза зажмуривает глаза.
4. Круговая мышца рта закрывает ротовое отвер-
стие, выдвигая губы вперед.
5. Мышца смеха растягивает рот, у некоторых людей
при этом образуются ямочки на щеках. *
Жевательные мышцы. Жевательные мышцы располагаются
на боковых отделах черепа по четыре с каждой стороны. Все
жевательные мышцы, прикрепляясь к нижнечелюстной кости,
приводят ее в движение при своем сокращении.
К жевательным мышцам относятся: жевательная и
височная мышцы, лежащие более поверхностно, и две
мышцы, глубже расположенные, — медиальная и ла-
теральная крыловидные мышцы (последниедве
начинаются от крыловидного отростка клиновидной кости,
что и определило их название).
Работа мышц. Утомление
' Электрические импульсы, приходящие по двигательным
нервным волокнам, вызывают в мышечных волокнах возбуж-
дение, проявляющееся в их сокращении. В скелетных мышцах
мышечные волокна изолированы друг от друга, поэтому воз-
буждение, возникающее в одном из них, не распространяется
127
на соседние. При этом возбужденное мышечное волокно
сокращается по закону «все или ничего», т. е. если нервнйе •
импульсы пришли к мышечному волокну и оказались способ- \
ними вызвать его возбуждение, то мышечное волокно сокра- •’
щается с максимально возможной для него силой. Сократить- <
ся в полсилы оно не может. Поэтому степень сокращения
мышцы зависит только от числа сократившихся в данный мо- ч
мент волокон. Сила мышц с продольным расположением во- .
локон зависит от их толщины. Мышцы, выполняющие общую
работу в одном и том же суставе, называют мышцами-синерги-
стами (от греч. sinergio — сотрудничество). Например, плече-
вая мышца и двуглавая мышца плеча сгибают руку в локтевом
суставе, средняя и малая ягодичные мышцы отводят ногу в та-
зобедренном суставе. Мышцы антагонисты, наоборот, при со-
кращении вызывают противоположные движения в одном и
том же суставе. Так, трехглавая мышца плеча разгибает руку в
локтевом суставе, а двуглавая мышца плеча вызывает ее сги-
бание. Портняжная мышца приводит к сгибанию в коленном '
суставе, а четырехглавая — к разгибанию. По отношению к та-
зобедренному суставу эти же две мышцы являются уже не ан-
тагонистами, а синергистами, так как совместно сгибают бед-
ро в тазобедренном суставе.
Длительная мышечная работа приводит к мышечному
утомлению. Утомление — временное снижение работоспо-
собности (клетки, органа или всего организма), наступающее
в результате работы и исчезающее после отдыха. Статическая
нагрузка быстрее приводит к утомлению, чем динамическая.
Динамическая нагрузка характеризуется быстрым измене-
нием во времени ее значения и приводит к чередованию со-
кращения и расслабления мышц. При статической работе,
например при удерживании груза, мышцы находятся в на-
пряженном состоянии длительное время. При этом импуль-
сы поступают к мышечным волокнам с большой частотой,
поэтому каждый очередной нервный импульс приходит к
мышце раньше, чем она успеет расслабиться после предыду-
щего импульса.
В развитии утомления, возникающего при мышечной ра-
боте, ведущую роль играет не усталость самих мышц (пери-
ферический механизм утомления), а особое состояние двига-
тельных нервных центров (центральный механизм утомления).
Отсюда становится ясно, почему физическая работоспособ-
ность так зависит от настроения. Если работа выполняется с
интересом, утомление наступает не так скоро. Убедительным
доказательством ведущей роли состояния нервных центров в
128
развитии утомления (центральный механизм утомления) явля-
ются результаты описываемого эксперимента. Человек, находя-
щийся под гипнозом, быстро устает и обливается потом, под-
нимая легкую корзинку, если ему внушили, что у него в руках
тяжелая гиря. И, наоборот, легко и долго ритмично поднимает
гирю, если, находясь под гипнозом, думает, что у него в руках
легкая корзинка.
С состоянием двигательных нервных центров связано и
другое явление. Когда одно и то же движение попеременно
выполняется правой и левой рукой, усталость возникает не так
скоро, как в случае, если то же движение и в том же ритм® осу-
ществляется только одной рукой. Поэтому для восстановле-
ния работоспособности какой-нибудь группы мышц более
благоприятен не полный покой, а интенсивная работа другой
мышечной группы. Великий русский физиолог Иван Михай-
лович Сеченов называл такой отдых «активным отдыхом».
Преимущество активного отдыха И. М. Сеченов рассматривал
как доказательство преобладания центральных механизмов в
развитии утомления над периферическими.
При динамической работе скорость утомления зависит от
двух показателей — физической нагрузки, падающей на мыш-
цу, и от ритма работы, т. е. от частоты мышечных сокращений.
При увеличении нагрузки или при учащении ритма мы-
шечных сокращений утомление наступает быстрее. Влияние
этих условий на объем выполненной работы изучал в начале
нашего века И. М. Сеченов. Оказалось, что если увеличивать
нагрузку, интенсивность выполняемой работы возрастает, но
только до определенного уровня, а затем снижается. Мышеч-
ная работа достигает максимального объема при средних на-
грузках и средних скоростях сокращения мышц. Таким обра-
зом, И. М. Сеченовым были заложены основы новой науки —
гигиены труда.
КРОВЕНОСНАЯ СИСТЕМА
КРОВЬ
Кровь, лимфа и тканевая жидкость составляют внутреннюю
среду организма.
Кровь является особым видом соединительной ткани, так
как образуется из соединительной ткани зародыша — мезен-
химы.
5 — 281
129
Состав крови и характериспнп ее иомюяятов
Кровь состоит из плазмы — жидкой части, на которую при-
ходится 42—45% массы крови, и форменных элементов (эрит- \
родитов, лейкоцитов, тромбоцитов), составляющих 55—
58%крови (рис. 5.19).
1. Плазма является сложной биологической средой, в состав
которой входит вода (90-92%) и важные для функций крови
органические и неорганические вещества, различные белки 'i
(альбумины 4,5—5%, глобулины а, Д, у 2—3%, фибриноген 0,2— |
0,4%, глюкоза 0,11%), аминокислоты, минеральные вещест- |
ва 0,90%, среди которых находятся ионы Na+, К+, Са2+, Mg2+, j-
СГ, НРО*4, HiCO‘> В плазме содержатся гормоны, витамины,
растворимые газы, а также присутствуют ферменты, участвую- >
щие в свертывании крови, в расщеплений пищевых веществ и •
окислительно-восстановительных процессах. Сывороткой кро- <
ви называется ее жидкая часть (плазма) без фибриногена.
2. Эритроциты — это красные кровяные тельца размером 7—
8 мкм, которые являются основным типом клеток крови, по-
Рис. 5.19. Свежий препарат крови человека:
1 — эритроциты; 2 — нейтрофильные лейкоциты; 3 — эозинофил; 4, 5, б —
лимфоциты; 7 — моноцит; 8 — кровяные пластинки
скольку их в 1000 раз больше, чем лейкоцитов. В 1 мм3 около
4—5 млн эритроцитов. По форме эритроциты представляют
собой двояковогнутый диск. Такая форма клетки увеличивает
поверхность для диффузии дыхательных газов, а также делает
эритроциты способными к обратимой деформации при про-
хождении через узкие изогнутые капилляры. Эритроциты —
специализированные безъядерные клетки. Они образуются в
красном костном мозге, разрушаются в селезенке и печени.
В начальных фазах своего развития эритроциты имеют
ядро и называются ретикулоцитами. По мере созревания ядро
замещается дыхательным пигментом — гемоглобином (Нв),
составляющим 90% сухого вещества эритроцитов. Гемоглобин
обладает свойством легко связывать и отщеплять О2. Присое-
диняя его, он становится оксигемоглобином (НвО8). Отдавая
О2 в местах с малым его содержанием, он превращается в вос-
становленный (редуцированный) гемоглобин. Оксигемогло-
бин несколько отличается по цвету от гемоглобина, и поэтому
венозная кровь, содержащая восстановленный гемоглобин,
имеет темно-вишневый цвет. В венозной крови также содер-
жится нестойкое соединение гемоглобина с СО2 — карбгемог-
лобин. Кроме О2 гемоглобин может соединяться в организме
человека с некоторыми другими газами, например, с угарным
газом СО. Эго соединение носит название карбоксигемогло-
бина (НвСО), которое в 300 раз прочнее, чем соединение ге-
моглобина с О2. Примесь 0,1% СО в. воздухе ведет к тому, что
80% гемоглобина оказывается связанным с СО. В этом случае
гемоглобин не присоединяет О2, что является очень опасным
для жизни. При отравлении угарным газом необходимо выне-
сти человека на свежий воздух. Карбоксигемоглобин в этих ус-
ловиях постепенно распадается. Прй уменьшении количества
гемоглобина в эритроцитах или уменьшении числа эритроци-
тов в крови возникает анемия.
При добавлении в кровь антикоагулянтов наблюдается осе-
дание эритроцитов. У здоровых мужчин скорость оседания
эритроцитов (СОЭ) колеблется от 3 до 9 мм/ч, у здоровых
женщин — от 1 до 12 мм/ч. Время циркуляции эритроцитов в
крови около 90 суток.
Функция эритроцитов — перенос кислорода от
легких к тканям и двуокиси углерода от тканей к органам ды-
хания.
3. Лейкоциты, или белые кровяные тельца, представляют
собой ядерные бесцветные клетки, способные к активному пе-
редвижению и изменению своей формы. Число лейкоцитов
составляет 4—9 тыс. в 1 мм3 крови. Они образуются в лимфати-
5* • ’?*
ческих узлах, селезенке и красном костном мозге. Лейкоциты д
делят на два главных типа, один из которых характеризуется
зернистой цитоплазмой (гранулоциты), а другой — незернис- t
той (агранулоциты).
Существуют три типа зернистых лейкоцитов, различаемых '*
по окраске их цитоплазматических гранул. Те клетки, специ-
фические гранулы которых интенсивно окрашиваются кислым
красителем эозином, называются эозинофилами. Лейкоциты, I
гранулы которых окрашиваются основными красителями, на-
зываются базофилами. Лейкоциты, гранулы которых при нор-
мальных значениях pH (7,0) не воспринимают ни основные, ?
ни кислые красители, называются нейтрофилами. Нейтрофи-
лы чаЪто называют полиморфно-ядерными лейкоцитами. Они
отличаются формами ядер, а не гранул. По этим качествам по-
лиморфно-ядерные лейкоциты различаются на юные, палоч-
коядерные и сегментированные. Юные и палочкоядерные яв-
ляются незрелыми формами нейтрофилов.
Среди агранулоцитов существуют два типа лейкоцитов. Бо-
лее многочисленные и мелкие называются лимфоцитами, так
как они обнаруживаются не только в крови, но и в лимфе, бо-
лее крупные и не столь многочисленные — моноциты.
Функции гранулоцитов:
эозинофилы составляют 1—4% всех лейкоцитов. Выраба-
тываются так же, как и нейтрофилы, в костном мозге, после
чего они несколько часов находятся в крови, а затем поки-
дают кровеносное русло и уходят в ткани, где остаются в те-
чение нескольких дней. Основным местом локализации яв-
ляется соединительная ткань кишечника, кожи и легких.
Эозинофилы выполняют следующие основные функции:
а) накапливаются в зонах аллергических реакций и уменьша-
ют повреждающее действие аллергена; б) играют важную роль
в нейтрализации глистных инвазий (мигрируют в места лока-
лизации паразита, вызывают путем экзоцитоза на его поверх-
ности свои лизосомальные ферменты, причиняя тем самым
прямой вред паразиту; в) способны фагоцитировать и унич-
тожать патогенные микроорганизмы, но в меньшей степени,:
чем нейтрофилы;
базофилы составляют всего 0,5% лейкоцитов в крови. Вы-
рабатываются в костном мозге, попадают в ток крови и очень
быстро расселяются в соединительной ткани различных час-
тей организма. Четкая роль базофилов не установлена, однако
они способны: а) фагоцитировать микроорганизмы; б) скапли-
ваться в местах! воспаления; в) участвовать в нейтрализации
воспалительных и аллергических реакций; г) продуцировать
132
гепарин и гистамин. Гепарин препятствует свертыванию кро-
ви в очаге воспаления, а гистамин расширяет капилляры, что
способствует рассасыванию и заживлению;
нейтрофилы составляют 50—70% от всех лейкоцитов. Раз-
виваются в костном мозге, где они проходят много стадий пре-
образования, прежде чем принимают свою зрелую форму и
выходят в кровоток. В кровяном русле они пребывают не более
8—12 ч, после чего выходят из мелких сосудов (венул) и попа-
дают в соединительную ткань. Нейтрофилы выполняют следу-
ющие основные функции: а) фагоцитируют бактерии и уча-
ствуют в их разрушении; б) участвуют в нейтрализации
вредных агентов в зонах воспаления. Гной состоит в основном
из нейтрофилов или их остатков. И. И. Мечников назвал та-
кие лейкоциты, фагоцитами, а само явление поглощения и
разрушения лейкоцитами чужеродных тел — фагоцитозом.
Функции агранулоцитов:
лимфоциты крови здорового человека занимают второе
место после нейтрофилов. Различают два типа лимфоцитов,
называемых В- и Т-лимфоцитами. Морфологически они не
различимы. Различить их можно лишь соответствующими
иммунологическими методами. У человека клетки В-типа
происходят из костного мозга и лимфоузлов, а Т-лимфоциты
из тимуса (зобной железы). Функционально эти два типа лим-
фоцитов участвуют в различных реакциях, таких, как образова-
ние антител и реакции клеточного иммунитета;
моноциты в норме составляют от 2 до 8% всех лейкоци-
тов. Образуются в костном мозге и являются предшественни-
ками макрофагов. После выхода моноцитов в русло крови
они, пробыв в ней около 3 сут, покидают кровоток и перехо-
дят в ткани, превращаясь там в макрофаги. Таким образом,
к основным функциям моноцитов можно отнести: а) фагоци-
тоз крупных частиц чужеродных веществ, попавших в орга-
низм человека; б) участие в иммунных реакциях с погло-
щением продуктов иммунного ответа антиген — антитело;
в) участие в формировании иммунного ответа в организме че-
ловека.
4. Тромбоциты являются небольшими фрагментами, кото-
рые образуются при распаде очень крупных клеток костного
мозга — мегакириоцитов. При этом каждая пластинка цели-
ком покрыта клеточной мембраной и не содержит компонен-
тов ядра. Число циркулирующих в крови тромбоцитов насчи-
тывает 250—350 клеток в 1 мм3.
К основным функциям тромбоцитов можно
отнести: а) участие в процессе свертывания крови; б) участие в
133
закупорке поврежденных отверстий сосудов. Этот процесс на- ;
зывается агрегацией пластинок.
Механизм свертывания крови
Механизм свертывания крови представлен на рис. 5.20. г
Кровяной тромбопластин включает в себя около 16 факте-
ров — это антигемофилические факторы крови. Например: |
а) 2-й антигемофилический глобулин крови, б) факторы 8-й, |
9-й и т. д., а также в) ноны кальция. Отсутствие хотя бы одного
из 16 факторов ведет к гемофилии.
Одним из белков плазмы крови является фибриноген.
В нормальных условиях он находится в растворе (т. е. он ра-
створим). Кроме того, в плазме содержится глобулин, называ- |
емый протромбином, который в обычных условиях не активен.
В месте повреждения происходит высвобождение тканевого f
тромбопластина, который наряду с кровяным тромбопласти- |
ном (антигемофилическими факторами крови и ионами киль- |
ция) осуществляет превращение протромбина в тромбин.
Тромбин вызывает полимеризацию растворимого фибриноге-
на в нерастворимые волокна фибрина. Фибрин обычно форми- >
руется в местах агрегации (прилипания) тромбоцитов. При >
повреждении целостности сосудов происходит прилипание з
тромбоцитов к поврежденной стенке сосудов при наличии
ионов кальция. У прилипших тромбоцитов происходит высво-
Фибриноген
(растворимый
белок)
Фибрин (тромб)
(нерастворимый белок)
Тромбин
(активный фермент)
Ca+t
Тканевый тромбопластин
(секрет разрушенных тромбоцитов)
Кровяной тромбопластин
Протромбин
(неактивный фермент)
Рис. 5»20. Механизм свертывания крови
вождение из секреторных гранул разнообразных веществ
(тромбина, коллагена) в окружающую среду. Это ведет к при-
липанию все новых и новых тромбоцитов к тем, которые уже
агрегировали. Так образуется пластиночный сгусток, который
затем пронизывается нитями фибрина. Закупорка поврежден-
ных сосудов или просвета сосудов называется тромбозом.
Скопление тромбоцитов на внутренней поверхности крове-
носного сосуда называют белым тромбом, потому что масса аг-
регирующих пластинок в свежем состоянии имеет белый цвет.
Белый тромб может образовываться только в текучей крови.
Он растет за счет вовлечения все новых и новых пластинок из
протекающей мимо крови. Белый тромб по своей природе от-
личается от красного, образующегося при свертывании непод-
вижной крови. Красный тромб в основном формируется из во-
локон фибрина, образующих сеть, в которой застревает много
эритроцитов.
Группы крови
Известна система АВО, включающая четыре группы крови.
В крови имеются особые белковые вещества; в эритроцитах —
агглютиногены (А и В), в плазме — агглютинины (а и р). Если
агглютинин а встречается с агглютиногеном А или агглюти-
нин р с агглютиногеном В, то происходит реакция агглютина-
ции (склеивание эритроцитов).
Таблица 5.1
Совместимость групп крови
Группа крови Наличие агглютиногенов в эритроцитах Наличие агглютининов в плазме
1(0)- ' Нет а, р
и (А) А р
Ш (В) В а
IV (АВ) АВ Нет
Людей с 1 группой крови называют универсальными доно-
рами, так как их кровь можно переливать всем четырем груп-
пам. Людей с IV группой называют универсальными реципи-
ентами, так как им можно переливать кровь любых групп.
Кровь II группы может быть перелита лицам с II и IV группа-
ми, кровь III группы — лицам, имеющим III и IV группы.
В настоящее время предпочитают переливать одногруппную
-кровь в небольших дозах.
135
Резус-фактор. У подавляющего большинства людей (до 85%)
эритроциты содержат особый антиген, который называют резус-
фактором. У других людей (15%) резус-фактор отсутствует. Лиц, в
эритроцитах которых содержится резус-фактор, называют резус-
положительными, а тех, у которых он отсутствует — резус-отри-
цательными. В плазме крови нет антител, комплементарных ан-
тигену резус-фактору. Но если резус-положительная кровь
попадет к человеку с резус-отрицательной кровью, то у него в
плазме образуются антитела и эритроциты начинают разрушать-
ся. При повторном попадании резус-положительной крови в
организм резус-отрицательного человека возникает быстрая ре-
акция по разрушению эритроцитов донорской крови. Это приво-
дит к резус-конфликту и созд ает угрозу для жизни человека.
Иммунитет
Иммунитет — способ защиты организма от генетически чу-
жеродных веществ и инфекционных агентов. Защитные реак-
ции организма обеспечиваются клетками-фагоцитами, а также
белками-антителами. Антитела вырабатывают плазматические
клетки, образующиеся из В-лимфоцитов в ответ на появление
в организме чужеродных белков — антигенов. Антитела связы7
ваются с антигенами, образуя комплекс «антиген — антитело»,
в котором антиген теряет свои патогенные свойства.
Различают естественный иммунитет, выработанный самим
организмом без искусственных вмешательств, и искусствен-
ный — возникающий при введении в организм специальных
веществ.
Естественный иммунитет может быть врожденным и приобре-
тенным. При врожденном организм получает иммунные тела
от матери через плаценту или с материнским молоком. При
приобретенном антитела в организме образуются после пере-
несенного заболевания.
Искусственный иммунитет может быть активным и пассив-
ным. Активный иммунитет вырабатывается при введении
вакцины, содержащей ослабленные или убитые возбудители
заболеваний или их токсины. Такой иммунитет сохраняется
долго. Принцип создания лечебных вакцин и введение их в
медицинскую практику принадлежит Л. Пастеру. Пассивный
иммунитет возникает при введении в организм лечебной сы-
воротки с уже готовыми антителами. Такой иммунитет сохра-
няется недолго — 4—6 нед. Сыворотку получают из крови жи-
вотных (чаще всего лошадей), которым вводят постепенно
возрастающие дозы микроорганизмов или их токсинов.
КРОВООБРАЩЕНИЕ.
СТРОЕНИЕ И РАБОТА СЕРДЦА
Общее представление о системе кровообращения
Кровообращением называют непрерывное движение крови
в организме. Сердце с кровеносными сосудами (артериями,
венами) и лимфатическая система составляют сосудистую сис-
тему организма. Кровь и лимфа в результате своего движения
по сосудам человеческого тела осуществляют доставку кисло-
рода и питательных веществ к клеткам, удаляют продукты обме-
на веществ и осуществляют гуморальную регуляцию деятельно-
сти органов и систем в организме. По характеру жидкости,
циркулирующей в сосудистой системе человека и позвоноч-
ных, последняя, подразделяется на два отдела: кровеносный:
представляющий собой систему сосудов, по которым циркули-
рует кровь (артерии, капилляры, вены и сердце), и лимфати-
ческий, по системе сосудов которого движется лимфа.
Кровеносная система состоит из центрального органа —
сердца и находящейся в соединении с ним замкнутой системы
трубок различного диаметра, называемых кровеносными сосу-
дами.
Сосуды, несущие кровь от сердца, называются а р т е р и я -
м и, а сосуды, по которым кровь течет к сердцу — венами.
Артерии, постепенно разветвляясь и уменьшаясь в диаметре,
переходят в капилляры. Капилляры — это мельчайшие со-
суды, по которым артериальная кровь подходит непосред-
ственно к тканям. Здесь происходит обмен газов между кровью
и тканями, кровь превращается в венозную и переходит в вену-
лы. Венулы, сливаясь, образуют сначала мелкие, а затем круп-
ные вены, по которым кровь направляется к сердцу. Кровь
циркулирует по единой замкнутой системе сосудов, в которой
различают большой и малый круги кровообращения.
Лимфатическая система в процессе эмбриогенеза вступает
в связь с кровеносной системой и является добавочным для ве-
нозных сосудов руслом.
Строение артерий, капилляров, вен
Стенка артерии имеет три слоя: наружный, средний и внут-
ренний. Наружный слой соединительно-тканный, средний —
гладкомышечный, содержащий много эластических волокон,
внутренний состоит из одного слоя плоских клеток эпителия.
Различают артерии эластического (ближайшие к сердцу круп-
137
ные сосуды) и мышечного типа (средние и более мелкие арте-
рии). которые регулируют приток крови к органу, а также арте-
риолы (мельчайшие артерии), которые, разветвляясь, перехо-
дят в капилляры.
Через стенки капилляров, состоящие из одного слоя плос-
ких эндотелиальных клеток и базальной мембраны, осуществ-
ляется обмен газами и питательными веществами.
Стенки вен, так же как и артерий, состоят из трех слоев, но
они тоньше и беднее мышечными и эластическими волокна-
ми (рис. 5.21), поэтому вены менее упруги и могут спадаться.
Большинство вен снабжены клапанами, которые препятству-
ют обратному току крови (рис. 5.22).
Строегие сердца
Сердце человека представляет собой полый мышечный
орган, который ритмически сокращается в течение всей жизни
организма. Сердце человека состоит из четырех отделов: двух
желудочков — правого и левого, и двух предсердий — правого
и левого. Форма сердца конусовидная, основание конуса обра-
щено назад, вправо и кверху, а верхушка вперед, влево и вниз.
Сердце располагается в средостении. Его ббльшая часть нахо-
дится в левой половине тела, а меньшая — в правой. Верхушка
сердца лежит на уровне 5-го левого межреберного промежутка
несколько кнутри от вертикальной линии, проходящей через
сосок и середину ключицы. Левая граница сердца идет наис-
кось к его верхушке от места соединения хряща и кости 3-го
ребра. Правая граница в виде изогнутой линии выступает на
1—2 см за правый край грудины на уровне 3-5-го ребер; а ниж-
няя проходит от хряща 5-го правого ребра к верхушке сердца
(рис. 5.23).
Сердце окружено околосердечной сумкой, или перикар-
дом, наружный слой которого состоит из нерастяжимой бе-
лой фиброзной ткани. Этот наружный слой перикарда
срастается с грудиной и сухожильным центром диафрагмы.
Рис. 5.23. Расположение сердца в грудной клетке
Внутренний слой перикарда состоит из двух листков: висце-
рального (эпикард) и париетального. Между двумя листками
серозного перикарда выделяется перикардиальная жидкость,
которая уменьшает трение между стенками сердца и окружа-
ющими тканями. Висцеральный листок (эпикард) срастается
с мышечным слоем сердца — миокардом, а париетальный ли-
сток — с фиброзной тканью (рис. 5.24). Неэластичный в це-
лом характер перикарда препятствует излишнему растяже-
нию сердца и переполнению его кровью.
Миокард — средний слой сердечной стенки, является наи-
более мощным. Образован он поперечно-полосатой мышеч-
ной тканью.
От скелетной мускулатуры мышечная ткань сердца отлича-
ется следующими особенностями:
1) ее сокращение происходит непроизвольно, каку гладкой
мышечной ткани;
2) клетки (кардиомиоциты) бывают двух видов: типичные
сократительные мышечные клетки и атипичные миоциты, со-
ставляющие проводящую систему сердца;
3) типичные кардиомиоциты в отличие от волокон попе-
речно-полосатой мышечной ткани имеют намного больше ми-
Рис. 5.24. Строение сердца:
1 — верхняя полая вена; 2 — дуга аорты; 3 — легочная артерия; 4 — легоч-
ная вена; 5 — правый желудочек; 6 — левый желудочек; 7 — нижняя полая
вена; 8 — створчатые клапаны; 9 — левое предсердие; 10 — правое предсер-
дие; 11 — венечные сосуды
140
тохондрий, что обеспечивает высокий уровень окислительных
процессов. Эндоплазматическая сеть (ЭПС) в кардиомиоци-
тах, наоборот, хуже развита, чем в волокнах скелетной мышеч-
ной ткани. Ядро кардиомиоцитов занимает центральное поло-
жение, миофибриллы располагаются по периферии клеток
строго параллельно;
4) атипические миоциты генерируют и проводят импульсы
внутри сердца. Это большие клетки, богатые гранулированной
ЭПС с малым числом миофибрилл и митохондрий, с крупным
ядром, не всегда расположенным в центре клетки, и иннерви-
рованные окончаниями нейронов.
Миокард желудочков имеет три слоя мышц, а миокард
предсердий — два слоя.
Эндокард — внутренний слой сердца, представляет собой
тонкую оболочку из эндотелиальных клеток, расположенных
на соединительно-тканном слое с эластическими волокнами и
гладкомышечными клетками.
Сердце состоит из двух несообщающихся между собой поло-
вин — правой и левой. В правую половину впадают вены, принсн
сящие венозную кровь, поэтому правую половину называют ве-
нозным сердцем. В левую половину сердца по легочным
венам поступает артериальная кровь, поэтому правую половину
сердца называют артериальным сердцем. Каждая по-
ловина сердца делится на две камеры — предсердие и желудочек.
Правое предсердие является полостью, которая спереди пе-
реходит в полый отросток — правое ушко. В полость правого
предсердия открываются верхняя и нижняя полые вены. Место
впадения полых вен называется венозным синусом, который у
человека является частью правого предсердия. В правое пред-
сердие также открываются небольшими отверстиями многочис-
ленные мелкие вены сердца. На границе правого предсердия и
правого желудочка находится правое предсердно-желудочковое
отверстие. Кровь из предсердия поступает в желудочек через
правое предсердно-желудочковое отверстие. По краю этого от-
верстия расположен тоехстворчатый клапан. Каждая его створ-
ка, состоящая из волокнистой ткани и покрытая эндотелием,
является складкой внутренней оболочки сердца (эндокарда).
К свободным краям створок прикрепляются сухожильные нити,
которые своими противоположными краями присоединены к
верхушкам сосочковых мышц. Сухожильные нити и сосочковые
мышцы не позволяют выворачиваться створкам клапана в пред-
сердие во время сокращения желудочка, когда под давлением
крови створчатый клапан захлопывается. Из правого желудочка
выходит крупный сосуд — легочный ствол. В устье легочного
141
ствола расположены три полулунных клапана, которые свобод-
но пропускают кровь из желудочка в легочный ствол, но пре-
пятствуют обратному току (рис. 5.25). *
Левое предсердие находится сзади и слева в основании
сердца. Его ушко выходит на переднюю поверхность сердца.
В левое предсердие впадает четыре легочные вены. На грани-
це левого предсердия и левого желудочка находится левое
предсердно-желудочковое отверстие. По краю левого пред-
сердно-желудочкового отверстия расположен двухстворчатый
клапан (митральный) (рис. 5.26). К свободным краям ство-
рок прикреплены сухожильные нити, которые своим проти-
воположным концом присоединены к двум сосочковым
мышцам. От левого желудочка начинается аорта. В устье аор-
ты, как и в устье легочного ствола, расположены три полулун-
ных клапана.
Рис. 5.25. Полулунный
клапан:
Г — полулунный клапан;
2 — линия продольного
разреза сосуда
Рис. 5.26. Створчатые клапаны:
/ — створчатые клапаны; 2 — гголусу-
хожильные нити
Свойства сердечной мьнпцы
Сердечная мышца обладает следующими свойствами:
1) возбудимостью; 2) проводимостью; 3) сократимостью и
4) ритмичной автоыатией.
Воэбуденоеть седая проявляется в возникновении возбуж-
дения при действии разных раздражителей. Сила раздражения
должна быть не менее пороговой; яри этом пороговые раздра-
жители вызывают сокращения сердечной мышцы максималь-
ной силы. Эта особенность возникновения возбуждения полу-
чила название закона «все или ничего», так как сердечная
мышца отвечает на раздражение либо возбуждением всех воло-
кон, либо не реагирует совсем.
Возбудимость сердечной мышцы непостоянна, она изменя-
ется по ходу возбуждения. В начальном его периоде сердечная
мышца не восприимчива (рефрактерна) к повторным раздраже-
ниям. Этот период называется фазой абсолютной рефрактерпос-
ти (невосприимчивости). У человека она длится 0,2-0,3 с, т. е.
совпадает со временем сокращения сердца. По окончании фазы
абсолютной рефрактерности возбудимость сердечной мышцы
постепенно восстанавливается и в течение короткого периода
становится исходной. Благодаря периоду абсолютной рефрак-
терности сердечная мышца не ъ$ожвт сокращаться в обычных
условиях по типу тетануса. Это очень важно для координа-
ции работы предсердий и желудочков.
Проводимость сердца обеспечивает распространение воз-
буждения от клеток водителя ритма по всему миокарду. Рас-
пространение возбуждения по сердцу осуществляется электри-
ческим путем. Потенциал действия, возникающий в одной
мышечной клетке, является раздражителем для других. Спо-
собность к провед ению возбуждения зависит как от структур-
ных особенностей мышечных клеток сердца, так и от темпера-
турного фактора, содержания в клетках гликогена и кислорода.
Сократимость сердечной мышцы обусловливает увеличение
напряжения или укорочение ее мышечных элементов при воз-
буждении.
Ритмичной автоматией сердца называют его способность
ритмически сокращаться без внешних раздражений под влия-
нием' импульсов, возникающих в нем самом. За автоматию
сердца отвечают узлы автоматии, которые представляют собой
скопление атипической мышечной ткани. В сердце два основ-
ных узла автоматии:
1) синоатриальный узел (синусно-предсердный), располо-
женный в месте впадения полых вея в правое предсердие;
2) атриовентрикулярный узел (предсердно-желудоч-
ковый), находящийся в правом предсердии в межпредсерд-
ной перегородке. От этого узла отходит пучок Гиса, кото-
рый идет по межжелудочковой перегрродке. Далее пучок
Гиса разделяется на две ножки, от которых отходят волокна
Пуркинье, иннервирующие мышечные клетки желудочков
(рис. 5.27).
Синоатриальный узел называют водителем ритма сердца
(пейсмекером), или центром автомашин первого порядка.
Именно в нем генерируются импульсы, вызывающие сокраще-
ния сердца.
Атриовентрикулярный узел является центром автоматии
второго порядка, так как он генерирует импульсы с меньшей
частотой, чем синусный узел, примерно в два раза. Пучок Гиса
и волокна Пуркинье тоже обладают автоматией, но их ритми-
ческая активность более низкая, чем у атриовентрикулярного
узла.
1
3 8
Рис. 5.27. Проводящая система сердца (схема):
V - верхняя полая вена; 2 — атриовентрикулярный узел; 3 — атриовен-
трикулярные клапаны; 4 — ножка пучка Гиса; 5 — левый желудочек; 6 —
перегородка между желудочками; 7 — сеть волокон Пуркинье; 8 — правый
желудочек; 9 — нижняя полая вена; 10 — пучок Гиса; 11 — венечный
синус; 12 — синоатриальный узел; 13 — левое предсердие; 14 — правое
предсердие
144
В физиологических условиях водителем ритма сердца явля-
ется синусный узел. Другие отделы проводящей системы функ-
ционируют лишь как проводники возбуждения. При угнетении
деятельности синусного узла ритмический поток импульсов к
атриовентрикулярному узлу прекращается, и в таких условиях
проявляется собственная автоматия этого отдела проводящей
системы сердца.
Большой круг кровообращения
Большой круг кровообращения (рис. 5.28) служит для дос-
тавки ко всем органам и тканям О2 и питательных веществ. Он
начинается в левом желудочке сердца, из которого выходит
Рис. 5.28. Большой и малый
круги кровообращения:
7 — аорта; 2 — легочные арте-
рии; 3 — легочные вены; 4 — ар-
терии внутренних органов; 5 —
капилляры; 6 — воротная вена
печени; 7 — верхняя и нижняя
полые вены
145
аорта, несущая артериальную кровь. Аорта разветвляется на $
артерии, которые идут ко всем органам и тканям тела. Через £
стенку капилляров из крови в ткани переходят питательные .
вещества и кислород, а из ткани в кровь — продукты обмена
веществ, в том числе и COj. В капиллярах кровь из артериаль- >
ной превращается в венозную, которая продвигается по венам ?
и поступает в правое предсердие по верхней и нижней полым |
венам. Из правого предсердия венозная кровь переходит в
правый желудочек. ?
Основные артерии большого круга кровообращения — это |
аорта, левая и правая общие подвздошные, плечеголовной $
ствол, левая и правая общие сонные, левая и правая подклю-
чичные. Основные вены — яремные, подключичные, под-
вздошные, верхняя и нижняя полые.
Малый круг кровообращения.
Малый круг кровообращения (см. рис. 5.28) начинается из
правого желудочка легочным стволом, который, разветвляясь
дает начало правой и левой легочным артериям. В легких они
разветвляются на множество артерий, переходящих в капилля-
ры. В капиллярных сетях, оплетающих альвеолы, венозная
кровь отдает СО2 и получает О2, превращаясь в артериальную.
Обогащенная кислородом артериальная кровь поступает из
капилляров в вены, которые, слившись в четыре легочные
вены (по д ве с каждой стороны), впадают в левое предсердие.
Кровоснабжение сердца
Кровоснабжение сердца начинается двумя венечными арте-
риями, которые отходят от начальной расширенной части
аорты (луковицы аорты). Они снабжают кровью стенки серд-
ца. Заканчивается «сердечный круг кровообращения» венами
сердца. Последние сливаются в венечный синус, впадающий в
правое предсердие; ряд вей открываются непосредственно в
полость пред сердия.
Работасердца
Сокращения отделов сердца называются их систолой, рас-
слабление —диастолой. Систола и диастола согласованы меж-
ду собой и составляют цикл работы сердца.
Рассмотрим пример сердечного цикла при частоте сердеч-
ных сокращений (ЧСС) 72 раза в минуту. Началом каждого
146
цикла является сокращение (систола) предсердий (0,1 с). По
окончании систолы предсердий начинается систола желудоч-
ков (0,3 с); предсердия в это время расслабляются. Затем на-
ступает диастола желудочков, во время которой расслаблены
одновременно и предсердия и желудочки (0,4 с).
Причинами одностороннего движения крови в сердце от
предсердий в желудочки и по сосудистой системе являются:
1) последовательное сокращение и расслабление мио-
карда;
2) изменение давления крови в отделах сердца;
3) деятельность клапанов сердца;
4) деятельность клапанов аорты и легочного ствола.
При систоле предсердий давление крови в них повышается,
и поэтому кровь поступает из предсердий в желудочки сердца,
где давление в это время меньше. Обратному току крови из
предсердий в полые и легочные вены в это время препятствует
сокращение кольцеобразных мцшц, наподобие
сфинктеров охватывающих отверстия вен.
По окончании систолы предсердий начинается систола
желудочков, которая делится на две фазы: фаза напряжения и
фаза изгнания крови. В первую фазу мускулатура желудочков
напрягается, ио объем их не меняется. Постепенно давление
крови в желудочках повышается, и створчатые клапаны смыка-
ются. Сосочковые мышцы желудочков сокращаются, сухо-
жильные хорды натягиваются и препятствуют выворачиванию
створчатых клапанов в предсердия.
Когда давление крови в желудочке становится выше, чем в
аорте и легочном стволе, наступает фаза изгнания. Полулунные
клапаны открываются, мышцы желудочков сокращаются и
кровь под большим давлением выбрасывается в аорту и легоч-
ный ствол.
После систолы желудочков вслед за фазой изгнания насту-
пает диастола желудочков. Благодаря расслаблению желудоч-
ков давление в аорте и легочном стволе становится больше,
чем в желудочках, и поэтому полулунные клапаны захлопыва-
ются. Одновременно открываются створчатые клапаны, и
кровь поступает из предсердий в желудочки.
В это время кровь свободно протекает из верхней и ниж-
ней полых вен в правое предсердие, а из легочных вен — в
левое предсердие. Так как створчатые клапаны открыты, то
кровь попадает в желудочки. Таким образом, большая часть
крови — примерно 70% ее объема — поступает в желудочки
самотеком во время общего расслабления
с е р д ц а, а во время систолы предсердий — только 30%.
147
В конце диастолы желудочков начинается систола предсер-
дий, т. е. начинается новый, очередной цикл работы сердца.
В покое ЧСС в среднем равна 60—80 раз в минуту. При фи-
зических нагрузках она возрастает до 150—200 раз в минуту.
Количество крови, выбрасываемой из сердца в среднем в
минуту, называют минутным объемом кровообращения. В со-
стоянии покоя он равен 4,5-5 л крови. Количество крови, ко-
торое выбрасывается во время одной систолы, называют сис-
толическим объемом. Его можно узнать, если разделить
минутный объем крови на число сердечных сокращений. Он в
среднем равен 65—75 мл крови.
Кровяное давление
Кровь движется благодаря разности давлений. Самое высо-
кое давление наблюдается в аорте и крупных сосудах. По мере
продвижения крови по артериям часть энергии давления ис-
пользуется на преодоление трения крови о стенки сосудов, и
поэтому величина его постепенно падает. Особенно значи-
тельное падение давления происходит в самых маленьких
артериях и капиллярах — они оказывают наибольшее сопро-
тивление движению крови. В венах давление продолжает сни-
жаться, и в полых венах оно равно атмосферному давлению .
или даже несколько ниже его.
Для медицинских целей артериальное давление определяют
обычно в плечевой артерии. У здоровых людей в возрасте от
15 до 50 лет максимальное давление равно 105—125 мм рт. ст.
У лиц старше 50 лет артериальное давление, как правило, повы-
шается, и к 60 годам оно достигает 135—140 мм рт. ст. У ново-
рожденных максимальное (систолическое^ или сердечное) арте-
риальное давление равно 50 мм рт. ст.
Минимальное (диастолическое, или сосудистое) давление
крови у людей среднего возраста в плечевой артерии в среднем
составляет 60—80 мм рт. ст.
Регуляция сердечной деятельности
Сердце имеет развитую парасимпатическую и симпатичес-
кую иннервацию (рис. 5.29).
Парасимпатическая иннервация осуществляется блуждаю-
щим нервом, который вызывает урежение ритма и умень-
шение силы сердечных сокращений. При достаточно силь-
ном раздражении блуждающего нерва происходит остановка
сердца.
148
Рис. 5.29. Иннервация сердца
(схема):
1 — кора мозга; 2 — продолгова-
тый мозг; 3 — спинной мозг; 4 —
симпатические волокна; 5 — шей-
ные симпатические узлы; б —
блуждающий нерв; 7— промежу-
точный мозг
Возбуждение симпатических нервов вызывает учащение и
усиление сердечных сокращений.
Центры, регулирующие деятельность сердца, находятся в
продолговатом и спинном мозге. Кроме того, имеются центры
регуляции сердечной деятельности в гипоталамусе и коре
больших полушарий. Изменение работы сердца происходит
рефлекторно в ответ на самые различные раздражения, дей-
ствующие на организм (тепло, холод, страх и т. д.).
Большую роль в регуляции сердечной деятельности играют
различные гуморальные влияния. Так, гормон надпочечника
адреналин учащает й усиливает работу сердца, а ацетил-
холин (медиатор) обладает противоположным эффектом.
149
ЛИМФА И ЛИМФООБРАЩЕНИЕ
Лшифа — это жидкая ткань, циркулирующая в сосудах
лимфатической системы. Среди ее форменных образований
преобладают лимфоциты и очень мало эритроцитов или по-
следние вообще отсутствуют. При повреждении кровеносных
капилляров и увеличении проницаемости их стенок, напри-
мер при действии ионизирующей радиации, число эритро-
цитов в лимфе возрастает. Химический состав лимфатичес-
кой жидкости близок к составу плазмы крови, но в ней
содержится меньше (в 3—4 раза) белков, поэтому лимфа обла-
дает небольшой вязкостью. В лимфе содержится фибрино-
ген, и она способна свертываться, но гораздо медленнее, чем
кровь.
Лимфообразование обеспечивается переходом жидкости
(воды и ряда растворенных в ней веществ) из кровеносных капил-
ляров в ткани и поступлением ее из межклеточного пространства
в капилляры лимфатической системы (рис. 5.30). Лимфа, оттека-
ющая от разных частей тела, имеет неодинаковый состав, и он
Рис. 5.30. Лимфатическая система:
1 — артерия; 2 — тканевая жидкость;
3— вена; 4— лимфатические капилляры;
5 — клетки
150
определяется специфической деятельностьк резных органов и
тканей.
В состав лимфатической системы входят: лимфатические
капилляры, лимфатические сосуды и лимфатические узлы.
Лимфатическая система выполняет следующие функции:
1) защитную (в лимфатических сосудах происходит размно-
жение лимфоцитов и фагоцитирование болезнетворных орга-
низмов, а также вырабатываются иммунные тела);
2) является дополнительной системой оттока жидкости от
органов;
3) участвует в обмене веществ (всасывание продуктов рас-
щепления жира).
Лимфатические сосуды распространены почти во всех тка-
нях и органах. Их ход совпадает с ходом вен. Началом лимфа-
тической системы являются замкнутые лимфатическое капил-
ляры, в них фильтруется тканевая (межклеточная) жидкость,
образуя лимфу. Из сетей лимфатических капилляров берут на-
чало более крупные лимфатические сосуды, снабженные клапа-
нами. По лимфатическим сосудам лимфа направляется в
грудной лимфатический п р о т о к. Она поступает
от всех органов, за исключением правой половины головы,
правой руки и правой части груди (рис. 5.31). Из этих участков
тела лимфа собирается в правый лимфатический
проток. Лимфатические протоки впадают в вены большого
круга кровообращения, соответственно в правый и левый ве-
нозные углы. Венозный угол образован слиянием внутренней
яремной и подключичной вен; справа — правый венозный
угол, слева — левый. По ходу лимфатических сосудов имеются
лимфатические узлы (рис. 5.32). В них лимфа обогащается лей-
коцитами, там же задерживаются и обезвреживаются микроор-
ганизмы. При попадании бактерий в лимфатические узлы
последние увеличиваются и становятся болезненными. Скоп-
ления лимфатических узлов отмеченола шее под нижней че-
люстью, в локтевых и коленных сгибах, в подмышечной впади-
не, в паху и других местах.
Лимфатические узлы представляют собой органы бобовид-
ной формы. В ворота узла входит артерия и выходят вены вме-
сте с выносящими лимфатическими сосудами. Снаружи орган
покрыт соединительно-тканной капсулой, от которой внутрь
органа отходят прослойки, или перекладины. Между перекла-
динами располагается ретикулярная ткань, образующая осно-
ву органа. В лимфатическом узле различают два слоя: располо-
женный на периферии темный корковый слой и занимающий
центральную часть органа светлый мозговой слой. В корковом
131
б
Рис. 5.31. Направление
тока лимфы
(схема).
. а — расположение
групп лимфатических
узлов;
б — расположение
областей, откуда
собирается лимфа
в грудной и правый
лимфатические протоки
(область последнего
заштрихована):
1 — шейные лимфатичес-
кие узлы; 2 — подкрыльцо-
вые узлы; 3 — паховые
узлы; 4 — локтевые узлы;
5 — правый поясничный
ствол; 6 — левый пояснич-
ный ствол; 7 — кишечный
ствол; 8 — грудной проток;
9 — место впадения груд-
ного .протока; 10 — место
впадения правого лимфати-
ческого протока
слое находятся лимфатические фолликулы, которые состоят
из ретикулярной ткани. В петлях ретикулярной ткани залега-
ют лимфоциты, их предшественники лимфобласты и другие
клетки. - , '
Лимфатические приносящие сосуды впадают в узел на его
выпуклой поверхности. Входя в него, эти сосуды теряют свою
стенку. Таким образом, лимфа, притекающая к лимфатичес-
кому узлу, вливается в орган и проходит между его тканевыми
элементами. Между фолликулами расположены светлые про-
странства — лимфатические синусы. Сливаясь вместе, они об-
разуют выносящие лимфатические сосуды, которые выходят
из органа через его ворота. В лимфатических фолликулах про-
исходит не только обогащение лимфы лимфоцитами, но и
152
Рис. 5.32. Лимфатические
узлы:
1 — верхняя полая вена; 2 —
грудной проток; 3 — лимфати-
ческие капилляры; 4 — лимфа-
тические узлы
обезвреживание чужеродных для организму веществ (защит-
ная функция).
Ток лимфы очень медленный, в крупных лимфатических
сосудах он равен 10,25-0,3 мм/мин. Механизм передвижения
лимфы сходен с механизмом передвижения крови в венах.
' Факторами движения лимфы по сосудам являются:
а) ритмическое сокращение стенок крупных лимфатичес-
ких сосудов (8—10 раз в минуту);
б) наличие клапанов в лимфатических сосудах (обусловли-
вает однонаправленное движение лимфы);
в) присасывающее действие расширенного грудного лимфа-
тического протока в момент увеличения объема грудной поло-
сти при вдохе и отрицательное давление в грудной полости
при выдохе;
г) сокращение скелетных мышц.
. з
ДЫХАНИЕ
ЭТАПЫ ПРОЦЕССА ДЫХАНИЯ
Дыхание в физиологии рассматривают как сложный биоло-1
гический процесс, обеспечивающий доставку кислорода тка-1
ням, использование его в процессе метаболизма и удаление |
образующегося СО2 из организма. I
В процессе дыхания выделяют следующие этапы: |
1. Внешнее дыхание — это газообмен между организмом и I
окружающим его атмосферным воздухом. *
2. Транспорт газов в кровь — это газообмен между альвео- j
лярным воздухом и кровью легочных капилляров. g
3. Транспорт газов кровью — это движение газов из легочных |
капилляре» к тканям и органам и от тканей и органов к клет-
кам. Кислород транспортируется в двух состояниях: химичес- '
кой связи с гемоглобином (оксигемоглобин) и в виде простого '
растворения в плазме крови. Углекислый газ транспортируется:
в виде солей угольной кислоты (бикарбонатов), в соединении с |
гемоглобином (карбгемоглобин) и в растворенном состоянии. |
4. Транспорт газов в ткани — это переход газов из кровенос-1
ных капилляров органа в его клетки; -
5. Тканевое дыхание, или внутреннее, — это дыхание, свя-
занное с потреблением О2 митохондриями при аэробном
окислении и высвобождение СО2 из клетки.
СТРОЕНИЕ ОРГАНОВ ДЫХАНИЯ
К дыхательной системе относят легкие и дыхательные пути,
по которым воздух проходит в легкие и обратно (рис. 5.33).
Дыхательные пути представлены носовой полостью, глот-
кой, гортанью, трахеей и бронхами. Обычно человек дышит«
через нос при сомкнутом рте, однако возможно дыхание и че-
рв’рот.
Полость носа
Полость носа образована лицевыми костями и хрящами и
поделена носовой перегородкой (сзади костной, спереди
хрящевой) на две симметричные половины, которые спереди
сообщаются с атмосферой через ноздри, а сзади — с глоткой.
В каждой половине носа имеются по три носовые ракови-
ны — верхняя, средняя и нижняя, которые соответственно
154
Рис. 5.33. Дыхательная система (схема):
7 — полость рта; 2— носовая часть глотки; 3 — мягкое небо; 4—язык; 5—
ротовая часть глотки; 6 — надгортанник; 7 — гортанная часть пкихи; 8 —
гортань; 9 — пищевод; Ю — трахея; 11 — верхушка легкого; 12 — левое
легкое; 13 — левый главный бронх; 14, 15 — альвеолы; 16 — правый глав-
ный бронх; 17 — правое легкое; 18 — подъязычная кость; 19 — нижняя
челюсть; 20 — преддверие рта; 21 — роговая щель; 22 — твердое небо; 23 —
носовая полость
образуют три носовых хода. Вдыхаемый воздух проникает в
полость носа через два передних отверстия — ноздри и,
пройдя по носовым хешам, выходит в носоглотку через хоаны.
В полость носа открывается носослезный канал, по которому
избыток слезной жидкости выводится в носовую полость.
Слизистая оболочка носа представлена многорядным мерца-
тельным эпителием. Оболочка содержит слизистые железы,
секрет которых обволакивает частички пыли и увлажняет воз-
дух; она богата кровеносными сосуд ами, что способствует со-
греванию воздуха.
155
Приспособления слизистой оболочки, способствующие
очистке и обогреванию воздуха, наиболее развиты в средних и
нижних носовых ходах, поэтому данная часть полости носа на-
зывается дыхательной. В области верхней носовой раковины
слизистая оболочка выстлана особым обонятельным эпители-
ем, содержащим рецепторные обонятельные клетки, являю-
щиеся окончаниями обонятельного нерва. Таким образом, по-
мимо дыхательной носовая полость выполняет функцию органа
обоняния.
Из полости носа вдыхаемый воздух через хоаны попадает в
носоглотку, далее в ротовую часть глотки и затем в гортань.
При дыхании через рот воздух не согревается и не очищается,
что может привести к заболеванию организма.
Гортань
Гортань является не только каналом для прохождения воз-
духа, но одновременно функционирует и как голосовой аппа-
рат. Гортань располагается на передней поверхности шеи, вы-
ступая между мышцами этой области на уровне 4—6-го
шейных позвонков. С помощью подъязычно-щитовидной
мембраны гортань соединяется с подъязычной костью и по-
этому следует за ее движениями, опускаясь и поднимаясь (на-
пример, при глотании).
Гортань имеет три парных и три непарных хряща. К непар-
ным относятся щитовидный, перстневидный и надгортанный
хрящи, к парным — черпаловидный, рожковидный и клино-
видный хрящи (рис. 5.34).
Наиболее крупным является щитовидный хрящ, состоя-
щий из двух пластинок, срастающихся спереди под прямым
или почти прямым углом. У взрослых мужчин эти пластинки
наиболее развиты и образуют спереди угловой выступ (адамо-
во яблоко, или кадык). Надгортанный хрящ, или надгортан-
ник, представляет собой листовидную пластинку, отделяю-
щую гортань от глотки и прикрывающую отверстие гортани.
Он построен из эластического хряща. Перстневидный хрящ
находится в основании гортани. Хрящи гортани могут изме-
нять свое положение относительно друг друга благодаря сус-
тавам и мышцам.
Полость гортани покрыта слизистой оболочкой, представ-
ленной многорядным мерцательным эпителием, за исключе-
нием поверхности голосовых связок и надгортанника.
Наиболее сложно устроена средняя часть гортани, где на
боковых стенках имеются две пары складок, образующих верх-
156
1
Рис. 5.34. Строение гортани:
1 — надгортанник; 2 — щитовой хрящ; 3 — голосовые связки; 4 — голосовая
щель; 5 — трахея
ние*и нижние голосовые связки. Верхние связки называют
ложными, нижние связки — истинными. Голосовые
связки натянуты между щитовидным и черпаловидным хряща-
ми и служат для воспроизведения звука. Пространство между
правыми И левыми связками называются голосовой щелью. При
сокращении определенных мышц гортани изменяется поло-
жение хрящей, между которыми натянуты связки. Вследствие
этого меняется ширина голосовой щели и натяжение связок.
Выдыхаемый воздух вызывает колебания голосовых связок, в
результате чего появляются звуки различного тона и силы.
В произношении звуков и членораздельной речи принимают
участие язык, губы, полость рта и носа, а также система орга-
нов дыхания.
Внизу гортань переходит в дыхательное горло, или трахею,
расположенную по средней линии шеи под кожей й окружен-
ную небольшим слоем мышц.
Гортань участвует в акте глотания, смещаясь при этом вверх
(за счет сокращения мышц, поднимающих подъязычную
кость). Вход в гортань закрывается в результате того, что над-
гортанник отклоняется назад под давлением на него языка и
отчасти за счет сокращения черпаловидно-надгортанных
мышц.
157
Трахея
Трахея представляет собой трубку длиной 10—15 см. «Ске- +
лет» трахеи представлен хрящевыми полукольцами, которые
сзади не замкнуты и соединены соединительно-тканной пере- $
понкой. Это обстоятельство имеет большое значение: позади |
расположен пищевод, который при прохождении комка пищи |
может расширяться, внедряясь в просвет трахеи. Верхним с во- |
им концом она прикрепляется к перстневидному хрящу с по- ]
мощью связки на уровне межпозвоночного хряща между б-м и ;
7-м шейными позвонками. На уровне 4—5-го грудного позвон- •
ка трахея переходит в бронхи. Ввиду того, что трахея распола-
гается и в области шеи я в грудной полости, ее принято делить ?
на два отдела: шейный и грудной.
Внутренняя поверхность трахеи покрыта слизистой оболочкой, .
которая имеет отдельные узелки лимфоидной ткани и выстлана
мерцательным эпителием. В шейном отделе впереди трахеи рас-
положены мышцы: грудино-подъязычная и грудино-щитовид-
ная, а на уровне 2-4-го колец трахеи — перешеек щитовид ной
железы. Желобок между трахеей и пищеводом служит местом 4
прохождения нервов и кровеносных сосудов. В грудной полости
трахея располагается в средостении. Здесь к ней спереди приле-
жат вилочковая железа, аорта и левая плечеголовная вена.
Бронхи
Трахея делится на д ва главных бронха: правый и левый. Пра-
вый бронх более короткий и толстый и отходит под меныпим уг-
лом, чем левый. Поэтому инородные тела чаще попадают в пра-
вый бронх. Каждый из главных бронхов входит в ворота
соименного легкого и разделяется по числу основных долей лег-
кого на три долевых бронха в правом легком и два — в левом.
В свою очередьэти бронхиальные ветви (бронхи II порадка), раз-
ветвляясь, дают начало сегментарным бронхам (брон-
хам Ш порядка). Последние продолжают делиться дихотомичес-
ки; образуется бронхиальное дерево. Крупные бронхи по
строению напоминают трахею. По мере уменьшения диаметра
бронхов вместо хрящевых полуколец в их стенке появляются хря-
щевые пластинки, которые совершенно исчезают в мельчайших
бронхах (концевых или терминальных бронхиолах диаметром
около 0,5 мм).
Легкие
Правое и левое легкие располагаются в грудной полости по
бокам от средостения. По форме они напоминают усеченный
158
конус, правое легкое короче и толще, чем левое. Легкие делят-
ся щелями на доли (рис. 5.35). Правое на три доли, левое — на
две. Доли подразделяются на бронхолеговдые сегменты. Пра-
вое легкое имеет 10 сегментов, а левое 9. Сегменты состоят из
первичных долек. Дольки легких отделены друг от друга про-
слойками соединительной ткани. Легкие пронизаны бронха-
ми, образующими бронхиальное дерево. Воздух, пройдя носо-
вую полость, гортань, трахею попадает в два бронха: правый и
левый.
На внутренней поверхности легкого находятся ворота, пред-
ставляющие собой углубления, через которые проходят: бронх,
кровеносные сосуды и лимфатические сосуды и нервы. Все это
составляет корень легкого.
Морфологической и функциональной единицей легкого
является ацинус (система разветвлений одной концевой брон-
хиолы). Ацинус (рис. 5.36) начинается бронхиолами, перехо-
дящими в разветвления альвеолярных ходов, которые заканчи-
ваются альвеолярными мешочками. Ацинусы отделены друг от
друга тонкими соединительно-тканными прослойками; 12—
18 ацинусов образуют легочную дольку.
Рис. 5.35. Трахея, Сфонхи и легкие (вид спереди):
J — верхушка легкого; 2 — верхняя доля; За и 36 — междолевые борозды;
4— нижняя доля; 5 — сердечная вырезка; 6 — средняя доля (правое лег-
кое); 7 — нижняя доля; 8 — бронхи; 9 — трахея
159
Рис. 5.36. Схема ацинуса:
1 — конечная (терминальная) брон-
хиола; 2 — респираторные бронхио- '
лы; 3 — альвеолярные ходы; 4 — аль-
веолы
На стенках альвеолярных ходов и мешочков располагаются
альвеолы. Альвеолы имеют вид открытого пузырька. Внутрен-
няя поверхность их выстлана плоским дыхательным эпители-
ем, находящимся на базальной мембране. Снаружи к базаль-
ной мембране подходит сеть кровеносных капилляров. Так как
альвеолы тесно прилегают друг к другу, то капилляры, оплета-
ющие их, одной своей поверхностью граничат с одной альвео-
лой, а другой поверхностью — с другой соседней альвеолой,
что обеспечивает оптимальные условия обмена газов между
воздухом альвеол и кровью капилляров. Число альвеол в лег-
ких человека равно 600—700 млн, а их общая поверхность при
вдохе колеблется в пределах 40—120 м2.
Каждое легкое погружено в серозный мешок, который на-
зывается плеврой. Серозная оболочка состоит из двух листков:
висцерального и пристеночного (париетального). Внутренний
висцеральный листок плотно прикрывает само легкое, заходит
в его борозды и таким образом отделяет доли легкого друг от
друга. Наружный пристеночный плевральный листок сраста-
ется со стенками грудной полости, а внутренней своей сторо-
ной обращен к висцеральному листку. Между двумя этими ли-
стками (висцеральным и пристеночным) имеется щелевидное
пространство, которое называется полостью плевры. Эта по-
лость шириной 5—10 мкм содержит серозную жидкость, сход-
ную по составу с лимфой, и благодаря которой уменьшается
трение между прилегающими друг к другу висцеральным и
пристеночным листками. Легкие постоянно находятся в рас-
тянутом состоянии, так как давление в межплевральной поло-
сти всегда чуть ниже атмосферного, поэтому его условно назы-
вают отрицательным, а атмосферное давление при этом
принимают за нулевое. Атмосферный воздух, растягивая лег-
кие, прижимает их к плевре и грудной стенке. Вследствие это-
го плевральная полость по форме представляет собой узкую
закрытую щель между двумя листками. Но легочная ткань
обладает большой эластичностью, противодействуя рас-
тяжению. Эта эластическая тяга легких уменьшает величину
160
давления воздуха (760 мм рт. ст.) на 4—9 мм рт. ст. (756—
751 ммрт. ст.). Чем сильнее растягиваются легкие, тем выше
становится их эластическая тяга и тем меньше меняется давле-
ние в плевральной полости. Во время вдоха оно равно —9 мм
рт. ст., во время выдоха -4 мм рт. ст.
Париетальная плевра'разделяется на реберную, диафраг-
мальную и медиастинальную. Реберная плевра покрывает реб-
ра, диафрагмальная — диафрагму, медиастинальная — средо-
стение.
Средостение — это комплекс органов, расположенных меж-
ду правым и левым листками медиастинальной плевры (серд-
це, пищевод, дуга аорты, вилочковая железа, нервы и пр.).
В месте перехода реберной плевры в диафрагмальную образу-
ется реберно-диафрагмальный синус — запасное пространство
для легких при их расширении.
МЕХАНИЗМ ВДОХА И ВЫДОХА
Акт вдоха обеспечивается сокращением межреберных
мышц и диафрагмы (рис. 5.37). Межреберные мышцы, сокра-
щаясь, приподнимают ребра,.несколько поворачивают их вок-
Рис. 5.37. Положение грудной клетки и диафрагмы при выдохе
(слева) и вдохе (справа):
1 — наружные межреберные мышцы; 2 — внутренние межреберные мыш-
цы; 3 — диафрагма
6-281
161
руг своей оси и отводят в стороны, а грудину — вперед, В ре-
зультате объем грудной полости увеличивается в переднезад-
нем и боковых направлениях. Одновременно сокращается диа-
фрагма; при этом меняется ее форма: из куполообразной она
становится плоской, за счет чего увеличивается объем грудной
полости в продольном направлении (см. рис. 5.37).
Поступление воздуха в легкие является пассивным процес-
сом и обусловлено разностью его давления в легких и окружа-
ющей среде. При вдохе объем грудной полости увеличивается,
давление в плевральной полости становится более отрицатель-
ным (9—8 мм рт. ст). Вследствие этого сопротивление в легких
к растяжению уменьшается и они растягиваются. Разность
давлений воздуха в атмосфере и в альвеолах легких является не-
посредственной причиной д вижения воздуха из окружающей
среды в легкие — происходит вдох (рис. 5.38, 5.39). В конце
вдоха эластическая тяга грудной клетки начинает противодей-
ствовать вдоху. Ее направление совпадает с направлением эла-
стической тяги легких, что препятствует продолжению вдоха.
Рис. 5.38. Плевральное давление на различных фазах дыхания.
Под влиянием эластической тяти легких, действующей по
направлению стрелок, в плевральной полости создается
«отрицательное» (по отношению к окружающей среде) давление
которое можно зарегистрировать с помощью манометра
162
J
Эластическая
тяга при
дыхательных
движениях
j Эластическая
у тяга в покое
Рис. 5.39. Схема, объясняющая изменения плеврального давления
(Рщ,) и альвеолярного давления (Ра) при вдохе (слева) и выдохе
(справа):
Рр — давление в полости рта; R — аэродинамическое сопротивление воз-
духоносных путей
Акт выд оха начинается с расслабления наружных дыхатель-
ных мышц и диафрагмы. Вследствие этого под действием элас-
тических сил легких и давления внутренних органов, а также
силы тяжести грудной клетки объем ее уменьшается. Эласти-
ческая тяга легких обусловлена тремя факторами: 1) поверхно-
стным натяжением пленки жидкости, покрывающей внутрен-
нюю поверхность альвеол; 2) упругостью ткани стенок альвеол
вследствие наличия в них эластических волокон; 3) тонусом
бронхиальных мышц. Внутренняя поверхность альвеол выст-
лана сурфактантом, имеющим низкое поверхностное на-
6*
163
тяжение. Это очень важно для стабилизации состояния альве-
ол. Основная функция сурфактанта — обеспечение расшире-
ния альвеол при вдохе й противодействие спаданию их при
выдохе. Кроме того, сурфактант обладает бактерицидными
свойствами, а также препятствует прохождению жидкости в
полость альвеол. В его образовании участвуют фосфолипиды и
гликопротеиды.
Уменьшение объема грудной полости обусловливает повы-
шение плеврального давления. Оно становится менее отрица-
тельным (5—3 мм рт. ст.). В результате этого и под действием
эластической тяги легких их объем уменьшается, давление
воздуха в них становится выше атмосферного (см. рис. 5.39), и
воздух удаляется в окружающую среду. Выдох заканчивается
тогда, когда эластическая тяга легких уравновешивается повы-
шающимся давлением в плевральной полости. Акт выдоха ста-
новится активным при сокращении внутренних межреберных
мышц.
Типы дыхания. В зависимости от того, какие мышцы пре-
имущественно участвуют в акте вдоха, различают грудной,
брюшной (диафрагмальный) и смешанный типы дыхания.
У мужчин тип дыхания брюшной, у женщин — грудной, хотя
типы дыхания могут меняться в зависимости от некоторых ус-
ловий (например, физическая работа).
Пневмоторакс. Попадание воздуха в плевральную полость
получило название пневмоторакса. Он .может произойти при
повреждении плевры, например, при ранениях стенки груд-
ной полости. При этом явлении легкое спадается, дыхание
может прекратиться. '
Пневмоторакс может быть и искусственным — его вызыва-
ют для лечения туберкулеза легких. В этом случае прокалыва-
ют грудную стенку иглой от шприца и впускают необходимое
количество воздуха в плевральную полость. Легкое до извест-
ной степени спадается, его движения ограничиваются, что
способствует закрытию и заживлению патологических поло-
стей (каверн). Через некоторое время воздух из плевральной
полости рассасывается и давление в ней вновь падает.
ВЕНТИЛЯЦИЯ ЛЕГКИХ И ДЫХАТЕЛЬНЫЙ АКТ
Количество воздуха, находящееся в легких после макси-
мального вдоха, составляет общую емкость легких. У взрослого
человека она колеблется от 4200 до 6000 мл. Общая емкость
легких состоит из жизненной емкости легких (ЖЕЛ) (3300—
164
4800 мл) и остаточного объема воздуха (’/з от ЖЕЛ) (НЮ-
1500 мл) (рис. 5.40).
. Жизненная емкость легких представляет собой то коли-
чество воздуха, которое можно выдохнуть после максимально-
го вдоха. ЖЕЛ составляют три легочных объема: 1) дыха-
тельный объем — это то количество воздуха, которое
вдыхается и выдыхается при каждом обычном спокойном вдо-
хе (400—500мл); 2) резервный объем вдоха (допол-
нительный воздух) — это то количество воздуха, которое мож-
но вдохнуть при максимальном вдохе после обычного вдоха
(1900—3200 мл); 3) резервный объем выдоха
(резервный воздух) — это количество воздуха, которое можно
выдохнуть при максимальном выдохе после обычного выдоха
(700-1000 мл).
Остаточный объем воздуха — это тот воздух, который оста-
ется в легких после максимального выдоха, поэтому легкие
полностью не спадаются.
СПИРОМЕТРИЯ
ИЗМЕНЕНИЕ ОБЪЕМА ВОЗДУХА В ЛЕГКИХ
ОТ ГЛУБОКОГО ВЫДОХА ДО ГЛУБОКОГО ВДОХА
Гпубокий ВЫДОХ
Покойный ВЫДОХ
Покойный ВДОХ
Гпубокий ВДОХ
1000 см3
1500 СМ3
2000 СМ3
3500 СМ3
Рис. 5.40. Определение жизненной емкости легких
165
При спокойном дыхании после выдоха в легких остается1
резервный объем выдоха и остаточный объем воздуха. Сумму
этих объемов называют функциональной остаточ-
ной. емкостью.
Дыхательным мертвым пространством на-
зывается тот объем дыхательных путей, в котором не происхо-
дит процесса газообмена между воздухом и кровью. У человека
среднего возраста объем дыхательного мертвого пространства
составляет 140—150 мл, или */з дыхательного объема при спо-
койном дыхании. В альвеолах к концу спокойного выдоха на-
ходится около 3000 мл воздуха (функциональная остаточная
емкость), поэтому при каждом спокойном вдохе обновляется
лишь */7 — '/io альвеолярного воздуха.
Показателями вентиляции легких являются:
а) минутный объем дыхания, определяемый как
произведение дыхательного объема на число дыханий в мину-
ту. У взрослого человека в спокойном состоянии минутный
объем дыхания составляет 6—8 л в минуту, при физической на-
грузке — от 30 до 100—250 л в минуту;
б) частота дыхательных движений в покое, состав-
ляющая 12—16 в минуту.
ДИФФУЗИЯ ГАЗОВ В КРОВЬ
Диффузия газов из окружающей среды в жидкость и из
жидкости в воздух подчиняется законам движения газов. Если
над жидкостью находится смесь газов, то каждый газ растворя-
ется в жидкости соответственно его парциальному давлению.
Парциальное давление — это то давление газа, которое прихо-
дится на его долю от общего давления смеси газов. Таким об-
разом, парциальное давление пропорционально содержанию
каждого газа в смеси. По законам движения газы из области с
высоким парциальным давлением переходят в область с низ-
ким парциальным давлением. Поэтому вдыхаемый (атмосфер-
ный) и выдыхаемый воздух отличаются по содержанию в нем
О2 и СО2 (табл. 5.2).
Направление и интенсивность перехода из альвеолярно-
го воздуха в кровь легочных микрососудов, а СО2 в обратном
направлении определяет разница между парциальным дав-
лением в альвеолярном воздухе и парциальным давлением
растворенного газа в крови. Так, для О2 парциальное давле-
ние в альвеолах составляет 102 мм рт. ст., а в крови, посту-
пающей в легкие, 40 мм рт. ст. Следовательно, О2 из альвео-
Таблица 5.2
Содержание газов, %, во вдыхаемом, выдыхаемом
и альвеолярном воздухе
Содержание газа, % Воздух
вдыхаемый выдыхаемый альвеолярный
Кислород 20,94 16,0 14,0
Углекислый газ 0,03 4,0 - 5,6
Азот* 79,03 80,0 80,4
. * Имеется в виду азот и все инертные газы.
лярного воздуха будет диффундировать в кровь. Для С€>г
парциальное давление составляет в альвеолярном воздухе
40 мм рт. ст., а в крови, притекающей к легким, 47 мм рт. ст.
Поэтому СО2 будет диффундировать из крови в альвеоляр-
ный воздух. Кислород в крови находится в растворенном
виде (менее 1%) и в соединении с Нв (99%). Каждый грамм
Нв способен связать 1,4 мл Ог и, следовательно (при содер-
жании Нв 150 г/л), каждые 100 мл крови мохуг переносить
около 21 мл Ог- Эта величина характеризует кислородную ем-
кость крови (КЕК).
При задержке дыхания при глубоком вдохе величина диф-
фузии газов оказывается наибольшей. За счет гравитационно-
го перераспределения кровотока и объема крови в легочных
капиллярах диффузная поверхность легких в положении лежа
больше, чем в положении сидя, а сидя — больше, чем в поло-
жении стоя. С возрастом диффузная способность легких сни-
жается.
ТРАНСПОРТ ГАЗОВ КРОВЬЮ
Из альвеолярного воздуха в кровь поступает кислород, а
из тканей — углекислый газ переносится кровью. Кислород
далее поступает в ткани, а углекислый газ в легкие. Основная
масса О2 переносится Нв. Нв образует с Ог непрочное, диссо-
циирующее соединение — оксигемоглобин Нв(О2)4. В этой
форме он транспортируется' тканям. В тканях оксигемогло-
бин легко отдает О2 (за счет разности парциального давле-
ния) и превращается в восстановленный Нв(ННв), который
вновь транспортируется к легким. СОг поступает из тканей.
В тканевых капиллярах растворяющийся в плазме СОг за счет
167
разности парциального давления диффундирует в эритроци-
ты, где под влиянием фермента карбоангидразы превращает-
ся в угольную кислоту (СО2 + Н2О = Н2СО3). Угольная кис-
лота диссоциирует в эритроцитах на ионы Н+ и НСОз", так
как мембрана эритроцитов проницаема для анионов, то
ионы НСОз" диффундируют в плазму. Вместо иона НСОз" в
эритроциты поступают ионы хлора. В плазме благодаря
переходу ионов хлора в эритроциты освобождаются ионы
натрия, с которыми соединяются ионы НСОз", образуя
NaHCO3. Освобождающийся Н+ ион угольной кислоты в
эритроцитах связывается с Нв и образует восстановленный
Нв(ННВ) (рис. 5.41).
Кроме того, СОг (около 10% от общего количества) соеди-
няется с Нв, образуя карбгемоглобин НвСО2. Выяснено, что
СО2 присоединяется к аминной группе Нв путем карбамин-
ной связи. Таким образом, СО2, поступающий в кровь из тка-
ней, переносится в кровь в виде бикарбоната, в виде карбге-
моглобина и свободно — в растворенном состоянии.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Регуляция дыхания осуществляется рефлекторным и гумо-
ральным путями. Оба этих механизма обеспечивают ритми-
ческий характер дыхания и изменение его интенсивности,
приспосабливая организм к различным условиям окружаю-
щей среды. Дыхательный центр расположен в продолговатом
мозге. Нервный центр — парный. От групп нервных клеток в
правой половине продолговатого мозга импульсы поступают к
дыхательным мышцам правой половины тела. От групп клеток
в левой половине — к мышцам левой половины тела. В свою
очередь дыхательный центр состоит из центра вдоха (инспира-
торного центра) и центра выдоха (экспираторного центра).
К дыхательному центру также относится группа клеток верх-
ней части варолиева моста. Это так называемый центр пневмо-
таксиса.
Дыхательному центру свойственна автоматия, т. е. способ-
ность генерировать ритмические импульсы без поступления в
него каких-либо возбуждений. Вдох и выдох обеспечивается
следующими нервными процессами. Афферентные импульсы
(к мозгу) дыхательный центр получает от механорецепторов
легких, дыхательных путей, дыхательных мышц, а также от хе-
морецепторов сосудистых рефлексогенных зон. Благодаря
автоматии и раздражающему воздействию афферентных им-
168
В тканях
Эритроцит Плазма Клетка
В легких
Рис. 5.41. Процессы, происходящие в эритроците при поглощении
или отдаче кровью кислорода и углекислого газа (схема)
пульсов происходит возбуждение центра вдоха, от которого
нервная импульсация распространяется1 по эфферентным пу-
тям. Это приводит к сокращению дыхательных мышц, в ре-
зультате чего происходит вдох. Растяжение мышц при вдохе
169
вызывает возбуждение механорецепторов, находящихся в стен-
ках альвеол. Импульсы от этих рецепторов распространяются
по чувствительным волокнам блуждающего нерва к нейронам
центра выдоха и возбуждают его. Одновременно импульсы от
инспираторных нейронов поступают вцентры пневмотаксиса,
а от него приходят к нейронам экспираторного центра и тоже
возбуждают его. Возникшее возбуждение центра выдоха тор-
мозит инспираторный центр, вследствие чего импульсация к
дыхательным мышцам прекращается. В результате легкие спа-'
даются — происходит выдох. Благодаря этому возбуждение
механорецепторов, подающих импульсацию в экспираторный
центр, прерывается. Затем прекращается тормозящее действие
нейронов центра выдоха на инспираторный центр. Это ведет к
новому возбуждению инспираторных нейронов и происходит
новый вдох.
Кроме того, рефлекторная регуляция дыхания осуществля-
ется под влиянием гуморальных факторов. Специфическим
гуморальным регулятором дыхательных движений является
СО2, который накапливается в крови и вызывает возбуждение
дыхательного центра, дыхание при Этом становится глубже.
Недостаток О2 в крови, омывающей нейроны дыхательного
центра, оказывает влияние на их активность и дыхание учаща-
ется.
ПИЩЕВАРЕНИЕ
ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОГО
ТРАКТА
Для жизнедеятельности организма необходима энергия.
Эту энергию организм получает с пищей. В пище содержатся
различные вещества, к которым относятся белки, жиры, угле-
воды, минеральные вещества, витамины. Минеральные веще-
ства (соли), вода и витамины усваиваются человеком в том
виде, в котором они находятся в пище. Белки, жиры, углеводы
(полисахариды), являясь высокомолекулярными соединения-
ми, не проходят через биологические мембраны и нуждаются
в дополнительном расщеплении на более низкомолекулярные
соединения.
Процесс расщепления высокомолекулярных соединений с
последующим всасыванием продуктов расщепления в кровь и
лимфу называется пищеварением (рис. 5.42).
170
Пища — 800-1000 г
Вода — 1,2-1,5 л
Рис. 5.42. Последовательность процессов, происходящих
в пищеварительном конвейере
171
Пищеварение происходит в пищеварительном тракте под
воздействием ферментов. Пищеварительный тракт осуществ-
ляет следующие основные функции: секреторную, моторную,
всасывательную и экскреторную.
1. Секреторная функция заключается в расщеплении белков,
жиров, углеводов под воздействием ферментов. Протеазы —
это ферменты, расщепляющие белки, липазы — расщепляю-
щие жиры, и амилазы — расщепляющие углеводы. Все три
группы ферментов относятся к гидролитическим ферментам,
под воздействием которых белки распадаются до аминокис-
лот, жиры — до глицерина и жирных кислот, углеводы — до
моносахаридов.
Ферменты обладают большой специфичностью —
каждый из них действует только на определенное химическое
соединение или продукты его распада. Большинство фермен-
тов являются белковыми веществами и для их проявления не-
обходимы: 1) оптимальная температура 36—37’ С; 2) опреде-
ленная реакция среды. Ферменты в весьма малых количествах
действуют наподобие катализаторов, ускоряя химические пре-
вращения, не меняя при этом свою химическую структуру.
2. Моторная, или двигательная, функция осуществляется
мускулатурой пищеварительного тракта и обеспечивает: 1) же-
вание; 2) глотание; 3) передвижение пищи по пищеваритель-
ному тракту; 4) выбрасывание непереваренных остатков.
3. Всасывательная функция осуществляется слизистой обо-
лочкой желудка, тонкого и толстого кишечника. Этот процесс
обеспечивает поступление переваренных органических веществ,
солей, витаминов и воды во внутреннюю среду организма.
4. Экскреторная функция проявляется в выведении из внут-
ренней среды веществ в просвет желудочно-кишечного тракта.
Железы пищеварительного тракта способны выделять азотис-
тые соединения (мочевину, мочевую кислоту и др.), соли,
воду, ядовитые и различные лекарственные вещества и продук-
ты их метаболизма.
В пищеварительной системе различают пищеварительный
тракт (канал) и крупные пищеварительные железы, открываю-
щиеся в него своими выводными протоками.
Пищеварительный тракт (рис. 5.43) начинается ротовым
отверстием, за которым следует полость рта, глотка, пищевод,
желудок, тонкая и толстая кишка. Стенка пищеварительного
тракта состоит из трех основных слоев: 1) наружного (соедини-
тельно-тканного); 2) среднего (мышечного) и 3) внутреннего
(слизистого), состоящего из эпителия и соединительно-ткан-
ной пластинки.
172
Рис. 5.43. Строение
пи щеварител ьного
канала (схема):
1 — глотка; 2 — пищевод;
3 — вход в желудок; 4 — ме-
сто перехода желудка в две-
надцатиперстную кишку;
5 — место перехода двенад-
цатиперстной кишки в то-
щую; 6 — тощая кишка;
7 - нисходящая ободочная
кишка; 8— сигмовидная
кишка; 9 — червеобразный
отросток; 10 — подвздош-
ная кишка; 11 — слепая
кйшка; 12— восходящая
ободочная кишка; 13 — ме-
сто перехода восходящей
ободочной кишки в попе-
речную ободочную кишку
(большая часть поперечной
ободочной кишки удалена);
14 — двенадцатиперстная
кишка
ЭТАПЫ ПРОЦЕССА ПИЩЕВАРЕНИЯ
Пищеварение в пищеварительном тракте осуществляется
следующим образом.
В полости рта происходит измельчение пищи, смачивание
ее слюной, частичное расщепление углеводов и формирование
пищевого комка.
На передней стенке ротовой полости (рис. 5.44) находится
ротовое отверстие, образованное губами. Верхняя стенка рото-
вой полости образована твердым и мягким небом. Твердое
небо отделяет носовую полость от ротовой, представляя для
первой дно, а для второй крышу. Мягкое небо состоит из по-
крытых слизистой оболочкой мышц. К ним относят мышцы,
173
Рис. 5.44. Полость рта (щеки разрезаны):
1 — верхняя губа; 2 — уздечка; 3 — десна; 4 — верхние зубы; 5 — твердое
небо; 6 — мягкое небо; 7 — небво-язычиая дужка; 8 — небно-глоточная
-дужка; 9 — небная миндалина; 10 — разрез щеки; 11 — нижние зубы; 12 —
десна; 13 — нижняя губа; 14 — уздечка нижней губы; 15 — язык (спинка
языка); К — зев; 77 — язычок
поднимающие и натягивающие мягкое небо, и непарная мыш-
ца — язычок. При сокращении этих мышц во время глотания
мягкое небо поднимается и герметически отделяет носовую
часть глотки — носоглотку—от остальной ее части.
В ротовой полости находятся зубы. В каждом зубе разли-
чают коронку, шейку и корень (рис. 5.45). С помощью корня
зуб укрепляется в челюсти. Выступающая в полость рта и со-
прикасающаяся с пищей часть зуба называется коронкой.
174
Верхние зубы
4
Нижние зубы
б
Рис. 5.45. Зубы человека.
а — расположение зубов разные
типов в челюсти и их внешнее
строение:
1 — резцы; 2 — клыки; 3 — малые
коренные зубы; 4 — большие ко-
ренные зубы;
б — внутреннее строение зубов:
I — эмаль; 2 — дентин; 3 — пульпа*
4 — артерия; 5 — вена; б — нерв
12 3 4
Зубная кость, которая составляет главную часть стенки зуба,
называется дентином. Коронка зуба поверх дентина покрыта
эмалью, а корень покрыт цементом. Цемент — это грубово-
локнистая кость, основное вещество которой пропитано со-
лями извести.
Зубная система человека гетеродонтная, т. е. в ней присут-
ствуют зубы различной формы: в каждой половине верхней и
нижней челюсти имеется по два резца, одному клыку, два ма-
лых коренных зуба, а у взрослых еще и три больших коренных
зуба.
175
В ротовой полости находится мышечный орган — язык.
Язык состоит из поперечно-полосатой мускулатуры и являет-
ся органом речи, участвует в оценке вкуса пищи (за счет вкусо-
вых рецепторов), помогает ее перемешиванию при жевании,
смачиванию ее слюной и глотанию.
В полость рта открываются и выделяют секрет слюнные желе-
зы. Они делятся на две группы: железы слизистой оболочки рта и
железы, расположенные за пределами слизистой оболочки.
Железы слизистой оболочки рта и языка получают свое на-
звание по месту своего расположения: небные, щечные, языч-
ные, губные, зубные.
Из второй группы самая крупная — околоушная железа. Ее
выводной проток открывается на уровне верхнего второго
большого коренного зуба.
Подчелюстная железа лежит в подчелюстной ямке под диа-
фрагмой рта. Выводной проток открывается под языком.
Подъязычная железа лежит под языком на диафрагме рта. Ее
выводной проток соединяется с протоком подчелюстной же-
лезы и открывается вместе с ним.
Слюна представляет собой вязкую бесцветную жидкость,
состоящую на 95,5—99% из воды и на 1—1,5% из органических
и неорганических веществ. К органическим веществам отно-
сятся: белковое слизистое вещество муцин, некоторое коли-
чество глобулинов, аминокислот, мочевина и др. В слюне со-
держится неорганических веществ (соли кальция, калия и др.)
в 2—3 раза меньше, чем органических. Реакция слюны — сла-
бощелочная. Муцин, как основной компонент слюны, уча-
ствует в формировании пищевого комка. Кроме того, в слюне
содержится лизоцим (бактерицидное вещество), фермен-
ты амилаза и мал ьтаза. Амилаза расщепляет крахмал
до мальтозы, а мальтаза — дисахарид мальтозу на две молеку-
лы глюкозы. Пища находится в ротовой полости 15—20 с и по-
этому здесь не происходит полного расщепления крахмала.
У человека в сутки отделяется в среднем около 1000—1200 мл
слюны. Пищевой комок движением языка и щек передвигается
на спинку языка и направляется к его корню. Затем комок при-
жимается к твердому небу и передвигается в глотку.
Глотка — мышечная трубка, расположенная впереди шей-
ных позвонков. В ней различают три части: носоглотку,
ротоглотку и гортанную ч а с т ь глотки. В ротовой
части перекрещиваются дыхательные и пищеварительные
пути. В глотку открываются хоаны (соединяют ее с носовой
полостью), отверстия евстахиев труб (связывают глотку с поло-
стью среднего уха). В ротовой части глотка через зев сообщает-
176
ся с полостью рта, а в гортанную часть открываются вход в гор-
тань и отверстие пищевода. Как только пищевой комок посту-
пил в глотку, сокращаются мышцы, суживающие глотку, выше
комка и он передвигается в пищевод.
Пищевод — мышечная трубка длиной 25—30 см. Верхняя
треть пищевода образована поперечно-полосатой мышечной
тканью, остальная часть — гладкой мышечной тканью. Пище-
вод проходит через отверстие в диафрагме в брюшную полость
и здесь переходит в желудок. Функция пищевода — активное
проведение пищевого комка перистальтическими сокращени-
ями мышечной оболочки. Таким образом из, пищевода пища
попадает в желудок (рис. 5.46).
Желудок представляет собой расширенную часть пищева-
рительной трубки. В нем различают место перехода пищевода
в желудок, т. е. вход в него (кардиа), и прилегающую кардиаль-
ную часть желудка. Верхняя часть желудка, выпуклая кверху,
называется сводом. Место перехода в двенадцатиперстную
Рис. 5.46. Расположение печени, желудка
и поджелудочной железы:
1 — пищевод; 2 — желудок; 3 — поджелудочная железа; 4 — тонкая кишка;
5 — прямая кишка; 6 — толстая кишка; 7 — двенадцатиперстная кишка; 8—
желчный пузырь; 9 — печень
177
кишку называется ь;: лвратником (пилорус). Часть желудка,
прилегающая к прнь^тнику, называется пилорической. В же-
лудке различают две поверхности — переднюю и заднюю, и
два изогнутых края — верхний вогнутый (малая кривизна) и
нижний выпуклый (большая кривизна). Большая часть желуд-
ка, кроме пилорической, называется телом желудка. Емкость
желудка составляет от одного до нескольких литров. Стенка
его состоит из соединительно-тканной, мышечной и слизи-
стой оболочек. В слизистой оболочке находятся трубчатые же-
лезы, вырабатывающие желудочный сок (2,0—2,5 л/сут). Раз-
личают три вида желудочных желез: собственные,
пилорические и кардиальные. Преобладают собственные же-
лезы желудка, которые расположены в области тела и дна же-
лудка. Эти железы построены из трех видов клеток: главных,
обкладочных и добавочных. В главных клетках образуются пи-
щеварительные ферменты, в обкладочных клетках вырабатыва-
ется соляная кислота, а в добавочных — мукоидный слизи-
стый секрет.
Желудочный сок имеет кислую реакцию среды. Соляная
кислота обладает бактерицидным действием; кроме того, она
активирует пепсин, вызывает денатурацию и набухание белков
и тем самым способствует их последующему расщеплению
пепсином. Когда пищевой комок попадает в желудок, в нем
действие амилазы и мальтазы продолжается до тех пор, пока
пищевой комок не пропитывается кислым желудочным соком,
в котором ферменты слюны теряют свою активность. В состав
желудочного сока входят следующие ферменты: пепсин, жела-
тиназа, липаза, химозин. Пепсин расщепляет белки до по-
липептидов; желатиназа гидролизует желатин; липаза
расщепляет эмульгированные жиры молока на глицерин и
жирные кислоты; химозин створаживает молоко.
И. П. Павлов изучал секрецию и состав желудочного сока в
опытах на собаках (рис. 5.47). Один из опытов И. П. Павлова
по изучению секреции желудочного сока состоял в перерезке
пищевода у собаки и выведении двух образовавшихся концов
на поверхность шеи, так что при кормлении собаки пища,
вместо того чтобы попадать в желудок, выходила наружу через
отверстие в шее. Хотя пища при этом не использовалась, такое
«мнимое кормление» приводило к выделению желудочного
сока в количестве, равном примерно четверти нормального ко-
личества (у человека при каждом приеме пищи в норме выде-
ляется от 400 до 800 мл желудочного сока). При перерезке не-
рвов, идущих к желудку, сокоотделение полностью
прекращалось. Когда пищу вкладывали в ту часть перерезан-
178
Рис. 5.47. Собака с эзофаготомией и фистулой желудка
кого пищевода, которая ведет в желудок, так, что собака не
могла видеть пищу и ощущать ее запах и вкус, поступление
пищи в желудок вызывало примерно половину нормального
количества сока. Это сокоотделение происходило даже при
перерезке идущих к желудку нервов, хотя при этом сока выде-
лялось меньше. Следовательно, сокоотделение зависит от-
части от нервной стимуляции желудочных желез импульсами
клеток слизистой желудка, отчасти — от действия гормона
гастрина. Существование и действие этого гормона было дока-
зано в опытах с перекрестным кровообращением. При этом
кровеносную систему одной собаки соединяли с помощью ре-
зиновых трубок с кровеносной системой другой собаки. Когда
в пилорическую область желудка одной собаки вводили пищу,
желудочные железы другой собаки начинали секретировать
желудочный сок. Поскольку между обеими собаками не было
никаких нервных связей, секреция желудочного сока у второй
собаки вызывалась веществом, переносимым кровью, т. е. га-
стрином.
При опыте с «мнимым кормлением» выделение желудоч-
ного сока не прекращается сразу же после окончания «мни-
мого кормления», а длится еще полтора-два часа. Но этот
опыт не может полностью воспроизвести условия и процес-
сы, происходящие в желудке в период пищеварения у нор-
мального животного, потому что во время опыта пищи в же-
лудке нет. И поэтому И. П. Павлов предложил еще одну
операцию, которая состояла в том, что из дна желудка выре-
зали й сшивали маленький, «павловский», желудочек. Нервы
и кровеносные сосуды при этом не перерезали. При кормле-
нии животного пища попадала только в большой желудок,
179
где и переваривалась, а в маленьком желудочке пиши не
было, но сок выделялся так же, как и в большом желудке. Сок
из маленького желудочка собирали через фистулу, и по его ?
выделению наблюдали за работой всего желудка. В результате
этого опыта было установлено, что желудочный сок выде-
ляется не полтора-два часа, как при «мнимом кормлении», а
в течение всего времени пока пища находится в желудке (от
четырех до десяти часов). Причиной столь длительного соко-
отделения являются вещества, которые образовались при
действии продуктов частичного расщепления пищи. Эти ве-
щества, попадая в кровь, транспортируются ею к желудочным
железам и стимулируют их работу.
Таким образом, регуляция желудочного сока состоит из
двух фаз: нервной и гуморальной. Первая начинается раздра-
жением зрительных и обонятельных рецепторов видом и запа-
хом пищи (условно-рефлекторное сокоотделение). Когда
пища попадает в рот и раздражает вкусовые рецепторы, проис-
ходит безусловно-рефлекторное выделение желудочного сока.
В ходе переваривания пищй в желудке образуются химические
вещества, которые вызывают дальнейшее соковыделение (гу-
моральная фаза). Без нервной фазы невозможны гуморальная
фаза и переваривание пищи в желудке. Поэтому И. П. Павлов
назвал первую (нервную) фазу аппетитным (запаль-
ным) сокоотделением.
Гладкая мускулатура стенок желудка обладает автома-
тией и обеспечивает двигательную функцию желудка. К гу-
моральным факторам, усиливающим сокращения желудка,
относятся гастрин, гистамин и ацетилхолин. Тормозят мо-
торику адреналин, норадреналин и энтерогастрон. Пища нахо-
дится в желудке от 5 до 10 часов в зависимости от состава
пищи. Дольше всего (до 10 часов) в желудке остается жир-
ная пища.
Далее жидкая или полужидкая пищевая масса, называемая
химусом, переходит в тонкий кишечник.
Тонкая кишка начинается у привратниковой части желудка
и оканчивается утачала толстой кишки. Ее длина у взрослого
человека 5—6 м. Тонкую кишку делят на три отдела: 1) двенад-
цатиперстная кишка (ближайший к желудку отдел длиной
25—30 см); 2) тощая кишка и 3) подвздошная.
Эвакуация пищи в двенадцатиперстную кишку происходит
порциями, благодаря периодическому расслаблению и сокра-
щению сфинктера привратника (пилорического сфинктера).
В двенадцатиперстной кишке реакция среды щелочная. Пере-
ход пищи в кишечник длится до тех пор, пока реакция в две-
180
надцатиперстной кишке не станет кислой. Тогда соляная кис-
лота раздражает рецепторы в слизистой оболочке кишки,
вследствие чего опять-таки рефлекторно мышцы сфинктера
сокращаются и закрывают проход из желудка в кишечник. Как
только реакция становится щелочной вследствие нейтрализа-
ции соками поджелудочной железы, желез кишечника и жел-
чи, мышцы сфинктера вновь расслабляются и новая порция
пищи переходит из желудка в кишечник. Закрытие выхода из
привратника при поступлении соляной кислоты в двенадца-
типерстную кишку получило название запирательного
пилорического рефлекса.
Слизистая оболочка тонкой кишки имеет складки и много-
численные ворсинки (от 18 до 40 на 1 мм2), что увеличивает ее
общую поверхность. В ворсинку входят нервы, кровеносные и
лимфатические сосуды (рис. 5.48).
Поджелудочная железа располагается позади желудка, у зад-
ней брюшной стенки, состоит из головки, тела и хвоста и яв-
ляется железой смешанной секреции. Она, с одной стороны,
относится к железам внешней секреции, так как ее железистые
клетки продуцируют поджелудочный с о к, который че-
рез систему выводных протоков поступает в двенадцатиперст-
ную кишку, и, с другой стороны, является железой внутренней
секреции, так как ее особые клетки, называемые островки
Лангерганса, выделяют гормоны инсулин и глюкагон,
поступающие непосредственно в кровь.
Рис. 5.48. Строение
кишечной ворсинки:
1 — артерия; 2 — вена; 3 —
гладкие мышцы; 4 — цент-
ральный лимфатический
сосуд; стрелками показано
направление тока крови
181
I «И Г I <-А'< KV ’ н
7'
ЧТ-! г; rj
Поджелудочный сок представляет собой бесцветную про-
зрачную жидкость щелочной реакции. В нем содержатся фер-
менты в неактивном состоянии — тоипсиноген. химотрипси-
ноген и проэластаза. И. П. Павловым установлено, что
тоипсиноген переходит в активное состояние, т. е. в трипсин.
под влиянием фермента энтерокиназы, содержащейся в ки-
шечном соке. Трипсин воздействует на химотрипсиноген и
проэластазу и переводит их в активные фермента химотрип-
син и панкреопептидазу соответственно.
Трипсин и химотрипсин расщепляют белки в двенадцатипер-
стной кишке до аминокислот. Таким образом, в двенадцатипер-
стной кишке происходит полное расщепление белков, начатое в
желудке под действием пепсина. Липаза, вырабатываемая под-
желудочной железой, расщепляет жиры на глицерин и жирные
кислота. Амилаза, мальтаза, лактаза, сахараза расщепляют угле-
воды. Желчь, выделяемая печенью, эмульгирует жиры, активизи-
рует работу ферментов, усиливает двигательную активность ки-
шечника, тормозит развитее гнилостных процессов.
Продукты расщепления сложных углеводов (глюкоза) и
белков (аминокислота) всасываются микроворсинками и по-
ступают в кровеносное русло. Глицерин и жирные кислоты,
образовавшиеся в результате расщепления жиров, в клетках
ворсинок вновь превращаются в жиры, которые поступают за-
тем в лимфатические капилляры. В тонком кишечнике проис-
ходит всасывание витаминов, воды, минеральных солей.
Два слоя гладких мышц в стенке тонкого кишечника, со-
кращаясь, обусловливают его маятникообразные движения,
что, в свою очередь, способствует перемешиванию пищевых
масс. Благод аря перистальтическим сокращениям кишечника
его содержимое проталкивается в сторону толстой кишки.
Толстый кишечник делят на слепую кишку счервеоб-
разнымотростком (аппендиксом), ободочную (восходя-
щую, поперечную, нисходящую, сигмовидную) и прямую.
Железы толстого кишечника вырабатывают сок, относительно
бедный пищеварительными ферментами. Зато здесь находит-
ся огромное количество бактерий, ферменты которых расщеп-
ляют сложный углевод — клетчатку. Бактерии синтезируют
также витамины К и В^. Хотя длина толстого кишечника от-
носительно невелика (примерно равна длине тела), поступаю-
щие сюда пищевые массы (химус) находятся здесь дольше, чем
в других отделах пищеварительного тракта. По мере продвиже-
ния химус превращается в плотные каловые массы. Этому спо-
собствует интенсивное всасывание воды. В нижней части пря-
мой кишки расположены две кольцевые мышцы. Одна из них
182
образована поперечно-полосатыми мышечными волокнами,
что д елает акт дефекации процессом произвольным.
Печень — самая крупная железа человека. Ее масса близка к
2 кг. Структурной и функциональной единицей печени явля-
ется печеночная долька, диаметр которой составляет 1,5 мм.
Она имеет форму призмы и состоит из клеток гепатоци-
тов. В отличие от всех других органов печень получает кровь из
двух источников: артериальную — из печеночной артерии и ве-
нозную — из воротной вены. Последняя собирает кровь от пи-
щеварительного канала, селезенки, поджелудочной железы.
В капиллярах печеночной дольки кровь смешивается. Крове-
носные сосуды, выходящие из дольки, укрупняясь, собираются
в несколько печеночных вен, впадающих в нижнюю полую вену.
Секрет печени — желчь— накапливается в желчном пу-
зыре и порциями поступает в двенадцатиперстную кишку. За
сутки у человека образуется 500—700 мл желчи. В состав желчи
входят вода (90%), органические (желчные кислоты и желчные
пигменты) и неорганические вещества (минеральные соли).
К желчным пигментам относится билирубин и биливердин,
образующиеся из гемоглобина разрушающихся эритроцитов.
Среди многочисленных функций печени следует отметить
ее защитную (барьерную) роль и участие в обмене белков, жи-
ров и углеводов. В печени синтезируется витамин А, фибрино-
ген, протромбин, холестерин. Здесь синтезируется и накапли-
вается гликоген, а также откладываются в запас витамины А, Д,
К, РР. В печени задерживаются микроорганизмы, попавшие в
организм, и осуществляется обезвреживание ядовитых веществ.
Например, из сильнодействующего яда — аммиака синтезиру-
ется менее ядовитое вещество—мочевина; подвергается распа-
ду фенол — промежуточный продукт белкового обмена.
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
ОБЩЕЕ ПРЕДСТАВЛЕНИЕ
ОБ ОБМЕНЕ ВЕЩЕСТВ И ЭНЕРГИИ
Между организмом и окружающей средой происходит об-
мен веществ и энергии. Этот обмен обеспечивает пластичес-
кие и энергетические потребности организма. Энергию для
поддержания жизнедеятельности организм получает из посту-
пивших в него питательных веществ.
183
Энергия используется: а) для синтеза белков, нуклеиновых
кислот и липидов; б) для построения клеточных мембран и
органелл клетки; в) для выполнения механической, химичес-
кой, осмотической и электрической работ в организме; г) для
транспорта ионов через клеточную мембрану и т. д.
В обмене веществ и энергии (метаболизме) выделяют два
взаимосвязанных, но разнонаправленных процесса: анабо-
лизм и катаболизм.
Анаболизм (от греч. «подъем»), или ассимиляция (лат.
«пластический обмен»), — это совокупность процессов био-
синтеза органических веществ, компонентов клетки и других
структур органов и тканей. Анаболизм обеспечивает рост, раз-
витие, обновление биологических структур и накопление
энергетических субстратов.
Катаболизм (от греч. «сбрасывание» или «разрушение»),
или диссимиляция (от лат. «несходный»), — это совокупность
процессов расщепления сложных молекул до простых веществ
(с использованием части из них в качестве предшественников
биосинтеза), а затем и до конечных продуктов распада с обра- ’
зованием макроэргических и восстановленных соединений
(АТФ, реатинфосфат, НАДФ и др.).
Процессы анаболизма и катаболизма находятся в организ-
ме в состоянии динамического равновесия.
Преобладание анаболических процессов над катаболичес- ,
кими приводит к росту, накоплению массы тканей, а преобла- |
дание катаболических процессов — к частичному разрушению 1
тканевых структур. Состояние равновесного или неравновес- i
ного анаболизма и катаболизма зависит от возраста, состоя-
ния здоровья, выполняемой организмом физической нагрузки
или от переживаемых эмоций.
ОБМЕН БЕЛКОВ
Функции белков:
1) структурная — образует каркас клетки;
2) ферментативная;
3) сократительная (за счет белков актина-миозина);
4) гормональная — некоторые гормоны являются пептидами;
. 5) энергетическая — при распаде выделяется энергия.
В организме здорового человека количество распавшегося
за сутки белка равно количеству вновь синтезированного. Че-
ловек и другие животные могут добывать азот только из ами-
нокислот, поступающих в организм с белковой пищей.
134
Аминокислоты делят на две большие группы: незамени-
мые — это десять из двадцати существующих аминокислот (ва-
лин, триптофан, метионин, лицин и т. д.) и заменимые (ала-
нин, цистеин, глутаминовая и аспарагиновая кислота и т. д.).
Незаменимые это те, которые поступают в организм только
с пищей. Заменимые аминокислоты могут синтезироваться в
организме человека из других аминокислот.
В зависимости от аминокислотного состава белки также де-
лятся на две группы: полноценные белки, которые содержат пол-
ный набор аминокислот, и неполноценные белки, которые не со-
держат полного набора аминокислот.
Скорость распада и обновления белков в организме различ-
на. В среднем белки организма человека обновляются за 80 су-
ток. О суммарном обмене белка судят по количеству азота, вы-
водимого из организма человека. Установлено, что выделение
организмом 1 г азота соответствует распаду 6,25 г белка. О по-
треблении же количества азота судят по составу пищи, посту-
пающей в организм.
В тех случаях, когда в организм поступает больше азота,
чем его выделяется из организма, говорят о положитель-
ном азотистом балансе. Он обычно наблюдается при
увеличении массы мышечной ткани, в период роста организ-
ма, беременности, выздоровлении после истощающего заболе-
вания. Состояние, при котором количество выводимого из
организма азота превышает его поступление в организм, назы-
вают отрицательным азотистым балансом.Оно
имеет место при питании неполноценными белками, белко-
вом голодании и сопровождает некоторые заболевания. Норма
потребления белка для здорового человека составляет 0,75 г на
1 кг массы в сутки. Для здорового человека массой 70 кг необ-
ходимо около 53 г белка в сутки. Белки используются в орга-
низме в первую очередь в качестве пластических веществ.
В процессе их разрушения освобождается энергия для синтеза
АТФ и образования тепла.
Установлено, что после потребления белковой пищи ин-
тенсивность обмена временно повышается, иногда на 30% по
сравнению с уровнем основного обмена; это сопряжено с до-
полнительным освобождением энергии, необходимой для
превращения части аминокислот в углеводы и жиры.
Такое специфическое динамическое действие белков зави-
сит от определенного воздействия печени на аминокислоты,
так как после скармливания белка животным с удаленной пе-
ченью интенсивность-обмена не повышается. Животное с уда-
ленной печенью может жить в течение 48 ч и более, если при
185
этом ему внутривенно вводить глюкозу. Однако в конце кон
нов оно погибает из-за накопления в организме аммиака.
У нормальных животных ферменты печеночных клеток пре-
вращают аммиак в мочевину.
ОБМЕН ЛИПИДОВ
Функции липидов:
1) структурная — входят в состав клеточных мембран;
2) энергетическая — запас энергии взрослого организма
обеспечивается в основном за счет жиров;
3) жиры являются источником образования эндогенной
воды. При окислении 100 г нейтрального жира в организме об-
разуется около 107 г воды.
Липиды — это главным образом нейтральные сложные
эфиры глицерина и высших жирных кислот — триглицериды,
фосфолипиды и стерины. Жирные кислоты делят на две боль-
шие группы: а) насыщенные жирные кислоты (стеарино-
вая); б) ненасыщенные жирные кислоты (арахидоновая,
линолевая, ланолиновая).
Некоторые из ненасыщенных жирных кислот по своим фи-
зиологическим свойствам близко стоят к витаминам. Каки бел-
ки, высшие жирные кислоты подразделяются на заменимые и
незаменимые, которые поступают только с пищей (арахидоно-
вая, линолевая, ланолиновая кислоты поступают с раститель-
ными маслами — оливковым, подсолнечным, конопляным).
Генетические особенности липвдов. Липиды живых организ-
мов обладают генетической специфичностью. Различают их
видовую специфичность и индивидуальную специфичность.
Поскольку в синтезе липидов принимают участие фермен-
ты, которые контролируются генетическим аппаратом орга-
низма, то специфичность липидов, определяется генетически-
ми особенностями белков и ферментов.
Если основную роль в удовлетворении энергетических по-
требностей организма играет нейтральная молекула жира —
триглицериды, то пластическую функцию липидов в организ-
ме осуществляют главным образом фосфолипиды, холестерин
и их производные. Эти липидные молекулы выполняют функ-
цию структурных компонентов клеточных мембран, липопро-
теидов, являющихся предшественниками синтеза стероидных
гормонов, желчных кислот и простагландинов. В отличие от
белков, которые не образуют специфических запасных форм,
служащих источником энергии, запасы нейтральных жиров
1«6
триглицеридов в жировых депо человека в среднем составляют
10—20% массы его тела. Изних около половины локализованы
в подкожной жировой клетчатке. Кроме того, значительные
запасы нейтральных жиров откладываются в большом сальни-
ке, околопочечной клетчатке и между мышцами.
Жиры и масла — наиболее концентрированные пищевые
продукты, так как они значительно превосходят по калорий-
ности углеводы и белки, но и содержат меньше воды, чем угле-
водные и белковые продукты.
Суточный пищевой рацион жира составляет 80-100 г. При
этом в пищевой рацион должны быть включены обязательно
растительные масла (20—25 г).
ОБМЕН УГЛЕВОДОВ
Функции углеводов:
1) энергетическая—основной источник энергии в организме;
2) структурная — составная часть гликопротеидов, входя-
щих в клеточные мембраны. Используются для синтеза липи-
дов и некоторых аминокислот.
Организм человека получает углеводы в виде сахарозы, глю-
козы, фруктозы, растительного полисахарида — крахмала и в
виде животного полисахарида — гликогена. В желудочно-ки-
шечном тракте осуществляется расщепление сложных углево-
дов до уровня моносахаридов (глюкозы, фруктозы, галактозы).
Моносахариды всасываются в кровь и через воротную вену по-
ступают в клетки печени. Здесь фруктоза и галактоза превра-
щаются в глюкозу. Внутриклеточная концентрация глюкозы в
гепатоцитах близка к ее концентрации в крови. При избыточ-
ном поступлении в печень глюкозы она фосфорилируется и
превращается в резервную форму ее хранения — гликоген. Ко-
личество гликогена может составить в организме человека
150—200 г. В случае ограничения потребления пищи или по
мере снижения уровня глюкозы в крови происходит расщепле-
ние гликогена и поступление глюкозы в кровь. С током крови
глюкоза доставляется в различные клетки организма. Здесь
она является важнейшим источником энергии.
ОБМЕН ВОДЫ И ЛИПИДОВ
Содержание воды в организме в протоплазме клеток взрос-
лого человека составляет в среднем 73%. Водный баланс в орга-
187
низме поддерживается за счет равенства объема потерь воды и
ее поступления в организм. Суточная потребность в воде удов-
летворяется за счет поступления ее при питье (1500 мл), с пи-
щей (1000 мл) и образования в организме в ходе обменных про-
цессов эндогенной воды (400-500 мл). Из организма вода
выводится с мочой (1400 мл), калом (100 мл), посредством ис-
парения с кожи и дыхательных путей (1000—1200 мл). Таким об-
разом, минимальная потребность в воде в сутки составляет
1700 мл. Потребность организма в воде зависит от характера пи-
тания. Пища, богатая белками, а также повышенный прием
соли обусловливают большую потребность в воде, которая необ-
ходима для экскреции с большим объемом мочи осмотически
активных веществ — мочевины и минеральных веществ.
Недостаточное поступление в организм воды или избыточная
ее потеря приводит к дегидратации, что сопровождается сгуще-
нием крови и нарушением гидродинамики. Недостаток^ орга-
низме воды в объеме 20% массы тела ведет к летальному исходу.
Избыточное поступление воды в организм или снижение ее
объемов, выводимых из организма, приводит к водяной интокси-
кации, что может сопровождаться мышечными судорогами.
Физиологическая роль минеральных ионов для организма
человека чрезвычайно велика (см. табл. 5.3).
Обмен веществ и обмен минеральных ионов в организме
тесно связаны, что обусловлено необходимостью поддержания
осмотического давления.
ВИТАМИНЫ
В 1880 г. Н. И. Лунин показал, что естественные пищевые
продукты содержат дополнительные, еще не известные факто-
ры питания. Именно это положение легло в основу создания
теории витаминов.
Витамины — это группа веществ разнообразной химичес-
кой природы, которая необходима для роста, выживания и
размножения организмов.
Витамины имеют следующие общебиологические свойства.
1. Биосинтез витаминов происходит в основном вне орга-
низма человека. Эндогенный биосинтез некоторых из них,
осуществленный кишечной микрофлорой, не в состоянии по-
крыть потребность организма, поэтому человек получает их
извне, с пищей.
2. Витамины не являются пластическим материалом и не
служат источником энергии.
188
Таблица 5.3.
Физиологическая роль минеральных ионов
Мине- ральные ионы Физиологическая роль Суточная потреб- ность Источник
Na Натрий Участвует в процессе возбужде- ния клетки, в поддержании ос- мотического давления и PH сре- ды, влияет на работу почек . 2-3 г Поваренная соль в составе животной и рас- тительной пищи
Са Кальций Структурный компонент в тка- нях зубов и костей. Необходим при свертывании крови, синап- тической передаче 0,8 г Молоко, овощи, зеленые листья
К Калий Необходим для возбуждения не- рвных клеток, проведения им- пульсов, сокращения мышц, в том числе мцокарда и других клеток 2-3 г Овощи, мясо, сухофрукты, орехи
С1 Хлор Участвует в поддержании PH желудочного сока, также играет роль в процессе возбуждения и торможения в нервных клетках 3-5 г Поваренная соль, расти- тельная и жи- вотная пища
Р Фосфор Структурный компонент костей и зубов, входит в состав АТФ и клеточных мембран в виде фос- фолипидов Ю,7-0,8 г Молоко, рыба, яйца, орехи, злаки
Fe Железо Структурный компонент гемо- глобина крови, находится в со- ставе ферментов окислительного фосфорилирования (цитохромы) 10-15 мг Мясо, печень, рыба, орехи
J Йод Необходим для нормальной дея- тельности щитовидной железы 0,15— 0,3 мг Морепродукты, овощи, рыбий жир
Си Медь Играет роль в процессе всасыва- ния железа, образовании гемо- глобина 2-5 мг Яйца, печень, почки, рыба, ви- ноград, шпинат
F Фтор Структурный компонент зубной ткани 1 мг С пищевыми продуктами
Mg Магний Входит в состав коферментов, для функционирования мышеч- ной, нервной и костной ткани * 250- 350 мг Мясо, молоко, зерна
S Сера Входит в состав аминокислот, белков (инсулин) и витаминов (В1.Н) * 1г Мясо, печень, рыба, яйца
Zn Цинк Важнейший компонент фермен- тов, необходимых для нормаль- ного роста 10-15 мг Крабы, мясо, бобы, желток
Со Кобальт В составе витамина В12 100- 200 мкг Печень
189
3. Витамины совершенно необходимы для всех жизненных
процессов и биологически активны уже в малых количествах.
4. При поступлении в организм витамины оказывают влия-
ние на биохимические процессы, протекающие в различных
тканях и органах.
5. Недостаточное поступление в организм отдельных вита-
минов или нарушение их усвоения ведет к развитию патологи-
ческих процессов в виде специфических гипо- и авитамино-
зов. Для лечения этих процессов используется специфическое
действие соответствующих витаминов.
б. В повышенных дозах витамины используются в лечебных
целях в качестве мощных неспецифических фармацевтических
средств.
В настоящее время известно несколько десятков витаминов,
но в лечебной практике используются лишь около 20 из них.
Витамины разделяют на две большие группы: водораство-
римые и жирорастворимые.
I. Водорастворимые витамины
Витамин Вi (тимин)
Авитаминоз. Болезнь называется бери-бери. Основ-
ным симптомом болезни является полиневрит (поражение не-
рвной-системы) й недостаточность сердечно-сосудистой сис-
темы (одышка, сердцебиение).
Роль в организме. 1. Участвует в углеводном и белко-
вом обмене. 2. Участвует в качестве кофермента декарбоксилаз
цикла Кребса (в расщеплении пировиноградной кислоты).
3. Является важным фактором в передаче нервных импуль-
сов — усиливает активность ацетилхолина как медиатора не-
рвной системы. 4. Участвует в синтезе нуклеиновых кислот.
Источник поступления. Животные и человек ви-
тамин В] не синтезируют, поэтому полностью зависят от
поступления его из внешней среды. Тимин синтезируется зе-
леными растениями и многочисленными видами микроорга-
низмов. Значительно содержание витамина в дрожжах, в заро-
дышах и оболочках зерновых злаков, а следовательно, и в хлебе
из муки грубого помола.
Суточная потребность. При обычной нагрузке на
смешанной диете составляет 2-3 мг. Поступающий с пищей
витамин В] всасывается в основном в двенадцатиперстной и
тощей кишке. Усвоение тиамина кишечником ограничено 6—
10 мг в сутки.
190
Витамин В2 (рибофлавин)
Авитаминоз. Недостаток поступления с пищей рибо-
флавина или плохое его усвоение в организме ведет к разви-
тию арибофлавиноза. Специфическими изменениями, свя-
занными с недостатком рибофлавина, являются поражения
слизистой оболочки рта (хейлозангулярный стоматит), губ
(трещины, «заеды»), кожи (шелушение) и глаз (светобоязнь,
слезоточение).
Роль в организме. 1.Участвуете окислительно-вос-
становительных процессах, т. е. образует часть окислительно-
восстановительной системы, действуя в качестве переносчика
водорода к кислороду. 2. Участвует в обмене аминокислот в
организме, в усвоении и синтезе жиров. -
Источник поступления. Наиболее богатыми ис-
точниками витамина В2 являются дрожжи, яичный белок, мо-
локо, печень, мясо, рыба и птица. В растительном жире и зер-
новых его мало. Рибофлавин устойчив к высокой температуре,
поэтому при кулинарной обработке разрушается незначитель-
но. Его потери увеличиваются в щелочной среде, а также при
стоянии пищи на свету.
Суточная потребность. Составляет для человека
2—3 мг. Она может возрастать при физической нагрузке, при
повышении калорийности пищи с увеличением в ней белков.
Витамин Bj (пантотеновая кислота)
Авитаминоз. У человека встречается довольно редко,
так как витамин В5 широко распространен в продуктах пита-
ния и образуется микробами кишечника. Длительная недоста-
точность в пантотеновой кислоте (апантотеноз) проявляется
общим угнетением, апатией и неустойчивостью сердечно-со-
судистой системы. К этому периоду отмечается развитие син-
дрома жжения ног, который чаще начинается постепенно на-
растающими покалываниями и онемением в пальцах нижних
конечностей.
Роль в организме. 1. Пантотеновая кислота входит в
состав многих ферментов, участвующих в обезвреживании в
организме ряда токсических веществ. 2. Входит в состав фер-
ментов, участвующих в образовании фосфолипидов. 3. Прини-
мает участие в окислении пировиноградной кислоты. 4. Явля-
ется катализатором в синтезе полипептидов. 5. Стимулирует
синтез ацетилхолина, медиатора нервной системы. 6. Входит в
состав кофермента А.
Источник поступления. Пантотеновая кислота
содержится во всех продуктах, а также синтезируется кишеч-
191
ной микрофлорой человека в количестве, достаточном для
Предупреждения гиповитаминоза.
Суточная потребность человека в пантотеновой
кислоте определяется в 10—12 мг.
Витамин Bj (витамин РР, или никотиновая кислота)
Авитаминоз. В результате скудного однообразного пи-
тания с нарушением белкового обмена возникает заболева-
ние — пеллагра. Диагностика пеллагры — классические «три
Д» (дерматит, диарея, дененция). Под дененцией понимают
нарушение психики.
Роль в организме. 1. Вместе с другими витаминами
принимает участие в окислительно-восстановительных про-
цессах, являясь незаменимым компонентом коферментов
НАД и НАДФ. 2. Участвует в обмене веществ в организме (осо-
бенно в белковом и азотистом обмене). 3. Влияет через авто-
номную (вегетативную) нервную систему на гемодинамичес-
кие реакции сердечно-сосудистой системы (например,
снижение артериального давления).
Источник поступления. Витамин синтезируется в
организме из аминокислоты триптофан, а также содержится в го-
товом виде (никотиновая кислота), содержится во многих про-
дуктах как животного, так и растительного происхождения. Ею
богаты печень, почки, зерновые культуры (ячмень, пшено, рис,
рожь и др.), фрукты и овощи бедны никотиновой кислотой.
Суточная потребность в никотиновой кислоте со-
ставляет 15—25 мг.
Витамин Bf (пиридоксин)
Авитаминоз. Явления витаминной недостаточности
могут развиться при повышенной потребности в этом витами-
не. Это бывает при употреблении большого количества белков
с пищей, при охлаждении, длительной физической нагрузке,
приеме серосодержащих лекарств, а также при беременности.
Недостаточность пиридоксина у человека вызывает сниже-
ние аппетита, появление тошноты, дерматит, стоматит, пси-
хические расстройства, полиневрит верхних и нижних конеч-
ностей. При недостаточности пиридоксина у детей могут
появляться судороги и развиться анемия.
Роль в организме. 1. Входит в состав различных фер-
ментов, участвующих в белковом обмене. 2. Необходим для
поддержания нормальной функции центральной нервной сис-
темы. 3. Влияет на обмен серосодержащих аминокислот.
4. Способствует образованию гемоглобина. 5. Принимает уча-
192
стие в обмене ненасыщенных жирных кислот (линолевой, ли-
ноленовой).
Источник поступления. Потребность в пиридок-
сине покрывается синтезом бактериальной флоры кишечника
человека. Витамин В6 хорошо сохраняется во время кулинар-
ной обработки пищи, а также при консервировании пищевых
продуктов. Однако жарение, копчение и тушение мяса ведет к
потере от 20 до 50% содержащегося.в нем пиридоксина. Про-
филактика авитаминоза состоит в назначении пиридоксина во
время лечения больных сульфаниламидами и антибиотиками.
Суточная потребноеть человека в пиридоксине
ориентировочно исчисляется в 2—4 мг.
Витамин BJ2 (цианкобаламин)
Авитаминоз. Заболевание носит название гиперхром-
ной анемии Аддисона—Бирмера. Антианемическое вещество,
эффективное при лечении анемии Аддисона—Бирмера, явля-
ется соединением внутреннего фактора и внешнего фактора.
Внешним фактором является витамин В12. Внутренний же
фактор образуется в желудке и содержится в желудочном соке
в виде гастромукопротеина. Поступающий с пищей витамин
В|2 связывается с внутренним фактором и в виде белково-вита-
мИнного комплекса поступает в кишечник, а затем всасывает-
ся в кровь. Снижение уровня витамина В12 чаще всего возника-
ет после резекции желудка. Это связано с потерей внутреннего
фактора за счет удаления части желудка и атрофии оставшейся
слизистой оболочки.
Роль в организме. 1. Участвует в созревании кровя-
ных элементов в костном мозгу. 2. Необходим для нормально-
го роста человека и животных вследствие его стимулирующего
воздействия на образование нуклеиновых кислот и на синтез
белка. 3. Вызывает липотропный эффект — понижение холе-
стерина у больных атеросклерозом. 4. Положительно влияет
на обменные процессы в сердечной мышце и щитовидной же-
лезе. 5. Способствует превращению каротина в витамин А.
Источник поступления. Основным поставщиком
витамина Вц являются продукты животного происхождения.
Зеленые растения цианкобаламина не синтезируют. Вита-
мин В12 нетоксичен. Его инъекции безболезненны.
В природе синтез витамина Bi2 осуществляется лишь мик-
роорганизмами (бактерии, грибы и т. д.). Синтез витамина В12
бактериями происходит и в толстом отделе кишечника, но от-
сутствие здесь внутреннего фактора резко ограничивает его
всасывание.
7-281
193
Суточная потребность человека в витамине В>
составляет 2 мкг. При парентеральном введении усвоение ви
тамина в несколько раз выше, чем при приеме вовнутрь.
Витамин В и (пангамовая кислота)
Авитаминоз. Недостатокпангамовой кислоты в орга-
низме человека привадит к угнетению утилизации кислорода
клетками, что ведет к интоксикации и даже анемии тканей.
Роль в организме. 1. Участвует в окислительных про-
цессах. 2. Способствует усилению и нормализации углеводно-
го и липидного обменов. 3. Ускоряет окисление этилового
спирта в печени и усиливает регенеративные процессы пече-
ночной ткани. 4. Оказывает благоприятное действие на тече-
ние сердечно-сосудистых заболеваний. 5. В настоящее время
признан одним из ведущих средств в профилактике и лечении
преждевременного старения.
Источник поступления. Пангамовая кислота на-
ходится в семенах большинства растений (название витамина
происходит от греческих слов «пан» — всюду и «гами» —
семя), а также в говяжьей печени, пивных дрожжах.
Суточная потребность организма в этом витами-
не равняется 2 мг. В лечебных целях пангамовая кислота на-
значается в виде пангамата кальция до 250 мг в день. Витамин
Ви токсическими свойствами не обладает.
Витамин Вс (фолиевая кислота, или фолацин)
Авитаминоз. Клинические проявления авитаминоза
фолиевой кислоты, как правило, развиваются постепенно. На-
рушается прежде всего созревание эритроцитов за счет тормо-
жения синтеза гемоглобина. Эритроциты обладают меньшей
стойкостью, что ведет к их быстрому разрушению в печени в
возникновению анемии. Вслед за анемией развиваются лейко-
немия (уменьшение лейкоцитов в крови) и патология органов
пищеварения (гастрит, стоматит, энтерит).
Роль в организме. 1. Фолиевая кислота действует на
кроветворение — ведет к повышению содержания гемоглоби-
на и росту числа эритроцитов, лейкоцитов и тромбоцитов.
2. Входит в состав ферментов, участвующих в синтезе белка и
аминокислот (метионина).
Источник поступления. Им являются дрожжи,
печень, грибы, шпинат, цветная капуста, зеленые листья.
Фолиевая кислота синтезируется некоторыми видами ки-
шечных бактерий, главным образом молочнокислыми и
дрожжами.
194
Суточная потребность человека в фолиевой кис-
лоте составляет 2—3 мг. Лечебная суточная доза — 20—30 мг.
Фолиевая кислота токсична (может вызывать гистаминопо-
добное действие).
Витамин Н (биотин)
Авитаминоз. Биотин-авитаминоз характеризуется по-
ражением кожи я изменениями со стороны органов пищеваре-
ния (исчезновение аппетита, тошнота, отечность языка).
Имеют место также мышечные боли вялость, депрессия. Не-
достаточность организма человека в биотине может возник-
нуть при подавлении синтеза этого витамина кишечной фло-
рой (сульфаниламиды, антибиотики) или при употреблении
большого количества яиц. Дело в том, что сырой яичный бе-
лок содержит токсически действующий альбумин — авизин.
В кишечнике авизин связывается с биотином, образуя авцзи-
ново-биотиновый комплекс. В таком виде биотин не способен
всасываться в кровь и организмом не используется.
Имеется описание хронической биотиновой недостаточно-
сти у человека, который длительное время принимал по 12 сы-
рых яиц в день.
Роль в организме. 1. Биотин участвует в нервно-тро-
фических процессах. 2. Принимает участие в окислении пирови-
ноградной кислоты. 3. Играет активную роль в жировом обмене.
Источник поступления. Биотин содержится во
всех продуктах, где находятся витамины группы В. Наиболее
богаты биотином печень, почки. Кроме того, у человека био-
тин синтезируется бактериями кишечной флоры.
Суточная потребность человека в биотине —око-
ло 0,01 мг. Она обеспечивается продуктами обычного питания.
Холин
Авитаминоз. Самым характерным признаком недоста-
точности в организме холина является жировая дистрофия пе-
чени. Накопления жира в печени в последующем ведет к не-
крозам и развитию соединительной ткани с нарушением всех
функций печени (цирроз).
Роль в организме. 1. Входит в состав фосфолипи-
дов — лецитина и сфингомиелина, которые принимают актив-
ное участие в обмене и транспорте жиров. 2. Является исход-
ным веществом для построения ацетилхолина (сложный эфир
холина) — одного из основных медиаторов возбуждения в не-
рвной системе. 3. Способствует удалению жира из печени.
4. Участвует в процессе всасывания жира из кишечника.
7
195
5. Участвует в окислительных реакциях и в процессе кроветво-
рения (вместе с витамином В12 и фолиевой кислотой).
Источник поступления. В организме человека хо-
лин синтезируется из аминокислоты метионина. Его источни-
ком являются: печень, мозг, белая мука. Кроме того, он имеет-
ся в яйцах, мясе, злаках и овощах.
Суточная потребность организма в холине связа-
на с составом пищевого рациона и колеблется от 1,5 до 3,0 г.
При обычном питании человек с пищей получает 1,5-4 г хо-
лина. Этот витамин малотоксичен.
Витамин N (липоевая, или тиоктоновая кислота)
Авитаминоз. Недостаток в организме липоевой кислоты
вызывает нарушение липоидного обмена и, как следствие, повы-
шение вероятности поражения печени и сосудов (атеросклероз).
.Роль в организме. 1. Входит в состав ферментов, уча-
ствующих в окислении пировиноградной и а-кетоглутаровой
кислот. 2. Вызывает снижение холестерина в крови и препят-
ствует развитию атеросклероза. 3. Используется как лечебное
средство при хронических гепатитах, циррозах печени, атеро-
склерозе и сердечно-сосудистых заболеваниях.
Источник поступления. Наибольшее количество
липоевой кислоты содержится в мясе (говядине), молоке; уме-
ренное — в рисе и наименьшее — в овощах.
Суточная потребность в витамине N составляет
1—2 мг. В лечебных дозах (10—20 мг в сутки) она не вызывает
токсических и других побочных явлений.
Витамин U (противоязвенный фактор, от лат. ulcus — «язва»)
Авитаминоз. Недостаток витамина в организме спо-
собствует развитию эрозивных процессов в слизистой оболоч-
ке желудка и двенадцатиперстной кишки.
Роль в организме. 1. Необходим для сохранения це-
лостности слизистой оболочки. 2. Вызывает детоксикацию ги-
стамина, способствующего возникновению язвы желудка и
двенадцатиперстной кишки. 3. Улучшает моторику кишечни-
ка и желчного пузыря, повышает их тонус.
Источник поступления. Противоязвенный фактор
является пищевым фактором, поступающим в организм с ово-
щами. Высокую витаминную ценность представляет сок све-
жих овощей, и особенно капустный сок.
Суточная потребность в витамине U точно не
установлена. В качестве лечебной дозы при язвенной болезни
используют до 300 мг витамина в сутки.
196
Витамин С (аскорбиновая кислота)
Авитаминоз. Цинга. Первые описания цинги как мас-
сового заболевания относятся к XIII в. Стадия развитой, «цве-
тущей» цинги характеризуется выраженной кровоточивостью
десен и кровоизлияниями в мышцы, подкожную клетчатку, в
кожу, суставы и надкостницу. Цинга нередко сопровождается
нарастающей анемией.
Роль в организме. 1. Стимулирует процесс образо-
вания коллагена. Этим он способствует поддержанию нор-
мальной проницаемости капилляров. 2. Принимает участие в
обменных процессах, являясь переносчиком водорода. 3. кон-
тролирует различные фазы белкового обмена. 4. Влияет на
скорость образования ДНК клеточного ядра. 5. Контролирует
сложные биохимические реакции в клетках центральной не-
рвной системы. 6. Оказывает стабилизирующее действие на
молекулы адреналина путем их защиты от окисления. 7. По-
вышает фагоцитарные свойства крови главным образом за счет
нарастания фагоцитарной активности лейкоцитов.
Источник поступления. Источником витамина С
являются растительные продукты: шиповник, помидоры, чер-
ная смородина, зеленый грецкий орех, капуста, цитрусовые,
картофель, болгарский перец и т. д.
Суточная потребность в витамине С в обычных
условиях составляет 70—80 мг. Противопоказаний к назначе-
нию аскорбиновой кислоты нет.
Витамин Р (биофлавоноиды—рутин, катехин)
Авитаминоз. Недостаточное содержание витамина Р в
пище приводит к нарушению проницаемости капилляров и
появление геморрагической сыпи. В отличие от больших ге-
моррагий при цинге кровоизлияния при Р-авитаминозе часто
ограничиваются только волосяными мешочками.
Роль в организме. 1. Оказывает влияние на прони-
цаемость сосудов, образуя ферментативную систему вместе с
витамином С, активно участвующую в процессах клеточного
дыхания в качестве окислительно-восстановительного ком-
плекса. 2. Способствует более быстрому повышению уровня
аскорбиновой кислоты в организме. 3. Повышает актив-
ность аскорбиновой кислоты и способствует ее накоплению
в организме. 4. Предохраняет от окисления аскорбиновую
кислоту, адреналин и другие гормоны надпочечников.
5. Влияет на желчеотделение. 6. Влияет на кроветворение и
нормализацию уровня кровяного давления при гипертони-
ческой болезни.
197
Источник поступления. Витамин Р широко рас-
пространен в цитрусовых, черной смородине, шиповнике, в
листьях чая и других пищевых продуктах.
Суточная потребность для человека исчисляется
в 50 мг. Лечебная доза — 300 мг в д ень.
II. Жирорастворимые витамины
Витамин А (ретинол)
Авитаминоз. Заболевание — «куриная слепота» — xaJ
растеризуется нарушением способности глаза приспосабли-
ваться к слабому освещению. Больной «куриной слепотой» с
наступлением сумерек, в ночное время и днем в затемненном
помещении плохо видит (или совсем не видит) и, следователь-
но, недостаточно ориентируется в обстановке. Для авитаминоза
витамина А характерно нарушение функций эпителия органов с
превращением различных видов эпителия в многослойный
плоский эпителий и усилением процесса ороговения.
Роль в организме. 1. Принимает участие в образова-
нии зрительного пигмента палочек сетчатки глаза — родопси-
на, а также зрительного пигмента колбочек — йодопсина.
2. Необходим для обеспечения нормальной дифференциров-
ки эпителиальной ткани. 3. Является антагонистом особого
вещества, которое вызывает кератинизацию эпителиальных
клеток. 4. Оказывает влияние на регуляцию клеточного деле-
ния. При низком содержании ретинола число митозов умень-
шается. 5. Является антагонистом тироксина (гормона щито-
видной железы), уменьшая его Действие в отношении
усиления обмена веществ. 6. Противодействует (антагонист)
токсическому действию витамина Д (витамин А задерживает
проявления цинги, а витамин Д их ускоряет). 7. Участвует в
минеральном обмене (обмене фосфора) и образовании холе-
стерина.
Источник поступления. Организм человека полу-
чает ретинол с пищей. Среди продуктов животного происхож-
дения наиболее богат витамином А жир печени морских жи-
вотных и рыб. В продуктах растительного происхождения
витамин А Находится в виде провитамина — каротина. В орга-
низме человека и животных (в стенке кишечника, в печени, в
крови и т. д.) при участии каротиназы и холина каротин пре-
вращается в витамин А.
Суточная потребность человека в витамине А рав-
на 1,5 мг для взрослого человека и 2 мгдля беременных и кор-
мящих.
198
Витамин Д (кальциферолы)
Авитаминоз. Заболевание детей рахитом известно уже
. давно. Недостаток в организме витамина Д ведет к изменению
фосфорно-кальциевого обмена с нарушением костеобразова-
ния, функций нервно-мышечного аппарата с проявлениями
расстройств центральной нервной системы. При этом наруша-
ются трофические процессы и в других органах и системах.
Роль в организме. 1.Регулируетфосфорно-кальцие-
вый обмен в организме и тем самым способствует процессу ко-
. стеобразования. 2. Повышает усвоение пищевого кальция в
кишечнике, поддерживает нормальный уровень кальция в кро-
ви. 3. Улучшает обеспечение организма фосфором за счет уси-
ления его реабсорбции в почках.
Источник получения. С целью предупреждения и
лечения рахита уже давно используют рыбий жир и солнечный
свет. Предшественники витамина Д (провитамины) находятся
в основном в коже человека и животных. Под действием ульт-
рафиолетовых лучей с длиной волны 255-313 нм из них обра-
зуются все разновидности витамина Д.
Суточная потребность человека в витамине Д со-
ставляет около 2,5 мкг при одновременном введении соответ-
ствующего количества кальция и фосфора.
Витамин Е (токоферолы)
Ав итаминоз. Е-авитаминоз у человека не описан. Не-
достаточность же витамина Е сопровождается гемолитической
анемией у недоношенных детей. Недостаток витамина Е в
организме взрослого человека проявляется в нарушении про-
цесса оплодотворения, увеличения непроизвольных абортов,
мышечной дистрофии.
Роль в организме. 1. Участвует в образовании гона-
дотропного гормона гипофиза. 2. Способствует накоплению в
организме витамина А и других жирорастворимых витаминов,
предохраняет ненасыщенные жирные кислоты от окисления,
участвует в фосфорилировании. 3. Способствует нормальному
содержанию гликогена печени, улучшает жировой, белковый и
минеральный обмен. 4. Входит в состав противосвертывающей
системы крови и участвует в предотвращении неестественной
коагуляции в сосудах.
Источник поступления. Токоферолы образуются
только в зеленых частях растения, и особенно в молодых рост-
ках злаков. Отсюда богаты витамином Е шпинат, салат, капус-
та и зародыши пшеницы. Животные не синтезируют токофе-
ролы. Однако витамин Е содержится в мясе, печени, яичном
199
желтке, масле и молоке (особенно летнем). Для усвоения орга-
низмом витамина Е необходимо^наличие желчи в кишечном
содержимом.
Суточная потребность человека в витамине Е
предположительно составляет 10—25 мг. Лечебная доза дости-
гает 1 г.
Витамин К (филлохинон, фарнохинон, викасол)
Авитаминоз. К-авитаминоз проявляется геморрагичес-
ким диатезом — кровоточивостью.
Роль в организме. 1. Участвует в процессе сверты-
ваемости крови за счет регуляции биосинтеза свертывающих
факторов в печени (протромбина и др.). 2. Повышает сократи-
тельную способность мышц в результате воздействия витами-
на К на миозин — сократительный белок мышечных волокон.
3. Усиливает регенерацию тканей и ускоряет зажйвление ран,
а также обладает болеутоляющим действием и повышает со-
противляемость организма к инфекциям.
Источник поступления. Витамин К образуется в
зеленых частях растений, а также синтезируется кишечной
микрофлорой.
Суточная потребность организма в витамине К
исчисляется равной 0,2—0,3 мг. При желтухе витамин К надо
вводить внутримышечно или внутривенно.
ВЫДЕЛЕНИЕ
ОБЩЕЕ ПРЕДСТАВЛЕНИЕ О СИСТЕМЕ ВЫДЕЛЕНИЯ
Выделение — это процесс удаления конечных продуктов
метаболизма, которые уже не могут быть использованы орга-
низмом, а также чужеродных веществ (нередко ядовитых), ко-
торые попали в организм извне. Все эти вещества выводятся
из организма с помощью органов выделения.
В процессах выделения участвуют почки, потовые железы,
легкие, кишечник.
Почками удаляются вода, ряд продуктов обмена веществ,
избыток солей, чужеродные и токсические вещества после
инактивирования в других органах. Например,-лекарственные
вещества и продукты их метаболизма.
Потовыми железами удаляются вода и соли. Они играют,
так же как и почки, важную роль в поддержании постоянства
Ж
осмотического давления, ионного состава и реакции внутрен-
ней среды.
Легкие принимают участие в выделительных процессах,
удаляя из организма углекислый газ, воду и некоторые летучие
вещества, например пары эфира, хлороформа и др.
Через слизистую оболочку кишечника удаляются из крови
некоторые продукты обмена, например, желчные пигменты и
соли тяжелых металлов.
СТРОЕНИЕ МОЧЕВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ.
ОБРАЗОВАНИЕ И СВОЙСТВА МОЧИ.
В мочевыделительную систему входят две почки, экскретом
которых является моча, и органы, служащие для скопления и
выведения мочи (мочеточники, мочевой пузырь, мочеиспус-
кательный канал).
Почки расположены на задней стенке брюшной полости по
бокам позвоночника на уровне последнего грудного и 1—2-го
поясничных позвонков. Правая почка лежит ниже левой в
среднем на 1—1,5 см.
Почка имеет темно-красный цвет и бобовидную форму.
На вогнутом крае имеются ворота, через которые в почку вхо-
дят почечная артерия и нервы, а выходят вена и мочеточник
(рис. 5.49).
Почка окружена собственной фиброзной оболочкой в виде
тонкой гладкой пластинки, непосредственно прилегающей к
веществу почки. В области ворот и по задней поверхности на-
ходится слой рыхлой жировой ткани, составляющей жировую
капсулу почки.
В почке различают корковое вещество и мозговое вещество.
Корковое вещество толщиной около 4 мм находится на пери-
ферии органа. Под ним располагается мозговое вещество. Оно
состоит.из образований конической формы, носящих .назва-
ние почечных пирамидок, разделенных прослойками корково-
го вещества. Верхушки пирамидок, называемые сосочками,
направлены в почечную лоханку. В почке в среднем имеется
12 сосочков. Каждый сосочек усеян маленькими отверстиями,
через которые моча выделяется в начальные части мочевых пу-
тей — малые чашечки. Малые чашечки (8-9 штук) впадают в
две большие чашечки. Две большие чашечки (верхняя и ниж-
няя) сливаются в одну почечную лоханку, которая охватывает
ворота позади почечных сосудов и, загибаясь вниз, переходят
в мочеточник (рис. 5.50).
201
Рис. 5.49. Почки
и мочеточники:
1 — правая почка; 2 — по-
чечная артерия; 3 — над-
почечники; 4 — нижняя
полая вена; 5 — аорта; 6—
левая почка; 7 — почеч-
ная вена; 8, 9— мочеточ-
ники
Таким образом, моча, выделяющаяся насосочках, на своем
пути до мочевого пузыря проходит через малые чашечки, по-
чечную лоханку и мочеточник.
Основной структурной и функциональной единицей почки яв-
ляется нефрон (рис. 5.51). У человека в почке их содержится около
миллиона. Нефрон включает в себя несколько отделов, сильно
различающихся по строению и физиологической функции:
• 1) почечное тельце (мальпигиево тельце), состоящее из
капсулы Боумена-Шумлянского и мальпигиева клубочка;
2) проксимальный извитой каналец (каналец 1-го поряд-
ка), направляющийся к мозговому веществу почки;
3) петля Генле;
4) дистальный извитой каналец (каналец 2-го порядка).
Каждый нефрон начинается микроскопической двухстен-
ной капсулой, имеющей форму чаши и называемой капсулой
Боумена-Шумлянского. Она расположена в корковом веще-
стве. Капсула охватывает сплетение капилляров, называемое
мальпигиевым клубочком. Вместе с клубочком капсула обра-
зует мальпигиево, или почечное, тельце. Между стенками кап-
сулы имеется узкая полость, от которой начинается просвет
почечного канальца. Внутренний листок капсулы образован
плоскими эпителиальными клетками, расположенными на
тонкой базальной мембране и плотно прилегающими со всех
Рис. 5.50. Почка
(в разрезе):
1 — большая чашечка; 2 —
малые чашечки; 3 — моз-
говое вещество; 4— сосоч-
ки; 5— корковое вещество;
6 — мочеточник; 7 — по-
чечная лоханка
Рис. 5.51.
Строение
нефрона (схема):
7 —клубочек; 2 —
дистальный из-
витой каналец;
3 — проксимальный
извитой каналец;
4 — собирательные
кружки; 5— нисхо-
дящая часть петли
Генле; 6 — восхо-
дящая часть петли
Генле. В кружках
изображено стро-
ение эпителия в
различных частях
нефрона
203
сторон к капиллярам клубочка. Между эпителиальными клет-
ками внутренней стенки капсулы имеются узкие щели. В свою
очередь между эндотелиальными клетками капилляров также
имеются отверстия диаметром около ОД мкм. Таким образом,
кровь, циркулирующая в капиллярах, по существу отделена от
полости капсулы лишь тонкой базальной мембраной.
От полости капсулы Боумена—Шумлянского отходит по-
чечный каналец извитой формы, так называемый извитой ка-
налец 1-го порядка (проксимальный извитой каналец), на-
правляющийся к мозговому слою почки. На границе между
корковым и мозговым слоями каналец суживается и выпрям-
ляется, образуя в мозговом слое петлю Генле, состоящую из
нисходящей и восходящей части. Восходящий прямой кана-
лец в мозговом слое переходит в извитой каналец 2-го порядка
(дистальный извитой каналец). Извитые канальцы 2-го поряд-
ка открываются в так называемые собирательные трубочки,
которые, начинаясь в корковом слое, переходят в мозговой,
доходят до вершин пирамидок и через сосочковые ходы от-
крываются^ почечные чашечки. Длина канальцев нефрона
различна — от 18 до 50 мм, а суммарная длина всех канальцев
почки очень велика — около 100 км.
Нефрон имеет ряд особенностей кровоснабжения. Артерия
почки, разветвляясь, образует артериолы, каждая из которых
входит в капсулу Боумена—Шумлянского и распадается на ка-
пиллярную сеть, образующую мальпигиев клубочек. Эти ка-
пилляры вновь собираются в артериолу, по которой артери-
альная кровь оттекает от мальпигиева клубочка. Артериола,
доставляющая кровь в клубочек, называется приносящей, а та,
по которой кровь оттекает, — отводящей. В капиллярах маль-
пигиева клубочка давление крови высокое — 70-80 мм рт. ст.
Это обусловлено следующим: 1) почечная артерия отходит не-
посредственно от брюшной аорты, и путь ее до почки корот-
кий; 2) диаметр приносящей артериолы клубочка примерно в
два раза больше, чем выносящей.
Выносящая артериола клубочка вновь разветвляется, обра-
зуя густую сеть капилляров, оплетающую извитые канальцы
1-го, 2-го порядка и петлю Генле. В этой сети благодаря газо-
обмену артериальная кровь, превращаясь в венозную, поступа-
ет в мелкие вены, которые, постепенно сливаясь, образуют по-
чечную вену, впадающую в нижнюю полую вену.
Около 80% всех нефронов имеют структуру и кровообраще-
ние, описанные выше.Юни называются корковыми нефрона-
ми. Остальные нефроны расположены так, что большая часть
их составных элементов лежит в мозговом слое.
Образование мочи складывается из трех процессов:
1) фильтрации;
2) реабсорбции;
3) канальцевой секреции.
Фильтрационная фаза: В эту фазу происходит фильтрация
плазмы крови из капилляров мальпигиева клубочка в полость
капсулы Боумена-Шумлянского. Давление в капиллярах
клубочка 70—80 мм рт. ст., что обеспечивает фильтрацию
плазмы в полость капсулы. Этой фильтрации препятствует
онкотическое давление крови (30 мм рт. ст.) и давление
фильтрата в полости капсулы и канальцев (внутрипочечное
давление — около 20 ммрт. ст.). Поэтому давление, обеспе-
чивающее фильтрацию, равно: 75 — (30 + 20) = 25 мм рт. ст.
Плазма фильтруется со всеми растворенными в ней вещества-
ми, кроме белков. Фильтрация плазмы происходит через
межэндотелиальные щели капилляров клубочка, базальную
мембрану и щели между клетками внутренней стенки капсу-
лы. Органические вещества плазмы с низкой молекулярной
массой (глюкоза, аминокислоты, мочевина, мочевая кислота
и др.), а также неорганические соли свободно диффундируют
через эти образования. Таким образом, состав фильтрата тот
же, что и плазмы крови, за исключением высокомолекуляр-
ных белков. Этот фильтрат называют первичной мо-
чой.
В сутки у человека образуется 150—180 л первичной мочи.
Такой большой объем первичной, мочи возникает благодаря
следующим причинам: 1) через почки проходит большое ко-
личество крови (1700—1800 л в сутки), и приблизительно из
каждых 10 л крови отфильтровывается 1 л первичной мочи;
2) интенсивной фильтрации способствует большая поверх-
ность капилляров мальпигиевых клубочков, достигающая 1,5—
2,0 м2, и высокое давление в них.
Реабсорбционная фаза. Первичная моча из капсулы Боуме-
на—Шумлянского поступает в почечные канальцы. Здесь про-
исходит обратное всасывание в кровь воды и ряда веществ, ко-
торые содержатся в первичной моче. Из первичной мочи
образуется только 1,0— 1,5л вторичной мочи. Она по-
ступает в почечные лоханки и выводится из организма. Экспе-
риментально установлено, что обратно в кровоток всасывается
в извитых канальцах 1-го порядка 40—45% воды, в петле Ген-
ле — 25—28%, в извитых канальцах 2-го порядка — 10% и в со-
бирательных трубочках — 20%. Так, только 1% воды, входящей
в состав первичной мочи, остается нереабсорбированной, и
она поступает в почечные лоханки. При этом невсасывающие-
205
ся вещества (мочевина, мочевая кислота, аммиак, креатинин,
сульфаты, фосфаты и др.) концентрируются. Таким образом,
эти продукты выводятся в сравнительно небольшом количе-
стве суточной мочи (1,0—1,5 л).
Реабсорбция веществ из первичной мочи зависит от их
концентрации в крови в данное время. Так, например, если в
крови глюкоза находится в избытке (свыше 0,15—0,18%), то
часть глюкозы из первичной мочи не всасывается обратно, а
выводится наружу с конечной мочой. Наоборот, при недостат-
ке, например, поваренной соли в пище, выведение ее с мочой
почти прекращается.
Для каждого вещества существует определенный предел их
концентрации в крови, выше которого они не могут полно-
стью реабсорбироваться в канальцах. Этот предел называют
порогом выведения. Однако некоторые вещества явля-
ются беспороговыми, так как они не подвергаются реабсорб-
ции даже при очень малом содержании в крови. К ним отно-
сится мочевина, сульфаты, креатинин.
Канальцевая секреция. Эпителий почечных канальцев спосо-
бен кроме своей основной (реабсорбционной) функции секре-
тировать (т. е. выделять) некоторые вещества. Так, например,
аммиак может образовываться и секретироваться канальцевым
эпителием. Некоторые лекарственные вещества (пенициллин)
выводятся преимущественно (на 80—90%) с помощью канальце-
вой секреции.
Свойства мочи. Относительная плотность мочи равна
1,012—1,020, pH мочи колеблется в пределах 4,7—6,5. Цвет
мочи слабо-желтый за счет присутствия пигментов уроби-
лина и у ро хром а. Эти пигменты образуются в кишечни-
ке и почках из пигментов желчи и, в свою очередь, происходят
из продуктов распада гемоглобина. Азотистые продукты выво-
дятся с мочевиной.
РЕГУЛЯЦИЯ ДЕЯТЕЛЬНОСТИ ПОЧЕК
Регуляция деятельности почек осуществляется нервно-реф-
лекторным и гуморальным механизмами.
1. Почки снабжены симпатическими и парасимпатическими
нервами. Раздражение симпатических нервов ведет к усилению
реабсорбции, что обусловливает снижение в моче хлоридов, а
также сужение почечных сосудов. Раздражение парасимпатичес-
ких нервов ведет к увеличению выделения хлоридов с мочой за
счет уменьшения их реабсорбции.
206
2. Регуляция деятельности почек осуществляется и гумо-
ральными механизмами. Это происходит под влиянием анти-
дауретического гормона (АДГ), или вазопрессина, который вы-
рабатывается в клетках гипоталамуса, накапливается в задней
доле гипофиза и отсюда поступает в кровь.
При поступлении в кровь избыточного количества солей
повышается се осмотическое давление. Вследствие этого раз-
дражаются осморецепторы, находящиеся в самом гипоталаму-
се. Сигналы от этих рецепторов вызывают повышенное выде-
ление ДЦГ гипоталамусом. Из задней доли гипофиза в кровь
поступает повышенное количество этого гормона, что приво-
дит к усилению обратного всасывания воды из мочи. Концен-
трация солей в конечной моче увеличивается, а в крови умень-
шается, что обусловливает падение осмотического давления в
последней. В том случае, если в организм поступает избыточ-
ное количество воды, то осмотическое давление в крови пада-
ет. При этом стимуляция осморецепторов ослабляется и сни-
жается секреция АДГ. Диурез увеличивается и организм
освобождается от воды. Благодаря этому процессу осмотичес-
кое давление крови нормализуется.
При нарушении деятельности гипоталамуса или задней
доли гипофиза выделение АДГ может прекращаться, вода пе-
рестает реабсорбироваться, вследствие чего резко возрастает
диурез (до 20—25 л в сутки). Это заболевание называется неса-
харным мочеизнурением.
На образование мочи оказывают влияние и другие гормо-
ны. Так, адреналин в больших дозах вызывает сужение
приносящих сосудов клубочков и тем самым снижает диурез.
Гормон щитовидной железы тироксин, наоборот, усилива-
ет диурез, ослабляя реабсорбцию воды в канальцах.
ВНУТРЕННЯЯ СЕКРЕЦИЯ
ОБЩЕЕ ПРЕДСТАВЛЕНИЕ
О ЖЕЛЕЗАХ ВНУТРЕННЕЙ СЕКРЕЦИИ
Изучением эндокринной системы занимается эндокрино-
логия — наука о гуморальной регуляции функций' организма,
ее общих принципах и закономерностях.
В организме существует два типа желез — экзокринные и
эндокринные.
207
Экзокринные железы (железы желудочно-кишечного трак-
та, потовые железы и др.) выделяют секрет через имеющиеся в
них выводные протоки в полость тела или на его
поверхность (внешняясекреция).
Эндокринные железы не имеют выводного протока и выде-
ляют секрет непосредственно в кровь и лимфу
(внутренняя секреция) (рис. 5.52). Секреты, вырабатываемые
в железах внутренней секреции (ЖВС), называют гормонами.
Гормоны, образуясь в клетках железы, поступают непосред-
ственно в кровь сосудов, снабжающих железы, разносятся ею
по организму, усиливая или ослабляя при этом деятельность
разнообразных органов.
Гормон — это вещество, выделяемое клетками в одной час-
ти тела и переходящее в какую-нибудь другую его часть, где
оно действует в очень малой концентрации, регулируя рост
или активность клеток. Таким образом, «гормон» — это не хи-
мическое понятие, так как этот термин не относится к какому-
либо определенному классу химических соединений, а скорее,
физиологическое понятие. Считается, что вещество может на-
зываться истинным гормоном, если оно обладает рядом при- t
сущих ему свойств. ,
1. Специфичность. Определенный гормон отличает-
ся от других гормонов тремя параметрами: строго определен-
Рис. 5.52. Расположение
желез внутренней
секреции:
1 — гипофиз; 2 — околощито-
видные железы; 3 — зобная
железа; 4 — поджелудочная
железа; 5 — половые железы;
6 — надпочечники; 7—щито-
видная железа; 8 — эпифиз
ным своеобразием химической структуры, местом его образо-
ваний (определенные клетки, определенные железы) и выпол-
няемойфункцией.
2. Высокая биологическая активность.Гор-
моны оказывают свое действие в чрезвычайно низких кон-
центрациях. Они действуют в основном на процессы, проис-
ходящие в клетках, в их цитоплазматических структурах и
неактивны в межклеточной среде.
3. Секретируемость железой. Казалось бы, этот
признак не требует пояснений. Вместе с тем следует отметить,
что ряд специфических биологически активных веществ может
образовываться в эндокринной железе в качестве промежуточ-
ных продуктов биосинтеза гормонов или их катаболитов, но
при этом не секретироваться в кровь в обычйых условиях.
4. Дистгнтность действия. Отдаленность места
действия гормонов от места их образования позволяет отли-
чить истинные гормоны от местных, или тканевых, гормонов
(гормоноидов).
Таким образом, перечисленные свойства гормонов позво-
ляют охарактеризовать их как особый класс биоорганических
соединений системного действия.
К железам внутренней секреции относят: гипофиз, щито-
видную, паращитовидную железы, надпочечники, эпифиз, ви-
лочковую железу. Кроме желез внешней и внутренней секре-
ции существуют железы смешанной секреции — половые и
поджелудочная.
Гормоны играют ведущую роль в гуморальной регу-
ляции функций организма. По химической природе гормо-
ны делят на три группы: полипептиды и белки (инсулин);
аминокислоты и их производные (тироксин, адреналин); сте-
роиды (половые гормоны). Гормоны циркулируют в крови в
свободном состоянии и в виде соединений с белками. Связан-
ные с белками гормоны, как правило, переходят в неактивную
форму.
ГИПОФИЗ
Размеры железы небольшие — от 1 до 1,5 см в разных плос-
костях. Гипофиз лежит в углублении костного выступа на вер-
хней стороне клиновидной кости, который назвали «турецким
седлом» за его форму. Гипофиз целиком «сидит» в седле, бла-
годаря чему защищен костными стенками спереди, снизу и
сзади. Надежность защиты гипофиза соответствует его значе-
209
нию для организма. Анатомически гипофиз связав ножкой с
дном третьего мозгового желудочка промежуточного мозга и
имеет здесь тесный контакт с гипоталамусом.
Гипофиз содержит три доли — переднюю, промежуточную
и заднюю. Переднюю и промежуточную доли объединяют под
названием аденогипофиза, а заднюю долю называют нейроги-
пофизом.
Передняя доля гипофиза—полиморфная эндокринная же-
леза, состоящая из функционально и структурно различных
клеток, выполняющих роль специфических регуляторов дру-
гих (периферических) эндокринных желез. Это так называе-
мые тропные гормоны. Принято считать, что в передней доле
гипофиза образуется семь различных гормонов.
1. Гормон роста (ГР), который называют соматотропным
(СТР),—регулирует рост. Гиперфункция в детском воз-
расте приводит к гигантизму, у взрослого человека возникает
акпомегалия (увеличение размеров носа, нижней челюсти, ки-
стей рук и стоп ног). При гипофункции в детском возра-
сте происходит задержка роста — каплнковость. Гипофизар-
ные карлики характеризуются правильными пропорциями
тела и нормальным развитием психики. При гипофункции у
взрослых происходят изменения обмена веществ: либо сдвиг к
ожирению, либо к общему похуданию.
2. Тиреотропный гормон (ПТ)—воздействует нарост и фун-
кцию щитовидной железы.
3. Адренокортикотропный гормон (АКТГ), называемый также
кортикотропином, оказывает воздействие на кору надпочеч-
ников, усиливая синтез ее гормонов.
4. Фомыкуяоетимуяирующий гормон — способствует росту
фолликула яичника.
5. Лютеинизирующий гормон—усиливает образование поло-
вых гормонов и рост половых клеток.
б. Лютеотроиный гормон—способствует образованию жел-
того тела и синтезу прогестерона.
7. Лактогенный гормон (пролактин) — стимулирует выра-
ботку молока молочными железами.
Последние четыре гормона относят к гонадотропным.
Промежуточная доля гипофиза у человека развита очень сла-
бо. Так же как у некоторых видов позвоночных (рыб, амфи-
бий), клетки промежуточной доли образуют меланоцитости-
мулирующий гормон (интермидин), который стимулирует
образование пигмента меланина и его внутриклеточную миг-
рацию в меланоциты. Также интермидин восстанавливает цвет
кожи, лишенной пигментации.
210
Залит доля гипофиза, или нейрогипофиз, выделяет два
нейрогормона, местом образования которых являются нейро-
секреторные клетки.
1. Антидиуретический гормон (АДГ) — вазопрессин, регули-
рует содержание воды в организме и обмен жидкости в крове-
носных сосудах. При обеднении организма водой (повышение
осмотического давления крови) секреция этого гормона уве-
личивается. Основное действие АДГ связано с канальцами
почек, в которых усиливается реабсорбция воды. Вследствие
этого диурез уменьшается. Гипофункция приводит к не-
сахаоному мочеизнурению. Выделяется большое количество
мочи, не содержащей сахарами возникает сильная жажда.
2. Окситоцин — способствует сокращению гладкой муску-
латуры матки в конце беременности и отделению молока во
время лактации.
ГИПОТАЛАМУС И ЕГО СВЯЗЬ С ГИПОФИЗОМ
Гипоталамус—это подбугорный отдел промежуточного моз-
га, являющийся одновременно нервным образованием и эндо-
кринным органом. Эта область, расположенная сразу над гипо-
физом и несколько сзади, осуществляет трансформацию
нервных импульсов в эндокринный процесс. Нейроэндокринные
клетки гипоталамуса вырабатывают вазопрессин, окситоцин и
резилинг-факторы. Одни из резияинг-факторов играют стимули-
рующую гипофизарную роль (либерины), другие — ингибитор-
ную (статины). Попадая по аксонам нейросекреторных клеток из
гипоталамуса в заднюю долю гипофиза, резилинг-факторы про-
никают в кровьи с ее током достигают передней доли, где при-
нимают участие в известных регуляторных функциях.
Таким образом, гипоталамус — место непосредственного
взаимодействия нервной и эндокринной систем и верховный
орган гормональной регуляции рядя эндокринных функций. Ги-
поталамус и гипофиз образуют единый структурно-функцио-
нальный комплекс.
ЭПИФИЗ {ШИШКОВИДНАЯ ЖЕЛЕЗА)
Энифиз — это конусообразная железа длиной 0,5—1 см, на-
поминающая по форме еловую шишку. Эпифиз развивается
из задних отделов крыши третьего желудочка и остается свя-
занным с ним, выступая кзади так, что оказывается располо-
211
женным дорзальнее среднего мозга. Эпифиз секретирует в
кровь два гормонам мелатонин и серотонин. В дневные часы
синтезируется серотонин, а в ночное время — мелатонин.
Роль мелатонина состоит в сдерживании развития репро-
дуктивной системы до достижения определенного возраста,
так как этот гормон угнетает секрецию гонадотропных гормо-
нов гипофиза. Действуя на гипофиз, мелатонин проходит не-
сколько стадий превращений до активной формы. Одна из
этих стадий — стадия превращения в серотонин. После удале-
ния эпифиза наступает преждевременное половое созревание.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа расположена на шее впереди гортани.
Ее масса составляет 30-40 г. В ней различают две доли и пе-
решеек. Железа покрыта двумя капсулами: наружной и внут-
ренней. Внутренняя образует в ткани железы перегородки,
содержащие в своей толще сосуды и нервы. Перегородки не
соединены друг с другом и не изолируют целиком одну доль-
ку от другой, поэтому щитовидная железа псевдодольчатая.
В дольках щитовидной железы человека насчитывается около
30 млн. фолликулов. Фолликул является морфофункциональ-
ной единицей щитовидной железы. В фолликулярных клет-
ках железы образуются три гормона: тироксин, трийодтиро-
нин и кальцитонин. Основными функциями этих гормонов
являются: стимуляция окислительных процессов в клетках,
регуляция водного, белкового, жирового, углеводного и ми-
нерального обменов, роста и развития организма. Они ока-
зывают действие на функции центральной нервной системы.
При гипофункции, проявляющейся в детском воз-
расте, возникает кретинизм (задержка роста, психического и
полового развития). У взрослых организмов развиваются
слизистые отеки тканей. Соединительная ткань утолщена
и уплотнена, наблюдаются некоторая степень ожирения и
склонность к облысению. Речь у таких больных замедлена,
мыслительная активность снижена.
При гиперфункции возникает базедова болезнь (ти-
реотоксикоз). Характеризуется увеличением массы щитовид-.
ной железы (проявляется припухлостью на шее — зобом), пу-
чеглазием, учащенным сердцебиением, раздражительностью,
бессонницей, потливостью, повышением основного обмена и
температуры тела, увеличением содержания йода в крови.
Больные потребляют большое количество пищи, но в то же
212
время быстро худеют. Все эти явления выражены в разной сте-
пени в зависимости от стадии заболевания. Считают, что при-
чина заболевания связана в основном с психическими травма-
ми. При тяжелой форме заболевания прибегают к частичному
или полному удалению щитовидной железы. При образова-
нии паренхиматозного (эндемического) зоба пища с низким
содержанием йода не обеспечивает необходимого количества
этого элемента для синтеза гормонов щитовидной железы,
поэтому содержание их в крови снижается. Передняя доля ги-
пофиза начинает усиленно выделять ТТГ (тиреотропный гор-
мон). Он стимулирует секреторную активности и рост фолли-
кулярных клеток. Происходит увеличение клеток паренхимы
железы, но содержимое их становится разжиженным и они те-
ряют активность.
ОКОЛОЩИТОВИДНАЯ ЖЕЛЕЗА
У человека имеются четыре околощитовидные железы, две
из которых располагаются на задней поверхности щитовидной
железы и две — у нижнего ее полюса, а иногда в самой ткани.
Общая масса четырех околощитовидных желез составляет все-
го около 100 мг, а их наибольший диаметр — около 0,5 см.
Эти железы вырабатывают паратиреоидный гормон (ПТГ) —
паратгормон, который оказывает прямое действие на повыше-
ние концентрации кальция и фосфора в крови; Если у живот-
ного удалить околощитовидные железы, то возникающие при-
ступы судорог всей скелетной мускулатуры (тетания) приводят
к смерти. Непосредственной причиной смерти является рас-
стройство дыхания вследствие судорог дыхательных мышц.
Тетания развивается в результате понижения содержания
кальция в кровй и в спинномозговой жидкости. Это подтверж-
дается тем, что введение солей кальция животным с удаленны-
ми околощитовидными железами предупреждает развитие те-
тании. ПТГ вызывает быстрый выход кальция из костного
вещества, воздействуя на остеокласты, которые оказывают ли-
тическое действие на прилежащее к ним межклеточное веще-
ство. Предполагается, что высвобождаемый таким образом
кальций поступает пакостным канальцам в кровеносные ка-
пилляры, что и ведет к повышению концентрации кальция в
крови. Кроме того, паратгормон усиливает всасывание каль-
ция в кишечнике и реабсорбцию его в почечных канальцах.
Все это обусловливает повышенное содержание кальция в
крови.
213
У человека при гипофункции околощитовидных желез
содержание кальция в крови падает и одновременно увеличива-
ется содержание в ней фосфатов. Вследствие этого резко повыша-
ется возбудимость центральной нервной системы, а у детей, кро-
ме того, нарушается рост костей, зубов и волос. С лечебной
целью в таких случаях применяют препараты из этих же желез.
ТИМУС (ВИЛОЧКОВАЯ ЖЕЛЕЗА)
Вилочковая, или зобная, железа помещается за грудиной.
Вверху она примыкает к трахее, внизу — к аорте. Железа состоит
из двух асимметричных частей — правой и левой долей, плотно
прилегающих друг к другу и соединенных рыхлой соединитель-
ной тканью. Сверху железа покрыта соединительной капсулой.
Проникающие внутрь железы соединительно-тканные тяжи де-
лят ее на дольки. Каждая долька состоит из коркового слоя и
мозгового вещества. Мозговая часть дольки содержит значитель-
но меньше лимфоцитов, чем корковая, и преимущественно со-
стоит из эпителиальных клеточных элементов различной формы
и величины. Основную массу клеток вилочковой железы пред-
ставляют лимфоциты, ихлорые составляют 90% всей клеточной
популяции. Гормональную функцию несут эпителиальные клет-
ки мозгового и коркового слоев, причем во многих местах они
формируют так называемые тельца Гассаля — очаги кистозной
дегенерации с эндокринной активностью.
Существует определенная взаимосвязь между деятельнос-
тью железы и возрастом. Абсолютная масса железы у новорож-
денных составляет 13 г. Наибольшей массы (25—37 г) железа
достигает в период роста — от б до 16 лет. Впоследствии она
уменьшается в массе и у взрослых ее значительная часть заме-
щается жировой тканью. Полагают, что до периода полового
созревания зобная железа усиленно функционирует и подав-
ляет функцию половых желез. С наступлением половой зрело-
сти ее масса уменьшается и действие ослабляется.
Вилочковая железа вырабатывает следующие гормоны:
1. Лимфоцитстимулирующий гормон (ЛСГ) — стимулирует -
лимфопоэз и развитие лимфоидной ткани. У облученных лю-
дей восстанавливает клеточную иммунную реактивность. Пре-
дотвращает развитие синдрома истощения (отставание в росте
по сравнению со сверстниками, малоподвижность и т. д.).
2. Тимозин—предотвращает синдром истощения, стимули-
рует реакции клеточного иммунитета, обладает противоопухо-
левым эффектом.
214
3. Тимин — ингибирует (блокирует) передачу в нервно-мы-
шечных синапсах и является стимулятором иммунных про-
цессов.
4. Тимостерин — стимулирует продукцию антител, облада-
ет противоопухолевым действием.
5. Гипокальциемический фактор — сходен с кальцитонином
паращитовидной и щитовидной желез, т. е. понижает уровень
кальция и фосфата в костной ткани и вызывает соответствую-
щее изменение их уровня в сыворотке крови.
Патологические повреждения вилочковой железы встреча-
ются как самостоятельные и как сопутствующие заболевания,
и их можно подразделить на две группы:
а) врожденные, являющиеся последствием эмбрио-
нального или постэмбрионального повреждения костного
мозга, продуцирующего стволовые клетки. Вследствие этого в
вилочковой железе затруднена нормальная дифференцировка
стволовых клеток в иммунокомпетентные Т-лимфоциты, что
ведет к угнетению синтеза антител и иммунных реакций кле-
точного типа. Эти состояния наблюдаются у новорожденных и
обычно заканчиваются смертью до конца 1-го года жизни;
б) вторичные иммунодефицитные состояния, развива-
ющиеся при ряде заболеваний и отражающиеся на состоянии
иммунной системы. К таковым относятся опухолевые и ряд
вирусных заболеваний (например, поражение лимфоидной
системы вирусом СПИДа).
НАДПОЧЕЧНИКИ
Это парные уплощенные образования желтого цвета, лежа-
щие, как это и следует из их названия, в непосредственной бли-
зости от верхнего полюса почек. Длина каждой из этих желез
около 6 см, ширина 3—4 см, толщина немного меньше 1 см. Каж-
дый надпочечник состоит из коркового и мозгового вещества.
Кора надпочечников вырабатывает следующие гормоны.
1. Глюкокортикоиды (гидрокортизон, кортизон и кортико-
стерон). Из коры надпочечников выделены многочисленные
кортикостероиды (производные холестерина), но только неко-
торые из них активны. Глюкокортикоиды стимулируют синтез
белков-ферментов, которые осуществляют следующие воздей-
ствия на организм:
а) влияют на белковый и углеводный обмен;
б) подавляют образование клеток-киллеров и изменяют
иммунный ответ;
215
в) замедляют рост и формирование различных видов соеди-
нительной ткани (костной, хрящевой и т. д.);_
г) подавляют аллергические и воспалительные реакции.
2. Минералокортикоиды, наибольшей активностью среди
которых обладает альдостерон, который участвует в под-
держании баланса натрия и калия в организме, усиливая реаб-
сорбцию натрия в почечных канальцах. Отсюда роль альдосте-
рона состоит в сохранении в организме натрия.
Недостаток минералокортикоидов ведет к уменьшению ре-
абсорбции натрия и хлора. Организм при этом теряет большое
количество натрия, что несовместимо с жизнью. Только введе-
ние минералокортикоидов в этом случае поможет избежать ле-
тального исхода.
3. Половые гормоны являются слабыми андрогенами и эстро-
генами, которые образуются у взрослого человека в небольшом
количестве и контролируют развитие вторичных половых при-
знаков. Соответственно в организме мужчин образуются жен-
ские (эстрогены), а в организме женщин мужские (андрогены)
половые гормоны.
Мозговое вещество надпочечников секретирует следующие
гормоны.
1. Адреналин усиливает и учащает сокращение сердца, повы-
шает возбудимость, сердечной мышцы, при этом он расширяет
сосуды сердца и мозга, а также вызывает расширение сосудов
работающих мышц. Под его воздействием сокращаются глад-
кие мышцы желчных и мочевыводящих путей, матки и влага-
лища, и мышца, расширяющая зрачок; ослабляется моторная
функция желудка и кишечника, расслабляются мышцы моче-
вого и желчного пузыря, бронхов и бронхиол. Поэтому этот
гормон используют при лечении больных бронхиальной аст-
мой. Адреналин вызывает сужение сосудов (артериол) брюш-
ных органов и кожи (кроме кожи лица). Он усиливает рас-
щепление гликогена в печени и в мышцах до глюкозы и
дальнейшее его окисление. Под влиянием адреналина повы-
шается возбудимость рецепторов сетчатки глаза, слухового и
вестибулярного аппаратов, а также ЦНС. Таким образом, ад-
реналин может вызвать экстренное изменение состояния орга-
низма, направленное на повышение работоспособности.
2. Норадреналин отличается от адреналина отсутствием од-
ной метильной группы, поэтому норадреналин непосредствен-
но предшествует образованию самого адреналина. Адреналин и
норадреналин сходны по своим свойствам и физиологическому
влиянию, но пороги их действия различны. Так, норадреналин
вызывает сужение всех кровеносных сосудов.
216
Образование адреналина и норадреналина в мозговом слое
надпочечников регулируется, симпатической нервной систе-
мой. Возбуждение симпатических нервов, проходящих в чрев-
ном нерве, или раздражение определенных зон гипоталамуса
усиливает образование этих гормонов. Во время сильных эмо-
ций (гнев, спортивные напряжения, сильная радость) поступ-
ление адреналина в кровь увеличивается. Страх вызывает пре-
имущественный выброс норадреналина. Это свидетельствует о
том, что центры высших отделов мозга оказывают влияние на
продукцию этих гормонов.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
Поджелудочная железа относится к железам смешанной
секреции. В ней наряду с образованием пищеварительного
сока, содержащего пищеварительные ферменты (внешняя сек-
реция), вырабатываются гормоны, поступающие непосред-
ственно в кровь (внутренняя секреция).
Гормоны поджелудочной железы образуются в особых
группах клеток, называемых островками Лангерганса. Подже-
лудочная железа вырабатывает следующие гормоны.
1. Инсулин — по химической природе является белком. При
введении инсулина в организм происходит усиленный переход
глюкозы из плазмы крови в клетки печени, скелетной муску-
латуры, сердечной мышцы и др. Это происходит из-за того,
что инсулин увеличивает проницаемость мембраны клеток
для глюкозы, а в меньшей степени и для аминокислот, что
способствует синтезу белка. В клетках из глюкозы синтезиру-
ется гликоген. Инсулин также увеличивает поступление глю-
козы в жировые клетки,'где из нее образуются жиры. Таким
образом, он способствует использованию глюкозы в качестве
энергетического, пластического и запасного материала. Не-
достаточное количество инсул ина ведет к на-
рушению углеводного, белкового и жирового обмена. Это за-
болевание называется сахарным диабетом. При этом
заболевании повышается количество сахара в крови (гипергли-
кемия), достигая 200—300—400 мг% (0,2—0,4%) вместо 80—
120 мг% (0,08—0,12%) в норме. Понижается способность тка-
ней использовать поступающую в организм глюкозу и откла-
дывать ее в клетках печени й мышц в виде гликогена. Глюкоза
из крови переходит в мочу и выводится с ней из организма,
что называется глюкозурией. Это происходит потому, что в
почечных канальцах понижается реабсорбция глюкозы из пер-
217
вичной мочи. Выделение сахара сопровождается возрастанием
диуреза (полиурия) до 4—5 л в сутки. При диабете в связи с на-
рушением углеводного обмена резко увеличивается расходова-
ние белков и жиров в качестве источников энергий. Но так как
нормальный ход обмена веществ нарушается, то накапливают-
ся продукты неполного окисления жиров, к которым относят-
ся кетоновые тела — оксимасляная и ацетоуксусная кислоты.
Значительная часть белков, поступающих в организм, превра-
щается в углеводы, что сопровождается образованием большо-
го количества промежуточных кислых продуктов. При этом
идут изменения реакции крови в кислую сторону, возникает
ацидоз. Все эти явления ведут к отравлению организма. При
тяжелых формах диабета возможен смертельный исход. Тяже-
лое состояние организма облегчают путем внутривенного вве-
дения глюкозы.
2. Глюкагон. Этот гормон поджелудочной железы действует
противоположно инсулину: а) усиливает расщепление глико-
гена в печени; б) повышает уровень сахара в крови; в) стимули-
рует расщепление жира в жировой ткани.
Секреция инсулина и глюкагона находится под влиянием
автономной нервной системы. Раздражение блуждающего нерва
стимулирует образование инсулина, а раздражение симпатичес-
ких нервов тормозит его. Повышение количества глюкозы в
крови во время пищеварения, при физической нагрузке приво-
дит к возрастанию секреции инсулина. Наоборот, понижение
концентрации глюкозы в крови тормозит секрецию инсулина,
но повышает образование и выделение глюкагона. Такие меха-
низмы обеспечивают постоянный уровень сахара в крови.
3. Липокаин — стимулирует образование лецитина и окис-
ление жирных кислот в печени, т. е. способствует использова-
нию организмом жиров.
4. Ваготонин — повышает активность нейронов блуждаю-
щих нервов и усиливает возбудимость парасимпатического от-
дела автономной нервной системы.
5. Центропнеин—стимулирует нейроны дыхательного цен-
тра и расширяет бронхи; его применяют при лечении бронхи-
альной астмы.
ТКАНЕВЫЕ ГОРМОНЫ
Эти гормоны (точнее — гормоноиды) вырабатываются в
специализированных клетках, расположенных в разных орга-
нах. Чаще всего они действуют местно, на небольшом расстоя-
218
нии от выделения, хотя могут поступать в кровь. Они регули-
руют разнообразные физиологические и обменные процессы.
К тканевым гормонам относятся следующие. '
1. Гормоны желудочно-кишечного тракта, которые выделя-
ются во время пищеварения: секретин, гастрин, холецистоки-
нин и др. (см. тему «Пищеварение»).
2. Гистамин — обладает широким спектром действия.
В области выделения расширяет артериолы, капилляры и ве-
нулы, увеличивает их проницаемость, вызывает зуд и боль,
стимулирует образование желудочного сока. Гистамин играет
роль в развитии воспалительных и аллергических процессов.
3. Серотонин — действует местно, суживая сосуды, снижает
проницаемость капилляров, способствует агрегации тромбо-
цитов. Гистамин и серотонин участвуют в передаче нервных
импульсов в центральной нервной системе.
4. Кинины. К представителям этой группы относится поли-
пептид брадикинин, который снижает артериальное давление,
расширяет капилляры и увеличивает их проницаемость, явля-
ясь одним из важных факторов развития воспалительного про-
цесса. Брадикинин стимулирует сокращения гладких мышц
бронхов, кишечника и матки.
5. Простагландины — это циклические продукты ненасы-
щенных органических кислот, содержащих 20 атомов углеро-
да. Образуются простагландины во всех тканях организма. Не-
которые из них имеют значение в развитии воспалительных и
аллергических реакций. Среди них есть гормоноиды, вызыва-
ющие сокращения гладких мышц (применяются при родах),
но есть и расслабляющие гладкие мышцы. Некоторые из дан-
ных гормонов снижают артериальное давление.
6. Кейлоны — это низкомолекулярные гликопротеиды, об-
ладающие способностью блокировать синтез ДНК в клетках и
их размножение.
РАЗМНОЖЕНИЕ
СПОСОБЫ РАЗМНОЖЕНИЯ
ЖИВЫХ ОРГАНИЗМОВ
Все живые организмы, от бактерий до млекопитающих, об-
ладают способностью к самовоспроизведению, или размноже-
нию. Различают половое и беспалое размножение.
219
К бесполому размножению относятся различные способы раз-
множения организмов, характеризующиеся отсутствием полово-
го процесса и осуществляющиеся без участия половых клеток.
В процессе эволюции возникло более сложное половое раз-
множение, свойственное всем живым организмам, у которых
образуются половые клетки. Человеку также свойственно по-
ловое размножение, осуществляющееся за счет специализиро-
ванных половых клеток — яйцеклеток, образующихся в жен-
ских половых железах, и сперматозоидов, образующихся в
мужских половых железах. Развитие нового организма проис-
ходит только после их слияния, т. е. оплодотворения.
МУЖСКАЯ ПОЛОВАЯ СИСТЕМА
• <•
Мужская половая система включает в себя внутренние и на-
ружные половые органы. К внутренним мужским половым орга-
нам относят семенники (или яички), семявыносящие протоки,
предстательную железу, бульбоуретральные (куперовы) железы
и семенные пузырьки. К наружным половым органам принад-
лежат мужской половой член и мошонка (рис. 5.53).
Семенники (яички) — это половые железы, образующие муж-
ские половые клетки — сперматозоиды (внешняя секреция) и
половые гормоны (внутренняя секреция). Семенники — парная
железа округлой формы размером около 3 х 4 х 2 см. Семенник
снаружи покрыт плотной соединительно-тканной (белочной)
оболочкой, от которой внутрь органа отходят перегородки и
делят его на множество (100—300) долек (рис. 5.54). В каждой
дольке расположено по одному или по два извитых семенных
канальца, являющихся структурной и функциональной едини-
цей мужской половой железы. Длина каждого канальца 50—
80 см, а общая их длина достигает 250 м. Стенки канальцев
выстланы сперматогенным эпителием (рис. 5.55), из которого
образуются сперматозоиды, и клетками Сертоли, которые
способствуют созреванию сперматозоидов, участвуют в мета-
болизме андрогенов (мужских половых гормонов) и захватыва-
ют из крови питательные вещества. В тканях между семенны-
ми канальцами находятся клетки Лейдиха, вырабатывающие
андрогены, являющиеся стероидами — производными холесте-
рина. 90% андрогенов составляет самый активный гормон те-
стостерон. Тестостерон обладает активным воздействием
на белковый обмен, что способствует увеличению роста, мы-
шечной массы и силы у мальчиков, достигших полового созре-
вания.
220
26
27
22
18
28
2
19
16
15
Рис. 5.53. Мочеполовой
аппарат мужчины
(полусхематично):
1 ~ левая почка; 2 — корковое
вещество; 3 — мозговое вещество;
4 — лоханка почки; 5 ~ левый мо-
четочник; 6 — верхушка мочевого
пузыря; 7 — пузырно-пупочная
связка (остаток мочевого протока
зародыша); 8 — тело мочевого пу-
зыря; 9 — губчатое тело полового
члена; 10 — пещеристое тело по-
лового члена; 11 — губчатая часть
мочеиспускательного канала; 12—
головка полового члена; 13 —
дольки яичка; 14 — яичко; 15 — придаток яичка; 16 — семявыносящий
проток; 17 — седалищно-пещеристая мышца; 18 — луковично-губчатая
мышца; 19 — луковица полового члена; 20 — бульбоуретральная железа
(железа Купера); 21 — перепончатая часть мочеиспускательного канала;
22 — предстательная железа; 23 — семенной пузырек; 24 — ампула семявы-
носящего протока; 25 — дно мочевого пузыря; 26 — правая почка; 27 —
ворота почки; 28 — почечная артерия; 29 — почечная вена
221
•X.
Рис. 5.54. Яичко (белочная
оболочка частично вскрыта):
1 — белочная оболочка; 2 — сеть
средостения яичка; 3 — дольки яич-
ка; 4 — выносящий каналец; 5 —
проток придатка яичка; 6 — семя-
выносящий проток
Рис. 5.55. Сперматогенный эпителий:
1 — формирующиеся сперматозоиды; 2 — сперматиды; 3 — сперматоциты
первого порядка; 4 — сперматогонии; 5 — поддерживающие клетки (клет-
ки Сертоли)
222
У новорожденного ребенка извитые канальцы не имеют про-
света. Он появляется только на 7—8-м году жизни. В это же время
среди клеток стенки канальцев выделяются гоноциты. Из гено-
цидов после их деления формируются сперматогокии. Спермато-
гонии, в свою очередь, являются клетками, исходными дая обра-
зования сперматозоидов. Процесс образования сперматозоидов
назван сперматогенезом (подробно описан в курсе об-
щей биологии в пособии «Генетика и онтогенез», с. 37).
Созревшие сперматозоиды заполняют систему выносящих
канальцев, которые, выходя иэ семенника, формируют прида-
ток семенника. Придаток плотно прирастает к заднему краю
половой железы. Он является не только хранилищем сперма-
тозоидов, но и местом, где половые клетки становятся способ-
ными к оплод отворению. Выносящие канальцы сливаются в
штопороббразно закрученный проток придатка, который, в
свою очередь, переходит в семявыносящий проток. Левый и
правый семявыносящие протоки направляются ко дну мочево-
го пузыря, где они сближаются. В конечном отделе каждого
семявыносящего протока образуется как бы вырост — семен-
ной пузырек (см. рис. 5.53). Он представляет собой сильно из-
витую трубочку (ее размеры 5x2x1 см), в которой образуется
густой желтоватый секрет. Этот секрет поступает в семявыно-
сящий проток и обеспечивает питание и активизацию сперма-
тозоидов. При слиянии протока семенного пузырька с семя-
выносящим протоком формируется семявыбрасывающий
проток, который проходит через предстательную железу и от-
крывается в мочеиспускательный канал.
Предстательная железа, массой примерно в 20 г и размерами
около 4x3x2 см, представляет собой железисто-мышечный
орган. В ней вырабатывается жидкий секрет, который выбрасы-
вается в момент семяизвержения через мельчайшие отверстия
выводных протоков в мочеиспускательный канал. Предстатель-
ная железа прилегает своим основанием к мочевому пузырю, а
ее задняя поверхность — к стенке прямой кишки.
В просвет мочеиспускательного канала поступает еще один
секрет, который предохраняет слизистую оболочку канала от
раздражающего действия мочи. Этот секрет, имеющий вязкую
консистенцию, вырабатывается в бульбоуретральной (куперо-
вой) железе. Эта железа парная, размером с горошину.
Сперматогенез у человека составляет примерно 64—70 дней,
а число сперматозоидов, образующихся в семенниках взрослого
мужчины, огромно. Так, в одном миллилитре семенной жидко-
сти их содержится до 100 млн, а во время одного семяизверже-
ния выделяется 200—400 млн.
223
ЖЕНСКАЯ ПОЛОВАЯ СИСТЕМА
К женской половой системе относят наружные половые
органы, расйЪлагающиеся на поверхности тела, и внутренние,
находящиеся в полости малого таза.
К внутренним женским половым органам относятся: влагали-
ще, матка, маточные (фаллопиевы) трубы и яичники (рис. 5.56).
Влагалище — эластическая, легко растяжимая трубка дли-
ной 10-12 см, соединяющая наружные половые органы с мат-
кой. Стенки влагалища образованы из двух оболочек: слизис-
той и мышечной. Поверхностные слои слизистой оболочки
ежемесячно отторгаются, в связи с менструальным циклом.
Матка — гладкомышечный орган грушевидной формы дли-
ной 7—9 см. Полость матки имеет треугольную форму, верхние
углы которой открываются в устье маточных труб, а нижний
угол — в канал шейки матки. Стенка матки состоит из трех
Рис. 5.56. Внутренние половые органы женщины (вид сзади).
Матка, влагалище, левые яичник и маточная труба в разрезе:
1 — яичник; 2 — бахромки; 3 — отверстие маточной трубы, открывающееся
в полость брюшины; 4 — маточная труба; 5 — отверстие маточной трубы,
открывающееся в полость матки; 6 — дно матки; 7 — полость матки; 8 —
собственная связка яичника; 9 — широкая связка матки; 10 — канал шейки
матки; 11 — влагалище; 12 — наружный маточный зев; 13 — шейка матки;
14 — тело матки; 15 — круглая связка матки; 16 — пузырчатый яичниковый
фолликул (граафов пузырек)
224
слоев: внутреннего — эндометрия, среднего — миометрия и
наружного — периметрия. Поверхностные слои эндометрия
(слизистой оболочки) также изменяются в зависимости от фаз
менструального цикла.
Маточные трубы, или яйцеводы, — парные протоки, по ко-
торым перемещаются яйцеклетки. Длина их 10—12 см, начи-
наются они от углов матки и заканчиваются бахромками, или
фимбриями (см. рис. 5.56).
Яичники — женские половые железы, где образуются яй-
цеклетки (внешняя секреция) и половые гормоны (внутрен-
няя секреция). Размеры яичника примерно 4 х 3 х 1,5 см и его
масса 5—6 г. Он покрыт соединительно-тканной оболочкой,
под которой располагается корковое и мозговое вещество.
В корковом веществе происходит образование половых клеток
и выработка овариальных гормонов. Мозговое вещество обра-
зовано соединительной тканью, в которой проходят сосуды и
нервы.
В корковом слое располагаются многочисленные первич-
ные фолликулы. Каждый первичный фолликул состоит из за-
чатковой яйцеклетки (овоцита), окруженной однослойным
фолликулярным эпителием. Среди первичных фолликул
встречаются созревающие, атрофирующиеся фолликулы, жел-
тые и беловатые тела (рис. 5.57).
Размножение женских половых клеток происходит во внут-
риутробном периоде. В результате образуются первичные
фолликулы, каждый из которых содержит незрелую женскую
половую клетку — овоцит. В двух яичниках новорожденной
девочки содержится до 800 тыс. первичных фолликулов. Их
число после рождения уменьшается благодаря их рассасыва-
нию.
Ко времени наступления половой зрелости насчитывается
лишь 400—500 шт. первичных фолликулов, которые преобра-
зуются в зрелые пузырчатые фолликулы яичника (граафовы
пузырьки). Рост фолликулов происходит циклически каждые
28 дней, под влиянием фолликулостимулирующего гормона
(ФСГ), вырабатываемого гипофизом. При этом клетки фолли-
кулярного эпителия интенсивно делятся и эпителий стано-
вится многослойным. Фолликулярные клетки начинают выра-
батывать жидкость фолликула, содержащую женские половые
гормоны — эстрогены.
Яйцеклетка, окруженная слоем фолликулярных клеток, от-
тесняется к одному из йЬлюсов фолликула (яйценосный хол-
мик). Так образуется пузырчатый (вторичный) фолликул (см.
рис. 5.56).
8-281
225
Рис. 5.57. Яичник. Схема развития фолликула:
1 — примордиальный фолликул; 2 — растущий фолликул; 3 — о&олачка
фолликула; 4 — фолликулярная жидкость; 5 — белочная оболочка; 6 —
зрелый пузырчатый фолликул (граафов пузырек); 7 — белое тело; 8 —
атретический фолликул; 9 — интерстициальная ткань; 10 — овуляция (раз-
рыв граафова пузырька); 11 — мозговая часть яичника; 12 — зачатковый
эпителий; 13 — желтое тело; 14 — регрессирующее желтое тело
После созревания пузырчатый фолликул разрывается, и
овоцит, окруженный 3-4 тыс. фолликулярных клеток, выхо-
дит в брюшную полость, откуда попадает в маточную трубу.
Процесс выхода овоцита носит название овуляции. Она
происходит у человека регулярно под действием лютеинизи-
226
рующего гормона (ЛГ) лютропина, вырабатываемого гипофи-
зом. Под влиянием этого же гормона на месте лопнувшего
пузырька в яичнике развивается желтое тело — времен-
ная железа внутренней секреции. Желтое тело продуцирует
гормон прогестерон, который готовит слизистую оболочку
матки для имплантации оплодотворенной яйцеклетки. Про-
гестерон, кроме того, угнетает развитие фолликулов. Если
оплодотворения не происходит, желтое тело функционирует
лишь 12—14 дней, и на его месте в яичнике остается соедини-
тельно-тканный рубец — беловатое тело. В случае оп-
лодотворения желтое тело сохраняется в течение всей бере-
менности. <
НЕРВНАЯ СИСТЕМА
ОБЩИЕ ПРИНЦИПЫ ОРГАНИЗАЦИИ
НЕРВНОЙ СИСТЕМЫ
Нервная система регулирует и координирует деятельность
всех органов и их систем, обусловливая целостность функцио-
нирования организма. Благодаря ей осуществляется связь
организма с внешней средой И его адаптация к постоянно ме-
няющимся условиям. Нервная система является материаль-
ной основой сознательной деятельности человека, его мышле-
ния, поведения, речи.
Нервная система в организме представлена группой орга-
нов, ведущая роль в образовании которых принадлежит не-
рвной ткани, которая, в свою очередь, состоит из нейронов и
глиальных клеток.
Нервная система имеет два вида деления. Функцио-
нально она подразделяется на соматическую и автономную
(вегетативную), анатомически — на центральную не-
рвную систему (ЦНС) и периферическую нервную систему.
Соматическая нервная система регулирует работу скелет-
ных мышц и обеспечивает чувствительность человеческого
тела. Автономная (вегетативная) нервная система регули-
рует обмен веществ, работу внутренних органов и гладких
мышц.
Центральная нервная система состоит из головного моз-
га, заключенного в мозговом отделе черепа, и спинного моз-
га, который проходит по каналу позвоночника и заканчива-
8*
227
ется на уровне границы между 1-м и 2-м поясничными по-
звонками.
Периферическая нервная система образована многочис-
ленными парными нервами, отходящими от головного и спин-
ного мозга. По ходу таких нервов расположены скопления тел
нервных клеток — ганглии, также относящиеся к периферичес-
кой нервной системе.
СТРУКТУРНЫЕ ЭЛЕМЕНТЫ
НЕРВНОЙ СИСТЕМЫ
Нейрон. Основным элементом нервной системы является
нервная клетка, называемая нейроном. В каждой из них мож-
но выделить тело клетки и отростки — длинные и короткие.
Нейроны классифицируются по числу их цитоплазматических
отростков (рис. 5.58):
1) биполярные нейроны (имеют два отростка);
2) мультиполярные нейроны (имеют более двух отростков);
3) униполярные нейроны (имеют один выраженный цито-
плазматический отросток). *
Мозг человека, как полагают, состоит из 2,5 х 1О10 нейро-
нов. Это приблизительно столько же, сколько звезд в нашей
Галактике. Все процессы химическбго синтеза протекают в
теле нейрона, откуда продукты поступают в различные отрост-
ки. Во взрослом организме не наблюдается признаков митоза
йейронов, что указывает на приобретение полного или по
крайней мере почти полного набора нервных клеток уже ко
времени рождения.
Нервное волокно. От тела нейрона отходят тонкие отрост-
ки, по которым распространяются нервные импульсы. Нейро-
ны имеют один (и только один) аксон (от греч. ахоп — ось),
названный так, вероятно, потому, что он часто остается нераз-
ветвленным. Таким образом, аксоном является только один из
двух отростков биполярного нейрона или один из многих у
мультиполярного. Другой отросток биполярного нейрона и
все остальные отростки мультиполярного нейрона называют
дендритами (от греч. dendron — дерево), так как многие из них
разветвляются наподобие дерева.
Нервные импульсы аксоны передают от тела нейрона к ме-
сту назначения, а дендриты ~ к телу клетки.
Чувствительные (сенсорные) нейроны передают информа-
цию^ расположенных на периферии рецепторов к ЦНС.
Двигательные (моторные) нейроны проводят нервные импуль-
228
Рис. 5.58. Нервные
клетки:
А — мультиполярный
нейрон; Б — униполяр-
ный нейрон; В — бипо-
лярный нейрон; 1 — ней-
рит; 2 — дендриты
сы от ЦНС к скелетным мышцам, к секреторным клеткам и
гладким мышцам внутренних органов и кожи.
Нервные клетки образуют специализированные контакты,
или синапсы, с постсинаптической клеткой (нервной, мышеч-
ной или железистой). Подавляющее большинство синапсов
ЦНС образовано окончаниями аксонов одних нейронов на
дендритах других. Специальная функция синапса заключается
в передаче импульса от клетки к клетке. Отдельные клетки об-
разуют до 2000 синапсов каждая.
С функциональной точки зрения важной частью крупного
нервного волокна является аксоплазма — тонкий тяж прото-
плазмы, продолжающий протоплазму тела клетки. Аксоплаз-
ма окружена плазматической мембраной и почти всегда по-
верх нее бывает одета дополнительными оболочками. Над
плазматической мембраной располагается прочная, неэлас-
тичная многоклеточная оболочка — неврилемма. Она образо-
вана особыми шванновскими клетками (леммоцитами).
229
Обычно часть шванновской клетки многократно наматывает-
ся на участок отростка нейрона, образуя подобие рулета.
Слои очень тонкие и состоят из жирового вещества мембран.
Таким образом формируется так называемая миелиновая обо-
лочка. В промежутке между двумя соседними шванновскими
клетками — «рулетами» — миелиновая оболочка нервного
волокна прерывается. Такая область именуется перехватом
Ранвье.
Различают две основные группы нервных волокон: - м и е -
л и н о в ы е (мякотные) (рис. 5.59) и безмиелиновые
(безмякотные).
Нервы построены из мякотных и безмякотных нервных во-
локон и соединительно-тканных оболочек. Мякотные нервные
волокна входят в состав чувствительных и двигательных нервов,
безмякотные нервные волокна в основном принадлежат авто-
номной нервной системе. Между нервными волокнами распо-
лагается тонким слоем соединительная ткань — эндонервий.
Снаружи нерв покрывает волокнистая соединительная ткань —
перинервий. Таким образом, крупные аксоны как бы заключены
в футляр из мембранных слоев шванновских клеток.
Клетки глии. Пространство между нейронами заполняют
глиальные клетки. На их долю приходится около 90% общего
числа клеток в нервной системе и примерно половина ее объе-
ма. Глия обеспечивает структурную и метаболическую опору
для сети нейронов, создает их над лежащее взаиморасположе-
ние. Среди них различают:
1) астроциты, находящиеся в головном и спинном мозгу.
Они расположены между кровеносными капиллярами и тела-
Рис. 5.59. Мякотное нервное волокно (продольный разрез):
1 — неврилемма; 2 — леммоцит (шванновская клетка); 3 — осевой цилиндр
с проходящими в нем нейрофибриллами; 4 — миелиновая оболочка; 5 —
перехват Ранвье
230
ми нейронои, где образуют астроцитарные ножки.
Астроциты периодически подвергаются дегенерации, однако
их популяция восстанавливается за счет деления клеток-пред-
шественников (астробластов);
2) олигодендроциты, тесно связанные в ЦНС с длинными
нервными путями, образованными пучками аксонов, а также с
нервами. Они выполняют функцию образования миелина, как
и шванновские клетки, однако образуют миелиновую оболоч-
ку волокна между перехватами Ранвье;
3) эпендимные клетки глии, в основном образующие непре-
рывную эпителиальную выстилку желудочков мозга —эпен-
диму. Они сохраняются и в центральном канале спинного
мозга. В некоторых участках желудочков эпендима впячивает-
ся, одевая сосудистые сплетения;
4) микроглию, состоящую из мелких клеток, разбросанных в
белом и сером веществе мозга. Клетки микроглии редки в не-
поврежденном мозгу, в области же повреждений ткани мозга
они всегда представлены в изобилии. Подобно макрофагам
они выполняют фагоцитарную функцию.
Мозговые оболочки. Головной и спинной мозг защищены
не только костным футляром (черепом и позвоночником), но
и тремя соединительно-тканными мозговыми оболочками.
Первая из них — мягкая (сосудистая) оболочка — является
внутренней. Она заходит во все щели И борозды, образована
соединительно-тканными эластическими волокнами и прони-
зана множеством кровеносных сосудов. Проникая в мозг, кро-
веносные сосуды обеспечивают его питание.
Паутинная оболочка составляет средний слой. Он отделен
от мягкой мозговой оболочки (и в то же время соединен с нею)
паутиноподобной сетью тонких соединительно-тканных пере-
городок.
Паутинная оболочка выстлана слоем уплощенных клеток.
Как и ее перегородки (трабекулы), она содержит коллагеновые
и эластические волокна и с наружной, и внутренней поверх-
ности. Между паутинной и мягкой оболочками имеется
пространство, заполненное цереброспинальной жидкостью и
называемое подпаутинным субарахноидальным про-
странством. Связь спинномозговой жидкости с жидкими тка-
нями организма осуществляется через отростки паутинной
оболочки, проникающие в лимфатические кровеносные ка-
пилляры.
Твердая мозговая оболочка — самая наружная. Она имеет же-
сткую консистенцию и состоит из плотной соединительной
ткани, богатой коллагеновыми и эластическими волокнами и
231
отличается повышенной прочностью. Пространство между
твердой и паутинной оболочками называется субдураль-
ным. Она обычно содержит небольшое количество жидкости,
которая не является цереброспинальной. Твердая мозговая
оболочка головного мозга срастается с внутренней надкостни-
цей костей основания черепа. Менее прочно она связана с ко-
стями свода черепа. В спинном мозге твердая мозговая обо-
лочка отделена от надкостницы, выстилающей позвоночный
канал, эпидуральным пространством — узкой щелью,
заполненной рыхлой соединительной тканью, богатой венами
и жировыми клетками.
Рецепторы. Центральная нервная система получает инфор-
мацию о состоянии окружающей среды от рецепторов. Каж-
дый рецептор воспринимает определенный раздражитель —
химический, электромагнитный (световые волны), механичес-
кий или температурный. Первичная реакция любого рецепто-
ра состоит в генерации рецепторного потенциала, возникаю-
щего в результате взаимодействия между раздражителем и
мембраной рецептора. Затем информация в виде электричес-
ких импульсов по чувствительным нервам поступает в сенсор-
ные структуры нервной системы, где подвергается декодирова-
нию и анализу.
Морфологически и физиологически каждый рецептор при-
способлен д ля восприятия строго определенного раздражителя,
т. е. раздражителя, к которому рецептор наиболее чувствителен.
По этому признаку все рецепторы обычно делят на пять групп:
фоторецепторы, воспринимающие свет;
механорецепторы, реагирующие на прикосновение, давле-
ние, звуковые волны;
хеморецепторы, чувствительные к уровню глюкозы в крови,
концентрации газов;
терморецепторы, чувствительные к холоду и теплу:
болевые рецепторы, реагирующие на повреждение ткани.
В зависимости от пространственного восприятия раздра-
жителя рецепторы подразделяют на четыре группы:
дистантные экстерорецепторы, реагирующие на отдален-
ные раздражители (зрительные, обонятельные и слуховые);
контактные экстерорецепторы, воспринимающие раздра-
жения поверхности тела (рецепторы прикосновения, давле-
ния, температурные и вкусовые);
интерорецепторы, воспринимающие раздражение от внут-
ренних органов и уровень химических веществ в крови;
проприорецепторы, сигнализирующие о положении тела в
пространстве (раздражение от суставов, сухожилий, мышц).
232
ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА
НЕРВНОГО ВОЛОКНА И НЕРВОВ
1. Возбудимость. В 1791 г. ученый из Болоньи Гальвани выд-
винул идею о существовании «животного электричества» в не-
рвах и мышцах. Его соотечественник Маттеучи в 40-х гг. XIX в.
получил первые доказательства электрической природы не-
рвного импульса, а Гельмгольц, ставший впоследствии знаме-
нитым физиком, в 1850 г. измерил скорость проведения не-
рвного импульса,, определив передачу его по нерву не как
физическое проведение, а как активный биологичес-
кий процесс. В связи с этим нервные импульсы были на-
званы потенциалом действия.
В результате получили широкое распространение пред-
ставления о том, что, во-первых, нейрон можно рассматривать
как клетку, предназначенную для выработки импульсов, и, во-
вторых, что эти импульсы служат одним из средств обмена
сигналами между нервными клетками.
2. Проводимость. Функция аксона заключается в проведе-
нии нервных импульсов. Проведение нервного импульса мож-
но уподобить распространению электрического тока. Обычно
потенцйал действия зарождается в начальном, ближайшем к
телу клетки сегменте аксона и пробегает по аксону к его окон-
чаниям.
За счет различных ионов (Na+, К+), которые постоянно пе-
ремещаются через мембрану живой клетки (вследствие диф-
фузии), на ее поверхности формируется заряд (мембранный
потенциал). В покое на внутренней стороне мембраны регист-
рируется отрицательный потенциал. Постоянный отрицатель-
ный потенциал, регистрируемый на нейронах, называют мем-
бранным потенциалом покоя, а данное явление — поляризацией.
Уменьшение степени поляризации (смещение потенциала к
нулю) называют деполяризацией, а увеличение — гиперполя-
ризацией.
После раздражения нейрона возникший потенциал дей-
ствия состоит из двух фаз:
а) фазы деполяризации мембраны, обусловленной времен-
ным повышением ее проницаемости для на-
тр и я. В этот момент открываются специфические натриевые
каналы, и Na+ лавинообразно устремляются в клетку.
Этот приток положительных ионов приводит к уменьше-
нию заряда на противоположной мембране;
б) фазы деполяризации потенциала действия, связанной с
закрытием натриевых и открытием калие-
233
вых каналов. По мере выхода К+ удаляются положитель-
ные заряды, и мембранный потенциал вновь достигает уровня
покоя, после чего наступает гиперполяризация. Заканчива-
ется эта фаза закрытием калиевых каналов, что приводит к
восстановлению прежнего уровня проницаемости для К+ и
Na+ и исходного уровня мембранного потенциала (обычно он
равен 70 мВ).
Возникновение потенциала действия имеет следующие
особенности:
все возбудимые клетки характеризуются, определенным
пороговым значением изменения мембранного по-
тенциала, при котором возникает потенциал действия;
потенциал действия создает короткую рефрактер-
но с т ь (т. е. невосприимчивость мембраны в этот момент к
новому раздражению);
не существует сильных или слабых импульсов: волокно или
включается полностью или не включается вовсе, нейроны ра-
ботают по принципу «в с е или ничего»;
Потенциал действия (импульс) характеризуется посто-
янством амплитуды при распространении на большие
расстояния.
Проведение потенциала действия по различным аксонам
нейронов не равнозначно. Так, в миелизированных волокнах
скорость прохождения импульса достигает 120 м/с, а в безмя-
котных она значительно ниже. Разница связана с тем, что в
миелиновых волокнах деполяризующий ток не протекает че-
рез участки, покрытые миелиновой оболочкой, а «прыгает»
лишь по небольшим обнаженным участкам аксона (перехва-
там Ранвье). Потенциал действия здесь распространяется
скачкообразно (сальтоторное проведение).
» 3. Целостность нервного волокна. Возбуждение распростра-
няется по нервному волокну лишь при сохранении его анато-
мической и физиологической целостности. Потеря структур-
ных и физиологических свойств в результате перевязки,
охлаждения или воздействия токсических веществ ведет к на-
рушению проводимости нервного волокна.
4. Двустороннее проведение возбуждения по нервному волок-
ну. Явление открыто русским ученым Р. И. Рабухиным, кото-
рый показал, что возбуждение, возникнув в какой-либо облас-
ти волокна, распространяется в обе стороны, независимо от
того, какое это волокно — центростремительное или центро-
бежное.
5. Свойство изолированного проведения нервного импульса.
Если возбуждение возникло в одном нервном волокне, то оно
234
не может перейти на соседнее нервное волокно, находящееся в
одном и том же нерве. Важное значение этого свойства прояв-
ляется в том, что большинство нервов являются смешанными,
состоящими из тысяч функционально различных нервных во-
локон.
6. Относительная неутомляемость нерва. Это свойство было
определено в 1884 г. Н. Е. Введенским, который показал, что
нерв сохраняет способность к проведению возбуждения даже
при длительном непрерывном его раздражении, т. е. нерв прак-
тически неутомляем. Лишь изменения морфофункциональных
свойств нерва постепенно подавляют его проводимость.
7. Функциональная лабильность нервной ткани. Понятие,
сформулированное Н. Е. Введенским в 1892 т., который обна-
ружил, что нерв может отвечать на данную частоту раздраже-
ния такой же частотой возбуждения лишь до определенного
предела. Отсюда мерой лабильности по Н. Е. Введенскому яв-
ляется наибольшее число возбуждений, которое ткайь может
воспроизвести в 1 секунду в полном соответствии с частотой
раздражений. Например, наибольшее число импульсов двига-
тельного нерва теплокровных составляет до 1000 в 1 секунду.
Возбудимая ткань в зависимости от функционального состоя-
ния способна изменять свою лабильность как в сторону ее по-
нижения, так и повышения. В этом случае возбудимая ткан»:
начинает усваивать новые, более высокие (или низкие), ранее
не доступные ей ритмы активности. Снижение функциональ-
ной лабильности в процессе жизнедеятельности ведет к т о р -
можению функции.
ПЕРЕДАЧА ВОЗБУЖДЕНИЯ С НЕРВА
НА ИННЕРВИРУЕМУЮ КЛЕТКУ
Структуры, обеспечивающие переход возбуждения с не-
рвного волокна на иннервируемую им клетку — нервную, мы-
шечную, секреторную — называют синапсами (от греч.
synapsis — соединение). В местах межклеточных контактов
мембраны клеток разделены синаптической щелью. Участок
мембраны нервного волокна называется пресинаптической
мембраной, а участок мембраны контактирующей с ним клет-
ки — постсинаптической мембраной. Выделяют химичес-
кие синапсы (с химической передачей импульса) и элект-
рические (с электрической передачей импульса). Первые
осуществляют проведение возбуждения только в одном на-
правлении, а вторые — в обоих направлениях. Последние у
235
млекопитающих встречаются редко. Химический синапс име-
ет сложное строение. В нервном окончании, покрытом преси-
наптической мембраной, располагаются особые пузырьки (ве-
зикулы), содержащие химические вещества, передающие
возбуждение от пресинаптической мембраны на постсинапти-
ческую мембрану и называемые медиаторами. Медиаторы
синтезируются в теле нейрона и током аксоплазмы транспор-
тируются к пресинаптической мембране.
Распространение потенциала действия по аксону вызывает
диффундирование микродоз нейромедиатора в синаптичес-
кую щель и образование временного контакта между клетка-
ми. Такой контакт вызывает возбуждение постсинаптической
мембраны и возникновение на ней постсинаптического по-
тенциала. Его величина зависит от количества медиатора, ко-
торый выделяется в синаптическую щель и от чувствительнос-
ти постсинаптической мембраны. Достижение критического
уровня йостсинаптического потенциала вызывает возникнове-
ние потенциала действия, распространяющегося по иннерви-
руемой клетке.
Наиболее распространенными медиаторами являются
ацетилхолин и норадреналин.
После прохождения импульсов через синапс с одной клетки
на другую в синаптическом пространстве медиатор подвергает-
ся расщеплению ферментами, которые находятся на мембра-
нах. Компоненты медиатора затем в значительном количестве
захватываются пресинаптическими окончаниями, транспорти-
руются в цитоплазму, где вновь используются для биосинтеза.
АНАТОМИЯ ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ
Анатомически нервную систему принято делить на цент-
ральную — головной и спинной мозг, и перифери-
ческую — нервы (нервные стволы) и нервные узлы
(ганглии).
К периферической нервной системе относятся нервные
стволы (из головного и спинного мозга), а также нервные
сплетения возле сосудов и в органах, нервные узлы и разнооб-
разные нервные окончания.
Нервные стволы (нервы) при выходе из мозга образуют на-
чальную часть нерва, называемую к о р е ш к о м. Из спинного
мозга нервы выходят двумя корешками — передним и задним.
От головного мозга отходят 12 пар нервов:
236
1) обонятельный нерв — чувствительный, идет от обонятель-
ного эпителия слизистой оболочки носовой полости к о б о -
нятельным луковицам переднего мозга;
2) зрительный нерв — чувствительный, берет начало от сет-
чатки глаза, вместе с выростами промежуточного мозга образу-
ет зрительный перекрест и затем составляет даль-
нейший ход проводящих путей зрительного анализатора;
3) глазодвигательный нерв — двигательный, иннервирует че-
тыре из шести наружных мышц глаза;
4) блоковый нерв — двигательный, иннервирует верхнюю ко-
сую мышцу глаза;
5) тройничный нерв — смешанный крупный нерв с тремя ос-
новными ветвями: глазной, верхнечелюстной и нижнечелюст-
ной;
6) отводящий нерв — двигательный, идет к задней прямой
мышце глаза, которая отводит глаз вбок или назад;
7) лицевой нерв — смешанный, иннервирует все мимичес-
кие мышцы, слезную и слюнные железы, включает и чувстви-
тельные волокна от вкусовых рецепторов языка;
8) слуховой, или преддверно-улитковый нерв — чувствитель-
ный, идет от внутреннего уха. Разветвляется на две части:
преддверную, несущую импульсы от вестибулярного ап-
парата, и улитковую, несущую импульсы от спирального
' органа;
9) языкоглоточный нерв — смешанный, двигательные волокна
иннервируют некоторые мышцы глотки, а чувствительные —
мышцы органа вкуса, зева, евстахиевой трубы и барабанной по-
лости;
10) блуждающий нерв — смешанный, кроме мускулатуры го-
ловы иннервирует многие внутренние органы — сердце, желу-
док, кишечник, печень, почки, поджелудочную железу;
11) добавочный нерв — двигательный, иннервирует две мыш-
цы — грудино-ключично-сосцевидную и трапециевидную;
12) подъязычный нерв — двигательный, является переходным
от спинномозговых нервов к черепным. Иннервирует мышцы
языка и некоторые мышцы шеи.
Спинномозговые нервы смешанные, их 31 пара, они распо-
ложены слева и справа в каждом из сегментов тела. Нервы бы-
вают различной толщины и длины. Чем крупнее нервный
ствол, тем большую область он иннервирует. Поэтому по тол-
щине, длине, положению и ветвлению нервного ствола можно
приблизительно судить об области, иннервируемой им. Са-
мые длинные — это нервные стволь^конечностей, особенно
нижних (седалищный нерв). Большинство нервов состоят из
237
множества нервных волокон. Чем толще нерв, тем больше
приходится нервных волокон на площадь его поперечного сре-
за. Особенно много нервных волокон приходится на мышцы
глазного яблока. Поэтому нервные стволы глазных мышц,
кожные нервы лица и пальцев относительно тайце, чем про-
чие нервы.
Главный ствол спинального нерва и его основные ветви
включают в свой состав как афферентные (от рецепторов
импульсы проводятся к спинному и головному мозгу), так и
эфферентные (от 1ЩС импульсы проводятся к мышеч-
ным и железистым структурам) волокна. Однако корешки у
человека имеют четкое разграничение функций. Нередкие ко-
решки — проводят импульсы центробежно по эфферентным
волокнам, тела которых лежат в спинном мозге. Задние кореш-
ки, напротив, включают афферентные волокна, проводящие
центростремительные импульсы. Тела соответствующих ней-
ронов лежат, как правило, в ганглии заднего корешка. Эти
клетки считаются униполярными, так как имеют отросток, ко-
торый почти сразу Т-образно разветвляется: одна его часть до-
ставляет импульсы от чувствительных структур, другая переда-
ет импульсы в ЦНС.
АНАТОМИЯ ЦЕНТРАЛЬНОЙ
НЕРВНОЙ СИСТЕМЫ
Спинной мозг расположен в позвоночном канале. Он начи-
нается от головного мозга и имеет вид белого, несколько
сплюснутого в спинно-брюшном направлении тяжа длиной
около 45 см. На уровне 1—2-го поясничных позвонков он за-
канчивается мозговым конусом, от которого идет концевая
нить, присоединяющаяся к надкостнице копчика. Внутри
спинного мозга находится узкий центральный канал, содержа-
щий спинномозговую жидкость. Этот канал является остатком
полости первичной нервной трубки. Вверху он сообщается с
полостью IV мозгового желудочка, а внизу заканчивается не-
большим расширением — концевым желудочком.
Спинной мозг имеет два утолщения, образованные скопле-
нием тел нервных клеток, иннервирующих конечности: шей-
ное и поясничное (рис. 5.60).
В спинном мозге различают серое и белое вещество.
Серое вещество, образованное преимущественно телами
нервных клеток, расположено вокруг центрального канала и на
поперечном срезе напоминает по форме букву «Н» (рис. 5.61).
238
8
Рис. 5.60. Спинной мозг.
а — вид спереди; б — вид
сзади:
/ — передняя срединная щель;
2 — задняя срединная борозда;
3 — шейное утолщение; 4 — по-
ясничное утолщение; 5 — моз-
говой конус; 6 — концевая нить;
7 — продолговатый мозг; 8 —
мост
а б
2
рис. 5.61.
Горизонтальный разрез
спинного мозга:
1 — задняя срединная бо-
розда; 2 — задние столбы
белого вещества спинного
мозга; 3 — задний корешок;
4 — боковые столбы белого
вещества спинного мозга;
5 — передний корешок; 6 —
передние столбы белого ве-
щества спинного мозга; 7—
передняя срединная щель;
8 — передний рог, 9 — бо-
ковой рог; Ю — задний рог
239
В нем различают передние, или вентральные, и задние, ши
дорзальные, рога.
В грудной и верхней части поясничного отделов выделяют
еще боковые рога серого вещества, образованные телами ней-
ронов симпатического отдела автономной (или вегетативной)
нервной системы. Вокруг серого вещества располагается белое
вещество, образованное нервными волокнами (длинными от-
ростками нервных клеток).
Нейроны функционально делятся на чувствительные
(сенсорные), двигательные, или моторные, и вставочные,
обеспечивающие связь первых со вторыми.
К задним рогам серого вещества спинного мозга подходят
аксоны чувствительных нейронов и образуют задние спинно-
мозговые корешки. От передних рогов спинного мозга отходят
аксоны двигательных нейронов и образуют передние корешки
спинномозговых нервов.
Белое вещество спинного мозга делят на передние, задние
и боковые столбы, в которых расположены проводящие пути.
По восходящим проводящим путям возбуждение проходит от
рецепторов к вставочным нейронам спинного мозга и далее к
вышележащим отдела'м головного мозга. По нисходящим про-
водящим путям электрические импульсы передаются от раз-
ных отделов головного мозга к рабочим органам. В этом про-
является проводниковая функция спинного мозга,
вторая функция — рефлекторная. Она заключается в
том, что спинной мозг обеспечивает осуществление простей-
ших рефлексов, таких как сгибание и разгибание конечностей,
коленный рефлекс, отдергивание руки, ритмические рефлексы
(например, процесс шагания).
Рефлекс — это ответная реакция организма на внешнее
или внутреннее раздражение, осуществленная с участием
ЦНС и под ее контролем. Путь, по которому осуществляется
рефлекс, называют рефлекторной дугой.
Рефлекторная дуга состоит из пяти основных звеньев
(рис. 5.62):
I — рецептор;
II — чувствительный путь;
III — центральная нервная система;
IV — двигательный путь;
V — рабочий орган.
На схеме изображена двух- и трехнейронная рефлекторная
дуга. Она начинается от рецептора 1, находящегося в коже,
дендрит чувствительного нейрона 2 которого составляет боль-
шую часть чувствительного пути (II). Тела чувствительных
240
Рис. 5.62. Рефлекторная дуга.
а — двухнейронная; б — трехнейронная:
1 — рецептор; 2 — чувствительный (центростремительный) нерв; 3 — чув-
ствительный нейрон в спинномозговом ганглии; 4 — аксон чувствительного
нейрона; 5 — задние корешки спинномозговых нервов; 6 — вставочный
нейрон; 7 — аксон вставочного нерва; 8 — двигательный нейрон в рогах
спинного мозга; 9 — спинной мозг; 10 — аксон двигательного (центробеж-
ного) нейрона; 11 — рабочий орган
/
нейронов 3 находятся всегда за пределами ЦНС и формируют
узлы на задних корешках 5 спинномозговых нервов. Аксон
чувствительного нейрона 4, войдя в ЦНС, передает электри-
ческие импульсы на вставочный нейрон 6. Тела вставочных
нейронов 6 слагают задние рога серого вещества спинного
мозга. Передние рога серого вещества образованы телами дви-
гательных нейронов 8. На их дендриты 7 поступает возбуж-
дение с аксона, вставочного нейрона 6. Аксон двигательного
нейрона'в составе спинномозгового нерва формирует чувстви-
тельный путь (IV) и заканчивается в мышце (V).
Еслд рефлекторная дуга двухнейронная (моносинаптичес-
кая), то в этом случае ее начало и окончание находится в од-
ном и том же органе. Примером моносинаптической рефлек-
торной дуги может служить коленный рефлекс.
Головной мозг располагается в полости черепа. Его масса у
взрослого человека составляет 1500 г (от 1100 до 2000 г). Коле-
бания в размерах мозга человека не являются критерием для
суждения о степени развития интеллекта. Так, масса мозга Бай-
рона 2238 г, Тургенева — 2012 г, Кромвеля — 2000 г, Шилле-
ра — 1871 г, химика Либиха — 1325 г, поэта Уитмена — 1282 г,
Анатоля Франса — 1017 г.
Головной мозг имеет следующие отделы: продолговатый
мозг, задний мозг (мост и мозжечок), средний мозг и пере-
241
дний мозг (конечный мозг, представляющий собой большие
полушария, и промежуточный мозг) (рис. 5.63).
Головной мозг имеет четыре полости, связанные друг с дру-
гом и называемые желудочками. Полость IV желудочка распо-
ложена на уровне продолговатого мозга и моста. III желудочек
локализован в промежуточном мозге. Эти полости соединены
водопроводом мозга — узким каналом, проходящем в среднем
мозге. I и II желудочки называют боковыми. Они симметрич-
но расположены в больших полушариях головного мозга.
Рассмотрим подробнее каждый из отделов головного мозга.
Продолговатый мозг является непосредственным продол-
жением спинного мозга. Длина его около 25 мм, форма при-
ближается к усеченному конусу, обращенному основанием
вверх.
Центры продолговатого мозга регулируют деятельность
многих органов грудной и брюшной полостей. Нейроны этого
отдела участвуют в регуляции дыхания, кровообращения, пи-
щеварения.
В продолговатом мозге замыкаются рефлекторные дуги за-
щитных рефлексов: кашля, чихания, рвоты.
7
8
1 2 3 4 5
Рис. 5.63. Продольный разрез головного мозга:
1 — продолговатый мозг; 2 — варолиев мост; 3 — средний мозг; 4 — гипофиз;
5 — промежуточный мозг; 6 — мозолистое тело; 7 — полушарие переднего
мозга; 8 — четверохолмие; 9 — червячок; 10 — полушарие мозжечка
242
Мост появляется лишь у млекопитающих. У человека он
достигает наибольшего развития. Проводящие дуги моста свя-
зывают кору больших полушарий со спинным мозгом и с ко-
рой полушарий мозжечка.
Нейроны моста воспринимают сигналы от кожи лица, от
языка и слизистой оболочки ротовой полости (вкусовая чув-
ствительность); сюда поступает информация от рецепторов
слухового и вестибулярного анализаторов. В нем находятся не-
рвные центры, регулирующие работу слюнных и слезных же-
лез, жевательных и мимических мышц.
Мозжечок расположен в затылочной части головного моз-
га над продолговатым мозгом и мостом. Мозжечок состоит
из коры, представленной серым веществом. Толщина коры
колеблется в пределах I—2,5 мм. Под ней в толще белого ве-
щества находятся ядра. В мозжечке различают два полуша-
рия и соединяющую их срединную часть — червь. Поверх-
ность полушарий и червя разделяют поперечные борозды,
увеличивающие ее площадь до 850 см2. Мозжечок тремя па-
рами мозжечковых ножек связан с продолговатым мозгом, ва-
ролиевым мостом и средним мозгом. Его основная роль за-
ключается в поддержании равновесия и координации
движений.
Средний мозг соединяет передний мозг с задним. Здесь рас-
положены центры поддержания напряжения (тонуса) скелет-
ных мышц, центры зрения и слуха. В среднем мозге замыка-
ются дуги ориентировочных рефлексов на зрительные и
слуховые раздражения. Ориентировочные рефлексы проявля-
ются в поворотах головы и тела в сторону раздражителя.
В промежуточный мозг электрические импульсы от всех ре-
цепторов тела и после частичной обработки в этом отделе на-
правляются к большим полушариям. Ядрами продолговатого
мозга осуществляется регуляция сложных двигательных реф-
лексов, таких как плавание, ходьба, бег. Отсюда координиру-
ется работа внутренних органов, регулируется обмен веществ,
потребление пищи и воды. В промежуточном мозге располо-
жены высшие центры автономной нервной системы. Нейроны
некоторых ядер этого отдела мозга вырабатывают биологичес-
ки активные вещества (БАВ), осуществляя гуморальную регу-
ляцию.
Большие полушария головного мозга достигают наибольшего
развития у человека. На них приходится 78% общей массы го-
ловного мозга. Поверхность больших полушарий образована
серым веществом — корой. Ее площадь около 2200 см2. Эго
достигается благодаря ее складчатому строению, образованию
243
борозд и извилин. Причем поверхность извилин более чем в
два раза меньше суммарной поверхности боковых и нижних
стенок многочисленных борозд.
В коре у человека насчитывается около 14 млрд нервных
клеток. Нейроны одинаковой структуры расположены в виде
отдельных слоев:
1-й слой — молекулярный. Представлен большим
числом отростков нейронов, тела которых залегают глубже, и
мелкими мультиполярными нейронами (имеют большое чис-
ло отростков);
2-й слой — наружный зернистый. Образован тела-
ми мелких нейронов;
3-й слой — пирамидный. Образован нейронами пира-
мидной формы, которые значительно крупнее нейронов вто-
рого слоя;
4-й слой — внутренний зернистый. Образован
мелкими нейронами звездчатой формы;
5-йслой— внутренний пирамидный. Внемзале-
гают очень крупные пирамидные клетки (до 125 млн);
6-йслой— полиморфные клетки. Образован ней-
ронами различной формы и размеров.
Скопления серого вещества находятся под корой в глубине
полушарий, в их белом веществе. Они образуют подкорковые
(базальные) ядра. К ним относятся полосатые тела, которые у
низших позвоночных составляют преобладающую массу ко-
нечного мозга.
Большие полушария отделены друг от друга продоль-
ной щелью, в глубине которой видно соединяющее их
мозолистое тело, образованное белым веществом. Каж-
дое полушарие разделено бороздами на лобную, теменную,
височную, островковую и затылочную доли.
Центральная борозда отделяет лобную долю от те-
менной, латеральная борозда — височную долю от
лобной и теменной (в глубине латеральной борозды). Кру-
говая борозда островка отделяет островковую долю
от других отделов полушария, теменно-затылочная
борозда разделяет теменную и затылочную доли. Более мел-
кие борозды делят доли на извилины.
В коре больших полушарий выделяют чувствительные, дви-
гательные и ассоциативные зоны.
В чувствительные зоны поступают импульсы от
органов чувств, кожи, внутренних органов, мышц, сухожилий.
При возбуждении нейронов чувствительных зон возникают
ощущения. Зрительная зона расположена в затылочной доле
244
коры, слуховая — в височной доле. В теменной доле находится
центр кожной и мышечно-суставной чувствительности. Здесь
особенно большая поверхность приходится на представитель-
ство рецепторов кисти рук, голосового аппарата и лица, значи-
тельно меньшая — туловища, бедра, голени. В этом отражены
различия в числе рецепторов, имеющихся в коже туловища и
наиболее чувствительных участков тела — губах, языке, поду-
шечках пальцев.
В каждое полушарие электрические импульсы поступают с
противоположной стороны тела. Двигательные зоны
коры расположены в задних областях лобных долей и ответ-
ственны за работу мышц также противоположных сторон тела.
Возникновения возбуждения в двигательной зоне вызывает
четкие координированные реакции.
В ассоциативные зоны коры приходят импульсы
от разных рецепторов. Например, значительная часть лобных
долей не отвечает на электрическое раздражение, а разруше-
ние их отдельных участков может либо остаться без видимых
последствий, либо, наоборот, привести к заметным психичес-
ким нарушениям.
Таким образом, наиболее четко фиксированы в коре чув-
ствительные и двигательные зоны, а ассоциативные функцио-
нируют как единое целое. Они связаны главным образом со
сложными проявлениями и высшей нервной деятельностью.
Лобные доли мозга ответственны за выработку программ
поведения и управления трудовой деятельности. Именно у че-
ловека они достигают особенного развития, в чем в значитель-
ной мере связан высокий уровень психических возможностей
человека по сравнению с животными. Их масса у человека со-
ставляет более 50% от общей массы головного мозга.
В составе человеческого мозга имеются структуры, которых
нет у животных, например речевые центры. Кроме того, у че-
ловека развита функциональная асимметрия мозга, т. е. спе-
циализация полушарий. Так, левое полушарие отвечает за аб-
страктно-логическое мышление. Правое полушарие руководит
процессами образного мышления и ответственно за музыкаль-
ное и художественное творчество.
В эволюции человека речь первоначально лишь использо-
вала, а затем развила и качественно преобразовала уже имею-
щуюся межполушарную асимметрию.
Правое полушарие быстрее, чем левое, обрабатывает посту-
пающую в него информацию. Согласно одной из концепций,
в процессе обучения правое полушарие работает по принципу
дедукции, т. е. сначала осуществляет синтез, а затем анализ;
245
левое же полушарие функционирует по принципу индукции,
сначала анализируя раздражители, а затем синтезируя их.
Человек с превалированием правополушарных функций тя-
готеет к конкретным видам деятельности, он медлителен и не-
разговорчив, но наделен способностью тонко чувствовать и пе-
реживать, он склонен к созерцательности и воспоминаниям.
«Левополушарный» человек тяготеет к теории, имеет большой
словарный запас и активно им пользуется, ему присуща двига-
тельная активность, целеустремленность, способность прогно-
зировать события.
АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА
По функциональному принципу нервную систему делят на
соматическую и автономную (вегетативную).
Отделы автономной нервной системы. Автономную нервную
систему делят на два отдела: симпатический и парасимпати-
ческий. В каждом отделе автономной нервной системы име-'
ются центральная и периферическая части. Центральную
часть образуют тела нейронов, лежащих в спинном и головном
мозге. Эти скопления нервных клеток получили название ве-
гетативных ядер (симпатических и парасимпатических). Отхо-
дящие от ядер волокна, а также узлы автономной нервной сис-
темы и нервные сплетения в стенках внутренних органов
образуют периферическую часть. Путь импульсов в автоном-
ной нервной системе состоит из двух нейронов. «Первые» на-
ходятся в ядрах ЦНС, а «вторые» на периферии в узлах или в
нервных сплетениях.
Симпатические ядра расположены в спинном мозге в облас-
ти боковых рогов серого вещества и образованы телами «не-
рвных» нейронов. Волокна, отходящие от ядер, идут в составе
передних спинномозговых корешков и заканчиваются в сим-
патических узлах. Симпатические узлы образованы телами
«вторых» нейронов, расположены попарно слева и справа от
позвоночного столба и соединены друг с другом. Образован-
ные ими парные пограничные симпатические стволы начина-
ются на уровне шейных позвонков и заканчиваются у копчика.
От узлов симпатической цепочки импульсы идут к рабочему
органу (коже, сосуду, железам, внутренним органам, органам
чувств). Часть тел «вторых» нейронов не входит в состав пар-
ных пограничных симпатических стволов. Они образуют узлы
нервных сплетений, расположенных вблизи органов (солнеч-
ное, сердечное, подчревное и другие сплетения). Путь от ЦНС
246
до нервного узла называют преганглионарным (ганглий — си-
ноним слова узел), а от узла рабочего органа — постганглио-
нарным. Особенность симпатического отдела автономной не-
рвной системы в том, что у него преганглионарный путь
короткий, постганглионарный — значительно длиннее.
Парасимпатические ящ>а лежат в среднем и продолговатом
мозге, а также в крестцовом отделе спинного мозга. Они обра-
зованы телами «нервных» нейронов автономного пути. Не-
рвные волокна от ядер среднего* мозга в составе глазодвига-
тельного нерва (III пара) идут к органам зрения. Поступающее
по ним возбуждение изменяет величину зрачка, меняет форму
хрусталика, приспосабливая глаз к ясному видению предметов
на различных, расстояниях.
Значительное число волокон от ядер продолговатого мозга
входит в состав блуждающего нерва (X пара), снабжающего па-
расимпатическими волокнами бблыпую часть органов трудной
и брюшной полости. От ядер крестцовой части спинного мозга
парасимпатические волокна идут к толстой кишке, мочевому
пузырю, половым органам. В парасимпатическом отделе не-
рвные узлы образованы телами «вторых» нейронов и разброса-
ны по всему телу; они находятся около или в самом иннервиру-
емом органе. В связи с этим в парасимпатическом отделе
автономной нервной системы преганглионарные волокна име-
ют большую длину, а постганглионарный путь очень короткий.
Функции автономной нервной системы. Симпатический и
парасимпатический отделы обладают различными свойства-
ми. Симпатическая нервная система регулирует функциони-
рование всех органов и тканей тела человека; ее волокна под-
ходят не только ко всем внутренним органам, но и скелетной
мускулатуре, коже. Волокна парасимпатического отдела не
подходят к гладкой мускулатуре кожи, к скелетным мышцам и
' к большинству кровеносных сосудов. Парасимпатическую ин-
нервацию имеют только сосуды половых органов, слюнных же-
лез и языка.
Парасимпатическая и симпатическая нервные системы ока-
зывают противоположное влияние на функции разных органов.
Симпатическая нервная система — это система «тревоги»,
защиты организма, мобилизации резервов, необходимых для
активного взаимодействия со средой.
Раздражение симпатического отдела увеличивает частоту и
силу сердечных сокращений, расширяет сосуды сердца, сужает
большинство других кровеносных сосудов, уменьшает образо-
вание мочи в почках, расширяет бронхи, расслабляет желчные
протоки и желчный пузырь, замедляет двигательную функцию
247
желудочно-кишечного тракта, усиливает обмен веществ, сти-
мулирует секрецию пота, расширяет зрачки и глазные щели.
Парасимпатическая нервная система оказывает успокаива-
ющее действие на организм. Она тормозит работу большин-
ства систем: замедляет ритм сердечных сокращений, уменьша-
ет их силу, сужает бронхи. Дыхание становится редким и
глубоким. Она не влияет на работу почек, селезенки, потовых
желез, на большинство кровеносных сосудов. Но она повыша-
ет тонус мочевого пузыря, способствует его опорожнению, ак-
тивизирует работу желудочно-кишечного тракта, усиливая его
двигательную функцию и повышая тонус, сокращает желчный
пузырь и расслабляет сфинктер, осуществляя изгнание желчи.
Под ее влиянием суживаются зрачки и глазные щелй.
Функции автономной нервной системы контролируют вы-
шележащие отделы головного мозга и коры больших полуша-
рий.
В свою очередь симпатическая нервная система влияет на
деятельность головного мозга.
ОРГАНЫ ЧУВСТВ
ОБЩИЕ ПРЕДСТАВЛЕНИЯ ОБ ОРГАНАХ ЧУВСТВ
Органами чувств названы органы восприятия раздражений,
идущих из окружающей и внутренней среды. К ним относят
органы осязания, зрения, слуха, вкуса и обоняния. В органах
чувств находятся скопления специфических рецепторов. В ре-
цепторе энергия внешнего возбуждения трансформируется в
нервные импульсы, а затем по чувствительным нервным путям
эти импульсы поступают в соответствующую зону коры боль-
ших полушарий головного мозга, где формируются специфи-
ческие ощущения. В зависимости от способа взаимодействия
с раздражителем различают рецепторы контактные (кожи, вку-
совые)^ дистантные (зрительные, слуховые, обонятельные).
И. П. Павлов всю совокупность нейронов, участвующих в
восприятии и проведении возбуждений, а также чувствитель-
ные нейроны коры головного мозга считал единой системой и
назвал ее анализатором. Рецепторы являются перифери-
ческим звеном анализатора, афферентные нейроны
и проводящие пути составляют проводниковый отдел
анализатора, участки головного мозга, которые восприни-
248
мают возбуждение, являются корковыми (централь-
ными) представительствами анализаторов.
Основные свойства рецепторов. Каждый рецептор приспо-
соблен к определенному виду раздражителя (адекватный раздра-
житель). К такому раздражителю рецептор обладает очень вы-
сокой чувствительностью. Но можно вызвать его возбуждение
другим, неадекватным по характеру раздражителем, в этом
случае сила раздражителя во много раз больше, чем для адек-
ватного. Например, механическое (удар) раздражение глаза
или уха может вызвать соответственно ощущение вспышки
света или звука.
Для каждого рецептора имеется определенный порог раз-
дражения, характеризующийся определенной силой и дли-
тельностью воздействия раздражителя.
Возбудимость рецепторов непостоянна. Она может менять-
ся при изменениях состояния самих рецепторов и централь-
ной нервной системы. Так, например, количество возбужден-
ных тепловых и холодовых рецепторов на единицу площади
кожного покрова изменяется в течение непродолжительных
промежутков времени (часы, минуты). Такое явление названо
функциональной лабильностью рецепторов.
Адаптация рецепторов (привыкание) проявляется в сниже-
нии чувствительности к постоянно или повторно действую-
щим раздражителям: табачный дым, холод, постоянный шум
на улице, звуки радио и т. д.
КОЖА
Кожа образует наружный покров тела, площадь которого
составляет 1,5-1,6 м2.
Функции кожи: 1) защитная (от вредных воздействий и
проникновения микроорганизмов); 2) терморегуляционная (че-
рез кровеносные сосуды кожи, потовые железы, подкожную
жировую клетчатку); 3) выделительная (благодаря потовым же-
лезам); 4) рецепторная (в коже находятся болевые, температур-
ные, тактильные рецепторы); 5) депо крови (в сосудах кожи де-
понируется до 1 л крови); 6) синтез витамина Д (в коже
содержится предшественник витамина Д, который под влия-
нием ультрафиолетовых лучей превращается в витамин Д).
Компоненты кожи. Кожа состоит из двух главных частей: тон-
кого наружного слоя — эпидермиса, в котором нет кровеносных
сосудов, и внутреннего, более толстого слоя, — дермы, богатой
кровеносными сосудами и нервными окончаниями (рис. 5.64).
249
Рис. 5.64. Кожа
человека.
а — поперечный срез:
1 — эпидермис; 2 — волос;
3 — сальная железа; 4 —
собственно кожа; 5 — эпи-
телиальное корневое вла-
галище; 6 — потовая же-
леза; 7 — кожная артерия;
8 — кожная вена; 9 — не-
рвное окончание; 10 —
жировая клетчатка;
8 7
a
б
б — схема структур,
видимых на срезе:
1 — зона эластических
и коллагеновых волокон;
2— мышца, поднимающая
волос; 3 — зернистый
слой; 4 — роговой слой;
5 — мальпигиев слой; 6 —
волосяной фолликул; 7 —
сальные железы; 8 — сек-
реторные клетки; 9 —
проток железы; 10 — во-
лосяной сосочек; 11 —
подкожные жировые клет-
ки; 12 —часть стержня во-
лоса; 13 — кровеносный
сосуд
250
Эпидермис представлен многослойным плоским орогове-
вающим эпителием, в котором выделяют пять слоев: ба-
зальный (здесь происходит размножение клеток, благода-
ря чему восполняются все слои эпидермиса, и здесь же
находятся пигментные клетки, содержащие в цитоплазме
пигмент -меланин, защищающий организм человека от уль-
трафиолетовых лучей); шиповатый (состоит из клеток,
соединенных между собой с помощью отростков); зернис-
тый (клетки содержат в цитоплазме гранулы специального
белка кератогиалина); блестящий (ядра клеток разруше-
ны, а цитоплазма пропитана веществом, образовавшимся из
кератогиалина); роговой (самый поверхностный, состоит
из ороговевших клеток, содержащих кератин; этот слой пол-
ностью обновляется за 5—30 суток). От количества пигмента,
содержащегося в клетках эпидермиса, зависит цвет кожи чело-
века.
В дерме различают два слоя: сосочковый и сетчатый.
Сосочковый слой состоит из рыхлой соединитель-
ной ткани. Он образует многочисленные сосочки, вдающиеся
в эпидермис. От него зависит рисунок на коже. В сосочковом
слое имеются гладкие мышечные клетки, кровеносные и лим-
фатические сосуды, нервные окончания. Сетчатый слой
образован плотной соединительной тканью. Пучки коллаге-
новых и эластических волокон образуют сеть и придают коже
прочность. В этом слое расположены потовые и сальные желе-
зы и корни волос.
За дермой следует подкожный слой жировой клетчатки.
Она состоит из рыхлой соединительной ткани, содержащей
жировые отложения.
Производными кожи являются: волосы, ногти, сальные, по-
товые и молочные железы.
Половые железы — трубчатые железы, сосредоточенные на
границе сетчатого слоя и подкожной жировой клетчатки. Вы-
водные протоки открываются на поверхности кожи порами.
Потовыми железами богата кожа ладоней, подошв ног, подмы-
шечных впадин. Состав пота: вода (98—99%), хлорид натрия,
мочевая кислота, аммиак, мочевина и др. При потении проис-
ходит теплоотдача.
Сальные железы расположены в сетчатом слое, на границе с
сосочковым. Йх протоки открываются в волосяную сумку. Они
выделяют секрет — кожное сало, которое смазывает волосы и
смягчает кожу, сохраняя ее эластичность.
Роговыми производными эпидермиса кожи являются воло-
сы и ногти.
251
Волос состоит из корня и стержня. Корень волоса заканчива-
ется расширением — волосяной луковицей, в которую снизу вда-
ется волосяной сосочек с сосудами и нервами. За счет деления
клеток волосяной луковицы происходит рост волоса. Корень во-
лоса снаружи окружен волосяной сумкой. К волосяной сумке
прикрепляется гладкая мышца, поднимающая волос. В месте пе-
рехода корня волоса в стержень образуется углубление — волося-
ная воронка, в которую открываются протоки сальных желез.
Стержень включает мозговое вещество (построено из оро-
говевших клеток, которые содержат зерна пигмента и мелкие
пузырьки воздуха) й корковое вещество (состоит из ороговев-
ших клеток, заполненных воздухом и гранулами меланина).
Снаружи волос покрыт кутикулой, представленной роговыми
чещуями. К старости в ороговевших клетках (роговых чешуях)
мозгового и коркового вещества снижается количество пиг-
мента и нарастает количество пузырьков воздуха — волосы се-
деют.
Ногти представляют собой выпуклые в поперечнике рого-
вые пластинки, лежащие на тыльной поверхности концевых
фаланг. Ноготь лежит в ложе, состоящем из росткового эпите-
лия и соединительной ткани. Кожа ногтевого ложа богата кро-
веносными сосудами и нервными окончаниями.
Кожа содержит рецепторы, воспринимающие раздражения
из внешней среды. Болевые раздражения воспринимаются
свободными нервными окончаниями, холодовые — более по-
верхностными (ближе к эпидермису), тепловые — находящи-
мися в глубоких слоях дермы и подкожном слое. Холодовые
рецепторы могут возбуждаться не только понижением темпе-
ратуры, но и действием тепла (выше 45°С). Наибольшей чув-
ствительностью обладает кожа губ, носа; наименьшей — кожа
спины, живота, подошв ног. Центральный отдел кожного ана-
лизатора находится в задней центральной извилине (темен-
ной доле коры больших полушарий головного мозга).
ОРГАН ЗРЕНИЯ
Органом зрения является глаз. Свет, возбуждая рецептор-
ный аппарат глаза, вызывает зрительные ощущения.
Строение глаза. Глаз состоит из глазного яблока и вспомога-
тельных элементов (глазные мышцы, веки и слезные железы).
В стенке глазного яблока различают три оболочки: 1) наружную
фиброзную, 2) срединную сосудистую и 3) внутреннюю сетча-
тую (рис. 5.65).
252
Рис. 5.65. Горизонтальный разрез глазного яблока
(схема):
1 — конъюнктива; 2 — реснитчатое тело; 3 — связка, с помощью кото-
рой хрусталик прикреплен к реснитчатому телу; 4 — роговица; 5 — хру-
сталик; 6 — передняя камера глаза; 7 — радужная оболочка; 8 — задняя
камера глаза; 9, 17 — мышца глазного яблока; 10 — склера; 11 — соб-
ственно сосудистая оболочка; 12 — сетчатая оболочка; 13 — желтое
пятно; 14 — зрительный нерв; 15 — диск зрительного нерва; 16 — стек-
ловидное тело
Наружная оболочка глазного яблока называется склерой, ко-
торая представляет собой непрозрачную жесткую сферическую
соединительно-тканную капсулу, защищающую внутренние
структуры и помогающую глазу сохранять свою форму. На пере-
дней поверхности глаза склера переходит в прозрачную, более
толстую роговицу, через которую и проникает свет в глаз.
Средняя оболочка называется сосудистой. В ней имеется
три отдела: 1) собственно сосудистая оболочка, 2) реснитчатое
тело и 3) радужная оболочка.
253
Собственно сосудистая оболочка (толщиной 0,1 мм) — это
задняя часть средней оболочки. Она содержит пигментный
слой, который поглощает свет.
В реснитчатом теле различают наружный отдел, образован-
ный реснитчатой мышцей, и внутренний отдел, образованный
отростками, которые окружают хрусталик.
Радужная оболочка представляет собой пластинку с отвер-
стием в центре — зрачком. Вблизи зрачкового края радуж-
ной оболочки в этом слое располагаются мышцы, суживающие
(кольцевые) и расширяющие (радиальные) зрачок. При этом
диаметр зрачка может меняться от 2 до 8 мм.
За радужной оболочкой располагается плотное тело —
хрусталик (9,0 х 4,4 мм), прикрепленный к реснитчатым
отросткам с помощью особой пинновой связки. Хрусталик
представляет собой эластичное чечевицеобразное тело. Оно
располагается сразу позади зрачка и преломляет входящие в
глаз лучи света, фокусируя их на сетчатке. В преломлении лу-
чей также участвует выпуклая поверхность роговицы, внутри-
глазные жидкости, стекловидное тело.
Между роговицей и радужной оболочкой глаза располага-
ется передняя камера глаза, сообщающаяся через зрачок с зад-
ней камерой глаза. Последняя ограничена задней поверхнос-
тью радужной оболочки и хрусталиком. Камеры глаза
заполнены водянистой жидкостью. Пространство за хрустали-
ком до сетчатки заполнено желатиноподобной массой —
стекловидным телом.
Внутренняя сетчатая оболочка (сетчатка) (рис. 5.66) пред-
ставляет собой светочувствительный слой, который связан со
зрительным нервом. В нем присутствует множество рецептор-
ных клеток, называемых палочками и колбочками.
В глазу всего около 125 млн палочек и 6—7 млн колбочек. На
месте выхода зрительного нерва из глазного яблока нет ни па-
лочек, ни колбочек. Поэтому это место не воспринимает свет
и его называют слепым пятном. Распределение палочек
и колбочек неравномерное. В середине сетчатки (на прямой,
проходящей через центры роговицы и хрусталика) находится
область наибольшей остроты зрения — центральная ямка.
Здесь сосредоточены колбочки, ответственные за «дневное»
зрение (зрение при ярком свете), за различение деталей и за
цветовое зрение. Существует три различных типа колбочек —
для красного, зеленого и синего цвета. Центральная ямка яв-
ляется центром желтого пятнан находится на расстоя-
нии около 4 мм несколько латеральнее от слепого пятна.
Палочки же являются рецепторами, ответственными за «суме-
254
Волокна зрительного нерва
Слои нейронов
Рис. 5.66. Строение сетчатки:
1 — палочка; 2 — колбочка
речное» зрение (при низких уровнях яркости). По направле-
нию к периферии сетчатки число колбочек уменьшается, а па-
лочек, отсутствующих в центре, увеличивается.
Каждая палочка содержит светочувствительный Пигмент
родопсин (зрительный пурпур) — соединение ретинена (рети-
наля) с белком (липопротеином) сетчатки опсином. Рети-
нен — это альдегидная форма витамина А. Ретинен может со-
единяться со специфическим опсином из палочек, образуя
активный пигмент родопсин, или с другим опсином из колбо-
чек, образуя активный пигмент йодопсин (зрительный фио-
летовый). Синтез .родопсина нарушается при недостатке ви-
тамина А, что приводит к резкому ухудшению сумеречного
зрения •— «куриной слепоте».
Глаз состоит из двух основных систем: 1) оптической сис-
темы светопреломляющих сред и 2) рецепторной системы сет-
чатки.
К светопреломляющим средам глаза относятся: роговица,
водянистая жидкость передней камеры глаза, хрусталик и
стекловидное тело.
К рецепторной системе относятся светочувствительные
клетки: колбочки и палочки, связанные с нейронами коры го-
ловного мозга.
Глаз имеет вспомогательные образования, которые выпол-
няют защитно-приспособительную роль. К ним относятся
веки, брови, ресницы и слезные железы. Основу век составляет
255
плотная соединительнб-тканная пластинка, снаружи покрытая
кожей, изнутри — соединительно-тканной оболочкой (конъ-
юнктивой). Конъюнктива с век переходит на переднюю по-
верхность глазного яблока. Здесь она покрыта многослойным
плоским эпителием. При coi/кнутых веках образуется узкое
пространство между конъюнктивой век и конъюнктивой глаз-
ного яблока — конъюнктивальный мешок. При закрывании и
открывании век наружная часть роговицы увлажняется и очи-
щается. Слезная железа располагается выше верхней части на-
ружного края глазницы. Ее протоки открываются в конъюнкти-
вальный мешок, который переходит в слезно-носовой канал.
Аккомодация глаза. Аккомодация — это способность глаза
ясно видеть предметы, находящиеся от него на различных рас-
стояниях. Она осуществляется за счет изменения кривизны
хрусталика. При расслаблении связочного аппарата хрусталик
становится более выпуклым, что ведет к усилению преломляю-
щей способности. Вследствие этого, в фокус на сетчатке сходят-
ся лучи от близко расположенных предметов. При сокращении
связочного аппарата хрусталик становится более плоским, что
способствует рассмотрению более отдаленных предметов.
Световая чувствительность. Минимальное количество свето-
вой энергии, достаточное для возникновения светового ощуще-
ния, определяет абсолютную чувствительность глаза. Предель-
ная чувствительность зрения у человека варьирует от 2 до
нескольких десятков квантов света при длительном пребывании
в полной темноте. Глаз человека максимально чувствителен к
желто-зеленой части спектра. Зрительное ощущение возникает
при продолжительности раздражения глаза менее 0,001 с.
Адаптация. Адаптация — это приспособление глаза к виде-
нию при разных уровнях освещенности, обусловленное наличи-
ем двух типов фоторецепторов — колбочек и палочек. Напри-
мер, при выходе из темного помещения на яркий солнечный
свет в первый момент мы ничего не различаем, так как требует-
ся некоторое время на включение колбочкового зрения. Затем
мы постепенно адаптируемся к свету и начинаем воспринимать
окружающие предметы — это световая адаптация глаза. При
быстром переходе от света к темноте тоже получается состояние
полной слепоты, которая постепенно исчезает благодаря тем-
новой адаптации, т. е. переходу к палочковому зрению. Время,
необходимое для адаптации, определяется временем протека-
ния химических реакций в фоторецепторах на свету или в тем-
ноте. Время световой адаптации намного меньше темновой.
Острота зрения. Острота зрения — это способность глаза раз-
личать две близко расположенные точки. Расстояние между точ-
256
ками определяется углом между лучами, идущими от двух точек
через центр зрачка глаза. Нормальный глаз при дневной осве-
щенности различает две точки под углом 60 угловых секунд (1').
Острота зрения зависит от уровня освещенности, что обусловле-
но работой либо колбочкового, либо палочкового аппарата. Кол-
бочки связаны нейронами поштучно (в центральной ямке), а па-
лочки — десятками и сотнями (на периферии сетчатки).
Бинокулярное зрение. Зрение человека осуществляется двумя
глазами, бинокулярно, что позволяет видеть окружающий мир
объемным и обеспечивает пространственную ориентацию. При
зрении двумя глазами на сетчатке каждого глаза получается
изображение рассматриваемого предмета, т. е. два изображе-
ния. Однако мы воспринимаем эти два изображения как одно и
происходит это потому, что изображений каждой точки предме-
та попадает на так называемые соответственные участки сетчат-
ки, равноудаленные от желтого пятна. Это происходит за счет
согласованного действия двух групп глазных мышц, ответствен-
ных за аккомодацию и поворот глазных яблрк. Расстройство би-
нокулярного зрения имеет место при косоглазии и большой
разнице рефракции левого и правого глаза.
Рефракция глаза и ее нарушения. Рефракция — преломляющая
способность глаза, которая зависит от двух основных факторов:
от оптической силы преломляющих сред глаза и размеров (дли-
ны) глазного яблока. Если параллельные лучи света, проникаю-
щие в глаз, преломляясь оптической системой глаз, сходятся в
фокус на сетчатке, то на ней получается четкое изображе-
ние рассматриваемого предмета, что обусловливает хорошую
видимость предмета. Такой глаз называется эмметропическим,
или нормальным (рис. 5.67).
Существуют следующие отклонения рефракции глаза от
нормы: близорукость, или миопия, дальнозоркость, или ги-
перметропия, и астигматизм.
Если параллельные лучи света, преломляясь оптической си-
стемой глаза, сходятся в фокус не на сетчатке, а перед ней, на
сетчатку падает пучок расходящихся лучей и изображение рас-
сматриваемого предмета становится расплывчатым; такой вид
рефракции носит название близорукости. Близорукость может
возникать, если сетчатка расположена дальше фокусного рас-
стояния хрусталика (см. рис. 5.67). Для коррекции близорукос-
ти используются очки с отрицательными линзами.
Если же фокус оптической системы глаза лежит поза-
ди сетчатки, ина ней при этом также получается нечет-
кое изображение предмета, то такой вид рефракции носит
название дальнозоркости (см. рис. 5.67). Дальнозоркость
9-281
257
Рис. 5.67. Схема рефракции
в нормальном (/),
дальнозорком (2 )
и близоруком (5) глазу
может возникнуть, если сетчатка расположена слишком
близко к хрусталику и фокусировка хороша только при рас-
сматривании далеко расположенных предметов. Для кор-
рекции дальнозоркости используются очки с положитель-
ными линзами.
Астигматизм вызван невозможностью схождения всех
лучей в одну точку, в один фокус вследствие неодинакового
преломления лучей в разных меридианах глаза. Астиг-
матизм зависит от различной кривизны роговицы и хруста-
лика в различных меридианах глаза. Поэтому лучи, пада-
ющие на один меридиан, преломляются сильнее и
фокусируются раньше, чем другие лучи, преломляющиеся
по другим меридианам. Для коррекции астигматизма ис-
пользуются очки со сфероцилиндрическими линзами, а в
настоящее время контактные линзы, которые, плавая в слез-
ной жидкости над роговицей, компенсируют ее отклонения
от правильной формы.
ОРГАН СЛУХА И РАВНОВЕСИЯ
Строение органа слуха. Орган слуха воспринимает механи-
ческую энергию колебаний воздуха. Он состоит из трех частей:
наружного, среднего и внутреннего уха.
Наружное ухо. Оно представлено ушной раковиной и на-
ружным слуховым проходом. Ушная раковина — это
свободно выступающая на поверхности головы складка кожи, в
основе которой располагается пластинка эластического хряща.
Наружный слуховой проход у взрослого имеет дли-
258
ну до 3,5 см, выстлан кожей, особенностью которой является
наличие желез, выделяющих ушную серу. Сера и волоски име-
ют защитное значение, так как препятствуют попаданию в
глубокие части наружного слухового прохода инородных тел.
На границе между наружным и средним ухом натянута бара-
банная перепонка. Ее толщина около 0,1 мм. Барабан-
ная перепонка представляет собой соединительно-тканную плас-
тинку овальной формы, диаметром 9 и И мм, наружная сторона
которой покрыта эпидермисом. Приблизительно в центре пере-
понка втянута внутрь среднего уха, так как в этом месте в ее ткань
вплетается конец рукоятки молоточка (рис. 5.68).
Среднее ухо. Представлено барабанной полостью, имеющей
неправильную форму, емкостью в 0,75 мл. Ее наружной стен-
кой является барабанная перепонка. В среднем ухе три косточ-
ки: 1) молоточек, 2) наковальня и 3) стремечко. Молото-
чек вплетен рукояткой в барабанную перепонку, а другой
Рис. 5.68. Строение уха (схема):
1 — полость среднего уха (барабанная область); 2 — барабанная перепонка;
3 — наружный слуховой проход; 4 — молоточек; 5 — наковальня; 6 —
стремечко; 7 — полукружные каналы; 8 — преддверие; 9 — овальное окно;
10 — вестибулярная лестница; 11 — барабанная лестница; 12 — слуховая
(евстахиева) труба
9* 25°
стороной сочленяется с наковальней. В свою очередь
длинный отросток наковальни сочленяется с головкой стре-
мечка. Стремечко посредством кольцевой связки соединяет-
ся с краем овального окна, расположенного на внутренней стен-
ке барабанной полости у основания вестибулярного канала и
упирается в затягивающую его перепонку. Кроме овального от-
верстия на внутренней стенке барабанной полости у основания
барабанного канала имеется круглое отверстие (круглое окно).
В улитке содержится жидкость, колебания которой, возникаю-
щие у овального окна и прошедшие по ходам улитки, доходят
до круглого окна, выпячивая его перепонку в полость среднего
уха. Если бы этого окна не было, то колебания были бы невоз-
можны вследствие несжимаемости жидкости.
Барабанная полость через отверстие в передней стенке со-
общается посредством евстахиевой (слуховой) тру-
бы с полостью глотки (длина слуховой трубы 3,5 см, а диа-
метр 2 мм). При акте глотания отверстие трубы открывается в
глотку, что ведет к выравниванию давления в среднем ухе с на-
ружным атмосферным давлением. Это устройство обеспечива-
ет одинаковое давление воздуха по обе стороны барабанной
перепонки.
Внутреннее ухо. Включает костный лабиринт,кото-
рый лежит в височной кости и представляет собой сложно уст-
роенные каналы. Внутри костного лабиринта находится пе-
репончатый лабиринт, повторяющий форму костного
лабиринта (рис. 5.69). Между костным и перепончатым лаби-
ринтом имеется узкое пространство, заполненное жидко-
стью — перилимфой. Перепончатый лабиринт — это замкну-
тая система полостей и каналов, также заполненная
жидкостью — эндолимфой. Костный лабиринт состоит из
трех отделов: преддверия, улитки и полукружных каналов.
Преддверие занимает центральное положение. Оно сооб-
щается с полукружными каналами сзади и с улиткой спереди.
Улитка (рис. 5.70) представляет собой широкий костный
канал, который у человека делает 2 3/4 оборота вокруг своей
оси. Перепончатый,канал улитки отделен от костного двумя
мембранами — более тонкой вестибулярной и более плотной
и упругой — основной. Эти мембраны делят костный лаби-
ринт на три узких хода — верхний, средний и нижний. Верх-
ний канал носит название вестибулярной лестницы. Он начи-
нается от овального окна и продолжается до вершины улитки,
где через отверстие сообщается с нижним каналом, называе-
мым барабанной лестницей. Она берет начало в об-
ласти круглого окна. Оба эти канала заполнены перилимфой и
Рис. 5.69. Поперечный разрез улитки
(схема):
1 — лестница преддверия; 2 — вестибулярная мембрана; 3 — проток улит-
ки; 4 — секреторный эпителий; 5 — покровная мембрана; 6 — спиральный
(кортиев) орган; 7 — спиральная связка; 8 — основная мембрана; 9 — бара-
банная лестница; 10 — спиральный ганглий; 11 — наружные волосковые
клетки; 12 — нервные волокна, подходящие к волосковым клеткам; 13 —
внутренние волосковые клетки
261
1
Рис. 5.70. Строение костного и перепончатого лабиринтов
(схема):
1 — преддверие; 2 — вещество височной кости; 3 — сообщение между
лестницей преддверия и барабанной лестницей; 4 — перепончатый канал
улитки; 5 — лестница преддверия; 6 — лестница барабанная; 7 — мешочек;
8 — круглое отверстие; 9 — овальное отверстие; 10 — барабанная полость;
11 — расширение (ампула) перепончатого полукружного канала; 12 — ма-
точка; 13 — костный полукружный канал; 14 — перепончатый полукруж-
ный канал
представляют собой единый канал, отделенный от воздушной
полости среднего уха мембранами овального и круглого окон.
Между верхним и нижним каналом находится средний
перепончатый канал, полость которого не сообщается с полос-
тью других каналов и заполнена эндолимфой. Внутри средне-
го канала на основной мембране располагается звуковоспри-
нимающий аппарат—кортие в орган. Он состоит из 3-
4 рядов рецепторных (волосковых) клеток. В кортиевом органе
24 000 волосковых клеток, которые тянутся рядами вдоль за-
витков улитки по всей ее длине. Каждая рецепторная клетка
одним концом фиксирована на основной (базилярной) мемб-
ране, второй ее полюс находится в полости перепончатого ка- '=
нала. На конце полюса рецепторной клетки находятся волос- !
ки (от 30 до 120 у каждой клетки), которые омываются ,
эндолимфой. Волоски контактируют с подвижной покров-
ной (текториальной) мембраной, расположенной
над волосковыми клетками; один ее конец свободный, дру-
гой — прикреплен к основной мембране.
Передача и восприятие звуковых колебаний. Ушная ракови-
на улавливает звуки и направляет их в наружный слуховой про-
ход. Звуковые волны вызывают колебания барабанной пере- ;
понки. Они передаются слуховым косточкам — молоточку,
наковальне и стремечку, а через мембрану овального окна рас-
262
пространяются на перилимфу верхнего (вестибулярной лест-
ницы) и нижнего (барабанной лестницы) канала. Так как вес-
тибулярная мембрана, разделяющая верхний и нижний канал,
очень тонкая, то колебания перилимфы легко передаются на
эндолимфу среднего (перепончатого) канала. Эти колебания
приводят в движение основную мембрану, где расположен
кортиев орган. При этом волоски рецепторных клеток касают-
ся покровной мембраны, деформируются и благодаря этому в
рецепторах возникает возбуждение. Возбуждение передается
волокнам слухового нерва, которые образуют синапсы на ре-
цепторных клетках. Импульсы, проходящие по волокнам слу-
хового нерва, передают информацию о действующих звуковых
волнах в центральную нервную систему. По восходящим не-
рвным путям возбуждение доходит до корковых нейронов.
В слуховой зоне коры мозга нервные импульсы трансформиру-
ются в слуховые ощущения.
Орган равновесия
(вестибулярный аппарат)
Вестибулярный аппарат является органом восприятия поло-
жения тела и сохранения равновесия. Любое изменение поло-
жения и движения тела в пространстве вызывает раздражение
рецепторов вестибулярного аппарата, в результате чего возни-
кают рефлексы, обеспечивающие сокращения скелетных
мышц для сохранения равновесия.
Вестибулярный аппарат включает преддверие костного лаби-
ринта и три полукружных канала (рис. 5.71), расположенных в
трех взаимно перпендикулярных плоскостях. Один конец каж-
дого канала расширен и называется ампулой. Перепончатый
лабиринт в области преддверия представлен двумя мешочка-
ми, одий из которых расположен ближе к улитке, другой —
ближе к полукружным каналам.
В мешочках находится так называемый отолитовый
аппарат, имеющий сложную структуру. В каждом мешочке
имеются возвышения, называемые пятнами. Это скопле-
ние рецепторных клеток. Клетки имеют цилиндрическую
форму, суженную на одном конце. Суженная часть имеет
один подвижный волосок и 50-80 склеенных неподвижных
волосков. Волоски рецепторных клеток погружены в студени-
стую волокнистую массу, покрывающую пятно. В этой массе
находится большое количество микроскопических кристал-
лических образований, состоящих из углекислого кальция.
Они называются отолитами исобраныв отолитовый
263
Рис. 5.71. Левый лабиринт
млекопитающего (схема):
1, 2, 3 — полукружные каналы;
4 — ампулы каналов; 5, 6 — пред-
дверие (vestibulum), которое раз-
деляется на два мешочка: utricutes
(3) и sacculus пунктиром отмече-
ны «слуховые пятна» — macula
sacculae; 7—.улитка
слой (мембрану). Этот слой оказывает при изменении по-
ложения головы механическое давление на волоски.
В перепончатых полукружных каналах рецепторные клетки
имеются только в ампулах, где они образуют скопления, полу-
чившие название гребешков. Клетки имеют длинные во-
лоски, изгибающиеся при движении эндолимфы.
К рецепторным клеткам вестибулярного аппарата подходят
отростки биополярных нейронов вестибулярного узла, распо-
ложенные во внутреннем слуховом проходе. Вторые отростки
этих клеток при выходе из узла образуют вестибулярную ветвь
VIII пары черепных нервов.
Разрушение преддверия и полукружных каналов вызывает
потерю чувства равновесия.
Во время изменения положения тела отолитовая мембрана
надавливает на волоски рецепторных клеток, что приводит к
их возбуждению, а в полукружных каналах рецепторные клет-
ки раздражаются движением эндолимфы. Возникающая сиг-
нализация информирует центральную нервную систему о по-
ложении тела. В результате происходит перераспределение
тонуса скелетных мышц, обеспечивающее выравнивание поло-
жения тела и сохранение равновесия.
При повышенной чувствительности вестибулярного аппа-
рата во время пользования различными видами транспорта
возникает состояние, называемое морской болезнью. При
264
этом ощущается головокружение, появляются рвота, головная
боль. Регулярными тренировками человек может снизить чув-
ствительность вестибулярного аппарата и избавиться от этой
болезни.
ОРГАНЫ ОБОНЯНИЯ И ВКУСА
Орган обоняния
Этот орган, обеспечивающий восприятие запаха, заложен в
слизистой оболочке верхней и нижней части средней носовой
раковины. Здесь располагается обонятельный эпителий, состоя-
щийиз рецепторных и опорных клеток.
Рецепторные клетки (нейроны) имеют два отростка: корот-
кий — периферический, и длинный — центральный. Корот-
кий отросток на своем округлом конце несет до 10—12 заост-
ренных подвижных ресничек, или волосков? У человека в
органе обоняния насчитывается до 40 млн рецепторных кле-
ток. При вдохе молекулы пахучих веществ оседают на обо-
нятельном эпителии, растворяются в слизи и раздражают
реснички рецепторных клеток. Импульсы от обонятельных ре-
цепторов поступают по обонятельному нерву в обонятельную
зону коры больших-полушарий.
Орган вкуса
Этот орган обеспечивает восприятие вкуса пищи. Рецепторы
вкуса представлены совокупностью вкусовых луковиц (печек), рас-
положенных в многослойном эпителии сосочков языка. Вкусо-
вые луковицы могут располагаться и на других органах: тубах, на-
ружной й внутренней поверхности надгортанника, голосовых
связках. У человека насчитывается около 2000 вкусовых луковиц.
Каждая вкусовая луковица имеет эллипсоидную форму и состоит
из веретенообразных вкусовых и опорных клеток.
На поверхности этих клеток насчитывается до 40—50 микровор-
синок, увеличивающих площадь воспринимающей мембраны.
На вершине луковицы находится отверстие — вкусовая
пора. Вкусовые вещества адсорбируются на поверхности мик-
роворсинок. Возбуждение от вкусовых луковиц передается в виде
сигналов по нервным путям в центральный отдел вкусового ана-
лизатора. На возникновение вкусовых ощущений оказывают вли-
яние не только возбуждение вкусовых рецепторов, но и обоня-
тельных, а также болевых и температурных рецепторов полости
265
рта. При насморке, когда обоняние подавлено, вкус пищи ощу-
щается плохо.
Рецепторы, специфичные к восприятию сладкого, располо-
жены на кончике языка, горького — на корне языка, кислого и
соленого — на боковых его поверхностях.
Центральные отделы вкусового и обонятельного анализато-
ров сосредоточены на внутренней поверхности височной и
лобной долей коры больших полушарий.
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА
Единство и целостность организма проявляются во взаимо-
зависимости и соподчинении отдельных его частей. Взаимо-
отношения между частями тела и органами человека регулиру-
ются деятельностью ЦНС и ЖВС.
Еще в прошлом веке решающую роль в обеспечении един-
ства организма с внешнйм миром И. М. Сеченов (1829-1905) —
основоположник отечественной физиологии, отводил нервной
системе при доминирующем влиянии ее высшего отдела — го-
ловного мозга. Его ученик И. П. Павлов (1849—1936) ввел в на-
уку понятие о высшей нервной деятельности (ВИД). Высшая
нервная деятельность высших отделов ЦНС обеспечивает слож-
ные отношения между организмом и внешним миром.
К высшим отделам ЦНС относят кору больших полушарий^
и ближайшие к ней подкорковые образования. В основе ВНД
лежит рефлекторная и интегративная деятельность мозга.
БЕЗУСЛОВНЫЕ И УСЛОВНЫЕ РЕФЛЕКСЫ
Термин «рефлекс» предложил чешский анатом и физиолог
Й. Прохаска (1749—1820). Представления о рефлекторном ха-
рактере деятельности головного мозга впервые были развиты
И. М. Сеченовым («Рефлексы головного мозга», 1866), обосно-
вавшим рефлекторную природу психической деятельности че-
ловека. Так впервые был поставлен вопрос об объективном,
физиологическом анализе психических процессов.
Идея И. М. Сеченова в дальнейшем развивалась И. П. Пав-
ловым, который ввел понятие о безусловных и условных рефлек-
сах, разделив всю совокупность рефлекторных реакций на две
группы (табл. 5.4). Наиболее тонкие приспособления организ-
46
Таблица 5.4
Сравнительная характеристика безусловных и условных рефлексов
Условные рефлексы
Безусловные рефлексы
1. Врожденные, наследственно
передающие реакции организма
2. Видовые, т. е. свойственные
всем представителям данного
вида
3. Появляются при первом же
воздействии раздражителя. Та-
кие раздражители называют
безусловными
4. Относительно постоянны
5. Рефлекторные дуги замыкают-
ся в спинном мозге или в под-
корковых структурах головного
мозга
1. Приобретенные организмом в
процессе индивидуального раз-
вития на основе «жизненного
опыта».
2. Индивидуальные: у одних пред-
ставителей одного и того же вида
они могут быть, а у других отсут-
ствовать
3. Для образования условного
рефлекса необходимо сочетание
во времени какого-либо изме-
нения в окружающей среде,
воспринятого корой больших
полушарий головного мозга, с
осуществлением того или иного
безусловного рефлекса. Такой
раздражитель, вызывающий об-
разование условного рефлекса,
называют условным раздражи-
телем. Условный рефлекс появ-
ляется после нескольких соче-
таний условного и безусловного
раздражителей
4. Непостоянны и в зависимости от
определенных условий могут вы-
рабатываться, закрепиться или
исчезнуть; это свойство отраже-
но в самом их названии
5. Рефлекторные дуги замыкаются
в коре больших полушарий го-
ловного мозга. В последнее вре-
мя показана в экспериментах на
животных с удаленной корой
большого мозга возможность
выработки условного рефлекса.
Однако образование рефлексов
происходит с большим трудом и
только в ответ на простые раз-
дражители. Таким образом,
структура условного рефлекса
чрезвычайно сложна, и выде-
лить чисто корковые и подкор-
ковые «функции» очень трудно.
В целом можно сказать, что в
поведенческих рефлексах веду-
щая роль принадлежит коре, в
образовании вегетативных ус-
ловных рефлексов (изменение
ЧСС, слюноотделение и т, д.)
не менее важна и роль подкор-
ковых структур
267
ма к окружающей среде достигаются при образовании и исчез-
новении различных условных рефлексов, которые и в процессе
эволюции и в онтогенезе особи усложняются и совершенству-
ются.
Совокупность сложных безусловных рефлексов, определя-
ющих некоторые формы поведения, называют инстинктом
(например, построение гнезда, забота о ребенке). В основе на-
выков — автоматизированных действий, выполняемых «почти»
без контроля сознания, лежит комплекс условных рефлексов
(например, чтение, письмо, управление автомашиной).
УСЛОВИЯ ОБРАЗОВАНИЯ УСЛОВНЫХ
РЕФЛЕКСОВ
1. Совпадение во времени действия индифферентного
(нейтрального) и безусловного раздражителя.
2. Начало действия условного (индифферентного) раздра-
жителя должно несколько опередить начало действия безус-
ловного раздражителя. Например, если сначала появляется за-
пах, затем кислота во рту, условный рефлекс появляется после
20 сочетаний. Если сначала кислота во рту, а через 10 секунд
появляется запах, условный рефлекс не вырабатывается даже
после 427 сочетаний.
3. Необходимо деятельное состояние коры. Так, у накорм-
ленного сонливого животного выработать пищевой рефлекс
невозможно.
ТОРМОЖЕНИЕ УСЛОВНЫХ
РЕФЛЕКСОВ
Возбуждение — это реакция на раздражение, выработан-
ная в процессе эволюции. Заключается в переходе оТ состоя-
ния относительного физиологического покоя к деятельно-
сти. В основе лежит изменение проницаемости мембран
заряда на мембране.
Торможение — активный нервный процесс, приводящий к
угнетению или предупреждению возбуждения. Выделяют два
типа торможения: безусловное и условное.
Безусловное торможение (внешнее по Павлову) относится к
врожденным свойствам нервной системы. Оно выявляется при
первом же предъявлении сильного постороннего раздражите-
ля. Например, громкий звонок вызывает торможение слюно-
268
отделительного рефлекса. Ситуация в жизни: представьте, Вам
одной фразой испортили аппетит. Вторая ситуация: малень-
кий ребенок хочет есть и поэтому плачет. Пища еще не готова.
Мама отвлекает его, давая игрушку. Плачь прекращается, так
как поведение, связанное с требованием пищи, оказывается
заторможенным.
Условное торможение бывает двух видов — угасание и диф-
ференцировка.
Угасание формируется постепенно при неподкрепле-
нии безусловным раздражителем (например, пища) условного
раздражителя (например, свет лампочки или бутылочка с со-
ской).
Дифференцировка — это способность отличить
один сигнал от других, похожих. Например, только хозяйка
дает кошке лакомства. Кошка бежит к открыванию двери, если
пришла хозяйка (слышит звук открываемого замка). На при-
ход других людей кошка не реагирует (замок открывают чуть-
чуть по-другому — «не те это сигналы»).
Внимание
Внимание — произвольный процесс, осуществляющийся
при участии процессов условного торможения. Характеризует-
ся: 1) устойчивостью — способностью сопротивляться утомле-
нию и отвлечению и 2) способностью быстро переключать
внимание.
Понятие о первой и второй
сигнальных системах
Многообразные условные рефлексы, возникающие на ос-
нове безусловных рефлексов, принадлежат к первой сигналь-
ной системе. Они свойственны человеку и животным.
У человека, кроме того, появилась и развилась новая, более
совершенная, вторая сигнальная система — сигнализация сло-
вом. Она находится в тесном взаимодействии с первой сиг-
нальной системой и при этом играет ведущую роль в созна-
тельной жизни человека.
Вторая сигнальная система — это система речевых сигна-
лов. Произносимое, слышимое и видимое слово может вы-
звать и вызывает у человека те же действия и реакции, что и
внешнее и внутреннее раздражение иного характера. Слоро,
по выражению И. П. Павлова, становится сигналом сигналов.
В слове и в совокупности слов (речи) обобщаются сигналы
269
первой сигнальной системы, т. е. предметы и явления окружа-
ющего мира.
Речь и, как результат, — вторая сигнальная система возник-
ли в процессе общественного труда и в процессе общения че-
ловека с другими людьми. Сигнальное значение слова опреде-
ляется его смысловым содержанием в отличие от слухового
или видимого раздражителей, на которые отвечает и мозг жи-
вотного. «Слово, — писал И. П. Павлов, — сделало нас людь-
ми». Способность человеческого мозга к анализу и синтезу ре-
чевых сигналов составляет «специально человеческое, высшее
мышление» (И. П. Павлов). Способность через слово к обоб-
щенному отражению происходящего во внешнем мире позво-
лило человеку создать науку и значительно повысить возмож-
ность ориентации в окружающем мире. При наличии второй
сигнальной системы человек может получать знания и накап-
ливать опыт без непосредственного контакта с окружающей
его действительностью.
Деятельность второй сигнальной системы условно-реф-
лекторная и подчиняется тем же згусонам, что и деятель-
ность первой. Установлено, что процесс обобщения словом
развивается как результат выработки системы условных
связей.
Развитие второй сигнальной системы — результат деятель-
ности всей коры больших полушарий.
Первая и вторая сигнальные системы — это разные уровни
единой ВИД человека. Они находятся в тесном взаимодей-
ствии. Ведущую роль в этом взаимодействии играет вторая
сигнальная система.
Соотношение между первой и второй сигнальной системой
меняется в процессе онтогенеза. На первых этапах постсигналь-
ного развития преобладающее развитие имеет первая сигналь-
ная система. С возрастом повышается роль второй сигнальной
системы. После незначительного ослабления в пубертатном пе-
риоде вторая сигнальная система вновь занимает ведущее поло-
жение и, постепенно развиваясь, сохраняет его у взрослого че-
ловека.
Типы высшей нервной
деятельности
Типы ВНД — это комплексы врожденных и приобретенных
индивидуальных особенностей нервной системы, которые оп-
ределяют различия в поведении разных людей и в их отноше-
нии к одним и тем же внешним воздействиям.
И. П. Павлов выделял четыре основных типа ВИД:
1. С«пьный неуравновешенный тип — сильная нервная си-
стема (высокий уровень работоспособности нервных кле-
ток) и преобладание процессов возбуждения над торможе-
нием.
2. Сильный уравновешенный подвижный тип — высокая под-
вижность сильных нервных процессов и уравновешенность
возбуждения и торможения.
3. Сильный уравновешенный инертный тип — при значитель-
ной силе нервных процессов их низкая подвижность (замкну-
тый переход из состояния возбуждения в торможение или на-
оборот).
4. Слабый тип — низкая работоспособность корковых кле-
ток (их неспособность выдерживать сильные или длительные
нагрузки) и отсюда — слабость нервных процессов.
Между указанными типами существует множество перехо-
дов, и люди образуют по силе нервной системы непрерывный
ряд.
И. П. Павлов усматривал совпадение между выделенными
типами нервных процессов и типами темпераментов у людей
(по классификации Гиппократа). Первому из приведенных
типов по И. П. Павлову соответствует холерический темпера-
мент, второму — сангвинический, третьему — флегматичес-
кий, а четвертому — меланхолический.
Память
Одним из основных свойств нервной системы является бо-
лее или менее длительное хранение информации о событиях
внешнего мира и ответной реакции организма на эти события.
Способность накопления информации о внешнем мире,, ее
хранение и последующее использование (воспроизведение
опыта) называют памятью.
Процесс фиксации информации, процесс ее хранения и
процесс ее воспроизведения — это три составных элемента па-
мяти. Память лежит в основе сознательной деятельности че-
ловека. Она является необходимым элементом процессов
мышления и связана с фиксацией условно-рефлекторных свя-
зей в мозге.
Объем памяти возрастал в ходе эволюции по мере увеличе-
ния числа нервных клеток, взаимосвязей между ними и с ус-
ложнением структуры мозга.
Период, связанный с хранением информации, может быть
коротким (краткосрочная память) и длительным (долгосроч-
271
ная память). Оба вида памяти в процессе мыслительной дея-
тельности человека тесно связаны. Возможен, например, пе-
реход информации из структур кратковременной памяти в не-
рвные структуры, обеспечивающие долговременную память в
процессе закрепления информации.
Распространено представление, что в основе кратко-
временной памяти лежат электрофизиологические механиз-
мы, связанные с «реверберацией импульсации» (циркуля-
цией нервных импульсов) по замкнутой системе нейронов
(П. К. Анохин, 1968). Долговременная память формируется
на основе синтеза макромолекул (нуклеиновых кислот, бел-
ков) и связана с активацией генетического аппарата. Допус-
кают, что в качестве субстрата индивидуальной памяти выс-
тупает ДНК.
Процессы долговременной памяти зависят также от уровня
метаболизма гамма-аминомасляной и глутаминовой кислот,
циклических нуклеотидов, а также ионов кальция.
Важное значение в длительности хранения информации
усматривают и в росте симпатических связей и самих синап-
сов. Подтверждением этому служит возникновение симпати-
ческих выростов-шипиков, число которых увеличивается в
процессе обучения и накопления информации.
Формирование механизмов памяти человека зависит от на-
следственных факторов и факторов окружающей среды. Те и
другие факторы находятся в тесном взаимодействии. Память
можно тренировать.
Сознание
Сознание — это высшая форма обобщенного отражения
человеком окружающей действительности и своего собствен-
ного мира (осознание собственного «Я»); с сознанием связа-
но целенаправленное поведение человека в окружающем
мире и предусмотрение результатов этого поведения. Созна-
ние является функцией мозга, которая обеспечивает наибо-
лее оптимальное приспособление организма к окружающей
среде. Сознание не является врожденным. Однако имеются
предпосылки его возникновения в онтогенезе, что определя-
ется особенностями нервной системы, ее структурой. Эта
возможность переходит в действительность в условиях обще-
ственной жизни.
Возникновение и развитие сознания исторически сопря-
жено с возникновением и развитием трудовой деятельности
людей. Материальной формой возникновения сознания явля-
ется язык (система условных звуков). Не исключено парал-
лельное использование человеком других символов, напри-
мер, математических, художественных образов и т. д.
Сознание определяет (Моунткастел, 1977, цит. по
р. Шмидт): 1) внимание и способность сосредоточиться;
2) способность порождать абстрактные мысли и оперировать
ими, а также выражать их словами или каким-либо другим
способом; 3) возможность оценить предстоящий по« :упок,
т. е. способность к ожиданию и прогнозированию; 4) осозна-
ние своего «я» и признание других индивидуумов; 5) наличие
эстетических ценностей.
Функциональными предпосылками сознания человека
является определенный уровень функционирования ЦНС
(активности нейронов). Формируется сознание в результате
взаимодействия между структурами коры и подкорки. Если
в реакцию вовлекается относительно небольшое число ней-
ронов коры и подкорки, то такие реакции протекают как
подсознательные; если реакции протекают при достаточно
выраженной («глобальной» — по выражению Бабского,
1985) активности ЦНС, нейронных «ансамблей» коры и
подкорковых структур, то она осуществляется с участием со-
знания. Как- слишком низкая (при наркозах, в состоянии
комы), так и чрезмерная нейронная активность (при элект-
рическом шоке, эпилептическом припадке) несовместимы с
сознанием.
Подсознание и сознание — это части единого целостного
процесса, два уровня, обеспечивающие непрерывность взаи-
модействия организма и среды. Высшим регулятором челове-
ческого поведения является сознание, в подчинении которого
находится информация, накопленная в процессе жизненного
опыта. Сознание определяет целенаправленное поведение че-
ловека с учетом накопленного опыта.
Мышление
При расширении жизненных задач в процессе приспособ-
ления человека к условиям жизни большое место отводится
мышлению — сложнейшему виду мозговой деятельности.
Мысль является обобщенным отражением действительности.
При этом источником подсознания мира являются не только
непосредственные ощущения; в процессе мыслительной дея-
тельности возникают образы с их свойствами на основании
логических построений и умозаключений. Будучи вполне ес-
тественным для человека процессом, мышление является
273
функцией мозга. Проявление полноты мышления человека
становится возможным при овладении языком, понятиями,
логикой. Все это является продуктом развития общественной
деятельности. Общественная деятельность является критери-
ем истинности мышления, а также является той основой, на
которой вырастают логические правила и законы.
раздел 6
ВОЛЮЦИЯ
и экология
ЭВОЛЮЦИЯ ЖИЗНИ НА ЗЕМЛЕ
КОСМИЧЕСКАЯ ЭВОЛЮЦИЯ МАТЕРИИ
Вселенная представляет собой уникальный всеобъемлющий
предмет для размышления и изучения.
Эволюцию материи во Вселенной мы можем представить в
виде трех ее составляющих — вещества, энергии, организации.
Если модуль материи, т. е. сумма вещества энергии и органи-
зации остается постоянным (по законам сохранения), то вся
эволюция материи составляет многократный и непрерывный
переход вещества в энергию, энергии в организацию и, наобо-
рот, организации в энергию, энергии в вещество (по законам
перехода).
В настоящее время наибольшее количество фактов в свою
пользу набрала теория «Большого взрыва», которая, несмотря
на все ее недостатки, позволяет рассуждать о процессе услож-
нения материи, опираясь на известные нам фундаментальные
законы.
Изучение любого процесса невозможно без расчленения его
на части, без рассмотрения этих частей отдельно с целью ана-
лиза различных его сторон. Рассматривая эволюцию началь-
ной стадии развития материи, астрофизики используют для
нового качественного этапа название «эра». И это несмотря на
то, что длительность эры может быть очень малой по нашим
понятиям.
ЭРА ПЕРВИЧНОГО ВЕЩЕСТВА
В самом НАЧАЛЕ — ноль-пункте времени — вещество и
энергия были сдавлены в единую гигантскую массу, которая
не имела ни пространства, ни времени. Поскольку наука не
обладает более ранним представлением о материи, начало
«Большого взрыва» иногда обозначают «точкой неопределен-
ности». Температура в «точке неопределенности» достигала ко-
лоссальной величины. Состояние ее являлось неустойчивым.
Произошел взрыв. Не такой взрыв, который мы можем наблю-
дать на Земле и который начинается из определенного центра и
276
затем распространяется, захватывая все большее и большее
пространство, а взрыв, который произошел одновременно вез-
де, организовав и заполнив с самого начала все пространство.
При этом каждая частица материи устремилась прочь от лю-
бой другой частицы, образуя вектор времени. На основании
приобретенных наукой знаний считается, что проследить эво-
люцию Вселенной от момента «Большого взрыва» возможно.
Для этого необходимо задать плотность трем сохраняющимся
величинам в исходный момент ранней Вселенной, которыми
являются: электрический заряд, барионное число, лептонное
число.
1. Электрический заряд. Возможно появление или исчезно-
вение пары частиц с равными по величине и противоположны-
ми по знаку электрическими зарядами, но полный электричес-
кий заряд никогда не меняется.
2. Барионное число. Барион — это объединяющий термин,
включающий ядерные частицы (протоны и нейтроны) вместе
с более тяжелыми нестабильными частицами, известными как
гипероны. Барионы и антибарионы могут рождаться и уничто-
жаться парами; кроме того, барионы могут распадаться на дру-
гие барионы, как в бета-распаде радиоактивных ядер, когда
нейтрон переходит в протон, и наоборот. Однако полное чис-
ло барионов минус число антибарионов (антипротонов, анти-
нейтронов, антигиперонов) никогда не меняется.
3. Лептонное число. Лептоны — это легкие, отрицательно за-
ряженные частицы — электрон и мюон, а также электрически
нейтральная частица нулевой массы, называемая нейтрино, и
их античастицы— позитрон, антимюон и антинейтрино. Не-
смотря на нулевые массы и заряд, нейтрино и антинейтрино
несут энергию и импульс, как и любые другие частицы. Пол-
ное число лептонов минус полное число антилептонов никогда
ре меняется.
Э?А ТЕПЛОВОГО РАВНОВЕСИЯ
В результате «Большого взрыва» Вселенная быстро расши-
ряется и остывает. Основную роль в пространстве, которое те-
перь занимает материя, играют мюоны, электроны, позитроны,
нейтрино и антинейтрино. Но Вселенная еще столь плотна, что
даже нейтрино, которые могут годами «путешествовать» сквозь
свинцовые бруски, не будучи рассеянными, удерживаются в
тепловом равновесии с электронами, позитронами и фотонами,
благодаря быстрым столкновениям с ними и друг с другом.
277
Температура Вселенной, равная 100 миллиардам градусов?
Кельвина (10й К), в момент взрыва резко падает и уже черед
две секунды от НАЧАЛА достигает 10 миллиардов градусов^
Кельвина (1О10 К). Время этой эры быстротечно и составило^
менее 10 с. Основным конечным результатом этой эры являет-
ся аннигиляция электронно-позитронных пар, превращение их
в фотоны, что приводит к заполнению Вселенной светом.
ЭРА НУКЛЕОСИНТЕЗА
Менее чем через 14 с от НАЧАЛА температура Вселенной
упала до 3 миллиардов градусов Кельвина (3-109 К). С этого мо-
мента, говоря о температуре Вселенной, подразумевают темпе-
ратуру фотонов. Теперь уже достаточно «прохладно» д ля того,
чтобы образовались различные стабильные ядра вроде гелия и
водорода. То есть большинство свободных электронов и пози-
тронов исчезло, перешло к электрическому типу взаимодей-
ствия, и главными составными частями Вселенной являются те-
перь фотоны, нейтрино и антинейтрино. Почти все нейтроны
также ушли на образование ядер гелия. Вселенная продолжала
расширяться и охлаждаться, и в течение 700 000 лет не про-
изошло ничего «интересного». После первых трех минут мате-
риал, из которого должны были образоваться звезды, состоял на
22—28% из гелия, а остальное — почти все из водорода. Таким
образом, Солнце и другие звезды начали жизнь, имея в своем
составе лишь водород и гелий.
ЗВЕЗДНАЯ ЭРА
Наша эра. Температура по истечении 700 000 лет стансу
вится ниже 3000 К. Время течет, и качественных скачков во
Вселенной не возникает. Вещество эволюционирует, образуя
галактики и звезды в результате сгущения. Начинают образо-
вываться атомы (рис. 6.1) — элементарные «кирпичики» веще-
ства, которые Д. И. Менделееву удалось привести в систему.
Большинство из этих элементов образуются в звездах и попа-
дают в пространство при взрывах звезд. В недрах звезд образу-
ются углерод, железо и т. п. Разлет сложных элементов при
взрыве звезд приводит их к новому сгущению и образованию
планетных систем.
Диаметр нашей Солнечной системы, ограниченной орбитой
Плутона, составляет 12 млрд км. Лучу света требуется более
278
• Протон
< Нейтрон
.. • Электрон
Н Не
1,0079 а.е.м. 4,0026 а.е.м.
Реакция слияния атомов водорода
4x1,0079 а.е.м. 4,0026 а.е.м. + энергия,
(4,0316 а.е.м.) эквивалентная
0,029 ае.м.
Рис. 6.1. Пример слияния атомов водорода с образованием
атома гелия. Один атом гелия легче, чем четыре атома водорода.
Дефект массы соответствует выделению энергии излучения
(а. е. м. — атомная единица массы)
11 ч для того, чтобы пройти это расстояние, в то время как до-
рогу от Земли до Луны он покрывает за 1,25 с. Возможно, что
Вселенная сейчас имеет конечную длину окружности, оцени-
ваемую примерно в 125 млрд световых лет (рис. 6.2).
ГЕОЛОГИЧЕСКАЯ ЭВОЛЮЦИЯ НА ЗЕМЛЕ
ОБОЛОЧКИ ЗЕМЛИ
Возраст Земли оценивается приблизительно в 6 млрд лет.
Согласно геофизическим данным, современная Земля состоит
из следующих разнородных слоев (оболочек) (рис. 6.3).
1. Ядро. Делится на внешний (жидкий) и внутренний (твер-
дый) слои. Ядро преимущественно состоит из железа. Толщина
внешнего расплавленного слоя составляет 2220 км, а диаметр
внутреннего слоя равен 2510 км. Наличие расплавленного слоя
обусловлено высокими температурой и давлением в глубине
планеты.
279
Рис. 6.2. Сопоставление орбит и размеров планет
Солнечной системы
2. Мантия. Состоит из обогащенных железом магмати-
ческих пород и простирается от поверхности на глубину до
2900 км.
3. Земная кора. Это верхний слой твердой земли. Кора обра-
зовалась из застывшей расплавленной магмы, а также путем
осаждения на дно водных бассейнов частиц пород, разрушен-
280
Кора
Рис. 6.3. Строение земных недр
Мантия
Внешнее ядро
(жидкое)
Внутреннее ядро
(твердое)
ных действием ветра, воды и других факторов, и перекристал-
лизаций изверженных и осадочных пород. Поверхностный
слой мантии вместе с земной корой называется литосферой.
4. Гидросфера. Составляет одну четырехтысячную долю
массы всей Земли. Около 94% массы гидросферы — это соле-
ные воды Мирового океана. Из остальных 6% гидросферы три
четверти приходится на подземные воды и четверть — на лед-
ники Антарктиды и Гренландии. Прочие ледники и озера со-
ставляют очень малую долю гидросферы.
5. Атмосфера. Составляет одну миллионную долю массы
всей Земли. Сухой воздух состоит из 78,08% азота, 20,95%
кислорода, 0,93% аргона, около 3% углекислого газа и малых
количеств благородных газов и водорода, а также небольшого
количества водяных паров, несущих различные активные при-
меси.
В зависимости от плотности газов, колебаний температуры
и других факторов в атмосфере различают:
а) тропосферу — нижний слой атмосферы, который прости-
рается до 10—12 км. Чем больше высота, тем ниже температура
и давление;
б) стратосферу — слой, простирающийся до 80—100 км от
поверхности Земли. Этот слой имеет относительно постоянную
температуру порядка —40° С. На высоте около 20 км в этом
слое регистрируется максимальное содержание озона;
в) ионосферу — верхний слой атмосферы, где солнечное из-
лучение производит ионизацию. Условно этот слой занимает
до 500 км от поверхности Земли (рис. 6.4).
281
70 -
60 -
50 “
40 |
30
’ 20
10
О
10
20
30
^Тропосфера
/ / ‘ .* _______1 " "" 11 : /
Литосфера —yj идросфера*
Рис. 6.4. Геосферы Земли
СТРОЕНИЕ И ЭВОЛЮЦИЯ ЗЕМНОЙ КОРЫ
Главными элементами рельефа поверхности нашей планеты
являются материки и океанические впадины. Поэтому земная
кора делится на два основных типа: материковую и океаническую
кору. Земная кора материков состоит из трех мощных слоев, От-
личающихся по своим свойствам и плотности. Верхний слой сло-
жен сравнительно неплотными осадочными породами, средний
называется гранитным, а нижний — базальтовым. Земная кора
под океанами отличается от материковой не только своей толщи-
ной, но и отсутствием гранитного слоя. Таким образом, под океа-
нами присутствуют лишь два слоя—осадочный и базальтовый.
Самыми крупными структурами земной коры материков
являются геосинклинальные складчатые пояса и древние плат-
формы.
Геосинклинальные складчатые пояса представляют собой
подвижные участки земной коры, геологическая история ко-
торых характеризовалась интенсивным осадконакоплением,
многократно проявлявшимися складкообразовательными про-
цессами и сильной вулканической деятельностью. В них на-
капливались мощные толщи осадочных пород, формировались
магматические породы, часто проявлялись землетрясения. Гео-
синклинальные пояса занимают обширные участки матери-
ков, располагаясь между древними платформами или по их
краям в виде широких полос.
Древние платформы — наиболее устойчивые и малоподвиж-
ные участки материков. В отличие от геосинклинальных поясов
282
древние платформы испытывали медленные колебательные дви-
жения, в их пределах накапливались осадочные породы обычно
небольшой толщи, отсутствовали складкообразовательные про-
цессы, редко проявлялись вулканизм и землетрясения; Древние
платформы образуют в составе континентов участки, являющие-
ся остовами всех материков.
Геологическая история нашей планеты, начиная примерно
с 3,5 млрд лет, делится на пять крупных естественных этапов
развития, получивших название архейской, протерозойской,
палеозойской, мезозойской и кайнозойской эры. В составе
трех последних выделяют периоды, которые делятся на эпохи,
состоящие, в свою очередь, из веков. В сумме эти естественные
отрезки геологической истории Земли образуют геохронологи-
ческую шкалу (рис. 6.5 и 6.6).
Время,
млн лет
Геологическая
эра
События
— 1000
— 2000
— 3000
— 4000
ЩИ Кайнозой111111
Мезозой
Микроископаемые (?)
Химическая эволюция
Образование Земли
Биологическая
(дарвиновская)
эволюция
Эволюция человека
Появление млекопитающих
Первые находки позвоночных
Первые находки
многоклеточных
— 5000
Возникновение Солнечной
системы
Рис. 6Л. Геохронологическая шкала
283
бремя от начала периода ДО наших М дней, млн лет Продолжи- тельность, млн лет Периоды а
2 ЧЕТВЕРТИЧНЫЙ Спкпммшыяа 1ПЯл эпоха Плейстоцен )S о
65 63 [третичный Плиоцен Миоцен ‘ Олигоцен Эоцен Палеоцен | К а й н о з
135 190 70 Мел I И о ’
55 Юра- ст О СО
ф
35 Триас S
225 280 55 Пермь )3
о
345 65 Карбон со
50 Девон о
390 35 Силур ф
430 500 70 Ордовик е;
се
570 70 Кембрий С
Рис. 6.6. Палеонтологическая летопись Земли
284
В архейскую эру (около 2 млрд лет назад) древние платфор-
мы образовывали единый монолитный материк Пангею. Ос-
новную массу его составляли только что возникшие древние
платформы, по окраинам которых располагались геосинкли-
нальные пояса. Пангея была окружена древними океанами.
Возможно, что уже существовал Тихий океан, являющийся
основным водным бассейном Земли. Клином между континен-
тами Северного и Южного полушарий вдавался гипотетический
океан — Тетис, получивший свое название в честь греческой
богини моря. В дальнейшем этот океан разделил Пангею на
континенты Северного и Южного полушарий-(рис. 6.7). Пер-
вый представлял суперконтинент Лавразию, а второй — су-
перконтинент Гондвану. Это были голые, гористые пустыни с
довольно разветвленными речными артериями. Северный Ле-
довитый океан выглядел небольшим внугриконтинентальным
морем; Атлантический и Индийский океаны еще тогда не заро-
дились.
Рис. 6.7. Разделение древней Пангеи.
Стрелками обозначено движение материков:
а — конец Перми, около 230 млн лет; б — конец Триаса, около 195 млн лег;
в — конец Юры, около 140 млн лет; г — конец Мела, около 70 млн лет
285
В протерозойскую эру развитие геосинклинали# и платформ
шло различными путями. Происходило регрессивное разрас-
тание платформ за счет сокращения территорий геосинклина-
ли#. Лавразия и Гондвана представляли собой ассоциации
платформ. Их типичным ландшафтом являлись пустынные
континентальные равнины, обрамленные горными массивами.
Океаны и моря были относительно мелководными, с большим
числом архипелагов. Важным событием позднепротерозойской
эры явилось появление в атмосфере Земли свободного кисло-
рода и сокращение содержания углекислоты. Наличие свобод-
ного кислорода послужило началом формирования озона (О3).
Постепенно возникает озоновый «экран», задерживаю-
щий до 97% коротковолновой ультрафиолетовой радиации.
Повышается соленость океанических вод, достигая современ-
ных значений. Все это предопределило бурное развитие орга-
нической жизни в последующие этапы геологической исто-
рии Земли.
В палеозойскую эру происходит значительное увеличение
территории платформ. Во многих местах геосинклинали# (осо-
бенно в девоне и карбоне) интенсивно проявляется вулканизм.
На месте многих геосинклинали# возникают горы, поднятия
испытывают и платформы.
В позднем палеозое наряду с областями теплового тропичес-
кого климата существовали полярные зоны. Развитие матери-
ковых ледников установлено на месте юга Африки, в Австра-
лии, Индии, Южной Америке, Антарктиде, т. е. центральные
области Гондваны (особенно в карбоне) были покрыты ледя-
ным панцирем.
В мезозойскую эру произошло изменение очертания матери-
ков. Единые супергигантские континенты — Лавразия и Гонд-
вана распадаются. Между отдельными блоками литосферы
Гондваны намечается отделение Африки, Индии, Австралии и
Антарктиды. Между ними закладывается морской бассейн,
превратившийся в д альнейшем в Индийский океан. Возникает
узкая коленообразная лента Атлантики при отделении от Лав-
разии Северной Америки.
В кайнозойскую эру продолжается «расползание* материков.
Интенсивно формируется Атлантика. Произошло разделение
Индии, Австралии и Антарктиды. Менее подвижной оказа-
лась Африка.
В антропогене мощное обледенение охватывает континенты
Северного полушария. В Европе центром оледенения являлись
Скандинавский полуостров и Альпы. Обширные оледенения
захватили также азиатскую часть бывшего СССР и Северную
286
Америку. Последние примерно 10 тыс. лет приходятся на меж-
ледниковую эпоху. Границы ледников в Северном полушарии
резко сократились. Материковые льды остались в Гренландии
и на некоторых островах Ледовитого океана.
На современных материках выделяют от 10 до 16 древних
платформ (наиболее крупными являются: Восточно-Европей-
ская, Сибирская, Северо-Американская, Африкано-Аравий-
ская, Индостанская, Австралийская и Антарктическая), а также
7 геосинклинальных поясов (Среднеземноморский, Тихооке-
анский, Атлантический, Урало-Монгольский, Арктический,
Бразильский и Внутриафриканский) (рис. 6.8).
Рис. 6.8. Схема расположения древних платформ
и геосинклинальных поясов:
I — древние платформы (цифры на карте): 1 — Восточно-Европейская; 2 —
Сибирская; 3 — Таримская; 4 — Китайско-Корейская; 5 — Южно-Китайская;
6 — Северо-Американская; 7 — Африкано-Аравийская; 8 — Индостанская;
9 — Австралийская; 10 — Южно-Американская; 11 — Восточно-Бразильская;
12 — Антарктическая;
II — малые геосинклинальные складчатые пояса (буквк на карте):*
Б — Бразильский, ВА — Внутриафриканский
287
ХИМИЧЕСКАЯ ЭВОЛЮЦИЯ
И ПРОИСХОЖДЕНИЕ ЖИЗНИ
ТЕОРИЯ ПРОИСХОЖДЕНИЯ ЖИЗНИ НА ЗЕМЛЕ
ОПАРИНА - ХОЛДЕЙНА
Примерно через миллиард лет после зарождения Земли
возникли одноклеточные организмы путем эволюции из орга-
нических веществ, образовавшихся абиогенно в несодержав-
шей свободного кислорода атмосфере. Эта основная идея
была высказана в ее современной форме в 1928 г. А. И. Опа-
риным и Дж. Холдейном. В 1959 г. первые эксперименты по
неорганическому синтезу «органических» веществ в восста-
новительной среде провел С. Миллер, бывший в ту пору сту-
дентом.
Миллер использовал очень простой прибор — колбу, в кото-
рой создавались электрические разряды (рис. 6.9). Прибор за-
полнялся водой и различными газами. В основном использова-
лись водород, метан и аммиак. Свободный кислород в колбу не
«допускался». В верхней части колбы непрерывно происходили
электрические разряды. Внизу кипела вода, создавая циркуля-
цию пара и воды через прибор. Аналогичные пробы были про-
Рис. 6.9. Аппарат для воздействия
Нагрев искровыми разрядами на смеси газов
288
ведены и другими учеными во всем мире. Опираясь на эти ис-
следования, было сделано предположение о том, что «живые
системы» появились на земной поверхности в результате взаи-
модействия «первичных источников энергии» с неким набором
«первичных молекул» (т. е. неживых молекул, имевшихся на
поверхности Земли).
Jjecb эволюционный процесс живой материи на Земле
был теоретически обоснован условиями начального со-
стояния.
1. Наличие бескислородной первичной атмосферы. Предпо-
лагается, что первичная атмосфера Земли содержала преиму-
щественно водород, метан, воду и аммиак.
2. Воздействие на атмосферу источника энергии. Оценить
роль источников энергии нелегко. Наиболее вероятными яв-
ляются: ультрафиолетовое излучение, вулканизм, энергия ме-
теоритных ударов. Среди них ультрафиолетовое излучение
представляет собой самый значительный источник энергии,
далеко превосходящий по величине все другие.
3. Наличие на поверхности Земли жидкой воды (таким обра-
зом колебания температуры в водоемах должны быть в преде-
лах от 0 до 100’ С).
4. Наличие в атмосфере, гидросфере и литосфере большого
разнообразия несвязанных в сложные структуры химических
элементов.
СТАДИИ БИОГЕНЕЗА
Биогенез — это процесс развития от неживого к живому.
Различают несколько его стадий.
1. Стадия неорганического фотосинтеза (т. е. образование
малых «органических» молекул в неорганических реакциях). '
Отсутствие кислородно-озонового «экрана» позволяло корот-
коволновому ультрафиолетовому излучению падать на поверх-
ность Земли. Оно вызывало неорганические фотохимические
реакции, т. е. элементы атмосферы и гидросферы после погло-
щения световых квантов образовывали органические молеку-
лярные соединения. Поскольку процесс этот основан на по-
глощении квантов света, идущих от Солнца, его можно назвать
фотосинтезом. Но это неорганический фотосинтез, так как
происходит без участия живых организмов. В водоемах Земли
создавалась концентрация простых соединений углерода и
других элементов. Часто эту смесь называют «первичным
бульоном».
10-281
289
2. Стадия коацервации. Взвешенное в виде частиц про-
стейшее «органическое» вещество в капле воды получило
название коацервата. Исследователи с давних времен заме-
чали, что растворы высокомолекулярных веществ при сто-
янии могут мутнеть, оттого что молекулы их слипаются в
комплексы, а комплексы образуют крошечные капли, взве-
шенные в жидкости. Внешне они похожи на капли молочно-
го жира в молоке.. Концентрация веществ в таких каплях
может быть в десятки и сотни раз выше, чем в окружающем
их растворе. Однако коацерватные системы нестабильны и
через некоторое время оседают на дно, образуя неводный
слой.
3. Стадия образования оболочек, или мембран. Стабилиза-
ция коацерватных систем была обеспечена в первую очередь
возникновением вокруг них внешнего плотного слоя, сходного
со слоем клеточных мембран. Полагают, что образование при-
митивных мембран вокруг коацерватных капель начинается с
выстраивания полярных молекул на поверхности раздела воз-
дух,—вода или липид — вода.
4. Стадия появления метаболита. В принципе это не что
иное, как поглощение и выделение с промежуточной реакцией
(или цепью реакций) некоторых других органических веществ.
В протоклеточной системе могли бы протекать реакции, обес-
печивающие высвобождение необходимой энергии, сходные с
современным фотосинтезом или дыханием. Примитивный ме-
таболизм также способствовал стабилизации и длительному
существованию коацерватных капель.
5. Стадия воспроизведения (редупликации). Последняя ста-
дия биогенеза — приобретение способности к воспроизведе-
нию, благодаря которому жизнь может продолжать сама себя.
В эпоху преджизни могли синтезироваться упорядоченные по-
лимеры, которые можно назвать самоупорядоченными. Для
этого не требуются живые организмы или нуклеиновые кисло-
ты. Возникновение самоупорядоченности, видимо, может быть
объяснено на основе трехмерной структуры и электронной
конфигурации молекул аминокислот. Со временем могли на-
чаться взаимодействия между неорганически синтезирован-
ным протеиноидом, или предбелком, и нуклеотидами того же
происхождения. Так постепенно нуклеиновые кислоты могли
приобрести свою главенствующую роль, свойственную им
теперь,
В результате биогенеза на Земле возникли прокариотичес-
кие клетки, не обладавшие оформленным клеточным ядром и
ядерной оболочкой.
290
АЛЬТЕРНАТИВНЫЕ ТЕОРИИ
ВОЗНИКНОВЕНИЯ ЖИЗНИ НА ЗЕМЛЕ
Теория панспермии. Согласно теории панспремии, завоевав-
шей популярность в XIX веке, жизнь могла распространяться
от одной Солнечной системы к другой в виде спор микроорга-
низмов. Ф. Крик и Л. Оргель высказали недавно еще более
смелое предположение. По их мнению, Земля и, возможно,
другие первоначально лишенные жизни планеты могли быть
«засеяны» намеренно какими-то разумными существами, оби-
тателями тех солнечных систем, которые в своем развитии опе-
редили нашу планету на миллиарды лет. По мнению Крик и
Оргель, такая «направленная панспермия» могла бы, напри-
мер, объяснить, почему молибден—довольно редкий на Земле
элемент — совершенно необходим живым существам как ко-
фактор образования многих важнейших ферментов. Однако
эта теория не отвечает на вопрос: каким образом могла воз-
никнуть и эволюционировать жизнь на планете, допустим,
сходной с нашей Землей?
Теория стационарного состояния. В этой теории, предложен-
ной в конце 40-х годов Г. Бонди, утверждается, что Вселенная
всегда была почти такой же, как сейчас. Отсюда жизнь, как
мы ее себе представляем, является ровесницей всего вещества
Вселенной. .
ПРОИСХОЖДЕНИЕ ЭУКАРИОТ
1. Гипотеза мембраногенеза Робертсона (1959). Согласно дан-
ной гипотезе, органеллы клетки (митохондрии и пластиды) и
все остальные мембранные устройства, включая ядро, возникли
в протоклетке из выпячивания и разрастания плазмолеммы. По
Робертсону, все процессы филогенетического развития цито-
плазматических органелл и ядра происходят в результате нали-
чия мембранных связей между органеллами и ядром, а также
формирования органелл (пикосом, фагосом) путем впячивания
плазмолеммы.
Однако трудно себе представить, что развитие одной специ-
ализированной структуры в цитоплазме осуществлялось, как
правило, за счет других специализированных структур, в част-
ности митохондрий, а также за счет эргастосом и ретикулосом.
2. Гипотеза симбиогенеза Мережковского (1910), согласно
которой эволюционное развитие клетки сводится к симбиоти-
ческому объединению предковых организмов бактериального
10*
291
типа. Однако явления фаго- и пиноцитоза у прокариотов не)
обнаружены, в силу чего пути проникновения в матрикс бакте-j
рий для других организмов сомнительны.
3. Гипотеза синбактериогенеза Студитского (1962, 1981) сво-
дится к эволюционному построению клетки из органелл, воз-
никших путем специализации из бактериоидных предков,
сформировавших клетку путем комплексирования. Гипотеза
основывается на фактах онтогенетической и функциональной
динамики клеточных структур, что позволяет видеть отражение
их исторического прошлого, которое, согласно синбактериоген-
ной гипотезе, включает доклеточное состояние в виде бактери-
оидных форм, ставших в ходе эволюции органеллами клетки.
С позиции данной гипотезы происхождения клетки получили
вполне правдоподобное объяснение такие явления, как размно-Ч
жение, активная подвижность, развитие безъядерных эритро-!
цитов и кровяных пластинок, и многие другие проявлениям
структурного динамизма клетки.
ЭВОЛЮЦИЯ ОРГАНИЗМОВ j
РАЗНООБРАЗИЕ ЖИВЫХ СУЩЕСТВ
** j
Органический мир состоит из животных и растений. Иско-
паемые и современные животные подразделяются на две груп-
пы: беспозвоночных и позвоночных. Беспозвоночные в свою
очередь делятся еще на две группы организмов: одноклеточных
(составляют один тип простейших) и многоклеточных (все ос-
тальные типы).
Первые одноклеточные существа, появившиеся в глубокой
древности, в дальнейшем развивались двумя различными путя-
ми: из одних произошли растения, из других — животные. Ос-
новным отличием растений от животных является принцип пи-
тания организмов. Клетки растений способны сами создавать
органические вещества на свету из веществ неорганических —
углекислого газа и воды. Строятся эти вещества с помощью
хлоропластов, наполненных хлорофиллом и плавающих в ци-
топлазме клеток растений. Такой тип питания, характерный для
растений, называется автотрофным. Животные таким способом
не могут вырабатывать органические вещества. В их клетках от-
сутствуют хлоропласты, и поэтому они потребляют уже готовые
органические вещества, созданные растениями. Такой тип пи-
тания называется гетеротрофным и свойствен животным орга-
292
низмам и грибам. Таким образом, животные питаются либо рас-
тениями, либо другими травоядными животными. Некоторые
простейшие одноклеточные организмы обладают и автотроф-
ным и гетеротрофным типом питания. Особенно интересны од-
ноклеточные, питающиеся одновременно двумя способами. Та-
ков жгутиконосец — эвглена зеленая. Это позволяет считать,
что такие простейшие стоят недалеко от общих истоков проис-
хождения как растительного, так и животного мира. Те потомки
простейших одноклеточных, у которых сохранился хлорофилл,
но не было приспособлений д ля поглощения чужих белков, д али
начало миру растений. Другие потомки приобрели способность
поглощать и переваривать чужие белки, углеводы и жиры, но в
их клетках хлорофилловые зерна не развивались. Такие орга-
низмы дали начало миру животных.
Под макроэволюцией подразумевают события, происхо-
дящие в геологическом масштабе времени и состоящие в
развитии признаков, по которым различаются такие груп-
I пы, как род, семейство, отряд, класс и тип. Макроэволюцию
нельзя изучать путем прямых наблюдений или эксперимен-
тов. Вскрыть эти закономерности помогают данные палеон-
тологии, сравнительной эмбриологии, а также молекуляр-
ной биологии и генетики.
Микроэволюция изучает изменения, происходящие внутри
вида и приводящие к его дифференцировке, завершаясь видо-
образованием.
Происхождение многоклеточных организмов
В настоящее время не вызывает сомнения, что многокле-
точные организмы произошли от одноклеточных. Из суще-
ствующих теорий происхождения многоклеточных основными
являются теория гастреи Э. Геккеля и теория паренхимеллы
И. И. Мечникова. Обе теории основываются на биогенетичес-
ком законе Геккеля—Мюллера: «Онтогенез является кратким
и быстрым повторением филогенеза, обусловленным наслед-
ственностью и приспособлением» (рис. 6.10).
Теория гастреи. Геккель обратил внимание на то, что заро-
дышевые стадии многоклеточных очень сходны с колониаль-
ными жгутиковыми и низшими многоклеточными. Так, все
многоклеточные проходят стадию однослойного шара — блас-
тулы, которая очень похожа на шарообразную колонию воль-
вокс. Следующая стадия — двухслойный зародыш, или гаст-
рула, — соответствует строению низших многоклеточных —
кишечнополостных. Переход однослойной стадии зародыша в
293
Рис. 6.10. Происхождение многоклеточных животных:
верхний ряд — фазы эмбрионального развития
многоклеточных животных:
1 — образование двух бластомеров; 2 — полное равномерное дробление;
3 — бластула; 4 — паренхимула; 5 — гаструла;
нижний ряд — животные, организация которых соответствует
определенной фазе индивидуального развития:
1 — делящаяся эвглена; 2 — колония эудорины; 3 — колония вольвокса;
4 — фагоцителла (гипотетический предок многоклеточных животных); 5 —
гидра
двухслойную происходит чаще всего путем впячивания одной
из сторон шара. На основании этого Геккель предположил,
что многоклеточные в своем зародышевом развитии повторя-
ют этапы перехода от одноклеточных к многоклеточным.
По мнению Геккеля, первые многоклеточные возникли
следующим образом. У шарообразной колонии типа вольвокс
произошло впячивание одной из сторон шара, и образовался
простейший двухслойный организм. На месте впячивания
формируется рот, внутренний слой выстилает кишку. После
того как образовалось два слоя, функции клеток разделились.
Наружный слой стал выполнять защитную функцию, а внут-
ренний — пищеварительную. Этот первичный многоклеточ-
ный организм Геккель назвал гастреей. Он правильно связал
происхождение многоклеточных с колониальными простей-
шими, но не мог объяснить причин, которые привели к образо-
ванйю двухслойности.
294
Теория пареняпаеллы. С точки зрения И. И. Мечникова, в
однослойных шаровидных колониях второй — внутренний —
слой образовался не путем впячивания, а путем выселения
клеток из наружного слоя внутрь (иммиграция). Отдельные
простейшие однослойной колонии после захвата пищи погру-
жались внутрь шара и образовывали там рыхлую массу (парен-
химу). Внутри они переваривали пищу, после чего вновь воз-
вращались на поверхность. Постепенно в процессе эволюции
часть клеток, переваривающих пищу, утратила способность
возвращаться на поверхность и образовала внутренний слой.
Сначала они получали пищу через клетки наружного слоя, а
затем развилось ротовое отверстие и полость кишки стала спо-
собной сообщаться с внешней средой. Как видно, теория
Мечникова объясняет образование простейшего двухслойного
организма вполне конкретными причинами — захватом и пе-
ревариванием пищи. Этот первичный многоклеточный орга-
низм Мечников назвал паренхимеллой, или фагоцителлой.
Современные прокариоты охватывают широкий круг орга-
низмов, отличающихся по химическому составу и физиологи-
ческим функциям. К ним относятся такие разнородные груп-
пы организмов, как бактерии и сине-зеленые водоросли.
Бактерии. Те из бактерий, которые не нуждаются для своего
питания ни в каких других животных и растениях и сущест-
вуют за счет химических реакций, происходящих в растворах
газообразных или минеральных веществ, называются хемо-
трофными. Считается, что хемотрофы были первичными орга-
низмами на Земле, когда на ней еще не было необходимых
условий для жизни растений и животных. Один из наиболее
древних остатков живых существ — это так называемые желе-
зобактерии железных руд в сланцах архейского отложения,
найденных в Северной Америке. В более ранних отложениях не
найдено пока никаких следов жизни.
Между тем среди хемотрофных бактерий железобактерии
являются далеко не простейшими по своему строению, и воз-
можно, что от появления на Земле первых живых существ и до
времени отложения слоев кристаллических архейских сланцев
прошел не один миллион лет.
Среди бактерий есть такие, которые произошли, по всей ве-
роятности, путем упрощения более сложных организмов. Так,
среди серных бактерий встречаются представители, очень сход-
ные по окраске с имеющими хлорофилл сине-зелеными водо-
295
;;ослями, а живущие в крови высших животных бактерии спи-
рохеты могут иметь животное происхождение — огпростейших.
Образ жизни бактерий-паразитов способствует упрощению
организации благодаря своему однообразию и делает сходными
организмы, различные по происхождению. Когда развилась ра-
стительная и животная жизнь, то легкость питания органичес-
кими веществами, естественно, способствовала развитию бак-
терий, питающихся за счет животных и растений.
Сине-зеленые водоросли — простейшая группа растительных
организмов. Наличие двух фотосистем (ФС1 и ФС2) у сине-зе-
леных водорослей свидетельствует о высоком уровне их эволю-
ционного развития. Однако качественный состав фотосистем у
сине-зеленых водорослей отличается от высших растений и зе-
леных водорослей. После изучения состава пигментов (хлоро-
филл А, ФС1, ФС2, фикобиллины) и их свойств у сине-зеленых
водорослей и бактерий (бактериохлорофилл, ФС2) стала оче-
видной безосновательность попыток выводить одинаковое про-
исхождение сине-зеленых водорослей и бактерий, как это было
принято. Сходство между ними скорее внешнее. Таким обра-
зом, вероятнее всего, что сине-зеленые водоросли представляют
собой самостоятельную ветвь от первоначальных прокариотов.
Однако поскольку эти водоросли — прокариоты, было предло-
жено назвать их цианобактериями, чтобы как-то отличать от
эукариотических водорослей.
Филогения зеленых водорослей
Зеленые водоросли — прогрессивная группа растительных
организмов по сравнению с сине-зелеными. Полагают, что зе-
леные водоросли возникли из прокариот, содержащих хлоро-
филл А и В.
В отличие от сине-зеленых в клетках зеленых одноклеточ-
ных водорослей содержатся вполне сформировавшиеся хло-
ропласты, или, как их называют у водорослей, хроматофоры.
В дальнейшем определяющим фактором в эволюции водорос-
лей явилась эволюция хроматофора. Наличие у многих пред-
ставителей водорослей осевого хроматофора позволяет обна-
ружить и изучить родственные связи зеленых водорослей и
проследить их историческое развитие.
Общие направления функциональной эволюции зеленых
водорослей можно сформулировать в следующих тезисах.
1. Массивный осевой хроматофор является первичным, бо-
лее примитивным, и лишь впоследствии, в процессе прогрес-
сивной эволюции, из него возник пристенный, функциональ-
296
но более совершенный и целесообразный для фотосинтетичес-
* кой деятельности. Осевой хроматофор, по мнению академика
* А. В. Топачевского, мог возникнуть лишь у амебовидных пред-
ставителей водорослей, так как при расположении в центре
клетки он не мешал образованию псевдоподий и не подвергал-
ся их влиянию. Таким образом, амебовидные формы следует
считать первичными среди зеленых водорослей. Другие типы
строения хроматофора — чашевидный, пластинчатый и осо-
бенно пристенный — более прогрессивные и преобладают у по-
давляющего большинства современных зеленых водорослей.
2. Жгутиковые формы водорослей, среди которых наиболее
простыми являются хламидомонады, имеют вторичное проис-
хождение от амебовидных организмов. Основной критерий
обоснования этого вывода — сложность жгутиков и относи-
тельная простота строения псевдоподий.
3. Из более простых хламидомонад путем утраты подвижности
возникли хлорококковые, а из них — нитчатые улотриксовые.
4. Появление многоядерных клеток положило начало разви-
тию кладофоровых.
5. Конъюгаты, включающие как одноклеточные, так и нит-
чатые формы (спирогира), представляют собой отдельную
группу водорослей, берущую начало от первичных амебовид-
ных форм. Поэтому в цикле их размножения отсутствуют жгу-
тиковые формы.
6. Диатомовые водоросли происходят от жгутиковых орга-
низмов, окрашенных в желтый цвет и способных отлагать в
своих оболочках небольшое количество кремнезема. Диатомо-
вые характеризуются разнообразием клеточных форм, приспо-
собленностью к свободноплавающему образу жизни или к об-
разу жизни организма, прикрепляющегося под водой к скалам
или другим твердым предметам. Они не переходят ни в какой
высший тип, являясь как бы законченным в своем развитии.
Филогения высших растений
Рассмотрим основные признаки, приобретенные зелеными
нитчатыми водорослями при переходе от одноклеточных форм
к многоклеточным, а затем к высшим растениям.
1. Сходство зеленых нитчатых водорослей с высшими рас-
тениями прежде всего в крупных размерах их тела (таллома).
Они широко распространены в тех же биотопах, что и высшие
растения. Возникновение крупных макроскопических талло-
мов способствовало захвату жизненного пространства в борьбе
за условия существования.
297
2. Наличие хорошо развитых вакуолей, связанных в основ-
ном с многоклеточностыо организмов, что наблюдается у нит- ,
чатых водорослей и низших растений. Установлено, что объем
протоплазмы,клеток зеленых нитчатых водорослей изменяется
в зависимости от их фотосинтетической активности. На про-
тяжении световой фазы протопласт увеличивается в размерах
за счет уменьшения объема вакуоли. Во время темновой фазы
уменьшается протопласт, а объем вакуоли увеличивается. Сле-
довательно, центральная вакуоль — это пространственный ре-
зерв для интенсивного роста в процессе фотосинтеза.
3. Появление фотопериодизма. Связь жизненных функций
со сменой дня и ночи у нитчатых водорослей аналогична выс-
шим растениям.
4. Предками первичных наземных растений могли быть зе-
леные нитчатые водоросли с многорядными нитями, поэтому
легко представить возникновение флоэмы. Ситовидная трубка
является аналогом и гомологом нити зеленых нитчатых водо-
рослей и выполняет ту же функцию проводящей системы.
Ксилемы у зеленых нитчатых водорослей нет. Однако процесс
ее закладки у высших растений свидетельствует о том, что эта
ткань возникла не сразу.
5. У зеленых водорослей впервые наблюдаются различные
сочетания ядерных фаз. Обычно у них преобладает гаплоидная
фаза (гаметофит) над диплоидной (спорофит), но у более слож-
ных нитчатых водорослей уже наблюдается обратное явление.
Поэтому с выходом на сушу дальнейшее сокращение гамето-
фита вплоть до половых клеток явилось одним из важнейших
факторов завоевания различных ареалов с более жесткими ус-
ловиями обитания, чем в водной среде.
Происхождение высших растешгй из нитчатых водорослей
является наиболее вероятнъш. Предками этих первичных на-
земных растений могли быть зеленые нитчатые водоросли с
многорядными нитями. Большинство из них прикреплялось к
субстрату (камням) с помощью специальных приспособле-
ний — ризоидов, которые являлись продолжением главной оси
растения (рис. 6.11).
Древнейшие остатки растений обнаружены в раннедевон-
ских отложениях. Эти растения относятся к классу риниопсида
(псилофиты). Для представителей этой группы древнейших
высших растений характерны дихотомическое (вильчатое) вет-
вление и отсутствие листьев. Вместо корней у них были ризо-
моиды, вместо корневых волосков — ризоиды. Корней у ри-
ниопсидов еще не было. Они возникли впоследствии из
побегов. У таких древнейших растений обнаружена микориза
2%
Атмосфера
Вода
Плазмодесмы
Зачаточные
межклеточники
Вторичные
ризоиды
Первичные
ризоиды
Твердый
субстрат
Ветвление
б
Рис. 6Л1. Зеленая нитчатая водоросль
как элементарное высшее растение:
а — кладофора; б — хетоморфа
(гриб-симбионт), что нашло отражение в дальнейшей эволю-
ции высших растений. Деление клеток и рост риниопсидов
верхушечный. Вторичного роста еще нет. Расположение спо-
рангиев тоже верхушечное. Устьичный аппарат лишь в зача-
точном состоянии. Для них характерно чередование гамето-
фита и спорофита с преобладанием последнего. Именно из
299
риниофитных растений в процессе эволюции возникло боль-
шое количество разнообразных наземных форм, в том числе
сосудистые растения (рис. 6.12).
Происхождение красных и бурых водорослей. Это своеобраз-
ные группы водорослей как по морфологическому строению
слоевища, так и по биохимическому составу. Прежде всего
следует подчеркнуть, что они — эукариоты. Главным отличием
этих водорослей является наличие в них кроме хлорофилла А
каротиноидов и дополнительных пигментов: в красных — фи-
коэритрина и фикоцианина, а в бурых — фукоксантина. Эти
типы растений имеют значительную филогенетическую обо-
собленность. У них не прослеживается переход от одноклеточ-
ной организации к многоклеточной. Вместе с тем в эволюци-
папоротники
Мезозойские
.семенные
папоротники
Водорослевые предки
высших растений
Рис. 6.12. Схема эволюции высших растений
300
онной цепи многоклеточные водоросли (бурые и багрянки)
зашли в тупик, достигнув значительного расцвета и высокой
степени дифференциации в пределах типа растительного цар-
ства. Можно лишь только предположить их эволюционное
родство с низшими представителями зеленых водорослей.
Происхождение мохообразных. Мохообразные происходят
от водорослей, у которых гаметофит преобладает над спорофи-
том. Гаметофит эволюционировал из простертой части много-
нитчатого таллома, а спорофит — из вертикальной. Данная
группа растений стоит особняком в филогенетическом отно-
шении. Это слепая ветвь эволюции. Возникновение мохооб-
разных происходило в силуре.
Происхождение плауновидных. Среди современных высших
растений плауновидные — одна из древнейших групп. Они воз-
никли в середине девона, отчленившись от растений, близких к
риниофитам, которые имели антеридии с двухжгутиковыми спер-
матозоидами. Процветали плауновидные в палеозое, когда суще-
ствовали их гигантские древовидные формы (лепидодендроны).
Происхождение хвощевидных. Появились хвощевидные в
девоне и процветали от карбона до триаса. Несмотря на чле-
нистость стеблевых органов и колосков, они по многим при-
знакам напоминают риниофиты. Вероятно, древние хвощи
произошли от растений, близких к риниофитам, имевших
многожгутиковые сперматозоиды в антеридиях. Древовидные
хвощи (каламиты) наряду с древними плауновидными образо-
вали леса каменноугольного периода.
Происхождение папоротникообразных. Древнейшие ископа-
емые формы известны с девона. В карбоне крупные древовид-
ные папоротники наравне с другими споровыми растениями
составляли обширные влажные леса, остатки которых уча-
ствовали в образовании залежей каменного угля. Наряду с воз-
никновением типичной сосудистой системы у папоротников
появились также листья и корни. По своему происхождению
(от древних риниофитов). папоротники имеют значительно
больше родства с хвощевидными, чем с плауновидными.
Происхождение голосеменных. Первыми голосеменными
растениями стали семенные папоротники, которые размно-
жались уже не спорами, а семенами. Важнейшая особенность
семени — наличие в нем зародыша, защищенного от небла-
гоприятных условий среды и мутагенных факторов, а также
запасных веществ — белков; углеводов и жиров, необходимых
для роста зародыша. Оплодотворение у семенных папорот-
ников происходило уже не в водной среде. Это дало им воз-
можность избежать массового вымирания сначала в пермском
301
периоде, а затем в триасовом периоде, когда климат стал бо-
лее сухим. Таким образом, расцвет голосеменных относится к
концу палеозоя и мезозою — эпохе горообразования, подня-
тия материков и проявления магматизма.
Происхождение покрытосеменных. В середине мелового пери-
ода (около 120 млн лет тому назад) по не вполне понятным при-
чинам наступила вспышка видообразования цветковых растений,
в результате чего к концу данного периода покрытосеменные за-
воевали сушу. Господство покрытосеменных продолжается в те-
чение всей кайнозойской эры до настоящего времени. Важней-
шая особенность покрытосеменных растений — возникновение
цветка, открывшего возможность перекрестного оплодотворения
путем переноса пыльцы сначала ветром, а затем и насекомыми.
Цветок в процессе эволюции возник из сильно укороченного ли-
стостебельного побега. Чашелистики, лепестки, тычинки и пес-
тик — это видоизмененные листья, которые у предков цветковых
растений окружали органы размножения. Крупным эволюцион-
ным достижением цветковых растений является двойное опло-
дотворение, открытое в 1989 г. С. Г. Навашиным. Вероятным
предком покрытосеменных считают семенные папоротники,
обитавшие в мезозойской эре.
Филогения животных
Наиболее известна науке история животных. Существует
точка зрения, что губки возникли от какого-то одного предка,
принадлежащего простейшим, а остальные многоклеточные
животные — от какого-то другого. Самые крупные категории,
на которые делятся животные, — это типы; на протяжении гео-
логического времени возникло по меньшей мере 35 типов, из
которых 26 существуют до сих пор, а 9 вымерли (рис. 6.13).
Происхождение простейших. Тип простейших произошел от
жгутиконосцев, давшим начало голым, а затем раковинным кор-
неножкам, а также, возможно, инфузориям. Что касается споро-
виков, то часть их происходит от амеб, а другие формы этого
класса непосредственно от жгутиконосцев. Одиночные жгутико-
вые дают начало колониальным формам, причем потомками ша-
рообразных колоний жгутиковых являются кишечнополостные,
а из других колониальных форм, жгуты которых окружены плаз-
матическими воротничками, развились губки (рис. 6.14).
В настоящее время традиционной точке зрения о происхож-
дении инфузорий от жгутиконосцев можно противопоставить
представление о возникновении инфузорий от многоклеточных
плоских червей. В ходе развития клеточные границы исчезли, а
302
20
1. PORIFERA (губки)
2. CNI DARIA (актинии, медувы,
кораллы)
3. CTENOPHORA (гребневики)
4. PLATYHELMiNTNES (плоские черви)
5. NEMERTINA (немертины)
8. ACANTHQCEPHALA (скребки)
7. ROTIFERA (коловратки)
8. GASTROTRICHA (брюхоресничные
черви)
9. KINORHYNCHA (хинориыхи)
10. NEMATODA (круглые черви)
11. PRIAPUUDA (приалулиды)
12. ENTOPROCTA (стебельчатые
животные, снабженные
щупальцами)
13. ANNELIDA (дождевые черви,
сабеллиды)
14. ARTHROPODA (насекомые,
ракообразные, морские жалуди)
15. SIPUNCUUDA (сипункулиды)
16. ECHIUROIDEA (ехиуриды)
17. MOLLUSCA (улитки, осьминоги,
кальмары)
18. PHORONIDA (форониды)
19. BRACHIOPODA (плеченогие)
20. ECTOPROCTA (мшанки)
21. CHAETOGNATHA (щетинкочелюстные)
22. POGONOPHORA
23. ECHINODERMATA (морские звезды,
морские ежи)
24. HEMICHORDATA (баданоглосс)
25. UROCHORDATA (асцидии)
26. CHORDATA (ланцетник, рыбы,
амфибии, рептилии, птицы.
млекопитающие)
Рис. 6.13. Ныне живущие типы животных (сгруппированы
в соответствии с планом их строения; для каждого типа
приведено изображение одного из его представителей)
соматические ядра слились в единый макронуклеус, обладаю-
щий сверхвысокой плоидностью. Гипотеза о происхождении
инфузорий от многоклеточных животных, утерявших клеточ-
ные границы, была высказана в 1924 г. И. И. Месяцевым.
В пользу данной гипотезы можно привести следующие ар-
гументы.
303
ЖГУТИКОНОСЦЫ
Рис. 6.14. Филогения простейших
1. Имеется некоторая аналогия между половым процессом
у инфузорий и плоских червей. Спаривание плоских червей
внешне напоминает конъюгацию инфузорий.
2. У мелких форм турбеллярий, как и у инфузорий, движе-
ние осуществляется преимущественно за счет ресничек. У ин-
фузорий неоднократно обнаруживали многоклеточность, а
также нервные узлы и нервные пучки, что является особеннос-
тью многоклеточных организмов.
Происхождение губок и кишечнополостных (рис. 6.15). Как
уже сказано, губки и одиночные гидроиды происходят от жгу-
тиконосцев. Одиночные гидроиды дают начало колониальным
формам, а от последних развиваются сцифоидные и гребневи-
ки. Гребневики образуют переход к ресничным червям, т. е.
к трехслойным животным. Об этом свидетельствует появление
впервые у гребневиков следующих прогрессивных признаков.
1. Развитие между экто- и энтодермой зачатка третьего за-
родышевого листка —мезодермы. За счет этого зачатка у
взрослых форм в студенистом веществе мезоглии развиваются
многочисленные мышечные элементы.
2. В связи с переходом от плавания к ползанию впервые
обособились вентральная и дорзальная стороны тела.
3. Впервые появляются элементы двусторонней (билате-
ральной) симметрии.
4. Нервные клетки начинают концентрироваться в нервные
узлы (ганглии).
Происхождение червей (рис. 6.16). Ресничные черви дают
начало плоским (сосальщикам и ленточным), круглым и
кольчатым червям. Из последних многощетинковые черви
304
КИШЕЧНОПОЛОСТНЫЕ
Жгутиконосцы
Рис. 6.15. Филогения губок и кишечнополостных
Рис. 6.16. Филогения червей
имеют наиболее примитивное устройство, и от них происхо-
дят более поздние и специализированные формы (многоще-
тинковые и пиявки). От многощетинковых кольчецов также
обособились два новых типа животных — членистоногие и
моллюски.
305
Происхождение моллюсков. Ближе всех к кольчецам — '•
класс боконервных моллюсков; более высокодифференциро-
ванные — брюхоногие и пластинчатожаберные; высший класс •
составляют головоногие (осьминоги и каракатицы).
Происхождение членистоногих (рис. 6.17). Весьма специа- -
лизированный тип членистоногих, происходя от кольчатых ;
червей, очень давно разделился на две ветви. Одна из них .
имеет исходной формой ископаемых — трилобитов, от кото- i
рых произошли, с одной стороны, ракообразные, с другой —
ракоскорпионы, давшие начало паукообразным. Другая ветвь
ведет к насекомым через древнюю форму первичнотрахей-
ных, отдельные представители которых сохранились до на-
шего времени, и через филогенетически более поздних мно-
гоножек (рис. 6.18).
Происхождение иглокожих (рис. 6.19). Предки иглокожих
были двусторонне симметричными животными, перешедши-
306
Рис. 6.18. Происхождение насекомых (схематично):
1 — кольчатые черви; П — параподии; Р — рот* а — головной конец; 2 —
смещение 1, 2, 3 сегментов-члеников кпереди, сегменты 1+2+3 образовали
сросшуюся воедино голову. Их придатки стали челюстями; 4 — насекомые;
позади головы обособляются 1, 2 и 3 членики, образуя грудь. Остальные
образовали брюшко. На нем остатки конечностей (Ц) и яйцеклад (Я), иногда
становящийся жалом
°ис. 6.19. Филогения иглокожих
307
ми потом к сидячему образу жизни; к этому времени (начало
палеозойской эры),относится и возникновение лучевой сим-
метрии. Современные иглокожие в большинстве случаев ведут
свободный образ жизни (кроме морских лилий) и сохраняют
лучевую симметрию (кроме сердцевидных морских ежей и го-
лотурий, у которых наблюдается частичный возврат к двусто-
ронней симметрии). Наиболее древними из известных пред-
ков иглокожих являются шаровики (начало палеозоя); от них
возникли морские бутоны (каменноугольный период), а от
близких к последним форм пошло постепенное усложнение
организации через морских лилий к морским звездам, офиу-
рам и морским ежам. Голотурии отделились от общего ствола
иглокожих очень рано, происходя непосредственно от шаро-
виков.
Филогения хордовых
Происхождение первых позвоночных (рис. 6.20). Предками
позвоночных были наиболее просто организованные беспоз-
воночные из всех существовавших в прошлое время. Это про-
стые, сидячие формы, напоминающие ныне существующих
глубоководных морских животных, относящихся к классу
крыложаберных. Хотя крыложаберных относят К типу хордо- ’
вых, тело этих животных, прикрепленное ко дну с помощью
стебелька, в основном состоит из пищеварительного тракта.
Вокруг рта находятся щупальца, служащие для захвата пище-
вых частиц, приносимых водой. Дальнейшее развитие этих
простейших существ, по-видимому, завершилось появлени-
ем жаберных щелей, которые представляют собой парные от-
верстия, открывающиеся по бокам тела и ведущие из глотки
наружу.
Первоначально жаберные щели служили фильтрами для
задержания пищевых частиц, в дальнейшем же они превра-
тились в аппарат для дыхания. На этой стадии эволюци-
онного процесса наши предки, должно быть, напоминали
современных оболочников. Оболочники, как сидячие, пита-
ющиеся за счет фильтрования воды формы, представляют
собой эволюционный тупик. Что же касается их личинок, то
в процессе эволюции неоднократно имело место явление
так называемого педоморфоза, когда взрослые особи элими-
нировались, а личиночные формы становились полово-,
зрелыми и способными к воспроизведению себе подобных.
Вполне вероятно, что в те древнейшие времена личинки
животных, напоминавших оболочников, имели жаберный
308
Рис. 6.20. Схема, показывающая возникновение первых
позвоночных из личиночной формы животных, напоминающих
оболочников, в результате педоморфоза
аппарат на уровне глотки; мускульный хвост, усиленный
толстой гибкой продольной струной; дорзальный нервный
тяж, который в головном отделе получал сенсорно инфор-
мацию от рудиментарных органов чувств. Первые хордовые,
возникшие путем педоморфоза, по всей вероятности, мало
отличались от современных ланцетников. Ископаемые ос-
татки небольших примитивных хордовых, не имевших щу-
палец и челюстей, обнаружены в силурских отложениях.
В девонском периоде уже в изобилии появляются первые
позвоночные, представленные рыбообразными бесчелюст-
ными животными.
Рыбы, давшие начало наземным позвоночным. Первым
большим достижением ранних позвоночных было развитие
челюстей. Однако и до наших дней дошли такие реликты,
как круглоротые (миноги, миксины). Основные пути эволю-
ции рыб сопровождались усовершенствованием челюстей с
современным развитием жаберных крышек, защищающих
жабры, и окостенением скелета. Так появился класс костис-
тых рыб.
В середине девона костные рыбы дивергируют в двух раз-
личных направлениях, различаясь главным образом строе-
нием плавников. У одних плавники не имели покрытых че-
309
шуей лопастей основания, у других основания плавников
напоминали мясистую лопасть. У последних передняя пара
плавников, расположенных на груди, могла легко трансфор-
мироваться в переднюю пару конечностей наземных. Зад-
няя пара плавников, расположенная позади средней части
туловища, также могла превратиться в заднюю пару конеч-
ностей. Совершенно очевидно, что только примитивные
костные рыбы девонского периода, обитавшие в мелких,
периодически пересыхавших внутриматериковых водоемах,
были существами достаточно «земноводными» для того, что-
бы дать начало наземным позвоночным (рис. б Л). В классе
рыб представители лопастеперых оказались, однако, не осо-
бенно преуспевающими. В настоящее время только три
рода двоякодышащих рыб, которые до сих пор обитают в
мелких пресных водоемах, и один вид латимерии, живущих
в океане, являются представителями этой эволюционной
ветви (рис. 6.22).
Происхождение амфибий. Первыми амфибиями, ведущими
свое происхождение от вида, близкого к костным рыбам, были
ихтиостеги. У них имелись уже хорошо развитые плечевой и
тазовый пояса, с помощью которых они управляли расчленен-
ными передними и задними конечностями. Такие конечности
вполне моти обеспечить передвижение животного по суше.
Развитие амфибий в каменноугольном и пермском периодах
было следующим.
1. Развитие прочной кожи, часто покрытой костными плас-
тинами, предохраняющими от потери влаги в результате испа-
рения.
2. Укрепление позвоночного столба вследствие формиро-
вания цельных позвонков, которые с помощью мышц и свя-
зок образуют прочный горизонтальный ствол, поддерживаю-
щий тело.
3. Усовершенствование дыхательной системы путем обра-
зования хорошо развитых носовых ходов, которые открыва-
ются ноздрями, расположенными на дор&альной поверхно-
сти черепа.
Одни из самых первых потомков ихтиостег, известных под
общим названием Эмболомеры, уже в раннем карбоне опреде-
лились в направлении развития как будущие рептилии. Тем
временем основная ветвь амфибий (так называемые Лабирин-
тодонты, среди них Рахитомии) развивались в направлении
современных, ныне живущих лягушек и жаб. Предки же хвос-
татых земноводных (тритонов, саламандр, протеев) оказались
совсем иного происхождения. С самого начала эволюции ам-
310
Рис. 6.21. Возникновение и развитие позвоночных
(снизу вверх):
а — кнстеперые рыбы, выходящие на сушу; €—первые
земноводные;# — первые пресмыкающиеся,
уже откладывающие яйца на суше
ЗН
Рис. 6.22. Деление девонских рыб на лучеперых
и лопастеперых
фибий существовала небольшая ветвь, которая резко отлича-
лась от основной ветви, получившей общее название Лепос-
пондилы (рис. 6.23). Современные хвостатые амфибии как раз
и представляют собой потомков древней группы Лепоспон-
дилов.
Появление высших позвоночных — рептилий и птиц. Во
время каменноугольного периода произошло крупнейшее со-
бытие в эволюции наземных позвоночных — животные при-
обрели ряд приспособлений к развитию яйца вне водной
312
Рис. 6.23. Филогения амфибий
среды. Первыми животными, у которых выработался ряд та-
ких приспособлений, быди рептилии. Так, благодаря появле-
нию яиц с зародышевыми оболочками отпала необходимость
в личиночной стадии в процессе онтогенетического разви-
тия. Все зародышевые структуры заключены в скорлупу, дос-
таточно прочную для того, чтобы защищать содержимое
яйца, и в то же время достаточно пористую для того, чтобы
обеспечить доступ кислорода в яйцо и выделение из него уг-
лекислого газа.
Превращение одной из наиболее древних лабиринтообраз-
ных амфибий {Эмболомеры) в рептилий {Котилозавров) про-
изошло в конце пермского периода. На первом этапе диверген-
313
ции потомки Котилозавров образовали пять подклассов, три
из которых нас интересуют особенно: Диапсида, Анапсида и
Синапсида. Из этих трех подклассов Диапсида была преуспева-
ющей ветвью рептилий (рис. 6.24). Они характеризовались на-
личием двух задних височных ям, разделенных заглазничной
и чешуйчатой костями.
В начале мезозойской эры (триас) ветвь подкласса Диапсид
представлена была так называемыми архозаврами. История
эволюции этой ветви рептилий, кульминацией которой было
появление динозавров, достаточно хорошо известна. Репти-
лии, принадлежащие к подклассам Анапсида и Синапсида,
занимали весьма скромное положение. Кроме динозавров ар-
хозавры дали также начало новому классу позвоночных — пти-
цам. Как класс птицы произошли от могущественной ветви
>5
О
О
W
ф
Котилозавры (первые рептилии)
I Амниоты
Эмболомеры (амфибии)
&
Q
Д
8
Рис. 6.24. Три подкласса рептилий и их потомки
314
архозавров, когда последние находились в зените своего рас-
цвета. Может показаться, что данный случай противоречит
общему правилу, согласно которому только примитивные
виды какого-либо класса делают большой скачок и дают нача-
ло новому, более совершенному классу. Следует сказать, одна-
ко, что процесс возникновения птиц из рептилий не сопро-
вождался появлением каких-либо крупных новшеств. Поэтому
не удивительно, что птиц часто называют пернатыми репти-
лиями 0$ис. 6.25).
Ныне живущие ящерицы и змеи, а также крокодилы и ал-
лигаторы произошли от рептилий, относящихся к подклассу
Диапсида. Напротив, черепахи являются потомками Анапси-
да. В отличие от Диапсида у Анапсида отсутствуют височные
ямы позади глаз.
Рис. 6.25. Происхождение птиц. Чешуя (Ч) превращается
.в перья и пух, сохраняясь на ногах:
/ — древние ящерицы; 2—3 — лазающие формы; 4 — археоптерикс; 5 —
птицы (правая сторона без перьев, чтобы показать остатки пальцев (П)
и укоротившийся хвост (Хв)
315
. Появление млекопитающих. Представители подкласса Си-
напсида так и не достигли господствующего положения как
рептилии. Непосредственно продолжая линию примитивных
рептилий, эта группа дала начало первым млекопитающим.
Именно они являются соединяющим звеном примитивных
рептилий, с одной стороны, и млекопитающих — с другой
(рис. 6.26).
Характерной чертой подкласса было наличие одной височ-
ной ямы, ограниченной заглазничной и чешуйчатой костями.
В отличие от Диапсида представители Синапсида на протяже-
нии всей своей эволюции сохранили горизонтальное положе-
ние тела и передвигались с помощью четырех конечностей.
Рис. 6.26. Происхождение млекопитающих (1—4)
от звероящеров до первых яйцекладущих млекопитающих:
В — расположение волос, растущих между чешуйками. Видно, как чешуя
постепенно заменялась волосяным покровом; М — расположение млечных
желез
316
Среди Синапсид наиболее близким к млекопитающим были
представители, известные под общим названием Териодонты.
Позднее появилась группа рептилий, которая служила проме-
жуточным звеном между прогрессивными Териодонтами и
примитивными млекопитающими. К таким рептилиям отно-
сились Ихтиозавры. Несмотря на то что эта группа рептилий
как бы заложила фундамент для последующего развития и
триумфа млекопитающих уже в раннем мезозое, прошли мно-
гие миллионы лет, прежде чем наступил век млекопитающих.
Ныне живущие млекопитающие принадлежат к трем под-
классам: Однопроходные (Яйцекладущие, или Клоачные),
Сумчатые и Плацентарные (рис. 6.27).
Рис. 6.27. Схема, иллюстрирующая происхождение
однопроходных, сумчатых и плацентарных млекопитающих
от звероподобных рептилий
317
Происхождение приматов (рис. 6.28). Самыми первыми и ,
наиболее примитивными плацентарными млекопитающими
были насекомоядные, и, очевидно, именно они были родо-
начальниками всех других плацентарных. От этой ветви при- '
митивных насекомоядных произошло 28 отрядов плацентар-
ных млекопитающих, 11 из которых к настоящему времени
вымерли.
От какого-то гипотетического насекомоядного предка,
обитавшего на деревьях и напоминавшего тупайю, приматы
развивались в трех главных направлениях. Первая радиация
примитивных приматов в палеоцене (50—70 млн лет назад)
привела к возникновению подотряда, к которому относятся
лемуры, лори и долгопяты.
В результате второй радиации в раннем эоцене (примерно
40 млн лет назад) появился другой подотряд. К нему принад-
лежат все обезьяны Старого и Нового Света. Третья радиация в
раннем олигоцене (приблизительно 35 млн лет назад) привела
3
2
х
а
л
о
® 3
©
Q
Лемуры
5"
С ъ
Питекантроп
(Homo erectus)
_ -1 млн
ч. Австралопитек
ДриоЛигек
-11 млн
Лимнрпитек
^Проконсул
Гоминиды
______" I -26 млн
З4 Парапитек Проплиопитек
Антропоиды
Насекомоядные
-36 млн
Первые приматы
Рис. 6.28. Схема родословного древа приматов
318
к образованию надсемейства Гоминид. Предок Гоминид про-
плиопитек обладал тенденцией к увеличению тела и мозга.
Следом за проплиопитеком появились лимнопитеки и про-
консул. Лимнопитеки, по-видимому, представляли собой бо-
ковую ветвь, которая дала начало современным гиббонам, тогда
как менее специализированный проконсул стал предком со-
временных Гоминид. Потомками проконсула были различные
дриопитеки. Эти обезьяны, вероятно, напоминавшие по
внешнему виду шимпанзе, были предками не только совре-
менных шимпанзе, горилл и орангутанов, но, возможно, и нас
самих.
АНТРОПОГЕНЕЗ
Современная антропология выделяет несколько этапов ант-
ропогенеза.
I этап — предшественники человека. В основании развития
линии приматов, приведшей к возникновению Человека, нахо-
дятся австралопитеки (южная обезьяна). Находки австрало-
питеков сделаны на территории главным образом Южной Аф-
рики.
Австралопитеки — достаточно крупные обезьяны (масса
приблизительно 20—50 кг, рост 120—150 см), ходили на двух
ногах при выпрямленном положении тела. Масса мозга была
близка к 550 г. Зубы расположены в виде широкой, как у чело-
века, а не узкой, как у обезьяны, дуги. Клыки у них были не-
большие. Австралопитеки — обитатели открытых пространств,
жившие в пещерах. Они широко использовали различные кос-
ти животных, в качестве ударных орудий применяли палки,
камни. Австралопитеки вымерли до окончания плейстоцена
(т. е. они жили в период от 9 млн до 750 000 лет тому назад).
Среди прогрессивных австралопитеков выделяется Человек
умелый (Homo habilis). Масса мозга у него была больше, чем у
среднего австралопитека (650 г). Первый палец стопы Челове-
ка умелого не был отведен в сторону, как у обезьяны; концевые
кисти фаланг были короткими и плоскими, как у настоящего
человеческого существа. Человек умелый уже мог изготовлять
грубые галечные орудия. Это произошло 2,6 млн лет тому на-
зад. Значение находки галечных орудий огромно, так как тру-
довая деятельность является границей, отделяющей Человека
от остальной природы.
II этап — древнейшие люди (архантропы). Внешне архантро-
пы были уже весьма похожи на современного человека. Масса
319
мозга у них составляла около 800—1000 г. Именно при такой!
массе мозга становится возможным овладение речью. У архан-|
тропов прогрессивно развивалась способность к производств^!
орудий труда. Средний рост мужчин был около 160 см, а жен*!!
щин — 150 см. С зоологической точки зрения эти формы, ви-
димо, относятся к одному большому виду — Человеку выпрями-
ленному (Homo erectus).
Занимая промежуточное положение между современным!
человеком, с одной стороны, и австралопитеком, с другой сто-'
роны, архантропы начали быстро расселяться по Африке, Сре- j
диземноморью, Южной, Центральной и Юго-Восточной;
Азии. Вероятно, в этот же период начал использоваться огонь. |
Расселяясь и попадая в новые условия существования, архант- •
ропы образовали отдельные изолированные формы. Сейчас •
изучено довольно значительное число форм древнейших лю-
дей. Наиболее известные: питекантроп (о. Ява), синантроп
(Китай), гейдельбергский человек (Средняя Европа), атлант-.
роп (Алжир), телантроп (Южная Африка), олдовайский пите- ’
кантроп (Центральная Африка).
III этап — древние люди (палеонтропы). Древние люди за-
нимают промежуточное положение между древнейшими
людьми — архантропами и ископаемыми формами Человека
разумного — как по строению тела, так и по развитию культу-
ры. По месту первого описания таких людей5 (долина Неан-
дерталь близ Дюссельдорфа) их часто называют неандерталь-
цами. Неандертальцы имели сравнительно небольшой рост
(155—165 см). Масса их мозга составляла около 1500 г., при-
чем сильное развитие получили отделы, связанные с логичес-
ким мышлением. Орудия неандертальцев — ручные рубила,
остроконечники и скребки — были более совершенными,
чем примитивные обработанные гальки.
Увеличение числа находок позволило выяснить, что неан-
дертальцы были весьма неоднородной группой. Они подразде-
лялись на две главные расы: поздние неандертальцы и ранние
неандертальцы. Поздние неандертальцы выглядели
звероподобными, однако масса их мозга превышала массу
мозга современного человека (1600 г). Среди них были развиты
внутригрупповые связи и на охоте, и при защите от врагов, и от
неблагоприятных погодных условий. Представители поздних
неандертальцев очень небольшими группами могли выживать
и побеждать в борьбе за существование. Ранние неан-
дертальцы оказались на совершенно ином эволюционном
пути — они шли к победе в борьбе за существование через
объединение сил отдельных индивидов, что привело 50—
320
40 тыс. лет назад к возникновению того вида, к которому при-
надлежим и мы с вами, — Человека разумного (Homo sapiens).
IV этап — современные люди (неантропы). Такие качества не-
адтропов (кроманьонцев), как развитие речи, интеллект, кол-
лективная трудовая деятельность, привели в процессе дальней-
шего развития к возникновению человеческого общества.
Существуют две основные точки зрения на происхождение
человека современного типа. Первая точка зрения — гипотеза
полицентризма, вторая — гипотеза моноцентризма. Согласно
первой гипотезе Человек разумный возник в нескольких мес-
тах планеты из разных предковых форм, принадлежащих к па-
леонтропам (или даже к архантропам), согласно второй — су-
ществовало единое место возникновения человечества из
какого-то общего предкового ствола. В последнее время полу-
чила всестороннюю аргументацию гипотеза широкого моно-
центризма Я. Я. Рогинского. Согласно этой гипотезе человек
возник в одном месте, после чего началось широкое и активное
расселение возникших неантропов по планете — они быстро
захватили Переднюю Азию, Европу, Центральную и Южную
Африку.
Дифференциация Человека разумного на расы. Все совре-
менное человечество принадлежит к единому полиморфному
виду — Человек разумный. Единство человечества основано на
общности происхождения, социально-психического развития,
на неограниченной способности к скрещиванию людей раз-
личных рас, а также на практически одинаковом уровне об-
щего физического и умственного развития представителей
всех рас.
Вид Человек разумный распадается на три большие расы —
европеоидную, монголоидную и негроидную. В пределах каждой
большой расы выделяют еще малые, или вторичны'е,
расы. Между большими расами существуют переходные
(рис. 6.29). Механизм образования рас человека таков же, как
и механизм возникновения внутривидовых группировок у жи-
вотных.
Происхождение и развитие рас представляет собой очень
сложный исторический процесс. Около 100 тыс. лет назад на
Азиатском континенте с прилегающими к нему областями Аф-
рики и Европы сформировались две расовые группы — юго-
западная и северо-восточная.
Эти группы разделялись горными цепями Гималаев и Гин-
дукуша и хребтом Индокитая. Первая из этих ветвей позднее
дала начало европеоидной и негроидной большим расам. Вто-
рая же сформировала монголоидную расу, распавшуюся на ряд
II **281
321
Рис. 6.29. Схема взаимоотношения
современных рас человека
мелких ветвей, представители одной из которых, американс-
кой, проникли в районе Берингова пролива из Азии в Север-
ную Америку и затем заселили весь Новый Свет.
Различия между расами касаются лишь второстепенных
признаков, обычно связанных с частными приспособлениями
к конкретным условиям существования. Сторонники расиз-
ма в своем утверждении о биологической неравноценности
рас исходят главным образом из происхождения их от разных
предков. Они считают, что европеоидная раса ведет свое на-
чало от кроманьонцев, тогда как все остальные связаны в |
своем происхождении с неандертальцами. Однако эта кон-
цепция основана на извращении научных данных. Палеонто-
логия все больше и больше накапливает факты, свидетель-
ствующие о том, что прародина современного человека зани-
мала обширную территорию, но без резко обозначенных изо-
лированных центров формирования рас, как это утверждает
теория полицентрического расообразования. Труды ученых
подтверждают, что расовые особенности не играют роли в
развитии психической деятельности человека. Далее история
учит, что человеческая культура не есть результат умственно-
го развития какой-нибудь одной расы, она возникла в про-
цессе многовековой трудовой деятельности всего человече-
ства.
Факторы антропогенеза
1. Прямохождение. Предпосылкой для превращения пред-
человека в человека было такое изменение образа его жизни,
при котором он начал отвыкать от помощи рук при ходьбе, ус-
ваивая прямую походку. Рука стала свободной и могла теперь
приобретать новые навыки.
2. Труд. Первое основное условие всей человеческой жизни.
Человек имеет орган труда — руку; у животных, в том числе
обезьян, их конечности служат лишь органами передвижения.
В процессе трудовой деятельности человек производит все не-
обходимое для своей жизни. Начало трудовой деятельности,
связанное с использованием передних конечностей, относится
к периоду 3—2,5 млн лет тому назад.
3. Открытие и использование огня. Это событие привело к
активному потреблению мясной пищи, без которой человек не
мог стать человеком. Дело в том, что человеку, как и другим
млекопитающим, нужны незаменимые аминокислоты, синте-
зировать которые его организм не может (например, метионин,
' триптофан). Содержание незаменимых аминокислот в расти-
тельной пище далеко не оптимально. Чтобы получить необхо-
димую норму, нужно съесть очень много растений. Жизнь в
этом случае превращается в непрерывное поедание, ни на что
другое времени не остается.
4. Стадный образ жизни. Способность выполнять несколько
важных функций. Члены стада объединяются для удержания
территории и защиты от хищников. Кроме того, что очень важ-
но, стадо создает ту среду, в которой происходит передача опы-
та друг другу и обучение детенышей другого рода жизненно
важным навыкам.
5. Возникновение речи. Потребность в передаче информации
и координации деятельности отдельных особей в стадах при-
вело к развитию языка. Обучение языкам требует не только
наличия природных интеллектуальных особенностей, но и ,
времени (особенно времени периода развития, предшествую- .
щего половой зрелости).
Человекообразные обезьяны, возможно, обладают способ-
ностями, необходимыми для того, чтобы выучиться основам
человеческого языка, но не могут реализовать эти способнос-
ти, так как слишком быстро достигают зрелости.
6. Развитие высшей нервной деятельности. Трудовая деятель-
ность и речь .вместе со стадным образом жизни за последние
несколько сотен тысяч лет привели человека к наращиванию ,
массы головного мозга. Развитие мозга повысило способность
к обучению и в свою очередь оказало влияние на трудовую дея-
тельность людей.
7. Развитие альтруистических наклонностей. Трудовая дея-
тельность человека требовала передачи накопленного опыта
своим современникам, объединяя усилия отдельных индиви-
дов в процессе труда, охоты и защиты. Происходила грандиоз-
ная переоценка ценностей.
Преимущества перед другими получили те племена, которые
«догадались» позаботиться о стариках и поддерживать физи-
чески не таких, сильных, но полезных своими умственными
способностями особей (Мастера по выделке орудий, хорошие ,
следопыты и наблюдатели и т. п.). Так возникла сложная
структура стада, племени, орды. Так возникло Человечество,
т. е. Человек вышел за рамки биологических законов, сделав
возможным освоение окружающей среды (приручение живот-
ных и окультуривание растений), а также создание искусства.
КУЛЬТУРНАЯ эволюция
Эволюция человека состоит из двух процессов: органичес-
кой эволюции и культурной эволюции. Направление органи-
ческой эволюции приводится в движение длительным отбо-
ром. Культурная эволюция имеет собственные движущие
силы, но, однако, в рамках естественного отбора. В эпоху
культурной эволюции естественный отбор продолжает дей-
ствовать и органическая эволюция не прекращается, только в
конкурентную борьбу (отбор) вступают такие признаки, как
интеллект, способность к обучаемости, восприятию культуры
и т. п.
К факторам культурной эволюции относятся:
а) общее накопление культурного наследия;
324
б) тенденции в развитии культуры, возникающие в резуль-
тате конкуренции между сообществами, различающимися в
культурном отношении (но не генетически).
Принято считать, что культурная эволюция совершается
путем постепенного накопления культурного наследия. Снача-
ла происходит приобретение полезных знаний, затем эти зна-
ния включаются в общую массу знаний древнего общества,
после чего передаются последующим поколениям путем обу-
чения. Каждое следующее поколение добавляет к этому запасу
знаний то, что ему удается открыть самому. Новые знания пе-
редаются дальше тем же путем. Следовательно, культурное на-
следие имеет кумулятивный характер.
ЭВОЛЮЦИОННОЕ УЧЕНИЕ
ЭТАПЫ РАЗВИТИЯ ЭВОЛЮЦИОННОГО УЧЕНИЯ
4
Развитие эволюционной идеи до XIX века
Мыслители древности, находясь еще под значительным
влиянием мифов, создавали теории происхождения и развития
жизни, в которых элементы фантазии переплетались с наблю-
дениями. Разумеется, их воззрения очень далеки от современ-
ных представлений. Коротко остановимся'на представлениях
некоторых древних ученых, касающихся вопроса о происхож-
дении и дальнейшей судьбе живых существ.
Фалес из Милета (V в. до н. э., Греция) был первым, кто пы-
тался найти общий источник возникновения всего существу-
ющего. Этим общим источником Фалес считал воду, что дало
повод объявить его предшественником современных иссле-
дователей, также полагающих, что жизнь зародилась на дне
Океана.
Анаксимандр. Согражданин и современник Фалеса. Пола-
гал, что твердь сначала была покрыта водой и только постепен-
но начала высыхать. К этому времени на ее поверхности по-
явились люди, которые ранее жили в воде и напоминали рыб.
Анаксимен. Ученик Анаксимандра. Считал, что все суще-
ствующее возникло из воздуха. В частности, из воздуха появи-
лась земля и вода, их смешение образовало ил, из которого под
влиянием солнечного тепла произошли путем самозарождения
растения, животные и люди.
325
Гераклит (конец VI — начало V в. до н. э.). Основой мира
считал движение. «Все течет, — говорил Гераклит, — и ничто
не остается постоянным». Эта основная идея могла бы лечь в
основу действительно исторического взгляда на природу, но
она осталась неразвитой Гераклитом.
Эмпедокл (483—423 до н. э.). Все существующее, по Эмпе-
доклу, имеет в основе четыре первых физических принципа:
огонь, воздух, воду и землю. Этими принципами управляют
две силы: объединяющая — любовь и разъединяющая — нена-
висть. Борьба этих двух сил приводила к появлению различ-
ных комбинаций живых существ в растительном и животном
царствах.
Демокрит (460—370 до н. э.). Подверг сравнению строение
человеческого тела с устройством других живых организмов.
Демокрит обратил внимание на приспособление отдельных ор-
ганов к выполнению определенных функций; однако объясне-
ния этой упорядоченности и целесообразной организации он
не дал.
Анаксагор. Современник Демокрита. Полагал, что целесо-
образное устройство живых существ обязано наличию разум-
ного плана, источником которого является всемогущий дух,
или разум.
Платон (V—IV вв. до н. э.). Крупнейший древнегреческий
философ. Высказал мысль о существовании двух миров: мира
вещей, воспринимаемых чувствами, и мира идей, воспринима-
емых разумом, мышлением. Платоновское учение об идеях
оказало влияние на биологию, оно легло в основу представле-
ния об идеальном плане строения живых существ, к которому
их действительное устройство только приближается.
Аристотель (384—322 до н. э.). Организмы, по мысли Арис-
тотеля, отличаются от тел неживой природы присутствием
души, которая является исчерпывающей причиной их жизнен-
ных свойств. Наиболее примитивная — питающая — душа
имеется у растений. Животные помимо питающей души обла-
дают еще чувствующей, в силу чего они проявляют волю и
способны к движению. Человек сверх питательной и чувству-
ющей души обладает еще мыслящей душой, способной обра-
зовывать понятия и строить обобщения. Отметив черты разли-
чия и черты сходства, Аристотель не увидел в этом сходстве
родства, общности происхождения.
Парацельс (1493—1541). Выразителем биологических пред-
ставлений эпохи Возрождения был врач Теофраст фон Гоген-
гейм, известный в истории под именем Парацельса. Наряду с
совершенно фантастическими представлениями о жизненных
326
соках, имеющих различный состав, и учением об архее — вер-
ховном начале, управляющем всеми химическими процессами
в организме, Парацельс высказал мысль о наследственных ве-
щах, переходящих к детям от отца и матери. Та же мысль о
комбинировании наследственных вещей легла у Парацельса в
основу представлений о возникновении разнообразных форм
растений и животных.
Лейбниц (1646—1716). Основная идея Лейбница заключа-
ется в утверждении, что все тела состоят из частиц — монад,
обладающих индивидуальностью и жизненными свойствами.
Важнейшее жизненное свойстве монад заключается в том, что
каждая из них наделена силой и поэтому может действовать
сама по себе, не нуждаясь в побуждении извне. Характеризую-
щая монаду сила подчиняется закону причинности, но в то же
самое время она действует целесообразно, осуществляя и меха-
нические движения тел и гармонично их устраивая. Одним из
важнейших в философии Лейбница является закон непрерыв-
ности, заключающийся в том, что все тела природы образуют
ряд непрерывных переходов от одного к другому, без пропусков
и повторений. Так как, по представлениям Лейбница, монады
не возникают и не уничтожаются, то и развитие организмов
должно мыслиться как развертывание (эволюция) предше-
ствующих свойств, а смерть — как свертывание (инволюция)
этих свойств, переход их в скрытое состояние.
Шарль Бонне (1720—1793). Известен развитием лейбници-
анской идеи о постепенных переходах от одного тела природы
к другому, выраженной в виде лестницы существ (лестницы
тел природы), которые располагаются по принципу убываю-
щей высоты организации (человек — четвероногие — расте-
ния — кристаллы и т. д.).
Карл Линней (1707—1778). Славу великого ученого Линней
завоевал небольшим по объему трудом, озаглавленным «Систе-
ма природы», который он опубликовал в 1735 г. в Лейдене
(Голландия). В этой работе Линней упорядочил классифика-
цию органического мира (класс — отряд — род — вид — разно-
видность). Каждый вид получил два латинских обозначения:
первое, написанное с прописной буквы, обозначает род, вто-
рое — вид.
Линней первым дал точное описание видов и родов расте-
ний (около 10 000). Впервые указал на необходимость созда-
ния естественной системы, построенной с учетом всей сово-
купности признаков организма.
Жорж Бюффон (1707—1778). Заслугой Бюффона является
установление связи между организмами и средой их обитания.
Он излагал, правда, в несколько фантастическом виде, исто-
рию Земли и пытался сформулировать положение об измене-
нии живых существ в зависимости от изменений окружающе-
го их ландшафта. Однако представление об изменчивости
организмов под влиянием условий существования не вылилось ;
у Бюффона в учение об историческом развитии живого мира. .
Эразм Дарвин (1731—1801). Дед великого Чарлза Дарвина.
Автор четырехтомного труда «Зоономия», в котором он помес-
тил теоретические суждения об изменчивости видов, приспо-
собляемости, остаточных органах и т. п. Основой развития он
считал внутренние причины, определяющие направление и ха-
рактер развития жизни.
Развитие эволюциошюй вдеи в XIX веке
Жан Батист Ламарк (1744—1829). Ламарк считается, по
справедливости, первым эволюционистом. Превосходный ис-
следователь природы, Ламарк заметил, что существовавшие в
его время типы и семейства животных можно расположить в
некий ряд — от простейших организмов до самых сложных с
постепенным переходом от одних к другим. Легко видеть, что
непрерывный ряд организмов, связанный с понятием града-
ции, родствен лейбницианской идее, развитой Бонне, о лестни-
це существ. Наряду с этим между Ламарком и Бонне суще-
ствует принципиальное отличие в том отношении, что ряд
живых существ, по Ламарку, исторически изменяется — низ-
шие формы, усложняясь, превращаются в высшие. Эту теорию
он сформулировал в 1803 г. в книге «Философия зоологии».
Здесь Ламарк привел многочисленные доказательства медлен-
ного изменения видов животных эволюционным путем, соста-
вил первые родословные отдельных классов животного мира
(включая человека) в виде ступеней прогрессивной эволюции.
В этом трактате Ламарк излагает принципы естественной сис-
темы животных и основы первой эволюционной теории. Впер-
вые в мире на основе построения естественной системы живот-
ных Ламарк высказывает гениальную мысль о том, что процесс
л исторического развития живых организмов носит наступателд-
цый характер.
Основными положениями эволюционной теории Ламарка '
были следующие.
1. Ламарк утверждал, что Творец Вселенной создал лишь
материю и законы, постояннодействующие в природе и при-
водящие ее в движение. Бог дал первый толчок природе, после
чего уже не вмешивался в ее дела.
12Я
'2. В отличие от лестницы тел природы ввел принцип града-
ции — рассматривал ступени как результат прогрессивной эво-
люции, усложнения форм жизни.
3. Пришел к заключению, что в природе видов нет, суще-
ствует только непрерывная цепь изменяющихся особей, кото-
рую лишь искусственно в целях удобства классификации
подраздедшацта виды. Этот вывод был неверным. Видысуще-
ств^ют^еально^\и это не мешает признавать нам йх ^Нменяе-г
млеть v V V ч. - . X-
ч/~ЗгОбратй]I внимание на фактор времени, необходимый для
эволюции. Рассматривал эволюцию как длительный истори-
ческий процесс. По мнению Ламарка, эволюция животных и
растений протекает настолько медленно, что человек не заме-
чает происходящих изменений.
5. Ламарк признавал два фактора эволюции: один из них —
активное влияние внешней среды, которая изменяет и модифи-
цирует организмы в пределах каждой из ступеней развития
природы, второй — тенденция к совершенствованию, которая
обуславливает прогрессивное развитие организмов.
6. Ламарк неоднократно подчеркивал, что живые существа
построены целесообразно и в такой же мере целесообразны
проявления их жизнедеятельности, их ответы, на внешнее воз-
действие. По Ламарку, целесообразность в органическом мире
есть свойство, изначально присущее живым существам. Ее
возникновение нельзя объяснить иначе, чем вмешательством
Божественного Разума.
Жоффруа Сент-Илер (1772—1844). Занимался в основном
сравнительной анатомией животных. Почти не осталось ана-
томических проблем, которые этот знаменитый исследователь
не пытался бы тщательно изучить. Обнаружил достаточно
много фактов, говорящих в пользу идеи единого плана строе-
ния организмов. Сформулировал положение о гомологичных
органах. Пришел к выводу, что атррфия_одного органа сопро-
вождается гипертрофией друго^го,^ обратно, т. е. в организме
господствует принцип равновесия органов. В остальном воз-
зрения Сент-Ил ера принципиально не отличались от воззре-
ний Ламарка.
Жорж Кювье (1769—1832). Кювье пользовался заслужен-
ным авторитетом в области систематической зоологии, срав-
нительной анатомии и палеонтологии, основы которой были в
значительной мере заложены его трудами. Убедившись в том,
что ископаемые формы очень резко отличаются от ныне жи-
вущих, и не находя между теми и другими переходных форм,
он пришел к выводу о неизменчивости животных. Смена на-
329
селения Земли происходила, по Кювье, не в результате эво-
люции, а в силу геологических катастроф, уничтожавших на-
селение той или иной части Земли, после чего опустошенная
местность заселялась новыми формами с других материков
или имело место повторное творение Богом новых форм жи-
вотных.
В1830 г. между Кювье и Сент-Илером в стенах Парижской
Академии наук возник знаменитый спор. Кювье защищал уче-
ние о нескольких (четырех) планах строения организмов,
Сент-Илер отстаивал иДею единого плана. Формально спор
шел не об эволюции, а о том, следует ли признавать четыре
плана строения или один. Однако по существу этот спор, не-
сомненно, относился к эволюционной идее. Противники оста-
лись при своих мнениях, хотя формально победа была одержа-
на Кювье.
В результате этого спора развитие эволюционной идеи в пе-
риод с 1830 по 1858 г. затормозилось. Взгляды большинства
ученых, высказывавшихся об эволюции, в тот период были
крайне осторожными и сдержанными.
Русские эволюционисты первой половины XIX века
В этот период в России выдвигается плеяда ученых, разви-
вающих трансформистские и эволюционные представления.
Среди них выделяются видный эмбриолог академик К. М. Бэр
(1792-1876), врач И. Е. Дядьковский (1784-1841), К. Ф. Рулье
(1814—1858) и его ученик Н. А. Северцов (1827—1885).
ЭВОЛЮЦИОННОЕ УЧЕНИЕ ЧАРЛЗА ДАРВИНА
Концепции возникновения и развития
живых организмов
Основным вопросом, подлежащим разрешению в каждой
теории, претендующей на истолкование эволюции, является
вопрос о происхождении целесообразного устройства орга-
низмов, их приспособленности к условиям существования.
На всех этапах своей истории биология имела ряд концеп-
ций, объясняющих возникновение органической целесооб-
разности.
1. Креационизм — представление о возникновении живого в
результате акта творения, о постоянстве и неизменности всего
существующего, об абсолютной целесообразности.
^30
2. Трансформизм — концепция, допускающая изменяе-
мость живого в пределах особей и не связывающая наблюдае-
мые в органическом мире изменения с поступательным харак-
тером развития и происхождением высших, более сложных
организованных форм, от низших, более примитивных. Целе-
сообразное строение считалось изначальным свойством орга-
низмов, и только в этом усматривалась мудрость Создателя
Вселенной.
3. Эволюционная теория Ламарка—для которой характерно
признание исторического развития живого, глубокое обосно-
вание Идеи эволюции органического мира и анализ ее факто-
ров. Однако по-прежнему органическая целесообразность рас-
сматривается как свойство, изначально присущее живым
существам.
4 Дарвинам — эволюционная теория, изучающая общие
закономерности развития органического мира, рассматриваю-
щая целесообразность в строении организмов как относитель-
ную и Зависящую от действия условий внешней среды.
Предпосылки возникновения дарвинизма
1. К середине XIX в. Англия (родина дарвинизма) занимала
первое место на мировом рынке среди капиталистических
стран. Здесь высокого уровня достигает как тяжелая, так и
легкая промышленность. Успешно развивающаяся текстиль-
ная промышленность потребовала большого количества каче-
ственного сырья. Это стимулировало широкое распростране-
ние и развитие овцеводства, вело к формированию новых
методов совершенствования существующих и выведения но-
вых пород овец. Одновременно практическая селекция начи-
нает в€е сильней проникать в различные отрасли животновод-
ства, а также в растениеводство. Английские селекционеры
добиваются больших успехов в создании новых пород живот-
ных и сортов .растений. Развиваясь стихийно, практическая
селекция нуждалась в глубоком теоретическом обобщении на-
копленных ею фактов, и именно это обобщение позволило раз-
решить некоторые проблемы начинающей складываться эво-
люционной теории.
2. В геологии Ч. Лайель (1797—1875) доказал несостоятель-
ность представлений Кювье о том, что изменения на Земле
происходят под действием каких-то особенных сил, и устано-
вил, что поверхность Земли меняется не в результате катаст-
роф, а постоянно. Знакомство с этой теорией способствовало
формированию эволюционных взглядов Дарвина.
331
3. В различных областях биологии (сравнительная анато-
мия, систематика, палеонтология, эмбриология и клеточная
теория) накопилось большое количество фактов, свидетель-
ствующих об изменчивости вида. Фактический материал этих
наук противоречил представлению о неизменности видов. Все
это послужило идейной предпосылкой возникновения теории
Дарвина.
* Жизнь и научное творчество
Чарлза Дарвина
Чарлз Роберт Дарвин родился 12 февраля 1809 г. в Шрюсбе-
ри. Его дед Эразм Дарвин был известен как ученый, медик и
поэт. Отец Дарвина также был врачом. В 1818 г. Чарлз посту-
пил в школу, где провел семь лет. В октябре 1826 г. он стал сту-
дентом медицинского факультета Эдинбургского университе-
та. Через два года Дарвин по настояцию отца перевелся на
богословский факультет Кембриджской университета. Кроме
богословия он увлекался естественной! историей. В 1831 г. по
окончании университета Дарвин в качестве натуралиста от-
правляется в кругосветное путешествие на корабле «Бигль»
(«Ищейка»). Во время путешествия у Дарвина зародилась но-
вая концепция, объясняющая законы развития жизни лучше,
чем какая-либо высказанная до него. Вернувшись в Англию,
Дарвин с семьей поселился в Доу, близ Лондона, где в течение
17 лет работал над теорией происхождения видов. 18 июня
1858 г. Дарвин получает от известного зоолога Альфреда Уол-
леса, знаменитого исследователя тропических стран, находив-
шегося в то время на Малайском архипелаге, небольшую ста-
тью, в которой излагались мысли, весьма сходные с
основными положениями его теории. В результате 1 июля
1858 г. на заседании Лондонского Линнеевского общества
были одновременно доложены записки Уоллеса и краткий
очерк теории Дарвина. После этого события 24 ноября 1859 г.
вышла в свет знаменитая книга Дарвина «Происхождение ви-
дов путем естественного отбора, или Сохранение благоприят-
ствуемых пород в борьбе за жизнь» — труд, который совершил
переворот в естествознании.
Все последующие работы Дарвина представляли -собой
дальнейшее развитие его теории, изложенной в «Происхожде-
нии видов».
Умер Дарвин 19 апреля 1882 г. По постановлению Палаты
общин он был погребен в Вестминстерском аббатстве. Его мо-
гила находится неподалеку от могилы Ньютона.
332
Основные положения теории
Чарлза Дарвина
Дарвин о происхождении культурных растений и домашних
животных. На примере культурных растений и домашних
животных Дарвин показал значительную пластичность орга-
низмов, т. е. утверждал, что все разнообразие пород и сор-
тов выведено человеком от одного или небольшого числа
диких предков. Например, все многообразие сортов капусты
происходит от одного вида дикой европейской капусты
(рис. 6.30).
Все без исключения сорта и породы служат для удовлетво-
рения каких-либо потребностей человека — экономических
или эстетических. Это* и доказывает, что они созданы челове-
ком. Породы и сорта отличаются друг от друга в первую оче-
редь по тем особенностям, которые интересуют человека. На-
пример, у различных сортов моркови листья, плоды и семена
весьма сходны, корни же различаются большим разнообрази-
ем по форме, цвету, содержанию сахара и т. д. То же относится
к свекле, редису и другим корнеплодам. Сорта капусты отли-
чаются друг от друга листьями, сорта роз — цветами, гороха —
семенами и т.-д. -
Анализируя методы работы селекционеров, Дарвин пришел
к заключению, что создание новых сортов и пород основано на
Рис. 6.30. Многообразие сортов капусты:
1 — джорастущая; 2 — белокочанная; 3 — краснокочанная; 4 — цветная;
5 — кольраби; 6 — брюссельская; 7 — савойская; 8 — кормовая
333
использовании человеком трех факторов — изменчивости, на-
следственное™ и отбора. J
Учение Дарвина об изменчивое™. Дарвин отмечал, что при '
выведении новых пород и сортов селекционеры опираются на ?•
несколько форм изменчивое™.
1. Определенная изменчивость. Эта форма изменчивости "
связана с прямым влиянием внешней среды. Особи, подвер- <
гавшиеся влиянию измененных условий, испытывают сход- ?
ные, т. е. определенные изменения. Так, дикие растения, пере-
саженные на хорошо удобренную почву садов и огородов, |
изменяются в сторону более сильного развития вегетативных';
органов. Животные же под влиянием обильного питания до- :
стигают более крупных размеров и меняют свое телосложение.
2. Изменчивость при упражнении и неупражнении органов.
Дарвин приводит многочисленные примеры, показывающие, '
что органы при увеличении физиологической нагрузки уси- j
ленно развиваются, а при ее недостатке остаются недоразви- '
тыми.
В настоящее время эти две формы изменчивое™ относятся >
к модификационной и представляют собой изменения,
происходящие в пределах нормы реакции.
3. Неопределенная изменчивость. Проявляется, как отмечал
Дарвин, лишь у отдельных особей и выражается в слабых, едва
заметных уклонениях в развитии породы или сорта. Накапли-
ваясь из поколения в поколение, они могут изменить любые
свойства' организма в любом желательном направлении. Не-
определенность этих изменений заключается в том, что особи,
обитающие в сходных условиях среды, уклоняются от своей
породы или сорта в самых различных направлениях. В отдель-
ных случаях неопределенная изменчивость возникает скачко-
образно. По современным представлениям это мутацион-
ная изменчивость.
4. Почковые вариации у растений. Под этим названием
Дарвин описывает случаи изменения отдельных ветвей, раз-
вивающихся из какой-либо одной почки. Так, дерево желтой
сливы дало однажды ветку, на которой росли красные сливы.
При размножении видоизмененных побегов путем прививки
они не теряли вновь приобретенных свойств. Это давало воз-
можность использовать почковые вариации для создания но-
вых сортов. С точки зрения генетики это соматические
мутации.
5. Скрещивание как источник изменчивости. Большинство
пород домашних животных было получено путем скрещивания
различных рас с последующим отбором особей с заранее наме-
уияыми свойствами. Таким образом, скрещивание служило
одним из способов получения измененных особей, т. е. благо-
даря комбинационной изменчивости потомство гибридов от-
личается значительным разнообразием.
6. Коррелятивная изменчивость. Она заключается в том, что
изменение какого-либо органа влечет за собой всегда или по-
чти всегда изменение других органов и их функций. В основе
коррелятивной изменчивости, как полагает современная гене-
тика, лежит плейотропное действие генов.
7. Компенсационная изменчивость. При такой изменчивости
проявляется обратная корреляция в развитии двух сопряжен-
ных признаков, т. е. с развитием одних органов и функций
происходит подавление других, которые недоразвиваются и
могут редуцироваться. Так, одни и те же сорта капусты не дают
одновременно обильной листвы и обильного количества семян.
Трудно добиться, чтобы корова давала много молока и в то же
время сильно жирела.
Учение Дарвина об искусственном отборе. В отличие от своих
предшественников Дарвин впервые заметил, что свойства со-
здаваемых человеком пород и сортов нельзя объяснить только
изменчивостью организмов. Особенностью домашних орга-
низмов является то, что у них всегда имеются признаки, полез-
ные для человека, т. е. удовлетворяющие какие-то его потреб-
ности. Человек по своему усмотрению накапливает изменения
путем отбора. Природа приносит последовательные изменения,
человек же суммирует их в известных полезных для него на-
правлениях. Таким образом, он приспосабливает животных и
растения к своим потребностям или прихотям. Отбор основы-
вается на изменчивости и наследственности, но эти свойства
сами по себе не могут объяснить возникновения новых форм с
желательными для человека качествами. Изменчивость дает
только материал для отбора, новые же формы создаются чело-
веком, накапливающим желательные для него изменения в
ряде поколений.
Итак, под искусственным отбором следует понимать процесс
создания новых пород животных и сортов культурных растений
путем систематического сохранения особей с определенными,
ценными для человека признаками и свойствами в ряде поколе-
ний при содействии их размножению. Дарвин различает две фор-
мы искусственного отбора.
1. Бессознательный искусственный отбор. Это наиболее
ранняя форма селекционного процесса. Сущность бессозна-
тельного отбора заключается в том, что человек не ставил пе-
ред собой цели улучшения своих одомашненных животных и
выведенных культурных растений, но стремился сохранить
для размножения лучших животных и лучшие семена; тем са-
мым уничтожались менее ценные животные и растения. На-
пример, когда человека интересовала яйценоскость, то в хо-
зяйстве сохранялись лучшие несушки, что в конце концов
приводило к появлению высокояйценосных пород кур; если
интересовала молочность, то сохранялись именно такие коро-
вы, и т. д. Таким образом возникли тонкорунные овцы, аро-
матные розы, высокомасличные сорта подсолнечника и мно-
гое другое.
2. Методический искусственный отбор заключается в том,
что селекционер сознательно ставит перед собой определенную
задачу и тщательно подбирает производителей, наиболее соот-
ветствующих намеченной цели, выбраковывая, уничтожая ос-
тальных. Методический отбор всегда являлся творческим про-
цессом, приводившим к образованию новых пород и сортов.
Пользуясь этим методом, селекционер, как скульптор, «лепит»
новые органические формы по заранее продуманному проекту.
Принципы искусственного отбора.
1. Изменчивость и наследственность. Без изменчивости
возникновение новых форм было бы невозможно, так как
только изменчивость организмов дает материал для отбора.
2. Отбор и размножение небольшого числа «лучших», наи-
более совершенных особей и устранение (элиминация) из
процесса воспроизведения всех менее совершенных форм.
3. Накопление положительных изменений в результате по-
вторяющихся в ряде поколений избирательного размножения
только наиболее совершенных форм.
Учение Дарвина о естественном отборе. Установив общие
принципы, объясняющие возникновение новых форм живот-
ных и растений в практике сельского хозяйства, Дарвин по-
ставил перед собой вопрос: не распространяются ли эти прин-
ципы и на возникновение новых форм в природе, т. е. не лежат
ли они в основе движущей силы эволюционного процесса.
Анализируя свои наблюдения и фактический материал,
описанный в литературе, Дарвин в своей работе «Происхожде-
ние видов путем естественного отбора» доказал, что изменчи-
вость и наследственность — общие свойства всех организмов.
Далее, им было показано, что материал для отбора наиболее
«лучших» (наиболее приспособленных) в природе всегда есть,
так как организмам свойственно интенсивное размножение, а
отбор «лучших» осуществлялся путем уничтожения (элимина-
ции) «худших». Тем самым естественный отбор функциониру-
ет при наличии двух факторов, которые дают возможность на
336
основании тех же принципов, что и при искусственном отбо-
ре, обеспечивать возникновение новых форм в животном и ра-
стительном мире. Это интенсивность размножения и борьба
за существование.
Интенсивность размножения. Нет сомнений, что в природе
рождается намного больше особей, чем может выжить. Дарвин
приводит следующий пример. Одуванчик, по приближенным
подсчетам, дает 100 семян. Из них на следующий год могло бы
вырасти 100 растений, каждое из которых снова дало бы по
100 семян. Значит, при беспрепятственном размножении чис-
ло потомков одного одуванчика можно было бы представить
геометрической прогрессией, а значит, для расселения потом-
ства одного одуванчика на десятый год потребовалась бы пло-
щадь, в 15 раз превышающая площадь всей суши Земли. Одна-
ко такого числа потомков никогда не бывает. Интенсивная
гибель организмов — результат борьбы за существование.
Борьба за существование. Дарвин многократно подчерки-
вал, что борьбу за существование не следует понимать в узком
смысле, упрощенно, как прямое физическое уничтожение од-
них организмов другими.
Борьба за существование — это совокупность многообраз-
ных и весьма сложных взаимоотношений, существующих
между организмами и окружающими их условиями природы.
Дарвин различает три формы борьбы за существование.
1. Межвидовая борьба. Примеры межвидовой
борьбы многочисленны. Это взаимоотношения между хищни-
ком и его жертвой, травоядными животными и растениями,
насекомыми и насекомоядными птицами; это конкурентная
борьба между культурными растениями и сорняками; взаимо-
отношения между деревьями различных видов в лесу и между
травами на лугу. Сюда же можно отнести и явление антагониз-
ма микробов.
2. Внутривидовая б о р ь б а. Это борьба, включаю-
щая взаимоотношения между особями одного вида. Так, на-
пример, рыжие тараканы часто полностью вытесняют черных,
серая крыса — черную, европейская пчела — австралийскую,
европейский дождевой червь — южноамериканского. Конку-
рентные внутривидовые отношения широко известны повсю-
ду. Птицы одного и того же вида конкурируют из-за мест гнез-
дования. Самцы многих видов млекопитающих и птиц в
брачный период вступают друг с другом в борьбу за обладание
самкой. Среди рыб широко распространен каннибализм —
поедание особей своего вида. Например, крупные щуки неред-
ко поедают своих меньших по размеру собратьев. Внутривидо-
337
вую борьбу Дарвин считал главнейшим видом борьбы за суще-
ствование.
3. Борьба с неблагоприятными условиями?
внешней среды. Огромную роль в процессе элиминация
особей в растительном и животном мире играют условия неор-
ганической внешней среды. Огромное число растений уничто-
жается почти ежегодно поздними заморозками, засухой, рез-у
кими климатическими колебаниями. Много растений гибнет-
во время суровых малоснежных зим. При недостатке в воде
кислорода в водоемах происходит гибель рыб. Масса семян
растений заносится ветром в неблагоприятные условия и гиб-1
нет. Этих примеров достаточно, чтобы показать большую ин-
тенсивность борьбы с условиями внешней среды.
Все виды борьбы за существование приводят к истребле-
нию огромного числа особей или, что не менее важно, к тоодЯ
что часть особей не оставляет после себя потомства.
Какие особи выживают в борьбе за существование и какие
гибнут?
Некоторые последователи Дарвина утверждали, что в борь-1
бе за существование выживают самые сильные, ловкие, быст-
рые животные. Это вульгарное представление искажает самый
смысл учения Дарвина. Он подчеркивал, что нет ни одного*
свойства, которое было бы полезно при всех условиях. Следо-
вательно, в результате борьбы за существование выживают осо-,
би, наиболее приспособленные к тем конкретным условиям
среды, в которых протекает жизнь данного вида, Таким обра-,
зом, в природе нет идеально, абсолютно приспособленных к,
условиям их жизни органических форм. Органы, первоначаль-:
но сформировавшиеся под влиянием естественного отбора,
сделавшись бесполезными, могут проявлять большую измен-
чивость под влиянием новых факторов среды. При изменении
условий, сформировавших данный признак, то, что было по-
лезным, станет вредным. Отсюда возникает и современное
представление об относительной целесообразности в органи-
ческой природе. Целесообразность — это взаимосвязь и соот-
ветствие всех органов в организме и приспособленность их
строения и функций к существованию в определенных услови-
ях среды. Целесообразность возникает в результате естествен-
ного отбора, и поэтому эволюционный процесс всегда носит
приспособительный характер.
Возникновение в природе целесообразных приспособлений.
Приведем несколько примеров.
1. Возникновение приспособлений к перекрестному опыле-
нию у растений связано с препятствием к самоопылению. Та-
338
кие препятствия возможны при формировании однополых
цветков, неодновременном созревании тычинок и рылец, по-
явлении разных форм цветков.
2. Возникновение покровительственных окрасок и форм у
животных чрезвычайно широко распространено в различных
систематических группах. Под покровительственными окрас-
ками и формами следует понимать любые окраски и формы,
которые помогают животным сохранять свою жизнь. Сюда от-
носятся: а) маскировка, включающая в себя все случаи окрас-
ки, при которых животные делаются незаметными на фоне ок-
ружающей их обстановки; б) демонстрация, когда благодаря
контрастному сочетанию цветов животное легко обнаружить.
Обычно подобная окраска сочетается с несъедобностью орга-
низма; в) подражательное сходство с определенными предме-
тами, не представляющими интереса для врага или жертвы,
помогающее животному достигнуть безопасности; г) мими-
крия — сходство в окраске и форме между двумя или несколь-
кими видами насекомых. Мимикрирующие виды всегда имеют
яркую демонстрационную окраску. При этом, как правило,
один из членов мимикрирующей пары оказывается несъедоб-
ным, в то время как другой лишен защитных свойств и только
«подражает» своей защищенной модели.
Приспособительные явления возникают не сразу; Есте-
ственный отбор, как и искусственный, базируется на различи-
ях между особями. Если те или другие, даже незначительные
изменения полезны и способны увеличивать вероятность вы-
живания, то они будут накапливаться из поколения в поколе-
ние, в результате чего особи приобретут значительные отли-
чия от своих родоначальников, которые в новых условиях
становятся целесообразными. Таким образом, Дарвин дока-
зал, что целесообразность в природе носит относительный ха-
рактер и является следствием отбора — выживания наиболее
приспособленных.
Процесс видообразования по Дарвину. Дивергенция. Изуче-
ние процесса естественного отбора привело Дарвина к созда-
нию монофилетической теории происхождения видов, сущ-
ность которой заключается в том, что каждая крупная группа
животных и растений происходит от единого корня. В процессе
эволюции за счет одного родоначального вида может возник-
нуть несколько новых. Такой процесс Дарвин объясняет рас-
хождением признаков, или дивергенцией (рис. 6.31). Хотя ди-
вергенция и является основным направлением эволюционного
процесса, однако в результате приспособления к одинаковым
Условиям среды может возникнуть конвергентное сходство у
339
Степень изоляции
Рис. 6.31. Стадии дивергенции
форм, неродственных или находящихся в очень отдаленном
филогенетическом родстве.
Так, червеобразная форма языка конвергентно развивалась
у ехидны (клоачные), муравьеда (неполнозубые) и у сумчатого
муравьеда (сумчатые) в связи с приспособлением к питанию
муравьями.
В основе теории происхождения видов, созданной Дарви-
ном, лежат следующие принципиально важные положения:
1. Филогенетическое изменение вида всегда вызывается из-
менением условий среды.
2. Образование нового вида начинается с формирования
новых внутривидовых форм, которые Дарвин называл разно-
видностями.
3. В результате естественного отбора происходит приспособ-'
ление новых органических форм к изменившимся условиям.
4. Образование видов является длительным историческим
процессом.
5. Основным направлением филогенеза является диверген-’
ция, но не исключена возможность образования и конвергентч
ных форм.
б. Каждая крупная группа животных или растений всегд а про-
исходит от одного корня (монофилетическое происхождение).
340
J
ЭВОЛЮЦИЯ КАК РЕАЛЬНОСТЬ
(ПРИЗНАКИ И ДОКАЗАТЕЛЬСТВА)
Понятия вада и популяции
Часто интуитивно мы считаем, что вид — это группа очень
сходных между собой организмов, например, людей, лошадей *
или морковок. Научное определение вида неоднократно изме-
нялось, и в настоящее время наиболее широко распространен-
ным является следующее: вид — это группа морфологически
сходных организмов, имеющих общее происхождение и потен-
циально способных к скрещиванию между собой в естествен-
ных условиях. Особи, из которых слагается вид, не всегда жи-
вут в непосредственном соседстве друг с другом. Обычно они
образуют более или менее изолированные группы, называемые
популяциями. Изучая процесс эволюции, мы обычно рассмат-
риваем составляющие вид популяции. Популяция — это сооб-
щество скрещивающихся между собой организмов. Особи
скрещиваются друг с другом в пределах данной популяции
либо могут покидать свою популяцию и скрещиваться с осо-
бями, принадлежащими другой популяции.
Критерии вида. 1. Морфологический критерий свидетельству-
ет о морфологической изоляции вида. Он является одним из
самых ранних видовых критериев. Йм пользовались система-
тики всех поколений, начиная с Линнея. Он наиболее прост и
удобен в практической работе. На его основе составлены опре-
делительные таблицы как в ботанике, так и в зоологии.
Ограниченность морфологического критерия выступает
особенно отчетливо на примере так называемых видов-двойни-
ков, или биологических видов. Виды-двойнйки можно
определить как морфологически сходные или идентичные,
репродуктивно изолированные природные популяции.
2. Физиологе-биохимический критерий. В основе этого
критерия заложено следующее понятие: виды различны,
потому что различны их химические структуры. Особенно-
сти вида передаются от поколения к поколению, так как в
гаметах закодированы процессы синтеза определенных, спе-
цифических белков. Отсюда следует, что для биохимичес-
кой оценки видовой специфики особый интерес представ-
ляет изучение особенностей ДНК вида. Изучение видовой
специфики ДНК естественно подводит нас к следующему
видовому критерию.
3. Генетический критерий. В основе его лежат два положе-
ния. Во-первых, виды различаются кариотипами, т. е. сово-
341
купностью признаков хромосомного комплекса. Во-вторых*
особи разных видов в естественных условиях, как правило, на
скрещиваются. Препятствием для скрещивания являются раз*
личные изолирующие механизмы (физиологические, морфоло-
гические, экологические), вырабатывающиеся в результате
эволюционного процесса. Однако для большинства животных
изолирующие механизмы, предотвращающие скрещивание'
близкородственных видов, до конца не изучены. Определение
вида, которое возникает из такой теоретической концепции»
гласит: виды — это группы скрещивающихся естественных
популяций, репродуктивно изолированные от других таких
групп.
Однако существуют виды, которые не изолированы друг от.
друга барьером стерильности. Например, кряква и шило-
хвость, два разных вида уток, в неволе способны скрещиваться
друг с другом и давать многочисленное потомство. Очевидно,
разобщенность этих двух видов поддерживается какими-то
другими факторами.
4. Эколого-географический критерий. Используется с целью
установления закономерностей географического распростра-
нения биологических форм и особенностей их взаимоотноше-
ния с факторами внешней среды. Каждый вид занимает опре-
деленную область распространения, т. е. ареал. Ареалы видов
одного рода могут быть полностью изолированными либо, со-
впадать частично или полностью. Виды, имеющие изолиро-
ванные ареалы,называются аллопатрическими.Виды,
у которых ареалы перекрываются или совпадают, имёнуются
симпатрическими. Видовая специфика каждого из них
вскрывается в результате их экологических особенностей. На-
пример, отсутствие гибридов между органическими формами в
местах перекрывания ареала.
Иерархическая классификация организмов
(таксономия)
Нетрудно видеть, что все рассмотренные нами критерии до-
полняют друг друга, помогая вскрывать специфику вида. По-
этому в практике описания видов животных и растений они
используются все вместе, в зависимости от того, насколько
изучены те или иные стороны морфологии, физиологии, гене-
тики и экологии изучаемой формы.
Виды, совпадающие по многим существенным признакам,
но все же репродуктивно изолированные друг от друга, объеди-
няются в роды. По мере ступенчатого расширения критериев
342
сходства близкие между собой роды соединяются в семейства,
семейства — в отряды, отряды — в классы, классы — в типы
(рис. 6.32). Иерархическое соподчинение этих таксонов долж-
но указывать на степень филогенетического родства. В постро-
енной таким образом естественной системе объединение видов
в таксон высшего ранга говорит о наличии у них общей исход-
ной формы в определенное геологическое время, причем чем
дальше от современности момент рождения родственных форм,
тем выше должен быть ранг охватывающей виды системати-
ческой категории.
Такие категории, как особи, скрещивающиеся популяции,
виды, г- это естественные, реально существующие объекты,
тогда как таксоны более высокого ранга представляют собой
абстрактные логические категории. Каждая из них — это
класс объектов, в который входят все категории более низкого
ранга, имеющие определенные общие признаки. Добавляя к
вышеназванным категориям префиксы под- и над- (в система-
тике животных), можно создавать промежуточные категории,
например, подвиды, надсемейства и т. п.
Сама возможность построения иерархической классифика-
ции организмов (таксономия) по различным уровням сходства
объяснима только тем, что категории этой системы отражают в
той или иной степени близкое или отдаленное филогенетичес-
кое «кровное родство». Именно в их выявлении состоит смысл
таксономических исследований.
Итак, таксономия, или систематика, дает весьма ценные
; доказательства в пользу эволюционного учения.
1. Независимо от эволюционного учения систематика об-
наруживает родственные связи между организмами.
Рис. 6.3.2. Диаграмма взаимоотношений между систематическими
категориями
343
Рис. 6.33. Единство плана
строения:
1 — скелет человека; 2 — скелет
шимпанзе; 3 — скелет лягушки
344
2. Независимо от эволюци- .
онного учения систематика i
приходит к выводу о необхо- :
димости изображать отноше- •
ния между естественными |
группами организмов в виде |
генеалогического древа. f
3. Систематика накаплива- I
ет ценный материал по вопро- |
сам изменчивости и изменяв- |
мости видов и дает основания |
считать справедливым взгляд, -|
согласно которому виды эво- |
люционируют. I
Доказательства эволюции |
в области сравнительной |
анатомии 1
'• 1
Сравнительная анатомия |
изучает строение животных f
(или растений), в результате |
чего удается установить черты |
сходства и различия и опреде- |
лить степень родства между I
ныне живущими организмами. |
В общей форме естествен- |
ное сходство выражается в 1
том, что у сравниваемых форм !
наблюдается сходный общий f
план организации, так назы- |
ваемое единство плана строе- |
ния. В узком смысле закон -
единства плана строения при- *
меним к отношениям, суще- •
ствующим в пределах каждого
отдельного типа животного J
царства. Например, сравнивая :
скелеты позвоночных живот- I
ных, легко установить в общей ;
организации, расположении, <
числе и форме частей этих*;
скелетов взаимное сходство •
(рис. 6.33). Факт замечатель- J
ного общего сходства в строении позвоночных естественнее
всего объясняется предположением, что все эти формы связа-
ны узами родства.
Возникает, однако, вопрос: возможно ли более широкое
толкование закона единства плана строения? Можно ли, на-
пример, применить это понятие не только к отношениям, су-
ществующим в пределах одного класса, но и к отношениям,
существующим между типами? На этот вопрос в ряде случаев
можно дать положительный ответ, в особенности если мы бу-
дем искать единство плана строения не только среди взрос-
лых, но и личиночных форм. При такой оговорке можно лег-
ко показать замечательное сходство, например, между
личиночными формами кольчатых червей и некоторых мол-
люсков, что позволяет ставить вопрос о родстве между этими
двумя типами.
В ряде случаев сходен не только общий план строения срав-
ниваемых форм, но и детали их организации. Это выражается,
в частности, в установлении гомологии органов, имеющей пер-
востепенное значение для выяснения филогенетического род-
ства между организациями.
Гомологичные органы — это такие органы разных животных
(растений), которые имеют сходное строение, одинаковое по-
ложение в теле и развиваются из тех же самых эмбриональных
закладок. Гомологичные органы могут выполнять у разных
животных одинаковые или разные функции. Например, иглы
ежа гомологичны волосам других млекопитающих. Усики го-
роха, служащие для прикрепления его стебля к другим расте-
ниям, являются гомологами листа.
Рудиментарные органы. Особым видом гомологичных орга-
нов являются рудименты (зачаток). Это органы, утерявшие в
филогенезе свое значение и функцию и оставшиеся в виде не-
доразвитых образований. Поэтому точнее было бы назвать их
не зачаточными, а. остаточными. В теле каждого животного
можно встретить много рудиментов. Немало рудиментов имеет-
ся и в теле человека (рис. 6.34).
Атавизм — это появление признаков, присущих далеким
предкам и в норме не встречающихся у современных форм. Ес-
тественно, что такие признаки говорят о филогенетическом
родстве между различными организмами: теми, у кого такие
признаки отсутствуют как норма, и теми, у кого они проявля-
ются лишь как атавизм. Примеры атавизма у человека — шей-
ная фистула (образование, гомологичное жаберной щели),
хвостовой придаток, обильная волосатость всего тела, наличие
более двух молочных желез и т. д. (рис. 6.35).
345
Рис. 6.34.
Рудиментарные органы
человека:
1 — волосяной покров на
голове пятимесячного
эмбриона; 2 — ушная ра-
ковина с Дарвиновым бу-
горком (а); 3 — полулунная
складка (п. с.)
Аналогичные органы — это
органы, выполняющие одинако-
вую функцию, но не имеющие,
единства строения и происхож-
дения. Например, у рыб органы
водного дыхания — жабры. У ра-
кообразных органами дыхания
тоже служат жабры, но они нахо-
дятся в конечностях. Тот факт,
что эти органы развиваются раз-
личным путем, указывает, что
морфологическое сходство меж-
ду ними чисто внешнее, не отра-
жающее родства между организ-
мами.
Конвергенция. Во многих слу-
чаях, когда неродственные орга-
низмы живут в одинаковых усло-
виях внешней среды и ведут
сходный образ жизни, у них под
влиянием естественного отбора
возникают аналогичные органы
и сходные формы тела. Такое
сходство можно видеть, сравни-
вая быстро плавающих морских
хищников — акулу (класс рыб),
ихтиозавра (класс пресмыкаю-
щихся) и дельфина (класс мле-
копитающих).
Учение о гомологии и анало-
гии органов уточнило методы
систематики (таксономии), по-
зволило избегать ошибочного
объединения животных в систе-
матические группы на основе
лишь внешнего сходства. Клас-
сификация, построенная на эволюционном принципе, дока-
зывает, что все систематические группы — типы, классы, отря-
ды — связаны друг с другом общностью происхождения.
Свидетельством этому служат так называемые сборные формы,
т е. животные, сочетающие признаки разных систематичес-
ких групп. Так, ланцетник связывает позвоночных с беспозво-
ночными. Кистеперые рыбы обладают и жабрами и легкими,
связывая тем самым рыб с амфибиями.
346
Рис. 6.35. Развитие атавистических признаков у людей:
1—2 — волосатые люди: Адриан Евтихеев (7) и мексиканская танцовщица
Юлия Пастрана (2 ); 3—десятилетний мальчик из Индии с мягким хвостом;
4 — случай многососковости
Доказательства эволюции на базе
эмбриологии
Сравнительный метод исследования фактов эмбрионально-
го развития дает ценные свидетельства в пользу эволюционно-
го учения. Между зародышами животных наблюдается необы-
чайное сходство, с предельной ясностью указывающее на
«кровное родство».
347
На основании сравнительно-эмбриологических исследо-
ваний К. Бэр сформулировал закон зародышевого сходства,
согласно которому черты, общие для большой группы род-
ственных животных, проявляются раньше частных призна-
ков. По Бэру, в зародышевом развитии животных наиболее
рано формируются признаки, отражающие принадлежность
к определенному классу, затем отряду, роду и, наконец,
виду. Закладка органов, их положение, форма, способы раз-
вития, отношение друг к другу необычайно сходны, напри-
мер, у зародыша рыбы, саламандры и черепахи, у эмбриона
крысы и человека (рис. 6.36). В дальнейшем развитии заро-
дышей позвоночных отмеченное сходство ослабевает. По-
степенно зародыши как бы «расходятся» и явственно приоб-
ретают черты, свойственные уже не только позвоночным
вообще, но и определенным классам, к которым они при-
надлежат.
Сопоставление организации низших и высших животных
с их зародышевыми стадиями показывает, что каждое высшее
животное проходит в своём индивидуальном развитии этапы,
соответствующие более низкоорганизованным животным.
Это представление позволило Э. Геккелю в 1866 г. сформули-
ровать основной биогенетический закон, в соответствии с ко-
торым онтогенез представляет собой краткое и быстрое повто-
рение филогенеза. Согласно Геккелю, новые признаки,
имеющие эволюционное значение, возникают во взрослом
состоянии. По мере усложнения организации взрослых форм
зародышевое развитие удлиняется за счет включения допол-
нительных стадий. Этим законом объясняется далеко идущее
сходство между эмбриональными стадиями животных. На-
пример, тот факт, что зародыши всех высших животных име-
ют жаберные карманы в глотке, находит свое наиболее есте-
ственное объяснение в утверждении, что все они генетически
родственны и произошли в конечном счете от предков, жив-
ших в воде.
Тот факт, что все многоклеточные организмы начинают
свое индивидуальное развитие с яйца (с одноклеточного со-
стояния) и проходят сходные стадии эмбрионального разви-
тия, дает право предлагать и единственно естественное
объяснение этого факта, а именно, что многоклеточные орга-
низмы произошли от одноклеточных предков, т. е. от одного
общего корня.
Начиная с Геккеля биологи широко использовали биогене-
тический закон в целях выявления филогении определенных
систематических групп. Однако результаты применения этого
348
I стадия
Рыба Саламандра Черепаха Крыса Человек
Рис. 6.36. Эмбрионы позвоночных животных
на ранних стадиях развития (I стадия) сходны друг с другом,
на более поздних (II и III стадии) похожи
на филогенетически родственные формы
закона к выявлению закономерностей эволюции не всегда ока-
зывались удовлетворительными. Сам Геккель был принужден
констатировать, что в ряде случаев в онтогенезе не происходит
рекапитуляции, т. е. повторения пройденного предками пути
исторического развития. В этой связи он выделил палингеие-
зы и ценогенезы.
349
X’
Под палингенезами он понимал повторение филогенетичес-
ких изменений в индивидуальном развитии.
Ценогенезы, по Геккелю, представляют собой эмбрио-
нальные нарушения, происходящие в результате приспо-J
собления зародышей или молодых особей к определенным;
условиям индивидуального развития. Геккель различал два *
типа ценогенезов — гетеротопии и гетерохронии. При г е - f
теротопии происходит сдвигание местоположения орга-
нов; гетерохронии представляют собой нарушения во
время закладки органов. Примером гетерохронии служит4;
более ранняя закладка нервной системы и запаздывание в;
формировании половой системы у высших позвоночных и
человека по сравнению с низшими. Примером гетеротопии^
является закладка легких, представляющих собой видоизме- .
нение задней пары жаберных мешков, расположенных по j
бокам кишечника, на его брюшной сторрне. Примером не- '
ногенеза можно считать появление провизорных органов у
зародышей наземных позвоночных (амниона, хориона, ал- ;
лантоиса).
Крупным шагом в развитии эволюционной эмбриологии
явилось учение о филэмбриогенезах, созданное А. Н. Север-
цевым.
Филэмбриогенезами кхзывахп новые наследственные свойства,
которые возникают в зародышевой жизни животных, сохраняют-
ся у взрослых форм и передаются последующим поколениям,
включаясь, таким образом, в филогенез. Филэмбриогенезы от
ценогенезов отличаются тем, что являются не приспособления-
ми зародыша, а новыми свойствами, остающимися у взрослых
животных.
Согласно Северцову, филэмбриогенезы могут возникнуть
на любой стадии эмбрионального развития. Основное поло-
жение теории заключается в том, что эволюционные измене-
ния состоят в изменении хода онтогенетического развития.
Различают три основных способа филэмбриогенезов: анабо-
лия, девиация, архаллаксис.
Под анаболией понимают такой способ изменений,
который возникает на последних фазах развития. Девиа-
циями хода онтогенеза называют изменения, происходя-
щие на средних фазах морфогенеза. Наконец, к архал-
лаксисам относят такие изменения, которые происходят
в самом начале эмбрионального развития органа и поэтому
резко изменяют весь ход морфогенеза. Примером всех трех
видов филэмбриогенезов могут служить изменения в разви-
тии придатков кожи пресмыкающихся, птиц и млекопитаю-
350
щих по сравнению с таковыми у хрящевых рыб, древние
виды которых были предками наземных позвоночных
(рис. 6.37).
Рис. 6.37. Схема филогенетического развития
костных (I) и роговых (П) чешуй, перьев (III)
и волос (IV) позвоночных:
1—6— развитие плакоидной чешуи акулы; 7—12—развитие чешуй ящерицы.
Ход их развития изменен на средних стациях (девиация); 13—15 — развитие
пера. Сперва развитие пера повторяет пути развития чешуи пресмыкающихся
(7—//)» но на последних стадиях (Н) происходит изменение (анаболия)
и развитие пера идет по новому пути (В—15); 16—21 — развитие волоса.
С наиболее ранних стаций направление развития волоса уклоняется от
направлений развития прочих кожных образований; повторения нет,
наблюдается архаллаксис
351
Доказательства эволюции из области -%
палеонтологии ,4
Изучение процесса эволюции на основании палеонтологи- •
ческих материалов привело к формулировке ряда законов,
часть которых в значительной мере правильно обобщает на- '
блюденйя, другие же требуют некоторых поправок. Приведем ••
наиболее известные законы. ??
1. Закон неспециализированное™. Согласно этому закону, к ;
эволюционному развитию способны малоспециализирован-
ные формы; прогрессирующая специализация отдельных вет-
вей ведет к вымиранию соответствующей группы организмов. |
Причину этого вымирания можно объяснить прогрессивно |
уменьшающейся изменчивостью специализированных форм. %
Вымирание специализированных ветвей в связи с пониженной й
изменчивостью не может, однако, считаться универсальной 1
закономерностью, так как даже среди очень крупных специа- 1
лизированных форм, например, бабочек, являющихся конеч- I
ной филогенетической ветвью насекомых, изменчивость чрез- '
вычайно велика.
2. Закон необратимости эволюции. Согласно этому закону, i
эволюция организмов, изменившихся в каком-нибудь направ-.
лении, никогда не приводит к появлению признаков, характер- :
ных для его предков. То есть эволюционный процесс не воз-
вращается вспять по старому пути. Так, лошадь, утратившая
все пальцы, кроме одного, ни при каких обстоятельствах не -
сможет развить свои рудиментарные пальцы.
На основании остатков организмов, находимых в разных •
слоях почвы, можно судить об их строении и отчасти об их
жизнедеятельности, о периоде жизни Земли, когда эти орга- •
низмы ее населяли, об их распространении и, что самое глав-
ное, об их генетической связи друг с другом и с современными
живыми существами.
Чтобы убедиться в этом, попытаемся рассмотреть общие '
направления эволюционного процесса и смену форм во време-
ни (табл. 6.1).
Таблица 6.1
Эра Периоды Характерные группы организмов i
Докемб- рийская Плохая сохранность палеонтологических документов. Случайные находки. Аольча- J тые черви (аннелиды), ракообразные (трило- J биты).
352
Продолжение табл. 6.1
Эра Периоды Характерные группы организмов
Палеозойская
Кембрийский Среди простейших уже имеются и совре- менные роды. Кишечнополостные', еще нет настоящих кораллов. Черви', высокодиффе- ренцированные аннелиды. Членистоногие'. низшие ракообразные. Иглокожие', древние формы.
Силурийский Кишечнополостные', появились настоящие кораллы. Иглокожие, древние группы в рас- цвете. Еще мало морских звезд и ежей. Мол- люски'. бросается в глаза множество при- митивных головоногих. Членистоногие'. трилобиты и ракоскорпионы. Первые члени- стоногие, дышащие воздухом, в том числе древние скорпионы. Позвоночные', низшие формы, так называемые бесчелюстные, в том числе так называемые панцирные рыбы. Настоящие рыбы: древние акуловые, двоя- кодышащие, кистеперые. Растения', kqjw- росли, некоторые папоротники.
Девонский Кишечнополостные', многие древние формы в упадке. Кораллы многочисленны. Иглоко- жие'. древние (силурийские) морские лилии. Морских ежей еще мало. Моллюски: среди головоногих все еще господствуют прими- тивные наутилиды. Позвоночные', древние рыбообразные панцирные формы достигают расцвета. Рыбы: акуловые, химеры, древ- ние кистеперые, примитивные осетровые, двоякодышащие. Земноводные', появились стегоцефалы. Растения', первые многочис- ленные наземные формы, хвощи, папорот- ники и др.
Каменноуголь- ный Кишечнополостные', много древних „корал- лов, ныне не существующих. Иглокожие'. многочисленные морские лилии, древние ежи, ныне не существующие. Моллюски'. среди брюхоногих моллюсков много легоч- ных (наземных) форм. Членистоногие', мно- гоножки, пауки с членистым брюшком, много насекомых, в том числе представите- ли отрядов: бескрылых, прямокрылых, сет- чатокрылых, полужесткокрылых (клопы). Ракоскорпионы в упадке. Трилобиты в упадке. Позвоночные', двоякодышащие и
353
П-281
Продолжение табл. 6.^.
Эра Периоды Характерные группы организмов
м Q 3S О кистеперые рыбы сокращаются в числе. Большим числом форм представлены при- митивные земноводные — стегоцефалы. В5 конце периода появляются первые пресмьь кающиеся. Растения: папоротники, хвощи; некоторые голосеменные. •
СП О О § с Пермский Моллюски: много аммонитов и наутилид. Чле^ нистоногие: трилобиты вымирают. Позвоноч- ные: прежние группы рыб, много настоящих^ акул. Земноводные: многочисленные и разной образные стегоцефалы. Пресмыкающиеся*. примитивные группы. Растения: развитием хвойных.
Мезозойская Триасовый Простейшие: много современных родов. Ки- шечнополостные: среди кораллов смена древ- них новыми, шестилучевыми. Иглокожие*. среди морских лилий многие вымерли, ио все же эта группа господствует. Морские ежи многочисленнее, древние вымерли, по- явились новые формы. Моллюски: в расцве- те, много новых групп. Мезозойские брюхо- ногие моллюски вымирают, наутилиды сокращаются в числе. Аммонитов много, в том числе новые формы. Белемниты еще редки. Членистоногие: трилобиты исчезли, появляются десятиногие раки. Позвоночные: появляются костистые рыбы. Земноводные: последние стегоцефалы. Пресмыкающиеся: динозавры, крокодилы, черепахи, плезио- завры, ихтиозавры и др. Появляются млеко- питающие: примитивные группы сборного и переходного типов. Растения: среди хвой- ных — араукария и сосна.
Юрский Кишечнополостные: пышный расцвет рифооб- разующих кораллов. Иглокожие: все еще мно- го морских лилий. Морские ежи еще много- численнее, чем в триасе. Моллюски: множество новых форм. Кораблики в упадке и представлены одним родом — Наутилус (кораблик). Белемниты в расцвете, новые аммониты. Членистоногие: среди насекомых первые представители двукрылых и пере- пончатокрылых. Позвоночные. Рыбы: харак- терны примитивные костистые. Пресмыка-
354
Окончание табл. 6.1
Эра Периоды Характерные группы организмов
Мезозойская ющиеся: в расцвете диплодок, стегозавр, игуанодон, черепахи, крокодилы, ихтиозав- ры, плезиозавры и др. Появляются птицы: археоптерикс, археоникс. Млекопитающие. еще далеко до расцвета, мелкие примитив- ные формы. Растения', хвощи, папоротни- ки, хвойные.
Меловой Иглокожие, морские лилии в упадке. Первое место переходит к морским ежам. Моллюски: множество аммонитов и белемнитов. Наути- лус (кораблик). Членистоногие: много десяти- ногих раков. Рыбы: массовое развитие костис- тых рыб (расцвет). Земноводные: появляются формы, близкие к современным. Пресмыкаю- щиеся в расцвете. Появляются настоящие ящерицы и крокодилы. Ихтиозавры вымира- ют. Зубастые птицы. Млекопитающие, те же древние мелкие формы, но появляются сум- чатые и неполнозубые. Растения: еще в ниж- нем мелу появляются однодольные и двудоль- ные, в том числе ива, тополь, дуб, береза, эвкалипт и др.
Кайнозойская Третичный Вымирают новые формы мелового периода. Иглокожие: много современных родов. Мол- люски: белемниты и аммониты приходят в упадок и вымирают. Позвоночные: высшие костистые рыбы. Земноводные. близки совре- менным. Вымирают все древиие пресмыкаю- щиеся., и остаются гаттерии, ящерицы, кроко- дилы и черепахи. Впервые появляются змеи. Среди птиц много близких современным. Происходит закладка современных отрядов млекопитающих..
Четвертичный Современные формы животного и раститель- ного мира. Человек.
Распределение зевотных
но зоогеографическим областям
Многообразный животный мир, или фауна, каждой зоогео-
графической области включает в своем составе специфические
Руководящие формы, определяющие границы области.
12»
355
Существуют следующие шесть областей: Палеарктическая^-
Неоарктическая, Восточная (Индо-Малайская, или Индийс-
кая), Неотропическая, Эфиопская, Австралийская. Каждая и^
них обладает особой фауной (рис. 6.38). ?
Перечислим некоторые формы, характеризующие отделы. ;
ные области.
Палеарктическая область. Эту область, охватывающую Ев-%
pony, Северную Африку и Северную и Среднюю Азию, харак- •
теризуют: зубр, серна, сайгак, каменный баран (архар), двугор-
бый верблюд, выхухоль, настоящие кроты; среди J
многочисленных птиц — усатые синицы, саджа, тетерев-косач,)
глухарь, фазаны (рис. 6.39). Эти формы свойственны только;
этой области. |
Неоарктическая область. Она охватывает Северную Америку, 8
Гренландию, прилегающие к Северной Америке острова, в том#
числе Бермудские и Антильские. Для Неоарктической области
характерны: бизон, мускусный бык (в Гренландии), американ-1
ская вонючка, американский енот, опоссум; среди птиц — ди- ]
кие индейки, степной тетерев, странствующий голубь, каролин-1
ский попугай; среди змей важно отметить ядовитую гремучую |
змею (рис. 6.40). - 5
1”~'| гттт'Н Г7"У1 EZZZZI
Австралии- Неотро- Эфиоп- Восточная Палеарк- Неоарк-
ская пическая ская тическая тическая
Рис. 638. Зоогеографические области
356
Восточная (Индо-Малайская, или Индийская) область.
Включает Индию, Индо-Китай, Южный Китай, острова, при-
легающие к Южной Азии, вплоть до границ Австралийской
области. Область характеризуют: две человекообразные обезь-
яны (орангутан и гиббон), лемуры (полуобезьяны), чепрачный
Рис. 6.39. Характерные представители
Палеарктической области:
1 — косуля; 2 — сайгак (сайга); 3 — лось; 4 — верблюд; 5 — серна; 6 —
сибирский горный козел; 7 — лошадь Пржевальского; 8 — выхухоль; 9 —
тетерев; 10 — горная индейка; 11 — усатая синица; 12 — кавказский фазан;
13 — саджа
357
тапир, индийский слон (короткоухий, с короткими бивнями)^
банкивские куры (родичи домашних кур), птица-носорог, фа^
зан-аргус; среди пресмыкающихся — гавиал (род крокодилов^
своеобразная ящерица — летающий дракон (рис. 6.41).
Неотропическая область географически соответствует Юж-s
ной и Центральной Америке. Характерными животными этой*
Рис. 6.40. Характерные представители |
Неоарктической области: 1
1 — вилорог; 2 — овцебык; 3 — йгот; 4 — древесный дикобраз; 5 — снежная j
коза; 6 — бизон; 7 — американская вонючка; 8 — опоссум; 9 — луговая j
собачка; 10— странствующий голубь; 11 — луговой тетерев; 12 — глазчатая
индейка; 13 — гремучая змея . -1
358
области являются: опоссумы — особое семейство сумчатых,
неполнозубые — броненосцы, муравьеды, а также водосвинка,
американские тапиры, наконец, широконосые обезьяны с их
примечательным, играющим роль дополнительного органа
хватания хвостом. Среди птиц необходимо назвать трехпалого
страуса нанду» замечательного гаоцина, птенцы которого ис-
Рис. 6.41. Характерные представители Восточной
(Индо-Малайской, или Индийской) области:
1 — гиббон; 2 — шестокрыл; 3 — лемур долгопят; 4— чепрачный тапир; 5—
орангутан; 6 — таял; 7 — индийский носорог; 8 — индийский слон; 9 —
дикий петух; 10 — яванский павлин; 11 — птица-носорог; 12 — летучий
дракон; 13— крокодил-гавиал
359
пользуют крылья как органы лазания по ветвям, группу колиб- .
ри — самых маленьких птиц мира, наконец, огромного кондо-
ра — представителя хищников (рис. 6.42). :
Эфиопская область. В нее входят Африка, кроме Северной
Африки, и Мадагаскар. Область характеризуется рядом свое-
образных форм: ящер, трубкозуб, златокрот, бегемот, жирафа,
ряд антилоп, в том числе антилопа гну, африканский слон ?:
Рис. 6.42. Характерные представители
Неотропической области:
1 — броненосец; 2 — муравьед; 3 — ленивец; 4 — обезьяна капуцин; 5 —
обезьяна уистити; 6 — водосвинка; 7 — тапир; 8 — гуанако; 9 — гоацин;
10 — колибри; 11 — тукан; 12 — кондор; 13 — нанду; 14 — игуана
360
(большие уши, длинные бивни), две человекообразные обезь-
яны (горилла и шимпанзе), полуобезьяны; среди птиц — сек-
ретарь (охотник на змей), двупалый страус, так называемые ба-
наноеды и др. (рис. 6.43).
Австралийская область. К ней относятся Австралия, Новая
Зеландия, Новая Гвинея, Полинезия, Сандвичевы острова,
Зондский архипелаг (часть). Для этой области характерны со-
Рис. 6.43. Характерные представители Эфиопской области:
I — даман; 2 — трубкозуб; 3 — ящер; 4 — шимпанзе; 5 — гиппопотам; 6 —
антилопа гну; 7 — черная антилопа; 8 — окапи; 9 — жираф; 10 — лемурвари;
11 — двурогий носорог; 12— африканский слон; 13 — страус; 14 — секретарь;
15 — шлемонская цесарка
361
временные сумчатые и однопроходные. Сумчатые дают ряд
групп — хищники, грызуны, травоядные. Однопроходные
представлены утконосом, ехиднами и проехиднами. Птицы: <
попугай-нестор, венценосный голубь, новозеландский киви я
птица-лира, райские птицы (рис. 6.44). Среди пресмыкаю-
Рис. 6.44. Характерные представители
Австралийской области:
1 — ехидна; 2 — утконос; 3 — гигантский кенгуру; 4 — древесный кенгуру;
5 — сумчатая летяга; 6 — вомбат; 7 — сумчатый вояк; 8 — сумчатый крот;
9 — венценосный голубь; 10 — какаду-ворон; 11 — казуар; 12—эму; 13 —
лирохвост; 14 — киви; 15 — совиный попугай
362
щихся следует отметить туатару (гаттерию) — представителя
вымершей группы первоящеров (Прозауриа).
Сравнительный обзор зоогеографических областей позво-
ляет обнаружить, что сходство между их фаунами тем больше,
чем крупнее сравниваемые систематические категории, и на-
оборот, тем меньше, чем меньше сравниваемые систематичес-
кие категории.
Эта закономерность справедлива для всех областей. Удов-
летворительное объяснение этому явлению дает только эволю-
ционная теория. Очевидно, что в геологическом прошлом наша
планета была заселена относительно менее дифференцирован-
ными формами животных. Обособившийся в разных областях
животный мир все более изменялся, причем в пределах каждой
области развился свой специфический видовой состав. Это все
тот же процесс расхождения, дивергенции, который можно
проследить в палеонтологических ветвях, в систематике и на
эмбриологическом материале.
БИОЛОГИЧЕСКИЕ ОСНОВЫ СОВРЕМЕННОЙ СЕЛЕКЦИИ
Учение Дарвина об отборе легло в основу такой науки, как
селекция. Предметом селекции является исследование специ-
фических закономерностей эволюции домашних животных и
возделываемых растений, направляемой человеком. Главная
цель и задачи селекции состоят в создании высокопродуктив-
ных пород животных и сортов растений, которые наилучшим
образом повышают производительность труда в животновод-
стве и растениеводстве. Порода и сорт входят в средства сель-
скохозяйственного производства. Породой и сортом называют
популяцию организмов, искусственно создаваемую человеком
и имеющую определенные наследственные особенности. Все
особи внутри породы и сорта имеют очень сходные, наслед-
ственно закрепленные показатели продуктивности, биологи-
ческие свойства и морфологические признаки.
Основные методы селекции
1. Генетические метод ы. Изменчивость исходного материала
является основой для создания новых пород животных и сор-
тов растений. При этом имеют значение такие типы изменчи-
вости, как а) мутационная', б) комбинационная; в) полиплоидия.
2. Методы скрещивания. Наличие наследственной изменчи-
вости позволяет путем различных систем скрещивания соче-
363
тать определенные наследственные признаки в одном орга-
низме, а также избавляться от нежелательных свойств. При
разведении применяют различные системы скрещиваний, ко-
торые делят на несколько типов:
а) родственное скрещивание, часто называемое инбридин-
гом. Родственным называют скрещивание особей, имеющих
близкую степень родства: брат—сестра, двоюродные братья
и сестры и т. д. Его смысл заключается в том, что генетичес-
кий состав популяции оценивается путем разложения ее на
генетически различные линии. При этом гены, находящие-
ся в гетерозиготном состоянии, переходят в гомозиготное
состояние;
б) неродственное скрещивание, когда у скрещиваемых особей
имеется отсутствие ближайших общих предков в 4—6 поколе-
ниях их родословных. Ценность этого скрещивания в том, что
помеси первого поколения оказываются более жизнеспособ-
ными, более устойчивыми к заболеваниям, имеют повышен-
ную плодовитость, т. е. проявляют гетерозис, или явление
гибридной мощности. Под гетерозисом понимают
свойство гибридов первого поколения при скрещивании раз-
личных рас, пород животных и сортов растений, а также ин-
бридных (чистых) линий превосходить по ряду признаков и
свойств лучший из исходных родительских организмов.
Дальнейшее скрещивание гибридов между собой ведет к зату-
ханию этого эффекта в следующих поколениях. О наличии
гетерозиса следует говорить лишь в том случае, когда межли-
нейный гибрид превосходит не только родителей (линии),
но и сорта или породы, от которых произошли эти линии.
В настоящее время гетерозис установлен для всех изученных
видов.
3. Методы отбора. Отбор является одним из основных мето-
дов селекции. В сочетании с генетическими методами он по-
зволяет создавать новые формы, сорта и плоды с определенны-
ми свойствами и признаками. Различают:
а) массовый отбор — т. е. отбор особей по внешним пока-
зателям (фенотипу), отвечающим принятому стандарту для
популяции сорта или породы без проверки генотипа отдель-
ных особей;
б) индивидуальный отбор — т. е. отбор, при котором прежде
всего оценивается потомство отдельного растения или живот-
ного в раду поколений. Вследствие этого становится возмож-
ным оценивать наследственные качества отдельных индивиду-
умов — способность передавать свои свойства и норму реакции
генотипа потомству.
364
4. Онтогенетический метод. Крупный вклад в развитие се-
лекции растений внес русский ученый И. В. Мичурин. Им
поднят совершенно новый для селекционно-генетической те-
ории вопрос, а именно проблема развития наследственных
признаков в онтогенезе. Он доказал, что сортовые признаки
растений, например их теплолюбивость или холодоустойчи-
вость, форма, размер и качество плодов, формируются на ран-
них стадиях онтогенеза под влиянием наследственности и ус-
ловий внешней среды. И. В. Мичурин создал стройную
систему селекционного процесса, в которой сочетаются три
основных звена: гибридизация, воздействие условиями внешней
среды на развивающиеся гибриды и отбор. Мичурин разрабо-
тал специальные методы преодоления нескрещиваемости у ра-
стений при отдаленной гибридизации. (Отдаленной гибриди-
зацией называется скрещивание форм, относящихся к разным
разновидностям, видам, родам и другим таксономическим
единицам более высокого порядка. Например, мул был полу-
чен при скрещивании лошади и осла; грейпфрут — лимона и
апельсина.)
К числу основных методов Мичурина относятся:
а) метод предварительного вегетативного сближения. За-
ключается в предварительной прививке скрещиваемых расте-
ний друг на друга (например, сеянец рябины гибридного про-
исхождения прививают в крону взрослого дерева груши для
дальнейшего преодоления нескрещиваемости между рябиной и
грушей);
б) метод посредника. Данный метод лучше всего рассмот-
реть на конкретном примере. Мичурин, чтобы продвинуть на
Север культуру персика, решил осуществить гибридизацию
этого культурного растения с диким монгольским миндалем
бобовником. Поскольку прямого скрещивания между ними
добиться не удалось, экспериментатор предварительно скрес-
тил миндаль с полукультурным персиком Давида. Получен-
ный от скрещивания гибрид Мичурин назвал посредником,
так как он уже сравнительно легко скрещивался с культурны-
ми сортами персика;
в) метод направленного воспитания сеянцев. Здесь особенно
известен метод «ментора». «Ментором» Мичурин называет
взрослое растение, которое служит воспитателем для молодого
сеянца;
г) метод опыления смесью пыльцы в целях преодоления не-
скрещиваемости растений. Так, при скрещивании яблони
(мать) с грушей (отец) к пыльце последней добавляется не-
большое количество пыльцы яблони. Это делает рыльце ябло-
365
ни восприимчивым и к пыльце груши, вследствие чего часть
семяпочек оплодотворяется материнской пыльцой, а часть — У
пыльцой чужого вида.
Селекция и учение Н. И. Вавилова
Процесс окультуривания растений имеет разную длитель- ;
посты у одних он продолжается несколько тысячелетий, а у
других начался не более двухсот лет назад. В решении пробле-
мы происхождения культурных растений ждущую роль сыгра-
ло учение академика Н. И. Вавилова о центрах происхождения
культурных растений. В результате его исследований было ус-
тановлено семь очагов земного шара, откуда ведет начало вся
мировая земледельческая культура (табл. 6.2, рис. 6.45).
Таблица 6.2 .
Центры происхождения культурных растений
Центры Растения
1. Южноазиатский Тропический рис, сахарный трост- ник, значительная часть зерновых бобовых, тропические плодовые — банан, кокосовая пальма
2. Восточноазиатский Просо, гречиха, ряд клубнеплодов и корнеплодов, многие, плодовые (груша, яблоня, слива и др.), неко- торые цитрусовые и декоративные растения
3. Юго-западаоазиатский i Мягкая пшеница, карликовая пшеница, круглозерная пшени- ца, горох, чечевица, чина, кон- ские бобы, нут, хлопчатник-гуза
4. Средиземноморский Маслина, рожковое дерево, ряд овощных (свекла и др.)
5. Абиссинский .Твердая пшеница, ячмень, ко- фейное дерево
6. Центральноамериканский Кукуруза, американские фасоли, тыквы, перец, какао, хлопчатник- упланд
7. Андийский (Южноамериканский) Картофель и ряд других корне- плодов, табак, ананас, земляной орех, хинное дерево
366
Рис. 6.45. Центры происхождения культурных растений
Детальное изучение изменчивости многих видов растений
дало возможность Н. И. Вавилову обобщить вбе известные
факты в форме общего закона, известного под названием зако-
на гомологичных рядов в наследственной изменчивости. Сущ-
ность этого закона заключается в следующем: «генетически
близкие виды и роды характеризуются сходными рядами на-
следственной изменчивости с такой правильностью, что, зная
ряд форм в пределах одного вида, можно предвидеть нахожде-
ние параллельных форм у других видов и родов».
Закон дает возможность обнаружить сходную направлен-
ность изменчивости, например, у злаковых (рис. 6.46), и пре-
дугадать существование форм, еще не известных науке.
ФАКТОРЫ ЭВОЛЮЦИИ
I. Изоляция как элементарный фактор эволюции. Изоля-
ция — возникновение любых барьеров» препятствующих скре-
щиванию данного вида с другими видами. Различают два вида
изоляции.
367
1 23 4 5 6 7 8 9 Й
•ф
Рис. 6.46. Гомологические ряды изменчивости
у видов пшеницы и ячменя по признаку остистости:
1—3 — формы мягких пшениц (42 хромосомы); 4— б — формы твердых Ц
пшениц (28 хромосом); 7— 9 — формы шестирунного ячменя (7, 4, 7 —
остистые формы; 2, 5, 8 — короткоостые формы; 3, 6, 9 — безостые формы)
1. Пространственная, или географическая, изоляция. Эта
изоляция может существовать в разных формах: водные барье-
ры разделяют «сухопутные» виды, барьеры суши изолируют
виды-гидробиоты, возвышенности — равнинные популяции, а
равнины — горные популяции и т. д.
2. Репродуктивная изоляция. Предотвращает межвидовые
скрещивания, а также уменьшает успешность межвидовых
скрещиваний. Это связано с отсутствием встреч между по-
тенциально брачными партнерами (сезонная, или б и о -
топическая, изоляция); с отсутствием спаривания при
встрече между потенциально брачными партнерами (этоло-
гическая изоляция); с отсутствием осеменения при
попытке к копуляции (механическая изоляция); с не-
жизнеспособностью оплодотворенных яиц либо неполноцен-
ностью последующих поколений (гибридологическая
изоляция).
II. Изменчивость как элементарный фактор эволюции. Без
изменения генотипического состава популяции невозможно
протекание любого эволюционного процесса. Генетическая ге-
терогенность популяции связана с качественными перестрой-
ками генофонда. С генетической точки зрения, эволюция —
368
это изменение генов в данной популяции. Частота какого-
либо гена (точнее, аллели) зависит от ряда различных факто-
ров, среди которых следует отметить:
1. Мутационный процесс. Мутация — это изменение, затра-
гивающее последовательность нуклеотидов или их число в мо-
лекуле ДНК и создающее тем самым новые аллели.
2. Рекомбинация. Процесс, с помощью которого имеющая-
ся в популяции аллель перемешивается (рекомбинирует) в
результате скрещивания между собой. Эта генетическая ком-
бинаторика многократно изменяет значение мутаций: они
входят в новые геномы, оказываются в новой генотипической
среде.
3. Поток генов. Частота генов в данной популяции может
изменяться в результате эмиграции или иммиграции особей.
Это обмен генами между популяциями, осуществляющийся в
форме происходящих время от времени спариваний между осо-
бями, принадлежащими к различным скрещивающимся попу-
ляциям, проникновения в данную популяцию животного, ро-
дившегося в другом месте, или же внесения чужих семян или
пыльцы. Непосредственный эффект такого события сходен с
эффектом мутации — в популяцию вносится новый источник
изменчивости.
4. Закон Харди — Вайнберга и поддержание изменчивости.
Английский математик Г. Харди и независимо от него немец-
кий генетик В. Вайнберг сформулировали положение, которое
сейчас именуется законом Харди — Вайнберга и описывает ча-
стоту встречаемости генов в равновесной популяции свободно
скрещивающихся организмов. Этот закон описывает некую
абстрактную «идеальную» популяцию со свойствами, для ре-
альных популяций невозможными:
а) численность такой популяции должна равняться беско- •
нечности;
б) скрещивание особей внутри популяции полностью сво-
бодно, т. е. осуществляется без всяких ограничений. Такие пот
пуляцииназываются панмиксными (свободносмешива-
ющимися);
в) популяция должна быть абсолютно изолированной.
Допустим, что частота встречаемости доминантного гена А в
популяции равна q, а рецессивного а соответственно 1 — q, тог-
да соотношение гомо- и гетерозигот будет следующим:
q2AA + 2^(1 — q)Aa + (1 - q)2aa.
Это известная из школьной алгебры формула квадрата сум-
мы двух чисел. Если q = 0,5, как в случае первого поколения
369
ж
менделевских гибридов, то во втором поколении мы имеем
следующее соотношение:
0,25.44 + 0,54а + 6,25оа,
а так как Аа в случае полной доминантности фенотипически
выражены как АА, мы получаем менделевское расщепление — Л:
0,75:0,25, т. е. 3 :1. f
5. Дрейф генов (генетико-автоматические процессы). Итак,
;:м
относительная численность аллелей в популяции абсолютно
точно описывалась бы уравнением Харди — Вайнберга, если f
бы численность самой популяции была бы бесконечной. Чем Ц
меньше популяция, тем больше отклонения, и если числен- J
ность размножающихся особей в ней окажется ниже 500, на-
пинаются любопытные явления. Их назвали генетико-автома- М
тическими процессами, или коротко, дрейфом генов. Суть 1
дрейфа генов в том, что из-за случайных колебаний один из ал- |
лелей может вообще выпасть из популяции, и она окажется по |
данному гену гомозиготной. В результате происходит гомози-
данному гену гомозиготной. В результате происходит гомози-
готизация особей и затухание изменчивости. Нетрудно понять,
что это открывает путь случайной дифференцировке популя-
ций вплоть до видового обособления.
III. Популяционные волны как элементарный эволюционный
фактор. Популяционными волнами, или «волнами жизни» (по
С. С. Четверикову), называют периодические или апериоди-
1
ческие колебания численности организмов в природных по- |
пуляциях. Изменение генофондов популяций происход ит как |
на подъеме, так и на спаде популяционной волны. При росте |
численности организмов наблюдаются слияние ранее разоб- |
щенных популяций и объединение их генофондов. При спаде |
численности наблюдается распад крупных популяций. Возни- |
кающие малочисленные популяции характеризуются изме- |
ненными генофондами. |
В условиях массовой гибели организмов редкие мутантные |
аллели могут быть генофондом вида. При сохранении редкого |
аллеля его концентрация в генофонде малочисленной популя- I
ции автоматически возрастает. Популяционные волны дей- |
ствуют на генофонды ненаправленно, поэтому они так же, как «
и мутационный процесс, только предоставляют эволюцион- .;
ный материал к действию естественного отбора. 1
IV. Естественный отбор — движущий фактор эволюции. Под |
естественным отбором нужно понимать избирательное (диф- |
ференциальное) воспроизведение генотипов (или генных ком- |
плексов). В процессе естественного отбора важны не столько ?
выживание или гибель особей, сколько их дифференциальное |
370
размножение. Успех в размножении разных особей может слу-
жить объективным генетико-эволюционным критерием есте-
ственного отбора.
В зависимости от результата различают направленную, ста-
билизирующую и дизруптивную формы естественного отбора
(рис. 6.47).
1. Направленный отбор (движущий) вызывает прогрессив-
ное или направленное в одну сторону изменение генетическо-
го состава популяций, что проявляется в сдвиге средних зниче-
ний отбираемых признаков в сторону их усиления или
ослабления. Он происходит в тех случаях, когда популяция
находится в процессе приспособления к новой среде или же
когда происходит постепенное изменение среды, а вслед за
ней и постепенное изменение популяции.
2. Стабилизирующий сохраняет в популяции средний
вариант фенотипа или признака. Действие отбора состоит в
элиминации плохо приспособленных и периферических вари-
аций, возникающих в результате мутационного процесса, по-
тока генов и рекомбинации. Это наиболее распространенный
тип отбора.
Направленный
ТИПЫ ОТБОРА
Дизрутмвный
3
ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Рис. 6.47. Воздействие отбора трех различных типов
на генетическую изменчивость в популяции
371
3. Дизруптивный отбор благоприятствует сохранению край?-
них типов и элиминации промежуточных. В результате оц
приводит к сохранению и усилению полиморфизма. i
Как и другие элементарные эволюционные факторы, есте-,
ственный отбор вызывает изменения в соотношении аллелей
в генофондах популяций. Особенность его действия состоим
в том, что эти изменения направленны. В эволюции есте---
ственному отбору принадлежит творческая роль. Исключая
из репродукции генотипы с малой приспособительной цен-4
ностью, сохраняя благоприятные генные комбинации разно-
го достоинства, он преобразует картину генотипической!
изменчивости, складывающуюся первоначально под действиЛ
ем случайных факторов, в биологически целесообразном на-|
правлении. 1
Плата за отбор. Генетический груз. Естественный отбор имеет|
как позитивный, так и негативный аспекты. Он влечет за собой!,
предпочтительное выживание и размножение одних генотипов и|
предпочтительную элиминацию и невозможность размножения
других. Невозможность размножения той или иной особи, обус-'
ловленная отбором в его негативном аспекте — проявляется ли *
она в форме избирательной смертности или неудавшегося раз- ?
множения как такового, известна под названием генетической^
гибели (или гибели, обусловленной отбором). Генетическая ги- |
бель снижает репродуктивный потенциал данной популяции. |
Популяция обладала бы более высокой численностью, если бы -
не потери за счет генетической гибели. Поэтому генетическая J
гибель представляет собой, по крайней мере потенциально, не- ?
кое бремя для популяции. Это бремя называется генетическим |
грузом. Различают несколько типов генетического груза, корре- I
лирующегося с разными типами отбора. |
1. Мутационный груз —неизбежный побочный про- !
дукт мутационного процесса. Этот процесс порождает вредные #
мутации, которые затем должны быть выметены стабилизиру- г
ющим отбором.
2. Сегрегационный груз существует в популяциях, |
которые используют преимущества, создаваемые превосход- >
ством гетерозигот. При этом в каждом поколении происходит ’
выщепление менее приспособленных гомозигот, понижающих '
среднюю приспособленность популяции. Гомозиготы пред- 5
ставляют собой плату за уравновешивающий отбор.
3. Субституционный г р у з, который порождает на- ;
правленный отбор. Замещение старой аллели превосходящей t
ее новой влечет за собой генетическую гибель носителей ста- 5
рой аллели. *
Пути и способы видообразования. Процесс образования ви- •
дов осуществляется в результате взаимодействия элементар-
ных эволюционных факторов. Различают следующие способы
видообразования.
1. Аллотропическое видообразование (географическое) — но-
вый вид может возникнуть из одной или группы смежных попу-
ляций, расположенных на периферии ареала исходного вида.
2. Симпатрическое видообразование — новый вид возникает
внутри ареала исходного вида.
3. Дивергентное видообразование — первоначальный единый
вид разделяется на два или более новых в связи с возникновени-
ем межпопуляционных изоляционных барьеров и углублением
различий под действием естественного отбора вплоть до генети-
ческой изоляции. Такой процесс, ведущий к увеличению коли-
чества видов, называется истинным видообразованием.
4. Филитическое видообразование — новый вид может воз-
никнуть посредством постепенного изменения одного и того же
вида во времени без какой-либо дивергенции исходных групп,
что наблюдается при изменении условий во всем ареале.
5. Гибридогенное видообразование — новый вид возникает
путем гибридизации с последующим удвоением числа хромо-
сом (аллополиплоидия). 1
ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИОННОГО ПРОЦЕССА
Изучение филогенеза животного мира показало, что эволю-
ция идет различными путями (рис. 6.48). Вопрос об основных
направлениях эволюционного процесса был разработан акаде-
миком А. Н. Северцовым.
Рис. 6.48. Схема основных направлений эволюционного процесса.
Две верхние плоскости (а, б) возникли путем ароморфоза,
нижняя (в) — путем морфологического регресса.
Ветвящиеся линии в пределах плоскости идиоадаптации
373
Ароморфоз — основной путь, прогрессивной эволюции,
В этом направлении шла эволюция от одноклеточных к мно-
гоклеточным, от двухслойных к трехслойным, от низших хор*
довыхк позвоночным. Ароморфозы — это подъем организа*
ции с одного уровня на другой, более высокий.
Идиоадаптация — путь узкоприспособительных изменении,
не связанных с повышением уровня развития. Путь идиоадан*,
тации объясняет возникновение в пределах каждого типа и
класса многочисленных видов, имеющих одинаковый уровень
организации, но приспособленных, например, к жизни в пре-
сной или в соленой воде.
Морфофизиологический регресс выражается в упрощении
строения. Этот путь эволюции, ведущий от сложного к более
простому, довольно широко распространен в природе и не при-
водит к вымиранию упростившихся форм, если они достаточ-
но приспособлены к среде обитания. Примером может служить
эволюция ленточных червей, утративших в процессе филогене-
за органы пищеварения.
ЖИЗНЬ И ВТОРОЙ ЗАКОН
ТЕРМОДИНАМИКИ
Жизнь на Земле зародилась около двух или четырёх милли- '
ардов лет назад. Толчком научного изучения мира живого по-
служило создание в прошлом веке трех главных научных сис-
тем, в которых вскрыты различные аспекты единства живой
материи.
1. Клеточная теория Шванна и Шейдена (1838).
2. Закон наследственности Менделя (1856).
3. Теория эволюции путем естественного отбора Дарвина и •
Уоллеса (1858).
КРИТЕРИИ ЖИВОЙ МАТЕРИИ
Между двумя состояниями материи, живым и неживым,
существуют некие различия, которые можно считать непре-
менными критериями живой материи. Перечислим важней-
шие из них.
1. Обмен веществ как между компонентами живой системы,
так и между нею и окружающей средой.
374 . J
2. Самовоспроизведение, т. е. система сама воспроизводит
себя и поддерживает свою целостность, используя для этого
элементы окружающей среды с более низкой упорядоченно-
стью.
3. Эволюционная непрерывность. Жизнь есть процесс, начи-
нающийся от некоторого простого уровня и спонтанно услож-
няющийся в результате последовательных событий, которые
мы называем эволюцией.
4. Пространственная компактность, которая достигается за
счет организации деталей живого механизма на молекулярном
уровне, так что невероятно высокая организация материи уме-
щается в очень сжатом объеме.
5. Энергетическая экономичность. КПД живых систем зна-
чительно превышает КПД машин, созданных человеком.
6. Универсальность углерода как материала для скелета био-
логических молекул.
7. Вода как универсальный растворитель для живой системы.
8. Нуклеиновые кислоты как стабильные хранители наслед-
ственной информации.
9. Белки как катализаторы и регуляторы в биологических сис-
темах.
10. Макроэргические фосфаты как аккумуляторы энергии.
Таким образом, можно перечислить еще много других кри-
териев живых систем, и все они будут необходимы, однако не-
достаточны для «полного» понимания такого явления, как
жизнь. Исходя из этого, мы можем дать лишь относительно
истинные определения понятию «жизнь».
Определение сущности жизни
Историческое определение сущности жизни (по Ф. Энгель-
су): «Жизнь — есть способ существования белковых тел, и этот
способ существования состоит по своему существу в постоян-
ном обновлении химических составных частей этих тел».
Определение сущности жизни с позлет физиологии: «Жизнь—
это совокупность процессов, совершающихся в энергетически
открытой и саморегулируемой системе, важнейшими компо-
нентами которой являются белок, нуклеиновые кислоты, фос-
форорганические соединения, а характернейшим способом су-
ществования является самообновление химического состава,
происходящее благодаря обмену веществ с окружающей средой,
обеспечивающее живому существу пластичность, относительную
устойчивость, а также самоохранение путем воспроизведения
себе подобных».
375
Определение сущности жизни с позиции генетики: «Система
может быть названа живой, если она обладает специфичное-?,
тью или информацией, которую можно передавать потомству,
кроме того, информация может претерпевать изменения, с тем
чтобы измененная информация или специфичность также пе-;
редавалась потомству».
Обобщенное определение сущности жизни: «Жизнь — это ак-
тивное, идущее с затратой энергии поддержание и воспроиз- |
ведение специфической структуры».
Упорядоченность живых систем |
4'
Из вышесказанного прослеживается одна из характерных *¥
черт живых систем как рабочих механизмов, заключающаяся 1
в том, что они тяготеют к локальному повышению упорядочен- |
ности, усваивая простые молекулы и создавая из них сложные |
соединения, имеющие упорядоченное строение. Однако орга- f
низмы берут извне не упорядоченность, а энергию: растения — |
в виде квантов света, животные — в виде малоокисленных со- J
единений, которые можно сжечь в процессе дыхания. За счет *
этой энергии они строят свою индивидуальную упорядочен- ь
ность, пренебрегая чужой.
Согласно второму закону термодинамики — одному из :
фундаментальных законов физической Вселенной — клетка ?
(организм) использует лишь часть общей энергии, содержа-
щейся в химических соединениях, так называемую свобод-
ную энергию, остальная же часть энергии рассеивается в виде -
тепла. Соотношение между этими величинами можно выра-
зить уравнением: Н= G + TS, из которого видно, что измене- ?
ние общей энергии (ДЯ) складывается из изменения свобод-
ной энергии (А(?) и изменения той части энергии, которая
рассеивается в виде тепла (в этом уравнении Т — температура
по Кельвину; S— энтропия системы). Итак, энтропия пред- '
ставляет часть общей энергии, которая не может быть ис-
пользована в данной системе. Здесь S служит мерой необра-
тимости реакции. Согласно второму закону термодинамики'
энтропия изолированной системы стремится к некоторому
максимальному значению, при котором достигается равнове-
сие и жизнедеятельность в биологических системах прекра-
щается. Если приложить энергию, чтобы перевести биологи-
ческие процессы с более низкого энергетического уровня на
более высокий, то энтропия уменьшается. Однако такого
рода процессы термодинамически невозможны, если они не
связаны е другой системой, в которой энтропия соответствен-
376
ро увеличивается, тем самым компенсируя ее уменьшение в
цервой системе.
Какими же свойствами должна обладать живая система для
поддержания упорядоченности, т. е. понижения энтропии?
На современном этапе развития науки такими сврйствами
считают:
1. Существование живых форм как открытых систем,
вследствие чего обеспечивается поступление извне опреде-
ленных химических соединений (т. е. дополнительной энер-
гии). Таким образом, живая* система способна функциониро-
вать за счет свободной энергии, поступающей из внешней
среды.
2. Живые формы являются слабо или сильно неравиовес-
нымисистемами. Отсюда следует, что живые системы совер-
шают постоянную работу против термодинамического равно-
весия за счет свободной энергии.
3. В живых системах все процессы периодические. В об-
щем смысле ритм выступает как тип связи происходящих
событий, интегрирующий и организующий отдельные ком-
поненты в единое, целое, функционально организованное
образование.
4. Живые системы — нелинейные системы. Нелинейность
означает, что имеется несколько степеней свободы в развитии
того или иного биологического процесса.
5. Живые системы подчиняются законам управления. Меха-
низм управления обеспечивает переход из одного состояния
организма в другое. Существует несколько уровней управле-
ния регуляции морфогенетических процессов в живых фор-
мах:
а) метаболический, к которому относятся все процессы, на-
ходящиеся под общим контролем функциональных белков;
б) оперонный, охватывающий системы управления, изменя-
ющие скорость синтеза биополимеров;
в) клеточный, представляющий генетические системы, у
которых продолжительность основных состояний сравнима с
продолжительностью клеточного цикла;
г) онтогенетический — организменный, представляемый
как совокупность клеточных генетических систем, взаимодей-
ствующих между собой на разных уровнях через эффекторы и
специализированные агенты;
д) психологический, осуществляемый за счет внушения.
У высших животных и человека этим органом управления яв-
ляется нервная система, и прежде всего головной мозг, его
большие полушария.
377
Основные биологические ' <
принципы ч"
В последнее время делаются попытки выявить и сформу»
лировать основные концепции и логические связи, характе-
ризующие живые системы в отличие от неживых, и рассмот-
реть вытекающие из них общефилософские представления, -,-
И хотя всеобъемлющая теория жизни в настоящее время по
существу отсутствует, имеются теоретические осмысления^
дающие аксиоматическую интерпретацию жизни. Заслу-;*
живает внимания система принщнюв (аксиом) биологии по^’
Б. М. Медникову. ,Д.
I принцип. Все живые организмы должны быть единством фе-"^
нотипа и программы для его построения (генотипа), передающе-^
гося по наследству из поколения в поколение. Ц
Согласно этому принципу по наследству передается не<
структура, а описание структуры и инструкции по ее изготов-1
лению.
П принцип. Наследственные молекулы синтезируются мат-$
ричным путем. В качестве матрицы, на которой строится ген$
будущего поколения, используется ген предыдущего поколения. • ,1
Различают следующие процессы матричного синтеза: |
1) репликацию (ДНК->ДНК); 2) транскрипцию (ДНК->РНК);|
3) трансляцию (РНК->белок). |
Ш принцип. В процессе передачи из поколения в поколение ге-
нетические программы в результате многих причин изменяются Д;
случайно и ненаправленно, и лишь случайно эти изменения оказы- %
ваются приспособительными.
Из этого принципа вытекает, что принципиально невоз- |
можно достаточно точно определить координаты и импульсы '
частиц, атакующих гены (по принципу Гейзенберга), а значит, ]
возникновение мутаций непредсказуемо и ненаправлено.
IV принцип. Случайные изменения генетических программ при
становлении фенотипов многократно усиливаются и подверга- ;
ются отбору условиями внешней среды. I
Принцип усиления означает, что размножение клетки —
носительницы мутации какого-нибудь гена, ведет последова- •
тельно к умножению гена, умножению мРНК, умножению !
специфического белка. Отбор, таким образом, действует не i
прямо на изменение генетических программ, а на фенотипы, в ;
которых каждое изменение в миллиарды миллиардов раз уси- >
ливается. Здесь отбор выступает в качестве «демона Дарвина», '
т. е. предотвращает повышение энтропии, сохраняя структур-
ную организацию живых систем.
378
ОСНОВЫ ЭКОЛОГИИ
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Жизнь растений и животных невозможна вне окружающей
их среды. Закономерностями, возникающими при взаимодей-
ствии организмов со средой, занимается биологическая на-
ука — экология. Среда — это окружающий нас материальный
мир, мир живой (органической) и неживой (неорганической)
природы. Элементы окружающей среды, оказывающие опре-
деленное воздействие на организмы, называют экологическими
факторами.
Многообразные экологические факторы подразделяют на
две большие группы — абиотических и биотических факторов.
К абиотическим (отгреч.«а» — отрицание,«биос»—
жизнь) отно'сят проявление свойств неживой природы, к
биотическим — влияние живой природы, в том числе и
воздействие человека (антропогенные факторы). Влия-
ние экологических факторов сказывается на всех процессах
жизнедеятельности организмов, и прежде всего на их обмене
веществ.
АБИОТИЧЕСКИЕ ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
В процессе эволюционного развития каждый вид организ-
мов приспособился к определенной географической области,
т. е. имеет свой ареал. Существование каждого вида организ-
мов ограничено определенным максимумом и минимумом ка-
ких-либо факторов, оказывающих ограничивающее действие
на его жизненно важные функции. К действию каждого факто-
ра у любого вида организмов существуют крайние границы
терпимости — ограничивающий фактор.
Ограничивающий фактор — фактор среды, выходящей за
пределы выносливости организма (за пределы допускаемого
максимума и минимума): света, влаги, температуры и т. д. Но
существует и оптимальный фактор, при котором развитие
организма происходит при наиболее благоприятной интен-
сивности экологического фактора (свет, температура, воздух
и т. д.). Так, крайние температурные границы, при которых
происходит развитие Яйца аскариды, — это 12—40“С; оптимум
составляет 25’С.
379
Для всех организмов существуют свои границы по каждому
экологическому фактору. Границы, за пределами которых не-
возможно существование организма (верхние слои атмосферы,
горячий источник), называются пределами выносли-
в о ст и.
Приспособление организмов к условиям среды может быть '
быстрым и обратимым или довольно медленным, что зависит
от глубины воздействия фактора. Организмы, которые могут \
жить в довольно разнообразных условиях внешней среды, на-
зываются эврибионтными (от греч. «эврос» — широ- д
кий). Например, медведь бурый живет в условиях холодного и S
теплого климата, в сухих и влажных районах, питается разно-
образной растительной и животной пищей. Стенобионт-|
н ы е организмы (от греч. «стенос» — узкий) приспособлены к у
существованию лишь в узком диапазоне условий среды. На- |
пример, форель может жить только в чистых холодных горных |
реках. Стенобионтами являются большинство внутренних и |
внешних паразитов, глубоководные рыбы и др. д
£
Свет •§
Это наиболее важный физический фактор на Земле, кото- j
рый оказывает огромное биологическое влияние на организмы. |
Биологическое действие солнечного света обуславливается его
спектральным составом, интенсивностью, суточной и сезон- ’
ной периодичностью. Разные участки солнечного спектра не- ?
равнозначны по биологическому действию.
Ультрафиолетовая часть характеризуется самой высокой
энергией квантов и высокой фотохимической активностью. "
С помощью ультрафиолетовых лучей в организме животных s
осуществляется биосинтез витамина D. Эти лучи лучше всего .
воспринимаются зрением многих насекомых, у растений они ;
способствуют синтезу некоторых биологически активных со- ;
единений (витаминов, пигментов).
Инфракрасные лучи несут основное количество тепловой
энергии. Это приводит к нагреванию всего организма, что име-
ет большое значение для холоднокровных животных (насеко-
мых, рептилий и др.). Важнейшими функциями инфракрасных
лучей по отношению к растениям являются осуществление
транспирации, с помощью которой из листьев отводится водя-
ными парами излишек тепла, а также создание оптимальных |
условий для вхождения углекислого газа через устьица.
Потребность организмов в периодической смене опреде- |
ленной продолжительности дня и ночи называется фотопе- |
380 I
риодизмом (от греч. «фотос» — свет). Регулируемая фотопери-
одизмом реакция организмов на изменение времен года (при
наступлении осеннего короткого дня опадают листья с дере-
вьев и готовятся к перезимовке животные; при наступлении
весеннего длинного дня пробуждаются растения и восста-
навливается жизненная активность животных) называется
сезонным ритмом. Реакция организмов в течение суток
на периоды света и темноты определенной длительности (по-
кой и активность у животных, суточные-ритмы движения
цветков и листьев у растений, ритмичность деления клеток,
процесса фотосинтеза и т. д.) называется биологичес-
кимичасами.
Температура
Температура является важным ограничивающим фактором.
У большинства организмов процессы жизнедеятельности лро-
текают в пределах от —4 до,40—45° С. Этим объясняется скуд-
ность форм в арктических областях и в условиях пустынь.
Температура, наиболее благоприятная для жизнедеятельности
и роста, называется оптимальной. Например, отдельные виды
бактерий и сине-зеленых водорослей могут существовать в го-
рячих источниках при температуре около 80’С. Полярные
воды с температурой от 0 до —2° С населены разными предста-
вителями животного и растительного мира — беспозвоночны-
ми, рыбами, водорослями.
В природных условиях температура очень редко держится
на уровне, благоприятном для жизни. Поэтому у растений и
животных возникают специальные приспособления, которые
ослабляют резкие колебания температуры.
1. Теплокровность — способность к терморегуляции орга-
низма у млекопитающих и птиц, одно из самых важных про-
грессивных приспособлений.
2. Зимняя спйчка — приспособление животных к перенесе-
нию зимнего периода (зимний сон).
3. Анабиоз — временное состояние организма, при котором
жизненные процессы замедлены до минимума и отсутствуют
все видимые признаки жизни (наблюдается у холоднокровных
животных зимой и в жаркий период года).
4. Морозостойкость — способность организмов выносить
низкие отрицательные температуры.
5. Состояние покоя. Зимний покой — приспособительное
свойство многолетнего растения, для которого характерно пре-
кращение видимого роста и жизнедеятельности, отмирание
381
надземных побегов у травянистых жизненных форм и опаде-'
ние листьев у древесных и кустарниковых форм.
Лепишй покой — приспособительное свойство раннецвету-
щих (тюльпан, шафран, пролеска), растений тропических рай--*'
онов, пустынь и полупустынь.
Состояние покоя разной длительности характерно также
для свежесобранных семян, плодов, клубней, луковиц, корне- ‘
плодов.
Влажность
Это один из наиболее важных факторов среды. Недостаток ,
воды является причиной ограничения жизнедеятельности и
географического распространения наземных организмов.
По приуроченности к местообитаниям с различными усло-
виями увлажнения выделяют следующие основные формы на-
земных растений: гигрофиты, мезофиты и ксерофиты.
Гигрофиты — растения влажных местообитаний. Для мно-
гих из них характерны тонкие цельные голые или с небольшим
опушением листья с малым числом устьиц. К гигрофитам от-
носятся: калужница болотная, дербенник иволистый, подма-
ренник болотный, рис, росянка, папирус, а также папоротники
и орхидные. Избыточная влага у гигрофитов выделяется с по-
мощью специальных водяных устьиц — гидатод, располагаю-
щихся по краям листьев.
Мезофиты — это растения, приспособленные к жизни в
условиях умеренного увлажнения. В основном это растений,
произрастающие на лугах и в лесах. К мезофитам относятся
эфемеры и эфемероиды, которые быстро заканчивают свой ве-
гетационный период (не более 4—б нед) и засушливое время
переживают в виде семян (эфемеры) либо в виде покоящихся
луковиц, клубней и корневищ (эфемероиды).
Ксерофиты — растения сухих местообитаний, способные
переносить недостаток влаги (почвенной,'атмосферной).
У многих ксерофитов выработались анатомо-морфологические
особенности, направленные на уменьшение испарения влаги
либо на добывание воды из глубоких почвенных горизонтов.
У многих ксерофитов уменьшена площадь поверхности листь-
ев, их опушение очень обильное (коровяк). Пустынные и по-
лупустынные ксерофиты обладают глубокими корневыми сис-
темами (верблюжья колючка, у которой главный корень
достигает до 15 м в глубину).
Интересные приспособления к недостатку питьевой воды
наблюдаются и у животных засушливых мест. Они максималь-
382
но используют так называемую метаболическую воду, которая
образуется при биохимических реакциях в организме, накап-
ливают жировые запасы, «водоотдача» которых самая большая
(из 100 г жира побочно образуется 105—107 г воды). Такие жи-
вотные обычно бывают хорошими бегунами и способны пре-
одолевать большие расстояния.
Соленость среды
Так как живые организмы возникли в океане, до сих пор
внутренние жидкие ткани многих эволюционно древнейших
видов с простым строением (акулы, скаты, ракообразные)
сохранили значительное сходство с морской водой по соот-
ношению основных ионов (натрия, калия, кальция, хлора).
Сухопутные животные, оставив морскую среду, которая
дала им жизнь, тоже сохраняют постоянным водно-электро-
литный состав своих тканей. Такая способность отдельных
клеток и организмов поддерживать внутреннюю среду в хи-
мически неизменном состоянии с помощью компактных
механизмов является одним из чудесных свойств живой
материи; Такие механизмы особенно четко и безошибочно
должны работать у мигрирующих рыб (лосось, кета, горбу-
ша), которые периодически переходят из морской воды в
пресную, и наоборот.
Растительные организмы в процессе эволюции также при-
способились к слишком высокой концентрации солей в среде.
У таких растений имеется повышенная концентрация вакуо-
лярного сока, которая по всасывающей силе выше почвенного
раствора, и они называются галофитами (представители се-
мейства мареновых, хлопчатник, томаты). Излишек солей не-
которые из них способны выделять наружу, где эти соли после
высыхания образуют твердые пленки, чешуйки или кристал-
лические скопления.
Кислород
Для большинства живых организмов кислород является
жизненно необходимым. В бескислородной среде могут разви-
ваться только анаэробные бактерии.
Кислород обеспечивает осуществление экзотермических
реакций, в ходе которых освобождается необходимая д ля жиз-
недеятельности энергия. Этот элемент является конечным ак-
цептором электрона, который отщепляется от атома водорода в
процессе энергетического обмена.
383
На Земле только зеленые растения являются продуцентами ’;
кислорода в процессе фотосинтеза. Поглощение организмами??
кислорода из внешней среды происходит:
а) всей поверхностью тела (простейшие черви);
б) трахеями (насекомые);
в) жабрами (рыбы);
г) легкими (позвоночные).
Кислород химически связывается и переносится по орга- f ;
низму специальными пигментами крови — гемоглобином (по- J
звоночные), гемоцианином (моллюски, ракообразные). При f
недостатке кислорода в среде у организмов выработались coot- Т
ветствующие приспособления: повышенная кислородная ем- ?
кость крови, более частые и глубокие дыхательные движения,
большой объем легких (у жителей высокогорья, птиц).
БИОТИЧЕСКИЕ ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Под биотическими факторами понимаются воздействия на
организм всех других живых существ того же и других видов.
В результате этих воздействий возникают межвидовые и внут-
ривидовые взаимоотношения, которые выражаются в пищевых
связях (цепи питания), конкуренции и т. д.
Цепи питания
Источником энергии, за счет которой существуют все орга-
низмы, является Солнце (рис. 6.49). Однако ими может усваи-
ваться и использоваться далеко не вся энергия солнечного излу-
чения. Большая часть солнечной энергии теряется в виде тепла.
Все организмы по типу питания можно разделить на три
функциональные группы. Зеленые растения — автотрофы.
Они способны аккумулировать солнечную энергию в процессе
фотосинтеза и синтезировать органические вещества. Автотро-
фы — это продуценты (от лат. «продуцентис» — производя-
щий), т. е. производители органического вещества, первая
функциональная группа организмов биоценоза.
Кроме того, существуют гетеротрофные организмы, кото-
рым для питания необходимы уже готовые органические веще-
ства. Такие организмы, являющиеся растительноядными и
плотоядными животными, называются консументами (от лат.
«консумо» — употреблять, расходовать). Отмершие тела и тру-
пы консументов разлагаются до минеральных элементов, кото-
рые опять поступают в окружающую среду.
384
Рис. 6.49. Роль организмов в круговороте веществ
в биосфере
Следующей функциональной группой являются сапрофа-
ги — микроорганизмы и грибы, которые разрушают останки
до углекислоты и воды. Эти организмы называются редуцента-
ми (от лат. «редуцере» — уменьшение, упрощение).
Перенос энергии от ее источника (растений) через ряд орга-
низмов называют пищевой цепью. Все живые организмы свя-
заны между собой энергетическими отношениями, поскольку
являются объектами питания других организмов. Травоядные
животные (потребители первого порядка) поедают растения,
первичные хищники (потребители второго порядка) поедают
травоядных, вторичные хищники (потребители третьего поряд-
ка) поедают хищников помельче. Таким образом, создаются
цепи питания из продуцентов и консументов, которые на раз-
ных этапах смыкаются с сообществом редуцентов.
Цепь питания — это цепь взаимосвязанных видов, последо-
вательно извлекающих органическое вещество и энергию из
исходного пищевого вещества; каждое предыдущее звено яв-
ляется пищей для последующего.
Примеры цепей питания можно наблюдать повсюду: листья
растений поедаются гусеницами, гусениц поедают птицы,
птиц поедают лисицы, на них паразитируют клещи и блохи, в
13 — 28,
385
кишечном тракте последних имеются простейшие, бактерии^
а отбросами этих животных и их трупами питаются разные
трупоядные насекомые, после которых минерализацию прово-
дят микроорганизмы. X
Возможность существования низкоорганизованных форм
обуславливается тем, что они занимают среду или потребляю^
продукты, не нужные другим (например, бактерии поедают
органические остатки, которые уже непригодны для потреблен
ния другими организмами). Следовательно, среда разделена на
сферы влияния, своеобразные экологические ниши.
Экологическая ниша — место в биогеоценозе, которое за-
нимает вид, не конкурируя с другими видами за источник!
энергии.
Формы биотических связей
Конкретные формы биотических связей организмов до-
вольно разнообразны. Самыми распространенными биологи^
ческйми взаимоотношениями на антагонистической основе;
являются хищничество, конкуренция и паразитизм, а положи-^
тельной основе — симбиоз и взаимопомощь.
Хищничество. Хищники — это организмы, которые ловят и-
поедают свою жертву (сразу или по частям). Это хищные пти-
цы и рыбы, некоторые рептилии, насекомые; из млекопитаю-
щих — это кошки, собаки, ласки, тюлени, скунсы, моржи.
Конкуренция. Межвидовой конкуренцией называют ак-
тивный поиск представителей двух или нескольких видов од- ;
них и тех же пищевых ресурсов среды обитания. То есть это
любое взаимодействие между двумя или более популяциями, >
которое вредно отражается на их росте и выживании. Конку-
рентные отношения наблюдаются, когда организмы являются
антагонистами в борьбе за пищу, место существования и дру-
гие необходимые для жизни факторы; например, это конку-
ренция между растениями за свет, питательные вещества
почвы.
Паразитизм. Это форма антагонистического сожительства
организмов разных видов, при котором один организм живет
за счет другого (хозяина), используя его в качестве среды
обитания и источника питания, нанося тем самым ему вред.
В зависимости от длительности контакта паразитов с орга-
низмом хозяина различают паразитизм временный и стацио-
нарный (постоянный), когда паразит пребывает в организме
хозяина в течение основного периода своего развития. При-
мерами временного паразитизма являются малярийный
386
длазмодий, паразитические амебы, а примерами стационар-
ного паразитизма — грибы-паразиты растений и животных,
плодожорка, аскариды и др.
Симбиоз (мутуализм, от лат. «мутус» — взаимный) — взаи-
мовыгодное сожительство двух или более организмов. Симби-
онтами могут быть растения, растения и животные или только
животные. Классическим примером симбиоза являются ли-
шайники, а также клубеньковые бактерии с бобовыми, мико-
риза некоторых грибов с корнями деревьев.
Синойкия (греч. «син» — вместе, «ойкос» — дом) — сожи-
тельство, при котором партнер использует другого только как
жилище. Комменсализм — сожительство с использованием
партнера в качестве жилища и в качестве источника питания,
однако без вреда для него. Пресноводная рыба горчак откла-
дывает икринки в мантийную полость двустворчатых моллюс-
ков. Икринки, защищенные раковиной моллюска, развивают-
ся, не нанося вред хозяину.
Некоторые морские полипы, относящиеся к типу кишечно-
полостных, поселяясь на очень крупных рыбах, в качестве
пищи используют их испражнения. В первом примере хозяин
предоставляет своему партнеру только место для жизни, во вто-
ром — и «квартиру», и питание.
Межвидовая взаимопомощь играет большую роль в борьбе
за существование. Примером могут быть птицы, которые
уничтожают личинок-паразитов под кожей у буйвола; птицы,
которые очищают пасть крокодилов от пиявок. В раститель-
ном мире это взаимосвязи между растениями и насекомыми-
опылителями.
Правило экологической пирамиды
Первое звено всякой цепи питания — превращение в про-
цессе фотосинтеза световой энергии в химическую и образова-
ние органических веществ. При этом лишь около 3% световой
энергии, падающей на растение, переходит в потенциальную
энергию органических веществ; остальная рассеивается в виде
тепла. Каждое из последующих звеньев способно использо-
вать лишь 5—12% энергии корма и превратить ее во вновь по-
строенное вещество своего тела. Остальная энергия или пре-
вращается в тепло и рассеивается, или (чаще всего) просто не
усваивается. В среднем коэффициент полезного действия каж-
дого звена цепи не превышает 10%, что неизбежно ведет к
Уменьшению количества образующегося органического веще-
ства на каждом последующем пищевом уровне. Поэтому ти-
13»
487
личная цепь питания состоит не более чем из четырех—шести
взаимно связанных пищевых уровней.
Например, для образования 1 кг массы дельфина нужно
около 10 кг съеденной рыбы, а этим 10 кг нужно 100 кг их кор-
ма — водных беспозвоночных, которым, в свою очередь, для
образования такой массы необходимо съесть 1000 кг водорос- У
лей и бактерий. Если в соответствующем масштабе изобразить
эти величины в порядке их зависимости, то действительно об- (
разуется своеобразная пирамида (рис. 6.50). Различают не- |
сколько категорий экологических пирамид. Пирамида чисел
отражает число особей на каждом уровне пищевой цепи; пира- |
мида биомассы — количественное соотношение органического
вещества; пирамида энергии — количество энергии в пище
каждого уровня.
юоо
Рис. 6.50. Правило экологической пирамиды
388
Антропогенные факторы
По сравнению с рассмотренными факторами среды влия-
ние антропогенных факторов, т. е. влияние человека на расти-
тельный и животный мир, особенно велико. Достаточно ука-
зать на роль человека в создании и распространении новых
видов, сортов, пород и т. д.
Воздействие человека на природу иногда оказывается для
нее губительным. Так, при непосредственном участии челове-
ка за последние 200 лет с лица Земли исчезло 150 видов жи-
вотных. Численность некоторых видов животных и растений
настолько уменьшилась, что возникла угроза их дальнейше-
му существованию. В связи с этим создана международная
«Красная, книга», куда занесены самые ценные виды, кото-
рые находятся под угрозой вымирания и поэтому требуют
тщательной охраны (белый медведь, уссурийский тигр, лан-
дыш майский и т. д.), а также создан «Черный список» —
международный список вымерших видов животных и рас-
тений.
СООБЩЕСТВА И БИОГЕОЦЕНОЗЫ
Разные растения взаимодействуют между собой и образуют
сообщества. От климата и типов растительности в свою оче-
редь зависит распространение животных. Каждое раститель-
ное сообщество, будь то естественное или искусственное,
обычно имеет более или менее определенный набор видов жи-
вотных, формирующийся в соответствии с условиями жизни в'
нем и составом видов растений. Сложные и тесные взаимодей-
ствия между климатом, живыми организмами и физическим
субстратом приводят к образованию характерных региональ-
ных сообществ. Эта совокупность растений, животных и усло-
вий окружающей среды на определенной территории называ-
ется биогеоценозом. Своеобразие любого биогеоценоза зависит
от состава его населения и существующих в нем связей между
популяциями.
Состав населения биогеоценоза определяет условия среды, в
которой живут организмы. В то же время организмы, взаимодей-
ствуя со средой, очень изменяют ее, создавая тем самым специ-
фические черты местообитания, неповторимые в иных условиях.
Обычно условия жизни в биогеоценозе называют биотопом, а'
его население — биоценозом. Биоценоз — это только комплекс
живых организмов в биогеоценозу, т. е. только его часть.
389
Биогеоценозы могут быть разных размеров. Кочка среди .
болота или пень в лесу, нора с ее населением, аквариум — при- .
меры микробиогеоценозов, или микроэкосистем; определен-
ную растительную ассоциацию со всеми ее компонентами (на- ;
пример, ельник-кисличник, ельник-брусничник и т. п.),
озеро можно назвать мезобиогеоценозами, или мезоэкосисте-
мами, а океан, суша и отдельные типы растительности (лес, :
степь, луг и т. п.) являются макробиогеоценозами, или макро-
экосистемами. Совокупность всех биогеоценозов (экосистем)
нашей планеты создает гигантскую глобальную экосистему,
называемую биосферой.
В структуре биогеоценоза можно выделить четыре, звена.
1. Абиотическое окружение, т. е. комплекс факторов нежи-
вой природы, из которой биогеоценоз черпает средства жизни и
куда выделяет продукты обмена.
2. Комплекс зеленых растений, обеспечивающий органи-
ческим веществом все живущее население. Это — продуценты,
или образователи, среди которых есть хемосинтезирующие
организмы.
3. Комплекс всевозможных организмов, живущих за счет
питательных веществ, созданных продуцентами. Это — консу-
менты, или потребители.
4. Комплекс организмов, разлагающих органические со-
единения до минерального состояния. Это — редуценты, или
разлагатели, представленные микроорганизмами — бактерия-
ми, грибками, простейшими.
Между всеми четырьмя звеньями существует тесная взаи-
мосвязь (рис. 6.51).
Местообитание
Местообитание (биотоп) — это не только территория, зани-
маемая сообществом, но и присущий ей комплекс экологичес-
ких условий среды. Различают следующие основные биотопы.
1. Тундра. Это холодная безлесная равнина, занимающая
огромные пространства в Евразии, Северной Америке и Грен-
ландии, опоясывающая Северную полярную область.
2. Хвойные леса. Непосредственно к югу от тундры лежит
огромная зона хвойного леса, проходящая по всему Северному
полушарию.
3. Лиственные леса. К югу от хвойных лесов лежит область
листопадных лесов умеренной зоны. Цивилизация достигла
наибольшего развития именно в этих районах, и в результате от
некогда огромных лесных массивов почти ничего не осталось.
390
Рис. 6.51. Структура биогеоценоза
4. Влажный тропический лес. Эта зона, опоясывающая Зем-
лю по экватору, имеет наиболее сложную структуру. Число
различных видов растений здесь необычайно велико.
5. Степные биомы. Зона постепенного перехода от лесного
биома к степному называется лесостепью. В тропиках наиболее
обширную область такого типа образуют африканские саванны.
Там, где есть реки, в саванну вклиниваются полосы леса (так на-
зываемые галерейные леса). Наиболее известные степные био-
мы — это степи Старого Света, североамериканские прерии, юж-
ноамериканские льяносы и пампа и африканские вельды.
6. Пустыни. Одну седьмую часть всей поверхности суши за-
нимают области, где дожди не восполняют потерю влаги в ре-
зультате испарения. Для пустынь характерен недостаток воды,
а в Центральной Сахаре имеются районы, где<совсем не бывает
дождей.
Между этими сообществами, или биомами, существуют пе- |
реходы, которые могут быть постепенными или сравнительно
резкими. В том и другом случае имеется переходная зона —
так называемый экотоп. Здесь встречаются виды, характерные
для обоих биомов, а иногда еще и виды, свойственные только
переходной области.
Вертикальная зональность
Как мы уже отмечали, климат изменяется не только в.зави-
симости от широты и местных условий, но и в зависимости от
высоты над уровнем моря. Поэтому можно ожидать,-что в го-
рах распространение живых организмов изменяется аналогич-
ным образом. Здесь может быть 4—5 узких растительных по-
ясов, причем последовательность их направления снизу вверх
соответствует последовательности широтных зон от экватора к
полюсу на одной и той же высоте (снизу вверх: пустыни — луга
и степи — карликовый хвойный лес — северный хвойный
лес — тундра — снега и ледники).
Биологические зоны океана
В морях и океанах на организмы оказывают влияние многие
факторы: глубина бассейна, соленость воды, освещенность, :
давление, насыщенность кислородом, температура, характер
морского дна и т. д. Из перечисленных факторов наибольшее
влияние на организм оказывает глубина и соленость воды. Ос-
новная масса морских организмов сосредоточена на глубине до
200 м, где наиболее благоприятны условия обитания.
Бентосные организмы населяют практически все биономи- :
ческие зоны моря (рис. 6.52). Эти зоны — литораль, сублито-
раль, эпибатиаль, батиаль и абиссаль — четко выделяются на у
дне океана в зависимости от его глубины. Первые три зоны час- I
то объединяются под названием неритовой области. Литоралью |
называется прибрежная зона приливов и отливов; живущие |
здесь организмы приспособились к периодическим осушениям i
и сильным волнениям воды. Далее до глубины 200 м следует J
сублитораль — наиболее благоприятная зона для обитания •
организмов. Ниже, от 200 до 500 м, находится эпибатиаль; здесь |
исчезают растения и резко уменьшается число животных. Бати- 1
аль расположена на континентальном склоне до глубины 1000— 1
3000 м. Еще глубже — абиссаль; эта зона охватывает огромные |
площади океанического дна. Органический мир батиали и I
абиссали весьма беден. |
392 I
Сукцессия и виды сообществ
Биогеоценоз обладает саморегуляцией, т. е. способностью к
восстановлению внутреннего равновесия после какого-либо
воздействия природного или антропогенного фактора. Под
влиянием природных факторов внешней среды происходит
смена биогеоценозов — сукцессия: на месте болот образуются
луга, на месте лугов — леса. Смена биогеоценозов может быть
вызвана также стихийными бедствиями (пожар, массовое раз-
множение вредителей) или влиянием человека (вырубка леса,
осушение или орошение земель). Восстановление биогеоценоза
может происходить естественным путем, что длится на протя-
жении нескольких этапов (после пожара или вырубки еловый
лес восстанавливается более 100 лет). Искусственное восстанов-
ление биоценоза происходит за счет комплекса мероприятий,
обеспечивающих возобновление прежнего биоценоза: посев се-
мян, посадка саженцев деревьев, возвращение исчезнувших жи-
вотных.
Фитоценоз (от греч. «фитон» — растение, «ценоз» — об-
щий) — растительное сообщество, исторически сложившееся
в результате сочетания взаимодействующих растений на од-
нородном участке территории. Его характеризуют определен-
ный видовой состав, жизненные формы, ярусность (надзем-
ная и подземная), обилие видов, размещение, внешний вид,
жизненность, сезонные изменения, развитие (смена сооб-
ществ).
Агроценоз (от греч. «агрос — поле, «ценоз» — общий). Это
искусственно созданный человеком биоценоз. Он не способен
393
длительно существовать без вмешательства человека, не обла-
дает саморегуляцией и в то же время характеризуется высокой
продуктивностью (урожайностью) одного или нескольких ви-
дов (сортов) растений или животных.
Биоценоз (от 1реч. «био» — жизнь, «ценоз» — общий) — со-
общество растений и животных, населяющих одну террито-
рию, взаимно связанных в цепи питания и влияющих друг на
друга.
БИОСФЕРА И УЧЕНИЕ В. И. ВЕРНАДСКОГО
Биосферой называется оболочка Земли, которая населена
и активно преобразуется живыми существами. Согласно
В. И. Вернадскому, биосфера — это такая оболочка, в кото-
рой существует или существовала в прошлом жизнь и кото-
рая подвергается воздействию живых организмов. Она вклю-
чает в себя:
1) живое вещество, образованное совокупностью организмов;
2) биогенное вещество, которое создается и перерабатывает-
ся в процессе жизнедеятельности организмов (газы атмосфе-
ры, каменный уголь, нефть, сланцы, известняки и др.);
3) южное вещество, которое образуется без участия живых
организмов (продукты тектонической деятельности, метеориты);
4) биокосное вещество, представляющее собой совместный
результат жизнедеятельности организмов и абиогенных про-
цессов (почвы).
В XX веке впервые в истории Земли человек узнал и охватил
всю биосферу, закончил географическую карту Земли, рассе-
лился по всей ее поверхности. Человечество стало единым це-
лым. На Земле нет ни одного места , где бы человек не мог
прожить, если бы это было ему нужно. Эволюция человека пе-
решла на новую качественную ступень, а именно: все человече-
ство как единый целый организм стало объектом эволюционного
процесса. В ходе геологического времени вектор эволюционно-
го процесса теперь связан с усовершенствованием централь-
ной нервной системы (мозга). Это крупное эмпирическое
обобщение получило название «принципа цефализации
Дана». Результат цефализации Дана отражается не столько в.
росте человеческого мозга, сколько в разумной деятельности
человека. Если поставить всех людей рядом, они не заняли бы
площади более нескольких десятков квадратных километров.
Таким образом, человечество представляет ничтожную массу
вещества планеты. Мощь его связана не столько с этим самым
живым веществом, сколько с его мозгом, с его разумом и с на-
правленным этим разумом его трудом. Исторический процесс
на наших глазах меняется коренным образом. Впервые в исто-
рии человечества интересы народных масс — и всех людей и
каждой свободной личности отдельно — определяют жизнь
человечества, являются материалом его представлений о спра-
ведливости. Человечество, взятое в целом, становится мощной
геологической силой, и перед его мыслью и трудом возникает
вопрос о перестройке биосферы в интересах свободно мысля-
щего человечестЬа как единого целого. Это новое состояние
биосферы и есть «ноосфера». Ноосфера — новое геологическое
явление на нашей планете. В йей впервые человек становится
крупнейшей геологической силой. Он может и должен корен-
ным образом перестраивать своим трудом и мыслью область
своей жизни. В геологической истории биосферы перед челове-
чеством открывается светлое будущее, но только в том случае,
если он осознает свою миссию и не будет направлять свой ра-
зум и труд на самоистребление.
ПРИМЕРНАЯ ПРОГРАММА
. ВСТУПИТЕЛЬНЫХ ЭКЗАМЕНОВ
(ИСПЫТАНИЙ) ПО БИОЛОГИИ
ОБЩИЕ УКАЗАНИЯ
На экзамене по биологии поступающий в высшее учебное
заведение должен показать:
— знание основных понятий, закономерностей и законов в
области строения, жизни и развития растительного, животного
организмов и человека, развития в целом органического мира;
— знание строения и жизни растений, животных, человека,
классификации растений и животных;
— умение обосновывать выводы, используя биологические
термины, объяснять явления природы, применять знания в
практической деятельности.
РАСТЕНИЯ
Общее знакомство с цветковыми растениями
Цветковое растение и его органы: корень и побег; строение
побега: стебель, листья, почки; цветок — видоизмененный по-
бег. Плоды и семена, приспособленность их к распростране-
нию в природе. Состав растений (органические и неорганичес-
кие вещества).
Клеточное строение растительного
организма
Знакомство с увеличительными приборами (лупа, микро-
скоп).
Клетка и ее строение: оболочка, цитоплазма, ядро, пластиды,
вакуоли. Жизнедеятельность клетки: движение цитоплазмы, по-
ступление веществ в клетку, ее рост и деление. Ткани.
Клеточное строение растений.
396
Корень
Корень. Виды корней. Типы корневых систем. Почва, ее зна-
чение для жизни растений. Охрана почв.
Внешнее и внутреннее строение корня. Зоны корня. Рост
корня. Основные функции корня: поглощение воды и мине-
ральных веществ, укрепление растения в почве. Дыхание кор-
ня. Удобрения. Значение обработки почвы, внесения удобре-
ний. Корнеплоды, их использование человеком.
Побег
Побег. Почка — зачаточный побег, ее строение. Развитие по-
бега из почки.
Лист. Внешнее строение листа. Жилкование. Листья про-
стые и сложные. Листорасположение.
Особенности микроскопического строения листа в связи с
его функциями: покровная ткань (кожица, устьица), основная,
проводящая и механическая ткани листа.
Фотосинтез.
Дыхание. Испарение воды листьями. Видоизменения листь-
ев. Листопад. Необходимость зашиты воздуха от загрязнения. -
Озеленение населенных пунктов и помещений.
Стебель. Рост стебля в длину, ветвление. Формирование кро-
ны. Внутреннее строение древесного стебля в связи с его функ-
циями. Рост стебля в толщину. Образование годичных колец.
Передвижение минеральных и органических веществ по стеб-
лю. Отложение запасных веществ.
Видоизмененные побеги: корневище, клубень, луковица, их
строение, биологическое и хозяйственное значение.
Размножение растений
Размножение и его значение. Способы размножения.
Вегетативное размножение. Размножение растений с помо-
щью вегетативных органов в природе и растениеводстве (видо-
измененными побегами, черенками, отводками, делением кус-
та, прививкой). Биологическое и хозяйственное значение веге-
тативного размножения. Размножение растений семенами.
Цветок — видоизмененный побег. Значение цветка в размноже-
нии растений. Строение околоцветника, тычинки, пестика.
Соцветия и их биологическое значение. Перекрестное опыле-
ние насекомыми, ветром. Самоопыление.
Оплодотворение. Образование семян и плодов. Значение
плодов и семян в природе и жизни человека. Вред, наноси-
397
мый природе массовым сбором дикорастущих растений.
Охрана цветковых растений.
Строение семян (на примере двудольных и однодольных ра-
стений), их химический состав. Условия прорастания семян.
Дыхание семян. Питание и рост проростков. Агротехника посе-
ва семян и выращивания растений.
Растения
и окружающая среда
Растение — целостный организм. Взаимосвязи клеток, тка-
ней и органов. Основные процессы жизнедеятельности расти-
тельного организма. Растительное сообщество. Экологические
факторы неживой и живой природы и связанные с деятельнос-
тью человека. Взаимосвязь растений и факторов неживой и жи-
вой природы на примере растений леса, луга и др. Приспособ-
ленность растений к совместной жизни в лесу, на лугу и т. д.
Роль растений в природе и жизни человека.
Влияние деятельности человека на жизнь растений леса, луга.
Охрана растений, защита среды их обитания, законы об охране
природы.
Отделы растений
Водоросли. Строение и жизнедеятельность одноклеточных
и многоклеточных водорослей. Размножение водорослей. Нит-
чатые водоросли. Морские водоросли. Роль водорослей в при-
роде и народном хозяйстве, их охрана.
Мхи. Строение и размножение (на примере местных видов).
Образованиеторфа, его значение.
Папоротники. Строение и размножение, роль в природе и
жизни человека. Хвощи. Плауны.
Голосеменные. Строение и размножение (на примере сосны,
ели и других хвойных). Распространение хвойных, их значение
в природе, народном хозяйстве.
Покрытосеменные (цветковые). Особенности строения и
жизнедеятельности покрытосеменных как наиболее высокоор-
ганизованной группы растений, их господство на Земле.
Покрытосеменные растения
Многообразие цветковых растений.
Класс Двудольные растения. Семейства: крестоцветные (ка-
пустные), розоцветные, бобовые, пасленовые, сложноцветные
(астровые), мальвовые, маревые, виноградные.
Класс Однодольные растения. Семейства: лилейные,
злаки.
398
Отличительные признаки растений: перечисленных се-
мейств, их биологические особенности, народнохозяйственное
значение.
Влияние деятельности человека на видовое многообразие
цветковых растений. Охрана редких видов цветковых растений.
Сельскохозяйственные растения
Происхождение культурных растений. Понятие сорта. До-
стижения науки в выведении новых сортов растений.
Важнейшие сельскохозяйственные растения (зерновые, пло-
дово-ягодные, овощные, масличные, технические и др.), био-
логические основы их выращивания.
Рштие растительного мира
Многообразие растений и их происхождение. Доказательства
исторического развития растений: ископаемые остатки.
Основные этапы в развитии растительного мира: возникно-
вение одноклеточных и многоклеточных водорослей; возник-
новение фотосинтеза; выход растений на сушу (псилофиты, мхи,
папоротники, голосеменные, покрытосеменные). Усложнение
растений в процессе исторического развития.
Господство покрытосеменных в настоящее время, их много-
образие и распространение на земном шаре. Влияние хозяй-
ственной деятельности человека на растительный мир. Охрана
растений.
БАКТЕРИИ. ГРИБЫ. ЛИШАЙНИКИ
Бактерии
Строение и жизнедеятельность бактерий. Размножение бак-
терий. Распространение бактерий в воздухе, почве, воде, живых
организмах. Роль бактерий в природе, промышленности, меди-
цине, сельском хозяйстве. Болезнетворные бактерии и борьба с
ними.
Грибы. Лишайники
Общая характеристика грибов. Шляпочные грибы, их строе-
ние, питание. Симбиоз грибов с растениями. Съедобные и ядо-
витые грибы. Правила сбора грибов и их охрана. Профилактика
отравления ядовитыми грибами. Плесневые грибы. Пеницилл,
его использование для получения антибиотиков. Дрожжи. Гри-
бы-паразиты, вызывающие болезни растений. Роль грибов в
природе и хозяйстве.
Строение лишайника. Симбиоз. Питание. Размножение.
Роль лишайников в природе и хозяйстве.
ЖИВОТНЫЕ
Общие сведения о животном мире
Многообразие животного мира. Основные отличия животных
от растений, черты их сходства. Систематика животных.
Одноклеточные животные
Обыкновенная амеба. Особенности строения клетки одно-
клеточного организма. Среда обитания. Передвижение. Пита-
ние. Дыхание. Выделение. Размножение. Образование цисты.
Многообразие одноклеточных животных. Зеленая эвглена, осо-
бенности ее строения и питания. Инфузория-туфелька. Раздра-
жимость. Малярийный паразит. Морские простейшие (форами-
ниферы, радиолярии).
Значение простейших в природе, жизни человека. Общая ха-
рактеристика простейших.
Тип Кишечнополостные
Пресноводная гидра. Среда обитания. Внешнее строение.
Лучевая симметрия. Двухслойность. Особенности строения
клетки многоклеточного животного организма. Покровные,
стрекательные, нервные клетки. Нервная система. Рефлекс.
Питание. Регенерация. Размножение.
Многообразие кишечнополостных (коралловые полипы и
медузы), их значение.
Общая характеристика типа.
Типы Плоские, Круглые,
Кольчатые черви
Тип Плоские черви, их многообразие.'Печеночный сосаль-
щик. Двусторонняя симметрия. Особенности строения и про-
цессов жизнедеятельности, обусловленные паразитическим
образом жизни; вред, наносимый животноводству, меры борь-
бы. Многообразие видов (бычий цепень, эхинококк, белая пла-
нария). Общая характеристика типа.
Тип Круглые черви. Человеческая аскарида — паразит чело-
века. Меры предупреждения заражения.
Тип Кольчатые черви, их многообразие. Дождевой червь, его
среда обитания, внешнее строение, передвижение. Ткани, орга-
400
ны, системы органов. Процессы жизнедеятельности. Регенера-
ция. Размножение. Роль дождевых червей в почвообразовании.
Общая характеристика типа.
Тип Моллюски
Беззубка. Среда обитания, особенности внешнего строения,
питания, дыхания, размножения.
Многообразие моллюсков (большой прудовик, виноградная
улитка, слизни, устрица, мидия}, их значение в природе, жиз-
ни человека.
Общая характеристика типа.
Тип Членистоногие
Класс Ракообразные. Среда обитания ракообразных. Осо-
бенности строения, жизнедеятельности; размножение, много-
образие ракообразных. Общая характеристика класса.
Класс Паукообразные. Особенности внешнего строения, пи-
тания, дыхания, поведения паука в связи с жизнью на суше.
Клещи. Внешнее строение. Клещи — вредители культурных
растений и меры борьбы с ними. Паразитические клещи — воз-
будители и переносчики опасных болезней. Меры защиты от
клещей. Общая характеристика класса.
Класс Насекомые. Особенности строения, процессов жиз-
недеятельности, насекомых на примере жука. Размножение.
Типы развития насекомых.
Основные отряды насекомых. Чешуекрылые. Черты приспо-
собленности к среде обитания во внешнем строении; размно-
жение и развитие бабочек. Тутовый шелкопряд. Шелководство.
Двукрылые. Комнатная муха — переносчик возбудителей
опасных заболеваний человека и меры борьбы с ней.
Перепончатокрылые. Медоносная пчела. Состав и жизнь
пчелиной семьи: танцы пчел, зимовка. Инстинкты — основы
поведения насекомых. Пчеловодство. Общая характеристика
класса.
Многообразие насекомых (колорадский жук, муравьи, на-
ездник), их роль в природе; практическое и эстетическое зна-
чение. Биологический способ борьбы с насекомыми — вре-
дителями сельскохозяйственных культур и его роль в сохра-
нении урожая. Охрана насекомых.
Общая характеристика типа.
- Тип Хордовые
Ланцетник. Среда обитания. Особенности строения ланцет-
ника как низшего хордового.
401
Класе Рыбы
Среда обитания рыб. Особенности внешнего строения, ске- 3
лета и мускулатуры. Полость тела. Особенности строения сис-
тем внутренних органов в связи с их функциями. Обмен ве-
ществ. Нервная система и органы чувств. Рефлексы. Поведе-
ние.
Размножение, нерест и развитие. Забота о потомстве. При-
способленность рыб к среде обитания. Миграции. Многообра-
зие рыб (отряды: сельдеобразные, кистеперые).
Общая характеристика класса.
Хозяйственное значение рыб. Искусственное разведение
рыб, прудоводство. Охрана рыб.
Класс Земноводные
Лягушка. Особенности строения, передвижения в связи со
средой обитания. Нервная система и органы чувств. Размно-
жение и развитие.
Многообразие земноводных (отряды: хвостатые, бесхвостые),
их происхождение, значение и охрана.
Общая характеристика класса.
Класс Пресмыкающиеся
Ящерица. Среда обитания, особенности строения, размно-
жения, поведения в связи с жизнью на суше. Регенерация.
Многообразие современных пресмыкающихся (отряды: че-
шуйчатые, черепахи, крокодилы), их практическое значение и
охрана. Происхождение пресмыкающихся. Древние пресмыка-
ющиеся: динозавры, зверозубые ящеры.
Общая характеристика класса.
Класс Птицы
Внешнее строение, скелет, мускулатура. Особенности внут-
реннего строения, обмена веществ птицы, связанные с поле-
том. Усложнение нервной системы, органов чувств; поведение
птиц.
Размножение и развитие птиц. Забота о потомстве.
Приспособленность птиц к сезонным явлениям в природе
(гнездование, кочевки, перелеты). Происхождение птиц.
Общая характеристика класса.
Птицы парков, садов, лугов, полей, лесов, болот; побережий,
водоемов, степей, пустынь, хищные птицы.
Роль птиц в природе и жизни человека, система мероприя-
тий по охране птиц.
Птицеводство. Происхождение домашних птиц, их породы.
Класс Млекопитающие
Особенности внешнего строения, скелета, мускулатуры, внут-
реннего строения, обмена веществ млекопитающего. Усложне-
ние нервной системы, органов чувств, поведения. Размножение
и развитие, забота о потомстве.
Общая характеристика класса.
Происхождение млекопитающих. Первозвери. Сумчатые.
Отрады Плацентарных. Насекомоядные и рукокрылые. Гры-
зуны. Зайцеобразные. Хищные. Ластоногие и китообразные. Ко-
пытные. Приматы.
Роль млекопитающих в природе и жизни человека, их
охрана.
Сельскохозяйственные животные класса млекопитающих.
Крупный рогатый скот, овцы, свиньи, лошади. Происхож-
дение домашних животных. Содержание, кормление и разве-
дение.
эволюция
ЖИВОТНОГО МИРА
Доказательства исторического развития животного мира:
сравнительно-анатомические, эмбриологические, палеонтоло-
гические.
Ч. Дарвин о причинах эволюции животного мира (наслед-
ственность, изменчивость, естественный отбор). Происхожде-
ние одноклеточных. Происхождение многоклеточных. Услож-
нение строения и жизнедеятельности позвоночных животных в
процессе исторического развития животного мира. Родство че-
ловека с животными.
Несостоятельность взглядов о неизменности животного
мира.
ПРИРОДНЫЕ СООБЩЕСТВА
Среда обитания организмов. Основные экологические
факторы среды, их влияние на растения и животных. При-
родные сообщества (на примере леса, луга, водоема). Роль
растений, животных, грибов и бактерий в природном сооб-
ществе. Взаимосвязи в природном сообществе. Цепи пита-
ния.
Значение природных сообществ в жизни человека. Вли-
яние деятельности человека на природные сообщества, их
охрана.
403
ЧЕЛОВЕК И ЕГО ЗДОРОВЬЕ
Введение. Общий обзор организма
человека
Значение знаний о строении, жизнедеятельности организ-
ма человека и гигиене для охраны его здоровья. Человек и
окружающая среда.
Органы и системы органов. Строение клетки (цитоплазма,
ядро, рибосомы, митохондрии, мембрана). Основные процес-
сы жизнедеятельности клетки (питание, дыхание, деление).
Краткие сведения о строении и функциях основных тканей.
Рефлексы. Нервная и гуморальная регуляция деятельности орга-
низма. Организм — единое целое.
Опорно-двигательная система
Значение опорно-двйгательнОй системы. Скелет человека,
сходство скелетов человека и животных. Особенности скелета ’
человека, связанные с трудовой деятельностью и прямохожде-
нием. Типы соединения костей. Состав, строение и свойства
костей, рост костей. Первая помощь при ушибах, растяжениях
связок, вывихах, переломах.
Мышцы, их функции. Основные группы мышц тела челове-
ка. Работа мышц. Статическая и динамическая нагрузки. Влия-
ние ритма и нагрузки на работу мышц.
Кровь и кровообращение
Внутренняя среда организма (кровь, межклеточная жид- 4
кость, лимфа) и ее относительное постоянство. Значение крови »'
и кровообращения. Состав крови. Плазма крови. Свертывание 4
крови как защитная реакция организма. Строение и функции |
эритроцитов и лейкоцитов. Иммунитет. Роль И. И. Мечни- |
кова в создании учения об иммунитете. Инфекционные за- |
болевания и борьба с ними. Предупредительные прививки. I
СПИД и борьба с ним. Группы крови. Переливание крови. До- |
норство. |
Органы кровообращения: сердце и сосуды (артерии, капил- 1
ляры, вены). Сердце, его строение и работа. Большой и малый |
круги кровообращения, лимфообращение. Движение крови по |
сосудам. Кровяное давление. Нервная и гуморальная регуляция 1
деятельности сердца и сосудов. Предупреждение сердечйо-со- |
судистых заболеваний. Первая помощь при кровотечениях. 1
Вредное влияние курения и употребления алкоголя на сердце и !
сосуды. J
404 1
Дыхание
Значение дыхания. Строение и функции органовдыхания.
Голосовой аппарат. Газообмен в легких и тканях. Дыхатель-
ные движения. Жизненная емкость легких. Нервная и гумо-
ральная регуляция дыхания. Искусственное дыхание. Инфек-
ционные болезни, передающиеся через воздух, предупрежде-
ние воздушно-капельных Инфекций, гигиенический режим во
время болезни. Гигиена органов дыхания. Вредное влияние
курения на органы дыхания. Охрана окружающей воздушной
среды.
Пищеварение
Значение пищеварения. Питательные вещества и пищевые
продукты. Строение и функции органов пищеварения. Зубы,
профилактика болезней зубов. Пищеварительные ферменты и
их значение. Роль Н.П. Павлова в изучении функций органов
пищеварения. Пищеварение. Печень и поджелудочная железа,
их роль в пищеварении. Всасывание. Регуляция процессов пи-
щеварения. Гигиенические условия нормального пищеварения.
Предупреждение глистных и желудочно-кишечных заболева-
ний, пищевых отравлений, первая доврачебная помощь при них.
Влияние курения и употребления алкоголя на пищеварение.
Обмен веществ и энергии.
Выделение
Общая характеристика обмена веществ и энергии. Пласти-
ческий обмен, энергетический обмен и их взаимосвязь. Значе-
ние для организма белков, жиров и углеводов, воды и минераль-
ных солей. Влияние алкоголя и токсических веществ, наркоти-
ков на обмен веществ. Витамины. Их роль в обмене веществ.
Основные гиповитаминозы. Пшервитаминозы.
Способы сохранения витаминов в пищевых продуктах. Нор-
мы питания. Рациональное питание. Режим питания школь-
ников.
Значение выделения.из организма конечных продуктов об-
мена веществ. Органы мочевыделительной системы, их функ-
ции, профилактика заболеваний.
Кожа
Строение и функции кожи. Роль кожи в теплорегуляции. За-
каливание организма. Гигиена кожи, гигиенические требования
к одежде и обуви. Профилактика и первая помощь при тепло-
вом и солнечном ударах, ожогах и обморожениях, электрошоке.
405
Железы внутренней секреции
Значение желез внутренней секреции для роста, развития
и регуляции функций организма. Гормоны. Внутрисекретор-
ная деятельность желез внутренней секреции. Роль половых
желез в развитии организма. Половое созревание. Гигиена
юноши и девушки.
Нервная система. Органы чувств. Высшая нервная
деятельность
Значение нервной системы в регуляции и согласованности
функций организма человека и взаимосвязи организма со сре-
дой. Центральная и периферическая нервная система.
Строение и функции спинного мозга и отделов головного
мозга. Роль вегетативной нервной системы в регуляции работы
внутренних органов.
Кора больших полушарий. Органы чувств, их значение.
Анализаторы. Строение, функции, гигиена.
Роль И.М. Сеченова и И.П. Павлова в создании учения о
высшей нервной деятельности.
Безусловныеи условные рефлексы. Биологическое значение
образования и торможения условных рефлексов. Особенности
высшей нервной деятельности человека. Речь и мышление. Со-
знание как функция мозга. Социальная обусловленность пове-
дения человека.
Сон, его значение и гигиена. Изменение работоспособности
в трудовом процессе. Гигиена умственного труда. Режим для
школьников. Вредное влияние никотина, алкоголя и наркоти-
ков на нервную систему.
Размножение и развитие
Система органов размножения.
Оплодотворение и внутриутробное развитие. Рождение ре-.
бенка. Рост и развитие ребенка. Гигиена грудных детей. Вредное
влияние алкоголя, никотина и других факторов на потомство.
Характеристика подросткового период а.
ОБЩАЯ БИОЛОГИЯ
Изучение общих биологических закономерностей — задача
заключительного раздела курса биологии. Уровни организации
живой природы: клеточный, организменный, видовой, биоце-
нотический, биосферный. Значение биологической науки для
сельского хозяйства, промышленности, медицины, гигиены,
охраны природы.
406
Эволюционное учете
Основные положения эволюционной теории Ч. Дарвина. Зна-
чение теории эволюции для развития естествознания.
Критерии вида. Популяция — единица вида и эволюции.
Понятие сорта растений и породы животных.
Движущие силы эволюции: наследственность, изменчивость,
борьба за существование, естественный отбор. Ведущая роль ес-
тественного отбора в эволюции.
Искусственный отбор и наследственная изменчивость—ос-
нова выведения пород домашних животных и сортов культур-
ных растений.
Возникновение приспособлений. Относительный характер
приспособленности.
Микроэволюция. Видообразование.
Результаты эволюции: приспособленность организмов, мно-
гообразие видов.
Развитие органического мира
Доказательства эволюции органического мира. Главные на-
правления эволюции. Ароморфоз, идиоадаптация. Соотноше-
ние различных направлений эволюции. Биологический про-
гресс и регресс.
Краткая история развития органического мира. Основные
ароморфозы в эволюции органического мира. Основные направ-
ления эволюции покрытосеменных, насекомых, птиц и млеко-
питающих в кайнозойскую эру
Происхождение человека
Ч. Дарвин о происхождении человека от животных.
Движущие силы антропогенеза: социальные и биологичес-
кие факторы. Ведущая роль законов общественной жизни в
социальном прогрессе человечества.
Древнейшие, древние и ископаемые люди современного
типа.
Человеческие расы, их происхождение и единство. Антина-
учная, реакционная сущность социального дарвинизма и ра-
сизма. .
Основы экологии
Предмет и задачи экологии. Экологические факторы: абио-
тические, биотические, антропогенный, их комплексное воз-
действие на организм.
Фотопериодизм. Среды жизни. Экологическая ниша.
407
Вид, его экологическая характеристика. Популяция, изме-
нение ее численности, способы регулирования численности. Ра-
циональное использование видов, сохранение их разнообразия.
Биогеоценоз. Разнообразие популяций в биогеоценозе и их
взаимосвязи. Цепи питания.
Основы учения о биосфере
Биосфера. В.И. Вернадский о возникновении биосферы. Гра-
ницы биосферы. Биомасса поверхности суши, Мирового океа-
на, почвы. Живое вещество и его функции. Круговорот веществ
и превращение энергии в биосфере. Ноосфера.
Основы цитологии
Основные положения клеточной теории. Клетка — струк-
турная и функциональная единица живого. Строение и функ-
ции ядра, оболочки, цитоплазмы и ее основных органоидов.
Особенности строения клеток прокариот, эукариот. •
Содержание химических элементов в клетке. Вода и другие
неорганические вещества, их роль в жизнедеятельности клетки.
Органические вещества:‘углеводы, липиды, белки, нуклеино-
вые кислоты, АТФ, биополимеры, их роль в клетке. Ферменты,
их роль в регуляции процессов жизнедеятельности. Самоудвое-
ниеДНК.
Обмен веществ и превращение энергии — основа жизнедея-
тельности клетки. Энергетический обмен в клетке и его сущ-
ность. Значение АТФ в энергетическом обмене.
Пластический обмен. Фотосинтез. Биосинтез белков. Ген
и его роль в биосинтезе. Код ДНК. Реакции матричного син-
теза. Взаимосвязь процессов пластического и энергетического
обмена.
Вирусы, особенности их строения и жизнедеятельности.
Вирус СПИДа.
Профилактика СЦИДа.
Размножение и индивидуальное развитие
организмов
Деление клетки — основа размножения и индивидуального
развития организмов. Подготовка клетки кделению. Удвоение
молекул ДНК. Синтез белка. Хромосомы, их гаплоидный и ди-
плоидный набор, постоянство числа и формы. Фазы деления
клетки. Значение деления клетки.
Половое и бесполое размножение организмов. Половые клет-
ки. Мейоз. Развитие яйцеклеток и сперматозоидов. .Оплодот-
ворение;
408
Развитие зародыша (на примере животных). Постэмбрио-
нальное развитие. Вредное влияние алкоголя и никотина на
развитие организма человека.
Возникновение жизни на Земле.
Основы генетики
Генетика — наука о наследственности и изменчивости орга-
низмов. Основные методы генетики. Моно- и дигибридное скре-
щивание. Анализ потомства.
Законы наследственности, установленные Г. Менделем. До-
минантные и рецессивные признаки. Аллельные гены. Фено-
тип и генотип. Гомозигота и гетерозигота. Единообразие пер-
вого поколения.
Промежуточный характер наследования. Закон расщепления
признаков. Статистический характер явлений расщепления.
Цитологические основы единообразия первого поколения и рас-
щепления признаков во втором поколении. Закон независимо-
го наследования и его цитологической основы.
Сцепленное наследование. Нарушение сцепления. Пере-
крест хромосом.
Генотип как целостная исторически сложившаяся система.
Генетика пола. Хромосомная теория наследственности.
Значение генетики для медицины и здравоохранения. Вред-
ное влияние никотина, алкоголя и наркотиков на наследствен-
ность человека.
Роль генотипа и условий внешней среды в формировании,
фенотипа. Модификационная изменчивость. Норма реакции.
Статистические закономерности модификационной изменчи-
вости.
Мутации, их причины. Закон гомологических рядов в на-
следственной изменчивости, сформулированный Н.И. Вавило-
вым. Экспериментальное получение мутаций. Мутации как
материал д ля искусственного и естественного отбора. Загрязне-
ние природной среды мутагенами и его последствия.
Генетика и теория эволюции. Генетика популяций. Формы
естественного отбора: движущий и стабилизирующий.
Основы селекции
Н.И. Вавилов о происхождении культурных растений.
Основные методы селекции: гибридизация и искусственный
отбор. Роль естественного отбора в селекции.
Селекция растений. Самоопыление перекрестноопыляемых
растений. Гетерозис. Полиплоидия и отдалённая гибридиза-
ция. Достижения селекции растений.
409
Селекция животных. Типы скрещивания и методы разведе-
ния. Метод анализа наследственных хозяйственно-ценных при-
знаков у животных-производителей. Отдаленная гибридизация
ломавших животных.
Биотехнология и ее основные направления: микробиологи-
ческий синтез, генная и клеточная инженерия. Значение био-
технологии для селекции.
СОДЕРЖАНИЕ
БОТАНИКА
Растительные ткани................. ......... 4
Корень..................................... ... 16
Стебель.............................. ........ 22
Лист........................................... 31
Раздражимость и движение растительных
организмов................................... 36
Таксономические категории и таксоны.
Бинарная номенклатура.’...................... 38
Надцарство доядерные организмы
(прокариоты). Царство дробянки................ 39
Надцарство ядерные организмы
(эукариоты)................................... 45
Царство грибы...................................45
Царство растения............................. 51
АНАТОМИЯ И ФИЗИОЛОГИЯ
Краткий исторический очерк развития
анатомии................................ 96
Опорно-двигательный аппарат...............100
Кровеносная система.............;....... 129
Дыхание 154
Пищеварение...............................170
Обмен веществ и энергии................. 183
Выделение............................... 200
Внутренняя секреция.......................207
Размножение............................. 219
Нервная система 227
Органы чувств 248
Высшая нервная деятельность человека 266
ЭВОЛЮЦИЯ И экология
ЭВОЛЮЦИЯ ЖИЗНИ НА ЗЕМЛЕ....................276
Космическая эволюция материи...............276
Геологическая эволюция на Земле ...........279
Химическая эволюция и происхождение жизни...288
Эволюция организмов........................292
Эволюционное учение...................... 325
411
Жизнь и второй закон термодинамики.....374
ОСНОВЫ ЭКОЛОГИИ........................379
Экологические факторы..................379
Биосфера и учение В. И. Вернадского....394
ПРИМЕРНАЯ ПРОГРАММА
ВСТУПИТЕЛЬНЬЕХ ЭКЗАМЕНОВ
(ИСПЫТАНИЙ) по биологии..................396
Учебное издание
Чебышев Николай Васильевич, Кузнецов Сергей Владимирович,
Зайчикова Светлана Геннадьевна, ^ленков Сергей Иванович
БИОЛОГИЯ
Том 2
Редактор Я. А. Литвина
Компьютерная верстка Я. В. Ивановой
Технический редактор Я. В. Яшкова
Корректор Я. С Хачанян
ИД № 03434 от 15.12.00.
ИД № 03433 от 15.12.00.
Подписано в печать 27.02.2002. Формат 84 х 108 732.
Бумага типографская. Печать высокая. Гарнитура Ньютон.
Усл. печ. л. 21,84. Уч.-изд. л. 23,22. Тираж 5 000 экз. Заказ № 281
ООО «Издательство Новая Волна».
107078, Москва, ул. Садовая-Спасская, 3.
Тел. (095) 208-38-92, факс (095) 207-04-11.
Интернет/Home page — www.newwave.msk.ru
Электронная почта/E-mail — sales@newwave.msk.ru
Издатель Умеренков.
Текст отпечатан с готовых диапозитивов
в ФГУП Владимирская книжная типография.
600000, Владимир, Октябрьский проспект, 7.
Издательство «Новая Волна»
осуществляет уникальную программу
издания учебных пособий для поступающих в вузы
Приобретая наши издания,
вы получаете великолепную возможность
воспользоваться знаниями и опытом
самых авторитетных авторов,
советы которых уже помогли стать
студентами лучших учебных, заведений
десяткам миллионов абитуриентов
По вопросам оптовых закупок и «книги-почтой»
обращаться в отдел реализации издательства:
101000, г. Москва, Главпочтамт, а/я 251
тел. (095) 208-38-92
тел ./факс (095) 207-04-11
e-mail: sales@newwave.msk.ru
http://www.newwave.msk.ru
М. Н. Зуев
«История России»
(пособие для поступающих в вузы) в 2 кн.
В пособии систематизированно
излагаются основные факты и собы-
тия истории России с древнейших
времен до наших дней. Изложенный
в нем материал соответствует про-
грамме вступительных экзаменов в
вузы и в то же время содержит мно-
го расширяющих кругозор дополни-
тельных сведений. В приложении
приводится развернутая хронология
российской истории.
Обложка. Формат 84 х 108 */ЗТ
448 стр. (кн. 1). 608 стр. (кн. 2).
Издательство «Новая Волна»
предлагает:
Г. П. Хомченко
«Пособие по химии для поступающих в вузы»
В пособии освещены все вопросы
приемных экзаменов по химии для
поступающих в вузы. Для лучшего
усвоения курса химии приведены
некоторые дополнительные сведе-
ния. После каждой главы даются ти-
повые задачи с решениями и задачи
для самостоятельной работы.
Обложка. Формат 84х1081/ЗТ 480 стр.
Г. П. Хомченко, И. Г. Хомченко
«Сборник задач по химии
для поступающих в вузы»
Сборник включает задачи по об-
щей, неорганической и органичес-
кой химии в соответствии с програм-
мой по химии для поступающих в
вузы. Последний раздел задачника
содержит задачи повышенной слож-
ности. В каждом разделе приводят-
ся типовые задачи с решениями.
Обложка. Формат 84 х 1081/ЗТ 288стр.
Издательство «Новая Волна»
предлагает:
И. В. Кондаков, Л. Я. Шнейберг
«Русская литература ХХвека»
(пособие для поступающих в вузы) в 2 кн.
В пособии дан анализ произве-
дений отечественной литературы
XX века, ставших событием не толь-
ко художественной, но и обществен-
ной жизни страны, раскрывается
идейное содержание, особенности
сюжета и композиции, своеобразие
образов и стиля произведений выда-
ющихся писателей прошлого века —
от Гумилева до Солженицына.
Обложка. Формат 84 х 1081/зх
576 стр. (кн. 1). 576 стр. (кн. 2).
Л. М. Малышева
«Пишем сочинение. Практический курс»
(пособие для поступающих в вузы)
В пособии рассмотрены не только
теоретические вопросы написания
сочинений, но и даны практические
рекомендации сдающему экзамен.
Издание содержит анализ творчества
русских классиков XIX — начала
XX веков и практические советы по
освещению любой заданной темы на
основе этого материала. Приведены
образцы сочинений, при написании
которых использованы практические
методы, предлагаемые автором.
Обложка. Формат 84 х 1081/зх 384стр.
Издательство «Новая Волна»
предлагает:
Л. А. Морозова
«Основы государства и права»
(пособие для поступающих в вузы)
В учебном пособии раскрывают-
ся важнейшие понятия теории госу-
дарства и права. Освещаются осно-
вы гражданского, семейного, трудо-
вого, уголовного и других отраслей
права с позиций новой, общедемо-
кратически ориентированной Кон-
ституции РФ 1993 г. В конце книги
имеются пояснительные схемы, по-
могающие успешному усвоению ос-
новных тем, и предметный указатель.
Обложка, Формат 84 х 108f/32. 352 стр.
Ю. Г« Павленко
«Физика»
(пособие для поступающих в вузы)
В пособии освещены все вопро-
сы приемных экзаменов по физике.
Для лучшего усвоения курса физики
приведены некоторые дополнитель-
ные сведения. В каждой главе дают-
ся типовые задачи с решениями и за-
дачи для самостоятельной работы,
охватывающие все разделы програм-
мы по физике и отражающие наибо-
лее существенные для каждой темы
понятия и законы.
Обложка. Формат 84 х 108 */& 640 стр.
v;
'иология
*?..-a-
пособие
для поступающих
в вузы